Область техники, к которой относится изобретение
Настоящее изобретение относится к получению мультиспецифических антител, прежде всего таких мультиспецифических антител, которые содержат кроссовер доменов в одной из их цепей. В способе, указанном в настоящем описании, уровень экспрессии рекомбинантной клетки млекопитающего, секретирующей мультиспецифическое антитело, повышают путем интродукции дополнительной кассеты экспрессии для цепи с обмененными доменами в уже трансфектированную или трансдуцированную клетку.
Предпосылки создания изобретения
В US №5958727 описан метод получения полипептида, включающий культивирование мутантной клетки в условиях, приводящих к производству полипептида, в котором мутантная клетка является родственной родительской клетке, содержащей первую ДНК-последовательность, которая кодирует полипептид, путем интродукции конструкции нуклеиновой кислоты в геном родительской клетки в локус, который не находится в первой ДНК-последовательности, не находится во второй ДНК-последовательности, кодирующей белок, который является отрицательным регулятором транскрипции, трансляции или секреции полипептида, и не находится в третьей ДНК-последовательности, которая кодирует протеазу, гидролизующую полипептид в данных условиях; и мутантная клетка продуцирует большее количество полипептида, чем родительская клетка при культивировании обоих клеток в данных условиях; и выделение полипептида.
Genzel Y. с соавторами установили, что замена глутамина на пируват снижает образование аммиака и ингибирует рост клеток млекопитающих (Biotechnol. Prog. 21, 2005, сс. 58-69). De la Cruz Edmonds M. С. с соавторами разработали протоколы трансфекции и высокопроизводительного скрининга для клеточной системы CHOK1 SV (Mol. Biotechnol. 34, 2006, сс. 179-190). В WO 2007/036291 описана улучшенная среда для культивирования клеток. В ЕР 1482031 описана бессывороточная среда для культивирования клеток млекопитающих и ее применение. Link Т. с соавторами разработали биологический процесс получения рекомбинантного слитого белка MUC1, экспрессируемого клетками СНО-K1 в безбелковой среде (J. Biotechnol. 110, 2004, сс. 51-62). В ЕР 0481791 описана культуральная среда для СНО-клеток и адаптированных клеток. В US 2007/161079 описаны рекомбинантные клеточные клоны, обладающие повышенной стабильностью, и методы их получения и применения. В ЕР 0659880 описан метод культивирования клеток животных или продуцирующих антитела клеток. У Butler М. с соавторами описана адаптация клеток млекопитающих к неаммониегенным средам (Cytotechnol. 15, 1994, сс. 87-94). Altamirano С.с соавторами описали улучшенный состав среды для культивирования СНО-клеток: а именно, одновременную замену глюкозы и глутамина (Biotechnol. Prog. 16, 2000, сс.69-75).
В ЕР 0569678 описаны двойные трансфектанты по генам ГКГС в качестве клеточных вакцин для иммунопрофилактики метастазов опухолей. В WO 97/08342 описан усовершенствованный метод измерения активности промоторной последовательности в клетке млекопитающего с использованием репортерного гена. Применение анти-RhoA и анти-RhoC siPHK для специфического ингибирования синтеза RhoA или RhoC описано в WO 2005/113770. Метод рекомбинантного производства или экспрессии эукариотического мутанта по щелочной фосфатазе в клетках дрожжей описан в US №7202072. В WO 2001/038557 описан метод скрининга многократно трансформированных клеток с помощью бицистронной экспрессии флуоресцентных белков. Метод получения рекомбинантных линий эукариотических клеток, которые экспрессируют множество представляющих интерес белков или РНК, описан в WO 1999/47647. Системы, включающие методы, композиции и наборы, для трансфекции клеток материалами для трансфекции с использованием кодируемых носителей описаны в WO 2003/076588. В US №5089397 описана экспрессионная система для рекомбинантного получения требуемого белка, включающая СНО-клетки, трансформированные ДНК-последовательностью, которая имеет кодирующую последовательность требуемого белка под контролем промотора человеческого металлотионеина-II. Метод получения рекомбинантных белков описан в US 2003/0040047. Lamango с соавторами (Lamango N.S. и др., Arch. Biochem. Biophys. 330, 1996, сс. 238-250) описали зависимость производства прогормона конвертазы 2 от присутствия нейроэндокринного полипептида 7 В2. Трансфекция экспрессионным вектором на основе BPV-1 клеток, несущих неинтегрированные реплицирующие BPV-1 геномы, описана у Waldenstroem М. и др., Gene 120, 1992, сс.175-181. В US №4912038 описаны методы и векторы для получения альвеолярного сурфактантного белка 32K собак и человека.
В WO 89/10959 описаны технологии рекомбинантной ДНК и экспрессия полипептидов млекопитающих в созданных с помощью генной инженерии эукариотических клетках. Повторный совместный перенос экспрессионного вектора для человеческого гормона роста и экспрессионного вектора для гена маркера селекции описан в DD 287531. В WO 93/01296 описано производство антител в инфицированных вирусом коровьей оспы клетках. В WO 95/17513 описана повторная трансформация нитчатых грибов. В WO 89/00999 описана модульная сборка генов антител, полученные с ее помощью антитела и их применение. В US 2003/096341 описана экспрессия щелочной фосфатазы в дрожжах.
В ЕР 1453966 описан метод получения рекомбинантного полипептида. В WO 03/046187 описан метод получения рекомбинантного полипептида. В US №5550036 описан метод совместной амплификации генов человеческого белка С в клетках человека. В ЕР 0921194 описано семейство генов лиганда TNF. В ЕР 0319206 описана амплификация генов. У Lin F.K. с соавторами описаны клонирование и экспрессия гена человеческого эритропоэтина (Proc. Natl. Acad. Sci. USA 82, 1985, сс. 7580-7584). В WO 00/28066 описаны клетки-хозяева, экспрессирующие рекомбинантный человеческий эритропоэтин. У Chen S. с соавторами описано производство рекомбинантных белков в клетках млекопитающих (в: Curr. Prot. Prot. Sci., 1998, 5.10.1-5.10.41).
В WO 89/00605 описаны трансфектированные клетки, содержащие векторы, которые имеют гены, ориентированные в противоположных направлениях, и методы их получения. В US №5420019 описаны стабильные бактерицидные/повышающие проникновение белковые продукты и содержащие их фармацевтические композиции. В US №5639275 описаны биосовместимые изолированные от воздействия иммунной системы капсулы, содержащие генетически измененные клетки для доставки биологически активных молекул.
Kemball-Cook G. с соавторами описали высокопродуктивное производство человеческих факторов коагуляции крови VII и XI с использованием нового экспрессионного вектора для клеток млекопитающих (Gene 139, 1994, сс. 275-279). В ЕР 1010758 описана система экспрессии для производства рекомбинантного человеческого эритропоэтина, метод очистки секретированного человеческого эритропоэтина и его применение.
У Mulligan R.C. и Berg Р. описана селекция клеток животных, которые экспрессируют ген Escherichia coli, кодирующий ксантин-
гуанинфосфорибозилтрансферазу (Proc. Natl. Acad. Sci. USA 78, 1981, сс. 2072-2076). У Colosimo А. с соавторами описан перенос и экспрессия чужеродного гена в клетках млекопитающих (BioTechniques 29, 2000, сс. 314-331). У Maruyama K. с соавторами описаны трансфекция культивируемых клеток млекопитающих экспрессионными векторами для клеток млекопитающих (Meth. Nucleic Acids Res. 1991, сс. 283-305). У Wang D.Z. с соавторами описано лечение пациентов с острым инсультом с помощью внутривенного введения tPA (Stroke 31, 2000, сс. 77-81). Sakamoto Т. с соавторами описали предупреждение артериальной повторной окклюзии после тромболиза с помощью активированного белка С (Circulation 90, 1994, сс. 427-432). Lee G.M. с соавторами разработали бессывороточную среду для получения эритропоэтина с помощью суспензионной культуры рекомбинантных клеток яичника китайского хомячка с использованием статистического конструирования (J. Biotechnol. 69, 1999, сс. 85-93). У Lusky М. и Botchan M.R. описана характеризация поддерживающих последовательностей вектора вируса бычьей папилломы (Cell 36, 1984, сс. 391-401).
В US 2014/0242079 описано соотношение векторов 1:2:1:1 для векторов одной кассеты экспрессии, предназначенной для кратковременной экспрессии в клетках HEK.
В WO 2015/052230 описаны мультиспецифические антитела с обмененными доменами в общей вариабельной легкой цепи.
В WO 2012/023053 описаны методы создания мультиспецифических и многовалентных антител.
В WO 2005/072112 описаны методы получения и идентификации мультиспецифических антител.
В WO 02/079255 описаны рекомбинантные антитела, совместно экспрессируемые с GnTIII.
В US 2002/06210 описан метод получения мультиспецифических антител, имеющих гетеромультимерные и общие компоненты.
В US 2013/045888 описана мультикопийная стратегия для получения высоких титров высокоочищенных мультисубъединичных белков, таких как антитела, в трансформированных микробах, таких как Pichia pastoris.
У Frenzel с соавторами в Front. Immunol. 4, 2013, статья 217 описана экспрессия рекомбинантных антител.
Wurm с соавторами описали получение терапевтических средств в виде рекомбинантных белков в культивируемых клетках млекопитающих (Nat. Biotechnol. 22, 2004, сс. 1393-1398).
Краткое изложение сущности изобретения
Установлено, что для создания клеточных линий, предназначенных для производства гетеродимерных, т.е. мультиспецифических антител, целесообразно применять для трансфекции экспрессионный вектор, который содержит в качестве единственной кассеты экспрессии для цепи антитела кассету экспрессии для легкой цепи. Указанный вектор можно применять вместе с другими экспрессионными векторами для совместной трансфекции или отдельно на второй последующей стадии трансфекции. С помощью указанного подхода к производству можно получать клеточную линию, которая продуцирует гетеродимерное антитело с улучшенным профилем продукта, т.е. с увеличенным количеством продукта и пониженным количеством родственных продукту примесей.
Одним из объектов настоящего изобретения является способ получения мультиспецифического антитела, которое содержит/состоит из/включает по меньшей мере три различных полипептида, который включает следующие стадии:
- культивирование клетки млекопитающего в среде для культивирования (в условиях, пригодных для экспрессии мультиспецифического антитела), где клетка млекопитающего создана путем
а) трансфекции клетки млекопитающего (не экспрессирующей антитело) первым экспрессионным вектором и одним, двумя или тремя дополнительными экспрессионными векторами,
при этом первый экспрессионный вектор содержит строго одну нуклеотидную последовательность, которая кодирует полипептид мультиспецифического антитела, и один, два или три дополнительных экспрессионных вектора, каждый из которых содержит по меньшей мере две нуклеотидные последовательности, каждая из которых кодирует различные полипептидные цепи мультиспецифического антитела,
где строго одна нуклеотидная последовательность первого экспрессионного вектора представляет собой нуклеотидную последовательность, которая кодирует полипептид легкой цепи мультиспецифического антитела, где трансфекцию первым экспрессионным вектором осуществляют либо одновременно, либо до, либо после трансфекции одним, двумя или тремя дополнительными экспрессионными векторами, и
б) отбор клетки (стабильно) трансфектированной на стадии а), которая растет в условиях селективного культивирования,
выделение мультиспецифического антитела из клетки или среды для культивирования,
и получение тем самым мультиспецифического антитела.
Одним из объектов настоящего изобретения является способ создания/продуцирования/получения клетки млекопитающего (обладающей способностью к (стабильной) экспрессии мультиспецифического антитела, которое содержит/состоит по меньшей мере из трех различных полипептидов, включающий следующую стадию:
а) трансфекция клетки млекопитающего (не экспрессирующей антитело) первым экспрессионным вектором и одним, двумя или тремя дополнительными экспрессионными векторами,
при этом первый экспрессионный вектор содержит строго одну нуклеотидную последовательность, которая кодирует полипептид мультиспецифического антитела, и один, два или три дополнительных экспрессионных вектора, каждый из которых содержит по меньшей мере две нуклеотидные последовательности, каждая из которых кодирует различные полипептидные цепи мультиспецифического антитела,
где строго одна нуклеотидная последовательность первого экспрессионного вектора представляет собой нуклеотидную последовательность, которая кодирует полипептид легкой цепи мультиспецифического антитела, где трансфекцию первым экспрессионным вектором осуществляют либо одновременно, либо до, либо после трансфекции одним, двумя или тремя дополнительными экспрессионными векторами, и
б) отбор клетки, трансфектированной на стадии а), которая растет в условиях селективного культивирования,
и тем самым создание/продуцирование/получение клетки млекопитающего, которая (стабильно) экспрессирует мультиспецифическое антитело.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, два из полипептидов мультиспецифического антитела содержат/имеют обмен (когнатными) доменами.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, строго одна нуклеиновая кислота первого экспрессионного вектора кодирует полипептид легкой цепи с обменом доменами мультиспецифического антитела.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, стадия а) включает: совместную трансфекцию клетки млекопитающего (не экспрессирующей антитело) первым экспрессионным вектором и одним, двумя или тремя дополнительными экспрессионными векторами.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, стадия а) включает следующие стадии: I) трансфекция (одновременная или последовательная) клетки млекопитающего одним, двумя или тремя дополнительными экспрессионными векторами, необязательно II) отбор (стабильно) трансфектированной клетки, III) трансфекция клетки, указанной в подпункте I) или II), первым экспрессионным вектором и необязательно IV) отбор (стабильно) трансфектированной клетки.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, отбор основан на уровне экспрессии и/или количестве родственных продукту побочных продуктов/примесей.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, отбирают (стабильно)
трансфектированную(ые) клетку(и), которая(ые) продуцирует(ют) наименьшее количество (фракцию) родственных продукту побочных продуктов/примесей.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, отбирают (стабильно)
трансфектированную(ые) клетку(и), которая(ые) продуцирует(ют) наименьшее количество (фракцию) родственных продукту побочных продуктов/примесей и характеризуется(ются) наибольшим выходом.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, клетка млекопитающего стабильно экспрессирует мультиспецифическое антитело.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, клетка млекопитающего представляет собой СНО-клетку.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, обмен доменами представляет собой CH1-CL-кроссовер или VH-VL-кроссовер.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, мультиспецифическое антитело представляет собой трехвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь полноразмерного антитела, которое специфически связывается с первым антигеном,
б) вторую тяжелую цепь полноразмерного антитела, которая при спаривании с первой легкой цепью специфически связывается с первым антигеном, и
в) Fab-фрагмент, который специфически связывается со вторым антигеном, слитый через пептидный линкер с С-концом одной из тяжелых цепей, указанных в подпункте а) или б), в котором константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
Одним из объектов настоящего изобретения является (стабильно трансфектированная) клетка млекопитающего, полученная с помощью способа, представленного в настоящем описании.
Одним из объектов настоящего изобретения является способ получения мультиспецифического антитела, который включает следующие стадии:
культивирование (стабильно трансфектированной) клетки, указанной в настоящем описании, в среде для культивирования (в условиях, пригодных для экспрессии мультиспецифического антитела),
выделение мультиспецифического антитела из клетки или среды для культивирования,
необязательно очистка выделенного антитела с помощью одной или нескольких стадии(й) хроматографии,
и получение тем самым мультиспецифического антитела.
Одним из объектов настоящего изобретения является способ получения препарата мультиспецифического антитела с низким/пониженным количеством родственных продукту примесей, который включает следующие стадии:
- получение/продуцирование/создание (стабильно трансфектированной) клетки млекопитающего, которая (стабильно) экспрессирует мультиспецифическое антитело, с помощью способа, представленного в настоящем описании,
- культивирование полученной/продуцированной/созданной клетки млекопитающего в среде для культивирования,
- выделение препарата антитела из клетки или среды для культивирования,
- необязательно очистка выделенного антитела с помощью одной или нескольких стадии(й) хроматографии,
и получение тем самым препарата мультиспецифического антитела с низким/пониженным количеством родственных продукту примесей.
Одним из объектов настоящего изобретения является применения способа, представленного в настоящем описании, для снижения количества родственных продукту примесей в препарате мультиспецифического антитела.
В настоящем описании представлен способ получения мультиспецифического антитела, которое содержит по меньшей мере одну цепь с кроссовером доменов, в рекомбинантной клетке млекопитающего. Способ обеспечивает улучшенный процесс, где улучшение, среди прочего, представляет собой снижение количества родственных продукту побочных продуктов и повышение количества правильно уложенного/правильно собранного мультиспецифического антитела.
Одним из объектов настоящего изобретения является способ получения мультиспецифического антитела (содержащего по меньшей мере одну полипептидную цепь с кроссовером доменов), который включает следующие стадии:
а) получение (стабильно трансфектированной) клетки млекопитающего, которая (стабильно) экспрессирует мультиспецифическое антитело,
б) трансфекция клетки млекопитающего, указанной на стадии а), с помощью кассеты экспрессии, кодирующей полипептидную цепь мультиспецифического антитела, которая имеет кроссовер доменов,
в) культивирование клетки, указанной на стадии б), и выделение антитела из клетки или среды для культивирования и получение тем самым мультиспецифического антитела,
г) необязательно очистка выделенного антитела с помощью одной или нескольких стадии(й) хроматографии.
В одном из вариантов осуществления всех объектов изобретения клетка млекопитающего, экспрессирующая мультиспецифическое антитело, стабильно экспрессирует мультиспецифическое антитело.
В одном из вариантов осуществления изобретения кассета экспрессии, указанная на стадии б), представляет собой экспрессионный вектор.
В одном из вариантов осуществления всех объектов изобретения полипептидная цепь мультиспецифического антитела, которая имеет кроссовер доменов, представляет собой легкую цепь антитела.
В одном из вариантов осуществления всех объектов изобретения кроссовер доменов представляет собой CH1-CL-кроссовер или VH-VL-кроссовер.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело или трехвалентное биспецифическое антитело, или четырехвалентное биспецифическое антитело.
В одном из вариантов осуществления всех объектов изобретения клетку млекопитающего, экспрессирующую мультиспецифическое антитело, получают путем трансфекции клетки млекопитающего одной или несколькими молекулой(ами) нуклеиновых(ой) кислоты(т), которая(ые) кодирует(ют) мультиспецифическое антитело, и отбирают стабильно трансфектированную клетку.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой триспецифическое или тетраспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь полноразмерного антитела, которое специфически связывается с первым антигеном, и
б) вторую (модифицированную) легкую цепь и вторую (модифицированную) тяжелую цепь полноразмерного антитела, которое специфически связывается со вторым антигеном, в котором вариабельные домены VL и VH заменены друг на друга и/или константные домены CL и СН1 заменены друг на друга, и
в) в котором один-два антигенсвязывающих пептидов, которые специфически связываются с одним или двумя дополнительными антигенами (т.е. с третьим и/или четвертым антигеном), слиты через пептидный линкер с С-или N-концом легких цепей или тяжелых цепей, указанных в подпунктах а) и/или б).
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой биспецифическое четырехвалентное антитело, которое содержит
а) две легкие цепи и две тяжелые цепи антитела, которое специфически связывается с первым антигеном (и содержит два Fab-фрагмента),
б) два дополнительных Fab-фрагмента антитела, которое специфически связывается со вторым антигеном, в котором указанные дополнительные Fab-фрагменты оба слиты через пептидный линкер либо с С-, либо с N-концами тяжелых цепей, указанных в подпункте а),
и
где в Fab-фрагментах осуществлены следующие модификации
I) в обоих Fab-фрагментах, указанных в подпункте а), или в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и/или константные домены CL и СН1 заменены друг на друга,
или
II) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменены друг на друга и константные домены CL и СН1 заменены друг на друга, и
в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга или константные домены CL и СН1 заменены друг на друга, или
III) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменены друг на друга или константные домены CL и СН1 заменены друг на друга, и
в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и константные домены CL и СН1 заменены друг на друга, или
IV) в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и в обоих Fab-фрагментах, указанных в подпункте б), константные домены CL и СН1 заменены друг на друга, или
V) в обоих Fab-фрагментах, указанных в подпункте а), константные домены CL и СН1 заменены друг на друга и в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга.
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществлены следующие модификации
I) в обоих Fab-фрагментах, указанных в подпункте а), или в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и/или
константные домены CL и СН1 заменены друг на друга.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой биспецифическое четырехвалентное антитело, которое содержит:
а) (модифицированную) тяжелую цепь первого антитела, которое специфически связывается с первым антигеном и содержит первую доменную пару VH-CH1, где С-конец указанной тяжелой цепи слит с N-концом второй доменной пары VH-CH1 указанного первого антитела через пептидный линкер,
б) две легкие цепи указанного первого антитела, указанного в подпункте а),
в) (модифицированную) тяжелую цепь второго антитела, которое специфически связывается со вторым антигеном и содержит первую доменную пару VH-CL, где С-конец указанной тяжелой цепи слит с N-концом второй доменной пары VH-CL указанного второго антитела через пептидный линкер, и
г) две (модифицированные) легкие цепи указанного второго антитела, указанного в подпункте в), каждая из которых содержит доменную пару CL-СН1.
Во всех объектах изобретения, представленных в настоящем описании, первая легкая цепь содержит VL-домен и CL-домен и первая тяжелая цепь содержит VH-домен, CH1-домен, шарнирную область, СН2-домен и СН3-домен.
В одном из вариантов осуществления всех объектов изобретения антитело, полученное способом, указанным в настоящем описании, представляет собой мультиспецифическое антитело, которое нуждается в гетеродимеризации по меньшей мере двух полипептидов тяжелых цепей.
В одном из вариантов осуществления изобретения полноразмерное антитело представляет собой
а) полноразмерное антитело человеческого IgG1-подкласса,
б) полноразмерное антитело человеческого IgG4-подкласса,
в) полноразмерное антитело человеческого IgG1-подкласса с мутациями L234A, L235A и P329G,
г) полноразмерное антитело человеческого IgG4-подкласса с мутациями S228P, L235E и P329G,
д) полноразмерное антитело человеческого IgG1-подкласса с мутациями L234A, L235A и P329G в обеих тяжелых цепях и мутациями T366W и S354C или Y349C в одной тяжелой цепи и мутациями T366S, L368A, Y407V и Y349C или S354C в соответствующей другой тяжелой цепи,
е) полноразмерное антитело человеческого IgG4-подкласса с мутациями S228P и P329G в обеих тяжелых цепях и мутациями T366W и S354C в одной тяжелой цепи и мутациями T366S, L368A, Y407V и Y349C в соответствующей другой тяжелой цепи,
ж) полноразмерное антитело человеческого IgG1-подкласса с мутациями L234A, L235A, P329G, 1253А, Н310А и Н435А в обеих тяжелых цепях и мутациями T366W и S354C в одной тяжелой цепи и мутациями T366S, L368A, Y407V и Y349C в соответствующей другой тяжелой цепи, или
з) полноразмерное антитело человеческого IgG1-подкласса с мутациями L234A, L235A, P329G, M252Y, S254T и Т256Е в обеих тяжелых цепях и мутациями T366W и S354C в одной тяжелой цепи и мутациями T366S, L368A, Y407V и Y349C в соответствующей другой тяжелой цепи.
Одним из объектов изобретения, указанных в настоящем описании, является клетка, которая содержит нуклеиновую кислоту, кодирующую биспецифическое антитело, которое получено с помощью способа, представленного в настоящем описании.
Одним из объектов изобретения, указанных в настоящем описании, является способ получения мультиспецифического антитела, представленного в настоящем описании, который включает следующие стадии:
а) культивирование клетки, указанной в настоящем описании, которая продуцирует /экспрессирует мультиспецифическое антитело, и
б) выделение мультиспецифического антитела из клетки или среды для культивирования,
и получение тем самым мультиспецифического антитела, представленного в настоящем описании.
Одним из объектов изобретения, указанных в настоящем описании, является антитело, которое получено способом, представленным в настоящем описании.
Одним из объектов изобретения, указанных в настоящем описании, является фармацевтическая композиция, которая содержит антитело, полученное способом, который представлен в настоящем описании, и фармацевтически приемлемый носитель.
Одним из объектов изобретения, указанных в настоящем описании, является антитело, полученное способом, который представлен в настоящем описании, для применения в качестве лекарственного средства.
Одним из объектов изобретения, указанных в настоящем описании, является применение биспецифического антитела, полученное способом, который представлен в настоящем описании, для приготовления лекарственного средства.
В одном из вариантов осуществления всех объектов изобретения биспецифическое антитело выбирают из группы биспецифических антител, которая состоит из антитела к белку А-бета/рецептору трансферрина, антитела к CD20/рецептору трансферрина, антитела к PD1/Tim3 и антитела к FAP/DR5.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой биспецифическое четырехвалентное антитело, которое содержит
а) две легкие цепи и две тяжелые цепи антитела, которое специфически связывается с первым антигеном (и содержит два Fab-фрагмента),
б) два дополнительных Fab-фрагмента антитела, которое специфически связывается со вторым антигеном, в котором каждый из указанных дополнительных Fab-фрагментов слит через пептидный линкер с индивидуальным С-концом одной из тяжелых цепей, указанных в подпункте а), и
в котором в дополнительных Fab-фрагментах осуществлены следующие модификации: в обоих дополнительных Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и/или константные домены CL и СН1 заменены друг на друга,
при этом I) первый антиген представляет собой DR5, а второй антиген представляет собой FAP или II) первый антиген представляет собой FAP, а второй антиген представляет собой DR5,
где две тяжелые цепи антитела, которое специфически связывается с первым антигеном, относятся к человеческому IgG1-подклассу с мутациями L234A, L235A и P329G.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, где вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга,
при этом I) первый антиген представляет собой PD1, а второй антиген представляет собой Tim3 или II) первый антиген представляет собой Tim3, а второй антиген представляет собой PD1,
где первая тяжелая цепь и вторая тяжелая цепь обе относятся к человеческому IgG1-подклассу с мутациями L234A, L235A и P329G и с мутацией T366W и необязательно S354C или Y349C в одной тяжелой цепи, и с мутациями T366S, L368A, Y407V и необязательно Y349C или S354C в соответствующей другой тяжелой цепи, при этом концевой глицин или глицин-лизиновый дипептид может отсутствовать,
в котором первая легкая цепь содержит в константном домене легкой цепи (CL) в положении 123 аминокислотный остаток аргинина (вместо присутствующего в молекуле дикого типа остатка глутаминовой кислоты; мутация E123R) и в положении 124 аминокислотный остаток лизина (вместо присутствующего в молекуле дикого типа остатка глутамина; мутация Q124K) (нумерация согласно Кэботу),
в котором первая тяжелая цепь содержит в первом константном домене тяжелой цепи (СН1) в положении 147 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа остатка лизина; мутация K147E) и в положении 213 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа аминокислотного остатка лизина; мутация K213E) (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой трехвалентное биспецифическое антитело, которое содержит
а) две легкие цепи и две тяжелые цепи антитела, которое специфически связывается с первым антигеном (и содержит два Fab-фрагмента),
б) один дополнительный Fab-фрагмент антитела, которое специфически связывается со вторым антигеном, в котором указанный дополнительный Fab-фрагмент слит через пептидный линкер с С-концом одной из тяжелых цепей, указанных в подпункте а),
и
в котором в дополнительном Fab-фрагменте осуществлены следующие модификации: вариабельные домены VL и VH заменены друг на друга и/или константные домены CL и СН1 заменены друг на друга, при этом I) первый антиген представляет собой А-бета, а второй антиген представляет собой рецептор трансферрина или II) первый антиген представляет собой CD20, а второй антиген представляет собой рецептор трансферрина.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой биспецифическое антитело, которое содержит
а) одно полноразмерное антитело, содержащее две пары цепей, каждая из которых состоит из легкой цепи полноразмерного антитела и тяжелой цепи полноразмерного антитела, где сайты связывания, образованные каждой из пар, состоящих из полноразмерной тяжелой цепи и полноразмерной легкой цепи, специфически связываются с первым антигеном, и
б) один дополнительный Fab-фрагмент, где дополнительный Fab-фрагмент слит с С-концом одной из тяжелых цепей полноразмерного антитела, при этом сайт связывания дополнительного Fab-фрагмента специфически связывается со вторым антигеном,
в котором каждая из легких цепей полноразмерного антитела содержит в константном домене легкой цепи (CL) в положении 123 аминокислотный остаток аргинина (вместо присутствующего в молекуле дикого типа остатка глутаминовой кислоты; мутация E123R) и в положении 124 аминокислотный остаток лизина (вместо присутствующего в молекуле дикого типа остатка глутамина; мутация Q124K) (нумерация согласно Кэботу),
в котором каждая из тяжелых цепей полноразмерного антитела содержит в первом константном домене тяжелой цепи (СН1) в положении 147 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа остатка лизина; мутация K147E) и в положении 213 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа аминокислотного остатка лизина; мутация K213E) (нумерация согласно EU-индексу Кэбота),
в котором дополнительный Fab-фрагмент, специфически связывающийся со вторым антигеном, содержит кроссовер доменов, такой как замена друг на друга константного домена легкой цепи (CL) и константного домена 1 тяжелой цепи (СН1), и
где первый антиген представляет собой человеческий белок А-бета, а второй антиген представляет собой рецептор человеческого трансферрина.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой биспецифическое антитело, которое содержит
а) одно полноразмерное антитело, содержащее две пары цепей, каждая из которых состоит из легкой цепи полноразмерного антитела и тяжелой цепи полноразмерного антитела, где связывания, образованные каждой из пар, состоящих из полноразмерной тяжелой цепи и полноразмерной легкой цепи, специфически связываются с первым антигеном, и
б) один дополнительный Fab-фрагмент, где дополнительный Fab-фрагмент слит с С-концом одной из тяжелых цепей полноразмерного антитела, при этом сайт связывания дополнительного Fab-фрагмента специфически связывается со вторым антигеном,
в котором каждая из легких цепей полноразмерного антитела содержит в константном домене легкой цепи (CL) в положении 123 аминокислотный остаток аргинина (вместо присутствующего в молекуле дикого типа остатка глутаминовой кислоты; мутация E123R) и в положении 124 аминокислотный остаток лизина (вместо присутствующего в молекуле дикого типа остатка глутамина; мутация Q124K) (нумерация согласно Кэботу),
в котором каждая из тяжелых цепей полноразмерного антитела содержит в первом константном домене тяжелой цепи (СН1) в положении 147 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа остатка лизина; мутация K147E) и в положении 213 остаток глутаминовой кислоты (вместо присутствующего в молекуле дикого типа аминокислотного остатка лизина; мутация K213E) (нумерация согласно EU-индексу Кэбота),
в котором дополнительный Fab-фрагмент, специфически связывающийся со вторым антигеном, содержит кроссовер доменов, такой как замена друг на друга константного домена легкой цепи (CL) и константного домена 1 тяжелой цепи (СН1), и
где первый антиген представляет собой человеческий CD20, а второй антиген представляет собой рецептор человеческого трансферрина.
В одном из вариантов осуществления всех объектов изобретения, указанных в настоящем описании, каждый полипептид находится в кассете экспрессии, каждая из которых содержит в направлении 5'→3' промотор, структурный ген, кодирующий полипептид, последовательность полиаденилирования и необязательно терминирующую последовательность. В одном из вариантов осуществления изобретения все кассеты экспрессии имеют одинаковый промотор, одинаковый сайт полиаденилирования и необязательно одинаковую терминирующую последовательность. В одном из вариантов осуществления изобретения промотор представляет собой промотор человеческого CMV (цитомегаловирус). В одном из вариантов осуществления изобретения промотор CMV содержит интрон А. В одном из вариантов осуществления изобретения сайт полиаденилирования представляет собой сайт полиаденилирования BGH (бычий гормон роста). В одном из вариантов осуществления изобретения терминатор присутствует и представляет собой HGT (терминатор человеческого гормона роста). В одном из вариантов осуществления изобретения промотор представляет собой промотор CMV, необязательно содержащий интрон А, а сайт полиаденилирования представляет собой сайт полиаденилирования BGH. В одном из вариантов осуществления изобретения промотор представляет собой промотор CMV, необязательно содержащий интрон А, сайт полиаденилирования представляет собой сайт полиаденилирования BGH и терминатор представляет собой HGT.
В одном из вариантов осуществления изобретения дополнительный экспрессионный вектор содержит или каждый из дополнительных экспрессионных векторов содержит по меньшей мере две нуклеотидные последовательности, каждая из которых кодирует различные поли пептидные цепи мультиспецифического антитела, при этом каждая нуклеиновая кислота присутствует/содержится строго один раз в соответствующем векторе.
Подробное описание изобретения
Модули для димеризации по типу «knobs-into-holes» («выступы во «впадины»») и их применение для создания антител описаны у Carter Р.; Ridgway J.В.В.; Presta L.G.: Immunotechnology, том 2, №1, февраль 1996 г., сс. 73-73(1).
Общая информация, касающаяся нуклеотидных последовательностей легких и тяжелых цепей человеческих иммуноглобулинов, представлена у: Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
В контексте настоящего описания аминокислотные положения всех константных областей и доменов тяжелой и легкой цепи пронумерованы согласно системе нумерации Кэбота, которая описана у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991, и обозначена в настоящем описании как «нумерация согласно Кэботу». В частности, систему нумерации Кэбота (см. сс. 647-660), описанную у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991, применяют для константного домена легкой цепи CL каппа-и лямбда-изотипа, а систему нумерации на основе EU-индекса Кэбота (см. сс. 661-723) применяют для константных доменов тяжелой цепи (СН1, шарнир, СН2 и СН3, которую в этом случае дополнительно уточняют с помощью обозначения «нумерация согласно EU-индексу Кэбота).
Пригодные для осуществления настоящего изобретения методы и технологии описаны, например, в: Current Protocols in Molecular Biology, т. I-III, под ред. Ausubel F.M., 1997; DNA Cloning: A Practical Approach, т. I и II, под ред. Glover N.D. и Hames B.D., изд-во Oxford University Press, 1985; Animal Cell Culture - a practical approach, под ред. Freshney R.I., изд-во IRL Press Limited, 1986; Watson J.D. и др., Recombinant DNA, 2-ое изд., изд-во CHSL Press, 1992; Winnacker E.L., From Genes to Clones; изд-во N.Y., VCH Publishers, 1987; Cell Biology, под ред. Celis J., 20-е изд., изд-во Academic Press, 1998; Freshney R.I., Culture of Animal Cells: A Manual of Basic Technique, 20-е изд., изд-во Alan R. Liss, Inc., N.Y., 1987.
Применение технологии рекомбинантной ДНК позволяет создавать производные нуклеиновой кислоты. Указанные производные, например, могут быть модифицированы в одном или нескольких нуклеотидных положениях путем замены, изменения, обмена, делеции или инсерции. Модификацию или дериватизацию можно, например, осуществлять методами сайтнаправленного мутагенеза. Указанные модификации может легко осуществлять специалист в данной области (см., например, Sambrook J. и др., Molecular Cloning: A laboratory manual, изд-во Cold Spring Harbor Laboratory Press, New York, USA, 1999; Hames B.D. и Higgins S.G., Nucleic acid hybridization - a practical approach, изд-во IRL Press, Oxford, England, 1985).
Определения
«Мультиспецифическое антитело» означает антитело, обладающее связывающими специфичностями в отношении по меньшей мере двух различных эпитопов на одном и том же антигене или на двух различных антигенах. Мультиспецифические антитела можно получать в виде полноразмерных антител или фрагментов антител (например, биспецифические антитела типа F(ab')2) или их комбинаций (например полноразмерное антитело плюс дополнительные scFv- или Fab-фрагменты). Описаны сконструированные антитела с двумя, тремя или большим количеством (например, четырьмя) функциональных антигенсвязывающих сайтов (см., например, US 2002/0004587 А1).
Понятие «правило уложенное /правильно собранное» в контексте настоящего описания означает, что антитело имеет правильную стехиометрию, т.е. содержит совместимое количество и копии индивидуальных /соответствующих легких и тяжелых цепей. Например, «нативное человеческое антитело IgG-типа» является» является правило уложенным/правильно собранным, когда выделенная молекула содержит два полипептида легких цепей и два полипептида тяжелых цепей. Например, если мультиспецифическое антитело представляет собой двухвалентное биспецифическое нативное человеческое антитело IgG-типа, то оно является правило уложенным/правильно собранным, когда выделенная молекула состоит из первой пары когнатной первой легкой цепи и когнатной первой тяжелой цепи, связывающейся с первым антигеном, и второй пары когнатной второй легкой цепи и когнатной второй тяжелой цепи, связывающейся со вторым антигеном, т.е. состоит из четырех различных полипептидов. Все антитела, которые не являются правило уложенными/правильно собранными, т.е. которые содержат меньшее или большее количество по сравнению с требуемым количеством цепей и/или содержат неправильно ассоциированные цепи, т.е. не образующие когнатную пару тяжелой и легкой цепи, обозначают как «родственные продукту побочные продукты».
Понятие «кроссовер доменов» в контексте настоящего описания означает, что в паре, состоящей из VH-CH1-фрагмента тяжелой цепи антитела и соответствующей когнатной легкой цепи антитела, т.е. в связывающем плече антитела (т.е в Fab-фрагменте), последовательность доменов отличается от встречающейся в естественных условиях последовательности тем, что по меньшей мере один домен тяжелой цепи заменен на соответствующий домен легкой цепи и наоборот. Существует три общих типа кроссовера доменов (I) кроссовер СН1- и CL-доменов, который приводит к кроссоверу доменов легкой цепи, которая содержит последовательность доменов VL-CH, и к кроссоверу доменов фрагмента тяжелой цепи, который содержит последовательность доменов VH-CL (или тяжелой цепи полноразмерного антитела, которая содержит последовательность доменов VH-CL-шарнир-СН2-СН3), (II) кроссовер доменов VH- и VL-доменов, который приводит к кроссоверу доменов легкой цепи, которая содержит последовательность доменов VH-CL, и к кроссоверу доменов фрагмента тяжелой цепи, который содержит последовательность доменов VL-CH1, и (III) кроссовер доменов полной легкой цепи (VL-CL) и фрагмента полной тяжелой цепи VH-CH1 («кроссовер, Cross-Fab»), который приводит к кроссоверу доменов легкой цепи, которая содержит последовательность доменов VH-CH1, и к кроссоверу доменов фрагмента тяжелой цепи, который содержит последовательность доменов VL-CL (все вышеуказанные последовательности доменов приведены в направлении от N-конца к С-концу).
В контексте настоящего описания понятие «заменены друг на друга» касательно соответствующих доменов тяжелых и легких цепей относится к вышеописанным кроссоверам доменов. Так, когда СН1- и CL-домены «заменены друг на друга», то это относится к кроссоверу доменов, указанному в подпункте (I) и образованным таким путем последовательностям доменов тяжелой и легкой цепей. Соответственно, когда VH и VL «заменены друг на друга», то это относится к кроссоверу доменов, указанному в подпункте (II); и когда СН1- и CL-домены «заменены друг на друга» и VH1- и VL-домены «заменены друг на друга», то это относится к кроссоверу доменов, указанному в подпункте (III). Биспецифические антитела, включающие кроссоверы доменов, описаны, например, в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254 и у Schaefer W. и др., Proc. Natl. Acad. Sci USA 108, 2011, сс. 11187-11192.
Мультиспецифическое антитело, полученное способом, представленным в настоящем описании, главным образом содержат Fab-фрагменты, включающие кроссовер доменов СН1- и CL-доменов, указанный выше в подпункте (I), или кроссовер доменов VH- и VL-доменов, указанный выше в подпункте (II). Fab-фрагменты, специфически связывающиеся с одним(ими) и тем(и) же антигеном(ами) конструируют так, чтобы они имели одинаковую последовательность доменов. Таким образом, том в случае, когда более одного Fab-фрагмента с кроссовером доменов содержится в мультиспецифическом антителе, указанный(ые) Fab-фрагмент(ы) специфически связывается(ются) с одним и тем же антигеном.
В контексте настоящего описания понятие «антитело» используется в его наиболее широком смысле, и оно относится к различным структурам антител, включая (но, не ограничиваясь только ими) моноклональные антитела, поликлональные антитела и мультиспецифические антитела (например, биспецифические антитела), при условии, что они обладают требуемой антигенсвязывающей активностью.
Понятие «фрагмент антитела» относится к молекуле, отличной от интактного антитела, которая содержит часть интактного антитела, обладающую способностью связываться с антигеном, с которым связывается интактное антитело. Примерами фрагментов антител являются (но, не ограничиваясь только ими) Fv, Fab, Fab', Fab'-SH, F(ab')2, димерные антитела (диабоди); линейные антитела; одноцепочечные молекулы антител (например, scFv); и мультиспецифические антитела, образованные из фрагментов антител.
Понятие «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из конкретного источника или конкретного вида, а остальная часть тяжелой и/или легкой цепи происходит из другого источника или другого вида.
Понятие «класс» антитела относится к типу константного домена или константной области, входящего/входящей в тяжелую цепь. Существует пять основных классов антител: IgA, IgD, IgE, IgG и IgM, а некоторые из них можно дополнительно подразделять на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелых цепей, соответствующие различным классам иммуноглобулинов, обозначают как α, δ, ε, γ и μ соответственно.
Понятие «Fc-рецептор» в контексте настоящего описания относится к активирующим рецепторам, отличающимся присутствие цитоплазматической ITAM-последовательности, ассоциированной с рецептором (см., например,. Ravetch J.V. и Bolland S., Annu. Rev. Immunol. 19, 2001, сс. 275-290). Указанные рецепторы представляют собой FcγRI, FcγRIIA и FcγRIIIA. Понятие «не связывающееся с FcγR» означает, что у антитела, полученного способом, представленным в настоящем описании, в концентрации 10 мкг/мл связывание с NK-клетками составляет 10% или менее от связывания антитела к OX40L LC.001, которое описано в WO 2006/029879.
В то время как для IgG4 характерно пониженное связывание с FcR, антитела из других подклассов IgG отличаются сильным связыванием. Однако Pro238, Asp265, Asp270, Asn297 (утрата слитого с Fc углевода), Pro329 и 234, 235, 236 и 237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435 представляют собой остатки, которые при их изменении также снижают способность связываться с FcR (Shields R.L. и др., J. Biol. Chem. 276, 2001, сс. 6591-6604; Lund J. и др., FASEB J. 9, 1995, сс. 115-119; Morgan А. и др., Immunology 86, 1995, сс. 319-324; и ЕР 0307434).
В одном из вариантов осуществления изобретения антитело, полученное способом, представленным в настоящем описании, относится к подклассу IgGl или IgG2 и содержит мутацию PVA236, GLPSS331 и/или L234A/L235A. В одном из вариантов осуществления изобретения антитело, полученное способом, представленным в настоящем описании, относится к подклассу IgG4 и содержит мутацию L235E. В одном из вариантов осуществления изобретения антитело дополнительно содержит мутацию S228P.
Понятие «Fc-область» в контексте настоящего описания относится к С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Понятие включает нативную последовательность Fc-областей и вариант Fc-областей. В одном из вариантов осуществления изобретения Fc-область тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Однако С-концевой лизин (Lys447) Fc-области может либо присутствовать, либо отсутствовать. Если специально не указано иное, то нумерация аминокислотных остатков в Fc-области или константном участке соответствует системе EU-нумерации, которую называют также EU-индексом, описанной у Kabat и др., Sequences of Proteins of Immunological Interest, 5-ое изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991. публикация NIH 91-3242.
Антитела, полученные способом, представленным в настоящем описании, содержат в качестве Fc-области в одном из вариантов осуществления изобретения Fc-область человеческого происхождения. В одном из вариантов осуществления изобретения Fc-область содержит все части человеческой константной области. Fc-область антитела принимает непосредственное участие в активации комплемента, связывании C1q, активации С3 и связывании Fc-рецептора. В то время как влияние антитела на систему комплемента зависит от определенных условий, связывание с C1q зависит от определенных сайтов связывания в Fc-области. Указанные сайты связывания известны в данной области и описаны у Lukas T.J. и др., J. Immunol. 127, 1981, сс. 2555-2560; Brunhouse R. и Cebra J.J., Mol. Immunol. 16, 1979, сс. 907-917; Burton D R. и др., Nature 288, 1980, сс. 338-344; Thommesen J.E. и др., Mol. Immunol. 37, 2000, сс. 995-1004; Idusogie E E. и др., J. Immunol. 164, 2000, сс. 4178-4184; Hezareh M. и др., J. Virol. 75, 2001, cc. 12161-12168; Morgan А. и др., Immunology 86, 1995, сс. 319-324; и ЕР 0307434. Указанные сайты связывания представляют собой, например, L234, L235, D270, N297, Е318, K320, K322, Р331 и Р329 (нумерация согласно EU-индексу Кэбота; если специально не указано иное, то нумерацию аминокислотных остатков в Fc-области или константной области осуществляют согласно системе EU-нумерации, которую обозначают также как EU-индекс, описанной у Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-oe изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991. публикация NIH 91-3242). Антитела подклассов IgGl, IgG2 и IgG3, как правило, обладают способностью активировать комплемент, связываться с C1q и активировать С3, в то время как IgG4 не активируют систему комплемента, не связываются с C1q и не активируют С3. «Fc-область антитела» представляет собой понятие, хорошо известно специалисту в данной области, и ее определяют на основе расщепления антител папаином. В одном из вариантов осуществления изобретения Fc-область представляет собой человеческую Fc-область. В одном из вариантов осуществления изобретения Fc-область представляет собой Fc-область человеческого IgG4-подкласса, содержащую мутации S228P и/или L235E (нумерация согласно EU-индексу Кэбота). В одном из вариантов осуществления изобретения Fc-область представляет собой Fc-область человеческого IgG1-подкласса, содержащую мутации L234A и L235A (нумерация согласно EU-индексу Кэбота).
Понятия «полноразмерное антитело», «интактное антитело» и «цельное антитело» в контексте настоящего описания используют взаимозаменяемо для обозначения антитела, имеющего структуру, практически сходную со структурой нативного антитела, или имеющего тяжелые цепи, которые содержат указанную в настоящем описании Fc-область. «Полноразмерное антитело» представляет собой антитело, которое содержит антигенсвязывающую вариабельную область, а также константный домен легкой цепи (CL) и константные домены тяжелой цепи CH1, СН2 и СН3. Константные домены могут иметь нативную последовательность константных доменов (например, константные домены, имеющие человеческую нативную последовательность) или варианты ее аминокислотной последовательности. Более конкретно полноразмерное антитело содержит две легкие цепи антитела (каждая из которых содержит вариабельный домен легкой цепи и константный домен легкой цепи) и две тяжелые цепи антитела (каждая из которых содержит вариабельный домен тяжелой цепи, шарнирную область и константные домены CH1, СН2 и СН3 тяжелой цепи). С-концевые аминокислотные остатки K или GK могут присутствовать или отсутствовать независимо друг от друга в двух тяжелых цепях полноразмерного антитела.
В контексте настоящего описания понятия «клетка-хозяин», «линия клеток-хозяев» и «культура клеток-хозяев» используются взаимозаменяемо, и они относятся к клеткам, в которые интродуцирована экзогенная нуклеиновая кислота, включая потомство указанных клеток. Клетки включают «трансформантов» и «трансформированные клетки», которые включают первичные трансформированные клетки, а также потомство, полученное из них, независимо от количества пересевов. Потомство может не быть строго идентичным родительской клетке по составу нуклеиновых кислот, а может содержать мутации. Под данное понятие подпадает мутантное потомство, которое обладает такой же функцией или биологической активностью, что и отобранная путем скрининга или селекции исходная трансформированная клетка.
Понятие «гуманизированное» антитело относится к химерному антителу, которое содержит аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых вариантах осуществления изобретения гуманизированное антитело может содержать практически все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или практически все HVR (например, CDR) соответствуют участкам нечеловеческого антитела, а все или практически все FR соответствуют участкам человеческого антитела. Гуманизированное антитело необязательно может содержать по меньшей мере часть константной области антитела, происходящей из человеческого антитела. Понятие «гуманизированная форма» антитела, например, нечеловеческого антитела, относится к антителу, которое подвергнуто гуманизации.
«Выделенное» антитело представляет собой антитело, которое отделено от компонента его естественного окружения. В некоторых вариантах осуществления изобретения антитело очищают до чистоты, превышающей 95% или 99% по данным, например, электрофоретических анализов (таких, например, как ДСН-ПААГ, изоэлектрическое фокусирование (ИЭФ), капиллярный электрофорез) или хроматографических анализов (таких, например, как ионообменная хроматография или ЖХВР с обращенной фазой). Обзор методов оценки чистоты антител см., например, у Flatman S. и др., J. Chrom. В 848, 2007, сс. 79-87.
Понятие «моноклональное антитело» в контексте настоящего описания относится к антителу, полученному из популяции практически гомогенных антител, т.е. индивидуальные антитела, входящие в популяцию, идентичны и/или связываются с одним и тем же эпитопом, за исключением возможных вариантов антител, например, содержащих встречающиеся в естественных условиях мутации или возникающих в процессе производства препарата моноклонального антитела, указанные варианты, как правило, присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые, как правило, включают различные антитела к различным детерминантам (эпитопам), каждое моноклональное антитело из препарата моноклонального антитела направлено против одной детерминанты антигена. Таким образом, прилагательное «моноклональный» определяет особенность антитела, характеризуя его как полученное из практически гомогенной популяции антител, а не сконструированное в соответствии с требованиями к получению антитела с помощью какого-либо конкретного метода. Например, моноклональные антитела, которые можно применять согласно настоящему изобретению, можно создавать с помощью различных технологий, включая (но, не ограничиваясь только ими) метод гибридом, методы рекомбинантной ДНК, методы фагового дисплея и методы на основе трансгенных животных, которые содержат все или часть локусов иммуноглобулина, указанные методы и другие, приведенные в качестве примера методы получения моноклональных антител, представлены в настоящем описании.
Понятие «нативные антитела» относятся к встречающимся в естественных условиях молекулам иммуноглобулинов с различными структурами. Например, нативные антитела IgG-типа представляют собой гетеротетрамерные гликопротеины массой примерно 150000 Да, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, связанных дисульфидными мостиками. Каждая тяжелая цепь в направлении от N- к С-концу содержит вариабельную область (VH), которую называют также вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которой расположены три константных домена (CH1, СН2 и СН3), которые называют также константной областью тяжелой цепи. Аналогично этому каждая легкая цепь в направлении от N- к С-концу содержит вариабельную область (VL), которую называют также вариабельным легким доменом или вариабельным доменом легкой цепи, за которой расположен константный легкий (CL) домен. Легкая цепь может относиться к одному из двух типов, обозначенных как каппа (κ) или лямбда (λ), на основе аминокислотной последовательности ее константного домена. Антитело, «подобное нативному», имеет такую же структуру, что и «нативное антитело», но отличается специфичностью связывания.
В контексте настоящего описания понятие «вектор» относится к молекуле нуклеиновой кислоты, которая обладает способностью амплифицировать другую нуклеиновую кислоту, с которой она связна. Понятие относится к вектору как самореплицирующейся содержащей нуклеиновую кислоту структуре, а также к вектору, встроенному в геном клетки-хозяина, в которую интродуцирован вектор. Некоторые векторы обладают способностью контролировать экспрессию нуклеиновых кислот, с которым они функционально связаны. Такие векторы в контексте настоящего описания обозначают как «экспрессионные векторы».
Понятие «кассета экспрессии» относится к конструкции, которая содержит необходимые регуляторные элементы, такие как промотор и сайт полиаденилирования, для экспрессии по меньшей мере содержащейся в ней нуклеиновой кислоты в клетке.
Понятие «экспрессионный вектор» означает нуклеиновую кислоту, содержащую все необходимые элементы для экспрессии содержащегося(ищся) в ней структурного(ых) гена(ов) в клетке. Как правило, экспрессионный вектор содержит плазмидную единицу для размножения прокариотического организма, например в Е. coli, содержащую сайт инициации репликации, маркер для селекции, эукариотический маркер для селекции и одну или несколько кассет экспрессии для экспрессии представляющего(их) интерес структурного(ых) гена(ов), каждая из которых содержит промоторную нуклеиновую кислоту, структурный ген и терминатор транскрипции, включающий сигнал полиаденилирования. Экспрессия гена, как правило, находится под контролем промоторной нуклеиновой кислоты, и такой структурный ген обозначают как «функционально связанный» с промоторной нуклеиновой кислотой. Аналогично этому, регуляторный элемент и коровая промоторная нуклеиновая кислота являются функционально связанными, если регуляторный элемент модулирует активность коровой промотрной нуклеиновой кислоты.
Понятие «функционально связанные» означает взаимное расположение двух или большего количества компонентов, при котором указанные компоненты находятся во взаимосвязи, обеспечивающей им требуемую функцию. Например, промотор и/или энхансер являются функционально связанными с кодирующей последовательностью, если действует(ют) в цис-направлении касательно контроля или модуляции транскрипции сцепленной последовательности. Как правило, но необязательно, ДНК-последовательности, которые являются «функционально связанными», являются смежными и, если требуется соединять две кодирующие белки области, такие как секреторный лидер и полипептид, смежными и расположенными в рамке (считывания). Однако, хотя функционально связанный промотор, как правило, локализован в обратном направлении относительно кодирующей последовательности, не является необходимым, чтобы он был смежный с ней. Энхансер является функционально связанным с кодирующей последовательностью, если энхансер усиливает транскрипцию кодирующей последовательности. Функционально связанные энхансеры могут быть локализованы в обратном направлении, внутри кодирующих последовательностей или в прямом направлении относительно кодирующих последовательностей и на существенном расстоянии от промотора. Сайт полиаденилирования является функционально связанным с кодирующей последовательностью, если он локализован на находящемся в прямом направлении конце кодирующей последовательности, так, чтобы транскрипция происходила через кодирующую последовательность в последовательности полиаденилирования. Стоп-кодон трансляции является функционально связанным с последовательностью экзонной нуклеиновой кислоты, если он локализован в находящемся в прямом направлении конце (3'-конец) кодирующей последовательности, так, чтобы трансляция проходила через кодирующую последовательность до стоп-кодона и прекращалась на нем. Связывание осуществляют с помощью методов рекомбинации, известных в данной области, например, с использованием ПЦР-методологии и/или путем лигирования в пригодных сайтах рестрикции. Если пригодные сайты рестрикции не существуют, то применяют синтетические адаптеры или линкеры в соответствии с принятой практикой.
Понятие «полипептид» означает полимер, состоящий из аминокислот, сцепленных пептидными связями, полученный либо в естественных условиях, либо путем синтеза. Полипептиды, состоящие менее чем из примерно 20 аминокислотных остатков, можно обозначать как «пептиды», в то время как молекулы, состоящие из двух или большего количества полипептидов или содержащие один полипептид, состоящий более чем из 100 аминокислотных остатков, можно обозначать как «белки». Полипептид может содержать также не относящиеся к аминокислотам компоненты, такие как углеводные группы, ионные металлов или эфиры карбоновых кислот.Не относящиеся к аминокислотам компоненты могут поставлять клеткой, в которой полипептид экспрессируется, и могут варьироваться в зависимости от типа клетки. В контексте настоящего описания полипептиды определяют в зависимости от структуры их аминокислотного каркаса или нуклеиновой кислоты, кодирующей их. Добавки, такие как углеводные группы, как правило, не указываются, но тем не менее они могут присутствовать.
Понятие «получение» означает экспрессию представляющего интерес структурного гена, встроенного в кассету экспрессии, в клетке. Понятие включает процессы транскрипции и трансляции нуклеиновой кислоты. Получение осуществляют в соответствующих прокариотических или эукариотических клетках, и экспрессированный, т.е. полученный полипептид, можно выделять из клеток после лизиса или из супернатанта культуры.
Понятие «нуклеиновая кислота-промотор, нуклеотидный промотор» означает полинуклеотидную последовательность, которая контролирует транскрипцию гена/структурного гена или нуклеотидной последовательности, с которой она функционально связана. Нуклеотидный промотор включает сигналы для связывания РНК-полимеразы и инициации транскрипции. Применяемый нуклеотидный промотор должен быть функциональным в клетке, в которой предполагается экспрессия выбранного структурного гена. В данной области хорошо известно большое количество нуклеотидных промоторов, включая конститутивные, индуцибельные и репрессируемые промоторы из широкого разнообразия различных источников (и их можно идентифицировать в базах данных, таких как GenBank), и они доступны в виде или в составе клонированных полинуклеотидов (например, находящихся в депозитариях, таких как АТСС, а также в других коммерческих или индивидуальных источниках).
Как правило, нуклеотидный промотор локализован в 5' некодирующей или нетранслируемой области гена, проксимально относительно сайта начала транскрипции структурного гена. Элементы последовательности в нуклеотидных промоторах, функция которых состоит в инициации транскрипции, часто характеризуются консенсусными нуклеотидными последовательностями. Эти элементы включают сайты связывания РНК-полимеразы, последовательности TATA, последовательности СААТ, специфические для дифференциации элементы (DSE), респондерные элементы циклического АМФ (CRE), сывороточные респондерные элементы (SRE), глюкокортикостероидные респондерные элементы (GRE) и сайты связывания других факторов транскрипции, таких как CRE/ATF, АР2, SP1, белок, связывающий респондерный элемент цАМФ (CREB) и октамерные факторы. Если нуклеотидный промотор представляет собой индуцибельный нуклеотидный промотор, то скорость транскрипции возрастает в ответ на индуцирующий агент, как в случае нуклеотидного промотора CMV, за которым следуют два сайта tet-оператора, нуклеотидных промоторов металлотионеина и белков теплового шока. Скорость транскрипции не регулируется индуцирующим агентом, если нуклеотидный промотор представляет собой конститутивно активный нуклеотидный промотор. Эукариотическими нуклеотидный промоторами, которые идентифицированы в качестве сильных нуклеотидных промоторов для экспрессии, являются нуклеотидный ранний промотор SV40, нуклеотидный главный поздний промотор аденовирусов, нуклеотидный промотор мышиного металлотионеина-I, длинный концевой повтор вируса саркомы Раусы, фактор элонгации 1 альфа китайского хомячка (CHEF-1), человеческий EF-1 альфа, убиквитин и главный немедленно-ранний нуклеотидный промотор человеческого цитомегаловируса (hCMV MIE).
Понятие «маркер для селекции (селектирующий маркер)» означает нуклеиновую кислоту, которая обеспечивает специфическую селекцию клеток, которые его несут, в отношении или против присутствия соответствующего селектирующего агента (культивирование в условиях селективного культивирования). Как правило, селектирующий маркер может придавать устойчивость к лекарственному средству или компенсировать метаболический или катаболический дефект в клетке, в которую он интродуцирован. Селектирующий маркер может быть позитивным, негативным или бифункциональным. Ценным позитивным селектирующим маркером является ген устойчивости к антибиотикам, обеспечивающий отбор клеток, трансформированный им, в присутствии соответствующего селектирующего агента, например, антибиотика. Нетрансформированная клетка не может расти или выживать в селективных условиях, т.е. в присутствии селектирующего агента. Негативные селектирующие маркеры обеспечивают избирательную элиминацию клеток, несущих маркер. Применяемые для эукариотических клеток селектирующие маркеры включают, например, структурные гены, кодирующие аминогликозидфосфотрансферазу (АРН), такие, например, как маркеры для селекции по признаку устойчивости к гигромицину (hyg), неомицину (neo) и G418, кодирующие дигидрофолатредуктазу (DHFR), тимидинкиназу (tk), глутаминсинтетазу (GS), аспарагинсинтетазу, триптофансинтетазу (селектирующий агент индол), гистидинолдегидрогеназу (селектирующий агент гистидинол D), и нуклеиновые кислоты, придающие устойчивость к пуромицину, блеомицину, флеомицину, хлорамфениколу, зеоцину и микофеноловой кислоте.
Способы
Настоящее изобретение основано по меньшей мере частично на открытии того, что для создания клеточных линий для производства гетеродимерных антител целесообразно применять для трансфекции экспрессионный вектор, который содержит в качестве единственной кодирующей (антитело) полипептид нуклеиновой кислоты нуклеиновую кислоту, которая кодирует полипептид легкой цепи, т.е. вектор, содержащий в качестве единственной кассеты экспрессии полипептида антитела кассету экспрессии легкой цепи. Указанный вектор применяют в сочетании с дополнительными экспрессионными векторами для котрансфекции или трансфекции отдельно на второй последующей стадии. С помощью указанного подхода можно получать линию клеток-продуцентов, которые продуцируют гетеродимерное антитело с улучшенным профилем продукта, т.е. с повышенным количеством продукта и пониженным количеством родственных продукту примесей.
Один из подходов к созданию мультиспецифических антител известен как «технология CrossMab». Указанный подход базируется на кроссовере доменов между тяжелой и легкой цепями, что приводит тем самым к созданию различных расположений доменов в тяжелых цепях и легких цепях различной специфичности (см., например, в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, у Schaefer W. и др., Proc. Natl. Acad. Sci. USA 108, 2011, сс. 11187-11192 данные, касающиеся двухвалентных биспецифических антител IgG-типа с кроссовером доменов; в WO 2010/145792 и WO 2010/145792 данные, касающиеся четырехвалентных антигенсвязывающих белков с кроссовером доменов).
Мультиспецифические антитела с заменой/обменом VH/VL в одном сайте связывания для предупреждения ошибочного спаривания легкой цепи (CrossMabVH-VL), которые описаны в WO 2009/080252 (см. также Schaefer W. и др., Proc. Natl. Acad. Sci. USA, 108, 2011, сс. 11187-11191, существенно снижали образование побочных продуктов, обусловленное ошибочным спариванием легкой цепи, которая связывается с первым антигеном, с «неправильной» тяжелой цепью, которая связывается со вторым антигеном (по сравнению с подходами, в которых не использован указанных обмен доменами). Однако их получение не полностью свободно от образования побочных продуктов. Основной побочный продукт образуется в результате взаимодействия бенс-джонсовского типа «неправильной» легкой цепи с тяжелой цепью с обмененными доменами (см. также Schaefer W. и др., Proc. Natl. Acad. Sci. USA, 108, 2011, сс.11187-11191; на фиг. S1I в приложении).
В WO 2015/101588 А1 описаны модули шаттлов для гематоэнцефалического барьера. В WO2015/101588 А1 упомянуты двухвалентные биспецифические антитела с кроссовером VH/VL-доменов в одном из связывающих плечей с мутациями в поверхности раздела CH1/CL. В WO 2015/101588 А1 не описан технический эффект указанных мутаций.
Известны различные методы создания клеточных линий для получения четырехцепочечных гомодимерных двухвалентных антител, т.е. антител типа нативных антител. Для повышения производительности (продуктивности) указанных клеточных линий некоторые из указанных методов основаны на так называемом подходе «супертрансфекции». При применении указанного подхода клетки трансфектируют по меньшей мере двукратно с селекцией промежуточной клеточной линии. Каждый из векторов, применяемых в указанном подходе супертрансфекции, как правило, содержит полную информацию для кодирования подлежащего экспрессии антитела, т.е. для легкой цепи и для тяжелой цепи. В некоторых специальных методах супертрансфекции применяют очень сходные или даже идентичные векторы, отличающиеся только селектирующим маркером для того, чтобы достигать тесной интеграции в геном в известной продуктивной области. Подобно методам амплификации генов с использованием DHFR методы супертрансфекции служат для повышения уровня (выхода) экспрессии путем повышения количества функциональных кассет экспрессии в клетках.
Для новых комплексных трехвалентных биспецифических форматов антител, содержащих гетеродимерную Fc-область и так называемый обмен доменами, которые оба интродуцированы для ограничения или даже исключения ошибочного спаривания цепей и тем самым повышения выхода правильно уложенного и собранного полученного мультиспецифического антитела, описана сложная процедура котрансфекции 3-4 векторами, каждый из который содержит одну кассету экспрессии, с использованием различных соотношений векторов (см., например, WO 2013/026833).
Настоящее изобретение основано, по меньшей мере частично, на открытии того факта, что уровень экспрессии мультиспецифического антитела в рекомбинантной клетке можно повышать, если клетку повторно трансфектировать кассетой экспрессии для экспрессии легкой цепи указанного мультиспецифического антитела. Это является особенно ценным, если мультиспецифическое антитело содержит вариант тяжелой и легкой цепей с кроссовером доменов.
Одним из объектов изобретения, представленных в настоящем описании, является способ получения мультиспецифического антитела (содержащего по меньшей мере один полипептид с кроссовером доменов), включающий следующие стадии:
а) получение клетки млекопитающего, которая экспрессирует антитело,
б) трансфекция клетки млекопитающего, указанной в подпункте а), экспрессионным вектором, который содержит кассету экспрессии, кодирующую полипептид антитела, которое имеет кроссовер доменов,
в) культивирование клетки, указанной в подпункте б), и выделение антитела из клетки или среды для культивирования и тем самым получение мультиспецифического антитела.
Модифицированные клетки, полученные с помощью способа, представленного в настоящем описании, «секретируют» в большем количестве мультиспецифическое антитело в правильно уложенной и собранной форме, и их обозначают в контексте настоящего описания как клетки, в которых количество правильно уложенного и правильно собранного мультиспецифического антитела, высвободившегося во внеклеточную среду, увеличено по сравнению с родительской клеткой. Анализ методом иммуноблоттинга, анализы биологической активности и методы физико-химического разделения можно применять для определения абсолютных количеств правильно уложенного и собранного мультиспецифического антитела, высвободившегося из модифицированной клетки по сравнению с родительской клеткой.
Одним из объектов изобретения, представленных в настоящем описании, является способ получения мультиспецифического антитела, включающий следующие стадии:
а) культивирование модифицированной клетки в условиях, пригодных /благоприятных для получения мультиспецифического антитела, при этом
I) модифицированная клетка является родственной родительской клетке, где родительская клетка содержит последовательности первой ДНК, кодирующей мультиспецифическое антитело, посредством интродукции нуклеиновой кислоты в геном родительской клетки в локус, который не находится в последовательностях первой ДНК; и
II) модифицированная клетка продуцирует большее количество мультиспецифического антитела, чем родительская клетка, когда обе клетки культивируют в одинаковых условиях; и
б) выделение полипептида.
Форматы антител с кроссовером доменов
Способ, представленный в настоящем описании, в целом пригоден для получения любого мультиспецифического антитела, которое содержит отдельно кодируемую тяжелую и легкую цепь.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга.
Антитело, указанное в подпункте а), не содержит модификацию, указанную в подпункте б), и тяжелая цепь и легкая цепь, указанные в подпункте а), представляют собой выделенные цепи.
В антителе, указанном в подпункте б),
в легкой цепи
вариабельный домен легкой цепи VL заменен вариабельным доменом тяжелой цепи VH указанного антитела, и
в тяжелой цепи
вариабельный домен тяжелой цепи VH заменен вариабельным доменом легкой цепи VL указанного антитела.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга и в котором константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
Антитело, указанное в подпункте а), не содержит модификацию, указанную в подпункте б), и тяжелая цепь и легкая цепь, указанные в подпункте а), представляют собой выделенные цепи.
В антителе, указанном в подпункте б),
в легкой цепи
вариабельный домен легкой цепи VL заменен вариабельным доменом тяжелой цепи VH указанного антитела, и константный домен легкой цепи CL замен константным доменом тяжелой цепи СН1 указанного антитела; и
в тяжелой цепи
вариабельный домен тяжелой цепи VH заменен вариабельным доменом легкой цепи VL указанного антитела, и константный домен тяжелой цепи СН1 заменен константным доменом легкой цепи CL указанного антитела.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
Антитело, указанное в подпункте а), не содержит модификацию, указанную в подпункте б), и тяжелая цепь и легкая цепь, указанные в подпункте а), представляют собой выделенные цепи.
В антителе, указанном в подпункте б),
в легкой цепи
константный домен легкой цепи CL заменен константным доменом тяжелой цепи СН1 указанного антитела; и
в тяжелой цепи
константный домен тяжелой цепи СН1 заменен константным доменом легкой цепи CL указанного антитела.
В одном из вариантов осуществления всех объектов изобретения мультиспецифическое антитело представляет собой триспецифическое или тетраспецифическое антитело, которое содержит
а) первую легкую цепь и первую тяжелую цепь полноразмерного антитела, которое специфически связывается с первым антигеном, и
б) вторую (модифицированную) легкую цепь и вторую (модифицированную) тяжелую цепь полноразмерного антитела, которое специфически связывается со вторым антигеном, в котором вариабельные домены VL и VH заменены друг на друга и/или в котором константные домены CL и СН1 заменены друг на друга, и
в) в котором 1-4 антигенсвязывающих пептида, которые специфически связываются с одним или двумя дополнительными антигенами (т.е. с третьим и/или четвертым антигеном), слиты через пептидный линкер с С- или N-концом легких цепей или тяжелых цепей, указанных в подпунктах а) и/или б).
Антитело, указанное в подпункте а), не содержит модификацию, указанную в подпункте б), и тяжелая цепь и легкая цепь, указанные в подпункте а), представляют собой выделенные цепи.
В одном из вариантов осуществления изобретения триспецифическое или тетраспецифическое антитело содержит один или два антигенсвязывающих пептида, указанных в подпункте в), которые специфически связываются с одним или двумя дополнительными антигенами.
В одном из вариантов осуществления изобретения антигенсвязывающие пептиды выбирают из группы, состоящей из scFv-фрагмента и scFab-фрагмента.
В одном из вариантов осуществления изобретения антигенсвязывающие пептиды представляют собой scFv-фрагменты.
В одном из вариантов осуществления изобретения антигенсвязывающие пептиды представляют собой scFab-фрагменты.
В одном из вариантов осуществления изобретения антигенсвязывающие пептиды слиты с С-концом тяжелых цепей, указанных в подпунктах а) и/или б).
В одном из вариантов осуществления изобретения триспецифическое или тетраспецифическое антитело содержит один или два, указанных в подпункте в), антигенсвязывающих пептида, которые специфически связываются с одним дополнительным антигеном.
В одном из вариантов осуществления изобретения триспецифическое или тетраспецифическое антитело содержит два идентичных антигенсвязывающих пептида, указанных в подпункте в), которые специфически связываются с третьим антигеном. В одном предпочтительном варианте осуществления изобретения два идентичных антигенсвязывающих пептида слиты оба через одинаковый пептидный линкер с С-концом тяжелых цепей, указанных в подпунктах а) и б). В одном предпочтительном варианте осуществления изобретения указанные два идентичных антигенсвязывающих пептида представляют собой либо scFv-фрагмент, либо scFab-фрагмент.
В одном из вариантов осуществления изобретения триспецифическое или тетраспецифическое антитело содержит два антигенсвязывающих пептида, указанных в подпункте в), которые специфически связываются с третьи и четвертым антигеном.
В одном из вариантов осуществления изобретения указанные два антигенсвязывающих пептида слиты оба через одинаковый пептидный коннектор с С-концом тяжелых цепей, указанных в подпунктах а) и б). В одном предпочтительном варианте осуществления изобретения указанные два антигенсвязывающих пептида представляют собой либо scFv-фрагмент, либо scFab-фрагмент.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое четырехвалентное антитело, которое содержит
а) две легкие цепи и две тяжелые цепи антитела, которое специфически связывается с первым антигеном (и содержит два Fab-фрагмента),
б) два дополнительных Fab-фрагмента антитела, которое специфически связывается со вторым антигеном, в котором указанные дополнительные Fab-фрагменты оба слиты через пептидный линкер либо с С-, либо с N-концами тяжелых цепей, указанных в подпункте а),
и
где в Fab-фрагментах осуществлены следующие модификации
I) в обоих Fab-фрагментах, указанных в подпункте а), или в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и/или константные домены CL и СН1 заменены друг на друга,
или
II) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменены друг на друга и константные домены CL и СН1 заменены друг на друга, и
в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга или константные домены CL и СН1 заменены друг на друга, или
III) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменены друг на друга или константные домены CL и СН1 заменены друг на друга, и
в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга и константные домены CL и СН1 заменены друг на друга, или
IV) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменены друг на друга и в обоих Fab-фрагментах, указанных в подпункте б), константные домены CL и СН1 заменены друг на друга,
или
V) в обоих Fab-фрагментах, указанных в подпункте а), константные домены CL и СН1 заменены друг на друга и в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменены друг на друга.
В одном из вариантов осуществления изобретения указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер либо с С-концами тяжелых цепей, указанных в подпункте а), либо с N-концами тяжелых цепей, указанных в подпункте а).
В одном из вариантов осуществления изобретения указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер с С-концами тяжелых цепей, указанных в подпункте а).
В одном из вариантов осуществления изобретения указанные дополнительные Fab-фрагменты слиты оба через пептидный коннектор с N-концами тяжелых цепей, указанных в подпункте а).
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществляют следующие модификации:
I) в обоих Fab-фрагментах, указанных в подпункте а), или в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменяют друг на друга,
и/или
константные домены CL и СН1 заменяют друг на друга.
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществляют следующие модификации:
I) в обоих Fab-фрагментах, указанных в подпункте а), вариабельные домены VL и VH заменяют друг на друга и/или
константные домены CL и СН1 заменяют друг на друга.
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществляют следующие модификации:
I) в обоих Fab-фрагментах, указанных в подпункте а), константные домены CL и СН1 заменяют друг на друга.
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществляют следующие модификации:
I) в обоих Fab-фрагментах, указанных в подпункте б), вариабельные домены VL и VH заменяют друг на друга и/или
константные домены CL и СН1 заменяют друг на друга.
В одном из вариантов осуществления изобретения в Fab-фрагментах осуществляют следующие модификации:
I) в обоих Fab-фрагментах, указанных в подпункте б), константные домены CL и СН1 заменяют друг на друга.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое четырехвалентное антитело, которое содержит:
а) (модифицированную) тяжелую цепь первого антитела, которое специфически связывается с первым антигеном и содержит первую доменную пару VH-CH1, в котором с С-концом указанной тяжелой цепи слит через пептидный линкер N-конец второй доменной пары VH-CH1 указанного первого антитела,
б) две легкие цепи указанного первого антитела, указанного в подпункте а),
в) (модифицированную) тяжелую цепь второго антитела, которое специфически связывается со вторым антигеном и содержит первую доменную пару VH-CL, в котором с С-концом указанной тяжелой цепи слит через пептидный линкер N-конец второй доменной пары VH-CL указанного второго антитела, и
г) две (модифицированные) легкие цепи второго антитела, указанного в подпункте в), каждая из которых содержит доменную пару CL-CH1.
В одном из вариантов осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое антитело, которое содержит:
а) тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с первым антигеном, и
в) тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается со вторым антигеном, в котором N-конец тяжелой цепи соединен с С-концом легкой цепи через пептидный линкер.
Антитело, указанное в подпункте а), не содержит модификацию, указанную в подпункте б), и тяжелая цепь и легкая цепь, указанные в подпункте а), представляют собой выделенные цепи.
Во всех объектах изобретения, представленных в настоящем описании, первая легкая цепь содержит VL-домен и CL-домен, а первая тяжелая цепь содержит VH-домен, CH1-домен, шарнирную область, СН2-домен и СН3-домен.
В одном из вариантов осуществления изобретения антитело, полученное способом, представленным в настоящем описании, представляет собой мультиспецифическое антитело, которое нуждается в гетеродимеризации по меньшей мере двух полипептидов тяжелых цепей.
Несколько подходов к СН3-модификациям, способствующих гетеродимеризации, описано, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954, WO 2013/096291, которые включены в настоящее описание в качестве ссылки. Как правило, в подходах, известных в данной области, СН3-домен первой тяжелой цепи и СН3-домен второй тяжелой цепи оба конструируют комплементарным образом, в результате чего тяжелая цепь, содержащая один из сконструированных СН3-доменов, не может более образовывать гомодимер с другой тяжелой цепью такой же структуры (например, первая тяжелая цепь со сконструированным СН3 не может более образовывать гомодимер с другой первой тяжелой цепью со сконструированным СН3; и вторая тяжелая цепь со сконструированным СН3 не может более образовывать гомодимер с другой второй тяжелой цепью со сконструированным СН3). Таким образом, у первой тяжелой цепи, которая содержит один сконструированный СН3-домен, усиливается способность к гетеродимеризации с другой тяжелой цепью, содержащей СН3-домен, сконструированный комплементарным образом. Согласно этому варианту осуществления изобретения СН3-домен первой тяжелой цепи и СН3-домен второй тяжелой цепи создают комплементарным образом с помощью аминокислотных замен, которые усиливают способность к гетеродимеризации первой тяжелой цепи и второй тяжелой цепи, в то время как первая тяжелая цепь и вторая тяжелая цепь более не могут образовывать гомодимер (например, из-за стерических препятствий).
В данной области известны различные подходы, способствующие гетеродимеризации тяжелых цепей, которые процитированы и описаны выше, и они рассматриваются в качестве различных альтернатив, применяемых в мультиспецифическом антителе, предлагаемом в изобретении, которое содержит «нескрещенную Fab-область» из первого антитела, которое специфически связывается с первым антигеном, и «скрещенную Fab-область» из второго антитела, которое специфически связывается со вторым антигеном, в сочетании с конкретными аминокислотными заменами, описанными выше в изобретении.
СН3-домены мультиспецифического антитела, полученного с помощью способа, представленного в настоящем описании, можно изменять с использованием технологии «knob-into-holes» (взаимодействие по типу «выступ-во-впадину»), которая описана подробно с помощью нескольких примеров, например, в WO 96/027011, у Ridgway J.B. и др., Protein Eng 9, 1996, сс. 617-621 и Merchant A.M. и др., Nat Biotechnol 16, 1998, сс. 677-681. При осуществлении этого метода поверхности взаимодействия двух СН3-доменов изменяют с целью повышения гетеродимеризации обеих тяжелых цепей, содержащих два указанных СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может содержать «выступ», а другой может содержать «впадину». Интродукция дисульфидного мостика дополнительно стабилизирует гетеродимеры (Merchant A.M. и др., Nature Biotech 16, 1998, сс. 677-681; Atwell S. и др., J. Mol. Biol. 270, 1997, сс. 26-35) и повышает выход.
В одном предпочтительном варианте осуществления изобретения мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутацию T366W в СН3-домене «цепи с «выступами» и мутации T366S, L368A, Y407V в СН3-домене «цепи с впадиной» (нумерация согласно EU-индексу Кэбота). Можно применять также дополнительный межцепочечный дисульфидный мостик между СН3-доменами (Merchant A.M. и др., Nature Biotech 16, 1998, сс. 677-681), например, путем интродукции мутации Y349C в СН3-домен «цепи с «выступами» и мутации Е356С или мутации S354C в СН3-домен «цепи с впадиной». Таким образом, согласно другому предпочтительному варианту осуществления изобретения мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации Е356С, T366S, L368A и Y407V во втором из двух СН3-доменов, или мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V во втором из двух СН3-доменов (дополнительная мутация Y349C в одном из СН3-доменов и дополнительная мутация Е356С или S354C в другом СН3-домене приводит к образованию межцепочечного дисульфидного мостика) (нумерация согласно EU-индексу Кэбота).
Однако в альтернативном или дополнительном варианте можно применять также другие варианты технологии «knobs-in-holes», представленные в ЕР 1870459А1. В одном из вариантов осуществления изобретения мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутации R409D и K370E в СН3-домене «цепи с «выступами» и мутации D399K и E357K в СН3-домене «цепи с впадиной» (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления изобретения мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутацию T366W в СН3-домене «цепи с «выступами» и мутации T366S, L368A и Y407V в СН3-домене «цепи с впадиной» (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления изобретения мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V во втором из двух СН3-доменов, или мультиспецифическое антитело, полученное с помощью способа, представленного в настоящем описании, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V во втором из двух СН3-доменов и дополнительно содержит мутации R409D и K370E в СН3-домене «цепи с «выступами» и мутации D399K и E357K в СН3-домене «цепи с впадиной» (нумерация согласно EU-индексу Кэбота).
Помимо технологии «knob-into-hole» в данной области известны другие технологии модификации СН3-доменов тяжелых цепей мультиспецифического антитела для усиления гетеродимеризации. Указанные технологии, прежде всего описанные в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954 и WO 2013/096291, рассматриваются в настоящем описании в качестве альтернативы технологии «knob-into-hole» касательно мультиспецифического антитела, полученного с помощью способа, указанного в настоящем описании.
В одном из вариантов всех объектов и вариантов осуществления изобретения, которые представлены в настоящем описании, мультиспецифическое антитело представляет собой биспецифическое антитело или триспецифическое антитело. В одном из предпочтительных вариантов осуществления изобретения мультиспецифическое антитело представляет собой биспецифическое антитело.
В одном из вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, антитело представляет собой двухвалентное или трехвалентное антитело. В одном из вариантов осуществления изобретения антитело представляет собой двухвалентное антитело.
В одном из вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело имеет структуру константного домена, соответствующую антителу IgG-типа. В другом варианте осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG1-подклассу или человеческому IgG1-подклассу с мутациями L234A и L235A. В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG2-подклассу. В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG3-подклассу. В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG4-подклассу или человеческому IgG4-подклассу с дополнительной мутацией S228P. В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG1-подклассу или человеческому IgG4-подклассу. В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG1-подклассу с мутациями L234A и L235A (нумерация согласно EU-индексу Кэбота). В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG1-подклассу мутациями L234A, L235A и P329G (нумерация согласно EU-индексу Кэбота). В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG4-подклассу с мутациями S228P и L235E (нумерация согласно EU-индексу Кэбота). В одном из дополнительных вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, мультиспецифическое антитело отличается тем, что указанное мультиспецифическое антитело относится к человеческому IgG4-подклассу с мутациями S228P, L235E и P329G (нумерация согласно EU-индексу Кэбота).
В одном из вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, антитело, которое содержит тяжелую цепь, включающую СН3-домен, указанный в настоящем описании, содержит дополнительный С-концевой дипептид глицин-лизин (G446 и K447, нумерация согласно EU-индексу Кэбота). В одном из вариантов осуществления всех объектов изобретения, которые представлены в настоящем описании, антитело, которое содержит тяжелую цепь, включающую СН3-домен, указанный в настоящем описании, содержит дополнительный С-концевой остаток глицина (G446, нумерация согласно EU-индексу Кэбота).
Биспецифическое трехвалентное антитело к человеческому А-бета/рецептору человеческого трансферрина
Указанное антитело представляет собой биспецифическое антитело, состоящее из полноразмерного базового антитела и слитое с Fab-фрагментом, в котором определенные домены «крестообразно» обменены. Таким образом, образовавшееся биспецифическое антитело является несимметричным. Для этой цели биспецифическое антитело получали с использованием технологии гетеродимеризации, обозначенной как «knobs-into-holes», с применением первой тяжелой цепи с мутациями, приводящими к образованию так называемых «выступов» (НС«выступ»), и второй тяжелой цепи с мутациями, приводящими к образованию так называемых впадин (НС«впадина»).
В этом примере применяли котрансфекцию.
Антитело 0012, антитело 0015, антитело 0020 и антитело 0024 описаны в WO 2017/055540 A1 (SEQ ID NO: 06-09, SEQ ID NO: 01-03 и SEQ ID NO: 10, SEQ ID NO: 11-13 и SEQ ID NO: 14-17 в WO 2017/055540 A1 соответственно).
Антитело 0012 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VL-домен, где в Fab VH- и VL-домены обменены (кроссовер доменов VH-VL). Оба Fab без кроссовера доменов полноразмерного антитела были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0015 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию «выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VH-домен Fab, где в Fab CH1 - и CL-домены обменены (кроссовер доменов CH-CL). Оба Fab без кроссовера доменов полноразмерного антитела были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0020 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию «выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VL-домен одноцепочечного Fab (отсутствие кроссовера доменов). Оба Fab без кроссовера доменов полноразмерного антитела были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0024 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию «выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VH-домен Fab, где в Fab CH1 - и CL-домены обменены (кроссовер доменов CH-CL).
Для рекомбинантного получения биспецифических антител применяли различное распределение/различную комбинацию соответствующих полипептидов в различных экспрессионных векторах, различные соотношения образовавшихся векторов и различные последовательности для трансфекции.
LC+НС«впадина»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «впадины», и легкую цепь.
LCcross+HC«выступ»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «выступа», и легкую цепь с кроссовером доменов.
LC: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи.
LCcross: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи с кроссовером доменов.
НС«выступ»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «выступа», и слитую с scFab.
Ниже в таблице представлены результаты, полученные с использованием клеток СНО-К1.

Биспецифические антитела получали в небольшом масштабе в клетках CHO-S и распределение побочного продукта анализировали после первой стадии очистки с использованием аффинной хроматографии на белке А и после второй стадии очистки с использованием препаративной гель-фильтрации. Результаты представлены ниже в таблице.
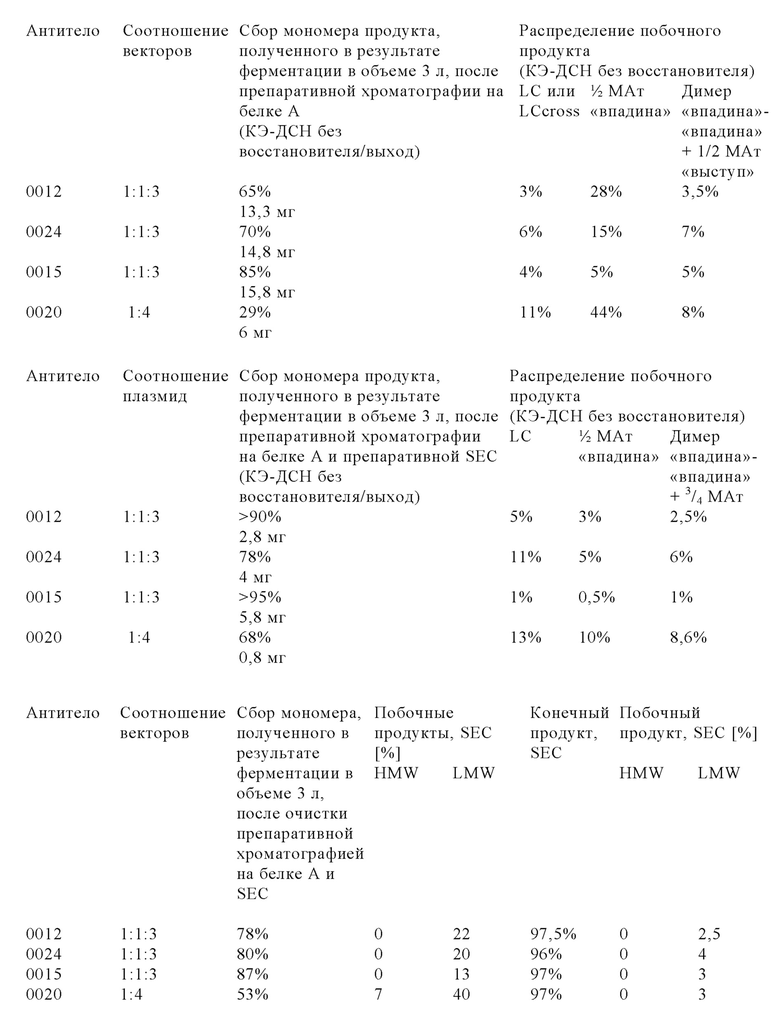
При применении указанного подхода можно получать линию клеток-продуцентов, которая продуцирует гетеродимерное антитело с улучшенным профилем продуктов, т.е. с повышенным количеством продукта и пониженным количеством родственных продукту примесей.
Для создания стабильных линий клеток-продуцентов применяли котрансфекцию экспрессионной плазмидой, содержащей кассету экспрессии только для легкой цепи антитела.
Клетки СНО-K1 трансфектировли с использованием соотношения плазмид 1(LC):1(LC+HC«впадина»):3(LCcross+HC«выступ»). Клетки со стабильной интегрированной в их геном чужеродной ДНК отбирали с помощью метотрексата. Стабильные клеточные линии выделяли и оценивали с использованием четырехдневной культуры с подпиткой в отношении качества продукта. Продукт выделяли с помощью аффинной хроматографии на белке А и анализировали с помощью КЭ-ДСН.
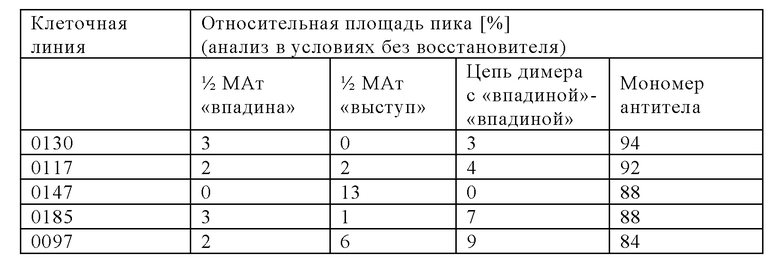
Биспецифическое трехвалентное антитело к человеческому CD20/ рецептору человеческого трансферрина
Указанное антитело представляет собой биспецифическое антитело, состоящее из полноразмерного базового антитела и слитое с Fab-фрагментом, в котором определенные домены «крестообразно» обменены. Таким образом, образовавшееся биспецифическое антитело является несимметричным. Для этой цели биспецифическое антитело получали с использованием технологии гетеродимеризации, обозначенной как «knobs-into-holes», с применением первой тяжелой цепи с мутациями, приводящими к образованию так называемых «выступов» (НС«выступ»), и второй тяжелой цепи с мутациями, приводящими к образованию так называемых впадин (НС«впадина»).
В этом примере применяли котрансфекцию.
Антитело 0039, антитело 0041, антитело 0040 и антитело 0042 описаны в WO 2017/055542 A1 (SEQ ID NO: 06-09, SEQ ID NO: 01-03 и SEQ ID NO: 10, SEQ ID NO: 11-13 и SEQ ID NO: 22 и SEQ ID NO: 14-17 в WO 2017/055542 A1 соответственно).
Антитело 0038 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер scFab. Оба плеча канонического Fab были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0039 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VL-домен Fab, где в Fab VH- и VL-домены обменены (кроссовер доменов VH-VL). Оба Fab с неизмененными доменами были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0041 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VH-домен, где в Fab CH1 - и CL-домены обменены (кроссовер доменов CH-CL). Обе пары тяжелых и легких цепей полноразмерного антитела, также как Fab, были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке. Оба Fab с неизмененными доменами были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Антитело 0040 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с С-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит через линкер VH Fab, где в Fab CH1 - и CL-домены обменены (кроссовер доменов CH-CL).
Антитело 0042 представляет собой полноразмерное антитело, содержащее одну тяжелую цепь с мутациями, приводящими к образованию «впадины», и одну тяжелую цепь с мутациями, приводящими к образованию ««выступа», в котором с N-концом тяжелой цепи с мутациями, приводящими к образованию «выступа», слит CH1 Fab, где в Fab VH- и VL-домены слитой тяжелой цепи обменены (кроссовер доменов VH-VL). Оба Fab с неизмененными доменами были модифицированы таким образом, чтобы они содержали заряды, что способствовало правильной сборке.
Для рекомбинантного получения биспецифических антител применяли различное распределение/различную комбинацию соответствующих полипептидов в различных экспрессионных векторах, различные соотношения образовавшихся векторов и различные последовательности для трансфекции.
LC+НС«впадина»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «впадины», и легкую цепь.
LCcross+HC«выступ»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «выступа», и легкую цепь с кроссовером доменов.
LC: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи.
LCcross: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи с кроссовером доменов.
НС«выступ»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «выступа», и слитую с scFab.
Для рекомбинантного получения биспецифических антител в HEK-клетках применяли различное распределение/различную комбинацию соответствующих полипептидов в различных экспрессионных векторах, различные соотношения образовавшихся векторов и различные последовательности для трансфекции. Результаты представлены ниже в таблице.

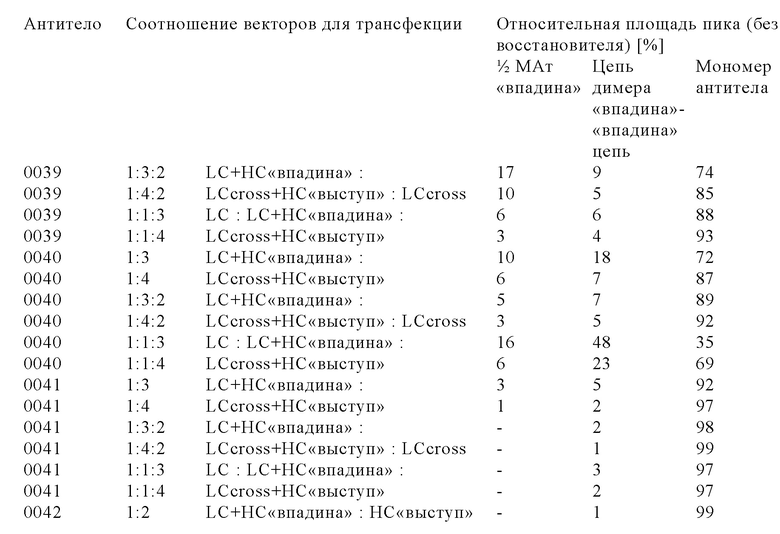
Для рекомбинантного получения биспецифических антител в клетках СНО-K1 применяли различное распределение/различную комбинацию соответствующих полипептидов в различных экспрессионных векторах, различные соотношения образовавшихся векторов и различные последовательности для трансфекции. Результаты представлены ниже в таблице.


Биспецифические антитела получали в небольшом масштабе в клетках CHO-S и распределение побочного продукта анализировали после первой стадии очистки с использованием аффинной хроматографии на белке А и после второй стадии очистки с использованием препаративной гель-фильтрации. Результаты представлены ниже в таблице.
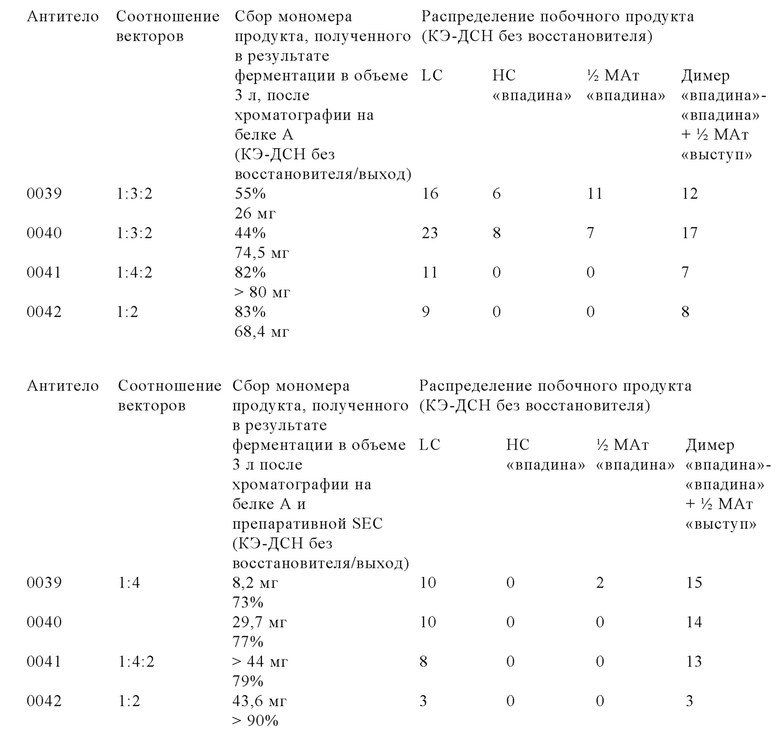
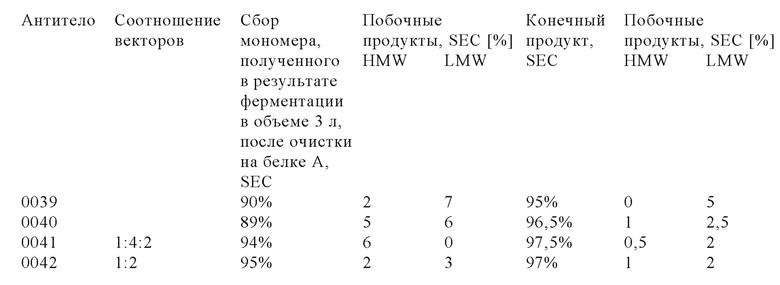
Биспецифические антитела получали в различных клеточных линиях. Результаты представлены ниже в таблице.

При применении указанного подхода можно получать линию клеток-продуцентов, которая продуцирует гетеродимерное антитело с улучшенным профилем продуктов, т.е. с повышенным количеством продукта и пониженным количеством родственных продукту примесей.
Биспецифическое двухвалентное антитело к человеческому PD1/человеческому Tim3
Указанное антитело представляет собой биспецифическое антитело, состоящее из полноразмерного антитела с мутациями «knob-into-hole» в Fc-области и искусственным дисульфидным мостиком между СН3-доменами, в котором в паре тяжелой и легкой цепи, образующей сайт связывания для PD1, VH- и VL-домены заменены друг на друга. Таким образом, образовавшееся биспецифическое антитело является несимметричным. Для этой цели биспецифическое антитело получали с использованием технологии гетеродимеризации, обозначенной как «knobs-into-holes», с использованием первой тяжелой цепи с мутациями, приводящими к образованию так называемых «выступов» (НС«выступ»), и второй тяжелой цепи с мутациями, приводящими к образованию так называемых впадин (НС«впадина»). Последовательности описаны в WO 2017/055404 А1
В этом примере применяли котрансфекцию.
Для этого объединяли несколько версий экспрессионных плазмид с получением клеточной линии, экспрессирующей указанное выше антитело. Эти подходы отличались по комбинации плазмид, но не отличались полученными антителами.
Для получения антитела 0516 осуществляли совместную трансфекцию в соотношении 1:1 применяемым для трансфекции вектором 1, который содержит кассеты экспрессии для первой легкой цепи (LC-1) и первой тяжелой цепи с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласса, и вектором 2, который содержит кассеты экспрессии для второй легкой цепи с обменом доменов (CrossLC-2), и второй тяжелой цепи с обменом доменов с мутациями, приводящими к образованию «выступа» (CrossHC-2-«выступ»), IgG1-подкласса.
Для получения антитела 0517 осуществляли совместную трансфекцию в соотношении 1:1:1 применяемым для трансфекции вектором 1, который содержит кассеты экспрессии для первой легкой цепи (LC-1) и первой тяжелой цепи с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласса, вектором 2, который содержит кассеты экспрессии для второй легкой цепи с обменом доменов (CrossLC-2), и второй тяжелой цепи с обменом доменов с мутациями, приводящими к образованию «выступа» (CrossHC-2-«выступ»), IgG1-подкласса, и вектором 3, который содержит кассету экспрессии для второй легкой цепи с обменом доменов (CrossLC-2).
Для получения антитела 0518 осуществляли совместную трансфекцию в соотношении 1:1 применяемым для трансфекции вектором 1, который содержит кассеты экспрессии для второй легкой цепи с обменом доменов (CrossLC-2) и первой тяжелой цепи с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласса, и вектором 2, который содержит кассеты экспрессии для первой легкой цепи (LC-1) и второй тяжелой цепи с обменом доменов с мутациями, приводящими к образованию «выступа» (CrossHC-2-«выступ»), IgG1-подкласса.
Для получения антитела 0519 осуществляли совместную трансфекцию в соотношении 1:1:1 применяемым для трансфекции вектором 1, который содержит кассеты экспрессии для второй легкой цепи с обменом доменов (CrossLC-2) и первой тяжелой цепи с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласса, вектором 2, который содержит кассеты экспрессии для первой легкой цепи (LC-1) и второй тяжелой цепи с обменом доменов с мутациями, приводящими к образованию «выступа» (CrossHC-2-«выступ»), IgG1-подкласса, и вектором 3, который содержит кассету экспрессии для второй легкой цепи с обменом доменов (CrossLC-2). Результаты представлены ниже в таблице.
Правильно собранное антитело имеет стехиометрию, соответствующую ABCD, где А обозначает вторую тяжелую цепь с мутациями, приводящими к образованию «выступа» (CrossHC-2-«выступ»), IgG1-подкласса, В обозначает первую тяжелую цепь с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласс, С обозначает вторую легкую цепь с обменом доменов (CrossLC-2), a D обозначает первую легкую цепь (LC-1).
В основном образовавшиеся родственные соединению побочные продукты представляли собой неправильно собранные антитела. Два основных побочных продукта оба представляли собой четырехцепочечные антитела. Первое представляло собой гетеро-«впадина»-«выступ»-НС-димер, в котором скрещенная легкая цепь заменена нескрещенной легкой цепью (ABD2). Второе представляло собой гомо-«впадина»-«впадина»-димер полуантитела (B2D2).
Установлено, что при применении для трансфекций дополнительной плазмиды, содержащей только кассету экспрессии для легкой цепи с обменом доменов, можно получать улучшенный результаты, т.е. снижение присутствия родственных продукту побочных продуктов (см. фиг. 1А-1Г).

Как продемонстрировано на фиг. 1, можно снижать количество родственных продукту побочных продуктов, прежде всего побочного продукта ABD2. Это одновременно уменьшает потерю продукта в процессе последующих стадий очистки. Например, либо можно уменьшать количество необходимых стадий очистки, либо можно снижать потерю продукта, связанную с перекрыванием пиков и фракционированием (пики в большей степени разделены и их можно «вырезать» с уменьшенной потерей продукта), либо могут иметь место оба этих явления. Таким образом, можно увеличивать получаемый выход.
При применении указанного подхода можно получать линию клеток-продуцентов, которая продуцирует гетеродимерное антитело с улучшенным профилем продуктов, т.е. с повышенным количеством продукта и пониженным количеством родственных продукту примесей.
Биспецифическое четырехвалентное антитело к человеческому FAP/DR5
Биспецифические антитела к FAP-DR5 создавали путем слияния FAP-связывающего домена с тяжелой цепь анти-DRS IgG на С-конце через линкер (G4S)4. Соответствующая антителу к DR5 часть состояла из вариабельной легкой цепи (VL) и вариабельной тяжелой цепи (VH) дрозитумаба (см. US 2007/003141401) или новых антител к DR5, созданных с помощью фагового дисплея. Для минимизации количества побочных продуктов в виде неправильно спаренных легких цепей применяли CrossMab-технологию для кроссовера доменов. FAP-связывающий модуль создавали в виде скрещенного Fab, в котором VH-домен слит с константной легкой цепью (CL) и VL-домен слит с CHI-доменом (константная тяжелая цепь 1). Последовательности описаны в WO 2016/055432.
В этом примере применяли котрансфекцию.
Соответствующие полипептидам кассеты экспрессии распределяли в различные экспрессионные векторы.
LC+НС«впадина»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «впадины», и легкую цепь.
LCcross+HC«выступ»: экспрессионный вектор, который содержит одну кассету экспрессии для тяжелой цепи с мутацией, приводящей к образованию «выступа», и легкую цепь с кроссовером доменов.
LC: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи.
LCcross: экспрессионный вектор, который содержит одну кассету экспрессии для легкой цепи с кроссовером доменов.
Клон 131 получали с помощью стандартной трансфекции двумя плазмидами, каждая из которых содержала две кассеты экспрессии для экспрессии биспецифического антитела (полноразмерное антитело с каждым одним из CH1/CL cross-Fab, присоединенным к каждому из С-концов тяжелых цепей).
Этот клон продуцировал продукты следующего состава.

Указанный клон применяли в качестве основного клона для второй трансфекции с использованием плазмиды, которая содержала только скрещенную легкую цепь FAP-связывающего сайта.
Ниже в таблице в качестве примера представлены характеристики некоторых полученных клонов.
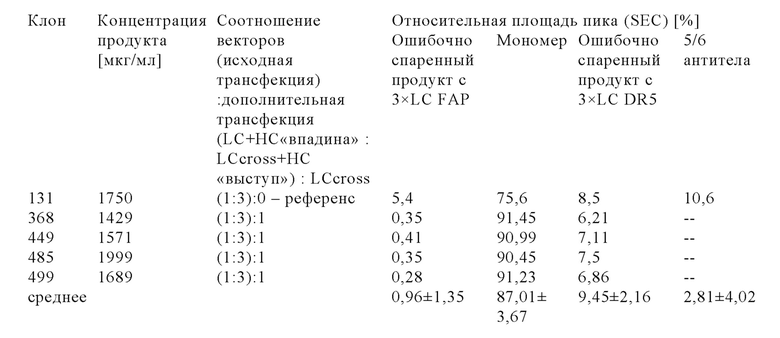
Результаты, полученные с помощью КЭ-ДСН (231=5/6 антитела; 242=мономер), представлены ниже в таблице и на фиг. 2.
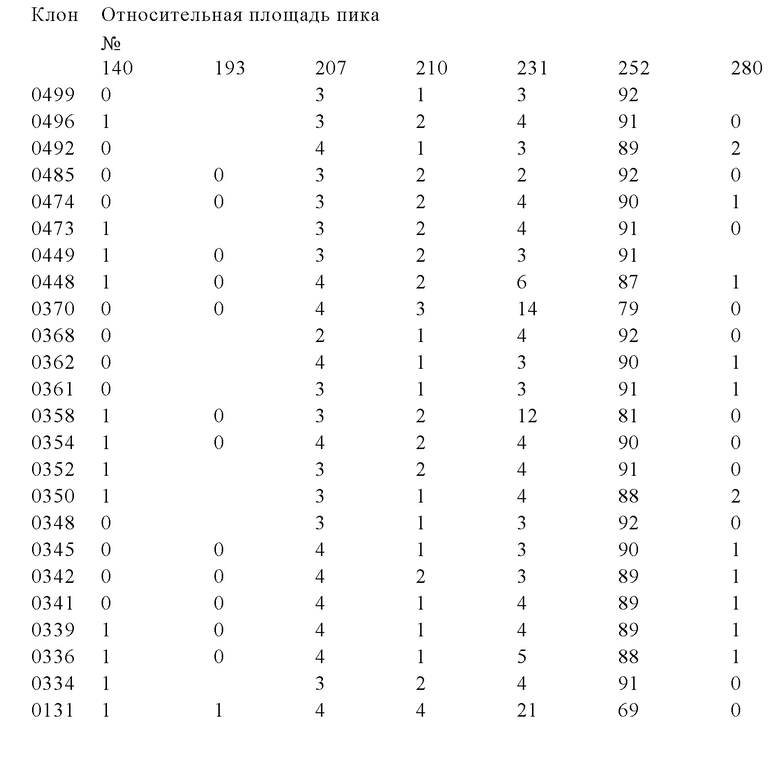
При применении указанного подхода можно получать линию клеток-продуцентов, которая продуцирует гетеродимерное антитело с улучшенным профилем продуктов, т.е. с повышенным количеством продукта и пониженным количеством родственных продукту примесей.
Общие методы рекомбинации и композиции, применяемые для получения антител
Антитела можно получать, используя методы рекомбинации и композиции, например, описанные в US №4816567. При осуществлении этих методов получают одну или несколько выделенную(ых) нуклеиновую(ых) кислоту(т), кодирующую(ие) антитело.
В случае нативного антитела или фрагмента нативного антитела требуется две нуклеиновые кислоты, она для легкой цепи или ее фрагмента и одна для тяжелой цепи или ее фрагмента. Указанная(ые) нуклеиновая(ые) кислота(ы) кодирует(ют) аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкой(их) и/или тяжелой(ых) цепи(ей) антитела). Эти нуклеиновые кислоты могут находиться в одном экспрессионном векторе или в различных экспрессионных векторах.
В случае биспецифического антитела с гетеродимерными тяжелыми цепями требуется четыре нуклеиновые кислоты, одна для первой легкой цепи, одна для второй легкой цепи, которая содержит первый гетеромономерный полипептид Fc-области, одна для второй легкой цепи и одна для второй тяжелой цепи, которая содержит второй гетеромономерный полипептид Fc-области. Например, одна гетеродимерная тяжелая цепь содержит так называемые «мутации, приводящие к образованию «выступов»» (T366W и необязательно одна из S354C или Y349C), а другая содержит так называемые «мутации, приводящие к образованию «впадины» (T366S, L368A и Y407V и необязательно Y349C или S354C) (см., например, Carter Р. и др., Immunotechnol. 2, 1996, с. 73). Указанная(ые) нуклеиновая(ые) кислота(ы) кодирует(ют) аминокислотную последовательность, которая содержит первый VL, и/или аминокислотную последовательность, которая содержит первый VH, включающий первую гетеромономерную Fc-область, и/или аминокислотную последовательность, которая содержит второй VL, и/или аминокислотную последовательность, которая содержит второй VH, включающий вторую гетеромономерную Fc-область антитела (например, первую и/или вторую легкую и/или первую и/или вторую тяжелые цепи антитела). Указанные нуклеиновые кислоты могут находиться в одном экспрессионном векторе или в различных экспрессионных векторах, как правило, эти нуклеиновые кислоты локализованы в двух или трех экспрессионных векторах, т.е. один вектор может содержать более одной из указанных нуклеиновых кислот.Примерами таких биспецифических антител являются CrossMab и Т-клеточные биспецифические антитела.
Одним из вариантов осуществления изобретения являются выделенные нуклеиновые кислоты, кодирующие антитело, которое применяют в способах, представленных в настоящем описании.
Следующим вариантом осуществления изобретения является(ются) один или несколько векторов (например, экспрессионных векторов), который(ые) содержит(ат) указанную(ые) нуклеиновую(ые) кислоту(ы).
Следующим вариантом осуществления изобретения является клетка-хозяин, содержащая указанную(ые) нуклеиновую(ые) кислоту(ы).
В одном из указанных вариантов осуществления изобретения клетка-хозяин содержит (например, трансформирована):
- в случае нативного антитела или фрагмента нативного антитела:
(1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или
(2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела;
- в случае биспецифического антитела с гетеродимерными тяжелыми цепями:
(1) первый вектор, содержащий первую пару нуклеиновых кислот, которые кодируют аминокислотные последовательности одной из них, содержащей первый VL, и другой, содержащей первый VH антитела, и второй вектор, содержащий вторую пару нуклеиновых кислот, которые кодируют аминокислотные последовательности одной из них, содержащей второй VL, и другой, содержащей второй VH антитела, или
(2) первый вектор, содержащий первую нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую один из вариабельных доменов (предпочтительно вариабельный домен легкой цепи), второй вектор, содержащий пару нуклеиновых кислот, которые кодируют аминокислотные последовательности одной из них, содержащей вариабельный домен легкой цепи, и другой, содержащей вариабельный домен первой тяжелой цепи, и третий вектор, содержащий пару нуклеиновых кислот, которые кодируют аминокислотные последовательности одной из них, содержащей соответствующий вариабельный домен легкой цепи, отличный от присутствующего во втором векторе, и другой их них, содержащей вариабельный домен второй тяжелой цепи, или
(3) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую первый VL антитела, второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую первый VH антитела, третий вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую второй VL антитела, и четвертый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую второй VH антитела.
В одном из вариантов осуществления изобретения клетка-хозяин является эукариотической, например, представляет собой клетку яичника китайского хомячка (СНО) или лимфоидную клетку (например, Y0-, NS0-,Sp20-клетку). Одним из вариантов осуществления изобретения является способ получения анти-[[PRO]] антитела, где способ включает культивирование клетки-хозяина, которая содержит нуклеиновые кислоты, кодирующие антитело, описанное выше, в условиях, пригодных для экспрессии антитела, и необязательно выделение антитела из клетки-хозяина (или среды для культивирования клетки-хозяина).
Для рекомбинантного получения анти-[[PRO]] антитела нуклеиновые кислоты, кодирующие антитело, например, описанное выше, выделяют и встраивают в один или несколько векторов для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Указанные нуклеиновые кислоты легко можно выделять и секвенировать с использованием общепринятых процедур (например, путем использования олигонуклеотидных зондов, которые обладают способностью специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела), или получать методами рекомбинации, или получать с помощью химического синтеза.
Приемлемые клетки-хозяева для клонирования или экспрессии кодирующих антитело векторов включают прокариотические или эукариотические клетки, представленные в настоящем описании. Например, антитела можно получать в бактериях, в частности, когда не требуется гликозилирование и связанная с Fc эффекторная функция. Сведения об экспрессии фрагментов антител и полипептидов в бактериях см. например, в US №5648237, US №5789199 и US №5840523 (см. также у Charlton K.А. в: Methods in Molecular Biology, под ред. Lo В.K.С, изд-во Humana Press, Totowa, NJ, т. 248, 2003, сс. 245-254 описание экспрессии фрагментов антител в Е. coli.) После экспрессии антитело можно выделять из пасты бактериальных клеток в виде растворимой фракции и можно дополнительно очищать.
Помимо прокариотических организмов в качестве хозяев, пригодных для клонирования или экспрессии векторов, которые кодируют антитела, можно использовать эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования которых были «гуманизированы», что позволяет получать антитело с частично или полностью человеческой схемой гликозилирования (см. Gerngross T.U., Nat. Biotech. 22, 2004, сс. 1409-1414 и Li Н. и др., Nat. Biotech. 24, 2006, сс. 210-215).
Клетки-хозяева, которые можно использовать для экспрессии гликозилированного антитела, получают также из многоклеточных организмов (беспозвоночных и позвоночных животных). Примерами клеток беспозвоночных являются клетки насекомых, а также можно применять клетки растений. Были выявлены многочисленные штаммы бакуловирусов и соответствующие пригодные для них в качестве хозяев клетки насекомых, прежде всего для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев можно применять также культуры растительных клеток (см., например, US №5959177, US №6040498, US №6420548, US №7125978 и US №6417429 (описание технологии PLANTIBODIES™ для получения антител в трансгенных растениях)).
В качестве хозяев можно применять также клетки позвоночных. Например, можно использовать клеточные линии млекопитающих, которые адаптированы к росту в суспензии. Другими примерами приемлемых линий клеток-хозяев млекопитающих являются линия клеток почки обезьяны CV1, трансформированная с помощью ОВ40 (COS-7); линия клеток почки эмбриона человека (НЕК 293-клетки или клетки линии 293, описанные, например, у Graham F.L. и др., J. Gen. Virol., 36, 1977, сс.59-74); клетки почки детеныша хомяка (ВНК); клетки Сертоли мыши (ТМ4-клетки, описанные, например, у Mather J.P., Biol. Reprod., 23, 1980, сс. 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76,); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени бычьей крысы (BRL 3А); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, описанные, например, у Mather J.Р. и др., Annals N.Y. Acad. Sci., 383, 1982, сс. 44-68); клетки MRC 5 и клетки FS4. Другими ценными линиями клеток-хозяев млекопитающих являются клетки яичника китайского хомячка (СНО), включая DHFR-CHO-клетки (Urlaub G. и др., Proc. Natl. Acad. Sci. USA, 77, 1980, сс. 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор конкретных линий клеток-хозяев млекопитающих, которые можно применять для производства антител, см., например, у Yazaki Р. и Wu A.M., в: Methods in Molecular Biology под ред. В.K.C. Lo, изд-во Humana Press, Totowa, NJ, т. 248, 2003, сс. 255-268.
Краткое описание чертежей
На чертежах показано:
на фиг.1 - результаты общей ионной ESI-MC после
дегликозилирования антител для следующих вариантов:
(A) трансфекция 0516;
(Б) трансфекция 0517;
(B) трансфекция 0518;
(Г) трансфекция 0519;
А: вторая тяжелая цепь с мутациями, приводящими к образованию
«выступа» (CrossHC-2-«выступ»), IgG1-подкласса; Б: первая тяжелая цепь с мутациями, приводящими к образованию «впадины» (НС-1-«впадина»), IgG1-подкласса; В: вторая легкая цепь с обменом доменов (CrossLC-2), Г: первая легкая цепь (LC-1);
на фиг. 2 - относительное содержание мономера (А) и побочного продукта в виде 5/6 антитела (Б) при применении референс-клона 0131 и клонов, полученных с помощью способа, представленного в настоящем описании, по данным, полученным с помощью КЭ-ДСН.
Примеры
Ниже представлены примеры способов и композиций, предлагаемых в изобретении. Как должно быть очевидно, можно воплощать на практике различные другие варианты осуществления изобретения с учетом представленного выше описания изобретения в целом.
Материалы и общие методы
Общую информацию, касающуюся нуклеотидных последовательностей легких и тяжелых цепей человеческих иммуноглобулинов, см. у: Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-е изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991. Аминокислоты цепей антител нумеровали и обозначали в соответствии с системой нумерации Кэбота (Kabat Е.А. и др., Sequences of Proteins of Immunological Interest, 5-е изд., изд-во Public Health Service, National Institutes of Health, Bethesda, MD, 1991)
Методы рекомбинантной ДНК
Для манипуляций с ДНК использовали стандартные методы, описанные у Sambrook J. и др., Molecular cloning: A laboratory manual; изд-во Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Реагенты для молекулярной биологии применяли согласно инструкциям производителей.
Синтез генов
Требуемые сегменты генов создавали из олигонуклеотидов, полученных с помощью химического синтеза. Длинные сегменты генов, фланкированные единичными сайтами, распознаваемыми рестрикционными эндонуклеазами, собирали путем отжига и лигирования олигонуклеотидов, включая ПЦР-амплификацию и последующее клонирование через указанные сайты рестрикции. Последовательности ДНК субклонированных фрагментов генов подтверждали ДНК-секвенированием. Синтез фрагментов генов заказывали в соответствии с данными спецификациями на фирме Geneart (Регенсбург, Германия).
Определение последовательности ДНК
Последовательности ДНК определяли путем секвенирования двух цепей, которое осуществляли на фирме MediGenomix GmbH (Мартинсрид, Германия) или на фирме SequiServe GmbH (Фатерштеттен, Германия).
Анализ последовательностей ДНК и белков и обработка данных о последовательностях
Пакет программ GCG (фирма Genetics Computer Group, Мэдисон, шт. Висконсин), версия 10.2 и Infomax's Vector NT1 Advance suite, версия 8.0 применяли для создания, картирования, анализа, аннотирования и иллюстрации последовательностей.
Экспрессионные векторы
Для экспрессии описанных биспецифических антител можно применять экспрессионные векторы для кратковременной экспрессии (например, в HEK293-клетках), основанные на структуре кДНК с промотором CMV, содержащим или несодержащим интрон А, или на геномной структуре с промотором CMV.
Помимо кассеты экспрессии для антитела векторы содержат:
- сайт инициации репликации, который обеспечивает репликацию этого вектора в Е. coli, и
- ген β-лактамазы, который придает устойчивость к ампициллину Е. coli.
Транскрипционная единица гена антитела состоит из следующих элементов:
- уникальный(ые) сайт(ы) рестрикции на 5'-конце,
- немедленно-ранний энхансер и промотор из человеческого цитомегаловируса,
- последовательность интрона А в случае структуры на основе кДНК,
- 5'-нетранслируемая область из гена человеческого антитела, сигнальная последовательность тяжелой цепи иммуноглобулина,
- нуклеиновая кислота, кодирующая соответствующую цепь антитела либо в виде структуры кДНК, либо геномной экзон-интронной структурой,
- 3'-нетранслируемая область с последовательностью сигнала полиаденилирования и
- уникальный(ые) сайт(ы) рестрикции на 3'-конце.
Слитые гены, кодирующие цепи антител, создавали с помощью ПЦР и/или синтеза генов и собирали с использованием известных методов и технологий рекомбинации путем соединения сегментов нуклеиновых кислот, например, используя уникальные сайты рестрикции в соответствующих векторах. Последовательности субклонированных нуклеиновых кислот подтверждали секвенированием ДНК. Для кратковременных трансфекций получали большие количества векторов с использованием препаратов векторов из трансформированных культур Е. coli (фирмы Nucleobond АХ, Macherey-Nagel).
Для всех конструкций применяли технологию гетеродимеризации «выступ-во-впадину» с типичной заменой, приводящей к образованию «выступа» (T366W) в первом СН3-домене, и соответствующими заменами, приводящими к образованию «впадины» (T366S, L368A и Y407V) во втором СН3-домене (а также с двумя интродуцированными остатками цистеина S354C/Y349'C) (которые находились в соответствующих последовательностях тяжелых цепей (НС), указанных выше).
Методики культивирования клеток
Применяли стандартные методики культивирования клеток, описанные в: Current Protocols in Cell Biology, под ред. Bonifacino J.S., Dasso M., Harford J.В., Lippincott-Schwartz J. и Yamada K.M., изд-во John Wiley & Sons, Inc., 2000.
Кратковременные трансфекции в системе HEK293-F
Биспецифические антитела получали путем кратковременной экспрессии. Для этой цели осуществляли трансфекцию соответствующими векторами, используя систему HEK293-F (фирма Invitrogen) согласно инструкции производителя. В целом, метод состоял в следующем: клетки HEK293-F (фирма Invitrogen), растущие в суспензии либо во встряхиваемой колбе, либо в ферментере с перемешиванием в бессывороточной среде для экспрессии FreeStyle™ 293 (фирма Invitrogen), трансфектировали смесью соответствующих экспрессионных векторов и 293fectin™ или фектина (фирма Invitrogen). При применении 2-литровой встряхиваемой колбы (фирма Corning) клетки HEK293-F высевали с плотностью 1,0×106 клеток/мл в 600 мл и инкубировали при 120 об/мин в атмосфере с 8% СО2. На следующий день клетки трансфектировали при плотности клеток примерно 1,5×106 клеток/мл, используя примерно 42 мл смеси А) 20 мл среды Opti-MEM (фирма Invitrogen), содержащей 600 мкг общей векторной ДНК (1 мкг/мл), и Б) 20 мл среды Opti-MEM, дополненной 1,2 мл 293fectin™ или фектина (2 мкл/мл). Из-за того, что имело место поглощение глюкозы, в процессе ферментации добавляли раствор глюкозы. Супернатант, содержащий секретированное антитело, собирали через 5-10 дней и антитела либо очищали непосредственно из супернатанта, либо супернатант замораживали и хранили. Определение белков
Концентрацию белка в очищенных антителах и производных определяли, измеряя оптическую плотность (ОП) при 280 нм, используя коэффициент молярной экстинкции, рассчитанный на основе аминокислотной последовательности, согласно Расе и др., Protein Science 4, 1995, сс. 2411-1423. Определение концентрации антител в супернатантах Концентрацию антител и их производных в супернатантах клеточных культур определяли с помощью иммунопреципитации с использованием белок А-агарозных гранул (фирма Roche Diagnostics GmbH, Маннгейм, Германия). Для этого 60 мкл белок А-агарозных гранул промывали трижды в TBS-NP40 (50 мМ Трис-буфер, рН 7,5, дополненный 150 мМ NaCl и 1% Nonidet-P40). Затем 1-15 мл супернатанта клеточной культуры наносили на белок А-агарозные гранулы, предварительно уравновешенные в TBS-NP40. После инкубации в течение 1 ч при комнатной температуре гранулы промывали на фильтрующей колонке Ultrafree-MC (фирма Amicon), однократно с использованием 0,5 мл TBS-NP40, дважды с использованием 0,5 мл 2-кратного забуференного фосфатом физиологического раствора (2×ЗФР, фирма Roche Diagnostics GmbH, Маннгейм, Германия) и кратковременно 4 раза с использованием 0,5 мл 100 мМ Na-цитратного буфера (рН 5,0). Связанное антитело элюировали, добавляя 35 мкл буфера для образца NuPAGE® LDS (фирма Invitrogen). Половину образца объединяли с восстановителем для образца NuPAGE® или оставляли без восстановления соответственно, и выдерживали в течение 10 мин при 70°С. Затем 5-30 мкл вносили в 4-12% NuPAGE® Бис-Трис-гель для ДСН-ПААГ (фирма Invitrogen) (с MOPS-буфером для ДСН-ПААГ без восстановителя и MES-буфером в качестве подвижного буфера для NuPAGE® с антиоксидантной добавкой (фирма Invitrogen) для ДСН-ПААГ с применением восстановителя) и окрашивали кумасси голубым.
Концентрацию антител в супернатантах клеточных культур оценивали количественно с помощью аффинной ЖХВР-хроматографии. В целом, метод стоял в следующем: супернатанты клеточных культур, содержащие антитела, которые связывались с белком А, наносили на колонку Applied Biosystems Poros А/20 в буфере, содержащем 200 мМ KH2PO4, 100 мМ цитрат натрия, рН 7,4, и элюировали с помощью буфера, содержащего 200 мМ NaCl, 100 мМ лимонную кислоту, рН 2,5, в системе Agilent HPLC 1100. Количественно оценивали концентрацию элюированного антитела с использованием УФ-абсорбции и интегрирования площадей пиков. Очищенное стандартное антитело IgG1-подкласса служило в качестве стандарта.
Альтернативно этому, концентрацию антител и их производных в супернатантах клеточных культур определяли с помощью сэндвич-IgG-ELISA. В целом, метод состоял в следующем: 96-луночные титрационные микропланшеты, отличающийся высокой способностью связываться со стрептавидином (StreptaWell High Bind Streptavidin А) (фирма Roche Diagnostics GmbH, Маннгейм, Германия), сенсибилизировали, используя 100 мкл/лунку молекулы биотинилированного захватывающего антитела к человеческому IgG F(ab')2<h-Fcγ>BI (фирма Dianova) в концентрации 0,1 мкг/мл, в течение 1 ч при комнатной температуре или в альтернативном варианте в течение ночи при 4°С и затем промывали трижды, используя 200 мкл/лунку ЗФР, 0,05% Твин (ЗФР-Т, фирма Sigma). Затем добавляли в лунки из расчета 100 мкл/лунку серийные разведения в ЗФР (фирма Sigma) содержащих соответствующее антитело супернатантов клеточных культур и инкубировали в течение 1-2 ч на шейкере при комнатной температуре. Лунки промывали трижды, используя 200 мкл/лунку ЗФР-Т, и осуществляли детекцию связанного антитела с помощью 100 мкл F(ab')2<hFcγ>POD (фирма Dianova) в концентрации 0,1 мкг/мл в качестве идентифицирующего антитела, путем инкубации в течение 1-2 ч на шейкере при комнатной температуре. Несвязанное идентифицирующее антитело удаляли путем трехкратной промывки с использованием 200 мкл/лунку ЗФР-Т. Связанное идентифицирующее антитело выявляли путем добавления 100 мкл ABTS/лунку с последующей инкубацией. Определение абсорбции осуществляли с помощью спектрометра Tecan Fluor, осуществляя измерения при длине волны 405 нм (длина референс-волны 492 нм).
Препаративная очистка антител
Антитела очищали из профильтрованных супернатантов клеточных культур согласно стандартному протоколу. В целом, метод состоял в следующем: антитела вносили в колонку с белок А-Сефарозой (фирма GE Healthcare) и промывали с помощью ЗФР. Элюцию белков осуществляли при значении рН 2,8, после чего немедленно проводили нейтрализацию образца. Агрегированные белки отделяли от мономерных антител с помощью гель-фильтрации (Супердекс 200, фирма GE Healthcare) в ЗФР или в 20 мМ гистидиновом буфере, содержащем 150 мМ NaCl (рН 6,0). Фракции мономерных антител объединяли, концентрировали (при необходимости), используя, например, центрифужный концентратор MILLIPORE Amicon Ultra (30 MWCO), замораживали и хранили при температуре -20°С или -80°С. Часть образцов использовали для последующего аналитического анализа белков и аналитической характеризации, например, с помощью ДСН-ПААГ, гель-фильтрации (SEC) или масс-спектрометрии.
ДСН-ПААГ
Гелевую систему NuPAGE® Pre-Cast (фирма Invitrogen) применяли согласно инструкции производителя. В частности, применяли 10% или 4-12% гели NuPAGE® Novex® Bis-TRIS Pre-Cast (рН 6,4) и подвижный буфер NuPAGE® MES (восстановленные гели с антиоксидантной добавкой для подвижного буфера NuPAGE®) или MOPS (невосстановленные гели).
КЭ-ДСН
Чистоту и целостность антител анализировали с помощью КЭ-ДСН с использованием микрофлюидной технологии Labchip (фирма PerkinElmer, США). Для этого 5 мкл раствора антитела подготавливали для КЭ-ДСН-анализа, используя набор реагентов для экспрессии белков НТ (Protein Express Reagent Kit) согласно инструкциям производителя, и анализировали с помощью системы LabChip GXII с использованием чипа НТ Protein Express. Данные анализировали с помощью программного обеспечения LabChip GX.
Аналитическая гель-фильтрация
Гель-фильтрацию (SEC) для определения агрегированного и олигомерного состояния антител осуществляли с помощью ЖХВР-хроматографии. В целом, метод состоял в следующем: очищенные на белке А антитела вносили в колонку Tosoh TSKgel G3000SW в буфере, содержащем 300 мМ NaCl, 50 мМ KH2PO4/K2HPO4 (рН 7,5), в системе Dionex Ultimate® (фирма Thermo Fischer Scientific) или в колонку Супердекс 200 (фирма GE Healthcare) в 2 × ЗФР в системе Dionex HPLC. Элюированные белки оценивали количественно с на основе УФ-абсорбции и интегрирования площадей пиков. В качестве стандарта служил BioRad Gel Filtration Standard 151-1901.
Масс-спектрометрия
В этом разделе описана характеризация биспецифических антител в отношении их правильной сборки. Ожидаемые первичные структуры анализировали с помощью масс-спектрометрии с использованием ионизация электроспреем (ESI-MC) дегликозилированного интактного антитела и в специальных вариантах дегликозилированного/ограничено расщепленного LysC антитела.
Антитела дегликозилировали с помощью N-гликозидазы F в фосфатном или Трис-буфере при 37°С в течение вплоть до 17 ч с использованием белка в концентрации 1 мг/мл. Ограниченные расщепления LysC (фирма Roche Diagnostics GmbH, Маннгейм, Германия) осуществляли с использованием 100 мкг дегликозилированного антитела в Трис-буфере (рН 8) при комнатной температуре в течение 120 ч или при 37°С в течение 40 мин соответственно. Перед осуществлением масс-спектрометрии образцы обессоливали посредством ЖХВР на колонке Сефадекс G25 (фирма GE Healthcare). Общую массу определяли с помощью ESI-MC с использованием МС-системы maXis 4G UHR-QTOF (фирма Bruker Daltonik), снабженной источником TriVersa NanoMate (фирма Advion).
Пример 1
Экспрессия и очистка
Биспецифические антитела получали согласно методу, описанному выше в разделе «Материалы и общие методы».
Биспецифические антитела очищали из супернатанта путем комбинации аффинной хроматографии на белке А и гель-фильтрации. Полученные продукты характеризовали в отношении их идентичности с помощью масс-спектрометрии и в отношении аналитических свойств, таких как чистота по данным КЭ-ДСН, содержание мономера и стабильность.
Ожидаемые первичные структуры анализировали с помощью масс-спектрометрии с использованием ионизация электроспреем (ESI-MC) дегликозилированного интактного антитела и дегликозилированного/ расщепленного плазмином или в альтернативном варианте дегликозилированного/ограничено расщепленного LysC антитела согласно методу, описанному в разделе «Общие методы».
Дополнительные аналитические методы (например, методы оценки термостабильности, масс-спектрометрию и функциональную оценку) применяли только после очистки с помощью белка А и SEC.
Пример 2
Определение связывания с фибриллами Аβ 1-40 in vitro с помощью ELISA Связывание биспецифических антител с фибриллярным Аβ оценивали с помощью анализа ELISA. В целом, метод состоял в следующем: планшеты Maxisorb сенсибилизировали с использованием Аβ (1-40) в концентрации 7 мкг/мл в ЗФР в течение 3 дней при 37°С с получением фибриллярного белка Абета, а затем сушили в течение 3 ч при КТ. Планшет блокировали с помощью 1% CroteinC и 0,1% RSA (крысиный сывороточный альбумин) в ЗФР (блокирующий буфер) в течение 1 ч при КТ, затем однократно промывали буфером для промывки. Биспецифические антитела или контроли добавляли в концентрациях вплоть до 100 нМ в блокирующем буфере и инкубировали при 4°С в течение ночи. После четырех стадий промывки конструкции выявляли, добавляя античеловеческий-IgG-HRP (фирма Jackson Immunoresearch) в разведении 1:10000 в блокирующем буфере (1 ч, КТ), после чего осуществляли 6 промывок и инкубировали в ТМВ (фирма Sigma). Абсорбцию определяли при 450 ни после прекращения цветовой реакции с помощью 1н. HCl.
Пример 3
Определение связывания с рецептором трансферрина in vitro Связывание биспецифических антител с рецептором мышиного трансферрина оценивали с помощью FACS-анализа с использованием клеток мышиной миеломы линии X63.AG8-563. Если в случае антител к Аβ установлена определенная тенденция к неспецифическому связыванию с Ag8-клетками, то специфическое связывание можно оценивать количественно путем совместной инкубации с 20-кратным избытком антитела к мышиному TfR. Клетки собирали путем центрифугирования, промывали однократно ЗФР и 5×104 клеток инкубировали с серийными разведениями от 1,5пМ до 10нМ полипептидных слияний с добавлением или без добавления 200нМ антитела к мышиному TfR в 100 мкл RPMI/10% FCS в течение 1,5 ч на льду. После двух промывок RPMI/10% FCS клетки инкубировали с козьим античеловеческим IgG, сшитым с фикоэритрином (фирма Jackson Immunoresearch), в разведении 1:600 в RPMI/19% FCS в течение 1,5 ч на льду. Клетки вновь промывали, ресуспендировали в RPMI/10% FCS и измеряли флуоресценцию фикоэритрина с помощью устройства FACS-Array (фирма Becton-Dickinson). Пример 4
Анализ связывания с помощью поверхностного плазмонного резонанса для оценки взаимодействия человеческий TfR-антитело
Эксперимент по связыванию осуществляли с использованием устройства BIAcore В 4000 (фирма GE Healthcare), снабженного сенсорным чипом C1 (фирма GE Healthcare, каталожный №. BR1005-35), предварительно обработанным антителом к человеческому Fab (фирма GE Healthcare, каталожный №28-9583-25), используя стандартную химию на основе аминного сочетания согласно руководству поставщика.
Для кинетических измерений образец антитела иммобилизовывали, осуществляя контакт в течение 60 с, и используя скорость потока 10 мкл/мин в забуференном фосфатом физиологическом растворе (рН 7,4), дополненном 0,05% Твин 20, при 25°С. Рекомбинантный меченный His6 рецептор человеческого трансферрина (фирма R&D systems, каталожный №2474-TR-050) наносили в возрастающих концентрациях и осуществляли мониторинг сигнала в течение времени. Установлено, что при скорости потока 30 мкл/мин средняя продолжительность времени ассоциации составляла 150 с, а времени диссоциации 600 с. Данные аппроксимировали с использование модели связывания 1:1 (изотерма Лэнгмюра).
Пример 5
Окрашивание нативных человеческих β-амилоидных бляшек в срезах головного мозга пациента с болезнью Альцгеймера с помощью непрямой иммунофлуоресценции с использованием биспецифического антитела, полученного с помощью способа, представленного в настоящем описании
Можно тестировать способность биспецифических антител окрашивать нативные человеческие β-амилоидные бляшки с помощью иммуногистохимического анализа с использованием непрямой иммунофлуоресценции. Удалось продемонстрировать специфическое и чувствительное окрашивание подлинных человеческих β-амилоидных бляшек. Криостатные срезы нефиксированной ткани из височной коры головного мозга, полученной после смерти пациентов с диагностированной болезнью Альцгеймера, метили для непрямой иммунофлуоресценции. Применяли двухстадийную инкубацию для обнаружения связанного биспецифического антитела, которое выявляли с помощью очищенного аффинной хроматографией козьего античеловеческого (GAH555) IgG (H+L), конъюгированного с красителем Alexa 555 (фирма Molecular Probes). Контроли включали неродственные человеческие антитела IgG1-подкласса (фирма Sigma) и индивидуально вторичное антитело, которые все дали отрицательные результаты.
Пример 6
In vivo выявление β-амилоидных бляшек биспецифическим антителом, полученным способом, представленным в настоящем описании, на мышиной модели болезни Альцгеймера
Биспецифическое антитело можно тестировать на двойной трансгенной мышиной модели APP/PS2, мышиной модели родственного AD амилоидоза (Richards J. Neuroscience, 23, 2003, сс. 8989-9003), в отношении способности осуществлять иммунное декорирование (выявление) β-амилоидных бляшек in vivo. Это позволяет оценивать степень проникновения в головной мозг и связывание с бляшками амилоида-β. Слитый полипептид можно вводить в различных дозах в сравнении с «голым» моноклональным антителом к Аβ и через 6 дней осуществлять перфузию животных забуренным фосфатом физиологическим раствором, замораживать образцы головного мозга на сухом льду и подготавливать для получения криосрезов.
Присутствие антител, связанных с β-амилоидными бляшками, можно оценивать с использованием нефиксированных криостатных срезов с помощью метода непрямой иммунофлуоресценции с одной меткой с применением козьего античеловеческого IgG (H+L), конъюгированного с красителем Alexa555 (GAH555) (фирма Molecular Probes), в концентрации 15 мкг/мл в течение 1 ч при комнатной температуре. Контрастное окрашивание амилоидных бляшек можно осуществлять путем инкубации с ВАР-2, мышиным моноклональным антителом против Аβ, конъюгированным с Alexa 488, в концентрации 0,5 мкг/мл в течение 1 ч при комнатной температуре. Микроскопические препараты заливали флуоресцентной поддерживающей средой (S3023, фирма Dako) и визуализировали с помощью конфокального лазерного микроскопа.
Хотя выше описаны некоторые детали изобретения, приведенные с целью иллюстрации и примера для простоты понимания, описание и примеры не должны рассматриваться как ограничивающие объем изобретения. Описание всей процитированной патентной и научной литературы специально полностью включены в настоящее описание в качестве ссылки.
Пример 7
Трансфекция стабильной клеточной линии, экспрессирующей биспецифическое анти-DR5/FAP антитело, экспрессионным вектором, содержащим кассету экспрессии для легкой цепи с обменом доменами
Клон клеток 0131 трансфектировали экспрессионным вектором для CrossLC, содержащим одну кассету экспрессии для легкой цепи с кроссовером доменов. Трансфекции осуществляли с использованием линеаризированной ДНК в среде определенного химического состава с использованием нуклеофекции (фирма Amaxa) и 0,6/1,2/2,4пМ (общей) плазмиды, с получением 2×3 клеточных пулов, которые подвергали дифференцированной селекции.
Пулы трансфектированных клонов отбирали в среде определенного химического состава, дополненной глутамином в концентрации 10 ммолей/л и 250нМ МТХ (для DHFR) плюс 500нМ и 700нМ гигромицином В. Через 3 недели пулы анализировали с помощью КЭ-ДСН и иммуногистохимии (ИГХ) в отношении уменьшения побочных пиков и повышения основного пика.
На основе полученных результатов для серийного разведения (LD) отобрано три пула, для селекции которых использовали 250нМ МТХ и 700нМ HygB (0314, 0316, 0318), и каждый из них высевали в 3 х 384-луночных планшета со средой определенного химического состава, дополненной глутамином в концентрации 10 ммолей/л и МТХ в концентрации 250 нмолей/л и 700нМ HygB.
Через 1 неделю супернатанты, полученные при использовании 3 × 384-луночных планшетов, тестировали в отношении связывания с DR5 и FAP с помощью методов ELISA и ELISA с (двойным) DR5-FAP-связыванием. Отобрано 158 клонов с высокими титрами и высокой реактивностью в отношении обоих антигенов и их размножали, начиная с использования 24-луночных планшетов, доходя до 6 лунок, в которых их оценивали в виде полученной в эксперименте четырехдневной партии («оценивали титры последовательно размножаемой культуры») в отношении связывания с мишенями с помощью ELISA и ELISA с двойным связыванием, рост, продуктивность и профиль побочных продуктов оценивали с помощью КЭ-ДСН. Из них 46 клонов с титрами, составляющими вплоть до 830 мкг/мл, и приемлемым качеством продукта дополнительно характеризовали в 14-дневных культурах с подпиткой с использованием системы Ambr15 и анализировали в отношении связывания с мишенями, особенностей роста и профиля побочных продуктов с помощью КЭ-ДСН и ИГХ. Из них 20 клонов отбирали и дополнительно оценивали с помощью масс-спектрометрии (МС). Из них 10 отобранных клонов культивировали во встряхиваемых колбах в среде определенного химического состава и хранили в виде PSB (бульон, содержащий пептон, сорбит и желчные соли).
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2780629C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2573914C2 |
| ТРИ- ИЛИ ТЕТРАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2010 |
|
RU2570633C2 |
| БИСПЕЦИФИЧЕСКИЕ ЧЕТЫРЕХВАЛЕНТНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ БЕЛКИ | 2010 |
|
RU2604189C2 |
| НОВЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ СПЕЦИФИЧЕСКИ СВЯЗЫВАТЬСЯ С CD40 И FAP | 2018 |
|
RU2766234C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |
| СПОСОБ ПОЛУЧЕНИЯ И ОТБОРА МОЛЕКУЛ, ВКЛЮЧАЮЩИХ ПО МЕНЬШЕЙ МЕРЕ ДВЕ РАЗЛИЧНЫЕ ГРУППИРОВКИ, И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2646159C2 |
| УСТОЙЧИВЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2018 |
|
RU2779602C2 |
| ДВУХВАЛЕНТНЫЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2008 |
|
RU2547615C2 |
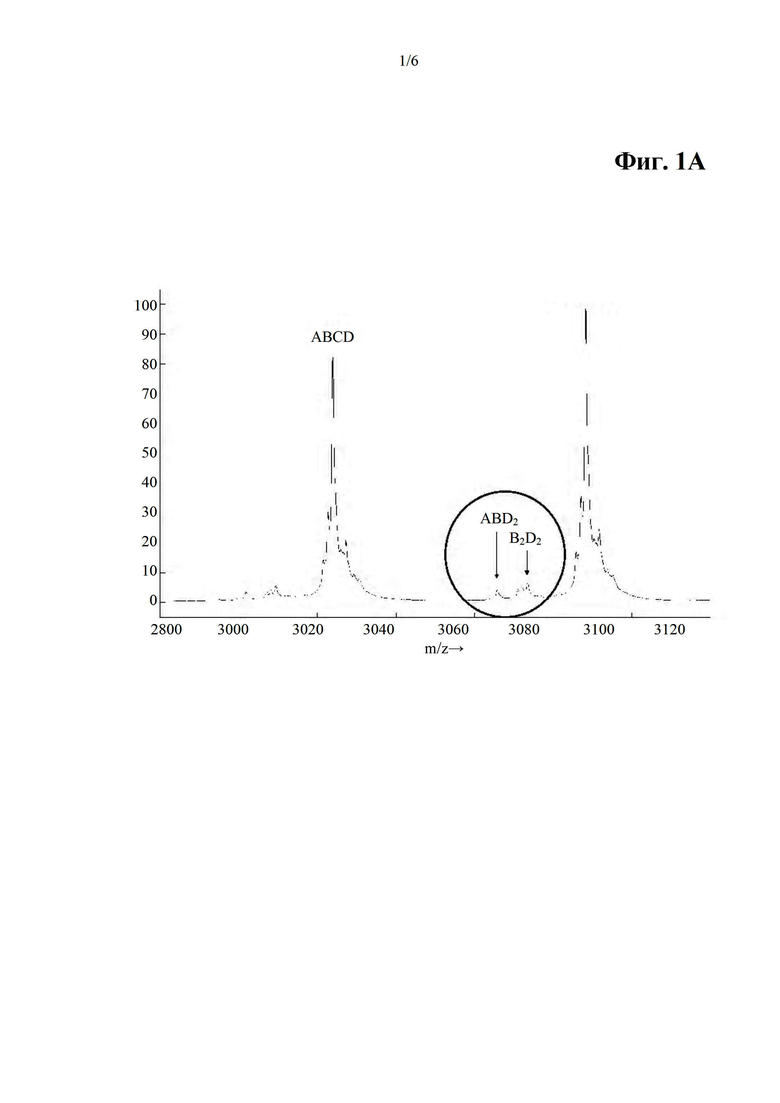
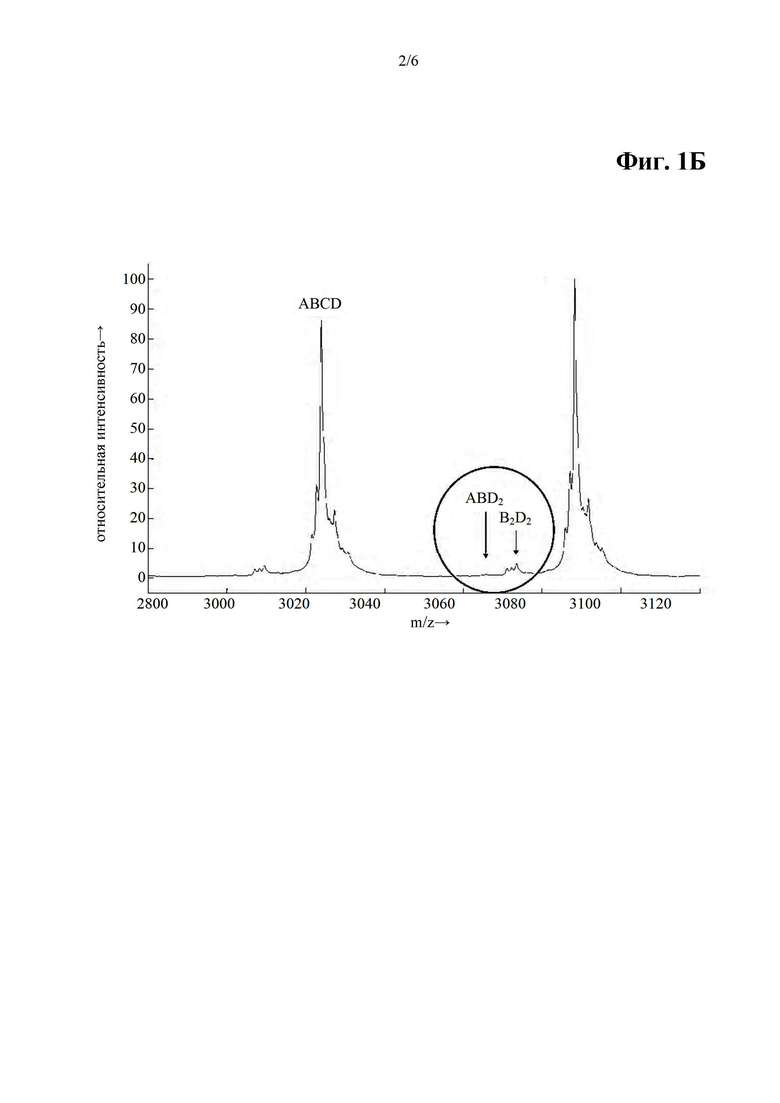
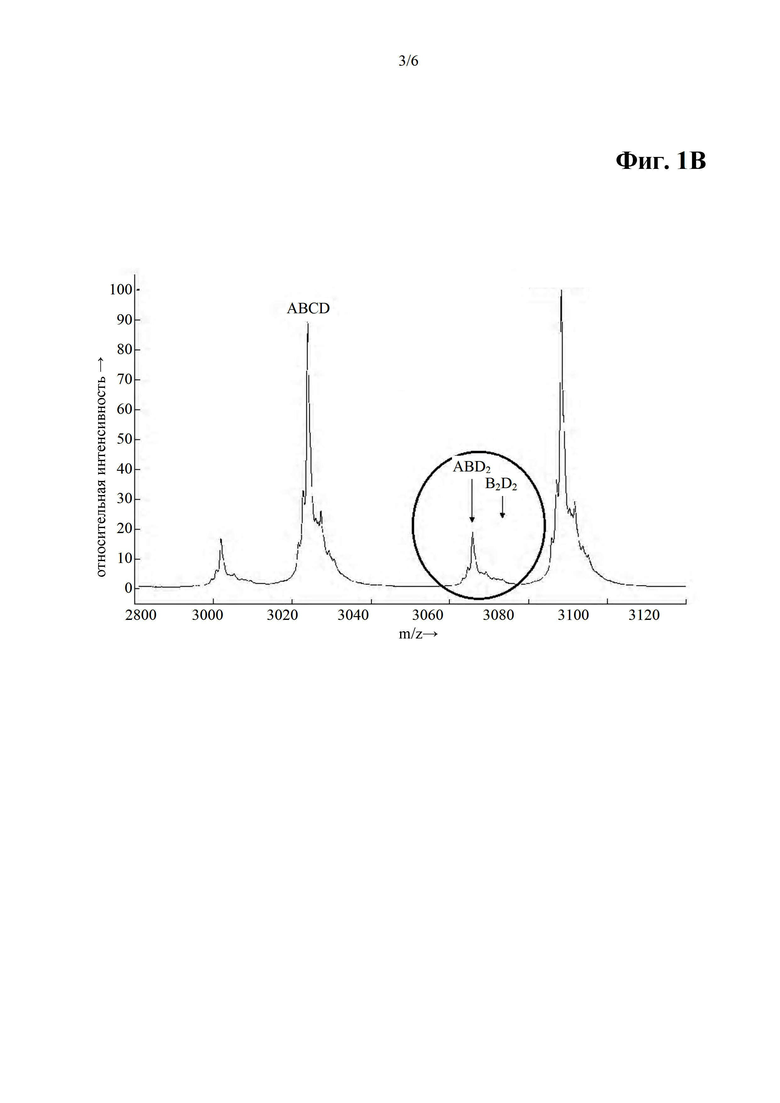
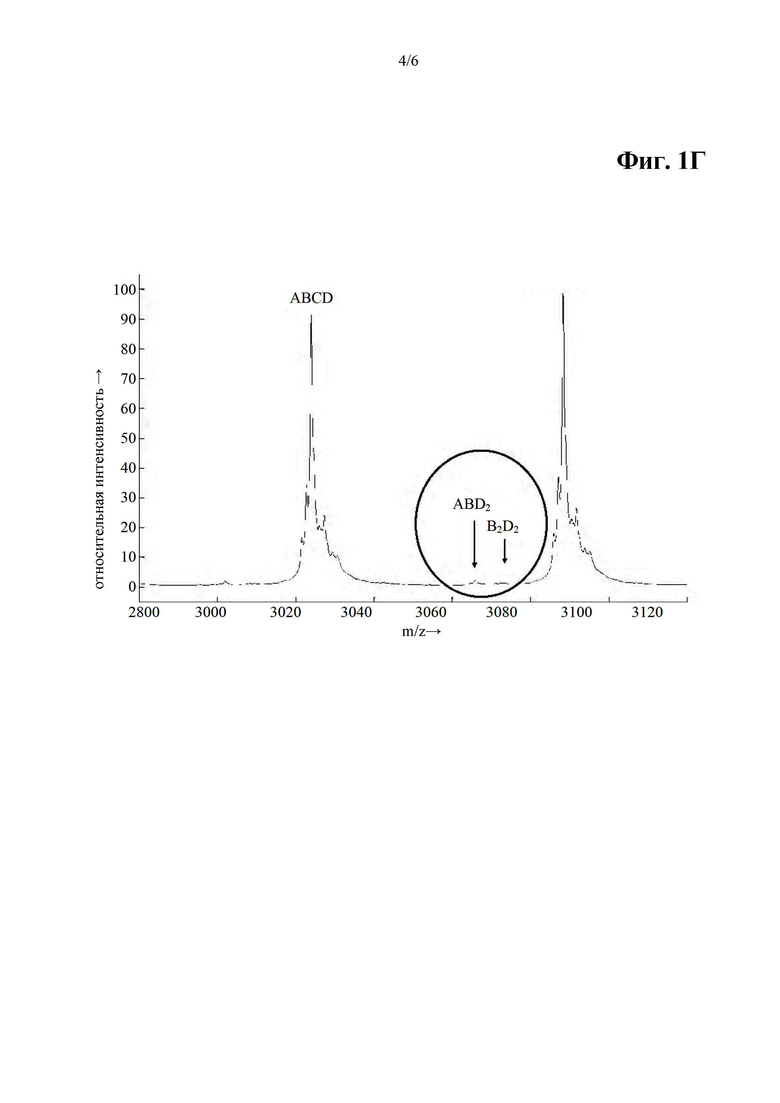
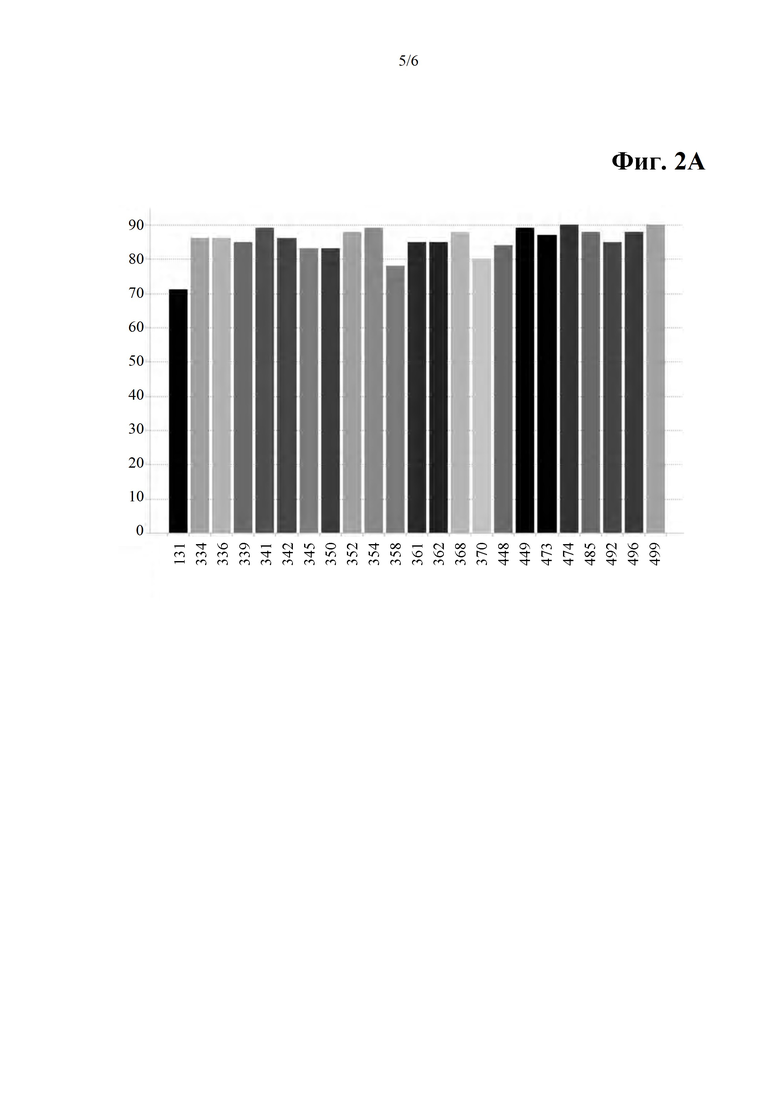
Настоящее изобретение относится к области иммунологии. Предложен способ получения мультиспецифического антитела. Настоящее изобретение может найти дальнейшее применение в продуцировании гетеродимерных антител с улучшенным профилем продукта. 11 з.п. ф-лы, 6 ил., 7 пр.
1. Способ получения мультиспецифического антитела, которое относится к подклассу человеческого IgG1, где мультиспецифическое антитело включает:
- первую легкую цепь,
- первую тяжелую цепь, содержащую первый гетеромономерный полипептид Fc-области,
- вторую легкую цепь и
- вторую тяжелую цепь, содержащую второй гетеромономерный полипептид Fc -области,
который включает следующие стадии:
- культивирование клетки млекопитающего в среде для культивирования, где клетка млекопитающего создана путем
a) трансфекции клетки млекопитающего первым экспрессионным вектором и одним, двумя или тремя дополнительными экспрессионными векторами,
при этом первый экспрессионный вектор содержит только последовательность нуклеиновых кислот, кодирующую цепь мультиспецифического антитела, причем эта последовательность нуклеиновых кислот кодирует легкую цепь мультиспецифического антитела,
где один, два или три дополнительных экспрессионных вектора, каждый из которых содержит последовательности нуклеиновых кислот, кодирующие цепи мультиспецифического антитела, причем по меньшей мере две последовательности нуклеиновых кислот, каждая кодирует различные цепи мультиспецифического антитела,
- выделение мультиспецифического антитела из клетки или среды для культивирования и
- получение тем самым мультиспецифического антитела.
2. Способ по п. 1, в котором две цепи мультиспецифического антитела содержат обмен доменами.
3. Способ по п. 2, в котором нуклеиновая кислота первого экспрессионного вектора кодирует легкую цепь с обменом доменами мультиспецифического антитела.
4. Способ по одному из пп. 1-3, в котором стадия a) включает совместную трансфекцию первым экспрессионным вектором и одним, двумя или тремя дополнительными экспрессионными векторами.
5. Способ по одному из пп. 1-3, в котором клетку млекопитающего трансфектируют сначала одним, двумя или тремя дополнительными экспрессионными векторами и затем трансфектируют первым экспрессионным вектором.
6. Способ по одному из пп. 1-5, в котором клетка млекопитающего стабильно экспрессирует мультиспецифическое антитело.
7. Способ по одному из пп. 1-6, в котором клетка млекопитающего представляет собой CHO-клетку.
8. Способ по одному из пп. 2-7, в котором обмен доменами представляет собой CH1-CL-кроссовер или VH-VL-кроссовер.
9. Способ по одному из пп. 1-8, в котором мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменены друг на друга.
10. Способ по одному из пп. 1-8, в котором мультиспецифическое антитело представляет собой двухвалентное биспецифическое антитело, которое содержит
a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антигеном, и
б) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антигеном, в котором константные домены CL и CH1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
11. Способ по одному из пп. 1-8, в котором мультиспецифическое антитело представляет собой трехвалентное биспецифическое антитело, которое содержит
a) первую легкую цепь и первую тяжелую цепь полноразмерного антитела, которое специфически связывается с первым антигеном,
б) вторую тяжелую цепь полноразмерного антитела, которая при спаривании с первой легкой цепью специфически связывается с первым антигеном, и
в) Fab-фрагмент, который специфически связывается со вторым антигеном, слитый через пептидный линкер с C-концом одной из тяжелых цепей, указанных в подпункте а) или б), в котором константные домены CL и CH1 второй легкой цепи и второй тяжелой цепи заменены друг на друга.
12. Способ по одному из пп. 1-11, где способ предназначен для получения препарата мультиспецифического антитела с низким/пониженным количеством родственных продукту примесей.
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |

