Перекрестная ссылка
Данная заявка испрашивает приоритет предварительной заявки на патент США №62/352698, поданной 21 июня 2016 года, предварительной заявки на патент США №62/394360, поданной 14 сентября 2016 года и предварительной заявки на патент США №62/491908, поданной 28 апреля 2017 года, которые включены в данный документ посредством ссылки в полном объеме.
Уровень техники
Иммунная система организма служит защитой от инфекции, травм и рака. Две отдельные, но взаимосвязанные системы, гуморальная и клеточная иммунные системы, работают вместе, чтобы защитить организм. Гуморальная система опосредуется растворимыми факторами, называемыми антителами, которые нейтрализуют продукты, признанные организмом чужеродными. Тогда как, клеточная система включает клетки, такие как Т-клетки и макрофаги, которые удаляют и нейтрализуют чужеродные вторжения.
Активация Т-клеток имеет решающее значение для стимуляции иммунных реакций. Т-клетки проявляют иммунологическую специфичность и направляют большинство клеточных иммунных ответов. Хотя Т-клетки не секретируют антитела, они необходимы для секреции антител В-лимфоцитами. Активация Т-клеток требует участия ряда молекул клеточной поверхности, таких как рецепторный комплекс Т-клеток, и молекулы CD4 или CD8. Антиген-специфический Т-клеточный рецептор (ТКР) состоит из дисульфид-связанного гетеродимера, мембранного гликопротеина с цепями, альфа и бета (α и β) или гамма и дельта (γ и δ). ТКР нековалентно связан с комплексом инвариантных белков, обозначенных CD3.
Известно, что Т-клетки оказывают сильное противоопухолевое действие в многочисленных экспериментальных условиях. Антитела, способные эффективно вовлекать Т-клетки против опухолевых клеток, были доступны в виде биспецифических антител, например, направленных на антигены, ассоциированные с опухолью (ТАА - tumor-associated antigen) и агонистические мембранные белки Т-клеток, такие как комплекс ТКР/CD3 и CD28. Эти биспецифические антитела способны активировать Т-клетки независимо от специфичности их ТКР, что приводит к специфическому лизису клеток, несущих соответствующие TAA.
Однако, хотя биспецифические антитела к CD3 могут перенаправлять лизис, опосредованный Т-клетками, на злокачественные клетки, клинические испытания с применением бсАт на основе CD3 показали высокую токсичность у пациентов. Неспецифическая активация Т-клеток от бсАт может происходить антиген-независимым образом из-за взаимодействия Fc/Fc-рецептора (FcR) или антиген-зависимым образом, в случае если антиген экспрессируется как на нормальных, так и на опухолевых клетках. Оба механизма могут быть ответственны за токсичность, наблюдаемую в предыдущих клинических исследованиях (См., например, Link и соавт. (1998 год) Int. J. Cancer 77(2):251-6; Durben и соавт. Molecular Therapy (2015 год); 23 4, 648-655). Из-за возникающего в результате синдрома высвобождения цитокинов имели место значительные ограничения в разработке этих антител для терапевтических целей.
Например, Mack и соавт. (1995 год), PNAS 92:7021-7025 сообщали о биспецифическом одноцепочечном (бсоц) - антителе с CD3-специфичностью, обозначаемом как биспецифический вовлекатель Т-клеток (BiTE - bispecific T-cell engager). Однако как и в более ранних исследованиях с различными биспецифическими антителами к CD3, чрезмерная активация Т-клеток и высвобождение цитокинов, индуцированные при применении in vivo, ограничивают безопасные применимые дозы до менее чем 100 мкг/день, что приводит к концентрации в сыворотке ниже 1 нг/мл.
CD3-специфические антитела и биспецифические антитела, полученные из них, обеспечиваются данным изобретением.
Публикации
Антитела к CD3 описаны, например, в патентах США% 5585097; 5929212; 5968509; 6706,265; 6750325; 7381803; 7728114. Биспецифические антитела со специфичностью связывания CD3 описаны, например, в патентах США №7262276; 7635472; 7862813; и 8236308, каждый из которых специально включен в данный документ посредством ссылки.
Сущность изобретения
Композиции и способы их применения предоставлены для семейства родственных антител, которые связываются и активируют передачу сигналов посредством CD3, например, активацией CD3+ T-клеток. Антитела в семействе содержат набор последовательностей CDR, как определено в данном документе. Семейство антител обеспечивает ряд преимуществ, которые способствуют полезности в качестве клинического лекарственного средства(в). Антитела в семействе включают членов с рядом аффинностей связывания, позволяющих выбирать конкретную последовательность с желаемой аффинностью. Способность точно настраивать аффинность имеет особое значение для управления уровнем активации CD3 у индивида, которого лечат, и, таким образом, для снижения токсичности. Члены семейства антител могут обладать аффинностью (KD) к CD3 в диапазоне от около 10-6 до около 10-11 M. Некоторые члены семейства антител по данному изобретению перекрестно реагируют с белком CD3 яванского макака и для такой перекрестной реакции идентифицируется необходимый специфический мотив, что позволяет отбирать антитела для доклинических или клинических испытаний на этой основе.
Антитела к CD3, которые имеют аффинность (KD) 50 нМ или более, 100 нМ или более, 500 нМ или более или 1 мкМ, или более, могут быть желательными для более точной имитации взаимодействия ТКР/ГКГС (главный комплекс гистосовместимости) и минимизации высвобождения токсических цитокинов при поддержании эффективного лизиса опухолевых клеток. В некоторых вариантах реализации изобретения антитела к CD3 характеризуются или отбираются для снижения склонности индуцировать высвобождение цитокинов при связывании с компетентной Т-клеткой, например, для высвобождения ИЛ-2 и ИФНγ. Антитела могут быть отобраны для терапевтического применения, которое оптимизирует уничтожение опухолевых клеток и сниженное высвобождение цитокинов, например антитело, в семействе последовательностей антител, описанных в данном документе, индуцирует высвобождение цитокинов, которое составляет менее чем около половины от максимума, наблюдаемого для члена семьи в сравнительном анализе, и может быть меньше, например, меньше и около 25% от максимума, наблюдаемого для члена семьи в сравнительном анализе.
В некоторых вариантах реализации изобретения предлагаются биспецифические или мультиспецифические антитела, которые содержат по меньшей мере вариабельную область тяжелой цепи из семейства антител по данному изобретению и могут содержать вариабельную область тяжелой и легкой цепи, представленную в данном документе. Биспецифические антитела содержат по меньшей мере вариабельную область тяжелой цепи антитела, специфичного к белку, отличному от CD3, и могут содержать вариабельную область тяжелой и легкой цепи. В некоторых таких вариантах реализации изобретения вторая специфичность антитела связывается с антигеном, ассоциированным с опухолью, нацеливающим антигеном, например, интегринами и тому подобным, патогенным антигеном, белком контрольной точки и тому подобным. Различные форматы биспецифических антител находятся в пределах объема изобретения, включая без ограничения одноцепочечные полипептиды, двухцепочечные полипептиды, трехцепочечные полипептиды, четырехцепочечные полипептиды и кратные им.
Семейство CD3-специфических антител по данному изобретению содержит домен VH, содержащий последовательности CDR1, CDR2 и CDR3 в каркасе VH человека. Последовательности CDR могут быть расположены, например, в области около аминокислотных остатков 26-35; 53-59; и 98-117 для CDR1, CDR2 и CDR3, соответственно, предоставленных типовых последовательностей вариабельной области, изложенных в SEQ ID NO: 1-68. Специалисту в данной области техники должно быть понятно, что последовательности CDR могут находиться в другом положении, в случае если выбрана другая каркасная последовательность, хотя обычно порядок последовательностей будет оставаться тем же.
Последовательности CDR для антитела по данному изобретению могут иметь следующие формулы последовательности. Х обозначает вариабельную аминокислоту, которая может представлять собой специфические аминокислоты, как указано ниже.
CDR1
при этом:
X5 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X5 представляет собой S или R;
X7 и X8 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X7 и X8 независимо представляют собой S или G. В некоторых вариантах реализации изобретения X7X8 представляют собой SS или GG;
X9 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X9 представляет собой H или Y; в некоторых вариантах реализации изобретения X9 представляет собой H;
X10 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X10 представляет собой Y или F; в некоторых вариантах реализации изобретения X10 представляет собой Y.
В некоторых вариантах реализации изобретения последовательность CDR1 имеет формулу: G G S I X5 S H H G Y, при этом X5 такой, как определено выше. В некоторых вариантах реализации изобретения последовательность CDR1 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 26-35.
CDR2
при этом:
X2' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X2' представляет собой S, Y или H;
X3' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X3' представляет собой Y, H или R;
X6' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X6' представляет собой S, N или I, или R;
X7' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X7' представляет собой T или P.
В некоторых вариантах реализации изобретения последовательность CDR2 имеет формулу: I X2' X3' S G S T; или I X2' X3' S G N P при этом X2’ и X3’ такие, как определено выше. В некоторых вариантах реализации изобретения последовательность CDR2 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 53-59.
CDR3
X1'' R2'' W3'' R4'' H5'' D6'' I7'' X8'' X9'' X10'' Y11'' P12'' Y13'' Y14'' Y15'' Y16'' G17'' M18'' D19'' V20''
при этом:
X1'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X1'' представляет собой A или G.
X8'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X8'' представляет собой L или F.
X9'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X9'' представляет собой T или A
X10'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X10'' представляет собой G, A или R.
В некоторых вариантах реализации изобретения X8'' X9'' X10'' представляют собой F A A, мотив которого соответствует антителам, которые перекрестно реагируют с белком CD3 яванского макака. В других вариантах реализации изобретения X8'' X9'' X10'' представляют собой L T A. В некоторых вариантах реализации изобретения последовательность CDR3 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 98-117.
В некоторых вариантах реализации изобретения CD3-связывающий домен VH связан с доменом вариабельной области легкой цепи. В некоторых таких вариантах реализации изобретения легкая цепь представляет собой фиксированную легкую цепь. В некоторых вариантах реализации изобретения легкая цепь содержит домен VL с последовательностями CDR1, CDR2 и CDR3 в каркасе VL человека. Последовательности CDR могут быть теми, которые содержатся в SEQ ID NO: 69. В некоторых вариантах реализации изобретения последовательность CDR содержит аминокислотные остатки 27-32; 50-52; 89-97 для CDR1, CDR2, CDR3 соответственно.
В некоторых вариантах реализации изобретения последовательности CDR антитела по данному изобретению представляют собой последовательность с по меньшей мере 85% идентичностью, по меньшей мере 90% идентичностью, по меньшей мере 95% идентичностью, по меньшей мере 99% идентичностью относительно последовательности CDR или набора последовательностей CDR в любой из SEQ ID NO: 1-69. В некоторых вариантах реализации изобретения последовательность CDR по данному изобретению содержит одну, две, три или более аминокислотных замен по отношению к последовательности CDR или набора последовательностей CDR в любой из SEQ ID NO:1-69. В некоторых вариантах реализации изобретения указанная аминокислотная замена(ы) представляет собой одну или более в положении 5 или 10 из CDR1, положении 2, 6 или 7 из CDR2, положении 1, 8, 9 или 10 из CDR3 относительно формул, представленных выше.
В некоторых вариантах реализации изобретения биспецифическое антитело по данному изобретению содержит CD3-связывающую вариабельную область, описанную в данном документе, спаренную с легкой цепью. В некоторых вариантах реализации изобретения легкая цепь содержит последовательность вариабельной области, изложенную в SEQ ID NO: 69, или вариабельную область, содержащую набор последовательностей CDR в SEQ ID NO: 69, и каркасные последовательности. Различные последовательности Fc находят применение, включая, без ограничения, человеческие IgG1, IgG2a, IgG2b, IgG3, IgG4 и тому подобные. В некоторых вариантах реализации изобретения второе плечо биспецифического антитела содержит вариабельную область, которая специфически связывается с антигеном, ассоциированным с опухолью. В некоторых вариантах реализации изобретения второе плечо биспецифического антитела содержит вариабельную область, которая специфически связывается с ВСМА. В некоторых вариантах реализации изобретения плечо к BCMA представляет собой одноцепочечную вариабельную область, например, как изображено на фиг 2В. В некоторых вариантах реализации изобретения плечо к BCMA содержит последовательность вариабельной области, изложенную в SEQ ID NO: 70; или тандемную последовательность вариабельной области, изложенную в SEQ ID NO: 71. Последовательность Fc плеча к BCMA может представлять собой, без ограничения, IgG1, IgG2a, IgG2b, IgG3, IgG4 человека и тому подобное. Последовательности CDR могут быть теми, которые содержатся в SEQ ID NO: 70. В некоторых вариантах реализации изобретения последовательность CDR содержит аминокислотные остатки 26-33; 51-58; 97-108 для CDR1, CDR2, CDR3 соответственно.
В других вариантах реализации изобретения предлагаются фармацевтические композиции, содержащие по меньшей мере CD3-связывающий домен VH по данному изобретению, например, моноспецифическое, биспецифическое и тому подобное антитело или антителоподобный белок, содержащий по меньшей мере CD3-связывающий домен VH по данному изобретению; и фармацевтически приемлемый наполнитель. Композиция может быть лиофилизирована, суспендирована в растворе и тому подобное. И может быть представлена в виде стандартной дозы.
В некоторых вариантах реализации изобретения предложен способ лечения рака, включающий введение индивиду, имеющему для этого показания, эффективной дозы моноспецифического, биспецифического и тому подобного антитела по данному изобретению. Когда антитело является биспецифическим, второй антигенсвязывающий участок может специфически связывать опухолевый антиген, белок контрольной точки и тому подобное. В различных вариантах реализации изобретения рак выбран из группы, состоящей из рака яичников, рака молочной железы, желудочно-кишечного тракта, рака головного мозга, рака головы и шеи, рака предстательной железы, рака толстой кишки, рака легких, лейкемии, лимфомы, саркомы, карциномы, опухолей нервных клеток, плоскоклеточного рака, эмбрионально-клеточных опухолей, метастазов, недифференцированных опухолей, семином, меланом, миелом, нейробластом, опухолей смешанных клеток и неоплазий, вызванных инфекционными агентами.
В некоторых вариантах реализации изобретения предложен способ лечения инфекционного заболевания, включающий введение индивидууму, имеющему для этого показания, эффективной дозы моноспецифического, биспецифического и тому подобного антитела по данному изобретению. Когда антитело является биспецифическим, второй антигенсвязывающий участок может специфически связывать патогенный антиген, например, бактерии, вирусы или паразиты.
В других вариантах реализации изобретения предложен способ получения биспецифического антитела по данному изобретению, включающий экспрессию последовательностей антитела, например, одной или более кодирующих последовательностей легкой цепи, одной или более кодирующих последовательностей тяжелой цепи в одной клетке-хозяине. В различных вариантах реализации изобретения клетка-хозяин может представлять собой прокариотическую или эукариотическую клетку, такую как клетка млекопитающего.
Краткое описание графических материалов
Изобретение лучше всего будет понятно из последующего подробного описания при чтении вместе с прилагаемыми графическими материалами. Файл патента или заявки содержит по меньшей мере одно графическое изображение, выполненное в цвете. Копии этого патента или патентной заявки с цветным графическим материалом(и) будут предоставлены патентным ведомством по запросу и после уплаты необходимого взноса. Следует подчеркнуть, что в соответствии со установившейся практикой согласно общепринятой практике, различные объекты графических материалов проиллюстрированы без соблюдения масштаба. Напротив, размеры различных объектов произвольно увеличены или уменьшены для ясности. В графические материалы включены следующие фигуры.
Фиг. 1A-1B. (FAM1_aCD3_CDR_seqalign). На фиг. 1A проиллюстрировано выравнивание областей CDR1, 2 и 3 всех членов семейства антител, распознающих CD3 человека. Последовательности CDR соответствуют аминокислотным остаткам 26-35; 53-59; и 98-117 для CDR1, CDR2 и CDR3 соответственно, из последовательностей вариабельной области, указанных в SEQ ID NO: 1-68. На фиг. 1B проиллюстрированы области CDR1, 2 и 3 фиксированной легкой цепи (SEQ ID NO: 69); и типовая последовательность к ВСМА (SEQ ID NO: 70 и SEQ ID NO: 71).
На фиг. 2A-2E проиллюстрированы схематические модели биспецифических антител человека. На фиг. 2A проиллюстрировано биспецифическое антитело к CD3:опухолевому антигену с общей легкой цепью (всего 3 уникальные цепи). На фиг. 2B проиллюстрировано биспецифическое антитело к CD3:опухолевому антигену с 2 уникальными легкими цепями (всего 4 уникальные цепи). На фиг. 2C проиллюстрировано биспецифическое антитело к CD3:опухолевому антигену с тяжелой цепью только опухолевого антигенсвязывающего домена (3 уникальные цепи). На фиг. 2D проиллюстрировано биспецифическое антитело к CD3:опухолевому антигену с опухолевым антигенсвязывающим доменом scFv (всего 3 уникальные цепи). На фиг. 2E проиллюстрировано биспецифическое антитело к CD3:опухолевому антигену с связывающим доменом scFv к CD3 (всего 3 уникальные цепи).
На фиг. 3 проиллюстрирована активация CD8+ T-клеток человека с помощью биспецифического FlicAb α-CD3/α-PD-L1. Очищенные CD8+ Т-клетки человека были сокультивированы с биспецифическими в указанных концентрациях и опухолевыми клетками. Ramos (В-клеточная лимфома) представляет собой клеточную линию, отрицательную по PD-L1, а HDLM2 (множественная миелома) представляет собой клеточную линию, положительную по PD-L1. CD69 представляет собой мембранную молекулу, которая активируется на активированных Т-клетках. Средняя интенсивность флуоресценции (СИФ) человеческих CD8+ Т-клеток, окрашенных флуоресцентно-меченным моноклональным антителом к CD69, коррелирует со степенью активации. Активация зависела от присутствия как опухолевых клеток, экспрессирующих PD-L1, так и биспецифических.
На фиг. 4 проиллюстрирован цитолиз опухолевых клеток с помощью биспецифического FlicAb α-CD3/α-PD-L1. Опухолевые клетки (HDLM2) инкубировали с очищенными человеческими CD8+ T-клетками и биспецифическими антителами. Клетки HDLM2 не экспрессируют CD20, и сокультура с биспецифическим FlicAb α-CD3/α-CD20 не приводила к гибели клеток HDLM2. Только сокультура человеческих CD8+ Т-клеток и HDLM2 с биспецифическим FlicAb α-CD3/α-PD-L1 приводит к значительному уничтожению. HDLM2 экспрессируют PD-L1 на своей поверхности.
На фиг. 5 проиллюстрирована таблица, обобщающая состояние антител к CD3 в моноспецифическом и биспецифическом формате. В столбце 1 показан идентификатор последовательности для последовательности VH к CD3. В столбце 2 показано значение СИФ для связывания клетками Jurkat исходного моноспецифического (антитела) к CD3. В столбце 3 показано значение СИФ для связывания T-клетками яванского макака исходного моноспецифического (антитела) к CD3. В столбце 4 показано название биспецифического антитела aCD3:aBCMA. В столбце 5 показаны пикограммы ИЛ-2, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 6 показаны пикограммы ИЛ-6, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 7 показаны пикограммы ИЛ-10, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 8 показаны пикограммы ИФН-γ, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 9 показаны пикограммы ФНОα, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 10 показана ЭК50 лизиса опухолевых клеток U266, опосредованного биспецифическими антителами, в присутствии пан-Т-клеток человека. В столбце 11 показан процент лизиса опухолевых клеток U266, в присутствии биспецифического антитела и пан-Т-клеток человека при дозе 333 нг/мл биспецифического антитела. В столбце 12 показана аффинность белкового связывания плеча к CD3 биспецифического антитела, измеренная с помощью Octet. В столбце 13 показано значение СИФ для связывания клетками Jurkat биспецифического антитела.
На фиг. 6 проиллюстрирован опосредованный биспецифическими антителами лизис опухолевых клеток. Четыре биспецифических антитела αCD3_fam1:aBCMA, каждое с уникальным плечом к CD3 и общим плечом к BCMA, были протестированы на способность убивать опухолевые клетки U266 BCMA+ посредством перенаправления активированных первичных Т-клеток. В этом эксперименте клетки U266, которые экспрессируют BCMA, смешивали с активированными пан-Т-клетками в соотношении E:T 10:1 наряду с добавлением биспецифического антитела. Ось x демонстрирует концентрацию применяемого антитела, а ось y демонстрирует % лизиса опухолевых клеток через 6 часов после добавления антитела.
На фиг. 7 проиллюстрирована биспецифическая активность уничтожения U266, коррелирующая с высвобождением ИЛ-2. Сравнение активности лизиса опухолевых клеток, опосредованной биспецифическими антителами, с высвобождением цитокина ИЛ-2 изображено на диаграмме рассеяния. Корреляция между производством ИЛ-2 и лизисом опухолевых клеток U266 составляет R2= 0,37.
На фиг. 8 проиллюстрирована биспецифическая активность уничтожения U266, коррелирующая с высвобождением ИФН-γ. Сравнение активности лизиса опухолевых клеток, опосредованной биспецифическими антителами, с высвобождением цитокина ИФН-γ изображено на диаграмме рассеяния. Корреляция между производством ИФН-γ и лизисом опухолевых клеток U266 составляет R2=0,53.
На фиг. 9 проиллюстрирована биспецифическая активность уничтожения U266, коррелирующая с аффинностью связывания с CD3. Сравнение активности лизиса опухолевых клеток U266, опосредованной биспецифическими антителами, с аффинностью связывания с CD3 изображено на диаграмме рассеяния. Корреляция между ЭК50 уничтожения U266 и аффинностью связывания белка составляет R2=0,93.
На фиг. 10A-10D проиллюстрирован опосредованный биспецифическими антителами лизис опухолевых клеток. Биспецифические антитела αCD3_F1F: αBCMA анализировали на способность уничтожать три различных линии опухолевых клеток ВСМА+ и одну линию ВСМА-негативных клеток путем перенаправления активированных исходных Т-клеток. Антитела состояли из плеча αCD3 (SEQ ID NO: 1 и SEQ ID NO: 69) и плеча αBCMA (SEQ ID NO: 70 или SEQ ID NO: 71). В этом эксперименте опухолевые клетки смешивали с активированными пан-Т-клетками в соотношении E:T 10:1 наряду с добавлением биспецифического антитела. На фиг. 10A проиллюстрировано уничтожение клеток RPMI-8226. На фиг. 10B проиллюстрировано уничтожение клеток NCI-H929, на фиг. 10C проиллюстрировано уничтожение клеток U-266 и на фиг. 10D проиллюстрировано уничтожение клеток K562, отрицательный контроль. Ось x демонстрирует концентрацию применяемого антитела, а ось y демонстрирует % лизиса опухолевых клеток через 6 часов после добавления антитела.
На фиг. 11A-11D проиллюстрировано опосредованное биспецифическими антителами высвобождение ИЛ-2. Уровень высвобождения цитокинов ИЛ-2 измеряли после культивирования Т-клеток человека в покое с различными линиями опухолевых клеток и увеличивающимися дозами биспецифического антитела aCD3_F1F:aBCMA (как проиллюстрировано на фиг. 10). На фиг. 11A проиллюстрировано высвобождение ИЛ-2, стимулированное клетками RPMI-8226, на фиг. 11B проиллюстрировано высвобождение ИЛ-2, стимулированное клетками NCI-H929, на фиг. 11C проиллюстрировано высвобождение ИЛ-2, стимулированное клетками U-266 и на фиг. 11D проиллюстрировано высвобождение ИЛ-2, стимулированное клетками K562, отрицательный контроль.
На фиг. 12A-12D проиллюстрировано опосредованное биспецифическими антителами высвобождение ИФН-γ. Уровень высвобождения цитокина ИФН-γ измеряли после культивирования Т-клеток человека в покое с различными линиями опухолевых клеток и увеличивающимися дозами биспецифического антитела aCD3_F1F:aBCMA (как проиллюстрировано на фиг. 10). На фиг. 12A проиллюстрировано высвобождение ИФН-γ, стимулированное клетками RPMI-8226, на фиг. 12B проиллюстрировано высвобождение ИФН-γ, стимулированное клетками NCI-H929, на фиг. 12C проиллюстрировано высвобождение ИФН-γ, стимулированное клетками U-266 и на фиг. 12D проиллюстрировано высвобождение ИФН-γ, стимулированное клетками K562, отрицательный контроль.
Подробное описание сущности изобретения
Чтобы облегчить понимание этого изобретения, ряд терминов определен ниже.
Перед описанием данных активных агентов и способов, следует понимать, что это изобретение не ограничивается конкретной описанной методологией, продуктами, устройствами и факторами, поскольку такие способы, устройства и составы, конечно, могут видоизменяться. Следует также понимать, что применяемая в данном документе терминология предназначена только для описания конкретных вариантов реализации изобретения и не предназначена для ограничения объема данного изобретения, которое будет ограничено только прилагаемой формулой изобретения.
Необходимо отметить, что в данном документе и в прилагаемой формуле изобретения формы единственного числа включают обозначаемые объекты во множественном числе, если иное непосредственно не следует из контекста. Таким образом, например, ссылка на "потенциальный лекарственный препарат" относится к одному или более таким потенциальным кандидатам, а ссылка на "способ" включает ссылки на эквивалентные этапы и способы, известные специалистам в данной области техники, и так далее.
Если не указано иное, все употребляемые в данном документе технические и научные термины имеют значения, обычно подразумеваемые специалистом в данной области техники, к которой принадлежит изобретение. Все публикации, упомянутые в данном документе, включены в данный документ посредством ссылки с целью описания и раскрытия устройств, составов и методов, которые описаны в публикации и которые могут применяться в связи с описанным выше изобретением.
В случае, когда предусмотрен диапазон значений, следует понимать, что каждое промежуточное значение, до десятой единицы нижнего предела, если из контекста явно не следует иное, между верхним и нижним пределом этого диапазона и любое другое указанное или промежуточное значение в этом указанном диапазоне охватываются в пределах изобретения. Верхний и нижний пределы этих меньших диапазонов могут быть независимо включены в меньшие диапазоны, а также включены в данное изобретение, с учетом любого конкретно исключенного предела в указанном диапазоне. В тех случаях, когда указанный диапазон включает одно или оба из этих пределов, в изобретение также включены диапазоны, исключающие любой или оба из этих включенных пределов.
В последующем описании изложены многочисленные конкретные подробности с целью обеспечения более полного понимания данного изобретения. Однако для специалиста в данной области техники будет очевидно, что данное изобретение может применяться на практике и без одной или более указанных конкретных подробностей. В других случаях общеизвестные особенности и процедуры, хорошо известные специалистам в данной области техники, не описываются с целью избегания затруднения понимания изобретения.
Обычно в данном изобретении применяются общепринятые способы синтеза белка, культуры рекомбинантных клеток и выделения белка и методы рекомбинантной ДНК, известные специалистам в данной области техники. Такие методы подробно описаны в литературе, см., например, Maniatis, Fritsch & Sambrook, Molecular Cloning:A Laboratory Manual (1982 год); Sambrook, Russell and Sambrook, Molecular Cloning:A Laboratory Manual (2001 год); Harlow, Lane and Harlow, Using Antibodies:A Laboratory Manual:Portable Protocol No. I, Cold Spring Harbor Laboratory (1998 год); и Harlow and Lane, Antibodies:A Laboratory Manual, Cold Spring Harbor Laboratory; (1988 год).
Определения терминов
Под термином "содержащий" подразумевается, что перечисленные элементы требуются для композиции/способа/набора, однако для формирования композиции/способа/набора и тому подобного в рамках формулы изобретения могут быть включены и другие элементы.
Под термином "состоящий по существу из” подразумевается ограничение рамок описанной композиции или способа указанными материалами или поэтапными действиями, не оказывающими существенного влияния на основную и новую характеристику (характеристики) объекта изобретения.
Под термином “состоящий из” подразумевается исключение любого элемента, поэтапного действия или ингредиента, не указанного в формуле изобретения, из композиции, способа или набора.
Термины “лекарственное средство”, “лечение” и тому подобное используются в данном документе для общего обозначения желательного фармакологического и/или физиологического эффекта. Эффект может быть профилактическим в смысле полного или частичного предотвращения заболевания или его симптома, и/или терапевтическим в смысле частичного или полного излечения заболевания и/или нежелательного эффекта, относящегося к указанному заболеванию. Термин “лечение” в данном документе охватывает любое лечение заболевания у млекопитающего, и включает: (a) профилактику возникновения заболевания у субъекта, который, возможно, предрасположен к данному заболеванию, однако которому не поставлен диагноз данного заболевания; (b) ингибирование заболевания, то есть, остановку его развития; или (c) облегчение заболевания, то есть, регрессию заболевания. Терапевтический агент можно вводить до, во время или после начала заболевания или травмы. Лечение текущего заболевания, при котором лечение стабилизирует или ослабляет нежелательные клинические симптомы у пациента, представляет особый интерес. Такое лечение желательно выполнять до полной потери функции пораженных тканей. Рассматриваемое лекарственное средство можно вводить во время симптоматической стадии заболевания, и, в некоторых случаях, после симптоматической стадии заболевания.
"Терапевтически эффективное количество" означает количество активного агента, необходимое для получения терапевтического благоприятного эффекта у субъекта. Например, “терапевтически эффективное количество” представляет собой количество, индуцирующее, облегчающее или иным образом вызывающее улучшение патологических симптомов, прогрессирования заболевания или физиологических состояний, ассоциированных с заболеванием или, или улучшающее устойчивость к расстройству.
Термины “субъект”, “индивид” и “пациент” в данном документе используются на равных основаниях и относятся к млекопитающему, подвергаемому оценке на предмет лечения и/или лечению. В одном варианте реализации млекопитающее является человеком. Термины “субъект”, “индивид” и “пациент” без ограничений охватывают индивидов с раком, аутоиммунными заболеваниями, патогенными инфекциями и тому подобным. Субъекты могут являться людьми, но включают и других млекопитающих, особенно пригодных для применения в качестве лабораторных моделей заболеваний человека, например, мышь, крысу и тому подобное.
Термины "рак", "новообразование" и "опухоль" применяются в данном документе взаимозаменяемо для обозначения клеток, которые демонстрируют автономный нерегулируемый рост таким образом, что они демонстрируют аберрантный фенотип роста, характеризующийся значительной потерей контроля над пролиферацией клеток. Клетки, представляющие интерес для обнаружения, анализа или лечения в данной заявке, включают предраковые (например, доброкачественные), злокачественные, предметастатические, метастатические и неметастатические клетки. Рак практически каждой ткани является известным. Фраза "раковая нагрузка" относится к количеству раковых клеток или объему рака у субъекта. Соответственно, снижение раковой нагрузки означает уменьшение количества раковых клеток или объема рака у субъекта. В данном контексте термин "раковая клетка" относится к любой клетке, которая представляет собой раковую клетку или получена из раковой клетки, например, клона раковой клетки. Специалистам в данной области техники известны многие типы раковых заболеваний, включая солидные опухоли, такие как карциномы, саркомы, глиобластомы, меланомы, лимфомы, миеломы и тому подобное, и циркулирующие злокачественные новообразования, такие как лейкемии, включая, в частности, В-клеточные лейкемии, Т-клеточные лейкемии и тому подобное. Примеры рака включают, но без ограничений, рак яичников, рак молочной железы, рак толстой кишки, рак легких, рак простаты, гепатоцеллюлярный рак, рак желудка, рак поджелудочной железы, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, рак мочевыводящих путей, рак щитовидной железы, рак почки, карциному, меланому, рак головы и шеи и рак головного мозга.
Термины "антителозависимая клеточноопосредованная цитотоксичность" и "АЗКЦ" относятся к реакции, опосредованной клетками, в которой неспецифические цитотоксические клетки, которые экспрессируют рецепторы Fc, такие как естественные клетки-киллеры, нейтрофилы и макрофаги, распознают связанное антитело на целевой клетке и вызывают лизис целевой клетки. Активность АЗКЦ может быть оценена с применением способов, таких как описанные в патенте США №5,821,337. АЗКЦ относится к антителозависимому клеточноопосредованному фагоцитозу.
"Эффекторные клетки" представляют собой лейкоциты, которые экспрессируют один или более рецепторов константной области и выполняют эффекторные функции.
"Цитокин" представляет собой белок, высвобождаемый одной клеткой для воздействия на другую клетку в качестве межклеточного медиатора.
Термин "неиммуногенный" относится к материалу, который не инициирует, не провоцирует и не усиливает иммунный ответ, при этом иммунный ответ включает в себя адаптивный и/или врожденный иммунный ответ.
Термин "выделенный" означает, что материал извлечен из своего исходного окружения (например, природной среды, если он встречается в естественных условиях). Например, природный полинуклеотид или полипептид, присутствующий в живом животном, не является выделенным, но тот же полинуклеотид или полипептид, отделенный от материалов, совместно с ним присутствующих в естественной системе, является выделенным. Такие нуклеотиды могут входить в состав вектора, и/или такие полинуклеотиды или полипептиды могут входить в состав композиции, и по-прежнему являться выделенными в том смысле, что такой вектор или композиция не являются частью их природного окружения.
Под термином "фармацевтически приемлемое вещество" подразумевают вспомогательное вещество, которое можно применять при получении фармацевтической композиции, и которое является в целом безопасным, нетоксичным и желательным; этот термин включает вспомогательные вещества, приемлемые как для ветеринарного использования, так и для фармацевтического использования для лечения людей. Такие вспомогательные вещества могут быть твердыми, жидкими, полужидкими или, в случае аэрозольной композиции, газообразными.
Под термином “фармацевтически приемлемые соли и сложные эфиры” подразумевают соли и сложные эфиры, являющиеся фармацевтически приемлемыми и обладающие желательными фармакологическими свойствами. Такие соли включают соли, которые могут образовываться при реакции кислых протонов, присутствующих в соединениях, с неорганическими или органическими основаниями. Подходящие неорганические соли включают соли, образованные щелочными металлами, например, натрием и калием, магнием, кальцием и алюминием. Подходящие органические соли включают соли, образованные органическими основаниями, например, аминными основаниями, например, этаноламином, диэтаноламином, триэтаноламином, трометамином, N-метилглюкамином и тому подобным. Такие соли также включают кислотно-аддитивные соли, образованные неорганическими кислотами (например, соляной и бромистоводородной кислотой) и органическими кислотами (например, уксусной кислотой, лимонной кислотой, малеиновой кислотой и алкан- и аренсульфоновыми кислотами, например, метансульфоновой кислотой и бензолсульфоновой кислотой). Фармацевтически приемлемые сложные эфиры включают эфиры, образованные из карбокси-, сульфонилокси- и фосфоноксигрупп, присутствующих в соединениях, например, C1-6 алкиловые эфиры. При наличии двух кислотных групп фармацевтически приемлемая соль или сложный эфир может представлять собой монокислоту-моносоль или сложный эфир или двойную соль, или сложный эфир, и, аналогичным образом, при наличии более чем двух кислотных групп, некоторые или все из таких групп могут находиться в виде соли или этерифицированном виде. Соединения, названия которых указаны в данном изобретении, могут присутствовать вне формы соли или сложного эфира, или в форме соли и/или сложного эфира, и подразумевается, что наименование таких соединений включают как исходное соединение (не соль и не сложный эфир), так и его фармацевтически приемлемые соли и сложные эфиры. Кроме того, некоторые соединения, названия которых указаны в данном изобретении, могут присутствовать в более чем одной стереоизомерной форме, и подразумевается, что наименование таких соединений включают все одиночные стереоизомеры и все смеси (рацемические или другие) таких стереоизомеров.
Термины "фармацевтически приемлемый", "физиологически переносимый" и их грамматические формы по отношению к композициям, носителям, разбавителям и реагентам применяются на равных основаниях и означают, что указанные материалы можно вводить или наносить на организм человека без нежелательных физиологических эффектов, которые могли бы препятствовать введению композиции.
"Гомологию" между двумя последовательностями определяют по идентичности последовательностей. Если две последовательности, которые должны сравниваться друг с другом, отличаются по длине, идентичность последовательности предпочтительно относится к проценту нуклеотидных остатков более короткой последовательности, которые идентичны нуклеотидным остаткам более длинной последовательности. Обычно идентичность последовательностей можно определять с помощью компьютерных программ, таких как программа Bestfit (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive Madison, Wis. 53711). Программа Bestfit применяет локальный алгоритм гомологии Смита-Уотермана (Smith и Waterman), Advances in Applied Mathematics 2 (1981 год), 482-489, для поиска сегмента с самой высокой идентичностью последовательности между двумя последовательностями. В случае применения Bestfit или другой программы выравнивания последовательностей с целью определения того, идентична ли конкретная последовательность, например, на 95% эталонной последовательности, предлагаемой в данном изобретении, параметры предпочтительно регулируют таким образом, чтобы процент идентичности рассчитывался для всей длины эталонной последовательности, и при этом, чтобы допускались гэпы в гомологии вплоть до 5% от общего количества нуклеотидов в эталонной последовательности. В случае применения Bestfit так называемые необязательные параметры предпочтительно оставляют в своих предустановленных ("по умолчанию") значениях. Отклонения, появляющиеся при сравнении данной последовательности и описанных выше последовательностей, предлагаемых в данном изобретении, могут быть вызваны, например, добавлением, делецией, заменой, вставкой или рекомбинацией. Такое сравнение последовательностей можно предпочтительно выполнять с помощью программы "fasta20u66" (версия 2. 0u66, сентябрь 1998 года, разработана William R. Pearson и University of Virginia; смотри также публикацию W.R. Pearson (1990 год), Methods in Enzymology 183, 63-98, прилагаемые примеры и http://workbench.sdsc.edu/). Для этой цели могут применять настройки параметров "по умолчанию".
Термин "вариант" относится к полипептидам, имеющим аминокислотные последовательности, которые в некоторой степени отличаются от полипептида с нативной последовательностью. Обычно варианты аминокислотных последовательностей будут обладать, по меньшей мере, около 80% идентичностью последовательности, более предпочтительно, по меньшей мере, на около 90% гомологичны по последовательности. Варианты аминокислотной последовательности могут иметь замены, делеции и/или вставки в определенных положениях в эталонной аминокислотной последовательности.
Термин "вектор" в контексте данного документа обозначает молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, с ней соединенную. Один тип вектора представляет собой "плазмиду", которая относится к циклической двухцепочечной петле ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другой тип вектора представляет собой вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в вирусный геном. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальную точку начала репликации, и эписомальные векторы млекопитающих). Другие векторы (например, не-эписомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при введении в клетку хозяина и, таким образом, реплицироваться вместе с геномом хозяина. Более того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы упоминаются в данном документе как "рекомбинантные экспрессионные векторы" (или просто, "рекомбинантные векторы"). В общем, векторы экспрессии, используемые в технологиях рекомбинантной ДНК, часто имеют форму плазмид. В данном описании изобретения "плазмида" и "вектор" могут быть взаимозаменяемы, поскольку плазмида является наиболее часто используемой формой вектора.
Применяемый в данном документе термин "клетка-хозяин" (или "рекомбинантная клетка-хозяин") предназначен для обозначения клетки, которая была генетически изменена или способна генетически изменяться путем введения экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. Следует понимать, что такие термины предназначены для обозначения не только конкретной исследуемой клетки, но и клеток, происходящих из нее. Поскольку некоторые модификации могут произойти в последующих поколениях из-за мутаций или воздействия окружающей среды, такое потомство не может, по сути, быть идентичным родительской клетке, но все еще включено в термин "клетка-хозяин", как применяется в данном документе.
"Аффинность связывания" обычно относится к силе суммарного количества нековалентных взаимодействий между одним сайтом связывания молекулы (например, антителом или другой связывающей молекулой) и ее партнером по связыванию (например, антигеном или рецептором). Аффинность молекулы X к ее партнеру Y, как правило, можно представить в виде константы диссоциации (Kd). Аффинность можно измерить с помощью общепринятых способов, известных в данной области техники, в том числе тех, которые описаны в данном документе. Низкоаффинные антитела слабо связывают антиген (или рецептор) и склонны легко диссоциировать, тогда как высокоаффинные антитела связывают антиген (или рецептор) более плотно и имеют тенденцию оставаться связанными дольше.
Если специально не указано иное, термин "конъюгат", как описано и заявлено в данном документе, определяется как гетерогенная молекула, образованная ковалентным присоединением одного или более фрагментов антитела к одной или более молекуле(ам) полимера, при этом гетерогенная молекула растворима в воде, то есть растворима в физиологических жидкостях, таких как кровь, и при этом гетерогенная молекула свободна от любого структурированного агрегата. Представляющий интерес конъюгат представляет собой ПЭГ. В контексте вышеприведенного определения термин "структурированный агрегат" относится к (1) любому агрегату молекул в водном растворе, имеющему структуру сфероида или сфероидной оболочки, так что гетерогенная молекула не находится в мицелле или другой эмульсионной структуре, и не фиксирована в липидном бислое, везикуле или липосоме; и (2) любому агрегату молекул в твердой или нерастворимой форме, такой как матрица хроматографических шариков, который не высвобождает гетерогенную молекулу в раствор при контакте с водной фазой. Соответственно, термин "конъюгат", как он определено в данном документе, охватывает вышеупомянутую гетерогенную молекулу как преципитат, осадок, биоразрушаемая матрица или другое твердое вещество, способное высвобождать гетерогенную молекулу в водный раствор при гидратации твердого вещества.
Слово "метка" при использовании в данном документе относится к детектируемому соединению или композиции, которое прямо или косвенно конъюгировано с антителом. Сама метка может быть выявляемой сама по себе (например, метки-радиоизотопы или флуоресцентные метки) или, в случае ферментной метки, может катализировать такие химические изменения субстратного соединения или композиции, которые можно выявить.
Под "твердой фазой" подразумевается безводная матрица, к которой может приставать антитело по данному изобретению. Примеры твердых фаз, охватываемых данным документом, включают те, которые сформированы частично или полностью из стекла (например, стекла с контролируемым размером пор), полисахаридов (например, агарозы), полиакриламидов, полистирола, поливинилового спирта и силиконов. В определенных вариантах реализации изобретения, в зависимости от контекста, твердая фаза может содержать лунку аналитического планшета; в других это колонка очистки (например, колонка аффинной хроматографии). Этот термин также включает в себя прерывистую твердую фазу из дискретных частиц, такую как описанная в патенте США № 4,275,149.
Антитела, также называемые иммуноглобулинами, обычно содержат по меньшей мере одну тяжелую цепь и одну легкую цепь, причем аминоконцевой домен тяжелой и легкой цепей является вариабельным по последовательности, и поэтому обычно называется доменом вариабельной области, или вариабельным доменом тяжелой цепи (VH) и вариабельным доменом легкой цепи (VH). Эти два домена обычно связываются друг с другом, образуя специфическую область связывания, хотя здесь также обсуждается, что специфическое связывание также можно получить с применением только вариабельных последовательностей тяжелой цепи, и в данной области техники известно и применяется большое количество неприродных конфигураций антител.
"Функциональное" или "биологически активное" антитело или антигенсвязывающая молекула (включая антитела, содержащие только тяжелые цепи, и биспецифичные трехцепочечные антителоподобные молекулы (ТСА - three-chain antibody-like molecules) в данном документе) представляет собой антитело или молекулу, способную проявлять одну или более из своих естественных активностей при структурных, регуляторных, биохимических или биофизических явлениях. Например, функциональное антитело или другая связывающая молекула, например, TCA, может обладать способностью специфически связывать антиген, и это связывание, в свою очередь, может вызывать или изменять клеточные, или молекулярные явления, например, передачу сигнала или ферментативную активность. Функциональное антитело или другая связывающая молекула, например, TCA, также может блокировать активацию лиганда или рецептора или действовать как агонист или антагонист. Способность антитела или другой связывающей молекулы, например, TCA, проявить одну или более своих природных активностей, зависит от нескольких факторов, включая правильный фолдинг и сборку полипептидных цепей.
Термин "антитело" в данном документе применяют в его наиболее широком смысле; он специфически охватывает моноклональные антитела, поликлональные антитела, мономеры, димеры, мультимеры, полиспецифические антитела (например, биспецифические антитела), антитела, содержащие только тяжелую цепь, трехцепочечные антитела, одноцепочечные Fv-фрагменты, нанотела и тому подобное, а также включает фрагменты антител, при условии, что они обладают желательной биологической активностью (Miller и соавт. (2003) Jour. of Immunology 170:4854-4861). Антитела могут являться антителами мыши, человека, гуманизированными, химерными антителами или могут быть происходить от других видов животных.
Термин антитело может относиться к полноразмерной тяжелой цепи, полноразмерной легкой цепи, интактной молекуле иммуноглобулина; или иммунологически активному фрагменту любого из этих полипептидов, то есть, полипептиду, содержащему антигенсвязывающий сайт, иммуноспецифически связывающий антиген или мишень, представляющую интерес, или их фрагмент, причем такие мишени включают раковую клетку или клетки, продуцирующие аутоиммунные антитела, ассоциированные с аутоиммунным заболеванием, но не ограничиваются ими. Описанный в данном документе иммуноглобулин может содержать любую подходящую область Fc, включая, без ограничения, IgG, IgE, IgM, IgD, IgA, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2 человека или других млекопитающих, например, яванских макак, или подкласс молекул иммуноглобулинов, включая сконструированные подклассы с измененными Fc-фрагментами, обеспечивающими ослабление или усиление активности эффекторных клеток. Иммуноглобулины могут происходить от любого вида животных. В одном аспекте иммуноглобулин имеет в основном человеческое происхождение.
Термин “вариабельный” относится к тому факту, что последовательности некоторых частей вариабельных доменов сильно различаются у разных антител и вносят вклад в связывание и специфичность каждого конкретного антитела по отношению к его конкретному антигену. В то же время изменчивость не является равномерно распределенной по вариабельным доменам антител. Она сосредоточена в трех сегментах вариабельных доменов как легкой, так и тяжелой цепи, называемых гипервариабельными областями. Более консервативные фрагменты вариабельных доменов называются каркасными областями (FR). Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит четыре FR, преимущественно принимающих конфигурацию бета-листа, соединенных тремя гипервариабельными участками, которые образуют петли, соединяющие структуру бета-листа, а в некоторых случаях - образующие часть структуры бета-листа. Гипервариабельные области каждой цепи объединены друг с другом в непосредственной близости от FR и, вместе с гипервариабельными областями другой цепи, участвуют в образовании антигенсвязывающего сайта антител (см. Kabat и соавт. (1991), Sequences of Proteins of Immunological Interest, 5-е Изд. Public Health Service, National Institutes of Health, Бетесда, штат Мэриленд). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, однако проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности (АЗКЦ).
Термин "гипервариабельная область", в случае если применяется в данном документе, относится к аминокислотным остаткам антитела, которые отвечают за связывания антигена. Гипервариабельная область может содержать аминокислотные остатки из "определяющей комплементарность области" или "CDR" и/или эти остатки из "гипервариабельной петли". Остатки "каркасной области" или "FR" представляют собой остатки вариабельного домена, отличные от остатков гипервариабельной области, как определено в данном документе.
Вариабельные области, представляющие интерес, включают, по меньшей мере, одну последовательность CDR из вариабельных областей, представленных в данном документе, обычно, по меньшей мере, 2 последовательности CDR и, более обычно, 3 последовательности CDR. Типовые обозначения CDR показаны в данном документе, однако специалист в данной области техники поймет, что обычно применяется ряд определений CDR, включая определение по Кабату (см. "Zhao и соавт., A germline knowledge based computational approach for determining antibody complementarity determining regions". Mol Immunol. 2010 год; 47:694-700), которое основано на изменчивости последовательности и является наиболее часто применяемым. Определение по Чотиа основано на расположении областей структурной петли (Chothia и соавт. "Conformations of immunoglobulin hypervariable regions." Nature. 1989 год; 342:877-883). Предоставляющие интерес альтернативные определения CDR включают, без ограничения, те, которые раскрыты в работах Honegger, "Yet another numbering scheme for immunoglobulin variable domains: an automatic modeling and analysis tool." J Mol Biol. 2001 год; 309:657-670; Ofran и соавт. "Automated identification of complementarity determining regions (CDRs) reveals peculiar characteristics of CDRs and B cell epitopes." J Immunol. 2008 год; 181:6230-6235; Almagro "Identification of differences in the specificity-determining residues of antibodies that recognize antigens of different size: implications for the rational design of antibody repertoires." J Mol Recognit. 2004 год; 17:132-143; и Padlanet и соавт. "Identification of specificity-determining residues in antibodies." Faseb J. 1995 год; 9:133-139., каждая из которых специально включена в данном документе посредством ссылки.
Термин "моноклональное антитело" в данном документе относится к антителу, полученному из популяции в значительной степени однородных антител, то есть, отдельные антитела в составе популяции являются идентичными, за исключением мутаций, происходящих по естественным причинам, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифичными, поскольку они направлены против одиночного антигенного сайта. Кроме того, по сравнению с препаратами поликлональных антител, которые содержат различные антитела против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одиночной детерминанты антигена. Кроме специфичности, моноклональные антитела обладают преимуществом, заключающимся в том, что их можно синтезировать в виде, не загрязненном другими антителами. Модификатор “моноклональное” указывает на то, что антитело получено из практически однородной популяции антител; его не следует интерпретировать как требование о продукции антитела посредством какого-либо конкретного способа.
Антитела в данном документе включают "химерные" антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям антител конкретного вида животных или антител, принадлежащих к конкретному классу или подклассу, в то время как остальная часть цепи(ей) идентична или гомологична соответствующим последовательностям антител другого вида животных или антител, принадлежащих к другому классу или подклассу, а также фрагментам таких антител, при условии, что они демонстрируют желательную биологическую активность (патент США №4,816,567; и Morrison и соавт. (1984 год) Proc. Natl. Acad. Sci. USA, 81:6851-6855). Представляющие интерес химерные антитела включают "приматизированные" антитела, содержащие антигенсвязывающие последовательности вариабельного домена, полученные от примата отличного от человека (например, мартышки Старого Света, обезьяны и тому подобное), и человеческие последовательности константной области.
"Интактная цепь антитела" в данном документе представляет собой цепь, содержащую полноразмерную вариабельную область и полноразмерную константную область. Интактное “стандартное” антитело содержит интактную легкую цепь и интактную тяжелую цепь, а также константный домен легкой цепи (CL) и константные домены тяжелой цепи - CH1, шарнир, CH2 и CH3 (для секретируемого IgG). Другие изотипы, например, IgM или IgA, могут содержать другие CH-домены. Константные домены могут представлять собой константные домены с нативной последовательностью (например, константные домены человека с нативной последовательностью) или варианты их аминокислотных последовательностей. Интактное антитело может обладать одной или более “эффекторными функциями”, которые относятся к его биологической активности, относимой на счет Fc-константной области (нативной последовательности Fc-области или аминокислотной последовательности вариантной Fc-области) антитела. Примеры эффекторных функций антител включают связывание C1q; комплемент-зависимую цитотоксичность; связывание с рецептором Fc; антитело-зависимую клеточно-опосредованную цитотоксичность (АЗКЦ); фагоцитоз; и подавление рецепторов поверхности клетки. Варианты константной области включают варианты, изменяющие эффекторный профиль, связывание с рецепторами Fc и тому подобным.
В зависимости от аминокислотных последовательностей константных доменов тяжелых цепей интактные антитела можно отнести к различным "классам". Существует пять основных классов интактных антител иммуноглобулина: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно подразделены на "подклассы" (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA, и IgA2. Константные домены тяжелой цепи, соответствующие различным классам антител, называются α, δ, ε, γ и μ, соответственно. Хорошо известны субъединичная структура и трехмерная конфигурация различных классов иммуноглобулинов. Формы Ig включают модификации шарнирной области или бесшарнирные формы (Roux и соавт. (1998 год) J. Immunol. 161:4083-4090; Lund и соавт. (2000 год) Eur. J. Biochem. 267:7246-7256; США 2005/0048572; США 2004/0229310). Легкие цепи антител любого вида позвоночных можно отнести к одному из двух четко различающихся типов, называемых κ- и λ-цепями, на основе аминокислотной последовательности их константных доменов.
"Функциональная область Fc" обладает "эффекторной функцией" нативной последовательности области Fc. Типовые эффекторные функции включают связывание C1q; КЗЦ (комплементзависимую цитотоксичность); Связывание с рецептором Fc; АЗКЦ; АЗКФ (антителозависимый клеточный фагоцитоз); подавление рецепторов клеточной поверхности (например, B-клеточного рецептора) и тому подобное. Такие эффекторные функции обычно требуют взаимодействия области Fc с рецептором, например, рецепторами FcγRI; FcγRIIA; FcγRIIB1; FcγRIIB2; FcγRIIIA; FcγRIIIB, и рецептором FcRn, обладающим низкой аффинностью; их оценку можно выполнить с помощью различных анализов, описанных, например, в определениях в данном документе. "Мертвая" область Fc представляет собой область Fc, мутированную с целью сохранения активности по отношению к, например, увеличению продолжительности периода полужизни сыворотки, однако не активирующую рецептор Fc, обладающий высокой аффинностью.
“Нативная последовательность Fc-области” содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, встречающейся в природе. Нативные последовательности Fc-областей человека включают нативную последовательность Fc-области IgG1 человека (отличных от А и А аллотипов); нативную последовательность Fc-области IgG2 человека; нативную последовательность Fc-области IgG3 человека; и нативную последовательность Fc-области IgG4 человека, а также их варианты, встречающиеся в природе.
“Вариантная Fc-область” содержит аминокислотную последовательность, отличающуюся от нативной последовательности Fc-области за счет модификации по меньшей мере одной аминокислоты, предпочтительно замены одной или более аминокислот. Предпочтительно, вариантная Fc-область содержит по меньшей мере одну аминокислотную замену по сравнению с нативной последовательностью Fc-области или Fc-области исходного полипептида, например, от приблизительно одной до приблизительно десяти аминокислотных замен, и предпочтительно от приблизительно одной до приблизительно пяти аминокислотных замен в нативной последовательности Fc-области или Fc-области исходного полипептида. Вариантная область Fc согласно данному изобретению предпочтительно обладает по меньшей мере около 80% гомологией по отношению к нативной последовательности области Fc и/или с области Fc исходного полипептида, и наиболее предпочтительно, по меньшей мере около 90% гомологией по отношению к ним, более предпочтительно, по меньшей мере около 95% гомологией по отношению к ним.
Вариантные последовательности Fc могут содержать три аминокислотных замены в области СН2 с целью ослабления связывания FcγRI в положениях 234, 235 и 237 согласно индексу ЕС (см., Duncan и соавт., (1988 год) Nature 332:563). Две аминокислотные замены в сайте связывания комплемента C1q в положениях 330 и 331 согласно индексу ЕС, ослабляют фиксацию комплемента (см., Tao и соавт., J. Exp. Med. 178:661 (1993 год) и Canfield and Morrison, J. Exp. Med. 173:1483 (1991 год)). Замена остатков в положениях 233-236 IgG1 или IgG2 человека и в положениях 327, 330 и 331 IgG4 значительно ослабляет АЗКЦ и КЗЦ (см., например, Armour KL. и соавт., 1999 год Eur J Immunol. 29(8):2613-24; и Shields RL. и соавт., 2001 год. J Biol Chem. 276(9):6591-604). Возможны другие варианты Fc-области, в том числе, без ограничений, вариант, в котором область, способная образовывать дисульфидную связь, удалена, или вариант, в котором удалены некоторые аминокислотные остатки на N-конце нативной Fc-формы или к ним добавлен остаток метионина. Таким образом, в одном варианте реализации данного изобретения один или более из Fc-фрагментов молекулы scFc может содержать одну или более мутацию в шарнирной области с целью устранения возможности образования дисульфидных связей. В еще одном варианте реализации шарнирная область Fc может быть полностью удалена. В еще одном варианте реализации молекула может содержать вариант Fc-области.
Кроме того, можно сконструировать вариант Fc-области, удалив или значительно ослабив эффекторные функции посредством замены, делеции или добавления аминокислотных остатков для влияния на связывание комплемента или рецептора Fc. В качестве неограничивающего примера, возможна делеция в сайте связывания комплемента, например, C1q-связывающем сайте. Методы получения таких производных последовательностей фрагмента Fc иммуноглобулинов описаны в международных патентных публикациях № WO 97/34631 и WO 96/32478. Кроме того, можно модифицировать домен Fc путем фосфорилирования, сульфатирования, ацилирования, гликозилирования, метилирования, фарнезилирования, ацетилирования, амидирования и тому подобного.
Fc-область может находиться в форме, содержащей нативные углеводные цепи, увеличенное количество углеводных цепей по сравнению с нативной формой или уменьшенное количество углеводных цепей по сравнению с нативной формой, или может находиться в агликозилированной или дегликозилированной форме. Увеличение, уменьшение, удаление или другие модификации углеводных цепей можно осуществить с помощью способов, часто применяемых в данной области техники, например, химического способа, ферментативного способа или посредством экспрессии в генетически сконструированной линии клеток-продуцентов. Такие линии клеток могут включать микроорганизмы, например, Pichia Pastoris и клеточную линию млекопитающих, например, клетки СНО, которые естественным образом экспрессируют гликозилирующие ферменты. Кроме того, можно сконструировать микроорганизмы или клетки для экспрессии гликозилирующих ферментов или сделать их неспособными к экспрессии ферментов гликозилирования (см., например, Hamilton и соавт., Science, 313:1441 (2006 год); Kanda, и соавт, J. Biotechnology, 130:300 (2007 год); Kitagawa и соавт., J. Biol. Chem., 269 (27):17872 (1994 год); Ujita-Lee и соавт., J. Biol. Chem., 264 (23):13848 (1989 год); Imai-Nishiya и соавт., BMC Biotechnology 7:84 (2007 год); и WO 07/055916). В качестве примера сконструированных клеток с модифицированной активностью сиалирования, в клетки яичника китайского хомячка и в клетки sf9 рекомбинантно внедрили ген альфа-2,6-сиалилтрансферазы 1. Антитела, экспрессируемые этими рекомбинантными клетками, подвергаются сиалированию продуктом экзогенного гена. Дополнительный способ получения молекул Fc с модифицированным количеством углеводных остатков по сравнению с множеством нативных молекул включает разделение указанного множества молекул на гликозилированную и негликозилированную фракции, например, с помощью аффинной хроматографии с лектином (см., например, WO 07/117505). Показано, что наличие конкретных гликозилирующих функциональных групп приводит к изменению функции иммуноглобулинов. Например, удаление углеводных цепей с молекулы Fc приводит к резкому снижению аффинности связывания с фрагментом C1q первого компонента комплемента C1 и ослаблению или потере антителозависимой клеточноопосредованной цитотоксичности (АЗКЦ) или комплементзависимой цитотоксичности (КЗЦ) и, следовательно, отсутствию индукции ненужных иммунных реакций in vivo. Дополнительные важные модификации включают сиалирование и фукозилирование: присутствие сиаловой кислоты в IgG коррелировало с противовоспалительной активностью (см., например, Kaneko, и соавт., Science 313:760 (2006 год)), в то время как удаление фукозы из IgG приводит к усилению активности АЗКЦ (см., например, Shoj-Hosaka, и соавт., J. Biochem., 140:777 (2006 год)).
В альтернативных вариантах реализации антитела по данному изобретению могут содержать последовательность Fc с усиленными эффекторными функциями, например, повышенной способностью к связыванию с FcγRIIIA и повышенной активностью АЗКЦ. Например, присоединение фукозы к N-связанному гликану при Asn-297 области Fc стерически затрудняет взаимодействие Fc с FcγRIIIA, а удаление фукозы посредством гликоинженерии может усилить связывание с FcγRIIIA, что приводит к > 50-кратному увеличению активности АЗКЦ по сравнению с контрольными IgG1 дикого типа. Белковая инженерия позволяет получить различные варианты с повышенной аффинностью связывания Fc с FcγRIIIA за счет мутаций аминокислот в области Fc IgG1. Примечательно, что тройной мутант с заменой на аланин S298A/E333A/K334A демонстрирует 2-кратное усиление связывания с FcγRIIIA и функцию АЗКЦ. Варианты S239D/I332E (2X) и S239D/I332E/A330L (3X) характеризуются значительным увеличением сродства связывания с FcγRIIIA и усилением АЗКЦ in vitro и in vivo. Другие варианты области Fc, выявленные посредством фагового дисплея, также демонстрировали улучшенное связывание с FcγRIIIA и усиленное уничтожение опухолевых клеток в моделях ксенотрансплантатов у мышей. См., например, Liu и соавт. (2014 год) JBC 289(6):3571-90, специально включенный в данный документ посредством ссылки.
Термин "антитело, содержащее область Fc" относится к антителу, содержащему область Fc. С-концевой остаток лизина (остаток 447 в соответствии с ЕС-системой нумерации) в Fc-области можно удалить, например, при очистке антитела или с помощью рекомбинантной инженерии нуклеиновой кислоты, кодирующей антитело. Соответственно, антитело, содержащее область Fc согласно данному изобретению, может включать антитело с K447 или без него.
"Fv" представляет собой минимальный фрагмент антитела, содержащий полный антигенраспознающий и антигенсвязывающий участок. Связывающие CD3 антитела по данному изобретению содержат димер одного домена вариабельной области тяжелой цепи и одного домена вариабельной области легкой цепи, соединенных жесткой нековалентной связью; однако дополнительные антитела, например, для применения в мультиспецифической конфигурации может содержать VH в отсутствие последовательности VL. Даже один вариабельный домен (или половина Fv, содержащего только три гипервариабельные области, специфичные для антигена) обладает способностью распознавать и связывать антиген, хотя аффинность может быть ниже, чем аффинность сайта связывания двух доменов.
Фрагмент Fab также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Фрагменты Fab' отличаются от фрагментов Fab по дополнительным нескольким остаткам на карбоксильном конце домена CH1 тяжелой цепи, включая один или более цистеинов из шарнирной области антитела. В данном описании Fab'-SH представляет собой обозначение Fab', в котором цистеиновый(е) остаток(и) константных доменов несет по меньшей мере одну свободную тиольную группу. Фрагменты F(ab')2 антитела первоначально получали в виде пар фрагментов Fab', между которыми находятся шарнирные остатки цистеина. Известны также другие варианты химического связывания фрагментов антител.
"Гуманизированные" формы антител нечеловеческого происхождения (например, грызунов), включая одноцепочечные антитела, представляют собой химерные антитела (включая одноцепочечные антитела), содержащие минимальные последовательности, происходящие от иммуноглобулина нечеловеческого происхождения. См., например, Jones и соавт., (1986 год) Nature 321:522-525; Chothia и соавт., (1989 год) Nature 342:877; Riechmann и соавт., (1992 год) J. Mol. Biol. 224, 487-499; Foote and Winter, (1992 год) J. Mol. Biol. 224:487-499; Presta и соавт., (1993 год) J. Immunol. 151, 2623-2632; Werther и соавт., (1996 год) J. Immunol. Methods 157:4986-4995; и Presta и соавт., (2001 год) Thromb. Haemost. 85:379-389. Дополнительную информацию см. в патентах США №5,225,539; 6,548,640; 6,982,321; 5,585,089; 5,693,761; 6,407,213; Jones и соавт., (1986 год) Nature, 321:522-525; и Riechmann и соавт., (1988 год) Nature 332:323-329.
В данном контексте термин "одноцепочечное антитело" означает одну полипептидную цепь, содержащую один или более антигенсвязывающих доменов, которые связывают эпитоп антигена, при этом такие домены получены из или имеют идентичность последовательности с вариабельной областью тяжелой или легкой цепи антитела. Части такой вариабельной области могут кодироваться генными сегментами VH или VL, генными сегментами D и JH или генными сегментами JL. Вариабельная область может кодироваться перестроенными генными сегментами VHDJH, VLDJH, VHJL или VLJL. Генные сегменты V-, D- и J- могут быть получены от людей и различных животных, включая птиц, рыб, акул, млекопитающих, грызунов, приматов, отличных от человека, верблюдов, лам, кроликов и тому подобных.
CD3-связывающие антитела по данному изобретению находят конкретное применение в мультиспецифических конфигурациях, которые включают, без ограничения, биспецифические антитела, трифункциональные антитела и тому подобное. Большое разнообразие способов и конфигураций белков известно и применяется в биспецифических моноклональных антителах (БсмАт), триспецифических антителах и тому подобных.
БсмАт первого поколения состояли из двух тяжелых и двух легких цепей, по одной от двух разных антител. Две области Fab направлены против двух антигенов. Область Fc состоит из двух тяжелых цепей и образует третий сайт связывания с рецептором Fc на иммунных клетках (см., например, Lindhofer и соавт., The Journal of Immunology, Том 155, стр. 219-225, 1995 год). Антитела могут быть из одного или разных видов. Например, клеточные линии, экспрессирующие антитела крысы и мыши, секретируют функциональное биспецифическое Ат из-за преимущественного, ограниченного видом, спаривания тяжелых и легких цепей. В других вариантах реализации изобретения области Fc сконструированы так, что они подходят друг другу только определенным образом.
Другие типы биспецифических антител включают химически связанные Fab, состоящие только из областей Fab. Два химически связанных фрагмента Fab или Fab2 образуют искусственное антитело, которое связывается с двумя различными антигенами, что делает его типом биспецифического антитела. Получаются антигенсвязывающие фрагменты (Fab или Fab2) двух разных моноклональных антител связанные химическими средствами, такими как тиоэфир (см., Glennie, M J и соавт., Journal of immunology 139, стр. 2367-75, 1987 год; Peter Borchmann и соавт., Blood, Том 100, № 9, стр. 3101-3107, 2002 год).
Различные другие способы получения поливалентных искусственных антител были разработаны путем рекомбинантного слияния вариабельных доменов двух антител. Одноцепочечный вариабельный фрагмент (scFv) представляет собой слитый белок вариабельных областей тяжелых (VH) и легких цепей (VL) иммуноглобулинов, связанных с коротким линкерным пептидом из 10-25 аминокислот. Обычно линкер насыщен глицином для гибкости, а также серином или треонином для растворимости и может соединять N-конец VH с C-концом VL, или наоборот. Биспецифические одноцепочечные вариабельные фрагменты (ди-scFvs, би-scFvs) могут быть сконструированы путем связывания двух scFv с различной специфичностью. Производится одна пептидная цепь с двумя областями VH и двумя областями VL, что приводит к получению двухвалентных scFv.
Биспецифические тандемные scFv также известны как привлекающие T-клетки биспецифические активаторы (BiTE). Биспецифические scFv могут быть созданы с помощью линкерных пептидов, которые являются слишком короткими для того, чтобы две вариабельные области складывались вместе (длиной около пяти аминокислот), заставляя scFvs димеризоваться. Этот тип известен как диатела (Adams и соавт., British journal of cancer 77, стр. 1405-12, 1998 год). Технология платформы двуафинного перенацеливания (DART - Dual-Affinity Re-Targeting) (Macrogenics, Роквилл, штат Мэриленд). Эта технология слияния белков применяет два одноцепочечных вариабельных фрагмента (scFv) различных антител на одной пептидной цепи длиной около 55 килодальтон. SCORPION Therapeutics (Emergent Biosolutions, Inc., Сиэтл, штат Вашингтон) объединяет два антигенсвязывающих домена в одноцепочечный белок. Один связывающий домен находится на С-конце, а второй связывающий домен находится на N-конце эффекторного домена на основе областей Fc иммуноглобулина.
Тетравалентные и биспецифические подобные антителам белки также включают DVD-Ig, которые сконструированы из двух моноклональных антител (Wu, C. и соавт., Nature Biotechnology, 25, стр. 1290-1297, 2007 год). Для конструирования молекулы DVD-Ig домены V двух мАт сливаются в тандеме с помощью короткого линкера (TVAAP) с вариабельным доменом легкой цепи (VL) первого антитела на N-конце, за которым следуют VL и Ck других антител для образования легкой цепи белка DVD-Ig. Аналогичным образом, вариабельные области тяжелой (VH) цепи двух мАт сливаются в тандеме с помощью короткого линкера (ASTKGP) с первым антителом на N-конце, за которым следует другое антитело и константные домены тяжелой цепи с образованием тяжелой цепи (VH1/VL1) белка DVD-Ig. Все константные домены легкой цепи и тяжелой цепи сохраняются в конструкции DVD-Ig, поскольку они являются критическими для образования дисульфид-связанной полной IgG-подобной молекулы. Котрансфекция клеток млекопитающих экспрессионными векторами, кодирующими легкую цепь и тяжелую цепь DVD-Ig, приводит к секреции IgG-подобной молекулы одного вида с молекулярной массой приблизительно 200 кДа. Эта молекула теперь имеет четыре сайта связывания, по 2 от каждого мАт.
Термин "биспецифическая трехцепочечная антитело-подобная молекула" или "ТСА" применяется в данном документе для обозначения антитело-подобных молекул, включающих, состоящих по существу или состоящих из трех полипептидных субъединиц, две из которых включают в себя, состоят по существу из или состоят из одной тяжелой и одной легкой цепи моноклонального антитела или функциональных антигенсвязывающих фрагментов таких цепей антитела, содержащих антигенсвязывающую область и по меньшей мере один домен СН. Эта пара тяжелая цепь/легкая цепь обладает специфичностью связывания для первого антигена. Третья полипептидная субъединица содержит, состоит по существу или состоит из антитела с только тяжелой цепью, содержащего участок Fc, содержащий домены СН2 и/или СН3, и/или СН4, в отсутствие домена СН1, и антигенсвязывающий домен, который связывает эпитоп второго антигена или другой эпитоп первого антигена, при этом такой связывающий домен происходит из или имеет идентичность последовательности с вариабельной областью тяжелой или легкой цепи антитела. Части такой вариабельной области могут кодироваться генными сегментами VH или VL, генными сегментами D и JH или генными сегментами JL. Вариабельная область может кодироваться перестроенными генными сегментами VHDJH, VLDJH, VHJL или VLJL.
Белок TCA использующий "антитело только с тяжелой цепью" или "антитело с тяжелой цепью" или "полипептид тяжелой цепи", в данном контексте означает одноцепочечное антитело, содержащее тяжелые цепи СН2 и/или СН3, и/или СН4, но не имеющее домена СН1. В одном варианте реализации изобретения антитело с тяжелой цепью состоит из антигенсвязывающего домена, по меньшей мере, части шарнирной области и доменов CH2 и CH3. В другом варианте реализации изобретения антитело с тяжелой цепью состоит из антигенсвязывающего домена, по меньшей мере, части шарнирной области и домена CH2. В дополнительном варианте реализации изобретения антитело с тяжелой цепью состоит из антигенсвязывающего домена, по меньшей мере, части шарнирной области и домена CH3. Антитела с тяжелой цепью, в которых домен СН2 и/или СН3 укорочен, также включены в данный документ. В дополнительном варианте реализации изобретения тяжелая цепь состоит из антигенсвязывающего домена и, по меньшей мере, одного домена СН (СН1, СН2, СН3 или СН4), но без шарнирной области. Антитело только с тяжелой цепью может быть в форме димера, в котором две тяжелые цепи дисульфидно связаны друг с другом или ковалентно или нековалентно связаны друг с другом. Антитело с тяжелой цепью может принадлежать к подклассу IgG, но антитела, принадлежащие к другим подклассам, таким как подкласс IgM, IgA, IgD и IgE, также включены в данный документ. В конкретном варианте реализации изобретения антитело с тяжелой цепью относится к подтипу IgG1, IgG2, IgG3 или IgG4, в частности подтипу IgG1.
Антитела с тяжелой цепью составляют около одной четвертой части антител IgG, продуцируемых верблюдовыми, например, верблюдами и ламами (Hamers-Casterman C., и соавт. Nature. 363, 446-448 (1993 год)). Эти антитела образованы двумя тяжелыми цепями, но лишены легких цепей. Как следствие, вариабельная антигенсвязывающая часть называется доменом VHH и представляет собой наименьший встречающийся в природе интактный антигенсвязывающий сайт длиной всего около 120 аминокислот (Desmyter A., и соавт. J. Biol. Chem. 276, 26285-26290 (2001 год)). Антитела с тяжелой цепью с высокой специфичностью и аффинностью могут производиться против различных антигенов посредством иммунизации (van der Linden, R. H., и соавт. Biochim. Biophys. Acta. 1431, 37-46 (1999 год)), а часть VHH может быть легко клонирована и экспрессирована в дрожжах (Frenken, L.G.J., и соавт. J. Biotechnol. 78, 11-21 (2000 год)). Их уровни экспрессии, растворимости и стабильности значительно выше, чем у классических фрагментов F(ab) или Fv (Ghahroudi, M. A. и соавт. FEBS Lett. 414, 521-526 (1997 год)). Также было показано, что акулы имеют один VH-подобный домен в своих антителах, называемый VNAR. (Nuttall и соавт. Eur. J. Biochem. 270, 3543-3554 (2003 год); Nuttall и соавт., Function and Bioinformatics 55, 187-197 (2004 год); Dooley и соавт., Molecular Immunology 40, 25-33 (2003 год)).
Антитело или антигенсвязывающая молекула, включая антитела, содержащие только тяжелую цепь, и биспецифические трехцепочечные антителоподобные молекулы (ТСА) в данном документе, "которые связывают" представляющий интерес антиген, представляют собой молекулу, которая связывает антиген с достаточной аффинностью, так что антитело или связывающая молекула полезна в качестве диагностического и/или терапевтического агента при нацеливании на антиген и не имеет значительного перекрестного взаимодействия с другими белками. В таких вариантах реализации изобретения степень связывания антитела или другой связывающей молекулы с нецелевым антигеном будет составлять не более 10%, как определено анализом сортировки клеток с активированной флуоресценцией (FACS - fluorescence activated cell sorting) или радиоиммунопреципитацией (РИА)
Белки по данному изобретению
Данное изобретение обеспечивает семейство родственных антител, которые связываются и активируют передачу сигналов посредством CD3, например, активацией CD3+ T-клеток. Антитела в семействе содержат набор последовательностей CDR, как определено в данном документе, и примерами которых являются предоставленные последовательности VH из SEQ ID NO: 1-68 и приведенная в качестве примера последовательность VL из SEQ ID NO: 69. Семейство антител обеспечивает ряд преимуществ, которые способствуют полезности в качестве клинического лекарственного средства(в). Антитела в семействе включают членов с рядом аффинностей связывания, позволяющих выбирать конкретную последовательность с желаемой аффинностью. Способность точно настраивать аффинность имеет особое значение для управления уровнем активации CD3 у индивида, которого лечат, и, таким образом, для снижения токсичности. Например, если целью являются опухолевые антигены, присутствующие в небольшом количестве (менее 10000 молекул на клетку), ожидается, что применяются высокоаффинные связыватели CD3 (<30 нМ). Если целью являются опухолевые антигены, присутствующие в большом количестве (более 50000 молекул на клетку), предпочтительными являются низкоаффинные связыватели CD3 (>50 нМ).
Подходящее антитело может быть выбрано из библиотеки для разработки и применения, включая без ограничения применение в качестве биспецифического антитела. Определение аффинности к белку-кандидату может быть выполнено с применением способов, известных в данной области техники, например, измерения Biacore и тому подобного. Члены семейства антител могут иметь аффинность к CD3 с Kd от около 10-6 до около 10-11, включая без ограничений: от около 10-6 до около 10-10; от около 10-6 до около 10-9; от около 10-6 до около 10-8; от около 10-8 до около 10-11; от около 10-8 до около 10-10; от около 10-8 до около 10-9; от около 10-9 до около 10-11; от около 10-9 до около 10-10; или любое значение в этих диапазонах. Выбор аффинности может быть подтвержден биологической оценкой активации Т-клеток, например, in vitro или в доклинической модели, и оценкой потенциальной токсичности.
Некоторые члены семейства антител по данному изобретению перекрестно реагируют с белком CD3 яванского макака и для такой перекрестной реакции идентифицируется необходимый специфический мотив, что позволяет отбирать антитела для доклинических или клинических испытаний на этой основе. Было обнаружено, что антитела, имеющие аминокислотный мотив FAA в CDR3 тяжелой цепи, особенно эффективны в перекрестной реакции с белками CD3 приматов, отличных от человека.
Вовлечение рецептора T-клетки (ТКР) путем связывания комплексов MH-пептид или антител к ТКР/CD3 инициирует активацию T-клеток. Примерами антител к ТКР/CD3, которые активируют Т-клетки, являются OKT3 и UCHT1. Эти антитела к CD3 перекрестно конкурируют за связывание с CD3 на Т-клетках и обычно применяются в анализах активации Т-клеток. Антитела к CD3 по данному изобретению перекрестно конкурируют с OKT3 за связывание с CD3 человека. В зависимости от аффинности связывания CD3 и эпитопа на CD3 антитела к CD3 активировали Т-клетки с различными функциональными результатами. Инкубация in vitro Т-клеток человека с низкоаффинными антителами к CD3 приводила к неполной активации Т-клеток, низкой продукции ИЛ-2 и ИЛ-10. Напротив, высокоаффинные связыватели CD3 активировали Т-клетки, чтобы продуцировать значительно больше ИЛ-2 и других цитокинов. Низкоаффинные антитела к CD3 считаются частичными агонистами, которые избирательно индуцируют некоторые эффекторные функции, сильное уничтожение опухоли и активацию CD69, но не вызывают других, таких как продукция ИЛ-2 и ИЛ-10. Высокоаффинные связыватели по данному изобретению являются полными агонистами, активирующими многие иммунные эффекторные функции Т-клеток. Сила взаимодействия с CD3 и узнаваемым эпитопом привела к качественно различной активации Т-клеток. Максимальная продукция цитокинов Т-клетками, активированными низкоаффинными антителами к CD3, была ниже, чем максимальная активация высокоаффинными антителами к CD3. В некоторых вариантах реализации антитело по данному изобретению приводит к более низкому высвобождению одного или обоих ИЛ-2 и ИЛ-10 при объединении с Т-клетками в анализе активации по сравнению с эталонным антителом к CD3 в том же анализе, при этом эталонное антитело может представлять собой ID 304703 (или антитело с эквивалентной аффинностью). Максимальное высвобождение ИЛ-2 и/или ИЛ-10 может составлять менее чем около 75% высвобождения с помощью эталонного антитела, менее чем около 50% высвобождения с помощью эталонного антитела, менее чем около 25% высвобождения с помощью эталонного антитела, и может составлять менее чем около 10% высвобождения с помощью эталонного антитела.
В некоторых вариантах реализации изобретения предлагаются биспецифические или мультиспецифические антитела, которые могут иметь любую из обсуждаемых в данном документе конфигураций, включая, без ограничения, биспецифическую трехцепочечную конфигурацию. Биспецифические антитела содержат по меньшей мере вариабельную область тяжелой цепи антитела, специфичного к белку, отличному от CD3, и могут содержать вариабельную область тяжелой и легкой цепи. В некоторых таких вариантах реализации изобретения вторая специфичность антитела связывается с антигеном, ассоциированным с опухолью, нацеливающим антигеном, например, интегринами и тому подобное, патогенным антигеном, белком контрольной точки и тому подобным. Различные форматы биспецифических антител находятся в пределах объема изобретения, включая без ограничения одноцепочечные полипептиды, двухцепочечные полипептиды, трехцепочечные полипептиды, четырехцепочечные полипептиды и кратные им.
Семейство CD3-специфических антител по данному изобретению содержит домен VH, содержащий последовательности CDR1, CDR2 и CDR3 в каркасе VH человека. Последовательности CDR могут быть расположены, например, в области аминокислотных остатков 26-35; 53-59; и 98-117 для CDR1, CDR2 и CDR3, соответственно, предоставленных типовых последовательностей вариабельной области, изложенных в SEQ ID NO: 1-68. Специалисту в данной области техники должно быть понятно, что последовательности CDR могут находиться в другом положении, в случае если выбрана другая каркасная последовательность, хотя обычно порядок последовательностей будет оставаться тем же.
Последовательности CDR для антитела по данному изобретению могут иметь следующие формулы последовательности. Х обозначает вариабельную аминокислоту, которая может представлять собой специфические аминокислоты, как указано ниже.
CDR1
при этом:
X5 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X5 представляет собой S или R;
X7 и X8 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X7 и X8 независимо представляют собой S или G. В некоторых вариантах реализации изобретения X7X8 представляют собой SS или GG;
X9 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X9 представляет собой H или Y; в некоторых вариантах реализации изобретения X9 представляет собой H;
X10 может быть любой аминокислотой; в некоторых вариантах реализации изобретения X10 представляет собой Y или F; в некоторых вариантах реализации изобретения X10 представляет собой Y.
В некоторых вариантах реализации изобретения последовательность CDR1 имеет формулу: G G S I X5'S H H G Y, при этом X5 такой, как определено выше. В некоторых вариантах реализации изобретения последовательность CDR1 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 26-35.
CDR2
при этом:
X2' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X2' представляет собой S, Y или H;
X3' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X3' представляет собой Y, H или R;
X6' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X6' представляет собой S, N или I, или R;
X7' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X7' представляет собой T или P.
В некоторых вариантах реализации изобретения последовательность CDR2 имеет формулу: I X2’ X3’ S G S T; или I X2’ X3’ S G N P при этом X2’ и X3’ такие, как определено выше. В некоторых вариантах реализации изобретения последовательность CDR2 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 53-59.
CDR3
X1'' R2'' W3'' R4'' H5'' D6'' I7'' X8'' X9'' X10'' Y11'' P12'' Y13'' Y14'' Y15'' Y16'' G17'' M18'' D19'' V20''
при этом:
X1'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X1' представляет собой A или G.
X8'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X8'' представляет собой L или F.
X9'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X9'' представляет собой T или A
X10'' может быть любой аминокислотой; в некоторых вариантах реализации изобретения X10'' представляет собой G, A или R.
В некоторых вариантах реализации изобретения X8'' X9'' X10'' представляют собой F A A, мотив которого соответствует антителам, которые перекрестно реагируют с белком CD3 яванского макака. В других вариантах реализации изобретения X8'' X9'' X10'' представляют собой L T A. В некоторых вариантах реализации изобретения последовательность CDR3 антитела к CD3 по данному изобретению содержит последовательность, изложенную в любой из SEQ ID NO: 1-68, остатки 98-117.
В некоторых вариантах реализации изобретения CD3-связывающий домен VH связан с доменом вариабельной области легкой цепи. В некоторых таких вариантах реализации изобретения легкая цепь представляет собой фиксированную легкую цепь. В некоторых вариантах реализации изобретения легкая цепь содержит домен VL с последовательностями CDR1, CDR2 и CDR3 в каркасе VL человека. Последовательности CDR могут быть последовательностями из SEQ ID NO: 69. В некоторых вариантах реализации изобретения последовательность CDR1 содержит аминокислотные остатки 27-32; 50-52; 89-97 для CDR1, CDR2, CDR3 соответственно.
В некоторых вариантах реализации изобретения последовательности CDR антитела по данному изобретению представляют собой последовательность с по меньшей мере 85% идентичностью, по меньшей мере 90% идентичностью, по меньшей мере 95% идентичностью, по меньшей мере 99% идентичностью относительно последовательности CDR или набора последовательностей CDR в любой из SEQ ID NO: 1-68. В некоторых вариантах реализации изобретения последовательность CDR по данному изобретению содержит одну, две, три или более аминокислотных замен по отношению к последовательности CDR или набора последовательностей CDR в любой из SEQ ID NO: 1-68. В некоторых вариантах реализации изобретения указанная аминокислотная замена(ы) представляет собой одну или более в положении 5 или 10 из CDR1, положении 2, 6 или 7 из CDR2, положении 1, 8, 9 или 10 из CDR3 относительно формул, представленных выше.
Когда белок по данному изобретению представляет собой биспецифическое антитело, один связывающий фрагмент, то есть комбинация VH/VL или только VH, является специфичным для CD3 человека, тогда как другое плече может быть специфичным для клеток-мишеней, включая раковые клетки, такие как клетки яичника, молочной железы, желудочно-кишечного тракта, головного мозга, головы и шеи, предстательной железы, толстой кишки и рак легкого и тому подобное, а также гематологические опухоли, такие как В-клеточные опухоли, включая лейкемии, лимфомы, саркомы, карциномы, опухоли нервных клеток, плоскоклеточные карциномы, эмбрионально-клеточные опухоли, метастазы, недифференцированные опухоли, семиномы, меланомы, миеломы, нейробластомы, опухоли смешанных клеток, новообразования, вызванные инфекционными агентами, и другие злокачественные новообразования, клетки, инфицированные патогеном, аутореактивные клетки, вызывающие воспаление и/или аутоиммунную реакцию. Часть, не содержащая CD3, также может быть специфичной для иммунорегуляторного белка, как будет описано в данном документе.
Опухолеассоциированные антигены (TAA) относительно ограничены опухолевыми клетками, тогда как опухолеспецифические антигены (TSA) являются уникальными для опухолевых клеток. TSA и TAA обычно представляют собой части внутриклеточных молекул, экспрессируемых на поверхности клетки как часть основного комплекса гистосовместимости.
Тканеспецифические дифференцировочные антигены представляют собой молекулы, присутствующие на опухолевых клетках и их нормальных клеточных аналогах. Опухолеассоциированные антигены, о которых известно, что они распознаются терапевтическими мАт, подразделяются на несколько различных категорий. Гематопоэтические дифференцировочные антигены представляют собой гликопротеины, которые обычно связаны с группами кластера дифференцировки (CD) и включают в себя CD20, CD30, CD33 и CD52. Дифференцировочные антигены клеточной поверхности представляют собой разнообразную группу гликопротеинов и углеводов, которые находятся на поверхности как нормальных, так и опухолевых клеток. Антигены, которые участвуют в передаче сигналов роста и дифференцировки, часто являются факторами роста и рецепторами факторов роста. Факторы роста, которые являются целями для антител у онкопациентов, включают CEA, рецептор эпидермального фактора роста (EGFR (epidermal growth factor receptor); также известный как ERBB1), ERBB2 (также известный как HER2), ERBB3, MET (также известный как HGFR), рецептор инсулиноподобного фактора роста 1 (IGF1R - insulin-like growth factor 1 receptor), рецептор эфрина A3 (EPHA3), рецептор ФНО-зависимого апоптоз-индуцирующего лиганда 1 (TRAILR1 (tumor necrosis factor (TNF)-related apoptosis-inducing ligand receptor 1); также известный как TNFRSF10A), TRAILR2 (также известный как TNFRSF10B) и лиганд рецептора-активатора ядерного фактора κВ (RANKL; также известный как TNFSF11). Антигенами, участвующими в ангиогенезе, обычно являются белки или факторы роста, которые поддерживают образование нового микроциркуляторного русла, включая фактор роста сосудистого эндотелия (VEGF - vascular endothelial growth factor), рецептор VEGF (VEGFR), интегрин αVβ3 и интегрин α5β1. Строма опухоли и внеклеточный матрикс являются необходимыми опорными структурами для опухоли. Антигены стромального и внеклеточного матрикса, которые являются терапевтическими мишенями, включают белок активации фибробластов (FAP - fibroblast activation protein) и тенасцин.
Примеры лекарственных средств на основе моноклональных антител, полезных в биспецифических конфигурациях, включают, без ограничения, ритуксимаб; Ибритумомаб; Тиуксетан; Тозитумомаб; Брентуксимаб; ведотин; Гемтузумаб; озогамицин; Алемтузумаб; IGN101; адекатумумаб; Лабетузумаб; huA33; Пемтумомаб; ореговомаб; CC49 (минретумомаб); cG250; J591; MOv18; MORAb-003 (фарлетузумаб); 3F8, ch14. 18; KW-2871; hu3S193; IgN311; Бевацизумаб; IM-2C6; CDP791; Етарацизумаб; Волоциксимаб; Цетуксимаб, панитумумаб, нимотузумаб; 806; Трастузумаб; пертузумаб; MM-121; AMG 102, METMAB; SCH 900105; AVE1642, IMC-A12, MK-0646, R1507; CP 751871; KB004; IIIA4; Мапатумумаб (HGS-ETR1); HGS-ETR2; CS-1008; Деносумаб; Сибротузумаб; F19; и 81C6.
Рецепторы иммунной контрольной точки, которые наиболее активно изучались в контексте клинической иммунотерапии рака, антиген 4 ассоциированный с цитотоксическими Т-лимфоцитами (CTLA4 - cytotoxic T-lymphocyte-associated antigen 4; также известный как CD152) и белок 1 запрограммированной смерти клетки (PD1 - programmed cell death protein 1; также известный как CD279) - оба представляют собой ингибирующие рецепторы. Клиническая активность антител, которые блокируют любой из этих рецепторов, подразумевает, что противоопухолевый иммунитет может быть усилен на нескольких уровнях и что комбинаторные стратегии могут быть разумно разработаны, руководствуясь механистическими соображениями и доклиническими моделями.
Двумя лигандами для PD1 являются лиганд 1 PD1 (PDL1; также известный как B7-H1 и CD274) и PDL2 (также известный как B7-DC и CD273). PDL1 экспрессируется на раковых клетках и посредством связывания с его рецептором PD1 на Т-клетках ингибирует активацию/функцию Т-клеток.
Ген 3 активации лимфоцитов (LAG3; также известный как CD223), 2B4 (также известный как CD244), аттенюатор (attenuator) В и Т-лимфоцитов (BTLA; также известный как CD272), белок 3 мембраны Т-клеток (TIM3; также известный как HAVcr2), рецептор аденозина A2a (A2aR) и семейство рецепторов подавления цитотоксичности, были связаны с ингибированием активности лимфоцитов и в некоторых случаях с индукцией анергии лимфоцитов. Нацеливание антител этих рецепторов можно применять в способах по данному изобретению.
Агенты, которые агонизируют иммунную костимулирующую молекулу, также полезны в способах по данному изобретению. Такие агенты включают агонисты или CD40 и OX40. CD40 представляет собой костимулирующий белок, обнаруженный в антигенпрезентирующих клетках (АПК) и необходимый для их активации. Эти АПК включают фагоциты (макрофаги и дендритные клетки) и B-клетки. CD40 является частью семейства рецепторов ФНО. Первичными активирующими сигнальными молекулами для CD40 являются ИФНγ и лиганд CD40 (CD40L). Стимуляция через CD40 активирует макрофаги.
Представляющие интерес антитела к CCR4 (CD194) включают в себя гуманизированные моноклональные антитела, направленные против C-C-хемокинового рецептора 4 (CCR4) с потенциальной противовоспалительной и противоопухолевой активностями. CCR2 экспрессируется на свободных макрофагах, которые могут быть обнаружены при различных воспалительных состояниях, например, ревматоидном артрите; и также были идентифицированы как экспрессирующиеся на макрофагах, стимулирующих опухоль. CCR2 также экспрессируется на регуляторных Т-клетках, а лиганд CCR2, CCL2, опосредует рекрутирование регуляторных Т-клеток в опухоли. Регуляторные Т-клетки подавляют ответ на противоопухолевые Т-клетки, и поэтому желательно их ингибирование или истощение их популяции.
Получение белков по данному изобретению
Хотя антитела могут быть получены с помощью химического синтеза, их обычно получают методами рекомбинантной ДНК, такими как коэкспрессия всех цепей, составляющих белок в одной рекомбинантной клетке-хозяине, или коэкспрессия полипептида тяжелой цепи и антитела, например, человеческого антитела. Кроме того, тяжелая и легкая цепи антитела также могут быть экспрессированы с применением одного полицистронного экспрессионного вектора. Очистка отдельных полипептидов достигается с применением стандартных методов очистки белков, таких как аффинная (белок А) хроматография, эксклюзионная хроматография и/или хроматография гидрофобного взаимодействия. Биспецифические достаточно различаются по размеру и гидрофобности, поэтому очистку можно проводить с применением стандартных процедур.
Количество антитела и полипептида тяжелой цепи, продуцируемых в одной клетке-хозяине, можно минимизировать путем конструирования константных областей антитела и тяжелой цепи, так что гомодимеризация является предпочтительнее гетеродимеризации, например, путем введения самокомплементарных взаимодействий (см., например, WO 98/50431 о возможностях, таких как стратегии "выпуклости-в-полость" (см., WO 96/27011)). Поэтому еще одним аспектом данного изобретения является предоставление способа получения биспецифического в рекомбинантном хозяине, способа, включающего этап: экспрессии в рекомбинантной клетке-хозяине последовательности нуклеиновой кислоты, кодирующей по меньшей мере два полипептида тяжелой цепи, при этом указанные полипептиды тяжелой цепи отличаются своими константным областям в достаточной степени, чтобы уменьшить или предотвратить образование гомодимера, но увеличить образование биспецифического.
Когда белок содержит три цепи, например, FlicAb, они могут быть получены путем коэкспрессии трех цепей (2 тяжелых цепи и одна легкая цепь), составляющих молекулу в одной рекомбинантной клетке-хозяине.
Для рекомбинантной продукции белков в данном документе применяют одну или более нуклеиновых кислот, кодирующих все цепи, например, 2, 3, 4 и тому подобное, выделяют и встраивают в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. Многие векторы являются доступными. Компоненты вектора, как правило, включают в себя, но не ограничиваются ими, один или более из следующих элементов: сигнальную последовательность, точку начала репликации, один или более маркерных генов и энхансерный элемент, промотор и последовательность терминации транскрипции.
В предпочтительном варианте реализации изобретения клетка-хозяин в соответствии со способом по данному изобретению способна к экспрессии человеческого иммуноглобулина на высоком уровне, то есть, по меньшей мере, 1 пг/клетка/день, предпочтительно, по меньшей мере, 10 пг/клетка/день и даже более предпочтительно по меньшей мере 20 пг/клетка/день или более без необходимости амплификации молекул нуклеиновой кислоты, кодирующих отдельные цепи в указанной клетке-хозяине.
Фармацевтическая композиция
Другим аспектом данного изобретения является предоставление фармацевтических композиций, содержащих один или более белков по данному изобретению в смеси с подходящим фармацевтически приемлемым носителем. Примеры фармацевтически приемлемых носителей, применяемых в данном документе, представляют собой, но без ограничений, адъюванты, твердые носители, воду, буферы или другие носители, применяемые в данной области техники для хранения терапевтических компонентов или их комбинаций.
Терапевтические составы белков, применяемые в соответствии с данным изобретением, готовят для хранения путем смешивания белков, имеющих желаемую степень чистоты, с необязательными фармацевтически приемлемыми носителями, наполнителями или стабилизаторами (см., например, Remington's Pharmaceutical Sciences 16-е издание, Osol, A. Ред. (1980 год)), например, в виде лиофилизированных составов или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях, и включают буферные вещества, например, фосфат, цитрат и другие органические соли; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (например, хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензэтония; фенол; бутиловый или бензиловый спирт; алкилпарабены, например, метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и m-крезол); низкомолекулярные (содержащие менее 10 остатков) полипептиды; белки, например, сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, например, поливинилпирролидон; аминокислоты, например, глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, например, ЭДТК; сахара, например, сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, например, натрия; металлсодержащие комплексы (например, комплексы Zn-белок) и/или неионогенные ПАВ, например, TWEEN™, PLURONICS™ или полиэтиленгликоль (ПЭГ).
Составы антител к CD3 описаны, например, в патентной публикации США №20070065437, полное описание которой включено в данный документ посредством ссылки. Подобные составы могут быть применены для белков по данному изобретению. Основными компонентами таких составов являются рН-буферное средство, эффективное в диапазоне от 3,0 до 6,2, соль, поверхностно-активное вещество и эффективное количество биспецифического со специфичностью к CD3.
Способы применения
Предлагаются способы лечения или снижения заболеваемости, включая, без ограничения, инфекцию, аутоиммунное заболевание, первичный или метастатический рак и тому подобное, в схеме приема лекарств, включающей контакт клеток-мишеней с антигенсвязывающей композицией по данному изобретению, особенно когда антигенсвязывающая композиция представляет собой мультиспецифическое антитело, подходящее для состояния, подвергаемого лечению, например, где один связывающий фрагмент специфически связывается с антигеном, ассоциированным с опухолью, для лечения соответствующих раковых клеток; связывающий фрагмент, специфичный для патогена, представляющего интерес, для лечения соответствующей инфекции, и тому подобное. Такие способы включают в себя введение субъекту, нуждающемуся в этом, терапевтически эффективного количества или эффективной дозы агентов по данному изобретению, включая без ограничения комбинации реагента с химиотерапевтическим лекарственным средством, лучевую терапию или хирургическое вмешательство.
Эффективные дозы композиций по данному изобретению для лечения заболевания варьируют в зависимости от многих разных факторов, в том числе способа введения, целевого сайта, физиологического состояния пациента, от того, представляет ли собой пациент человека или животное, других вводимых медикаментов, и того, является ли лечение профилактическим или терапевтическим. Обычно пациентом является человек, но млекопитающие, отличные от человека, также могут подвергаться лечению, например, домашние животные, такие как собаки, кошки, лошади и тому подобное, лабораторные млекопитающие, такие как кролики, мыши, крысы и тому подобное, и тому подобное. Дозировки для лечения можно подбирать для оптимизации безопасности и эффективности.
Уровни дозировки могут быть легко определены квалифицированным лечащим врачом и могут быть изменены при необходимости, например, при необходимости изменения реакции субъекта на терапию. Количество активного ингредиента, которое можно комбинировать с веществами-носителями для получения разовой лекарственной формы, изменяется в зависимости от получающего лечение индивида и конкретного способа введения. Единичные лекарственные формы обычно содержат от около 1 мг до около 500 мг активного ингредиента.
В некоторых вариантах реализации изобретения терапевтическая дозировка агента может составлять от около 0,0001 до 100 мг/кг и более, обычно от 0,01 до 5 мг/кг от массы тела хозяина. Например, дозировки могут составлять 1 мг/кг массы тела или 10 мг/кг массы тела или в пределах 1-10 мг/кг. Типовая схема лечения включает в себя введение один раз каждые две недели или один раз в месяц или один раз каждые 3-6 месяцев. Терапевтические препараты по данному изобретению обычно вводят несколько раз. Интервалы между введением каждой дозы могут быть еженедельными, ежемесячными или ежегодными. Интервалы также могут быть неравномерными, на что указывает измерение уровня терапевтического препарат в крови пациента. В альтернативном варианте, терапевтические препараты по данному изобретению могут быть введены в виде состава для продолжительного высвобождения, в этом случае необходимо менее частое введение. Дозировка и частота варьируют в зависимости от времени полужизни полипептида у пациента.
В профилактических целях относительно небольшая доза может вводиться через относительно длительные интервалы на протяжении продолжительного периода времени. Некоторые пациенты продолжают получать лечение на протяжении всей жизни. В других терапевтических применениях иногда требуется относительно высокая дозировка с относительно короткими интервалами, до тех пор, пока прогрессирование заболевания не уменьшится или не прекратится, и предпочтительно, до тех пор, пока пациент не покажет частичное или полное улучшение симптомов заболевания. После этого пациенту может быть назначен профилактический режим.
В еще других вариантах реализации способы по данному изобретению включают в себя лечение, уменьшение или предотвращение роста опухоли, метастазирования опухоли или инвазии опухоли раков, включая карциномы, гематологические злокачественные опухоли, такие как лейкемии и лимфомы, меланомы, саркомы, глиомы, и тому подобное. Для профилактического применения фармацевтические композиции или лекарственные препараты вводят пациенту, подверженному риску заболевания или иным образом восприимчивому к заболеванию, в количестве, достаточном для устранения или уменьшения риска, уменьшения степени тяжести или задержки начала заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, присутствующие в ходе развития заболевания.
Композиции для лечения заболевания могут вводиться парентеральным, местным, внутривенным, внутриопухолевым, пероральным, подкожным, внутриартериальным, внутричерепным, внутрибрюшинным, интраназальным или внутримышечным способом. Типичным путем введения является внутривенный или внутриопухолевый, хотя другие пути могут быть одинаково эффективными.
Обычно композиции готовят в виде инъекционных растворов, либо в виде жидких растворов, либо суспензий; также могут быть приготовлены твердые формы, подходящие для растворения или суспендирования в жидких носителях перед инъекцией. Препарат также может быть эмульгирован или инкапсулирован в липосомы или микрочастицы, такие как полилактид, полигликолид или сополимер, для усиления адъювантного эффекта, как обсуждалось выше. Langer, Science 249:1527, 1990 год и Hanes, Advanced Drug Delivery Reviews 28:97-119, 1997 год. Агенты по данному изобретению можно вводить в форме инъекции веществ замедленного всасывания или имплантата препарата, который может быть составлен таким образом, чтобы обеспечить длительное или пульсирующее высвобождение активного ингредиента. Фармацевтические композиции, как правило, составлены как стерильные, по существу изотонические и полностью соответствующие всем нормам Надлежащей Производственной Практики (GMP - Good Manufacturing Practice) США. Управление по контролю качества пищевых продуктов и лекарственных средств США.
Токсичность белков, описанных в данном документе, может быть определена стандартными фармацевтическими процедурами на клеточных культурах или экспериментальных животных, например, путем определения ЛД50 (доза, летальная для 50% популяции) или ЛД100 (доза, летальная для 100% популяции). Соотношение доз между токсическим и терапевтическим эффектом, представляет собой терапевтический индекс. Данные, полученные из этих анализов клеточных культур и исследований на животных, могут быть применены при определении диапазона доз, который не токсичен для применения у человека. Дозировка белков, описанных в данном документе, предпочтительно находится в диапазоне значений концентраций в кровообращении, которые включают эффективную дозу с небольшой токсичностью или без нее. Дозировка может меняться в этом диапазоне в зависимости от применяемой формы дозировки и используемого пути введения. Точный состав, способ введения и дозировка могут быть выбраны отдельным врачом с учетом состояния пациента.
Фармацевтические композиции можно вводить в различных стандартных лекарственных формах в зависимости от способа введения. Например, стандартные лекарственные формы, подходящие для перорального введения, включают, но без ограничения, порошок, таблетки, пилюли, капсулы и пастилки для рассасывания. Общепризнано, что композиции по данному изобретению при пероральном введении должны быть защищены от воздействия пищеварения. Это обычно достигается либо путем образования комплексов молекул с композицией для придания им устойчивости к кислотному и ферментативному гидролизу, либо путем упаковки молекул в подходяще устойчивый носитель, такой как липосома или защитный барьер. Средства защиты агентов от системы пищеварения хорошо известны в данной области техники.
Композиции для введения обычно содержат антитело или другой абляционный агент, растворенный в фармацевтически приемлемом носителе, предпочтительно в водном носителе. Могут применяться различные водные носители, например, забуференный физиологический раствор и тому подобное. Эти растворы являются стерильными и, как правило, не содержат нежелательных веществ. Эти композиции могут быть стерилизованы обычными, хорошо известными методами стерилизации. Композиции могут содержать фармацевтически приемлемые вспомогательные вещества, которые необходимы для соответствия физиологическим условиям, такие как средства регулирующие рН и буферные средства, средства, регулирующие токсичность и тому подобное, например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция, лактат натрия и тому подобное. Концентрация активного агента в этих составах может варьироваться в широких пределах и будет выбираться в первую очередь на основе объемов жидкости, вязкости, массы тела и тому подобного в соответствии с конкретным выбранным путем введения и потребностями пациента (например, Remington's Pharmaceutical Science (15-е изд., 1980 год) и Goodman & Gillman, The Pharmacological Basis of Therapeutics (Hardman и соавт., ред., 1996 год)).
Также в объем данного изобретения входят наборы, содержащие активные агенты и их составы, по данному изобретению и инструкции по применению. Набор может дополнительно содержать по меньшей мере один дополнительный реагент, например, химиотерапевтическое лекарственное средство и тому подобное. Наборы обычно включают в себя этикетку, указывающую на предполагаемое применение содержимого набора. Термин "этикетка" включает в себя любые письменные или записанные материалы, поставляемые в комплекте или вместе с ним, или иным образом сопровождающие набор.
Композиции можно вводить для терапевтического лечения. Композиции вводят пациенту в количестве, достаточном для существенного удаления клеток-мишеней, как описано выше. Количество, достаточное для достижения этого, определяется как "терапевтически эффективная доза", которая может обеспечить улучшение общей выживаемости. Композиции можно вводить однократно или многократно в зависимости от требуемой и переносимой пациентом дозировки и частоты. Конкретная доза, необходимая для лечения, будет зависеть от медицинского диагноза и истории заболевания млекопитающего, а также от других факторов, таких как возраст, вес, пол, путь введения, эффективность и тому подобного.
Теперь, когда изобретение полностью описано, для специалиста в данной области техники будет очевидно, что различные изменения и модификации могут быть сделаны без отклонения от сущности или объема изобретения.
Примеры
Пример 1
Генетически сконструированные крысы, экспрессирующие антитела, содержащие только тяжелые цепи
Человеческий локус IgH был сконструирован и собран из нескольких частей, которые включали модификацию и объединение генов С-области крысы, которые затем соединялись по ходу транскрипции области VH6-D-JH человека. Два BAC с отдельными кластерами человеческих генов VH были затем совместно введены с BAC, кодирующим собранный (человеческий VH6-D-JH-крысиный C) фрагмент.
Были получены трансгенные крысы, несущие искусственные локусы иммуноглобулинов тяжелой цепи в неупорядоченной конфигурации. Включенные гены константной области кодируют IgM, IgD, IgG2b, IgE, IgA и 3'-энхансер. ОТ-ПЦР и анализ сыворотки (ИФА) трансгенных крыс выявили продуктивную перестройку локусов трансгенных иммуноглобулинов и экспрессию в сыворотке различных изотипов антител только с тяжелой цепью. Трансгенные крысы перекрестно скрещивались с крысами с мутированными эндогенными локусами тяжелой цепи и легкой цепи, ранее описанными в публикации патента США 2009/0098134 А1. Анализ таких животных продемонстрировал инактивацию экспрессии тяжелой и легкой цепи иммуноглобулина крысы и высокий уровень экспрессии антител тяжелой цепи с вариабельными областями, кодируемыми генами V, D и J человека. Иммунизация трансгенных крыс приводила к получению сывороточных ответов с высоким титром антигенспецифических антител с тяжелой цепью. Эти трансгенные крысы, экспрессирующие антитела с тяжелой цепью с областью VDJ человека, были названы UniRats.
Пример 2
Генетически сконструированные крысы, экспрессирующие антитела с фиксированной легкой цепью
Наборы трансгенных человеческих антител были получены из Н-цепей с разнообразной (VH-D-JH)n перестройкой в сочетании с уникальной L-цепью. Для этого перестроенная L-цепь, человеческий Vk-Jk1-Ck, была интегрирована в зародышевую линию крысы посредством микроинъекции ДНК, и полученные трансгенные животные были скрещены с ранее описанной линией крысы, которая естественным образом экспрессирует набор человеческой H-цепи (Osborn и соавт., 2013 год). Эта новая линия крыс была названа OmniFlic.
Иммунизация крыс OmniFlic с применением множества различных антигенов продуцировала высокие уровни антигенспецифического IgG, сходного с другими трансгенными крысами, несущими тот же локус IgH. Анализ набора методом ОТ-ПЦР позволил выявить сильно вариабельные перестройки гена VH при высоких уровнях транскрипта и белка. Кроме того, был идентифицирован только один продукт L-цепи, также экспрессирующийся на высоком уровне.
Антигенспецифические связыватели из OmniFlic были получены с помощью сиквенирования нового поколения (NGS) и отобраны из библиотек кДНК (дрожжей, E. coli, фагов), которые после сиквенирования идентифицировали разнообразные транскрипты H-цепи. Для экспрессии в клетках млекопитающих гипермутированные конструкции H-цепи трансфицировали в комбинации с исходной трансгенной последовательностью Igκ. В этом перестроенном Vk-Jk1-Ck не допускались мутационные изменения, и всегда одна и та же L-цепь экспрессировалась с различными продуктами H-цепи для генерации моноклонального человеческого IgG.
Пример 3
Получение антигенспецифических антител у трансгенных крыс.
Для получения антигенспецифических антител с тяжелой цепью у крыс, экспрессирующих генетически сконструированных крыс, иммунизировали двумя способами.
Иммунизация рекомбинантными внеклеточными доменами PD-L1 и BCMA. Рекомбинантные внеклеточные домены PD-L1 и BCMA были приобретены у R&D Systems, разбавлены стерильным физиологическим раствором и объединены с адъювантом. Иммуногены либо комбинировали с полным адъювантом Фрейнда (CFA) и неполным адъювантом Фрейнда (IFA), либо с адъювантами Titermax и Ribi. Первую иммунизацию (примирование) иммуногеном в CFA или Titermax вводили в левую и правую ноги. После первой иммунизации иммуногенами в CFA в каждую ногу вводили еще две иммунизации в IFA (бустер-инъекции) или еще 4 иммунизации в Ribi и еще одну в Titermax. Эта последовательность иммунизаций приводит к развитию В-клеток, продуцирующих высокоаффинные антитела. Концентрации иммуногена составляли 10 мкг на ногу. Сыворотку собирали у крыс при последнем кровопускании для определения титров сыворотки.
Для получения антител к CD3δε человека генетически сконструированных крыс иммунизировали с применением протоколов иммунизации на основе ДНК.
Крыс OmniFlic иммунизировали конструкциями CD3-эпсилон/дельта человека и яванской макаки в Aldevron, Inc. (Фарго, штат Северная Дакота) с применением технологии антител GENOVAC. Дренирующие лимфатические узлы собирали после окончательной иммунизации и выделяли РНК. После синтеза кДНК набор антител тяжелой цепи IgH был охарактеризован с помощью сиквенирования нового поколения и нашего собственного программного обеспечения для внутреннего использования. Были отобраны кандидатные антигенспецифические последовательности VH, демонстрирующие доказательства антигенспецифического положительного отбора. Несколько сотен последовательностей VH, кодирующих FlicAb, были отобраны для сборки генов и клонированы в экспрессионный вектор. Впоследствии полностью человеческие антитела FlicAb IgG1 экспрессировали в клетках HEK для анализа с помощью Flow и ИФА. Человеческие FlicAb тестировали на связывание с первичными человеческими Т-клетками и клетками Jurkat с помощью потока. Кроме того, человеческие FlicAb были протестированы с применением рекомбинантных белков CD3δε в ИФА. Все FlicAb с положительным связыванием с Т-клетками человека перечислены на фиг. 1. Выбранные последовательности были дополнительно охарактеризованы в анализах активации Т-клеток.
Пример 4. Характеристика антител OmniFlic к CD3
Антитела, полученные в результате кампании, как описано выше, были дополнительно охарактеризованы по их способности активировать Т-клетки для продукции цитокинов и активировать CD69 на их поверхности. Для количественного определения ИЛ-2, продуцируемого клетками Jurkat или лимфоцитами периферической крови, применяли набор ИФА max BioLegend. Результаты эксперимента с применением Т-клеток периферической крови человека и различных концентраций антител OmniFlic проиллюстрированы на фиг. 3.
Пример 5. Характеристика биспецифических антител
FlicAb к CD3 ID 304703 (SEQ ID NO: 39) был выбран для дальнейшей разработки в биспецифических. Этот FlicAb перекрестно реагирует с CD3 яванской макаки и существенно стимулирует Т-клетки человека. Было получено два типа биспецифических (см. фиг. 2 для схематического представления). Для создания биспецифичных FlicAb применялась технология "выступ-во-впадину" (Protein Engineering том. 9, № 7, стр. 617-621, 1996 год, 'Knobs-into-holes' engineering of antibody CH3 domains for heavy chain heterodimerization. John B. B. Ridgway, Leonard G. Presta и Paul Carter). С-конец тяжелой цепи с помощью выступа был помечен С-меткой, а гетеродимерные антитела были очищены с применением CaptureSelect C-Tag Affinity Matrix (Thermo Fischer Scientific). Было произведено биспецифическое FlicAb с одним плечом, реагирующим с CD3 (ID 304703), а другим - с человеческим PD-L1, и было показано, что оно активирует человеческие CD8+ T-клетки только в присутствии PD-L1-позитивных опухолевых клеток (фиг. 3). HDLM2 представляет собой клеточную линию множественной миеломы, которая экспрессирует PD-L1 на поверхности. Ramos представляет собой клеточную линию лимфомы Беркитта, которая отрицательна по PD-L1. Экспрессия CD69 применялась как регистрируемый показатель. Биспецифическое антитело применяли в указанных концентрациях.
Как проиллюстрировано на фиг. 4, опухолевые клетки (HDLM2, которые экспрессируют PD-L1 на поверхности клетки) инкубировали с очищенными T-клетками CD8 + человека и биспецифическими антителами. Клетки HDLM2 не экспрессируют CD20, и сокультура с биспецифическим FlicAb α-CD3/α-CD20 не приводила к гибели клеток HDLM2. Только сокультура человеческих CD8+ Т-клеток и HDLM2 с биспецифическим FlicAb α-CD3/α-PD-L1 приводила к значительному уничтожению.
На фиг. 5 обобщены данные для антител в моноспецифическом и биспецифическом формате. В столбце 1 показан идентификатор последовательности для VH к CD3 идентификатор последовательности ID NO: 304703 (SEQ ID NO: 39), 314171 (SEQ ID NO: 13), 313306 (SEQ ID NO: 1), 313329 (SEQ ID NO: 6) и 313283 (SEQ ID NO: 18). В столбце 2 показано значение СИФ для связывания клетками Jurkat исходного моноспецифического (антитела) к CD3. В столбце 3 показано значение СИФ для связывания T-клетками яванского макака исходного моноспецифического (антитела) к CD3. В столбце 5 показаны пикограммы ИЛ-2, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 6 показаны пикограммы ИЛ-6, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 7 показаны пикограммы ИЛ-10, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 8 показаны пикограммы ИФН-γ, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 9 показаны пикограммы ФНОα, высвобождаемого пан-Т-клетками, стимулированными биспецифическим антителом, связывающим белок ВСМА, помещенным на пластике, в указанной дозе. В столбце 10 показана ЭК50 лизиса опухолевых клеток U266, опосредованного биспецифическими антителами, в присутствии пан-Т-клеток человека. В столбце 11 показан процент лизиса опухолевых клеток U266, в присутствии биспецифического антитела и пан-Т-клеток человека при дозе 333 нг/мл биспецифического антитела. В столбце 12 показана аффинность белкового связывания плеча к CD3 биспецифического антитела, измеренная с помощью Octet. В столбце 13 показано значение СИФ для связывания клетками Jurkat биспецифического антитела.
Пример 6
Корреляция активности биспецифического уничтожения и высвобождения цитокинов.
Как проиллюстрировано на фиг. 6, четыре биспецифических антитела αCD3_fam1:aBCMA, каждое с уникальным плечом к CD3 (как показано) и общим плечом к BCMA, были протестированы на способность убивать опухолевые клетки U266 BCMA+ посредством перенаправления активированных первичных Т-клеток. В этом эксперименте клетки U266, которые экспрессируют BCMA, смешивали с активированными пан-Т-клетками в соотношении E:T 10:1 наряду с добавлением биспецифического антитела. Ось x демонстрирует концентрацию применяемого антитела, а ось y демонстрирует % лизиса опухолевых клеток через 6 часов после добавления антитела. Активность уничтожения коррелировала с высвобождением ИЛ-2 (фиг. 7); с высвобождением ИФН-γ (фиг. 8) и с аффинностью связывания CD3 (фиг. 9). Корреляция между производством ИЛ-2 и лизисом опухолевых клеток U266 составляет R2=0,37. Корреляция между производством ИФН-γ и лизисом опухолевых клеток U266 составляет R2=0,53. Корреляция между ЭК50 уничтожения U266 и аффинностью связывания белка составляет R2=0,93.
Биспецифические антитела αCD3_F1F:αBCMA анализировали на способность уничтожать три различных линии опухолевых клеток ВСМА+ и одну линию ВСМА-негативных клеток путем перенаправления активированных исходных Т-клеток. Антитела состояли из плеча αCD3 (SEQ ID NO: 1 и SEQ ID NO: 69) и плеча αBCMA (SEQ ID NO: 70 или SEQ ID NO: 71) (проиллюстрировано на фиг. 10-12). В этом эксперименте опухолевые клетки смешивали с активированными пан-Т-клетками в соотношении E:T 10:1 наряду с добавлением биспецифического антитела. На фиг. 10A проиллюстрировано уничтожение клеток RPMI-8226, на фиг. 10B проиллюстрировано уничтожение клеток NCI-H929, на панели C проиллюстрировано уничтожение клеток U-266 и на фиг. 10D проиллюстрировано уничтожение клеток K562, отрицательный контроль. Ось x демонстрирует концентрацию применяемого антитела, а ось y демонстрирует% лизиса опухолевых клеток через 6 часов после добавления антитела.
На фиг. 11 проиллюстрирован уровень высвобождения цитокинов ИЛ-2 измеренный после культивирования Т-клеток человека в покое с различными линиями опухолевых клеток и увеличивающимися дозами биспецифического антитела aCD3_F1F:aBCMA (как проиллюстрировано на фиг. 10). На фиг. 11A проиллюстрировано высвобождение ИЛ-2, стимулированное клетками RPMI-8226, на фиг. 11B проиллюстрировано высвобождение ИЛ-2, стимулированное клетками NCI-H929, на фиг. 11C проиллюстрировано высвобождение ИЛ-2, стимулированное клетками U-266 и на фиг. 11D проиллюстрировано высвобождение ИЛ-2, стимулированное клетками K562, отрицательный контроль.
Уровень высвобождения цитокина ИФН-γ измеряли после культивирования Т-клеток человека в покое с различными линиями опухолевых клеток и увеличивающимися дозами биспецифического антитела aCD3_F1F:aBCMA (как проиллюстрировано на фиг. 10). На фиг. 12A проиллюстрировано высвобождение ИФН-γ, стимулированное клетками RPMI-8226, на фиг. 12B проиллюстрировано высвобождение ИФН-γ, стимулированное клетками NCI-H929, на фиг. 12C проиллюстрировано высвобождение ИФН-γ, стимулированное клетками U-266 и на фиг. 12D проиллюстрировано высвобождение ИФН-γ, стимулированное клетками K562, отрицательный контроль.
Примеры указаны таким образом, чтобы обеспечить обычным специалистам в этой области техники полное раскрытие и описание того, как изготавливать и применять данное изобретение, и не предназначены для ограничения объема того, что авторы изобретения считают своим изобретением, и они не предназначены для представления того, что эксперименты, приведенные ниже, это все эксперименты, которые проводились. Были предприняты меры для обеспечения точности в отношении применяемых чисел (например, количеств, температуры и тому подобного), но некоторые экспериментальные ошибки и отклонения должны учитываться. Если не указано иное, части представляют собой части по весу, молекулярная масса представляет собой среднюю молекулярную массу, температура представлена в градусах Цельсия, а давление равно или близко к атмосферному.
В то время как данное изобретение было описано со ссылками на конкретные варианты его реализации, специалистам в данной области техники следует понимать, что могут быть проведены различные изменения и эквиваленты могут быть заменены без отхода от истинной сути и объема данного изобретения. Кроме того, многие модификации могут быть произведены для того, чтобы адаптировать конкретную ситуацию, материал, состав вещества, процесс, технологические стадии или стадию к задаче, сути и объему данного изобретения. Предполагается, что все указанные модификации находятся в пределах объема прилагаемой формулы изобретения.
--->
Перечень последовательностей
<110> TENEOBIO, INC.
<120> CD3 BINDING ANTIBODIES
<130> 60792.00027US01 (TNO-0010-US1)
<140> 16/312,743
<141> 2019-07-22
<150> PCT/US2017/038373
<151> 2017-06-20
<150> 62/491,908
<151> 2017-04-28
<150> 62/394,360
<151> 2016-09-14
<150> 62/352,698
<151> 2016-06-21
<160> 105
<170> PatentIn version 3.5
<210> 1
<211> 128
<212> PRT
<213> Homo sapiens
<400> 1
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Thr Thr Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 2
<211> 128
<212> PRT
<213> Homo sapiens
<400> 2
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Glu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 3
<211> 128
<212> PRT
<213> Homo sapiens
<400> 3
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Glu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 4
<211> 128
<212> PRT
<213> Homo sapiens
<400> 4
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser His Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 5
<211> 128
<212> PRT
<213> Homo sapiens
<400> 5
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 6
<211> 128
<212> PRT
<213> Homo sapiens
<400> 6
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Ile Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Val Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 7
<211> 128
<212> PRT
<213> Homo sapiens
<400> 7
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Met Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 8
<211> 128
<212> PRT
<213> Homo sapiens
<400> 8
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 9
<211> 128
<212> PRT
<213> Homo sapiens
<400> 9
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Arg Gly Leu Glu
35 40 45
Trp Val Gly Tyr Ile Ser Tyr Ser Gly Arg Thr Tyr Tyr Asp Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 10
<211> 128
<212> PRT
<213> Homo sapiens
<400> 10
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Asp
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Thr Thr Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 11
<211> 128
<212> PRT
<213> Homo sapiens
<400> 11
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Glu Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 12
<211> 128
<212> PRT
<213> Homo sapiens
<400> 12
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 128
<212> PRT
<213> Homo sapiens
<400> 13
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 14
<211> 128
<212> PRT
<213> Homo sapiens
<400> 14
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 15
<211> 128
<212> PRT
<213> Homo sapiens
<400> 15
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 16
<211> 128
<212> PRT
<213> Homo sapiens
<400> 16
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 17
<211> 128
<212> PRT
<213> Homo sapiens
<400> 17
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Met Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 18
<211> 128
<212> PRT
<213> Homo sapiens
<400> 18
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 19
<211> 128
<212> PRT
<213> Homo sapiens
<400> 19
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 20
<211> 128
<212> PRT
<213> Homo sapiens
<400> 20
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 21
<211> 128
<212> PRT
<213> Homo sapiens
<400> 21
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Asp Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Thr Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 22
<211> 128
<212> PRT
<213> Homo sapiens
<400> 22
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Leu
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Arg Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 23
<211> 128
<212> PRT
<213> Homo sapiens
<400> 23
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Asn Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 24
<211> 128
<212> PRT
<213> Homo sapiens
<400> 24
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Ile Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 25
<211> 128
<212> PRT
<213> Homo sapiens
<400> 25
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 26
<211> 128
<212> PRT
<213> Homo sapiens
<400> 26
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 27
<211> 128
<212> PRT
<213> Homo sapiens
<400> 27
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Val Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 28
<211> 128
<212> PRT
<213> Homo sapiens
<400> 28
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Ile Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Leu
65 70 75 80
Ser Leu Lys Val Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 29
<211> 128
<212> PRT
<213> Homo sapiens
<400> 29
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Met Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 30
<211> 128
<212> PRT
<213> Homo sapiens
<400> 30
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly Tyr Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 31
<211> 128
<212> PRT
<213> Homo sapiens
<400> 31
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Phe Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 32
<211> 128
<212> PRT
<213> Homo sapiens
<400> 32
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Phe Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 33
<211> 128
<212> PRT
<213> Homo sapiens
<400> 33
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Phe Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 34
<211> 128
<212> PRT
<213> Homo sapiens
<400> 34
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Asp
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 35
<211> 128
<212> PRT
<213> Homo sapiens
<400> 35
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ser Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 36
<211> 128
<212> PRT
<213> Homo sapiens
<400> 36
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 37
<211> 128
<212> PRT
<213> Homo sapiens
<400> 37
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ser Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 38
<211> 128
<212> PRT
<213> Homo sapiens
<400> 38
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 39
<211> 128
<212> PRT
<213> Homo sapiens
<400> 39
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 40
<211> 128
<212> PRT
<213> Homo sapiens
<400> 40
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr His Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Met Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 41
<211> 128
<212> PRT
<213> Homo sapiens
<400> 41
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 42
<211> 128
<212> PRT
<213> Homo sapiens
<400> 42
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ile Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 43
<211> 128
<212> PRT
<213> Homo sapiens
<400> 43
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Asn Thr Tyr Tyr Lys Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Met Asn Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 44
<211> 128
<212> PRT
<213> Homo sapiens
<400> 44
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 45
<211> 128
<212> PRT
<213> Homo sapiens
<400> 45
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 46
<211> 128
<212> PRT
<213> Homo sapiens
<400> 46
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 47
<211> 128
<212> PRT
<213> Homo sapiens
<400> 47
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ser Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 48
<211> 128
<212> PRT
<213> Homo sapiens
<400> 48
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 49
<211> 128
<212> PRT
<213> Homo sapiens
<400> 49
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 50
<211> 128
<212> PRT
<213> Homo sapiens
<400> 50
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ser Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Gly Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 51
<211> 128
<212> PRT
<213> Homo sapiens
<400> 51
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 52
<211> 128
<212> PRT
<213> Homo sapiens
<400> 52
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 53
<211> 128
<212> PRT
<213> Homo sapiens
<400> 53
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Gln Ser Arg Val Phe Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Asn Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 54
<211> 128
<212> PRT
<213> Homo sapiens
<400> 54
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 55
<211> 128
<212> PRT
<213> Homo sapiens
<400> 55
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Arg Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 56
<211> 128
<212> PRT
<213> Homo sapiens
<400> 56
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Ser Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Met Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Arg Leu Asn Ser Val Thr Ala Val Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 57
<211> 128
<212> PRT
<213> Homo sapiens
<400> 57
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 58
<211> 128
<212> PRT
<213> Homo sapiens
<400> 58
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 59
<211> 128
<212> PRT
<213> Homo sapiens
<400> 59
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Val Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 60
<211> 128
<212> PRT
<213> Homo sapiens
<400> 60
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Asn Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 61
<211> 128
<212> PRT
<213> Homo sapiens
<400> 61
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 62
<211> 128
<212> PRT
<213> Homo sapiens
<400> 62
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 63
<211> 128
<212> PRT
<213> Homo sapiens
<400> 63
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Ile Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 64
<211> 128
<212> PRT
<213> Homo sapiens
<400> 64
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Asn Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 65
<211> 128
<212> PRT
<213> Homo sapiens
<400> 65
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 66
<211> 128
<212> PRT
<213> Homo sapiens
<400> 66
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln Tyr Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile His Arg Ser Gly Asn Pro Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Leu Leu Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Arg Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 67
<211> 128
<212> PRT
<213> Homo sapiens
<400> 67
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Ser Val Ser Gly Gly Ser Ile Arg Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Tyr Ser Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Ser Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 68
<211> 128
<212> PRT
<213> Homo sapiens
<400> 68
Arg Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Gly
20 25 30
Gly His Tyr Trp Ser Trp Ile Arg Gln His Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Tyr Ile Tyr His Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr
100 105 110
Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 69
<211> 107
<212> PRT
<213> Homo sapiens
<400> 69
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Asn Trp Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 70
<211> 243
<212> PRT
<213> Homo sapiens
<400> 70
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile Arg Gly Ser Asp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gln Gly Glu Asn Asp Gly Pro Phe Asp His Arg Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Tyr Gly Met Ser Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val Ser Gly Ile Arg
165 170 175
Gly Ser Asp Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe
180 185 190
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Lys Gln Gly
210 215 220
Glu Asn Asp Gly Pro Phe Asp His Arg Gly Gln Gly Thr Leu Val Thr
225 230 235 240
Val Ser Ser
<210> 71
<211> 119
<212> PRT
<213> Homo sapiens
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Gly Ile Arg Gly Ser Asp Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gln Gly Glu Asn Asp Gly Pro Phe Asp His Arg Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 72
<211> 10
<212> PRT
<213> Homo sapiens
<400> 72
Gly Gly Ser Ile Arg Ser Gly Gly His Tyr
1 5 10
<210> 73
<211> 7
<212> PRT
<213> Homo sapiens
<400> 73
Ile Ser Tyr Ser Gly Ser Thr
1 5
<210> 74
<211> 20
<212> PRT
<213> Homo sapiens
<400> 74
Ala Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 75
<211> 7
<212> PRT
<213> Homo sapiens
<400> 75
Ile His His Ser Gly Ser Thr
1 5
<210> 76
<211> 20
<212> PRT
<213> Homo sapiens
<400> 76
Ala Arg Trp Arg His Asp Ile Phe Thr Thr Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 77
<211> 10
<212> PRT
<213> Homo sapiens
<400> 77
Gly Gly Ser Ile Ser Ser Gly Gly His Tyr
1 5 10
<210> 78
<211> 7
<212> PRT
<213> Homo sapiens
<400> 78
Ile His Tyr Ser Gly Ser Thr
1 5
<210> 79
<211> 20
<212> PRT
<213> Homo sapiens
<400> 79
Ala Arg Trp Arg His Asp Ile Phe Ala Ala Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 80
<211> 7
<212> PRT
<213> Homo sapiens
<400> 80
Ile His Arg Ser Gly Asn Pro
1 5
<210> 81
<211> 6
<212> PRT
<213> Homo sapiens
<400> 81
Gln Ser Val Ser Ser Asn
1 5
<210> 82
<211> 9
<212> PRT
<213> Homo sapiens
<400> 82
Gln Gln Tyr Asn Asn Trp Pro Trp Thr
1 5
<210> 83
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (7)..(10)
<223> Any amino acid
<220>
<223> See specification as filed for detailed description of
substitutions and preferred embodiments
<400> 83
Gly Gly Ser Ile Xaa Ser Xaa Xaa Xaa Xaa
1 5 10
<210> 84
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<220>
<221> MOD_RES
<222> (1)..(1)
<223> Any amino acid
<220>
<221> MOD_RES
<222> (8)..(10)
<223> Any amino acid
<220>
<223> See specification as filed for detailed description of
substitutions and preferred embodiments
<400> 84
Xaa Arg Trp Arg His Asp Ile Xaa Xaa Xaa Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 85
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 85
Thr Val Ala Ala Pro
1 5
<210> 86
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<400> 86
Ala Ser Thr Lys Gly Pro
1 5
<210> 87
<211> 10
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<220>
<221> MOD_RES
<222> (5)..(5)
<223> Any amino acid
<220>
<223> See specification as filed for detailed description of
substitutions and preferred embodiments
<400> 87
Gly Gly Ser Ile Xaa Ser His His Gly Tyr
1 5 10
<210> 88
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<220>
<221> MOD_RES
<222> (2)..(3)
<223> Any amino acid
<220>
<223> See specification as filed for detailed description of
substitutions and preferred embodiments
<400> 88
Ile Xaa Xaa Ser Gly Ser Thr
1 5
<210> 89
<211> 7
<212> PRT
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
peptide
<220>
<221> MOD_RES
<222> (2)..(3)
<223> Any amino acid
<220>
<223> See specification as filed for detailed description of
substitutions and preferred embodiments
<400> 89
Ile Xaa Xaa Ser Gly Asn Pro
1 5
<210> 90
<211> 10
<212> PRT
<213> Homo sapiens
<400> 90
Gly Gly Ser Ile Ser Ser Ser Ser His Tyr
1 5 10
<210> 91
<211> 10
<212> PRT
<213> Homo sapiens
<400> 91
Gly Gly Ser Ile Arg Ser Gly Gly Tyr Tyr
1 5 10
<210> 92
<211> 10
<212> PRT
<213> Homo sapiens
<400> 92
Gly Gly Ser Ile Ser Ser Gly Gly His Phe
1 5 10
<210> 93
<211> 7
<212> PRT
<213> Homo sapiens
<400> 93
Ile Tyr His Ser Gly Ser Thr
1 5
<210> 94
<211> 7
<212> PRT
<213> Homo sapiens
<400> 94
Ile Tyr Tyr Ser Gly Ser Thr
1 5
<210> 95
<211> 7
<212> PRT
<213> Homo sapiens
<400> 95
Ile Ser Tyr Ser Gly Arg Thr
1 5
<210> 96
<211> 7
<212> PRT
<213> Homo sapiens
<400> 96
Ile Tyr His Ser Gly Ile Thr
1 5
<210> 97
<211> 7
<212> PRT
<213> Homo sapiens
<400> 97
Ile Tyr His Ser Gly Asn Thr
1 5
<210> 98
<211> 20
<212> PRT
<213> Homo sapiens
<400> 98
Gly Arg Trp Arg His Asp Ile Leu Thr Ala Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 99
<211> 20
<212> PRT
<213> Homo sapiens
<400> 99
Ala Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 100
<211> 20
<212> PRT
<213> Homo sapiens
<400> 100
Gly Arg Trp Arg His Asp Ile Leu Thr Gly Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 101
<211> 20
<212> PRT
<213> Homo sapiens
<400> 101
Ala Arg Trp Arg His Asp Ile Leu Thr Arg Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 102
<211> 20
<212> PRT
<213> Homo sapiens
<400> 102
Ala Arg Trp Arg His Asp Ile Leu Thr Val Tyr Pro Tyr Tyr Tyr Tyr
1 5 10 15
Gly Met Asp Val
20
<210> 103
<211> 8
<212> PRT
<213> Homo sapiens
<400> 103
Gly Phe Thr Val Ser Ser Tyr Gly
1 5
<210> 104
<211> 8
<212> PRT
<213> Homo sapiens
<400> 104
Ile Arg Gly Ser Asp Gly Ser Thr
1 5
<210> 105
<211> 12
<212> PRT
<213> Homo sapiens
<400> 105
Ala Lys Gln Gly Glu Asn Asp Gly Pro Phe Asp His
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| НОВЫЕ МОЛЕКУЛЫ АГОНИСТИЧЕСКИХ АНТИ-TNFR2 АНТИТЕЛ | 2019 |
|
RU2840049C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CTLA-4, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2756100C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2790288C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2793167C2 |


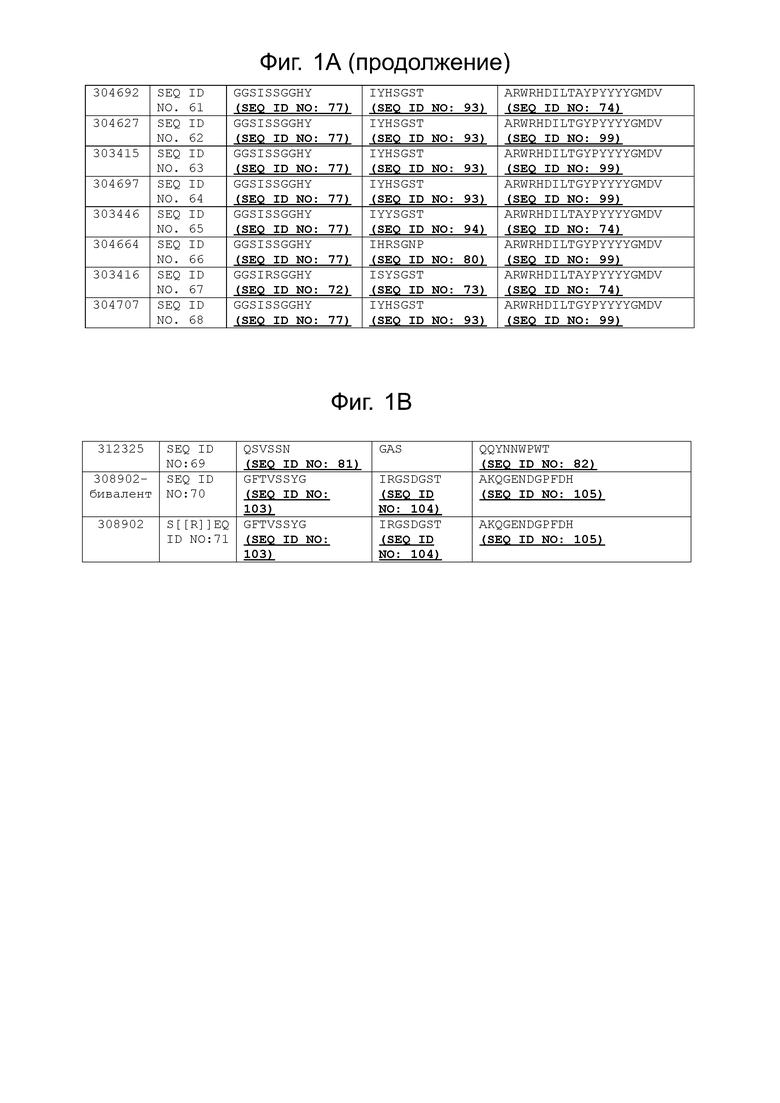
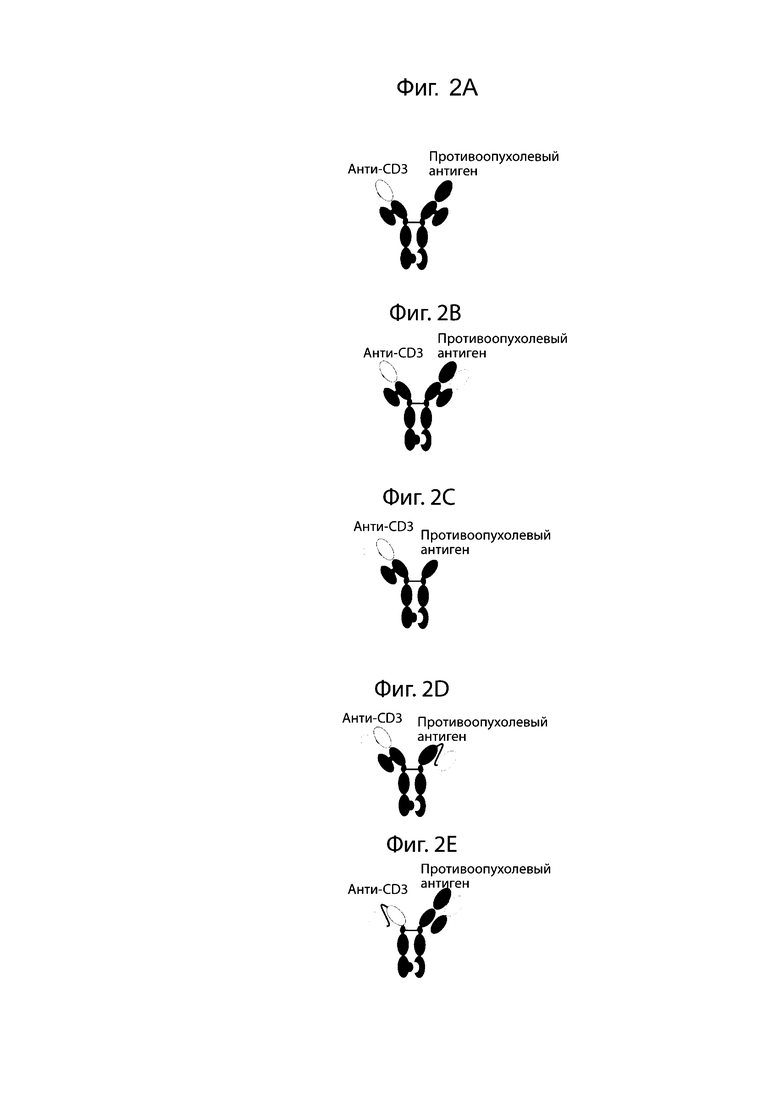
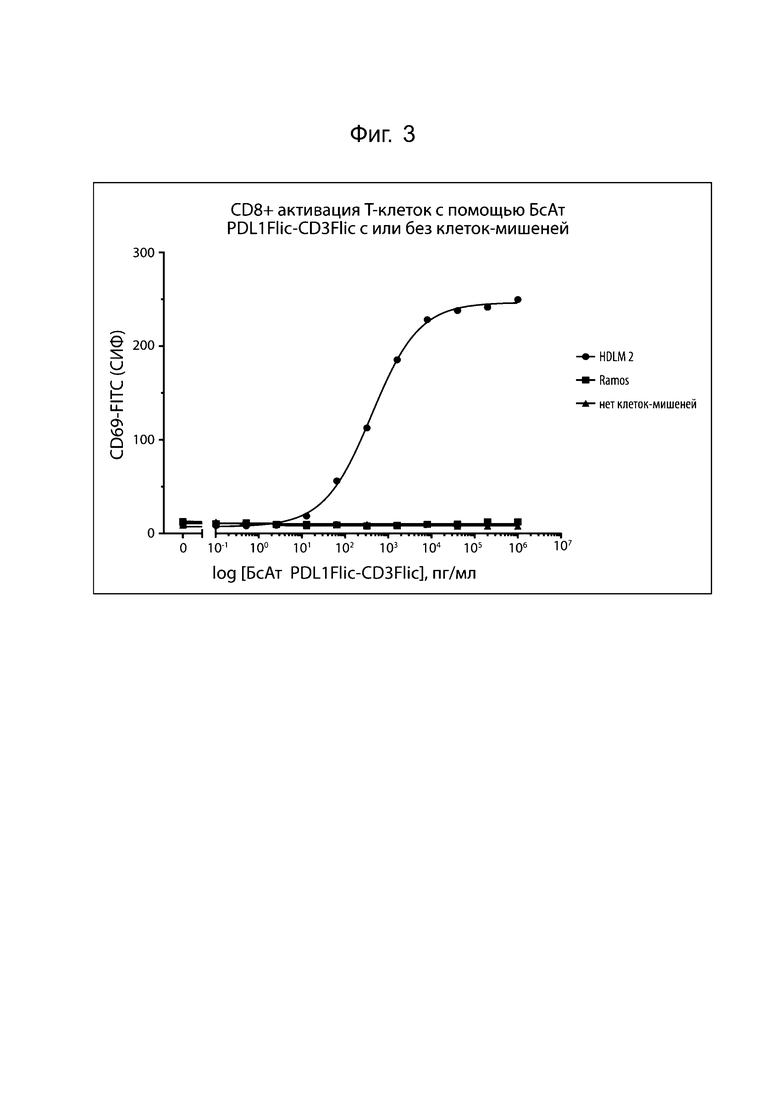
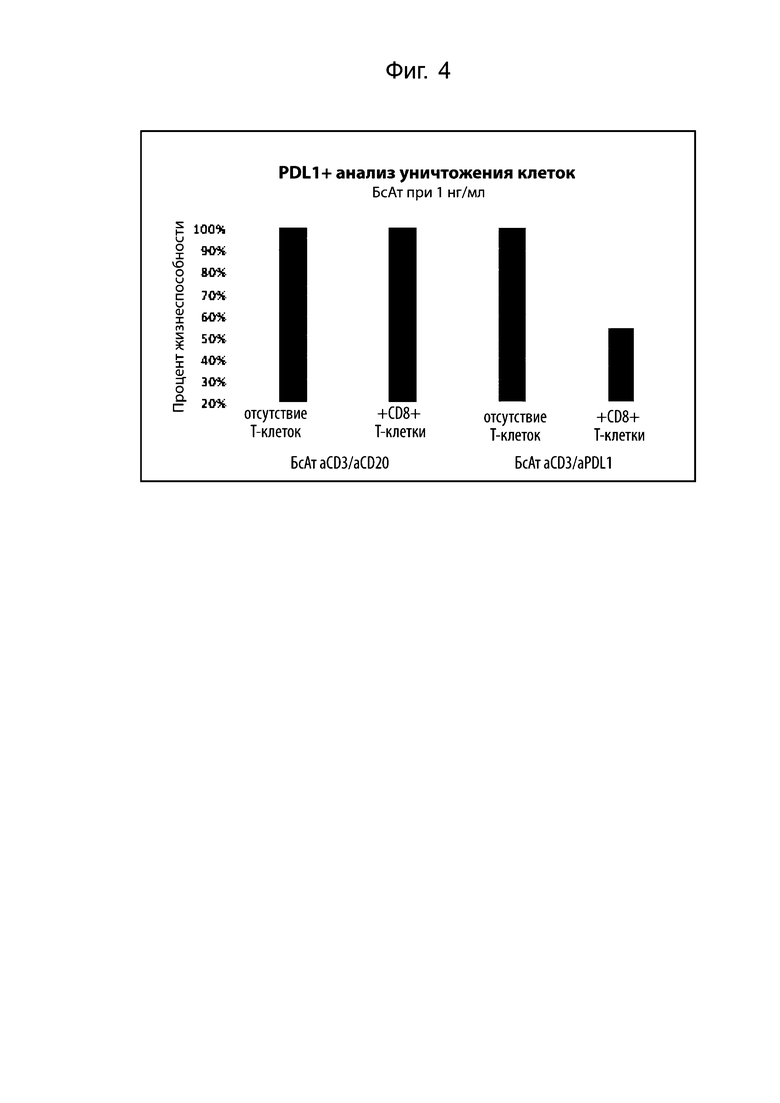
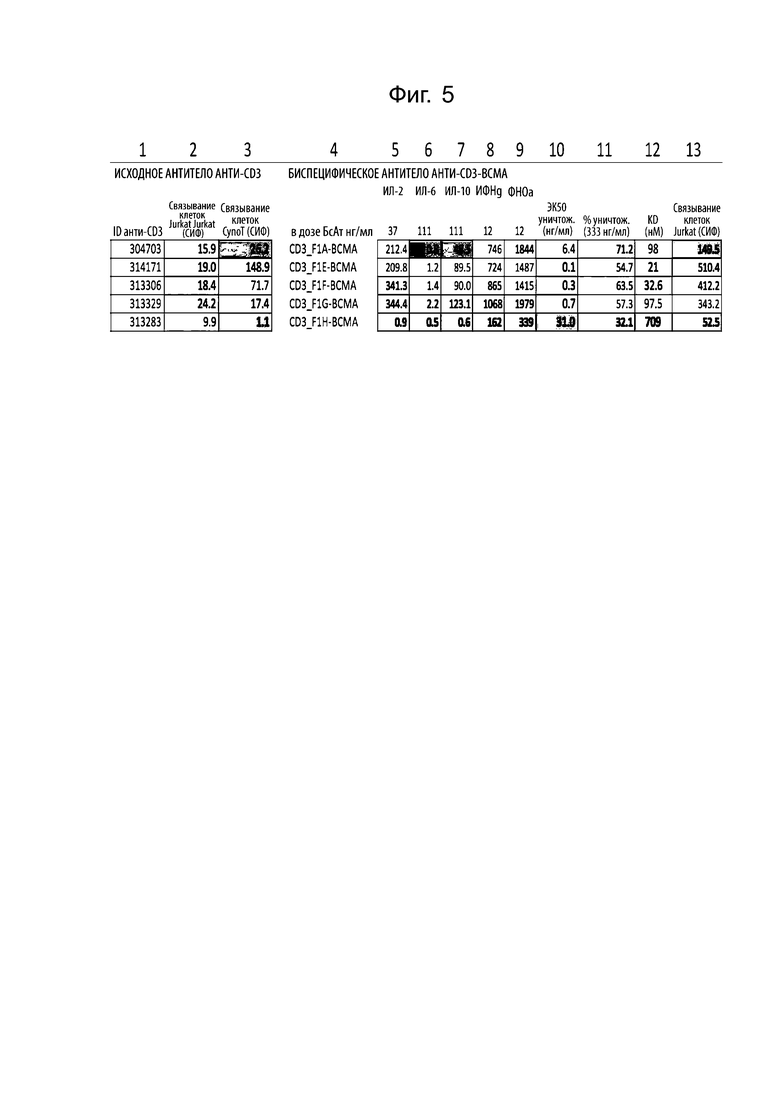

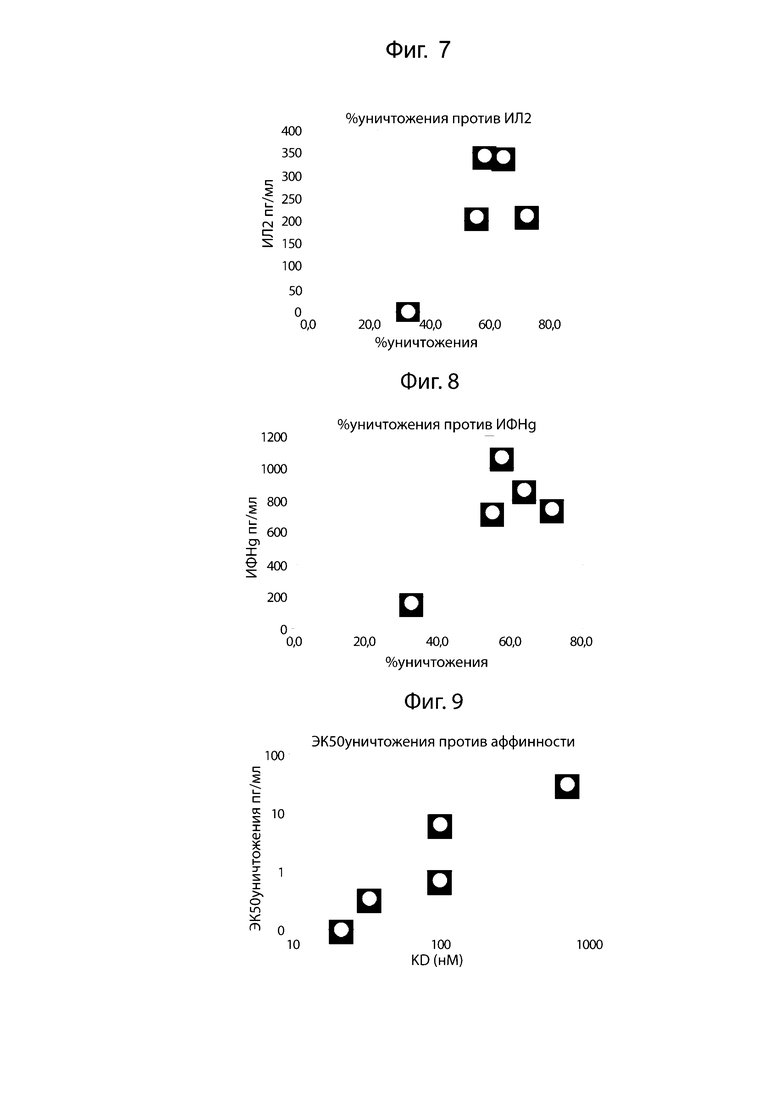
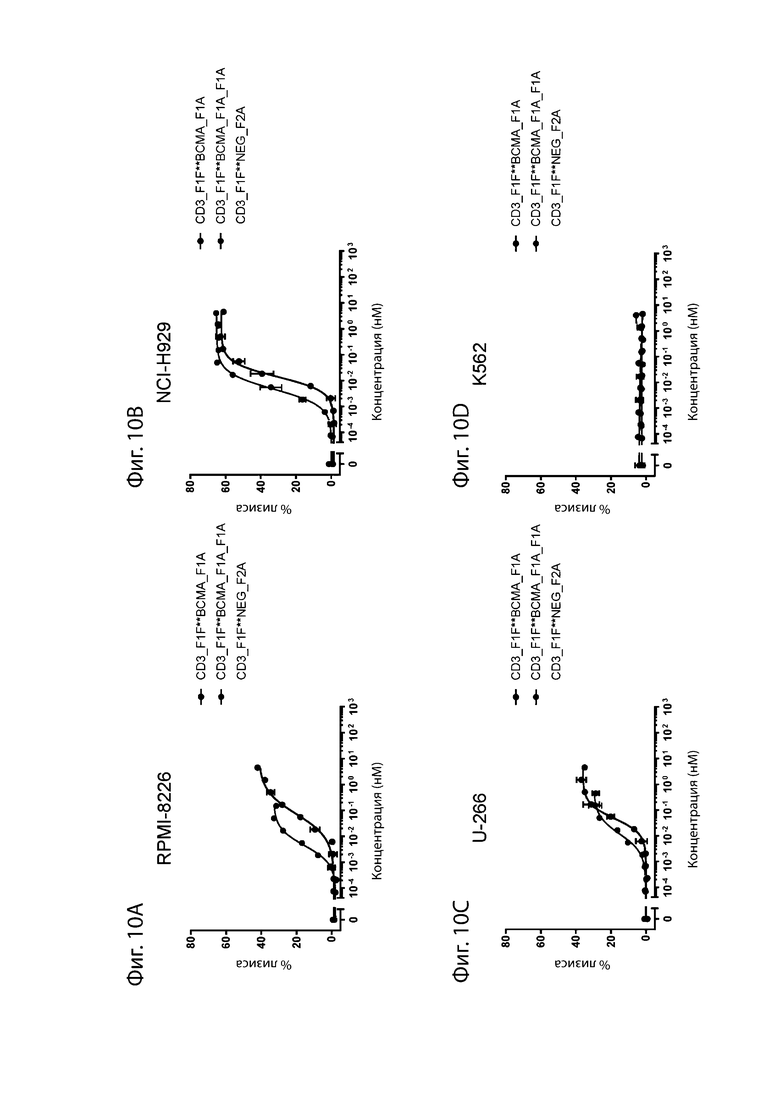
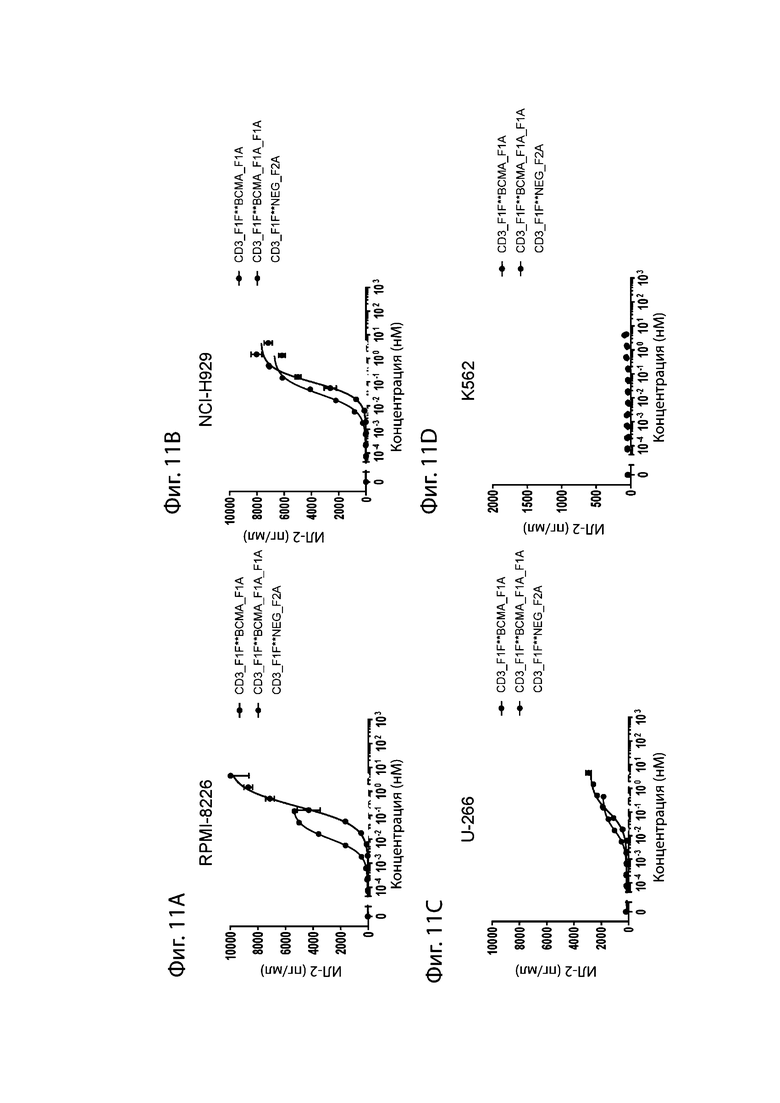
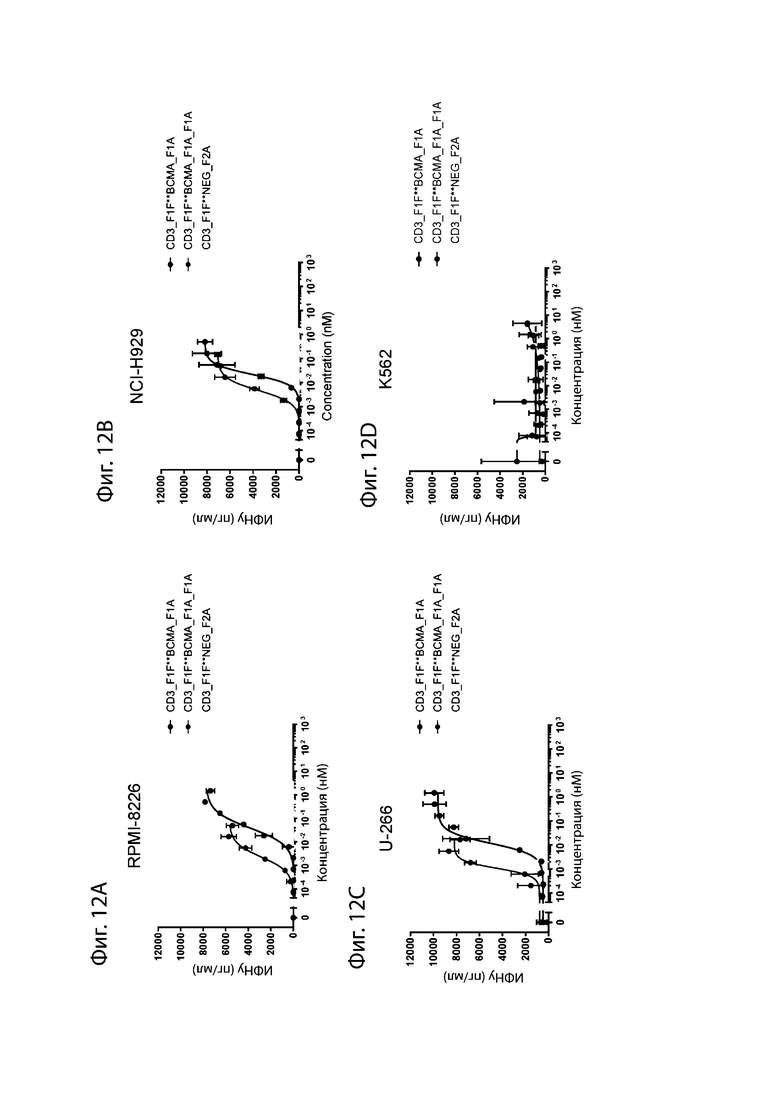
Изобретение относится к области биотехнологии, в частности к мультиспецифичному антителу, специфичному к CD3δε человека и антигену патогенна, содержащей его композиции, а также к способу получения указанного антитела. Также раскрыт полинуклеотид, кодирующий вышеуказанное антитело, а также вектор и клетка, его содержащие. Изобретение эффективно для лечения инфекции. 9 н. и 22 з.п. ф-лы, 6 пр., 12 ил.
1. Мультиспецифичное антитело, специфичное к CD3δε человека и антигену патогена, содержащее первый связывающий фрагмент, который специфично связывается с CD3δε человека, и второй связывающий фрагмент, который специфично связывается с антигеном патогена, отличающееся тем, что этот первый связывающий фрагмент содержит:
первый вариабельный домен тяжелой цепи, содержащий:
(а) CDR1 последовательности GGSIRSGGHY, CDR2 последовательности ISYSGST и CDR3 последовательности ARWRHDILTAYPYYYYGMDV; или
(b) CDR1 последовательности GGSIRSGGHY, CDR2 последовательности IHHSGST и CDR3 последовательности ARWRHDIFTTYPYYYYGMDV; или же
(c) CDR1 последовательности GGSISSGGHY, CDR2 последовательности IHYSGST и CDR3 последовательности ARWRHDIFAAYPYYYYGMDV;
(d) CDR1 последовательности GGSIRSGGHY, CDR2 последовательности IHRSGNP и CDR3 последовательности ARWRHDIFAAYPYYYYGMDV; или же
(e) CDR1 последовательности GGSISSGGHY, CDR2 последовательности IHHSGST и CDR3 последовательности ARWRHDIFAAYPYYYYGMDV; и
вариабельный домен легкой цепи, содержащий последовательность CDR1, содержащую QSVSSN, последовательность CDR2, содержащую GAS, и последовательность CDR3, содержащую QQYNNWPWT в каркасе VL человека.
2. Мультиспецифичное антитело по п. 1, в котором последовательности CDR1, CDR2 и CDR3 указанного первого вариабельного домена тяжелой цепи представлены в каркасе VH человека.
3. Мультиспецифичное антитело по любому одному из пп. 1, 2, отличающееся тем, что вариабельный домен тяжелой цепи содержит аминокислотную последовательность, изложенную в любой из SEQ ID NO: 18, 1, 39, 6 и 13.
4. Мультиспецифичное антитело по любому одному из пп. 1-3, отличающееся тем, что вариабельный домен легкой цепи содержит аминокислотную последовательность SEQ ID NO: 69.
5. Мультиспецифичное антитело по любому одному из пп. 1-4, отличающееся тем, что при контакте с Т-клетками в анализах активации мультиспецифичное антитело индуцирует высвобождение пониженных уровней одного или обоих из ИЛ-2 и ИЛ-10 относительно эталонного антитела к CD3.
6. Мультиспецифичное антитело по любому одному из пп. 1-5, содержащее область Fc.
7. Мультиспецифичное антитело по п. 6, отличающееся тем, что область Fc была сконструирована для снижения эффекторных функций.
8. Мультиспецифичное антитело по любому одному из пп. 1-7, отличающееся тем, что аффинность (KD) белка к CD3 дельта эпсилон человека составляет от около 10-6 M до около 10-11 M.
9. Мультиспецифичное антитело по любому одному из пп. 1-7, отличающееся тем, что перекрестно реагирует с белком CD3 человека и яванского макака.
10. Мультиспецифичное антитело по любому одному из пп. 1-9, отличающееся тем, что содержит одиночную цепь.
11. Мультиспецифичное антитело по любому одному из пп. 1-9, отличающееся тем, что содержит две цепи или множество цепей.
12. Мультиспецифичное антитело по любому одному из пп. 1-9, отличающееся тем, что содержит три цепи.
13. Мультиспецифичное антитело по любому одному из пп. 1-9, отличающееся тем, что содержит три цепи и два антигенсвязывающих плеча, причем первое антигенсвязывающее плечо содержит тяжелую цепь антитела и легкую цепь антитела, а второе антигенсвязывающее плечо содержит тяжелую цепь антитела, которая не содержит домен CH1.
14. Мультиспецифичное антитело по любому одному из пп. 1-13, отличающееся тем, что при контакте с Т-клетками в анализах активации указанное мультиспецифичное антитело индуцирует высвобождение пониженных уровней одного или обоих из ИЛ-2 и ИЛ-10 относительно эталонного антитела к CD3; и
индуцирует более 30% цитотоксичности к опухоли в стандартных анализах in vitro с применением опухолевых клеток и человеческих Т-клеток.
15. Мультиспецифичное антитело по п. 14, отличающееся тем, что второй связывающий фрагмент, который специфично связывается с антигеном патогена, представляет собой домен только тяжелой цепи.
16. Мультиспецифичное антитело по п. 15, отличающееся тем, что второй связывающий фрагмент, который специфично связывается с антигеном патогена, содержит вариабельный домен легкой цепи.
17. Мультиспецифичное антитело по п. 15, отличающееся тем, что вариабельный домен легкой цепи является таким же, что и вариабельный домен легкой цепи CD3-связывающей области.
18. Направленная против CD3 фармацевтическая композиция, содержащая терапевтически эффективное количество мультиспецифичного антитела по любому одному из пп. 1-17.
19. Фармацевтическая композиция по п. 18 в составе на одну дозу.
20. Полинуклеотид, кодирующий мультиспецифичное антитело по любому одному из пп. 1-17.
21. Экспрессионный вектор, содержащий полинуклеотид по п. 20.
22. Рекомбинантная клетка-хозяин для продукции мультиспецифичного антитела по любому из пп. 1-17, содержащая вектор по п. 21.
23. Способ получения мультиспецифичного антитела по любому одному из пп. 1-17, включающий в себя выращивание рекомбинантной клетки-хозяина по п. 22 в условиях, допускающих экспрессию мультиспецифичного антитела, и выделение мультиспецифичного антитела из клеток.
24. Способ лечения инфекции, включающий в себя введение индивидуальной эффективной дозы мультиспецифичного антитела по любому одному из пп. 1-17 или фармацевтической композиции по п.18.
25. Применение мультиспецифичного антитела по любому из пп. 1-17 для приготовления лекарственного средства для лечения инфекции.
26. Способ по любому из пп. 23 и 24 или применение по п. 25, где индивидом является человек.
27. Биспецифичное антитело, специфичное к CD3δε человека и антигену патогена, содержащее:
(i) CD3-связывающий вариабельный домен тяжелой цепи в каркасной области VH человека;
(ii) CD3-связывающий вариабельный домен легкой цепи, содержащий последовательность CDR1, содержащую QSVSSN, последовательность CDR2, содержащую GAS, и последовательность CDR3, содержащую QQYNNWPWT, в каркасной области VL человека; а также
(iii) одноцепочечный вариабельный домен в одноцепочечной или тандемной конфигурации, который специфично связывается с указанным антигеном патогена,
где биспецифичное антитело отличается тем, что CD3-связывающий вариабельный домен тяжелой цепи включает:
(а) последовательность CDR1 GGSIRSGGHY, последовательность CDR2 ISYSGST и последовательность CDR3 ARWRHDILTAYPYYYYGMDV; или же
(b) последовательность CDR1 GGSIRSGGHY, последовательность CDR2 IHHSGST и последовательность CDR3 ARWRHDIFTTYPYYYYGMDV; или
(c) последовательность CDR1 GGSISSGGHY, последовательность CDR2 IHYSGST и последовательность CDR3 ARWRHDIFAAYPYYYYGMDV;
(d) последовательность CDR1 GGSIRSGGHY, последовательность CDR2 IHRSGNP и последовательность CDR3 ARWRHDIFAAYPYYYYGMDV; или
(e) последовательность CDR1 GGSISSGGHY, последовательность CDR2 IHHSGST и последовательность CDR3 ARWRHDIFAAYPYYYYGMDV.
28. Биспецифичное антитело по п. 27, содержащее область Fc.
29. Биспецифичное антитело по п. 28, где CD3-связывающий вариабельный домен тяжелой цепи содержит последовательность SEQ ID NO: 18, а вариабельный домен легкой цепи содержит последовательность SEQ ID NO: 69.
30. Биспецифичное антитело по п. 27, содержащее CD3-связывающий вариабельный домен тяжелой цепи, содержащий последовательности CDR1, CDR2 и CDR3 SEQ ID NO: 18 в каркасной области VH человека.
31. Биспецифичное антитело по п. 27, содержащее вариабельный домен легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3 SEQ ID NO: 69 в каркасной области VL человека.
| WO2016079177 A1, 26.05.2016 | |||
| WO2015095412 A1, 25.06.2015 | |||
| WO2014022540 A1, 06.02.2014 | |||
| US2013273055 A1, 17.10.2013 | |||
| ВЫДЕЛЕННОЕ АНТИ-CD30 АНТИТЕЛО (ВАРИАНТЫ), ХОЗЯЙСКАЯ КЛЕТКА, СПОСОБ ПОЛУЧЕНИЯ ХИМЕРНОГО ИЛИ ГУМАНИЗИРОВАННОГО ВАРИАНТА АНТИ-CD30 АНТИТЕЛ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ РОСТА КЛЕТОК CD30+ И СПОСОБ ИНГИБИРОВАНИЯ РОСТА ОПУХОЛЕВЫХ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ CD30 | 2006 |
|
RU2492186C2 |

