ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к антителу к PD-L1 (лиганд 1 программируемой гибели) или к его антигенсвязывающему фрагменту, к кодирующей его нуклеиновой кислоте, вектору, включающему данную нуклеиновую кислоту, к клетке, трансфицированной данным вектором, к способу получения данного антитела или его антигенсвязывающего фрагмента, к содержащей его композиции для предупреждения или лечения рака и к включающей его композиции для комбинированной терапии для предупреждения или лечения рака.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
PD-L1 представляет собой трансмембранный белок типа 1, имеющий два lg-подобных домена в пределах внеклеточной области, трансмембранный домен и короткий цитоплазматический домен. Цитоплазматический домен не имеет известного мотива трансдукции сигнала, указывая то, что PD-L1 не имеет сигнализации для взаимодействия с его рецептором. PD-L1 имеет молекулярную массу 40 кДа (290 аминокислот) и кодируется геном CD274 на мышиной хромосоме 19 и человеческой хромосоме 9. PD-L1 является членом семейства белка В7 и имеет примерно 20%-ную идентичность аминокислотной последовательности с В7.1 и В7.2. Человеческий PD-L1 имеет идентичность аминокислот 70% и 93% с PD-L1 мышиного ортолога и ортолога яванского макака соответственно.
PD-L1 связывается с PD-1, который представляет собой его рецептор, с аффинностью (KD) 770 нМ. PD-1 экспрессируется на активированных Т-клетках, В-клетках и клетках костного мозга, и модулирует активацию или подавление клеточных иммунных ответов. Экспрессия PD-L1 в клетках может опосредовать защиту против гибели от цитотоксических Т-лимфоцитов (CTL), что является регуляторным механизмом, который притупляет хронические иммунные ответы во время вирусной инфекции. Раковые заболевания, такие как хронические и провоспалительные заболевания, подрывают иммунозащитный путь через повышающую регуляцию экспрессии PD-L1 для того, чтобы, таким образом, ускользнуть от иммунного ответа хозяина. При активном иммунном ответе IFNγ также осуществляет повышающую регуляцию экспрессии PD-L1.
PD-L1 также опосредует иммуносупрессию через взаимодействие с другим белком В7.1 (также известным как CD80), таким образом, блокируя способность передавать один из вторичных сигналов активации к Т-клеткам через CD28. Ввиду экспрессии PD-L1 на опухолевых клетках и занятия В7.1, релевантность данного специфичного взаимодействия в иммунорезистентности опухоли остается неясной.
Иммунная функция человеческого организма регулирует общую функцию Т-лимфоцитов через контроль костимулирующих и коингибирующих сигналов в то же самое время, что и распознавание антигена. Данный регуляторный механизм называется иммунологической контрольной точкой. Иммунная функция человеческого организма выявляет опухолеспецифичные неоантигены, экспрессируемые из-за таких изменений, как мутации, происходящие в опухолевых клетках, и удаляет посредством этого опухолевые клетки или источники вирусной инфекции.
Однако некоторые опухолевые клетки подавляют иммунную функцию посредством изменения микроокружения опухоли для того, чтобы избегать такой иммунной атаки или стимулировать иммунное ускользание через иммунотолерантность или иммуноредактирование Т-клеток.
В качестве одной из данных стратегий ускользания подавляется функция опухолеспецифичных Т-лимфоцитов через изменения в функции иммунологической контрольной точки. В частности, посредством активации ингибирующей иммунологической контрольной точки в опухолевых клетках избегается атака опухолеспецифичных Т-лимфоцитов. В данном отношении противоопухолевый эффект может быть получен посредством усиления подавленных ингибированием их функции клеточной активности и эффекта опухолеспецифичных Т-лимфоцитов с использованием моноклонального антитела к PD-1 или лиганда PD-L1.
Относительно данного технического уровня авторы настоящей заявки попытались разработать антитело, которое специфично связывается с PD-L1. В результате, авторы настоящего изобретения разработали антитело против PD-L1, которое связывается с PD-L1 с высокой аффинностью, и установили то, что данное антитело против PD-L1 может ингибировать образование комплекса PD-1/PD-L1 и, таким образом, может желательно функционировать в качестве иммуноонкологического лекарственного средства, посредством этого достигая цели настоящего изобретения.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предложение нового антитела к PD-L1 или его антигенсвязывающего фрагмента.
Другой целью настоящего изобретения является предложение нуклеиновой кислоты, кодирующей антитело или его антигенсвязывающий фрагмент.
Еще одной другой целью настоящего изобретения является предложение вектора, включающего нуклеиновую кислоту, клетки, трансфицированной данным вектором, и способа их получения.
Еще одной другой целью настоящего изобретения является предложение композиции для предупреждения или лечения рака или инфекционного заболевания, содержащей данное антитело или его антигенсвязывающий фрагмент.
Еще одной целью настоящего изобретения является предложение композиции для комбинированной терапии для предупреждения или лечения рака посредством введения антитела или его антигенсвязывающего фрагмента в комбинации с другим противораковым средством.
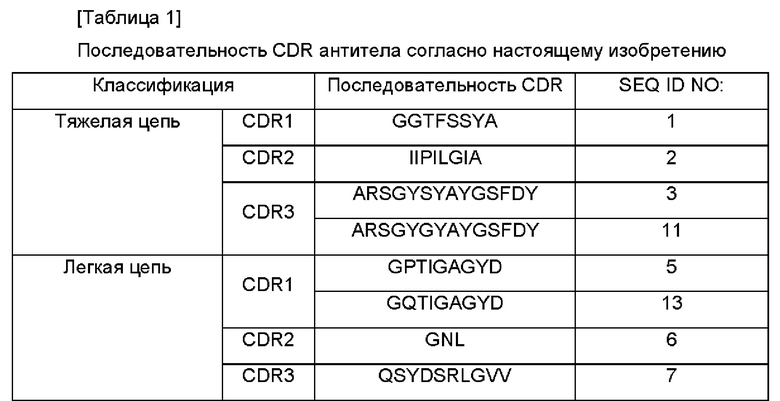
Для того чтобы осуществить приведенные выше цели согласно настоящему изобретению предложено антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент, которое содержит CDR1 (определяющая комплементарность область 1) тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 1, CDR2 тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 2, CDR3 тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 3 или SEQ ID NO: 11, CDR1 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 5 или SEQ ID NO: 13, CDR2 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 6, и CDR3 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 7.
Кроме того, согласно настоящему изобретению предложена нуклеиновая кислота, кодирующая данное антитело или его антигенсвязывающий фрагмент.
Кроме того, согласно настоящему изобретению предложен вектор, содержащий данную нуклеиновую кислоту.
Кроме того, согласно настоящему изобретению предложена клетка, трансфицированная данным вектором.
Кроме того, согласно настоящему изобретению предложен способ получения антитела или его антигенсвязывающего фрагмента, включающий (а) культивирование клетки, описанной выше, и (b) очистку антитела или его антигенсвязывающего фрагмента из данной культивируемой клетки.
Кроме того, согласно настоящему изобретению предложена композиция для предупреждения или лечения рака, содержащая антитело или его антигенсвязывающий фрагмент в качестве активного ингредиента.
Кроме того, согласно настоящему изобретению предложена композиция для комбинированной терапии для предупреждения или лечения рака посредством введения данного антитела или его антигенсвязывающего фрагмента в комбинации с другим противораковым средством.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На ФИГ. 1 показаны результаты реакции ELISA (твердофазный иммуноферментный анализ) для подтверждения связывания индивидуальных клонов антитела (примерно 1400), полученных посредством эксперимента по фаговому дисплею, на белке-антигене PD-L1.
На ФИГ. 2 показаны результаты эксперимента FACS (флуоресцентная сортировка клеток) по репрезентативным клонам антитела на клетках 293Т, в которых на их поверхности искусственно экспрессируется продукт человеческого гена PD-L1, для того, чтобы проанализировать связывание клонов антитела, связывающихся с PD-L1, с белком-антигеном на поверхности клетки.
На ФИГ. 3 показаны результаты подтверждения изменения сенсограммы SPR (поверхностный плазмонный резонанс) при совместном включении образца антитела по сравнению с условиями раствора (синяя линия), содержащего только белок-антиген PD-L1, посредством анализа оценки эффективности на основе SPR для подтверждения эффективности ингибирования связывания PD-1/PD-L1 9 репрезентативных клонов антител.
На ФИГ. 4 показаны результаты анализа связывания антител-кандидатов типа IgG, нацеленных на PD-L1, с мышиным белком PD-L1 с использованием линии клеток МС38 рака толстой кишки мыши, обработанной mIFN-γ (мышиный интерферон-гамма).
На ФИГ. 5 показаны результаты, подтверждающие эффективность ингибирования связывания между белками PD-1/PD-L1 in vitro в анализе подтверждения эффективности антител-кандидатов на основе клеток.
На ФИГ. 6 показаны результаты анализа противораковой эффективности in vivo антител-кандидатов с использованием линии клеток МС38 рака толстой кишки мышиного происхождения и модели сингенных мышей C57BL/6.
На ФИГ. 7 показаны результаты, подтверждающие способность выбранного антитела-кандидата (KL001) увеличивать активность иммунных клеток;
На ФИГ. 8 показаны результаты, подтверждающие способность антитела-кандидата KL001-13, отобранного после созревания аффинности антител-кандидатов, увеличивать активность иммунных клеток.
На ФИГ. 9 показаны результаты, подтверждающие итоговую картину увеличения, в зависимости от порядка пэннинга эксперимента по оптимизации улучшения эффективности антител-кандидатов.
На ФИГ. 10 показаны результаты реакции ELISA для отбора антител с улучшением аффинности.
На ФИГ. 11 показаны результаты измерения способности отобранного антитела к связыванию с человеческим PD-L1 и мышиным PD-L1 с использованием прибора Biacore;
На ФИГ. 12 показаны результаты оценки эффективности ингибирования роста опухоли при в.б. (внутрибрюшинное) введении антитела с оптимизированной эффективностью «KL001 -13», предоптимизированного антитела «KL001» и сравнительного антитела MPDL3280A в подкожной модели сингенной мыши МС38;
На ФИГ. 13 показаны результаты измерения значения IC50 (полумаксимальная ингибирующая концентрация) отобранного антитела с использованием in vitro анализа на основе клеток;
На ФИГ. 14 показаны результаты подтверждения противоракового эффекта in vivo по комбинированной обработке IL-2 (интерлейкин-2) и антителом KL001-13 или сравнительным антителом MPDL3280A.
На ФИГ. 15 показаны результаты подтверждения противоракового эффекта in vivo по комбинированной обработке антителом против CTLA-4 (9D9) и антителом KL001-13 или сравнительным антителом MPDL3280A.
На ФИГ. 16 показаны последовательности CDR антитела по настоящему изобретению при использовании разных схем нумерации, помимо схемы нумерации IMGT; и
На ФИГ. 17 показана перекрестная реактивность антитела по изобретению в отношении людей, мышей, обезьян и собак.
СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Если не определено иначе, все технические и научные термины, использованные в данном документе, имеют такие же значения, как и значения, типично понятные специалистам в области, к которой принадлежит настоящее изобретение. В общем, номенклатура, использованная в данном документе, является хорошо известной в данной области и типичной.
В одном аспекте настоящее изобретение относится к антителу, связывающемуся с PD-L1, или к его антигенсвязывающему фрагменту, которое содержит CDR1 тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 1, CDR2 тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 2, CDR3 тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 3 или SEQ ID NO: 11, CDR1 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 5 или SEQ ID NO: 13, CDR2 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 6, и CDR3 легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 7.
«PD-L1» в том виде, в котором он используется в данном документе, представляет собой лиганд иммуносупрессивного рецептора «рецептора программируемой гибели 1 (PD-1)», главным образом экспрессируемого на активированных Т- и В-клетках, и при связывании PD-1 и лигандов PD-L1 и/или PD-L2 сигнализация рецептора антигена может подвергаться негативной регуляции. Лиганды для PD-1 (PD-L1 и PD-L2) могут конститутивно экспрессироваться или индуцироваться во многих типах клеток, включающих негематопоэтические ткани и разные типы опухолей. PD-L1 экспрессируется на В-клетках, Т-клетках, клетках костного мозга и дендритных клетках (DC), и также экспрессируется на периферических клетках, аналогичных эндотелиальных клетках микрососудов и в нелимфатических органах, таких как сердце, легкие и т.д. В отличие от этого, PD-L2 обнаруживается только на макрофагах и дендритных клетках. Картина экспрессии лиганда PD-1 может отражать роль PD-1 в поддержании периферической толерантности и может способствовать регуляции ответов аутореактивных Т-клеток и В-клеток на периферии. PD-L1 и PD-L2 представляют собой трансмембранные рецепторы типа 1, которые содержат и IgV- и IgC-подобные домены в пределах внеклеточной области. Оба лиганда содержат короткие цитоплазматические домены, имеющие неизвестные мотивы сигнализации.
Многочисленные исследования выявили, что взаимодействие PD-1 с его лигандом ингибируют пролиферацию лимфоцитов in vitro и in vivo. Известно, что блокада взаимодействия PD-1/PD-L1 увеличивает пролиферацию Т-клеток и продукцию цитокинов, и блокирует прохождение клеточного цикла. Блокада взаимодействия PD-1/PD-L1 может индуцировать усиленный опухолеспецифичный Т-клеточный иммунитет, таким образом, помогая иммунной системе устранять опухолевые клетки. Кроме того, при хронической ВИЧ-инфекции ВИЧ-специфичные Т-клетки CD8+ функционально ослабевают, и снижаются способности к продукции цитокинов и эффекторных молекул, и к их пролиферации. PD-1 высоко экспрессируется в ВИЧ-специфичных Т-клетках CD8+ ВИЧ-инфицированных индивидов, и блокада взаимодействия PD-1/PD-L1 способна улучшать способность к пролиферации ВИЧ-специфичных Т-клеток и к продукции цитокинов в ответ на стимуляцию пептидом ВИЧ, усиливая, посредством этого, активность Т-клеток или противовирусный иммунный ответ.
Термин «антитело» в том виде, в котором он используется в данном документе, относится к антителу против PD-L1, которое специфично связывается с PD-L1. В объем настоящего изобретения включаются полное антитело, которое специфично связывается с PD-L1 и также антигенсвязывающий фрагмент молекулы данного антитела.
Полное антитело имеет структуру, имеющую две полноразмерные легкие цепи и две полноразмерные тяжелые цепи, причем данные легкие цепи, соответственно, связываются с тяжелыми цепями посредством связывания дисульфидными связыми. Константная область тяжелой цепи имеет гамма (γ), мю (μ) альфа (α), дельта (δ) и эпсилон (ε) типы, и также имеет гамма 1 (γ1), гамма 2 (γ2), гамма 3 (γ3), гамма 4 (γ4), альфа 1 (α1) и альфа 2 (α2) подклассы. Константная область легкой цепи имеет каппа (κ) и лямбда (λ) типы.
Антигенсвязывающий фрагмент антитела или фрагмент антитела представляет собой фрагмент, имеющий антигенсвязывающую функцию, и он включает Fab, F(ab'), F(ab')2, Fv и тому подобные. Среди данных фрагментов антитела Fab имеет структуру, имеющую вариабельные области легкой цепи и тяжелой цепи, константную область легкой цепи, первую константную область тяжелой цепи (СН1) и имеет один антигенсвязывающий сайт. Fab' отличается от Fab тем, что Fab' имеет шарнирную область, содержащую по меньшей мере один остаток цистеина на С-конце домена СН1 тяжелой цепи.
F(ab')2 фрагмент анитела создается дисульфидной связью между остатками цистеина в шарнирной области Fab'. Fv представляет собой минимальный фрагмент антитела, имеющий только вариабельную область тяжелой цепи и вариабельную область легкой цепи. Двухцепочечный Fv представляет собой фрагмент, в котором вариабельная область тяжелой цепи и вариабельная область легкой цепи связаны нековалентной связью, и одноцепочечный Fv (scFv) представляет собой фрагмент, в котором вариабельная область тяжелой цепи и вариабельная область легкой цепи обычно связаны ковалентной связью через пептидный линкер между ними или прямо связываются на С-конце, образуя димерную структуру, подобную двухцепочечному Fv. Такие фрагменты антитела могут быть получены с использованием протеаз (например, Fab может быть получен рестрикционным расщеплением полного антитела папаином, и фрагмент F(ab')2 может быть получен рестрикционным расщеплением полного антитела пепсином) или может быть получен посредством технологии генной инженерии.
В одном воплощении антитело по данному изобретению находится в виде Fv (например, scFv) или в виде полного антитела. Также константная область тяжелой цепи может представлять собой любую константную область, выбранную среди таких изотипов, как гамма (γ), мю (μ), альфа (α), дельта (δ) и эпсилон (ε). Например, константная область может представлять собой гамма 1 (IgG1), гамма 3 (IgG3) или гамма 4 (IgG4). Константная область легкой цепи может быть типа каппа или лямбда.
Термин «тяжелая цепь» в том виде, в котором он используется в данном документе, относится к полноразмерной тяжелой цепи, содержащей домен VH вариабельной области, содержащий аминокислотную последовательность, имеющую достаточную последовательность вариабельной области для придания специфичности к антигену, и три домена константной области СН1, СН2 и СН3, и ее фрагменту. Также термин «легкая цепь» в том виде, в котором он используется в данном документе, относится к полноразмерной легкой цепи, содержащей домен VL вариабельной области, содержащий аминокислотную последовательность, имеющую достаточную последовательность вариабельной области для придания специфичности к антигену, и домен CL константной области, и ее фрагменту.
Примеры антитела по настоящему изобретению включают моноклональные антитела, мультиспецифичные антитела, человеческие антитела, гуманизированные антитела, химерные антитела, одноцепочечные Fv (scFv), одноцепочечные антитела, фрагменты Fab, фрагменты F(ab'), Fv, связанные дисульфидом (sdFv), антиидиотипические антитела (анти-Id), эпитопсвязывающие фрагменты таких антител и тому подобные, но не ограничиваются ими.
Моноклональное антитело представляет собой антитело, полученное из популяции по существу гомогенных антител, в которой отдельные антитела, которые составляют данную популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в малых количествах. Моноклональное антитело является высокоспецифичным и индуцируется против одного антигенного сайта. В отличие от типичных препаратов (поликлональных) антител, которые типично включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене.
Термин «эпитоп» относится к белковой детерминанте, с которой может специфично связываться антитело. Данный эпитоп обычно состоит из группы химически активных поверхностных молекул, например, аминокислот или боковых цепей сахаров, и обычно имеет специфические трехмерные структурные характеристики и специфические характеристики заряда. Стерические и нестерические эпитопы различаются тем, что связывание с первыми, но не с последними, теряется в присутствии денатурирующего растворителя.
Антитело, не являющееся человеческим, в «гуманизированной» форме представляет собой химерное антитело, которое содержит минимальную последовательность, происходящую из иммуноглобулина, не являющегося человеческим (например, мышиного). В большинстве случаев данное гуманизированное антитело представляет собой человеческий иммуноглобулин (антитело-акцептор), в котором остаток из гипервариабельной области акцептора заменяется остатком из гипервариабельной области вида, не являющегося человеком (антитело-донор), имеющего желательную специфичность, аффинность и эффективность, например, мышей, крыс, кроликов или приматов, не являющихся человеком.
«Человеческое антитело» представляет собой молекулу, происходящую из человеческого иммуноглобулина, и данный термин означает то, что все аминокислотные последовательности составляющие антитело, содержащие область, определяющую комплементарность, и структурную область, входят в состав человеческого иммуноглобулина.
Часть тяжелой цепи и/или легкой цепи является идентичной или гомологичной соответствующей последовательности в антителе, происходящем из конкретного вида или принадлежащем к конкретному классу или подклассу антитела, тогда как остальная(ные) цепь(пи) включает «химерное» антитело (иммуноглобулин), который является идентичным или гомологичным соответствующей последовательности в антителе, происходящем из другого вида или принадлежащем к другому классу или подклассу антитела, а также фрагментам таких антител, которые демонстрируют желательную биологическую активность.
Термин «вариабельный домен антитела» в том виде, в котором он используется в данном документе, относится к частям легкой цепи и тяжелой цепи молекулы антитела, содержащим аминокислотные последовательности области, определяющей комплементарность (CDR; т.е. CDR1, CDR2 и CDR3), и к каркасной области (FR). VH относится к вариабельному домену тяжелой цепи, a VL относится к вариабельному домену легкой цепи.
Термин «область, определяющая комплементарность» (CDR; т.е. CDR1, CDR2 и CDR3) относится к аминокислотному остатку вариабельного домена антитела, который является необходимым для связывания антигена. Каждый вариабельный домен типично имеет три CDR, идентифицированные как CDR1, CDR2 и CDR3.
Настоящее изобретение относится к антителу, связывающемуся с PD-L1 или к его антигенсвязывающему фрагменту, содержащему CDR1 тяжелой цепи, содержащей последовательность SEQ ID NO: 1, CDR2 тяжелой цепи, содержащей последовательность SEQ ID NO: 2, CDR3 тяжелой цепи, содержащей последовательность SEQ ID NO: 3 или SEQ ID NO: 11, CDR1 легкой цепи, содержащей последовательность SEQ ID NO: 5 или SEQ ID NO: 13, CDR2 легкой цепи, содержащей последовательность SEQ ID NO: 6, и CDR3 легкой цепи, содержащей последовательность SEQ ID NO: 7 (схема нумерации CDR, используемая в настоящем патенте, осуществляется согласно «нумерации IMGT»).
Согласно настоящему изобретению антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может включать вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с SEQ ID NO: 1, CDR2 тяжелой цепи с SEQ ID NO: 2 и CDR3 тяжелой цепи с SEQ ID NO: 3, или вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с SEQ ID NO: 1, CDR2 тяжелой цепи с SEQ ID NO: 2 и CDR3 тяжелой цепи с SEQ ID NO: 11.
В настоящем изобретении антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может содержать вариабельную область легкой цепи, содержащую CDR1 легкой цепи с SEQ ID NO: 5, CDR2 легкой цепи с SEQ ID NO: 6 и CDR3 легкой цепи с SEQ ID NO: 7, или вариабельную область легкой цепи, содержащую CDR1 легкой цепи с SEQ ID NO: 13, CDR2 легкой цепи с SEQ ID NO: 6 и CDR3 легкой цепи с SEQ ID NO: 7.
В частности, в настоящем изобретении антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может содержать: вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с SEQ ID NO: 1, CDR2 тяжелой цепи с SEQ ID NO: 2 и CDR3 тяжелой цепи с SEQ ID NO: 3, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи с SEQ ID NO: 5, CDR2 легкой цепи с SEQ ID NO: 6 и CDR3 легкой цепи с SEQ ID NO: 7; или
вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с SEQ ID NO: 1, CDR2 тяжелой цепи с SEQ ID NO: 2 и CDR3 тяжелой цепи с SEQ ID NO: 11, и вариабельную область легкой цепи, содержащую CDR1 легкой цепи с SEQ ID NO: 13, CDR2 легкой цепи с SEQ ID NO: 6 и CDR3 легкой цепи с SEQ ID NO: 7.
«Каркасная область» (FR) представляет собой остаток вариабельного домена, отличный от остатка CDR. Каждый вариабельный домен типично имеет четыре FR, идентифицированных как FR1, FR2, FR3 и FR4.
Антитело против PD-L1 является одновалентным или двухвалентным и содержит одну цепь или две цепи. Функционально аффинность связывания антитела против PD-L1 попадает в диапазон от 10-5 М до 10-12 М. Например, аффинность связывания антитела против PD-L1 может составлять от 10-6 М до 10-12 М, от 10-7 М до 10-12 М, от 10-8 М до 10-12 М, от 10-9 М до 10-12 М, от 10-10 М до 10-12 М, от 10-11 М до 10-12 М, от 10-5 М до 10-11 М, от 10-6 М до 10-11 М, от 10-7 М до 10-11 М, от 10-8 М до 10-11 М, от 10-9 М до 10-11 М, от 10-10 М до 10-11 М, от 10-5 М до 10-10 М, от 10-8 М до 10-10 М, от 10-7 М до 10-10 М, от 10-8 М до 10-10 М, от 10-9 М до 10-10 М, от 10-5 М до 10-9 М, от 10-6 М до 10-9 М, от 10-7 М до 10-9 М, от 10-8 М до 10-9 М, от 10-5 М до 10-8 М, от 10-6 М до 10-8 М, от 10-7 М до 10-8 М, от 10-5 М до 10-7 М, от 10-6 М до 10-7 М или от 10-5 М до 10-6 М.
Антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может содержать вариабельную область тяжелой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательносью SEQ ID NO: 4 или SEQ ID NO: 12. Антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может содержать вариабельную область тяжелой цепи с SEQ ID NO: 4 или SEQ ID NO: 12. Также антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент может содержать вариабельную область легкой цепи, содержащую последовательность, имеющую по меньшей мере 90%-ную гомологию последовательности с последовательностью SEQ ID NO: 8 или SEQ ID NO: 14.
В конкретном воплощении согласно настоящему изобретению может содержаться вариабельная область тяжелой цепи с SEQ ID NO: 4 и вариабельная область легкой цепи с SEQ ID NO: 8, или может содержаться вариабельная область тяжелой цепи с SEQ ID NO: 12 и вариабельная область легкой цепи с SEQ ID NO: 14.
При этом определение последовательности (области) CDR может слегка отличаться для антитела, имеющего ту же самую вариабельную область, в зависимости от его схемы нумерации.
В частности, как показано в Таблице 2 ниже, даже в той же самой вариабельной области последовательность CDR может быть определена по-разному, в зависимости от схемы нумерации.
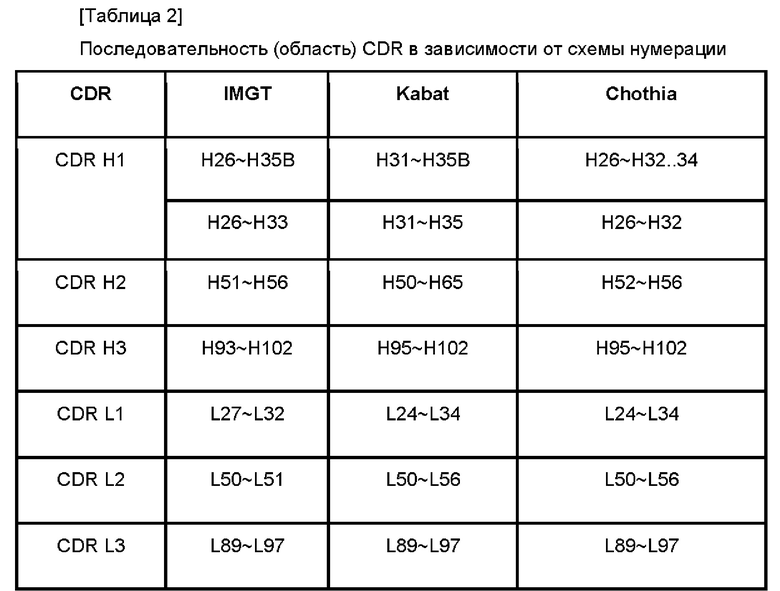
Таким образом, последовательность CDR антитела по настоящему изобретению имеет последовательность, показанную в Таблице 3 ниже, при использовании разных схем нумерации, помимо схемы нумерации IMGT (ФИГ. 16).


В одном воплощении антитело по данному изобретению демонстрирует перекрестную реактивность в отношении человека и мышей, а также у человека и млекопитающих, отличных от человека. В частности, млекопитающие, отличные от человека, могут представлять собой обезьян или собак (ФИГ. 17). Термин «фаговый дисплей» в том виде, в котором он используется в данном документе, представляет собой методику для осуществления дисплея варианта полипептида в виде слитого белка с по меньшей мере частью белка оболочки на поверхности фага, например, частицы нитчатого фага. Польза фагового дисплея состоит в том, что он может быстро и эффективно классифицировать последовательности, которые связываются с антигенами-мишенями с высокой аффинностью в больших библиотеках рандомизированных вариантов белка. Осуществление дисплея библиотек пептидов и белков на фагах использовали для скрининга миллионов полипептидов для того, чтобы идентифицировать полипептиды, имеющие свойства специфичного связывания.
Методика фагового дисплея оказалась мощным средством для получения и отбора новых белков, которые связываются со специфичными лигандами (например, антигенами). С использованием методики фагового дисплея могут быть получены большие библиотеки вариантов белков, и могут быть быстро классифицированы последовательности, которые связываются с антигенами-мишенями с высокой аффинностью. Нуклеиновую кислоту, кодирующую вариант полипептида, сливают с последовательностью нуклеиновой кислоты, кодирующей белок оболочки вируса, например, с геном III белка или с геном VIII белка. Была разработана система одновалентного фагового дисплея, в которой последовательность нуклеиновой кислоты, кодирующую белок или полипептид, сливают с последовательностью нуклеиновой кислоты, кодирующей часть гена III белка. В данной системе одновалентного фагового дисплея слитый ген экспрессируется на низком уровне, и также экспрессируется ген III белка дикого типа, таким образом, что поддерживается инфективность частицы.
Осуществление демонстрации экспрессии пептидов на поверхности нитчатого фага и экспрессии функциональных фрагментов антитела в периплазме Е. coli является важным при разработке библиотек фагового дисплея антител. Библиотеки антител или антигенсвязывающих полипептидов были получены разными способами, например, посредством способов изменения одного гена посредством вставки случайной последовательности ДНК или клонирования родственных последовательностей гена. Библиотеки могут подвергаться скринингу на экспрессию антител или антигенсвязывающих белков, имеющих желательные характеристики.
Методика фагового дисплея имеет несколько преимуществ в сравнении с типичными способами гибридомы и рекомбинантными способами получения антител, имеющих желательные характеистики. Данная методика обеспечивает продукцию больших библиотек антител, имеющих разные последовательности, в пределах короткого времени без применения животных. Продукция гибридом или гуманизированных антител может требовать производственного периода в несколько месяцев. Кроме того, поскольку не требуется иммунитет, библиотеки фагового дисплея могут продуцировать антитела против антигенов, которые являются токсичными или имеют низкую антигенность. Библиотеки фагового дисплея могут использоваться для продукции и идентификации новых терапевтических антител.
С использованием библиотек фагового дисплея можно применять методики получения человеческих антител от иммунизированных или неиммунизированных людей, последовательностей зародышевой линии или репертуаров lg наивных В-клеток. Можно использовать множество лимфоидных тканей для получения несенсибилизированных или неиммуногенных антигенсвязывающих библиотек.
Технология, способная идентифицировать и выделять высокоаффинные антитела из библиотек фагового дисплея, является важной для выделения новых терапевтических антител. Выделение высокоаффинных антител из данных библиотек может зависеть от размера библиотек, эффективности продукции в бактериальных клетках и разнообразия библиотек. Размер библиотек уменьшается неправильным свертыванием антитела или антигенсвязывающего белка и неэффективной продукцией из-за присутствия терминатора. Экспрессия в бактериальных клетках может ингибироваться, когда антитело или антигенсвязывающий домен не сворачивается правильно. Экспрессия может быть улучшена посредством альтернативного мутирования остатков на поверхности контакта вариабельных/константных доменов или в выбранных остатках CDR. Последовательность каркасной области является элементом для обеспечения правильного сворачивания при продукции фаговых библиотек антител в бактериальных клетках.
Для выделения высокоаффинных антител важной является продукция разнообразных библиотек антител или антигенсвязывающих белков. Часто обнаруживали то, что область CDR3 участвует в связывании антигена. Поскольку область CDR3 на тяжелой цепи значительно варьирует по размеру, последовательности и структурной конформации, с ее использованием можно получить целый ряд библиотек.
Кроме того, разнообразие может быть создано рандомизацией CDR вариабельных тяжелых и легких цепей с использованием всех 20 аминокислот в каждом положении. Применение всех 20 аминокислот может приводить к высокоразнообразным последовательностям вариантов антител и может увеличивать вероятности идентификации новых антител.
Антитело или фрагмент антитела по настоящему изобретению может включать не только последовательность антитела против PD-L1 по настоящему изобретению, но также его биологические эквиваленты в пределах интервала, который обеспечивает специфичное распознавание PD-L1. Например, можно делать дополнительные модификации аминокислотной последовательности антитела для того, чтобы дополнительно улучшать аффинность связывания и/или другие биологические свойства антитела. Такие модификации включают, например, делецию, вставку и/или замену остатков аминокислотной последовательности антитела. Вариации аминокислот основываются на относительном сходстве заместителей боковой цепи аминокислот, например, гидрофобности, гидрофильности, заряде, размере и тому подобном. На основе анализа размера, формы и типа заместителей боковой цепи аминокислот все из аргинина, лизина и гистидина являются положительно заряженными остатками, аланин, глицин и серии имеют аналогичные размеры, и фенилаланин, триптофан и тирозин имеют аналогичные формы. Следовательно, на основе данных соображений аргинин, лизин и гистидин могут рассматриваться как биологически функциональные эквиваленты, аланин, глицин и серии могут рассматриваться как биологически функциональные эквиваленты, и фенилаланин, триптофан и тирозин могут рассматриваться как биологически функциональные эквиваленты.
Принимая во внимание вышеописанные вариации, имеющие эквивалентную биологическую активность, антитело по настоящему изобретению или кодирующая его молекула нуклеиновой кислоты истолковывается как содержащая последовательность, демонстрирующую существенную идентичность с последоваельностью, изложенной в номере последовательности. При выравнивании последовательности по настоящему изобретению и любых других последовательностей таким образом, чтобы они в максимальной степени соответствовали друг другу, и анализе выровненной последовательности с использованием алгоритма, обычно используемого в данной области, существенная идентичность относится к последовательности, демонстрирующей по меньшей мере 90%-ную гомологию, предпочтительно по меньшей мере 95%-ную гомологию, по меньшей мере 96%-ную гомологию, по меньшей мере 97%-ную гомологию, по меньшей мере 98%-ную гомологию или по меньшей мере 99%-ную гомологию. Способы выравнивания для сравнения последовательностей хорошо известны в данной области. Базовый инструмент поиска местного выравнивания (BLAST) от NCBI доступен посредством NBCI и т.д., и может использоваться в сочетании с программами секвенирования, такими как blastp, blastm, blastx, tblastn и tblastx в интернете. BLAST доступен на сайте www.ncbi.nlm.nih.gov/BLAST/. Способ сравнения гомологии последовательности с использованием данной прораммы можно найти на сайте www.ncbi.nlm.nih.gov/BLAST/blast_help.html.
Основываясь на этом, антитело по настоящему изобретению или его антигенсвязывающий фрагмент может иметь 90%-ную, 91%-ную, 92%-ную, 93%-ную, 94%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную или более высокую гомологию с определенной последовательностью или со всеми последовательностями, описанными в данном описании изобретения. Такая гомология может быть определена посредством сравнения и/или выравнивания последовательностей с использованием способов, известных в данной области. Например, процентная доля гомологии последовательности нуклеиновой кислоты или белка по настоящему изобретению может быть определена с использованием алгоритма сравнения последовательностей (т.е. BLAST или BLAST 2.0), выравнивания вручную или визуальной проверки.
В другом аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей антитело или его антигенсвязывающий фрагмент.
Антитело или его антигенсвязывающий фрагмент может быть получено рекомбинантно посредством выделения нуклеиновой кислоты, кодирующей антитело по настоящему изобретению или его антигенсвязывающий фрагмент.Данная нуклеиновая кислота выделяется и вставляется в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или дальнейшей экспрессии. В еще одном другом аспекте, на его основе, настоящее изобретение относится к вектору, содержащему нуклеиновую кислоту.
Термин «нуклеиновая кислота» в том виде, в котором он используется в данном документе, имеет значение, полностью охватывающее молекулы ДНК (гДНК (геномная ДНК) и кДНК (комплементарная ДНК)) и РНК, а нуклеотиды, которые являются основными строительными блоками нуклеиновых кислот, включают природные нуклеотиды, а также аналоги, в которых области сахара или основания являются модифицированными. Последовательность нуклеиновой кислоты, кодирующая вариабельные области тяжелой цепи и легкой цепи, по настоящему изобретению может быть модифицирована. Такие модификации включают присоединение, делецию или неконсервативную, или консервативную замену нуклеотидов.
ДНК, кодирующая антитело, легко выделяется или синтезируется с использованием типичных методик (например, с использованием олигонуклеотидного зонда, способного к специфичному связыванию с ДНК, кодирующей тяжелую и легкую цепи антитела). Многие векторы доступны в продаже. Компонент вектора обычно включает, но не ограничивается, по меньшей мере одним компонентом, выбранным среди сигнальной последовательности, репликатора, по меньшей мере одного гена-маркера, энхансерного элемента, промотора и последовательности терминации транскрипции.
Термин «вектор» в том виде, в котором он используется в данном документе, относится к средству для осуществления экспрессии гена-мишени в клетке-хозяине, включая плазмидный вектор, космидный вектор, вирусный вектор, такой как бактериофага вый вектор, аденовирусный вектор, ретровирусный вектор или вектор на основе аденосателлитного вируса и тому подобные. В данном векторе нуклеиновая кислота, кодирующая антитело, связана функциональным образом с промотором.
Термин «связанный функциональным образом» в том виде, в котором он используется в данном документе, означает функциональную связь между последовательностью контроля экспрессии нуклеиновой кислоты (например, промотор, сигнальная последовательность или группа сайтов связывания регулятора транскрипции) и другой последовательностью нуклеиновой кислоты, при этом последовательность контроля служит для контроля транскрипции и/или трансляции последовательности другой нуклеиновой кислоты.
При использовании прокариотической клетки в качестве хозяина обычно включают сильный промотор, способный продвигать транскрипцию (например, промотор tac, промотор lac, промотор lacUV5, промотор lpp, промотор pLλ, промотор pRλ, промотор rac5, промотор amp, промотор recA, промотор SP6, промотор trp или промотор Т7), сайт связывания рибосомы для инициации трансляции и последовательность терминации транскрипции/трансляции. Кроме того, например, при использовании в качестве хозяина эукариотической клетки, можно использовать промотор, происходящий из генома клетки млекопитающего (например, промотор металлотионина, промотор β-актина, промотор человеческого гемоглобина или промотор креатинина человеческих мышц), или промотор, происходящий из вирусов млекопитающих (например, поздний промотор аденовируса, промотор вируса осповакцины 7.5K, промотор SV40, промотор цитомегаловируса (CMV), tk промотор HSV (вирус простого герпеса), промотор мышиного вируса опухоли молочной железы (MMTV), промотор LTR ВИЧ, промотор вируса Молони, промотор вируса Эпштейна-Барр (EBV) или промотор вируса саркомы Рауса (RSV), и обычно он имеет последовательность полиаденилирования в качестве последовательности терминации транскрипции.
В некоторых случаях данный вектор может быть слит с другой последовательностью для того, чтобы облегчать очистку антитела, экспрессируемого от него. Примеры последовательности, которая сливается, включают глутатион-S-трансферазу (Pharmacia, США), мальтозосвязывающий белок (NEB, США), FLAG (IBI, США) и 6х His (гексагистидин; Qiagen, США).
Данный вектор содержит, в качестве селективного маркера, ген устойчивости к антибиотику, который обычно используется в данной области, например, ген, придающий устойчивость к ампициллину, гентамицину, карбенициллину, хлорамфениколу, стрептомицину, канамицину, генетицину, неомицину или тетрациклину.
В еще одном другом аспекте настоящее изобретение относится к клетке, трансфицированной вектором, описанным выше. Примеры клетки, используемой для продукции антитела по настоящему изобретению, могут включать прокариотические клетки, дрожжевые клеткми и клетки высших эукариотов, но не ограничиваются ими.
Можно использовать штаммы, принадлежащие к роду Bacillus, такие как Escherichia coli, Bacillus subtilis и Bacillus thuringiensis, и прокариотические клетки-хозяева, такие как Streptomyces, Pseudomonas (например, Pseudomonas putida), Proteus mirabilis и Staphylococcus (например, Staphylococcus carnosus).
Здесь клетки животных представляют наибольший интрес, и примеры полезных линий клеток-хозяев могут включать COS-7, BHK, СНО, CHO-S, CHOK1, GS-CHO, DXB-11, DG-44, CHO/-DHFR, CV1, HEK293, ТМ4, VERO, HELA, MDCK, BRL 3А, W138, Hep G2, SK-Hep, ММТ, TRI, MRC 5, FS4, 3Т3, RIN, А549, РС12, K562, PER.C6, SP2/0, NS-0, U20S и НТ1080, но не ограничиваются ими.
В еще одном другом аспекте настоящее изобретение относится к способу получение антитела или его антигенсвязывающего фрагмента, включающему (а) культивирование клеток, описанных выше, и (b) очистку антитела или его антигенсвязывающего фрагмента из культивируемых клеток.
Данные клетки могут культивироваться в разных средах. В качестве культуральной среды можно использовать любую имеющуюся в продаже среду без ограничения. Все другие важные добавки, известные специалистам в данной области, могут содержаться в подходящих концентрациях. Условия культивирования, такие как температура, рН и т.д., представляют собой условия, уже используемые для клеток-хозяев, отобранных для экспрессии, как будет очевидно специалистам в данной области.
Для очистки антитела или его антигенсвязывающего фрагмента можно удалять примеси, например, посредством центрифугирования или ультрафильтрации, и образующийся продукт можно очищать с использованием, например, аффинной хроматографии. Можно использовать другие дополнительные методики очистки, такие как анионо- или катионообменная хроматография, хроматография на основе гидрофобных взаимодействий, хроматография на гидроксиапатите и тому подобные.
В другом аспекте настоящее изобретение относится к композиции для предупреждения или лечения рака, содержащей описанное выше антитело в качестве активного ингредиента.
В настоящем изобретении может рассматриваться, например, фармацевтическая композиция для предупреждения или лечения рака, содержащая (а) фармацевтически эффективное количество антитела к PD-L1 или его антигенсвязывающего фрагмента по настоящему изобретению и (b) фармацевтически приемлемый носитель. Кроме того, настоящее изобретение относится к способу предупреждения или лечения рака, включающему введение антитела к PD-L1 или его антигенсвязывающего фрагмента по настоящему изобретению в эффективном количестве, требующемся для пациента.
Поскольку в данной композиции используется вышеупомянутое антитело против PD-L1 или его антигенсвязывающий фрагмент согласно настоящему изобретению в качестве активного ингредиета, описание общих компонентов между ними опущено.
Связывание PD-L1 с PD-1 негативно модулирует антигенспецифичные ответы Т-клеток, важные для толерантности и предупреждения аутоиммунных заболеваний и иммунопатологии. Однако избыточные взаимодействия PD-L1/PD-1, которые можно индуцировать хронической антигенной стимуляцией, могут приводить к подавлению антигенспецифичных ответов Т-клеток и потере Т-клеток, что является характеристиками исчерпания Т-клеток. Исчерпание Т-клеток является состоянием дисфункции Т-клеток, которое может происходить при хронических инфекциях и раковых заболеваниях. Оно определяется плохой эффекторной функцией, поддерживаемой экспрессией ингибирующих рецепторов и отличным транскрипционным состоянием от состояния функциональных эффекторных Т-клеток или Т-клеток памяти. Исчерпание препятствует контролю инфекции и прогрессирования опухоли.
Как продемонстрировано позднее со ссылкой на примеры, антитело или его антигенсвязывающий фрагмент по настоящему изобретению связывается с PD-L1 с высокой аффинностью и, таким образом, ингибирует образование комплекса PD-1/PD-L1 и, посредством этого, может с пользой использоваться для лечения рака, который индуцирует исчерпание Т-клеток, который ускользает от противоопухолевой активности Т-клеток.
Настоящее изобретение также относится к композиции для комбинированной терапии для предупреждения или лечения рака посредством введения антитела или его антигенсвязывающего фрагмента согласно настоящему изобретению в комбинации с другим противораковым средством.
В настоящем изобретении, например, может рассматриваться композиция для комбинированной терапии для предупреждения или лечения рака, содержащая (а) фармацевтически эффективное количество антитела к PD-L1 или его антигенсвязывающего фрагмента согласно настоящему изобретению и (b) фармацевтически приемлемый носитель. Настоящее изобретение также относится к способу комбинированной терапии для предупреждения или лечения рака, включающему введение антитела к PD-L1 или его антигенсвязывающего фрагмента согласно настоящему изобретению в эффективном количестве, требующемся для пациента.
Поскольку в композиции используется вышеупомянутое антитело против PD-L1 или его антигенсвязывающий фрагмент согласно настоящему изобретению в качестве активного ингредиента, описание общих компонентов между ними опускается.
Как описано выше, антитело согласно настоящему изобретению используется в комбинации с другим противораковым средством, таким образом, делая возможным эффективное нацеливание на опухолевые клетки, сверхэкспрессирующие PD-L1, и увеличение противоопухолевой активности Т-клеток, увеличивая, посредством этого, иммунный ответ, нацеленный на опухолевые клетки.
Можно использовать другие антинеопластические или иммуногенные средства [(например, ослабленные раковые клетки, опухолевые антигены (включающие молекулы рекомбинантных белков, пептидов и углеводов), антигенпрезентирующие клетки, например, дендритные клетки, подвергнутые импульсному воздействию происходящих из опухоли антигенов или нуклеиновых кислот, иммуностимулирущие цитокины (например, IL-2 (интерлейкин-2), IFNα2 (интерферон-альфа2), GM-CSF (гранулоцитарно-макрофагальный
колониестимулирующий фактор) и клетки, трансфицированные генами, кодирующими иммуностимулирующие цитокины (которые включают, например, GM-CSF, но не ограничиваются им)], клетки, содержащие антитело, нацеленное на раковый антиген (например, CAR-T (Т-клетки с химерным рецептором антигена), CAR-NK (клетки-природные киллеры с химерным рецептором антигена)), иммунодепрессивные ингибиторы микроокружения опухоли (например, ингибитор IDO), онколитический вирус, стандартную противораковую терапию (например, химиотерапию, лучевую терапию или хирургию) или другие ассоциированные с раком антигены.
Кроме того, антитело или его антигенсвязывающий фрагмент по настоящему изобретению можно использовать наряду с другим антителом (примеры другого антитела, чем антитело к PD-L1 могут включать антитела k VEGF (фактор роста эндотелия сосудов), EGFR (рецептор эпидермального фактора роста), Her2/neu, рецептору VEGF, другим рецепторам факторов роста, CD20, CD40, CTLA-4 (антиген 4 цитотоксических Т-лимфоцитов), ОХ-40, 4-1 ВВ и ICOS).
В частности, антитело, которое может использоваться, наряду с антителом или его антигенсвязывающим фрагментом по настоящему изобретению, может использоваться без ограничения, при условии, что оно представляет собой антитело, способное к специфичному связыванию с антигеном, ассоциированным с раком или аутоиммунным заболеванием, и примеры такого антигена могут включать 4-1ВВ, интегрин, ангиопоэтин, аналог 3 ангиопоэтина, фактор, активирующий В-клетки (BAFF), В7-Н3, CCR4, CD3, CD4, CD6, CD11a, CD19, CD20, CD22, CD30, CD33, CD38, CD40, CD52, CD62, CD79b, CD80, CGRP, ОХ-40, ICOS, клаудин-18, CTLA4, DLL3, рецептор EGF, рецептор Fc, FGF23, рецептор фолата, GD2, GM-CSF, HER2, Her2/neu, HER3, VEGF, рецептор VEGF, рецептор интерферона, интерферон-гамма, IgE, рецептор IGF-1, интерлейкин 1, рецептор интерлейкина 2, рецептор интерлейкина 4, интерлейкин 5, рецептор интерлейкина 5, интерлейкин 6, рецептор интерлейкина 6, интерлейкин 7, интерлейкин 12/23, интерлейкин 13, интерлейкин 17А, рецептор А интерлейкина 17, рецептор интерлейкина 31, рецептор интерлейкина 36, LAG3, LFA3, NGF (фактор роста нервов), PVSK9, PD-1, PD-L1, RANK-L, SLAMF7 и тканевой фактор, но не ограничиваются ими.
Более конкретно, примеры антитела, которое связывается с антигеном, описанным выше, включают следующие:
антитела к 4-1 ВВ, такие как утомилумаб;
антитела к интегрину, такие как натализумаб, этролизумаб, ведолизумаб и бимагрумаб;
антитела к амилоиду-бета, такие как бапинеузумаб, кренезумаб, соланезумаб, адуканумаб и гантенерумаб;
антитела к ангиопоэтину, такие как AMG 780;
антитела к аналогу 3 ангиопоэтина, такие как эвинакумаб;
антитела к фактору, активирующему В-клетки (BAFF), такие как табалумаб, ланалумаб и белимумаб;
антитела к В7-Н3, такие как омбуртамаб;
антитела к CCR4, такие как могамулизумаб;
антитела к CD3, такие как отеликсизумаб, теплизумаб, муромонаб, тебентафусп, которое представляет собой биспецифичное антитело к GP100 и CD3, блинатумомаб, которое представляет собой биспецифичное антитело к CD19 и REGN1979, которое представляет собой биспецифичное антитело к CD20 и CD3;
антитела к CD4, такие как ибализумаб и занолимумаб; антитела к CD6, такие как итолизумаб; антитела к CD11 а, такие как эфализумаб;
антитела к CD19, такие как инэбилизумаб, тафаситамаб и лонкастуксимаб тесирин в качестве ADC (конъюгат антитело-лекарственное средство);
антитела к CD20, такие как окрелизумаб, ублитуксимаб, обинутузумаб, офатумумаб, ритуксимаб, тозитумомаб и ибритумомаб тиуксетан в качестве ADC;
антитела к CD22, такие как эпратузумаб и, в качестве ADC, инотузумаб озогамицин и моксетумомаб пасудотокс;
ADC к CD30, такие как брентуксимаб ведотин;
ADC к CD33, такие как вадастуксимаб талирин и гемтузумаб озогамицин;
антитела KCD38, такие как даратумумаб и изатуксимаб;
антитела к CD52, такие как алемтузумаб;
антитела к CD62, такие как кризанлизумаб;
ADC к CD79b, такие как полатузумаб ведотин;
антитела к CD80, такие как галиксимаб;
антитела к CGRP, такие как эптинезумаб, фреманезумаб, галканезумаб и эренумаб;
антитела к клаудину-18, такие как золбетуксимаб;
антитела к CTLA4, такие как тремелимумаб, залифрелимаб и ипилимумаб; ADC к DLL3, такие как ровалпитузумаб тезирин;
антитела к рецептору EGF, такие как цетуксимаб, депатуксизумаб, залутумумаб, нецитумумаб и панитумумаб;
антитела к рецептору Fc, такие как нипокалимаб и розаноликсизумаб; антитела к FGF23, такие как бурозумаб;
антитела к рецептору фолата, такие как фарлетузумаб и мирветуксимаб соравтазин в качестве ADC;
антитела к GD2, такие как динутуксимаб и накситамаб; антитела к GM-CSF, такие как отилимаб;
антитела к HER2, такие как маргетуксимаб, пертузумаб, трастузумаб и, в качестве ADC, трастузумаб дерукстекан, трастузумаб эмтанзин и трастузумаб дуокармазин;
антитела к HER3, такие как патритумаб;
антитела к рецептору интерферона, такие как анифролумаб;
антитела к интерферону-гамма, такие как эмапалумаб;
антитела к IgE, такие как лигелизумаб и омализумаб;
антитела к рецептору IGF-1, такие как далотузумаб, фигитумумаб и тепротумумаб;
антитела к интерлейкину 1, такие как гевокизумаб и канакинумаб;
антитела к рецептору интерлейкина 2, такие как даклизумаб и базиликсимаб;
антитела к рецептору интерлейкина 4, такие как дупилумаб; антитела к интерлейкину 5, такие как меполизумаб и реслизумаб; антитела к рецептору интерлейкина 5, такие как бенрализумаб; антитела к интерлейкину 6, такие как клазакизумаб, олокизумаб, сирукумаб и силтуксимаб;
антитела к рецептору интерлейкина 6, такие как сарилумаб, сатрализумаб, тоцилизумаб и REGN88;
антитела к интерлейкину 7, такие как секукинумаб;
антитела к интерлейкину 12/23, такие как устекинумаб и бриакинумаб;
антитела к интерлейкину 13, такие как лебрикизумаб и тралокинумаб;
антитела к интерлейкину 17А, такие как иксекизумаб и бимекизумаб;
антитела к рецептору А интерлейкина 17, такие как бродалумаб;
антитела к интерлейкину 23, такие как бразикумаб, гузелкумаб, ризанкизумаб, тилдракизумаб и мирикизумаб;
антитела к рецептору интерлейкина 31, такие как немолизумаб;
антитела к рецептору интерлейкина 36, такие как спесолимаб;
антитела к LAG3, такие как релатлимаб;
антитела к NASP2, такие как нарсоплимаб;
антитела к NGF, такие как фазинумаб и танезумаб;
антитела к PVSK9, такие как алирокумаб, эволокумаб и бокоцизумаб;
антитела к PD-1, такие как ламбролизумаб, балстилимаб, камрелизумаб, цемиплимаб, достарлимаб, пролголимаб, синтилимаб, спартализумаб, тислелизумаб, пембролизумаб и ниволумаб;
антитела к PD-L1, такие как атезолизумаб, авелумаб, энвафолимаб, дурвалумаб и бинтрафусп альфа, который представляет собой биспецифичное антитело к TGF-бета и PD-L1;
антитела к RANK-L, такие как деносумаб;
антитела к SLAMF7, такие как элотузумаб;
антитела к тканевому фактору, такие как концизумаб и марстацимаб;
антитела к TNF (фактор некроза опухолей), в частности, к TNFα, такие как инфликсимаб, адалимумаб, голимумаб, цертолизумаб пегол, который представляет собой фрагмент антитела, и озорализумаб, который представляет собой биспецифичное антитело k TNF и альбумину;
антитела к VEGF, такие как бролуцизумаб, ранибизумаб, бевацизумаб и фарицимаб, которое представляет собой биспецифичное антитело к VEGF и Ang2; и
антитела к рецептору VEGF, такие как рамуцирумаб, но не ограничиваются ими.
Рак, который представляет собой заболевание, против которого применяется данная композиция, типично включает рак, который отвечает на иммунотерапию, и рак, который ранее не связывали с иммунотерапией. Неограничивающие примеры рака, подлежащего лечению, могут включать следующие: меланома (например, метастатическая злокачественная меланома), рак почки (например, светлоклеточная карцинома), рак предстательной железы (например, аденокарцинома предстательной железы, трудно поддающаяся лечению гормонами), аденокарцинома поджелудочной железы, рак груди, рак толстой кишки, рак легкого (например, немелкоклеточный рак легкого), рак пищевода, плоскоклеточная карцинома головы и шеи, рак печени, рак яичника, рак шейки матки, рак щитовидной железы, глиобластома, глиома, лейкоз, лимфома и другие новообразования. Кроме того, настоящее изобретение охватывает трудно поддающийся лечению или рецидивирующий рак, рост которого может ингибироваться с использованием антитела по настоящему изобретению.
Антитело или фрагмент антитела по настоящему изобретению также может использоваться одно или в комбинации с вакциной для стимуляции иммунного ответа на патогены, токсины и собственные антигены. Данное антитело или его антигенсвязывающий фрагмент может использоваться для стимуляции иммунного ответа на вирусы, которые инфицируют человека, включающие, например, вирус иммунодефицита человека, вирус гепатита классов А, В и С, вирус Эпштейна-Барр, человеческий цитомегаловирус, человеческий вирус папилломы и вирус герпеса, но не ограничивающиеся ими. Антитело или его антигенсвязывающий фрагмент можно использовать для стимуляции иммунного ответа на инфекцию бактериальными или грибковыми паразитами и другими патогенами.
Настоящее изобретение также относится к композиции, содержащей антитело или его антигенсвязывающий фрагмент согласно настоящему изобретению и полезные бактерии. Данные полезные бактерии используются в качестве бактерий или пробиотиков, имеющих противораковую эффективность, и их примеры могут включать Anaerococcus, Anaerostipes, Alistipes, Akkermansia, Bacillus, Bacteroides, Bifidobacterium, Blautia, Capnocytophaga, Clostridium, Collinsella, Desulfovibrio, Dorea, Enterococcus, Escherichia, Eubacterium, Faecaiibacterium, Fusobacterium, Gardnerella, Gemmiger, Klebsiella, Lactobacillus, Leuconostoc, Moryella, Paraprevotella, Parabacteroides, Phascolarctobacterium, Porphyromonas, Prevotella, Pseudobutyrivibrio, Roseburia, Rothia, Ruminococcus, Shigella, Streptococcus, Veillonella, Weissella и тому подобные, но не ограничиваются ими.
Фармацевтически приемлемый носитель, содержащийся в композиции по настоящему изобретению, может включать носители, обычно используемые в препаратах, например, лактозу, декстрозу, сахарозу, сорбит, маннит, крахмал, аравийскую камедь, фосфат кальция, альгинат, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, воду, сироп, метилцеллюлозу, метилгидроксибензоат, пропилгидроксибензоат, тальк, стеарат магния и минеральное масло, но не ограничивается ими. Композиция по настоящему изобретению может дополнительно включать смазку, увлажнитель, подсластитель, корригент, эмульгатор, суспендирующее средство, консервант и тому подобные, помимо приведенных выше компонентов.
Фармацевтическая композиция по настоящему изобретению может вводиться перорально или парентерально, и примеры парентерального введения могут включать внутривенную инъекцию, подкожную инъекцию, внутримышечную инъекцию, внутрибрюшинную инъекцию, эндотелиальное введение, местное введение, интраназальное введение, внутрилегочное введение, интраректальное введение и т.д.
При пероральном введении белок или пептид является перевариваемым, и, следовательно, пероральная композиция должна быть приготовлена таким образом, чтобы активное средство было покрыто или защищено от распада в желудке. Также данная фармацевтическая композиция может вводиться с использованием любого устройства, способного транспортировать активное средство в клетки-мишени.
Подходящая доза композиции по настоящему изобретению может варьировать, в зависимости от таких факторов, как способ приготовления, способ введения, возраст пациента, масса и пол, заболеваемость, пища, время введения, путь введения, скорость выведения и чувствительность ответа, и доза, которая является эффективной для желательного лечения или предупреждения, может быть легко определена и прописана врачами обычной квалификации. Например, ежесуточная доза фармацевтической композиции по настоящему изобретению составляет 0,0001-100 мг/кг. Термин «фармацевтически эффективное количество» в том виде, в котором он используется в данном документе, относится к достаточному количеству для предупреждения или лечения рака.
Фармацевтическая композиция по настоящему изобретению может быть приготовлена в стандартной лекарственной форме или может быть приготовлена в многодозовом контейнере с использованием фармацевтически приемлемого носителя и/или эксципиента согласно способу, который может легко осуществляться обычным специалистом в области, к которой принадлежит настоящее изобретение. Здесь данный препарат может принимать форму раствора, суспензии или эмульсии в масляной или водной среде, или может принимать форму экстракта, порошка, суппозитория, порошка, гранулы, таблетки или капсулы, и может дополнительно включать диспергент или стабилизатор.
Композиция по настоящему изобретению может вводиться одна в качестве терапевтического средства или может вводиться в комбинации с другим терапевтическим средством и может вводиться последовательно или одновременно с традиционными терапевтическими средствами.
В еще одном другом аспекте согласно настоящему изобретению предложен конъюгат антитело-лекарственное средство, в котором лекарственное средство конъюгировано с антителом против PD-L1 или его антигенсвязывающим фрагментом согласно настоящему изобретению, и содержащая его фармацевтическая композиция. Кроме того, согласно настоящему изобретению предложен способ лечения опухоли с использованием конъюгата антитело-лекарственное средство, в котором лекарственное средство конъюгировано с антителом против PD-L1 или его антигенсвязывающим фрагментом, и содержащая их фармацевтическая композиция.
Антитело против PD-L1 или его антигенсвязывающий фрагмент может быть связано с лекарственным средством через линкер. Данный линкер представляет собой сайт, связывающий антитело против PD-L1 или его антигенсвязывающий фрагмент с лекарственным средством. Например, данный линкер является расщепляемым при внутриклеточных условиях, и, в частности, лекарственное средство может высвобождаться от антитела посредством расщепления линкера во внутриклеточной среде.
Данный линкер может расщепляться расщепляющим агентом, присутствующим во внутриклеточной среде, например, в лизосоме или эндосоме, и может представлять собой, например, пептидный линкер, который может расщепляться внутриклеточным пептидазным или протеазным ферментом, таким как лизосомальная или эндосомальная протеаза. Как правило, пептидный линкер имеет длину по меньшей мере в две аминокислоты. Расщепляющий агент может включать катепсин В, катепсин D и плазмин, и он способен гидролизовать пептид с высвобождением лекарственного средства в клетки-мишени.
Пептидный линкер может расщепляться тиолзависимой протеазой катепсином-В, который сверхэкспрессируется в раковых тканях, и, например, может использоваться линкер Phe-Leu или Gly-Phe-Leu-Gly. Кроме того, данный пептидный линкер может расщепляться, например, внутриклеточной протеазой и может представлять собой линкер Val-Cit или линкер Phe-Lys.
В одном воплощении расщепляемый линкер является рН-чувствительным и может быть чувствительным к гидролизу при определенном значении рН. В общем, рН-чувствительные линкеры могут гидролизоваться при кислотных условиях. Примеры кислотолабильных линкеров, способных к гидролизу в лизосоме, могут включать гидразоны, семикарбазоны, тиосемикарбазоны, цис-аконитовые амиды, ортоэфиры, ацетали, кетали и тому подобное.
В другом воплощении линкер может расщепляться при восстанавливающих условиях и может включать, например, дисульфидный линкер. Разные дисульфидные связи могут образоваться с использованием SATA (N-сукцинимидил-S-ацетилтиоацетат), SPDP (N-сукцинимидил-3-(2-пиридилдитио)пропионат), SPDB (N-сукцинимидил-3-(2-пиридилдитио)бутират) и SMPT (N-сукцинимидил-оксикарбонил-альфа-метил-альфа-(2-пиридил-дитио)толуол).
Лекарственное средство и/или лекарственное средство-линкер может случайным образом быть конъюгирован через лизин антитела или через цистеин, экспонированный при восстановлении цепи дисульфидной связи. В некоторых случаях линкер-лекарственное средство может связываться через генетически модифицированную метку, например, цистеин, присутствующий в пептиде или белке. Данная генетически модифицированная метка, например, пептид или белок, может включать аминокислотный мотив, который может распознаваться, например, изопреноидтрансферазой. Данный пептид или белок может иметь делецию на карбокси конце пептида или белка, или может иметь присоединение посредством ковалентной связи спейсерного элемента к карбоксильному (С) концу данного пептида или белка.
Также линкер может представлять собой, например, нерасщепляемый линкер, и лекарственное средство может высвобождаться посредством только одного этапа гидролиза антитела с получением, таким образом, например, комплеса аминокислота/линкер/лекарственное средство. Данный тип линкера может представлять собой тиоэфирную группу или малеимидокапроильную группу и может оставаться стабильным в крови.
Лекарственное средство в конъюгате антитело-лекарственное средство может представлять собой средство, демонстрирующее фармакологический эффект, и может связываться с антителом, и их конкретные примеры могут включать химиотерапевтическое средство, токсин, микроРНК (миРНК), миРНК (малая интерферирующая РНК), кшРНК (короткая шпилечная РНК) и радиоактивный изотоп. Химиотерапевтическое средство может представлять собой, например, цитотоксическое средство или иммуносупрессивное средство. В частности, оно может включать ингибитор микротрубочек, ингибитор митоза, ингибитор топоизомеразы или химиотерапевтическое средство, способное функционировать в качестве интеркалятора ДНК. Оно также может включать иммуномодулирующее соединение, противораковое средство, противовирусное средство или их комбинации.
Такое лекарственное средство может представлять собой по меньшей мере одно, выбранное из группы, состоящей из, например, следующих: майтансиноид, ауристатин, аминоптерин, актиномицин, блеомицин, талисомицин, камптотецин, N8-ацетилспермидин, 1-(2-хлорэтил)-1,2-диметилсульфонилгидразид, эсперамицин, этопозид, 6-меркаптопурин, доластатин, трихотецен, калихеамицин, таксол, таксан, паклитаксел, доцетаксел, метотрексат, винкристин, винбластин, доксорубицин, мелфалан, митомицин А, митомицин С, хлорамбуцил, дуокармицин, L-аспарагиназа, меркаптопурин, тиогуанин, гидроксимочевина, цитарабин, циклофосфамид, ифосфамид, нитрозомочевина, цисплатин, карбоплатин, митомицин, дакарбазин, прокарбазин, топотекан, азотистый иприт, цитоксан, 5-фторурацил, CNU (бисхлорэтилнитрозомочевина), иринотекан, идарубицин, даунорубицин, дактиномицин, пликамицин, митоксантрон, аспарагиназа, винорелбин, хлорамбуцил, мелфалан, кармустин, ломустин, бусульфан, треосульфан, дакарбазин, тенипозид, топотекан, 9-аминокамптотецин, криснатол, триметрексат, микофеноловая кислота, тиазофурин, рибавирин, EICAR (5-этинил-1-бета-дрибофуранозилимидазол-4-карбоксамид), гидроксимочевина, дефероксамин, флоксуридин, доксифлуридин, ралтитрексед, цитарабин (ara С), цитозина арабинозид, флударабин, тамоксифен, ралоксифен, мегестрол, гозерелин, лейпролида ацетат, флутамид, бикалутамид, ЕВ1089, СВ1093, KH1060, вертепорфин, фталоцианин, фотосенсибилизатор Ре4, диметокси-гипокреллин А, интерферон-α, интерферон-γ, фактор некроза опухолей, гемцитабин, велкад, ревлимид, талидомид, ловастатин, ион 1-метил-4-фенилпиридиния, стауриспорин, актиномицин D, дактиномицин, блеомицин А2, блеомицин В2, пепломицин, эпирубицин, пирарубицин, зорубицин, митоксантрон, верапамил, тапсигаргин, нуклеазы и токсины, происходящие из бактерий или животных и растений, но не ограничиваясь ими.
В еще одном другом аспекте согласно настоящему изобретению предложено биспецифичное антитело, в котором антитело против PD-L1 или его антигенсвязывающий фрагмент по настоящему изобретению связывается с антителом, которое связывается с другим антигеном.
Антитело, образующее биспецифичное антитело, наряду с антителом против PD-L1 или его антигенсвязывающим фрагментом по настоящему изобретению, может использоваться без ограничения, при условии, что оно представляет собой антитело, способное к специфичному связыванию с антигеном, ассоциированным с раком или аутоиммунным заболеванием. Примеры такого антигена могут включать 4-1ВВ, интегрин, ангиопоэтин, аналог 3 ангиопоэтина, фактор, активирующий В-клетки (BAFF), В7-Н3, CCR4, CD3, CD4, CD6, CD11a, CD19, CD20, CD22, CD30, CD33, CD38, CD40, CD52, CD62, CD79b, CD80, CGRP, OX-40, ICOS, клаудин-18, CTLA4, DLL3, рецептор EGF, рецептор Fc, FGF23, рецептор фолата, GD2, GM-CSF, HER2, Her2/neu, HER3, VEGF, рецептор VEGF, рецептор интерферона, интерферон-гамма, IgE, рецептор IGF-1, интерлейкин 1, рецептор интерлейкина 2, рецептор интерлейкина 4, интерлейкин 5, рецептор интерлейкина 5, интерлейкин 6, рецептор интерлейкина 6, интерлейкин 7, интерлейкин 12/23, интерлейкин 13, интерлейкин 17А, рецептор А интерлейкина 17, рецептор интерлейкина 31, рецептор интерлейкина 36, LAG3, LFA3, NGF, PVSK9, PD-1, PD-L1, RANK-L, SLAMF7, тканевой фактор, TGF-β и тому подобное.
Более конкретно, примеры антитела, которое связывается с антигеном, описанным выше, включают:
антитела к 4-1 ВВ, такие как утомилумаб;
антитела к интегрину, такие как натализумаб, этролизумаб, ведолизумаб и бимагрумаб;
антитела к амилоиду-бета, такие как бапинеузумаб, кренезумаб, соланезумаб, адуканумаб и гантенерумаб;
антитела к ангиопоэтину, такие как AMG 780;
антитела к аналогу ангиопоэтина 3, такие как эвинакумаб;
антитела к фактору, активирующему В-клетки (BAFF), такие как табалумаб, ланалумаб и белимумаб;
антитела к В7-Н3, такие как омбуртамаб;
антитела к CCR4, такие как могамулизумаб;
антитела к CD3, такие как отеликсизумаб, теплизумаб, муромонаб, тебентафусп, которое представляет собой биспецифичное антитело к GP100 и CD3, блинатумомаб, который представляет собой биспецифичное антитело KCD19 и CD3, и REGN1979, который представляет собой биспецифичное антитело к CD20 и CD3;
антитела к CD4, такие как ибализумаб и занолимумаб; антитела к CD6, такие как итолизумаб; антитела к CD11 а, такие как эфализумаб;
антитела к CD19, такие как инэбилизумаб, тафаситамаб и лонкастуксимаб тесирин в качестве ADC;
антитела к CD20, такие как окрелизумаб, ублитуксимаб, обинутузумаб, офатумумаб, ритуксимаб, тозитумомаб и ибритумомаб тиуксетан в качестве ADC;
антитела к CD22, такие как эпратузумаб и, в качестве ADC, инотузумаб озогамицин и моксетумомаб пасудотокс;
ADC к CD30, такие как брентуксимаб ведотин;
ADC к CD33, такие как вадастуксимаб талирин и гемтузумаб озогамицин;
антитела к CD38, такие как даратумумаб и изатуксимаб;
антитела к CD52, такие как алемтузумаб;
антитела к CD62, такие как кризанлизумаб;
ADC к CD79b, такие как полатузумаб ведотин;
антитела к CD80, такие как галиксимаб;
антитела к CGRP, такие как эптинезумаб, фреманезумаб, галканезумаб и эренумаб;
антитела к клаудину-18, такие как золбетуксимаб;
антитела к CTLA4, такие как тремелимумаб, залифрелимаб и ипилимумаб; ADC к DLL3, такие как ровалпитузумаб тезирин;
антитела к рецептору EGF, такие как цетуксимаб, депатуксизумаб, залутумумаб, нецитумумаб и панитумумаб;
антитела к рецептору Fc, такие как нипокалимаб и розаноликсизумаб; антитела к FGF23, такие как бурозумаб;
антитела к рецептору фолата, такие как фарлетузумаб и мирветуксимаб соравтазин в качестве ADC;
антитела к GD2, такие как динутуксимаб и накситамаб; антитела к GM-CSF, такие как отилимаб;
антитела к HER2, такие как маргетуксимаб, пертузумаб, трастузумаб и, в качестве ADC, трастузумаб дерукстекан, трастузумаб эмтанзин и трастузумаб дуокармазин;
антитела к HER3, такие как патритумаб;
антитела к рецептору интерферона, такие как анифролумаб;
антитела к интерферону-гамма, такие как эмапалумаб;
антитела к IgE, такие как лигелизумаб и омализумаб;
антитела к рецептору IGF-1, такие как далотузумаб, фигитумумаб и тепротумумаб;
антитела к интерлейкину 1, такие как гевокизумаб и канакинумаб; антитела к рецептору интерлейкина 2, такие как даклизумаб и базиликсимаб;
антитела к рецептору интерлейкина 4, такие как дупилумаб; антитела к интерлейкину 5, такие как меполизумаб и реслизумаб; антитела к рецептору интерлейкина 5, такие как бенрализумаб; антитела к интерлейкину 6, такие как клазакизумаб, олокизумаб, сирукумаб и силтуксимаб;
антитела к рецептору интерлейкина 6, такие как сарилумаб, сатрализумаб, тоцилизумаб и REGN88;
антитела к интерлейкину 7, такие как секукинумаб;
антитела к интерлейкину 12/23, такие как устекинумаб и бриакинумаб;
антитела к интерлейкину 13, такие как лебрикизумаб и тралокинумаб;
антитела к интерлейкину 17А, такие как иксекизумаб и бимекизумаб;
антитела к рецептору А интерлейкина 17, такие как бродалумаб;
антитела к интерлейкину 23, такие как бразикумаб, гузелкумаб, ризанкизумаб, тилдракизумаб и мирикизумаб;
антитела к рецептору интерлейкина 31, такие как немолизумаб;
антитела к рецептору интерлейкина 36, такие как спесолимаб;
антитела к LAG3, такие как релатлимаб;
антитела к NASP2, такие как нарсоплимаб;
антитела к NGF, такие как фазинумаб и танезумаб;
антитела к PVSK9, такие как алирокумаб, эволокумаб и бокоцизумаб;
антитела к PD-1, такие как ламбролизумаб, балстилимаб, камрелизумаб, цемиплимаб, достарлимаб, пролголимаб, синтилимаб, спартализумаб, тислелизумаб, пембролизумаб и ниволумаб;
антитела к PD-L1, такие как атезолизумаб, авелумаб, энвафолимаб, и дурвалумаб;
антитела к RANK-L, такие как деносумаб; антитела к SLAMF7, такие как элотузумаб;
антитела к тканевому фактору, такие как концизумаб и марстацимаб;
антитела k TNF, в частности, k TNFα, такие как инфликсимаб, адалимумаб, голимумаб, цертолизумаб пегол, который представляет собой фрагмент антитела, и озорализумаб, который представляет собой биспецифичное антитело к TNF и альбумину;
антитела к VEGF, такие как бролуцизумаб, ранибизумаб, бевацизумаб и фарицимаб, которое представляет собой биспецифичное антитело к VEGF и Ang2; и
антитела к рецептору VEGF, такие как рамуцирумаб, но не ограничиваются ими.
Лучшее понимание настоящего изобретения может быть получено посредством следующих примеров. Данные примеры просто излагаются для иллюстрации настоящего изобретения, и их не следует истолковывать как ограничивающие объем настоящего изобретения, как будет очевидно обычным специалистам в данной области.
ПРИМЕРЫ
Пример 1. Экспрессия и очистка антигена PD-L1
С использованием вектора (Sino Biological), содержащего ген кДНК PD-L1 в качестве матрицы, фрагмент, содержащий внеклеточную область (Met1-Thr239) из сигнальной последовательности на N-конце гена PD-L1, подвергали ПЦР(полимеразная цепная реакция)-амплификации и вставляли в сайты рестрикционных ферментов NheI и SfiI вектора pCEP4-Fc в качестве вектора для слитой экспрессии человеческого фрагмента Fc, таким образом, конструируя вектор "pCEP4-PDL1-Fc".
Затем экспрессионный вектор PD-L1 вводили в клетки Freestyle 293-F с использованием FectoPRO (Polyplus), который представляет собой реактив трансфекции клеток, согласно способу трансфекции, предоставленному изготовителем, и культуру осуществляли в течение примерно от 6 до 10 суток таким образом, что белок-антиген высвобождался из клеток.
Белок-антиген PD-L1, высвобожденный в культуральную среду клеток, очищали посредством способа очистки с открытой колонкой с использованием смолы с белком A (Amicogen), элюировали лимонной кислотой и затем нейтрализовали 1 М раствором Tris при рН 9,4. Нейтрализованный белок-антиген очищали посредством способа диализа 5 раз с использованием 1×буфера PBS и использовали для последующих экспериментов.
Пример 2. Повторная амплификация библиотеки фагов
1) Для отбора человеческих антител, связывающихся с PD-L1, использовали методику «фагового дисплея». Для отбора человеческих антител-кандидатов посредством фагового дисплея использовали три типа библиотек человеческих антител, и они представляли собой: 1) библиотеку наивных scFv, описанную в 'PNAS, 1999 vol.96, pp. 6953-6958', 2) библиотеку синтетических scFv, описанную в 'Molecules and Cells, 2009, vol. 27, pp.225-235', и 3) 'библиотеку наивных scFv LG', разработанную LG Life Sciences.
2) Для фагового дисплея фаги повторно амплифицировали из Е, coli для того, чтобы, таким образом, продуцировать фаги, осуществляющие дисплей scFv. Данный способ был следующим: библиотеку «1)» получали с использованием способа «спасения фага, осуществляющего дисплей scFv» в данной статье, библиотеку «2)» получали согласно способу «спасения библиотеки», упомянутому в данной статье, и библиотеку «3)» получали посредством отдельного способа, и его подробное описание было следующим:
3) 1 флакон клеток, содержащих фаги, оттаивали, высевали в 2 л 2×YT (содержащей 1% глюкозы, 34 мкг/мл хлорамфеникола и 5 мМ MgCl2), культивировали во встряхиваемом инкубаторе при 37°С до достижения ОП600 (оптическая плотность при длине волны 600 нм) 0,5, смешивали с хелперным фагом VCSM13 при значении MOI (множественность заражения) 20 и культивировали в инкубаторе при 37°С в течение 45 минут без встряхивания для обеспечения полной инфекции индивидуальных клеток хелперным фагом.
4) Инфицированные клетки переносили в 500 мл пробирку Sorvall и центрифугировали при 5000 об./мин в течение 15 минут, после чего клеточный осадок ресуспендировали в 2 л 2×YT (содержащей 1 мМ IPTG (изопропилтиогалактозид), 34 мкг/мл хлорамфеникола, 70 мкг/мл канамицина и 5 мМ MgCl2) и затем культивировали в течение ночи во встряхиваемом инкубаторе (220 об./мин) при 30°С, таким образом, продуцируя фаги.
5) После культивирования клеток в течение ночи (220 об./мин, 30°С) среду клеточной культуры центрифугировали при 4°С и 8000 об./мин в течение 20 минут, осадок клеточной культуры отбрасывали, и в супернатант добавляли раствор PEG (полиэтиленгликоль)/NaCl (20% масс/об. PEG 6000, 0,25 М NaCl) в объеме, соответствующем 1/5 объема супернатанта, и инкубировали на льду в течение примерно 1 часа для обеспечения агломерации и оседания на дно фагов, с последующим центрифугированием при 8000 об./мин при 4°С в течение 20 минут с получением осадка фага. Супернатант декантировали, осадок фага ресуспендировали примерно в 80 мл PBS (фосфатно-солевой буферный раствор) с получением образца раствора фага, и примеси в образце раствора фага удаляли с использованием 0,45 мкм шприцевого фильтра, завершая, посредством этого, получение образца фаговой библиотеки.
Пример 3. Фаговый пэннинг
1) С использованием эпоксишариков Dynabeads™ М-270, доступных от Dynal, примерно 10 мкг белка-антигена PD-L1 связывали с 12,5 мкл шариков согласно экспериментальному способу, предоставленному изготовителем, для получения магнитных шариков, с которыми был связан белок-антиген, с последующей реакцией блокирования посредством инкубации при комнатной температуре в течение 1 часа в буфере PBS, содержащем 3% BSA.
2) Примерно 1013 человеческого антитела, экспрессированного на поверхности фагов, очищенного посредством осаждения PEG/NaCl с использованием шприцевого фильтра, суспендировали в растворе PBS, содержащем 1% BSA (бычий сывороточный альбумин) и 0,05% Tween, и помещали в иммунопробирку (Nunc), покрытую 12,5 мкг/мл ранее полученного образца человеческого антитела (Green Cross, 10% в.в. (внутривенная) инъекция глобулина SN), с последующей реакцией вычитания для удаления фагов, связанных с областью Fc (при комнатной температуре в течение 2 часов).
3) После завершения вычитания образующийся раствор фага смешивали с магнитными шариками, с которыми был связан белок-антиген, перемешивали при комнатной температуре в течение примерно 2 часов, таким образом, что обеспечивалось связывание с магнитными шариками фагов с антителом, способных к связыванию с белком-антигеном, и затем помещали в магнитизированную колонку MACS (Miltenyi Biotech) для обеспечения связывания антител с магнитными шариками, с которыми был связан белок-антиген.
4) Во время пэннинга жесткость условий корректировали таким образом, что могли быть отобраны многие высокоаффинные антитела при увеличении числа процессов промывки, например, сначала промывая один раз 6 мл PBS, второй и третий раз промывая два раза 6 мл PBS, четвертый раз промывая три раза 7 мл PBS и тому подобное.
5) После промывки буфером PBS в колонку MACS инъецировали 700 мкл раствора 0,25% трипсина для заполнения данной колонки раствором трипсина, колонку культивировали в инкубаторе при 37°С в течение 30 минут для индукции элюции фага, и данную колонку центрифугировали при 1000 об./мин в течение 1 минуты для получения элюированных фагов, которые затем смешивали со свежеприготовленной Е. coil ER2738 (NEB), культивируемой до уровня ОП600 примерно 0,7-0,8 для индукции инфекции Е. coli элюированными фагами.
6) Е. coli, инфицированные фагами, распределяли на чашки с твердой средой LB (содержащей 1% глюкозы), включающей соответствующие антибиотики, и культивировали в течение ночи в инкубаторе при 30°С таким образом, что Е. coli могли вырастать до состояния лужайки. После завершения роста Е. coli, покрывающие чашку, отбирали с использованием скребка, и фаговые частицы повторно амплифицировали посредством способа, использованного в Примере 2.
Пример 4. Отбор антитела, связывающегося с белком-антигеном PD-L1, посредством экспериментов ELISA (твердофазный иммуноферментный анализ) и FACS (флуоресцентная сортировка клеток).
1) Для того, чтобы подтвердить связывание с белком-антигеном PD-L1, получали периплазматические фракции, содержащие фрагменты антитела, из примерно 1700 бактериальных штаммов, экспрессирующих индивидуальные антитела, и осуществляли эксперименты по отбору ELISA и FACS.
2) Для того, чтобы получить фрагмент белка антитела от каждого штамма Е. coli, единичные колонии бактерий высевали в 96-глубоколуночный планшет, в котором распределяли примерно 1 мл среды LB, культивировали во встряхиваемом инкубаторе при 37°С и затем в нее добавляли 1 мМ IPTG при достижении значения ОП600 примерно 0,7-0,8, с последующей инкубацией со встряхиванием в течение ночи при 30°С для индукции экспрессии фрагмента антитела в периплазме Е. coli.
3) На следующие сутки, для того, чтобы экстрагировать периплазматический компонент, культивируемую Е. coli центрифугировали (4000 об./мин, 15 минут, 4°С), супернатант декантировали, и Е. coli полностью ресуспендировали в 80 мкл ледяного 1×буфера TES (20% масс/об. сахарозы, 50 мМ Tris, 1 мМ EDTA (этилендиаминтетрауксусная кислота), рН 8,0), давали постоять на льду в течение 30 минут, дополнительно добавляли 120 мкл ледяного 0,2 × буфера TES, перемешивали и давали постоять в течение еще примерно 30 минут для индукции экстракции периплазматического компонента с использованием осмотического давления. Затем клетки удаляли посредством центрифугирования, и супернатант использовали для экспериментов по подтверждению ELISA и FACS в качестве периплазматического компонента, содержащего фрагменты антитела.
4) Для эксперимента по подтверждению ELISA для подтверждения связывания белка-антигена PD-L1 примерно 100 нг/лунку белка антигена растворяли в 100 мкл буфера PBS и инкубировали в течение ночи при 4°С для того, чтобы таким образом покрыть им половину размера лунок планшета ELISA (Corning, 3690). На следующие сутки буфер заменяли примерно 170 мкл буфера PBS, содержащего 3% BSA, с последующей реакцией блокирования посредством инкубации при 37°С в течение примерно 1 часа.
5) 50 мкл супернананта, образующегося в результате экстракции периплазматического компонента, и 50 мкл PBS инкубировали при комнатной температуре в течение примерно 1 часа таким образом, что фрагменты антитела могли связываться с покрывающим белком-антигеном, с последующей промывкой 2-3 раза. Распределяли буфер PBS, содержащий вторичное антитело против myc, конъюгированное с HRP (пероксидаза хрена), или антитело против НА, конъюгированное с HRP, с последующей инкубацией при комнатной температуре в течение примерно 1 часа и затем промывая 2-3 раза буфером PBS.
6) 100 мкл реакционного раствора ELISA, содержащего тетраметилбензидин (ТМВ), который является субстратом HRP, распределяли каждый раз, и связывание фрагмента антитела с антигеном PD-L1 определяли посредством наблюдения цветной реакции.
7) Эксперимент FACS проводили для того, чтобы подтвердить связывание отобранных фрагментов антитела с природной конформацией PD-L1, экспрессируемого на поверхности клеток. Для этого белок-антиген PD-L1 экспрессировали в клетках 293Т посредством трансфекции, после чего определяли связывание фрагментов антитела с экспрессируемым на поверхности антигеном PD-L1 посредством проточной цитометрии, и экспериментальную процедуру проводили согласно общему экспериментальному способу FACS.
8) Клетки 293Т, в которые был введен экспрессионный вектор белка PD-L1, делили таким образом, чтобы получать примерно от 5×105 до 1 х106 клеток/100 мкл, распределяли в 96-луночном планшете с V-образным дном, подвергали реакции блокирования и давали реагировать со 100 мкл смешанного раствора периплазматического компонента, содержащего фрагмент антитела против PD-L1 с буфером PBS 1:1 при 4°С (или на льду) в течение примерно 30-60 минут, таким образом, индуцируя связывание фрагмента антитела с белком-антигеном. Затем несвязанные компоненты удаляли посредством центрифугирования (1000 об./мин в течение 1 минуты при 4°С), проводили промывку буфером FACS, и образующиеся клетки ресуспендировали в буфере FACS, содержащем разведение 1:400 вторичных антител против метки, конъюгированных с Alexa Fluor 488, с последующей реакцией при 4°С в течение еще 30-60 минут. Затем несвязанные компоненты удаляли посредством центрифугирования (1000 об./мин, 1 минута, 4°С), флуоресцентно меченые клетки ресуспендировали в 400-700 мкл буфера FACS, и присутствие или отсутствие флуоресцентного окрашивания клеток анализировали с использованием проточного цитометра FACSCalibur (Becton Dickinson).
Как показано на ФИГ. 1, периплазматический компонент получали из примерно 1800 колоний, полученных из 3-ей и 4-ой элюций эксперимента по пэннингу белка PD-L1, реакцию ELISA проводили для того, чтобы подтвердить его связывание с белком-антигеном PD-L1, которым был покрыт 96-луночный планшет, и клоны, демонстрирующие цветную реакцию, подвергали секвенированию. В результате получали примерно 72 индивидуальных клонов антител.
Как показано на ФИГ. 2, на основе результатов FACS на клетках 293Т с введением белка PD-L1 для того, чтобы подтвердить способность нативной конформации данных 72 индивидуальных клонов к связыванию с белком PD-L1, обнаружили, что целый ряд антител связывается с поверхностью клеток, на которых экспрессируется белок PD-L1.
Пример 5. Анализ ингибирования связывания PD-1/PD-L1 на основе технологии BLI
1) Эксперимент для отбора функционального антитела проводили на основе принципа «интерферометрии биослоя (BLI)» с использованием прибора Octet, доступного у Pall. В частности, проводили гидратацию биосенсоров AR2G, активацию ЕОС/сульфо-NHS и инактивацию непрореагировавших функциональных групп с использованием 1 М этаноламина, при покрытии 5 мкг/мл белка PD-1 в 10 мМ ацетате натрия при рН 5, и инкубацию в 1×подвижном буфере, приводя к уравновешиванию.
2) Для того чтобы определять влияние интерференции на взаимодействие между антигеном PD-L1 и белком PD-1 из-за связывания белка-антигена PD-L1 и фрагмента антитела, биосенсор, покрытый белком PD-1, погружали в лунку, в которой были смешаны белок-антиген PD-L1 и антитело, связывающееся с PD-L1 (или сравнительное антитело), и измеряли изменение массы на поверхности биосенсора, приписываемое связыванию белка-антигена PD-L1 в растворе.
3) Экспериментальные результаты анализировали с использованием программы для анализа данных, предоставленной для применения с прибором Octet.
Как показано на ФИГ. 3, оценивали, ингибировалось ли связывание белка-антигена PD-L1 в растворе с белком PD-1, которым был покрыт биосенсор, посредством обеспечения связывания отобранного антитела с PD-L1 через последовательную реакцию отобранного антитела scFv и PD-L1-Fc после связывания белка PD-1-Fc с биосенсором Octet.
Эффективность ингибирования связывания отдельных антител-кандидатов, имеющих разные вариабельные области тяжелой цепи и легкой цепи, измеряли с использованием прибора Octet и, таким образом, при совместной инкубации фрагментов антитела идентифицировали примерно 50 или больше антител-кандидатов (среди всего 72 антител-кандидатов), демонстрирующих такую характеристику, что увеличение картины сенсограммы из-за снижения связывания PD-L1 было снижено.
Пример 6: превращение IgG
1) Вариабельные области VH и VL примерно 58 отдельно взятых антител подвергали ПЦР-амплификации с использованием набора праймеров человеческих антител для осуществления амплификации областей VH и VL отдельных антител, идентифицированных посредством секвенирования отдельных клонов антител, с последующей очисткой.
2) Фрагменты VH обрабатывали рестрикционными ферментами Kpnl или BamHI и Nhel, очищали и вставляли в соответствующий сайт рестрикционного фермента вектора pCEP4-VH для экспрессии тяжелой цепи, а фрагменты VL обрабатывали рестрикционными ферментами Kpnl или BamHI и BsiWI, очищали и вставляли в соответствующий сайт рестрикционного фермента вектора pCEP4-VL для экспрессии легкой цепи, полученные таким образом векторы способны экспрессировать индивидуальные антитела в клетках животных (58 типов экспрессионных векторов каждой тяжелой цепи и легкой цепи).
3) Каждый вектор очищали с использованием набора DNA Maxi-prep, доступного от Qiagen, и клетки Freestyle 293-F трансфицировали им с использованием трансфекционного реактива FectoPRO, доступного от Polyplus, таким образом, экспрессируя индивидуальные антитела. После культивирования образующийся супернатант подвергали способу аффинной очистки с использованием смолы с белком А для очистки белка антитела.
4) Для того, чтобы служить в качестве контрольного антитела, в соответствующих патентах идентифицировали аминокислоты вариабельной области тяжелой цепи и легкой цепи МК3475, которое представляет собой нацеленное на PD-1 антитело, разработанное Merck, и MPDL3280A, которое представляет собой нацеленное на PD-L1 антитело, разработанное Genentech, синтезировали кДНК и вставляли в вектор pCEP4-VH и в вектор pCEP4-VL для получения экспрессионных векторов, и осуществляли такие же процедуры, как и для антител-кандидатов, при этом получали два антитела, очищали и использовали в качестве контрольных антител.

Пример 7. Анализ FACS связывания мышиного PD-L1
1) Клетки "МС38" (5×106 клеток/10 мл) из линии клеток мышиного рака толстой кишки, экспрессирующие мышиный PD-L1, обрабатывали 100 нг/мл IFN-γ и культивировали в течение примерно 48 часов для повышения уровня экспрессии PD-L1 на поверхности данной линии раковых клеток.
2) После культивирования клетки МС38 делили таким образом, чтобы получать примерно от 5×105 до 1×106 клеток/100 мкл и распределяли в 96-луночном планшете с V-образным дном, с последующей реакцией блокирования согласно общему экспериментальному способу FACS, после чего давали реагировать очищенным антителам против PD-L1 при концентрации 10 мкг/мл при 4°С (или на льду) в течение примерно 30-60 минут, после чего несвязавшиеся антитела отмывали и удаляли посредством центрифугирования с использованием промывочного буфера FACS (1000 об./мин в течение 1 минуты при 4°С).
3) Клетки со связавшимся антителом ресуспендировали в буфере FACS, содержащем разведение 1:400 вторичных антител козы против человеческих антител, конъюгированных с Alexa Fluor 488, и затем давали реагировать при 4°С в течение примерно 30-60 минут.Затем несвязавшиеся вторичные антитела отмывали промывочным буфером FACS посредством центрифугирования (1000 об./мин в течение 1 минуты при 4°С).
4) Клетки ресуспендировали в 400-700 мкл буфера FACS, после чего присутствие или отсутствие флуоресцентного окрашивания клеток анализировали с использованием проточного цитометра FACSCalibur (Becton Dickinson), таким образом, что была подтверждена способность первичных антител связываться с поверхностью мышиного белка PD-L1.
Как показано на ФИГ. 4, на основе результатов, подтверждающих способность к связыванию с поверхностью клеток МС38, экспрессирующих мышиный PD-L1, с использованием антител-кандидатов, экспрессированных и очищенных в виде IgG посредством FACS, обнаружили большое число антител, демонстрирующих способность к связыванию с мышиным PD-L1. В частности, подтвердили то, что антитела KL001 (PL110) и PL112 имели очень сильную способность к связыванию.
Пример 8. Анализ блокирования связывания PD-1/PD-L1
1) Известны разные экспериментальные способы в качестве способов подтверждения эффективности на основе клеток in vitro для подтверждения эффективности ингибирования связывания белка иммунологической контрольной точки PD-1 и белка PD-L1 в качестве его лиганда, но эффективность ингибирующей способности PD-L1 отобранных клонов антител была подтверждена с использованием «биоанализа блокировки PD-1/PD-L1», который представляет собой набор для анализа на основе клеток, предоставленный Promega, и в наборе Promega интенсивность значения люминисцеции, измеренная для клеток, была пропорциональна интенсивности эффективности ингибирования связывания PD-1/PD-L1.
2) Проводили эксперимент согласно протоколу, предоставленному в наборе Promega, и он кратко обобщается ниже.
3) За одни сутки до анализа «клетки PD-L1 оттаять и использовать» были оттаяны в водяной бане при 37°С, добавлены в среду для извлечения клеток и смешаны с легкий перемешиванием, после чего примерно 100 мкл их каждый раз распределяли в планшете для анализа с белым дном с использованием многоканальной пипетки, с последующей инкубацией в течение ночи.
4) В сутки анализа супернатант декантировали, и образцы для оценки эффективности, а именно: образцы серийно разведенного раствора, сравнительного антитела и антитела-кандидата, добавляли в каждую лунку в объеме раствора 40 мкл, после чего забирали из морозильника эффекторные клетки PD1 (CS187105) «оттаивай и используй», оттаивали в водяной бане при 37°С, тщательно суспендировали в буфере для анализа, и 40 мкл его распределяли в каждую лунку, обработанную образцом антитела.
5) Обеспечивали протекание реакции активации-ингибирования клеток Jurkat (эффекторные клетки PD-1) в инкубаторе с CO2 при 37°С в течение 6 часов, и помещали в каждую лунку 80 мкл раствора анализа люциферазы Bio-Glo, содержащего субстрат люциферазы, который нагревали, пока он не становился теплым, и значение люминисценции измеряли с использованием многометочного планшет-ридера VICTOR (Perkin Elmer).
Как показано на ФИГ. 5, на основе результатов биоанализа блокировки PD-1/PD-L1 in vitro от Promega на 58 антителах, связывающихся с PD-L1, очищенных в виде IgG, подтвердили то, что были открыты клоны-кандидаты, имеющие сильную эффективность ингибирования связывания PD-1/PD-L1, аналогичную антителу, нацеленному на PD-1 (антитело MK3475), и антителу, нацеленному на PD-L1 (антитело MPDL3280A), используемых в качестве контрольных антител.
В качестве значения принимали эффективность ингибирования связывания PD-1/PD-L1 антител-кандидатов относительно 100%-ной эффективности ингибирования антитела MK3475, и клон №50, и клон KL001 демонстрировали эффективность ингибирования 91% и 89% соответственно. Клон KL001 демонстрировал перекрестную реактивность с антигеном PD-L1 человека-мыши, и клон №50 демонстрировал характеристику связывания только с человеческим PD-L1, и данные клоны, соответственно, обозначали кодовыми наименованиями KL001 и KL002 с разработкой кандидатов.
Пример 9. Модель исследования сингенных мышей MC38/C57BL/6 in vivo
1) Клетки мышиного рака толстой кишки МС38 культивировали в среде, дополненной 10% фетальной телячьей сыворотки, при 37°С и 5% CO2 в течение примерно 1 недели, и проверяли их морфологию, жизнеспособность, время удвоения и загрязнение микоплазмой. Получали незагрязненные раковые клетки в нормальном состоянии для применения в экспериментах на животных, и при инъекции экспериментальным животным использовали клетки с жизнеспособностью клеток 95% или больше.
2) Клетки МС38 отбирали с использованием трипсина-EDTA и суспендировали в концентрации 1×107 клеток/мл в холодном PBS для пересадки, и полученную таким образом суспензию клеток подкожно инъецировали в количестве 5×105 клеток/50 мкл в правую заднюю ногу мышей C57BL/6 с использованием шприца для пересадки. Затем периодически наблюдали образование и рост опухоли, и объем опухоли рассчитывали подстановкой измеренных размеров опухоли в следующее уравнение.
[Объем опухоли=(a2b)/2; а - короткое измерение опухоли, b - длинное измерение опухоли]
3) При достижении размера опухоли 80 мм3 плюс/минус 20 мышей переклассифицировали, в зависимости от данного объема и группировали в контрольные группы (группа обработки PBS и группа обработки сравнительным антителом) и 4 типа групп обработки антителом, нацеленным на PD-L1, примерно 200 мкг/200 мкл образца антитела вводили посредством внутрибрюшинной инъекции, и устанавливали цикл инъекций 3 раза (1 сутки, 4 сутки и 7 суток) с интервалом 3 суток после первой инъекции.
4) Рост раковой ткани измеряли каждые 3 суток до начала обработки образцом антитела и каждые 2 суток после начала обработки, создавали кривую роста опухоли с объемом опухоли с исходной даты обработки образцом антитела до конца эксперимента (в течение примено 2 недель) и получали для применения в качестве подтверждающих данных для оценки эффективности антитела. При достижении объемом опухоли предела, определенного Институциональным комитетом по уходу и применению животных (IACUC), соответствующих животных умерщвляли. В конце эксперимента мышей умерщвляли, опухоль вырезали, опухоль взвешивали, и фотографировали форму опухоли.
5) Для того чтобы сравнить противораковую эффективность in vivo антител, демонстрирущих высокую ингибирующую эффективность, и антител, демонстрирующих среднюю эффективность в анализе оценки эффективности in vitro, проводили эксперимент по оценки эффективности in vivo с использованием четырех типов антител: №50 (KL002), KL001 (PL110), №8, №61 и контрольного антитела MPDL3280A.
Как показано на ФИГ. 6, на основе результатов анализа противораковой эффективности in vivo антител-кандидатов с использованием линии клеток МС38 рака толстой кишки мышиного происхождения и модели сингенных мышей C57BL/6, антитело-кандидат KL001 (PL110), имеющее перекрестную реактивность человек-мышь, демонстрировало превосходную противораковую эффективность, и антитело №50 (KL002), связывающееся только с человеческим PD-L1, не демонстрировало противораковой эффективности.
Также для клонов №8 и №61, которые не демонстрировали высокой эффективности в эксперименте по эффективности in vitro, противораковая эффективность in vivo не была сравнительно высокой.
Пример 10. Анализ иммунорегулирующей активности ex-vivo антитела KL001
1) Для того чтобы подтвердить эффективность ex-vivo антител-кандидатов при взаимодействии между иммунными клетками в крови посредством подтверждения того, может ли увеличиваться иммунная активность посредством связывания антитела-кандидата, нацеленного на PD-L1, в явлении ограничения иммунной активности из-за связывания PD-1/PD-L1, индуцированного в условиях иммунной активации между иммунными клетками в человеческом организме, из человеческой крови выделяли одноядерные клетки периферической крови (РВМС), РВМС, стимулированные суперантигенами - SEA или SEB - обрабатывали иммунными противораковыми антителами, и измеряли изменения активности между иммунными клетками.
2) После получения одобрения от Институционального экспертного совета (IRB) штаб-квартиры по гемотрансфузиологии Корейского красного креста приобретали концентрат эритроцитов (cRBC) в Gangwon Blood Center, разводили 1:1 с использованием PBS и центрифугировали на Ficoll-Paque таким образом, что разделяли эритроциты (RBC), одноядерные клетки периферической крови (РВМС) и плазму, в зависимости от удельной плотности клеток, и часть в виде РВМС экстрагировали и использовали для эксперимета.
3) Отделенные одноядерные клетки периферической крови (РВМС) распределяли в количестве 1×105 клеток/100 мкл в 96 лунок (u-образное дно), после чего каждую лунку обрабатывали 0,003 мкг/100 мкл SEB (стафилококковый энтеротоксин В - суперантиген, Toxin Technology), и затем 10 мкг/мл каждого антитела из экспериментальной группы и контрольной группы с созданием условий для осуществления активации иммунных клеток. Через 96 часов количественно определяли количество IL-2, генерированного из-за активации иммунных клеток в среде культуры клеток с использованием набора ELISA для IL-2, подтверждая, посредством этого, иммунорегулирующую активность антител-кандидатов против человеческих иммунных клеток.
Как показано на ФИГ. 7 и 8, в анализе активации иммунных клеток посредством анализа SEB подтвердили то, что происходило случайное поперечное связывание между АРС (антигенпрезентирующие клетки) и Т-клетками во время обработки SEB, и что активность иммунных клеток возрастала, и, таким образом, увеличивалась секреция IL-2. Подтвердили способность к активации иммунных клеток отобранного ранее антитела-кандидата (KL001) (на данной Фиг. показано среднее значение пяти экспериментов), и также подтвердили то, что антитело-кандидат KL001-13, отобранное после созревания аффинности антител-кандидатов, демонстрирует аналогичную способность регулировать активность иммунных клеток.

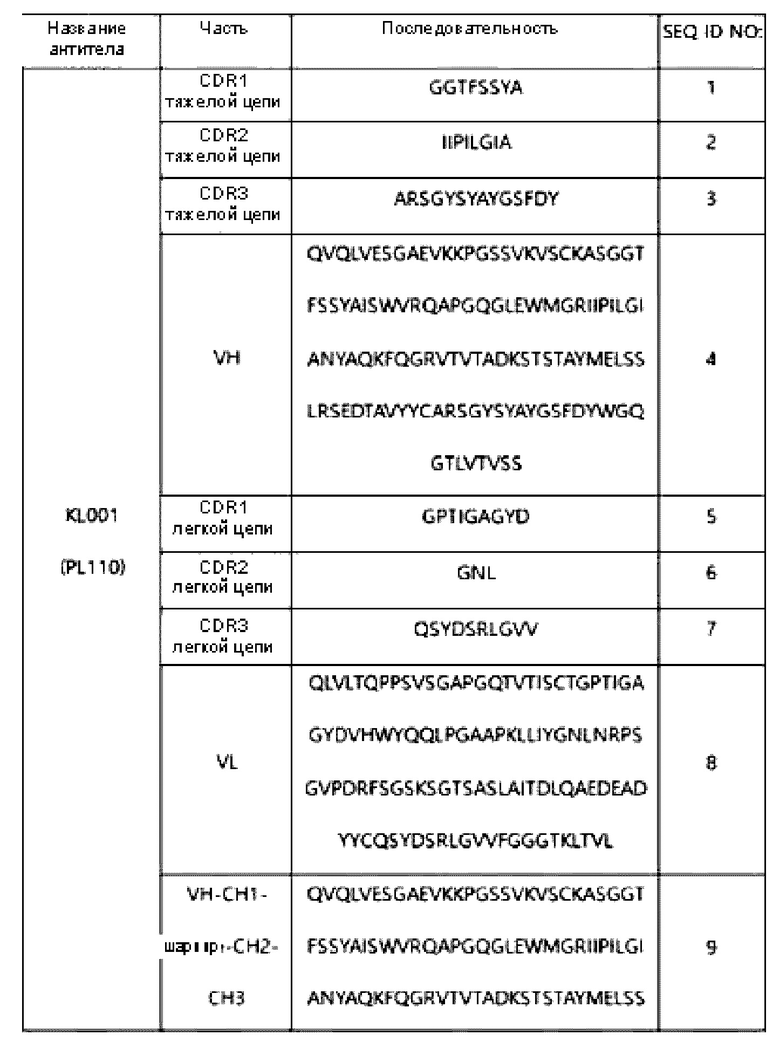


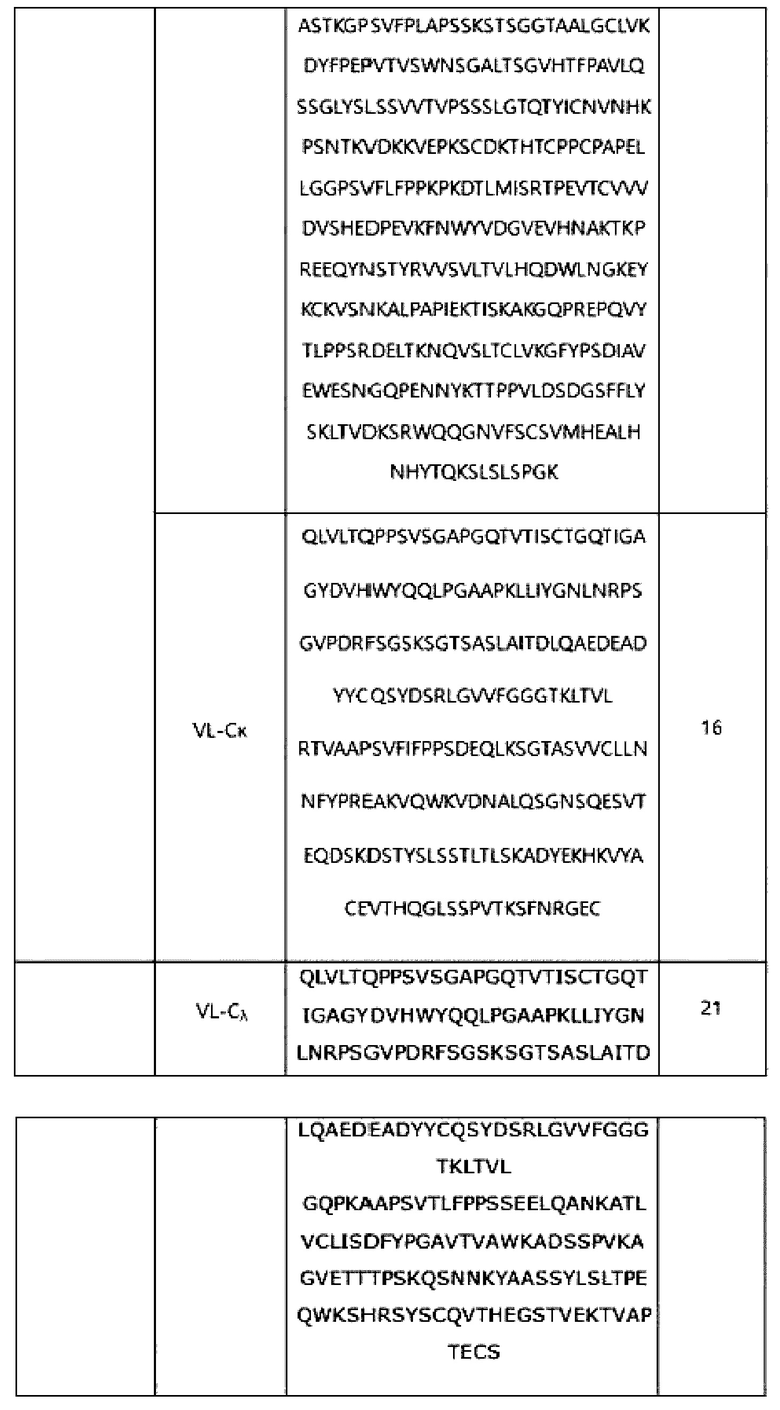
Пример 11. Эксперимент по созреванию аффинности
1) Конструировали библиотеку направленного мутагенеза посредством интенсивного введения мутаций в области CDR2 и 3 тяжелой цепи, и области CDR1 и 3 легкой цепи в базовом остове антитела KL001.
2) Фрагменты, в которых мутации были введены в области VH и VL клона KL001 были амплифицированы в качестве вставок для конструирования библиотеки с использованием ПЦР с перекрывающимися праймерами, данный фрагмент KL001 обрабатывали рестрикционным ферментом для клонирования в вектор фагового дисплея «pComb3XSS», и проводили реакцию лигирования на векторе, обработанном таким же рестрикционным ферментом.
3) Лигированные вектор фагового дисплея pComb3X и вставку трансформировали в компетентные для электропорации клетки, полученные с использованием клеток ER2738, для конструирования библиотеки дисплея мутантного антитела KL001.
4) С использованием хелперного фага VCSM13 получали фаги, осуществляющие дисплей мутантных антител KL001, и затем очищали посредством способа осаждения с использованием PEG (полиэтиленгликоль) и NaCl.
5) С использованием биотинилированного антигена PD-L1-Fc проводили 3-4 цикла пэннинга, и колонии, полученные от 3-го и 4-го циклов пэннинга, использовали для реакции ELISA для отбора антител на улучшение аффинности.
6) Отобранные фрагменты антител экспрессировали в периплазме с использованием IPTG, периплазматические фракции получали с использованием сахарозы, и затем отбирали антитела-кандидаты, для которых ожидалось то, что они продемонстрируют повышенную способность к связыванию, посредством ELISA с использованием планшета, покрытого антигеном PD-L1.
7) Отбор антител-кандидатов, имеющих повышенную способность к связыванию, завершали с использованием конкурентного ELISA.
Как показано на ФИГ. 9, фрагменты ДНК направленного мутагенеза, образующиеся в результате введения сайт-направленных мутаций в области CDR2 и 3 тяжелой цепи и области CDR1 и 3 легкой цепи антитела KL001, связывали посредством ПЦР с перекрывающимися праймерами с получением остова антитела Fab-типа, который затем вставляли в сайт рестрикционного фермента Sfil вектора фагового дисплея «pComb3XSS» посредством лигирования и трансформировали в клетки Е, coli ER2738 для фагового дисплея посредством электропорации, конструируя, посредством этого, библиотеку дисплея мутантного антитела KL001 размера «1,3×109». Для того чтобы обогатить клоны антител, имеющих сильную способность к связыванию, с использованием биотинилированного антигена PD-L1-Fc осуществляли 3-4 цикла пэннинга при жестких экспериментальных условиях таким образом, что количество антигена снижалось. Здесь подтвердили от результатов измерения пэннинга на входе и на выходе то, что количество сильных связывателей эффективно увеличивалось посредством пэннинга.
Как показано на ФИГ. 10, периплазматические фракции получали из примерно 1600 индивидуальных колоний (от набора 1 до набора 16), образующихся в результате 3-го и 4-го циклов пэннинга, проводили реакцию ELISA для отбора антител с улучшенной аффинностью, и первично отбирали клоны-кандидаты, демонстрирующие положительный сигнал (примерно 130 клонов).
Проводили конкурентный ELISA для того, чтобы определять, улучшалась ли способность к связыванию антигена (или улучшалось ли значение Koff), инкубацию проводили в течение еще примерно 4 часов в растворе, содержащем белок-антиген PD-L1, и подтверждали клоны антител, сохраняющие связывание, таким образом, что идентифицировали примерно 27 типов антител-кандидатов, которые были сильно связаны с PD-L1, покрывающим планшет, по сравнению с KL001 (исходное антитело KL001, которое представляет собой сравнительную группу, демонстрировало значение ELISA «0,278», и антитела с улучшенной аффинностью были идентифицированы как клоны, представляющие более высокие значения, чем данное значение).
Как очевидно из Таблицы 6, приведенной ниже, аминокислотные последовательности антител, в которые были введены разные мутации, были идентифицированы посредством секвенирования всех антител, для которых ожидается то, что они продемонстрируют повышенную способность к связыванию.
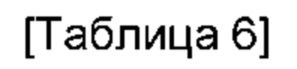
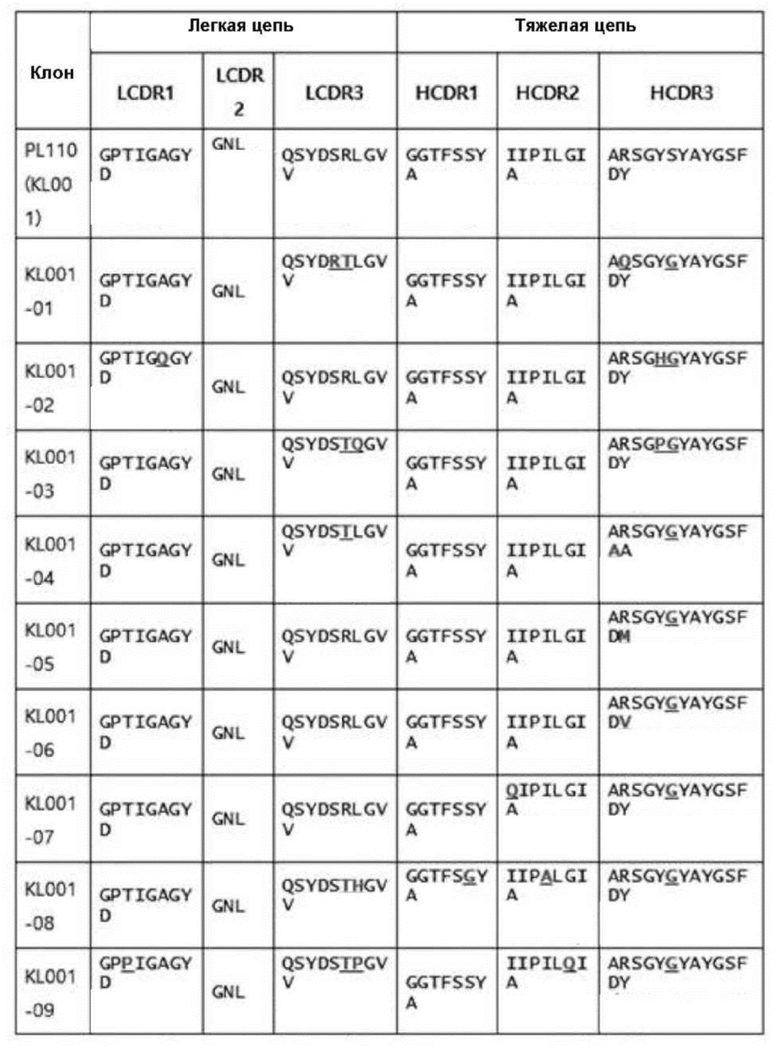


Для того чтобы измерить способность антител с улучшенной аффинностью препятствовать взаимодействию между антигеном PD-L1 и белком PD-1 из-за связывания белка-антигена PD-L1 и фрагмента антитела, биосенсор, покрытый белком PD-1, погружали в лунку, в которой были смешаны белок-антиген PD-L1 и антитело, связывающееся с PD-L1 (или сравнительное антитело), и, таким образом, измеряли изменение массы на поверхности биосенсора, приписываемое связыванию белка-антигена PD-L1 в растворе, с использованием прибора Octet на основе BLI, упомянутого в Примере 5.
В Таблице 7 ниже показаны результаты способности к связыванию антител с улучшенной аффинностью, проанализированных с использованием программы для анализа данных, конкретно предоставленной для применения с прибором Octet.
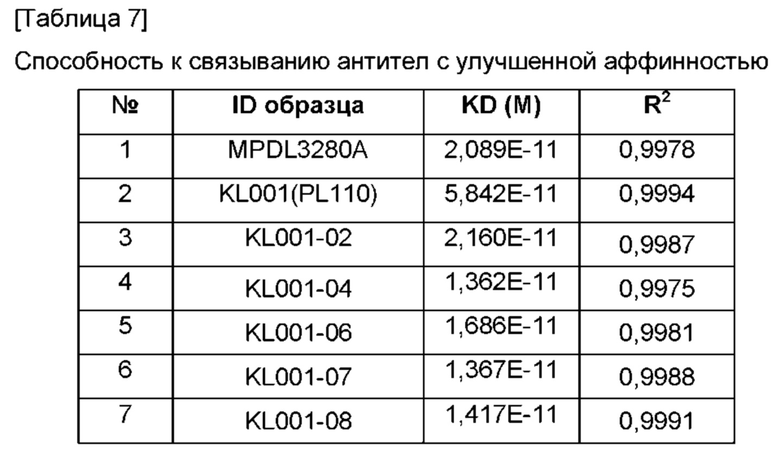
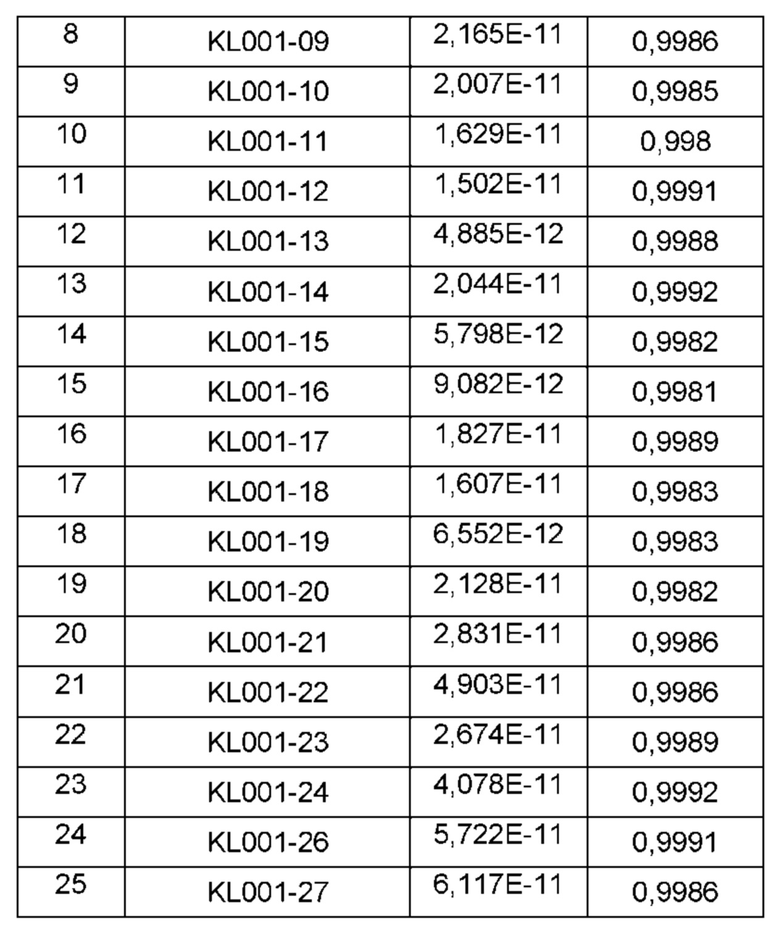
Пример 12. Анализ Biacore
1) Для того чтобы измерить способность антитела-кандидата к связыванию с белком-антигеном PD-L1, измеряли силу связывания с использованием прибора Biacore T200 на основе принципа поверхностного плазмонного резонанса (SPR).
2) В анализе аффинности связывания антитела против PD-L1 с использованием способа захвата антитела сначала на поверхности чипа СМ5 иммобилизовали антитело против человеческого IgE (Fc) с использованием набора для захвата человеческих антител согласно руководству для данного экспериментального способа, предоставленному изготовителем, и антитела-кандидаты KL001-8 и KL001-13 разводили в подвижном буфере PBS-P, инъецировали при скорости тока 10 мкл/мин в течение 30 секунд и захватывали на уровне от 300 до 330 RU, после чего человеческий белок-антиген PD-L1 (Sino Biological) подвергали серийному 2-кратному разведению от 5 нМ, и наблюдали реакции ассоциации и диссоциации при скорости тока 30 мкл/мин (было установлено соответствующее им время реакции 4 и 7 минут). При измерении способности к связыванию с мышиным белком-антигеном PD-L1 способность к связыванию измеряли посредством серийного 2-кратного разведения от 10 нМ, а ассоциацию и диссоциацию - в течение 4 мин и 3 мин, соответственно, при скорости тока 30 мкл/мин.
3) Для того чтобы удалить связавшееся антитело в каждом цикле, инъецировали регенерирующий раствор (3 М MgCl2) при скорости тока 30 мкл/мин в течение 30 секунд, и экспериментальные данные были поучены с использованием модели связывания 1:1 программы BIA evaluation версии 1.0.
4) В эксперименте для анализа аффинности связывания антитела против PD-L1 с использованием способа захвата антигена антитело против His иммобилизовали на поверхности чипа CM5 с использованием набора для захвата His согласно инструкциям изготовителя, захватывали белок-антиген PD-L1, антитело-кандидат подвергали серийному 2-кратному разведению от 2,5 нМ, и его способность к связыванию измеряли посредством ассоциации и диссоциации в течение 4 мин и 7 мин, соответственно, при скорости тока 30 мкл/мин.
Как показано на ФИГ. 11, антитело против человеческого IgG (Fc) иммобилизовали в проточных ячейках №3 и 4. Иммобилизацию проводили согласно инструкциям для набора для захвата человеческих антител, указывающим иммобилизацию в пределах интервала примерно от 11000 до 13000 RU в каждой проточной ячейке. Антитела KL001-8 и KL001-13 захватывали с использованием подвижного буфера (PBS-P), PD-L1 подвергали серийному разведению и инъецировали, и измеряли кинетику, на основе чего подтверждали то, что антитело KL001-8 демонстрировало способность к связыванию с человеческим PD-L1 с KD 6,714×10-11 M и с мышиным PD-L1 с KD 6,401×10-9 M, и антитело KL001-13 демонстрировало способность к связыванию с человеческим PD-L1 примерно с KD 6,320×10-11 M и с мышиным PD-L1 - 2,797×10-9 M.
Пример 13. Оценка противораковой эффективности посредством одиночной обработки с использованием модели сингенных мышей in vivo
Противораковую эффективность антител-кандидатов подтверждали с использованием модели сингенных мышей in vivo
Как показано на ФИГ. 12, два типа антител с оптимизированной эффективностью и антитело MPDL3280A в.б. вводили в подкожную модель сингенных мышей МС38, и оценивали их эффективность по ингибированию роста опухоли. Относительно контрольной группы, которой вводили PBS, группа MPDL3280A демонстрировала ингибирование 94,3%, группа KL001 (PL110) демонстрировала ингибирование 97,5%, группа антитела KL001-13 с улучшенной аффинностью связывания демонстрировала ингибирование 98,9%, и не наблюдали изменения массы тела из-за лекарственного средства.

Таблица, демонстрирующая результаты TGI (% ингибирования роста опухоли, TGI%=(1-(T/C))×100)
Пример 14. Измерение IC50 для подтверждения эффективности in vitro
1) Для того чтобы подтвердить эффект улучшения эффективности in vitro антител-кандидатов, полученных посредством исследований для улучшения аффинности и физических свойств, в отличие от сравнительного антитела, значение IC50 для ингибирующего эффекта подтверждали посредством измерения способности ингибирования связывания PD-1/PD-L1 серийно разведенных образцов антитела-кандидата и сравнительного антитела с использованием набора для «биопробы блокады PD-1/PD-L1», предоставленного Promega.
2) Данный эксперимент проводили согласно протоколу, предоставленному в наборе Promega, и он обобщается ниже.
3) За одни сутки до анализа "клетки PD-L1 оттаивай и используй» оттаивали в водяной бане при 37°С, добавляли в среду для восстановления клеток, смешивали легким помешиванием, и примерно 100 мкл их распределяли каждый раз в планшет для анализа с белым дном с использованием многоканальной пипетки, с последующей инкубацией в течение ночи.
4) В сутки анализа супернатант декантировали, и образцы для оценки эффективности, а именно: образцы серийно разведенных раствора, сравнительного антитела и антитела-кандидата добавляли в каждую лунку в объеме раствора 40 мкл, после чего эффекторные клетки PD1 «оттаивай и используй» (CS187105) забирали из морозильника, оттаивали в водяной бане при 37°С, аккуратно суспендировали в буфере для анализа, и 40 мкл их распределяли в каждую лунку, обработанную образцом антитела.
5) Обеспечивали прохождение реакции активации-ингибирования с клетками Jurkat в инкубаторе с СО2 при 37°С в течение 6 часов, добавляли в каждую лунку 80 мкл раствора для анализа люциферазы Bio-Glo, содержащего субстрат люциферазы, который нагревали, пока он не становился теплым, и значение люминисценции измеряли с использованием многометочного планшет-ридера VICTOR (Perkin Elmer).
Как показано на ФИГ. 13, на основе результатов, подтверждающих уровень эффективности отобранных антител посредством измерения значения IC50 отобранного антитела с использованием анализа на основе клеток in vitro, большинство антител-кандидатов с улучшенной аффинностью имело низкие значения концентрации IC50 по сравнению со сравнительным антителом MPDL3280A, указывая на улучшенную способность регулировать иммунную активность.
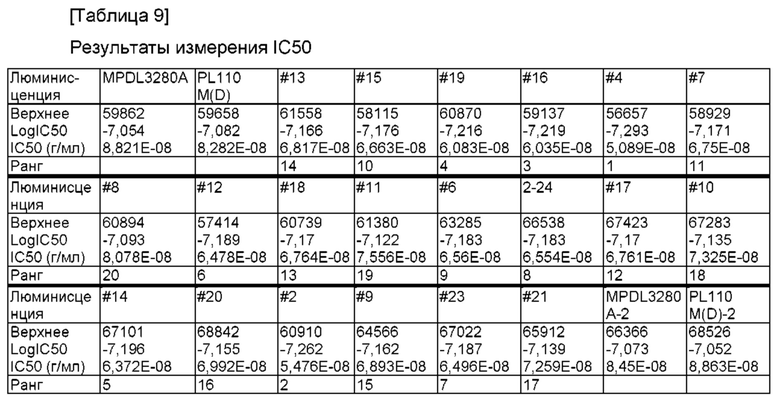
Пример 15. Оценка противоракового эффекта посредством комбинированного лечения in vivo
1) Для того чтобы подтвердить, наблюдался ли синергический эффект в животной модели оценки противораковой эффективности, при использовании иммунного противоракового актитела-кандидата к PD-L1 в комбинации с другими противораковыми средствами, проводили эксперимент для подтверждения эффективности репрезентативного белка иммуноактивации, а именно: рекомбинантного IL-2, антитела антитела против CTLA-4 (клон 9D9, BioXcell) и антитела-кандидата, нацеленного на PD-L1 (KL001 -13), при использовании в комбинации.
2) Для разработки комбинированной терапии с использованием антитела KL001-13 проводили эксперимент по комбинированной обработке с использованием линии клеток мышиного рака толстой кишки МС38 и мышиной модели 6-недельных самцов  C57BL/6, причем данные группы делили всего на 8 групп, включая четыре группы, которым вводили одни PBS, IL-2, CTLA-4 и KL001-13, и четыре группы с совместным введением IL-2 плюс KL001-13, CTLA-4 плюс KL001-13, IL-2 плюс MPDL3280A и CTLA-4 плюс MPDL3280A с 8 мышами в каждой группе (п равен 8).
C57BL/6, причем данные группы делили всего на 8 групп, включая четыре группы, которым вводили одни PBS, IL-2, CTLA-4 и KL001-13, и четыре группы с совместным введением IL-2 плюс KL001-13, CTLA-4 плюс KL001-13, IL-2 плюс MPDL3280A и CTLA-4 плюс MPDL3280A с 8 мышами в каждой группе (п равен 8).
3) Линию клеток мышиного рака толстой кишки МС38 культивировали при плотности клеток 70-90%, обрабатывали 0,25% трипсином-EDTA, собирали и ресуспендировали в среде DMEM (среда Игла, модифицированная по Дульбекко) (бессывороточная) при концентрации клеток 1×106 клеток/100 мкл с получением образца клеток МС38 с жизнеспособностью клеток 95% или больше, который затем подкожно инъецировали в количестве 1×106 клеток/100 мкл в левую сторону тела мышей C57BL/6, анестезированных 2,5%-ным авертином, и давали им расти в течение примерно 7-10 суток в экспериментальных животных - мышах, пока размер опухоли инъецированной линии клеток МС38 не достигал 100 мм3. Здесь размер опухоли рассчитывали следующим образом.
[Объем опухоли=(a2b)/2; а - короткое измерение опухоли, b - длинное измерение опухоли]
4) При достижении размера опухоли 100 мм3, внутрибрюшинно инъецировали приготовленные PBS, антитело-кандидат, сравнительное антитело и цитокин IL-2 либо одиночно, либо в комбинации. Здесь антитела KL001-13, MPDL3280A и CTLA-4 вводили всего 4 раза одни или в комбинации в концентрации 10 мг/кг (200 мкг/100 мкл) с интервалом 3 суток, и цитокин IL-2 вводили всего 5 раз один или в комбинации в концентрации 1×104 IU(международные едницы)/100 мкл с интервалом 2 суток. Размер опухоли (мм3) измеряли 3 раза в неделю, и наблюдали степень роста опухоли. В анализе для подтверждения объединенной эффективности IL-2 и антитела KL001-13 IL-2 обычно вводится в экспериментах на животных ежесуточно из-за короткого периода полувыведения IL-2 у животных, но в настоящем эксперимента IL-2 вводился в малом количестве каждые 2 суток для того, чтобы определить, демонстрировал ли IL-2 в качестве стимулятора иммунной активности синергический эффект при применении в комбинации с антителом, нацеленным на PD-L1.
Как показано на ФИГ. 14, при введении IL-2 и антитела KL001-13 в комбинации могли наблюдать, что противораковая эффективность значимо возрастала по сравнению с тем, когда IL-2 и антитело KL001-13 вводились одиночно, и также, что их противораковый эффект превосходил объединенный эффект IL-2 и антитела MPDL3280A, доступного от Genentech, которым обрабатывали сравнительную группу.
Как показано на ФИГ. 15, на основе результатов подтверждения эффективности введения антитела, нацеленного на белок CTLA-4 иммунологической контрольной точки (клон 9D9, BioXcell), и антитела KL001-13 либо одних, либо в комбинации, наблюдали хороший противораковый эффект в группе, которой вводили одно антитело против PD-L1 по сравнению с группой, которой вводили одно антитело против CTLA-4, и при введении двух данных антител в комбинации был продемонстрирован выдающийся синергический эффект в такой степени, что наблюдали много случаев, при которых раковые клетки были полностью убиты. В частности, наблюдали, что комбинированный эффект антитела KL001-13 превосходил комбинированную эффективность антитела против CTLA-4 и антитела MPDL3280A, доступного от Genentech, использованного в качестве сравнительного антитела.
Промышленная применимость
Согласно настоящему изобретению может быть подтверждено то, что антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент демонстрирует перекрестную реактивность человек-мышь, человек-обезьяна и человек-собака, связывается с PD-L1 с очень высокой аффинностью и ингибирует образование комплекса PD-1/PD-L1. Кроме того, превосходные эффекты могут быть продемонстрированы в анализах на основе клеток in vitro, в экспериментах по эффективности in vivo и в экспериментах по комбинированной терапии. Посредством этого, антитело, связывающееся с PD-L1, или его антигенсвязывающий фрагмент по настоящему изобретению может эффективно использоваться для предупреждения или лечения рака, как это желательно, и может демонстрировать синергический эффект при использовании в комбинации с другим противораковым средством.
Хотя конкретные воплощения настоящего изобретения и были подробно раскрыты, как описано выше, обычным специалистам в данной области будет очевидно то, что данное описание представляет собой просто предпочтительные типичные воплощения, и его не следует истолковывать как ограничивающее объем настоящего изобретения. Следовательно, существенный объем настоящего изобретения будет определен приложенной формулой изобретения и ее эквивалентами.
[Перечень последовательностей в развернутой форме]
Прикреплен электронный файл.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SCRIPPS KOREA ANTIBODY INSTITUTE
<120> АНТИТЕЛА ПРОТИВ ЛИГАНДА 1 ПРОГРАММИРУЕМОЙ ГИБЕЛИ И ИХ ПРИМЕНЕНИЯ
<130> PF-B2653
<140> PCT/KR2020/004854
<141> 2020-04-09
<150> KR 10-2019-0042501
<151> 2019-04-11
<160> 85
<170> PatentIn version 3.5
<210> 1
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 1
Gly Gly Thr Phe Ser Ser Tyr Ala
1 5
<210> 2
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 2
Ile Ile Pro Ile Leu Gly Ile Ala
1 5
<210> 3
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 3
Ala Arg Ser Gly Tyr Ser Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 4
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 4
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Val Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Ser Tyr Ala Tyr Gly Ser Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 5
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 5
Gly Pro Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 6
<211> 3
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 6
Gly Asn Leu
1
<210> 7
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 7
Gln Ser Tyr Asp Ser Arg Leu Gly Val Val
1 5 10
<210> 8
<211> 108
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 8
Gln Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Pro Thr Ile Gly Ala Gly Tyr Asp
20 25 30
Val His Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Asn Leu Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Asp Leu Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu Gly
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 9
<211> 451
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 9
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Val Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Ser Tyr Ala Tyr Gly Ser Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 10
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 10
Gln Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Pro Thr Ile Gly Ala Gly Tyr Asp
20 25 30
Val His Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Asn Leu Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Asp Leu Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu Gly
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 11
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 11
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 12
<211> 121
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 12
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Val Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 13
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 13
Gly Gln Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 14
<211> 108
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 14
Gln Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Gln Thr Ile Gly Ala Gly Tyr Asp
20 25 30
Val His Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Asn Leu Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Asp Leu Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu Gly
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 15
<211> 451
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 15
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Val Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 16
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 16
Gln Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Gln Thr Ile Gly Ala Gly Tyr Asp
20 25 30
Val His Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Asn Leu Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Asp Leu Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu Gly
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 17
<211> 448
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 18
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 19
<211> 450
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Tyr Met Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Lys Phe
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Phe Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 20
<211> 218
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 20
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Gly Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Leu Ala Ser Tyr Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Ser Arg
85 90 95
Asp Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 21
<211> 214
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 21
Gln Leu Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Thr Val Thr Ile Ser Cys Thr Gly Gln Thr Ile Gly Ala Gly Tyr Asp
20 25 30
Val His Trp Tyr Gln Gln Leu Pro Gly Ala Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Asn Leu Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Asp Leu Gln Ala
65 70 75 80
Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Arg Leu Gly
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 22
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 22
Ser Tyr Ala Ile Ser
1 5
<210> 23
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 23
Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala
1 5 10
<210> 24
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 24
Ser Gly Tyr Ser Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 25
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 25
Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 26
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 26
Thr Gly Pro Thr Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 27
<211> 12
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 27
Thr Gly Gln Thr Ile Gly Ala Gly Tyr Asp Val His
1 5 10
<210> 28
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 28
Gly Asn Leu Asn Arg Pro Ser
1 5
<210> 29
<211> 7
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 29
Gly Gly Thr Phe Ser Ser Tyr
1 5
<210> 30
<211> 6
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 30
Ile Pro Ile Leu Gly Ile
1 5
<210> 31
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 31
Gly Pro Thr Ile Gly Gln Gly Tyr Asp
1 5
<210> 32
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 32
Gly Pro Pro Ile Gly Ala Gly Tyr Asp
1 5
<210> 33
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 33
Gly Gln Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 34
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 34
Gly Pro Gln Ile Gly Ala Gly Tyr Asp
1 5
<210> 35
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 35
Gly Pro Thr Ile Gln Ala Gly Tyr Asp
1 5
<210> 36
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 36
Gly Pro Val Ile Gly Ala Gly Tyr Asp
1 5
<210> 37
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 37
Gly Gln Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 38
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 38
Gly Pro Thr Ile Gly Ala Gly Phe Asp
1 5
<210> 39
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 39
Gln Pro Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 40
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 40
Gly Pro Thr Ile Gly Gln Gly Tyr Asp
1 5
<210> 41
<211> 9
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 41
Gly Gln Thr Ile Gly Ala Gly Tyr Asp
1 5
<210> 42
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 42
Gln Ser Tyr Asp Arg Thr Leu Gly Val Val
1 5 10
<210> 43
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 43
Gln Ser Tyr Asp Ser Thr Gln Gly Val Val
1 5 10
<210> 44
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 44
Gln Ser Tyr Asp Ser Thr Leu Gly Val Val
1 5 10
<210> 45
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 45
Gln Ser Tyr Asp Ser Thr His Gly Val Val
1 5 10
<210> 46
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 46
Gln Ser Tyr Asp Ser Thr Pro Gly Val Val
1 5 10
<210> 47
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 47
Gln Ser Tyr Asp Gln Thr Leu Gly Val Val
1 5 10
<210> 48
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 48
Gln Ser Tyr Asp Arg Thr Leu Gly Val Val
1 5 10
<210> 49
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 49
Gln Ser Tyr Asp Ser Ala Leu Gly Val Val
1 5 10
<210> 50
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 50
Gln Ser Tyr Asp Ser Thr Glu Gly Val Val
1 5 10
<210> 51
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 51
Gln Ser Tyr Asp Ser Thr Val Gly Val Val
1 5 10
<210> 52
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 52
Gln Ala Tyr Asp Ser Thr Leu Gly Val Val
1 5 10
<210> 53
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 53
Gln Ser Met Asp Ser Thr Leu Gly Val Val
1 5 10
<210> 54
<211> 10
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 54
Gln Ser Tyr Asp Ser Thr Asp Gly Val Val
1 5 10
<210> 55
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 55
Gly Gly Thr Phe Ser Gly Tyr Ala
1 5
<210> 56
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 56
Gln Ile Pro Ile Leu Gly Ile Ala
1 5
<210> 57
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 57
Ile Ile Pro Ala Leu Gly Ile Ala
1 5
<210> 58
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 58
Ile Ile Pro Ile Leu Gln Ile Ala
1 5
<210> 59
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 59
Ile Ile Pro Ile Met Gly Ile Ala
1 5
<210> 60
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 60
Ile Ile Pro Val Leu Gly Ile Ala
1 5
<210> 61
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 61
Ile Ser Pro Ile Leu Gly Ile Ala
1 5
<210> 62
<211> 8
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 62
Ile Ile Pro Ile Leu Gly Gln Ala
1 5
<210> 63
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 63
Ala Gln Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 64
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 64
Ala Arg Ser Gly His Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 65
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 65
Ala Arg Ser Gly Pro Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 66
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 66
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Ala Ala
1 5 10
<210> 67
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 67
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Met
1 5 10
<210> 68
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 68
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Val
1 5 10
<210> 69
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 69
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 70
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 70
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 71
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 71
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 72
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 72
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 73
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 73
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 74
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 74
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 75
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 75
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 76
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 76
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 77
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 77
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 78
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 78
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 79
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 79
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 80
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 80
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 81
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 81
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 82
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 82
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 83
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 83
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 84
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 84
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Ser Phe Asp Tyr
1 5 10
<210> 85
<211> 14
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 85
Ala Arg Ser Gly Tyr Gly Tyr Ala Tyr Gly Val Phe Asp His
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| АНТИ-PD-L1 АНТИТЕЛА | 2015 |
|
RU2722212C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ РЕЦЕПТОР TGF-БЕТА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2776204C1 |
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714232C2 |
| АНТИТЕЛО ПРОТИВ VSIG4 ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2826236C1 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
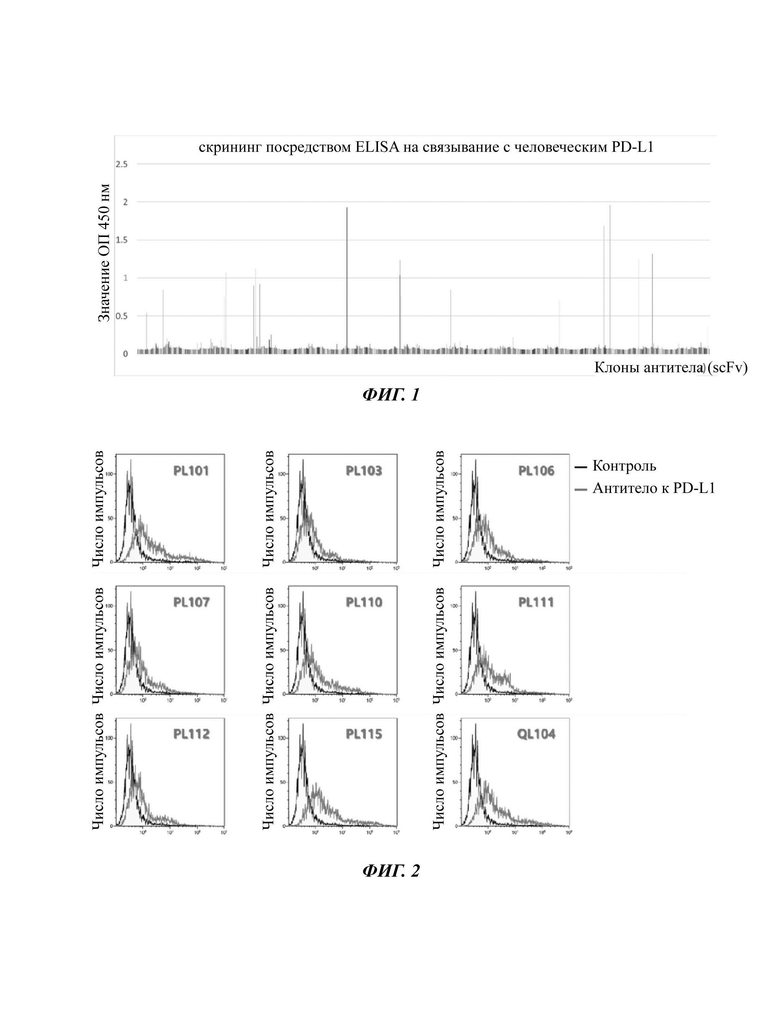
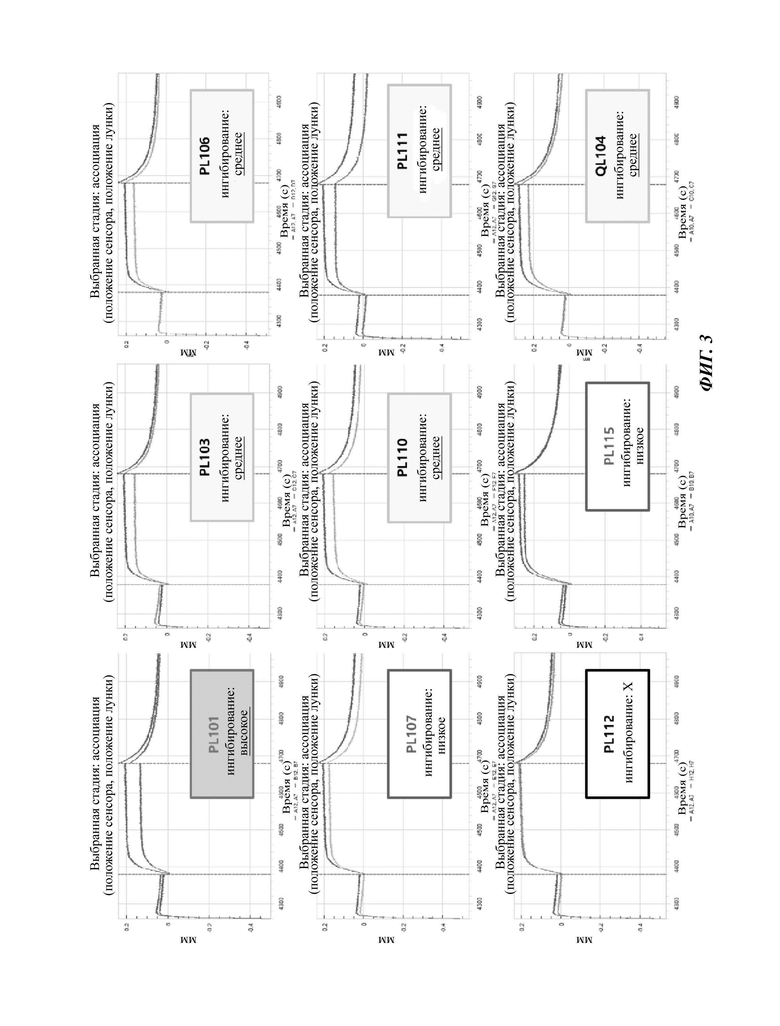

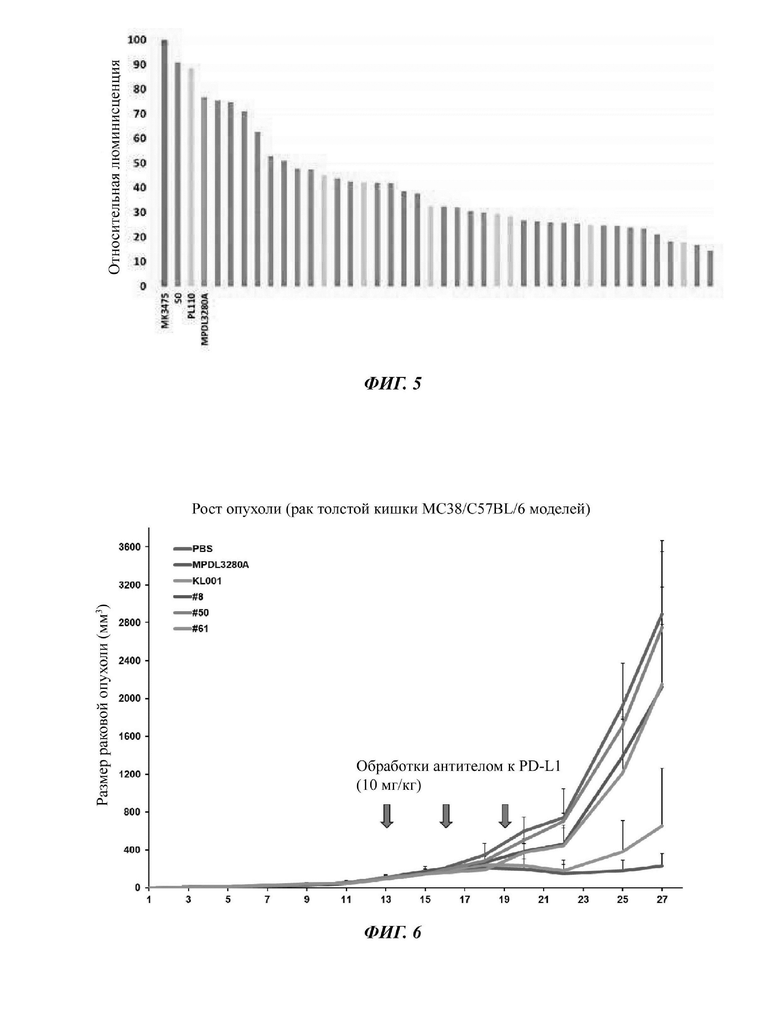
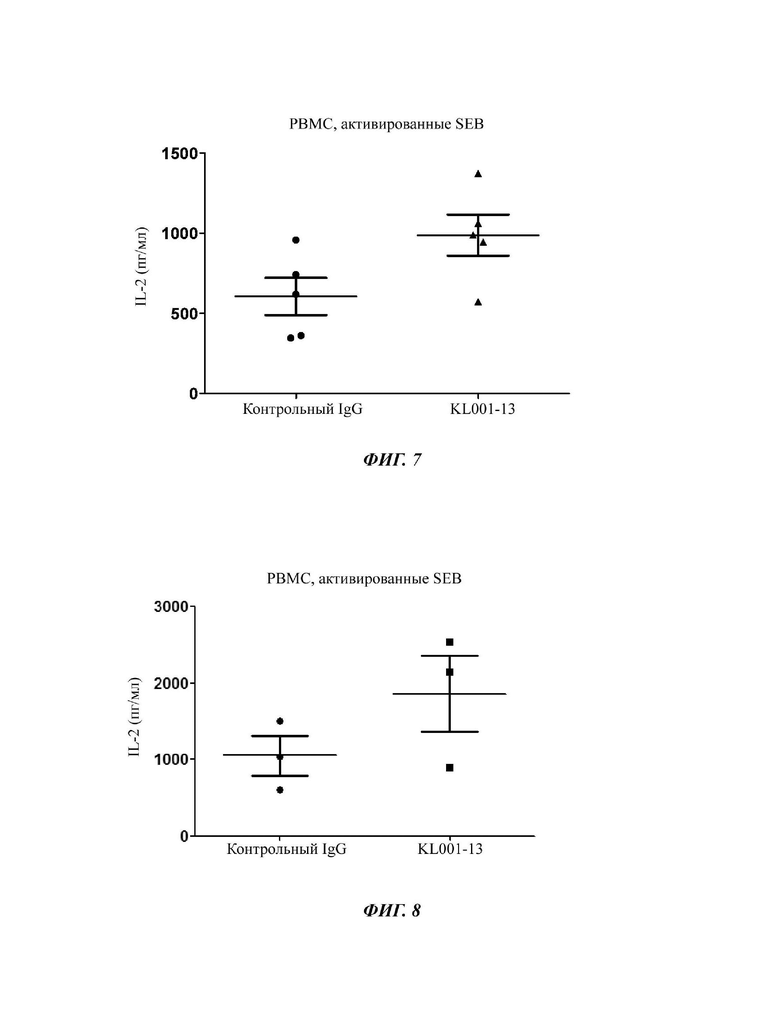
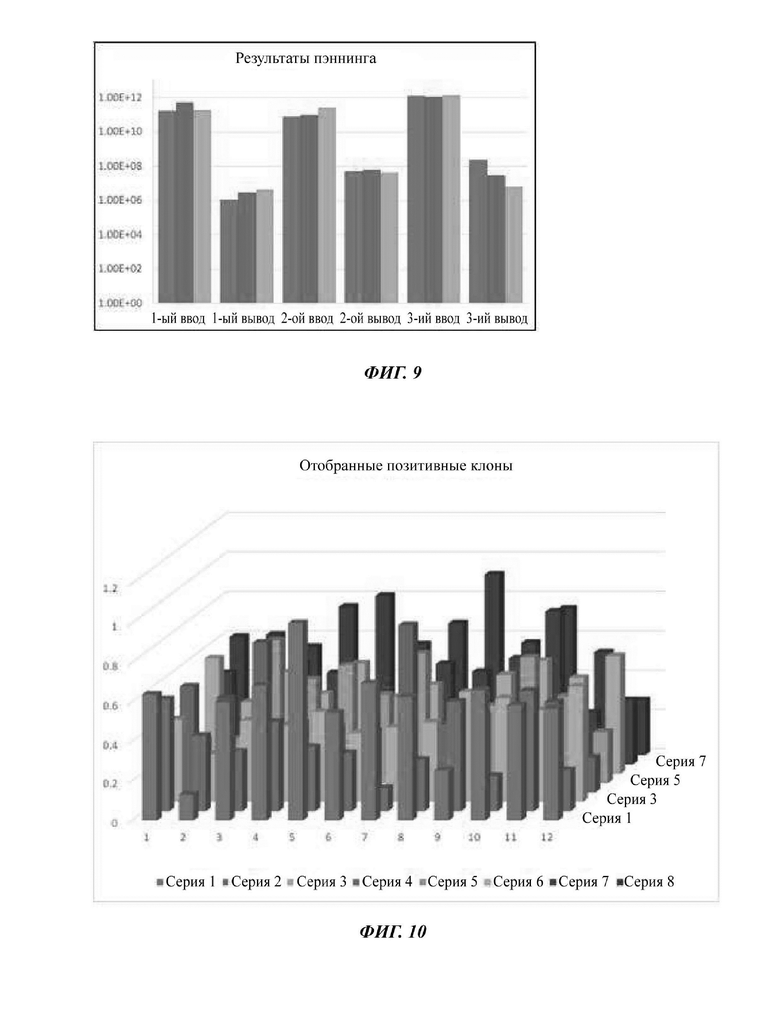
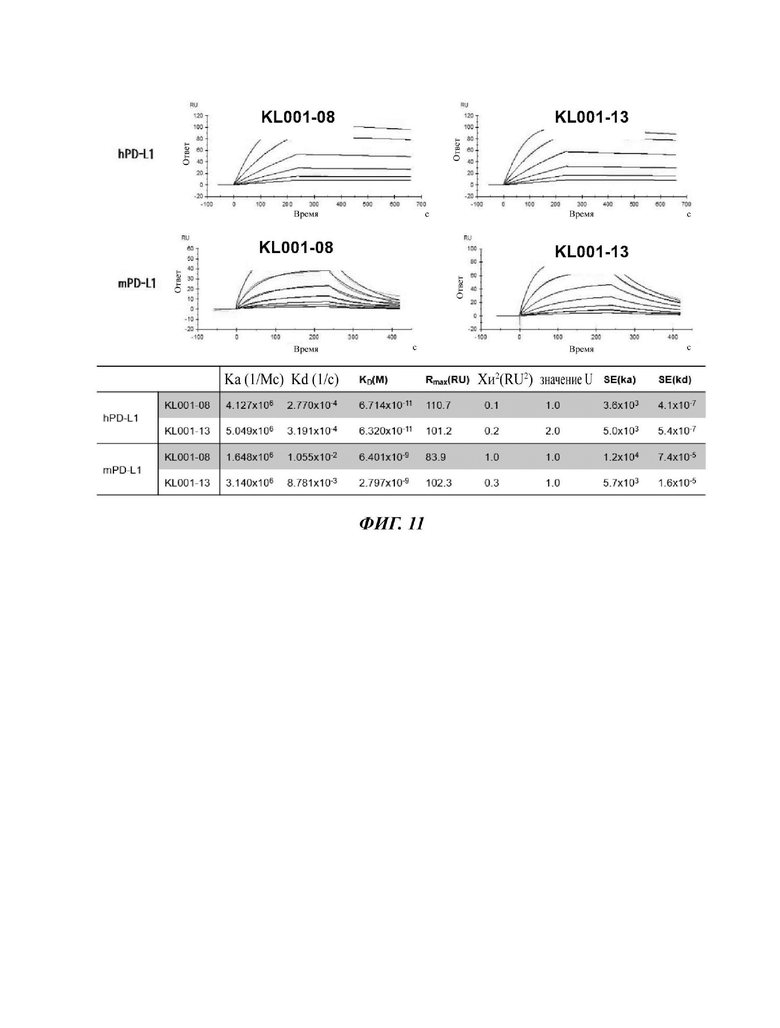
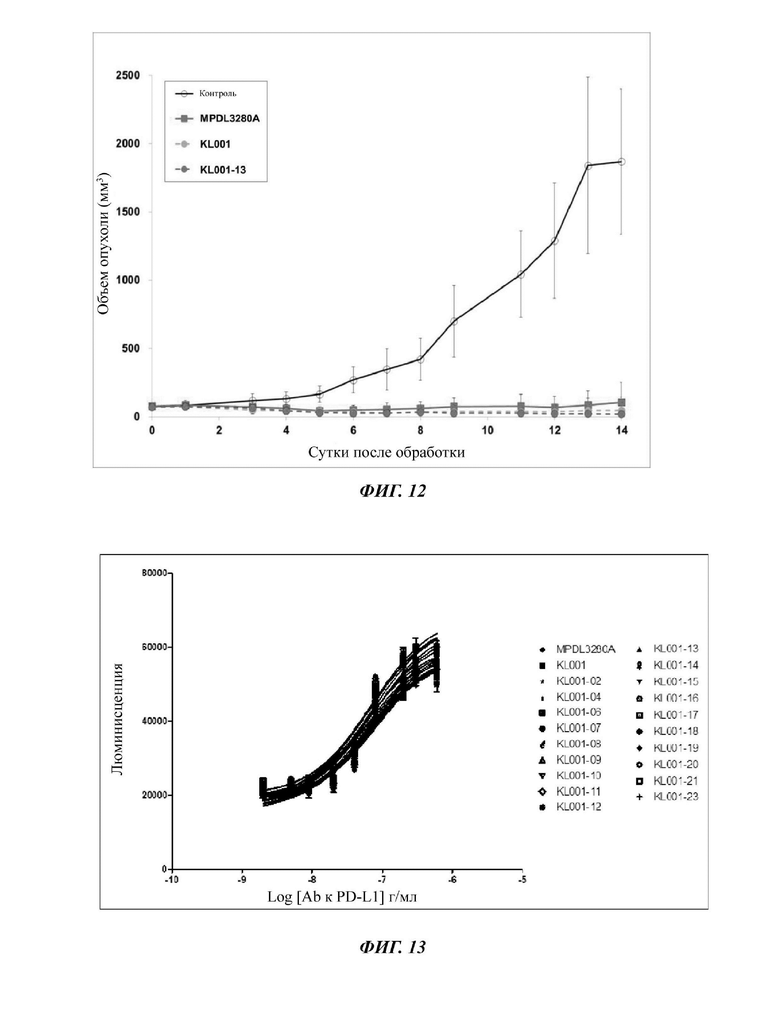


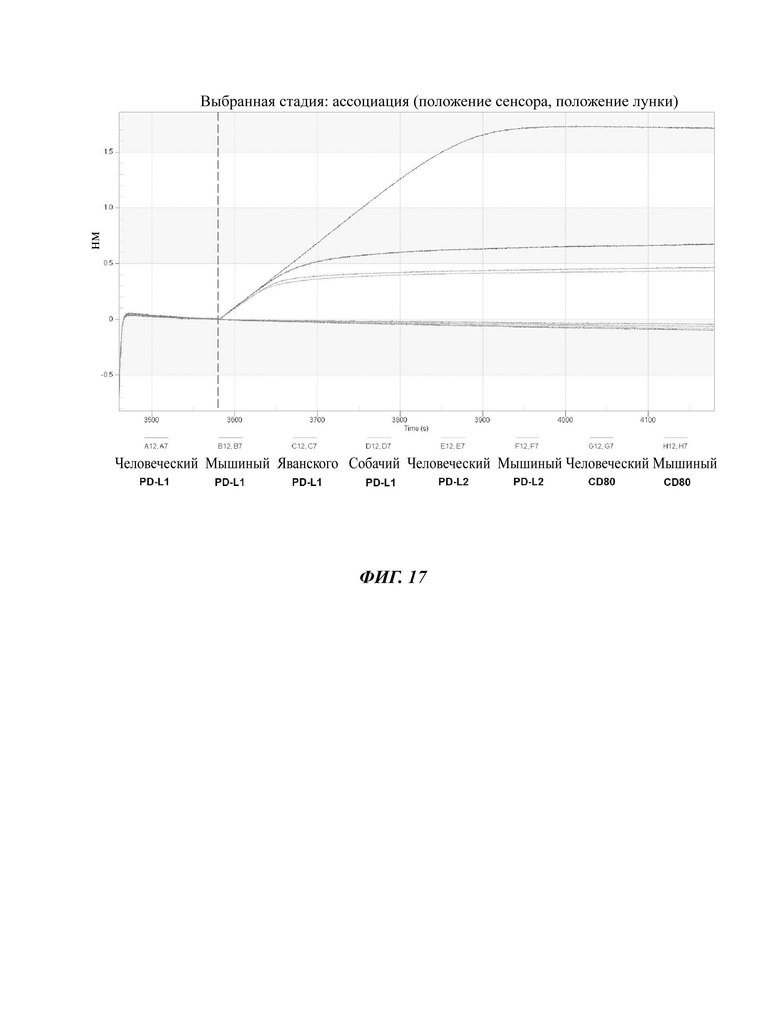
Группа изобретений относится к иммунологии. Предложено антитело к PD-L1 и антигенсвязывающий фрагмент, охарактеризованные аминокислотными последовательностями определяющих комплементарность областей (CDRs) с SEQ ID NO: 1-3 в составе вариабельных областей тяжелой цепи и с SEQ ID NO: 5-7 в составе вариабельных областей легкой цепи или с SEQ ID NO: 1,2,11 в составе вариабельных областей тяжелой цепи и с SEQ ID NO: 13,6,7 в составе вариабельных областей легкой цепи. Кроме того, предложена нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент; экспрессионный вектор; клетка для получения антитела или его антигенсвязывающего фрагмента; способ получения антитела или его антигенсвязывающего фрагмента; композиция, содержащая в качестве активного ингредиента антитело или его антигенсвязывающий фрагмент, для предупреждения или лечения рака. Изобретение позволяет получить новое антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с PD-L1, которые эффективны для предупреждения или лечения рака. 6 н. и 2 з.п. ф-лы, 17 ил., 9 табл., 15 пр.
1. Антитело, специфически связывающееся с PD-L1, или его антигенсвязывающий фрагмент, содержащее: вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с последовательностью SEQ ID NO: 1, CDR2 тяжелой цепи с последовательностью SEQ ID NO: 2 и CDR3 тяжелой цепи с последовательностью SEQ ID NO: 3, вариабельную область легкой цепи, содержащую CDR1 легкой цепи с последовательностью SEQ ID NO: 5, CDR2 легкой цепи с последовательностью SEQ ID NO: 6 и CDR3 легкой цепи с последовательностью SEQ ID NO: 7, или вариабельную область тяжелой цепи, содержащую CDR1 тяжелой цепи с последовательностью SEQ ID NO: 1, CDR2 тяжелой цепи с последовательностью SEQ ID NO: 2 и CDR3 тяжелой цепи с последовательностью SEQ ID NO: 11, вариабельную область легкой цепи, содержащую CDR1 легкой цепи с последовательностью SEQ ID NO: 13, CDR2 легкой цепи с последовательностью SEQ ID NO: 6 и CDR3 легкой цепи с последовательностью SEQ ID NO: 7.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO: 4 или SEQ ID NO: 12.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, содержащее вариабельную область легкой цепи, содержащую последовательность SEQ ID NO: 8 или SEQ ID NO: 14.
4. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3.
5. Экспрессионный вектор, содержащий нуклеиновую кислоту по п. 4.
6. Клетка для получения антитела или его антигенсвязывающего фрагмента по любому из пп. 1-3, которая трансфицирована экспрессионным вектором по п. 5.
7. Способ получения антитела, специфически связывающегося с PD-L1, или его антигенсвязывающего фрагмента, включающий:
(а) культивирование клетки по п. 6; и
(b) очистку антитела или его антигенсвязывающего фрагмента из культивированной клетки.
8. Композиция для предупреждения или лечения рака, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3 в качестве активного ингредиента.
| US 2017319690 A1, 09.11.2017 | |||
| WO 2018195226 A1, 25.10.2018 | |||
| US 8715743 B2, 06.05.2014 | |||
| KR 20180016321 A, 14.02.2018 | |||
| АНТИТЕЛА К PD-L1 И ИХ ПРИМЕНЕНИЕ ДЛЯ УСИЛЕНИЯ ФУНКЦИИ Т-КЛЕТОК | 2009 |
|
RU2636023C2 |
| SHARPE A.H | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| NAT IMMUNOL, 2007, v.8, no.3, p.239-45 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |

