Изобретение относится к области биотехнологии и генной инженерии. Изобретение описывает получение нового интеграционного вектора pPIG-1, имеющего модульную конструкцию и позволяющего осуществлять интеграцию и экспрессию гетерологичных генов в Pichia pastoris. Интеграционный вектор содержит селективный маркер для отбора трансформантов в клетках Е.coli, сайт начала репликации, сайт интеграции и экспрессионную кассету, в состав которой входят промотор, терминатор транскрипции и кодирующую полную последовательность 18S рРНК. Особенность данного вектора заключается в использовании полноразмерной последовательности гена 18S рРНК для гомологичной рекомбинации вектора, что позволяет производить интеграцию гена вставки без выщепления нативного гена. Использование данного вектора для трансформации позволяет увеличить продуктивность штамма продуцента растительной хитиназы из Drosera capensis.
Изобретение относится к биотехнологии, генетической инженерии и представляет собой технологию получения вектора для мультикопийной интеграции и экспрессии различных генов в штаммах Pichia pastoris.
Дрожжи Pichia pastoris являются хорошо известными продуцентами, способными производить широкий спектр различных ферментов, антител и пептидов, продуцируя их как внутриклеточно так и в культуральную среду. Традиционно для получения штаммов продуцентов используют вектора серии pPIC (Thermo Fisher Scientific cat. №. V19520) имеющие фланкируюие участки для рекомбинации, гомологичные нативному промотору алкоголь оксидазы (AOX1), либо селективному гену (обычно His4), что позволяет получать штаммы с жестким контролем экспрессии, но при этом частота интеграции целевого гена не велика и редко превышает 5 копий на геном [Vogl et al., 2018]. Стандартным решением для получения рекомбиантных штаммов Pichia pastoris является использование векторов серии pPICZα, имеющих удобную систему селекции на антибиотике зеоцин и обеспечивающий прогнозируемый результат при трансформации.
Несмотря на то, что AOX1 промотор является одним из самых сильных известных промоторов в P. pastoris, он не всегда удобен для применения в промышленных и лабораторных условиях. Для регуляции данного промотора необходим метанол в ростовой среде. Опасность метанола и повышенные санитарно-гигиенические требования к организации работ с метанолом являются серьезным препятствием для реализации биотехнологических производств на основе индуцибельных систем экспрессии, использующих AOX1 промотор. В связи с этим, представляется важным найти альтернативные пути, которые позволили бы использовать метилотрофные дрожжи для продукции целевого белка без использования метанола для индукции. С другой стороны, необходимо сохранить эффективность экспрессии сопоставимой с индуцибельными промоторами.
Задачей предлагаемого изобретения является расширение арсенала плазмидных векторов, пригодных для конструирования на их основе штаммов-продуцентов промышленно значимых белков.
Для решения поставленной задачи была предложена идея универсального интегративного вектора, обеспечивающего высокую частоту интеграции (мультикопийность) целевого гена.
Из коммерческих векторов, известен pGAPZa (Thermo Fisher Scientific cat. №. V20020), который позволяет получать штаммы-продуценты рекомбинантных белков, в которых экспрессия происходит конститутивно. Однако из-за конструктивных особенностей вектора, события инсерции генов в локусе промотора GAP возникают в результате единичного кроссинговера между локусом и областью GAP промотора на векторах pGAPZ или pGAPZa. Это приводит к вставке одной копий экспрессионной кассеты с целевым геном в 3`- область от локуса GAP промотора, при этом множественная встройка целевого гена хоть и возможна, но вероятность данного события крайне мала. Еще одним минусом данного подходя является высокая вероятность нарушения структуры гена GAP. В качестве селекционного маркера в данной серии плазмид используется ген BleoR под контролем промотора TEF1.
Существенным отличием предлагаемого к патентованию вектора pPIG-1 от коммерчески доступных векторов является использование полноразмерной последовательности 18S рРНК для интеграции. В геноме P. pastoris содержится 8 копий данного гена, присутствующего на всех 4 хромосомах [Schutter De и др., 2009], тогда как обычно используемые сайты интеграции (AOX1 промотор, His4 и др.) представлены одной копией на весь геном. При этом при многокопийной интеграции большинство генов вставки будет сосредоточена в одном месте генома, что может негативно сказаться на эффективности транскрипции.
В качестве прототипа данного вектора используется конструкция из патента RU 2388823 где успешно реализовали идею гомологичной рекомбинации по 18s рРНК. Однако в качестве плечей для рекомбинации используются не полная последовательность 18S рРНК, а два фрагмента: 5' 18S рРНК длиной 269 п.о. и фрагмент 3' 18S рРНК длиной 689 п.о., что может приводить к нарушению нативной структуры гена при рекомбинации и не является оптимальным. В работе [Näätsaari L, et al, 2012] продемонстрированно, что наибольшую частоту интеграции вставки в хромосому обеспечивают одинаковые по размеру плечи длиной более 650 п.о. Так же, в предлагаемом изобретении осуществлена модификация нуклеотидной последовательности пре-про области α-фактора Saccharomyces cerevisiae и введен сайт рестрикции HindIII, что позволяет вставлять последовательность целевого гена без добавления промежуточных аминокислот на N -концевой участок секретируемого белка.
Заявляемый мультикопийный интеграционный вектор pPIG-1 для экспрессии генов в дрожжах конструируют с использованием стандартных генно-инженерных методик: ПЦР, overlap-ПЦР и сборка полученных фрагментов методом Гибсона [Gibson et al., 2009].
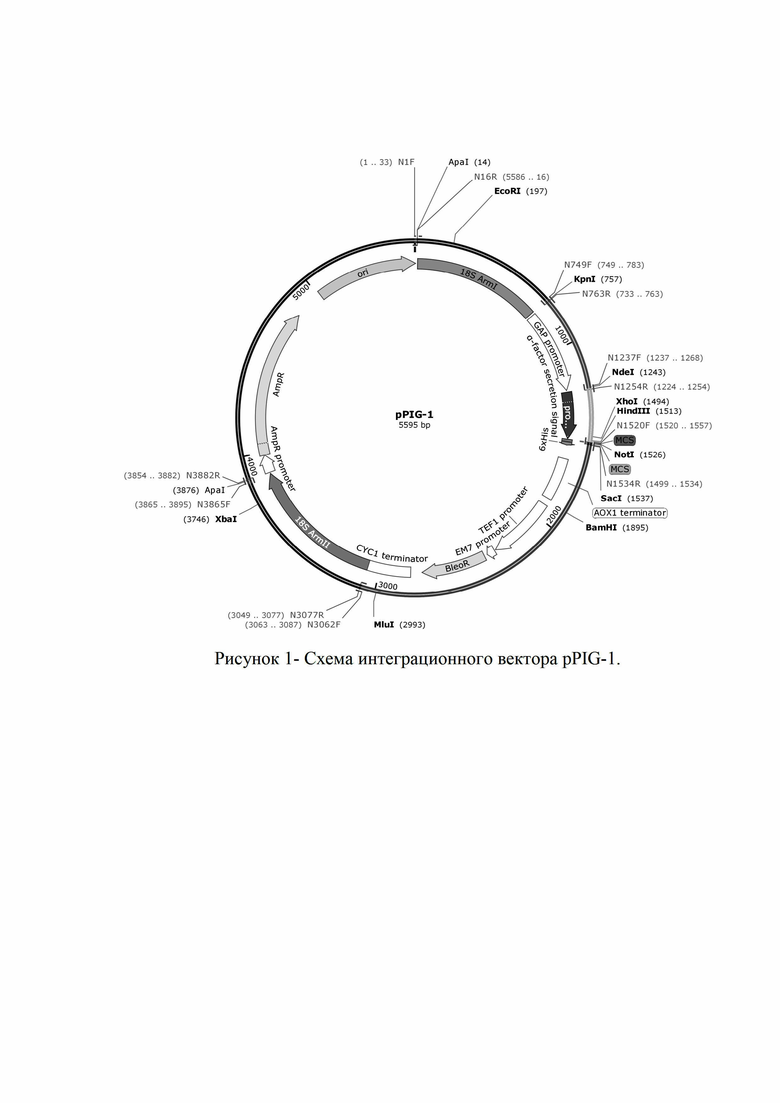
Предложенная генетическая конструкция состоит из 3’ фрагмента 18s рРНК (18-750 п.о), GAP промотора (764-1240 п.о.), модифицированный сигнал секреции гена MF из S.cerevisiae (1251-1517 п.о.), сайта множественного клонирования (1518-1537 п.о.), AOX1 терминатора (1635-1881 п.о.), TEF1 промотора (1921-2307 п.о.), EM7 промотора (2315-2362 п.о.), гена устойчивости к зеоцину BleoR (2381-2755 п.о.), CYC1 терминатора (2821-3068 п.о.) 5’ фрагмента 18s рРНК (3069-3870), гена устойчивости к ампициллину AmpR (3837-4842 п.о.) и точки начала репликации ori (5013-6 п.о.). Плечи для интеграции образуют полноразмерный ген 18S рРНК. Вектор имеет модульную структуру. Перед последовательностью GAP промотора введен сайт рестрикции KpnI. Между GAP промотором и пре-про сегментом α-фактора введен сайт рестрикции NdeI, что позволяет свободно менять сигнальные пептиды и промоторы, получая на основе интегративного вектора pPIG-1 новые конструкции. Кассета, содержащая AmpR и ori фланкированы сайтами ApaI, которые выщепляются при линеаризации вектора перед трансформацией, что позволяет избегать вставки бактериальных генов в геном дрожжей.
Именно такое сочетание элементов вектора дает возможность эффективной трансформации Pichia pastoris и множественной интеграции в геном целевой последовательности.
Для демонстрации эффективности нового вектора pPIG-1 были проведены эксперименты по клонированию в него гена chi19, кодирующего растительную хитиназу 19-го семейства гликозилгидролаз [AN GeneBank: MK093978.1]. Данный фермент был выбран в связи с его потенциальной промышленной значимостью для сельского хозяйства и невозможностью его получения в других экспрессионных системах из-за либо нерастворимости (в клетках E.coli), либо токсичности для хитинсодержащей клеточной стенки (в случае экспрессии в мицелиальных грибах) [Синельников и др., 2020].
Изобретение иллюстрируется следующими изображениями:
Рис. 1 - карта вектора pPIG-1
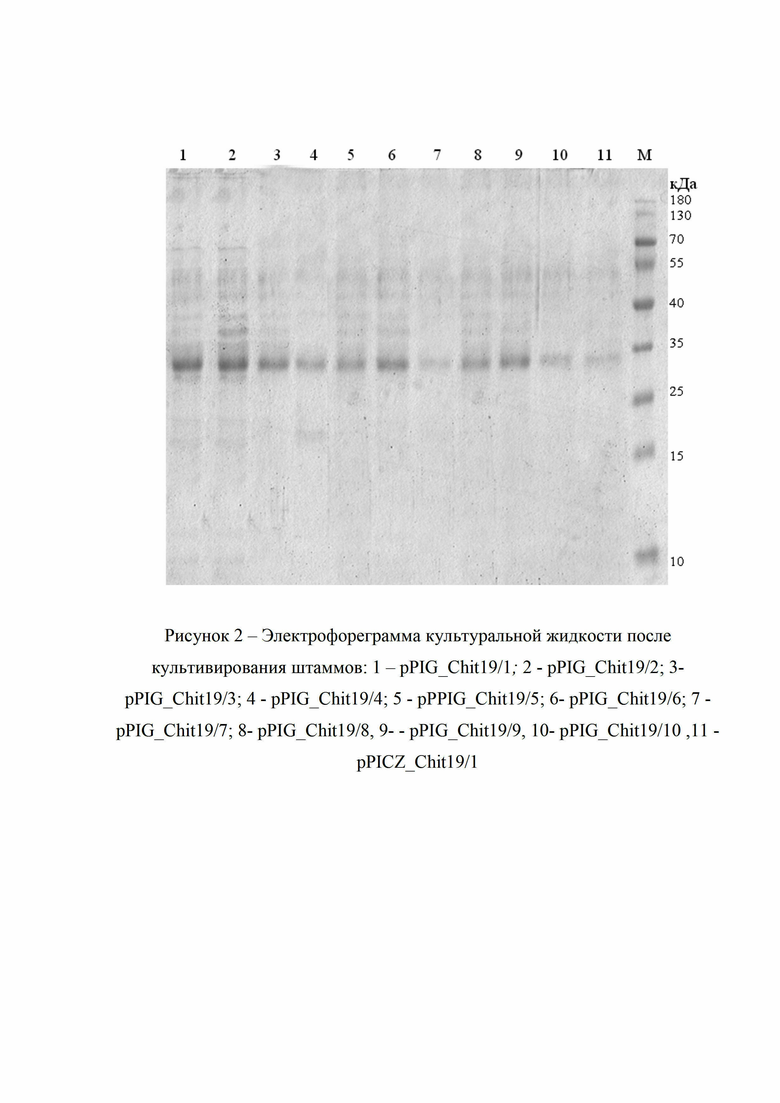
Рис. 2 - электрофореграмма культуральной жидкости pPIG_Chit19 и pPICZ_Chit19
Изобретение сопровождается двумя таблицами:
Таблица 1 - Структура праймеров, использованных при получении интеграционного вектора pPIG-1
Таблица 2 - Количество копий гена chi19 в геноме продуцентов хитиназы 19 семейства pPIG_Chit19 и pPICZ_Chit19
Пример 1 Получение интеграционного вектора pPIG-1
Поставленная задача решена путем конструирования инеграционного вектора pPIG-1 для экспрессии в дрожжах Pichia pastoris. Сконструированный вектор имеет размер 5595 п.о. (SEQ ID NO 1) (рис. 1)
Фрагмент ДНК размером 1747 п.о. (SEQ ID NO 2), кодирующий последовательность ori и гена устойчивости к ампицилину bla получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы (Thermo Fisher Scientific) и праймеров N3865F и N16R. На концах олигонуклеотидных праймеров добавлены последовательности, кодирующие сайты рестрикции ApaI. В качестве матрицы для ПЦР используют плазмиду pUC19 [https://www.addgene.org/50005/].
Фрагмент ДНК (SEQ ID NO 3), кодирующий кассету обеспечивающую устойчивость трансформантов к Зеоцину, который состоит из: фрагмента множественного сайта клонирования, 6His метки, AOX1 терминатора, дрожжевого TEF1 промотора, бактериального промотора EM7, гена ble, CYC1 терминатор размером 1558 п.о. При помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы и праймеров N749F и N1254R. В качестве матрицы для ПЦР используют коммерческую плазмиду pPICZAα (Thermo Fisher Scientific Каталожный номер: V19020). Фрагмент получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы и праймеров N1520F и N3077R.
Фрагмент ДНК, кодирующий сигнал секреции α-фактора и часть множественного сайта клонирования длиной 298 п.о. (SEQ ID NO 4) получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы (Thermo Fisher Scientific) и праймеров N1237F и N1534R. В качестве матрицы для ПЦР используют коммерческую плазмиду pPICZAα.
Для получения нижеперечисленных элементов конструкции, из штамма штамма Pichia pastoris GS115 (Thermo Fisher Scientific cat. № C18100) выделяют хромосомную ДНК [Lõoke, et al 2011], которая служит матрицей для синтеза GAP промотора, и фрагментов гена 18S рРНК.
Фрагмент ДНК кодирующий промотор глицеральдегид-3-фосфат - дегидрогеназы (GAP промотор) [GenBank NC_012964.1] длиной 506 п.о. фланкированный сайтами рестрикции KpnI и NdeI (SEQ ID NO 5),. получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы и праймеров N749F и N1254R. В качестве матрицы для ПЦР используют геномную ДНК штамма Pichia pastoris GS115
Фрагмент, кодирующий 3’ конец 18S рРНК (SEQ ID NO 6) получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы и праймеров N1F и N763R. В качестве матрицы для ПЦР используют геномную ДНК штамма Pichia pastoris GS115.
Фрагмент, кодирующий 5’-конец 18S рРНК (SEQ ID NO 7) получают при помощи ПЦР с использованием Phusion™ High-Fidelity DNA полимеразы и праймеров N3062F и N3882R. В качестве матрицы для ПЦР используют геномную ДНК штамма Pichia pastoris GS115.
Праймеры, использованные при получении интеграционного вектора представлены в Таблице 1.
Таблица 1 - Структура праймеров, использованных при получении интеграционного вектора pPIG-1
Амплифицированные фрагменты ДНК очищают из геля, используя с этой целью набор Qiagen (Qiagen, cat. №28706)
Далее очищенные фрагменты 2, 3, 4, 5, 6 и 7 смешивались в эквимолярных количествах и лигировались по методу Гибсона с применением Gibson Assembly® Master Mix (NEB cat. № E2611S). Полученной лигазной смесью трансформируют компетентные клетки Escherichia coli One Shot™ Mach1™ (Thermo Fisher Scientific cat. № C862003), после стандартной процедуры трансформации [Маниатис, 1984] клетки разводят в три раза средой LB и инкубируют при 37°С 1 час, после чего высевают на агар LB, содержащий ампициллин в концентрации 100 мкг/мл. Посевы инкубируют 24 часа при 37°С, после чего выросшие ампициллин-резистентные колонии выращиваются в 5 мл среды LB, после чего выделяют плазмидную ДНК при помощи набора QIAGEN Plasmid Mini Kit (QIAGEN Cat No.12123). Правильность сборки всех элементов проверяется путем секвенирования по методу Сэнгера. При расщеплении рестриктазами: ApaI, KpnI, HindIII и NdeI вектор pPIG-1 дает фрагменты массой 2363, 1733, 743 и 270 п.о., что позволяет надежно идентифицировать данную конструкцию.
Схема интеграционного вектора pPIG-1 приведено на Рисунке 1.
Пример 2 Создание штамма продуцента растительной хитиназы 19-ого семейства (pPIG_Chit19) на основе вектора pPIG-1
Для проверки применимости, полученного в примере 1, интеграционного вектора для создания штаммов продуцентов, был создан штамм-продуцент растительной хитиназы на основе штамма Pichia pastoris GS115.
Синтетический ген растительной хитиназы 19 семейства (SEQ ID NO 8) амплифицируют при помощи Phusion™ High-Fidelity полимеразы и праймеров ChitF (5`-AAGCTTGTCCAGTGTGGTAGCGAAGTCG-3`) и ChitR (5`-ATGCGGCCGCTCAGCTGAACGGACGTTGATTG-3`). Полученный ПЦР-фрагмент обрабатывают рестриктазами HindIII и NotI и очищают из геля, используя набор Qiagen (Qiagen, cat. № 28706).
Вектор pPIG-1 обрабатывают рестриктазами HindIII и NotI, после чего очищают из геля, используя набор Qiagen (Qiagen, cat. № 28706).
Вектор и вставка смешиваются в соотношении 1 к 5 и лигируются Т4 лигазой (Thermo Fisher Scientific cat. № EL0011). Лигазной смесью трансформируют компетентные клетки Escherichia coli One Shot™ Mach1™ (Thermo Fisher Scientific cat. № C862003), после стандартной процедуры трансформации [Маниатис, 1984] клетки разводят в три раза средой LB и инкубируют при 37°С 1 час, после чего высевают на агар LB, содержащий ампициллин в концентрации 100 мкг/мл. Посевы инкубируют 24 часа при 37°С, после чего выросшие ампициллинрезистентные колонии выращиваются в 5 мл среды LB, после чего выделяют плазмидную ДНК при помощи набора QIAGEN Plasmid Mini Kit (QIAGEN Cat No. 12123). Правильность сборки плазмидной ДНК проверяют путем секвенирования по методу Сэнгера в обоих направлениях. В результате получена конструкция pPIG_Chit19.
Для проведения трансформации pPIG_Chit19 препаративно выделили из клеток Ε.coli и линеаризовали с помощью рестриктазы ApaI. Трансформацию линеаризованной pPIG_Chit19 проводили с использованием электропорации и последующим отбором трансформантов на среде, содержащей 300 мкг/мл зеоцина (Thermo Fisher Scientific, cat. № R25005). Полученные штаммы-трансформанты серии pPIG_Chit19 пересевали на стандартную среду YPD. В результате был получен штамм продуцент pPIG_Chit19.
Продуктивность полученного штамма определяют путем скрининга 10 выбранных клонов путем анализа культуральной жидкости на содержание целевого белка. Штаммы культивировали в 10 мл стандартной среды YP c добавлением 2% глицерина в течении 36 часов и анализировали культуральную жидкость методом электрофореза в полиакриламидном геле в денатурирующих условиях (рис. 2).
Таким образом за 36 часов культивирования после скрининга определен выход целевой хитиназы в культуральной жидкости каждого штамма, который составил от 20 до 150 мг/л. Выход белка определялся количественно методом Лоури [Lowry и др., 1951]
Для сравнения эффективности были создан штамм продуцент pPICZ_Chit19, представляющий собой штамм Pichia pastoris GS115, который был трансформирован плазмидой, представляющей собой коммерческий вектором pPICZAα (Invitrogen, США), трансформированный синтетическим геном хитиназы 19 семейства под контролем AOX1 промотора. Выбранные трансформанты были ферментированы согласно рекомендациям производителя [Thermo Fisher Scientific MAN00000034]. Что позволило получить выходы от 10 до 30 мг/л chit19 на 4 сутки после индукции метанолом.
В данном примере показано, что при использовании плазмид на основе pPIG-1 удается получать целевой белок (в данном случае растительную хитиназу 19-ой семьи гликозилгидролаз) с эффективностью, сопоставимой с сильным индуцибельным промотором pAOX1, при этом не требуется использование сложных сред и добавление метанола в качестве индукции.
Пример 3 Определение частоты интеграции структурных элементов вектора pPIG-1 c геном chi19
Для измерения частоты интеграции конструкции pPIG_Chit19 в геном Pichia Pastoris используют ПЦР в реальном времени. В трех наиболее продуктивных штаммах pPIG_Chit19 и pPICZ_Chit19 (из примера 2) анализировали количество последовательностей pAOX1 (промотор алкогольоксидазы 1) chi19 (хитиназа D.capensis), pGAP (промотор глицеральдегид-3-фосфат - дегидрогеназы) и act (актин) в геноме трансформантов, показавших наибольшую продуктивность. В качестве контроля и образца для построения калибровочных кривых использовали родительский штамм Pichia pastoris GS115. Результаты представлены в Таблице 2.
Таблица 2 - Количество копий структурных элементов в геноме продуцентов хитиназы 19 семейства pPIG_Chit19 и pPICZ_Chit19.
Для проверки точности и стабильности количественного определения были построены стандартные кривые для всех генов. Значения коэффициента вариации (CV) для данных последовательностей составили от 0,55% до 6,15%, а значения стандартного отклонения (SD) варьировалось от 0,03 до 0,16. Эти данные показали, что значения CV и SD незначительно различались в экспериментах, указывая на то, что системы РТ-ПЦР функционировали стабильно и надежно.
Максимальное количество копий гена Chit 19 в геном дрожжей составило 12 для штамма продуцента pPPIG_chit19/1, что в 4 раза выше, чем максимальное число интеграций при использовании конструкций на основе коммерческого вектора pPICZAα. Количество GAP промоторов увеличивается пропорционально количеству интеграций гена Chit 19, что говорит о введении полной экспрессионной кассеты, а не отдельных ее элементов.
Таким образом, полученный интеграционный вектор pPIG-1 повышает эффективность интеграции экспрессионной кассеты в геном P. pastoris GS115 в 4~12 раз по сравнению с коммерческим вектором pPICZAα.
Список используемых источников
1. Gibson D.G., Young L., Venter C., Hutchinson C., Smith H. Enzymatic assembly of DNA molecules up to several hundred kilobases // Nat. Methods. 2009. Т. 6. № 5. С. 343–345.
2. RU 2388823 (10.05.2010). Интегративный плазмидный вектор для экспрессии генов в дрожжах.
3. Lõoke M., Kristjuhan K., Kristjuhan A. Extraction of genomic DNA from yeasts for PCR-based applications // Biotechniques. 2011. Т. 50. № 5. С. 325–328.
4. Методы генетической инженерии. Молекулярное клонирование'; Маниатис, Т.И; Фрич, Э.; Сэмбрук, Дж.; Изд-во: М.: Мир, 1984 г
5. Lowry O.H. Protein measurement with the Folin phenol reagent. // J. Biol. Chem. 1951. Т. 193. № 1. С. 265–275.
6. Näätsaari L, Mistlberger B, Ruth C, Hajek T, Hartner FS, Glieder A. Deletion of the Pichia pastoris KU70 homologue facilitates platform strain generation for gene expression and synthetic biology// PLoS One. 2012;7(6):e39720.
7. Schutter K. De, Lin Y-C., Van Hecke A., Glinka S., Weber-Lehmann J., Rouze P., Peer Y., Cakkewaert N. Genome sequence of the recombinant protein production host Pichia pastoris // Nat. Biotechnol. 2009. Т. 27. № 6. С. 561–566.
8. Vogl T., Gebbie L., Palfreyman R., Speight, R. Effect of plasmid design and type of integration event on recombinant protein expression in Pichia pastoris // Appl. Environ. Microbiol. 2018. Т. 84. № 6.
9. Синельников И.Г. и др. Клонирование и экспрессия новой хитиназы из хищных растений Drosera Capensis // Вестник Московского университета. Серия 2 Химия. 2020. Т. 61. № 5. С. 361–368.
SEQ ID NO 1
TGGAAAAACGGGCCCGCAGGTTCACCTACGGAAACCTTGTTACGACTTTTACTTCCTCTAAATGACCAAGTTTGTCCAAGTTCAGGCTCGCGCCCTCCCAAAGCCTCACTAAACCATTCAATCGGTAGTAGCGACGGGCGGTGTGTACAAAGGGCAGGGACGTAATCAGCGCGAGCTGATGACTCGCGCTTACTAGGAATTCCTCGTTGAAGCGCCTCTTGCAAAGCGCTATCCCCAGCACGACGGAGTCTAAGATTCCCCGGCCATCTCTGGCAAGGACTCGCTGCCTCCGTCAGTGTAGCGCGCGTGCGGCCCAGAACGTCTAAGGGCATCACAGACCTGTTATTGCCTCGCTTCCGCTGGCTTGCGCCAGTTGTCCTTCTAAGAAGATCCCCCAGCAATGCCAGGTAACCTAGTTAAAAGCCAAGGTCTCGTTCGTTATCGCAATTAAGCAGACAAATCACTCCACCAACTAAGAACGGCCATGCACCACCACCCACAAAATCAAGAAAGTGCTCTCATCCTGTCAATCCTCATTGTGTCTGGACCTGGTGAGTTTCCCCGTGTTGAGTCAAATTAAGCCGCAGGCTCCACTCCTGGTGGTGCCCTTCCGTCAATTCCTTTAAGTTTCAGCCTTGCGACCATACTCCCCCCAGAACCCAAAGACTTTGATTTCTCGTAAGGTGCCGGGGAAGGCTATTCCCCGATCCCTAGTCGGCATCGTTTATGGTTAAGACTACGACGGTATCTGGGGTACCATCCTTTTTTGTAGAAATGTCTTGGTGTCCTCGTCCAATCAGGTAGCCATCTCTGAAATATCTGGCTCCGTTGCAACTCCGAACGACCTGCTGGCAACGTAAAATTCTCCGGGGTAAAACTTAAATGTGGAGTAATGGAACCAGAAACGTCTCTTCCCTTCTCTCTCCTTCCACCGCCCGTTACCGTCCCTAGGAAATTTTACTCTGCTGGAGAGCTTCTTCTACGGCCCCCTTGCAGCAATGCTCTTCCCAGCATTACGTTGCGGGTAAAACGGAAGTCGTGTACCCGACCTAGCAGCCCAGGGATGGAAAAGTCCCGGCCGTCGCTGGCAATAATAGCGGGCGGACGCATGTCATGAGATTATTGGAAACCACCAGAATCGAATATAAAAGGCGAACACCTTTCCCAATTTTGGTTTCTCCTGACCCAAAGACTTTAAATTTAATTTATTTGTCCCTATTTCAATCAATTGAACAACTATCCATATGGAAATGAGATTTCCTTCAATTTTTACTGCTGTTTTATTCGCAGCATCCTCCGCATTAGCTGCTCCAGTCAACACTACAACAGAAGATGAAACGGCACAAATTCCGGCTGAAGCTGTCATCGGTTACTCAGATTTAGAAGGGGATTTCGATGTTGCTGTTTTGCCATTTTCCAACAGCACAAATAACGGGTTATTGTTTATAAATACTACTATTGCCAGCATTGCTGCTAAAGAAGAAGGGGTATCTCTCGAGAAAAGAGAGGCTGAAGCTTCTGCAGGCGGCCGCGAGCTCATCATCATCATCATCATTGAGTTTGTAGCCTTAGACATGACTGTTCCTCAGTTCAAGTTGGGCACTTACGAGAAGACCGGTCTTGCTAGATTCTAATCAAGAGGATGTCAGAATGCCATTTGCCTGAGAGATGCAGGCTTCATTTTTGATACTTTTTTATTTGTAACCTATATAGTATAGGATTTTTTTTGTCATTTTGTTTCTTCTCGTACGAGCTTGCTCCTGATCAGCCTATCTCGCAGCTGATGAATATCTTGTGGTAGGGGTTTGGGAAAATCATTCGAGTTTGATGTTTTTCTTGGTATTTCCCACTCCTCTTCAGAGTACAGAAGATTAAGTGAGACCTTCGTTTGTGCGGATCCCCCACACACCATAGCTTCAAAATGTTTCTACTCCTTTTTTACTCTTCCAGATTTTCTCGGACTCCGCGCATCGCCGTACCACTTCAAAACACCCAAGCACAGCATACTAAATTTTCCCTCTTTCTTCCTCTAGGGTGTCGTTAATTACCCGTACTAAAGGTTTGGAAAAGAAAAAAGAGACCGCCTCGTTTCTTTTTCTTCGTCGAAAAAGGCAATAAAAATTTTTATCACGTTTCTTTTTCTTGAAATTTTTTTTTTTAGTTTTTTTCTCTTTCAGTGACCTCCATTGATATTTAAGTTAATAAACGGTCTTCAATTTCTCAAGTTTCAGTTTCATTTTTCTTGTTCTATTACAACTTTTTTTACTTCTTGTTCATTAGAAAGAAAGCATAGCAATCTAATCTAAGGGGCGGTGTTGACAATTAATCATCGGCATAGTATATCGGCATAGTATAATACGACAAGGTGAGGAACTAAACCATGGCCAAGTTGACCAGTGCCGTTCCGGTGCTCACCGCGCGCGACGTCGCCGGAGCGGTCGAGTTCTGGACCGACCGGCTCGGGTTCTCCCGGGACTTCGTGGAGGACGACTTCGCCGGTGTGGTCCGGGACGACGTGACCCTGTTCATCAGCGCGGTCCAGGACCAGGTGGTGCCGGACAACACCCTGGCCTGGGTGTGGGTGCGCGGCCTGGACGAGCTGTACGCCGAGTGGTCGGAGGTCGTGTCCACGAACTTCCGGGACGCCTCCGGGCCGGCCATGACCGAGATCGGCGAGCAGCCGTGGGGGCGGGAGTTCGCCCTGCGCGACCCGGCCGGCAACTGCGTGCACTTCGTGGCCGAGGAGCAGGACTGACACGTCCGACGGCGGCCCACGGGTCCCAGGCCTCGGAGATCCGTCCCCCTTTTCCTTTGTCGATATCATGTAATTAGTTATGTCACGCTTACATTCACGCCCTCCCCCCACATCCGCTCTAACCGAAAAGGAAGGAGTTAGACAACCTGAAGTCTAGGTCCCTATTTATTTTTTTATAGTTATGTTAGTATTAAGAACGTTATTTATATTTCAAATTTTTCTTTTTTTTCTGTACAGACGCGTGTACGCATGTAACATTATACTGAAAACCTTGCTTGAGAAGGTTTTGGGACGCTCGAAGGCTTTAATTTGCTCATCTTCGATCCCCTAACTTTCGTTCTTGATTAATGAAAACGTCCTTGGCGAATGCTTTCGCAGTAGTTAGTCTTGGGGCGATCCAAGAATTTCACCTCTGACGCCCCAATACTGACGCCCCCGACCGTCCCTGTTAATCATTACGCGGCCCCGAACCAACAAAAGAACCGTATCCTCTTCTGTTATTCCATGCTAATATATTCAACTACTGCCTTGAACACTCTAATTTCCTCAAAGTAACGTCCGTTCAACTACGAGCTTTTTAACTGCAACAACTTTAATATACGCTATTGGAGCTGGAATTACCGCGGCTGCTGGCACCAGACTTGCCCTCCAATTGTTCCTCGTTAAGGTATTTACGTTGTACTCATTCCAATTACAAGACCAAAGGCCCTGTATCGTTATTTATTGTCACTACCTCCCTGTGTCAGGATTGGGTAATTTGCGCGCCTGCTGCCTTCCTTGGATGTGGTAGCCGTCTCTCAGGCTCCCTCTCCGGAATCGAACCCTTATTCCCCGTTACCCGTAGAAACCATGGTAGGCCTCTATCCTACCATCGAAAGTTGATAGGGCAGAAATTTGAATGAACCATCCTAAGATTCGAAAAGTTATTATGAATCACCAAAACGAAGGTTTTATCTAATAAATACGCCCGAGGGCTGATCAAGTATTAGCTCTAGAATTACCACGGTTATCCTTGTAGCAACACTATCAAATAAACGATAACTGATTTAATGAGCCATTCGCAGTTTCACCGTATAATGCTATACTTAGACATGCATGGCTTAATCTTTGAGACAAGGGCCCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTATTGACGCCGGGCAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGCGGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCACGATGCCTGTAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAATAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATATATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTA
SEQ ID NO 2
GAGACAAGGGCCCGCGGAACCCCTATTtgtTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTATTGACGCCGGGCAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGCGGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCACGATGCCTGTAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAATAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATATATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTTCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACgggcccg
SEQ ID NO 3
TGCAGGCGGCCGCGAGCTCATCATCATCATCATCATTGAGTTTGTAGCCTTAGACATGACTGTTCCTCAGTTCAAGTTGGGCACTTACGAGAAGACCGGTCTTGCTAGATTCTAATCAAGAGGATGTCAGAATGCCATTTGCCTGAGAGATGCAGGCTTCATTTTTGATACTTTTTTATTTGTAACCTATATAGTATAGGATTTTTTTTGTCATTTTGTTTCTTCTCGTACGAGCTTGCTCCTGATCAGCCTATCTCGCAGCTGATGAATATCTTGTGGTAGGGGTTTGGGAAAATCATTCGAGTTTGATGTTTTTCTTGGTATTTCCCACTCCTCTTCAGAGTACAGAAGATTAAGTGAGACCTTCGTTTGTGCGGATCCCCCACACACCATAGCTTCAAAATGTTTCTACTCCTTTTTTACTCTTCCAGATTTTCTCGGACTCCGCGCATCGCCGTACCACTTCAAAACACCCAAGCACAGCATACTAAATTTTCCCTCTTTCTTCCTCTAGGGTGTCGTTAATTACCCGTACTAAAGGTTTGGAAAAGAAAAAAGAGACCGCCTCGTTTCTTTTTCTTCGTCGAAAAAGGCAATAAAAATTTTTATCACGTTTCTTTTTCTTGAAATTTTTTTTTTTAGTTTTTTTCTCTTTCAGTGACCTCCATTGATATTTAAGTTAATAAACGGTCTTCAATTTCTCAAGTTTCAGTTTCATTTTTCTTGTTCTATTACAACTTTTTTTACTTCTTGTTCATTAGAAAGAAAGCATAGCAATCTAATCTAAGGGGCGGTGTTGACAATTAATCATCGGCATAGTATATCGGCATAGTATAATACGACAAGGTGAGGAACTAAACCATGGCCAAGTTGACCAGTGCCGTTCCGGTGCTCACCGCGCGCGACGTCGCCGGAGCGGTCGAGTTCTGGACCGACCGGCTCGGGTTCTCCCGGGACTTCGTGGAGGACGACTTCGCCGGTGTGGTCCGGGACGACGTGACCCTGTTCATCAGCGCGGTCCAGGACCAGGTGGTGCCGGACAACACCCTGGCCTGGGTGTGGGTGCGCGGCCTGGACGAGCTGTACGCCGAGTGGTCGGAGGTCGTGTCCACGAACTTCCGGGACGCCTCCGGGCCGGCCATGACCGAGATCGGCGAGCAGCCGTGGGGGCGGGAGTTCGCCCTGCGCGACCCGGCCGGCAACTGCGTGCACTTCGTGGCCGAGGAGCAGGACTGACACGTCCGACGGCGGCCCACGGGTCCCAGGCCTCGGAGATCCGTCCCCCTTTTCCTTTGTCGATATCATGTAATTAGTTATGTCACGCTTACATTCACGCCCTCCCCCCACATCCGCTCTAACCGAAAAGGAAGGAGTTAGACAACCTGAAGTCTAGGTCCCTATTTATTTTTTTATAGTTATGTTAGTATTAAGAACGTTATTTATATTTCAAATTTTTCTTTTTTTTCTGTACAGACGCGTGTACGCATGTAACATTATACTGAAAACCTTGCTTGAGAAGGTTTTGGGACGCTCGAAGGCTTTAATTTGCtcatcttcg
SEQ ID NO 4
ctatcCATATGGaaATGAGATTTCCTTCAATTTTTACTGCTGTTTTATTCGCAGCATCCTCCGCATTAGCTGCTCCAGTCAACACTACAACAGAAGATGAAACGGCACAAATTCCGGCTGAAGCTGTCATCGGTTACTCAGATTTAGAAGGGGATTTCGATGTTGCTGTTTTGCCATTTTCCAACAGCACAAATAACGGGTTATTGTTTATAAATACTACTATTGCCAGCATTGCTGCTAAAGAAGAAGGGGTATCTCTCGAGAAAAGAGAGGCTGAAGCTTCTGCAGGCGGCCGCGA
SEQ ID NO 5
CTGGGGTACCATCCTTTTTTGTAGAAATGTCTTGGTGTCCTCGTCCAATCAGGTAGCCATCTCTGAAATATCTGGCTCCGTTGCAACTCCGAACGACCTGCTGGCAACGTAAAATTCTCCGGGGTAAAACTTAAATGTGGAGTAATGGAACCAGAAACGTCTCTTCCCTTCTCTCTCCTTCCACCGCCCGTTACCGTCCCTAGGAAATTTTACTCTGCTGGAGAGCTTCTTCTACGGCCCCCTTGCAGCAATGCTCTTCCCAGCATTACGTTGCGGGTAAAACGGAAGTCGTGTACCCGACCTAGCAGCCCAGGGATGGAAAAGTCCCGGCCGTCGCTGGCAATAATAGCGGGCGGACGCATGTCATGAGATTATTGGAAACCACCAGAATCGAATATAAAAGGCGAACACCTTTCCCAATTTTGGTTTCTCCTGACCCAAAGACTTTAAATTTAATTTATTTGTCCCTATTTCAATCAATTGAACAACTATCCATATGGAAATGA
SEQ ID NO 6
TGGAAAAACGGGCCCGCAGGTTCACCTACGGAAACCTTGTTACGACTTTTACTTCCTCTAAATGACCAAGTTTGTCCAAGTTCAGGCTCGCGCCCTCCCAAAGCCTCACTAAACCATTCAATCGGTAGTAGCGACGGGCGGTGTGTACAAAGGGCAGGGACGTAATCAGCGCGAGCTGATGACTCGCGCTTACTAGGAATTCCTCGTTGAAGCGCCTCTTGCAAAGCGCTATCCCCAGCACGACGGAGTCTAAGATTCCCCGGCCATCTCTGGCAAGGACTCGCTGCCTCCGTCAGTGTAGCGCGCGTGCGGCCCAGAACGTCTAAGGGCATCACAGACCTGTTATTGCCTCGCTTCCGCTGGCTTGCGCCAGTTGTCCTTCTAAGAAGATCCCCCAGCAATGCCAGGTAACCTAGTTAAAAGCCAAGGTCTCGTTCGTTATCGCAATTAAGCAGACAAATCACTCCACCAACTAAGAACGGCCATGCACCACCACCCACAAAATCAAGAAAGTGCTCTCATCCTGTCAATCCTCATTGTGTCTGGACCTGGTGAGTTTCCCCGTGTTGAGTCAAATTAAGCCGCAGGCTCCACTCCTGGTGGTGCCCTTCCGTCAATTCCTTTAAGTTTCAGCCTTGCGACCATACTCCCCCCAGAACCCAAAGACTTTGATTTCTCGTAAGGTGCCGGGGAAGGCTATTCCCCGATCCCTAGTCGGCATCGTTTATGGTTAAGACTACGACGGTATCTGGGGTACCATCCT
SEQ ID NO 7
ATTTGCTCATCTTCGATCCCCTAACTTTCGTTCTTGATTAATGAAAACGTCCTTGGCGAATGCTTTCGCAGTAGTTAGTCTTGGGGCGATCCAAGAATTTCACCTCTGACGCCCCAATACTGACGCCCCCGACCGTCCCTGTTAATCATTACGCGGCCCCGAACCAACAAAAGAACCGTATCCTCTTCTGTTATTCCATGCTAATATATTCAACTACTGCCTTGAACACTCTAATTTCCTCAAAGTAACGTCCGTTCAACTACGAGCTTTTTAACTGCAACAACTTTAATATACGCTATTGGAGCTGGAATTACCGCGGCTGCTGGCACCAGACTTGCCCTCCAATTGTTCCTCGTTAAGGTATTTACGTTGTACTCATTCCAATTACAAGACCAAAGGCCCTGTATCGTTATTTATTGTCACTACCTCCCTGTGTCAGGATTGGGTAATTTGCGCGCCTGCTGCCTTCCTTGGATGTGGTAGCCGTCTCTCAGGCTCCCTCTCCGGAATCGAACCCTTATTCCCCGTTACCCGTAGAAACCATGGTAGGCCTCTATCCTACCATCGAAAGTTGATAGGGCAGAAATTTGAATGAACCATCCTAAGATTCGAAAAGTTATTATGAATCACCAAAACGAAGGTTTTATCTAATAAATACGCCCGAGGGCTGATCAAGTATTAGCTCTAGAATTACCACGGTTATCCTTGTAGCAACACTATCAAATAAACGATAACTGATTTAATGAGCCATTCGCAGTTTCACCGTATAATGCTATACTTAGACATGCATGGCTTAATCTTTGAGACAAGGGCCCGCGGA
SEQ ID NO 8
atgCGCATCACGGTCCTGTTGCTCTTGTGTGTCGCTCCTCTGTTGTCTGGCACGTATGCTGTCCAGTGTGGTAGCGAAGTCGGTGGAGCTCTGTGTCCGAATGGTCTGTGTTGCAGCAAGTATGGCTACTGTGGCACTACGTCTGCCTACTGTGGTCCGGGCTGTCAGAGCCAGTGTGGTGGTTCCTCTCCTCCGCCTGCTCCTCCCAGCCCGACTCCGAGTCCTCCGTCTCCCTCTGGAGGTGGTGATGTGTCCAGCATCATCACCTCCCAGATCTTCAATCAGATGCTGCTCCATCGCAATGACAATGCCTGTCCTGCCCATGGCTTCTACAGCTATCAAGCCTTCTTGGATGCTGCACGCAAGTTTACTGGTTTCGGTACGACTGGTGACATCAACACTCGCAAACGTGAACTGGCTGCCTTCTTTGGTCAGACGAGCCACGAGACCACTGGTGGCTGGCCCACTGCTCCTGATGGTCCGTATGCCTGGGGCTACTGCTTCAAACAGGAACAAGGCAATCCTGGTGACTACTGTGTCCAGTCTTCCACGTATCCCTGTGCCCCTGGCAAGAAGTACTATGGTCGTGGACCGATTCAGATCTCCTACAACTACAACTATGGTCAGTGTGGAGCCGCCATTAATCAACCCCTGCTGAGCAATCCGGATCTGGTCGCGTCCAATGCCGATGTGTCCTTCGAGACTGCCATCTGGTTCTGGATGACTCCTCAAGGTAGCAAACCCTCCTGTCATGCCGTCGCCACTGGTCAGTGGACTCCGTCCGTCGCCGATCAAGCTGCTGGACGTGTTCCTGGCTATGGTGTCATTACGAACATCATCAATGGAGGTGTCGAGTGTGGCAAAGGCACGGTCCCGCAAGTTGCCGATCGCATTGGCTTCTATCAACGCTACTGCTCCATCATGGGTATTGCGCCTGGTGGCAATCTTGGCTGCTACAATCAACGTCCGTTCAGCGCGGCCGCat
--->
Перечень последовательностей
<110> Федеральное государственное учреждение «Федеральный
Исследовательский центр «Фундаментальные основы биотехнологии»
Российской академии наук»
<120> Интеграционный вектор для многокопийной интеграции в 18S рРНК
дрожжей Pichia pastoris
<160> 7
<210> 1
<211> 5595
<212> plasmid DNA
<213> synthetic DNA construct
<221> misc_feature
<222> (18)...(750)
<223> 18S ArmI
<221> promoter
<222> (764)...(1240)
<223> GAP promoter. Pichia pastoris promoter
glyceraldehyde-3-phosphate dehydrogenase
<221> CDS
<222> (1251)...(1517)
<223> N-terminal secretion signal from S. cerevisiae
MF-alpha-factor-1
<221> misc_feature
<222> (1514)...(1537)
<223> multiple cloning site
<221> CDS
<222> (1538)...(1555)
<223> 6xHis affinity tag
<221> terminator
<222> (1635)...(1881)
<223> Pichia pastoris AOX1 terminator
<221> promoter
<222> (1921)...(2307)
<223> TEF1 promoter from S. cerevisiae
<221> promoter
<222> (2315)...(2362)
<223> synthetic bacterial promoter EM7
<221> CDS
<222> (2381)...(2755)
<223> Sh ble gene product from Streptoalloteichus hindustanus
<221> terminator
<222> (2821)...(3068)
<223> S. cerevisiae CYC1 terminator
<221> misc_feature
<222> (3069)...(3870)
<223> 18S ArmII
<221> promoter
<222> (3878)...(3981)
<223> AmpR promoter
<221> CDS
<222> (3982)...(4842)
<223> beta-lactamase
<221> rep_origin
<222> (5013)...(6)
<223> high-copy-number ColE1/pMB1/pBR322/pUC origin of replication
<400> 1
1 tggaaaaacg ggcccgcagg ttcacctacg gaaaccttgt tacgactttt acttcctcta
61 aatgaccaag tttgtccaag ttcaggctcg cgccctccca aagcctcact aaaccattca
121 atcggtagta gcgacgggcg gtgtgtacaa agggcaggga cgtaatcagc gcgagctgat
181 gactcgcgct tactaggaat tcctcgttga agcgcctctt gcaaagcgct atccccagca
241 cgacggagtc taagattccc cggccatctc tggcaaggac tcgctgcctc cgtcagtgta
301 gcgcgcgtgc ggcccagaac gtctaagggc atcacagacc tgttattgcc tcgcttccgc
361 tggcttgcgc cagttgtcct tctaagaaga tcccccagca atgccaggta acctagttaa
421 aagccaaggt ctcgttcgtt atcgcaatta agcagacaaa tcactccacc aactaagaac
481 ggccatgcac caccacccac aaaatcaaga aagtgctctc atcctgtcaa tcctcattgt
541 gtctggacct ggtgagtttc cccgtgttga gtcaaattaa gccgcaggct ccactcctgg
601 tggtgccctt ccgtcaattc ctttaagttt cagccttgcg accatactcc ccccagaacc
661 caaagacttt gatttctcgt aaggtgccgg ggaaggctat tccccgatcc ctagtcggca
721 tcgtttatgg ttaagactac gacggtatct ggggtaccat ccttttttgt agaaatgtct
781 tggtgtcctc gtccaatcag gtagccatct ctgaaatatc tggctccgtt gcaactccga
841 acgacctgct ggcaacgtaa aattctccgg ggtaaaactt aaatgtggag taatggaacc
901 agaaacgtct cttcccttct ctctccttcc accgcccgtt accgtcccta ggaaatttta
961 ctctgctgga gagcttcttc tacggccccc ttgcagcaat gctcttccca gcattacgtt
1021 gcgggtaaaa cggaagtcgt gtacccgacc tagcagccca gggatggaaa agtcccggcc
1081 gtcgctggca ataatagcgg gcggacgcat gtcatgagat tattggaaac caccagaatc
1141 gaatataaaa ggcgaacacc tttcccaatt ttggtttctc ctgacccaaa gactttaaat
1201 ttaatttatt tgtccctatt tcaatcaatt gaacaactat ccatatggaa atgagatttc
1261 cttcaatttt tactgctgtt ttattcgcag catcctccgc attagctgct ccagtcaaca
1321 ctacaacaga agatgaaacg gcacaaattc cggctgaagc tgtcatcggt tactcagatt
1381 tagaagggga tttcgatgtt gctgttttgc cattttccaa cagcacaaat aacgggttat
1441 tgtttataaa tactactatt gccagcattg ctgctaaaga agaaggggta tctctcgaga
1501 aaagagaggc tgaagcttct gcaggcggcc gcgagctcat catcatcatc atcattgagt
1561 ttgtagcctt agacatgact gttcctcagt tcaagttggg cacttacgag aagaccggtc
1621 ttgctagatt ctaatcaaga ggatgtcaga atgccatttg cctgagagat gcaggcttca
1681 tttttgatac ttttttattt gtaacctata tagtatagga ttttttttgt cattttgttt
1741 cttctcgtac gagcttgctc ctgatcagcc tatctcgcag ctgatgaata tcttgtggta
1801 ggggtttggg aaaatcattc gagtttgatg tttttcttgg tatttcccac tcctcttcag
1861 agtacagaag attaagtgag accttcgttt gtgcggatcc cccacacacc atagcttcaa
1921 aatgtttcta ctcctttttt actcttccag attttctcgg actccgcgca tcgccgtacc
1981 acttcaaaac acccaagcac agcatactaa attttccctc tttcttcctc tagggtgtcg
2041 ttaattaccc gtactaaagg tttggaaaag aaaaaagaga ccgcctcgtt tctttttctt
2101 cgtcgaaaaa ggcaataaaa atttttatca cgtttctttt tcttgaaatt ttttttttta
2161 gtttttttct ctttcagtga cctccattga tatttaagtt aataaacggt cttcaatttc
2221 tcaagtttca gtttcatttt tcttgttcta ttacaacttt ttttacttct tgttcattag
2281 aaagaaagca tagcaatcta atctaagggg cggtgttgac aattaatcat cggcatagta
2341 tatcggcata gtataatacg acaaggtgag gaactaaacc atggccaagt tgaccagtgc
2401 cgttccggtg ctcaccgcgc gcgacgtcgc cggagcggtc gagttctgga ccgaccggct
2461 cgggttctcc cgggacttcg tggaggacga cttcgccggt gtggtccggg acgacgtgac
2521 cctgttcatc agcgcggtcc aggaccaggt ggtgccggac aacaccctgg cctgggtgtg
2581 ggtgcgcggc ctggacgagc tgtacgccga gtggtcggag gtcgtgtcca cgaacttccg
2641 ggacgcctcc gggccggcca tgaccgagat cggcgagcag ccgtgggggc gggagttcgc
2701 cctgcgcgac ccggccggca actgcgtgca cttcgtggcc gaggagcagg actgacacgt
2761 ccgacggcgg cccacgggtc ccaggcctcg gagatccgtc ccccttttcc tttgtcgata
2821 tcatgtaatt agttatgtca cgcttacatt cacgccctcc ccccacatcc gctctaaccg
2881 aaaaggaagg agttagacaa cctgaagtct aggtccctat ttattttttt atagttatgt
2941 tagtattaag aacgttattt atatttcaaa tttttctttt ttttctgtac agacgcgtgt
3001 acgcatgtaa cattatactg aaaaccttgc ttgagaaggt tttgggacgc tcgaaggctt
3061 taatttgctc atcttcgatc ccctaacttt cgttcttgat taatgaaaac gtccttggcg
3121 aatgctttcg cagtagttag tcttggggcg atccaagaat ttcacctctg acgccccaat
3181 actgacgccc ccgaccgtcc ctgttaatca ttacgcggcc ccgaaccaac aaaagaaccg
3241 tatcctcttc tgttattcca tgctaatata ttcaactact gccttgaaca ctctaatttc
3301 ctcaaagtaa cgtccgttca actacgagct ttttaactgc aacaacttta atatacgcta
3361 ttggagctgg aattaccgcg gctgctggca ccagacttgc cctccaattg ttcctcgtta
3421 aggtatttac gttgtactca ttccaattac aagaccaaag gccctgtatc gttatttatt
3481 gtcactacct ccctgtgtca ggattgggta atttgcgcgc ctgctgcctt ccttggatgt
3541 ggtagccgtc tctcaggctc cctctccgga atcgaaccct tattccccgt tacccgtaga
3601 aaccatggta ggcctctatc ctaccatcga aagttgatag ggcagaaatt tgaatgaacc
3661 atcctaagat tcgaaaagtt attatgaatc accaaaacga aggttttatc taataaatac
3721 gcccgagggc tgatcaagta ttagctctag aattaccacg gttatccttg tagcaacact
3781 atcaaataaa cgataactga tttaatgagc cattcgcagt ttcaccgtat aatgctatac
3841 ttagacatgc atggcttaat ctttgagaca agggcccgcg gaacccctat ttgtttattt
3901 ttctaaatac attcaaatat gtatccgctc atgagacaat aaccctgata aatgcttcaa
3961 taatattgaa aaaggaagag tatgagtatt caacatttcc gtgtcgccct tattcccttt
4021 tttgcggcat tttgccttcc tgtttttgct cacccagaaa cgctggtgaa agtaaaagat
4081 gctgaagatc agttgggtgc acgagtgggt tacatcgaac tggatctcaa cagcggtaag
4141 atccttgaga gttttcgccc cgaagaacgt tttccaatga tgagcacttt taaagttctg
4201 ctatgtggcg cggtattatc ccgtattgac gccgggcaag agcaactcgg tcgccgcata
4261 cactattctc agaatgactt ggttgagtac tcaccagtca cagaaaagca tcttacggat
4321 ggcatgacag taagagaatt atgcagtgct gccataacca tgagtgataa cactgcggcc
4381 aacttacttc tgacaacgat cggaggaccg aaggagctaa ccgctttttt gcacaacatg
4441 ggggatcatg taactcgcct tgatcgttgg gaaccggagc tgaatgaagc cataccaaac
4501 gacgagcgtg acaccacgat gcctgtagca atggcaacaa cgttgcgcaa actattaact
4561 ggcgaactac ttactctagc ttcccggcaa caattaatag actggatgga ggcggataaa
4621 gttgcaggac cacttctgcg ctcggccctt ccggctggct ggtttattgc tgataaatct
4681 ggagccggtg agcgtgggtc tcgcggtatc attgcagcac tggggccaga tggtaagccc
4741 tcccgtatcg tagttatcta cacgacgggg agtcaggcaa ctatggatga acgaaataga
4801 cagatcgctg agataggtgc ctcactgatt aagcattggt aactgtcaga ccaagtttac
4861 tcatatatac tttagattga tttaaaactt catttttaat ttaaaaggat ctaggtgaag
4921 atcctttttg ataatctcat gaccaaaatc ccttaacgtg agttttcgtt ccactgagcg
4981 tcagaccccg tagaaaagat caaaggatct tcttgagatc ctttttttct gcgcgtaatc
5041 tgctgcttgc aaacaaaaaa accaccgcta ccagcggtgg tttgtttgcc ggatcaagag
5101 ctaccaactc tttttccgaa ggtaactggc ttcagcagag cgcagatacc aaatactgtt
5161 cttctagtgt agccgtagtt aggccaccac ttcaagaact ctgtagcacc gcctacatac
5221 ctcgctctgc taatcctgtt accagtggct gctgccagtg gcgataagtc gtgtcttacc
5281 gggttggact caagacgata gttaccggat aaggcgcagc ggtcgggctg aacggggggt
5341 tcgtgcacac agcccagctt ggagcgaacg acctacaccg aactgagata cctacagcgt
5401 gagctatgag aaagcgccac gcttcccgaa gggagaaagg cggacaggta tccggtaagc
5461 ggcagggtcg gaacaggaga gcgcacgagg gagcttccag ggggaaacgc ctggtatctt
5521 tatagtcctg tcgggtttcg ccacctctga cttgagcgtc gatttttgtg atgctcgtca
5581 ggggggcgga gccta
<210> 2
<211> 1747
<212> DNA
<213> The sequence encodes part of vector pPIG-1 responsible for
replication in E.coli
<221> misc_feature
<222> (1)...(15)
<223> sequence encodes 5 'end for Gibson assembly
<221> promoter
<222> (13)...(117)
<223> AmpR promoter
<221> CDS
<222> (118)...(978)
<223> beta-lactamase
<221> rep_origin
<222> (1149)...(1737)
<223> high-copy-number ColE1/pMB1/pBR322/pUC origin of replication
<221> misc_feature
<222> (1730)...(1747)
<223> sequence encodes 3 'end for Gibson assembly
<400> 1
1 gagacaaggg cccgcggaac ccctatttgt ttatttttct aaatacattc aaatatgtat
61 ccgctcatga gacaataacc ctgataaatg cttcaataat attgaaaaag gaagagtatg
121 agtattcaac atttccgtgt cgcccttatt cccttttttg cggcattttg ccttcctgtt
181 tttgctcacc cagaaacgct ggtgaaagta aaagatgctg aagatcagtt gggtgcacga
241 gtgggttaca tcgaactgga tctcaacagc ggtaagatcc ttgagagttt tcgccccgaa
301 gaacgttttc caatgatgag cacttttaaa gttctgctat gtggcgcggt attatcccgt
361 attgacgccg ggcaagagca actcggtcgc cgcatacact attctcagaa tgacttggtt
421 gagtactcac cagtcacaga aaagcatctt acggatggca tgacagtaag agaattatgc
481 agtgctgcca taaccatgag tgataacact gcggccaact tacttctgac aacgatcgga
541 ggaccgaagg agctaaccgc ttttttgcac aacatggggg atcatgtaac tcgccttgat
601 cgttgggaac cggagctgaa tgaagccata ccaaacgacg agcgtgacac cacgatgcct
661 gtagcaatgg caacaacgtt gcgcaaacta ttaactggcg aactacttac tctagcttcc
721 cggcaacaat taatagactg gatggaggcg gataaagttg caggaccact tctgcgctcg
781 gcccttccgg ctggctggtt tattgctgat aaatctggag ccggtgagcg tgggtctcgc
841 ggtatcattg cagcactggg gccagatggt aagccctccc gtatcgtagt tatctacacg
901 acggggagtc aggcaactat ggatgaacga aatagacaga tcgctgagat aggtgcctca
961 ctgattaagc attggtaact gtcagaccaa gtttactcat atatacttta gattgattta
1021 aaacttcatt tttaatttaa aaggatctag gtgaagatcc tttttgataa tctcatgacc
1081 aaaatccctt aacgtgagtt ttcgttccac tgagcgtcag accccgtaga aaagatcaaa
1141 ggatcttctt gagatccttt ttttctgcgc gtaatctgct gcttgcaaac aaaaaaacca
1201 ccgctaccag cggtggtttg tttgccggat caagagctac caactctttt tccgaaggta
1261 actggcttca gcagagcgca gataccaaat actgttcttc tagtgtagcc gtagttaggc
1321 caccacttca agaactctgt agcaccgcct acatacctcg ctctgctaat cctgttacca
1381 gtggctgctg ccagtggcga taagtcgtgt cttaccgggt tggactcaag acgatagtta
1441 ccggataagg cgcagcggtc gggctgaacg gggggttcgt gcacacagcc cagcttggag
1501 cgaacgacct acaccgaact gagataccta cagcgtgagc tatgagaaag cgccacgctt
1561 cccgaaggga gaaaggcgga caggtatccg gtaagcggca gggtcggaac aggagagcgc
1621 acgagggagc ttccaggggg aaacgcctgg tatctttata gtcctgtcgg gtttcgccac
1681 ctctgacttg agcgtcgatt tttgtgatgc tcgtcagggg ggcggagcct atggaaaaac
1741 gggcccg
<210> 3
<211> 1558
<212> DNA
<213> The sequence encodes part of vector pPIG-1: MCS, 6xHis-tag, AOX1
terminator and cassette required for
zeocin selection
<221> misc_feature
<222> (1)...(15)
<223> sequence encodes 5 'end for Gibson assembly
<221> CDS
<222> (19)...(39)
<223> 6xHis affinity tag
<221> terminator
<222> (116)...(362)
<223> Pichia pastoris AOX1 terminator
<221> promoter
<222> (402)...(788)
<223> TEF1 promoter from S. cerevisiae
<221> promoter
<222> (796)...(843)
<223> synthetic bacterial promoter EM7
<221> CDS
<222> (862)...(1236)
<223> Sh ble gene product from Streptoalloteichus hindustanus
<221> terminator
<222> (1302)...(1549)
<223> S. cerevisiae CYC1 terminator
<221> misc_feature
<222> (1544)...(1558)
<223> sequence encodes 3 'end for Gibson assembly
<400> 1
1 tgcaggcggc cgcgagctca tcatcatcat catcattgag tttgtagcct tagacatgac
61 tgttcctcag ttcaagttgg gcacttacga gaagaccggt cttgctagat tctaatcaag
121 aggatgtcag aatgccattt gcctgagaga tgcaggcttc atttttgata cttttttatt
181 tgtaacctat atagtatagg attttttttg tcattttgtt tcttctcgta cgagcttgct
241 cctgatcagc ctatctcgca gctgatgaat atcttgtggt aggggtttgg gaaaatcatt
301 cgagtttgat gtttttcttg gtatttccca ctcctcttca gagtacagaa gattaagtga
361 gaccttcgtt tgtgcggatc ccccacacac catagcttca aaatgtttct actccttttt
421 tactcttcca gattttctcg gactccgcgc atcgccgtac cacttcaaaa cacccaagca
481 cagcatacta aattttccct ctttcttcct ctagggtgtc gttaattacc cgtactaaag
541 gtttggaaaa gaaaaaagag accgcctcgt ttctttttct tcgtcgaaaa aggcaataaa
601 aatttttatc acgtttcttt ttcttgaaat tttttttttt agtttttttc tctttcagtg
661 acctccattg atatttaagt taataaacgg tcttcaattt ctcaagtttc agtttcattt
721 ttcttgttct attacaactt tttttacttc ttgttcatta gaaagaaagc atagcaatct
781 aatctaaggg gcggtgttga caattaatca tcggcatagt atatcggcat agtataatac
841 gacaaggtga ggaactaaac catggccaag ttgaccagtg ccgttccggt gctcaccgcg
901 cgcgacgtcg ccggagcggt cgagttctgg accgaccggc tcgggttctc ccgggacttc
961 gtggaggacg acttcgccgg tgtggtccgg gacgacgtga ccctgttcat cagcgcggtc
1021 caggaccagg tggtgccgga caacaccctg gcctgggtgt gggtgcgcgg cctggacgag
1081 ctgtacgccg agtggtcgga ggtcgtgtcc acgaacttcc gggacgcctc cgggccggcc
1141 atgaccgaga tcggcgagca gccgtggggg cgggagttcg ccctgcgcga cccggccggc
1201 aactgcgtgc acttcgtggc cgaggagcag gactgacacg tccgacggcg gcccacgggt
1261 cccaggcctc ggagatccgt cccccttttc ctttgtcgat atcatgtaat tagttatgtc
1321 acgcttacat tcacgccctc cccccacatc cgctctaacc gaaaaggaag gagttagaca
1381 acctgaagtc taggtcccta tttatttttt tatagttatg ttagtattaa gaacgttatt
1441 tatatttcaa atttttcttt tttttctgta cagacgcgtg tacgcatgta acattatact
1501 gaaaaccttg cttgagaagg ttttgggacg ctcgaaggct ttaatttgct catcttcg
<210> 4
<211> 298
<212> DNA
<213> The sequence encodes part of vector pPIG-1: N-terminal
secretion signal from S. cerevisiae MF-alpha-factor-1 and MCS
<221> misc_feature
<222> (1)...(18)
<223> sequence encodes 5 'end for Gibson assembly
<221> CDS
<222> (15)...(281)
<223> N-terminal secretion signal from S. cerevisiae MF-alpha-factor-1
<221> misc_feature
<222> (277)...(298)
<223> multiple cloning site
<221> misc_feature
<222> (284)...(298)
<223> sequence encodes 3 'end for Gibson assembly
<400> 1
1 ctatccatat ggaaatgaga tttccttcaa tttttactgc tgttttattc gcagcatcct
61 ccgcattagc tgctccagtc aacactacaa cagaagatga aacggcacaa attccggctg
121 aagctgtcat cggttactca gatttagaag gggatttcga tgttgctgtt ttgccatttt
181 ccaacagcac aaataacggg ttattgttta taaatactac tattgccagc attgctgcta
241 aagaagaagg ggtatctctc gagaaaagag aggctgaagc ttctgcaggc ggccgcga
<210> 5
<211> 506
<212> DNA
<213> The sequence encodes part of vector pPIG-1: GAP promoter
<221> misc_feature
<222> (1)...(15)
<223> sequence encodes 5 'end for Gibson assembly
<221> promoter
<222> (16)...(492)
<223> GAP promoter. Pichia pastoris promoter
glyceraldehyde-3-phosphate dehydrogenase
<221> misc_feature
<222> (489)...(506)
<223> sequence encodes 3 'end for Gibson assembly
<400> 1
1 ctggggtacc atcctttttt gtagaaatgt cttggtgtcc tcgtccaatc aggtagccat
61 ctctgaaata tctggctccg ttgcaactcc gaacgacctg ctggcaacgt aaaattctcc
121 ggggtaaaac ttaaatgtgg agtaatggaa ccagaaacgt ctcttccctt ctctctcctt
181 ccaccgcccg ttaccgtccc taggaaattt tactctgctg gagagcttct tctacggccc
241 ccttgcagca atgctcttcc cagcattacg ttgcgggtaa aacggaagtc gtgtacccga
301 cctagcagcc cagggatgga aaagtcccgg ccgtcgctgg caataatagc gggcggacgc
361 atgtcatgag attattggaa accaccagaa tcgaatataa aaggcgaaca cctttcccaa
421 ttttggtttc tcctgaccca aagactttaa atttaattta tttgtcccta tttcaatcaa
481 ttgaacaact atccatatgg aaatga
<210> 6
<211> 763
<212> DNA
<213> The sequence encodes 18s rRNA from Pichia pastoris
<221> misc_feature
<222> (1)...(16)
<223> sequence encodes 5 'end for Gibson assembly
<221> misc_feature
<222> (18)...(750)
<223> Pichia pastoris 18s rRNA
<221> misc_feature
<222> (749)...(763)
<223> sequence encodes 3 'end for Gibson assembly
<210> 7
<211> 820
<212> DNA
<213> The sequence encodes 18s rRNA from Pichia pastoris
<221> misc_feature
<222> (1)...(15)
<223> sequence encodes 5 'end for Gibson assembly
<221> misc_feature
<222> (7)...(808)
<223> Pichia pastoris 18s rRNA
<221> misc_feature
<222> (803)...(820)
<223> sequence encodes 3 'end for Gibson assembly
<210> 8
<211> 975
<212> DNA
<213> The sequence encodes of chitinase 19 family frome Drosera capensis
<221> CDS
<222> (1)...(975)
<223> chitinase 19 family frome Drosera capensis
<400> 1
ATG CGC ATC ACG GTC CTG TTG CTC TTG TGT GTC GCT CCT CTG TTG TCT GGC ACG TAT GCT
M R I T V L L L L C V A P L L S G T Y A
GTC CAG TGT GGT AGC GAA GTC GGT GGA GCT
V Q C G S E V G G A
CTG TGT CCG AAT GGT CTG TGT TGC AGC AAG TAT GGC TAC TGT GGC ACT ACG TCT GCC TAC
L C P N G L C C S K Y G Y C G T T S A Y
TGT GGT CCG GGC TGT CAG AGC CAG TGT GGT
C G P G C Q S Q C G
GGT TCC TCT CCT CCG CCT GCT CCT CCC AGC CCG ACT CCG AGT CCT CCG TCT CCC TCT GGA
G S S P P P A P P S P T P S P P S P S G
GGT GGT GAT GTG TCC AGC ATC ATC ACC TCC
G G D V S S I I T S
CAG ATC TTC AAT CAG ATG CTG CTC CAT CGC AAT GAC AAT GCC TGT CCT GCC CAT GGC TTC
Q I F N Q M L L H R N D N A C P A H G F
TAC AGC TAT CAA GCC TTC TTG GAT GCT GCA
Y S Y Q A F L D A A
CGC AAG TTT ACT GGT TTC GGT ACG ACT GGT GAC ATC AAC ACA CGC AAA CGT GAA CTG GCT
R K F T G F G T T G D I N T R K R E L A
GCC TTC TTT GGT CAG ACG AGC CAC GAG ACC
A F F G Q T S H E T
ACT GGT GGC TGG CCC ACT GCT CCT GAT GGT CCG TAT GCC TGG GGC TAC TGC TTC AAA CAG
T G G W P T A P D G P Y A W G Y C F K Q
GAA CAA GGC AAT CCT GGT GAC TAC TGT GTC
E Q G N P G D Y C V
CAG TCT TCC ACG TAT CCC TGT GCC CCT GGC AAG AAG TAC TAT GGT CGT GGA CCG ATT CAG
Q S S T Y P C A P G K K Y Y G R G P I Q
ATC TCC TAC AAC TAC AAC TAT GGT CAG TGT
I S Y N Y N Y G Q C
GGA GCC GCC ATT AAT CAA CCC CTG CTG AGC AAT CCG GAT CTG GTC GCG TCC AAT GCC GAT
G A A I N Q P L L S N P D L V A S N A D
GTG TCC TTC GAG ACT GCC ATC TGG TTC TGG
V S F E T A I W F W
ATG ACT CCT CAA GGT AGC AAA CCC TCC TGT CAT GCC GTC GCC ACT GGT CAG TGG ACT CCG
M T P Q G S K P S C H A V A T G Q W T P
TCC GTC GCC GAT CAA GCT GCT GGA CGT GTT
S V A D Q A A G R V
CCT GGC TAT GGT GTC ATT ACG AAC ATC ATC AAT GGA GGT GTC GAG TGT GGC AAA GGC ACG
P G Y G V I T N I I N G G V E C G K G T
GTC CCG CAA GTT GCC GAT CGC ATT GGC TTC
V P Q V A D R I G F
TAT CAA CGC TAC TGC TCC ATC ATG GGT ATT GCG CCT GGT GGC AAT CTT GGC TGC TAC AAT
Y Q R Y C S I M G I A P G G N L G C Y N
CAA CGT CCG TTC AGC TGA
Q R P F S *
<---
Изобретение относится к области биотехнологии и генной инженерии. Предложен интеграционный вектор pPIG-1 для осуществления экспрессии рекомбинантных белков в дрожжах P. pastoris, имеющий размер 5595 п.о. и характеризующийся нуклеотидной последовательностью, приведенной в SEQ ID NO 1. Изобретение обеспечивает высокую частоту интеграции целевого гена. 3 пр., 2 табл., 2 ил.
Интеграционный вектор pPIG-1 для осуществления экспрессии рекомбинантных белков в дрожжах P. pastoris, имеющий размер 5595 п.о. и характеризующийся нуклеотидной последовательностью, приведенной в SEQ ID NO 1.
| ИНТЕГРАТИВНЫЙ ПЛАЗМИДНЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ ГЕНОВ В ДРОЖЖАХ | 2008 |
|
RU2388823C1 |
| pGAPZα A Plasmid | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

