В данной заявке испрашивается приоритет заявки на патент Китая №202010790877.0 с названием "РЕКОМБИНАНТНЫЙ ШТАММ, ПРОДУЦИРУЮЩИЙ L-ЛИЗИН, СПОСОБЫ ЕГО КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ", поданной 7 августа 2020 года, и заявки на патент Китая №202010514023.X с названием "РЕКОМБИНАНТНЫЙ ШТАММ С МОДИФИЦИРОВАННЫМ ГЕНОМ DAPB, СПОСОБЫ ЕГО КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ", поданной 8 июня 2020 года; обе заявки включены в данный документ во всей полноте путем ссылки.
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области генной инженерии и микроорганизмам и касается рекомбинантного штамма с усиленной способностью продуцировать L-лизин и способов его конструирования и его применения.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
L-Лизин выполняет такие физиологические функции, как стимуляция развития, усиление иммунитета и улучшение функции центральной нервной ткани. Это одна из восьми незаменимых аминокислот, которые люди и животные не способны синтезировать самостоятельно и нуждаются в них для роста. В настоящее время L-лизин является второй наиболее распространенной аминокислотой в мире и производится в основном посредством ферментации. Наиболее важным штаммом, используемым для производства аминокислот, является Corynebacterium, включая Corynebacterium glutamicum, C. flavum, C. crenatum, Corynebacterium pekinense и т.д. Среди них важным продуцирующим лизин штаммом является Corynebacterium glutamicum. Приблизительно 90% L-лизина, произведенного промышленным путем, применяют в качестве пищевой добавки в кормовой промышленности, а 10% применяют в качестве ароматизатора и подсластителя в пищевой промышленности, а также в качестве фармацевтического промежуточного соединения в фармацевтической промышленности.
Улучшение способов ферментации для производства L-лизина может затрагивать технологии ферментации, такие как перемешивание и подача кислорода, или затрагивать состав питательной среды, такой как концентрация сахара во время ферментации, или затрагивать процессирование культуральной жидкости в продукт подходящей формы, например, путем высушивания культуральной жидкости и пеллетирования или путем ионообменной хроматографии, или может затрагивать характеристики и свойства, присущие соответствующим микроорганизмам.
Способы улучшения характеристик и свойств указанных микроорганизмов включают мутагенез, отбор и скрининг мутантов. Штаммы, полученные таким путем, устойчивы к метаболитам или являются ауксотрофами по метаболитам, имеющим регуляторное значение, и способны продуцировать L-лизин.
Если взять в качестве примера Corynebacterium glutamicum, для синтеза 1 моль L-лизина в биосинтетическом пути С. glutamicum необходимо 4 моль NADPH. Таким образом, для повышения накопления L-лизина в биосинтетическом пути С. glutamicum, очень важной стратегией является увеличение количества NADPH в метаболическом пути С. glutamicum или уменьшение количества NADPH, требующегося в пути синтеза L-лизина.
Дигидродипиколинатредуктаза (DHDPR) является вторым ключевым ферментом в биосинтезе диаминопимелиновой кислоты и L-лизина у бактерий и высших растений и катализирует реакцию NAD(P)H-зависимого восстановления дигидродипиколината с образованием гексагидродипиколината. Данный фермент играет ключевую роль в формировании клеточной стенки. DHDPR использует в качестве кофактора либо NADH, либо NADPH. DHDPR различных бактерий имеет разную аффинность к различным кофакторам. Например, для участия в синтезе L-лизина DHDPR Е. coli предпочитает NADH, тогда как DHDPR С. glutamicum в основном использует в качестве кофактора NADPH. Как и DHDPR, обнаруженные в других организмах, DHDPR С. glutamicum кодируется геном dapB, и ее синтетическая активность не регулируется конечными продуктами пути синтеза, но ингибируется 2,6-пиридиндикарбоновой кислотой (2,6-PDC).
Лизин связан с ферментативной активностью в биосинтетическом пути, которая обычно может усиливаться в результате амплификации одного или более генов в пути биосинтеза лизина или за счет применения модифицированных промоторов генов.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В изобретении предложен микроорганизм или рекомбинантный штамм, продуцирующий L-лизин, где экспрессия полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3, повышена и/или основания в положениях -49, -51 и от -54 до -58 в промоторной области, показанной в SEQ ID NO: 29, мутированы. В изобретении также предложен способ получения L-лизина с использованием микроорганизма или рекомбинантного штамма.
В первом аспекте изобретения предложен микроорганизм или рекомбинантный штамм, продуцирующий L-лизин, относящийся к роду Corynebacterium, который имеет повышенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3. Согласно изобретению, повышенная экспрессия относится к тому, что экспрессия полинуклеотида усилена, или полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, имеет точечную мутацию, или полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, имеет точечную мутацию и усиленную экспрессию.
Аминокислотная последовательность SEQ ID NO: 3 представляет собой аминокислотную последовательность белка, кодируемого геном NCg12176.
По сравнению со штаммом дикого типа или родительским штаммом, микроорганизм или рекомбинантный штамм имеет усиленную способность продуцировать L-лизин.
Полинуклеотид может кодировать аминокислотную последовательность, обладающую гомологией приблизительно 90% или выше, приблизительно 92% или выше, приблизительно 95% или выше, приблизительно 97% или выше, приблизительно 98% или выше, приблизительно 99% или выше с аминокислотной последовательностью SEQ ID NO: 3. В данном документе термин «гомология» относится к проценту идентичности между двумя полинуклеотидами или двумя полипептидными модулями. Гомология последовательностей между одним модулем и другим может быть установлена способами, известными в данной области техники. Например, такая гомология последовательностей может быть установлена алгоритмом BLAST.
Экспрессия полинуклеотида может быть усилена путем замены или мутации последовательности, регулирующей экспрессию, введения мутации в полинуклеотидную последовательность, увеличения числа копий полинуклеотида посредством включения в хромосому или введения вектора или их комбинации и тому подобного.
Последовательность, регулирующая экспрессию полинуклеотида, может быть модифицирована. Последовательность, регулирующая экспрессию полинуклеотида, контролирует экспрессию полинуклеотида, с которым она функционально связана, и может включать промотор, терминатор, энхансер, сайленсер и т.д. Полинуклеотид может иметь замену в стартовом кодоне. Полинуклеотид может быть встроен в хромосому в специфическом сайте, тем самым увеличивая число копий. В изобретении специфический сайт включает сайт транспозона, межгенный сайт и т.д. Кроме того, полинуклеотид можно встраивать в экспрессионный вектор и затем этот экспрессионный вектор вводить в клетку-хозяина, тем самым увеличивая число копий.
В одном воплощении изобретения полинуклеотид или полинуклеотид с точечной мутацией встраивают в хромосому микроорганизма в специфическом сайте, тем самым увеличивая число копий.
В одном воплощении изобретения полинуклеотид, несущий последовательность промотора, или полинуклеотид с точечной мутацией, несущий последовательность промотора, встраивают в хромосому микроорганизма в специфическом сайте, тем самым вызывая сверхэкспрессию нуклеотидной последовательности.
В одном воплощении изобретения полинуклеотид или полинуклеотид с точечной мутацией встраивают в экспрессионный вектор и затем этот экспрессионный вектор вводят в клетку-хозяина, тем самым увеличивая число копий.
В одном воплощении изобретения полинуклеотид, несущий последовательность промотора, или полинуклеотид с точечной мутацией, несущий последовательность промотора, встраивают в экспрессионный вектор и затем этот экспрессионный вектор вводят в клетку-хозяина, тем самым вызывая сверхэкспрессию нуклеотидной последовательности.
В конкретном воплощении изобретения полинуклеотид может содержать нуклеотидную последовательность SEQ ID NO: 1.
В одном воплощении изобретения полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, имеет точечную мутацию, так что остаток лизина в положении 176 аминокислотной последовательности SEQ ID NO: 3 заменен на другой аминокислотный остаток.
Согласно изобретению предпочтительно, чтобы остаток лизина в положении 176 был заменен на остаток аспарагина.
Согласно изобретению, аминокислотная последовательность после замены остатка лизина (K) в положении 176 аминокислотной последовательности, показанной в SEQ ID NO: 3, на остаток аспарагина (N), показана в SEQ ID NO: 4.
В одном воплощении изобретения полинуклеотидная последовательность с точечной мутацией образована вследствие мутации основания в положении 528 полинуклеотидной последовательности, показанной в SEQ ID NO: 1.
Согласно изобретению, мутация включает мутацию основания аденин (А) на цитозин (С) в положении 528 полинуклеотидной последовательности, показанной в SEQ ID NO: 1.
В одном воплощении изобретения полинуклеотидная последовательность с точечной мутацией содержит полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
В данном описании термин «функционально связана» относится к функциональной связи между регуляторной последовательностью и полинуклеотидной последовательностью, где регуляторная последовательность контролирует транскрипцию и/или трансляцию полинуклеотидной последовательности. Регуляторная последовательность может быть сильным промотором, способным повышать уровень экспрессии полинуклеотида. Регуляторная последовательность может быть промотором, происходящим из микроорганизмов, принадлежащих к роду Corynebacterium, или может быть промотором, происходящим из других микроорганизмов. Например, промотор может представлять собой промотор trc, промотор gap, промотор tac, промотор Т7, промотор lac, промотор trp, промотор araBAD или промотор cj7.
В конкретном воплощении изобретения промотор представляет собой промотор полинуклеотида (ген NCg12176), кодирующий аминокислотную последовательность SEQ ID NO: 3.
В данном описании термин «вектор» относится к полинуклеотидной конструкции, содержащей последовательность гена и его регуляторную последовательность, и сконструирован с возможностью экспрессии целевого гена в подходящей клетке-хозяине. Альтернативно, вектор может относиться к полинуклеотидной конструкции, содержащей последовательности, полезные для гомологичной рекомбинации, где регуляторная последовательность эндогенного гена в геноме клетки-хозяина может быть изменена, или целевой ген, который может экспрессироваться, встраивают в специфический сайт в геноме хозяина после введения вектора в клетку-хозяина. В связи с этим, вектор, который может найти применение в изобретении, может дополнительно содержать маркер селекции для определения введения вектора в клетку-хозяина или встраивания вектора в хромосому клетки-хозяина. Маркер селекции может включать маркер, придающий селектируемый фенотип, такой как устойчивость к лекарствам, ауксотрофия, устойчивость к цитотоксическим агентам или экспрессия поверхностных белков. В контексте применения таких селективных агентов можно отбирать трансформированные клетки, поскольку только клетки, экспрессирующие маркер селекции, могут выживать или демонстрировать различные фенотипические признаки.
В некоторых конкретных воплощениях изобретения используемый вектор представляет собой плазмиду pK18mobsacB или плазмиду pXMJ19.
В данном документе термин «трансформация» относится к введению полинуклеотида в клетку-хозяин, так что полинуклеотид может реплицироваться как внегеномный элемент или как элемент, встроенный в геном клетки-хозяина. Способ трансформации вектором, используемым в изобретении, может включать способ введения молекулы нуклеиновой кислоты в клетку. Кроме того, как известно в данной области техники, в зависимости от клетки-хозяина можно применять электрический импульс.
Согласно изобретению, микроорганизм или рекомбинантный штамм, относящийся к роду Corynebacterium, может представлять собой Corynebacterium glutamicum, Brevibacterium flavum, Brevibacterium lactofermentum, Corynebacterium ammoniagenes, Corynebacterium pekinense.
В одном воплощении изобретения микроорганизм, относящийся к роду Corynebacterium, представляет собой Corynebacterium glutamicum YP97158, депонированный 16 августа 2016 в Главном центре коллекций микробиологических культур Китая (адрес: No. 1 West Beichen Road, Chaoyang District, Beijing, телефон: 010-64807355) с номером депонирования CGMCC №12856 и указанный в заявке на патент Китая CN 106367432 A (дата подачи: 1 сентября 2016; дата публикации: 1 февраля 2017).
Согласно изобретению, микроорганизм или рекомбинантный штамм может также иметь другие преимущества, связанные с повышенным продуцированием L-лизина, например, повышенную или пониженную экспрессию генов, связанных с продуцированием NADPH (таких как ген, кодирующий глюкозодегидрогеназу ген, кодирующий глюконаткиназу ген, кодирующий глицеральдегид-3-фосфатдегидрогеназу ген, кодирующий глюкозо-6-фосфатдегидрогеназу, или ген, кодирующий 6-фосфоглюконатдегидрогеназу), и/или других генов, участвующих в биосинтезе или секреции L-лизина (таких как ген, кодирующий аспартатаминотрансферазу ген, кодирующий аспартаткиназу, ген, кодирующий аспартат-полуальдегиддегидрогеназу ген, кодирующий дигидродипиколинатсинтазу ген, кодирующий дигидродипиколинатредуктазу, ген, кодирующий м-диаминопимелатдегидрогеназу ген, кодирующий диаминопимелатдекарбоксилазу, lysE) или замену генов чужеродным геном.
Во втором аспекте изобретения предложена полинуклеотидная последовательность, аминокислотная последовательность, кодируемая полинуклеотидной последовательностью, рекомбинантный вектор, содержащий полинуклеотидную последовательность, и рекомбинантный штамм, содержащий полинуклеотидную последовательность.
Согласно изобретению, полинуклеотидная последовательность содержит полинуклеотид, кодирующий пептид, содержащий аминокислотную последовательность SEQ ID NO: 3, где остаток лизина в положении 176 заменен на другой аминокислотный остаток.
Согласно изобретению, предпочтительно, чтобы остаток лизина в положении 176 был заменен на остаток аспарагина.
Согласно изобретению, аминокислотная последовательность после замены остатка лизина (K) в положении 176 аминокислотной последовательности, показанной в SEQ ID NO: 3, на остаток аспарагина (N), показана в SEQ ID NO: 4.
Согласно изобретению, предпочтительно, чтобы полинуклеотидная последовательность, кодирующая полипептид, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 3, содержала полинуклеотидную последовательность, показанную в SEQ ID NO: 1.
В одном воплощении изобретения полинуклеотидная последовательность образована вследствие мутации нуклеотида в положении 528 полинуклеотидной последовательности, показанной в SEQ ID NO: 1.
Согласно изобретению, мутация относится к замене основания/нуклеотида сайта, и способ мутации может быть выбран по меньшей мере из способов мутагенеза, сайт-направленного мутагенеза посредством ПЦР и гомологичной рекомбинации. В изобретении предпочтительно применяют сайт-направленный мутагенез посредством ПЦР и/или гомологичную рекомбинацию.
Согласно изобретению, мутация включает мутацию основания аденин (А) на цитозин (С) в положении 528 полинуклеотидной последовательности, показанной в SEQ ID NO: 1.
В одном воплощении изобретения полинуклеотидная последовательность содержит полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
Согласно изобретению, аминокислотная последовательность содержит аминокислотную последовательность, показанную в SEQ ID NO: 4.
Согласно изобретению, рекомбинантный вектор сконструирован посредством введения полинуклеотидной последовательности в плазмиду.
В одном воплощении изобретения плазмида представляет собой плазмиду pK18mobsacB.
В другом воплощении изобретения плазмида представляет собой плазмиду pXMJ19.
В частности, рекомбинантный вектор может быть сконструирован из полинуклеотидной последовательности и плазмиды с использованием системы рекомбинации NEBuider.
Согласно изобретению, рекомбинантный штамм содержит полинуклеотидную последовательность.
В одном воплощении изобретения исходным штаммом для рекомбинантного штамма является YP97158.
В третьем аспекте изобретения также предложен способ конструирования рекомбинантного штамма Corynebacterium glutamicum.
Согласно изобретению, способ конструирования содержит стадию:
модификации полинуклеотидной последовательности гена NCg12176 дикого типа, показанной в SEQ ID NO: 1, в штамме-хозяине для мутации основания в положении 528 с получением рекомбинантного штамма рода Corynebacterium, содержащего мутированный кодирующий ген NCg12176.
Согласно способу конструирования по изобретению, способ модификации может представлять собой по меньшей мере один из способов мутагенеза, сайт-направленного мутагенеза посредством ПЦР и гомологичной рекомбинации.
Согласно способу конструирования по изобретению, мутация относится к мутации аденина (А) на цитозин (С) в положении 528 в SEQ ID NO: 1; в частности, полинуклеотидная последовательность содержит мутированный кодирующий ген NCg12176, показанный в SEQ ID NO: 2.
Кроме того, способ конструирования включает следующие стадии:
(1) модификация полинуклеотидной последовательности гена NCg12176 дикого типа, показанной в SEQ ID NO: 1, для мутирования нуклеотида в положении 528 с получением полинуклеотидной последовательности мутированного гена NCg12176;
(2) лигирование мутированной полинуклеотидной последовательности с плазмидой для конструирования рекомбинантного вектора;
(3) введение рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма Corynebacterium:, содержащего мутированный кодирующий ген NCg 12176.
Согласно способу конструирования по изобретению, стадия (1) включает стадию конструирования гена NCg12176 с точечной мутацией: согласно геномной последовательности Corynebacterium glutamicum, синтез двух пары праймеров, Р1 и Р2, а также Р3 и Р4, для амплификации фрагмента гена NCg12176, введение точечной мутации в ген дикого типа NCg12176, как показано в SEQ ID NO: 1, посредством сайт-направленного мутагенеза, с получением гена NCg12176 с точечной мутацией, имеющего нуклеотидную последовательность SEQ ID NO: 2, которая обозначена как NCg12176A528C.
В одном воплощении изобретения геном Corynebacterium glutamicum может иметь происхождение из штамма АТСС13032, геномная последовательность которого доступна на сайте NCBI.
В одном воплощении изобретения праймеры на стадии (1) являются следующими:
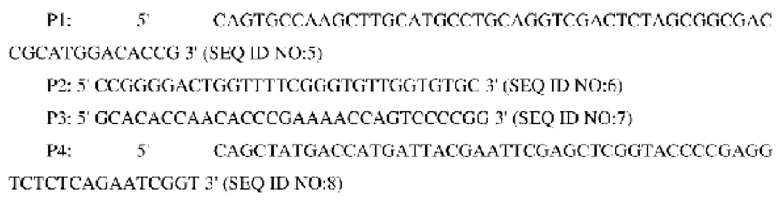
В одном воплощении изобретения процедура амплификации методом ПЦР является следующей: 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 40 с.
В одном воплощении изобретения процедура ПЦР-амплификации методом перекрывания-удлинения является следующей: 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 90 с.
Согласно способу конструирования по изобретению, стадия (2) содержит стадию конструирования рекомбинантной плазмиды, включающую: сборку выделенного и очищенного NCg12176A528C и плазмиды pK18mobsacB при помощи системы рекомбинации NEBuider с получением рекомбинантной плазмиды pK18-NCg12176A528C.
Согласно способу конструирования по изобретению, стадия (3) содержит стадию конструирования рекомбинантного штамма, включающую: трансформацию штамма-хозяина рекомбинантной плазмидой pK18-NCg12176A528C с получением рекомбинантного штамма.
В одном воплощении изобретения способ трансформации на стадии (3) представляет собой способ электротрансформации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения рекомбинация достигается гомологичной рекомбинацией.
В четвертом аспекте изобретения также предложен способ конструирования рекомбинантного штамма Corynebacterium.
Согласно изобретению, способ конструирования включает следующие стадии:
амплификацию фрагментов гомологических участков, расположенных в направлении против хода транскрипции и по ходу транскрипции относительно гена NCg12176, кодирующей области гена NCg12176 и последовательности его промоторной области, или кодирующей области гена NCg12176A528C и последовательности его промоторной области, и введение гена NCg12176 или NCg12176A528C в геном штамма-хозяина посредством гомологичной рекомбинации для реализации сверхэкспрессии гена NCg12176 или NCg12176 А528С в штамме.
В одном воплощении изобретения праймеры для амплификации фрагментов гомологических участков, расположенных в направлении против хода транскрипции, являются следующими:
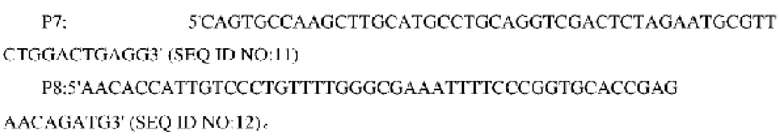
В одном воплощении изобретения праймеры для амплификации фрагментов гомологических участков, расположенных в направлении по ходу транскрипции, являются следующими:

В одном воплощении изобретения праймеры для амплификации кодирующей области гена и последовательности его промоторной области являются следующими:
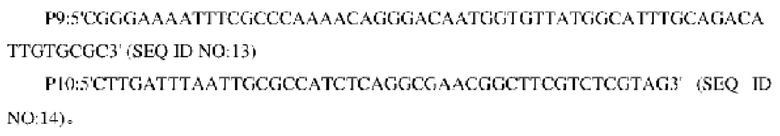
В одном воплощении изобретения проводили ПЦР с использованием вышеуказанных Р7 и Р12 в качестве праймеров и смеси трех фрагментов гомологического фрагмента, расположенного в направлении против хода транскрипции, и гена NCg12176 или NCg12176A528C с его собственным промотором в качестве матрицы для получения и объединенного фрагмента гомологического участка.
В одном воплощении изобретения используемая система ПЦР является следующей: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл; процедура ПЦР-амплификации является следующей: первоначальная денатурация при 94°С в течение 5 мин, 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 120 с; и финальная элонгация при 72°С в течение 10 мин.
В одном воплощении изобретения для сборки челночной плазмиды PK18mobsacB и объединенного фрагмента гомологичного участка с получением объединяющей плазмиды используют систему рекомбинации NEBuider.
В одном воплощении изобретения объединяющей плазмидой трансфицируют штамм-хозяин и вводят ген NCg12176 или NCg12176A528C в геном штамма-хозяина посредством гомологичной рекомбинации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQID NO: 2.
В пятом аспекте изобретения также предложен способ конструирования рекомбинантного штамма Corynebacterium.
Согласно изобретению, способ конструирования включает следующие стадии:
амплификацию кодирующей области гена NCg12176 и последовательности его промоторной области, или кодирующей области гена NCg12176A528C и последовательности его промоторной области, конструирование сверхэкспрессирующего плазмидного вектора и введение вектора в штамм-хозяин для реализации сверхэкспрессии гена NCg12176 или NCg12176A528C в штамме.
В одном воплощении изобретения праймеры для амплификации кодирующей области гена и последовательности его промоторной области являются следующими:
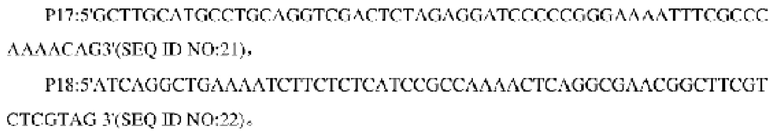
В одном воплощении изобретения использованная система ПЦР является следующей: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл; процедура ПЦР-амплификации является следующей: первоначальная денатурация при 94°С в течение 5 мин, 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 90 с; и финальная элонгация при 72°С в течение 10 мин.
В одном воплощении изобретения для сборки челночной плазмиды pXMJ19 и фрагмента NCg12176 или NCg12T76A528C с его собственным промотором с получением сверхэкспрессирующей плазмиды используют систему рекомбинации NEBuider.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQID NO: 2.
Рекомбинантный штамм, полученный по изобретению, может быть использован в отдельности для получения L-лизина посредством ферментации или может быть смешан с другими продуцирующими L-лизин бактериями для получения L-лизина посредством ферментации.
В дополнительном аспекте изобретения предложена нуклеотидная последовательность промотора, содержащая нуклеотидную последовательность, полученную путем мутации оснований в положениях -49, -51 и от -54 до -58 в промоторной области, показанной в SEQ ID NO: 29.
Согласно изобретению, нуклеотид в положении -49 промоторной области, показанной в SEQ ID NO: 29, мутирован с цитозина (С) в аденин (А), нуклеотид в положении -51 мутирован с гуанина (G) в тимин (Т), и нуклеотидная последовательность в положениях от -54 до -58 мутирована с CTGCA в GGTGT.
Согласно изобретению, нуклеотидная последовательность промотора является следующей:
(а) нуклеотидная последовательность, показанная в SEQ ID NO: 30; или
(б) нуклеотидная последовательность, обладающая идентичностью по меньшей мере 90%, предпочтительно идентичностью по меньшей мере 95% или по меньшей мере 98%, с нуклеотидной последовательностью, приведенной в SEQ ID NO: 30 и сохраняющая активность усиления промотора в (а), где нуклеотид в положении -49 сохранен как аденин (А), нуклеотид в положении -51 сохранен как тимин (Т), и нуклеотидная последовательность в положениях от -54 до -58 сохранена как GGTGT.
В изобретении также предложена экспрессионная кассета, содержащая вышеупомянутый промотор, где экспрессионная кассета содержит промотор и кодирующую последовательность, функционально связанную с промотором. В одном воплощении изобретения кодирующая последовательность представляет собой кодирующую последовательность гена dapB.
В изобретении также предложен рекомбинантный вектор, содержащий нуклеотидную последовательность промотора по изобретению.
Согласно изобретению, рекомбинантный вектор сконструирован посредством лигирования нуклеотидной последовательности промотора по изобретению с челночной плазмидой; в одном воплощении изобретения челночная плазмида представляет собой плазмиду pK18mobsacB.
В изобретении также предложен рекомбинантный штамм, содержащий вышеупомянутую нуклеотидную последовательность промотора или вышеупомянутого рекомбинантного вектора.
Согласно изобретению, рекомбинантный штамм содержит нуклеотидную последовательность, показанную в SEQ ID NO: 30. Нуклеотидная последовательность, показанная в SEQ ID NO: 30, представляет собой промоторную область гена dapB. Кроме того, нуклеотидная последовательность, показанная в SEQ ID NO: 30, связана с кодирующей последовательностью гена dapB. В частности, рекомбинантный штамм может содержать вышеупомянутую экспрессионную кассету или рекомбинантный вектор по изобретению. В частности, рекомбинантный штамм по изобретению получен путем трансформации экспрессионной кассетой или рекомбинантным вектором. Согласно изобретению, рекомбинантный штамм образован путем введения мутированной нуклеотидной последовательности промотора в штамм-хозяин для рекомбинации; штамм-хозяин может быть выбран из известных в данной области техники штаммов, продуцирующих L-лизин, например, выбран из по меньшей мере одного из штаммов Corynebacterium, где штамм Corynebacterium может представлять собой Corynebacterium glutamicum, Brevibacterium flavum, Corynebacterium crenatum, Corynebacterium pekinense; предпочтительно, Corynebacterium glutamicum. В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
Согласно рекомбинантному штамму по изобретению, в качестве вектора используют плазмиду pK18mobsacB.
Согласно изобретению, рекомбинантный штамм может дополнительно содержать другие модификации.
В изобретении также предложен способ конструирования рекомбинантного штамма, продуцирующего L-лизин, включающий следующие стадии:
(1) модификация промоторной области, показанной в SEQ ID NO: 29, для мутации оснований в положениях -49, -51 и от -54 до -58 с получением нуклеотидной последовательности, содержащей мутированную промоторную область.
Согласно изобретению, мутация относится к тому, что нуклеотид в положении -49 промоторной области, показанной в SEQ ID NO: 29, мутирован с цитозина (С) в аденин (А), нуклеотид в положении -51 мутирован с гуанина (G) в тимин (Т), и нуклеотидная последовательность в положениях от -54 до -58 мутирована с CTGCA в GGTGT. В частности, нуклеотидная последовательность мутированной промоторной области представлена в SEQ ID NO: 30. Кроме того, способ конструирования также включает следующие стадии:
(2) лигирование полинуклеотидной последовательности мутированной промоторной области с плазмидой для конструирования рекомбинантного вектора;
(3) введение рекомбинантного вектора в штамм-хозяин с получением продуцирующего L-лизин рекомбинантного штамма, содержащего мутированную промоторную область.
Согласно изобретению, на стадии (1) способ мутации включает мутагенез, сайт-направленный мутагенез посредством ПЦР или гомологичную рекомбинацию, предпочтительно, сайт-направленный мутагенез посредством ПЦР
Согласно изобретению, стадия (1) содержит: конструирование двух пар праймеров для амплификации промоторной области гена dapB, а затем выполнение ПЦР с получением нуклеотидной последовательности мутированной промоторной области.
В одном воплощении изобретения праймеры, используемые на стадии (1), являются следующими:
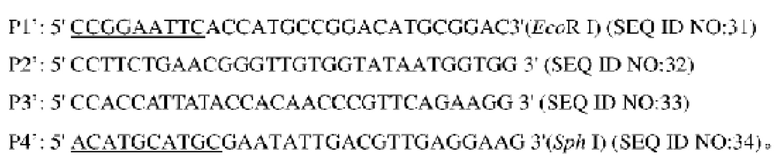
В одном воплощении изобретения стадия (1) включает использование Corynebactsrium glutamicum ATCC13032 в качестве матрицы, осуществление ПЦР-амплификации с праймерами Р1' и Р2', Р3' и Р4', соответственно, с получением двух фрагментов ДНК, содержащих точечные мутации; использование двух фрагментов ДНК в качестве матрицы, осуществление ПЦР-амплификации методом перекрывания-удлинения с праймерами Р1' и Р4', с получением фрагмента ДНК, содержащего нуклеотидную последовательность (SEQ ID NO: 30) промоторной области по изобретению.
Согласно изобретению, на стадии (1) в результате ПЦР-амплификации методом перекрывания-удлинения на двух концах полученного фрагмента ДНЕ находятся рестрикционные сайты BcoRI и Sph I, соответственно.
Согласно изобретению, стадия (2) включает: выделение и очистку продукта, амплифицированного в ходе ПЦР-реакции методом перекрывания-удлинения, лигирование расщепленного двумя рестриктазами (EcoR. I/Spk I) фрагмента с расщепленной этими же двумя рестриктазами (EcoR I/Sph I) челночной плазмидой с получением рекомбинантного вектора посредством аллельной замены.
Согласно изобретению, челночная плазмида представляет собой плазмиду pK18mobsacB, сконструированный рекомбинантный вектор представляет собой pK18-PdapB(C(-49)A,G(-51)T, CTGCA(-54--58)GG TGT).
В одном воплощении изобретения рекомбинантная плазмида имеет маркер устойчивости к канамицину.
В одном воплощении изобретения способ трансформации на стадии (3) представляет собой способ электро-трансформации; в качестве примера, на стадии (3) рекомбинантной плазмидой трансформируют штамм YP97158.
В изобретении также предложено применение вышеупомянутого микроорганизма или рекомбинантного штамма по изобретению для получения L-лизина; или способ повышения выхода ферментации L-лизина; или способ получения L-лизина.
Согласно применению и способу по изобретению он включает ферментирование микроорганизма или рекомбинантного штамма и выделение L-лизина из культуры с получением L-лизина. Согласно применению и способу по изобретению, рекомбинантный штамм по изобретению может быть использован в отдельности или в смеси с другими продуцирующими L-лизин бактериями.
Микроорганизм можно культивировать в подходящей среде в условиях культивирования, известных в данной области техники. Среда может содержать: источники углерода, источники азота, микроэлементы и их комбинации. В ходе процесса культивирования можно доводить рН культуры. Кроме того, в ходе процесса культивирования может быть включена стадия предупреждения образования пузырьков воздуха, например, с использованием пеногасителя. Кроме того, в ходе процесса культивирования может быть включена стадия инжектирования газа в культуру. Газ может включать любой газ, способный поддерживать аэробные условия культивирования. В ходе процесса культивирования температура может составлять от 20°С до 45°С. Полученный L-лизин можно выделять из культуры путем обработки культуры серной кислотой или соляной кислотой и затем можно применять комбинацию способов, таких как анионообменная хроматография, концентрирование, кристаллизация и изоэлектрическая преципитация.
В данном изобретении:
SEQ ID NO 1: Последовательность открытой рамки считывания (ORF) NCg12176 дикого типа

SEQ ID NO 2: Последовательность (ORF) NCg12176A528C

SEQ ID NO 3: аминокислотная последовательность белка, кодируемого геном NCg 12176 дикого типа

SEQ ID NO 4: аминокислотная последовательность белка, кодируемого геном NCg12176K176N

SEQ ID NO 29: последовательность промотора дикого типа

SEQ ID NO 29: мутированная последовательность промотора

Б данном изобретении, осуществляя подавление или нокаут гена NCg12176 обнаружили, что продукт, кодируемый данным геном, оказывает влияние на способность продуцировать L-лизин, и в результате введения точечной мутации в кодирующую последовательность или увеличения числа копий гена или сверхэкспрессии гена получили рекомбинантный штамм, который обладает преимуществами для производства L-лизина в высоких концентрациях по сравнению с немодифицированным штаммом.
Кроме того, рекомбинантный штамм получен путем введения точечных мутаций в промоторную область гена dapB. По сравнению с не мутированным штаммом полученный штамм также значительно увеличивал продуцирование L-лизина, дополнительно улучшал эффективность продуцирования и снижал стоимость продуцирования. Таким образом, изобретение подходит для внедрения в производство и применения.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее будут более подробно описаны технические решения изобретения с отсылкой к конкретным примерам. Следует понимать, что приведенные ниже Примеры служат только в иллюстративных целях для объяснения изобретения и не должны рассматриваться как ограничивающие объем изобретения. Использованные методики основаны на вышеупомянутом содержании изобретения и входят в заявленный объем правовой защиты изобретения. Если не указано иное, исходные материалы и реагенты, использованные в следующих ниже Примерах, доступны для приобретения или могут быть получены известными способами, все выполненные операции известны в данной области техники или осуществляются согласно инструкциям производителя доступных для приобретения продуктов.
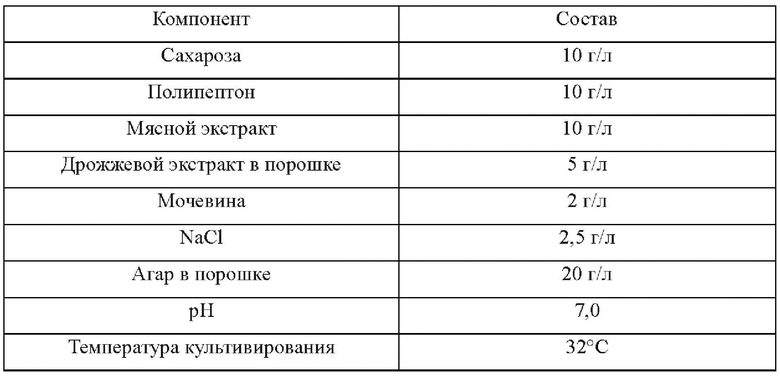
В следующих ниже Примерах композиции базовых сред, использованных для культивирования штаммов, идентичны, и в эту базовую среду можно добавлять, соответственно, необходимую сахарозу, канамицин или хлорамфеникол и т.д. Базовая среда имеет следующий состав:
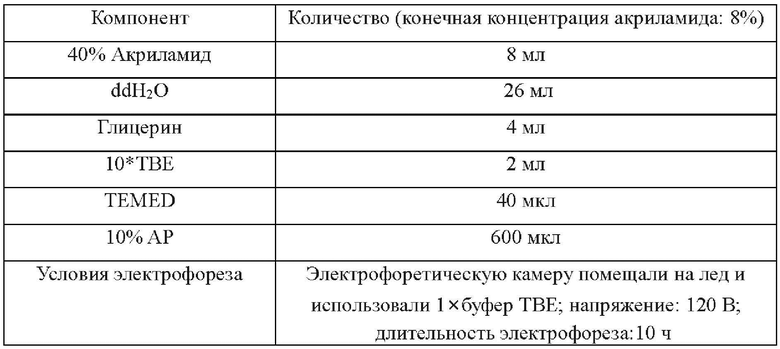
Пропись и условия проведения SSCP-PAGE в следующих Примерах являются следующими:
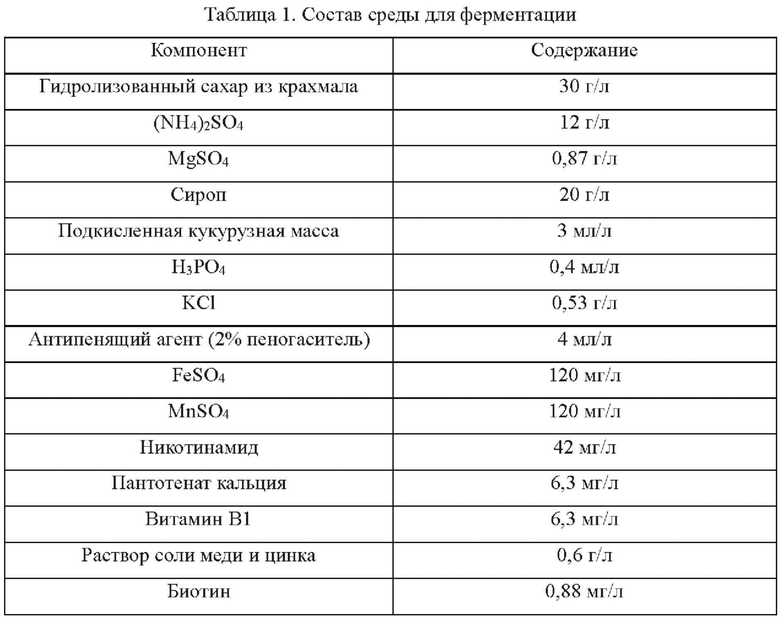
Состав среды для ферментации и контролируемый процесс ферментации L-лизина в следующих Примерах являются следующими:
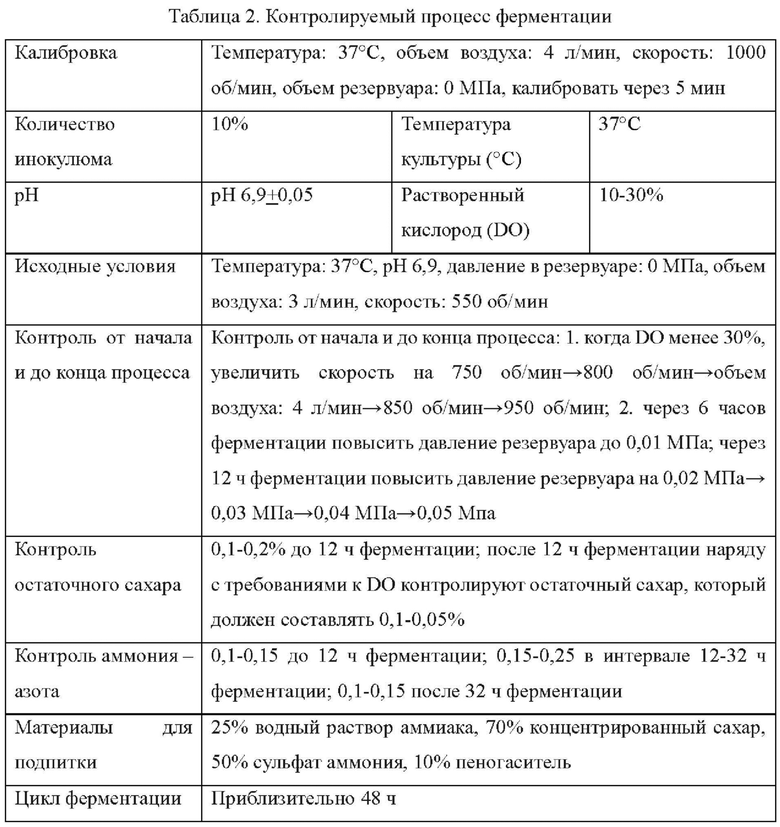
Пример 1. Конструирование вектора для трансформации pK18-NCg12176A528C, содержащего кодирующую область гена NCg12176 с точечной мутацией
Согласно геномной последовательности Corynebacterium glutamicum АТСС13032 дикого типа, опубликованной в NCBI, конструировали и синтезировали две пары праймеров для амплификации последовательности кодирующей области гена NCg12176 и вводили точечную мутацию в кодирующую область гена NCg12176 (SEQ ID NO: 1) (кодируемый им белок имел аминокислотную последовательность, приведенную в SEQ ID NO: 3) штамма YP97158, депонированного 16 августа 2016 года в Главном центре коллекций микробиологических культур Китая (адрес: No. 1 West Beichen Road, Chaoyang District, Beijing, телефон: 010-64807355) с номером депонирования CGMCC №12865 и указанного в заявке на патент Китая CN106367432A (дата подачи: 1 сентября 2016 года, дата публикации: 1 февраля 2017 года) путем аллель ной замены, так что основание в положении 528 нуклеотидной последовательности гена NCg12176 изменялось с аденина (А) на цитозин (С) (SEQ ID NO: 2: NCg12176A528C), и аминокислотный остаток в положении 176 аминокислотной последовательности соответствующего кодируемого белка изменялся с остатка лизина на остаток аспарагина (SEQIDNO: 4: NCg12176K176N).
Конструировали следующие праймеры (синтезировали в компании Invitrogen, Shanghai):
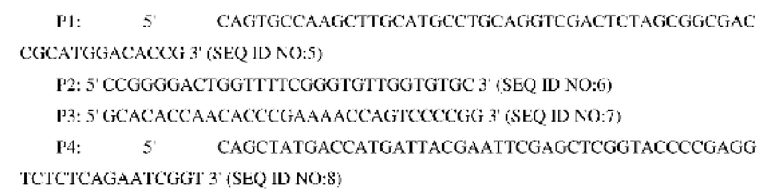
Способ конструирования: используя геномную ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы, осуществляли ПЦР-амплификацию с праймерами Р1 и Р2, а также праймерами Р3 и Р4, соответственно.
ПЦР-система: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 40 с и финальная элонгация при 72°С в течение 10 мин. Получали два фрагмента ДНК, содержащих кодирующую область гена NCg12176, NCg12176 верх и NCg12176 низ, размером 796 пн и 786 пн, соответственно.
Два вышеупомянутых фрагмента выделяли и очищали посредством электрофореза в агарозном гене и затем, используя два очищенных фрагмента ДНК в качестве матрицы, осуществляли ПЦР-амплификацию методом перекрывания-удлинения с праймерами Р1 и Р4 с получением фрагмента приблизительно 1552 пн.
ПЦР-система: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 90 с и финальная элонгация при 72°С в течение 10 мин.
Указанный фрагмент ДНК (NCg12176A528C) вызывал замену основания в положении 528 кодирующей области гена NCg12176 YP97158 с аденина (А) на цитозин (С) и затем замену аминокислотного остатка в положении 176 кодируемого белка с лизина (K) на остаток аспарагина (N).
Плазмиду pK18mobsacB (Addgene) расщепляли с помощью Xba I. NCg12176A528C и линеаризованную плазмиду pK18mobsacB выделяли и очищали путем электрофорез а в агарозном геле и затем подвергали сборке с использованием системы рекомбинации NEBuider с получен нем вектора pK18-NCg12176A528C, содержащего маркер устойчивости к канамицину. Вектор pE18-NCg12176A528C отправляли на секвенирование и правильный вектор pK18-NCg12176A528C, содержащий точечную мутацию (А-С), хранили для дальнейшего применения.
Пример 2. Конструирование генно-модифицированного штамма, содержащего ген NCg12176A528C с точечной мутацией
Способ конструирования: плазмидой pK18-NCg12176A528C для аллельной замены трансформировали продуцирующий L-лизин штамм YP97158 (способ его конструирования описан в WO 2014121669 А1; путем секвенирования подтверждали, что кодирующая область гена NCg12176 дикого типа сохранялась в хромосоме данного штамма) посредством электропорации; одиночные колонии, полученные после культивирования, анализировали с использованием праймера Р1 и универсального праймера M13R, соответственно, и штамм, у которого удавалось выявить полосу приблизительно 1559 пн, считали положительным штаммом. Этот положительный штамм культивировали в среде, содержащей 15% сахарозы, и одиночные колонии, полученные после культивирования, выращивали в среде, содержащей канамицин и без канамицина в одно и то же время; выявляли штамм, росший на среде без канамицина и не росший на среде, содержащей канамицин, при помощи ПЦР с использованием следующих праймеров (синтезированных в компании Invitrogen, Shanghai):
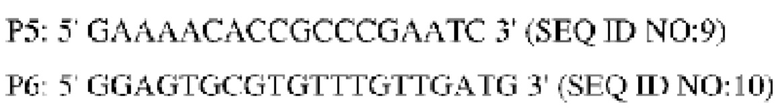
После денатурации при высокой температуре и помещения в ледяную баню вышеупомянутый продукт ПЦР-амплификации подвергали электрофорезу для анализа одноцепочечного конформационного полиморфизма (sscp-электрофорезу) (амплифицированный фрагмент плазмиды pK18-NCg12176A528C использовали в качестве положительного контроля, амплифицированный фрагмент YP97158 использовали в качестве отрицательного контроля, а воду использовали в качестве холостой пробы). Из-за различий в структуре фрагменты имели разное положение на электрофореграмме. Штамм, у которого расположение фрагмента не совпадало с фрагментом, служившим отрицательным контролем, но было таким же, как у фрагмента, служившего положительным контролем, имел успешную аллельную замену. Вновь выполняли ПЦР-амплификацию с праймерами Р5 и Р6 для амплификации целевого фрагмента штамма с успешной аллельной заменой и амплифицированный фрагмент лигировали с вектором PMD19-T для секвенирования. Штамм с успешной аллельной заменой верифицировали путем исследования последовательности с мутированным основанием и обозначили его YPL-4-011.
Пример 3. Конструирование генно-модифицированного штамма, сверхэкспрессирующего ген NCg12176 или NCg12176A528C в геноме
Согласно геномной последовательности Corynebacterium glutamicum дикого типа АТСС13032, опубликованной в NCBI, конструировали и синтезировали три пары праймеров для амплификации фрагментов гомологических участков, расположенных в направлении против хода транскрипции и по ходу транскрипции и кодирующей области гена NCg12176 или NCg12176A528C и последовательность промоторной области. Ген NCg12176 или NCg12176A528C вводили в штамм YP97158 посредством гомологичной рекомбинации.
Конструировали следующие праймеры (синтезировали в компании Invitrogen, Shanghai):

Способ конструирования: используя в качестве матрицы геномную ДНК Corynebacterium glutamicum АТСС13032 или YPL-4-011, соответственно, осуществляли ПЦР-амплификацию с праймерами Р7 и Р8, Р9 и Р10, а также Р11 и Р12 с получением расположенного в направлении против хода транскрипции фрагмента гомологического участка размером приблизительно 720 пн, гена NCg12176 и фрагмента его промотора приблизительно 1092 пн, гена NCg12176A528C и фрагмента его промотора приблизительно 1092 пн и расположенного в направлении по ходу транскрипции фрагмента гомологического участка размером приблизительно 653 пн. Используя смесь вышеупомянутых трех амплифицированных фрагментов (то есть расположенного в направлении против хода транскрипции фрагмента гомологического участка, гена NCg12176 и фрагмента его промотора, расположенного в направлении по ходу транскрипции фрагмента гомологического участка; или расположенного в направлении против хода транскрипции фрагмента гомологического участка, гена NCg12176A528C и фрагмента его промотора, расположенного в направлении по ходу транскрипции фрагмента гомологического участка) в качестве матрицы, осуществляли амплификацию с праймерами Р7 и Р12 с получением объединенного фрагмента гомологического участка.
После ПЦР-реакции амплифицированные продукты выделяли путем алектрофореза и выделяли необходимые фрагменты ДНК размером приблизительно 2504 пн с использованием набора колонок для выделения ДНК из геля (TLANGEN). Фрагменты лигировали с плазмидой PK18mobsacB, расщепленной с помощью Xba I, с использованием системы рекомбинации NEBuider с получением объединяющих плазмид PK18mobsacB-NCg12176 и PK18mobsacB-NCg12176A528C, соответственно. Эти две плазмиды содержали маркер устойчивости к канамицину, и рекомбинанты с интегрированными в геномы плазмидами можно было получить посредством скрининга на устойчивость к канамицину.
ПЦР-система: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 120 с и финальная элонгация при 72°С в течение 10 мин.
Продуцирующий L-лизин штамм YP97158 подвергали электро -трансформации двумя объединяющими плазмидами, соответственно, и полученные после культивирования одиночные колонии анализировали при помощи ПЦР с праймерами Р13 и Р14. Из положительных штаммов удалось амплифицировать фрагмент приблизительно 1609 пн, а из исходных штаммов амплифицировать фрагмент было невозможно. Проводили скрининг положительных штаммов с 15% сахарозой и культивировали в среде, содержащей канамицин и без канамицина в то же время. Штамм, росший на среде без канамицина, и не росший на среде, содержащей канамицин, далее анализировали при помощи ПЦР с использованием праймеров Р15 и Р16, и штамм, из которого удалось амплифицировать фрагмент приблизительно 1123 пн, представлял собой штамм с геномом YP97158, в который был интегрирован ген NCg12176 или NCg12176A528C, обозначенный YPL-4-012 (без точечной мутации) и YPL-4-013 (с точечной мутацией), соответственно.
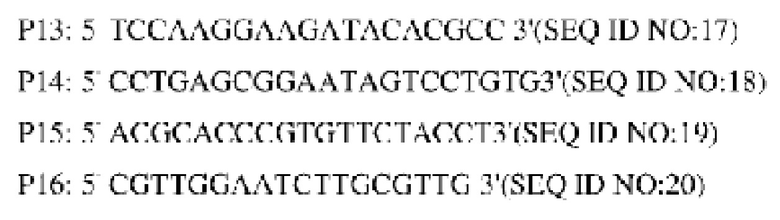
Пример 4. Конструирование генно-модифицированного штамма, сверхэкспрессирующего ген NCg12176 или NCg12176A528C на плазмиде
Согласно геномной последовательности Corynebacterium glutamicum дикого типа АТСС13032, опубликованной в NCBI, конструировали и синтезировали три пары праймеров для амплификации кодирующих участков гена NCg12176 или NCg12176A528C и последовательности промоторной области. Конструировали следующие праймеры (синтезировали в компании Invitrogen, Shanghai):
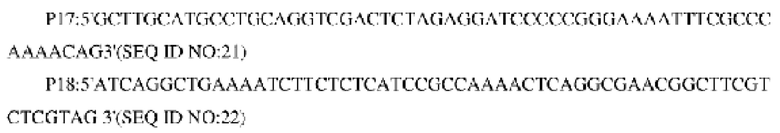
Способ конструирования: используя в качестве матрицы геномную ДНК Corynehacterium glutamicum дикого типа АТСС13032 или YPL-4-011, соответственно, осуществляли ПЦР-амплификацию с праймерами Р17 и Р18 с получением гена NCg12176 и или NCg12176A528C и фрагмента его промотора размером приблизительно 1140 пн. Амплифицированные продукты выделяли путем электрофореза и выделяли необходимые фрагменты ДНК размером приблизительно 1140 пн с использованием набора колонок для выделения ДНК из геля (TIANGEN). Фрагменты лигировали с расщепленной с помощью EcoR I челночной плазмидой pXMJ19 с использованием системы рекомбинации NEBuider с получением сверхэкспрессирующих плазмид pXMJ19-NCg12176 и pXbH19-NCg12176A528C, соответственно. Эти две плазмиды содержали маркеры устойчивости к хлорамфениколу, и штаммы, трансформированные плазмидами, можно было получить посредством скрининга на устойчивость к хлорамфениколу.
ПЦР-система: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 90 с; и финальная элонгация при 72°С в течение 10 мин.
Продуцирующий L-лизин штамм YP97158 подвергали электро-трансформации двумя плазмидами, соответственно, и после культивирования анализировали одиночные колонии при помощи ПНР с праймерами M13R(-48) и P18. Штаммы, из которых при помощи ПЦР удалось амплифицировать фрагмент размером приблизительно 1147 пн, представляли собой штаммы, трансформированные плазмидами, которые обозначили YPL-4-014 (без точечной мутации) и YPL-4-015 (с точечной мутацией), соответственно.
Пример 5. Конструирование генно-модифицированного штамма с геномом, лишенным гена NCg12176
Согласно геномной последовательности Corynebacterium glutamicum ATCC13032, опубликованной в NCBI, синтезировали две пары праймеров для амплификации фрагментов двух концов кодирующей области гена NCg12176 и амплифицированные при помощи ПЦР фрагменты использовали в качестве фрагментов гомологических участков, расположенных в направлении против хода транскрипции и по ходу транскрипции. Конструировали следующие праймеры (синтезировали в компании Invitrogen, Shanghai): 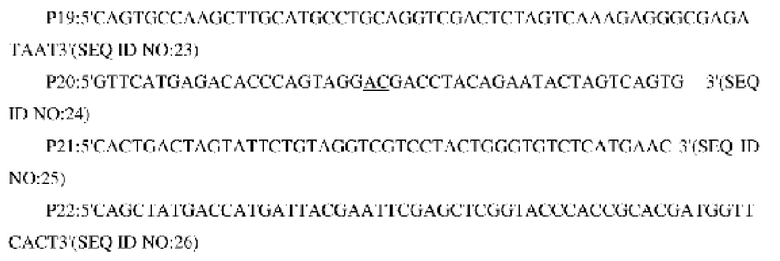
Используя в качестве матрицы геномную ДНК Corynebacterium glutamicum АТСС13032, осуществляли ПЦР-амплификацию с праймерами Р19 и Р20, а также Р21 и Р22, соответственно, с получением расположенного в направлении против хода транскрипции фрагмента гомологического участка размером приблизительно 852 пн и расположенного в направлении по ходу транскрипции фрагмента гомологического участка размером приблизительно 787 пн и затем осуществляли ПЦР методом перекрывания-удлинения с праймерами Р19 и Р22 с получением всего фрагмента гомологического участка размером 1639 пн. После ПЦР-реакции амплифицированный продукт выделяли путем электрофореза и выделяли необходимый фрагмент ДНК размером приблизительно 1639 пн с использованием набора колонок для выделения ДНК из геля (TIANGEN). Фрагмент лигировали с расщепленной с помощью Xba I челночной плазмидой pXMJ19 с использованием системы рекомбинации NEBuider с получением плазмиды с нокаутом Плазмида содержала маркер устойчивости к канамицину.
Продуцирующий лизин штамм YP97158 подвергали электро-трансформации плазмидой, приводящей к нокауту, и полученные после культивирования одиночные колонии анализировали при помощи ПЦР со следующими праймерами (синтезированы в компании Invitrogen, Shanghai):
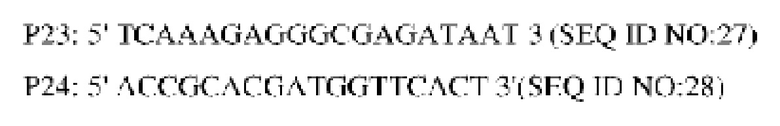
Штаммы, из которых при помощи вышеупомянутой ПЦР удалось амплифицировать фрагменты размером 1521 пн и 2556 пн, были положительными штаммами, а штаммы, из которых удалось амплифицировать только фрагмент размером 2556 пн, были исходными штаммами. Проводили скрининг положительных штаммов на среде с 15% сахарозы и культивировали в среде, содержащей канамицин и без канамицина, соответственно. Штамм, росший на среде без канамицина, и не росший на среде, содержащей канамицин, далее анализировали при помощи ПЦР с праймерами Р23 и Р24, и штамм, из которого удалось амплифицировать фрагмент размером 1521 пн, представлял собой генетически модифицированный штамм с нокаутом кодирующей области гена NCg12176, который обозначили YPL-4-016.
Пример 6. Эксперимент с ферментацией L-лизина
Осуществляли ферментацию культуральной среды, приведенной в Таблице 1, с ипсользованием штаммов, сконструированных в Примерах 2-5, и исходного штамма YP97158 в ферментаторе модели BLBIO-5GC-4-H (приобретенном в Shanghai Bailun Biotechnology Co., Ltd.) согласно контролируемому процессу, приведенному в Таблице 2 для экспериментов с ферментацией. С каждым штаммом проводили эксперимент в трех повторностях. Результаты показаны в Таблице 3.
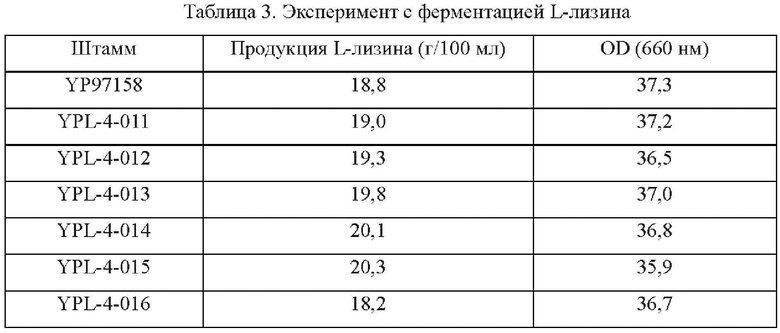
Результаты показаны в Таблице 3. Сверхэкспрессия гена NCg12176 или точечная мутация в кодирующей области гена NCg12176, т.е. NCg12176A528C, и сверхэкспрессия в Corynebacterium glutamicum способствуют улучшению продуцирования L-лизина, тогда как подавление или нокаут гена не приводят к накоплению лизина.
Пример 7. Конструирование трансформирующего вектора pK18-PdapB(C(-49)A, G(-51)T, CTGCA(-54--58)GGTGT), содержащего промоторную область гена dapB с точечной мутацией
Согласно геномной последовательности дикого типа Corynebacterium glutamicum АТСС13032, опубликованной в NCBI, конструировали и синтезировали две пары праймеров для амплификации последовательности промоторной области гена dapB и вводили точечные мутации в промоторную область гена dap В (SEQ II) NO: 29) в штамм YP97158 посредством аллельной замены, так что нуклеотид в положении -49 нуклеотидной последовательности промоторной области гена dapB был мутирован с цитозина (С) в аденин (А), нуклеотид в положении -51 мутирован с гуанина (G) в тимин (Т), а нуклеотидная последовательность в положениях от -54 до -58 мутирована с CTGCA в GGTGT.
Конструировали следующие праймеры (синтезировали в компании Invitrogen, Shanghai):
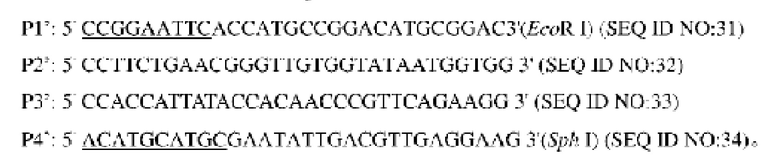
Способ конструирования: используя геномную ДНК Corynebacterium glutamicum АТСС13032 в качестве матрицы, осуществляли ПЦР-амплификацию с праймерами Р1' и Р2', а также праймерами Р3' и Р4', соответственно.
ПЦР-система: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 60 с и финальная элонгация при 72°С в течение 10 мин.
Получали два фрагмента ДНК, содержащих точечную мутацию, dapB-верх и dapB-низ, размером 665 пн и 664 пн, соответственно. Два вышеупомянутых фрагмента выделяли и очищали посредством электрофореза в агарозном гене и затем, используя два очищенных фрагмента ДНК в качестве матрицы, осуществляли ПЦР-амплификацию методом перекрывания-удлинения с праймерами Р1' и Р4' с получением фрагмента верх-низ размером приблизительно 1279 пн.
Система ПЦР методом перекрывания-удлинения: 10× кратный буфер Ex Taq 5 мкл, смесь дНТФ (по 2,5 ммоль каждый) 4 мкл, Mg2+ (25 ммоль) 4 мкл, праймеры (10 пмоль) по 2 мкл каждый, Ex Taq (5 Ед/мкл) 0,25 мкл, общий объем 50 мкл.
Процедура ПЦР-амплификации методом перекрывания-удлинения: первоначальная денатурация при 94°С в течение 5 мин; 30 циклов денатурации при 94°С в течение 30 с, отжиг при 52°С в течение 30 с и элонгация при 72°С в течение 90 с и финальная элонгация при 72°С в течение 10 мин.
Указанный фрагмент верх-низ выделяли и очищали посредством электрофореза в агарозном геле. Фрагмент содержал промоторную область гена dapB и расположенные в направлении против хода транскрипции и по ходу транскрипции от него последовательности, и на двух концах фрагмента находились сайты рестрикции EcoR I и Sph I, соответственно. Указанный фрагмент ДНК приводил к замене нуклеотида С в положении -49 промоторной области гена dapB YP97158 на А и нуклеотида G в положении -51 на Т и нуклеотидной последовательности CTGCA в положениях с -54 до -58 на GGTGT.
Фрагмент расщепляли двумя рестрикционными ферментами EcoR I и Sph I и затем лигировали с челночной плазмидой pK18mobsacB (Addgene), которую расщепляли теми же двумя рестрикционными ферментами EcoR I и Sph I, с получением плазмиды pK18-VdapB(C(-49)A,G(-51)T, CTGCA(-54--58)GGTGT) для аллельной замены, содержащей маркер устойчивости к канамицину. Вектор pK18-PdapB(C(-49)A,G(-51)T, CTGCA(-54--58)GGTGT) отправляли на секвенирование и правильный вектор pK18-PdapB(C(-49)A,G(-51)T, CTGCA(-54--58)GGTGT), содержащий точечные мутации, хранили для дальнейшего использования.
Пример 8. Конструирование генно-модифицированного штамма, содержащего вектор pK18-PdapB(C(-49)A,G(-51)T,CTGCA(-54--58)GGTGT) с точечной мутацией
Продуцирующий L-лизин штамм YP97158 подвергали электро-трансформации плазмидой pK18-PdapB(C(-49)A,G(-51)T,CTGCA(-54--58)GGTGT) для аллельной замены (путем секвенирования подтверждали, что промоторная область гена dapB дикого типа сохранялась в хромосоме данного штамма), и одиночные колонии, полученные после культивирования, анализировали с использованием праймера Р1' и универсального праймера M13R, соответственно. Из положительных штаммов удалось амплифицировать фрагмент размером приблизительно 1350 пн. Положительные штаммы культивировали на среде, содержащей 15% сахарозы, и полученные после культивирования одиночные колонии, культивировали в среде, содержащей канамицин и без канамицина, соответственно.
Штамм, росший на среде без канамицина и не росший на среде, содержащей канамицин, далее анализировали при помощи ПЦР с использованием следующих праймеров (синтезированных в компании Invitrogen, Shanghai):
После денатурации при высокой температуре и помещения в ледяную баню вышеупомянутый продукт ПЦР-амплификации подвергали sscp-электрофорезу (амплифицированный фрагмент плазмиды pK18-PdapB(C(-49)A,G(-51)T,CTGCA(-54--58)GGTGT) использовали в качестве положительного контроля, амплифицированный фрагмент YP97158 использовали в качестве отрицательного контроля, а воду использовали в качестве холостой пробы). Из-за различий в структуре фрагменты имели разное положение на электрофореграмме. Штамм, у которого расположение фрагмента не совпадало с фрагментом, служившим отрицательным контролем, но было таким же, как у фрагмента, служившего положительным контролем, имел успешную аллельную замену. Вновь выполняли ПЦР-амплификацию для амплификации целевого фрагмента положительного штамма и амплифицированный фрагмент лигировали с вектором PMD19-T для секвенирования. Штамм с успешной аллельной заменой верифицировали путем исследования последовательности с мутированным основанием и обозначили его YPL-4-010.
Пример 9. Эксперимент с ферментацией L-лизина
Осуществляли ферментацию культуральной среды, приведенной в Таблице 1, с использованием штамма YPL-4-010, сконструированного в Примере 8, и исходного штамма YP97158 в ферментаторе модели BLBIO-5GCM-H (приобретенном в Shanghai Bailun Biotechnology Co., Ltd) согласно контролируемому процессу, приведенному в Таблице 2 для экспериментов с ферментацией. С каждым штаммом проводили эксперимент в трех повтори остях. Результаты показаны в Таблице 4.
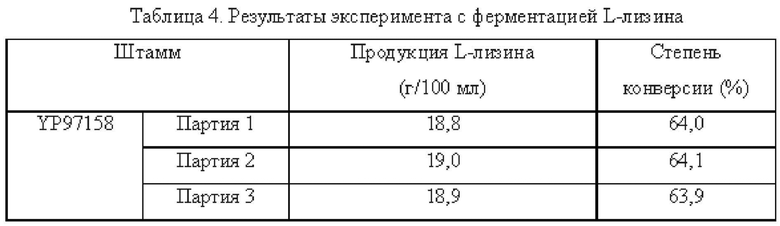
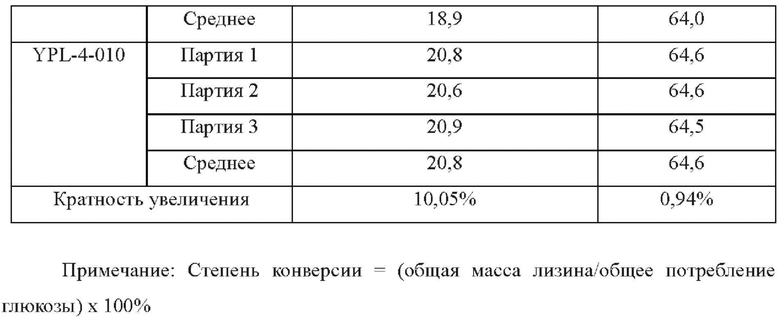
Результаты показаны в Таблице 4, точечная мутация в промоторе гена dapB С. glutamicum, PdapB(C(-49)A,G(-51)T, CTGCA(-54--58)GGTGT), способствовала улучшению продуцирования L-лизина.
Воплощения изобретения описаны выше. Однако изобретение не ограничивается описанными выше воплощениями. Любые модификации, эквивалентные замены, улучшения и так далее, сделанные в соответствии с идеей и принципами изобретения, входят в объем правовой защиты изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> HEILONGJIANG EPPEN BIOTECH CO., LTD.
<120> РЕКОМБИНАНТНЫЙ ШТАММ, ПРОИЗВОДЯЩИЙ L-ЛИЗИН, И СПОСОБЫ ЕГО
КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ
<130>
<160> 36
<170> PatentIn version 3.5
<210> 1
<211> 1035
<212> DNA
<213> Corynebacterium glutamicum
<400> 1
atggcatttg cagacattgt gcgcagcgtc gaaaaccgca ccaacgcagc gaccctcaac 60
tggtccatca aaaatggctg gaagcccgaa gtcaccggat tttccgggta cggctccggg 120
cgtcgagtgc gcgtccttgc gcgcgtgctc atgtccaacc ccgaaaattt gcttgtcgac 180
gccccctccc aatcaattac ccaacaagca cagcgcggtt ggcgccagtt cttcaccatc 240
caagtgccca acctgccagt aactgtcacc gttggtggga aaacagttac ctcatccacc 300
aacgacaacg gctacgttga cctcctggtg gaagaccaca accttgaccc cggctggcac 360
accatccaga tccaagccga aggttccacc cccgccgaag cccgcgtcct catcgtggaa 420
aacaccgccc gaatcggact catctccgac atcgacgaca ccatcatggt cacctggctt 480
ccccgagcac tcctcgccgc atggaactcg tgggttttgc acaccaaaac ccgaaaacca 540
gtccccggaa tgaaccgctt ctacgaagaa ctcctcaaag accaccccga cgcacccgtg 600
ttctacctct ccaccggcgc atggaacacc tttgaaaccc tccaagagtt catcaacaaa 660
cacgcactcc ccgacggccc catgctgctc accgactggg gaccaacccc cacaggacta 720
ttccgctcag gtcaagagca caagaaagtc caactgcgca acctgtttat cgaatacccc 780
gacatgaaat ggatcctcgt cggcgacgat ggccaacacg atcccctcat ctacggcgaa 840
gcagtcgaag aacaccccaa ccgcatcgca ggcgttgcaa tccgtgagct ctcccccggc 900
gaacatgtgc tctcccacgg aacaactgcg tcactgtcca ccatcacgac caacgggggc 960
caaggagtcc cagtagttca cggccgcgat ggatatgagt tgctgcagcg ctacgagacg 1020
aagccgttcg cctga 1035
<210> 2
<211> 1035
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 2
atggcatttg cagacattgt gcgcagcgtc gaaaaccgca ccaacgcagc gaccctcaac 60
tggtccatca aaaatggctg gaagcccgaa gtcaccggat tttccgggta cggctccggg 120
cgtcgagtgc gcgtccttgc gcgcgtgctc atgtccaacc ccgaaaattt gcttgtcgac 180
gccccctccc aatcaattac ccaacaagca cagcgcggtt ggcgccagtt cttcaccatc 240
caagtgccca acctgccagt aactgtcacc gttggtggga aaacagttac ctcatccacc 300
aacgacaacg gctacgttga cctcctggtg gaagaccaca accttgaccc cggctggcac 360
accatccaga tccaagccga aggttccacc cccgccgaag cccgcgtcct catcgtggaa 420
aacaccgccc gaatcggact catctccgac atcgacgaca ccatcatggt cacctggctt 480
ccccgagcac tcctcgccgc atggaactcg tgggttttgc acaccaacac ccgaaaacca 540
gtccccggaa tgaaccgctt ctacgaagaa ctcctcaaag accaccccga cgcacccgtg 600
ttctacctct ccaccggcgc atggaacacc tttgaaaccc tccaagagtt catcaacaaa 660
cacgcactcc ccgacggccc catgctgctc accgactggg gaccaacccc cacaggacta 720
ttccgctcag gtcaagagca caagaaagtc caactgcgca acctgtttat cgaatacccc 780
gacatgaaat ggatcctcgt cggcgacgat ggccaacacg atcccctcat ctacggcgaa 840
gcagtcgaag aacaccccaa ccgcatcgca ggcgttgcaa tccgtgagct ctcccccggc 900
gaacatgtgc tctcccacgg aacaactgcg tcactgtcca ccatcacgac caacgggggc 960
caaggagtcc cagtagttca cggccgcgat ggatatgagt tgctgcagcg ctacgagacg 1020
aagccgttcg cctga 1035
<210> 3
<211> 344
<212> PRT
<213> Corynebacterium glutamicum
<400> 3
Met Ala Phe Ala Asp Ile Val Arg Ser Val Glu Asn Arg Thr Asn Ala
1 5 10 15
Ala Thr Leu Asn Trp Ser Ile Lys Asn Gly Trp Lys Pro Glu Val Thr
20 25 30
Gly Phe Ser Gly Tyr Gly Ser Gly Arg Arg Val Arg Val Leu Ala Arg
35 40 45
Val Leu Met Ser Asn Pro Glu Asn Leu Leu Val Asp Ala Pro Ser Gln
50 55 60
Ser Ile Thr Gln Gln Ala Gln Arg Gly Trp Arg Gln Phe Phe Thr Ile
65 70 75 80
Gln Val Pro Asn Leu Pro Val Thr Val Thr Val Gly Gly Lys Thr Val
85 90 95
Thr Ser Ser Thr Asn Asp Asn Gly Tyr Val Asp Leu Leu Val Glu Asp
100 105 110
His Asn Leu Asp Pro Gly Trp His Thr Ile Gln Ile Gln Ala Glu Gly
115 120 125
Ser Thr Pro Ala Glu Ala Arg Val Leu Ile Val Glu Asn Thr Ala Arg
130 135 140
Ile Gly Leu Ile Ser Asp Ile Asp Asp Thr Ile Met Val Thr Trp Leu
145 150 155 160
Pro Arg Ala Leu Leu Ala Ala Trp Asn Ser Trp Val Leu His Thr Lys
165 170 175
Thr Arg Lys Pro Val Pro Gly Met Asn Arg Phe Tyr Glu Glu Leu Leu
180 185 190
Lys Asp His Pro Asp Ala Pro Val Phe Tyr Leu Ser Thr Gly Ala Trp
195 200 205
Asn Thr Phe Glu Thr Leu Gln Glu Phe Ile Asn Lys His Ala Leu Pro
210 215 220
Asp Gly Pro Met Leu Leu Thr Asp Trp Gly Pro Thr Pro Thr Gly Leu
225 230 235 240
Phe Arg Ser Gly Gln Glu His Lys Lys Val Gln Leu Arg Asn Leu Phe
245 250 255
Ile Glu Tyr Pro Asp Met Lys Trp Ile Leu Val Gly Asp Asp Gly Gln
260 265 270
His Asp Pro Leu Ile Tyr Gly Glu Ala Val Glu Glu His Pro Asn Arg
275 280 285
Ile Ala Gly Val Ala Ile Arg Glu Leu Ser Pro Gly Glu His Val Leu
290 295 300
Ser His Gly Thr Thr Ala Ser Leu Ser Thr Ile Thr Thr Asn Gly Gly
305 310 315 320
Gln Gly Val Pro Val Val His Gly Arg Asp Gly Tyr Glu Leu Leu Gln
325 330 335
Arg Tyr Glu Thr Lys Pro Phe Ala
340
<210> 4
<211> 344
<212> PRT
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 4
Met Ala Phe Ala Asp Ile Val Arg Ser Val Glu Asn Arg Thr Asn Ala
1 5 10 15
Ala Thr Leu Asn Trp Ser Ile Lys Asn Gly Trp Lys Pro Glu Val Thr
20 25 30
Gly Phe Ser Gly Tyr Gly Ser Gly Arg Arg Val Arg Val Leu Ala Arg
35 40 45
Val Leu Met Ser Asn Pro Glu Asn Leu Leu Val Asp Ala Pro Ser Gln
50 55 60
Ser Ile Thr Gln Gln Ala Gln Arg Gly Trp Arg Gln Phe Phe Thr Ile
65 70 75 80
Gln Val Pro Asn Leu Pro Val Thr Val Thr Val Gly Gly Lys Thr Val
85 90 95
Thr Ser Ser Thr Asn Asp Asn Gly Tyr Val Asp Leu Leu Val Glu Asp
100 105 110
His Asn Leu Asp Pro Gly Trp His Thr Ile Gln Ile Gln Ala Glu Gly
115 120 125
Ser Thr Pro Ala Glu Ala Arg Val Leu Ile Val Glu Asn Thr Ala Arg
130 135 140
Ile Gly Leu Ile Ser Asp Ile Asp Asp Thr Ile Met Val Thr Trp Leu
145 150 155 160
Pro Arg Ala Leu Leu Ala Ala Trp Asn Ser Trp Val Leu His Thr Asn
165 170 175
Thr Arg Lys Pro Val Pro Gly Met Asn Arg Phe Tyr Glu Glu Leu Leu
180 185 190
Lys Asp His Pro Asp Ala Pro Val Phe Tyr Leu Ser Thr Gly Ala Trp
195 200 205
Asn Thr Phe Glu Thr Leu Gln Glu Phe Ile Asn Lys His Ala Leu Pro
210 215 220
Asp Gly Pro Met Leu Leu Thr Asp Trp Gly Pro Thr Pro Thr Gly Leu
225 230 235 240
Phe Arg Ser Gly Gln Glu His Lys Lys Val Gln Leu Arg Asn Leu Phe
245 250 255
Ile Glu Tyr Pro Asp Met Lys Trp Ile Leu Val Gly Asp Asp Gly Gln
260 265 270
His Asp Pro Leu Ile Tyr Gly Glu Ala Val Glu Glu His Pro Asn Arg
275 280 285
Ile Ala Gly Val Ala Ile Arg Glu Leu Ser Pro Gly Glu His Val Leu
290 295 300
Ser His Gly Thr Thr Ala Ser Leu Ser Thr Ile Thr Thr Asn Gly Gly
305 310 315 320
Gln Gly Val Pro Val Val His Gly Arg Asp Gly Tyr Glu Leu Leu Gln
325 330 335
Arg Tyr Glu Thr Lys Pro Phe Ala
340
<210> 5
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 5
cagtgccaag cttgcatgcc tgcaggtcga ctctagcggc gaccgcatgg acaccg 56
<210> 6
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 6
ccggggactg gttttcgggt gttggtgtgc 30
<210> 7
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 7
gcacaccaac acccgaaaac cagtccccgg 30
<210> 8
<211> 58
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 8
cagctatgac catgattacg aattcgagct cggtaccccg aggtctctca gaatcggt 58
<210> 9
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 9
gaaaacaccg cccgaatc 18
<210> 10
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 10
ggagtgcgtg tttgttgatg 20
<210> 11
<211> 55
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 11
cagtgccaag cttgcatgcc tgcaggtcga ctctagaatg cgttctggac tgagg 55
<210> 12
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 12
aacaccattg tccctgtttt gggcgaaatt ttcccggtgc accgagaaca gatg 54
<210> 13
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 13
cgggaaaatt tcgcccaaaa cagggacaat ggtgttatgg catttgcaga cattgtgcgc 60
<210> 14
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 14
cttgatttaa ttgcgccatc tcaggcgaac ggcttcgtct cgtag 45
<210> 15
<211> 45
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 15
ctacgagacg aagccgttcg cctgagatgg cgcaattaaa tcaag 45
<210> 16
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 16
cagctatgac catgattacg aattcgagct cggtacccgc tatgacacct tcaacggatc 60
<210> 17
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 17
tccaaggaag atacacgcc 19
<210> 18
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 18
cctgagcgga atagtcctgt g 21
<210> 19
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 19
acgcacccgt gttctacct 19
<210> 20
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 20
cgttggaatc ttgcgttg 18
<210> 21
<211> 59
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 21
gcttgcatgc ctgcaggtcg actctagagg atcccccggg aaaatttcgc ccaaaacag 59
<210> 22
<211> 59
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 22
atcaggctga aaatcttctc tcatccgcca aaactcaggc gaacggcttc gtctcgtag 59
<210> 23
<211> 55
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 23
cagtgccaag cttgcatgcc tgcaggtcga ctctagtcaa agagggcgag ataat 55
<210> 24
<211> 46
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 24
gttcatgaga cacccagtag gacgacctac agaatactag tcagtg 46
<210> 25
<211> 46
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 25
cactgactag tattctgtag gtcgtcctac tgggtgtctc atgaac 46
<210> 26
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 26
cagctatgac catgattacg aattcgagct cggtacccac cgcacgatgg ttcact 56
<210> 27
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 27
tcaaagaggg cgagataat 19
<210> 28
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 28
accgcacgat ggttcact 18
<210> 29
<211> 192
<212> DNA
<213> Corynebacterium glutamicum
<400> 29
cttaagtctc atatttcaaa catagttcca cctgtgtgat taatccctag aacggaacaa 60
actgatgaac aatcgttaac aacacagacc aaaacggtca gttaggtatg gatatcagca 120
ccttctgaac gggtacgtct agactggtgg gcgtttgaaa aactcttcgc cccacgaaaa 180
tgaaggagca ta 192
<210> 30
<211> 192
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 30
cttaagtctc atatttcaaa catagttcca cctgtgtgat taatccctag aacggaacaa 60
actgatgaac aatcgttaac aacacagacc aaaacggtca gttaggtatg gatatcagca 120
ccttctgaac gggttgtggt ataatggtgg gcgtttgaaa aactcttcgc cccacgaaaa 180
tgaaggagca ta 192
<210> 31
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 31
ccggaattca ccatgccgga catgcggac 29
<210> 32
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 32
ccttctgaac gggttgtggt ataatggtgg 30
<210> 33
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 33
ccaccattat accacaaccc gttcagaagg 30
<210> 34
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 34
acatgcatgc gaatattgac gttgaggaag 30
<210> 35
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 35
agatcgtcgg actcattgac 20
<210> 36
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Искусственная последовательность
<400> 36
caaacatagt tccacctgtg 20
<---
Изобретение относится к биотехнологии и представляет собой микроорганизм, продуцирующий L-лизин, относящийся к роду Corynebacterium, который имеет повышенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3; или где основания в положениях -49, -51 и от -54 до -58 в промоторной области, показанной в SEQ ID NO: 29, мутированы; или где указанный микроорганизм имеет экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 4. Использование указанного микроорганизма позволяет повысить выход ферментации L-лизина, продуцируемого штаммом с такими мутациями. 6 н. и 9 з.п. ф-лы, 4 табл., 7 пр.
1. Микроорганизм, продуцирующий L-лизин, относящийся к роду Corynebacterium, который имеет повышенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3; или где основания в положениях -49, -51 и от -54 до -58 в промоторной области, показанной в SEQ ID NO: 29, мутированы; или
где указанный микроорганизм имеет экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 4.
2. Микроорганизм по п. 1, где полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, содержит нуклеотидную последовательность SEQ ID NO: 1.
3. Микроорганизм по п. 1 или 2, где полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 4, содержит нуклеотидную последовательность SEQ ID NO: 2.
4. Микроорганизм по любому из пп. 1-3, где нуклеотид в положении -49 промоторной области, показанной в SEQ ID NO: 29, мутирован с цитозина (C) в аденин (A), нуклеотид в положении -51 мутирован с гуанина (G) в тимин (T), и нуклеотидная последовательность в положениях от -54 до -58 мутирована с CTGCA в GGTGT; предпочтительно, нуклеотидная последовательность промотора является следующей:
(а) нуклеотидная последовательность, показанная в SEQ ID NO: 30; или
(б) нуклеотидная последовательность, обладающая идентичностью по меньшей мере 90%, предпочтительно идентичностью по меньшей мере 95% или по меньшей мере 98%, с нуклеотидной последовательностью, показанной в SEQ ID NO: 30, и сохраняющая активность усиления промотора в (a), где нуклеотид в положении -49 сохранен как аденин (A), нуклеотид в положении -51 сохранен как тимин (T), и нуклеотидная последовательность в положениях от -54 до -58 сохранена как GGTGT.
5. Микроорганизм по любому из пп. 1-4, где микроорганизм представляет собой Corynebacterium glutamicum, предпочтительно YP97158.
6. Промотор, содержащий нуклеотидную последовательность, полученную путем мутации оснований в положениях -49, -51 и от -54 до -58 в промоторной области, показанной в SEQ ID NO: 29.
7. Промотор по п. 6, где нуклеотид в положении -49 промоторной области, показанной в SEQ ID NO: 29, мутирован с цитозина (C) в аденин (A), нуклеотид в положении -51 мутирован с гуанина (G) в тимин (T), и нуклеотидная последовательность в положениях от -54 до -58 мутирована с CTGCA в GGTGT.
8. Промотор по п. 6 или 7, где нуклеотидная последовательность промотора является следующей:
(а) нуклеотидная последовательность, показанная в SEQ ID NO: 30; или
(б) нуклеотидная последовательность, обладающая идентичностью по меньшей мере 90%, предпочтительно идентичностью по меньшей мере 95% или по меньшей мере 98%, с нуклеотидной последовательностью, показанной в SEQ ID NO: 30, и сохраняющая активность усиления промотора в (a), где нуклеотид в положении -49 сохранен как аденин (A), нуклеотид в положении -51 сохранен как тимин (T), и нуклеотидная последовательность в положениях от -54 до -58 сохранена как GGTGT.
9. Экспрессионная кассета промотора, содержащая нуклеотидную последовательность промотора по любому из пп. 6-8 и кодирующую последовательность, функционально связанную с промотором.
10. Экспрессионная кассета промотора по п. 9, где кодирующая последовательность представляет собой кодирующую последовательность гена dapB.
11. Экспрессионный рекомбинантный вектор, содержащий нуклеотидную последовательность промотора по любому из пп. 6-8.
12. Экспрессионный рекомбинантный вектор по п. 11, где указанный рекомбинантный вектор сконструирован посредством лигирования нуклеотидной последовательности промотора с плазмидой pK18mobsacB.
13. Продуцирующая L-лизин экспрессионная рекомбинантная клетка-хозяин, содержащая нуклеотидную последовательность промотора по любому из пп. 6-8 или экспрессионную кассету по п. 9 или 10 или рекомбинантный вектор по п. 11 или 12.
14. Рекомбинантная клетка-хозяин по п. 13, где штамм-хозяин указанной рекомбинантной клетки-хозяина представляет собой YP97158.
15. Способ получения L-лизина, включающий культивирование микроорганизма по любому из пп. 1-5 или рекомбинантной клетки-хозяина по п. 13 или 14 и выделение L-лизина из культуры.
| CN 109721658 A, 07.05.2019 | |||
| CN 111197021 A, 26.05.2020 | |||
| CN 109721658 A, 07.05.2019 | |||
| Микроорганизм, обладающий способностью продуцировать L-лизин, и способ получения L-лизина с использованием этого микроорганизма | 2015 |
|
RU2694729C2 |
| CN 111197021 A, 26.05.2020 | |||
| УСТРОЙСТВО ЗАЩИТЫ СИСТЕМЫ ПИТАНИЯ ПРИ ГАЗОПЛАМЕННОЙ ОБРАБОТКЕ МАТЕРИАЛОВ | 2000 |
|
RU2169063C1 |
| EP 3153586 A1, 12.04.2017. | |||

