ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается преимущество и приоритет по предварительной заявке на патент США № 62/457780, поданной 10 февраля 2017 г., полное содержание которой включено в настоящее описание в виде ссылки для всех целей.
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит списки последовательностей, которые представлены в электронном виде в формате ASCII и настоящим полностью включены в виде ссылки. Указанная копия ASCII, созданная 8 февраля 2018 года, называется DFY–003PC_SL.txt и имеет размер 91310 байтов. Данная заявка посредством ссылки полностью включает машиночитаемую форму (CRF) списке последовательностей в текстовом формате ASCII, поданную через EFS-Web. Текстовый файл списка последовательностей, поданный через EFS-Web, названный 14247-424-999_SUB_SEQ_LISTING.txt, был создан 12 июля 2021 г. и имеет размер 98150 байт.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с антигеном созревания В–клеток (ВСМА), рецептором NKG2D и CD16.
УРОВЕНЬ ТЕХНИКИ
[0004] Рак продолжает оставаться серьезной проблемой для здоровья, несмотря на значительные исследовательские усилия и научные достижения в лечении этого заболевания, описанные в литературе. Рак крови и костного мозга являются часто диагностируемыми видами рака, включая множественные миеломы, лейкемию и лимфомы. Существующие способы лечения этих видов рака не являются эффективными для всех пациентов и/или могут иметь существенные побочные эффекты. Другие виды рака также не поддаются лечению с помощью современных методов лечения.
[0005] Иммунотерапия рака является желательной благодаря своей высокой специфичности и способствует разрушению раковых клеток при участии иммунной системы самого пациента. Слитые белки, такие как биспецифические рекрутеры T–клеток, представляют собой описанные в литературе иммунотерапевтические агенты для лечения рака, которые связываются с опухолевыми клетками и T–клетками, способствуя разрушению опухолевых клеток. В литературе описаны антитела, которые связываются с определенными антигенами, ассоциированными с опухолями, и с определенными иммунными клетками. См., например, WO 2016/134371 и WO 2015/095412.
[0006] Натуральные киллерные (NK) клетки (натуральные киллеры) являются компонентом врожденной иммунной системы и составляют примерно 15% циркулирующих лимфоцитов. NK–клетки проникают практически во все ткани и отличаются главным образом своей способностью эффективно убивать опухолевые клетки без необходимости в предварительной сенсибилизации. Активированные NK–клетки убивают клетки–мишени с помощью средств, аналогичных цитотоксическим Т–клеткам, т.е. посредством цитолитических гранул, которые содержат перфорин и гранзимы, а также через рецепторные пути гибели клеток. Активированные NK–клетки также секретируют воспалительные цитокины, такие как IFN–гамма и хемокины, которые рекрутируют другие лейкоциты в ткани–мишени.
[0007] NK–клетки реагируют на сигналы посредством множества активирующих и ингибирующих рецепторов на своей поверхности. Например, когда NK–клетки встречают здоровые аутоклетки, их активность подавляется путем активации иммуноглобулин–подобных рецепторов (KIR) киллерных клеток. Альтернативно, когда NK–клетки сталкиваются с чужеродными или раковыми клетками, они активируются посредством своих активирующих рецепторов (например, NKG2D, NCR, DNAM1). NK–клетки также активируются константной областью некоторых иммуноглобулинов через рецепторы CD16 на их поверхности. Общая чувствительность NK–клеток к активации зависит от суммы стимулирующих и ингибирующих сигналов.
[0008] ВСМА представляет собой трансмембранный белок, принадлежащий к суперсемейству рецепторов TNF. Он специфически связывается с суперсемейством факторов (лигандов) некроза опухолей, членом 13b (TSF13B/TALL–1/BAFF), что приводит к активации NF–κB и MAPK8/JNK. Его экспрессия ограничена линией B–клеток и, как было показано, важна для развития B–клеток и аутоиммунного ответа. BCMA также связывается с различными членами семейства TRAF и, таким образом, может трансдуцировать сигналы выживания и пролиферации клеток. ВСМА задействован в различных раковых заболеваниях, таких как множественная миелома, лимфома и лейкоз. Настоящее изобретение обеспечивает определенные преимущества для улучшения лечения рака экспрессирующего ВСМА.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0009] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с ВСМА на раковой клетке и рецептором NKG2D и рецептором CD16 на натуральных киллерных клетках. Такие белки могут взаимодействовать с более чем одним типом рецепторов, активирующих NK, и могут блокировать связывание природных лигандов с NKG2D. В определенных вариантах осуществления белки могут оказывать агонистическое воздействие на NK–клетки у людей и у других видов, таких как грызуны и яванские макаки. Различные аспекты и варианты осуществления изобретения более подробно описаны ниже.
[0010] Соответственно, один из аспектов изобретения относится к белку, который включает первый антигенсвязывающий участок, который связывается с NKG2D; второй антигенсвязывающий участок, который связывается с ВСМА; и FС–домен антитела, его часть, достаточную для связывания CD16, или третий антигенсвязывающий участок, который связывается с CD16. Каждый антигенсвязывающий участок может включать вариабельный домен тяжелой цепи антитела и вариабельный домен легкой цепи антитела (например, расположенный как в антителе или слитый с scFv), или один или более антигенсвязывающих участков могут представлять собой однодоменное антитело, такое как VHH антитело, такое как верблюжье антитело, или VNAR антитело, такое как антитело, обнаруженное у хрящевых рыб.
[0011] Первый антигенсвязывающий участок, который связывается с NKG2D, в одном из вариантов осуществления может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 1, такой как имеющий аминокислотную последовательность, по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичную последовательности SEQ ID NO: 1 и/или включающий аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 64), CDR2 (SEQ ID NO: 65) и CDR3 (SEQ ID NO: 66) последовательности SEQ ID NO: 1. Альтернативно, первый антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 41, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 42. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичным SEQ ID NO: 41 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 67), CDR2 (SEQ ID NO: 68) и CDR3 (SEQ ID NO: 69) последовательности SEQ ID NO: 41. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичным SEQ ID NO: 42 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 70), CDR2 (SEQ ID NO: 71) и CDR3 (SEQ ID NO: 72) последовательности SEQ ID NO: 42. В других вариантах осуществления первый антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 43, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 44. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 43 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 73), CDR2 (SEQ ID NO: 74) и CDR3 (SEQ ID NO: 75) последовательности SEQ ID NO: 43. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 44 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 76), CDR2 (SEQ ID NO: 77) и CDR3 (SEQ ID NO: 78) последовательности SEQ ID NO: 44.
[0012] Альтернативно, первый антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 45, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 46, такие как имеющие аминокислотные последовательности, по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичные SEQ ID NO: 45 и SEQ ID NO: 46, соответственно. В другом варианте осуществления первый антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 47, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 48, такие как имеющие аминокислотные последовательности по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичные SEQ ID NO: 47 и SEQ ID NO: 48, соответственно.
[0013] Второй антигенсвязывающий участок необязательно может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 49, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 53 или SEQ ID NO: 54. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичным SEQ ID NO: 49 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 50), CDR2 (SEQ ID NO: 51) и CDR3 (SEQ ID NO: 52) последовательности SEQ ID NO: 49. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 53 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 55), CDR2 (SEQ ID NO: 56) и CDR3 (SEQ ID NO: 57) последовательности SEQ ID NO: 53. Альтернативно, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичным SEQ ID NO: 54 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 55), CDR2 (SEQ ID NO: 56) и CDR3 (SEQ ID NO: 58) последовательности SEQ ID NO: 54.
[0014] Альтернативно, второй антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 59, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 60. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 59 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 79), CDR2 (SEQ ID NO: 80) и CDR3 (SEQ ID NO: 81) последовательности SEQ ID NO: 59. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 60 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 82), CDR2 (SEQ ID NO: 83) и CDR3 (SEQ ID NO: 84) последовательности SEQ ID NO: 60.
[0015] В другом варианте осуществления второй антигенсвязывающий участок может включать вариабельный домен тяжелой цепи, относящийся к SEQ ID NO: 61, и вариабельный домен легкой цепи, относящийся к SEQ ID NO: 62. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или 100% идентичным SEQ ID NO: 61 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 85), CDR2 (SEQ ID NO: 86) и CDR3 (SEQ ID NO: 87) последовательности SEQ ID NO: 61. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего участка может быть по меньшей мере на 90%, по меньшей мере на 95% или на 100% идентичным SEQ ID NO: 62 и/или включать аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 88), CDR2 (SEQ ID NO: 89) и CDR3 (SEQ ID NO: 90) последовательности SEQ ID NO: 62.
[0016] В некоторых вариантах осуществления второй антигенсвязывающий участок включает вариабельный домен легкой цепи, имеющий аминокислотную последовательность, идентичную аминокислотной последовательности вариабельного домена легкой цепи, присутствующей в первом антигенсвязывающем участке.
[0017] В некоторых вариантах осуществления белок включает часть FС–домена антитела, достаточную для связывания CD16, причем FС–домен антитела содержит шарнирный и CH2–домены и/или аминокислотные последовательности по меньшей мере на 90% идентичные аминокислотной последовательности 234–332 человеческого антитела IgG.
[0018] Также предлагаются составы, содержащие один из этих белков; клетки, содержащие одну или более нуклеиновых кислот, экспрессирующих эти белки, и способы ускорения гибели опухолевых клеток с помощью этих белков.
[0019] В другом аспекте изобретения предлагается способ лечения рака у пациента. Способ включает введение нуждающемуся в этом пациенту терапевтически эффективного количества описанного в настоящей заявке полиспецифического связывающего белка. Типичные виды рака, подходящие для лечения с помощью полиспецифических связывающих белков включают, например, множественную миелому, острый миеломоноцитарный лейкоз, Т–клеточную лимфому, острый моноцитарный лейкоз и фолликулярную лимфому.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0020] На фиг. 1 представлено гетеродимерное полиспецифическое антитело. NKG2D–связывающий домен (правое плечо); домен, связывающий опухолевый антиген (левое плечо). Легкие цепи, которые являются общими, представлены на чертеже одинаковым оттенком или узором.
[0021] На фиг. 2 представлено гетеродимерное полиспецифическое антитело. NKG2D–связывающий домен – scFv (правое плечо); домен, связывающий опухолевый антиген (левое плечо).
[0022] На фиг. 3 представлено TriNKET в форме Triomab, которое представляет собой трифункциональное биспецифическое антитело, сохранившее IgG–подобную форму. Эта химера состоит из двух полу–антител, каждое из которых содержит одну легкую и одну тяжелую цепь, полученные из двух родительских антител. Форма Triomab может представлять собой гетеродимерную конструкцию, содержащую 1/2 антитела крысы и 1/2 антитела мыши.
[0023] На фиг. Фиг.4 представлено TriNKET в форме KiH с общей легкой цепью (Common Light Chain (LC)), полученное с помощью технологии «выступ–во–впадину» (KIH). KiH представляет собой гетеродимер, содержащий 2 Fab, связывающихся с мишенями 1 и 2, и FС, стабилизированный обеспечивающими гетеродимеризацию мутациями. TriNKET в формате KiH может представлять собой гетеродимерную конструкцию с 2 Fab, связывающимися с мишенью 1 и мишенью 2, содержащую две разные тяжелые цепи и общую легкую цепь, которая спаривается с обеими тяжелыми цепями.
[0024] На фиг. 5 представлено TriNKET в форме иммуноглобулина с двумя вариабельными доменами (DVD–IgТМ), в котором мишень–связывающие домены двух моноклональных антител объединены посредством гибких встречающихся в природе линкеров, образуя четырехвалентную IgG–подобную молекулу. DVD–IgТМ представляет собой гомодимерную конструкцию, в которой вариабельный домен, нацеленный на антиген 2, слит с N–концом вариабельного домена Fab, нацеленного на антиген 1. Конструкт содержит нормальный FС.
[0025] На фиг. Фиг.6 представлено TriNKET в форме ортогонального Fab интерфейса (Ortho–Fab), который представляет собой гетеродимерную конструкцию, содержащую 2 Fab–фрагмента, связывающихся с мишенью 1 и мишенью 2, слитых с FС. Спаривание LC–HC обеспечивается ортогональным интерфейсом. Гетеродимеризация обеспечивается мутациями в FС.
[0026] На фиг. 7 представлено TrinKET в формате 2–в–1 Ig.
[0027] На фиг. 8 представлено TriNKET в форме ES, представляющую собой гетеродимерную конструкцию, содержащую два разных Fab–фрагмента, связывающихся с мишенью 1 и мишенью 2, слитых с FС. Гетеродимеризация обеспечивается мутациями, усиливающими электростатические взаимодействия в FС.
[0028] На фиг. 9 представлено TriNKET в форме обмена Fab–плечами: антитела, в которых произошел обмен Fab–плечами путем замены тяжелой цепи и присоединения легкой цепи (полу–молекулы) на пару тяжелая–легкая цепи из другой молекулы, что привело к образованию биспецифических антител. Форма обмена Fab–плечами (Fab Arm Exchange form) (cFae) представляет собой гетеродимер, содержащий 2 Fab, связывающихся с мишенью 1 и 2, и FС, стабилизированный обеспечивающими гетеродимеризацию мутациями.
[0029] На фиг. 10 представлено TriNKET в форме тела SEED, которое представляет собой гетеродимер, содержащий 2 Fab, связывающихся с мишенью 1 и 2, и FС, стабилизированный обеспечивающими гетеродимеризацию мутациями.
[0030] На фиг. 11 представлено TriNKET в форме LuZ–Y, в котором для инициации гетеродимеризации двух различных HC используется лейциновая молния. Форма LuZ–Y представляет собой гетеродимер, содержащий два разных слитых с FС scFab, связывающихся с мишенями 1 и 2. Гетеродимеризация обеспечивается мотивами лейциновой молнии, слитыми с С–концом FС.
[0031] На фиг. 12 представлено TriNKET в форме Cov–X–тела.
[0032] На фиг. 13A–13B представлены TriNKET в формах ĸλ–тела, которые представляют собой гетеродимерные конструкции с двумя разными Fab, слитыми с FС, стабилизированным обеспечивающими гетеродимеризацию мутациями: Fab1, нацеленный на антиген 1, содержит каппа–LC, тогда как второй Fab, нацеленный на антиген 2, содержит лямбда–LC. На фиг. 13А показан примерный внешний вид одной из форм ĸλ–тела; на фиг. 13B показан примерный внешний вид другого ĸλ–тела.
[0033] На фиг. 14 показаны линейные графики, демонстрирующие аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с человеческим рекомбинантным NKG2D в анализе ИФА.
[0034] На фиг. 15 показаны линейные графики, демонстрирующие аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с рекомбинантным NKG2D яванского макака в анализе ИФА.
[0035] На фиг. 16 показаны линейные графики, демонстрирующие аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с рекомбинантным NKG2D мыши в анализе ИФА.
[0036] На фиг. 17 показана гистограмма, демонстрирующая связывание NKG2D–связывающих доменов (перечисленных в виде клонов) с клетками EL4, экспрессирующими NKG2D человека, полученная методом проточной цитометрии, которая отражает увеличение средней интенсивности флуоресценции (СИФ) относительно фона.
[0037] На фиг. 18 представлена гистограмма, демонстрирующая связывание NKG2D–связывающих доменов (перечисленных в виде клонов) с клетками EL4, экспрессирующими мышиный NKG2D, полученная методом проточной цитометрии, которая отражает увеличение средней интенсивности флуоресценции (СИФ) относительно фона.
[0038] На фиг. 19 представлены линейные графики, демонстрирующие специфическую аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с рекомбинантным человеческим NKG2D–FС в условиях конкуренции с природным лигандом ULBP–6.
[0039] На фиг. 20 представлены линейные графики, демонстрирующие специфическую аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с рекомбинантным человеческим NKG2D–FС в условиях конкуренции с природным лигандом MICA.
[0040] На фиг. 21 представлены линейные графики, демонстрирующие специфическую аффинность связывания NKG2D–связывающих доменов (перечисленных в виде клонов) с рекомбинантным мышиным NKG2D–FС в условиях конкуренции с природным лигандом дельта Rae–1.
[0041] На фиг. 22 представлена гистограмма, демонстрирующая активацию человеческого NKG2D с помощью NKG2D–связывающих доменов (перечисленных в виде клонов), полученная путем количественного определения процентного содержания T–альфа–положительных клеток, экспрессирующих человеческие NKG2D–CD3 дзета слитые белки.
[0042] На фиг. 23 представлена гистограмма, демонстрирующая активацию мышиного NKG2D с помощью NKG2D–связывающих доменов (перечисленных в виде клонов), полученная путем количественного определения процента T–альфа–положительных клеток, экспрессирующих мышиные NKG2D–CD3 дзета слитые белки.
[0043] На фиг. 24 представлена гистограмма, демонстрирующая активацию человеческих NK–клеток с помощью NKG2D–связывающх доменов (перечисленных в виде клонов).
[0044] На фиг. 25 представлена гистограмма, демонстрирующая активацию человеческих NK–клеток с помощью NKG2D–связывающих доменов (перечисленных в виде клонов).
[0045] На фиг. 26 представлена гистограмма, демонстрирующая активацию мышиных NK–клеток с помощью NKG2D–связывающих доменов (перечисленных в виде клонов).
[0046] На фиг. 27 представлена гистограмма, демонстрирующая активацию мышиных NK–клеток с помощью NKG2D–связывающих доменов (перечисленных в виде клонов).
[0047] На фиг. 28 представлена гистограмма, демонстрирующая цитотоксическое действие NKG2D–связывающих доменов (перечисленных в виде клонов) на опухолевые клетки.
[0048] На фиг. 29 представлена гистограмма, демонстрирующая температуру плавления NKG2D–связывающих доменов (перечисленных в виде клонов), измеренную методом дифференциальной сканирующей флуориметрии.
[0049] На фиг. 30 представлены линейные графики, показывающие профиль связывания нацеленных на BCMA TriNKET с NKG2D, экспрессируемыми клетками EL4.
[0050] На фиг. 31 представлены линейные графики, показывающие профиль связывания нацеленных на BCMA TriNKET с BCMA, экспрессируемыми клетками миеломы человека MM.1S.
[0051] На фиг. 32 представлена гистограмма, демонстрирующая активацию человеческих NK в культуре с BCMA–положительными клетками миеломы человека ММ.1S.
[0052] На фиг. 33 представлены линейные графики, показывающие, что нацеленные на BCMA TriNKET с разными NKG2D–связывающими доменами усиливают лизис человеческих NK–клеток в клетках миеломы KMS12–PE.
[0053] На фиг. 34A–34C представлены гистограммы синергетической активации NK–клеток с помощью CD16 и NKG2D. На фиг. 34А показаны уровни CD107a; на фиг. 34B показаны уровни IFNγ; на фиг. 34C показаны уровни CD107a и IFNγ. На графиках показано среднее значение (n=2) ± SD. Данные получены в пяти независимых экспериментах с участием пяти разных здоровых доноров.
[0054] На фиг. 35 представлена гетеродимерная конструкция Oasc–Fab, которая включает Fab, связывающийся с мишенью 1, и scFab, связывающийся с мишенью 2, слитые с FС. Гетеродимеризация обеспечивается мутациями в FС.
[0055] На фиг. 36 представлено DuetMab, которое представляет собой гетеродимерную конструкцию, содержащую два разных Fab, связывающихся с антигенами 1 и 2, и FС, стабилизированный обеспечивающими гетеродимеризацию мутациями. Fab 1 и 2 содержат разные S–S мосты, обеспечивающие правильное спаривание легкой цепи (LC) с тяжелой цепью (HC).
[0056] На фиг. 37 представлено CrossmAb, которое представляет собой гетеродимерную конструкцию с двумя разными Fab, связывающимися с мишенями 1 и 2, которые слиты с FС, стабилизированным гетеродимеризацией. Домены CL и CH1 и домены VH и VL подвергнуты переключению, например, CH1 является линейно слитым с VL, в то время как CL является линейно слитым с VH.
[0057] На фиг. 38 представлено Fit–Ig, которое представляет собой гомодимерную конструкцию, в которой Fab, связывающийся с антигеном 2, слит с N–концом HC Fab, который связывается с антигеном 1. Конструкция содержит FС дикого типа.
[0058] На фиг. 39 представлен график, показывающий, что TriNKET усиливают лизис клеток миеломы KMS12–PE, обеспечиваемый человеческими NK–клетками.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0059] Согласно изобретению предоставляются полиспецифические связывающие белки, которые связывают ВСМА на раковой клетке и рецептор NKG2D и рецептор CD16 на натуральных киллерных клетках, для активации натуральных киллерных клеток, фармацевтические композиции, содержащие такие полиспецифические связывающие белки, и терапевтические способы, в которых используются такие полиспецифические белки и фармацевтические композиции, в том числе для лечения рака. Различные аспекты изобретения изложены в приведенных ниже разделах; однако аспекты изобретения, описанные в одном из конкретных разделов, никоим образом не ограничены никаким другим конкретным разделом.
[0060] Для гарантии понимания настоящего изобретения ниже приведено определение ряда терминов и фраз.
[0061] Используемое в настоящем описании единственное число означает «один или более» и включают множественное число, если из контекста в явном виде не следует иное.
[0062] Используемый в настоящем описании термин «антигенсвязывающий участок» относится к части молекулы иммуноглобулина, которая участвует в связывании антигена. В человеческих антителах антигенсвязывающий участок образован аминокислотными остатками N–концевых вариабельных ("V") областей тяжелой ("H") и легкой ("L") цепей. Три сильно различающиеся участка в V–областях тяжелой и легкой цепей называются «гипервариабельными областями», расположенными между более консервативными фланкирующими участками, известными как «каркасные области» или «FR». Таким образом, термин «FR» относится к аминокислотным последовательностям, которые в природе расположены в иммуноглобулинах между и рядом с гипервариабельными областями. В молекуле человеческого антитела три гипервариабельные области легкой цепи и три гипервариабельные области тяжелой цепи расположены относительно друг друга в трехмерном пространстве, формируя антигенсвязывающую поверхность. Антигенсвязывающая поверхность является комплементарной трехмерной поверхности связанного антигена, и три гипервариабельные области каждой из тяжелых и легких цепей называются «определяющими комплементарность областями» или «CDR». У некоторых животных, таких как верблюды и хрящевые рыбы, антигенсвязывающий участок образован одной цепью антитела, обеспечивающей «однодоменное антитело». Антигенсвязывающие участки могут существовать в интактном антителе, в антигенсвязывающем фрагменте антитела, в котором сохранена антигенсвязывающая поверхность, или в рекомбинантном полипептиде, таком как scFv, в котором для соединения в одном полипептиде вариабельного домена тяжелой цепи с вариабельным доменом легкой цепи используется пептидный линкер.
[0063] Используемый в настоящем описании термин «ассоциированный с опухолью антиген» означает любой антиген, включая, без ограничения, белок, гликопротеин, ганглиозид, углевод, липид, ассоциированный с раком. Такой антиген может экспрессироваться в злокачественных клетках или в микроокружении опухоли, например в кровеносных сосудах, ассоциированных с опухолью, внеклеточном матриксе, мезенхимальной строме или иммунных инфильтратах.
[0064] Используемые в настоящем описании термины «субъект» и «пациент» относятся к организму, подверженному лечению способами и композициями, описанными в настоящем описании. Такие организмы предпочтительно включают, без ограничения, млекопитающих (например, мышей, обезьян, лошадей, крупного рогатого скота, свиней, собак, кошек и т.п.) и более предпочтительно включают людей.
[0065] Используемый в настоящем описании термин «эффективное количество» относится к количеству соединения (например, соединения по настоящему изобретению), достаточному для достижения полезных или требуемых результатов. Эффективное количество может быть введено в виде одного или более введений, применений или дозировок и не ограничено конкретным препаратом или путем введения. Используемый в настоящем описании термин «лечение» включает любой эффект, например снижение, уменьшение, модуляцию, ослабление или устранение, который приводит к улучшению состояния, ослаблению заболевания, расстройства и т.п., или купированию его симптома.
[0066] Используемый в настоящем описании термин «фармацевтическая композиция» относится к комбинации активного агента с носителем, инертным или активным, делающим композицию особенно подходящей для диагностического или терапевтического применения in vivo или ex vivo.
[0067] Используемый в настоящем описании термин «фармацевтически приемлемый носитель» относится к любому стандартному фармацевтическому носителю, такому как забуференный фосфатом физиологический раствор, вода, эмульсии (например, такие как эмульсия масло/вода или вода/масло), и различные типы смачивающих агентов. Композиции также могут включать стабилизаторы и консерванты. Примеры носителей, стабилизаторов и адъювантов см., например, Martin, Remington's Pharmaceutical Sciences, 15th Ed., Mack Publ. Co., Easton, PA [1975].
[0068] Используемый в настоящем описании термин «фармацевтически приемлемая соль» относится к любой фармацевтически приемлемой соли (например, кислоте или основанию) соединения по настоящему изобретению, которая при введении субъекту способна обеспечить образование соединения по настоящему изобретению или его активный метаболит или остаток. Как известно специалистам в данной области техники, «соли» соединений по настоящему изобретению могут быть получены из неорганических или органических кислот и оснований. Примеры кислот включают, без ограничения, соляную, бромистоводородную, серную, азотную, хлорную, фумаровую, малеиновую, фосфорную, гликолевую, молочную, салициловую, янтарную, толуол–п–сульфоновую, винную, уксусную, лимонную, метансульфоновую, этансульфоновую, муравьиную, бензойную, малоновую, нафталин–2–сульфоновую, бензолсульфоновую кислоту и т.п. Другие кислоты, такие как щавелевая кислота, которые сами по себе хотя и не являются фармацевтически приемлемыми, могут быть использованы для получения солей, полезных в качестве промежуточных соединений при получении соединений по изобретению и их фармацевтически приемлемых аддуктов кислот.
[0069] Типичные основания включают, без ограничения, гидроксиды щелочных металлов (например, натрия), гидроксиды щелочноземельных металлов (например, магния), аммиак и соединения формулы NW4+, где W представляет собой C1–4 алкил и т.п.
[0070] Примеры солей включают, без ограничения: ацетат, адипат, альгинат, аспартат, бензоат, бензолсульфонат, бисульфат, бутират, цитрат, камфорат, камфорсульфонат, циклопентан–пропионат, диглюконат, додецилсульфат, этансульфонат, фумарат, адипафлукогептаноат, глицерофосфат, гемисульфат, гептаноат, гексаноат, гидрохлорид, гидробромид, гидройодид, 2–гидроксиэтансульфонат, лактат, малеат, метансульфонат, 2–нафталинсульфонат, никотинат, оксалат, пальмоат, пектинат, персульфат, фенилпропионат, пикрат, пивалат, пропионат, сукцинат, тартрат, тиоционат, тозилат, ундеканоат и т.п. Другие примеры солей включают анионы соединений по настоящему изобретению, соединенные с подходящим катионом, таким как Na+, NH4+ и NW4+ (где W представляет собой C1–4алкильную группу), и т.п.
[0071] Для терапевтического применения рассматриваются соли соединений по настоящему изобретению, которые являются фармацевтически приемлемыми. Однако соли кислот и оснований, которые не являются фармацевтически приемлемыми, также могут найти применение, например, при получении или очистке фармацевтически приемлемого соединения.
[0072] В описании в местах, относящихся к композициям, раскрытым как имеющие, включающие или содержащие конкретные компоненты, или относящихся к процессам и способам, раскрытым как имеющие, включающие или содержащие конкретные этапы, предполагается, что также существуют композиции по настоящему изобретению, которые по существу состоят или состоят из перечисленных компонентов, и процессы и способы по настоящему изобретению, которые по существу состоят или состоят из перечисленных этапов обработки.
[0073] Как правило, процент, указанный для композиции, является весовым, если не указано иное. Кроме того, если переменная не сопровождается определением, то применимо предыдущее определение этой переменной.
I. БЕЛКИ
[0074] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с ВСМА на раковой клетке и рецептором NKG2D и рецептором CD16 на натуральных киллерных клетках, активируя таким образом натуральные киллерные клетки. Полиспецифические связывающие белки используются в описанных в настоящей заявке фармацевтических композициях и терапевтических способах. Связывание полиспецифического связывающего белка с рецептором NKG2D и рецептором CD16 на натуральной киллерной клетке усиливает активность натуральной киллерной клетки, которая направлена на разрушение раковой клетки. Связывание полиспецифического связывающего белка с ВСМА на раковой клетке способствует сближению раковой клетки с натуральной киллерной клеткой, что облегчает прямое и опосредованное разрушение раковой клетки натуральной киллерной клеткой. Дальнейшее описание примеров полиспецифических связывающих белков приведено ниже.
[0075] Первый компонент полиспецифических связывающих белков связывается с экспрессирующими рецептор NKG2D клетками, включающими, без ограничения, NK–клетки, γδ T–клетки и CD8+ αβ T–клетки. При связывании NKG2D полиспецифические связывающие белки могут блокировать связывание NKG2D с природными лигандами, такими как ULBP6 и MICA.
[0076] Второй компонент полиспецифических связывающих белков связывается с экспрессирующими BCMA клетками, которые могут включать, без ограничения, клетки множественной миеломы, острого миеломоноцитарного лейкоза, Т–клеточной лимфомы, острого моноцитарного лейкоза и фолликулярной лимфомы.
[0077] Третий компонент полиспецифических связывающих белков связывается с клетками, экспрессирующими CD16, FС–рецептор на поверхности лейкоцитов, включающими натуральные киллерные клетки, макрофаги, нейтрофилы, эозинофилы, тучные клетки и фолликулярные дендритные клетки.
[0078] Полиспецифические связывающие белки могут принимать несколько форматов, как показано, без ограничения, в приведенных ниже примерах. Один из форматов представляет собой гетеродимерное полиспецифическое антитело, которое включает первую тяжелую цепь иммуноглобулина, вторую тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина. Первая тяжелая цепь иммуноглобулина включает первый FС–домен (шарнир–CH2–CH3), первый вариабельный домен тяжелой цепи и необязательно первый домен CH1 тяжелой цепи. Легкая цепь иммуноглобулина включает вариабельный домен легкой цепи и константный домен легкой цепи; вместе с первой тяжелой цепью иммуноглобулина легкая цепь иммуноглобулина образует антигенсвязывающий участок, который связывает NKG2D. Вторая тяжелая цепь иммуноглобулина содержит второй FС–домен (ширнир–CH2–CH3), второй вариабельный домен тяжелой цепи и второй домен CH1 тяжелой цепи, которые могут спариваться с легкой цепью иммуноглобулина, идентичной той, которая спаривается с первой тяжелой цепью иммуноглобулина, за исключением того, что когда легкая цепь иммуноглобулина спарена со второй тяжелой цепью иммуноглобулина, полученный антигенсвязывающий участок связывается с ВСМА. Первый FС–домен и второй FС–домен вместе могут связываться с CD16 (фиг.1).
[0079] Другой приведенный в качестве примера формат включает гетеродимерное полиспецифическое антитело, которое включает первую тяжелую цепь иммуноглобулина, легкую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина. Первая тяжелая цепь иммуноглобулина включает первый FС–домен (шарнир–CH2–CH3), слитый либо через линкер, либо шарнирную область антитела с одноцепочечным Fv (scFv), который связывается с NKG2D. Для связывания scFv с первым FС–доменом или с самим scFv могут быть использованы различные линкеры. Кроме того, для стабилизации общей структуры scFv последний может включать мутации, способствующие образованию дисульфидной связи. ScFv также может включать мутации для модификации изоэлектрической точки всей первой тяжелой цепи иммуноглобулина и/или для облегчения последующей очистки. Вторая тяжелая цепь иммуноглобулина включает второй FС–домен (шарнир–CH2–CH3) и второй вариабельный домен тяжелой цепи, и необязательно второй домен CH1 тяжелой цепи. Легкая цепь иммуноглобулина включает вариабельный домен легкой цепи и константной домен легкой цепи. Вторая тяжелая цепь иммуноглобулина спаривается с легкой цепью иммуноглобулина и связывается с ВСМА. Первый FС–домен и второй FС–домен могут вместе связываться с CD16 (фиг.2).
[0080] Один или более дополнительных мотивов связывания могут быть слиты с С–концом домена СН3 константной области, необязательно, через линкерную последовательность. В некоторых вариантах осуществления антигенсвязывающий участок может представлять собой одноцепочечную или стабилизированную дисульфидной связью вариабельную область (scFv) или может образовывать четырехвалентную или трехвалентную молекулу.
[0081] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Triomab, которое представляет собой трифункциональное биспецифическое антитело, сохранившее IgG–подобную форму. Эта химера состоит из двух полу–антител, каждое из которых содержит одну легкую и одну тяжелую цепи, происходящие от двух родительских антител.
[0082] В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой форму KiH с общей легкой цепью (Common Light Chain) (LC), полученную с помощью технологии «выступ–во–впадину» (KIH). Технология KIH включает конструирование доменов CH3 для создания «выступа» или «впадины» в каждой тяжелой цепи, обеспечивающих возможность гетеродимеризации. Концепция получения FС с помощью технологии «выступ–во–впадину (KiH)» заключается в образовании «выступа» в одном домене CH3 (CH3A) путем замены небольшого остатка на объемный остаток (т.е. T366WCH3A согласно нумерации ЕС). Для получения «выступа» на другом домене CH3 (CH3B) создают комплементарную поверхность со «впадиной» путем замены соседних остатков, ближайших к выступу, меньшими по размеру (т.е. T366S/L368A/Y407VCH3B). Мутация «впадина» была оптимизирована посредством структурно–управляемого скрининга фаговых библиотек (Atwell S, Ridgway JB, Wells JA, Carter P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. J. Mol. Biol. (1997) 270(1):26–35). Рентгеновские кристаллические структуры KiH вариантов FС (Elliott JM, Ultsch M, Lee J, Tong R, Takeda K, Spiess C., et al., Antiparallel coormation of knob and hole aglycosylated half–antobody homodimers is mediated by a CH2–CH3 hydrophobic interaction, J. Mol. Biol. (2014) 426(9):1947–57; Mimoto F, Kadono S, Katada H, Igawa T, Kamikawa T, Hattori K. Crystal structure of a novel assymetrically engineered Fc variant with improved affinity for FcgammaRs. Mol Immunol (2014) 58(1):132–8) показали, что гетеродимеризация является выгодной с точки зрения термодинамики за счет гидрофобных взаимодействий, возникающих благодаря пространственному соответствию (комплементарности) в интерфейсе между CH3 доменами в ядре, тогда как интерфейсы выступ–выступ и впадина–впадина не способствуют гомодимеризации из–за пространственных затруднений и, соответственно, нарушения благоприятных взаимодействий.
[0083] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме иммуноглобулина с двумя вариабельными доменами (DVD–IgTM), в котором мишень–связывающие домены двух моноклональных антител объединены посредством гибких встречающихся в природе линкеров, образуя четырехвалентную IgG–подобную молекулу.
[0084] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме ортогонального Fab интерфейса (ortho–Fab). В подходе ortho–Fab IgG (Lewis SM, Wu X, Pustilnik A, Sereno A, Huang F, Rick HL, et al. Generation of bispecific IgG antibodies by structure–based design of an orthogonal Fab interface. Nat. Biotechnol. (2014) 32(2):191–8), при конструировании на основе локальной структуры в интерфейс между LC и HCVH–CH1 вводят комплементарные мутации только в один Fab без каких–либо изменений другого Fab.
[0085] В некоторых вариантах осуществления полиспецифический связывающий белок находится в формате Ig 2–в–1. В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме ES, которая представляет собой гетеродимерную конструкцию, содержащую два разных слитых с FС Fab–фрагмента, связывающихся с мишенью 1 и мишенью 2. Гетеродимеризация обеспечивается мутацией, усиливающей электростатические взаимодействия в FС. В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме ĸλ–тела, которое представляет собой гетеродимерные конструкции с двумя разными Fab, слитыми с FС, стабилизированными обеспечивающими гетеродимеризацию мутациями: Fab1, нацеленный на антиген 1, содержит каппа–LC, тогда как второй Fab, нацеленный на антиген 2, содержит лямбда LC. На фиг. 13А показан пример одной формы ĸλ–тела; на фиг. 13B показан пример другого ĸλ–тела.
[0086] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме обмена Fab–плечами (антитела, в которых произошел обмен Fab–плечами путем замены тяжелой цепи и присоединенной легкой цепи (полу–молекулы) на пару тяжелая–легкая цепи из другой молекулы, что приводит к образованию биспецифических антител). В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме тела SEED. Платформа, позволяющая создавать домен путем обмена цепей, (SEED) была разработана для генерации асимметричных и биспецифических антителоподобных молекул, что расширяет возможности применения природных антител в терапевтических целях. Эта платформа, позволяющая конструировать белки, основана на обмене структурно родственных последовательностей иммуноглобулина в консервативных доменах СН3. Конструкция SEED позволяет эффективно генерировать гетеродимеры AG/GA, в то же время препятствуя гомодимеризации AG и GA в доменах CH3 в SEED. (Muda M. et al., Protein Eng. Des. Sel. (2011, 24 (5):447–54)). В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме LuZ–Y, в которой лейциновая молния используется для инициации гетеродимеризации двух разных HC. (Wranik, BJ. Et al., J. Biol. Chem. (2012), 287:43331–9).
[0087] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Cov–X–тела. В биспецифических CovX–телах два разных пептида соединены вместе посредством разветвленного азетидинового линкера и слиты с каркасным антителом в мягких условиях сайт–специфическим образом. В то время как фармакофоры отвечают за функциональную активность, каркас антитела обеспечивает длительный период полураспада и Ig–подобное распределение. Фармакофоры могут быть химически оптимизированы или заменены другими фармакофорами для получения оптимизированных или уникальных биспецифических антител. (Doppalapudi VR et al., PNAS (2010), 107(52); 22611–22616).
[0088] В некоторых вариантах осуществления полиспецифический связывающий белок находится в гетеродимерной форме Oasc–Fab, которая включает Fab, связывающийся с мишенью 1, и scFab, связывающийся с мишенью 2, оба слитые с FС. Гетеродимеризация обеспечивается мутациями в FС.
[0089] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме DuetMab, которая представляет собой гетеродимерную конструкцию, содержащую два разных Fab, связывающихся с антигенами 1 и 2, и FС, стабилизированный мутациями, обеспечивающими гетеродимеризацию. Fab 1 и 2 содержат разные мосты S–S, которые обеспечивают правильное спаривание LC и HC.
[0090] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме CrossmAb, которая представляет собой гетеродимерную конструкцию с двумя разными Fab, связывающимися с мишенями 1 и 2, которые слиты с FС, стабилизированным гетеродимеризацией. Домены CL и CH1 и домены VH и VL подвергнуты переключению, например, CH1 является линейно слитым с VL, в то время как CL является линейно слитым с VH.
[0091] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Fit–Ig, которая представляет собой гомодимерные конструкции, в которых Fab, связывающийся с антигеном 2, слит с N–концом HC Fab, который связывается с антигеном 1. Конструкция содержит FС дикого типа.
[0092] В таблице 1 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с NKG2D.
(SEQ ID NO:1)
CDR1 (SEQ ID NO:64) – GSFSGYYWS
CDR2 (SEQ ID NO:65) –
EIDHSGSTNYNPSLKS
CDR3 (SEQ ID NO:66) –
ARARGPWSFDP
(SEQ ID NO:2)
(SEQ ID NO:3)
(SEQ ID NO:4)
(A40)
(SEQ ID NO:5)
(SEQ ID NO:6)
(SEQ ID NO:7)
(SEQ ID NO:8)
(SEQ ID NO:9)
(SEQ ID NO:10)
(SEQ ID NO:11)
(SEQ ID NO:12)
(C26)
(SEQ ID NO:13)
(SEQ ID NO:14)
(SEQ ID NO:15)
(SEQ ID NO:16)
(SEQ ID NO:17)
(SEQ ID NO:18)
(SEQ ID NO:19)
(SEQ ID NO:20)
(SEQ ID NO:21)
(SEQ ID NO:22)
(SEQ ID NO:23)
(SEQ ID NO:24)
(SEQ ID NO:25)
(SEQ ID NO:26)
(SEQ ID NO:27)
(SEQ ID NO:28)
(SEQ ID NO:29)
(SEQ ID NO:30)
(SEQ ID NO:31)
(SEQ ID NO:32)
(SEQ ID NO:33)
(SEQ ID NO:34)
(SEQ ID NO:35)
(SEQ ID NO:36)
(SEQ ID NO:37)
(SEQ ID NO:38)
(F47)
(SEQ ID NO:39)
(SEQ ID NO:40)
(SEQ ID NO:41)
CDR1 (SEQ ID NO:67) –
GTFSSYAIS
CDR2 (SEQ ID NO:68) –
GIIPIFGTANYAQKFQG
CDR3 (SEQ ID NO:69) –
ARGDSSIRHAYYYYGMDV
(SEQ ID NO:42)
CDR1 (SEQ ID NO:70) –
KSSQSVLYSSNNKNYLA
CDR2 (SEQ ID NO:71) –
WASTRES
CDR3 (SEQ ID NO:72) –
QQYYSTPIT
(F43)
(SEQ ID NO:43)
CDR1 (SEQ ID NO:73) –
GSISSSSYYWG
CDR2 (SEQ ID NO:74) –
SIYYSGSTYYNPSLKS
CDR3 (SEQ ID NO:75) –
ARGSDRFHPYFDY
(SEQ ID NO:44)
CDR1 (SEQ ID NO:76) –
RASQSVSRYLA
CDR2 (SEQ ID NO:77) –
DASNRAT
CDR3 (SEQ ID NO:78) –
QQFDTWPPT
(F04)
(SEQ ID NO:95)
(SEQ ID NO:96)
(SEQ ID NO:97)
(SEQ ID NO:98)
(A44)
(SEQ ID NO:99)
CDR1 (SEQ ID NO:105) – FTFSSYAMS
CDR2 (SEQ ID NO:106) – AISGSGGSTYYADSVKG
CDR3 (SEQ ID NO:107) – AKDGGYYDSGAGDY
(SEQ ID NO:100)
CDR1 (SEQ ID NO:108) – RASQGIDSWLA
CDR2 (SEQ ID NO:109) – AASSLQS
CDR3 (SEQ ID NO:110) – QQGVSYPRT
(A49)
(SEQ ID NO:101)
CDR1 (SEQ ID NO:111) – FTFSSYSMN
CDR2 (SEQ ID NO:112) – SISSSSSYIYYADSVKG
CDR3 (SEQ ID NO:113) – ARGAPMGAAAGWFDP
(SEQ ID NO:102)
CDR1 (SEQ ID NO:114) – RASQGISSWLA
CDR2 (SEQ ID NO:115) – AASSLQS
CDR3 (SEQ ID NO:116) – QQGVSFPRT
(F63)
(SEQ ID NO:103)
CDR1 (SEQ ID NO:117) – YTFTGYYMH
CDR2 (SEQ ID NO:118) – WINPNSGGTNYAQKFQG
CDR3 (SEQ ID NO:119) – ARDTGEYYDTDDHGMDV
(SEQ ID NO:104)
CDR1 (SEQ ID NO:120) – RASQSVSSNLA
CDR2 (SEQ ID NO:121) – GASTRAT
CDR3 (SEQ ID NO:122) – QQDDYWPPT
[0093] Альтернативно, вариабельный домен тяжелой цепи, определенный SEQ ID NO: 45, может быть спарен с вариабельным доменом легкой цепи, определенным SEQ ID NO: 46, образуя антигенсвязывающий участок, который может связываться с NKG2D, как показано в US 9273136.
QVQLVESGGGLVKPGGSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAFIRYDGSNKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDRGLGDGTYFDYWGQGTTVTVSS (SEQ ID: 45)
QSALTQPASVSGSPGQSITISCSGSSSNIGNNAVNWYQQLPGKAPKLLIYYDDLLPSGVSDRFSGSKSGTSAFLAISGLQSEDEADYYCAAWDDSLNGPVFGGGTKLTVL (SEQ ID NO: 46)
[0094] Альтернативно, вариабельный домен тяжелой цепи, определенный SEQ ID NO: 47, может быть спарен с вариабельным доменом легкой цепи, определенным SEQ ID NO: 48, с образованием антигенсвязывающего участка, который может связываться с NKG2D, как показано в US 7879985.
QVHLQESGPGLVKPSETLSLTCTVSDDSISSYYWSWIRQPPGKGLEWIGHISYSGSANYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCANWDDAFNIWGQGTMVTVSS (SEQ ID NO: 47)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPWTFGQGTKVEIK (SEQ ID NO: 48)
[0095] В таблице 2 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с ВСМА.
(SEQ ID NO:49)
CDR1(SEQ ID NO:50) – DYYIN
CDR2 (SEQ ID NO:51) –WIYFASGNSEYNQKFTG
CDR3 (SEQ ID NO:52) – LYDYDWYFDV
или DIVMTQTPLSLSVTPGQPASISCKSSQSLVHSNGNTYLHWYLQKPGQSPQLLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDVGIYYCSQSSIYPWTFGQGTKLEIK
(SEQ ID NO:54)
CDR1(SEQ ID NO:55) – KSSQSLVHSNGNTYLH
CDR2 (SEQ ID NO:56) – KVSNRFS
CDR3 – AETSHVPWT (SEQ ID NO:57) или SQSSIYPWT (SEQ ID NO:58)
(PCT/US15/64269)
(SEQ ID NO:59)
CDR1 (SEQ ID NO:79) – DYSIN
CDR2 (SEQ ID NO:80) – WINTETREPAYAYDFR
CDR3 (SEQ ID NO:81) – DYSYAMDY
(SEQ ID NO:60)
CDR1 (SEQ ID NO:82) – RASESVTILGSHLIH
CDR2 (SEQ ID NO:83) – LASNVQT
CDR3 (SEQ ID NO:84) – LQSRTIPRT
(US14,122,391)
(SEQ ID NO:61)
CDR1 (SEQ ID NO:85) – NYWMH
CDR2 (SEQ ID NO:86) – ATYRGHSDTYYNQKFKG
CDR3 (SEQ ID NO:87) – GAIYNGYDVLDN
(SEQ ID NO:62)
CDR1 (SEQ ID NO:88) – SASQDISNYLN
CDR2 (SEQ ID NO:89) – YTSNLHS
CDR3 (SEQ ID NO:90) – QQYRKLPWT
(US20170051068)
CDR1: SSSYFWG (SEQ ID NO:125)
CDR2: SIYYSGITYYNPSLKS (SEQ ID NO:126)
CDR3: HDGATAGLFDY (SEQ ID NO:127)
CDR1: GGNNIGSKSVH (SEQ ID NO:128)
CDR2: DDSDRPS (SEQ ID NO:129)
CDR3: QVWDSSSDHVV (SEQ ID NO:130)
(WO2017021450)
CDR1: DNAMG (SEQ ID NO:131) или GFTFSDNAMG (SEQ ID:143)
CDR2: AISGPGSSTYYADSVK (SEQ ID NO:132)
CDR3: VLGWFDY (SEQ ID NO:133)
CDR1: RASQSVSDEYLS (SEQ ID NO:134)
CDR2: SASTRAT (SEQ ID NO:135)
CDR3: QQYGYPPDFT (SEQ ID NO:136)
(WO2017021450)
CDR1: SYAMS (SEQ ID NO:137)
CDR2: AISGSGGSTYYADSVKG (SEQ ID NO:138)
CDR3: VLGWFDY (SEQ ID NO:139)
RASQSVSSSYLA (SEQ ID NO:140)
GASSRAT (SEQ ID NO:141)
QQYGYPPDFT (SEQ ID NO:142)
[0096] Альтернативно, новые антигенсвязывающие участки, которые могут связываться с ВСМА, могут быть идентифицированы с помощью скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 63.
SEQ ID NO: 63
MLQMAGQCSQNEYFDSLLHACIPCQLRCSSNTPPLTCQRYCNASVTNSVKGTNAILWTCLGLSLIISLAVFVLMFLLRKINSEPLKDEFKNTGSGLLGMANIDLEKSRTGDEIILPRGLEYTVEECTCEDCIKSKPKVDSDHCFPLPAMEEGATILVTTKTNDYCKSLPAALSATEIEKSISAR
[0097] В пределах FС–домена связывание CD16 опосредовано шарнирной областью и доменом CH2. Например, в человеческом IgG1 взаимодействие с CD16 главным образом сосредоточено на аминокислотных остатках Asp 265 – Glu 269, Asn 297 – Thr 299, Ala 327 – Ile 332, Leu 234 – Ser 239 и углеводном остатке N–ацетил–D–глюкозамина в домене СН2 (см. Sondermann et al., Nature, 406 (6793):267–273). Для усиления или уменьшения аффинности связывания с CD16 мутации могут быть выбраны, исходя из известных доменов, например, путем использования библиотек фагового дисплея или дрожжевого дисплея для скрининга кДНК, или могут быть разработаны, исходя из известной трехмерной структуры взаимодействия.
[0098] Сборка тяжелых цепей гетеродимерных антител может осуществляться путем экспрессии двух разных последовательностей тяжелых цепей антител в одной и той же клетке, что может привести к образованию гомодимеров каждой тяжелой цепи антитела, а также гетеродимеров. Преимущественная сборка гетеродимеров может быть получена путем включения различных мутаций в домен СН3 каждой константной области тяжелой цепи антитела, как показано в US13/494870, US16/028850, US11/533709, US12/875015, US13/289934, US14/773418, US12/811207, US13/866756, US14/647480 и US14/830336. Например, мутации могут быть введены в домен CH3 человеческого IgG1 и включать различные пары аминокислотных замен в первом полипептиде и втором полипептиде, которые позволяют осуществить селективную гетеродимеризацию этих двух цепей друг с другом. Положения аминокислотных замен, показанные ниже, пронумерованы в соответствии с индексом ЕС, как в Kabat.
[0099] В одном из сценариев в результате аминокислотной замены в первом полипептиде исходная аминокислота заменяется более крупной аминокислотой, выбранной из аргинина (R), фенилаланина (F), тирозина (Y) или триптофана (W), и в результате по меньшей мере одной аминокислотной замены во втором полипептиде исходная аминокислота(–ы) заменяется аминокислотой(–ами) меньшего размера, выбранными из аланина (A), серина (S), треонина (T) или валина (V) таким образом, чтобы замена на более крупную аминокислоту (выпуклость) соответствовала поверхности с заменами на более мелкие аминокислоты (впадина). Например, один полипептид может включать замену T366W, а другой может включать три замены, включая T366S, L368A и Y407V.
[0100] Вариабельный домен тяжелой цепи антитела по изобретению необязательно может быть связан с аминокислотной последовательностью, по меньшей мере на 90% идентичной константной области антитела, такой как константная область IgG, содержащая шарнирную область, домены СН2 и СН3 и необязательно домен СН1. В некоторых вариантах осуществления аминокислотная последовательность константной области по меньшей мере на 90% идентична константной области человеческого антитела, такой как константная область человеческого IgG1, константная область IgG2, константная область IgG3 или константная область IgG4. В некоторых других вариантах осуществления аминокислотная последовательность константной области по меньшей мере на 90% идентична константной области антитела другого млекопитающего, такого как кролик, собака, кошка, мышь или лошадь. Константная область может содержать одну или более мутаций относительно константной области человеческого IgG1, например, в Q347, Y349, L351, S354, E356, E357, K360, Q362, S364, T366, L368, K370, N390, K392, T394, D399, S400, D401, F405, Y407, K409, T411 и/или K439. Типичные замены включают, например, Q347E, Q347R, Y349S, Y349K, Y349T, Y349D, Y349E, Y349C, T350V, L351K, L351D, L351Y, S354C, E356K, E357Q, E357L, E3636K, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364, S364H, S364D, T366V, T366I, T366L, T366M, T366K, T366W, T366S, L368E, L368A, L368D, K370S, N390D, N390E, K392L, K392M, K392, K392, K392D, T396, T39F T3F, D399V, S400K, S400R, D401K, F405A, F405T, Y407A, Y407I, Y407V, K409F, K409W, K409D, T411D, T411E, K439D и K439E.
[00101] В определенных вариантах осуществления мутации, которые могут быть включены в CH1 константной области человеческого IgG1, могут находиться в аминокислотах V125, F126, P127, T135, T139, A140, F170, P171 и/или V173. В некоторых вариантах осуществления мутации, которые могут быть включены в Cκ константной области человеческого IgG1, могут находиться в аминокислотах E123, F116, S176, V163, S174 и/или T164.
[00102] Аминокислотные замены могут быть выбраны из следующих наборов замен, показанных в таблице 3.
[00103] Альтернативно, аминокислотные замены могут быть выбраны из следующих наборов замен, показанных в таблице 4.
[00104] Альтернативно, аминокислотные замены могут быть выбраны из следующего набора замен, показанного в таблице 5.
[00105] Альтернативно, по меньшей мере, одна аминокислотная замена в каждой полипептидной цепи может быть выбрана из Таблицы 6.
[00106] Альтернативно, по меньшей мере, одна аминокислотная замена может быть выбрана из следующего набора замен, показанного в Таблице 7, где положение(я), указанное(ые) в столбце «Первый полипептид», заменено любой известной отрицательно заряженной аминокислотой, и положение(я), указанное(ые) в столбце «Второй полипептид», заменено(ы) любой известной положительно заряженной аминокислотой.
[00107] Альтернативно, по меньшей мере, одна аминокислотная замена может быть выбрана из следующего набора, показанного в Таблице 8, где положение(я), указанное(ые) в столбце «Первый полипептид», заменено(ы) любой известной положительно заряженной аминокислотой, и положение(я), указанное(ые) столбце «Второй полипептид», заменено(ы) любой известной отрицательно заряженной аминокислотой.
[00108] В качестве альтернативы или в дополнение структурная стабильность гетеромультимерного белка может быть увеличена путем введения S354C в первую или вторую полипептидную цепь и Y349C в противоположную полипептидную цепь с образованием таким образом искусственного дисульфидного мостика в интерфейсе между двумя полипептидами.
[00109] Полиспецифические белки, описанные выше, могут быть получены методом рекомбинантной ДНК, хорошо известным специалисту в данной области. Например, первая последовательность нуклеиновой кислоты, кодирующая первую тяжелую цепь иммуноглобулина, может быть клонирована в первый вектор экспрессии; вторая последовательность нуклеиновой кислоты, кодирующая вторую тяжелую цепь иммуноглобулина, может быть клонирована во второй вектор экспрессии; третью последовательность нуклеиновой кислоты, кодирующую легкую цепь иммуноглобулина, можно клонировать в третий вектор экспрессии; первый, второй и третий векторы экспрессии вместе могут быть стабильно трансфицированы в клетки–хозяева для продуцирования мультимерных белков.
[00110] Для получения максимального выхода полиспецифического белка можно исследовать различные соотношения первого, второго и третьего векторов экспрессии с тем, чтобы определить оптимальное соотношение для трансфекции в клетки–хозяева. После трансфекции могут быть выделены отдельные клоны для создания банка клеток методами, известными в данной области, такими как ограниченное разведение, ИФА, FACS, микроскопия или Clonepix.
[00111] Клоны можно культивировать в условиях, подходящих для роста в биореакторе и поддержания экспрессии полиспецифического белка. Полиспецифические белки могут быть выделены и очищены способами, известными в данной области техники, включая центрифугирование, глубинную фильтрацию, лизис клеток, гомогенизацию, замораживание–оттаивание, аффинную очистку, гель–фильтрацию, ионообменную хроматографию, обменную хроматографию гидрофобного взаимодействия и хроматографию в смешанном режиме.
II. Характеристика TriNKET
[00112] В определенных вариантах осуществления описанные в настоящей заявке TriNKET, которые включают NKG2D–связывающий домен и домен, связывающий ассоциированный с опухолью антиген, связываются с клетками, экспрессирующими человеческий NKG2D. В некоторых вариантах осуществления TriNKET, которые включают NKG2D–связывающий домен и домен, связывающий ассоциированный с опухолью антиген, связываются с ассоциированным с опухолью антигеном на уровне, сопоставимом с таковым для моноклонального антитела, имеющего такой же домен, связывающий ассоциированный с опухолью антиген.
[00113] TriNKET, описанные в настоящей заявке, более эффективно снижают рост опухоли и уничтожают раковые клетки.
[00114] В определенных вариантах осуществления описанные в настоящей заявке TriNKET, которые включают NKG2D–связывающий домен и домен, связывающий ассоциированный с опухолью антиген, активируют первичные человеческие NK–клетки в условиях культивирования с опухолевыми клетками, экспрессирующими этот антиген. Активация NK–клеток характеризуется увеличением дегрануляции CD107a и продуцирования цитокинов IFNγ. Кроме того, по сравнению с моноклональным антителом, которое включает домен, связывающий ассоциированный с опухолью антиген, TriNKET демонстрируют превосходную активацию человеческих NK–клеток в присутствии опухолевых клеток, экспрессирующих этот антиген. Например, по сравнению с анти–BCMA моноклональным антителом TriNKET по настоящему изобретению, имеющие BCMA–связывающий домен, демонстрируют превосходную активацию человеческих NK–клеток в присутствии BCMA–экспрессирующих раковых клеток.
[00115] В определенных вариантах осуществления описанные в настоящей заявке TriNKET, включающие NKG2D–связывающий домен и домен, связывающий ассоциированный с опухолью антиген, усиливают активность покоящихся и IL–2–активированных человеческих NK–клеток в присутствии опухолевых клеток, экспрессирующих этот антиген. Покоящиеся NK–клетки показали более слабое фоновое продуцирование IFNγ и дегрануляцию CD107a, чем IL–2–активированные NK–клетки. В некоторых вариантах осуществления покоящиеся NK–клетки демонстрируют более сильное изменение продуцирования IFNγ и дегрануляции CD107a по сравнению с IL–2–активированными NK–клетками. В некоторых вариантах осуществления IL–2–активированные NK–клетки показывают более высокий процент клеток, ставших IFNγ+; CD107a+ после стимуляции с помощью TriNKET.
[00116] В определенных вариантах осуществления описанные в настоящей заявке TriNKET, которые включают NKG2D–связывающий домен и домен, связывающий ассоциированный с опухолью антиген BCMA, усиливают цитотоксическую активность покоящихся и IL–2–активированных человеческих NK–клеток в присутствии опухолевых клеток, экспрессирующих этот антиген. Кроме того, TriNKET (например, A40–TriNKET, A44–TriNKET, A49–TriNKET, C26–TriNKET, F04–TriNKET, F43–TriNKET, F47–TriNKET и F63–TriNKET), которые включают домен, связывающий ассоциированный с опухолью антиген BCMA, более мощно направляют ответ активированных и покоящихся NK–клеток против клеток опухоли по сравнению с моноклональным антителом, которое включает такой же участок связывания с ассоциированным с опухолью антигеном. В некоторых вариантах осуществления TriNKET имеют преимущество в борьбе с опухолевыми клетками, экспрессирующими средний и низкий уровни ВСМА, по сравнению с моноклональными антителами, которые включают ВСМА–связывающий участок. Следовательно, терапия, включающая TriNKET, может быто более эффективной чем терапия анти–BCMA моноклональными антителами.
[00117] В определенных вариантах осуществления, описанные в настоящей заявке TriNKET (например, A40–TriNKET, A44–TriNKET, A49–TriNKET, C26–TriNKET, F04–TriNKET, F43–TriNKET, F47–TriNKET и F63–TriNKET), которые включают домен, связывающий ассоциированный с опухолью антиген ВСМА, имеют преимущество относительно моноклональных антител в лечении видов рака, для которых характерна высокая экспрессия FС–рецептора (FСR), или видов рака в микроокружении опухолей с высокими уровнями FСR. Моноклональные антитела оказывают влияние на рост опухоли посредством множества механизмов, включая, среди прочего, ADCC, CDC, фагоцитоз и сигнальную блокаду. Среди FcγR CD16 имеет самое низкое сродство к FС IgG; FcγRI (CD64) является высокоаффинным FСR, который связывается с FС IgG примерно в 1000 раз сильнее, чем CD16. CD64 обычно экспрессируется во многих гематопоэтических линиях, таких как миелоидная линия, и может экспрессироваться в опухолях, происходящих из этих типов клеток, таких как острый миелолейкоз (ОМЛ). Иммунные клетки, проникающие в опухоль, такие как MDSC и моноциты, также экспрессируют CD64 и, как известно, проникают в микроокружение опухоли. Экспрессия CD64 опухолью или в микроокружении опухоли может оказывать негативное влияние на терапию моноклональными антителами. Экспрессия CD64 в микроокружении опухоли затрудняет взаимодействие этих антител с CD16 на поверхности NK–клеток, поскольку антитела предпочитают связываться с рецептором, имеющим высокое сродство. TriNKET, за счет направленного воздействия двух активирующих рецепторов на поверхность NK–клеток, могут преодолеть пагубное влияние экспрессии CD64 (либо в опухоли, либо в микроокружении опухоли) на терапию моноклональными антителами. Независимо от экспрессии CD64 в опухолевых клетках, TriNKET способны опосредовать ответы человеческих NK–клеток против всех опухолевых клеток, поскольку двойное нацеливание двух активирующих рецепторов на NK–клетки обеспечивает более сильное специфическое связывание с NK–клетками.
[00118] В некоторых вариантах осуществления описанные в настоящей заявке TriNKET (например, A40–TriNKET, A44–TriNKET, A49–TriNKET, C26–TriNKET, F04–TriNKET, F43–TriNKET, F47–TriNKET и F63–TriNKET), которые включают домен, связывающий ассоциированный с опухолью антиген BCMA, обеспечивают улучшенный профиль безопасности благодаря уменьшению внеопухолевых побочных эффектов. Натуральные киллерные клетки и CD8 T–клетки способны непосредственно лизировать опухолевые клетки, хотя механизмы распознавания нормальных аутоклеток и опухолевых клеток у NK–клеток и CD8 T–клеток отличаются. Активность NK–клеток регулируется балансом сигналов, поступающих от активирующих (NCR, NKG2D, CD16 и т.д.) и ингибирующих (KIR, NKG2A и т.д.) рецепторов. Баланс этих активирующих и ингибирующих сигналов позволяет NK–клеткам отличать здоровые аутоклетки от подвергнутых стрессу, инфицированных вирусом или трансформированных аутоклеток. Этот «встроенный» механизм аутотолерантности позволяет защищать нормальную здоровую ткань от ответов NK–клеток. Для расширения этого принципа, исходя из аутотолерантности NK–клеток, можно с помощью TriNKET оказывать направленное воздействие на антигены, экспрессируемые как в самих этих клетках, так и в опухолях, не вызывая при этом внеопухолевых побочных эффектов или с увеличенным терапевтическим окном. В отличие от натуральных киллерных клеток для Т–клеток необходимо распознавание специфического пептида, презентированного молекулами МНС, для активации и осуществления эффекторных функций. Т–клетки были основной целью иммунотерапии, и было разработано много стратегий для перенаправления Т–клеточных ответов против опухоли. Биспецифические Т–клетки, ингибиторы контрольных точек и CAR–T–клетки были одобрены FDA, но их недостатком часто оказывалась токсичность, которая является причиной ограничения дозы. Биспецифические Т–клетки и клетки CAR–T работают на уровне системы распознавания TCR–MHC, используя связывающие домены для нацеливания антигенов на поверхность опухолевых клеток и сконструированные сигнальные домены для передачи сигналов активации в эффекторную клетку. Несмотря на то, что эти методы лечения эффективны для вызова противоопухолевого иммунного ответа, они часто сопровождаются синдромом высвобождения цитокинов (CRS) и внеопухолевыми побочными эффектами. TriNKET уникальны в этом контексте, так как они не «перекрывают» природные системы активации и ингибирования NK–клеток. Вместо этого TriNKET предназначены для изменения баланса и обеспечения дополнительных сигналов активации NK–клеток, сохраняя при этом NK–толерантность к здоровым клеткам.
[00119] В некоторых вариантах осуществления описанные в настоящей заявке TriNKET, включающие NKG2D–связывающий домен (например, A40–TriNKET, A44–TriNKET, A49–TriNKET, C26–TriNKET, F04–TriNKET, F43–TriNKET, F47–TriNKET и F63–TriNKET) и домен, связывающий ассоциированный с опухолью антиген BCMA, задерживают прогрессирование опухоли более эффективно, чем моноклональные антитела, которые включают такой же домен, связывающий опухолевый антиген. В некоторых вариантах осуществления TriNKET, включающие NKG2D–связывающий домен и домен, связывающий опухолевый антиген BCMA, являются более эффективными против метастазов рака, чем моноклональные антитела, которые включают анти–ВСМА–связывающий домен.
III. Терапевтическое применение
[00120] Изобретение относится к способам лечения рака с помощью полиспецифического связывающего белка, описанного в настоящей заявке, и/или фармацевтической композиции, описанной в настоящей заявке. Способы могут быть использованы для лечения различных видов рака, которые экспрессируют BCMA, путем введения нуждающемуся в этом пациенту терапевтически эффективного количества полиспецифического связывающего белка, описанного в настоящей заявке.
[00121] Терапевтический способ можно охарактеризовать в зависимости от рака, подлежащего лечению. Например, в определенных вариантах осуществления злокачественные новообразования происходят из клеток крови или костного мозга. Типичные примеры включают множественную миелому, острый миеломоноцитарный лейкоз, Т–клеточную лимфому, острый моноцитарный лейкоз и фолликулярную лимфому. Т–клеточные лимфомы могут включать лимфобластные лимфомы из Т–клеток–предшественников, периферические Т–клеточные лимфомы, кожные Т–клеточные лимфомы, ангиоиммунобластные Т–клеточные лимфомы, экстранодальные NK/Т–клеточные лимфомы, Т–клеточные лимфомы, ассоциированные с энтеропатией, подкожную панникулит–подобную Т–клеточную лимфому, анапластическую крупноклеточную лимфому или периферическую Т–клеточную лимфому.
[00122] В некоторых вариантах осуществления рак представляет собой В–клеточную лимфому, такую как диффузная В–клеточная крупноклеточная лимфома, первичную средостенную В–клеточную лимфому, фолликулярную лимфому, малую лимфоцитарную лимфому, мантийноклеточную лимфому, В–клеточную лимфому маргинальной зоны, экстранодальную B–клеточную лимфому маргинальной зоны, нодальную B–клеточную лимфому маргинальной зоны, B–клеточную лимфому маргинальной зоны селезенки, лимфому Беркитта, лимфоплазмоцитарную лимфому, волосатоклеточный лейкоз или первичную лимфому центральной нервной системы (ЦНС).
[00123] В некоторых других вариантах осуществления рак представляет собой солидную опухоль, такую как рак головного мозга, рак мочевого пузыря, рак молочной железы, рак шейки матки, рак толстой кишки, рак ободочной и прямой кишки, рак эндометрия, рак пищевода, лейкоз, рак легких, рак печени, меланому, рак яичников, рак поджелудочной железы, рак предстательной железы, рак прямой кишки, рак почки, рак желудка, рак яичка или рак матки. В других вариантах осуществления рак представляет собой васкуляризованную опухоль, плоскоклеточный рак, аденокарциному, мелкоклеточный рак, меланому, глиому, нейробластому, саркому (например, ангиосаркому или хондросаркому), рак гортани, рак околоушной железы, рак желчных путей, рак щитовидной железы, акральную лентигинозную меланому, актинический кератоз, острый лимфоцитарный лейкоз, острый миелоидный лейкоз, аденоидно–кистозную карциному, аденомы, аденосаркому, аденосквамозную карциному, рак анального канала, анальный рак, аноректальный рак, астроцитарную опухоль, карциному бартолиновой железы, базально–клеточную карциному, рак желчных протоков, рак костей, рак костного мозга, рак бронхов, рак бронхиальной железы, карциноид, холангиокарциному, хондосаркому, папиллому/карциному хориоидного сплетения, хронический лимфолейкоз, хроническую миелоидную лейкемию, светлоклеточный рак, рак соединительной ткани, цистаденому, рак пищеварительной системы, рак двенадцатиперстной кишки, рак эндокринной системы, опухоль эндодермального синуса, гиперплазию эндометрия, эндометриальную стромальную саркому, эндометриоидную аденокарциному, эндотелиальный рак, эпендимальный рак, эпителиально–клеточный рак, саркому Юинга, рак глаз и орбиты, рак женских половых органов, фокальную узловую гиперплазию, рак желчного пузыря, рак антрума желудка, рак дна желудка, гастриному, глиобластому, глюкогоному, рак сердца, гемангиобластому, гемангиоэндотелиому, гемангиомы, печеночную аденому, печеночный аденоматоз, рак гепатобилиарной системы, гепатоцеллюлярную карциному, болезнь Ходжкина, рак подвздошной кишки, инсулиному, интраэпителиальную неоплазию, плоскоклеточную интраэпителиальную неоплазию, рак внутрипеченочных желчных протоков, инвазивную плоскоклеточную карциному, рак тощей кишки, рак суставов, саркому Капоши, рак таза, крупноклеточную карциному, рак задней кишки, лейомиосаркому, злокачественную лентиго–меланому, лимфому, рак мужских половых органов, злокачественную меланому, злокачественные мезотелиальные опухоли, медуллобластому, медуллоэпителиому, менингеальный рак, мезотелиальный рак, метастатический рак, рак рта, мукоэпидермоидный рак, множественную миелому, рак мышц, рак носового канала, рак нервной системы, нейроэпителиальную аденокарциному, узловую меланому, неэпителиальный рак кожи, неходжкинскую лимфому, овсяно–клеточный рак, олигодендроглиальный рак, рак ротовой полости, остеосаркому, папиллярную серозную аденокарциному, рак полового члена, рак глотки, опухоли гипофиза, плазмоцитому, псевдосаркому, легочную бластому, рак прямой кишки, почечно–клеточный рак, рак дыхательной системы, ретинобластому, рабдомиосаркому, саркому, серозную карциному, рак пазухи, рак кожи, мелкоклеточный рак, рак тонкой кишки, рак гладких мышц, рак мягких тканей, соматостатин–секретирующую опухоль, рак позвоночника, плоскоклеточную карциному, рак поперечно–полосатой мускулатуры, субмезотелиальный рак, поверхностно распространяющуюся меланому, Т–клеточный лейкоз, рак языка, недифференцированный рак, рак мочеточника, рак мочеиспускательного канала, рак мочевого пузыря, рак мочевой системы, рак шейки матки, рак тела матки, увеальную меланому, рак влагалища, веррукозную карциному, ВИПому, рак вульвы, хорошо дифференцированную карциному или опухоль Вильмса.
[00124] Рак, подлежащий лечению, может быть охарактеризован в соответствии с наличием конкретного антигена, экспрессируемого на поверхности раковой клетки. В некоторых вариантах осуществления раковая клетка может экспрессировать, дополнительно к BCMA, одно или более из следующего: CD2, CD19, CD20, CD30, CD38, CD40, CD52, CD70, EGFR/ERBB1, IGF1R, HER3/ERBB3, HER4/ERBB4, MUC1, cMET, SLAMF7, PSCA, MICA, MICB, TRAILR1, TRAILR2, MAGE–A3, B7.1, B7.2, CTLA4 и PD1.
IV. Комбинированная терапия
[00125] Другой аспект изобретения относится к комбинированной терапии. Полиспецифические связывающие белки, описанные в настоящей заявке, используются в комбинации с дополнительными терапевтическими агентами для лечения рака.
[00126] Примеры терапевтических агентов, которые могут быть использованы в качестве части комбинированной терапии при лечении рака, включают, например, облучение, митомицин, третиноин, рибомустин, гемцитабин, винкристин, этопозид, кладрибин, митобронитол, метотрексат, доксорубицин, карбоквон, пентостатин, нитракрин, зиностатин, цетрореликс, летрозол, ралтитрексид, даунорубицин, фадрозол, фотемустин, тималфазин, собузоксан, недаплатины, цитарабин, бикалутамид, винорелбина, веснаринон, аминоглютетимид, амсакрин, проглумид, эллиптиния ацетат, кетансерин, доксифлуридин, этретинат, изотретиноин, стрептозоцин, нимустин, виндезин, флутамид, дрогенил, бутоцин, кармофур, разоксан, сизофилан, карбоплатины, митолактол, тегафур, ифосфамид, преднимустин, пицибанил, левамизол, тенипозид, импросульфан, эноцитабин, лизурид, оксиметолон, тамоксифен, прогестерон, мепитиостан, эпитиостанол, форместан, интерферон–альфа, интерферон–2 альфа, интерферон–бета, интерферон–гамма, колониестимулирующий фактор–1 колониестимулирующий фактор–2, денилейкин дифитокс, интерлейкин–2, фактор высвобождения лютеинизирующего гормона и варианты вышеупомянутых агентов, которые могут дифференциально связываться со своим родственным рецептором, а также увеличивать или уменьшать период полувыведения из сыворотки.
[00127] Дополнительным классом агентов, которые могут быть использованы в качестве части комбинированной терапии при лечении рака, являются ингибиторы иммунной контрольной точки. Типичные ингибиторы иммунной контрольной точки включают агенты, которые ингибируют одно или более из следующего: (i) цитотоксический T–лимфоцит–ассоциированный антиген 4 (CTLA4), (ii) белок 1 запрограммированной смерти клеток (PD1), (iii) PDL1, (iv) LAG3, (v) B7–H3, (vi) B7–H4 и (vii) TIM3. Ипилимумаб, ингибитор CTLA4, был одобрен Управлением по контролю за продуктами и лекарствами США для лечения меланомы.
[00128] Другими агентами, которые могут быть использованы в качестве части комбинированной терапии при лечении рака, представляют собой моноклональные антитела, которые нацелены на мишени, не являющиеся контрольными точками (например, герцептин), и нецитотоксические агенты (например, ингибиторы тирозинкиназы).
[00129] Другие категории противораковых агентов включают, например: (i) ингибитор, выбранный из ингибитора ALK, ингибитора ATR, антагониста А2А, ингибитора эксцизионной репарации оснований, ингибитора Bcr–Abl–тирозинкиназы, ингибитора тирозинкиназы Брутона, ингибитора CDC7, ингибитора CHK1, ингибитора циклинзависимой киназы, ингибитора ДНК–ПК, ингибитора ДНК–ПК и mTOR, ингибитора DNMT1, ингибитора DNMT1 плюс 2–хлор–деоксиаденозина, ингибитора HDAC, ингибитора сигнального пути Hedgehog, ингибитора IDO, ингибитора JAK, ингибитора mTOR, ингибитора MEK, ингибитора MELK, ингибитора MTH1, ингибитора PARP, ингибитора фосфоинозитид–3–киназы, ингибитора PARP1 и DHODH, ингибитора протеасом, ингибитора топоизомеразы–II, ингибитора тирозинкиназы, ингибитора VEGFR и ингибитора WEE1; (ii) агонист OX40, CD137, CD40, GITR, CD27, HVEM, TRSF25 или ICOS; и (iii) цитокин, выбранный из IL–12, IL–15, GM–CSF и G–CSF.
[00130] Белки по изобретению также можно использовать в качестве дополнения к хирургическому удалению первичного поражения.
[00131] Количество полиспецифического связывающего белка и дополнительного терапевтического агента и относительные сроки введения могут быть выбраны таким образом, чтобы был достигнут желаемый комбинированный терапевтический эффект. Например, при назначении комбинированной терапии нуждающемуся в этом пациенту терапевтические агенты в комбинации или фармацевтическую композицию или композиции, содержащие терапевтические агенты, можно вводить в любом порядке, например, последовательно, параллельно, вместе, одновременно и т.п. Кроме того, например, полиспецифический связывающий белок может быть введен в то время, когда дополнительный терапевтический агент(ы) оказывает свое профилактическое или терапевтическое действие, или наоборот.
V. Фармацевтические композиции
[00132] Настоящее раскрытие также относится к фармацевтическим композициям, которые содержат терапевтически эффективное количество белка, описанного в настоящей заявке. Композиция может быть приготовлена для использования в различных системах доставки лекарственного вещества. Для получения соответствующего состава в композицию также могут быть включены один или более физиологически приемлемых наполнителей или носителей. Подходящие составы для использования в настоящем раскрытии описаны в Remington's Pharmaceutical Sciences, Mack Publishing Company, Philadelphia, PA, 17th ed., 1985. Краткий обзор способов доставки лекарственных веществ см., например, у Langer (Science 249: 1527–1533, 1990).
[00133] Состав для внутривенной доставки лекарственного вещества по настоящему изобретению может находиться в пакете, ручке или шприце. В определенных вариантах осуществления пакет может быть соединен с каналом, содержащим трубку и/или иглу. В определенных вариантах осуществления состав может представлять собой лиофилизированный состав или жидкий состав. В определенных вариантах осуществления состав может быть сублимированным (лиофилизированным) и содержаться примерно в 12–60 флаконах. В некоторых вариантах осуществления состав может быть сублимирован, и 45 мг суьлимированного состава может содержаться в одном флаконе. В некоторых вариантах осуществления примерно 40–100 мг сублимированного состава может содержаться в одном флаконе. В некоторых вариантах осуществления для получения терапевтической дозы белка в лекарственном составе для внутривенного введения объединяют сублимированный состав из 12, 27 или 45 флаконов. В определенных вариантах осуществления состав может представлять собой жидкий состав и храниться в количестве от примерно 250 мг/флакон до примерно 1000 мг/флакон. В определенных вариантах осуществления состав может представлять собой жидкий состав и храниться в количестве примерно 600 мг/флакон. В определенных вариантах осуществления состав может представлять собой жидкий состав и храниться в количестве примерно 250 мг/флакон.
[00134] Состав по настоящему изобретению может находиться в виде жидкого водного фармацевтического состава, включающего терапевтически эффективное количество белка в забуференном растворе, образующем состав.
[00135] Эти композиции могут быть стерилизованы обычными методами стерилизации или могут быть стерильно отфильтрованы. Полученные водные растворы могут быть упакованы для применения в том виде, в каком они находятся, или могут быть лиофилизированы, причем лиофилизированный препарат объединяют со стерильным водным носителем перед введением. Значение рН препаратов обычно находится в диапазоне от 3 до 11, более предпочтительно от 5 до 9 или от 6 до 8, и наиболее предпочтительно от 7 до 8, например от 7 до 7,5. Полученные композиции в твердой форме могут быть упакованы в несколько емкостей однократной дозы, каждая из которых содержит фиксированное количество вышеупомянутого агента или агентов. Композиция в твердой форме также может быть упакована в контейнер переменного объема.
[00136] В определенных вариантах осуществления настоящего изобретения предоставляется состав с увеличенным сроком хранения, включающий белок по настоящему раскрытию в комбинации с маннитом, моногидратом лимонной кислоты, цитратом натрия, дигидратом динатрийфосфата, дигидратом дигидрофосфата натрия, хлоридом натрия, полисорбатом 80, водой и гидроксидом натрия.
[00137] В определенных вариантах осуществления готовят водный состав, включающий белок по настоящему изобретению в рН–забуференном растворе. Буфер по настоящему изобретения может иметь рН в диапазоне от примерно 4 до примерно 8, например, от примерно 4,5 до примерно 6,0 или от примерно 4,8 до примерно 5,5, или может иметь рН от примерно 5,0 до примерно 5,2. Диапазоны, промежуточные по отношению к указанным выше значениям pH, также являются частью настоящего изобретения. Например, диапазоны значений, включающие комбинацию любого из приведенных выше значений в качестве верхнего и/или нижнего пределов, также включены в настоящее изобретение. Примеры буферов, позволяющие регулировать рН в этом диапазоне, включают ацетатные (например, ацетат натрия), сукцинатные (такие как сукцинат натрия), глюконатные, гистидиновые, цитратные и другие буферы органических кислот.
[00138] В определенных вариантах осуществления состав включает буферную систему, которая содержит цитрат и фосфат для поддержания pH в диапазоне от примерно 4 до примерно 8. В некоторых вариантах осуществления диапазон pH может составлять от примерно 4,5 до примерно 6,0 или от примерно рН от 4,8 до примерно 5,5 или в диапазоне рН от примерно 5,0 до примерно 5,2. В некоторых вариантах осуществления буферная система включает моногидрат лимонной кислоты, цитрат натрия, дигидрат динатрийфосфата и/или дигидрат дигидрофосфата натрия. В некоторых вариантах осуществления буферная система включает примерно 1,3 мг/мл лимонной кислоты (например, 1,305 мг/мл), примерно 0,3 мг/мл цитрата натрия (например, 0,305 мг/мл), примерно 1,5 мг/мл дигидрата динатрийфосфата (например, 1,53 мг/мл), примерно 0,9 мг/мл дигидрата дигидрофосфата натрия (например, 0,86) и примерно 6,2 мг/мл хлорида натрия (например, 6,165 мг/мл). В некоторых вариантах осуществления буферная система включает 1–1,5 мг/мл лимонной кислоты, от 0,25 до 0,5 мг мл цитрата натрия, от 1,25 до 1,75 мг/мл дигидрата динатрийфосфата, от 0,7 до 1,1 мг/мл дигидрата дигидрофосфата натрия и от 6,0 до 6,4 мг/мл хлорида натрия. В определенных вариантах осуществления pH состава корректируют с помощью гидроксида натрия.
[00139] Полиол, который действует как регулятор тоничности и может стабилизировать антитело, также может быть включен в состав. Полиол добавляют в состав в количестве, которое может меняться в зависимости от требуемой изотоничности состава. В определенных вариантах осуществления водный состав может быть изотоническим. Количество добавляемого полиола также может меняться зависимости от молекулярной массы полиола. Например, может быть добавлено меньшее количество моносахарида (например, маннита) по сравнению с дисахаридом (таким как трегалоза). В некоторых вариантах осуществления полиол, который может быть использован в составе в качестве регулятора тоничности, представляет собой маннит. В определенных вариантах осуществления концентрация маннита может составлять от примерно 5 до примерно 20 мг/мл. В определенных вариантах осуществления концентрация маннита может составлять примерно 7,5–15 мг/мл. В определенных вариантах концентрация маннита может составлять примерно 10–14 мг/мл. В определенных вариантах осуществления концентрация маннита может составлять примерно 12 мг/мл. В определенных вариантах осуществления в качестве полиола в состав может быть включен сорбит.
[00140] В состав также может быть добавлен детергент или поверхностно–активное вещество. Типичные детергенты включают неионогенные детергенты, такие как полисорбаты (например, полисорбаты 20, 80 и т.д.) или полоксамеры (например, полоксамер 188). Количество добавляемого детергента является таким, которое позволяет уменьшить агрегацию находящегося в составе антитела и/или минимизировать образование частиц в составе и/или уменьшить адсорбцию. В определенных вариантах осуществления состав может включать поверхностно–активное вещество, которое представляет собой полисорбат. В определенных вариантах осуществления состав может содержать детергент, такой как полисорбат 80 или Твин 80. Твин 80 – это термин, используемый для описания полиоксиэтилен (20) сорбитан моноолеата (см. Fiedler, Lexikon der Hifsstoffe, Editio Cantor Verlag Aulendorf, 4th ed., 1996). В определенных вариантах осуществления состав может содержать от примерно 0,1 до примерно 10 мг/мл полисорбата 80 или от примерно 0,5 до примерно 5 мг/мл. В определенных вариантах осуществления в состав может добавлено примерно 0,1% полисорбата 80.
[00141] В вариантах осуществления белковый продукт по настоящему изобретению готовят в виде жидкого состава. Жидкий состав может быть представлен в концентрации 10 мг/мл в любом флаконе USP/Ph Eur типа I 50R, закрытом резиновой пробкой и запечатанном алюминиевой обжимной крышкой. Пробка может быть изготовлена из эластомера, соответствующего USP и Ph Eur. В некоторых вариантах осуществления флаконы могут быть заполнены 61,2 мл раствора белкового продукта, обеспечивающего экстрагируемый объем 60 мл. В определенных вариантах осуществления жидкий состав может быть разбавлен 0,9% солевым раствором.
[00142] В определенных вариантах осуществления жидкий состав по настоящему изобретению может быть приготовлен в виде раствора с концентрацией 10 мг/мл в комбинации со стабилизирующими уровнями сахара. В определенных вариантах осуществления жидкий состав может быть приготовлен в водном носителе. В определенных вариантах осуществления стабилизатор может быть добавлен в количестве, не превышающем количество, которое может привести к вязкости, нежелательной или непригодной для внутривенного введения. В некоторых вариантах осуществления сахар может представлять собой дисахариды, например сахарозу. В определенных вариантах осуществления жидкий состав также может включать одно или более из следующего: буферный агент, поверхностно–активное вещество и консервант.
[00143] В определенных вариантах осуществления рН жидкой композиции можно регулировать путем добавления фармацевтически приемлемой кислоты и/или основания. В определенных вариантах осуществления фармацевтически приемлемая кислота может представлять собой соляную кислоту. В определенных вариантах осуществления основание может представлять собой гидроксид натрия.
[00144] В дополнение к агрегации, дезамидирование является распространенным вариантом пептидного и белкового продукта, которое может происходить во время ферментации, сбора/осветления клеточной культуры, очистки, хранения лекарственного вещества/лекарственного продукта и во время анализа образцов. Дезамидирование представляет собой потерю NH3 белком с образованием промежуточного сукцинимидного соединения, которое может подвергаться гидролизу. Содержание промежуточного сукцинимидного соединения приводит к снижению массы исходного пептида на 17 дальтон. Последующий гидролиз приводит к увеличению массы на 18 дальтон. Выделение промежуточного сукцинимидного соединения затруднено из–за его нестабильности в водных условиях. Таким образом, дезамидирование обычно определяют по увеличению массы на 1 дальтон. Дезамидирование аспарагина приводит к образованию аспарагиновой или изоаспарагиновой кислоты. Параметры, влияющие на скорость дезамидирования, включают pH, температуру, диэлектрическую проницаемость растворителя, ионную силу, первичную последовательность, локальную конформацию полипептида и третичную структуру. Аминокислотные остатки, расположенные рядом с Asn в пептидной цепи, влияют на скорости дезамидирования. Gly и Ser, следующие за Asn в белковых последовательностях, приводят к более высокой восприимчивости к дезамидированию.
[00145] В определенных вариантах осуществления жидкий состав по настоящему изобретению может храниться в условиях pH и влажности, предотвращающих дезаминирование белкового продукта.
[00146] Представляющий интерес водный носитель является фармацевтически приемлемым (безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкого состава. Примеры носителя включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), pH–забуференный раствор (например, физиологический раствор с фосфатным буфером), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00147] Для уменьшения воздействия бактерий в составы по настоящему изобретению необязательно может быть добавлен консервант. Добавление консерванта может, например, способствовать получению состава для многоразового (многократного) применения.
[00148] Внутривенные (IV) составы могут быть предпочтительным путем введения в конкретных случаях, например, когда пациент находится в больнице после трансплантации и получает все лекарственные средства через IV введение. В некоторых вариантах осуществления жидкий состав перед введением разбавляют 0,9% раствором хлорида натрия. В определенных вариантах осуществления разбавленный лекарственный продукт для инъекций является изотоническим и пригодным для введения путем внутривенной инфузии.
[00149] В определенных вариантах осуществления, соль или буферные компоненты могут быть добавлены в количестве от 10 мМ до 200 мМ. Соли и/или буферы являются фармацевтически приемлемыми и полученными из различных известных кислот (неорганических и органических) с «образующими основание» металлами или аминами. В определенных вариантах осуществления буфер может представлять собой фосфатный буфер. В определенных вариантах осуществления буфер может представлять собой глицинатный, карбонатный, цитратный буферы, и в этом случае в качестве противоиона могут служить ионы натрия, калия или аммония.
[00150] Для уменьшения воздействия бактерий в составы по настоящему изобретению необязательно может быть добавлен консервант. Добавление консерванта может, например, способствовать получению состава для многоразового (многократного) применения.
[00151] Представляющий интерес водный носитель является фармацевтически приемлемым (безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкого состава. Иллюстративные носители включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), pH–забуференный раствор (например, физиологический раствор с фосфатным буфером), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00152] Состав по настоящему изобретению может существовать в виде лиофилизированного состава, включающего белки и лиопротектор. Лиопротектором может быть сахар, например дисахариды. В определенных вариантах осуществления лиопротектор может представлять собой сахарозу или мальтозу. Лиофилизированный состав также может включать одно или более из следующего: буферный агент, поверхностно–активное вещество, образующий объем агент и/или консервант.
[00153] Количество сахарозы или мальтозы, используемое для стабилизации лиофилизированного лекарственного продукта, может находиться в массовом соотношении белка к сахарозе или мальтозе, равном по меньшей мере 1:2. В некоторых вариантах осуществления массовое соотношение белок/сахароза или мальтоза может составлять от 1:2 до 1:5.
[00154] В определенных вариантах осуществления значение pH состава перед лиофилизацией может быть отрегулировано путем добавления фармацевтически приемлемой кислоты и/или основания. В определенных вариантах осуществления фармацевтически приемлемая кислота может представлять собой соляную кислоту. В определенных вариантах осуществления фармацевтически приемлемым основанием может быть гидроксид натрия.
[00155] Перед лиофилизацией значение pH раствора, содержащего белок по настоящему изобретению, можно регулировать таким образом, чтобы оно находилось в диапазоне от 6 до 8. В некоторых вариантах осуществления диапазон pH для лиофилизированного лекарственного продукта может составлять от 7 до 8.
[00156] В определенных вариантах осуществления соль или буферные компоненты могут быть добавлены в количестве от 10 мМ до 200 мМ. Соли и/или буферы являются фармацевтически приемлемыми и полученными из различных известных кислот (неорганических и органических) с «образующими основание» металлами или аминами. В определенных вариантах осуществления буфер может представлять собой фосфатный буфер. В некоторых вариантах осуществления буфер может представлять собой глицинатный, карбонатный, цитратный буферы, и в этом случае в качестве противоиона могут служить ионы натрия, калия или аммония.
[00157] В определенных вариантах осуществления может быть добавлен «образующий объем агент». «Образующий объем агент» представляет собой соединение, которое увеличивает массу лиофилизированной смеси и обеспечивает физическую структуру лиофилизированной лепешки (например, облегчает получение по существу однородной лиофилизированной лепешки, которая способна сохранять структуру с открытыми порами). Примеры образующего объем агента включают маннит, глицин, полиэтиленгликоль и сорбит. Лиофилизированные составы по настоящему изобретению могут содержать такие образующие объем агенты.
[00158] Для уменьшения воздействия бактерий в составы по настоящему изобретению необязательно может быть добавлен консервант. Добавление консерванта может, например, способствовать получению состава для многоразового (многократного) применения.
[00159] В определенных вариантах осуществления лиофилизированный лекарственный продукт может быть приготовлен с помощью водного носителя. Представляющий интерес водный носитель является фармацевтически приемлемым (например, безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкого состава после лиофилизации. Примеры разбавителя включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), pH–забуференный раствор (например, физиологический раствор с фосфатным буфером), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00160] В определенных вариантах осуществления лиофилизированный лекарственный продукт по настоящему изобретению восстанавливают либо с помощью стерильной воды для инъекций, USP (SWFI), либо 0,9% инъекцией хлорида натрия, USP. Во время восстановления лиофилизированный порошок растворяют в растворе.
[00161] В определенных вариантах осуществления лиофилизированный белковый продукт по настоящему изобретению состоит из примерно 4,5 мл воды для инъекций и разбавлен 0,9% солевым раствором (раствором хлорида натрия).
[00162] Фактические уровни дозирования активных ингредиентов в фармацевтических композициях по настоящему изобретению можно менять для получения количества активного ингредиента, которое является эффективным для достижения требуемого терапевтического ответа для конкретного пациента, композиции и способа введения, и при этом не является токсичным для пациента.
[00163] Конкретная доза может представлять собой унифицированную дозу для каждого пациента, например, 50–5000 мг белка. Альтернативно, доза может быть адаптирована относительно примерной массы тела пациента или площади поверхности пациента. Другие факторы определения подходящей дозы могут включать заболевание или состояние, которое подлежит лечению или профилактике, тяжесть заболевания, способ введения, а также возраст, пол и состояние здоровья пациента. Специалисты в данной области обычно проводят дополнительные уточняющие расчеты, необходимые для определения подходящей дозы для лечения, особенно с учетом информации о дозировании и результатов анализа, раскрытых в настоявшем описании. Доза также может быть определена с помощью известных анализов для определения доз, используемых в комбинации с соответствующими данными доза–ответ. Дозу для отдельного пациента можно корректировать по мере наблюдения за развитием заболевания. Для определения, необходима ли корректировка дозы для достижения или поддержания эффективной концентрации, в крови у пациента могут быть измерены уровни нацеливаемой конструкции или комплекса. Для определения того, какие нацеливаемые конструкции и/или комплексы и их дозы с наибольшей вероятностью окажутся эффективными для данного индивидуума, может быть использована фармакогеномика (Schmitz et al., Clinica Chimica Acta 308:43–53, 2001; Steimer et al., Clinica Chimica Acta 308:33–41, 2001).
[00164] Как правило, дозировки, основанные на массе тела, составляют от примерно 0,01 мкг до примерно 100 мг на кг массы тела, например от примерно 0,01 мкг до примерно 100 мг/кг массы тела, от примерно 0,01 мкг до примерно 50 мг/кг массы тела, от примерно 0,01 до примерно 10 мг/кг массы тела, от примерно 0,01 до примерно 1 мг/кг массы тела, от примерно 0,01 до примерно 100 мкг/кг массы тела, от примерно 0,01 до примерно 50 мкг/кг массы тела, от примерно 0,01 до примерно 10 мкг/кг массы тела, от примерно 0,01 до примерно 1 мкг/кг массы тела, от примерно 0,01 до примерно 0,1 мкг/кг массы тела, от примерно 0,1 мкг до примерно 100 мг/кг массы тела, от примерно 0,1 мкг до примерно 50 мг/кг массы тела, от примерно 0,1 мкг до примерно 10 мг/кг массы тела, от примерно 0,1 мкг до примерно 1 мг/кг массы тела, от примерно 0,1 мкг до примерно 100 мкг/кг массы тела, от примерно 0,1 мкг до примерно 10 мкг/кг массы тела, от примерно 0,1 мкг до примерно 1 мкг/кг массы тела, от примерно 1 мкг до примерно 100 мг/кг массы тела, от примерно 1 мкг до примерно 50 мг/кг массы тела, от примерно 1 мкг до примерно 10 мг/кг массы тела, от примерно 1 мкг до примерно 1 мг/кг массы тела, от примерно 1 мкг до примерно 100 мкг/кг массы тела, от примерно 1 мкг до примерно 50 мкг кг массы тела, от примерно 1 мкг до примерно 10 мкг/кг массы тела, от примерно 10 мкг до примерно 100 мг/кг массы тела, от примерно 10 мкг до примерно 50 мг/кг массы тела, от примерно 10 мкг до примерно 10 мг/кг массы тела, от примерно 10 мкг до примерно 1 мг/кг массы тела, от примерно 10 мкг до примерно 100 мкг/кг массы тела, от примерно 10 мкг до примерно 50 мкг/кг массы тела, от примерно 50 мкг до примерно 100 мг/кг массы тела, от примерно 50 мкг до примерно 50 мг/кг массы тела, от примерно 50 мкг до примерно 10 мг/кг массы тела, от примерно 50 мкг до примерно 1 мг/кг массы тела, от примерно 50 мкг до примерно 100 мкг/кг массы тела, от примерно 100 мкг до примерно 100 мг/кг массы тела, от примерно 100 мкг до примерно 50 мг/кг массы тела, от примерно 100 мкг до примерно 10 мг/кг массы тела, от примерно 100 мкг до примерно 1 мг/кг массы тела, от примерно 1 мг до примерно 100 мг/кг массы тела, от примерно 1 мг до примерно 50 мг/кг массы тела, от примерно 1 мг до примерно 10 мг/кг массы тела, от примерно 10 мг до примерно 100 мг/кг массы тела, от примерно 10 мг до примерно 50 мг/кг массы тела, от примерно 50 мг до примерно 100 мг/кг массы тела.
[00165] Дозы могут вводиться один или более раз в сутки, еженедельно, ежемесячно или ежегодно, или даже один раз каждые 2–20 лет. Специалисты в данной области техники могут легко оценить частоту введения доз, исходя из результатов измерения времени пребывания и концентрации нацеливаемой конструкции или комплекса в жидкостях или тканях организма. Введение по настоящему изобретению может быть внутривенным, внутриартериальным, внутрибрюшинным, внутримышечным, подкожным, внутриплевральным, интратекальным, внутриполостным, путем перфузии через катетер или путем прямой внутриклеточной инъекции. Введение может осуществляться один или более раз в сутки, один или более раз в неделю, один или более раз в месяц и один или более раз в год.
ПРИМЕРЫ
[00166] Изобретение, описанное в целом, будет более понятным со ссылкой на приведенные ниже примеры, которые включены исключительно с целью иллюстрации определенных аспектов и вариантов осуществления настоящего изобретения и не предназначены для ограничения изобретения.
Пример 1. NKG2D–связывающие домены связываются с NKG2D
NKG2D–связывающие домены связываются с очищенным рекомбинантным NKG2D
[00167] Выполняли слияние последовательности нуклеиновых кислот эктодоменов NKG2D человека, мыши или яванского макака с последовательностями нуклеиновых кислот, кодирующими FС–домены IgG1 человека, и вводили в клетки млекопитающих для экспрессии. После очистки слитые белки NKG2D–FС адсорбировали в лунках микропланшетов. После блокирования лунок бычьим сывороточным альбумином для предотвращения неспецифического связывания, NKG2D–связывающие домены титровали и добавляли в лунки с предварительно адсорбированными NKG2D–FС слитыми белками. Для определения связывания первичного антитела использовали вторичное антитело, конъюгированное с пероксидазой хрена, которое специфически распознает человеческую легкую цепь каппа, что позволяет избежать проблем с перекрестной реактивностью с FС–фрагментом. 3,3',5,5'–Тетраметилбензидин (TMB), субстрат для пероксидазы хрена, добавляли в лунки для визуализации сигнала связывания, поглощение которого измеряли при 450 нМ и корректировали при 540 нМ. В каждую лунку добавляли клон NKG2D–связывающего домена, контроль изотипа или положительный контроль (выбранный из SEQ ID NO: 45–48 или антимышиных клонов NKG2D MI–6 и CX–5, доступных в eBioscience).
[00168] Контроль изотипа показал минимальное связывание с NKG2D–FС рекомбинантными белками, в то время как положительный контроль наиболее сильно связывался с рекомбинантными антигенами. NKG2D–связывающие домены, продуцируемые всеми клонами, показали связывание с NKG2D–FС рекомбинантными белками человека (фиг. 14), мыши (фиг. 16) и яванского макака (фиг. 15), хотя с разным сродством в зависимости от клона. Как правило, каждый анти–NKG2D–клон связывался с рекомбинантным NKG2D–FС человека (фиг. 14) и яванского макака (фиг. 15) с аналогичной аффинностью, но с рекомбинантным NKG2D–FС мыши с более низкой аффинностью (фиг. 16).
NKG2D–связывающие домены связываются с клетками, экспрессирующими NKG2D
[00169] Для экспрессии NKG2D–сигнальный домен CD3–дзета химерного антигенного рецептора человека или мыши создавали линию клеток мышиной лимфомы EL4. NKG2D–связывающий клон, контроль изотипа или положительный контроль использовали в концентрации 100 нМ для окрашивания внеклеточного NKG2D, экспрессированного в клетках EL4. Для определения связывания антител использовали вторичные антитела против человеческого IgG, конъюгированные с флуорофором. Клетки анализировали методом проточной цитометрии и рассчитывали увеличение относительно фона (FOB) средней интенсивности флуоресценции (СИФ) NKG2D–экспрессирующих клеток по сравнению с родительскими клетками EL4.
[00170] NKG2D–связывающие домены, продуцированные всеми клонами, связывались с клетками EL4, экспрессирующими NKG2D человека и мыши. Антитела положительного контроля (выбранные из SEQ ID NO: 45–48 или клоны MI–6 и CX–5 анти–мышиных NKG2D, доступные на eBioscience) показали самый лучший сигнал связывания согласно FOB. Аффинность связывания каждого из клонов с NKG2D была аналогичной между клетками, экспрессирующими человеческий NKG2D (фиг. 17) и мышиный NKG2D (фиг. 18).
Пример 2. NKG2D–связывающие домены блокируют связывание природного лиганда с NKG2D
Конкуренция с ULBP–6
[00171] Человеческие NKG2D–FС рекомбинантные белки адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для снижения неспецифического связывания. Насыщенную концентрацию ULBP–6–His–биотина добавляли в лунки с последующим добавлением клонов NKG2D–связывающего домена. После 2–часовой инкубации лунки промывали, и ULBP–6–His–биотин, который оставался связанным в лунках, покрытых NKG2D–FС, детектировали, используя стрептавидин, конъюгированный с пероксидазой хрена и субстратом TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона вычисляли специфическое связывание NKG2D–связывающих доменов с NKG2D–FС белками по процентному количеству ULBP–6–His–биотина, который блокировался при связывании с NKG2D–FС белками в лунках. Антитело положительного контроля (выбранное из SEQ ID NO: 45–48) и различные NKG2D–связывающие домены блокировали связывание ULBP–6 с NKG2D, в то время как контроль изотипа показал слабую конкуренцию с ULBP–6 (фиг. 19).
Конкуренция с MICA
[00172] Человеческие MICA–FС рекомбинантные белки адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для уменьшения неспецифического связывания. NKG2D–FС–биотин добавляли в лунки, после чего добавляли NKG2D–связывающие домены. После инкубации и промывки NKG2D–FС–биотин, который оставался связанным в лунках, покрытых MICA–FС, детектировали с помощью субстрата стрептавидин–HRP и TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона специфическое связывание NKG2D–связывающих доменов с белками NKG2D–FС вычисляли, исходя из процента NKG2D–FС–биотина, который блокировался при связывании с лунками, покрытыми MICA–FС. Антитело положительного контроля (выбранное из SEQ ID NO: 45–48) и различные NKG2D–связывающие домены блокировали связывание MICA с NKG2D, в то время как контроль изотипа показал слабую конкуренцию с MICA (фиг. 20).
Конкуренция с Rae–1 дельта
[00173] Мышиный рекомбинантный Rae–1дельта–FС (приобретенный у компании R&D Systems) адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для уменьшения неспецифического связывания. Мышиный NKG2D–FС–биотин добавляли в лунки с последующими добавлением NKG2D–связывающих доменов. После инкубации и промывки NKG2D–FС–биотин, который оставался связанным в лунках, покрытых Rae–1дельта–FС, детектировали с помощью субстрата стрептавидин–HRP и TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона специфическое связывание NKG2D–связывающих доменов с белками NKG2D–FС вычисляли, исходя из процентного содержания NKG2D–FС–биотина, который блокировался при связывании с лунками, покрытыми Rae–1дельта–FС. Положительный контроль (выбранный из SEQ ID NO: 45–48 или анти–мышиных NKG2D–клонов MI–6 и CX–5, доступных на eBioscience) и различные клоны NKG2D–связывающего домена блокировали связывание Rae–1дельта с мышиным NKG2D, в то время как изотипное контрольное антитело демонстрировало слабую конкуренцию с Rae–1дельта (фиг. 21).
Пример 3. Клоны NKG2D–связывающего домена активируют NKG2D
[00174] Выполняли слияние последовательностей нуклеиновых кислот NKG2D человека и мыши с последовательностями нуклеиновых кислот, кодирующими сигнальный домен CD3–дзета, для получения конструкций химерного антигенного рецептора (CAR). Затем конструкции NKG2D–CAR клонировали в ретровирусный вектор с помощью сборки методом Гибсона и трансфицировали в клетки expi293 для получения ретровируса. Клетки EL4 инфицировали вирусами, содержащими NKG2D–CAR вместе с 8 мкг/мл полибрена. Через 24 часа после заражения уровни экспрессии NKG2D–CAR в клетках EL4 анализировали методом проточной цитометрии, и отбирали клоны, которые экспрессировали высокие уровни NKG2D–CAR на клеточной поверхности.
[00175] Для определения, активируют ли NKG2D–связывающие домены NKG2D, их адсорбировали в лунках микропланшета, и клетки NKG2D–CAR EL4 культивировали в лунках, покрытых фрагментом антитела, в течение 4 часов в присутствии брефельдина–A и монензина. Внутриклеточное продуцирование T–альфа, индикатора активации NKG2D, анализировали методом проточной цитометрии. Процент TNF–альфа–положительных клеток нормализовали по отношению к клеткам, обработанным положительным контролем. Все NKG2D–связывающие домены активировали как человеческие NKG2D (фиг. 22), так и мышиные NKG2D (фиг. 23).
Пример 4. NKG2D–связывающие домены активируют NK–клетки
Первичные человеческие NK–клетки
[00176] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкоцитарной оболочки периферической крови центрифугированием в градиенте плотности. NK–клетки (CD3–CD56+) выделяли из PBMC посредством отрицательного отбора с помощью магнитных бусин, при этом чистота выделенных NK–клеток обычно превышала 95%. Затем выделенные NK–клетки культивировали в среде, содержащей 100 нг/мл IL–2, в течение 24–48 часов, после чего их переносили в лунки микропланшета, на котором были адсорбированы NKG2D–связывающие домены, и культивировали в среде, содержащей конъюгированное с флуорофором анти–CD107a антитело, брефелдин–А и монензин. После культивирования NK–клетки анализировали методом проточной цитометрии, используя конъюгированные с флуорофором антитела против CD3, CD56 и IFN–гамма. Для оценки активации NK–клеток анализировали окрашивание CD107a и IFN–гамма в клетках CD3–CD56+. Увеличение количества CD107a/IFN–гамма–двойных положительных клеток свидетельствовало о более высокой активации NK–клеток, обеспечиваемой двумя, а не одним, активирующими рецепторами. NKG2D–связывающие домены и положительный контроль (выбранный из SEQ ID NO: 45–48) показали более высокий процент NK–клеток, которые стали CD107a+ и IFN–гамма+, по сравнению с контролем изотипа (на фиг. 24 и фиг. 25 приведены данные из двух независимых экспериментов, в каждом из которых использовали PBMC разных доноров для приготовления NK–клеток).
Первичные мышиные NK–клетки
[00177] У мышей C57Bl/6 извлекали селезенки, которые измельчали через 70 мкм сито для клеток для получения суспензии отдельных клеток. Клетки осаждали и ресуспендировали в лизирующем буфере ACK (приобретенном у Thermo Fisher Scientific # A1049201; 155 мМ хлорида аммония, 10 мМ бикарбоната калия, 0,01 мМ EDTA) для удаления эритроцитов. Оставшиеся клетки культивировали с 100 нг/мл hIL–2 в течение 72 часов, а затем собирали и готовили для выделения NK–клеток. Затем из клеток селезенки выделяли NK–клетки (CD3–NK1.1+) с чистотой, превышающей 90%, методом отрицательного истощения, используя магнитные бусины. Очищенные NK–клетки культивировали в среде, содержащей 100 нг/мл mIL–15, в течение 48 часов, после чего их переносили в лунки микропланшета, на котором были адсорбированы NKG2D–связывающие домены, и культивировали в среде, содержащей конъюгированное с флуорофором анти–CD107a антитело, брефелдин–А и монензин. После культивирования в лунках, покрытых NKG2D–связывающим доменом, NK–клетки анализировали методом проточной цитометрии с помощью конъюгированных с флуорофором антител против CD3, NK1.1 и IFN–гамма. Для оценки активации NK–клеток анализировали окрашивание CD107a и IFN–гамма в клетках CD3–NK1.1+. Увеличение количества CD107a/IFN–гамма–двойных положительных клеток свидетельствовало о более высокой активации NK–клеток, обеспечиваемой двумя, а не одним, активирующими рецепторами. NKG2D–связывающие домены и положительный контроль (выбранный из анти–мышиных NKG2D–клонов MI–6 и CX–5, доступных в eBioscience) показали более высокий процент NK–клеток, которые стали CD107a+ и IFN–гамма+, по сравнению с контролем изотипа (на фиг. 26 и фиг. 27 представлены данные двух независимых экспериментов, в каждом из которых использовали для приготовления NK–клеток разных мышей).
Пример 5. NKG2D–связывающие домены обеспечивают цитотоксичность опухолевых клеток–мишеней
[00178] Результаты анализа активации первичных человеческих и мышиных NK–клеток показали увеличенный уровень маркеров цитотоксичности на NK–клетках после инкубации с NKG2D–связывающими доменами. Для определения, приводит ли это к усилению лизиса опухолевых клеток, использовали клеточный анализ, в котором каждый NKG2D–связывающий домен превращали в моноспецифическое антитело. FС–область использовали в качестве одного плеча направленного воздействия, тогда как область Fab (NKG2D–связывающий домен) действовал в качестве другого плеча направленного воздействия для активации NK–клеток. Клетки THP–1, которые имеют человеческое происхождение и экспрессируют высокие уровни FС–рецепторов, использовали в качестве опухоли–мишени, и использовали набор для определения цитотоксичности Perkin Elmer DELFIA. Клетки THP–1 метили реагентом BATDA и ресуспендировали в 105/мл в культуральной среде. Затем меченые клетки THP–1 объединяли с антителами NKG2D и выделенными мышиными NK–клетками в лунках микротитровального планшета при 37°С в течение 3 часов. После инкубации извлекали 20 мкл культурального супернатанта, смешивали с 200 мкл раствора европия и инкубировали при встряхивании в течение 15 минут в темноте. Флуоресценцию измеряли с течением времени с помощью планшетного ридера PheraStar, оборудованного модулем флуоресценции с временным разрешением (возбуждение при 337 нм, излучение при 620 нм), и в соответствии с прилагаемыми к набору инструкциями рассчитывали показатель специфического лизиса.
[00179] Положительный контроль, ULBP–6, являющийся природным лигандом для NKG2D, показал увеличенный уровень специфического лизиса клеток–мишеней THP–1 мышиными NK–клетками. Анти–NKG2D антитела также увеличивали специфический лизис клеток–мишеней THP–1, тогда как антитело к контролю изотипа показало сниженный уровень специфического лизиса. Пунктирная линия указывает на специфический лизис клеток THP–1 мышиными NK–клетками без добавления антител (фиг. 28).
Пример 6. Анти–NKG2D антитела показывают высокую термостабильность
[00180] Температуры плавления NKG2D–связывающих доменов анализировали методом дифференциальной сканирующей флуориметрии. Экстраполированные значения кажущейся температуры плавления оказались высокими по сравнению с типичными антителами IgG1 (фиг. 29).
Пример 7. Полиспецифические связывающие белки связываются с NKG2D
[00181] Для экспрессии человеческого NKG2D создавали линии клеток EL4 мышиных лимфом. Триспецифические связывающие белки (TriNKET), каждый из которых содержал NKG2D–связывающий домен, ассоциированный с опухолью антигенсвязывающий домен (BCMA–связывающий домен) и FС–домен, который связывается с CD16, как показано на фиг. 1, тестировали на сродство к внеклеточному NKG2D, экспрессированному в клетках EL4. Связывание полиспецифических связывающих белков с NKG2D детектировали с использованием вторичных антител против человеческого IgG, конъюгированных с флуорофором. Клетки анализировали методом проточной цитометрии и вычисляли увеличение относительно фона (FOB) средней интенсивности флуоресценции (СИФ) NKG2D–экспрессирующих клеток по сравнению с родительскими клетками EL4.
[00182] Протестированные TriNKET включают BCMA–TriNKET–C26 (ADI–28226 и BCMA–связывающий домен), BCMA–TriNKET–F04 (ADI–29404 и BCMA–связывающий домен), BCMA–TriNKET–F43 (ADI–29443 и BCMA–связывающий домен) и BCMA–TriNKET–F47 (ADI–29447 и BCMA–связывающий домен).
[00183] BCMA–связывающий домен, использованный в тестируемых молекулах, состоял из вариабельного домена тяжелой цепи и вариабельного домена легкой цепи, как указано ниже.
[00184] Вариабельный домен тяжелой цепи EM–801 (SEQ ID NO: 91):

[00185] Вариабельный домен легкой цепи EM–801 (SEQ ID NO: 92):

[00186] Вариабельный домен тяжелой цепи EM–901 (SEQ ID NO: 93)

[00187] Вариабельный домен легкой цепи EM–901 (SEQ ID NO: 94)

Пример 8. Полиспецифические связывающие белки связываются с опухолевыми антигенами человека.
Триспецифические связывающие белки связываются с ВСМА
[00188] Клетки миеломы человека MM.1S, экспрессирующие ВСМА, использовали для анализа связывания TriNKET с антигеном ВСМА, ассоциированным с опухолью. TriNKET разбавляли и инкубировали с соответствующими клетками. TriNKET и, необязательно, родительское анти–ВСМА моноклональное антитело (ЕМ–801) инкубировали с клетками, и связывание детектировали с помощью конъюгированных с флуорофором вторичных антител против человеческого IgG. Клетки анализировали методом проточной цитометрии, и увеличение относительно фона (FOB) вычисляли по средней интенсивности флуоресценции (СИФ) для TriNKET и EM–801, нормализованной относительно контрольных вторичных антител. C26–TriNKET–BCMA, F04–TriNKET–BCMA, F43–TriNKET–BCMA и F47–TriNKET–BCMA показали сопоставимые с EM–801 уровни связывания с BCMA, экспрессируемыми в клетках MM.1S, (фиг. 31).
Пример 9. Полиспецифические связывающие белки активируют NK–клетки
Первичные человеческие NK–клетки активируются TriNKET при совместном культивировании с линиями раковых клеток человека, экспрессирующих мишень
[00189] Совместное культивирование первичных человеческих NK–клеток с BCMA–положительными клетками миеломы MM.1S приводило к TriNKET–опосредованной активации первичных человеческих NK–клеток. TriNKET, нацеленные на BCMA (например, C26–TriNKET–BCMA и F04–TriNKET–BCMA), опосредовали активацию человеческих NK–клеток, совместно культивированных с клетками миеломы MM.1S, о чем свидетельствовало увеличение дегрануляции CD107a и продуцирования цитокинов IFNγ (фиг. 32). По сравнению с TriNKET изотипом, TriNKET, нацеленные на BCMA (например, A44–TriNKET–BCMA, A49–TriNKET–BCMA, C26–TriNKET–BCMA, F04–TriNKET–BCMA, F43–TriNKET–BCMA, F43–TriNKET–BCMA, F47–TriNK–BCMA и F63–TriNKET–BCMA), показали увеличение активности NK–клеток (фиг. 32).
Пример 10. Триспецифические связывающие белки обеспечивают цитотоксичность раковых клеток–мишеней.
[00190] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкоцитарной оболочки периферической крови центрифугированием в градиенте плотности. NK–клетки (CD3–CD56+) выделяли из PBMC посредством отрицательного отбора с помощью магнитных бусин, при этом чистота выделенных NK–клеток обычно превышала 90%. Выделенные NK–клетки культивировали в среде, содержащей 100 нг/мл IL–2, для активации или оставляли на ночь без цитокина. IL–2–активированные или покоящиеся NK–клетки использовали на следующий день в анализах на цитотоксичность.
DELFIA анализ цитотоксичности:
[00191] Линии раковых клеток человека, экспрессирующие представляющую интерес мишень, собирали из культуры, клетки промывали PBS и ресуспендировали в ростовой среде при 106 мл/мл для мечения реагентом BATDA (Perkin Elmer AD0116). Мечение клеток–мишеней выполняли согласно инструкциям производителя. После мечения клетки промывали 3 раза PBS и ресуспендировали при 0,5–1,0×105/мл в культуральной среде. Для приготовления фоновых лунок брали аликвоту меченых клеток, и клетки извлекали из среды. В лунки осторожно, чтобы не повредить гранулированные клетки, добавляли 100 мкл среды в трех экземплярах. 100 мкл BATDA–меченных клеток добавляли в каждую лунку 96–луночного планшета. Часть лунок оставляли для спонтанного высвобождения из клеток–мишеней, и часть лунок готовили для максимального лизиса клеток–мишеней путем добавления 1% Triton–X. Моноклональные антитела или TriNKET к представляющей интерес опухоли–мишени разводили в культуральной среде, в каждую лунку добавляли 50 мкл разведенного mAb или TriNKET. Покоящиеся и/или активированные NK–клетки собирали из культуры, клетки промывали и ресуспендировали при 105–2,0×106/мл в культуральной среде в зависимости от требуемого соотношения E:T. 50 мкл NK–клеток добавляли в каждую лунку планшета, получая общий объем культуры, равный 200 мкл. До проведения анализа планшет инкубировали при 37ºC с 5% CO2 в течение 2–3 часов.
[00192] После культивирования в течение 2–3 часов планшет извлекали из инкубатора, и клетки осаждали центрифугированием при 200g в течение 5 минут. 20 мкл культурального супернатанта переносили в чистый микропланшет, предоставленный производителем, и в каждую лунку добавляли 200 мкл раствора европия при комнатной температуре. Планшет защищали от света и инкубировали на шейкере при 250 об/мин в течение 15 минут. Планшет считывали на приборе Victor 3 или SpectraMax i3X. Процент специфического лизиса рассчитывали следующим образом: % специфического лизиса = ((Экспериментальное высвобождение – спонтанное высвобождение)/(Максимальное высвобождение – спонтанное высвобождение))*100%.
[00193] Анализировали TriNKET–опосредованный лизис BCMA–положительных клеток миеломы. На фиг. 39 показан TriNKET–опосредованный лизис BCMA–положительных клеток миеломы KMS12–PE в присутствии покоящихся эффекторных NK–клеток человека. Два TriNKET (cFAE–A49.801 и cFAE–A49.901), содержащие один и тот же NKG2D–связывающий домен (A49), но разные домены, направленные на BCMA, тестировали относительно их in vitro эффективности. Оба TriNKET в одинаковой степени усиливали опосредованный NK–клетками лизис клеток KMS12–PE, но TriNKET, содержащий домен EM–901 направленного воздействия, обеспечивал повышенную активность (фиг. 39).
[00194] На фиг. 33 показана цитотоксическая активность нескольких TriNKET, содержащих разные NKG2D–связывающие домены (A40, A44, A49, C26 и F47), но один и тот же домен, нацеленный на BCMA. Изменение NKG2D–связывающего домена в TriMAKET, нацеленном на BCMA, влияло на максимальный уровень гибели клеток, а также на эффективности TriNKET. Для всех TriNKET был продемонстрирован увеличенный уровень гибели клеток–мишеней KMS12–PE по сравнению с моноклональным антителом EM–901 (фиг. 33).
Пример 11
[00195] Исследовали синергетическую активацию человеческих NK–клеток путем сшивания NKG2D и CD16.
Первичный анализ активации человеческих NK–клеток
[00196] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкоцитарной оболочки периферической крови человека центрифугированием в градиенте плотности. NK–клетки очищали от РВМС с помощью отрицательных магнитных бусин (StemCell # 17955). Согласно результатам проточной цитометрии NK–клетки были CD3–CD56+ на >90%. Затем перед использованием в анализах активации клетки размножали в течение 48 часов в среде, содержащей 100 нг/мл hIL–2 (Peprotech # 200–02). Антитела наносили в 96–луночный планшет с плоским дном с концентрацией 2 мкг/мл (анти–CD16, Biolegend # 302013) и 5 мкг/мл (анти–NKG2D, R&D # MAB139) в 100 мкл стерильного PBS в течение ночи при 4°С с последующей тщательной промывкой лунок для удаления избытка антител. Для оценки дегрануляции IL–2–активированные NK–клетки ресуспендировали при 5×105 клеток/мл в культуральной среде, дополненной 100 нг/мл hIL2 и 1 мкг/мл APC–конъюгированного анти–CD107a mAb (Biolegend # 328619). Затем в планшеты, покрытые антителами, добавляли 1×105 клеток/лунку. Ингибиторы транспорта белка Брефелдин А (BFA, Biolegend # 420601) и монензин (Biolegend # 420701) добавляли с конечным разведением 1:1000 и 1:270 соответственно. Планшеты с клетками инкубировали в течение 4 часов при 37°С в 5% СО2. Для внутриклеточного окрашивания IFN–γ NK–клетки метили анти–CD3 (Biolegend # 300452) и анти–CD56 mAb (Biolegend # 318328), а затем фиксировали и пермеабилизовали, и метили анти–IFN–γ mAb (Biolegend # 506507). NK–клетки анализировали на экспрессию CD107a и IFN–γ методом проточной цитометрии после стробирования на живые CD56+CD3– клетки.
[00197] Для изучения относительной активности комбинации рецепторов выполняли связывание NKG2D или CD16 и совместное связывание обоих рецепторов путем стимуляции связывания на планшете. Как показано на фиг. 34 (фиг. 34A–3C), комбинированная стимуляция CD16 и NKG2D приводила к сильно увеличенным уровням CD107a (дегрануляция) (фиг. 3A) и/или продуцирования IFN–γ (фиг. 34B). Пунктирные линии представляют аддитивный эффект отдельных стимуляций каждого рецептора.
[00198] Уровни CD107a и внутриклеточного продуцирования IFN–γ у IL–2–активированных NK–клеток анализировали через 4 часа после начала стимуляции связывания на планшете с помощью анти–CD16, анти–NKG2D или комбинации обоих моноклональных антител. На графиках показано среднее значение (n=2) ± SD. На фиг. 34А показаны уровни CD107a; на фиг. 34B показаны уровни IFNγ; на фиг. 34C показаны уровни CD107a. Данные, показанные на фиг. 34A–34C, получены в пяти независимых экспериментах с участием пяти здоровых доноров.
ВКЛЮЧЕНИЕ В ВИДЕ ССЫЛКИ
[00199] Полное раскрытие каждого из патентных документов и научных статей, упомянутых в настоящем описании, включено в виде ссылки для всех целей.
ЭКВИВАЛЕНТЫ
[00200] Изобретение может быть реализовано в других конкретных формах без отклонения от сущности или его существенных характеристик. Таким образом, приведенные выше варианты осуществления следует рассматривать исключительно как иллюстративные, а не ограничивающие изобретение, описанное в настоящей заявке. Таким образом, объем изобретения обозначен прилагаемой формулой изобретения, а не предшествующим описанием, и все изменения, которые охвачены значением и диапазоном эквивалентов формулы изобретения, включены в объем изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> DRAGONFLY THERAPEUTICS, INC.
<120> БЕЛКИ, СВЯЗЫВАЮЩИЕ BCMA, NKG2D И CD16
<130> 14247-424-999
<140> US 16/484,936
<141> 2019-08-09
<150> PCT/US2018/017653
<151> 2018-02-09
<150> 62/457,780
<151> 2017-02-10
<160> 143
<170> PatentIn version 3.5
<210> 1
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 1
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 2
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 3
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 3
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 4
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 4
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Ile Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 5
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 5
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 6
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr His Ser Phe Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 7
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 7
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 8
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Ser Tyr Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 9
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 9
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 10
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 10
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 11
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 11
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Gly Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 12
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 12
Glu Leu Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asp Ile Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 13
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 13
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 14
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ser Phe Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 15
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 15
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 16
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Lys Glu Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 17
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 18
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 19
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 19
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 20
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 20
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ile Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 21
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 21
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 22
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 23
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 23
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 24
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 25
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 25
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 26
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 27
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 27
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 28
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Phe Ser Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 29
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 29
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 30
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Ser Tyr Ser Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 31
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 31
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 32
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 33
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 33
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 34
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 34
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 35
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 35
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 36
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr His Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 37
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 37
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 38
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Leu Tyr Ser Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 39
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 39
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 40
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Thr Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 41
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 41
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Ser Ser Ile Arg His Ala Tyr Tyr Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 42
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 42
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Thr Pro Ile Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 43
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 43
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Gly Ser Asp Arg Phe His Pro Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 44
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 44
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Thr Trp Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 45
<211> 121
<212> Белок
<213> Homo sapiens
<400> 45
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Gly Leu Gly Asp Gly Thr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 46
<211> 110
<212> Белок
<213> Homo sapiens
<400> 46
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Asp Leu Leu Pro Ser Gly Val Ser Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Phe Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 47
<211> 115
<212> Белок
<213> Homo sapiens
<400> 47
Gln Val His Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Asp Asp Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly His Ile Ser Tyr Ser Gly Ser Ala Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Asn Trp Asp Asp Ala Phe Asn Ile Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 48
<211> 108
<212> Белок
<213> Homo sapiens
<400> 48
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 49
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 49
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Pro Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Tyr Phe Ala Ser Gly Asn Ser Glu Tyr Asn Gln Lys Phe
50 55 60
Thr Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Asp Tyr Asp Trp Tyr Phe Asp Val Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser
115
<210> 50
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 50
Asp Tyr Tyr Ile Asn
1 5
<210> 51
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 51
Trp Ile Tyr Phe Ala Ser Gly Asn Ser Glu Tyr Asn Gln Lys Phe Thr
1 5 10 15
Gly
<210> 52
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 52
Leu Tyr Asp Tyr Asp Trp Tyr Phe Asp Val
1 5 10
<210> 53
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 53
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Ala Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ala Glu Thr
85 90 95
Ser His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 54
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 54
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Ile Tyr Tyr Cys Ser Gln Ser
85 90 95
Ser Ile Tyr Pro Trp Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 55
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 55
Lys Ser Ser Gln Ser Leu Val His Ser Asn Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 56
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 56
Lys Val Ser Asn Arg Phe Ser
1 5
<210> 57
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 57
Ala Glu Thr Ser His Val Pro Trp Thr
1 5
<210> 58
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 58
Ser Gln Ser Ser Ile Tyr Pro Trp Thr
1 5
<210> 59
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 59
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Lys Arg Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 60
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 60
Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu Ala Met Ser Leu Gly
1 5 10 15
Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Thr Ile Leu
20 25 30
Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln Thr Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe Thr Leu Thr Ile Asp
65 70 75 80
Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr Cys Leu Gln Ser Arg
85 90 95
Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 61
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Asn Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ala Thr Tyr Arg Gly His Ser Asp Thr Tyr Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Ile Tyr Asn Gly Tyr Asp Val Leu Asp Asn Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 62
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Asn Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Arg Lys Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 63
<211> 184
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 63
Met Leu Gln Met Ala Gly Gln Cys Ser Gln Asn Glu Tyr Phe Asp Ser
1 5 10 15
Leu Leu His Ala Cys Ile Pro Cys Gln Leu Arg Cys Ser Ser Asn Thr
20 25 30
Pro Pro Leu Thr Cys Gln Arg Tyr Cys Asn Ala Ser Val Thr Asn Ser
35 40 45
Val Lys Gly Thr Asn Ala Ile Leu Trp Thr Cys Leu Gly Leu Ser Leu
50 55 60
Ile Ile Ser Leu Ala Val Phe Val Leu Met Phe Leu Leu Arg Lys Ile
65 70 75 80
Asn Ser Glu Pro Leu Lys Asp Glu Phe Lys Asn Thr Gly Ser Gly Leu
85 90 95
Leu Gly Met Ala Asn Ile Asp Leu Glu Lys Ser Arg Thr Gly Asp Glu
100 105 110
Ile Ile Leu Pro Arg Gly Leu Glu Tyr Thr Val Glu Glu Cys Thr Cys
115 120 125
Glu Asp Cys Ile Lys Ser Lys Pro Lys Val Asp Ser Asp His Cys Phe
130 135 140
Pro Leu Pro Ala Met Glu Glu Gly Ala Thr Ile Leu Val Thr Thr Lys
145 150 155 160
Thr Asn Asp Tyr Cys Lys Ser Leu Pro Ala Ala Leu Ser Ala Thr Glu
165 170 175
Ile Glu Lys Ser Ile Ser Ala Arg
180
<210> 64
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 64
Gly Ser Phe Ser Gly Tyr Tyr Trp Ser
1 5
<210> 65
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 65
Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 66
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 66
Ala Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro
1 5 10
<210> 67
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 67
Gly Thr Phe Ser Ser Tyr Ala Ile Ser
1 5
<210> 68
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 68
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 69
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 69
Ala Arg Gly Asp Ser Ser Ile Arg His Ala Tyr Tyr Tyr Tyr Gly Met
1 5 10 15
Asp Val
<210> 70
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 70
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 71
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 71
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 72
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 72
Gln Gln Tyr Tyr Ser Thr Pro Ile Thr
1 5
<210> 73
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 73
Gly Ser Ile Ser Ser Ser Ser Tyr Tyr Trp Gly
1 5 10
<210> 74
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 74
Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 75
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 75
Ala Arg Gly Ser Asp Arg Phe His Pro Tyr Phe Asp Tyr
1 5 10
<210> 76
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 76
Arg Ala Ser Gln Ser Val Ser Arg Tyr Leu Ala
1 5 10
<210> 77
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 77
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 78
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 78
Gln Gln Phe Asp Thr Trp Pro Pro Thr
1 5
<210> 79
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 79
Asp Tyr Ser Ile Asn
1 5
<210> 80
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 80
Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe Arg
1 5 10 15
<210> 81
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 81
Asp Tyr Ser Tyr Ala Met Asp Tyr
1 5
<210> 82
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 82
Arg Ala Ser Glu Ser Val Thr Ile Leu Gly Ser His Leu Ile His
1 5 10 15
<210> 83
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 83
Leu Ala Ser Asn Val Gln Thr
1 5
<210> 84
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 84
Leu Gln Ser Arg Thr Ile Pro Arg Thr
1 5
<210> 85
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 85
Asn Tyr Trp Met His
1 5
<210> 86
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 86
Ala Thr Tyr Arg Gly His Ser Asp Thr Tyr Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 87
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 87
Gly Ala Ile Tyr Asn Gly Tyr Asp Val Leu Asp Asn
1 5 10
<210> 88
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 88
Ser Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 89
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 89
Tyr Thr Ser Asn Leu His Ser
1 5
<210> 90
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 90
Gln Gln Tyr Arg Lys Leu Pro Trp Thr
1 5
<210> 91
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 91
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Leu Gly Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 92
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 92
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Tyr Pro Pro
85 90 95
Asp Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 93
<211> 116
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 93
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Asn
20 25 30
Ala Met Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Pro Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Val Leu Gly Trp Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 94
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 94
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Asp Glu
20 25 30
Tyr Leu Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile His Ser Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Ala Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Tyr Pro Pro
85 90 95
Asp Phe Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 95
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 95
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 96
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 96
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Glu Gln Tyr Asp Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 97
<211> 126
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 97
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Arg Lys Ala Ser Gly Ser Phe Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 98
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 98
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Glu Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Pro Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 99
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 99
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Gly Gly Tyr Tyr Asp Ser Gly Ala Gly Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 100
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 100
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Asp Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Val Ser Tyr Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 101
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 101
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Met Gly Ala Ala Ala Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 102
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 102
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Val Ser Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 103
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 103
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Thr Gly Glu Tyr Tyr Asp Thr Asp Asp His Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 104
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
полипептид
<400> 104
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asp Asp Tyr Trp Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 105
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 105
Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5
<210> 106
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 106
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 107
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 107
Ala Lys Asp Gly Gly Tyr Tyr Asp Ser Gly Ala Gly Asp Tyr
1 5 10
<210> 108
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 108
Arg Ala Ser Gln Gly Ile Asp Ser Trp Leu Ala
1 5 10
<210> 109
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 109
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 110
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 110
Gln Gln Gly Val Ser Tyr Pro Arg Thr
1 5
<210> 111
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 111
Phe Thr Phe Ser Ser Tyr Ser Met Asn
1 5
<210> 112
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 112
Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 113
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 113
Ala Arg Gly Ala Pro Met Gly Ala Ala Ala Gly Trp Phe Asp Pro
1 5 10 15
<210> 114
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 114
Arg Ala Ser Gln Gly Ile Ser Ser Trp Leu Ala
1 5 10
<210> 115
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 115
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 116
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 116
Gln Gln Gly Val Ser Phe Pro Arg Thr
1 5
<210> 117
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 117
Tyr Thr Phe Thr Gly Tyr Tyr Met His
1 5
<210> 118
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 118
Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 119
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 119
Ala Arg Asp Thr Gly Glu Tyr Tyr Asp Thr Asp Asp His Gly Met Asp
1 5 10 15
Val
<210> 120
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 120
Arg Ala Ser Gln Ser Val Ser Ser Asn Leu Ala
1 5 10
<210> 121
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 121
Gly Ala Ser Thr Arg Ala Thr
1 5
<210> 122
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 122
Gln Gln Asp Asp Tyr Trp Pro Pro Thr
1 5
<210> 123
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 123
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Phe Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ile Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg His Asp Gly Ala Thr Ala Gly Leu Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 124
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 124
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Ile Gly Ser Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Pro Pro Gly Gln Ala Pro Val Val Val Val Tyr
35 40 45
Asp Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Val Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ser Asp His
85 90 95
Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 125
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 125
Ser Ser Ser Tyr Phe Trp Gly
1 5
<210> 126
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 126
Ser Ile Tyr Tyr Ser Gly Ile Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 127
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 127
His Asp Gly Ala Thr Ala Gly Leu Phe Asp Tyr
1 5 10
<210> 128
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 128
Gly Gly Asn Asn Ile Gly Ser Lys Ser Val His
1 5 10
<210> 129
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 129
Asp Asp Ser Asp Arg Pro Ser
1 5
<210> 130
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 130
Gln Val Trp Asp Ser Ser Ser Asp His Val Val
1 5 10
<210> 131
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 131
Asp Asn Ala Met Gly
1 5
<210> 132
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 132
Ala Ile Ser Gly Pro Gly Ser Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
<210> 133
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 133
Val Leu Gly Trp Phe Asp Tyr
1 5
<210> 134
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 134
Arg Ala Ser Gln Ser Val Ser Asp Glu Tyr Leu Ser
1 5 10
<210> 135
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 135
Ser Ala Ser Thr Arg Ala Thr
1 5
<210> 136
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 136
Gln Gln Tyr Gly Tyr Pro Pro Asp Phe Thr
1 5 10
<210> 137
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 137
Ser Tyr Ala Met Ser
1 5
<210> 138
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 138
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 139
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 139
Val Leu Gly Trp Phe Asp Tyr
1 5
<210> 140
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 140
Arg Ala Ser Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 141
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 141
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 142
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический пептид
<400> 142
Gln Gln Tyr Gly Tyr Pro Pro Asp Phe Thr
1 5 10
<210> 143
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: синтетический
пептид
<400> 143
Gly Phe Thr Phe Ser Asp Asn Ala Met Gly
1 5 10
<---

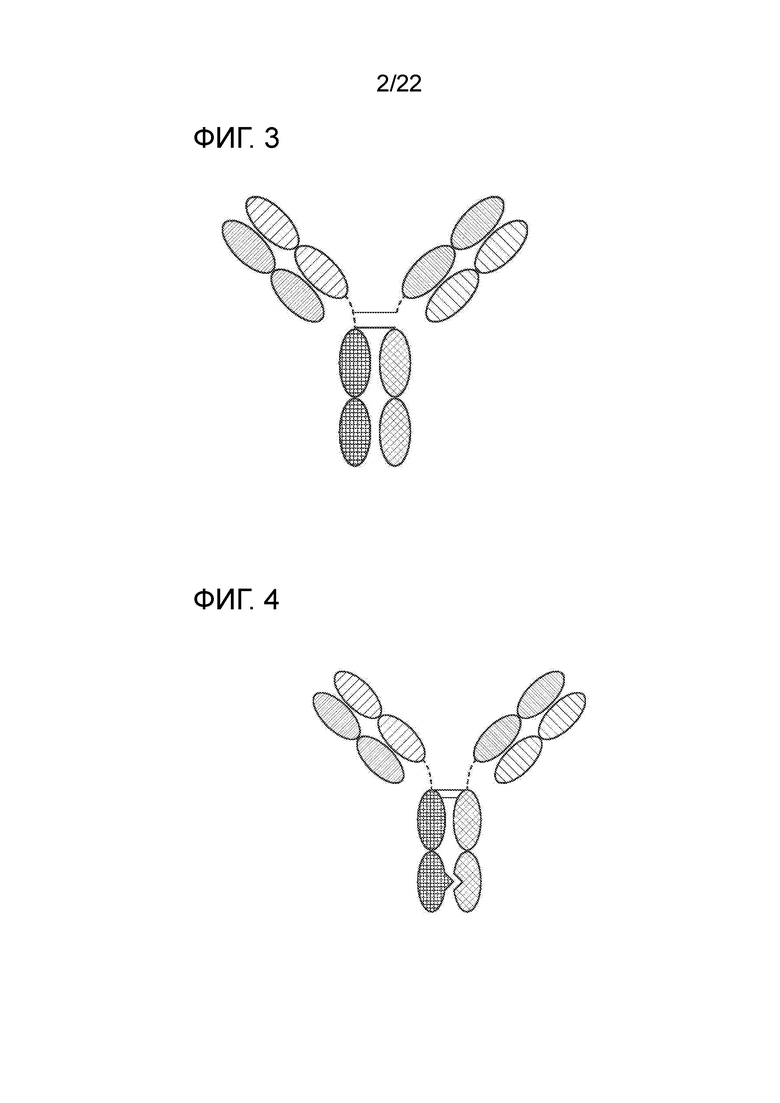
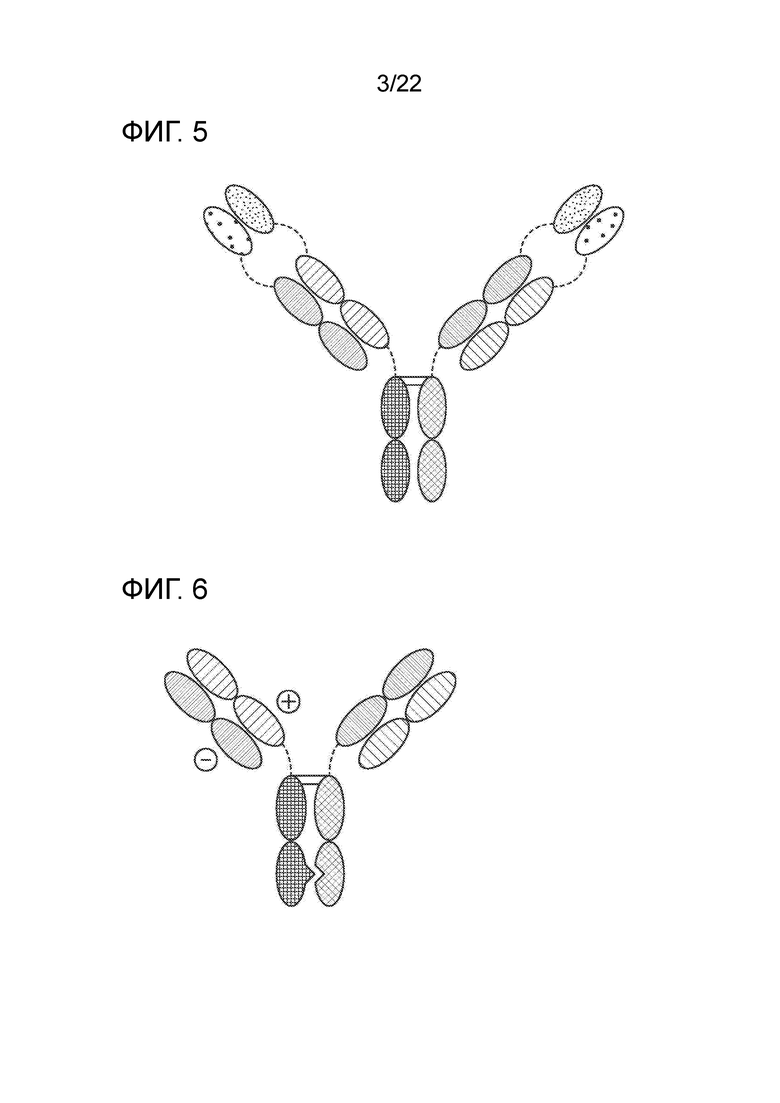
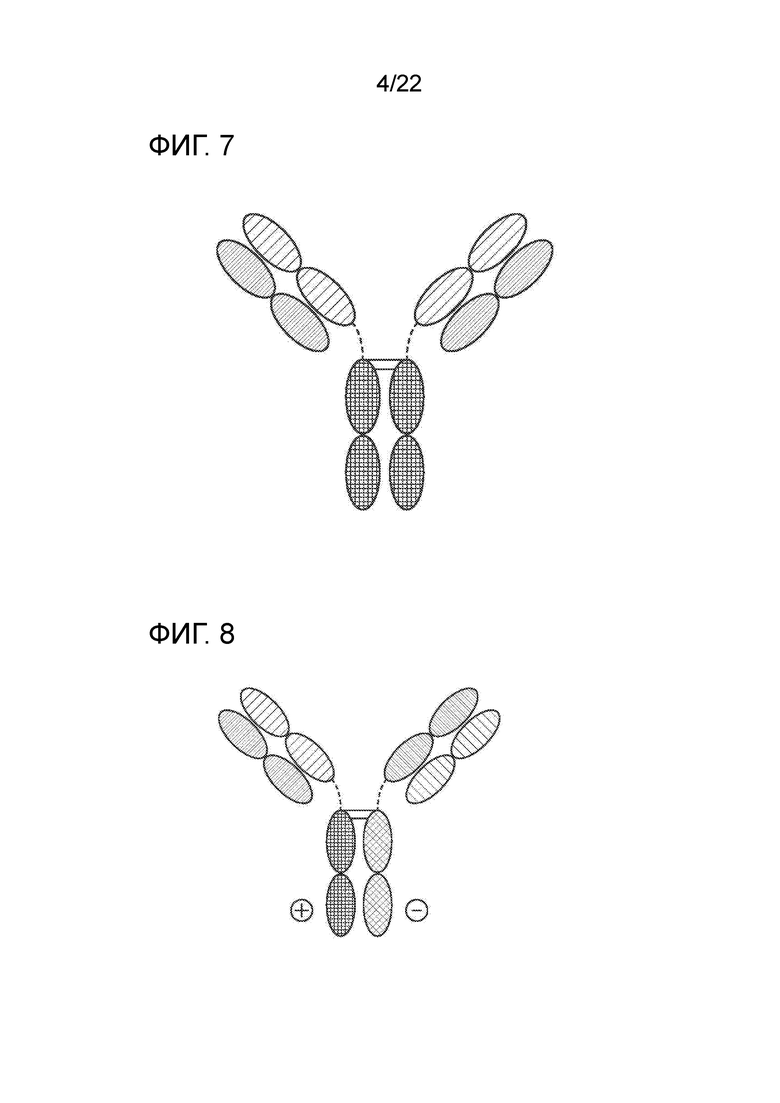
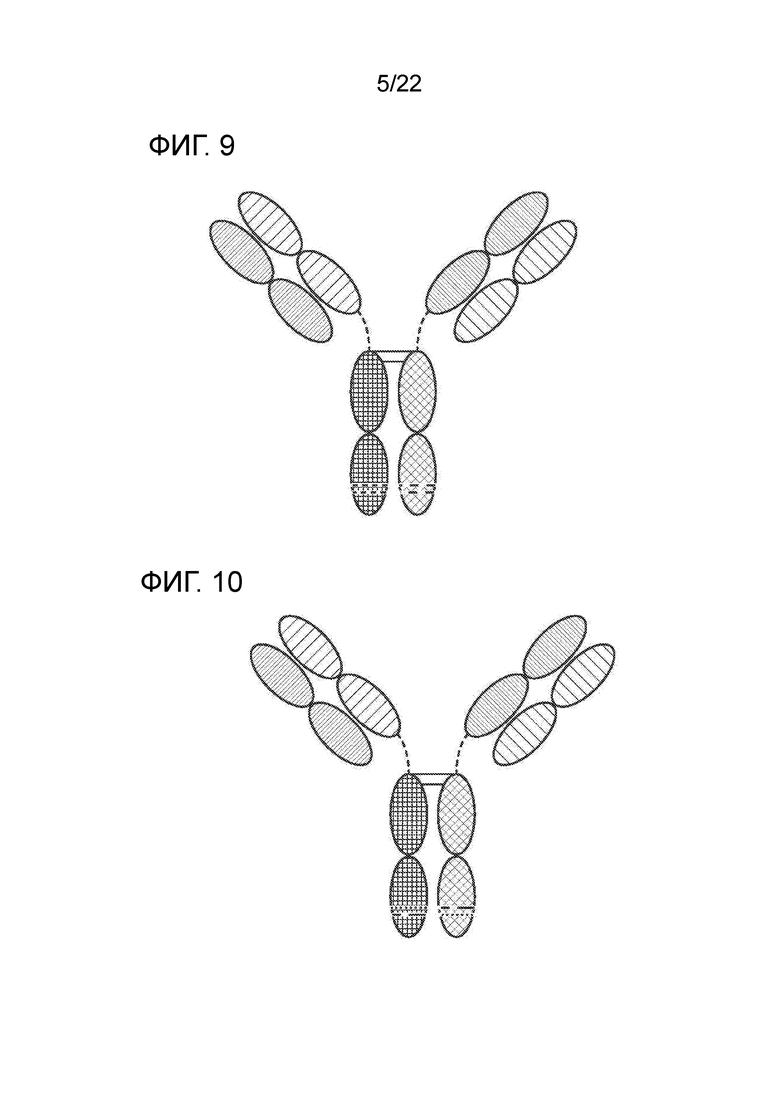
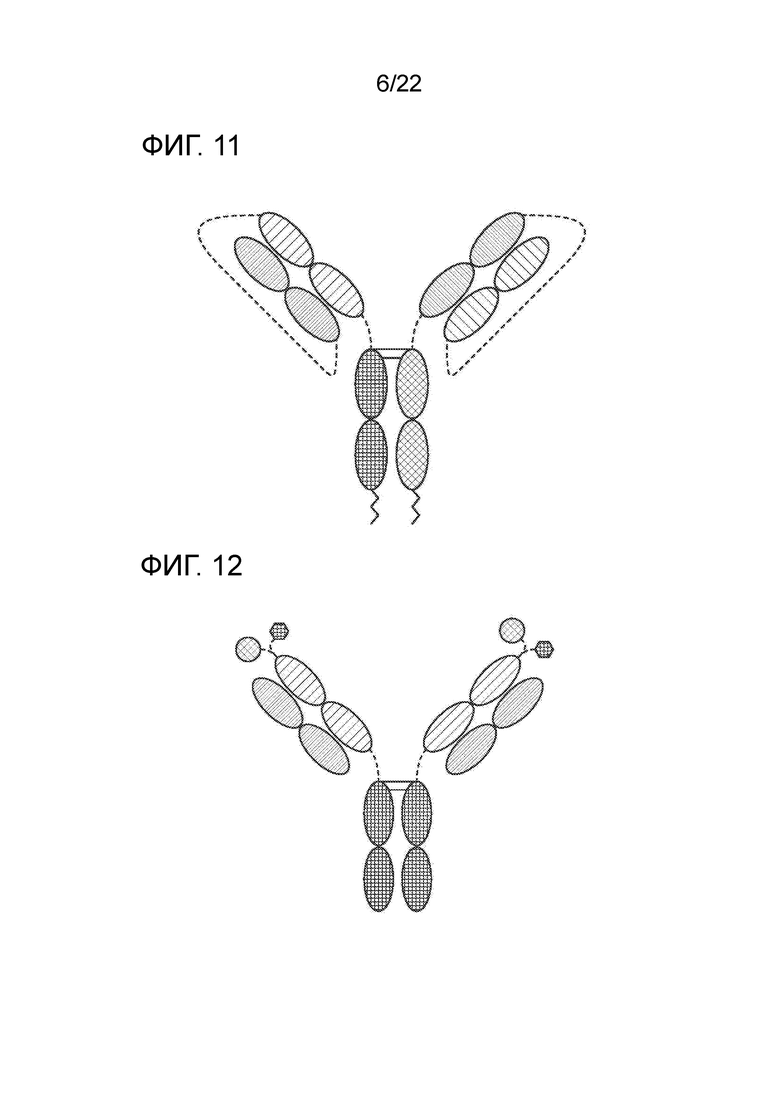
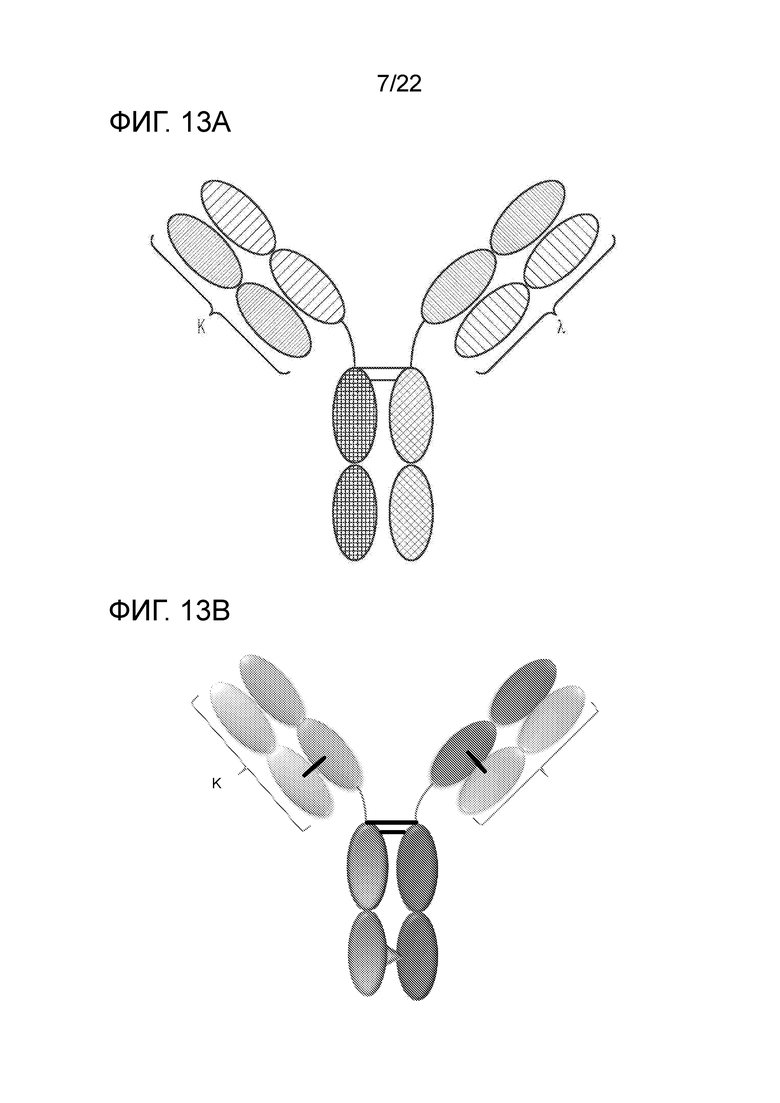

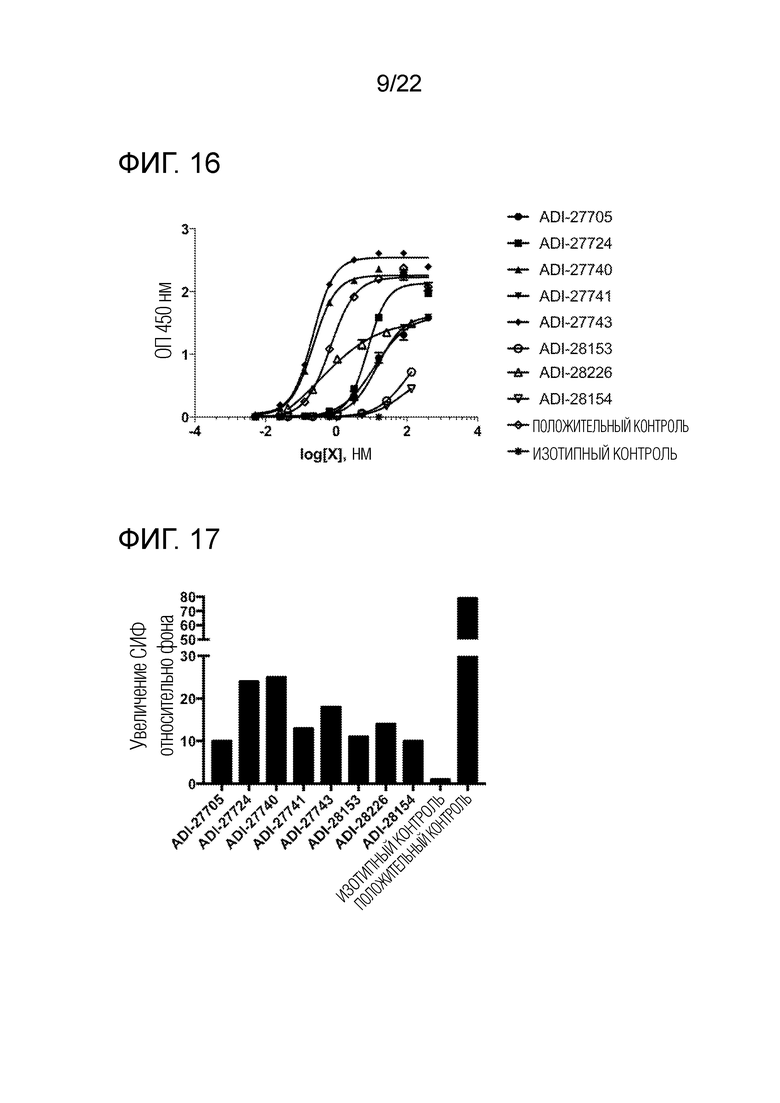
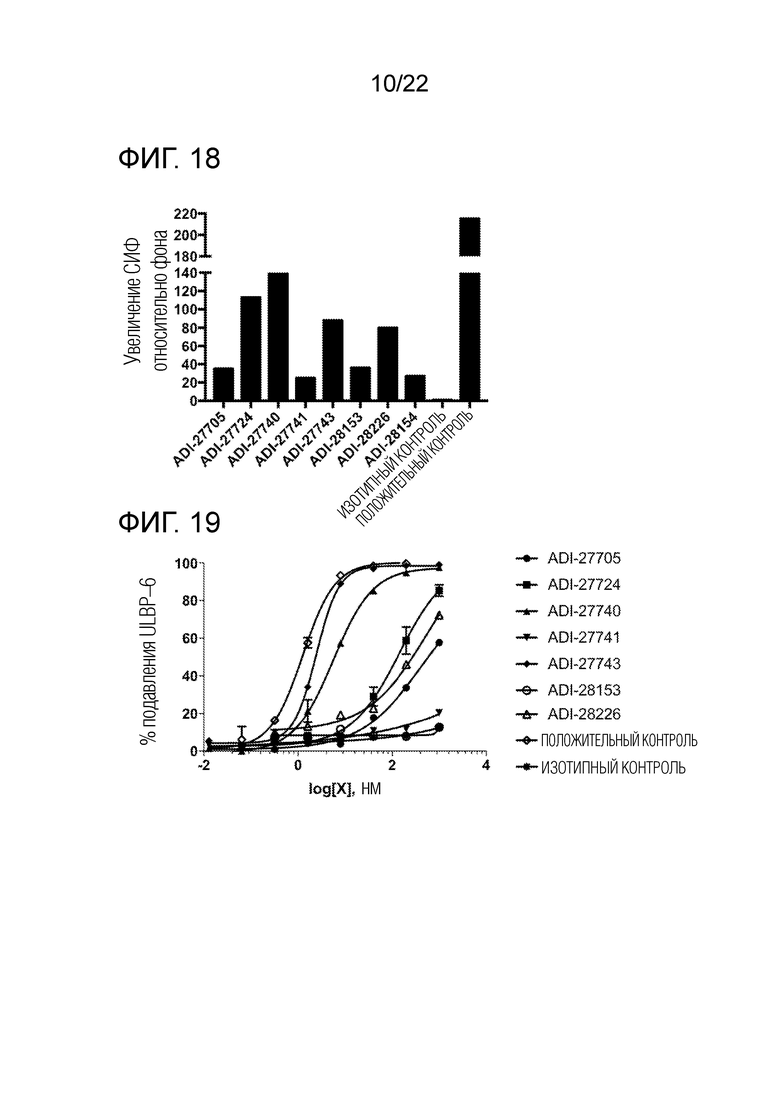

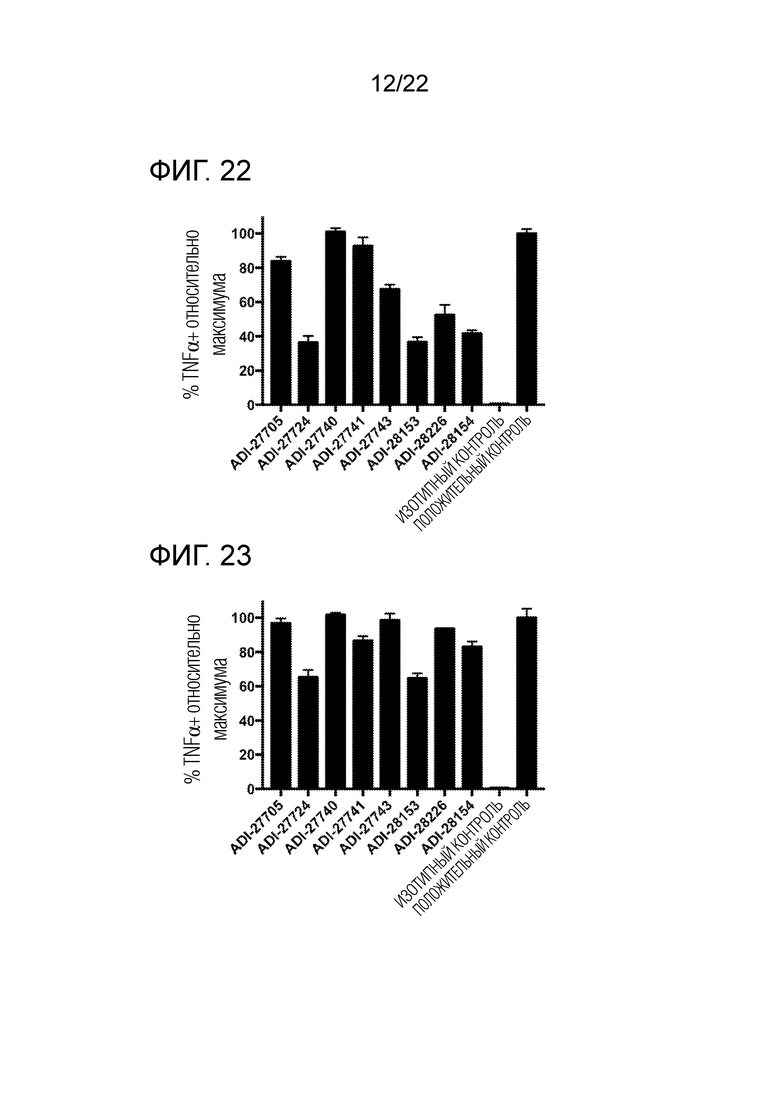
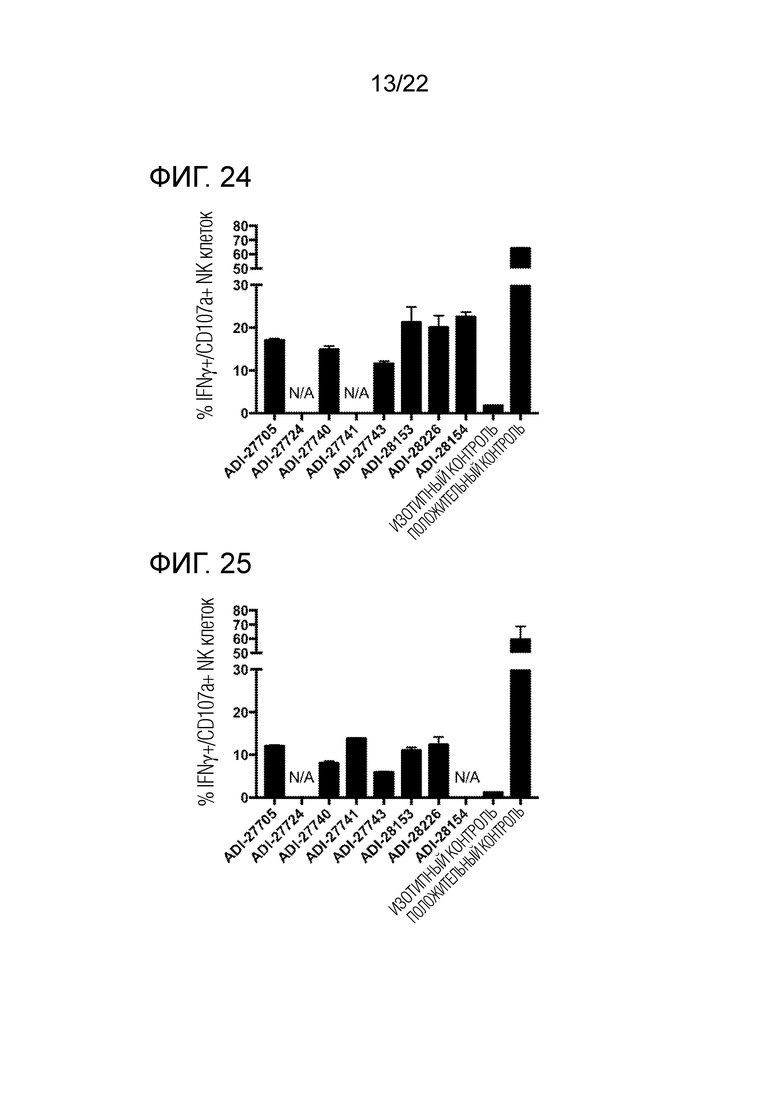
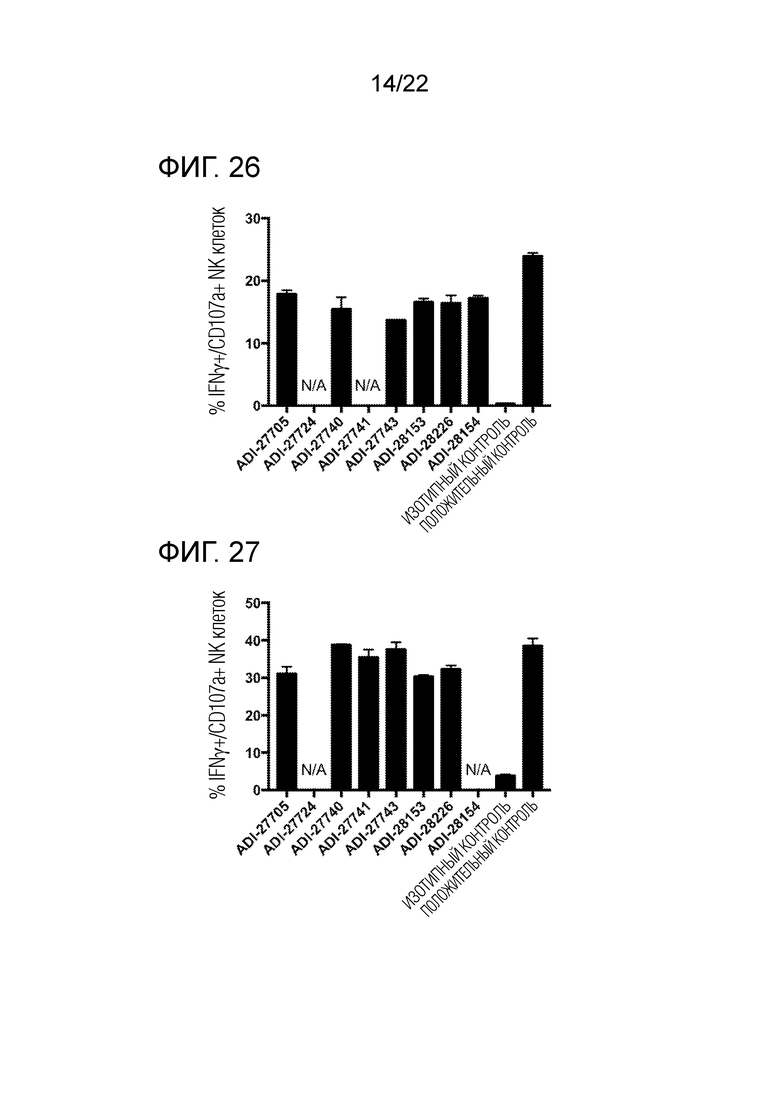
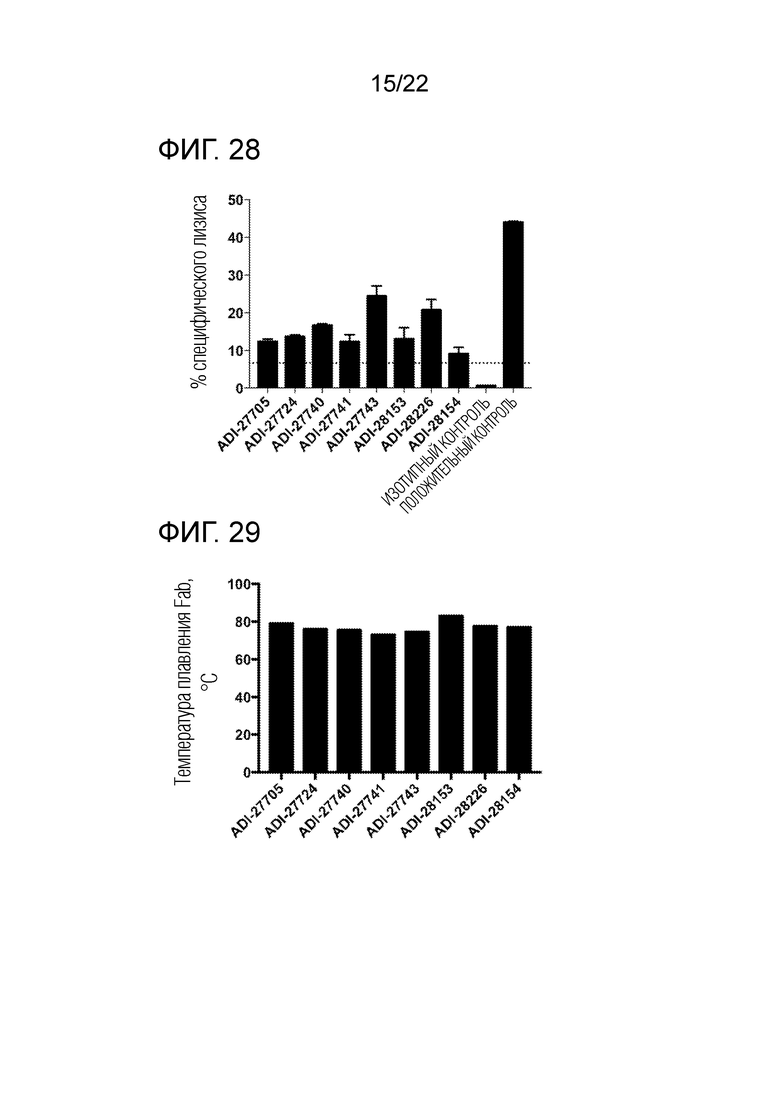

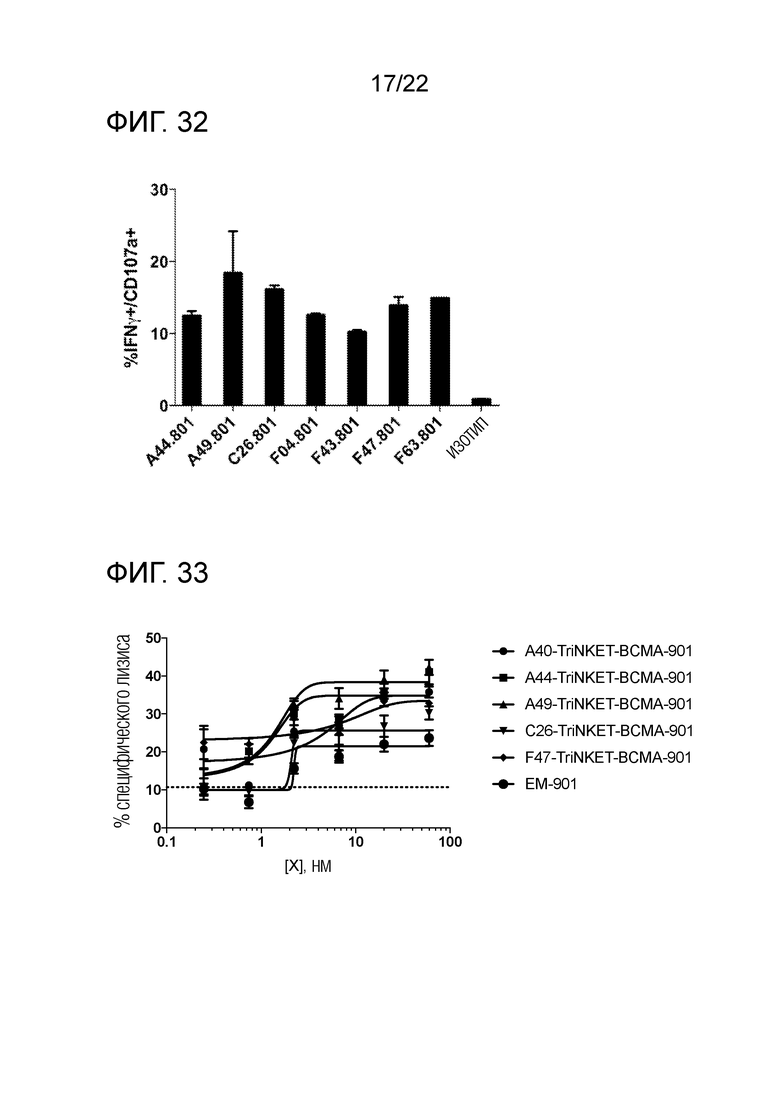
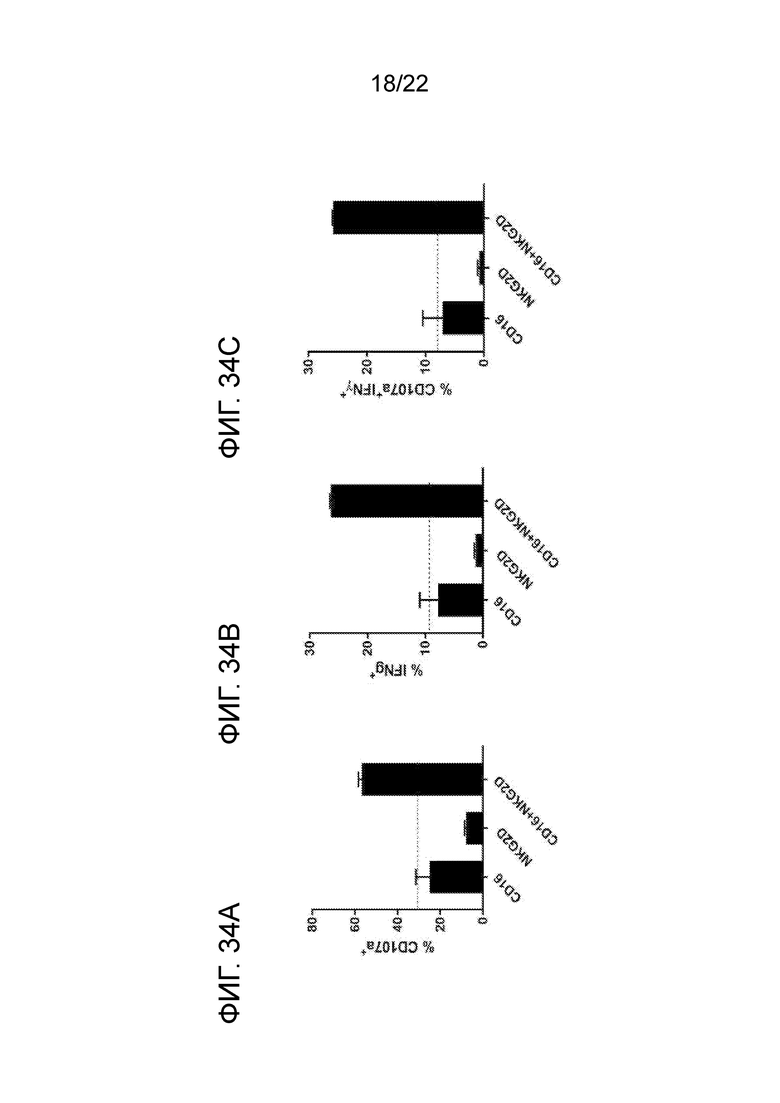
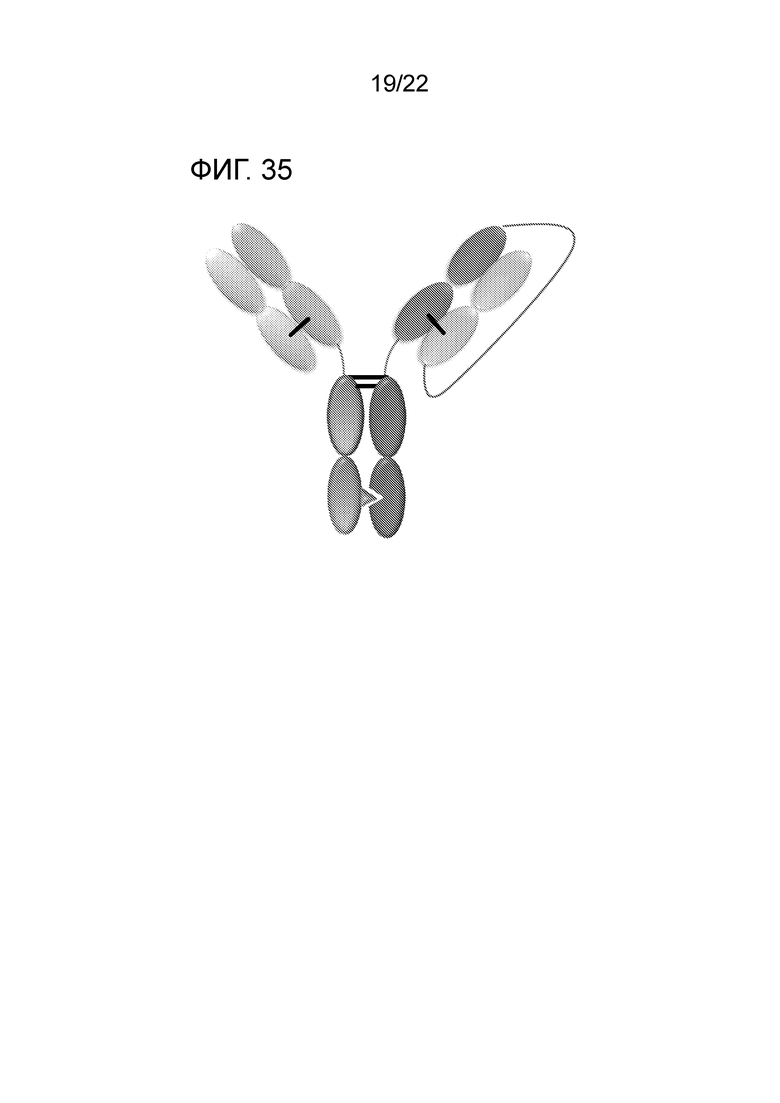
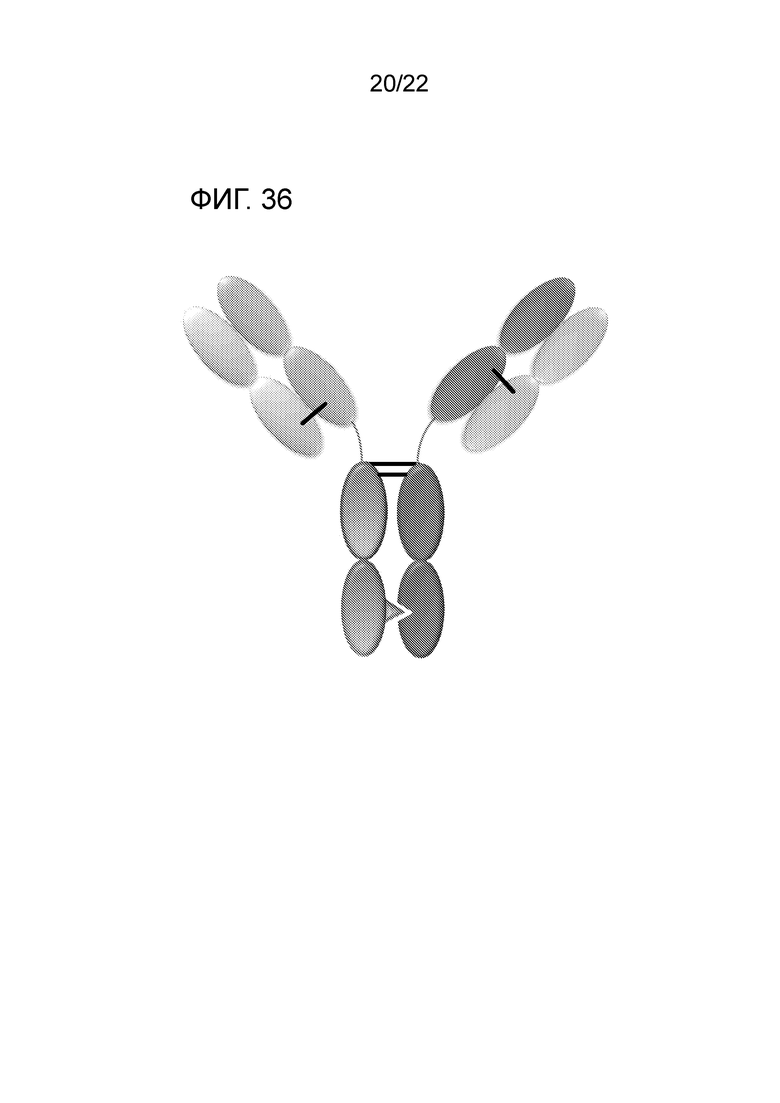
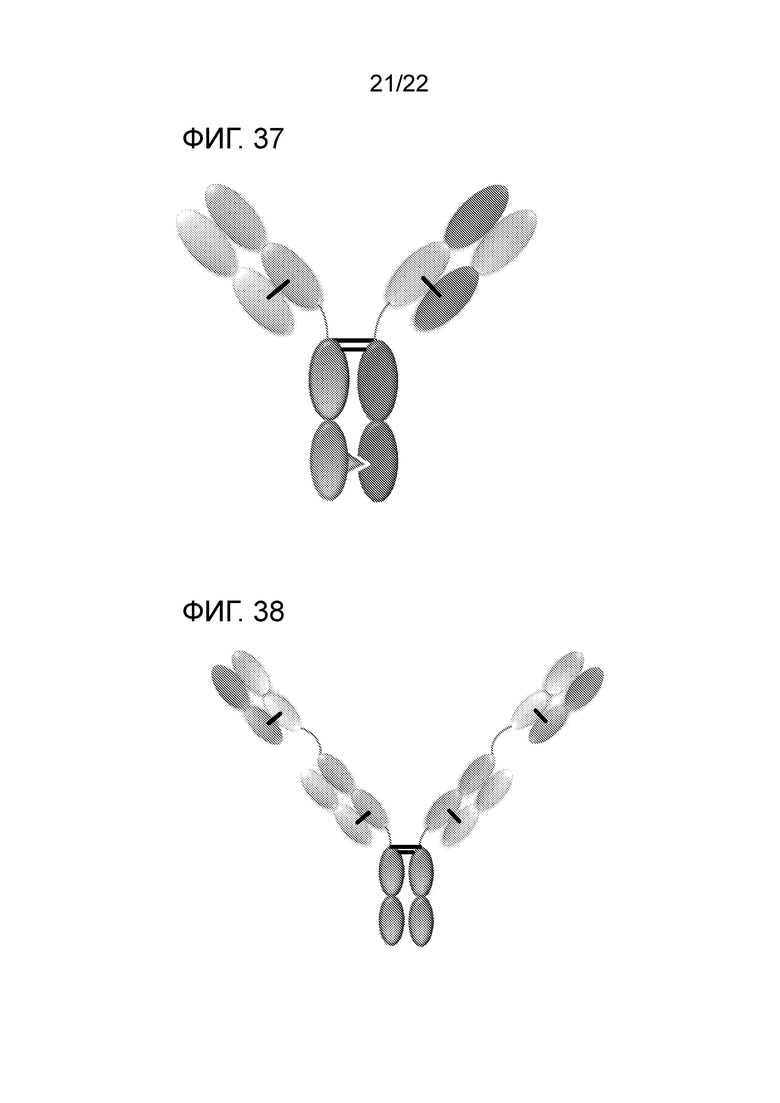
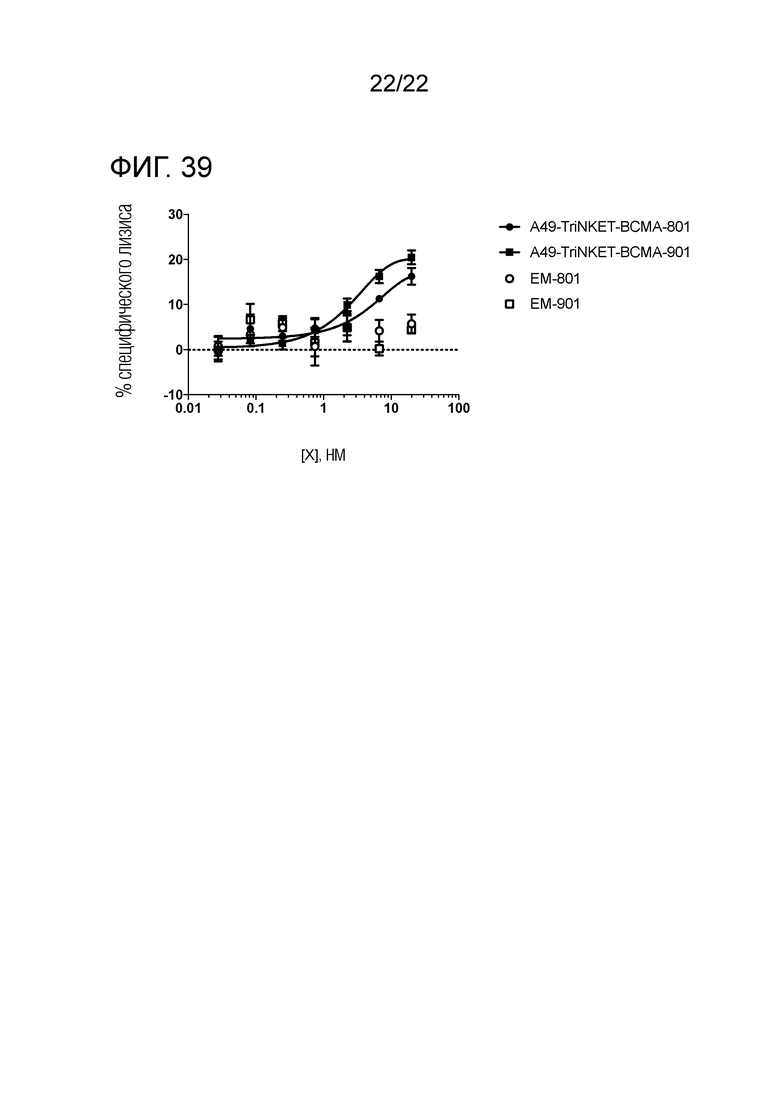
Изобретение относится к биохимии и медицине, в частности к полиспецифическим связывающим белкам, которые связываются с BCMA, рецептором NKG2D и CD16, а также фармацевтическим композициям и терапевтическим способам, полезным для лечения рака. Настоящее изобретение обеспечивает определенные преимущества для улучшения лечения рака, экспрессирующего ВСМА. 5 н. и 22 з.п. ф-лы, 39 ил., 8 табл., 11 пр.
1. Полиспецифический связывающий белок, который связывается с NKG2D, BCMA и CD16, содержащий:
(а) первый антигенсвязывающий участок молекулы иммуноглобулина, который связывается с NKG2D, где указанный первый антигенсвязывающий участок содержит вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL);
(b) второй антигенсвязывающий участок молекулы иммуноглобулина, который связывается с ВСМА, где указанный второй антигенсвязывающий участок содержит VH и VL; и
(c) домен Fc антитела или его часть, достаточную для связывания CD16, где указанный домен Fc антитела содержит первый домен Fc и второй домен Fc, и
где указанные VH или VL первого антигенсвязывающего участка связаны с указанным первым доменом Fc и указанные VH или VL второго антигенсвязывающего участка связаны с указанным вторым доменом Fc, и
где указанный второй антигенсвязывающий участок содержит:
(i) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 131 или SEQ ID NO: 143, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 132, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 133, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 134, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 135, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 136;
(ii) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 50, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 51, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 52, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 55, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 56, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 57 или SEQ ID NO: 58;
(iii) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 79, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 80, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 81, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 82, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 83, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 84;
(iv) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 85, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 86, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 87, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 88, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 89, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 90;
(v) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 125, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 126, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 127, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 128, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 129, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 130; или
(vi) последовательность CDR1 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 137, последовательность CDR2 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 138, последовательность CDR3 тяжелой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 139, последовательность CDR1 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 140, последовательность CDR2 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 141, и последовательность CDR3 легкой цепи, идентичной аминокислотной последовательности, представленной в SEQ ID NO: 142.
2. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок связывается с NKG2D у людей, приматов, не являющихся людьми, и грызунов.
3. Полиспецифический связывающий белок по п.1, в котором указанный VH первого антигенсвязывающего участка и указанный VL первого антигенсвязывающего участка находятся в одном и том же полипептиде.
4. Полиспецифический связывающий белок по п.1, в котором указанный VH второго антигенсвязывающего участка и указанный VL второго антигенсвязывающего участка находятся в одном и том же полипептиде.
5. Полиспецифический связывающий белок по п.1, в котором указанный VL первого антигенсвязывающего участка имеет аминокислотную последовательность, идентичную аминокислотной последовательности указанного VL второго антигенсвязывающего участка.
6. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 1.
7. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 101, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную SEQ ID NO: 102.
8. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 41, и аминокислотную последовательность VL, по меньшей мере на 95% идентичный SEQ ID NO: 42.
9. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 43, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную SEQ ID NO: 44.
10. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 45, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную SEQ ID NO: 46.
11. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 47, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную SEQ ID NO: 48.
12. Полиспецифический связывающий белок по п.1, в котором первый антигенсвязывающий участок содержит:
(i) последовательность CDR1 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 111, последовательность CDR2 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 112, последовательность CDR3 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 113, последовательность CDR1 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 114, последовательность CDR2 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 115, и последовательность CDR3 легкой цепи, идентичную к аминокислотной последовательности, представленной в SEQ ID NO: 116;
(ii) последовательность CDR1 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 64, последовательность CDR2 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 65, и последовательность CDR3 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 66;
(iii) последовательность CDR1 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 67, последовательность CDR2 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 68, последовательность CDR3 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 69, последовательность CDR1 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 70, последовательность CDR2 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 71, и последовательность CDR3 легкой цепи, идентичную к аминокислотной последовательности, представленной в SEQ ID NO: 72; или
(iv) последовательность CDR1 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 73, последовательность CDR2 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 74, последовательность CDR3 тяжелой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 75, последовательность CDR1 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 76, последовательность CDR2 легкой цепи, идентичную аминокислотной последовательности, представленной в SEQ ID NO: 77, и последовательность CDR3 легкой цепи, идентичную к аминокислотной последовательности, представленной в SEQ ID NO: 78.
13. Полиспецифический связывающий белок по п.1, где второй антигенсвязывающий сайт содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 93, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную SEQ ID NO: 94.
14. Полиспецифический связывающий белок по п.1, в котором второй антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 49, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную последовательности SEQ ID NO: 53 или SEQ ID NO: 54.
15. Полиспецифический связывающий белок по п.1, в котором второй антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 59, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную последовательности SEQ ID NO: 60.
16. Полиспецифический связывающий белок по п.1, в котором второй антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 61, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную последовательности SEQ ID NO: 62.
17. Полиспецифический связывающий белок по п.1, в котором второй антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 123, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную последовательности SEQ ID NO: 124.
18. Полиспецифический связывающий белок по п.1, в котором второй антигенсвязывающий участок содержит аминокислотную последовательность VH, по меньшей мере на 95% идентичную SEQ ID NO: 91, и аминокислотную последовательность VL, по меньшей мере на 95% идентичную последовательности SEQ ID NO: 92.
19. Полиспецифический связывающий белок по п.1, где указанный полиспецифический белок содержит указанную часть домена Fc антитела, достаточную для связывания CD16, причем указанный домен Fc антитела содержит шарнирный и CH2 домены.
20. Полиспецифический связывающий белок по п.19, в котором указанный домен Fc антитела содержит шарнирный и СН2 домены человеческого антитела IgG1.
21. Полиспецифический связывающий белок по п.19, в котором указанный домен Fc антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную аминокислотам 234–332 человеческого IgG1 антитела.
22. Полиспецифический связывающий белок по п.19, в котором указанный домен Fc антитела содержит аминокислотную последовательность, по меньшей мере на 95% идентичную домену Fc человеческого IgG1, и отличается в одном или более положений, выбранных из группы, состоящей из Q347, Y349, L351, S354, E356, E357, K360, Q362, S364, T366, L368, K370, N390, K392, T394, D399, S400, D401, F405, Y407, K409, T411 и K439.
23. Фармацевтический состав для лечения рака, экспрессирующего ВСМА, содержащий эффективное количество полиспецифического связывающего белка по любому из предшествующих пунктов и фармацевтически приемлемый носитель.
24. Клетка-хозяин для получения полиспецифического связывающего белка по любому из пп.1–22, содержащая один или более экспрессионных векторов, содержащих одну или более нуклеиновых кислот, кодирующих полиспецифический связывающий белок по любому из пп.1–22.
25. Способ усиления гибели клеток опухоли, экспрессирующей BCMA, включающий приведение клетки опухоли, экспрессирующей BCMA, и натуральной киллерной клетки в контакт с полиспецифическим связывающим белком по любому из пп.1–22.
26. Способ лечения рака, экспрессирующего ВСМА, включающий введение пациенту эффективного количества полиспецифического связывающего белка по любому из пп.1–22.
27. Способ по п.26, в котором рак, экспрессирующий ВСМА, выбирают из группы, состоящей из множественной миеломы, острого миеломоноцитарного лейкоза, Т–клеточной лимфомы, острого моноцитарного лейкоза и фолликулярной лимфомы.
| WO 2016166629 A1, 20.10.2016 | |||
| COLMAN P | |||
| M., Effects of amino acid sequence changes on antibody-antigen interactions, Research in Immunology, 1994, V | |||
| Заслонка для русской печи | 1919 |
|
SU145A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| SAFDARI Y | |||
| et al., Antibody humanization methods-a review and update, Biotechnology and Genetic Engineering Reviews, 2013, V | |||
| Солесос | 1922 |
|
SU29A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| SHEN J | |||
| et al., Single variable | |||

