Ссылка на список последовательностей, предоставленный в электронном виде
[0001] Содержание предоставленного в электронном виде списка последовательностей в текстовом файле ASCII (название 2943_083STR0_SeqListing_ST25.txt; размер: 143686 байт и дата создания: 3 марта 2016 года), поданного с заявкой, полностью включено в настоящее описание посредством ссылки.
Уровень техники, предшествующий изобретению
[0002] Плазмацитоидные дендритные клетки (pDC) представляют собой отдельную популяцию дендритных клеток (DC) в периферической крови и вторичных лимфоидных органах, которые составляют только приблизительно от 0,1 до 0,5% мононуклеарных клеток периферической крови (PBMC). Однако эти клетки являются особенно важными регуляторами иммунной системы, т.к. они являются основным источником интерферона I типа (IFN). IFN I типа поддерживают функцию NK клеток, B-клеток, T-клеток и миелоидных дендритных клеток. Эти IFN являются важными при первичных иммунных ответах и обладают противовирусной и противоопухолевой активностью. Однако полагают, что pDC и IFN I типа также играют роль в развитии аутоиммунных заболеваний, таких как системная красная волчанка, хронический ревматизм и псориаз. Таким образом, понимание того, как регулировать молекулярные пути, участвующие в высвобождении IFN, является пригодным для подавления иммунных ответов и лечения и профилактики заболевания.
[0003] pDC высвобождают IFN в ответ на нуклеиновые кислоты, которые распознаются Toll-подобными рецепторами (TLR) TLR7 и TLR9, экспрессируемые на поверхности pDC. Индуцированные TLR ответ регулируется рецепторами, содержащими иммунорецепторные тирозиновые активирующие мотивы (ITAM). Иммуноглобулиноподобный транскрипт 7 (ILT7), также называемый LIRA4, LILRA4 или CD85G, является одним из таких рецепторов.
[0004] ILT7 является представителем семейства генов иммуноглобулиноподобного транскрипта (ILT) или лейкоцитарного иммуноглобулиноподобного рецептора (LIR). ILT7 селективно экспрессируется на поверхности плазмацитоидных дендритных клеток человека (pDC) и не экспрессируется на миелоидных дендритных клетках или других лейкоцитах периферической крови. Cao et al., J. Exp. Medicine 6:1399-1405 (2006). ILT7 содержит четыре иммуноглобулиноподобных внеклеточных домена и трансмембранный домен. Внеклеточная часть является важной для взаимодействия с лигандом ILT7, антигеном стромальных клеток костного мозга 2 (BST2), и трансмембранный домен ILT7 содержит положительно заряженный остаток, который позволяет ему образовывать комплекс с FcεRIγ. Предполагалось, что взаимодействие BST2-ILT7 отрицательно регулирует естественную иммунную функцию pDC, возможно, как механизм отрицательной обратной связи. Кроме того, было показано, что антитело, сшивающее ILT7, in vitro отрицательно регулирует продукцию IFN-альфа и TNF-альфа посредством pDC. Таким образом, необходимыми являются антитела и другие связывающие ILT7 молекулы, которые являются пригодными для нейтрализующей ILT7 и регулирующей pDC активности и высвобождения IFN, например, для лечения и профилактики заболеваний, таких как аутоиммунные заболевания.
Область техники, к которой относится изобретение
[0005] Изобретение относится к связывающим ILT7 молекулам, например, к антителам к ILT7 и их антигенсвязывающим фрагментам, вариантам или производным, способам использования антител и фрагментов, и способам лечения или профилактики аутоиммунных заболеваний и состояний, связанных с экспрессирующими ILT7 клетками.
Краткое описание сущности изобретения
[0006] В настоящее описании предоставлены связывающие ILT7 молекулы, например, антитела к ILT7 и их антигенсвязывающие фрагменты.
[0007] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, который может связываться с тем же эпитопом ILT7 то и антитело, содержащее вариабельную область тяжелой цепи (VH) SEQ ID NO:202 и вариабельную область легкой цепи (VL) SEQ ID NO:207.
[0008] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, который конкурентно ингибирует связывание с ILT7 антитела, содержащего VH SEQ ID NO:202 и VL SEQ ID NO:207.
[0009] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, содержащий определяющие комплементарность области (CDR) HCDR1, HDR2, HCDR3, LCDR1, LCDR2 и LCDR3, содержащие последовательности SEQ ID NO: 203, 204, 205, 208, 209 и 210, соответственно.
[0010] В одном из вариантов осуществления связывающий ILT7 белок содержит VH по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный SEQ ID NO:202 и/или VL по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичный SEQ ID NO:207.
[0011] В одном из вариантов осуществления связывающий ILT7 белок содержит VH, содержащую SEQ ID NO:202, и VL, содержащую SEQ ID NO:207.
[0012] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, содержащий VH, содержащую SEQ ID NO:202.
[0013] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, содержащий VH, содержащую SEQ ID NO:207.
[0014] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, который может связываться с тем же эпитопом ILT7, что и антитело, содержащее VH и VL, выбранные из группы, состоящей из SEQ ID NO:12 и SEQ ID NO:17, соответственно SEQ ID NO:22 и SEQ ID NO:27, соответственно; SEQ ID NO:32 и SEQ ID NO:37, соответственно; SEQ ID NO:42 и SEQ ID NO:47, соответственно; SEQ ID NO:52 и SEQ ID NO:57, соответственно; SEQ ID NO:62 и SEQ ID NO:67, соответственно; SEQ ID NO:72 и SEQ ID NO:77, соответственно; SEQ ID NO:82 и SEQ ID NO:87, соответственно; SEQ ID NO:92 и SEQ ID NO:97, соответственно; SEQ ID NO:102 и SEQ ID NO:107, соответственно; SEQ ID NO:112 и SEQ ID NO:117, соответственно; SEQ ID NO:122 и SEQ ID NO:127, соответственно; SEQ ID NO:132 и SEQ ID NO:137, соответственно; SEQ ID NO:142 и SEQ ID NO:147, соответственно; SEQ ID NO:152 и SEQ ID NO:157, соответственно; SEQ ID NO:162 и SEQ ID NO:167, соответственно; SEQ ID NO:172 и SEQ ID NO:177, соответственно; SEQ ID NO:182 и SEQ ID NO:187, соответственно; SEQ ID NO:192 и SEQ ID NO:197, соответственно; SEQ ID NO:212 и SEQ ID NO:217, соответственно; SEQ ID NO:222 и SEQ ID NO:227, соответственно; SEQ ID NO:232 и SEQ ID NO:237, соответственно и SEQ ID NO:242 и SEQ ID NO:247, соответственно.
[0015] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, который конкурентно ингибирует связывание с ILT7 антитела, содержащего VH и VL, выбранные из группы, состоящей из SEQ ID NO:12 и SEQ ID NO:17, соответственно SEQ ID NO:22 и SEQ ID NO:27, соответственно; SEQ ID NO:32 и SEQ ID NO:37, соответственно; SEQ ID NO:42 и SEQ ID NO:47, соответственно; SEQ ID NO:52 и SEQ ID NO:57, соответственно; SEQ ID NO:62 и SEQ ID NO:67, соответственно; SEQ ID NO:72 и SEQ ID NO:77, соответственно; SEQ ID NO:82 и SEQ ID NO:87, соответственно; SEQ ID NO:92 и SEQ ID NO:97, соответственно; SEQ ID NO:102 и SEQ ID NO:107, соответственно; SEQ ID NO:112 и SEQ ID NO:117, соответственно; SEQ ID NO:122 и SEQ ID NO:127, соответственно; SEQ ID NO:132 и SEQ ID NO:137, соответственно; SEQ ID NO:142 и SEQ ID NO:147, соответственно; SEQ ID NO:152 и SEQ ID NO:157, соответственно; SEQ ID NO:162 и SEQ ID NO:167, соответственно; SEQ ID NO:172 и SEQ ID NO:177, соответственно; SEQ ID NO:182 и SEQ ID NO:187, соответственно; SEQ ID NO:192 и SEQ ID NO:197, соответственно; SEQ ID NO:212 и SEQ ID NO:217, соответственно; SEQ ID NO:222 и SEQ ID NO:227, соответственно; SEQ ID NO:232 и SEQ ID NO:237, соответственно и SEQ ID NO:242 и SEQ ID NO:247, соответственно.
[0016] В одном из вариантов осуществления выделенный связывающий ILT7 белок представляет собой связывающий ILT7 белок, содержащий CDR: HCDR1, HDR2, HCDR3, LCDR1, LCDR2 и LCDR3, выбранные из группы, состоящей из SEQ ID NO: 13, 14, 15, 18, 19 и 20, соответственно; SEQ ID NO: 23, 24, 25, 28, 29 и 30, соответственно; SEQ ID NO: 33, 34, 35, 38, 39 и 40, соответственно; SEQ ID NO: 103, 104, 105, 108, 109 и 110, соответственно; SEQ ID NO: 213, 214, 215, 218, 219 и 220, соответственно; SEQ ID NO: 223, 224, 225, 228, 229 и 230, соответственно; SEQ ID NO: 233, 234, 235, 238, 239 и 240, соответственно и SEQ ID NO: 243, 244, 245, 248, 249 и 250, соответственно.
[0017] В одном из вариантов осуществления связывающий ILT7 белок содержит VH и VL по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичные: SEQ ID NO:12 и SEQ ID NO:17, соответственно; SEQ ID NO:22 и SEQ ID NO:27, соответственно; SEQ ID NO:32 и SEQ ID NO:37, соответственно; SEQ ID NO:42 и SEQ ID NO:47, соответственно; SEQ ID NO:52 и SEQ ID NO:57, соответственно; SEQ ID NO:62 и SEQ ID NO:67, соответственно; SEQ ID NO:72 и SEQ ID NO:77, соответственно; SEQ ID NO:82 и SEQ ID NO:87, соответственно; SEQ ID NO:92 и SEQ ID NO:97, соответственно; SEQ ID NO:102 и SEQ ID NO:107, соответственно; SEQ ID NO:112 и SEQ ID NO:117, соответственно; SEQ ID NO:122 и SEQ ID NO:127, соответственно; SEQ ID NO:132 и SEQ ID NO:137, соответственно; SEQ ID NO:142 и SEQ ID NO:147, соответственно; SEQ ID NO:152 и SEQ ID NO:157, соответственно; SEQ ID NO:162 и SEQ ID NO:167, соответственно; SEQ ID NO:172 и SEQ ID NO:177, соответственно; SEQ ID NO:182 и SEQ ID NO:187, соответственно; SEQ ID NO:192 и SEQ ID NO:197, соответственно; SEQ ID NO:212 и SEQ ID NO:217, соответственно; SEQ ID NO:222 и SEQ ID NO:227, соответственно; SEQ ID NO:232 и SEQ ID NO:237, соответственно, или SEQ ID NO:242 и SEQ ID NO:247, соответственно.
[0018] В одном из вариантов осуществления VH и VL содержат SEQ ID NO:12 и SEQ ID NO:17, соответственно; SEQ ID NO:22 и SEQ ID NO:27, соответственно; SEQ ID NO:32 и SEQ ID NO:37, соответственно; SEQ ID NO:42 и SEQ ID NO:47, соответственно; SEQ ID NO:52 и SEQ ID NO:57, соответственно; SEQ ID NO:62 и SEQ ID NO:67, соответственно; SEQ ID NO:72 и SEQ ID NO:77, соответственно; SEQ ID NO:82 и SEQ ID NO:87, соответственно; SEQ ID NO:92 и SEQ ID NO:97, соответственно; SEQ ID NO:102 и SEQ ID NO:107, соответственно; SEQ ID NO:112 и SEQ ID NO:117, соответственно; SEQ ID NO:122 и SEQ ID NO:127, соответственно; SEQ ID NO:132 и SEQ ID NO:137, соответственно; SEQ ID NO:142 и SEQ ID NO:147, соответственно; SEQ ID NO:152 и SEQ ID NO:157, соответственно; SEQ ID NO:162 и SEQ ID NO:167, соответственно; SEQ ID NO:172 и SEQ ID NO:177, соответственно; SEQ ID NO:182 и SEQ ID NO:187, соответственно; SEQ ID NO:192 и SEQ ID NO:197, соответственно; SEQ ID NO:212 и SEQ ID NO:217, соответственно; SEQ ID NO:222 и SEQ ID NO:227, соответственно; SEQ ID NO:232 и SEQ ID NO:237, соответственно, или SEQ ID NO:242 и SEQ ID NO:247, соответственно.
[0019] В одном из вариантов осуществления выделенный связывающий ILT7 белок содержит VH, содержащую SEQ ID NO:12, 22, 32, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 212, 222, 232 или 242.
[0020] В одном из вариантов осуществления выделенный связывающий ILT7 белок содержит VL, содержащую SEQ ID NO:17, 27, 37, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187, 197, 217, 227, 237 или 247.
[0021] В одном из вариантов осуществления связывающий ILT7 белок содержит антитело или его антигенсвязывающий фрагмент. В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент являются афукозилированными.
[0022] В одном из вариантов осуществления связывающий ILT7 белок связывается с областью Ig1 ILT7. В одном из вариантов осуществления связывающий ILT7 белок связывается с область Ig2 ILT7.
[0023] В одном из вариантов осуществления связывающий ILT7 белок связывается с ILT7 человека и яванского макака.
[0024] В одном из вариантов осуществления связывающий ILT7 белок подавляет высвобождение интерферона (IFN) альфа из мононуклеарных клеток периферической крови (PBMC). В одном из вариантов осуществления связывающий ILT7 белок обладает активностью ADCC против плазмацитоидных дендритных клеток (pDC) в PMBC.
[0025] В одном из вариантов осуществления связывающий ILT7 белок содержит антитело мыши, антитело человека, химерное, гуманизированное антитело или антитело с измеренной поверхностью или его антигенсвязывающий фрагмент.
[0026] В одном из вариантов осуществления связывающий ILT7 белок содержит антитело, Fab, Fab', F(ab')2, Fd, одноцепочечный Fv или scFv, связанный дисульфидными связями Fv, домен V-NAR, IgNar, интраантитело, IgGΔCH2, миниантитело, F(ab')3, тетратело, триотело, диатело, однодоменное антитело, DVD-Ig, Fcab, mAb2, (scFv)2 или scFv-Fc.
[0027] В одном из вариантов осуществления связывающий ILT7 белок содержит моноклональное антитело или его антигенсвязывающий фрагмент.
[0028] В одном из вариантов осуществления связывающий ILT7 белок содержит константный домен тяжелой цепи иммуноглобулина, выбранный из группы, состоящей из: (a) константного домена IgA; (b) константного домена IgD; (c) константного домена IgE; (d) константного домена IgG1; (e) константного домена IgG2; (f) константного домена IgG3; (g) константного домена IgG4 и (h) константного домена IgM.
[0029] В одном из вариантов осуществления связывающий ILT7 белок содержит константный домен легкой цепи иммуноглобулина, выбранный из группы, состоящей из: (a) константного домена каппа-цепи Ig и (b) константного домена лямбда-цепи Ig.
[0030] В одном из вариантов осуществления связывающий ILT7 белок содержит константный домен IgG1 человека и константный домен лямбда-цепи человека.
[0031] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой клетку-хозяина, продуцирующую связывающую ILT7 молекулу.
[0032] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VH, где VH содержит аминокислотную последовательность по меньшей мере на 85%, 90%, 95% идентичную или идентичную VH SEQ ID NO:202, 12, 22, 32, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 212, 222, 232 или 242. В одном из вариантов осуществления полинуклеотид содержит последовательность по меньшей мере на 85%, 90%, 95% идентичную или идентичную SEQ ID NO:201, 11, 21, 31, 41, 51, 61, 71, 81, 91, 101, 111, 121, 131, 141, 151, 161, 171, 181, 191, 211, 221, 231 или 241.
[0033] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой выделенный полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VL, где VL содержит аминокислотную последовательность по меньшей мере на 85%, 90%, 95% идентичную, или идентичную VL 207, 17, 27, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187, 197, 217, 227, 237 или 247. В одном из вариантов осуществления полинуклеотид содержит последовательность по меньшей мере на 85%, 90%, 95% идентичную или идентичную SEQ ID NO:206, 16, 26, 36, 46, 56, 66, 76, 86, 96, 106, 116, 126, 136, 146, 156, 166, 176, 186, 196, 216, 226, 236 или 246.
[0034] В одном из вариантов осуществления нуклеиновая кислота является функционально связанной с контрольной последовательностью. В одном из вариантов осуществления антитело или его антигенсвязывающий фрагмент, содержащий VH или VL, кодируемые нуклеиновой кислотой, может специфически связываться с ILT7.
[0035] В одном из вариантов осуществления полинуклеотид, кодирующей связывающую ILT7 молекулу, приведен в настоящем описании.
[0036] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой вектор, содержащий полинуклеотид.
[0037] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой полипептид, кодируемый полинуклеотидом.
[0038] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой клетку-хозяина, трансформированную полинуклеотидом, предоставленным в настоящем описании (например, полинуклеотидом, содержащим нуклеиновую кислоту, кодирующую VH, и полинуклеотидом, содержащий нуклеиновую кислоту, кодирующую VL).
[0039] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой клетку-хозяина, содержащую полинуклеотид, предоставленный в настоящем описании (например, полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VH, и полинуклеотид, содержащий нуклеиновую кислоту, кодирующую VL), вектор, предоставленный в настоящем описании, или полипептид, предоставленный в настоящем описании. В одном из вариантов осуществления клетка-хозяин является клеткой-хозяином млекопитающих. В одном из вариантов осуществления клетка-хозяин представляет собой миеломные клетки мыши NS0, клетке человека PER.C6® или клетки яичника китайского хомяка (CHO). В одном из вариантов осуществления клетка-хозяин не содержит фермент α-1,6-фукозилтрансферазу.
[0040] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ получения связывающей молекулы против ILT7, включающий культивирование клетки-хозяина, предоставленной в настоящем описании, и выделение указанной связывающей молекулы. Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой связывающую молекулу против ILT7, получаемую способом.
[0041] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ детекции экспрессии ILT7 в образце, включающий (a) приведение образца в контакт со связывающей ILT7 молекулой, предоставленной в настоящем описании, и (b) детекцию связывания связывающей молекулы в образце.
[0042] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ детекции плазмацитоидных дендритных клеток, включающий (a) приведение образца, содержащего клетки, в контакт со связывающей ILT7 молекулой, предоставленной в настоящем описании, и (b) детекцию связывания связывающей молекулы в образце.
[0043] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой фармацевтическую композицию, содержащую (a) связывающую ILT7 молекулу, предоставленную в настоящем описании, полинуклеотид, предоставленный в настоящем описании, вектор, предоставленный в настоящем описании, полипептид, предоставленный в настоящем описании, или клетку-хозяина, предоставленную в настоящем описании, и (b) носитель.
[0044] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ снижения высвобождения IFN-альфа из плазмацитоидной дендритной клетки, включающий приведение плазмацитоидной дендритной клетки в контакт со связывающей ILT7 молекулой, представленной в настоящем описании, полинуклеотидом, предоставленном в настоящем описании, вектором, предоставленном в настоящем описании, полипептидом, предоставленном в настоящем описании, клеткой-хозяином, предоставленной в настоящем описании, или фармацевтической композицией, предоставленной в настоящем описании.
[0045] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ лечения являющегося человеком индивидуума с аутоиммунным заболеванием, включающий введение индивидууму эффективного количества связывающей ILT7 молекулы, предоставленной в настоящем описании, полинуклеотидом, предоставленным в настоящем описании, вектором, предоставленным в настоящем описании, полипептидом, предоставленным в настоящем описании, клеткой-хозяином, предоставленным в настоящем описании, или фармацевтической композицией, предоставленной в настоящем описании.
[0046] Один из вариантов осуществления, предоставленный в настоящем описании, представляет собой способ профилактики аутоиммунного заболевания у являющегося человеком индивидуума, включающий введение индивидууму эффективного количества связывающей ILT7 молекулы, предоставленной в настоящем описании, полинуклеотида, предоставленного в настоящем описании, вектора, предоставленного в настоящем описании, полипептида, предоставленного в настоящем описании, клетки-хозяина, предоставленной в настоящем описании, или фармацевтической композиции, предоставленной в настоящем описании. В одном из вариантов осуществления аутоиммунное заболевание представляет собой системную красную волчанку. В одном из вариантов осуществления аутоиммунное заболевание представляет собой хронический ревматизм.
Краткое описание чертежей/фигур
[0047] Фигуры 1A и 1B: демонстрируют выравнивания последовательностей вариабельной области тяжелой цепи (1A) и вариабельной области легкой цепи (1B) антител SBI28 (№28), 10D10 и 7C7. Затенение указывает на последовательности CDR. Открытые прямоугольники представляют мутации, вводимые в 10D10 с получением 7C7.
[0048] Фигура 2: демонстрирует связывание антител к ILT7 и антитела отрицательного контроля (R437) с клетками CT-550, экспрессирующими ILT7 человека, как определяют проточной цитометрией. SBI33 относится к антителу к ILT7 ILT7№33, как приведено в опубликованной заявке США № 2009/0280128.
[0049] Фигура 3: демонстрирует связывание антител к ILT7 и антитела отрицательного контроля (R437) с клетками CT-125, экспрессирующими ILT7 яванского макака, как определяют проточной цитометрией.
[0050] Фигура 4: демонстрирует активность ADCC антител к ILT7 и антитела отрицательного контроля (R437) против экспрессирующих ILT7 человека клеток.
[0051] Фигура 5: демонстрирует активность ADCC антител к ILT7 и антитела отрицательного контроля (R437) против экспрессирующих ILT7 яванского макака клеток.
[0052] Фигуры 6A и 6B: демонстрируют связывание антител к ILT7 и антитела отрицательного контроля (R437) с плазмацитоидными дендритными клетками (pDC) в мононуклеарных клетках периферической крови (PBMC).
[0053] Фигура 7: демонстрирует связывание афукозилированных антител к ILT7 и их исходных антител с клетками CT-550, экспрессирующими ILT7 человека (левая панель) и ILT7 яванского макака (правая панель) ILT7, как определяют проточной цитометрией.
[0054] Фигура 8: демонстрирует активность ADCC афукозилированных антител к ILT7 и их исходных антител против экспрессирующих ILT7 человека (левая панель) и ILT7 яванского макака (правая панель) клеток.
[0055] Фигуры 9A и 9B: демонстрируют выравнивания последовательностей вариабельной области тяжелой цепи (9A) и вариабельной области легкой цепи (9B) семи вариантов ILT70080. В выравнивании также продемонстрированы наиболее близкие последовательности зародышевой линии (IGHV1-69*01 и IGLV3-21*01).
[0056] Фигуры 10A и 10B: демонстрируют выравнивания последовательностей вариабельной области тяжелой цепи (10A) и вариабельной области легкой цепи (10B) девяти вариантов ILT70083. В выравнивании также продемонстрированы наиболее близкие последовательности зародышевой линии (IGHV3-23*01 и IGLV1-51*01).
[0057] Фигура 11: демонстрирует связывание вариантов ILT70080 с клетками экспрессирующими ILT7 человека (CT-550; верхняя панель) и клетками, экспрессирующими ILT7 яванского макака (CT-125; нижняя панель).
[0058] Фигура 12: демонстрирует связывание вариантов ILT70083 с клетками, экспрессирующими ILT7 человека (верхняя панель) или ILT7 яванского макака (нижняя панель).
[0059] Фигура 13: демонстрирует активность ADCC антител вариантов ILT70080 против экспрессирующих ILT7 человека клеток.
[0060] Фигура 14: демонстрирует активность ADCC антител вариантов ILT70083 против экспрессирующих ILT7 человека клеток.
[0061] Фигура 15: демонстрирует связывание афукозилированных антител к ILT70080.6 и ILT70083 с экспрессирующими ILT7 человек (левая панель) и яванского макака клетками (правая панель).
[0062] Фигура 16: демонстрирует активность ADCC афукозилированных антител к ILT70080.6 и ILT70083 на экспрессирующие ILT7 человека (левая панель) и ILT7 яванского макака (правая панель) клетки.
[0063] Фигура 17: демонстрирует цитотоксичность (слева) и секрецию IFN-α (сперва) PBMC человека, подвергаемых действию афукозилированных антител к ILT70080.6 и ILT70083.
[0064] Фигура 18: демонстрирует связывание афукозилированного ILT70137 с клетками, экспрессирующими ILT7 человека (левая панель) или ILT7 яванского макака (правая панель). Кружки означают афукозилированный ILT70137, и треугольники означают контроль.
[0065] Фигура 19: демонстрирует активность ADCC афукозилированного ILT70137 на клетки, экспрессирующие ILT7 человека (левая панель) или ILT7 яванского макака (правая панель). Треугольники означают афукозилированный ILT70137, и кружки означают контроль.
[0066] Фигура 20: демонстрирует активность ADCC афукозилированного ILT70137 путем измерения ингибирования продукции IFN-альфа как непрямая оценка способности антитела индуцировать ADCC мононуклеарных клеток периферической крови (PBMC) in vitro.
[0067] Фигура 21: демонстрирует связывание афукозилированного ILT70137 с первичными плазмацитоидными дендритными клетками (pDC) человека.
[0068] Фигура 22: демонстрирует истощение pDC у яванских макак, обрабатываемых афукозилированным 7C7 или афукозилированным ILT70137. Стрелки внизу графика указывают на моменты времени введения антитела.
[0069] Фигура 23: демонстрирует продукцию IFNα после обработки афукозилированным 7C7 или афукозилированным ILT70137. Стрелки внизу графика указывают на моменты времени введения антитела.
Подробное описание изобретения
I. Определения
[0070] Следует отметить, что термин в единственном числе относится к одному или более объектам; например, понимают, что "антитело к ILT7" представляет собой один или более антител к ILT7. Таким образом, в настоящем описании термины в единственном числе, "один или более" и "по меньшей мере один" могут использоваться взаимозаменяемо.
[0071] Как используют в настоящем описании, термин "полипептид" предназначен включать отдельный "полипептид", а также многие "полипептиды" и относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также известными как пептидные связи). Термин "полипептид" относится к любой цепи или цепям двух или более аминокислот и не относится к определенной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды, "белок", "аминокислотная цепь" или любой другой термин, используемый для обозначения цепи или цепей двух или более аминокислот, входят в определение "полипептида", и термин "полипептид" можно использовать вместо или взаимозаменяемо по отношению к любому из этих терминов. Термин "полипептид" также предназначен обозначать продукты постэкспрессионной модификаций полипептида, включая без ограничения, гликозилирование, ацетилирование, фосфорилирование, амидирование, дериватизацию известными защитными/блокирующими группами, протеолитическое расщепление или модификацию неприродными аминокислотами. Полипептид можно получать из естественного биологического источника или получать рекомбинантной технологией, но он не обязательно транслируется из указанной последовательности нуклеиновой кислоты. Его можно получать любым способом, включая химический синтез.
[0072] Размер полипептида по изобретению может составлять приблизительно 3 или более, 5 или более, 10 или более, 20 или более, 25 или более, 50 или более, 75 или более, 100 или более, 200 или более, 500 или более, 1000 или более, или 2000 или более аминокислот. Полипептиды могут иметь определенную трехмерную структуру, хотя они не обязательно имеют такую структуру. Полипептиды определенной трехмерной структурой обозначают как свернутые, и полипептиды, не обладают определенной трехмерной структурой, но могут принимать большое число различных конформаций, обозначают как несвернутый. Как используют в настоящем описании, термин гликопротеин относится к белку, связанному по меньшей мере с одной углеводной группой, которая присоединена к белку через содержащую кислород или содержащую азот боковую цепь аминокислотного остатка, например, остатка серина или остатка аспарагина.
[0073] Под "выделенным" полипептидом или его фрагментом, вариантом или производным понимают полипептид, который не находится в своем природном окружение. Не требуется конкретные уровни очистки. Например, выделенные полипептид можно удалять его нативного или природного окружения. Рекомбинантно продуцируемые полипептиды и белки, экспрессируемые в клетках-хозяевах считаются изолированными для целей изобретения, а также являются нативными или рекомбинантными полипептидами, которые отделяли, фракционировали или частично или в значительной степени очищали любым подходящим способом.
[0074] Также в понятие полипептиды по настоящему изобретению входят фрагменты, производные, аналоги или варианты указанных выше полипептидов и любое их сочетание. Термины "фрагмент," "вариант", "производное" и "аналог" по отношению к антителам к ILT7 или полипептидам антител по настоящему изобретению включают любые полипептиды, которые сохраняют по меньшей мере определенные антигенсвязывающие свойства соответствующего антитела или полипептида антитела по изобретению. Фрагменты полипептидов по настоящему изобретению включают протеолитические фрагменты, а также фрагменты делеции, в дополнение к конкретным фрагментам антител, описываемым в другом месте в настоящем описании. Варианты антител к ILT7 и полипептидов антител по настоящему изобретению включают фрагменты, как описано выше, а также полипептиды с измененными аминокислотными последовательностями вследствие замены, делеций или вставок аминокислот. Варианты могут возникать естественным образом или являться неприродными. Неприродные варианты можно получать с использованием известных техник мутагенеза. Полипептиды-варианты могут содержать консервативные или неконсервативные замены, делеции или добавления аминокислот. Полипептиды-варианты также могут быть обозначены в настоящем описании как "аналоги полипептидов". Как используют в настоящем описании, "производное" антитела к ILT7 или полипептида антитела означает отдельный полипептид, содержащий один или более остатков подвергнутых химической дериватизации в результате взаимодействия функциональной группы боковой цепи. Также в качестве "производных" включают такие пептиды, которые содержат одно или более природных производных аминокислот двадцати стандартных аминокислот. Например, 4-гидроксипролин можно замещать пролином; 5-гидроксилизин можно замещать лизином; 3-метилгистидин можно замещать гистидином; гомосерин можно замещать серином, и орнитин можно замещать лизином. Производные антител к ILT7 и полипептиды антител по настоящему изобретению могут включать полипептиды, которые изменяли таким образом, чтобы они обладали дополнительными свойствами, которые не встречаются у эталонного антитела или полипептида антитела по изобретению.
[0075] Термин "полинуклеотид" предназначен включать отдельную нуклеиновую кислоту, а также совокупность нуклеиновых кислот и относится к выделенной молекуле нуклеиновой кислоты или конструкции, например, информационной РНК (иРНК) или плазмидной ДНК (пДНК). Полинуклеотид может содержать общепринятую фосфодиэфирнйю связь или необщепринятую связь (например, амидную связь, такую как встречающуюся пептидных нуклеиновых кислотах (ПНК)). Термин "нуклеиновая кислота" относится к любому один или более сегментов нуклеиновой кислоты, например, фрагментам ДНК или РНК, содержащимся в полинуклеотиде. "Выделенная" нуклеиновая кислота или полинуклеотид предназначен включать молекулу нуклеиновой кислоты, ДНК или РНК, которую удалили из ее природного окружения. Например, рекомбинантный полинуклеотид, кодирующий связывающую молекулу против ILT7, например, антитело или его антигенсвязывающий фрагмент, содержащийся в векторе, считают выделенным для целей настоящего изобретения. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах, или очищенные (частично или по существу) полинуклеотиды в растворе. Выделенные молекулы РНК включают РНК транскрипты in vivo или in vitro полинуклеотидов по настоящему изобретению. Выделенные полинуклеотиды или нуклеиновые кислоты по настоящему изобретению дополнительно включают такие молекулы, получаемые синтетически. Кроме того, полинуклеотид или нуклеиновая кислота может представлять собой или может включать регуляторный элемент, такой как промотор, участок связывания рибосомы или терминатор транскрипции.
[0076] Как используют в настоящем описании, "кодирующая область" представляет собой участок нуклеиновой кислоты, который состоит из кодонов, транслируемых в аминокислоты. Несмотря на то, что "стоп-кодон" (TAG, TGA или TAA) не транслируется в аминокислоту, его можно считать частью кодирующей области, но какие-либо фланкирующие последовательности, например, промоторы, участки связывания рибосомы, терминаторы трансляции, интроны и т.п., не являются частью кодирующей области. Два или более кодирующие области по настоящему изобретению могут содержаться в одной полинуклеотидной конструкции, например, в одном векторе или в отдельных полинуклеотидных конструкциях, например, в отдельных (различных) векторах. Кроме того, любой вектор может содержать одну кодирующую область или может содержать две или более кодирующие области, например, один вектор может отдельно кодировать вариабельную область тяжелой цепи иммуноглобулина и вариабельную область легкой цепи иммуноглобулина. Кроме того, вектор, полинуклеотид или нуклеиновая кислота по изобретению может кодировать гетерологичные кодирующие области, слитые или не слитые с нуклеиновой кислотой, кодирующие антитело к ILT7 или его фрагмент, вариант или производное. Гетерологичные кодирующие области включают без ограничения специализированные элементы или мотивы, такие как секреторный сигнальный пептид или гетерологичный функциональный домен.
[0077] В определенных вариантах осуществления полинуклеотид или нуклеиновая кислота представляет собой ДНК. В случае ДНК полинуклеотид, содержащий нуклеиновую кислоту, кодирующую полипептид, как правило, может содержать промотор и/или другие элементы контроля транскрипции или трансляции, функционально связанные с одной или более кодирующими областями. Функционально связанный означает, в случае кодирующей области продукта гена, например, полипептида, является связанной с одной или более регуляторных последовательностей таким образом, чтобы помещать экспрессию продукта гена под влияние или контроль регуляторной последовательности(ей). Два фрагмента ДНК (такие как кодирующая область полипептида и промотор, связанный с ней) являются "функционально связанными", если введение функции промотора приводит к транскрипции иРНК, кодирующей желаемый продукт гена, и если природа связи между двумя фрагментами ДНК не нарушает способности регулирующих экспрессию последовательностей управлять экспрессией продукта гена или не нарушает способность ДНК-матрицы транскрибироваться. Таким образом, промоторная область функционально связана с нуклеиновой кислотой, кодирующей полипептид, если промотор мог осуществлять транскрипцию этой нуклеиновой кислоты. Промотор может представлять собой клеткоспецифичный промотор, который направляет по существу транскрипцию ДНК только в определенных клетках. Другие элементы контроля транскрипции, наряду с промотором, например, энхансеры, операторы, репрессоры и сигналы терминации транскрипции, можно функционально связывать с полинуклеотидом для направления клеткоспецифичной транскрипцией. Подходящие промоторы и другие области контроля транскрипции описаны в настоящем описании.
[0078] Специалистам в данной области известны различные области контроля транскрипции. Они включают без ограничения области контроля транскрипции, которые функционируют в клетках позвоночных, такие как, но, не ограничиваясь ими, промоторные и энхансерные сегменты из цитомегаловирусов (предранний промотор в сочетании с интроном A), вируса обезьян 40 (ранний промотор) и ретровирусов (таких как вирус саркомы Рауса). Другие области контроля транскрипции включат области контроля транскрипции, получаемые из генов позвоночных, такие как актин, белок теплового шока, бычий гормон роста и β-глобин кролика, а также другие последовательности, способные регулировать экспрессию гена в эукариотических клетках. Дополнительные подходящие области контроля транскрипции включают тканеспецифические промоторы и энхансеры, а также индуцибельные лимфокином промоторы (например, промоторы, индуцируемые интерферонами или интерлейкинами).
[0079] Подобным образом, специалистам в данной области известны различные элементы контроля трансляции. Они включают, но не ограничиваются ими, участки связывания рибосомы, кодоны инициации трансляции и терминации трансляции, и элементы, получаемые из пикорнавирусов (в частности, внутренний участок связывания рибосомы или IRES, также обозначаемый как последовательность CITE).
[0080] В других вариантах осуществления полинуклеотид по настоящему изобретению представляет собой РНК, например, в форме информационной РНК (иРНК).
[0081] Полинуклеотид и нуклеиновая кислота, кодирующие области по настоящему изобретению, можно связывать с дополнительными кодирующими областями, которые кодируют секреторные или сигнальные пептиды, которые направляют секрецию полипептида, кодируемого полинуклеотидом по настоящему изобретению. Согласно сигнальной гипотезе белки, секретируемые клетками млекопитающих, содержат сигнальный пептид или секреторную лидерную последовательность, которая отщепляются от зрелого белка после того, как началось выведение растущей белковой цепи через шероховатый эндоплазматический ретикулум. Специалистам в данной области известно, что полипептиды, секретируемые клетками позвоночных, как правило, содержат сигнальный пептид, слитый с N-концом полипептида, который отщепляется от полного или "полноразмерного" полипептида с получением секретируемой или "зрелой" формы полипептида. В определенных вариантах осуществления нативный сигнальный пептид, например, используют сигнальный пептид тяжелой цепи или легкой цепи иммуноглобулина или функциональное производное такой последовательности, которое сохраняет способность направлять секрецию полипептида, который является функционально связанным с ним. Альтернативно, можно использовать гетерологичный сигнальный пептид млекопитающего или его функциональное производное. Например, лидерную последовательность дикого типа можно замещать лидерной последовательностью активатора тканевого плазминогена человека(TPA) или β-глюкуронидазы мыши.
[0082] "Связывающая молекула" или "антигенсвязывающая молекула" по настоящему изобретению относится в широком смысле к молекуле, которая специфически связывается с антигенной детерминантой. В одном из вариантов осуществления связывающая молекула специфически связывается с ILT7, например, полноразмерным ILT7 или зрелым ILT7. В другом варианте осуществления связывающая молекула по изобретению представляет собой антитело или его антигенсвязывающий фрагмент. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере одну тяжелую или легкую цепь CDR молекула эталонного антитела. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере две CDR из одной или более молекул эталонного антитела. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере три CDR из одной или более молекул эталонного антитела. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере четыре CDR из одной или более молекул эталонного антитела. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере пять CDR из одной или более молекул эталонного антитела. В другом варианте осуществления связывающая молекула по изобретению содержит по меньшей мере шесть CDR из одной или более молекул эталонного антитела. В определенных вариантах осуществления эталонная молекула антител представляет собой 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052.
[0083] Настоящее изобретение относится к определенным антителам к ILT7 или их антигенсвязывающим фрагментам, вариантам или производным. Термин "антитело" означает молекулу иммуноглобулина, которая распознает и специфически связывается с мишенью, такой как белок, полипептид, пептид, углевод, полинуклеотид, липид или сочетания указанных выше через по меньшей мере один антиген участок распознавания в вариабельной области молекулы иммуноглобулина. Как используют в настоящем описании, термин "антитело" включает интактные поликлональные антитела, интактные моноклональные антитела, химерные антитела, гуманизированные антитела, антитела человека, слитые белки, содержащие антитело, и любую другую модифицированную молекулу иммуноглобулина при условии, что антитела обалдеют желаемой биологической активностью. Антитело может принадлежать к любому из пяти основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM или их подклассов (изотипов) (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2), на основании идентичности их константных доменов тяжелых цепей, обозначаемых как альфа, дельта, эпсилон, гамма и мю, соответственно. Различные классы иммуноглобулинов обладают различными и хорошо известными структурами субъединиц и трехмерными конфигурациями. Антитела могут являться "голыми" или конъюгированными с другими молекулами, такими как токсины, радиоактивные изотопы и т.д.
[0084] Термин "фрагмент антитела" или "его фрагмент антитело" относится к части интактного антитела. "Антигенсвязывающий фрагмент" или "его антигенсвязывающий фрагмент" относится к части интактного антитела, которая связывается с антигеном. Антигенсвязывающий фрагмент может содержать антигенопределяющие вариабельные области интактного антитела. Примеры фрагментов антител включают, но не ограничиваются ими, Fab, Fab', F(ab')2 и фрагменты Fv, линейные антитела, scFv и одноцепочечные антитела.
[0085] Как используют в настоящем описании, антитела "человека" или "полностью принадлежащие человеку" антитела включают антитела, содержащие аминокислотную последовательность иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулина человека или от животных, трансгенных по одному или более иммуноглобулинам человека, и у которых не экспрессируются эндогенные иммуноглобулины, как описано ниже и, например, в патенте США № 5939598 на имя Kucherlapati et al. Полностью принадлежащие человеку антитела являются особенно желательными для терапевтического лечения являющегося человеком пациента.
[0086] Антитела человека можно получать различными известными в данной области способами, включая способы фагового дисплея с использованием библиотек антител, получаемых из последовательностей иммуноглобулина человека, как описано у Vaughan et al., Nat. Biotech. 14:309-314 (1996), Sheets et al., Proc. Nat'l. Acad. Sci. 95:6157-6162 (1998), Hoogenboom and Winter, J. Mol. Biol. 227:381 (1992) и Marks et al., J. Mol. Biol. 222:581 (1991)). Дополнительные примеры способов фагового дисплея, которые можно использовать для получения и использования антител, включают способы, описываемые у Rothe et al., J. Mol. Biol., 376:1182 (2008), Brinkman et al., J. Immunol. Methods 182:41-50 (1995); Ames et al., J. Immunol. Methods 184:177-186 (1995); Kettleborough et al., Eur. J. Immunol. 24:952-958 (1994); Persic et al., Gene 187:9-18 (1997); Burton et al., Advances in Immunology 57:191-280 (1994); заявке PCT № PCT/GB91/01134; публикациях PCT WO 90/02809, WO 91/10737, WO 92/01047, WO 92/18619, WO 93/11236, WO 95/15982, WO 95/20401 и патентах США № 6172197, 5885793, 6521404, 6544731, 6555313, 6582915, 6593081, 6300064, 6653068, 6706484, 7264963, 5698426, 5223409, 5403484, 5580717, 5427908, 5750753, 5821047, 5571698, 5427908, 5516637, 5780225, 5658727, 5733743 и 5969108, каждый из которых полностью включен в настоящее описание посредством ссылки.
[0087] Кроме того, как известно в данной области, антитела человека можно получать с использованием трансгенных мышей, у которых не могут экспрессироваться функциональные эндогенные иммуноглобулины, но у которых могут экспрессировать гены иммуноглобулинов человека. Для обзора этой технологии см. Lonberg and Huszar, Int. Rev. Immunol. 13:65-93 (1995).
[0088] Дополнительные способы, доступные в данной области конструирования антител, позволили выделять антитела человека или их фрагменты. Например, гибридомы человека можно получать так, как описано Kontermann and Sefan. Antibody Engineering, Springer Laboratory Manuals (2001). Полностью принадлежащие человеку антитела можно, аналогично, получать различными технологиями дисплея, например, фагового дисплея или другими системами на основе вирусного дисплея. В способах фагового дисплея функциональные домены антитела предоставляют на поверхности фаговых частиц, которые несут полинуклеотидные последовательности, кодирующие их. Например, последовательности ДНК, кодирующие области VH и VL амплифицируют из библиотек кДНК животного (например, библиотек кДНК из лимфоидной ткани человека или мыши) или библиотек синтетической кДНК. В определенных вариантах осуществления ДНК, кодирующие области VH и VL, соединяют друг с другом через линкер scFv посредством ПЦР и клонируют в фагмидный вектор (например, p CANTAB 6 или pComb 3 HSS). Вектор электропорируют в E. coli и инфицируют E. coli фагом-помощником. Используемый в этих способах фаг, как правило, представляет собой нитевидный фаг, включая fd и M13, и области VH или VL, как правило, рекомбинантно сливают с геном III или геном VIII фага. Фаг, экспрессирующий антигенсвязывающий домен, который связывается с представляющим интерес антигеном (т.е. ILT7) можно выбирать или идентифицировать с антигеном, например, с использованием меченого антигена или антигена, связанного или захваченного на твердой поверхности или грануле.
[0089] Антитела "человека" или "полностью принадлежащие человеку" антитела также включают антитела, содержащие по меньшей мере вариабельный домен тяжелой цепи или по меньшей мере вариабельные домены тяжелой цепи и легкой цепи, где вариабельный домен(ы) содержат аминокислотную последовательность вариабельного домена(ов) иммуноглобулина человека.
[0090] Антитела "человека" или "полностью принадлежащие человеку" антитела также включают антитела "человека" или "полностью принадлежащие человеку" антитела, как описано выше, которые содержат, по существу состоят из или состоят из вариантов (включая производные) молекул антител (например, области VH и/или области VL), описываемых в настоящем описании, где антитела или их антигенсвязывающие фрагменты, варианты или производные иммуноспецифически связываются с полипептидом ILT7 или его фрагментом или вариантом. Стандартные способы, известных специалистам в данной области можно использовать для введения мутаций в нуклеотидную последовательность, кодирующую антитело человека к ILT7, включая, но, не ограничиваясь ими, сайт-направленный мутагенез и опосредованный ПЦР мутагенез, которые приводят к заменам аминокислот. Варианты (включая производные) могут кодироваться менее чем 50 заменами аминокислот, менее чем 40 заменами аминокислот, менее чем 30 заменами аминокислот, менее чем 25 заменами аминокислот, менее чем 20 заменами аминокислот, менее чем 15 заменами аминокислот, менее чем 10 заменами аминокислот, менее чем 5 заменами аминокислот, менее чем 4 заменами аминокислот, менее чем 3 заменами аминокислот или менее чем 2 заменами аминокислот относительно эталонной области VH, VHCDR1, VHCDR2, VHCDR3, области VL, VLCDR1, VLCDR2 или VLCDR3.
[0091] В определенных вариантах осуществления замены аминокислот являются консервативными заменами аминокислот, дополнительно описанными ниже. Альтернативно, мутации можно вводить в случайном порядке по всей длине или в части кодирующей последовательности, таким образом, как посредством насыщающего мутагенеза, и можно проводить скрининг получившихся мутантов на биологическую активность для идентификации мутантов, которые сохраняют активность (например, способность связываться с полипептидом ILT7, например, человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши). Такие варианты (или их производные) антител "человека" или "полностью принадлежащих человеку" антител также могут быть обозначены как антитела человека или полностью принадлежащие человеку антитела, которые являются "оптимизированными" или "оптимизированными по связыванию антигена", и включают антитела, которые обладают улучшенной аффинностью к антигену.
[0092] Основные структуры иммуноглобулина в позвоночных организмах относительно хорошо известны. См., например, Harlow et al. (1988) Antibodies: A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory Press).
[0093] Как описано более подробно ниже, термин "иммуноглобулин" включает различные обширные классы полипептидов, которые можно различать на биохимическом уровне. Специалистам в данной области понятно, что тяжелые цепи классифицируют как гамма, мю, альфа, дельта или эпсилон, (γ, μ, α, δ, ε) с некоторыми подклассами среди них (например, γ1-γ4). Именно природа этой цепи определяет "класс" антитела как IgG, IgM, IgA IgG или IgE, соответственно. Подклассы иммуноглобулинов (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и т.д. хорошо охарактеризованы и, как известно, обеспечивают функциональную специализацию. Специалист в данной области может легко различать модифицированные варианты каждого из этих классов и изотипов, принимая во внимание настоящее описание, и, таким образом, они входят в объем настоящего изобретения. Несмотря на то, что следующее ниже описание, как правило, относится к молекулам иммуноглобулинов класса IgG, все классы иммуноглобулинов полностью входят в объем настоящего изобретения. Касательно IgG, стандартная молекула иммуноглобулина содержит два идентичных полипептида легкой цепи молекулярной массы приблизительно 23000 Дальтон, и два идентичных полипептида тяжелой цепи молекулярной массой 53000-70000. Четыре цепи, как правило, соединены дисульфидными связями в конфигурации "Y", где легкие цепи закрепляют тяжелые цепи, начинающиеся в месте начала "Y" и продолжающиеся в вариабельной области.
[0094] Легкие цепи классифицируют как каппа или лямбда (κ, λ). Каждый класс тяжелых цепей может встречаться с легкой каппа- или лямбда-цепью. В основном, легкие и тяжелые цепи являются ковалентно связанными друг с другом, и "хвостовые" участки двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями, когда иммуноглобулины получают посредством гибридом, B-клеток или сконструированными с помощью генетической инженерии клетками-хозяевами. В тяжелой цепи аминокислотные последовательности идет от N-конца на раздвоенном конце Y-конфигурации к C-концу на конце каждой цепи.
[0095] Основание антитела "Y" называется Fc-область (кристаллизующийся фрагмент) и состоит из двух тяжелых цепей, которые предоставляют два или три константных домена в зависимости от класса антитела. Таким образом, Fc-область связывается с конкретным классом Fc-рецепторов и другими иммунными молекулами, такими как комплементные белки. Легкие и тяжелые цепи разделяют на области структурной и функциональной гомологии. Термины "константный" и "вариабельный" используют в отношении функции. В отношении этого, следует понимать, что вариабельные домены участков легких (VL или VK) и тяжелых (VH) цепей определяют распознавание антигена и специфичность. В противоположность этому, константные домены легкой цепи (CL) и тяжелой цепи (CH1, CH2 или CH3) обеспечивают важные биологические свойства, такие как секреция, трансплацентарный перенос, связывание Fc-рецептора, связывание комплемента и т.п. По соглашению нумерация доменов константной области увеличивается по мере того, как они отдаляются от антигенсвязывающего участка или N-конца антитела. N-концевой участок представляет собой вариабельную область, и на C-концевой участке располагается константная область; домены CH3 и CL фактически содержат C-конец тяжелых и легких цепей, соответственно.
[0096] Как указано выше, вариабельная область позволяет антителу селективно распознавать и специфически связывать эпитопы на антигенах. Таким образом, домен VL и домен VH или подгруппа определяющих комплементарность областей (CDR) в этих вариабельных доменах антитела объединяется, образуя вариабельную область, которая определяет трехмерный антигенсвязывающий участок. Такая четвертичная структура антитела образует антигенсвязывающий участок, присутствующий на конце каждого плеча Y. Более конкретно, антигенсвязывающий участок определяют три CDR на каждой из цепей VH и VL. В некоторых случаях, например, определенные молекулы иммуноглобулина получают от видов верблюдовых или конструируют на основании иммуноглобулинов верблюдовых, полная молекула иммуноглобулина может состоять только из тяжелых цепей без легких цепей. См., например, Hamers-Casterman et al., Nature 363:446-448 (1993).
[0097] В природных антителах шесть "определяющих комплементарность областей" или "CDR", содержащиеся на каждом антигенсвязывающем домене, представляют собой короткие, непрерывные последовательности аминокислот, которые располагаются особым образом, образуя антигенсвязывающий домен, т.к. антитело принимает свою трехмерную конфигурацию в водной среде. Оставшаяся часть аминокислот в антигенсвязывающих доменах, обозначаемые как "каркасные" области, обладают меньшей межмолекулярной изменчивостью. Каркасные области главным образом принимают конформацию β-листа, и CDR образуют петли, которые соединяют и в некоторых случаях образуют часть структуры β-листа. Таким образом, каркасные области функционируют с образованием каркаса, который обеспечивает расположение CDR в правильной ориентации в результате межцепочечных нековалентных взаимодействий. Антигенсвязывающий домен, образованный расположенными в особом порядке CDR, определяет поверхность, комплементарную эпитопу на иммунореактивном антигене. Такая комплементарная поверхность способствует нековалентному связыванию антитела с его когнатным эпитопом. Специалист в данной области может легко идентифицировать аминокислоты, содержащие CDR и каркасные области, соответственно, для любого данного вариабельного домена тяжелой или легкой цепи, т.к. они были точно определены (см. ниже).
[0098] В случае, когда имеются два или более определений термина, которые используются и/или приняты в данной области, то используемое в настоящем описании определение термина предназначено включать все такие значения, если явно не указано иное. Конкретным примером является использование термина "определяющая комплементарность область" ("CDR") для описания несмежных антигенсвязывающих участков, встречающихся в вариабельной области полипептидов тяжелой и легкой цепей. Эта конкретная область описана у Kabat et al. (1983) U.S. Dept. of Health and Human Services, "Sequences of Proteins of Immunological Interest" и Chothia and Lesk, J Mol. Biol. 196:901-917 (1987), которые включены в настоящее описание посредством ссылки, где определения включают перекрывающиеся или подгруппы аминокислотных остатков при сравнении друг с другом. Однако применение любого определения для ссылки на CDR антитела или его вариантов предназначено входить в объем термина, как определено и как используют в настоящем описании. IMGT (ImMunoGeneTics) также предоставляет систему нумерации вариабельных областей иммуноглобулинов, содержащих CDR. См., например, Lefranc M.P. et al., Dev. Comp. Immunol. 27: 55-77(2003), который включен в настоящее описание посредством ссылки. Система нумерации IMGT была основана на выравнивании более чем 5000 последовательностей, данных о структуре и характеристике гипервариабельных петель и позволяет легко сравнивать вариабельные области и области CDR для всех видов. Подходящие аминокислотные остатки, которые включают CDR, как определяют посредством каждой из указанных выше ссылок, указаны ниже в таблице 1 как сравнение. Точная нумерация остатков, которые включают конкретную CDR, может изменяться в зависимости от последовательности и размера CDR. Специалисты в данной области могут общепринято определять, какие остатки содержат конкретную CDR с учетом аминокислотной последовательности вариабельной области антитела.
Таблица 1. Определения CDR1
1Нумерация все определений CDR в таблице 1 соответствует системам нумераций, указанных Kabat et al. (см. ниже).
[0099] Kabat et al. также определили систему нумерации для последовательностей вариабельных доменов, которая является применимой к любому антителу. Специалист в данной области может однозначно применять данную систему "нумерации по Kabat" к любой последовательности вариабельного домена без учета каких-либо экспериментальных данных за пределами самой последовательности. Как используют в настоящем описании, термин "нумерация по Kabat" относится к системе нумерации, изложенной у Kabat et al. (1983) U.S. Dept. of Health and Human Services, "Sequence of Proteins of Immunological Interest".
[00100] Антитела или их антигенсвязывающие фрагменты, варианты или производные по изобретению включают без ограничения поликлональные, моноклональные, антитела мыши, антитела человека, гуманизированные, приматизированные или химерные антитела, одноцепочечные антитела, связывающие эпитоп фрагменты, например, Fab, Fab' и F(ab')2, Fd, Fv, одноцепочечные Fv (scFv), связанные дисульфидными связями Fv (sdFv), фрагменты, содержащие домен VL или VH, фрагменты, получаемые посредством expression библиотеки экспрессии Fab, и анти-идиотипические антитела (антитела Id) (включая, например, антитела Id к антителу к ILT7, описываемому в настоящем описании). Молекулы scFv известны в данной области и описаны, например, в патенте США № 5892019. Молекулы иммуноглобулина или антитела по изобретению могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2, и т.д.) или подкласса молекулы иммуноглобулина.
[00101] Как используют в настоящем описании, термин "часть тяжелой цепи" включает аминокислотные последовательности, получаемые из тяжелой цепи иммуноглобулина. Полипептид, содержащий часть тяжелой цепи содержит по меньшей мере один из: домена CH1, шарнирного домена (например, верхней, средней и/или нижней шарнирной области), домена СН2, домена СН3 или их варианта или фрагмента. Например, связывающий полипептид для применения в изобретении может содержать полипептидную цепь, содержащую домен СН1; полипептидную цепь, содержащую домен СН1, по меньшей мере часть шарнирного домена и домен СН2; полипептидную цепь, содержащую домен СН1 и домен СН3; полипептидную цепь, содержащую домен СН1, по меньшей мере часть шарнирного домена, и домен СН3 или полипептидную цепь, содержащую домен СН1, по меньшей мере часть шарнирного домена, домен СН2 и домен СН3. В другом варианте осуществления полипептид по изобретению содержит полипептидную цепь, содержащую домен СН3. Кроме того, связывающий полипептид для применения в изобретении может не содержать по меньшей мере часть домена СН2 (например, весь или часть домена СН2). Как указано выше, специалисту в данной области должно быть понятно, что эти домены (например, части тяжелой цепи) можно модифицировать таким образом, чтобы они отличались аминокислотной последовательностью от природной молекулы иммуноглобулина.
[00102] В определенных антителах к ILT7 или их антигенсвязывающих фрагментах, вариантах или производных, описываемых в настоящем описании, части тяжелой цепи одной полипептидной цепи мультимера являются идентичными частям на второй полипептидной цепи мультимера. Альтернативно, содержащие часть тяжелой цепи мономеры по изобретению не являются идентичными.
[00103] Части тяжелой цепи связывающей молекулы для применения в способах диагностики и лечения, описанных в настоящем описании, можно получать из различных молекул иммуноглобулина. Например, часть тяжелой цепи полипептида может содержать домен CH1, получаемый из молекулы IgG1, и шарнир, получаемый из молекулы IgG3. В другом примере часть тяжелой цепи может содержать шарнирную часть, получаемую частично из молекулы IgG1 и частично из молекулы IgG3. В другом примере часть тяжелой цепи может содержать химерный шарнир, получаемый частично из молекулы IgG1 и частично из молекулы IgG4.
[00104] Как используют в настоящем описании, термин "часть легкой цепи" включает аминокислотные последовательности, получаемые из легкой цепи иммуноглобулина, например, легкой каппа- или лямбда-цепи. Часть легкой цепи может содержать по меньшей мере один из домена VL или CL.
[00105] Антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные, описываемые в настоящем описании, можно описывать или определять в отношении эпитопа(ов) или части (частей) антигена, например, описываемого в настоящем изобретении целевого полипептида (например, полноразмерного или зрелого ILT7), который они распознают или специфически связывают. Часть полипептида-мишени, которая специфически взаимодействует с антигенсвязывающим доменом антитела, представляет собой "эпитоп" или "антигенную детерминанту". Полипептид-мишень может содержать один эпитоп, но, как правило, содержит по меньшей мере два эпитопа и может включать любое число эпитопов в зависимости от размера, конформации и типа антигена. Кроме того, следует отметить, что "эпитоп" на полипептиде-мишени может представлять собой или может содержать не полипептидные элементы, например, эпитоп может содержать углеводородную боковую цепь.
[00106] Считается, что минимальный размер пептидного или полипептидного эпитопа для антитела составляет приблизительно от четырех до пяти аминокислот. Пептидные или полипептидные эпитопы могут содержать по меньшей мере семь, предпочтительнее по меньшей мере девять или по меньшей мере приблизительно от 15 до приблизительно 30 аминокислот. Поскольку CDR может распознавать антигенный пептид или полипептид в его третичной форме, аминокислоты, содержащие эпитоп, не должны быть непрерывными, и в некоторых случаях, могут даже не находиться на одной и той же пептидной цепи. Пептидный или полипептидный эпитоп, распознаваемый антителами к ILT7 по настоящему изобретению, может содержать последовательность по меньшей мере из 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, предпочтительнее по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25 или от приблизительно 15 до приблизительно 30 смежных или непрерывных аминокислот ILT7.
[00107] Под "специфически связывается" в целом подразумевается, что антитело связывается с эпитопом через его антигенсвязывающий домен, и что связывание влечет за собой некоторую комплементарность между антигенсвязывающим доменом и эпитопом. В соответствии с этим определением говорят, что антитело "специфически связывается" с эпитопом, когда оно связывается с таким эпитопом через свой антигенсвязывающий домен легче, чем оно связывалось бы со случайным, неродственным эпитопом. Термин "специфичность" используется в настоящем описании для определения относительной аффинности, с которой определенное антитело связывается с определенным эпитопом. Например, можно считать, что антитело "А" имеет более высокую специфичность к данному эпитопу, чем антитело "В", или можно сказать, что антитело "А" связывается с эпитопом "С" с большей специфичностью, чем оно имеет специфичность к родственному эпитопу "D".
[00108] Под "предпочтительно связывается" подразумевают, что антитело специфически связывается с эпитопом легче, чем оно бы связывалось с родственным, подобным, гомологичным или аналогичным эпитопом. Таким образом, антитело, которое "предпочтительно связывается" с данным эпитопом, более вероятно связывалось бы с этим эпитопом, чем с родственным эпитопом, даже хотя такое антитело может перекрестно реагировать с родственным эпитопом.
[00109] В другом неограничивающем примере, можно считать, что антитело предпочтительно связывается с первым антигеном, если оно связывается с первым эпитопом с аффинностью, которая по меньшей мере на один порядок величины меньше, чем KD антитела ко второму эпитопу. В другом неограничивающем примере, можно считать, что антитело предпочтительно связывается с первым эпитопом, если оно связывается с первым эпитопом с аффинностью, которая является по меньшей мере на два порядка величины меньше, чем KD антитела ко второму эпитопу.
[00110] В другом неограничивающем примере можно считать, что антитело предпочтительно связывает первый эпитоп, если оно связывается с первым эпитопом со скоростью обратной реакции (k(off)), которая является меньше, чем k(off) антитела k(off) ко второму эпитопу. В другом неограничивающем пример, можно сказать, что антитело предпочтительно связывается с первым эпитопом, если оно связывается с первым эпитопом с аффинностью, которая по меньшей мере на один порядок величины меньше, чем k(off) антитела ко второму эпитопу. В другом неограничивающем примере можно считать, что антитело предпочтительно связывается с первым эпитопом, если оно связывается с первым эпитопом с аффинностью, которая по меньшей мере на два порядка величины меньше, чем k(off) антитела ко второму эпитопу. Можно сказать, что антитело или его антигенсвязывающий фрагмент, вариант или производное, описываемое в настоящем описании, связывается с описываемым в настоящем описании полипептидом-мишенью (например, ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши) или его фрагментом или вариантом со скоростью обратной реакции (k (off)) меньшей или равной 5×10-2 с-1, 10-2 с-1, 5×10-3 с-1 или 10-3 с-1. Можно сказать, что антитело связывается с описываемым в настоящем описании полипептидом-мишенью (например, ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши) или его фрагментом или вариантом со скоростью обратной реакции (k(off)) меньшей или равной 5×10-4 с-1, 10-4 с-1, 5×10-5 с-1 или 10-5 с-1, 5×10-6 с-1, 10-6 с-1, 5×10-7 с -1 или 10-7 с-1.
[00111] Можно сказать, что антитело или его антигенсвязывающий фрагмент, вариант или производное, описываемое в настоящем описании, связывается с полипептидом-мишенью, описываемым в настоящем описании, (например, ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши) или его фрагментом или вариантом со скоростью прямой реакции (k(on)) большей или равной 103 M-1 с-1, 5×103 M-1 с-1, 104 M-1 с-1 или 5×104 M-1 с-1. Антитело по изобретению может связываться с полипептидом-мишенью, описываемым в настоящем описании, (например, ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примат и мыши) или его фрагментом или вариантом со скоростью прямой реакции (k(on)) большей или равной 105 M-1 с-1, 5×105 M-1 с-1, 106 M-1 с-1 или 5×106 M-1 с-1, или 107 M-1 с-1.
[00112] Считают, что антитело конкурентно ингибирует связывание эталонного антитела с данным эпитопом, если предпочтительно связывается с этим эпитопом или перекрывающимся эпитопом в тех случаях, когда оно блокирует до некоторой степени связывание эталонного антитела с эпитопом. Конкурентное ингибирование можно определять любым известным в данной области способом, например, анализами конкурентного ELISA. Можно сказать, что антитело конкурентно ингибирует связывание эталонного антитела с данным эпитопом по меньшей мере на 90%, по меньшей мере на 80%, по меньшей мере на 70%, по меньшей мере на 60% или по меньшей мере на 50%.
[00113] Как используют в настоящем описании, термин "аффинность" относится к мере силы связывания отдельного эпитопа с CDR молекулы иммуноглобулина. См., например, Harlow et al. (1988) Антитела: Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory Press, 2nd ed.) pages 27-28. Как используют в настоящем описании, термин "авидность" относится к общей стабильности комплекса популяции иммуноглобулинов с антигеном, т.е. прочности функционального соединения смеси иммуноглобулинов с антигеном. См., например, Harlow на страницах 29-34. Авидность относится к аффинности отдельных молекул иммуноглобулина в популяции с конкретными эпитопами, а также с валентностями иммуноглобулинов и антигена. Например, взаимодействие двухвалентного моноклонального антитела и антигена с высокоповторяющейся структурой эпитопа, таким как полимер, было бы взаимодействием с высокой авидностью.
[00114] Антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные по изобретению также описывают или определяют с точки зрения их перекрестной реактивности. Как используют в настоящем описании, термин "перекрестная реактивность" относится к способности антитела, специфичного к одному антигену, взаимодействовать со вторым антигеном; показателем родства между двумя различными антигенными веществами. Таким образом, антитело дает перекрестную реакцию, если оно связывается с эпитопом, отличным от эпитопа, который индуцирует его образование. Перекрестно реагирующий эпитоп, как правило, содержит многие из тех же комплементарных структурных признаков, что и индуцирующий эпитоп, и в некоторых случаях, может фактически лучше подходить, чем оригинальный.
[00115] Например, определенные антитела обладают некоторой степенью перекрестной реактивности, которая заключается в том, что они связываются с родственными, но неидентичными эпитопами, например, эпитопами по меньшей мере с 95%, по меньшей мере с 90%, по меньшей мере с 85%, по меньшей мере с 80%, по меньшей мере с 75%, по меньшей мере с 70%, по меньшей мере с 65%, по меньшей мере с 60%, по меньшей мере с 55% и по меньшей мере с 50% идентичностью (как рассчитывают известными в данной области способами и описываемыми в настоящем описании) с эталонным эпитопом. Можно сказать, что антитело обладает небольшой перекрестной реактивностью или не обладает ей, если оно не связывается с эпитопами с идентичностью менее 95%, менее 90%, менее 85%, менее 80%, менее 75%, менее 70%, менее 65%, менее 60%, менее 55% и менее 50% (как рассчитывают известными в данной области способами и описываемыми в настоящем описании) с эталонным эпитопом. Антитело можно считать "высокоспецифичным" к определенному эпитопу, если оно не связывается с каким-либо другим аналогом, ортологом или гомологом этого эпитопа.
[00116] Связывающие молекулы против ILT7, например, антитела или их антигенсвязывающие фрагменты, варианты или производные по изобретению также можно описывать или определять в отношении их аффинности связывания с полипептидом по изобретению, например, ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши. Пригодные аффинности связывания включают аффинности связывания с константой диссоциации или Kd менее чем 5×10-2 M, 10-2 M, 5×10-3 M, 10-3 M, 5×10-4 M, 10-4 M, 5×10-5 M, 10-5 M, 5×10-6 M, 10-6 M, 5×10-7 M, 10-7 M, 5×10-8 M, 10-8 M, 5×10-9 M, 10-9 M, 5×10-10 M, 10-10 M, 5×10-11 M, 10-11 M, 5×10-12 M, 10-12 M, 5×10-13 M, 10-13 M, 5×10-14 M, 10-14 M, 5×10-15 M или 10-15 M.
[00117] В определенных вариантах осуществления антитело связывается с человек ILT7 с константой диссоциации или Kd менее 1 нМ. В определенных вариантах осуществления антитело связывается с ILT7 яванского макака с константой диссоциации или Kd менее 5 нМ. В определенных вариантах осуществления антитело связывается с человек ILT7 с константой диссоциации или Kd менее 1 нМ и связывается с ILT7 яванского макака с константой диссоциации или Kd менее 5 нМ.
[00118] Как указано выше, структуры субъединицы и трехмерная конфигурация константных областей различных классов иммуноглобулинов являются хорошо известными. Как используют в настоящем описании, термин "домен VH" включает вариабельный домен на N-конце тяжелой цепи иммуноглобулина, и термин "домен CH1" включает первый (в большинстве случаев на N-конце) домен константной области тяжелой цепи иммуноглобулина. Домен CH1 является смежным с доменом VH и располагается на N-конце по отношению к шарнирной области тяжелой цепи молекулы иммуноглобулина.
[00119] Как используют в настоящем описании, термин "домен CH2" включает часть молекулы тяжелой цепи, которая располагается, например, приблизительно от остатка 244 до остатка 360 антитела с использованием общепринятых систем нумерации (остатки от 244 до 360 система нумерации по Kabat; и остатки 231-340 система нумерации EU; см. Kabat EA et al.). Домен CH2 является уникальным тем, что он не находится в тесной связи с другим доменом. Наоборот, две N-связанные разветвленные углеводные цепи располагаются между двумя доменами CH2 интактной нативной молекулы IgG. Также документально подтверждено, что домен CH3 простирается от домена CH2 до C-конца молекулы IgG и содержит приблизительно 108 остатков.
[00120] Как используют в настоящем описании, термин "шарнирная область" включает часть молекулы тяжелой цепи, которая соединяет домен CH1 с доменом CH2. Такая шарнирная область содержит приблизительно 25 остатков и является гибкой, таким образом, обеспечивая возможность двум N-концевым антигенсвязывающим областям двигаться независимо. Шарнирные области можно разделить на три отдельных домена: верхний, средний и нижний домены шпильки (Roux et al., J. Immunol. 161:4083 (1998)).
[00121] Как используют в настоящем описании термин, "дисульфидная связь" включает ковалентную связь, образуемую между двумя атомами серы. Аминокислота цистеин содержит тиольную группу, которая может образовывать дисульфидную связь или мостик со второй тиольной группой. Во многих природных молекулах IgG, области CH1 и CL являются связанными дисульфидной связью, и две тяжелые цепи являются связанными двумя дисульфидными связями в положениях, соответствующих 239 и 242 с использованием система нумерации по Kabat (положение 226 или 229 система нумерации EU).
[00122] Как используют в настоящем описании, термин "химерное антитело" будет означать любое антитело, где иммунореактивную область или участок получают или выделяют от первого вида, а константную область (которая может быть интактной, частичной или модифицированной в соответствии с настоящим изобретением) получают от второго вида. В определенных вариантах осуществления связывающая область-мишень или участок является из не принадлежащего человеку источника (например, мыши или примата), и константная область принадлежит человеку.
[00123] Как используют в настоящем описании, термин "сконструированное антитело" относится к антителу, в котором вариабельный домен в тяжелой или легкой цепи или в обеих изменяют по меньшей мере частичной заменой одной или более CDR из антитела с известной специфичностью и при необходимости частичной заменой каркасной области и изменением последовательности. Хотя CDR можно выделять из антитела того же класса или даже подкласса, что и антитело, из которого получают каркасные области, предусмотрено, что CDR выделяют из антитела другого класса или из антитела от другого вида. Сконструированное антитело, в котором одну или более "донорных" CDR из не принадлежащего человеку антитела известной специфичности пересаживают в каркасную область тяжелой или легкой цепи человека, обозначают в настоящем описании как "гуманизированное антитело". Может необязательно являться необходимой замена всех CDR полностью CDR из донорного вариабельного домена для передачи антигенсвязывающей способности одного вариабельного домена другому. Наоборот, необходимым может являться только перенос этих остатков, которые являются необходимыми для сохранения активности целевого участка связывания.
[00124] Кроме того, понятно, что каркасные области в пределах вариабельного домена в тяжелой или легкой цепи или обеих гуманизированного антитела могут содержать только остатки человеческого происхождения, в этом случае такие каркасные области гуманизированного антитела обозначают как "полностью принадлежащие человеку каркасные области". Альтернативно, один или более остатков каркасной области(ей) донорного вариабельного домена можно конструировать в соответствующем положении принадлежащей человеку каркасной области(й) вариабельного домена в тяжелой или легкой цепи, или обеих гуманизированного антитела, если необходимо сохранять соответствующее связывание или повышать связывание с антигеном ILT7. Принадлежащую человеку каркасную область, которую конструировали, таким образом, чтобы она содержит смесь принадлежащих человеку и донору каркасных остатков, и обозначают ее в настоящем описании как "частично принадлежащую человеку каркасную область.
[00125] Например, гумманизацию антитела к ILT7 можно по существу проводить следующим ниже способом Winter и соавторами (Jones et al., Nature 321:522-525 (1986); Riechmann et al., Nature 332:323-327 (1988); Verhoeyen et al., Science 239:1534-1536 (1988)), заменой CDR антитело грызуна к ILT7 или мутантного антитела грызуна к ILT7 или последовательностей CDR соответствующими последовательностями человека. Также см. патенты США № 5225539; 5585089; 5693761; 5693762; 5859205; включенные в настоящее описание посредством ссылки. Получаемое гуманизированное антитело к ILT7 будет содержать по меньшей мере одну CDR грызуна или мутантную CDR грызуна в полностью принадлежащих человека каркасных областях вариабельного домена тяжелой и/или легкой цепи гуманизированного антитела. В некоторых случаях остатки в каркасных областях одного или более вариабельных доменов гуманизированного антитела к ILT7 заменяют соответствующими не принадлежащими человеку (например, принадлежащими грызуна) остатками (см., например, патенты США № 5585089; 5693761; 5693762 и 6180370), в случае чего получаемое гуманизированное антитело к ILT7 содержит частично каркасные области человека в вариабельном домене тяжелой и/или легкой цепи.
[00126] Кроме того, гуманизированные антитела могут содержать остатки, которые не встречаются в реципиентном антителе или в донорном антителе. Эти модификации проводят для дополнительного улучшения характеристик антитела (например, для получения желаемой аффинности). В основном, гуманизированное антитело содержит по существу все и по меньшей мере из одного и, как правило, двух вариабельных доменов, в которых все или по существу все CDR соответствуют всем CDR не принадлежащего человеку иммуноглобулина, и все или по существу все каркасные области являются каркасными областями последовательности иммуноглобулина человека. Гуманизированное антитело также необязательно содержат по меньшей мере часть константной области иммуноглобулина (Fc), как правило, константной области иммуноглобулина человека. Более подробно см. Jones et al., Nature 331:522-525 (1986); Riechmann et al., Nature 332:323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2:593-596 (1992); включенные в настоящее описание посредством ссылки. Таким образом, такие "гуманизированные" антитела могут включать антитела, где по существу менее интактный вариабельный домен человека заменяли соответствующей последовательностью от не являющихся человеком видов. На практике, гуманизированные антитела, как правило, представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, некоторые каркасные остатки заменяют остатками участков-аналогов в антителах грызунов. См., например, патенты США № 5225539, 5585089, 5693761, 5693762, 5859205. Также см. патент США № 6180370 и международную публикацию № WO 01/27160, где описаны гуманизированные антитела и техники получение гуманизированных антител с улучшенной аффинностью к предопределенным антигенам.
[00127] Как используют в настоящем описании, термины "связанный", "слитый" или "слияния" используют взаимозаменяемо. Эти термины относятся к соединению друг с другом двух других элементов или компонентов, любыми способами, включая химическую конъюгацию или рекомбинантные способы. Слияние "в рамке считывания" относится к соединению двух или более открытых рамок считывания (ORF) полинуклеотида с образованием непрерывной большей ORF, таким образом, чтобы сохранять правильную рамку считывания трансляции исходных ORF. Таким образом, рекомбинантный слитый белок представляет собой один белок, содержащий два или более сегмента, которые соответствуют полипептидам, кодируемым оригинальными ORF (где сегменты обычно не соединены таким образом в природе). Несмотря на то, что рамка считывания, получаемая таким образом, является непрерывной на всем протяжении слитых сегментов, сегменты могут являться физически или пространственное разделенными, например, линкерной последовательностью в рамке считывания. Например, полинуклеотиды, кодирующие CDR вариабельной области иммуноглобулина могут являться слитыми в рамке считывания, но разделенными полинуклеотидом, кодирующим по меньшей мере одну каркасную область или дополнительные области CDR иммуноглобулина, при условии, что "слитые" CDR котранслируются как часть непрерывного полипептида.
[00128] В отношении полипептидов "линейная последовательность" или "последовательность" представляет собой порядок аминокислот в полипептиде в направлении от N- к C-концу, в котором остатки, которые располагаются по соседству друг с другом в последовательности, являются смежными в первичной структуре полипептида.
[00129] Термин "экспрессия", как используют в настоящем описании, относится к процессу, в результате которого ген продуцирует биохимическое соединение, например, полипептид. Процесс включает любое проявление функционального присутствия гена в клетке, включая без ограничения, нокдаун гена, а также транзиторную экспрессию и стабильную экспрессию. Он включает без ограничения транскрипцию гена в информационную РНК (иРНК) и трансляцию такой иРНК в полипептид(ы). Если конечный желаемый продукт является биохимическим соединением, экспрессия включает образование этого биохимического соединения и любых предшественников. Экспрессия гена привод к образованию "продукта гена". Как используют в настоящем описании, продукт гена может представлять собой нуклеиновую кислоту, например, информационную РНК, продуцируемую в результате транскрипции гена, или полипептид, который транслируется из транскрипта. Продукты генов, описываемые в настоящем описании, дополнительно включают нуклеиновые кислоты с посттранскрипционными модификациями, например, полиаденилированием, или полипептиды с посттрансляционными модификациями, например, метилированием, гликозилированием, добавлением липидов, связью с другими белковыми субъединицами, протеолитическим расщеплением и т.п.
[00130] Как используют в настоящем описании, термины "лечить" или "лечение" относятся к терапевтическому лечению и профилактическим или превентивным мерам, где целью является предотвращение или замедление (уменьшение) нежелательного физиологического изменения или нарушение, такого как прогрессирование аутоиммунного состояния. Благоприятные или желаемые клинические результаты включают, но не ограничиваются ими, ослабление симптомов, уменьшение тяжести заболевания, стабилизированное (т.е. не ухудшающееся) состояние заболевания, отсрочивание или замедление прогрессирования заболевания, улучшение состояния или облегчение состояния болезни и ремиссию (независимо от того, являются ли она частичной или полной), независимо от того, являются ли они детектируемыми или недетектируемыми. "Лечение" также может означать увеличение выживаемости по сравнению с ожидаемым выживанием, в случае отсутствия лечения. Те, кто нуждается в лечении, включают те, которые уже страдают состоянием или нарушением, а также тех, кто предрасположен к состоянию или нарушению, или тех, у кого следует проводить профилактику состояния или нарушения.
[00131] Под "объектом" или "индивидуумом", или "животным", или "пациентом", или "млекопитающим" подразумевают любого индивидуума, в частности являющегося млекопитающим индивидуума, у которого желательным является проведение диагностики, прогноза или терапии. Являющиеся млекопитающими индивидуумы включают людей, домашних животных, сельскохозяйственных животных и животных из зоопарка, участвующих в спорте животных или домашних животных, таких как собаки, кошки, морские свинки, кролики, крысы, мыши, лошади, крупный рогатый скот, коровы и т.д.
[00132] Как используют в настоящем описании, фразы, такие как "индивидуум, для которого введение антитела к ILT7 будет являться эффективным", и "нуждающееся в лечении животное" включает индивидуумов, таких как являющиеся млекопитающими индивидуумы, для которых эффективным будет введение антитела к ILT7, используемого, например, для детекции антитела к полипептиду ILT7 (например, для диагностической процедуры) и/или для лечения, т.е. облегчения или профилактики заболевания антителом к ILT7.
II. ILT7
[00133] Как используют в настоящем описании, термины "ILT7" и "полипептид ILT7" используют взаимозаменяемо. В определенных вариантах осуществления ILT7 является полноразмерным. В другом варианте осуществления ILT7 представляет собой зрелый ILT7 (аминокислоты 24-499). В других вариантах осуществления ILT7 может включать полноразмерный ILT7, его фрагмент или вариант полипептида ILT7, где фрагмент ILT7 или вариант полипептида ILT7 сохраняет некоторые или все функциональные свойства активного ILT7.
[00134] Полноразмерный ILT7 человека представляет собой белок длиной 499 аминокислоты (номер доступа P59901), содержащий сигнальный пептид (аминокислоты 1-23), внеклеточный домен (аминокислоты 24-446), трансмембранный домен (аминокислоты 447-467) и цитоплазматический домен (аминокислоты 468-499). Внеклеточный домен содержит четыре иммуноглобулиноподобных домена C2 (аминокислоты 24-118, 123-213, 224-313 и 324-413). ILT7 является представителем семейства генов иммуноглобулиноподобного транскрипта (ILT) или лейкоцитарного иммуноглобулиноподобного рецептора (LIR). Последовательность ILT7 яванского макака предоставлена как SEQ ID NO:292:
[00135] PRTHMQAENLLKPILWAEPGPVIIWKKPVTIWCQGTLEAQEYRLDKEGNSISRHMLKTLESENKAKFSIPSMMWEHAGRYHCYYQSPAGWSEPSDPLELVVTAYSRPSLSALPSPVVTSGVNVTLRCASRLGLGRFTLIEEGDHRLSWTLDSHQHNHGKFQALFPVGPLTFSNRGTFRCYGYENNTPYVWSEPSDPLQLLVSGVSRKPSLLTLQGPVVAPGDNLTLQCGSDVGYIRYALYKEGGDGLPQRPGQQSQAGLSQASFTLNPVRGSHGGQYRCYGAHNVSSKWSAPSDPLDILIAGQIPDRPSLSVQLGPTVASGEKVTLLCQSWGPMFTFLLAKEGAAHPPLRLRSTYRAQQYQAEFPMSPVTSAHAGTYRCYGSRSSDPYLLSHSSEPLELVVSEATETLNPAQNKSDSKTAPHLQDYTVENLIRMGIAGLVLVFLGILLFEAQQSQRSPTRCSQEVNSREDNAPFRVVEPWEQI (SEQ ID NO:292).
[00136] ILT7 селективно экспрессируется на подгруппе мононуклеарных клеток периферической крови (PBMC), называемых плазмацитоидными дендритными клетками (pDC). pDCs являются основным источником иммуномодулирующей молекулы интерферона (IFN)-альфа, и ILT7 играет роль в регуляции высвобождения IFN-альфа из этих клеток.
III. Связывающие молекулы против ILT7
[00137] В определенных вариантах осуществления связывающие ILT-7 молекулы, предоставленные в настоящем описании, представляют собой антитела или их антигенсвязывающие фрагменты, которые содержат последовательности и/или обладают свойствами связывающими ILT7 антител, предоставленных в настоящем описании. SEQ ID NO последовательностей антител к ILT7 приведены в таблице 2.
Таблица 2: SEQ ID NO последовательности антитела к ILT7
[00138] В определенных вариантах осуществления связывание молекул, например, антител или их антигенсвязывающих фрагментов, вариантов или производных по изобретению, например, антител 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 и ILT70052 связываются с ILT7 и ингибируют высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00139] В определенных вариантах осуществления антитела по изобретению включают антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные, которые связываются с ILT7, например, антитела 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 и ILT70052. В определенных вариантах осуществления антитела к ILT7 связываются с ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примат и мыши.
[00140] В одном из вариантов осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с тем же эпитопом ILT7, как и антитело 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052. В другом варианте осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с тем же эпитопом ILT7, как и антитело, содержащее VH и VL 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052. В другом варианте осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с тем же эпитопом ILT7, как и антитело, содержащее VH или VL 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052.
[00141] В другом варианте осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с ILT7 и конкурентно ингибирует специфическое связывание антитела 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052 с ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши. В другом варианте осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с ILT7 и конкурентно ингибирует специфическое связывание антитела, содержащего VH и VL 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052 с ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши. В другом варианте осуществления настоящее изобретение относится к выделенной связывающей молекуле, например, антителу или его антигенсвязывающему фрагменту, варианту или производному, которое специфически связывается с ILT7 и конкурентно ингибирует специфическое связывание антитела, содержащего VH или VL 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052, с ILT7, например, ILT7 человека, примата, мыши или любым сочетанием ILT7 человека, примата и мыши.
[00142] В определенных вариантах осуществления связывающая молекула по изобретению содержит аминокислотную последовательность, которая обладает по меньшей мере 80%, 85%, 88%, 89%, 90%, 91%, 92%, 93%, 94% или 95% идентичностью последовательности с аминокислотной последовательностью молекулы эталонного антитела к ILT7. В дополнительном варианте осуществления связывающая молекула обладает по меньшей мере 96%, 97%, 98%, 99% или 100% идентичность последовательности с эталонным антителом. В определенных вариантах осуществления эталонное антитело представляет собой 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052.
[00143] В другом варианте осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, варианту или производному, содержащему, состоящему по существу из или состоящему из домена VH, который содержит аминокислотную последовательность, которая является по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности VH SEQ ID NO: 22, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 202, 212, 222, 232, 242, 252 и 262, где антитело или его антигенсвязывающий фрагмент, вариант или производное, содержащее домен VH, специфически или преимущественно связывается с ILT7. В дополнительном варианте осуществления антитело или его антигенсвязывающий фрагмент, вариант или производное ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00144] В дополнительном варианте осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, варианту или производному, содержащему, состоящему по существу из или состоящему из домена VL, который содержит аминокислотную последовательность, которая является по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной аминокислотной последовательности VL SEQ ID NO: 27, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187, 197, 207, 217, 227, 237, 247, 257 или 267, где антитело или его антигенсвязывающий фрагмент, вариант или производное, содержащее домен VL, специфически или преимущественно связывается с ILT7. В дополнительном варианте осуществления и антитело или его антигенсвязывающий фрагмент, вариант или производное ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00145] В дополнительном варианте осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, варианту или производному, содержащему, состоящему по существу из или состоящему из домена VH и домена VL, которые содержат аминокислотные последовательности, которые являются по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичными последовательностям VH и VL SEQ ID NO: 22 и 27; 42 и 47; 52 и 57; 62 и 67; 72 и 77; 82 и 87; 92 и 97; 102 и 107; 112 и 117; 122 и 127; 132 и 137; 142 и 147; 152 и 157; 162 и 167; 172 и 177; 182 и 187; 192 и 197; 202 и 207; 212 и 217; 222 и 227; 232 и 237; 242 и 247; 252 и 257 или 262 и 267, соответственно, где антитело или его антигенсвязывающий фрагмент, вариант или производное, содержащее домены VH и VL, специфически или преимущественно связывается с ILT7. В дополнительном варианте осуществления и антитело или его антигенсвязывающий фрагмент, вариант или производное ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00146] В дополнительном варианте осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, варианту или производному, содержащему, состоящему по существу из или состоящему из домена VH и домена VL, которые содержат последовательности VH-CDR1, VH-CDR2, VH-CDR3, VL-CDR1, VL-CDR2 и VL-CDR3 SEQ ID NO: 23, 24, 25, 28, 29 и 30; 43, 44, 45, 48, 49 и 50; 53, 54, 55, 58, 59 и 60; 63, 64, 65, 68, 69 и 70; 73, 74, 74, 78, 79 и 80; 83, 84, 85, 88, 89 и 90; 93, 94, 95, 98, 99 и 100; 103, 104, 105, 108, 109 и 110; 113, 114, 115, 118, 119 и 120; 123, 124, 125, 128, 129 и 130; 133, 134, 135, 138, 139 и 140; 143, 144, 145, 148, 149 и 150; 153, 154, 155, 158, 159 и 160; 163, 164, 165, 168, 169 и 170; 173, 174, 175, 178, 179 и 180; 183, 184, 185, 188, 189 и 190; 193, 194, 195, 198, 199 и 200; 203, 204, 205, 208, 209 и 210; 213, 214, 215, 218, 219 и 220; 223, 224, 225, 228, 229 и 230; 233, 234, 235, 238, 239 и 240; 243, 244, 245, 248, 249 и 250; 253, 254, 255, 258, 259 и 260; 263, 264, 265, 268, 269 и 270, соответственно, где антитело или его антигенсвязывающий фрагмент, вариант или производное, содержащее домены VH и VL, специфически или преимущественно связывается с ILT7. В дополнительном варианте осуществления и антитело или его антигенсвязывающий фрагмент, вариант или производное ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00147] Подходящие биологически активные варианты антител к ILT7 по изобретению можно использовать в способах по настоящему изобретению. Такие варианты сохраняют желаемые свойства связывания родительского антитела к ILT7. Способы получения вариантов антител, как правило, доступны в данной области.
[00148] Способы мутагенеза и изменений нуклеотидной последовательности хорошо известны в данной области. См., например, Walker and Gaastra, eds. (1983) Techniques in Molecular Biology (MacMillan Publishing Company, New York); Kunkel, Proc. Natl. Acad. Sci. USA 82:488-492 (1985); Kunkel et al., Methods Enzymol. 154:367-382 (1987); Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual (Cold Spring Harbor, N.Y.); патент США № 4873192 и цитируемые в них ссылки, включенные в настоящее описание посредством ссылки. Руководство относительно соответствующих замены аминокислот, которые не влияют на биологическую активность представляющего интерес полипептида можно найти в модели у Dayhoff et al. (1978) в Atlas of Protein Sequence and Structure (Natl. Biomed. Res. Found., Washington, D.C.), pp. 345-352, полностью включенном в настоящее описание посредством ссылки. В модели Dayhoff et al. используют матрицу подобия закрепившейся точечной мутации (PAM) аминокислоты (матрица PAM 250) для определения подходящих консервативных замен аминокислот. Консервативные замены, такие как замена одной аминокислоты другой, обладающей сходными свойствами, могут являться полезными. Примеры консервативных замен аминокислот, как указано в матрице PAM 250 модели Dayhoff et al. включают, но не ограничиваются ими, Gly↔Ala, Val↔Ile↔Leu, Asp↔Glu, Lys↔Arg, Asn↔Gln и Phe↔Trp↔Tyr.
[00149] При конструировании вариантов связывающих молекул против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного делают модификации таким образом, что варианты сохраняют требуемые свойства, например, являются способными специфически связываться с ILT7, и в определенных вариантах осуществления являются способными ингибировать высвобождение IFN-альфа. Очевидно, что любые мутации, проводимые в ДНК, кодирующей вариант полипептида, не должны выносить последовательность за рамку считывания. В определенном варианте осуществления любые мутации, проводимые в ДНК, не будут создавать комплементарные области, которые могут образовывать вторичную структуру иРНК.
[00150] Способы измерения специфичности связывания антитела связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, включают, но не ограничиваются ими, стандартные анализы конкурентного связывания, анализы цитотоксичности, анализы высвобождения IFN, анализы ELISA и т.п.
[00151] При обсуждении в настоящем описании независимо от того, является ли какой-либо конкретный полипептид, включая константные области, CDR, домены VH или домены VL, описываемые в настоящем описании, по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, или даже 100% идентичным другому полипептиду, % идентичности можно определять с применением способов и компьютерных программ/программного обеспечения, известных в данной области, такие как, но не ограничиваясь ими, программа BESTFIT (Wisconsin Sequence Analysis Package, Version 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Madison, Wis. 53711). В BESTFIT используют алгоритм локальной гомологии Smith and Waterman (1981) Adv. Appl. Math. 2:482-489, для поиска лучшего сегмента гомологии между двумя последовательностями. При использовании BESTFIT или любой другой программы для выравнивания последовательностей для определения, является ли конкретная последовательность, например, на 95% идентичной эталонной последовательности по настоящему изобретению, параметры устанавливают, как известно, таким образом, чтобы процент идентичности рассчитывать по всей длине последовательности эталонного полипептида, и чтобы допускать пропуски в гомологии до 5% от общего числа аминокислот в эталонной последовательности.
[00152] Для целей по настоящему изобретению процент идентичность последовательности можно определять с использованием алгоритма поиска гомологии Смита-Уотермана с использованием поиска аффинного пропуска со штрафом за создание пропуска 12 и штрафом за продление пропуска 2, матрицы BLOSUM 62. Алгоритм поиска гомологии Смита-Уотермана описан у Smith and Waterman (1981) Adv. Appl. Math. 2:482-489. Вариант, например, может отличаться от эталонного антитела к ILT7 (например, 7C7, ILT70080, ILT70080.1-ILT70080.7, ILT70083, ILT70083.1-ILT70083.9, ILT70089, ILT70100, ILT70137, ILT70142, ILT70144 или ILT70052) только на 1-15 аминокислотных остатков, только на 1-10 аминокислотных остатков, таких как 6-10, только на 5, только на 4, 3, 2, или даже на 1 аминокислотный остаток.
[00153] Точная химическая структура полипептида, способного специфически связываться с ILT7 и сохранять желаемую активность зависит от ряда факторов. Вследствие того, что в молекуле присутствуют ионизируемые амино- и карбоксильные группы, конкретный полипептид можно получать как кислую или основную соль, или в нейтральной форме. Все такие препараты, которые сохраняют свою биологическую активность при помещении в подходящие условия окружающей среды, входят в определение антител к ILT7, как используют в настоящем описании. Кроме того, первичную аминокислотную последовательность полипептида можно увеличивать посредством дериватизации с использованием фрагментов сахаров (гликозилирование) или посредством других дополнительных молекул, таких как липиды, фосфатные, ацетильные группы и т.д. ЕЕ также можно увеличивать посредством конъюгации с сахаридами. Определенные аспекты такого увеличения проводят через системы посттрансляционного процессинга продуцирующего хозяина; другие такие модификации можно вводить in vitro. В любом случае такие модификации входят в определение антитела к ILT7, используемое в настоящем описании, при условии, что желаемые свойства антитела к ILT7 не нарушаются. Ожидают, что такие модификации могут количественно или качественно влиять на активность, путем увеличения или уменьшения активности полипептида в различных анализах. Кроме того, отдельные аминокислотные остатки в цепи можно модифицировать путем окисления, восстановления или другой дериватизации, и полипептид можно расщеплять с получением фрагментов, которые сохраняют активность. Такие изменения, которые не нарушают желаемые свойства (например, специфичность связывания к ILT7, аффинность связывания и сопутствующую активность, например, способность ингибировать направляемое ILT7 высвобождение цитокинов тучными клетками, эндотелиальными клетками и пролиферацию клеток TF-1) не исключает последовательность полипептида из определения представляющих интерес антител к ILT7, как используют в настоящем описании.
[00154] Уровень техники предоставляет основное руководство в отношении получения и использования вариантов полипептидов. При получении вариантов связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, специалист в данной области может легко определять, какие модификации в нативной нуклеотидной или аминокислотной последовательности белка приведут к варианту, который является пригодным для использования в качестве терапевтически активного компонента фармацевтической композиции, используемой в способах по настоящему изобретению.
[00155] Константную область антитела к ILT7 можно подвергать мутации для изменения эффекторной функция несколькими способами. Например, см. патент США № 6737056B1 и публикацию патентной заявки США № 2004/0132101A1, в которых описаны мутации Fc, которые оптимизируют связывание антитела с Fc-рецепторами.
[00156] В определенных антителах к ILT7 участок Fc можно подвергать мутации для повышения эффекторной функции с использованием техник, известных из уровня техники. Например, делеция или инактивация (через точечные мутации или другими способами) домена константной области может уменьшать can reduce связывание Fc-рецептора циркулирующего модифицированного антитела. В других случаях может происходить следующее: модификации константной области в соответствии с настоящим изобретением ослабляют связывание комплемента и, таким образом, уменьшают время полувыведения из сыворотки и неспецифическое связывание конъюгированного цитотоксина. Еще одни другие модификации константной области можно использовать для модификации дисульфидных связей или олигосахаридных фрагментов, что обеспечит возможность повышенной локализации вследствие увеличенной антигенной специфичности или гибкости антитела. Получаемый физиологический профиль, биодоступность и другие биохимические эффекты модификаций, такие как биораспределение и время полувыведения из сыворотки, можно легко измерять и количественно оценивать с использованием хорошо известных иммунологических техник без излишнего экспериментирования.
[00157] Определенные антитела к ILT7, предоставленные в настоящем описании, являются афукозилированными. Антитела, не содержащие коровую фукозу остатки из N-гликанов Fc, проявляют сильную ADCC при более низких концентрациях с гораздо более высокой эффективностью по сравнению с фукозилированными аналогами, и они могут ускользать от ингибирующего действия иммуноглобулина G (IgG) сыворотки в отношении ADCC в результате своего сильного связывания с гамма-рецептором IIIa (Fc FcγRIIIa).
[00158] Антитела к ILT7 по изобретению также включают производные, которые модифицируют, например, ковалентным присоединением любого типа молекулы с антителом, таким образом, что ковалентное присоединение не нарушает специфическое связывание антитела с его когнатным эпитопом. Например, но не в качестве ограничения, производные антител включают антитела, которые модифицировали, например, гликозилированием, ацетилированием, пегилированием, фосфорилированием, амидированием, дериватизацикй известными защитными/блокирующими группами, протеолитическим расщеплением, связыванием с клеточным лигандом или другим белком и т.д. Любую из многих химических модификаций можно проводить известными способами, включая, но, не ограничиваясь ими, специфическое химическое расщепление, ацетилирование, формилирование и т.д. Дополнительно производное может содержать одну или более неклассических аминокислот.
[00159] "Консервативная замена аминокислоты" представляет собой замену, при которой аминокислотный остаток заменяют аминокислотным остатком, содержащим боковую цепь с аналогичным зарядом. Семейства аминокислотных остатков с боковыми цепями с аналогичными зарядами были определены в уровне технике. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагинов кислота, глутаминовую кислоту), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Альтернативно, мутации можно вводить в случайном порядке по всей длине или части кодирующей последовательности, таким образом, как посредством насыщающего мутагенеза и много проводить скрининг получаемых мутантов на биологическую активность для идентификации мутантов, которые сохраняют активность (например, способность связываться с антителом к полипептиду ILT7).
[00160] Например, можно вводить мутации только в каркасные области или только в области CDR молекулы антитела. Вводимые мутации могут быть молчащими или нейтральными бессмысленными мутациями, т.е. не оказывают или оказывают незначительный эффект на способность антитело связываться с антигеном. Эти типы мутаций могут являться пригодными для оптимизации частоты использования кодонов или улучшения продукции антитела гибридомы. Альтернативно, ненейтральные бессмысленные мутации могут изменять способность антитела связываться с антигеном. Локализация наиболее молчащих и нейтральных бессмысленных мутаций, вероятнее всего, приходится на каркасные области, тогда как локализации наиболее ненейтральных бессмысленных мутаций вероятнее всего, приходится на CDR, хотя это не является абсолютным требованием. Специалист в данной области способен конструировать и тестировать мутантные молекулы с желаемыми свойствами, такими как отсутствие изменения антигенсвязывающей активности или изменение активности связывания (например, улучшения антигенсвязывающей активности или изменение специфичности антитела). После мутагенеза кодируемый белок можно общепринято экспрессировать и функциональную и/или биологическую активность кодируемого белка, (например, способность иммуноспецифически связываться по меньшей мере с одним эпитопом полипептида ILT7) можно определять с использованием техник, описываемых в настоящем описании, или общепринятыми модифицирующими техниками, известными в данной области.
[00161] В определенных вариантах осуществления антитела к ILT7 по изобретению содержат по меньшей мере одну оптимизированную определяющую комплементарность область (CDR). Под "оптимизированной CDR" подразумевают, что CDR модифицировали и оптимизированные последовательности отбирали на основании устойчивой или улучшенной аффинности связывания и/или активности антитела к ILT7, которая передается антителу к ILT7, содержащему оптимизированную CDR. "Активность антитела к ILT7" может включать, например, активность, которая модулирует одну или более следующих ниже видов активности, ассоциированных с ILT7, например, управляемое ILT7 высвобождение интерферона плазмацитоидные дендритные клетки, цитотоксичность к экспрессирующим ILT7 клеткам, или любая другая активность, связанная с ILT7. Активность антитела к ILT7 также может быть связана со снижением частоты возникновения или тяжести заболеваний, связанных с экспрессией ILT7, включая, но, не ограничиваясь ими, определенные типы аутоиммунных состояний, например, системную красную волчанку, хронический ревматизм и псориаз. Модификации могут улучшать замену аминокислотных остатков в CDR, таким образом, что антитело к ILT7 сохраняет специфичность к антигену ILT7 и обладает улучшенной аффинностью связывания и/или улучшенной активностью антитела к ILT7.
IV. Полинуклеотиды, кодирующие антитела к ILT7
[00162] Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующей антитела к ILT7 по изобретению или их антигенсвязывающие фрагменты, варианты или производные.
[00163] В дополнительном варианте осуществления настоящее изобретение относится к выделенному полинуклеотиду, содержащему, состоящему по существу или состоящему из нуклеиновой кислоты, кодирующей домен VH, который содержит аминокислотную последовательность, которая является по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной эталонной последовательности домена VH полипептида, содержащего SEQ ID NO: 12, 22, 32, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 202, 212, 222, 232 или 242, где антитело к ILT7, содержащее кодируемый домен VH, специфически или преимущественно связывается с ILT7. В определенных вариантах осуществления полинуклеотид кодирует антитело или его антигенсвязывающий фрагмент, вариант или производное, которое ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00164] В дополнительном варианте осуществления настоящее изобретение относится к выделенному полинуклеотиду, содержащему, состоящему по существу или состоящему из нуклеиновой кислоты, кодирующей домен VL, который содержит аминокислотную последовательность, которая является по меньшей мере на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичной эталонной последовательности домена VL полипептида, содержащего SEQ ID NO: 17, 27, 37, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187, 197, 207, 217, 227, 237 или 247, где антитело к ILT7, содержащее кодируемый домен VL, специфически или преимущественно связывается с ILT7. В определенных вариантах осуществления полинуклеотид кодирует антитело или его антигенсвязывающий фрагмент, вариант или производное, которое ингибирует высвобождение IFN-альфа плазмацитоидными дендритными клетками.
[00165] Любой из описанных выше полинуклеотидов может дополнительно содержать дополнительные нуклеиновые кислоты, кодирующие, например, сигнальный пептид для направления секреции кодируемого полипептида, константных областей антитела, как описано в настоящем описании, или других гетерологичных полипептидов, как описано в настоящем описании. Также, как описано более подробно в другом месте в настоящем описании, настоящее изобретение относится к композициям, содержащим один или более полинуклеотидов, описанных выше.
[00166] В одном из вариантов осуществления изобретение включает композиции, содержащие первый полинуклеотид и второй полинуклеотид, где указанный первый полинуклеотид кодирует домен VH, как описано в настоящем описании, и где указанный второй полинуклеотид кодирует домен VL, как описано в настоящем описании. В частности, композиция может содержать, состоять по существу или состоять из кодирующего домен VH полинуклеотида, как указано в SEQ ID NO:11, 21, 31, 41, 51, 61, 71, 81, 91, 101, 111, 121, 131, 141, 151, 161, 171, 181, 191, 201, 211, 221, 231 или 241, и кодирующего домен VL полинуклеотида, как указано в SEQ ID NO:16, 26, 36, 46, 56, 66, 76, 86, 96, 106, 116, 126, 136, 146, 156, 166, 176, 186, 196, 206, 216, 226, 236 или 246. Композиция также может содержать, состоять по существу или состоять из кодирующего домен VH полинуклеотида, кодирующего последовательность, указанную в SEQ ID NO:12, 22, 32, 42, 52, 62, 72, 82, 92, 102, 112, 122, 132, 142, 152, 162, 172, 182, 192, 202, 212, 222, 232 или 242, и кодирующего домен VL полинуклеотида, кодирующего последовательность, указанную в SEQ ID NO:17, 27, 37, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187, 197, 207, 217, 227, 237, 247, 257 или 267. В определенных вариантах осуществления кодирующий домен VH полипептид и кодирующий домен VL полипептид находятся в одном и том же векторе. В определенных вариантах осуществления кодирующий домен VH полипептид и кодирующий домен VL полипептид находятся в разных векторах.
[00167] Настоящее изобретение также относится к фрагментам полинуклеотидов по изобретению, как описано где-либо в другом месте. Дополнительно полинуклеотиды, которые кодируют слитые полипептиды, фрагменты Fab и другие производные, как описано в настоящем описании, также предусмотрены изобретением.
[00168] Полинуклеотиды можно получать или производить любым известным в данной области способом. Например, если известна нуклеотидная последовательность антитела, полинуклеотид, кодирующий антитело, можно собирать из химически синтезированных олигонуклеотидов (например, как описано у Kutmeier et al., Bio Techniques 17:242 (1994)), что в кратком изложении включает синтез перекрывающихся олигонуклеотидов, содержащих участки последовательностей, кодирующих антитело, выравнивание и лигирование таких олигонуклеотидов, а затем амплификацию лигированных олигонуклеотидов с помощью ПЦР.
[00169] Альтернативно, полинуклеотид, кодирующий антитело к ILT7 или его антигенсвязывающий фрагмент, вариант или производное по изобретению, можно получать из нуклеиновой кислоты из подходящего источника. Если клон, содержащий нуклеиновую кислоту, кодирующую конкретное антитело, недоступен, но известна последовательность молекулы антитела, нуклеиновую кислоту, кодирующую антитело, можно химически синтезировать или получать из подходящего источника (например, библиотеки кДНК антител или библиотеки кДНК, получаемой из любой ткани или клеток, экспрессирующих антитело или другое антитело к ILT7, или нуклеиновой кислоты, например, поли A+РНК, выделенной из любой ткани или клеток, экспрессирующих антитело или другое антитело к ILT7, таких как гибридомные клетки, отобранные для экспрессии антитела) амплификацией ПЦР с использованием синтетических праймеров, гибридизуемых на 3'- и 5'-концах последовательностей, или клонированием с использованием олигонуклеотидного зонда, специфического к последовательности конкретного гена для идентификации, например, лона кДНК библиотеки кДНК, кодирующей антитело или другое антитело к ILT7. Амплифицированные нуклеиновые кислоты, получаемые ПЦР, можно затем клонировать в реплицирующиеся клонирующие векторы любым хорошо известным в данной области способом.
[00170] После того, как определяют нуклеотидную последовательность и соответствующую аминокислотную последовательность антитела к ILT7 или его антигенсвязывающего фрагмента, варианта или производного, его нуклеотидную последовательность можно подвергать манипуляции с применением хорошо известных в данной области способов манипуляции нуклеотидными последовательностями, например, технологии рекомбинантной ДНК, сайт-направленный мутагенез, ПЦР и т.д. (см., например, техники, описанные у Sambrook et al. (1990) Molecular Cloning, A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.) and Ausubel et al., eds. (1998) Current Protocols in Molecular Biology (John Wiley & Sons, NY), которые полностью включены посредством ссылки в настоящем описании), для получения антител с различной аминокислотной последовательностью, например, для проведения замен аминокислот, делеций и/или вставок.
[00171] Полинуклеотид, кодирующий связывающая молекула против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, может состоять из любого полирибонуклеотида или полидезоксирибонуклеотида, который может предоставлять собой немодифицированную РНК или ДНК, или модифицированную РНК или ДНК. Например, полинуклеотид, кодирующий антитело к ILT7, или его антигенсвязывающий фрагмент, вариант или производное может состоять из одноцепочечной и двухцепочечной ДНК, ДНК, которая является смесью одноцепочечных и двухцепочечных областей, одноцепочечной и двухцепочечной РНК, и РНК, которая является смесью одноцепочечных и двухцепочечных областей, гибридных молекул, содержащих ДНК и РНК, которые могут быть одноцепочечными или, что более характерно, двухцепочечными или смесью одноцепочечных и двухцепочечных областей. Кроме того, полинуклеотид, кодирующий связывающая молекула против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное может состоять из трехцепопченых областей, содержащих РНК или ДНК или как РНК, так и ДНК. Полинуклеотид, кодирующий связывающая молекула против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, также может содержать одно или более модифицированных оснований или остовов ДНК или РНК, модифицированных для стабильности или по другим причинам. "Модифицированные" основания включают, например, тритилированные основания и необычные основания, такие как инозин. Ряд модификаций можно вносить в ДНК и РНК; таким образом, "полинуклеотид" включает химически, ферментативно или метаболически модифицированные формы.
[00172] Выделенный полинуклеотид, кодирующий неприродный вариант полипептида, происходящий из иммуноглобулина (например, части тяжелой цепи или части легкой цепи иммуноглобулина), можно получать путем внесения одной или нескольких нуклеотидных замен, вставок или делеций в нуклеотидную последовательность иммуноглобулина, так чтобы вносить в кодируемый белок одну или более аминокислотных замен, вставок или делеций. Мутации можно вводить стандартными способами, такими как сайт-специфический мутагенез и опосредованный ПЦР мутагенез. Консервативные аминокислотные замены можно получать в одном или более не относящихся к незаменимым аминокислотных остатков.
V. Слитые белки и конъюгаты антитела
[00173] Как более подробно описано в другом месте в настоящем описании, связывающие молекулы против ILT7, например, антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные, можно дполнительно подвергать рекомбинантному слиянию с гетерологичным полипептидом на N-или С-конце или химически конъюгировать (включая ковалентную и нековалентную конъюгации) с полипептидами или другими композициями. Например, антитела к ILT7 можно подвергать рекомбинантному слиянию или конъюгировать с молекулами, пригодными в качестве меток в анализах для детекции и эффекторными молекулами, такими как гетерологичные полипептиды, лекарственные средства, радионуклиды или токсины. См., например, публикации РСТ WO 92/08495; WO 91/14438; WO 89/12624; патент США № 5314995 и ЕР 396387.
[00174] Антитела к ILT7 по изобретению или их антигенсвязывающие фрагменты, варианты или производные могут включать производные, которые модифицировали, т.е. путем ковалентного присоединения любого типа молекулы к антителу, таким образом, чтобы ковалентное присоединение не препятствовало связыванию антитела с ILT7. Например, но, не ограничиваясь этим, производные антител включают антитела, которые модифицировали, например, путем гликозилирования, ацетилирования, пегилирования, фосфорилирования, амидирования, дериватизация с помощью известных защитных/блокирующих групп, протеолитического расщепления, связывания с клеточным лигандом или другим белком и т.д. Любые из многих химических модификаций можно проводить известными способами, включая, но не ограничиваясь ими, специфическое химическое расщепление, ацетилирование, формилирование и т.д. Кроме того, производное может содержать одну или несколько неклассических аминокислот.
[00175] Связывающие молекулы против ILT7, например антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные могут состоять из аминокислот, связанных друг с другом пептидными связями или модифицированными пептидными связями, т.е. пептидных изостер, и могут содержать аминокислоты, отличные от кодируемых генами 20 аминокислот. Например, антитела к ILT7 можно модифицировать с помощью природных процессов, таких как посттрансляционный процессинг, или способами химической модификации, которые хорошо известны в данной области. Такие модификации широко описаны в основных руководствах и в более подробных монографиях, а также в обширной научной литературе. Модификации могут происходить в любой части связывающей молекулы против ILT7, включая пептидный остов, боковые цепи аминокислот и N- или С-концы, или в таких молекулах, как углеводы. Следует понимать, что один и тот же тип модификации может присутствовать в одинаковой или в различной степени в нескольких участках в данной связывающей молекуле против ILT7. Также данная связывающая молекула против ILT7 может содержать многие типы модификаций. Связывающие молекулы против ILT7 могут быть разветвленными, например, в результате убиквитинилирования, и они могут быть циклическими с ветвлением или без него. Циклическая, разветвленная и разветвленная циклическая связывающая молекула против ILT7 может являться результатом посттрансляционных природных процессов, или ее можно получать синтетическими способами. Модификации включают ацетилирование, ацилирование, ADP-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение группы гема, ковалентное присоединение нуклеотида или производного нуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфотидилинозитола, сшивание, циклизацию, образование дисульфидных связей, деметилирование, образование ковалентных сшивок, образование цистеина, образование пироглутамата, формилирование, гамма-карбоксилирование, гликозилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, пегилирование, протеолитический процессинг, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, опосредуемое транспортной РНК добавление аминокислот к белкам, такое как аргинилирование и убиквитинилирование (см., например, Proteins - Structure and Molecular Properties, T.E. Creighton, W.H. Freeman and Company, New York 2nd Ed. (1993); Posttranslational Covalent Modification Of Proteins, В.С. Johnson, Ed. (1983), Academic Press, New York, p. 1-12; Scifter et al., Meth Enzymol 182:626-646 (1990); Rattan et al., Ann NY Acad Sci 663:48-62 (1992)).
[00176] Настоящее изобретение также относится к слитым белкам, содержащим антитело к ILT7 или его антигенсвязывающий фрагмент, вариант или производное и гетерологичный полипептид. Гетерологичный полипептид, с которым сливают антитело, может являться пригодным для функции или пригодным для направленной доставки к клеткам, экспрессирующим полипептид против ILT7.
[00177] В одном из вариантов осуществления слитый белок по изобретению содержит, состоит по существу или состоит из полипептида, содержащего аминокислотную последовательность любого одного или более доменов VH антитела по изобретению или аминокислотную последовательность любого одного или более доменов VL антитело по изобретению или ее фрагменты, варианты или производные и гетерологичную последовательность полипептида.
[00178] В другом варианте осуществления слитый белок для применения в способах диагностики и лечения, описываемых в настоящем описании, содержит, состоит по существу или состоит из полипептида, содержащего аминокислотную последовательность любой одной, двух, трех CDR домена VH антитела к ILT7 или его фрагментов, вариантов или производных, или аминокислотную последовательность любой одной, двух, трех CDR домена VL антитела к ILT7 или его фрагментов, вариантов или производных, и последовательность гетерологичного полипептида. В одном из вариантов осуществления слитый белок содержит полипептид, содержащий аминокислотную последовательность по меньшей мере одного домена VH антитела к ILT7 по изобретению и аминокислотную последовательность по меньшей мере одного домена VL антитела к ILT7 по изобретению или его фрагментов, производных или вариантов, и последовательность гетерологичного полипептида. В определенных вариантах осуществления домены VH и VL слитого белка соответствуют антителу из одного источника (или фрагменту scFv или Fab), которое специфично связывается по меньшей мере с одним эпитопом ILT7. В еще одном другом варианте осуществления слитый белок для применения в способах диагностики и лечения, описываемых в настоящем описании, содержит полипептид, содержащий аминокислотную последовательность любой одной, двух, трех или более CDR домена VH антитела к ILT7 и аминокислотную последовательность любой одной, двух, трех или более CDR домена VL антитела к ILT7 или его фрагментов или вариантов, и последовательность гетерологичного полипептида. В определенных вариантах осуществления две, три, четыре, пять, шесть или более CDR домена VH или домена VL соответствуют антителу из одного источника (или фрагменту scFv или Fab) по изобретению. Также изобретение охватывает молекулы нуклеиновых кислот, кодирующие такие слитые белки.
[00179] Иллюстративные слитые белки, описанные в литературе, включают слияния T-клеточного рецептора (Gascoigne et al., Proc Natl Acad Sci USA 54:2936-2940 (1987)); CD4 (Capon et al., Nature 337:525531 (1989); Traunecker et al., Nature 339:68-70 (1989); Zettmeissl et al., DNA Cell Biol. USA 9:347-353 (1990); и Byrn et al., Nature 344:667-670 (1990)); L-селектина (рецептор хоминга) (Watson et al., J. Cell Biol. 110:2221-2229 (1990); и Watson et al., Nature 349:164-167 (1991)); CD44 (Aruffo et al., Cell (57:13031313 (1990)); CD28 и B7 (Linsley et al., J. Exp Med. 773:721-730 (1991)); CTLA-4 (Lisley et al., J. Exp Med. 174:561-569 (1991)); CD22 (Stamenkovic et al., Cell 66: 1133-1144 (1991)); рецептора TNF (Ashkenazi et al., Proc Natl Acad Sa USA 55:10535-10539 (1991); Lesslauer et al., Eur J. Immunol. 27:2883-2886 (1991); и Pep-pel et al., J. Exp Med. 774: 1483-1489 (1991)); и рецептора IgE (Ridgway and Gorman, J. Cell Biol Vol. 115, Abstract № 1448 (1991)).
[00180] Как обсуждалось в других разделах настоящего описания, связывающие молекулы против ILT7, например, антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные можно сливать с гетерологичными полипептидами для увеличения времени полужизни in vivo полипептидов или для применения в иммунологических анализах с применением известных в данном уровне техники способов. Например, в одном из вариантов осуществления PEG можно конъюгировать с антителами к ILT7 по изобретению для увеличения их времени полужизни in vivo. См. Leong et al., Cytokine 16:106 (2001); Adv. in Drug Deliv. Rev. 54:531 (2002); or Weir et al., Biochem. Soc. Transactions 30:512 (2002).
[00181] Кроме того, связывающие молекулы против ILT7, например антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные можно сливать с маркерными последовательностями, такими как пептид, для облегчения их очистки или детекции. В определенных вариантах осуществления маркерная аминокислотная последовательность представляет собой гекса-гистидиновый пептид, такой как маркер, предоставленный в векторе pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, Calif., 91311), наряду с другими, многие из которых являются коммерчески доступными. Как описано у Gentz et al., Proc. Natl. Acad. Sci. USA 55:821-824 (1989), например, гексагистидин обеспечивает удобную очистку слитого белка. Другие пептидные метки, пригодные для очистки, включают, но не ограничиваются ими, метку "НА", которая соответствует эпитопу, происходящему из белка гемагглютинина вируса гриппа (Wilson et al., Cell 37:161 (1984)) и метку "flag".
[00182] Слитые белки можно получать с применением способов, которые хорошо известны в данной области (см. например патенты США № 5116964 и 5225538). Точный участок, в котором можно проводить слияние, можно выбирать эмпирически для оптимизации секреции или характеристик связывания слитого белка. Затем ДНК, кодирующую слитый белок, трансфицируют в клетку-хозяина для экспрессии.
[00183] Связывающие молекулы против ILT7, например, антитела по настоящему изобретению или их антигенсвязывающие фрагменты, варианты или производные можно использовать в неконъюгированной форме или их можно конъюгировать по меньшей мере с одним типом молекул, например, для улучшения терапевтических свойств молекулы, для облегчения детекции мишени или для визуализации и терапии пациента. Связывающие молекулы против ILT7, например, антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные можно метить или конъюгировать либо до очистки, либо после нее или когда проводят очистку.
[00184] В частности, антитела к ILT7 по изобретению или их антигенсвязывающие фрагменты, варианты или производные можно конъюгировать с терапевтическими средствами, пролекарствами, пептидами, белками, ферментами, вирусами, липидами, модификаторами биологического ответа, фармацевтическими средствами или PEG.
[00185] Специалистам в данной области понятно, что конъюгаты также можно формировать с применением различных способов в зависимости от выбранного средства, подлежащего конъюгации. Например, конъюгаты с биотином получают, например, посредством реакции связывающего полипептида с активированным сложным эфиром биотина, таким как N-гидроксисукцинимидный эфир биотина. Подобным образом, конъюгаты с флуоресцентным маркером можно получать в присутствии присоединяющего средства, например средств, перечисленных в настоящем описании, или посредством реакции с изотиоцианатом, таким как флуоресцеин-изотиоцианат. Конъюгаты антител к ILT7 по изобретению или их антигенсвязывающие фрагменты, варианты или производные получают аналогичным образом.
[00186] Настоящее изобретение дополнительно охватывает связывающие молекулы против ILT7, например, антитела по изобретению или их антигенсвязывающие фрагменты, варианты или производные, конъюгированные с диагностическим или терапевтическим средством. Антитела к ILT7, включая их антигенсвязывающие фрагменты, варианты или производные, можно использовать в диагностических целях, например, для мониторинга развития или прогрессирования заболевания в качестве части процедуры клинического исследования, например, для определения эффективности данной схемы лечения и/или профилактики. Например, детекцию можно облегчать связыванием антитела к ILT7 или его антигенсвязывающего фрагмента, варианта или производного с детектируемым веществом. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества, радиоактивные вещества, позитронно-активные металлы при использовании различных способов позитронно-эмиссионной томографии и нерадиоактивных ионов парамагнитных металлов. См., например, патент США № 4741900 для ионов металлов, которые можно конъюгировать с антителами для применения в качестве диагностических средств в соответствии с настоящим изобретением. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных веществ включают умбеллиферон, флуоресцеин, флуоресцеина изотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, дансилхлорид или фикоэритрин; пример люминесцентного вещества включает люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин, и примеры пригодного радиоактивного веещства включают 125I, 131I, 111In, 90Y или 99Tc.
[00187] Связывающую молекулу против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное также можно метить детектируемой меткой путем присоединения его к хемилюминесцентному соединению. Затем присутствие меченной хемилюминесцентным соединением связывающей молекулы против ILT7 определяют детекцией наличия люминесценции, которая появляется в ходе химической реакции. Примерами особенно пригодных хемилюминесцентных метящих соединений являются люминол, изолюминол, тероматический сложный эфир акридиния, имидазол, соль акридиния и сложный эфир щавелевой кислоты.
[00188] Одним из путей, которым антитело к ILT7 или его антигенсвязывающий фрагмент, вариант или производное можно детектируемо метить, является его связывание с ферментом и использование связанного продукта в ферментном иммуноанализе (EIA) (Voller, A., "The Enzyme Linked Immunosorbent Assay (ELISA)" Microbiological Associates Quarterly Publication, Walkersville, Md.; Diagnostic Horizons 2:1-7 (1978); Voller et al., J. Clin. Pathol. 37:507-520 (1978); Butler, Meth. Enzymol. 73:482-523 (1981); Maggio, ed. (1980), Enzyme Immunoassay, CRC Press, Boca Raton, Fla.; Ishikawa et al., eds. (1981), Enzyme Immunoassay (Kgaku Shorn, Tokyo). Фермент, который связывают с антителом к ILT7, взаимодействует с соответствующим субстратом, таким как хромогенным субстратом, таким образом, чтобы образовывалась химическая группа, которую можно детектировать, например, спектрофотометрическими, флуориметрическими или визуальными способами. Ферменты, которые можно использовать для детектируемого мечения антитела, включают, но не ограничиваются ими, малатдегидрогеназу, стафилококковую нуклеазу, дельта-5-стероидизомеразу, алкогольдегидрогеназу дрожжей, альфа-глицерофосфат, дегидрогеназу, триозофосфатизомеразу, пероксидазу хрена, щелочную фосфатазу, аспарагиназу, глюкозооксидазу, бета-галактозидазу, рибонуклеазу, уреазу, каталазу, глюкоза-6-фосфатдегидрогеназу, глюкоамилазу и ацетилхолинэстеразу. Кроме того, детекцию можно проводить колориметрическими способами, в которых применяют хромогенный субстрат для фермента. Также детекцию можно проводить путем визуального сравнения степени ферментативной реакции субстрата по сравнению со сходным образом полученными стандартами.
[00189] Детекцию также можно проводить с использованием любого из ряда других иммунологических анализов. Например, путем радиоактивного мечения связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, возможно проводить детекцию связывающей молекулы с использованием радиоиммунологического анализа (RIA) (см., например, Weintraub (March, 1986), Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques (The Endocrine Society), включенной в настоящее описание посредством ссылки). Радиоактивн изотоп можно детектировать с помощью средств, включающих, но не ограничивающихся ими, гамма-счетчик, сцинтилляционный счетчик или радиоавтографию.
[00190] Связывающую молекулу против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное также можно детектируемо метить с использованием испускающих флуоресценцию металлов, таких как 152Eu или другие металлы серии лантаноидов. Эти металлы можно присоединять к связывающей молекуле с использованием таких хелатирующих металлы групп, как диэтилентриаминпентауксусная кислота (DTPA) или этилендиаминтетрауксусная кислота (EDTA).
[00191] Способы конъюгации различных молекул с антителом (например, антителом к ILT7) или его антигенсвязывающим фрагментом, вариантом или производным хорошо известны, см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), p. 243-56 (Alan R. Liss, Inc. (1985); Hellstrom et al., "Antibodies For Drug Delivery", Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), Marcel Dekker, Inc., p. 623-53 (1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al. (eds.), p. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds), Academic Press p. 303-16 (1985), и Thorpe et al. (1982), "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev. 62:11958.
VI. Экспрессия полипептидов антител
[00192] Последовательности ДНК, которые кодируют легкую и тяжелую цепи антитела, можно получать одновременно или по отдельности с использованием обратной транскриптазы и ДНК-полимеразы в соответствии с хорошо известными способами. ПЦР можно инициировать с помощью праймеров для консенсусной константной области или с помощью более специфических праймеров на основе опубликованных последовательностей ДНК и аминокислотных последовательностей тяжелых и легких цепей. Как указано выше, ПЦР также можно использовать для выделения клонов ДНК, кодирующих легкие и тяжелые цепи антитела. В этом случае библиотеки можно подвергать скринингу с помощью консенсусных праймеров или более крупных гомологичных зондов, таких как зонды константной области мыши.
[00193] ДНК, как правило, плазмидную ДНК, можно выделять из клеток с применением известных в данной области способов, подвергать рестрикционному картированию и секвенировать в соответствии со стандартными хорошо известными способами, представленными в деталях, например, в указанных выше ссылках, касающихся технологий рекомбинантных ДНК. Как известно, ДНК может быть синтетической в соответствии с настоящим изобретением в любой момент в процессе выделения или при последующем анализе.
[00194] После манипуляции выделенным генетическим материалом для получения антител к ILT7 или их антигенсвязывающих фрагментов, вариантов или производных по изобретению полинуклеотиды, кодирующие антитела к ILT7, как правило, встраивают в экспрессирующий вектор для введения в клетки-хозяева, которые можно использовать для получения желаемого количества антитела к ILT7.
[00195] Для рекомбинантной экспрессии антитела или его фрагмента, варианта или производного, например тяжелой или легкой цепи антитела, которое связывается с молекулой-мишенью, описываемой в настоящем описании, например, ILT7, требуется конструкции экспрессирующего вектора, содержащего полинуклеотид, кодирующий антитело. После получения полинуклеотида, кодирующего молекулу антитела или тяжелую или легкую цепь антитела, или его части (например, содержащей вариабельный домен тяжелой или легкой цепи) по изобретению, можно получать вектор для продукции молекулы антитела технологией рекомбинантных ДНК с использованием способов, хорошо известных в данной области. Таким образом, способы получения белка посредством экспрессии полинуклеотида, содержащего кодирующую антитело нуклеотидную последовательность, описаны в настоящем описании. Способы, которые хорошо известны специалистам в данной области, можно использовать для конструкции экспрессирующих векторов, содержащих кодирующие антитело последовательности и подходящие сигналы контроля транскрипции и трансляции. Эти способы включают, например, способы рекомбинантных ДНК in vitro, синтетические способы и генетическую рекомбинацию in vivo. Таким образом, изобретение относится к реплицирующимся векторам, содержащим нуклеотидную последовательность, кодирующую молекулу антитела по изобретению или его тяжелую или легкую цепь, или вариабельный домен тяжелой или легкой цепи, функционально связанный с промотором. Такие векторы могут включать нуклеотидную последовательность, кодирующую константную область молекулы антитела (см., например, публикацию РСТ WO 86/05807; публикацию РСТ WO 89/01036 и патент США № 5122464) и в такой вектор можно клонировать вариабельный домен антитела для экспрессии всей тяжелой или легкой цепи.
[00196] Термин "вектор" или "экспрессирующий вектор" используют в настоящем описании для обозначения векторов, используемых в соответствии с настоящим изобретением в качестве носителей для введения в клетку-хозяина и экспрессии в клетке-хозяине желаемого гена. Как известно специалистам в данной области, такие векторы можно легко выбирать из группы, состоящей из плазмид, фагов, вирусов и ретровирусов. Как правило, векторы, совместимые с настоящим изобретением, содержат селективный маркер, подходящие участки рестрикции для облегчения клонирования желаемого гена и способности проникать и/или реплицироваться в эукариотических или прокариотических клетках.
[00197] Для целей этого изобретения можно применять ряд систем экспрессирующих векторов. Например, в одном классе векторов используют элементы ДНК, которые происходят из вирусов животных, таких как вирус папилломы крупного рогатого скота, вирус полиомы, аденовирус, вирус осповакцины, бакуловирус, ретровирусы (RSV, MMTV или MOMLV) или вирус SV40. Другие векторы предполагают использование полицистронных систем с внутренними участками связывания рибосом. Кроме того, клетки, в хромосомы которых встроилась ДНК, можно отбирать путем введения одного или нескольких маркеров, которые позволяют селекцию трансфицированных клеток-хозяев. Маркер может обеспечивать фототрофность для ауксотрофного хозяина, устойчивость к биоцидам (например, антибиотикам) или устойчивость к тяжелым металлам, таким как медь. Ген селективного маркера можно прямо связывать с последовательностями ДНК, которые необходимо экспрессировать, или вводить в ту же клетку котрансформацией. Также для оптимального синтеза мРНК необходимыми могут являться дополнительные элементы. Такие элементы включают сигнальные последовательности, сигналы сплайсинга, а также промоторы транскрипции, энхансеры и сигналы терминации.
[00198] В определенных вариантах осуществления клонированные гены вариабельной области встраивают в экспрессирующий вектор совместно с генами константной области тяжелой и легкой цепей (например, человека), синтезированными как указано выше. Как известно, в настоящем изобретении можно использовать любой экспрессирующий вектор, который способен обеспечивать экспрессию в эукариотических клетках. Примеры подходящих векторов включают, но не ограничиваются ими, плазмиды pcDNA3, pHCMV/Zeo, pCR3.1, pEF 1/His, pIND/GS, pRc/HCMV2, pSV40/Zeo2, pTRACER-HCMV, pUB6/V5-His, pVAX1 и pZeoSV2 (доступные от Invitrogen, San Diego, Calif.) и плазмиду pCI (доступную от Promega, Madison, Wis.). Как правило, скрининг больших количеств трансформированных клеток в отношении клеток, которые соответственно экспрессируют высокие уровни тяжелой и легкой цепей иммуноглобулинов, является общепринятым экспериментированием, которое можно проводить, например, с помощью роботизированных систем.
[00199] В более общем смысле, после получения вектора или последовательности ДНК, кодирующей мономерную субъединицу антитела к ILT7, экспрессирующий вектор можно вводить в подходящую клетку-хозяина. Введение плазмиды в клетку-хозяина можно проводить различными способами, хорошо известными специалистам в данной области. Они включают, но не ограничиваются ими, трансфекцию (включая электрофорез и электропорацию), слияние протопластов, осаждение фосфатом кальция, слияние клеток с заключенной в оболочку ДНК, микроинъекцию и инфекцию неизмененным вирусом. См. Ridgway (1988), "Mammalian Expression vectors", Vectors, ed. Rodriguez and Denhardt, Eds. (Butterworths, Boston, Mass.), Chapter 24.2, p. 470-472 (1988). Как правило, плазмиду вводят в хозяина электропорацией. Клетки-хозяева, несущие экспрессирующую конструкцию, выращивают в условиях, подходящих для продукции легких цепей и тяжелых цепей, и анализируют в отношении синтеза белков тяжелой и/или легкой цепей. Иллюстративные способы анализа включают твердофазный иммуноферментный анализ (ELISA), радиоиммунный анализ (RIA) или анализ посредством активированной флуоресценцией сортировки клеток (FACS), иммуногистохимию и т.п.
[00200] Экспрессирующий вектор переносят в клетку-хозяина общепринятыми способами, а затем трансфицированные клетки культивируют общепринятыми способами с получением антитела для применения в способах, описываемых в настоящем описании. Таким образом, изобретение включает клетки-хозяева, содержащие полинуклеотид, кодирующий антитело по изобретению или его тяжелую или легкую цепь, функционально связанные с гетерологичным промотором. В определенных вариантах осуществления для экспрессии двухцепочечных антител векторы, кодирующие как тяжелые, так и легкие цепи, можно коэкспрессировать в клетке-хозяине для экспрессии полной молекулы иммуноглобулина, как подробно описано ниже.
[00201] Как используют в настоящем описании, термин "клетки-хозяева" относится к клеткам, которые содержат векторы, сконструированные с использованием технологий рекомбинантных ДНК и кодирующие по меньшей мере один гетерологичный ген. В описаниях способов выделения антител из рекомбинантных хозяев термины "клетка" и "клеточная культура" используют взаимозаменяемо для обозначения источника антитела, если явно не указано иное. Другими словами, выделение полипептида из "клеток" может означать выделение либо из осажденных целых клеток, либо из клеточной культуры, содержащей как среду, так и суспендированные клетки.
[00202] Для экспрессии молекул антител для применения в способах, описываемых в настоящем описании, можно использовать ряд систем хозяин-экспрессирующий вектор. Такие системы хозяин-экспрессирующий вектор представляют собой носители, посредством которых можно получать и затем очищать представляющие интерес кодирующие последовательности, а также они представляют собой клетки, которые при трансформации или трансфекции подходящими нуклеотидными кодирующими последовательностями могут экспрессировать молекулу антитела по изобретению in situ. Они включают, но не ограничиваются ими, микроорганизмы, такие как бактерии (например, Е. coli, В. subtilis), трансформированные рекомбинантными экспрессирующими векторами с бактерофаговой ДНК, плазмидной ДНК или космидной ДНК, содержащими кодирующие антитело последовательности; дрожжи (например, Saccharomyces, Pichia), трансформированные рекомбинантными экспрессирующими векторами дрожжей, содержащими кодирующие антитело последовательности; системы клеток насекомых, инфицированные рекомбинантными вирусными экспресирующими векторами (например, бакуловирусом), содержащими кодирующие антитело последовательности; системы клеток растений, инфицированных рекомбинантными вирусными экспрессирующими векторами (например, вирусом мозаики цветной капусты, CaMV; вирусом табачной мозаики, TMV) или трансформированных рекомбинантными плазмидными экспрессирующими векторами (например, плазмидой Ti), содержащими кодирующие антитело последовательности; или системы клеток млекопитающих (например, клетки COS, CHO, BLK, 293, 3Т3), несущие рекомбинантные экспрессирующие конструкции, содержащие промоторы, происходящие из генома клеток млекопитающих (например, промотор металлотионеина) или из вирусов млекопитающих (например, поздний промотор аденовируса; промотор 7.5К вируса осповакцины). Предпочтительно для экспрессии рекомбинантной молекулы антитела используют бактериальные клетки, такие как Escherichia coli, или эукариотические клетки, особенно для экспрессии целой рекомбинантной молекулы антитела. Например, эффективной экспрессирующей системой для антител являются клетки млекопитающих, такие как клетки яичника китайского хомяка (СНО), в сочетании с векторами, такими как основной средне-ранний промоторный элемент из цитомегаловируса (Foecking et al., Gene 5:101 (1986); Cockett et al., Bio/Technology 8:2 (1990)).
[00203] Линия клеток-хозяев, используемая для экспрессии белков, часто происходит из млекопитающих; специалисты в данной области способны определять конкретные линии клеток-хозяев, которые наилучшим образом подходят для экспрессии в них желаемого продукта гена. Иллюстративные линии клеток-хозяев включают, но не ограничиваются ими, клетки СНО (китайский хомяк), DG44 и DUXBII (линии яичника китайского хомка, DHFR-минус), HELA (карцинома шейки матки человека), CVI (линия почки обезьяны), COS (производное CVI с Т-антигеном SV40), VERY, BHK (почки детенышей хомячка), MDCK, 293, WI38, R1610 (фибробласты китайского хомяка) BALBC/3T3 (фибробласты мыши), НАК (линия почки хомяка), SP2/0 (миелома мыши), P3.times.63-Ag3.653 (миелома мыши), BFA-1c 1BPT (эндотелиальные клетки крупного рогатого скота), RAJI (лимфоциты человека) и 293 (почка человека). Линии клеток-хозяев, как правило, являются доступными от коммерческих организаций, American Tissue Culture Collection или из опубликованной литературы.
[00204] Кроме того, можно выбирать линию клеток-хозяев, которая модулирует экспрессию встроенных последовательностей или модифицирует и процессирует продукт гена определенным желаемым образом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важны для функции белка. Различные клетки-хозяева обладают характерными и конкретными механизмами посттрансляционного процессинга и модификации белков и продуктов генов. Для обеспечения правильной модификации и процессинга чужеродного экспрессируемого белка можно выбирать подходящие линии клеток или системы хозяев. С этой целью, можно использовать эукариотические клетки-хозяева, которые обладают клеточным аппаратом для правильного процессинга первичного транскрипта, гликозилирования и фосфорилирования продукта гена.
[00205] Для длительной продукции рекомбинантных белков с высоким выходом предпочтительной является стабильная экспрессия. Например, можно конструировать линии клеток, которые стабильно экспрессируют молекулу антитела. Вместо применения экспрессирующих векторов, которые содержат вирусные участки начала репликации, клетки-хозяева можно трансформировать ДНК, контролируемой соответствующими элементами контроля экспрессии (например, промоторной, энхансерной последовательностями, терминаторами транскрипции, участками полиаденилирования и т.д.), и с селективным маркером. После введения чужеродной ДНК сконструированные клетки можно оставлять расти в течение 1-2 суток в обогащенной среде, а затем их можно переключать на селективные среды. Селективный маркер в рекомбинантной плазмиде придает устойчивость при селекции и позволяет клеткам стабильно встраивать плазмиду в их хромосомы и расти с образованием очагов, которые, в свою очередь, можно клонировать и размножать в клеточные линии. Этот способ можно преимущественно использовать для конструирования линий клеток, которые стабильно экспрессируют молекулу антитела.
[00206] Можно использовать ряд систем для селекции, включая, но, не ограничиваясь ими, гены тимидинкиназы вируса простого герпеса (Wigler et al., Cell 11: 223 (1977)), гипоксантин-гуанин-фосфорибозилтрансферазы (Szybalska & Szybalski, Proc. Natl. Acad. Sci. USA 48:202 (1992)) и аденинфосфорибозилтрансферазы (Lowy et al., Cell 22:817, 1980) в tk-, hgprt- или aprt-клетках соответственно. Также можно использовать устойчивость к антиметаболитам в качестве основы для селекции для следующих генов: dhfr, который придает устойчивость к метотрексату (Wigler et al., Natl. Acad. Sci. USA 77:357 (1980); O'Hare et al., Proc. Natl. Acad. Sci. USA 78: 1527 (1981)); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan & Berg, Proc. Natl. Acad. Sci. USA 78:2072 (1981)); neo, который придает устойчивость к аминогликозиду G-418, Clinical Pharmacy 12:488-505; Wu and Wu, Biotherapy 3:87-95 (1991); Tolstoshev, Ann. Rev. Pharmacol. Toxicol. 32:573-596 (1993); Mulligan, Science 260:926-932 (1993); и Morgan and Anderson, Ann. Rev. Biochem. (52:191-217 (1993); TIB TECH 11(5): 155-215 (May, 1993), и hygro, который придает устойчивость к гигромицину (Santerre et al., Gene 30:147 (1984)). Способы, широко известные в области технологии рекомбинантных ДНК, которые можно использовать, описаны у Ausubel et al. (1993), Current Protocols in Molecular Biology, John Wiley & Sons, NY; Kriegler (1990), Gene Transfer and Expression, A Laboratory Manual, Stockton Press, NY; и в главах 12 и 13, Dracopoli et al. (eds) (1994), Current Protocols in Human Genetics. John Wiley & Sons, NY; Colberre-Garapin et al. (1981), J. Mol. Biol. 150:1, которые полностью включены в настоящее описание посредством ссылки в полном объеме.
[00207] Уровни экспрессии молекулы антитела можно увеличивать посредством амплификации вектора (для обзора см. Bebbington and Hentschel (1987), "The Use Of Vectors Based On Gene Amplification For The Expression Of Cloned Genes In Mammalian Cells In DNA Cloning". (Academic Press, New York), Vol. 3). Когда маркер в векторной системе, экспрессирующей антитело, является амплифицируемым, увеличение уровня ингибитора, присутствующее в культуре клеток-хозяев, приведет к увеличению количества копий маркерного гена. Поскольку амплифицированная область связана с геном антитела, продукция антитела также будет возрастать (Grouse et al., Mol. Cell. Biol. 3:257 (1983)).
[00208] Продукция in vitro позволяет увеличивать масштаб с получением больших количеств желаемых полипептидов. Способы культивирования клеток млекопитающих в условиях культивирования тканей известны в данной области и включают гомогенную суспензионную культуру, например, в эрлифтном ферментере или в реакторе с непрерывным перемешиванием, или иммобилизованную или заключенную клеточную культуру, например в полых волокнах, микрокапсулах, на агарозных микрогранулах или керамических кассетах. Если необходимо и/или желательно, растворы полипептидов можно очищать стандартными способами хроматографии, например гель-фильтрацией, ионообменной хроматографией, хроматографией над DEAE-целлюлозой или (иммуно)аффинной хроматографией, например, после избирательного биосинтеза синтетического полипептида шарнирной области или перед или после этапа HIC-хроматографии, описываемой в настоящем описании.
[00209] Гены, кодирующие антитела к CD100 или их антигенсвязывающие фрагменты, варианты или производные по изобретению, также можно экспрессировать в клетках не принадлежащих млекопитающим, таких как клетки бактерий или дрожжей, или растений. Бактерии, которые легко захватывают нуклеиновые кислоты, включают представителей enterobacteriaceae, таких как штаммы Escherichia coli или Salmonella; Bacillaceae, такие как Bacillus subtilis; Pneumococcus; Streptococcus и Haemophilus influenzae. Кроме того, понятно, что при экспрессии в бактериях гетерологичные полипептиды, как правило, становятся частью телец включения. Гетерологичные полипептиды необходимо выделять, очищать, а затем собирать в функциональные молекулы. В случае, когда четырехвалентные формы антител являются желательными, то субъединицы затем будут подвергаться самосборке в четырехвалентные антитела (WO 02/096948A2).
[00210] В бактериальных системах ряд экспрессирующих векторов можно преимущественно выбирать в зависимости от предполагаемого применения молекулы антитела, которую экспрессируют. Например, когда необходимо получать большое количество такого белка для получения фармацевтических композиций молекулы антитела, необходимым могут являться векторы, которые направляют экспрессию высоких уровней слитых белковых продуктов, очистку которых легко проводить. Такие векторы включают, но не ограничиваются ими, экспрессирующий вектор Е. coli pUR278 (Ruther et al., EMBO J. 2: 1791 (1983)), в котором кодирующая антитело последовательность можно лигировать индивидуально в вектор в рамке считывания с кодирующей lacZ областью, таким образом, чтобы получать слитый белок; векторы pIN (Inouye & Inouye, Nucleic Acids Res. 73: 3101-3109 (1985); Van Heeke and Schuster, J. Biol Chem. 24:5503-5509 (1989)) и т.п. Также можно использовать векторы pGEX для экспрессии чужеродных полипептидов в виде слитых белков с глутатион S-трансферазой (GST). Как правило, такие слитые белки являются растворимыми, и их можно легко выделять из лизированных клеток адсорбцией и связыванием с гранулами на основе матрицы из глутатион-агарозы с последующим элюированием в присутствии свободного глутатиона. Векторы pGEX конструируют так, чтобы они включали участки расщепления протеазы тромбина или фактора Ха, так чтобы продукт клонированного гена-мишени мог высвобождаться из группы GST.
[00211] В дополнение к прокариотам также можно использовать эукариотические микроорганизмы. Saccharomyces cerevisiae или обычные пекарские дрожжи наиболее часто используются среди эукариотических микроорганизмов, хотя широко доступен ряд других штаммов, например, Pichia pastoris.
[00213] В системе насекомых в качестве вектора для экспрессии чужеродных, как правило, генов используют вирус ядерного полиэдроза Autographa californica (AcNPV). Этот вирус растет в клетках Spodoptera frugiperda. Кодирующую антитело последовательность можно клонировать индивидуально в не являющиеся необходимыми области (например, ген полиэдрина) вируса и помещать под контроль промотора AcNPV (например, промотора полиэдрина).
[00214] После рекомбинантной экспрессии связывающая молекула по изобретению ее можно очищать любым известным в данной области способом очистки молекулы иммуноглобулина, например, хроматографией (например, ионообменной, аффинной в частности на основе аффинности к специфическому антигену после хроматографии с белком А и колоночной хроматографии с распределением по размеру), центрифугирования, дифференциальной растворимости или любым другим стандартным способом очистки белков. Альтернативно, предпочтительный способ увеличения аффинности антител по изобретению описан в публикации патентной заявки США № 2002-0123057 А1.
VII. Способы лечения с использованием терапевтических связывающих молекул против ILT7
[00215] Способы по изобретению относятся к применению связывающих молекул против ILT7, например, антител, включая их антигенсвязывающие фрагменты, варианты и производные, для лечения пациентов, страдающих заболеванием, связанным с экспрессией ILT7 или экспрессирующими ILT7 клетками. Под "экспрессирующей ILT7 клеткой" подразумевают клетки, экспрессирующие антиген ILT7. Способы детекции экспрессии ILT7 в клетках хорошо известны в данной области и включают, но не ограничиваются ими, способы ПЦР, иммуногистохимию, проточную цитометрию, вестерн-блоттинг, ELISA и т.п.
[00216] Хотя следующее ниже обсуждение относится к способам диагностики и лечения различных заболеваний и нарушений антителом к ILT7 по изобретению, способы, описываемые в настоящем описании, также применимы для антигенсвязывающих фрагментов, вариантов и производных этих антител к ILT7, которые сохраняют желаемые свойства антител к ILT7 по изобретению, например, способны специфично связывать ILT7 и обладают нейтрализующей активностью в отношении ILT7.
[00217] В одном варианте осуществления лечение включает применение или введение связывающая молекула против ILT7, например, антитела или его антигенсвязывающего фрагмента по настоящему изобретению пациенту или применение или введение связывающей молекулы против CD100 в выделенную ткань или линию клеток от пациента, где пациент страдает заболеванием, имеет симптомом заболевания или предрасположенность к заболеванию. В другом варианте осуществления лечение также включает применение или введение фармацевтической композиции, содержащей связывающую молекулу против ILT7, например, антитело или его антигенсвязывающий фрагмент по настоящему изобретению, пациенту или применение или введение фармацевтической композиции, содержащей связывающую молекулу против ILT7, в выделенную ткань или линию клеток от пациента, который страдает заболевание, имеет симптомом заболевания или предрасположенность к заболеванию.
[00218] Связывающие молекулы против ILT7, например, антитела или их антигенсвязывающие фрагменты, варианты или производные по настоящему изобретению являются пригодными для лечения различных аутоиммунных состояний. Например, терапия по меньшей мере одним антителом к ILT7 вызывает физиологическую реакцию, например, снижение интерферона, которое является благоприятным для лечении состояний болезни, связанных с экспрессирующими ILT7 клетками у человека.
[00219] В одном из вариантов осуществления изобретение относится к связывающим молекулам против ILT7, например, антителам или их антигенсвязывающим фрагментам, вариантам или производным для применения в качестве лекарственного средства, в частности для применения в лечении или профилактики аутоиммунного состояния или заболевания. Примеры аутоиммунных заболеваний включают, но не ограничиваются ими: миозит, диабет, болезнь Хашимото, аутоиммунную надпочечниковую недостаточность, эритроцитарную анемию, рассеянный склероз, ревматоидный кардит, системную красную волчанку, псориаз, ревматоидный артрит, хроническое воспаление, синдром Шегрена, полимиозит, дерматомиозит, миозит с тельцами включения, юношеский миозит и склеродермию.
[00220] В соответствии со способами по настоящему изобретению по меньшей мере одну связывающую молекулу против ILT7, например, антитело или его антигенсвязывающий фрагмент, вариант или производное, как определено в другом месте в настоящем описании, используют для стимуляции положительного терапевтического ответа в отношении аутоиммунного ответа. Под "положительным терапевтическим ответом" по отношению к лечению аутоиммунного заболевания подразумевают улучшение состояния заболевания в связи с активностью этих связывающих молекул, например, антител или их антигенсвязывающих фрагментов, вариантов или производных и/или улучшение симптомов, ассоциированных с заболеванием. Таким образом, можно наблюдать понижение уровней интерферона-альфа, снижение количества или активности плазмацитоидных дендритных клеток или улучшение одного или более симптомов, ассоциированных с заболеванием. Таким образом, например, улучшение состояния заболевания можно характеризовать как полный ответ. Под "полным ответом" подразумевают отсутствие клинически детектируемого заболевания с нормализацией любых полученных ранее результатов исследования. Такой ответ может сохраняться в течение по меньшей мере одного месяца после лечения способами по изобретению. Альтернативно улучшение состояния заболевания может быть классифицировано как частичный ответ.
[00221] Связывающие молекулы против ILT7, например, антитела или их антигенсвязывающие фрагменты, варианты или производные, описываемые в настоящем описании, также могут являться пригонными при лечении аутоиммунных заболеваний и дефицитных состояний или нарушений иммунной системы, которые ассоциированы с экспрессирующими ILT7 клетками. Аутоиммунные заболевания характеризуются повреждением на уровне клеток, тканей и/или органов, вызываемым иммунологической реакцией индивидуума на свои собственные клетки, ткани и/или органы. В одном из вариантов осуществления аутоиммунное заболевание представляет собой системную красную волчанку.
[00222] Клинический ответ можно оценивать с использованием способов скрининга, таких как сканирование магнитно-резонансной томографией (MRI), рентгеновскую радиографическую визуализацию, сканирование компьютерной томографией (СТ), проточную цитометрию или анализ активированной флуоресценцией сортировкой клеток (FACS), гистологию, макроскопическое исследование патологии и химию крови, включая, но не ограничиваясь ими, изменения, детектируемые посредством ELISA, RIA, хроматографии и т.п. В дополнение к этим положительным терапевтическим ответам, индивидуум, подвергающийся лечению связывающей молекулой против ILT7, например, антителом или его антигенсвязывающим фрагментом, может испытывать благоприятный эффект улучшения симптомов, ассоциированных с заболеванием.
[00223] Дополнительным вариантом осуществления изобретения является применение связывающей молекулы против ILT7, например, антитела или его антигенсвязывающих фрагментов, вариантов или производных для диагностического мониторинга уровней белков в ткани в качестве части процедуры клинического тестирования, например, для определения эффективности данной схемы лечения. Например, детекцию можно облегчать связыванием антитела с детектируемым веществом. Примеры детектируемых веществ включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества и радиоактивные вещества. Примеры подходящий ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных вещетсв включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; пример люминесцентного вещества включает люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин, и примеры подходящего пригодного радиоактивного вещества включают 125I, 131I, 35S или 3H.
VIII. Фармацевтические композиции и способы введения
[00224] Способы получения и введения связывающих молекул против ILT7, например, антител или их антигенсвязывающих фрагментов, вариантов или производных, предоставленных в настоящем описании, нуждающемуся в этом индивидууму хорошо известны специалистам в данной области, или специалистам в данной области легко их определяют.
[00225] Как описано в настоящем описании, связывающие молекулы против ILT7, например, антитела или их антигенсвязывающие фрагменты, варианты или производные, предоставленные в настоящем описании можно вводить в фармацевтически эффективном количестве для лечения in vivo опосредуемых экспрессирующими ILT7 клетками заболеваний, таких как определенные типы аутоиммунных заболеваний. В отношении этого, следует понимать, что описанные связывающие молекулы по изобретению можно формулировать так, чтобы облегчать введение и обеспечивать стабильность активного вещества. Фармацевтические композиции в соответствии с настоящим изобретением могут содержать фармацевтически приемлемый нетоксичный стерильный носитель. Для целей настоящей заявки фармацевтически эффективное количество связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, конъюгированного или неконъюгированного, должно означать количество, достаточное для достижения эффективного связывания с мишенью и для положительного эффекта, например, для улучшения состояния симптомов заболевания или состояния или детекции вещества или клетки.
[00226] Фармацевтические композиции, подходящие инъекций, должны быть стерильными и должны быть жидкими в той степени, чтобы существовала возможность введения в шприц. Они должны быть стабильными в условиях производства и хранения и надежно защищены от заражающего действия микроорганизмов, таких как бактерии и грибы. Предупреждение действия микроорганизмов можно обеспечивать различными антибактериальными и противогрибковыми средствами. Подходящие составы для применения в терапевтических способах, описываемых в настоящем описании, описаны в Remington's Pharmaceutical Sciences (Mack Publishing Co.) 16th ed. (1980).
[00227] В соответствии с объемом настоящего изобретения антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные по изобретению можно вводить человеку или другому животному в соответствии с указанными выше способами лечения в количестве, достаточном для оказания терапевтического действия. Антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные по изобретению можно вводить такому человеку или другому животному в общепринятой лекарственной форме, получаемой объединением антитела или его антигенсвязывающего фрагмента, варианта или производного по изобретению с общепринятым фармацевтически приемлемым носителем или разбавителем в соответствии с известными способами. Специалисту в данной области будет понятно, что форма и характер фармацевтически приемлемого носителя или разбавителя зависит от количества активного ингредиента, с которым его объединяют, пути введения и других хорошо известных переменных. Специалисты в данной области также будет понятно, что смесь, содержащая один или более типов связывающих молекул против ILT7, например, антител или их антигенсвязывающих фрагментов, вариантов или производных по изобретению может оказаться особенно эффективной.
[00228] Под "терапевтически эффективной дозой или количеством" или "эффективным количеством" подразумевают количество связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного, которое при введении приводит к положительному терапевтическому ответу в отношении лечения пациента с заболеванием или состоянием, подлежащим лечению.
[00229] Терапевтически эффективные дозы композиций по настоящему изобретению для лечения опосредованных экспрессирующими ILT7 клетками заболеваний, таких как определенные типы аутоиммунных заболеваний, включая, например, системную красную волчанку, изменяются в зависимости от многих различных факторов, включая пути введение, участок-мишень, физиологическое состояние пациента, является ли пациент человеком или животным, других вводимых лекарственных препаратов, и является ли лечение профилактическим или терапевтическим. Как правило, пациент является человеком, но не принадлежащие человеку млекопитающие, включая трансгенных млекопитающих также можно лечить. Лечебные дозы можно ститровать для оптимизации показателей безопасности и эффективности.
[00230] Настоящее изобретение также относится к использованию связывающей молекулы против ILT7, например, антитела или его антигенсвязывающего фрагмента, варианта или производного в производстве лекарственного средства для лечения аутоиммунное заболевание, включая, например, системную красную волчанку.
IX. Диагностика
[00231] Изобретение дополнительно относится к способу диагностики, пригодному во время диагностики опосредованных экспрессирующими ILT7 клетками заболеваний, таких как определенные типы аутоиммунных заболеваний, включая, например, системную красную волчанку, которые включают измерение уровня экспрессии белка или трансркипта ILT7 в ткани или других клетках или жидкостях организма от индивидуума и сравнение измеряемого уровня экспрессии со стандартным уровнем экспрессии ILT7 в нормальной ткани или жидкости организма, тем самым увеличение уровня экспрессии по сравнению со стандартом является показателем нарушения.
[00232] Антитела к ILT7 по изобретению и их антигенсвязывающие фрагменты, варианты и производные можно использовать для анализа уровней белка ILT7 в биологическом образце, включая применение классических иммуногистологических способов, известных специалистам в данной области (например, см. Jalkanen, et al., J. Cell. Biol. 101:976-985 (1985); Jalkanen et al., J. Cell Biol. 105:3087-3096 (1987)). Другие способы на основе антител, пригодные для детекции экспрессия белка ILT7, включают иммунологические анализы, такие как твердофазный иммуноферментный анализ (ELISA), иммунопреципитацию или вестерн-блоттинг. Подходящие анализы более подробно описаны в другом месте в настоящем описании.
[00233] Под "анализом уровня экспрессии полипептида ILT7" подразумевают качественное или количественное измерение или оценку уровня полипептида ILT7 в первом биологическом образце либо прямо (например, путем определения или оценки абсолютного уровня белка), либо относительно (например, путем сравнения с ассоциированным с заболеванием уровнем полипептида во втором биологическом образце). Уровень экспрессии полипептида ILT7 в первом биологическом образце можно измерять или оценивать и сравнивать со стандартным уровнем полипептида ILT7, где стандарт берут из второго биологического образца, получаемого от индивидуума, не страдающего нарушением, или определяют по усредненным уровням для популяции индивидуумов, не страдающих нарушением. Как будет понятно в данной области, после того, как "стандартный" уровень полипептида ILT7 известен, его можно повторно использовать в качестве стандарта для сравнения.
[00234] Под "биологическим образцом" понимают любой биологический образец, получаемый от индивидуума, из линии клеток, тканевой культуры или другого источника клеток, потенциально экспрессирующих ILT7. Способы получения биопсий тканей и жидкостей организма от млекопитающих хорошо известны в данной области.
X. Иммунологические анализы
[00235] Связывающие молекулы против ILT7 например, антитела или их антигенсвязывающие фрагменты, варианты или производные по изобретению можно анализировать в отношении иммуноспецифического связывания любым способом, известным в данной области. Иммуноанализы, которые можно использовать, включают, но не ограничиваются ими, системы конкурентного и неконкурентного анализа с применением способов, таких как вестерн-блоттинг, радиоиммунологические анализы, ELISA (твердофазный иммуноферментный анализ), радиоиммунологические анализы формата "сэндвич", анализы иммунопреципитации, реакции с преципитином, реакции с преципитином с диффузией в геле, анализы иммунодиффузии, анализы агглютинации, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммунологические анализы, иммунологические анализы с белком А, наряду с другими. Такие анализы являются общепринятыми и хорошо известны специалисту в данной области (см., например, Ausubel et al., eds (1994), Current Protocols in Molecular Biology (John Wiley & Sons, Inc., NY), Vol. 1, которая полностью включена в настоящее описание посредством ссылки). Иллюстративные иммунологические анализы описаны в кратком изложении ниже (но не предназначены для ограничения).
[00236] Антитела к ILT7 или их антигенсвязывающие фрагменты, варианты или производные по изобретению дополнительно можно применять гистологически, как в иммунофлуоресценции, иммуноэлектронной микроскопии или неиммунологических анализах для детекции in situ белка ILT7 или его консервативных вариантов или пептидных фрагментов. Детекцию in situ можно проводить путем извлечения гистологического образца из пациента и нанесения на него меченого антитела к ILT7 или его антигенсвязывающего фрагмента, варианта или производного, например, наносимых наслоением меченого антитела (или фрагмента) на биологический образец. С использованием такой процедуры можно определить не только присутствие белка ILT7 или консервативных вариантов или пептидных фрагментов, а также его распределение в исследуемой ткани. С использованием настоящего изобретения специалист в данной области может легко понять, что любой из широкого спектра гистологических способов (таких как процедуры окрашивания) можно модифицировать для проведения такой детекции in situ.
[00237] Иммуноанализы и неиммунные анализы продуктов гена ILT7 или их консервативных вариантов или пептидных фрагментов, как правило, включают инкубацию образца, такого как биологическая жидкость, экстракт ткани, свежесобранные клетки или лизаты клеток, которые инкубировали в клеточной культуре, в присутствии меченого поддающейся детекции меткой антитела, способного связываться с ILT7 или его консервативными вариантами или пептидными фрагментами, и детекцию связанного антитела любым из ряда хорошо известных в данной области способов.
[00238] Биологический образец можно приводить в контакт или иммобилизовывать на твердофазной подложке или носителе, таких как нитроцеллюлоза или другая твердая подложка, на которой можно иммобилизовывать клетки, клеточные частицы или растворимые белки. Затем подложку можно промывать подходящими буферами с последующей обработкой меченным поддающейся детекции меткой антителом к ILT7 или его антигенсвязывающим фрагментом, вариантом или производным. Затем твердофазную подложку можно промывать буфером второй раз для удаления несвязанного антитела. Затем антитело необязательно метят. Затем можно проводить детекцию количества связанной метки на твердой подложке общепринятыми способами.
[00239] Под "твердофазной подложкой или носителем" подразумевают любую подложку, способную связывать антиген или антитело. Хорошо известные подложки или носители включают стекло, полистирол, полипропилен, полиэтилен, декстран, нейлон, амилазы, нейтральные и модифицированные целлюлозы, полиакриламиды, габбро и магнетит. Для целей настоящего изобретения характер носителя может быть либо растворимым до некоторой степени, либо нерастворимым. Материал подложки может иметь практически любую возможную структурную конфигурацию, при условии, что связанная с ним молекула способна связываться с антигеном или антителом. Таким образом, конфигурация подложки может быть сферической, как в гранулах, или цилиндрической, как во внутренней поверхности тестовой пробирки или внешняя поверхность стержня. Альтернативно, поверхность может быть плоской, такой как лист, тест-полоска и т.д. Иллюстративные подложки включают полистироловые гранулы. Специалистам в данной области известны многие другие пригодные носители для связывания антитела или антигена, или они способны определить их с использованием общепринятого экспериментирования.
[00240] Активность связывания данной партии антител к ILT7 или их антигенсвязывающего фрагмента, варианта или производного можно хорошо определять известными способами. Специалисты в данной области способны определять действующие и оптимальные условия анализа для каждого определения с использованием общепринятого экспериментирования.
[00241] Аффинность связывания антитела с антигеном и скорость обратной реакции взаимодействия антитело-антиген можно определять анализами конкурентного связывания. Один из примеров анализа конкурентного связывания представляет собой радиоиммунологический анализ, включающий инкубацию меченого антиген (например, 3H или 125I) с представляющим интерес антителом в присутствии увеличивающихся количеств немеченого антигена и детекцию антитела, связанного с меченым антигеном. Аффинность представляющего интерес антитела к конкретному антигену и скорости обратных реакций связывание можно определять на основании данных посредством анализа графика Скэтчарда. Также можно определять конкуренцию со вторым антителом с использованием радиоиммунологических анализов. В этом случае антиген инкубируют с представляющим интерес антителом, конъюгированным с меченым соединением (например, 3H или 125I) в присутствии увеличивающихся количеств немеченого второго антитела.
[00242] Доступно множество способов измерения аффинности взаимодействия антитело-антиген, но относительно мало для определения констант скоростей реакций. Большинство способов основаны на мечении антитела или антигена, что неизбежно осложняет повседневное измерения и вносит неточность в измеренные количества.
[00243] Поверхностный плазмонный резонанс (SPR), как проводимый на BIACORE®, обеспечивает ряд преимуществ над общепринятыми способами измерения аффинности взаимодействий антитело-антиген: (i) не требуется мечение ни антитела, ни антигена; (ii) не требуется предварительная очистка антител, можно использовать непосредственно супернатант культуры клеток; (iii) для многих целей оценки возможны и достаточны измерения в реальном времени, позволяющие быстрое полуколичественное сравнение различных взаимодействий моноклональных антител; (iv) биоспецифическую поверхность можно регенерировать так, чтобы серии различных моноклональных антител можно было легко сравнивать в идентичных условиях; (v) аналитические процедуры являются полностью автоматизированными и можно проводить обширные серии измерений без вмешательства пользователя. BIAapplications Handbook, version АВ (новое издание в 1998 году), BIACORE® code № BR-1001-86; BIAtechnology Handbook, version AB (reprinted 1998), BIACORE® code № BR-1001-84. Основанные на SPR исследования связывания требуют, чтобы один представитель связывающей пары был иммобилизован на поверхности сенсора. Иммобилизованный партнер по связыванию обозначают как лиганд. Партнер по связыванию в растворе обозначают как анализируемое соединение. В некоторых случаях лиганд связан непрямо с поверхностью через связывание с другой иммобилизованной молекулой, которую обозначают как улавливающая молекула. Ответ SPR отражает изменение в весовой концентрации на поверхности детектора по мере связывания или диссоциации анализируемых соединений.
[00244] На основе SPR способы измерения BIACORE® в реальном времени осуществляют мониторинг взаимодействий, непосредственно когда они происходят. Способ хорошо подходит для определения кинетических параметров. Ранжирование сравнительной аффинности проводить просто, и кинетические константы и константы аффинности можно устанавливать из данных сенсограмм.
[00245] Когда анализируемое соединение инъецируют отдельным импульсом над поверхностью лиганда, полученную сенсограмму можно подразделить на три существенных фазы: (i) ассоциация анализируемого соединения с лигандом в процессе инъекции образца; (ii) состояние равновесия или устойчивое состояние в процессе инъекции образца, где скорость связывания анализируемого соединения уравновешивается диссоциацией из комплекса; (iii) диссоциация анализируемого соединения с поверхности во время течения буфера.
[00246] Фазы ассоциации и диссоциации обеспечивают информацию о кинетике взаимодействия анализируемое соединение-лиганд (ka и kd, скорости образования комплекса и диссоциации kd/ka=KD). Фаза равновесия обеспечивает информацию об аффинности взаимодействия анализируемое соединение-лиганд (KD).
[00247] Программное обеспечение BIAevaluation обеспечивает обширные средства для аппроксимации кривых с использованием алгоритмов как численного интегрирования, так и глобальной аппроксимации. При подходящем анализе данных из простых исследований BIACORE® можно получать отдельные константы скорости и аффинности для взаимодействия. Диапазон аффинности, поддающейся измерению этим способом, является очень широким, от мМ до пМ.
[00248] Эпитопная специфичность является важной характеристикой моноклонального антитела. Картирование эпитопа с помощью BIACORE® в отличие от общепринятых способов с использованием радиоиммунологического анализа, ELISA или других способов поверхностной адсорбции не требует мечения или очистки антител и позволяет тесты специфичности множества участков с использованием последовательности из нескольких моноклональных антител. Кроме того, большие количества анализов можно проводить автоматически.
[00249] Эксперименты попарного связывания тестируют способность двух MAb связываться одновременно с одним и тем же антигеном. MAb, направленные против отдельных эпитопов, связываются независимо, в то время как MAb, направленные против идентичных или близкородственных эпитопов, будут препятствовать связыванию друг друга. Эти эксперименты по связыванию с использованием BIACORE® легко выполнять.
[00250] Например, можно использовать улавливающую молекулу для связывания первого Mab с последующим добавлением антигена и второго MAb последовательно. Сенсограммы выявляют: (1) сколько антигена связывается с первым Mab, (2) в какой степени второе MAb связывается со связанным с поверхностью антигеном, (3) если второе MAb не связывается, изменит ли результаты обратный порядок попарного теста.
[00251] Пептидное ингибирование является другим способом, используемым для картирования эпитопов. Этот способ может дополнять исследования попарного связывания антител и может соотносить функциональные эпитопы со структурными признаками, когда первичная последовательность антигена известна. Пептиды или антигенные фрагменты тестируют в отношении ингибирования связывания различных MAb с иммобилизованным антигеном. Пептиды, которые могут препятствовать связыванию данного MAb, считают структурно родственными эпитопу, определенному с помощью этого MAb.
[00252] В практическом осуществлении настоящего изобретения используют, если не указано иное, общепринятые способы клеточной биологии, клеточной культуры, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантных ДНК и иммунологии, которые известны специалистам в данной области. Такие способы подробно объяснены в литературе. См., например, Sambrook et al., ed. (1989), Molecular Cloning A Laboratory Manual (2nd ed.; Cold Spring Harbor Laboratory Press); Sambrook et al., ed. (1992), Molecular Cloning: A Laboratory Manual, (Cold Springs Harbor Laboratory, NY); D. N. Glover ed. (1985), DNA Cloning, Volumes I and II; Gait, ed. (1984), Oligonucleotide Synthesis; Mullis et al. патент США № 4683195; Hames and Higgins, eds. (1984), Nucleic Acid Hybridization; Hames and Higgins, eds. (1984), Transcription And Translation; Freshney (1987), Culture Of Animal Cells (Alan R. Liss, Inc.); Immobilized Cells And Enzymes (IRL Press) (1986); Perbal (1984), A Practical Guide To Molecular Cloning; the treatise, Methods In Enzymology (Academic Press, Inc., N.Y.); Miller and Calos eds. (1987), Gene Transfer Vectors For Mammalian Cells, (Cold Spring Harbor Laboratory); Wu et al., eds., Methods In Enzymology, Vol. 154 и 155; Mayer and Walker, eds. (1987), Immunochemical Methods In Cell And Molecular Biology (Academic Press, London); Weir and Blackwell, eds. (1986), Handbook Of Experimental Immunology, Volumes I-IV; Manipulating the Mouse Embryo, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., (1986); и Ausubel et al. (1989), Current Protocols in Molecular Biology (John Wiley and Sons, Baltimore, Md.).
[00253] Общие принципы конструирования антител указаны у Borrebaeck, ed. (1995), Antibody Engineering (2nd ed.; Oxford Univ. Press). Общие принципы инженерии белков указаны в Rickwood et al., eds. (1995), Protein Engineering, A Practical Approach (IRL Press at Oxford Univ. Press, Oxford, Eng.). Общие принципы антител и связывания антитело-гептен указаны в: Nisonoff (1984), Molecular Immunology (2nd ed.; Sinauer Associates, Sunderland, Mass.); и Steward (1984), Antibodies, Their Structure and Function (Chapman and Hall, New York, N.Y.). Кроме того, как правило, следуют стандартным способам в иммунологии, известным в данной области, и не описанным конкретно, как в Current Protocols in Immunology, John Wiley & Sons, New York; Stites et al., eds. (1994), Basic and Clinical Immunology (8th ed; Appleton & Lange, Norwalk, Conn.) и Mishell and Shiigi (eds) (1980), Selected Methods in Cellular Immunology (W.H. Freeman and Co., NY).
[00254] Стандартные справочные издания, в которых указаны общие принципы иммунологии, включают Current Protocols in Immunology, John Wiley & Sons, New York; Klein (1982), J., Immunology: The Science of Self-Nonself Discrimination (John Wiley & Sons, NY); Kennett et al., eds. (1980), Monoclonal Antibodies, Hybridomas: A New Dimension in Biological Analyses (Plenum Press, NY); Campbell (1984), "Monoclonal Antibody Technology", Laboratory Techniques in Biochemistry and Molecular Biology, ed. Burden et al. (El-severe, Amsterdam); Goldsby et al., eds. (2000), Kuby Immunnology (4th ed.; H. Freemand & Co.); Roitt et al. (2001), Immunology (6th ed.; London: Mosby); Abbas et al. (2005), Cellular and Molecular Immunology (5th ed.; Elsevier Health Sciences Division); Kontermann and Dubel (2001), Antibody Engineering (Springer Verlan); Sambrook and Russell (2001), Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press); Lewin (2003), Genes VIII (Prentice Hall2003); Harlow and Lane (1988), Antibodies: A Laboratory Manual (Cold Spring Harbor Press); Dieffenbach and Dveksler (2003), PCR Primer (Cold Spring Harbor Press).
[00255] Все цитированные выше ссылки, а также все цитируемые в настоящем описании ссылки полностью включены в настоящее описание посредством ссылки.
[00256] Следующие ниже примеры предоставлены в качестве иллюстрации и не для ограничения.
ПРИМЕРЫ
Материалы и способы
Биологические образцы
[00257] Периферическую кровь человека от нормальных здоровых добровольцев получали через программу MedImmune Blood Donor с письменным информированным согласием и одобрением от IRB. Мононуклеарные клетки периферической крови (PBMC) выделяли из свежей цельной крови с использованием пробирок для получения клеток Vacutainer CPT с цитратом натрия (Becton Dickinson Biosciences, NJ, USA). Пробирки центрифугировали при 17000g в течение 25 мин, 22°C с минимальным торможением. После центрифугирования удаляли сыворотку и переносили клеточный лейкоцитарный слой в конические пробирки 50 мл (BD Biosciences). Очищенные клетки дважды отмывали стерильным фосфатно-солевым буфером (PBS) (Invitrogen Life Technologies, CA, USA) при 350g в течение 10 минут при 22°C. Клетки ресуспендировали в PBS или среде RPMI 1640, дополненной 10% эмбриональной телячьей сывороткой (Invitrogen) и фильтровали с использованием пробирок BD Falcon 5 мл с крышками с фильтром для клеток (BD Biosciences). Плотности клетка определяли с использованием счетчика клеток Vi-Клетка XR® (Beckman Coulter, CA, USA).
[00258] Периферическую кровь яванских макак от здоровых животных получали от Bioqual (Bioqual, Inc. MD, USA) в соответствии руководствами National Institutes of Health for care and use of primates. PBMC яванских макак выделяли с использованием пробирок для получения клеток Vacutainer CPT с цитратом натрия (как описано выше) или с Histopaque 10771 (Sigma-Aldrich, MO, USA). В кратком изложении, свежую цельную кровь доводили до 50× начального объема крови стерильным PBS. Затем 25 мл разбавленной крови наносили на 10 мл 90% Histopaque 10771 (Sigma Aldrich) и центрифугировали образцы при 400g в течение 20 мин при комнатной температуре с минимальным торможением. Кольцо клеток удаляли и переносили в новую 50 мл коническую пробирку. Очищенные клетки дважды отмывали стерильным PBS при 350g в течение 10 минут при 22°C. Клетки ресуспендировали в PBS или среде RPMI 1640, дополненной 10% эмбриональной телячьей сывороткой, фильтровали и подсчитывали, как описано выше.
Клетки
[00259] Клетки CT-125 и CT-550 получали от Dr. Yong-Jun Liu (University of Texas M.D. Anderson Cancer Center, Houston, TX, USA). Клетки CT-125 получали трансдукцией T-клеточной гибридомы 2B4 мыши немеченым FcεR1γ мыши и репортерный ген NFAT-GFP, и клетки CT-550 получали трансдукцией клеток CT-125 меченным HA ILT7 человека (Ohtsuka M. et al., PNAS 101: 8126-8131 (2004); Cao W. et al., JEM 203: pp 1399-1405 (2006)). Стабильную линию клеток CT-125 Cyno ILT7 получали франсфекцией клеток CT-125 геном ILT7 яванского макака, клонированным в плазмидный вектор pME18X. клетки CT культивировали в RPMI 1640, дополненной 10% эмбриональной телячьей сывороткой (FBS) и 1× пенициллином/стрептомицином (все реактивы от Invitrogen Life Technologies).
[00260] Клетки KC1333 получали от Biowa (Biowa, NJ, USA). Клетки KC1333 культивировали в Advance RPMI 1640, дополненной 10% FBS, 4 мМ L-глутамина, 0,2 мкг/мл генетицина (все реактивы от Invitrogen) и 18,3 пг/мл рекомбинантного IL-2 человека (PeproTech, NJ, USA).
Антитела и реагенты
[00261] Гуманизированные варианты антитело к ILT7, 7C7 к ILT7 (7C7), и гуманизированный изотипический контроль R347 получали в рамках MedImmune. Гуманизированные варианты конъюгированного с аллофикоцианином (APC) антитела к ILT7, 7C7 и изотипический контроль R347 получали с использованием наборов APC monoclonal antibody labeling (Thermo Fisher Scientific, IL, USA). Меченное R-фикоэритрином (PE) и FITC антитело к BDCA-2 человеку (клон AC144), антитело-R-PE к BDCA-4 человека (клон AD5-17F6) и FcR человека блокирующий реагент были от Miltenyi Biotech, CA, USA. антитело к CD123 человека (клон 7G3), конъюгированное с R-PE, FITC или APC, меченное Alexa Fluor 488 антитело к CD8 человека (клон RPA-T8), меченное Alexa Fluor 488 антитело к CD3 человека (клон SP34-2), меченное FITC антитело к CD14 человека (клон M5E2), меченое FITC антитело к CD20 человека (клон 2H7) и меченое PerCP-Cy 5,5 anti-человек HLA-DR (клон G46-6) получали от BD Biosciences. Pacific Blue антитело к человек CD56 (клон MEM-188) получали от BioLegend, CA, USA. DyLight 649 антитело к IgG человека и полостью принадлежащий человеку IgG получали от Jackson Immunoresearch, PA, USA.
[00262] Окрашивание цельной крови проводили с использованием лизирующего раствора BD FACS (BD Biosciences). 7-AAD получали от Invitrogen. Плазму крови группы AB человека мужского пола получали от Sigma-Aldrich. Рекомбинантный IL-2 человека получали от R&D Systems, MN, USA, и рекомбинантный интерферон β человека (IFN-β) получали от PBL Biomedical, NJ, USA. CpG A ODN 2216 получали от InvivoGen, CA, USA.
Мечение рекомбинантного ILT7 человека и яванского макака
[00263] Белки подвергали биотинилированию через свободные амины с использованием EZ-link Sulfo-NHS-LC-Biotin (Thermo/Pierce, product: 21335). Реагент растворяли в безводном диметилформамиде и растворы белков на основе PBS доводили до pH~8 1 M NaHCO3 в D-PBS.
[00264] Встраивания метки оценивали масс-спектрометрией MALDI-TOF во всех случаях, и непрореагировавшие реагенты удаляли заменой буфера с использованием уравновешенных D-PBS одноразовых колонок Sephadex G25. Для биотинилирования конечные концентрации белков определяли по оптической плотности при 280 нм с использованием коэффициентов экстинкции, рассчитываемых из аминокислотных последовательностей.
Анализ связывания ELISA
[00265] Одноцепочечные фрагменты Fv предоставляли на фаговых частицах и тестировали в анализе связывания для определения перекрестной реактивности и специфичности к панели рекомбинантных антигенов. Образцы супернатанта предоставленных на фагах scFv получали в 96-луночных планшетах с глубокими лунками так, как указано ниже. 5 мл культуры из каждой лунки 96-луночного исходного планшета переносили в планшет для культивирования Greiner с глубокими лунками, содержащий 500 мл среды 2TYAG (2TY+100 мг/мл ампициллина+2% глюкозы) и инкубировали в течение 5 часов при 37°C, 280 об./мин. Затем добавляли 100 мл/лунку фага-помощника K07 M13 (разбавленного до 1,5×1011 БОЕ/мл в 2TYAG) и инкубировали планшет при 37°C, 150 об./мин. для обеспечения инфекции. Затем планшет центрифугировали при 3200 об./мин. в течение 10 минут и удаляли супернатант. Бактериальные осадки ресуспендировали в 500 мл/лунка 2TYAK (2TY+100 мг/мл ампициллина+50 мг/мл канамицина) и инкубировали планшет в течение ночи при 25°C, 280 об./мин. Утром в каждую лунку добавляли 500 мл 6% (масс./об.) обезжиренное порошковое молоко в 2× PBS и инкубировали планшет в течение 1 часа при комнатной температуре. Затем планшет центрифугировали при 3200 об./мин. в течение 10 минут и использовали блокированные супернатанты предоставляемых фагом scFv непосредственно в экспериментах ELISA.
[00266] Для определений EC50, как правило, очищенные IgG разбавляли в 3 раза в 3% (масс./об.) порошковым молоком в PBS (PBS-M) с получением 11 точек концентраций. Для разбавления препарата использовали 96-луночные полипропиленовые планшеты Greiner (Greiner, 650201). Как правило, каждое разбавление получали в двух повторениях. Перед использованием непосредственно в экспериментах ELISA разбавленные IgG оставляли в PBS-M в течение 1 часа при комнатной температуре для блокирования.
[00267] Анализы связывания IL-T7 проводили в планшетах для ELISA по существу так, как указано ниже. В каждом эксперименте не использовали все антигены, а, как правило, тестировали антиген IL-T7 человека, мыши и яванского макака. Для тестирования неспецифического связывания также использовали соответствующие контрольные антигены (бычий инсулин плюс IL-4Rα FLAG®His,при необходимости). За исключением бычьего инсулина все антигены подвергали биотинилированию и их всех получали с использованием бактериальной экспрессии. Антигены IL-T7 подвергали биотинилированию через свободные сульфгидрильные группы с использованием EZ link Biotin-BMCC (Perbio/Pierce 21900). Способ получения IL-4Rα FLAG®His, который использовали в качестве контрольного антигена, описан в WO/2010/070346. IL-4Rα FLAG®His подвергали биотинилированию через свободные амины с использованием EZ link Sulfo-NHS-LC-Biotin (Perbio/Pierce, 21335).
[00268] Планшеты со стрептавидином (Thermo Scientific, AB-1226) покрывали биотинилированным антигеном при 0,5мкг/мл в PBS и инкубировали в течение ночи при 4°C. Планшеты отмывали 3× PBS и блокировали 300 мкл/лунку блокирующим буфером (PBS-M) в течение 1 часа. Планшеты отмывали 1× PBS и добавляли блокированные образцы 50 мкл/лунку в течение 1 часа при комнатной температуре. Планшеты отмывали 3× PBS-T (PBS+1% (об./об.) Tween-20) и добавляли реагенты для детекции [антитело к IgG человека HRP (Sigma, A0170) или антитело к M13-HRP (Amersham, 27-9421-01) для детекции IgG или предоставленные фагом scFv, соответственно] в разведении 1:5000 по 50 мкл/лунку в PBS-M в течение 1 часа при комнатной температуре. Планшеты отмывали 3× PBS-T и проявляли TMB 50 мкл/лунку (Sigma, T0440). Реакцию гасили 0,1M H2SO4 50 мкл/лунку перед чтением на EnVision™ планшетном спектрофотометре или аналогичном оборудовании при 450 нм.
[00269] Кривые доза-ответ графически отображали для титрований IgG с использованием программного обеспечения для подбора кривой Prism (Graphpad). Считали, что предоставляемые фагом scFv связываются с антигеном IL-T7, если оптическая плотность 450 нм составляла >0,5 и <0,1-0,2 для того же образца на контролях (инсулин и IL-4Rα Flag®His). Одноцепочечные фрагменты Fv предоставляли на фаговых частицах и тестировали как неочищенные препараты в одноточечном скрининге ELISA.
Анализ связывания клеток с использованием технологии флуоресцентного анализа в микрообъеме (FMAT)
[00270] Этот гомогенный анализ оценивает связывание образцов супернатанта неочищенных scFv или очищенного IgG с клетками китайского хомяка (CHO), экспрессирующими ILT7 человек или яванского макака, в 384-луночном (Costar 3655) формате. Связывание ScFv или Ab с клетками детектировали с использованием антитела мыши к смеси меченных His/Alexafluor®-647 антител козы против коза антитела мыши (Molecular Probes A21236) или антитело козы к меченному антителу человека Alexafluor®-647 (Molecular Probes A21445), соответственно. Планшеты читали на устройстве для чтения планшетов Applied Biosystems Cellular Detection system 8200. Гелий-неоновый лазер фокусировали на глубине 100 мкм от дна лунки, сканировали площадь 1 мм2. Клетки, оседающие на дне лунки, и при возбуждении лазером при 633 нм, те гранулы со связанным флуорофором (где локальная концентрация флуорофора является относительно высокой по сравнению с несвязанным флуорофором) испускали сигнал при 650-685 нм, который измеряли с использованием фотоэлектронного умножителя 1 (PMT1). Несвязанный флуорофор в растворе находился за пределами глубины возбуждения или в относительно низкой локальной концентрации и, таким образом, не испускал существенного сигнала. Присутствие образцов scFv или IgG, связывающихся с клетками на дне лунки, вызывало увеличение детекции меченного Alexafluor антитела в глубине возбуждения. Это измеряли как увеличение флуоресценции.
[00271] В этих экспериментах буфер для анализа представлял собой PBS (Gibco 14190-094), содержащий 0,1% BSA (Sigma A9576-50 мл), 0,1% Tween-20 (Sigma P2287) и 0,01% азид натрия. Для получения смеси для детекции ScFv антитела мыши к His и антитела к AF647 мыши смешивали при 1 мкг/мл и 2 мкг/мл соответственно в буфере для анализа. Для получения смеси для детекции IgG получали антитело к AF647 человека в концентрации 2 мкг/мл в буфере для анализа.
[00272] Используемые клетки представляли собой клетки CHO-K1, экспрессирующие ILT7 человека или яванского макака, которые культивировали стандартными техниками культивирования тканей. Клетки выращивали приблизительно до 80% конфлюэнтности в F-10 (Gibco, 22390-025)+10% FCS (SAFC Biosciences, 13068C)+0,5 мг/мл зеоцина В (Invitrogen, R250-01), промывали PBS, открепляли с помощью аккутазы (PAA, L11-007) и ресуспендировали в PBS 1,5×105 клеток/мл.
[00273] образцы супернатанта неочищенного scFv получали в 96-луночных планшетах в глубокими лунками. 5 мкл культуры из каждой лунки 96-луночного исходного планшета переносили в планшет для культивирования Greiner с глубокими лунками, содержащий 900 мкл среды 2TY (1,6% триптон, 1% дрожжевого экстракта, 0,5% NaCl, pH 7,0)+ 100 мкг/мл ампициллина+0,1% глюкозы и инкубировали в течение 5 часов при 37°C, 280 об./мин. Затем добавляли 100 мкл/лунку 10 мМ IPTG в TY и инкубировали блок в течение ночи при 30°C, 280 об./мин. Утром блок центрифугировали при 3200 об./мин. в течение 15 минут. Для высокопроизводительного скрининга супернатанты scFv из блока с глубокими лунками переносили непосредственно в планшет для анализа для необходимого разбавления 20%.
[00274] В тестируемые лунки 384-луночного черного планшета Costar с прозрачном дном из несвязывающей поверхности добавляли следующее: 10 мкл образца (IgG или ScFv), 10 мкл антитела для детекции или смеси антител и 30 мкл клеток. Отрицательные контроли, используемые в этих экспериментах, как правило, включали добавление изотипических (IgG) или нерелевантных (ScFv) контролей, или буфера для анализа вместо экспериментального образца. Планшеты герметизировали и инкубировали в течение четырех часов при комнатной температуре в темноте, а затем читали на устройстве для чтения планшетов Applied Biosystems Cellular Detection system 8200. Как правило, данные анализировали с использованием алгоритма по скорости с гейтированием, устанавливаемым как отношение цветов <0,4, размер 15-30 и минимальное число событий 20. Попадания от образцов супернатанта неочищенного scFv определяли как демонстрирующие 50% или большее ингибирование сигнала по сравнению с лунками общего контрольного связывания. Кривые доза-ответ графически отображали для титрований IgG с использованием программного обеспечения для подбора кривой Prism (Graphpad).
[00275] Для определений IC50, как правило, очищенные IgG разбавляли в 2 раза в буфере для анализа от 500 нМ с получением 11 точек концентраций. Для разбавления препаратов использовали 96-луночные полипропиленовые планшеты Greiner(Greiner, 650201). Как правило, каждое разведение получали в двух повторениях. Альтернативно, тестирование IgG проводили с одной концентрацией в диапазоне концентраций от 500 нМ до 0,2 нМ.
Оценка связывания антитела на линиях клеток проточной цитометрией
[00276] Связывание вариантов антител к ILT7 и изотипических контролей с ILT7 человека и яванского макака оценивали анализом проточной цитометрии с использованием клеток CT-550 и cynoILT7 CT-125, соответственно. Клетки CT-125 использовали в качестве контроля. Клетки ресуспендировали в блокирующим буфере (PBS, дополненной 10% FBS) в концентрации 5 миллионов клеток на мл и переносили в круглодонные 96-луночные планшеты (BD FalconTM Clear Microtest Plate, BD Biosciences) 100 мкл на лунку. варианты антител к ILT7 и контроль добавляли к клеткам в течение 30 мин при 4°C на встряхивателе для планшетов. Клетки отмывали три раза PBS и ресуспендировали в блокирующем буфере (100 мкл/лунку). Связывание IgG человека на клеточной поверхности детектировали с использованием вторичного антитела к IgG человека конъюгированного с DyLight 649 (1 в 1000 разбавлении). Клетки инкубировали в темноте в течение 30 мин при 4°C на встряхивателе для планшетов. Клетки отмывали три раза PBS и регистрировали флуоресценцию на поверхности с использованием системы для проточной цитометрии LSRII и программного обеспечения FACSDiva (оба от BD Biosciences).
Assessment связывания антителе на цельной крови и PBMC проточной цитометрией
[00277] Связывание меченных APC антител к ILT7 и изотипических контролей на цельной крови человека и яванского макака оценивали анализом проточной цитометрии. Цельную кровь переносили в 50 мл конические пробирки, 1 мл на пробирку. Меченные APC антитела добавляли непосредственно в цельную кровь. Антитела к BDCA-2-PE и антитело к CD123-PE использовали в качестве специфических маркеров плазмацитоидных дендритных клеток (pDC) при окрашивании цельной крови человека и окрашивании цельной крови яванского макака, соответственно. Цельную кровь инкубировали с антителами в течение 30 мин при 4°C в темноте на встряхивателе для планшетов. Кровь обрабатывали лизирующим раствором BD FACS по инструкциям производителя. Клетки отмывали и оценивали связывание антитела проточной цитометрией с использованием системы проточной цитометрии LSRII и программного обеспечения FACSDiva.
[00278] Для окрашивания PBMC сначала PBMC промывали PBS и ресуспендировали в холодном блокирующем буфере на основе PBS, содержащем 50% плазмы человека мужского пола с группой крови AB, 20 мкг/мл IgG человека и 200 мкл/мл блокирующего FcR человека реагента в течение 15 мин при 4°C на встряхивателе для планшетов. Через 15 мин варианты меченных APC антител к ILT7 или меченного APC изотипического контроля добавляли непосредственно в блокирующий раствор. Антитела к BDCA-2-PE и антитела к BDCA-4-PE альтернативно использовали в качестве специфического к pDC маркера для окрашивания PBMC человека. В PBMC яванского макака pDC определяли как HLA-DR+, Lineage-, CD11c- и CD123high (Malleret et al., Immunology, 124: 223-233 (2008)). Таким образом, антитела к HLA-DR PerCP-Cy5,5, Lineage-FITC (антитела к CD3, CD8, CD20 и CD14) и антитела к CD123-PE использовали в качестве специфического к pDC маркера для окрашивания PBMC яванского макака. PBMC инкубировали в течение 30 мин 4°C в темноте на встряхивателе для планшетов. Клетки отмывали и оценивали связывание антитела проточной цитометрией с использованием системы проточной цитометрии LSRII и программного обеспечения FACSDiva.
Оценка активности антитела анализом антителозависимой клеточной цитотоксичности (ADCC) с использованием линий клеток
[00279] Активность антител к ILT7 определяли с использованием анализа на основе ADCC клеток in vitro. Клетки KC1333 (эффекторы) и клетки CT (мишени) совместно культивировали в отношении 5:1 (2,5×105 KC1333 на 0,5×105 клеток CT) в круглодонных 96-луночных планшетах. Клетки совместно культивировали в присутствии антител к ILT7 или изотипического контроля в течение 16 часов в среде для культивирования RPMI 1640, дополненной 10% FBS, при 37°C, 5% CO2. Затем клетки отмывали и переносили в блокирующий буфер (PBS-10% FBS). Клетки KC1333 детектировали с использованием Pacific-Blue-антитела к CD56. Мертвые клетки детектировали с использованием 7-AAD. Жизнеспособность клеток-мишеней оценивали проточной цитометрией с использованием системы проточной цитометрии LSRII и программного обеспечения FACSDiva. Процент цитотоксичности получали с применением следующей ниже формулы: процент цитотоксичности=100-(количество живых мишеней/количество живых мишеней на исходном уровне)×100.
Оценка активности антитела анализом ADCC с использованием PBMC человека
[00280] PBMC человека отмывали PBS и ресуспендировали в среде RPMI, дополненной 10% FBS и 200 нг/мл рекомбинантного IL-2 человека в концентрации 5,0×106 клеток на мл. PBMC высевали в двух повторениях в круглодонные 96-луночные планшеты, 100 мкл на лунку. Получали 10-кратные серийные разведения антител к ILT7 и контрольных антител и в соответствующие лунки добавляли 100 мкл растворов антител добавляли до конечных концентраций 33,85 нМ-3,385 фМ. Клетки инкубировали в течение 6 часов при 37°C, 5% CO2. После инкубаций клетки дважды отмывали в 250 мкл холодного PBS. Клетки ресуспендировали в 1000 мкл холодного блокирующего буфера на основе PBS, содержащего 50% плазмы человека мужского пола с группой крови, 20 мкг/мл IgG человека и 200 мкл/мл блокирующего FcR человека реагента в течение 15 мин при 4°C. После этапа блокирования 100 мкл холодного блокирующего буфера, содержащего FITC-антитела к BDCA2 человека и APC-антитела к CD123 человека, добавляли в соответствующие лунки. Планшеты инкубировали, аккуратно встряхивали в течение 30 минут при 4°C. После инкубации клетки дважды отмывали в 250 мкл холодного PBS с конечным ресуспендированием в 200 мкл холодного PBS. Во все лунки добавляли 50 мкл холодного раствора 7-AAD (Invitrogen) и положительные по 7-AAD плазмацитоидные дендритные клетки оценивали с использованием системы проточной цитометрии LSRII и программного обеспечения FACSDiva.
Анализы секреции IFNα с PBMC человека
[00281] PBMC человека отмывали PBS и высевали в двух повторениях в круглодонные 96-луночные планшеты в конечной плотности 150000-156000 клеток на лунку в среде RPMI, дополненной 10% FBS и 200нг/мл рекомбинантного IL-2 человека. Получали 10-кратные серийные разведения антител к ILT7 и контрольных антител и в соответствующие лунки добавляли 100О мкл растворов антител до конечных концентраций 6,77 нМ-0,677 фМ. Клетки и антитела инкубировали в течение 9,5-10 часов при 37°C, 5% CO2. После инкубаций 50 мкл ODN2216 (Invitrogen™) добавляли в соответствующие лунки до конечной концентрации 0,5 мкМ и дополнительно инкубировали планшеты еще 16 часов при 37°C, 5% CO2. После инкубаций планшеты центрифугировали при 350g в течение 10 минут, осторожно удаляли супернатанты и количественно определяли IFNα с использованием набора IFNα Multi-Subtype ELISA (PBL Biomedical).
Анализы секреции IFNα с использованием PBMC яванского макака
[00282] PBMC яванского макака отмывали PBS и ресуспендировали в RPMI 1640, дополненной 10% FBS, 220 нг/мл рекомбинантного IL-2 человека и 500 МЕд./мл рекомбинантного IFN-β человека. В соответствующие лунки добавляли максимальные количества клеток в плотностях в диапазоне от 314000 до 818000 клеток на лунку. Получали 10 кратные серийные разведения антител к ILT7 и контрольных антител и в соответствующие лунки добавляли 100 мкл растворов антител до конечных концентраций 33,85 нМ-3,385 фМ. Клетки и антитела инкубировали в течение 9,5-10 часов при 37°C, 5% CO2. После инкубаций в соответствующие лунки добавляли 50 мкл ODN2216 (Invitrogen™) до конечной концентрации 0,5 мкМ и дополнительно инкубировали планшеты еще 16 часов при 37°C, 5% CO2. После инкубаций планшеты центрифугировали при 350g в течение 10 минут, аккуратно удаляли супернатанты и количественно определяли IFNα в супернатанте с использованием набора Rhesus/Cynomolgus IFNα ELISA (PBL Biomedical).
Статистический анализ
[00283] Кривые EC50 и IC50 для анализов связывания, ADCC и секреция цитокинов анализы получали с использованием программного обеспечения GraphPad Prims 5 (GraphPad Software, CA, USA).
Пример 1
Получение гуманизированных антител к ILT7 из антитела мыши SB128
[00284] mAb мыши SBI28 (SBI28 относится к антителу к ILT7 ILT7№28, предоставленному в опубликованной заявке США № 2009/0280128) гуманизировали путем перестановки каркаса (Dall'Acqua et al., Methods 36:43-60 (2005)). С применением этого способа mAb мыши SBI28 гуманизировали с помощью синтезированной комбинаторной библиотеки, состоящей из шести CDR, слитых в рамке чтения с совокупность отдельных каркасов зародышевой области человека. Гены каркаса человека выбрали из общедоступной совокупности генов зародышевой линии антитела. Эти универсальные праймерные совокупности каркасов включают 46 генов каппа-цепи зародышевой линии человека, 5 последовательностей Jk зародышевой линии человека, 44 генов тяжелой цепи зародышевой линии человека и 6 последовательностей JH зародышевой линии человека. Конструировали банк праймеров так, чтобы кодировать каждый каркас каждого гена зародышевой линии. Также синтезировали праймеры CDR, специфические к антителу, с вырожденными концами, которые перекрываются с совокупностями каркасов. Конструировали библиотеку с перестановкой каркаса SBI28 путем объединения подбиблиотеки с перестановкой каркаса вариабельная область тяжелой цепи с подбиблиотекой с перестановкой каркаса вариабельная область легкой цепи. Подбиблиотеки с перестановкой каркаса собирали последовательно с использованием ПЦР путем перекрывающейся достройки цепей. Первую "ПЦР-слияние" проводили для синтеза каждого отдельного каркаса зародышевой линии человека, слитого в рамке чтения с участком соответствующих CDR. Затем проводили вторую "ПЦР-сборку" с использованием продукта ПЦР-слияния в качестве матрицы для амплификации подбиблиотек полноразмерных VH и VL. Библиотеку с перестановкой каркаса SBI28 клонировали в вектор экспрессии Fab на основе M13 с использованием способа гибридизационного мутагенеза по Кункелю. Приблизительно 1300 клонов из библиотеки с перестановкой каркаса SBI28 подвергали скринингу на клетках CHO, экспрессирующих рекомбинантный ILT7CHO-клетка, с использованием анализа MesoScale Discovery (MSD). Один гуманизированный вариант, 10D10, связывался с ILT7 человека с аффинностью в 3 раза ниже по сравнению с его химерным родителем ("SBI28ch"), как измеряли поверхностным плазмонным резонансом (SPR) на ProteOn. SBI28ch относится к антителу к ILT7 ILT7№28, как предоставлено в опубликованной заявке США № 2009/0280128, которая полностью включена в настоящее описание посредством ссылки.
[00285] Оптимизацию аффинности 10D10 начали проводить для улучшения его аффинности связывания с ILT7 человека и яванского макака. Сначала 10D10 клонировали в вектор для экспрессии ScFv на основе M13 для экономного мутагенеза. В этом способе каждую отдельную аминокислоту всех 6 CDR подвергали случайной мутации с использованием двух отдельных библиотек (NSS и NWS) на положение остатка. Конструировали всего 12 независимых библиотек для 6 CDR способа гибридизационного мутагенеза по Кункелю. (Kunkel T. A., et al., Methods Enzymol. 154:367 (1987)). Скрининг синтезированных библиотек состоял из одноточечного ELISA, сконструированного с возможностью захвата ограниченных концентраций секретируемого ScFv из бактериальных сред для культивирования для нормализации концентрации scFv в каждой лунке. Меченый антиген ILT7 связывался с захваченным ScFv, и сила сигнала от этого взаимодействия коррелировала с относительной аффинностью связывания. Приблизительно от 2000 до 3000 клонов подвергали скринингу. Чтобы дополнительно сконструировать вариант с улучшенной аффинностью, все благоприятные единичные замены аминокислот совместно кодировали с получением небольшой и слитой комбинаторной библиотеки. На этом этапе 14 отдельных положительных попаданий в 9 положениях в 6 CDR кодировали одновременно с получением комбинаторной библиотеки scFv. В кратком изложении, конструировали вырожденные праймеры, которые кодировали все благоприятные аминокислотные замены, а также родительский остаток в том же положении. Эту комбинаторную библиотеку подвергали скринингу с использованием ELISA с возможностью захвата одной точки, как описано ранее. Скринингу подвергали приблизительно 1200 клонов. Вариабельные области варианта с улучшенной аффинностью 7C7 индивидуально клонировали в вектор млекопитающих экспрессии pOE и транзиторно экспрессировали в клетки HEK293. Секретируемые растворимые IgG человека выделяли непосредственно из кондиционированных сред. Очищенные IgG анализировали на связывание с rILT7 с использованием ProteOn и FACS. В эксперименте ProteOn демонстрировали 60-кратное увеличение KD аффинности оптимизированного антитела 7C7 по сравнению с SBI28ch. Посредством FAC, в котором измеряли связывание с рекомбинантным ILT7 человек и яванского макака, экспрессируемого клетками CHO, для 7C7 демонстрировали в 2,2 раза и в 14 раз большее EC50 для ILT7 человека и яванского макака, соответственно, по сравнению с SBI28ch. Выравнивания последовательностей VH и VL SBI28, 10D10 и 7C7 приведены на фигурах 1A и 1B, соответственно.
Пример 2
Получение антител к ILT7 человека из принадлежащей человеку библиотеки
[00286] В дополнение к гуманизации антитела мыши к ILT7 (как описано выше в примере 1) получали антитела человека с использованием библиотеки последовательностей человека. Применение нескольких стратегий получения антител к ILT7 максимально увеличивает шансы получения антитела к ILT7 с различными признаками, таким образом, что можно выбрать идеальное антитело для конкретной цели.
2.1 Селекция
[00287] Для селекции использовали большую библиотеку антител человека на основе одноцепочечного Fv (scFv), получаемую с использованием индивидуальных вариабельной области тяжелой цепи и вариабельных областей легкой цепи, выделяемых из костного мозга у взрослых не получавших ранее лечение доноров, которые клонировали в фагмидный вектор на основе нитевидного фага M13 (Hutchings C., "Generation of Naïve Human Antibody Libraries" in Antibody Engineering, Dubel. Berlin, Springer Laboratory Manuals: p. 93 (2001); Lloyd et al., Protein Eng. Des. Sel. 22(3):159-68 (2009)). Антитела со специфическим к ILT7 scFv выделяли из библиотек фагового дисплея в серии повторных циклов селекции рекомбинантного ILT7 человека и/или яванского макака по существу, как описано ранее у Vaughan et al. (Nat. Biotechnol. 14(3):309-14 (1996)). В кратком изложении, scFv-фаговые частицы инкубировали с биотинилированным рекомбинантным ILT7 в растворе (биотинилированным через свободные амины с использованием EZ link Sulfo-NHS-LC-Biotin (Thermo/Pierce, продукт: 21335)). Как правило, scFv-фагов частицы инкубировали с 100 нМ биотинилированного рекомбинантного ILT7 в течение 1 часа. Затем ScFv, связанный с антигеном улавливали на покрытые стрептавидином парамагнитные гранулы (Dynabeads® M-280), следуя рекомендациям производителя. Несвязанный фаг отмывали в серии циклов промывания с использованием PBS-Tween. Фаговые частицы, удерживаемые на антигене, элюировали, вводили путем инфекции в бактерии и спасали для следующего раунда селекции. Как правило, таким образом проводили три раунда селекции.
2.2 Идентификация специфических к ILT7 связывающих средств посредством фагового ELISA
[00288] scFvs предоставляли на фаговых частицах и тестировали в анализе связывания для определения перекрестной реактивности и специфичности к рекомбинантным антигенам. Подробное описание способа анализа приведено в разделе "материалы и способы". Приблизительно 2100 отдельных точек данных получали из анализа связывания и идентифицировали попадания, т.е. клоны scFv, для которых демонстрировали связывание с рекомбинантным ILT7, подвергали секвенированию ДНК (Osbourn et al., Immunotechnology 2(3):181-96 (1996); Vaughan et al., Nat. Biotechnol. 14(3):309-14 (1996)).
2.3 Идентификация связывающих ILT7 средств посредством FMAT
[00289] Уникальные scFv экспрессировали в периплазме бактерий и подвергали скринингу на их активность связывания в анализе связывания на основе технологии флуоресцентного анализа в микрообъеме (FMAT). Связывание scFv с ILT7, экспрессируемыми на поверхности клеток детектировали с использованием антитела козы к меченному Alexafluor®-647 антителу мыши. Подробное описание способа анализа приведено в разделе "материалы и способы".
2.4 Переформатирование scFv в IgG1
[00290] Наиболее эффективные scFv связывающие средства преобразовывали в формат полного иммуноглобулин G1 (антитело IgG1), по существу как описано Persic et al. (Gene 187(1):9-18 (1997)), со следующими модификациями. Фрагмент OriP включали в экспрессирующие векторы для облегчения использования с транзиторными клетками CHO и для обеспечения эписомальной репликации. Домен VH клонировали в вектор (pEU1.3), содержащий константные домены человека тяжелой цепи и регуляторные элементы для экспрессии полной тяжелой цепи IgG1 в клетках млекопитающих. Подобным образом, домен VL клонировали в вектор (pEU4.4) для экспрессии константных доменов легкой цепи (лямбда) человека и регуляторных элементов для экспрессии легкой полной цепи IgG в клетках млекопитающих. Для получения IgG, векторы, экспрессирующие тяжелые и легкие цепи IgG, трансфицировали в CHO-транзиторные клетки млекопитающих. IgG экспрессировали, и он секретировался в среду. Перед очисткой сборы объединяли и фильтровали. Затем IgG очищали с использованием хроматографии с белком A. Супернатанты культур нагружали на керамическую колонку подходящего размера с белком A (BioSepra) и промывали 50 мМ Tris-HCl pH 8,0, 250 мМ NaCl. Связанный IgG элюировали из колонки с использованием 0,1 M цитрата натрия (pH 3,0) и нейтрализовали добавлением Tris-HCl (pH 9,0). В элюированном веществе заменяли буфер на PBS с использованием колонок Nap10 (Amersham, №17-0854-02) и определяли концентрацию IgG спектрофотометрически с использованием коэффициента экстинкции на основе аминокислотной последовательности IgG (Mach et al., Anal. Biochem. 200(1):74-80 (1992)).
2.5 Анализ связывания IgG
[00291] Виды перекрестной реактивности антител к ILT7 определяли с использованием анализа связывания FMAT. Подробное описание способа анализа приведено в разделе "материалы и способы". Следующие 11 антител идентифицировали как антитела, которые эффективно связывались с ILT7 человека и яванского макака в скрининговом анализе FMAT: ILT70019, ILT70028, ILT70052, ILT70076, ILT70080, ILT70083, ILT70089, ILT70100, ILT70137, ILT70142 и ILT70144.
Пример 3
Антитела к ILT7 связываются с экспрессирующими ILT7 клетками
[00292] Для определения связывания EC50 ILT70019, ILT70028, ILT70052, ILT70076, ILT70080, ILT70083, ILT70089, ILT70100, ILT70137, ILT70142 и ILT70144 с клетками, экспрессирующими ILT7 человека, кандидатов подвергали скринингу на связывание с клетками CT-550 проточной цитометрией. ILT70080 (EC50=0,28 нМ), ILT70083 (EC50=0,37 нМ), ILT70137 (EC50=0,41 нМ), ILT70144, ILT70142, ILT70052 и ILT70100 связывались с экспрессирующими ILT7 человека клетками. Кандидаты ILT70019, ILT70028 и ILT70076 не связывались с экспрессирующими ILT7 человека клетками. Антитела к ILT7 7C7 (7C7 описано выше в примере 1) и SBI33 (SBI33 относится к антителу к ILT7 ILT7№33, предоставленному опубликованной заявке США № 2009/0280128) использовали в качестве положительных контролей. Изотипический контроль R347 использовали в качестве отрицательного контроля и не демонстрировали для него какого-либо связывание с экспрессирующими ILT7 клетками. На графике, представленном на фигуре 2, приведено среднее значение двух независимых экспериментов, и в таблице, представленной на фигуре 2, продемонстрирована средняя EC50.
[00293] Для определения EC50 связывания вариантов с клетками, экспрессирующими ILT7 яванского макака, антитела подвергали скринингу на связывание с клетками CynoILT7 CT-125 проточной цитометрией. ILT70052 (EC50=0,35 нМ), ILT70080 (EC50=0,44 нМ), ILT70083 (EC50=1,37 нМ), ILT70137 (EC50=1,40 нМ), ILT70100 (EC50=1,63 нМ) и ILT70144 (EC50=7,81 нМ), ILT70142 и ILT70089 давали положительный результат в анализе связывания с ILT7 человека. ILT70019, ILT70028 и ILT70076 не вызвались с экспрессирующими ILT7 яванского макака клетками. Для изотипического контроля R347 не демонстрировали какого-либо связывания с экспрессирующими ILT7 клетками. На графике, представленном на фигуре 3, приведено среднее значение двух независимых экспериментов, и таблице, представленной на фигуре 3, продемонстрировано средняя EC50.
[00294] Таким образом, все ILT70052, ILT70080, ILT70083, ILT70100, ILT70137, ILT70142 и ILT70144 связывались с клетками, экспрессирующими ILT7 яванского макака или ILT7 человека. Особенно низкие значения EC50 получали с клетками, экспрессирующими ILT7 как яванского макака, так и человека, с использованием ILT70080, ILT70083 и ILT70137.
Пример 4
Активность ADCC антител к ILT7
[00295] Антитела к ILT7 тестировали на активность ADCC против экспрессирующих ILT7 человека линий клеток с использованием клеточного анализа in vitro. Клетки, экспрессирующие ILT7 человека, (клетки-мишени) высевали в соотношении 1:5 с линией клеток естественных киллеров (NK) KC1333 (эффекторные клетки) в присутствии вариантов антител к ILT7 или изотипического контроля в течение 18 часов. Во время анализа проточной цитометрии клетки KC1333 выделяли с использованием маркера NK CD56 (Biolegend №304624) и использовали 7-AAD, чтобы отличать живые клетки от мертвых. Процент живых клеток-мишеней рассчитывали с применением этого способа и сравневали с исходных уровнем (контроль без антитела). Цитотоксичность рассчитывали по следующей ниже формуле:
Процент цитотоксичности=100-(количество живых мишеней/количество живых мишеней в контроле без антител)×100
[00296] ILT70080 обладал самой высокой ADCC активностью по отношению к экспрессирующим ILT7 человека клеткам (EC50=0,022 нМ), далее следовали ILT70137 (EC50=0,044 нМ) и ILT70083 (EC50=0,094 нМ). ILT70142, ILT70052, ILT70100 и ILT70144 также проявляли активность ADCC (фигура 4). Изотипический контроль R347 и афукозилированный вариант R347 ("Afuc R347") не проявляли какой-либо активности ADCC с экспрессирующими ILT7 человека клетками.
[00297] Также тестировали антитела к ILT7 на ADCC активность по отношению к экспрессирующим ILT7 яванского макака клеткам с помощью активности на основе клеток in vitro. ILT70080 обалдел самой высокой ADCC активностью по отношению к экспрессирующим ILT7 яванского макака клеткам (EC50=0,008 нМ), далее следовали ILT70137 (EC50=0,015 нМ), ILT70142 (EC50=0,058 нМ), ILT70052 (EC50=0,073 нМ), ILT70144 (EC50=0,123), ILT70100 (EC50=0,188 нМ) и ILT70083 (EC50=0,433 нМ). ILT70089 также проявлял активность ADCC. Положительный контроль 7C7 проявлял ADCC, и изотипический (отрицательный) контроль R347 не проявлял какой-либо ADCC с экспрессирующими ILT7 яванского макака клетками. На графике и в таблице на фигуре 5 приведены два независимых эксперимента.
[00298] Таким образом, ILT70080 и ILT70137 обладали самой высокой активностью ADCC в клетках, экспрессирующих ILT-7 яванского макака, и клетках, экспрессирующих ILT-7 человек.
Пример 5
Связывание антитела к ILT7 с PBMC
[00299] Связывание антител к ILT7 ILT70080, ILT70083 и ILT70137 с PBMC человека оценивали проточной цитометрией с использованием концентрации антитела 2,5 мкг/мл. ITL70080, ILT70083 и ILT70137 специфически связывались с pDC (клетки BDCA-4+) (фигуры 6A и B). Связывание не происходило с изотипическим контролем R347.
[00300] Связывание антител к ILT7 ILT70080, ILT70083 и ILT70137 с PBMC яванского макака также оценивали проточной цитометрией. ITL70080 и ILT70083 специфически связывались с pDC (клетки HLA-DR+, Lineage-, CD123high).
Пример 6
Эффект антител к ILT7 на секрецию IFN-альфа
[00301] Варианты антител к ILT7 тестировали на ADCC активность на PBMC человека и яванского макака, как описано выше. Секрецию IFNα в супернатанте PBMC, культивированных с антителами к ILT7 и CpG-A, измеряли посредством ELISA. Все из ILT70080, ILT70083 и ILT70137 подавляли реакцию IFNα на CpG-A в PBMC человека и яванского макака. ILT70080 обладал наибольшим подавляющим эффектом в отношении реакции IFNα.
Пример 7
Афукозилирование антител ILT70080 и ILT70083
[00302] Антитела IgG1 содержат два участка N-связанных олигосахаридов в Fc-области, и эти участки являются в значительной степени фукозилированными в антителах человека. Антителозависимая клеточная цитотоксичность (ADCC) опосредована связыванием лимфоцитарных рецепторов с Fc-областями антитела, и на это влияет степень фукозилирования. Увеличение ADCC наблюдали при пониженном фукозилировании. Таким образом, получали и анализировали афукозилированные варианты ILT7.
7.1 Получение афукозилированного варианта антител к ILT7
[00303] ILT70080 и ILT70083 IgG1 экспрессировали в линии клеток CHO, которые не содержат фермент α-1,6-фукозилтрансферазу. Экспрессия в этой линии клеток приводит к антителу, которое не содержит молекулу α-1,6-фукозы на N-гликане в Asn-297 тяжелой цепи.
7.2 Тестирование афукозилированных антител к ILT7 ILT70080 и ILT70083
[00304] Анализ связывания с афукозилированными и родительскими антителами ILT70080 и ILT70083 на экспрессирующих ILT7 клетках проводили для оценки, влияло ли афукозилирование на EC50 связывания антител. Для родительских и афукозилированных антител демонстрировали сходное связывание с клетками, экспрессирующими ILT7 человека, и клетками, экспрессирующими ILT7 яванского макака (фигура 7).
[00305] ADCC активность афукозилированных антител ILT70080 и ILT70083 тестировали на экспрессирующих ILT7 человека и яванского макака клетках с использованием клеточного анализ in vitro, описанного выше (Пример 3). Афукозилирование повышало ADCC активность у всех тестируемых кандидатов (фигура 8). Наблюдали увеличение активность в десять раз для антитела ILT70080 при афукозилировании в анализах как для человека, так и для яванского макака (от EC50=0,013 нМ до EC50=0,001 нМ и от EC50=0,006 нМ до EC50=0,00051 нМ, соответственно), при этом наблюдали увеличение в 6-7 раз для ILT70083 (от EC50=0,089 нМ до EC50=0,0105 нМ и от EC50=0,36 нМ до EC50=0,057 нМ, соответственно). Афукозилированный изотипический контроль R347 не проявлял какой-либо ADCC по отношению к экспрессирующим ILT7 клеткам.
[00306] Связывание афукозилированных антитела к ILT7 ILT70080 и ILT70083 с PBMC человека оценивали проточной цитометрией. Афукозилированные варианты ITL70080 и ILT70083 специфически связывались с pDC (клетки BDCA-2+). Связывание отсутствовало с изотипическим контролем R347.
[00307] Связывание афукозилированных вариантов антител к ILT7 ILT70080 и ILT70083 с PBMC яванского макака также оценивали проточной цитометрией. Афукозилированные варианты ITL70080 и ILT70083 специфически связывались с pDC (HLA-DR+ Lineage- CD123high). Связывание отсутствовало с изотипическим контролем R347.
Пример 8
Конструирование антител ILT70080 и ILT70083
8.1 Конструирование ILT70080
[00308] Аминокислотные последовательности VH и VL ILT70080 выравнивали с известными последовательностями зародышевой линии человека в базе данных VBASE (Althaus H-H, Müller W and Tomlinson I: V BASE; http://vbase.mrc-cpe.cam.ac.uk/) и идентифицировали наиболее близкую последовательность зародышевой линии по сходству последовательности. Для домена VH она представляла собой VH1-69 (DP-10), и для домена VL она представляла собой Vlambda3-h. Выбирали семь остатков в каркасах (FW) каждого домена VH (A13K, T16S, L69I*, S70T, L80M, Y84S и D85E) и домена VL (E3V, K20R, S22T, M46L*, M48I*, F50Y* и I66N*) для преобразования в наиболее близкую последовательность зародышевой линии. Мутации, отмеченные звездочкой, находятся в положениях, классифицированных как остатки Вернье (Foote J. et al. J. Mol. Biol. 224: 487 (1992)), и их, как правило, не изменяли. Однако на основании анализа как классификации CDR по Kabat (Wu T. T. and Kabat E. A. J. Exp. Med. 132:211-250 (1970)) и IMGT (Lefranc, M.-P. et al. Dev. Comp. Immunol. 27: 55-77 (2003)) полагают, что эти положения обеспечивают дополнительную возможность дополнительно уменьшать иммуногенность с небольшим риском изменения свойств связывания родительского антитела. Кроме того, мутагенез тяжелой цепи N64Q проводили в последовательности CDR2 VH (как определено по Kabat) для устранения возможного участка дезамидирования (NG) в этом положении. Мутагенез проводили на последовательности scFv ILT70080 в pCantab6 (McCafferty et al., Appl Biochem Biotech 47:157 (1994)) стандартными способами молекулярной биологии. В реакция множественного мутагенеза использовали различные комбинации мутагенных олигонуклеотидов для получения библиотек последовательностей, содержащих различные комбинации мутаций FW. Затем тестировали панели вариантов scFv ILT70080 на сохранение связывания с ILT7 человека в виде неочищенных периплазматических экстрактов в клеточном анализе связывания FMAT, как описано выше.
[00309] Получали семь ILT70080 вариантов в виде IgG. См. фигуры 9A и 9B для выравнивания последовательностей VH и VL, соответственно.
8.2 Конструирование ILT70083
[00310] Также осуществляли приведение ILT70083 к зародышевой линии. Наиболее близкими идентифицированными последовательностями зародышевой линии являлись VH3-23 (DP-47) и Vlambda1-b (DPL-5) для последовательностей VH и VL, соответственно. Один остаток FW выбирали для мутагенеза в домене VH (W66R) и восемь остатков FW выбирали в домене VL, снова включая выбранные положения Вернье (V4L*, R42T, A64G*, I66K*, S68G*, A72T, A74G и E81G). Варианты ILT70083, содержащие различные комбинации мутаций, получали непосредственно в векторах pEU, содержащих отдельные цепи VH и VL, стандартными способами молекулярной биологии. Затем совместно трансфицировали цепи VH и VL ILT70083 в различные комбинации с получением девяти вариантов ILT70083 IgG1. См. фигуры 10A и 10B для выравниваний VH и VL, соответственно.
8.3 Тестирование сконструированных антител
[00311] Получаемые IgG1 тестировали для подтверждения того, что изменения последовательности вводили в ILT70080 и ILT70083, и они не оказывали отрицательного влияния на связывание родительского антитела с клетками, экспрессирующими ILT7 человека (клетки CT-550) или ILT7 яванского макака (клетки CT-125). Варианты подвергали скринингу на связывание проточной цитометрией. Все варианты ILT70080 обладали связыванием с ILT7 человека и яванского макака аналогичным родительскому антителу ILT70080 (EC50=0,213 нМ и 0,547 нМ, соответственно). См. фигуру 11. Связывание вариантов ILT70083 с ILT7 человека также являлось аналогичным родительскому антителу (EC50=0,464 нМ). См. фигуру 12. Однако пять вариантов ILT70083 (ILT70083.4, ILT70083.9, ILT70083.3, ILT70083.6 и ILT70083.8) обладали улучшенной связывающей способностью по сравнению с родительским антителом с ILT7 яванского макака. См. фигуру 12.
[00312] Сконструированные антитела ILT70080 и ILT70083 тестировали на ADCC активность по отношению к экспрессирующим ILT7 человека линиям клеток с использованием клеточного анализ in vitro. Все варианты ILT70080 обладали повышенной ADCC активностью по сравнению с родительским антителом (EC50<14,1 пМ). См. фигуру 13. Два кандидата с самыми низкими EC50 представляли собой ILT70080.6 (EC50=6,9 пМ) и ILT70080.1 (EC50=8,0 пМ). Значения EC50 для других вариантов ILT70080 являлись таким, как указано ниже: ILT70080.1 EC50=10,0 пМ; ILT70080.3 EC50=11,0 пМ; ILT70080.4 EC50=11,9 пМ; ILT70080.5 EC50=8,6 пМ и ILT70080.7 EC50=7,8 пМ. Было выявлено, что все варианты ILT70083 обладают повышенной активностью по сравнению с родительским антителом (EC50>89,0 пМ). См. фигуру 14.
Пример 9
Афукозилирование сконструированных антител ILT70080 и ILT70083
[00313] Получали афукозилированный вариант ILT70080.6. Афукозилирование антитела ILT70080.6 не влияло на его связывание с клетками, экспрессирующими ILT7 человека или яванского макака. EC50 связывания афукозилированного ILT70080.6 с экспрессирующими ILT7 человека и яванского макака клетками составляло 152,3 пМ и 366,2 пМ, соответственно. См. фигуру 15. В таблицах на фигуре 15 приведены средние результаты трех независимых экспериментов связывания, в которых измеряли среднюю интенсивность флуоресценции (MFI).
[00314] Также оценивали ADCC активность афукозилированных вариантов ILT70080.6 и ILT70083 (см. пример 7 выше). Афукозилирование ILT70080.6 улучшало его ADCC активность по отношению как к экспрессирующим ILT7 человека клеткам, так и к экспрессирующим ILT7 яванского макака клеткам приблизительно в 10 раз. См. фигуру 16. EC50 афукозилированного ILT0080.6 составляло 1,12 пМ против экспрессирующих ILT7 человека клеток и 0,44 пМ против экспрессирующих ILT7 яванского макака клеток. В таблицах на фигуре 16 приведены средние результаты трех независимых анализов ADCC, в которых измеряли цитотоксичность.
[00315] Афукозилированные ILT70080.6 и ILT70083 тестировали на ADCC активность на PBMC человека. Цитотоксичность антител оценивали проточной цитометрией и измеряли в супернатанте опосредованную CpG A секрецию IFNα с помощью ELISA. Результаты приведены на фигуре 17. В PBMC яванского макака значения EC50 для секреции IFNα с использованием афукозилированных антител ILT70080.6 и ILT70083 составляли 58 пМ и 5216 пМ, соответственно.
[00316] Было выявлено, что в цельной крови и PBMC человека афукозилированные антитела ILT70080.6 и ILT70083 специфически связывались с BDCA-2-полоижтельными клетками. Связывание антител ограничивалось pDC человека для всех тестируемых концентраций (0,1-5,0 мкг/мл).
[00317] Было выявлено, что в цельной крови яванского макака афукозилированные антитела ILT70080.6 и ILT70083 связывались с pDC (клетки HLA-DR+ Lineage- CD123high) во всех тестируемых концентрациях (0,5-2,5 мкг/мл).
Пример 10
Афукозилирование антитела ILT70137
[00318] Афукозилированный вариант антитела ILT70137 получали, как описано выше в примере 7 для антител ILT70080 и ILT70083.
10.1 Связывание с растворимым рекомбинантным ILT7 человека
[00319] Использовали BIAcore (поверхностный плазмонный резонанс) для измерения кинетических констант скорости (kon, koff) для связывания афукозилированного IgG1 ILT70137 с белком ILT7 человека с использованием формата анализа захвата IgG. Связывание каждой концентрации двухкратной серии разбавлений белка ILT7 регистрировали после первого захвата IgG на поверхности сенсорного чипа, за которым следовал белок ILT7 или буфер устройства. Между каждой парой инъекций поверхность захвата IgG регенерировали. Затем рассчитывали индивидуальные константы скорости ассоциации и диссоциации на основании получаемых кривых связывания с использованием программного обеспечения Biaevaluation доступного через программное обеспечение поставщика, использующее модель аппроксимации 1:1, которая включала термин для корректировки на ограниченное массопереносом связывание, если его будут детектировать. Из графика данных BIAcore с высоким разрешением определяли, что константа скорости ассоциации и константы скорости диссоциации для связывания белка ILT7 с афукозилированным IgG1 ILT70137 составляет 1,855×105 M-1 с-1. Этот же график также использовали для определения соответствующей константы скорости диссоциации для таких взаимодействий, которую измеряли как 3,175×10-2с-1. Из этих констант скоростей реакций затем рассчитывали KD из коэффициента koff/kon равного 171 нМ. Эти результаты обобщены в таблице 3 ниже. Ошибки наблюдателя для kon и koff являлись низкими, и полная совместимость с данными являлась хорошей, как судили по значениям ChI2 ~1% от рассчитанного Rmax (максимального ответа). В своей совокупности это позволяет предположить, что использование модели односайтового связывания для аппроксимации данных являлось оправданным. Оценка не указывала на то, что связывание являлось ограниченным массоперенос, таким образом, измеряемые константы скорости ассоциации считали верными.
Таблица 3: Обобщение кинетической константы скорости реакции и данных KD для связывания белка ILT7 человека с афукозилированным IgG1 ILT70137
Kon=константа скорости ассоциации; Koff=константа скорости диссоциации; KD=равновесная константа связывания.
10.2 Связывание с линиями клеток, экспрессирующими ILT7
[00320] Связывание афукозилированного ILT70137 с ILT7 определяли с использованием линий клеток, стабильно экспрессирующих ILT7 человека или яванского макака. Среднюю интенсивность флуоресценции связанных с клетками антител оценивали проточной цитометрией. Клетки инкубировали с тестируемым антителом в увеличивающихся концентрациях в диапазоне от 0,004 до 333,3 нМ в течение 30 минут при 4°C. После инкубации клетки отмывали холодным PBS и инкубировали в течение 30 минут при 4°C с антителом к человек-Alexa Fluor 647. Затем определяли интенсивность флуоресценции посредством FACS и рассчитывали значения EC50 с использованием нелинейного эмпирического уравнения в программном обеспечении GraphPad Prism 6.
[00321] Результаты представлены на фигуре 18. Было выявлено, что афукозилированное ILT70137 связывалось с клетками, экспрессирующими рекомбинантным ILT7 человека и ILT7 яванского макака зависимым от дозы способом. Не наблюдали значимого связывания с изотипическими контролями. Средняя половина максимальной эффективной концентрации (EC50) афукозилированного ILT70137 составляла 0,303 нМ для связывания с клетками, экспрессирующими ILT7 человека, и 2,148 нМ для клеток, экспрессирующих ILT7 яванского макака.
10.3 ADCC активность на линиях клеток, экспрессирующих ILT7
[00322] Способность афукозилированного ILT70137 индуцировать ADCC измеряли посредством анализа активируемой флуоресценцией сортировки клеток (FACS) на клетках-мишенях, которые экспрессировали ILT7 человека или ILT7 яванского макака. Клетки-мишени культивировали совместно с линией эффекторных NK клеток KC1333 в отношении 1:5 в присутствии увеличивающихся концентраций афукозилированного ILT70137 или изотипического контроля (в диапазоне от 0 до 6,66×10-9 M). Для оценки жизнеспособности клеток-мишеней проточной цитометрией KC1333 выделяли с использованием CD56 и мертвые клетки выделяли с использованием витального красителя 7-амино-актиномицина D (7-AAD). Живые клетки-мишени определяли как отрицательные по CD56, отрицательные по 7-AAD. Процент цитотоксичности рассчитывали по следующей ниже формуле: % цитотоксичности=100- (процент живых клеток-мишеней/процент живых мишеней в контролях без антител)×100. Значения полумаксимальной эффективной концентрации (EC50) рассчитывали с использованием нелинейного эмпирического уравнения в программном обеспечении GraphPad Prism. Ось x: концентрация антитела.
[00323] Результаты представлены на фигуре 19. Афукозилированное ILT70137 индуцировало ADCC на клетках, экспрессирующих ILT7, зависящим от дозы образом с EC50 4,19 пМ на клетках, экспрессирующих ILT-7 человека, и 1,89 пМ для клеток, экспрессирующих ILT7 яванского макака.
10.4 ADCC активность на первичных плазмацитоидных дендритных клетках
[00324] Секреция IFN-α в ответ на агонист Toll-подобного рецептора 9 (TLR9) во многом обусловлена плазмацитоидными дендритными клетками (pDC) в препарате мононуклеарных клеток периферической крови (PBMC). Таким образом, непосредственно определяли возможность афукозилирования ILT70137 индуцировать ADCC первичных pDC путем оценки его способности блокировать секрецию IFN-α в PBMC. В этих анализах очищенные PBMC высевали в 96-луночные круглодонные планшеты в среде, дополненной 10% эмбриональной телячьей сывороткой и 200 нг/мл рекомбинантного IL-2 человека. В соответствующие лунки добавляли серийные разведения афукозилированного ILT70137 и контрольных антител в двух повторениях и инкубировали в течение 9,5 часов. После инкубации в каждую лунку добавляли 50 мкл TLR9 агониста ODN2216 до конечной концентрации 0,5 мкМ. IFN-α количественно определяли в супернатантах с использованием набора MultiSubtype IFN-α ELISA, и его значение приведено в виде пг/мл супернатанта на фигуре 20. IC50 ADCC рассчитывали с использованием нелинейного эмпирического уравнения в программном обеспечении GraphPad Prism v5.01.
[00325] Афукозилированный ILT70137 снижал опосредованную TLR9 секрецию IFN-α в PBMC зависящим от дозы образом с полумаксимальной ингибирующей концентрацией (IC50) 0,048 нМ. Эти результаты указывают на то, что афукозилированный ILT70137 эффективно истощал природные первичные pDC человека в PBMC.
10.5 Связывание с первичными плазмацитоидными дендритными клетками
[00326] В мононуклеарных клетках периферической крови (PBMC) оценивали специфичность афукозилированного ILT70137 к первичным плазмацитоидным дендритным клеткам (pDC) человека посредством FACS. PBMC выделяли от являющихся людьми доноров. Для правильной идентификации этой подпопуляции дендритных клеток сначала использовали маркеры CD123 (экспрессируемые pDC и базофилами) и CD304 (уникальный для pDC). pDC явяллись CD123+CD304+ двойными положительным, и окрашивание CD304 было достаточным для идентификации pDC. См. фигуру 21 (верхняя панель). Афукозилированный ILT70137 связывался только с CD304-положительными клетками, что указывает на то, что оно связывалось только с pDC. См. фигуру 21 (нижняя правая панель). Не наблюдали значимого связывания с этой популяцией с афукозилированным контрольным антителом R3-47 изотипом IgG1 человека. См. фигуру 21 (нижняя левая панель).
Пример 11
Активность in vivo антител к ILT7
[00327] Три антитела к ILT7 вводили самцам яванского макака: афукозилированное 7C7, афукозилированное ILT70080.6 и афукозилированное IgG1 ILT70137. Все три антитела активно истощали плазмацитоидные дендритные клетки (pDC).
[00328] Введение афукозилированного ILT70080.6, как правило, хорошо переносилось. Однако наблюдали следующие результаты патологического исследования: уменьшение числа нейтрофилов, сосудистый лейкоцитоз, увеличение гломерулярного матрикса и васкулярного/периваскулярного воспаления. Кроме того, появление антител к афукозилированному антителу ILT70080.6 (антитела к лекарственному средству) было ассоциировано с увеличенным клиренсом афукозилированного ILT70080.6.
[00329] В другом исследовании изучали токсикокинетику афукозилированного 7C7 и афукозилированного ILT70137. В настоящем исследовании 5 эквивалентных доз антител вводили яванским макакам посредством инфузии. После введения экспозиция являлась сопоставимой между афукозилированным 7C7 и афукозилированным ILT70137 в равновесном состоянии. Кроме того, как продемонстрировано на фигуре 22, специфическое и обратимое истощение pDC получали с использованием любого антитела. Истощение pDC приводило к ингибированию ex vivo продукции IFNα. См. фигуру 23.
[00330] Однако патология животных, которых обрабатывали афукозилированным 7C7 и афукозилированным ILT70137, была различной. У некоторых животных, которых обрабатывали афукозилированным 7C7 наблюдали увеличение массы селезенки. Также наблюдали микроскопические данные у некоторых животных, которых обрабатывали афукозилированным 7C7. В частности, в селезенке наблюдали красную пульпу и гипертрофию/гиперплазию макрофагов. В печени наблюдали гипертрофию/гиперплазию клеток Купфера. Кроме того, иммуногистохимия показала содержащие IgG человека/7C7 и IgG обезьяны гранулярные отложения, связанные с гипертрофированными/гиперплазированными клетками Купфера в печени и макрофагами красной пульпы в селезенке. Эти наблюдения соответствуют чрезмерному физиологическому клиренсу иммунных комплексов, содержащих лекарственное средство (7C7) и антитела к лекарственному средству (ADA). В противоположность этому с афукозилированным ILT70137 не наблюдали изменений массы органов и макроскопических или микроскопических данных.
[00331] Кроме того, несмотря на то, что количество нейтрофилов падало ниже 1 E3/мкл у двух обезьян, которых обрабатывали афукозилированным антителом 7C7, не наблюдали значимых изменений количества нейтрофилов у обезьян, которых обрабатывали контролем или афукозилированным ILT70137.
[00332] Таким образом, несмотря на то, что все три антитела истощали pDC in vivo превосходные показатели безопасности и отсутствие антител к лекарственному средству после введения афукозилированного ILT70137 являлись неожиданным преимуществом.
Пример 12
Картирование эпитопов
[00333] Для определения эпитопа, связываемого антителами к ILT7, конструировали химерные полипептиды, содержащие полипептиды ILT7 и ILT1, и тестировали связывание антител к ILT7 с этими конструкциями. ILT1 (номер доступа Q8N149) выбирали для конструирования химерных вариантов, т.к. он обладает такой же модульной структурой как и ILT7 и обладает 65% идентичностью с ILT7, но не распознается моноклональными антителами ILT7. Химерные полипептиды получали заменой внеклеточных доменов Ig ILT7 аналогами ILT1. Все эти конструкции содержали N-концевую метку Flag. Результаты демонстрировали, что ILT70080 и ILT70083 связываются с доменом Ig1 ILT7. В противоположность этому, антитело 7C7 связывается с доменом Ig2 ILT7.
[00334] Указанное выше описание конкретных вариантов осуществления полностью откроет общий характер изобретения, который специалисты в данной области, применяя знания в данной области, могут легко модифицировать и/или адаптировать для различных применений, таких как конкретные варианты осуществления, без излишнего экспериментирования, не выходя за рамки общей концепции настоящего изобретения. Таким образом, такие адаптации и модификации предназначены входить в значение и диапазон эквивалентов раскрываемых вариантов осуществления, на основании указаний и руководства, предоставленных в настоящем описании. Следует понимать, что формулировки или терминологию в настоящем описании используют для цели описания, а не ограничения, таким образом, что специалист в данной области интерпретирует терминологию или формулировки настоящего описания в свете указаний и руководства.
[00335] Объем и сущность настоящего изобретения не должны быть ограничены каким-либо из описанных выше иллюстративных вариантов осуществления, а следует их определять только в соответствии со следующей ниже формулой изобретения и ее эквивалентами.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MedImmune
<120> СВЯЗЫВАЮЩИЕ ILT7 МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130>
<160> 292
<170> PatentIn version 3.5
<210> 1
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> SBI28 VH
<400> 1
caggttcagc tgcagcagtc tggggctgag ctggtgaagc ctggggcctc agtgaagatg 60
tcctgcaagg cttttggcta caccttcact acctatccaa tagagtggat gaagcagaat 120
catgggaaga gcctagagtg gattggaaat tttcatcctt acaatgatga tactaagtac 180
aatgaaaaat tcaagggcaa ggccaaattg actgtagaaa aatcctctag cacagtctac 240
ttggagctca gccgattaac atctgatgac tctgctgttt attactgtgc aaggggggat 300
gattacggga tggactactg gggtcaagga acctcagtca ccgtctcctc a 351
<210> 2
<211> 117
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VH
<400> 2
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Phe Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Pro Ile Glu Trp Met Lys Gln Asn His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asn Phe His Pro Tyr Asn Asp Asp Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Lys Leu Thr Val Glu Lys Ser Ser Ser Thr Val Tyr
65 70 75 80
Leu Glu Leu Ser Arg Leu Thr Ser Asp Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Asp Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 3
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VHCDR1
<400> 3
Thr Tyr Pro Ile Glu
1 5
<210> 4
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VHCDR2
<400> 4
Asn Phe His Pro Tyr Asn Asp Asp Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 5
<211> 8
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VHCDR3
<400> 5
Gly Asp Asp Tyr Gly Met Asp Tyr
1 5
<210> 6
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> SBI28 VL
<400> 6
gacattgtga tgacccagtc tcaaaaattc atgtccacat cagtaggaga cagggtcagc 60
atcacctgca aggccagtca gaatgttcgt actgctgtag cctggtatca acagaaacca 120
gggcagtctc ctaaagcact gatttacttg gcatccaacc ggcacactgg agtccctgat 180
cgcttcacag gcagtggatc tgggacagat ttcactctca ccattagcaa tgtgcaatct 240
gaagacctgg cagattattt ctgtctgcaa cattggaatt atccattcac gttcggctcg 300
gggacaaagt tggaaataaa a 321
<210> 7
<211> 107
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VL
<400> 7
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Arg Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Ala Leu Ile
35 40 45
Tyr Leu Ala Ser Asn Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Leu Gln His Trp Asn Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 8
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VLCDR1
<400> 8
Lys Ala Ser Gln Asn Val Arg Thr Ala Val Ala
1 5 10
<210> 9
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VLCDR2
<400> 9
Leu Ala Ser Asn Arg His Thr
1 5
<210> 10
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> SBI28 VLCDR3
<400> 10
Leu Gln His Trp Asn Tyr Pro Phe Thr
1 5
<210> 11
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 7C7 VH
<400> 11
gaggtgcagc tggtggagtc tgggggaggc gtagtacagc ctgggagatc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc atctacccca tcgagtgggt gcgacaggct 120
cctggacagg gcctggaatg gatcggcaac ttccacccct acaacgacga caccaagtac 180
aacgagaagt tcaagggcag agtcaccatg accacagaca catccacgag cacagtgtac 240
atggagctga gcagcctgag atctgaggac acggccgtgt attactgtac gagaggcgac 300
gactacggcc tggactattg gggccagggc accctcgtga ccgtgtcctc t 351
<210> 12
<211> 117
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VH
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ile Tyr
20 25 30
Pro Ile Glu Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Phe His Pro Tyr Asn Asp Asp Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Gly Asp Asp Tyr Gly Leu Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 13
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VHCDR1
<400> 13
Ile Tyr Pro Ile Glu
1 5
<210> 14
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VHCDR2
<400> 14
Asn Phe His Pro Tyr Asn Asp Asp Thr Lys Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 15
<211> 8
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VHCDR3
<400> 15
Gly Asp Asp Tyr Gly Leu Asp Tyr
1 5
<210> 16
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 7C7 VL
<400> 16
aatattcaga tgacccagag cccgagcagc ctgagcgcaa gcgttggtga tcgtgttacc 60
attacctgtg acgccagcca gaatgttcgt accgcagttg catggtatca gcagaaaccg 120
ggtaaagcac cgaaacgtct gatttatctg gcaagtaatc gtcataccgg tgttccgagc 180
cgttttagcg gtagcggttc tggcaccgat tttaccctga ccattagcag cctgcagagc 240
gaagattttg ccacctatta ttgtctgcag cattggaatt atccgtttac ctttggtccg 300
ggtacaaaac tggaaattaa a 321
<210> 17
<211> 107
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VL
<400> 17
Asn Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Asp Ala Ser Gln Asn Val Arg Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile
35 40 45
Tyr Leu Ala Ser Asn Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln His Trp Asn Tyr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 18
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VLCDR1
<400> 18
Asp Ala Ser Gln Asn Val Arg Thr Ala Val Ala
1 5 10
<210> 19
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VLCDR2
<400> 19
Leu Ala Ser Asn Arg His Thr
1 5
<210> 20
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> 7C7 VLCDR3
<400> 20
Leu Gln His Trp Asn Tyr Pro Phe Thr
1 5
<210> 21
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080 VH
<400> 21
caggtgcagc tggtgcaatc tggggctgag gtgaaggcgc ctgggacttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tcaacggcag agtcaccctt tccgcggacg aatccacgag cacagcctac 240
ttggaactga gcagtctaag atatgacgac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cgagt 375
<210> 22
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VH
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Ala Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Asn Gly Arg Val Thr Leu Ser Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Glu Leu Ser Ser Leu Arg Tyr Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 23
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VHCDR1
<400> 23
Asn Tyr Ala Val Ser
1 5
<210> 24
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VHCDR2
<400> 24
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Asn
1 5 10 15
Gly
<210> 25
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VHCDR3
<400> 25
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 26
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080 VL
<400> 26
tcctatgagc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaagatt 60
tcctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccatctctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 27
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VL
<400> 27
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Lys Ile Ser Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ile Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 28
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VLCDR1
<400> 28
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 29
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VLCDR2
<400> 29
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 30
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080 VLCDR3
<400> 30
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 31
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VH
<400> 31
caggtgcagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt accgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 32
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VH
<400> 32
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 33
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VHCDR1
<400> 33
Asn Tyr Ala Val Ser
1 5
<210> 34
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VHCDR2
<400> 34
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 35
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VHCDR3
<400> 35
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 36
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VL
<400> 36
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccatctctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 37
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VL
<400> 37
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ile Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 38
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VLCDR1
<400> 38
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 39
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VLCDR2
<400> 39
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 40
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.1 VLCDR3
<400> 40
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 41
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VH
<400> 41
caggtgcagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt tccgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 42
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VH
<400> 42
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Ser Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 43
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VHCDR1
<400> 43
Asn Tyr Ala Val Ser
1 5
<210> 44
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VHCDR2
<400> 44
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 45
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VHCDR3
<400> 45
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 46
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VL
<400> 46
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccatctctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 47
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VL
<400> 47
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ile Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 48
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VLCDR1
<400> 48
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 49
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VLCDR2
<400> 49
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 50
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.2 VLCDR3
<400> 50
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 51
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VH
<400> 51
caggtgcagc tggtgcaatc tggggctgag gtgaaggcgc ctgggacttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt tccgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 52
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VH
<400> 52
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Ala Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Ser Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 53
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VHCDR1
<400> 53
Asn Tyr Ala Val Ser
1 5
<210> 54
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VHCDR2
<400> 54
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 55
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VHCDR3
<400> 55
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 56
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VL
<400> 56
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccatctctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 57
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VL
<400> 57
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ile Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 58
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VLCDR1
<400> 58
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 59
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VLCDR2
<400> 59
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 60
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.3 VLCDR3
<400> 60
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 61
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VH
<400> 61
caggtgcagc tggtgcaatc tggggctgag gtgaaggcgc ctgggacttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt tccgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 62
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VH
<400> 62
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Ala Pro Gly Thr
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Ser Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 63
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VHCDR1
<400> 63
Asn Tyr Ala Val Ser
1 5
<210> 64
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VHCDR2
<400> 64
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 65
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VHCDR3
<400> 65
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 66
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VL
<400> 66
tcctatgagc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaagatt 60
tcctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccatctctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 67
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VL
<400> 67
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Lys Ile Ser Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Ile Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 68
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VLCDR1
<400> 68
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 69
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VLCDR2
<400> 69
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 70
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.4 VLCDR3
<400> 70
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 71
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VH
<400> 71
caggtgcagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt accgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 72
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VH
<400> 72
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 73
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VHCDR1
<400> 73
Asn Tyr Ala Val Ser
1 5
<210> 74
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VHCDR2
<400> 74
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 75
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VHCDR3
<400> 75
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 76
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VL
<400> 76
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgatggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 77
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VL
<400> 77
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Met Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 78
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VLCDR1
<400> 78
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 79
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VLCDR2
<400> 79
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 80
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.5 VLCDR3
<400> 80
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 81
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VH
<400> 81
caggtgcagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt accgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 82
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VH
<400> 82
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 83
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VHCDR1
<400> 83
Asn Tyr Ala Val Ser
1 5
<210> 84
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VHCDR2
<400> 84
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 85
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VHCDR3
<400> 85
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 86
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VL
<400> 86
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgctggtcat ttattataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 87
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VL
<400> 87
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 88
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VLCDR1
<400> 88
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 89
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VLCDR2
<400> 89
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 90
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.6 VLCDR3
<400> 90
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 91
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VH
<400> 91
caggtgcagc tggtgcaatc tggggctgag gtgaagaagc ctgggtcttc ggtgaaagtc 60
tcctgcaagg catctggaga cagcttcagg aactatgctg tcagttgggt gcgacaggcc 120
ccaggacaag gtcttgagtg gatgggagcg atgatgccta gttttggaac aagggagcac 180
tcacagcagt tccagggcag agtcaccctt accgcggacg aatccacgag cacagcctac 240
atggaactga gcagtctaag atctgaggac acggccgtct attactgtgc gggtagtcgg 300
gactacaatg cttaccattt ttggactggt ccccctgact tctggggccg aggaaccctg 360
gtcaccgtct cctca 375
<210> 92
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VH
<400> 92
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Asp Ser Phe Arg Asn Tyr
20 25 30
Ala Val Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe
50 55 60
Gln Gly Arg Val Thr Leu Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro
100 105 110
Asp Phe Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 93
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VHCDR1
<400> 93
Asn Tyr Ala Val Ser
1 5
<210> 94
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VHCDR2
<400> 94
Ala Met Met Pro Ser Phe Gly Thr Arg Glu His Ser Gln Gln Phe Gln
1 5 10 15
Gly
<210> 95
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VHCDR3
<400> 95
Ser Arg Asp Tyr Asn Ala Tyr His Phe Trp Thr Gly Pro Pro Asp Phe
1 5 10 15
<210> 96
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VL
<400> 96
tcctatgtgc tgactcagcc accctcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacagcgt tggcagtaca agtgtccact ggtaccagca gaagccgggc 120
caggcccctg tgctggtcat gttttataac agtgaccggc cctcagggat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg actattactg tcaggtgtgg gatactagta gtgatcatcc ggagttcggc 300
ggagggacca agctgaccgt ccta 324
<210> 97
<211> 108
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VL
<400> 97
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Ser Val Gly Ser Thr Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Met Phe
35 40 45
Tyr Asn Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Thr Ser Ser Asp His
85 90 95
Pro Glu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 98
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VLCDR1
<400> 98
Gly Gly Asp Ser Val Gly Ser Thr Ser Val His
1 5 10
<210> 99
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VLCDR2
<400> 99
Tyr Asn Ser Asp Arg Pro Ser
1 5
<210> 100
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70080.7 VLCDR3
<400> 100
Gln Val Trp Asp Thr Ser Ser Asp His Pro Glu
1 5 10
<210> 101
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083 VH
<400> 101
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggctg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cgagt 375
<210> 102
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VH
<400> 102
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Trp Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 103
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VHCDR1
<400> 103
Ser Tyr Ala Met Ser
1 5
<210> 104
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VHCDR2
<400> 104
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 105
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VHCDR3
<400> 105
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 106
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083 VL
<400> 106
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggagag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccgccc tggccatcac cggactccag 240
actgaggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 107
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VL
<400> 107
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Arg Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Ala Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 108
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VLCDR1
<400> 108
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 109
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VLCDR2
<400> 109
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 110
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083 VLCDR3
<400> 110
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 111
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VH
<400> 111
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggctg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cgagt 375
<210> 112
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VH
<400> 112
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Trp Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 113
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VHCDR1
<400> 113
Ser Tyr Ala Met Ser
1 5
<210> 114
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VHCDR2
<400> 114
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 115
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VHCDR3
<400> 115
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 116
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VL
<400> 116
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 117
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VL
<400> 117
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 118
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VLCDR1
<400> 118
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 119
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VLCDR2
<400> 119
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 120
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.1 VLCDR3
<400> 120
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 121
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VH
<400> 121
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggctg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cgagt 375
<210> 122
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VH
<400> 122
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Trp Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 123
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VHCDR1
<400> 123
Ser Tyr Ala Met Ser
1 5
<210> 124
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VHCDR2
<400> 124
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 125
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VHCDR3
<400> 125
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 126
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VL
<400> 126
cagtctgtcc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 127
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VL
<400> 127
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 128
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VLCDR1
<400> 128
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 129
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VLCDR2
<400> 129
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 130
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.2 VLCDR3
<400> 130
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 131
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VH
<400> 131
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggctg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cgagt 375
<210> 132
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VH
<400> 132
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Trp Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 133
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VHCDR1
<400> 133
Ser Tyr Ala Met Ser
1 5
<210> 134
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VHCDR2
<400> 134
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 135
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VHCDR3
<400> 135
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 136
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VL
<400> 136
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctggctccaa atctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 137
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VL
<400> 137
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 138
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VLCDR1
<400> 138
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 139
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VLCDR2
<400> 139
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 140
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.3 VLCDR3
<400> 140
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 141
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VH
<400> 141
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggctg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cgagt 375
<210> 142
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VH
<400> 142
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Trp Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 143
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VHCDR1
<400> 143
Ser Tyr Ala Met Ser
1 5
<210> 144
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VHCDR2
<400> 144
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 145
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VHCDR3
<400> 145
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 146
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VL
<400> 146
cagtctgtcc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctggctccaa gtctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 147
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VL
<400> 147
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 148
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VLCDR1
<400> 148
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 149
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VLCDR2
<400> 149
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 150
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.4 VLCDR3
<400> 150
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 151
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VH
<400> 151
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cctca 375
<210> 152
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VH
<400> 152
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 153
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VHCDR1
<400> 153
Ser Tyr Ala Met Ser
1 5
<210> 154
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VHCDR2
<400> 154
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 155
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VHCDR3
<400> 155
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 156
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VL
<400> 156
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggagag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccgccc tggccatcac cggactccag 240
actgaggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 157
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VL
<400> 157
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Arg Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Ala Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 158
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VLCDR1
<400> 158
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 159
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VLCDR2
<400> 159
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 160
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.5 VLCDR3
<400> 160
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 161
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VH
<400> 161
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cctca 375
<210> 162
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VH
<400> 162
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 163
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VHCDR1
<400> 163
Ser Tyr Ala Met Ser
1 5
<210> 164
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VHCDR2
<400> 164
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 165
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VHCDR3
<400> 165
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 166
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VL
<400> 166
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 167
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VL
<400> 167
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 168
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VLCDR1
<400> 168
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 169
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VLCDR2
<400> 169
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 170
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.6 VLCDR3
<400> 170
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 171
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VH
<400> 171
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cctca 375
<210> 172
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VH
<400> 172
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 173
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VHCDR1
<400> 173
Ser Tyr Ala Met Ser
1 5
<210> 174
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VHCDR2
<400> 174
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 175
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VHCDR3
<400> 175
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 176
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VL
<400> 176
cagtctgtcc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctgcctccat ctcttccacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 177
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VL
<400> 177
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Ala Ser Ile Ser Ser Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 178
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VLCDR1
<400> 178
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 179
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VLCDR2
<400> 179
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 180
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.7 VLCDR3
<400> 180
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 181
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VH
<400> 181
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cctca 375
<210> 182
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VH
<400> 182
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 183
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VHCDR1
<400> 183
Ser Tyr Ala Met Ser
1 5
<210> 184
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VHCDR2
<400> 184
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 185
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VHCDR3
<400> 185
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 186
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VL
<400> 186
cagtctgtcg tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctggctccaa atctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 187
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VL
<400> 187
Gln Ser Val Val Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 188
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VLCDR1
<400> 188
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 189
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VLCDR2
<400> 189
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 190
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.8 VLCDR3
<400> 190
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 191
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VH
<400> 191
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcggaca 300
tattactatg atagtgatgg tcactcggat gtttttgata tttggggccg gggcaccctg 360
gtcaccgtct cctca 375
<210> 192
<211> 125
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VH
<400> 192
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe
100 105 110
Asp Ile Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 193
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VHCDR1
<400> 193
Ser Tyr Ala Met Ser
1 5
<210> 194
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VHCDR2
<400> 194
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 195
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VHCDR3
<400> 195
Arg Thr Tyr Tyr Tyr Asp Ser Asp Gly His Ser Asp Val Phe Asp Ile
1 5 10 15
<210> 196
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VL
<400> 196
cagtctgtcc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgttctg gaagcgcctc caatattggg agtaattttg tgtcctggta ccaacaactc 120
cccgggacag cccccaaact cctcatttat gacaatgata aacgagactt agggattcct 180
gaccgcttct ctggctccaa gtctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggaca ccagtctgac tgttggggtt 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 197
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VL
<400> 197
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn
20 25 30
Phe Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asp Lys Arg Asp Leu Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Thr Ser Leu
85 90 95
Thr Val Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 198
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VLCDR1
<400> 198
Ser Gly Ser Ala Ser Asn Ile Gly Ser Asn Phe Val Ser
1 5 10
<210> 199
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VLCDR2
<400> 199
Asp Asn Asp Lys Arg Asp Leu
1 5
<210> 200
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70083.9 VLCDR3
<400> 200
Gly Thr Trp Asp Thr Ser Leu Thr Val Gly Val
1 5 10
<210> 201
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70137 VH
<400> 201
caggtacagc tgcagcagtc aggagctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggtta cacctttacc agctatggta tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggatgg atcagcgctt acaatggtaa cacaaactat 180
gcacagaagc tccagggcag agtcaccatg accacagaca catccacgag cacagcctac 240
atggagctga ggagcctgag atctgacgac acggccgtgt attactgtgc gagaaatggc 300
ctctgggggt gggactctga tgcttttgat atctggggcc gaggaaccct ggtcaccgtc 360
tcctca 366
<210> 202
<211> 122
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VH
<400> 202
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 203
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VHCDR1
<400> 203
Ser Tyr Gly Ile Ser
1 5
<210> 204
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VHCDR2
<400> 204
Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu Gln
1 5 10 15
Gly
<210> 205
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VHCDR3
<400> 205
Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile
1 5 10
<210> 206
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70137 VL
<400> 206
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ggttataact atgtctcctg gtaccaacag 120
cacccaggca aagcccccaa actcatgatt tatgatgtca gtaatcggcc ctcaggggtt 180
tctaatcgct tctctggctc caagtctggc aacacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agctcatata caagcagcag cactgtggta 300
ttcggcggag ggaccaaggt caccgtccta 330
<210> 207
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VL
<400> 207
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Val Val Phe Gly Gly Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 208
<211> 14
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VLCDR1
<400> 208
Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr Asn Tyr Val Ser
1 5 10
<210> 209
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VLCDR2
<400> 209
Asp Val Ser Asn Arg Pro Ser
1 5
<210> 210
<211> 10
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70137 VLCDR3
<400> 210
Ser Ser Tyr Thr Ser Ser Ser Thr Val Val
1 5 10
<210> 211
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70052 VH
<400> 211
gaggtgcagc tggtggagtc tgggggaggc ttggtcaagc ctggagggtc cctgagactc 60
tcctgtgaag cctccggatt caccttcact gactactaca tgaactggat ccgccaggct 120
ccagggaagg ggctggagtg gatttcatac attagtcctg ggggtactac cgtatactac 180
gcagactctg tgaagggccg attcaccatc tccagggaca acgccaagaa ctcattatat 240
ctccaaatga acagcctgag agtcgaggac acggccgtct attactgtgc gagagacatt 300
agtggttatg gcgactggtt cgacccctgg ggcaagggca ccctggtcac cgtctcgagt 360
<210> 212
<211> 120
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VH
<400> 212
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Glu Ala Ser Gly Phe Thr Phe Thr Asp Tyr
20 25 30
Tyr Met Asn Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Tyr Ile Ser Pro Gly Gly Thr Thr Val Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Val Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Ile Ser Gly Tyr Gly Asp Trp Phe Asp Pro Trp Gly Lys
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 213
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VHCDR1
<400> 213
Asp Tyr Tyr Met Asn
1 5
<210> 214
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VHCDR2
<400> 214
Tyr Ile Ser Pro Gly Gly Thr Thr Val Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 215
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VHCDR3
<400> 215
Asp Ile Ser Gly Tyr Gly Asp Trp Phe Asp Pro
1 5 10
<210> 216
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70052 VL
<400> 216
cagtctgtcg tgacgcagcc gccctcagcg tctgggaccc ccgggcagag ggtcaccatc 60
tcttgttctg gaagcagctc caacatcgga agtaataccg taaactggta ccagctgctc 120
ccagctacgg cccccagaat cctcatctat aataataatc agcggccgtc aggggtccca 180
gaccgattcg ctggctccaa gtctggcacc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgga gcatgggatg acagcctgaa tagtccggtg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 217
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VL
<400> 217
Gln Ser Val Val Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Leu Leu Pro Ala Thr Ala Pro Arg Ile Leu
35 40 45
Ile Tyr Asn Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ala
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Gly Ala Trp Asp Asp Ser Leu
85 90 95
Asn Ser Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 218
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VLCDR1
<400> 218
Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn Thr Val Asn
1 5 10
<210> 219
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VLCDR2
<400> 219
Asn Asn Asn Gln Arg Pro Ser
1 5
<210> 220
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70052 VLCDR3
<400> 220
Gly Ala Trp Asp Asp Ser Leu Asn Ser Pro Val
1 5 10
<210> 221
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70100 VH
<400> 221
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaggcatagg 300
ggccggtggc taccgtactt cgatctctgg ggccagggga caatggtcac cgtctcgagt 360
<210> 222
<211> 120
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VH
<400> 222
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Arg Gly Arg Trp Leu Pro Tyr Phe Asp Leu Trp Gly Gln
100 105 110
Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 223
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VHCDR1
<400> 223
Ser Tyr Ala Met Ser
1 5
<210> 224
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VHCDR2
<400> 224
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 225
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VHCDR3
<400> 225
His Arg Gly Arg Trp Leu Pro Tyr Phe Asp Leu
1 5 10
<210> 226
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70100 VH
<400> 226
gacatccagt tgacccagtc tccatcctcc ctgtctgcat ctgtgggaga gagagtcacc 60
atcacttgcc aggcgagtca ggacattcgc agctatttaa attggtatca gcaaaaacca 120
gggaaagccc ctaaactcct gatctatgat gtatccaatt tggaaaccgg ggtcccatca 180
cggttcagtg gaagtggatc tgggacagag tatactttca gcatcagcag cctgcagcct 240
gaagattttg caacatatta ttgtcaacag tatgataaag ccccgcttac tttcggcgga 300
gggaccaagg tggaaatcaa a 321
<210> 227
<211> 107
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VL
<400> 227
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Arg Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Val Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Tyr Thr Phe Ser Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Lys Ala Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 228
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VLCDR1
<400> 228
Gln Ala Ser Gln Asp Ile Arg Ser Tyr Leu Asn
1 5 10
<210> 229
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VLCDR2
<400> 229
Asp Val Ser Asn Leu Glu Thr
1 5
<210> 230
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70100 VLCDR3
<400> 230
Gln Gln Tyr Asp Lys Ala Pro Leu Thr
1 5
<210> 231
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70142 VH
<400> 231
caggtccagc tggtacagtc tggagctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggtta cacctttacc agctatggta tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggatgg atcagcgctt acaatggtaa cacaaactat 180
gcacagaagc tccagggcag agtcaccatg accagggaca cgtccatcag cacagcctac 240
atggagctga gcaggctgag atctgacgac acggccgtgt attactgtgc gagaaatggc 300
ctctgggggt gggactctga tgcttttgat atctgggggc aagggaccac ggtcaccgtc 360
tcgagt 366
<210> 232
<211> 122
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VH
<400> 232
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 233
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VHCDR1
<400> 233
Ser Tyr Gly Ile Ser
1 5
<210> 234
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VHCDR2
<400> 234
Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu Gln
1 5 10 15
Gly
<210> 235
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VHCDR3
<400> 235
Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile
1 5 10
<210> 236
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70142 VL
<400> 236
cagtctgtgc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgctctg gaagcagctc caacattggg aataattatg tatcctggta ccagcagctc 120
ccaggaacag cccccaaact cctcatttat gacaataata agcgaccctc agggattcct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggata gcagcctgag tgctggcgtc 300
ttcggaactg ggacccagct caccgtttta 330
<210> 237
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VL
<400> 237
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Ser Leu
85 90 95
Ser Ala Gly Val Phe Gly Thr Gly Thr Gln Leu Thr Val Leu
100 105 110
<210> 238
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VLCDR1
<400> 238
Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn Tyr Val Ser
1 5 10
<210> 239
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VLCDR2
<400> 239
Asp Asn Asn Lys Arg Pro Ser
1 5
<210> 240
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70142 VLCDR3
<400> 240
Gly Thr Trp Asp Ser Ser Leu Ser Ala Gly Val
1 5 10
<210> 241
<211> 366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70144 VH
<400> 241
caggtccagc tggtacagtc tggagctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggtta cacctttacc agctatggta tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggatgg atcagcgctt acaatggtaa cacaaactat 180
gcacagaagc tccagggcag agtcaccatg accagggaca cgtccatcag cacagcctac 240
atggagctga gcaggctgag atctgacgac acggccgtgt attactgtgc gagaaatggc 300
ctctgggggt gggactctga tgcttttgat atctgggggc aagggaccac ggtcaccgtc 360
tcgagt 366
<210> 242
<211> 122
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VH
<400> 242
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 243
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VHCDR1
<400> 243
Ser Tyr Gly Ile Ser
1 5
<210> 244
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VHCDR2
<400> 244
Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu Gln
1 5 10 15
Gly
<210> 245
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VHCDR3
<400> 245
Asn Gly Leu Trp Gly Trp Asp Ser Asp Ala Phe Asp Ile
1 5 10
<210> 246
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70144 VL
<400> 246
cagtctgtgc tgacgcagcc gccctcagtg tctgcggccc caggacagaa ggtcaccatc 60
tcctgctctg gaagcagctc caacattggg aataattatg tatcctggta ccagcagctc 120
ccaggaacag cccccaaact cctcatttat gacaataata agcgaccctc agggattcct 180
gaccgattct ctggctccaa gtctggcacg tcagccaccc tgggcatcac cggactccag 240
actggggacg aggccgatta ttactgcgga acatgggata gcagcctgag tgctggcgtc 300
ttcggaactg ggacccagct caccgtttta 330
<210> 247
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VL
<400> 247
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Ser Leu
85 90 95
Ser Ala Gly Val Phe Gly Thr Gly Thr Gln Leu Thr Val Leu
100 105 110
<210> 248
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VLCDR1
<400> 248
Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn Tyr Val Ser
1 5 10
<210> 249
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VLCDR2
<400> 249
Asp Asn Asn Lys Arg Pro Ser
1 5
<210> 250
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70144 VLCDR3
<400> 250
Gly Thr Trp Asp Ser Ser Leu Ser Ala Gly Val
1 5 10
<210> 251
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70019 VH
<400> 251
caggtccagc tggtgcagtc tgggggaggc ctggtcaagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt agctatagca tgaactgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcatcc attagtagta gtagttacat atactacgca 180
gactcagtga agggccgatt caccatctcc cgagacaacg ccaagaactc actgtatctg 240
caaatgaaca gcctgagagc cgaggacacg gctgtgtatt actgtgcgag agaggcacca 300
aatggtctaa ttgactaccg gggccggggg acaatggtca ccgtctcttc a 351
<210> 252
<211> 117
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VH
<400> 252
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Ala Pro Asn Gly Leu Ile Asp Tyr Arg Gly Arg Gly Thr Met
100 105 110
Val Thr Val Ser Ser
115
<210> 253
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VHCDR1
<400> 253
Ser Tyr Ser Met Asn
1 5
<210> 254
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VHCDR2
<400> 254
Ser Ile Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val Lys Gly
1 5 10 15
<210> 255
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VHCDR3
<400> 255
Glu Ala Pro Asn Gly Leu Ile Asp Tyr
1 5
<210> 256
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70019 VL
<400> 256
gacatcgtga tgacccagtc tccttccacc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggccagtca gggtattagt agctggttgg cctggtatca gcagaaacca 120
gggagagccc ctaaggtctt gatctataag gcatctactt tagaaagtgg ggtcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag agttacagta ccccgtggac gttcggccaa 300
gggaccaagc tggagatcaa a 321
<210> 257
<211> 107
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VL
<400> 257
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Arg Ala Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Thr Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 258
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VLCDR1
<400> 258
Arg Ala Ser Gln Gly Ile Ser Ser Trp Leu Ala
1 5 10
<210> 259
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VLCDR2
<400> 259
Lys Ala Ser Thr Leu Glu Ser
1 5
<210> 260
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70019 VLCDR3
<400> 260
Gln Gln Ser Tyr Ser Thr Pro Trp Thr
1 5
<210> 261
<211> 351
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70028 VH
<400> 261
cagctgcagc tgcaggagtc gggcgcaaga atgttgaagc cttcggagac cctgtccctc 60
acgtgcgacg tccagaatgg gtccttcagt actcactact ggacttggat ccgccagccc 120
ccagggaagg ggctggagtg gattggagac atcaatcaaa ggggaagtac caaccacaat 180
ccgtccctca ggagtcgact caccatatca gtagacccgt ccaagaatca gttctcagtg 240
aggttgactt ctgtgaccgc cgcggacacg gctgtctatt actgtgcgag attgaacgac 300
gatgatgctt ttgatatctg gggccggggg acaatggtca ccgtctcgag t 351
<210> 262
<211> 117
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VH
<400> 262
Gln Leu Gln Leu Gln Glu Ser Gly Ala Arg Met Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Asp Val Gln Asn Gly Ser Phe Ser Thr His
20 25 30
Tyr Trp Thr Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn Gln Arg Gly Ser Thr Asn His Asn Pro Ser Leu Arg
50 55 60
Ser Arg Leu Thr Ile Ser Val Asp Pro Ser Lys Asn Gln Phe Ser Val
65 70 75 80
Arg Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Leu Asn Asp Asp Asp Ala Phe Asp Ile Trp Gly Arg Gly Thr Met
100 105 110
Val Thr Val Ser Ser
115
<210> 263
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VHCDR1
<400> 263
Thr His Tyr Trp Thr
1 5
<210> 264
<211> 16
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VHCDR2
<400> 264
Asp Ile Asn Gln Arg Gly Ser Thr Asn His Asn Pro Ser Leu Arg Ser
1 5 10 15
<210> 265
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VHCDR3
<400> 265
Leu Asn Asp Asp Asp Ala Phe Asp Ile
1 5
<210> 266
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70028 VL
<400> 266
cagtctgtgc tgacgcagcc gccctcagcg tctgggaccc ccgggcaggg ggtcaccatc 60
tcgtgttctg gaagcagctc caacatcggg agtcatactg taaattggta ccaacatctc 120
gcaggaacgg cccccaaact cctcatctat agtaataatc agcggccctc aggggtccct 180
gaccgattct ctggctccaa gtctgcctcc tcagcctccc tggccatcag tgggctccag 240
tctgaggatg aggctgatta ttactgtgca gcctgggatg ccggtgtcga tggttatgtc 300
ttcggaaccg ggaccaaggt caccgtccta 330
<210> 267
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VL
<400> 267
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Gly Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser His
20 25 30
Thr Val Asn Trp Tyr Gln His Leu Ala Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Ala Ser Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Ala Gly Val
85 90 95
Asp Gly Tyr Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu
100 105 110
<210> 268
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VLCDR1
<400> 268
Ser Gly Ser Ser Ser Asn Ile Gly Ser His Thr Val Asn
1 5 10
<210> 269
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VLCDR2
<400> 269
Ser Asn Asn Gln Arg Pro Ser
1 5
<210> 270
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70028 VLCDR3
<400> 270
Ala Ala Trp Asp Ala Gly Val Asp Gly Tyr Val
1 5 10
<210> 271
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70076 VH
<400> 271
gaggtgcagc tggtggagtc cggagctgag gtgaagaagc ctggggcctc agtgaaggtc 60
tcctgcaagg cttctggtta cacctttacc agctacggta tcagctgggt gcgacaggcc 120
cctggacaag ggcttgagtg gatgggatgg atcagcgctt acaatggtaa cacaaactat 180
gcacagaagc tccagggcag agtcaccatg accacagaca catccacgag cacagcctac 240
atggagctga ggagcctgag atctgacgac acggccgtgt attactgtgc gagaggtccg 300
gggtggctgg gctttgacta ctggggccgg ggcaccctgg tcaccgtctc gagt 354
<210> 272
<211> 118
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VH
<400> 272
Glu Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Pro Gly Trp Leu Gly Phe Asp Tyr Trp Gly Arg Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 273
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR1
<400> 273
Ser Tyr Gly Ile Ser
1 5
<210> 274
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR2
<400> 274
Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu Gln
1 5 10 15
Gly
<210> 275
<211> 9
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR3
<400> 275
Gly Pro Gly Trp Leu Gly Phe Asp Tyr
1 5
<210> 276
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70076 VL
<400> 276
cagtctgccc tgactcagcc tgcctccgtg tctgggtctc ctggacagtc gatcaccatc 60
tcctgcactg gaaccagcag tgacgttggt ggttataact atgtctcttg gtaccaacaa 120
gacccaggca aggcccccaa actcatgatt tataatgtca gtaatcggcc ctcaggggtt 180
tctaatcgct tctctggctc caagtctggc aatacggcct ccctgaccat ctctgggctc 240
caggctgagg acgaggctga ttattactgc agttcatata caagcagcag cacttatgtc 300
ttcggaactg ggaccaagct gaccgtccta 330
<210> 277
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VL
<400> 277
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Asp Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asn Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Tyr Val Phe Gly Thr Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 278
<211> 14
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR1
<400> 278
Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr Asn Tyr Val Ser
1 5 10
<210> 279
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR2
<400> 279
Asn Val Ser Asn Arg Pro Ser
1 5
<210> 280
<211> 10
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70076 VHCDR3
<400> 280
Ser Ser Tyr Thr Ser Ser Ser Thr Tyr Val
1 5 10
<210> 281
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70089 VH
<400> 281
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gctcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gaaagattta 300
ggatatactt tagatacaag tgacttctgg ggccggggca ccctggtcac cgtctcgagt 360
<210> 282
<211> 120
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VH
<400> 282
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Leu Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Leu Gly Tyr Thr Leu Asp Thr Ser Asp Phe Trp Gly Arg
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 283
<211> 5
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VHCDR1
<400> 283
Ser Tyr Ala Met Ser
1 5
<210> 284
<211> 17
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VHCDR2
<400> 284
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 285
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VHCDR3
<400> 285
Asp Leu Gly Tyr Thr Leu Asp Thr Ser Asp Phe
1 5 10
<210> 286
<211> 330
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ILT70089 VL
<400> 286
tcctatgtgc tgactcagcc accctcagcg tctgggtccc ccgggcaggg ggtcaccatc 60
tcttgttctg gaagcagctc caacatcgga actaatactg taaactggta cctgcacgtt 120
ccaggaacgg cccccaaact cctcacttat actaataatc agcggccctc aggggtccaa 180
gatcgattct ctggctccaa gtctgacacg tcagcctccc tggccatcag tgggctccag 240
gctgaggatg agggtgatta ttactgtgcg acatgggatg acagcctgaa tggtctagcg 300
ttcggcggag ggaccaagct gaccgtccta 330
<210> 287
<211> 110
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VL
<400> 287
Ser Tyr Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Gly Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Thr Asn
20 25 30
Thr Val Asn Trp Tyr Leu His Val Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Thr Tyr Thr Asn Asn Gln Arg Pro Ser Gly Val Gln Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Asp Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Gly Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Gly Leu Ala Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 288
<211> 13
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VLCDR1
<400> 288
Ser Gly Ser Ser Ser Asn Ile Gly Thr Asn Thr Val Asn
1 5 10
<210> 289
<211> 7
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VLCDR2
<400> 289
Thr Asn Asn Gln Arg Pro Ser
1 5
<210> 290
<211> 11
<212> белок
<213> Искусственная последовательность
<220>
<223> ILT70089 VLCDR3
<400> 290
Ala Thr Trp Asp Asp Ser Leu Asn Gly Leu Ala
1 5 10
<210> 291
<211> 499
<212> белок
<213> Homo sapiens
<400> 291
Met Thr Leu Ile Leu Thr Ser Leu Leu Phe Phe Gly Leu Ser Leu Gly
1 5 10 15
Pro Arg Thr Arg Val Gln Ala Glu Asn Leu Pro Lys Pro Ile Leu Trp
20 25 30
Ala Glu Pro Gly Pro Val Ile Thr Trp His Asn Pro Val Thr Ile Trp
35 40 45
Cys Gln Gly Thr Leu Glu Ala Gln Gly Tyr Arg Leu Asp Lys Glu Gly
50 55 60
Asn Ser Met Ser Arg His Ile Leu Lys Thr Leu Glu Ser Glu Asn Lys
65 70 75 80
Val Lys Leu Ser Ile Pro Ser Met Met Trp Glu His Ala Gly Arg Tyr
85 90 95
His Cys Tyr Tyr Gln Ser Pro Ala Gly Trp Ser Glu Pro Ser Asp Pro
100 105 110
Leu Glu Leu Val Val Thr Ala Tyr Ser Arg Pro Thr Leu Ser Ala Leu
115 120 125
Pro Ser Pro Val Val Thr Ser Gly Val Asn Val Thr Leu Arg Cys Ala
130 135 140
Ser Arg Leu Gly Leu Gly Arg Phe Thr Leu Ile Glu Glu Gly Asp His
145 150 155 160
Arg Leu Ser Trp Thr Leu Asn Ser His Gln His Asn His Gly Lys Phe
165 170 175
Gln Ala Leu Phe Pro Met Gly Pro Leu Thr Phe Ser Asn Arg Gly Thr
180 185 190
Phe Arg Cys Tyr Gly Tyr Glu Asn Asn Thr Pro Tyr Val Trp Ser Glu
195 200 205
Pro Ser Asp Pro Leu Gln Leu Leu Val Ser Gly Val Ser Arg Lys Pro
210 215 220
Ser Leu Leu Thr Leu Gln Gly Pro Val Val Thr Pro Gly Glu Asn Leu
225 230 235 240
Thr Leu Gln Cys Gly Ser Asp Val Gly Tyr Ile Arg Tyr Thr Leu Tyr
245 250 255
Lys Glu Gly Ala Asp Gly Leu Pro Gln Arg Pro Gly Arg Gln Pro Gln
260 265 270
Ala Gly Leu Ser Gln Ala Asn Phe Thr Leu Ser Pro Val Ser Arg Ser
275 280 285
Tyr Gly Gly Gln Tyr Arg Cys Tyr Gly Ala His Asn Val Ser Ser Glu
290 295 300
Trp Ser Ala Pro Ser Asp Pro Leu Asp Ile Leu Ile Ala Gly Gln Ile
305 310 315 320
Ser Asp Arg Pro Ser Leu Ser Val Gln Pro Gly Pro Thr Val Thr Ser
325 330 335
Gly Glu Lys Val Thr Leu Leu Cys Gln Ser Trp Asp Pro Met Phe Thr
340 345 350
Phe Leu Leu Thr Lys Glu Gly Ala Ala His Pro Pro Leu Arg Leu Arg
355 360 365
Ser Met Tyr Gly Ala His Lys Tyr Gln Ala Glu Phe Pro Met Ser Pro
370 375 380
Val Thr Ser Ala His Ala Gly Thr Tyr Arg Cys Tyr Gly Ser Arg Ser
385 390 395 400
Ser Asn Pro Tyr Leu Leu Ser His Pro Ser Glu Pro Leu Glu Leu Val
405 410 415
Val Ser Gly Ala Thr Glu Thr Leu Asn Pro Ala Gln Lys Lys Ser Asp
420 425 430
Ser Lys Thr Ala Pro His Leu Gln Asp Tyr Thr Val Glu Asn Leu Ile
435 440 445
Arg Met Gly Val Ala Gly Leu Val Leu Leu Phe Leu Gly Ile Leu Leu
450 455 460
Phe Glu Ala Gln His Ser Gln Arg Ser Pro Pro Arg Cys Ser Gln Glu
465 470 475 480
Ala Asn Ser Arg Lys Asp Asn Ala Pro Phe Arg Val Val Glu Pro Trp
485 490 495
Glu Gln Ile
<210> 292
<211> 483
<212> белок
<213> Яванский макак
<400> 292
Pro Arg Thr His Met Gln Ala Glu Asn Leu Leu Lys Pro Ile Leu Trp
1 5 10 15
Ala Glu Pro Gly Pro Val Ile Ile Trp Lys Lys Pro Val Thr Ile Trp
20 25 30
Cys Gln Gly Thr Leu Glu Ala Gln Glu Tyr Arg Leu Asp Lys Glu Gly
35 40 45
Asn Ser Ile Ser Arg His Met Leu Lys Thr Leu Glu Ser Glu Asn Lys
50 55 60
Ala Lys Phe Ser Ile Pro Ser Met Met Trp Glu His Ala Gly Arg Tyr
65 70 75 80
His Cys Tyr Tyr Gln Ser Pro Ala Gly Trp Ser Glu Pro Ser Asp Pro
85 90 95
Leu Glu Leu Val Val Thr Ala Tyr Ser Arg Pro Ser Leu Ser Ala Leu
100 105 110
Pro Ser Pro Val Val Thr Ser Gly Val Asn Val Thr Leu Arg Cys Ala
115 120 125
Ser Arg Leu Gly Leu Gly Arg Phe Thr Leu Ile Glu Glu Gly Asp His
130 135 140
Arg Leu Ser Trp Thr Leu Asp Ser His Gln His Asn His Gly Lys Phe
145 150 155 160
Gln Ala Leu Phe Pro Val Gly Pro Leu Thr Phe Ser Asn Arg Gly Thr
165 170 175
Phe Arg Cys Tyr Gly Tyr Glu Asn Asn Thr Pro Tyr Val Trp Ser Glu
180 185 190
Pro Ser Asp Pro Leu Gln Leu Leu Val Ser Gly Val Ser Arg Lys Pro
195 200 205
Ser Leu Leu Thr Leu Gln Gly Pro Val Val Ala Pro Gly Asp Asn Leu
210 215 220
Thr Leu Gln Cys Gly Ser Asp Val Gly Tyr Ile Arg Tyr Ala Leu Tyr
225 230 235 240
Lys Glu Gly Gly Asp Gly Leu Pro Gln Arg Pro Gly Gln Gln Ser Gln
245 250 255
Ala Gly Leu Ser Gln Ala Ser Phe Thr Leu Asn Pro Val Arg Gly Ser
260 265 270
His Gly Gly Gln Tyr Arg Cys Tyr Gly Ala His Asn Val Ser Ser Lys
275 280 285
Trp Ser Ala Pro Ser Asp Pro Leu Asp Ile Leu Ile Ala Gly Gln Ile
290 295 300
Pro Asp Arg Pro Ser Leu Ser Val Gln Leu Gly Pro Thr Val Ala Ser
305 310 315 320
Gly Glu Lys Val Thr Leu Leu Cys Gln Ser Trp Gly Pro Met Phe Thr
325 330 335
Phe Leu Leu Ala Lys Glu Gly Ala Ala His Pro Pro Leu Arg Leu Arg
340 345 350
Ser Thr Tyr Arg Ala Gln Gln Tyr Gln Ala Glu Phe Pro Met Ser Pro
355 360 365
Val Thr Ser Ala His Ala Gly Thr Tyr Arg Cys Tyr Gly Ser Arg Ser
370 375 380
Ser Asp Pro Tyr Leu Leu Ser His Ser Ser Glu Pro Leu Glu Leu Val
385 390 395 400
Val Ser Glu Ala Thr Glu Thr Leu Asn Pro Ala Gln Asn Lys Ser Asp
405 410 415
Ser Lys Thr Ala Pro His Leu Gln Asp Tyr Thr Val Glu Asn Leu Ile
420 425 430
Arg Met Gly Ile Ala Gly Leu Val Leu Val Phe Leu Gly Ile Leu Leu
435 440 445
Phe Glu Ala Gln Gln Ser Gln Arg Ser Pro Thr Arg Cys Ser Gln Glu
450 455 460
Val Asn Ser Arg Glu Asp Asn Ala Pro Phe Arg Val Val Glu Pro Trp
465 470 475 480
Glu Gln Ile
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
| МОНОВАЛЕНТНЫЕ АНТИТЕЛА К ПРОПЕРДИНУ И ФРАГМЕНТЫ АНТИТЕЛ | 2018 |
|
RU2790103C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА CAIX, ANO1, МЕЗОТЕЛИН, TROP2, СEA ИЛИ КЛАУДИН-18.2 | 2018 |
|
RU2792671C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА NKG2D, CD16 И TROP2 | 2018 |
|
RU2820629C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ ВСМА, NKG2D И CD16 | 2018 |
|
RU2805254C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ CD33, NKG2D И CD16 | 2018 |
|
RU2820603C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ ДЛЯ АКТИВАЦИИ КЛЕТОК-НАТУРАЛЬНЫХ КИЛЛЕРОВ И ИХ ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2018 |
|
RU2809125C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |






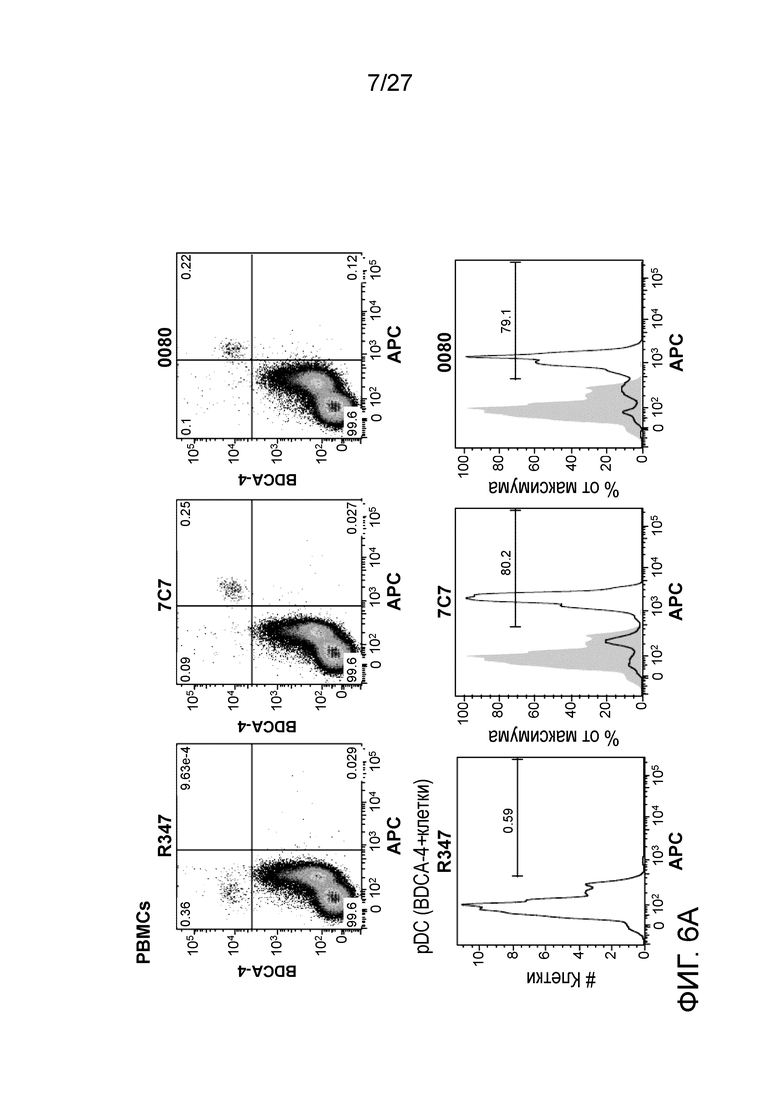




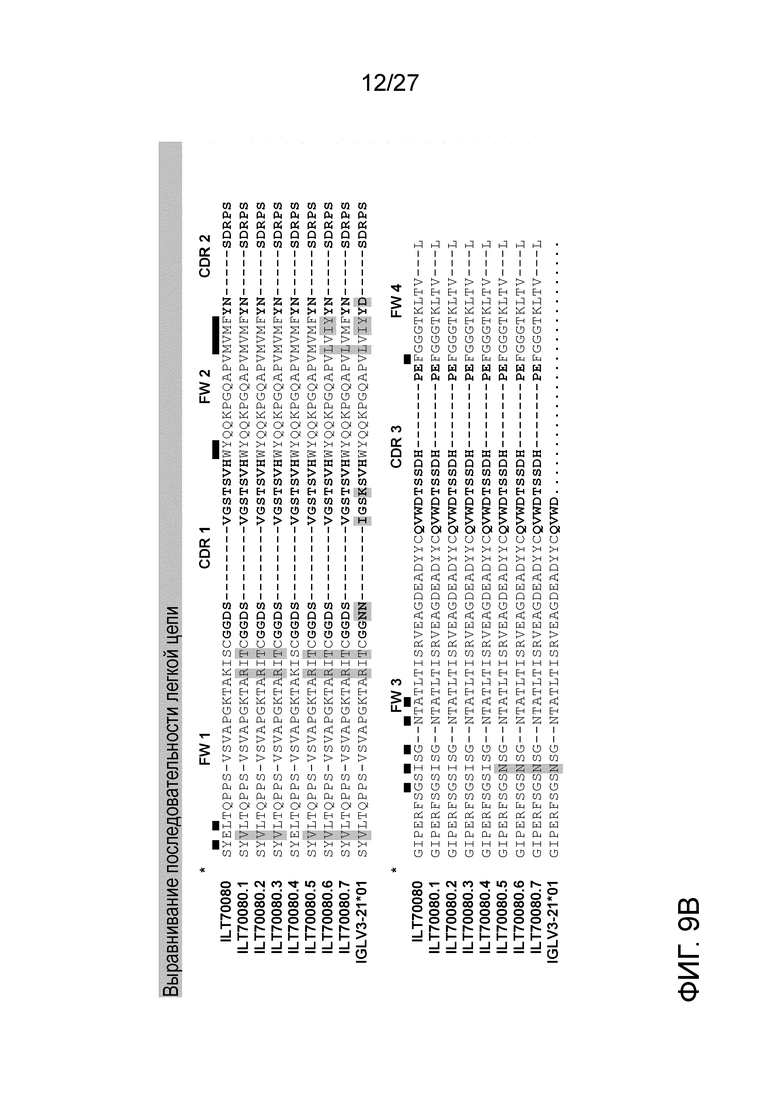


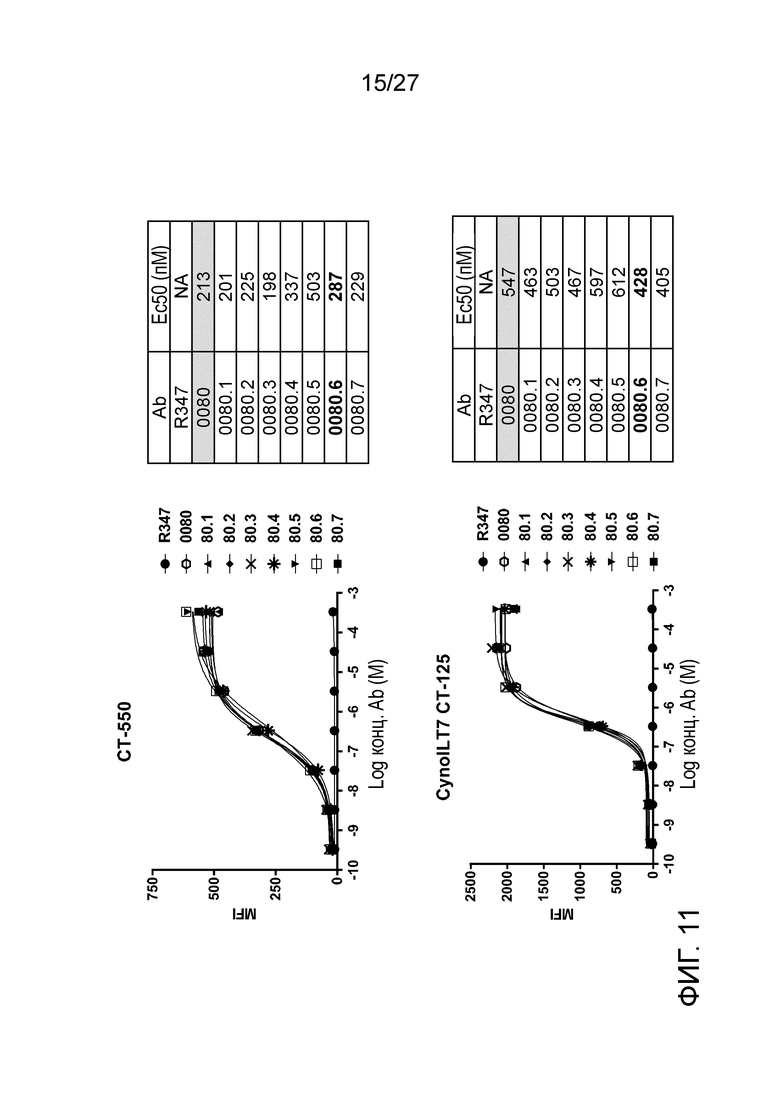

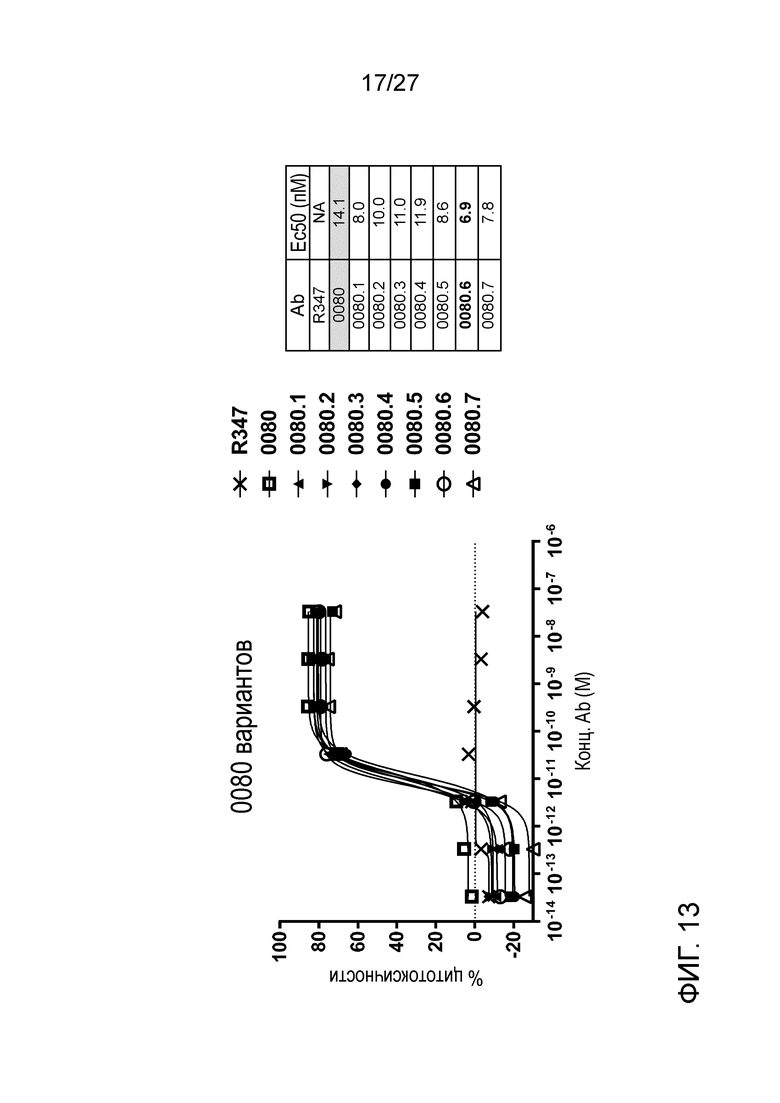
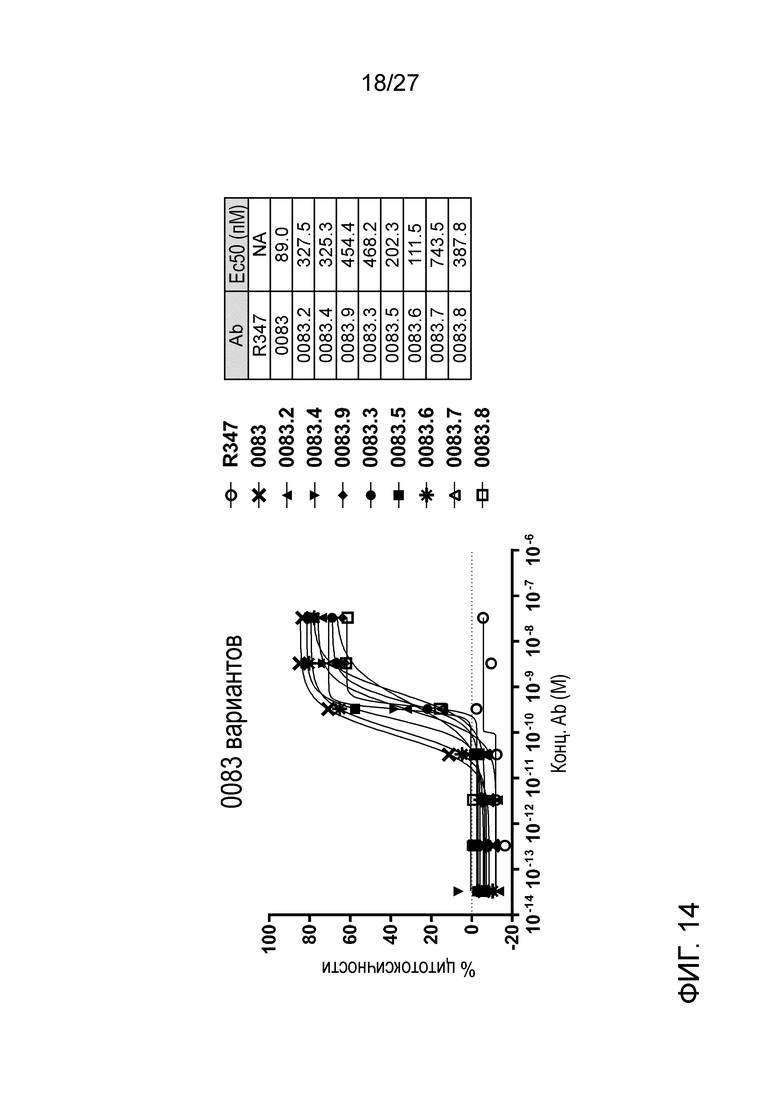
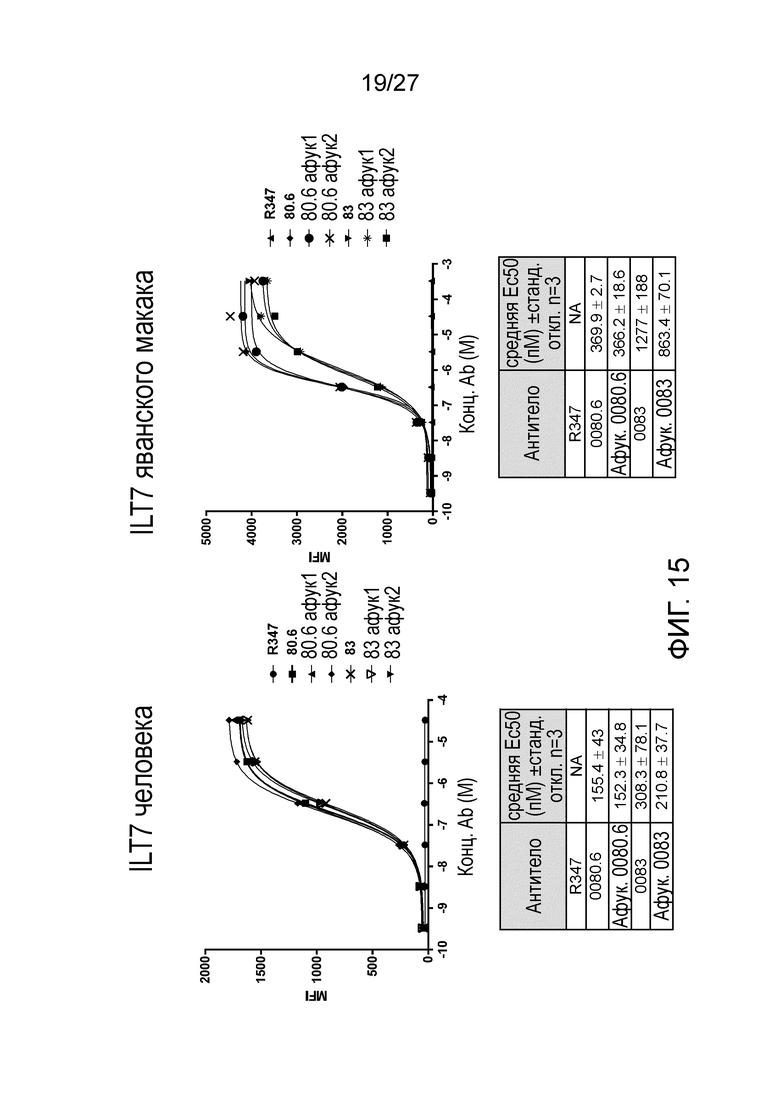

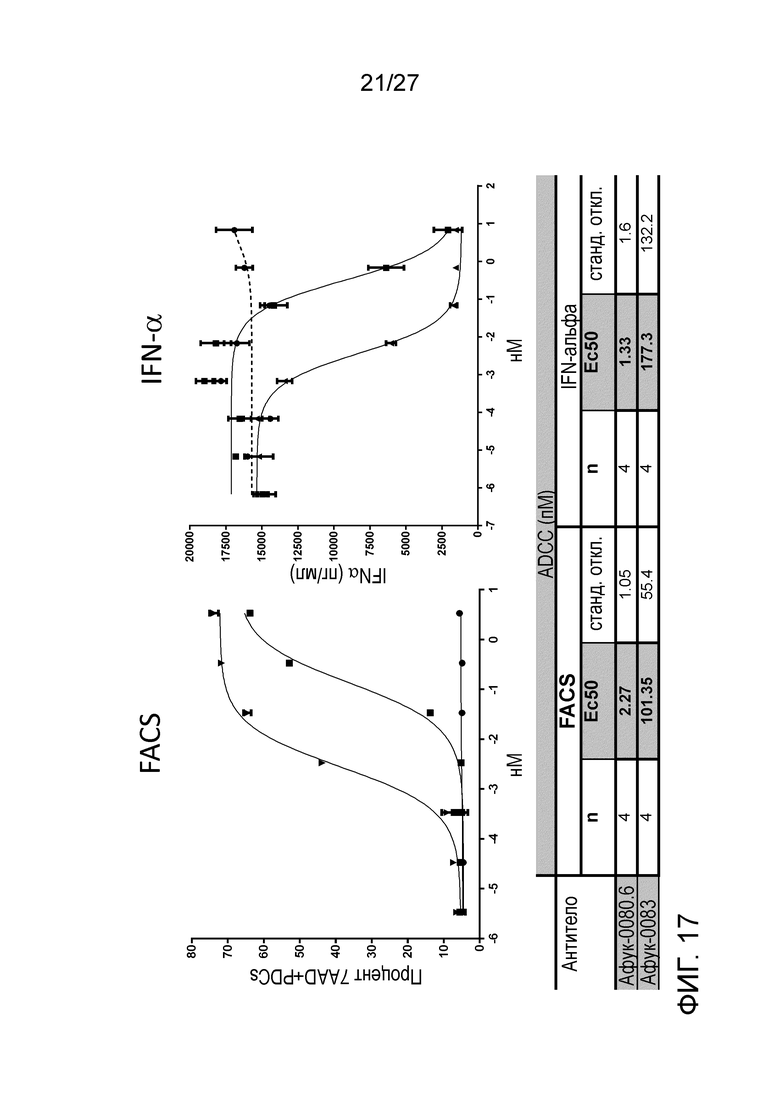
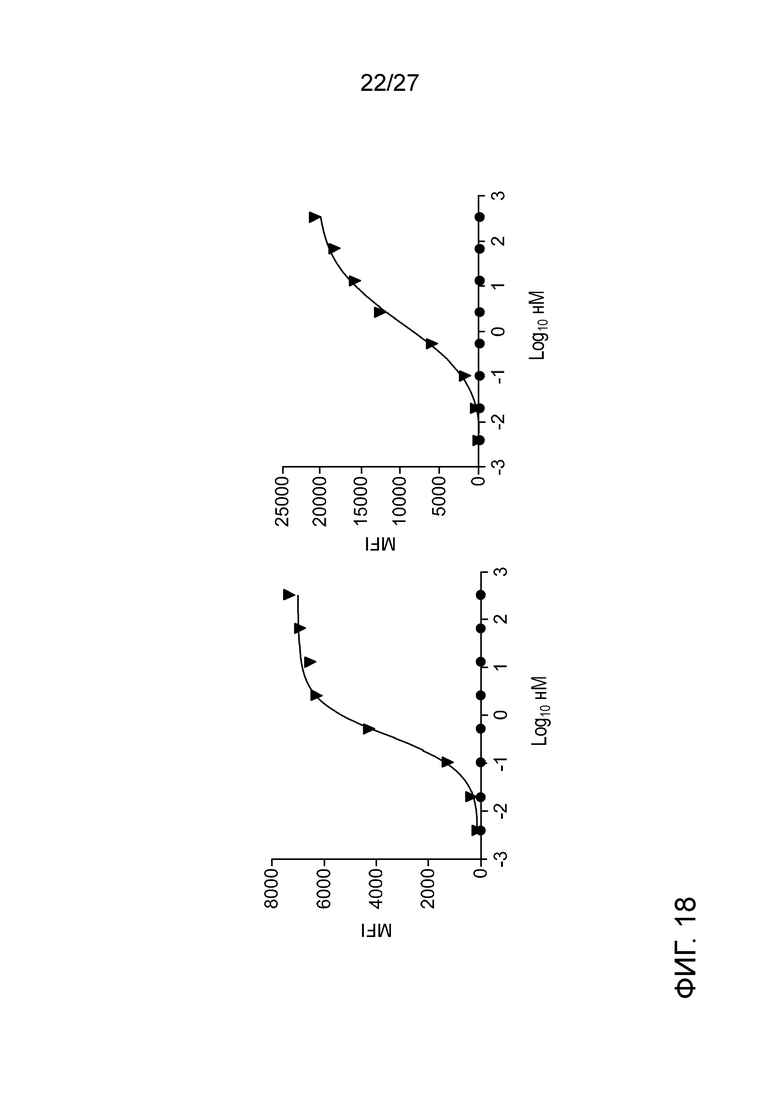
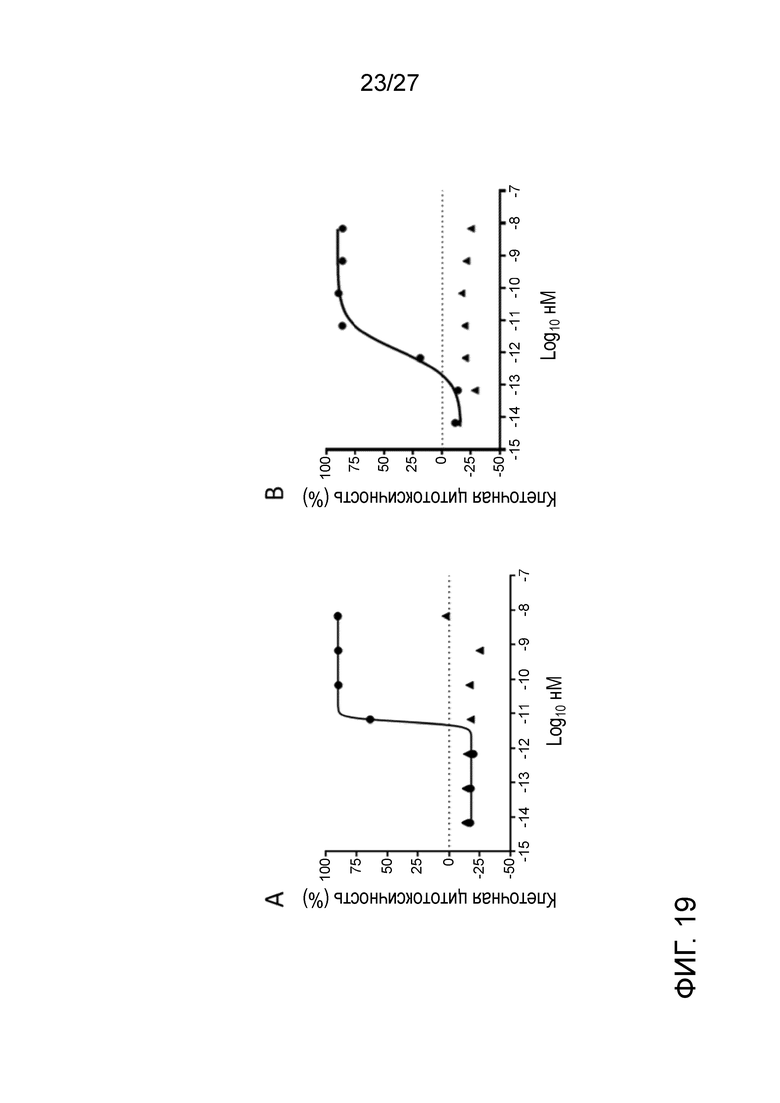


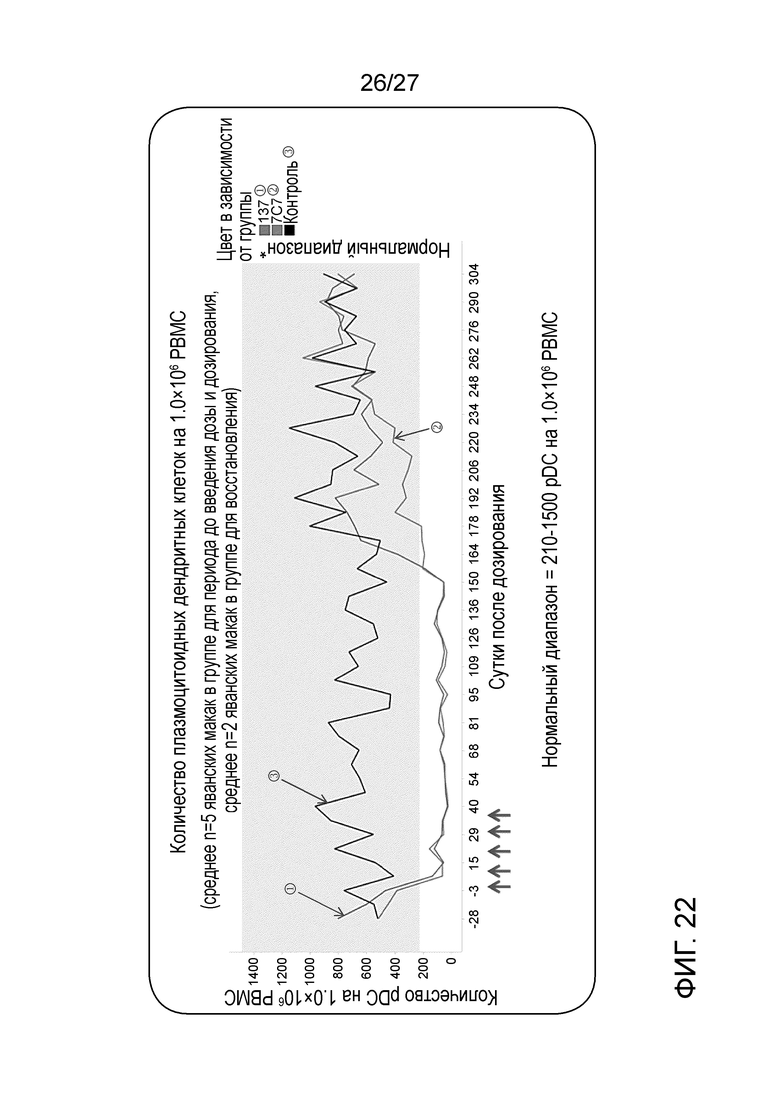

Изобретение относится к области биотехнологии, конкретно к антителу, которое специфично связывается с иммуноглобулиноподобным транскриптом 7 (ILT7), и может быть использовано в медицине в лечении и профилактике аутоиммунного заболевания. Изобретение обеспечивает уменьшение высвобождения интерферона (IFN)-альфа плазмацитоидной дендритной клеткой (pDC). 11 н. и 11 з.п. ф-лы, 23 ил., 3 табл., 12 пр.
1. Антитело, которое специфично связывается с иммуноглобулиноподобным транскриптом 7 (ILT7), содержащее определяющие комплементарность области (CDR) HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3, где указанные HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 содержат аминокислотные последовательности SEQ ID NO: 203, 204, 205, 208, 209 и 210 соответственно.
2. Антитело по п. 1, где указанное антитело содержит вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где указанные области VH и VL содержат аминокислотную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 202 и SEQ ID NO: 207 соответственно.
3. Антитело по п. 2, где указанные области VH и VL содержат аминокислотную последовательность SEQ ID NO: 202 и SEQ ID NO: 207 соответственно.
4. Антитело по п. 1, где указанное антитело содержит константный домен тяжелой цепи иммуноглобулина, выбранный из группы, состоящей из константного домена IgA, константного домена IgE, константного домена IgG1, константного домена IgG2, константного домена IgG3, константного домена IgG4 и константного домена IgM.
5. Антитело по п. 4, где указанное антитело содержит константный домен IgG1.
6. Антитело по п. 1, где указанное антитело содержит константный домен легкой цепи иммуноглобулина, выбранный из группы, состоящей из константного домена каппа-цепи Ig и константного домена лямбда-цепи Ig.
7. Антитело по п. 6, где указанное антитело содержит константный домен лямбда.
8. Антитело по п. 1, где указанное антитело представляет собой моноклональное антитело.
9. Антитело по п. 1, где указанное антитело представляет собой антитело человека, химерное антитело или антитело с измененной поверхностью.
10. Антитело по п. 9, где указанное антитело представляет собой антитело человека.
11. Антитело по п. 1, где указанное антитело является афукозилированным.
12. Антитело по п. 1, где указанное антитело специфично связывается с ILT7 человека и яванского макака.
13. Афукозилированное человеческое моноклональное антитело, которое специфично связывается с ILT7, содержащее:
i) определяющие комплементарность области (CDR) HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3;
ii) константный домен IgG1 человека; и
iii) константный домен лямбда человека,
где указанные HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 содержат аминокислотные последовательности SEQ ID NO: 203, 204, 205, 208, 209 и 210 соответственно.
14. Полинуклеотид, кодирующий аминокислотную последовательность антитела по любому из пп. 1-13.
15. Экспрессионный вектор, содержащий полинуклеотид по п. 14.
16. Клетка-хозяин для получения антитела, которое специфично связывается с ILT7, содержащая вектор по п. 15, где указанная клетка-хозяин представляет собой миеломную клетку мыши NS0, клетку человека PER.C6, клетку яичника китайского хомяка, клетку карциномы шейки матки человека HELA, клетку CVI обезьяны, COS, VERY, клетку почки детенышей хомячка, MDCK, WI38, клетку фибробласта китайского хомяка R1610, клетку фибробласта мыши BALBC/3T3, клетку почки хомяка НАК, клетку миеломы мыши SP2/0, клетку миеломы мыши P3x63-Ag3.653, эндотелиальную клетку крупного рогатого скота BFA-1c1BPT или клетку клеточной линии лимфоцита человека RAJI.
17. Способ получения антитела, которое специфично связывается с ILT7, включающий культивирование клетки-хозяина по п. 16 и выделение указанного антитела.
18. Способ детекции экспрессии белка ILT7 в образце, включающий (a) приведение указанного образца в контакт с антителом по любому из пп. 1-13 и (b) детекцию связывания указанного антитела с белком ILT7 в указанном образце.
19. Способ уменьшения высвобождения интерферона (IFN)-альфа плазмацитоидной дендритной клеткой (pDC), включающий приведение pDC в контакт с антителом по любому из пп. 1-13.
20. Применение антитела, специфичного к ILT7, в лечении субъекта-человека с аутоиммунным заболеванием,
где указанное антитело содержит определяющие комплементарность области тяжелой цепи (HCDR) HCDR1, HCDR2, HCDR3 и определяющие комплементарность области легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, где указанные HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 содержат аминокислотные последовательности SEQ ID NO: 203, 204, 205, 208, 209 и 210 соответственно.
21. Применение антитела, специфичного к ILT7, в профилактике аутоиммунного заболевания у субъекта-человека,
где указанное антитело содержит определяющие комплементарность области тяжелой цепи (HCDR) HCDR1, HCDR2, HCDR3 и определяющие комплементарность области легкой цепи (LCDR) LCDR1, LCDR2 и LCDR3, где указанные HCDR1, HCDR2, HCDR3, LCDR1, LCDR2 и LCDR3 содержат аминокислотные последовательности SEQ ID NO: 203, 204, 205, 208, 209 и 210 соответственно.
22. Афукозилированное человеческое моноклональное антитело IgG1, которое специфично связывается с ILT7, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), где указанная вариабельная область тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 202 и указанная вариабельная область легкой цепи содержит аминокислотную последовательность SEQ ID NO: 207.
| US 2009280128 А1, 12.11.2009 | |||
| CHO M | |||
| ET AL., SAGE library screening reveals ILT7 as a specific plasmacytoid dendritic cell marker that regulates type I IFN production, INTERNATIONAL IMMUNOLOGY, 2008, v.20, n.1, p.155 - 164 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ ИНТЕРФЕРОНА-АЛЬФА И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2431638C2 |
| DASHIVETS T | |||
| еt al., Oxidation in the complementarity-determining regions differentially | |||

