Область техники, к которой относится изобретение
Изобретение относится к моноклональным анти-CTLA-4-антителам, нуклеиновым кислотам, кодирующим антитела, экспрессирующим векторам и рекомбинантным клеткам, включающим нуклеиновые кислоты, и фармацевтическим композициям, включающим антитела. Также предложены способы получения антител и способы применения антител для лечения заболеваний, включая рак и аутоиммунные заболевания.
Предшествующий уровень техники
Иммунотерапия рака, недавний прорыв в лечении онкологических заболеваний, использует собственную иммунную систему пациента для атаки на опухолевые клетки. Содействие устойчивому CD8 Т-клеточно-зависимому цитотоксическому ответу в микроокружении опухоли важно для генерации эффективного противоопухолевого иммунного ответа. Тем не менее, опухоль имеет тенденцию избегать иммунного надзора, используя механизм подавления Т-клеток. Истощение инфильтрирующих опухоль лимфоцитов (TIL) приводит к анергии цитотоксических Т-клеток и выходу опухолевых клеток из-под надзора (Wherry and Kurachi, 2015, Nat Rev Immunol., 2015, 15: 486-499; Dyck and Mills, 2017, Eur., J. Immunol., 47(5): 765-779).
Ингибиторы иммунных белков контрольной точки обладают потенциалом для лечения различных опухолей, таких как метастатическая меланома, рак легкого, рак молочной железы и почечно-клеточный рак. CTLA-4 (CD152) представляет собой такую ингибирующую молекулу контрольной точки на поверхности Т-клеток. Первоначально она был идентифицирована путем дифференциального скрининга библиотеки кДНК мышиных цитолитических Т-клеток (Brunet et al., 1987, Nature, 328: 267-270). Предполагается, что CTLA-4 может функционировать в качестве отрицательного регулятора активации Т-клеток (Walunas et al., 1994, Immunity, 1: 405-413). CTLA-4 экспрессируется конститутивно на поверхности регуляторных Т-клеток, но его количество относительно низкое. Она активируется при активации Т-клеток. После активации CTLA-4 взаимодействует с CD80 (B7.1) и CD86 (B7.2), которые также являются лигандами для CD28, с гораздо более высокой аффинностью связывания, чем CD28 (van der Merwe et al.,1997, J Exp Med. 185:393-403; Alegre et al., 2001, Nat Rev Immunol, 1: 220-228). Передача сигналов CD28 способствует активации Т-клеток, в то время как взаимодействие CTLA-4 с его лигандами B7.1 и B7.2 предотвращает дальнейшую активацию Т-клеток.
Антагонисты CTLA-4 привлекательны, поскольку блокада CTLA-4 с помощью антагонистов была показана как эффективная противоопухолевая терапия (пат. США № 6984720). Ингибирование этого поверхностного рецептора с использованием антагониста, такого как mAb против CTLA-4, усиливало эффекторные ответы CD4 и CD8 Т-клеток и снижало супрессирующую функцию клеток Treg. В последние годы лечение на основе антагонистов CTLA-4 быстро прогрессировало. Ипилимумаб (VERYOR®) является гуманизированным антителом и блокирует эффекты CTLA-4, которые усиливают ответы Т-клеток на опухолевые клетки. Ипилимумаб был первым лекарственным средством, продемонстрировавшим улучшение общей выживаемости пациентов с метастатической меланомой в рандомизированном контролируемом исследовании фазы 3. Он имеет управляемый профиль безопасности в дозе 3 мг/кг в качестве монотерапии у пациентов, ранее получавших другие виды терапии, и в дозе 10 мг/кг в сочетании с дакарбазином у пациентов, не получавших лечение. Помимо злокачественной меланомы, ипилимумаб также находится в стадии исследования для лечения рака предстательной железы и немелкоклеточного рака легкого.
Однако, несмотря на прогресс, упомянутый выше, желательны антагонисты CTLA-4 с улучшенной аффинностью, специфичностью и способностью к совершенствованию. Кроме того, также необходимы более эффективные терапевтические средства, включающие анти-CTLA-4 антитела, которые эффективно ингибируют сигнальную активность CTLA-4, в то же время вызывая минимальные неблагоприятные побочные эффекты у людей.
Сущность изобретения
Раскрыто выделенное моноклональное антитело, содержащее CD152-связывающий домен, где CD152-связывающий домен включает вариабельную область тяжелой цепи иммуноглобулина, содержащую CDR1, CDR2 и CDR3, где CDR1, CDR2 и CDR3 содержат аминокислотные последовательности, имеющие, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с (1) SEQ ID NO: 4, 5 и 6, соответственно; (2) SEQ ID NO: 10, 11 и 12, соответственно; (3) SEQ ID NO: 16, 17 и 18, соответственно; (4) SEQ ID NO: 22, 23 и 24, соответственно; (5) SEQ ID NO: 28, 29 и 30, соответственно; (6) SEQ ID NO: 34, 35 и 36, соответственно; (7) SEQ ID NO: 40, 41 и 42, соответственно; (8) SEQ ID NO: 46, 47 и 48, соответственно; (9) SEQ ID NO: 52, 53 и 54, соответственно; (10) SEQ ID NO: 58, 59 и 60, соответственно; (11) SEQ ID NO: 64, 65 и 66, соответственно; (12) SEQ ID NO: 70, 71 и 72, соответственно; (13) SEQ ID NO: 76, 77 и 78, соответственно; (14) SEQ ID NO: 82, 83 и 84, соответственно; (15) SEQ ID NO: 88, 89 и 90, соответственно; (16) SEQ ID NO: 94, 95 и 96, соответственно; (17) SEQ ID NO: 100, 101 и 102, соответственно; (18) SEQ ID NO: 106, 107 и 108, соответственно; (19) SEQ ID NO: 112, 113 и 114, соответственно; (20) SEQ ID NO: 118, 119 и 120, соответственно; (21) SEQ ID NO: 124, 125 и 126, соответственно; (22) SEQ ID NO: 130, 131 и 132, соответственно; (23) SEQ ID NO: 136, 137 и 138, соответственно; (24) SEQ ID NO: 142, 143 и 144, соответственно; (25) SEQ ID NO: 148, 149 и 150, соответственно; (26) SEQ ID NO: 154, 155 и 156, соответственно; (27) SEQ ID NO: 160, 161 и 162, соответственно; (28) SEQ ID NO: 166, 167 и 168, соответственно; (29) SEQ ID NO: 172, 173 и 174, соответственно; (30) SEQ ID NO: 178, 179 и 180, соответственно; (31) SEQ ID NO: 184, 185 и 186, соответственно; или (32) SEQ ID NO: 190, 191 и 192, соответственно.
В некоторых случаях антитело представляет собой антитело, содержащее только тяжелые цепи. В некоторых случаях антитело не содержит легкой цепи иммуноглобулина. В некоторых случаях антитело содержит одну тяжелую цепь иммуноглобулина. В некоторых случаях антитело содержит две тяжелые цепи иммуноглобулина. В некоторых случаях антитело состоит из двух тяжелых цепей иммуноглобулина. В некоторых случаях, по меньшей мере, одна из двух тяжелых цепей иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 2, 8, 14, 20, 26, 32, 38, 44, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128, 134, 140, 146, 152, 158, 164, 170, 176, 182 или 188.
В некоторых случаях вариабельная область тяжелой цепи иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, на 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 3, 9, 15, 21, 27, 33, 39, 45, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183 или 189.
В некоторых случаях антитело специфически связывается с CD152 человека. В некоторых случаях антитело связывается с CD152 человека с высокой аффинностью. В некоторых случаях антитело связывается с CD152 человека с аффинностью выше, чем у аналога ипилимумаба. В некоторых случаях антитело связывается с CD152 человека с аффинностью, которая, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 30 раз, по меньшей мере, в 50 раз или, по меньшей мере, в 100 раз выше, чем у аналога ипилимумаба.
В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-7 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-8 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-9 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-10 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-11 М или менее. В некоторых случаях Kd составляет 6,0*10-11 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd, который ниже, чем у аналога ипилимумаба. В некоторых случаях антитело диссоциирует от CD152 человека с Kd, который, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 30 раз, по меньшей мере, в 50 раз или, по меньшей мере, в 100 раз ниже, чем у аналога ипилимумаба. В некоторых случаях Kd определяется поверхностным плазмонным резонансом.
В некоторых случаях антитело специфически связывается с CD152 обезьяны. В некоторых случаях антитело специфически не связывается с CD152 обезьяны. В некоторых случаях антитело блокирует связывание CD152 с CD80, CD86 или с обеими. В некоторых случаях антитело способствует секреции IL-2 иммунными клетками. В некоторых случаях антитело индуцирует активацию Т-клеток. В некоторых случаях антитело стимулирует противоопухолевый иммунный ответ иммунных клеток. В некоторых случаях антитело представляет собой человеческое, гуманизированное или химерное антитело.
В другом аспекте в данном документе раскрыто выделенное моноклональное антитело, содержащее только тяжелые цепи, включающее CD152-связывающий домен, где антитело специфически связывается с CD152 человека. В некоторых случаях антитело не содержит легкой цепи иммуноглобулина. В некоторых случаях антитело содержит одну тяжелую цепь иммуноглобулина. В некоторых случаях антитело содержит две тяжелые цепи иммуноглобулина. В некоторых случаях антитело состоит из двух тяжелых цепей иммуноглобулина.
В некоторых случаях антитело связывается с CD152 человека с высокой аффинностью. В некоторых случаях антитело связывается с CD152 человека с аффинностью выше, чем у аналога ипилимумаба. В некоторых случаях антитело связывается с CD152 человека с аффинностью, которая, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 30 раз, по меньшей мере, в 50 раз или, по меньшей мере, в 100 раз выше, чем у аналога ипилимумаба.
В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-7 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-8 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-9 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-10 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd 1,0*10-11 М или менее. В некоторых случаях Kd составляет 6,0*10-11 М или менее. В некоторых случаях антитело диссоциирует от человеческого CD152 с Kd, который ниже, чем у аналога ипилимумаба. В некоторых случаях антитело диссоциирует от CD152 человека с Kd, который, по меньшей мере, в 2 раза, по меньшей мере, в 5 раз, по меньшей мере, в 10 раз, по меньшей мере, в 20 раз, по меньшей мере, в 30 раз, по меньшей мере, в 50 раз или, по меньшей мере, в 100 раз ниже, чем у аналога ипилимумаба. В некоторых случаях Kd определяется поверхностным плазмонным резонансом.
В некоторых случаях антитело специфически связывается с CD152 обезьяны. В некоторых случаях антитело специфически не связывается с CD152 обезьяны. В некоторых случаях CD152-связывающий домен включает вариабельную область тяжелой цепи иммуноглобулина, содержащую CDR1, CDR2 и CDR3, где CDR1, CDR2 и CDR3 содержат аминокислотные последовательности, имеющие, по меньшей мере, 80%, 85%, 88%, 90% 92%, 95%, 97%, 98%, 99% или 100% идентичности с (1) SEQ ID NO: 4, 5 и 6, соответственно; (2) SEQ ID NO: 10, 11 и 12, соответственно; (3) SEQ ID NO: 16, 17 и 18, соответственно; (4) SEQ ID NO: 22, 23 и 24, соответственно; (5) SEQ ID NO: 28, 29 и 30, соответственно; (6) SEQ ID NO: 34, 35 и 36, соответственно; (7) SEQ ID NO: 40, 41 и 42, соответственно; (8) SEQ ID NO: 46, 47 и 48, соответственно; (9) SEQ ID NO: 52, 53 и 54, соответственно; (10) SEQ ID NO: 58, 59 и 60, соответственно; (11) SEQ ID NO: 64, 65 и 66, соответственно; (12) SEQ ID NO: 70, 71 и 72, соответственно; (13) SEQ ID NO: 76, 77 и 78, соответственно; (14) SEQ ID NO: 82, 83 и 84, соответственно; (15) SEQ ID NO: 88, 89 и 90, соответственно; (16) SEQ ID NO: 94, 95 и 96, соответственно; (17) SEQ ID NO: 100, 101 и 102, соответственно; (18) SEQ ID NO: 106, 107 и 108, соответственно; (19) SEQ ID NO: 112, 113 и 114, соответственно; (20) SEQ ID NO: 118, 119 и 120, соответственно; (21) SEQ ID NO: 124, 125 и 126, соответственно; (22) SEQ ID NO: 130, 131 и 132, соответственно; (23) SEQ ID NO: 136, 137 и 138, соответственно; (24) SEQ ID NO: 142, 143 и 144, соответственно; (25) SEQ ID NO: 148, 149 и 150, соответственно; (26) SEQ ID NO: 154, 155 и 156, соответственно; (27) SEQ ID NO: 160, 161 и 162, соответственно; (28) SEQ ID NO: 166, 167 и 168, соответственно; (29) SEQ ID NO: 172, 173 и 174, соответственно; (30) SEQ ID NO: 178, 179 и 180, соответственно; (31) SEQ ID NO: 184, 185 и 186, соответственно; или (32) SEQ ID NO: 190, 191 и 192, соответственно.
В некоторых случаях CD152-связывающий домен включает, по меньшей мере, одну тяжелую цепь иммуноглобулина, где тяжелая цепь иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 2, 8, 14, 20, 26, 32, 38, 44, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128, 134, 140, 146, 152, 158, 164, 170, 176, 182 или 188. В некоторых случаях вариабельная область тяжелой цепи иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 3, 9, 15, 21, 27, 33, 39, 45, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183 или 189.
В некоторых случаях антитело блокирует связывание CD152 с CD80, CD86 или с обеими. В некоторых случаях антитело способствует секреции IL-2 иммунными клетками. В некоторых случаях антитело индуцирует активацию Т-клеток. В некоторых случаях антитело стимулирует противоопухолевый иммунный ответ иммунных клеток. В некоторых случаях антитело представляет собой человеческое, гуманизированное или химерное антитело.
В другом аспекте в данном документе раскрыта фармацевтическая композиция, включающая любое антитело, раскрытое в данном документе, и его фармацевтически приемлемое вспомогательное вещество. В некоторых случаях фармацевтически приемлемое вспомогательное вещество выбирают из группы, состоящей из носителей, поверхностно-активных агентов, загустителей или эмульгаторов, твердых связующих, дисперсионных или суспендирующих добавок, солюбилизаторов, красителей, ароматизаторов, покрытий, агентов для улучшения распадаемости, смазывающих веществ, подсластителей, консервантов, изотонических агентов и их комбинаций. В некоторых случаях фармацевтическая композиция дополнительно содержит второе антитело, где второе антитело представляет собой иммуностимулирующее антитело или костимулирующее антитело. В некоторых случаях иммуностимулирующее антитело выбирают из группы, состоящей из анти-PD-1 антитела, анти-PD-L1 антитела, анти-LAG-3 антитела, анти-TIM 3 антитела, анти-STAT3 антитела и анти-ROR1 антитела. В некоторых случаях костимулирующее антитело представляет собой анти-CD137 антитело или анти-GITR антитело.
В другом аспекте в данном документе раскрыта выделенная молекула нуклеиновой кислоты, кодирующая любое антитело, раскрытое в данном документе. В некоторых случаях молекула нуклеиновой кислоты включает нуклеотидную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 92%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO.: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187. В некоторых случаях молекула нуклеиновой кислоты включает нуклеотидную последовательность, представленную в SEQ ID NO: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187.
В другом аспекте в данном документе раскрыт экспрессирующий вектор, содержащий сегмент нуклеиновой кислоты, кодирующий любое антитело, раскрытое в данном документе, где сегмент нуклеиновой кислоты функционально связан с регуляторными последовательностями, подходящими для экспрессии сегмента нуклеиновой кислоты в клетке-хозяине. В некоторых случаях сегмент нуклеиновой кислоты включает нуклеотидную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 92%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO : 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187. В некоторых случаях сегмент нуклеиновой кислоты включает нуклеотидную последовательность, представленную в SEQ ID NO: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187.
В другом аспекте в данном документе раскрыта клетка-хозяин, содержащая любой экспрессирующий вектор, раскрытый в данном документе.
В другом аспекте в данном документе раскрыт способ получения CD152-связывающего моноклонального антитела, причем способ включает: культивирование клетки-хозяина, содержащей любой экспрессирующий вектор, в условиях, в которых экспрессируется сегмент нуклеиновой кислоты, с получением, таким образом, CD152-связывающего моноклонального антитела. В некоторых случаях клетка-хозяин представляет собой линию клеток-хозяев CHO, HEK293 или COS. В некоторых случаях клеточная линия-хозяин CHO представляет собой клеточную линию CHO-K1. В некоторых случаях способ дополнительно включает извлечение CD152-связывающего моноклонального антитела.
В другом аспекте в данном документе раскрыт способ индукции антителозависимой клеточно-опосредованной цитотоксичности (ADCC) против клетки, экспрессирующей антиген, ассоциированный с опухолью, причем способ включает: контакт Т-клетки с любым антителом, раскрытым в данном документе, где указанный контакт происходит в условиях, при которых индуцируется ADCC против клетки, экспрессирующей ассоциированный с опухолью антиген.
В другом аспекте в данном документе раскрыт способ лечения расстройства у объекта, включающий введение объекту терапевтически эффективного количества любого антитела, раскрытого в данном документе, или любой фармацевтической композиции, раскрытой в данном документе. В некоторых случаях расстройство является раком. В некоторых случаях рак выбирают из группы, состоящей из лейкоза, лимфомы, CLL, малой лимфоцитарной лимфомы, B-клеточной лимфомы из клеток маргинальной зоны, лимфомы Беркетта, почечно-клеточного рака, рака толстой кишки, колоректального рака, рака молочной железы, эпителиального плоскоклеточного рака, меланомы, миеломы, рака желудка, рака головного мозга, рака легкого, рака поджелудочной железы, рака шейки матки, рака яичников, рака печени, рака мочевого пузыря, рака предстательной железы, рака яичка, рака щитовидной железы и рака головы и шеи. В некоторых случаях способ дополнительно включает терапевтический агент. В некоторых случаях терапевтическое средство представляет собой противораковое лекарственное средство. В некоторых случаях терапевтическое средство представляет собой ипилимумаб или его биоподобный продукт. В некоторых случаях расстройство является аутоиммунным заболеванием.
В другом аспекте в данном документе раскрыто применение любой фармацевтической композиции, раскрытой в данном документе, для приготовления лекарственного средства для лечения расстройства. В некоторых случаях расстройство является раком. В некоторых случаях рак выбирают из группы, состоящей из лейкоза, лимфомы, CLL, малой лимфоцитарной лимфомы, B-клеточной лимфомы из клеток маргинальной зоны, лимфомы Беркетта, почечно-клеточного рака, рака толстой кишки, колоректального рака, рака молочной железы, эпителиального плоскоклеточного рака, меланомы, миеломы, рака желудка, рака головного мозга, рака легкого, рака поджелудочной железы, рака шейки матки, рака яичников, рака печени, рака мочевого пузыря, рака предстательной железы, рака яичка, рака щитовидной железы и рака головы и шеи. В некоторых случаях расстройство является аутоиммунным заболеванием.
В другом аспекте изобретение относится к выделенному моноклональному антителу или его антигенсвязывающей части, имеющему вариабельную область тяжелой цепи, которая включает область CDR1, область CDR2 и область CDR3, где область CDR1, область CDR2 и область CDR3 содержит аминокислотные последовательности, имеющие, по меньшей мере, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичности с (1) SEQ ID NO: 4, 5 и 6, соответственно; (2) SEQ ID NO: 10, 11 и 12, соответственно; (3) SEQ ID NO: 16, 17 и 18, соответственно; (4) SEQ ID NO: 22, 23 и 24, соответственно; (5) SEQ ID NO: 28, 29 и 30, соответственно; (6) SEQ ID NO: 34, 35 и 36, соответственно; (7) SEQ ID NO: 40, 41 и 42, соответственно; (8) SEQ ID NO: 46, 47 и 48, соответственно; (9) SEQ ID NO: 52, 53 и 54, соответственно; (10) SEQ ID NO: 58, 59 и 60, соответственно; (11) SEQ ID NO: 64, 65 и 66, соответственно; (12) SEQ ID NO: 70, 71 и 72, соответственно; (13) SEQ ID NO: 76, 77 и 78, соответственно; (14) SEQ ID NO: 82, 83 и 84, соответственно; (15) SEQ ID NO: 88, 89 и 90, соответственно; (16) SEQ ID NO: 94, 95 и 96, соответственно; (17) SEQ ID NO: 100, 101 и 102, соответственно; (18) SEQ ID NO: 106, 107 и 108, соответственно; (19) SEQ ID NO: 112, 113 и 114, соответственно; (20) SEQ ID NO: 118, 119 и 120, соответственно; (21) SEQ ID NO: 124, 125 и 126, соответственно; (22) SEQ ID NO: 130, 131 и 132, соответственно; (23) SEQ ID NO: 136, 137 и 138, соответственно; (24) SEQ ID NO: 142, 143 и 144, соответственно; (25) SEQ ID NO: 148, 149 и 150, соответственно; (26) SEQ ID NO: 154, 155 и 156, соответственно; (27) SEQ ID NO: 160, 161 и 162, соответственно; (28) SEQ ID NO: 166, 167 и 168, соответственно; (29) SEQ ID NO: 172, 173 и 174, соответственно; (30) SEQ ID NO: 178, 179 и 180, соответственно; (31) SEQ ID NO: 184, 185 и 186, соответственно; или (32) SEQ ID NO: 190, 191 и 192, соответственно; где антитело или его антигенсвязывающий фрагмент связывают CTLA-4.
В другом аспекте выделенное моноклональное антитело или его антигенсвязывающая часть по настоящему изобретению включает вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 3, 9, 15, 21, 27, 33, 39, 45, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183 или 189.
В одном воплощении антитело по настоящему изобретению включает тяжелую цепь, содержащую аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичности с SEQ ID NO: 2, 8, 14, 20, 26, 32, 38, 44, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128, 134, 140, 146, 152, 158, 164, 170, 176, 182 или 188, которые могут кодироваться последовательностью нуклеиновой кислоты, имеющей, по меньшей мере, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентичности с SEQ ID No: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 157, 163, 169, 175, 181 или 187.
В некоторых воплощениях антитело по настоящему изобретению состоит по существу или состоит из двух тяжелых цепей, описанных выше.
В другом аспекте изобретения антитело или его антигенсвязывающая часть является частью иммуноконъюгата, который содержит терапевтический агент, например цитотоксин или радиоактивный изотоп, связанный с антителом. В другом аспекте антитело является частью биспецифической молекулы, которая содержит второй функциональный фрагмент (например, второе антитело), обладающий специфичностью связывания, отличной от указанного антитела или его антигенсвязывающей части. В другом аспекте антитело или его антигенсвязывающие части по настоящему изобретению могут быть превращены в часть химерного антигенного рецептора (CAR) или сконструированного Т-клеточного рецептора. Антитело или его антигенсвязывающие части по настоящему изобретению также могут кодироваться или применяться в сочетании с онколитическим вирусом.
Также предлагается фармацевтическая композиция, содержащая антитело или его антигенсвязывающую часть, иммуноконъюгат, биспецифическую молекулу, химерный антигенный рецептор, сконструированный Т-клеточный рецептор или онколитический вирус по изобретению, необязательно составленный в фармацевтически приемлемом носителе.
Фармацевтическая композиция может дополнительно включать другие противораков агенты. Фармацевтическая композиция может дополнительно включать, по меньшей мере, одно дополнительное иммуностимулирующее антитело, выбранное из группы, состоящей из анти-PD-1 антитела, анти-PD-L1 антитела, анти-LAG-3 антитела, анти-TIM 3 антитела, анти-STAT3-антитела и анти-ROR1-антитела и/или костимулирующего антитела, которое может быть анти-CD137-антителом или анти-GITR-антителом.
Также предлагается молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающую часть (например, вариабельные области и/или CDR) по изобретению, а также экспрессирующий вектор, включающий нуклеиновую кислоту и клетку-хозяина, содержащую экспрессирующий вектор. Также предлагается способ получения анти-CTLA-4 антитела с использованием клетки-хозяина, содержащей экспрессирующий вектор, и включающий стадии (i) экспрессии антитела в клетке-хозяине и (ii) выделения антитела из клетки-хозяина.
В еще одном воплощении изобретение относится к способу ингибирования роста опухолевых клеток у объекта, включающему введение объекту терапевтически эффективного количества антитела или его антигенсвязывающей части по изобретению. Опухоль может представлять собой солидную или несолидную опухоль, выбранную из группы, состоящей из лейкоза, лимфомы, CLL, малой лимфоцитарной лимфомы, В-клеточной лимфомы маргинальных клеток, лимфомы Беркетта, почечно-клеточного рака, рака толстой кишки, колоректального рака, рака молочной железы, эпителиального плоскоклеточного рака, меланомы, миеломы, рака желудка, рака головного мозга, рака легкого, рака поджелудочной железы, рака шейки матки, рака яичников, рака печени, рака мочевого пузыря, рака предстательной железы, рака яичка, рака щитовидной железы и рака головы и шеи. Предпочтительно, опухоль представляет собой меланому, рак предстательной железы или немелкоклеточный рак легкого. В еще одном воплощении изобретение относится к способу лечения аутоиммунного заболевания у объекта, включающему введение объекту терапевтически эффективного количества антитела или его антигенсвязывающей части по изобретению. В еще одном воплощении изобретение относится к способу лечения вирусной инфекции у объекта, включающему введение объекту терапевтически эффективного количества антитела или его антигенсвязывающей части по изобретению. В другом воплощении способ включает введение фармацевтической композиции, биспецифического агента или иммуноконъюгата по изобретению.
В другом аспекте изобретение относится к анти-CTLA-4 антителу и композиции по изобретению для применения в вышеупомянутых способах или для получения лекарственного средства для применения в вышеупомянутых способах (например, для лечения).
Другие особенности и преимущества настоящего раскрытия будут очевидны из следующего подробного описания и примеров, которые не должны рассматриваться как ограничивающие. Содержание всех источников, записей Genbank, патентов и опубликованных патентных заявок, процитированных по всей данной заявке, прямо включено в данный документ ссылкой.
Краткое описание чертежей
Вышеизложенное краткое изложение, а также последующее подробное описание изобретения будет лучше поняты при рассмотрении вместе с чертежами. Следует понимать, что изобретение не ограничено воплощениями, представленными на чертежах.
На чертежах:
На фиг. 1 показана активность связывания анти-CTLA-4 антител по настоящему изобретению с CTLA-4 человека, экспрессированной на клетках 293.
На фиг. 2 показана активность связывания анти-CTLA-4-антител по настоящему изобретению с CTLA-4 яванского макака, экспрессированной на клетках 293.
На фиг. 3 показана способность анти-CTLA-4 антител по настоящему изобретению блокировать взаимодействие между CTLA-4 и B7.1.
На фиг. 4А и 4В показано влияние анти-CTLA-4 антител CL3, CL5, CL11 и CL22 (A) и CL24, CL25 и CL30 (B) на секрецию IL-2 в анализе стимуляции SEB-зависимых Т-лимфоцитов с использованием PBMC от донора 1.
На фиг. 5А и 5В показана способность к связыванию анти-CTLA-4 антител экспрессированный CL5, CL11, CL22, CL24 и CL25 (A) и CL5 (B) с CTLA-4 человека или CTLA-4 мыши.
На фиг. 6А и 6В показано влияние анти-CTLA-4 антител CL5, CL11 и их мутантов (A) и CL22, CL25 и их мутантов (B) на секрецию IL-2 в анализе стимуляции SEB-зависимых Т-лимфоцитов с использованием PBMC от донора 2.
На фиг. 7 показана активность связывания мутантов анти-CTLA-4-антитела с CTLA-4 человека.
На фиг. 8 показана активность мутантов анти-CTLA-4-антител в отношении блокирующего взаимодействия между CTAL-4 и меченным биотином B7.1.
На фиг. 9 показано влияние мутантов анти-CTLA-4-антител на секрецию IL-2 в анализе стимуляции SEB-зависимых Т-лимфоцитов с использованием PBMC от донора 3.
На фиг. 10 показана активность связывания анти-CTLA-4 антитела CL5 и его мутантов с CTLA-4 человека или яванского макака, измеренная с помощью BiaCore.
На фиг. 11 показана активность ADCC in vitro анти-CTLA-4-антитела CL5 и его мутанта на клетках CHO K1-CTLA-4.
На фиг. 12 показан профиль зависимости сывороточной концентрации от времени анти-CTLA-4-антитела CL5 у мышей-самцов C57BL/6.
На фиг. 13 показано соотношение опухоли к сыворотке концентрации анти-CTLA4 HCAb.
На фиг. 14А, 14В и 14С показана противоопухолевая активность анти-CTLA-4 антител у мышей с опухолями МС38. (А) Кривые роста опухоли в разных группах. (B) Кривая выживания времени до конечной точки Каплана-Мейера. (C) Массы тела мышей оставались относительно постоянными вместе с обработкой человеческим анти-CTLA-4 антителом. Данные были выражены как Среднее + SEM, N = 9.
На фиг. 15А и 15В показано ингибирование роста опухоли человеческими анти-CTLA-4 антителами у мышей, несущих МС38. (А) Кривая роста опухоли. (B) Масса тела мышей оставалась относительно постоянной на фоне обработки человеческим анти-CTLA-4 антителом. Данные были выражены как Среднее + SEM, N = 6.
Подробное описание изобретения
Общий обзор
В раскрытии предлагаются антитела, которые специфически связываются с CD152. Эти связывающие молекулы могут специфически связываться с CD152 и с другой мишенью. Введение терапевтически эффективного количества CD152-связывающего антитела нуждающемуся в этом пациенту полезно при лечении определенных расстройств, включая определенные виды рака. Связывание антитела с Т-клеткой, экспрессирующей CD152, индуцирует антителозависимую клеточно-опосредованную цитотоксичность в отношении клетки, экспрессирующей антиген, ассоциированный с опухолью. Терапевтические средства, связывающиеся с CD152, по настоящему изобретению, предлагают различные преимущества при лечении пациентов, например, эффективное связывание с CD152, эффективную индукцию антителозависимой клеточно-опосредованной цитотоксичности и/или более низкий риск неблагоприятных событий (например, токсичности). В определенных аспектах CD152-связывающие антитела связываются с CD152 более эффективно в определенных форматах (например, антитело, содержащее только тяжелые цепи, по сравнению с типичным полноразмерным антителом), что приводит к более высокой эффективности и улучшенной применимости при лечении расстройств, связанных с CD152.
Определения
Используемые в данном документе заголовки разделов предназначены только для организационных целей и не должны истолковываться как ограничивающие описанный объект изобретения. Все документы или части документов, цитируемых в данном документе, включая, без ограничения указанным, патенты, патентные заявки, статьи, книги и трактаты, настоящим прямо включены ссылкой в полном объеме для любых целей. В случае, если один или несколько из включенных документов или частей документов определяют термин, который противоречит определению этого термина в заявке, определение, которое появляется в этой заявке, имеет преимущественную силу. Однако упоминание любых источника, статьи, публикации, патента, публикации патента и заявки на патент, цитируемых в данном документе, не является и не должно восприниматься как подтверждение или какая-либо форма предположения о том, что они представляют собой действующий предшествующий уровень техники или являются частью широко распространёнными общими знаниями в любой стране мира.
В настоящем описании любой диапазон концентрации, процентный диапазон, диапазон отношения или целочисленный диапазон следует понимать как включающий значение любого целого числа в указанном диапазоне и, при необходимости, его доли (например, одну десятую и одну сотую целого числа), если не указано иное. Следует понимать, что термины, приведенные в единственном числе, относятся к «одному или нескольким» из перечисленных компонентов, если не указано иное. Использование альтернативы (например, «или») следует понимать как означающее либо одну, обе или любую комбинацию альтернатив. Используемые в данном документе термины «включать» и «содержать» используются как синонимы. Кроме того, следует понимать, что полипептиды, содержащие различные комбинации компонентов (например, доменов или областей) и заместителей, описанных в данном документе, раскрыты в настоящей заявке в той же степени, как если бы каждый полипептид был изложен индивидуально. Таким образом, выбор конкретных компонентов отдельных полипептидов находится в пределах объема настоящего раскрытия.
Термин «около» и его грамматические эквиваленты по отношению к референсному числовому значению и его грамматические эквиваленты, используемые в данном документе, могут включать диапазон значений плюс или минус 10% от этого значения, такой как диапазон значений плюс или минус 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% от этого значения. Например, сумма «около 10» включает суммы от 9 до 11.
При использовании в данном документе «полипептид» или «полипептидная цепь» представляет собой единое, линейное и непрерывное расположение ковалентно связанных аминокислот. Он не включает две полипептидные цепи, которые связаны друг с другом нелинейным образом, например, через дисульфидную связь между цепями (например, половинная молекула иммуноглобулина, в которой легкая цепь связана с тяжелой цепью через дисульфидную связь). Полипептиды могут иметь или образовывать одну или несколько внутрицепочечных дисульфидных связей. Что касается полипептидов, как описано в данном документе, ссылка на аминокислотные остатки, соответствующие остаткам, указанным в SEQ ID NO, включает посттрансляционные модификации таких остатков.
«Белок» представляет собой макромолекулу, содержащую одну или несколько полипептидных цепей. Белок также может содержать непептидные компоненты, такие как углеводные группы. Углеводы и другие непептидные заместители могут быть добавлены к белку клеткой, в которой производится белок, и будут варьировать в зависимости от типа клетки. Белки определены в данном документе с точки зрения их структуры аминокислотной цепи; заместители, такие как углеводные группы, обычно не указаны, но, тем не менее, могут присутствовать.
Термины «антитело», «иммуноглобулин» или «Ig» могут использоваться в данном документе взаимозаменяемо и означают молекулу иммуноглобулина, которая распознает и специфически связывается с мишенью, такой как белок, полипептид, пептид, углевод, полинуклеотид, липид или комбинации вышеуказанных через, по меньшей мере, один сайт распознавания антигена в вариабельной области молекулы иммуноглобулина. При использовании в данном документе термин «антитело» охватывает интактные поликлональные антитела, интактные моноклональные антитела, фрагменты антител (такие как фрагменты Fab, Fab′, F(ab′)2 и Fμ), одноцепочечные мутанты Fμ (scFv), мультиспецифичные антитела, такие как биспецифичные антитела (включая антитела с двойным связыванием), химерные антитела, гуманизированные антитела, человеческие антитела, гибридные белки, содержащие часть определения антигена из антитела, и любую другую модифицированную молекулу иммуноглобулина, содержащую сайт распознавания антигена, при условии, что антитела демонстрируют искомую биологическую активность. Термин «антитело» может также относиться к Y-образному гликопротеину с молекулярной массой приблизительно 150 кДа, который состоит из четырех полипептидных цепей: двух легких (L) цепей и двух тяжелых (H) цепей. Существует пять типов изотипов тяжелых цепей млекопитающих Ig, обозначаемых греческими буквами альфа (α), дельта (δ), эпсилон (ε), гамма (γ) и мю (μ). Тип тяжелой цепи определяет класс антител, то есть IgA, IgD, IgE, IgG и IgM, соответственно. Классы γ и α дополнительно подразделяются на подклассы на основе различий в последовательности и функции константного домена, например, IgG1, IgG2A, IgG2B, IgG3, IgG4, IgA1 и IgA2. У млекопитающих есть два типа легких цепей иммуноглобулина, λ и κ.
При использовании в данном документе термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, то есть отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций и/или модификаций после трансляции (например, изомеризации, амидиации), которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифичными и направлены против одного антигенного сайта. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты на антигене. В дополнение к их специфичности, моноклональные антитела имеют то преимущество, что они синтезируются культурой гибридомы, не загрязненной другими иммуноглобулинами. Модификатор «моноклональный» указывает на характер антитела, получаемого по существу из гомогенной популяции антител, и его не следует истолковывать как требующее получения антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены различными способами, включая, например, гибридомный способ (например, Kohler and Milstein, Nature, 256:495-97 (1975); Hongo et al., Hybridoma, 14 (3): 253-260 (1995), Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981)), способы рекомбинантной ДНК (см., например, Пат. США 4816567), технологии фагового дисплея (см., например, Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol.340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284 (1-2): 119-132 (2004) и технологии получения антител человека или антител, подобных антителам человека в животных, которые имеют части или все локусы иммуноглобулина человека или гены, кодирующие последовательности иммуноглобулина человека (см. например, WO 1998/24893; WO 1996/34096; WO 1996/33735; WO 1991/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA 90: 2551 (1993); Jakobovits et al., Nature 362: 255-258 (1993); Bruggemann et al., Year in Immunol. 7:33 (1993); Пат. США No. 5545807; 5545806; 5569825; 5625126; 5633425; и 5661016; Marks et al., Bio/Technology 10: 779-783 (1992); Lonberg et al., Nature 368: 856-859 (1994); Morrison, Nature 368: 812-813 (1994); Fishwild et al., Nature Biotechnol. 14: 845-851 (1996); Neuberger, Nature Biotechnol. 14: 826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol. 13: 65-93 (1995).
При использовании в данном документе термин «антитело, содержащее только тяжелые цепи» (HCAb) относится к антителу, которое состоит только из двух тяжелых цепей и не содержит двух легких цепей, обычно присутствующих в полноразмерных антителах.
Термин «выделенное антитело», при использовании для описания различных антител, раскрытых в данном документе, означает антитело, которое было идентифицировано и отделено и/или извлечено из клетки или клеточной культуры, из которой оно было экспрессировано. Загрязняющие компоненты его естественной среды представляют собой материалы, которые обычно мешают диагностическому или терапевтическому применению полипептида и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых воплощениях антитело очищают до чистоты более 95% или 99%, что определяется, например, электрофоретическим способом (например, SDS-PAGE, изоэлектрическое фокусирование (IEF), капиллярный электрофорез) или хроматографическим способом (например, ионно-обменная или обратно-фазная ВЭЖХ). Для обзора способов оценки чистоты антител см., например, Flatman et al., J. Chromatogr. B 848: 79-87 (2007). В предпочтительных воплощениях антитело будет очищено (1) до степени, достаточной для получения, по меньшей мере, 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатора с вращающимся стаканом, или (2) до гомогенности с помощью SDS-PAGE при невосстанавливающих или восстанавливающих условия с использованием кумасси синего или, предпочтительно, серебряного красителя. Выделенное антитело включает антитела in situ в рекомбинантных клетках, поскольку, по меньшей мере, один компонент полипептидной природной среды не будет присутствовать. Обычно, однако, выделенный полипептид будет получен, по меньшей мере, на одной стадии очистки.
Термин «выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее естественной среды. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует внехромосомно или в локации в хромосоме расположении, которая отличается от ее естественного расположения в хромосоме.
Термины «вариабельная область легкой цепи» (также называемая «вариабельный домен легкой цепи» или «VL» или VL) и «вариабельная область тяжелой цепи» (также называемая «вариабельный домен тяжелой цепи» или «VH» или VH) относятся к вариабельной области связывания антитела легкой и тяжелой цепи, соответственно. Вариабельные области связывания состоят из дискретных, четко определенных субобластей, известных как «области, определяющие комплементарность» (CDR) и «каркасные области» (FR), обычно включающие FR1-CDR1-FR2-CDR2-FR3-CDR3 -FR4 от аминоконца до карбоксильного конца. В одном воплощении FR являются гуманизированными. Термин «CL» относится к «константной области легкой цепи иммуноглобулина» или «константной области легкой цепи», то есть константной области легкой цепи антитела. Термин «СН» относится к «константной области тяжелой цепи иммуноглобулина» или «константной области тяжелой цепи», которая дополнительно делится в зависимости от изотипа антитела на СН1, СН2 и СН3 (IgA, IgD, IgG) или домены CH1, CH2, CH3 и CH4 (IgE, IgM). «Fab» (фрагмент, связывающий антиген) представляет собой часть антитела, которая связывается с антигенами и включает вариабельную область и домен CH1 тяжелой цепи, связанный с легкой цепью через межцепную дисульфидную связь.
При использовании в данном документе термин «связывающий домен» или «связывающая область» относится к домену, области, части или сайту белка, полипептида, олигопептида или пептида или антитела или связывающего домена, полученного из антитела, которые обладают способностью специфически распознавать и связывать молекулу-мишень, такую как антиген, лиганд, рецептор, субстрат или ингибитор (например, CD152). Типичные связывающие домены включают вариабельные области одноцепочечных антител (например, доменные антитела, sFv, scFv, scFab), рецепторные эктодомены и лиганды (например, цитокины, хемокины). В некоторых воплощениях связывающий домен включает или состоит из антигенсвязывающего сайта (например, содержащего вариабельную последовательность тяжелой цепи и вариабельную последовательность легкой цепи или три области, определяющие комплементарность из легкой цепи (CDR) и три CDR из тяжелой цепи антитела, помещенных в альтернативный вариант каркасной области (FR) (например, FR человека, необязательно содержащей одну или несколько аминокислотных замен). Известно множество анализов для идентификации доменов связывания по настоящему изобретению, которые специфически связывают конкретную мишень, включая вестерн-блот, ELISA, скрининг библиотеки фагового дисплея и анализ взаимодействия BIACORE®. При использовании в данном документе, «CD152-связывающий домен» может иметь вариабельные области тяжелой цепи иммуноглобулина, содержащие три CDR тяжелой цепи: CDR1, CDR2 и CDR3.
Антитело или связывающий домен «специфически связывает» мишень, если оно связывает мишень с аффинностью или Ka (то есть константой равновесной ассоциации конкретного связывающего взаимодействия с единицами 1/М), равной или превышающей 105 М-1 и при этом незначительно связывает другие компоненты, присутствующие в тестовом образце. Антитела или связывающие домены могут быть классифицированы как «высокоаффинные» антитела или связывающие домены и «низкоаффинные» антитела или связывающие домены. «Высокоаффинные» антитела или связывающие домены относятся к тем антителам или связывающим доменам с Ka, по меньшей мере, 107 М-1, по меньшей мере, 108 М-1, по меньшей мере, 109 М-1, по меньшей мере, 1010 М-1, по меньшей мере, 1011 М-1, по меньшей мере, 1012 М-1 или, по меньшей мере, 1013 М-1. «Низкоаффинные» антитела или связывающие домены относятся к тем антителам или связывающим доменам с Ka до 107 М-1, до 106 М-1, до 105 М-1. В ином случае, аффинность может быть определена как равновесная константа диссоциации (Kd) конкретного связывающего взаимодействия с единицами М (например, 10-5 M - 10-13 M). В случае связывания антитела с антигеном Ka = 1/Kd. Аффинность антител или связывающих доменов в соответствии с настоящим изобретением может быть легко определена с использованием обычных методов, таких как поверхностный плазмонный резонанс (см., например, Scatchard et al. (1949) Ann. N.Y. Acad. Sci. 51:660; и Пат. США № 5283173, 5468614 или эквивалент).
При использовании в данном документе «CD152» относится к кластеру дифференцировки 152, который также известен как белок 4, ассоциированный с цитотоксическими Т-лимфоцитами (CTLA-4). Термины «CD152», «CTLA-4» и «CTLA4» используются в данном документе взаимозаменяемо. Аналогично, «анти-CD152», «анти-CTLA-4», «анти-CTLA4» также используются в данном документе взаимозаменяемо.
При использовании в данном документе «CD80» относится к кластеру дифференцировки 80, который представляет собой белок, обнаруженный в дендритных клетках, активированных В-клетках и моноцитах, который обеспечивает костимулирующий сигнал, необходимый для активации и выживания Т-клеток. Термины «CD80», «B7-1» и «B7.1» используются в данном документе взаимозаменяемо.
При использовании в данном документе «CD86» относится к кластеру дифференцировки 86, который представляет собой белок, экспрессируемый на антигенпрезентирующих клетках, который обеспечивает костимулирующие сигналы, необходимые для активации и выживания Т-клеток. Термины «CD86», «B7-2» и «B7.2» используются в данном документе взаимозаменяемо.
При использовании в данном документе «консервативная замена» в данной области техники известна как замена одной аминокислоты другой аминокислотой, которая имеет сходные свойства. Типичные консервативные замены хорошо известны в данной области (см., например, WO 97/09433, стр. 10, опубликовано 13 марта 1997 г.; Lehninger, Biochemistry, Second Edition; Worth Publishers, Inc. NY:NY (1975), pp.71-77; Lewin, Genes IV, Oxford University Press, NY and Cell Press, Cambridge, MA (1990), p. 8). В некоторых воплощениях консервативная замена включает замену лейцина на серин.
При использовании в данном документе «аналог ипилимумаба» относится к моноклональному антителу, которое специфически связывается с CTLA-4, содержащему тяжелую цепь с аминокислотной последовательностью SEQ ID NO.: 199 и легкую цепь с аминокислотной последовательностью SEQ ID № 200.
При использовании в данном документе, если не указано иное, любое непатентованное или общее название биологического продукта включает биологический продукт и любой его биоподобный продукт. Например, непатентованное название, ипилимумаб, относится к биологическому продукту, продаваемому под торговой маркой YERVOY; это также включает любой биоподобный продукт биологического продукта.
При использовании в данном документе, если не указано иное, термин «биоподобный продукт» относится к 1) биологическому продукту, имеющему аминокислотную последовательность, которая идентична референсному продукту; 2) биологический продукт, имеющий аминокислотную последовательность, отличную (например, укорочения на N- или C-конце) от референсного продукта; или 3) биологический продукт, имеющий посттрансляционную модификацию, отличающуюся от (например, гликозилированием или фосфорилированием) от референсного продукта, где биоподобный продукт и референсный продукт используют один и тот же механизм или механизмы действия для профилактики, обработки или лечения заболевания или состояния.
При использовании в данном документе термин «производное» относится к модификации одного или нескольких аминокислотных остатков пептида химическими или биологическими средствами, либо с ферментом, либо без него, например, путем гликозилирования, алкилирования, ацилирования, образования сложного эфира или амида. образование.
При использовании в данном документе указание того, что полипептид или аминокислотная последовательность, «получены из» указанного полипептида или белка, относится к происхождению полипептида. В некоторых воплощениях полипептидная или аминокислотная последовательность, которая получена из конкретной последовательности (иногда называемой «исходной» или «родителем» или «родительской» последовательностью), имеет аминокислотную последовательность, которая по существу идентична исходной последовательности или ее части, причем часть состоит, по меньшей мере, из 10-20 аминокислот, по меньшей мере, 20-30 аминокислот, или, по меньшей мере, 30-50 аминокислот, или, по меньшей мере, 50-150 аминокислот, или которые иным образом идентифицируемы обычным специалистом в данной области техники, как происходящие из исходной последовательности. Например, связывающий домен может быть получен из антитела, например, Fab, F(ab’)2, Fab’, scFv, однодоменного антитела (sdAb) и т.д.
Полипептиды, полученные из другого полипептида, могут иметь одну или несколько мутаций относительно исходного полипептида, например, один или несколько аминокислотных остатков, которые были замещены другим аминокислотным остатком или которые имеют одну или несколько вставок или делеций аминокислотных остатков. Полипептид может содержать аминокислотную последовательность, которая не встречается в природе. Такие вариации обязательно имеют менее чем 100% идентичности или сходства последовательности с исходным полипептидом. В одном воплощении вариант будет иметь аминокислотную последовательность от около 60% до менее чем 100% идентичности или сходства аминокислотной последовательности с аминокислотной последовательностью исходного полипептида. В другом воплощении вариант будет иметь аминокислотную последовательность от около 75% до менее чем 100%, от около 80% до менее чем 100%, от около 85% до менее чем 100%, от около 90% до менее чем 100 %, от около 95% до менее чем 100% идентичности или сходства аминокислотной последовательности с аминокислотной последовательностью исходного полипептида.
При использовании в данном документе, если не указано иное, положение аминокислотного остатка в вариабельной области молекулы иммуноглобулина нумеруется в соответствии с системой нумерации IMGT (Brochet, X, et al., Nucl. Acids Res. (2008) 36, W503-508), а положение аминокислотного остатка в константной области молекулы иммуноглобулина пронумеровано в соответствии с номенклатурой EU (Ward et al., 1995 Therap. Immunol. 2:77-94). В данной области известны другие системы нумерации (например, система нумерации по Kabat (Kabat, Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, MD: Public Health Service, National Institutes of Health (1991)).
При использовании в данном документе термин «человеческое» антитело относится к антителу человеческого происхождения или гуманизированному антителу.
При использовании в данном документе термин «гуманизированный» относится к процессу создания с помощью методов генной инженерии антител или иммуноглобулиновых связывающих белков и полипептидов, полученных от не являющегося человеком вида (например, мыши или крысы), менее иммуногенными для человека, с сохранением при этом антигенсвязывающих свойств оригинального антитела. В некоторых воплощениях связывающий домен(домены) антитела или иммуноглобулиновых связывающих белков и полипептидов (например, вариабельные области легкой и тяжелой цепи, Fab, scFv) являются гуманизированными. Нечеловеческие связывающие домены могут быть гуманизированы с использованием методов, известных как прививка CDR (Jones et al., Nature 321: 522 (1986)) и их вариантов, включая «решейпинг» (Verhoeyen, et al., 1988 Science 239:1534-1536; Riechmann, et al., 1988 Nature 332:323-337; Tempest, et al., Bio/Technol 1991 9:266-271), «гиперхимеризация» (Queen et al., 1989 Proc Natl Acad Sci USA 86: 10029-10033; Co, et al., 1991 Proc Natl Acad Sci USA 88: 2869-2873; Co, et al., 1992 J. Immunol. 148: 1149-1154) и «венирование» (Mark, et al., "Derivation of therapeutically active humanized and veneered anti-CD18 antibodies.” In: Metcalf BW, Dalton BJ, eds. Cellular adhesion: molecular definition to therapeutic potential. New York: Plenum Press, 1994: 291-312). Если они получены из не являющегося человеком источника, другие области антитела или иммуноглобулиновые связывающие белки и полипептиды, такие как шарнирная область и домены константной области, также могут быть гуманизированы.
При использовании в данном документе термин «нуждающийся пациент» или «нуждающийся объект» относится к пациенту или объекту, подверженному риску заболевания или расстройства или состояния, которое поддается лечению или улучшению с помощью CD152-связывающего антитела или его композиции, представленных в данном документе.
При использовании в данном документе термин «фармацевтически приемлемый» относится к молекулярным веществам и композициям, которые обычно не вызывают аллергических или других серьезных побочных реакций при введении с использованием путей, хорошо известных в данной области. Молекулярные объекты и композиции, одобренные регулирующим органом федерального правительства или правительства штата или перечисленные в Фармакопее США или в других общепризнанных фармакопеях для применения на животных, и, более конкретно, на людях, считаются «фармацевтически приемлемыми».
При использовании в данном документе термин «лечение», «лечить» или «улучшать состояние» относится либо к терапевтическому лечению, либо к профилактическому/превентивному лечению. Лечение является терапевтическим, если, по меньшей мере, один симптом заболевания у индивидуума, получающего лечение, улучшается или лечение может отсрочить ухудшение прогрессирующего заболевания у индивидуума или предотвратить возникновение дополнительных сопутствующих заболеваний.
При использовании в данном документе термин «терапевтически эффективное количество (или доза)» или «эффективное количество (или доза)» конкретной связывающей молекулы или соединения относится к такому количеству соединения, которое достаточно, чтобы привести к ослаблению одного или нескольких симптомов заболевания, подвергаемого лечению, статистически значимым образом или статистически значимому улучшению функции органов. При ссылке на отдельный активный ингредиент, вводимый отдельно, терапевтически эффективная доза относится только к этому ингредиенту. Применительно к комбинации терапевтически эффективная доза относится к объединенным количествам активных ингредиентов, которые приводят к терапевтическому эффекту, вне зависимости вводятся ли они последовательно или одновременно (в одной и той же композиции или одновременно в отдельных композициях).
При использовании в данном документе термины «антитело-зависимая клеточно-опосредованная цитотоксичность» и «ADCC» относятся к клеточно-опосредованному процессу, в котором неспецифические цитотоксические клетки, которые экспрессируют FcγR (например, моноцитарные клетки, такие как клетки-натуральные киллеры (NK) и макрофаги) распознают связанное антитело (или другой белок, способный связывать FcγR) на клетке-мишени и впоследствии вызывают лизис клетки-мишени. В принципе, любая эффекторная клетка с активирующим FcγR может быть иницирована для опосредования ADCC. Первичными клетками для опосредования ADCC являются NK-клетки, которые экспрессируют только FcγRIII, тогда как моноциты, в зависимости от их состояния активации, локализации или дифференцировки, могут экспрессировать FcγRI, FcγRII и FcγRIII. Обзор экспрессии FcγR на гемопоэтических клетках см., например, Ravetch et al., 1991, Annu. Rev. Immunol., 9: 457-92.
При использовании в данном документе термин «промотор» относится к области ДНК, вовлеченной в связывание РНК-полимеразы для инициации транскрипции.
При использовании в данном документе термины «нуклеиновая кислота», «молекула нуклеиновой кислоты» или «полинуклеотид» относятся к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам в одно- или двухцепочечной форме. За исключением случаев конкретного ограничения, термины охватывают нуклеиновые кислоты, содержащие аналоги природных нуклеотидов, которые обладают свойствами связывания, сходными с эталонной нуклеиновой кислотой, и метаболизируются способом, подобным встречающимся в природе нуклеотидам. Если не указано иное, конкретная последовательность нуклеиновой кислоты также неявно включает ее консервативно модифицированные варианты (например, вырожденные замены кодонов) и комплементарные последовательности, а также явно указанную последовательность. В частности, вырожденные замены кодонов могут быть достигнуты путем получения последовательностей, в которых третье положение одного или нескольких выбранных (или всех) кодонов замещено остатками смешанного основания и/или дезоксиинозина (Batzer et al. (1991) Nucleic Acid Res. 19:5081; Ohtsuka et al. (1985) J. Biol. Chem. 260:2605-2608; Cassol et al. (1992); Rossolini et al. (1994) Mol. Cell. Probes 8:91-98). Термин нуклеиновая кислота используется взаимозаменяемо с геном, кДНК и мРНК, кодируемой геном. При использовании в данном документе термины «нуклеиновая кислота», «молекула нуклеиновой кислоты» или «полинуклеотид» предназначены для включения молекул ДНК (например, кДНК или геномной ДНК), молекул РНК (например, мРНК), аналогов ДНК или РНК, полученных с использованием нуклеотидных аналогов и их производных, фрагментов и гомологов.
Термин «экспрессия» относится к биосинтезу продукта, кодируемого нуклеиновой кислотой. Например, в случае сегмента нуклеиновой кислоты, кодирующего представляющий интерес полипептид, экспрессия включает транскрипцию сегмента нуклеиновой кислоты в мРНК и трансляцию мРНК в один или несколько полипептидов.
Термины «единица экспрессии» и «кассета экспрессии» используются в данном документе взаимозаменяемо и обозначают сегмент нуклеиновой кислоты, кодирующий представляющий интерес полипептид и способный обеспечивать экспрессию сегмента нуклеиновой кислоты в клетке-хозяине. Единица экспрессии обычно включает промотор транскрипции, открытую рамку считывания, кодирующую представляющий интерес полипептид, и терминатор транскрипции, все в функциональной конфигурации. В дополнение к транскрипционному промотору и терминатору, единица экспрессии может дополнительно включать другие сегменты нуклеиновой кислоты, такие как, например, энхансер или сигнал полиаденилирования.
При использовании в данном документе термин «экспрессирующий вектор» относится к молекуле нуклеиновой кислоты, линейной или кольцевой, содержащей одну или несколько единиц экспрессии. В дополнение к одной или нескольким единицам экспрессии экспрессирующий вектор может также включать дополнительные сегменты нуклеиновой кислоты, такие как, например, одну или несколько точек начала репликации или один или несколько селектируемых маркеров. Экспрессирующие векторы обычно происходят из плазмидной или вирусной ДНК или могут включать элементы обоих.
При использовании в данном документе термин «идентичность последовательности» относится к взаимосвязи между двумя или более полинуклеотидными последовательностями или между двумя или более полипептидными последовательностями. Когда положение в одной последовательности занято одним и тем же основанием нуклеиновой кислоты или аминокислотным остатком в соответствующем положении сопоставленной последовательности, говорят, что последовательности «идентичны» в этом положении. Процент «идентичности последовательности» рассчитывают путем определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, чтобы получить количество «идентичных» положений. Количество «идентичных» положений затем делится на общее количество положений в окне сравнения и умножается на 100, чтобы получить процент «идентичности последовательности». Процент «идентичности последовательности» определяется путем сравнения двух оптимально выровненных последовательностей в окне сравнения. Окно сравнения для последовательностей нуклеиновых кислот может составлять, например, по меньшей мере, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 или более нуклеиновых кислот в длину. Окно сравнения для полипептидных последовательностей может быть, например, по меньшей мере, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 300 или более аминокислот в длину. Чтобы оптимально выровнять последовательности для сравнения, часть полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции, называемые пробелами, в то время как референсная последовательность поддерживается постоянной. Оптимальное выравнивание - это такое выравнивание, которое, даже с пропусками, создает максимально возможное количество «идентичных» положений между референсной последовательностью и последовательностью сравнения. Процент «идентичности последовательностей» между двумя последовательностями можно определить с помощью версии программы «BLAST 2 Sequences», которая была доступна в Национальном центре биотехнологической информации по состоянию на 1 сентября 2004 года, и эта программа включает программы BLASTN (для сравнения нуклеотидных последовательностей) и BLASTP (для сравнения полипептидных последовательностей), программы которых основаны на алгоритме Karlin и Altschul (Proc. Natl. Acad. Sci. USA 90 (12): 5873-5877, 1993). При использовании «BLAST 2 Sequences» параметры, которые были параметрами по умолчанию на 1 сентября 2004 года, могут использоваться для размера «слова» (3), штрафа за открытый разрыв (11), штрафа за продолжение разрыва (1), прекращение разрыва (50), ожидаемое значение (10) и любой другой обязательный параметр, включая, без ограничения, опцию матрицы. Считается, что две нуклеотидные или аминокислотные последовательности имеют «по существу сходную идентичность последовательности» или «существенную идентичность последовательности», если две последовательности имеют, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 96 %, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичности последовательностей относительно друг друга.
Антитела
В данном документе раскрыты человеческие моноклональные антитела, содержащие CD152-связывающий домен. Антитела могут быть антителами только на основе тяжелых цепей. Антитела могут состоять только из двух тяжелых цепей. Антитела могут не содержать легких цепей. Антитела могут специфически связываться с CD152. Антитела могут быть выделенными моноклональными антителами, которые специфически связываются с CD152 с высокой аффинностью.
Описанные в данном документе анти-CD152-антитела могут специфически связываться с CD152 человека. В некоторых случаях анти-CD152 антитела могут связываться с CD152 человека с высокой аффинностью (например, KD <6,0*10-11M). Анти-CD152 антитела могут иметь сравнимую или более высокую аффинность к CTLA-4 по сравнению с аналогом ипилимумабом. Анти-CD152 антитела также могут блокировать связывание CD152 с его лигандами B7.1. Анти-CD152 антитела могут иметь повышенное соотношение опухоль/периферическая сыворотка, чем аналог ипилимумаб. Анти-CD152 антитела могут индуцировать более высокую ADCC, например, антитела могут индуцировать, по меньшей мере, около 2-кратное, по меньшей мере, около 3-кратное, по меньшей мере, около 4-кратное, по меньшей мере, около 5-кратное, по меньшей мере, около 6-кратное, по меньшей мере, около 7-кратное, по меньшей мере, около 8-кратное, по меньшей мере, около 9-кратное, по меньшей мере, около 10-кратное, по меньшей мере, около 15 -кратное или, по меньшей мере, около 20-кратное, увеличение лизисной активности NK-клеток.
Анти-CD152 антитела могут содержать CD152-связывающий домен, который включает вариабельную область тяжелой цепи иммуноглобулина, содержащую CDR1, CDR2 и CDR3. Анти-CD152 антитела также могут содержать CDR1, CDR2 и CDR3, которые отличаются от этих областей из анти-CD152 антител, раскрытых в данном документе, одной или несколькими консервативными модификациями. В данной области понятно, что могут быть выполнены определенные модификации консервативных последовательностей, которые не удаляют связывание антигена. См., например, Brummell et al., 1993, Biochem 32: 1180-8; de Wildt et al., 1997, Prot. Eng. 10:835-41; Komissarov et al., 1997, J. Biol. Chem. 272:26864-26870; Hall et al., 1992, J. Immunol. 149:1605-12; Kelley and O'Connell, 1993, Biochem.32:6862-35; Adib-Conquy et al., 1998, Int. Immunol.10:341-6 и Beers et al., 2000, Clin. Can. Res. 6:2835-43.
Фармацевтические композиции и составы
Фармацевтическая композиция может содержать одно или несколько анти-CD152 антител, описанных в данном документе, в составе вместе с фармацевтически приемлемым вспомогательным веществом. Говорят, что эксципиент является «фармацевтически приемлемым эксципиентом», если его пациент может переносить его прием. Эксципиенты, которые могут быть использованы, включают носители, поверхностно-активные вещества, загущающие или эмульгирующие агенты, твердые связующие, дисперсионные или суспендирующие добавки, солюбилизаторы, красители, ароматизаторы, покрытия, разрыхлители, смазывающие вещества, подсластители, консерванты, изотонические агенты и их комбинации. Выбор и применение подходящих эксципиентов изучается в Gennaro, ed., Remington: The Science and Practice of Pharmacy, 20th Ed. (Lippincott Williams & Wilkins 2003), и в Gennaro, ed., Remington's Pharmaceutical Sciences (Mack Publishing Company, 19th ed. 1995). Стерильный фосфатно-солевой буферный раствор является одним из примеров фармацевтически приемлемого эксципиента. Композиции могут дополнительно включать один или несколько носителей, разбавителей, консервантов, солюбилизаторов, буферных агентов, альбумина для предотвращения потери белка на поверхностях сосудов и т.д.
Количество активного ингредиента, которое можно комбинировать с материалом-носителем для получения единичной дозированной формы, может варьировать в зависимости от объекта, которого лечат, и конкретного способа введения, и, как правило, может быть таким количеством фармацевтической композиции, которое оказывает терапевтический эффект. Как правило, количество активного ингредиента может составлять от около 0,01% до около 99% (масс./масс.) композиции, например, может составлять около 0,1% -1%, около 0,1% -5%, около 0,1-10%. около 0,1% -20%, около 0,5% -1%, около 0,5% -5%, около 0,5% -10%, около 0,5% -20%, около 1% -5%, около 1% -10%, около 1% -20%, около 5% -10%, около 5% -20%, около 10% -20%, около 10% -30%, около 20% -30%, около 20% -40%, около 30% -40%, около 30% -50%, около 40% -50%, около 40% -60%, около 50% -60%, около 50% -70%, около 60% -70%, около 60 % -80%, около 70% -80%, около 70% -90%, около 80% -90%, около 80% -95% или 95% -99% фармацевтической композиции. Предпочтительно количество активного ингредиента может составлять от около 0,1% до около 70% и наиболее предпочтительно от около 1% до около 30% фармацевтической композиции.
Фармацевтическая композиция может быть подходящей для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активный ингредиент может быть покрыт материалом, защищающим его от действия кислот и других природных условий, которые могут его инактивировать. Словосочетание «парентеральное введение» используемое в данном документе, означает режимы введения отличные от энтерального или местного введения, обычно инъекцией, и включает, без ограничений указанным, внутривенную, внутримышечную, внутриартериальную, внутритрахеальную, внутрикапсулярную, внутриорбитальную, внутрикардиальную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, внутрисуставную, субкапсулярную, субарахноидальную, внутрипозвоночную, эпидуральную и надчревную инъекцию или инфузию. В ином случае, антитело по изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или слизистый путь введения, например, интраназально, перорально, вагинально, ректально, сублингвально или местно. Фармацевтическая композиция может быть в форме стерильных водных растворов или дисперсий. Фармацевтическая композиция также может быть составлена в виде микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства.
Фармацевтическая композиция может быть составлена в дозированной форме, выбранной из группы, состоящей из: пероральной дозированной лекарственной формы, внутривенной дозированной лекарственной формы, интраназальной дозированной лекарственной формы, дозированной лекарственной формы для суппозиториев, внутрикожной дозированной лекарственной формы, внутримышечной дозированной лекарственной формы, внутрибрюшинной дозированной лекарственной формы, подкожной дозированной лекарственной формы, эпидуральная дозированной лекарственной формы, сублингвальной дозированной лекарственной формы и внутримозговой дозированной лекарственной формы. Дозированная лекарственная форма для перорального применения может быть выбрана из группы, состоящей из таблеток, пилюль, пеллет, капсул, порошков, пастилок, гранул, растворов, суспензий, эмульсий, сиропов, эликсиров, составов с замедленным высвобождением, аэрозолей и спреев.
Фармацевтическая композиция может представлять собой композицию с контролируемым высвобождением, включая имплантаты, трансдермальные пластыри и микрокапсулированные системы доставки. Могут быть использованы биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Описанные в данном документе моноклональные антитела могут быть составлены для обеспечения надлежащего распределения in vivo. Например, чтобы гарантировать, что терапевтические антитела по изобретению проникают через гематоэнцефалический барьер, они могут быть включены в липосомы, которые могут дополнительно содержать нацеливающие фрагменты для усиления селективного транспорта к конкретным клеткам или органам. См., например, пат. США No. 4522811; 5374548; 5416016; и 5399331; V. V. Ranade, 1989, J. Clin.Pharmacol.29:685; Umezawa et al., 1988, Biophys. Res. Commun. 153:1038; Bloeman et al., 1995, FEBS Lett.357:140; M. Owais et al., 1995, Antimicrob. Agents Chemother. 39:180; Briscoe et al., 1995, Am. J. Physiol. 1233:134; Schreier et al., 1994, J. Biol. Chem. 269:9090; Keinanen and Laukkanen, 1994, FEBS Lett. 346:123; и Killion and Fidler, 1994, Immunomethods 4:273.
Фармацевтическая композиция может необязательно включать один или несколько дополнительных фармацевтически активных ингредиентов, таких как другое антитело или лекарственное средство. Фармацевтические композиции по изобретению также можно вводить в комбинированной терапии, например, с другим противоопухолевым средством, другим противовоспалительным средством или вакциной.
Фармацевтические композиции могут поставляться в виде набора, включающего контейнер, который содержит фармацевтическую композицию, как описано в данном документе. Фармацевтическая композиция может быть предоставлена, например, в форме раствора для инъекций для однократной или многократных доз или в виде стерильного порошка, который будет восстановлен перед инъекцией. В ином случае, такой набор может включать устройство, диспергирующее сухой порошок, генератор жидкого аэрозоля или распылитель для введения фармацевтической композиции. Такой набор может дополнительно включать письменную информацию о показаниях и применении фармацевтической композиции.
Способы лечения
Кроме того, в настоящем описании раскрыт способ лечения расстройства путем введения объекту терапевтически эффективного количества антитела или фармацевтической композиции, раскрытой в данном документе. Анти-CD152 антитела, описанные в данном документе, могут быть использованы в способе лечения объекта (например, человека или примата, не являющегося человеком) или для изготовления лекарственного средства для лечения объекта. Обычно такие способы включают введение объекту, нуждающемуся в таком лечении, анти-CD152-антител, описанных в данном документе.
Анти-CD152 антитела, описанные в данном документе, могут быть использованы в способе лечения объекта (например, человека или примата, не являющегося человеком) или для изготовления лекарственного средства для лечения объекта. Как правило, такие способы включают введение объекту, нуждающемуся в таком лечении, анти-CD152 антитела, описанного в данном документе. В некоторых воплощениях анти-CD152 антитело включает, по меньшей мере, одну эффекторную функцию, выбранную из антителозависимой клеточно-опосредованной цитотоксичности (ADCC) и/или комплементзависимой цитотоксичности (CDC), так что анти-CD152 антитело индуцирует ADCC и/или CDC против CD152-экспрессирующих клеток у объекта.
Также в данном документе раскрыт способ лечения расстройства, характеризующегося сверхэкспрессией опухолевого антигена, такого как рак. Примеры опухолевых антигенов, которые могут распознаваться биспецифическим анти-CD152-антителом, могут включать PSMA, CD19, CD20, CD37, CD38, CD123, Her2, ROR1, RON, антиген гликопротеина A33 (gpA33) и CEA. Обычно такие способы включают введение объекту, нуждающемуся в таком лечении, терапевтически эффективного количества анти-CD152 антитела, содержащего второй связывающий домен, который связывает опухолевый антиген, как описано в данном документе. Анти-CD152 антитело может индуцировать перенаправленную T-клеточную цитотоксичность (RTCC) против опухолевых антиген-экспрессирующих клеток у объекта.
Способ может быть использован для лечения рака, такого как рак предстательной железы, колоректальный рак, почечно-клеточный рак, рак мочевого пузыря, рак слюнной железы, рак поджелудочной железы, рак яичников, немелкоклеточный рак легкого, меланома, рак молочной железы (например, тройной негативный рак молочной железы), рак надпочечника, лимфома мантийных клеток, острый лимфобластный лейкоз, хронический лимфолейкоз, неходжкинская лимфома, острый миелоидный лейкоз (AML), В-лимфолейкоз, бластная плазмацитоидная дендритоклеточная опухоль (BPDCN), волосатоклеточный лейкоз.
Также в данном документе раскрыт способ лечения аутоиммунного расстройства, включающий введение терапевтически эффективного количества фармацевтических композиций или анти-CD152 антитела, описанного в данном документе, пациенту, нуждающемуся в этом.
Объекты для введения анти-CD152-антител, как описано в данном документе, включают пациентов с высоким риском развития конкретного расстройства, а также пациентов с имеющимся таким расстройством. Как правило, у объекта диагностируют расстройство, для которого требуется лечение. Кроме того, объекты могут в течение курса лечения наблюдаться на предмет любого изменения расстройства (например, увеличения или уменьшения клинических симптомов расстройства). Кроме того, в некоторых вариантах объект не страдает от другого расстройства, требующего лечения, которое включает нацеливание на клетки, экспрессирующие CD152.
В профилактических целях фармацевтические композиции можно вводить пациенту, подверженному или иным образом подверженному риску конкретного расстройства, в количестве, достаточном для устранения или уменьшения риска или задержки возникновения расстройства. В терапевтических применениях композиции можно вводить пациенту, подозреваемому или уже страдающему таким расстройством, в количестве, достаточном для лечения или, по меньшей мере, частичной остановки симптомов расстройства и его осложнений. Количество, достаточное для достижения этого, называется терапевтически эффективной дозой или количеством. Как в профилактических, так и в терапевтических режимах агенты можно вводить в нескольких дозировках, пока не будет достигнут достаточный ответ. Как правило, ответ контролируется, и повторные дозы назначаются, если искомый ответ начинает исчезать.
Для идентификации пациентов, подлежащих лечению, в соответствии со способами раскрытия, могут быть использованы принятые способы скрининга для определения факторов риска, связанных с конкретными расстройствами, или для определения состояния существующего расстройства, выявленного у объекта. Такие способы могут включать, например, определение того, есть ли у человека родственники, которым был поставлен диагноз определенного заболевания. Способы скрининга могут также включать, например, обычные обследования для определения семейного статуса для конкретного заболевания, которое, как известно, имеет наследственный компонент. Например, известно, что различные виды раков имеют определенные наследуемые компоненты. Наследуемые компоненты рака включают, например, мутации во множественных генах, которые являются трансформирующими (например, Ras, Raf, EGFR, cMet и др.), наличие или отсутствие определенных молекул HLA и рецептора подавления цитотоксичности (KIR) или механизмы, с помощью которых раковые клетки способны модулировать иммуносупрессию таких клеток, как NK-клетки и Т-клетки, прямо или косвенно (см., например, Ljunggren and Malmberg, Nature Rev. Immunol. 7:329-339, 2007; Boyton and Altmann, Clin. Exp. Immunol. 149:1-8, 2007). С этой целью нуклеотидные зонды могут обычно использоваться для идентификации лиц, несущих генетические маркеры, связанные с конкретным представляющим интерес расстройством. Кроме того, в данной области техники известно множество разнообразных иммунологических способов, которые полезны для идентификации маркеров специфического расстройства. Например, в данной области техники доступны и хорошо известны различные способы иммуноферментного анализа ELISA, в которых используются зонды моноклональных антител для выявления антигенов, связанных со специфическими опухолями. Скрининг может быть осуществлен в соответствии с известными симптомами пациента, возрастными факторами, связанными факторами риска и т.д. Эти способы позволяют врачу регулярно отбирать пациентов, нуждающихся в способах, описанных в данном документе, для лечения. В соответствии с этими способами нацеливание на патологические опухолевые антиген-экспрессирующие клетки может быть реализовано в виде независимой программы лечения или в качестве последующей, вспомогательной или координирующей схемы лечения по сравнению с другими видами лечения.
Определение эффективных дозировок в этом контексте обычно основывается на модельных исследованиях на животных, за которыми следуют клинические испытания на людях, и руководствуется определением эффективных дозировок и протоколов введения, которые значительно уменьшают возникновение или серьезность расстройства у модельных объектов. Эффективные дозы композиций по настоящему изобретению варьируют в зависимости от многих различных факторов, включая способы введения, целевой участок, физиологическое состояние пациента, является ли пациент человеком или животным, вводятся другие лекарственные средства, является ли лечение профилактическим или терапевтическим, а также конкретная активность самой композиции и ее способность вызывать искомый ответ у индивидуума. Обычно пациент является человеком, но при некоторых заболеваниях пациент может быть млекопитающим, не являющимся человеком. Как правило, режимы дозирования корректируются для обеспечения оптимального терапевтического ответа, то есть для оптимизации безопасности и эффективности. Соответственно, терапевтически эффективное количество также представляет собой количество, в котором любые нежелательные побочные эффекты перевешиваются полезными эффектами введения анти-CD152 антитела, как описано в данном документе. Для введения анти-CD152 антитела дозировка может варьировать от около 0,1 мкг до 100 мг/кг или от 1 мкг/кг до около 50 мг/кг, а чаще от 10 мкг до 5 мг/кг массы тела объекта. В более конкретных воплощениях эффективное количество агента составляет от около 1 мкг/кг до около 20 мг/кг, от около 10 мкг/кг до около 10 мг/кг или от около 0,1 мг/кг до около 5 мг/кг. Дозировки в этом диапазоне могут быть достигнуты путем однократного или многократного введения, включая, например, многократное введение в день или ежедневное, еженедельное, двухнедельное или ежемесячное введение. Например, в определенных вариациях режим состоит из первоначального введения, за которым следуют множественные последующие введения с еженедельными или двухнедельными интервалами. Другой режим состоит из начального введения с последующим многократным, последующим введением с месячными или двухмесячными интервалами. В качестве альтернативы, введение может осуществляться нерегулярно, на основании мониторинга клинических симптомов расстройства.
Дозировка фармацевтической композиции может быть изменена лечащим врачом для поддержания искомой концентрации в целевом участке. Например, если выбран внутривенный способ доставки, локальная концентрация агента в кровотоке в ткани-мишени может составлять около от 0,01-50 наномолей композиции на литр, иногда от около 1,0 наномоля на литр до 10, 15 или 25 наномолей на литр в зависимости от состояния объекта и прогнозируемого измеренного ответа. Более высокие или более низкие концентрации могут быть выбраны на основе способа доставки, например, трансэпидермальной доставки по сравнению с доставкой на поверхность слизистой оболочки. Дозировка также должна регулироваться на основе скорости высвобождения введенной композиции, например, назального спрея в сравнении с порошком, пероральных или инъецированных частиц с замедленным высвобождением, трансдермальных композиций и т.д. Для достижения того же уровня концентрации в сыворотке, например, частицы с медленным высвобождением со скоростью высвобождения 5 нМ (в стандартных условиях) должны вводиться в дозе, примерно вдвое превышающей дозу частиц со скоростью высвобождения 10 нМ.
Анти-CD152 терапевтическое средство (например, анти-CD152 антитело) также можно вводить в суточной дозе от около 0,001 до около 10 миллиграммов (мг) на килограмм (mpk) массы тела, предпочтительно, в виде однократной суточной дозы или в разделенных дозах от около двух до шести раз в день. Для введения взрослому пациенту-человеку терапевтически эффективное количество можно вводить в дозах в диапазоне от 0,2 до 800 мг на дозу, включая, без ограничения указанным, 0,2 мг на дозу, 0,5 мг на дозу, 1 мг на дозу, 5 мг на дозу, 10 мг на дозу, 25 мг на дозу, 100 мг на дозу, 200 мг на дозу и 400 мг на дозу, и в ходе лечения можно вводить многократные, обычно последовательные ежедневные дозы. Анти-CD152 терапевтическое средство можно вводить в разное время суток. В одном воплощении оптимальная терапевтическая доза может быть введена вечером. В другом воплощении оптимальная терапевтическая доза может быть введена утром. Общая суточная доза анти-CD152 терапевтического средства, таким образом, в одном воплощении может составлять от около 1 мг до около 2 г, и часто варьирует от около 100 мг до около 1,5 г, и наиболее часто варьирует от около 200 мг до около 1200 мг. В случае типичного взрослого человека массой 70 кг общая суточная доза анти-CD152 терапевтического средства может составлять от около 2 мг до около 1200 мг и часто будет варьировать, как отмечено выше, от около 0,2 мг до около 800 мг.
Режимы дозирования также можно регулировать для обеспечения оптимального искомого ответа (например, терапевтического ответа). Например, может быть введен один болюс, несколько разделенных доз могут вводиться с течением времени, или доза может быть пропорционально уменьшена или увеличена, как указано в зависимости от терапевтической ситуации. Особенно выгодно составлять парентеральные композиции в дозированной форме для простоты введения и однородности дозировки. Используемая в данном документе дозированная лекарственная форма относится к физически дискретным единицам, подходящим в качестве единичных доз для объектов, подлежащих лечению; каждая единица содержит заранее определенное количество активного ингредиента, рассчитанное для получения искомого терапевтического эффекта, в сочетании с необходимым фармацевтическим носителем. В ином случае, антитело может быть введено в виде состава с замедленным высвобождением, в случае которого требуется менее частое введение.
Для введения антитела дозировка может составлять от около 0,0001 до 100 мг/кг и более обычно от 0,01 до 5 мг/кг массы тела хозяина. Например, дозировки могут составлять 0,3 мг/кг массы тела, 1мг/кг массы тела, 3 мг/кг массы тела, 5 мг/кг массы тела или 10 мг/кг массы тела в пределах диапазона 1-10 мг/кг. Примерный режим лечения включает введение раз в неделю, раз каждые две недели, раз каждые три недели, раз каждые четыре недели, раз в месяц, раз каждые три месяца или раз каждые три-шесть месяцев. Предпочтительные режимы дозирования для анти-CD152 антитела по изобретению включают 1мг/кг массы тела или 3 мг/кг массы тела посредством внутривенного введения, с антителом, которое дается одним из следующих режимов дозирования: (i) каждые четыре недели для шести доз, затем каждые три месяца; (ii) каждые три недели; (iii) 3 мг/кг массы тела однократно с последующим 1 мг/кг массы тела каждые три недели. В некоторых способах дозу корректируют до достижения концентрации антител в плазме около 1-1000 мкг/мл, а в некоторых способах около 25-300 мкг/мл.
«Терапевтически эффективная дозировка» анти-CD152 антитела по изобретению предпочтительно приводит к уменьшению выраженности симптомов заболевания, увеличению частоты и продолжительности периодов без симптомов заболевания или предотвращению нарушения или нетрудоспособности вследствие поражения заболеванием. Например, для лечения объектов, имеющих опухоль, «терапевтически эффективная доза» предпочтительно ингибирует рост опухоли, по меньшей мере, на около 20%, более предпочтительно, по меньшей мере, на около 40%, еще более предпочтительно, по меньшей мере, на около 60%, и еще более предпочтительно, по меньшей мере, около на 80% по сравнению с необработанными объектами. Терапевтически эффективное количество терапевтического антитела может уменьшить размер опухоли или иным образом ослабить симптомы у объекта, который обычно является человеком или может быть другим млекопитающим.
Что касается лечения солидных опухолей, протоколы для оценки конечных точек и противоопухолевой активности хорошо известны в данной области. Хотя каждый протокол может определять оценки ответа опухоли по-разному, критерии RECIST (Критерии оценки ответа при солидных опухолях) в настоящее время считаются рекомендованными руководящими принципами для оценки ответа опухоли Национального института онкологии США (см. Therasse et al., J. Natl. Cancer Inst. 92: 205-216, 2000). В соответствии с критериями RECIST опухолевый ответ означает уменьшение или устранение всех измеримых повреждений или метастазов. Заболевание, как правило, считается измеримым, если оно включает поражения, которые можно точно измерить, по меньшей мере, в одном измерении, например,> 20 мм с помощью традиционных методов или> 10 мм с помощью спиральной компьютерной томографии с четко определенными полями с помощью медицинской фотографии или рентгеновской компьютерной осевой томографии (КТ), магнитно-резонансной визуализации (МРТ) или клинического обследования (если поражения поверхностные). Неизмеримое заболевание означает, что заболевание включает поражения <20 мм, измеренные с помощью традиционных методов или <10 мм с помощью спиральной компьютерной томографии, и действительно неизмеримые поражения (слишком маленькие, чтобы их можно было точно измерить). К неизмеримым заболеваниям относятся плевральные выпоты, асциты и заболевание, подтвержденное косвенными данными.
Критерии объективного статуса необходимы для протоколов, чтобы оценить реакцию солидной опухоли. Типичные критерии включают следующее: (1) полный ответ (CR), определяемый как полное исчезновение всех измеримых заболеваний; нет новых поражений; нет связанных с болезнью симптомов; нет доказательств неизмеримых заболеваний; (2) Частичный ответ (PR), определяемый как уменьшение суммы самого длинного диаметра целевых поражений на 30% (3) Прогрессирующее заболевание (PD), определяемое как увеличение суммы наибольшего диаметра целевых поражений на 20% или появление любого нового поражения; (4) Стабильное состояние или отсутствие ответа, определяемое как не отвечающее критериям CR, PR или прогрессирующего заболевания (См. Therasse et al., выше.).
Дополнительные конечные точки, которые принимаются в области онкологии, включают общую выживаемость (OS), выживаемость без заболеваний (DFS), частоту объективных ответов (ORR), время до прогрессирования (TTP) и выживаемость без прогрессирования (PFS) (см. Guidance for Industry: Clinical Trial Endpoints for the Approval of Cancer Drugs and Biologics, April 2005, Center for Drug Evaluation and Research, FDA, Rockville, MD.)
Анти-CD152 антитела могут быть использованы для подавления опосредованных CTLA-4 сигнальных путей, которые отрицательно регулируют иммунные ответы, и, следовательно, для усиления опухолеспецифических иммунных ответов, либо в виде монотерапии, либо в сочетании с моноклональными антителами против PD-L1 или другими противоопухолевыми средствами.
Способы приготовления антител
Антитела, раскрытые в данном документе, могут представлять собой антитело, состоящее только из тяжелых цепей человека (HCAb), полученное от гуманизированных мышей Harbour (пат. США No. 9353179, 9346877 и 8921522 и европейские патенты No. 1776383 и 1864998). Молекулы, продуцируемые мышами HCAb, могут быть растворимыми и могут иметь аффинность, разнообразие и/или физико-химические свойства, сравнимые с традиционными человеческими антителами IgG.
Получение HCAb в HCAb-мышах может способствовать образованию растворимых доменов VH человека, минимальной иммуноглобулиновой единицы распознавания и, таким образом, конструированию новых многофункциональных молекул, содержащих либо несколько доменов VH, либо домен(домены) VH, связанные с другими молекулами, например, биспецифическими, конъюгаты антитело-лекарственное средство или диагностические или терапевтические молекулы, полученные из домена VH.
Анти-CD152 антитела также могут быть получены с использованием антитела, имеющего одну или несколько последовательностей VH анти-CD152 антитела, раскрытого в данном документе, в качестве исходного материала для конструирования модифицированного антитела. Антитело может быть сконструировано путем модификации одного или нескольких остатков в вариабельных областях (то есть VH и/или VL), например, в пределах одной или нескольких областей CDR и/или в одной или нескольких каркасных областях. Дополнительно или альтернативно, антитело может быть сконструировано путем модификации остатков в константной области (областях), например, для изменения эффекторной функции (функций) антитела.
Полинуклеотидные молекулы, содержащие искомую полинуклеотидную последовательность, могут размножаться путем помещения молекулы в вектор. Могут использоваться вирусные и невирусные векторы, включая плазмиды. Выбор плазмиды будет зависеть от типа клетки, в которой желательно размножение, и цели размножения. Определенные векторы полезны для амплификации и получения больших количеств искомой последовательности ДНК. Другие векторы пригодны для экспрессии в клетках в культуре. Еще другие векторы пригодны для переноса и экспрессии в клетках у целого животного или индивидуума. Выбор подходящего вектора находится в пределах квалификации в данной области. Многие такие векторы доступны на рынке. Частичный или полноразмерный полинуклеотид встраивается в вектор обычно посредством присоединения ДНК-лигазы к расщепленному сайту рестриктазы в векторе. В ином случае, искомая нуклеотидная последовательность может быть вставлена путем гомологичной рекомбинации in vivo. Обычно это достигается путем присоединения областей гомологии к вектору на боковых сторонах искомой нуклеотидной последовательности. Области гомологии добавляют путем лигирования олигонуклеотидов или с помощью полимеразной цепной реакции с использованием, например, праймеров, включающих как область гомологии, так и часть искомой нуклеотидной последовательности.
Для экспрессии можно использовать кассету или систему экспрессии. Для экспрессии нуклеиновой кислоты, кодирующей полипептид, раскрытый в данном документе, молекулу нуклеиновой кислоты, кодирующую полипептид, функционально связанную с регуляторными последовательностями, которые контролируют транскрипционную экспрессию в экспрессирующем векторе, вводят в клетку-хозяина. В дополнение к транскрипционным регуляторным последовательностям, таким как промоторы и энхансеры, экспрессирующие векторы могут включать последовательности регуляции трансляции и маркерный ген, который подходит для отбора клеток, несущих экспрессирующий вектор. Генный продукт, кодируемый полинуклеотидом по изобретению, экспрессируется в любой удобной системе экспрессии, включая, например, системы на основе бактерий, дрожжей, насекомых, амфибий и млекопитающих. В экспрессирующем векторе полипептид-кодирующий полинуклеотид связан с регуляторной последовательностью, если это необходимо, для получения искомых свойств экспрессии. Они могут включать промоторы, энхансеры, терминаторы, операторы, репрессоры и индукторы. Промоторы могут регулируемыми (например, промотор из вектора pIND, индуцируемый стероидами (Invitrogen)) или конститутивными (например, промоторы из последовательностей CMV, SV40, фактора элонгации или LTR). Они связаны с искомой нуклеотидной последовательностью, с помощью способов, описанных выше для связывания с векторами. Могут быть использованы любые методы, известные в данной области техники. Соответственно, экспрессирующий вектор обычно обеспечивает область инициации транскрипции и трансляции, которая может быть индуцибельной или конститутивной, где кодирующая область функционально связана под транскрипционным контролем области инициации транскрипции и области терминации транскрипции и трансляции.
Экспрессирующая кассета может быть введена в различные векторы, например, плазмиду, ВАС, YAC, бактериофаг, такой как лямбда, Р1, М13 и т.д., вирусные векторы растений или животных (например, векторы на основе ретровирусов, аденовирусные векторы) и т.п., где векторы обычно характеризуются способностью обеспечивать отбор клеток, содержащих экспрессирующие векторы. Векторы могут обеспечивать внехромосомное поддержание, особенно в виде плазмид или вирусов, или интеграцию в хромосому хозяина. Если требуется внехромосомное поддержание, для репликации плазмиды предоставляется последовательность точки начала репликации, которая может обеспечивать низкое или высокое число копий. Для выбора доступно большое разнообразие маркеров, особенно тех, которые защищают от токсинов, более конкретно от антибиотиков. Конкретный маркер, который выбран, выбирается в соответствии с природой хозяина, где в некоторых случаях может использоваться комплементация ауксотрофных хозяев. Для введения конструкции ДНК можно использовать любой удобный способ, включая, например, конъюгацию, бактериальную трансформацию, осажденную кальцием ДНК, электропорацию, слияние, трансфекцию, заражение вирусными векторами, биолистику и тому подобное. Раскрытие относится к экспрессирующему вектору, включающему сегмент нуклеиновой кислоты, где указанный сегмент нуклеиновой кислоты может содержать нуклеотидную последовательность, представленную в SEQ ID NO: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187.
Соответственно, белки для применения в настоящем раскрытии могут быть получены в генетически сконструированных клетках-хозяевах в соответствии с традиционными методами. Подходящими клетками-хозяевами являются те типы клеток, которые могут быть трансформированы или трансфицированы экзогенной ДНК и выращены в культуре и включают бактерии, грибные клетки и культивируемые клетки высших эукариот (включая культивируемые клетки многоклеточных организмов), в частности культивируемые клетки млекопитающих. Методы обработки клонированных молекул ДНК и введения экзогенной ДНК в различные клетки-хозяева раскрыты в Sambrook and Russell, Molecular Cloning: A Laboratory Manual (3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 2001), и Ausubel et al., Short Protocols in Molecular Biology (4th ed., John Wiley & Sons, 1999).
Для направления рекомбинантного белка в секреторный путь клетки-хозяина, в экспрессирующем векторе предоставляется секреторная сигнальная последовательность (также известная как лидерная последовательность). Секреторная сигнальная последовательность может быть последовательностью нативной формы рекомбинантного белка или может быть получена из другого секретируемого белка или синтезирована de novo. Секреторная сигнальная последовательность функционально связана с кодирующей полипептид ДНК-последовательностью, то есть две последовательности соединены в правильной рамке считывания и расположены так, чтобы направлять вновь синтезированный полипептид в секреторный путь клетки-хозяина. Секреторные сигнальные последовательности обычно расположены 5' относительно последовательности ДНК, кодирующей представляющий интерес полипептид, хотя некоторые сигнальные последовательности могут быть расположены в другом месте представляющей интерес последовательности ДНК (см., например, Welch et al., Пат. США № 5037743; Holland et al., Пат. США № 5143830).
Культивируемые клетки млекопитающих являются подходящими хозяевами для продуцирования рекомбинантных белков для применения в настоящем описании. Способы введения экзогенной ДНК в клетки-хозяева млекопитающих включают трансфекцию, опосредованную фосфатом кальция (Wigler et al., Cell 14: 725, 1978; Corsaro и Pearson, Somatic Cell Genetics 7: 603, 1981: Graham and Van der Eb, Virology 52: 456, 1973), электропорацию (Neumann et al., EMBO J. 1: 841-845, 1982), трансфекцию, опосредованную DEAE-декстраном (Ausubel et al., выше), и трансфекцию, опосредованную липосомами (Hawley-Nelson et al., Focus 15:73, 1993; Ciccarone et al., Focus 15:80, 1993). Получение рекомбинантных полипептидов в культивируемых клетках млекопитающих описано, например, Levinson et al., пат. США No. 4713339; Hagen et al., пат. США № 4784950; Palmiter et al., пат. США № 4579821; и Ringold, пат. США № 4656134. Примеры подходящих клеток-хозяев млекопитающих включают клетки почки африканской зеленой мартышки (Vero; ATCC CRL 1587), клетки почки эмбриона человека (293-HEK; ATCC CRL 1573), клетки почки детеныша хомяка (BHK-21, BHK-570; ATCC CRL 8544)., ATCC CRL 10314), клетки почки собаки (MDCK; ATCC CCL 34), клетки яичника китайского хомяка (CHO-K1; ATCC CCL61; CHO DG44; CHO DXB11 (Hyclone, Logan, UT); см. также, например, Chasin et al., Som. Cell. Molec. Genet. 12: 555, 1986)), клетки гипофиза крысы (GH1; ATCC CCL82), клетки HeLa S3 (ATCC CCL2.2), клетки гепатомы крысы (H-4-II-E; ATCC CRL 1548) SV40-трансформированные клетки почки обезьяны (COS-1; ATCC CRL 1650) и мышиные эмбриональные клетки (NIH-3T3; ATCC CRL 1658). Дополнительные подходящие клеточные линии известны в данной области и доступны в общественных депозиториях, таких как Американская коллекция типовых культур, Манассас, Вирджиния. Могут быть использованы сильные промоторы транскрипции, такие как промоторы из SV-40 или цитомегаловирус. См., например, пат. США № 4956288. Другие подходящие промоторы включают промоторы из генов металлотионеина (пат. США No. 4579821 и 4601978) и основной поздний промотор аденовируса.
Отбор лекарственными средствами обычно используется для отбора культивируемых клеток млекопитающих, в которые была вставлена чужеродная ДНК. Такие клетки обычно называют «трансфектантами». Клетки, которые были культивированы в присутствии селективного агента и способны передавать представляющий интерес ген своему потомству, называются «стабильными трансфектантами». Типичные селектируемые маркеры включают ген, кодирующий устойчивость к антибиотику неомицину, который позволяет проводить селекцию в присутствии лекарственного средства типа неомицина, такого как G-418 или тому подобного; ген gpt для ксантин-гуанин-фосфорибозилтрансферазы, который обеспечивает рост клеток-хозяев в присутствии микофенольной кислоты/ксантина; и маркеры, которые обеспечивают устойчивость к зеоцину, блеомицину, бластоцидину и гигромицину (см., например, Gatignol et al., Mol. Gen. Genet. 207: 342, 1987; Drocourt et al., Nucl. Acids Res. 18: 4009, 1990). Системы отбора могут также использоваться для повышения уровня экспрессии представляющего интерес гена, в процессе, называемом «амплификацией». Амплификацию проводят путем культивирования трансфектантов в присутствии низкого уровня селективного агента и последующего увеличения количества селективного агента для отбора клеток, которые продуцируют высокие уровни продуктов введенных генов. Типичным амплифицируемым селектируемым маркером является дигидрофолатредуктаза, которая придает устойчивость к метотрексату. Также могут быть использованы другие гены устойчивости к лекарственным средствам (например, устойчивость к гигромицину, устойчивость к нескольким лекарственным средствам, пуромицин-ацетилтрансфераза).
Другие высшие эукариотические клетки также могут быть использованы в качестве хозяев, включая клетки насекомых, клетки растений и клетки птиц. Использование Agrobacterium rhizogenes в качестве вектора для экспрессии генов в клетках растений было рассмотрено Sinkar et al., J. Biosci. (Bangalore) 11:47-58, 1987. Трансформация клеток насекомых и продуцирование в них чужеродных полипептидов раскрыта в Guarino et al., US 5162222 и WO 94/06463.
Клетки насекомых могут быть инфицированы рекомбинантным бакуловирусом, обычно полученным из вируса ядерного полиэдроза Autographa californica (AcNPV). См. King and Possee, The Baculovirus Expression System: A Laboratory Guide (Chapman & Hall, London); O’Reilly et al., Baculovirus Expression Vectors: A Laboratory Manual (Oxford University Press., New York 1994); и Baculovirus Expression Protocols. Methods in Molecular Biology (Richardson ed., Humana Press, Totowa, NJ, 1995). Рекомбинантный бакуловирус также может быть получен путем использования системы на основе транспозона, описанной Luckow et al. (J. Virol. 67: 4566-4579, 1993). Эта система, которая использует трансферные векторы, коммерчески доступна в форме набора (набор BAC-TO-BAC; Life Technologies, Гейтерсберг, Мэриленд). Трансферный вектор (например, PFASTBAC1; Life Technologies) содержит транспозон Tn7 для перемещения ДНК, кодирующей представляющий интерес белок, в геном бакуловируса, который содержится в E.coli, в виде большой плазмиды, называемой «бакмидой». См. Hill-Perkins and Possee, J. Gen. Virol. 71: 971-976, 1990; Bonning et al., J. Gen. Virol. 75: 1551-1556, 1994; и Chazenbalk and Rapoport, J. Biol. Chem. 270: 1543-1549, 1995. Кроме того, трансферные векторы могут включать слияние в рамке с ДНК, кодирующей удлинение полипептида или аффинную метку, как описано выше. С использованием методов, известных в данной области, трансферный вектор, содержащий последовательность ДНК, кодирующую белок, трансформируют в клетки-хозяева E.coli, и клетки подвергают скринингу на наличие бактерий, которые содержат прерванный ген lacZ, указывающий на рекомбинантный бакуловирус. Бакмидную ДНК, содержащую рекомбинантный бакуловирусный геном, выделяют, используя обычные методы, и используют для трансфекции клеток Spodoptera frugiperda, таких как клетки Sf9. Затем получают рекомбинантный вирус, который экспрессирует представляющий интерес белок. Рекомбинантные вирусные стоки получают способами, обычно используемыми в данной области.
Для продуцирования белка рекомбинантный вирус используется для инфицирования клеток-хозяев, как правило, клеточной линии, происходящей от совки кукурузной листовой, Spodoptera frugiperda (например, клетки Sf9 или Sf21) или Trichoplusia ni (например, клетки HIGH FIVE; Invitrogen, Карлсбад, Калифорния). См. в общем плане Glick and Pasternak, Molecular Biotechnology, Principles & Applications of Recombinant DNA (ASM Press, Washington, D.C., 1994). См. также пат. США № 5300435. Бессывороточные среды используются для роста и поддержания клеток. Подходящие составы сред известны в данной области и могут быть получены от коммерческих поставщиков. Клетки выращивают до плотности инокуляции приблизительно от 2-5 × 105 клеток до 1-2 × 106 клеток, и в этот момент добавляют рекомбинантный вирусный сток при множественности инфекции (MOI) от 0,1 до 10, более типично около 3. Используемые процедуры обычно описаны в доступных лабораторных руководствах (см., например, King and Possee, выше; O’Reilly et al., Выше; Richardson, выше).
Грибные клетки, включая дрожжевые клетки, также могут быть использованы в настоящем описании. В этом отношении виды дрожжей включают, например, Saccharomyces cerevisiae, Pichia pastoris и Pichia methanolica. Способы трансформации клеток S. cerevisiae экзогенной ДНК и получения из них рекомбинантных полипептидов описаны, например, в Kawasaki, пат. США № 4599311; Kawasaki et al., пат. США № 4931373; Brake, пат. США № 4870008; Welch et al., пат. США № 5,037,743; и Murray et al., пат. США № 4845075. Трансформированные клетки отбирают по фенотипу, определяемому селектируемым маркером, обычно лекарственной устойчивостью или способностью расти в отсутствие определенного питательного вещества (например, лейцина). Примерной векторной системой для применения в Saccharomyces cerevisiae является векторная система POT1, раскрытая Kawasaki et al. (пат. США № 4931373), которая позволяет выбирать трансформированные клетки путем роста в среде, содержащей глюкозу. Подходящие промоторы и терминаторы для применения в дрожжах включают промоторы из генов гликолитического фермента (см., например, Kawasaki, пат. США № 4599311; Kingsman et al., пат. США № 4615974; и Bitter, пат. США № 4979092) и генов алкогольдегидрогеназы. См. также пат. США No. 4990446; 5063154; 5139936; и 4661454. В данной области известны системы трансформации для других дрожжей, включая Hansenula polymorpha, Schizosaccharomyces pombe, Kluyveromyces lactis, Kluyveromyces fragilis, Ustilago maydis, Pichia pastoris, Pichia methanolica, Pichia guillermondii и Candida maltosa. См., например, Gleeson et al., J. Gen. Microbiol. 132:3459-3465, 1986; Cregg, пат. США № 4882279; и Raymond et al., Yeast 14: 11-23, 1998. Клетки Aspergillus могут быть использованы в соответствии со способами McKnight et al., пат. США № 4935349. Способы трансформации Acremonium chrysogenum раскрыты в Sumino et al., пат. США № 5162228. Способы трансформации Neurospora раскрыты Lambowitz, пат. США № 4486533. Продуцирование рекомбинантных белков в Pichia methanolica описано в пат. США No. 5716808; 5736383; 5854039; и 5888768.
Прокариотические клетки-хозяева, включая штаммы бактерий Escherichia coli, Bacillus и других родов, также являются полезными клетками-хозяевами в рамках настоящего изобретения. Способы трансформации этих хозяев и экспрессии клонированных в них чужеродных последовательностей ДНК хорошо известны в данной области (см., например, Sambrook and Russell, выше). При экспрессии рекомбинантного белка в бактериях, таких как E.coli, белок может удерживаться в цитоплазме, обычно в виде нерастворимых гранул, или может направляться в периплазматическое пространство с помощью последовательности бактериальной секреции. В первом случае клетки лизируют, а гранулы извлекают и денатурируют, используя, например, изотиоцианат гуанидина или мочевину. Затем денатурированный белок может быть повторно свернут и димеризован путем разбавления денатуранта, например, диализом против раствора мочевины и комбинации восстановленного и окисленного глутатиона, с последующим диализом против буферного солевого раствора. В ином случае, белок может быть извлечен из цитоплазмы в растворимой форме и выделен без использования денатурирующих агентов. Белок извлекается из клетки в виде водного экстракта, например, в фосфатно-солевом буферном растворе. Для захвата представляющего интерес белка экстракт наносят непосредственно на хроматографическую среду, такую как иммобилизованное антитело или гепарин-сефарозная колонка. Секретируемые белки могут быть извлечены из периплазматического пространства в растворимой и функциональной форме путем разрушения клеток (например, путем обработки ультразвуком или осмотическим шоком), чтобы высвободить содержимое периплазматического пространства и восстановления белка, тем самым устраняя необходимость в денатурации и рефолдинге. Антитела, включая одноцепочечные антитела, можно продуцировать в бактериальных клетках-хозяевах в соответствии с известными способами. См., например, Bird et al., Science 242: 423-426, 1988; Huston et al., Proc. Natl. Acad. Sci. США 85: 5879-5883, 1988; и Pantoliano et al., Biochem. 30:10117-10125, 1991.
Трансформированные или трансфицированные клетки-хозяева культивируют в соответствии с традиционными процедурами в культуральной среде, содержащей питательные вещества и другие компоненты, необходимые для роста выбранных клеток-хозяев. Различные подходящие среды, включая определенные среды и сложные среды, известны в данной области и обычно включают источник углерода, источник азота, незаменимые аминокислоты, витамины и минералы. Среды по необходимости могут также содержать такие компоненты, как факторы роста или сыворотка. Среда для роста, как правило, отбирает клетки, содержащие экзогенно добавленную ДНК, например, путем отбора на лекарственном средстве или с помощью дефицита в существенном питательном веществе, которое дополняется селектируемым маркером, который переносится на экспрессирующем векторе или совместно трансфицируется в клетку-хозяина.
Анти-CD152 антитела могут быть очищены обычными способами очистки белка, обычно с помощью комбинации хроматографических методов. См. в общем плане Affinity Chromatography: Principles & Methods (Pharmacia LKB Biotechnology, Uppsala, Sweden, 1988); Scopes, Protein Purification: Principles and Practice (Springer-Verlag, New York 1994). Белки, содержащие Fc-область иммуноглобулина, могут быть очищены аффинной хроматографией на иммобилизованном протеине А или протеине G. Дополнительные стадии очистки, такие как гель-фильтрация, могут быть использованы для получения искомого уровня чистоты или для обессоливания, замены буфера и т.п.
Следующие примеры изобретения служат для дополнительной иллюстрации сущности изобретения. Следует понимать, что следующие примеры не ограничивают изобретение и что объем изобретения должен определяться прилагаемой формулой изобретения.
Примеры
Пример 1 Получение анти-CTLA-4 антител
Человеческий белок CTLA-4-ECD (Acro Bio) использовали в качестве иммуногена для получения анти-CTLA-4 антител. Использование технологии трансгенных по человеческому иммуноглобулину мышей для разработки и получения человеческих антител было впервые описано Abgenix (ксеномышь и Medarex (HuMab «мышь»); Lonberg et al., 1994, Nature, 368: 856-859; Lonberg and Huszar, 1995, Internal Rev. Immunol., 13: 65-93; Harding and Lonberg, 1995, Ann. N.Y. Acad. Sci., 764: 536-546).
Мышей HCAb иммунизировали белком CTLA-4-ECD человека в дозе 20 мг/мышь каждые две недели три раза, и шесть из них иммунизировали еще пять раз по 44 мг/мышь. За исключением первой инъекции, где в качестве адъюванта использовался Stimune (Prionics), все бустерные введения проводили с адъювантом Ribi (адъювант Sigma S6322-1VL). После иммунизации суспензии отдельных клеток выделяли из костного мозга, селезенки и лимфатических узлов мыши. Затем плазматические клетки мыши выделяли с использованием набора для выделения плазматических клеток (Miltenyi, Cat.No.130-092-530). Вкратце, была получена суммарная РНК из плазматических клеток мыши и обратно транскрибирована в кДНК в большом пуле. VH-области человека амплифицировали из кДНК с использованием праймеров следующим образом.
Прямой праймер:
lib-3-23/53-S: 5’-GTGTCCAGTGTGAGGTGCAGCTG (SEQ ID NO: 193) и
lib-3-11-S: 5’-GTGTCCAGTGTCAGGTGCAGCTG (SEQ ID NO: 194)
Обратный праймер:
mG1hrv: 5’-ggcttacaaccacaatccctgggc (SEQ ID NO: 195)
Все амплифицированные фрагменты ПЦР, содержащие домен VH, клонировали в экспрессирующий вектор млекопитающих pTT5. Полученные плазмиды трансформировали в бактерии (DH5α) путем электропорации. Плазмиды дублировали и очищали, а затем трансфицировали в клетки НЕК 293 для получения антител.
Клетки 293 инкубировали в среде 293 FreeStyle (12338018, Thermo) в течение 10 дней, и надосадочные жидкости подвергали скринингу с помощью анализа ELISA. Рекомбинантные человеческие белки CTLA4-his (Acro Bio) разводили в PBS с концентрацией 2 мкг/мл, и 100 мкл разведенных белков CTLA-4-his добавляли на лунку к микропланшетам ELISA, которые инкубировали в течение ночи при 4°С для покрытия планшетов рекомбинантными белками. Затем планшеты блокировали блокирующим раствором для ELISA (включающим 2% BSA, 0,05% (об./об.) Tween-20, буфер PBS pH 7,4, масс./об.) при 37°C в течение двух часов и затем инкубировали с надосадочными жидкостями в течение 1 часа при 37°С. Планшеты промывали и инкубировали с козьим антителом против человеческого IgG (H + L), конъюгированным с пероксидазой хрена (HRP) (A18805, Life technologies), при 37°C в течение одного часа. Добавляли 100 мкл тетраметилбензидина (TMB) и планшеты инкубировали при комнатной температуре в течение 15 минут. Для прекращения реакции добавляли 50 мкл 1 Н HCl. Тридцать пять положительных клонов со значительным окрашиванием отбирали для дальнейших испытаний.
35 клонов секвенировали, и 9 клонов из 35 отобрали с уникальными последовательностями CDR3. Последовательности нуклеиновых кислот и аминокислот этих 9 анти-CTLA-4 антител суммированы в таблице 1. Антитела HCAb содержали только две тяжелые цепи.
Таблица 1. Последовательности нуклеиновых кислот и аминокислотные последовательности антител против CTLA-4 человека
тяжелая цепь
нк: нуклеиновая кислота; ак: аминокислота
Пример 2. Получение и очистка анти-CTLA-4 антител
Стадия 1. Получение клеток HEK 293F со сверхэкспрессией hCTLA-4
Нуклеотидная последовательность, кодирующая CTLA-4 человека (SEQ ID NO: 196, кодирующая аминокислотную последовательность SEQ ID NO: 197), была субклонирована в вектор pcDNA3.1 (Clontech) для получения плазмиды. Клетки HEK293 и CHO-K1 (Invitrogen) транзиторно трансфицировали плазмидами с использованием PEI, и трансформанты культивировали в культуральной среде DMEM, содержащей 0,5 г/мл пенициллина/стрептомицина и 10% (масс./масс.) фетальной бычьей сыворотки (FBS) в течение 2 недель. Осуществляли лимитирующее разведение в 96-луночном культуральном планшете, и планшет инкубировали при 37°С с 5% (об./об.) СО2 в течение приблизительно 2 недель. Моноклоны наращивали в 6-луночных планшетах, и нарощенные клоны подвергали скринингу с помощью проточной цитометрии с использованием коммерчески доступных анти-hCTLA-4 антител (R & D Systems). Клоны, демонстрирующие более высокие скорости роста и более высокую интенсивность флуоресценции, как измерено с помощью FACS, дополнительно наращивали и подвергали криоконсервации в жидком азоте.
Стадия 2. Определение активности связывания анти-CTLA-4 антител в клеточной среде HEK 293F с помощью ELISA и анализа связывания FACS на клеточной основе
Рекомбинантные человеческие белки CTLA4-his (Acro Bio) разводили в PBS с концентрацией 2 мкг/мл, и 100 мкл разведенных белков CTLA-4-his добавляли на лунку к микропланшетам для ELISA, которые инкубировали в течение ночи при 4°С для покрытия планшетов рекомбинантными белками. Затем планшеты блокировали блокирующим раствором для ELISA (содержащим 2% BSA, 0,05% (об./об.) Tween-20, буфер PBS pH 7,4, об./об.) при 37°C в течение двух часов и затем инкубировали с клеточной средой 293F, содержащей анти-CTLA-4 антитела (см. Пример 1) в течение 1 часа при 37°С. Планшеты промывали и инкубировали с козьим антителом против человеческого IgG (H + L), конъюгированным с пероксидазой хрена (HRP) (A18805, Life technologies), при 37°C в течение одного часа. Добавляли 100 мкл тетраметилбензидина (TMB) и планшеты инкубировали при комнатной температуре в течение 15 минут. 50 мкл 1 Н HCl добавляли для прекращения реакции, и OD 450 нм определяли с помощью ридера для ELISA-планшетов.
При этом клетки 293-hCTLA-4, полученные на стадии 1, культивировали и использовали для измерения активности связывания антител. Клетки обрабатывали бесферментным раствором для диссоциации клеток (раствор Versene, Invitrogen) и затем собирали. BSA добавляли к суспензии клеток до конечной концентрации 1%, и клетки блокировали в течение 30 минут на льду и затем дважды промывали HBSS. Клетки собирали после центрифугирования и ресуспендировали в буфере FACS (HBSS + 1% BSA, об./об.) при 2 × 106 клеток/мл. Затем в каждую лунку 96-луночного планшета добавляли 100 мкл клеточной суспензии. 100 мкл клеточной среды 293F, содержащей анти- CTLA-4 антитела (см. Пример 1), добавляли в каждую лунку 96-луночного планшета и инкубировали в течение 1 часа на льду. Клетки дважды промывали буфером FACS, и добавляли 100 мкл Alexa 488-меченного антитела против белка человека (H + L) (Invitrogen) в 96-луночный планшет и инкубировали в течение 1 часа на льду. Образцы трижды промывали буфером FACS, и в каждую лунку добавляли 100 мкл буфера для фиксации (4% параформальдегидного об./об.) и инкубировали в течение 10 минут. Затем клетки дважды промывали буфером FACS и ресуспендировали в 100 мкл буфера FACS. Среднюю интенсивность флуоресценции (MFI) определяли с использованием FACS Calibur (BD).
Стадия 3. Продуцирование и очистка ведущих антител-кандидатов
Концентрация антител из клеток HEK293 составляла около 1-10 мкг/мл и широко варьировала. Кроме того, FBS и компоненты культуральной среды могут мешать анализу. Следовательно, необходимо было осуществить препаративное получение (1-5 мг) и очистку антитела.
Конструкции, содержащие нуклеотидные последовательности, кодирующие анти- CTLA-4 антитела (как указано в таблице 1), вводили в клетки 293. Надосадочную жидкость, содержащую антитела-мишени, собирали через 6-7 дней после трансфекции центрифугированием и фильтрацией. Моноклональные антитела очищали, пропуская их через 2 мл колонки с Протеином G (GE Healthcare). Колонки с Протеином G сначала уравновешивали буфером PBS (рН 7,2), а затем надосадочные жидкости гибридомной культуры наносили на уравновешенные колонки с Протеином G с постоянной скоростью потока 3 мл/мин. Затем каждую колонку промывали PBS-буфером, объем которого в 3 раза превышал объем колонки. Анти- CTLA-4 антитела затем элюировали элюирующим буфером (0,1 М ацетатный буфер, pH 2,5), и УФ-поглощение элюатов контролировали с использованием УФ-детектора (пик поглощения УФ-излучения A280). 10% 1 М трис-HCL-буфера добавляли к элюатам для нейтрализации рН, и образцы подвергали стерильной фильтрации, пропуская их через фильтры 0,22 мкм. Были получены очищенные в стерильных условиях анти- CTLA-4 антитела.
Концентрации очищенных анти-CTLA-4 антител определяли по УФ-поглощению (A280/1.4) и измеряли чистоту и уровень эндотоксина (набор Lonza). Очищенные анти-CTLA-4 антитела имели концентрацию эндотоксина менее чем 1 EU/мг.
Пример 3. Характеризация ведущих антител-кандидатов
Клетки 293 стабильно трансфицировали плазмидами pTT5, содержащими последовательность нуклеиновой кислоты, кодирующую CTLA-4 человека (SEQ ID NO: 196), для получения клеток 293F, стабильно экспрессирующих человеческие CTLA-4 (в настоящем документе называемые клетками 293-hCTLA-4). Дополнительные клетки 293 были стабильно трансфицированы плазмидами pIRES, включающими последовательность нуклеиновой кислоты, кодирующую полноразмерный CTLA-4 яванского макака (SEQ ID NO: 198), для генерации клеток 293, стабильно экспрессирующих CTLA-4 яванского макака (в настоящем документе называемых клетками 293-cynoCTLA-4). Клетки 293-hCTLA-4 и 293-cynoCTLA-4 культивировали и наращивали в колбах для культивирования T-75 до конфлуентности 90%. Культуральную среду аспирировали и клетки дважды промывали HBSS (сбалансированный солевой раствор Хэнкса, Invitrogen). Клетки обрабатывали бесферментным раствором для диссоциации клеток (раствор Versene, Invitrogen) и собирали. Затем клетки дважды промывали HBSS, определяли количество клеток и ресуспендировали клетки с использованием HBSS при 2 × 106 клеток/мл. BSA добавляли к суспензии клеток до конечной концентрации 1%, и клетки блокировали в течение 30 минут на льду и затем дважды промывали HBSS. Клетки собирали после центрифугирования и ресуспендировали в буфере FACS (HBSS + 1% BSA, об./об.) при 2 × 106 клеток/мл. Затем в каждую лунку 96-луночного планшета добавляли 100 мкл клеточной суспензии. 100 мкл очищенных анти-CTLA-4 антител из Примера 2 или контрольных антител добавляли в каждую лунку 96-луночного планшета и инкубировали в течение 1 часа на льду, где тяжелая цепь и легкая цепь аналога ипилимумаба имели аминокислотные последовательности SEQ ID NO: 199 и SEQ ID NO: 200, соответственно. Клетки дважды промывали буфером FACS, и добавляли 100 мкл Alexa 488-меченного антитела против белка человека (H + L) (Invitrogen) в 96-луночный планшет и инкубировали в течение 1 часа на льду. Образцы трижды промывали буфером FACS и в каждую лунку добавляли 100 мкл буфера для фиксации (содержащего 4% параформальдегида, об./об.) и инкубировали в течение 10 минут. Затем клетки дважды промывали буфером FACS и ресуспендировали в 100 мкл буфера FACS.
Среднюю интенсивность флуоресценции (MFI) определяли с использованием FACS Calibur (BD), и результаты представлены на фиг. 1 и фиг. 2. Антитела из Примера 2 обладали активностью связывания с CTLA человека или яванского макака на клетках 293F, сравнимой с аналогом ипилимумабом.
Пример 4. Определение способности антител против CTLA-4 блокировать связывание CTLA-4 с B7.1.
Анализ связывания рецептор-лиганд на основе клеток проводили для определения способности антител против CTLA-4 блокировать связывание CTLA-4 с его лигандами B7.1. Рекомбинантный белок B7.1ECD-Fc (B71-H5259, Acro Bio) биотинилировали с использованием EZ-LINK NHS-PEG12-Biotin (Thermo Scientific # 21312) в соответствии с инструкцией производителя. Биотинилированный белок B7.1ECD-Fc концентрировали и удаляли свободную метку с использованием центрифужного фильтра Amicon (отсечка 10 кДа). Внеклеточный домен B7.1 соответствовал аминокислотам Val35-Asn242 белка базы данных Uniprot P33681.
Клетки 293-hCTLA-4, полученные в Примере 3, культивировали и наращивали в колбах для культивирования Т-75 до конфлуентности 60-80%. Культуральную среду аспирировали и клетки дважды промывали PBS. Клетки обрабатывали бесферментным раствором для диссоциации клеток (раствор Versene, Invitrogen) и собирали. Раствор диссоциации нейтрализовали добавлением 8 мл культуральной среды и определяли количество клеток. Клетки центрифугировали при 300 g в течение 5 минут и ресуспендировали в блокирующем буфере (содержащем 2% BSA, буфер PBS pH 7,4, масс./об.) при 1 × 106 клеток/мл. Клетки блокировали в течение 15 минут при 37°С. Тем временем лунки 96-луночных круглодонных планшетов блокировали 200 мкл блокирующего буфера в течение 1 часа при 37°С. Блокирующий буфер отбрасывали, и 200 мкл клеток распределяли в каждую лунку 96-луночного планшета (2 × 105 клеток/лунку). Планшеты центрифугировали при 500 g в течение 5 минут и надосадочные жидкости отбрасывали. Клетки ресуспендировали в 100 мкл анти-CTLA-4 антител, приготовленных в блокирующем буфере с различными концентрациями. 100 мкл биотинилированного ECD-Fc B7.1 (60 мкг/мл в блокирующем буфере) добавляли в каждую лунку 96-луночного планшета и осторожно перемешивали при встряхивании. Планшеты инкубировали при 4°С в течение 90 минут и дважды промывали 200 мкл блокирующего буфера. Блокирующий буфер отбрасывали и клетки ресуспендировали в 100 мкл раствора стрептавидина-Alexa 488 (Invitrogen, 1: 500 в блокирующем буфере) и инкубировали при 4°С в течение 1 часа. Планшеты трижды промывали блокирующим буфером и добавляли 200 мкл блокирующего буфера.
Среднюю интенсивность флуоресценции (MFI) определяли с использованием FACS Calibur (BD). Результаты, как показано на фиг. 3, продемонстрировали, что анти- CTLA-4 антитела могут блокировать связывание экспрессируемого клеткой CTLA-4 с его лигандом B7.1 на уровне, сопоставимом с аналогом ипилимумабом.
Пример 5. Антитела против CLTA-4 HCAb, способствующие высвобождению IL-20
стадия 1. Тест стимуляции РВМС
100 мкл PBMC (содержащих 1 × 105 клеток) добавляли в лунки 96-луночного планшета, и затем добавляли 50 мкл каждого тестируемого антитела, имеющего различные концентрации, в 96-луночный планшет и инкубировали в течение 15 минут при комнатной температуре. 50 мкл 100 нг/мл SEB добавляли в каждую лунку и культивировали при 37°C, 5% CO2 в течение 72 часов. Надосадочные жидкости собирали и хранили при -20°С до анализа.
Стадия 2. Детекция секреции интерлейкина IL-2 методом ELISA
Количественное определение уровня IL-2 в культуральной надосадочной жидкости проводили с использованием набора ELISA для определения IL-2 человека Quantikine (DY202, R & D Systems), следуя инструкциям, предоставленным производителем. Вкратце, поликлональные антитела против IL-2 наносили на микропланшеты для ELISA, и 100 мкл культуральной надосадочной жидкости, а также стандарта добавляли в каждую лунку и инкубировали при комнатной температуре в течение 2 часов. Планшеты 4 раза промывали промывочным буфером с последующим добавлением конъюгированных с HRP антител против IL-2 человека и инкубировали при комнатной температуре в течение 2 часов. После промывки добавляли хромогенный субстрат (5120-0077, SeraCare) и инкубировали в темноте при комнатной температуре в течение 30 минут, и реакцию останавливали добавлением стоп-раствора (E661006-0200, BBI Life Sciences).
Поглощение при 450 нм определяли с использованием ридера для ELISA-планшетов, и результаты, как показано на фиг. 4A и 4B, продемонстрировали, что некоторые из анти-CTLA-4 антител могут увеличивать секрецию IL-2 при низких концентрациях по сравнению с человеческим IgG (AB170090 (Crown Bio) или аналогом ипилимумабом.
Пример 6. Анти-CTLA-4 антитела HCAb, связанные с CTLA-4 человека, но не с CTLA-4 мыши в анализе ELISA
Рекомбинантный человеческий или мышиный белок CTLA4-his (CT4-H5229 для белка человека, CT4-M52H5 для белка мыши, Acro Bio) разводили в PBS до концентрации 2 мкг/мл и 100 мкл разведенного белка CTLA-4-his на лунку добавляли к микропланшетам для ELISA, которые инкубировали в течение ночи при 4°С для покрытия планшетов рекомбинантными белками. Затем планшеты блокировали раствором для ELISA (включающим 2% BSA, 0,05% (об./об.) Tween-20, буфер PBS pH 7,4, об./об.) при 37°C в течение двух часов и затем инкубировали с анти-CTLA-4 антителами при различных концентрациях в течение 1 часа при 37°С. Планшеты промывали и инкубировали с козьим антителом против человеческого IgG (H + L), конъюгированным с пероксидазой хрена (HRP) (A18805, Life technologies), при 37°C в течение одного часа. Добавляли 100 мкл тетраметилбензидина (TMB) и планшеты инкубировали при комнатной температуре в течение 15 минут. 50 мкл 1 Н HCl добавляли для прекращения реакции, и OD 450 нм определяли с помощью ридера для ELISA-планшетов.
Данные показаны на фиг. 5A и фиг. 5B, предполагая, что все клоны связываются с CTLA-4 человека, но почти не связываются с CTLA-4 мыши.
Пример 7. Мутанты анти-CTLA-4 антител, способствующие высвобождению IL-20
Был проведен анализ стимуляции Т-клеток для некоторых антител, упомянутых выше, и их мутантов для изучения влияния этих антител на стимуляцию Т-клеток путем блокирования связывания CTLA-4 с его лигандами B7.1 и B7.2.
Мутанты антител получали путем применения мутаций S239D и I332E в константном домене Fc клона 5 HCAb, клона 11, клона 22, клона 25 и клона 30 с помощью ПЦР. Мутированные антитела экспрессировали в клетках НЕК293 и очищали, как описано в Примере 2. Нуклеотидные и аминокислотные последовательности мутантов антител, определенных с использованием стандартных методов молекулярной биологии, представлены в таблице 2.
Таблица 2. Нуклеотидные и аминокислотные последовательности мутантов анти-CTLA-4 антител
тяжелая цепь
нк: нуклеиновая кислота; ак: аминокислота
Тест стимуляции PBMC и обнаружение секреции интерлейкина IL-2 с помощью ELISA выполняли, как в Примере 5.
Результаты на фиг. 6A и 6B продемонстрировали, что мутанты антител против CTLA-4 усиливали секрецию IL-2 по сравнению с изотипическим контрольным IgG1 человека и их родительскими клонами.
Пример 8. Варианты анти-CTLA-4 антител с удаленным PTM
Аминокислотные последовательности 7 клонов HCAb (CL22, CL25, CL5, CL3, CL11, CL30, CL24) выравнивали с геном IGHV3-53*01, и они представлены в таблице 3. Различия в гене зародышевой линии и участках PTM были выделены. Последовательности были пронумерованы согласно схеме нумерации Chothia.
Ген зародышевой линии IGHV3-53 изначально не имеет мотива N-гликозилирования, и эти мотивы N-гликозилирования, присутствующие в 7 клонах HCAb, были образованы соматическими мутациями. Следовательно, один из подходов к удалению этого мотива состоял в том, чтобы заменить его соответствующими аналогичными остатками в зародышевой линии, например, заменить мотив NVS в CDR1 CL11 на TVS зародышевой линии. Альтернативный подход был также изучен путем объединения удаления PTM с «зародышевой линией» на основе концепции CDR-трансплантации, используемой для гуманизации. Во втором подходе CDR каждого HCAb были трансплантированы к каркасам IGHV3-53 зародышевой линии, и ключевые остатки каркаса из родительского HCAb также были сохранены. Для некоторых антител были восстановлены аминокислотные мутации, расположенные в специальных остатках, которые могут влиять на активность связывания между CTLA-4 и антителами. Последовательности нуклеиновых кислот и аминокислот вариантов человеческих анти-CTLA-4 антител HCAb с удалением PTM перечислены в таблице 4.
Таблица 3. Различия аминокислотной последовательности HCAb по сравнению с последовательностью зародышевой линии
Таблица 4. Последовательности нуклеиновых кислот и аминокислотные последовательности вариантов антител против CTLA-4 с удалением PTM
тяжелая цепь
нк: нуклеиновая кислота; ак: аминокислота
Пример 9. Связывающая активность на основе клеток вариантов анти-CTLA-4 антител с удалением PTM
Анализ связывания на основе клеток проводили для определения способности связывания вариантов антител против CTLA-4 с удалением PTM в отношении человеческого CTLA-4. Процедура анализа была аналогична описанной в Примере 3. Вкратце, клетки 293F-hCTLA-4 собирали с использованием бесферментного раствора для диссоциации клеток (раствор Versene, Invitrogen), а затем нейтрализовали культуральной средой и определяли количество клеток. Клетки центрифугировали и блокировали в блокирующем буфере (содержащем 2% BSA, буфер PBS pH 7,4, масс/объем) при 1 × 106 клеток/мл в течение 15 минут при 37°С. 200 мкл клеток распределяли в каждую лунку 96-луночного планшета (2 × 105 клеток/на лунку). Планшеты центрифугировали и надосадочные жидкости отбрасывали. Клетки ресуспендировали в 100 мкл анти-CTLA-4 антител, приготовленных в блокирующем буфере. Планшеты инкубировали при 4°С в течение 90 минут и дважды промывали. После того как блокирующий буфер удаляли, клетки ресуспендировали в 100 мкл меченного Alexa 488 антитела против белка человека (H + L) (1: 500, Invitrogen) и инкубировали при 4°C в течение 1 часа. Клетки промывали и ресуспендировали в 200 мкл блокирующего буфера. Среднюю интенсивность флуоресценции (MFI) определяли с использованием FACS Calibur (BD).
Результаты, как показано на фиг. 7, продемонстрировали, что варианты анти-CTLA-4 антител с удалением PTM связывались с человеческим CTLA-4, экспрессируемым клетками.
Пример 10. Блокирующая активность вариантов анти-CTLA-4 антител с удалением PTM при взаимодействии CTLA-4-B7.1 и B7.2
Анализ связывания лиганда с рецептора на основе клеток проводили, как описано в Примере 4, для определения способности вариантов анти-CTLA-4 антител с удалением PTM блокировать связывание CTLA-4 с его лигандами B7.1.
Результаты, как показано на фиг. 8, продемонстрировали, что варианты анти-CTLA-4 антител с удалением PTM блокировали связывание экспрессируемого клетками CTLA-4 с его лигандом B7.1 и B7.2 (данные не показаны) на уровне, сопоставимым с аналогом ипилимумабом.
Пример 11. Варианты анти-CTLA антител 4 с удалением PTM, способствующие высвобождению IL-20
Стимуляцию РВМС и количественную оценку уровня IL-20 проводили, как описано в Примере 5.
Результаты, как показано на фиг. 9, продемонстрировали, что варианты анти-CTLA-4 антител с удалением PTM все еще могут стимулировать секрецию IL-2. Анти-CTLA-4 антитела с мутациями S239D и I332E и удалением PTM индуцировали повышенную секрецию IL-2, по сравнению с антителами с мутациями S239D и I332E, но без удаления PTM.
Пример 12. Константа аффинности связывания и диссоциации вариантов анти-CTLA-4 антител с удалением PTM
Константы диссоциации определяли с помощью Biacore T200 (GE Healthcare) в соответствии со спецификациями прибора, предоставленными производителем. Вкратце, разведенные 1 мкг/мл анти-CTLA-4 антитела в 10 мМ NaOAc (pH 5, Sigma) иммобилизовали на проточной ячейке сенсорного чипа CM5 Series S. Оставшиеся активные эфирные группы блокировали 1 М этаноламином (рН 8,5). С HBS-EP + в качестве рабочего буфера, рекомбинантные белки CTLA-4-his человека (CT4-H5229, AcroBio) и CTLA-4-his яванского макака (CT4-C5227, AcroBio) с пятью последовательно разведенными концентрациями инъецировали в проточные клетки при 30 мкл/мин со временем ассоциации 180 с. Буферный поток поддерживали для диссоциации в течение 600 с. Значение KD для каждого взаимодействия между антителом и антигеном оценивали, используя оценочное программное обеспечение Biacore T200 1.0 и подходящую модель связывания 1:1.
Результаты показаны на фиг. 10 и в таблице 5. Аффинность связывания двух клонов была аналогична таковой у аналога ипилимумаба.
Таблица 5. Кинетика и аффинность связывания человеческих антител против CTLA-4 с белком CTLA-4ECD-his человека и белком cynoCTLA-4ECD-his яванского макака, как определено Biacore T200
Пример 13. Анализ функции ADCC in vitro
Чтобы подтвердить предполагаемую NK-зависимую цитотоксическую активность человеческих антител против CTLA-4, был проведен анализ антителозависимой клеточно-опосредованной цитотоксичности (ADCC) как на CTLA-4-экспрессирующих клетках CHO-K1, так и на CTLA-4-экспрессирующих in vitro стимулированных клетках Treg.
Клетки CHO-K1, экспрессирующие CTLA-4, как описано в Примере 2, стадия 1, доводили до концентрации 2 × 105 клеток/мл в среде ADCC (содержащей RPMI 1640 без фенолового красного, 10% FBS и 1% пенициллин/стрептомицин). 50 мкл клеточных суспензий (1 × 104 жизнеспособных клеток) добавляли в каждую лунку 96-луночного планшета с v-образным дном. Тестируемые антитела последовательно разводили в среде ADCC (без фенолового красного) и 50 мкл каждого полученного раствора добавляли в лунки в трех экземплярах. Конечные концентрации антител составили: 0,087 мкМ, 0,44 мкМ, 2,2 мкМ, 10,9 мкМ, 54 мкМ, 272 мкМ, 1,36 нМ и 6,8 нМ. Планшет инкубировали при 37°С в течение 30 минут. Клетки NK92, стабильно трансфицированные FcγR158V, доводили средой ADCC (без фенолового красного) так, чтобы путем добавления 100 мкл клеток NK92, стабильно трансфицированных FcγR158V, к клеткам-мишеням соотношение эффекторных клеток и клеток-мишеней составляло 5:1. Затем планшет инкубировали при 37°С в течение 6 часов. После 6 часов инкубации планшет центрифугировали и 50 мкл каждого супернатанта переносили в новый планшет. Надосадочную жидкость инкубировали с 50 мкл буфера для обнаружения LDH при комнатной температуре в течение 30 минут и измеряли оптическую плотность при 490 нм. Для максимального контроля клеточного лизиса добавляли 50 мкл CTLA-4-экспрессирующих клеток CHO-K1, 50 мкл среды ADCC и 100 мкл 1% буфера тритон-X100 для обнаружения LDH. Для контроля минимального клеточного лизиса добавляли 50 мкл CTLA-4-экспрессирующих клеток CHO-K1 и 150 мкл среды ADCC для обнаружения LDH. Измеряли поглощение при 492/650 нм. Процент лизиса клеток рассчитывали как 100 * (поглощение образцов - поглощение фона)/(поглощение максимального высвобождения - поглощение минимального высвобождения). Все проценты значений клеточного лизиса рассчитывали с использованием GraphPad Prism 5.0.
Результаты, как показано на фиг. 11, показали, что антитело клона CL5-dPTM' индуцировало эффект ADCC на CTLA-4-экспрессирующих клетках CHO-K1, а клон CL5-eA-dTPM' с дополнительными мутациями S239D и I332E показал более высокую активность ADCC, чем аналог ипилимумаб.
CTLA-4-экспрессирующие in vitro стимулированные Treg-клетки получали in vitro из выделенных наивных CD4+ T-клеток. Сначала наивные CD4 + Т-клетки выделяли из первичных РВМС в соответствии с инструкцией производителя (Miltenyi, 130-094-131). Затем наивные CD4 + Т-клетки активировали путем инкубации с гранулами с активатором человеческих T-клеток CD3/CD28 (Dynabeads human T-activator CD3/CD28) (1:1) (Thermo, 11131D), 10 нг/мл IL-2 (PeproTech, 200-02-B) и 20 нг/мл TGF-β1 (PeproTech, 100-21) в течение трех дней. В день эксперимента по опосредованному ADCC уничтожению стимулированные Treg-клетки доводили до концентрации 1 × 106 клеток/мл в среде ADCC (содержащей RPMI 1640 без фенолового красного, 10% FBS и 1% пенициллин/стрептомицин). Клетки 1 × 106 окрашивали 5 мкл кальцеина AM (Therom, C34851, исходный материал, приготовленный как 50 мкг на 50 мкл ДМСО) в течение 1 ч при 37°С. Treg-клетки трижды промывали средой ADCC. 50 мкл клеточных суспензий Treg (5 × 103 жизнеспособных клеток) добавляли в каждую лунку 96-луночного планшета с v-образным дном. Тестируемые антитела последовательно разводили в среде ADCC (без фенолового красного) и 50 мкл каждого полученного раствора добавляли в лунки в трех экземплярах. Конечные концентрации антител составляли 1 мкМ, 10 мкМ, 100 мкМ, 1 нМ, 10 нМ и 100 нМ. Планшет инкубировали при комнатной температуре в течение 30 минут. Свежие PBMC (Miaotong) доводили до 5 × 106 клеток/мл средой ADCC (без фенолового красного). При добавлении 50 мкл свежей РВМС к окрашенным клеткам Treg соотношение эффекторных клеток и клеток-мишеней составляло 50: 1. Затем планшет инкубировали при 37°С в течение 2 часов. После 2 часов инкубации планшет центрифугировали и 100 мкл каждой надосадочной жидкости переносили в новый планшет. Надосадочную жидкость измеряли прибором Enspire. Для контроля максимального клеточного лизиса добавляли 50 мкл окрашенных кальцеином AM клеток Treg, 50 мкл среды ADCC и 100 мкл 1% буфера тритон-X100 для обнаружения высвобожденного кальцеина AM. Для контроля минимального клеточного лизиса добавляли 50 мкл клеток, окрашенных кальцеином AM, и 150 мкл среды ADCC для обнаружения высвобожденного кальцеина AM. Измеряли поглощение при 520/650 нм. Процент удельного уничтожения рассчитывается как 100 * (поглощение образцов - поглощение фона)/(поглощение максимального высвобождения - поглощение минимального высвобождения). Все проценты значений удельного уничтожения рассчитывали с использованием GraphPad Prism 5.0.
Результаты показали, что человеческие анти-CTLA-4 антитела индуцировали эффект ADCC в отношении CTLA-4-экспрессирующих клеток Treg, а антитело с мутациями S239D и I332E показало более высокую активность ADCC, чем антитело без мутаций.
Пример 14. Фармакокинетическое исследование анти-CTLA-4 антител
Стадия 1. Обработка однократной дозой анти- CTLA-4 антитела у мышей
Самцам мышей C57BL/6 вводили 3 мг/кг анти- CTLA-4 антитела через хвостовую вену для измерения сывороточной концентрации антител против CTLA-4. Животных сдерживали вручную и приблизительно 100 мкл крови/в момент времени собирали с помощью ретроорбитальной пункции, момент времени соответствовал моменту до введения дозы, 0,167, 1, 4, 8, 24 ч, 2, 4, 7, 14 дней после инъекции антитела. Терминальный забор осуществляли посредством пункцию сердца. Образцы крови оставляли при комнатной температуре и центрифугировали при 2000 × g в течение 5 минут при 4°С для получения образцов сыворотки.
Стадия 2. Концентрация анти-CTLA-4 антитела в сыворотке, измеренная методом ELISA
Все образцы сыворотки сначала разбавляли в 20 раз в аналитическом разбавителе. Дополнительное разведение было сделано в 5% мышиной сыворотке (PBS, об./об.). Рекомбинантный человеческий белок CTLA4-his (Acro Bio) разводили в PBS до концентрации 0,5 мкг/мл, и 50 мкл разведенного образца белка CTLA-4-his добавляли на лунку в микропланшеты для ELISA, которые инкубировали в течение ночи при 4°С для покрытия планшетов рекомбинантными белками. Затем планшеты блокировали блокирующим раствором для ELISA (включающим 2% BSA, 0,05% (об./об.) Tween-20, буфер PBS pH 7,4, об./об.) при 37°C в течение двух часов. Блокирующий буфер удаляли и планшеты инкубировали с разведенными образцами сыворотки в течение 1 часа при 37°С. Планшеты трижды промывали промывочным буфером (PBS + 0,01% (об./об.) Tween 20) и инкубировали с козьим антителом против человеческого IgG (Fc), конъюгированным с пероксидазой хрена (A0170, Sigma), при 37°С в течение одного часа. Добавляли 100 мкл тетраметилбензидина (TMB) и планшеты инкубировали при комнатной температуре в течение 15 минут. 100 мкл 0,1 Н HCl добавляли в каждую лунку для остановки реакции. Поглощение при 450 нм измеряли с помощью ридера для ELISA-планшетов (SpectraMax M2).
Соответствующая сывороточная концентрация анти-CTLA-4 антитела показана на фиг. 12, а подробные данные приведены в таблице 6.
Таблица 6. Средняя концентрация антител в сыворотке после внутривенного введения (в.в.) в дозе 3 мг/кг для мышей
мл/кг
Кроме того, отношение опухоли к сыворотке концентрации антитела против CTLA4 HCAb измеряли у мышей C57BL/6 с опухолями MC38. Мышам вводили 3 мг/кг анти-CTLA-4 антитела через хвостовую вену для измерения концентрации анти-CTLA-4 антител в сыворотке и опухоли. Животных удерживали вручную и приблизительно 100 мкл крови/момент времени собирали с помощью ретроорбитальной пункции, момент времени составлял 8 и 24 часа после инъекции. Конечный сбор осуществляли посредством пункцию сердца. Образцы крови хранили при комнатной температуре и центрифугировали при 2000 × g в течение 5 минут при 4°С для получения образцов сыворотки. Концентрации анти-CTLA-4 антитела в сыворотке и опухоли также тестировали с помощью ELISA, как на стадии 2.
Отношение опухоли к сыворотке концентрации антитела против CTLA4 HCAb в группе CL5-eA-dPTM' составляло примерно в 1 раз выше, чем в группе аналогов Ipi, как показано на фиг. 13, что может быть связано с уникальным признаком антител HCAb, содержащих только тяжелую цепь. Более высокое распределение опухоли к сыворотке может привести к более высокому проникновению в опухолевую ткань антител HCAb.
Пример 15. Мыши, несущие опухоль MC38, лучше выживали с человеческими анти-CTLA-4 антителами
Криоконсервированная клеточная линия карциномы толстой кишки мыши MC-38 была выделена и культивирована в среде DMEM, содержащей 10% эмбриональной бычьей сыворотки (FBS) и 1% пенициллина, стрептомицина при 37 °C, чтобы получить достаточно клеток для имплантации опухоли. Культивируемые клетки МС-38 собирали, ресуспендировали в PBS с плотностью 1 × 107 клеток/мл с жизнеспособностью> 90% и подкожно имплантировали в правый бок 120 hCTLA-4-нокаутных мышей (GempharmaTech). Через пять дней после инокуляции опухоли отобрали 81 мышь с размером опухоли в диапазоне от 26 до 64 мм3 (средний размер опухоли составляла 40 мм3) и распределили на 9 групп с использованием стратифицированной рандомизации с 9 мышами на группу в зависимости от их объема опухоли. Обработку начинали со дня рандомизации (определяемого как D0). Группу 1 обрабатывали hlgG1 в.б. (внутрибрюшинно) при 10 мг/кг в D0, D3, D6, D10, D13, D16; группу 2 обрабатывали CL20’-еА (последовательности в таблице 7) в.б. при 5,4 мг/кг в D0, D3, D6, D10, D13, D16; группу 3 обрабатывали аналогом ипилимумаба, в.б. при 10 мг/кг в D0, D3, D6, D10, D13, D16; группу 4 обрабатывали аналогом ипилимумаба в.б. при 1 мг/кг в D0, D3, D6, D10, D13, D16; группу 5 обрабатывали CL5'-dPTM' в.б. при 5,4 мг/кг в D0, D3, D6, D10, D13, D16; группу 6 обрабатывали CL5'-dPTM' в.б. при 0,54 мг/кг в D0, D3, D6, D10, D13, D16; группу 7 обрабатывали CL5'-eA-dPTM' в.б. при 5,4 мг/кг в D0, D3, D6, D10, D13, D16; группу 8 обрабатывали CL5'-eA-dPTM' в.б. при 1,5 мг/кг в D0, D3, D6, D10, D13, D16 и группу 9 обрабатывали CL5'-eA-dPTM' в.б. при 0,54 мг/кг в D0, D3, D6, D10, D13, D16.
Таблица 7. Нуклеотидные и аминокислотные последовательности клона CL20’-eA
тяжелая цепь
нк: нуклеиновая кислота; ак: аминокислота
Размеры опухоли измеряли три раза в неделю во время обработки. Когда отдельное животное достигло конечной точки (TV> 2000 мм3), его умерщвляли. Время от начала обработки до прекращения считалось временем его выживания. Кривая выживания построена по методу Каплана-Мейера. Среднее время выживания (MST) рассчитывали для каждой группы. Увеличение продолжительности жизни (ILS) рассчитывалось по следующей формуле:
ILS (%) = (MSTобработка-MSTноситель)/MSTноситель x 100%).
ILS (%)> 25% будет считаться биологически значимым преимуществом выживания в соответствии с критериями Национального института рака.
Относительное изменение массы тела (RCBW) каждой мыши рассчитывали по следующей формуле: RCBW (%) = (BWi - BW0)/BW0 × 100%, где BWi относится к средней массе тела в день i, а BW0 относится к средней массе в день 0.
Объемы опухолей (TV) рассчитывали по следующей формуле: объем опухоли = (длина × ширина2)/2.
Степень ингибирования роста опухоли (TGI%) для каждой группы, получавшей дозу, рассчитывали по следующей формуле: TGI% = [1-TVi/TVvi] * 100%, где TVi относится к среднему объему опухоли в группе, получающей дозу, в день i, и TVvi относится к среднему объему опухоли в группе, получавшей носитель, в день i.
Среднее значение и стандартная ошибка среднего (SEM) массы тела мышей, RCBW и объема опухоли в каждой группе были рассчитаны с использованием Microsoft Excel 2007. Фигуры массы тела, относительного изменения массы тела, кривая роста опухоли и ингибирования роста опухоли были построены с использованием GraphPad Prism 5. Рост опухоли между различными группами анализировали с использованием двухстороннего РМ ANOVA. Кривые выживаемости Каплана-Мейера были проанализированы с использованием логрангового критерия. P-значения <0,05 рассматривали, как статистически значимые.
Все исследование было прекращено на D65. Индивидуальные кривые роста опухолей каждой группы показаны на фиг. 14А. Кривые выживаемости животных по времени до конечной точки Каплана-Мейера показаны на фиг. 14В. Все процедуры переносились без каких-либо побочных эффектов, как показано на фиг. 14C. В большинстве этих групп мышей умерщвляли, когда объем опухоли достигал 2000 мм3. В группе 7 3 из 9 мышей, получавших CL5’-eA-dPTM’ при 5,4 мг/кг, выжили без опухоли на D65. В группе 9 2 из 9 мышей, которым вводили CL5’-eA-dPTM’ в дозе 0,54 мг/кг, выжили без опухоли на D65.
MST рассчитывали для каждой группы, и они представлены в таблице 8. MST группы, получавшей носитель, hIgG1 при 5,4 мг/кг и CL20’-eA при 5,4 мг/кг составили 14 дней. MST групп обработки аналогом ипилимумаба при 10 мг/кг и 1 мг/кг, CL5'-dPTM' при 5,4 мг/кг и 0,54 мг/кг, CL5'-eA-dPTM' при 5,4 мг/кг, 1,5 мг/кг и 0,54 мг/кг составили 14, 23, 19, 21 14, 40, 21 и 28 дней, соответственно. Также можно видеть в таблице 8, ILS в группах обработки составляли 64,3%, 35,7%, 50%, 0%, 185,7%, 50% и 100% по сравнению с группой hIgG1, получавшей носитель. Аналог ипилимумаб при 10 мг/кг, CL5'-dPTM' при 5,4 мг/кг, CL5’-eA-dPTM' при 5,4 мг/кг, 1,5 мг/кг и 0,54 мг/кг значительно увеличил среднее время выживания. Мыши, получавшие CL5’-eA-dPTM при 5,4 мг/кг, продемонстрировали лучшее ILS, чем мыши с аналогом ипилимумабом при 10 мг/кг.
Таблица 8. Анализ выживания
Примечание: ILS = (MSTGn – MSTG1)/MSTG1 * 100%; P- значением являются все группы по сравнению с группой G1.
Пример 16. Ингибирование роста опухоли MC38 у hCTLA-4-нокаутных мышей с помощью анти- CTLA-4 антитела HCAb.
Криоконсервированная клеточная линия карциномы толстой кишки мыши MC-38 была выделена и культивирована в среде DMEM, содержащей 10% эмбриональной бычьей сыворотки (FBS) и 1% пенициллина, стрептомицина при 37°C, чтобы получить достаточно клеток для имплантации опухоли. Культивируемые клетки МС-38 собирали, ресуспендировали в PBS с плотностью 5 × 106 клеток/мл с жизнеспособностью> 90% и подкожно имплантировали в правый бок 60 hCTLA-4-нокаутных мышей. Через семь дней после инокуляции опухоли были отобраны 30 мышей с опухолями со средним размером 102 мм3, которые были рандомизированы на 5 групп (n = 6) в зависимости от размера опухоли. Обработку начинали в день рандомизации (определяемый как D0). Включенных в исследование мышей обрабатывали hIgG1 (0,5 мг/кг), аналогом ипилимумаба (0,5 и 0,2 мг/кг) и CL5'-eA-dPTM' (0,27 и 0,1 мг/кг), соответственно, внутрибрюшинно (ip) в D0, D6, D9, D13, D16 и D19. Мышей контролировали ежедневно, а массу тела регистрировали в рабочие дни. Размеры опухоли измеряли два раза в неделю во время обработки.
Относительное изменение массы тела (RCBW), объемов опухоли (TV) и степени ингибирования роста опухоли (TGI%) рассчитывали, как в Примере 16. Среднее значение, стандартная ошибка среднего (SEM) массы тела мышей, RCBW и объема опухоли в каждой группе рассчитывали с использованием Microsoft Excel 2007. Фигуры массы тела, относительного изменения массы тела, кривая роста опухоли и ингибирования роста опухоли были построены с использованием GraphPad Prism 5. Рост опухоли между различными группами анализировали с использованием двухстороннего РМ ANOVA. Кривые выживаемости Каплана-Мейера были проанализированы с использованием логрангового критерия. P-значения <0,05 рассматривали, как статистически значимые.
Кривые роста опухоли показаны на фиг. 15А. Все виды обработок переносились без каких-либо побочных эффектов, как показано на фиг. 15B. Мыши в группах CL5’-eA-dPTM’ при 0,1 мг/кг и 0,27 мг/кг, аналог ипилимумаба при 0,5 мг/кг показали значительное ингибирование роста опухоли от D20 до D30 по сравнению с мышами в группе hIgG1 при 0,2 мг/кг. В то время как обработка аналогом ипилимумабом в дозе 0,2 мг/кг не показала значительного ингибирования роста опухоли по сравнению с hIgG1 при 0,2 мг/кг. CL5’-eA-dPTM ’способствует лучшему ингибированию роста опухоли, чем аналог ипилимумаб в той же дозе.
Хотя изобретение было описано подробно и со ссылкой на его конкретные воплощения, для специалиста в данной области техники будет очевидно, что в него могут быть внесены различные изменения и модификации, не выходящие за пределы сущности и объема изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Harbour Biomed (Shanghai) Co., Ltd
<120> ANTIBODIES BINDING CTLA-4 AND USES THEREOF
<130> 55543 00001
<150> US62/607917
<151> 2017-12-20
<160> 200
<170> PatentIn version 3.5
<210> 1
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 1
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gattggagga ggcgagatcc agccgggggg gtccctgaga 120
ctctcgtgtg cagcctctgg gttcaacgtc agtaagaact acatgagttg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtaatctata gtggtggtgg tacagaatac 240
gcagcctccg tgaagggccg atttaacatc tccagagaca gttccaagaa tatgttgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attattgtgc gagggccgtc 360
cccaggcccc atgggactga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 2
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 2
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ile Gly Gly Gly Glu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Asn Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Gly Thr Glu Tyr
65 70 75 80
Ala Ala Ser Val Lys Gly Arg Phe Asn Ile Ser Arg Asp Ser Ser Lys
85 90 95
Asn Met Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro Arg Pro His Gly Thr Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 3
<211> 118
<212> PRT
<213> Homo Sapiens
<400> 3
Glu Val Gln Leu Val Glu Ile Gly Gly Gly Glu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Gly Thr Glu Tyr Ala Ala Ser Val Lys
50 55 60
Gly Arg Phe Asn Ile Ser Arg Asp Ser Ser Lys Asn Met Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Arg Pro His Gly Thr Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 4
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 4
Gly Phe Asn Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 5
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 5
Val Ile Tyr Ser Gly Gly Gly Thr Glu Tyr Ala Ala Ser Val Lys Gly
1 5 10 15
<210> 6
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 6
Ala Val Pro Arg Pro His Gly Thr Asp Ile
1 5 10
<210> 7
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 7
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggcgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaatgtc agtaagaatt acatgagctg ggtccgccag 180
gttccaggga aggggctgga gtgggtcgca gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcatcctc tccagagaca attccgacaa cacactgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 8
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 8
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Asn Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 9
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 9
Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Asn Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 10
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 10
Gly Phe Asn Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 11
<211> 16
<212> PRT
<213> Homo Sapiens)
<400> 11
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 12
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 12
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 13
<211> 1116
<212> DNA
<213> Homo Sapiens)
<400> 13
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaacgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgttt 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccaagtgccc ctagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 14
<211> 372
<212> PRT
<213> Homo Sapiens)
<400> 14
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Asn Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Phe Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 15
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 15
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asn Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Phe Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 16
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 16
Gly Phe Asn Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 17
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 17
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 18
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 18
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 19
<211> 1119
<212> DNA
<213> Homo Sapiens
<400> 19
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gtccggggga ggcttagttc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg attcaccttc agtaactact ggatgcactg ggtccgccaa 180
gctccaggga aggggctggt gtgggtctca cgtattaata gtgatgggag taacacaacc 240
tacgcggact ccgtgaaggg ccgattcacc atctccagag acaacgccaa gaacacgctg 300
tatctgcaaa tgaacagtct gagagccgag gacacggctg tgtattactg tgcaagatgg 360
ttcggggagc actactacgg aatggacgtc tggggccaag ggaccacggt caccgtctct 420
tcagagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 480
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 540
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 600
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag 660
cagtacaaca gcacgtaccg ggtggtcagc gtcctcaccg tcctgcacca ggactggctg 720
aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa 780
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 840
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 900
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 960
cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag 1020
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1080
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1119
<210> 20
<211> 373
<212> PRT
<213> Homo Sapiens
<400> 20
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Phe Ser Asn Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Val Trp Val Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr
65 70 75 80
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
85 90 95
Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met
115 120 125
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Glu Pro Lys
130 135 140
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
145 150 155 160
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
165 170 175
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
180 185 190
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
195 200 205
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
210 215 220
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
225 230 235 240
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
245 250 255
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
260 265 270
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
275 280 285
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
290 295 300
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
305 310 315 320
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
325 330 335
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
340 345 350
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
355 360 365
Leu Ser Pro Gly Lys
370
<210> 21
<211> 120
<212> PRT
<213> Homo Sapiens
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 22
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 22
Gly Phe Thr Phe Ser Asn Tyr Trp Met His
1 5 10
<210> 23
<211> 17
<212> PRT
<213> Homo Sapiens
<400> 23
Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 24
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 24
Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val
1 5 10
<210> 25
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 25
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga gggctgatcc agccgggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agaaacaact acatgagctg ggcccgccag 180
gttccaggga agggactgga gtgggtctca gttatttata gcggtgggag tataacctac 240
gcagactccg tgaagggccg attcatcatc tccagagaca attccaacaa cacgctatat 300
cttcaaatga acggtctgag agccgaggac acggccgtat attactgtgc gagagccgtc 360
ccacatgccc caagtgcttt tgacatctgg ggccagggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 26
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 26
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Arg Asn Asn Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Asn
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 27
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 27
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Arg Asn Asn
20 25 30
Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Asn Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 28
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 28
Gly Phe Thr Val Arg Asn Asn Tyr Met Ser
1 5 10
<210> 29
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 29
Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 30
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 30
Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile
1 5 10
<210> 31
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 31
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga ggcttaatcc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg gctcaatgtc agtaggcact acttaagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtcgca gtaatttata gcggtggtgg tatagagtac 240
gtagactccg tgaagggccg attcaccatc tccagagaca atgccaacaa cacgctgtat 300
cttcaaatga gaagcctgag aaccgaagac tcggccgtat attactgtgc gcgggccatt 360
cccagacccc atggttcgga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 32
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 32
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu
35 40 45
Asn Val Ser Arg His Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ala Val Ile Tyr Ser Gly Gly Gly Ile Glu Tyr
65 70 75 80
Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Asn
85 90 95
Asn Thr Leu Tyr Leu Gln Met Arg Ser Leu Arg Thr Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Ile Pro Arg Pro His Gly Ser Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 33
<211> 118
<212> PRT
<213> Homo Sapiens
<400> 33
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Asn Val Ser Arg His
20 25 30
Tyr Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Tyr Ser Gly Gly Gly Ile Glu Tyr Val Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Asn Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Arg Ser Leu Arg Thr Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Ile Pro Arg Pro His Gly Ser Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 34
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 34
Gly Leu Asn Val Ser Arg His Tyr Leu Ser
1 5 10
<210> 35
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 35
Val Ile Tyr Ser Gly Gly Gly Ile Glu Tyr Val Asp Ser Val Lys Gly
1 5 10 15
<210> 36
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 36
Ala Ile Pro Arg Pro His Gly Ser Asp Ile
1 5 10
<210> 37
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 37
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg gttcaccgtc agtagcaact acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gttatttata gcggtggtag cacatactac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 300
cttcaaatga gcaacctgac cgccgaggac acggccgtgt attactgtgc gagagccgtt 360
ccccactccc ccagtgcttt tgataggtgg ggccagggaa ccctggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 38
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 38
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Ser Asn Leu Thr Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp
115 120 125
Arg Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 39
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 39
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Ser Asn Leu Thr Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 40
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 40
Gly Phe Thr Val Ser Ser Asn Tyr Met Ser
1 5 10
<210> 41
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 41
Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 42
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 42
Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg
1 5 10
<210> 43
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 43
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaacgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgtat 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccacatgccc ctagtaattt tgataaatgg ggccaaggga caagggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 44
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 44
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Asn Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp
115 120 125
Lys Trp Gly Gln Gly Thr Arg Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 45
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 45
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asn Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys Trp Gly Gln Gly
100 105 110
Thr Arg Val Thr Val Ser Ser
115
<210> 46
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 46
Gly Phe Asn Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 47
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 47
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 48
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 48
Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys
1 5 10
<210> 49
<211> 1125
<212> DNA
<213> Homo Sapiens
<400> 49
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gtctggggga ggcttggttc agccgggggg gtccctgcga 120
ctctcctgtg tagcctctgg attcaccttc agtaactatt ggatgcactg ggtccgccaa 180
attccaggga aggggatggt gtggatctca gaaattaaca gtgacgggaa taggacaggc 240
tatgcggact ccgtaaaggg ccgattcacc atctccagag acaacgccaa gaagatgttg 300
tatctgcaaa tgaacagtct gagagccgag gacacggctg tttattattg tgcaagaaac 360
tggggatcgg gcccccatca taccgctttt gatggttggg gccaagggac aatggtcacc 420
gtctcttcag agcccaaatc ttgtgacaaa actcacacat gcccaccgtg cccagcacct 480
gaactcctgg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 540
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 600
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 660
gaggagcagt acaacagcac gtaccgggtg gtcagcgtcc tcaccgtcct gcaccaggac 720
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 780
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 840
ccatcccggg aggagatgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 900
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 960
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 1020
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 1080
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaa 1125
<210> 50
<211> 375
<212> PRT
<213> Homo Sapiens
<400> 50
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe
35 40 45
Thr Phe Ser Asn Tyr Trp Met His Trp Val Arg Gln Ile Pro Gly Lys
50 55 60
Gly Met Val Trp Ile Ser Glu Ile Asn Ser Asp Gly Asn Arg Thr Gly
65 70 75 80
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
85 90 95
Lys Lys Met Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr
100 105 110
Ala Val Tyr Tyr Cys Ala Arg Asn Trp Gly Ser Gly Pro His His Thr
115 120 125
Ala Phe Asp Gly Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu
130 135 140
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
145 150 155 160
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
165 170 175
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
180 185 190
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
195 200 205
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
210 215 220
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
225 230 235 240
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
245 250 255
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
260 265 270
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
275 280 285
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
290 295 300
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
305 310 315 320
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
325 330 335
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
340 345 350
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
355 360 365
Leu Ser Leu Ser Pro Gly Lys
370 375
<210> 51
<211> 122
<212> PRT
<213> Homo Sapiens
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ile Pro Gly Lys Gly Met Val Trp Ile
35 40 45
Ser Glu Ile Asn Ser Asp Gly Asn Arg Thr Gly Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Lys Met Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Trp Gly Ser Gly Pro His His Thr Ala Phe Asp Gly Trp
100 105 110
Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 52
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 52
Gly Phe Thr Phe Ser Asn Tyr Trp Met His
1 5 10
<210> 53
<211> 17
<212> PRT
<213> Homo Sapiens
<400> 53
Glu Ile Asn Ser Asp Gly Asn Arg Thr Gly Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 54
<211> 13
<212> PRT
<213> Homo Sapiens
<400> 54
Asn Trp Gly Ser Gly Pro His His Thr Ala Phe Asp Gly
1 5 10
<210> 55
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 55
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaccgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgttt 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccaagtgccc ctagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 56
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 56
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Thr Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Phe Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 57
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 57
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Phe Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 58
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 58
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 59
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 59
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 60
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 60
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 61
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 61
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gagcggagga ggcttgatcc agccgggggg gtccctgaga 120
ctctcctgtg ccgcctctgg gtttaccgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaagggccg atttaccatc tccagagaca gttccaagaa cacggtgtac 300
cttcaaatga acagcctgag agccgaggac accgccgtgt attactgtgc gagagccgtc 360
ccaagtgccc ctagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 62
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 62
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys
85 90 95
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 63
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 63
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 64
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 64
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 65
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 65
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 66
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 66
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 67
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 67
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga gggctgatcc agccgggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agaaacaact acatgagctg ggcccgccag 180
gttccaggga agggactgga gtgggtctca gttatttata gcggtgggag tataacctac 240
gcagactccg tgaagggccg attcatcatc tccagagaca attccaagaa cacgctatat 300
cttcaaatga acggtctgag agccgaggac acggccgtat attactgtgc gagagccgtc 360
ccacatgccc caagtgcttt tgacatctgg ggccagggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 68
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 68
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Arg Asn Asn Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 69
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 69
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Arg Asn Asn
20 25 30
Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 70
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 70
Gly Phe Thr Val Arg Asn Asn Tyr Met Ser
1 5 10
<210> 71
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 71
Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 72
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 72
Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile
1 5 10
<210> 73
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 73
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gagcggagga gggctgatcc agccgggggg gtccctgaga 120
ctctcctgtg cagcctctgg gttcaccgtc agaaacaact acatgagctg ggtgcgccag 180
gccccaggga agggactgga gtgggtctca gttatttata gcggtgggag tataacctac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctatat 300
cttcaaatga acagcctgag agccgaggac acggccgtat attactgtgc gagagccgtc 360
ccacatgccc caagtgcttt tgacatctgg ggccagggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 74
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 74
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Arg Asn Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 75
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Arg Asn Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 76
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 76
Gly Phe Thr Val Arg Asn Asn Tyr Met Ser
1 5 10
<210> 77
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 77
Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 78
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 78
Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile
1 5 10
<210> 79
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 79
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg gttcaccgtc agtagcaact acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gttatttata gcggtggtag cacatactac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 300
cttcaaatga gcagcctgag ggccgaggac acggccgtgt attactgtgc gagagccgtt 360
ccccactccc ccagtgcttt tgataggtgg ggccagggaa ccctggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 80
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 80
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp
115 120 125
Arg Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 81
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 81
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Ser Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 82
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 82
Gly Phe Thr Val Ser Ser Asn Tyr Met Ser
1 5 10
<210> 83
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 83
Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 84
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 84
Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg
1 5 10
<210> 85
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 85
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gagcggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg gttcaccgtc agtagcaact acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gttatttata gcggtggtag cacatactac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 300
cttcaaatga acagcctgag ggccgaggac acggccgtgt attactgtgc gagagccgtt 360
ccccactccc ccagtgcttt tgataggtgg ggccagggaa ccctggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 86
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 86
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp
115 120 125
Arg Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 87
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 87
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 88
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 88
Gly Phe Thr Val Ser Ser Asn Tyr Met Ser
1 5 10
<210> 89
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 89
Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 90
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 90
Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg
1 5 10
<210> 91
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 91
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaccgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgtat 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccacatgccc ctagtaattt tgataaatgg ggccaaggga caagggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 92
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 92
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Thr Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp
115 120 125
Lys Trp Gly Gln Gly Thr Arg Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 93
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 93
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys Trp Gly Gln Gly
100 105 110
Thr Arg Val Thr Val Ser Ser
115
<210> 94
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 94
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 95
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 95
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 96
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 96
Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys
1 5 10
<210> 97
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 97
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggcgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agtaagaatt acatgagctg ggtccgccag 180
gttccaggga aggggctgga gtgggtcgca gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcatcctc tccagagaca attccgacaa cacactgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 98
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 98
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys
85 90 95
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp
115 120 125
Lys Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 99
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 99
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 100
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 100
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 101
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 101
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 102
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 102
Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys
1 5 10
<210> 103
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 103
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggcgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agtaagaatt acatgagctg ggtccgccag 180
gttccaggga aggggctgga gtgggtcgca gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcatcctc tccagagaca attccgacaa cacactgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 104
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 104
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 105
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 105
Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 106
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 106
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 107
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 107
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 108
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 108
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 109
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 109
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gagcggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agtaagaatt acatgagctg ggtccgccag 180
gccccaggga aggggctgga gtgggtcagc gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacactgtat 300
cttcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 110
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 110
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 111
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 112
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 112
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 113
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 113
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 114
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 114
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 115
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 115
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaacgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgttt 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccaagtgccc ctagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 116
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 116
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Asn Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Phe Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 117
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 117
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asn Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Phe Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 118
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 118
Gly Phe Asn Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 119
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 119
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 120
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 120
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 121
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 121
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga gggctgatcc agccgggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agaaacaact acatgagctg ggcccgccag 180
gttccaggga agggactgga gtgggtctca gttatttata gcggtgggag tataacctac 240
gcagactccg tgaagggccg attcatcatc tccagagaca attccaacaa cacgctatat 300
cttcaaatga acggtctgag agccgaggac acggccgtat attactgtgc gagagccgtc 360
ccacatgccc caagtgcttt tgacatctgg ggccagggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 122
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 122
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Arg Asn Asn Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Asn
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 123
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 123
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Arg Asn Asn
20 25 30
Tyr Met Ser Trp Ala Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Ile Ser Arg Asp Asn Ser Asn Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 124
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 124
Gly Phe Thr Val Arg Asn Asn Tyr Met Ser
1 5 10
<210> 125
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 125
Val Ile Tyr Ser Gly Gly Ser Ile Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 126
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 126
Ala Val Pro His Ala Pro Ser Ala Phe Asp Ile
1 5 10
<210> 127
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 127
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagcctctgg gttcaccgtc agtagcaact acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gttatttata gcggtggtag cacatactac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 300
cttcaaatga gcaacctgac cgccgaggac acggccgtgt attactgtgc gagagccgtt 360
ccccactccc ccagtgcttt tgataggtgg ggccagggaa ccctggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 128
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 128
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
35 40 45
Thr Val Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Ser Asn Leu Thr Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp
115 120 125
Arg Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 129
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 129
Glu Val Gln Leu Val Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Ser Asn Leu Thr Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 130
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 130
Gly Phe Thr Val Ser Ser Asn Tyr Met Ser
1 5 10
<210> 131
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 131
Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 132
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 132
Ala Val Pro His Ser Pro Ser Ala Phe Asp Arg
1 5 10
<210> 133
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 133
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggagga gactggagga gacttgatcc agccgggggg gtccctgaga 120
ctctcctgta cagcctctgg gtttaacgtc agtagaatct acatgagctg ggtccgccag 180
gctccaggga aggggctgga gtgggtctca gtgatttata gcggtggaag tatatattac 240
gcagattccg tgaggggccg atttaccatc tccagagaca gttccacgaa cacggtgtat 300
cttcaaatga acagcctgag agccgaggac tcggccgtgt attactgtgc gagagccgtc 360
ccacatgccc ctagtaattt tgataaatgg ggccaaggga caagggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 134
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 134
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe
35 40 45
Asn Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr
65 70 75 80
Ala Asp Ser Val Arg Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr
85 90 95
Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp
115 120 125
Lys Trp Gly Gln Gly Thr Arg Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 135
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 135
Glu Val Gln Leu Glu Glu Thr Gly Gly Asp Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Phe Asn Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Thr Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Ser Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys Trp Gly Gln Gly
100 105 110
Thr Arg Val Thr Val Ser Ser
115
<210> 136
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 136
Gly Phe Asn Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 137
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 137
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Arg Gly
1 5 10 15
<210> 138
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 138
Ala Val Pro His Ala Pro Ser Asn Phe Asp Lys
1 5 10
<210> 139
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 139
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggcgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaatgtc agtaagaatt acatgagctg ggtccgccag 180
gttccaggga aggggctgga gtgggtcgca gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcatcctc tccagagaca attccgacaa cacactgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 140
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 140
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Asn Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 141
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 141
Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Asn Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 142
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 142
Gly Phe Asn Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 143
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 143
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 144
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 144
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 145
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 145
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggcgga gactggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agtaagaatt acatgagctg ggtccgccag 180
gttccaggga aggggctgga gtgggtcgca gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcatcctc tccagagaca attccgacaa cacactgtat 300
cttcaaatga acaacctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 146
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 146
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 147
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 147
Glu Val Gln Leu Ala Glu Thr Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Ile Leu Ser Arg Asp Asn Ser Asp Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 148
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 148
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 149
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 149
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 150
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 150
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 151
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 151
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
tctgaggtgc agctggtgga gagcggagga ggcttgatcc agcctggggg gtccctgaga 120
ctctcctgtg cagtctctgg gttcaccgtc agtaagaatt acatgagctg ggtccgccag 180
gccccaggga aggggctgga gtgggtcagc gttgtttata gcggtggtag caaaacctac 240
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacactgtat 300
cttcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagccgtt 360
cctcattccc ccagttcttt tgatatctgg ggccaaggga caatggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 152
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 152
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu
20 25 30
Ile Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe
35 40 45
Thr Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys
50 55 60
Gly Leu Glu Trp Val Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp
115 120 125
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 153
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 153
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 154
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 154
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 155
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 155
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 156
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 156
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 157
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 157
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagtc cgggggaggc ttagttcagc ctggggggtc cctgagactc 120
tcctgtgcag cctctggatt caccttcagt aactactgga tgcactgggt ccgccaagct 180
ccagggaagg ggctggtgtg ggtctcacgt attaatagtg atgggagtaa cacaacctac 240
gcggactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 300
ctgcaaatga acagtctgag agccgaggac acggctgtgt attactgtgc aagatggttc 360
ggggagcact actacggaat ggacgtctgg ggccaaggga ccacggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 158
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 158
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Phe Ser Asn Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Val Trp Val Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met Asp
115 120 125
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 159
<211> 120
<212> PRT
<213> Homo Sapiens
<400> 159
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 160
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 160
Gly Phe Thr Phe Ser Asn Tyr Trp Met His
1 5 10
<210> 161
<211> 17
<212> PRT
<213> Homo Sapiens
<400> 161
Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 162
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 162
Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val
1 5 10
<210> 163
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 163
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagag cggaggaggc ttgatccagc cgggggggtc cctgagactc 120
tcctgtgccg cctctgggtt taccgtcagt agaatctaca tgagctgggt ccgccaggct 180
ccagggaagg ggctggagtg ggtctcagtg atttatagcg gtggaagtat atattacgca 240
gattccgtga agggccgatt taccatctcc agagacagtt ccaagaacac ggtgtacctt 300
caaatgaaca gcctgagagc cgaggacacc gccgtgtatt actgtgcgag agccgtccca 360
agtgccccta gttcttttga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 164
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 164
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn
85 90 95
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 165
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 165
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 166
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 166
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 167
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 167
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 168
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 168
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 169
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 169
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagag cggaggaggc ttgatccagc ctggggggtc cctgagactc 120
tcctgtgcag tctctgggtt caccgtcagt aagaattaca tgagctgggt ccgccaggcc 180
ccagggaagg ggctggagtg ggtcagcgtt gtttatagcg gtggtagcaa aacctacgca 240
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac actgtatctt 300
caaatgaaca gcctgagagc cgaggacacg gccgtgtatt actgtgcgag agccgttcct 360
cattccccca gttcttttga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 170
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 170
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr
35 40 45
Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 171
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 171
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 172
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 172
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 173
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 173
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 174
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 174
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 175
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 175
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagag cggaggaggc ttgatccagc cgggggggtc cctgagactc 120
tcctgtgccg cctctgggtt taccgtcagt agaatctaca tgagctgggt ccgccaggct 180
ccagggaagg ggctggagtg ggtctcagtg atttatagcg gtggaagtat atattacgca 240
gattccgtga agggccgatt taccatctcc agagacagtt ccaagaacac ggtgtacctt 300
caaatgaaca gcctgagagc cgaggacacc gccgtgtatt actgtgcgag agccgtccca 360
agtgccccta gttcttttga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccggacg tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccgagga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 176
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 176
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Val Ser Arg Ile Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn
85 90 95
Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Glu
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 177
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 177
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Arg Ile
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Ser Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 178
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 178
Gly Phe Thr Val Ser Arg Ile Tyr Met Ser
1 5 10
<210> 179
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 179
Val Ile Tyr Ser Gly Gly Ser Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 180
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 180
Ala Val Pro Ser Ala Pro Ser Ser Phe Asp Ile
1 5 10
<210> 181
<211> 1113
<212> DNA
<213> Homo Sapiens
<400> 181
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagag cggaggaggc ttgatccagc ctggggggtc cctgagactc 120
tcctgtgcag tctctgggtt caccgtcagt aagaattaca tgagctgggt ccgccaggcc 180
ccagggaagg ggctggagtg ggtcagcgtt gtttatagcg gtggtagcaa aacctacgca 240
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac actgtatctt 300
caaatgaaca gcctgagagc cgaggacacg gccgtgtatt actgtgcgag agccgttcct 360
cattccccca gttcttttga tatctggggc caagggacaa tggtcaccgt ctcttcagag 420
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 480
ggaccggacg tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 540
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 600
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 660
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 720
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccgagga gaaaaccatc 780
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag 840
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 900
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 960
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1020
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1080
acgcagaaga gcctctccct gtctccgggt aaa 1113
<210> 182
<211> 371
<212> PRT
<213> Homo Sapiens
<400> 182
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr
35 40 45
Val Ser Lys Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Glu Trp Val Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
115 120 125
Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Glu Pro Lys Ser Cys
130 135 140
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
145 150 155 160
Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
165 170 175
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
180 185 190
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
195 200 205
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
210 215 220
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
225 230 235 240
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Glu
245 250 255
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
260 265 270
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
275 280 285
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
290 295 300
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
305 310 315 320
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
325 330 335
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
340 345 350
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
355 360 365
Pro Gly Lys
370
<210> 183
<211> 119
<212> PRT
<213> Homo Sapiens
<400> 183
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Ile Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Phe Thr Val Ser Lys Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 184
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 184
Gly Phe Thr Val Ser Lys Asn Tyr Met Ser
1 5 10
<210> 185
<211> 16
<212> PRT
<213> Homo Sapiens
<400> 185
Val Val Tyr Ser Gly Gly Ser Lys Thr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 186
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 186
Ala Val Pro His Ser Pro Ser Ser Phe Asp Ile
1 5 10
<210> 187
<211> 1116
<212> DNA
<213> Homo Sapiens
<400> 187
atggagaccg acaccctgct gctgtgggtg ctgctgctgt gggtgcccgg atccaccggc 60
gaggtgcagc tggtggagtc cgggggaggc ttagttcagc ctggggggtc cctgagactc 120
tcctgtgcag cctctggatt caccttcagt aactactgga tgcactgggt ccgccaagct 180
ccagggaagg ggctggtgtg ggtctcacgt attaatagtg atgggagtaa cacaacctac 240
gcggactccg tgaagggccg attcaccatc tccagagaca acgccaagaa cacgctgtat 300
ctgcaaatga acagtctgag agccgaggac acggctgtgt attactgtgc aagatggttc 360
ggggagcact actacggaat ggacgtctgg ggccaaggga ccacggtcac cgtctcttca 420
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 480
gggggaccgg acgtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 540
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 600
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 660
tacaacagca cgtaccgggt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 720
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccga ggagaaaacc 780
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 840
gaggagatga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 900
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 960
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1020
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1080
tacacgcaga agagcctctc cctgtctccg ggtaaa 1116
<210> 188
<211> 372
<212> PRT
<213> Homo Sapiens
<400> 188
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val
20 25 30
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
35 40 45
Phe Ser Asn Tyr Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly
50 55 60
Leu Val Trp Val Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr
65 70 75 80
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys
85 90 95
Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala
100 105 110
Val Tyr Tyr Cys Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met Asp
115 120 125
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Glu Pro Lys Ser
130 135 140
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
145 150 155 160
Gly Gly Pro Asp Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
165 170 175
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
180 185 190
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
195 200 205
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
210 215 220
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
225 230 235 240
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
245 250 255
Glu Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
260 265 270
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
275 280 285
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
290 295 300
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
305 310 315 320
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
325 330 335
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
340 345 350
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
355 360 365
Ser Pro Gly Lys
370
<210> 189
<211> 120
<212> PRT
<213> Homo Sapiens
<400> 189
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Val Trp Val
35 40 45
Ser Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 190
<211> 10
<212> PRT
<213> Homo Sapiens
<400> 190
Gly Phe Thr Phe Ser Asn Tyr Trp Met His
1 5 10
<210> 191
<211> 17
<212> PRT
<213> Homo Sapiens
<400> 191
Arg Ile Asn Ser Asp Gly Ser Asn Thr Thr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 192
<211> 11
<212> PRT
<213> Homo Sapiens
<400> 192
Trp Phe Gly Glu His Tyr Tyr Gly Met Asp Val
1 5 10
<210> 193
<211> 23
<212> DNA
<213> Artificial sequence
<220>
<223> Primer
<400> 193
gtgtccagtg tgaggtgcag ctg 23
<210> 194
<211> 23
<212> DNA
<213> Artificial sequence
<220>
<223> Primer
<400> 194
gtgtccagtg tcaggtgcag ctg 23
<210> 195
<211> 24
<212> DNA
<213> Artificial sequence
<220>
<223> Primer
<400> 195
ggcttacaac cacaatccct gggc 24
<210> 196
<211> 672
<212> DNA
<213> Homo Sapiens
<400> 196
atggcttgcc ttggatttca gcggcacaag gctcagctga acctggctac caggacctgg 60
ccctgcactc tcctgttttt tcttctcttc atccctgtct tctgcaaagc aatgcacgtg 120
gcccagcctg ctgtggtact ggccagcagc cgaggcatcg ccagctttgt gtgtgagtat 180
gcatctccag gcaaagccac tgaggtccgg gtgacagtgc ttcggcaggc tgacagccag 240
gtgactgaag tctgtgcggc aacctacatg atggggaatg agttgacctt cctagatgat 300
tccatctgca cgggcacctc cagtggaaat caagtgaacc tcactatcca aggactgagg 360
gccatggaca cgggactcta catctgcaag gtggagctca tgtacccacc gccatactac 420
ctgggcatag gcaacggaac ccagatttat gtaattgatc cagaaccgtg cccagattct 480
gacttcctcc tctggatcct tgcagcagtt agttcggggt tgttttttta tagctttctc 540
ctcacagctg tttctttgag caaaatgcta aagaaaagaa gccctcttac aacaggggtc 600
tatgtgaaaa tgcccccaac agagccagaa tgtgaaaagc aatttcagcc ttattttatt 660
cccatcaatt ga 672
<210> 197
<211> 223
<212> PRT
<213> Homo Sapiens
<400> 197
Met Ala Cys Leu Gly Phe Gln Arg His Lys Ala Gln Leu Asn Leu Ala
1 5 10 15
Thr Arg Thr Trp Pro Cys Thr Leu Leu Phe Phe Leu Leu Phe Ile Pro
20 25 30
Val Phe Cys Lys Ala Met His Val Ala Gln Pro Ala Val Val Leu Ala
35 40 45
Ser Ser Arg Gly Ile Ala Ser Phe Val Cys Glu Tyr Ala Ser Pro Gly
50 55 60
Lys Ala Thr Glu Val Arg Val Thr Val Leu Arg Gln Ala Asp Ser Gln
65 70 75 80
Val Thr Glu Val Cys Ala Ala Thr Tyr Met Met Gly Asn Glu Leu Thr
85 90 95
Phe Leu Asp Asp Ser Ile Cys Thr Gly Thr Ser Ser Gly Asn Gln Val
100 105 110
Asn Leu Thr Ile Gln Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile
115 120 125
Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Leu Gly Ile Gly
130 135 140
Asn Gly Thr Gln Ile Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser
145 150 155 160
Asp Phe Leu Leu Trp Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe
165 170 175
Tyr Ser Phe Leu Leu Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys
180 185 190
Arg Ser Pro Leu Thr Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu
195 200 205
Pro Glu Cys Glu Lys Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
210 215 220
<210> 198
<211> 672
<212> DNA
<213> Macaca mulatta
<400> 198
atggcttgcc ttggatttca gcggcacaag gctcggctca acctggctac caggacccgg 60
ccctacactc tcctgttttc tcttctcttc atccctgtct tctccaaagc aatgcacgtg 120
gcccagcctg ctgtggtgct ggccaacagc cgagggatcg ccagctttgt gtgtgagtat 180
gcatctccag gcaaagccac tgaggtccgg gtgacagtgc ttcggcaggc cgacagccag 240
gtgactgaag tctgtgcggc aacatacatg atggggaatg agttgacctt cctagatgat 300
tccatctgca cgggcacctc cagtggaaat caagtgaacc tcactatcca aggactgagg 360
gctatggaca caggactcta catctgcaag gtggagctca tgtacccacc accatactac 420
atgggcatag gcaatggaac ccagatttat gtaattgatc cagaaccgtg cccagattct 480
gacttcctcc tctggatcct tgcagcagtt agttcggggt tgttttttta tagctttctc 540
ctcacagctg tttctttgag caaaatgcta aagaaaagaa gccctctcac aacaggggtc 600
tatgtgaaaa tgcccccaac agagccagaa tgtgaaaagc aatttcagcc ttattttatt 660
cccatcaatt ga 672
<210> 199
<211> 448
<212> PRT
<213> Artificial sequence
<220>
<223> heavy chain of Ipilimumab analogue
<400> 199
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Thr Phe Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Trp Leu Gly Pro Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 200
<211> 215
<212> PRT
<213> Artificial sequence
<220>
<223> light chain of Ipilimumab analogue
<400> 200
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Phe Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЧАСТИЦЫ, КОНЪЮГИРОВАННЫЕ С ПЕПТИДОМ | 2014 |
|
RU2813163C2 |
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| СПЕЦИФИЧЕСКИЕ АНТИТЕЛА К PD-L1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2749109C2 |
| НОВЫЕ МОЛЕКУЛЫ АГОНИСТИЧЕСКИХ АНТИ-TNFR2 АНТИТЕЛ | 2019 |
|
RU2840049C2 |
| АНТИ-PRE-S1 HBV АНТИТЕЛА | 2016 |
|
RU2739955C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2779489C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ КОМПОЗИЦИЙ T-КЛЕТОК И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2835034C2 |
| СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РАКОВОГО МЕТАСТАЗА И РАЗРЕЖЕНИЯ КОСТИ, СВЯЗАННОГО С РАКОВЫМ МЕТАСТАЗОМ | 2007 |
|
RU2470665C2 |
| БИОЛОГИЧЕСКИЕ МАТЕРИАЛЫ ДЛЯ МОДУЛЯЦИИ ИММУННЫХ ОТВЕТОВ | 2017 |
|
RU2770060C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
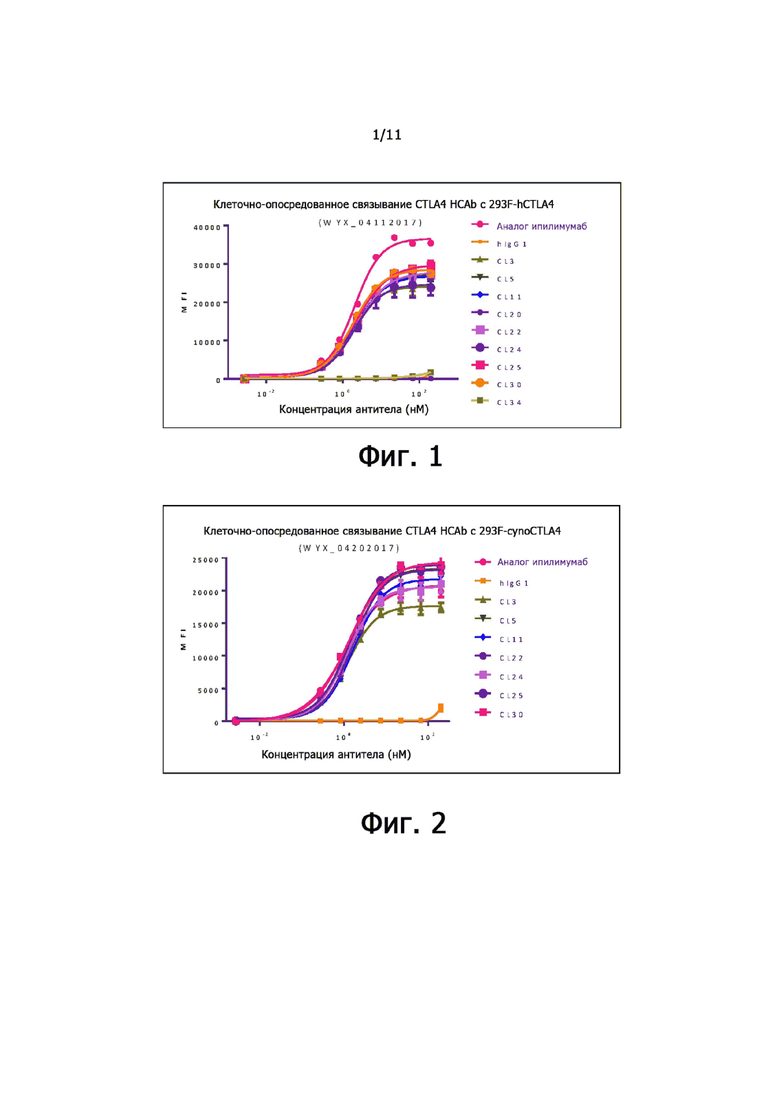
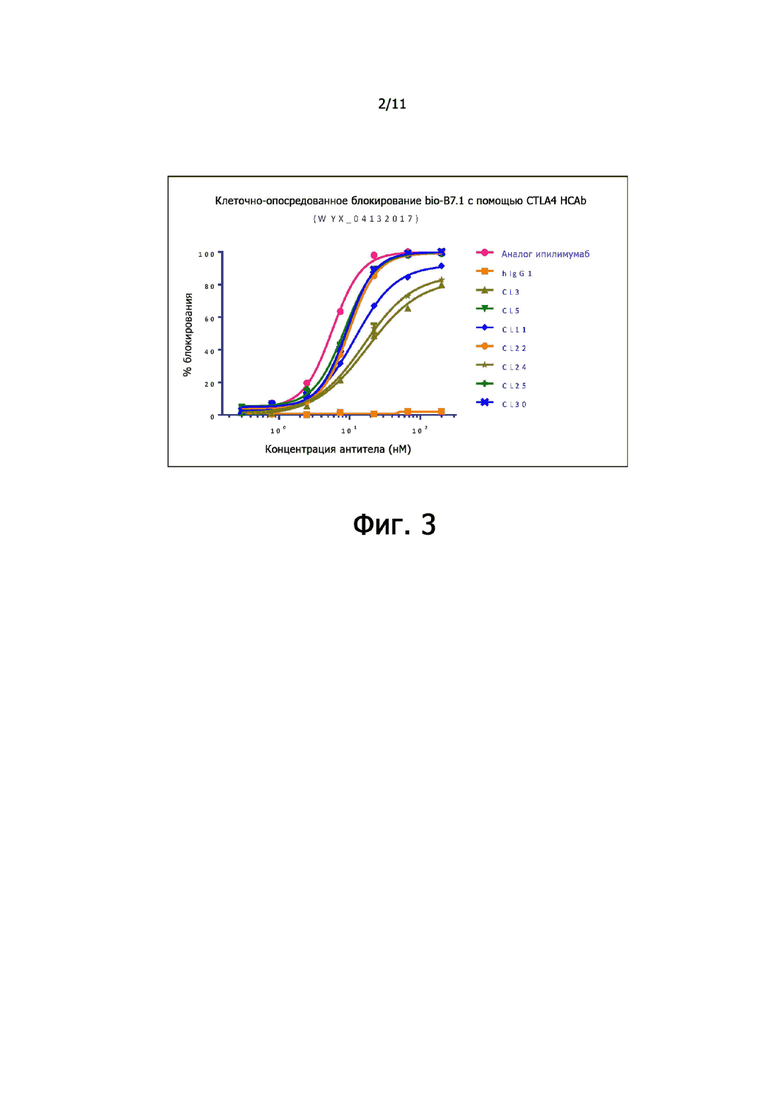

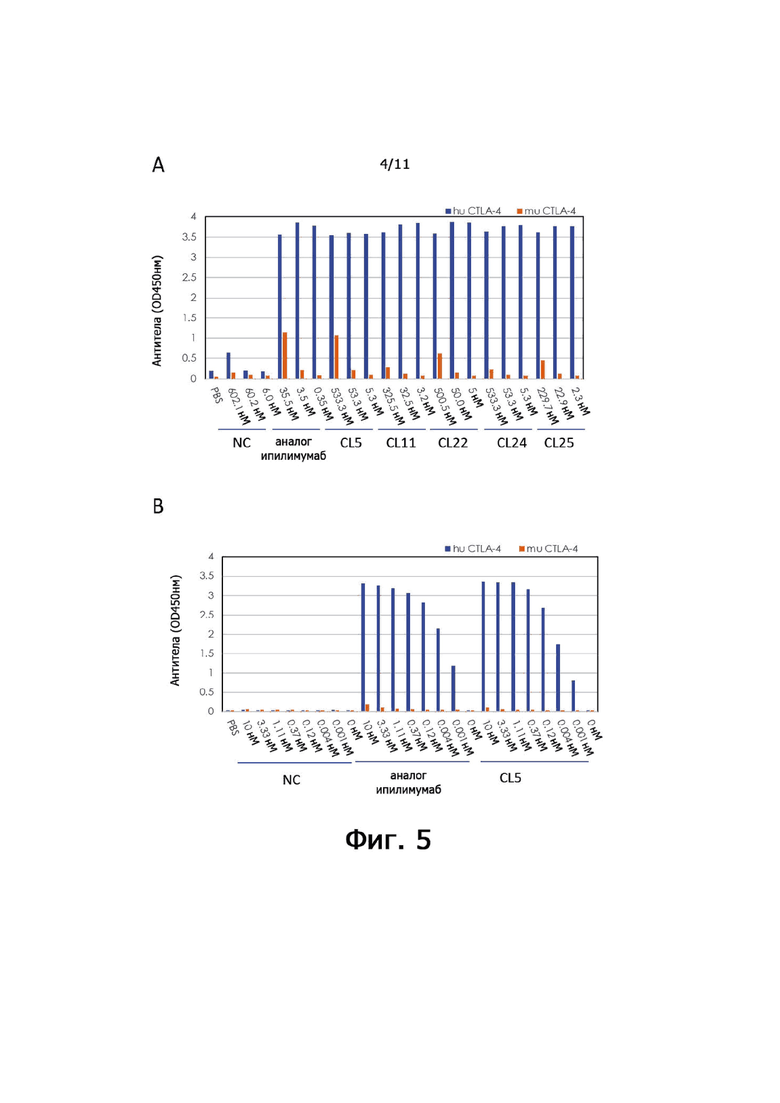
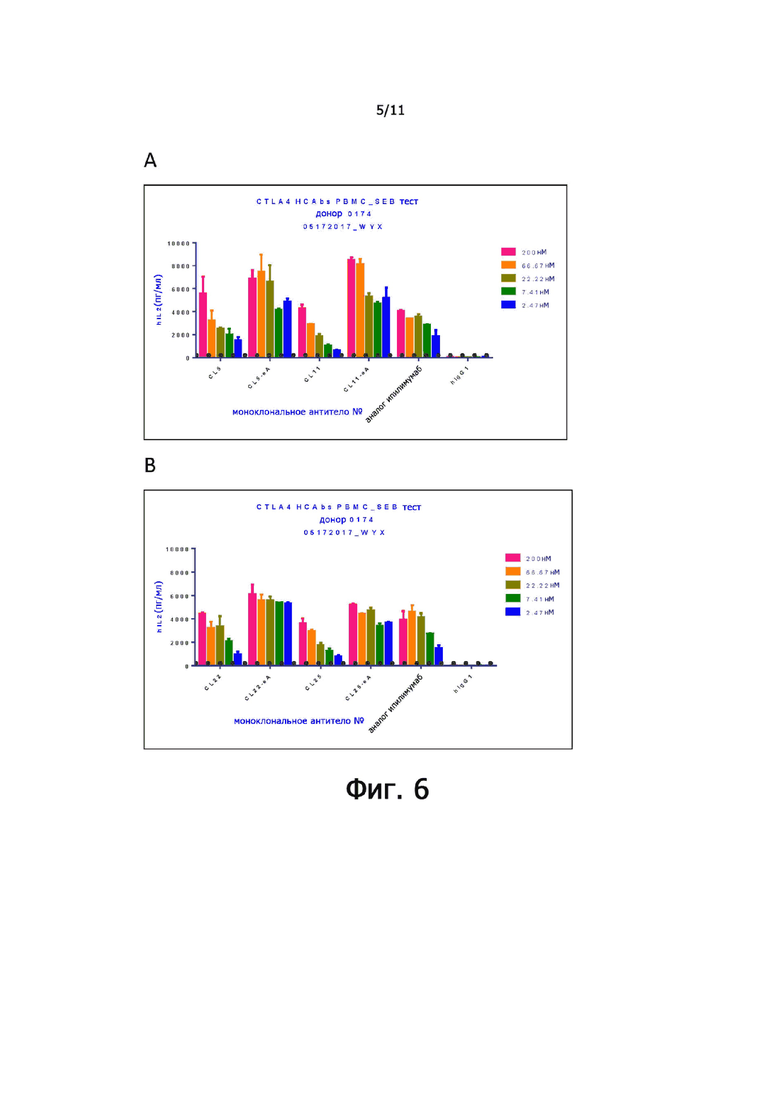
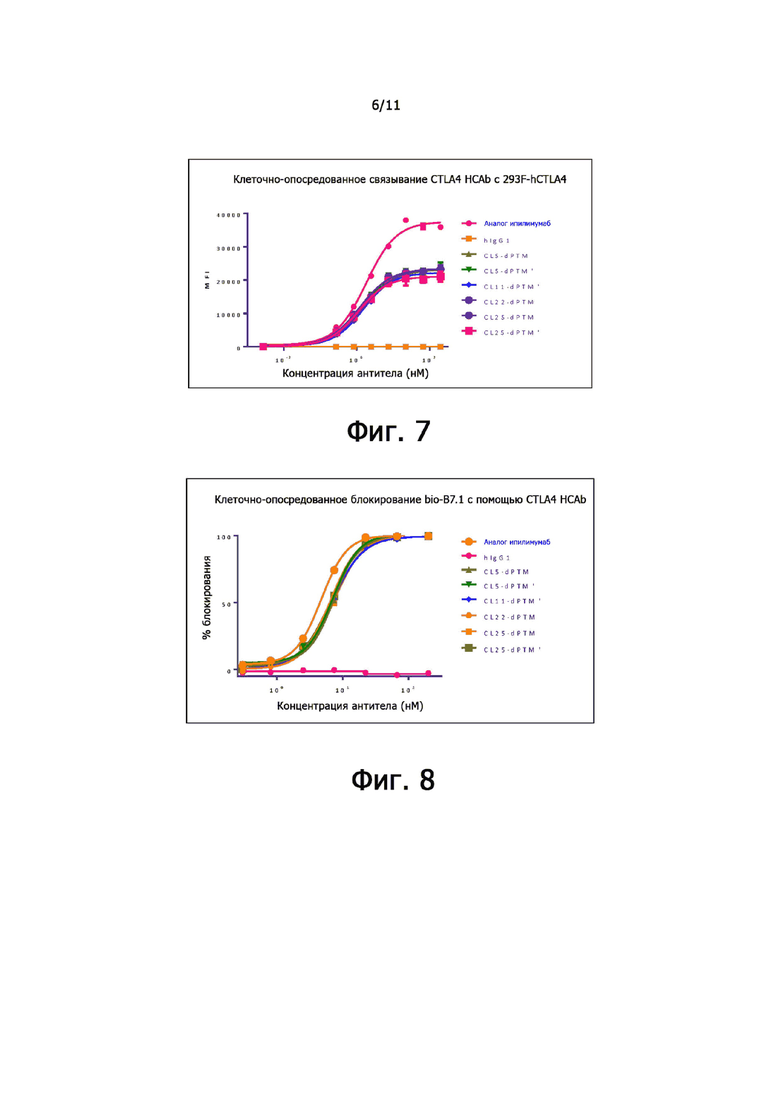
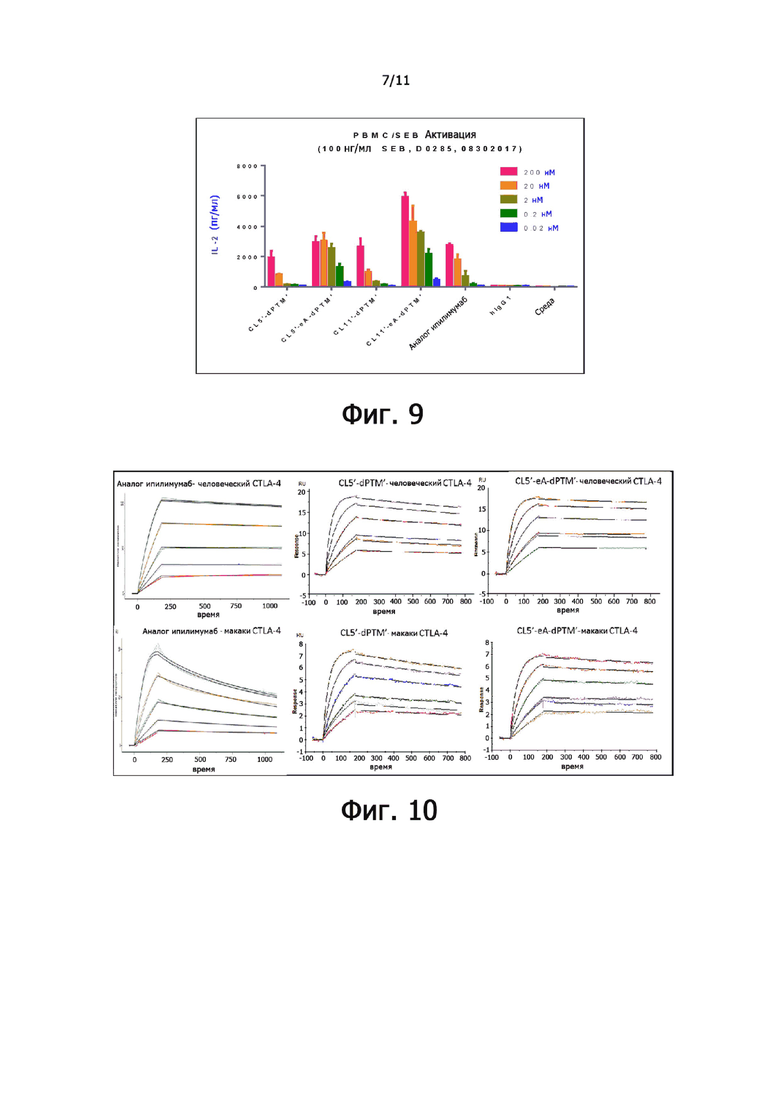

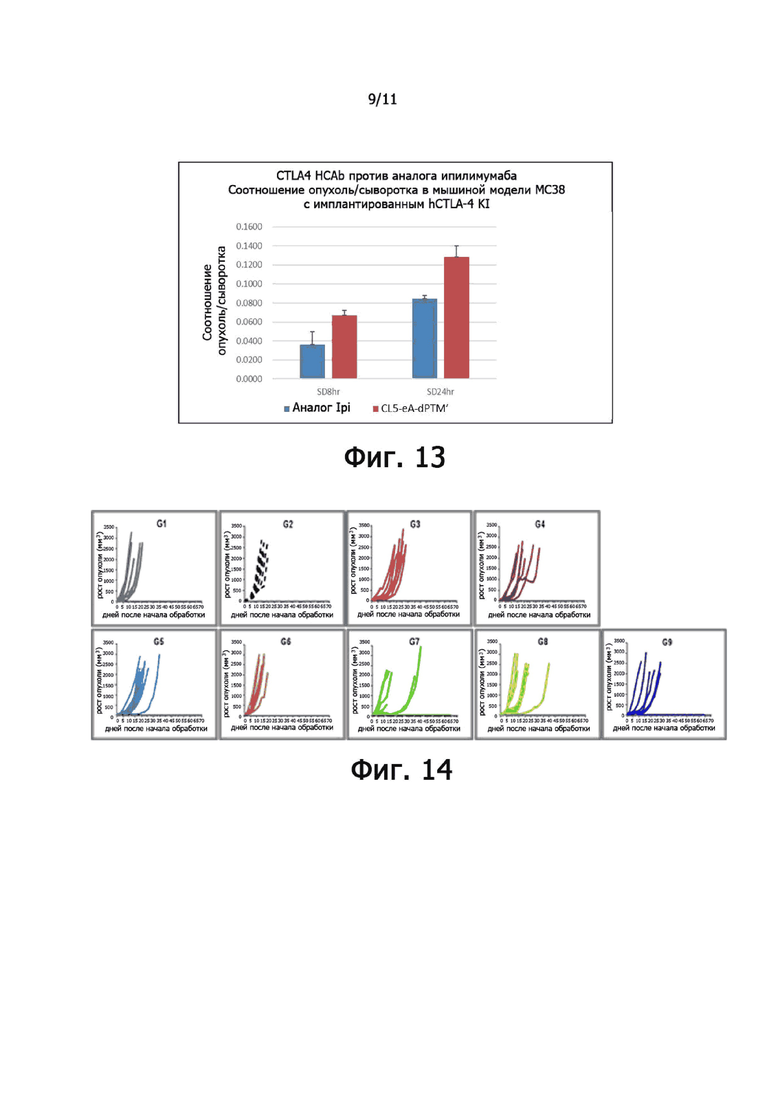
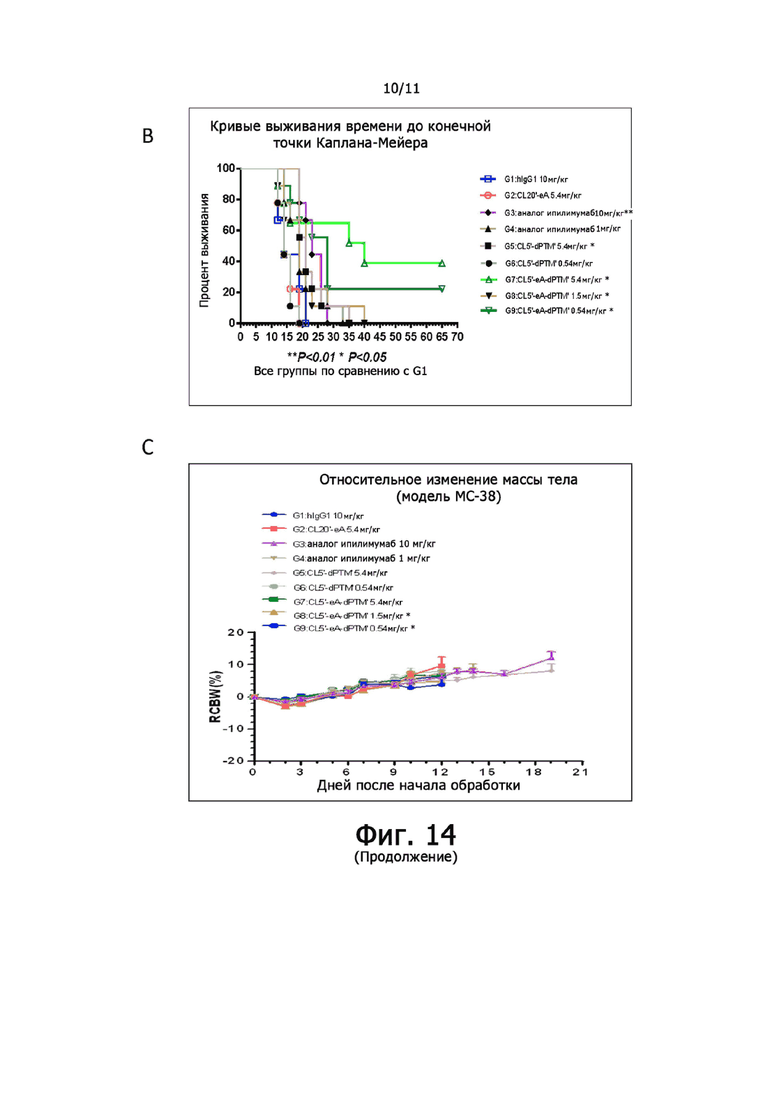
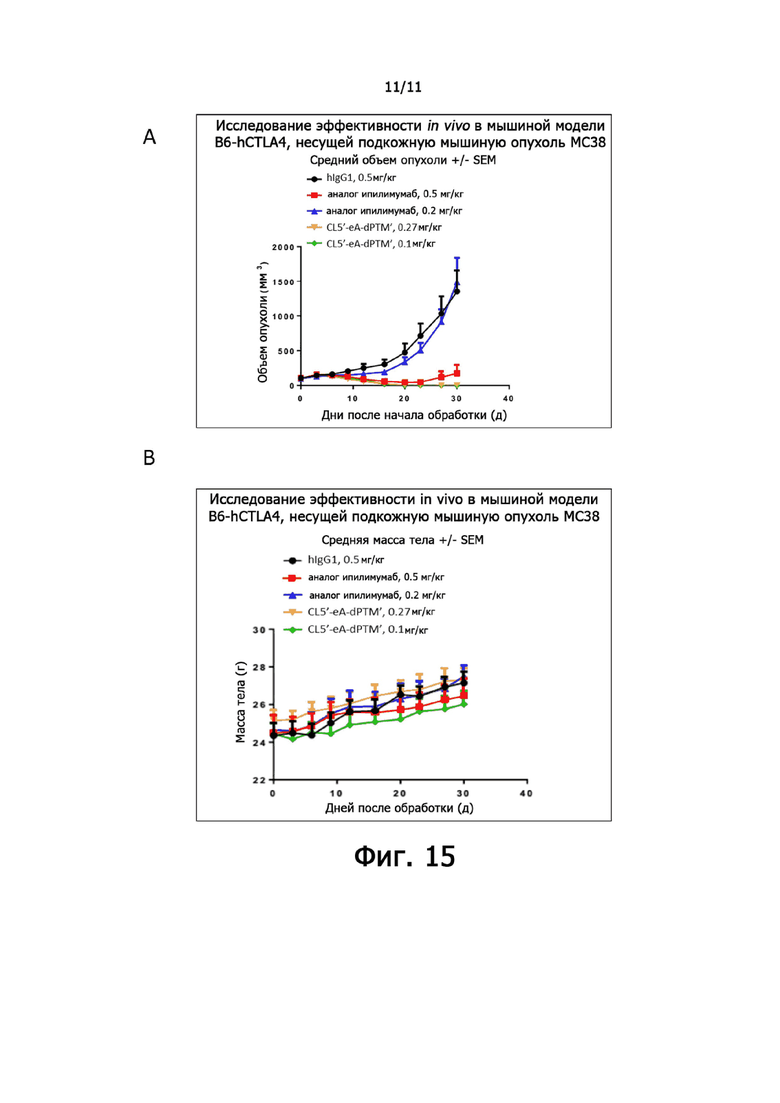
Изобретение относится к области биотехнологии. Предложено специфически связывающее CD152 антитело, включающее только тяжелые цепи. Также изобретение относится к нуклеиновой кислоте, экспрессирующему вектору и клетке-хозяину для получения антитела и к применению антитела для лечения рака и аутоиммунного заболевания. Изобретение обеспечивает эффективное ингибирование сигнальной активности CD152 с минимальными неблагоприятными побочными эффектами. 7 н. и 20 з.п. ф-лы, 15 ил., 8 табл., 16 пр.
1. Выделенное моноклональное антитело, специфически связывающее CD152, включающее CD152-связывающий домен, где CD152-связывающий домен включает вариабельную область тяжелой цепи иммуноглобулина, содержащую CDR1, CDR2 и CDR3, где CDR1, CDR2 и CDR3 содержат аминокислотные последовательности:
(1) SEQ ID NO: 4, 5 и 6, соответственно;
(2) SEQ ID NO: 10, 11 и 12, соответственно;
(3) SEQ ID NO: 16, 17 и 18, соответственно;
(4) SEQ ID NO: 22, 23 и 24, соответственно;
(5) SEQ ID NO: 28, 29 и 30, соответственно;
(6) SEQ ID NO: 34, 35 и 36, соответственно;
(7) SEQ ID NO: 40, 41 и 42, соответственно;
(8) SEQ ID NO: 46, 47 и 48, соответственно;
(9) SEQ ID NO: 52, 53 и 54, соответственно;
(10) SEQ ID NO: 58, 59 и 60, соответственно;
(11) SEQ ID NO: 64, 65 и 66, соответственно;
(12) SEQ ID NO: 70, 71 и 72, соответственно;
(13) SEQ ID NO: 76, 77 и 78, соответственно;
(14) SEQ ID NO: 82, 83 и 84, соответственно;
(15) SEQ ID NO: 88, 89 и 90, соответственно;
(16) SEQ ID NO: 94, 95 и 96, соответственно;
(17) SEQ ID NO: 100, 101 и 102, соответственно;
(18) SEQ ID NO: 106, 107 и 108, соответственно;
(19) SEQ ID NO: 112, 113 и 114, соответственно;
(20) SEQ ID NO: 118, 119 и 120, соответственно;
(21) SEQ ID NO: 124, 125 и 126, соответственно;
(22) SEQ ID NO: 130, 131 и 132, соответственно;
(23) SEQ ID NO: 136, 137 и 138, соответственно;
(24) SEQ ID NO: 142, 143 и 144, соответственно;
(25) SEQ ID NO: 148, 149 и 150, соответственно;
(26) SEQ ID NO: 154, 155 и 156, соответственно;
(27) SEQ ID NO: 160, 161 и 162, соответственно;
(28) SEQ ID NO: 166, 167 и 168, соответственно;
(29) SEQ ID NO: 172, 173 и 174, соответственно;
(30) SEQ ID NO: 178, 179 и 180, соответственно;
(31) SEQ ID NO: 184, 185 и 186, соответственно; или
(32) SEQ ID NO: 190, 191 и 192, соответственно.
2. Выделенное моноклональное антитело по п. 1, отличающееся тем, что антитело представляет собой антитело, включающее только тяжелые цепи.
3. Выделенное моноклональное антитело по п. 1 или 2, отличающееся тем, что антитело включает две тяжелые цепи иммуноглобулина.
4. Выделенное моноклональное антитело по любому из пп. 1-3, отличающееся тем, что антитело состоит из двух тяжелых цепей иммуноглобулина.
5. Выделенное моноклональное антитело по любому из пп. 1-4, отличающееся тем, что вариабельная область тяжелой цепи иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NOs: 3, 9, 15, 21, 27, 33, 39, 45, 51, 57, 63, 69, 75, 81, 87, 93, 99, 105, 111, 117, 123, 129, 135, 141, 147, 153, 159, 165, 171, 177, 183 или 189.
6. Выделенное моноклональное антитело по любому из пп. 3, 4, отличающееся тем, что, по меньшей мере, одна из двух тяжелых цепей иммуноглобулина включает аминокислотную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100% идентичности с SEQ ID NO: 2, 8, 14, 20, 26, 32, 38, 44, 50, 56, 62, 68, 74, 80, 86, 92, 98, 104, 110, 116, 122, 128, 134, 140, 146, 152, 158, 164, 170, 176, 182 или 188.
7. Выделенное моноклональное антитело по любому из пп. 1-6, которое проявляет одно или комбинацию следующих свойств: (a) связывание с CD152 человека; (b) специфически связывается с CD152 обезьяны; (c) не связывается с CD152 мыши; (d) блокирование связывания CD152 с CD80, CD86 или с обоими; (е) стимулирует секрецию IL-2 иммунными клетками; (f) индуцирует активацию Т-клеток; (g) стимулирует противоопухолевый иммунный ответ иммунными клетками.
8. Выделенное моноклональное антитело по любому из пп. 1-7, которое проявляет одно или комбинацию следующих свойств: (а) связывание с CD152 человека с аффинностью, более высокой, чем у аналога ипилимумаба; (b) связывание с CD152 человека с аффинностью, которая, по меньшей мере, в 2 раза выше, по меньшей мере, в 5 раз выше, по меньшей мере, в 10 раз выше, по меньшей мере, в 20 раз выше, по меньшей мере, в 30 раз выше, по меньшей мере, в 50 раз выше или, по меньшей мере, в 100 раз выше, чем у аналога ипилимумаба; (c) диссоциация от CD152 человека с Kd 1,0*10-9 М или менее; (d) диссоциация от CD152 человека с Kd, которая меньше, чем у аналога ипилимумаба; (e) диссоциация от CD152 человека с Kd, которая, по меньшей мере, в 2 раза меньше, по меньшей мере, в 5 раз меньше, по меньшей мере, в 10 раз меньше, по меньшей мере, в 20 раз меньше, по меньшей мере, в 30 раз меньше, по меньшей мере, в 50 раз меньше или, по меньшей мере, в 100 раз меньше, чем у аналога ипилимумаба.
9. Выделенное моноклональное антитело по любому из пп. 1-8, отличающееся тем, что антитело диссоциирует от CD152 человека с Kd 6,0*10-11 М или менее.
10. Выделенное моноклональное антитело по любому из пп. 1-9, отличающееся тем, что антитело представляет собой человеческое, гуманизированное или химерное антитело.
11. Фармацевтическая композиция для лечения рака, содержащая эффективное количество выделенного моноклонального антитела по любому из пп. 1-10 и фармацевтически приемлемое вспомогательное вещество.
12. Фармацевтическая композиция по п. 11, отличающаяся тем, что фармацевтически приемлемое вспомогательное вещество выбрано из группы, состоящей из носителей, поверхностно-активных агентов, загущающих или эмульгирующих агентов, твердых связующих веществ, дисперсионных или суспендирующих добавок, солюбилизаторов, красителей, ароматизаторов, покрытий, дезинтегрирующих агентов, смазывающих веществ, подсластителей, консервантов, изотонических агентов и их комбинации.
13. Фармацевтическая композиция по п. 11 или 12, дополнительно содержащая второе антитело, где второе антитело представляет собой иммуностимулирующее антитело или костимулирующее антитело.
14. Фармацевтическая композиция по п. 13, отличающаяся тем, что иммуностимулирующее антитело выбрано из группы, состоящей из анти-PD-1 антитела, анти-PD-L1 антитела, анти-LAG-3 антитела, анти-TIM 3 антитела, анти-STAT3 антитела и анти-ROR1 антитела.
15. Фармацевтическая композиция по п. 13 или 14, отличающаяся тем, что костимулирующее антитело представляет собой анти-CD137 антитело или анти-GITR антитело.
16. Молекула нуклеиновой кислоты, кодирующая выделенное антитело по любому из пп. 1-10.
17. Молекула нуклеиновой кислоты по п. 16, отличающаяся тем, что молекула нуклеиновой кислоты включает нуклеотидную последовательность, имеющую, по меньшей мере, 80%, 85%, 88%, 90%, 92%, 95%, 97%, 98%, 99% или 100%. идентичности с SEQ ID NO: 1, 7, 13, 19, 25, 31, 37, 43, 49, 55, 61, 67, 73, 79, 85, 91, 97, 103, 109, 115, 121, 127, 133, 139, 145, 151, 163, 169, 175, 181 или 187.
18. Экспрессирующий вектор, включающий молекулу нуклеиновой кислоты по п. 16 или 17, отличающийся тем, что молекула нуклеиновой кислоты функционально связана с регуляторными последовательностями, подходящими для экспрессии сегмента нуклеиновой кислоты в клетке-хозяине.
19. Клетка-хозяин для продуцирования антитела по любому из пп.1-10, содержащая экспрессирующий вектор по п. 18.
20. Способ индукции антителозависимой клеточно-опосредованной цитотоксичности (ADCC) против экспрессирующей CD152 клетки, причем указанный способ включает контакт NK-клетки с выделенным антителом по любому из пп. 1-10, где указанный контакт происходит в условиях, посредством чего индуцируется ADCC против экспрессирующей CD152 клетки.
21. Применение выделенного моноклонального антитела по любому из пп. 1-10 или фармацевтической композиции по любому из пп. 11-15 в приготовлении лекарственного средства для лечения расстройства, где расстройством является рак или аутоиммунное заболевание.
22. Применение по п. 21, отличающееся тем, что расстройство представляет собой рак.
23. Применение по п. 22, отличающееся тем, что рак выбран из группы, состоящей из лейкоза, лимфомы, CLL, малой лимфоцитарной лимфомы, В-клеточной лимфомы маргинальных клеток, лимфомы Беркетта, почечно-клеточного рака, рака толстой кишки, колоректального рака, рака молочной железы, эпителиального плоскоклеточного рака, меланомы, миеломы, рака желудка, рака головного мозга, рака легкого, рака поджелудочной железы, рака шейки матки, рака яичников, рака печени, рака мочевого пузыря, рака предстательной железы, рака яичка, рака щитовидной железы и рака головы и шеи.
24. Применение по любому из пп. 21-23, отличающееся тем, что лекарственное средство дополнительно включает дополнительный терапевтический агент.
25. Применение по п. 24, отличающееся тем, что дополнительный терапевтический агент представляет собой противоопухолевый агент.
26. Применение по п. 24, отличающееся тем, что дополнительным терапевтическим агентом является ипилимумаб и его биоподобный продукт.
27. Применение по п. 21, отличающееся тем, что расстройство представляет собой аутоиммунное заболевание.
| CN 106220732 A, 14.12.2016 | |||
| CN 106046164 A, 26.10.2016 | |||
| ПРИМЕНЕНИЕ АНТИТЕЛ К CTLA-4 | 2005 |
|
RU2346702C2 |
| КОМПОЗИЦИЯ АНТИТЕЛ ПРОТИВ CTLA-4 | 2006 |
|
RU2356579C1 |
| CALLAHAN M | |||
| K | |||
| et al., Anti-CTLA-4 Antibody Therapy: Immune Monitoring During Clinical Development of a Novel Immunotherapy, Seminars in Oncology, Vol.37, No.5, 2010, pp.473-484. | |||

