Область техники, к которой относится изобретение
Настоящее изобретение касается новых антител, специфически связывающихся с иммуносупрессором опухолей TIGIT (T-клеточный иммунорецептор с иммуноглобулиновым доменом и ингибирующим мотивом на основе тирозина [ITIM]), либо их антигенсвязывающих фрагментов, нуклеиновых кислот, кодирующих эти антитела либо их антигенсвязывающие фрагменты, векторов и клеток хозяина, содержащих такие нуклеиновые кислоты, способа получения таких антител либо их антигенсвязывающих фрагментов, фармацевтических композиций, содержащих эти антитела либо их антигенсвязывающие фрагменты в качестве активного ингредиента, и применения таких фармацевтических композиций.
Антитела либо их антигенсвязывающие фрагменты, специфически связывающиеся с TIGIT, и фармацевтические композиции, содержащие их в качестве активного ингредиента, предпочтительно применяются для лечения рака или опухолей, но настоящее изобретение не ограничивается этим.
Уровень техники
Иммунная система человека работает для защиты человеческого организма, атакуя патогены или вирусы (антигены), которые проникают извне, и аномальные клетки типа раковых клеток. Таким образом, главная функция иммунной системы человека состоит в том, чтобы отличать нормальные клетки организма от внешних захватчиков и аномальных клеток типа раковых клеток и определять, нужно ли атаковать эти клетки. Репрезентативными иммунными клетками, которые могут различать раковые клетки в иммунной системе человека, являются T-клетки, и здоровые люди могут эффективно уничтожать раковые клетки посредством иммунных реакций, хотя раковые клетки и растут в организме. Соответственно, прогрессирование рака означает, что иммунная система ненормальна.
В иммунной системе человека имеется иммунная система обнаружения для подавления гипериммунных реакций, вызванных гиперпролиферацией T-клеток. Такая иммунная система обнаружения именуется “контрольной точкой иммунитета”, а белки, задействованные в контрольных точках иммунитета, именуются “белками контрольных точек иммунитета”.
По сути дела, функция контрольных точек иммунитета заключается в подавлении гипериммунных реакций при гиперактивации и/или гиперпролиферации T-клеток, но раковые клетки злоупотребляют контрольными точками иммунитета, чтобы предотвратить нападение T-клеток на раковые клетки, что в конечном итоге приводит к прогрессированию рака.
В данной области уже известно, что заболевания типа раковых можно лечить с помощью ингибиторов таких контрольных точек иммунитета. В настоящее время коммерчески доступны препараты антител, направленных на белки контрольных точек иммунитета, и проводится разработка различных ингибиторов контрольных точек иммунитета.
Первым разработанным терапевтическим средством типа ингибитора контрольных точек иммунитета был ипилимумаб, который представляет собой моноклональное антитело, специфичное к CTLA-4 (связанному с цитотоксическими T-лимфоцитами антигену-4), рецептору типа контрольной точки иммунитета, и оказался эффективным против метастатической злокачественной меланомы. Впоследствии были разработаны моноклональные антитела, специфичные к PD-1 (белку-1 запрограммированной смерти клеток) и PD-L1 (лиганду-1 запрограммированной смерти), который является лигандом для PD-1. Репрезентативные примеры таковых включают ниволумаб, пембролизумаб, авелумаб, атезолизумаб и дурвалумаб. Ингибиторы PD-1 или PD-L1 эффективны при злокачественных меланомах, а также при различных опухолях.
TIGIT (T-клеточный иммунорецептор с иммуноглобулиновым доменом и ингибирующим мотивом на основе тирозина [ITIM]) представляет собой рецептор, который экспрессируется главным образом в активированных T-клетках и NK-клетках (природных киллерах) и относится к белкам контрольных точек иммунитета в широком смысле.
TIGIT связывается с такими лигандами, как CD155 и CD112, на поверхности раковых клеток, подавляя активацию иммунных клеток. Сообщалось, что антитела против TIGIT индуцируют активацию T-клеток CD8+ вместе с антителами, блокирующими PD-1/PD-L1, и тем самым эффективно устраняют опухоли или вирусы.
На сегодняшний день сообщалось о нескольких антителах против TIGIT (как-то US 9713641 B, US 2016/0176963 A, US 9,499,596 B), но исследования их конкретных механизмов недостаточны, и антитела, обладающие эффективностью, практически применимой к терапевтическим средствам, еще не разработаны. Таким образом, все еще существует насущная потребность в TIGIT-специфичных антителах, обладающих высокой эффективностью.
Соответственно, в результате интенсивных усилий по разработке новых антител, специфически связывающихся с TIGIT, авторы настоящего изобретения изобрели новые антитела против TIGIT, обладающие высоким сродством к TIGIT, который гиперэкспрессируется в раковых клетках, и установили потенциальную возможность того, что антитела либо их антигенсвязывающие фрагменты по настоящему изобретению будут эффективными противораковыми средствами и тем самым завершили настоящее изобретение.
Документы предшествующего уровня техники
US Patent No. 9,713,641 (2017.7.25).
US Publication No. 2016/0176963 (2016.6.23).
US Patent No. 9,499,596 (2016.11.22).
Сущность изобретения
Техническая проблема
Итак, одной из целей настоящего изобретения является получение новых антител против TIGIT либо их антигенсвязывающих фрагментов, специфически связывающихся с TIGIT.
Другой целью настоящего изобретения является получение фармацевтических композиций, в частности, фармацевтических композиций для иммунологических противораковых средств (иммуноонкологических средств), содержащих антитела против TIGIT либо их антигенсвязывающие фрагменты в качестве активного ингредиента.
Еще одной целью настоящего изобретения является получение способа лечения рака или опухолей, включающего введение антител против TIGIT либо их антигенсвязывающих фрагментов, применение антител против TIGIT либо их антигенсвязывающих фрагментов для лечения рака или опухолей и применение антител против TIGIT либо их антигенсвязывающих фрагментов для приготовления лекарственных средств для лечения рака или опухолей.
Еще одной целью настоящего изобретения является получение композиций для совместного введения при лечении рака или опухолей, содержащих антитела против TIGIT либо их антигенсвязывающие фрагменты и другое терапевтическое средство для лечения рака.
Еще одной целью настоящего изобретения является получение нуклеиновых кислот, кодирующих антитела против TIGIT либо их антигенсвязывающих фрагментов, векторов и клеток хозяина, содержащих такие нуклеиновые кислоты, и способа получения антител против TIGIT либо их антигенсвязывающих фрагментов с их помощью.
Техническое решение
В соответствии с настоящим изобретением, вышеуказанные и другие цели могут быть достигнуты путем получения антител против TIGIT либо их антигенсвязывающих фрагментов, включающих вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 1 или 2, CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 3 или 4, и CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 5 или 6, и вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 7 или 8, CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 9 или 10, и CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 11 или 12.
Антитела против TIGIT либо их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 13 или 14, и вариабельную область легкой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 15 или 16.
Краткое описание фигур
Вышеуказанные и другие цели, особенности и другие преимущества настоящего изобретения станут более понятными из следующего подробного описания в сочетании с прилагаемыми фигурами, где
на фиг. 1 представлены результаты ELISA по идентификации связывания антител против TIGIT с антигенами TIGIT человека, мыши и макаки-резуса с целью определения межвидовой перекрестной реактивности при первоначальном скрининге антител против TIGIT;
на фиг. 2 представлен график, показывающий результаты ELISA по идентификации связывания антител против TIGIT с TIGIT, CD96, CD155, CD112, CD113 и CD226 с целью определения специфичности по суперсемейству TIGIT при первоначальном отборе антител против TIGIT;
на фиг. 3 представлены результаты измерений с помощью флуоресцентного проточного цитометра по идентификации связывания выбранных при первоначальном отборе антител против TIGIT с антигенами TIGIT, экспрессированными на поверхности клеток;
на фиг. 4 представлены результаты измерения степени связывания антител против TIGIT с белками TIGIT на клеточной поверхности с помощью флуоресцентного проточного цитометра;
на фиг. 5 представлены результаты анализа блокирования при определении степени ингибирования связывания между TIGIT и CD155 при обработке антителами против TIGIT;
на фиг. 6 представлено количество IFN-γ, секретируемого клетками линии NK92 при обработке антителами против TIGIT в совместной культуре клеток линии NK92, гиперэкспрессирующих TIGIT, и клеток линии HeLa, гиперэкспрессирующих PVR;
на фиг. 7 представлены результаты измерения экспрессии NKG2D в клетках линии NK92 при обработке антителами против TIGIT с помощью флуоресцентного проточного цитометра в условиях совместного культивирования клеток линии NK92, гиперэкспрессирующих TIGIT, и клеток линии HeLa, гиперэкспрессирующих PVR;
на фиг. 8 представлен график, показывающий объем опухолей в последний день тестирования по оценке эффективности in vivo антител против TIGIT в соответствии с одним воплощением, результаты которого свидетельствуют об эффективности на модели опухолей CT26; и
на фиг. 9 представлен график, показывающий объем опухолей в последний день тестирования по оценке эффективности антител против TIGIT в соответствии с одним воплощением по каждой дозе in vivo при введении по отдельности и в комбинации с антителом против PD-L1, результаты которого свидетельствуют об эффективности на модели опухолей CT26.
Раскрытие сущности изобретения
Если не указано иначе, все используемые здесь технические и научные термины имеют такие значения, которые понятны специалистам в той области, к которой относится настоящее изобретение. В целом используемая здесь номенклатура хорошо известна в данной области и обычно она и применяется.
В одном аспекте настоящего изобретения предусмотрены антитела против TIGIT либо их антигенсвязывающие фрагменты, включающие:
вариабельную область тяжелой цепи, содержащую:
CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 1 или 2,
CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 3 или 4, и
CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 5 или 6, и
вариабельную область легкой цепи, содержащую:
CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 7 или 8,
CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 9 или 10, и
CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 11 или 12.
Кроме того, антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат: вариабельную область тяжелой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 13 или 14; и вариабельную область легкой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 15 или 16.
Предпочтительно антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат:
(1) вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 1; CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 3; и CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 5; и
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 7; CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 9; и CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 11; или
(2) вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 2; CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 4; и CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 6; и
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 8; CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO:10; и CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 12; или
(3) вариабельную область тяжелой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 13; и
вариабельную область легкой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 15; или
(4) вариабельную область тяжелой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 14; и
вариабельную область легкой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 16.
TIGIT, который экспрессируется на поверхности таких иммунных клеток, как T-клетки, NK-клетки и дендритные клетки, связывается с PVR (рецептором полиовируса, CD155) на поверхности раковых клеток, подавляя активность иммунных клеток. Антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению специфически связываются с сайтом связывания CD155 у TIGIT и ингибируют передачу сигналов при взаимодействии TIGIT/CD155, индуцируя активацию иммунных клеток и подавляя рост опухолевых клеток. CD155 экспрессируется на клеточной поверхности различных млекопитающих типа людей, обезьян, мышей и крыс и передает сигналы, которые ингибируют активацию иммунных клеток при связывании с TIGIT.
То есть антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению ингибируют передачу сигналов при взаимодействии TIGIT/CD155, ослабляя сигналы ингибирования иммунных клеток от раковых клеток, вызывая реактивацию иммунного ответа для эффективной атаки на раковые клетки и тем самым оказывая противораковое действие. В конечном счете, антитела против TIGIT либо их антигенсвязывающие фрагменты можно использовать для иммунологической противораковой терапии, нацеленной на TIGIT, иммуносупрессор опухолей. В частности, антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению снижают либо ингибируют экспрессию или активность TIGIT у субъектов, страдающих раком, и индуцируют непрерывный противораковый ответ T-клеток или NK-клеток, тем самым обеспечивая эффект лечения рака.
Белок TIGIT, действующий в качестве антигена для антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению и тесно связанный с ингибированием активности иммунных клеток, представляет собой мембранный белок, который находится на поверхности иммунных клеток и служит субингибиторным рецептором для иммунных клеток. TIGIT может происходить из млекопитающих типа приматов, в том числе людей и обезьян, и грызунов, включая мышей и крыс.
В настоящем изобретении термин “TIGIT” является общим термином для любых вариантов, изоформ или видовых гомологов TIGIT, которые естественно экспрессируются в клетках, предпочтительно это TIGIT человека, но настоящее изобретение не ограничивается этим и включает TIGIT из других животных и т.п.
Антитела против TIGIT по настоящему изобретению предпочтительно специфически связываются с сайтом связывания CD155 или ингибирующим связывание CD155 сайтом TIGIT человека (hTIGIT; SEQ ID NO: 21; № доступа NCBI: NP_776160), но настоящее изобретение не ограничивается этим.
Аминокислотные последовательности и последовательности участков CDR тяжелой цепи и CDR легкой цепи вариабельной области антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению представлены в табл. 1-4.
Таблица 1. Аминокислотные последовательности участков CDR тяжелой цепи антител против TIGIT по изобретению
Таблица 2. Аминокислотные последовательности участков CDR легкой цепи антител против TIGIT по изобретению
Таблица 3. Аминокислотные последовательности вариабельных областей тяжелой цепи антител против TIGIT по настоящему изобретению (участки CDR подчеркнуты)

Таблица 4. Аминокислотные последовательности вариабельных областей легкой цепи антител против TIGIT по настоящему изобретению (участки CDR подчеркнуты)

В то же время антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, последовательность которой на 80% или больше, предпочтительно на 90% или больше, более предпочтительно на 99% или больше идентична каждому из участков CDR1 тяжелой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 1 или 2; CDR2 тяжелой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 3 или 4; и CDR3 тяжелой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 5 или 6, причем антитела либо их антигенсвязывающие фрагменты, имеющие такие же характеристики, как и TIGIT по настоящему изобретению, тоже входят в рамки антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению.
Антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, последовательность которой на 80% или больше, предпочтительно на 90% или больше, более предпочтительно на 99% или больше идентична вариабельной области тяжелой цепи, включающей аминокислотную последовательность, приведенную в SEQ ID NO: 13 или 14, причем антитела либо их антигенсвязывающие фрагменты, имеющие такие же характеристики, как и TIGIT по настоящему изобретению, тоже входят в рамки антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению.
Кроме того, антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, последовательность которой на 80% или больше, предпочтительно на 90% или больше, более предпочтительно на 99% или больше идентична каждому из участков CDR1 легкой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 7 или 8; CDR2 легкой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 9 или 10; и CDR3 легкой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 11 или 12, причем антитела либо их антигенсвязывающие фрагменты, имеющие такие же характеристики, как и TIGIT по настоящему изобретению, тоже входят в рамки антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению.
Кроме того, антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, последовательность которой на 80% или больше, предпочтительно на 90% или больше, более предпочтительно на 99% или больше идентична каждой из вариабельных областей легкой цепи, включающих аминокислотную последовательность, приведенную в SEQ ID NO: 15 или 16, причем антитела либо их антигенсвязывающие фрагменты, имеющие такие же характеристики, как и TIGIT по настоящему изобретению, тоже входят в рамки антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению.
Кроме того, антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению могут включать и такие антитела, либо их антигенсвязывающие фрагменты, у которых часть аминокислотной последовательности антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению подверглась замене посредством консервативных замен.
В настоящем изобретении термин “консервативная замена” означает такую модификацию полипептида, которая включает замену одной или нескольких аминокислот на одну или несколько аминокислот с близкими биологическими или биохимическими свойствами, которые не вызывают потери биологических или биохимических функций полипептида. Термин “консервативная замена аминокислоты” означает замену аминокислотного остатка на аминокислотный остаток с аналогичной боковой цепью. Классы аминокислотных остатков со сходными боковыми цепями установлены и хорошо известны в данной области. Такие классы включают аминокислоты с основными боковыми цепями (напр., лизин, аргинин, гистидин), аминокислоты с кислотными боковыми цепями (напр., аспарагиновая кислота, глутаминовая кислота), аминокислоты с незаряженными полярными боковыми цепями (напр., глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), аминокислоты с неполярными боковыми цепями (напр., аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), аминокислоты с разветвленными боковыми цепями (напр., треонин, валин, изолейцин) и аминокислоты с ароматическими боковыми цепями (напр., тирозин, фенилаланин, триптофан, гистидин). Предполагается, что антитела по настоящему изобретению содержат консервативные замены аминокислот и все-таки сохраняют активность.
В настоящем изобретении термин “TIGIT-специфичное антитело” означает такое антитело, которое связывается с TIGIT и ингибирует биологическую активность TIGIT, причем он применяется взаимозаменяемо с “антителом против TIGIT”.
В настоящем изобретении термин “антитело против TIGIT” включает и поликлональные антитела, и моноклональные антитела, а предпочтительно это моноклональные антитела и может быть целые антитела. Целое антитело представляет собой структуру с двумя полноразмерными легкими цепями и двумя полноразмерными тяжелыми цепями, включающими константную область, причем каждая легкая цепь соединяется с соответствующей тяжелой цепью дисульфидной связью.
Целые антитела против TIGIT по настоящему изобретению включают формы IgA, IgD, IgE, IgM и IgG, а IgG включают подтипы IgG1, IgG2, IgG3 и IgG4.
Антитела против TIGIT по настоящему изобретению предпочтительно представляют собой полностью человеческие антитела, отобранные из библиотек человеческих антител, но настоящее изобретение не ограничивается этим.
В настоящем изобретении термин “антигенсвязывающий фрагмент” антител против TIGIT означает такой фрагмент, который способен связываться с антигеном антител против TIGIT, то есть с TIGIT, и охватывает Fab, Fab', F(ab')2, scFv, (scFv)2, scFv-Fc, Fv и др., причем он применяется взаимозаменяемо с “фрагментом антител”.
Fab включает в себя вариабельную область каждой из тяжелой цепи и легкой цепи, константную область легкой цепи и первый константный участок (домен CH1) тяжелой цепи, каждая из которых содержит антигенсвязывающий сайт. Fab' отличается от Fab тем, что он также содержит шарнирную область, содержащую по меньшей мере один остаток цистеина на C-конце домена CH1 тяжелой цепи. F(ab')2 образуется посредством дисульфидной связи между остатками цистеина в шарнирной области Fab'.
Fv (вариабельный фрагмент), включающий в себя вариабельные области тяжелой цепи и легкой цепи, представляет собой минимальный фрагмент антитела, обладающий исходной специфичностью исходного иммуноглобулина. Двухцепочечный Fv (dsFv, стабилизованный дисульфидом Fv) образуется при связывании вариабельной области легкой цепи с вариабельной областью тяжелой цепи через дисульфидную связь. Одноцепочечный Fv (scFv) представляет собой такой Fv, в котором соответствующие вариабельные области тяжелой цепи и легкой цепи ковалентно связаны через пептидный линкер. Эти фрагменты антител могут быть получены обработкой целых антител протеазой (напр., Fab можно получить путем ограниченного расщепления целых антител папаином, а фрагмент F(ab')2 можно получить путем ограниченного расщепления целых антител пепсином), а предпочтительно их конструируют по технологии генетической рекомбинации (например, путем амплификации ДНК, кодирующей тяжелую цепь антитела или ее вариабельную область, либо ДНК, кодирующей легкую цепь или ее вариабельную область, в качестве матрицы методом ПЦР (полимеразной цепной реакции) с помощью пары праймеров, и амплификации с помощью комбинации пар праймеров для соединения ДНК, кодирующей пептидный линкер, по каждому из его концов с тяжелой цепью или ее вариабельной областью и легкой цепью или ее вариабельной областью).
В другом аспекте настоящего изобретения предусмотрены нуклеиновые кислоты, кодирующие антитела против TIGIT по изобретению. При этом нуклеиновые кислоты могут находиться в клетках или в клеточном лизате, в частично очищенном виде или в практически чистом виде. Нуклеиновые кислоты могут быть “выделенными” или “практически чистыми”, если они очищены от других клеточных компонентов или других примесей, к примеру, нуклеиновых кислот или белков из других клеток, стандартными методами, включающими, к примеру, щелочную обработку с SDS, зональное центрифугирование в CsCl, колоночную хроматографию, электрофорез в агарозном геле и другие, хорошо известные в данной области. Нуклеиновая кислота по настоящему изобретению может представлять собой, к примеру, ДНК или РНК и может включать или не включать в себя последовательности интронов.
В другом аспекте настоящего изобретения предусмотрены векторы, содержащие нуклеиновые кислоты. Для экспрессии антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению получают ДНК, кодирующую частичные или полноразмерные легкие и тяжелые цепи, стандартными методами молекулярной биологии (напр., ПЦР-амплификации или клонирования кДНК с помощью гибридом, экспрессирующих искомое антитело), причем ДНК может быть “функционально связана” с контролирующими транскрипцию и трансляцию последовательностями, которые вставляются в экспрессирующий вектор.
В настоящем изобретении термин “функционально связанный” может означать то, что гены, кодирующие антитела, лигированы в векторе с тем, чтобы контролирующие транскрипцию и трансляцию последовательности могли выполнять предназначенную им функцию регулирования транскрипции и трансляции генов антител.
Выбирают такие экспрессирующие векторы и контролирующие транскрипцию и трансляцию последовательности, которые совместимы с используемыми для экспрессии клетками хозяина. Гены легкой цепи антител и гены тяжелой цепи антител вставляют в отдельные векторы или же оба гена вставляют в один и тот же экспрессирующий вектор. Антитела вставляют в экспрессирующие векторы стандартными методами (напр., лигирования фрагментов генов антител и соответствующих сайтов рестрикционных ферментов в векторы либо лигирования по тупым концам, когда нет сайтов рестрикционных ферментов). В некоторых случаях рекомбинантные экспрессирующие векторы могут кодировать сигнальные пептиды, которые облегчают секрецию цепей антител из клеток-хозяев. Гены цепей антител можно клонировать в векторы таким образом, чтобы сигнальные пептиды присоединялись к N-концам генов цепей антител в соответствии с рамкой. Сигнальные пептиды могут представлять собой сигнальные пептиды иммуноглобулинов или гетерологичные сигнальные пептиды (т.е. сигнальные пептиды, полученные из других белков, помимо иммуноглобулина). Кроме того, рекомбинантные экспрессирующие векторы содержат регуляторные последовательности, контролирующие экспрессию генов цепей антител в клетках хозяина. “Регуляторные последовательности” могут включать промоторы, энхансеры и другие контролирующие экспрессию элементы (напр., сигналы полиаденилирования), которые контролируют транскрипцию или трансляцию генов цепей антител. Специалистам должно быть известно, что конструкция экспрессирующих векторов может варьироваться путем выбора различных регуляторных последовательностей в зависимости от таких факторов, как выбор трансформируемых клеток хозяина и уровень экспрессии белка.
В другом аспекте настоящего изобретения предусмотрены клетки хозяина, содержащие нуклеиновые кислоты или векторы. Клетки хозяина по настоящему изобретению предпочтительно выбирают из группы, состоящей из клеток животных, растительных, дрожжевых клеток, Escherichia coli и клеток насекомых, но настоящее изобретение не ограничивается этим.
В частности, клетки хозяина по настоящему изобретению могут представлять собой прокариотические клетки типа Escherichia coli, Bacillus subtilis, Streptomyces sp., Pseudomonas sp., Proteus mirabilis или Staphylococcus sp. Кроме того, клетки хозяина могут быть выбраны из грибков типа Aspergillus sp., дрожжей типа Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces sp. или Neurospora crassa и других эукариотических клеток, включая клетки низших эукариот и клетки высших эукариот, полученные из насекомых.
Клетки хозяина также могут происходить из растений или млекопитающих. Предпочтительно клетки хозяина выбирают из группы, состоящей из клеток почек обезьян (COS7), клеток NSO, SP2/0, клеток яичников китайского хомячка (CHO), W138, клеток почек детенышей хомячка (BHK), MDCK, клеток миеломы, клеток HuT78 и клеток HEK 293, но настоящее изобретение не ограничивается этим. Особенно предпочтительны клетки СНО.
Нуклеиновой кислотой или вектором трансформируют или трансфицируют клетки хозяина. При “трансформации” или “трансфекции” для введения чужеродной нуклеиновой кислоты (ДНК или РНК) в прокариотические или эукариотические клетки хозяина широко применяются различные методы, включая электрофорез, осаждение фосфатом кальция, трансфекцию DEAE-декстраном, липофекцию и т.п. Для экспрессии антител против TIGIT по изобретению можно использовать различные экспрессионные комбинации хозяин/вектор. Подходящие экспрессирующие векторы для эукариотических хозяев включают, без ограничения, регулирующие экспрессию последовательности, полученные из SV40, коровьего папилломавируса, аденовирусов, аденоассоциированных вирусов, цитомегаловирусов и ретровирусов. Экспрессирующие векторы для бактериальных хозяев включают бактериальные плазмиды, полученные из Escherichia coli, как-то pET, pRSET, pBluescript, pGEX2T, вектор pUC, col E1, pCR1, pBR322, pMB9 и их производные, плазмиды с более широким кругом хозяев типа RP4, фаговые ДНК, к примеру, различные производные фага лямбда типа λgt10, λgt11 и NM989 и других ДНК-фагов типа ДНК-фага M13 и нитчатых фагов с одноцепочечной ДНК. Экспрессирующие векторы, пригодные для дрожжевых клеток - это плазмиды 2°C и их производные. Пригодный для клеток насекомых вектор - pVL941.
В другом аспекте настоящего изобретения предусмотрен способ получения антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению, который включает культивирование клеток хозяина для экспрессии антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению.
При введении в клетки млекопитающих рекомбинантного экспрессирующего вектора, способного экспрессировать антитело против TIGIT или его антигенсвязывающий фрагмент, антитело можно получить при инкубации в течение времени, достаточного для того, чтобы антитело экспрессировалось в клетках хозяина, более предпочтительно в течение времени, достаточного для того, чтобы антитело секретировалось в культуральную среду.
В некоторых случаях экспрессируемое антитело можно отделить от клеток хозяина и очистить до гомогенности. Разделение или очистка антител может проводиться методами разделения и очистки, которые обычно применяются для белков, напр., методом хроматографии. Хроматография может включать, к примеру, аффинную хроматографию, включающую колонку с белком A и колонку с белком G, ионообменную хроматографию или гидрофобную хроматографию. Наряду с хроматографией, антитела можно отделять и очищать комбинированием фильтрования, ультрафильтрации, высаливания, диализа и т.п.
В другом аспекте настоящего изобретения предусмотрены фармацевтические композиции для лечения рака или опухолей, содержащие антитела против TIGIT либо их антигенсвязывающие фрагменты в качестве активного ингредиента.
В другом аспекте настоящего изобретения предусмотрен способ лечения рака или опухолей, включающий введение антител против TIGIT либо их антигенсвязывающих фрагментов пациентам, нуждающимся в профилактике или лечении.
В другом аспекте настоящего изобретения предусмотрено применение антител против TIGIT либо их антигенсвязывающих фрагментов для лечения рака или опухолей.
В другом аспекте настоящего изобретения предусмотрено применение антител против TIGIT либо их антигенсвязывающих фрагментов для приготовления лекарственных средств для лечения рака или опухолей.
Термин “рак” или “опухоль” относится или означает физиологическое состояние, которое обычно характеризуется неконтролируемым ростом/пролиферацией клеток у млекопитающих.
Раковые заболевания, которые можно лечить с помощью композиций по настоящему изобретению, не имеют особых ограничений и включают как солидный рак, так и рак крови. Примеры таких раковых заболеваний включают рак кожи типа меланомы, рак печени, гепатоцеллюлярную карциному, рак желудка, рак молочной железы, рак легких, рак яичников, рак бронхов, рак носоглотки, рак гортани, рак поджелудочной железы, рак мочевого пузыря, колоректальный рак, рак толстой кишки, рак шейки матки, рак мозга, рак простаты, рак костей, рак щитовидной железы, рак паращитовидной железы, рак почек, рак пищевода, холангиокарциному, рак яичек, рак прямой кишки, рак головы и шеи, рак шейки матки, рак мочеточников, остеосаркому, нейробластому, фибросаркому, рабдомиосаркому, астроцитому, нейробластому и глиому, но не ограничиваются этим. Предпочтительно рак, который можно лечить композициями настоящего изобретения, выбирают из группы, состоящей из рака толстой кишки, рака молочной железы, рака легких и рака почек.
Настоящим изобретением предусмотрены фармацевтические композиции, содержащие терапевтически эффективное количество антител против TIGIT либо их антигенсвязывающих фрагментов и фармацевтически приемлемый носитель. Термин “фармацевтически приемлемый носитель” относится к веществам, которые могут быть добавлены к активному ингредиенту для облегчения составления или стабилизации лекарственной формы и не оказывают значительных вредных токсических эффектов на пациентов.
Носитель означает такой носитель или разбавитель, который не раздражает пациентов и не влияет на биологическую активность и свойства вводимого соединения. Фармацевтические носители, приемлемые для композиций, составляемых в виде жидких растворов, включают стерильные биосовместимые ингредиенты, а их примеры включают физраствор, стерильную воду, раствор Рингера, забуференный физраствор, растворы альбумина для инъекций, растворы декстрозы, растворы мальтодекстрина, глицерин, этанол и их смеси. При необходимости могут быть добавлены другие стандартные добавки, как-то антиоксиданты, буферы и бактериостатические средства. Кроме того, также могут быть добавлены и разбавители, диспергаторы, поверхностно-активные вещества, связующие и смазывающие вещества для приготовления растворов для инъекций типа водных растворов, суспензий и эмульсий, пилюль, капсул, гранул или таблеток. Описаны и другие носители, к примеру, в Remington's Pharmaceutical Sciences (E.W. Martin). Такие композиции могут содержать терапевтически эффективное количество по меньшей мере одного антитела против TIGIT или его антигенсвязывающего фрагмента.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных растворов или дисперсий для инъекций ex temporo. Применение таких сред и средств для фармацевтически активных ингредиентов хорошо известно в данной области. Композиции предпочтительно предназначены для парентерального введения. Композиции могут быть приготовлены в виде растворов, микроэмульсий, липосом или других упорядоченных структур, подходящих для высоких концентраций лекарственных средств. Носителем может быть, к примеру, растворитель или дисперсионная среда, содержащая воду, этанол, полиол (как-то глицерин, пропиленгликоль и жидкий полиэтиленгликоль) и подходящие смеси таковых. В некоторых случаях композиции могут содержать и изотонические средства, например, сахара, многоатомные спирты типа маннита, сорбита или хлорид натрия. Стерильные растворы для инъекций можно приготовить путем включения необходимого количества активного ингредиента, необязательно вместе с одним или с комбинацией ингредиентов, описанных выше, в соответствующий растворитель с последующей стерилизацией микрофильтрованием. Дисперсии обычно получают путем включения активного соединения в стерильный носитель, содержащий основную дисперсионную среду и другие необходимые ингредиенты из числа описанных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций некоторые способы получения включают вакуумную сушку и сублимационную сушку (лиофилизацию) для получения порошка активного ингредиента и любых дополнительных требуемых ингредиентов из их предварительно стерилизованных и фильтрованных растворов.
Дозировка фармацевтических композиций по настоящему изобретению не имеет особых ограничений, но может варьироваться в зависимости от различных факторов, включая состояние здоровья и вес пациентов, тяжесть заболевания, тип препарата, способ введения и время введения. Фармацевтические композиции по настоящему изобретению можно вводить в виде одной или нескольких доз в день таким млекопитающим, как крысы, мыши, домашние животные и люди, стандартным приемлемым способом, включая, без ограничения, внутрибрюшинное введение, внутривенное введение, внутримышечное введение, подкожное введение, внутрикожное введение, пероральное введение, местное введение, интраназальное введение, внутрилегочное введение или интраректальное введение.
Фармацевтические композиции по настоящему изобретению можно вводить пациентам болюсом или путем непрерывной инъекции, если нужно. Например, введение болюсом антител против TIGIT либо их антигенсвязывающих фрагментов по настоящему изобретению, представленных в виде Fab-фрагментов, может проводиться в дозе от 0,01 мкг/кг массы тела до 100 мг/кг массы тела, предпочтительно от 1 мкг/кг массы тела до 10 мг/кг массы тела.
В настоящем изобретении термин “терапевтически эффективное количество” означает такое количество комбинации антитела против TIGIT или его антигенсвязывающего фрагмента, которое требуется для получения измеримых преимуществ in vivo у нуждающихся в лечении пациентов. Точное количество будет зависеть от ряда факторов, включая, без ограничения, ингредиенты и физические свойства терапевтической композиции, популяцию предполагаемых пациентов и соображения соответствующих пациентов, и может быть легко определено специалистами в данной области. При полном учете этих факторов нужно вводить минимальное количество, достаточное для достижения максимального эффекта и не вызывающее побочных эффектов, и специалист в данной области может легко определить такую дозу.
В другом аспекте настоящего изобретения предусмотрен способ лечения рака и торможения разрастания рака путем введения нуждающимся в лечении субъектам антител против TIGIT либо их антигенсвязывающих фрагментов или содержащих их фармацевтических композиций.
Антитела против TIGIT либо их антигенсвязывающие фрагменты или содержащие их фармацевтические композиции по настоящему изобретению для лечения раковых клеток либо их метастазов или же для подавления разрастания рака можно вводить в фармацевтически эффективном количестве.
Фармацевтически эффективное количество может зависеть от типа рака, возраста и веса пациентов, природы и тяжести симптомов, типа текущего лечения, количества курсов лечения, формы введения и способа введения, и его легко может определить специалист в данной области. Композиции по настоящему изобретению можно вводить одновременно или последовательно с вышеприведенными фармакологическими или физиологическими ингредиентами, а также в комбинации со стандартными терапевтическими средствами, последовательно или одновременно со стандартным терапевтическим средством. Такое введение может быть однократным или многократным. Нужно вводить минимальное количество, способное оказывать максимальный эффект и не вызывать побочных эффектов с учетом всех вышеприведенных факторов, и специалисты в данной области могут легко его определить.
В настоящем изобретении термин “субъект” служит для обозначения таких млекопитающих, предпочтительно людей, которые страдают или предрасположены к тому заболеванию, которое можно облегчить, подавлять или лечить путем введения антител против TIGIT либо их антигенсвязывающих фрагментов или же содержащих их фармацевтических композиций.
Антитела против TIGIT либо их антигенсвязывающие фрагменты и содержащие их фармацевтические композиции могут применяться в комбинации со стандартными терапевтическими средствами.
Соответственно, в другом аспекте настоящего изобретения предусмотрены композиции для совместного введения при лечении рака или опухолей, содержащие антитела против TIGIT либо их антигенсвязывающие фрагменты и другое терапевтическое средство против рака, а также способ лечения рака или опухолей с их помощью.
Другое терапевтическое средство против рака означает такое терапевтическое средство, которое может применяться для лечения рака в дополнение к антителам против TIGIT либо их антигенсвязывающим фрагментам по настоящему изобретению.
В настоящем изобретении терапевтическое средство против рака может быть ингибитором контрольной точки иммунитета, но настоящее изобретение не ограничивается этим.
В настоящем изобретении ингибиторы контрольных точек иммунитета также называются “ингибиторами контрольных точек”, причем ими могут быть антитела против CTLA-4, антитела против PD-1 или антитела против PD-L1, но настоящее изобретение не ограничивается этим. В частности, ингибитором контрольной точки иммунитета может быть ипилимумаб, ниволумаб, пембролизумаб, атезолизумаб, авелумаб, дурвалумаб и т.п., но настоящее изобретение не ограничивается этим.
Термин “применяется в комбинации (при совместном введении)” означает то, что антитело против TIGIT или его антигенсвязывающий фрагмент и каждое из других терапевтических средств против рака можно вводить одновременно, последовательно или в обратном порядке, а также в виде комбинации соответствующих эффективных количеств в том объеме, который может быть установлен специалистами в данной области.
В одном воплощении настоящего изобретения было подтверждено, что введение в комбинации (совместное введение) антитела против PD-L1 и антитела против TIGIT по настоящему изобретению дополнительно ингибирует рост опухолей.
Композиции для совместного введения содержат антитела против TIGIT, причем связанные с ними конфигурации являются такими же, как те, что содержатся в композициях для профилактики или лечения рака, описанных выше, так что описание каждой конфигурации в равной степени применимо к композициям для совместного введения.
В другом аспекте настоящего изобретения предусмотрены конъюгаты антитело-лекарственное средство, содержащие лекарственное средство, конъюгированное с антителом против TIGIT или его антигенсвязывающим фрагментом по настоящему изобретению, а также фармацевтические композиции, содержащие конъюгаты антитело-лекарственное средство. Настоящим изобретением также предусмотрен способ лечения опухолей с помощью конъюгатов антитело-лекарственное средство, содержащих лекарственное средство, конъюгированное с антителом против TIGIT или его антигенсвязывающим фрагментом по настоящему изобретению, и фармацевтических композиций, содержащих конъюгаты антитело-лекарственное средство.
Антитело против TIGIT или его антигенсвязывающий фрагмент может связываться с препаратом через линкер. Линкер представляет собой участок, соединяющий антитело против TIGIT или его антигенсвязывающий фрагмент с препаратом. Например, линкер позволяет высвобождение препарата из антитела при расщеплении линкера в присутствии такого реагента, который может расщепляться во внутриклеточных условиях, то есть во внутриклеточной среде.
Линкер может расщепляться расщепляющим агентом, присутствующим во внутриклеточной среде типа лизосом или эндосом, и может представлять собой, к примеру, пептидный линкер, который может расщепляться внутриклеточной пептидазой или протеазой типа лизосомной или эндосомной протеазы. Обычно пептидный линкер имеет длину по меньшей мере в две аминокислоты. Расщепляющий агент может включать катепсин B и катепсин D или плазмин и может гидролизировать пептид с высвобождением препарата в клетки мишени.
Пептидный линкер может расщепляться тиол-зависимой протеазой, катепсином B, который гиперэкспрессируется в раковых тканях, а линкер представляет собой, к примеру, линкер Phe-Leu или Gly-Phe-Leu-Gly. Кроме того, пептидный линкер может расщепляться, к примеру, внутриклеточной протеазой, а линкер представляет собой линкер Val-Cit или линкер Phe-Lys.
В одном воплощении расщепляемый линкер может быть чувствительным к pH и подверженным гидролизу при определенном значении pH. Обычно pH-чувствительные линкеры могут гидролизироваться в кислых условиях. Примеры кислотолабильных линкеров, которые могут гидролизироваться в лизосомах, включают гидразоны, семикарбазоны, тиосемикарбазоны, цис-аконитовые амиды, ортоэфиры, ацетали, кетали и т.п.
В другом воплощении линкер может расщепляться в восстановительных условиях, примером чего является дисульфидный линкер. Можно образовывать различные дисульфидные связи с помощью N-сукцинимидил-S-ацетилтиоацетата (SATA), N-сукцинимидил-3-(2-пиридилдитио)пропионата (SPDP), N-сукцинимидил-3-(2-пиридилдитио)бутирата (SPDB) и N-сукцинимидилоксикарбонил-α-метил-α-(2-пиридилдитио)толуола (SMPT).
Препарат и/или препарат с линкером можно конъюгировать случайным образом через лизин антитела или же конъюгировать через экспонированный цистеин при восстановлении цепи с дисульфидной связью. В некоторых случаях препарат с линкером можно конъюгировать через цистеин, находящийся в генно-инженерной метке, напр., в пептиде или белке. Генно-инженерная метка, напр., пептид или белок, может содержать аминокислотный мотив, который может распознаваться, к примеру, изопреноидтрансферазой. Пептид или белок содержит делецию на карбоксильном конце пептида или белка либо добавление спейсерного звена через ковалентную связь на карбоксильном (C) конце пептида или белка.
Кроме того, линкер может представлять собой, к примеру, нерасщепляемый линкер, и препарат может высвобождаться только за одну стадию гидролиза антитела с образованием, к примеру, комплекса аминокислота-линкер-препарат. Этот тип линкера может представлять собой тиоэфирную или малеимидокапроильную группу и оставаться стабильным в крови.
Препарат в конъюгате антитело-лекарственное средство может быть связан с антителом в качестве агента, оказывающего фармакологический эффект, в частности, это может быть химиотерапевтическое средство, токсин, микроРНК (миРНК), киРНК, кшРНК или радиоизотоп. Химиотерапевтическое средство может представлять собой, к примеру, цитотоксическое средство или иммуносупрессивное средство. В частности, препарат может содержать ингибитор микротубулина, ингибитор митоза, ингибитор топоизомеразы или же химиотерапевтическое средство, способное служить в качестве интеркалятора ДНК. Препарат также может содержать иммуномодулирующее соединение, противораковое средство или противовирусное средство либо их комбинацию.
Такие препараты могут включать одно или несколько выбранных из группы, состоящей из майтанзиноида, ауристатина, аминоптерина, актиномицина, блеомицина, талидомида, камптотецина, N8-ацетилспермидина, 1-(2-хлорэтил)-1,2-диметилсульфонилгидразида, эсперамицина, этопозида, 6-меркаптопурина, доластатина, трихотецена, калихеамицина, таксола, таксана, паклитакселя, доцетакселя, метотрексата, винкристина, винбластина, доксорубицина, мелфалана, хлорамбуцила, дуокармицина, L-аспарагиназы, меркаптопурина, тиогуанина, гидроксимочевины, цитарабина, циклофосфамида, ифосфамида, нитрозомочевины, цисплатина, карбоплатина, митомицина (митомицина A, митомицина C), дакарбазина, прокарбазина, топотекана, азотистого иприта, цитоксана, этопозида, 5-фторурацила, CNU (бисхлорэтилнитрозомочевины), иринотекана, камптотецина, блеомицина, идарубицина, даунорубицина, дактиномицина, пликамицина, митоксантрона, аспарагиназы, винорелбина, хлорамбуцила, мелфалана, кармустина, ломустина, бусульфана, треосульфана, дакарбазина, этопозида, тенипозида, топотекана, 9-аминокамптотецина, криснатола, триметрексата, микофеноловой кислоты, тиазофурина, рибавирина, EICAR (5-этинил-1-бета-D-рибофуранозилимидазол-4-карбоксамида), гидроксимочевины, дефероксамина, флоксуридина, доксифлуридина, ралтитрекседа, цитарабина (ara C), цитозин-арабинозида, флударабина, тамоксифена, ралоксифена, мегестрола, госерелина, лейпролида ацетата, флутамида, бикалутамида, EB1089, CB1093, KH1060, вертепорфина, фталоцианина, Pe4 (фотосенсибилизатор), деметоксигипокреллина A, интерферона-α, интерферона-γ, фактора некроза опухолей, гемцитабина, Velcade, ревлимида, таломида, ловастатина, 1-метил-4-фенилпиридиния, стауроспорина, актиномицина D, дактиномицина, блеомицина A2, блеомицина B2, пепломицина, эпирубицина, пирарубицина, зорубицина, верапамила, тапсигаргина, нуклеаз и токсинов, полученных из бактерий либо растений или животных, но не ограничиваются этим.
Далее настоящее изобретение будет описано более подробно на следующих примерах. Однако специалистам в данной области должно быть ясно, что эти примеры приводятся только для иллюстрации настоящего изобретения и не должны рассматриваться как ограничивающие объем настоящего изобретения.
ПРИМЕРЫ
Пример 1. Скрининг антител против TIGIT
1.1. Скрининг клонов scFv и Fab антител человека против TIGIT
Антитела, специфически связывающиеся с TIGIT, отбирали путем скрининга методом фагового дисплея, используя библиотеки scFv и Fab антител человека. Библиотека scFv была получена согласно описанию в “Construction of a Large Synthetic Human scFv Library with Six Diversified CDRs and High Functional Diversity (Yang HY et al., Molecules and Cells 27, 225-235), а библиотека Fab была получена согласно описанию в Корейском патенте №1694832. Скрининг методом фагового дисплея проводили вплоть до четвертого раунда в целом, а по мере возрастания количества раундов уменьшали количество антигена и повышали количество промываний. Применяли метод перекрестных антигенов, используя антигены TIGIT-ECD-Fc человека для первого и третьего раунда скрининга и антигены TIGIT-ECD-Fc мыши для второго и четвертого раунда скрининга. В иммунопробирки вносили по 20 мкг антигена TIGIT, разведенного в буфере PBS, и инкубировали при 4°C в течение ночи для фиксации антигена TIGIT на поверхности иммунопробирки. Покрытые антигеном TIGIT иммунопробирки блокировали в растворе PBS-T/BSA (5%) в течение 1 часа при комнатной температуре, туда же добавляли фаги из библиотек scFv или Fab антител человека в количестве 4,7×1012 или 1,2×1013, соответственно, и инкубировали смесь при комнатной температуре в течение 2 часов для связывания фагов из библиотек антител человека с антигенами TIGIT. Фаги, не связавшиеся с антигенами TIGIT, удаляли промывкой раствором PBST (pH 7,4). Остаток элюировали 0,1 М раствором глицина (pH 3,0) и нейтрализовали 1 М раствором трис-HCl (pH 8,0). Элюированными фагами инфицировали E. coli ER2537 (OD600 = 0,5) при 37°C, амплифицировали с помощью хелперных фагов VCSM13 и использовали в следующем раунде скрининга. Результаты определения количества фагов при каждом раунде скрининга посредством пэннинга и соотношения общего количества фагов к количеству элюированных фагов показали, что количество фагов, связавшихся с антигенами TIGIT, возрастало с повышением числа раундов пэннинга.
Клонами фагов, полученных из продуктов каждого раунда скрининга, инфицировали штаммы E. coli ER2537 и высеивали на чашки с ампициллином для получения колоний. Специфичность связывания колоний с антигенами TIGIT определяли следующим методом ELISA, используя периплазматические экстракты. Отобранные колонии высеивали в 96-луночный планшет, в который вносили по 120 мкл среды SB/карбенициллин (50 мкг/мл), и культивировали на качалке для планшетов при 37°C (2-я скорость) до достижения OD600 = 0,6. Добавляли 30 мкл среды SB/карбенициллин (50 мкг/мл) с 5 мМ IPTG и инкубировали в течение ночи на качалке для планшетов при 37°C (2-я скорость). Образцы центрифугировали при 3000 об/мин в течение 10 мин для удаления супернатантов. Полученные осадки тщательно ресуспендировали (диссоциировали) в 100 мкл раствора BBS (200 мМ борной кислоты, 150 мМ NaCl, 1 мМ EDTA) и инкубировали при 4°C в течение 1 часа. После центрифугирования при 3000 об/мин в течение 20 минут выделяли только супернатанты, получая тем самым периплазматические экстракты. Смешивали 80 мкл периплазматического экстракта с 80 мкл TBST (5% BSA), а затем блокировали при комнатной температуре в течение 1 часа. Блокированные периплазматические экстракты вносили в дозе 80 мкл на лунку в 96-луночный планшет, покрытый антигенами IgG человека, TIGIT-Fc человека и TIGIT-Fc мыши, и инкубировали при комнатной температуре в течение 1 часа для связывания антител с антигенами. После трехкратной отмывки TBST в лунки добавляли раствор TBST (5% BSA) с разведенным 1:3000 антителом против HA-HRP (Roche) в дозе 30 мкл на лунку и инкубировали при комнатной температуре в течение 1 часа. После трехкратной отмывки TBST добавляли раствор TMB в дозе 30 мкл на лунку для развития окраски. После остановки реакции с помощью 1N H2SO4 измеряли поглощение при 450 нм. При скрининге клонов антител, обладающих сродством связывания с антигенами TIGIT, методом ELISA с использованием периплазматических экстрактов антител отобрали 14 различных клонов антител (S02, S03, S04, S05, S06, S11, S12, S14, S19, S32, S39, S43, S62, S64) из библиотеки scFv и 4 различных клона антител (F01, F02, F03, F04) из библиотеки Fab.
1.2. Клонирование отобранных клонов scFv и Fab в виде IgG и получение и очистка антител
Для получения антител типа IgG из отобранных клонов scFv и Fab каждый ген вариабельной области подвергали клонированию с использованием экспрессирующего вектора, содержащего ген константной области антитела IgG1. Гены вариабельной области ПЦР-амплифицированных тяжелых и легких цепей из клонов scFv и Fab подвергали клонированию с помощью рестрикционных ферментов ClaI (NEB), NheI (NEB) или комбинации ClaI и BsiWI (NEB), получая векторы, которые могут экспрессироваться в виде IgG. Для тяжелых цепей использовали векторы pcDNA3.3 (Invitrogen), а для легких цепей - векторы pOptiVEC (Invitrogen). Получение антител типа IgG проводили путем краткосрочной трансфекции клеток линии 293F (Invitrogen). Клетки 293F трансфицировали ДНК векторов pcDNA3.3 и pOptiVEC, клонированных в виде IgG, и на 6-й день собирали культуру клеток и использовали для очистки. Очистку Fc проводили на смоле с протеином A для очистки антител из культуры антител. Культуру антител пропускали со скоростью 1 мл/мин через смолу MabSelect SuRe Protein A (GE Healthcare), уравновешенную с 1xPBS (pH 7,4) для индукции связывания. По завершении связывания антител смолу сначала промывали 1xPBS (pH 7,4), а затем еще 0,1 М раствором глицина (pH 5,5). Для получения конечных антител их элюировали с помощью 0,1 М раствора глицина (pH 3,5) и нейтрализовали 1 М раствором трис-HCl (pH 8,0).
Для определения межвидовой перекрестной реактивности антител против TIGIT, отобранных при скрининге методом фагового дисплея, проверяли методом ELISA, будут ли антитела против TIGIT связываться с антигенами TIGIT человека (R&D Systems), TIGIT мыши (R&D Systems) и TIGIT макаки-резуса. При этом антигены TIGIT макаки-резуса экспрессировали и очищали в слитом с Fc виде путем синтеза генов в соответствии с последовательностью гена TIGIT макаки-резуса (№ доступа NCBI: XP_014985303.1). Каждый из трех видов антигенов TIGIT разводили до концентрации 1 мг/мл в PBS, вносили по 30 мкл на лунку в 96-луночный планшет и инкубировали в течение ночи при 4°C, чтобы их фиксировать. Затем проводили связывание по 30 нг отобранных антител с антигеном TIGIT, а затем в каждую лунку добавляли 30 мкл разведенного 1:3000 вторичного антитела в TBST (5% BSA) и инкубировали при комнатной температуре в течение 1 часа. После трехкратной отмывки TBST в каждую лунку добавляли 30 мкл раствора TMB для развития окраски. После остановки реакции с помощью 1N H2SO4 измеряли окраску по поглощению при 450 нм. Межвидовую перекрестную реактивность отобранных антител проверяли по связыванию с 6 типами антигенов (фиг. 1). Результаты показали, что большинство антител, использовавшихся при проверке, связывались как с TIGIT человека, так и с TIGIT мыши.
Для определения специфичности к TIGIT у антител против TIGIT, отобранных при скрининге методом фагового дисплея, проверяли методом ELISA их связывание с TIGIT из различных суперсемейств, как-то CD96 (Sinobiological), CD155 (Sinobiological), CD112 (R&D Systems), CD113 (R&D Systems) и CD226 (R&D Systems). Шесть типов антигенов, включая TIGIT, наносили по 100 нг на лунку в 96-луночный планшет, добавляли по 30 нг отобранных антител и инкубировали при комнатной температуре в течение 1 часа. После трехкратной отмывки TBST в каждую лунку добавляли 30 мкл разведенного 1:3000 вторичного антитела против Fab-HRP человека (Jackson) в TBST (5% BSA) и инкубировали при комнатной температуре в течение 1 часа. После трехкратной отмывки TBST в каждую лунку добавляли 30 мкл раствора TMB. После остановки реакции с помощью 1N H2SO4 определяли окраску по поглощению при 450 нм. Проверяли связывание выбранных антител с шестью антигенами. В результате оказалось, что все антитела специфически связываются только с антигеном TIGIT (фиг. 2).
Наконец, проводили FACS-анализ на клетках линии CHO-S (CHO-hTIGIT), гиперэкспрессирующих белки TIGIT человека, с тем, чтобы проверить связывание отобранных антител с антигенами TIGIT, экспрессированными на поверхности клеток. Клетки линии CHO-S (CHO-hTIGIT) получали путем трансдукции клеток CHO-S полноразмерным геном TIGIT человека с помощью лентивирусного вектора и последующего отбора только клеток CHO, гиперэкспрессирующих TIGIT человека, с помощью антибиотика бластицидина. Полученные клетки линии CHO-hTIGIT промывали ледяным PBS, вносили по 5×104 клеток в пробирки, добавляли 1 мкг каждого из полученных IgG-антител и инкубировали на льду в течение 1 часа. После этого к клеткам добавляли 1 мкг вторичного антитела против IgG человека с FITC (Invitrogen) и инкубировали на льду в течение 1 часа. Клетки промывали ледяным PBS, а затем подвергали FACS-анализу для определения связывания антител с антигенами TIGIT человека, экспрессированными на поверхности клеток (фиг. 3). В качестве положительного контроля использовали полученное из хомяка антитело 10A7 против антител человека, допускающее перекрестное связывание с мышиными, а вариабельная область была получена авторами настоящего изобретения путем получения генов в виде антитела, содержащего константную область IgG2a мыши, на основе последовательности, описанной в US Patent No. 2015/0216970. Антитело 10A7 связывается с антигеном TIGIT человека, но не связывается с ниволумабом (антителом против PD1), используемым в качестве отрицательного контроля. Это значит, что были получены нормальные клетки CHO-hTIGIT. Было установлено, что отобранные антитела специфически связываются с клетками CHO-hTIGIT.
Пример 2. Оптимизация антител против TIGIT
2.1. Оптимизация антител F04 и S64
В описанном выше процессе скрининга антител в конечном счете были отобраны два типа клонов, F04 и S64, в качестве антител человека против TIGIT. Для улучшения стабильности этих антител составляли суббиблиотеки на основе аминокислотных последовательностей антител F04 и S64 и проводили скрининг для повышения стабильности при высокой температуре методом усиленной отмывки. Получали суббиблиотеку антител F04 при одновременной перетасовке CDRH1 и CDRH2 путем наложения одного на другой. Получали три типа суббиблиотек, включая библиотеку, полученную при одновременной перетасовке CDRH1 и CDRH2, библиотеку, полученную при одновременной перетасовке CDRL1, CDRL2 и CDRL3, и библиотеку, полученную при перетасовке других CDR, за исключением CDRH3, методом перекрывающихся реакций ПЦР. Участки CDR, обладающие разнообразием аминокислотных последовательностей для клональной оптимизации F04 и S64, представлены ниже в табл. 5 (участки CDR подчеркнуты).
Таблица 5. Последовательности вариабельной области F04 и S64

Из суббиблиотеки, составленной для отбора более устойчивых клонов, чем исходные клоны, извлекали фаги, а затем нагревали перед связыванием с антигенами, чтобы удалить неустойчивые клоны. В первом и втором раунде скрининга методом фагового дисплея фаги обрабатывали при 60°C в течение 10 мин, а в третьем и вплоть до шестого раунда скрининга фаги обрабатывали при 80°C в течение 10 мин. Кроме того, при ELISA проводили скрининг при остаточном соотношении, составленном для различения клонов с улучшенной стабильностью от исходных клонов при увеличении времени отмывки до 2 часов и повышении температуры до 37°C. Исходя из этого отбирали клоны с улучшенной стабильностью и анализировали их последовательности (табл. 6 и 7).
Таблица 6. Последовательности CDR антител при скрининге на основе клонов F04
Таблица 7. Последовательности CDR антител при скрининге на основе клонов scFv
Наконец, проводили отбор клонов F04-10 путем скрининга суббиблиотеки F04, а отбор клонов S64-39 проводили путем скрининга суббиблиотеки S64.
Антитела на основе клонов F04-10 включают в себя аминокислотную последовательность, приведенную в SEQ ID NO: 13 в качестве вариабельной области тяжелой цепи, и аминокислотную последовательность, приведенную в SEQ ID NO: 15 в качестве вариабельной области легкой цепи, а антитела на основе клонов S64-39 включают в себя аминокислотную последовательность, приведенную в SEQ ID NO: 14 в качестве вариабельной области тяжелой цепи, и аминокислотную последовательность, приведенную в SEQ ID NO: 16 в качестве вариабельной области легкой цепи.
Кроме того, антитела на основе клонов F04-10 включают: вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 1; CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 3; и CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 5; и вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 7; CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 9; и CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 11.
Кроме того, антитела на основе клонов S64-39 включают: вариабельную область тяжелой цепи, содержащую участок CDR1 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 2; CDR2 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 4; и CDR3 тяжелой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 6; и вариабельную область легкой цепи, содержащую участок CDR1 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 6; CDR2 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 10; и CDR3 легкой цепи, включающий аминокислотную последовательность, приведенную в SEQ ID NO: 12.
2.2. Клонирование генов антител против TIGIT и очистка антител
Экспрессирующие векторы антител F04-10-IgG1 и S64-39-IgG1 получали таким же образом, как в Примере 1.2. Экспрессирующие векторы тяжелой цепи антител S64-39-IgG4 получали следующим образом. Из экспрессирующих векторов S64-39-IgG1 удаляли гены, соответствующие константной области IgG1, с помощью рестрикционных ферментов NheI и XhoI (NEB), а туда вставляли гены, в которые была субклонирована константная область тяжелой цепи антитела ниволумаб против PD-1, обработанная NheI и XhoI, получая такие экспрессирующие векторы тяжелых цепей, чтобы тяжелые цепи S64-39 в конечном счете экспрессировались в виде IgG4. Вариабельные области легкой цепи антител F04-10 и S64-39 субклонировали в векторы pOptiVEC с помощью рестрикционных ферментов ClaI и XhoI, соответственно.
Получение и очистку трех типов антител, получаемых в виде IgG, проводили методом краткосрочной экспрессии, используя клетки линии 293F и смолу с белком A MabSelect SuRe таким же образом, как в Примере 1.2 выше.
Пример 3. Тест для выявления связывания антител против TIGIT с TIGIT на клеточной поверхности
Для проверки способности трех типов антител против TIGIT, полученных в Примере 2, связываться с TIGIT, экспрессированным на поверхности клеток, гиперэкспрессирующие TIGIT клетки линии СНО (далее именуются клетками линии CHO-TIGIT) обрабатывали антителами против TIGIT, а затем выявляли антитела против TIGIT, связавшиеся с TIGIT на поверхности клеток, с помощью флуоресцентного проточного цитометра.
В частности, клетки линии CHO-TIGIT культивировали в инкубаторе с 5% CO2 при 37°C в течение 48-72 часов со средой определенного химического состава (CD FortiCHO Chemically Defined Medium + 8 мМ L-глутамина + 20 мкг/мл бластицидина + 1% средства против слипания). Культивируемые клетки линии CHO-TIGIT собирали центрифугированием, разбавляли раствором FACS (PBS + 5% FBS) и вносили при плотности 1×105 клеток на лунку в 96-луночный круглодонный планшет (Corning). Затем, чтобы полностью удалить среду определенного состава, оставшуюся на поверхности клеток, добавляли раствор FACS, центрифугировали 3 мин при 2000 об/мин и трижды промывали для удаления супернатанта. Промытые клетки линии CHO-TIGIT ресуспендировали добавлением 100 мкл раствора FACS. В лунки 96-луночного круглодонного планшета с клетками линии CHO-TIGIT добавляли по 100 мкл антител против TIGIT при каждой концентрации, которые были разбавлены до 2-кратной конечной концентрации раствором FACS, и инкубировали при 4°C в течение 1 часа. Затем, чтобы удалить антитела против TIGIT, остающиеся в супернатанте и не связавшиеся с TIGIT на поверхности клеток, в каждую лунку добавляли раствор FACS, центрифугировали 3 мин при 2000 об/мин и трижды промывали для удаления супернатанта. Затем для выявления антител против TIGIT, связавшихся с TIGIT на поверхности клеток, добавляли 100 мкл перекрестно абсорбированного вторичного козьего антитела против IgG человека (H+L) (Invitrogen), разведенного до 10 мкг/мл раствором FACS, и инкубировали при 4°C в течение 1 часа. После трехкратной отмывки каждый образец переносили в пробирку 12×75 мм (BD Biosciences) и анализировали с помощью флуоресцентного проточного цитометра. В результате было установлено, что связывающая способность (EC50) антител против TIGIT к TIGIT на поверхности клеток составляла 16,41 нг/мл для F04-10-IgG1, 69,01 нг/мл для S64-39-IgG1, 80,78 нг/мл для S64-39-IgG4 и 16,54 нг/мл для 10A7 (фиг. 4).
Пример 4. Тест на ингибирование антителами против TIGIT связывания TIGIT с CD155
Для проверки активности трех типов антител против TIGIT, полученных выше в Примере 2, тестировали ингибирование связывания между TIGIT и CD155.
В частности, в этом тесте проверяли ингибирующее действие антител против TIGIT на связывание между TIGIT и CD155 в системе совместного культивирования экспрессирующих CD155 клеток линии aAPC/CHO-K1 и экспрессирующих TIGIT клеток эффекторной линии с помощью набора для анализа блокирования TIGIT/CD155 (Promega).
Экспрессирующие CD155 клетки линии aAPC/CHO-K1 растворяли и разбавляли в 14,5 мл основной среды (среда F-12, содержащая 10% FBS) и вносили по 100 мкл полученных клеток в 96-луночный планшет (Costar) и хранили в инкубаторе с CO2 от 16 до 24 часов. Для F04-10-IgG1 концентрат, содержащий антитело против TIGIT, путем 4-кратного серийного разведения доводили до 2 мг/мл, что вдвое выше, чем концентрация при обработке, используя аналитическую среду (среда RPMI 1640, содержащая 10% FBS), а S64-39-IgG1 и S64-39-IgG4 доводили до 3 мг/мл путем 2-кратного серийного разведения. Используемое в качестве контроля антитело 10A7 доводили до 2,4 мг/мл путем 3-кратного серийного разведения. Из 96-луночного планшета, содержащего экспрессирующие CD155 клетки линии aAPC/CHO-K1, полностью удаляли среду и вносили полученные антитела против TIGIT по 40 мкл в каждую лунку, чтобы получить следующие фактические концентрации: для F04-10-IgG1 образец 4-кратного серийного разведения использовался для обработки при 1 мг/мл, а для S64-39-IgG1 и S64-39 IgG4 образцы 2-кратных серийных разведений использовались для обработки при 1 мг/мл. Для контрольного антитела 10А7 образец 3-кратного серийного разведения использовался для обработки при 1,2 мг/мл. Затем растворяли и разбавляли экспрессирующие TIGIT клетки эффекторной линии в 6 мл аналитической среды, вносили в каждую лунку по 40 мкл разбавленных клеток, а затем инкубировали при 37°C в инкубаторе с 5% CO2 в течение 6 часов. В каждую лунку добавляли 80 мкл реагента Bio-Glo™, приготовленного путем добавления буфера Bio-Glo™ к субстрату Bio-Glo™, и проводили реакцию при комнатной температуре в течение 10 мин. Измеряли величину сигнала (в относительных единицах люминесценции, RLU) на считывающем люминесценцию устройстве (Molecular Devices, SpectraMax L) и рассчитывали соотношение уровней сигналов (кратность сигнала = RLUс разведенным Ab/RLUконтроль без антитела), то есть отношение величины сигнала при обработке антителом против TIGIT к величине сигнала без обработки антителом против TIGIT. Получили следующие результаты: ингибирование связывания (EC50) антителами против TIGIT составило 23,89 нМ для F04-10-IgG1, 0,581 мкМ для S64-39-IgG1, 1,08 мкМ для S64-39-IgG4 и 0,752 мкМ для 10A7 (фиг. 5).
Пример 5. Тест на активацию NK-клеток антителами против TIGIT
Для того чтобы определить активность клеток линии NK92 после обработки тремя типами антител против TIGIT, полученных в Примере 2, измеряли уровень секреции IFN-γ в клетках линии NK92 и проводили тест на наличие экспрессии NKG2D с помощью флуоресцентного проточного цитометра в условиях совместного культивирования гиперэкспрессирующих TIGIT клеток линии NK92 и гиперэкспрессирующих PVR клеток раковой линии яичников человека (линии клеток HeLa).
В частности, гиперэкспрессирующие TIGIT клетки линии NK92 разводили до концентрации 2×105/мл в полной среде (alpha-MEM + 12,5% FBS + 12,5% лошадиной сыворотки + 0,1 мМ 2-меркаптоэтанола + 100 ед./мл IL-2), а затем культивировали в колбе T25 (Corning) в объеме 5 мл при 37°C в инкубаторе с 5% CO2 в течение 16-24 часов. К клеткам добавляли 25 мкг/мл антитела против TIGIT для ингибирования гиперэкспрессии TIGIT в культуре клеток линии NK92 и культивировали при 37°C в инкубаторе с 5% CO2 в течение 72 часов. Во время культивирования клеток линии NK92 с антителом против TIGIT разводили гиперэкспрессирующие PVL клетки линии HeLa и культивировали в полной среде (среда RPMI 1640 с добавлением 10% FBS) в концентрации 3×105/мл в объеме 15 мл в колбе T75 (Corning) и культивировали в инкубаторе с 5% CO2 при 37°C в течение 24-48 часов. Затем клетки линии NK92 и клетки линии HeLa культивировали совместно в течение 4-6 часов при 37°C в инкубаторе с 5% CO2 в объеме 1 мл в 12-луночном планшете (Corning) при соотношении 1:10 (1×105 NK92 : 1×106 HeLa). По завершении совместного культивирования получали культуральный супернатант и хранили его при -20°C для теста ELISA на IFN-γ, а культивированные клетки разбавляли PBS (Gibco) для препаратов.
Использовали культуральный супернатант и клетки, полученные по описанной выше методике.
Сначала проводили тест ELISA для определения количества IFN-γ, секретируемого из клеток линии NK92, используя культуральный супернатант. Этот тест проводили с помощью набора Qantikine для анализа IFN-γ человека методом ELISA (R&D Systems). Из результатов анализа количества IFN-γ, секретируемого клетками NK92 после обработки антителом против TIGIT, видно, что группы, обработанные антителами F04-10-IgG1 и S64-39-IgG1, проявляли значительное повышение секреции IFN-γ по сравнению с группой, обработанной контрольным антителом, а группа, обработанная антителом S64-39-IgG4, проявляла уровень секреции IFN-γ, эквивалентный таковому в группе, обработанной контрольным антителом (фиг. 6).
После этого авторы изобретения проводили анализ с помощью флуоресцентного проточного цитометра на наличие экспрессии NKG2D, одного из белков-маркеров активации NK-клеток, у клеток, полученных при вышеизложенном совместном культивировании. Для иммуноокрашивания клеток их сначала разводили до концентрации 1×106/мл в буфере для окрашивания клеток (Biolegend) и проводили окрашивание с помощью антитела против CD56 с eFluor (eBioscience) и антитела против NKG2D с PE (BD Biosciences) при 4°C в течение 20 минут в отсутствие света, чтобы специфически выделялись только клетки NK92. Затем, чтобы удалить красители, добавляли 1 мл буфера для окрашивания клеток и центрифугировали при 2000 об/мин в течение 5 мин, повторяя эту операцию три раза. Затем каждый образец переносили в пробирку 12×75 мм для флуоресцентной проточной цитометрии (BD Biosciences) и определяли профиль экспрессии NKG2D в клетках экспрессирующей CD56 линии с помощью флуоресцентного проточного цитометра. В результате было установлено, что экспрессия NKG2D значительно повышается в группах, обработанных антителами F04-10-IgG1, S64-39-IgG1 и S64-39-IgG4, по сравнению с группой, обработанной контрольным антителом (фиг. 7).
Пример 6. Тест для измерения сродства антител против TIGIT к антигенам TIGIT
Для измерения способности трех типов антител против TIGIT, полученных в Примере 2, связываться с TIGIT человека (rhTIGIT-Fc) и TIGIT мыши (rmTIGIT-Fc), использовали метод поверхностного плазмонного резонанса (SPR) на приборе BIAcore T200 (GE Healthcare). Метод SPR основан на том принципе, что показатель преломления света, проходящего через сенсорный чип, изменяется в зависимости от состояния вещества, нанесенного на сенсорный чип. При пропускании антигена или антитела по чипу, покрытому антигеном или антителом, показатель преломления изменяется вследствие связывания между ними, и по измеренным значениям рассчитывается сродство (KD).
Антитела против TIGIT иммобилизировали на сенсорном чипе CM5 Series S (GE Healthcare) до уровня 500 RU с помощью 10 мМ раствора ацетата (pH 4,0) и набора для конъюгирования аминов (GE Healthcare). Делали 2-кратные серийные разведения белков TIGIT-Fc человека или мыши (R&D Systems) от концентрации 40 нМ в буфере HBS-EP (0,01 М HEPES, pH 7,4, 0,15 М NaCl, 3 мМ EDTA, 0,005% детергента P20, GE Healthcare) и пропускали каждое из них для измерения сродства антиген-антитело, вызывая ассоциацию, диссоциацию и регенерацию с антителом, иммобилизованным на сенсорном чипе. Ассоциацию белков TIGIT-Fc измеряли при скорости 30 мкл/мин в течение 600 сек, диссоциацию измеряли в течение 2000 сек, а регенерацию проводили в 10 мМ растворе глицина (pH 1,5) при скорости 100 мкл/мин в течение 25 сек. Результаты измерения способности к связыванию с rhTIGIT-Fc и rmTIGIT-Fc представлены ниже в табл. 8 и 9.
Таблица 8. Результаты измерения способности к связыванию с rhTIGIT-Fc
Таблица 9. Результаты измерения способности к связыванию с rmTIGIT-Fc
Пример 7. Ингибирующее действие антител против TIGIT на рост опухолей
Для оценки активности антител против TIGIT in vivo была поставлена модель опухолей у мышей (сингенная модель колоректальной карциномы CT26 на мышах BALB/c). При этом антитело положительного контроля (10A7) и три вида антител (F04-10-IgG1, S64-39-IgG1, S64-39-IgG4), полученных в Примере 2, вводили по отдельности или в комбинации с антителом против PD-L1 (10F.9G2 типа IgG2b крыс) и проводили сравнительную оценку эффектов ингибирования ими роста опухолей.
Сначала, чтобы поставить модель опухолей у мышей, культивируемые опухолевые клетки CT26 имплантировали подкожно (день 0) путем инъекции в дозе 100 мкл (1×106 клеток) на мышь и давали опухолям вырасти до определенного размера. Через 8 дней, когда объем опухолей достигал 119 мм3 (день 8, день начала введения), внутрибрюшинно вводили отрицательный контроль (IgG2b крыс в дозе 10 мг/кг) и 4 исследуемых вещества (в дозах 25 мг/кг) по три раза с интервалами в 3 дня в сочетании с антителом против PD-L1 (в дозе 10 мг/кг). Затем с интервалами в 2 недели измеряли объем опухолей и вес тела. Ингибирующее действие на рост опухолей выражали в виде TGI, который рассчитывали по объемам опухолей, измеренным в последний день тестирования in vivo (день 28), по следующей формуле:
степень TGI (%) = 100 × (1-ΔT/ΔC),
где: ΔT = средний объем опухолей в группе, получавшей исследуемое вещество, при измерении в последний день - средний объем опухолей в группе, получавшей исследуемое вещество, при измерении в день начала введения;
ΔC = средний объем опухолей в группе, получавшей отрицательный контроль, при измерении в последний день - средний объем опухолей в группе, получавшей отрицательный контроль, при измерении в день начала введения.
Объем опухолей возрастал примерно в 23 раза на последний день в группе, получавшей отрицательный контроль, по сравнению с днем начала введения. По сравнению с отрицательным контролем, антитело положительного контроля и антитело против PD-L1 проявляли умеренное противоопухолевое действие при введении по отдельности, а введение комбинации двух антител давало более сильный и значительный эффект ингибирования опухолей (фиг. 8). В случае антител против TIGIT, два вида антител типа IgG1 (S64-39-IgG1, F04-10-IgG1) проявляли действие, аналогичное эффекту антитела положительного контроля при введении по отдельности, а S64-39-IgG1 проявлял эквивалентный уровень эффекта при введении в комбинации. Антитело S64-39-IgG4 оказывало несколько меньший эффект, чем S64-39-IgG1, при введении отдельно и в комбинации.
Итак, три вида антител (F04-10-IgG1, S64-39-IgG1, S64-39-IgG4), полученных в Примере 2 выше, проявляли значительный противоопухолевый эффект по сравнению с группой, получавшей отрицательный контроль, при введении по отдельности или в комбинации с антителом против PD-L1 (F04-10-IgG1, S64-39-IgG1). В частности, при введении по отдельности два вида антител (F04-10-IgG1, S64-39-IgG1) давали эквивалентный противоопухолевый эффект при сравнении с группой, получавшей отрицательный контроль, а при введении в комбинации один вид антител (S64-39-IgG1) давал эквивалентный противоопухолевый эффект в сравнении с группой, получавшей отрицательный контроль.
Статистический анализ, приведенный на фиг. 8, проводили по критерию множественных сравнений Даннета с помощью GraphPad Prism 5, а статистическую значимость отличий при сравнении с группой, получавшей отрицательный контроль, выражали следующим образом: * p <0,05; ** p <0,01; и *** p <0,001.
Пример 9. Ингибирующее рост опухолей действие антитела против TIGIT в зависимости от дозы и применения в комбинации
Для оценки активности антитела против TIGIT in vivo в зависимости от дозы и применения в комбинации была поставлена модель опухолей у мышей (сингенная модель колоректальной карциномы CT26 на мышах BALB/c). Полученное в примере 2 антитело F04-10 вводили мышам на модели опухолей отдельно или в комбинации с антителом против PD-L1 (10F.9G2 типа IgG2b крыс) для сравнительной оценки эффекта ингибирования роста опухолей.
Сначала, чтобы поставить модель опухолей у мышей, культивируемые опухолевые клетки CT26 имплантировали подкожно (день 0) путем инъекции в дозе 100 мкл (1×106 клеток) на мышь и давали опухолям вырасти до определенного размера. Через 7 дней, когда объем опухолей достигал 80 мм3 (день 7, день начала введения), испытуемых мышей случайным образом разделяли на группы с одинаковым средним объемом опухолей, а затем внутрибрюшинно вводили отрицательный контроль (комбинация по 10 мг/кг IgG2b крыс и 25 мг/кг IgG1 человека) либо F04-10 (5, 10, 25 мг/кг) по отдельности или в сочетании с антителом против PD-L1 по три раза с интервалами в 3 дня. Затем с интервалами в 2 недели измеряли объем опухолей и вес тела. Ингибирующее действие на рост опухолей выражали в виде TGI, который рассчитывали по объемам опухолей, измеренным в последний день тестирования in vivo (день 24), по следующей формуле:
степень TGI (%) = 100 × (1-ΔT/ΔC),
где: ΔT = средний объем опухолей в группе, получавшей исследуемое вещество, при измерении в последний день - средний объем опухолей в группе, получавшей исследуемое вещество, при измерении в день начала введения;
ΔC = средний объем опухолей в группе, получавшей отрицательный контроль, при измерении в последний день - средний объем опухолей в группе, получавшей отрицательный контроль, при измерении в день начала введения.
Объем опухолей возрастал примерно в 16 раз на последний день в группе, получавшей отрицательный контроль, по сравнению с днем начала введения. С другой стороны, группа, получавшая только F04-10, проявляла небольшой противоопухолевый эффект при дозах 3 и 10 мг/кг и максимальный эффект при дозе 25 мг/кг, то есть проявлялось сильное ингибирующее действие на рост опухолей. Введение 10 мг/кг одного лишь антитела против PD-L1 давало умеренный эффект ингибирования опухолей. При введении антитела против PD-L1 в сочетании с F04-10 в дозах 3 и 10 мг/кг ингибирующий эффект на рост опухолей был сильнее, чем при введении такой же дозы одного F04-10. С другой стороны, при 25 мг/кг F04-10, что давало наибольший эффект, не проявлялось усиления эффектов при комбинации с антителом против PD-L1 (фиг. 9).
Итак, антитело F04-10, полученное в Примере 2, проявляло значительное максимальное ингибирующее действие при 25 мг/кг при введении по отдельности, тогда как антитело F04-10 при введении в дозах 3 и 10 мг/кг, т.е. менее 25 мг/кг, проявляло улучшение эффектов при введении в комбинации с антителом против PD-L1 по сравнению с введением по отдельности.
Статистический анализ, приведенный на фиг. 9, проводили по критерию множественных сравнений Даннета с помощью GraphPad Prism 5, а статистическую значимость отличий при сравнении с группой, получавшей отрицательный контроль, выражали следующим образом: * p <0,05.
Промышленная применимость
Антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению, как оказалось, очень специфически и сильно связываются с TIGIT и проявляют отличную терапевтическую эффективность по сравнению с обычными антителами против TIGIT. Соответственно, фармацевтические композиции, содержащие антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению в качестве активного ингредиента, могут применяться в качестве противораковых иммунотерапевтических средств на основе активации иммунных клеток.
Кроме того, фармацевтические композиции, содержащие антитела против TIGIT либо их антигенсвязывающие фрагменты по настоящему изобретению в качестве активного ингредиента, могут применяться в комбинированной терапии с химическими препаратами и другими химиотерапевтическими средствами.
Хотя конкретные конфигурации настоящего изобретения были описаны подробно, специалистам в данной области должно быть понятно, что данное описание приводится в виде предпочтительных воплощений для иллюстративных целей и не должно рассматриваться как ограничивающее объем настоящего изобретения. Поэтому по существу объем настоящего изобретения определяется прилагаемой формулой изобретения и ее эквивалентами.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> YUHAN CORPORATION
<120> Антитела против TIGIT т их применение
<130> PF-B2449
<140> PCT/KR2019/002440
<141> 2019-02-28
<150> 10-2018-0024822
<151> 2018-02-28
<160> 21
<170> PatentIn версии 3.5
<210> 1
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 1 вариабельной области тяжелой цепи
<400> 1
Ser Tyr Tyr Met Ser
1 5
<210> 2
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 1 вариабельной области тяжелой цепи
<400> 2
Asn Tyr Ala Met Ser
1 5
<210> 3
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 2 вариабельной области тяжелой цепи
<400> 3
Ser Ile Gly Ser Gly Ser Pro Ser Ser Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly
<210> 4
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 2 вариабельной области тяжелой цепи
<400> 4
Gly Ile Ser Pro Ser Gly Ser Ser Ile Tyr Tyr Ala Asp Ser Val Gln
1 5 10 15
Gly
<210> 5
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 3 вариабельной области тяжелой цепи
<400> 5
Ser Ser Tyr Ser Gly Gly Asn Gly Tyr Tyr Tyr Tyr Ala Tyr Ala Phe
1 5 10 15
Asp Tyr
<210> 6
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 3 вариабельной области тяжелой цепи
<400> 6
Ala Ile Arg Thr Cys Ser Leu Ser His Cys Tyr Tyr Tyr Tyr Gly Met
1 5 10 15
Asp Val
<210> 7
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 1 вариабельной области легкой цепи
<400> 7
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 8
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 1 вариабельной области легкой цепи
<400> 8
Ser Ser Ser Ser Ser Asn Ile Gly Ser Asn Ala Val Asn
1 5 10
<210> 9
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 2 вариабельной области легкой цепи
<400> 9
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 10
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 2 вариабельной области легкой цепи
<400> 10
Tyr Asp Asn Gln Arg Pro Ser
1 5
<210> 11
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_CDR 3 вариабельной области легкой цепи
<400> 11
Gln Gln Gly Tyr His Arg Tyr Ala Thr
1 5
<210> 12
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_CDR 3 вариабельной области легкой цепи
<400> 12
Ala Thr Trp Asp Tyr Ser Leu Ser Gly Tyr Val
1 5 10
<210> 13
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_Вариабельная область тяжелой цепи
<400> 13
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Gly Ser Gly Ser Pro Ser Ser Thr Tyr Tyr Ala Asp Ser
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
65 70 75 80
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Ser Ser Tyr Ser Gly Gly Asn Gly Tyr Tyr Tyr Tyr Ala
100 105 110
Tyr Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 14
<211> 127
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_Вариабельная область тяжелой цепи
<400> 14
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Pro Ser Gly Ser Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ala Ile Arg Thr Cys Ser Leu Ser His Cys Tyr Tyr Tyr Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 15
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04-10_Вариабельная область легкой цепи
<400> 15
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Tyr His Arg Tyr
85 90 95
Ala Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 16
<211> 110
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64-39_Вариабельная область легкой цепи
<400> 16
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Ser Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Tyr Ser Leu
85 90 95
Ser Gly Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 17
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04_Вариабельная область тяжелой цепи
<400> 17
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Gly Ser Tyr Tyr Thr Tyr Tyr Ala Asp Ser Val Lys Gly
50 55 60
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
65 70 75 80
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
85 90 95
Ser Ser Tyr Ser Gly Gly Asn Gly Tyr Tyr Tyr Tyr Ala Tyr Ala Phe
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 18
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон F04_Вариабельная область легкой цепи
<400> 18
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Tyr His Arg Tyr
85 90 95
Ala Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 19
<211> 127
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64_Вариабельная область тяжелой цепи
<400> 19
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Tyr Pro Gly Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Ala Ile Arg Thr Cys Ser Leu Ser His Cys Tyr Tyr Tyr Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 20
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Клон S64_Вариабельная область легкой цепи
<400> 20
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Cys Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ser Trp Asp Tyr Ser Leu
85 90 95
Ser Ala Tyr Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 21
<211> 244
<212> Белок
<213> Искусственная последовательность
<220>
<223> Предшественник T-клеточного иммунорецептора с Ig-доменом и ITIM
[Homo
sapiens]
<400> 21
Met Arg Trp Cys Leu Leu Leu Ile Trp Ala Gln Gly Leu Arg Gln Ala
1 5 10 15
Pro Leu Ala Ser Gly Met Met Thr Gly Thr Ile Glu Thr Thr Gly Asn
20 25 30
Ile Ser Ala Glu Lys Gly Gly Ser Ile Ile Leu Gln Cys His Leu Ser
35 40 45
Ser Thr Thr Ala Gln Val Thr Gln Val Asn Trp Glu Gln Gln Asp Gln
50 55 60
Leu Leu Ala Ile Cys Asn Ala Asp Leu Gly Trp His Ile Ser Pro Ser
65 70 75 80
Phe Lys Asp Arg Val Ala Pro Gly Pro Gly Leu Gly Leu Thr Leu Gln
85 90 95
Ser Leu Thr Val Asn Asp Thr Gly Glu Tyr Phe Cys Ile Tyr His Thr
100 105 110
Tyr Pro Asp Gly Thr Tyr Thr Gly Arg Ile Phe Leu Glu Val Leu Glu
115 120 125
Ser Ser Val Ala Glu His Gly Ala Arg Phe Gln Ile Pro Leu Leu Gly
130 135 140
Ala Met Ala Ala Thr Leu Val Val Ile Cys Thr Ala Val Ile Val Val
145 150 155 160
Val Ala Leu Thr Arg Lys Lys Lys Ala Leu Arg Ile His Ser Val Glu
165 170 175
Gly Asp Leu Arg Arg Lys Ser Ala Gly Gln Glu Glu Trp Ser Pro Ser
180 185 190
Ala Pro Ser Pro Pro Gly Ser Cys Val Gln Ala Glu Ala Ala Pro Ala
195 200 205
Gly Leu Cys Gly Glu Gln Arg Gly Glu Asp Cys Ala Glu Leu His Asp
210 215 220
Tyr Phe Asn Val Leu Ser Tyr Arg Ser Leu Gly Asn Cys Ser Phe Phe
225 230 235 240
Thr Glu Thr Gly
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО АНТИ-EGFR/АНТИ-4-1ВВ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2831836C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
| КОНЬЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО, ВКЛЮЧАЮЩИЕ АНТИТЕЛО ПРОТИВ ЧЕЛОВЕЧЕСКОГО DLK1, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2801630C2 |
| АНТИТЕЛА ПРОТИВ VISTA И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750723C1 |
| АНТИТЕЛО ПРОТИВ ANG2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2788088C1 |
| АНТИТЕЛО ПРОТИВ TIE-2 И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2795455C2 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| АНТИТЕЛО ПРОТИВ VSIG4 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826236C1 |
| Эпитоп тиоредоксина-1, специфично связывающийся с моноклональным антителом | 2018 |
|
RU2804772C2 |
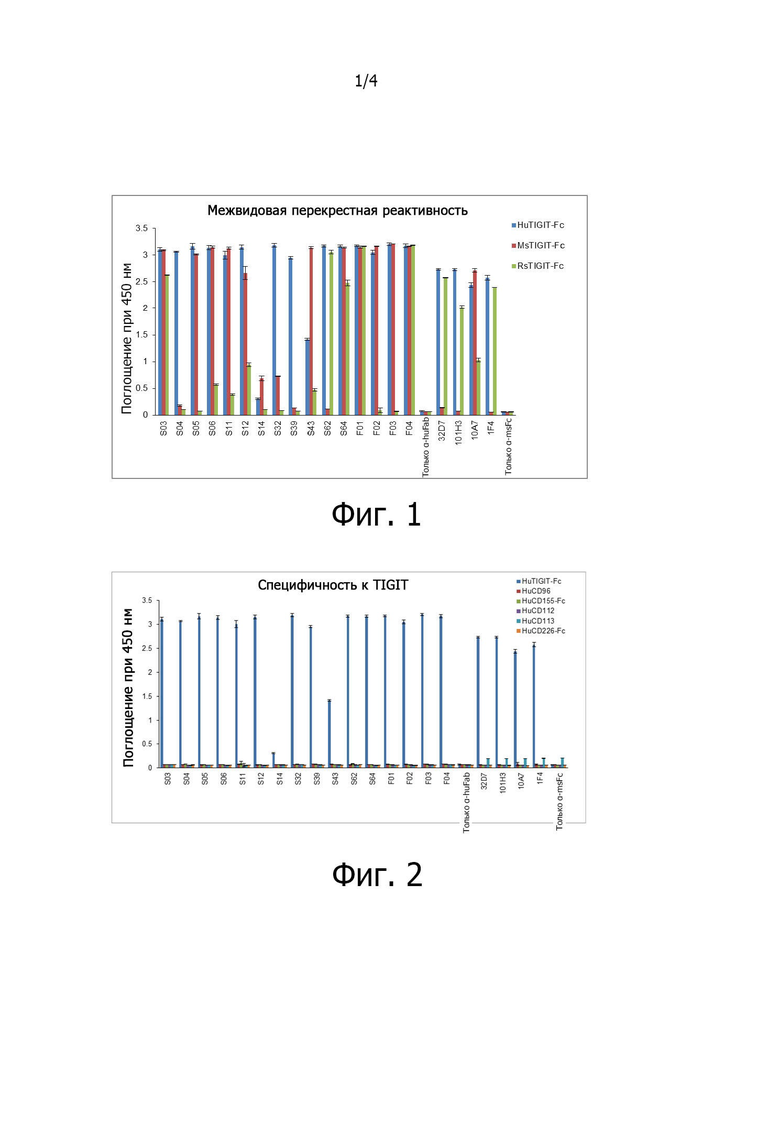
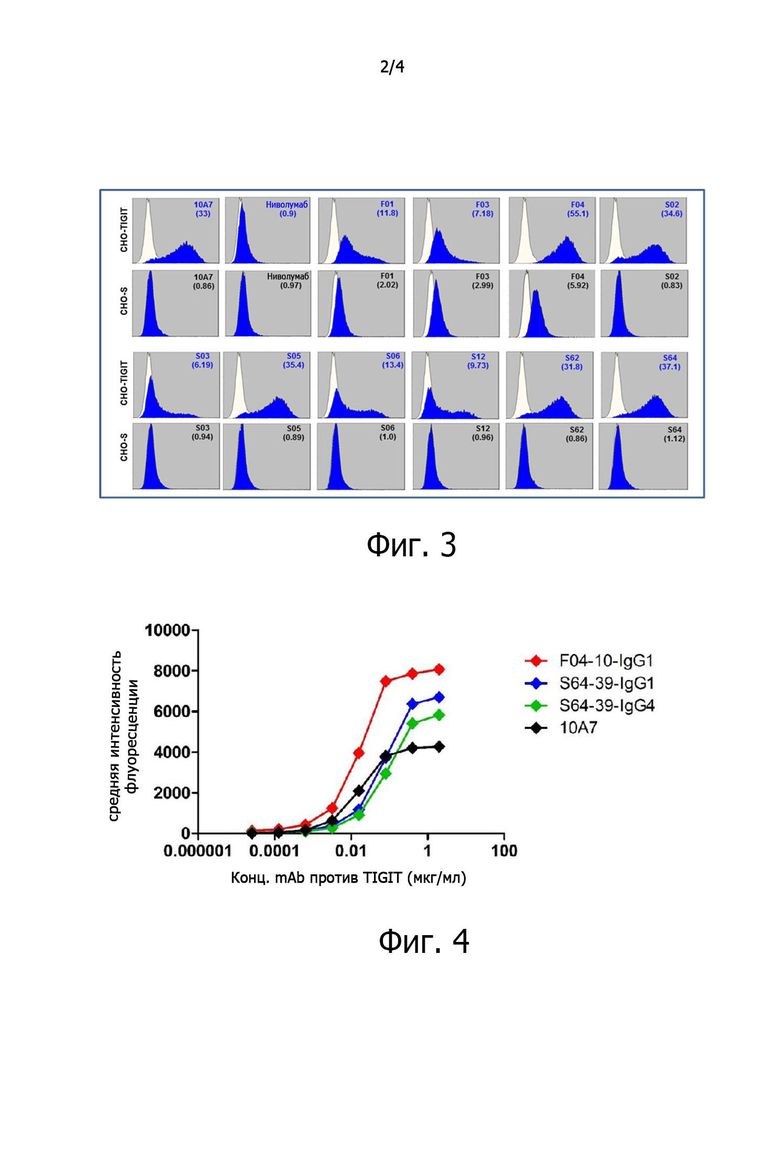
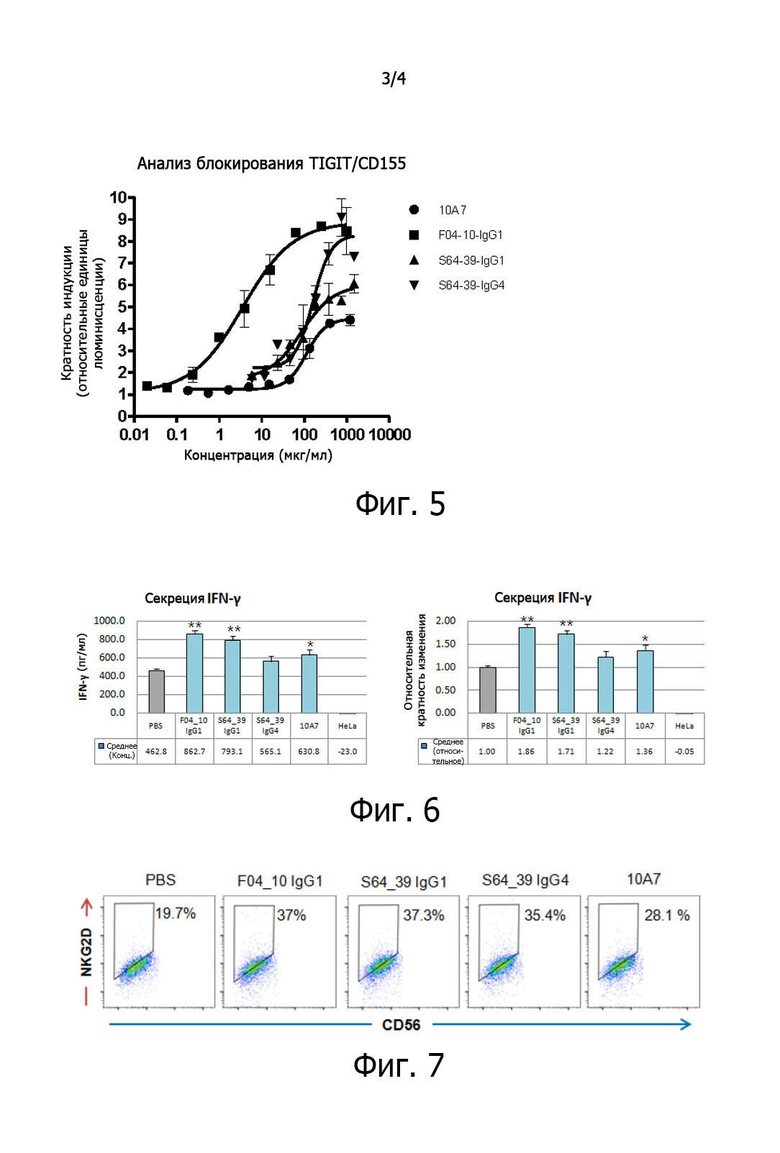
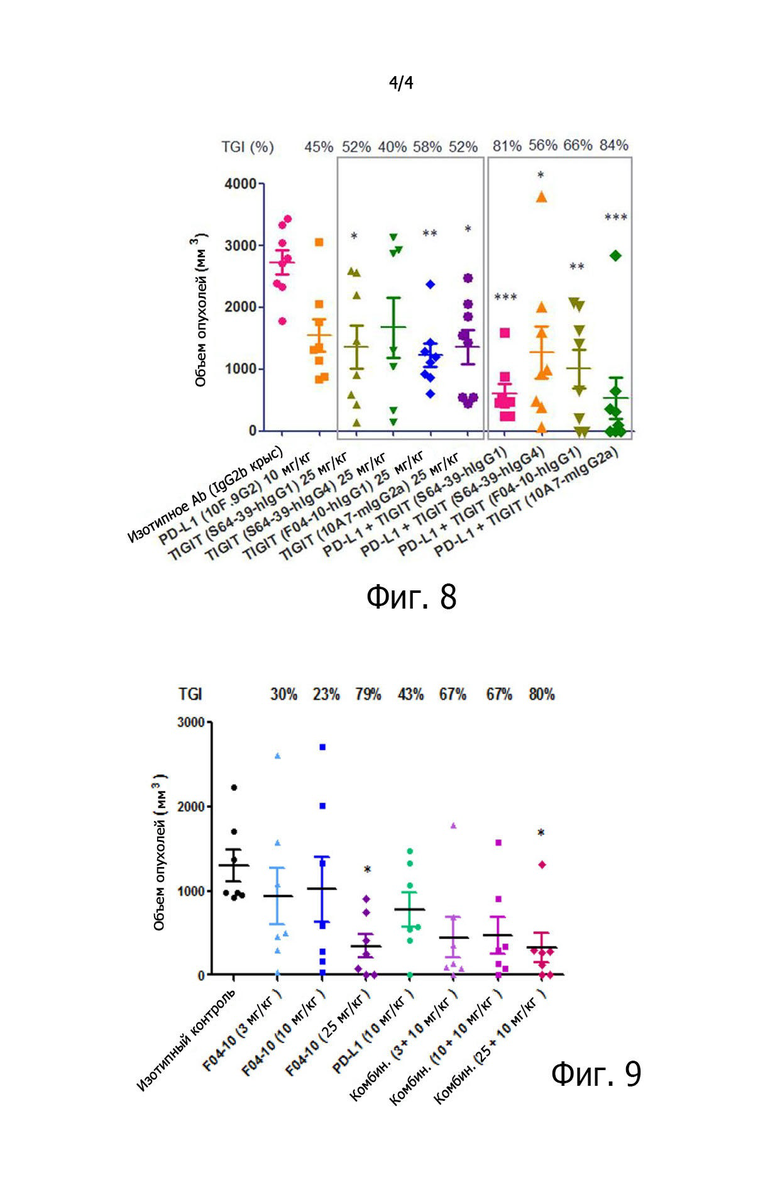
Группа изобретений относится к биотехнологии. Представлены новые антитело, специфически связывающееся с иммуносупрессором опухолей TIGIT (T-клеточный иммунорецептор с иммуноглобулиновым доменом и ингибирующим мотивом на основе тирозина [ITIM]), либо его антигенсвязывающий фрагмент, нуклеиновая кислота, кодирующая это антитело, вектор и клетка-хозяина, содержащая такую нуклеиновую кислоту, способ получения такого антитела либо его антигенсвязывающего фрагмента, фармацевтическая композиция, содержащая это антитело, либо его антигенсвязывающий фрагмент в качестве активного ингредиента. Данная группа изобретений применяется для эффективного лечения рака или опухолей. 9 н. и 8 з.п. ф-лы, 9 табл., 9 пр., 4 ил.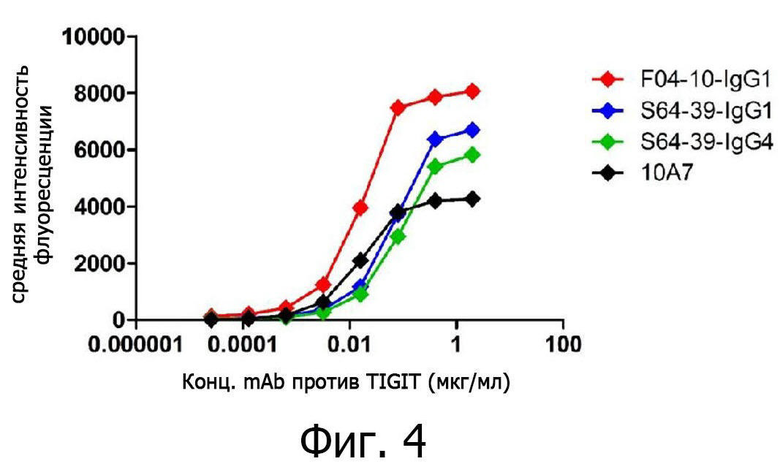
1. Антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с T-клеточным иммунорецептором с иммуноглобулиновым доменом и ингибирующим мотивом на основе тирозина [ITIM] (TIGIT), содержащие:
(1) вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 1, CDR2 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 3, и CDR3 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 5; и
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 7, CDR2 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 9, и CDR3 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 11; или
(2) вариабельный участок тяжелой цепи, содержащий CDR1 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 2, CDR2 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 4, и CDR3 тяжелой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 6; и
вариабельную область легкой цепи, содержащую CDR1 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 8, CDR2 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 10, и CDR3 легкой цепи, содержащий аминокислотную последовательность, указанную в SEQ ID NO: 12.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащие:
вариабельную область тяжелой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 13 или 14; и
вариабельную область легкой цепи, включающую аминокислотную последовательность, приведенную в SEQ ID NO: 15 или 16.
3. Антитело или его антигенсвязывающий фрагмент по п. 1 или 2, при этом антитело против TIGIT представляет собой моноклональное антитело.
4. Антитело или его антигенсвязывающий фрагмент по п. 1 или 2, при этом антигенсвязывающий фрагмент выбран из группы, состоящей из scFv, (scFv)2, scFv-Fc, Fab, Fab′ и F(ab′)2 антитела против TIGIT.
5. Фармацевтическая композиция для лечения рака или опухолей, содержащая эффективное количество антитела или его антигенсвязывающего фрагмента по п. 1 или 2 в качестве активного ингредиента.
6. Фармацевтическая композиция по п. 5, при этом рак или опухоль выбраны из рака кожи, рака печени, гепатоцеллюлярной карциномы, рака желудка, рака молочной железы, рака легких, рака яичников, рака бронхов, рака носоглотки, рака гортани, рака поджелудочной железы, рака мочевого пузыря, колоректального рака, рака толстой кишки, рака шейки матки, рака мозга, рака простаты, рака костей, рака щитовидной железы, рака паращитовидной железы, рака почек, рака пищевода, холангиокарциномы, рака яичек, рака прямой кишки, рака головы и шеи, рака шейки матки, рака мочеточников, остеосаркомы, нейробластомы, фибросаркомы, рабдомиосаркомы, астроцитомы, нейробластомы и глиомы.
7. Композиция для совместного введения при лечении рака или опухолей, содержащая антитело или его антигенсвязывающий фрагмент по п. 1 или 2 и другое терапевтическое средство против рака.
8. Композиция по п. 7, при этом другое терапевтическое средство против рака представляет собой ингибитор контрольной точки иммунитета.
9. Композиция по п. 8, при этом ингибитор контрольной точки иммунитета представляет собой антитело против CTLA-4, антитело против PD-1 или антитело против PD-L1.
10. Конъюгат антитело-лекарственное средство для лечения рака или опухоли, включающий антитело или его антигенсвязывающий фрагмент по п. 1 или 2.
11. Композиция для лечения рака или опухолей, содержащая эффективное количество конъюгата антитело-лекарственное средство по п. 10.
12. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по п. 1 или 2.
13. Рекомбинантный экспрессирующий вектор, содержащий нуклеиновую кислоту по п. 12.
14. Клетка-хозяин для продукции антитела или его антигенсвязывающего фрагмента по п.1 или 2, которая трансформирована рекомбинантным экспрессирующим вектором по п. 13.
15. Клетка-хозяин по п. 14, при этом клетка-хозяин выбрана из группы, состоящей из клеток животных, растительных клеток, дрожжей, Escherichia coli и клеток насекомых.
16. Клетка-хозяин по п. 15, при этом клетка-хозяин выбрана из группы, состоящей из клеток почек обезьян (COS7), клеток NSO, клеток SP2/0, клеток яичников китайского хомячка (CHO), W138, клеток почек детенышей хомячка (BHK), MDCK, клеток миеломы, клеток HuT78, клеток HEK293, Escherichia coli, Bacillus subtilis, Streptomyces sp., Pseudomonas sp., Proteus mirabilis, Staphylococcus sp., Aspergillus sp., Pichia pastoris, Saccharomyces cerevisiae, Schizosaccharomyces sp. и Neurospora crassa.
17. Способ получения антитела или его антигенсвязывающего фрагмента по п.1 или 2, включающий культивирование клетки-хозяина по любому из пп. 14-16.
| WO 2016106302 A1, 30.06.2016; | |||
| US 20170165366 A1, 15.06.2017 | |||
| US 20170198042 A1, 13.07.2017 | |||
| US 20170037133 A1, 09.02.2017 | |||
| RU 2015123032 A, 11.01.2017. |

