Область изобретения
Настоящее изобретение относится к области генной инженерии или биологической ферментации, а именно к способу эффективного биопревращения сбраживаемого углевода в 1,3-пропандиол с использованием одного рекомбинантного микроорганизма без добавления витамина B12.
Уровень техники
1,3-пропандиол является важным химическим сырьем и может быть использован в качестве органического растворителя во многих отраслях промышленности, таких как производство чернил, полиграфическая промышленность и производство красителей, лакокрасочная промышленность, производство смазочных материалов и/или антифризов. Главной особенностью 1,3-пропандиола является то, что он может быть использован в качестве мономера для синтеза полиэфира и полиуретана. В частности, 1,3-пропандиол может быть использован для получения политриметилентерефталата (РТТ) посредством реакции полимеризации с терефталевой кислотой. По сравнению с полиэтилентерефталатом (PЕТ) или полибутилентерефталатом (PBT), PTT обладает лучшими свойствами по множеству аспектов, таких как повышенная резистентность к загрязнению, ударная вязкость, упругое восстановление после деформации и устойчивость к ультрафиолетовому излучению, а также износостойкость, низкое водопоглощение и/или вырабатывание низкого статического электричества. Таким образом, PTT рассматривается как продукт PЕТ улучшенного качества и имеет широкие коммерческие перспективы.
В настоящее время, методами получения 1,3-пропандиола, в основном, являются химические и биологические методы. В химических методах, 1,3-пропандиол может быть синтезирован посредством сложного каталитического процесса с использованием пропиленоксида или пропилена в качестве исходного материала. Недостатками методов химического синтеза являются избыточное количество побочных продуктов, низкая селективность, условия работы, требующие высоких температур и высокого давления, огромные инвестиции в оборудование и невозобновляемость сырьевых ресурсов. Поэтому, от химических методов производства 1,3-пропандиола пришлось отказаться.
Биологические методы получения 1,3-пропандиола включают два основных технических протокола, один из которых заключается в получении 1,3-пропандиола с помощью природного микроорганизма с использованием глицерина в качестве сырья, а второй заключвется в получении 1,3-пропандиола с помощью рекомбинантного микроорганизма с использованием глюкозы в качестве сырья. Оба эти метода описаны ниже.
В одном техническом протоколе, 1,3-пропандиол получают с использованием глицерина качестве сырья с помощью природного микроорганизма, такого как Klebsiella pneumoniae, Clostridium butyricum или Citrobacter freundii, которые могут превращать глицерин в 1,3-пропандиол в анаэробных или микроаэробных условиях. Основными недостатками этого технического способа являются: 1. Строгий контроль за биологической безопасностью, который необходимо соблюдать при получении 1,3-пропандиола, поскольку бактерия Klebsiella pneumoniae, которая обычно используется, представляет собой кондиционированный патоген; 2. Синтез большого количества побочных продуктов, таких как уксусная кислота, молочная кислота, янтарная кислота и 2,3-бутандиол, который в значительной степени усложняет все последующие стадии экстракции; 3. Сильные колебания стоимости глицерина на коммерческом рынке.
Другим известным методом является получение 1,3-пропандиола с использованием глюкозы в качестве сырья с помощью рекомбинантных микроорганизмов. Так, например, специалисты DuPont осуществляют одну стадию превращения глюкозы в 1,3-пропандиол посредством экзогенной экспрессии пути синтеза глицерина (глицерин-3-фосфат-дегидрогеназы и глицерин-3-фосфатазы) из Saccharomyces cerevisiae, а также глицерин-дегидратазы и его активирующих факторов, происходящих от Klebsiella pneumoniae, в E.coli вместе с NADPH-зависимой алкогольдегидрогеназой YqhD, происходящей от E. coli (CN 200380104657.2). Недостаток этого способа заключается в том, что глицериндегидратаза требует наличия кофермента B12 в качестве кофактора, который не может синтезироваться E.coli, а поэтому в процессе ферментации необходимо добавлять дорогостоящий витамин B12, что значительно увеличивет расходы на производство и является невыгодным для промышленного и крупномасштабного производства.
Раскрытие изобретения
Целью настоящего изобретения является разработка нового способа прямого превращения сбраживаемого углевода в 1,3-пропандиол с использованием рекомбинантного микроорганизма, где указанный способ имеет низкую себестоимость и не требует добавления дорогостоящего витамина B12.
Во-первых, настоящее изобретение относится к рекомбинантному микроорганизму, способному сверхэкспрессировать:
(1) гены ацетил-CoA-карбоксилазы accBC и accDA;
(2) ген малонил-СоА-синтетазы mcr;
(3) ген 3-гидроксипропионил-CoA-синтетазы pcs;
(4) ген 3-гидроксипропионил-СоА-редуктазы pduP; и
(5) ген 1,3-пропандиолоксидоредуктазы yqhD.
Специалистам в данной области известно, что микроорганизмы согласно изобретению могут представлять собой стандартные микроорганизмы-модели, включая, но не ограничиваясь ими, E. coli, Corynebacterium glutamicum, Bacillus subtilis или Saccharomyces cerevisiae.
Рекомбинантный микроорганизм согласно изобретению может экспрессировать гены ацетил-СоА-карбоксилазы accBC и accDA, нуклеотидные последовательности которых представлены в SEQ ID NO: 1 и 2, соответственно.
В примерах настоящего изобретения, гены ацетил-СоА-карбоксилазы accBC и accDA происходят от Corynebacterium glutamicum.
Кроме того, рекомбинантный микроорганизм согласно изобретению может экспрессировать ген малонил-СоА-синтетазы mcr, нуклеотидная последовательность которого представлена в SEQ ID NO.3. В примерах настоящего изобретения, ген малонил-СоА-синтетазы mcr происходит от Chloroflexus aurantiacus.
Кроме того, рекомбинантный микроорганизм согласно изобретению может экспрессировать ген 3-гидроксипропионил-СоА-синтетазы pcs, ген 3-гидроксипропионил-CoA-редуктазы pduP и ген 1,3-пропандиолоксидоредуктазы yqhD. В примерах настоящего изобретения, ген 3-гидроксипропионил-СоА-синтетазы pcs происходит от Metallosphaera sedula, нуклеотидная последовательность которого представлена в SEQ ID NO: 4; ген 3-гидроксипропионил-CoA-редуктазы pduP происходит от Klebsiella pneumoniae, нуклеотидная последовательность которого представлена в SEQ ID NO: 5, а ген 1,3-пропандиолоксидоредуктазы yqhD происходит от E.coli, нуклеотидная последовательность которого представлена в SEQ ID NO: 6. Специалистам в данной области очевидно, что описание настоящего изобретения не ограничивается ферментами штаммов, описанных с Примерах, и что любой фермент, происходящий от других источников, имеющих такую же функцию, может также давать аналогичные технические эффекты.
В примерах настоящего изобретения, рекомбинантный микроорганизм был получен следующими способами:
(1) присоединения гена accBC, последовательность которого представлена в SEQ ID NO: 1, и гена accDA, последовательность которого представлена в SEQ ID NO: 2, к плазмиде pACYCDuet с получением рекомбинантной плазмиды pACYC-accDABC;
(2) присоединения фрагмента mcr размером 3,7 т.п.о., полученного путем ПЦР-амплификации с использованием нуклеотидной последовательности SEQ ID NO: 3 в качестве матрицы и с использованием серии праймеров, представленных в SEQ ID NO: 11-12, к плазмиде pACYC-accDABC с получением рекомбинантной плазмиды pACYC-accDABC-mcr; а затем переноса этой рекомбинантной плазмиды в E. coli с получением рекомбинантного штамма E.coli/pACYC-accDABC-mcr после скрининга;
(3) присоединения фрагмента pcs размером 2,0 т.п.о., полученного путем ПЦР-амплификации с использованием нуклеотидной последовательности SEQ ID NO: 4 в качестве матрицы и с использованием серии праймеров, представленных в SEQ ID NO: 13-14, фрагмента pduP SEQ ID NO: 5 и фрагмента yqhD SEQ ID NO: 6 к плазмиде pET28 с получением рекомбинантной плазмиды pET-pcs-pduP-yqhD;
(4) переноса pET-pcs-pduP-yqhD в рекомбинантный штамм E.coli/pACYC-accDABC-mcr, полученный в стадии (2), с образованием рекомбинантного штамма E.coli/pACYC-accDABC-mcr/pET-pcs-pduP-yqhD после скрининга, где указанный штамм представляет собой рекомбинантный микроорганизм согласно изобретению.
Настоящее изобретение относится к применению рекомбинантного микроорганизма для получения 1,3-пропандиола.
Настоящее изобретение относится к способу получения 1,3-пропандиола путем ферментации с использованием рекомбинантного микроорганизма, где указанный способ включает стадии:
(1) конструирования рекомбинантного микроорганизма, способного сверхэкспрессировать гены ацетил-СоА-карбоксилазы accBC и accDA, ген малонил-СоА-синтетазы mcr, ген 3-гидроксипропионил-СоА-синтетазы pcs, ген 3-гидроксипропионил-CoA-редуктазы pduP и ген 1,3-пропандиолоксидоредуктазы yqhD; и
(2) проведения аэробной ферментации с использованием исходного материала, содержащего сбраживаемый углевод в качестве субстрата, без необходимости добавления витамина B12.
Сырьем, содержащим сбраживаемый углевод на стадии (2), является меласса, сахароза, глюкоза, гидролизат крахмала, кукурузный сироп, ксилоза, манноза или глицерин.
Условия ферментации на стадии (2): 28-37°C, значение рН в пределах от 5 до 8, и значение растворенного кислорода более, чем 10%.
Предпочтительные условия ферментации на стадии (2) составляют: 30-37°C, значение рН в пределах от 6 до 7, и значение растворенного кислорода более, чем 10%.
Субстрат для ферментации на стадии (2) также содержит Na2HPO4, KH2PO4, MgSO4, NaCl, дрожжевой экстракт, NH4Cl, гидрохлорид тиамина и биотин.
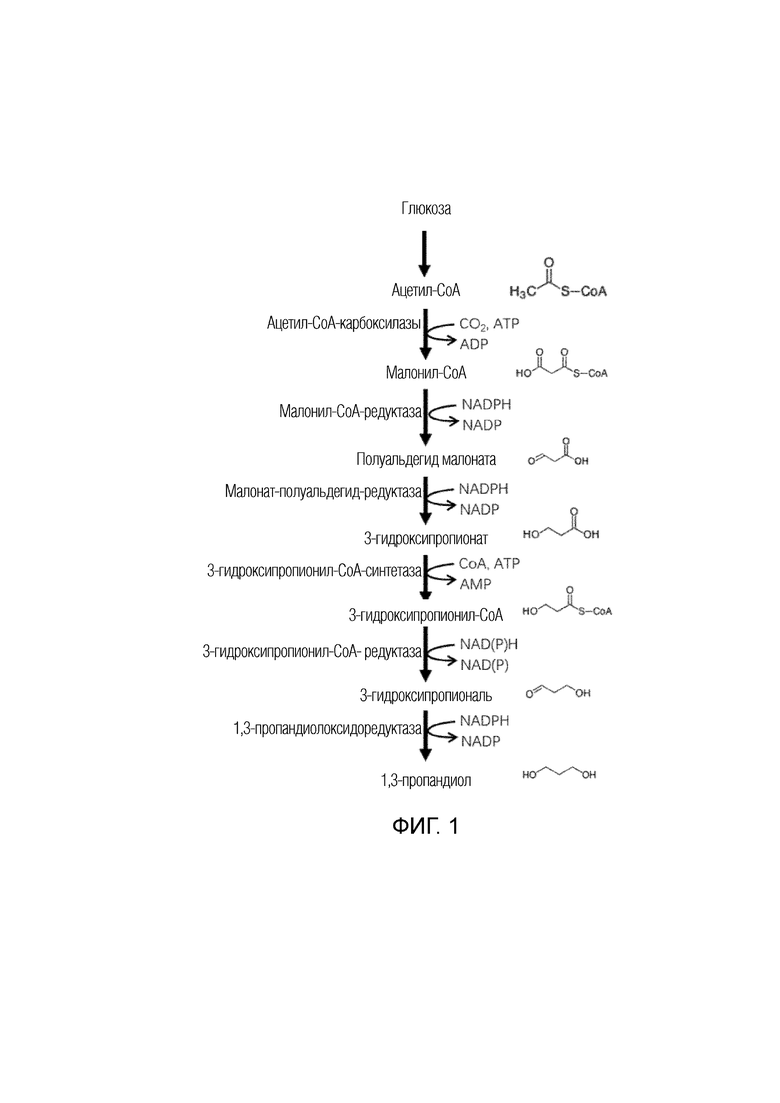
Настоящее изобретение относится к пути синтеза 1,3-пропандиола, как показано на фиг. 1: сначала, ацетил-СоА продуцируется из глюкозы (или из другого сбраживаемого углевода) по пути гликолиза в самом микроорганизме; ацетил-CoA продуцирует малонил-CoA в результате катализа ацетил-CoA-карбоксилазы; малонил-CoA продуцирует полуальдегид малоната в результате катализа малонил-CoA-редуктазы; полуальдегид малоната продуцирует 3-гидроксипропионат в результате катализа малонат-полуальдегид-редуктазы; 3-гидроксипропионат продуцирует 3-гидроксипропионил-CoA в результате катализа 3-гидроксипропионил- кофермент А-синтазы; 3-гидроксипропионил-CoA продуцирует 3-гидроксипропаналь в результате катализа 3-гидроксипропионил-CoA-редуктазы, а 3-гидроксипропаналь продуцирует 1,3-пропандиол в результате катализа 1,3-пропандиолоксидоредуктазы.
В соответствии с настоящим изобретением, 1,3-пропандиол получают путем ферментации в колбе или ферментере с использованием глюкозы в качестве субстрата с помощью рекомбинантного микроорганизма, способного сврехэкспрессировать гены ацетил-СоА-карбоксилазы accBC и accDA, ген малонил-СоА-синтетазы mcr, ген 3-гидроксипропионил-CoA-синтетазы pcs, ген 3-гидроксипропионил-CoA-редуктазы pduP и ген 1,3-пропандиолоксидоредуктазы yqhD. Во время ферментации, рекомбинантный микроорганизм согласно изобретению может утилизовать дешевую глюкозу в качестве сырья, при этом отсутствует необходимость в добавлении дорогостоящего витамина B12, что позволяет значительно снизить стоимость производства такого продукта и открывает широкие рыночные перспективы. Способ согласно изобретению является простым, не требует значительных затрат, позволяет получить 1,3-пропандиол с высоким выходом и минимальным числом побочных продуктов, и является подходящим для дальнейшего упрощения процесса выделения 1,3-пропандиола.
Краткое описание чертежей
На фиг. 1 представлена блок-схема способа получения 1,3-пропандиола путем ферментации глюкозы с использованием рекомбинантного микроорганизма согласно изобретению.
Подробное описание изобретения
Нижеследующие примеры используются для иллюстрации настоящего изобретения, но не ограничивают объема настоящего изобретения. В способы, стадии или условия изобретения могут быть внесены любые исправления и замены, не выходящие за рамки существа и объема изобретения, и эти исправления и замены также входят в объем настоящего изобретения.
Если это не оговорено особо, то все химические реагенты, используемые в примерах, являются обычными коммерчески доступными реагентами, а методы, используемые в примерах, являются стандартными методами, известными специалистам в данной области.
Пример 1. Конструирование рекомбинантной плазмиды для сверхэкспрессии генов ацетил-CoA-карбоксилазы accBC и accDA.
Ген accBC (нуклеотидная последовательность которого представлена в SEQ ID NО:1) размером приблизительно 1,8 т.п.о. получали путем ПЦР-амплификации с использованием генома Corynebacterium glutamicum ATCC 13032 в качестве матрицы и праймеров accBC-F (tagcgcagtaaAAGGAGATATACCatgtcagtcgagactaggaaga) и accBC-R (CTGCAGGCGCGCCGAGCTCGttacttgatctcgaggagaacaacgсс) и очищали.
Ген accDA (нуклеотидная последовательность которого представлена в SEQ ID NО:2) размером приблизительно 1,5 т.п.о. получали путем ПЦР-амплификации с использованием генома Corynebacterium glutamicum ATCC 13032 в качестве матрицы и праймеров accDA-F (GTTTAACTTTAATAAGGAGATATACatggtgtggggcatggaac) и accDA-R (TATATCTCCTTttactgcgctaaacgctcaaatcg) и очищали. Плазмиду pACYCDuet (Novagen) расщепляли ферментами NcoI и EcoRI, а очищенные фрагменты accBC и accDA подвергали одностадийному лигированию с pACYCDuet с использованием набора для сборки Гибсона (NEB) с получением рекомбинантной плазмиды, обозначенной pACYC-accDABC.
Пример 2. Конструирование рекомбинантной плазмиды для сверхэкспрессии гена малонил-СоА-синтетазы mcr.
Исходя из аминокислотной последовательности малонил-СоА-синтетазы, происходящей от Chloroflexus aurantiacus, оптимизированную последовательность нуклеиновой кислоты гена искусственно конструировали (последовательность гена представлена в SEQ ID NO.3) и синтезировали в Qinglan Biotech. Inc. Фрагмент mcr размером приблизительно 3,7 т.п.о. получали с помощью ПЦР с использованием генного фрагмента в качестве матрицы и с использованием праймеров mcr-F (GCGATCGCTGACGTCGGTACAAGGAGATATACATATGTCGGGCACTG) и mcr-R (TTTACCAGACTCGAGGGTACTTAAACGGTGATTGCGCGTCC) и очищали. Плазмиду pACYC-accDABC, полученную как описано в примере 1, расщепляли ферментом KpnI, и фрагмент mcr подвергали одностадийному лигированию с pACYC-accDABC с использованием набора для сборки Гибсона (NEB) с получением рекомбинантной плазмиды pACYC-accDABC-mcr. После переноса PACYC-accDABC-mcr в E.coli BL21 (DE3) методом тепловой трансформации, рекомбинантный микроорганизм отбирали в планшете со средой LB, содержащей 25 мг/л хлорамфеникола, и этот микроорганизм был обозначен E.coli/pACYC-accDABC-mcr.
Пример 3. Конструирование рекомбинантной плазмиды для сверхэкспрессии гена 3-гидроксипропионил-СоА-синтетазы pcs, гена 3-гидроксипропионил-CoA-редуктазы pduP и гена 1,3-пропандиолоксидоредуктазы yqhD.
Исходя из аминокислотной последовательности 3-гидроксипропионил-СоА-синтетазы Metallosphaera sedula, оптимизированную последовательность нуклеиновой кислоты гена искусственно конструировали (последовательность гена представлена в SEQ ID NO: 4) и синтезировали в Qinglan Biotech. Inc. Фрагмент pcs размером приблизительно 2,0 т.п.о. получали с помощью ПЦР с использованием генного фрагмента в качестве матрицы и с использованием праймеров pcs-F (CTTTAAGAAGGAGATATACCaggaggaaacagaaccATGTTTATGCGC) и pcs-R (acgttaatggTTAGGAAGTCTTTAATTCCTTCTTCAGTTCTTCCAC) и очищали. Ген pduP размером приблизительно 1,5 т.п.о. (нуклеотидная последовательность которого представлена в SEQ ID NО: 5) получали с помощью ПЦР с использованием генома Klebsiella pneumoniae DSM2026 в качестве матрицы, и с использованием праймеров pduP-F (GACTTCCTAAccattaacgtgagaactcatcaatgaatacag) и pduP-R (atATGTATATCTCCTTCTTAAAGTTttagcgaatggaaaaaaccgttggt), а затем очищали. Ген yqhD-F размером приблизительно 1,2 т.п.о. (нуклеотидная последовательность которого представлена в SEQ ID NО: 6) получали с помощью ПЦР с использованием генома E.coli W3110 в качестве матрицы, и праймеров yqhD-F (TAAGAAGGAGATATACATatgAACAACTTTAATCTGCACACC) и yqhD-R (CAAGCTTGTCGACGGAGCTCGCGGGCGGCTTCGTATATACG) и очищали. Плазмиду pET32a (Novagen) расщепяли ферментами NcoI и EcoRI, и фрагмент pcs, фрагмент pduP и фрагмент yqhD подвергали одностадийному лигированию с pET28a с получением рекомбинантной плазмиды, обозначенной pET-pcs-pduP-yqhD. После переноса pET-pcs-pduP-yqhD в E.coli/pACYC-accDABC-mcr, полученную как описано в Примере 2, методом тепловой трансформации, рекомбинантный микроорганизм отбирали в планшете со средой LB, содержащей 25 мг/л канамицина и 25 мг/л хлорамфеникола, и этот микроорганизм был обозначен E.coli/pACYC-accDABC-mcr/pET-pcs-pduP-yqhD.
Пример 4: Получение 1,3-пропандиола путем ферментации с помощью рекомбинантной E.coli
После культивирования штамма E.coli/pACYC-accDABC-mcr/pET-pcs-pduP-yqhD, полученного как описано в Примере 3, в планшете со средой LB, содержащей канамицин (25 мг/л) и хлорамфеникол (25 мг/л) в течение ночи, одну колонию, взятую из свежего планшета, инокулировали в 250 мл-колбу с перегородкой, содержащую 30 мл среды для посевной культуры, и инкубировали в течение 16 часов при 30°C и 200 об/мин.
Состав среды посевной культуры включает (г/л): глюкозу, 20; дрожжевой экстракт, 5,0; пептон, 10; NaCl, 5,0; хлорамфеникол, 0,025; и канамицин, 0,025.
Посевной бульон инокулировали в 1000 мл-колбу с перегородкой, содержащую 100 мл среды для сбраживания культуры в инокулируемом количестве 10% и инкубировали при 30°C и 200 об/мин. Через 6 часов после ферментации добавляли IPTG (0,5 мМ) и ферментацию продолжали еще 48 часов.
Состав среды для сбраживания культуры включает (г/л): глюкозу, 20; Na2HPO4 · 7H2O, 12,8; K2HPO4, 3,0; MgSO4, 0,5,; NaCl 0,5; NH4Cl, 0,5; дрожжевой экстракт, 10; биотин, 0,001; гидрохлорид тиамина, 0,001; хлорамфеникол, 0,025 и канамицин, 0,025.
Штамм E.coli/pACYC-accDABC-mcr/pET-pcs-pduP-yqhD, полученный как описано в Примере 3, тестировали через 24 часа после ферментации на Ферментативные активности ацетил-CoA-карбоксилазы, малонил-CoA-синтетазы, 3-гидроксипропионил-CoA- синтетазы, 3-гидроксипропионил-CoA-редуктазы и 1,3-пропандиолоксидоредуктазы, которые составляли 0,24 Ед/мг, 0,12 Ед/мг, 0,38 Ед/мг, 0,97 Ед/мг и 1,72 Ед/мг, соответственно, что указывает на то, что каждый рекомбинантный фермент экспрессировался нормально. В контрольном штамме E. coli BL21 (DE3) дикого типа каких-либо значимых соответствующих ферментативных активностей не обнаруживалось.
Через 48 часов после ферментации, штамм E.coli/pACYC-accDABC-mcr/pET-pcs-pduP-yqhD, полученный как описано в Примере 3, мог продуцировать 2,1 г/л 1,3-пропандиола со скоростью превращения массы 0,105 г/г глюкозы, что указывает на то, что сконструированный рекомбинантный штамм может превратить глюкозу непосредственно в 1,3-пропандиол без добавления кофермента B12. В контрольном эксперименте, проведенном в тех же условиях, ни штамм E.coli BL21 (DE3) дикого типа, ни штамм E.coli/pACYC-accDABC-mcr, полученный как описано в Примере 2, не могли продуцировать 1,3-пропандиол.
Хотя настоящее изобретение было подробно описано в общем описании и в его вышеуказанных вариантах, однако, специалистам в данной области очевидно, что в него могут быть внесены модификации или изменения исходя из раскрытия настоящего изобретения. Таким образом, эти модификации или изменения, не выходящие за рамки существа изобретения, входят в объем настоящего изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> TSINGHUA UNIVERSITY
<120> СПОСОБ ПОЛУЧЕНИЯ 1,3-ПРОПАНДИОЛА ПУТЕМ ФЕРМЕНТАЦИИ
РЕКОМБИНАНТНОГО МИКРООРГАНИЗМА
<130> P2018TC532
<150> CN 201711405440.5
<151> 2017-12-22
<160> 18
<170> PatentIn version 3.5
<210> 1
<211> 1776
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген accBC
<400> 1
atgtcagtcg agactaggaa gatcaccaag gttcttgtcg ctaaccgtgg tgagattgca 60
atccgcgtgt tccgtgcagc tcgagatgaa ggcatcggat ctgtcgccgt ctacgcagag 120
ccagatgcag atgcaccatt cgtgtcatat gcagacgagg cttttgccct cggtggccaa 180
acatccgctg agtcctacct tgtcattgac aagatcatcg atgcggcccg caagtccggc 240
gccgacgcca tccaccccgg ctacggcttc ctcgcagaaa acgctgactt cgcagaagca 300
gtcatcaacg aaggcctgat ctggattgga ccttcacctg agtccatccg ctccctcggc 360
gacaaggtca ccgctcgcca catcgcagat accgccaagg ctccaatggc tcctggcacc 420
aaggaaccag taaaagacgc agcagaagtt gtggctttcg ctgaagaatt cggtctccca 480
atcgccatca aggcagcttt cggtggcggc ggacgtggca tgaaggttgc ctacaagatg 540
gaagaagtcg ctgacctctt cgagtccgca acccgtgaag caaccgcagc gttcggccgc 600
ggcgagtgct tcgtggagcg ctacctggac aaggcacgcc acgttgaggc tcaggtcatc 660
gccgataagc acggcaacgt tgttgtcgcc ggaacccgtg actgctccct gcagcgccgt 720
ttccagaagc tcgtcgaaga agcaccagca ccattcctca ccgatgacca gcgcgagcgt 780
ctccactcct ccgcgaaggc tatctgtaag gaagctggct actacggtgc aggcaccgtt 840
gagtacctcg ttggctccga cggcctgatc tccttcctcg aggtcaacac ccgcctccag 900
gtggaacacc cagtcaccga agagaccacc ggcatcgacc tggtccgcga aatgttccgc 960
atcgcagaag gccacgagct ctccatcaag gaagatccag ctccacgcgg ccacgcattc 1020
gagttccgca tcaacggcga agacgctggc tccaacttca tgcctgcacc aggcaagatc 1080
accagctacc gcgagccaca gggcccaggc gtccgcatgg actccggtgt cgttgaaggt 1140
tccgaaatct ccggacagtt cgactccatg ctggcaaagc tgatcgtttg gggcgacacc 1200
cgcgagcagg ctctccagcg ctcccgccgt gcacttgcag agtacgttgt cgagggcatg 1260
ccaaccgtta tcccattcca ccagcacatc gtggaaaacc cagcattcgt gggcaacgac 1320
gaaggcttcg agatctacac caagtggatc gaagaggttt gggataaccc aatcgcacct 1380
tacgttgacg cttccgagct cgacgaagat gaggacaaga ccccagcaca gaaggttgtt 1440
gtggagatca acggccgtcg cgttgaggtt gcactcccag gcgatctggc actcggtggc 1500
accgctggtc ctaagaagaa ggccaagaag cgtcgcgcag gtggtgcaaa ggctggcgta 1560
tccggcgatg cagtggcagc tccaatgcag ggcactgtca tcaaggtcaa cgtcgaagaa 1620
ggcgctgaag tcaacgaagg cgacaccgtt gttgtcctcg aggctatgaa gatggaaaac 1680
cctgtgaagg ctcataagtc cggaaccgta accggcctta ctgtcgctgc aggcgagggt 1740
gtcaacaagg gcgttgttct cctcgagatc aagtaa 1776
<210> 2
<211> 1455
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген accDA
<400> 2
atggtgtggg gcatggaaca cacttcagca ttgacgctca tagactcggt tttggaccct 60
gacagcttca tttcttggaa tgaaactccc caatatgaca acctcaatca aggctatgca 120
gagaccttgg agcgggctcg aagcaaggcc aaatgcgatg aatcggtaat tactggagaa 180
ggcaccgtgg agggcattcc ggtagccgtt attttgtccg atttttcctt cctcggcggt 240
tctttgggca cggtcgcgtc ggtgcgcatc atgaaggcga ttcaccgcgc cacagagctg 300
aaactcccac tgctggtctc ccctgcttcc ggtggtgcgc gcatgcagga agacaatcga 360
gcttttgtca tgatggtgtc cataaccgcg gctgtgcagc gtcaccgcga ggcgcatttg 420
ccgttcctgg tgtatttgcg caatcccacg atgggtggcg ccatggcctc gtggggttca 480
tctgggcatc tcacttttgc ggaacccggc gcgcagatag gtttcctggg tcctcgcgtg 540
gtggagttaa ccactgggca tgcgcttcca gacggtgtgc agcaggcgga gaatttggtg 600
aaaactggtg tgattgatgg aattgtgtcg ccactccaat tgcgtgcagc ggtggcaaaa 660
accctcaagg ttattcagcc ggtagaggca acggatcgtt tttctccaac aactcctggc 720
gtggcacttc cggtgatgga ggcgattgcg cgttctcgtg acccgcagag gcctggaatc 780
ggggagatta tggaaacgtt gggggcagac gtcgtcaagc tttctggtgc gcgtgctggc 840
gcattgagcc cggctgtgcg cgttgccctg gcgcgcatcg ggggccggcc cgtggtgctg 900
attgggcagg atcgccgctt cacgcttggg ccgcaggagc tgcgttttgc gcgtcgtggc 960
atttcgctgg cgcgcgagct aaacctgccg atcgtgtcca tcatcgacac ctccggcgcc 1020
gaattgtcgc aggcggctga ggagctcggc atcgcaagct cgattgcgcg caccttgtcc 1080
aagcttatcg acgctcccct ccccaccgtt tcggtcatta ttggtcaggg cgttggcggt 1140
ggcgcgctgg ccatgctgcc cgccgatctg gtctacgcgg ccgaaaacgc gtggctgtcc 1200
gcattgccac cagagggcgc ctcggccatc ctcttccgcg acaccaacca cgccgcggaa 1260
atcatagagc gacaaggcgt gcaggcgcac gcacttttaa gccaagggct tatcgacggg 1320
atcgtcgccg aaaccgagca ctttgttgaa gaaattctcg gcacaatcag caacgccctc 1380
tccgaattgg ataacaatcc ggagagggcg ggacgcgaca gtcgcttcac acgatttgag 1440
cgtttagcgc agtaa 1455
<210> 3
<211> 3663
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген mcr
<400> 3
atgtcgggca ctgggcgttt agccggtaaa atcgcgttga tcacgggcgg agcgggtaac 60
attggttctg aattgacgcg tcgctttttg gcggagggcg cgacggtcat catttctgga 120
cgcaaccgcg ccaaactgac ggcgttggct gagcgcatgc aagccgaagc aggagtgcct 180
gctaaacgca tcgatctgga agtcatggac ggttcggatc ccgtggcagt acgcgcgggt 240
atcgaagcaa ttgtagcacg ccacggacag attgatattt tagtcaacaa cgctggttcg 300
gcgggggcgc agcgccgtct ggcagaaatc ccattaactg aggcagagtt agggcccggt 360
gcagaagaga ccttacatgc gtccattgct aacctgcttg ggatgggctg gcacttgatg 420
cgcatcgcgg ctccacacat gcctgtaggc tctgccgtga ttaatgtaag cacaatcttt 480
tcacgcgccg agtactatgg tcgtatccct tatgtaacgc caaaggcagc ccttaatgca 540
cttagtcagc ttgcggcgcg cgagttaggt gctcgtggca tccgtgtgaa tacgattttt 600
ccagggccaa ttgagtccga tcgcattcgt acggtgtttc agcgcatgga tcagttgaag 660
ggccgcccgg aaggagacac agcacaccac tttctgaata ctatgcgcct ttgtcgtgcg 720
aacgaccaag gagctttgga acgccgtttc ccgagcgtcg gagacgtggc agatgctgcc 780
gtgtttttgg ccagtgccga gagtgctgcg ctttcgggag agactatcga ggtcactcac 840
ggtatggaac ttccagcgtg ctccgagaca tcacttctgg cacgtactga cttgcgtact 900
attgacgcgt caggccgtac taccctgatt tgcgcgggag atcaaattga agaggtcatg 960
gcgttgaccg gcatgctgcg cacatgtggg agtgaagtaa tcatcggatt ccgttccgca 1020
gcagcgttgg cgcagtttga acaggctgtt aacgaatccc gccgtttggc aggggctgac 1080
ttcactcctc ctatcgccct gccattagat ccacgtgatc cggctacaat cgatgcagtg 1140
ttcgattggg gtgcaggaga aaacacgggc gggatccacg ctgctgtaat cttaccggcg 1200
accagtcacg agcccgctcc ctgtgtcatc gaggtcgatg acgagcgtgt ccttaatttt 1260
ctggctgatg aaattacagg gaccattgta atcgcatctc gccttgcccg ttattggcaa 1320
agtcagcgtt tgacgcctgg tgcgcgtgcg cgcgggccac gtgtaatttt tctgtcgaat 1380
ggcgctgacc aaaatggcaa tgtgtacgga cgcattcaga gcgcagcaat tgggcaactt 1440
atccgtgtgt ggcgtcatga ggctgaattg gattatcagc gtgcaagtgc tgcgggggat 1500
cacgttttac ctcccgtgtg ggcaaaccag attgttcgct ttgccaaccg tagtctggag 1560
ggtctggagt ttgcctgtgc ttggacggcg caacttctgc actcacagcg ccacattaac 1620
gagattactc tgaacatccc tgctaacatt tccgcgacca ccggcgcccg ttcggcttcg 1680
gtggggtggg cggaatcatt aatcgggctg catcttggta aggtggcgtt aattactgga 1740
ggtagcgccg gcattggagg gcaaattggg cgcctgcttg ctttatctgg ggcccgtgtg 1800
atgttggcgg cgcgcgaccg tcataagctg gaacagatgc aggctatgat tcagagcgag 1860
ttagccgaag tagggtatac cgacgttgaa gatcgcgtcc acattgcacc gggttgcgat 1920
gtatcaagcg aagctcaatt agccgattta gttgagcgca ccttgtccgc atttggtacc 1980
gtcgattatt taatcaataa tgcgggcatt gcgggcgtag aggagatggt tatcgacatg 2040
ccagtcgaag gctggcgcca cacgcttttt gcgaatctga tcagtaatta tagtttgatg 2100
cgcaaattag ctccgttaat gaagaagcaa ggatccggct acatcctgaa tgtatcatct 2160
tatttcggcg gagaaaaaga tgcggcgatt ccatacccga accgtgcgga ttacgctgta 2220
tcgaaggctg gtcaacgcgc tatggccgag gtatttgccc gtttcttagg tccagagatt 2280
cagatcaatg ccattgcacc aggccccgtt gagggcgatc gcttacgcgg gactggggag 2340
cgcccagggt tgtttgcgcg ccgcgcccgc cttatccttg agaataaacg cttaaacgaa 2400
ttgcacgcag ctcttatcgc cgctgcccgc actgacgagc gtagcatgca cgagttagtg 2460
gaactgttgt taccaaacga tgttgctgcg ctggagcaga atcctgccgc tcccactgcc 2520
ttgcgtgaat tagcacgccg ttttcgtagc gaaggtgacc ctgctgcctc gtcgtcatcc 2580
gcccttctta accgctccat tgctgccaag ttgctggccc gtttacacaa cggcgggtat 2640
gtgctgcccg ctgacatctt cgcgaatctg ccgaatccac ccgatccctt cttcacccgt 2700
gctcaaatcg accgcgaagc gcgcaaagtt cgcgatggta tcatgggcat gctgtacttg 2760
cagcgcatgc cgactgaatt tgatgtagcc atggcgaccg tgtactattt ggctgatcgc 2820
aacgtcagcg gagagacctt tcacccgtcc gggggattac gttatgagcg tactcctacg 2880
gggggagaat tgtttggcct gccttcacca gagcgccttg ccgaacttgt gggttcgact 2940
gtatacttaa tcggagaaca cttaactgaa catctgaatc tgctggcacg cgcatatctt 3000
gagcgttatg gagcgcgtca agtggtcatg attgtcgaaa ctgagaccgg tgccgagacg 3060
atgcgtcgtt tattacacga ccacgtggaa gccggacgct taatgacaat cgtagctggg 3120
gatcaaatcg aagcagcaat cgaccaagcg attacccgtt acggtcgccc ggggccggtc 3180
gtgtgcaccc cttttcgtcc tcttcccact gtccctttag tagggcgtaa ggacagtgac 3240
tggagtaccg tgctttcaga agccgagttt gctgagttgt gcgagcatca attaacacat 3300
catttccgcg tagcccgtaa gatcgcgtta tcagatggtg catctttggc tcttgttact 3360
ccagaaacaa ctgctacctc tacgaccgag cagtttgctt tggccaactt tattaagaca 3420
acattacatg ccttcacggc caccatcggc gttgagagcg aacgcaccgc tcagcgtatt 3480
ctgattaatc aggtggactt gactcgccgt gcgcgtgcgg aagagccgcg tgatccacat 3540
gagcgtcaac aagaattgga acgttttatt gaagcggttt tacttgttac agctcctctt 3600
cccccagagg ctgatactcg ctacgcgggt cgcattcacc gcggacgcgc aatcaccgtt 3660
taa 3663
<210> 4
<211> 1986
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген pcs
<400> 4
atgtttatgc gctacatcat ggtggaagag cagactttga aaactggaag ccaagagctt 60
gaagaaaaag ctgattataa tatgcgttac tacgcacacc ttatgaagct gtctaaagaa 120
aagcctgccg aattttgggg ctctttggcg caagatcttc tggattggta tgagccctgg 180
aaggaaacga tgcgccaaga ggaccccatg acgcgctggt tcattggcgg gaagattaat 240
gcgtcctaca atgctgtgga ccgtcatttg aacggccccc gcaaattcaa agctgcggta 300
atctgggaga gtgagttggg ggaacgtaaa atcgttactt accaagacat gttctacgaa 360
gtcaaccgct gggcaaacgc cttacgcagt cttggcgtag gtaaaggaga tcgcgtgacc 420
atttatatgc cgctgacccc agaaggaatc gcggccatgc tggcgtcagc tcgtattgga 480
gcgattcatt cagttatctt cgccggattt ggatcgcaag ccatcgccga ccgcgttgag 540
gacgccaagg caaaagtcgt aatcaccgct gacgcatatc ctcgtcgtgg taaagtagtt 600
gaactgaaaa agacagtgga cgaggccctt aattctttag gtgaacgtag tccagtgcaa 660
cacgtcctgg tctatcgccg catgaagaca gacgttaaca tgaaagaagg gcgcgacgtt 720
ttcttcgacg aagtgggcaa ataccgctat gtggagcctg aacgtatgga ttctaacgac 780
cccttattta ttctgtacac gtcaggtact acaggtaaac caaaggggat tatgcactcg 840
actggaggat atttgaccgg gacggcggtt atgctgcttt ggtcatatgg actgtcgcag 900
gagaatgatg tattattcaa tacttcagac atcggctgga tcgtgggtca ctcttacatc 960
acttactcgc cgttaatcat gggtcgcacg gtggtgatct atgagtctgc ccccgattac 1020
ccatatcccg ataagtgggc ggagatcatt gagcgttatc gcgctacaac ttttggtact 1080
tcagccacgg cgttgcgcta tttcatgaaa tacggggacg aatatgttaa gaatcatgat 1140
ttaagttcta ttcgtattat cgttacgaat ggcgaagttt tgaactatag cccctggaaa 1200
tggggcttgg aagtgcttgg tggcggcaag gtattcatgt cgcatcaatg gtggcaaacg 1260
gagaccggag cacctaattt aggctacctg cctggcatta tttatatgcc gatgaaatct 1320
gggccagcgt ctggctttcc tctgcctgga aatttcgttg aagtccttga cgagaatggt 1380
aacccgagtg cgcctcgtgt acgtggctac cttgtaatgc gtccaccctt ccccccaaac 1440
atgatgatgg gcatgtggaa tgacaacgga gagcgcctga agaaaactta tttctcaaaa 1500
tttggaagct tatactaccc cggagacttc gctatggttg atgaagacgg ttatatctgg 1560
gtactgggtc gtgcggacga gacgttaaag atcgcggcac atcgtatcgg tgccggcgag 1620
gtcgaaagcg caatcacctc tcacccatca gttgcagaag ccgcagttat tggagttccg 1680
gattcagtga agggcgagga ggtgcatgcc ttcgtggttc tgaaacaagg gtacgctccg 1740
tcctctgagc ttgcaaagga cattcaaagt catgtccgta aagtcatggg ccctattgta 1800
tcgccacaga ttcactttgt tgataaactt cctaagactc gctcgggtaa ggtgatgcgc 1860
cgtgtcatta aggccgtgat gatgggatcg tcggcaggcg accttaccac gattgaagac 1920
gaagcaagca tggatgaaat caagaaggcg gtggaagaac tgaagaagga attaaagact 1980
tcctaa 1986
<210> 5
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген pduP
<400> 5
atgaatacag cagaactgga aacccttatc cgcaccatcc tcagtgaaaa gctcgcgccg 60
acgccccctg cccctcagca agagcagggc attttctgcg atgtcggcag cgccatcgac 120
gccgctcatc aggcttttct ccgctatcag cagtgtccgc taaaaacccg cagcgccatt 180
atcagcgccc tgcgggagac gctggccccc gagctggcga cgctggcgga agagagcgcc 240
acggaaaccg gcatgggcaa caaagaagat aaatatctga aaaataaagc cgctcttgaa 300
aacacgccgg gcatagagga tctcactacc agcgccctca ccggcgatgg cgggatggtg 360
ctgtttgagt actcgccgtt cggggttatt ggcgccgtgg cgcccagcac caacccaacg 420
gaaaccatta tcaacaacag tatcagcatg ctggcggcgg gtaacagcgt ctatttcagc 480
ccccatcccg gcgcgaaaaa ggtctcgttg aagcttatcg ccaggatcga agagatcgcc 540
taccgctgca gcgggatccg taacctggtg gtgaccgttg ccgagccgac ctttgaagcc 600
acccagcaaa tgatgtccca cccgctgatt gccgttctgg ctatcaccgg cggccctggc 660
attgtggcga tgggcatgaa aagcggtaaa aaagtgatcg gcgctggcgc cggcaatccg 720
ccgtgcatcg ttgatgaaac cgccgatctc gtcaaagccg ccgaagatat catcagcggc 780
gccgccttcg attacaacct gccctgtatc gccgaaaaaa gcctgatcgt cgtcgcctcc 840
gtcgctgacc gcctgatcca gcagatgcag gattttgacg cgctgctgtt gagccgacag 900
gaggccgata ccctgcgtgc cgtctgcctg cccgacggcg cggcgaataa aaaactggtc 960
ggtaaaagcc cggctgcgct gctggcggcg gcgggtctcg ccgttccgcc tcgcccccct 1020
cgcctgctga tagccgaggt ggaggcgaac gacccctggg tgacctgcga gcagctgatg 1080
ccggtgctgc cgatcgtcag ggtcgccgac tttgacagcg ccctggcgct ggccctgcgc 1140
gttgaggagg gtctgcacca caccgccatt atgcactcgc agaatgtctc gcggctcaat 1200
ctggcggcac gcacgctgca gacctccatt tttgtcaaaa atggcccgtc ttacgcggga 1260
atcggcgtcg gcggcgaagg gtttaccacc ttcaccatcg ccacgccaac cggagaaggc 1320
accacctccg cgcggacgtt cgcccgcctg cggcgctgcg tgttgaccaa cggtttttcc 1380
attcgctaa 1389
<210> 6
<211> 1164
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ген yqhD
<400> 6
atgaacaact ttaatctgca caccccaacc cgcattctgt ttggtaaagg cgcaatcgct 60
ggtttacgcg aacaaattcc tcacgatgct cgcgtattga ttacctacgg cggcggcagc 120
gtgaaaaaaa ccggcgttct cgatcaagtt ctggatgccc tgaaaggcat ggacgtgctg 180
gaatttggcg gtattgagcc aaacccggct tatgaaacgc tgatgaacgc cgtgaaactg 240
gttcgcgaac agaaagtgac tttcctgctg gcggttggcg gcggttctgt actggacggc 300
accaaattta tcgccgcagc ggctaactat ccggaaaata tcgatccgtg gcacattctg 360
caaacgggcg gtaaagagat taaaagcgcc atcccgatgg gctgtgtgct gacgctgcca 420
gcaaccggtt cagaatccaa cgcaggcgcg gtgatctccc gtaaaaccac aggcgacaag 480
caggcgttcc attctgccca tgttcagccg gtatttgccg tgctcgatcc ggtttatacc 540
tacaccctgc cgccgcgtca ggtggctaac ggcgtagtgg acgcctttgt acacaccgtg 600
gaacagtatg ttaccaaacc ggttgatgcc aaaattcagg accgtttcgc agaaggcatt 660
ttgctgacgc taatcgaaga tggtccgaaa gccctgaaag agccagaaaa ctacgatgtg 720
cgcgccaacg tcatgtgggc ggcgactcag gcgctgaacg gtttgattgg cgctggcgta 780
ccgcaggact gggcaacgca tatgctgggc cacgaactga ctgcgatgca cggtctggat 840
cacgcgcaaa cactggctat cgtcctgcct gcactgtgga atgaaaaacg cgataccaag 900
cgcgctaagc tgctgcaata tgctgaacgc gtctggaaca tcactgaagg ttccgatgat 960
gagcgtattg acgccgcgat tgccgcaacc cgcaatttct ttgagcaatt aggcgtgccg 1020
acccacctct ccgactacgg tctggacggc agctccatcc cggctttgct gaaaaaactg 1080
gaagagcacg gcatgaccca actgggcgaa aatcatgaca ttacgttgga tgtcagccgc 1140
cgtatatacg aagccgcccg ctaa 1164
<210> 7
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер accBC-F
<400> 7
tagcgcagta aaaggagata taccatgtca gtcgagacta ggaaga 46
<210> 8
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер accBC-R
<400> 8
ctgcaggcgc gccgagctcg ttacttgatc tcgaggagaa caacgcc 47
<210> 9
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер accDA-F
<400> 9
gtttaacttt aataaggaga tatacatggt gtggggcatg gaac 44
<210> 10
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер accDA-R
<400> 10
tatatctcct tttactgcgc taaacgctca aatcg 35
<210> 11
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер mcr-F
<400> 11
gcgatcgctg acgtcggtac aaggagatat acatatgtcg ggcactg 47
<210> 12
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер mcr-R
<400> 12
tttaccagac tcgagggtac ttaaacggtg attgcgcgtc c 41
<210> 13
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер pcs-F
<400> 13
ctttaagaag gagatatacc aggaggaaac agaaccatgt ttatgcgc 48
<210> 14
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер pcs-R
<400> 14
acgttaatgg ttaggaagtc tttaattcct tcttcagttc ttccac 46
<210> 15
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер pduP-F
<400> 15
gacttcctaa ccattaacgt gagaactcat caatgaatac ag 42
<210> 16
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер pduP-R
<400> 16
atatgtatat ctccttctta aagttttagc gaatggaaaa accgttggt 49
<210> 17
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер yqhD-F
<400> 17
taagaaggag atatacatat gaacaacttt aatctgcaca cc 42
<210> 18
<211> 41
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер yqhD-R
<400> 18
caagcttgtc gacggagctc gcgggcggct tcgtatatac g 41
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ ПОЛУЧЕНИЯ МЕТАКРИЛОВОЙ КИСЛОТЫ ИЛИ СЛОЖНЫХ ЭФИРОВ МEТАКРИЛОВОЙ КИСЛОТЫ | 2008 |
|
RU2491346C9 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2009 |
|
RU2521502C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ ИЗОПРЕНА | 2008 |
|
RU2545699C2 |
| Новая аденилосукцинат-синтетаза и способ получения нуклеотидов пурина с ее использованием | 2018 |
|
RU2770464C1 |
| Рекомбинантный штамм, продуцирующий L-лизин, способы его конструирования и его применение | 2020 |
|
RU2832108C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-ЦИСТЕИНА, L-ЦИСТИНА, S-СУЛЬФОЦИСТЕИНА ИЛИ ТИАЗОЛИДИНОВОГО ПРОИЗВОДНОГО L-ЦИСТЕИНА, ИЛИ ИХ СМЕСИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2458982C2 |
| НОВАЯ АЦЕТИЛ-COA-КАРБОКСИЛАЗА | 2010 |
|
RU2551779C2 |
Изобретение относится к биотехнологии и представляет собой способ получения 1,3-пропандиола посредством ферментации рекомбинантного микроорганизма, который может сверхэкспрессировать гены ацетил-СоА-карбоксилазы accBC и accDA; ген малонил-СоА-синтетазы mcr; ген 3-гидроксипропионил-СоА-синтетазы pcs, ген 3-гидроксипропионил-СоА-редуктазы pduP и ген 1,3-пропандиолоксидоредуктазы yqhD. Рекомбинантный микроорганизм подвергают ферментации в сбраживаемой культуре в колбе или в ферментере с использованием глюкозы в качестве сырья с получением 1,3-пропандиола. Рекомбинантный микроорганизм может утилизовать глюкозу, сахарозу, мелассу, ксилозу и т.п. в качестве сырья в процессе ферментации без добавления дорогостоящего витамина B12. Изобретение позволяет получать 1,3-пропандиол с высокой степенью эффективности. 3 н. и 7 з.п. ф-лы, 1 ил., 4 пр.
1. Рекомбинантный микроорганизм для продукции 1,3-пропандиола, отличающийся тем, что рекомбинантный микроорганизм может сверхэкспрессировать:
(1) гены ацетил-СоА-карбоксилазы accBC и accDA;
(2) ген малонил-СоА-синтетазы mcr;
(3) ген 3-гидроксипропионил-CoA-синтетазы pcs;
(4) ген 3-гидроксипропионил-CoA-редуктазы pduP и
(5) ген 1,3-пропандиолоксидоредуктазы yqhD.
2. Рекомбинантный микроорганизм по п. 1, где указанный микроорганизм представляет собой E. coli, Corynebacterium glutamicum, Bacillus subtilis или Saccharomyces cerevisiae.
3. Рекомбинантный микроорганизм по п. 1 или 2, где нуклеотидные последовательности accBC и accDA представлены в SEQ ID NO: 1-2.
4. Рекомбинантный микроорганизм по п. 1 или 2, где нуклеотидная последовательность гена малонил-СоА-синтетазы mcr представлена в SEQ ID NO: 3.
5. Рекомбинантный микроорганизм по п. 1 или 2, где нуклеотидная последовательность гена 3-гидроксипропионил-CoA-синтетазы pcs представлена в SEQ ID NO: 4.
6. Рекомбинантный микроорганизм по п. 1 или 2, где нуклеотидная последовательность гена 3-гидроксипропионил-СоА-редуктазы pduP представлена в SEQ ID NO: 5.
7. Рекомбинантный микроорганизм по п. 1 или 2, где нуклеотидная последовательность гена 1,3-пропандиолоксидоредуктазы yqhD представлена в SEQ ID NO: 6.
8. Применение рекомбинантного микроорганизма по любому из пп. 1-7 для получения 1,3-пропандиола путем ферментации углевода.
9. Способ получения 1,3-пропандиола с использованием рекомбинантного микроорганизма по любому из пп. 1-7, где указанный способ включает стадии:
(1) конструирования рекомбинантного микроорганизма, способного сверхэкспрессировать гены ацетил-СоА-карбоксилазы accBC и accDA; ген малонил-СоА-синтетазы mcr; ген 3-гидроксипропионил-СоА-синтетазы pcs; ген 3-гидроксипропионил-CoA-редуктазы pduP и ген 1,3-пропандиолоксидоредуктазы yqhD; и
(2) проведения аэробной ферментации с использованием исходного материала, содержащего сбраживаемый углевод в качестве субстрата, без необходимости добавления кофермента витамина B12.
10. Способ по п. 9, где сырье, содержащее сбраживаемый углевод стадии (2), представляет собой мелассу, сахарозу, глюкозу, гидролизат крахмала, кукурузный сироп, ксилозу, маннозу или глицерин, а условиями ферментации являются: 28-37°C, величина рН в пределах от 5 до 8, а количество растворенного кислорода составляет более 10%.
| ОДНОСТАДИЙНЫЙ СПОСОБ ПОЛУЧЕНИЯ 1,3-ПРОПАНДИОЛА ПУТЕМ ГИДРОФОРМИЛИРОВАНИЯ И ГИДРИРОВАНИЯ | 2002 |
|
RU2286330C2 |
| KR 1020120099315 A, 10.09.2012 | |||
| US 20040197881 A1, 07.10.2004. | |||

