ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителам против VSIG4 (V-подобный Ig домен-содержащий 4 белок) или антигенсвязывающим фрагментам и их применению.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Механизм иммунной эвазии раковых клеток достигается за счет инактивации цитотоксических Т-клеток, которые обладают цитотоксической активностью, при связывании с белком иммунной контрольной точки, присутствующим на поверхности Т-клеток. Это дает теоретическое обоснование ингибитора иммунной контрольной точки, с помощью которого инфицированные вирусом клетки или раковые клетки могут быть уничтожены путем восстановления функции Т-клеток с использованием иммунной контрольной точки в качестве мишени для усиления его активации.
Ингибиторы иммунных контрольных точек в качестве противораковых иммунотерапевтических агентов третьего поколения были впервые одобрены в 2010 году Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов и, начиная с клинического лечения меланомы, идет непрерывный поток результатов исследований, демонстрирующих превосходные терапевтические эффекты в противораковой терапии рака легких, рака печени и тому подобного. За последние 10 лет тема ингибиторов иммунных контрольных точек обрела всемирную важность. Поскольку противораковый иммунотерапевтический агент представляет собой антитело, которое вырабатывается таким образом, что раковые клетки атакуются Т-клетками, есть отчеты по результатам исследований, демонстрирующим, что превосходный эффект проявляется даже при комбинированной терапии с обычными противораковыми агентами. На сегодняшний день известны различные белки иммунных контрольных точек, включая CTLA-4 (цитотоксический Т-лимфоцит-ассоциированный белок 4), PD-1 (белок запрограммированной гибели клеток 1), TIM-3 (белок 3, содержащий домены Т-клеточного иммуноглобулина и муцина), LAG-3 (ген активации лимфоцитов 3), TIGIT (Т-клеточный иммунорецептор с иммуноглобулином и доменом ингибирующего мотива иммунорецептора на основе тирозина) и VISTA (v-домен Ig-содержащий супрессор активации Т-клеток).
V-подобный Ig домен-содержащий 4 белок (VSIG4, CRIg или Z39Ig) является белком иммунной контрольной точки, который изучается в последние годы, и представляет собой белок семейства, связанного с В7. Известно, что VSIG4 высокоэкспрессируется в печени, дендритных клетках, нейтрофилах и дремлющих макрофагах, однако в других органах, включая легкие, сердце, селезенку и лимфатические узлы, он экспрессируется в небольших количествах, при этом он не экспрессируется в Т-клетках и В-клетках. Белки семейства VSIG4 и В7 имеют общую консервативную аминокислотную последовательность, a VSIG4 имеет один полный домен IgV типа и расщепленный домен IgC типа (Vogt L. et al., J Clin Invest. (2006) 116: 2817-2826; Helmy KY. et al., Cell (2006) 124: 915-927). Известно, что VSIG4 ингибирует альтернативный путь активации комплемента, связываясь с субъединицей C3b конвертазы. Кроме того, сообщается, что, при связывании с неизвестным Т-клеточным рецептором, VSIG4 может ингибировать пролиферацию CD4+ и CD8+ Т-клеток. VSIG4 был изучен на предмет возникновения аутоиммунных и/или воспалительных расстройств, поскольку было показано, что растворимый слитый белок VSIG4-Fc, по-видимому, защищает от развития экспериментального аутоиммунного артрита, увеоретинита и гепатита (Не et al., Mol. Immunol. (2008) Molecular Immunology 45(16): 4041-4047). Однако недавно также стало известно, что экспрессия VSIG4 связана с регуляцией противоопухолевого иммунитета, такого как развитие рака легкого и плохой прогноз глиомы высокой степени злокачественности или тому подобное (Liao Y. et al., Lab Invest. (2014) 94: 706-715; Xu T. et al., Am J Transl Res. (2015) 7: 1172-1180). Кроме того, согласно исследованиям Jung et al. (Hepatology (2012) 56 (5):1838-48), существует разница в месте связывания VSIG4 между противовоспалительным и Т-клеточным ингибированием.
Антитела, направленные против VSIG4, были описаны ранее (см., например, WO 2020/069507). Однако эти антитела связывают только одну из двух форм белка, тем самым опосредуя лишь частичное ингибирование его активности.
Таким образом, по-прежнему существует необходимость в предложении новых антител против VSIG4, которые могут установить оптимальный противоопухолевый иммунитет.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ЦЕЛЬ
Цель настоящего раскрытия - предложить новое антитело против VSIG4 или его антигенсвязывающий фрагмент.
Дополнительной целью настоящего изобретения, таким образом, является предоставление композиции для лечения рака, содержащей вышеупомянутые антитела или антигенсвязывающие фрагменты.
ТЕХНИЧЕСКИЕ СПОСОБЫ ДОСТИЖЕНИЯ ВЫШЕУКАЗАННОЙ ЦЕЛИ
Для достижения вышеуказанной цели в соответствии с настоящим изобретением предложено моноклональное антитело, специфически связывающееся с VSIG4, или его антигенсвязывающий фрагмент. Описанное в данном документе антитело связывает как длинную, так и короткую формы VSIG4, что приводит к эффективному подавлению противовоспалительных сигналов, опосредованных VSIG4. Антитело против VSIG4, раскрытое в настоящем документе, таким образом, активирует иммунный ответ у пациента, нуждающегося в этом, тем самым придавая пациенту защитный противоопухолевый иммунитет.
Настоящее изобретение относится, в частности, к моноклональному антителу против VSIG4 или его антигенсвязывающему фрагменту, содержащему три CDR тяжелой цепи и три CDR легкой цепи, где последовательности CDR выбраны из группы последовательностей, изложенных в SEQ ID NO: 3-58. Более конкретно, раскрытое в настоящем документе антитело содержит три CDR тяжелой цепи и три CDR легкой цепи, как изложено в таблице 2.
Настоящее изобретение также относится к моноклональному антителу против VSIG4 или его антигенсвязывающему фрагменту, содержащему любую одну вариабельную область тяжелой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 129, 131, 133, 135, 137, 139, 141, 143, 145, 147 и 149; и любую одну вариабельную область легкой цепи, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO: 130, 132, 134, 136, 138, 140, 142, 144, 146, 148 и 150, и антигенсвязывающему фрагменту моноклонального антитела.
Кроме того, настоящее изобретение дополнительно относится к полинуклеотиду, кодирующему вариабельную область тяжелой цепи и вариабельную область легкой цепи моноклонального антитела или его антигенсвязывающего фрагмента.
Кроме того, настоящее изобретение дополнительно относится к экспрессионному вектору, содержащему полинуклеотид.
Кроме того, настоящее изобретение дополнительно относится к трансформанту, трансформированному экспрессионным вектором.
Кроме того, настоящее изобретение дополнительно относится к способу получения моноклонального антитела, специфически связывающегося с VSIG4, или его антигенсвязывающего фрагмента, путем культивирования указанного трансформанта.
Кроме того, настоящее изобретение относится к композиции для стимуляции иммунного ответа, содержащей в качестве эффективного ингредиента моноклональное антитело, специфически связывающееся с VSIG4, или его антигенсвязывающий фрагмент.
Кроме того, настоящее изобретение дополнительно относится к фармацевтической композиции для лечения рака, содержащей в качестве эффективного ингредиента моноклональное антитело, специфически связывающееся с VSIG4, или его антигенсвязывающий фрагмент.
Кроме того, настоящее изобретение дополнительно относится к способу лечения рака, включающему введение индивидууму фармацевтической композиции для лечения рака.
Кроме того, настоящее изобретение дополнительно относится к конъюгату антитело-лекарственное средство, содержащему лекарственное средство, связанное с моноклональным антителом, специфически связывающимся с VSIG4, или его антигенсвязывающий фрагмент.
Кроме того, настоящее изобретение дополнительно относится к белку CAR (химерный антигенный рецептор), включающему i) вышеуказанные антитела; ii) трансмембранный домен и iii) CAR (химерный антигенный рецептор) с доменом внутриклеточной сигнализации, характеризующийся тем, что он вызывает активацию Т-клеток в соответствии со связыванием указанного выше i) антитела с антигеном.
Кроме того, настоящее изобретение дополнительно относится к мультиспецифическому антителу, содержащему моноклональное антитело, специфически связывающееся с VSIG4 или его антигенсвязывающим фрагментом.
ПОЛЬЗА ИЗОБРЕТЕНИЯ
Поскольку новое антитело по настоящему изобретению связывается с VSIG4, и его антигенсвязывающий фрагмент может связываться с VSIG4 для ингибирования активности VSIG4, ожидается, что они могут быть преимущественно использованы для разработки различных иммунотерапевтических средств для лечения расстройства, связанного с VSIG4.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На фиг. 1 показаны структура и экспрессия hVSIG4(S) и hVSIG4(L). (А) диаграмма, иллюстрирующая структуру двух форм белка VSIG4 (по материалам Small et al., Swiss Med Wkly. (2016) 146:w14301). (В) Результат вестерн-блоттинга для тестирования экспрессии hVSIG4(L) и hVSIG4(S) в макрофагах: rechVSIG4: рекомбинантный hVSIG4 (длинный и короткий); 264, 265 и 266: доноры, у которых были выделены PBMS. AF4646: поликлональное антитело против VSIG4 (R&D Systems, Миннеаполис, Миннесота, США).
На фиг. 2 показано, что активация CD4+Т-клеток ингибируется hVSIG4(S) и hVSIG4(L). CD4+ клетки приводили в контакт с антителом против CD3 ОКТ3 (BioxCell ref ВЕ0001-2, клон ОКТ3) в присутствии рекомбинантных белков (hVSIG4(L)-Fc, hVGIG4(S)-Fc, PDL1-Fc (R&D Systems 156-B7) или изотипического контроля hIgG1 (c9G4)). Пролиферацию CD4+ Т-клеток (А) и высвобождение IFNγ (В) определяли с помощью проточной цитометрии.
Фиг. 3 представляет собой диаграмму, иллюстрирующую раскрытый в настоящем описании способ скрининга моноклонального антитела, специфически связывающегося с VSIG4.
Фиг. 4 представляет собой диаграмму, иллюстрирующую вектор экспрессии белка-антигена VSIG4.
На фиг. 5 показан результат ДНС-ПААГ-электрофореза очищенного белка-антигена VSIG4.
На фиг. 6 показан результат проведения ИФА полифагов для тестирования специфичности положительного пула поли scFv-фаговых антител, который был получен посредством пэннинга каждого раунда (т.е. первого, второго и третьего раунда), в отношении антигена.
На фиг. 7 показан результат проведения ИФА для отбора положительных фагов с превосходными связующими свойствами в отношении антигена VSIG4.
На фиг. 8 показан результат проведения ДНС-ПААГ анализа 11 одиночных человеческих антител к рекомбинантному VSIG4.
На фиг. 9 показаны результаты проведения FACS (анализ сортировки клеток с активированной флуоресценцией) анализа трансформированных клеток, сверхэкспрессирующих человеческий VSIG4, с использованием антитела против VSIG4 человека, связанного с флуоресцентным материалом аллофикоцианин (АРС).
На фиг. 10 показаны результаты FACS-анализа специфичности связывания для клеток, сверхэкспрессирующих человеческий VSIG4, с 11 антителами против VSIG4 человека. (А) HEK293E: тест на неспецифическое связывание. (В) hVSIG4/HEK293E: специфическое связывание CVSIG4 на клеточной поверхности.
На фиг. 11 показано связывание 11 человеческих моноклональных антител против VSIG4 с hVSIG4(S) и hVSIG4(L). (А) Связывание с hVSIG4(S) и hVSIG4(L) анализировали с помощью ИФА с использованием исходных scFv версий 11 человеческих антител против VSIG4. (В) Связывание с hVSIG4(S) и hVSIG4(L) анализировали вестерн-блоттингом с использованием 11 полноразмерных человеческих антител против VSIG4. NRH: нередуцированная форма, нагреваемая; RH: редуцированная форма, нагреваемая.
На фиг. 12 показано, что мышиный т6Н8 и его гуманизированная версия hz6H8-A2 связываются с hVSIG4(L), но не с hVSIG4(S). (А) Вестерн-блоттинг: rechVSIG4: рекомбинантный hVSIG4 (длинный и короткий); 264, 265 и 266: доноры, у которых были выделены PBMS. AF4646: поликлональное антитело против VSIG4 (R&D Systems, Миннеаполис, Миннесота, США). (В) ИФА с hVSIG4-His (короткая форма) и hVSIG4 Fc (длинная форма): m9G4: изотипический контроль, козий IgG контроль: отрицательный контроль.
На фиг. 13 показан результат проведения ELISA для картирования эпитопов 11 scFv человеческих монокпональных антител против VSIG4 с определенными 8 группами эпитопов. Нумерация групп не связана ни с положением в последовательности, ни с трехмерной структурой антигена.
Фиг. 14 представляет собой диаграмму, иллюстрирующую раскрытый в данном документе способ тестирования 11 полноразмерных человеческих монокпональных антител против VSIG4 в анализе на воспаление.
Фиг. 15 представляет собой диаграмму, иллюстрирующую раскрытый в данном документе способ тестирования 11 полноразмерных человеческих монокпональных антител против VSIG4 в анализе иммуносупрессии.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение станет более полно понятым из подробного описания, приведенного в настоящем документе, и из сопроводительных чертежей, которые приведены только в качестве иллюстрации и не ограничивают представленный объем изобретения.
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимает специалист в области химии, биохимии, клеточной биологии, молекулярной биологии и медицинских наук.
Термин «примерно» или «приблизительно» относится к нормальному диапазону погрешности для данного значения или к диапазону, известному специалисту в данной области. Обычно это означает в пределах 20%, например, в пределах 10% или в пределах 5% (или 1% или менее) заданного значения или диапазона.
В контексте данного документа, «вводить» или «введение» относится к инъекции или иной физической доставке вещества в том виде, в котором оно существует вне тела (например, антитела против VSIG4, предложенного в данном документе) пациенту, например, например, через слизистую оболочку, путем внутрикожной, внутривенной, внутримышечной доставки и/или любого другого способа физической доставки, описанного в данном документе или известного в данной области техники. Когда лечат заболевание или его симптом, введение вещества как правило проводят после наступления заболевания или его симптомов. Когда проводят профилактику заболевания или его симптомов, введение вещества обычно осуществляют до наступления заболевания или его симптомов. Путь введения композиции по настоящему изобретению может представлять собой любой из различных путей, включая пероральный и парентеральный пути, при условии, что они позволяют доставить композицию в ткань-мишень. В частности, введение можно осуществлять общепринятым способом пероральным, колоректальным, местным, внутривенным, внутрибрюшинным, внутримышечным, внутриартериальным, трансдермальным, интраназальным, ингаляционным, внутриглазным или интрадермальным путем.
Термины «антитело» и «иммуноглобулин» или «Ig» используются в контексте данного документа взаимозаменяемо. Эти термины используются в данном документе в самом широком смысле и конкретно охватывают моноклональные антитела (включая полноразмерные моноклональные антитела) любого изотипа, такие как IgG, IgM, IgA, IgD и IgE, поликлональные антитела, мультиспецифические антитела, химерные антитела и фрагменты антител, при условии, что указанные фрагменты сохраняют требуемую биологическую функцию. Эти термины предназначены для включения полипептидного продукта В-клеток в полипептиды класса иммуноглобулинов, способные связываться со специфичным молекулярным антигеном и состоят из двух идентичных пар полипептидных цепей, соединенных между собой дисульфидными связями, где каждая пара имеет одну тяжелую цепь (около 50-70 кДа) и одну легкую цепь (около 25 кДа), и каждая аминоконцевая часть каждой цепи включает вариабельную область из примерно 100 - 130 или более аминокислот, и каждая карбоксиконцевая часть каждой цепи включает константную область (см. Borrebaeck (ed.) (1995) Antibody Engineering, Second Ed., Oxford University Press.; Kuby (1997) Immunology, Third Ed., W.H. Freeman and Company, New York). Каждая вариабельная область каждой тяжелой и легкой цепи состоит из трех областей, определяющих комплементарность (CDR), которые также известны как гипервариабельные области, и четырех каркасных областей (FR), более высококонсервативных частей вариабельных доменов, расположенных от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействуюет с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями-хозяевами или факторами, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента. В некоторых вариантах осуществления конкретный молекулярный антиген, который может связываться с антителом, предложенным в настоящем документе, включает целевой полипептид VSIG4, фрагмент или эпитоп. Антитело, реагирующее с конкретным антигеном, может быть получено с помощью рекомбинантных способов, таких как отбор библиотек рекомбинантных антител в фаговых или аналогичных векторах, или путем иммунизации животного антигеном или нуклеиновой кислотой, кодирующей антиген.
Антитела также включают, но не ограничиваются ими, синтетические антитела, моноклональные антитела, антитела, полученные рекомбинантным способом, мультиспецифические антитела (включая биспецифические антитела), человеческие антитела, гуманизированные антитела, верблюжьи антитела, химерные антитела, интратела, антиидиотипические (анти-Id) антитела и функциональные фрагменты любого из вышеперечисленного, которые относятся к части тяжелой или легкой цепи антитела, которая сохраняет некоторые или все биологические функции антитела, из которого был получен фрагмент. Антитела, предложенные в данном документе, могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), любого класса (например, IgG1, IgG2, IgG3, IgG4 IgA1 и IgA2) или любого подкласса (например, IgG2a и IgG2b) молекулы иммуноглобулина.
Термины «антитела против VSIG4», «антитела, которые связываются с VSIG4», «антитела, которые связываются с эпитопом VSIG4» и аналогичные термины используются взаимозаменяемо в контексте данного документа и означают антитела, которые связываются с полипептидом VSIG4, таким как антиген или эпитоп VSIG4. Такие антитела включают поликлональные и моноклональные антитела, в том числе химерные, гуманизированные и человеческие антитела. Антитело, которое связывается с антигеном VSIG4, может перекрестно реагировать с родственными антигенами. В некоторых вариантах осуществления антитело, которое связывается с VSIG4, не вступает в перекрестную реакцию с другими антигенами, такими как, например, другие пептиды или полипептиды, принадлежащие к суперсемейству В7. Антитело, которое связывается с VSIG4, может быть идентифицировано, например, с помощью иммунологического анализа, BIAcore или других методик, известных специалистам в данной области техники. Антитело связывается с VSIG4, например, когда оно связывается с VSIG4 с более высокой аффинностью, чем с любым кросс-реактивным антигеном, что определяется с использованием экспериментальных методик, таких как радиоиммуноанализы (RIA) и твердофазные иммуноферментные анализы (ИФА), например, антитело, которое специфически связывается с VSIG4. Как правило, специфичная или селективная реакция будет по меньшей мере в два раза больше фонового сигнала или шума и может быть более чем в 10 раз больше фонового сигнала. См., например Paul, ed., 1989, Fundamental Immunology Second Edition, Raven Press, New York на стр. 332-336 для обсуждения специфичности антител. В некоторых вариантах осуществления антитело, «которое связывает» целевой антиген, представляет собой антитело, которое связывает антиген с достаточной аффинностью, такой, что антитело пригодно в качестве диагностического и/или терапевтического агента при нацеливании на клетку или ткань, экспрессирующую антиген, и не вступает в значительной степени в перекрестную реакцию с другими белками. В таких вариантах осуществления степень связывания антитела с «нецелевым» белком будет составлять менее примерно 10% от связывания антитела с его конкретным белком-мишенью, что определяется сортировкой клеток с активированной флуоресценцией (FACS) или радиоиммунопреципитацией (RIPA). Что касается связывания антитела с молекулой-мишенью, то термин «специфическое связывание» или «специфично связывается с» или является «специфичным в отношении» конкретного полипептида или эпитопа у конкретного полипептида-мишени означает связывание, которое измеримо отличается от неспецифесконо взаимодействия. Специфическое связывание может быть измерено, например, посредством определения связывания молекулы по сравнению со связыванием контрольной молекулы, которая, как правило, представляет собой молекулу с аналогичной структурой, не обладающую активностью связывания. Например, специфическое связывание может быть определено по конкурированию с контрольной молекулой, которая схожа с мишенью, например, избытком немеченой мишени. В данном случае индикатором специфичного связывания является конкурентное ингибирование связывания меченой мишени с зондом избытком немеченой мишени. Термин «специфическое связывание» или «специфично связывается с» или является «специфичным в отношении» конкретного полипептида или эпитопа на конкретном полипептиде-мишени, используемый в контексте данного документа, может быть представлен, например, молекулой, имеющей KD в отношении мишени по меньшей мере приблизительно 10-4 М, в альтернативном варианте по меньшей мере примерно 10-5 М, в альтернативном варианте по меньшей мере примерно 10-6 М, в альтернативном варианте по меньшей мере примерно 10-7 М, в альтернативном варианте по меньшей мере примерно 10-8 М, в альтернативном варианте по меньшей мере примерно 10-9 М, в альтернативном варианте по меньшей мере примерно 10--10 М, в альтернативном варианте по меньшей мере примерно 10-11 М, в альтернативном варианте по меньшей мере примерно 10--12 М или более. В некоторых вариантах осуществления термин «специфическое связывание» относится к связыванию, когда молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде без связывания по существу с любым другим полипептидом или эпитопом полипептида. В некоторых вариантах осуществления антитело, которое связывается с VSIG4, имеет константу диссоциации (KD) ≤ 1 мкМ, ≤100 нМ, ≤10 нМ, ≤1 нМ или ≤0,1 нМ.
Используемый в контексте данного документа термин «антиген» относится к заданному антигену, с которым антитело может избирательно связываться. Антиген-мишень может представлять собой полипептид, углевод, нуклеиновую кислоту, липид, гаптен или другое природное или синтетическое соединение. В некоторых вариантах осуществления антиген-мишень представляет собой полипептид, включающий, например, полипептид VSIG4.
Термины «антигенсвязывающий фрагмент», «антигенсвязывающий домен», «антигенсвязывающая область» и аналогичные термины обозначают ту часть антитела, которая содержит аминокислотные остатки, которые взаимодействуют с антигеном и придают связующему агенту его специфичность и аффинность к антигену (например, области, определяющие комплементарность (CDR)). Под выражением «антигенсвязывающий фрагмент» антитела подразумевается указание на любой пептид, полипептид или белок, сохраняющий способность связываться с мишенью (также обычно обозначаемой антигеном) указанного антитела, как правило, с тем же эпитопом, и содержащий аминокислотную последовательность из по меньшей мере из 5 последовательных аминокислотных остатков, по меньшей мере 10 последовательных аминокислотных остатков, по меньшей мере 15 последовательных аминокислотных остатков, по меньшей мере 20 последовательных аминокислотных остатков, по меньшей мере 25 последовательных аминокислотных остатков, по меньшей мере 40 последовательных аминокислотных остатков, по меньшей мере 50 последовательных аминокислотных остатков, по меньшей мере 60 последовательных аминокислотных остатков, по меньшей мере 70 последовательных аминокислотных остатков, по меньшей мере 80 последовательных аминокислотных остатков, по меньшей мере 90 последовательных аминокислотных остатков, по меньшей мере 100 последовательных аминокислотных остатков, по меньшей мере 125 последовательных аминокислотных остатков, по меньшей мере 150 последовательных аминокислотных остатков, по меньшей мере 175 последовательных аминокислотных остатков или по меньшей мере 200 последовательных аминокислотных остатков аминокислотной последовательности антитела. В конкретном варианте осуществления указанный антигенсвязывающий фрагмент содержит по меньшей мере одну CDR антитела, из которого он получен. При этом в предпочтительном варианте осуществления указанный антигенсвязывающий фрагмент содержит 2, 3, 4 или 5 CDR, более предпочтительно 6 CDR антитела, из которого он получен.
«Антигенсвязывающие фрагменты» могут быть выбраны, без ограничений, в группе, состоящей из фрагментов Fab, Fab', (Fab')2, Fv, scFv (sc для одной цепи), бис-scFv, scFv-Fc, Fab2, Fab3, минител, диател, триател, тетрател и нанотел, а также слитых белков с неупорядоченными пептидами, такими как XTEN (удлиненный рекомбинантный полипептид) или мотивы PAS, и любого фрагмента, время полужизни которого может быть увеличено путем химической модификации, такой как добавление поли(алкилен)гликоля, такого как поли(этилен)гликоль («ПЭГилирование») (пэгилированные фрагменты, называемые Fv-PEG, scFv-PEG, Fab-PEG, F(ab')2-PEG или Fab'-PEG) («PEG» для поли(этилен)гликоля), или путем включения в липосому, причем указанные фрагменты имеют по меньшей мере один из типичных CDR антитела по изобретению. Среди фрагментов антител Fab имеет структуру, включающую вариабельные области легкой цепи и тяжелой цепи, константную область легкой цепи и первую константную область тяжелой цепи (СН1), и имеет один антигенсвязывающий сайт. Fab' отличается от Fab тем, что он имеет шарнирную область, включающую один или несколько цистеиновых остатков на С- конце домена СН1 тяжелой цепи. F(ab')2 антитело образуется, когда цистеиновые остатки шарнирной области Fab' образуют дисульфидную связь. Fv представляет собой минимальный фрагмент антитела, который имеет только вариабельную область тяжелой цепи и вариабельную область легкой цепи, а рекомбинантный способ получения Fv фрагмента описан в международной публикации WO 88/10649 или тому подобном. В двухцепочечном Fv (dsFv) вариабельная область тяжелой цепи и вариабельная область легкой цепи связаны друг с другом через дисульфидную связь, а в одноцепочечном Fv (scFv) вариабельная область тяжелой цепи и вариабельная область легкой цепи ковалентно связаны друг с другом как правило через пептидный линкер. Данные фрагменты антител могут быть получены с использованием протеиназы (например, Fab может быть получен путем рестрикционного расщепления целого антитела папаином, а фрагмент F(ab')2 может быть получен путем рестрикционного расщепления пепсином), и предпочтительно могут быть получены методами генной инженерии. Предпочтительно, указанные «антигенсвязывающие фрагменты» будут представлять собой или будут содержать частичную последовательность вариабельной области тяжелой или легкой цепи антитела, из которого они получены, причем указанная частичная последовательность будет достаточной для сохранения той же специфичности связывания, что и специфичность антитела, из которого они получены, и достаточной аффинности, предпочтительно, по меньшей мере, равной 1/100, более предпочтительно, по меньшей мере 1/10 аффинности антитела, из которого они получены, относительно мишени. Такие фрагменты антител описаны, например, в Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York (1989); Myers (ed.), Molec. Biology and Biotechnology: A Comprehensive Desk Reference, New York: VCH Publisher, Inc.; Huston et al., Cell Biophysics, 22:189-224 (1993); Plückthun and Skerra, Meth. Enzymol., 178:497-515 (1989) and in Day, E.D., Advanced Immunochemistry, Second Ed., Wiley-Liss, Inc., New York, NY (1990).
Термины «связывает» или «связывание», используемые в контексте данного документа, обозначают взаимодействие между молекулами с образованием комплекса, который в физиологических условиях является относительно стабильным. Взаимодействия могут быть, например, нековалентными взаимодействиями, включая водородные связи, ионные связи, гидрофобные взаимодействия и/или Ван-дер-Ваальсовы взаимодействия. Комплекс может также включать связывание двух или более молекул, удерживаемых вместе ковалентными или нековалентными связями, взаимодействиями или силами. Сила всех нековалентных взаимодействий между одним антигенсвязывающим сайтом на антителе и одним эпитопом молекулы-мишени, такой как VSIG4, представляет собой аффинность антитела или функционального фрагмента к этому эпитопу. Отношение ассоциации (k1) к диссоциации (k-1) антитела к моновалентному антигену (k1/k-1) представляет собой константу ассоциации K, которая является мерой аффинности. Значение K варьируется для разных комплексов антитело-антиген и зависит как от k1 так и от k-1. Константа ассоциации K для антитела, предложенного в настоящем документе, может быть определена с использованием любого способа, предложенного в настоящем документе, или любого другого способа, хорошо известного специалистам в данной области техники. Аффинность на одном сайте связывания не всегда отражает истинную силу взаимодействия между антителом и антигеном. Когда комплексные антигены, содержащие множественные, повторяющиеся антигенные детерминанты, такие как поливалентный VSIG4, вступают в контакт с антителами, содержащими множество сайтов связывания, взаимодействие антитела с антигеном на одном сайте повысит вероятность реакции на втором сайте. Сила таких множественных взаимодействий между поливалентным антителом и антигеном называется авидностью. Авидность антитела может быть лучшей мерой его связывающей способности, чем аффинность его отдельных сайтов связывания. Например, высокая авидность может компенсировать низкую аффинность, как это иногда обнаруживается для пентамерных антител IgM, которые могут иметь более низкую аффинность, чем IgG, но высокая авидность IgM, возникающая в результате его поливалентности, позволяет ему эффективно связывать антиген. Способы определения того, происходит ли связывание двух молекул, хорошо известны в данной области техники и включают, например, равновесный диализ, поверхностный плазмонный резонанс и тому подобное. В конкретном варианте осуществления указанное антитело или его антигенсвязывающий фрагмент связывается с VSIG4 с аффинностью, которая по меньшей мере в два раза превышает его аффинность при связывании с неспецифичной молекулой, такой как BSA или казеин. В более конкретном варианте осуществления указанное антитело или его антигенсвязывающий фрагмент связывается только с VSIG4.
Используемый в контексте данного документа термин «биологический образец» или «образец» означает образец, который был получен из биологического источника, такого как пациент или субъект. «Биологический образец», используемый в контексте денного документа, относится, в частности, к целому организму или подмножеству его тканей, клеток или составных частей (например, кровеносный сосуд, включая артерии, вены и капилляры, жидкости организма, включая, но не ограничиваясь ими, кровь, сыворотку, слизь, лимфатическую жидкость, синовиальную жидкость, спинномозговую жидкость, слюну, околоплодную жидкость, околоплодными пуповинную кровь, мочу, вагинальную жидкость и сперму). «Биологический образец» также относится к гомогенату, лизату или экстракту, полученному из целого организма или подмножества его тканей, клеток или составных частей, или их фракции либо части. Наконец, «биологический образец» относится к среде, такой как питательный бульон или гель, в которой размножился микроорганизм, которая содержит клеточные компоненты, такие как белки или молекулы нуклеиновых кислот.
В контексте данного документа термин «биопэннинг» означает процесс отбора из библиотеки фагов, экспонирующих пептид на фаговой оболочке, только тех фагов, которые экспонируют на поверхности пептид, обладающий свойством связываться с молекулой-мишенью (например, антитело, фермент и рецептор клеточной поверхности). В одном варианте осуществления биопэннинг, используемый в настоящем документе, включает четыре стадии, где первая стадия представляет собой стадию получения библиотеки фагов, вторая стадия представляет собой стадию захвата, включающую приведение в контакт фаговой библиотеки с молекулой-мишенью, третья стадия - стадия промывки, включающая удаление фагов, которые не связываются с молекулой-мишенью, и четвертая стадия - элюирование, посредством которой извлекаются целевые фаги. Пример биопэннинга представлен в примерах настоящего изобретения.
Термин «блок» или его грамматический эквивалент при использовании в контексте антитела относится к антителу, которое предотвращает или останавливает биологическую активность антигена, с которым связывается данное антитело. Блокирующее антитело включает антитело, которое объединяется с антигеном, не вызывая реакции, но которое блокирует другой белок от последующего объединения или образования комплекса с данным антигеном. Блокирующий эффект антитела может быть таким, который приводит к измеримому изменению биологической активности антигена.
Термины «клеточное пролиферативное расстройство» и «пролиферативное расстройство» относятся к расстройствам, которые ассоциированы с некоторой степенью аномальной пролиферации клеток. В некоторых вариантах осуществления клеточное пролиферативное расстройство представляет собой опухоль или рак. Термин «опухоль», используемый в настоящем документе, относится к росту и пролиферации всех неопластических клеток, злокачественных или доброкачественных, а также ко всем предраковым и раковым клеткам и тканям. Термины «рак», «злокачественный», «клеточное пролиферативное расстройство», «пролиферативное расстройство» и «опухоль» не являются взаимоисключающими в контексте настоящего документа. Термины «рак» и «раковый» относятся или описывают физиологическое состояние у млекопитающих, которое как правило характеризуется нерегулируемым ростом клеток.
Используемый в контексте данного документа термин «рак» представляет собой любое злокачественное новообразование, возникающее в результате нежелательного роста, инвазии и, при определенных условиях, метастазирования пораженных клеток в организме. Клетки, вызывающие рак, имеют генетические нарушения и обычно теряют свою способность контролировать клеточное деление, миграцию клеток, статус дифференцировки и/или механизм гибели клеток. При большинстве видов рака образуется опухоль, однако при некоторых видах рака кроветворной системы, таких как лейкемия, этого не происходит. Таким образом, термин «рак», используемый в контексте данного документа, может включать как доброкачественные, так и злокачественные виды рака. Термин «рак», используемый в данном документе, относится, в частности, к любому раку, который можно лечить человеческим антителом по настоящему изобретению без каких-либо ограничений. Примеры рака включают рак печени, рак молочной железы, рак почки, опухоль головного мозга, рак желчных протоков, рак пищевода, рак желудка, рак толстой кишки, колоректальный рак, рак носоглотки, рак гортани, рак легкого, рак восходящего отдела толстой кишки, рак шейки матки, рак щитовидной железы, лейкемию, болезнь Ходжкина, лимфому и множественную миелому, рак крови, но не ограничиваются ими.
«Химиотерапевтический агент» представляет собой химический или биологический агент (например, агент, включающий низкомолекулярное лекарственное средство или биологический препарат, такой как антитело или клетка), пригодный для лечения рака, независимо от механизма действия. Химиотерапевтические агенты включают соединения, используемые в таргетной терапии и обычной химиотерапии. Химиотерапевтические агенты включают, но не ограничиваются ими, алкилирующие агенты, антиметаболиты, противоопухолевые антибиотики, митотические ингибиторы, ингибиторы функции хроматина, противоангиогенезные агенты, антиэстрогены, антиандрогены или иммуномодуляторы.
Термин «химерное» антитело означает антитело, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида. В одном из вариантов осуществления «химерное антитело» представляет собой антитело, в котором константная область или ее часть изменена, замещена или заменена так, что вариабельная область связана с константной областью другого вида или принадлежит к другому классу или подклассу антител. В другом варианте осуществления «химерное антитело» относится к антителу, в котором вариабельная область или ее часть изменена, замещена или заменена так, что константная область связана с вариабельной областью другого вида или принадлежит к другому классу или подклассу антител.
Используемый в контексте данного документа термин «CDR» означает одну из трех гипервариабельных областей (Н1, Н2 или Н3) в некаркасной области VH β-листовой структуры иммуноглобулина (Ig или антитела) или одну из трех гипервариабельных областей (L1, L2 или L3) в некаркасной области VL β-листовой структуры антитела. Соответственно, CDR представляют собой последовательности вариабельной области, вставленные между последовательностями каркасной области. Области CDR хорошо известны специалистам в данной области техники и были определены, например, Kabat как области наибольшей гипервариантности в вариабельных доменах (V) антител (Kabat et al., J. Biol. Chem. 252:6609-6616 (1977); Kabat, Adv. Prot. Chem. 32:1-75 (1978). CDR по Kabat основаны на вариабельности последовательностей и являются наиболее часто используемыми (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Вместо этого Chothia обращается к расположению структурных петель (Chothia и Lesk J Mol. Bioi. 196:901-917 (1987)). Последовательности областей CDR также были структурно определены Chothia как те остатки, которые не являются частью консервативной структуры β-листа и, таким образом, способны принимать различные конформации (Chothia and Lesk, J. Mol. Bioi. 196:901-917 (1987)). Конец петли CDR-H1 по Chothia при нумерации с использованием нумерации Kabat варьируется между Н32 и Н34 в зависимости от длины петли (это связано с тем, что схема нумерации Kabat помещает вставки в Н35А и Н35В; если нет ни 35А, ни 35В, петля заканчивается на 32; если присутствует только 35А, петля заканчивается на 33; если присутствуют и 35А, и 35В, петля заканчивается на 34). Обе терминологии хорошо известны в данной области техники. Последовательности областей CDR также были охарактеризованы AbM, Contact и IMGT. Гипервариабельные области по AbM представляют собой компромисс между CDR по Kabat и структурными петлями по Chothia и используются в программном обеспечении AbM от Oxford Molecular для моделирования антител. «Контактные» гипервариабельные области основаны на анализе имеющихся сложных кристаллических структур. Недавно была разработана и получила широкое распространение универсальная система нумерации ImMunoGeneTics (IMGT) (Lafranc et al., Дев. Комп. Иммунол. 27(1):55-77 (2003)). Универсальная нумерация IMGT была описана для сравнения вариабельных доменов независимо от рецептора антигена, типа цепи или вида. [Lefranc М.-Р., Immunology Today 18, 509 (1997) / Lefranc М.-Р., The Immunologist, 7, 132-136 (1999)]. В универсальной нумерации IMGT консервативные аминокислоты всегда имеют одно и то же положение, например, цистеин 23 (1-й CYS), триптофан 41 (консервативный TRP), гидрофобная аминокислота 89, цистеин 104 (2-й CYS), фенилаланин или триптофан 118 (J-PHE или J-TRP). Универсальная нумерация IMGT обеспечивает стандартизированное разграничение каркасных областей (FR1-IMGT: положения 1-26, FR2-IMGT: 39-55, FR3-IMGT: 66-104 и FR4-IMGT: 118-128) и областей, определяющих комплементарность: CDR1-IMGT: 27-38, CDR2-IMGT: 56-65 и CDR3-IMGT: 105-117. Поскольку гэпы представляют собой незанятые положения, длины CDR-IMGT (показаны в скобках и разделены точками, например, [8.8.13]) становятся важной информацией. Универсальная нумерация IMGT используется в 2D графических представлениях, обозначенных как IMGT Colliers de Perles [Ruiz, M. and Lefranc, M.-P., Immunogenetics, 53, 857-883 (2002) / Kaas, Q. and Lefranc, M.-P., Current Bioinformatics, 2, 21-30 (2007)], и в 3D-структурах в IMGT/структура-DB [Kaas, Q., Ruiz, M. and Lefranc, M.-P., T cell receptor and MHC structural data. Nucl. Acids. Res., 32, D208-D210 (2004)]. Положения CDR в вариабельном домене эталонного антитела были определены путем сравнения многочисленных структур (Al-Lazikani et al., J. Mol. Biol. 273:927-948 (1997); Morea et al., Methods 20:267-279 (2000)). Поскольку количество остатков в гипервариабельной области варьирует в разных антителах, дополнительные остатки относительно канонических положений обычно нумеруются буквами а, b, с и т.д. рядом с номером остатка в схеме нумерации канонических вариабельных доменов. (Al-Lazikani et al., см выше (1997)). Такая номенклатура также хорошо известна специалистам в данной области техники.
Гипервариабельные области могут содержать «удлиненные гипервариабельные области»: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 или 89-96 (L3) в VL и 26-35 или 26-35А (Н1), 50-65 или 49-65 (Н2) и 93-102, 94-102 или 95-102 (Н3) в VH. Остатки вариабельного домена имеют номер 25 в соответствии с Kabat et al., supra, для каждого из этих определений. Используемые в настоящем документе термины «HVR» и «CDR» используются взаимозаменяемо.
Используемый в настоящем документе термин «ингибитор контрольной точки» относится к молекуле, такой как, например, малая молекула, растворимый рецептор или антитело, которое нацелено на иммунную контрольную точку и блокирует функцию указанной иммунной контрольной точки. Более конкретно, «ингибитор контрольной точки», используемый в контексте данного документа, представляет собой молекулу, такую как, например, малая молекула, растворимый рецептор или антитело, которое способно ингибировать или иным образом снижать одну или более биологических активностей иммунной контрольной точки. В некоторых вариантах осуществления ингибитор белка-иммунной контрольной точки (например, антагонистическое антитело, предложенное в настоящем документе) может, например, действовать путем ингибирования или иного уменьшения путей активации и/или клеточной сигнализации клетки, экспрессирующей указанный белок-иммунную контрольную точку (например, Т-клетки), тем самым ингибируя биологическую активность указанной клетки в сравнении с биологической активностью в отсутствие антагониста. Примеры ингибиторов иммунных контрольных точек включают низкомолекулярные лекарственные средства, растворимые рецепторы и антитела.
Термин «константная область» или «константный домен» относится к карбоксиконцевому участку легкой и тяжелой цепи, который непосредственно не участвует в связывании антитела с антигеном, но проявляет различные эффекторные функции, такие как взаимодействие с Fc рецептором. Данные термины означают часть молекулы иммуноглобулина, имеющую более консервативную аминокислотную последовательность по сравнению с другой частью иммуноглобулина, вариабельным доменом, который содержит антигенсвязывающий сайт. Константный домен содержит домены СН1, СН2 и СН3 тяжелой цепи и домен CL легкой цепи.
Используемый в контексте данного документа термин «цитотоксический агент» означает агент, который при введении субъекту лечит или предотвращает развитие клеточной пролиферации, предпочтительно развитие рака в организме субъекта, путем ингибирования или предотвращения клеточной функции и/или вызывая гибель клеток. Цитотоксический агент, который может быть использован в настоящем конъюгате антитело-лекарственное средство, включает любой агент, его часть или остаток, обладающий цитотоксическим эффектом или ингибирующим действием на клеточную пролиферацию. Примеры таких агентов включают (i) химиотерапевтический агент, способный функционировать в качестве ингибитора микротубуллина, митотического ингибитора, ингибитора топоизомеразы или интеркалятора ДНК; (ii) белковый токсин, способный функционировать ферментативным путем; и iii) радиоизотопы (радионуклиды). Цитотоксический агент может быть конъюгирован с антителом, таким как, например, антитело против VSIG4, с образованием иммуноконъюгата. Предпочтительно, цитотоксический агент высвобождается из антитела в специфических условиях, например, в кислых условиях, тем самым терапевтически воздействуя на клетки-мишени, например, предотвращая их пролиферацию или проявляя цитотоксический эффект.
Термин «сниженный», используемый в настоящем описании, относится к уровню биомаркера, например, VSIG4, у субъекта, по меньшей мере, в 1 раз (например, в 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 1000, 10000 раз или более) ниже его эталонного значения. Термин «сниженный», поскольку он относится к уровню биомаркера, например, VSIG4, у субъекта, означает также сниженный по меньшей мере на 5% (например, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 95%), 99%) или 100%) по сравнению с уровнем в эталонном образце или относительно эталонного значения для указанного маркера.
Термин «обнаружение», используемый в настоящем документе, охватывает количественное или качественное обнаружение.
Термин «обнаруживаемый зонд» или «обнаруживаемый агент», используемый в контексте данного документа, относится к композиции, которая дает обнаруживаемый сигнал. Термин относится к веществу, которое может быть использовано для установления существования или присутствия требуемой молекулы, такой как антитело, предложенное в настоящем документе, в образце или субъекте. Обнаруживаемый агент может представлять собой вещество, которое можно визуализировать, или вещество, которое можно определить и/или измерить иным образом (например, путем количественного определения). Данный термин включает, без ограничения, любой флуорофор, хромофор, радиоактивную метку, фермент, антитело или фрагмент антитела и т.п., которые обеспечивают детектируемый сигнал посредством своей активности.
Используемый в контексте данного документа термин «диагноз» или «идентификация субъекта, имеющего» относится к процессу идентификации заболевания, состояния или травмы по его признакам и симптомам. Диагноз - это, в частности, процесс определения того, страдает ли человек заболеванием или недугом (например, раком). Рак диагностируется, например, путем обнаружения наличия маркера, связанного с раком, такого как, например, VSIG4.
Термин «кодировать» или его грамматические эквиваленты, используемый в отношении молекулы нуклеиновой кислоты, означает молекулу нуклеиновой кислоты в ее нативном состоянии или в измененном способами, хорошо известными специалистам в данной области техники, которая может быть транскрибирована для получения мРНК, которая затем транслируется в полипептид и/или его фрагмент. Антисмысловая цепь является комплементом такой молекулы нуклеиновой кислоты, и из нее можно вывести кодирующую последовательность.
«Эффективное количество» или «терапевтически эффективное количество» агента, например, фармацевтической композиции, относится к количеству, эффективному в дозировках и в течение периодов времени, необходимых для получения требуемого биологического ответа у субъекта. Такой ответ включает облегчение симптомов заболевания или расстройства, подлежащего лечению, предотвращение, ингибирование или отсрочку рецидива симптома заболевания или самого заболевания, увеличение продолжительности жизни субъекта по сравнению с продолжительностью жизни при отсутствии лечения, или предотвращение, ингибирование или отсрочку прогрессирования симптома заболевания или самого заболевания. «Эффективное количество» представляет собой, в частности, количество агента, эффективное для достижения требуемого терапевтического или профилактического результата. Более конкретно, «эффективное количество», используемое в контексте данного документа, представляет собой количество агента, которое обеспечивает терапевтический эффект. Терапевтически эффективное количество также представляет собой такое количество, при котором терапевтически полезные эффекты превосходят любые токсические или вредные эффекты агента.
Эффективное количество может быть введено за одно или более введений, нанесений или дозировок. Такая доставка зависит от ряда переменных, включая период времени, в течение которого должна использоваться отдельная единица дозирования, биодоступность агента, способ введения и т.д. В некоторых вариантах осуществления эффективное количество также означает количество антитела (например, антитела против VSIG4), предложенного в настоящем документе, для достижения конкретного результата (например, ингибирования биологической активности иммунной контрольной точки, например модулирование Т-клеточной активации). В некоторых вариантах осуществления данный термин означает количество терапии (например, ингибитора иммунной контрольной точки, такого как, например, анти-VSIG4 антитело), которое является достаточным для уменьшения и/или ослабления тяжести и/или продолжительности данного заболевания, расстройства или состояния и/или связанного с ним симптома. Данный термин также охватывает количество, необходимое для снижения или ослабления развития или прогрессирования данного заболевания, расстройства или состояния, снижения или ослабления рецидива, развития или начала данного заболевания, расстройства или состояния и/или для улучшения или усиления профилактического или терапевтического эффекта (эффектов) другой терапии (например, терапия, отличная от указанного ингибитора иммунной контрольной точки). В контексте противораковой терапии терапевтический эффект означает, например, любое улучшение состояния при раке, в том числе любое из них или их комбинацию следующего: остановка или замедление прогрессирования рака (например, от одной стадии рака к другой), остановку или отсрочку обострения или ухудшения симптомов или признаков рака, уменьшение тяжести рака, индуцирование ремиссии рака, ингибирование пролиферации опухолевых клеток, размера опухоли или количества опухолей или снижение уровней биомаркеров, указывающих на рак. В некоторых вариантах осуществления эффективное количество антитела, предложенного в данном документе, составляет от примерно 0,1 мг/кг (мг антитела на кг массы субъекта) до примерно 100 мг/кг. В некоторых вариантах осуществления эффективное количество антитела, предложенного в данном документе, составляет примерно 0,1 мг/кг, примерно 0,5 мг/кг, примерно 1 мг/кг, 3 мг/кг, 5 мг/кг, примерно 10 мг/кг, примерно 15 мг/кг, примерно 20 мг/кг, примерно 25 мг/кг, примерно 30 мг/кг, примерно 35 мг/кг, примерно 40 мг/кг, примерно 45 мг/кг, примерно 50 мг/кг, примерно 60 мг/кг, примерно 70 мг/кг, примерно 80 мг/кг примерно 90 мг/кг или примерно 100 мг/кг (или диапазон, представленный здесь).
Термин «эпитоп», используемый в контексте данного документа, относится к области антигена, такой как полипептид VSIG4 или фрагмент полипептида VSIG4, с которым связывается антитело. Предпочтительно эпитоп, используемый в контексте данного документа, представляет собой локализованную область на поверхности антигена, такого как полипептид VSIG4 или фрагмент полипептида VSIG4, способную связываться с одной или более антигенсвязывающими областями антитела и который обладает антигенной или иммуногенной активностью у животного, такого как млекопитающее (например, человек), которая способна вызвать иммунный ответ.Эпитоп, обладающий иммуногенной активностью, представляет собой часть полипептида, которая вызывает иммунный ответ у животного. Эпитоп, обладающий антигенной активностью, представляет собой часть полипептида, с которой связывается антитело, что определяется любым способом, хорошо известным в данной области техники, например, иммуноанализом. Антигенные эпитопы необязательно должны быть иммуногенными. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как аминокислоты, и имеют специфические трехмерные структурные характеристики, а также специфические характеристики заряда. В некоторых вариантах осуществления эпитопы могут включать детерминанты, которые представляют собой химически активные поверхностные группировки молекул, такие как боковые цепи сахаров, фосфорильные группы или сульфониловые группы, и, в некоторых вариантах осуществления, могут иметь специфические трехмерные структурные характеристики и/или специфические характеристики заряда. Эпитоп может образовываться из смежных остатков или из несмежных остатков, сближенных в результате фолдинга антигенного белка. Эпитопы, образованные смежными аминокислотами, обычно сохраняются при воздействии денатурирующих растворителей, тогда как эпитопы, образованные несмежными аминокислотами, обычно разрушаются при данном воздействии. Как правило, антиген имеет несколько или много различных эпитопов и вступает в реакцию со многими различными антителами. Определение эпитопа, связанного антителом, может быть выполнено с помощью любой технологии эпитопного картирования, известной специалисту в данной области техники.
Термины «полноразмерное антитело», «интактное антитело» или «целое антитело» используются взаимозаменяемо для обозначения антитела в его по существу интактной форме, в отличие от фрагмента антитела. В частности, «полноразмерные антитела», используемые в контексте данного документа, включают антитела с тяжелыми и легкими цепями, включая Fc-область. Константные домены могут представлять собой константные домены с нативной последовательностью (например, константные домены с нативной последовательностью человека) или их аминокилотные варианты. В некоторых случаях интактное антитело может иметь одну или несколько эффекторных функций.
Термин «гликозилирование», описанный в контексте данного документа, означает способ процессинга для доставки гликозильной группы к белку. Гликозилирование осуществляется путем связывания гликозильной группы с остатком серина, треонина, аспарагина или гидроксилизина белка-мишени, опосредованного гликозилтрансферазой. Гликозилированный белок может не только использоваться в качестве структурного материала живой ткани, но и играет важную роль в распознавании клетками на клеточной поверхности. Таким образом, согласно настоящему изобретению, путем изменения гликозилирования или модели гликозилирования моноклонального антитела по настоящему изобретению или его антигенсвязывающего фрагмента можно достичь улучшенного эффекта антитела.
Термин «тяжелая цепь» при использовании в отношении антитела относится к полипептидной цепи, составляющей примерно 50-70 кДа, при этом аминоконцевая часть включает вариабельную область из примерно 120-130 или более аминокислот, а карбоксиконцевая часть включает константную область. Константная область может представлять собой один из пяти разных типов, обозначаемых как альфа (α), дельта (δ), эпсилон (ε), гамма (γ) и мю (μ), в зависимости от аминокислотной последовательности константной области тяжелой цепи. Разные тяжелые цепи различаются по размеру: α, δ и γ содержат примерно 450 аминокислот, в то время как μ и ε содержат примерно 550 аминокислот. В сочетании с легкой цепью данные разные типы тяжелых цепей образуют пять хорошо известных классов антител, IgA, IgD, IgE, IgG и IgM, соответственно, включая четыре подкласса IgG, а именно IgG1, IgG2, IgG3 и IgG4. Тяжелая цепь может представлять собой тяжелую цепь человека.
Термины «клетка-хозяин», «линия клетки-хозяина» и «культура клетки-хозяина» используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформанты» и «трансформированные клетки», которые включают первичную трансформированную клетку и полученное из нее потомство независимо от количества пассажей. Потомство может быть не полностью идентичным по содержанию нуклеиновых кислот родительской клетке, и может содержать мутации. Мутантное потомство, обладающее той же функцией или биологической активностью, что и подвергаемая скринингу или выбранная в исходно трансформированной клетке, включено в настоящий документ.
«Человеческое антитело» представляет собой антитело, которое обладает аминокислотной последовательностью, соответствующей последовательности антитела, продуцируемого человеком, и/или полученное с использованием любого из способов получения человеческих антител, раскрытых в настоящем документе. Данное определение человеческого антитела, в частности, исключает гуманизированное антитело, содержащее нечеловеческие антигенсвязывающие остатки. Человеческие антитела могут быть получены с использованием различных способов, известных в данной области техники, включая библиотеки фаговых дисплеев, как раскрыто в настоящем описании. Hoogenboom and Winter, J, Mol, Biol., 227:381 (1991); Marks et al., J, Mol, Biol., 222:581 (1991). Также для получения человеческих моноклональных антител доступны способы, описанные в Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al., J. Immunol., 147(1):86-95 (1991). См. также van Dijk and van de Winkel, Cur., Opin. Pharmacol., 5: 368-74 (2001). Человеческие антитела также могут быть получены путем введения антигена трансгенному животному, которое было модифицировано для продуцирования таких антител в ответ на антигенный вызов, но чьи эндогенные локусы были отключены, например, иммунизированной ксеномыши (см., например, патенты США №6075181 и 6150584 относительно технологии XENOMOUSE™). См. также, например, Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006) относительно человеческих антител, получаемых с помощью технологии гибридомы В-клеток человека.
«Гуманизированное» антитело относится к химерному антителу, которое содержит минимальную последовательность, полученную из нечеловеческого иммуноглобулина. В одном варианте осуществления гуманизированное антитело представляет собой человеческий иммуноглобулин (реципиентное антитело), в котором остатки от CDR реципиента заменены остатками от CDR вида, не принадлежащего человеку (донорское антитело), такого как мышь, крыса, кролик или нечеловеческий примат, обладающего требуемой специфичностью, аффинностью и/или способностью. В некоторых случаях некоторые остатки сегментов скелета (называемые FR для каркаса) могут быть модифицированы для сохранения аффинности связывания в соответствии со способами, известными специалисту в данной области (Jones et al., Nature, 321:522-525, 1986). В некоторых вариантах осуществления FR-остатки иммуноглобулина человека заменены соответствующими остатками нечеловеческого происхождения. В некоторых вариантах осуществления гуманизированное антитело будет содержать по существу все из меньшей мере одного, и как правило двух вариабельных доментов, в которых все или по существу все CDR соответствуют CDR нечеловеческого антитела, и все или по существу все FR соответствуют FR человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела (Fc), как правило, иммуноглобулина человека. «Гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подверглось гуманизации. Целью гуманизации является снижение иммуногенности ксеногенного антитела, такого как мышиное антитело, для введения в человека при сохранении полной антигенсвязывающей аффинности и специфичности антитела. Для получения более подробной информации см., например, Jones et al, Nature 321: 522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992). См. также, например, Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op. Biotech. 5:428-433 (1994); и патенты США №6982321 и 7087409.
Используемый в данном документе термин «идентификация» в отношении субъекта, имеющего какое-либо состояние, относится к процессу оценки субъекта и определения того, что субъект имеет какое-либо состояние, например, страдает раком.
Используемые в контексте данного документа термины «иммунная контрольная точка» или «белок иммунной контрольной точки» относятся к определенным белкам, продуцируемым некоторыми типами клеток иммунной системы, такими как Т-клетки, и некоторыми раковыми клетками. Такие белки регулируют функцию Т-кпеток в иммунной системе. Примечательно, что они помогают держать иммунные реакции под контролем и могут препятствовать уничтожению раковых клеток Т-клетками. Указанные белки иммунных контрольных точек достигают этого результата, взаимодействуя со специфическими лигандами, которые посылают сигнал в Т-клетку и по существу отключают или ингибируют функцию Т-клеток. Ингибирование этих белков приводит к восстановлению функции Т-клеток и иммунному ответу против раковых клеток. Примеры белков контрольных точек включают, но не ограничиваются ими, CTLA-4, PDL1, PDL2, PD1, В7-Н3, В7-Н4, BTLA, HVEM, TIM3, GAL9, LAG3, VISTA, KIR, 2В4 (принадлежит к семейству молекул CD2 и экспрессируется на всех NK, γδ и CD8+ (αβ) Т-клетках памяти), CD 160 (также называемый BY55), CGEN-15049, СНK1 и CHK2 киназы, IDO1, A2aR и различные лиганды семейства В7.
Термин «повышенный», используемый в настоящем описании, относится к уровню биомаркера, например, VSIG4, субъекта, по меньшей мере, однократно (например, в 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 1000, 10000 раз или более) превышающему его эталонное значение. «Повышенный» в отношении уровня биомаркера, например, VSIG4, субъекта, означает также по меньшей мере на 5% больше (например, 5%, 6%, 7%, 8%, 9%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%), 95%), 99%) или 100%) уровня в эталонном образце или в сравнении с эталонным значением для указанного маркера.
Используемый в контексте данного документа термин «ингибитор» или «антагонист» относится к молекуле, которая способна ингибировать или иным образом снижать одну или более биологических активностей белка-мишени, такого как любой из белков иммунных контрольных точек, описанных выше.
«Выделенное» антитело представляет собой антитело, которое было отделено от компонента его естественной среды. В некоторых вариантах осуществления изобретения антитело очищают до чистоты более 95% или 99%, определяемой, например, с помощью электрофореза (например, ДСН-ПААГ-электрофореза, изоэлектрического фокусирования (ИЭФ), капиллярного электрофореза) или хроматографии (например, ионообменной или обращенно-фазовой ВЭЖХ). Обзор способов оценки чистоты антител см., например, в Flatman et al., J. Chromatogr. В 848:79-87 (2007).
«Выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественной среды. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но данная молекула нуклеиновой кислоты присутствует вне хромосом или в хромосомной локации, которая отличается от ее естественной хромосомной локации
Термин «KD», используемый в контексте данного документа, означает константу диссоциации специфичного взаимодействия антитело-антиген и используется в качестве индикатора для измерения аффинности антитела к антигену. Более низкая KD означает более высокую аффинность антитела к антигену.
Как предполагается в настоящем документе, «уровень» биомаркера, например, VSIG4 заключается в количественном значении биомаркера в образце, например, в образце, взятом у больного раком. В некоторых вариантах осуществления количественное значение состоит не из абсолютного значения, которое измеряют фактически, а состоит из конечного значения, полученного в результате учета отношения сигнал-шум, возникающего при используемом формате анализа, и/или с учетом калибровочных эталонных значений, которые используются для повышения воспроизводимости измерений уровня маркера рака от анализа к анализу. В некоторых вариантах осуществления «уровень» биомаркера, например, VSIG4 выражается в произвольных единицах измерения, поскольку важным является то, что одинаковые произвольные единицы сравниваются (i) от анализа к анализу, или (ii) от одного пациента, страдающего раком, к другим, или (iii) между анализами, проводимыми в разные периоды времени для одного и того же пациента, или (iv) между уровнем биомаркера, измеренным в образце пациента, и заданным эталонным значением (которое также может называться здесь «пороговым» значением).
Термин «легкая цепь» при использовании в отношении антитела относится к полипептидной цепи приблизительно 25 кДа, где аминоконцевая часть включает вариабельную область от примерно 100 до примерно 110 или более аминокислот и карбоксиконцевая часть включает константную область. Приблизительная длина легкой цепи составляет от 211 до 217 аминокислот. Существует два различных типа, обозначаемых как каппа (κ) и лямбда (λ), исходя из аминокислотной последовательности константных доменов. Аминокислотные последовательности легкой цепи хорошо известны в данной области техники. Легкая цепь может представлять собой человеческую легкую цепь.
Используемый в контексте данного документа термин «моноклональное антитело» обозначает антитело, происходящее из почти однородной популяции антител, где популяция содержит идентичные антитела, за исключением нескольких возможных естественных мутаций, которые можно обнаружить в минимальных долях. Моноклональное антитело возникает в результате роста одноклеточного клона, такого как гибридома, и характеризуется тяжелыми цепями одного класса и подкласса и легкими цепями одного типа. Используемое в настоящем описании моноклональное антитело демонстрирует специфическое связывание с одним антигенным сайтом (т.е. одним эпитопом), когда ему презентировано антитело. Моноклональное антитело может быть получено различными способами, которые хорошо известны в соответствующей области техники.
Как описано в настоящем описании, термин «пэгилирование» означает способ обработки для увеличения времени удерживания антитела в крови путем введения полиэтиленгликоля в вышеупомянутое моноклональное антитело или его антигенсвязывающий фрагмент. В частности, при пегилировании полимерных наночастиц полиэтиленгликолем повышается гидрофильность на поверхности наночастиц и, соответственно, быстрый распад в живом организме можно предотвратить за счет так называемого эффекта незаметности, препятствующего распознаванию иммунной активностью, в том числе макрофагами в организме человека, чтобы вызвать фагоцитоз и расщепление патогенов, продуктов жизнедеятельности и инородных тел, попавших извне. Таким образом, время удерживания антитела в крови может быть увеличено путем пэгилирования. Пэгилирование, используемое в настоящем описании, может быть осуществлено способом, с помощью которого образуется амидная группа на основе связи между карбоксильной группой гиалуроновой кислоты и аминогруппой полиэтиленгликоля, но не ограничивается этим, и пэгилирование может осуществляться различными способами. При этом, что касается полиэтиленгликоля, предназначенного для использования, предпочтительно используется полиэтиленгликоль, имеющий молекулярную массу от 100 до 1000 и линейную или разветвленную структуру, хотя он конкретно этим не ограничивается.
Используемый в контексте данного документа термин «процент идентичности» или «% идентичности» между двумя последовательностями нуклеиновых кислот или аминокислот относится к проценту идентичных нуклеотидов или аминокислотных остатков между двумя сравниваемыми последовательностями, полученному после оптимального выравнивания, причем этот процент является чисто статистическим, а различия между двумя последовательностями распределяются случайным образом по их длине. Сравнение двух последовательностей нуклеиновых кислот или аминокислот как правило проводят путем сравнения последовательностей после их оптимального выравнивания, при этом указанное сравнение можно проводить по сегментам или с использованием «окна выравнивания». Оптимальное выравнивание последовательностей для сравнения может быть осуществлено, помимо сравнения вручную, с помощью способов, известных специалисту в данной области.
Для аминокислотной последовательности, проявляющей по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, или по меньшей мере 99% идентичности с эталонной аминокислотной последовательностью, предпочтительные примеры включают те, которые содержат эталонную последовательность, определенные модификации, в частности удаление, добавление или замену по меньшей мере одной аминокислоты, усечение или удлинение. В случае замены одной или более последовательных или непоследовательных аминокислот предпочтительны замены, при которых замещенные аминокислоты заменяются «эквивалентными» аминокислотами. В данном документе выражение «эквивалентные аминокислоты» предназначено для обозначения любых аминокислот, которые могут быть заменены одной из структурных аминокислот без изменения биологической активности соответствующих антител и конкретных примеров антител, определенных ниже. Эквивалентные аминокислоты могут быть определены либо по их структурной гомологии с аминокислотами, которые они заменяют, либо по результатам сравнительных тестов биологической активности между различными антителами, которые могут быть сгенерированы.
В качестве неограничивающего примера в таблице 1 ниже обобщены возможные замены, которые, вероятно, будут осуществлены без значительной модификации биологической активности соответствующего модифицированного антигенсвязывающего белка; обратные замены, естественно, возможны при тех же условиях.
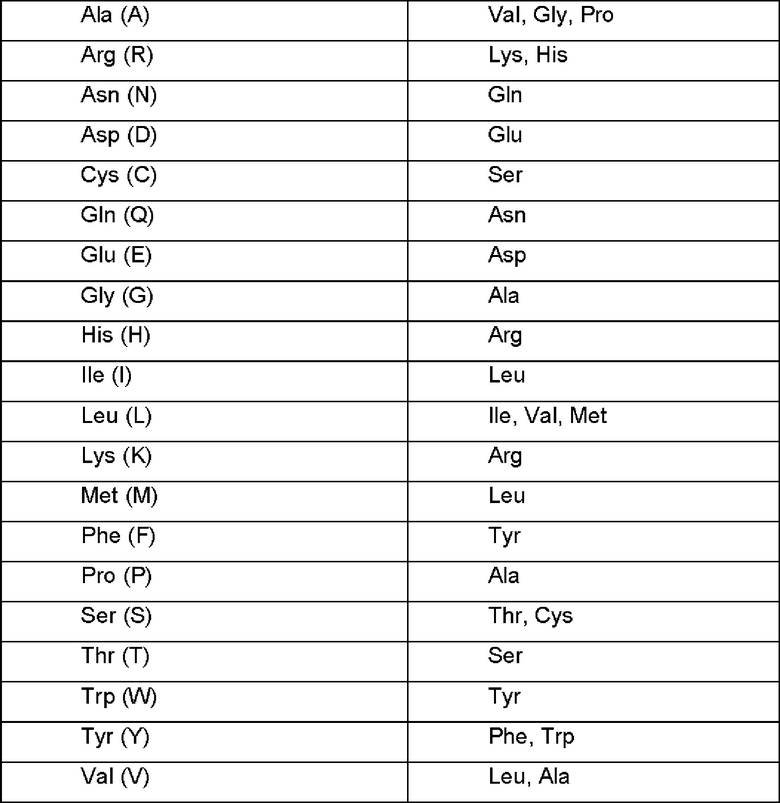
Термин «фармацевтически приемлемый», используемый в настоящем документе, означает одобренный регулирующим органом федерального правительства или правительства штата или включенный в фармакопею США, Европейскую фармакопею или другую общепризнанную фармакопею для использования среди животных, и особенно среди людей. Более конкретно, когда речь идет о носителе, выражение «фармацевтически приемлемый» означает, что носитель (носители) совместим с другим ингредиентом (ингредиентами) композиции и не является вредным для его получателя. Соответственно, используемое в контексте данного документа выражение «фармацевтически приемлемый носитель» означает носитель или разбавитель, который не ингибирует биологическую активность и свойства соединения для введения без стимуляции живого организма. Тип носителя может быть выбран, исходя из предполагаемого пути введения. Количество каждого используемого носителя может варьироваться в пределах диапазонов, общепринятых в данной области техники. В качестве фармацевтически приемлемого носителя в композиции, которую готовят в виде жидкого раствора, в качестве стерилизованного носителя, подходящего для живого организма можно использовать физиологический раствор, стерилизованную воду, буферного солевой раствор, инъекционный раствор альбумина, раствор декстрозы, раствор мальтодекстрина, глицерина и смеси одного или более из них. При необходимости могут быть добавлены общие добавки, такие как антиоксидант, буферный раствор и бактериостат. Кроме того, путем дополнительного добавления разбавителя, диспергатора, поверхностно-активного вещества, связующего вещества или смазки композиция может быть приготовлена в форме препарата для инъекций, такого как водный раствор, суспензия и эмульсия, пилюля, капсула, гранула или таблетка.
Используемый в контексте данного документа термин «поликлональное антитело» относится к антителу, которое было продуцировано среди или в присутствии одного или более других, неидентичных антител. Как правило, поликлональные антитела вырабатываются из В-лимфоцита в присутствии нескольких других В-лимфоцитов, продуцирующих неидентичные антитела. Обычно поликлональные антитела получают непосредственно от иммунизированного животного.
Термин «эталонное значение», используемый в контексте данного документа, относится к уровню экспрессии рассматриваемого биомаркера (например, VSIG4) в эталонном образце. «Эталонный образец», используемый в контексте данного документа, означает образец, полученный от субъектов, предпочтительно двух или более субъектов, которые не имеют заболевание или, в альтернативном варианте, от общей популяции. Подходящие эталонные уровни экспрессии биомаркера могут быть определены путем измерения уровней экспрессии указанного биомаркера у нескольких подходящих субъектов, и такие эталонные уровни могут быть скорректированы с учетом конкретных популяций субъектов. Эталонное значение или эталонный уровень может представлять собой абсолютное значение; относительное значение; значение, имеющее верхний или нижний предел; диапазон значений; среднее значение; медианное значение, среднее значение или значение по сравнению с конкретным контрольным или базовым значением. Эталонное значение может быть основано на отдельном выборочном значении, таком как, например, значение, полученное из образца испытуемого, но в более ранний момент времени. Эталонное значение может быть основано на большом количестве образцов, например, на популяции субъектов хронологической возрастной группы, или на основе пула образцов, включая или исключая образец, подлежащий тестированию.
«Субъект», который может быть подвергнут описанной в настоящем документе методике, может быть любым из млекопитающих, включая человека, собаку, кошку, крупный рогатый скот, козу, свинью, свиней, овец и обезьян. Известно, что пациентом может быть человек. В одном варианте осуществления «субъект» или «нуждающийся субъект» относится к млекопитающему, которое страдает раком или у которого подозревают рак, или у которого диагностирован рак. Используемый в контексте данного документа термин «субъект, страдающий раком», относится к млекопитающему, которое страдает раком или у которого был диагностирован рак. «Контрольный субъект» относится к млекопитающему, которое не страдает раком и у него не подозревают рак.
Используемый в контексте данного документа термин «лечение» заболевания у субъекта или «лечение» субъекта, имеющего заболевание, относится к подверганию субъекта фармацевтическому лечению, например, введению лекарственного средства, так что степень заболевания снижается или заболевание предотвращают. Например, лечение приводит к уменьшению по меньшей мере одного признака или симптома заболевания или состояния. Лечение включает (но не ограничивается) введение композиции, такой как фармацевтическая композиция, и может быть выполнено как профилактически, так и после наступления патологического события. Лечение может потребовать введения агента и/или обработку более одного раза.
«Вариабельная область» или «вариабельный домен» антитела относится к амино-концевым доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может обозначаться как «VH». Вариабельный домен легкой цепи может обозначаться как «VL». Эти домены, как правило, являются наиболее вариабельными частями антитела и содержат антигенсвязывающие сайты.
Термин «вектор» относится к веществу, которое используется для введения молекулы нуклеиновой кислоты в клетку-хозяина. В частности, «вектор», используемый в контексте данного документа, представляет собой молекулу нуклеиновой кислоты, способную передавать другую молекулу нуклеиновой кислоты, с которой она связана. Одним из примеров вектора является «плазмида», которая относится к кольцевой двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим примером вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальное происхождение репликации и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. Термин «вектор», таким образом, включает вектор как само реплицирующуюся структуру нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он был введен Векторы, пригодные для использования, включают, например, экспрессионные векторы, плазмиды, фаговые векторы, вирусные векторы, эписомы и искусственные хромосомы, которые могут включать селекционные последовательности или маркеры, обеспечивающие стабильную интеграцию в хромосому клетки-хозяина.
Определенные векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы в настоящем документе обозначаются как «рекомбинантные экспрессионные векторы» (или просто «экспрессионные векторы»). Как правило, экспрессионные векторы, применяемые в методиках рекомбинантных ДНК, находятся в форме плазмид. В настоящем описании «плазмида» и «вектор» могут использоваться взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако предполагается, что данное изобретение включает такие формы экспрессионных векторов, как бактериальные плазмиды, YAC, космиды, ретровирусы, эписомы, происходящие из EBV, и все другие векторы, которые известны специалисту в данной области техники как подходящие для обеспечения экспрессии тяжелых и/или легких цепей интересующего антитела (например, антитела против VSIG4). Специалисту в данной области будет понятно, что полинуклеотиды, кодирующие тяжелую и легкую цепи, можно клонировать в разные векторы или в один и тот же вектор.
Векторы могут включать один или более селектируемых маркерных генов и соответствующие последовательности контроля экспрессии. Селектируемые маркерные гены, которые могут быть включены, например, обеспечивают устойчивость к антибиотикам или токсинам, восполняют ауксотрофный дефицит или поставляют необходимые питательные вещества, не содержащиеся в культуральной среде. Последовательности контроля экспрессии могут включать конститутивные и индуцируемые промоторы, энхансеры транскрипции, терминаторы транскрипции и т.п., которые хорошо известны в данной области техники. Когда две или более молекул нуклеиновых кислот подлежат совместной экспрессии (например, тяжелые и легкие цепи антитела), обе молекулы нуклеиновых кислот могут быть вставлены, например, в один экспрессионный вектор или в отдельные экспрессионные векторы. Для экспрессии одного вектора кодирующие нуклеиновые кислоты могут быть функционально связаны с одной общей последовательностью контроля экспрессии или с разными последовательностями контроля экспрессии, такими как один индуцируемый промотор и один конститутивный промотор. Введение молекул нуклеиновых кислот в клетку-хозяина может быть подтверждено с использованием способов, хорошо известных в данной области техники. Такие методы включают, например, анализ нуклеиновых кислот, такой как Нозерн-блоттинг или полимеразная цепная реакция (ПЦР), амплификация мРНК, или иммуноблоттинг для экспрессии генных продуктов, или другие подходящие аналитические способы для проверки экспрессии введенной последовательности нуклеиновой кислоты или ее соответствующего генного продукта. Специалистам в данной области техники понятно, что молекула нуклеиновой кислоты экспрессируется в достаточном количестве для получения требуемого продукта (например, антитело против VSIG4, предложенное в настоящем документе), и также понятно, что уровни экспрессии могут быть оптимизированы для получения достаточной экспрессии с использованием способов, хорошо известных в данной области техники.
Термин «VSIG4» или «полипептид VSIG4» и аналогичные термины относятся к полипептиду («полипептид», «пептид» и «белок» используются взаимозаменяемо в контексте данного документа), кодируемому геном V-подобный Ig домен-содержащего 4 белка (VIG4), который расположен в перицентромерной области Х-хромосомы человека, а также известен в данной области техники как белок суперсемейства иммуноглобулинов Z39IG, Z39IG, рецептор комплемента суперсемейства иммуноглобулинов, CRIg. Последовательность гена VSIG4 может быть, например, представлена последовательностью, имеющей номер доступа GenBank, такой как, например, No. NM_007268.2, NM_001100431.1, NM_001184831.1, NM_001184830.1 или NM_001257403.1.
VSIG4 (V-подобный Ig домен-содержащий 4) представляет собой V-подобный и Ig домен-содержащий 4 белок, который структурно связан с семейством иммунных регуляторных белков В7. У людей существуют две различные формы белка VSIG4. Длинная форма содержит как константный (С2-тип), так и вариабельный (V-тип) домен иммуноглобулина, в то время как короткая форма содержит только домен иммуноглобулина V-типа, без С2-типа. Эти две формы проиллюстрированы на фиг. 1А. В одном варианте осуществления человеческий белок VSIG4 имеет последовательность, представленную последовательностью в Uniprot с номером доступа Q9Y279. В одном варианте осуществления длинная форма человеческого белка VSIG4 имеет последовательность, представленную последовательностью в Uniprot с номером доступа Q9Y279-1. Предпочтительно, длинная форма VSIG4 имеет последовательность, изложенную в SEQ ID NO: 1. В одном варианте осуществления короткая форма человеческого белка VSIG4 имеет последовательность, представленную последовательностью Uniprot с номером доступа Q9Y279-3. Предпочтительно, короткая форма VSIG4 имеет последовательность, изложенную в SEQ ID NO: 2.
VSIG4 функционирует как рецептор комплемента, функционально ингибируя активность комплемента путем связывания с сегментами комплемента iC3b и C3b, тем самым опосредуя клиренс C3b-опсонизированных патогенов. Было обнаружено, что экспрессия VSIG4 ограничена тканевыми макрофагами, и было показано, что она подавляется в ответ на липополисахарид (LPS) (Vogt et al. (2006) J. of Clin. Invest. 116:2817).
VSIG4 является белком иммунной контрольной точки, обладающим противовоспалительными и иммуносупрессивными свойствами. Растворимый слитый белок VSIG4 ингибирует воспаление (Small et al., Swiss Med Wkly. (2016) 146:w14301), тогда как недостаток VSIG4 инициирует воспаление, опосредованное макрофагами (Liao et al. (2014) Lab. Invest. 94: 706). Данное ингибирование активации макрофагов посредством VSIG4, по-видимому, является C3b-независимым (Li et al. (2017) Nat Commun. 8(1):1322). VSIG4 выполняет регуляторную функцию в активации Т-клеток (Vogt et al. (2006) J. of Clin. Invest. 116:2817; Xu et al. (2010) Immunol Lett. 18; 128(1):46-50; Jung et al. (2012) Hepatology. 56(5): 1838-48; Jung et al. (2015) Immunol Lett. 165(2):78-83; Munawara et al. (2019) Front Immunol. 10;10:2892). Примечательно, что VSIG4 является сильным отрицательным регулятором Т-клеточной пролиферации и продукции IL-2 путем связывания неидентифицируемого Т-клеточного лиганда рецептора (Vogt et al. (2006) J. of Clin. Invest. 116:2817).
Как и в случае со многими белками иммунных контрольных точек, активность VSIG4 способствует росту опухоли, активируя иммунную толерантность. У мышей с дефицитом VSIG4 растут меньшие по размеру опухоли, чем у мышей дикого типа, что дает основание предполагать, что отсутствие VSIG4 активирует иммунный ответ, который предотвращает рост опухоли. Массивные инфильтраты VSIG4-экспрессирующих макрофагов в микроокружении опухоли наблюдались у пациентов с диагнозом немелкоклеточного рака легкого (Liao et al. (2014) Lab. Invest. 94:706). Ген VSIG4 сверхэкспрессируется на нескольких видах раковых клеток, таких как рак легкого, рак яичников, рак молочной железы, гепатома и множественная меланома, и действует как онкоген, который подавляет иммунные реакции и способствует прогрессированию опухоли. Высокий уровень экспрессии VSIG4 действительно коррелирует с глиомой высокой степени злокачественности и плохим прогнозом пациента (Xu et al. (2015) Am. J. Transl. Res. 7: 1172).
Антитела против VSIG4
Иммунные контрольные точки играют крайне важную роль в поддержании аутотолерантности и ограничении иммунноопосредованного повреждения тканей в физиологических условиях. VSIG4 представляет собой трансмембранный белок типа I, принадлежащий к суперсемейству иммуноглобулинов, родственных В7, который экспрессируется на покоящихся макрофагах. VSIG4 представляет собой коингибирующий лиганд, который осуществляет негативную регуляцию Т-клеточной активации посредством ингибирования пролиферации Т-кпеток CD4+ и CD8+ и продукции IL-2. Известны две формы VSIG4, длинная форма (huVSIG4(L)) и короткая форма (huVSIG4(S)), которые отличаются наличием мембранного проксимального домена, представляющего собой иммуноглобулиновый домен IgC-типа в длинной форме.
Авторы настоящего изобретения показали, что обе формы экспрессируются в макрофагах. Кроме того, обе формы функциональны: растворимые версии либо huVSIG4(L), либо huVSIG4(S) ингибируют активацию Т-клеток CD4+ человека, о чем свидетельствует ингибирование Т-клеточной пролиферации и продукции IFNγ. Таким образом, как длинная, так и короткая формы VSIG4 участвуют в регуляторной активности белка, а это означает, что обе формы должны быть ингибированы для ослабления иммуносупрессии.
В настоящем изобретении предложены новые моноклональные антитела, специфически связывающиеся с VSIG4 человека. Более конкретно, в настоящем изобретении предложены новые моноклональные антитела, способные связываться как с длинной, так и с короткой формой белка. Кроме того, описанные в данном документе антитела индуцируют интернализацию в дополнение к связыванию с VSIG4, тем самым способствуя удалению рецептора с клеточной поверхности. Это отличается от антител предшествующего уровня техники, таких как, например, антитела, описанные в WO 2020/069507, которые способны связываться только с длинной формой VSIG4 человека и не могут вызывать интернализацию.
Авторы настоящего изобретения обнаружили, что эффективная блокада VSIG4 достигается с использованием антител против VSIG4, раскрытых в настоящем документе. Действительно, данные антитела модулируют противовоспалительные функции и ингибируют иммуносупрессивные свойства VSIG4, о чем свидетельствует их способность запускать высвобождение провоспалительных цитокинов и блокировать секрецию противовоспалительных цитокинов макрофагами и способствовать активации Т-клеток. Таким образом, антитела против VSIG4, раскрытые в настоящем в настоящем документе, полезны для генерации противоопухолевого иммунного ответа у пациентов с раком.
В первом аспекте настоящего изобретения предложено моноклональное антитело или его антигенсвязывающий фрагмент, способное специфически связываться с человеческим VSIG4. В одном из вариантов осуществления указанное антитело способно связывать как длинную форму человеческого VSIG4, так и короткую форму VSIG4. В одном из вариантов осуществления длинная форма человеческого белка VSIG4 имеет последовательность, изложенную в SEQ ID NO: 1. В одном из вариантов осуществления короткая форма человеческого белка VSIG4 имеет последовательность, изложенную в SEQ ID NO: 2.
В одном из вариантов осуществления антитело против VSIG4 индуцирует интернализацию при связывании с VSIG4.
В одном из вариантов осуществления интернализация антитела по изобретению может быть оценена иммунофлуоресценцией (как показано ниже в настоящей заявке) или любым способом или процессом, известным специалисту в данной области техники, специфичной для механизма интернализации.
Комплекс VSIG4/антитело интернализуется после связывания антитела с внеклеточным доменом (ECD) VSIG4, тем самым индуцируя снижение количества VSIG4 на поверхности клеток. Данное снижение можно количественно определить любым способом, известным специалисту в данной области техники, таким как, в качестве неограничивающих примеров, вестерн-блоттинг, анализ сортировки клеток с активированной флуоресценцией (FACS), иммунофлуоресценция и тому подобное.
В одном из вариантов осуществления это снижение, отражая таким образом интернализацию, измеряется посредством FACS и выражается как разница или дельта между средней интенсивностью флуоресценции (MFI), измеренной при 4°С, и MFI, измеренной при 37°С, в обоих случаях после инкубации клеток с антителом в течение 4 часов.
Данное значение дельта может быть, например, определено на основе значений MFI, полученных с использованием необработанных клеток и клеток, обработанных антителом с использованием i) VSIG4-трансфицированных клеток НЕK293 после 4-часовой инкубации с антителом, описанным в настоящем документе, и ii) вторичного антитела, меченного А1еха488. Данный параметр определяют по следующей формуле: Δ(MFI4°C-MFI37°C).
Эта разница между MFI отражает понижающую регуляцию VSIG4, поскольку MFI пропорциональны экспрессии VSIG4 на клеточной поверхности.
Предпочтительно, данные антитела или любой их антигенсвязывающий фрагмент, описанные в настоящем документе, представляют собой моноклональные антитела, дающие A(MFI4°C-MFI37°C) на НЕK293, трансфицированных VSIG4, составляющую по меньшей мере 280, предпочтительно по меньшей мере 370.
Более подробно, вышеупомянутая дельта может быть измерена в соответствии со следующим способом, который нужно рассматривать в качестве иллюстративного, но не ограничивающего примера:
a) приведение в контакт целевых клеток с антителом по изобретению либо в холодной (4°С), либо в теплой (37°С) полной культуральной среде;
b) приведение в контакт клеток стадии а) и, параллельно, необработанных клеток со вторичным антителом;
c) измерение MFI (репрезентативной для количества VSIG4, присутствующего на поверхности) для обработанных и необработанных клеток с использованием вторичного меченого антитела, способного связываться с антителом по изобретению; и
d) вычисление дельты как разности между MFI, полученной для обработанных клеток, и MFI, полученной для необработанных клеток.
Исходя из этой дельты по MFI, процент интернализации можно определить следующим образом:
100×(MFU4°C-MFI37°C) / MFI4°C
Антитела или любой их антигенсвязывающий фрагмент, согласно изобретению, присутствуют на VSIG4-трансфицированных HEK293, процент интернализации составляет от 60% до 99%, предпочтительно от 61% до 96%.
В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент способен связываться с VSIG4 с ЕС50, составляющей от 10×10-10 до 1×10-9 М.
Используемый в контексте данного документа термин «ЕС50» относится к 50% эффективной концентрации. Более конкретно, термин «полумаксимальная эффективная концентрация» (ЕС50) соответствует концентрации лекарственного средства, антитела или токсина, которая индуцирует ответ на полпути между исходным уровнем и максимумом после некоторого конкретного времени воздействия. Он обычно используется в качестве меры эффективности лекарственного средства. Таким образом, ЕС50 кривой зависимости от возрастающей дозы представляет собой концентрацию соединения, где наблюдается 50% его максимального эффекта. ЕС50 кривой зависимости от дозы представляет собой концентрацию соединения, где 50% населения демонстрирует ответ после заданной продолжительности воздействия. Показатели концентрации обычно дают сигмоидальную кривую, быстро увеличиваясь при относительно небольшом изменении концентрации. Это может быть определено математически путем построения линии наибольшего соответствия.
В качестве предпочтительного варианта осуществления определенная в данном документе ЕС50 характеризует эффективность связывания антитела с внеклеточным доменом (ECD) VSIG4, экспонированным на клетках HEK293 человека. Параметр ЕС50 определяют с помощью анализа FACS. Параметр ЕС50 отражает концентрацию антитела, при которой получают 50% максимального связывания с человеческим IGF-1R, экспрессируемым на опухолевых клетках человека. Каждое значение ЕС50 рассчитывали как середину кривой зависимости доза-эффект с использованием четырехпараметрической логистической модели нелинейной регрессии (Prism Software). Этот параметр был выбран как репрезентативный для физиологических/патологических состояний.
Моноклональные антитела против VSIG4, используемые в контексте данного документа, включают, без ограничений, синтетические антитела, рекомбинантно продуцируемые антитела, мультиспецифические антитела (включая биспецифические антитела), человеческие антитела, гуманизированные антитела, верблюжьи антитела, химерные антитела, интратела, антиидиотипические (анти-Id) антитела и функциональные фрагменты любого из вышеперечисленных. Моноклональные антитела против VSIG4 могут быть человеческого или нечеловеческого происхождения. Примеры антител против VSIG4 нечеловеческого происхождения включают, но не ограничиваются ими, антитела млекопитающих (например, обезьян, грызунов, коз и кроликов). Поскольку каждая структура человеческого антитела происходит от человека, существует лишь небольшая вероятность наличия иммунного ответа по сравнению с обычным гуманизированным антителом или мышиным антителом, и, таким образом, оно имеет преимущество, заключающееся в том, что оно не вызывает какого-либо нежелательного иммунного ответа при введении человеку. Поэтому его с большим преимуществом можно использовать в качестве антитела для лечения. Соответственно, моноклональные антитела против VSIG4 для терапевтического использования у людей являются предпочтительно гуманизированными или полностью человеческими. Более предпочтительно, они являются полностью человеческими.
Согласно одному варианту осуществления настоящего изобретения, антитело, описанное в настоящем документе, является человеческим антителом, специфически связывающимся с VSIG4, полученным авторами настоящего изобретения на основании биопэннинга наивной библиотеки человеческих одноцепочечных Fv способом фагового дисплея.
В способах фагового дисплея функциональные домены антител отображаются на поверхности фаговых частиц, которые несут кодирующие их полинуклеотидные последовательности. В частности, последовательности ДНК, кодирующие домены VH и VL, амплифицируются из библиотек кДНК животных (например, человеческие или мышиные библиотеки кДНК пораженных тканей). ДНК, кодирующие домены VH и VL, рекомбинируют вместе с линкером scFv с помощью ПЦР и клонируют в фагемидный вектор. Вектор электропорируют в Е. coli, а Е. coli инфицируют хелперным фагом. Фаг, используемый в этих методах, обычно представляет собой нитевидный фаг, включая fd и М13, а домены VH и VL обычно подвергают рекомбинантному слиянию либо с геном III фага, либо с геном VIII. Фаг, экспрессирующий антигенсвязывающий домен, который связывается с определенным антигеном, может быть отобран или идентифицирован с использованием антигена, например, с использованием меченого антигена или антигена, связанного или захваченного твердой поверхностью или шариком. Примеры способов фагового дисплея, которые могут быть использованы для получения антител, предложенных в настоящем документе, включают те, которые раскрыты в Brinkman et al., 1995, J. Immunol. Methods 182:41-50; Ames et al, 1995, J. Immunol. Methods 184:177-186; Kettleborough et al., 1994, Eur. J. Immunol. 24:952-958; Persic et al., 1997, Gene 187:9-18; Burton et al., 1994, Advances in Immunology 57:191-280; PCT/GB91/01134; WO 90/02809, WO 91/10737, WO 92/01047, WO 92/18619, WO 93/1 1236, WO 95/15982, WO 95/20401, и W097/13844; и патенты США №5698426, 5223409, 5403484, 5580717, 5427908, 5750753, 5821047, 5571698, 5427908, 5516637, 5780225, 5658727, 5733743 и 5969108.
Как описано в приведенных выше ссылках, после фаговой селекции, области, кодирующие антитело, могут быть выделены из фага и использованы для получения полных антител, включая человеческие антитела или любой другой желаемый антигенсвязывающий фрагмент, и экспрессированы в любом желаемом хозяине, включая клетки млекопитающих, клетки насекомых, клетки растений, дрожжи и бактерии, например, как описано ниже. Методики рекомбинантного получения Fab-, Fab'- и F(ab')2-фрагментов также можно применять посредством способов, известных в данной области, таких как раскрытые в РСТ публикации № WO 92/22324; Mullinax et al., 1992, BioTechniques 12(6):864-869; Sawai et al., 1995, AJRI 34:26-34; и Better et al., 1988, Science 240:1041-1043.
Для получения полных антител, с целью амплификации последовательностей VH или VL в клонах scFv можно использовать ПЦР-праймеры, включающие нуклеотидные последовательности VH или VL, сайт рестрикции и фланкирующую последовательность для защиты сайта рестрикции. Используя методики клонирования, известные специалистам в данной области, амплифицированные посредством ПЦР VH-домены можно клонировать в векторы, экспрессирующие константную область VH, например, константную область гамма-4 человека, а амплифицированные посредством ПЦР VL-домены можно клонировать в векторы, экспрессирующие константную область VL, например, константные области каппа или лямбда человека. VH и VL домены также можно клонировать в один вектор, экспрессирующий необходимые константные области. Затем векторы для конверсии тяжелой цепи и векторы для конверсии легкой цепи совместно трансфицируют в клеточные линии для получения стабильных или переходных клеточных линий, которые экспрессируют полноразмерные антитела, например, IgG, с использованием методик, известных специалистам в данной области.
Антитела, получаемые по вышеуказанному способу, представляют собой антитела с повышенной аффинностью к антигену. Термин «аффинность» указывает на свойство специфически распознавать и связываться с сайтом конкретного антигена, и, вместе со специфичностью антитела к антигену, высокая аффинность является важным фактором иммунной реакции. В настоящем изобретении клетки библиотеки с гуманизированными тяжелыми цепями получают посредством случайной мутации вариабельной области тяжелых цепей, и для клеток библиотеки был проведен анализ колониеобразования для отбора первых вариантных клонов, обладающих высокими свойствами связывания антигена. Путем проведения конкурентного ELISA для отобранных клонов исследовали аффинность каждого клона. Помимо данного способа можно использовать различные способы измерения аффинности к антигену, и одним из примеров таких способов является технология поверхностного плазмонного резонанса.
В одном из вариантов осуществления моноклональное антитело против VSIG4, раскрытое в настоящем документе, специфически связывается с эпитопом в белке VSIG4. В частности, эпитоп, связанный настоящим антителом, может быть идентифицирован путем определения того, какие остатки VSIG4 прекращают связывание с антителом при мутации. В одном варианте осуществления VSIG4 является длинным вариантом. В другом варианте осуществления VSIG4 является коротким вариантом.
Предпочтительно антитело, раскрытое в настоящем документе, представляет собой антитело, которое связывается по меньшей мере с одной аминокислотой в одном или более эпитопах, причем эпитоп выбран в группе, состоящей из:
a) эпитоп М1, содержащий остатки Е24, V25, Е27, V29 и/или Т30 последовательности, изложенной в SEQ ID No: 2;
b) эпитоп М2, содержащий остатки D36, N38, L39 и/или Т42 последовательности, изложенной в SEQ ID No: 2;
c) эпитоп М3, содержащий остатки Q59, G61, S62, D63 и/или V65 последовательности, изложенной в SEQ ID No: 2;
d) эпитоп М4, содержащий остатки I77, А80, Y82 и/или Q83 последовательности, изложенной в SEQ No 2;
e) эпитоп М5, содержащий остатки Н87, Н90, K91 и/или V92 последовательности, изложенной в SEQ ID No: 2;
f) эпитоп М6, содержащий остатки S97, Q99, S101 и/или Т102 последовательности, изложенной в SEQ ID No: 2;
g) эпитоп М7, содержащий остатки R108, S109, Н110, Т112 и/или Е114 последовательности, изложенной в SEQ ID No: 2;
h) эпитоп М8, содержащий остатки Т119, Р120, D121, N123, Q124 и/или V125 последовательности, изложенной в SEQ ID No: 2.
Более предпочтительно, антитело, раскрытое в настоящем документе, представляет собой антитело, которое связывает:
a) по меньшей мере одну из аминокислот в М1;
b) по меньшей мере одну из аминокислот в М4 и, необязательно, по меньшей мере, один из остатков М3;
c) по меньшей мере одну из аминокислот в М7;
d) по меньшей мере одну из аминокислот в М8;
e) по меньшей мере одну из аминокислот в М7 и по меньшей мере одну из аминокислот в М8; или
f) по меньшей мере одну из аминокислот в М3, по меньшей мере одну из аминокислот в М7 и по меньшей мере одну из аминокислот в М8 и, необязательно, по меньшей мере, один из остатков М2 и/или по меньшей мере один из остатков М4.
Определение связывания антитела против VSIG4 с эпитопом можно выполнить любым способом или методом, известным специалисту в данной области техники, таким как, без ограничения, радиоактивность, Biacore, ELISA, проточная цитометрия и т.д., или в соответствии со способом, таким как описано в настоящем описании.
В одном варианте осуществления раскрытое в настоящем документе моноклональное антитело против VSIG4 содержит три CDR тяжелой цепи и три CDR легкой цепи. Предпочтительно антитело содержит тяжелую цепь и легкую цепь, причем тяжелая цепь содержит три CDR тяжелой цепи, а легкая цепь содержит три CDR легкой цепи.
Предпочтительно раскрытое в данном документе антитело содержит три CDR тяжелой цепи и три CDR легкой цепи, где последовательность каждой CDR выбрана из группы последовательностей, представленных в SEQ ID NO: 3-58.
В одном из вариантов осуществления антитело против VSIG4 содержит три CDR тяжелой цепи, содержащие последовательности, выбранные из группы, состоящей из SEQ ID NO: 3, 4, 5, 9, 10, 11, 12, 13, 17, 18, 19, 23, 24, 26, 27, 28, 32, 33, 34, 37, 38, 39, 43, 44, 45, 49, 50, 51 и 55.
В одном из вариантов осуществления антитело против VSIG4 содержит три CDR легкой цепи, содержащие последовательность, выбранную из группы, состоящей из SEQ ID NO: 6, 7, 8, 14, 15, 16, 20, 21, 22, 25, 29, 30, 31, 35, 36, 40, 41, 42, 46, 47, 48, 52, 53, 54, 56, 57 и 58.
В одном из предпочтительны вариантов осуществления предложено антитело против VSIG4, имеющее тяжелую цепь, содержащую три CDR тяжелой цепи, содержащие последовательности, выбранные из группы, состоящей из SEQ ID NO: 3, 4, 5, 9, 10, 11, 12, 13, 17, 18, 19, 23, 24, 26, 27, 28, 32, 33, 34, 37, 38, 39, 43, 44, 45, 49, 50, 51 и 55.
В другом предпочтительном варианте осуществления предложено антитело против VSIG4, имеющее легкую цепь, содержащую три CDR легкой цепи, содержащие последовательности, выбранные из группы, состоящей из SEQ ID NO: 6, 7, 8, 14, 15, 16, 20, 21, 22, 25, 29, 30, 31, 35, 36, 40, 41, 42, 46, 47, 48, 52, 53, 54, 56, 57 и 58.
В другом предпочтительном варианте осуществления антитело против VSIG4 содержит три CDR тяжелой цепи, при этом указанные CDR тяжелой цепи содержат последовательности, выбранные из группы, состоящей из SEQ ID NO: 3, 4, 5, 9, 10, 11, 12, 13, 17, 18, 19, 23, 24, 26, 27, 28, 32, 33, 34, 37, 38, 39, 43, 44, 45, 49, 50, 51 и 55; и три CDR легкой цепи, причем указанные CDR легкой цепи содержат последовательности, выбранные из группы, состоящей из SEQ ID NO: 6, 7, 8, 14, 15, 16, 20, 21, 22, 25, 29, 30, 31, 35, 36, 40, 41, 42, 46, 47, 48, 52, 53, 54, 56, 57 и 58.
В еще одном предпочтительном варианте осуществления антитело против VSIG4 содержит тяжелую цепь, причем указанная тяжелая цепь содержит три CDR тяжелой цепи, при этом указанные CDR тяжелой цепи содержат последовательности, выбранные из группы, состоящей из SEQ ID NO: 3, 4, 5, 9, 10, 11, 12, 13, 17, 18, 19, 23, 24, 26, 27, 28, 32, 33, 34, 37, 38, 39, 43, 44, 45, 49, 50, 51 и 55; и легкую цепь, причем указанная легкая цепь содержит три CDR легкой цепи, где указанные CDR легкой цепи содержат последовательности, выбранные из группы, состоящей из SEQ ID NO: 6, 7, 8, 14, 15, 16, 20, 21, 22, 25, 29, 30, 31, 35, 36, 40, 41, 42, 46, 47, 48, 52, 53, 54, 56, 57 и 58.
Более предпочтительно, описанное в контексте данного документа антитело выбрано из группы, состоящей из:
а) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
b) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 9, 10 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
c) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 11, 12 и 13 и три CDR легкой цепи с последовательностями SEQ ID NO: 14, 15 и 16;
d) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 19 и три CDR легкой цепи с последовательностями SEQ ID NO: 20, 21 и 22;
e) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 23, 24 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 25;
f) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 26, 27 и 28 и три CDR легкой цепи с последовательностями SEQ ID NO: 29, 30 и 31;
g) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 32, 33 и 34 и три CDR легкой цепи с последовательностями SEQ ID NO: 35, 36 и 16;
h) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 37, 38 и 39 и три CDR легкой цепи с последовательностями SEQ ID NO: 40, 41 и 42;
i) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 43, 44 и 45 и три CDR легкой цепи с последовательностями SEQ ID NO: 46, 47 и 48;
j) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 49, 50 и 51 и три CDR легкой цепи с последовательностями SEQ ID NO: 52, 53 и 54;
k) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 55 и три CDR легкой цепи с последовательностями SEQ ID NO: 56, 57 и 58.
В предпочтительном, но неограничивающем варианте осуществления антитело по изобретению выбирают в группе, состоящей из следующего:
a) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 129 или любой последовательностью, проявляющей по меньшей мере 80% идентичность с SEQ ID NO: 129, и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
b) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 131 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 131, и трех CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
c) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 133 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 133, и трех CDR легкой цепи с последовательностями SEQ ID NO: 14, 15 и 16;
d) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 135 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 135, и трех CDR легкой цепи с последовательностями SEQ ID NO: 20, 21 и 22;
e) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 137 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 137, и трех CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 25;
f) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 139 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 139, и трех CDR легкой цепи с последовательностями SEQ ID NO: 29, 30 и 31;
g) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 141 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 141, и трех CDR легкой цепи с последовательностями SEQ ID NO: 35, 36 и 16;
h) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 143 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 143, и трех CDR легкой цепи с последовательностями SEQ ID NO: 40, 41 и 42;
i) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 145 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 145, и трех CDR легкой цепи с последовательностями SEQ ID NO: 46, 47 и 48;
j) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 147 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 147, и трех CDR легкой цепи с последовательностями SEQ ID NO: 52, 53 и 54;
k) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 149 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 149, и трех CDR легкой цепи с последовательностями SEQ ID NO: 56, 57 и 58.
Под «любой последовательностью, проявляющей по меньшей мере 80%, предпочтительно 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 129», подразумевается последовательность, имеющая три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5 и, кроме того, проявляющая по меньшей мере 80%, предпочтительно 85%, 90%, 95% или 98% идентичность с полной последовательностью SEQ ID NO: 129 вне последовательностей, соответствующих CDR (т.е. SEQ ID №3, 4 и 5), где «вне последовательностей, соответствующих CDR» означает «исключение последовательностей, соответствующих CDR».
В другом предпочтительном, но не ограничительном варианте осуществления антитело по изобретению выбирают в группе, состоящей из следующего:
a) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 130 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 130 и три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5;
b) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 132 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 132 и три CDR тяжелой цепи с последовательностями SEQ ID NO: 9, 10 и 5;
c) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 134 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 134 и три CDR тяжелой цепи с последовательностями SEQ ID NO: 11, 12 и 13;
d) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 136 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 136 и три CDR тяжелой цепи с последовательностями SEQ ID No: 17, 18 и 19;
e) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 138 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 138 и три CDR тяжелой цепи с последовательностями SEQ ID No: 23, 24 и 5;
f) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 140 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 140 и три CDR тяжелой цепи с последовательностями SEQ ID No: 26, 27 и 28;
g) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 142 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 142 и три CDR тяжелой цепи с последовательностями SEQ ID No: 32, 33 и 34;
h) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 144 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 144 и три CDR тяжелой цепи с последовательностями SEQ ID No 37, 38 и 39;
i) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 146 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 146 и три CDR тяжелой цепи с последовательностями SEQ ID No: 43, 44 и 45;
j) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 148 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 148 и три CDR тяжелой цепи с последовательностями SEQ ID No: 49, 50 и 51; и
k) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 150 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 150 и три CDR тяжелой цепи с последовательностями SEQ ID No 17, 18 и 55.
Под «любой последовательностью, проявляющей по меньшей мере 80%, предпочтительно 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 130», подразумевается последовательность, имеющая три CDR легкой цепи с SEQ ID NO: 6, 7 и 8 и, кроме того, проявляющая по меньшей мере 80%, предпочтительно 85%, 90%, 95% или 98% идентичность с полной последовательностью SEQ ID NO: 130 за пределами последовательностей, соответствующих CDR (т.е. SEQ ID №6, 7 и 8).
Вариант осуществления изобретения относится к антителу, распознающему VSIG4 и выбранному в группе, состоящей из следующего:
a) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 129 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 129, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 130 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 130;
b) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 131 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 131, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 132 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 132;
c) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 133 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 133, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 134 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 134;
d) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 135 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 135, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 136 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с с SEQ ID No: 136;
e) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 137 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 137, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 138 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 138;
f) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 139 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 139, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 140 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 140;
g) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 141 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 141, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 142 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 142;
h) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 143 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 143, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 144 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 144;
i) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 145 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 145, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 146 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 146;
j) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 147 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 147, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 148 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 148; и
k) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 149 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 149, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 150 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 150.
Моноклональное антитело, специфически связывающееся с VSIG4 согласно одному варианту осуществления настоящего изобретения, или его антигенсвязывающий фрагмент предпочтительно представляет собой антитело, выбранное в группе, состоящей из следующего:
a) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 129, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 130;
b) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 131, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 132;
c) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 133, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 134;
d) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 135, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 136;
e) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 137, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 138;
f) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 139, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 140;
g) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 141, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 142;
h) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 143, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 144;
i) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 145, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 146;
j) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 147, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 148; и
k) антитело, содержащее вариабельную область тяжелой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 149, и вариабельную область легкой цепи, характеризующуюся аминокислотной последовательностью SEQ ID NO: 150.
Для большей ясности в следующей таблице 2 показаны последовательности (CDR, каркасные области, VH и VL) предпочтительных антител и эпитопов, связанных каждым из этих антител.
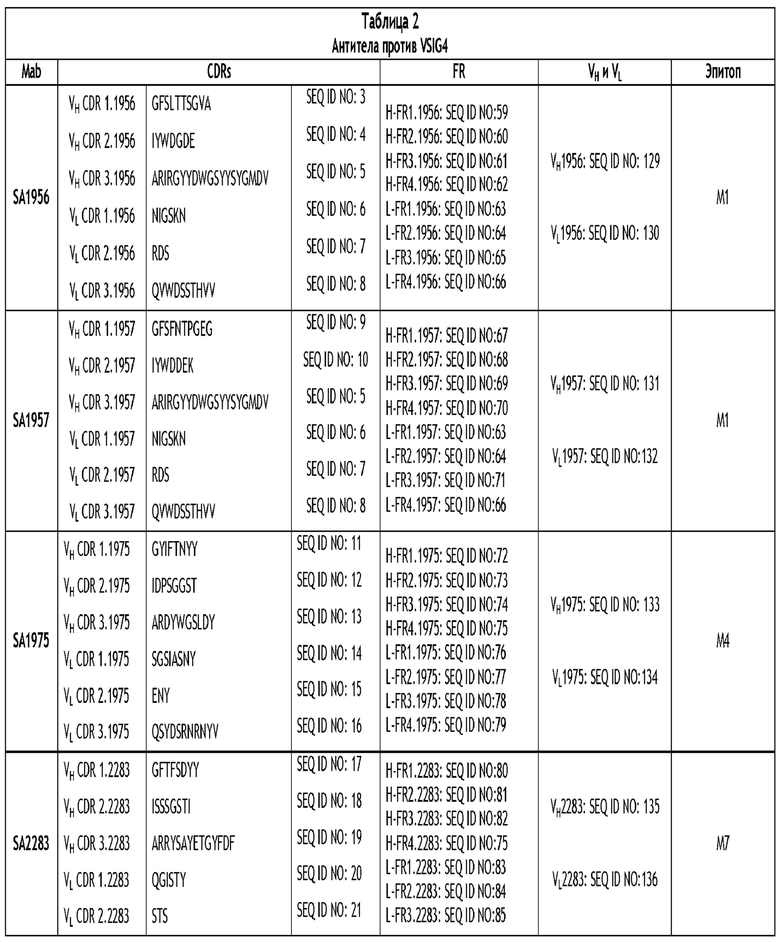
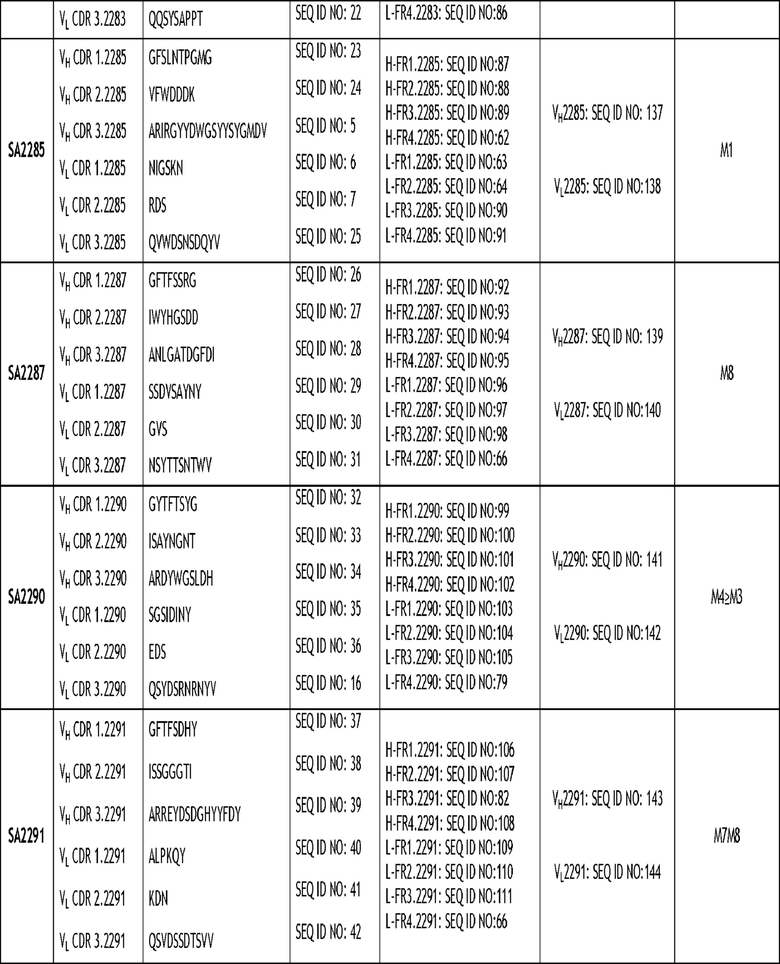
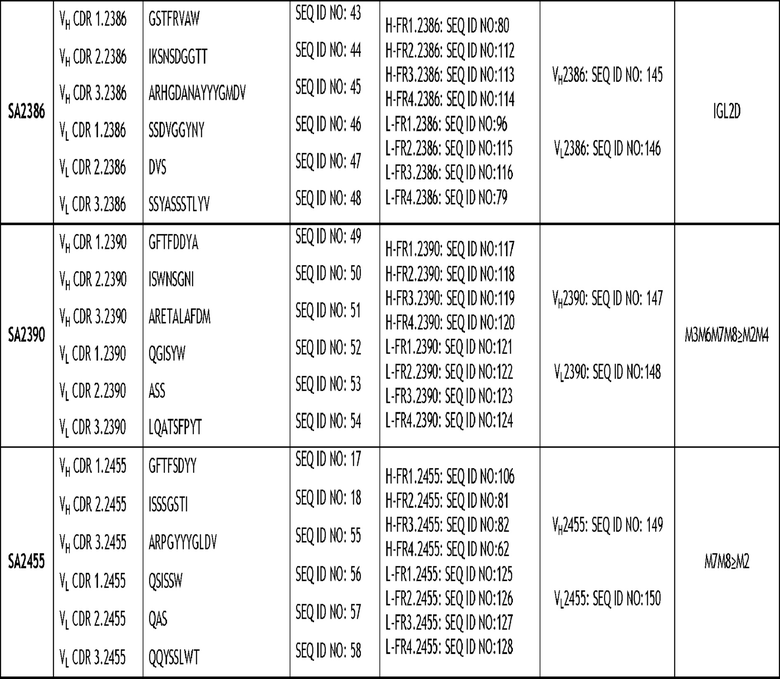
В диапазоне, в котором VSIG4 может быть специфически распознан, моноклональное антитело по настоящему изобретению или его антигенсвязывающий фрагмент может включать не только последовательность антитела против VSIG4 по настоящему изобретению, которая описана в настоящем описании, но и ее биологический эквивалент. Например, для дополнительного улучшения аффинности связывания и/или других биологических характеристик антитела могут быть внесены дополнительные изменения в аминокислотную последовательность антитела. Данные модификации включают, например, делецию, вставку и/или замену аминокислотной последовательности антитела. Данные аминокислотные модификации производятся на основе относительного сходства между заместителями боковой цепи аминокислоты, например, гидрофобности, гидрофильности, заряда, размера и т.п. На основе анализа размера, формы и типа заместителей боковой цепи аминокислоты установлено, что все аргинин, лизин и гистидин являются остатками с положительным зарядом; аланин, глицин и серин имеют схожие размеры; фенилаланин, триптофан и тирозин имеют схожую форму. Соответственно, исходя из этих соображений можно сказать, что биологически аргинин, лизин и гистидин; аланин, глицин и серин; фенилаланин, триптофан и тирозин являются функциональными эквивалентами.
В одном из вариантов осуществления моноклональные антитела против VSIG4, описанные в настоящем документе, могут быть в виде полноразмерных антител, многоцепочечных или одноцепочечных антител, фрагментов таких антител, которые избирательно связывают Р VSIG4 (включая, но не ограничиваясь Fab', (Fab')2, Fv и scFv), суррогатных антител (включая конструкцию суррогатной легкой цепи), однодоменных антител, гуманизированных антител, верблюжьих антител и тому подобного. Они также могут представлять собой или происходить из любого изотипа, включая, например, IgA (например, IgA1 или IgA2), IgD, IgE, IgG (например, IgG1, IgG2, IgG3 или IgG4) или IgM. В некоторых вариантах осуществления антитело против VSIG4 представляет собой IgG (например, IgG1, IgG2, IgG3 или IgG4). В одном из вариантов осуществления антитело дополнительно содержит константную область человека. В дополнительном варианте осуществления константную область человеческую выбирают из группы, состоящей из IgG1, IgG2, IgG2, IgG3, IgG4. В еще одном конкретном варианте осуществления константная область человека представляет собой IgG1. Кроме того, константная область тяжелой цепи имеет типы гамма (γ), мю (μ), альфа (α), дельта (δ) и эпсилон (ε), и, в качестве подкласса, она имеет гамма1 (γ1), гамма2 (γ2), гамма3 (γ3), гамма4 (γ4), альфа1 (α1) и альфа2 (α2). Константнаяобласть легкой цепи имеет каппа (κ) и лямбда (λ) типы.
Антитела против VSIG4 включают меченые антитела, пригодные в диагностических областях применения. Антитела могут быть использованы в диагностических целях, например, для обнаружения экспрессии интересующей мишени в конкретных клетках, тканях или сыворотке; или для мониторинга развития или прогрессирования иммунологического ответа в рамках процедуры клинических испытаний, чтобы, например, определить эффективность заданной схемы лечения. Обнаружение может быть облегчено путем соединения антитела с обнаруживаемым веществом или «меткой». Метка может быть конъюгирована напрямую или опосредованно с антителом против VSIG4 по изобретению. Сама метка может быть обнаруживаемой (например, радиоизотопные этикетки, изотопные этикетки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическое изменение субстратного соединения или композиции, которая является обнаруживаемой. Примеры обнаруживаемых веществ включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества, радиоактивные вещества, позитрон-активные металлы с использованием позитронно-эмиссионной томографии, и ионы нерадиоактивных парамагнитных металлов. Обнаруживаемое вещество может быть связано или конъюгировано либо непосредственно с антителом (или его фрагментом), либо опосредованно, через промежуточное соединение (такое как, например, линкер, известный в данной области техники) с использованием способов, известных в данной области техники. Примеры ферментативных меток включают люциферазы (например, люциферазу светлячка и бактериальную люциферазу; патент США No 4,737,456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уреазу, пероксидазу, такую как пероксидаза из хрена (HRPO), щелочную фосфатазу, β-галактозидазу, ацетилхолинэстеразу, глюкоамилазу, лизоцим, сахаридоксидазы (например, глюкозооксидаза, галактозооксидаза и глюкозо-6-фосфатдегидрогеназа), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидаза, микропероксидаза и тому подобное. Примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных веществ включают умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид, диметиламин-1-нафталинсульфонилхлорид или фикоэритрин и т.д.; пример люминесцентного вещества включает люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин; примеры подходящих изотопных веществ включают 13С, 15N и дейтерий; а примеры подходящего радиоактивного вещества включают 125I, 131I, 111 In или 99Тс.
Биспецифические антитела
Кроме того, в соответствии с настоящим изобретением предложено мультиспецифическое антитело, включая моноклональное антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент.
Вышеуказанное мультиспецифическое антитело в настоящем изобретении предпочтительно может быть биспецифическим антителом, но не ограничено им.
Мультиспецифическое антитело согласно настоящему изобретению предпочтительно имеет форму, при которой антитело против VSIG4, описанное в настоящем документе, связано с антителом, обладающим свойством связывания с молекулой-мишенью, специфичной для иммуноэффекторных клеток, или его фрагмент. Молекула-мишень, специфичная для иммуноэффекторных клеток, предпочтительно представляет собой иммунную контрольную точку, но она не ограничивается ею. Примеры молекул-мишеней, специфичных для иммуноэффекторных клеток, включают, например, PD-1, PD-L1, CTLA-4, TIM-3, TIGIT, BTLA, KIR, A2aR, VISTA, В7-Н3, TCR/CD3, CD16 (FcγRIIIa) CD44, Cd56, CD69, CD64 (FcγRI), CD89 и CD11b/CD18 (CR3).
Мультиспецифическое антитело представляет собой антитело, которое может одновременно распознавать разные множественные (два или более) эпитопы одного и того же антигена или двух или более отдельных антигенов, и антитела, принадлежащие к мультиспецифическим антителам, можно разделить на следующие: антитело на основе scFv, антитело на основе Fab, антитело на основе IgG и т.п. В случае мультиспецифического, например, биспецифического антитела два сигнала могут быть одновременно подавлены или усилены, и, таким образом, это может быть более эффективным, чем случай, когда происходит подавление/усиление одного сигнала. По сравнению со случаем, когда каждый сигнал обрабатывается сигнальным ингибитором для каждого из них, можно достигнуть низкодозированного введения, и два сигнала могут быть одновременно подавлены/усилены в одном и том же пространстве.
Способы получения биспецифического антитела широко известны. Как правило, рекомбинантное получение биспецифического антитела основано на совместной экспрессии пары тяжелая/легкая цепь двух иммуноглобулинов в условиях, при которых две тяжелые цепи обладают разной специфичностью.
В случае биспецифического антитела на основе scFv, комбинируя VL и VH различных scFv получают гибрид на основе scFv в гетеродимерной форме с образованием диатела (Holliger et al., Proc. Natl. Acad. Sci. U.S.A.,90:6444, 1993), и, соединяя разные scFv друг с другом можно получить тандемные ScFv. Экспрессируя СН1 и CL Fab на конце каждого scFv, можно получить гетеродимерное мини-антитело (Muller et al., FEBS lett., 432:45, 1998). Кроме того, путем замены части аминокислот СН3 домена гомодимерным доменом Fc происходит структурное изменение на форму «выступ-во-впадину» с получением гетеродимерной структуры, при этом указанные модифицированные СН3-домены экспрессируются на концах каждого отличного scFv, и таким образом можно получить минитело в гетеродимерной scFv форме. (Merchant et al., Nat. Biotechnol., 16:677, 1998).
В случае биспецифического антитела на основе Fab, при комбинации отдельных Fab' для конкретного антигена путем использования дисульфидной связи или медиатора, антитело можно получить в гетеродимерной Fab-форме и, посредством экспрессии scFv для другого антигена на конце тяжелой цепи или легкой цепи конкретного Fab, можно получить валентность антигена 2. Кроме того, при наличии шарнирной области между Fab и scFv, можно получить валентность антигена 4 в гомодимерной форме. Кроме того, в соответствующей области техники известен способ получения следующего: битело двойной направленности, с помощью которого получают валентность антигена 3 на основании слияния scFv для отличного антигена на конце легкой цепи и конце тяжелой цепи Fab, битело тройной направленности, с помощью которого получают валентность антигена 3 на основании слияния разных scFV с концом легкой цепи и концом тяжелой цепи Fab, и антитело тройной направленности F(ab')3 в простой форме, которое получают путем химического слияния трех различных Fab.
В случае биспецифического антитела на основе IgG способ получения биспецифического антитела путем получения гибридной гибридомы, так называемой квадромы, основанный на регибридизации гибридом мышей и крыс, известен из Trion Pharma. Кроме того, известен способ получения биспецифического антитела в так называемой форме «Дыры и ручка», в котором частичные аминокислоты гомодимерного домена СН3 Fc в различных тяжелых цепях модифицируются при совместном использовании легкой цепной части (Merchant et al., Nat. Biotechnol., 16:677, 1998), и, кроме биспецифического антитела в гетеродимерной форме, известен способ получения (scFv)4-IgG в гомодимерной форме по слиянию двух различных scFV с постоянной областью легкой цепи и тяжелой цепи IgG вместо переменной области с последующим выражением. Кроме того, ImClone Systems сообщила, что, основываясь на IMC-1C11 в качестве химерного моноклонального антитела для человеческого VEGFR-2, только один переменный домен для рецептора фактора роста мыши, полученного из тромбоцитов, а, сливается с аминокончиной легкой цепи антитела, чтобы продуцировать биспецифическое антитело. Кроме того, Россия и др. сообщили об антителе, имеющем высокую валентность антигена для CD20, на основе так называемого метода «стыковки и блокировки (DNL)» с использованием домена димеризации и стыковки (DDD) субъединицы протеинкиназы А (PKA) R и анкерного домена PKA (Rossi et al., Proc. Natl. Acad. Sci. США, 103:6841, 2006).
Производные антител
Антитела против VSIG4 по настоящему изобретению могут быть дополнительно модифицированы для того, чтобы содержать дополнительные небелковые фрагменты, которые известны в данной области техники и легкодоступны. В частности, в настоящий документ включены моноклональные антитела против VSIG4, которые дериватизированы, ковалентно-модифицированы или конъюгированы с другими молекулами, для использования в диагностических и терапевтических областях применения. Например, но не в качестве ограничения, дериватизированные антитела включают антитела, которые были модифицированы, например, путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидирования, дериватизации известными защитными/блокирующими группами, протеолитического расщепления, связывания с клеточным лигандом или другим белком и т.д. Любая из многочисленных химических модификаций может быть выполнена известными методиками, включая, но не ограничиваясь, специфическое химическое расщепление, ацетилирование, формилирование, метаболический синтез туникамицина и т.д. Дополнительно производное может содержать одну или несколько неклассических аминокислот.
В частности, моноклональное антитело по настоящему изобретению или его антигенсвязывающий фрагмент могут быть подвергнуты дериватизации, как описано выше, в частности, путем, например, гликозилирования и/или пэгилирования, с целью увеличения времени пребывания в живом организме, в который вводят антитело.
Что касается гликозилирования и/или пэгилирования, то различные паттерны гликозилирования и/или пэгилирования могут быть модифицированы способом, хорошо известным в данной области техники, при условии сохранения функции антитела по настоящему изобретению, при этом в антитело по настоящему изобретению включено вариантное моноклональное антитело, в котором модифицированы различные паттерны гликозилирования и/или пэгилирования, или его антигенсвязывающий фрагмент.
Предпочтительно, фрагменты, пригодные для дериватизации антитела, представляют собой водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, но не ограничиваются ими, полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля и пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена и малеинового ангидрида, полиаминоациды (гомополимеры или статические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропропиленгликоля, сополимеры полипропиленоксида и этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Количество полимеров, прикрепленных к антителу, может варьироваться, и если присоединено более одного полимера, они могут быть одними и теми же или разными молекулами. В общем, количество и/или тип полимеров, используемых для дериватизации, могут быть определены, исходя из соображений, включая, в том числе, конкретные свойства или функции антитела, подлежащего улучшению, будет ли производное антитела использоваться в терапии при определенных условиях и т.д.
В конкретном примере антитела против VSIG4 по настоящему изобретению могут быть присоединены к фрагментам поли(этиленгликоля) (ПЭГ). В конкретном варианте осуществления антитело представляет собой фрагмент антитела, и фрагменты ПЭГ присоединяются через боковую цепь любой доступной аминокислоты или концевую аминокислотную функциональную группу, расположенную во фрагменте антитела, например, любую свободную амино-, имино-тиоловую, гидроксильную или карбоксильную группу. Такие аминокислоты могут встречаться в природе во фрагменте антитела или могут быть сконструированы во фрагменте с использованием методов рекомбинантной ДНК. См., например, патент США No 5219996. Для присоединения двух или более молекул ПЭГ могут быть использованы несколько сайтов. Фрагменты ПЭГ могут быть ковалентно присоединены через тиольную группу по меньшей мере одного цистеинового остатка, расположенного во фрагменте антитела. В тех случаях, когда тиольная группа используется в качестве точки присоединения, могут быть использованы соответственно активированные эффекторные фрагменты, например, селективные производные тиола, такие как малеимиды и производные цистеина.
В конкретном примере конъюгат антитела против VSIG4 представляет собой модифицированный Fab' фрагмент, который пэгилирован, т.е. содержит ПЭГ (поли(этиленгликоль)) ковалентно присоединенный к нему, например, в соответствии со способом, раскрытым в ЕР0948544. См. также Poly(ethyleneglycol) Chemistry, Biotechnical and Biomedical Applications, (J. Milton Harris (ed.), Plenum Press, New York, 1992); Poly(ethyleneglycol) Chemistry and Biological Applications, (J. Milton Harris and S. Zalipsky, eds., American Chemical Society, Washington D.C., 1997); и Bioconjugation Protein Coupling Techniques for the Biomedical Sciences, (M. Aslam and A. Dent, eds., Grove Publishers, New York, 1998); и Chapman, 2002, Advanced Drug Delivery Reviews 54:531-545. ПЭГ может быть прикреплен к цистеину в шарнирной области. В одном из примеров модифицированный ПЭГ Fab' фрагмент содержит малеимидную группу, ковалентно связанную с одной тиольной группой в модифицированной шарнирной области. Остаток лизина может быть ковалентно связан с малеимидной группой, и к каждой из аминогрупп на остатке лизина может быть присоединен метоксиполи(этиленгликоль) полимер, имеющий молекулярную массу приблизительно 20000 Да. Таким образом, общая молекулярная масса ПЭГ, прикрепленного к Fab' фрагменту, может составлять приблизительно 40000 Да.
В другом варианте осуществления предложены конъюгаты антитела и небелкового фрагмента, которые могут быть избирательно нагреты воздействием излучения. В одном варианте осуществления небелковый фрагмент представляет собой углеродную нанотрубку (Kam et al, Proc. Natl. Acad. Sci. США 102: 11600-11605 (2005)). Излучение может быть любой длины волны и включает, без ограничения, длины волн, которые не наносят вреда обычным клеткам, но которые нагревают небелковый фрагмент до температуры, при которой клетки, находящиеся вблизи фрагмента антитело-белок, погибают.
Иммуноконъюгаты
В другом аспекте настоящего изобретения предложен иммуноконъюгат (взаимозаменяемо обозначаемый как «конъюгаты антитело-лекарственное средство» или «ADC»), содержащий антитело против VSIG4, как описано в настоящем описании, причем указанное антитело конъюгировано с цитотоксическим агентом.
Было выделено или синтезировано множество цитотоксических агентов, и они позволяют ингибировать пролиферацию клеток или уничтожать или снижать количество опухолевых клеток если не окончательно, то, по крайней мере, в значительной степени. Однако токсическая активность этих агентов не ограничивается опухолевыми клетками, и неопухолевые клетки также подвергаются воздействию и могут быть уничтожены. В частности, побочные эффекты наблюдаются на быстро обновляющихся клетках, таких как гемопоэтические клетки или клетки эпителия, в частности слизистых оболочек. Чтобы ограничить побочные эффекты на нормальные клетки при сохранении высокой цитотоксичности на опухолевых клетках, иммуноконъюгаты были использованы для местной доставки цитотоксических агентов при лечении рака (Lambert, J. (2005) Curr. Opinion in Pharmacology 5:543-549; Wu et al (2005) Nature Biotechnology 23(9): 1137-1146; Payne, G. (2003) i 3:207-212; Syrigos and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drug Deliv. Rev. 26:151-172; U.S. Pat. No. 4,975,278). Иммуноконъюгаты обеспечивают адресную доставку лекарственного фрагмента (т.е. цитотоксического агента) в опухоль и внутриклеточное накопление в ней, при этом системное введение неконъюгированных лекарственных средств может привести к неприемлемым уровням токсичности для нормальных клеток, а также опухолевых клеток, которые требуется уничтожить (Baldwin et al, Lancet (Mar. 15, 1986) стр. 603-05; Thorpe (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review," in Monoclonal Antibodies '84: Biological And Clinical Applications (A. Pinchera et al., eds) стр. 475-506. Сообщалось, что, как поликлональные, так и моноклональные антитела применимы в данных методиках (Rowland et al., (1986) Cancer Immunol. Immunother. 21: 183-87).
Цитотоксический агент, используемый в иммуноконъюгатах, раскрытых в настоящем описании, может представлять собой, без ограничения, лекарственное средство (т.е. «конъюгат антитело-лекарственное средство»), токсин (т.е. «иммунотоксин» или «конъюгат антитело-токсин»), радиоизотоп (т.е. «радиоиммуноконъюгат» или «конъюгат антитело-радиоизотоп») и т.д.
Предпочтительно иммуноконъюгат представляет собой связывающий белок, связанный по меньшей мере с лекарственным средством или лекарственным препаратом. Такой иммуноконъюгат обычно называют конъюгатом антитело-лекарство (или «ADC»), когда связывающий белок представляет собой антитело или его антигенсвязывающий фрагмент.
В первом варианте осуществления такие лекарственные средства могут быть описаны в отношении их механизма действия. В качестве неограничивающих примеров можно упомянуть алкилирующие агенты, такие как азотистый иприт, алкилсульфонаты, нитрозомочевину, оксазофорины, азиридины или иминэтилены, антиметаболиты, противоопухолевые антибиотики, ингибиторы митоза, ингибиторы функции хроматина, средства против ангиогенеза, антиэстрогены, антиандрогены, хелатирующие агенты, стимуляторы всасывания железа, ингибиторы циклооксигеназы, ингибиторы фосфодиэстеразы, ингибиторы ДНК, ингибиторы синтеза ДНК, стимуляторы апоптоза, ингибиторы тимидилата, ингибиторы Т-клеток, агонисты интерферона, ингибиторы рибонуклеозидтрифосфатредуктазы, ингибиторы ароматазы, антагонисты эстрогеновых рецепторов, ингибиторы тирозинкиназы, ингибиторы клеточного цикла, таксан, ингибиторы тубулина, ингибиторы ангиогенеза, стимуляторы макрофагов, антагонисты нейрокининовых рецепторов, агонисты каннабиноидных рецепторов, агонисты дофаминовых рецепторов, агонисты гранулоцитарного стимулирующего фактора, агонисты рецепторов эритропоэтина, агонисты рецепторов соматостатина, агонисты РФЛГ, сенсибилизаторы кальция, антагонисты рецепторов VEGF, антагонисты рецепторов интерлейкина, ингибиторы остеокластов, стимуляторы образования радикалов, антагонисты рецепторов эндотелина, алкалоид барвинка, антигормоны или иммуномодуляторы или любые другие новые лекарственные средства, удовлетворяющие критериям активности цитотоксина или токсина.
Такие лекарственные средства, например, приведены в справочнике Видаль 2010 на странице, посвященной соединениям, изложенным в колонке «Цитотоксические средства», посвященной онкологии и гематологии онкологии, причем данные цитотоксические соединения, приведенные со ссылкой на этот документ, изложены в данном документе как предпочтительные цитотоксические средства.
В частности, согласно изобретению, без ограничения, предпочтительными являются следующие лекарственные средства: мехлорэтамин, хлорамбукол, мелфален, хлоргидрат, пипобромен, преднимустин, динатрийфосфат, эстрамустин, циклофосфамид, альтретамин, трофосфамид, сульфофосфамид, ифосфамид, тиотепа, триэтиленамин, альтетрамин, кармустин, стрептозоцин, фотемустин, ломустин, бусульфан, треосульфан, импросульфан, дакарбазин, цисплатин, оксалиплатин, лобаплатин, гептаплатин, мириплатин гидрат, карбоплатин, метотрексат, пеметрексед, 5-фторурацил, флоксуридин, 5-фтордезоксиуридин, капецитабин, цитарабин, флударабин, цитозинарабинозид, 6-меркаптопурин (6-МП), неларабин, 6-тиогуанин (6-ТГ), хлордезоксиаденозин, 5-азацитидин, гемцитабин, кладрибин, дезоксикоформицин, тегафур, пентостатин, доксорубицин, даунорубицин, идарубицин, валрубицин, митоксантрон, дактиномицин, митрамицин, пликамицин, митомицин С, блеомицин, прокарбазин, паклитаксел, доцетаксел, винбластин, винкристин, виндезин, винорелбин, топотекан, иринотекан, этопозид, валрубицин, гидрохлорид амрубицина, пирарубицин, ацетат эллиптина, зорубицин, эпирубицин, идарубицин и тенипозид, разоксин, маримастат, батимастат, приномастат, таномастат, иломастат, CGS-27023A, галофугинон, COL-3, неовастат, талидомид, CDC 501, DMXAA, L-651582, скваламин, эндостатин, SU5416, SU6668, интерферон-альфа, EMD121974, интерлейкин-12, IM862, ангиостатин, тамоксифен, торемифен, ралоксифен, дролоксифен, йодоксифен, анастрозол, летрозол, эксеместан, флутамид, нилутамид, сприронолактон, ацетат ципротерона, финастерид, цимитидин, бортезомид, велкейд, бикалутамид, ципротерон, флутамид, фулвестран, экземестан, дазатиниб, эрлотиниб, гефитиниб, иматиниб, лапатиниб, нилотиниб, сорафениб, сунитиниб, ретиноид, рексиноид, метоксален, метиламинолевулинат, альдеслейкин, ОСТ-43, денилейкин дифтитокс, интерлейкин-2, тазонермин, лентинан, сизофилан, рокинимекс, пидотимод, пегадемаза, тимопентин, поли I:C, прокодазол, Tic BCG, corynebacterium parvum, NOV-002, украин, левамизол, 1311-chTNT, Н-101, целмолейкин, интерферон альфа-2а, интерферон альфа-2b, интерферон гамма-1а, интерлейкин-2, мобенакин, рексин-G, тецелейкин, акларубицин, актиномицин, арглабин, аспарагиназа, карзинофилин, хромомицин, дауномицин, лейковорин, мазопрокол, неокарзиностатин, пепломицин, саркомицин, соламаргин, трабектедин, стрептозоцин, тестостерон, кунекатехины, синекатехины, алитретиноин, белотекана гидрохлорид, калустерон, дромостанолон, эллиптиниум ацетат, этинилэстрадиол, этопозид, флуоксиместерон, форместан, фосфетрол, гозерелина ацетат, гексиламинолевулинат, гистрелин, гидроксипрогестерон, иксабепилон, лейпролид, медроксипрогестерона ацетат, мегестрола ацетат, метилпреднизолон, метилтестостерон, милтефозин, митобронитол, надролона фенилпропионат, норэтиндрона ацетат, преднизолон, преднизон, темсиролимус, тестолактон, триамконолон, трипторелин, вапреотида ацетат, зиностатин стималамер, амсакрин, триоксид мышьяка, бисантрена гидрохлорид, хлорамбуцил, хлортрианизен, цис-диамминдихлорплатина, циклофосфамид, диэтилстильбестрол, гексаметилмеламин, гидроксимочевина, леналидомид, лонидамин, мехлорэтанамин, митотан, недаплатин, нимустина гидрохлорид, памидронат, пипоброман, порфимер натрия, ранимустин, разоксан, семустин, собузоксан, мезилат, триэтиленмеламин, золедроновая кислота, камостата мезилат, фадрозола гидрохлорид, нафоксидин, аминоглютетимид, кармофур, клофарабин, цитозинарабинозид, децитабин, доксифлуридин, эноцитабин, флударабина фосфат, фторурацил, фторафур, урамустин, абареликс, бексаротен, ралтитерксед, тамибаротен, темозоломид, вориностат, мегастрол, клодронат динатрия, левамизол, ферумокситол, изомальтозид железа, целекоксиб, ибудиласт, бендамустин, альтретамин, митолактол, темсиролимус, пралатрексат, TS-1, децитабин, бикалутамид, флутамид, летрозол, клодронат динатрия, дегареликс, торемифен цитрат, гистамина дигидрохлорид, DW-166HC, нитракрин, децитабин, иринотекана гидрохлорид, амсакрин, ромидепсин, третиноин, кабазитаксел, вандетаниб, леналидомид, ибандроновая кислота, милтефозин, витеспен, мифамуртид, надропарин, гранисетрон, ондансетрон, трописетрон, ализаприд, рамосетрон, доласетрона мезилат, фосапрепитанта димеглюмин, набилон, апрепитант, дронабинол, TY-10721, лизурида гидроген малеат, эпицерам, дефибротид, дабигатрана этексилат, филграстим, пегфилграстим, редитукс, эпоэтин, молграмостим, опрелвекин, сипулеуцел-Т, М-Вакс, ацетил L-карнитин, донепезила гидрохлорид, 5-аминолевулиновая кислота, метиламинолевулинат, цетрореликса ацетат, икодекстрин, лейпрорелин, метилфенидат, октреотид, амлексанокс, плериксафор, менатетренон, анетолдитиолетион, доксеркальциферол, цинакалцета гидрохлорид, алефацепт, ромиплостим, тимоглобулин, тималфазин, убенимекс, имиквимод, эверолимус, сиролимус, Н-1011, лазофоксифен, трилостан, инкадронат, ганглиозиды, пегаптаниб октанатрий, вертопорфин, минодроновая кислота, золедроновая кислота, нитрат галлия, алендронат натрия, этидронат динатрия, памидронат динатрия, дутастерид, стибоглюконат натрия, армодафинил, дексразоксан, амифостин, WF-10, темопорфин, дарбэпоэтин альфа, анцестим, сарграмостим, палифермин, R-744, непидермин, опрелвекин, денилейкин дифтитокс, крисантаспаза, бусерелин, дезлорелин, ланреотид, октреотид, пилокарпин, бозентан, калихимицин, майтанзиноиды и циклоникат.
Для более подробной информации специалист в данной области техники может обратиться к руководству, изданному Association Francaise des Enseignants de Chimie Therapeutique под заголовком Traité de chimie therapeutique, vol. 6, Médicaments antitumouraux et perspectives dans le traitement des cancers, edition TEC & DOC, 2003"
В альтернативном варианте иммуноконъюгат может содержать связывающий белок, соединенный по меньшей мере с радиоизотопом. Такой иммуноконъюгат обычно называют конъюгатом антитело-радиоизотоп (или «ARC»), где связывающим белком является антитело или его антигенсвязывающий фрагмент.
Для избирательного разрушения опухоли антитело может содержать высокорадиоактивный атом. Для производства ARC доступны различные радиоактивные изотопы, такие как, без ограничений, At211, С13, N15, О17, FI19, I123, I131, I125, In111, Y90, Re186, Re188, Sm153, tc99m, Bi212, P32, Pb212, радиоактивные изотопы Lu, гадолиния, марганца или железа.
Любые способы или процессы, известные специалистам в данной области, могут быть использованы для включения такого радиоизотопа в ARC (см., например, "Monoclonal Antibodies in Immunoscintigraphy", Chatal, CRC Press 1989). В качестве неограничивающих примеров Tc99m или I123, Re186, Re188 и In111 могут быть присоединены через цистеиновый остаток. Y90 может быть присоединен через лизиновый остаток. I123 может быть присоединен с использованием метода IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80: 49-57).
Можно привести несколько примеров для иллюстрации знаний специалиста в данной области техники в сфере ARC, такие как Zevalin®, который представляет собой ARC, состоящий из моноклонального антитела против CD20 и радиоизотопа In111 или Y90, связанных тиомочевинным линкером-хелатором (Wiseman et al. (2000) Eur.Jour.Nucl.Med.27(7):766-77 Wiseman et al. (2002) Blood 99(12):4336-42 Witzig et al.(2002) J.CIin.Oncol. 20(10):2453-63, Witzig et al (2002) J.Clin.Oncol. 20(15):3262-69); или Mylotarg®, который состоит из антитела против CD33, связанного с калихеамицином (патенты США №4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Затем можно также упомянуть ADC, именуемый Адсетрис (соответствует брентуксимаб ведотину), который недавно был одобрен FDA для лечения лимфомы Ходжкина (Nature, vol. 476, стр 380-381, 25 августа 2011 г.).
В еще одном варианте осуществления изобретения иммуноконъюгат может содержать связывающий белок, соединенный с токсином. Такой иммуноконъюгат обычно обозначают как конъюгат антитело-токсин (или «АТС»), где связывающим белком является антитело или его антигенсвязывающий фрагмент.
Токсины представляют собой эффективные и специфические яды, продуцируемые живыми организмами. Обычно они состоят из аминокислотной цепи, молекулярная масса которой может варьироваться от нескольких сотен (пептиды) до ста тысяч дальтон (белки). Они также могут представлять собой низкомолекулярные органические соединения. Токсины продуцируются многочисленными организмами, например бактериями, грибами, водорослями и растениями. Многие из них чрезвычайно ядовиты, их токсичность на несколько порядков выше, чем у нервно-паралитических веществ.
Токсины, используемые в АТС, могут включать, без ограничений, все виды токсинов, которые могут оказывать свое цитотоксическое действие посредством механизмов, включающих связывание тубулина, связывание ДНК или ингибирование топоизомеразы.
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают цепь А дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, цепь А экзотоксина (из Pseudomonas aeruginosa), цепь А рицина, цепь А абрина, цепь А модекцина, альфа-сарцин, белки Aleurites fordii, диантиновые белки, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены.
В настоящем документе также представлены низкомолекулярные токсины, такие как доластатины, ауристатины, трихотецен и СС1065, и производные этих токсинов, обладающие активностью токсинов. Было показано, что доластатины и ауристатины препятствуют развитию микротрубочек, гидролизу GTP, ядерному и клеточному делению и обладают противораковой и противогрибковой активностью.
Описанные в данном документе иммуноконъюгаты могут дополнительно содержать линкер.
«Линкер», «линкерное звено» или «связь» означает химическую группу, содержащую ковалентную связь или цепь атомов, которая ковалентно присоединяет связывающий белок к по меньшей мере одному цитотоксическому агенту.
Линкеры могут быть получены с использованием различных бифункциональных агентов, связывающих белки таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (ИТ), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), бис-диазоний производные (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и биактивные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Меченая углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (MX-DTPA) является типичным хелатирующим агентом для конъюгации цитотоксических агентов с адресной системой. Другими сшивающими реагентами могут быть BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые имеются в продаже (например, от Pierce Biotechnology, Inc., Рокфорд, Иллинойс, США).
Линкер может быть «нерасщепляемым» или «расщепляемым» линкером.
Предпочтительно линкер представляет собой «расщепляемый линкер», облегчающий высвобождение цитотоксического агента в клетке. Например, можно использовать кислотолабильный линкер, пептидазочувствительный линкер, фотолабильный линкер, диметиловый линкер или дисульфид-содержащий линкер. Линкер предпочтительно расщепляется во внутриклеточных условиях, так что расщепление линкера высвобождает цитотоксический агент из связывающего белка во внутриклеточной среде.
Например, в некоторых вариантах осуществления линкер может быть расщеплен расщепляющим агентом, присутствующим во внутриклеточной среде (например, в лизосоме, эндосоме или кавеоле). Линкер может быть, например, пептидильным линкером, который расщепляется внутриклеточной пептидазой или ферментом протеазой, включая, в том числе, лизосомальную или эндосомальную протеазу. Как правило, длина пептидильного линкера составляет по меньшей мере две аминокислоты или по меньшей мере три аминокислоты. Расщепляющие агенты могут включать катепсины В и D и плазмин, каждый из которых, как известно, гидролизует дипептидные лекарственные производные, что приводит к высвобождению активного лекарственного средства внутри клеток-мишеней. Например, можно использовать пептидильный линкер, который расщепляется тиол-зависимой протеазой катепсин-В, которая высокоэкспрессируется в раковой ткани (например, линкер Phe-Leu или Gly-Phe-Leu-Gly). В конкретных вариантах осуществления пептидильный линкер, расщепляемый внутриклеточной протеазой, представляет собой линкер Val-Cit или линкер Phe-Lys. Одним из преимуществ использования внутриклеточного протеолитического высвобождения цитотоксического агента является то, что агент обычно ослабляется при конъюгировании, а стабильность конъюгатов в сыворотке обычно высока.
В других вариантах осуществления расщепляемый линкер является рН-чувствительным, т.е. чувствительным к гидролизу при определенных значениях рН. Как правило, рН-чувствительный линкер гидролизуется в кислых условиях. Например, можно использовать кислотолабильный линкер, который гидролизуется в лизосоме (например, гидразон, семикарбазон, тиосемикарбазон, цис-аконитамид, ортоэфир, ацеталь, кеталь и т.п.). Такие линкеры относительно стабильны в условиях нейтрального рН, например, в крови, но нестабильны при рН ниже 5,5 или 5,0, примерном рН лизосомы. В некоторых вариантах осуществления гидролизуемый линкер представляет собой тиоэфирный линкер (такой как, например, тиоэфир, присоединенный к терапевтическому агенту посредством ацилгидразоновой связи).
В других вариантах осуществления линкер может быть расщеплен в восстанавливающих условиях (например, дисульфидный линкер). В данной области известны различные дисульфидные линкеры, включая, например, те, которые могут быть образованы с использованием SATA (N-сукцинимидил-S-ацетилтиоацетат), SPDP (N-сукцинимидил-3-(2-пиридилдитио)пропионат), SPDB (N-сукцинимидил-3-(2-пиридилдитио)бутират) и SMPT (N-сукцинимидил-оксикарбонил-альфа-метил-альфа-(2-пиридилдитио)толуол)-, SPDB и SMPT.
У нерасщепляемых линкеров, напротив, не имеется очевидного механизма высвобождения лекарственного средства. Иммуноконъюгаты, содержащие такие нерасщепляемые линкеры, основаны на полной лизосомальной протеолитической деградации антитела, которое высвобождает цитотоксический агент после интернализации.
В качестве примера иммуноконъюгата, содержащего нерасщепляемый линкер, можно упомянуть иммуноконъюгат трастузумаб-эмтанзин- (TDM1), в котором трастузумаб комбинируется со связанным химиотерапевтическим агентом майтанзином (Cancer Research 2008; 68: (22). 15 ноября, 2008).
В предпочтительном варианте осуществления описанный в данном документе иммуноконъюгат может быть получен любым способом, известным специалисту в данной области, таким как, без ограничения, i) реакция нуклеофильной группы антигенсвязывающего белка с двухвалентным линкерным реагентом с последующей реакцией с цитотоксическим агентом или ii) реакция нуклеофильной группы цитотоксического агента с двухвалентным линкерным реагентом с последующей реакцией с нуклеофильной группой антигенсвязывающего белка.
Нуклеофильные группы на антигенсвязывающем белке включают, без ограничения, N-концевые аминогруппы, аминогруппы боковой цепи, например лизин, тиоловые группы боковой цепи и гидроксильные или аминогруппы сахара, когда антигенсвязывающий белок гликозилирован. Аминогруппы, тиоловые и гидроксильные группы являются нуклеофильными и способны реагировать с образованием ковалентных связей с электрофильными группами на линкерных фрагментах и линкерных реагентах, включая, без ограничения, активные сложные эфиры, такие как сложные эфиры NHS, сложные эфиры HOBt, галогенформиаты и галогенангидриды; алкил- и бензилгалогениды, такие как галогенацетамиды; альдегиды, кетоны, карбоксильные и малеимидные группы. Антигенсвязывающий белок может иметь восстанавливаемые межцепочечные дисульфиды, т.е. цистеиновые мостики. Антигенсвязывающие белки можно сделать реакционноспособными для конъюгации с линкерными реагентами путем обработки восстанавливающим агентом, таким как DTT (дитиотреитол). Таким образом, каждый цистеиновый мостик теоретически образует два реакционноспособных тиоловых нуклеофила. Дополнительные нуклеофильные группы могут быть введены в антигенсвязывающий белок с помощью любой реакции, известной специалисту в данной области. В качестве неограничивающего примера реакционноспособные тиоловые группы могут быть введены в антигенсвязывающий белок путем введения одного или более остатков цистеина.
Иммуноконъюгаты также можно получить путем модификации антигенсвязывающего белка для введения электрофильных фрагментов, которые могут реагировать с нуклеофильными заместителями линкерного реагента или цитотоксического агента. Сахара гликозилированного антигенсвязывающего белка могут быть подвергнуты окислению с образованием альдегидных или кетоновых групп, которые могут вступать в реакцию с аминогруппой линкерных реагентов или цитотоксического агента. Образовавшиеся в результате группы иминных оснований Шиффа могут образовывать стабильную связь или могут быть восстановлены с образованием стабильных аминных связей. В одном из вариантов осуществления реакция углеводной части гликозилированного антигенсвязывающего белка либо с галактозидазой, либо с метапериодатом натрия может давать карбонильные (альдегидные и кетоновые) группы в белке, которые могут реагировать с соответствующими группами лекарственного средства. В другом варианте осуществления белки, содержащие N-концевые остатки серина или треонина, могут реагировать с метапериодатом натрия, что приводит к образованию альдегида вместо первой аминокислоты.
Химерные антигенные рецепторы
В соответствии с настоящим изобретением дополнительно предложен белок CAR (химерный антигенный рецептор), включающий i) антитело по настоящему изобретению; ii) трансмембранный домен и; iii) внутриклеточный сигнальный домен, характеризующийся тем, что он вызывает активацию Т-клеток при связывании антитела, указанного выше i), с антигеном.
В настоящем изобретении белок CAR характеризуется тем, что он состоит из моноклонального антитела по настоящему изобретению, общеизвестного трансмембранного домена и внутриклеточного сигнального домена.
Как описано в настоящем документе, термин «CAR (химерный антигенный рецептор)» относится к рецептору неприродного происхождения, способному придавать иммуноэффекторной клетке специфичность в отношении конкретного антигена. В целом, CAR обозначает рецептор, который используется для придания специфичности моноклонального антитела Т-клеткам. CAR обычно состоит из внеклеточного домена, трансмембранного домена и внутриклеточного домена. Внеклеточный домен включает область распознавания антигена, и в настоящем изобретении сайт распознавания антигена представляет собой VSIG4-специфичное антитело. VSIG4-специфичное антитело является таким, как описано выше, и данное антитело, используемое в CAR, предпочтительно находится в форме фрагмента антитела. Более предпочтительно оно находится в форме Fab или scFv, но не ограничивается ими.
Далее, трансмембранный домен CAR имеет форму, в которой он связан с внеклеточным доменом, и он может происходить как из природной, так и из синтетической формы. Когда он происходит из природной формы, он может происходить из мембраносвязанного или трансмембранного белка, и он может представлять собой компонент, происходящий из трансмембранных доменов различных белков, таких как альфа-, бета- или дзета-цепь Т-клеточного рецептора, CD28, CD3-эпсилон., CD45, CD4, CD5, CDS, CD9, CD16, CD22, CD33, CD37, CD64, CD80, CD86, CD134, CD137, CD154 или CD8. Последовательности этих трансмембранных доменов можно получить из документов, хорошо известных в данной области, где в достаточной степени описан трансмембранный домен трансмембранного белка, но не ограничивается ими.
В CAR по настоящему изобретению внутриклеточный домен CAR является его частью, и он соединен с трансмембранным доменом. Внутриклеточный домен по настоящему изобретению может включать внутриклеточный сигнальный домен, который характеризуется свойством вызывать активацию Т-клеток, предпочтительно пролиферацию Т-клеток при связывании антигена с сайтом распознавания антигена CAR. Внутриклеточный сигнальный домен не имеет конкретных ограничений в части его типа при условии, что он может вызывать активацию Т-клеток при связывании антигена с сайтом распознавания антигена CAR, присутствующим вне клетки, и можно использовать различные виды внутриклеточных сигнальных доменов. Его примеры включают мотив активации иммунорецептора на основе тирозина (ITAM), при этом ITAM может включать домены, которые происходят из CD3 дзета (ξ), FcR гамма, FcR бета, CD3 гамма, CD3 дельта, CD3 эпсилон, CDS, CD22, CD79a, CD79b, CD66d или FcεRIγ, но не ограничивается ими.
Кроме того, предпочтительно, чтобы внутриклеточный домен CAR по настоящему изобретению дополнительно содержал костимулирующий домен с внутриклеточным сигнальным доменом, но не ограничивался этим. Костимулирующий домен представляет собой часть, которая содержится в CAR по настоящему изобретению и выполняет роль передачи сигнала Т-клеткам в дополнение к сигналу от внутриклеточного сигнального домена, и обозначает внутриклеточную часть CAR, включая внутриклеточный домен костимулирующей молекулы.
Костимулирующая молекула означает молекулу клеточной поверхности, необходимую для достаточной реакции лимфоцитов на антиген, и ее примеры включают среди прочего CD27, CD28, 4-1ВВ, ОХ40, CD30, CD40, PD-1, ICOS, LFA -1 (антиген-1, ассоциированный с функцией лимфоцитов), CD2, CD7, LIGHT, NKG2C и В7-Н3. Костимулирующий домен может представлять собой внутриклеточную часть молекулы, выбранной из группы, состоящей из этих костимулирующих молекул и их комбинации.
Кроме того, короткий олигопептидный или полипептидный линкер может избирательно связывать внутриклеточный домен и трансмембранный домен CAR. Хотя этот линкер может быть включен в CAR по настоящему изобретению, он не имеет конкретных ограничений в отношении длины линкера при условии, что он может индуцировать активацию Т-клеток посредством связывания внутриклеточного домена антигена с внеклеточным антителом.
Нуклеиновые кислоты и системы экспрессии
Настоящее изобретение охватывает полинуклеотиды, кодирующие последовательности легкой и тяжелой цепей иммуноглобулина антител, особенно антител против VSIG4, векторы, содержащие такие нуклеиновые кислоты, и клетки-хозяева, способные продуцировать антитела согласно настоящему изобретению. В настоящем документе также предложены полинуклеотиды, которые гибридизуются в условиях гибридизации высокой, промежуточной или более низкой жесткости, например, как определено выше, с полинуклеотидами, кодирующими антитело или модифицированное антитело, предложенное в настоящем документе.
В первом аспекте настоящее изобретение относится к одному или более полинуклеотидам, кодирующим антитело, в частности антитело, способное специфически связываться с VSIG4, или его фрагмент, как описано выше. В соответствии с настоящим изобретением, в частности, предложен полинуклеотид, кодирующий вариабельную область тяжелой цепи и/или вариабельную область легкой цепи моноклонального антитела или его антигенсвязывающего фрагмента. Более конкретно, в некоторых вариантах осуществления молекулы нуклеиновой кислоты, предложенные в настоящем документе, содержат или состоят из последовательности нуклеиновой кислоты, кодирующей вариабельную область тяжелой цепи и вариабельную область легкой цепи, раскрытые в настоящем документе, или любую их комбинацию (например, в виде нуклеотидной последовательности, кодирующей антитело, предложенное в настоящем документе, такое как, например, полноразмерное антитело, тяжелая и/или легкая цепь антитела или одноцепочечное антитело, предложенное в настоящем документе).
В одном из вариантов осуществления полинуклеотид кодирует три CDR тяжелой цепи антитела против VSIG4, описанного в настоящем документе. В одном из вариантов осуществления полинуклеотид кодирует три CDR легкой цепи антитела против VSIG4, описанного в настоящем документе. Водном из вариантов осуществления полинуклеотид кодирует три CDR тяжелой цепи и три CDR легкой цепи антитела против VSIG4, описанного в настоящем документе. В другом варианте осуществления предложена пара полинуклеотидов, где первый полинуклеотид кодирует три CDR тяжелой цепи антитела против VSIG4, описанного в настоящем документе; а второй полинуклеотид кодирует три CDR легкой цепи того же антитела против VSIG4, описанного в настоящем документе.
В одном из вариантов осуществления полинуклеотид кодирует вариабельную область тяжелой цепи антитела против VSIG4, описанного в настоящем документе. В одном из вариантов осуществления полинуклеотид кодирует вариабельную область легкой цепи антитела против VSIG4, описанного в настоящем документе. В одном из вариантов осуществления полинуклеотид кодирует вариабельную область тяжелой цепи и вариабельную область легкой цепи антитела против VSIG4, описанного в настоящем документе. В другом варианте осуществления предложена пара полинуклеотидов, где первый полинуклеотид кодирует вариабельную область тяжелой цепи описанного в данном документе антитела против VSIG4; а второй полинуклеотид кодирует вариабельную область легкой цепи того же самого антитела против VSIG4, описанного в настоящем документе.
В одном из вариантов осуществления полинуклеотид кодирует тяжелую цепь антитела против VSIG4, описанного в настоящем документе. В одном из вариантов осуществления полинуклеотид кодирует легкую цепь антитела против VSIG4, описанного в настоящем документе. В одном из вариантов осуществления полинуклеотид кодирует тяжелую цепь и легкую цепь описанного в данном документе антитела против VSIG4. В другом варианте осуществления предложена пара полинуклеотидов, где первый полинуклеотид кодирует тяжелую цепь антитела против VSIG4, описанного в настоящем документе; а второй полинуклеотид кодирует легкую цепь того же самого антитела против VSIG4, описанного в настоящем документе.
В одном из вариантов осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA1956 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 129.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA1956 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностью SEQ ID NO: 6, 7 и 8. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 130.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA1957 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 9, 10 и 5. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 131.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA1957 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 132.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA1975 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 11, 12 и 13. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 133.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA1975 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 14, 15 и 16. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 134.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2283 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 19. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 135.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2283 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 20, 21 и 22. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 136.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2285 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 23, 24 и 3. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 137.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2285 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностью SEQ ID NO: 6, 7 и 25. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 138.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2287 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 26, 27 и 28. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 139.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2287 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 29, 30 и 31. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 140.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2290 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 32, 33 и 34. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 141.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2290 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 35, 36 и 16. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 142.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2291 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 37, 38 и 39. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 143.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2291 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 40, 41 и 42. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 144.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2386 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 43, 44 и 45. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 145.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2386 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 46, 47 и 48. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 146.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2390 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 49, 50 и 51. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 147.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2390 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 52, 53 и 54. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 148.
В другом варианте осуществления предложен полинуклеотид, кодирующий тяжелую цепь антитела SA2455 против VSIG4, описанного выше. Предпочтительно указанная тяжелая цепь содержит три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 55. Более предпочтительно, указанная тяжелая цепь включает тяжелую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 149.
В другом варианте осуществления полинуклеотид кодирует легкую цепь антитела SA2455 против VSIG4, описанного выше. Предпочтительно указанная легкая цепь содержит три CDR легкой цепи с последовательностями SEQ ID NO: 56, 57 и 58. Более предпочтительно, указанная легкая цепь включает легкую цепь, содержащую вариабельную область с последовательностью SEQ ID NO: 150.
Из-за вырожденности кодона или с учетом кодона, предпочтительного в организме, в котором должны экспрессироваться легкая и тяжелая цепи человеческого антитела или его фрагмента, полинуклеотид, кодирующий легкую и тяжелую цепи моноклонального антитела по настоящему изобретению или его антигенсвязывающий фрагмент, может иметь различные вариации в кодирующей области в диапазоне, в котором аминокислотная последовательность легкой цепи и тяжелой цепи антитела, экспрессируемого из кодирующей области, не изменяется, и даже в области, отличной от кодирующей области, различные изменения или модификации могут быть сделаны в пределах диапазона, в котором они не влияют на экспрессию гена. Специалист в данной области без труда поймет, что эти вариантные гены также подпадают под объем настоящего изобретения. А именно, при условии, что белок, обладающий эквивалентной активностью, кодируется полинуклеотидом по настоящему изобретению, одно или более оснований нуклеиновой кислоты могут быть изменены путем замены, делеции, вставки или их комбинации, и они также входят в объем настоящего изобретения. Последовательность полинуклеотида может быть одноцепочечной или двухцепочечной, и это может быть либо молекула ДНК, либо молекула РНК (мРНК).
Согласно настоящему изобретению для экспрессии антитела по изобретению можно использовать различные системы экспрессии. В одном аспекте такие системы экспрессии представляют собой носители, с помощью которых можно получить и впоследствии очистить требуемые кодирующие последовательности, а также представляют собой клетки, которые могут при временной трансфекции соответствующими нуклеотидными кодирующими последовательностями экспрессировать антитело IgG in situ.
В соответствии с настоящим изобретением предложены векторы, содержащие полинуклеотиды, описанные выше. В одном варианте осуществления вектор содержит полинуклеотид, кодирующий тяжелую цепь целевого антитела (например, антитела против VSIG4). В другом варианте осуществления полинуклеотид кодирует легкую цепь целевого антитела (например, антитела против VSIG4). В другом варианте осуществления полинуклеотид кодирует тяжелую цепь и легкую цепь целевого антитела (например, антитела против VSIG4). В еще одном варианте осуществления предложена пара полинуклеотидов, где первый полинуклеотид кодирует тяжелую цепь целевого антитела (например, антитела против VSIG4), а второй полинуклеотид кодирует легкую цепь того же целевого антитела (например, антитела против VSIG4).
В соответствии с настоящим изобретением также предложены векторы, содержащие молекулы полинуклеотидов, кодирующие слитые белки, модифицированные антитела, фрагменты антител и зонды на их основе.
Для экспрессии тяжелой и/или легкой цепи целевого антитела (например, антитела против VSIG4) полинуклеотиды, кодирующие указанные тяжелые и/или легкие цепи, встраивают в экспрессионные векторы таким образом, чтобы гены были функционально связаны с транскрипционными и трансляционными последовательностями. В предпочтительном варианте осуществления эти полинуклеотиды клонируют в два вектора.
«Функционально связанные» последовательности включают как последовательности контроля экспрессии, смежные с целевым геном, так и последовательности контроля экспрессии, которые действуют в транс-положении или на расстоянии для контроля целевого гена. Используемый в контексте данного документа термин «последовательность контроля экспрессии» относится к полинуклеотидным последовательностям, которые необходимы для воздействия на экспрессию и процессинг кодирующих последовательностей, с которыми они лигированы. Последовательности контроля экспрессии включают соответствующие последовательности инициации, терминации транскрипции, промоторные последовательности и энхансерные последовательности; сигналы эффективного процессинга РНК, такие как сигналы сплайсинга и полиаденилирования; последовательности, стабилизирующие цитоплазматическую мРНК; последовательности, повышающие эффективность трансляции (например, консенсусная последовательность Козака); последовательности, повышающие стабильность белка; и, когда требуется, последовательности, которые усиливают секрецию белка. Природа таких контрольных последовательностей различается в зависимости от организма-хозяина; у прокариот такие контрольные последовательности обычно включают промотор, сайт связывания рибосомы и последовательность терминации транскрипции; у эукариот, как правило, такие контрольные последовательности включают промоторы и последовательность терминации транскрипции. Предполагается, что термин «контрольные последовательности» включает, как минимум, все компоненты, присутствие которых необходимо для экспрессии и процессинга, а также может включать дополнительные компоненты, присутствие которых предпочтительно, например, лидерные последовательности и партнерные последовательности по слиянию.
Полинуклеотиды по изобретению и векторы, содержащие эти молекулы, можно использовать для трансформации подходящей клетки-хозяина. Термин «клетка-хозяин», используемый в контексте данного документа, предназначен для обозначения клетки, в которую был введен рекомбинантный экспрессионный вектор для экспрессии целевого антитела (например, антитела против VSIG4). Следует понимать, что такие термины предназначены для обозначения не только конкретной рассматриваемой клетки, но также и потомства такой клетки. Поскольку определенные модификации могут возникать в последующих поколениях либо из-за мутации, либо из-за влияния окружающей среды, такое потомство может фактически не быть идентичным родительской клетке, но все же включено в объем термина «клетка-хозяин», используемый в настоящем документе.
Трансформацию можно проводить любым известным способом введения полинуклеотидов в клетку-хозяина. Такие способы хорошо известны специалистам в данной области и включают опосредованную декстраном трансформацию, осаждение фосфатом кальция, опосредованную полибреном трансфекцию, слияние протопластов, электропорацию, инкапсулирование полинуклеотида в липосомы, биолистическую инъекцию и прямую микроинъекцию ДНК в ядра.
Клетка-хозяин может быть совместно трансфицирована одним или более экспрессионными кеторами. Например, клетка-хозяин может быть трансфицирована вектором, кодирующим как тяжелую цепь, так и легкую цепь целевого антитела (например, антитела против VSIG4), как описано выше. В альтернативном варианте клетка-хозяин может быть трансформирована первым вектором, кодирующим тяжелую цепь целевого антитела (например, антитела против VSIG4), и вторым вектором, кодирующим легкую цепь указанного антитела. Клетки млекопитающих обычно используют для экспрессии рекомбинантных терапевтических иммуноглобулинов, особенно для экспрессии цельных рекомбинантных антител. Например, клетки млекопитающих, такие как клетки HEK293 или СНО, в сочетании с вектором, содержащим сигнал экспрессии, таким как вектор, несущий главный промоторный элемент гена раннего ответа цитомегаловируса человека, представляют собой эффективную систему экспрессии гуманизированного антитела против VSIG4 по изобретению (Foecking et al., 1986, Gene 45:101; Cockett et al., 1990, Bio/Technology 8: 2).
Кроме того, может быть выбрана клетка-хозяин, которая модулирует экспрессию вставленных последовательностей или модифицирует и процессирует генный продукт желаемым конкретным образом. Такие модификации (например, гликозилирование) и процессинг белковых продуктов могут быть важны для функции белка. Различные клетки-хозяева имеют особенности и специфические механизмы посттрансляционного процессинга и модификации белков и генных продуктов. Подходящие клеточные линии или системы-хозяева отбирают для обеспечения правильной модификации и процессинга целевого экспрессируемого антитела. Следовательно, эукариотические клетки-хозяева, обладающие клеточным механизмом для надлежащего процессинга первичного транскрипта, могут быть использованы для гликозилирования генного продукта. Такие клетки-хозяева млекопитающих включают, но не ограничиваются ими, клетки СНО, COS, HEK293, NS/0, BHK, Y2/0, 3Т3 или клетки миеломы (все эти клеточные линии доступны в общедоступных хранилищах, таких как Национальная коллекция культур микроорганизмов, Париж, Франция, или Американская коллекция типовых культур, Манассас, Вирджиния, США).
Для долгосрочного высокопродуктивного производства рекомбинантных белков предпочтительна стабильная экспрессия. В одном варианте осуществления изобретения могут быть сконструированы клеточные линии, которые стабильно экспрессируют антитело. Вместо использования экспрессионных векторов, которые содержат точки начала репликации вируса, клетки-хозяева трансформируют с использованием ДНК под контролем соответствующих регуляторных элементов экспрессии, включая промоторы, энхансеры, терминаторы транскрипции, сайты полиаденилирования и другие соответствующие последовательности, известные специалисту в данной области, и селектируемого маркера. После введения чужеродной ДНК сконструированным клеткам можно дать возможность расти в течение одних-двух суток в обогащенной среде, а затем перенести в селективную среду. Селектируемый маркер на рекомбинантной плазмиде придает устойчивость к селекции и позволяет клеткам стабильно интегрировать плазмиду в хромосому и размножаться в клеточной линии. В данной области известны другие способы конструирования стабильных клеточных линий. В частности, были разработаны способы сайт-специфической интеграции. В соответствии с данными способами трансформированная ДНК под контролем соответствующих регуляторных элементов экспрессии, включая промоторы, энхансеры, терминаторы транскрипции, сайты полиаденилирования и другие соответствующие последовательности, интегрируется в геном клетки-хозяина на конкретном сайте-мишени, который предварительно был расщеплен (Moele et al., Proc. Natl. Acad. Sci. U.S.A., 104(9): 3055-3060; US 5792632; US 5830729; US 6238924; WO 2009/054985; WO 03/025183; WO 2004/067753).
Согласно изобретению можно использовать ряд систем селекции, включая, в том числе, тимидинкиназу вируса простого герпеса (Wigler et al., Cell 11:223, 1977), гипоксантин-гуанинфосфорибозилтрансферазу (Szybalska et al., Proc Natl Acad Sci USA 48:202, 1992), отбор глутаматсинтазы в присутствии метионинсульфоксимида (Adv Drug Del Rev, 58:671, 2006, и веб-сайт или литература Lonza Group Ltd.) и гены аденин-фосфорибозилтрансферазы (Lowy et al., Cell 22: 817, 1980), в tk, hgprt или aprt клетках соответственно. Кроме того, резистентность к антиметаболитам можно использовать как основу для селекции следующих генов: dhfr, который придает резистентность к метотрексату (Wigler et al., Proc Natl Acad Sci USA 77: 357, 1980); gpt, придающий устойчивость к микофеноловой кислоте (Mulligan et al., Proc Natl Acad Sci USA 78: 2072, 1981); neo, придающий резистентность к аминогликозиду, G-418 (Wu et al., Biotherapy 3: 87, 1991); и hygro, придающий резистентность к гигромицину (Santerre et al., Gene 30: 147, 1984). Способы, известные в области технологии рекомбинантных ДНК, могут быть обычным образом применены для отбора требуемого рекомбинантного клона, и такие способы описаны, например, в Ausubel et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons (1993). Уровни экспрессии антитела можно повысить путем векторной амплификации. Когда маркер в векторной системе, экспрессирующей антитело, является амплифицируемым, повышение уровня ингибитора, присутствующего в культуре, будет увеличивать количество копий маркерного гена. Поскольку амплифицируемая область ассоциирована с геном, кодирующим антитело IgG по изобретению, продукция указанного антитела также будет увеличиваться (Crouse et al., Mol Cell Biol 3: 257, 1983). Существуют альтернативные способы экспрессии гена по изобретению и они известны специалистам в данной области. Например, можно сконструировать модифицированный белок «цинковые пальцы», который способен связывать регуляторные элементы экспрессии выше гена по изобретению; экспрессия указанного сконструированного белка «цинковые пальцы» (ZFN) в клетке-хозяине по изобретению приводит к увеличению продукции белка (см., например, Reik et al., Biotechnol. Bioeng., 97(5): 1180-1189, 2006). Более того, ZFN может стимулировать интеграцию ДНК в заранее определенную геномную локацию, что приводит к высокоэффективному сайт-специфическому добавлению гена (Moehle et al., Proc Natl Acad Sci USA, 104: 3055, 2007).
Целевое антитело (например, антитело против VSIG4) можно получить путем выращивания культуры трансформированных клеток-хозяев в условиях культивирования, необходимых для экспрессии требуемого антитела. Полученное в результате экспрессированное антитело можно затем очистить от культуральной среды или клеточных экстрактов. Растворимые формы целевого антитела (например, антитела против VSIG4) можно выделить из культурального супернатанта. Затем его можно очистить любым способом, известным в данной области техники, для очистки молекулы иммуноглобулина, например, хроматографией (например, ионообменная, аффинная, в частности, посредством аффинности белка А к Fc и т.д.), центрифугированием, дифференциальной растворимостью или любой другой стандартной методикой очистки белков. Подходящие способы очистки будут очевидны специалисту в данной области техники.
Таким образом, другой аспект изобретения относится к способу получения антитела (например, антитела против VSIG4), описанного в настоящем документе, причем указанный способ включает стадии:
a) выращивание описанной выше клетки-хозяина в культуральной среде в подходящих условиях культивирования; а также
b) выделение антитела (например, антитела против VSIG4) из культуральной среды или из указанных культивированных клеток.
Антитело, полученное культивированием трансформанта, можно использовать в неочищенном состоянии. Примеси могут быть удалены с помощью дополнительных различных общепринятых методов, таких как центрифугирование или ультрафильтрация, и полученный продукт может быть подвергнут диализу, осаждению солей, хроматографии и т.п., при этом метод может использоваться либо отдельно, либо в комбинации. Среди них наиболее широко используется аффинная хроматография, включая ионообменную хроматографию, эксклюзионную хроматографию, хроматографию гидрофобного взаимодействия, хроматографию на гидроксиапатите и т.п.
Фармацевтические композиции
В другом аспекте настоящее изобретение относится к композициям, содержащим антитело против VSIG4 или его антигенсвязывающий фрагмент, такое как, например, любое из антител против VSIG4, описанных в настоящем документе, или их конъюгат, т.е. иммуноконъюгат, содержащий одно из антител против VSIG4, описанных в настоящем документе.
Эти композиции особенно полезны, например, для стимулирования иммунного ответа у субъекта. Антитело по настоящему изобретению, которое специфически связывается с VSIG4, индуцирует активацию Т-клеток путем связывания с белком VSIG4, что ингибирует активацию Т-клеток, и, таким образом, антитело может стимулировать иммунный ответ.
Описанные в данном документе композиции также пригодны для лечения рака. Защитный противоопухолевый иммунитет может быть создан введением таких композиций, содержащих антитело против VSIG4, его антигенсвязывающие фрагменты или их конъюгаты, которые раскрыты в настоящем документе.
Необязательно композиции могут содержать одно или более дополнительных терапевтических средств, таких как ингибиторы иммунных контрольных точек, описанные ниже. Композиции обычно поставляются как часть стерильной фармацевтической композиции, которая обычно включает фармацевтически приемлемый носитель и/или вспомогательное вещество. Таким образом, в другом аспекте в соответствии с настоящим изобретением предложена фармацевтическая композиция, содержащая антитело против VSIG4 или его конъюгат и фармацевтически приемлемый носитель и/или вспомогательное вещество.
Эта композиция может находиться в любой подходящей форме (в зависимости от желаемого способа ее введения пациенту). Композиции, используемые в описанных в данном документе способах, можно вводить, например, интравитреально (в стекловидное тело) (например, интравитреальной инъекцией), посредством глазных капель, внутримышечно, внутривенно, внутрикожно, чрескожно, внутриартериально, внутрибрюшинно, внутрь очага поражения, внутрь черепа, внутрь сустава, внутрь простаты, интраплеврально, интратрахеально, интратекально, интраназально, интравагинально, интраректально, местно, внутриопухолево, перитонеально, подкожно, субконъюнктивально, интравезикулярно, мукозально, интраперикардиально, внутрипузырно, интраокулярно, интраорбитально, перорально, местно, чрескожно, путем ингаляций, путем инъекций, путем имплантации, путем инфузии, путем непрерывной инфузии, путем локализованной перфузии, непосредственно омывающей клетки-мишени, с помощью катетера, путем лаважа, кремов или липидных композиций. Композиции, используемые в описанных здесь способах, также можно вводить системно или местно. Способ введения может варьироваться в зависимости от различных факторов (например, вводимого соединения или композиции и тяжести состояния, заболевания или расстройства, подлежащего лечению). Наиболее подходящий путь введения в каждом конкретном случае будет зависеть от конкретного антитела, субъекта, характера и тяжести заболевания и физического состояния субъекта. Антитело против VSIG4, его антигенсвязывающий фрагмент или его конъюгат могут быть приготовлены в виде водного раствора и введены путем подкожной инъекции.
Фармацевтические композиции могут быть в целях удобства представлены в виде единичных дозированных форм, содержащих заданное количество антитела против VSIG4, его антигенсвязывающего фрагмента или его конъюгата на дозу. Такая единица может содержать, например, но без ограничения, от 5 мг до 5 г, например, от 10 мг до 1 г или от 20 до 50 мг. Фармацевтически приемлемые носители для использования в настоящем изобретении могут принимать самые разнообразные формы в зависимости, например, от состояния, подлежащего лечению, или пути введения.
Фармацевтические композиции по настоящему изобретению могут быть приготовлены для хранения в виде лиофилизированных составов или водных растворов путем смешивания антитела, имеющего требуемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами, обычно используемыми в данной области (все они упоминаются здесь как «носители»), а именно, буферными агентами, стабилизаторами, консервантами, изотонирующими добавками, неионогенными детергентами, антиоксидантами и другими различными добавками. См. Remington's Pharmaceutical Sciences, 16-е издание (Osol, ed. 1980). Такие добавки должны быть нетоксичными для реципиентов в применяемых дозировках и концентрациях.
Буферные агенты помогают поддерживать рН в диапазоне, близком к физиологическим условиям. Они могут присутствовать в диапазоне концентраций от примерно 2 мМ до примерно 50 мМ. Подходящие буферные агенты для использования в соответствии с настоящим изобретением включают как органические, так и неорганические кислоты и их соли, такие как цитратные буферы (например, смесь цитрата натрия и цитрата динатрия, смесь лимонной кислоты и тринатриевого цитрата, смесь лимонной кислоты и цитрата натрия и т.д.), сукцинатные буферы (например, смесь янтарной кислоты и сукцината натрия, смесь янтарной кислоты и гидроксида натрия, смесь янтарной кислоты и динатрийсукцината и т.д.), тартратные буферы (например, смесь винной кислоты и тартрата натрия, смесь винной кислоты и тартрата калия, смесь винной кислоты и гидроксида натрия и др.), фумаратные буферы (например, смесь фумаровой кислоты и фумарата натрия, смесь фумаровой кислоты и фумарата динатрия, смесь фумарата натрия и фумарата динатрия и т.д.), глюконатные буферы (например, смесь глюконовой кислоты и глюконата натрия, смесь глюконовой кислоты и гидроксида натрия, смесь глюконовой кислоты и глюконата калия и т.д.), оксалатный буфер (например, смесь щавелевой кислоты и оксалата натрия, смесь щавелевой кислоты и гидроксида натрия, смесь щавелевой кислоты и оксалата калия и т.д.), лактатные буферы (например, смесь молочной кислоты и лактата натрия, смесь молочной кислоты и гидроксида натрия, смесь молочной кислоты и лактата калия и т.д.) и ацетатные буферы (например, смесь уксусной кислоты и ацетата натрия, смесь уксусной кислоты и гидроксида натрия и т.д.). Кроме того, можно использовать фосфатные буферы, гистидиновые буферы и соли триметиламина, такие как Трис.
Консерванты могут быть добавлены для замедления роста микробов в количестве от 0,2% до 1% (масса/объем). Подходящие консерванты для использования в соответствии с настоящим изобретением включают фенол, бензиловый спирт, метакрезол, метилпарабен, пропилпарабен, хлорид октадецилдиметилбензиламмония, галогениды бензалкония (например, хлорид, бромид и йодид), хлорид гексаметония и алкилпарабены, такие как метилпарабены или пропилпарабен, катехол, резорцин, циклогексанол и 3-пентанол.
Изотонические вещества, иногда известные как «стабилизаторы», могут быть добавлены для обеспечения изотоничности жидких композиций по настоящему изобретению и включают многоатомные сахарные спирты, например, трехатомные или высшие сахарные спирты, такие как глицерин, эритрит, арабит, ксилит, сорбит и маннит. Стабилизаторы относятся к широкой категории вспомогательных веществ, функции которых могут варьироваться от наполнителя до добавки, которая солюбилизирует терапевтическое средство (например, антитело против VSIG4, его антигенсвязывающий фрагмент или его конъюгат) или помогает предотвратить денатурацию или прилипание к стенке контейнера.
Типичными стабилизаторами могут быть многоатомные сахароспирты (перечислены выше); аминокислоты, такие как аргинин, лизин, глицин, глутамин, аспарагин, гистидин, аланин, орнитин, L-лейцин, 2-фенилаланин, глутаминовая кислота, треонин и т.д., органические сахара или сахарные спирты, такие как лактоза, трегалоза, стахиоза, маннит, сорбит, ксилит, рибит, миоинизит, галактит, глицерин и т.п., включая циклиты, такие как инозитол; полиэтиленгликоль; полимеры аминокислот; серосодержащие восстановители, такие как мочевина, глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, α-монотиоглицерин и тиосульфат натрия; низкомолекулярные полипептиды (например, пептиды из 10 остатков или менее); белки, такие как человеческий сывороточный альбумин, бычий сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидоновые моносахариды, такие как ксилоза, манноза, фруктоза, глюкоза; дисахариды, такие как лактоза, мальтоза, сахароза и трисахариды, такие как раффиноза; и полисахариды, такие как декстран. Стабилизаторы могут присутствовать в количестве от 0,1 до 10000 масс на часть массы активного белка (например, антитела против VSIG4 или конъюгата, содержащего такое антитело).
Неионогенные поверхностно-активные вещества или детергенты (также известные как «смачивающие агенты») могут быть добавлены для способствования солюбилизации антитела против VSIG4 (или его конъюгата), а также для защиты терапевтического белка от агрегации, вызванной перемешиванием, что также дает возможность подвергать препарат поверхностному напряжению сдвига, не вызывая денатурации белка. Подходящие неионогенные поверхностно-активные вещества включают полисорбаты (20, 80 и т.д.), полиоксамеры (184, 188 и т.д.), плюроновые полиолы, моноэфиры полиоксиэтиленсорбитана (TWEEN®-20, TWEEN®-80 и т.д.). Неионогенные поверхностно-активные вещества могут присутствовать в диапазоне от примерно 0,05 мг/мл до примерно 1,0 мг/мл, например, от примерно 0,07 мг/мл до примерно 0,2 мг/мл.
Дополнительные различные вспомогательные вещества включают наполнители (например, крахмал), хелатирующие агенты (например, ЭДТА), антиоксиданты (например, аскорбиновая кислота, метионин, витамин Е) и сорастворители.
В соответствии с настоящим изобретением также предложена фармацевтическая композиция, содержащая по меньшей мере:
i) антитело против VSIG4, его антигенсвязывающий фрагмент или его конъюгат, как раскрыто в настоящем документе, и
ii) второй терапевтический агент, например, ингибитор иммунной контрольной точки, как описано ниже,
в виде комбинированных продуктов для одновременного, раздельного или последовательного применения.
«Одновременное применение» в контексте данного документа относится к введению двух соединений композиции по изобретению в одной и идентичной фармацевтической форме.
Используемый в контексте данного документа термин «раздельное применение» относится к одновременному введению двух соединений композиции по изобретению в разных фармацевтических формах.
«Последовательное применение» в контексте настоящего документа относится к последовательному введению двух соединений композиции по изобретению, каждого в отдельной фармацевтической форме.
Композиции антител против VSIG4 (или их антигенсвязывающих фрагментов или их конъюгатов) и вторых терапевтических агентов, таких как, например, ингибиторы иммунных контрольных точек, можно вводить однократно, в виде смесей одного или более антител против VSIG4 (или их антигенсвязывающих фрагментов или их конъюгатов) и/или одного или более вторых терапевтических агентов (например, ингибитор иммунных контрольных точек, как описано ниже), в смеси или комбинации с другими агентами, пригодными для лечения рака или в качестве дополнения к другой терапии рака. Ниже приведены примеры подходящей комбинированной и вспомогательной терапии.
Настоящее изобретение охватывает фармацевтические наборы, содержащие антитела против VSIG4 (или их антигенсвязывающие фрагменты или их конъюгаты) и описанные в настоящем документе. Фармацевтический набор представляет собой упаковку, содержащую антитело против VSIG4 (например, либо в лиофилизированной форме, либо в форме водного раствора) и один или более из следующих компонентов:
• Второй терапевтический агент, например ингибитор иммунной контрольной точки, как описано ниже;
• Устройство для введения антитела против VSIG4, например ручка, игла и/или шприц; а также
• Вода или буфер фармацевтической степени чистоты для ресуспендирования антитела, если ингибитор находится в форме антитела.
Каждая однократная доза антитела против VSIG4 (или его антигенсвязывающих фрагментов или его конъюгатов) может быть упакована отдельно, а набор может содержать одну или более однократных доз (например, две однократные дозы, три однократные дозы, четыре однократные дозы, пять однократных доз, восемь однократных доз, десять однократных доз или более. В конкретном варианте осуществления каждая из одной или более однократных доз находится в шприце или шприц-ручке.
Эффективные количества
Антитела против VSIG4, их антигенсвязывающие фрагменты и их конъюгаты, необязательно в комбинации с ингибиторами иммунных контрольных точек, обычно используют в количестве, эффективном для достижения желаемого результата, например, в количестве, эффективном для лечения рака у субъекта, нуждающегося в этом. Фармацевтические композиции, содержащие антитела против VSIG4 (или их антигенсвязывающие фрагменты или их конъюгаты) и/или ингибиторы иммунных контрольных точек можно вводить пациентам (например, людям) в терапевтически эффективных дозах.
Определение эффективного количества находится в пределах компетенции специалистов в данной области техники, особенно в свете подробного описания, представленного в настоящем документе. Токсичность и терапевтическую эффективность соединения или конъюгата можно определить стандартными фармацевтическими методиками на клеточных культурах и на экспериментальных животных. Эффективное количество настоящей комбинации или другого терапевтического средства для введения субъекту будет зависеть от стадии, категории и статуса заболевания (например, рака) и характеристик субъекта, таких как общее состояние здоровья, возраст, пол, масса тела и толерантность к лекарствам. Эффективное количество вводимого терапевтического средства или комбинации по настоящему изобретению также будет зависеть от способа введения и лекарственной формы. Величину дозы и интервал можно регулировать индивидуально для обеспечения уровней активного соединения в плазме, достаточных для поддержания требуемых терапевтических эффектов.
Количество вводимого антитела против VSIG4 или его антигенсвязывающего фрагмента или их конъюгатов будет зависеть от множества факторов, включая природу и стадию заболевания, которое лечат (например, рак), форму, путь и место введения, терапевтический режим (например, используется ли терапевтическое средство в комбинации с ингибиторами иммунных контрольных точек), возраст и состояние конкретного субъекта, подвергающегося лечению, восприимчивость пациента, которого лечат антителами или конъюгатами.
Подходящая доза может без труда быть определена специалистом в данной области. В конечном счете, врач определит соответствующие дозы, предназначенные для использования. Эту дозу можно применять повторно так часто, как это необходимо. Если развиваются побочные эффекты, количество и/или частота дозы могут быть изменены или уменьшены в соответствии с обычной клинической практикой. Надлежащая доза и схема лечения могут быть установлены путем наблюдения за ходом терапии с использованием обычных методов, известных специалистам в данной области.
Произвести начальную оценку эффективных доз можно, исходя из in vitro анализов. Например, начальную дозу для применения у животных можно составить таким образом, чтобы достичь концентрации антитела против VSIG4 в циркулирующей крови или сыворотке, которая равна или превышает аффинность связывания антитела против VSIG4, измеренную in vitro. Расчет доз для достижения таких концентраций в циркулирующей крови или сыворотке с учетом биодоступности конкретного антитела находится в пределах способностей квалифицированных специалистов. В качестве руководства читатель может обратиться к Fingl & Woodbury, «General Principles» в Goodman and Gilman's The Pharmaceutical Basis of Therapeutics, глава 1, последнее издание, Pagamonon Press, а также к приведенным в них ссылкам. Произвести оценку начальных доз можно, исходя из данных in vivo, например, животных моделей. Животные модели, пригодные для тестирования эффективности соединений для лечения конкретных заболеваний, таких как рак, обычно хорошо известны в данной области. Специалисты в данной области могут согласно обычной практике адаптировать такую информацию для того, чтобы определить дозы, подходящие для введения человеку.
Эффективная доза антитела против VSIG4, описанного в настоящем документе, может находиться в диапазоне от примерно 0,001 до примерно 75 мг/кг при однократном (например, струйном) введении, многократных введениях или непрерывном введении, или для достижения концентрации в сыворотке 0,01-5000 мкг/мл при однократном (например, струйном) введении, многократных введениях или непрерывном введении, или любом эффективном диапазоне или значении в зависимости от состояния, подвергаемого лечению, пути введения и возраста, веса и состояния субъекта. В конкретном варианте осуществления каждая доза может составлять от около 0,5 мкг до около 50 мкг на килограмм массы тела, например, от около 3 мкг до около 30 мкг на килограмм массы тела.
Количество, частота и продолжительность введения будут зависеть от множества факторов, таких как возраст пациента, вес и степень заболевания. Терапевтический режим введения может длиться от 2 недель до неопределенного срока, от 2 недель до 6 месяцев, от 3 месяцев до 5 лет, от 6 месяцев до 1 или 2 лет, от 8 месяцев до 18 месяцев или т.п. Необязательно терапевтический режим предусматривает повторное введение, например, один раз в сутки, два раза в сутки, каждые двое суток, трое суток, пять суток, каждую неделю, две недели или каждый месяц. Повторное введение может осуществляться в той же дозе или в другой дозе. Введение можно повторить один раз, два раза, три раза, четыре раза, пять раз, шесть раз, семь раз, восемь раз, девять раз, десять раз или более. Терапевтически эффективное количество антитела против VSIG4 или его конъюгата (необязательно в комбинации с ингибиторами иммунных контрольных точек) можно вводить в виде однократной дозы или на протяжении курса лечения, например, не протяжении недели, двух недель, трех недель, одного месяца, трех месяцев, шести месяцев, одного года или дольше.
Способы лечения
Способность антител против VSIG4 по настоящему изобретению индуцировать иммунный ответ, например, путем стимуляции дифференцировки М2-макрофагов и ингибирования VSIG4-опосредованной иммуносупрессии, делает их пригодными для лечения различных состояний, опосредованных VSIG4, включая рак. Таким образом, терапевтическое воздействие на путь ингибирования VSIG4 является многообещающим подходом к модулированию воспаления и Т-кпеточного иммунитета для лечения широкого спектра видов рака.
Таким образом, антитело против VSIG4, его антигенсвязывающий фрагмент или конъюгат, описанные в настоящем документе, можно использовать в способах лечения рака, индукции высвобождения провоспалительных цитокинов макрофагами, индукции пролиферации CD4+ Т-клеток, индукции пролиферации CD8+ Т-клеток, индукции продуцирования цитокинов CD4+ Т-клетками и индукции продуцирования цитокинов CD8+ Т-клетками, где указанные способы включают введение эффективного количества антитела против VSIG4, его антигенсвязывающего фрагмента или конъюгата субъекту, нуждающемуся в этом. Терапевтические способы, описанные в настоящем документе, могут включать введение антител, специфически связывающихся с VSIG4, описанных в настоящем документе, или их антигенсвязывающих фрагментов, или конъюгатов, содержащих данные антитела, как раскрыто в настоящем документе, пациенту, нуждающемуся в этом. Таким образом, антитела против VSIG4, их антигенсвязывающие фрагменты и их конъюгаты, раскрытые в настоящем документе, полезны при регуляции иммунитета, в частности Т-клеточного иммунитета, для лечения рака.
Соответственно, один из аспектов настоящего изобретения относится к антителу против VSIG4 или его антигенсвязывающему фрагменту или его конъюгату для применения при лечении рака у пациента. Кроме того, в настоящем документе предложен способ лечения рака у нуждающегося в этом субъекта, причем указанный способ включает введение пациенту антитела против VSIG4, его антигенсвязывающего фрагмента или конъюгата, раскрытых в настоящем документе. Настоящее изобретение также относится к применению антитела против VSIG4 или его антигенсвязывающего фрагмента или его конъюгата для изготовления лекарственного средства для лечения рака.
В одном из вариантов осуществления изобретение относится к композиции, содержащей антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент или конъюгат, для применения при лечении рака у пациента. Кроме того, в настоящем документе также предложен способ лечения рака у субъекта, нуждающегося в этом, причем указанный способ включает введение пациенту композиции, содержащей антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент или конъюгат. Настоящее изобретение также относится к применению композиции, содержащей раскрытое в данном документе антитело против VSIG4, или его антигенсвязывающий фрагмент, или конъюгат, для изготовления лекарственного средства для лечения рака.
В некоторых вариантах осуществления рак выбран из рака мочевого пузыря, рака молочной железы, рака шейки матки, рака толстой кишки, рака эндометрия, рака пищевода, рака фаллопиевых труб, рака желчного пузыря, рака желудочно-кишечного тракта, рака головы и шеи, гематологического рака (например, лейкемии, лимфомы или миеломы), рака гортани, рака печени, рак легких, лимфомы, меланомы, мезотелиомы, рака яичников, первичного рака брюшины, рака слюнных желез, саркомы, рака желудка, рака щитовидной железы, рака поджелудочной железы, почечно-кпеточной карциномы, глиобластомы и рака предстательной железы.
В одном из вариантов осуществления предложено антитело против VSIG4 или его антигенсвязывающий фрагмент, или его конъюгат для применения в индукции иммунного ответа у пациента с раком. В настоящем документе также предложен способ индукции иммунного ответа у пациента с раком, нуждающегося в этом, причем указанный способ включает введение пациенту антитела против VSIG4, его антигенсвязывающего фрагмента или конъюгата, раскрытых в настоящем документе. Настоящее изобретение также относится к применению антитела против VSIG4 или его антигенсвязывающего фрагмента или его конъюгата для изготовления лекарственного средства для индукции иммунного ответа у пациента с раком.
В одном из вариантов осуществления изобретение относится к композиции, содержащей антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент или конъюгат для применения при индукции иммунного ответа у пациента с раком. В настоящем документе также предложен способ иммунного ответа у пациента с раком, нуждающегося в этом, причем указанный способ включает введение пациенту композиции, содержащей антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент или конъюгат. Настоящее изобретение также относится к применению композиции, содержащей антитело против VSIG4, раскрытое в настоящем документе, или его антигенсвязывающий фрагмент или конъюгат, для изготовления лекарственного средства для индукции иммунного ответа у пациента с раком.
Таким образом, иммунный ответ, генерируемый антителом, раскрытым в данном документе, включает, без ограничения, индукцию высвобождения провоспалительных цитокинов макрофагами, индукцию пролиферации CD4+ Т-клеток, индукцию пролиферации CD8+ Т-клеток, индукцию продукции цитокинов CD4+ Т-клетками и индукцию продукции цитокинов CD8+ Т-клетками.
Антитело против VSIG4 или его антигенсвязывающий фрагмент или конъюгат можно смешивать с дополнительными химиотерапевтическими агентами, цитотоксическими агентами, антителами, лимфокинами или гемопоэтическим фактором роста. Примечательно, что терапевтические способы, описанные в настоящем документе, могут включать введение ингибитора иммунных контрольных точек вместе с антителом против VSIG4 или его антигенсвязывающим фрагментом или конъюгатом. Ингибитор иммунных контрольных точек и антитело против VSIG4 или его антигенсвязывающий фрагмент или конъюгат можно вводить одновременно, раздельно или последовательно.
Используемый в контексте данного документа термин «ингибитор контрольной точки» относится к молекуле, такой как, например, малая молекула, растворимый рецептор или антитело, которая нацеливается на иммунную контрольную точку и блокирует функцию указанной иммунной контрольной точки. Более конкретно, «ингибитор контрольной точки», используемый в данном документе, представляет собой молекулу, такую как, например, малая молекула, растворимый рецептор или антитело, которая блокирует определенные белки, вырабатываемые некоторыми типами клеток иммунной системы, такими как Т-клетки, и некоторые раковые клетки.
В первом варианте осуществления ингибитор иммунной контрольной точки представляет собой ингибитор любого из CTLA-4, PDL1, PDL2, PD1, В7-Н3, В7-Н4, BTLA, HVEM, TIM3, GAL9, LAG3, VISTA, KIR, 2В4 (принадлежит к семейству молекул CD2 и экспрессируется на всех NK, γδ и CD8+ (αβ) Т-клетках памяти), CD160 (также обозначаемый BY55), CGEN-15049, киназ CHK1 и CHK2, IDO1, A2aR и любых различных лигандов семейства В-7.
Типичные ингибиторы иммунных контрольных точек включают антитело против CTLA-4 (например, ипилимумаб), антитело против LAG-3 (например, BMS-986016), антитело против В7-Н3, антитело против В7-Н4, антитело против Tim3 (например, TSR-022, MBG453), антитело против BTLA, антитело против KIR, антитело против A2aR, антитело против CD200, антитело против PD-1 (например, пембролизумаб, ниволумаб, цемиплимаб, пидилизумаб), антитело против PD-L1 (например, атезолизумаб, авелумаб, дурвалумаб, BMS 936559), антитело против VISTA (например, JNJ 61610588), антитело против CD28, антитело против CD80 или -CD86, антитело против B7RP1, антитело против В7-Н3, антитело против HVEM, антитело против CD137 (например, урелумаб), антитело против CD137L, антитело против ОХ40 (например, 9 В12, PF-04518600, MEDI6469), антитело против OX40L, антитело против CD40 или -CD40L, антитело против GAL9, антитело против IL-10, слитый белок внеклеточного домена лиганда PD-1, например PDL-1 или PD-L2 и IgG1 (например, АМР-224), слитый белок внеклеточного домена лиганда ОХ40, например OX40L и IgG1 (например, MEDI6383), ингибитор IDO1 (например, эпакадостат) и лекарство, нацеленное на A2aR. Ряд ингибиторов иммунных контрольных точек был одобрен или в настоящее время проходит клинические испытания. К таким ингибиторам относятся ипилимумаб, пембролизумаб, ниволумаб, цемиплимаб, пидилизумаб, атезолизумаб, авелумаб, дурвалумаб, BMS 936559, JNJ 61610588, urelumab, 9 В12, PF-04518600, BMS-986016, TSR-022, MBG453, MEDI6469, MEDI6383 и эпакадостат.
Примеры ингибиторов иммунных контрольных точек приведены, в частности, в Marin-Acevedo et al., Journal of Hematology & Oncology 11:8, 2018; Kavecansky and Pavlick, AJHO 13(2): 9-20, 2017; Wei et al., Cancer Discov8(9): 1069-86, 2018.
Предпочтительно ингибитор иммунной контрольной точки представляет собой ингибитор CTLA-4, LAG-3, Tim3, PD-1, PD-L1, VISTA, CD137, ОХ40, или IDO1.
Способы диагностики
VSIG4 сверхэкспрессируется при различных видах рака, что указывает на то, что VSIG4 является надежным биомаркером для диагностики рака. Таким образом, реагенты, такие как предложенные в настоящем документе меченые антитела, которые связываются с белком VSIG4, можно использовать в диагностических целях для выявления, диагностики или мониторинга клеточно-пролиферативного заболевания, расстройства или состояния, такого как, например, рак.
Антитела против VSIG4, предложенные в настоящем документе, можно использовать для выявления VSIG4 или анализа уровней VSIG4 в биологическом образце с использованием классических иммуногистологических способов, описанных в настоящем документе или известных специалистам в данной области (например, см. Jalkanen et al., 1985, J. Cell Biol., 101:976-985 и Jalkanen et al., 1987, J.Cell.Biol., 105:3087-3096). Другие способы, основанные на антителах, пригодные для выявления экспрессии генов белков, включают иммунологические анализы, такие как твердофазный иммуноферментный анализ (ELISA) и радиоиммуноанализ (RIA). Подходящие метки для анализа на основе антител известны в данной области и включают ферментные метки, такие как глюкозооксидаза; радиоизотопы, такие как йод (125I, 121I), углерод (14С), сера (35S), тритий (3Н), индий (121In) и технеций (99Тс); люминесцентные метки, такие как люминол; и флуоресцентные метки, такие как флуоресцеин и родамин, и биотин.
Таким образом, в первом аспекте настоящее изобретение относится к способу in vitro обнаружения рака, экспрессирующего VSIG4, у субъекта, причем указанный способ включает следующие стадии:
a) приведение биологического образца указанного субъекта в контакт с антителом против VSIG4, раскрытым в данном документе, или его антигенсвязывающим фрагментом; и
b) обнаружение связывания указанного реагента с указанным биологическим образцом.
В соответствии с настоящим способом связывание VSIG4 указывает на наличие рака, экспрессирующего VSIG4. Предпочтительно связывание антитела против VSIG4 в иммунных инфильтратах микроокружения опухоли указывает на наличие рака, экспрессирующего VSIG4.
Изобретение также относится к способу in vitro обнаружения рака, экспрессирующего VSIG4, у субъекта, причем указанный способ включает следующие стадии:
a) приведение в контакт биологического образца указанного субъекта с антителом против VSIG4 или его антигенсвязывающим фрагментом; и
b) количественное определение связывания указанного реагента с указанным биологическим образцом.
Согласно настоящему способу связывание VSIG4 указывает на наличие рака, экспрессирующего VSIG4. Предпочтительно связывание антитела против VSIG4-B иммунных инфильтратах микроокружения опухоли указывает на наличие рака, экспрессирующего VSIG4.
Как будет очевидно специалисту в данной области техники, уровень связывания антитела с VSIG4 можно количественно определить любым способом, известным специалисту в данной области, как подробно описано ниже. Предпочтительные способы включают использование иммуноферментных анализов, таких как ELISA или ELISPOT, иммунофлуоресценции, иммуногистохимии (ИГХ), радиоиммуноанализа (РИА) или FACS.
Количественное определение на стадии b) настоящего способа является прямым отражением уровня экспрессии VSIG4 в образце, особенно в иммунных инфильтратах микроокружения опухоли. Таким образом, настоящий способ обеспечивает возможность идентифицировать рак, экспрессирующий VSIG4, путем определения уровня экспрессии VSIG4, как описано выше. В предпочтительном варианте осуществления уровень экспрессии VSIG4 в указанном образце, особенно в иммунных инфильтратах микроокружения опухоли, сравнивают с эталонным уровнем.
В соответствии с еще одним предпочтительным вариантом осуществления изобретение относится к способу обнаружения рака in vitro, экспрессирующего VSIG4, у субъекта, причем указанный способ включает следующие стадии:
а) определение уровня экспрессии VSIG4 в биологическом образце указанного субъекта; и
б) сравнение уровня экспрессии на стадии а) с эталонным уровнем;
при этом повышение анализируемого уровня VSIG4 на стадии а) по сравнению с эталонным уровнем свидетельствует о раке, экспрессирующем VSIG4.
Настоящее изобретение также относится к способу in vitro диагностики рака, экспрессирующего VSIG4, у субъекта, причем указанный способ включает следующие стадии:
а) определение уровня экспрессии VSIG4 в биологическом образце указанного субъекта; и
б) сравнение уровня экспрессии на стадии а) с эталонным уровнем;
при этом повышение анализируемого уровня VSIG4 на стадии а) по сравнению с эталонным уровнем свидетельствует о раке, экспрессирующем VSIG4.
Уровень экспрессии VSIG4 предпочтительно сравнивают или измеряют относительно уровней в контрольной клетке или образце, также называемых «эталонным уровнем» или «эталонным уровнем экспрессии». «Эталонный уровень», «эталонный уровень экспрессии», «контрольный уровень» и «контроль» используются в данном описании взаимозаменяемо. «Контрольный уровень» означает отдельно взятый исходный уровень, измеренный в сравниваемой контрольной клетке, в которой как правило не имеется заболеваний или рака. Указанная контрольная клетка может быть получена от того же индивидуума, поскольку даже у пациента с раком ткань, являющаяся местом расположения опухоли, все еще содержит неопухолевую здоровую ткань. Он также может происходить от другого индивидуума, который является нормальным или не страдает тем же заболеванием, что и индивидуум, от которого получен пораженный заболеванием или исследуемый образец. В контексте настоящего изобретения термин «эталонный уровень» относится к «контрольному уровню» экспрессии VSIG4, используемому для оценки тестируемого уровня экспрессии VSIG4 в образце пациента, содержащем раковые клетки. Например, когда уровень VSIG4 в биологическом образце пациента выше эталонного уровня VSIG4, считается, что клетки имеют высокий уровень экспрессии или сверхэкспрессию VSIG4. Эталонный уровень может можно определить множеством способов. Таким образом, уровни экспрессии могут выявлять клетки, несущие VSIG4, или, в альтернативном варианте, уровень экспрессии VSIG4 вне зависимости от количества клеток, экспрессирующих VSIG4. Таким образом, эталонный уровень для каждого пациента может быть задан с помощью эталонного соотношения VSIG4, при этом указанное эталонное соотношение может быть определено любым из способов определения эталонных уровней, описанных в настоящем документе.
Например, контроль может представлять собой заданное значение, которое может принимать различные формы. Это может быть одно пороговое значение, например медиана или среднее значение. «Эталонный уровень» может быть одним числом, одинаково применимым к каждому отдельно взятому пациенту, или эталонный уровень может варьироваться в зависимости от конкретных подгрупп пациентов. Так, например, пожилые мужчины могут иметь эталонный уровень, отличный от эталонного уровня молодых мужчин с тем же раковым заболеванием, а женщины могут иметь эталонный уровень, отличный от эталонного уровня мужчин с тем же раковым заболеванием. В альтернативном варианте «эталонный уровень» можно определить путем измерения уровня экспрессии VSIG4 в неонкогенных раковых клетках из той же ткани, что и ткань тестируемых неопластических клеток. Кроме того, «эталонный уровень» может представлять собой определенное отношение VSIG4 в неопластических клетках пациента к уровням VSIG4 в неопухолевых клетках у того же пациента. «Эталонный уровень» также может представлять собой уровень VSIG4 в клетках, культивируемых in vitro, которым можно манипулировать для имитации опухолевых клеток, или можно манипулировать любым другим способом, который дает экспрессионные уровни, в точности определяющие эталонный уровень. С другой стороны, «эталонный уровень» может быть установлен на основе сравнительных групп, таких как группы без повышенных уровней VSIG4 и группы с повышенными уровнями VSIG4. Другим примером сравнительных групп могут быть группы, имеющие конкретное заболевание, состояние или симптомы, и группы без заболевания. Заданное значение может быть установлено, например, когда испытуемая популяция делится поровну (или неравномерно) на группы, такие как группа низкого риска, группа среднего риска и группа высокого риска.
Эталонный уровень также можно определить путем сравнения уровня VSIG4 в популяциях пациентов с одним и тем же раком. Этого можно добиться, например, с помощью анализа гистограмм, в котором графически представлена вся группа пациентов, где первая ось обозначает уровень VSIG4, а вторая ось обозначает количество пациентов в группе, опухолевые клетки которых экспрессируют VSIG4 на заданном уровне. Две или более отдельные группы пациентов могут быть определены путем идентификации субпопуляций группы, которые имеют одинаковые или близкие уровни VSIG4. Затем можно определить эталонный уровень, исходя из уровня, который наилучшим образом различает данные отдельные группы. Эталонный уровень также может представлять уровни двух или более маркеров, одним из которых является VSIG4. Два или более маркера могут быть представлены, например, соотношением значений для уровней каждого маркера.
Аналогичным образом, клинически здоровая популяция будет иметь «нормальный» диапазон, отличный от диапазона популяции, о которой известно, что у нее имеется состояние, связанное с экспрессией VSIG4. Соответственно, выбранное заданное значение может учитывать категорию, в которую попадает индивидуум. Соответствующие диапазоны и категории могут быть выбраны специалистом в данной области не более чем путем рутинных экспериментов. Под «повышенным» подразумевается высокий уровень по сравнению с выбранным контролем. Как правило, контроль будет основываться на клинически здоровых нормальных индивидуумах в соответствующей возрастной группе.
Также следует понимать, что контролями согласно изобретению могут быть, в дополнение к заданным значениям, образцы материалов, тестируемых параллельно с экспериментальными материалами. Примеры включают ткани или клетки, полученные одновременно от одного и того же субъекта, например, части одной биопсии или части одного клеточного образца от субъекта.
Предпочтительно эталонный уровень VSIG4 представляет собой уровень экспрессии VSIG4 в образцах нормальной ткани (например,от пациента, не имеющего рака, экспрессирующего VSIG4, или от того же пациента до начала заболевания).
Более точный диагноз рака, экспрессирующего VSIG4, может позволить медицинским работникам применять профилактические меры или агрессивное лечение раньше, тем самым предотвращая развитие или дальнейшее прогрессирование рака, экспрессирующего VSIG4.
Ниже настоящее изобретение подробно поясняется с учетом примеров. Однако следующие примеры даны только для иллюстрации настоящего изобретения, и очевидно, что настоящее изобретение не ограничивается следующими примерами.
ПРИМЕРЫ
Пример 1: свойства длинных и коротких форм VSIG4
1-1. Экспрессия длинных и коротких форм VSIG4 на макрофагах
Известно, что VSIG4 экспрессируется макрофагами. Чтобы проверить, есть ли разница в экспрессии, в экстрактах макрофагов М1 и М2 искали каждую из двух форм VSIG4, т.е. VSIG4(L) и VSIG4(S).
50 нг/мл IFN-y (285-IF, R&D) добавляли к GM-CSF-дифференцированным МО-макрофагам для поляризации в провоспалительные М1-макрофаги. К M-CSF дифференцированным М0-макрофагам добавляли по 20 нг/мл каждого из следующих цитокинов IL-4 (130.093.922, Miltenyi Biotec), IL-10 (217-IL/CF, R&D) и TGF-|3 (130.095.066, Miltenyi Biotec) для поляризации в иммуносупрессивные М2-макрофаги. Дифференцированные МО-макрофаги инкубировали с цитокинами при 37°С, 5% CO2 в течение 2 суток. Поляризованные макрофаги М1 и М2 были получены на 8-е сутки. Поляризованные макрофаги активировали с использованием LPS в концентрации 100 нг/мл (L4516, Sigma) в течение 4 часов при 37°С, 5% CO2. Затем макрофаги собирали и промывали в культуральной среде. Связывание целевых антител с поляризованными М1-и М2-макрофагами оценивали с помощью проточной цитометрии после LPS активации.
15 мкг белковых экстрактов М1 и М2 вместе со 100 нг hVSIG4(L)-hFc и hVSIG4(S)-hFc помещали на ДСН-ПААГ гель, переносили на мембрану и зондировали либо поликлональным антителом, специфичным к VSIG4 (AF4646, R&D Systems, Миннеаполис, Миннесота, США) либо козьим изотипическим контролем.
В экстрактах М2 макрофагов были видны две полосы ожидаемых размеров (рис. 1В). Этот результат подтверждает предыдущие данные по экспрессии VSIG4 в макрофагах. Кроме того, он показывает, что как hVSIG4(S), так и hVSIG4(L) экспрессируются в макрофагах.
1-2. Экспрессия длинной и короткой форм VSIG4 в опухолях.
Исследовали экспрессию обеих форм VSIG4 в опухолях. Ген VSIG4 расположен на Х-хромосоме, и на генной модели изображено 7 экзонов. Это способствует образованию 2 информационных РНК, получаемых путем альтернативного сплайсинга. Одна длинная форма, Long-VSIG4 (uc004dwh.2) и одна короткая форма, Short-VSIG4 (uc004dwi.2), которые дают hVSIG4(L) и hVSIG4(S) соответственно. Атлас ракового генома (TCGA) содержит данные, полученные в результате характеристики более 20000 первичных раковых заболеваний и соответствующих нормальных образцов, охватывающих 33 типа рака. Данные по экспрессии опухоли TCGA (Tumor TCGA RNASeq) использовали для определения паттернов экспрессии данных двух изоформ с помощью ISOexpresso (Yang et al., BMC Genomics (2016) 17: 631; http://wiki.tqilab.org/ISOexpresso/).
Результаты представлены в Таблице 3.

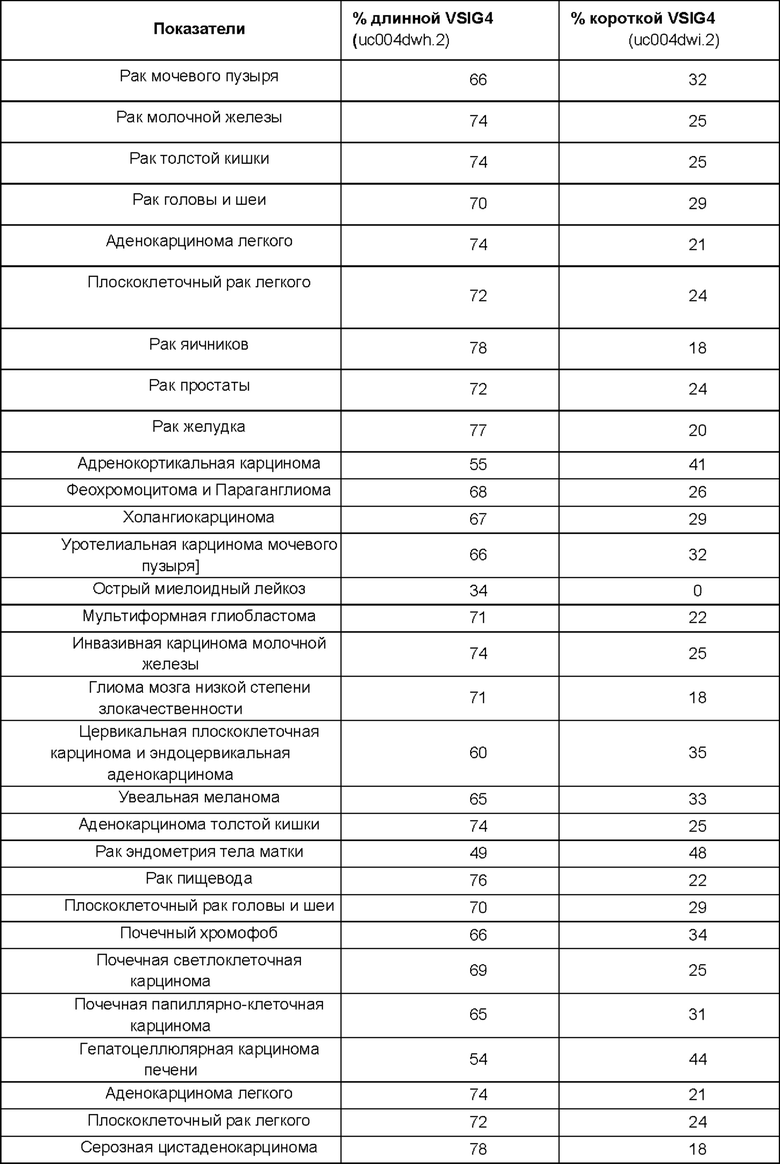
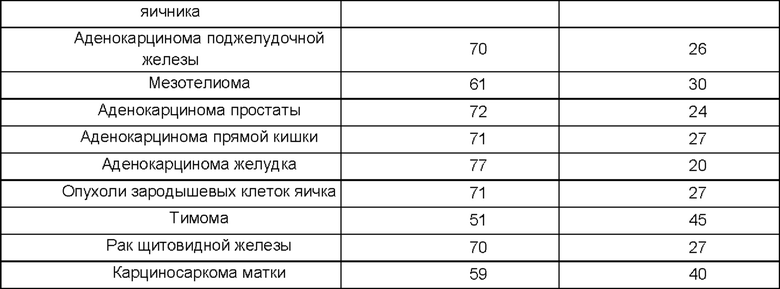
В опухолях экспрессируются как короткие, так и длинные изоформы VSIG4.
1-3. Ингибирование активации CD4+ Т-кпеток посредством hVSIG4(S) и hVSIG4(L)
96-луночные планшеты покрывали анти-CD3 антителом ОКТЗ в концентрации 2,5 мкг/мл (BioxCell ref ВЕ0001-2 клон ОКТ3) в количестве 100 мкл/лунка на 4 часа при 37°С, дважды промывали PBS и покрывали рекомбинантными белками (VSIG4(L)-Fc (SEQ ID NO: 183), +6+2 (SEQ ID NO: 184), PDL1-Fc (R&D Systems 156-B7) в концентрации 10 мкг/мл или изотипическим контролем hIgG1 (c9G4)) и инкубировали в течение ночи при 4°С. Лунки дважды промывали PBS и в каждую лунку, в 200 мкл культуральной среды, добавляли 200000 CD4+ Т-клеток, подвергнутых «негативной» очистке, от здорового донора и меченных CFSE.
После трех суток культивирования супернатанты переносили в новый планшет и анализировали с помощью MSD на высвобождение IFNγ. Кроме того, клетки анализировали с помощью проточной цитометрии для оценки скорости их пролиферации.
На фиг. 2А показано, что обе формы VSIG4 (VSIG4(S) и VSIG4(L)) ингибируют пролиферацию CD4+ Т-клеток. Также обе формы ингибируют высвобождение IFNγ CD4+ T-клетками (рис. 2В).
Пример 2. Получение и очистка антигена VSIG4
2-1. Конструирование вектора для экспрессии антигенного белка VSIG4
Для клонирования белка VSIG4 амплификацию проводили с использованием библиотеки кДНК клеток Jurkat (Stratagene, США) методом полимеразной цепной реакции (ПЦР) с использованием праймеров для VSIG4 (табл.4), которые включают сайты рестриктазы Sfi I в положениях 5' и 3' для получения только внеклеточного домена (20Arg - Ser281). Амплифицированный продукт ПЦР подвергали слиянию по карбокси-концу с человеческим Fc (hFc) или мышиным Fc (mFc) с использованием вектора N293F (фиг. 4).

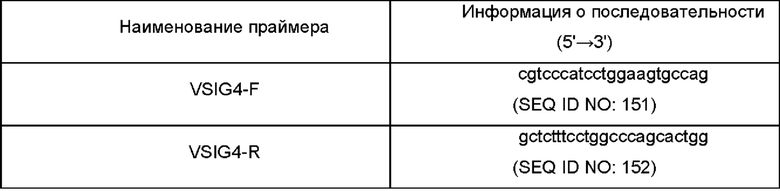
2-2. Экспрессия и очистка антигенного белка VSIG4
С помощью ПЭИ (полиэтиленимин: #23966, Polysciences, США) клетки HEK293F (Invitrogen, США) трансфицировали полученной антигенной плазмидой VSIG4. После этого клетки культивировали в течение 7 суток в среде для экспрессии Freestyle 293 (№ AG 100009, Thermo Fisher Scientific, США), которая не содержит сыворотки. Клеточную культуру, содержащую антиген VSIG4, собирали и центрифугировали в течение 10 мин при 5000 об/мин и удаляли остаточные клетки и плавающие вещества с помощью фильтра с бутылочным верхом с размером пор 0,22 мкм (Millipore, США). На основе аффинной хроматографии с использованием агарозной смолы с А-белком была проведена первая очистка антигена. Белок, полученный после первой очистки, подвергали второй очистке с использованием гель-фильтрационной хроматографии Superdex 200 (1,5 см × 100 см).
Чистоту очищенного белка определяли с помощью SDS-PAGE (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) в восстанавливающих условиях. В результате, как показано на фиг. 5, чистота очищенных белков VSIG4-hFc и VSIG4-mFc оказалась равной 95% или более.
Пример 3. Отбор человеческого антитела против VSIG
3-1. Биопэннинг
VSIG4-hFc и VSIG4-mFc, полученные в примере 2, и белковый антиген VSIG4-his (12163-Н08Н), который был приобретен у Sino Biological Inc., и ITGA6-Fc, используемый в качестве индикатора неспецифического связывания, наносили (50 мкг) на пробирку для иммуносорбента с последующим блокированием.
Что касается фагов из библиотеки человеческих антител, бактерии инфицировали библиотекой scFv (одноцепочечных вариабельных фрагментов) человека с вариативностью 2,7×1010, и затем культивировали в течение 16 часов при 30°С. После культивирования проводили центрифугирование для концентрирования супернатанта с использованием ПЭГ (полиэтиленгликоль, Sigma), и полученное вещество растворяли в фосфатно-солевом буферном растворе для получения библиотеки человеческих антител. Фаг из библиотеки добавляли в пробирку с иммуносорбентом и оставляли для протекания реакции на 2 часа при комнатной температуре. Затем после промывания 1Х PBS-Tween20 (PBS-T) и 1X PBS элюировали только scFv-фаги, специфически связанные с антигеном.
Посредством пэннинга, где бактерии повторно инфицируют элюированными фагами для амплификации, получали пул положительных фагов. С использованием фагов, амплифицированных в первом раунде, второй и третий раунды пэнинга проводили также, как и первый раунд, за исключением того, что количество стадий промывки PBS-T было увеличено. В результате, как показано в таблице 5, было подтверждено, что количество фагов, связанных с антигеном во время третьего раунда пэннинга, немного увеличилось на выходе относительно количества на входе.

3-2. ELISA полифагов
Для исследования антигенной специфичности положительного пула поли-scFv-фаговых антител, который был получен в процессе пэннинга каждого раунда примера 3-1, был проведен ELISA (иммуноферментный анализ) полифагов.
Банк клеток, замороженных после каждого пэнинга раундов 1-3, добавляли в среду, содержащую 2Х YTCM (10 г дрожжевого экстракта, 17 г триптона, 5 г NaCl, 34 мкг/мл хлорамфеникола), 2% глюкозы и 5 мМ хлорида магния (MgCl2) таким образом, чтобы OD600 составляла 0,1, а затем культивировали в течение 2-3 часов при 37°С (OD600=0,5-0,7). Затем, после инфицирования хелперным фагом М1, проводили культивирование в течение 16 часов при 30°С в среде, содержащей 2Х YTCMK (2Х YTCM, канамицин 35 мкг/мл), 5 мМ хлорида магния и 1 мМ IPTG. Культивируемые клетки центрифугировали (4500 об/мин, 15 минут, 4°С) и супернатант переносили в новую пробирку.
На 96-луночный иммунопланшет (#439454, NUNC, США) наносили каждый из двух антигенов в количестве 100 нг на лунку при 4°С в течение 16 часов с использованием посадочного буфера, а затем каждую лунку блокировали посредством 4% обезжиренного молока, растворенного в PBS. После этого каждую лунку промывали 0,2 мл PBS-T и в каждую лунку добавляли поли scFv-фаг из раундов 1-3 пэннинга, каждый в количестве 100 мкл, с последующей реакцией в течение 2 часов при комнатной температуре. Затем снова каждую лунку промывали 4 раза 0,2 мл PBS-T и после разбавления анти-M13-HRP (Amersham 27-9421-01) в качестве вторичного антитела в соотношении 1:2000 проводили реакцию с антителом в течение 1 часа при комнатной температуре. После промывания PBS-T OPD (о-фенилендиамина дигидрохлорид) таблетку (Sigma. 8787-TAB) готовили в PC буфере (0,1 M Na2HPO4, 0,005 М Na-цитрата, рН 5,0) и добавляли в лунку (100 мкл на лунку) для проявления окрашивания в течение 10 минут. Затем измеряли оптическую плотность при 490 нм с помощью спектрофотометра (Molecular Device, США).
Результат показан на фиг 6, посредством ELISA было подтверждено, что связующая способность в отношении антигена была улучшена в поли-scFv-фагах третьего раунда для указанных двух антигенов VSIG4.
3-3. Отбор положительного фага
Колонии, полученные из группы поликлональных фаговых антител, обладающих высокой связывающей способностью (третий пэнинг), культивировали в течение 16 часов при 37°С в 96-луночном планшете с глубокими лунками (#90030, Bioneer, Корея) с использованием 1 мл среды, содержащей 2Х YTCM, 2% глюкозы и 5 мМ хлорида магния. Из культивируемых клеток собирали от 100 до 200 мкл, так что OD600 составляла 0,1, затем вносили в среду, содержащую 1 мл 2Х YTCM, 2% глюкозы и 5 мМ хлорида магния, и культивировали в течение 2-3 часов при 37°С в 96-луночном планшете с глубокими лунками, так что OD600 составляла от 0,5 до 0,7. Инфицирование М1 хелперным фагом проводили до получения значения множественности заражения (MOI) 1:20, а затем культивировали в течение 16 часов при 30°С в среде, содержащей 2Х YTCMK, 5 мМ хлорида магния и 1 мМ IPTG.
На 96-луночный иммунопланшет наносили антиген VSIG4 в количестве 100 нг на лунку при 4°С в течение 16 часов, а затем каждую лунку блокировали с помощью 4% обезжиренного молока, растворенного в PBS. После этого каждую лунку промывали 0,2 мл PBS-T и вносили в каждую лунку один scFv-фаговый клон (соответственно 100 scFv-фагов) в количестве 100 мкл, культивируемый в течение 16 часов, для проведения реакции в течение 2 часов при комнатной температуре. Затем снова каждую лунку промывали 4 раза 0,2 мл PBS-T и после разбавления анти-M13-HRP в качестве вторичного антитела в соотношении 1:2000 проводили реакцию с антителом в течение 1 часа при комнатной температуре. После промывки PBS-T (0,2 мл) давали возможность проявиться окраске и измеряли оптическую плотность при 490 нм.
Результат, показанный на фиг. 7, относится к клонам одного фага, обладающим высокой связывающей способностью в отношении каждого антигена, всего было получено несколько десятков клонов одного фага для VSIG4.
3-4. Нуклеотидное секвенирование положительного фагового антитела
Для одного клона, который был отобран, как описано выше, для получения ДНК была проведена подготовка ДНК с использованием набора для очистки ДНК (Qiagen, Германия). Компании Macrogen, Корея, было предложено провести секвенирование ДНК. С учетом результатов секвенирования определяли сайт CDR вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL) отобранного антитела. Затем исследовали сходство между этими антителами и группой антител зародышевой линии с помощью программы Ig BLAST, представленной на веб-странице NCBI (http://www.ncbi.nlm.nih.gov/igblast/). В результате было получено 39 типов VSIG4-специфического фагового антитела, 11 из которых были охарактеризованы более конкретно. Они представлены в Таблице 2.
Пример 4. Продукция человеческого антитела против VSIG4
4-1. Превращение scFv формы в IgG форму
Для полного превращения 39 типов отобранного одного клонируемого фагового антитела против VSIG4 в форму IgG, последовательность ДНК, соответствующая вариабельным областям тяжелой и легкой цепей, была подвергнута ПЦР (iCycler iQ, BIO-RAD, США) с использованием праймеров, в которые включены сайты рестриктазы для SfiI/NheI и SfiI/BgllI соответственно. Продукты ПЦР тяжелой цепи и легкой цепи расщепляли с использованием каждого экспрессионного вектора, имеющего соответствующий сайт рестриктазы, и ДНК очищали с помощью набора для гель-экстракции ДНК (Qiagen). Для лигирования смешивали вектор (1 мкл, 10 нг), тяжелую или легкую цепь (от 100 до 200 нг, 15 мкл), буфер 10Х (2 мкл), лигазу (1 ед/мкл, 1 мкл) и воду, выдерживали от 1 до 2 часов при комнатной температуре и добавляли к клеткам для трансформации (компетентные клетки, XL1-blue). Полученную смесь выдерживали на льду в течение 5 минут, а затем подвергали тепловому шоку при 42°С в течение 90 секунд. После теплового шока к клеткам добавляли 1 мл среды и культивировали в течение 1 часа при 37°С с последующим нанесением на планшет LB-Amp и культивировали в течение 16 часов при 37°С. Полученные таким образом колонии собирали и инокулировали 5 мл среды LB Amp (Лизогенный бульон (LB), содержащий антибиотик ампициллин). После культивирования в течение 16 часов при 37°С проводили подготовку ДНК с использованием набора для подготовки ДНК (Nuclogen). Было запрошено секвенирование ДНК полученной таким образом ДНК (Macrogen, Корея).
В результате было подтверждено, что каждая тяжелая цепь и легкая цепь 11 типов клона антитела к VSIG4, которые были преобразованы в полноразмерные IgG, соответствует последовательности фагового антитела. После этого плазмидные ДНК тяжелой и легкой цепей с идентифицированной последовательностью использовали для получения антитела.
4-2. Получение человеческого антитела
Полученный экспрессионный вектор, содержащий тяжелую цепь и легкую цепь, подвергали совместной трансфекции в клетки HEK-293F в соотношении 6:4. Через семь суток после совместной трансфекции супернатант собирали, а клетки и плавающие материалы удаляли центрифугой и с помощью фильтра с бутылочным верхом с размером пор 0,22 мкм. Супернатант собирали и подвергали аффинной хроматографии с А-белком для очистки антитела IgG. После очистки антитело отделяли с использованием лизинового буфера и производили замену буфера таким образом, чтобы конечным ресуспендирующим буфером был PBS. Проводили количественный анализ антитела с помощью ВСА и Nano-drop для определения количества продукта. Затем антитело подвергали электрофорезу в полиакриламидном геле в присутствии додецилсульфата натрия с нагрузкой 5 мкг для каждого из восстанавливающих условий и невосстанавливающих условий. Соответственно, определяли степень чистоты и подвижности очищенного белка.
Результат представлен на фиг. 8, было выявлено 11 типов одного человеческого антитела против VSIG4 размером по меньшей мере 150 кДа в невосстанавливающих условиях, а количество получаемого варьировалось, т.е. от низкого 5 мг/л до очень большого 142,6 мг/л.
Пример 5. Свойства связывания VSIG4 антитела против VSIG4
5-1. Специфичность связывания антитела к VSIG4 на клеточной поверхности
Для получения трансформированного пула клеток, в которых сверхэкспрессируется человеческий VSIG4, НЕК293Е трансфицировали плазмидой pcDNA3.1, содержащей человеческий VSIG4, а затем проводили селекцию в среде для селекции, содержащей 400 мкг/мл зеоцина (#R25001, Thermo Fisher Scientific). После селекции пул клеток, в которых сверхэкспрессируется VSIG4, отделяли путем определения уровня экспрессии с помощью анализа FACS (сортировка клеток с активированной флуоресценцией) с использованием антитела против VSIG4 человека, связанного с флуоресцентным веществом АРС (аллофикоцианин) (#17-5757-42, ebioscience, США) (фиг. 9В), и после определения отсутствия базовой экспрессии VSIG4 с помощью анализа FACS с использованием антитела против VSIG4 человека, связанного с флуоресцентным веществом АРС, в клетках HEK293E, которые используются в качестве материнских клеток в клеточном пуле HEK293E, в котором сверхэкспрессируется человеческий VSIG4 (фиг. 9А), оценку свойств антитела проводили для 11 типов антитела против VSIG4 человека с использованием этих двух типов клеток.
Для подтверждения связывания клеток с антителом к VSIG4 для каждого образца готовили 0,5×106 клеток, которым давали прореагировать с антителом в концентрации 0,08 мкг/мл, 0,4 мкг/мл или 2 мкг/мл в течение 30 минут при 4°С. После этого клетки промывали 3 раза буфером, содержащим 2% PBS, и после реакции в течение 20 минут при 4°С с антителом против IgG человека (#FI-3000, Vectorlabs), связанным с флуоресцентным веществом FITC (изотиоцианат флуоресцеина), клетки промывали тем же способом, что и указанный выше, с последующим суспендированием в 0,5 мл PBS, содержащего 2% FPS. Затем клетки анализировали на проточном цитометре FACSCanto II. В результате было обнаружено, что все 11 типов одного антитела против VSIG4 человека хорошо связываются с клетками, сверхэкспрессирующими человеческий VSIG4, зависимым от концентрации образом (фиг. 10В). Однако в клетках HEK293E, не имеющих базовой экспрессии VSIG4, связывание отсутствовало (фиг. 10А). Этот результат указывает на то, что 11 типов одного антитела против VSIG4 человека специфически связываются с человеческим антигеном VSIG4.
5.2 Связывание человеческого антитела против VSIG4 с нативным человеческим VSIG4 посредством анализа FACS
Связывающие свойства ряда антител против VSIG4 оценивали с помощью анализа FACS на HEK293E, экспрессирующих человеческий VSIG4, с использованием возрастающих концентраций антитела. Аналогичный эксперимент был проведен с m6H8, мышиным моноклональным антителом, распознающим VSIG4 и описанным в WO 2020/069507. Для этой цели клетки (1×106 клеток/мл) инкубировали с каждым из 11 полных Ig, антителами против VSIG4 или m6H8, в течение 20 минут при 4°С в FACS-буфере (PBS, 0,1% BSA, 0,01% NaN3). Затем их промывали 3 раза и инкубировали с соответствующим вторичным антителом, связанным с Alexa 488, в течение 20 дополнительных минут при 4°С в темноте перед 3-кратной промывкой в FACS буфере. Связывание антител против VSIG4 или т6Н8 немедленно проводили на жизнеспособных клетках, которые идентифицировали с использованием йодида пропидия (окрашивающего мертвые клетки). Максимальную интенсивность сигнала, полученную от каждого антитела, обозначали как Bmax и выражали через среднее значение интенсивности флуоресценции (MFI). ЕС50 связывания, выраженную в молярности (М), рассчитывали с использованием нелинейного регрессионного анализа (GraphPad Prims 4.0).
Кривая титрования каждого мышиного или химерного антитела продемонстрировала, что все полученные антитела способны распознавать нативную форму VSIG4 с типичным профилем насыщения. ЕС50 связывания каждого антитела определяли с помощью нелинейного регрессионного анализа. ЕС50 варьировались от 1,2×10-9 до 9,3×10-10. Значения ЕС50 приведены в таблице 6.
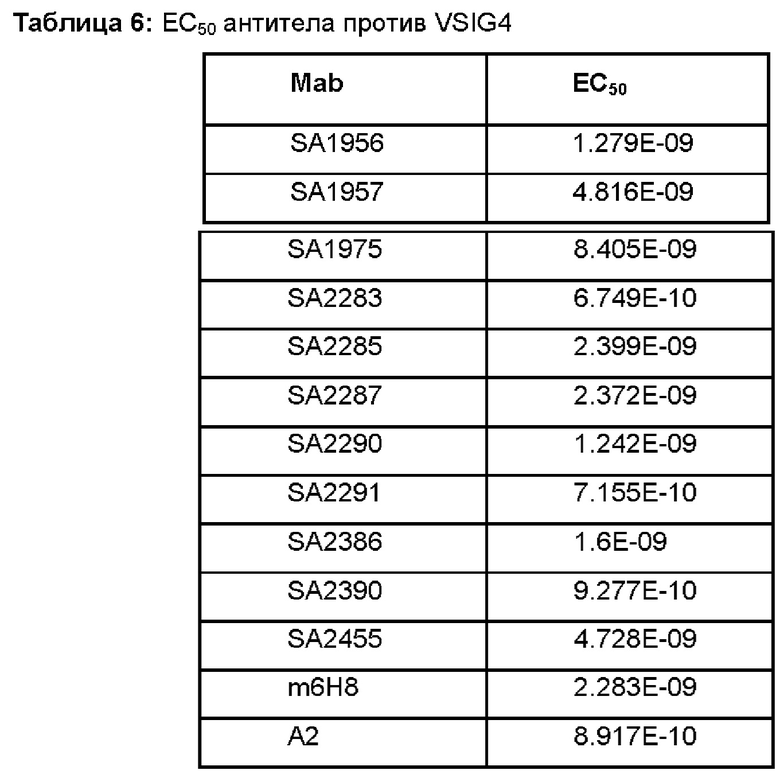
5.3 Связывание антитела против VSIG4 человека с длинной и короткой формами VSIG4
В первой серии экспериментов тестировали связывание 11 scFv-антител против VSIG4 с каждой из длинной и короткой форм VSIG4 посредством ELISA. В этих условиях было обнаружено специфическое связывание каждого из 11 протестированных антител с обеими формами (фиг. 11А).
Для подтверждения данного результата связывание 11 антител против VSIG4 с каждой из длинной и короткой форм VSIG4 анализировали посредством вестерн-блоттинга. В каждом случае 100 нг длинной формы hFc hVSIG4 (L) и короткой формы hFc hVSIG4 (S) зондировали специфическим антителом против VSIG4. Как показано на фиг. 11В, две полосы ожидаемого размера наблюдались для каждого из 11 антител против VSIG4. Напротив, m6Н8, мышиное моноклональное антитело, распознающее VSIG4 и описанное в WO 2020/069507, связывает только длинную форму VSIG4 (фиг. 12А). Данный результат был подтвержден в анализе ELISA (фиг. 12В).
5-4. Эпитопное картирование антител против VSIG4
Чтобы определить эпитоп, распознаваемый каждым из 11 антител против VSIG4 человека, их способность связывать ряд растворимых белков VSIG4, несущих конкретные мутации, анализировали с помощью ELISA. Используемые конструкции подробно описаны в таблице 7.
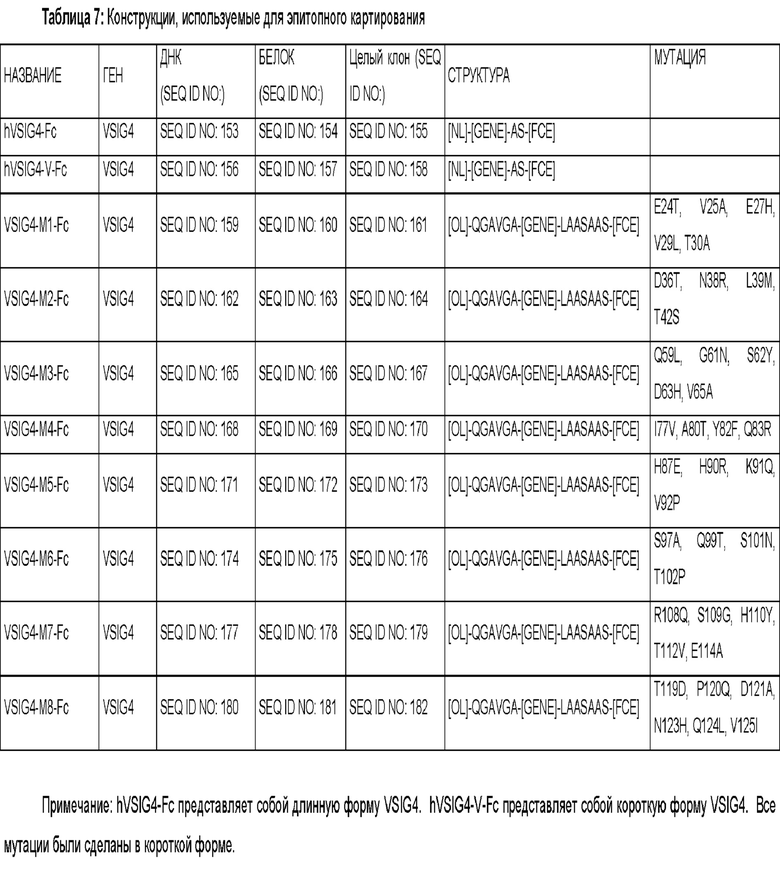
На 96-луночный иммунопланшет наносили различные антигены в количестве 100 нг на лунку при 4°С в течение 16 часов, а затем каждую лунку блокировали с использованием 4% обезжиренного молока, растворенного в PBS. После этого каждую лунку промывали 0,2 мл PBS-T и вносили в каждую лунку по 100 мкл каждого из 11 scFv-фагов, культивируемых в течение 16 часов, для получения реакции на 2 часа при комнатной температуре. Затем снова каждую лунку промывали 4 раза 0,2 мл PBS-T и после разбавления анти-M13-HRP в качестве вторичного антитела в соотношении 1:2000 проводили реакцию с антителом в течение 1 часа при комнатной температуре. После промывки PBS-T (0,2 мл) давали возможность проявиться окраске и измеряли оптическую плотность при 490 нм.
Результаты анализа представлены на фиг. 13
Эпитоп, распознаваемый антителами SA1956, SA1957 и SA2285, содержит по меньшей мере одну из аминокислот Е24, V25, Е27, V29 и Т30.
Эпитоп, распознаваемый антителами SA1975 и SA2290, содержит по меньшей мере одну из аминокислот I77, А80, Y82 и Q83. По крайней мере один из остатков Q59, G61, S62, D63 и V65 также может способствовать связыванию SA2290 с VSIG4.
Эпитоп, распознаваемый антителом SA2283, содержит по меньшей мере одну из аминокислот R108, S109, Н110, Т112 и Е114.
Эпитоп, распознаваемый антителом SA2287, содержит по меньшей мере одну из аминокислот Т119, Р120, D121, N123, Q124 и V125.
Эпитоп, распознаваемый антителом SA2291, содержит по меньшей мере одну из аминокислот R108, S109, Н110, Т112 и Е114, по меньшей мере одну из аминокислот Т119, Р120, D121, N123, Q124 и V125.
Эпитоп, распознаваемый антителом SA2390, содержит по меньшей мере одну из аминокислот Q59, G61, S62, D63 и V65, по крайней мере одну из аминокислот S97, Q99, S101 и Т102, по крайней мере одну из аминокислот R108, S109, Н110, Т112 и Е114 и по меньшей мере одну из аминокислот Т119, Р120, D121, N123, Q124 и V125. Кроме того, по меньшей мере один из остатков D36T, N38R, L39M, Т42 и по меньшей один из остатков I77, А80, Y82 и Q83 также может способствовать связыванию SA2390 с VSIG4.
Эпитоп, распознаваемый антителом SA2455, содержит по меньшей мере одну из аминокислот Q59, G61, S62, D63 и V65, по меньшей мере одну из аминокислот S97, Q99, S101 и Т102, по меньшей мере одну из аминокислот R108, S109, Н110, Т112 и Е114 и по меньшей мере одну из аминокислот Т119, Р120, D121, N123, Q124 и V125. Кроме того, по меньшей мере один из остатков D36T, N38R, L39M, Т42 также может способствовать связыванию SA2390 с VSIG4.
Пример 6: интернализация человеческого антитела против VSIG4
11 полноразмерных lg, антител против VSIG4, оценивали в анализе интернализации.
Для этого анализа клетки HEK-VSIG4 (т.е. клетки НЕК293, трансфицированные и экспрессирующие VSIG4 на поверхности) при конфлюэнтности 80% разделяли в трипсине и подсчитывали в счетчике ViCells. 100000 клеток HEK-VSIG4 инкубировали в течение 20 минут при 4°С в присутствии 10 мкг/мл каждого антитела в 100 мкл холодной культуральной среды: антитело против VSIG4, антитело m6H8 и А2, его гуманизированная версия (WO 2020/069507) или анти-IGFIR (Hz208F2-4). Затем клетки центрифугировали при 2000 об/мин и дважды промывали 200 мкл холодной среды.
Время Т0: клетки инкубировали непосредственно с 200 мкл вторичного козьего антитела против Alexa 488 человека, разбавленного 1/500, в течение 20 минут при 4°С. Затем их дважды промывали в холодной среде и анализировали с помощью FACS.
Время 4 ч: клетки инкубировали в 100 мкл холодной среды (4°С) или теплой среды (37°С) в течение 4 часов. Каждую партию клеток вращали при 2000 об/мин, дважды промывали 200 мкл холодной среды и инкубировали со вторичным козьим антителом Alexa 488 против человеческого антитела в течение 20 минут при 4°С в холодной среде. Затем клетки дважды промывали в холодной среде и анализировали с помощью FACS.
Уровень интернализации определяли путем расчета дельта MFI, соответствующей (MFI (при 4°С) - MFI (при 37°С)), а процент интернализации определяли путем расчета процента снижения MFI между 4°С и 37°С. (все MFI рассчитываются после вычета значения MFI изотипа).
Интернализирующее антитело против IGF-1R, Hz208F2-4 (WO 2015/162292), использовали в качестве положительного контроля в этом анализе, поскольку клетки HEK экспрессируют IGF-1R на клеточной поверхности.
Настоящие антитела против VSIG4 демонстрируют различные уровни интернализации (см. таблицу 8). С другой стороны, антитело m6H8 и его гуманизированная версия А2 не индуцировали какую-либо интернализацию.
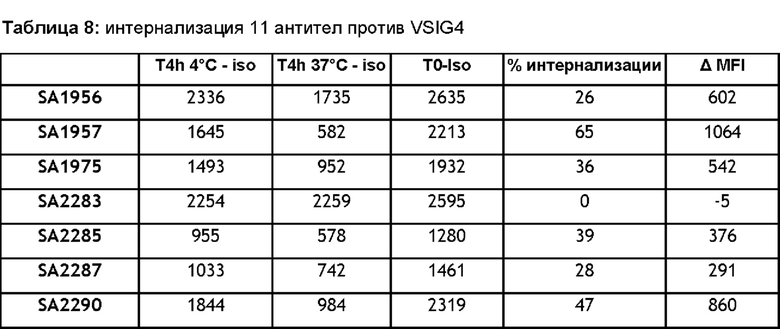
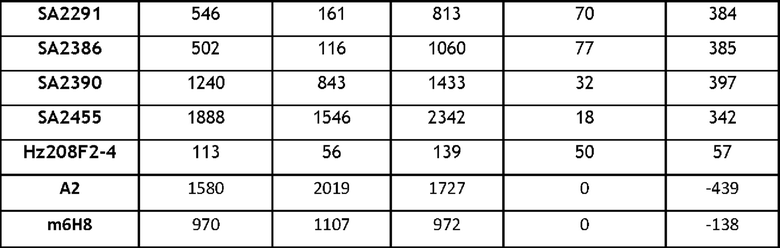
Пример 7: Ингибирование противовоспалительной и иммуносупрессивной функций VSIG4 человеческим антителом против VSIG4
7-1. Анализ воспаления
Для того, чтобы оценить способность антител против VSIG4 модулировать воспалительный фенотип макрофагов, проводили анализ высвобождения цитокинов на макрофагах, обработанных каждым из полных Ig, антител против VSIG4 человека. Схема эксперимента представлена на фиг. 14.
Мононуклеарные клетки периферической крови (РВМС) выделяли из крови человека путем центрифугирования в градиенте плотности из цитаферезного кольца, предоставленного EFS (Etablissement Francais du Sang). Затем моноциты очищали от РВМС с помощью позитивной иммуномагнитной клеточной селекции в соответствии с инструкциями производителя (130-050-201, Miltenyi Biotec).
Свежие моноциты высевали в 96-луночные обработанные культуральные планшеты с плоским дном (353072, Falcon) в культуральную среду (среда RPMI 1640 + 1% пенициллин-стрептомицин + 1% пируват натрия + 1% L-глутамин + 10% фетальная телячья сыворотка), содержащую 50 нг/мл M-CSF (130-096-492, Miltenyi Biotec). Их инкубировали при 37°С, 5% CO2 в течение 6 суток для дифференцировки в макрофаги. Дифференцированные М0-макрофаги были получены на 6-е сутки.
Связывание целевых антител с дифференцированными М0-макрофагами оценивали с помощью проточной цитометрии на 6-е сутки. К дифференцированным М0-макрофагам добавляли LPS (L4516, Sigma) в конечной концентрации 100 нг/мл. Тестируемые антитела или соответствующие изотипы добавляли к дифференцированным М0-макрофагам в трех концентрациях (2,5 мкг/мл, 5 мкг/мл и 10 мкг/мл). В данном анализе также тестировали мышиное антитело m6H8 и его гуманизированную форму, А2 (оба описаны в WO 2020/069507). В качестве контроля использовали контрольное антитело (R&D, Ref МАВ2078, клон 287219, mIgG2a), известное своей способностью имитировать высвобождение цитокинов из макрофагов М0 в направлении провоспалительного фенотипа, в конечной концентрации 5 мкг/мл. Для SA1956, SA2386, SA2390 и SA2455 в культуральную среду добавляли 50 мкг/мл C3b (А114, Complement Technology).
Дифференцированные МО-макрофаги инкубировали с LPS и тестируемыми антителами в течение 24 часов при 37°С, 5% CO2. Супернатанты клеточных культур собирали на 7-е сутки и переносили в новые 96-луночные планшеты с V-образным дном для анализа цитокинов. Измеряли концентрации IL-10, IL-6, IL-1β, IL-12/23р40 и TNF-α. Количественный анализ проводили с использованием технологии Meso Scale Discovery в соответствии с инструкциями производителя (K15UQK-4 и К151АОН-4, Meso Scale Discovery).
Для учета неоднородности среди здоровых доноров была проведена оценка по меньшей мере 5 доноров. Каждое экспериментальное условие было выполнено в трехкратной повторности и в одном эксперименте.
Результаты проведения анализа представлены в таблице 9.
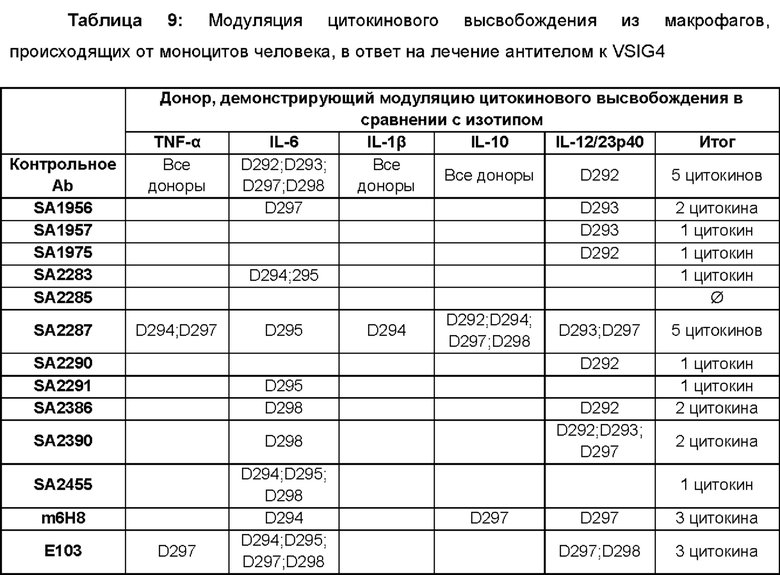
Все антитела против VSIG4, за исключением SA2285, приводят к повышенному уровню высвобождения провоспалительных цитокинов и/или к снижению секреции противовоспалительных цитокинов макрофагами.
Таким образом, эти антитела способны модулировать фенотип макрофагов человека.
7-2. Анализ иммуносупрессии
Мононуклеарные клетки периферической крови (РВМС) выделяли из крови человека путем центрифугирования в градиенте плотности из цитаферезного кольца, предоставленного EFS (Etablissement Francais du Sang). Затем моноциты и CD4+ Т-клетки очищали от РВМС от одного того же донора: моноциты очищали с помощью позитивной иммуномагнитной клеточной селекции в соответствии с инструкциями производителя (130-050-201, Miltenyi Biotec), в то время как CD4+ Т-клетки выделяли из непозитивной фракции от очистки моноцитов посредством негативной иммуномагнитной селекции клеток в соответствии с инструкциями производителя (19052, STEMCELL Technologies). CD4+ Т-клетки замораживали по 15×106 клеток на криопробирку в 1 мл среды для замораживания (07930, STEMCELL Technologies) для дальнейшего использования в совместном культивировании.
Свежие моноциты высевали в 96-луночные обработанные культуральные планшеты с плоским дном (353072, Falcon) в культуральную среду (среда RPMI 1640 + 1% пенициллин-стрептомицин + 1% пируват натрия + 1% L-глутамин + 10% фетальная телячья сыворотка), содержащую 50 нг/мл M-CSF (130-096-492, Miltenyi Biotec) для дальнейшей поляризации М2- макрофагов, или 50 нг/мл GM-CSF (130-093-866, Miltenyi Biotec) для дальнейшей поляризации М1- макрофагов. Их инкубировали при 37°С, 5% CO2 в течение 6 суток для дифференцировки в макрофаги. Дифференцированные МО-макрофаги были получены на 6-е сутки.
50 нг/мл IFN-γ (285-IF, R&D) добавляли к GM-CSF-дифференцированным МО-макрофагам для поляризации в провоспалительные М1-макрофаги. К M-CSF-дифференцированным МО-макрофагам для поляризации в иммуносупрессивные М2-макрофаги добавляли по 20 нг/мл каждого из следующих цитокинов: IL-4 (130.093.922, Miltenyi Biotec), IL-10 (217-IL/CF, R&D) и TGF-β (130.095.066, Miltenyi Biotec). Дифференцированные МО-макрофаги инкубировали с цитокинами при 37°С, 5% С02 в течение 2 сут. Поляризованные макрофаги М1 и М2 были получены на 8-е сутки. Поляризованные макрофаги активировали 100 нг/мл LPS (L4516, Sigma) в течение 4 часов при 37°С, 5% CO2. Затем макрофаги собирали и промывали в культуральной среде. Связывание целевых антител с поляризованными М1- и М2-макрофагами оценивали с помощью проточной цитометрии после LPS- активации.
М1- и М2-макрофаги высевали в классические 96-луночные планшеты с плоским дном по 20000 клеток на лунку в культуральную среду. Их инкубировали при 37°С, 5% CO2 в течение 24 часов. К макрофагам добавляли CD4+ Т-клетки в соотношении 1 макрофаг: 5 CD4 Т-клеток. В совместную культуру добавляли шарики CD3/CD28 (111-32D, Gibco) для активации CD4+ Т-кпеток в соотношении 1 шарик на 32 клетки.
Тестируемые антитела или соответствующие изотипы вносили в совместную культуру в конечной концентрации 10 мкг/мл. Авелумаб, моноклональное антитело против PD-L1, использовали в качестве положительного контроля. Для SA2386 в культуральную среду добавляли 50 мкг/мл C3b (А114, Complement Technology). Макрофаги и CD4+Т-клетки в совместной культуре инкубировали при 37°С, 5% CO2 в течение 5 суток. Супернатанты клеточных культур собирали на 14-е сутки и переносили в новые 96-луночные планшеты с V-образным дном для анализа цитокинов. Измеряли концентрацию IFN-γ. Проводили количественную оценку с использованием технологии Meso Scale Discovery в соответствии с инструкциями производителя (K151AEB-4, Meso Scale Discovery).
Было оценено по меньшей мере 5 доноров, чтобы принять во внимание неоднородность здоровых доноров. Каждое условие эксперимента было выполнено в трехкратной повторности и в одном эксперименте.
Результаты проведения анализа представлены в таблице 10.
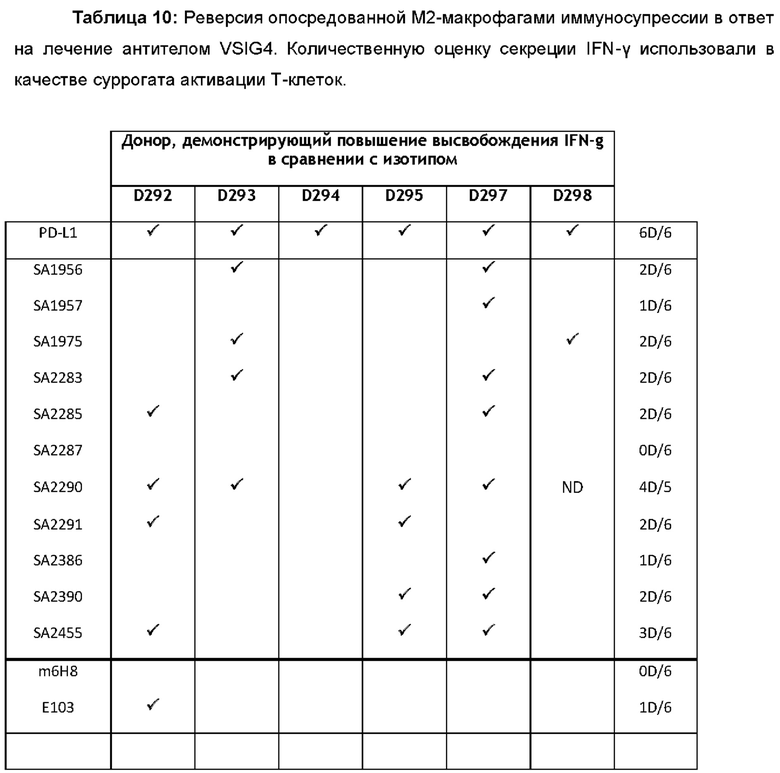
Все антитела против VSIG4, за исключением SA2285, индуцируют высвобождение IFN-γ CD4+ Т-клетками, что указывает на то, что они запускают активацию Т-клеток. Таким образом, эти антитела способны ингибировать иммуносупрессивную функцию VSIG4.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Уай-БИОЛОДЖИКС ИНК.
ПЬЕР ФАБР МЕДИКАМЕНТ
<120> АНТИТЕЛО ПРОТИВ VSIG4 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ
И ИХ ПРИМЕНЕНИЕ
<130> B378459PCTD40837
<150> KR10-2019-0109365
<151> 2019-09-04
<160> 184
<170> PatentIn version 3.5
<210> 1
<211> 399
<212> ПРТ
<213> Homo sapiens
<400> 1
Met Gly Ile Leu Leu Gly Leu Leu Leu Leu Gly His Leu Thr Val Asp
1 5 10 15
Thr Tyr Gly Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp
115 120 125
Lys Ile Thr Glu Leu Arg Val Gln Lys Leu Ser Val Ser Lys Pro Thr
130 135 140
Val Thr Thr Gly Ser Gly Tyr Gly Phe Thr Val Pro Gln Gly Met Arg
145 150 155 160
Ile Ser Leu Gln Cys Gln Ala Arg Gly Ser Pro Pro Ile Ser Tyr Ile
165 170 175
Trp Tyr Lys Gln Gln Thr Asn Asn Gln Glu Pro Ile Lys Val Ala Thr
180 185 190
Leu Ser Thr Leu Leu Phe Lys Pro Ala Val Ile Ala Asp Ser Gly Ser
195 200 205
Tyr Phe Cys Thr Ala Lys Gly Gln Val Gly Ser Glu Gln His Ser Asp
210 215 220
Ile Val Lys Phe Val Val Lys Asp Ser Ser Lys Leu Leu Lys Thr Lys
225 230 235 240
Thr Glu Ala Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr Ser Thr
245 250 255
Val Lys Gln Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr Leu Gly
260 265 270
Glu Thr Ser Ala Gly Pro Gly Lys Ser Leu Pro Val Phe Ala Ile Ile
275 280 285
Leu Ile Ile Ser Leu Cys Cys Met Val Val Phe Thr Met Ala Tyr Ile
290 295 300
Met Leu Cys Arg Lys Thr Ser Gln Gln Glu His Val Tyr Glu Ala Ala
305 310 315 320
Arg Ala His Ala Arg Glu Ala Asn Asp Ser Gly Glu Thr Met Arg Val
325 330 335
Ala Ile Phe Ala Ser Gly Cys Ser Ser Asp Glu Pro Thr Ser Gln Asn
340 345 350
Leu Gly Asn Asn Tyr Ser Asp Glu Pro Cys Ile Gly Gln Glu Tyr Gln
355 360 365
Ile Ile Ala Gln Ile Asn Gly Asn Tyr Ala Arg Leu Leu Asp Thr Val
370 375 380
Pro Leu Asp Tyr Glu Phe Leu Ala Thr Glu Gly Lys Ser Val Cys
385 390 395
<210> 2
<211> 305
<212> ПРТ
<213> Homo sapiens
<400> 2
Met Gly Ile Leu Leu Gly Leu Leu Leu Leu Gly His Leu Thr Val Asp
1 5 10 15
Thr Tyr Gly Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp
115 120 125
Lys Ile Thr Glu Leu Arg Val Gln Lys His Ser Ser Lys Leu Leu Lys
130 135 140
Thr Lys Thr Glu Ala Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr
145 150 155 160
Ser Thr Val Lys Gln Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr
165 170 175
Leu Gly Glu Thr Ser Ala Gly Pro Gly Lys Ser Leu Pro Val Phe Ala
180 185 190
Ile Ile Leu Ile Ile Ser Leu Cys Cys Met Val Val Phe Thr Met Ala
195 200 205
Tyr Ile Met Leu Cys Arg Lys Thr Ser Gln Gln Glu His Val Tyr Glu
210 215 220
Ala Ala Arg Ala His Ala Arg Glu Ala Asn Asp Ser Gly Glu Thr Met
225 230 235 240
Arg Val Ala Ile Phe Ala Ser Gly Cys Ser Ser Asp Glu Pro Thr Ser
245 250 255
Gln Asn Leu Gly Asn Asn Tyr Ser Asp Glu Pro Cys Ile Gly Gln Glu
260 265 270
Tyr Gln Ile Ile Ala Gln Ile Asn Gly Asn Tyr Ala Arg Leu Leu Asp
275 280 285
Thr Val Pro Leu Asp Tyr Glu Phe Leu Ala Thr Glu Gly Lys Ser Val
290 295 300
Cys
305
<210> 3
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 3
Gly Phe Ser Leu Thr Thr Ser Gly Val Ala
1 5 10
<210> 4
<211> 7
<212> ПРТ
<213> Homo sapiens
<400> 4
Ile Tyr Trp Asp Gly Asp Glu
1 5
<210> 5
<211> 19
<212> ПРТ
<213> Homo sapiens
<400> 5
Ala Arg Ile Arg Gly Tyr Tyr Asp Trp Gly Ser Tyr Tyr Ser Tyr Gly
1 5 10 15
Met Asp Val
<210> 6
<211> 6
<212> ПРТ
<213> Homo sapiens
<400> 6
Asn Ile Gly Ser Lys Asn
1 5
<210> 7
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 7
Arg Asp Ser
1
<210> 8
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 8
Gln Val Trp Asp Ser Ser Thr His Val Val
1 5 10
<210> 9
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 9
Gly Phe Ser Phe Asn Thr Pro Gly Glu Gly
1 5 10
<210> 10
<211> 7
<212> ПРТ
<213> Homo sapiens
<400> 10
Ile Tyr Trp Asp Asp Glu Lys
1 5
<210> 11
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 11
Gly Tyr Ile Phe Thr Asn Tyr Tyr
1 5
<210> 12
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 12
Ile Asp Pro Ser Gly Gly Ser Thr
1 5
<210> 13
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 13
Ala Arg Asp Tyr Trp Gly Ser Leu Asp Tyr
1 5 10
<210> 14
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 14
Ser Gly Ser Ile Ala Ser Asn Tyr
1 5
<210> 15
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 15
Glu Asn Tyr
1
<210> 16
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 16
Gln Ser Tyr Asp Ser Arg Asn Arg Asn Tyr Val
1 5 10
<210> 17
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 17
Gly Phe Thr Phe Ser Asp Tyr Tyr
1 5
<210> 18
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 18
Ile Ser Ser Ser Gly Ser Thr Ile
1 5
<210> 19
<211> 14
<212> ПРТ
<213> Homo sapiens
<400> 19
Ala Arg Arg Tyr Ser Ala Tyr Glu Thr Gly Tyr Phe Asp Phe
1 5 10
<210> 20
<211> 6
<212> ПРТ
<213> Homo sapiens
<400> 20
Gln Gly Ile Ser Thr Tyr
1 5
<210> 21
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 21
Ser Thr Ser
1
<210> 22
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 22
Gln Gln Ser Tyr Ser Ala Pro Pro Thr
1 5
<210> 23
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 23
Gly Phe Ser Leu Asn Thr Pro Gly Met Gly
1 5 10
<210> 24
<211> 7
<212> ПРТ
<213> Homo sapiens
<400> 24
Val Phe Trp Asp Asp Asp Lys
1 5
<210> 25
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 25
Gln Val Trp Asp Ser Asn Ser Asp Gln Tyr Val
1 5 10
<210> 26
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 26
Gly Phe Thr Phe Ser Ser Arg Gly
1 5
<210> 27
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 27
Ile Trp Tyr His Gly Ser Asp Asp
1 5
<210> 28
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 28
Ala Asn Leu Gly Ala Thr Asp Gly Phe Asp Ile
1 5 10
<210> 29
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 29
Ser Ser Asp Val Ser Ala Tyr Asn Tyr
1 5
<210> 30
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 30
Gly Val Ser
1
<210> 31
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 31
Asn Ser Tyr Thr Thr Ser Asn Thr Trp Val
1 5 10
<210> 32
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 32
Gly Tyr Thr Phe Thr Ser Tyr Gly
1 5
<210> 33
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 33
Ile Ser Ala Tyr Asn Gly Asn Thr
1 5
<210> 34
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 34
Ala Arg Asp Tyr Trp Gly Ser Leu Asp His
1 5 10
<210> 35
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 35
Ser Gly Ser Ile Asp Ile Asn Tyr
1 5
<210> 36
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 36
Glu Asp Ser
1
<210> 37
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 37
Gly Phe Thr Phe Ser Asp His Tyr
1 5
<210> 38
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 38
Ile Ser Ser Gly Gly Gly Thr Ile
1 5
<210> 39
<211> 15
<212> ПРТ
<213> Homo sapiens
<400> 39
Ala Arg Arg Glu Tyr Asp Ser Asp Gly His Tyr Tyr Phe Asp Tyr
1 5 10 15
<210> 40
<211> 6
<212> ПРТ
<213> Homo sapiens
<400> 40
Ala Leu Pro Lys Gln Tyr
1 5
<210> 41
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 41
Lys Asp Asn
1
<210> 42
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 42
Gln Ser Val Asp Ser Ser Asp Thr Ser Val Val
1 5 10
<210> 43
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 43
Gly Ser Thr Phe Arg Val Ala Trp
1 5
<210> 44
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 44
Ile Lys Ser Asn Ser Asp Gly Gly Thr Thr
1 5 10
<210> 45
<211> 15
<212> ПРТ
<213> Homo sapiens
<400> 45
Ala Arg His Gly Asp Ala Asn Ala Tyr Tyr Tyr Gly Met Asp Val
1 5 10 15
<210> 46
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 46
Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1 5
<210> 47
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 47
Asp Val Ser
1
<210> 48
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 48
Ser Ser Tyr Ala Ser Ser Ser Thr Leu Tyr Val
1 5 10
<210> 49
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 49
Gly Phe Thr Phe Asp Asp Tyr Ala
1 5
<210> 50
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 50
Ile Ser Trp Asn Ser Gly Asn Ile
1 5
<210> 51
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 51
Ala Arg Glu Thr Ala Leu Ala Phe Asp Met
1 5 10
<210> 52
<211> 6
<212> ПРТ
<213> Homo sapiens
<400> 52
Gln Gly Ile Ser Tyr Trp
1 5
<210> 53
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 53
Ala Ser Ser
1
<210> 54
<211> 9
<212> ПРТ
<213> Homo sapiens
<400> 54
Leu Gln Ala Thr Ser Phe Pro Tyr Thr
1 5
<210> 55
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 55
Ala Arg Pro Gly Tyr Tyr Tyr Gly Leu Asp Val
1 5 10
<210> 56
<211> 6
<212> ПРТ
<213> Homo sapiens
<400> 56
Gln Ser Ile Ser Ser Trp
1 5
<210> 57
<211> 3
<212> ПРТ
<213> Homo sapiens
<400> 57
Gln Ala Ser
1
<210> 58
<211> 8
<212> ПРТ
<213> Homo sapiens
<400> 58
Gln Gln Tyr Ser Ser Leu Trp Thr
1 5
<210> 59
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 59
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser
20 25
<210> 60
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 60
Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala
1 5 10 15
Leu
<210> 61
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 61
Arg Tyr Ser Ser Ser Leu Lys Asn Arg Leu Thr Ile Thr Lys Asp Thr
1 5 10 15
Ser Lys Ser Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp
20 25 30
Thr Ala Thr Tyr Tyr Cys
35
<210> 62
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 62
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
1 5 10
<210> 63
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 63
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn
20 25
<210> 64
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 64
Val His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile
1 5 10 15
Tyr
<210> 65
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 65
Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
1 5 10 15
Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly Asp Glu Ala
20 25 30
Asp Tyr Tyr Cys
35
<210> 66
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 66
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 67
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 67
Gln Val Thr Leu Arg Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser
20 25
<210> 68
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 68
Val Thr Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala
1 5 10 15
Leu
<210> 69
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 69
His Tyr Asn Pro Ser Leu Asn Gly Arg Leu Thr Ile Thr Lys Asp Thr
1 5 10 15
Ser Lys Ser Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp
20 25 30
Thr Ala Thr Tyr Tyr Cys
35
<210> 70
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 70
Trp Gly Gln Gly Thr Thr Ile Thr Val Ser Ser
1 5 10
<210> 71
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 71
Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
1 5 10 15
Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly Asp Glu Ala
20 25 30
Asp Tyr Tyr Cys
35
<210> 72
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 72
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Ile Lys Lys Pro Gly Ala
1 5 10 15
Ser Met Asn Val Ser Cys Lys Ala Ser
20 25
<210> 73
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 73
Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
1 5 10 15
Ile
<210> 74
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 74
Thr Tyr Ala Gln Lys Phe Gln His Arg Val Thr Met Thr Ser Asp Thr
1 5 10 15
Ser Thr Ser Thr Val Tyr Met Glu Leu Ile Gly Leu Gly Ser Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 75
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 75
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 76
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 76
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser
20 25
<210> 77
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 77
Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Ile Val Ile
1 5 10 15
Tyr
<210> 78
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 78
Leu Arg Pro Ser Glu Val Pro Asp Arg Phe Ser Gly Ser Ile Asp Ser
1 5 10 15
Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly Leu Lys Thr Glu Asp
20 25 30
Glu Gly Asp Tyr Tyr Cys
35
<210> 79
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 79
Phe Gly Thr Gly Thr Lys Val Thr Val Leu
1 5 10
<210> 80
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 80
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 81
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 81
Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10 15
Tyr
<210> 82
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 82
Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 83
<211> 26
<212> ПРТ
<213> Homo sapiens
<400> 83
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
20 25
<210> 84
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 84
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
1 5 10 15
Tyr
<210> 85
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 85
Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly Gly Gly Ser Gly
1 5 10 15
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
20 25 30
Thr Tyr Tyr Cys
35
<210> 86
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 86
Phe Gly Pro Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 87
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 87
Gln Val Thr Leu Arg Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Arg
20 25
<210> 88
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 88
Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Gly
1 5 10 15
Leu
<210> 89
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 89
Thr Tyr Arg Pro Ser Leu Arg Asn Arg Leu Thr Ile Thr Lys Glu Thr
1 5 10 15
Ser Lys Ser Gln Val Val Leu Thr Met Thr Asn Met Asp Pro Val Asp
20 25 30
Thr Ala Thr Tyr Tyr Cys
35
<210> 90
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 90
Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
1 5 10 15
Asn Thr Ala Thr Leu Thr Ile Ser Gly Val Glu Ala Gly Asp Glu Ala
20 25 30
Asp Tyr Tyr Cys
35
<210> 91
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 91
Phe Gly Thr Gly Ser Lys Val Thr Val Leu
1 5 10
<210> 92
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 92
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 93
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 93
Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ala
1 5 10 15
Leu
<210> 94
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 94
Ser Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ser Lys Asn Met Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 95
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 95
Trp Gly Gln Gly Thr Val Val Thr Val Ser Ser
1 5 10
<210> 96
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 96
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr
20 25
<210> 97
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 97
Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu Met Ile
1 5 10 15
Tyr
<210> 98
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 98
Asp Arg Pro Ser Gly Val Ser Phe Arg Phe Ser Gly Ser Lys Ser Gly
1 5 10 15
Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu Ala
20 25 30
Tyr Tyr Tyr Cys
35
<210> 99
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 99
Gln Met Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser
20 25
<210> 100
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 100
Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
1 5 10 15
Trp
<210> 101
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 101
Asn Tyr Ala Gln Lys Leu Gln Gly Arg Val Thr Met Thr Thr Asp Thr
1 5 10 15
Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 102
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 102
Trp Gly Gln Gly Ala Leu Val Thr Val Ser Ser
1 5 10
<210> 103
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 103
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Gly Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser
20 25
<210> 104
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 104
Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Ile Ile Ile
1 5 10 15
Tyr
<210> 105
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 105
Gln Arg Thr Ser Gly Val Pro Asp Arg Phe Ser Gly Ser Ile Asp Arg
1 5 10 15
Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly Leu Lys Thr Glu Asp
20 25 30
Glu Gly Asp Tyr Tyr Cys
35
<210> 106
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 106
Gln Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 107
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 107
Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
1 5 10 15
Ser
<210> 108
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 108
Trp Gly Gln Gly Ile Leu Val Thr Val Ser Ser
1 5 10
<210> 109
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 109
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Leu Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp
20 25
<210> 110
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 110
Ala Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Met
1 5 10 15
Tyr
<210> 111
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 111
Val Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Ser Ser Gly
1 5 10 15
Thr Thr Val Thr Leu Thr Ile Ser Arg Val Gln Ala Glu Asp Glu Ala
20 25 30
Asp Tyr Tyr Cys
35
<210> 112
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 112
Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Gly
1 5 10 15
Arg
<210> 113
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 113
Asp Tyr Ala Thr Pro Met Lys Asp Arg Phe Ile Ile Ser Arg Asp Asp
1 5 10 15
Ser Lys Asn Thr Val Phe Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 114
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 114
Trp Gly Gln Gly Thr Lys Val Thr Val Ser Ser
1 5 10
<210> 115
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 115
Ile Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Arg Leu Leu Ile
1 5 10 15
Tyr
<210> 116
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 116
Asp Arg Pro Ser Gly Val Ser Asn Arg Phe Ser Gly Ser Lys Ser Ala
1 5 10 15
Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu Ala
20 25 30
Asp Tyr Tyr Cys
35
<210> 117
<211> 25
<212> ПРТ
<213> Homo sapiens
<400> 117
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 118
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 118
Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Ser
1 5 10 15
Gly
<210> 119
<211> 38
<212> ПРТ
<213> Homo sapiens
<400> 119
Val Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
1 5 10 15
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 120
<211> 11
<212> ПРТ
<213> Homo sapiens
<400> 120
Trp Gly Pro Gly Thr Met Val Thr Val Ser Ser
1 5 10
<210> 121
<211> 26
<212> ПРТ
<213> Homo sapiens
<400> 121
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
20 25
<210> 122
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 122
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Ile Ala Pro Lys Leu Leu Ile
1 5 10 15
Phe
<210> 123
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 123
Arg Leu His Ser Gly Val Pro Ser Arg Phe Asn Gly Ser Gly Ser Gly
1 5 10 15
Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro Glu Asp Phe Ala
20 25 30
Thr Tyr Tyr Cys
35
<210> 124
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 124
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 125
<211> 26
<212> ПРТ
<213> Homo sapiens
<400> 125
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
20 25
<210> 126
<211> 17
<212> ПРТ
<213> Homo sapiens
<400> 126
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Ile Ala Pro Lys Leu Leu Ile
1 5 10 15
Tyr
<210> 127
<211> 36
<212> ПРТ
<213> Homo sapiens
<400> 127
Ile Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
1 5 10 15
Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala
20 25 30
Thr Tyr Tyr Cys
35
<210> 128
<211> 10
<212> ПРТ
<213> Homo sapiens
<400> 128
Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
1 5 10
<210> 129
<211> 127
<212> ПРТ
<213> Homo sapiens
<400> 129
Gln Val Thr Leu Lys Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Leu Thr Thr Ser
20 25 30
Gly Val Ala Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Tyr Trp Asp Gly Asp Glu Arg Tyr Ser Ser Ser
50 55 60
Leu Lys Asn Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Arg Gly Tyr Tyr Asp Trp Gly Ser Tyr Tyr Ser Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 130
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 130
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr His Val
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 131
<211> 127
<212> ПРТ
<213> Homo sapiens
<400> 131
Gln Val Thr Leu Arg Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Ser Gly Phe Ser Phe Asn Thr Pro
20 25 30
Gly Glu Gly Val Thr Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Ala Leu Ile Tyr Trp Asp Asp Glu Lys His Tyr Asn Pro Ser
50 55 60
Leu Asn Gly Arg Leu Thr Ile Thr Lys Asp Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Arg Gly Tyr Tyr Asp Trp Gly Ser Tyr Tyr Ser Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Thr Ile Thr Val Ser Ser
115 120 125
<210> 132
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 132
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Thr His Val
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 133
<211> 117
<212> ПРТ
<213> Homo sapiens
<400> 133
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Ile Lys Lys Pro Gly Ala
1 5 10 15
Ser Met Asn Val Ser Cys Lys Ala Ser Gly Tyr Ile Phe Thr Asn Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Asp Pro Ser Gly Gly Ser Thr Thr Tyr Ala Gln Lys Phe
50 55 60
Gln His Arg Val Thr Met Thr Ser Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ile Gly Leu Gly Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Trp Gly Ser Leu Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 134
<211> 112
<212> ПРТ
<213> Homo sapiens
<400> 134
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Glu Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Ala Ser Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Ile Val
35 40 45
Ile Tyr Glu Asn Tyr Leu Arg Pro Ser Glu Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Ser Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser
85 90 95
Arg Asn Arg Asn Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 135
<211> 121
<212> ПРТ
<213> Homo sapiens
<400> 135
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Tyr Ser Ala Tyr Glu Thr Gly Tyr Phe Asp Phe Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 136
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 136
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Thr Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Thr Ser Thr Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Gly Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Ala Pro Pro
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys
100 105
<210> 137
<211> 127
<212> ПРТ
<213> Homo sapiens
<400> 137
Gln Val Thr Leu Arg Glu Ser Gly Pro Thr Leu Val Lys Pro Thr Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Phe Arg Gly Phe Ser Leu Asn Thr Pro
20 25 30
Gly Met Gly Val Ala Trp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu
35 40 45
Trp Leu Gly Leu Val Phe Trp Asp Asp Asp Lys Thr Tyr Arg Pro Ser
50 55 60
Leu Arg Asn Arg Leu Thr Ile Thr Lys Glu Thr Ser Lys Ser Gln Val
65 70 75 80
Val Leu Thr Met Thr Asn Met Asp Pro Val Asp Thr Ala Thr Tyr Tyr
85 90 95
Cys Ala Arg Ile Arg Gly Tyr Tyr Asp Trp Gly Ser Tyr Tyr Ser Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 138
<211> 108
<212> ПРТ
<213> Homo sapiens
<400> 138
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Asn Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Arg Asp Ser Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp Gln
85 90 95
Tyr Val Phe Gly Thr Gly Ser Lys Val Thr Val Leu
100 105
<210> 139
<211> 118
<212> ПРТ
<213> Homo sapiens
<400> 139
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Arg
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr His Gly Ser Asp Asp Ser Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Met Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Asn Leu Gly Ala Thr Asp Gly Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Val Val Thr Val Ser Ser
115
<210> 140
<211> 110
<212> ПРТ
<213> Homo sapiens
<400> 140
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Ser Ala Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gly Val Ser Asp Arg Pro Ser Gly Val Ser Phe Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Tyr Tyr Tyr Cys Asn Ser Tyr Thr Thr Ser
85 90 95
Asn Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 141
<211> 117
<212> ПРТ
<213> Homo sapiens
<400> 141
Gln Met Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Trp Gly Ser Leu Asp His Trp Gly Gln Gly Ala Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 142
<211> 112
<212> ПРТ
<213> Homo sapiens
<400> 142
Asn Phe Met Leu Thr Gln Pro His Ser Val Ser Gly Ser Pro Gly Lys
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Arg Ser Ser Gly Ser Ile Asp Ile Asn
20 25 30
Tyr Val Gln Trp Tyr Gln Gln Arg Pro Gly Ser Ala Pro Thr Ile Ile
35 40 45
Ile Tyr Glu Asp Ser Gln Arg Thr Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Ile Asp Arg Ser Ser Asn Ser Ala Ser Leu Thr Ile Ser Gly
65 70 75 80
Leu Lys Thr Glu Asp Glu Gly Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser
85 90 95
Arg Asn Arg Asn Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 143
<211> 122
<212> ПРТ
<213> Homo sapiens
<400> 143
Gln Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp His
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Gly Gly Gly Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Glu Tyr Asp Ser Asp Gly His Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Ile Leu Val Thr Val Ser Ser
115 120
<210> 144
<211> 108
<212> ПРТ
<213> Homo sapiens
<400> 144
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Leu Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Ala Leu Pro Lys Gln Tyr Ala
20 25 30
Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Met Tyr
35 40 45
Lys Asp Asn Val Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ser Ser Gly Thr Thr Val Thr Leu Thr Ile Ser Arg Val Gln Ala Glu
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Val Asp Ser Ser Asp Thr Ser
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 145
<211> 124
<212> ПРТ
<213> Homo sapiens
<400> 145
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Phe Arg Val Ala
20 25 30
Trp Val Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Ile Lys Ser Asn Ser Asp Gly Gly Thr Thr Asp Tyr Ala Thr
50 55 60
Pro Met Lys Asp Arg Phe Ile Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Val Phe Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg His Gly Asp Ala Asn Ala Tyr Tyr Tyr Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Lys Val Thr Val Ser Ser
115 120
<210> 146
<211> 111
<212> ПРТ
<213> Homo sapiens
<400> 146
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Ile Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Arg Leu
35 40 45
Leu Ile Tyr Asp Val Ser Asp Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Ala Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Ser Ser
85 90 95
Ser Thr Leu Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 147
<211> 117
<212> ПРТ
<213> Homo sapiens
<400> 147
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Gly Ile Ser Trp Asn Ser Gly Asn Ile Val Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Thr Ala Leu Ala Phe Asp Met Trp Gly Pro Gly Thr Met
100 105 110
Val Thr Val Ser Ser
115
<210> 148
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 148
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Tyr Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Ile Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Ser Ser Arg Leu His Ser Gly Val Pro Ser Arg Phe Asn Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Ala Thr Ser Phe Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 149
<211> 118
<212> ПРТ
<213> Homo sapiens
<400> 149
Gln Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Tyr Tyr Tyr Gly Leu Asp Val Trp Gly Gln Gly Thr
100 105 110
Thr Val Thr Val Ser Ser
115
<210> 150
<211> 106
<212> ПРТ
<213> Homo sapiens
<400> 150
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Ile Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gln Ala Ser Ile Leu Asp Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Leu Trp Thr
85 90 95
Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 151
<211> 22
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 151
cgtcccatcc tggaagtgcc ag 22
<210> 152
<211> 23
<212> ДНК
<213> Искусственная
<220>
<223> Праймер
<400> 152
gctctttcct ggcccagcac tgg 23
<210> 153
<211> 792
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(L)-Fc)
<400> 153
cgtcccatcc tggaagtgcc agagagtgta acaggacctt ggaaagggga tgtgaatctt 60
ccctgcacct atgaccccct gcaaggctac acccaagtct tggtgaagtg gctggtacaa 120
cgtggctcag accctgtcac catctttcta cgtgactctt ctggagacca tatccagcag 180
gcaaagtacc agggccgcct gcatgtgagc cacaaggttc caggagatgt atccctccaa 240
ttgagcaccc tggagatgga tgaccggagc cactacacgt gtgaagtcac ctggcagact 300
cctgatggca accaagtcgt gagagataag attactgagc tccgtgtcca gaaactctct 360
gtctccaagc ccacagtgac aactggcagc ggttatggct tcacggtgcc ccagggaatg 420
aggattagcc ttcaatgcca ggctcggggt tctcctccca tcagttatat ttggtataag 480
caacagacta ataaccagga acccatcaaa gtagcaaccc taagtacctt actcttcaag 540
cctgcggtga tagccgactc aggctcctat ttctgcactg ccaagggcca ggttggctct 600
gagcagcaca gcgacattgt gaagtttgtg gtcaaagact cctcaaagct actcaagacc 660
aagactgagg cacctacaac catgacatac cccttgaaag caacatctac agtgaagcag 720
tcctgggact ggaccactga catggatggc taccttggag agaccagtgc tgggccagga 780
aagagcctgc ct 792
<210> 154
<211> 264
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(L)-Fc)
<400> 154
Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly
1 5 10 15
Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln
20 25 30
Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile
35 40 45
Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln
50 55 60
Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln
65 70 75 80
Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val
85 90 95
Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr
100 105 110
Glu Leu Arg Val Gln Lys Leu Ser Val Ser Lys Pro Thr Val Thr Thr
115 120 125
Gly Ser Gly Tyr Gly Phe Thr Val Pro Gln Gly Met Arg Ile Ser Leu
130 135 140
Gln Cys Gln Ala Arg Gly Ser Pro Pro Ile Ser Tyr Ile Trp Tyr Lys
145 150 155 160
Gln Gln Thr Asn Asn Gln Glu Pro Ile Lys Val Ala Thr Leu Ser Thr
165 170 175
Leu Leu Phe Lys Pro Ala Val Ile Ala Asp Ser Gly Ser Tyr Phe Cys
180 185 190
Thr Ala Lys Gly Gln Val Gly Ser Glu Gln His Ser Asp Ile Val Lys
195 200 205
Phe Val Val Lys Asp Ser Ser Lys Leu Leu Lys Thr Lys Thr Glu Ala
210 215 220
Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr Ser Thr Val Lys Gln
225 230 235 240
Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr Leu Gly Glu Thr Ser
245 250 255
Ala Gly Pro Gly Lys Ser Leu Pro
260
<210> 155
<211> 517
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(L)-Fc)
<400> 155
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Ala Asp
1 5 10 15
Val His Ser Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp
115 120 125
Lys Ile Thr Glu Leu Arg Val Gln Lys Leu Ser Val Ser Lys Pro Thr
130 135 140
Val Thr Thr Gly Ser Gly Tyr Gly Phe Thr Val Pro Gln Gly Met Arg
145 150 155 160
Ile Ser Leu Gln Cys Gln Ala Arg Gly Ser Pro Pro Ile Ser Tyr Ile
165 170 175
Trp Tyr Lys Gln Gln Thr Asn Asn Gln Glu Pro Ile Lys Val Ala Thr
180 185 190
Leu Ser Thr Leu Leu Phe Lys Pro Ala Val Ile Ala Asp Ser Gly Ser
195 200 205
Tyr Phe Cys Thr Ala Lys Gly Gln Val Gly Ser Glu Gln His Ser Asp
210 215 220
Ile Val Lys Phe Val Val Lys Asp Ser Ser Lys Leu Leu Lys Thr Lys
225 230 235 240
Thr Glu Ala Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr Ser Thr
245 250 255
Val Lys Gln Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr Leu Gly
260 265 270
Glu Thr Ser Ala Gly Pro Gly Lys Ser Leu Pro Ala Ser Glu Pro Lys
275 280 285
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
290 295 300
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
305 310 315 320
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
325 330 335
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
340 345 350
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
355 360 365
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
370 375 380
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
385 390 395 400
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
405 410 415
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
420 425 430
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
435 440 445
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
450 455 460
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
465 470 475 480
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
485 490 495
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
500 505 510
Leu Ser Pro Gly Lys
515
<210> 156
<211> 318
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-V-Fc)
<400> 156
cgtcccatcc tggaagtgcc agagagtgta acaggacctt ggaaagggga tgtgaatctt 60
ccctgcacct atgaccccct gcaaggctac acccaagtct tggtgaagtg gctggtacaa 120
cgtggctcag accctgtcac catctttcta cgtgactctt ctggagacca tatccagcag 180
gcaaagtacc agggccgcct gcatgtgagc cacaaggttc caggagatgt atccctccaa 240
ttgagcaccc tggagatgga tgaccggagc cactacacgt gtgaagtcac ctggcagact 300
cctgatggca accaagtc 318
<210> 157
<211> 106
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-V-Fc)
<400> 157
Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly
1 5 10 15
Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln
20 25 30
Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile
35 40 45
Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln
50 55 60
Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln
65 70 75 80
Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val
85 90 95
Thr Trp Gln Thr Pro Asp Gly Asn Gln Val
100 105
<210> 158
<211> 359
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-V-Fc)
<400> 158
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Ala Asp
1 5 10 15
Val His Ser Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Ala Ser Glu
115 120 125
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
130 135 140
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
145 150 155 160
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
165 170 175
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
180 185 190
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
195 200 205
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
210 215 220
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
225 230 235 240
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
245 250 255
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
260 265 270
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
275 280 285
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
290 295 300
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
305 310 315 320
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
325 330 335
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
340 345 350
Leu Ser Leu Ser Pro Gly Lys
355
<210> 159
<211> 348
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M1-Fc)
<400> 159
cgtcccatcc tgaccgcccc ccacagcctg gccggacctt ggaaagggga tgtgaatctt 60
ccctgcacct atgaccccct gcaaggctac acccaagtct tggtgaagtg gctggtacaa 120
cgtggctcag accctgtcac catctttcta cgtgactctt ctggagacca tatccagcag 180
gcaaagtacc agggccgcct gcatgtgagc cacaaggttc caggagatgt atccctccaa 240
ttgagcaccc tggagatgga tgaccggagc cactacacgt gtgaagtcac ctggcagact 300
cctgatggca accaagtcgt gagagataag attactgagc tccgtgtt 348
<210> 160
<211> 116
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M1-Fc)
<400> 160
Arg Pro Ile Leu Thr Ala Pro His Ser Leu Ala Gly Pro Trp Lys Gly
1 5 10 15
Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln
20 25 30
Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile
35 40 45
Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln
50 55 60
Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln
65 70 75 80
Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val
85 90 95
Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr
100 105 110
Glu Leu Arg Val
115
<210> 161
<211> 380
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M1-Fc)
<400> 161
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Arg Pro Ile Leu Thr Ala Pro
20 25 30
His Ser Leu Ala Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr
35 40 45
Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val
50 55 60
Gln Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly
65 70 75 80
Asp His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His
85 90 95
Lys Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp
100 105 110
Asp Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly
115 120 125
Asn Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala
130 135 140
Ser Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
145 150 155 160
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
165 170 175
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
180 185 190
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
195 200 205
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
210 215 220
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
225 230 235 240
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
245 250 255
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
260 265 270
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
275 280 285
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
290 295 300
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
305 310 315 320
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
325 330 335
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
340 345 350
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
355 360 365
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375 380
<210> 162
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M2-Fc)
<400> 162
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aagggaccgt gcggatgccc 60
tgcagctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgca tgtgagccac aaggttccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtt 345
<210> 163
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M2-Fc)
<400> 163
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Thr
1 5 10 15
Val Arg Met Pro Cys Ser Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 164
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M2-Fc)
<400> 164
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Thr Val Arg Met Pro Cys Ser Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 165
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M3-Fc)
<400> 165
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtactgcgg 120
aactaccacc ccgccaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgca tgtgagccac aaggttccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtc 345
<210> 166
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M3-Fc)
<400> 166
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Leu Arg Asn Tyr His Pro Ala Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 167
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M3-Fc)
<400> 167
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Leu
50 55 60
Arg Asn Tyr His Pro Ala Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 168
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M4-Fc)
<400> 168
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatgt gcagcagacc 180
aagttccggg gccgcctgca tgtgagccac aaggttccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtt 345
<210> 169
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M4-Fc)
<400> 169
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Val Gln Gln Thr Lys Phe Arg Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 170
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M4-Fc)
<400> 170
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Val Gln Gln Thr Lys Phe Arg Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 171
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M5-Fc)
<400> 171
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgga ggtgagccgg cagcccccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtt 345
<210> 172
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M5-Fc)
<400> 172
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu Glu Val Ser Arg Gln Pro Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 173
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M5-Fc)
<400> 173
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu Glu Val Ser Arg Gln
85 90 95
Pro Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 174
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M6-Fc)
<400> 174
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgca tgtgagccac aaggttccag gagatgtagc cctgaccctg 240
aaccccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtt 345
<210> 175
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M6-Fc)
<400> 175
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ala Leu Thr Leu
65 70 75 80
Asn Pro Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 176
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M6-Fc)
<400> 176
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ala Leu Thr Leu Asn Pro Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 177
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M7-Fc)
<400> 177
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgca tgtgagccac aaggttccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccagggctac tacgtgtgcg ccgtcacctg gcagactcct 300
gatggcaacc aagtcgtgag agataagatt actgagctcc gtgtt 345
<210> 178
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M7-Fc)
<400> 178
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Gln Gly Tyr Tyr Val Cys Ala Val Thr
85 90 95
Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 179
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M7-Fc)
<400> 179
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Gln Gly Tyr Tyr Val Cys Ala Val Thr Trp Gln Thr Pro Asp Gly Asn
115 120 125
Gln Val Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 180
<211> 345
<212> ДНК
<213> Искусственная
<220>
<223> Слитый белок (hVSIG(S)4-M8-Fc)
<400> 180
cccatcctgg aagtgccaga gagtgtaaca ggaccttgga aaggggatgt gaatcttccc 60
tgcacctatg accccctgca aggctacacc caagtcttgg tgaagtggct ggtacaacgt 120
ggctcagacc ctgtcaccat ctttctacgt gactcttctg gagaccatat ccagcaggca 180
aagtaccagg gccgcctgca tgtgagccac aaggttccag gagatgtatc cctccaattg 240
agcaccctgg agatggatga ccggagccac tacacgtgtg aagtcacctg gcaggaccag 300
gccggccacc tgatcgtgag agataagatt actgagctcc gtgtt 345
<210> 181
<211> 115
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (hVSIG4(S)-M8-Fc)
<400> 181
Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro Trp Lys Gly Asp
1 5 10 15
Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly Tyr Thr Gln Val
20 25 30
Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro Val Thr Ile Phe
35 40 45
Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala Lys Tyr Gln Gly
50 55 60
Arg Leu His Val Ser His Lys Val Pro Gly Asp Val Ser Leu Gln Leu
65 70 75 80
Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr Cys Glu Val Thr
85 90 95
Trp Gln Asp Gln Ala Gly His Leu Ile Val Arg Asp Lys Ile Thr Glu
100 105 110
Leu Arg Val
115
<210> 182
<211> 379
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок + Сигнальная последовательность (NLS-hVSIG4(S)-M8-Fc)
<400> 182
Met Gly Trp Ser Tyr Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Asp
1 5 10 15
Val His Ser Gln Gly Ala Val Gly Ala Pro Ile Leu Glu Val Pro Glu
20 25 30
Ser Val Thr Gly Pro Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr
35 40 45
Asp Pro Leu Gln Gly Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln
50 55 60
Arg Gly Ser Asp Pro Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp
65 70 75 80
His Ile Gln Gln Ala Lys Tyr Gln Gly Arg Leu His Val Ser His Lys
85 90 95
Val Pro Gly Asp Val Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp
100 105 110
Arg Ser His Tyr Thr Cys Glu Val Thr Trp Gln Asp Gln Ala Gly His
115 120 125
Leu Ile Val Arg Asp Lys Ile Thr Glu Leu Arg Val Leu Ala Ala Ser
130 135 140
Ala Ala Ser Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
145 150 155 160
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
165 170 175
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
180 185 190
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
195 200 205
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
210 215 220
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
225 230 235 240
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
245 250 255
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
260 265 270
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
275 280 285
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
290 295 300
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
305 310 315 320
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
325 330 335
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
340 345 350
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
355 360 365
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 183
<211> 513
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (сигнальный пептид-hVSIG4(L)-Fc)
<400> 183
Gln Gly Ile Leu Leu Gly Leu Leu Leu Leu Gly His Leu Thr Val Asp
1 5 10 15
Thr Tyr Gly Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp
115 120 125
Lys Ile Thr Glu Leu Arg Val Gln Lys Leu Ser Val Ser Lys Pro Thr
130 135 140
Val Thr Thr Gly Ser Gly Tyr Gly Phe Thr Val Pro Gln Gly Met Arg
145 150 155 160
Ile Ser Leu Gln Cys Gln Ala Arg Gly Ser Pro Pro Ile Ser Tyr Ile
165 170 175
Trp Tyr Lys Gln Gln Thr Asn Asn Gln Glu Pro Ile Lys Val Ala Thr
180 185 190
Leu Ser Thr Leu Leu Phe Lys Pro Ala Val Ile Ala Asp Ser Gly Ser
195 200 205
Tyr Phe Cys Thr Ala Lys Gly Gln Val Gly Ser Glu Gln His Ser Asp
210 215 220
Ile Val Lys Phe Val Val Lys Asp Ser Ser Lys Leu Leu Lys Thr Lys
225 230 235 240
Thr Glu Ala Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr Ser Thr
245 250 255
Val Lys Gln Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr Leu Gly
260 265 270
Glu Thr Ser Ala Gly Pro Gly Lys Ser Leu Pro Ala Ala Ala Asp Lys
275 280 285
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
290 295 300
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
305 310 315 320
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
325 330 335
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
340 345 350
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
355 360 365
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
370 375 380
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
385 390 395 400
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
405 410 415
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
420 425 430
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
435 440 445
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
450 455 460
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
465 470 475 480
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
485 490 495
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
500 505 510
Lys
<210> 184
<211> 419
<212> ПРТ
<213> Искусственная
<220>
<223> Слитый белок (сигнальный пептид-hVSIG4(S)-Fc)
<400> 184
Met Gly Ile Leu Leu Gly Leu Leu Leu Leu Gly His Leu Thr Val Asp
1 5 10 15
Thr Tyr Gly Arg Pro Ile Leu Glu Val Pro Glu Ser Val Thr Gly Pro
20 25 30
Trp Lys Gly Asp Val Asn Leu Pro Cys Thr Tyr Asp Pro Leu Gln Gly
35 40 45
Tyr Thr Gln Val Leu Val Lys Trp Leu Val Gln Arg Gly Ser Asp Pro
50 55 60
Val Thr Ile Phe Leu Arg Asp Ser Ser Gly Asp His Ile Gln Gln Ala
65 70 75 80
Lys Tyr Gln Gly Arg Leu His Val Ser His Lys Val Pro Gly Asp Val
85 90 95
Ser Leu Gln Leu Ser Thr Leu Glu Met Asp Asp Arg Ser His Tyr Thr
100 105 110
Cys Glu Val Thr Trp Gln Thr Pro Asp Gly Asn Gln Val Val Arg Asp
115 120 125
Lys Ile Thr Glu Leu Arg Val Gln Lys His Ser Ser Lys Leu Leu Lys
130 135 140
Thr Lys Thr Glu Ala Pro Thr Thr Met Thr Tyr Pro Leu Lys Ala Thr
145 150 155 160
Ser Thr Val Lys Gln Ser Trp Asp Trp Thr Thr Asp Met Asp Gly Tyr
165 170 175
Leu Gly Glu Thr Ser Ala Gly Pro Gly Lys Ser Leu Pro Ala Ala Ala
180 185 190
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
195 200 205
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
210 215 220
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
225 230 235 240
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
245 250 255
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
260 265 270
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
275 280 285
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
290 295 300
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
305 310 315 320
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
325 330 335
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
340 345 350
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
355 360 365
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
370 375 380
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
385 390 395 400
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
405 410 415
Pro Gly Lys
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714232C2 |
| АНТИТЕЛА ПРОТИВ VSIG4 ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2776638C1 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2819627C1 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| Композиция для предупреждения и лечения заболевания кожи, содержащая вещество, специфично связывающееся с пептидом, имеющим происхождение из виментина | 2018 |
|
RU2751486C1 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| КОНЪЮГАТ АНТИТЕЛА ПРОТИВ CEA И АНАЛОГА ЭКСАТЕКАНА И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2020 |
|
RU2833323C1 |
| АНТИ-PCSK9 АНТИТЕЛО, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2016 |
|
RU2739208C2 |
| Антитело против FXI/FXIA, его антигенсвязывающий фрагмент и фармацевтическое применение | 2021 |
|
RU2838408C1 |
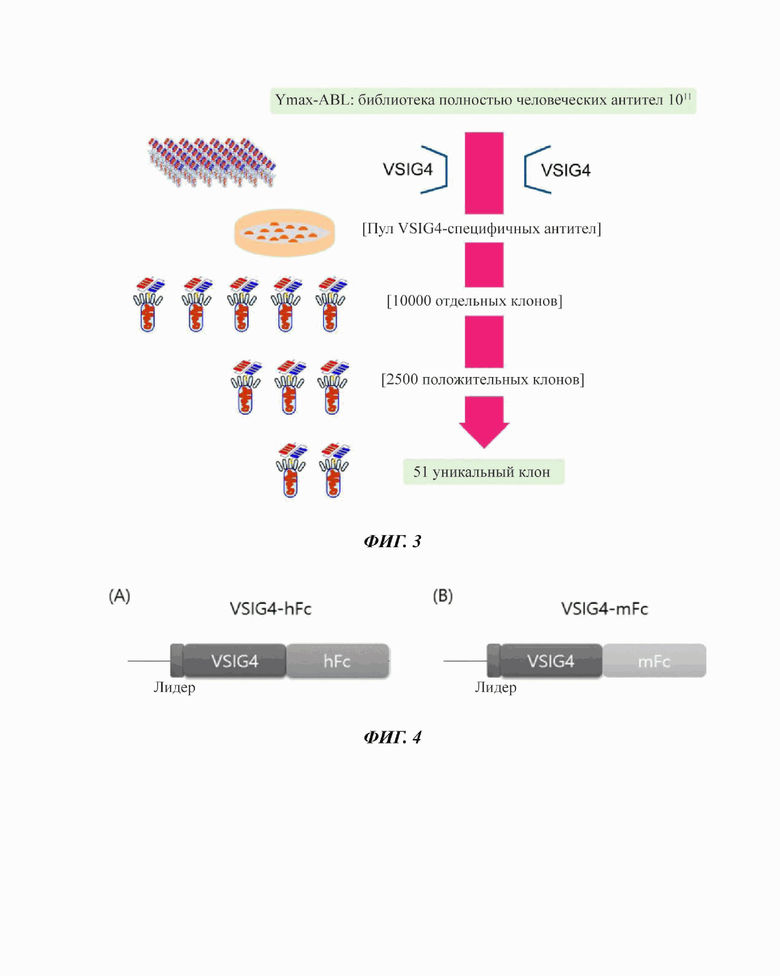
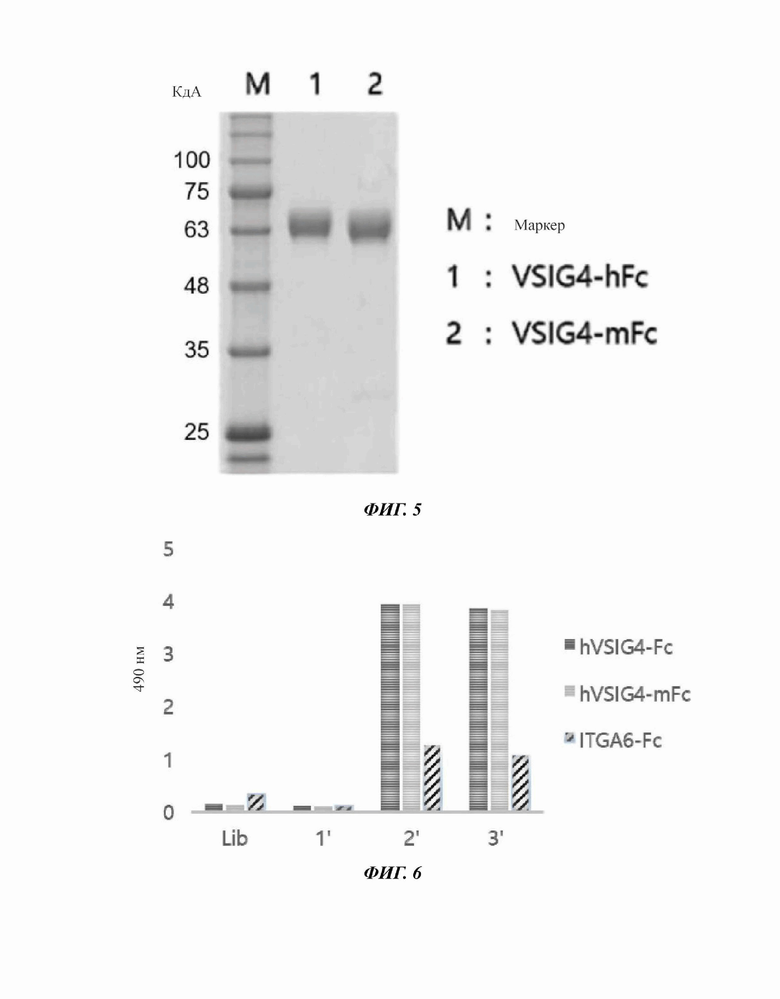
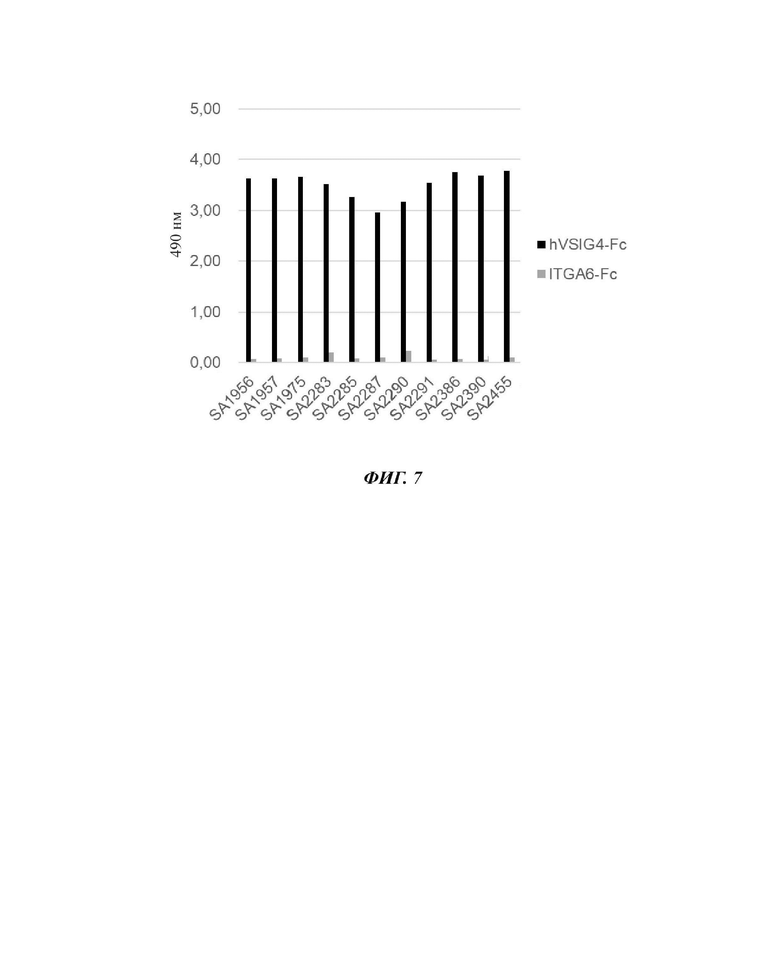
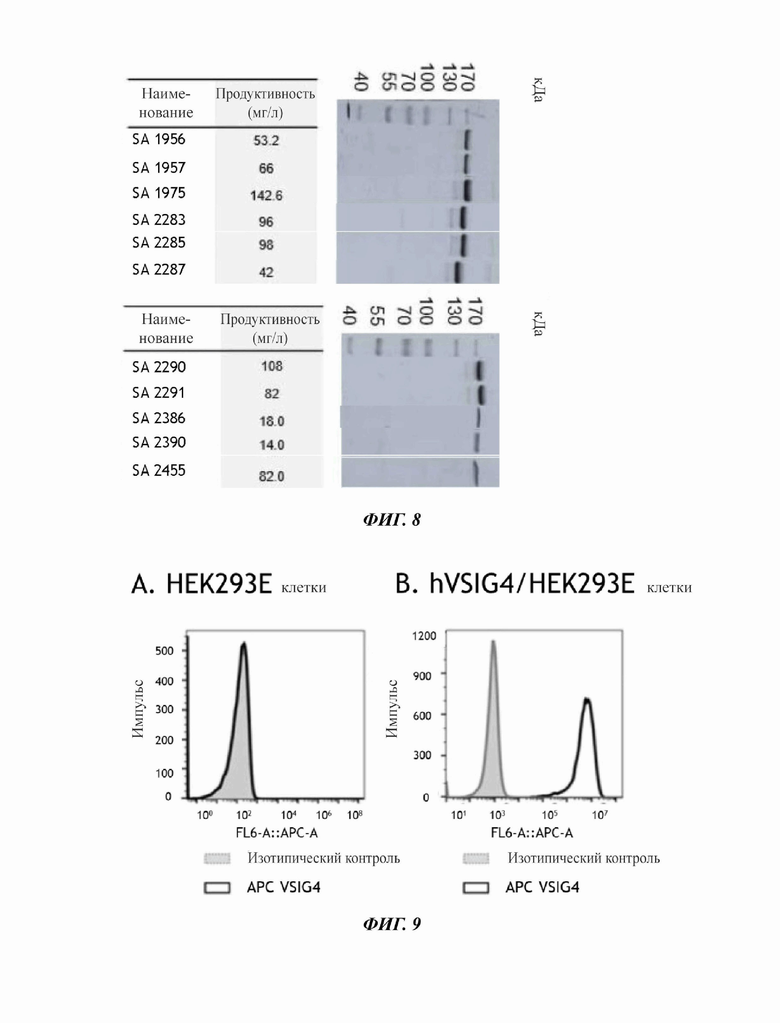
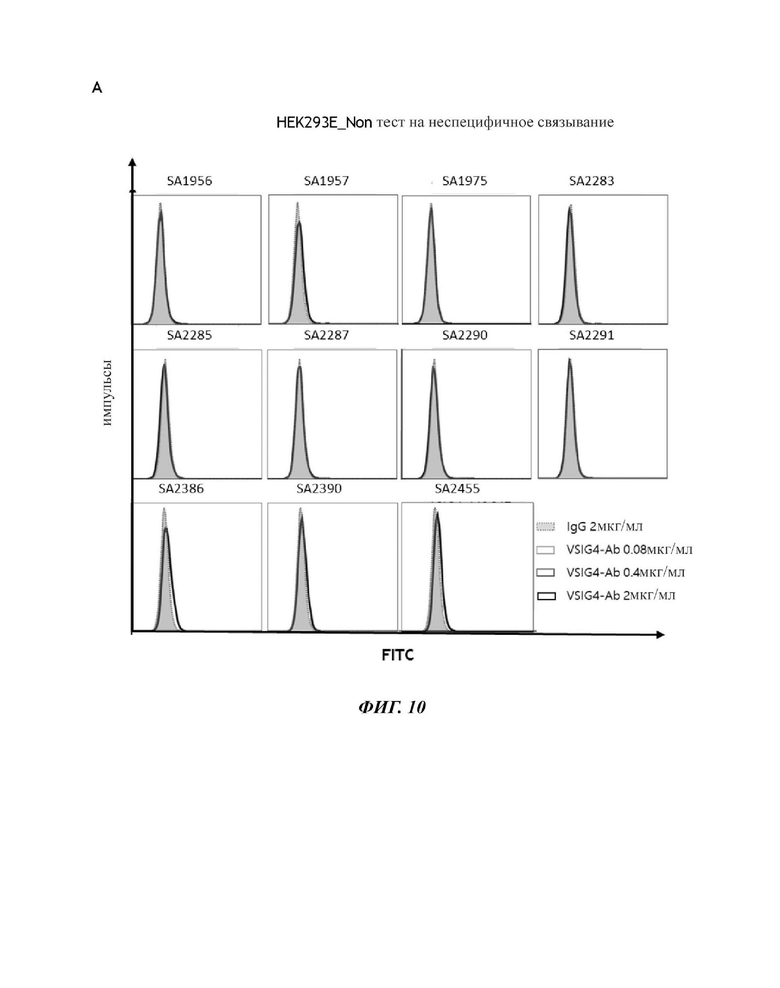
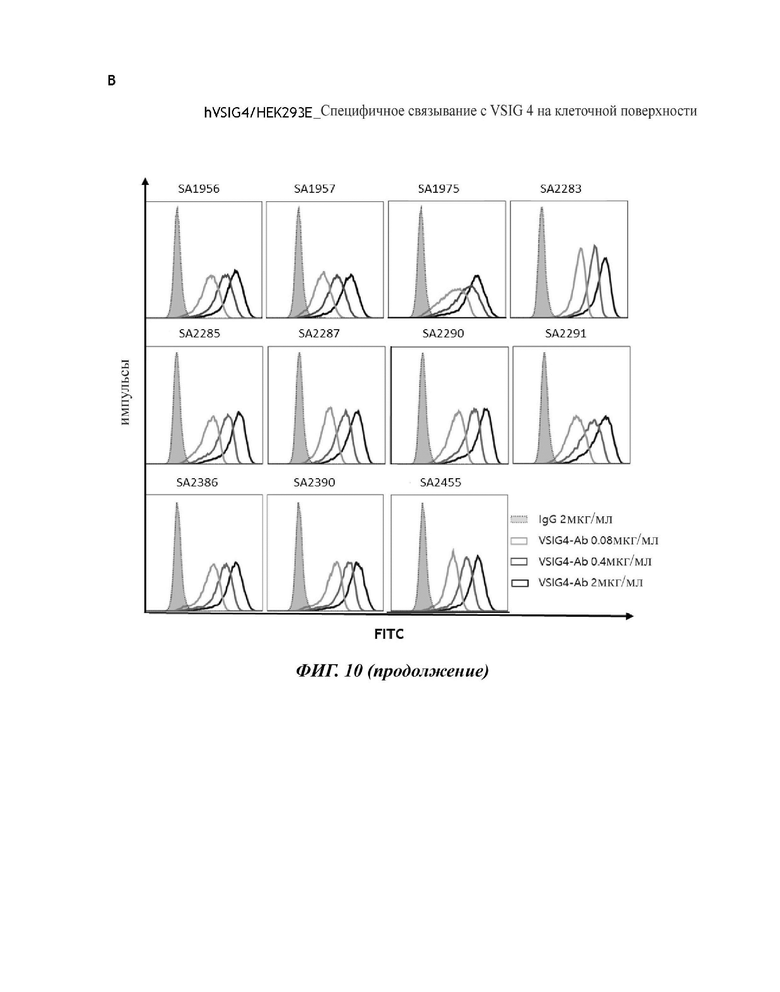

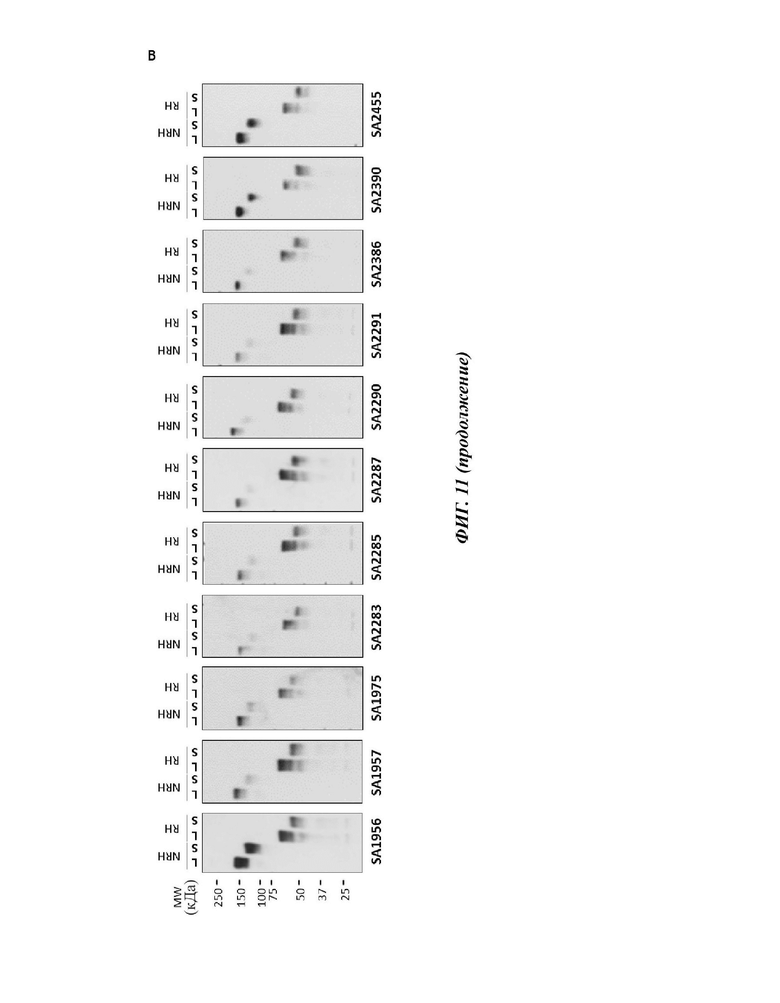
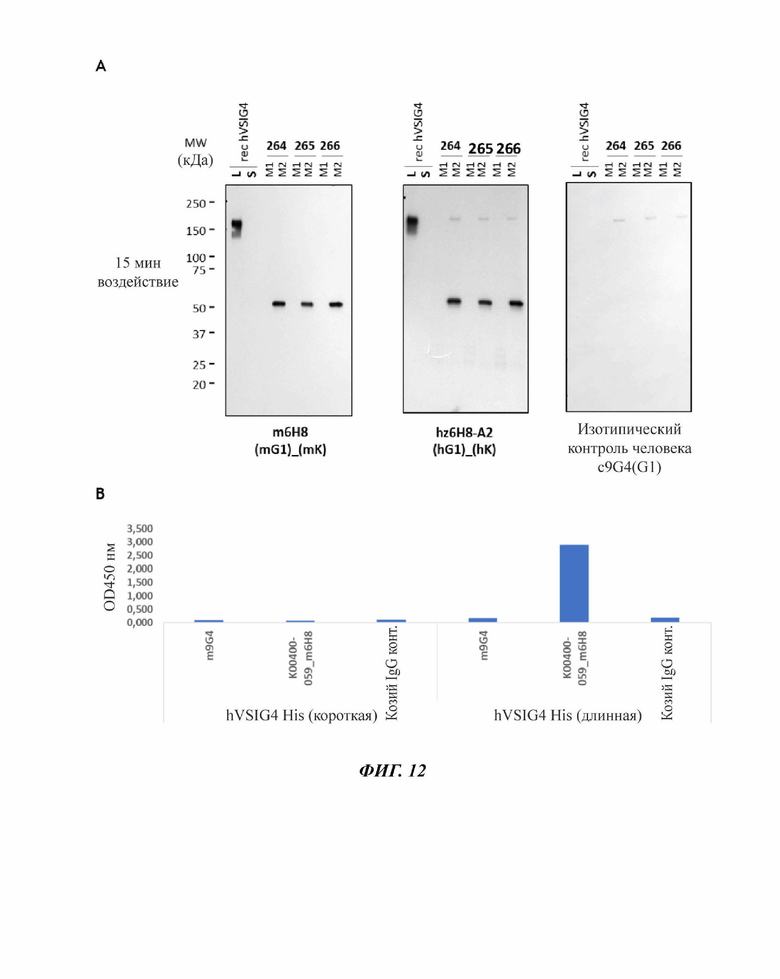
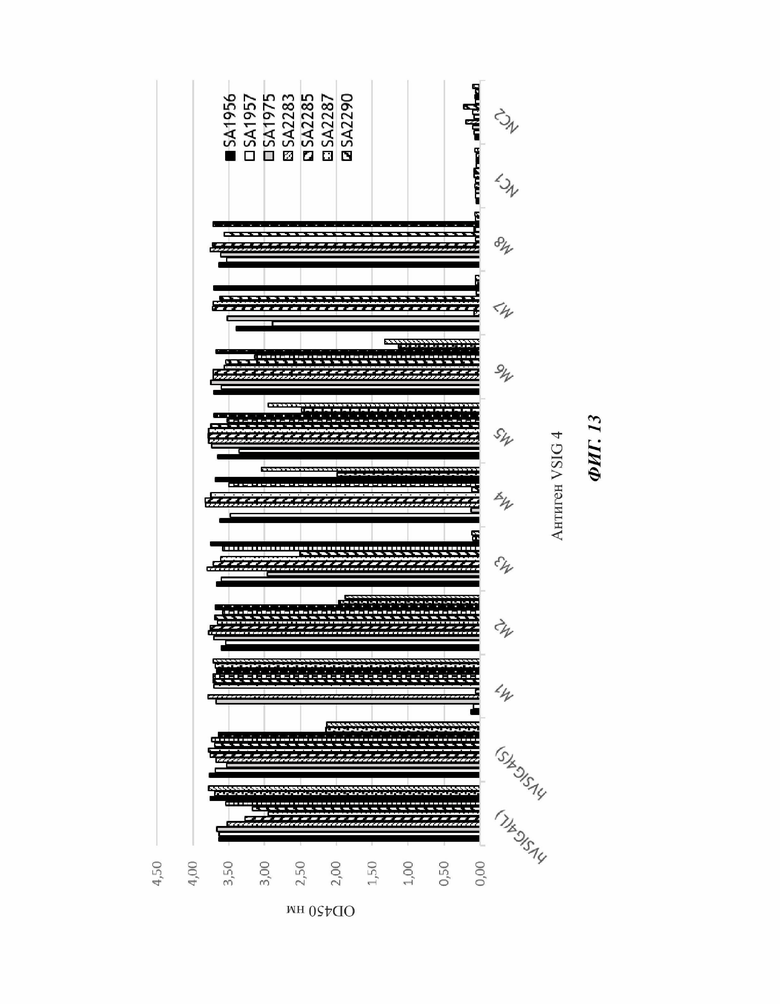
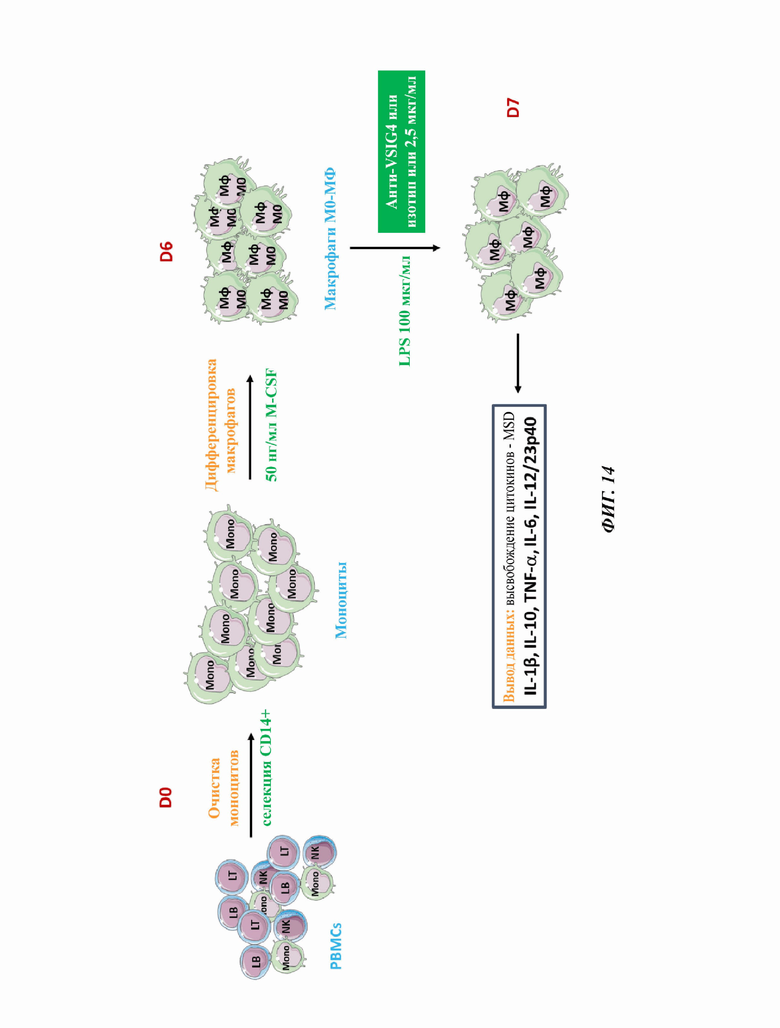
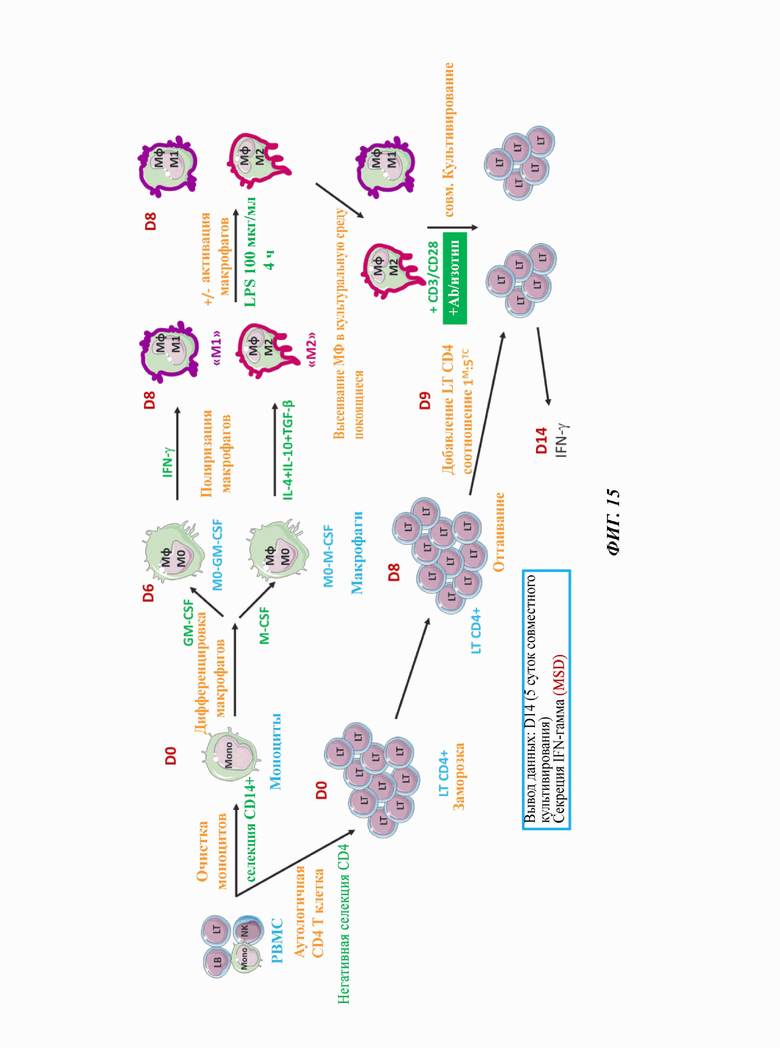
Изобретение относится к области биотехнологии, в частности к моноклональному антителу против V-подобного Ig домен-содержащего 4 белка (VSIG4) или его антигенсвязывающему фрагменту, а также к композиции и иммуноконъюгату, его содержащим. Также раскрыты полинуклеотид, кодирующей вышеуказанное антитело или его фрагмент, а также клетка и вектор, его содержащие. Изобретение также относится к способу получения вышеуказанного антитела или его фрагмента, а также к способу in vitro обнаружения рака, экспрессирующего VSIG4, используя вышеуказанное антитело или его фрагмент. Изобретение эффективно для стимуляции иммунного ответа у пациента, страдающего раком. 8 н. и 20 з.п. ф-лы, 15 ил., 10 табл., 7 пр.
1. Моноклональное антитело против V-подобного Ig домен-содержащего 4 белка (VSIG4) или его антигенсвязывающий фрагмент, причем указанное антитело выбрано из группы, состоящей из следующего:
a) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
b) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 9, 10 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
c) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 11, 12 и 13 и три CDR легкой цепи с последовательностями SEQ ID NO: 14, 15 и 16;
d) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 19 и три CDR легкой цепи с последовательностями SEQ ID NO: 20, 21 и 22;
e) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 23, 24 и 5 и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 25;
f) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 26, 27 и 28 и три CDR легкой цепи с последовательностями SEQ ID NO: 29, 30 и 31;
g) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 32, 33 и 34 и три CDR легкой цепи с последовательностями SEQ ID NO: 35, 36 и 16;
h) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 37, 38 и 39 и три CDR легкой цепи с последовательностями SEQ ID NO: 40, 41 и 42;
i) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 43, 44 и 45 и три CDR легкой цепи с последовательностями SEQ ID NO: 46, 47 и 48;
j) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 49, 50 и 51 и три CDR легкой цепи с последовательностями SEQ ID NO: 52, 53 и 54;
k) антитело, содержащее три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 55 и три CDR легкой цепи с последовательностями SEQ ID NO: 56, 57 и 58.
2. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по п. 1, где указанное антитело выбрано из одноцепочечных антител, верблюжьих антител, химерных антител, гуманизированных антител и человеческих антител.
3. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1 или 2, где указанное антитело представляет собой человеческое антитело.
4. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-3, где указанное антитело выбрано из антител IgA1, антител IgA2, антител IgD, антител IgE, антител IgG1, антител IgG2, антител IgG3, антител IgG4 и антител IgM.
5. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-3, где указанный антигенсвязывающий фрагмент выбран из группы, состоящей из фрагментов Fab, Fab', (Fab')2, Fv, scFv (sc для одной цепи), бис-scFv, scFv-Fc, Fab2, Fab3, мини-тел, диател, триател, тетрател и нанотел.
6. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-5, где указанный антигенсвязывающий фрагмент представляет собой scFv.
7. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-6, причем указанное антитело выбрано из группы, состоящей из следующего:
a) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 129 или любой последовательностью, проявляющей по меньшей мере 80% идентичность с SEQ ID NO: 129, и три CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
b) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 131 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 131, и трех CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 8;
c) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 133 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 133, и трех CDR легкой цепи с последовательностями SEQ ID NO: 14, 15 и 16;
d) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 135 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 135, и трех CDR легкой цепи с последовательностями SEQ ID NO: 20, 21 и 22;
e) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 137 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 137, и трех CDR легкой цепи с последовательностями SEQ ID NO: 6, 7 и 25;
f) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 139 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 139, и трех CDR легкой цепи с последовательностями SEQ ID NO: 29, 30 и 31;
g) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 141 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 141, и трех CDR легкой цепи с последовательностями SEQ ID NO: 35, 36 и 16;
h) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 143 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 143, и трех CDR легкой цепи с последовательностями SEQ ID NO: 40, 41 и 42;
i) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 145 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 145, и трех CDR легкой цепи с последовательностями SEQ ID NO: 46, 47 и 48;
j) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 147 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 147, и трех CDR легкой цепи с последовательностями SEQ ID NO: 52, 53 и 54;
k) антитело, содержащее или состоящее из вариабельного домена тяжелой цепи с последовательностью SEQ ID NO: 149 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 149, и трех CDR легкой цепи с последовательностями SEQ ID NO: 56, 57 и 58.
8. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-6, причем указанное антитело выбрано из группы, состоящей из следующего:
a) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 130 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 130, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 3, 4 и 5;
b) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 132 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 132, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 9, 10 и 5;
c) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 134 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 134, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 11, 12 и 13;
d) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 136 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 136, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 19;
e) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 138 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 138, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 23, 24 и 5;
f) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 140 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 140, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 26, 27 и 28;
g) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 142 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 142, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 32, 33 и 34;
h) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 144 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 144, и три CDR тяжелой цепи с последовательностями SEQ ID NO 37, 38 и 39;
i) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 146 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 146, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 43, 44 и 45;
j) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 148 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 148, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 49, 50 и 51; и
k) антитело, содержащее вариабельный домен легкой цепи с последовательностью SEQ ID NO: 150 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 150, и три CDR тяжелой цепи с последовательностями SEQ ID NO: 17, 18 и 55.
9. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-8, причем указанное антитело выбрано из группы, состоящей из следующего:
a) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 129 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 129, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 130 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 130;
b) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 131 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 131, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 132 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 132;
c) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 133 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 133, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 134 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 134;
d) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 135 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 135, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 136 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 136;
e) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 137 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 137, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 138 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 138;
f) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 139 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 139, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 140 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 140;
g) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 141 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 141, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 142 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 142;
h) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 143 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 143, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 144 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID No: 144;
i) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 145 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 145, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 146 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 146;
j) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 147 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 147, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 148 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 148; и
k) антитело, содержащее вариабельный домен тяжелой цепи с последовательностью SEQ ID NO: 149 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 149, и вариабельный домен легкой цепи с последовательностью SEQ ID NO: 150 или любой последовательностью, проявляющей по меньшей мере 80%, 85%, 90%, 95% или 98% идентичность с SEQ ID NO: 150.
10. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-9, где указанное антитело представляет собой интернализируемое антитело.
11. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-10, причем указанное антитело представляет собой антитело, которое связывается по меньшей мере с одной аминокислотой в одном или более эпитопах, при этом эпитоп выбран из группы, состоящей из следующего:
a) первый эпитоп, имеющий последовательность, содержащую остатки Е24, V25, Е27, V29 и/или Т30 последовательности, изложенной в SEQ ID NO: 2;
b) второй эпитоп, имеющий последовательность, содержащую остатки D36, N38, L39 и/или Т42 последовательности, изложенной в SEQ ID NO: 2;
c) третий эпитоп, имеющий последовательность, содержащую остатки Q59, G61, S62, D63 и/или V65 последовательности, изложенной в SEQ ID NO: 2;
d) четвертый эпитоп, имеющий последовательность, содержащую остатки I77, А80, Y82 и/или Q83 последовательности, изложенной в SEQ ID NO 2;
e) пятый эпитоп, имеющий последовательность, содержащую остатки Н87, Н90, K91 и/или V92 последовательности, изложенной в SEQ ID NO: 2;
f) шестой эпитоп, имеющий последовательность, содержащую остатки S97, Q99, S101 и/или Т102 последовательности, изложенной в SEQ ID NO: 2;
g) седьмой эпитоп, имеющий последовательность, содержащую остатки R108, S109, Н110, Т112 и/или Е114 последовательности, изложенной в SEQ ID NO: 2;
h) восьмой эпитоп, имеющий последовательность, содержащий остатки Т119, Р120, D121, N123, Q124 и/или V125 последовательности, изложенной в SEQ ID NO: 2.
12. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по п. 11, где указанное антитело связывает:
a) по меньшей мере одну из аминокислот в первом эпитопе;
b) по меньшей мере одну из аминокислот в четвертом эпитопе и, необязательно, по меньшей мере один из остатков в третьем эпитопе;
c) по меньшей мере одну из аминокислот в седьмом эпитопе;
d) по меньшей мере одну из аминокислот в восьмом эпитопе;
e) по меньшей мере одну из аминокислот в седьмом эпитопе и по меньшей мере одну из аминокислот в восьмом эпитопе; или
f) по меньшей мере одну из аминокислот в третьем эпитопе, по меньшей мере одну из аминокислот в седьмом эпитопе и по меньшей мере одну из аминокислот в восьмом эпитопе и, необязательно, по меньшей мере один из остатков второго эпитопа и/или по меньшей мере один из остатков четвертого эпитопа.
13. Иммуноконъюгат для стимуляции иммунного ответа у пациента, страдающего раком, содержащий моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12, где указанное антитело конъюгировано с цитотоксическим агентом.
14. Полинуклеотид, кодирующий моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12.
15. Экспрессионный вектор для получения моноклонального антитела против VSIG4 или его антигенсвязывающего фрагмента по любому из пп. 1-12, содержащий полинуклеотид по п. 14.
16. Клетка-хозяин для получения моноклонального антитела против VSIG4 или его антигенсвязывающего фрагмента по любому из пп. 1-12, трансформированная экспрессионным вектором по п. 15.
17. Способ получения моноклонального антитела против VSIG4 или его антигенсвязывающего фрагмента по любому из пп. 1-12, включающий:
a) культивирование клетки-хозяина по п. 16 в подходящих условиях и
b) выделение моноклонального антитела против VSIG4 или его антигенсвязывающего фрагмента из культуральной среды или из культивируемых клеток.
18. Фармацевтическая композиция для стимуляции иммунного ответа у пациента, страдающего раком, содержащая моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12 или иммуноконъюгат по п. 13 и фармацевтически приемлемый носитель и/или вспомогательное вещество.
19. Фармацевтическая композиция по п. 18, дополнительно содержащая ингибитор иммунной контрольной точки.
20. Фармацевтическая композиция по п. 19, где указанный ингибитор иммунной контрольной точки представляет собой ингибитор любого из CTLA-4, PDL1, PDL2, PD1, В7-Н3, В7-Н4, BTLA, HVEM, TIM3, GAL9, LAG3, VISTA, KIR, 2В4, CD 160, CGEN-15049, киназ CHK1 и CHK2, IDO1, A2aR и любого из различных лигандов семейства В-7.
21. Фармацевтическая композиция по любому из пп. 19 или 20, где указанный ингибитор иммунной контрольной точки выбран из группы, состоящей из ипилимумаба, пембролизумаба, ниволумаба, цемиплимаба, пидилизумаба, атезолизумаба, авелумаба, дурвалумаба, BMS 936559, JNJ 61610588, урелумаба, 9В12, PF-04518600, BMS-986016, TSR-022, MBG453, MEDI6469, MEDI6383 и эпакадостата.
22. Фармацевтическая композиция по любому из пп. 18 или 19 для одновременного, раздельного или последовательного применения.
23. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12, или иммуноконъюгат по п. 13, или фармацевтическая композиция по любому из пп. 18-22 для применения в лечении рака у пациента.
24. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12, или иммуноконъюгат по п. 13, или фармацевтическая композиция по любому из пп. 18-22 для применения в индукции иммунного ответа у пациента с раком.
25. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12, или иммуноконъюгат по п. 13, или фармацевтическая композиция по любому из пп. 18-22 для применения по п. 23, где иммунный ответ включает индукцию высвобождения провоспалительных цитокинов макрофагами, индукцию пролиферации CD4+ Т-клеток, индукцию пролиферации CD8+ Т-клеток, индукцию продукции цитокинов CD4+ Т-клетками и индукцию продукции цитокинов CD8+ Т-клетками.
26. Моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент по любому из пп. 1-12, или иммуноконъюгат по п. 13, или фармацевтическая композиция по любому из пп. 18-22 для применения по любому из пп. 23-25, где рак выбран из рака мочевого пузыря, рака молочной железы, рака шейки матки, рака толстой кишки, рака эндометрия, рака пищевода, рака фаллопиевых труб, рака желчного пузыря, рака желудочно-кишечного тракта, рака головы и шеи, гематологического рака, рака гортани, рака печени, рака легких, лимфомы, меланомы, мезотелиомы, рака яичников, первичного рака брюшины, рака слюнных желез, саркомы, рака желудка, рака щитовидной железы, рака поджелудочной железы, почечно-клеточной карциномы, глиобластомы и рака предстательной железы.
27. Способ in vitro обнаружения рака, экспрессирующего VSIG4, у субъекта, причем указанный способ включает следующие стадии:
а) приведение биологического образца указанного субъекта в контакт с антителом против VSIG4 или его антигенсвязывающим фрагментом по любому из пп. 1-12; и
b) обнаружение связывания указанного реагента с указанным биологическим образцом,
при этом связывание VSIG4 указывает на наличие рака, экспрессирующего VSIG4.
28. Способ по п. 27, где моноклональное антитело против VSIG4 или его антигенсвязывающий фрагмент мечено обнаруживаемой меткой.
| WO 2019005817 A2, 03.01.2019 | |||
| YURONG WEN et al., Structural evaluation of a nanobody targeting complement receptor Vsig4 and its cross reactivity, Immunobiology, 2017, 222(6), pp.807-813, doi:10.1016/j.imbio.2016.11.008 | |||
| YUNMEI LIAO et al., VSIG4 expression on macrophages facilitates lung cancer development, Laboratory Investigation, 2014, vol |

