ПЕРЕКРЕСТНАЯ ССЫЛКА НА СВЯЗАННЫЕ ЗАЯВКИ
[0001] Эта заявка испрашивает преимущество и приоритет заявки США № 62/464 341, поданной 27 февраля 2017 г., заявки США № 62/464 344, поданной 27 февраля 2017 г., заявки США № 62/464 347, поданной 27 февраля 2017 г., заявки США № 62/467 557, поданной 6 марта 2017 г., заявки США № 62/473 652, поданной 20 марта 2017 г., и заявки США № 62/473 659, поданной 20 марта 2017г., полное содержание каждой из которых настоящим включено в качестве ссылки в полном объеме для всех целей.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит список последовательностей, который представлен в электронном виде в формате ASCII и включен в настоящее описание в качестве ссылки в полном объеме. Указанная копия ASCII, созданная 27 февраля 2018 года, имеет имя DFY-012WO_ST25.txt и имеет размер 212 480 байтов.
ОБЛАСТЬ ТЕХНИКИ
[0003] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с рецептором NKG2D, CD16, и с опухолеспецифическим антигеном, выбранным из карбоангидразы 9 (CAIX), аноктамина-1 (ANO1), мезотелина, антигена клеточной поверхности трофобласта (TROP2), карциноэмбрионального антигена (СЕА) и клаудина-18.2.
Уровень техники
[0004] Злокачественное новообразование продолжает оставаться серьезной проблемой для здоровья, несмотря на значительные исследовательские усилия и научные достижения для лечения этого заболевания, о которых сообщается в литературе. Некоторые из наиболее часто диагностируемых видов злокачественных новообразований включают рак предстательной железы, рак молочной железы и рак легкого. Рак предстательной железы является наиболее распространенной формой злокачественных новообразований у мужчин. Рак молочной железы остается основной причиной смерти у женщин. Мелкоклеточный рак легкого остается одним из самых смертоносных злокачественных новообразований. Существующие методы лечения этих видов злокачественных новообразований не эффективны для всех пациентов и/или могут иметь существенные побочные эффекты. Другие виды злокачественных новообразований также остаются сложными для лечения с использованием существующих методов лечения.
[0005] Противоопухолевая иммунотерапия желательна, потому что она высокоспецифична и может способствовать разрушению опухолевых клеток с использованием собственной иммунной системы пациента. Слитые белки, такие как биспецифичные белки, рекрутирующие T-клетки, представляют собой противоопухолевые иммунотерапевтические средства, описанные в литературе, которые связываются с опухолевыми клетками и T-клетками для облегчения разрушения опухолевых клеток. Антитела, которые связываются с определенными опухолеспецифическими антигенами и с определенными иммунными клетками, описаны в литературе. См., например, WO 2016/134371 и WO 2015/095412.
[0006] Естественные клетки-киллеры (NK) являются компонентом врожденной иммунной системы и составляют приблизительно 15% циркулирующих лимфоцитов. NK-клетки проникают практически во все ткани, и первоначально характеризовались своей способностью эффективно уничтожать опухолевые клетки без необходимости предварительной сенсибилизации. Активированные NK-клетки уничтожают клетки-мишени с помощью средств, аналогичных цитотоксическим Т-клеткам, то есть с помощью цитолитических гранул, которые содержат перфорин и гранзимы, а также посредством путей рецепторов клеточной смерти. Активированные NK-клетки также секретируют воспалительные цитокины, такие как IFN-γ и хемокины, которые способствуют рекрутингу других лейкоцитов в ткани-мишени.
[0007] NK-клетки реагируют на сигналы через различные активирующие и ингибирующие рецепторы на своей поверхности. Например, когда NK-клетки встречают здоровые аутоклетки, их активность ингибируется активацией иммуноглобулино-подобных рецепторов (KIR) клеток-киллеров. Альтернативно, когда NK-клетки встречают чужеродные клетки или опухолевые клетки, они активируются через свои активирующие рецепторы (например, NKG2D, NCR, DNAM1). NK-клетки также активируются константной областью некоторых иммуноглобулинов через рецепторы CD16 на их поверхности. Общая чувствительность NK-клеток к активации зависит от суммы стимулирующих и ингибирующих сигналов.
[0008] Карбоангидраза 9 (CAIX) является металлоферментом цинка и относится к семейству ферментов, которые участвуют в обратимой гидратации углекислого газа с образованием бикарбоната и ионов водорода. CAIX имеет ограниченную экспрессию в нормальных тканях, но сверхэкспрессируется в широком спектре солидных опухолей и играет важную роль в развитии опухоли. CAIX, индуцированная гипоксическим опухолевым микроокружением, функционирует для борьбы с вредными эффектами высокого уровня гликолитического метаболизма в опухолевых клетках, поддерживая внутриклеточный рН, благоприятный для роста и выживания опухолевых клеток, в то же время создавая кислое внеклеточное пространство, облегчающее инвазивность опухолевых клеток. До настоящего времени сверхэкспрессия CAIX была обнаружена при почечно-клеточной карциноме, раке молочной железы, глиобластоме, раке головы и шеи, раке желудка, раке мочевого пузыря, раке яичников, а также при распространенных эпителиальных злокачественных новообразованиях, таких как карциномы пищевода, легкого, толстой кишки, почки, шейки матки, и при немелкоклеточной карциноме легкого.
[0009] Мезотелин представляет собой гликозилфосфатидилинозитол (GPI), закрепленный на клеточной поверхности белка, и экспрессируется в мезотелиальных клетках плевры, брюшины и перикарда. В дополнение к своей нормальной экспрессии, мезотелин сверхэкспрессируется при различных видах злокачественных новообразований, включающих мезотелиому, рак яичников, рак поджелудочной железы, немелкоклеточный рак легкого, рак молочной железы, холангиокарциному, рак желудка, серозную карциному матки, карциному тимуса и острый миелоидный лейкоз. Предполагается, что мезотелин может играть важную роль в развитии опухоли, включая клеточную адгезию, выживание/пролиферацию клеток и химиорезистентность.
[0010] Аноктамин-1 (ANO1) представляет собой чувствительный к напряжению кальций-активированный хлоридный канал. Он также известен под другими названиями, такими как TMEM16, TAOS2, ORAOV2 и DOG-1. Кальций-активированные хлоридные каналы функционируют во многих физиологических процессах, включая трансэпителиальную секрецию, возбуждение сердца и нейронов, сенсорную трансдукцию, сокращение гладких мышц и фертилизацию. ANO1 потенциально участвует в секреции эпителиальной жидкости, обонятельной и фототрансдукции, возбудимости нейронов и сердца и регуляции тонуса сосудов, включая моторику кишечника. ANO1 также высоко экспрессируется при некоторых злокачественных новообразованиях, где считается, что он увеличивает пролиферацию клеток, миграцию клеток и метастазирование, например, при плоскоклеточном раке пищевода (ESCC), стромальной опухоли желудочно-кишечного тракта (GIST), плоскоклеточной карциноме головы и шеи (HNSCC), раке поджелудочной железы, раке молочной железы, раке предстательной железы и саркоме. Обнаружено, что ~85% опухолей HNSCC являются ANO1-положительными, а у пациентов с HNSCC с высоким уровнем опухолевой экспрессии ANO1 общая выживаемость снижается; также установлено, что сверхэкспрессия ANO1 тесно связана со злокачественной опухолью рака предстательной железы и плохим прогнозом при раке молочной железы.
[0011] TROP2 представляет собой трансмембранный гликопротеин и трансдуктор сигнала Ca2+. В дополнение к его экспрессии в некоторых нормальных тканях, TROP2 сверхэкспрессируется при множестве злокачественных новообразований, включая рак молочной железы, легких, желудка, колоректальный рак, рак поджелудочной железы, предстательной железы, шейки матки, рак головы и шеи, карцинома носоглотки и карцинома яичника. Он передает сигналы опухолевым клеткам о пролиферации, инвазии и выживании.
[0012] Карциноэмбриональный антиген (СЕА), также известный как CD66e или CEACAM5, является членом суперсемейства иммуноглобулинов. Это гликопротеин с большой клеточной поверхностью, который в основном служит молекулой клеточной адгезии, опосредующей межклеточный контакт. Помимо функций клеточной адгезии и миграции, CEA сверхэкспрессируется при высоком проценте злокачественных новообразований человека, включая 90% рака желудочно-кишечного тракта, колоректального рака и поджелудочной железы, 70% немелкоклеточного рака легкого и 50% рака молочной железы. Было показано, что сверхэкспрессия СЕА коррелирует с онкогенностью и повышенной инвазивностью опухоли.
[0013] Клаудин-18.2 является одной из двух изоформ Клаудина-18, основным компонентом плотных контактов, которые регулируют проницаемость, барьерную функцию и полярность эпителиальных слоев. Claudin-18.2 экспрессируется только в короткоживущих дифференцированных клетках слизистой оболочки желудка в здоровых тканях. Тем не менее, он сверхэкспрессируется в до 80% желудочно-кишечных аденокарцином и 60% опухолей поджелудочной железы. Кроме того, его экспрессия наблюдалась у отдельных групп людей с раком пищевода, немелкоклеточной карциномой легкого, раком яичника, раком толстой кишки и некоторыми формами карциномы желчных протоков.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0014] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2 на опухолевой клетке и с рецептором NKG2D и рецептором CD16 на естественных клетках-киллерах. Такие белки могут взаимодействовать более чем с одним типом рецепторов, активирующих NK, и могут блокировать связывание природных лигандов с NKG2D. В некоторых вариантах осуществления белок может быть агонистом NK-клеток у людей и у других видов, таких как грызуны и яванские макаки. Различные аспекты и варианты осуществления изобретения описаны более подробно ниже.
[0015] Соответственно, один аспект изобретения относится к белку, который включает первый антигенсвязывающий сайт, который связывается с NKG2D; второй антигенсвязывающий сайт, который связывается с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2; и Fc-домен антитела, его часть, достаточную для связывания CD16, или третий антигенсвязывающий сайт, который связывается с CD16. Каждый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи антитела и вариабельный домен легкой цепи антитела (например, расположенный как в антителе или слитый вместе с scFv), или один или более антигенсвязывающих сайтов могут быть однодоменным антителом, таким как антитело VHH, такое как верблюжье антитело, или антитело VNAR, подобное тем, которые обнаружены у хрящевых рыб.
[0016] В одном варианте осуществления первый антигенсвязывающий сайт, который связывается с NKG2D, включает вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 1, как например, благодаря присутствию аминокислотной последовательности по меньшей мере на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%), идентичной SEQ ID NO: 1, и/или включая аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 51), CDR2 (SEQ ID NO: 52) и CDR3 (SEQ ID NO: 53) SEQ ID NO: 1. Вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 1, может быть связан с различными вариабельными доменами легкой цепи с образованием сайта связывания NKG2D. Например, первый антигенсвязывающий сайт, который включает вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 1, может дополнительно включать вариабельный домен легкой цепи, выбранный из любой из последовательностей, связанных с SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36 и 40. Например, первый антигенсвязывающий сайт включает вариабельный домен тяжелой цепи с аминокислотными последовательностями, по меньшей мере, на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичными SEQ ID NO: 1, и вариабельный домен легкой цепи с аминокислотными последовательностями, по меньшей мере, на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичными любой из последовательностей, выбранных из SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 22, 24, 26, 28, 30, 32, 34, 36 и 40. Альтернативно, первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 41, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 42. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 41 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 54), CDR2 (SEQ ID NO: 55) и CDR3 (SEQ ID NO: 56) SEQ ID NO: 41. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 42 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 57), CDR2 (SEQ ID NO: 58) и CDR3 (SEQ ID NO: 59) SEQ ID NO: 42. В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 43, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 44. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 43 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 60), CDR2 (SEQ ID NO: 61) и CDR3 (SEQ ID NO: 62) SEQ ID NO: 43. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 44 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 63), CDR2 (SEQ ID NO: 64) и CDR3 (SEQ ID NO: 65) SEQ ID NO: 44. В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 45, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 46. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 45 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 66), CDR2 (SEQ ID NO: 67) и CDR3 (SEQ ID NO: 68) SEQ ID NO: 45. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 46 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 69), CDR2 (SEQ ID NO: 70) и CDR3 (SEQ ID NO: 71) SEQ ID NO: 46.
[0017] Альтернативно, первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 47, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 48, например, путем наличия аминокислотных последовательностей, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичных SEQ ID NO: 47 и SEQ ID NO: 48, соответственно. В другом варианте осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 49, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 50, например, благодаря наличию аминокислотных последовательностей, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичных SEQ ID NO: 49 и SEQ ID NO: 50, соответственно.
[0018] В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 132, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 133. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 132 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 134), CDR2 (SEQ ID NO: 135) и CDR3 (SEQ ID NO: 136) SEQ ID NO: 132. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 133 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 137), CDR2 (SEQ ID NO: 138) и CDR3 (SEQ ID NO: 139) последовательности SEQ ID NO: 133.
[0019] В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 140, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 141. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 140 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 142), CDR2 (SEQ ID NO: 143) и CDR3 (SEQ ID NO: 144) SEQ ID NO: 140. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 141 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 145), CDR2 (SEQ ID NO: 146) и CDR3 (SEQ ID NO: 147) последовательности SEQ ID NO: 141.
[0020] В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 148, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 149. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 148 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 150), CDR2 (SEQ ID NO: 151) и CDR3 (SEQ ID NO: 152) SEQ ID NO: 148. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 149 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 153), CDR2 (SEQ ID NO: 154) и CDR3 (SEQ ID NO: 155) SEQ ID NO: 149.
[0021] В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 156, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 157. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 156 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 158), CDR2 (SEQ ID NO: 159) и CDR3 (SEQ ID NO: 160) SEQ ID NO: 156. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 157 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 161), CDR2 (SEQ ID NO: 162) и CDR3 (SEQ ID NO: 163) SEQ ID NO: 157.
[0022] В других вариантах осуществления первый антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 164, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 165. Например, вариабельный домен тяжелой цепи первого антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 164 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 166), CDR2 (SEQ ID NO: 167) и CDR3 (SEQ ID NO: 168) SEQ ID NO: 164. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 165 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 169), CDR2 (SEQ ID NO: 170) и CDR3 (SEQ ID NO: 171) SEQ ID NO: 165.
[0023] Второй антигенсвязывающий сайт может связываться с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2. В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с CAIX и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут быть связаны с образованием сайта связывания CAIX. Например, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 72, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 73. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 72 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 74), CDR2 (SEQ ID NO: 75) и CDR3 (SEQ ID NO: 76) SEQ ID NO: 72. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 73 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 77), CDR2 (SEQ ID NO: 78) и CDR3 (SEQ ID NO: 79) последовательности SEQ ID NO: 73.
[0024] Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 80, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 81. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 80 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 82), CDR2 (SEQ ID NO: 83) и CDR3 (SEQ ID NO: 84) последовательности SEQ ID NO: 80. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 81 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 85), CDR2 (SEQ ID NO: 86) и CDR3 (SEQ ID NO: 87) последовательности SEQ ID NO: 81.
[0025] Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 88, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 89. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 88 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 90), CDR2 (SEQ ID NO: 91) и CDR3 (SEQ ID NO: 92) последовательности SEQ ID NO: 88. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 89 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 93), CDR2 (SEQ ID NO: 94) и CDR3 (SEQ ID NO: 95) последовательности SEQ ID NO: 89.
[0026] В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с ANO1 и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут связываться с образованием сайта связывания ANO1. Например, второй антигенсвязывающий сайт может включать комплекс вариабельного домена тяжелой цепи, связанного с SEQ ID NO: 97, и вариабельного домена легкой цепи, связанного с SEQ ID NO: 98. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 97 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 99), CDR2 (SEQ ID NO: 100) и CDR3 (SEQ ID NO: 101) последовательности SEQ ID NO: 97. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 98 и/или включает аминокислотные последовательности, идентичные последовательностям CDR1 (SEQ ID NO: 102), CDR2 (SEQ ID NO: 103) и CDR3 (SEQ ID NO: 104) последовательности SEQ ID NO: 98.
[0027] В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с мезотелином и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут быть связаны с образованием сайта связывания мезотелина. Например, второй антигенсвязывающий сайт может включать набор вариабельного домена тяжелой цепи, связанного с SEQ ID NO: 106, и вариабельного домена легкой цепи, связанного с SEQ ID NO: 107. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 106 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 108), CDR2 (SEQ ID NO: 109) и CDR3 (SEQ ID NO: 110) последовательности SEQ ID NO: 106. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 107 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 111), CDR2 (SEQ ID NO: 112) и CDR3 (SEQ ID NO: 113) последовательности SEQ ID NO: 107. Альтернативно, второй антигенсвязывающий сайт может включать набор вариабельного домена тяжелой цепи, связанного с SEQ ID NO: 114, и вариабельного домена легкой цепи, связанного с SEQ ID NO: 115. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 114 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 116), CDR2 (SEQ ID NO: 117) и CDR3 (SEQ ID NO: 118) последовательности SEQ ID NO: 115. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 115 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 119), CDR2 (SEQ ID NO: 120) и CDR3 (SEQ ID NO: 121) последовательности SEQ ID NO: 115. Альтернативно, второй антигенсвязывающий сайт может включать набор вариабельного домена тяжелой цепи, связанного с SEQ ID NO: 122, и вариабельного домена легкой цепи, связанного с SEQ ID NO: 123. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 122 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 124), CDR2 (SEQ ID NO: 125) и CDR3 (SEQ ID NO: 126) последовательности SEQ ID NO: 122. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 123 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 127), CDR2 (SEQ ID NO: 128) и CDR3 (SEQ ID NO: 129) последовательности SEQ ID NO: 123.
[0028] В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с TROP2 и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут связываться с образованием сайта связывания TROP2. Например, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 172, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 173. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 172 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 174), CDR2 (SEQ ID NO: 175) и CDR3 (SEQ ID NO: 176) последовательности SEQ ID NO: 172. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 173 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 177), CDR2 (SEQ ID NO: 178) и CDR3 (SEQ ID NO: 179) последовательности SEQ ID NO: 173. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 180, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 181. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 180 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 182), CDR2 (SEQ ID NO: 183) и CDR3 (SEQ ID NO: 184) последовательности SEQ ID NO: 180. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 181 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 185), CDR2 (SEQ ID NO: 186) и CDR3 (SEQ ID NO: 187) последовательности SEQ ID NO: 181. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 188, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 189. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 188, и вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть по меньшей мере на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%), идентичен SEQ ID NO: 189. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 190, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 191. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 190, и вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть по меньшей мере на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%), идентичен SEQ ID NO: 191. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 192, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 193. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 192, и вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть по меньшей мере на 90% (например, на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 193.
[0029] В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с СЕА и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут быть связаны с образованием сайта связывания СЕА. Например, второй сайт связывания антигена может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 195, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 196. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 195 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 197), CDR2 (SEQ ID NO: 198) и CDR3 (SEQ ID NO: 199) последовательности SEQ ID NO: 195. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 196 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 200), CDR2 (SEQ ID NO: 201) и CDR3 (SEQ ID NO: 202) последовательности SEQ ID NO: 196. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 203, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 204. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 203 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 205), CDR2 (SEQ ID NO: 206) и CDR3 (SEQ ID NO: 207) последовательности SEQ ID NO: 203. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 204 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 208), CDR2 (SEQ ID NO: 209) и CDR3 (SEQ ID NO: 210) последовательности SEQ ID NO: 204. Альтернативно, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 211, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 212. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 211 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 213), CDR2 (SEQ ID NO: 214) и CDR3 (SEQ ID NO: 215) последовательности SEQ ID NO: 212. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 196 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 216), CDR2 (SEQ ID NO: 217) и CDR3 (SEQ ID NO: 218) последовательности SEQ ID NO: 212.
[0030] В некоторых вариантах осуществления второй антигенсвязывающий сайт связывается с Клаудином-18.2 и включает вариабельный домен тяжелой цепи и вариабельный домен легкой цепи, которые могут быть связаны с образованием сайта связывания Клаудина-18.2. Например, второй антигенсвязывающий сайт может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 220, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 221. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 220 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 222), CDR2 (SEQ ID NO: 223) и CDR3 (SEQ ID NO: 224) последовательности SEQ ID NO: 220. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 221 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 225), CDR2 (SEQ ID NO: 226) и CDR3 (SEQ ID NO: 227) последовательности SEQ ID NO: 221. Например, второй сайт связывания антигена может включать вариабельный домен тяжелой цепи, связанный с SEQ ID NO: 228, и вариабельный домен легкой цепи, связанный с SEQ ID NO: 229. Например, вариабельный домен тяжелой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 228 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 230), CDR2 (SEQ ID NO: 231) и CDR3 (SEQ ID NO: 232) последовательности SEQ ID NO: 228. Аналогично, вариабельный домен легкой цепи второго антигенсвязывающего сайта может быть, по меньшей мере, на 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%) идентичен SEQ ID NO: 229 и/или включает аминокислотные последовательности, идентичные CDR1 (SEQ ID NO: 233), CDR2 (SEQ ID NO: 234) и CDR3 (SEQ ID NO: 235) последовательности SEQ ID NO: 229.
[0031] В некоторых вариантах осуществления второй антигенсвязывающий сайт включает вариабельный домен легкой цепи, имеющий аминокислотную последовательность, идентичную аминокислотной последовательности вариабельного домена легкой цепи, присутствующей в первом антигенсвязывающем сайте.
[0032] В некоторых вариантах осуществления белок включает часть Fc-домена антитела, достаточную для связывания CD16, где Fc-домен антитела содержит шарнирные и CH2-домены и/или аминокислотные последовательности, по меньшей мере, на 90% идентичные аминокислотной последовательности 234-332 последовательности человеческого антитела IgG.
[0033] Предложены составы, содержащие один из этих белков; клетки, содержащие одну или более нуклеиновых кислот, экспрессирующих эти белки, и способы усиления гибели опухолевых клеток с использованием этих белков.
[0034] Другой аспект изобретения относится к способу лечения злокачественного новообразования у пациента. Способ включает введение пациенту, нуждающемуся в этом, терапевтически эффективного количества описанного в настоящем документе полиспецифичного связывающего белка. Злокачественные новообразования, подлежащие лечению с использованием CAIX-нацеленных полиспецифических связывающих белков, включают, например, почечно-клеточную карциному, рак молочной железы, глиобластому, рак головы и шеи, рак желудка, рак мочевого пузыря, рак яичников, опухоли пищевода, легких, толстой кишки, рак почки, шейки матки и немелкоклеточную карциному легкого. Злокачественные новообразования, подлежащие лечению с использованием мезотелин-нацеленных полиспецифических связывающих белков, включают, например, мезотелиому, рак яичников, рак поджелудочной железы, немелкоклеточный рак легкого, рак молочной железы, холангиокарциному, рак желудка, серозную карциному матки, карциному тимуса и острый миелоидный лейкоз. Злокачественные новообразования, подлежащие лечению с использованием ANO1-нацеленных полиспецифических связывающих белков, включают, например, плоскоклеточный рак пищевода (ESCC), желудочно-кишечную стромальную опухоль (GIST), плоскоклеточную карциному головы и шеи (HNSCC), рак поджелудочной железы, рак молочной железы, рак предстательной железы и саркому. Злокачественные новообразования, подлежащие лечению с использованием TROP2-нацеленных полиспецифических связывающих белков, включают, например, рак молочной железы, легких, желудка, колоректальный рак, рак поджелудочной железы, предстательной железы, шейки матки, рак головы и шеи, карциному носоглотки и карциному яичника. Злокачественные новообразования, подлежащие лечению с использованием CEA-нацеленных полиспецифических связывающих белков, включают, например, рак желудочно-кишечного тракта, колоректальный рак, рак поджелудочной железы, немелкоклеточный рак легкого и рак молочной железы. Злокачественные новообразования, подлежащие лечению с использованием клаудин-18.2-нацеленных полиспецифических связывающих белков, включают, например, рак пищевода, немелкоклеточную карциному легкого, рак яичника, рак толстой кишки и несколько форм карциномы желчных протоков.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0035] ФИГ. 1 представляет гетеродимерное полиспецифическое антитело. Каждое плечо может представлять собой либо NKG2D-связывающий домен, либо один из опухолеспецифических антигенсвязывающих доменов, выбранный из доменов, связывающихся с CAIX, ANO1, мезотелином, TROP2, CEA и Клаудином-18.2. В некоторых вариантах осуществления NKG2D- и опухолеспецифические антигенсвязывающие домены могут иметь общую легкую цепь.
[0036] ФИГ. 2 представляет гетеродимерное полиспецифическое антитело. Либо NKG2D-, либо опухолеспецтфтческий антигенсвязывающий домен, выбранный из доменов, связывающихся с CAIX, ANO1, мезотелином, TROP2, CEA и Клаудином-18.2, может принимать формат scFv (правое плечо).
[0037] ФИГ. 3 представляет собой линейные графики, демонстрирующие аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным NKG2D человека в анализе ИФА.
[0038] ФИГ. 4 представляет собой линейные графики, демонстрирующие аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным NKG2D яванской макаки в анализе ИФА.
[0039] ФИГ. 5 представляет собой линейные графики, демонстрирующие аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным NKG2D мыши в анализе ИФА.
[0040] ФИГ. 6 представляет собой гистограммы, демонстрирующие с помощью проточной цитометрии связывание NKG2D-связывающих доменов (перечисленных в виде клонов) с клетками EL4, экспрессирующими NKG2D человека, демонстрируя кратное превышение средней интенсивности флуоресценции (MFI) относительно фона.
[0041] ФИГ. 7 представляет собой гистограммы, демонстрирующие с помощью проточной цитометрии связывание NKG2D-связывающих доменов (перечисленных в виде клонов) с клетками EL4, экспрессирующими мышиный NKG2D, демонстрируя кратное превышение средней интенсивности флуоресценции (MFI) относительно фона.
[0042] ФИГ. 8 представляет собой линейные графики, демонстрирующие специфическую аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным человеческим NKG2D-Fc путем конкуренции с природным лигандом ULBP-6.
[0043] ФИГ. 9 представляет собой линейные графики, демонстрирующие специфическую аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным человеческим NKG2D-Fc путем конкуренции с природным лигандом MICA.
[0044] ФИГ. 10 представляет собой линейные графики, демонстрирующие специфическую аффинность связывания NKG2D-связывающих доменов (перечисленных в виде клонов) с рекомбинантным мышиным NKG2D-Fc путем конкуренции с природным лигандом Rae-1 дельта.
[0045] ФИГ. 11 представляет собой гистограммы, показывающие активацию человеческого NKG2D посредством NKG2D-связывающих доменов (перечисленных в виде клонов) путем количественного определения процента TNF-альфа-положительных клеток, которые экспрессируют слитые белки человеческий NKG2D-CD3 дзета.
[0046] ФИГ. 12 представляет собой гистограммы, показывающие активацию мышиного NKG2D с помощью NKG2D-связывающих доменов (перечисленных в виде клонов) путем количественного определения процента TNF-альфа-положительных клеток, которые экспрессируют слитые белки мышиный NKG2D-CD3 дзета.
[0047] ФИГ. 13 представляет собой гистограммы, показывающие активацию человеческих NK-клеток NKG2D-связывающими доменами (перечислены в виде клонов).
[0048] ФИГ. 14 представляет собой гистограммы, показывающие активацию человеческих NK-клеток NKG2D-связывающими доменами (перечислены в виде клонов).
[0049] ФИГ. 15 представляет собой гистограммы, показывающие активацию мышиных NK-клеток NKG2D-связывающими доменами (перечислены в виде клонов).
[0050] ФИГ. 16 представляет собой гистограммы, показывающие активацию мышиных NK-клеток NKG2D-связывающими доменами (перечислены в виде клонов).
[0051] ФИГ. 17 представляет собой гистограммы, показывающие цитотоксический эффект NKG2D-связывающих доменов (перечисленных в виде клонов) в отношении опухолевых клеток.
[0052] ФИГ. 18 представляет собой гистограммы, показывающие температуру плавления NKG2D-связывающих доменов (перечисленных в виде клонов), измеренную с помощью дифференциальной сканирующей флуориметрии.
[0053] ФИГ. 19А-19С представляет собой гистограммы синергетической активации NK-клеток с использованием связывания CD16 и NKG2D. ФИГ. 19А демонстрирует уровень CD107a; ФИГ. 19В демонстрирует уровень IFN-γ; ФИГ 19С демонстрирует уровень продуцирования CD107a и IFN-γ. Графики показывают среднее (n=2) ± SD. Данные представляют пять независимых экспериментов с использованием пяти разных здоровых доноров.
[0054] ФИГ. 20 представляет собой TriNKET в форме Triomab, которое представляет собой трифункциональное биспецифичное антитело, которое поддерживает IgG-подобную форму. Эта химера состоит из двух половинных антител, каждое с одной легкой и одной тяжелой цепью, которые происходят от двух родительских антител. Форма Triomab может представлять собой гетеродимерную конструкцию, содержащую 1/2 антитела крысы и 1/2 антитела мыши.
[0055] ФИГ. 21 представляет собой TriNKET в форме общей легкой цепи (LC) KiH, которая включает технологию «выступы-во-впадины» (KIH). KiH представляет собой гетеродимер, содержащий 2 Fab, связывающихся с мишенью 1 и 2, и Fc, стабилизированный мутациями гетеродимеризации. TriNKET в формате KiH может представлять собой гетеродимерную конструкцию с 2 Fab, связывающимися с мишенью 1 и мишенью 2, и содержит 2 различных тяжелых цепи и общую легкую цепь, которая спаривается с обеими тяжелыми цепями.
[0056] ФИГ. 22 представляет собой TriNKET в форме иммуноглобулина с двумя вариабельными доменами (DVD-Ig™), который объединяет домены, связывающиеся с мишенью двух моноклональных антител через гибкие встречающиеся в природе линкеры с получением четырехвалентной IgG-подобной молекулы. DVD-Ig™ представляет собой гомодимерную конструкцию, в которой вариабельный домен, нацеленный на антиген 2, слит с N-концом вариабельного домена Fab, нацеленного на антиген 1. Конструкция содержит нормальный Fc.
[0057] ФИГ. 23 представляет собой TriNKET в форме интерфейса ортогонального Fab (Орто-Fab), который представляет собой гетеродимерную конструкцию, которая содержит 2 Fab-фрагмента, связывающихся с мишенью 1 и мишенью 2, слитых с Fc. Спаривание LC-HC обеспечивается ортогональным интерфейсом. Гетеродимеризация обеспечивается мутациями в Fc.
[0058] ФИГ. 24 представляет собой TrinKET в формате 2-в-1 Ig.
[0059] ФИГ. 25 представляет собой TriNKET в форме ES, которая представляет собой гетеродимерную конструкцию, содержащую 2 различных Fab-фрагмента, связывающихся с мишенью 1 и мишенью 2, слитых с Fc. Гетеродимеризация обеспечивается мутациями электростатической регуляции в Fc.
[0060] ФИГ. 26 представляет собой TriNKET в форме с обменом Fab-фрагментами: антитела, которые обмениваются Fab-фрагментами путем замены тяжелой цепи и присоединенной легкой цепи (полу-молекулы) с парой тяжелых-легких цепей из другой молекулы, с получением биспецифичных антител. KiH представляет собой гетеродимер, содержащий 2 Fab, связывающихся с мишенью 1 и 2, и Fc, стабилизированный мутациями гетеродимеризации.
[0061] ФИГ. 27 представляет собой TriNKET в форме SEED Body, который представляет собой гетеродимер, содержащий 2 Fab-фрагмента, связывающихся с мишенью 1 и 2, и FC, стабилизированный мутациями гетеродимеризации.
[0062] ФИГ. 28 представляет собой TriNKET в форме LuZ-Y, в котором домен лейциновой застежки используется для индуцирования гетеродимеризации двух различных HC. LuZ-Y форма представляет собой гетеродимер, содержащий 2 разных scFab, связывающихся с мишенью 1 и 2, слитых с Fc. Гетеродимеризация обеспечивается благодаря мотивам лейциновой застежки, слитым с С-концом Fc.
[0063] ФИГ. 29 представляет собой TriNKET в форме Cov-X-Body.
[0064] ФИГ. 30A-30B - представляют собой TriNKET в формах ĸλ-Body, которые представляют собой гетеродимерные конструкции с двумя различными Fab, слитыми с Fc, стабилизированными мутациями гетеродимеризации: Fab 1, нацеленный на антиген 1, содержит каппа LC, а второй Fab, нацеленный на антиген 2, содержит лямбда LC. ФИГ. 30А является иллюстративным изображением одной формы ĸλ-Body; ФИГ. 30B является иллюстративным изображением другого ĸλ-Body.
[0065] ФИГ. 31 представляет собой гетеродимерную конструкцию Oasc-Fab, которая включает Fab, связывающийся с мишенью 1, и scFab, связывающийся с мишенью 2, слитые с Fc. Гетеродимеризация обеспечивается мутациями в Fc.
[0066] ФИГ. 32 представляет собой DuetMab, который представляет собой гетеродимерную конструкцию, содержащую 2 разных Fab, связывающихся с антигеном 1 и 2, и Fc, стабилизированный мутациями гетеродимеризации. Fab 1 и 2 содержат различные SS-мостики, которые обеспечивают правильное спаривание легкой цепи (LC) и тяжелой цепи (HC).
[0067] ФИГ. 33 представляет собой CrossmAb, который представляет собой гетеродимерную конструкцию с 2 различными Fab, связывающимися с мишенью 1 и 2, слитыми с Fc, стабилизированным гетеродимеризацией. Домены CL и CH1 и домены VH и VL переключаются, например, CH1 сливается в одной линии с VL, в то время как CL сливается в одной линии с VH.
[0068] ФИГ. 34 представляет собой Fit-Ig, который представляет собой гомодимерные конструкции, в которых Fab, связывающийся с антигеном 2, слит с N-концом HC Fab, который связывается с антигеном 1. Конструкция содержит Fc дикого типа.
[0069] ФИГ. 35 показывает связывание TriNKET, которые включают CAIX-связывающий домен и NKG2D-связывающий домен с NKG2D, экспрессируемым на клетках EL4.
[0070] ФИГ. 36 показывает связывание TriNKET, которые включают CAIX-связывающий домен и NKG2D-связывающий домен, с CAIX, экспрессируемым на клетках почечной клеточной карциномы человека A498.
[0071] ФИГ. 37A показывает TriNKET-опосредованную цитотоксичность эффекторных клеток KHYG-1 по отношению к CAIX-положительным клеткам почечно-клеточной карциномы A498. Пунктирная линия указывает на фоновое уничтожение клеток A498 во время совместного культивирования с покоящимися клетками KHYG-1 без добавления TriNKET. ФИГ. 37B показывает TriNKET-опосредованную цитотоксичность покоящихся NK-клеток в отношении CAIX-положительных клеток почечной карциномы A498.
ПОДРОБНОЕ ОПИСАНИЕ
[0072] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2 на опухолевой клетке и с рецептором NKG2D и рецептором CD16 на естественных клетках-киллерах, чтобы активировать естественные клетки-киллеры, к фармацевтическим композициям, содержащим такие полиспецифические связывающие белки, и к терапевтическим способам с использованием таких полиспецифических белков и фармацевтических композиций, в том числе для лечения злокачественного новообразования. Различные аспекты изобретения изложены ниже в разделах; однако аспекты изобретения, описанные в одном конкретном разделе, не должны быть ограничены каким-либо конкретным разделом.
[0073] Чтобы облегчить понимание настоящего изобретения, ряд терминов и выражений определены ниже.
[0074] Термин в единственном числе, используемый в настоящем описании, означает «один или более» и включают множественное число, если контекст не является неподходящим. Используемый в настоящем описании термин «антигенсвязывающий сайт» относится к части молекулы иммуноглобулина, которая участвует в связывании антигена. В антителах человека антигенсвязывающий сайт образуется аминокислотными остатками N-концевых вариабельных («V») областей тяжелой («H») и легкой («L») цепей. Три сильно расходящихся участка в V-областях тяжелой и легкой цепей называются «гипервариабельными областями», которые расположены между более консервативными фланкирующими участками, известными как «каркасные области» или «FR». Таким образом, термин «FR» относится к аминокислотным последовательностям, которые естественным образом находятся между и по соседству с гипервариабельными областями в иммуноглобулинах. В молекуле человеческого антитела три гипервариабельные области легкой цепи и три гипервариабельные области тяжелой цепи располагаются относительно друг друга в трехмерном пространстве с образованием антигенсвязывающей поверхности. Антигенсвязывающая поверхность комплементарна трехмерной поверхности связанного антигена, и три гипервариабельные области каждой из тяжелых и легких цепей обозначаются как «области, определяющие комплементарность» или «CDR». У некоторых животных, таких как верблюды и хрящевые рыбы, антигенсвязывающий сайт образован одной цепью антитела, образующей «однодоменное антитело». Антигенсвязывающие сайты могут существовать в интактном антителе, в антигенсвязывающем фрагменте антитела, который сохраняет антигенсвязывающую поверхность, или в рекомбинантном полипептиде, таком как scFv, с использованием пептидного линкера для соединения вариабельного домена тяжелой цепи с вариабельным доменом легкой цепи в одном полипептиде.
[0075] Используемый в настоящем описании термин «опухолеспецифический антиген» означает любой антиген, включая, но не ограничиваясь этим, белок, гликопротеин, ганглиозид, углевод, липид, который ассоциирован со злокачественным новообразованием. Такой антиген может экспрессироваться на злокачественных клетках или в микроокружении опухоли, как например, на кровеносных сосудах, ассоциированных с опухолью, внеклеточном матриксе, мезенхимальной строме или иммунных инфильтратах.
[0076] Используемые в настоящем описании термины «субъект» и «пациент» относятся к организму, подлежащему лечению способами и композициями, описанными в настоящем документе. Такие организмы предпочтительно включают, но не ограничиваются ими, млекопитающих (например, мышей, обезьян, лошадей, крупный рогатый скот, свиней, собак, кошек и тому подобное) и, более предпочтительно, включают человека.
[0077] Используемый в настоящем описании термин «эффективное количество» относится к количеству соединения (например, соединения по настоящему изобретению), достаточному для достижения полезных или желаемых результатов. Эффективное количество может быть введено за одно или более введений, применений или дозировок и не предназначено для ограничения конкретным составом или путем введения. Используемый в настоящем описании термин «лечение» включает любой эффект, например уменьшение, снижение, модуляцию, улучшение или устранение, которое приводит к улучшению состояния, заболевания, расстройства и т.п. или ослаблению его симптома.
[0078] Используемый в настоящем описании термин «фармацевтическая композиция» относится к комбинации активного агента с носителем, инертным или активным, что делает композицию особенно подходящей для диагностического или терапевтического применения in vivo или ex vivo.
[0079] Используемый в настоящем описании термин «фармацевтически приемлемый носитель» относится к любому из стандартных фармацевтических носителей, таких как фосфатно-буферный солевой раствор, вода, эмульсии (например, такие как эмульсии масло/вода или вода/масло) и различные типы смачивающих агентов. Композиции также могут включать стабилизаторы и консерванты. Для примеров носителей, стабилизаторов и адъювантов см., например, Martin, Remington's Pharmaceutical Sciences, 15th Ed., Mack Publ. Co., Easton, PA [1975].
[0080] Используемый в настоящем описании термин «фармацевтически приемлемая соль» относится к любой фармацевтически приемлемой соли (например, кислоты или основания) соединения по настоящему изобретению, которая при введении пациенту способна обеспечить соединение по настоящему изобретению или активный метаболит или его остаток. Как известно специалистам в данной области техники, «соли» соединений по настоящему изобретению могут быть получены из неорганических или органических кислот и оснований. Примеры кислот включают, но не ограничиваются ими, соляную, бромводородную, серную, азотную, хлорную, фумаровую, малеиновую, фосфорную, гликолевую, молочную, салициловую, янтарную, толуол-п-сульфоновую, винную, уксусную, лимонную, метансульфоновую, этансульфоновую, муравьиную, бензойную, малоновую, нафталин-2-сульфоновую, бензолсульфоновую кислота и тому подобное. Другие кислоты, такие как щавелевая кислота, хотя сами по себе они не являются фармацевтически приемлемыми, могут быть использованы при получении солей, полезных в качестве промежуточных соединений при получении соединений по изобретению и их фармацевтически приемлемых кислотно-аддитивных солей.
[0081] Примеры оснований включают, но не ограничиваются ими, гидроксиды щелочных металлов (например, натрия), гидроксиды щелочноземельных металлов (например, магния), аммиак и соединения формулы NW4+, где W представляет собой C1-4алкил и тому подобное.
[0082] Примеры солей включают, но не ограничиваются ими: ацетат, адипат, альгинат, аспартат, бензоат, бензолсульфонат, бисульфат, бутират, цитрат, камфорат, камфорсульфонат, циклопентанпропионат, диглюконат, додецилсульфат, этансульфонат, фумарат, флукогептаноат, глицерофосфат, гемисульфат, гептаноат, гексаноат, гидрохлорид, гидробромид, гидройодид, 2-гидроксиэтансульфонат, лактат, малеат, метансульфонат, 2-нафталинсульфонат, никотинат, оксалат, пальмоат, пектинат, персульфат, фенилпропионат, пикрат, пивалат, пропионат, сукцинат, тартрат, тиоцианат, тозилат, ундеканоат и тому подобное. Другие примеры солей включают анионы соединений по настоящему изобретению, соединенные с подходящим катионом, таким как Na+, NH4+ и NW4+ (где W представляет собой C1-4алкильную группу), и тому подобное.
[0083] Для терапевтического применения соли соединений по настоящему изобретению рассматриваются как фармацевтически приемлемые. Однако соли кислот и оснований, которые не являются фармацевтически приемлемыми, также могут найти применение, например, при получении или очистке фармацевтически приемлемого соединения.
[0084] Во всем описании, где композиции описаны как имеющие, включающие или содержащие конкретные компоненты, или где процессы и способы описаны как имеющие, включающие или содержащие конкретные стадии, предполагается, что дополнительно существуют композиции по настоящему изобретению, которые состоят по существу или состоят из перечисленных компонентов и что существуют процессы и способы согласно настоящему изобретению, которые состоят по существу или состоят из перечисленных стадий обработки.
[0085] Как правило, если в композиции указан процент, то это процент по массе, если не указано иное. Кроме того, если переменная не сопровождается определением, то предыдущее определение переменной является приоритетным.
I. Белки
[0086] Изобретение относится к полиспецифическим связывающим белкам, которые связываются с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2 на опухолевой клетке и рецептором NKG2D и рецептором CD16 на естественных клетках-киллерах, чтобы активировать естественную клетку-киллера. Полиспецифические связывающие белки пригодны в фармацевтических композициях и терапевтических способах, описанных в настоящем документе. Связывание полиспецифического связывающего белка с рецептором NKG2D и рецептором CD16 на естественной клетке-киллере усиливает активность естественной клетки-киллера в отношении разрушения опухолевой клетки. Связывание полиспецифического связывающего белка с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2 на опухолевой клетке приводит к тому, что опухолевая клетка приближается к естественной клетке-киллеру, что облегчает прямое и косвенное разрушение опухолевой клетки естественной клеткой-киллером. Дальнейшее описание примеров полиспецифических связывающих белков приведено ниже.
[0087] Первый компонент полиспецифических связывающих белков связывается с клетками, экспрессирующими рецептор NKG2D, которые могут включать, но не ограничиваются ими, NK-клетки, γδ T-клетки и CD8+ αβ T-клетки. При NKG2D-связывании полиспецифические связывающие белки могут блокировать природные лиганды, такие как ULBP6 и MICA, от связывания с NKG2D и активации рецепторов NKG2D.
[0088] Второй компонент полиспецифических связывающих белков связывается с клетками, экспрессирующими CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2. CAIX-экспрессирующие клетки включают, но не ограничиваются ими, почечно-клеточную карциному, рак молочной железы, глиобластому, рак головы и шеи, рак желудка, рак мочевого пузыря, рак яичников, опухоли пищевода, легких, толстой кишки, почек, шейки матки и немелкоклеточную карциному легких. Мезотелин-экспрессирующие клетки включают, но не ограничиваются ими, мезотелиому, рак яичника, рак поджелудочной железы, немелкоклеточный рак легкого, рак молочной железы, холангиокарциному, рак желудка, серозную карциному матки, карциному тимуса и острый миелоидный лейкоз. ANO1-экспрессирующие клетки включают, но не ограничиваются ими, плоскоклеточный рак пищевода (ESCC), желудочно-кишечную стромальную опухоль (GIST), плоскоклеточную карциному головы и шеи (HNSCC), рак поджелудочной железы, рак молочной железы, рак предстательной железы и саркому. TROP2-экспрессирующие клетки включают, но не ограничиваются ими, рак молочной железы, легких, желудка, колоректального, поджелудочной железы, предстательной железы, шейки матки, рак головы и шеи, карциному носоглотки и карциному яичника. CEA-экспрессирующие клетки включают, но не ограничиваются ими, рак желудочно-кишечного тракта, колоректальный рак, рак поджелудочной железы, немелкоклеточный рак легкого и рак молочной железы. Клаудин-18.2-экспрессирующие клетки включают, но не ограничиваются ими, рак пищевода, немелкоклеточную карциному легкого, рак яичников, рак толстой кишки и несколько форм карциномы желчных протоков.
[0089] Третий компонент для полиспецифических связывающих белков связывается с клетками, экспрессирующими CD16, Fc-рецептор на поверхности лейкоцитов, включая естественные клетки-киллеры, макрофаги, нейтрофилы, эозинофилы, тучные клетки и фолликулярные дендритные клетки.
[0090] Описанные в настоящем документе полиспецифические связывающие белки могут принимать различные форматы. Например, один формат представляет собой гетеродимерное полиспецифическое антитело, которое включает первую тяжелую цепь иммуноглобулина, первую легкую цепь иммуноглобулина и вторую тяжелую цепь иммуноглобулина и вторую легкую цепь иммуноглобулина (ФИГ. 1). Первая тяжелая цепь иммуноглобулина включает первый Fc-домен (шарнир-CH2-CH3), первый вариабельный домен тяжелой цепи и необязательный первый домен CH1 тяжелой цепи. Первая легкая цепь иммуноглобулина включает первый вариабельный домен легкой цепи и первый константный домен легкой цепи. Первая легкая цепь иммуноглобулина вместе с первой тяжелой цепью иммуноглобулина образует антигенсвязывающий сайт, который связывается с NKG2D. Вторая тяжелая цепь иммуноглобулина содержит второй Fc-домен (шарнир-CH2-CH3), второй вариабельный домен тяжелой цепи и, необязательно, второй домен CH1 тяжелой цепи. Вторая легкая цепь иммуноглобулина включает второй вариабельный домен легкой цепи и второй константный домен легкой цепи. Вторая легкая цепь иммуноглобулина вместе со второй тяжелой цепью иммуноглобулина образует антигенсвязывающий сайт, который связывается с CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2. Первый Fc-домен и второй Fc-домен вместе могут связываться с CD16 (ФИГ.1). В некоторых вариантах осуществления первая легкая цепь иммуноглобулина идентична второй легкой цепи иммуноглобулина.
[0091] Другой примерный формат включает гетеродимерное полиспецифическое антитело, включающее первую тяжелую цепь иммуноглобулина, вторую тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина (ФИГ. 2). Первая тяжелая цепь иммуноглобулина включает первый Fc-домен (шарнир-CH2-CH3), слитый либо через линкер, либо за счет шарнира антитела с одноцепочечным вариабельным фрагментом (scFv), состоящим из вариабельного домена тяжелой цепи и вариабельного домена легкой цепи, которые спариваются и связываются с NKG2D или связываются с одним из опухолеспецифических антигенов, выбранных из CAIX, ANO1, мезотелина, TROP2, CEA и Клаудина-18.2. Вторая тяжелая цепь иммуноглобулина включает второй Fc-домен (шарнир-CH2-CH3), второй вариабельный домен тяжелой цепи и, необязательно, домен тяжелой цепи CH1. Легкая цепь иммуноглобулина включает вариабельный домен легкой цепи и константный домен легкой цепи. Вторая тяжелая цепь иммуноглобулина соединяется с легкой цепью иммуноглобулина и связывается с NKG2D или связывается с одним из опухолеспецифических антигенов, выбранных из CAIX, ANO1, мезотелина, TROP2, CEA и Клаудина-18.2. Первый Fc-домен и второй Fc-домен вместе могут связываться с CD16 (ФИГ. 2).
[0092] Один или более дополнительных мотивов связывания могут быть слиты с С-концом домена СН3 константной области, необязательно через линкерную последовательность. В некоторых вариантах осуществления антигенсвязывающий сайт может представлять собой одноцепочечную или стабилизированную дисульфидной цепью вариабельную область (scFv), или может образовывать четырехвалентную или трехвалентную молекулу.
[0093] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Triomab, который представляет собой трифункциональное биспецифичное антитело, которое поддерживает IgG-подобную форму. Эта химера состоит из двух половинных антител, каждое с одной легкой и одной тяжелой цепью, которые происходят от двух родительских антител.
[0094] В некоторых вариантах осуществления полиспецифический связывающий белок представляет собой форму с общей легкой цепью (LC) KiH, в которой используется технология «выступы во впадины» (KIH). KIH включает сконструированные домены CH3 для создания либо «выступа», либо «впадины» в каждой тяжелой цепи для содействия гетеродимеризации. Концепция технологии Fc «выступы-во-впадины (KiH)» заключалась в том, чтобы ввести «выступ» в одном домене CH3 (CH3A) путем замены небольшого остатка на объемный (например, T366WCH3A в нумерации EU). Чтобы приспособить «выступ», на другом домене CH3 (CH3B) была создана комплементарная поверхность «впадины» путем замены ближайших соседних к выступу остатков на меньшие (т.е. T366S/L368A/Y407VCH3B). Мутация «впадины» была оптимизирована путем структурно-управляемого скрининга фаговой библиотеки (Atwell S, Ridgway JB, Wells JA, Carter P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. Biol. (1997) 270(1):26-35). Рентгенокристаллические структуры вариантов KiH Fc (Elliott JM, Ultsch M, Lee J, Tong R, Takeda K, Spiess C, et al., Antiparallel conformation of knob and hole aglycosylated half-antibody homodimers is mediated by a CH2-CH3 hydrophobic interaction. J. Mol. Biol. (2014) 426(9):1947-57; Mimoto F, Kadono S, Katada H, Igawa T, Kamikawa T, Hattori K. Crystal structure of a novel asymmetrically engineered Fc variant with improved affinity for FcgammaRs. Mol. Immunol. (2014) 58 (1): 132-8) продемонстрировали, что гетеродимеризация термодинамически поддерживается гидрофобными взаимодействиями, обусловленными стерической комплементарностью в коровом интерфейсе между CH3-доменами, тогда как интерфейсы выступ-выступ и впадина-впадина не благоприятствуют гомодимеризация вследствие стерических помех и нарушения благоприятных взаимодействий, соответственно.
[0095] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме иммуноглобулина с двумя вариабельными доменами (DVD-Ig™), который объединяет домены связывания мишеней двух моноклональных антител через гибкие природные линкеры с получением четырехвалентной IgG-подобной молекулы.
[0096] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме ортогонального Fab-интерфейса (Орто-Fab). В подходе ortho-Fab IgG (Lewis SM, Wu X, Pustilnik A, Sereno A, Huang F, Rick HL, et al., Generation of bispecific IgG antibodies by structure-based design of an orthogonal Fab interface. Nat. Biotechnol. (2014) 32 (2): 191-8), региональный дизайн на основе структуры вводит комплементарные мутации в интерфейсе LC и HCVH-CH1 только в одном Fab без каких-либо изменений в другом Fab.
[0097] В некоторых вариантах осуществления полиспецифический связывающий белок имеет формат 2-в-1 Ig. В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме ES, которая представляет собой гетеродимерную конструкцию, содержащую 2 различных Fab, связывающихся с мишенью 1 и мишенью 2, слитых с Fc. Гетеродимеризация обеспечивается мутациями электростатического взаимодействия в Fc.
[0098] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме κλ-Body, которая представляет собой гетеродимерные конструкции с 2 различными Fab, слитыми с Fc, стабилизированными мутациями гетеродимеризации: Fab1, нацеленный на антиген 1, содержит каппа-LC, тогда как второй Fab, нацеленный на антиген 2, содержит лямбда LC. ФИГ. 30А является примерным представлением одной формы ĸλ-Body; ФИГ. 30B является примерным представлением другого ĸλ-Body.
[0099] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме с обменом Fab-фрагментов (антитела, которые обмениваются Fab-фрагментами путем замены тяжелой цепи и присоединенной легкой цепи (полу-молекулы) на пару тяжелых-легких цепей из другой молекулы, что приводит к получению биспецифичных антител).
[00100] В некоторых вариантах осуществления полиспецифический связывающий белок имеет форму SEED Body. Сконструированная доменная платформа с обменом цепей (SEED) была разработана для генерирования асимметричных и биспецифичных антителоподобных молекул, что позволяет расширять терапевтические применения природных антител. Эта платформа, сконструированная белками, основана на обмене структурно родственных последовательностей иммуноглобулина в консервативных доменах СН3. Конструкция SEED позволяет эффективно генерировать гетеродимеры AG/GA, в то же время препятствуя гомодимеризации доменов AG и GA SEED CH3. (Muda M. et al., Protein Eng. Des. Sel. (2011, 24 (5): 447-54).
[00101] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме LuZ-Y, в которой домен лейциновой застежки используется для индуцирования гетеродимеризации двух разных HC. (Wranik, BJ. Et al., J. Biol. Chem. (2012), 287: 43331-9).
[00102] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Cov-X-Body. В биспецифических CovX-Body два разных пептида соединяются вместе с использованием разветвленного азетидинонового линкера и сливаются с каркасным антителом в мягких условиях сайтспецифичным образом. В то время как фармакофоры отвечают за функциональную активность, каркас антител обеспечивает длительный период полужизни и Ig-подобное распределение. Фармакофоры могут быть химически оптимизированы или заменены другими фармакофорами для получения оптимизированных или уникальных биспецифичных антител. (Doppalapudi VR et al., PNAS (2010), 107 (52); 22611-22616).
[00103] В некоторых вариантах осуществления полиспецифический связывающий белок находится в гетеродимерной форме Oasc-Fab, которая включает Fab, связывающийся с мишенью 1, и scFab, связывающийся с мишенью 2, слитые с Fc. Гетеродимеризация обеспечивается мутациями в Fc.
[00104] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме DuetMab, которая представляет собой гетеродимерную конструкцию, содержащую 2 различных Fab, связывающихся с антигеном 1 и 2, и Fc, стабилизированный мутациями гетеродимеризации. Fab 1 и 2 содержат дифференциальные SS-мостики, которые обеспечивают правильное спаривание LC и HC.
[00105] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме CrossmAb, которая представляет собой гетеродимерную конструкцию с 2 различными Fab, связывающимися с мишенью 1 и 2, слитыми с Fc, стабилизированными гетеродимеризацией. Домены CL и CH1 и домены VH и VL переключаются, например, CH1 сливается в одной линии с VL, в то время как CL сливается в одной линии с VH.
[00106] В некоторых вариантах осуществления полиспецифический связывающий белок находится в форме Fit-Ig, которая представляет собой гомодимерные конструкции, где Fab, связывающийся с антигеном 2, слит с N-концом HC Fab, который связывается с антигеном 1. Конструкция содержит Fc дикого типа.
[00107] В Таблице 1 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с NKG2D. NKG2D-связывающие домены могут различаться по своей аффинности связывания с NKG2D, тем не менее, все они активируют человеческие NKG2D и NK-клетки.
(SEQ ID NO:1)
CDR1 (SEQ ID NO:51) - GSFSGYYWS
CDR2 (SEQ ID NO:52) -
EIDHSGSTNYNPSLKS
CDR3 (SEQ ID NO:53) -
ARARGPWSFDP
(SEQ ID NO:2)
(SEQ ID NO:3)
(SEQ ID NO:4)
(SEQ ID NO:5)
(SEQ ID NO:6)
(SEQ ID NO:7)
(SEQ ID NO:8)
(SEQ ID NO:9)
(SEQ ID NO:10)
(SEQ ID NO:11)
(SEQ ID NO:12)
(SEQ ID NO:13)
(SEQ ID NO:14)
(SEQ ID NO:15)
(SEQ ID NO:16)
(SEQ ID NO:17)
(SEQ ID NO:18)
(SEQ ID NO:19)
(SEQ ID NO:20)
(SEQ ID NO:21)
(SEQ ID NO:22)
(SEQ ID NO:23)
(SEQ ID NO:24)
(SEQ ID NO:25)
(SEQ ID NO:26)
(SEQ ID NO:27)
(SEQ ID NO:28)
(SEQ ID NO:29)
(SEQ ID NO:30)
(SEQ ID NO:31)
(SEQ ID NO:32)
(SEQ ID NO:33)
(SEQ ID NO:34)
(SEQ ID NO:35)
(SEQ ID NO:36)
(SEQ ID NO:37)
(SEQ ID NO:38)
(SEQ ID NO:39)
(SEQ ID NO:40)
(SEQ ID NO:41)
CDR1 (SEQ ID NO:54) -
GTFSSYAIS
CDR2 (SEQ ID NO:55) -
GIIPIFGTANYAQKFQG
CDR3 (SEQ ID NO:56) -
ARGDSSIRHAYYYYGMDV
(SEQ ID NO:42)
CDR1 (SEQ ID NO:57) -
KSSQSVLYSSNNKNYLA
CDR2 (SEQ ID NO:58) -
WASTRES
CDR3 (SEQ ID NO:59) -
QQYYSTPIT
(SEQ ID NO:43)
CDR1 (SEQ ID NO:60) -
GSISSSSYYWG
CDR2 (SEQ ID NO:61) -
SIYYSGSTYYNPSLKS
CDR3 (SEQ ID NO:62) -
ARGSDRFHPYFDY
(SEQ ID NO:44)
CDR1 (SEQ ID NO:63) -
RASQSVSRYLA
CDR2 (SEQ ID NO:64) -
DASNRAT
CDR3 (SEQ ID NO:65) -
QQFDTWPPT
(SEQ ID NO:45)
CDR1 (SEQ ID NO:66) - GTFSSYAIS
CDR2 (SEQ ID NO:67) - GIIPIFGTANYAQKFQG
CDR3 (SEQ ID NO:68) - ARRGRKASGSFYYYYGMDV
(SEQ ID NO:46)
CDR1 (SEQ ID NO:69) - ESSQSLLNSGNQKNYLT
CDR2 (SEQ ID NO:70) - WASTRES
CDR3 (SEQ ID NO:71) - QNDYSYPYT
(A44)
(SEQ ID NO:132)
CDR1 (SEQ ID NO:134) - FTFSSYAMS
CDR2 (SEQ ID NO:135) - AISGSGGSTYYADSVKG
CDR3 (SEQ ID NO:136) - AKDGGYYDSGAGDY
(SEQ ID NO:133)
CDR1 (SEQ ID NO:137) - RASQGIDSWLA
CDR2 (SEQ ID NO:138) - AASSLQS
CDR3 (SEQ ID NO:139) - QQGVSYPRT
(A49)
(SEQ ID NO:140)
CDR1 (SEQ ID NO:142) - FTFSSYSMN
CDR2 (SEQ ID NO:143) - SISSSSSYIYYADSVKG
CDR3 (SEQ ID NO:144) - ARGAPMGAAAGWFDP
(SEQ ID NO:141)
CDR1 (SEQ ID NO:145) - RASQGISSWLA
CDR2 (SEQ ID NO:146) - AASSLQS
CDR3 (SEQ ID NO:147) - QQGVSFPRT
(F63)
(SEQ ID NO:148)
CDR1 (SEQ ID NO:150) - YTFTGYYMH
CDR2 (SEQ ID NO:151) - WINPNSGGTNYAQKFQG
CDR3 (SEQ ID NO:152) - ARDTGEYYDTDDHGMDV
(SEQ ID NO:149)
CDR1 (SEQ ID NO:153) - RASQSVSSNLA
CDR2 (SEQ ID NO:154) - GASTRAT
CDR3 (SEQ ID NO:155) - QQDDYWPPT
(E79)
(SEQ ID NO:156)
CDR1 (SEQ ID NO:158) - YTFTSYYMH
CDR2 (SEQ ID NO:159) - IINPSGGSTSYAQKFQG
CDR3 (SEQ ID NO:160) - ARGAPNYGDTTHDYYYMDV
(SEQ ID NO:157)
CDR1 (SEQ ID NO:161) - RASQSVSSNLA
CDR2 (SEQ ID NO:162) - GASTRAT
CDR3 (SEQ ID NO:163) - QQYDDWPFT
(F47)
(SEQ ID NO:164)
CDR1 (SEQ ID NO:166) - GSFSGYYWS
CDR2 (SEQ ID NO:167) - EIDHSGSTNYNPSLKS
CDR3 (SEQ ID NO:168) - ARARGPWSFDP
(SEQ ID NO:165)
CDR1 (SEQ ID NO:169) - RASQSISSWLA
CDR2 (SEQ ID NO:170) - KASSLES
CDR3 (SEQ ID NO:171) - QQYDTFIT
[00108] Альтернативно, вариабельный домен тяжелой цепи, определенный SEQ ID NO: 47, может спариваться с вариабельным доменом легкой цепи, определенным SEQ ID NO: 48, с образованием антигенсвязывающего сайта, который может связываться с NKG2D, как проиллюстрировано в патенте США 9273136.
SEQ ID NO:47 QVQLVESGGGLVKPGGSLRLSCAASGFTFSSYGMHWVRQAPGKGLEWVAFIRYDGSN KYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKDRGLGDGTYFDYWGQGTTVTVSS
SEQ ID NO:75:
QSALTQPASVSGSPGQSITISCSGSSSNIGNNAVNWYQQLPGKAPKLLIYYDDLLPSGVSDRFSGSKSGTSAFLAISGLQSEDEADYYCAAWDDSLNGPVFGGGTKLTVL
[00109] Альтернативно, вариабельный домен тяжелой цепи, определенный SEQ ID NO: 49, может спариваться с вариабельным доменом легкой цепи, определенным SEQ ID NO: 50, с образованием антигенсвязывающего сайта, который может связываться с NKG2D, как проиллюстрировано в патенте США 7879985.
SEQ ID NO:49
QVHLQESGPGLVKPSETLSLTCTVSDDSISSYYWSWIRQPPGKGLEWIGHISYSGS ANYNPSLKSRVTISVDTSKNQFSLKLSSVTAADTAVYYCANWDDAFNIWGQGTMVTVSS
SEQ ID NO:50
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRA TGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYGSSPWTFGQGTKVEIK
[00110] В Таблице 2 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с CAIX.
(SEQ ID NO:72)
CDR1 (SEQ ID NO:74) - GFTFSNY
CDR2 (SEQ ID NO:75) - NSDGGI
CDR3 (SEQ ID NO:76) - HRSGYFSMDY
(SEQ ID NO:73)
CDR1(SEQ ID NO:77) - QNVVSAVA
CDR2 (SEQ ID NO:78) - SASNRYT
CDR3 (SEQ ID NO:79) - QQYSNYPWT
(US 8168758)
CDR1 (SEQ ID NO:82) - GFTFSSYGMS
CDR2 (SEQ ID NO:83) - GISSLGSTTYYADSVKG
CDR3 (SEQ ID NO:84) - TGSPGTFMHGDH
(SEQ ID NO:81)
CDR1 (SEQ ID NO:85) - RASQDINNYLS
CDR2 (SEQ ID NO:86) - YGASNLQS
CDR3 (SEQ ID NO:87) - QQYYGRPT
(US 12/016884)
(SEQ ID NO:88)
CDR1 (SEQ ID NO:90) - GFTFSNY
CDR2 (SEQ ID NO:91) - NSDGGI
CDR3 (SEQ ID NO:92) - HRSGYFSMDY
(SEQ ID NO:89)
CDR1 (SEQ ID NO:93) - QNVVSAVA
CDR2 (SEQ ID NO:94) - SASNRYT
CDR3 (SEQ ID NO:95) - QQYSNYPWT
[00111] Альтернативно, новые антигенсвязывающие сайты, которые связываются с CAIX, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 96.
SEQ ID NO:96
MAPLCPSPWLPLLIPAPAPGLTVQLLLSLLLLVPVHPQRLPRMQEDSPLGGGSSGE DDPLGEEDLPSEEDSPREEDPPGEEDLPGEEDLPGEEDLPEVKPKSEEEGSLKLEDLPTVE APGDPQEPQNNAHRDKEGDDQSHWRYGGDPPWPRVSPACAGRFQSPVDIRPQLAAFCP ALRPLELLGFQLPPLPELRLRNNGHSVQLTLPPGLEMALGPGREYRALQLHLHWGAAGR PGSEHTVEGHRFPAEIHVVHLSTAFARVDEALGRPGGLAVLAAFLEEGPEENSAYEQLLS RLEEIAEEGSETQVPGLDISALLPSDFSRYFQYEGSLTTPPCAQGVIWTVFNQTVMLSAK QLHTLSDTLWGPGDSRLQLNFRATQPLNGRVIEASFPAGVDSSPRAAEPVQLNSCLAAG DILALVFGLLFAVTSVAFLVQMRRQHRRGTKGGVSYRPAEVAETGA
[00112] В Таблице 3 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с ANO1.
(SEQ ID NO:97)
CDR1 (SEQ ID NO:99) - NYDMH
CDR2 (SEQ ID NO:100) - VIWGNGKTQYNSGLTS
CDR3 (SEQ ID NO:101) - SGYYYDGSYYSLFDY
(SEQ ID NO:98)
CDR1(SEQ ID NO:102) - RSSQSLLHSNGNTYLN
CDR2 (SEQ ID NO:103) - LVSRLES
CDR3 (SEQ ID NO:104) - VQSTHAPA
[00113] Альтернативно, новые антигенсвязывающие сайты, которые связываются с ANO1, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 105.
SEQ ID NO:105
MRVNEKYSTLPAEDRSVHIINICAIEDIGYLPSEGTLLNSLSVDPDAECKYGLYFR DGRRKVDYILVYHHKRPSGNRTLVRRVQHSDTPSGARSVKQDHPLPGKGASLDAGSGEPPMDYHEDDKRFRREEYEGNLLEAGLELERDEDTKIHGVGFVKIHAPWNVLCREAEFLKLKMPTKKMYHINETRGLLKKINSVLQKITDPIQPKVAEHRPQTMKRLSYPFSREKQHLFDLSDKDSFFDSKTRSTIVYEILKRTTCTKAKYSMGITSLLANGVYAAAYPLHDGDYNGENVEFNDRKLLYEEWARYGVFYKYQPIDLVRKYFGEKIGLYFAWLGVYTQMLIPASIVGIIVFLYGCATMDENIPSMEMCDQRHNITMCPLCDKTCSYWKMSSACATARASHLFDNPATVFFSVFMALWAATFMEHWKRKQMRLNYRWDLTGFEEEEEAVKDHPRAEYEARVLEKSLKKESRNKEKRRHIPEESTNKWKQRVKTAMAGVKLTDKVKLTWRDRFPAYLTNLVSIIFMIAVTFAIVLGVIIYRISMAAALAMNSSPSVRSNIRVTVTATAVIINLVVIILLDEVYGCIARWLTKIEVPKTEKSFEERLIFKAFLLKFVNSYTPIFYVAFFKGRFVGRPGDYVYIFRSFRMEECAPGGCLMELCIQLSIIMLGKQLIQNNLFEIGIPKMKKLIRYLKLKQQSPPDHEECVKRKQRYEVDYNLEPFAGLTPEYMEMIIQFGFVTLFVASFPLAPLFALLNNIIEIRLDAKKFVTELRRPVAVRAKDIGIWYNILRGIGKLAVIINAFVISFTSDFIPRLVYLYMYSKNGTMHGFVNHTLSSFNVSDFQNGTAPNDPLDLGYEVQICRYKDYREPPWSENKYDISKDFWAVLAARLAFVIVFQNLVMFMSDFVDWVIPDIPKDISQQIHKEKVLMVELFMREEQDKQQLLETWMEKERQKDEPPCNHHNTKACPDSLGSPAPSHAYHGGVL
[00114] В Таблице 4 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с мезотелином.
(SEQ ID NO:106)
CDR1 (SEQ ID NO:108) - GYSFTGY
CDR2 (SEQ ID NO:109) - TPYNGA
CDR3 (SEQ ID NO:110) - GGYDGRGFDY
(SEQ ID NO:107)
CDR1(SEQ ID NO:111) - SSVSYMH
CDR2 (SEQ ID NO:112) - DTSKLAS
CDR3 (SEQ ID NO:113) - QQWSKHPLT
(US 8425904)
(SEQ ID NO:114)
CDR1 (SEQ ID NO:116) - GFTFSRY
CDR2 (SEQ ID NO:117) - KQAGSE
CDR3 (SEQ ID NO:118) - EGAYYYDSASYYPYYYYYSMDV
(SEQ ID NO:115)
CDR1 (SEQ ID NO:119) - QSVSSSYLA
CDR2 (SEQ ID NO:120) - GASSRAT
CDR3 (SEQ ID NO:121) - QQYGSSQYT
(US 9023351)
(SEQ ID NO:122)
CDR1 (SEQ ID NO:124) - GYSFTSY
CDR2 (SEQ ID NO:125) - DPGDSR
CDR3 (SEQ ID NO:126) - GQLYGGTYMDG
(SEQ ID NO:123)
CDR1 (SEQ ID NO:127) - SSDIGGYNSVS
CDR2 (SEQ ID NO:128) - GVNNRPS
CDR3 (SEQ ID NO:129) - SSYDIESATPV
[00115] Альтернативно, новые антигенсвязывающие сайты, которые связываются с мезотелином, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 130.
SEQ ID NO:130
MALPTARPLLGSCGTPALGSLLFLLFSLGWVQPSRTLAGETGQEAAPLDGVLANP PNISSLSPRQLLGFPCAEVSGLSTERVRELAVALAQKNVKLSTEQLRCLAHRLSEPPEDLDALPLDLLLFLNPDAFSGPQACTRFFSRITKANVDLLPRGAPERQRLLPAALACWGVRGSLLSEADVRALGGLACDLPGRFVAESAEVLLPRLVSCPGPLDQDQQEAARAALQGGGPPYGPPSTWSVSTMDALRGLLPVLGQPIIRSIPQGIVAAWRQRSSRDPSWRQPERTILRPRFRREVEKTACPSGKKAREIDESLIFYKKWELEACVDAALLATQMDRVNAIPFTYEQLDVLKHKLDELYPQGYPESVIQHLGYLFLKMSPEDIRKWNVTSLETLKALLEVNKGHEMSPQAPRRPLPQVATLIDRFVKGRGQLDKDTLDTLTAFYPGYLCSLSPEELSSVPPSSIWAVRPQDLDTCDPRQLDVLYPKARLAFQNMNGSEYFVKIQSFLGGAPTEDLKALSQQNVSMDLATFMKLRTDAVLPLTVAEVQKLLGPHVEGLKAEERHRPVRDWILRQRQDDLDTLGLGLQGGIPNGYLVLDLSMQEALSGTPCLLGPGPVLTVLALLLASTLA
[00116] В Таблице 5 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с TROP2.
(SEQ ID NO:172)
CDR1 (SEQ ID NO:174) - GYTFTNY
CDR2 (SEQ ID NO:175) - NTYTGE
CDR3 (SEQ ID NO:176) - GGFGSSYWYFDV
(SEQ ID NO:173)
CDR1(SEQ ID NO:177) - QDVSIAVA
CDR2 (SEQ ID NO:178) - SASYRYT
CDR3 (SEQ ID NO:179) - QQHYITPLT
(PCT/CA08/000979)
(SEQ ID NO:180)
CDR1 (SEQ ID NO:182) - GYTFTNY
CDR2 (SEQ ID NO:183) - NTKTGE
CDR3 (SEQ ID NO:184) - GGYGSSYWYFDV
(SEQ ID NO:181)
CDR1 (SEQ ID NO:185) - QDVSIAVA
CDR2 (SEQ ID NO:186) - SASYRYT
CDR3 (SEQ ID NO:187) - QQHYITPLT
(US15/187179)
(SEQ ID NO:188)
QVQLVQSGAEVKKPGASVKVSCKASGYTFTTAGMQWVRQAPGQGLEWMGWINTHSGVPKYAEDFKGRVTISLDTSTSTAYLQLSSLKSEDTAVYYCARSGFGSSYWYFDVWGQGTLVTVSS
(SEQ ID NO:190)
QIQLVQSGAEVKKPGESVKVSCKASGYTFTTAGMQWVQQMPGKGLEWMGWINTHSGVPKYAEDFKGRVTFSLDTSTSTAYLQLSSLKSEDTAVYYCARSGFGSSYWYFDVWGQGTLVTVSS
(SEQ ID NO:192)
(SEQ ID NO:189)
DIVMTQSPSSLSASVGDRVTITCKASQDVSTAVAWYQQKPGKAPKLLIYSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFAVYYCQQHYITPLTFGQGTKLEIKRT
(SEQ ID NO:191)
DIVMTQSPSSLSASVGDRVTITCKASQDVSTAVAWYQQKPGKQPKLLIYSASYRYTGVPSRFSGSGSGTDFTLTISSLQPEDFAVYYCQQHYITPLTFGQGTKLEIKR
(SEQ ID NO:193)
[00117] Альтернативно, новые антигенсвязывающие сайты, которые связываются с TROP2, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 194.
SEQ ID NO:194
MARGPGLAPPPLRLPLLLLVLAAVTGHTAAQDNCTCPTNKMTVCSPDGPGGRCQCRALGSGMAVDCSTLTSKCLLLKARMSAPKNARTLVRPSEHALVDNDGLYDPDCDPEGRFKARQCNQTSVCWCVNSVGVRRTDKGDLSLRCDELVRTHHILIDLRHRPTAGAFNHSDLDAELRRLFRERYRLHPKFVAAVHYEQPTIQIELRQNTSQKAAGDVDIGDAAYYFERDIKGESLFQGRGGLDLRVRGEPLQVERTLIYYLDEIPPKFSMKRLTAGLIAVIVVVVVALVAGMAVLVITNRRKSGKYKKVEIKELGELRKEPSL
[00118] В Таблице 6 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с CEA.
(SEQ ID NO:195)
CDR1 (SEQ ID NO:197) - GFDFTTY
CDR2 (SEQ ID NO:198) - HPDSST
CDR3 (SEQ ID NO:199) - LYFGFPWFAY
(SEQ ID NO:196)
CDR1(SEQ ID NO:200) - QDVGTSVA
CDR2 (SEQ ID NO:201) - WTSTRHT
CDR3 (SEQ ID NO:202) - QQYSLYRS
(SEQ ID NO:203)
CDR1 (SEQ ID NO:205) - GYTFTEF
CDR2 (SEQ ID NO:206) - NTKTGE
CDR3 (SEQ ID NO:207) - WDFAYYVEAMDY
(SEQ ID NO:204)
CDR1 (SEQ ID NO:208) - AAVGTYVA
CDR2 (SEQ ID NO:209) - SASYRKR
CDR3 (SEQ ID NO:210) - HQYYTYPLFT
(US 9056911)
(SEQ ID NO:211)
CDR1 (SEQ ID NO:213) - GFNIKHY
CDR2 (SEQ ID NO:214) - NPENVD
CDR3 (SEQ ID NO:215) - YRYAGGGALDY
(SEQ ID NO:212)
CDR1 (SEQ ID NO:216) - SSVSYIH
CDR2 (SEQ ID NO:217) - DTSKLAS
CDR3 (SEQ ID NO:218) - QQWNNNPYS
[00119] Альтернативно, новые антигенсвязывающие сайты, которые связываются с CEA, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 219.
SEQ ID NO:219
MESPSAPPHRWCIPWQRLLLTASLLTFWNPPTTAKLTIESTPFNVAEGKEVLLLVHNLPQHLFGYSWYKGERVDGNRQIIGYVIGTQQATPGPAYSGREIIYPNASLLIQNIIQNDTGFYTLHVIKSDLVNEEATGQFRVYPELPKPSISSNNSKPVEDKDAVAFTCEPETQDATYLWWVNNQSLPVSPRLQLSNGNRTLTLFNVTRNDTASYKCETQNPVSARRSDSVILNVLYGPDAPTISPLNTSYRSGENLNLSCHAASNPPAQYSWFVNGTFQQSTQELFIPNITVNNSGSYTCQAHNSDTGLNRTTVTTITVYAEPPKPFITSNNSNPVEDEDAVALTCEPEIQNTTYLWWVNNQSLPVSPRLQLSNDNRTLTLLSVTRNDVGPYECGIQNKLSVDHSDPVILNVLYGPDDPTISPSYTYYRPGVNLSLSCHAASNPPAQYSWLIDGNIQQHTQELFISNITEKNSGLYTCQANNSASGHSRTTVKTITVSAELPKPSISSNNSKPVEDKDAVAFTCEPEAQNTTYLWWVNGQSLPVSPRLQLSNGNRTLTLFNVTRNDARAYVCGIQNSVSANRSDPVTLDVLYGPDTPIISPPDSSYLSGANLNLSCHSASNPSPQYSWRINGIPQQHTQVLFIAKITPNNNGTYACFVSNLATGRNNSIVKSITVSASGTSPGLSAGATVGIMIGVLVGVALI
[00120] В Таблице 7 перечислены пептидные последовательности вариабельных доменов тяжелой цепи и вариабельных доменов легкой цепи, которые в комбинации могут связываться с Клаудином-18.2.
AEEFKGRFAFSLETSASTAYLQINNLKNEDTATYFCARLGFGNAMDYWGQGTSVTVSS
(SEQ ID NO:220)
CDR1 (SEQ ID NO:222) - GYTFTNYG
CDR2 (SEQ ID NO:223) - INTNTGEP
CDR3 (SEQ ID NO:224) - ARLGFGNAMDY
ESGVPDRFTGSGSGTDFTLTISSVQAEDLAVYYCQNDYSYPLTFGAGTKLELK
(SEQ ID NO:221)
CDR1(SEQ ID NO:225) - QSLLNSGNQKNY
CDR2 (SEQ ID NO:226) - WAS
CDR3 (SEQ ID NO:227) - QNDYSYPLT
NQKFKDKATLTVDKSSSTAYMQLSSPTSEDSAVYYCTRSWRGNSFDYWGQGTTLTVSS
(SEQ ID NO:228)
CDR1 (SEQ ID NO:230) - GYTFTSYW
CDR2 (SEQ ID NO:231) - IYPSDSYT
CDR3 (SEQ ID NO:232) - TRSWRGNSFDY
ESGVPDRFTGSGSGTDFTLTISSVQAEDLAVYYCQNDYSYPFTFGSGTKLEIK
(SEQ ID NO:229)
CDR1 (SEQ ID NO:233) - QSLLNSGNQKNY
CDR2 (SEQ ID NO:234) - WAS
CDR3 (SEQ ID NO:235) - QNDYSYPFT
[00121] Альтернативно, новые антигенсвязывающие сайты, которые связываются с Клаудином-18.2, могут быть идентифицированы путем скрининга на связывание с аминокислотной последовательностью, определенной SEQ ID NO: 236.
SEQ ID NO:236
MAVTACQGLGFVVSLIGIAGIIAATCMDQWSTQDLYNNPVTAVFNYQGLWRSCVRESSGFTECRGYFTLLGLPAMLQAVRALMIVGIVLGAIGLLVSIFALKCIRIGSMEDSAKANMTLTSGIMFIVSGLCAIAGVSVFANMLVTNFWMSTANMYTGMGGMVQTVQTRYTFGAALFVGWVAGGLTLIGGVMMCIACRGLAPEETNYKAVSYHASGHSVAYKPGGFKASTGFGSNTKNKKIYDGGARTEDEVQSYPSKHDYV
[00122] Внутри Fc-домена связывание CD16 опосредуется шарнирной областью и доменом CH2. Например, в человеческом IgG1 взаимодействие с CD16, главным образом, сосредоточено на аминокислотных остатках Asp 265 - Glu 269, Asn 297 - Thr 299, Ala 327 - Ile 332, Leu 234 - Ser 239 и углеводном остатке N-ацетил-D -глюкозамин в домене СН2 (см. Sondermann et al., Nature, 406 (6793): 267-273). На основе известных доменов мутации могут быть выбраны для усиления или уменьшения аффинности связывания с CD16, например, с использованием представленных на фаге библиотек или библиотек кДНК, представленных на поверхности дрожжей, или могут быть сконструированы на основе известной трехмерной структуры взаимодействия.
[00123] Сборка гетеродимерных тяжелых цепей антител может быть осуществлена путем экспрессии двух разных последовательностей тяжелых цепей антител в одной и той же клетке, что может привести к сборке гомодимеров каждой тяжелой цепи антитела, а также сборке гетеродимеров. Стимулирование преимущественной сборки гетеродимеров может быть достигнуто путем включения различных мутаций в домен СН3 каждой константной области тяжелой цепи антитела, как показано в US13/494870, US16/028850, US11/533709, US12/875015, US13/289934, US14/773418, US12/811207, US13/866756, US14/647480, US14/830336. Например, мутации могут быть сделаны в домене CH3 на основе человеческого IgG1 и включают различные пары аминокислотных замен в первом полипептиде и втором полипептиде, которые позволяют этим двум цепям селективно гетеродимеризоваться друг с другом. Положения аминокислотных замен, показанные ниже, пронумерованы в соответствии с индексом EU, как в Kabat.
[00124] В одном сценарии аминокислотная замена в первом полипептиде заменяет исходную аминокислоту более крупной аминокислотой, выбранной из аргинина (R), фенилаланина (F), тирозина (Y) или триптофана (W) и, по меньшей мере, одна аминокислотная замена во втором полипептиде заменяет исходную аминокислоту (аминокислоты) меньшей аминокислотой(аминокислотами), выбранной из аланина (A), серина (S), треонина (T) или валина (V), так что более крупная аминокислотная замена (выпуклость) вписывается в поверхность более мелких аминокислотных замен (полость). Например, один полипептид может включать замену T366W, а другой может включать три замены, включая T366S, L368A и Y407V.
[00125] Вариабельный домен тяжелой цепи антитела по изобретению необязательно может быть связан с аминокислотной последовательностью, по меньшей мере, на 90% идентичной константной области антитела, такой как константная область IgG, включая шарнирные домены, домены СН2 и СН3 с доменом СН1 или без него. В некоторых вариантах осуществления аминокислотная последовательность константной области, по меньшей мере, на 90% идентична константной области человеческого антитела, такой как константная область человеческого IgG1, константная область IgG2, константная область IgG3 или константная область IgG4. В некоторых других вариантах осуществления аминокислотная последовательность константной области, по меньшей мере, на 90% идентична константной области антитела другого млекопитающего, такого как кролик, собака, кошка, мышь или лошадь. Одна или более мутаций могут быть включены в константную область по сравнению с константной областью человеческого IgG1, например, в Q347, Y349, L351, S354, E356, E357, K360, Q362, S364, T366, L368, K370, N390, K392, T394, D399, S400, D401, F405, Y407, K409, T411 и/или K439. Типичные замены включают, например, Q347E, Q347R, Y349S, Y349K, Y349T, Y349D, Y349E, Y349C, T350V, L351K, L351D, L351Y, S354C, E356K, E357Q, E357L, E357W, K360E, K360W, Q362E, S364K, S364E, S364H, S364D, T366V, T366I, T366L, T366M, T366K, T366W, T366S, L368E, L368A, L368D, K370S, N390D, N390E, K392L, K392M, K392V, K392F, K392D, K392E, T394F, T394W, D399R, D399K, D399V, S400K, S400R, D401K, F405A, F405T, Y407A, Y407I , Y407V, K409F, K409W, K409D, T411D, T411E, K439D, и K439E.
[00126] В некоторых вариантах осуществления мутации, которые могут быть включены в CH1 константной области человеческого IgG1, могут находиться в аминокислоте V125, F126, P127, T135, T139, A140, F170, P171 и/или V173. В некоторых вариантах осуществления мутации, которые могут быть включены в Cκ константной области человеческого IgG1, могут находиться в аминокислоте E123, F116, S176, V163, S174 и/или T164.
[00127] Альтернативно, аминокислотные замены могут быть выбраны из следующих наборов замен, показанных в Таблице 8.
[00128] Альтернативно, аминокислотные замены могут быть выбраны из следующих наборов замен, показанных в Таблице 9.
[00129] Альтернативно, аминокислотные замены могут быть выбраны из следующего набора замен, показанного в Таблице 10.
[00130] Альтернативно, по меньшей мере, одна аминокислотная замена в каждой полипептидной цепи может быть выбрана из Таблицы 11.
[00131] Альтернативно, по меньшей мере, одна аминокислотная замена может быть выбрана из следующего набора замен в Таблице 12, где положение(я), указанное в столбце «Первый полипептид», заменено любой известной отрицательно заряженной аминокислотой, и положение(я), указанное в столбце «Второй полипептид», заменено любой известной положительно заряженной аминокислотой.
[00132] Альтернативно, по меньшей мере, одна аминокислотная замена может быть выбрана из следующего набора в Таблице 13, где положение(я), указанное в столбце «Первый полипептид», заменено любой известной положительно заряженной аминокислотой, и указанное положение(я), указанное в столбце «Второй полипептид», заменено любой известная отрицательно заряженной аминокислотой.
[00133] Альтернативно, аминокислотные замены могут быть выбраны из следующего набора в Таблице 14.
[00134] Альтернативно или дополнительно, структурная стабильность гетеромультимерного белка может быть повышена путем введения S354C либо на первой, либо на второй полипептидной цепи, и Y349C на противоположной полипептидной цепи, которая образует искусственный дисульфидный мостик внутри интерфейса двух полипептидов.
[00135] Полиспецифические белки, описанные выше, могут быть получены с использованием технологии рекомбинантных ДНК, хорошо известных специалисту в данной области. Например, первая последовательность нуклеиновой кислоты, кодирующая первую тяжелую цепь иммуноглобулина, может быть клонирована в первый экспрессирующий вектор; вторая последовательность нуклеиновой кислоты, кодирующая вторую тяжелую цепь иммуноглобулина, может быть клонирована во второй экспрессирующий вектор; третью последовательность нуклеиновой кислоты, кодирующую легкую цепь иммуноглобулина, можно клонировать в третий экспрессирующий вектор; первый, второй и третий экспрессирующие векторы могут стабильно трансфецироваться вместе в клетки-хозяева для продуцирования мультимерных белков.
[00136] Для достижения наибольшего выхода полиспецифического белка можно изучить различные соотношения первого, второго и третьего экспрессирующего вектора, чтобы определить оптимальное соотношение для трансфекции в клетки-хозяева. После трансфекции отдельные клоны могут быть выделены для генерации банка клеток с использованием методов, известных в данной области, таких как лимитированное разведение, ИФА, FACS, микроскопия или Clonepix.
[00137] Клоны можно культивировать в условиях, подходящих для масштабирования для биореактора и поддержания экспрессии полиспецифического белка. Полиспецифические белки могут быть выделены и очищены с использованием методов, известных в данной области, включая центрифугирование, глубинную фильтрацию, лизис клеток, гомогенизацию, замораживание-оттаивание, аффинную очистку, гель-фильтрацию, ионообменную хроматографию, обменную хроматографию гидрофобного взаимодействия и смешанный режим хроматографии.
II. Характеристика полиспецифических белков
[00138] В некоторых вариантах осуществления полиспецифические белки, описанные в настоящем документе, которые включают в себя NKG2D-связывающий домен и связывающий домен для CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2, связываются с клетками, экспрессирующими NKG2D человека. В некоторых вариантах осуществления полиспецифические белки связываются с опухолеспецифическим антигеном CAIX, ANO1, мезотелином, TROP2, CEA или Клаудином-18.2 на уровне, сопоставимом с уровнем моноклонального антитела, имеющего такой же соответствующий связывающий домен для CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2. Однако описанные в настоящем документе полиспецифические белки могут быть более эффективными в снижении роста опухолей и уничтожении опухолевых клеток, экспрессирующих CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2, чем моноклональные антитела, соответствующие CAIX, ANO1, мезотелину, TROP2, CEA или Клаудину- 18,2.
[00139] В некоторых вариантах осуществления описанные в настоящем документе полиспецифические белки, которые включают в себя NKG2D-связывающий домен и связывающий домен для CAIX, ANO1, мезотелина, TROP2, CEA или Клауидина-18.2, могут активировать первичные NK-клетки человека при культивировании с опухолевыми клетками, экспрессирующими антиген CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2. Активация NK-клеток отмечена увеличением дегрануляции CD107a и продуцированием цитокинов IFNγ. Кроме того, по сравнению с моноклональным антителом, которое включает в себя тот же CAIX-, ANO1-, мезотелин-, TROP2-, CEA- или Клаудин-18.2-связывающий домен, полиспецифические белки демонстрируют превосходную активацию человеческих NK-клеток в присутствии опухолевых клеток, экспрессирующих антиген CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2.
[00140] В некоторых вариантах осуществления описанные в настоящем документе полиспецифические белки, которые включают в себя NKG2D-связывающий домен и связывающий домен для CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2, могут усиливать активность покоящихся и IL-2-активированных NK-клеток человека в присутствии опухолевых клеток, экспрессирующих антиген CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2.
[00141] В некоторых вариантах осуществления описанные в настоящем документе полиспецифические белки, которые включают в себя NKG2D-связывающий домен и связывающий домен для опухолеспецифического антигена CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2, могут усиливать цитотоксическую активность покоящихся и IL-2-активированных NK-клеток человека в присутствии опухолевых клеток, экспрессирующих антиген CAIX, ANO1, мезотелин, TROP2, CEA или Клауидин-18.2. В некоторых вариантах осуществления по сравнению с соответствующими моноклональными антителами полиспецифические белки могут давать преимущество против опухолевых клеток, экспрессирующих на среднем и низком уровне CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2.
[00142] В некоторых вариантах осуществления полиспецифические белки, описанные в настоящем документе, могут быть полезны при лечении злокачественного новообразования с высокой экспрессией рецептора Fc (FcR) или злокачественного новообразования, находящегося в микроокружении опухоли с высоким уровнем FcR, по сравнению с соответствующими моноклональными антителами CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2. Моноклональные антитела оказывают свое влияние на рост опухоли посредством множества механизмов, включая среди прочего ADCC, CDC, фагоцитоз и сигнальную блокаду. Среди FcγR CD16 имеет самую низкую аффинность к Fc IgG; FcγRI (CD64) является высокоаффинным FcR, который примерно в 1000 раз сильнее связывается с Fc IgG, чем CD16. CD64 обычно экспрессируется на многих клетках гемопоэтических линий, таких как миелоидная линия, и может экспрессироваться на опухолях, происходящих из этих типов клеток, таких как острый миелолейкоз (AML). Иммунные клетки, инфильтрующиеся в опухоль, такие как MDSC и моноциты, также экспрессируют CD64 и, как известно, инфильтруются в микроокружение опухоли. Экспрессия CD64 опухолью или в микроокружении опухоли может оказывать вредное влияние на терапию моноклональными антителами. Экспрессия CD64 в микроокружении опухоли затрудняет взаимодействие этих антител с CD16 на поверхности NK-клеток, поскольку антитела предпочитают связываться с высокоаффинным рецептором. Полиспецифические белки, направленно воздействуя на два активирующих рецептора на поверхности NK-клеток, могут преодолеть пагубное влияние экспрессии CD64 (либо на опухоль, либо на микроокружение опухоли) на терапию моноклональными антителами. Независимо от экспрессии CD64 на опухолевых клетках, полиспецифические белки способны опосредовать ответы человеческих NK-клеток против всех опухолевых клеток, поскольку двойное нацеливание двух активирующих рецепторов на NK-клетки обеспечивает более сильное специфическое связывание с NK-клетками.
[00143] В некоторых вариантах осуществления полиспецифические белки, описанные в настоящем документе, могут обеспечить лучший профиль безопасности за счет уменьшения ошибочного нацеливания вне опухоли. Естественные клетки-киллеры и CD8 T-клетки способны непосредственно лизировать опухолевые клетки, хотя механизмы, посредством которых NK-клетки и CD8 T-клетки отличают собственные нормальные клетки от опухолевых клеток, различаются. Активность NK-клеток регулируется балансом сигналов от активирующих (NCR, NKG2D, CD16 и т.д.) и ингибирующих (KIR, NKG2A и т.д.) рецепторов. Баланс этих активирующих и ингибирующих сигналов позволяет NK-клеткам определять здоровые собственные клетки из стрессированных, инфицированных вирусом или трансформированных собственных клеток. Этот «встроенный» механизм аутотолерантности поможет защитить нормальную здоровую ткань от ответов NK-клеток. Чтобы расширить этот принцип, аутотолерантность NK-клеток позволит TriNKET нацеливаться как на антигены, экспрессируемые собственными клетками, так и опухолью, без побочных эффектов, связанных с ошибочным нацеливанием, или с увеличенным терапевтическим окном. В отличие от естественных клеток-киллеров, Т-клетки требуют распознавания специфического пептида, презентированного молекулами МНС для активации и эффекторных функций. Т-клетки были основной целью иммунотерапии, и было разработано много стратегий для перенаправления Т-клеточных ответов против опухоли. Биспецифические Т-клетки, ингибиторы контрольных точек и CAR-T-клетки были одобрены FDA, но часто страдают от дозолимитирующей токсичности. Биспецифические Т-клетки и CAR-T-клетки функционируют вокруг системы распознавания TCR-MHC, используя связывающие домены для нацеливания на антигены на поверхности опухолевых клеток и используя сконструированные сигнальные домены для передачи сигналов активации в эффекторную клетку. Несмотря на то, что эти методы лечения эффективны для выявления противоопухолевого иммунного ответа, они часто сочетаются с синдромом высвобождения цитокинов (CRS) и побочными эффектами, связанными с ошибочным нацеливанием вне опухоли. Полиспецифические белки уникальны в этом контексте, поскольку они не будут «перекрывать» природные системы активации и ингибирования NK-клеток. Вместо этого, полиспецифические белки предназначены для поддержания баланса и обеспечения дополнительных сигналов активации для NK-клеток, сохраняя при этом толерантность NK к собственным клеткам.
[00144] В некоторых вариантах осуществления полиспецифические белки, описанные в настоящем документе, могут более эффективно задерживать прогрессирование опухоли, чем соответствующие моноклональные антитела CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2, которые включают один и тот же антигенсвязывающий домен. В некоторых вариантах осуществления полиспецифические белки, описанные в настоящем документе, могут быть более эффективными против метастазов рака, чем соответствующие моноклональные антитела CAIX, ANO1, мезотелина, TROP2, CEA или Клаудина-18.2, которые включают в себя один и тот же антигенсвязывающий домен.
III. Терапевтические применения
[00145] Изобретение относится к способам лечения злокачественного новообразования с использованием полиспецифического связывающего белка, описанного в настоящем документе, и/или к фармацевтической композиции, описанной в настоящем документе. Способы могут быть использованы для лечения различных видов злокачественных новообразований, клетки которых экспрессируют CAIX, ANO1, мезотелин, TROP2, CEA или Клаудин-18.2, путем введения пациенту, нуждающемуся в этом, терапевтически эффективного количества полиспецифического связывающего белка, описанного в настоящем документе.
[00146] Терапевтический способ может быть охарактеризован в зависимости от злокачественного новообразования, подлежащего лечению. Рак, подлежащий лечению с использованием CAIX-нацеленных полиспецифических связывающих белков, включает, например, почечно-клеточную карциному, рак молочной железы, глиобластому, рак головы и шеи, рак желудка, рак мочевого пузыря, рак яичников, опухоли пищевода, легких, толстой кишки, рак почки, шейки матки и немелкоклеточную карциному легкого. Злокачественные новообразования, подлежащие лечению с использованием мезотелин-нацеленных полиспецифических связывающих белков, включают, например, мезотелиому, рак яичников, рак поджелудочной железы, немелкоклеточный рак легкого, рак молочной железы, холангиокарциному, рак желудка, серозную карциному матки, карциному тимуса и острый миелоидный лейкоз. Злокачественные новообразования, подлежащие лечению с использованием ANO1-нацеленных полиспецифических связывающих белков, включают, например, плоскоклеточный рак пищевода (ESCC), желудочно-кишечную стромальную опухоль (GIST), плоскоклеточную карциному головы и шеи (HNSCC), рак поджелудочной железы, рак молочной железы, рак предстательной железы и саркому. Злокачественные новообразования, подлежащие лечению с использованием TROP2-нацеленных полиспецифических связывающих белков, включают, например, рак молочной железы, легких, желудка, колоректальный рак, рак поджелудочной железы, предстательной железы, шейки матки, рак головы и шеи, карциному носоглотки и карциному яичника. Злокачественные новообразования, подлежащие лечению с использованием CEA-нацеленных полиспецифических связывающих белков, включают, например, рак желудочно-кишечного тракта, колоректальный рак, рак поджелудочной железы, немелкоклеточный рак легкого и рак молочной железы. Злокачественные новообразования, подлежащие лечению с использованием Клаудин-18.2-нацеленных полиспецифических связывающих белков, включают, например, рак пищевода, немелкоклеточную карциному легкого, рак яичника, рак толстой кишки и несколько форм карциномы желчных протоков.
[00147] В некоторых других вариантах осуществления злокачественное новообразование представляет собой рак мозга, рак шейки матки, рак толстой кишки, колоректальный рак, рак эндометрия, рак пищевода, лейкоз, рак легкого, рак печени, меланому, рак яичников, рак поджелудочной железы, ректальный рак, рак почки, рак желудка, рак яичка или рак матки. Еще в других вариантах осуществления злокачественное новообразование представляет собой плоскоклеточную карциному, аденокарциному, мелкоклеточную карциному, меланому, нейробластому, саркому (например, ангиосаркому или хондросаркому), рак гортани, рак околоушной железы, рак желчных путей, рак щитовидной железы, акрально-лентигинозную меланому, актинический кератоз, острый лимфоцитарный лейкоз, острый миелоидный лейкоз, аденоидно-кистозную карциному, аденому, аденосаркому, железисто-плоскоклеточную карциному, рак анального канала, анальный рак, ректальный рак, астроцитарную опухоль, карциному бартолиновой железы, базальноклеточную карциному, рак печени, рак костей, рак костного мозга, рак бронхов, карциному бронхиальной железы, карциноид, холангиокарциному, хондосаркому, хориодпапиллому/карциному, хронический лимфолейкоз, хронический миелоидный лейкоз, светлоклеточную карциному, рак соединительной ткани, цистаденому, рак пищеварительной системы, рак двенадцатиперстной кишки, рак эндокринной системы, опухоль эндодермальной пазухи, гиперплазию эндометрия, стромальную саркому эндометрия, эндометриоидную аденокарциному, эндотелиальный рак, эпендимальный рак, эпителиально-клеточный рак, саркому Юинга, рак глаз и глазной орбиты, рак женских половых органов, фокальную нодулярную гиперплазию, рак желчного пузыря, рак полости желудка, рак дна желудка, гастриному, глиобластому, глюкагоному, рак сердца, гемангибластомы, гемангиоэндотелиому, гемангиомы, печеночную аденому, печеночный аденоматоз, гепатобилиарный рак, гепатоцеллюлярную карциному, болезнь Ходжкина, рак подвздошной кишки, инсулиному, интраэпителиальную неоплазию, интраэпителиальную плоскоклеточную неоплазию, рак внутрипеченочных желчных протоков, инвазивную плоскоклеточную карциному, рак тонкой кишки, рак соединительной ткани, саркому Капоши, рак малого таза, крупноклеточный рак, рак толстой кишки, лейомиосаркому, лентиго-меланому, лимфому, рак мужских половых органов, злокачественную меланому, злокачественные мезотелиальные опухоли, медуллобластому, медуллоэпителиому, менингеальный рак, мезотелиальный рак, метастатический рак, рак полости рта, мукоэпидермоидную карциному, множественную миелому, рак мышц, рак носового канала, рак нервной системы, нейроэпителиальную аденокарциному, нодулярную меланому, неэпителиальный рак кожи, неходжкинскую лимфому, овсяно-клеточный рак, олигодендроглиальный рак, рак полости рта, остеосаркому, папиллярную серозную аденокарциному, опухоли гипофиза, плазмоцитому, псевдосаркому, легочную бластому, рак прямой кишки, почечно-клеточный рак, рак дыхательной системы, ретинобластому, рабдомиосаркому, саркому, серозную карциному, рак пазухи, рак кожи, мелкоклеточный рак, рак тонкого кишечника, рак мягких тканей, соматостатин-секретирующую опухоль, рак позвоночника, плоскоклеточную карциному, рак поперечно-полосатых мышц, субмезотелиальный рак, меланому поверхностного распространения, Т-клеточный лейкоз, рак языка, недифференцированный рак, рак мочеточника, рак мочеиспускательного канала, рак мочевого пузыря, рак мочеполовой системы, рак шейки матки, рак тела матки, увеальную меланому, рак влагалища, бородавчатый рак, Випому, рак вульвы, хорошо дифференцированную карциному или опухоль Вильмса.
[00148] В некоторых других вариантах осуществления злокачественное новобразование представляет собой неходжкинскую лимфому, такую как В-клеточная лимфома или Т-клеточная лимфома. В некоторых вариантах осуществления неходжкинская лимфома представляет собой В-клеточную лимфому, такую как диффузная крупноклеточная В-клеточная лимфома, первичная средостенная В-клеточная лимфома, фолликулярная лимфома, малкоклеточная лимфоцитарная лимфома, лимфома мантийных клеток, В-клеточная лимфома маргинальной зоны, B-клеточная лимфома экстранодальной маргинальной зоны, B-клеточная лимфома узловой маргинальной зоны, B-клеточная лимфома маргинальной зоны селезенки, лимфома Беркитта, лимфоплазмоцитарная лимфома, волосатоклеточный лейкоз или лимфома первичной центральной нервной системы (ЦНС). В некоторых других вариантах осуществления неходжкинская лимфома представляет собой Т-клеточную лимфому, такую как лимфобластная лимфома предшественников Т-клеток, периферическая Т-клеточная лимфома, кожная Т-клеточная лимфома, ангиоиммунобластная Т-клеточная лимфома, экстранодальная лимфома естественных клеток-киллеров/Т-клеток, узловая Т-клеточная лимфома, кожная Т-клеточная лимфома, Т-клеточная лимфома энтеропатического типа, Т-клеточная лимфома типа подкожного панникулита, анапластическая крупноклеточная лимфома или узловая Т-клеточная лимфома.
[00149] Злокачественное новообразование, подлежащее лечению, можно охарактеризовать в соответствии с наличием определенного антигена, экспрессируемого на поверхности опухолевой клетки. В некоторых вариантах осуществления опухолевая клетка может экспрессировать один или более из следующих: CD2, CD19, CD20, CD30, CD38, CD40, CD52, CD70, EGFR/ERBB1, IGF1R, HER3/ERBB3, HER4/ERBB4, MUC1, cMET, SLAMF7, PSCA, MICA, MICB, TRAILR1, TRAILR2, MAGE-A3, B7.1, B7.2, CTLA4 и PD1.
IV. Комбинированная терапия
[00150] Еще один аспект изобретения предусматривает комбинированную терапию. Полиспецифические связывающие белки, описанные в настоящем документе, используются в комбинации с дополнительными терапевтическими агентами для лечения злокачественного новообразования.
[00151] Типичные терапевтические агенты, которые могут быть использованы в качестве части комбинированной терапии при лечении злокачественного новообразования, включают, например, облучение, митомицин, третиноин, рибомустин, гемцитабин, винкристин, этопозид, кладрибин, митобронит, метотрексат, доксорубицин, карбоквон, пентостатин, нитракрин, зиностатин, цетрореликс, летрозол, ралтитрексед, даунорубицин, фадрозол, фотемустин, тималфазин, собузоксан, недаплатин, цитарабин, бикалутамид, винорелбина, веснаринон, аминоглютетимид, амсакрин, проглумид, эллиптиния ацетат, кетансерин, доксифлуридин, этретинат, изотретиноин, стрептозоцин, нимустин, виндезины, флутамид, дрогенил, бутоцин, кармофур, разоксан, сизофилан, карбоплатин, митолактол, тегафур, ифосфамид, преднимустин, пицибанил, левамизол, тенипозид, импросульфан, эноцитабин, лизурид, оксимететолон, тамоксифен, прогестерон, мепитиостан, эпитиостанол, форместан, интерферон альфа, интерферон-2 альфа, интерферон-бета, интерферон-гамма, колониестимулирующий фактор-1, колониестимулирующий фактор-2, денилейкин дифитокс, интерлейкин-2, фактор высвобождения лютеинизирующего гормона и вариации вышеупомянутых агентов, которые могут проявлять дифференциальное связывание с их родственным рецептором, а также увеличивать или уменьшать период полужизни в сыворотке.
[00152] Дополнительный класс агентов, которые могут быть использованы в качестве части комбинированной терапии при лечении злокачественного новообразования, относится к ингибиторам иммунной контрольной точки. Типичные ингибиторы иммунной контрольной точки включают агенты, которые ингибируют один или более из (i) антигена 4 (CTLA-4), ассоциированного с цитотоксическими Т-лимфоцитами, (ii) белка 1 программированной клеточной смерти (PD1), (iii) PDL1, (iv) LAG-3, (v) B7-H3, (vi) B7-H4 и (vii) TIM3. Ингибитор CTLA4 ипилимумаб был одобрен Управлением по контролю за продуктами и лекарствами США для лечения меланомы.
[00153] Другие агенты, которые могут быть использованы в качестве части комбинированной терапии при лечении злокачественного новообразования, представляют собой агенты моноклональных антител, которые нацелены на мишени, отличные от контрольных точек (например, герцептин), и нецитотоксические агенты (например, ингибиторы тирозинкиназы).
[00154] Другие категории противоопухолевых агентов включают, например: (i) ингибитор, выбранный из ингибитора ALK, ингибитора ATR, антагониста A2A, ингибитора репарации исключения оснований, ингибитора Bcr-Abl тирозинкиназы, ингибитора тирозинкиназы Брутона, ингибитора CDC7, ингибитора CHK1, ингибитора циклин-зависимой киназы, ингибитора DNA-PK, ингибитора как DNA-PK, так и mTOR, ингибитора DNMT1, ингибитора DNMT1 плюс 2-хлор-дезоксиаденозин, ингибитора HDAC , ингибитора сигнального пути Hedgehog, ингибитора IDO, ингибитора JAK, ингибитора mTOR, ингибитора MEK, ингибитора MELK, ингибитора MTH1, ингибитора PARP, ингибитора фосфоинозитид-3-киназы, ингибитора обоих PARP1 и DHODH, протеосомного ингибитора, ингибитора топоизомеразы-II, ингибитора тирозинкиназы, ингибитора VEGFR и ингибитора WEE1; (ii) агонист OX40, CD137, CD40, GITR, CD27, HVEM, TNFRSF25 или ICOS; и (iii) цитокин, выбранный из IL-12, IL-15, GM-CSF и G-CSF.
[00155] Белки по изобретению также можно использовать в качестве дополнения к хирургическому удалению первичного поражения.
[00156] Количество полиспецифического связывающего белка и дополнительного терапевтического агента и относительные сроки введения могут быть выбраны для достижения желаемого комбинированного терапевтического эффекта. Например, при назначении комбинированной терапии пациенту, нуждающемуся в таком введении, терапевтические агенты в комбинации или фармацевтическую композицию или композиции, содержащие терапевтические агенты, можно вводить в любом порядке, таком как, например, последовательно, одновременно, вместе, одновременно и тому подобное. Кроме того, например, полиспецифический связывающий белок может быть введен в течение времени, когда дополнительный терапевтический агент(ы) оказывает свое профилактическое или терапевтическое действие, или наоборот.
V. Фармацевтические композиции
[00157] Настоящее раскрытие также относится к фармацевтическим композициям, которые содержат терапевтически эффективное количество белка, описанного в настоящем документе. Композиция может быть приготовлена для использования в различных системах доставки лекарственных средств. Одно или более физиологически приемлемых вспомогательных веществ или носителей также могут быть включены в композицию для подходящего состава. Подходящие составы для использования в настоящем раскрытии есть в Remington's Pharmaceutical Sciences, Mack Publishing Company, Philadelphia, PA, 17th ed., 1985. Для краткого обзора способов доставки лекарственных средств см., например, Langer (Science 249: 1527-1533, 1990).
[00158] Композиция для внутривенной доставки лекарственного средства по настоящему изобретению может содержаться в пакете, ручке или шприце. В некоторых вариантах осуществления пакет может быть соединен с каналом, содержащим трубку и/или иглу. В определенных вариантах осуществления состав может представлять собой лиофилизированный состав или жидкий состав. В определенных вариантах осуществления состав может быть заморожен-высушен (лиофилизирован) и может содержаться примерно в 12-60 флаконах. В некоторых вариантах осуществления композиция может быть лиофилизирована, и 45 мг лиофилизированной композиции может содержаться в одном флаконе. В некоторых вариантах осуществления примерно 40-100 мг лиофилизированной композиции может содержаться в одном флаконе. В некоторых вариантах осуществления лиофилизированный состав из 12, 27 или 45 флаконов объединяют для получения терапевтической дозы белка в составе внутривенного лекарственного средства. В некоторых вариантах осуществления композиция может представлять собой жидкую композицию и храниться в концентрации примерно от 250 мг/флакон примерно до 1000 мг/флакон. В определенных вариантах осуществления композиция может представлять собой жидкую композицию и храниться в концентрации примерно 600 мг/флакон. В некоторых вариантах осуществления композиция может представлять собой жидкую композицию и храниться в концентрации примерно 250 мг/флакон.
[00159] Настоящее раскрытие может существовать в жидкой водной фармацевтической композиции, включающей терапевтически эффективное количество белка в буферном растворе с образованием композиции.
[00160] Эти композиции могут быть стерилизованы обычными методами стерилизации или могут быть стерильно отфильтрованы. Полученные водные растворы могут быть упакованы для использования как есть или лиофилизированы, причем лиофилизированный препарат объединяют со стерильным водным носителем перед введением. РН препаратов обычно составляет от 3 до 11, более предпочтительно от 5 до 9 или от 6 до 8, и наиболее предпочтительно от 7 до 8, например от 7 до 7,5. Полученные композиции в твердой форме могут быть упакованы в виде нескольких однодозовых единиц, каждая из которых содержит фиксированное количество вышеупомянутого агента или агентов. Композиция в твердой форме также может быть упакована в контейнер для гибкого количества.
[00161] В некоторых вариантах осуществления настоящее раскрытие относится к составу с увеличенным сроком хранения, включающему белок по настоящему изобретению, в сочетании с маннитом, моногидратом лимонной кислоты, цитратом натрия, дигидратом динатрийфосфата, дигидратом дигидрофосфата натрия, хлоридом натрия, полисорбатом 80, вода и гидроксидом натрия.
[00162] В некоторых вариантах осуществления готовят водный состав, включающий белок по настоящему изобретению в рН-буферном растворе. Буфер данного изобретения может иметь рН в диапазоне примерно от 4 примерно до 8, например, примерно от 4,5 примерно до 6 или примерно от 4,8 примерно до 5,5, или может иметь рН примерно от 5 примерно до 5,2. Диапазоны, промежуточные по отношению к указанным выше значениям pH, также должны быть частью этого раскрытия. Например, диапазоны значений, использующие комбинацию любого из приведенных выше значений в качестве верхнего и/или нижнего пределов, предназначены для включения. Примеры буферов, которые будут контролировать рН в этом диапазоне, включают ацетатные (например, ацетат натрия), сукцинатные (такие как сукцинат натрия), глюконатные, гистидиновые, цитратные и другие буферы органических кислот.
[00163] В определенных вариантах осуществления композиция включает буферную систему, которая содержит цитрат и фосфат для поддержания рН в диапазоне примерно от 4 примерно до 8. В некоторых вариантах осуществления диапазон рН может составлять примерно от 4,5 примерно до 6, или примерно от рН 4,8 примерно до 5,5, или в диапазоне рН примерно от 5 примерно до 5,2. В некоторых вариантах осуществления буферная система включает моногидрат лимонной кислоты, цитрат натрия, дигидрат динатрийфосфата и/или дигидрат дигидрофосфата натрия. В некоторых вариантах осуществления буферная система включает в себя примерно 1,3 мг/мл лимонной кислоты (например, 1,305 мг/мл), примерно 0,3 мг/мл цитрата натрия (например, 0,305 мг/мл), примерно 1,5 мг/мл динатрийфосфата. дигидрат (например, 1,53 мг/мл), примерно 0,9 мг/мл дигидрата дигидрофосфата натрия (например, 0,86) и примерно 6,2 мг/мл хлорида натрия (например, 6,165 мг/мл). В некоторых вариантах осуществления буферная система включает от 1 до 1,5 мг/мл лимонной кислоты, от 0,25 до 0,5 мг/мл цитрата натрия, от 1,25 до 1,75 мг/мл дигидрата динатрийфосфата, от 0,7 до 1,1 мг/мл дигидрата дигидрофосфата натрия, и от 6 до 6,4 мг/мл хлорида натрия. В некоторых вариантах осуществления pH композиции доводят с помощью гидроксида натрия.
[00164] Полиол, который действует как тонизирующий агент и может стабилизировать антитело, также может быть включен в состав. Полиол добавляют к составу в количестве, которое может варьироваться в зависимости от желаемой изотоничности состава. В некоторых вариантах осуществления водная композиция может быть изотонической. Количество добавляемого полиола также может быть изменено по отношению к молекулярной массе полиола. Например, может быть добавлено меньшее количество моносахарида (например, маннита) по сравнению с дисахаридом (таким как трегалоза). В некоторых вариантах осуществления полиол, который может быть использован в составе в качестве средства для повышения тоничности, представляет собой маннит. В определенных вариантах осуществления концентрация маннита может составлять примерно от 5 примерно до 20 мг/мл. В некоторых вариантах осуществления концентрация маннита может составлять примерно 7,5-15 мг/мл. В некоторых вариантах концентрация маннита может составлять примерно 10-14 мг/мл. В определенных вариантах концентрация маннита может составлять примерно 12 мг/мл. В некоторых вариантах осуществления полиол сорбит может быть включен в состав.
[00165] Детергент или поверхностно-активное вещество также могут быть добавлены в состав. Типичные детергенты включают неионогенные детергенты, такие как полисорбаты (например, полисорбаты 20, 80 и т.д.) или полоксамеры (например, полоксамер 188). Количество добавляемого детергента таково, что оно уменьшает агрегацию антитела в составе и/или сводит к минимуму образование частиц в составе, и/или уменьшает адсорбцию. В определенных вариантах осуществления состав может включать поверхностно-активное вещество, которое представляет собой полисорбат. В некоторых вариантах осуществления композиция может содержать детергент полисорбат 80 или Tween 80. Tween 80 - это термин, используемый для описания полиоксиэтилен (20) сорбитанмоноолеата (см. Fiedler, Lexikon der Hifsstoffe, Editio Cantor Verlag Aulendorf, 4th ed., 1996). В некоторых вариантах осуществления композиция может содержать примерно от 0,1 примерно до 10 мг/мл полисорбата 80 или примерно от 0,5 примерно до 5 мг/мл. В некоторых вариантах осуществления в композицию можно добавлять примерно 0,1% полисорбата 80.
[00166] В некоторых вариантах осуществления белковый продукт по настоящему изобретению приготовлен в виде жидкой композиции. Жидкий состав может быть представлен в концентрации 10 мг/мл во флаконе любого типа USP/Ph Eur типа I 50R, закрытом резиновой пробкой и запечатанном с использованием алюминиевого обжимного уплотнения. Пробка может быть изготовлена из эластомера, соответствующего USP и Ph Eur. В некоторых вариантах осуществления флаконы могут быть заполнены 61,2 мл раствора белкового продукта, чтобы обеспечить экстрагируемый объем 60 мл. В некоторых вариантах осуществления жидкая композиция может быть разбавлена 0,9% солевым раствором.
[00167] В некоторых вариантах осуществления жидкая композиция по изобретению может быть приготовлена в виде раствора с концентрацией 10 мг/мл в комбинации с сахаром на стабилизирующем уровне. В некоторых вариантах осуществления жидкая композиция может быть приготовлена в водном носителе. В некоторых вариантах осуществления стабилизатор может быть добавлен в количестве, не превышающем того, которое может привести к вязкости, нежелательной или непригодной для внутривенного введения. В некоторых вариантах осуществления сахар может представлять собой дисахариды, например сахарозу. В некоторых вариантах осуществления жидкая композиция также может включать один или более из буферного агента, поверхностно-активного вещества и консерванта.
[00168] В некоторых вариантах осуществления pH жидкого состава может быть установлено путем добавления фармацевтически приемлемой кислоты и/или основания. В некоторых вариантах осуществления фармацевтически приемлемая кислота может представлять собой соляную кислоту. В некоторых вариантах осуществления основание может представлять собой гидроксид натрия.
[00169] В дополнение к агрегации, дезамидирование является распространенным вариантом продукта пептидов и белков, который может возникать во время ферментации, сбора/осветления клеток, очистки, хранения лекарственного вещества/лекарственного продукта и во время анализа образца. Дезамидирование представляет собой потерю NH3 из белка с образованием промежуточного сукцинимида, который может подвергаться гидролизу. Промежуточный сукцинимид приводит к снижению массы исходного пептида на 17 дальтон. Последующий гидролиз приводит к увеличению массы на 18 дальтон. Выделение промежуточного сукцинимида затруднено из-за нестабильности в водных условиях. По существу, дезамидирование обычно определяется как увеличение массы на 1 дальтон. Дезамидирование аспарагина приводит к получению аспарагиновой или изоаспарагиновой кислоты. Параметры, влияющие на скорость дезамидирования, включают pH, температуру, диэлектрическую постоянную растворителя, ионную силу, первичную последовательность, локальную конформацию полипептида и третичную структуру. Аминокислотные остатки, соседствующие с Asn в пептидной цепи, влияют на скорости дезамидирования. Gly и Ser, следующие за Asn в белковых последовательностях, приводят к более высокой чувствительности к дезамидированию.
[00170] В некоторых вариантах осуществления жидкая композиция по настоящему изобретению может быть сохранена в условиях pH и влажности для предотвращения дезаминирования белкового продукта.
[00171] Представляющий интерес водный носитель в настоящем описании является фармацевтически приемлемым (безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкой композиции. Иллюстративные носители включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), рН-буферный раствор (например, фосфатно-буферный солевой раствор), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00172] Консервант необязательно может быть добавлен в составы для уменьшения бактериального действия. Добавление консерванта может, например, способствовать получению многоразового (многодозового) состава.
[00173] Внутривенные (IV) составы могут быть предпочтительным путем введения в конкретных случаях, например, когда пациент находится в больнице после трансплантации, получая все лекарственные средства посредством IV пути. В некоторых вариантах осуществления жидкую композицию разбавляют 0,9% раствором хлорида натрия перед введением. В некоторых вариантах осуществления разбавленный лекарственный продукт для инъекций является изотоническим и пригодным для введения путем внутривенной инфузии.
[00174] В некоторых вариантах осуществления соли или буферные компоненты могут быть добавлены в количестве 10-200 мМ. Соли и/или буферы являются фармацевтически приемлемыми и получены из различных известных кислот (неорганических и органических) с металлами или аминами, образующими основание. В некоторых вариантах осуществления буфер может представлять собой фосфатный буфер. В некоторых вариантах осуществления буфер может представлять собой глицинатный, карбонатный, цитратный буферы, и в этом случае ионы натрия, калия или аммония могут служить в качестве противоиона.
[00175] Консервант может быть необязательно добавлен в составы для уменьшения бактериального действия. Добавление консерванта может, например, способствовать получению многоразового (многодозового) состава.
[00176] Представляющий интерес водный носитель в настоящем описании является фармацевтически приемлемым (безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкой композиции. Иллюстративные носители включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), рН-буферный раствор (например, фосфатно-буферный солевой раствор), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00177] Настоящее раскрытие может существовать в лиофилизированной композиции, включающей белки и лиопротектор. Лиопротектором может быть сахар, например дисахариды. В некоторых вариантах осуществления ликопротектор может представлять собой сахарозу или мальтозу. Лиофилизированный состав может также включать один или более из буферного агента, поверхностно-активного вещества, наполнителя и/или консерванта.
[00178] Количество сахарозы или мальтозы, полезное для стабилизации лиофилизированного лекарственного продукта, может быть в массовом соотношении по меньшей мере 1:2 белка к сахарозе или мальтозе. В некоторых вариантах осуществления массовое соотношение белок/сахароза или мальтоза может составлять от 1:2 до 1:5.
[00179] В некоторых вариантах осуществления pH композиции перед лиофилизацией может быть установлено путем добавления фармацевтически приемлемой кислоты и/или основания. В некоторых вариантах осуществления фармацевтически приемлемая кислота может представлять собой соляную кислоту. В некоторых вариантах осуществления фармацевтически приемлемым основанием может быть гидроксид натрия.
[00180] Перед лиофилизацией pH раствора, содержащего белок по настоящему изобретению, можно доводить в диапазоне от 6 до 8. В некоторых вариантах осуществления диапазон pH для лиофилизированного лекарственного продукта может составлять от 7 до 8.
[00181] В некоторых вариантах осуществления соли или буферные компоненты могут быть добавлены в количестве 10-200 мМ. Соли и/или буферы являются фармацевтически приемлемыми и получены из различных известных кислот (неорганических и органических) с металлами или аминами, образующими основание. В некоторых вариантах осуществления буфер может представлять собой фосфатный буфер. В некоторых вариантах осуществления буфер может представлять собой глицинатный, карбонатный, цитратный буферы, и в этом случае ионы натрия, калия или аммония могут служить в качестве противоиона.
[00182] В некоторых вариантах осуществления может быть добавлен «наполнитель». «Наполнитель» представляет собой соединение, которое добавляет массу к лиофилизированной смеси и способствует физической структуре лиофилизированной таблетки (например, облегчает производство по существу однородной лиофилизированной таблетки, которая поддерживает структуру с открытыми порами). Иллюстративные наполнители включают маннит, глицин, полиэтиленгликоль и сорбит. Лиофилизированные составы по настоящему изобретению могут содержать такие наполнители.
[00183] Консервант может быть необязательно добавлен в составы для уменьшения бактериального действия. Добавление консерванта может, например, способствовать получению многоразового (многодозового) состава.
[00184] В некоторых вариантах осуществления лиофилизированный лекарственный продукт может состоять из водного носителя. Представляющий интерес водный носитель настоящего описания является фармацевтически приемлемым (например, безопасным и нетоксичным для введения человеку) и полезным для приготовления жидкой композиции после лиофилизации. Иллюстративные разбавители включают стерильную воду для инъекций (SWFI), бактериостатическую воду для инъекций (BWFI), рН-буферный раствор (например, фосфатно-буферный солевой раствор), стерильный физиологический раствор, раствор Рингера или раствор декстрозы.
[00185] В некоторых вариантах осуществления лиофилизированный лекарственный продукт по настоящему изобретению восстанавливается либо стерильной водой для инъекций, USP (SWFI), либо 0,9% инъекцией хлорида натрия, USP. Во время восстановления лиофилизированный порошок растворяется в растворе.
[00186] В некоторых вариантах осуществления лиофилизированный белковый продукт по настоящему изобретению состоит из примерно 4,5 мл воды для инъекций и разбавляется 0,9% солевым раствором (раствор хлорида натрия).
[00187] Фактические уровни дозировки активных ингредиентов в фармацевтических композициях могут варьироваться так, чтобы получить количество активного ингредиента, которое эффективно для достижения искомого терапевтического ответа для конкретного пациента, композиции и режима введения, без токсичности для пациента.
[00188] Конкретная доза может представлять собой унифицированную дозу для каждого пациента, например, 50-5000 мг белка. Альтернативно, доза для пациента может быть адаптирована к приблизительной массе тела или площади поверхности пациента. Другие факторы в определении подходящей дозировки могут включать заболевание или состояние, которое подлежит лечению или предотвращению, тяжесть заболевания, способ введения, а также возраст, пол и состояние здоровья пациента. Специалисты в данной области обычно проводят дальнейшее уточнение расчетов, необходимых для определения подходящей дозировки для лечения, особенно в свете информации о дозировке и анализах, раскрытых в настоящем документе. Дозировка также может быть определена путем использования известных анализов для определения доз, используемых в сочетании с соответствующими данными доза-ответ. Дозировка индивидуального пациента может быть скорректирована по мере наблюдения за развитием заболевания. Уровни в крови нацеливающей конструкции или комплекса у пациента могут быть измерены, чтобы увидеть, нужно ли корректировать дозировку для достижения или поддержания эффективной концентрации. Фармакогеномика может быть использована для определения того, какие нацеливающие конструкции и/или комплексы и их дозы наиболее вероятно будут эффективны для данного индивидуума (Schmitz et al., Clinica Chimica Acta 308: 43-53, 2001; Steimer et al., Clinica Chimica Acta 308: 33-41, 2001).
[00189] Как правило, дозы на основе массы тела, составляют примерно от 0,01 мкг примерно до 100 мг на кг массы тела, например, примерно от 0,01 мкг примерно до 100 мг/кг массы тела, примерно от 0,01 мкг примерно до 50 мг/кг массы тела, примерно от 0,01 мкг примерно до 10 мг/кг массы тела, примерно от 0,01 мкг примерно до 1 мг/кг массы тела, примерно от 0,01 мкг примерно до 100 мкг/кг массы тела, примерно от 0,01 мкг примерно до 50 мкг/кг массы тела, примерно от 0,01 мкг примерно до 10 мкг/кг массы тела, примерно от 0,01 мкг примерно до 1 мкг/кг массы тела, примерно от 0,01 мкг примерно до 0,1 мкг/кг массы тела, примерно от 0,1 мкг примерно до 100 мг/кг массы тела, примерно от 0,1 мкг примерно до 50 мг/кг массы тела, примерно от 0,1 мкг примерно до 10 мг/кг массы тела, примерно от 0,1 мкг примерно до 1 мг/кг массы тела, примерно от 0,1 мкг примерно до 100 мкг/кг массы тела, примерно от 0,1 мкг примерно до 50 мкг/кг массы тела, примерно от 0,1 мкг примерно до 10 мкг/кг массы тела, примерно от 0,1 мкг примерно до 1 мкг/кг массы тела, примерно от 1 мкг примерно до 100 мг/кг массы тела, примерно от 1 мкг примерно до 50 мг/кг массы тела, примерно от 1 мкг примерно до 10 мг/кг массы тела, примерно от 1 мкг примерно до 1 мг/кг массы тела, примерно от 1 мкг примерно до 100 мкг/кг массы тела, примерно от 1 мкг примерно до 50 мкг/кг массы тела, примерно от 1 мкг примерно до 10 мкг/кг массы тела, примерно от 10 мкг примерно до 100 мг/кг массы тела, примерно от 10 мкг примерно до 50 мг/кг массы тела, примерно от 10 мкг примерно до 10 мг/кг массы тела, примерно от 10 мкг примерно до 1 мг/кг массы тела, примерно от 10 мкг примерно до 100 мкг/кг массы тела, примерно от 10 мкг примерно до 50 мкг/кг массы тела, примерно от 50 мкг примерно до 100 мг/кг массы тела, примерно от 50 мкг примерно до 50 мг/кг массы тела, примерно от 50 мкг примерно до 10 мг/кг массы тела, примерно от 50 мкг примерно до 1 мг/кг массы тела, примерно от 50 мкг примерно до 100 мкг/кг массы тела, примерно от 100 мкг примерно до 100 мг/кг массы тела, примерно от 100 мкг примерно до 50 мг/кг массы тела, примерно от 100 мкг примерно до 10 мг/кг массы тела, примерно от 100 мкг примерно до 1 мг/кг массы тела, примерно от 1 мг примерно до 100 мг/кг массы тела, примерно от 1 мг примерно до 50 мг/кг массы тела, примерно от 1 мг примерно до 10 мг/кг массы тела, примерно от 10 мг примерно до 100 мг/кг массы тела, примерно от 10 мг примерно до 50 мг/кг массы тела, примерно от 50 мг примерно до 100 мг/кг массы тела.
[00190] Дозы могут вводиться один или более раз в день, еженедельно, ежемесячно или ежегодно, или даже один раз каждые 2-20 лет. Специалисты в данной области техники могут легко оценить частоту повторения для дозирования на основании измеренного времени удерживания и концентраций нацеливающей конструкции или комплекса в жидкостях или тканях организма. Введение по настоящему изобретению может быть внутривенным, внутриартериальным, внутрибрюшинным, внутримышечным, подкожным, внутриплевральным, интратекальным, внутриполостным, путем перфузии через катетер или путем прямой внутриклеточной инъекции. Это может осуществляться один или более раз в день, один или более раз в неделю, один или более раз в месяц и один или более раз в год.
[00191] Вышеприведенное описание описывает множество аспектов и вариантов осуществления изобретения. В заявке на патент конкретно рассматриваются все комбинации и перестановки аспектов и вариантов осуществления.
ПРИМЕРЫ
[00192] Настоящее изобретение теперь в целом описано, оно будет более понятно с помощью ссылок на следующие примеры, которые включены только в целях иллюстрации некоторых аспектов и вариантов осуществления настоящего изобретения и не предназначены для ограничения изобретения каким-либо образом.
Пример 1. NKG2D-связывающие домены связываются с NKG2D
NKG2D-связывающие домены связываются с очищенным рекомбинантным NKG2D
[00193] Последовательности нуклеиновых кислот эктодоменов NKG2D человека, мыши или яванской макаки были слиты с последовательностями нуклеиновых кислот, кодирующими Fc-домены IgG1 человека, и введены в клетки млекопитающих для экспрессии. После очистки слитые белки NKG2D-Fc были адсорбированы в лунках микропланшетов. После блокирования лунок бычьим сывороточным альбумином для предотвращения неспецифического связывания, NKG2D-связывающие домены титровали и добавляли в лунки, на которые предварительно адсорбировали слитые белки NKG2D-Fc. Связывание первичного антитела определяли с использованием вторичного антитела, которое конъюгировано с пероксидазой хрена и специфически распознает легкую цепь каппа человека, чтобы избежать перекрестной реактивности Fc. 3,3',5,5'-тетраметилбензидин (TMB), субстрат для пероксидазы хрена, добавляли в лунки для визуализации сигнала связывания, поглощение которого измеряли при 450 нМ и корректировали при 540 нМ. Клон NKG2D-связывающего домена, контроль изотипа или положительный контроль (выбранный из SEQ ID NO: 47-50 или антимышиные клоны NKG2D MI-6 и CX-5, доступные в eBioscience) добавляли в каждую лунку.
[00194] Контроль изотипа показал минимальное связывание с рекомбинантными белками NKG2D-Fc, в то время как положительный контроль наиболее сильно связывался с рекомбинантными антигенами. NKG2D-связывающие домены, продуцируемые всеми клонами, демонстрировали связывание с рекомбинантными белками NKG2D-Fc человека, мыши и яванской макаки, хотя с разными аффинностями от клона к клону. Как правило, каждый анти-NKG2D-клон связывается с рекомбинантным NKG2D-Fc человека (ФИГ. 3) и яванской макаки (ФИГ. 4) с аналогичной аффинностью, но с более низкой аффинностью к рекомбинантному NKG2D-Fc мыши (ФИГ. 5).
NKG2D-связывающие домены связываются с клетками, экспрессирующими NKG2D
[00195] Клеточные линии мышиной лимфомы EL4 были сконструированы для экспрессии химерного антигенного рецептора NKG2D и сигнального домена CD3-дзета человека или мыши. NKG2D-связывающий клон, изотипический контроль или положительный контроль использовали в концентрации 100 нМ для окрашивания внеклеточного NKG2D, экспрессированного на клетках EL4. Связывание антител определяли с использованием вторичных антител против человеческого IgG, конъюгированных с флуорофором. Клетки анализировали с помощью проточной цитометрии и рассчитывали превышение относительно фона (fold-over-background) (FOB) с использованием средней интенсивности флуоресценции (MFI) NKG2D-экспрессирующих клеток по сравнению с родительскими клетками EL4.
[00196] NKG2D-связывающие домены, продуцируемые всеми клонами, связывались с клетками EL4, экспрессирующими NKG2D человека и мыши. Антитела положительного контроля (выбранные из SEQ ID NO: 45-48 или анти-мышиные клоны NKG2D MI-6 и CX-5, доступные на eBioscience) давали лучший сигнал связывания FOB.Аффинность связывания NKG2D для каждого клона была сходной между клетками, экспрессирующими NKG2D человека (ФИГ. 6) и мыши (ФИГ. 7).
Пример 2. NKG2D-связывающие домены блокируют связывание природного лиганда с NKG2D
Конкуренция с ULBP-6
[00197] Рекомбинантные белки NKG2D-Fc человека адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для снижения неспецифического связывания. Насыщенную концентрацию ULBP-6-His-биотин добавляли в лунки с последующим добавлением клонов NKG2D-связывающего домена. После 2-часовой инкубации лунки промывали и ULBP-6-His-биотин, который оставался связанным с лунками, покрытыми NKG2D-Fc, детектировали стрептавидином, конъюгированным с пероксидазой хрена и субстратом TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона специфическое связывание NKG2D-связывающих доменов с белками NKG2D-Fc рассчитывали по проценту ULBP-6-His-биотина, который блокировался от связывания с белками NKG2D-Fc в лунках. Антитело положительного контроля (выбранное из SEQ ID NO: 47-50) и различные NKG2D-связывающие домены блокировали связывание ULBP-6 с NKG2D, в то время как изотипический контроль показал небольшую конкуренцию с ULBP-6 (ФИГ. 8).
[00198] Последовательность ULBP-6 представлена SEQ ID NO: 131
MAAAAIPALLLCLPLLFLLFGWSRARRDDPHSLCYDITVIPKFRPGPRWCAVQGQVDEKTFLHYDCGNKTVTPVSPLGKKLNVTMAWKAQNPVLREVVDILTEQLLDIQLENYTPKEPLTLQARMSCEQKAEGHSSGSWQFSIDGQTFLLFDSEKRMWTTVHPGARKMKEKWENDKDVAMSFHYISMGDCIGWLEDFLMGMDSTLEPSAGAPLAMSSGTTQLRATATTLILCCLLIILPCFILPGI (SEQ ID NO: 131)
Конкуренция с MICA
[00199] Рекомбинантные белки MICA-Fc человека адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для уменьшения неспецифического связывания. NKG2D-Fc-биотин добавляли в лунки, за которыми следовало добавление NKG2D-связывающих доменов. После инкубации и промывания NKG2D-Fc-биотин, который оставался связанным с лунками, покрытыми MICA-Fc, детектировали с использованием субстрата стрептавидин-HRP и TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона специфическое связывание NKG2D-связывающих доменов с белками NKG2D-Fc рассчитывали по проценту NKG2D-Fc-биотина, который блокировался от связывания с MICA-Fc в лунках. Антитело позитивного контроля (выбранное из SEQ ID NO: 47-50) и различные NKG2D-связывающие домены блокировали связывание MICA с NKG2D, в то время как изотипический контроль показал небольшую конкуренцию с MICA (ФИГ. 9).
Конкуренция с Rae-1 дельта
[00200] Рекомбинантный мышиный Rae-1delta-Fc (приобретенный у R & D Systems) адсорбировали в лунках микропланшета, и лунки блокировали бычьим сывороточным альбумином для снижения неспецифического связывания. Мышиный NKG2D-Fc-биотин добавляли в лунки с последующим добавлением NKG2D-связывающих доменов. После инкубации и промывания NKG2D-Fc-биотин, который оставался связанным с лунками, покрытыми Rae-1delta-Fc, детектировали с использованием субстрата стрептавидин-HRP и TMB. Поглощение измеряли при 450 нМ и корректировали при 540 нМ. После вычитания фона специфическое связывание NKG2D-связывающих доменов с белками NKG2D-Fc рассчитывали по проценту NKG2D-Fc-биотина, который блокировался от связывания с лунками, покрытыми Rae-1delta-Fc. Положительный контроль (выбранный из SEQ ID NO: 47-50 или анти-мышиных клонов NKG2D MI-6 и CX-5, доступных на eBioscience) и различные клоны NKG2D-связывающего домена блокировали связывание Rae-1-дельта с мышиным NKG2D, в то время как антитело изотипического контроля демонстрировало небольшую конкуренцию с Rae-1delta (ФИГ. 10).
Пример 3. Клоны NKG2D-связывающего домена активируют NKG2D
[00201] Последовательности нуклеиновой кислоты NKG2D человека и мыши были слиты с последовательностями нуклеиновой кислоты, кодирующими сигнальный домен CD3 дзета, с получением конструкций химерного антигенного рецептора (CAR). Конструкции NKG2D-CAR затем клонировали в ретровирусный вектор с использованием сборки Гибсона и трансфицировали в клетки expi293 с получением ретровируса. Клетки EL4 инфицировали вирусами, содержащими NKG2D-CAR вместе с 8 мкг/мл полибрена. Через 24 часа после инфекции уровни экспрессии NKG2D-CAR в клетках EL4 анализировали проточной цитометрией и отбирали клоны, которые экспрессируют на высоком уровне NKG2D-CAR на клеточной поверхности.
[00202] Чтобы определить, активируют ли NKG2D-связывающие домены NKG2D, их адсорбировали в лунки микропланшета, и клетки NKG2D-CAR EL4 культивировали в лунках, покрытых фрагментом антитела, в течение 4 часов в присутствии брефельдина-A и монензина. Внутриклеточное продуцирование TNF-альфа, индикатора активации NKG2D, анализировали с помощью проточной цитометрии. Процент TNF-альфа-положительных клеток был нормализован по отношению к клеткам, обработанным положительным контролем. Все NKG2D-связывающие домены активировали как NKG2D человека (ФИГ. 11), так и мыши (ФИГ. 12).
Пример 4. NKG2D-связывающие домены активируют NK-клетки
Первичные человеческие NK-клетки
[00203] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкотромбоцитарного слоя периферической крови с использованием центрифугирования в градиенте плотности. NK-клетки (CD3-CD56+) выделяли с использованием отрицательного отбора с помощью магнитных гранул из PBMC, и чистота выделенных NK-клеток обычно составляла> 95%. Затем выделенные NK-клетки культивировали в среде, содержащей 100 нг/мл IL-2, в течение 24-48 часов, после чего их переносили в лунки микропланшета, на котором были адсорбированы NKG2D-связывающие домены, и культивировали в среде, содержащей конъюгированный с флуорофором антитело против CD107a, брефелдин-А и монензин. После культивирования NK-клетки анализировали проточной цитометрией с использованием антител, конъюгированных с флуорофором, против CD3, CD56 и IFN-γ. Окрашивание CD107a и IFN-γ анализировали в клетках CD3-CD56+ для оценки активации NK-клеток. Увеличение количества CD107a/IFN-γ-двойных положительных клеток свидетельствует о лучшей активации NK-клеток за счет рекрутирования двух активирующих рецепторов, а не одного рецептора. NKG2D-связывающие домены и положительный контроль (выбранный из SEQ ID NO: 45-48) показали более высокий процент NK-клеток, становящихся CD107a+ и IFN-γ+, чем изотипический контроль (ФИГ. 13 и ФИГ. 14 представляют два независимых эксперимента, в каждом из которых использовали разных доноров PBMC для приготовления NK-клеток).
Первичные мышиные NK-клетки
[00204] Селезенки получали от мышей C57Bl/6 и измельчали через сито для клеток 70 мкм для получения суспензии отдельных клеток. Клетки осаждали и ресуспендировали в лизирующем буфере ACK (приобретенном у Thermo Fisher Scientific # A1049201; 155 мМхлорида аммония, 10 мМбикарбоната калия, 0,01 мМEDTA) для удаления эритроцитов. Оставшиеся клетки культивировали с 100 нг/мл hIL-2 в течение 72 часов, а затем собирали и готовили для выделения NK-клеток. NK-клетки (CD3-NK1.1+) затем выделяли из клеток селезенки с использованием метода отрицательного истощения с использованием магнитных гранул с чистотой> 90%. Очищенные NK-клетки культивировали в среде, содержащей 100 нг/мл mIL-15, в течение 48 часов, после чего их переносили в лунки микропланшета, на котором были адсорбированы NKG2D-связывающие домены, и культивировали в среде, содержащей конъюгированный с флуорофором антитело против CD107a, брефелдин-А и монензин. После культивирования в лунках, покрытых NKG2D-связывающим доменом, NK-клетки анализировали проточной цитометрией с использованием антител, конъюгированных с флуорофором, против CD3, NK1.1 и IFN-γ. Окрашивание CD107a и IFN-γ анализировали в клетках CD3-NK1.1+ для оценки активации NK-клеток. Увеличение количества CD107a/IFN-γ-двойных положительных клеток свидетельствует о лучшей активации NK-клеток за счет вовлечения двух активирующих рецепторов, а не одного рецептора. NKG2D-связывающие домены и положительный контроль (выбранный из анти-мышиных NKG2D-клонов MI-6 и CX-5, доступных в eBioscience) показали более высокий процент NK-клеток, становящихся CD107a+ и IFN-γ+, чем изотипический контроль (ФИГ. 15 и ФИГ. 16 представляют два независимых эксперимента, в каждом из которых использовали разных мышей для приготовления NK-клеток).
Пример 5. NKG2D-связывающие домены обеспечивают цитотоксичность опухолевых клеток-мишеней
[00205] Анализы активации первичных NK-клеток человека и мыши демонстрируют повышенный уровень маркеров цитотоксичности для NK-клеток после инкубации с NKG2D-связывающими доменами. Чтобы определить, приводит ли это к усилению лизиса опухолевых клеток, был использован клеточный анализ, в котором каждый NKG2D-связывающий домен превращался в моноспецифическое антитело. Fc-область использовали в качестве одного нацеливающего фрагмента, тогда как Fab-область (NKG2D-связывающий домен) действовала как другой нацеливающий фрагмент для активации NK-клеток. Клетки THP-1, которые имеют человеческое происхождение и экспрессируют на высоком уровне Fc-рецепторы, использовали в качестве опухолевой мишени и использовали набор для цитотоксичности Perkin Elmer DELFIA. Клетки THP-1 метили реагентом BATDA и ресуспендировали в количестве 105/мл в культуральной среде. Меченые клетки THP-1 затем объединяли с антителами NKG2D и выделенными мышиными NK-клетками в лунках микропланшета при 37°С в течение 3 часов. После инкубации удаляли 20 мкл культурального супернатанта, смешивали с 200 мкл раствора европия и инкубировали при встряхивании в течение 15 минут в темноте. Флуоресценцию измеряли во времени с помощью планшет-ридера PheraStar, оборудованного модулем флуоресценции с разрешением по времени (возбуждение 337 нм, излучение 620 нм), и специфический лизис рассчитывали в соответствии с инструкциями набора.
[00206] Положительный контроль, ULBP-6 - естественный лиганд для NKG2D, показал повышенный специфический лизис клеток-мишеней THP-1 мышиными NK-клетками. Антитела NKG2D также увеличивали специфический лизис клеток-мишеней THP-1, тогда как антитело изотипического контроля показало снижение специфического лизиса. Пунктирная линия указывает на специфический лизис клеток THP-1 мышиными NK-клетками без добавления антител (ФИГ. 17).
Пример 6. Антитела NKG2D показывают высокую термостабильность
[00207] Температуру плавления NKG2D-связывающих доменов анализировали с использованием дифференциальной сканирующей флуориметрии. Экстраполированные кажущиеся температуры плавления являются высокими по сравнению с типичными антителами IgG1 (ФИГ. 18).
Пример 7. Синергическая активация человеческих NK-клеток путем перекрестного связывания NKG2D и CD16.
Анализ активации первичных человеческих NK-клеток
[00208] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкотромбоцитарного слоя периферической крови с использованием центрифугирования в градиенте плотности. NK-клетки очищали из РВМС с использованием отрицательных магнитных гранул (StemCell # 17955). NK-клетки были> 90% CD3-CD56+, как определено проточной цитометрией. Затем клетки наращивали 48 часов в среде, содержащей 100 нг/мл hIL-2 (Peprotech # 200-02), перед использованием в анализах активации. Антитела наносили на 96-луночный планшет с плоским дном в концентрации 2 мкг/мл (анти-CD16, Biolegend # 302013) и 5 мкг/мл (анти-NKG2D, R & D # MAB139) в 100 мкл стерильного PBS в течение ночи при 4°С с последующим тщательным промыванием лунок для удаления избытка антител. Для оценки дегрануляции IL-2-активированные NK-клетки ресуспендировали в количестве 5х105 клеток/мл в культуральной среде, дополненной 100 нг/мл hIL2 и 1 мкг/мл APC-конъюгированного анти-CD107a mAb (Biolegend # 328619). Затем на планшеты, покрытые антителами, добавляли 1х105 клеток/лунку. Ингибиторы транспорта белка Брефельдин А (BFA, Biolegend # 420601) и Монензин (Biolegend # 420701) добавляли в конечном разведении 1:1000 и 1:270, соответственно. Посеянные клетки инкубировали в течение 4 часов при 37°С в 5% СО2. Для внутриклеточного окрашивания IFN-γ NK-клетки метили анти-CD3 (Biolegend # 300452) и анти-CD56 mAb (Biolegend # 318328), а затем фиксировали и пермеабилизовали и метили анти-IFN-γ mAb (Biolegend # 506507). NK-клетки анализировали на экспрессию CD107a и IFN-γ методом проточной цитометрии после гейтинга живых CD56+CD3-клеток.
[00209] Чтобы исследовать относительную эффективность комбинации рецепторов, проводили перекрестное связывание NKG2D или CD16 и совместное связывание обоих рецепторов посредством стимулирования в состоянии, связанном с планшетом. Как показано на Фигуре. 19 (ФИГ. 19A-19C), комбинированное стимулирование CD16 и NKG2D приводило к сильно повышенным уровням CD107a (дегрануляция) (ФИГ. 19A) и/или к продуцированию IFN-γ (ФИГ. 19B). Пунктирные линии представляют аддитивный эффект отдельных стимулирований каждого рецептора.
[00210] Уровни CD107a и внутриклеточное продуцирование IFN-γ активированных IL-2 NK-клеток анализировали через 4 часа стимулирования, связанного с планшетом, с помощью анти-CD16, анти-NKG2D или комбинацией обоих моноклональных антител. Графики показывают среднее (n=2) ± SD. ФИГ. 19А демонстрирует уровень CD107a; ФИГ. 19B демонстрирует уровень IFNγ; ФИГ.19C демонстрирует уровень CD107a и IFNγ. Данные, показанные на ФИГ. 19А-19С представляют пять независимых экспериментов с использованием пяти различных здоровых доноров.
Пример 8. Полиспецифические связывающие белки связываются с NKG2D
[00211] Клеточные линии мышиной лимфомы EL4 были сконструированы для экспрессии NKG2D человека. Триспецифические связывающие белки (TriNKET), каждый из которых содержит NKG2D-связывающий домен, опухолеспецифический антигенсвязывающий домен (CAIX-связывающий домен) и Fc-домен, который связывается с CD16, как показано на ФИГ. 1, тестировали на их аффинность к внеклеточному NKG2D, экспрессированному на клетках EL4. Связывание полиспецифических связывающих белков с NKG2D определяли, используя конъюгированные с флуорофором вторичные антитела против человеческого IgG. Клетки анализировали с помощью проточной цитометрии и рассчитывали превышение относительно фона (fold-over-background) (FOB) с использованием средней интенсивности флуоресценции (MFI) NKG2D-экспрессирующих клеток по сравнению с родительскими клетками EL4.
[00212] Тестированные TriNKET включают A44-TriNKET-CAIX (ADI-27744 и CAIX-связывающий домен), F63-TriNKET-CAIX (ADI-29404 и CD33-связывающий домен), E79-TriNKET-CAIX (ADI-29379 и CAIX- связывающий домен), F47-TriNKET-CAIX (ADI-29447 и CAIX-связывающий домен) и A49-TriNKET-CAIX (ADI-27749 и CAIX-связывающий домен). CAIX-связывающий домен состоял из вариабельного домена тяжелой цепи SEQ ID NO: 80 и вариабельного домена легкой цепи SEQ ID NO: 81.
[00213] ФИГ. 35 показывает, что TriNKET, каждый из которых включает CAIX-связывающий домен и другой NKG2D-связывающий домен, связываются с NKG2D, экспрессируемым на клетках EL4.
Пример 9. Полиспецифические связывающие белки связываются с опухолевыми антигенами человека
Триспецифические связывающие белки связываются с CAIX
[00214] Линию почечно-клеточной карциномы человека A498, экспрессирующую CAIX, использовали для анализа связывания TriNKET с опухолеспецифическим антигеном CAIX. TriNKET и, необязательно, родительское моноклональное антитело против CAIX инкубировали с клетками и связывание детектировали с использованием вторичных антител против человеческого IgG, конъюгированных с флуорофором. Клетки анализировали с помощью проточной цитометрии и рассчитывали превышение относительно фона (FOB), используя среднюю интенсивность флуоресценции (MFI) от TriNKET и родительского моноклонального антитела против CAIX, нормализованного к контролям вторичных антител. TriNKET, нацеленные на CAIX, демонстрируют сопоставимые уровни связывания с CAIX на клетках A498 по сравнению с родительским антителом против CAIX (ФИГ. 36). Общий сигнал связывания сопоставим среди TriNKET, поскольку они содержат один и тот же домен связывания CAIX.
Пример 10. Триспецифические связывающиеся белки обеспечивают цитотоксичность опухолевых клеток-мишеней.
[00215] Мононуклеарные клетки периферической крови (РВМС) выделяли из лейкотромбоцитарного слоя периферической крови с использованием центрифугирования в градиенте плотности. NK-клетки (CD3-CD56+) выделяли с использованием отрицательного отбора с помощью магнитных гранул из PBMC, и чистота выделенных NK-клеток обычно составляла> 90%. Выделенные NK-клетки культивировали в среде, содержащей 100 нг/мл IL-2 для активации, или оставляли на ночь без цитокина. IL-2-активированные или покоящиеся NK-клетки использовали на следующий день в анализах на цитотоксичность.
[00216] NK/NK Т-клеточные линии KHYG-1 поддерживали в 10% HI-FBS-RPMI-1640 с 10 нг/мл IL-2. За день до использования в качестве эффекторных клеток клетки KHYG-1 собирали из культуры и клетки отмывали от среды, содержащей IL-2. После промывки клетки KHYG-1 ресуспендировали в 10% HI-FBS-RPMI-1640 и оставляли на ночь без цитокина.
Анализ цитотоксичности DELFIA:
[00217] Опухолевые клетки A498 человека, экспрессирующие CAIX, собирали из культуры; клетки промывали PBS и ресуспендировали в ростовой среде в количестве 106/мл для мечения реагентом BATDA (Perkin Elmer AD0116). Следовали инструкциям производителя по мечению клеток-мишеней. После мечения клетки промывали 3 раза PBS и ресуспендировали в количестве 0,5-1,0х105/мл в культуральной среде. Для приготовления фоновых лунок отбирали аликвоту меченых клеток, и клетки извлекали из среды. 100 мкл среды осторожно добавляли в лунки в трех экземплярах, чтобы не повредить осажденные клетки. 100 мкл BATDA-меченных клеток добавляли в каждую лунку 96-луночного планшета. Лунки предохраняли для спонтанного высвобождения из клеток-мишеней, и лунки были подготовлены для максимального лизиса клеток-мишеней путем добавления 1% Triton-X. TriNKET против интересующей опухоли-мишени разводили в культуральной среде, 50 мкл разведенного TriNKET добавляли в каждую лунку. Покоящиеся NK-клетки, активированные NK-клетки или клетки KHYG-1 собирали из культуры, промывали и ресуспендировали в количестве 105-2,0х106/мл в культуральной среде в зависимости от желаемого соотношения эффектора к мишени (Е:Т). 50 мкл NK-клеток добавляли в каждую лунку планшета, чтобы получить в общей сложности 200 мкл культурального объема. Планшет инкубировали при 37°C с 5% CO2 в течение 2-3 часов, прежде чем проводить анализ.
[00218] После культивирования в течение 2-3 часов планшет извлекали из инкубатора, и клетки осаждали центрифугированием при 200g в течение 5 минут. 20 мкл культурального супернатанта переносили в чистый микропланшет, предоставленный производителем, и 200 мкл раствора европия при комнатной температуре добавляли в каждую лунку. Планшет защищали от света и инкубировали на шейкере для планшетов при 250 об/мин в течение 15 минут. Планшет считывался с использованием инструментов Victor 3 или SpectraMax i3X. % специфического лизиса рассчитывали следующим образом: %Специфического лизиса = ((Экспериментальное высвобождение - спонтанное высвобождение)/(Максимальное высвобождение - спонтанное высвобождение)) * 100%.
[00219] TriNKET опосредуют цитотоксичность NK-клеток человека по отношению к CAIX-положительным опухолевым клеткам человека. Совместное культивирование покоящихся клеток KHYG-1 с клетками-мишенями A498 приводило к ~38% специфического лизиса опухоли (показано пунктирной линией на ФИГ. 37A). Добавление трех различных TriNKET, нацеленных на CAIX (A49-TriNKET-CAIX; E79-TriNKET-CAIX; и A44-TriNKET-CAIX), к совместной культуре усиливало специфический лизис опухолевых клеток примерно на 20% для всех трех NKG2D-связывающих доменов (ФИГ. 37А).
[00220] Покоящиеся человеческие NK-клетки смешивали с опухолевыми клетками A498. Покоящиеся человеческие NK-клетки не обладали цитотоксической активностью в отношении клеток A498 без добавления CAIX mAb или TriNKET. Моноклональное антитело против CAIX (вариабельные домены BAY79-4620 с hIgG1 в качестве mAb (гомодимер) смогли увеличить специфический лизис примерно на 9%. TriNKET (например, A49-TriNKET-CAIX, E79-TriNKET-CAIX и A44-TriNKET-CAIX) усиливали цитотоксическую активность покоящихся человеческих NK-клеток в отношении опухолевых клеток по сравнению с моноклональным антителом (ФИГ. 37B).
ВКЛЮЧЕНИЕ С ПОМОЩЬЮ ССЫЛКИ
[00221] Полное раскрытие каждого из патентных документов и научных статей, упомянутых в настоящем документе, включено в качестве ссылки для всех целей.
ЭКВИВАЛЕНТЫ
[00222] Изобретение может быть воплощено в других конкретных формах без отклонения от его сущности или существенных характеристик. Таким образом, вышеприведенные варианты осуществления следует рассматривать во всех отношениях как иллюстративные, а не ограничивающие изобретение, описанное в настоящем документе. Таким образом, объем изобретения обозначен прилагаемой формулой изобретения, а не предшествующим описанием, и все изменения, которые входят в значение и диапазон эквивалентности формулы изобретения, предназначены для включения в нее.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ADIMAB, LLC.
<120> ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА CAIX, ANO1,
МЕЗОТЕЛИН, TROP2, СЕА ИЛИ КЛАУДИН-18.2
<130> DFY-012WO
<140>
<141>
<150> 62/473,659
<151> 2017-03-20
<150> 62/473,652
<151> 2017-03-20
<150> 62/467,557
<151> 2017-03-06
<150> 62/464,347
<151> 2017-02-27
<150> 62/464,344
<151> 2017-02-27
<150> 62/464,341
<151> 2017-02-27
<160> 236
<170> PatentIn version 3.5
<210> 1
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 1
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 2
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 3
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 3
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 4
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 4
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Ile Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 5
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 5
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 6
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr His Ser Phe Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 7
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 7
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 8
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Asn Ser Tyr Tyr Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 9
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 9
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 10
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 10
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 11
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 11
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Gly Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 12
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 12
Glu Leu Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Thr Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ser Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Asp Ile Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 13
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 13
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 14
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ser Phe Pro Ile
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 15
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 15
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 16
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 16
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Lys Glu Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 17
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 17
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 18
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 19
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 19
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 20
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 20
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ile Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 21
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 21
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 22
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 23
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 23
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 24
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gly Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 25
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 25
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 26
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 27
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 27
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 28
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Ser Phe Ser Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 29
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 29
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 30
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Ser Tyr Ser Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 31
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 31
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 32
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 33
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 33
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 34
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 34
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Gln Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 35
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 35
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 36
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 36
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr His Ser Phe Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 37
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 37
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 38
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 38
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Glu Leu Tyr Ser Tyr
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 39
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 39
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 40
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 40
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Thr Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 41
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 41
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Asp Ser Ser Ile Arg His Ala Tyr Tyr Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 42
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 42
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Thr Pro Ile Thr Phe Gly Gly Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 43
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 43
Gln Leu Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Ile Ser Ser Ser
20 25 30
Ser Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr
85 90 95
Cys Ala Arg Gly Ser Asp Arg Phe His Pro Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 44
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 44
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Phe Asp Thr Trp Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 45
<211> 126
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 45
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Arg Lys Ala Ser Gly Ser Phe Tyr Tyr Tyr Tyr Gly
100 105 110
Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 46
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 46
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Glu Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Pro Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 47
<211> 121
<212> Белок
<213> Homo sapiens
<400> 47
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Arg Gly Leu Gly Asp Gly Thr Tyr Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 48
<211> 110
<212> Белок
<213> Homo sapiens
<400> 48
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Ala Val Asn Trp Tyr Gln Gln Leu Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asp Asp Leu Leu Pro Ser Gly Val Ser Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Phe Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Ala Trp Asp Asp Ser Leu
85 90 95
Asn Gly Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 49
<211> 115
<212> Белок
<213> Homo sapiens
<400> 49
Gln Val His Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Asp Asp Ser Ile Ser Ser Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly His Ile Ser Tyr Ser Gly Ser Ala Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Asn Trp Asp Asp Ala Phe Asn Ile Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 50
<211> 108
<212> Белок
<213> Homo sapiens
<400> 50
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 51
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 51
Gly Ser Phe Ser Gly Tyr Tyr Trp Ser
1 5
<210> 52
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 52
Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 53
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 53
Ala Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro
1 5 10
<210> 54
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 54
Gly Thr Phe Ser Ser Tyr Ala Ile Ser
1 5
<210> 55
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 55
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 56
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 56
Ala Arg Gly Asp Ser Ser Ile Arg His Ala Tyr Tyr Tyr Tyr Gly Met
1 5 10 15
Asp Val
<210> 57
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 57
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Asn Lys Asn Tyr Leu
1 5 10 15
Ala
<210> 58
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 58
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 59
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 59
Gln Gln Tyr Tyr Ser Thr Pro Ile Thr
1 5
<210> 60
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 60
Gly Ser Ile Ser Ser Ser Ser Tyr Tyr Trp Gly
1 5 10
<210> 61
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 61
Ser Ile Tyr Tyr Ser Gly Ser Thr Tyr Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 62
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 62
Ala Arg Gly Ser Asp Arg Phe His Pro Tyr Phe Asp Tyr
1 5 10
<210> 63
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 63
Arg Ala Ser Gln Ser Val Ser Arg Tyr Leu Ala
1 5 10
<210> 64
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 64
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 65
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 65
Gln Gln Phe Asp Thr Trp Pro Pro Thr
1 5
<210> 66
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 66
Gly Thr Phe Ser Ser Tyr Ala Ile Ser
1 5
<210> 67
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 67
Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 68
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 68
Ala Arg Arg Gly Arg Lys Ala Ser Gly Ser Phe Tyr Tyr Tyr Tyr Gly
1 5 10 15
Met Asp Val
<210> 69
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 69
Glu Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu
1 5 10 15
Thr
<210> 70
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 70
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 71
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 71
Gln Asn Asp Tyr Ser Tyr Pro Tyr Thr
1 5
<210> 72
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 72
Asp Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Lys Leu Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Leu Val
35 40 45
Ala Ala Ile Asn Ser Asp Gly Gly Ile Thr Tyr Tyr Leu Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg His Arg Ser Gly Tyr Phe Ser Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 73
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 73
Asp Ile Val Met Thr Gln Ser Gln Arg Phe Met Ser Thr Thr Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Val Ser Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Met Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Phe Phe Cys Gln Gln Tyr Ser Asn Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 74
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 74
Gly Phe Thr Phe Ser Asn Tyr
1 5
<210> 75
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 75
Asn Ser Asp Gly Gly Ile
1 5
<210> 76
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 76
His Arg Ser Gly Tyr Phe Ser Met Asp Tyr
1 5 10
<210> 77
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 77
Gln Asn Val Val Ser Ala Val Ala
1 5
<210> 78
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 78
Ser Ala Ser Asn Arg Tyr Thr
1 5
<210> 79
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 79
Gln Gln Tyr Ser Asn Tyr Pro Trp Thr
1 5
<210> 80
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 80
Gln Val Glu Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Ser Ser Leu Gly Ser Thr Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Thr Gly Ser Pro Gly Thr Phe Met His Gly Asp His Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 81
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Asn Asn Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Gly Arg Pro Thr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 82
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 82
Gly Phe Thr Phe Ser Ser Tyr Gly Met Ser
1 5 10
<210> 83
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 83
Gly Ile Ser Ser Leu Gly Ser Thr Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 84
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 84
Thr Gly Ser Pro Gly Thr Phe Met His Gly Asp His
1 5 10
<210> 85
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 85
Arg Ala Ser Gln Asp Ile Asn Asn Tyr Leu Ser
1 5 10
<210> 86
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 86
Tyr Gly Ala Ser Asn Leu Gln Ser
1 5
<210> 87
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 87
Gln Gln Tyr Tyr Gly Arg Pro Thr
1 5
<210> 88
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 88
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Gln Arg Leu Glu Leu Val
35 40 45
Ser Ala Ile Asn Ser Asp Gly Gly Ile Thr Tyr Tyr Leu Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Gly Ser Leu Arg Ala Glu Asp Met Ala Val Tyr Tyr Cys
85 90 95
Ala Arg His Arg Ser Gly Tyr Phe Ser Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 89
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 89
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asn Val Val Ser Ala
20 25 30
Val Ala Trp Tyr Leu Gln Lys Pro Gly Lys Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Tyr Ser Asn Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 90
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 90
Gly Phe Thr Phe Ser Asn Tyr
1 5
<210> 91
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 91
Asn Ser Asp Gly Gly Ile
1 5
<210> 92
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 92
His Arg Ser Gly Tyr Phe Ser Met Asp Tyr
1 5 10
<210> 93
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 93
Gln Asn Val Val Ser Ala Val Ala
1 5
<210> 94
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 94
Ser Ala Ser Asn Arg Tyr Thr
1 5
<210> 95
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 95
Gln Gln Tyr Ser Asn Tyr Pro Trp Thr
1 5
<210> 96
<211> 459
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
CAIX полипептид
<400> 96
Met Ala Pro Leu Cys Pro Ser Pro Trp Leu Pro Leu Leu Ile Pro Ala
1 5 10 15
Pro Ala Pro Gly Leu Thr Val Gln Leu Leu Leu Ser Leu Leu Leu Leu
20 25 30
Val Pro Val His Pro Gln Arg Leu Pro Arg Met Gln Glu Asp Ser Pro
35 40 45
Leu Gly Gly Gly Ser Ser Gly Glu Asp Asp Pro Leu Gly Glu Glu Asp
50 55 60
Leu Pro Ser Glu Glu Asp Ser Pro Arg Glu Glu Asp Pro Pro Gly Glu
65 70 75 80
Glu Asp Leu Pro Gly Glu Glu Asp Leu Pro Gly Glu Glu Asp Leu Pro
85 90 95
Glu Val Lys Pro Lys Ser Glu Glu Glu Gly Ser Leu Lys Leu Glu Asp
100 105 110
Leu Pro Thr Val Glu Ala Pro Gly Asp Pro Gln Glu Pro Gln Asn Asn
115 120 125
Ala His Arg Asp Lys Glu Gly Asp Asp Gln Ser His Trp Arg Tyr Gly
130 135 140
Gly Asp Pro Pro Trp Pro Arg Val Ser Pro Ala Cys Ala Gly Arg Phe
145 150 155 160
Gln Ser Pro Val Asp Ile Arg Pro Gln Leu Ala Ala Phe Cys Pro Ala
165 170 175
Leu Arg Pro Leu Glu Leu Leu Gly Phe Gln Leu Pro Pro Leu Pro Glu
180 185 190
Leu Arg Leu Arg Asn Asn Gly His Ser Val Gln Leu Thr Leu Pro Pro
195 200 205
Gly Leu Glu Met Ala Leu Gly Pro Gly Arg Glu Tyr Arg Ala Leu Gln
210 215 220
Leu His Leu His Trp Gly Ala Ala Gly Arg Pro Gly Ser Glu His Thr
225 230 235 240
Val Glu Gly His Arg Phe Pro Ala Glu Ile His Val Val His Leu Ser
245 250 255
Thr Ala Phe Ala Arg Val Asp Glu Ala Leu Gly Arg Pro Gly Gly Leu
260 265 270
Ala Val Leu Ala Ala Phe Leu Glu Glu Gly Pro Glu Glu Asn Ser Ala
275 280 285
Tyr Glu Gln Leu Leu Ser Arg Leu Glu Glu Ile Ala Glu Glu Gly Ser
290 295 300
Glu Thr Gln Val Pro Gly Leu Asp Ile Ser Ala Leu Leu Pro Ser Asp
305 310 315 320
Phe Ser Arg Tyr Phe Gln Tyr Glu Gly Ser Leu Thr Thr Pro Pro Cys
325 330 335
Ala Gln Gly Val Ile Trp Thr Val Phe Asn Gln Thr Val Met Leu Ser
340 345 350
Ala Lys Gln Leu His Thr Leu Ser Asp Thr Leu Trp Gly Pro Gly Asp
355 360 365
Ser Arg Leu Gln Leu Asn Phe Arg Ala Thr Gln Pro Leu Asn Gly Arg
370 375 380
Val Ile Glu Ala Ser Phe Pro Ala Gly Val Asp Ser Ser Pro Arg Ala
385 390 395 400
Ala Glu Pro Val Gln Leu Asn Ser Cys Leu Ala Ala Gly Asp Ile Leu
405 410 415
Ala Leu Val Phe Gly Leu Leu Phe Ala Val Thr Ser Val Ala Phe Leu
420 425 430
Val Gln Met Arg Arg Gln His Arg Arg Gly Thr Lys Gly Gly Val Ser
435 440 445
Tyr Arg Pro Ala Glu Val Ala Glu Thr Gly Ala
450 455
<210> 97
<211> 123
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 97
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Asn Tyr
20 25 30
Asp Met His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Asp Trp Met
35 40 45
Gly Val Ile Trp Gly Asn Gly Lys Thr Gln Tyr Asn Ser Gly Leu Thr
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Glu Asp Thr Ala Ile Tyr Phe Cys Thr
85 90 95
Arg Ser Gly Tyr Tyr Tyr Asp Gly Ser Tyr Tyr Ser Leu Phe Asp Tyr
100 105 110
Trp Gly Gln Gly Val Met Val Thr Val Ser Ser
115 120
<210> 98
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 98
Asp Val Val Leu Thr Gln Thr Pro Pro Thr Leu Ser Ala Thr Ile Gly
1 5 10 15
Gln Ser Val Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Asn Trp Leu Leu Gln Arg Pro Gly Gln Pro
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Arg Leu Glu Ser Gly Val Pro
50 55 60
Asn Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Gly Val Glu Ala Glu Asp Leu Gly Val Tyr Tyr Cys Val Gln Ser
85 90 95
Thr His Ala Pro Ala Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys
100 105 110
<210> 99
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 99
Asn Tyr Asp Met His
1 5
<210> 100
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 100
Val Ile Trp Gly Asn Gly Lys Thr Gln Tyr Asn Ser Gly Leu Thr Ser
1 5 10 15
<210> 101
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 101
Ser Gly Tyr Tyr Tyr Asp Gly Ser Tyr Tyr Ser Leu Phe Asp Tyr
1 5 10 15
<210> 102
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 102
Arg Ser Ser Gln Ser Leu Leu His Ser Asn Gly Asn Thr Tyr Leu Asn
1 5 10 15
<210> 103
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 103
Leu Val Ser Arg Leu Glu Ser
1 5
<210> 104
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 104
Val Gln Ser Thr His Ala Pro Ala
1 5
<210> 105
<211> 986
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
ANO1 полипептид
<400> 105
Met Arg Val Asn Glu Lys Tyr Ser Thr Leu Pro Ala Glu Asp Arg Ser
1 5 10 15
Val His Ile Ile Asn Ile Cys Ala Ile Glu Asp Ile Gly Tyr Leu Pro
20 25 30
Ser Glu Gly Thr Leu Leu Asn Ser Leu Ser Val Asp Pro Asp Ala Glu
35 40 45
Cys Lys Tyr Gly Leu Tyr Phe Arg Asp Gly Arg Arg Lys Val Asp Tyr
50 55 60
Ile Leu Val Tyr His His Lys Arg Pro Ser Gly Asn Arg Thr Leu Val
65 70 75 80
Arg Arg Val Gln His Ser Asp Thr Pro Ser Gly Ala Arg Ser Val Lys
85 90 95
Gln Asp His Pro Leu Pro Gly Lys Gly Ala Ser Leu Asp Ala Gly Ser
100 105 110
Gly Glu Pro Pro Met Asp Tyr His Glu Asp Asp Lys Arg Phe Arg Arg
115 120 125
Glu Glu Tyr Glu Gly Asn Leu Leu Glu Ala Gly Leu Glu Leu Glu Arg
130 135 140
Asp Glu Asp Thr Lys Ile His Gly Val Gly Phe Val Lys Ile His Ala
145 150 155 160
Pro Trp Asn Val Leu Cys Arg Glu Ala Glu Phe Leu Lys Leu Lys Met
165 170 175
Pro Thr Lys Lys Met Tyr His Ile Asn Glu Thr Arg Gly Leu Leu Lys
180 185 190
Lys Ile Asn Ser Val Leu Gln Lys Ile Thr Asp Pro Ile Gln Pro Lys
195 200 205
Val Ala Glu His Arg Pro Gln Thr Met Lys Arg Leu Ser Tyr Pro Phe
210 215 220
Ser Arg Glu Lys Gln His Leu Phe Asp Leu Ser Asp Lys Asp Ser Phe
225 230 235 240
Phe Asp Ser Lys Thr Arg Ser Thr Ile Val Tyr Glu Ile Leu Lys Arg
245 250 255
Thr Thr Cys Thr Lys Ala Lys Tyr Ser Met Gly Ile Thr Ser Leu Leu
260 265 270
Ala Asn Gly Val Tyr Ala Ala Ala Tyr Pro Leu His Asp Gly Asp Tyr
275 280 285
Asn Gly Glu Asn Val Glu Phe Asn Asp Arg Lys Leu Leu Tyr Glu Glu
290 295 300
Trp Ala Arg Tyr Gly Val Phe Tyr Lys Tyr Gln Pro Ile Asp Leu Val
305 310 315 320
Arg Lys Tyr Phe Gly Glu Lys Ile Gly Leu Tyr Phe Ala Trp Leu Gly
325 330 335
Val Tyr Thr Gln Met Leu Ile Pro Ala Ser Ile Val Gly Ile Ile Val
340 345 350
Phe Leu Tyr Gly Cys Ala Thr Met Asp Glu Asn Ile Pro Ser Met Glu
355 360 365
Met Cys Asp Gln Arg His Asn Ile Thr Met Cys Pro Leu Cys Asp Lys
370 375 380
Thr Cys Ser Tyr Trp Lys Met Ser Ser Ala Cys Ala Thr Ala Arg Ala
385 390 395 400
Ser His Leu Phe Asp Asn Pro Ala Thr Val Phe Phe Ser Val Phe Met
405 410 415
Ala Leu Trp Ala Ala Thr Phe Met Glu His Trp Lys Arg Lys Gln Met
420 425 430
Arg Leu Asn Tyr Arg Trp Asp Leu Thr Gly Phe Glu Glu Glu Glu Glu
435 440 445
Ala Val Lys Asp His Pro Arg Ala Glu Tyr Glu Ala Arg Val Leu Glu
450 455 460
Lys Ser Leu Lys Lys Glu Ser Arg Asn Lys Glu Lys Arg Arg His Ile
465 470 475 480
Pro Glu Glu Ser Thr Asn Lys Trp Lys Gln Arg Val Lys Thr Ala Met
485 490 495
Ala Gly Val Lys Leu Thr Asp Lys Val Lys Leu Thr Trp Arg Asp Arg
500 505 510
Phe Pro Ala Tyr Leu Thr Asn Leu Val Ser Ile Ile Phe Met Ile Ala
515 520 525
Val Thr Phe Ala Ile Val Leu Gly Val Ile Ile Tyr Arg Ile Ser Met
530 535 540
Ala Ala Ala Leu Ala Met Asn Ser Ser Pro Ser Val Arg Ser Asn Ile
545 550 555 560
Arg Val Thr Val Thr Ala Thr Ala Val Ile Ile Asn Leu Val Val Ile
565 570 575
Ile Leu Leu Asp Glu Val Tyr Gly Cys Ile Ala Arg Trp Leu Thr Lys
580 585 590
Ile Glu Val Pro Lys Thr Glu Lys Ser Phe Glu Glu Arg Leu Ile Phe
595 600 605
Lys Ala Phe Leu Leu Lys Phe Val Asn Ser Tyr Thr Pro Ile Phe Tyr
610 615 620
Val Ala Phe Phe Lys Gly Arg Phe Val Gly Arg Pro Gly Asp Tyr Val
625 630 635 640
Tyr Ile Phe Arg Ser Phe Arg Met Glu Glu Cys Ala Pro Gly Gly Cys
645 650 655
Leu Met Glu Leu Cys Ile Gln Leu Ser Ile Ile Met Leu Gly Lys Gln
660 665 670
Leu Ile Gln Asn Asn Leu Phe Glu Ile Gly Ile Pro Lys Met Lys Lys
675 680 685
Leu Ile Arg Tyr Leu Lys Leu Lys Gln Gln Ser Pro Pro Asp His Glu
690 695 700
Glu Cys Val Lys Arg Lys Gln Arg Tyr Glu Val Asp Tyr Asn Leu Glu
705 710 715 720
Pro Phe Ala Gly Leu Thr Pro Glu Tyr Met Glu Met Ile Ile Gln Phe
725 730 735
Gly Phe Val Thr Leu Phe Val Ala Ser Phe Pro Leu Ala Pro Leu Phe
740 745 750
Ala Leu Leu Asn Asn Ile Ile Glu Ile Arg Leu Asp Ala Lys Lys Phe
755 760 765
Val Thr Glu Leu Arg Arg Pro Val Ala Val Arg Ala Lys Asp Ile Gly
770 775 780
Ile Trp Tyr Asn Ile Leu Arg Gly Ile Gly Lys Leu Ala Val Ile Ile
785 790 795 800
Asn Ala Phe Val Ile Ser Phe Thr Ser Asp Phe Ile Pro Arg Leu Val
805 810 815
Tyr Leu Tyr Met Tyr Ser Lys Asn Gly Thr Met His Gly Phe Val Asn
820 825 830
His Thr Leu Ser Ser Phe Asn Val Ser Asp Phe Gln Asn Gly Thr Ala
835 840 845
Pro Asn Asp Pro Leu Asp Leu Gly Tyr Glu Val Gln Ile Cys Arg Tyr
850 855 860
Lys Asp Tyr Arg Glu Pro Pro Trp Ser Glu Asn Lys Tyr Asp Ile Ser
865 870 875 880
Lys Asp Phe Trp Ala Val Leu Ala Ala Arg Leu Ala Phe Val Ile Val
885 890 895
Phe Gln Asn Leu Val Met Phe Met Ser Asp Phe Val Asp Trp Val Ile
900 905 910
Pro Asp Ile Pro Lys Asp Ile Ser Gln Gln Ile His Lys Glu Lys Val
915 920 925
Leu Met Val Glu Leu Phe Met Arg Glu Glu Gln Asp Lys Gln Gln Leu
930 935 940
Leu Glu Thr Trp Met Glu Lys Glu Arg Gln Lys Asp Glu Pro Pro Cys
945 950 955 960
Asn His His Asn Thr Lys Ala Cys Pro Asp Ser Leu Gly Ser Pro Ala
965 970 975
Pro Ser His Ala Tyr His Gly Gly Val Leu
980 985
<210> 106
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 106
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Glu Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Leu Ile Thr Pro Tyr Asn Gly Ala Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr Trp Gly Ser Gly
100 105 110
Thr Pro Val Thr Val Ser Ser Ala
115 120
<210> 107
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 107
Asp Ile Glu Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Gly Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Asn Ser Tyr Ser Leu Thr Ile Ser Ser Val Glu Ala Glu
65 70 75 80
Asp Asp Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Lys His Pro Leu Thr
85 90 95
Phe Gly Ser Gly Thr Lys Val Glu Ile Lys Arg
100 105
<210> 108
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 108
Gly Tyr Ser Phe Thr Gly Tyr
1 5
<210> 109
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 109
Thr Pro Tyr Asn Gly Ala
1 5
<210> 110
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 110
Gly Gly Tyr Asp Gly Arg Gly Phe Asp Tyr
1 5 10
<210> 111
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 111
Ser Ser Val Ser Tyr Met His
1 5
<210> 112
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 112
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 113
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 113
Gln Gln Trp Ser Lys His Pro Leu Thr
1 5
<210> 114
<211> 131
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 114
Glu Val His Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Gln Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Lys Gln Ala Gly Ser Glu Lys Thr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ala Tyr Tyr Tyr Asp Ser Ala Ser Tyr Tyr Pro Tyr
100 105 110
Tyr Tyr Tyr Tyr Ser Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr
115 120 125
Val Ser Ser
130
<210> 115
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 115
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Ser Gln
85 90 95
Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 116
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 116
Gly Phe Thr Phe Ser Arg Tyr
1 5
<210> 117
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 117
Lys Gln Ala Gly Ser Glu
1 5
<210> 118
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 118
Glu Gly Ala Tyr Tyr Tyr Asp Ser Ala Ser Tyr Tyr Pro Tyr Tyr Tyr
1 5 10 15
Tyr Tyr Ser Met Asp Val
20
<210> 119
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 119
Gln Ser Val Ser Ser Ser Tyr Leu Ala
1 5
<210> 120
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 120
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 121
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 121
Gln Gln Tyr Gly Ser Ser Gln Tyr Thr
1 5
<210> 122
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 122
Gln Val Glu Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Trp Ile Gly Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asp Pro Gly Asp Ser Arg Thr Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Gln Leu Tyr Gly Gly Thr Tyr Met Asp Gly Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala
115 120
<210> 123
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 123
Asp Ile Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Ile Gly Gly Tyr
20 25 30
Asn Ser Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Gly Val Asn Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Asp Ile Glu
85 90 95
Ser Ala Thr Pro Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
<210> 124
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 124
Gly Tyr Ser Phe Thr Ser Tyr
1 5
<210> 125
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 125
Asp Pro Gly Asp Ser Arg
1 5
<210> 126
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 126
Gly Gln Leu Tyr Gly Gly Thr Tyr Met Asp Gly
1 5 10
<210> 127
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 127
Ser Ser Asp Ile Gly Gly Tyr Asn Ser Val Ser
1 5 10
<210> 128
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 128
Gly Val Asn Asn Arg Pro Ser
1 5
<210> 129
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 129
Ser Ser Tyr Asp Ile Glu Ser Ala Thr Pro Val
1 5 10
<210> 130
<211> 630
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
полипептид мезотелина
<400> 130
Met Ala Leu Pro Thr Ala Arg Pro Leu Leu Gly Ser Cys Gly Thr Pro
1 5 10 15
Ala Leu Gly Ser Leu Leu Phe Leu Leu Phe Ser Leu Gly Trp Val Gln
20 25 30
Pro Ser Arg Thr Leu Ala Gly Glu Thr Gly Gln Glu Ala Ala Pro Leu
35 40 45
Asp Gly Val Leu Ala Asn Pro Pro Asn Ile Ser Ser Leu Ser Pro Arg
50 55 60
Gln Leu Leu Gly Phe Pro Cys Ala Glu Val Ser Gly Leu Ser Thr Glu
65 70 75 80
Arg Val Arg Glu Leu Ala Val Ala Leu Ala Gln Lys Asn Val Lys Leu
85 90 95
Ser Thr Glu Gln Leu Arg Cys Leu Ala His Arg Leu Ser Glu Pro Pro
100 105 110
Glu Asp Leu Asp Ala Leu Pro Leu Asp Leu Leu Leu Phe Leu Asn Pro
115 120 125
Asp Ala Phe Ser Gly Pro Gln Ala Cys Thr Arg Phe Phe Ser Arg Ile
130 135 140
Thr Lys Ala Asn Val Asp Leu Leu Pro Arg Gly Ala Pro Glu Arg Gln
145 150 155 160
Arg Leu Leu Pro Ala Ala Leu Ala Cys Trp Gly Val Arg Gly Ser Leu
165 170 175
Leu Ser Glu Ala Asp Val Arg Ala Leu Gly Gly Leu Ala Cys Asp Leu
180 185 190
Pro Gly Arg Phe Val Ala Glu Ser Ala Glu Val Leu Leu Pro Arg Leu
195 200 205
Val Ser Cys Pro Gly Pro Leu Asp Gln Asp Gln Gln Glu Ala Ala Arg
210 215 220
Ala Ala Leu Gln Gly Gly Gly Pro Pro Tyr Gly Pro Pro Ser Thr Trp
225 230 235 240
Ser Val Ser Thr Met Asp Ala Leu Arg Gly Leu Leu Pro Val Leu Gly
245 250 255
Gln Pro Ile Ile Arg Ser Ile Pro Gln Gly Ile Val Ala Ala Trp Arg
260 265 270
Gln Arg Ser Ser Arg Asp Pro Ser Trp Arg Gln Pro Glu Arg Thr Ile
275 280 285
Leu Arg Pro Arg Phe Arg Arg Glu Val Glu Lys Thr Ala Cys Pro Ser
290 295 300
Gly Lys Lys Ala Arg Glu Ile Asp Glu Ser Leu Ile Phe Tyr Lys Lys
305 310 315 320
Trp Glu Leu Glu Ala Cys Val Asp Ala Ala Leu Leu Ala Thr Gln Met
325 330 335
Asp Arg Val Asn Ala Ile Pro Phe Thr Tyr Glu Gln Leu Asp Val Leu
340 345 350
Lys His Lys Leu Asp Glu Leu Tyr Pro Gln Gly Tyr Pro Glu Ser Val
355 360 365
Ile Gln His Leu Gly Tyr Leu Phe Leu Lys Met Ser Pro Glu Asp Ile
370 375 380
Arg Lys Trp Asn Val Thr Ser Leu Glu Thr Leu Lys Ala Leu Leu Glu
385 390 395 400
Val Asn Lys Gly His Glu Met Ser Pro Gln Ala Pro Arg Arg Pro Leu
405 410 415
Pro Gln Val Ala Thr Leu Ile Asp Arg Phe Val Lys Gly Arg Gly Gln
420 425 430
Leu Asp Lys Asp Thr Leu Asp Thr Leu Thr Ala Phe Tyr Pro Gly Tyr
435 440 445
Leu Cys Ser Leu Ser Pro Glu Glu Leu Ser Ser Val Pro Pro Ser Ser
450 455 460
Ile Trp Ala Val Arg Pro Gln Asp Leu Asp Thr Cys Asp Pro Arg Gln
465 470 475 480
Leu Asp Val Leu Tyr Pro Lys Ala Arg Leu Ala Phe Gln Asn Met Asn
485 490 495
Gly Ser Glu Tyr Phe Val Lys Ile Gln Ser Phe Leu Gly Gly Ala Pro
500 505 510
Thr Glu Asp Leu Lys Ala Leu Ser Gln Gln Asn Val Ser Met Asp Leu
515 520 525
Ala Thr Phe Met Lys Leu Arg Thr Asp Ala Val Leu Pro Leu Thr Val
530 535 540
Ala Glu Val Gln Lys Leu Leu Gly Pro His Val Glu Gly Leu Lys Ala
545 550 555 560
Glu Glu Arg His Arg Pro Val Arg Asp Trp Ile Leu Arg Gln Arg Gln
565 570 575
Asp Asp Leu Asp Thr Leu Gly Leu Gly Leu Gln Gly Gly Ile Pro Asn
580 585 590
Gly Tyr Leu Val Leu Asp Leu Ser Met Gln Glu Ala Leu Ser Gly Thr
595 600 605
Pro Cys Leu Leu Gly Pro Gly Pro Val Leu Thr Val Leu Ala Leu Leu
610 615 620
Leu Ala Ser Thr Leu Ala
625 630
<210> 131
<211> 246
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
ULBP-6 полипептид
<400> 131
Met Ala Ala Ala Ala Ile Pro Ala Leu Leu Leu Cys Leu Pro Leu Leu
1 5 10 15
Phe Leu Leu Phe Gly Trp Ser Arg Ala Arg Arg Asp Asp Pro His Ser
20 25 30
Leu Cys Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg Pro Gly Pro Arg
35 40 45
Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr Phe Leu His Tyr
50 55 60
Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro Leu Gly Lys Lys
65 70 75 80
Leu Asn Val Thr Met Ala Trp Lys Ala Gln Asn Pro Val Leu Arg Glu
85 90 95
Val Val Asp Ile Leu Thr Glu Gln Leu Leu Asp Ile Gln Leu Glu Asn
100 105 110
Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg Met Ser Cys Glu
115 120 125
Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln Phe Ser Ile Asp
130 135 140
Gly Gln Thr Phe Leu Leu Phe Asp Ser Glu Lys Arg Met Trp Thr Thr
145 150 155 160
Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp Glu Asn Asp Lys
165 170 175
Asp Val Ala Met Ser Phe His Tyr Ile Ser Met Gly Asp Cys Ile Gly
180 185 190
Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr Leu Glu Pro Ser
195 200 205
Ala Gly Ala Pro Leu Ala Met Ser Ser Gly Thr Thr Gln Leu Arg Ala
210 215 220
Thr Ala Thr Thr Leu Ile Leu Cys Cys Leu Leu Ile Ile Leu Pro Cys
225 230 235 240
Phe Ile Leu Pro Gly Ile
245
<210> 132
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 132
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Asp Gly Gly Tyr Tyr Asp Ser Gly Ala Gly Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 133
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 133
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Asp Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Val Ser Tyr Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 134
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 134
Phe Thr Phe Ser Ser Tyr Ala Met Ser
1 5
<210> 135
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 135
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 136
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 136
Ala Lys Asp Gly Gly Tyr Tyr Asp Ser Gly Ala Gly Asp Tyr
1 5 10
<210> 137
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 137
Arg Ala Ser Gln Gly Ile Asp Ser Trp Leu Ala
1 5 10
<210> 138
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 138
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 139
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 139
Gln Gln Gly Val Ser Tyr Pro Arg Thr
1 5
<210> 140
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 140
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Met Gly Ala Ala Ala Gly Trp Phe Asp Pro Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 141
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 141
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Val Ser Phe Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 142
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 142
Phe Thr Phe Ser Ser Tyr Ser Met Asn
1 5
<210> 143
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 143
Ser Ile Ser Ser Ser Ser Ser Tyr Ile Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 144
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 144
Ala Arg Gly Ala Pro Met Gly Ala Ala Ala Gly Trp Phe Asp Pro
1 5 10 15
<210> 145
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 145
Arg Ala Ser Gln Gly Ile Ser Ser Trp Leu Ala
1 5 10
<210> 146
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 146
Ala Ala Ser Ser Leu Gln Ser
1 5
<210> 147
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 147
Gln Gln Gly Val Ser Phe Pro Arg Thr
1 5
<210> 148
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 148
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Thr Gly Glu Tyr Tyr Asp Thr Asp Asp His Gly Met Asp
100 105 110
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 149
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 149
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Asp Asp Tyr Trp Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 150
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 150
Tyr Thr Phe Thr Gly Tyr Tyr Met His
1 5
<210> 151
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 151
Trp Ile Asn Pro Asn Ser Gly Gly Thr Asn Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 152
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 152
Ala Arg Asp Thr Gly Glu Tyr Tyr Asp Thr Asp Asp His Gly Met Asp
1 5 10 15
Val
<210> 153
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 153
Arg Ala Ser Gln Ser Val Ser Ser Asn Leu Ala
1 5 10
<210> 154
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 154
Gly Ala Ser Thr Arg Ala Thr
1 5
<210> 155
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 155
Gln Gln Asp Asp Tyr Trp Pro Pro Thr
1 5
<210> 156
<211> 126
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 156
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Pro Asn Tyr Gly Asp Thr Thr His Asp Tyr Tyr Tyr
100 105 110
Met Asp Val Trp Gly Lys Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 157
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 157
Glu Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asp Asp Trp Pro Phe
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 158
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 158
Tyr Thr Phe Thr Ser Tyr Tyr Met His
1 5
<210> 159
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 159
Ile Ile Asn Pro Ser Gly Gly Ser Thr Ser Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 160
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 160
Ala Arg Gly Ala Pro Asn Tyr Gly Asp Thr Thr His Asp Tyr Tyr Tyr
1 5 10 15
Met Asp Val
<210> 161
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 161
Arg Ala Ser Gln Ser Val Ser Ser Asn Leu Ala
1 5 10
<210> 162
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 162
Gly Ala Ser Thr Arg Ala Thr
1 5
<210> 163
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 163
Gln Gln Tyr Asp Asp Trp Pro Phe Thr
1 5
<210> 164
<211> 117
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 164
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 165
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 165
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp Thr Phe Ile Thr
85 90 95
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 166
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 166
Gly Ser Phe Ser Gly Tyr Tyr Trp Ser
1 5
<210> 167
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 167
Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser
1 5 10 15
<210> 168
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 168
Ala Arg Ala Arg Gly Pro Trp Ser Phe Asp Pro
1 5 10
<210> 169
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 169
Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala
1 5 10
<210> 170
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 170
Lys Ala Ser Ser Leu Glu Ser
1 5
<210> 171
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 171
Gln Gln Tyr Asp Thr Phe Ile Thr
1 5
<210> 172
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 172
Ser Val Gln Leu Gln Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Thr Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Asp Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Phe Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Ser Leu Val Thr Val Ser Ser
115 120
<210> 173
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 173
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Tyr Ile Thr Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Val Glu Ile Lys Arg
100 105
<210> 174
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 174
Gly Tyr Thr Phe Thr Asn Tyr
1 5
<210> 175
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 175
Asn Thr Tyr Thr Gly Glu
1 5
<210> 176
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 176
Gly Gly Phe Gly Ser Ser Tyr Trp Tyr Phe Asp Val
1 5 10
<210> 177
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 177
Gln Asp Val Ser Ile Ala Val Ala
1 5
<210> 178
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 178
Ser Ala Ser Tyr Arg Tyr Thr
1 5
<210> 179
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 179
Gln Gln His Tyr Ile Thr Pro Leu Thr
1 5
<210> 180
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 180
Gln Ile Gln Leu Val Gln Ser Gly His Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe
50 55 60
Lys Gly Arg Phe Val Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Met Tyr Phe Cys
85 90 95
Gly Arg Gly Gly Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 181
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 181
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Ile Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Val Tyr Tyr Cys Gln Gln His Tyr Ile Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 182
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 182
Gly Tyr Thr Phe Thr Asn Tyr
1 5
<210> 183
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 183
Asn Thr Lys Thr Gly Glu
1 5
<210> 184
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 184
Gly Gly Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp Val
1 5 10
<210> 185
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 185
Gln Asp Val Ser Ile Ala Val Ala
1 5
<210> 186
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 186
Ser Ala Ser Tyr Arg Tyr Thr
1 5
<210> 187
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 187
Gln Gln His Tyr Ile Thr Pro Leu Thr
1 5
<210> 188
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 188
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Ala
20 25 30
Gly Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr His Ser Gly Val Pro Lys Tyr Ala Glu Asp Phe
50 55 60
Lys Gly Arg Val Thr Ile Ser Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Phe Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 189
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 189
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Tyr Ile Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105
<210> 190
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 190
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Ala
20 25 30
Gly Met Gln Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr His Ser Gly Val Pro Lys Tyr Ala Glu Asp Phe
50 55 60
Lys Gly Arg Val Thr Ile Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Phe Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 191
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 191
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Tyr Ile Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr
100 105
<210> 192
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 192
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Ala
20 25 30
Gly Met Gln Trp Val Gln Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr His Ser Gly Val Pro Lys Tyr Ala Glu Asp Phe
50 55 60
Lys Gly Arg Val Thr Phe Ser Leu Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Lys Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Phe Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 193
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 193
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Gln Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Tyr Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Tyr Ile Thr Pro Leu
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 194
<211> 323
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
TROP2 полипептид
<400> 194
Met Ala Arg Gly Pro Gly Leu Ala Pro Pro Pro Leu Arg Leu Pro Leu
1 5 10 15
Leu Leu Leu Val Leu Ala Ala Val Thr Gly His Thr Ala Ala Gln Asp
20 25 30
Asn Cys Thr Cys Pro Thr Asn Lys Met Thr Val Cys Ser Pro Asp Gly
35 40 45
Pro Gly Gly Arg Cys Gln Cys Arg Ala Leu Gly Ser Gly Met Ala Val
50 55 60
Asp Cys Ser Thr Leu Thr Ser Lys Cys Leu Leu Leu Lys Ala Arg Met
65 70 75 80
Ser Ala Pro Lys Asn Ala Arg Thr Leu Val Arg Pro Ser Glu His Ala
85 90 95
Leu Val Asp Asn Asp Gly Leu Tyr Asp Pro Asp Cys Asp Pro Glu Gly
100 105 110
Arg Phe Lys Ala Arg Gln Cys Asn Gln Thr Ser Val Cys Trp Cys Val
115 120 125
Asn Ser Val Gly Val Arg Arg Thr Asp Lys Gly Asp Leu Ser Leu Arg
130 135 140
Cys Asp Glu Leu Val Arg Thr His His Ile Leu Ile Asp Leu Arg His
145 150 155 160
Arg Pro Thr Ala Gly Ala Phe Asn His Ser Asp Leu Asp Ala Glu Leu
165 170 175
Arg Arg Leu Phe Arg Glu Arg Tyr Arg Leu His Pro Lys Phe Val Ala
180 185 190
Ala Val His Tyr Glu Gln Pro Thr Ile Gln Ile Glu Leu Arg Gln Asn
195 200 205
Thr Ser Gln Lys Ala Ala Gly Asp Val Asp Ile Gly Asp Ala Ala Tyr
210 215 220
Tyr Phe Glu Arg Asp Ile Lys Gly Glu Ser Leu Phe Gln Gly Arg Gly
225 230 235 240
Gly Leu Asp Leu Arg Val Arg Gly Glu Pro Leu Gln Val Glu Arg Thr
245 250 255
Leu Ile Tyr Tyr Leu Asp Glu Ile Pro Pro Lys Phe Ser Met Lys Arg
260 265 270
Leu Thr Ala Gly Leu Ile Ala Val Ile Val Val Val Val Val Ala Leu
275 280 285
Val Ala Gly Met Ala Val Leu Val Ile Thr Asn Arg Arg Lys Ser Gly
290 295 300
Lys Tyr Lys Lys Val Glu Ile Lys Glu Leu Gly Glu Leu Arg Lys Glu
305 310 315 320
Pro Ser Leu
<210> 195
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 195
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Ala Ser Gly Phe Asp Phe Thr Thr Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile His Pro Asp Ser Ser Thr Ile Asn Tyr Ala Pro Ser Leu
50 55 60
Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys
85 90 95
Ala Ser Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Pro Val Thr Val Ser Ser
115
<210> 196
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 196
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asp Val Gly Thr Ser
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Trp Thr Ser Thr Arg His Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Leu Tyr Arg Ser
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 197
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 197
Gly Phe Asp Phe Thr Thr Tyr
1 5
<210> 198
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 198
His Pro Asp Ser Ser Thr
1 5
<210> 199
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 199
Leu Tyr Phe Gly Phe Pro Trp Phe Ala Tyr
1 5 10
<210> 200
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 200
Gln Asp Val Gly Thr Ser Val Ala
1 5
<210> 201
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 201
Trp Thr Ser Thr Arg His Thr
1 5
<210> 202
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 202
Gln Gln Tyr Ser Leu Tyr Arg Ser
1 5
<210> 203
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 203
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Glu Phe
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Lys Thr Gly Glu Ala Thr Tyr Val Glu Glu Phe
50 55 60
Lys Gly Arg Val Thr Phe Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala
115 120
<210> 204
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 204
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Ala Ala Val Gly Thr Tyr
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Tyr Arg Lys Arg Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Tyr Tyr Thr Tyr Pro Leu
85 90 95
Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 205
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 205
Gly Tyr Thr Phe Thr Glu Phe
1 5
<210> 206
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 206
Asn Thr Lys Thr Gly Glu
1 5
<210> 207
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 207
Trp Asp Phe Ala Tyr Tyr Val Glu Ala Met Asp Tyr
1 5 10
<210> 208
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 208
Ala Ala Val Gly Thr Tyr Val Ala
1 5
<210> 209
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 209
Ser Ala Ser Tyr Arg Lys Arg
1 5
<210> 210
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 210
His Gln Tyr Tyr Thr Tyr Pro Leu Phe Thr
1 5 10
<210> 211
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 211
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg Ser Gly Ala
1 5 10 15
Ser Ile Lys Leu Ser Cys Thr Ala Ser Gly Phe Asn Ile Lys His Tyr
20 25 30
Tyr Met His Trp Val Lys Gln Arg Pro Glu Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Asn Pro Glu Asn Val Asp Thr Glu Tyr Ala Pro Lys Phe
50 55 60
Gln Gly Lys Ala Thr Met Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Asn His Tyr Arg Tyr Ala Gly Gly Gly Ala Leu Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser Ala
115 120
<210> 212
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 212
Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Val Tyr
35 40 45
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Thr Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Asn Asn Asn Pro Tyr Ser
85 90 95
Phe Gly Gly Gly Thr Lys Val Val Ile Lys Thr Val
100 105
<210> 213
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 213
Gly Phe Asn Ile Lys His Tyr
1 5
<210> 214
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 214
Asn Pro Glu Asn Val Asp
1 5
<210> 215
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 215
Tyr Arg Tyr Ala Gly Gly Gly Ala Leu Asp Tyr
1 5 10
<210> 216
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 216
Ser Ser Val Ser Tyr Ile His
1 5
<210> 217
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 217
Asp Thr Ser Lys Leu Ala Ser
1 5
<210> 218
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 218
Gln Gln Trp Asn Asn Asn Pro Tyr Ser
1 5
<210> 219
<211> 702
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
CEA полипептид
<400> 219
Met Glu Ser Pro Ser Ala Pro Pro His Arg Trp Cys Ile Pro Trp Gln
1 5 10 15
Arg Leu Leu Leu Thr Ala Ser Leu Leu Thr Phe Trp Asn Pro Pro Thr
20 25 30
Thr Ala Lys Leu Thr Ile Glu Ser Thr Pro Phe Asn Val Ala Glu Gly
35 40 45
Lys Glu Val Leu Leu Leu Val His Asn Leu Pro Gln His Leu Phe Gly
50 55 60
Tyr Ser Trp Tyr Lys Gly Glu Arg Val Asp Gly Asn Arg Gln Ile Ile
65 70 75 80
Gly Tyr Val Ile Gly Thr Gln Gln Ala Thr Pro Gly Pro Ala Tyr Ser
85 90 95
Gly Arg Glu Ile Ile Tyr Pro Asn Ala Ser Leu Leu Ile Gln Asn Ile
100 105 110
Ile Gln Asn Asp Thr Gly Phe Tyr Thr Leu His Val Ile Lys Ser Asp
115 120 125
Leu Val Asn Glu Glu Ala Thr Gly Gln Phe Arg Val Tyr Pro Glu Leu
130 135 140
Pro Lys Pro Ser Ile Ser Ser Asn Asn Ser Lys Pro Val Glu Asp Lys
145 150 155 160
Asp Ala Val Ala Phe Thr Cys Glu Pro Glu Thr Gln Asp Ala Thr Tyr
165 170 175
Leu Trp Trp Val Asn Asn Gln Ser Leu Pro Val Ser Pro Arg Leu Gln
180 185 190
Leu Ser Asn Gly Asn Arg Thr Leu Thr Leu Phe Asn Val Thr Arg Asn
195 200 205
Asp Thr Ala Ser Tyr Lys Cys Glu Thr Gln Asn Pro Val Ser Ala Arg
210 215 220
Arg Ser Asp Ser Val Ile Leu Asn Val Leu Tyr Gly Pro Asp Ala Pro
225 230 235 240
Thr Ile Ser Pro Leu Asn Thr Ser Tyr Arg Ser Gly Glu Asn Leu Asn
245 250 255
Leu Ser Cys His Ala Ala Ser Asn Pro Pro Ala Gln Tyr Ser Trp Phe
260 265 270
Val Asn Gly Thr Phe Gln Gln Ser Thr Gln Glu Leu Phe Ile Pro Asn
275 280 285
Ile Thr Val Asn Asn Ser Gly Ser Tyr Thr Cys Gln Ala His Asn Ser
290 295 300
Asp Thr Gly Leu Asn Arg Thr Thr Val Thr Thr Ile Thr Val Tyr Ala
305 310 315 320
Glu Pro Pro Lys Pro Phe Ile Thr Ser Asn Asn Ser Asn Pro Val Glu
325 330 335
Asp Glu Asp Ala Val Ala Leu Thr Cys Glu Pro Glu Ile Gln Asn Thr
340 345 350
Thr Tyr Leu Trp Trp Val Asn Asn Gln Ser Leu Pro Val Ser Pro Arg
355 360 365
Leu Gln Leu Ser Asn Asp Asn Arg Thr Leu Thr Leu Leu Ser Val Thr
370 375 380
Arg Asn Asp Val Gly Pro Tyr Glu Cys Gly Ile Gln Asn Lys Leu Ser
385 390 395 400
Val Asp His Ser Asp Pro Val Ile Leu Asn Val Leu Tyr Gly Pro Asp
405 410 415
Asp Pro Thr Ile Ser Pro Ser Tyr Thr Tyr Tyr Arg Pro Gly Val Asn
420 425 430
Leu Ser Leu Ser Cys His Ala Ala Ser Asn Pro Pro Ala Gln Tyr Ser
435 440 445
Trp Leu Ile Asp Gly Asn Ile Gln Gln His Thr Gln Glu Leu Phe Ile
450 455 460
Ser Asn Ile Thr Glu Lys Asn Ser Gly Leu Tyr Thr Cys Gln Ala Asn
465 470 475 480
Asn Ser Ala Ser Gly His Ser Arg Thr Thr Val Lys Thr Ile Thr Val
485 490 495
Ser Ala Glu Leu Pro Lys Pro Ser Ile Ser Ser Asn Asn Ser Lys Pro
500 505 510
Val Glu Asp Lys Asp Ala Val Ala Phe Thr Cys Glu Pro Glu Ala Gln
515 520 525
Asn Thr Thr Tyr Leu Trp Trp Val Asn Gly Gln Ser Leu Pro Val Ser
530 535 540
Pro Arg Leu Gln Leu Ser Asn Gly Asn Arg Thr Leu Thr Leu Phe Asn
545 550 555 560
Val Thr Arg Asn Asp Ala Arg Ala Tyr Val Cys Gly Ile Gln Asn Ser
565 570 575
Val Ser Ala Asn Arg Ser Asp Pro Val Thr Leu Asp Val Leu Tyr Gly
580 585 590
Pro Asp Thr Pro Ile Ile Ser Pro Pro Asp Ser Ser Tyr Leu Ser Gly
595 600 605
Ala Asn Leu Asn Leu Ser Cys His Ser Ala Ser Asn Pro Ser Pro Gln
610 615 620
Tyr Ser Trp Arg Ile Asn Gly Ile Pro Gln Gln His Thr Gln Val Leu
625 630 635 640
Phe Ile Ala Lys Ile Thr Pro Asn Asn Asn Gly Thr Tyr Ala Cys Phe
645 650 655
Val Ser Asn Leu Ala Thr Gly Arg Asn Asn Ser Ile Val Lys Ser Ile
660 665 670
Thr Val Ser Ala Ser Gly Thr Ser Pro Gly Leu Ser Ala Gly Ala Thr
675 680 685
Val Gly Ile Met Ile Gly Val Leu Val Gly Val Ala Leu Ile
690 695 700
<210> 220
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 220
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 221
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 221
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 222
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 222
Gly Tyr Thr Phe Thr Asn Tyr Gly
1 5
<210> 223
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 223
Ile Asn Thr Asn Thr Gly Glu Pro
1 5
<210> 224
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 224
Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr
1 5 10
<210> 225
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 225
Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr
1 5 10
<210> 226
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 226
Trp Ala Ser
1
<210> 227
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 227
Gln Asn Asp Tyr Ser Tyr Pro Leu Thr
1 5
<210> 228
<211> 118
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 228
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 229
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 229
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 230
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 230
Gly Tyr Thr Phe Thr Ser Tyr Trp
1 5
<210> 231
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 231
Ile Tyr Pro Ser Asp Ser Tyr Thr
1 5
<210> 232
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 232
Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr
1 5 10
<210> 233
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 233
Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr
1 5 10
<210> 234
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 234
Trp Ala Ser
1
<210> 235
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 235
Gln Asn Asp Tyr Ser Tyr Pro Phe Thr
1 5
<210> 236
<211> 261
<212> Белок
<213> Неизвестный
<220>
<223> Описание неизвестного:
Клаудин-18.2 полипептид
<400> 236
Met Ala Val Thr Ala Cys Gln Gly Leu Gly Phe Val Val Ser Leu Ile
1 5 10 15
Gly Ile Ala Gly Ile Ile Ala Ala Thr Cys Met Asp Gln Trp Ser Thr
20 25 30
Gln Asp Leu Tyr Asn Asn Pro Val Thr Ala Val Phe Asn Tyr Gln Gly
35 40 45
Leu Trp Arg Ser Cys Val Arg Glu Ser Ser Gly Phe Thr Glu Cys Arg
50 55 60
Gly Tyr Phe Thr Leu Leu Gly Leu Pro Ala Met Leu Gln Ala Val Arg
65 70 75 80
Ala Leu Met Ile Val Gly Ile Val Leu Gly Ala Ile Gly Leu Leu Val
85 90 95
Ser Ile Phe Ala Leu Lys Cys Ile Arg Ile Gly Ser Met Glu Asp Ser
100 105 110
Ala Lys Ala Asn Met Thr Leu Thr Ser Gly Ile Met Phe Ile Val Ser
115 120 125
Gly Leu Cys Ala Ile Ala Gly Val Ser Val Phe Ala Asn Met Leu Val
130 135 140
Thr Asn Phe Trp Met Ser Thr Ala Asn Met Tyr Thr Gly Met Gly Gly
145 150 155 160
Met Val Gln Thr Val Gln Thr Arg Tyr Thr Phe Gly Ala Ala Leu Phe
165 170 175
Val Gly Trp Val Ala Gly Gly Leu Thr Leu Ile Gly Gly Val Met Met
180 185 190
Cys Ile Ala Cys Arg Gly Leu Ala Pro Glu Glu Thr Asn Tyr Lys Ala
195 200 205
Val Ser Tyr His Ala Ser Gly His Ser Val Ala Tyr Lys Pro Gly Gly
210 215 220
Phe Lys Ala Ser Thr Gly Phe Gly Ser Asn Thr Lys Asn Lys Lys Ile
225 230 235 240
Tyr Asp Gly Gly Ala Arg Thr Glu Asp Glu Val Gln Ser Tyr Pro Ser
245 250 255
Lys His Asp Tyr Val
260
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА NKG2D, CD16 И TROP2 | 2018 |
|
RU2820629C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ ВСМА, NKG2D И CD16 | 2018 |
|
RU2805254C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ CD33, NKG2D И CD16 | 2018 |
|
RU2820603C2 |
| МОДИФИЦИРОВАННЫЕ НЕПРИРОДНЫЕ ЛИГАНДЫ NKG2D, КОТОРЫЕ ИЗБИРАТЕЛЬНО ДОСТАВЛЯЮТ ПРИСОЕДИНЕННЫЕ ГЕТЕРОЛОГИЧНЫЕ МОЛЕКУЛЫ К НЕПРИРОДНЫМ РЕЦЕПТОРАМ NKG2D НА CAR-КЛЕТКАХ | 2020 |
|
RU2823728C2 |
| МОДИФИЦИРОВАННЫЕ ДОМЕНЫ α1-α2 НЕПРИРОДНЫХ ЛИГАНДОВ NKG2D, КОТОРЫЕ СВЯЗЫВАЮТСЯ С НЕПРИРОДНЫМИ РЕЦЕПТОРАМИ NKG2D | 2019 |
|
RU2815278C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ ДЛЯ АКТИВАЦИИ КЛЕТОК-НАТУРАЛЬНЫХ КИЛЛЕРОВ И ИХ ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2018 |
|
RU2809125C2 |
| УСЕЧЕННЫЕ ХИМЕРНЫЕ РЕЦЕПТОРЫ NKG2D И ПУТИ ИХ ПРИМЕНЕНИЯ В ИММУНОТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ | 2018 |
|
RU2830556C2 |

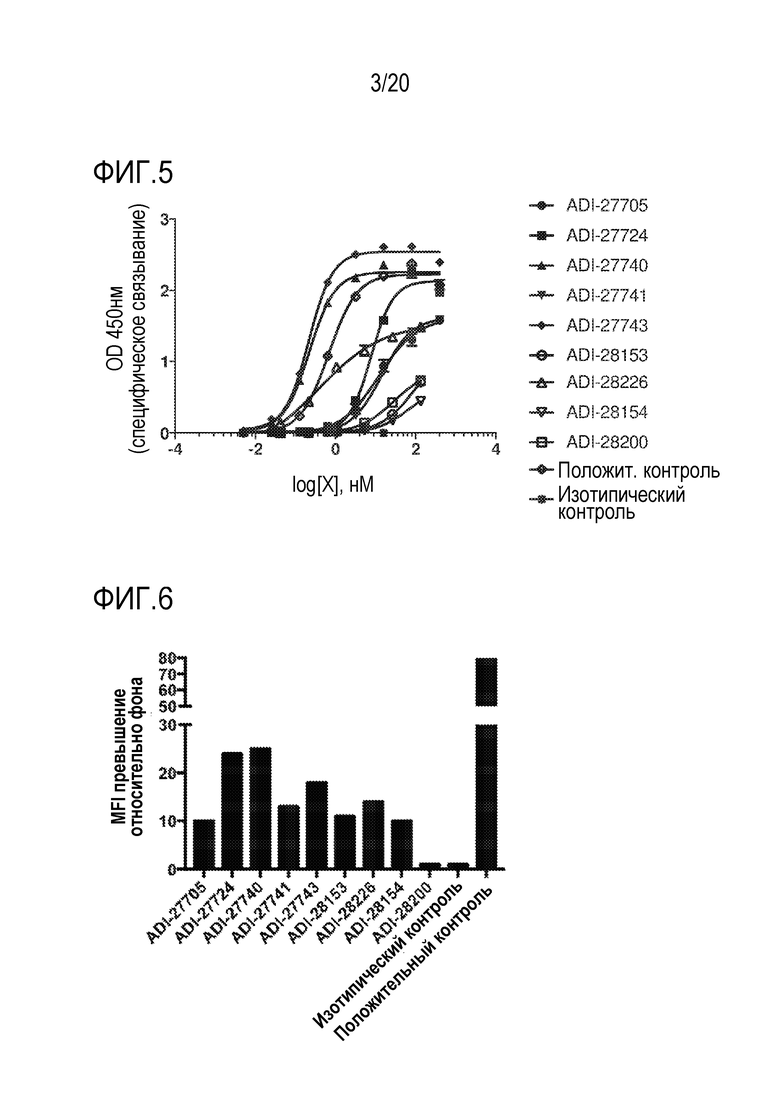
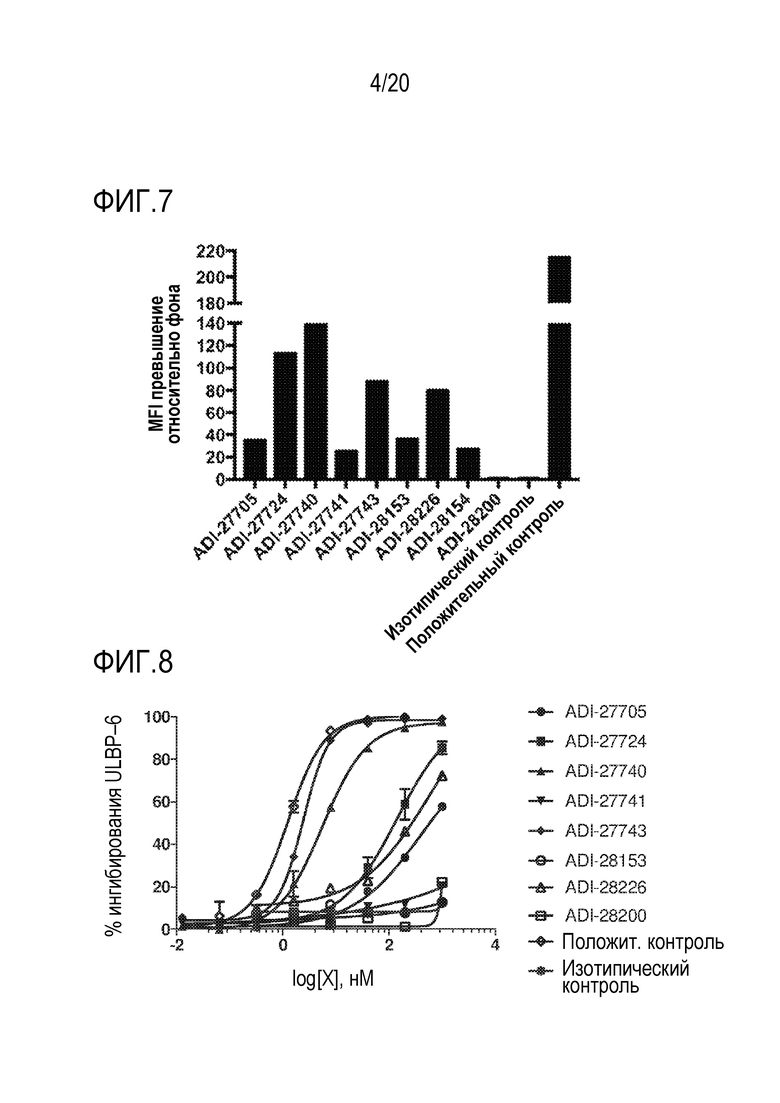

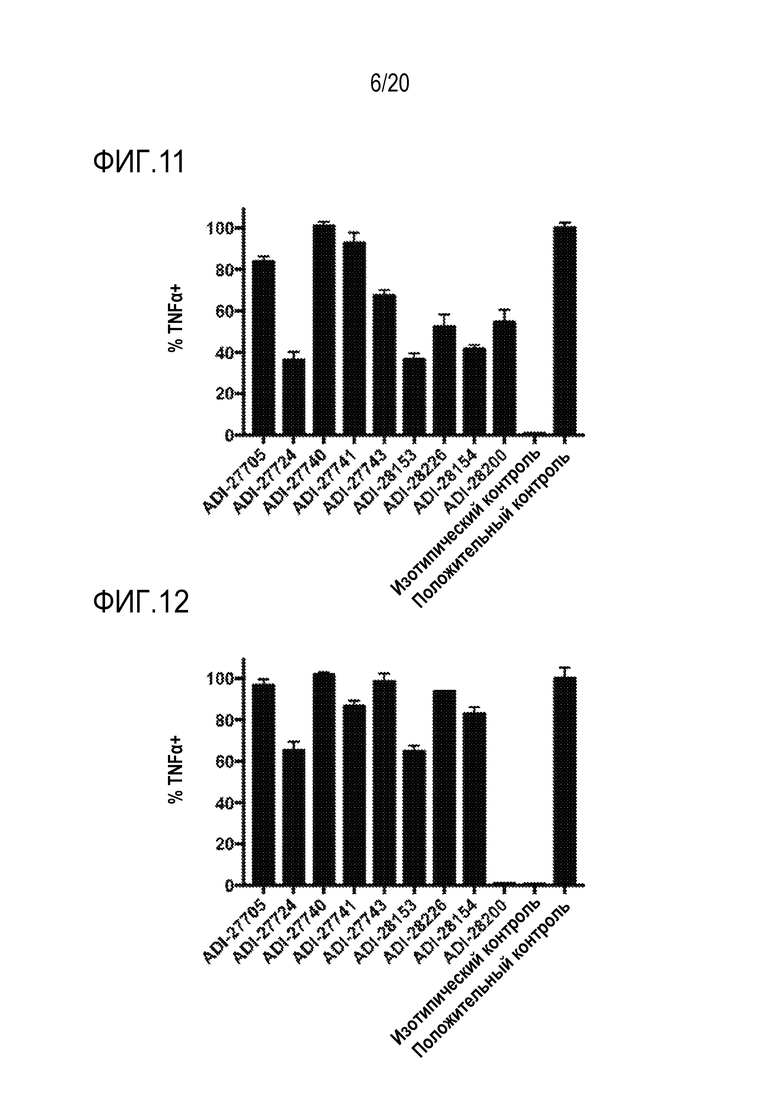

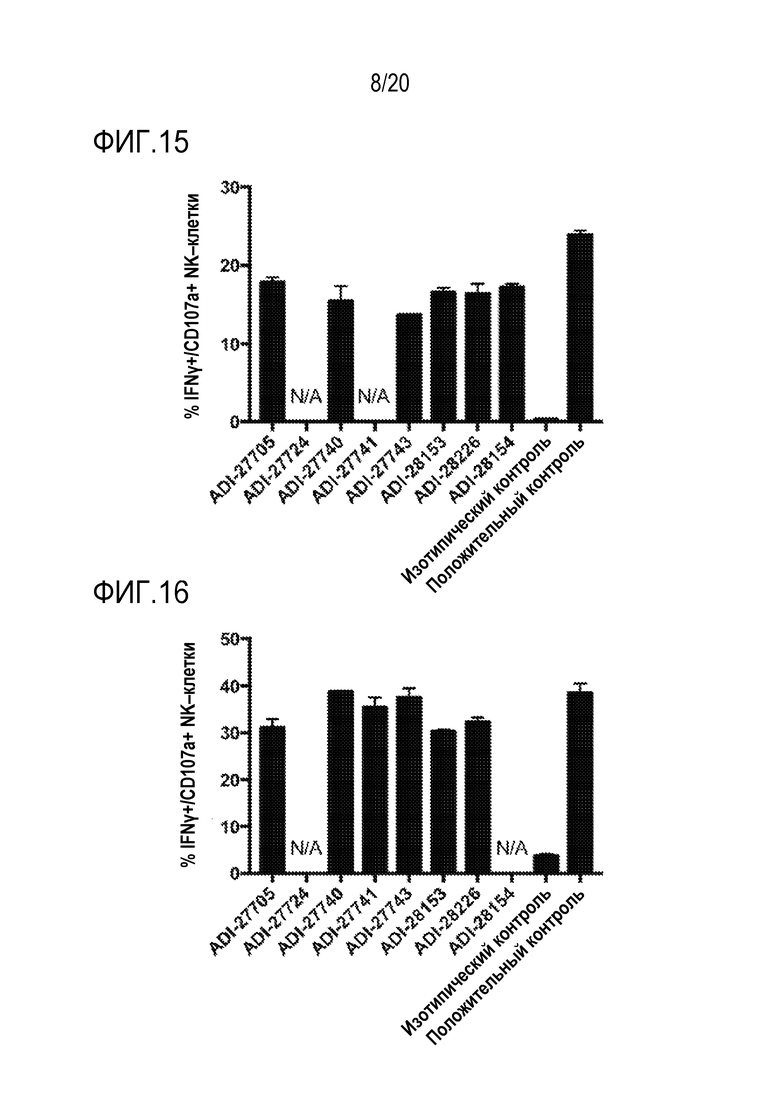
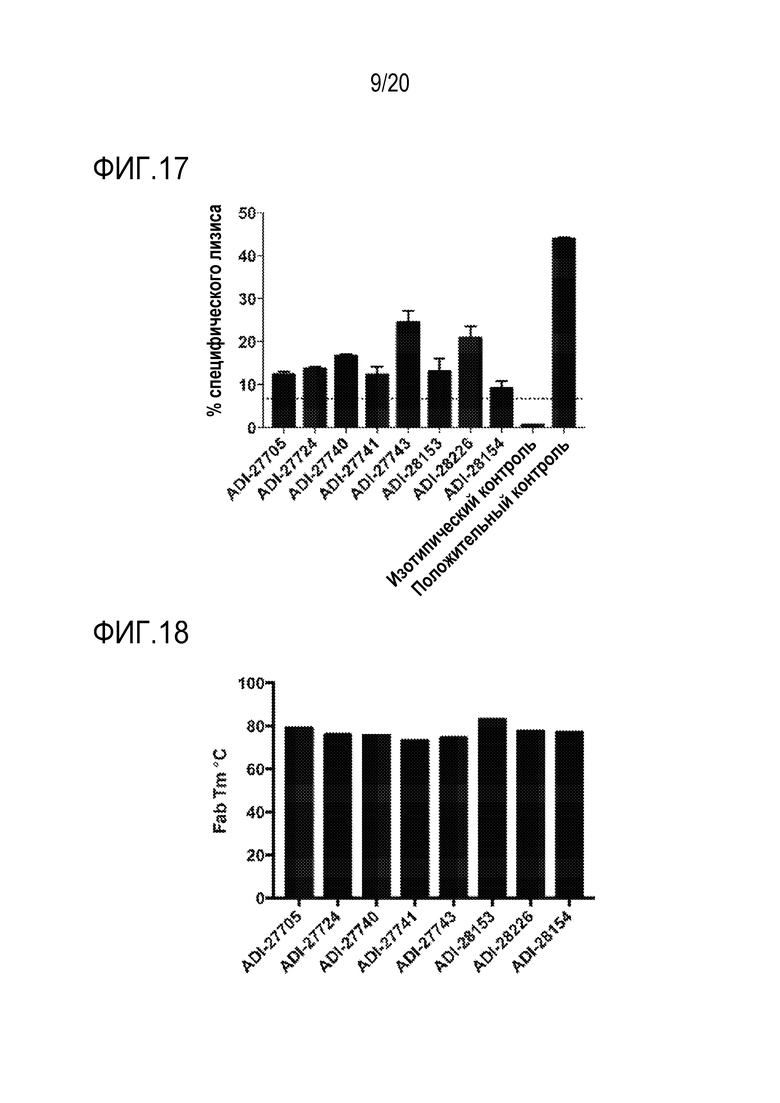

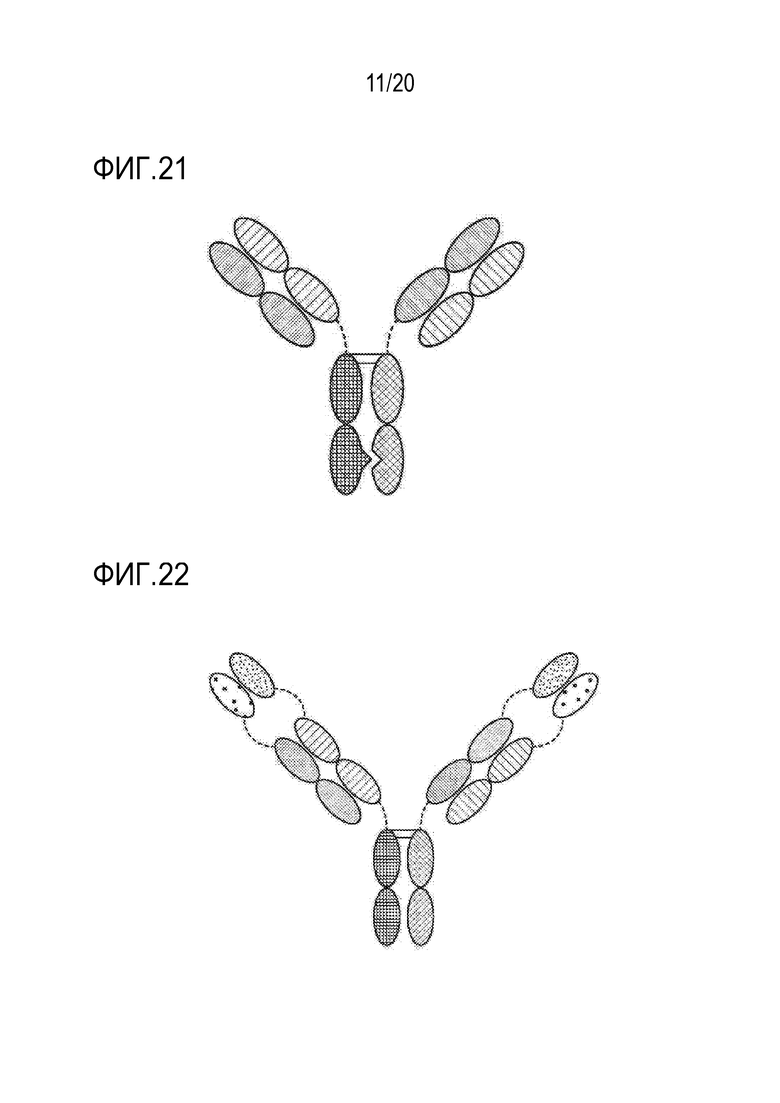
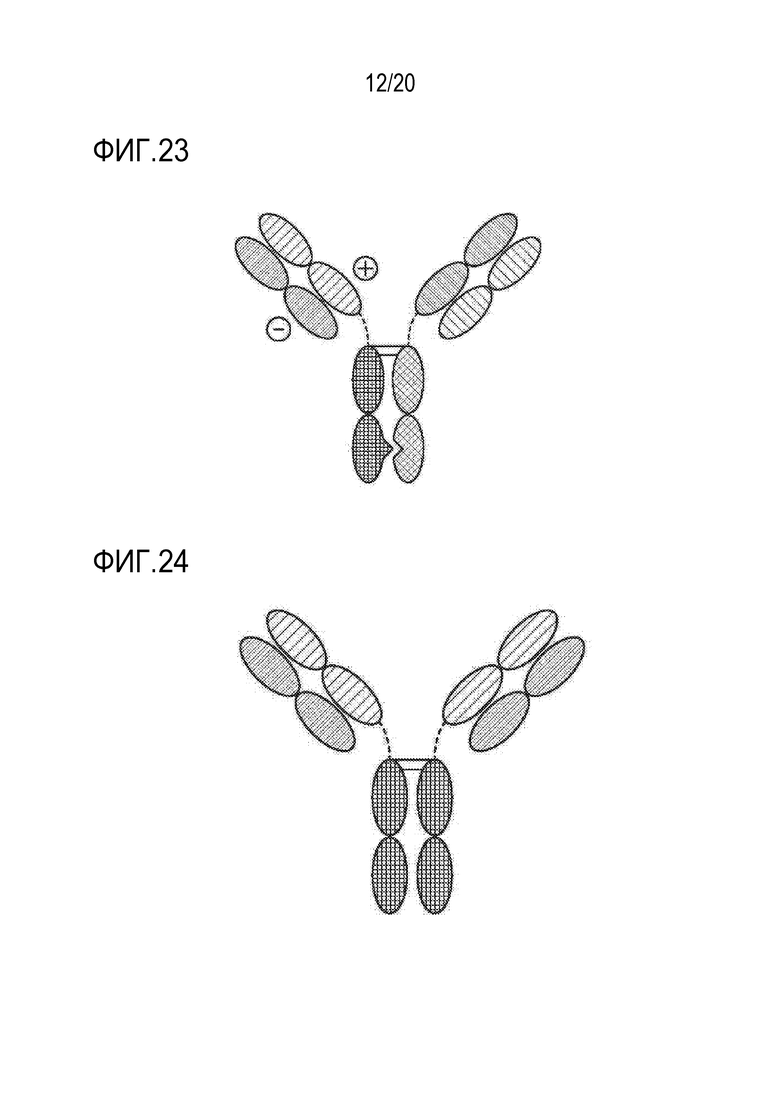



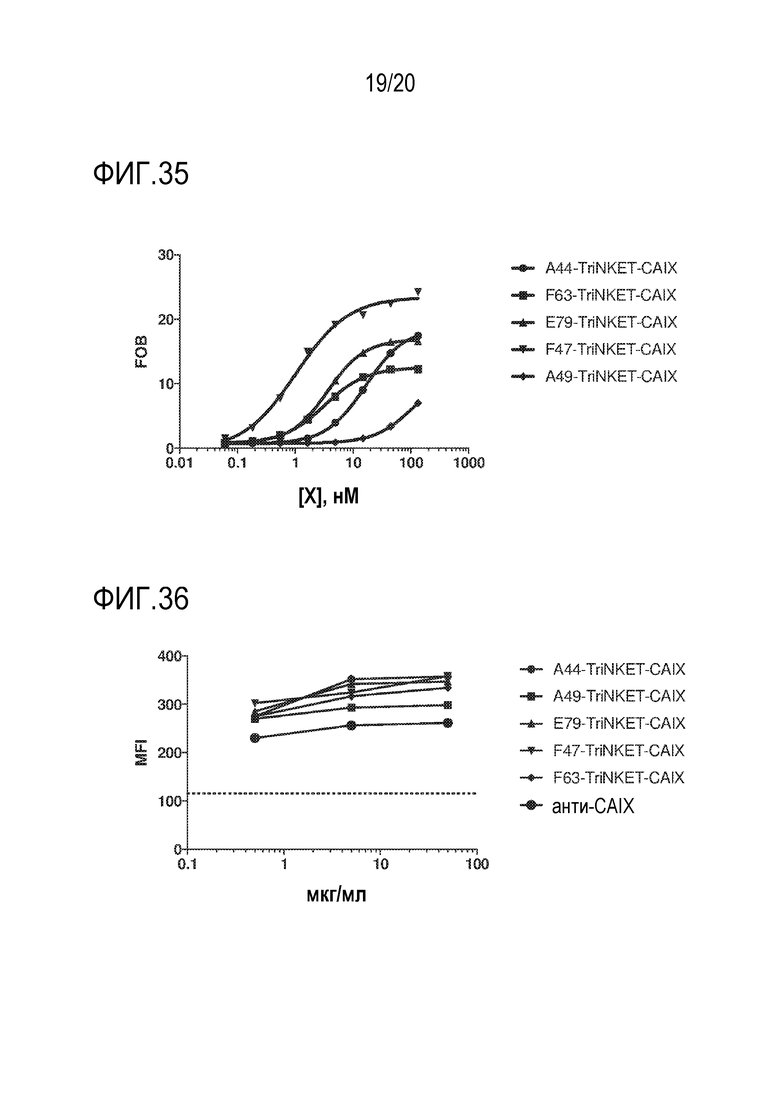
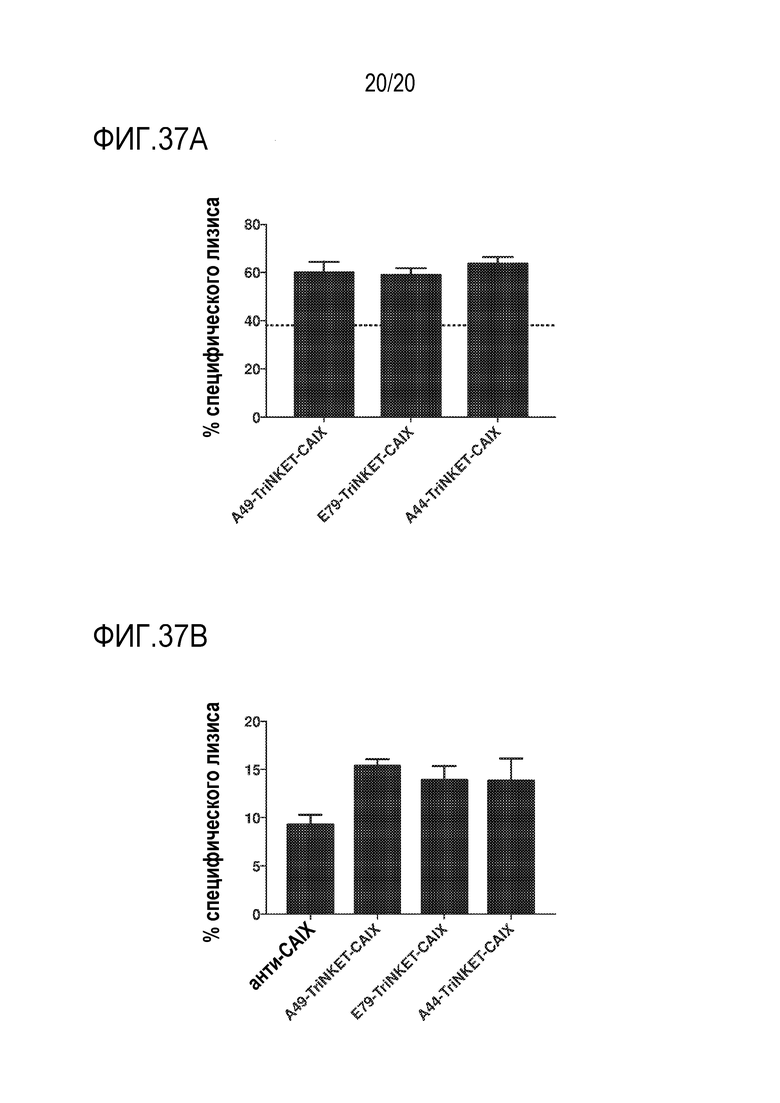
Настоящее изобретение относится к области биотехнологии, в частности к полиспецифическим антителам, и может быть использовано в медицине. Изобретение раскрывает новые белки, которые специфически связываются с рецептором NKG2D, CD16 и опухолеспецифическим антигеном CEA. Изобретение может быть применимо в медицинской практике в качестве терапевтического средства при лечении ряда злокачественных новообразований, например рака желудочно–кишечного тракта, колоректального рака, рака поджелудочной железы, немелкоклеточного рака легкого и рака молочной железы. 5 н. и 19 з.п. ф-лы, 37 ил., 14 табл., 10 пр.
1. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, содержащий:
(а) первый антигенсвязывающий сайт, содержащий вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) антитела к NKG2D;
(b) второй антигенсвязывающий сайт, содержащий VH и VL антитела к CEA; и
(c) первый полипептид Fc–домена антитела или его часть; и
(d) второй полипептид Fc–домена антитела или его часть,
где указанные первый полипептид Fc–домена антитела или его часть и указанные второй полипептид Fc–домена антитела или его часть вместе связывают CD16, и
где указанный VH или VL антитела к NKG2D слит с N-концом указанных первого полипептида Fc–домена антитела или его части, и указанный VH или VL антитела к CEA слит с N-концом указанных второго полипептида Fc–домена антитела или его части.
2. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.1, где VH и VL первого антигенсвязывающего сайта связываются с NKG2D у человека, приматов, отличных от человека, и грызунов.
3. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.1 или 2, где VH и VL первого антигенсвязывающего сайта присутствуют в одном и том же полипептиде.
4. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.1, где VH и VL второго антигенсвязывающего сайта присутствуют в одном и том же полипептиде.
5. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.4, где VL первого антигенсвязывающего сайта имеет аминокислотную последовательность, идентичную аминокислотной последовательности VL второго антигенсвязывающего сайта.
6. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 1.
7. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 41, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 42.
8. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 43, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 44.
9. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 45, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 46.
10. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 47, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 48.
11. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 49, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 50.
12. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–5, где VH первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 132, и VL первого антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 133.
13. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–12, где VH второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 195, и VL второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 196.
14. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–12, где VH второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 203, и VL второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 204.
15. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1–12, где VH второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 211, и VL второго антигенсвязывающего сайта содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную SEQ ID NO: 212.
16. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.1-15, где Fc–домен антитела содержит шарнирный и CH2–домены.
17. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.16, где Fc–домен антитела содержит шарнирный и СН2–домены человеческого антитела IgG1.
18. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по п.16 или 17, где Fc–домен антитела содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную аминокислотам 234–332 человеческого антитела IgG1.
19. Полиспецифический связывающий белок, который связывает NKG2D, CEA и CD16, по любому из пп.16-18, где Fc–домен антитела содержит аминокислотную последовательность, по меньшей мере, на 90% идентичную Fc–домену человеческого IgG1 и отличается в одном или более положениях, выбранных из группы, состоящей из Q347, Y349, L351, S354, E356, E357, K360, Q362, S364, T366, L368, K370, N390, K392, T394, D399, S400, D401, F405, Y407, K409, T411 и K439.
20. Фармацевтическая композиция для лечения злокачественного новообразования, содержащая эффективное количество полиспецифического связывающего белка по любому из пп.1-19 и фармацевтически приемлемый носитель.
21. Клетка-хозяин для получения полиспецифического связывающего белка, который связывает NKG2D, CEA и CD16, содержащая одну или более нуклеиновых кислот, экспрессирующих полиспецифический связывающий белок по любому из пп.1–19.
22. Способ прямого или косвенного усиления гибели опухолевых клеток, включающий воздействие полиспецифического связывающего белка по любому из пп.1–19 на опухоль и на естественные клетки–киллеры.
23. Способ лечения злокачественного новообразования, где указанный способ включает введение пациенту полиспецифического связывающего белка по любому из пп.1–19 или фармацевтической композиции по п.20.
24. Способ по п.23, где злокачественное новообразование, подлежащее лечению, выбирают из группы, состоящей из рака желудочно–кишечного тракта, колоректального рака, рака поджелудочной железы, немелкоклеточного рака легкого и рака молочной железы.
| US 20150056206 A1, 26.02.2015 | |||
| US 20160077105 A1, 17.03.2016 | |||
| US 20150166636 A1, 18.06.2015 | |||
| US 20150079088 A1, 19.03.2015 | |||
| RAAB S | |||
| et al., "Fc-optimized NKG2D-Fc constructs induce NK cell antibody-dependent cellular cytotoxicity against breast cancer cells independently of HER2/neu expression status", J | |||
| Immunol., 2014, v | |||
| Приспособление для градации давления в воздухопроводе воздушных тормозов | 1921 |
|
SU193A1 |

