Область техники, к которой относится изобретение
Данное изобретение относится к новым агонистическим антителам, которые специфически связываются с рецептором 2 фактора некроза опухоли (TNFR2) (tumor necrosis factor receptor 2), но не блокируют лиганд TNF-α от связывания с TNFR2. Изобретение также относится к их применению в медицине, например, для лечения рака или хронических воспалительных заболеваний.
Предпосылки к созданию изобретения
Рецептор 2 фактора некроза опухоли (TNF) (TNFR2, TNFR-2 или TNFRII), также известный как член суперсемейства рецепторов фактора некроза опухоли 1B (TNFRSF1B) и CD120b, представляет собой рецептор мембраны, который связывает фактор некроза опухоли альфа (TNF-α или TNFα). Он обнаруживается, например, на поверхности Т-клеток, моноцитов и макрофагов, и может активировать пролиферацию клеток, экспрессирующих рецептор TNFR2, через ядерный фактор каппа B (NF-κB). Примечательно, что TNFR2 в высокой степени повышающе регулируется при раке и, в частности, на инфильтрирующих опухоль иммунных клетках, например, регуляторных Т-клетках (Treg), цитотоксических эффекторных Т-клетках CD8+ и различных субпопуляциях миелоидных клеток
TNFR2 обсуждался как многообещающая мишень для иммунотерапии рака, и было описано, что он сильно экспрессируется на поверхности среди прочего внутриопухолевых Treg и многих опухолевых клеток человека (Williams GS et al, Oncotarget. 2016; 7(42): 68278–68291; Vanamee ES et al, Trends in Molecular Medicine, 2017, vol. 23, issue 11, 1037-1046; Frontiers in Immunology , November 2017 | Volume 8 | Article 1482, Sci Signal. 2018 Jan 2;11(511)).
Регуляторные Т-клетки (которые также могут называться Treg-клетками, Treg или Treg, и которые раньше были известны как супрессорные Т-клетки или супрессивные регуляторные Т-клетки) составляют субпопуляцию Т-клеток, способных подавлять другие иммунные клетки в нормальных и патологических иммунных условиях. Treg представляют собой CD4-положительные клетки (CD4+ клетки). Есть и другие CD4+ Т-клетки, которые не являются Treg; однако Treg могут быть отделены от не-Treg CD4+ клеток тем, что Treg также являются FOXP3-положительными (FOXP3+), тогда как не-Treg CD4+ являются FOXP3-отрицательными (FOXP3-). Treg также могут быть отделены от не-Treg клеток CD4+, в том смысле, что Treg также являются CD25+CD127отр/низкий, в то время как не-Treg клетки CD4+, являются либо CD25-CD127+, либо CD25+CD127+.
TNFR2 также обсуждался в связи с аутоиммунными заболеваниями (Faustman DL et al, Front Immunol. 2013; 4: 478, Clin Transl Immunology. 2016 Jan 8; 5(1): J Neurosci. 2016 May 4; 36(18):5128-43) и воспалительными заболеваниями (Ait-Ali D et al, Endocrinology. 2008 Jun; 149(6):2840-52, Sci Rep. 2016 Sep 7; 6:32834).
Анти-TNFR2 антитела разных типов и с различными характеристиками также были описаны ранее. Например, Williams et al (Oncotarget. 2016 Oct 18; 7(42):68278-68291) описывает как блокирующие лиганд, так и неблокирующие лиганд агонистические антитела.
WO 2014/124134 описывает применение агониста TNFR2, такого как агонистическое анти-TNFR2 антитело и/или активатор NF-κB, для получения in vitro композиции, обогащенной в CD4+CD25hi Treg. Утверждается, что композиция пригодна для лечения иммунологических нарушений или инфекционных заболеваний у пациентов.
WO 2017/040312 описывает анти-TNFR2 антитела и, в частности, агонистические анти-TNFR2 антитела, которые способны стимулировать передачу сигналов TNFR2 и оказывать влияние на распространение или пролиферацию Treg. WO 2017/040312 описывает антитела, которые специфически связываются с эпитопом, содержащим последовательность KCSPG, но не с эпитопом, содержащим последовательность KCRPG, таким образом исключая антитела из патента США 9821010, описанные выше, или, альтернативно, не связываются с другим членом суперсемейства TNFR. Утверждается, что агонистические антитела пригодны при лечении иммунологических заболеваний. WO 2017/040312 дополнительно описывает полную последовательность TNFR2 человека.
В WO 2017/083525 обсуждаются фармакологические композиции, содержащие анти-TNFR2 антитела, и их применение при лечении расстройств, связанных с TNF-α и/или TNFR2, например, рака. В WO 2017/083525 дополнительно обсуждаются антитела, содержащие Fc-домен IgG1 человека, который является нулевым для связывания с рецептором Fcγ, а также для подавления экспансии Treg.
Кроме того, анти-TNFR2 антитела, которые способны действовать как агонисты TNFR2, описаны в Galloway et al. (Eur. J. Immunol. 22:3045-3048, 1992), Tartaglia et al. (J. Biol. Chem.268:18542-18548, 1993), Tartaglia et al. (J. Immunol. 151:4637-4641, 1993), Smith et al. (J. Biol. Chem.269:9898-9905, 1994), и Amrani et al. (Am. J. Respir. Cell. Mol. Biol. 15:55-63, 1996).
Однако ни один из этих документов не описывает и не предлагает агонистических антител к TNFR2, которые специфически связываются с TNFR2, но не блокируют лиганд TNF-α от связывания с тем же TNFR2.
Fc-рецепторы представляют собой мембранные белки, которые обнаруживаются на клеточной поверхности иммунных эффекторных клеток, включая моноциты, макрофаги, дендритные клетки, нейтрофилы, тучные клетки, базофилы, эозинофилы, а также клетки натуральные киллеры и B-лимфоциты. Название происходит от специфичности их связывания с Fc-участком антител. Рецепторы Fc находятся на клеточной мембране, также известной как плазматическая мембрана или цитоплазматическая мембрана. Рецепторы Fc можно подразделить на активирующий FcγR и ингибирующий FcγR, которые, как известно, координируют клеточную активацию посредством связывания агрегированных Fc иммуноглобулина G и передачи активирующих или ингибирующих сигналов в клетку через внутриклеточные мотивы ITAM или ITIM. Связывание FcR агрегированного иммуноглобулина или иммунных комплексов может опосредовать интернализацию антитела в клетку и может приводить к антитело-опосредованному фагоцитозу, антителозависимой клеточно-опосредованной цитотоксичности или презентации антигена или перекрестной презентации антигена. Также известно, что FcR опосредуют или усиливают перекрестное связывание рецепторов на клеточной поверхности, связанных с антителами. Такое перекрестное связывание, как известно, требуется для некоторой (Li et al 2011. 'Inhibitory Fcgamma receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies', Science, 333: 1030-4.; White et al. 2011. 'Interaction with FcgammaRIIB is critical for the agonistic activity of anti-CD40 monoclonal antibody', J Immunol, 187: 1754-63 ) но не всей (Richman et al 2014. 'Anti-human CD40 monoclonal antibody therapy is potent without FcR crosslinking', Oncoimmunology, 3: e28610) способности антител активировать передачу сигналов в клетках-мишенях и может потребоваться, а может и не потребоваться для достижения терапевтических эффектов.
Подгруппой рецепторов Fc являются рецепторы Fcγ (Fc-гамма-рецепторы, FcgammaR, FcγR), которые специфичны для антител IgG. Существует два типа рецепторов Fcγ: активирующие рецепторы Fcγ (также обозначаемые как активационные рецепторы Fcγ) и ингибирующие рецепторы Fcγ. Активирующие и ингибирующие рецепторы передают свои сигналы через активационные тирозинсодержащие мотивы иммунорецепторов (ITAM) или ингибирующие тирозинсодержащие мотивы иммунорецепторов (ITIM) соответственно. У людей FcγRIIb (CD32b) является ингибирующим рецептором Fcγ, тогда как FcγRI (CD64), FcγRIIa (CD32a), FcγRIIc (CD32c) и FcγRIIIa (CD16a) активируют рецепторы Fcγ. FcγgRIIIb является GPI-связанным рецептором, экспрессируемым на нейтрофилах, у которого отсутствует мотив ITAM, но благодаря его способности сшивать липидные рафты и взаимодействовать с другими рецепторами он также считается активационным. У мышей активирующими рецепторами являются FcγRI, FcγRIII и FcγRIV.
Хорошо известно, что антитела могут модулировать активность иммунных клеток посредством взаимодействия с рецепторами Fcγ. В частности, то, как иммунные комплексы антител модулируют активацию иммунных клеток, определяется их относительным связыванием активирующих и ингибирующих рецепторов Fcγ. Различные изотипы антител связываются с активирующим и ингибирующим рецепторам Fcγ с различным сродством, что приводит к различным соотношениям A : I (соотношения активации к ингибированию) (Nimmerjahn et al; Science. 2005 Dec 2;310(5753):1510-2).
Связываясь с ингибирующими рецепторами Fcγ, антитело может ингибировать, блокировать и/или снижать функции эффекторных клеток. Связываясь с ингибирующим FcγR, антитела могут дополнительно стимулировать активацию клеток за счет агрегации нацеленных на антитела сигнальных рецепторов на клетке-мишени (Li et al. 2011. 'Inhibitory Fcgamma receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies', Science, 333: 1030-4; White et al 2011. 'Interaction with FcgammaRIIB is critical for the agonistic activity of anti-CD40 monoclonal antibody', J Immunol, 187: 1754-63; White et al 2014. 'Fcgamma receptor dependency of agonistic CD40 antibody in lymphoma therapy can be overcome through antibody multimerization', J Immunol, 193: 1828-35).
Связываясь с активирующим рецептором Fcγ, антитело может активировать функции эффекторных клеток и тем самым запускать такие механизмы, как антителозависимая клеточная цитотоксичность (АЗКЦ), антителозависимый клеточный фагоцитоз (ADCP - antibody dependent cellular phagocytosis), высвобождение цитокинов и/или антителозависимый эндоцитоз, а также нетоз (т.е. активация и высвобождение нейтрофилов, нейтрофильных внеклеточных ловушек) в случае нейтрофилов. Связывание антител с активирующим рецептором Fcγ также может приводить к увеличению некоторых маркеров активации, таких как CD40, MHCII, CD38, CD80 и/или CD86.
Недавние данные, опубликованные среди прочего авторами изобретения, демонстрируют критическую и дифференциальную зависимость агониста Т-клеток CD8 и Treg-истощающих анти-4-1BB антител от связывания с активирующими и ингибирующими FcγR соответственно, на терапевтическую эффективность (Buchan et al., 'Antibodies to Costimulatory Receptor 4-1BB Enhance Anti-tumor Immunity via T Regulatory Cell Depletion and Promotion of CD8 T Cell Effector Function', Immunity 2018 49(5):958-970). Более того, что важно, одновременное введение агониста Т-клеток CD8 и истощающих Treg анти-4-1BB антител, оптимизированных для связывания с активирующим и ингибирующим FcγR, соответственно, нарушения терапевтической активности. Эти данные демонстрируют критическую важность разработки антител с соответствующим и индивидуализированным участием активирующих и ингибирующих FcγR для максимизации терапевтической активности антител с различным механизмом действия. В то же время они демонстрируют, что неоптимальное взаимодействие активирующих и ингибирующих FcγR может серьезно снизить терапевтическую эффективность.
Эти данные были неожиданными, поскольку они контрастировали с данными об антителах с другим членам TNFSR, особенно с иммуностимулирующими анти-CD40 антителам, которые демонстрируют обязательную потребность в задействовании ингибирующего, но не активирующего FcγR (Li et al. 2011. 'Inhibitory Fcgamma receptor engagement drives adjuvant and anti-tumor activities of agonistic CD40 antibodies', Science, 333: 1030-4; White et al. 2011. 'Interaction with FcgammaRIIB is critical for the agonistic activity of anti-CD40 monoclonal antibody', J Immunol, 187: 1754-63). Взятые вместе, эти результаты демонстрируют, что FcγR-зависимость может варьироваться между антителами к разным мишеням одного и того же суперсемейства рецепторов, и даже между разными типами антител к одной и той же мишени таким образом, который трудно предсказать, но может иметь решающее значение для понимания и использования при разработке антител для терапевтического применения.
Сущность изобретения
В ходе работы, приведшей к данному изобретению, а также к параллельному изобретению, были идентифицированы две основные разные группы анти-TNFR2 антител с мощными терапевтическими эффектами и различными характеристиками и механизмами действия.
Авторы изобретения впервые определили мощную терапевтическую активность антагонистических анти-TNFR2 антител, которые блокируют TNF-α связывание с рецептором TNFR2. Было продемонстрировано, что активность таких антител зависит от FcγR-взаимодействий и, в частности, от связывания с активирующим FcγR для терапевтической активности in vivo. Было обнаружено, что эта группа или категория мощных терапевтических анти-TNFR2 реагентов характеризуется 1) выраженным блокированием и ингибированием TNF-α-индуцированной передачи сигналов TNFR2, и 2) активность, зависящая от FcγR-взаимодействия, наиболее сильная выгода от взаимодействия с активацией по сравнению с ингибирующими FcγR.
Затем изобретатели идентифицировали отдельную группу анти-TNFR2 антител со столь же мощной терапевтической активностью in vivo, но характеристики которых во многих отношениях противоположны характеристикам антагонистических, блокирующих типов антител TNFR2, составляющих первую группу. Анти-TNFR антитела этой второй группы не зависят от TNF-α блокады или ингибирования передачи сигналов TNFR2 для терапевтической активности, а скорее характеризуются сильной активацией передачи сигналов TNFR2. Кроме того, в отличие от блокирующих антител первой группы, агонистические антитела второй группы не проявляют облигатной зависимости от антитела:FcγR-взаимодействия, даже несмотря на то, что их активность улучшается с FcγR:взаимодействующими вариантами антитела. Кроме того, в отличие от антагонистических блокирующих антител первой группы, агонистические антитела второй группы проявляют наибольшую активность в вариантах антител с улучшенным связыванием с ингибирующим по сравнению с активирующим FcγR.
Данное изобретение относится ко второй группе анти-TNFR2 антител, то есть к молекулам агонистических антител, которые специфически связываются с TNFR2, но не блокируют лиганд TNF-α от связывания с TNFR2. Такие антитела являются мощными терапевтическими реагентами и пригодны в медицине.
Антагонистические блокирующие антитела, принадлежащие к первой группе, применяются в приведенных ниже. Примерах для сравнения с молекулами агонистических неблокирующих антител TNFR2 по данному изобретению. В примерах для сравнения используются также другие антитела с некоторыми характеристиками, аналогичными характеристикам первой или второй группы, или обеих, как дополнительно поясняется ниже.
Таким образом, данное изобретение относится к молекулам агонистических антител, которые специфически связываются с TNFR2 на клетке-мишени и не блокируют TNF-α связывание лиганда с TNFR2.
Данное изобретение также относится к конкретным примерам таких новых молекул агонистического антитела TNFR2.
Данное изобретение также относится к изолированным нуклеотидным последовательностям, кодирующим по меньшей мере одну из вышеуказанных молекул антител.
Данное изобретение также относится к плазмидам, содержащим по меньшей мере одну из указанных выше нуклеотидных последовательностей.
Данное изобретение также относится к вирусам, содержащим по меньшей мере одну из указанных выше нуклеотидных последовательностей или плазмид.
Данное изобретение также относится к клеткам, содержащим по меньшей мере одну из указанных выше нуклеотидных последовательностей, или по меньшей мере одну из указанных выше плазмид, или по меньшей мере один из указанных выше вирусов.
Данное изобретение также относится к указанным выше молекулам антител, нуклеотидным последовательностям, плазмидам, вирусам и/или клеткам для применения в медицине.
Данное изобретение также относится к указанным выше молекулам антител, нуклеотидным последовательностям, плазмидам, вирусам и/или клеткам для применения при лечении рака или хронического воспалительного заболевания.
Данное изобретение также относится к применению указанных выше молекул антител, нуклеотидным последовательностям, плазмидам, вирусам и/или клеткам для применения при лечении рака или хронического воспалительного заболевания.
Данное изобретение также относится к фармацевтическим композициям, содержащим или состоящим из по меньшей мере одной из указанных выше молекул антител, нуклеотидные последовательности, плазмиды, вирусы и/или клетки и, необязательно, фармацевтически приемлемый разбавитель, носитель или наполнитель. Такая фармацевтическая композиция может использоваться при лечении рака или хронического воспалительного заболевания.
Кроме того, данное изобретение относится к способам лечения рака у субъекта, включающим введение субъекту терапевтически эффективного количества по меньшей мере одной из вышеуказанных молекул антител, нуклеотидных последовательностей, плазмид, вирусов и/или клеток.
Данное изобретение также относится к молекулам антител, молекулам антител для применения, изолированным нуклеотидным последовательностям, изолированным нуклеотидным последовательностям для применения, плазмидам, плазмидам для применения, вирусам, вирусам для применения, клеткам, клеткам для применения, применениям, фармацевтическим композициям и способам лечения, как описано в данном документе, со ссылкой на сопроводительное описание, примеры и/или фигуры.
Подробное описание изобретения
Таким образом, данное изобретение касается молекул агонистического антитела TNFR2, которые специфически связываются с TNFR2, но не блокируют лиганд TNF-α от связывания с тем же TNFR2. Предпочтительно молекулы антител обладают внутренней агонистической активностью.
Описанные в данном документе молекулы агонистических антител не блокируют TNF-α от связывания с TNFR2 и, кроме того, они не блокируют передачу сигналов TNFR2. Было ясно продемонстрировано, что TNF-α опосредованная передача сигналов через TNFR2 запускает сигнальный каскад, который заканчивается активацией фактора ядерной транскрипции NFkB (Thommesen et al. “Distinct differences between TNF receptor 1- and TNF receptor 2-mediated activation of NFkappaB”. J Biochem Mol Biol. 2005 May 31;38(3):281-9; Yang et al. “Role of TNF-TNF Receptor 2 Signal in Regulatory T Cells and Its Therapeutic Implications”. Front Immunol. 2018 Apr 19; 9:784). Это, в свою очередь, приводит к активации клетки и синтезу нескольких провоспалительных факторов, одним из которых является ИФН-γ в НК-клетках (Liu et al. “NF-κB signaling in inflammation”. Signal Transduct Target Ther. 2017; 2. pii: 17023; Tato et al. “Opposing roles of NF-kappaB family members in the regulation of NK cell proliferation and production of IFN-gamma”. Int Immunol. 2006 Apr;18(4):505-13). В данном документе термины передача сигнала TNFR2 и активация TNFR2 используются взаимозаменяемо. Молекулы антител специфически связываются с TNFR2. Хорошо известно, что антитело специфически связывается или взаимодействует с определенной молекулой-мишенью или антигеном, и что это означает, что антитело предпочтительно и избирательно связывает свою мишень, а не молекулу, которая не является мишенью. Под «молекулой антитела, которая специфически связывает TNFR2» или «молекулой специфического антитела TNFR2» мы подразумеваем антитело, которое связывает белок TNFR2 дозозависимым образом, но не с неродственным белком. Кроме того, это же антитело связывает клетки, которые эндогенно экспрессируют TNFR2, и это связывание можно заблокировать путем предварительной инкубации тех же клеток с коммерчески доступным реагентом поликлональных антител к TNFR2, что показывает, что неспецифическое связывание не может быть обнаружено, когда TNFR2 маскируется поликлональным реагентом. Это показано в Примере 2.
Молекула антитела, которая специфически связывает TNFR2 (или молекулу анти-TNFR2 антитела), относится к молекуле антитела, которая специфически связывается по меньшей мере с одним эпитопом во внеклеточном домене TNFR2. Поверхностный клеточный антиген и эпитоп – это термины, которые может легко понять специалист в области иммунологии или клеточной биологии.
Способы оценки связывания белков известны специалистам в области биохимии и иммунологии. Было бы желательно, чтобы специалисты могли применять эти методы для оценки связывания антитела с мишенью и/или связывания участка Fc антитела с рецептором Fc, а также для оценки относительной силы или специфичности, или ингибирования, или уменьшения этих взаимодействий, или препятствования этим взаимодействиям. Примерами способов, которые можно применять для оценки связывания с белками, являются, например, иммуноанализы, BIAcore, вестерн-блоттинг, радиоиммуноанализ (РИА) и иммуноферментный анализ (ИФА) и проточная цитометрия (FACS). См. второе издание Fundamental Immunology, Raven Press, New York, стр. 332-336 (1989), где обсуждается специфичность антител.
Клетками-мишенями, экспрессирующими TNFR2, с которыми связывается агонистическое антитело в соответствии с данным изобретением, могут быть любые иммунные клетки, экспрессирующие TNFR2, такие как CD8-положительные клетки и миелоидные клетки.
Эффект связывания молекул агонистических антител согласно изобретению, с TNFR2 может выражаться в активации Т-клеток и/или миелоидных клеток; и/или инфильтрации Т-клеток и/или миелоидных клеток в пораженную ткань; и/или изменение состава Т-клеток и/или миелоидных клеток в пораженной ткани. Изменение в составе Т-клеток и/или миелоидных клеток означает в данном документе различные абсолютные или относительные количества различных субпопуляций клеток, таких как Treg, CD8-положительные клетки, опухолевые макрофаги (ТАМ - tumor associated macrophage) (включая их различные субпопуляции), клетки-супрессоры миелоидного происхождения (MDSC) и/или провоспалительные макрофаги.
В данном контексте «пораженная ткань» означает либо опухолевую ткань (т.е. все клетки в микроокружении опухоли, включая опухолевые клетки, иммунные клетки, эндотелиальные клетки и стромальные клетки), либо ткань, пораженную хроническим воспалительным заболеванием.
Чтобы решить, блокирует ли молекула антитела - или, скорее, в контексте данного изобретения не блокирует - связывание лиганда с TNFR2, можно использовать анализ ИФА, определяющий количество связанного TNF-α лиганда к иммобилизованному рецептору TNFR2 в присутствии антител, специфичных к TNFR2. Неблокирующее антитело не препятствует связыванию лиганда TNF-α с иммобилизованным рецептором TNFR2. Это продемонстрировано и объяснено более подробно в Примере 3 ниже. Более конкретно, молекула неблокирующего антитела TNFR2 согласно данному изобретению представляет собой молекулу антитела, которая снижает связывание TNF-α с TNFR2 менее чем на 50 % по сравнению со связыванием TNF-α в присутствии только молекулы антитела изотипического контроля. В некоторых вариантах осуществления данного изобретения, это определяется в высокодозном одноточечном ИФА или ИФА с титрованием дозы, как показано в Примере 3 и на Фиг. 6 и Фиг. 7.
Напротив, молекула блокирующего антагонистического антитела является полным блокатором, который, кроме того, способен противодействовать передаче сигналов TNFR2. Такие молекулы антител используются для сравнения в приведенных ниже примерах. Полный блокатор определяется в данном документе как молекула антитела, которая снижает TNF-α связывание с TNFR2 более чем на 98 %, т.е. до 100 %, по сравнению с TNF-α связыванием в присутствии только молекулы антитела изотипического контроля. Антитело изотипического контроля представляет собой антитело, продуцируемое против белка или другой структуры, которая не присутствует ни в какой форме в анализе при исследовании. Изотипический контроль в идеале имеет ту же структуру, но, по меньшей мере, ту же часть Fc, что и сравниваемые антитела. Это хорошо известно специалисту в данной области техники. В примерах, описанных в данном документе, изотипический контроль имел ту же основу, ту же часть Fc и был специфичен для Флуоресцеина изотиоцианата (FITC). В некоторых вариантах осуществления данного изобретения, полный блокатор снижает TNF-α связывание более чем на 99,5 %. Другие типы блокаторов - это частичные блокаторы и слабые блокаторы. В данном контексте частичный блокатор представляет собой молекулу антитела, которая снижает TNF-α связывание с TNFR2 на 60-98 % по сравнению с TNF-α связыванием в присутствии только молекулы антитела изотипического контроля, а слабый блокатор представляет собой молекулу антитела, которая снижает TNF-αсвязывание с TNFR2 менее чем на 60 %, например, 50-59,9 %, по сравнению с TNF-αсвязыванием в присутствии только молекулы антитела изотипического контроля.
Молекулы полных блокирующих антагонистических антител, молекулы частично блокирующих антител и молекулы слабо блокирующих антител применяются в примерах для сравнения с молекулами агонистических неблокирующих антител по данному изобретению.
Некоторые свойства и особенности могут лежать в основе и (совместно)-определять биологическую активность антител. Помимо способности блокировать или не блокировать связывание лиганда с рецептором, такие важные свойства включают способность молекул антител модулировать передачу сигналов рецептора, т.е. агонизировать или антагонизировать передачу сигналов рецептора, и зависимость антител от взаимодействий FcγR для придания терапевтической активности.
Мы сначала охарактеризовали способность антител полного блокирования, частичного блокирования и неблокирования модулировать передачу сигналов TNFR2. Были выявлены две крайности.
В первом случае мы идентифицировали антитела, которые полностью блокировали связывание лиганда с TNFR2, что блокировало TNF-α индуцированную передачу сигналов TNFR2 и которые сами по себе не индуцировали передачу сигналов при связывании с клеточно-эндогенно экспрессируемым TNFR2. Эта группа лиганд-блокирующих, антагонистических антител составляет отдельное изобретение и включена в данный документ для сравнения.
С другой стороны, мы идентифицировали антитела, которые не блокируют связывание лиганда с TNFR2, но при связывании с TNFR2 эндогенно экспрессирующие клетки агонизируют рецептор. Эта вторая группа антител составляет основу данного изобретения.
В данном контексте неблокирующее антитело представляет собой молекулу антитела, которая снижает связывание TNF-α с TNFR2 0-50 % (например, 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 %, включая все целые числа и все десятичные числа между ними) по сравнению со связыванием TNF-α в присутствии только молекулы антитела изотипического контроля.
Антитела и категории, определенные по частичному блокирующему агонисту, частичному блокирующему неагонисту и полному блокирующему неантагонисту, были дополнительно идентифицированы, демонстрируя сложную биологию и большую гетерогенность анти-TNFR2 антител, ясно демонстрируя, что антитела по данному изобретению образуют уникальную группу.
Чтобы определить, обладает ли антитело агонистической или антагонистической активностью, можно применять анализ клеток натуральных киллеров (НК), как описано в Примере 4. Вкратце описано, что НК-клетки отвечают на стимулы ИЛ-2 и ИЛ-12 секрецией ИФН-γ. Растворимый TNF-α продуцируется эндогенно и присутствует в устойчивых, но субоптимальных концентрациях (~100 пг/мл) для передачи сигналов TNFR2, что означает, что ИФН-γ может как увеличиваться, так и уменьшаться посредством модуляции передачи сигналов TNFR2. Следовательно, экзогенное добавление TNF-α в оптимальной концентрации при передаче сигнала TNFR2 увеличивает концентрации ИФН-γ в этом анализе, как и инкубация с агонистическим анти-TNFR2 антителом. Напротив, совместная инкубация с анти-TNF-α антителом или блокирующим лиганд антагонистическими антителами, описанными в данном документе для сравнения, снижают высвобождение ИФН-γ в этом анализе. Таким образом, этот анализ можно применять для идентификации агонистической или антагонистической активности или отсутствия таковой анти-TNFR2 антител. (TNFα Augments Cytokine-Induced NK Cell IFNγ Production through TNFR2. Almishri W.et al. J Innate Immun. 2016;8:617-629)
Следовательно, способность антител агонизировать, т.е. индуцировать передачу сигналов TNFR2, можно контролировать с применением этой экспериментальной установки. Способность антител самим индуцировать передачу сигналов при связывании с TNFR2 в том же анализе клеток натуральных киллеров (НК) может быть оценена путем мониторинга и сравнения увеличения высвобождения ИФН-γ с тем, которое наблюдается после культивирования в присутствии или в отсутствие экзогенного TNF-α, добавленного при оптимальных для передачи сигналов концентрациях, как описано в Примере 4. Следовательно, агонистическое антитело TNFR2 можно определить как антитело, которое усиливает высвобождение ИФН-γ НК-клетками в этом анализе. Антитело с внутренней агонистической способностью усиливает высвобождение ИФН-γ НК-клетками способом, который не зависит от перекрестного связывания антитела или взаимодействия с рецепторами Fc-гамма, и не зависит от присутствия растворимого TNF-α лиганда. Следовательно, внутреннюю агонистическую активность можно оценить с применением форматов антител, которые не взаимодействуют продуктивно с FcγR, например, агликозилированных антител, несущих мутацию N297A в Fc-домене, или в аналитических системах/клетках, лишенных FcγR. НК-клетки хорошо известны специалистам в данной области техники (Binyamin, L., et al (2008). Journal of immunology 180, 6392-6401; Blocking NK cell inhibitory self-recognition promotes antibody-dependent cellular cytotoxicity in a model of anti-lymphoma therapy.). Применяя этот анализ, агонистическое антитело определяется как антитело, приводящее к увеличению высвобождения ИФН-γ на > 100 % (> 2 раза). Поскольку в этом анализе применяются первичные клетки от доноров МКПК, необходимо включить по меньшей мере 4 донора и рассчитать средние значения для всех доноров. Клетки от каждого донора, которые должны быть включены в расчет среднего, должны отвечать на обработку положительным контролем (растворимый TNF-α) со > 100 % (> 2 раза) повышением уровней ИФН-γ по сравнению с обработкой изотипическим контролем.
Описанные в данном документе молекулы агонистических антител обладают внутренней агонистической активностью, как объяснено выше.
В некоторых вариантах осуществления данного изобретения предпочтительно, чтобы антитело увеличивало высвобождение ИФН-γ НК-клетками в описанном выше анализе по меньшей мере на 100 %.
В некоторых вариантах осуществления данного изобретения, агонистическая активность может быть улучшена молекулой антитела, связанной с рецептором Fcγ в дополнение к связыванию с TNFR2. В некоторых таких вариантах осуществления данного изобретения, молекулы агонистического неблокирующего TNFR2 антитела связываются с более высокой аффинностью с ингибирующими рецепторами Fcγ, чем с активирующими рецепторами Fcγ. Обладая более высокой аффинностью к ингибирующим рецепторам Fcγ, чем к активирующим рецепторам Fcγ, мы включаем значения вариантов, которые связываются с более высокой аффинностью с ингибирующими рецепторами Fcγ по сравнению с индивидуальными активирующими рецепторами Fcγ, например, по сравнению с любым из FcγRIIA, FcγRIIIA и FcγRI.
Относительно высокая гомология между системами FcγR мыши и человека объясняет многие общие аспекты консервативных опосредованных FcγR механизмов между видами. Однако подклассы IgG мыши и человека различаются по своей аффинности с родственными FcγR, это важно при переводе FcγR-опосредованных наблюдений в мышиной системе в терапевтические средства на основе IgG человека для выбора антитела, подкласса антител и/или сконструированного варианта подкласса, которые показывают соответствующее связывание с человеческими активирующими против ингибирующих FcγR. Аффинность и/или авидность молекул антител человека к индивидуальным FcγR человека можно определить с помощью поверхностного плазмонного резонанса (SPR - surface plasmon resonance).
В некоторых вариантах осуществления данного изобретения, связывание с рецептором Fc происходит посредством нормального взаимодействия между Fc-участком молекулы агонистического антитела и рецептором Fc. В некоторых таких вариантах осуществления данного изобретения, молекула антитела представляет собой IgG, который имеет Fc-участок, связывающийся с рецептором Fcγ. В некоторых таких вариантах осуществления данного изобретения, анти-TNFRII антитело представляет собой изотип IgG2 человека, которое имеет аналогичную промежуточную аффинность к человеческому ингибирующему FcγRIIB и человеческому активирующему FcγRIIA и FcγRIIIA, но не взаимодействует продуктивно с человеческим активирующим FcγRI. В некоторых вариантах осуществления данного изобретения, анти-TNFRII антитело относится к изотипу IgG1 человека, которое связывает FcγRIIB с более высокой аффинностью по сравнению с IgG2, но также связывает активирующие человеческие активирующие FcγRIIA, FcγRIIIA с более высокой аффинностью и дополнительно связывает активирующий FcγRI с высокой аффинностью. В других вариантах осуществления данного изобретения, анти-TNFRII антитело представляет собой IgG человека, сконструированный для усиленного связывания с FcγRIIB, например, с мутацией «SELF» (Chu et al. “Inhibition of B cell receptor-mediated activation of primary human B cells by coengagement of CD19 and FcgammaRIIb with Fc-engineered antibodies.” Mol Immunol. 2008 Sep; 45 (15): 3926-33) и/или сконструированы для относительного повышенного связывания с FcγRIIB по сравнению с активацией FcγR, например, мутаций V9 или V11 (Mimoto et al. “Engineered antibody Fc variant with selectively enhanced FcγRIIb binding over both FcγRIIaR131 and FcγRIIaH131”. Protein Eng Des Sel. 2013 Oct; 26(10): 589–598). Было показано, что такие варианты IgG, сконструированные для усиленного связывания с ингибирующим FcγRIIB или специфически повышенной аффинности связывания с ингибирующим FcγRIIB, но не активирующим FcγRIIA, увеличивают активность агониста in vivo, и терапевтическую активность агонистического антитела CD40 СР-870893 у гуманизированных животных для активации и ингибирования FcγR (Dahan et al. 2016. 'Therapeutic Activity of Agonistic, Human Anti-CD40 Monoclonal Antibodies Requires Selective FcgammaR Engagement', Cancer Cell, 29: 820-31).
Рецептор Fc, с которым молекула агонистического антитела может связываться в дополнение к TNFR2, представляет собой рецептор, обнаруженный на поверхности клеток миелоидного происхождения, таких как макрофаги, моноциты, MDCS, нейтрофилы, тучные клетки, базофилы или дендритные клетки, или на поверхности лимфоцитов, таких как НК-клетки, B-клетки или определенные T-клетки.
Как упоминалось выше, молекулы антител часто связываются с рецепторами Fc через свои Fc-участки. Поскольку описанные в данном документе молекулы агонистических антител обладают внутренней агонистической активностью, им не нужно связываться с рецепторами Fc, чтобы агонизировать TNFR2. Это означает, что в некоторых вариантах осуществления данного изобретения можно применять молекулы антитела, которые не зависят от связывания рецептора Fc через его Fc-участок, и фактически можно применять молекулы антитела, которые не имеют Fc-участка. В некоторых таких вариантах осуществления данного изобретения, молекула антитела может представлять собой Fab'2 или его пегилированную версию. В некоторых вариантах осуществления данного изобретения, молекулы антитела могут представлять собой двухвалентную или мультивалентную молекулу антитела, содержащую одноцепочечные антитела, Fab, Fv, scFv, Fab и/или (Fab')2. В других вариантах осуществления данного изобретения, молекулы антитела могут содержать модифицированный Fc-участок, такой как агликозилированный вариант молекулы антитела IgG1. Такое агликозилирование может быть достигнуто, например, путем замещения аминокислоты, аспарагина, в позиции 297 (N297X) в цепи этого антитела. Замещение может быть глютамином (N297Q) или аланином (N297A), или глицином (N297G), или аспарагином (N297D), или серином (N297S). Другие замены, например, описаны Jacobsen FW et al., JBC 2017, 292, 1865-1875 (см., например, Таблицу 1); такие дополнительные замены включают L242C, V259C, A287C, R292C, V302C, L306C, V323C, I332C и/или K334C.
В некоторых вариантах осуществления данного изобретения, молекула агонистического антитела TNFR2 представляет собой молекулу антитела IgG1, IgG3 или IgG4.
В некоторых вариантах осуществления данного изобретения, молекула агонистического антитела TNFR2 представляет собой молекулу антитела IgG, демонстрирующую улучшенное связывание с одним или более активирующими рецепторами Fc и/или сконструированная для улучшенного связывания с одним или более активирующими рецепторами Fcγ и/или сконструированная для улучшения относительного связывания с активирующими рецепторами над ингибирующими рецепторами Fcγ. В некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой Fc-сконструированное антитело IgG1 человека. Примеры таких сконструированных вариантов антител включают афукозилированные антитела с селективным улучшенным связыванием антител с FcγRIIIA и антитела, сконструированные путем направленной, мутационной или другой замены аминокислот, приводящей к улучшенному связыванию с одним или более активирующими рецепторами Fcγ по сравнению с ингибирующим FcγRIIB. (Richards et al. 2008. 'Optimization of antibody binding to FcgammaRIIa enhances macrophage phagocytosis of tumor cells', Mol Cancer Ther, 7: 2517-27; Lazar et al. 2006. 'Engineered antibody Fc variants with enhanced effector function', Proc Natl Acad Sci U S A, 103: 4005-10).
В некоторых вариантах осуществления данного изобретения, антитело IgG человека, которое сконструировано для улучшенного связывания с активирующими Fc гамма-рецепторами, может быть антителом IgG человека, несущим две мутации S239D и I332E или три мутации S239D, I332E и A330L, и/или мутации G236A в его Fc-участке. В некоторых вариантах осуществления данного изобретения, антитело IgG человека, которое сконструировано для улучшенного связывания с активирующими Fc гамма-рецепторами, может быть афукозилированным антителом IgG человека.
Как объяснялось выше, то, что молекулы антител являются внутренними агонистами, означает, что они являются агонистами как в отсутствие, так и в присутствии TNF-α. В некоторых вариантах осуществления данного изобретения, антитело является агонистическим в отсутствие TNF-α. В некоторых вариантах осуществления данного изобретения, антитело является агонистическим в присутствии TNF-α.
Клетка-мишень, экспрессирующая TNFR2, с которой связывается агонистическое антитело, в соответствии с данным изобретением, может быть выбрана из группы, состоящей из иммунных клеток, экспрессирующих TNFR2, или раковых клеток.
Антитела хорошо известны специалистам в области иммунологии и молекулярной биологии. Как правило, антитело состоит из двух тяжелых (H) цепей и двух легких (L) цепей. В данном документе, иногда эта полная молекула антитела называется полноразмерным антителом или антителом полной длины. Тяжелая цепь антитела содержит один вариабельный домен (VH) и три константных домена (CH1, CH2 и CH3), а легкая цепь молекулы антитела содержит один вариабельный домен (VL) и один константный домен (CL). Вариабельные домены (иногда вместе именуемые как участок FV) связываются с мишенью антитела или антигеном. Каждый вариабельный домен состоит из трех петель, называемых участками, определяющими комплементарность (CDR), которые отвечают за связывание с мишенью. Константные домены не участвуют непосредственно в связывании антитела с антигеном, но демонстрируют различные эффекторные функции. Антитела или иммуноглобулины могут быть отнесены к разным классам в зависимости от аминокислотной последовательности константного участка их тяжелых цепей. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, а у человека некоторые из этих иммуноглобулинов дополнительно делятся на подклассы (изотипы), например, IgG1, IgG2, IgG3 и IgG4; IgA1 и IgA2.
Другой частью антитела является участок Fc (также известный как кристаллизующийся фрагмент домена), который состоит из двух константных доменов каждой из тяжелых цепей антитела. Как уже упоминалось выше, Fc-участок отвечает за взаимодействие между антителом и рецептором Fc.
Термин «молекула антитела», употребляющийся в данной заявке, относится к полноразмерным антителам или антителам полной длины, а также к функциональным фрагментам полноразмерных антител и производным молекул таких антител.
Функциональные фрагменты полноразмерного антитела имеют те же характеристики связывания антигена, что и соответствующее полноразмерное антитело, и включают либо те же вариабельные домены (т.е. последовательности VH и VL), и/или те же последовательности CDR, что и соответствующее полноразмерное антитело. Функциональный фрагмент не всегда содержит все шесть CDR соответствующего полноразмерного антитела. Следует отметить, что молекулы, содержащие три или меньшее количество участков CDR (в некоторых случаях даже только один участок CDR или его часть), способны сохранять антигенсвязывающую активность антитела, производными которого являются CDR. Например, в работе Gao et al., 1994, J. Biol. Chem., 269: 32389-93 указано, что вся цепь VL (включая все три участка CDR) имеет высокое сродство к своему субстрату.
Молекулы, содержащие два участка CDR, описаны, например, в работе Vaughan & Sollazzo 2001, Combinatorial Chemistry & High Proposal Screening, 4: 417-430. На странице 418 (правая колонка – 3 «Наша стратегия проектирования») описано миниантитело, включающее только гипервариабельные участки H1 и H2 CDR, перемежающиеся внутри каркасных участков. Это миниантитело описывают, как способное связываться с мишенью. На работы Pessi et al., 1993, Nature, 362: 367-9, и Bianchi et al., 1994, J. Mol. Biol., 236: 649-59, ссылаются Vaughan и Sollazzo и более подробно описывают миниантитела H1 и H2 и их свойства. В работе Qiu et al., 2007, Nature Biotechnology, 25:921-9 показано, что молекула, состоящая из двух связанных участков CDR, способна связывать антиген. Quiocho 1993, Nature, 362: 293-4, дает краткое описание технологии «миниантитела». Ladner 2007, Nature Biotechnology, 25:875-7, комментирует, что молекулы, содержащие два участка CDR, способны сохранять антигенсвязывающую активность.
Молекулы антител, содержащие один участок CDR, описаны, например, в работе Laune et al., 1997, JBC, 272: 30937-44, в которой показано, что ряд гексапептидов, полученных из участка CDR, демонстрируют антигенсвязывающую активность, и отмечается, что синтетические пептиды полного единичного участка CDR демонстрируют сильную связывающую активность. В работе Monnet et al., 1999, JBC, 274: 3789-96, показано, что ряд 12-мерных пептидов и связанных с ними каркасных участков обладают антигенсвязывающей активностью, и прокомментировано, что один только CDR3-подобный пептид способен связывать антиген. В работе Heap et al., 2005, J. Gen. Virol., 86: 1791-1800, сообщается, что “микроантитело” (молекула, содержащая один участок CDR) способно связывать антиген, и показано, что циклический пептид из анти-ВИЧ антитела обладает антигенсвязывающей активностью и функцией. В работе Nicaise et al., 2004, Protein Science, 13: 1882-91, показано, что один участок CDR может придавать антигенсвязывающую активность и аффинность к своему лизоцимному антигену.
Таким образом, молекулы антител, имеющие пять, четыре, три или меньшее количество участков CDR способны сохранять антигенсвязывающие свойства полноразмерных антител, производными которых они являются.
Такая молекула антитела может также быть производной полноразмерного антитела или фрагментом такого антитела. При использовании производного оно должно обладать теми же антигенсвязывающими характеристиками, что и соответствующее полноразмерное антитело, в том смысле, что оно связывается с тем же эпитопом на мишени, что и полноразмерное антитело.
Таким образом, под термином «молекула антитела», который употребляется в настоящей заявке, понимаются все типы молекул антител и их функциональные фрагменты, а также их производные, включая: моноклональные антитела, поликлональные антитела, синтетические антитела, рекомбинантно полученные антитела, мультиспецифичные антитела, биспецифичные антитела, человеческие антитела, антитела человеческого происхождения, гуманизированные антитела, химерные антитела, одноцепочечные Fvs (scFv), фрагменты Fab, фрагменты F(ab')2, фрагменты F(ab'), Fvs (sdFv) с дисульфидной связью, тяжелые цепи антител, легкие цепи антител, гомодимеры тяжелых цепей антител, гомодимеры легких цепей антител, гетеродимеры тяжелых цепей антител, гетеродимеры легких цепей антител, антигенсвязывающие функциональные фрагменты таких гомо- и гетеродимеров.
Кроме того, термин «молекула антитела», употребляемый в данной заявке, включает все классы молекул антител и функциональных фрагментов, включая: IgG, IgG1, IgG2, IgG3, IgG4, IgA, IgM, IgD и IgE, если не указано иное.
В некоторых вариантах осуществления данного изобретения, молекула антитела представляет собой молекулу антитела человека, молекулу гуманизированного антитела или молекулу антитела человеческого происхождения. В некоторых таких вариантах осуществления данного изобретения, молекула антитела представляет собой антитело IgG. Известно, что оптимальная совместная стимуляция рецепторов агонистов суперсемейства TNFR, таких как TNFR2, зависит от взаимодействия антител с ингибирующим FcγRII. У мышей изотип IgG1, который связывается преимущественно с ингибирующим Fc-гамма-рецептором (FcγRIIB) и лишь слабо с активирующими Fc-гамма-рецепторами, как известно, оптимален для совместной стимулирующей активности моноклональных антител, направленных на суперсемейство TNFR. Хотя прямого эквивалента изотипа мышиного IgG1 у человека не было описано, антитела могут быть сконструированы так, чтобы демонстрировать аналогичное усиление связывания с ингибирующими, а не активирующими Fc-гамма-рецепторами человека. Такие сконструированные нацеленные на суперсемейство TNFR антитела также обладают улучшенной костимулирующей активностью in vivo у трансгенных мышей, сконструированных для экспрессии активирующих и ингибирующих Fc-гамма-рецепторов человека (Dahan et al, 2016, Therapeutic Activity of Agonistic, Human Anti-CD40 Monoclonal Antibodies Requires Selective FcγR Engagement. Cancer Cell. 29(6):820-31). Таким образом, в некоторых вариантах осуществления данного изобретения, молекула антитела имеет изотип, который оптимальным образом задействует ингибирующие рецепторы Fc. В некоторых вариантах осуществления данного изобретения, молекула антитела представляет собой антитело IgG2.
В некоторых вариантах осуществления данного изобретения, молекула агонистического антитела, которая специфично связывает TNFR2, может представлять собой антитело ламы и, в частности, антитело hcIgG ламы. Как и все млекопитающие, животное, относящееся к семейству верблюдовых, производят обычные антитела, состоящие из двух тяжелых цепей и двух легких цепей, связанных вместе дисульфидными связями в форме Y (IgG1). Однако они также продуцируют два уникальных подкласса иммуноглобулина G, IgG2 и IgG3, также известных как тяжелые цепи IgG (hcIgG). Эти антитела состоят только из двух тяжелых цепей, которые не имеют участка CH1, но все же имеют антигенсвязывающий домен на своем N-конце, называемом VнH. Обычный Ig требует ассоциации вариабельных участков, как тяжелой, так и легкой цепей, чтобы обеспечить большое разнообразие взаимодействий антиген-антитело. Хотя изолированные тяжелые и легкие цепи все же демонстрируют эту способность, они проявляют очень низкую аффинность по сравнению с парными тяжелыми и легкими цепями. Уникальной особенностью hcIgG является способность их мономерных антигенсвязывающих участков связывать антигены со специфичностью, аффинностью и особенно разнообразием, которые сопоставимы с обычными антителами, без необходимости соединения с другим участком.
Как указывается выше, данное изобретение включает в себя различные типы и формы молекул антител, которые должны быть известны специалисту в области иммунологии. Хорошо известно, что антитела, применяющиеся в терапевтических целях, часто модифицируются дополнительными компонентами, которые изменяют свойства молекулы антитела.
Соответственно, мы подразумеваем, что молекула антитела, описанная в данном документе, или молекула антитела, применяемая, как описано в данном документе (например, молекула моноклонального антитела, и/или молекула поликлонального антитела, и/или молекула биспецифичного антитела), содержат обнаруживаемый фрагмент и/или цитотоксический фрагмент.
Под «обнаруживаемым фрагментом молекулы» нами подразумевается одна или большее количество групп, состоящих из: фермента; радиоактивного атома; флуоресцентного фрагмента молекулы; хемилюминесцентного фрагмента молекулы; биолюминесцентного фрагмента молекулы. Обнаруживаемый фрагмент молекулы позволяет визуализировать молекулу антитела in vitro и/или in vivo, и/или ex vivo.
Под «цитотоксическим фрагментом молекулы» мы подразумеваем радиоактивную часть и/или фермент, например, причем фермент является каспазой; и/или токсин, например, причем токсин является бактериальным токсином или ядом; причем цитотоксический фрагмент способен индуцировать лизис клеток.
Также в данном изобретении подразумевается, что молекула антитела может быть в изолированной форме и/или очищенной форме, и/или может быть ПЕГилирована. Пегилирование – это метод, с помощью которого полимеры полиэтиленгликоля добавляются к молекуле, такой как молекула антитела или производное, с целью изменить ее поведение, например, продлить период полураспада путем увеличения ее гидродинамического размера, препятствуя выведению почками.
Как уже обсуждалось выше, CDR антитела связываются с антителом-мишенью. Назначение аминокислот каждому участку CDR, описанному в настоящей заявке, соответствует определениям, согласно работе Kabat EA et al. 1991, в "Sequences of Proteins of Immunological Interest" (последовательности белков, представляющих иммунологический интерес), пятое издание, публикация NIH No. 91-3242, pp xv-xvii.
Как может быть известно специалисту в данной области техники, существуют и другие способы назначения аминокислот каждому участку CDR. Например, Международная информационная система иммуногенетики (IMGT(R)) (http://www.imgt.org/ и Lefranc и Lefranc "The Immunoglobulin FactsBook" опубликованные издательством Academic Press, 2001).
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфично связывает TNFR2, является человеческим антителом.
В некоторых вариантах реализации изобретения молекула антитела, которая специфично связывает TNFR2, является антителом человеческого происхождения, т. е. первоначально человеческим антителом, которое было модифицировано, как описано в данной заявке.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфично связывает TNFR2, является гуманизированным антителом, т. е. первоначально нечеловеческим антителом, которое было модифицировано для увеличения его подобия с человеческим антителом. Гуманизированные антитела могут быть, например, мышиными антителами или антителами ламы.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфично связывает TNFR2, представляет собой молекулу антитела IgG2 человека.
В некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой антитело в форме антитела IgG2 человека, демонстрирующее улучшенное связывание с одним или несколькими ингибирующими Fc рецепторами и/или сконструированное для улучшения связывания с одним или несколькими ингибирующими Fc рецепторами; соответственно, в некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой антитело IgG2 человека, сконструированное с помощью Fc.
В некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой мышиное или гуманизированное мышиное антитело IgG3.
В некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой моноклональное антитело.
В некоторых вариантах осуществления данного изобретения, анти-TNFR2 антитело представляет собой поликлональное антитело.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, представляет собой молекулу антитела IgG1 человека, которая соответствует или мышиному IgG2a.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VH-CDR1, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VH-CDR2, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VH-CDR3, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VL-CDR1, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VL-CDR2, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула антитела, которая специфически связывает TNFR2, содержит одну из последовательностей VL-CDR3, перечисленных в Таблице 1 ниже.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, выбранную из группы, состоящей из молекул антитела, содержащих 6 CDR, выбранных из группы, состоящей из:
SEQ ID NO: 1, 2, 3, 4, 5 и 6;
SEQ ID NO: 9, 10, 11, 12, 13 и 14;
SEQ ID NO: 17, 18, 19, 20, 21 и 22;
SEQ ID NO: 25, 26, 27, 28, 29 и 30;
SEQ ID NO: 33, 34, 35, 36, 37 и 38;
SEQ ID NO: 41, 42, 43, 44, 45 и 46;
SEQ ID NO: 49, 50, 51, 52, 53 и 54;
SEQ ID NO: 57, 58, 59, 60, 61 и 62;
SEQ ID NO: 65, 66, 67, 68, 69 и 70;
SEQ ID NO: 73, 74, 75, 76, 77 и 78;
SEQ ID NO: 81, 82, 83, 84, 85 и 86;
SEQ ID NO: 89, 90, 91, 92, 93 и 94; и
SEQ ID NO: 97, 98, 99, 100, 101 и 102.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, содержащую 6 CDR, имеющих последовательности SEQ ID NO: 1, 2, 3, 4, 5 и 6; или молекула антитела, содержащая 6 CDR, имеющих SEQ ID NO: 9, 10, 11, 12, 13 и 14; или молекула антитела, содержащая 6 CDR, имеющих SEQ ID NO: 17, 18, 19, 20, 21 и 22; или молекула антитела, содержащая 6 CDR, имеющих SEQ ID NO: 25, 26, 27, 28, 29 и 30; или молекула антитела, содержащая 6 CDR, имеющих SEQ ID NO: 33, 34, 35, 36, 37 и 38; или молекула антитела, содержащая 6 CDR, имеющих SEQ ID NO: 41, 42, 43, 44, 45 и 46.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, содержащую 6 CDR, имеющих последовательности SEQ ID NO: 1, 2, 3, 4, 5 и 6.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, выбранную из группы, состоящей из молекул антитела, содержащих VH, выбранный из группы, состоящей из последовательностей SEQ ID NO: 7, 15, 23, 31, 39, 47, 55, 63, 71, 79, 87, 95 и 103.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, выбранную из группы, состоящей из молекул антитела, содержащих VL, выбранный из группы, состоящей из последовательностей SEQ ID NO: 8, 16, 24, 32, 40, 48, 56, 64, 72, 80, 88, 96 и 104.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, содержащую VH с SEQ ID NO: 7, 15, 23, 31, 39 или 47.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела представляет собой молекулу антитела, содержащую VH с SEQ ID NO: 7.
В некоторых вариантах осуществления данного изобретения, предпочтительно, чтобы молекула анти-TNFR2 антитела представляла собой молекулу антитела, содержащую VL, имеющий SEQ ID NO: 8, 16, 24, 32, 40 или 48.
В некоторых вариантах осуществления данного изобретения, более предпочтительно, чтобы молекула анти-TNFR2 антитела представляла собой молекулу антитела, содержащую VL, имеющий SEQ ID NO: 8.
В некоторых вариантах осуществления данного изобретения, предпочтительно, чтобы молекула анти-TNFR2 антитела содержала VH, имеющий SEQ ID NO: 7 и VH, имеющий SEQ ID NO: 8.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела содержит CH, имеющий SEQ ID NO: 217.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела содержит CL, имеющий SEQ ID NO: 218.
В некоторых вариантах осуществления данного изобретения, молекула анти-TNFR2 антитела содержит VH, имеющий SEQ ID NO: 7, VH, имеющий SEQ ID NO: 8, CH, имеющий SEQ ID NO: 217 и CL, имеющий SEQ ID NO: 218.
Таблица 1. Специфичные последовательности молекул агонистических антител TNFR2, которые не блокируют TNF-α связывание с TNFR2, как описано в данном документе (в последовательностях VH и VL, последовательности CDR выделены жирным шрифтом)
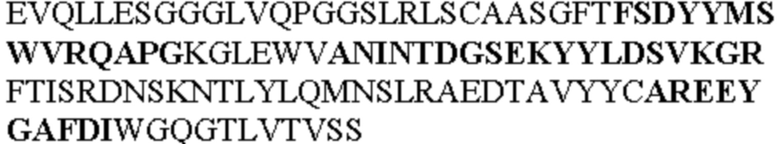
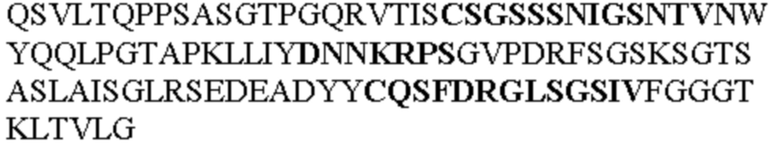
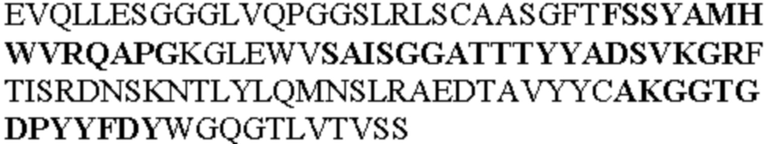
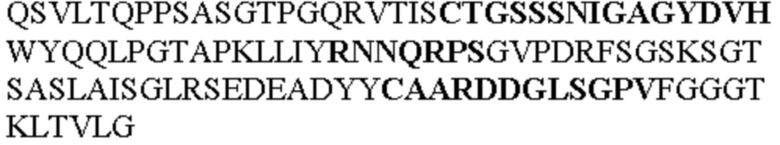
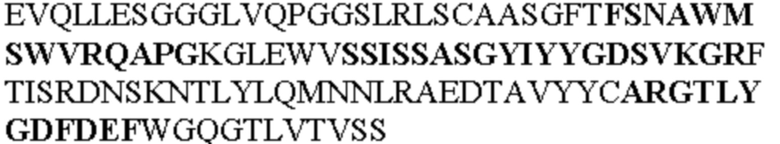
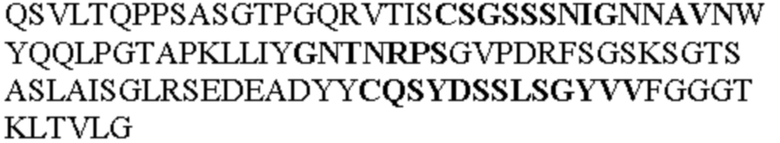
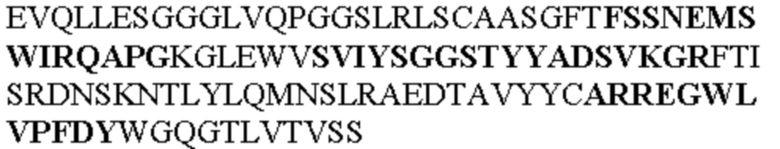
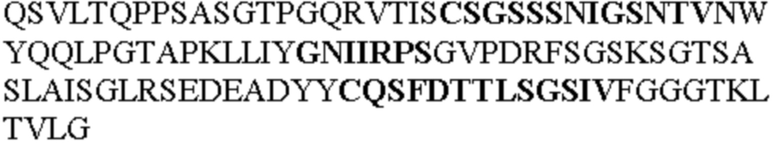


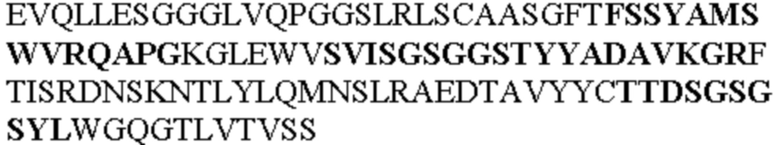
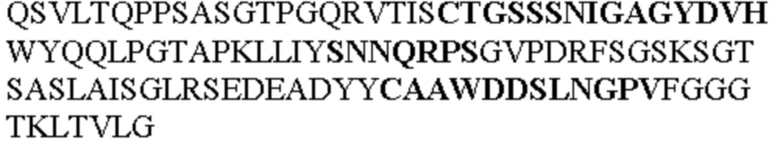
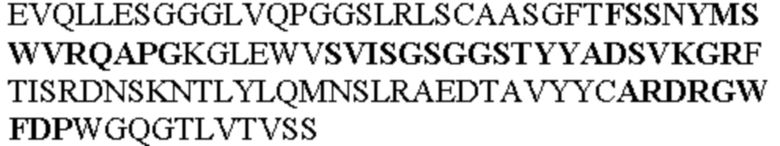

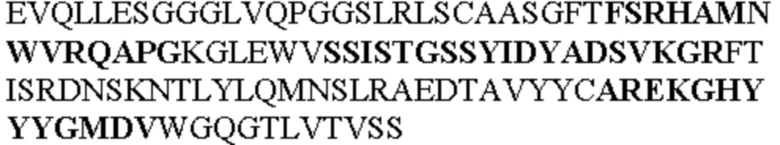
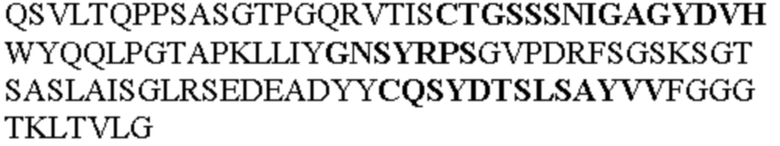
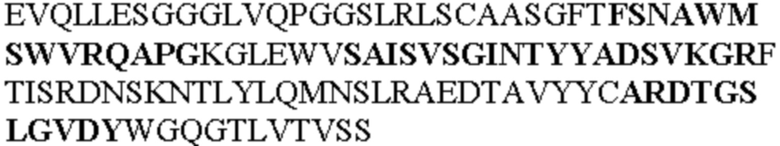
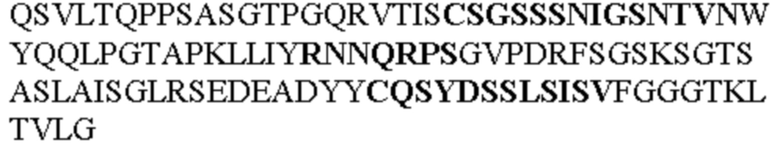
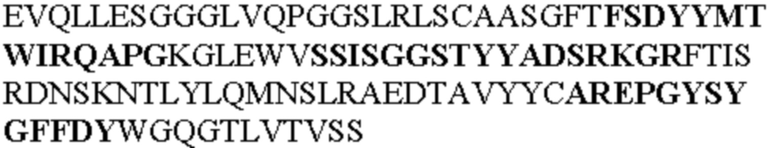
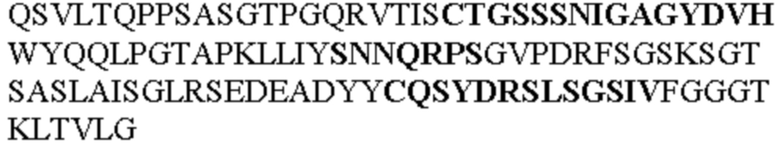
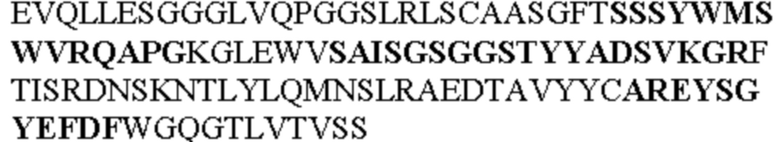
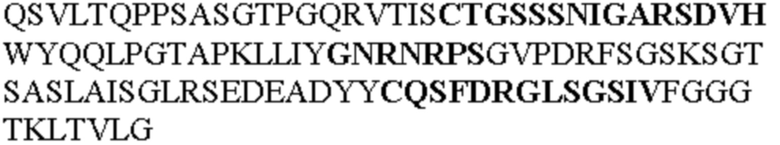
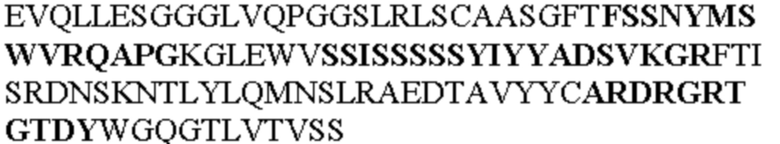
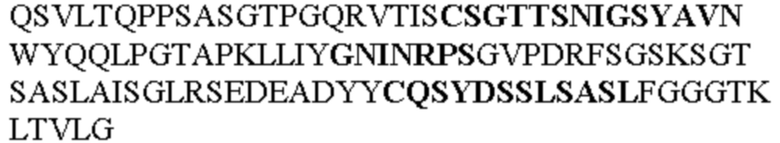
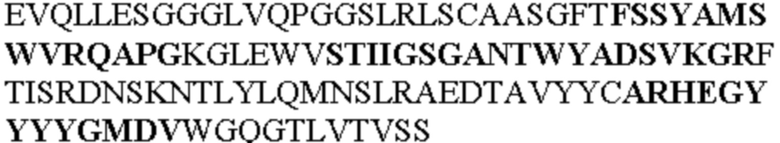
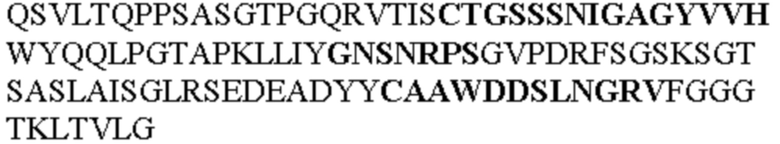
Чтобы определить или продемонстрировать особенности молекул антител по данному изобретению, их сравнивали с молекулами антител, которые блокируют TNF-α от связывания с TNFR2. Такие антитела показаны в Таблице 2.
Таблица 2. Специфичные последовательности молекул блокирующих антител TNFR2, упомянутые в данном документе в качестве эталонных антител (в последовательностях VH и VL, последовательности CDR выделены жирным шрифтом)
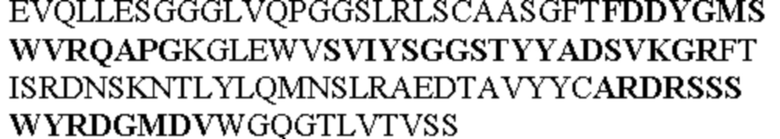
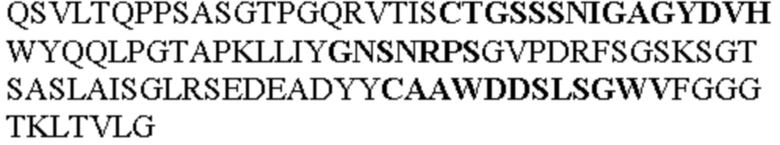
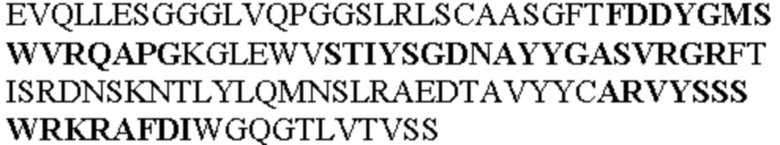
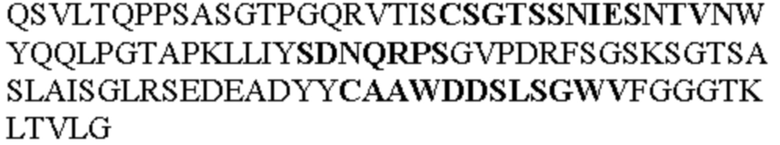

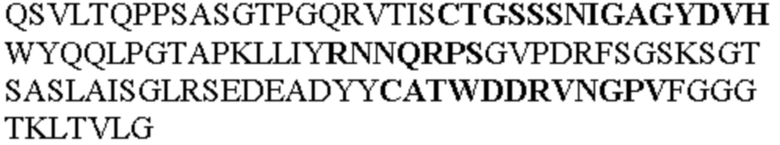
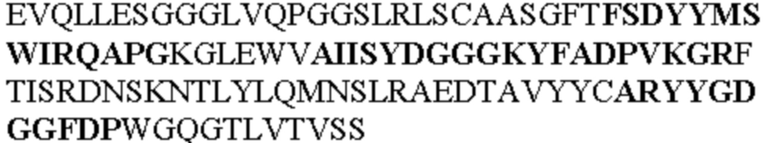
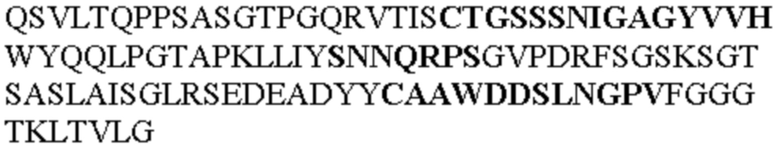
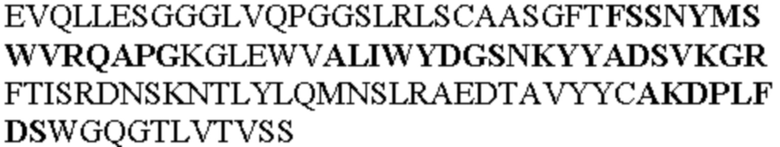
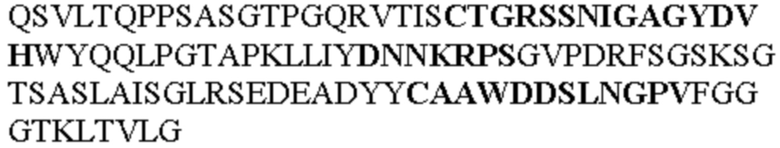
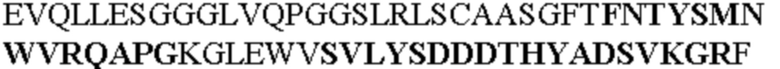
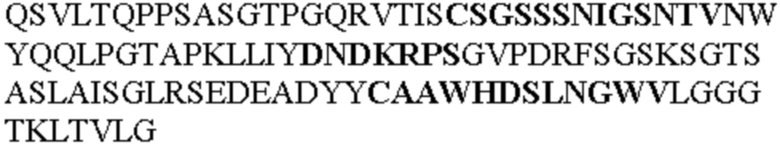
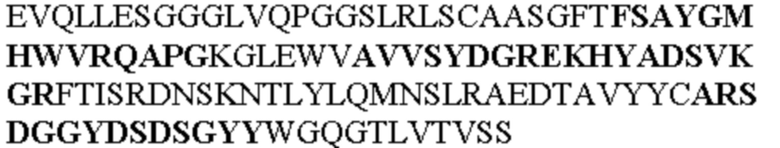
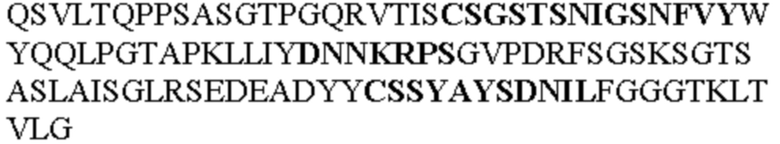
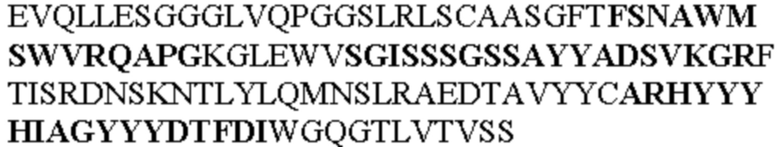
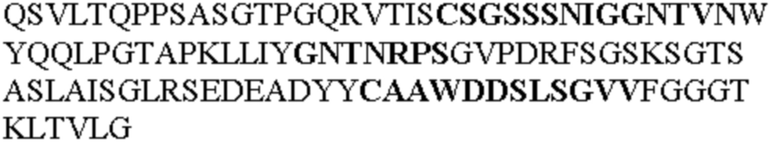
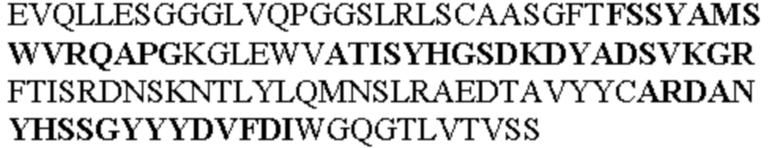
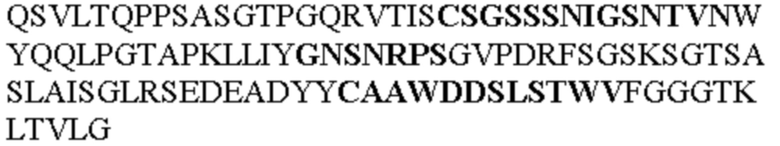
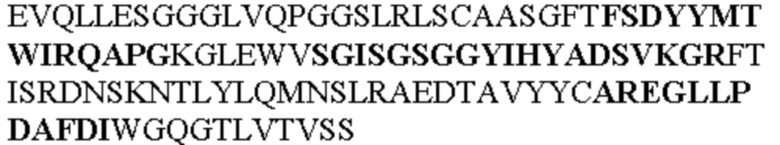

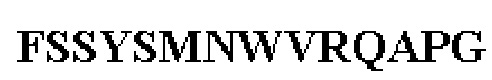

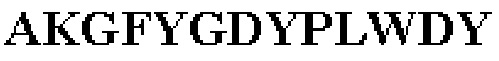
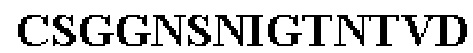
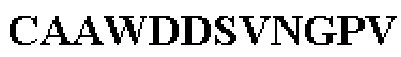
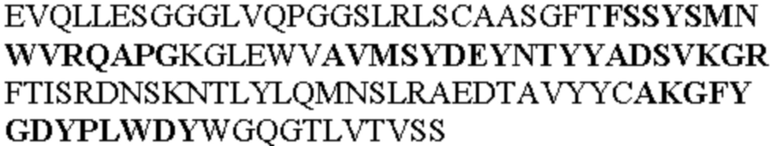
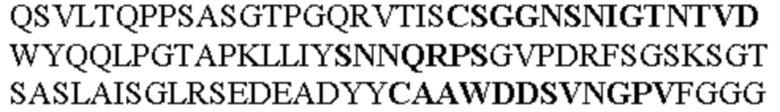
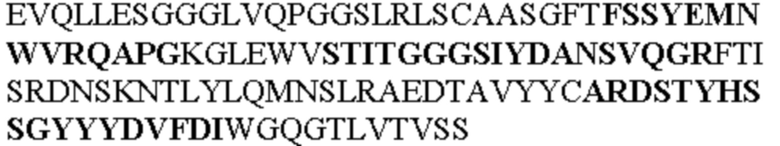
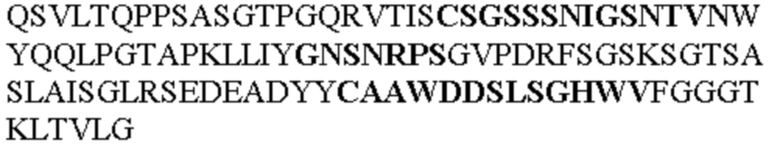
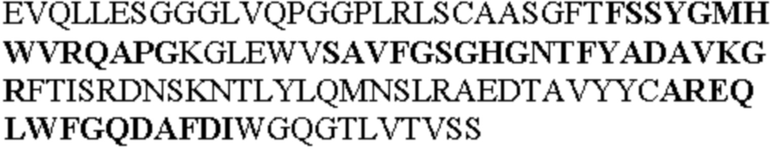
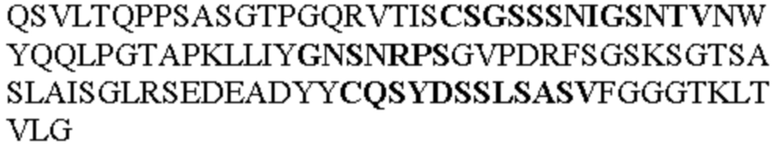
Все последовательности в Таблицах 1 и 2 выше имеют человеческое происхождение и происходят из библиотеки n-CoDeR®, как подробно объяснено в Примере 1.
В некоторых вариантах осуществления данного изобретения, молекулы антител, которые специфически связывают TNFR2, описанные в данном документе, также могут содержать один или обе константных участка (CH и/или CL), перечисленные в Таблице 3 ниже.
Таблица 3
Первый CH (SEQ ID NO: 217) и первый CL (SEQ ID NO: 218) в приведенной выше Таблице 3 имеют человеческое происхождение. Второй CH (SEQ ID NO: 219) и третий CH (SEQ ID NO: 220) в Таблице 3 оба получены из мышиного IgG2a, с той разницей, что третья последовательность CH (SEQ ID NO: 220) содержит мутацию N297A. Вторая последовательность CL (SEQ ID NO: 221) происходит из константного участка легкой цепи лямбда мыши. Эти мышиные последовательности применяются в примерах для суррогатных антител.
В некоторых вариантах осуществления данного изобретения, молекула антитела не связывает TNFR2 человека (hTNFR2). В некоторых вариантах осуществления данного изобретения, предпочтительно, чтобы молекулы агонистических антител прочно связывались с TNFR2 человека, т.е. чтобы они имели низкое значение EC50. Это продемонстрировано далее в Примере 2.
В некоторых вариантах осуществления данного изобретения, предпочтительно, чтобы молекула антитела связывалась как с hTNFR2, так и с TNFR2 яванского макака (cmTNFR2 или cynoTNFR2). Перекрестная реактивность с TNFR2, экспрессируемым на клетках яванского макака, также называемого крабоядной макакой или Macaca fascicularis, может быть выгодным, так как это позволяет тестировать молекулу антитела на животных без необходимости применять суррогатное антитело, уделяя особое внимание переносимости.
В некоторых вариантах осуществления данного изобретения, необходимо применять суррогатное антитело для тестирования функциональной активности молекулы антитела на соответствующих in vivo моделях на мышах. Чтобы гарантировать сопоставимость между эффектом молекулы антитела у людей и результатами in vivo для суррогатного антитела у мышей, важно выбрать функционально эквивалентное суррогатное антитело, имеющее такие же характеристики in vitro, что и молекула антитела человека. В некоторых вариантах осуществления данного изобретения, молекула антитела не связывается специфически с эпитопом TNFR2, содержащим последовательность KCSPG или состоящим из нее.
В некоторых вариантах осуществления данного изобретения, молекула антитела по данному изобретению или применяемая в соответствии с изобретением представляет собой молекулу антитела, которая способна конкурировать со специфическими антителами, представленными в данном документе, например, способная конкурировать с молекулами антитела, содержащими VH, выбранный из группы, состоящей из SEQ ID NO: 7, 15, 23, 31, 39, 47, 55, 63, 71, 79, 87, 95 и 103; и/или VL, выбранный из группы, состоящей из SEQ ID NO: 8, 16, 24, 32, 40, 48, 56, 64, 72, 80, 88, 96 и 104, для связывания с TNFR2.
Под «способное конкурировать за» подразумевается то, что конкурирующее антитело способно ингибировать или иным образом вмешиваться, по меньшей мере частично, в связывание молекулы антитела, как определено в данной заявке, с конкретной мишенью TNFR2.
Например, такая молекула конкурирующего антитела может быть способна ингибировать связывание описанной в данном документе молекулы антитела с TNRF2 по меньшей мере около на 10 %; например, по меньшей мере, около 20 % или, по меньшей мере, около 30 %, по меньшей мере, около 40 %, по меньшей мере, около 50 %, по меньшей мере, около 60 %, по меньшей мере, около 70 %, по меньшей мере, около 80 %, по меньшей мере, около 90 %, по меньшей мере, около 95 % или около 100 %.
Конкурентное связывание может быть определено способами, хорошо известными специалистам в данной области техники, такими как иммуноферментный анализ (ИФА).
Анализы ИФА можно применять для оценки эпитоп-модифицирующих или блокирующих антител. Дополнительные способы, пригодные для выявления конкурирующего антитела, описаны в лабораторном руководстве Antibodies: A Laboratory Manual, Harlow & Lane («Антитела: Лабораторное руководство», авторы – Харлоу и Лейн), которое включено в данную заявку посредством ссылки (например, см. стр. 567 – 569, 574 – 576, 583 и 590 – 612, 1988, CSHL, NY, ISBN 0-87969-314-2).
В некоторых вариантах осуществления данного изобретения, представляет интерес применение не самой молекулы антитела, а нуклеотидной последовательности, кодирующей такую молекулу антитела. Таким образом, данное изобретение охватывает нуклеотидные последовательности, кодирующие вышеуказанные молекулы агонистического неблокирующего TNFR-2 антитела.
Вышеописанные молекулы агонистических неблокирующих антител и нуклеотидные последовательности могут быть применены в медицине, а затем такая молекула антитела и/или нуклеотидная последовательность могут быть включены в фармацевтическую композицию, как дополнительно обсуждается ниже.
Вышеописанные молекулы агонистических неблокирующих антител, нуклеотидные последовательности и/или фармацевтические композиции можно применять при лечении рака, как дополнительно обсуждается ниже.
Вышеописанные молекулы агонистических неблокирующих антител, нуклеотидные последовательности и/или фармацевтические композиции могут применяться для лечения хронического воспалительного заболевания, как дополнительно обсуждается ниже.
Вышеописанные молекулы агонистических неблокирующих антител и/или нуклеотидные последовательности можно применять при производстве фармацевтической композиции для лечения рака.
Вышеописанные молекулы агонистических неблокирующих антител и/или нуклеотидные последовательности можно применять при производстве фармацевтической композиции для лечения хронического воспалительного заболевания.
Вышеописанные агонистические неблокирующие молекулы антител и/или фармацевтические композиции можно применять в способе лечения рака у пациента, в котором субъекту вводят терапевтически эффективное количество молекулы антитела или фармацевтической композиции.
Вышеописанные молекулы агонистических неблокирующих антител и/или фармацевтические композиции могут быть применены в способе лечения хронического воспалительного заболевания у пациента, в котором пациенту вводят терапевтически эффективное количество молекулы антитела или фармацевтической композиции.
В некоторых вариантах осуществления данного изобретения, относящихся к лечению рака, рак представляет собой солидный рак или лейкемический рак. Солидная опухоль - это аномальная масса ткани, которая обычно не содержит кист или жидких участков. Солидные опухоли могут быть доброкачественными (не рак) или злокачественными (рак). Злокачественные солидные опухоли в данном документе обозначают солидный рак. Различные типы солидных опухолей или рака названы в честь типа клеток, которые их образуют. Примерами солидных опухолей являются саркомы, карциномы и лимфомы.
Более специфичными примерами солидного рака являются рак легких, рак головы и шеи, рак желудка, рак молочной железы, колоректальный рак, рак простаты, рак мочевого пузыря, рак яичников, рак эндометрия, рак почки, рак печени, рак поджелудочной железы, рак щитовидной железы, рак мозга, рак центральной нервной системы, меланома, нейробластома, лимфома, опухоль Вильмса, рабдомиосаркома, ретинобластома и рак костей.
Более специфичными примерами лейкозного рака являются острый лимфоцитарный лейкоз, хроническое миелопролиферативное заболевание, острый нелимфоцитарный лейкоз, острый В-клеточный лимфоцитарный лейкоз, хронический лимфолейкоз, Т-клеточный острый лимфоцитарный лейкоз, неходжкинские лимфопролиферативные заболевания. В некоторых вариантах осуществления данного изобретения, описанные выше молекулы агонистического неблокирующего антитела можно применять в комбинации с молекулой антитела, которая специфически связывается с ингибитором контрольной точки. Альтернативно, обсуждаемые выше нуклеотидные последовательности, кодирующие молекулу агонистического неблокирующего TNFR2 антитела, можно применять в комбинации с молекулой антитела, которая специфически связывается с ингибитором контрольной точки или совместно стимулирующим агонистическим антителом. Примерами антител к ингибиторам контрольных точек являются антитела, нацеленные на CTLA4, PD1, PD-L1, VISTA, TIGIT, CD200, CD200R, BTLA, LAG3, TIM3, B7-H3, B7-H4, B7-H7. Примерами совместно стимулирующих агонистических антител являются антитела, нацеленные на OX40, 41BB, OX40L, 41BBL, GITR, ICOS, DR3, DR4, DR5, CD40, CD27, RANK, HVEM, LIGHT и B7-H6. Альтернативно, описанные выше молекулы агонистического неблокирующего TNFR2 антитела можно применять в комбинации с нуклеотидной последовательностью, кодирующей молекулу антитела, которая специфически связывается с ингибитором контрольной точки или совместно стимулирующим агонистом. Альтернативно, обсуждаемые выше нуклеотидные последовательности, кодирующие молекулу агонистического неблокирующего TNFR2 антитела, можно применять в комбинации с нуклеотидной последовательностью, которая кодирует молекулу антитела, которая специфически связывается с ингибитором контрольной точки или совместно стимулирующим агонистом. В некоторых таких вариантах осуществления данного изобретения, молекула антитела, которая специфически связывается с ингибитором контрольной точки, представляет собой анти-PD-1 антитело. Считается, что антитела к PD1 блокируют ингибирующий сигнал, опосредованный PD-L1, прежде всего в CD8+ Т-клетках; и тем самым допуская усиление опосредованного Т-клетками противоопухолевого ответа. Эти способы лечения могут взаимодействовать друг с другом. То же самое верно и для других ингибиторов контрольных точек и агонистических совместно стимулирующих антител.
Кроме того, описанные выше молекулы агонистического неблокирующего TNFR2 антитела можно применять в сочетании с другими противораковыми препаратами, такими как химиотерапия (например, но не ограничиваясь ими, доксорубицин, параплатин, циклофосфамид, паклитаксел, гемцитабин, 5-фторурацил, доцетаксел, винкристин, Митоксантрон, мутамицин, эпирубицин и метотрексат), низкомолекулярные ингибиторы тирозинкиназы или серин/треонинкиназы (например, но не ограничиваясь ими, ибрутиниб, иматиниб, сунтиниб, регорафениб, сорафениб, дазатиниб, эрлотиниб, вандетаниб, мидостаурин, вемурафениб, дабрафениб, палбоциклиб, рибоциклиб, Траметиниб или алектиниб), ингибиторы, нацеленные на рецепторы факторов роста (например, но не ограничиваясь ими, лекарственные средства, нацеленные на EGFR/HER1/ErbB1, EGFR2/HER2/ErbB2, EGFR3/HER3/ErbB3, VEGFR, PDGFR HGFR, RET, инсулиноподобный фактор роста IGFR, FGFR), антиангиогенные агенты (например, но не ограничиваясь ими, бевацизумаб, эверолимус, леналидомид, талидомид, зив-афлиберцепт) или облучение. Как правило, все вышеупомянутые противораковые препараты вызывают гибель раковых клеток, что приводит к воздействию неоантигенов и воспалению. Во время воздействия неоантигенов и притока воспалительных клеток в опухоль могут возникать синергические эффекты противоракового препарата.
Специалисту в области медицины может быть известно, что лекарства могут быть модифицированы с помощью различных добавок, например, чтобы изменить скорость всасывания лекарства в организме; и могут быть модифицированы в различных формах, например, чтобы обеспечить определенный путь введения в организм.
Соответственно, данная заявка включает молекулы агонистических неблокирующих антител, нуклеотидные последовательности, плазмиды, вирусы и/или клетки, описанные в данном документе, могут быть объединены с фармацевтически приемлемым наполнителем, носителем, разбавителем, носителем и/или адъювантом в фармацевтическую композицию. В этом контексте термин фармацевтическая композиция может применяться взаимозаменяемо с терминами фармацевтический препарат, фармацевтический состав, терапевтическая композиция, терапевтический препарат, терапевтический состав и терапевтический объект.
Описанные в данном документе фармацевтические композиции могут содержать или в некоторых вариантах осуществления данного изобретения состоять из молекул антител, нуклеотидных последовательностей, плазмид, вирусов или клеток.
Описанные в данном документе фармацевтические композиции могут в некоторых вариантах осуществления данного изобретения состоять или содержать плазмиды, содержащие нуклеотидные последовательности, кодирующие описанные выше молекулы антител или содержащие описанные выше нуклеотидные последовательности.
В некоторых вариантах осуществления данного изобретения, фармацевтические композиции могут содержать нуклеотидные последовательности, кодирующие части или полную молекулу антитела, описанную в данном документе, интегрированную в клеточный или вирусный геном или в вириом. Затем фармацевтическая композиция может содержать клетку или вирус в качестве носителя для доставки антитела по данному изобретению (или носителя для доставки нуклеотидной последовательности, кодирующей антитело по данному изобретению). Например, в варианте осуществления данного изобретения, вирус может быть в форме терапевтического онколитического вируса, содержащего нуклеотидные последовательности, кодирующие как минимум одну из молекул антител, описанных в данном документе. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие полноразмерное антитело IgG человека. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие молекулу антитела scFv, Fab или F(ab')2.
Как описано в прилагаемой формуле изобретения, некоторые из вариантов осуществления данного изобретения относятся к вирусу, содержащему нуклеотидную последовательность по данному изобретению или плазмиду по данному изобретению. Предпочтительно вирус представляет собой онколитический вирус, такой как терапевтический онколитический вирус. Такие онколитические вирусы известны специалистам в области медицины и вирусологии.
В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие аминокислотную последовательность, имеющую по меньшей мере 80 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит аминокислотную последовательность, имеющую по меньшей мере 85 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит аминокислотную последовательность, имеющую по меньшей мере 90 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит аминокислотную последовательность, имеющую по меньшей мере 95 % идентичности с последовательностью, указанной в Таблице 1 выше.
В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 7 и ID NO: 8. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 15 и ID NO: 16. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 23 и ID NO: 24. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 31 и ID NO: 32. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 39 и ID NO: 40. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ. ID NO: 47 и ID NO: 48. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 55 и ID NO: 56. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 63 и ID NO: 64. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 71 и ID NO: 72. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 79 и ID NO: 80. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 87 и ID NO: 88. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 95 и ID NO: 96. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие SEQ ID NO: 103 и ID NO: 104.
В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие аминокислотные последовательности, имеющие по меньшей мере 80 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие аминокислотные последовательности, имеющие по меньшей мере 85 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие аминокислотные последовательности, имеющие по меньшей мере 90 % идентичности с последовательностью, указанной в Таблице 1 выше. В некоторых вариантах осуществления данного изобретения, такой онколитический вирус содержит нуклеотидные последовательности, кодирующие аминокислотные последовательности, имеющие по меньшей мере 95 % идентичности с последовательностью, указанной в Таблице 1 выше.
В качестве примера, нуклеотидная последовательность, кодирующая антитело 001-F02, может быть такой, как представлена в Таблице 4.
Таблица 4. Пример нуклеотидных последовательностей, кодирующих антитело 001-F02 - части последовательностей, которые подчеркнуты в таблице, кодируют последовательности VH и VL, соответственно, 001-F02

CCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA
CCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTATCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATGA
Некоторые онколитические вирусы способны принимать достаточно большие вставки ДНК, чтобы обеспечить интеграцию полноразмерных последовательностей антител человека. Аттенуированные вирусы осповакцины и вирусы простого герпеса являются примерами терапевтических онколитических вирусов, чей геном достаточно велик для интеграции полноразмерных последовательностей антител IgG (Chan, W.M. et al 2014 Annu Rev Virol 1(1):119-141; Bommareddy, P.K., et al. 2018 Nat Rev Immunol 18(8):498-513). Полноразмерные антитела IgG были успешно интегрированы в онколитический Vaccinia вирус, что привело к экспрессии и внеклеточному высвобождению (продукции) полноразмерных антител IgG при инфицировании чувствительных к вирусу клеток-хозяев, например, раковых клеток (Kleinpeter, P., et al. 2016, Oncoimmunology 5(10): e1220467). Аденовирусы также могут быть сконструированы для кодирования полноразмерных антител IgG, которые функционально продуцируются и секретируются при клеточной инфекции (Marino, N., et al. 2017 J Clin Invest 123(6):2447-2463).
Изобретение также охватывает фармацевтические композиции, содержащие вирус, такой как онколитический вирус, как обсуждалось выше, и фармацевтически приемлемый разбавитель, носитель и/или адъювант.
Фармацевтическая композиция в некоторых вариантах осуществления данного изобретения может быть в форме CAR-T-клетки, несущей части или полные последовательности антитела, описанные в данном документе, как часть последовательности, кодирующей его химерный антиген Т-клеточного рецептора.
Изобретение также охватывает фармацевтические композиции, содержащие CAR-T-клетку, как обсуждалось выше, и фармацевтически приемлемый разбавитель, носитель и/или адъювант.
Изобретение также включает другие терапевтические методики или «формы» лекарственных средств, такие как конъюгаты антитело-лекарство, слитые белки и т.д., и фармацевтическую композицию, содержащую такие терапевтические методики.
Молекулы антител, нуклеотидные последовательности, плазмиды, вирусы, клетки и/или фармацевтические композиции, описанные в данном документе, могут быть пригодны для парентерального введения, включая водные и/или неводные стерильные растворы для инъекций, которые могут содержать антиоксиданты, и/или буферы, и/или бактериостатики, и/или растворенные вещества, которые делают композицию изотоничной крови предполагаемого реципиента; и/или водные и/или неводные стерильные суспензии, которые могут включать суспендирующие агенты и/или загустители. Молекулы антител, нуклеотидные последовательности, плазмиды, клетки и/или фармацевтические композиции, описанные в данном документе, могут быть представлены в контейнерах для однократной или многократной дозы, например, в запечатанных ампулах и флаконах, и могут храниться в высушенном сублимацией (т.е. лиофилизированных) состоянии, требующем добавления только стерильного жидкого носителя, например, воды для инъекций, непосредственно перед применением.
Экстемпоральные инъекционные растворы и суспензии можно приготовить из стерильных порошков и/или гранул, и/или таблеток ранее описанного вида.
При парентеральном введении пациентам-людям уровень суточной дозы молекулы анти- TNFR2 антитела обычно составляет от1 мг/кг веса тела пациента до 20 мг/кг, а в некоторых случаях даже до 100 мг/кг вводится в разовой или разделенной дозе. Более низкие дозы можно применять в особых обстоятельствах, например, в сочетании с продолжительным введением. В любом случае врач определит фактическую дозировку, которая будет наиболее подходящей для каждого отдельного пациента, и которая будет варьировать в зависимости от возраста, веса и реакции конкретного пациента. Приведенные выше дозы являются типичными для среднего случая. Конечно, могут быть отдельные случаи, когда оправданы более высокие или более низкие диапазоны дозировок, и такие случаи относятся к сфере применения данного изобретения.
Как правило, описанная в данном документе фармацевтическая композиция (или лекарственное средство), содержащая молекулу антитела, будет содержать молекулу анти- TNFR2 антитела в концентрации от между приблизительно 2 мг/мл и 150 мг/мл или от между приблизительно 2 мг/мл и 200 мг/мл.
Обычно для людей пероральное или парентеральное введение молекулы антител, нуклеотидных последовательностей, плазмид, вирусов, клеток и/или фармацевтических композиций, описанные в данном документе, является предпочтительным и наиболее удобным путем. Для ветеринарного применения молекулы антител, нуклеотидные последовательности, плазмиды, вирусы, клетки и/или фармацевтические композиции, описанные в данном документе, вводятся в виде подходящего приемлемого состава в соответствии с обычной ветеринарной практикой, и ветеринарный хирург определит режим дозирования и способ введения, который будет наиболее подходящем для конкретного животного. Таким образом, данное изобретение относится к фармацевтической композиции, содержащую количество молекулы антитела, нуклеотидной последовательности, плазмиды, вируса и/или клетки по данному изобретению, эффективное для лечения различных состояний (как описано выше и далее ниже). Желательно, чтобы молекулы антител, нуклеотидные последовательности, плазмиды, вирусы, клетки и/или фармацевтические композиции, описанные в данном документе, адаптированы для доставки с помощью пути, выбранном из группы, включающей: внутривенный (IV или в/в); внутриопухолевый (IM или i.m.); внутримышечный (SC или в/м) или подкожный.
Данное изобретение также включает молекулы антител, нуклеотидные последовательности, плазмиды, вирусы, клетки и/или фармацевтические композиции, описанные в данном документе, включающие фармацевтически приемлемые кислые аддитивные соли или основанные аддитивные соли целевых связывающих молекул или их частей по данному изобретения. Кислоты, которые применяются для получения фармацевтически приемлемых кислых аддитивных солей вышеупомянутых оснóвных соединений, полезных в настоящем изобретении, являются кислотами, которые образуют нетоксичные кислые аддитивные соли, т. е. соли, содержащие фармакологически приемлемые анионы, в частности, такие как соли гидрохлорид, гидробромид, гидройодид, нитрат, сульфат, бисульфат, фосфат, кислый фосфат, ацетат, лактат, цитрат, кислый цитрат, тартрат, битартрат, сукцинат, малеат, фумарат, глюконат, сахарат, бензоат, метансульфонат, этансульфонат, бензолсульфонат, п-толуолсульфонат и памоат [т. е. 1,1' - метилен-бис-(2-гидрокси-3 нафтоат)]. Фармацевтически приемлемые оснóвные аддитивные соли также можно применять для получения фармацевтически приемлемых солевых форм агентов согласно настоящему изобретению. Химические основания, которые можно применять в качестве реактивов для получения фармацевтически приемлемых оснóвных солей настоящих агентов, которые являются кислотными по своей природе, являются химические основания, которые образуют с такими соединениями нетоксичные оснóвные соли. Такие нетоксичные оснóвные соли включают, но не ограничиваются ими, нетоксичные оснóвные соли, являющиеся производными таких фармакологически приемлемых катионов, как катионы щелочных металлов (например, калия и натрия) и катионы щелочноземельных металлов (например, кальция и магния). кальций и магний), аммоний или водорастворимые аминовые аддитивные соли, такие как N-метилглюкамин-(меглюмин), а также в том числе низший алканоламмоний и другие оснóвные соли фармацевтически приемлемых органических аминов. Молекулы антител, нуклеотидные последовательности, плазмиды, вирусы и/или клетки, описанные в данном документе, могут быть лиофилизированы для хранения и восстановлены в подходящем носителе перед применением. Можно применять любой подходящий метод лиофилизации (например, распылительная сушка, сушка осадка) и/или методы ресуспензирования. Специалисты в данной области техники примут во внимание то, что лиофилизация и ресуспензирование могут привести к потере организмом в различной степени активности антител (например, при применении обычных иммуноглобулинов антитела IgM имеют тенденцию к большей потере активности, чем антитела IgG) и что для компенсации этого уровни применения могут быть скорректированы в сторону увеличения. В одном варианте осуществления данного изобретения лиофилизированный (сублимированный) полипептидсвязывающий фрагмент теряет не более чем около 20 %, или не более чем около 25 %, или не более чем около 30 %, или не более чем около 35 %, или не более чем около 40 %, или не более чем около 45 %, или не более чем около 50 % своей активности (до лиофилизации) при регидратации.
Молекулы анти-TNFR2 антитела, нуклеотидные последовательности и фармацевтические композиции, описанные в данном документе, могут быть применены для лечения рака у субъекта или пациента. В данном документе термины субъект и пациент используются как синонимы.
Термин «пациент» (или субъект), используемый в данном документе, относится к животному, включая человека, которому был поставлен диагноз: рак и/или проявляющие симптомы определенного заболевания.
В некоторых вариантах осуществления данного изобретения, пациент (или субъект) представляет собой животное, включая человека, у которого диагностирован рак.
В некоторых вариантах осуществления данного изобретения, пациент (или субъект) представляет собой животное, включая человека, у которого диагностировано хроническое воспалительное заболевание и/или проявляются симптомы хронического воспалительного заболевания. Хронические воспалительные заболевания в контексте данного описания включают аутоиммунные заболевания. Как описано выше, несколько разных иммунных клеток могут экспрессировать TNFR2, и в зависимости от заболевания и контекста относительные уровни экспрессии могут варьироваться. Хорошо известно, что, например, регуляторные Т-клетки могут экспрессировать высокие уровни TNFR2, и что они могут быть увеличены агонистами TNFR2. Регуляторные Т-клетки составляют субпопуляцию Т-клеток, способных подавлять другие иммунные клетки в нормальных и патологических иммунных условиях, и считаются инструментами предотвращения аутоиммунных атак на собственные ткани. Следовательно, стимуляция активности регуляторных Т-клеток может быть очень важной при лечении аутоиммунных заболеваний (Sharabi et al. “Regulatory T cells in the treatment of disease”. Nat Rev Drug Discov. 2018 Oct 12). Примерами хронических воспалительных заболеваний, помимо аутоиммунных заболеваний, являются остеоартрит и целиакия. Примерами аутоиммунных заболеваний являются ревматоидный артрит (РА), рассеянный склероз (MS - multiple sclerosis), сахарный диабет I типа, системная красная волчанка (СКВ), псориаз, воспалительное заболевание кишечника (ВЗК) или тяжелая миастения (MG - Myasthenia gravis).
В некоторых вариантах осуществления данного изобретения, пациент (или субъект) представляет собой пациента с высокой экспрессией TNFR2 в пораженной ткани. В этом контексте, высокая экспрессия означает более высокий уровень экспрессии TNFR2 по сравнению с соответствующей здоровой тканью. Обычно здоровая ткань, используемая для такого сравнения, представляет собой эталонную ткань (или стандартный эталон), собранную из здоровой ткани у одного или нескольких здоровых индивидуумов. Уровень экспрессии можно измерить стандартными методами, такими как иммуногистохимия (IHC - immunohistochemistry), сортировка клеток с активацией флуоресценции (FACS - fluorescence-activated cell sorting) или измерения экспрессии мРНК.
В данной заявке предполагается, что пациент может быть млекопитающим или не млекопитающим. Предпочтительно пациент-млекопитающее представляет собой человека, лошадь, корову, овцу, свинью, верблюда, собаку или кошку. Более всего предпочтительно, чтобы пациент-млекопитающее представлял собой человека.
Под «демонстрировать» в данной заявке подразумевается, что пациент демонстрирует симптом рака и/или диагностический маркер рака; и/или симптом рака и/или диагностический маркер рака можно измерить и/или оценить, и/или количественно определить.
Специалист в области медицины сразу поймет, каковы симптомы рака и диагностические маркеры рака, а также, как измерить и/или оценить, и/или количественно определить, происходит ли уменьшение или увеличение тяжести симптомов рака или уменьшение, или увеличение диагностических маркеров рака; а также как эти симптомы рака и/или диагностические маркеры рака можно использовать для формирования прогноза течения рака.
Лечение рака часто назначают в виде курса лечения, то есть лекарственный препарат вводят в течение определенного периода времени. Продолжительность курса лечения будет зависеть от ряда факторов, которые среди прочего могут включать тип вводимого лекарственного препарата; тип рака, который подвергается лечению; тяжесть рака, который подвергается лечению; возраст и состояние здоровья пациента.
Под «во время лечения» в данной заявке подразумевается то, что пациент в настоящее время проходит курс лечения и/или получает лекарственный препарат, и/или получает курс лекарственного препарата.
В некоторых вариантах осуществления данного изобретения рак, который подвергается лечению в соответствии с данным изобретением, является солидной опухолью.
Каждый из вышеописанных видов рака хорошо известен, а симптомы и диагностические маркеры рака хорошо описаны, как и лекарственные препараты, применяющиеся для лечения этих видов рака. Соответственно, симптомы, диагностические маркеры рака и лекарственные препараты, применяющиеся для лечения вышеуказанных типов рака, известны специалистам в области медицины.
Клинические определения диагноза, прогноза и прогрессирования большого количества раковых заболеваний основаны на определенных классификациях, известных как определение стадии рака. Эти системы определения стадии рака действуют для сопоставления ряда различных диагностических маркеров рака и симптомов рака, для обеспечения формулировки диагноза и/или прогноза, и/или прогрессирования рака. Специалисту в области онкологии будет известно, как оценить диагноз, и/или прогноз, и/или прогрессирование рака, используя систему определения стадии рака, и какие диагностические маркеры рака и симптомы рака следует применять для этого.
Под «определением стадии рака» в данной заявке подразумевается определение стадии рака по классификации Rai, которая включает в себя стадию 0, стадию I, стадию II, стадию III и стадию IV, и/или по классификации Binet, которая включает в себя стадию А, стадию B и стадию C, и/или по классификации Ann Arbour, которая включает стадию I, стадию II, стадию III и стадию IV.
Известно, что рак может вызывать аномалии в морфологии клеток. Эти аномалии часто повторно встречаются при определенных видах рака, что означает то, что исследование этих изменений в морфологии (также известное как гистологическое исследование) можно применять в диагностике или прогнозировании рака. Методы визуализации образцов для изучения морфологии клеток и подготовки образцов для визуализации хорошо известны в данной области техники; например, световая микроскопия или конфокальная микроскопия.
Под «гистологическим исследованием» в данной заявке подразумевается наличие мелких зрелых лимфоцитов и/или наличие мелких зрелых лимфоцитов с узкой каймой цитоплазмы, наличие мелких зрелых лимфоцитов с плотным ядром, лишенным отчетливо видимых ядрышек, и/или наличие мелких зрелых лимфоцитов с узкой каймой цитоплазмы, и/или с плотным ядром, лишенным отчетливо видимых ядрышек, и/или наличие атипичных клеток, и/или расщепленных клеток, и/или пролимфоцитов.
Хорошо известно, что рак является результатом мутаций в ДНК клетки, которые могут привести к тому, что клетка избегает клеточной гибели или бесконтрольно размножается. Поэтому исследование этих мутаций (также известное как цитогенетическое исследование) может быть полезным инструментом для оценки диагноза и/или прогноза рака. Примером этого является делеция хромосомной локации 13q14.1, которая характерна для хронической лимфоцитарной лейкемии. Методы исследования мутаций в клетках хорошо известны в данной области техники; например, флуоресцентная гибридизация in situ (FISH).
Под «цитогенетическим исследованием» в данной заявке подразумевается исследование ДНК в клетке и, в частности, в хромосоме. Цитогенетическое исследование можно применять для выявления изменений в ДНК, которые могут быть связаны с наличием рефрактерного рака и/или рецидивирующего рака. Такие изменения ДНК могут включать: делеции в длинном плече хромосомы 13 и/или делеции локуса 13q14.1 хромосомы, и/или трисомию хромосомы 12, и/или делеции в длинном плече хромосомы 12, и/или делеции в длинном плече хромосомы 11, и/или делецию 11q, и/или делеции в длинном плече хромосомы 6, и/или делецию 6q, и/или делеции в коротком плече хромосомы 17, и/или делецию в 17p, и/или транслокацию t(11:14), и/или транслокацию (q13:q32), и/или перестройку рецептора гена антигена, и/или перестройки BCL2, и/или перестройки BCL6, и/или транслокации t(14:18), и/или транслокации t(11:14), и/или транслокации (q13:q32), и/или транслокации (3:v), и/или транслокации (8:14), и/или транслокации (8:v), и/или транслокации t(11:14) и (q13:q32).
Известно, что больные раком проявляют определенные физикальные симптомы, которые часто являются результатом бремени рака на организм. Эти симптомы часто повторяются при одном и том же раке, и поэтому могут быть характерными для диагноза, и/или прогноза, и/или прогрессирования заболевания. Специалист в области медицины должен понимать, какие физикальные симптомы связаны с какими видами рака, и как оценка этих физикальных систем может коррелировать с диагнозом, и/или прогнозом, и/или прогрессированием заболевания. Под «физикальными симптомами» в данной заявке подразумевается гепатомегалия и/или спленомегалия.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
В приведенных ниже примерах делается ссылка на следующие фигуры:
На Фиг. 1 показано, что антитела по данному изобретению связывают TNFR2. Фиг.1 A-D: С помощью ИФА было показано, что человеческие антитела связываются с человеческим белком TNFR2 дозозависимым образом, генерируя различные значения EC50. Фиг. 1E: Мышиные антитела 3-F10 и 5-A05 связываются с mTNFR2 с аналогичной аффинностью.
На Фиг. 2 показано связывание специфичных к TNFR2 антител n-CoDeR® с in vitro активированными CD4+ Т-клетками. CD4+ Т-клетки, полученные из крови человека (Фиг. 2 AD), и CD4+ T-клетки селезенки мыши (Фиг. 2 E) были активированы ИЛ-2 и CD3/CD28 Dynabeads®. Аффинность TNFR2-специфических антител n-CoDeR® к активированным клеткам анализировали с помощью FACS в концентрациях от 0,002-267 нМ (человек) и 0,00003-133 нМ (мышь). Кривые показывают MFI после вычитания фона изотипического контроля (Фиг. 2 A (полные и частичные блокаторы), Фиг. 2 B (частичные блокаторы), Фиг. 2 C и D (неблокаторы), Фиг. 2 E (суррогатный полный блокатор мыши (3-F10) и неблокатор (5-A05)).
В то время как человеческие антитела TNFR2 связываются с различной аффинностью с in vitro активированными CD4+ Т-клетками (значения EC50 от 0,59 до 53 нМ), антитела TNFR2 мыши связываются с аналогичной аффинностью (значения EC50 в диапазоне от 0,072 до 0,11 нМ).
На Фиг. 3 показано, что антитела TNFR2 n-CoDeR® специфически связываются с TNFR2. CD4+ Т-клетки, полученные из крови человека (Фиг. 3 A), и CD4+ T-клетки селезенки мыши (Фиг. 3 B) активировали в течение 3 дней с помощью рекомбинантных активирующих гранул ИЛ-2 и CD3/CD28. Активированные in vitro клетки блокировали поликлональным антителом против TNFR2 (серая линия) или оставляли в ФСБ (черная линия) на 30 минут перед окрашиванием субоптимальной концентрацией различных антител TNFR2 n-CoDeR® или изотипического контроля (пунктирная линия) на 15 минут. Затем клетки промывали и инкубировали с вторичным антителом, конъюгированным с АПК, в течение 30 минут перед анализом с помощью проточной цитометрии.
Связывание всех антител может быть заблокировано поликлональным антителом TNFR2, следовательно, показано, что антитела TNFR2 n-CoDeR® (человека и мыши) специфичны к TNFR2.
На Фиг. 4 показана перекрестная реактивность человеческих TNFR2-специфических антител n-CoDeR® к Cynomolgus.
CD4+ Т-клетки выделяли из крови яванского макака и стимулировали PMA и иономицином. Через 2 дня клетки метили 0,1, 1 или 10 мкг/мл TNFR2-специфическими антителами n-CoDeR® или изотипическим контролем с последующей инкубацией со вторичным α-антителом человека, конъюгированным с АПК. Клетки анализировали способом проточной цитометрии. На фигуре показано процентное содержание TNFR2+ Т-клеток для индивидуальных антител по сравнению с изотипическим контролем. Результаты представляют собой среднее значение и стандартное отклонение (SD) из 2-3 отдельных экспериментов.
Большинство антител к TNFR2 демонстрируют перекрестно-реактивное связывание с клетками Cynomolgus.
Фиг. 5. Активированные клетки блокировали 40 мкг/мл антитела MR2-1 (Фиг. 5 A, черные столбцы) или оставляли в ФСБ (Фиг. 5 A, серые столбцы) на 30 минут, затем добавляли TNFR2-специфические антитела n-CoDeR®/поликлональные TNFR2 (pTNFR2) и клетки инкубировали 15 мин. Процент связанных антител TNFR2 n-CoDeR® анализировали с помощью FACS после инкубации со вторичными антителами, конъюгированными с АПК. На Фиг. 5B активированные CD4+ Т-клетки блокировали 40 мкг/мл TNFR2-специфических антител n-CoDeR®/pTNFR2 (черные столбцы) или оставляли в ФСБ (серая полоса), а затем инкубировали 15 мин с PE-конъюгированными антителами MR2-1. Затем клетки анализировали с помощью FACS.
Антитело MR2-1 не препятствовало связыванию TNFR2 специфических антител n-CoDeR®, а антитела n-CoDeR® не влияли на связывание MR2-1 с активированными клетками, показывая, что все антитела n-CoDeR® связывают другие домены белка TNFR2, чем антитело MR2-1. Это показывает, что все специфические TNFr2 антитела n-CoDeR® связывают другие эпитопы на белке TNFR2, чем клон MR2-1 TNFR2.
CD4+ Т-клетки, полученные из крови человека, стимулировали активационными гранулами rhIL-2 и CD3/CD28 в течение 2-3 дней. Активированные клетки блокировали 40 мкг/мл антитела MR2-1 (Фиг. 5 A, черные столбцы) или оставляли в ФСБ (Фиг. 5 A, серые столбцы) на 30 минут, затем добавляли TNFR2-специфические антитела n-CoDeR®/поликлональные TNFR2 (pTNFR2) и клетки инкубировали 15 мин. Процент связанных антител TNFR2 n-CoDeR® анализировали с помощью FACS после инкубации со вторичными антителами, конъюгированными с АПК. На Фиг. 5B активированные CD4+ Т-клетки блокировали 40 мкг/мл TNFR2-специфических антител n-CoDeR®/pTNFR2 (черные столбцы) или оставляли в ФСБ (серая полоса), а затем инкубировали 15 мин с PE-конъюгированными антителами MR2-1. Затем клетки анализировали с помощью FACS.
Антитело MR2-1 не препятствовало связыванию TNFR2 специфических антител n-CoDeR®, а антитела n-CoDeR® не влияли на связывание MR2-1 с активированными клетками, показывая, что все антитела n-CoDeR® связывают другие домены белка TNFR2, чем антитело MR2-1.
На Фиг. 6 показаны данные для антител, блокирующих лиганд. Блокирующий ИФА выполняли с мАт n-CoDeR®, специфическими для hTNFRII, для оценки характеристик блокирования лиганда. Фиг. 6 A) Все антитела инкубировали при 10 мкг/мл. Затем дозировали все антитела, снижающие сигнал, достигаемый с помощью изотипического контроля, более чем на 50 % (обозначено пунктирной линией) для дальнейшего изучения потенциала блокирования лиганда. Фиг. 6 B) показывает полные блокирующие мАт, Фиг. 6 C) и D) показывают частично блокирующие мАт, а Фиг. 6 E) показывает слабые блокирующие мАт. Все остальные мАт считаются неблокирующими мАт.
На Фиг. 7 показаны данные для антител, блокирующих лиганд. Блокирующий ИФА выполняли с мАт n-CoDeR®, специфическими для mTNFRII, для оценки характеристик блокирования лиганда. Фиг. 7 A) Все антитела инкубировали при 10 мк мкг/мл. Затем дозировали все антитела, снижающие сигнал, достигаемый с помощью изотипического контроля, более чем на 50 % (обозначено пунктирной линией) для дальнейшего изучения потенциала блокирования лиганда. Фиг. 7 B) показывает полные блокирующие мАт, Фиг. 7 C) и D) показывают частично блокирующие мАт, а Фиг. 7E) показывает слабые блокирующие мАт. Все остальные мАт считаются неблокирующими мАт.
На Фиг. 8 показано, что агонистические неблокирующие, но не антагонистические блокирующие TNFR2 специфические антитела n-CoDeR® усиливают продукцию ИФН-γ в НК-клетках, стимулированных ИЛ-2 и ИЛ-12. Фиг. 8А: НК-клетки, полученные из крови человека, стимулировали 20 нг/мл rhIL-2 и 20 нг/мл rhIL-12 с добавлением 10 мкг/мл TNFR2-специфических антител n-CoDeR®, изотипического контроля или 100 нг/мл rhTNF-α на 24 ч. Количество ИФН-γ в супернатантах культур измеряли с помощью MSD. Количество ИФН-γ нормализовано к изотипическому контролю и показано на Фиг. 8 A. Человеческие антитела, которые имели более высокое значение EC50, чем 25 нМ, к активированным in vitro CD4+ Т-клеткам не были включены в анализ. НК-клетки человека также продуцируют TNF-α в этих культурах (см. Фиг. 8D ниже). Результаты ИФН-γ представляют собой среднее значение для 3 доноров в 2 независимых экспериментах. Результаты показывают, что неблокирующие TNFR2 антитела являются агонистами и увеличивают продукцию ИФН-γ из НК-клеток, стимулированных цитокинами, в то время как блокирующие антитела являются скорее антагонистическими и снижают продукцию ИФН-γ НК-клетками. Кроме того, на Фиг. 8 BD показано, что агонистические антитела обладают внутренней агонистической активностью даже в отсутствие измеряемого TNF-α лиганда. Как показано на Фиг. 8B, добавление блокирующего TNF-α антитела снижает количество высвобождения ИФН-γ, но соотношение по сравнению с изотипическим контролем (Фиг. 8C) все еще сохраняется даже при полном отсутствии измеримого TNF-α в супернатанте (Фиг. 8D). На Фиг. 8D показано среднее значение уровней TNF-α у 2 доноров в присутствии или в отсутствие TNF-α нейтрализующиго антитела
На Фиг. 9 показано, что агонистические неблокирующие, но не антагонистические блокирующие TNFR2 специфические антитела n-CoDeR® активируют популяцию CD4+ Т-клеток памяти, что проявляется в увеличении доли CD25+ клеток. CD4+ Т-клетки, полученные из крови человека (Фиг. 9 A), и CD4+ T-клетки селезенки мыши (Фиг. 9 B) были активированы рекомбинантными ИЛ-2 и TNFR2-специфическими антителами n-CoDeR®, изотипическим контролем или рекомбинантным TNF-α. После 3 дней культивирования клетки окрашивали на CD25 и CD45RO (человек)/CD44 и CD62L (мышь) и анализировали с помощью проточной цитометрии. Результаты показывают процент клеток, экспрессирующих CD25, в популяции памяти (клетки CD45RO+ (человек)/CD44+ CD62L- (мышь)) по сравнению с процентом клеток CD25+, выделенных в культурах с изотипическим контролем. Результаты представляют собой среднее значение и SEM для 7 доноров (Фиг. 9 A, человек) и 3 мышей (в 2 независимых экспериментах) (Фиг. 9 B). В культурах как человека, так и мышей неблокирующие TNFR2 антитела индуцировали процент клеток памяти CD25+, в то время как блокирующие антитела не оказывали такого влияния на популяцию памяти. Как для культур человека (Фиг. 9 A), так и для мышей (Фиг. 9 B) добавление экзогенного TNF-α увеличить популяцию Т-клеток памяти CD25+. * = p <0,05, как рассчитано с помощью однофакторного дисперсионного анализа.
На Фиг. 10 показано, что анти-TNFR2 мАт модулируют супрессивную функцию производных миелоидных супрессивных клеток (MDSC - Myeloid-Derived Suppressive Cells). Для Фиг. 10 A и B MDSC предварительно инкубировали с анти-TNFR2 мАт человека в течение 30 мин. Затем без промывания клеток титруемые количества MDSC инкубировали с CD3+ T-клетками, меченными CFSE, в присутствии CD3/CD28 Dynabeads®. Фиг. 10А: % активированных CD25+ Т-клеток определяли через 3 дня с помощью FACS. Для нормализации между различными анализами фон изотипического контроля вычитали из всех точек данных. На Фиг. показана сумма 6 различных доноров в 3 независимых экспериментах при соотношении MDSC к Т-клеткам 1:4. Фиг. 10В: количество секретируемых цитокинов в супернатанте оценивали с помощью MSD и рассчитывали соотношение между высвобождением ИФН-γ и ИЛ-10 по отношению к изотипическому контролю. Данные были объединены из анализа супернатантов культур, полученных в экспериментах с 5 разными донорами.
Для Фиг. 10 были выделены миелоидные клетки CD11b+ из мышиной опухоли CT26 и предварительно инкубированы с мышиным анти-TNFR2 мАт в течение 30 мин. Титрование количества миелоидных клеток совместно культивировали с CD3+ Т-клетками, меченными CFSE, которые были очищены из наивных селезенок Balb/C. Клетки стимулировали CD3/CD28 Dynabeads® в течение 3 дней, а затем % пролиферирующих CFSEнизкий клеток анализировали с помощью FACS. (n = 4 независимых эксперимента). Было показано, что только агонистические/неблокирующие анти-TNFR2 мАт отменяют супрессивную функцию MDSC в анализах совместного культивирования Т-клеток/MDSC.
На Фиг. 10 D MDSC предварительно инкубировали в течение 30 минут с двумя разными анти-TNFR2 мАт человека (одно блокирующее = 1H10) и одно неблокирующее (= 1F02) в 3 разных изотипах, включая дефектный формат Fc, положение 297 аминокислоты было переключено, что привело к потере гликозилирования и, таким образом, уменьшению связывания с FcγR (в данном документе обозначено как N297Q). Затем без промывания клеток титруемые количества MDSC инкубировали с CD3+ Т-клетками, меченными CFSE, в присутствии CD3/CD28 Dynabeads®, и % активированных CD25+ Т-клеток определяли через 3 дня с помощью FACS. Для нормализации между различными анализами фон изотипического контроля вычитали из всех точек данных. На фигуре показано, что агонистическая активность неблокирующего антитела 1-F02 не зависит от изотипа антитела и связывания FcγR. На Фиг. показана сумма 4 разных доноров в 2 независимых экспериментах при соотношении MDSC к Т-клеткам 1:4.
Фиг. 11. Для Фиг. 11 мышам Balb/c подкожно вводили 1 × 106 клеток CT26. Через 8 дней при среднем размере опухоли 3x3 мм мышей дважды в неделю лечили 10 мг/кг антитела внутрибрюшинно, как показано на фигурах. Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли. Верхняя фигура показывает рост опухоли у мышей, получавших изотипический контроль, затем две фигуры ниже, на левой панели, показывают лиганд-блокирующее антагонистическое антитело (средняя фигура) и неблокирующее лиганд агонистическое антитело (нижняя фигура), в дефектном формате FcγR Ig. На средней панели показаны те же антитела в формате мышиного IgG2a, задействующие в первую очередь активирующие FcγR, а на правой панели - антитела в формате мышиного IgG1, задействованные в основном с ингибирующим FcγRIIb. На Фиг. 11В за выжившими мышами наблюдали в течение 70 дней. Как видно на фигурах, неблокирующее агонистическое антитело является наиболее эффективным при лечении опухолей в формате IgG1, в котором задействован в первую очередь ингибирующий FcγR. Кроме того, агонистическое антитело обладает независимым от FcγR противоопухолевым действием, что видно с применением формата N297A. С другой стороны, блокирующее антагонистическое антитело наиболее эффективно при лечении опухолей в формате IgG2a, в котором задействованы в первую очередь активирующие FcγR, и не оказывает никакого эффекта в дефектном формате связывания FcγR. *** = p <0,001 по сравнению с изотипическим контролем, рассчитанным с помощью лог-рангового критерия Мантела-Кокса.
Фиг. 12. Мышам C57/BL6 подкожно вводили 1 × 106 клеток MC38. При среднем размере опухоли 3x3 мм мышей дважды в неделю лечили 10 мг/кг антитела внутрибрюшинно, как показано на фигурах. На фиг. показаны кривые роста опухоли у отдельных мышей. Фиг. 12 A: изотипический контроль, Фиг. 12 B: нацеленное антитело к PD-1, Фиг. 12 C: антитело 5A05 (суррогатное антитело, лиганд неблокатор, агонист), Фиг. 12D: комбинация 5A05 и PD1. Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли. На Фиг. 12Е показаны кривые выживаемости для четырех различных групп лечения, * = p <0,05, *** = p <0,001 по сравнению с изотипическим контролем, рассчитанным с помощью лог-рангового критерия Мантела-Кокса.
На Фиг. 13 показано, что блокирующие лиганд агонистические антитела, эффективны в качестве противоопухолевого лечения в сочетании с анти-PD-L1. Мышам C57/BL6 подкожно вводили 1х106 клеток MC38. При среднем размере опухоли 5x5 мм мышей дважды обрабатывали изотипическим контролем антитела или 5A05 (день 1 и 4), или четыре дня подряд анти-PD-L1 с последующей пятой инъекцией через два дня (всего пять инъекций в день 1, 2, 3, 4 и 7), или их комбинацией. Все антитела вводили в дозе 10 мг/кг внутрибрюшинно. На Фиг. 13 показан средний рост опухоли +/- SEM, n = 10/группа. * = p <0,05, *** = p <0,001, как рассчитано с использованием однофакторного дисперсионного анализа.
Фиг. 14. Мышам C57/BL6 подкожно вводили 1 × 106 клеток B16. Через 3 дня мышам дважды в неделю вводили 10 мг/кг антитела внутрибрюшинно. Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли. Фиг. 14 A: изотипический контроль, Фиг. 14 B: антитело 5A05 (суррогатное антитело, неблокатор лиганда, агонист). На Фиг. 14C показаны кривые выживаемости для двух различных групп лечения. * = p <0,05 по сравнению с изотипическим контролем, рассчитанным с помощью лог-рангового критерия Мантела-Кокса.
Фиг. 15. Неблокирующее лиганд агонистическое суррогатное антитело 5A05 изменяет состав иммунных клеток в опухолях. Мышам инокулировали опухолевые клетки CT26, как описано, и вводили антитела, как указано, после того, как опухоли достигли размера приблизительно 7x7 мм после 3 инъекций на 8 день после начала лечения мышей умерщвляли и собирали опухоли. Суспензии единичных клеток опухоли анализировали на содержание иммунных клеток с помощью FACS. A) Неблокирующее лиганд агонистическое суррогатное антитело 5A05 вызывает истощение Treg и B) приток или рост CD8+ Т-клеток. Это вызывает сдвиг в соотношении Treg/CD8+ T-клеток, как показано на C) Фиг. D) показывает, что не только T-клетки, но также и миелоидные клетки, здесь количество связанных с опухолью макрофагов (TAM, определяемых как CD11b+, но отрицательных для обоих Ly6G и Ly6C) очень значительно уменьшено. Блокирующее лиганд агонистическое суррогатное антитело 3F10 также модулирует количество ТАМ, но все же значительно отличается от 5A05.
Фиг. 16. Мышам NOG в/в вводили 15-20 × 106 клеток МКПК. Через 10-12 дней у мышей удаляли селезенки, готовили суспензию единичных клеток и оценивали экспрессию TNFR2 с помощью FACS. Ранее экспрессию TNFR2 оценивали на Т-клетках, полученных из крови и образцов опухоли от 3 или 9 больных раком соответственно. Как показано на фигуре, экспрессия TNFR2 на Treg и CD8+ Т-клетки очень сопоставимы между Т-клетками человека, выращенными и активированными in vivo у мышей NOG, и Т-клетками из опухолей человека.
Фиг. 17. Мышам NOG в/в вводили 15-20 × 106 клеток МКПК. Через 10-12 дней у мышей удаляли селезенки, готовили суспензию отдельных клеток и анализировали клетки с помощью FACS. Фиг. 17A показывает средний процент окрашенных Treg, определенных как CD45+CD3+CD4+CD25+CD127низкий/отр, от общего числа клеток в селезенке. Фиг. 17B показывает средний процент Т-клеток (CD3+) от общего числа клеток в селезенке. Как показано на Фиг. 17, неблокирующее лиганд агонистическое антитело 1F02 увеличивает количество Treg (экспрессирующих самые высокие уровни TNFR2), а также общее количество Т-клеток в этой модели. Для сравнения, блокирующее лиганд антагонистическое антитело 1H10, не имеет такого эффекта.
Фиг. 18. Высвобождение ИФН-γ, индуцированное различными специфическими антителами к TNFR2, измеряли в трех различных системах in vitro. В качестве положительного контроля использовали анти-CD3 антитело - OKT3 или Муромонаб-CD3, анти-CD52 антитело - алемтузумаб и анти-CD28 антитело. Изотипический контроль применяли в качестве отрицательного контроля. Каждая точка представляет собой МКПК от одного донора-человека. На Фиг. 18А показаны результаты для клеточных культур высокой плотности, когда МКПК культивировали при концентрации 1 × 107 клеток/мл. Через 48 часов добавляли 10 мкг/мл антитела и инкубировали в течение 24 часов. Как видно на фигуре, и алемтузумаб, и OKT3 индуцировали значительное высвобождение ИФН-γ, но не индуцировали какие-либо специфические антитела к TNFR2. На Фиг. 18В показаны твердофазные культуры in vitro, полученные путем покрытия лунок 96-луночного планшета антителами перед добавлением МКПК. И снова, и алемтузумаб, и OKT3 индуцировали значительное высвобождение ИФН-γ вместе с некоторыми специфическими антителами TNFR2, особенно у одного из доноров. На Фиг. 18С показана стимуляция цельной крови антителом, и здесь алемтузумаб индуцировал значительное высвобождение ИФН-γ, но не каких-либо специфических антител к TNFR2.
Фиг. 19: NOG мыши инъецировали в/в 25-х106 МКПК клеток. Через 14 дней, когда было показано, что кровь мышей состоит приблизительно из 40 % Т-клеток человека, мышей обрабатывали 10 мкг антитела. Температуру тела измеряли через 1 час после инъекции (Фиг. 19 A). Эксперимент был прекращен через 5 часов после инъекции, и кровь была проанализирована на содержание ИФН-γ (Фиг. 19 B) или TNF-α (Фиг. 19 C). **** = p <0,0001 и ** = p <0,01, как рассчитано с помощью однофакторного дисперсионного анализа
На Фиг. 20 показано связывание с вариантами TNFR2, лишенными отдельных доменов. Связывание антител с вариантами TNFR2, экспрессируемыми на клетках HEK, тестировали методом проточной цитометрии. Отсутствие доменов 1 и 2 существенно не влияет на связывание (Фиг. 20 A и B), тогда как 3 и частично 4 полностью отменяют взаимодействие между антителом и TNFR2 (Фиг. 20 C и D). Точно так же отсутствие домена 1 + 3 полностью предотвращает связывание всех антител (кроме 1F06) (Фиг. 20 E), в то время как отсутствие домена 2 + 4 полностью отменяет связывание агонистических антител (1F02, 1F06, 4E08) и значительно снижает связывание также с антагонистами (1H10, 4H02, 5B08) (Фиг. 20 F). Темно-серый цвет указывает на положительный контроль, а белый - на антитело отрицательного контроля.
На Фиг. 21 показано сравнение аминокислотной последовательности домена 3 TNFR2 человека (H-D3) и мыши (M-D3). Сходные аминокислоты отмечены белым цветом, а различия - серым. Приведенные ниже пять последовательностей представляют 5 различных конструкций, против которых тестируются антитела. Обмены последовательностью от человека к мыши подчеркнуты, тогда как немеченная последовательность является полностью человеческой. Домены 1, 2 и 4 принадлежат человеку и не содержат замен или мутаций.
Фиг. 22. Связывание с TNFR2 дикого типа человека и мыши показано на левой панели. Мутировавшие конструкции hTNFR2 (m1, m2, m3 и m4) использовали для сужения сайта связывания для различных анти-hTNFR2 антител. Анализ проточной цитометрии показал, что мутации в аминокислотах 119-132 не влияют на связывание антител, но в аминокислотах 151-160 полностью отменяют связывание всех антител. Мутации в 134-144 нарушают связывание только блокирующих и антагонистических антител, но не оказывают значительного влияния на агонистические антитела. Темно-серые полосы указывают на положительный контроль и белые на отрицательные контрольные антитела. Пунктирная линия указывает на уровень антитела отрицательного контроля.
ПРИМЕРЫ
Теперь будут описаны конкретные примеры, не имеющие ограничительного характера, которые содержат в себе определенные аспекты данного изобретения.
Во многих примерах, в частности в примерах in vivo, применялось антитело 5-А05. Это мышиное антитело, которое является суррогатным антителом к человеческим антителам, описанным в данном документе. Оно было выбрано в качестве суррогатного антитела на основании его характеристик, аналогичных характеристикам неблокирующего лиганд агонистического антитела с хорошим значением EC50.
В некоторых примерах и на фигурах используется несколько иное название клонов антител, например, клон 001-F02 иногда сокращается до 1-F02 или 1F02, 005-B08 иногда сокращается до 5-B08 или 5B08 и т. д.
Пример 1 - Получение антител, специфичных к TNFR2
(См. также Фиг. 1 и приведенное выше описание этой фигуры).
Выделение фрагментов scFv антител
Библиотека н-Coder® scFv (BioInvent; Söderlind E, et al. Nat Biotechnol. 2000;18 (8):852-6) была использована для выделения фрагментов scFv антител, распознающих TNFR2 человека или мыши.
Библиотеку фагов использовали в трех последовательных пэннингах против рекомбинантного белка человека или мыши (Sino Biological). После инкубации фага клетки промывали для удаления не связавшихся фагов. Связывающие фаги элюировали трипсином и амплифицировали в E.coli. Полученный исходный раствор фага конвертировали в формат scFv. E.coli трансформировалась с плазмидами, несущими scFv, и были экспрессированы отдельные клоны scFv.
Идентификация уникального TNFR2 связывающего scFv
Конвертированные scFv из третьего пэннинга анализировали с применением гомогенного анализа FMAT (Applied Biosystems, Карлсбад, Калифорния, США) для связывания с клетками 293 FT, трансфицированными для экспрессии TNFR2 человека или мыши, или неродственного белка.
Вкратце, трансфицированные клетки добавляли в планшеты с прозрачным дном вместе с scFv-содержащим супернатантом из экспрессионных планшетов (разведенные 1:7), мышиное анти-His Tag антитело (0 4 мкг/мл; R&D Systems) и АПК-конъюгированные козьи антимышиные антитела (0,2 мкг / мл; кат. номер 115-136-146, Jackson Immunoresearch). Планшеты FMAT инкубировали при комнатной температуре в течение 9 ч перед считыванием. Бактериальные клоны, связывающие клетки, трансфицированные TNFR2, но не клетки, трансфицированные неродственным белком, классифицировали как активные и выборочно собирали в 96-луночный планшет.
Связывание IgG с TNFR2 в ИФА
96-луночные планшеты (планшет Lumitrac 600 LIA, Greiner) покрывали в течение ночи при 4°C с рекомбинантным белком TNFR2-Fc человека или мыши (Sino Biological) в концентрации 1 пмоль/лунку. После промывки титрованным дозам анти-TNFR2 мАт от 20 мкг/мл до 0,1 нг/мл (от 133 нМ до 1 пМ) давали возможность связываться в течение 1 часа. Затем планшеты снова промывали, и связанные антитела выявляли вторичным анти-человеческим-F(ab)-HRP антителом (Jackson ImmunoResearch), разведенным в 50 нг/мл. В качестве субстрата применяли Super Signal ELISA Pico (Thermo Scientific), и планшеты анализировали с применением ридера Tecan Ultra Microplate.
Данные, которые показаны в Таблице 5 и на Фиг. 1 AD, показывают, что все анти-TNFR2 антитела человека связываются с белком TNFR2 человека. Значения EC50 находятся в диапазоне от 0,082 нМ для 1-C08 до 4,4 нМ для 1-A09.
Кроме того, суррогатные клоны мышиных антител 3-F10 и 5-A05 также связываются с белком mTNFR2. Эти два клона связываются с очень похожей аффинностью (Таблица 5 и Фиг. 1E).
Таблица 5. Значения ЕС50 связывания антител с белком TNFR2 (человеческий белок, за исключением клона 3F10 и 5A05)
Пример 2 - Специфичность антител
(См. также Фиг. 2-5 и приведенное выше описание этих фигур).
Выделение CD4+ Т-клеток
МКПК из лейкоцитарной пленки человека и цельной крови яванского макака (M. fascicularis) были выделены с использованием градиентов Ficoll-Paque PLUS (GE Healthcare). CD4+ Т-клетки выделяли из МКПК с помощью магнитной сортировки клеток с применением набора для выделения CD4+ Т-клеток (человека) или CD4 MicroBeads, приматов, отличных человека (яванский макак) оба от Miltenyi. CD4+ Т-клетки мыши выделяли из селезенки с применением набора для выделения CD4+ Т-клеток (мыши) от Miltenyi.
Титрование TNFR2-специфических антител n-CoDeR®
Способность и аффинность антител TNFR2 n-CoDeR® связывать TNFR2, экспрессированный на клетках, были получены с применением in vitro активированных CD4+ Т-клеток. CD4+ Т-клетки человека стимулировали с rhIL-2 (50 нг/мл) (R&D systems) и Dynabeads® T-активатор CD3/CD28 для увеличения и активации Т-клеток (Gibco) 2-3 дня при 37 градусах. Клетки, активированные in vitro, метили возрастающим количеством антител n-CoDeR®, специфичных к TNFR2 или изотипическому контролю, в диапазоне 0,002-267 нМ. Затем клетки инкубировали со вторичным антителом a-человеческого IgG, конъюгированным с АПК (Jackson), с последующим анализом проточной цитометрией (FACSVerse, BD). Полученные кривые титрования показаны на Фиг. 3 A-D. CD4+ Т-клетки мыши стимулировали 135 ед/мл rmIL-2 (R&D systems) и Dynabeads® T-активатором CD3/CD28 для увеличения и активации Т-клеток (Gibco) 2-3 дня при 37 градусах. Активированные in vitro клетки метили возрастающим количеством антител n-CoDeR®, специфичных к TNFR2 или изотипическому контролю, в диапазоне от 0,00003-133 нМ. Затем клетки инкубировали со вторичным антителом a-мышиного IgG, конъюгированным с АПК (Jackson), с последующим анализом проточной цитометрией (FACSVerse, BD). Кривые титрования показаны на Фиг. 3E. Значения ЕС50 для кривых титрования были рассчитаны в Microsoft Excel и показаны в Таблице 6. Для человеческих антител значения ЕС50 различались от 0,6 нМ (4-H02) до 52,7 нМ (1-CO3). Антитела мыши связывались с активированными in vitro клетками со сходной аффинностью (0,072 нМ (3-F10) и 0,11 нМ (5-A05)).
Специфичность антител TNFR2 n-CoDeR®
Специфичность антител TNFR2 к TNFR2 была получена в экспериментах по блокированию FACS с коммерческими поликлональными антителами TNFR2 (R&D systems). CD4+ Т-клетки (мыши и человека), стимулированные 2-3 дня с помощью 50 нг/мл rhИЛ-2 (R&D systems) (человек)/135 ед/мл rmИЛ-2 (R&D systems) (мышь) и Dynabeads® T-активатором CD3/CD28 для увеличения и активации Т-клеток (Gibco), блокировали 40 мкг/мл поликлональным антителом TNFR2 (системы R&D) в течение 30 минут с последующей 15-минутной инкубацией с антителами TNFR2 n-CoDeR® или изотипическим контролем. Концентрация используемых антител n-CoDeR® была основана на кривых титрования для отдельных антител n-CoDeR® TNFR2, и была выбрана субоптимальная концентрация для каждого антитела. Затем клетки промывали и инкубировали 30 мин со вторичным антителом, конъюгированным с АПК (Jackson). Клетки анализировали проточной цитометрией (FACSVerse, BD). Все связывание специфичных к TNFR2 антител n-CoDeR® (как человеческих, так и мышиных) может быть блокировано поликлональным антителом против TNFR2, как показано на Фиг. 4. Эти результаты подтверждают, что антитела TNFR2 n-CoDeR® специфически связывают TNFR2 на CD4+ T-клетках, активированных in vitro.
Картирование эпитопа TNFR2-специфичных антител n-CoDeR® против клона MR2-1 антитела TNFR2
Клон MR2-1 антитела TNFR2 (Invitrogen) связывает специфический домен белка TNFR2. Если специфические TNFR2 антитела n-CoDeR® связаны с тем же доменом, что и MR2-1, их проверяли с помощью экспериментов по блокированию FACS.
CD4+ Т-клетки человека стимулировали 2-3 дня с помощью 50 нг/мл rhИЛ-2 (R&D systems) и Dynabeads® T-активатором CD3/CD28 для расширения и активации Т-клеток (Gibco). Активированные клетки блокировали 40 мкг/мл MR2-1 (черные столбцы на Фиг. 5A), TNFR2-специфическим антителом n-CoDeR®/поликлональным TNFR2 (R&D systems) (черные столбцы на Фиг. 6B) или ФСБ (серые столбцы на Фиг. 5). После 30 мин инкубации клетки немедленно окрашивали на TNFR2-специфические антитела n-CoDeR®/pTNFR2 (Фиг. 5A) или MR2-1 (Фиг. 5B) в течение 15 мин. Клетки на Фиг. 5A также инкубировали со вторичным реактивом IgG человека, конъюгированным с АПК (Jackson). Все клетки анализировали методом проточной цитометрии (FACSVerse, BD). Поскольку процент клеток MR2-1+ был таким же, как для блокированных n-CoDeR®, так и для неблокированных клеток (Фиг. 5B), и связывание антител n-CoDeR® было таким же с блокатором MR2-1 или без него (Фиг. 5A), эти антитела n-CoDeR®, вероятно, связывают другие эпитопы белка TNFR2, чем антитело MR2-1.
Связывание антител TNFR2 n-CoDeR® с яванским макаком
Для подтверждения перекрестной реактивности антител TNFR2 к яванскому макаку, T-клетки яванского макака CD4+ стимулировали 2 дня 50 нг/мл PMA (Sigma) и 100 нг/мл иономицина (Sigma). Клетки инкубировали со специфическими к TNFR2 антителами n-CoDeR® в 3 различных концентрациях (0,1, 1 и 10 мкг/мл), а затем инкубировали с реактивом вторичного a-человеческого IgG, конъюгированного с АПК (Jackson). Клетки были проанализированы с помощью проточной цитометрии (FACSVerse, BD), и результат показал, что большинство антител n-CoDeR®, специфичных к TNFR2 человека, могли связывать TNFR2 яванского макака, результаты для отдельных антител представлены на Фиг. 4.
Таким образом, данные в примере 2 показывают, что антитела человека специфически связываются сTNFR2 эндогенно экспрессируемого на иммунных клетках человека. Кроме того, данные показывают, что это связывание можно заблокировать путем добавления поликлонального коммерчески доступного антитела против TNFR2, что указывает на очень высокую специфичность к TNFR2. То же верно и для суррогатных клонов 3F10 и 5A05 в отношении мышиных клеток, экспрессирующих мышиный TNFR2. Кроме того, связывание человеческих клонов не зависит от антител MR2-1, проявляющих другую эпитопную специфичность по сравнению с MR2-1. Т
Таблица 6. Значения ЕС50, рассчитанные при титровании антител, специфичных к TNFR2, к активированным in vitro CD4+ Т-клеткам
Пример 3 - Проверка характеристик блокирования лиганда
(См. также Фиг. 6-7 и приведенное выше описание этих фигур).
ИФА метод
96-луночные планшеты покрывали hTNFRII (Синобиологический кат. nr10414-H08H) или mTNFRII (Синобиологический кат. No. 50128 M08H) в концентрации 2,5 пмоль/лунка в буфере для покрытия ИФА (0,1 M карбонат натрия, pH 9,5) и инкубировали в течение ночи при 4°С. После промывки в промывочном буфере для ИФА (ФСБ с 0,05 % Tween20), планшеты инкубировали при медленном перемешивании в течение 1 ч при комнатной температуре с моноклональными антителами n-CoDeR® в концентрации 10 мкг/мл (однодозовый ИФА) или 33 нМ, а затем 1:2 разведения (титрование ИФА) в блокирующем буфере, содержащем 0,45 % рыбьего желатина. Впоследствии рекомбинантный hTNF-α-bio (R&D, кат. № BT210) или mTNF-α (Gibco кат. No. PMC3014) добавляли в конечной концентрации 5 нМ и 2 нМ соответственно и оставляли для инкубирования еще 15 мин. После этого планшеты были промыты. Для ИФА человека добавляли стрептавидин-HRP (Jackson кат. № 016-030-084), разведенный 1:2000 в блокирующем буфере, и снова инкубировали в течение 1 ч при комнатной температуре с последующими промываниями сначала в буфере для ИФА, а затем в трис-буфере (pH 9,8). Субстрат (Super Signal ELISA Pico от Thermo Scientific кат. No. 37069) был затем разбавлен в соответствии с инструкциями производителя, добавлены в лунки и инкубированы в темноте в течение 10 минут перед считыванием на Tecan Ultra. Для ИФА мыши кроличьи анти-mTNF-α (Синобиологический кат. № 50349-RP02), разведенные до 1 мкг/мл, и оставляли инкубироваться в течение 1 часа при комнатной температуре. После промывания добавляли анти-кроличий-HRP, разведенный 1:10 000 в блокирующем буфере, и снова инкубировали в течение 1 ч при комнатной температуре. Добавление субстрата и считывание выполняли, как указано выше.
Данные для анти-человеческих антител и анти-мышиных антител представлены в Таблицах 7 и 8, соответственно, ниже и на Фиг. 6 и 7.
Таблица 7. Значения ЕС50 человеческих антител, блокирующих лиганд: антитела титровали и рассчитывали значения ЕС50
Таблица 8. Значения ЕС50 мышиных антител, блокирующих лиганд: антитела титровали и рассчитывали значения ЕС50
Определения блокаторов
- Полные блокаторы определяются как снижающие связывание TNF-α с более чем 98%
- Частичные блокаторы определяются как снижающие связывание TNF- α с 60-98%
- Слабые блокаторы определяются как снижающие связывание TNF- α менее чем 60%
- Неблокирующие антитела определяются как не достигающие блокирования более чем 50 % в высокодозном одноточечном ИФА, как показано на Фиг. 6A и 7A.
Данные, представленные в этом примере, показывают, что были получены различные антитела, начиная с антител, которые полностью ингибируют лиганд TNF-α от связывания с антителами, которые вообще не ингибируют блокирование лиганда. Это верно, как для антител человека, так и для суррогатов мыши.
Пример 4 - In vitro функциональность антител
(См. также Фиг. 8-10 и приведенное выше описание этих фигур).
Неблокирующие TNF-α антитела к TNFR2 усиливают выработку ИФН-γ, стимулированную цитокинами, НК-клетками, в отличие от блокирующих TNF-α антител TNFR2
Агонистические/антагонистические характеристики антител, специфичных к TNFR2, оценивали с применением анализа НК-клеток, описанного Almishri et al. (TNFα Augments Cytokine-Induced NK Cell IFNγ Production through TNFR2. Almishri W.et al. J Innate Immun. 2016;8:617-629).
Вкратце, НК-клетки человека выделяли из МКПК человека с помощью MACS с применением «набора для выделения НК» (Miltenyi). 100 мкл НК-клеток (1 × 106 клеток/мл) культивировали с 20 нг/мл rhИЛ-2 (R&D systems) и 20 нг/мл rhIL-12 (R&D systems) вместе с 10 мкг/мл TNFR2-специфичных антител, 10 мкг/мл изотипического контроля или 100 нг/мл TNF-α (R&D systems) в планшетах с U-образным дном (96-луночные микропланшеты Corning®, обработанные TC, Sigma-Aldrich). Супернатанты собирали через 24 часа, и количество продуцируемого ИФН-γ оценивали с помощью MSD. В качестве контроля анти-TNF-α антитела, нейтрализующие TNF-α (кат. № AF-210-NA, R&D systems). Как видно на фиг. 8D, доза 1 мкг/мл полностью нейтрализовала растворимый TNF-α и эта доза также снизила высвобождение ИФН-γ.
Неблокирующие TNFR2-антитела человека заметно усиливали выработку ИФН-γ стимулированными ИЛ-2 и ИЛ-12 НК-клетками (в 2-3 раза больше ИФН-γ, чем изотипический контроль), в то время как антагонистические антитела (здесь показаны с помощью полных блокаторов) проявляли антагонистические эффекты на выработку ИФН-γ НК-клетками (Фиг. 8А).
Этот тест считался нерепрезентативным для проведения с суррогатными антителами мыши из-за отсутствия эндогенно вырабатываемого TNF-α в мышиных культурах и экспрессии ингибирующего FcγR на мышиных НК-клетках и только активации FcγR на человеческом аналоге. Вместо этого для изучения агонистических или антагонистических свойств суррогатных антител мышей применяли анализ активации Т-клеток памяти (индукция CD25), а также анализ подавления супрессивных клеток миелоидного происхождения (MDSC) (оба описаны ниже).
Неблокирующие лиганд, но не блокирующие антитела TNFR2 индуцируют экспрессию CD25 на CD4+ Т-клетках памяти
Для дальнейшего понимания агонистических/антагонистических характеристик антител TNFR2 была оценена их способность увеличивать долю CD25, экспрессирующих Т-клетки памяти CD4+.
Вкратце, CD4+ Т-клетки человека выделяли из МКПК с помощью MACS с использованием «набора для выделения CD4+ Т-клеток» от Miltenyi. CD4+ Т-клетки культивировали с 10 нг/мл rhИЛ-2 (R&D systems) и 10 мкг/мл специфических антител к TNFR2 или с указанным количеством rhTNF-α (R&D systems). Через 3 дня экспрессию CD25 на клетках памяти (CD45RO+ клетки) анализировали с помощью FACS (Фиг. 9A).
Аналогичным образом CD4+ Т-клетки мыши были выделены из селезенки с помощью MACS с применением «набора для выделения CD4+ Т-клеток» (Miltenyi) и культивированы с 10 нг/мл rmИЛ-2 (R&D systems) и 10 мкг/мл антитела, специфичного к TNFR2, или по указанному количеству rmTNF-α (Системы НИОКР). Экспрессию CD25 на клетках памяти (CD44+ CD62L- клетки) анализировали с помощью FACS через 3 дня (Фиг. 9B).
Процент клеток, экспрессирующих CD25, увеличивался в культурах клеток памяти, стимулированных неблокирующим TNFR2, что снова демонстрирует агонистическую активность как у человека, так и у мыши. Однако стимуляция блокирующими антителами не увеличивала экспрессию CD25 в этих культурах.
Неблокирующее лиганд, но не блокирующее анти-TNFR2 мАт, подавляющая функция обратных миелоидных супрессивных клеток (MDSC) в анализах совместного культивирования Т-клеток/MDSC
Влияние анти-TNFR2 мАт человека на супрессивную функцию MDSC оценивали в анализах совместного культивирования Т-клеток/MDSC. Вкратце, MDSC были получены путем культивирования MACS-выделенных CD14+ моноцитов человека в 50 % асцитной жидкости, выделенной от больных раком, и 50 % R-10 в течение 3 дней. Затем MDSC промывали и предварительно инкубировали с 10 мкг/мл анти-TNFR2 мАт в течение 30 мин. Без промывания клеток титруемое количество MDSC культивировали совместно с MACS-выделенными, CFSE-меченными CD3+ Т-клетками в присутствии CD3/CD28 Dynabeads®. Через 72 часа процент активированных CD25+ Т-клеток оценивали с помощью FACS. Секрецию ИФН-γ и ИЛ-10 измеряли с помощью MSD в соответствии с инструкциями производителя.
В отличие от блокирующих анти-TNFR2 мАт (темно-серые столбцы), неблокирующие анти-TNFR2 мАт (светло-серые столбцы), как было показано, увеличивали процент активированных CD25+ в совместной культуре (Фиг. 10A). Путем измерения количества секретируемого гамма-интерферона (ИФН-γ) и интерлейкина-10 в супернатанте соотношение двух цитокинов использовали для оценки баланса Th1/Th2. Как показано на Фиг. 10B, отношение ИФН-γ к ИЛ-10 было значительно повышено неблокирующими анти-TNFR2 антителами по сравнению с блокирующими антагонистическими антителами, что указывает на то, что неблокирующие антитела вызывают сдвиг в сторону пути Th1.
Что касается человеческих антител, действие мышиных анти-TNFR2 антител было проверено в аналогичном анализе подавления. Здесь миелоидные клетки CD11b+ непосредственно MACS-выделяли из опухолей мыши CT26 и предварительно инкубировали в течение 30 мин с анти-TNFR2 мАт. CD3+-респондерные Т-клетки были очищены из наивных селезенок Balb/C. Миелоидные супрессорные клетки и CFSE-меченные Т-клетки, совместно культивировали в различных соотношениях в течение 3 дней. Процент пролиферирующих CFSEнизкий T-клеток определяли с помощью FACS. Подобно результатам анализов на людях, процент пролиферирующих респондерных клеток значительно ниже после инкубации с блокирующими, чем с неблокирующими антителами (Фиг. 10C).
В супрессорных культурах MDCS миелоидные клетки экспрессируют высокие уровни различных FcγR. Чтобы проверить, являются ли наблюдаемые агонистические и антагонистические эффекты зависимыми от Fc, неблокирующее лиганд агонистическое антитело 1F02 было протестировано в нескольких форматах, hIgG1, хорошее связывание с активирующими FcγR, hIgG2, хорошее связывание также с ингибирующим дефектный формат FcγRIIB и Fc с сильно уменьшенным связыванием со всеми FcγR. Результаты на Фиг. 10D показывают, что агонистическая активность неблокирующего антитела 1-F02 не зависит от изотипа антитела и связывания FcγR.
Таким образом, данные в примере 4 показывают, что неблокирующие лиганд антитела являются агонистическими, как измерено несколькими способами in vitro: опосредованное НК-клетками высвобождение ИФН-γ, активация клеток памяти CD4+, измеренная по экспрессии CD25 и пролиферации Т-клеток и CD25 экспрессия в анализе совместного культивирования MDCS. Кроме того, данные показывают, что это внутренняя характеристика, независимая от присутствия лиганда, а также от изотипа антитела. Мы также показываем, что суррогатные мышиные антитела 5A05 обладают сходными характеристиками агонизма. Антитела, стимулирующие Т-клетки или НК-клетки, могут быть применены при лечении рака и могут вызывать эндогенный иммунный ответ, в конечном итоге разрушая злокачественные клетки.
В соответствии с изобретением показаны неблокирующие лиганд агонистические антитела, тогда как блокирующие лиганд антагонистический антитела включены для сравнения.
Пример 5 - Суррогатное неблокирующее лиганд агонистическое анти-мышиное TNFR2 мАт имеет противоопухолевый эффект in vivo
(См. также Фиг. 11-17 и приведенное выше описание этих фигур).
Терапевтический эффект на разных моделях опухолей
Чтобы оценить in vivo противоопухолевый эффект неблокирующих лиганд агонистических моноклональных анти-TNFR2 антител, суррогат мыши, названный 5-A05, был исследован in vivo на различных моделях опухолей с применением различных форматов изотипов и отдельно или в комбинации с анти-PD-1, как описано ниже.
Мышей разводили и содержали на местных объектах в соответствии с рекомендациями домашнего офиса. Шести-восьминедельные самки BalbC и C57/BL6 мышей были предоставлены Taconic (Bomholt, Дания) и содержались в местных помещениях для животных. Клетки CT26, MC38 и B16 клетки (ATCC) выращивали в забуференной глутамаксом среде RPMI с добавлением 10 % FCS. Когда клетки были полуконфлюэнтными, их отделяли трипсином и ресуспендировали в стерильном ФСБ при 10 × 106 клеток/мл. Мышам п/к инъецировали 100 мкл клеточной суспензии, что соответствует 1 × 106 клеток/мышь. Через 3-8 дней после инъекции, в зависимости от модели, мышей дважды в неделю обрабатывали 10 мг/кг антитела внутрибрюшинно. (изотипический контроль, 3-F10 или 5-A05) и как показано на фигурах. Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли.
Неблокирующие лиганд агонистические анти-мышиные TNFR2 мАт 5-A05 проявляют терапевтический противоопухолевый эффект на трех различных моделях опухолей (Фиг. 11-14) с лечебным эффектом в более чувствительном к лечению CT26 (Фиг. 11) и эффектом ингибирования роста опухоли у более устойчивых к лечению MC38 и B16 (Фиг. 12-14).
Противоопухолевый эффект неблокирующего лиганд агонистического анти-мышиного TNFR2 мАт не требует связывания FcγR, но усиливается за счет такого связывания
Чтобы оценить важность взаимодействия Fc-FcγR на противоопухолевый эффект in vivo неблокирующего лиганд агонистического анти-TNFR2 мышиного суррогатного мАт 5A05, различные форматы Fc этого антитела исследовали in vivo на модели опухоли CT26, как описано ниже.
Мышей разводили и содержали, как описано выше. Клетки CT26 (ATCC) выращивали и инъецировали, как описано выше. Когда опухоли достигли 3х3 мм, мышам дважды в неделю вводили 10 мг/кг антитела внутрибрюшинно. (изотипический контроль, 5A05 IgG1, 5A05 IgG2a или 5-A05-N297A (дефектный Fc). Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли.
Fc-дефектное 5-A05-N297A демонстрирует четкую терапевтическую активность по сравнению с изотипическим контролем, что указывает на то, что вовлечение Fc не является полностью обязательным для терапевтической эффективности этого неблокирующего лиганд агонистического анти-мышиного TNFR2 мАт (Фиг. 11 A и B). Кроме того, оба формата IgG1 и IgG2a демонстрируют повышенную терапевтическую эффективность. Однако формат IgG1, предпочтительно связывающийся с ингибирующими Fcγ-рецепторами, демонстрирует превосходный терапевтический эффект, указывающий на агонизм, являющийся одним из важных механизмов действия этого анти-мышиного TNFR2 мАт (Фиг. 11 A-B). Это контрастирует с блокирующим лиганд антагонистическим суррогатным антителом 3-F10, которое не проявляет активности в дефектном формате Fc и проявляет лучшую активность в формате мышиного IgG2a, о котором известно, что оно предпочтительно связывает активирующий FcγR.
Неблокирующее лиганд агонистическое антитело (5-A05) в соответствии с данным изобретением, и блокирующее лиганд антагонистическое антитело (3-F10), включено в качестве эталона.
Комбинированный эффект с анти-PD-1 мАт
Чтобы оценить комбинированный in vivo противоопухолевый эффект неблокирующих лиганд агонистических моноклональных анти-TNFR2 суррогатных мышиных мАт (5-A05) с анти-PD-1, комбинацию лечения исследовали in vivo на модели опухоли MC38, как описано ниже.
Мышей разводили и содержали, как описано выше. Клетки MC38 (ATCC) выращивали и инъецировали, как описано выше. Через восемь дней после инъекции, мышам дважды в неделю вводили 10 мг/кг антитела внутрибрюшинно. (изотипический контроль, анти-мышиный PD-1, 5-A05 или комбинация анти-мышиного PD-1 и 5-A05) и как показано на Фиг. 12 A-E. Опухоли измеряли два раза в неделю, пока они не достигли диаметра 15 мм, после чего мышей умерщвляли.
Оба анти-мышиное PD-1 и неблокирующее лиганд агонистическое анти-мышиное TNFR2 мАт 5-A05 демонстрируют терапевтический эффект ингибирования роста опухоли в модели MC38 (Фиг. 12 A-E). Когда анти-PD1 и 5-A05 комбинируются, опухоли излечиваются в устойчивом к лечению MC38 (Фиг. 12 D-E).
Комбинированный эффект с анти-PD-L1 мАт
Чтобы оценить комбинированный противоопухолевый эффект in vivo агонистических анти-TNFR2 мАт, мы дополнительно объединили мышиный суррогатную (5A05) с анти-PD-L1 для лечения в модели опухоли MC38, как описано ниже.
Мышей разводили и содержали, как описано выше. Клетки MC38 (полученные от доктора М. Крэгга, Саутгемптонский университет) выращивали и вводили, как описано выше. Через шесть дней после инъекции, мышей дважды обрабатывали изотипическим контролем антитела или 3F10 (день 1 и 4), или четыре дня подряд анти-PD-L1 (клон 10F.9G2, Bioxcell) с последующей пятой инъекцией через два дня (всего пять инъекций на 1, 2, 3, 4 и 7 день) или их комбинацией. Все антитела вводили в дозе 10 мг/кг внутрибрюшинно. Опухоли измеряли штангенциркулем дважды в неделю до тех пор, пока они не достигли объема 2000 мм3, после чего мышей умерщвляли.
Оба анти-мышиное PD-L1 и агонистическое анти-мышиное TNFR2 мАт 5A05 демонстрируют терапевтический эффект ингибирования роста опухоли в модели MC38 (Фиг. 13). Когда объединяют анти-PD-L1 и антагонистическое анти-мышиной TNFR2 мАт 5A05, противоопухолевый эффект еще больше усиливается (Фиг. 13).
Модуляция иммунных клеток in vivo
Чтобы исследовать влияние иммунных клеток на опухоль in vivo, мышей BalbC инокулировали клетками CT26, как описано выше. После того, как опухоли достигли примерно 7x7 мм, мышей обрабатывали антителами в дозе 10 мг/кг, вводимыми внутрибрюшинно, как показано на Фиг. 15. Мышей обрабатывали на 1, 4 и 7 день и прекращали на 8 день. Опухоли иссекали, механически разделяли на мелкие кусочки и расщепляли с использованием смеси коллагеназы 100 мкг/мл либеразы и 100 мкг/мл ДНКазы при 37 °C в течение 2х5 минут с Vortex между ними. После фильтрации через фильтр 70 мкм, клеточную суспензию промывали (400 g в течение 10 мин) ФСБ, содержащим 10 % FBS. После этого клетки ресуспендировали в буфере MACS и окрашивали панелью антител, окрашивающей CD45, CD3, CD8, CD4 и CD25 или панель окрашивания антител MHCII, F4/80, Ly6C, CD11b и Ly6G. Перед окрашиванием клетки блокировали для неспецифического связывания с применением 100г/мл IVIG (очищенные иммуноглобулины для внутривенного введения). Клетки анализировали в FACS Verse. Мышиные Treg были количественно определены как CD45+CD3+CD4+CD25+, а TAM как CD11b+Ly6G-Ly6C-F4/80+MHCII+.
Как видно на Фиг. 15, лечение агонистическим антителом TNFR2 приводит к увеличению притока CD8+ Т-клеток в опухоль. Также наблюдается более слабая тенденция к истощению Treg. Вместе это приводит к значительному улучшению соотношения CD8+ T-клеток к Treg (Фиг. 15 C). Кроме того, агонистическое антитело модулирует миелоидный компартмент, уменьшая количество связанных с опухолью макрофагов.
Модель МКПК-NOG/SCID
Чтобы подтвердить полученные in vitro данные об агонистической/Т-клеточной пролиферативной активности неблокирующего лиганд агонистического анти-TNFR2 мАт 1-F02, мы проанализировали способность этого мАт индуцировать пролиферацию Т-клеток на модели МКПК-NOG в vivo, как описано ниже.
Мышей разводили и содержали на местных объектах в соответствии с рекомендациями домашнего офиса. Восьминедельных самок мышей NOG поставляла компания Taconic (Bomholt, Дания) и содержала в местных помещениях для животных. Для модели МКПК-NOG (первичный ксенотрансплантат человека) МКПК человека выделяли с применением Ficoll Paque PLUS и после промывки клетки ресуспендировали в стерильном ФСБ при 75 × 106 клеток/мл. Мышам NOG в/в вводили 200 мкл клеточной суспензии, что соответствует 15 × 106 клеток/мышь. Через две недели после инъекции мышей дважды (с интервалом в два дня) обрабатывали 10 мг/кг любого изотипического контроля, Ервой, анти-CD25, Campath, 1-F02 или 1-H10 (блокирующее лиганд антагонистическое анти-TNFR2 мАт). Селезенки мышей собирали через 2 дня после последней инъекции. Подмножества Т-клеток человека были идентифицированы и количественно определены с помощью FACS с применением следующих маркеров: CD45, CD3, CD4, CD8, CD25, CD127 (все от BD Biosciences). В отдельном эксперименте селезенки из необработанных МКПК человека были умерщвлены для определения экспрессии TNFR2 на Т-клетках человека с помощью FACS (Фиг. 16). Эти данные FACS показали, что экспрессия TNFR2 на Treg и CD8+ Т-клетках очень сопоставимы между Т-клетками человека, выращенными и активированными in vivo у мышей NOG, и Т-клетками из опухолей человека.
1-F02 индуцировал пролиферацию Т-клеток в соответствии с тем, что наблюдали in vitro и in vivo с неблокирующим лиганд агонистическим антимышиным суррогатным TNFR2 мАт, в то время как антагонистическое 1-H10 не индуцировало пролиферацию (Фиг. 17).
Таким образом, пример 5 показывает, что:
1. Неблокирующие лиганд агонистические антитела могут оказывать сильное противоопухолевое действие на нескольких моделях опухолей.
2. Этот эффект можно усилить за счет комбинации с анти-PD1 антителами.
3. Неблокирующие лиганд агонистические антитела не проявляют облигатной зависимости FcγR от противоопухолевого эффекта, но эффект усиливается за счет вовлечения, в первую очередь, ингибирующих, FcγR.
4. Неблокирующие лиганд агонистические антитела увеличивают приток CD8-положительных Т-клеток и уменьшают количество Treg в опухолях. Кроме того, они уменьшают количество ТАМ в опухолях, тем самым изменяя состав как Т-клеток, так и миелоидных клеток в опухоли.
5. В опухолях человека Т-клетки экспрессируют TNFR2.
6. В модели ксенотрансплантата человека, где экспрессия TNFR2 опухоли имитируется на Т-клетках, неблокирующее лиганд агонистическое антитело 1-F02 увеличивает количество Т-клеток.
Пример 6 - Неблокирующие лиганд агонистические антитела не индуцируют большие количества провоспалительных цитокинов
(См. также Фиг. 18-19 и приведенное выше описание этих фигур).
Высвобождение большого количества провоспалительных цитокинов является одним из возможных побочных эффектов иммуномодулирующих антител, применяемых для лечения пациентов. Следовательно, в данном документе мы измеряли высвобождение цитокинов, индуцированное неблокирующими лиганд агонистическими антителами, применяя два разных способа. Первый основан на стимуляции антителами в культурах in vitro, а второй основан на ксенотрансплантации иммунных клеток человека иммунодефицитным мышам. Было показано, что для in vitro создание культуры в значительной степени влияет на высвобождение цитокинов (Vessillier et al., J Immunol Methods. 2015 Sep; 424: 43–52). Чтобы учесть различия в методологиях, в соответствии с последними публикациями были применены три различных установки культивирования in vitro.
Для анализа высвобождения цитокинов (CRA - Cytokine Release Assays) на культуре клеток высокой плотности (HDC - High Density Cell Culture) МКПК культивировали с концентрацией 1 × 107 клеток/мл в бессывороточной среде CTL-Test (Cell Technology Limited) с добавлением 2 мМ глутамина, 1 мМ пирувата, 100 МЕ/мл пенициллина и стрептомицина. 2 мл клеточной культуры помещали в 12-луночный планшет. Через 48 часов 10 мкг/мл антитела добавляли к 1 × 105 предварительно инкубированных МКПК в 96-луночном планшете с плоским дном и инкубировали в течение 24 часов.
CRA МКПК твердой фазы (SP - Solid Phase) выполняли путем покрытия лунок 96-луночного планшета 1 мкг/мл антитела в течение 1 часа. После промывания планшета ФСБ добавляли 1 × 105 МКПК в 200 мкл полной среды на лунку и инкубировали в течение 48 часов.
Высвобождение цитокинов также измеряли после стимуляции 200 мкл цельной крови 5 мкг/мл антитела в течение 48 часов.
В конце периода инкубации планшеты центрифугировали, супернатант культуры отбирали и хранили при -20°C. Концентрации ИФН-γ, ИЛ-2, ИЛ-4, ИЛ-6, ИЛ-10, ИЛ-8 и TNF-α были измерены с применением изготовленных на заказ планшетов MSD в соответствии с инструкциями производителя (Meso Scale Discovery, США).
Таким образом, неблокирующие лиганд агонистические антитела индуцировали любое значительное высвобождение цитокинов, помимо ИФН-γ, только в любых условиях in vitro (данные не показаны). Антитела положительного контроля, алемтузумаб и OKT3 действительно индуцировали цитокины, наиболее выраженным из всех был ИФН-γ. Как видно на Фиг. 18, неблокирующие агонистические антитела не индуцировали ИФН-γ в двух из трех условий in vitro; кроме того, антитело 1-F02 не индуцировало ИФН-γ ни в одном из условий in vitro.
Модель переносимости МКПК-NOG
Чтобы исследовать переносимость неблокирующего лиганд агонистического мАт 1-F02 против TNFR2 человека, мы проанализировали высвобождение цитокинов in vivo в модели МКПК-NOG, как описано ниже.
Мышей разводили и содержали на местных объектах в соответствии с рекомендациями домашнего офиса. Восьминедельных самок мышей NOG поставляла компания Taconic (Bomholt, Дания) и содержала в местных помещениях для животных. Для модели МКПК-NOG (первичный ксенотрансплантат человека) МКПК человека выделяли с применением Ficoll Paque PLUS и после промывки клетки ресуспендировали в стерильном ФСБ при 125 × 106 клеток/мл. Мышам NOG в/в вводили 200 мкл клеточной суспензии, что соответствует 25 × 106 клеток/мышь. Через 2 недели после инъекции были взяты образцы крови для анализа уровня «гуманизации», означающего количество человеческих клеток в крови мышей NOG. Когда кровь состояла из прим. 40 % Т-клеток человека, мышей считали гуманизированными. Затем мышей обрабатывали 10 мкг либо Ервой, анти-CD3 (OKT-3), 1-F02, либо изотипического контрольного мАт. Температуру тела измеряли до инъекции антитела и через 1 час после инъекции (Фиг. 19 A). Как видно на Фиг. 19A, положительное контрольное антитело OKT3 вызывало резкое снижение температуры тела, как было опубликовано ранее, и в соответствии с токсичностью, наблюдаемой в клинике с этим антителом. Напротив, 1-F02 не оказало никакого влияния на температуру тела. Через пять часов после инъекции антител эксперименты были прекращены и кровь была собрана для анализа высвобождения цитокинов (MSD). Измеряемые цитокины представляли собой ИФН-γ, TNF-α, ИЛ-6 и ИЛ1β человека. Из них ИФН-γ и TNF-α были количественно определены на достаточно высоком уровне, чтобы быть надежными. Как видно на Фиг. 19 B и C, положительное контрольное антитело OKT3 индуцировало как значимые ИФН-γ, так и TNF-α высвобождение (в соответствии с токсичностью, наблюдаемой в клинике с этим антителом), тогда как у мышей, обработанных 1-F02, не было значительного высвобождения ИФН-γ. Однако наблюдалась тенденция к увеличению высвобождения TNF-α, хоть и не значительного и не такого драматичного, как для ОКТ3.
Таким образом, пример 6 показывает, что неблокирующие лиганд агонистические антитела TNFR2, в данном случае проиллюстрированные антителом, называемым 1-F02, не индуцируют существенных уровней высвобождения цитокинов, как измерено несколькими ранее опубликованными способами. Поскольку высвобождение цитокинов является ограничивающим фактором для некоторых иммуномодулирующих антител, это указывает на приемлемый профиль безопасности в этом отношении.
Пример 7 - Эпитопы генерированных нацеленных на TNFR2 антител
Нокауты в конструкции домена
В первой серии экспериментов использовали конструкции ДНК, кодирующие различные варианты TNFR2, в которых отсутствует один или более из 4 внеклеточных доменов, описанных в таблице 9. Во второй серии экспериментов использовали конструкции ДНК, кодирующие варианты TNFR2, в которых различные части домена 3 были заменены на соответствующую мышиную часть, как описано в таблице 10. Последнее возможно, поскольку ни одно из антител не обладает перекрестной реактивностью с мышиным TNFR. В обоих случаях конструкции были приобретены у GeneArt (ThermoFisher). Конструкции клонировали в вектор экспрессии, содержащий CMV-промотор и ориджин репликации плазмиды OriP, и временно экспрессировали в адаптированных к суспензии клетках HEK293-EBNA.
Таблица 9. Конструкции TNFR2, используемые для трансфекции, где один или несколько доменов были удалены
Таблица 10. Конструкции TNFR2, используемые для трансфекции, заменяли различные части домена 3 на соответствующие мышиные последовательности
Анализ связывания на основе проточной цитометрии
Клетки HEK-293-E трансфицировали соответствующими плазмидами кДНК вариантов TNFR2 с применением липофектамина 2000. Через 48 часов после трансфекции клетки собирали и окрашивали указанными антителами в течение 30 минут. После 2 стадий промывки ФСБ поверхностно-связанные антитела окрашивали вторичным анти-IgG, связанным с АПК. Перед анализом проточной цитометрии на проточном цитометре BD-Verse клетки промывали и окрашивали на наличие живых/мертвых клеток.
Эксперименты по связыванию трансфицированных клеток HEK 293 на основе проточной цитометрии четко показали, что домен 1 и домен 2 либо не влияют на связывание (домен 1), либо лишь незначительно влияют на связывание (домен 2) любого из антител к этим клеткам. В качестве положительного контроля использовали поликлональное антитело против TNFR2 человека. Антитело положительного контроля показало высокое связывание со всеми тестируемыми конструкциями, тогда как отрицательное антитело не показало связывания (Фиг. 20). Все протестированные антитела показали полную потерю связывания с TNFR2, лишенным домена 3. Точно так же большинство антител не могло связываться с TNFR2, если домен 4 отсутствовал. Все антагонистические антитела (1H10, 4H02 и 5B08) показали резко сниженное связывание с TNFR2 Δ4 более чем на 50 % по сравнению со связыванием с TNFR2 Δ1 и TNFR2 Δ2. Аналогичным образом, удаление двух доменов из TNFR2 ясно показало, что отсутствие домена 3 или 4 значительно отменяет связывание всех тестируемых антител с TNFR2, за возможным исключением агонистического антитела 1F06, в то время как отсутствие домена 4 отменяет связывание агонистических антител и снижает связывание антагонистических антител значительно. (Фиг. 20 E и F).
Связывание с химерным мышь-человек TNFR2
Для дальнейшего сужения сайта связывания и определения эпитопов части домена 3 TNFR2 человека были заменены соответствующей мышиной последовательностью. Поскольку все антитела проявляют очень небольшую перекрестную реактивность с TNFR2 мыши, потеря связывания с определенными конструкциями позволит улучшить связывающий эпитоп. На Фиг. 21 показаны различные химерные конструкции TNFR2 мышь-человек. Было произведено четыре различных замены, заменяющих 14 (m1), 12 (m2), 10 (m3) или 16 (m4) аминокислот из последовательности человека на соответствующую последовательность мыши. Остальные три домена (1, 2, 4) содержат исключительно человеческие последовательности.
Эти конструкции (домены 1-4 TNFR2 с мутациями в 3) затем трансфицировали в клетки HEK293, и антитела тестировали на связывание с применением подхода проточной цитометрии. В качестве положительного контроля использовали поликлональные антитела против TNFR2 мыши, а также против TNFR2 человека. Как и ожидалось, из-за сходства последовательностей оба поликлональных контрольных антитела показали значительную перекрестную реактивность и распознали TNFR2 как человека, так и мыши. Очевидно, что наилучшие сигналы были получены при сопоставлении антител с намеченной мишенью.
Наши моноклональные антитела показали сильное связывание с TNFR2 человека, но не связывались с TNFR2 мыши или имели очень слабое связывание (Фиг. 22, левая панель). Подобное связывание с очень небольшим восстановлением наблюдали для всех клонов конструкции hTNFR2 m1 с мутациями в аминокислотах 119-132, что указывает на то, что ни одно из антител не связывается с эпитопом в этом участке. Однако мутации в аминокислотах 134-144 (конструкция hTNFR2 m2) полностью аннулировали связывание для половины протестированных антител, соответствующих антагонистическим блокирующим антителам 1-H10, 4-H02 и 5-B08, что указывает на то, что антитела связываются, по меньшей мере, частично в пределах этого участка. 1-G10 представляет собой частичный блокиратор, также сильно пострадавший от этой замены. Примечательно, что агонистические антитела (1-F02, 1-F06 и 4-E08) сохраняли связывание с применением конструкции 2, что убедительно свидетельствует о другом эпитопе по сравнению с антагонистическими антителами. Интересно, что все антитела потеряли связывание с конструкцией hTNFR2 m3 с мутациями в аминокислотах 151–160. Это указывает на то, что все антитела, как агонисты, так и антагонисты, имеют частичный эпитоп в этой последовательности. Тестирование конструкции hTNFR2 m4 немного большего размера с мутациями в аминокислотах 130-144 показало такое же связывание, как и в случае конструкции hTNFR2 m2.
Выводы о связывании эпитопов
Группируя антитела по их функциональной роли, агонистические антитела (1-F02, 1-F06 и 4-E08), по-видимому, связывают очень дистальную С-концевую часть домена 3, охватывающую аминокислоты 151-160 домена 4, тогда как эпитоп для антагонистов (1-H10, 5-B08 и 4-H02) смещен больше к центру домена 3, охватывая аминокислоты 134-160 и, вероятно, покрывая меньшую часть домена 4. Однако, несмотря на это, их эпитопы, по-видимому, до некоторой степени перекрываются.
Ни одно из антител не связывается с N-концевой частью домена 3, аминокислоты 119-134. Сайты связывания с доменом 4 вполне вероятны для всех антител, но полностью не идентифицированы.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> БИОИНВЕНТ ИНТЕРНЭШНЛ АБ
<120> НОВЫЕ МОЛЕКУЛЫ АГОНИСТИЧЕСКИХ АНТИ-TNFR2 АНТИТЕЛ
<130> BI85 PCT
<150> EP 18203996.6
<151> 2018-11-01
<160> 223
<170> PatentIn version 3.5
<210> 1
<211> 14
<212> PRT
<213> Homo sapiens
<400> 1
Phe Ser Asp Tyr Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 2
<211> 19
<212> PRT
<213> Homo sapiens
<400> 2
Ala Asn Ile Asn Thr Asp Gly Ser Glu Lys Tyr Tyr Leu Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 3
<211> 10
<212> PRT
<213> Homo sapiens
<400> 3
Ala Arg Glu Glu Tyr Gly Ala Phe Asp Ile
1 5 10
<210> 4
<211> 14
<212> PRT
<213> Homo sapiens
<400> 4
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 5
<211> 7
<212> PRT
<213> Homo sapiens
<400> 5
Asp Asn Asn Lys Arg Pro Ser
1 5
<210> 6
<211> 13
<212> PRT
<213> Homo sapiens
<400> 6
Cys Gln Ser Phe Asp Arg Gly Leu Ser Gly Ser Ile Val
1 5 10
<210> 7
<211> 117
<212> PRT
<213> Homo sapiens
<400> 7
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Asn Thr Asp Gly Ser Glu Lys Tyr Tyr Leu Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Glu Tyr Gly Ala Phe Asp Ile Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 8
<211> 112
<212> PRT
<213> Homo sapiens
<400> 8
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Arg Gly Leu
85 90 95
Ser Gly Ser Ile Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 9
<211> 14
<212> PRT
<213> Homo sapiens
<400> 9
Phe Ser Ser Tyr Ala Met His Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 10
<211> 19
<212> PRT
<213> Homo sapiens
<400> 10
Ser Ala Ile Ser Gly Gly Ala Thr Thr Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 11
<211> 13
<212> PRT
<213> Homo sapiens
<400> 11
Ala Lys Gly Gly Thr Gly Asp Pro Tyr Tyr Phe Asp Tyr
1 5 10
<210> 12
<211> 15
<212> PRT
<213> Homo sapiens
<400> 12
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 13
<211> 7
<212> PRT
<213> Homo sapiens
<400> 13
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 14
<211> 12
<212> PRT
<213> Homo sapiens
<400> 14
Cys Ala Ala Arg Asp Asp Gly Leu Ser Gly Pro Val
1 5 10
<210> 15
<211> 120
<212> PRT
<213> Homo sapiens
<400> 15
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Gly Ala Thr Thr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Gly Thr Gly Asp Pro Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 16
<211> 112
<212> PRT
<213> Homo sapiens
<400> 16
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Arg Asp Asp Gly
85 90 95
Leu Ser Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 17
<211> 14
<212> PRT
<213> Homo sapiens
<400> 17
Phe Ser Asn Ala Trp Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 18
<211> 19
<212> PRT
<213> Homo sapiens
<400> 18
Ser Ser Ile Ser Ser Ala Ser Gly Tyr Ile Tyr Tyr Gly Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 19
<211> 12
<212> PRT
<213> Homo sapiens
<400> 19
Ala Arg Gly Thr Leu Tyr Gly Asp Phe Asp Glu Phe
1 5 10
<210> 20
<211> 14
<212> PRT
<213> Homo sapiens
<400> 20
Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn Ala Val Asn
1 5 10
<210> 21
<211> 7
<212> PRT
<213> Homo sapiens
<400> 21
Gly Asn Thr Asn Arg Pro Ser
1 5
<210> 22
<211> 13
<212> PRT
<213> Homo sapiens
<400> 22
Cys Gln Ser Tyr Asp Ser Ser Leu Ser Gly Tyr Val Val
1 5 10
<210> 23
<211> 119
<212> PRT
<213> Homo sapiens
<400> 23
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ala Ser Gly Tyr Ile Tyr Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Asn Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Thr Leu Tyr Gly Asp Phe Asp Glu Phe Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 24
<211> 112
<212> PRT
<213> Homo sapiens
<400> 24
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Thr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser Leu
85 90 95
Ser Gly Tyr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 25
<211> 14
<212> PRT
<213> Homo sapiens
<400> 25
Phe Ser Ser Asn Glu Met Ser Trp Ile Arg Gln Ala Pro Gly
1 5 10
<210> 26
<211> 18
<212> PRT
<213> Homo sapiens
<400> 26
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly Arg
<210> 27
<211> 12
<212> PRT
<213> Homo sapiens
<400> 27
Ala Arg Arg Glu Gly Trp Leu Val Pro Phe Asp Tyr
1 5 10
<210> 28
<211> 14
<212> PRT
<213> Homo sapiens
<400> 28
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 29
<211> 7
<212> PRT
<213> Homo sapiens
<400> 29
Gly Asn Ile Ile Arg Pro Ser
1 5
<210> 30
<211> 13
<212> PRT
<213> Homo sapiens
<400> 30
Cys Gln Ser Phe Asp Thr Thr Leu Ser Gly Ser Ile Val
1 5 10
<210> 31
<211> 118
<212> PRT
<213> Homo sapiens
<400> 31
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Glu Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Arg Glu Gly Trp Leu Val Pro Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 32
<211> 112
<212> PRT
<213> Homo sapiens
<400> 32
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Ile Ile Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Thr Thr Leu
85 90 95
Ser Gly Ser Ile Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 33
<211> 14
<212> PRT
<213> Homo sapiens
<400> 33
Phe Ser Arg Tyr Trp Met His Trp Val Arg Gln Val Pro Gly
1 5 10
<210> 34
<211> 19
<212> PRT
<213> Homo sapiens
<400> 34
Ser Gly Ile Ser Asp Ser Gly Val Val Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 35
<211> 10
<212> PRT
<213> Homo sapiens
<400> 35
Ala Arg Ala Gln Ser Val Ala Phe Asp Ile
1 5 10
<210> 36
<211> 15
<212> PRT
<213> Homo sapiens
<400> 36
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ala Gly His Asp Val His
1 5 10 15
<210> 37
<211> 7
<212> PRT
<213> Homo sapiens
<400> 37
Tyr Asp Asp Leu Leu Pro Ser
1 5
<210> 38
<211> 12
<212> PRT
<213> Homo sapiens
<400> 38
Cys Ala Ala Trp Asp Asp Ser Leu Ser Gly Trp Val
1 5 10
<210> 39
<211> 117
<212> PRT
<213> Homo sapiens
<400> 39
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met His Trp Val Arg Gln Val Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Asp Ser Gly Val Val Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ala Gln Ser Val Ala Phe Asp Ile Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 40
<211> 112
<212> PRT
<213> Homo sapiens
<400> 40
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
His Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Tyr Asp Asp Leu Leu Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Ser Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 41
<211> 14
<212> PRT
<213> Homo sapiens
<400> 41
Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 42
<211> 19
<212> PRT
<213> Homo sapiens
<400> 42
Ser Val Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ala Val
1 5 10 15
Lys Gly Arg
<210> 43
<211> 10
<212> PRT
<213> Homo sapiens
<400> 43
Thr Thr Asp Ser Gly Ser Gly Ser Tyr Leu
1 5 10
<210> 44
<211> 15
<212> PRT
<213> Homo sapiens
<400> 44
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 45
<211> 7
<212> PRT
<213> Homo sapiens
<400> 45
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 46
<211> 12
<212> PRT
<213> Homo sapiens
<400> 46
Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Pro Val
1 5 10
<210> 47
<211> 117
<212> PRT
<213> Homo sapiens
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ala Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Thr Asp Ser Gly Ser Gly Ser Tyr Leu Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 48
<211> 112
<212> PRT
<213> Homo sapiens
<400> 48
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 49
<211> 14
<212> PRT
<213> Homo sapiens
<400> 49
Phe Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 50
<211> 19
<212> PRT
<213> Homo sapiens
<400> 50
Ser Val Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 51
<211> 9
<212> PRT
<213> Homo sapiens
<400> 51
Ala Arg Asp Arg Gly Trp Phe Asp Pro
1 5
<210> 52
<211> 14
<212> PRT
<213> Homo sapiens
<400> 52
Cys Ser Gly Ser Arg Ser Asn Ile Asp Asn Ser Tyr Val Ser
1 5 10
<210> 53
<211> 7
<212> PRT
<213> Homo sapiens
<400> 53
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 54
<211> 12
<212> PRT
<213> Homo sapiens
<400> 54
Cys Ala Thr Trp Asp Asp Ser Leu Ser Gly Pro Val
1 5 10
<210> 55
<211> 116
<212> PRT
<213> Homo sapiens
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Gly Trp Phe Asp Pro Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 56
<211> 111
<212> PRT
<213> Homo sapiens
<400> 56
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Arg Ser Asn Ile Asp Asn Ser
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Ser Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 57
<211> 14
<212> PRT
<213> Homo sapiens
<400> 57
Phe Ser Arg His Ala Met Asn Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 58
<211> 19
<212> PRT
<213> Homo sapiens
<400> 58
Ser Ser Ile Ser Thr Gly Ser Ser Tyr Ile Asp Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 59
<211> 13
<212> PRT
<213> Homo sapiens
<400> 59
Ala Arg Glu Lys Gly His Tyr Tyr Tyr Gly Met Asp Val
1 5 10
<210> 60
<211> 15
<212> PRT
<213> Homo sapiens
<400> 60
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 61
<211> 7
<212> PRT
<213> Homo sapiens
<400> 61
Gly Asn Ser Tyr Arg Pro Ser
1 5
<210> 62
<211> 13
<212> PRT
<213> Homo sapiens
<400> 62
Cys Gln Ser Tyr Asp Thr Ser Leu Ser Ala Tyr Val Val
1 5 10
<210> 63
<211> 120
<212> PRT
<213> Homo sapiens
<400> 63
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg His
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Thr Gly Ser Ser Tyr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Lys Gly His Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 64
<211> 113
<212> PRT
<213> Homo sapiens
<400> 64
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Tyr Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Thr Ser
85 90 95
Leu Ser Ala Tyr Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 65
<211> 14
<212> PRT
<213> Homo sapiens
<400> 65
Phe Ser Asn Ala Trp Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 66
<211> 19
<212> PRT
<213> Homo sapiens
<400> 66
Ser Ala Ile Ser Val Ser Gly Ile Asn Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 67
<211> 11
<212> PRT
<213> Homo sapiens
<400> 67
Ala Arg Asp Thr Gly Ser Leu Gly Val Asp Tyr
1 5 10
<210> 68
<211> 14
<212> PRT
<213> Homo sapiens
<400> 68
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 69
<211> 7
<212> PRT
<213> Homo sapiens
<400> 69
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 70
<211> 12
<212> PRT
<213> Homo sapiens
<400> 70
Cys Gln Ser Tyr Asp Ser Ser Leu Ser Ile Ser Val
1 5 10
<210> 71
<211> 118
<212> PRT
<213> Homo sapiens
<400> 71
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Val Ser Gly Ile Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Thr Gly Ser Leu Gly Val Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 72
<211> 111
<212> PRT
<213> Homo sapiens
<400> 72
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser Leu
85 90 95
Ser Ile Ser Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 73
<211> 14
<212> PRT
<213> Homo sapiens
<400> 73
Ser Ser Ser Tyr Trp Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 74
<211> 19
<212> PRT
<213> Homo sapiens
<400> 74
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 75
<211> 11
<212> PRT
<213> Homo sapiens
<400> 75
Ala Arg Glu Tyr Ser Gly Tyr Glu Phe Asp Phe
1 5 10
<210> 76
<211> 15
<212> PRT
<213> Homo sapiens
<400> 76
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Arg Ser Asp Val His
1 5 10 15
<210> 77
<211> 7
<212> PRT
<213> Homo sapiens
<400> 77
Gly Asn Arg Asn Arg Pro Ser
1 5
<210> 78
<211> 13
<212> PRT
<213> Homo sapiens
<400> 78
Cys Gln Ser Phe Asp Arg Gly Leu Ser Gly Ser Ile Val
1 5 10
<210> 79
<211> 118
<212> PRT
<213> Homo sapiens
<400> 79
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ser Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Ser Gly Tyr Glu Phe Asp Phe Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 80
<211> 113
<212> PRT
<213> Homo sapiens
<400> 80
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Arg
20 25 30
Ser Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Arg Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Arg Gly
85 90 95
Leu Ser Gly Ser Ile Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 81
<211> 14
<212> PRT
<213> Homo sapiens
<400> 81
Ser Ser Ser Tyr Trp Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 82
<211> 19
<212> PRT
<213> Homo sapiens
<400> 82
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 83
<211> 11
<212> PRT
<213> Homo sapiens
<400> 83
Ala Arg Glu Tyr Ser Gly Tyr Glu Phe Asp Phe
1 5 10
<210> 84
<211> 15
<212> PRT
<213> Homo sapiens
<400> 84
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Arg Ser Asp Val His
1 5 10 15
<210> 85
<211> 7
<212> PRT
<213> Homo sapiens
<400> 85
Gly Asn Arg Asn Arg Pro Ser
1 5
<210> 86
<211> 13
<212> PRT
<213> Homo sapiens
<400> 86
Cys Gln Ser Phe Asp Arg Gly Leu Ser Gly Ser Ile Val
1 5 10
<210> 87
<211> 118
<212> PRT
<213> Homo sapiens
<400> 87
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Ser Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Tyr Ser Gly Tyr Glu Phe Asp Phe Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 88
<211> 113
<212> PRT
<213> Homo sapiens
<400> 88
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Arg
20 25 30
Ser Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Arg Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Phe Asp Arg Gly
85 90 95
Leu Ser Gly Ser Ile Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
Gly
<210> 89
<211> 14
<212> PRT
<213> Homo sapiens
<400> 89
Phe Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 90
<211> 19
<212> PRT
<213> Homo sapiens
<400> 90
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 91
<211> 11
<212> PRT
<213> Homo sapiens
<400> 91
Ala Arg Asp Arg Gly Arg Thr Gly Thr Asp Tyr
1 5 10
<210> 92
<211> 14
<212> PRT
<213> Homo sapiens
<400> 92
Cys Ser Gly Thr Thr Ser Asn Ile Gly Ser Tyr Ala Val Asn
1 5 10
<210> 93
<211> 7
<212> PRT
<213> Homo sapiens
<400> 93
Gly Asn Ile Asn Arg Pro Ser
1 5
<210> 94
<211> 12
<212> PRT
<213> Homo sapiens
<400> 94
Cys Gln Ser Tyr Asp Ser Ser Leu Ser Ala Ser Leu
1 5 10
<210> 95
<211> 118
<212> PRT
<213> Homo sapiens
<400> 95
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Gly Arg Thr Gly Thr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 96
<211> 111
<212> PRT
<213> Homo sapiens
<400> 96
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Thr Thr Ser Asn Ile Gly Ser Tyr
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Ile Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser Leu
85 90 95
Ser Ala Ser Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 97
<211> 14
<212> PRT
<213> Homo sapiens
<400> 97
Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 98
<211> 19
<212> PRT
<213> Homo sapiens
<400> 98
Ser Thr Ile Ile Gly Ser Gly Ala Asn Thr Trp Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 99
<211> 13
<212> PRT
<213> Homo sapiens
<400> 99
Ala Arg His Glu Gly Tyr Tyr Tyr Tyr Gly Met Asp Val
1 5 10
<210> 100
<211> 15
<212> PRT
<213> Homo sapiens
<400> 100
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Val Val His
1 5 10 15
<210> 101
<211> 7
<212> PRT
<213> Homo sapiens
<400> 101
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 102
<211> 12
<212> PRT
<213> Homo sapiens
<400> 102
Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Arg Val
1 5 10
<210> 103
<211> 120
<212> PRT
<213> Homo sapiens
<400> 103
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ile Gly Ser Gly Ala Asn Thr Trp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Glu Gly Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 104
<211> 112
<212> PRT
<213> Homo sapiens
<400> 104
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Val Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Asn Gly Arg Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 105
<211> 14
<212> PRT
<213> Homo sapiens
<400> 105
Phe Asp Asp Tyr Gly Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 106
<211> 18
<212> PRT
<213> Homo sapiens
<400> 106
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly Arg
<210> 107
<211> 16
<212> PRT
<213> Homo sapiens
<400> 107
Cys Ala Arg Asp Arg Ser Ser Ser Trp Tyr Arg Asp Gly Met Asp Val
1 5 10 15
<210> 108
<211> 15
<212> PRT
<213> Homo sapiens
<400> 108
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 109
<211> 7
<212> PRT
<213> Homo sapiens
<400> 109
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 110
<211> 12
<212> PRT
<213> Homo sapiens
<400> 110
Cys Ala Ala Trp Asp Asp Ser Leu Ser Gly Trp Val
1 5 10
<210> 111
<211> 121
<212> PRT
<213> Homo sapiens
<400> 111
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Arg Ser Ser Ser Trp Tyr Arg Asp Gly Met Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 112
<211> 112
<212> PRT
<213> Homo sapiens
<400> 112
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Ser Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 113
<211> 14
<212> PRT
<213> Homo sapiens
<400> 113
Phe Asp Asp Tyr Gly Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 114
<211> 18
<212> PRT
<213> Homo sapiens
<400> 114
Ser Thr Ile Tyr Ser Gly Asp Asn Ala Tyr Tyr Gly Ala Ser Val Arg
1 5 10 15
Gly Arg
<210> 115
<211> 15
<212> PRT
<213> Homo sapiens
<400> 115
Ala Arg Val Tyr Ser Ser Ser Trp Arg Lys Arg Ala Phe Asp Ile
1 5 10 15
<210> 116
<211> 14
<212> PRT
<213> Homo sapiens
<400> 116
Cys Ser Gly Thr Ser Ser Asn Ile Glu Ser Asn Thr Val Asn
1 5 10
<210> 117
<211> 7
<212> PRT
<213> Homo sapiens
<400> 117
Ser Asp Asn Gln Arg Pro Ser
1 5
<210> 118
<211> 12
<212> PRT
<213> Homo sapiens
<400> 118
Cys Ala Ala Trp Asp Asp Ser Leu Ser Gly Trp Val
1 5 10
<210> 119
<211> 121
<212> PRT
<213> Homo sapiens
<400> 119
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Tyr Ser Gly Asp Asn Ala Tyr Tyr Gly Ala Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Val Tyr Ser Ser Ser Trp Arg Lys Arg Ala Phe Asp Ile Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 120
<211> 111
<212> PRT
<213> Homo sapiens
<400> 120
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Thr Ser Ser Asn Ile Glu Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asp Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 121
<211> 14
<212> PRT
<213> Homo sapiens
<400> 121
Phe Ser Asp Tyr Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly
1 5 10
<210> 122
<211> 19
<212> PRT
<213> Homo sapiens
<400> 122
Ala Leu Ile Trp Tyr Asp Gly Gly Asn Glu Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 123
<211> 11
<212> PRT
<213> Homo sapiens
<400> 123
Val Arg Glu Thr Gly Asn Tyr Gly Met Asp Val
1 5 10
<210> 124
<211> 15
<212> PRT
<213> Homo sapiens
<400> 124
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 125
<211> 7
<212> PRT
<213> Homo sapiens
<400> 125
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 126
<211> 12
<212> PRT
<213> Homo sapiens
<400> 126
Cys Ala Thr Trp Asp Asp Arg Val Asn Gly Pro Val
1 5 10
<210> 127
<211> 118
<212> PRT
<213> Homo sapiens
<400> 127
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Gly Asn Glu Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Glu Thr Gly Asn Tyr Gly Met Asp Val Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 128
<211> 112
<212> PRT
<213> Homo sapiens
<400> 128
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Arg
85 90 95
Val Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 129
<211> 14
<212> PRT
<213> Homo sapiens
<400> 129
Phe Ser Asp Tyr Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly
1 5 10
<210> 130
<211> 19
<212> PRT
<213> Homo sapiens
<400> 130
Ala Leu Ile Trp Tyr Asp Gly Gly Asn Glu Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 131
<211> 11
<212> PRT
<213> Homo sapiens
<400> 131
Ala Arg Tyr Tyr Gly Asp Gly Gly Phe Asp Pro
1 5 10
<210> 132
<211> 15
<212> PRT
<213> Homo sapiens
<400> 132
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Val Val His
1 5 10 15
<210> 133
<211> 7
<212> PRT
<213> Homo sapiens
<400> 133
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 134
<211> 12
<212> PRT
<213> Homo sapiens
<400> 134
Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Pro Val
1 5 10
<210> 135
<211> 118
<212> PRT
<213> Homo sapiens
<400> 135
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Ser Tyr Asp Gly Gly Gly Lys Tyr Phe Ala Asp Pro Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Gly Asp Gly Gly Phe Asp Pro Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 136
<211> 112
<212> PRT
<213> Homo sapiens
<400> 136
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Val Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 137
<211> 14
<212> PRT
<213> Homo sapiens
<400> 137
Phe Ser Ser Asn Tyr Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 138
<211> 19
<212> PRT
<213> Homo sapiens
<400> 138
Ala Leu Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 139
<211> 8
<212> PRT
<213> Homo sapiens
<400> 139
Ala Lys Asp Pro Leu Phe Asp Ser
1 5
<210> 140
<211> 15
<212> PRT
<213> Homo sapiens
<400> 140
Cys Thr Gly Arg Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 141
<211> 7
<212> PRT
<213> Homo sapiens
<400> 141
Asp Asn Asn Lys Arg Pro Ser
1 5
<210> 142
<211> 12
<212> PRT
<213> Homo sapiens
<400> 142
Cys Ala Ala Trp Asp Asp Ser Leu Asn Gly Pro Val
1 5 10
<210> 143
<211> 115
<212> PRT
<213> Homo sapiens
<400> 143
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Pro Leu Phe Asp Ser Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 144
<211> 112
<212> PRT
<213> Homo sapiens
<400> 144
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Arg Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser
85 90 95
Leu Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 145
<211> 14
<212> PRT
<213> Homo sapiens
<400> 145
Phe Asn Thr Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 146
<211> 18
<212> PRT
<213> Homo sapiens
<400> 146
Ser Val Leu Tyr Ser Asp Asp Asp Thr His Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly Arg
<210> 147
<211> 17
<212> PRT
<213> Homo sapiens
<400> 147
Ala Arg Asp Cys Gly Gly Asp Cys His Ser Gly Asp Asp Ala Phe Asp
1 5 10 15
Ile
<210> 148
<211> 14
<212> PRT
<213> Homo sapiens
<400> 148
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 149
<211> 7
<212> PRT
<213> Homo sapiens
<400> 149
Asp Asn Asp Lys Arg Pro Ser
1 5
<210> 150
<211> 12
<212> PRT
<213> Homo sapiens
<400> 150
Cys Ala Ala Trp His Asp Ser Leu Asn Gly Trp Val
1 5 10
<210> 151
<211> 123
<212> PRT
<213> Homo sapiens
<400> 151
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asn Thr Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Leu Tyr Ser Asp Asp Asp Thr His Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Cys Gly Gly Asp Cys His Ser Gly Asp Asp Ala Phe Asp Ile
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 152
<211> 111
<212> PRT
<213> Homo sapiens
<400> 152
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp His Asp Ser Leu
85 90 95
Asn Gly Trp Val Leu Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 153
<211> 14
<212> PRT
<213> Homo sapiens
<400> 153
Phe Ser Ala Tyr Gly Met His Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 154
<211> 19
<212> PRT
<213> Homo sapiens
<400> 154
Ala Val Val Ser Tyr Asp Gly Arg Glu Lys His Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 155
<211> 14
<212> PRT
<213> Homo sapiens
<400> 155
Ala Arg Ser Asp Gly Gly Tyr Asp Ser Asp Ser Gly Tyr Tyr
1 5 10
<210> 156
<211> 14
<212> PRT
<213> Homo sapiens
<400> 156
Cys Ser Gly Ser Thr Ser Asn Ile Gly Ser Asn Phe Val Tyr
1 5 10
<210> 157
<211> 7
<212> PRT
<213> Homo sapiens
<400> 157
Asp Asn Asn Lys Arg Pro Ser
1 5
<210> 158
<211> 11
<212> PRT
<213> Homo sapiens
<400> 158
Cys Ser Ser Tyr Ala Tyr Ser Asp Asn Ile Leu
1 5 10
<210> 159
<211> 121
<212> PRT
<213> Homo sapiens
<400> 159
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Val Ser Tyr Asp Gly Arg Glu Lys His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Asp Gly Gly Tyr Asp Ser Asp Ser Gly Tyr Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 160
<211> 110
<212> PRT
<213> Homo sapiens
<400> 160
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Thr Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Tyr Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Tyr Ser Asp
85 90 95
Asn Ile Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 161
<211> 14
<212> PRT
<213> Homo sapiens
<400> 161
Phe Ser Asn Ala Trp Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 162
<211> 19
<212> PRT
<213> Homo sapiens
<400> 162
Ser Gly Ile Ser Ser Ser Gly Ser Ser Ala Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 163
<211> 18
<212> PRT
<213> Homo sapiens
<400> 163
Ala Arg His Tyr Tyr Tyr His Ile Ala Gly Tyr Tyr Tyr Asp Thr Phe
1 5 10 15
Asp Ile
<210> 164
<211> 14
<212> PRT
<213> Homo sapiens
<400> 164
Cys Ser Gly Ser Ser Ser Asn Ile Gly Gly Asn Thr Val Asn
1 5 10
<210> 165
<211> 7
<212> PRT
<213> Homo sapiens
<400> 165
Gly Asn Thr Asn Arg Pro Ser
1 5
<210> 166
<211> 12
<212> PRT
<213> Homo sapiens
<400> 166
Cys Ala Ala Trp Asp Asp Ser Leu Ser Gly Val Val
1 5 10
<210> 167
<211> 125
<212> PRT
<213> Homo sapiens
<400> 167
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Ala
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Ser Ser Gly Ser Ser Ala Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Tyr Tyr Tyr His Ile Ala Gly Tyr Tyr Tyr Asp Thr Phe
100 105 110
Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 168
<211> 111
<212> PRT
<213> Homo sapiens
<400> 168
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Gly Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Thr Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 169
<211> 14
<212> PRT
<213> Homo sapiens
<400> 169
Phe Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 170
<211> 19
<212> PRT
<213> Homo sapiens
<400> 170
Ala Thr Ile Ser Tyr His Gly Ser Asp Lys Asp Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 171
<211> 18
<212> PRT
<213> Homo sapiens
<400> 171
Ala Arg Asp Ala Asn Tyr His Ser Ser Gly Tyr Tyr Tyr Asp Val Phe
1 5 10 15
Asp Ile
<210> 172
<211> 14
<212> PRT
<213> Homo sapiens
<400> 172
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 173
<211> 7
<212> PRT
<213> Homo sapiens
<400> 173
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 174
<211> 12
<212> PRT
<213> Homo sapiens
<400> 174
Cys Ala Ala Trp Asp Asp Ser Leu Ser Thr Trp Val
1 5 10
<210> 175
<211> 125
<212> PRT
<213> Homo sapiens
<400> 175
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Tyr His Gly Ser Asp Lys Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ala Asn Tyr His Ser Ser Gly Tyr Tyr Tyr Asp Val Phe
100 105 110
Asp Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 176
<211> 111
<212> PRT
<213> Homo sapiens
<400> 176
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Thr Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 177
<211> 14
<212> PRT
<213> Homo sapiens
<400> 177
Phe Ser Asp Tyr Tyr Met Thr Trp Ile Arg Gln Ala Pro Gly
1 5 10
<210> 178
<211> 19
<212> PRT
<213> Homo sapiens
<400> 178
Ser Gly Ile Ser Gly Ser Gly Gly Tyr Ile His Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 179
<211> 11
<212> PRT
<213> Homo sapiens
<400> 179
Ala Arg Glu Gly Leu Leu Pro Asp Ala Phe Asp
1 5 10
<210> 180
<211> 14
<212> PRT
<213> Homo sapiens
<400> 180
Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn Tyr Val Ser
1 5 10
<210> 181
<211> 7
<212> PRT
<213> Homo sapiens
<400> 181
Arg Asn Asn Gln Arg Pro Ser
1 5
<210> 182
<211> 12
<212> PRT
<213> Homo sapiens
<400> 182
Cys Ala Ala Trp Asp Asp Ser Val Ser Gly Trp Val
1 5 10
<210> 183
<211> 119
<212> PRT
<213> Homo sapiens
<400> 183
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Thr Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Gly Ser Gly Gly Tyr Ile His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Leu Leu Pro Asp Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 184
<211> 111
<212> PRT
<213> Homo sapiens
<400> 184
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Arg Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Val
85 90 95
Ser Gly Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 185
<211> 14
<212> PRT
<213> Homo sapiens
<400> 185
Phe Ser Ser Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 186
<211> 19
<212> PRT
<213> Homo sapiens
<400> 186
Ala Val Met Ser Tyr Asp Glu Tyr Asn Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 187
<211> 13
<212> PRT
<213> Homo sapiens
<400> 187
Ala Lys Gly Phe Tyr Gly Asp Tyr Pro Leu Trp Asp Tyr
1 5 10
<210> 188
<211> 14
<212> PRT
<213> Homo sapiens
<400> 188
Cys Ser Gly Gly Asn Ser Asn Ile Gly Thr Asn Thr Val Asp
1 5 10
<210> 189
<211> 7
<212> PRT
<213> Homo sapiens
<400> 189
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 190
<211> 12
<212> PRT
<213> Homo sapiens
<400> 190
Cys Ala Ala Trp Asp Asp Ser Val Asn Gly Pro Val
1 5 10
<210> 191
<211> 120
<212> PRT
<213> Homo sapiens
<400> 191
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Met Ser Tyr Asp Glu Tyr Asn Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Gly Phe Tyr Gly Asp Tyr Pro Leu Trp Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 192
<211> 111
<212> PRT
<213> Homo sapiens
<400> 192
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Gly Asn Ser Asn Ile Gly Thr Asn
20 25 30
Thr Val Asp Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Val
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 193
<211> 14
<212> PRT
<213> Homo sapiens
<400> 193
Phe Ser Ser Tyr Glu Met Asn Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 194
<211> 18
<212> PRT
<213> Homo sapiens
<400> 194
Ser Thr Ile Thr Gly Gly Gly Ser Ile Tyr Asp Ala Asn Ser Val Gln
1 5 10 15
Gly Arg
<210> 195
<211> 18
<212> PRT
<213> Homo sapiens
<400> 195
Ala Arg Asp Ser Thr Tyr His Ser Ser Gly Tyr Tyr Tyr Asp Val Phe
1 5 10 15
Asp Ile
<210> 196
<211> 14
<212> PRT
<213> Homo sapiens
<400> 196
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 197
<211> 7
<212> PRT
<213> Homo sapiens
<400> 197
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 198
<211> 13
<212> PRT
<213> Homo sapiens
<400> 198
Cys Ala Ala Trp Asp Asp Ser Leu Ser Gly His Trp Val
1 5 10
<210> 199
<211> 124
<212> PRT
<213> Homo sapiens
<400> 199
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Glu Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Thr Gly Gly Gly Ser Ile Tyr Asp Ala Asn Ser Val Gln
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Ser Thr Tyr His Ser Ser Gly Tyr Tyr Tyr Asp Val Phe Asp
100 105 110
Ile Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 200
<211> 112
<212> PRT
<213> Homo sapiens
<400> 200
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Ser Gly His Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 201
<211> 14
<212> PRT
<213> Homo sapiens
<400> 201
Phe Ser Ser Tyr Gly Met His Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 202
<211> 20
<212> PRT
<213> Homo sapiens
<400> 202
Ser Ala Val Phe Gly Ser Gly His Gly Asn Thr Phe Tyr Ala Asp Ala
1 5 10 15
Val Lys Gly Arg
20
<210> 203
<211> 14
<212> PRT
<213> Homo sapiens
<400> 203
Ala Arg Glu Gln Leu Trp Phe Gly Gln Asp Ala Phe Asp Ile
1 5 10
<210> 204
<211> 14
<212> PRT
<213> Homo sapiens
<400> 204
Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 205
<211> 7
<212> PRT
<213> Homo sapiens
<400> 205
Gly Asn Ser Asn Arg Pro Ser
1 5
<210> 206
<211> 12
<212> PRT
<213> Homo sapiens
<400> 206
Cys Gln Ser Tyr Asp Ser Ser Leu Ser Ala Ser Val
1 5 10
<210> 207
<211> 122
<212> PRT
<213> Homo sapiens
<400> 207
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Pro Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Val Phe Gly Ser Gly His Gly Asn Thr Phe Tyr Ala Asp Ala
50 55 60
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu
65 70 75 80
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Glu Gln Leu Trp Phe Gly Gln Asp Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 208
<211> 111
<212> PRT
<213> Homo sapiens
<400> 208
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Arg
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Ser Ser Leu
85 90 95
Ser Ala Ser Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 209
<211> 14
<212> PRT
<213> Homo sapiens
<400> 209
Phe Ser Asp Ala Trp Met Thr Trp Val Arg Gln Ala Pro Gly
1 5 10
<210> 210
<211> 19
<212> PRT
<213> Homo sapiens
<400> 210
Ser Asp Leu Ser Asp Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
1 5 10 15
Lys Gly Arg
<210> 211
<211> 11
<212> PRT
<213> Homo sapiens
<400> 211
Gly Arg Leu Ala Ala Gly Gly Pro Val Asp Tyr
1 5 10
<210> 212
<211> 15
<212> PRT
<213> Homo sapiens
<400> 212
Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly Tyr Asp Val His
1 5 10 15
<210> 213
<211> 7
<212> PRT
<213> Homo sapiens
<400> 213
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 214
<211> 12
<212> PRT
<213> Homo sapiens
<400> 214
Cys Ser Val Trp Asp Asp Ser Leu Asn Ser Trp Val
1 5 10
<210> 215
<211> 118
<212> PRT
<213> Homo sapiens
<400> 215
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ala
20 25 30
Trp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Leu Ser Asp Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Gly Arg Leu Ala Ala Gly Gly Pro Val Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 216
<211> 112
<212> PRT
<213> Homo sapiens
<400> 216
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn Ile Gly Ala Gly
20 25 30
Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu
35 40 45
Leu Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu
65 70 75 80
Arg Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Val Trp Asp Asp Ser
85 90 95
Leu Asn Ser Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 217
<211> 330
<212> PRT
<213> Homo sapiens
<400> 217
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 218
<211> 105
<212> PRT
<213> Homo sapiens
<400> 218
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 219
<211> 330
<212> PRT
<213> Mus musculus
<400> 219
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 220
<211> 330
<212> PRT
<213> Mus musculus
<400> 220
Ala Lys Thr Thr Ala Pro Ser Val Tyr Pro Leu Ala Pro Val Cys Gly
1 5 10 15
Asp Thr Thr Gly Ser Ser Val Thr Leu Gly Cys Leu Val Lys Gly Tyr
20 25 30
Phe Pro Glu Pro Val Thr Leu Thr Trp Asn Ser Gly Ser Leu Ser Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu
50 55 60
Ser Ser Ser Val Thr Val Thr Ser Ser Thr Trp Pro Ser Gln Ser Ile
65 70 75 80
Thr Cys Asn Val Ala His Pro Ala Ser Ser Thr Lys Val Asp Lys Lys
85 90 95
Ile Glu Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
100 105 110
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
115 120 125
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
130 135 140
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
145 150 155 160
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
165 170 175
Glu Asp Tyr Ala Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
180 185 190
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
195 200 205
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
210 215 220
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
225 230 235 240
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
245 250 255
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
260 265 270
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
275 280 285
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
290 295 300
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
305 310 315 320
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
325 330
<210> 221
<211> 105
<212> PRT
<213> Mus musculus
<400> 221
Gln Pro Lys Ser Ser Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Glu Thr Asn Lys Ala Thr Leu Val Cys Thr Ile Thr Asp Phe
20 25 30
Tyr Pro Gly Val Val Thr Val Asp Trp Lys Val Asp Gly Thr Pro Val
35 40 45
Thr Gln Gly Met Glu Thr Thr Gln Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Met Ala Ser Ser Tyr Leu Thr Leu Thr Ala Arg Ala Trp Glu Arg
65 70 75 80
His Ser Ser Tyr Ser Cys Gln Val Thr His Glu Gly His Thr Val Glu
85 90 95
Lys Ser Leu Ser Arg Ala Asp Cys Ser
100 105
<210> 222
<211> 1356
<212> ДНК
<213> Homo sapiens
<400> 222
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttgat gattatggca tgagctgggt ccgccaagct 120
ccagggaagg ggctggagtg ggtctcagtt atttatagcg gtggtagtac atattacgca 180
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac gctgtatctg 240
caaatgaaca gcctgagagc cgaggacact gccgtgtatt actgtgcgag agatcgaagc 300
agcagctggt accgcgatgg tatggacgtc tggggccaag gtacactggt caccgtgagc 360
tcagcctcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct 420
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 480
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 540
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 600
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 660
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 720
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 780
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 840
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 900
aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 960
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 1020
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggat 1080
gagctgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 1140
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 1200
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1260
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1320
acgcagaaga gcctctccct gtctccgggt aaatga 1356
<210> 223
<211> 654
<212> ДНК
<213> Homo sapiens
<400> 223
cagtctgtgc tgactcagcc accctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcctgcactg ggagcagctc caacatcggg gcaggttatg atgtacactg gtatcagcag 120
ctcccaggaa cggcccccaa actcctcatc tatggtaaca gcaatcggcc ctcaggggtc 180
cctgaccgat tctctggctc caagtctggc acctcagcct ccctggccat cagtgggctc 240
cggtccgagg atgaggctga ttattactgt gcagcatggg atgacagcct gagtggttgg 300
gtgttcggcg gaggaaccaa gctgacggtc ctaggtcagc ccaaggctgc cccctcggtc 360
actctgttcc cgccctcctc tgaggagctt caagccaaca aggccacact ggtgtgtctc 420
ataagtgact tctacccggg agccgtgaca gtggcctgga aggcagatag cagccccgtc 480
aaggcgggag tggagaccac cacaccctcc aaacaaagca acaacaagta cgcggccagc 540
agctatctga gcctgacgcc tgagcagtgg aagtcccaca gaagctacag ctgccaggtc 600
acgcatgaag ggagcaccgt ggagaagaca gtggccccta cagaatgttc atga 654
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-PRE-S1 HBV АНТИТЕЛА | 2016 |
|
RU2739955C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CTLA-4, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2756100C1 |
| ЧАСТИЦЫ, КОНЪЮГИРОВАННЫЕ С ПЕПТИДОМ | 2014 |
|
RU2813163C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ CD3 | 2017 |
|
RU2779489C2 |
| CD3-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2020 |
|
RU2826453C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ КОМПОЗИЦИЙ T-КЛЕТОК И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2835034C2 |
| СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ РАКОВОГО МЕТАСТАЗА И РАЗРЕЖЕНИЯ КОСТИ, СВЯЗАННОГО С РАКОВЫМ МЕТАСТАЗОМ | 2007 |
|
RU2470665C2 |
| НОВЫЕ КОНСТРУКЦИИ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА, СОДЕРЖАЩИЕ ДОМЕНЫ TNFR2 | 2019 |
|
RU2808254C2 |
| СПЕЦИФИЧЕСКИЕ АНТИТЕЛА К PD-L1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2749109C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
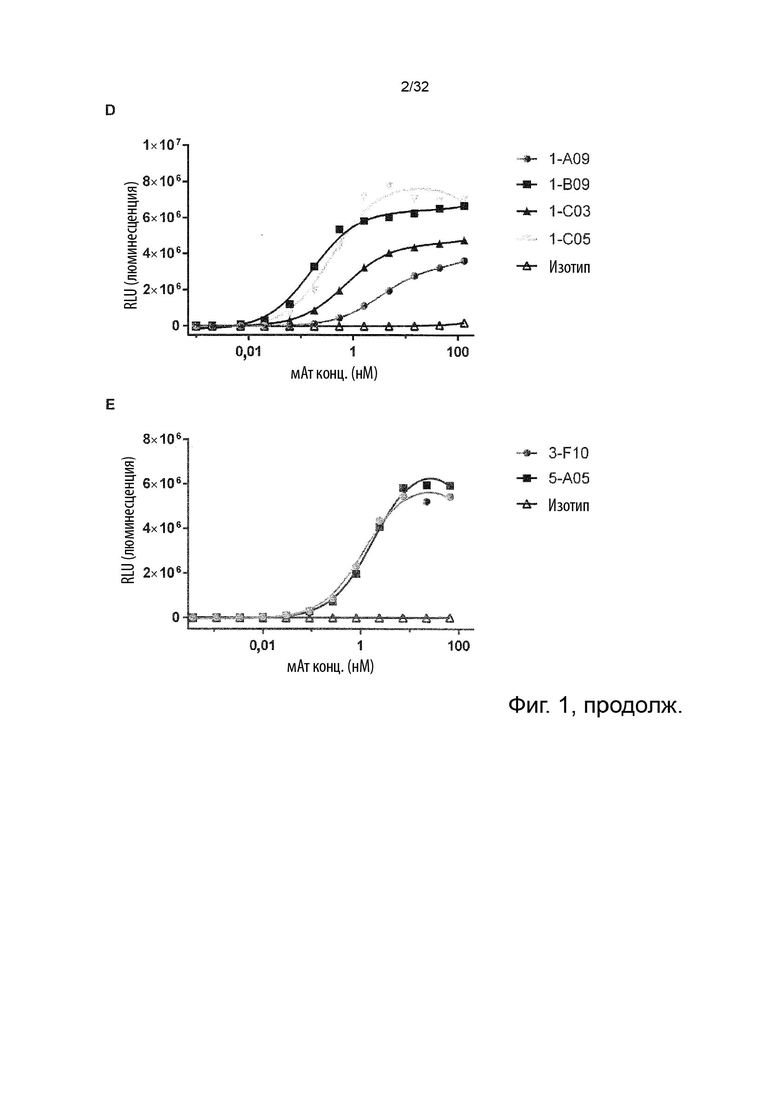
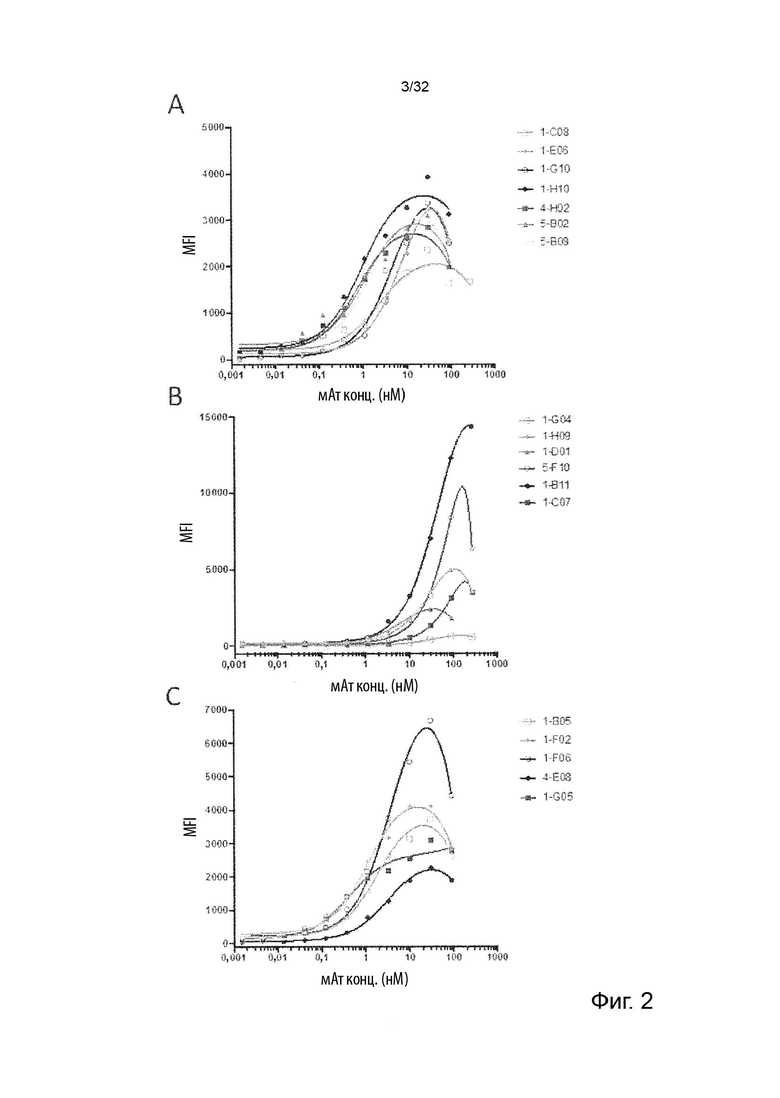
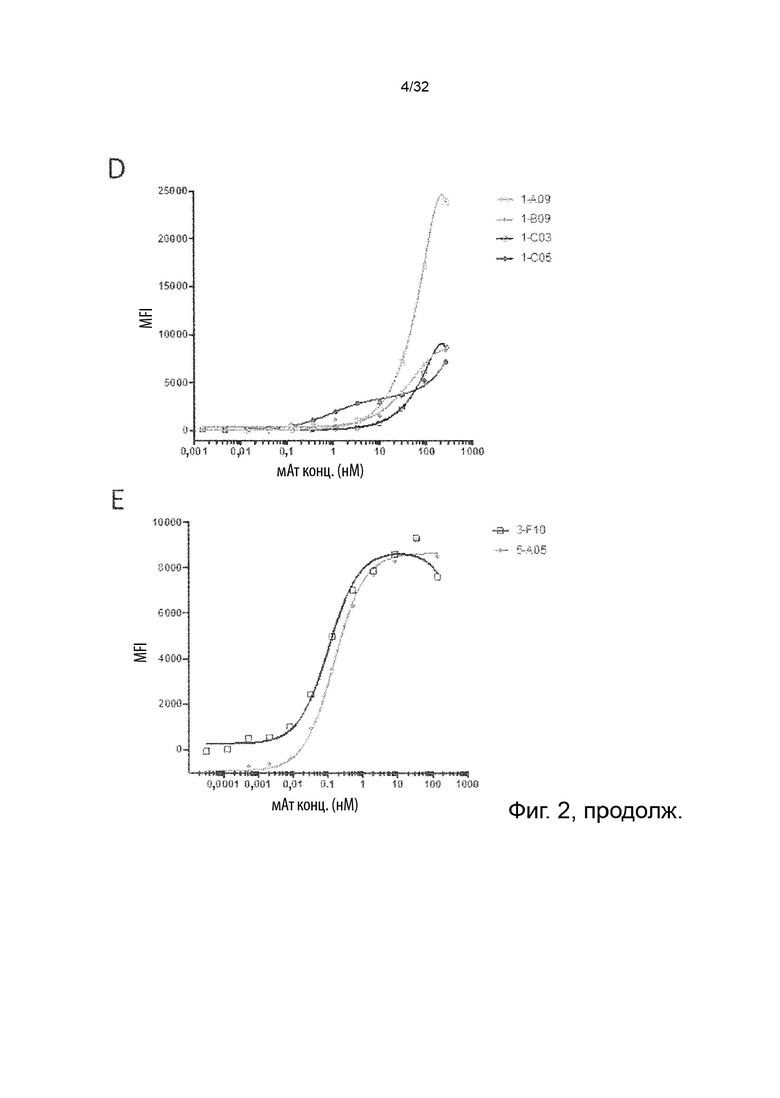
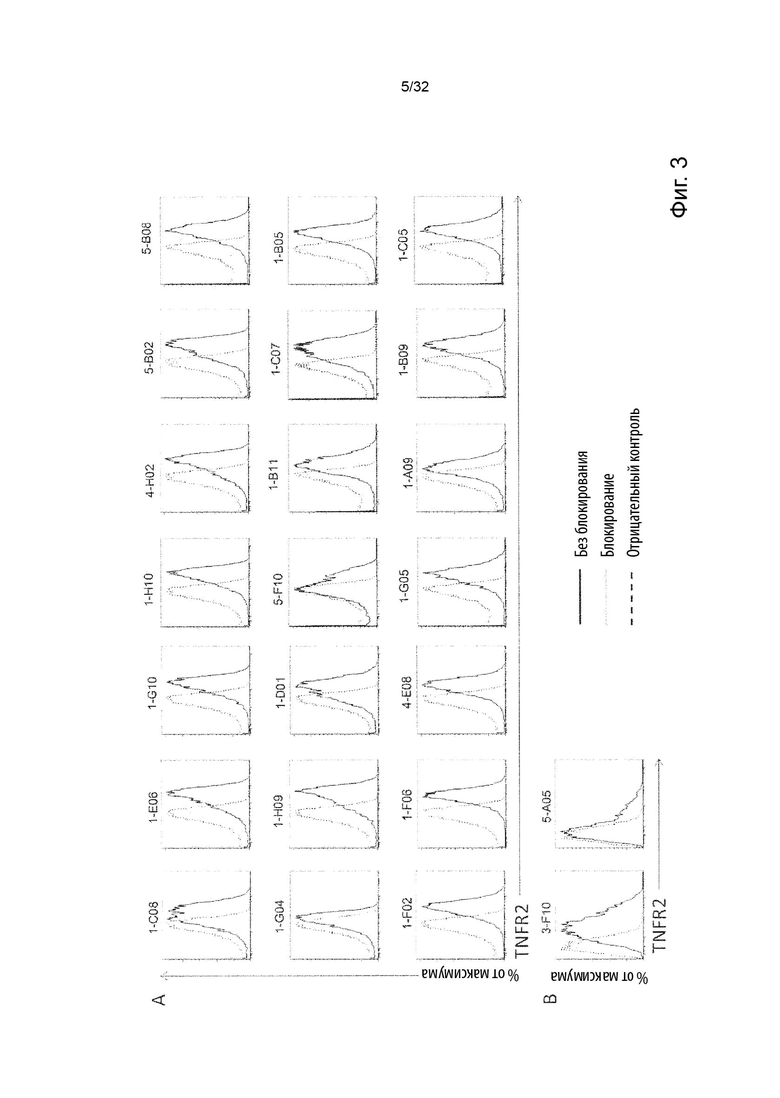
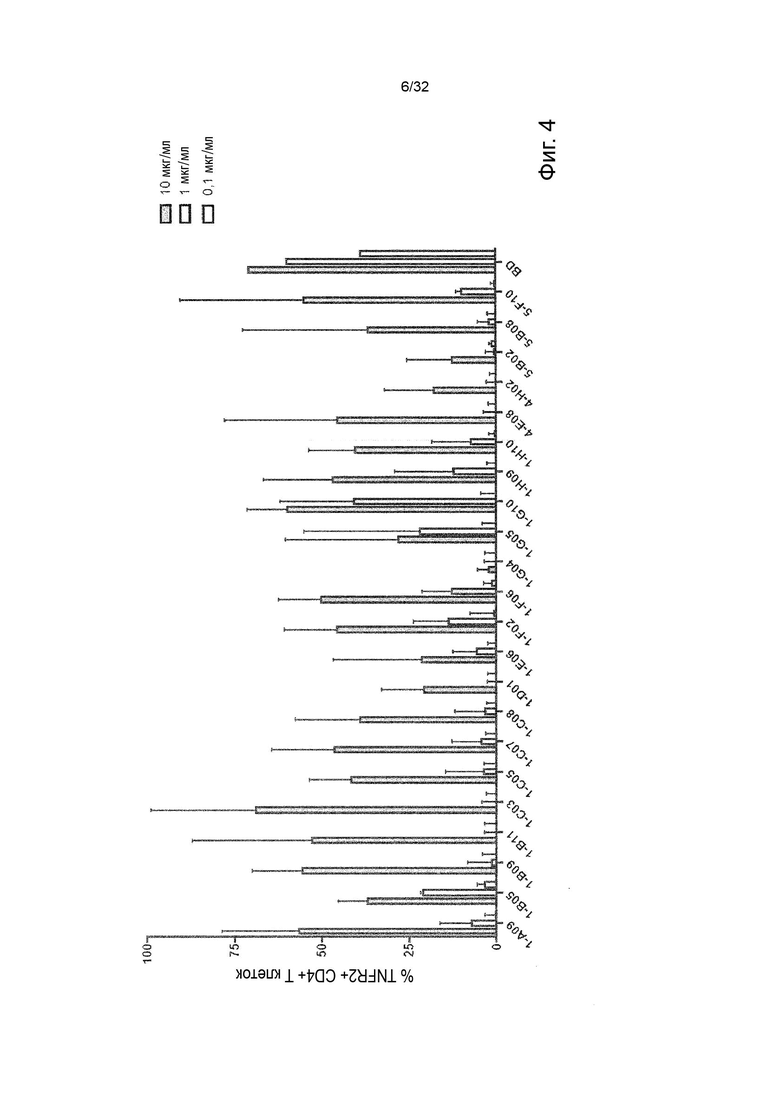
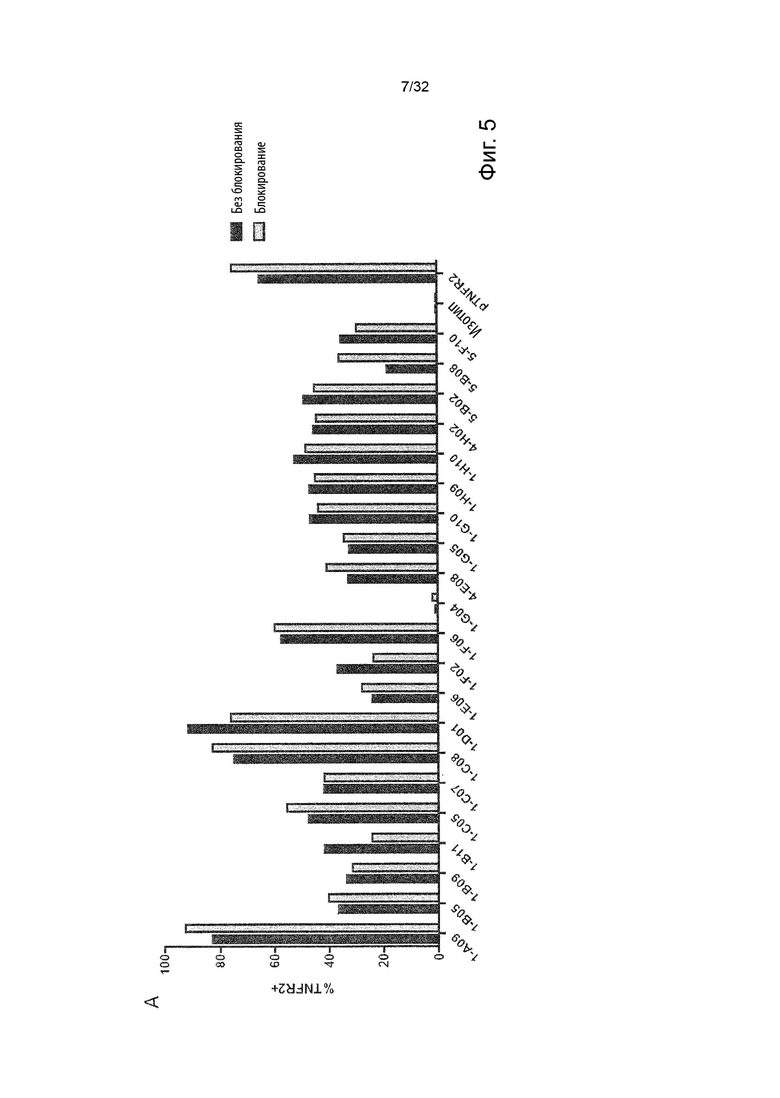
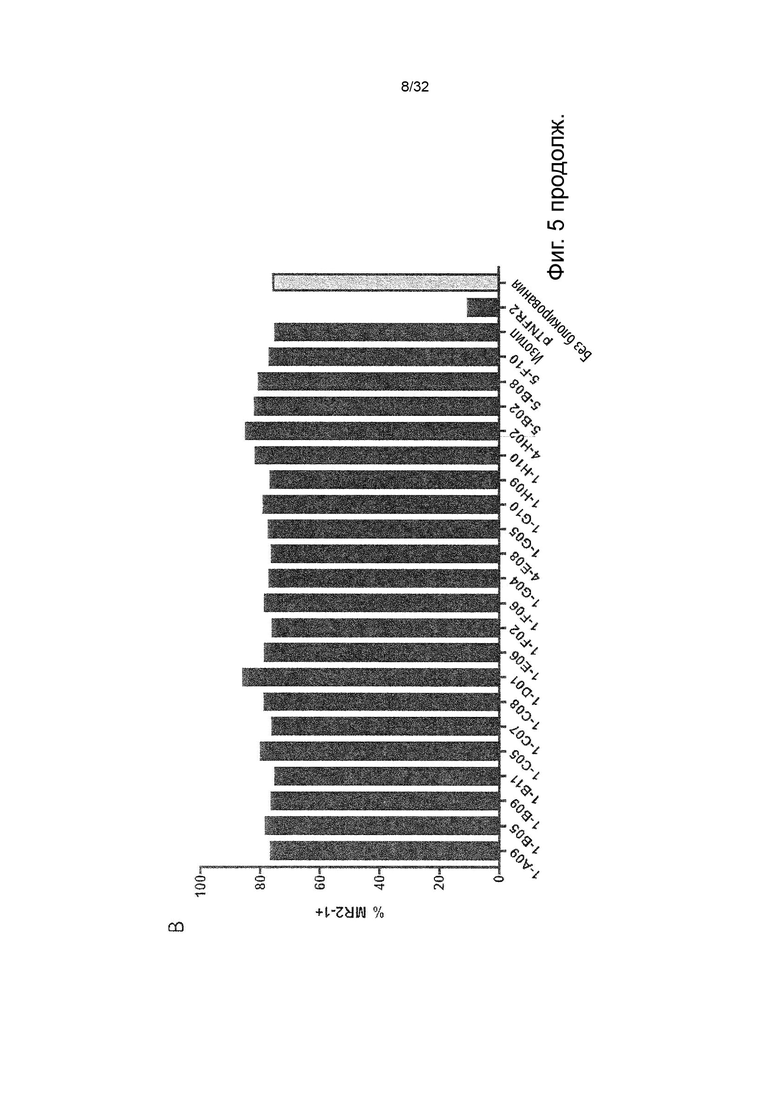
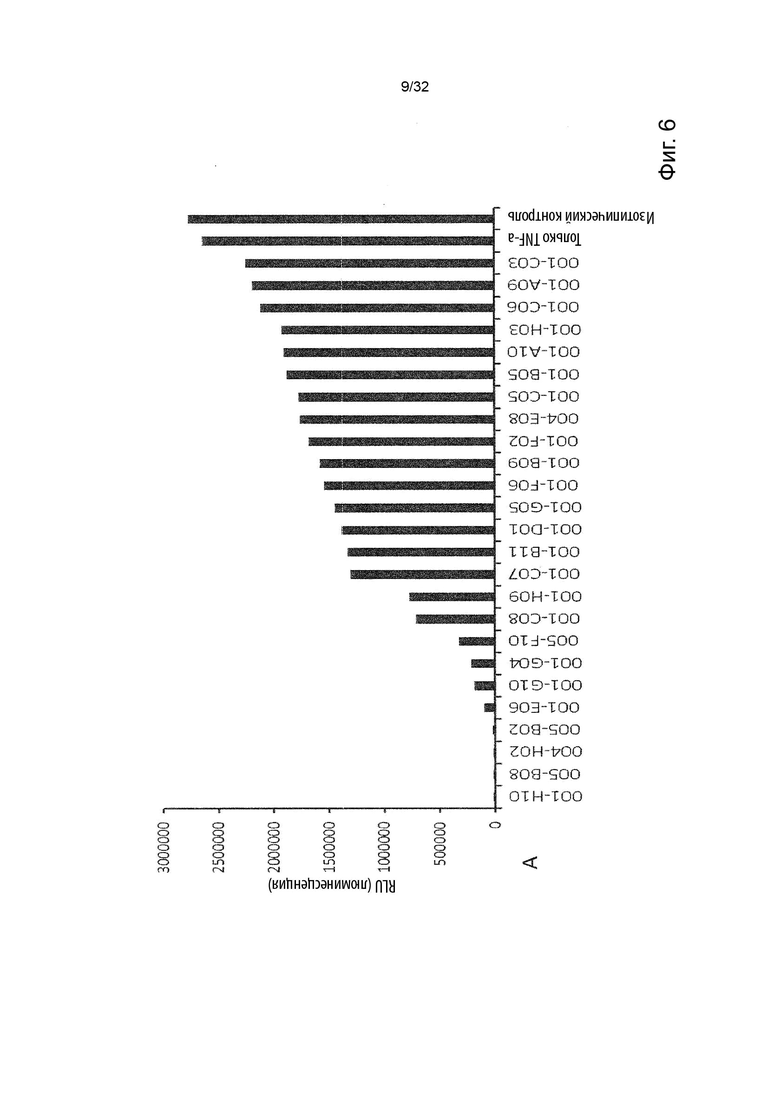
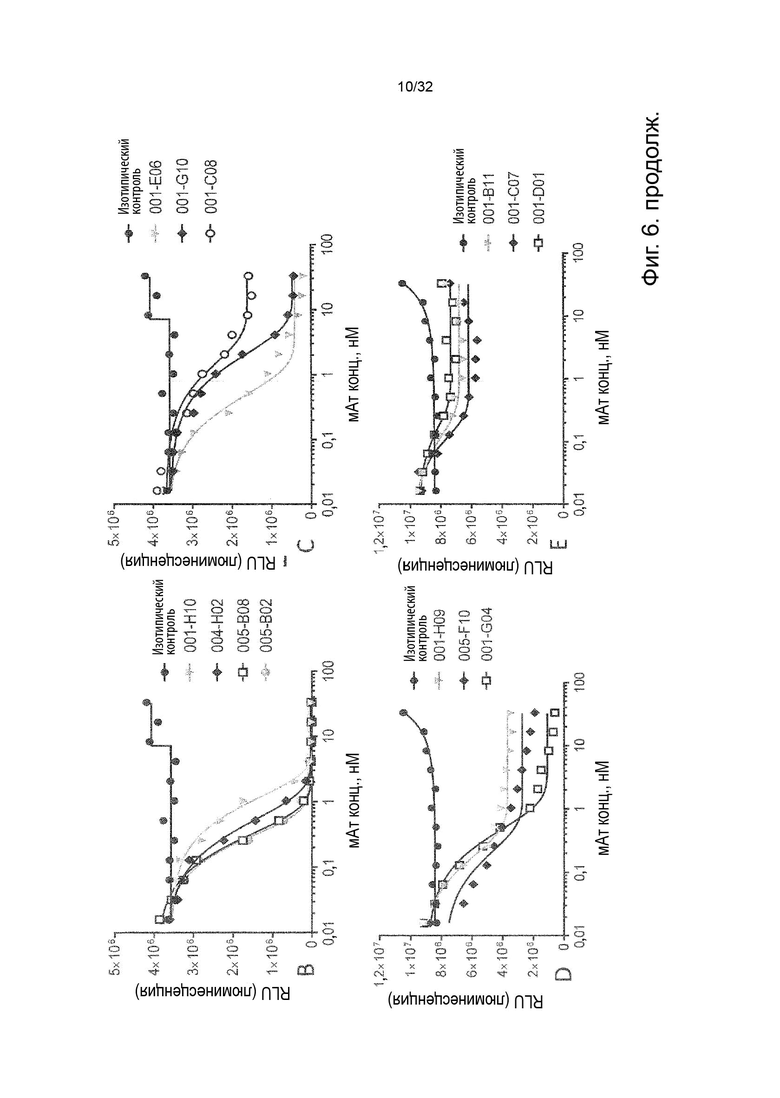
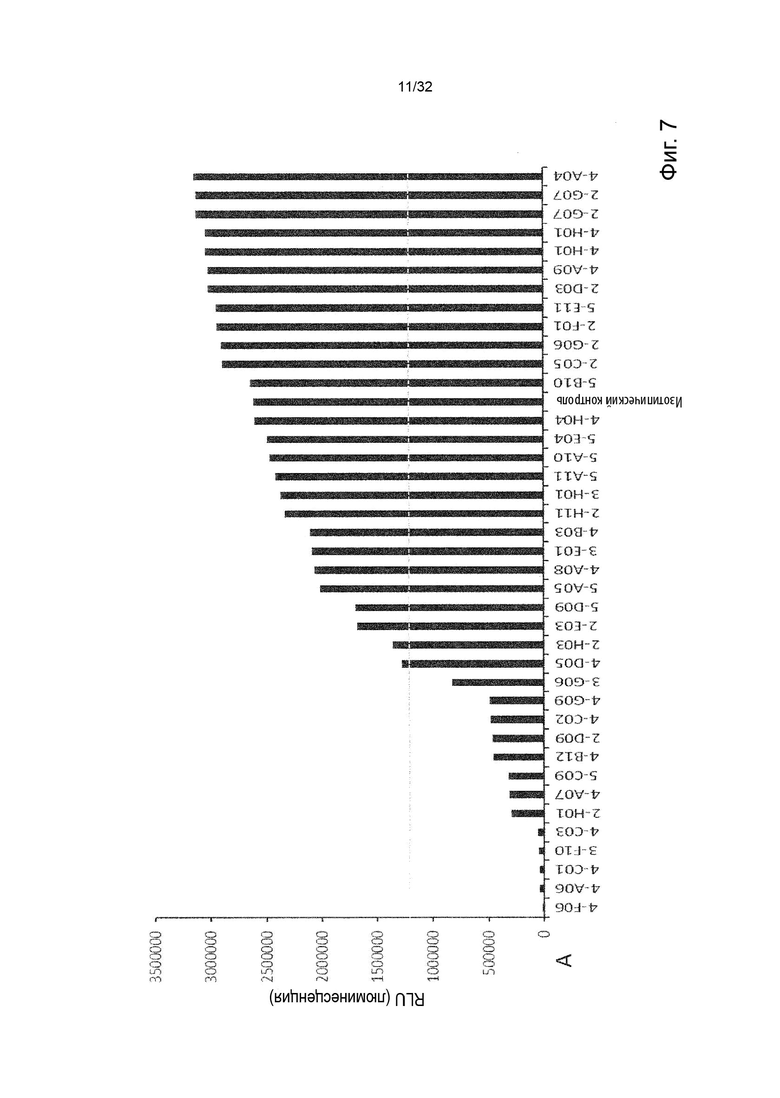
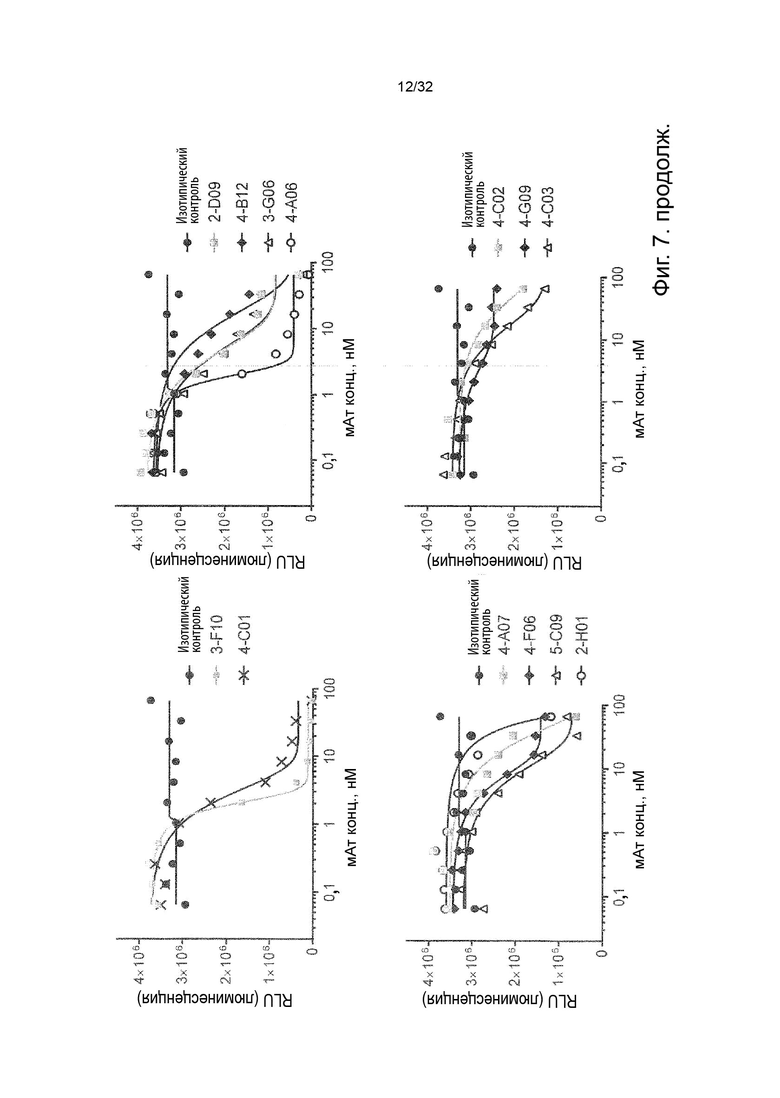
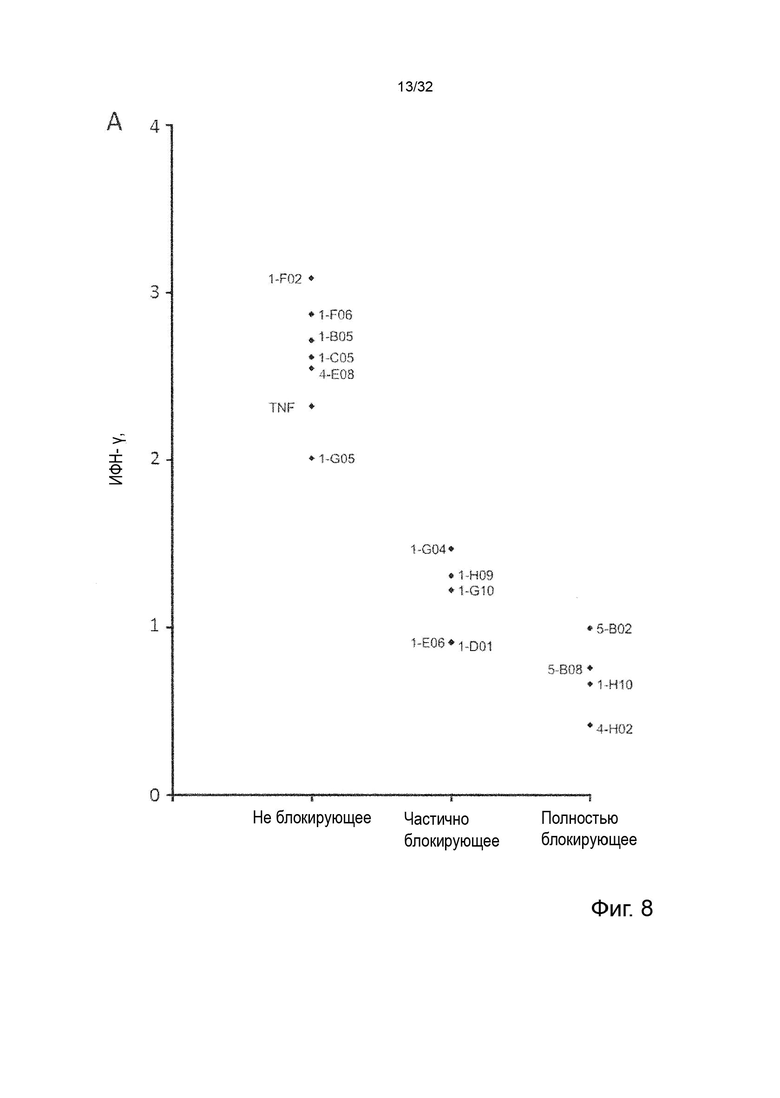
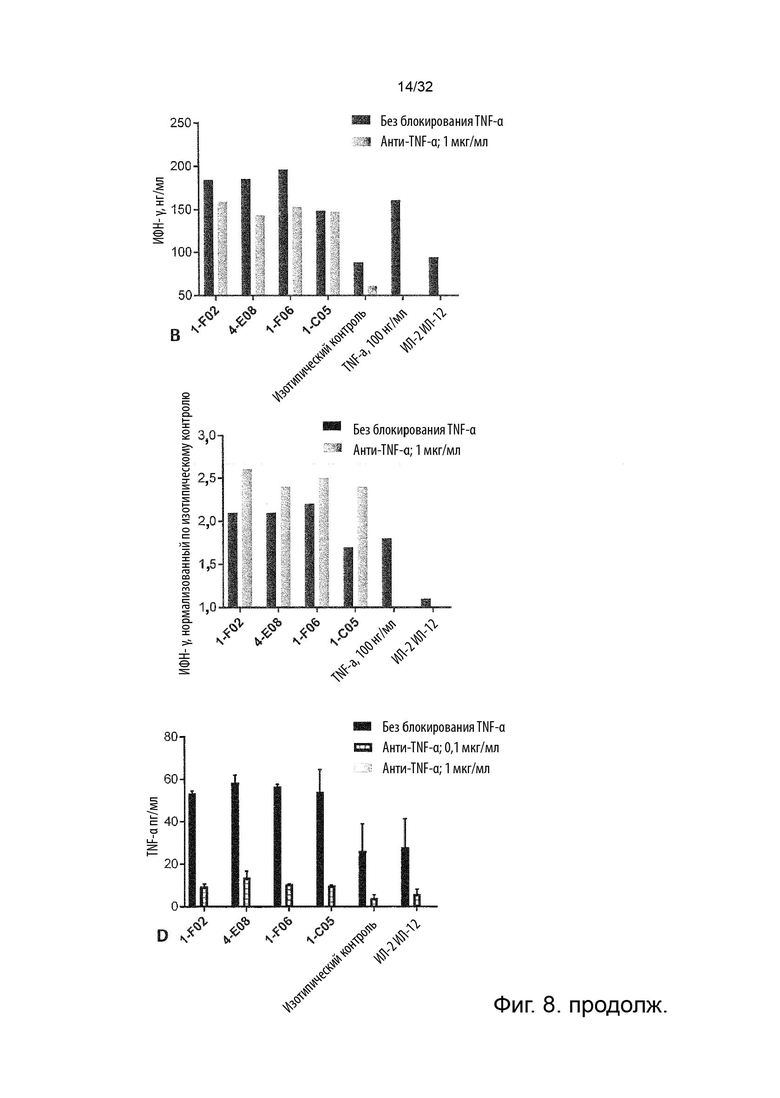
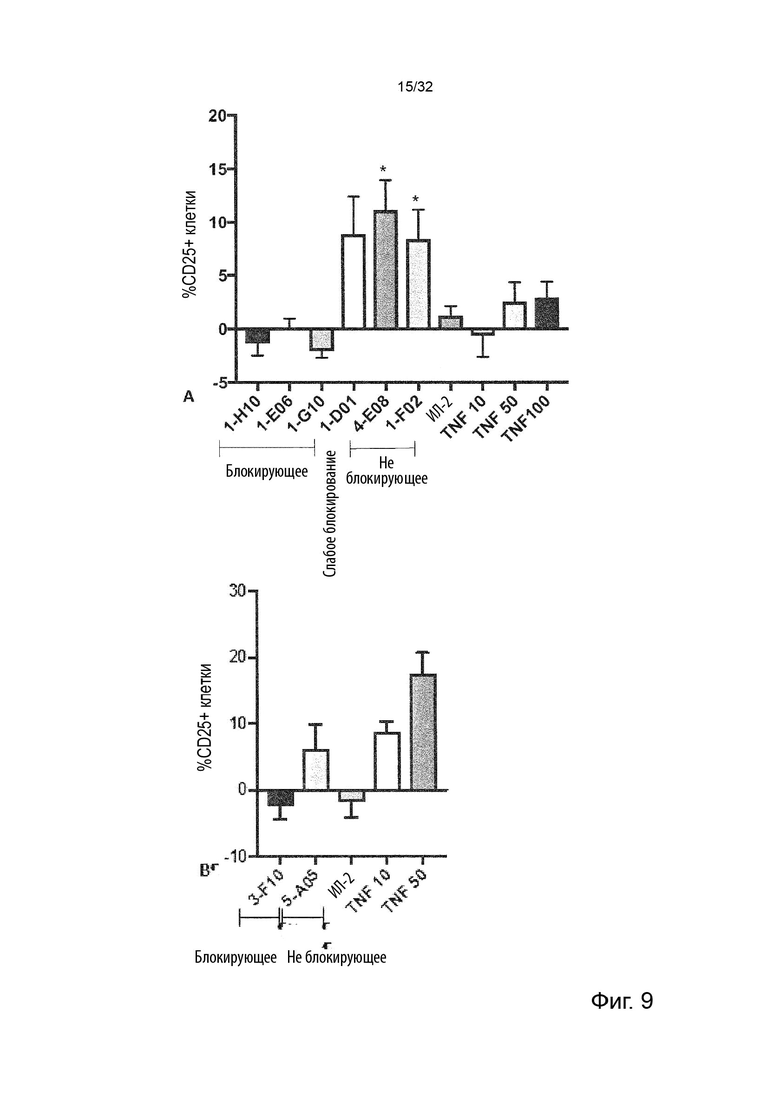
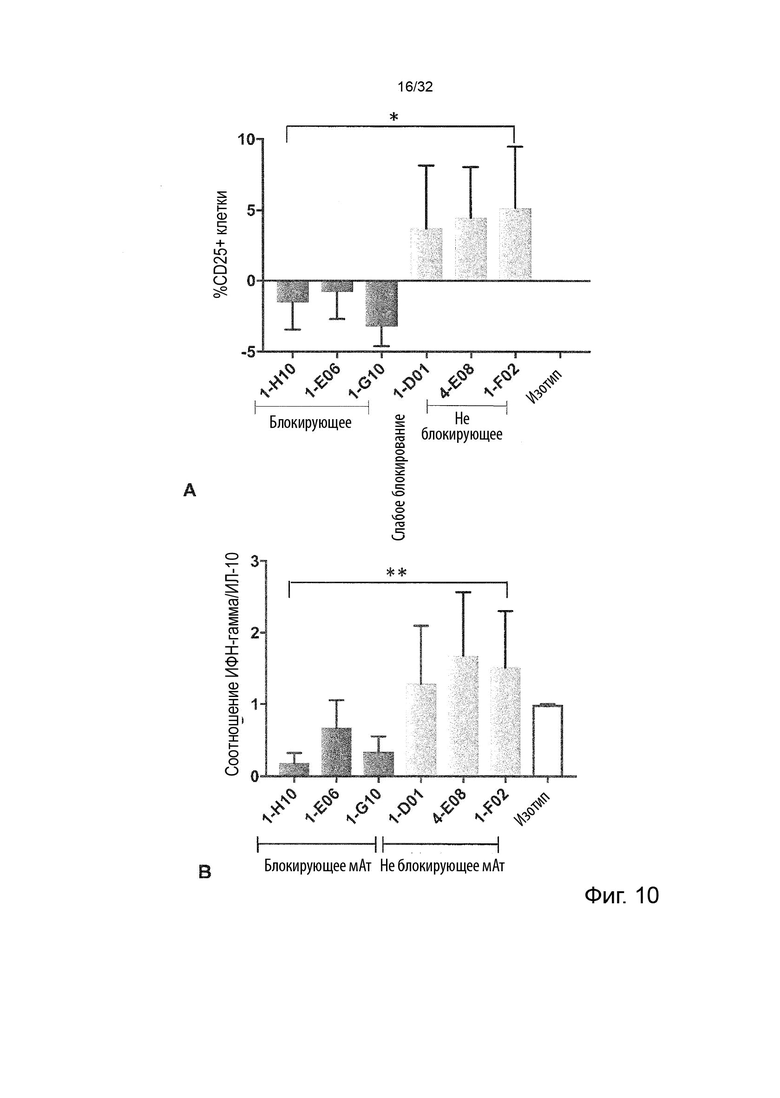
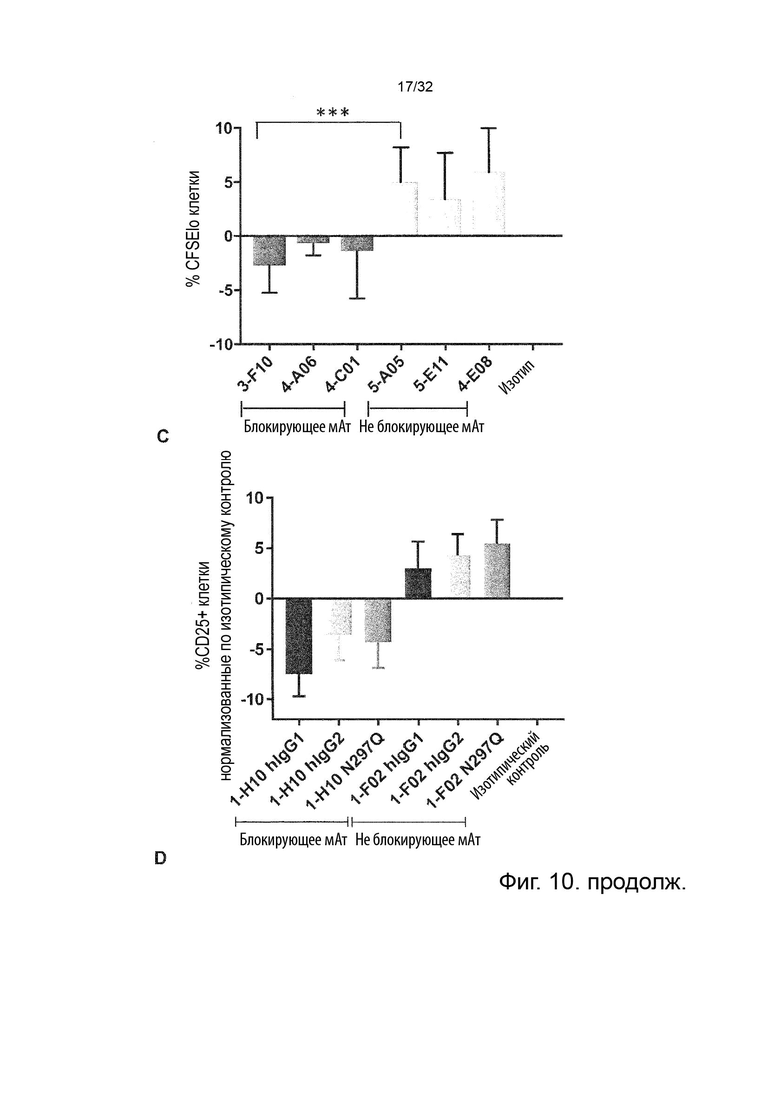
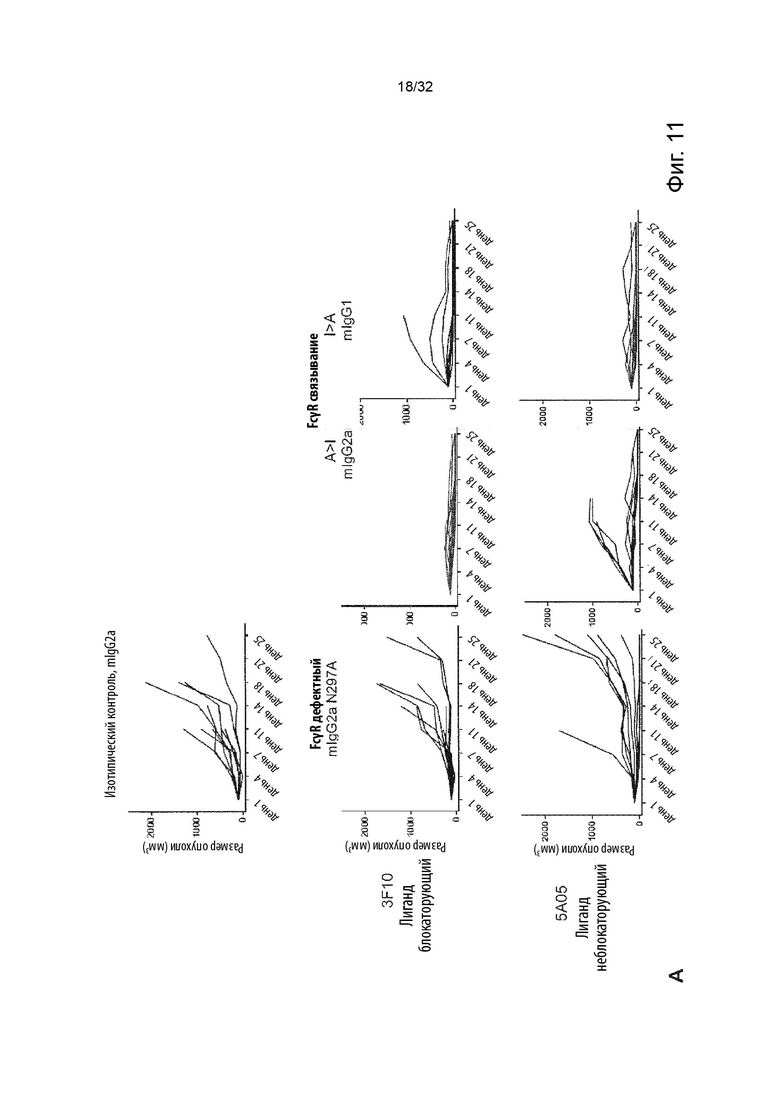
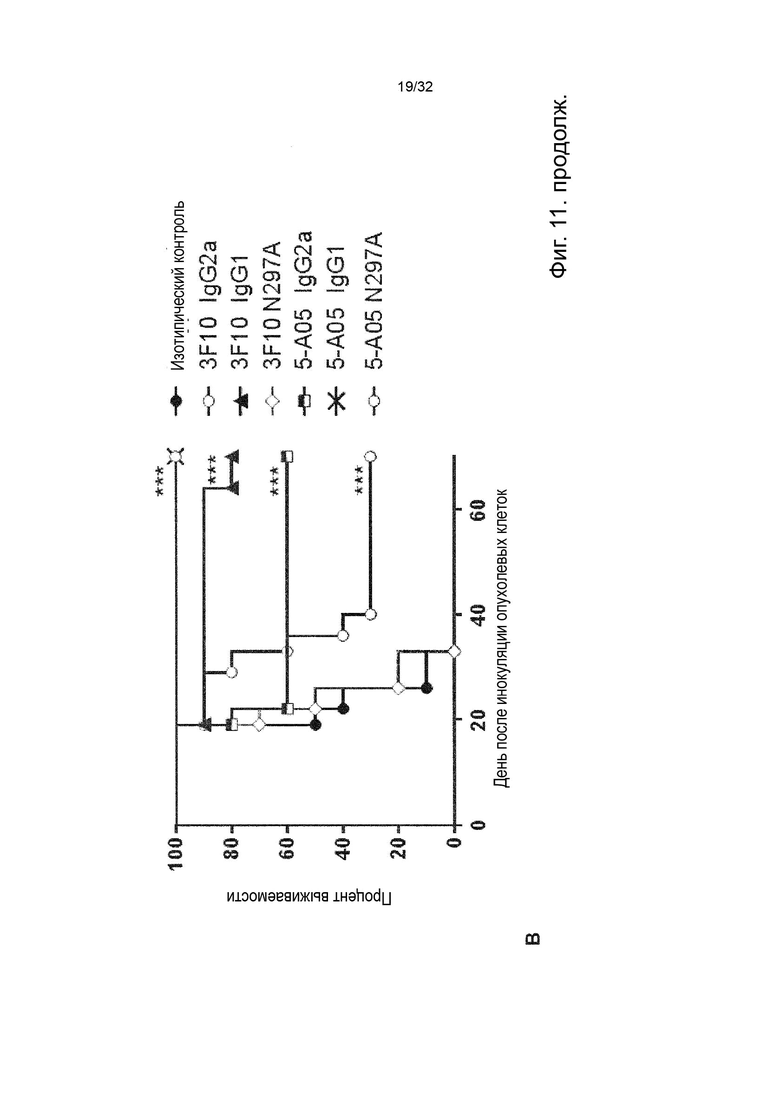
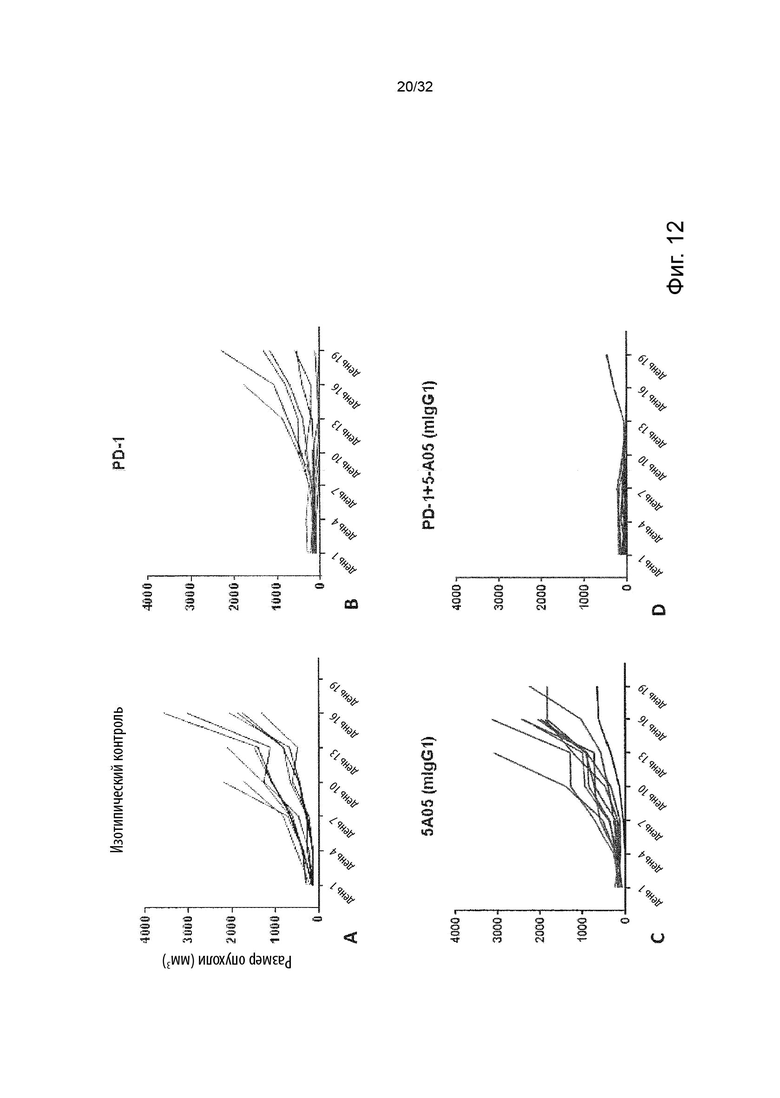
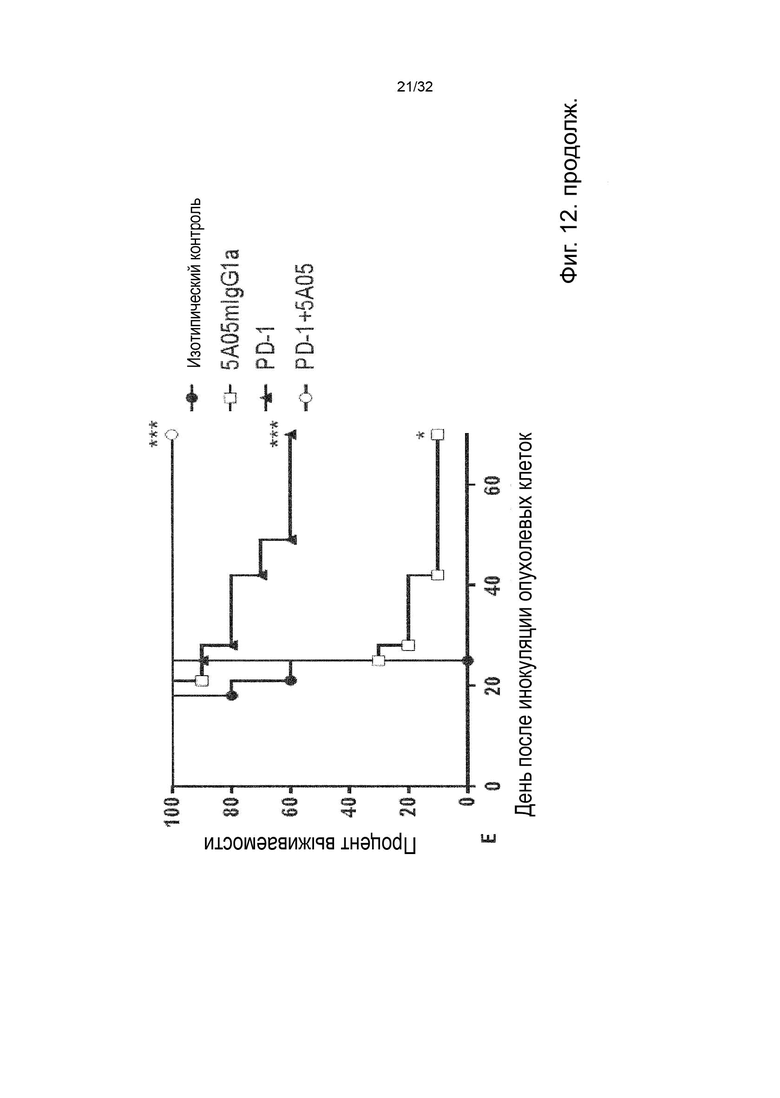
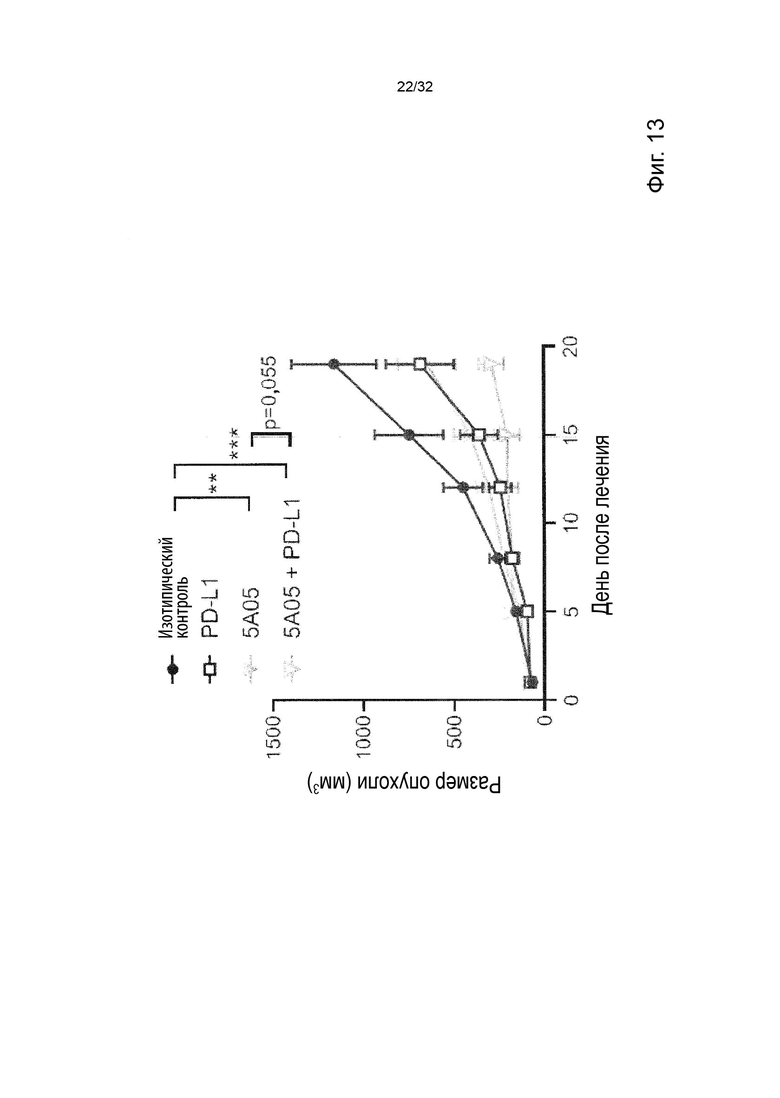
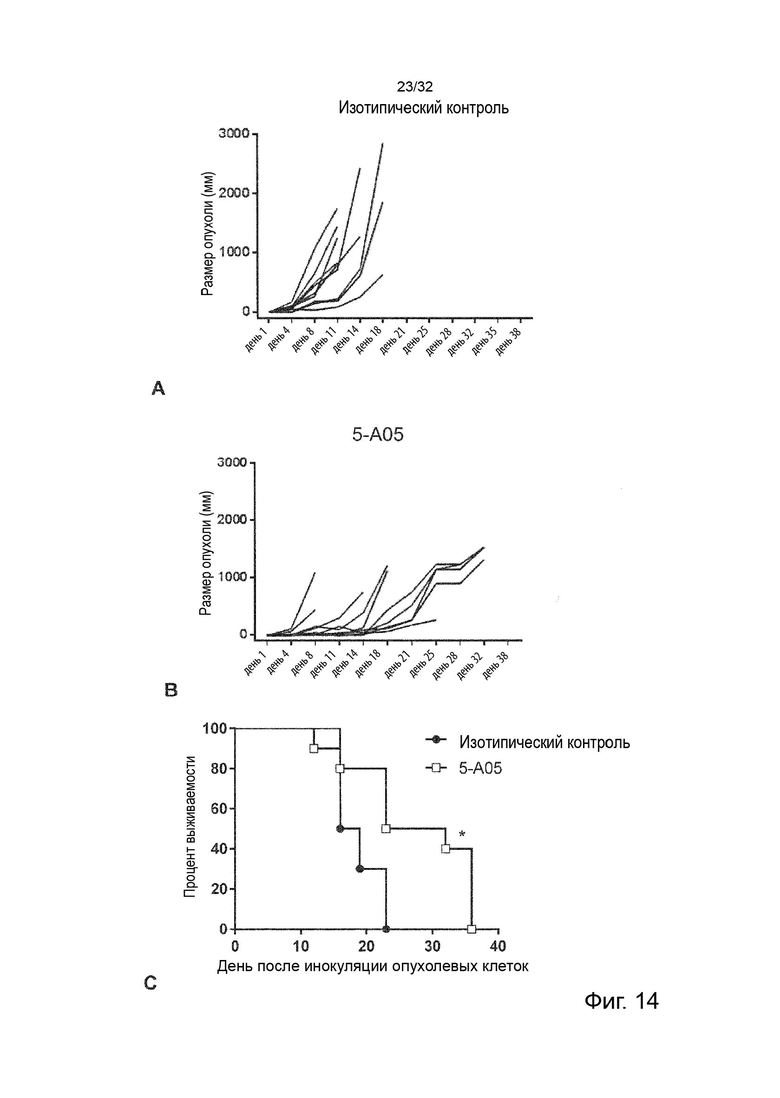
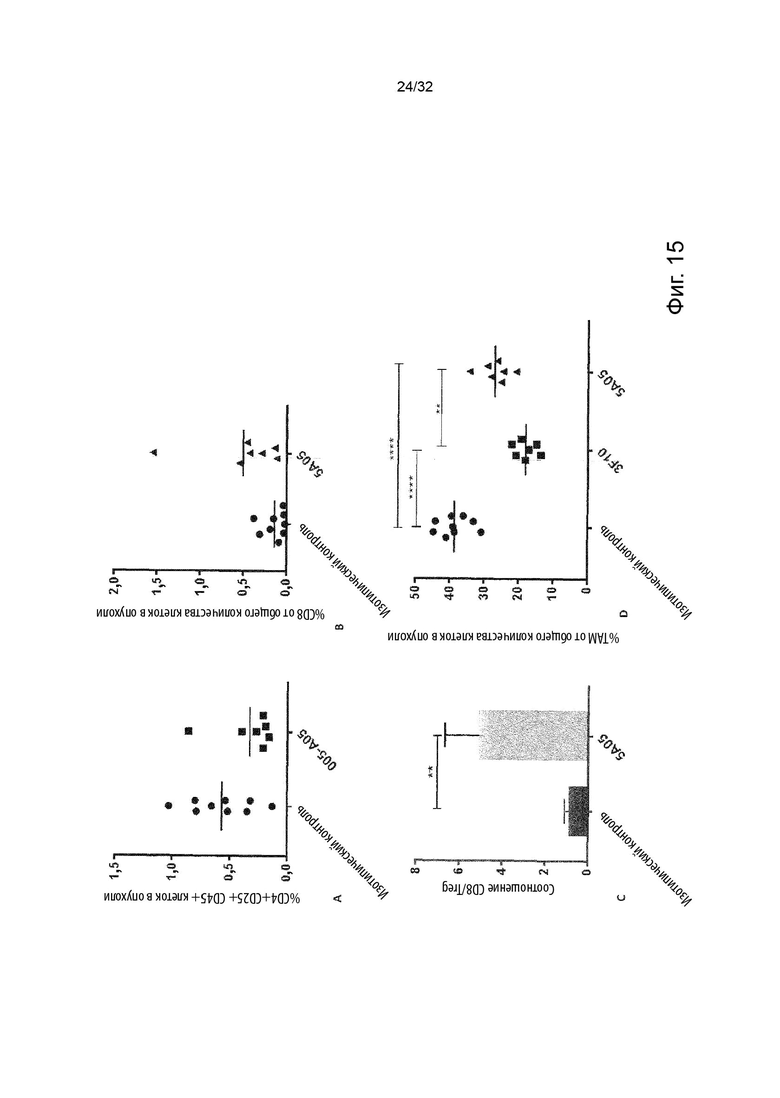
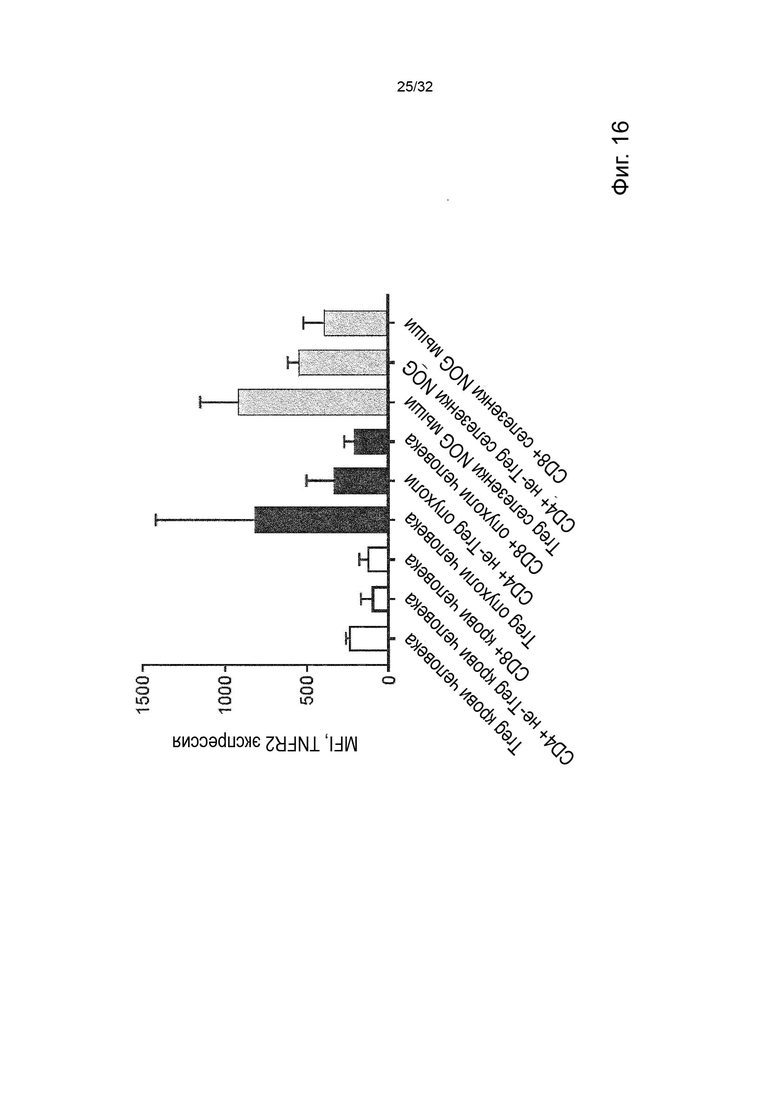
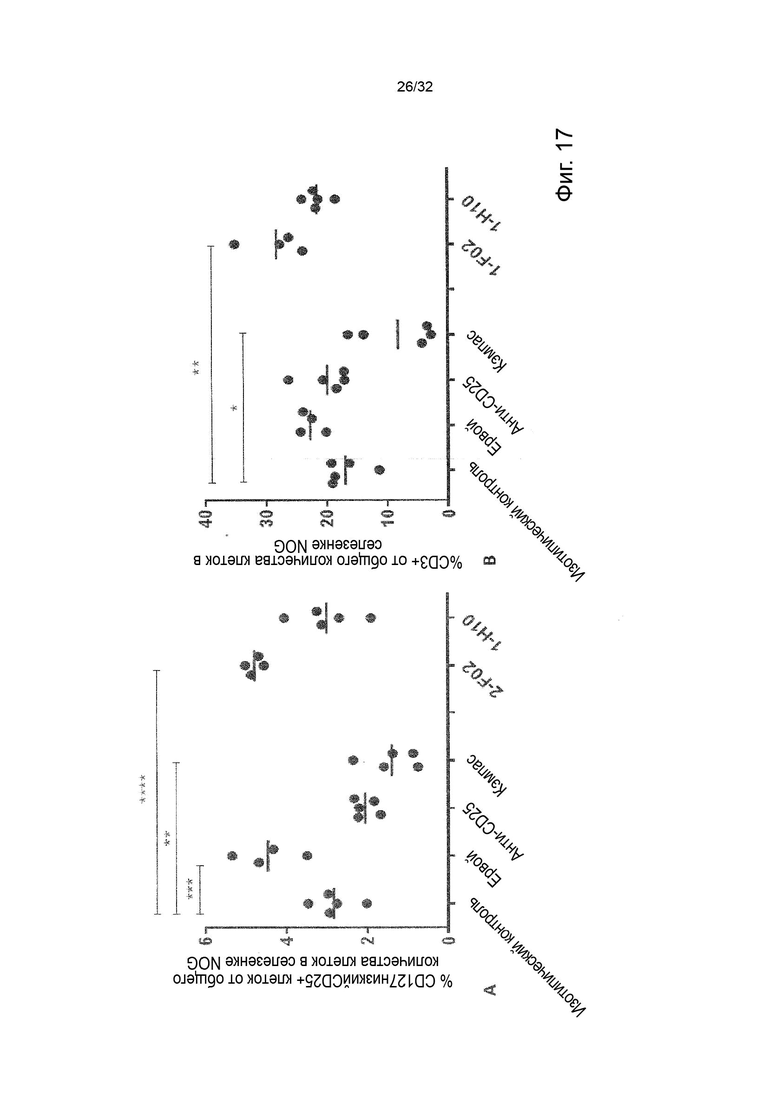
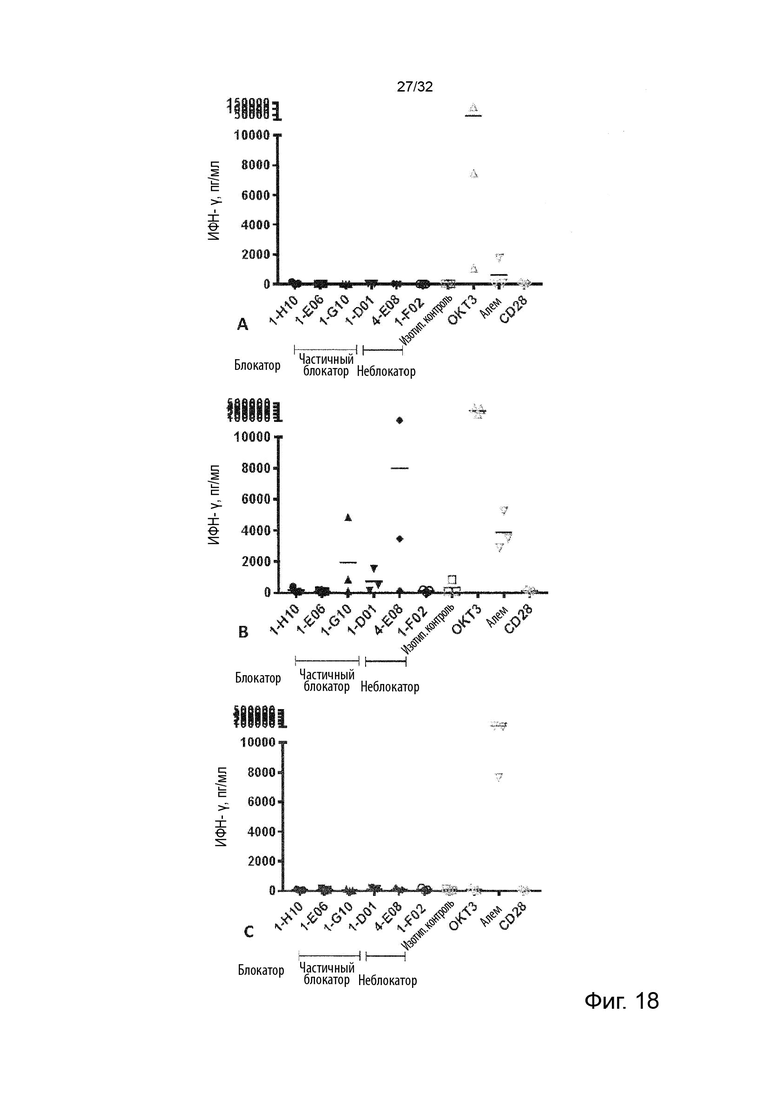
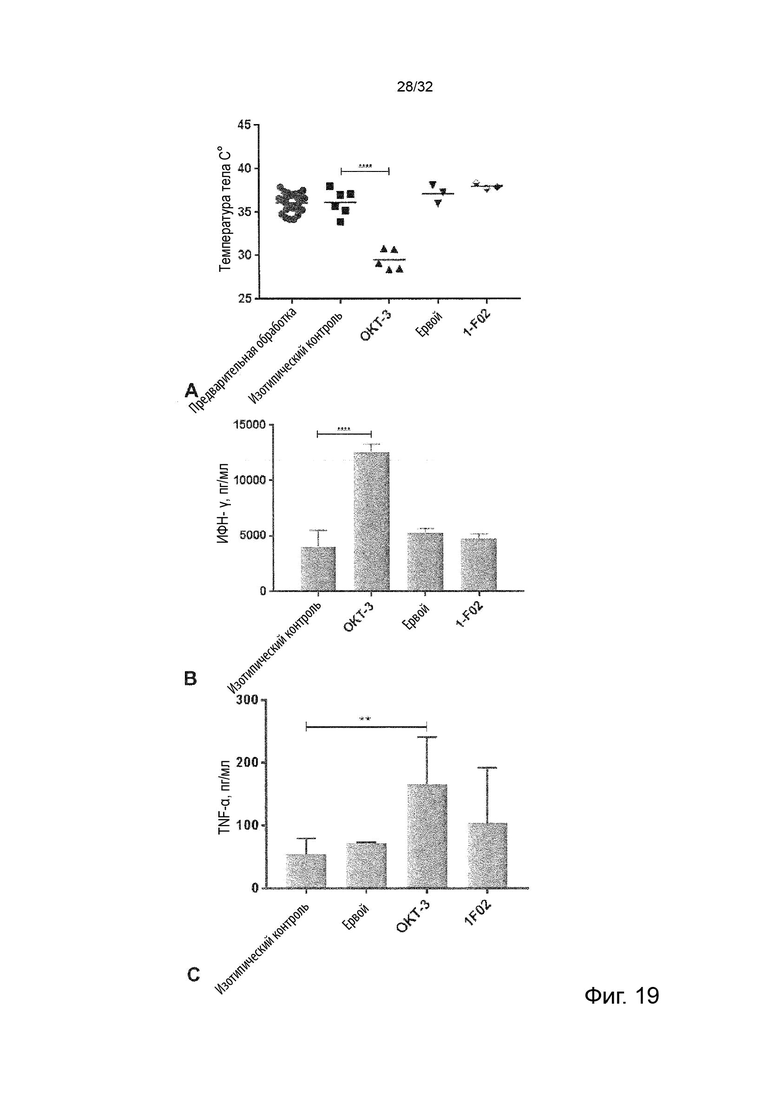
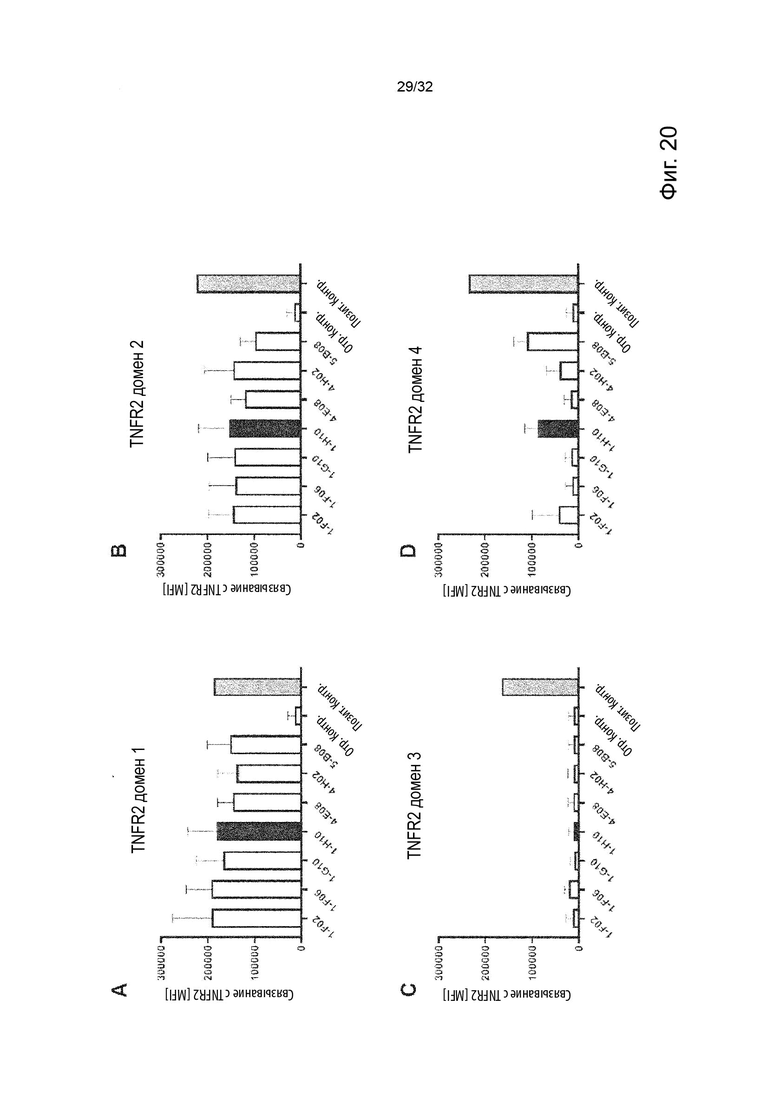
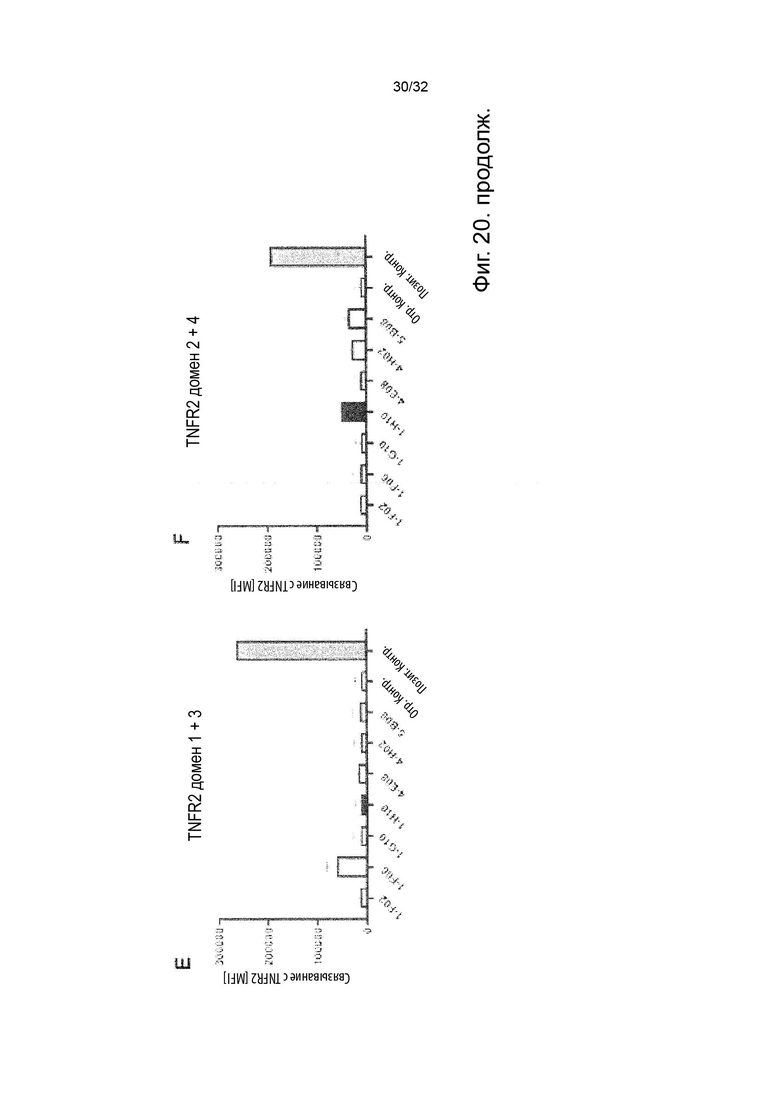

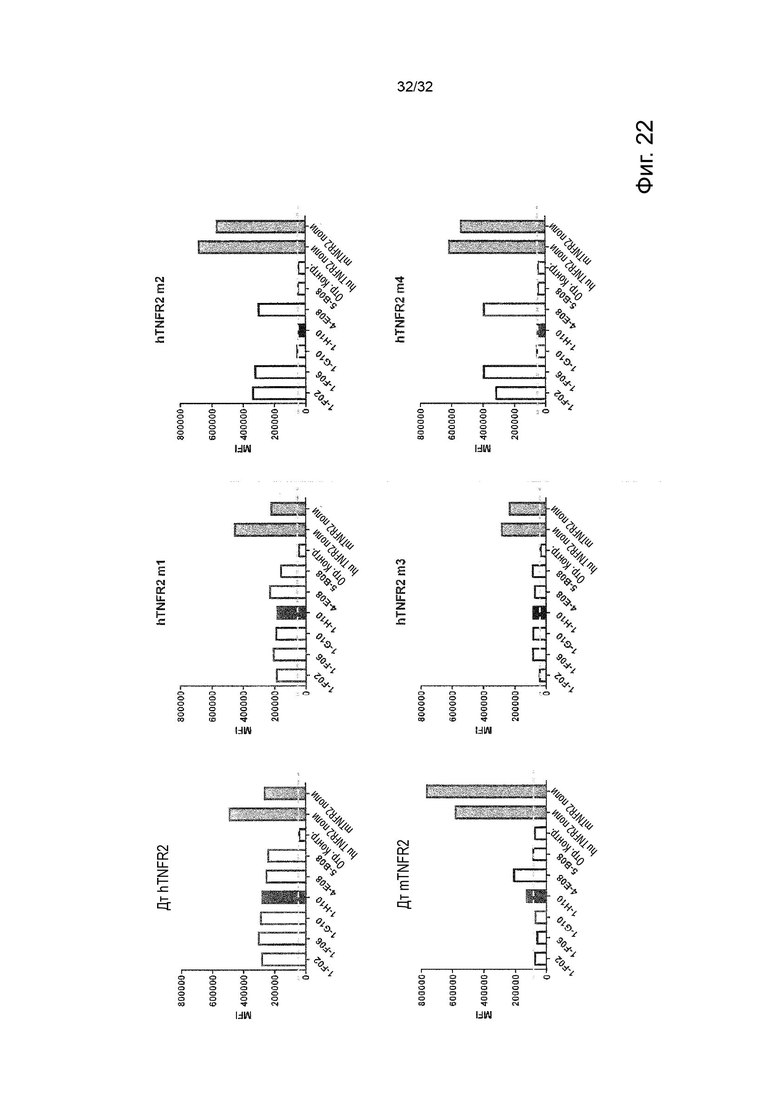
Изобретение относится к молекулярной биологии и медицине, в частности к новым молекулам антител, которые специфически связываются с TNFR2 на клетке-мишени и тем самым являются агонистами к TNFR2, но не блокируют лиганд TNF-α от связывания с TNFR2. Также описано применение таких молекул антител в медицине, например, при лечении рака или хронических воспалительных заболеваний. Антитела по настоящему изобретению являются мощными терапевтическими реагентами и пригодны в медицине. 10 н. и 27 з.п. ф-лы, 22 ил., 10 табл., 7 пр.
1. Молекула агонистического антитела или ее антигенсвязывающий фрагмент, специфически связывающиеся с TNFR2 на клетке-мишени, при этом указанная молекула антитела или ее антигенсвязывающий фрагмент не блокирует связывание лиганда TNF-α с TNFR2, где антитело или ее антигенсвязывающий фрагмент характеризуются внутренней агонистической активностью, где молекула антитела или ее антигенсвязывающий фрагмент содержат:
(a) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:1, SEQ ID NO:2 и SEQ ID NO:3, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:4, SEQ ID NO:5 и SEQ ID NO:6; или
(b) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:9, SEQ ID NO:10 и SEQ ID NO:11, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:12, SEQ ID NO:13 и SEQ ID NO:14; или
(c) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:17, SEQ ID NO:18 и SEQ ID NO:19, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:20, SEQ ID NO:21 и SEQ ID NO:22; или
(d) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:25, SEQ ID NO:26 и SEQ ID NO:27, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:28, SEQ ID NO:29 и SEQ ID NO:30; или
(e) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:33, SEQ ID NO:34 и SEQ ID NO:35, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:36, SEQ ID NO:37 и SEQ ID NO:38; или
(f) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:41, SEQ ID NO:42 и SEQ ID NO:43, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:44, SEQ ID NO:45 и SEQ ID NO:46; или
(g) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:49, SEQ ID NO:50 и SEQ ID NO:51, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:52, SEQ ID NO:53 и SEQ ID NO:54; или
(h) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:57, SEQ ID NO:58 и SEQ ID NO:59, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:60, SEQ ID NO:61 и SEQ ID NO:62; или
(i) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:65, SEQ ID NO:66 и SEQ ID NO:67, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:68, SEQ ID NO:69 и SEQ ID NO:70; или
(j) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:73, SEQ ID NO:74 и SEQ ID NO:75, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:76, SEQ ID NO:77 и SEQ ID NO:78; или
(k) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:81, SEQ ID NO:82 и SEQ ID NO:83, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:84, SEQ ID NO:85 и SEQ ID NO:86; или
(l) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:89, SEQ ID NO:90 и SEQ ID NO:91, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:92, SEQ ID NO:93 и SEQ ID NO:94; или
(m) вариабельную тяжелую цепь (VH), содержащую следующие CDR: SEQ ID NO:97, SEQ ID NO:98 и SEQ ID NO:99, и вариабельную легкую цепь (VL), содержащую следующие CDR: SEQ ID NO:100, SEQ ID NO:101 и SEQ ID NO:102.
2. Молекула антитела или ее антигенсвязывающий фрагмент по п. 1, отличающиеся тем, что также связываются с рецептором Fcγ.
3. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1, 2, отличающиеся тем, что связываются с более высокой аффинностью с ингибирующими рецепторами Fcγ, чем с активирующими рецепторами Fcγ.
4. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-3, отличающиеся тем, что их связывание с TNFR2 приводит к изменению в пораженной ткани количества и/или частоты клеток, экспрессирующих TNFR2.
5. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-4, отличающиеся тем, что их связывание с TNFR2 приводит к инфильтрации Т-клеток и/или миелоидных клеток в пораженную ткань, и/или изменению в составе Т-клеток и/или миелоидных клеток в пораженной ткани.
6. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-5, отличающиеся тем, что выбраны из группы, состоящей из полноразмерного антитела, химерного антитела, молекулы двухвалентного или поливалентного антитела, содержащего одноцепочечные антитела, Fab, Fv, scFv, Fab' и/или (Fab')2.
7. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-6, связывающиеся с TNFR2 человека (hTNFR2) и/или TNFR2 яванского макака (cmTNFR2).
8. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-7, где молекула антитела выбрана из группы, состоящей из молекулы антитела IgG человека, молекулы гуманизированного антитела IgG и молекулы антитела IgG человеческого происхождения.
9. Молекула антитела или ее антигенсвязывающий фрагмент по п. 8, где молекула антитела представляет собой антитело IgG2 человека.
10. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-9, где молекула антитела представляет собой моноклональное антитело.
11. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-10, отличающиеся тем, что не связываются специфически с эпитопом, содержащим или состоящим из последовательности KCSPG.
12. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-11, отличающиеся тем, что молекула антитела или его антигенсвязывающий фрагмент содержат:
(a) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:7 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:8; или
(b) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:15 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:16; или
(c) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:23 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:24; или
(d) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:31 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:32; или
(e) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:39 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:40; или
(f) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:47 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:48; или
(g) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:55 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:56; или
(h) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:63 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:64; или
(i) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:71 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:72; или
(j) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:79 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:80; или
(k) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:87 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:88; или
(l) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:95 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:96; или
(m) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:103 и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:104.
13. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-12, отличающиеся тем, что молекула антитела или его антигенсвязывающий фрагмент содержат:
(i) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:7, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:8; или
(ii) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:15, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:16; или
(iii) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:23, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:24; или
(iv) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:31, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:32; или
(v) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:39, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:40; или
(vi) аминокислотную последовательность вариабельной тяжелой цепи (VH) SEQ ID NO:47, и аминокислотную последовательность вариабельной легкой цепи (VL) SEQ ID NO:48.
14. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-11, где молекула антитела представляет собой молекулу антитела, которая способна конкурировать в связывании с TNFR2 с молекулой антитела, заявленной по любому из пп. 12, 13.
15. Изолированная нуклеиновая кислота, кодирующая молекулу антитела или ее антигенсвязывающий фрагмент, заявленные по любому из пп. 1-14, где указанная нуклеиновая кислота содержит нуклеотидную последовательность, кодирующую аминокислотную последовательность этого антитела или ее антигенсвязывающего фрагмента.
16. Экспрессионная плазмида, содержащая нуклеотидную последовательность по п. 15.
17. Вирус для доставки молекулы антитела или ее антигенсвязывающего фрагмента, содержащий нуклеиновую кислоту, кодирующую указанное антитело или его антигенсвязывающий фрагмент, где вирус характеризуется тем, что указанная нуклеиновая кислота представляет собой нуклеиновую кислоту по п. 15, а указанное антитело представляет собой антитело или его антигенсвязывающий фрагмент как определено в любом из пп. 1-14.
18. Вирус по п. 17, дополнительно содержащий нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
19. Клетка для доставки молекулы антитела или ее антигенсвязывающего фрагмента, заявленных по любому из пп. 1-14, содержащая нуклеиновую кислоту по п. 15.
20. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновая кислота по п. 15, плазмида по п. 16, вирус по любому из пп. 17, 18 и/или клетка по п. 19 для применения в лечении аутоиммунного заболевания, характеризующегося экспрессией TNF2 в иммунных клетках.
21. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновая кислота по п. 15, плазмида по п. 16, вирус по любому из пп. 17, 18 и/или клетка по п. 19 для применения в лечении рака у пациента, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью.
22. Молекула антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновая кислота по п. 15, плазмида по п. 16, вирус по п. 17 и/или клетка по п. 19 для применения в лечении рака у пациента, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью, в сочетании с:
молекулой антитела, которая специфически связывается с ингибитором контрольной точки;
нуклеотидной последовательностью, кодирующей молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки; и/или
клеткой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, или вирусом, содержащим нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
23. Применение молекулы антитела или ее антигенсвязывающего фрагмента по любому из пп. 1-14, нуклеиновой кислоты по п. 15, плазмиды по п. 16, вируса по п. 17 и/или клетки по п. 19 для получения фармацевтической композиции, применяемой в лечении рака у пациента, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью.
24. Применение по любому из пп. 22, 23, отличающееся тем, что указанную фармацевтическую композицию вводят в сочетании с:
молекулой антитела, которая специфически связывается с ингибитором контрольной точки;
нуклеотидной последовательностью, кодирующей молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки; и/или
клеткой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, или вирусом, содержащим нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
25. Применение молекулы антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновой кислоты по п. 15, плазмиды по п. 16, вируса по п. 17 и/или клетки по п. 19 для получения фармацевтической композиции, применяемой в лечении аутоиммунного заболевания, характеризующееся экспрессией TNF2 в иммунных клетках.
26. Применение по п. 25, отличающееся тем, что фармацевтическая композиция предназначена для применения при лечении аутоиммунного заболевания у пациента, имеющего высокую экспрессию TNFR2 в пораженной ткани по сравнению с соответствующей здоровой тканью.
27. Применение по п. 25 или 26, отличающееся тем, что указанную фармацевтическую композицию вводят в сочетании с:
молекулой антитела, которая специфически связывается с ингибитором контрольной точки;
нуклеотидной последовательностью, кодирующей молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки; и/или
клеткой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, плазмидой, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, или вирусом, содержащим нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
28. Фармацевтическая композиция для индуцирования передачи сигналов TNF2, содержащая молекулу антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновую кислоту по п. 15, плазмиду по п. 16, вирус по п. 17 и/или клетку по п. 19.
29. Фармацевтическая композиция по п. 28, где композиция содержит фармацевтически приемлемый разбавитель, носитель, наполнитель и/или эксципиент.
30. Фармацевтическая композиция по п. 28 для применения в лечении рака у пациента, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью, или аутоиммунного заболевания, характеризующегося экспрессией TNF2 в иммунных клетках.
31. Фармацевтическая композиция по п. 30 для применения в лечении рака у пациента, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью, в сочетании с фармацевтической композицией, содержащей:
молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
плазмиду, содержащую нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки; и/или
клетку, содержащую нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, плазмиду, содержащую нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, или вирус, содержащий нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
32. Способ лечения рака у субъекта, имеющего высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью, включающий введение субъекту терапевтически эффективного количества молекулы антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновой кислоты по п. 15, плазмиды по п. 16, вируса по п. 17, клетки по п. 19 или фармацевтической композиции по п. 28 или 29.
33. Способ лечения аутоиммунного заболевания, характеризующегося экспрессией TNF2 в иммунных клетках у субъекта, включающий введение субъекту терапевтически эффективного количества молекулы антитела или ее антигенсвязывающий фрагмент по любому из пп. 1-14, нуклеиновой кислоты по п. 15, плазмиды по п. 16, вируса по п. 17, клетки по п. 19 или фармацевтической композиции по п. 28 или 29.
34. Способ лечения рака по п. 32, отличающийся тем, что субъекту, имеющему высокую экспрессию TNF2 в пораженной ткани по сравнению с соответствующей здоровой тканью, также вводят терапевтически эффективное количество:
молекулы антитела, которая специфически связывается с ингибитором контрольной точки;
нуклеотидной последовательности, кодирующей молекулу антитела, которая специфически связывается с ингибитором контрольной точки;
плазмиды, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки; и/или
клетки, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, плазмиды, содержащей нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки, или вируса, содержащего нуклеотидную последовательность, кодирующую молекулу антитела, которая специфически связывается с ингибитором контрольной точки.
35. Молекула антитела или ее антигенсвязывающий фрагмент для применения по п. 22, нуклеиновая кислота для применения по п. 22, плазмида для применения по п. 22, вирус для применения по п. 22, клетка для применения по п. 22, применение по п. 24 или 27, фармацевтическая композиция по п. 31 или способ по п. 34, отличающиеся тем, что ингибитор контрольной точки представляет собой PD-1.
36. Молекула антитела или ее антигенсвязывающий фрагмент для применения по п. 22, нуклеиновая кислота для применения по п. 22, плазмида для применения по п. 22, вирус для применения по п. 22, клетка для применения по п. 22, применение по п. 24 или 27, фармацевтическая композиция по п. 31 или способ по п. 34, отличающиеся тем, что ингибитор контрольной точки представляет собой PD-L1.
37. Молекула антитела или ее антигенсвязывающий фрагмент для применения по пп. 21, 22 или 35, 36, нуклеиновая кислота для применения по пп. 21, 22 или 38, 39, плазмида для применения по пп. 21, 22 или 35, 36, вирус для применения по пп. 21, 22 или 35, 36, клетка для применения по пп. 21, 22 или пп. 35, 36, применение по пп. 23-27 или 35, 36, фармацевтическая композиция по пп. 28, 29, 31 или 35, 36 или способ по пп. 32, 33-36, отличающиеся тем, что рак представляет собой солидный рак.
| WILLIAMS G.S | |||
| et al., Phenotypic screening reveals TNFR2 as a promising target for cancer immunotherapy, Oncotarget, 2016, v | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Устройство для усиления микрофонного тока с применением самоиндукции | 1920 |
|
SU42A1 |
| Мотальный механизм конической и т.п. мотки | 1946 |
|
SU68278A2 |
| HE X | |||
| et al., A TNFR2-Agonist Facilitates High Purity Expansion of Human Low Purity Treg Cells, PLoS One, 2016, v | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| ANDO D | |||
| et al., Creation of mouse TNFR2-selective agonistic | |||

