Область техники, к которой относится изобретение
Настоящее изобретение относится к способам и фармацевтическим композициям для лечения рака почки.
Уровень техники
Рак почки составляет около 3% всех солидных опухолей. Около 85% почечных опухолей классифицируются как почечно-клеточный рак (ПКР). Приблизительно 80% диагностированных ПКР происходят из эпителиальных клеток, выстилающих проксимальные части мочеобразующих протоков почек, канальцы. Из-за своего вида под микроскопом этот тип рака известен как светлоклеточный почечно-клеточный рак (СКПКР, 65%) или папиллярный почечно-клеточный рак (ППКР, 15%). Почечно-клеточный рак (ПКР) является восьмым наиболее распространенным злокачественным новообразованием в Соединенных Штатах, для которого в 2016 году было зарегистрировано 62 700 новых случаев и 14 240 предполагаемых случаев смерти. За последнее десятилетие лучшее понимание генетической и метаболической основы ПКР привело к разработке нескольких новых направленных способов лечения метастатического ПКР (мПКР). В условиях метастатического заболевания последовательное использование ингибиторов тирозинкиназы (ИТК), которые нацелены на ангиогенез, и/или ингибиторов мишени рапамицина в клетках млекопитающих (mTOR) может привести к длительной выживаемости без прогрессирования заболевания и общей выживаемости в диапазоне 40 месяцев. Несмотря на этот прогресс, длительные ответы на эти препараты чрезвычайно редки. Таким образом, необходимо найти альтернативную стратегию лечения. Elabela (ELA), также известный как Toddler или Apela, представляет собой пептидный гормон, который недавно был идентифицирован как второй лиганд APJ, рецептора апелина. Полученный как предшественник из 32 аминокислот (aa), ELA также обнаруживается в виде 21 аминокислоты и 11 аминокислот. ELA ограниченно экспрессируется в плюрипотентных стволовых клетках человека, а также в почках и простате взрослых.
Сущность изобретения
Настоящее изобретение относится к способам и фармацевтическим композициям для лечения рака почки. В частности, настоящее изобретение определяется формулой изобретения.
Подробное описание изобретения
Авторы изобретения показывают, что, хотя Elabela (ELA) в основном экспрессируется в почках, его экспрессия снижается при раке почки человека. На модели ксенотрансплантата на животных (подкожная или субкапсулярная инъекция) Ela ингибирует прогрессирование опухоли. Это открытие идентифицируют Ela как новый ген-супрессор опухоли в почках.
Соответственно, первый объект настоящего изобретения относится к способу лечения рака почки у субъекта, нуждающегося в этом, включающему введение субъекту терапевтически эффективного количества полипептида ELA, содержащего аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с SEQ. ID NO: 1 (QRPVNLTMRRKLRKHNCLQRRCMPLHSRVPFP), где остаток аргинина (R) в положении 9, 10, 20 или 21 при необходимости мутирован.
Используемый в настоящей заявке термин «рак почки» имеет общее значение в данной области техники и относится к раку, возникшему из почки. В некоторых вариантах осуществления рак почки является почечно-клеточным раком. Используемый в настоящей заявке термин «почечно-клеточный рак» или «почечно-клеточная карцинома» (ПКР) относится к раку, который возникает в эпителии проксимальных извитых канальцев. Более конкретно, ПКР охватывает несколько относительно общих гистологических подтипов: светлоклеточный почечно-клеточный рак, папиллярный (хромофильный) рак, хромофобный рак, рак собирающих протоков и медуллярный рак. Светлоклеточный почечно-клеточный рак (скПКР) является наиболее распространенным подтипом ПКР.
Используемый в настоящей заявке термин «лечение» или «лечить» относится как к профилактическому или превентивному лечению, так и к лечебному или модифицирующему заболевание лечению, включая лечение пациента с риском развития заболевания или с подозрением на развитие заболевания, а также пациентов, которые больны или были диагностированы как страдающие от заболевания или медицинского состояния, и включает подавление клинического рецидива. Лечение может быть назначено субъекту, имеющему медицинское расстройство, или который в конечном счете может приобрести расстройство, чтобы предотвратить, вылечить, отсрочить начало, уменьшить тяжесть, или ослабить один или несколько симптомов расстройства или рецидив расстройства, или для того, чтобы продлить выживаемость субъекта сверх ожидаемого в отсутствие такого лечения. Под «терапевтическим режимом» понимают схему лечения заболевания, например, схему лечения, используемую во время терапии. Терапевтический режим может включать индукционный режим и поддерживающий режим. Термин «индукционный режим» или «индукционный период» относится к терапевтическому режиму (или части терапевтического режима), который используется для первоначального лечения заболевания. Общая цель индукционного режима состоит в том, чтобы обеспечить пациенту высокий уровень лекарственного средства в течение начального периода лечения. Индукционный режим может использовать (частично или полностью) «режим загрузки», который может включать введение большей дозы препарата, чем врач использовал бы во время поддерживающего режима, более частое введение препарата, чем врач использовал бы во время поддерживающего режима, или и то, и другое. Термин «поддерживающий режим» или «поддерживающий период» относится к терапевтическому режиму (или части терапевтического режима), который используется для поддержки пациента во время лечения заболевания, например, для поддержки пациента в состоянии ремиссии в течение длительного периода времени (месяцы или годы). В поддерживающей схеме может применяться непрерывная терапия (например, введение препарата через регулярные интервалы, например, еженедельно, ежемесячно, ежегодно и т.д.) или прерывистая терапия (например, прерывистое лечение, периодическое лечение, лечение при рецидиве или лечение после достижения определенного заранее заданного критерия (например, проявление заболевания и т. д.)).
Согласно изобретению первая аминокислотная последовательность, имеющая по меньшей мере 90% идентичности со второй аминокислотной последовательностью, означает, что первая последовательность имеет 90; 91; 92; 93; 94; 95; 96; 97; 98; 99 или 100% идентичности со второй аминокислотной последовательностью. Идентичность последовательности часто измеряют с точки зрения процентной идентичности (или сходства или гомологии); чем выше процент, тем более похожи две последовательности. Методы выравнивания последовательностей для сравнения хорошо известны в данной области техники. Различные программы и алгоритмы выравнивания описаны в: Smith and Waterman, Adv. Appl. Math., 2:482, 1981; Needleman and Wunsch, J. Mol. Biol., 48:443, 1970; Pearson and Lipman, Proc. Natl. Acad. Sci. U.S.A., 85:2444, 1988; Higgins and Sharp, Gene, 73:237-244, 1988; Higgins and Sharp, CABIOS, 5:151-153, 1989; Corpet et al. Nuc. Acids Res., 16:10881-10890, 1988; Huang et al., Comp. Appls Biosci., 8:155-165, 1992; and Pearson et al., Meth. Mol. Biol., 24:307-31, 1994). Altschul et al., Nat. Genet., 6:119-129, 1994, где представлено подробное рассмотрение методов выравнивания последовательностей и расчетов гомологии. Например, инструменты выравнивания ALIGN (Myers and Miller, CABIOS 4: 11-17, 1989) или LFASTA (Pearson and Lipman, 1988) могут использоваться для сравнения последовательностей (Internet Program® 1996, WR Pearson и Университет Вирджинии, fasta20u63 версия 2.0u63, дата выпуска декабрь 1996 г.). ALIGN сравнивает целые последовательности друг с другом, в то время как LFASTA сравнивает области локального сходства. Эти инструменты выравнивания и соответствующие учебные пособия доступны, например, в Интернете на веб-сайте NCSA. В качестве альтернативы, для сравнения аминокислотных последовательностей, содержащих более 30 аминокислот, можно использовать функцию последовательностей Blast 2, используя матрицу BLOSUM62 по умолчанию, установленную с параметрами по умолчанию (штраф за открытие гэпа 11 и штраф за продление гэпа 1). При выравнивании коротких пептидов (менее чем около 30 аминокислот) выравнивание следует выполнять с использованием функции последовательностей Blast 2, используя матрицу PAM30, установленную на параметры по умолчанию (штраф за открытие гэпа 9, штраф за продление гэпа 1). Система сравнения последовательностей BLAST доступна, например, на веб-сайте NCBI; см. также Altschul et al., J. Mol. Biol., 215:403-410, 1990; Gish. & States, Nature Genet., 3:266-272, 1993; Madden et al. Meth. Enzymol., 266:131-141, 1996; Altschul et al., Nucleic Acids Res., 25:3389-3402, 1997; и Zhang & Madden, Genome Res., 7:649-656, 1997.
Используемый в настоящей заявке термин «мутация» имеет общее значение в данной области техники, и относится к замене, делеции или вставке. Термин «замена» означает, что определенный аминокислотный остаток в определенной позиции удален, а другой аминокислотный остаток вставлен в ту же позицию. Термин «делеция» означает, что определенный аминокислотный остаток удален. Термин «вставка» означает, что один или несколько аминокислотных остатков вставлены до или после определенного аминокислотного остатка, более конкретно, что один или более, предпочтительно один или несколько аминокислотных остатков связаны с карбоксильной группой или аминогруппой определенного аминокислотного остатка.
В некоторых вариантах осуществления остаток аргинина в положении 9, 10, 20 или 21 мутирован, замещен таким образом, что заряд боковой цепи при pH = 7,4 обращен (например, отрицательный на положительный заряд) или становится нейтральным (например, отрицательный заменяется на нейтральный заряд). В некоторых вариантах осуществления масса и индекс гидрофобности остаются в одном и том же диапазоне.
В некоторых вариантах осуществления остаток аргинина (R) в положении 9, 10, 20 или 21 замещен аминокислотным остатком, выбранным из группы, состоящей из аланина (A) и/или серина (S).
Согласно изобретению, полипептид ELA по изобретению получают обычными автоматическими способами синтеза пептидов или рекомбинантной экспрессией. Общие принципы конструирования и получения белков хорошо известны специалистам в данной области техники. Полипептиды ELA по изобретению могут быть синтезированы в растворе или на твердой подложке в соответствии с общепринятыми методиками. Различные автоматические синтезаторы являются коммерчески доступными и могут использоваться в соответствии с известными протоколами, как описано в Stewart and Young; Tam et al., 1983; Merrifield, 1986 and Barany and Merrifield, Gross and Meienhofer, 1979. Полипептиды ELA по изобретению также можно синтезировать с помощью твердофазной технологии с использованием примерного пептидного синтезатора, такого как Модель 433A от Applied Biosystems Inc. Чистота любого данного белка, полученного с помощью автоматического пептидного синтеза или с помощью рекомбинантных способов, может быть определена с помощью анализа обращенно-фазовой ВЭЖХ. Химическая идентичность каждого пептида может быть установлена любым способом, хорошо известным специалистам в данной области техники. В качестве альтернативы автоматизированному синтезу пептидов может быть использована технология рекомбинантной ДНК, в которой нуклеотидную последовательность, которая кодирует выбранный белок, вставляют в вектор экспрессии, трансформируют или трансфицируют в подходящую клетку-хозяин и культивируют в условиях, подходящих для экспрессии, как описано здесь ниже. Рекомбинантные способы особенно предпочтительны для получения более длинных полипептидов. Разнообразные системы векторов экспрессии/хозяина могут быть использованы для содержания и экспрессии последовательности, кодирующей пептид или белок. Они включают микроорганизмы, такие как бактерии, трансформированные векторами экспрессии ДНК рекомбинантного бактериофага, плазмиды или космиды; дрожжи, трансформированные векторами экспрессии дрожжей (Giga-Hama et al., 1999); системы клеток насекомых, инфицированные вирусными векторами экспрессии (например, бакуловирусными, см. Ghosh et al., 2002); системы растительных клеток, трансфицированные вирусными векторами экспрессии (например, вирусом мозаики цветной капусты, CaMV; вирусом табачной мозаики, TMV) или трансформированные бактериальными векторами экспрессии (например, плазмидой Ti или pBR322; см., например, Babe et al., 2000); или системы клеток животных, но не ограничиваются ими. Специалистам в данной области известны различные способы оптимизации экспрессии белков у млекопитающих, см., например, Kaufman, 2000; Colosimo et al., 2000. Клетки млекопитающих, которые пригодны для продукции рекомбинантных белков, включают клетки VERO, клетки HeLa, клеточные линии яичника китайского хомячка (CHO), клетки COS (такие как COS-7), W138, BHK, HepG2, 3T3, RIN, MDCK, A549, PC12, K562 и 293 клетки, но не ограничиваются ими. Примерные протоколы рекомбинантной экспрессии пептидных субстратов или гибридных полипептидов в бактериях, дрожжах и других клетках беспозвоночных известны специалистам в данной области техники и кратко описаны здесь ниже. Системы млекопитающих хозяев для экспрессии рекомбинантных белков также хорошо известны специалистам в данной области техники. Штаммы клетки-хозяина могут быть выбраны по конкретной способности к процессингу экспрессированного белка или получению определенных посттрансляционных модификаций, которые будут полезны для обеспечения активности белка. Такие модификации полипептида включают ацетилирование, карбоксилирование, гликозилирование, фосфорилирование, липидирование и ацилирование, но не ограничиваются ими. Посттрансляционный процессинг, который расщепляет «препро» форму белка, также может быть важен для правильной вставки, свертывания и/или функционирования. Различные клетки-хозяева, такие как CHO, HeLa, MDCK, 293, WI38 и тому подобные, имеют специфические клеточные структуры и характерные механизмы для таких посттрансляционных активностей, и могут быть выбраны для обеспечения правильной модификации и процессинга введенного чужеродного белка.
В некоторых вариантах осуществления предполагается, что полипептиды ELA по изобретению, используемые в терапевтических способах по настоящему изобретению, могут быть модифицированы для улучшения их терапевтической эффективности. Такая модификация терапевтических соединений может использоваться для снижения токсичности, увеличения времени присутствия в циркуляции или изменения биораспределения. Например, токсичность потенциально важных терапевтических соединений может быть значительно снижена в сочетании с различными носителями лекарственного средства, которые модифицируют биораспределение. Стратегия повышения жизнеспособности лекарственного средства заключается в использовании водорастворимых полимеров. Было показано, что различные водорастворимые полимеры модифицируют биораспределение, улучшают режим клеточного поглощения, изменяют проницаемость через физиологические барьеры, и изменяют скорость клиренса из организма. Для достижения эффекта нацеливания или пролонгированного высвобождения были синтезированы водорастворимые полимеры, которые содержат компоненты лекарственного средства в качестве концевых групп, в качестве части каркаса или в качестве боковых групп в полимерной цепи. Полиэтиленгликоль (ПЭГ) широко используется в качестве носителя лекарственного средства, учитывая его высокую степень биосовместимости и простоту модификации. Было показано, что прикрепление к различным лекарствам, белкам и липосомам улучшает время пребывания и снижает токсичность. ПЭГ может быть связан с активными агентами через гидроксильные группы на концах цепи и другими химическими методами; однако сам ПЭГ ограничен максимум двумя активными агентами на молекулу. В другом подходе сополимеры ПЭГ и аминокислот были исследованы как новые биоматериалы, которые сохраняли бы свойства биосовместимости ПЭГ, но которые имели бы дополнительное преимущество в виде многочисленных точек присоединения на молекулу (обеспечивая большую нагрузку лекарственного средства) и которые могли бы быть синтетически разработаны для соответствия разнообразным применениям.
Второй объект настоящего изобретения относится к способу лечения рака почки у субъекта, нуждающегося в этом, включающему введение субъекту терапевтически эффективного количества молекулы нуклеиновой кислоты, которая кодирует полипептид ELA по настоящему изобретению.
Используемый в настоящей заявке термин «молекула нуклеиновой кислоты» имеет общее значение в данной области техники и относится к молекуле ДНК или РНК. Тем не менее, термин охватывает последовательности, которые включают любые из известных базовых аналогов ДНК и РНК, такие как 4-ацетилцитозин, 8-гидрокси-N6-метиладенозин, азиридинилцитозин, псевдоизоцитозин, 5-(карбоксигидроксилметил)-урацил, 5- фторурацил, 5-бромурацил, 5-карбоксиметиламинометил-2-тиоурацил, 5-карбоксиметиламинометилурацил, дигидроурацил, инозин, N6-изопентениладенин, 1-метиладенин, 1-метилпсевдоурацил, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-метиладенин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиамино-метил-2-тиоурацил, бета-D-маннозилквеозин, 5'-метоксикарбонилметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусная кислота, оксибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, урацил-5-оксиуксусной кислоты метиловый эфир, урацил-5-оксиуксусная кислота, псевдоурацил, квеозин, 2-тиоцитозин и 2,6-диаминопурин, но не ограничиваясь этим.
В некоторых вариантах осуществления молекула нуклеиновой кислоты по настоящему изобретению включена в подходящий вектор, такой как плазмида, космида, эписома, искусственная хромосома, фаг или вирусный вектор. Таким образом, еще один объект изобретения относится к вектору, содержащему нуклеиновую кислоту, кодирующую полипептид ELA по изобретению. Как правило, вектор является вирусным вектором, который представляет собой аденоассоциированный вирус (AAV), ретровирус, вирус бычьей папилломы, аденовирусный вектор, лентивирусный вектор, вирус коровьей оспы, вирус полиомы или инфекционный вирус. В некоторых вариантах осуществления вектор представляет собой вектор AAV. Используемый в настоящей заявке термин «AAV вектор» означает вектор, полученный из аденоассоциированного вирусного серотипа, включая AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9 и их мутантные формы, но не ограничиваясь ими. AAV векторы могут иметь один или несколько генов AAV дикого типа, полностью или частично удаленных, предпочтительно гены rep и/или cap, но сохраняют функциональные фланкирующие ITR последовательности. Ретровирусы могут быть выбраны в качестве векторов доставки генов благодаря их способности интегрировать свои гены в геном хозяина, переносить большое количество чужеродного генетического материала, инфицировать широкий спектр видов и типов клеток и быть упакованными в специальные клеточные линии. Чтобы создать ретровирусный вектор, нуклеиновую кислоту, кодирующую интересующий ген, встраивают в вирусный геном вместо определенных вирусных последовательностей, чтобы получить вирус, дефектный по репликации. Для получения вирионов конструируют пакующую клеточную линию, содержащую гены gag, pol и/или env, но без LTR и/или компонентов упаковки. Когда рекомбинантную плазмиду, содержащую кДНК, вместе с ретровирусным LTR и пакующими последовательностями, вводят в эту клеточную линию (например, посредством осаждения фосфатом кальция), пакующая последовательность позволяет упаковывать РНК-транскрипт рекомбинантной плазмиды в вирусные частицы, которые затем секретируются в культуральную среду. Среду, содержащую рекомбинантные ретровирусы, затем собирают, при необходимости концентрируют и используют для переноса гена. Ретровирусные векторы способны инфицировать самые разные типы клеток. Лентивирусы являются сложными ретровирусами, которые помимо обычных ретровирусных генов gag, pol и env содержат другие гены с регуляторной или структурной функцией. Более высокая сложность позволяет вирусу модулировать свой жизненный цикл, как при латентной инфекции. Некоторые примеры лентивирусов включают вирусы иммунодефицита человека (ВИЧ 1, ВИЧ 2) и вирус иммунодефицита обезьян (SIV). Лентивирусные векторы были получены путем многократного ослабления генов вирулентности ВИЧ, например, генов env, vif, vpr, vpu и nef удалены, что делает вектор биологически безопасным. В данной области известны лентивирусные векторы, см., например, патенты США № 6013516 и № 5994136, которые оба включены в настоящее описание в качестве ссылки. Как правило, векторы основаны на плазмидах или вирусах и сконструированы для переноса необходимых последовательностей для включения чужеродной нуклеиновой кислоты, для отбора и для переноса нуклеиновой кислоты в клетку-хозяина. Гены gag, pol и env интересующих векторов также известны в данной области техники. Таким образом, соответствующие гены клонируют в выбранный вектор и затем используют для трансформации интересующей клетки-мишени. Рекомбинантный лентивирус, способный инфицировать не-делящую клетку, где подходящая клетка-хозяин трансфицируется двумя или более векторами, несущими функции упаковки, а именно gag, pol и env, а также rev и tat, описаны в патенте США No. 5,994,136, включенном в настоящий документ посредством ссылки. Здесь описан первый вектор, который может обеспечить нуклеиновую кислоту, кодирующую вирусный gag и pol ген, и другой вектор, который может обеспечить нуклеиновую кислоту, кодирующую вирусный env, для получения пакующей клетки. Введение вектора, обеспечивающего гетерологичный ген, в эту пакующую клетку дает клетку-продуцент, которая высвобождает инфекционные вирусные частицы, несущие интересующий чужеродный ген. Env предпочтительно представляет собой белок амфотропной оболочки, который обеспечивает трансдукцию клетки человека и других видов. Как правило, молекула нуклеиновой кислоты или вектор по настоящему изобретению включают «контрольные последовательности», которые в совокупности относятся к промоторным последовательностям, сигналам полиаденилирования, последовательностям терминации транскрипции, вышележащим регуляторным доменам, точкам начала репликации, внутренним участкам посадки рибосомы («IRES»), энхансерам и тому подобным, которые совместно обеспечивают репликацию, транскрипцию и трансляцию кодирующей последовательности в клетке-реципиенте. Не все эти контрольные последовательности должны присутствовать всегда, если выбранная кодирующая последовательность способна реплицироваться, транскрибироваться и транслироваться в соответствующей клетке-хозяине. Другая последовательность нуклеиновой кислоты представляет собой «промоторную» последовательность, которая используется здесь в ее обычном смысле для обозначения нуклеотидной области, содержащей регуляторную последовательность ДНК, где регуляторная последовательность происходит от гена, который способен связывать РНК-полимеразу и инициировать транскрипцию нижележащей (в 3'-направлении) кодирующей последовательности. Промоторы транскрипции могут включать «индуцибельные промоторы» (где экспрессия полинуклеотидной последовательности, функционально связанной с промотором, индуцируется анализируемым веществом, кофактором, регуляторным белком и т.д.), «репрессируемые промоторы» (где экспрессия полинуклеотидной последовательности, функционально связанной с промотором, индуцируется анализируемым веществом, кофактором, регуляторным белком и т.д.), и «конститутивные промоторы».
Под «терапевтически эффективным количеством» подразумевается достаточное количество полипептида ELA или кодирующей его молекулы нуклеиновой кислоты для использования в способе лечения заболевания (например, рака почки) при разумном соотношении польза/риск, применимом к любому медицинскому лечению. Понятно, что общее ежедневное использование соединений и композиций по настоящему изобретению будет определяться лечащим врачом в рамках здравого медицинского заключения. Конкретный уровень терапевтически эффективной дозы для любого конкретного субъекта будет зависеть от множества факторов, включая возраст, массу тела, общее состояние здоровья, пол и диету субъекта; время введения, способ введения и скорость выведения конкретного используемого соединения; продолжительность лечения; лекарственные средства, используемые в комбинации или одновременно с конкретным используемым полипептидом; и подобные факторы, хорошо известные в медицине. Например, специалистам в данной области хорошо известно, что дозы соединения следует начинать с уровней, меньших, чем те, которые требуются для достижения необходимого терапевтического эффекта, и постепенно увеличивать дозировку до тех пор, пока не будет достигнут желаемый эффект. Однако суточная доза продуктов может находиться в широком диапазоне от 0,01 до 1000 мг на взрослого человека в сутки. Предпочтительно композиции содержат 0,01; 0,05; 0,1; 0,5; 1,0; 2,5; 5,0; 10,0; 15,0; 25,0; 50,0; 100; 250 и 500 мг активного ингредиента для симптоматической корректировки дозировки для субъекта, подлежащего лечению. Лекарственное средство обычно содержит примерно от 0,01 до 500 мг активного ингредиента, предпочтительно примерно от 1 до 100 мг активного ингредиента. Эффективное количество лекарственного средства обычно вводят в дозе от 0,0002 до 20 мг/кг массы тела в сутки, особенно примерно от 0,001 до 7 мг/кг массы тела в сутки.
Согласно изобретению, полипептид ELA или молекулу нуклеиновой кислоты (встроенную или не встроенную в вектор) по настоящему изобретению вводят субъекту в форме фармацевтической композиции. Как правило, полипептид ELA или молекула нуклеиновой кислоты (встроенная или не встроенная в вектор) по настоящему изобретению может быть объединена с фармацевтически приемлемыми вспомогательными веществами и при необходимости с матриксами для замедленного высвобождения, такими как биодеградируемые полимеры, для образования фармацевтических композиций. «Фармацевтически» или «фармацевтически приемлемый» относятся к молекулярным веществам и композициям, которые не вызывают нежелательную, аллергическую или другую неблагоприятную реакцию при введении млекопитающему, особенно человеку, в зависимости от ситуации. Фармацевтически приемлемый носитель или наполнитель относится к нетоксичному твердому, полутвердому или жидкому наполнителю, разбавителю, инкапсулирующему материалу или вспомогательному препарату любого типа. В фармацевтических композициях по настоящему изобретению для перорального, сублингвального, подкожного, внутримышечного, внутривенного, трансдермального, местного или ректального применения активное вещество по отдельности или в сочетании с другим активным веществом может вводиться в лекарственной форме в виде смеси с обычными фармацевтическими носителями животным и людям. Подходящие лекарственные формы включают формы для перорального введения, такие как таблетки, гелевые капсулы, порошки, гранулы и пероральные суспензии или растворы, формы для сублингвального и буккального введения, аэрозоли, имплантаты, подкожные, трансдермальные, местные, интраперитонеальные, внутримышечные, внутривенные, подкожные, трансдермальные, интратекальные и интраназальные формы, и формы для ректального введения. Как правило, фармацевтические композиции содержат носители, которые являются фармацевтически приемлемыми для препарата, который можно вводить посредством инъекции. Это могут быть, в частности, изотонические, стерильные солевые растворы (фосфат натрия или динатрия, хлорид натрия, калия, кальция или магния и т.п., или смеси таких солей) или сухие, особенно лиофилизированные композиции, которые после добавления в зависимости от случая, стерилизованной воды или физиологического раствора, обеспечивают получение растворов для инъекций. Фармацевтические формы, подходящие для инъекционного применения, включают стерильные водные растворы или дисперсии; составы, включающие кунжутное масло, арахисовое масло или водный пропиленгликоль; и стерильные порошки для немедленного приготовления стерильных инъекционных растворов или дисперсий. Во всех случаях форма должна быть стерильной и должна быть жидкой до такой степени, чтобы ее можно было легко вводить шприцем. Она должна быть стабильной в условиях производства и хранения, и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибки. Растворы, содержащие соединения по изобретению в виде свободного основания или фармакологически приемлемых солей, могут быть приготовлены в воде, подходящим образом смешанной с сурфактантом, таким как гидроксипропилцеллюлоза. Дисперсии также могут быть приготовлены в глицерине, жидких полиэтиленгликолях и их смесях и в маслах. В обычных условиях хранения и использования эти препараты содержат консервант для предотвращения роста микроорганизмов. Полипептид ELA или молекула нуклеиновой кислоты (встроенная или не встроенная в вектор) по настоящему изобретению может быть составлена в виде композиции в нейтральной или солевой форме. Фармацевтически приемлемые соли включают соли добавления кислоты (образованные с помощью свободных аминогрупп белка), и те, которые образованы с неорганическими кислотами, такими как, например, соляная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, миндальная, и тому подобное. Соли, образованные со свободными карбоксильными группами, также могут быть получены из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и тому подобное. Носителем также может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное), их подходящие смеси и растительные масла. Надлежащая текучесть может поддерживаться, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем использования сурфактантов. Предотвращение развития микроорганизмов может быть обеспечено различными антибактериальными и противогрибковыми агентами, например, парабенами, хлорбутанолом, фенолом, сорбиновой кислотой, мертиолятом и тому подобным. Во многих случаях предпочтительно включать изотонические агенты, например, сахара или хлорид натрия. Пролонгированная абсорбция композиций для инъекций может быть достигнута путем использования в композициях агентов, замедляющих абсорбцию, например, моностеарата алюминия и желатина. Стерильные растворы для инъекций готовят путем включения активных соединений в необходимом количестве в соответствующем растворителе с несколькими другими ингредиентами, перечисленными выше, по мере необходимости, с последующей стерилизующей фильтрацией. Обычно дисперсии готовят путем включения различных стерилизованных активных ингредиентов в стерильный носитель, который содержит основную дисперсионную среду и необходимые другие ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций типичными способами приготовления являются технологии вакуумной сушки и лиофильной сушки, которые дают порошок из активного ингредиента плюс любого дополнительного необходимого ингредиента из его предварительно стерильно отфильтрованного раствора. Также предполагается получение большего количества или высококонцентрированных растворов для прямой инъекции, когда предполагается, что использование ДМСО в качестве растворителя приводит к чрезвычайно быстрому проникновению, доставляя высокие концентрации активных агентов в небольшую область опухоли. После приготовления растворы вводят способом, совместимым с дозированной композицией, и в таком количестве, которое является терапевтически эффективным. Составы легко вводятся в различных лекарственных формах, таких как тип инъецируемых растворов, описанных выше, но также могут быть использованы капсулы для высвобождения лекарственного средства и тому подобное. Например, для парентерального введения в водном растворе раствор должен быть подходящим образом забуферен, если необходимо, и жидкий разбавитель сначала должен быть изотоническим с достаточным количеством физиологического раствора или глюкозы. Эти конкретные водные растворы особенно подходят для внутривенного, внутримышечного, подкожного и интраперитонеального введения. В связи с этим стерильные водные среды, которые могут быть использованы, будут известны специалистам в данной области техники в свете настоящего раскрытия. Некоторое изменение в дозировке обязательно произойдет в зависимости от состояния субъекта, которого лечат. Лицо, ответственное за введение, в любом случае определит подходящую дозу для индивидуального субъекта.
Еще один объект настоящего изобретения относится к способу диагностики рака почки у субъекта, включающему этапы:
(i) измерения уровня экспрессии Elabela (ELA) в образце, полученном от указанного субъекта;
(ii) сравнения экспрессии, измеренной на этапе (i), с её заранее заданным эталонным значением;
(iii) получения заключения о том, что субъект страдает от рака почки, когда уровень экспрессии Elabela (ELA) ниже, чем его предварительно определенное эталонное значение, или заключения о том, что субъект не страдает от рака почки, когда уровень экспрессии Elabela (ELA) выше, чем его предварительно определенное эталонное значение.
Используемый в настоящей заявке термин «диагностика» означает оценку того, страдает ли субъект от рака почки или нет.
Изобретение будет дополнительно проиллюстрировано следующими фигурами и примерами. Однако эти примеры и фигуры никоим образом не должны интерпретироваться как ограничивающие объем настоящего изобретения.
Краткое описание фигур
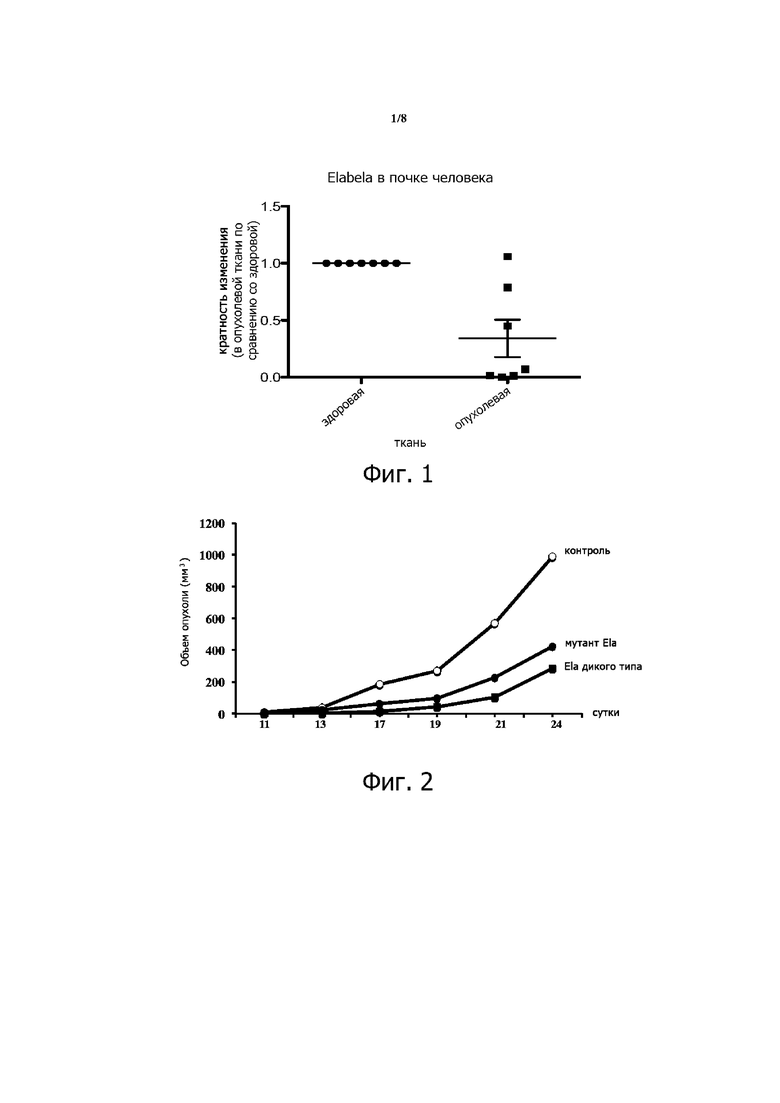
Фигура 1. Снижение экспрессии Elabela в ткани рака почки человека. ПЦР-анализ в режиме реального времени выявил понижающую регуляцию Еlabela в тканях опухоли почки по сравнению с псевдо-нормальными тканями, полученными от того же пациента.
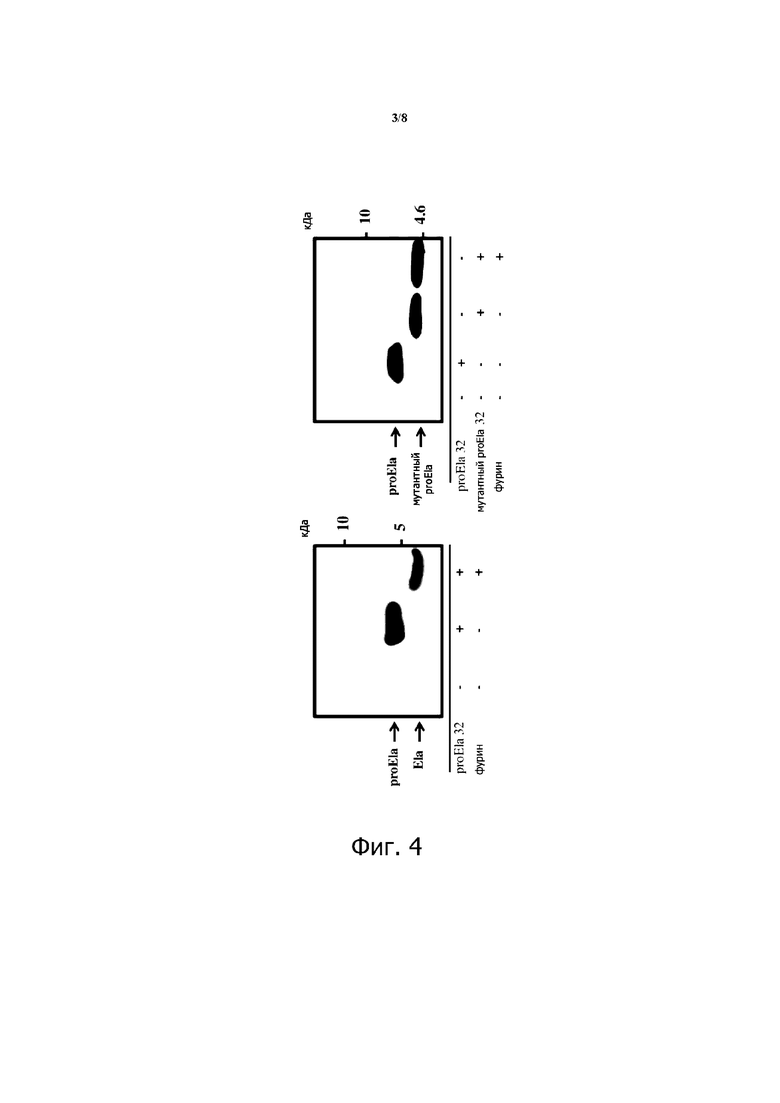
Фигура 2. Влияние экспрессии Elabela в клетках опухоли почки (RenCa) на прогрессирование подкожной опухоли. Elabela дикого типа (ela wt), мутант (Ela mut) или контрольный лентивирус были использованы для экспрессии Elabela, ela mut в клетках RenCa перед их инокуляцией сингенным мышам.
Фигура 3. Влияние экспрессии Elabela в клетках опухоли почки (RenCa) на прогрессирование опухоли почки. Для экспрессии ela wt в клетках RenCa до их субкапсулярной инокуляции у сингенных мышей использовали дикий тип Elabela (ela wt) или контрольный лентивирус.
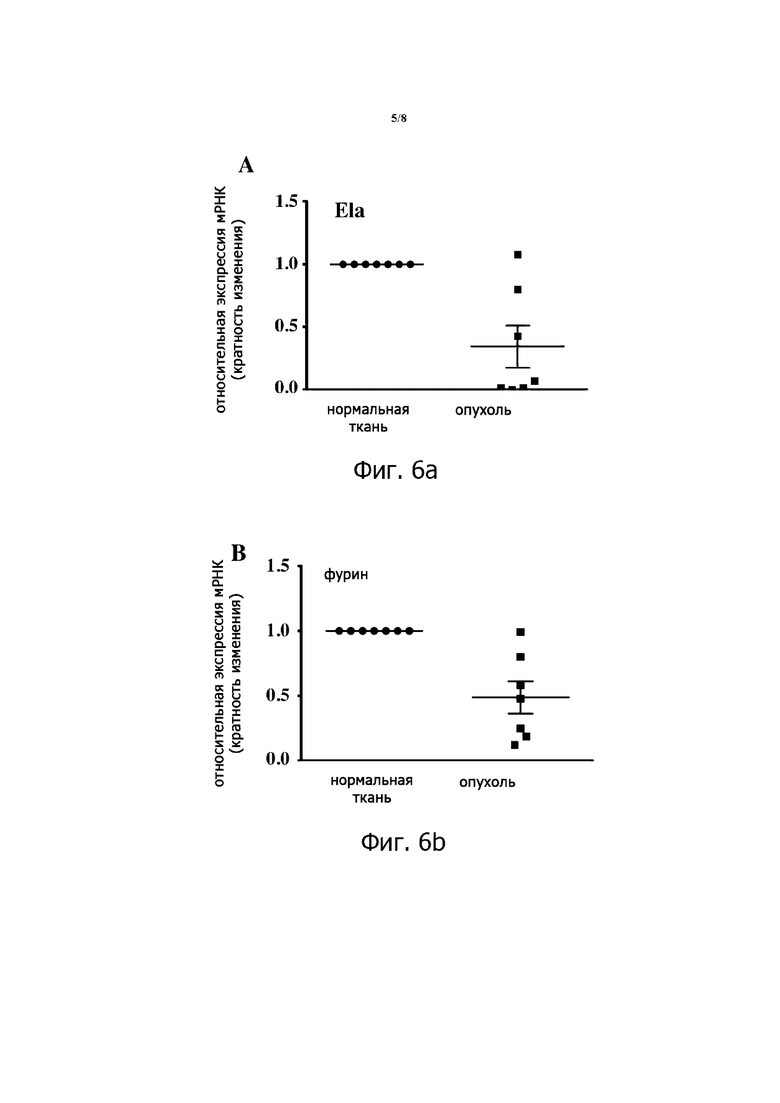
Фигура 4. Процессинг ProEla. Пептид дикого типа proEla 32 и mut proEla 32, в котором два сайта расщепления R31/R32 и R42/R43 были заменены на S31/S32 и S42/S43 соответственно, были синтезированы и инкубированы с фурином (0,2 × 10-4 Ед.) в течение 4 часов. Согласно оценке посредством вестерн-блоттинга с использованием антитела против Ela, фурин осуществляет процессинг proEla 32 дикого типа в соответствующих участках физиологического расщепления. Инкубация этого пептида с фурином генерирует только Ela 11 aa, что позволяет предположить, что proEla 32 эффективно расщепляется в двух участках расщепления. Инкубация mut proEla 32 с фурином не давала каких-либо продуктов.
Фигура 5. Экспресс-анализ Ela, фурина и APJ в тканях взрослой мыши. Общую РНК выделяли из указанных тканей, и анализ ПЦР в режиме реального времени проводили с использованием специфических праймеров для Ela (A) мыши, фурина (B) и APJ (C). Экспрессию конститутивных генов, которую оценивали в каждом образце, использовали в качестве эндогенного контроля в условиях, описанных в тексте. Показанные результаты являются примером от 3 экспериментов. Для сравнения печени (Ela и APJ) или кишечнику (фурин) было назначено значение 1 в зависимости от уровня экспрессии анализируемого гена. Данные представляют собой среднее значение ± стандартное отклонение (n = 3 на группу).
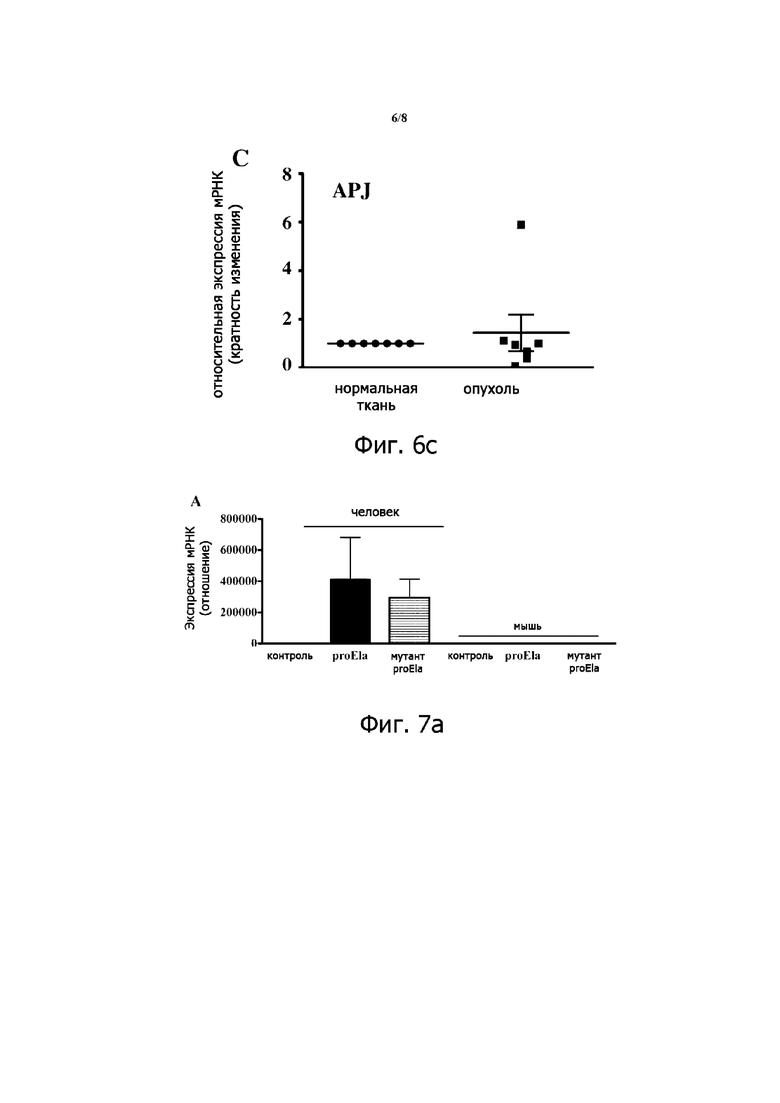
Фигура 6. Экспрессия Ela, фурина и APJ у пациентов с раком почки. Общую РНК экстрагировали из опухоли и окружающей неопухолевой ткани почечных клинических образцов от n=7 пациентов, и ПЦР-анализ в режиме реального времени выполняли с использованием специфических праймеров для человеческого Ela, фурина и APJ. Экспрессию конститутивных генов, которую оценивали в каждом образце, использовали в качестве эндогенного контроля в условиях, описанных в тексте. Графики показывают кратную разницу в экспрессии указанных транскриптов со значением «1», присвоенным нормальным тканям. Отмечается снижение экспрессии мРНК Ela в опухолевых тканях по сравнению с их нормальными аналогами.
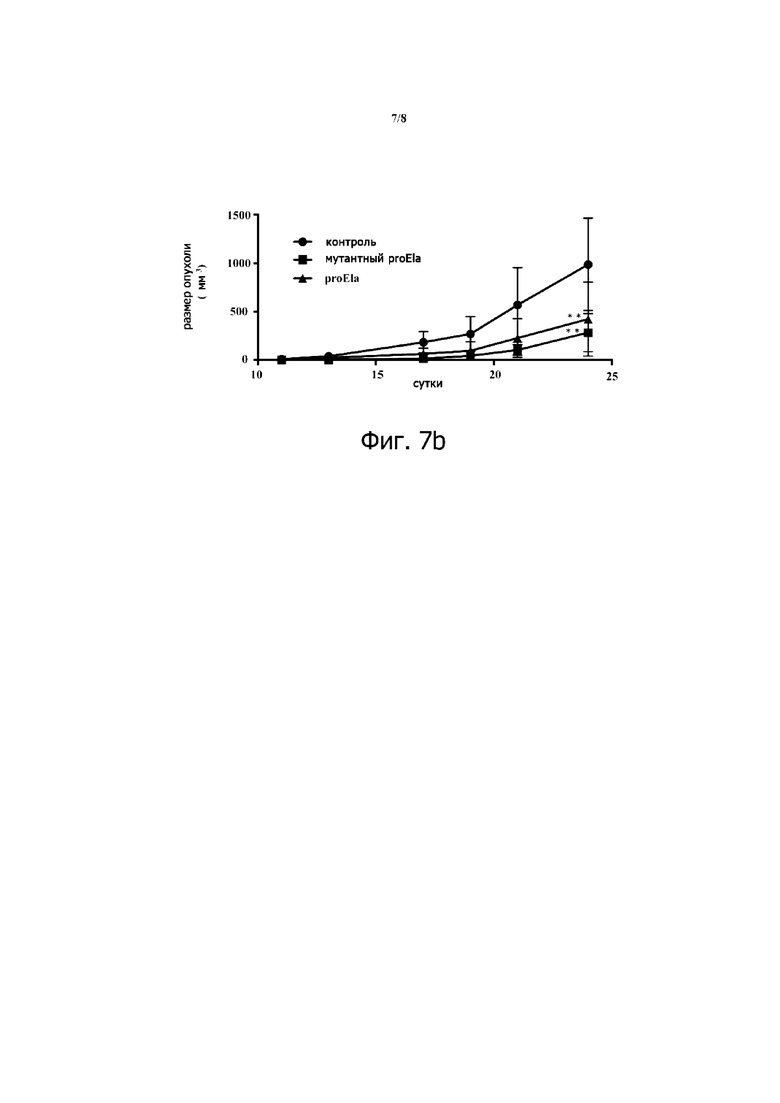
Фигура 7. Ингибирование роста опухоли proEla дикого типа и mut proEla. Раковые клетки почки RenCa (лишенные экспрессии Ela) были стабильно инфицированы пустым лентивирусным вектором (контроль) или тем же вектором, который содержит человеческую конструкцию proEla дикого типа или мутантный proEla. Экспрессия человеческого proEla дикого типа и mut proEla в этих клетках была подтверждена на уровне ARN с использованием ПЦР в режиме реального времени и человеческих праймеров. Использование мышиных праймеров Ela дано для сравнения (А). В трех группах самцов сингенных мышей Balb/c инокулировали подкожно 1×105 контрольных клеток RenCa, и те же самые клетки экспрессировали proEla 32 дикого типа или мутантный proEla 32. У животных наблюдали образование опухоли в течение указанных периодов. Отмечается меньший размер опухолей, индуцированных опухолевыми клетками, экспрессирующими proEla дикого типа или мутантный ProEla. ProEla 32 дикого типа более эффективно ингибировал рост опухоли по сравнению с мутантным pro Ela 32. Результаты представляют 2 эксперимента, выполненных с клетками RenCa, независимо инфицированными указанными лентивирусными векторами. Значения являются средними ± SEM (n = 6 на группу). *** Р <0,001.
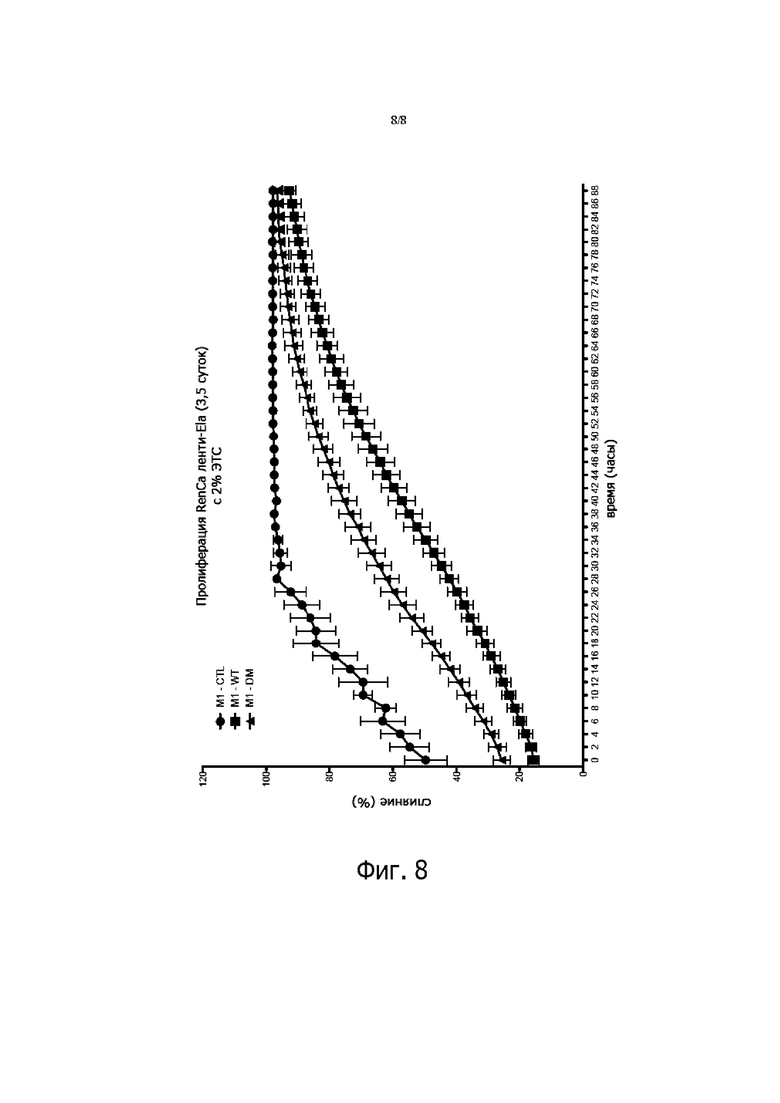
Фигура 8. Клетки RenCa (20000 клеток/лунку) высевали в 96-луночные планшеты, анализ пролиферации проводили с использованием системы визуализации живых клеток IncuCyte ZOOM™ (Essen BioScience, Мичиган, США). Эта система измеряет плотность клеток. Клетки размножали стандартным способом в среде RPMI 1640 (Life Technologies) в 2% эмбриональной телячьей сыворотке при 37°C. Клетки регулярно проверяли на микоплазмы (АТСС). Планшет помещали в устройство IncuCyte ZOOM ™, и изображения общего распределения клеток регистрировали каждые 2 часа общей продолжительностью 50 часов.
Пример 1
Elabela (ELA), также известный как Toddler или Apela, представляет собой пептидный гормон, который недавно был идентифицирован как второй лиганд APJ, рецептора апелина. Произведенный в качестве предшественника из 32 аминокислот (аа), ELA также обнаружен как 21 аа и 11 аа. Наши результаты показывают, что Ela в основном экспрессируется в почках, а его экспрессия снижается при раке почки человека (фиг. 1). На модели у животных с ксенотрансплантатом (подкожная или субкапсулярная инъекция) Ela ингибирует прогрессирование опухоли (Фиг.2 и 3). Кроме того, мы создали мутантный полипептид ELA, в котором остатки аргинина в положении 9, 10, 20 и 21 были замещены остатком серина. Мы показали, что мутантный полипептид ELA также способен ингибировать прогрессирование опухоли (фиг. 2). Эти находки идентифицируют Ela как новый ген-супрессор опухоли в почках.
Пример 2
Материал и методы
Образцы пациентов
Свежие образцы и соответствующие им нормальные ткани были получены из опухолей почек человека. Все пациенты дали письменное информированное согласие. Материал пациента деидентифицирован, и национальный комитет по этике исследований Франции утвердил протокол исследования. После операции образцы ткани немедленно переносили на лед и быстро замораживали в жидком азоте до тех пор, пока их не использовали для экстракции РНК.
Производство лентивирусных векторов, инфицирование и культивирование клеток
ProEla дикого типа и мутантные proEla (участки процессинга R31/R32 и R42/R43 были заменены на S31/S32 и S42/S43 aa) были клонированы в мультицистронный самоинактивирующийся лентивирусный вектор, содержащий репортерный ген tdTomato (pRRLsin-MND-hPGK-tdTomato-WPRE), под контролем энхансера вируса миелопролиферативной саркомы. Все конструкции были проверены секвенированием. Конструирование и продукцию лентивирусных векторов осуществляли на установке «Vect'UB» TMB-Core Университета Бордо. Псевдотипированные VSV-G лентивекторы получали тройной транзиторной трансфекцией в клетках HEK293T и концентрировали ультрафильтрацией (Vivaspin 20, Sartorius Biotech SA, США). Вирусные титры pLV-лентивекторов определяли путем трансформации клеток HEK293T последовательными разведениями вирусной надосадочной жидкости, а экспрессию tdTomato определяли через 5 дней с помощью анализа проточной цитометрией. В день инфицирования клетки аденокарциномы почки мыши RenCa (5×104 клеток/лунку) высевали в 24-луночный планшет с полибреном при 8 мкг/мл. Лентивирус, кодирующий proEla дикого типа, мутантный proEla или только tdTomato, добавляли в среду при MOI 10 (множественность инфекции). Уровни клеточной инфекции наблюдали через 72 часа с использованием флуоресцентного микроскопа. Клетки RenCa поддерживали в среде RPMI1640 с добавлением 10% ЭТС, 100 единиц/мл пенициллина/стрептомицина и 2 мМ L-глутамина.
Анализ полимеразной цепной реакции в режиме реального времени
Общую РНК из образцов человека экстрагировали с использованием набора для РНК NucleoSpin (Macherey-Nagel) в соответствии с инструкциями производителя. Общую РНК из образцов мышей экстрагировали с использованием реагента TRI (MRC Inc., США) в соответствии с инструкциями производителя. Один мкг общей РНК подвергали синтезу кДНК с использованием набора для обратной транскрипции кДНК высокой емкости (Applied Biosystems, Куртабёф, Франция). Качество РНК для образцов человека проверяли с использованием набора Agilent RNA 6000 Nano в соответствии с инструкциями производителя (Agilent). Относительную количественную оценку специфических мРНК проводили с помощью ПЦР в режиме реального времени с использованием ПЦР-системы StepOnePlusTM в режиме реального времени (Applied Biosystems, Куртабёф, Франция), Master Mix PCR (Eurogentec) и специфических праймеров, в соответствии с инструкциями производителя. Условия для реакции были следующими для SYBR Green кПЦР: 10 минут при 95°C, 40 циклов по 15 секунд при 95°C, 60 секунд при 60°C и затем 15 минут при 95°C, 60 секунд при 60°C и 15 минут при 95°С; и для Taqman кПЦР: 2 минуты при 50°C, 10 минут при 95°C, 40 циклов по 15 секунд при 95°C и 60 секунд при 60°C. Гены GAPDH, HPRT1 или S16 конститутивные гены использовали в качестве эндогенного контроля для клеток и тканей человека или мыши, как описано ранее (Scamuffa et al., 2008).
Синтез пептидов и ферментативное расщепление in vitro.
Пептиды Ela 11, proEla 32 дикого типа и мутантные пептиды proEla 32 были синтезированы Clinisciences. Пептиды Ela расщепляли фурином в течение 4 часов, как описано ранее (Sfaxi et al., 2014., Scamuffa et al., 2008), и подвергали вестерн-блоттингу.
Анализ посредством вестерн-блоттинга
Полученные продукты ферментативного расщепления in vitro подвергали электрофорезу в ДСН-полиакриламидном геле в 13% гелях. В качестве первичного антитела использовали анти-Ela 11 (Eurogentec). Вторичное антитело, конъюгированное с пероксидазой хрена, и усиленная хемилюминесценция (ECL + Plus, Amersham) были использованы для выявления первичных антител в соответствии с инструкциями производителей с использованием системы визуализации хемилюминесценции (GeneGnome, Syngene) (Sfaxi et al., 2014., Scamuffa et al., 2008).
Анализ интернализации рецепторов
Клетки HEK293A, стабильно экспрессирующие гибридный белок GFP-APJ человека, культивировали в бессывороточной среде в течение 24 часов и обрабатывали в течение 30 минут 1 мкМ Ela 11, proEla 32 дикого типа или мутантным пептидом proEla 32. Клетки фиксировали 4% параформальдегидом в течение 10 минут при комнатной температуре, и интернализацию рецептора APJ анализировали с использованием эпифлуоресцентной микроскопии Nikon.
Анализ активации ERK, AKT и P70
Клетки HEK293A, сверхэкспрессирующие человеческие гибридные белки GFP-APJ, поддерживали в бессывороточной среде в течение 24 часов и инкубировали с (1 мкМ) или без Ela 11, proEla дикого типа или мутантным proEla в течение 5, 15 и 30 минут при 37°С. Клетки лизировали в буфере RIPA (150 мМ NaCl, 50 мМ Трис, 1 мМ ЭДТА, 1% NP40, 0,25% дезоксихолата натрия, pH 8) и подвергали ДСН-ПАГЭ на 12% гелях. Клеточные лизаты анализировали вестерн-блоттингом на фосфорилирование ERK, AKT и P70 с использованием анти-фосфо-ERK; анти-фосфо-АКТ и анти-фосфо-P70 (Cell Signaling), соответственно. Пятна белков выделяли и повторно обрабатывали ERK, AKT или P70 (Cell Signaling) для нормализации данных. Первичные антитела визуализировали с использованием вторичных антител, конъюгированных с пероксидазой хрена (Amersham), и сигналы определяли с использованием хемилюминесцентной системы ECLPlus в соответствии с инструкциями производителей (Amersham).
Сигнальный путь mTOR
Клетки RenCa со сверхэкспрессией лентивируса, кодирующего proEla дикого типа, мутантный proEla или только tdTomato, содержали в бессывороточной среде в течение различного времени. Клетки лизировали в буфере RIPA (150 мМ NaCl, 50 мМ Трис, 1 мМ ЭДТА, 1% NP40, 0,25% дезоксихолата натрия, pH 8) и подвергали ДСН-ПАГЭ на 12% гелях. Клеточные лизаты анализировали вестерн-блоттингом на LC3, NFkB, Erk1/2, AKT, S6K или актин с использованием анти-фосфо-NFkB, анти-фосфо-Erk, анти-фосфо-Akt, анти-фосфо S6K. Пятна белка очищали и повторно обрабатывали ERK, AKT или P70 (Cell Signaling) для нормализации данных. Первичные антитела визуализировали с использованием вторичных антител, конъюгированных с пероксидазой хрена (Amersham), и сигналы определяли с использованием хемилюминесцентной системы ECLPlus в соответствии с инструкциями производителей (Amersham).
Анализ канцерогенности
7-8-недельных мышей Balb/c от Janvier Laboratories содержали в вентилируемых карусельных клетках и снабжали их стерильной пищей и питьевой водой. Совет по содержанию и экспериментам на животных правительства Франции одобрил все эксперименты на мышах, о которых здесь сообщалось. Чтобы оценить влияние экспрессии proEla дикого типа и мутантного proEla на способность клеток RenCa индуцировать рост опухоли, 1×105 клеток RenCa или тех же самых клеток, стабильно экспрессирующих proEla дикого типа или мутантный proEla, вводили подкожно сингенным мышам Balb/c. Образование опухолей контролировали каждые 2-3 дня, и мышей умерщвляли в конце экспериментов. Объем опухоли рассчитывали, как описано ранее (Sfaxi et al., 2014).
Анализ заживления ран
Клетки RenCa культивировали до субконфлюентной культуры. Царапину наносили наконечником на каждую лунку, и заживление раны наблюдали через 8 или 24 часа после культивирования в бессывороточной среде.
Статистика
Все данные были выражены как среднее значение ± стандартное отклонение (SD), и статистический анализ был выполнен с использованием Graphpad Prism 5.0 (GraphPad Software Inc., Сан-Диего, Калифорния). Значения р <0,05 считались значимыми.
Результаты
Процессинг рroEla 32 пропротеин-конвертазой фурином.
Структура кДНК человеческого ELA прогнозирует pre-proEla из 54 аминокислот (аа). После удаления 22 аминокислот сигнального пептида гормон высвобождается в виде proEla из 32 аминокислот. Наличие двух основных аминокислотных мотивов R31/R32 и R42/R43 в proEla 32 предполагает участие ПК в его обработке (данные не показаны). Посредством баз данных Genbank мы обнаружили, что последовательность proEla высоко консервативна, особенно вокруг ПК-подобных участков расщепления RX(K/R)RQ (данные не показаны). Чтобы изучить важность протеолитического созревания preEla 32 с помощью ПК для обеспечения его функции, мы сначала оценили экспериментальную обработку proEla 32 фурином с использованием анализа расщепления in vitro. Для этого мы синтезировали proEla 32aa дикого типа, который содержит участки процессинга proEla (R31/R32 и R42/R43), и мутантный пептид proEla 32, участки которого были мутированы в S31/S32 и S42/S43, соответственно. Как показано на фигуре 4, инкубация proEla дикого типа с рекомбинантным человеческим фурином (0,2 × 10-4 Ед.) генерировала преимущественно зрелый Ela 11aa, что свидетельствует о том, что участки R31/R32 и R42/R43 эффективно и одновременно расщепляются фурином, избегая генерации промежуточной формы ELA 22aa в этих условиях. Напротив, инкубация мутантного пептида ProEla 32 (S31/S32 и S42/S43) с фурином не позволила генерировать какие-либо зрелые продукты Ela, поддерживая специфический процессинг proEla посредством ПК на участках физиологического расщепления R31/R32 и R42/R43.
Анализ экспрессии Ela, фурина и APJ в тканях взрослых мышей и пациентов, страдающих раком почки.
ПЦР-анализ в режиме реального времени различных тканей взрослых мышей показал, что, хотя фурин и APJ экспрессируются во всех анализируемых тканях, Ela в основном экспрессируется в почках (фиг.5). Скоординированная экспрессия Ela, его рецептора APJ и его превращающего фермента фурина в нормальном эпителии почек мышей позволила нам исследовать уровень экспрессии этих генов при раке почки человека и оценить роль Ela и его процессинга в этом заболевании. Таким образом, с помощью ПЦР-анализа в режиме реального времени тканей, полученных от пациентов с раком почки, выявлена повсеместная экспрессия Ela, APJ и фурина (фиг. 6). По сравнению с нормальными тканями, в анализируемых тканях пациентов с раком почки была обнаружена пониженная экспрессия Ela и фурина. Из тканей 7 проанализированных пациентов экспрессия Ela была подавлена у 6 пациентов.
Интернализация APJ и Ela пептиды
Индуцированная лигандом интернализация рецепторов представляет собой клеточный ответ APJ на связывание лиганда и его активацию. Чтобы исследовать, будет ли зрелый ELA 11, дикий тип и мутантный про Ela 32 индуцировать интернализацию APJ, мы стабильно экспрессировали APJ в виде гибридного белка с усиленным зеленым флуоресцентным белком (GFP-APJ, данные не показаны) посредством лентивирусной инфекции в клетках HEK293 и исследовали его внутриклеточную локализацию в ответ на указанные пептиды Ela. На базовом уровне гибридный белок в основном локализован на клеточной поверхности. После обработки Ela пептидом через 30 мин в цитоплазме формировались большие везикулы, что позволяет предположить, что все формы Ela пептида способны активировать рецептор APJ и опосредовать его интернализацию. Подобным образом, обработка клеток Ela 11 и mut proEla 32 или proEla дикого типа и mut proEla также индуцировала интернализацию APJ.
Анализ активации ERK, AKT и p70
Чтобы оценить важность процессинга proEla в передаче сигналов ERK, AKT и p70, мы обработали клетки HEK293, экспрессирующие APJ, Ela дикого типа, mut proEla 32 или зрелым Ela 11. Все эти пептиды Ela (1 мкМ) были способны индуцировать фосфорилирование ERK в течение 5 мин после обработки (данные не показаны). Этот эффект был уменьшен через 15 минут, как показал вестерн-блоттинг анализ. Интересно, что в тех же условиях влияние proEla дикого типа на активацию ERK было выше по сравнению с эффектом mut proEla и Ela 11 (данные не показаны). Анализ активации AKT показал, что все протестированные пептиды индуцировали более низкую активацию AKT по сравнению с их влиянием на активацию ERK. Через 5 минут наблюдалось слабое видимое фосфорилирование AKT, которое достигало максимума примерно через 15 минут и затем уменьшалось (данные не показаны). Аналогичным образом, анализ AKT нижестоящего эффектора P70s6K показал, что применение 1 мкМ всех тестируемых пептидов не вызывало значительного эффекта в тех же условиях (данные не представлены).
Сигнальный путь mTOR
Чтобы оценить роль Elabela в прогрессировании рака почки, мы использовали клетки RenCa, экспрессирующие elabela или elabela, мутированные в участке фурина. Клетки культивировали в бессывороточной среде в течение 1, 3, 6, 12 или 24 часов, и влияние Elabela на путь mTOR оценивали с помощью анализа фосфорилирования nfkB, Erk1/2, akt или S6K.
Экспрессия версий ELABELA дикого типа или мутантных в клетках RenCa во время культивирования в бессывороточной среде вызывала:
1. Блокирование индукции аутофагии, оцениваемой по пониженным уровням LC3II.
2. Устойчивую активацию пути mTORC1, что определяется устойчивым фосфорилированием S6K и S6.
3. Усиленное ингибирование передачи сигналов ERK, что определяется увеличением фосфорилирования ERK.
4. Не влияла на передачу сигналов mTORC2, как определено P(473)AKT.
5. Не влияла на передачу сигналов PI3K, как определено P(308)AKT.
6. Не влияла на передачу сигналов NFkB.
Роль процессинга Ela и proEla в канцерогенезе.
Чтобы исследовать роль дикого типа и мутанта proEla 32 в развитии опухоли, мы воспользовались раковыми клетками почки RenCa мыши, в которых отсутствует экспрессия Ela, и использовали лентивирусные векторы pRRLsin-MND-hPGK-tdTomato-WPRE для доставки и стабильной экспрессии дикого типа и мутантного proEla 32 в этих клетках. Перед анализом раковые клетки RenCa, стабильно экспрессирующие proEla дикого типа и мутантные человеческие proEla, оценивали на экспрессию этих конструкций с использованием ПЦР в режиме реального времени (фиг.7А) и по наличию флуоресцентного белка tdTomato (данные не показаны). Трем группам самцов сингенных мышей Balb/c инокулировали подкожно 1 × 105 контрольных клеток, и тех же клеток, экспрессирующих proEla 32 дикого типа или мутантный proEla 32. Как показано на фиг.7B, экспрессия proEla дикого типа или мутантного proEla в этих опухолевых клетках значительно снижала рост опухоли по сравнению с контрольными клетками, экспрессирующими пустой вектор. Экспрессия proEla 32 дикого типа, по-видимому, ингибирует рост опухоли более эффективно, по сравнению с мутантом proEla 32. Эти данные подтверждаются экспериментом по заживлению ран (данные не показаны). Elabela ингибирует заживление ран клеток RenCa in vitro, а также на фигуре 8 в анализе пролиферации клеток. Elabela ингибирует пролиферацию клеток RenCa.
Обсуждение
Повсеместная экспрессия фурина и присутствие двухосновного мотива расщепления в предшественнике Ela (proEla 32) позволяет предположить, что ПК являются потенциальными протеазами для процессинга proEla (данные не показаны). В настоящем исследовании мы продемонстрировали, что ПК (фурин) участвуют в протеолитическом процессинге proEla в двух участках расщепления, а именно R31/R32 и R42/R43. В нашей модели участки расщепления proEla были подтверждены мутагенезом и ферментативным расщеплением in vitro пептидов дикого типа и мутантных пептидов proEla (фиг.4). Однако, хотя расщепление proEla (32aa) в этих двух участках расщепления предполагает образование пептидных форм Ela 22 и Ela 11, в этих условиях была обнаружена только форма Ela 11, что указывает на возможное быстрое превращение Ela 22 в Ela 11 в присутствии фурина (фиг.4). Анализ различных тканей взрослых мышей показал, что ко-экспрессия мРНК Ela, его рецепторов APJ и фурина (фиг.5) усиливает функциональную связь между ними. Эти данные дополнительно подтверждаются поразительной временной корреляцией между экспрессией Ela, APJ и фурина во время эмбрионального развития (Scamuffa et al., 2006., Helker et al., 2015), что свидетельствует о ключевой роли этих генов во время этих процессов. Соответственно, в предыдущих сообщениях указывалось, что мутантный фенотип ELA сходен с мутантами APJ у рыбок данио, а инактивация локуса fur путем гомологичной рекомбинации у мышей вызывает эмбриональную гибель вскоре после е10.5 (Scamuffa et al., 2006). Следовательно, предполагается, что ELA, его конвертирующая протеаза фурин и рецептор APJ участвуют в биологических функциях, необходимых для нормального развития эмбриона. Тем не менее, в этом исследовании мы обнаружили, что ингибирование процессинга предшественника proEla не оказывает существенного влияния на его биологическую функцию. Действительно, ELA первоначально был обнаружен высококонсервативным у позвоночных животных (Pauli et al., 2014., Chng et al., 2013), особенно вокруг участков расщепления ПК (данные не показаны), и описан как специфический лиганд для APJ, предполагающий функциональную консервацию пути ELA-APJ у млекопитающих. В последнее время сообщалось, что Ela опосредует другие функции посредством своего взаимодействия с гетерогенным ядерным рибонуклеопротеином L (hnRNPL), ингибирующим регулятором p53 (Li et al., 2015), поднимая вопросы о механизме (механизмах), которые определяют различные эффекты Ela in vitro и in vivo. Чтобы исследовать важность процессинга и функциональности proEla и его связь с передачей сигналов APJ, мы создали клетки HEK, стабильно экспрессирующие GFP-APJ. Мы продемонстрировали, что синтетические зрелые человеческие пептиды ELA, proEla дикого типа и мутантные пептиды proEla вызывают интернализацию человеческого APJ (данные не показаны), и что активация APJ этими пептидами приводит к значительной активации ERK (данные не показаны), таким образом, устанавливая, что процессированные или непроцессированные ELA могут активировать APJ при этих условиях. По сравнению с активацией ERK все протестированные пептиды Ela не смогли индуцировать значительную активацию AKT и активацию последующего эффектора P70 (данные не показаны). Ранее избыточную экспрессию APJ и фурина связывали с различными видами рака и метастазирующими опухолями. Анализ уровней экспрессии Ela в образцах нормальной и опухолевой ткани от 7 пациентов с раком почки показал, что экспрессия Ela была подавлена у 6/7 пациентов, что указывает на потенциальное супрессорное действие Ela в отношении опухоли (фиг.6). Таким образом, чтобы непосредственно исследовать биологическую роль процессинга Ela и proEla в росте опухоли, мы воспользовались преимуществами клеток RenCa мышиной почки, которые не экспрессируют Ela и вызывают опухоли у мышей Balb/c. Мы обнаружили, что экспрессия proEla дикого типа или мутантного proEla в этих клетках ингибировала способность опухолевых клеток к индукции роста опухоли у мышей (фиг.7B). Как Ela участвует в подавлении роста опухоли, в настоящее время неясно, но некоторые механизмы могут быть постулированы. Различия в способности Ela пептидов индуцировать активацию AKT и ERK могут быть фактором, вносящим вклад. Ранее было установлено, что результат активации ERK зависит от его уровней экспрессии и активности. В то время как в нормальных клетках высокий уровень активации ERK индуцирует клеточное старение, было обнаружено, что сниженная активность ERK спасает клетки от старения и облегчает их трансформацию онкогеном ras, что указывает на роль супрессора опухолей для передачи сигналов ERK (Deschênes-Simard et al., 2013). Сообщалось, что ERK-опосредованное старение включает деградацию протеасомой различных белков, необходимых для различных биологических функций, включая пролиферацию и миграцию клеток, метаболизм РНК и передачу сигналов клетками. Соответственно, уровни фосфо-ERK являются очень низкими при различных онкологических заболеваниях человека, включая рак молочной железы (Milde-Langosch et al., 2005), опухоль головного мозга (Mawrin et al., 2003., Mawrin et al., 2005), рак простаты (Malik et al. al., 2002), рак поджелудочной железы (Yip-Schneider et al., 2001) и опухоли почки (Lee et al., 2009., Svensson et al., 2009). Аналогично, пациенты с высокими уровнями ERK, коррелирующими с хорошим прогнозом, менее агрессивным фенотипом (Milde-Langosch et al., 2005; Lee et al., 2009., Svensson et al., 2009), имели лучшую выживаемость и лучше реагировали на лечение (Chadha et al., 2006). В противоположность этому, было обнаружено, что некоторые виды прогрессирующего рака коррелируют с низким уровнем фосфо-ERK и высоким уровнем AKT (Malik et al., 2002., Deng et al., 2015). В самом деле; путь AKT является наиболее часто нарушаемым сигнальным путем при раке человека (Millis et al., 2016), и аберрации пути AKT были выявлены в 40% всех типов опухолей. Многочисленные соединения, которые ингибируют путь AKT на всех уровнях, в настоящее время находятся в клинической разработке, включая те, которые непосредственно нацелены на AKT (Yap et al., 2008). Эти исследования предполагают, что некоторые опухолевые супрессорные функции пути ERK, но не AKT, могут быть реактивированы у больных раком. Основываясь на способности Ela индуцировать высокую активность ERK и низкую активность AKT, можно составить потенциальную стратегию лечения рака почки. В заключение, мы идентифицируем Ela как потенциальный новый ген-супрессор опухоли и описываем вероятный механизм, связывающий способность Ela с активацией ERK и ингибированием роста опухоли.
В клетках RenCa ELABELA (как дикого типа, так и мутантный) индуцировал активацию mTORC1 путем ингибирования фосфорилирования S6K, участвующего в пути mTORC1, и поскольку mTORC1 является прямым ингибитором mTOR, этот эффект действует также на путь ERK (данные не показаны).
Мы пришли к выводу, что Elabela действует через mTOR как супрессор опухолей.
Ссылки
На протяжении этой заявки различные ссылки описывают уровень техники, к которой относится это изобретение. Раскрытия этих ссылок включены посредством ссылки в настоящее описание.


Группа изобретений относится к области медицины и фармацевтики, а именно к применению полипептида ELA или кодирующей его нуклеиновой кислоты, где полипептид ELA содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с SEQ ID NO: 1 (QRPVNLTMRRKLRKHNCLQRRCMPLHSRVPFP), где остаток аргинина (R) в положениях 9, 10, 20 или 21 при необходимости замещен остатком аланина (A) и/или остатком серина (S) для лечения рака почки; и к применению полипептида ELA или кодирующей его нуклеиновой кислоты, где полипептид ELA содержит SEQ ID NO: 1 (QRPVNLTMRRKLRKHNCLQRRCMPLHSRVPFP), где остаток аргинина (R) в положениях 9, 10, 20 или 21 при необходимости замещен остатком аланина (A) и/или остатком серина (S) для лечения рака почки. Группа изобретений обеспечивает новые эффективные подходы к лечению рака почки. 2 н. и 4 з.п. ф-лы, 11 ил., 2 пр.
1. Применение полипептида ELA или кодирующей его нуклеиновой кислоты, где полипептид ELA содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичности с SEQ ID NO: 1 (QRPVNLTMRRKLRKHNCLQRRCMPLHSRVPFP), где остаток аргинина (R) в положениях 9, 10, 20 или 21 при необходимости замещен остатком аланина (A) и/или остатком серина (S) для лечения рака почки.
2. Применение по п.1, в котором субъект страдает почечно-клеточным раком.
3. Применение по п.2, где почечно-клеточный рак выбран из светлоклеточного почечно-клеточного рака, папиллярного почечно-клеточного рака, хромофобного почечно-клеточного рака, рака собирающих протоков и медуллярного рака.
4. Применение по п.3, где почечно-клеточный рак является светлоклеточным почечно-клеточным раком.
5. Применение по п.1, в котором молекула нуклеиновой кислоты включена в подходящий вектор, такой как плазмида, космида, эписома, искусственная хромосома, фаг или вирусный вектор.
6. Применение полипептида ELA или кодирующей его нуклеиновой кислоты, где полипептид ELA содержит SEQ ID NO: 1 (QRPVNLTMRRKLRKHNCLQRRCMPLHSRVPFP), где остаток аргинина (R) в положениях 9, 10, 20 или 21 при необходимости замещен остатком аланина (A) и/или остатком серина (S) для лечения рака почки.
| WO 2015084264 A1, 11.06.2015 | |||
| ЯКУБКЕ Х.-Д и др | |||
| Аминокислоты, пептиды, белки | |||
| - М.: Мир, 1985 | |||
| Подвижной рельс для пересечений железнодорожных путей | 1922 |
|
SU456A1 |
| WO 2016061141 A1, 21.04.2016 | |||
| RU 2188026 C1, 27.08.2002. | |||

