ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная заявка испрашивает приоритет предварительной заявки на патент США № 62/162720, поданной 16 мая 2015 года, содержание которой включено в данный документ с помощью ссылки во всей своей полноте для всех целей.
ПРЕДСТАВЛЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ТЕКСТОВОМ ФАЙЛЕ ASCII
[0002] Содержание нижеследующего представленного текстового файла ASCII включено в данный документ с помощью ссылки во всей своей полноте: машиночитаемая форма (CRF) перечня последовательностей (название файла: 159792013440SEQLIST.TXT, дата составления: 14 апреля 2016 г., размер: 49 кБ).
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Настоящее изобретение относится к способам лечения заболевания или нарушения, связанных с глубокой интронной мутацией, c использованием сконструированной не встречающейся в природе системы на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), а также к композициям, наборам и связанным с ними вирусным частицам.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[0004] Мутации в некодирующих последовательностях, таких как интроны, вовлечены в широкий спектр заболеваний. Геном человека содержит более высокую долю более длинных интронов, чем другие организмы, такие как черви и мухи; более 90% интронов человека составляют более 100 нуклеотидов в длину, причем более трети всех интронов составляют 2000 нуклеотидов или больше в длину (Molecular Biology of the Cell, 6th ed. (Alberts, B. et al. eds., 2014). Кроме того, глубокие интронные мутации были идентифицированы как важная и потенциально игнорируемая причина заболевания человека, при этом большая часть усилий была направлена на выявление связанных с заболеваниями мутаций, сосредоточенных в кодирующей последовательности (Homolova, K. et al. (2010) Hum. Mutat. 31:437-444). Из-за сложности сплайсинга mRNA у человека эти глубокие интронные мутации потенциально могут вызывать множество патологических состояний из-за механизмов, включающих, inter alia, дестабилизацию, деградацию и ошибочный сплайсинг mRNA (например, создание криптического сайта сплайсинга). В действительности, по некоторым оценкам до 5% менделевских заболеваний у людей могут быть связаны с глубокой интронной мутацией (Cooper, D.N. et al. (2010) Hum. Mutat. 31:631-655).
[0005] В качестве иллюстративного примера нарушения, которое в некоторых случаях было связано с глубокой интронной мутацией, врожденный амавроз Лебера (LCA) является самой тяжелой формой наследственной дистрофии сетчатки с возникновением симптомов на первом году жизни (Leber, T.K.G.v. (1869) Graefe's Archive for Clinical and Experimental Ophthalmology 15:1-25). Острота зрения у пациентов с LCA редко бывает лучше чем 20/400 (Cremers, F.P. et al. (2002) Human molecular genetics 11:1169-1176). LCA поражает приблизительно 1 на 30000 человек в общей популяции и составляет 5% всех наследственных форм дистрофии сетчатки (Koenekoop, R.K. (2004) Survey of ophthalmology 49:379-398). Наиболее частой генетической причиной LCA, составляющей примерно 15% всех случаев LCA в европейских странах и в Соединенных Штатах, является глубокая интронная мутация c.2991+1655A>G в интроне 26 гена CEP290, которая создает криптический сайт донора сплайсинга, приводящий к включению аберрантного экзона, содержащего преждевременный стоп-кодон (p.C998X), в mRNA CEP290 (denHollander,A.I.etal.(2006)Am.J.Hum.Genet.79:556-561;Perrault,I.etal.(2007)Hum.Mutat.28:416;Stone,E.M.(2007)Am.J.Ophthalmol.144:791-811;Wiszniewski,W.etal.(2011)Hum.Genet.129:319-327). Заболевание LCA, вызванное мутацией CEP290, известно как LCA10. Альтернативный сплайсинг криптического экзона в mRNA CEP290 происходит в некоторых, но не во всех транскриптах mRNA у гомозиготно пораженных индивидуумов (denHollander,A.I.etal.(2006)Am.J.Hum.Genet.79:556-561), подчеркивая гипоморфный характер этой интронной мутации.
[0006] Ген CEP290 человека охватывает 54 экзона, которые кодируют белок из 2479 аминокислот. CEP290 представляет собой центросомальный белок, который играет важную роль как в сборке цилий, так и в миграции цилиарных белков (Barbelanne, M. et al. (2013) Hum. Mol. Genet. 22:2482-2494; Craige, B. et al. (2010) J. Cell Biol. 190:927-940). В фоторецепторах, клетках сетчатки, наиболее подверженным мутациям по CEP290, CEP290 расположена в соединительной цилии (Chang, B. et al. (2006) Hum. Mol. Genet. 15:1847-1857), которая соединяет внутренний и внешний сегмент фоторецепторов.
[0007] В настоящее время не существует лечения LCA, вызванного мутацией CEP290. Два доклинических подхода в отношении данного заболевания представляют собой увеличение гена и антисмысловые олигонуклеотиды (AON). Размер комплементарной ДНК (кДНК) CEP290 человека превышает полный размер (~4,8 т.о.) рекомбинантных аденоассоциированных вирусов (rAAV). Лентивирусная векторная система способна вместить полноразмерную кДНК CEP290; однако она не способна точно контролировать уровень экспрессии CEP290. Предыдущие сообщения продемонстрировали, что фоторецепторы чувствительны к уровню экспрессии трансгена, и сверхэкспрессия CEP290 является цитотоксической (Burnight, E.R. et al. (2014) Gene Ther. 21:662-672; Tan, E. et al. (2001) Invest. Ophthalmol. Vis. Sci. 42:589-600). Альтернативной стратегий является применение AON для препятствования аберрантному сплайсингу CEP290 (Collin, R.W. et al. (2012) Mol. Ther. Nucleic Acids 1:e14; Gerard, X. et al. (2012) Mol. Ther. Nucleic Acids 1:e29). Однако данный подход требует еженедельных или ежемесячных субретинальных инъекций ретинологом в течение нескольких лет.
[0008] Соответственно, существует насущная необходимость в усовершенствованных терапевтических подходах для лечения нарушений, связанных с глубокими интронными мутациями, таких как LCA10, вызванный мутацией по CEP290.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0009] Настоящее изобретение предусматривает композиции для лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержащие сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает композиции для лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, которые содержат нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0010] В некоторых аспектах настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, которые содержат нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержащие сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2.
[0011] В некоторых вариантах осуществления вышеприведенных вариантов осуществления глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0012] В некоторых вариантах осуществления вышеприведенных вариантов осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0013] В некоторых вариантах осуществления вышеприведенных вариантов осуществления Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes (SEQ ID NO:40), белок Cas9 Staphylococcus aureus (SEQ ID NO: 55), белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку человека.
[0014] В некоторых вариантах осуществления вышеприведенных вариантов осуществления система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS). В некоторых вариантах осуществления Cas-белок содержит один или несколько NLS. В некоторых вариантах осуществления NLS представляет собой С-концевую последовательность в большом T-антигене SV40. В некоторых вариантах осуществления NLS содержит последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27).
[0015] В некоторых вариантах осуществления вышеприведенных вариантов осуществления содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0016] В некоторых вариантах осуществления вышеприведенных вариантов осуществления система CRISPR-Cas (например, первая направляющая РНК, вторая направляющая РНК и Cas-белок) находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты и/или белка клетками.
[0017] В некоторых вариантах осуществления вышеприведенных вариантов осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0018] В некоторых вариантах осуществления вышеприведенных вариантов осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых вариантах осуществления вектор находится в комплексе с системой доставки. В некоторых вариантах осуществления вектор находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты клетками.
[0019] В некоторых вариантах осуществления вышеприведенных вариантов осуществления вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV). В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5.
[0020] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0021] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вектор на основе rHSV получен из rHSV-1 или rHSV-2.
[0022] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV. В некоторых вариантах осуществления ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши. В некоторых вариантах осуществления ITR AAV представляют собой ITR AAV2. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор.
[0023] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид от аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0024] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0025] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
[0026] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид серотипа AAV из клад A-F. В некоторых вариантах осуществления вирусная частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота или AAV мыши, капсид rAAV2/HBoV1. В некоторых вариантах осуществления ITR и капсид вирусной частицы rAAV получены из AAV одного и того же серотипа. В некоторых вариантах осуществления ITR и капсид вирусных частиц rAAV получены из разных серотипов AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV2.
[0027] В некоторых аспектах настоящее изобретение предусматривает способы лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, включающие введение индивидууму терапевтически эффективного количества композиции, содержащей сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает способы лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, включающие введение индивидууму терапевтически эффективного количества композиции, содержащей нуклеиновую кислоту, которая кодирует сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0028] В некоторых аспектах настоящее изобретение предусматривает способы лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, включающие введение индивидууму терапевтически эффективного количества композиции, содержащей сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает способы лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, включающие введение индивидууму терапевтически эффективного количества композиции, содержащей нуклеиновую кислоту, которая кодирует сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2.
[0029] В некоторых вариантах осуществления вышеприведенных способов индивидуум является человеком. В некоторых вариантах осуществления млекопитающее является человеком. В некоторых вариантах осуществления композицию вводят в глаз индивидуума. В некоторых вариантах осуществления введение является субретинальным или интравитреальным.
[0030] В некоторых вариантах осуществления вышеприведенных способов глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0031] В некоторых вариантах осуществления вышеприведенных способов глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0032] В некоторых вариантах осуществления вышеприведенных способов Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes, белок Cas9 Staphylococcus aureus, белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку человека.
[0033] В некоторых вариантах осуществления вышеприведенных способов система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS). В некоторых вариантах осуществления Cas-белок содержит один или несколько NLS. В некоторых вариантах осуществления NLS представляет собой С-концевую последовательность в большом T-антигене SV40. В некоторых вариантах осуществления NLS содержит последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27).
[0034] В некоторых вариантах осуществления вышеприведенных способов содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0035] В некоторых вариантах осуществления вышеприведенных способов система CRISPR-Cas (например, первая направляющая РНК, вторая направляющая РНК и Cas-белок) находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты и/или белка клетками.
[0036] В некоторых вариантах осуществления вышеприведенных способов нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0037] В некоторых вариантах осуществления вышеприведенных способов нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых вариантах осуществления вектор находится в комплексе с системой доставки. В некоторых вариантах осуществления вектор находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты клетками.
[0038] В некоторых вариантах осуществления вышеприведенных вариантов осуществления вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV). В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5.
[0039] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0040] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вектор на основе rHSV получен из rHSV-1 или rHSV-2.
[0041] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV. В некоторых вариантах осуществления ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши. В некоторых вариантах осуществления ITR AAV представляют собой ITR AAV2. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор.
[0042] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид от аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0043] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0044] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
[0045] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид серотипа AAV из клад A-F. В некоторых вариантах осуществления вирусная частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота или AAV мыши, капсид rAAV2/HBoV1. В некоторых вариантах осуществления ITR и капсид вирусной частицы rAAV получены из AAV одного и того же серотипа. В некоторых вариантах осуществления ITR и капсид вирусных частиц rAAV получены из разных серотипов AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV2.
[0046] В некоторых вариантах осуществления вышеприведенных способов композиция представляет собой фармацевтическую композицию.
[0047] В некоторых аспектах настоящее изобретение предусматривает применение композиции согласно любому из вышеприведенных вариантов осуществления для лечения нарушения, связанного с глубокой интронной мутацией в гене индивидуума. В некоторых аспектах настоящее изобретение предусматривает применение композиции согласно любому из вышеприведенных вариантов осуществления в изготовлении лекарственного препарата для лечения нарушения, связанного с глубокой интронной мутацией в гене индивидуума. В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1. В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляет собой глазное заболевание. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2. В некоторых вариантах осуществления индивидуум является млекопитающим. В некоторых вариантах осуществления млекопитающее является человеком. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера.
[0048] В некоторых вариантах осуществления вышеприведенных применений композицию составляют для введения в глаз индивидуума. В некоторых вариантах осуществления введение, для которого получен состав, представляет собой субретинальное или интравитреальное введение.
[0049] В некоторых вариантах осуществления вышеприведенных путей применения последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0050] В некоторых вариантах осуществления вышеприведенных путей применения глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0051] В некоторых аспектах настоящее изобретение предусматривает наборы, содержащие композицию согласно любому из вышеприведенных вариантов осуществления. В некоторых вариантах осуществления набор содержит композицию согласно любому из вышеприведенных вариантов осуществления для применения в любом из способов, описанных в данном документе. В некоторых вариантах осуществления набор дополнительно содержит инструкции для применения.
[0052] В некоторых аспектах настоящее изобретение предусматривает вирусные частицы, содержащие вирусный вектор, где вирусный вектор содержит нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию в гене индивидуума, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глубокая интронная мутация в гене индивидуума ассоциирована с афибриногенемией, синдромом Альпорта, амиотрофическим латеральным склерозом, атаксией-телеангиэктазией, аутосомно-рецессивным поликистозным заболеванием почек, синдромом Барта, бета-талассемией, врожденной афибриногенемией, синдромом врожденной катаракты, лицевого дисморфизма и невропатии, врожденным нарушением гликозилирования Ia типа, врожденным нарушением гликозилирования II типа, кистозным фиброзом, дефицитом дигидроптеридинредуктазы, заболеванием Фабри, семейным тромбоцитарным нарушением с предрасположенностью к острому миелогенному лейкозу, анемией Фанкони, синдромом Гительмана, нечувствительностью к гормону роста, атаксией Фридрейха, гемофилией A, наследственной мегалобластной анемией 1, синдромом Германски-Пудлака, гомоцистинурией, заболеванием мочи с запахом кленового сиропа, синдромом Марфана, дефицитом метионинсинтазы, метилмалоновой ацидемией, дефицитом митохондриального трифункционального белка, мукополисахаридозом II типа, доброкачественной врожденной миопатией, мышечной дистрофией, нейрофиброматозом I типа, заболеванием Ниманна-Пика типа С, глазным альбинизмом I типа, дефицитом орнитин-дельта-аминотрансферазы, предрасположенностью к системной красной волчанке, пропионовой ацидемией, рабдоидными опухолями, синдромом Шварца-Джампеля, синдромом Стиклера, системной красной волчанкой, туберозным склерозом, синдромом Вернера, Х-связанной гипериммуноглобулинемией М или Х-сцепленной гипофосфатемией. В некоторых вариантах осуществления глубокая интронная мутация в гене индивидуума представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0053] В некоторых вариантах осуществления вирусная частица применяется для лечения индивидуума, у которого имеется афибриногенемия, синдром Альпорта, амиотрофический латеральный склероз, атаксия-телеангиэктазия, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемия, врожденная афибриногенемия, синдром врожденной катаракты, лицевой дисморфизм и невропатия, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемия Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксия Фридрейха, гемофилия A, наследственная мегалобластная анемия 1, синдром Германски-Пудлака, гомоцистинурия, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновая ацидемия, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественная врожденная миопатия, мышечная дистрофия, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизмом I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовая ацидемия, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системная красная волчанка, туберозный склероз, синдром Вернера, Х-сцепленная гипериммуноглобулинемия М или Х-сцепленная гипофосфатемия.
[0054] В некоторых аспектах настоящее изобретение предусматривает вирусные частицы, содержащие вирусный вектор, где вирусный вектор содержит нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию в гене индивидуума, ассоциированную с глазным заболеванием или нарушением, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глазное заболевание или нарушение представляют собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2. В некоторых вариантах осуществления вирусная частица применяется для лечения врожденного амавроза Лебера, атрофии зрительного нерва, пигментного ретинита, ретинобластомы, заболевания Штаргардта, синдрома Ашера или Х-сцепленного пигментного ретинита.
[0055] В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0056] В некоторых вариантах осуществления вышеприведенных вирусных частиц глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0057] В некоторых вариантах осуществления вышеприведенных вирусных частиц Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes, белок Cas9 Staphylococcus aureus, белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку человека.
[0058] В некоторых вариантах осуществления вышеприведенных вирусных частиц система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS). В некоторых вариантах осуществления Cas-белок содержит один или несколько NLS. В некоторых вариантах осуществления NLS представляет собой С-концевую последовательность в большом T-антигене SV40. В некоторых вариантах осуществления NLS содержит последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27).
[0059] В некоторых вариантах осуществления вышеприведенных вирусных частиц содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0060] В некоторых вариантах осуществления вышеприведенных вирусных частиц нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0061] В некоторых вариантах осуществления вышеприведенных вирусных частиц нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых вариантах осуществления вектор находится в комплексе с системой доставки. В некоторых вариантах осуществления вектор находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты клетками.
[0062] В некоторых вариантах осуществления вышеприведенных вирусных частиц вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV). В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид от аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0063] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0064] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вектор на основе rHSV получен из rHSV-1 или rHSV-2. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
[0065] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV. В некоторых вариантах осуществления ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши. В некоторых вариантах осуществления ITR AAV представляют собой ITR AAV2. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид серотипа AAV из клад A-F. В некоторых вариантах осуществления вирусная частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота или AAV мыши, капсид rAAV2/HBoV1. В некоторых вариантах осуществления ITR и капсид вирусной частицы rAAV получены из AAV одного и того же серотипа. В некоторых вариантах осуществления ITR и капсид вирусных частиц rAAV получены из разных серотипов AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV2.
[0066] В некоторых вариантах осуществления вышеуказанных вирусных частиц вирусная частица находится в фармацевтической композиции.
[0067] В некоторых аспектах настоящее изобретение предусматривает способы получения in vitro модели глазного заболевания, связанного с глубокой интронной мутацией в гене, включающие a) введение в эукариотические клетки нуклеиновой кислоты, кодирующей систему CRISPR-Cas, где система CRISPR-Cas содержит i) единую направляющую РНК, которая гибридизуется с противоположной нитью последовательности ДНК-мишени интрона в гене, ii) нуклеотидную последовательность, кодирующую Cas-белок, iii) однонитевой олигонуклеотид, содержащий матрицу для репарации, направляемой гомологией (HDR), содержащую гомологичные "плечи", фланкирующие требуемую интронную мутацию, и прилегающий к протоспейсеру мотив (PAM); и b) выделение клеток, которые содержат мутацию, включенную в ген.
[0068] В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0069] В некоторых вариантах осуществления PAM содержит мутацию, позволяющую избежать расщепления однонитевого олигонуклеотида экспрессированным Cas-белком в клетках.
[0070] В некоторых вариантах осуществления Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes, белок Cas9 Staphylococcus aureus, белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотических клетках. В некоторых вариантах осуществления эукариотические клетки представляют собой клетки млекопитающего. В некоторых вариантах осуществления эукариотические клетки представляют собой клетки человека. В некоторых вариантах осуществления эукариотические клетки представляют собой клетки глаза. В некоторых вариантах осуществления клетки глаза представляют собой клетки сетчатки.
[0071] В некоторых вариантах осуществления система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS). В некоторых вариантах осуществления Cas-белок содержит один или несколько NLS. В некоторых вариантах осуществления NLS представляет собой С-концевую последовательность в большом T-антигене SV40. В некоторых вариантах осуществления NLS содержит последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27).
[0072] В некоторых вариантах осуществления содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0073] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0074] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из единой направляющей РНК, Cas-белка или однонитевого олигонуклеотида, расположена на одном и том же или на разных векторах системы.
[0075] В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2. В некоторых вариантах осуществления вирусная частица применяется для лечения врожденного амавроза Лебера, атрофии зрительного нерва, пигментного ретинита, ретинобластомы, заболевания Штаргардта, синдрома Ашера или Х-сцепленного пигментного ретинита.
[0076] В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0077] В некоторых аспектах настоящее изобретение предусматривает способ расщепления нуклеиновой кислоты-мишени в клетке, включающий доставку в клетку эффективного количества композиции, содержащей: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых аспектах настоящее изобретение предусматривает способ лечения заболевания или нарушения, связанных с мутацией в нуклеиновой кислоте индивидуума, предусматривающий введение индивидууму терапевтически эффективного количества композиции, содержащей: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых аспектах настоящее изобретение предусматривает способ лечения глазного заболевания или нарушения, связанных с мутацией в нуклеиновой кислоте индивидуума, предусматривающий введение индивидууму терапевтически эффективного количества композиции, содержащей: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК, где Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию белка Cas по сравнению с экспрессией белка Cas до расщепления кассеты экспрессии Cas.
[0078] В некоторых вариантах осуществления вышеприведенных способов первая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК и целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления вторая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК и целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления первая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК, и вторая направляющая РНК гибридизуется с целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит последовательность полиаденилирования (polyA), функционально связанную с нуклеотидной последовательностью, кодирующей Cas-белок. В некоторых вариантах осуществления последовательность polyA представляет собой последовательность polyA из SV40. В некоторых вариантах осуществления расщепление целевого сайта первой или второй направляющей РНК Cas-белком разрывает функциональную связь между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA. В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, функционально связана с нуклеотидной последовательностью, кодирующей один или несколько сигналов внутриядерной локализации (NLS), так что Cas-белок, экспрессируемый с кассеты экспрессии Cas, слит внутри рамки считывания с одним или несколькими NLS. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая один или несколько NLS, находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью полиаденилирования (polyA). В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей один или несколько NLS, и последовательностью polyA. В некоторых вариантах осуществления один или несколько NLS содержат С-концевую последовательность большого T-антигена SV40. В некоторых вариантах осуществления один или несколько NLS содержат последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая систему CRISPR-Cas и/или кассету экспрессии Cas, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, функционально связана с промотором. В некоторых вариантах осуществления расщепление целевого сайта первой или второй направляющей РНК Cas-белком разрывает функциональную связь между регуляторным контрольным элементом и нуклеотидной последовательностью, кодирующей Cas-белок. В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между промотором и нуклеотидной последовательностью, кодирующей Cas-белок.
[0079] В некоторых вариантах осуществления вышеприведенных способов кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК, и где целевой сайт второй направляющей РНК примыкает к прилегающему к протоспейсеру мотиву (PAM), специфичному для Cas-белка; где расщепление целевого сайта первой направляющей РНК Cas-белком разрывает функциональную связь между регуляторным контрольным элементом и нуклеотидной последовательностью, кодирующей Cas-белок; где расщепление целевого сайта второй направляющей РНК Cas-белком разрывает функциональную связь между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA; и где после экспрессии Cas-белка и расщепления последовательностей ДНК-мишени Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК; где целевой сайт первой направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и промотором, функционально связанным с нуклеотидной последовательностью, кодирующей Cas-белок; где целевой сайт второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA, функционально связанной с нуклеотидной последовательностью, кодирующей Cas-белок; и где Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas.
[0080] В некоторых вариантах осуществления вышеприведенных способов заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0081] В некоторых аспектах настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, которые содержат нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержащие сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2.
[0082] В некоторых вариантах осуществления вышеприведенных вариантов осуществления глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0083] В некоторых вариантах осуществления вышеприведенных вариантов осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0084] В некоторых вариантах осуществления вышеприведенных вариантов осуществления Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes (SEQ ID NO:40), белок Cas9 Staphylococcus aureus (SEQ ID NO: 55), белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку человека.
[0085] В некоторых вариантах осуществления вышеприведенных вариантов осуществления содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0086] В некоторых вариантах осуществления вышеприведенных вариантов осуществления система CRISPR-Cas (например, первая направляющая РНК, вторая направляющая РНК и Cas-белок) находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты и/или белка клетками.
[0087] В некоторых вариантах осуществления вышеприведенных вариантов осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0088] В некоторых вариантах осуществления вышеприведенных вариантов осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых вариантах осуществления вектор находится в комплексе с системой доставки. В некоторых вариантах осуществления вектор находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты клетками.
[0089] В некоторых вариантах осуществления вышеприведенных вариантов осуществления вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV). В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5.
[0090] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0091] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вектор на основе rHSV получен из rHSV-1 или rHSV-2.
[0092] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV. В некоторых вариантах осуществления ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши. В некоторых вариантах осуществления ITR AAV представляют собой ITR AAV2. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор.
[0093] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид от аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0094] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0095] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
[0096] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид серотипа AAV из клад A-F. В некоторых вариантах осуществления вирусная частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота или AAV мыши, капсид rAAV2/HBoV1. В некоторых вариантах осуществления ITR и капсид вирусной частицы rAAV получены из AAV одного и того же серотипа. В некоторых вариантах осуществления ITR и капсид вирусных частиц rAAV получены из разных серотипов AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV2.
[0097] В некоторых аспектах настоящее изобретение предусматривает композицию для расщепления нуклеиновой кислоты-мишени в клетке, содержащую: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых аспектах настоящее изобретение предусматривает композиции для лечения заболевания или нарушения, связанных с мутацией в нуклеиновой кислоте индивидуума, содержащие: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых аспектах настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с мутацией в нуклеиновой кислоте индивидуума, содержащие: a) нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, и b) кассету экспрессии Cas, содержащую i) нуклеотидную последовательность, кодирующую Cas-белок, и ii) целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом первой направляющей РНК; где Cas-белок экспрессируется из кассеты экспрессии Cas; где Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию; и где Cas-белок расщепляет кассету экспрессии Cas по целевому сайту первой направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК, где Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию белка Cas по сравнению с экспрессией белка Cas до расщепления кассеты экспрессии Cas.
[0098] В некоторых вариантах осуществления вышеприведенных композиций первая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК и целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления вторая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК и целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления первая направляющая РНК гибридизуется с целевым сайтом первой направляющей РНК, и вторая направляющая РНК гибридизуется с целевым сайтом второй направляющей РНК. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит последовательность полиаденилирования (polyA), функционально связанную с нуклеотидной последовательностью, кодирующей Cas-белок. В некоторых вариантах осуществления последовательность polyA представляет собой последовательность polyA из SV40. В некоторых вариантах осуществления расщепление целевого сайта первой или второй направляющей РНК Cas-белком разрывает функциональную связь между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA. В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, функционально связана с нуклеотидной последовательностью, кодирующей один или несколько сигналов внутриядерной локализации (NLS), так что Cas-белок, экспрессируемый с кассеты экспрессии Cas, слит внутри рамки считывания с одним или несколькими NLS. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая один или несколько NLS, находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью полиаденилирования (polyA). В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей один или несколько NLS, и последовательностью polyA. В некоторых вариантах осуществления один или несколько NLS содержат С-концевую последовательность большого T-антигена SV40. В некоторых вариантах осуществления один или несколько NLS содержат последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая систему CRISPR-Cas и/или кассету экспрессии Cas, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, функционально связана с промотором. В некоторых вариантах осуществления расщепление целевого сайта первой или второй направляющей РНК Cas-белком разрывает функциональную связь между регуляторным контрольным элементом и нуклеотидной последовательностью, кодирующей Cas-белок. В некоторых вариантах осуществления целевой сайт первой или второй направляющей РНК находится между промотором и нуклеотидной последовательностью, кодирующей Cas-белок.
[0099] В некоторых вариантах осуществления вышеприведенных композиций кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК, и где целевой сайт второй направляющей РНК примыкает к прилегающему к протоспейсеру мотиву (PAM), специфичному для Cas-белка; где расщепление целевого сайта первой направляющей РНК Cas-белком разрывает функциональную связь между регуляторным контрольным элементом и нуклеотидной последовательностью, кодирующей Cas-белок; где расщепление целевого сайта второй направляющей РНК Cas-белком разрывает функциональную связь между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA; и где после экспрессии Cas-белка и расщепления последовательностей ДНК-мишени Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas. В некоторых вариантах осуществления кассета экспрессии Cas дополнительно содержит iii) целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК; где целевой сайт первой направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и промотором, функционально связанным с нуклеотидной последовательностью, кодирующей Cas-белок; где целевой сайт второй направляющей РНК находится между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью polyA, функционально связанной с нуклеотидной последовательностью, кодирующей Cas-белок; и где Cas-белок расщепляет кассету экспрессии Cas по целевым сайтам первой и второй направляющей РНК, уменьшая таким образом экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas.
[0100] В некоторых вариантах осуществления вышеприведенных композиций заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0101] В некоторых аспектах настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, которые содержат нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления настоящее изобретение предусматривает композиции для лечения глазного заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержащие сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2.
[0102] В некоторых вариантах осуществления вышеприведенных композиций глазное заболевание представляет собой врожденный амавроз Лебера. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В некоторых вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления первая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления вторая направляющая РНК кодируется с помощью ДНК, содержащей последовательность под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека. В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0103] В некоторых вариантах осуществления вышеприведенных композиций глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления посредством глубокой интронной мутации в ген вводят сайт донора сплайсинга или сайт акцептора сплайсинга.
[0104] В некоторых вариантах осуществления вышеприведенных композиций Cas-белок представляет собой белок Cas9. В некоторых вариантах осуществления белок Cas9 представляет собой белок Cas9 Streptococcus pyogenes (SEQ ID NO:40), белок Cas9 Staphylococcus aureus (SEQ ID NO: 55), белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola. В некоторых вариантах осуществления белок Cas 9 кодон-оптимизирован для экспрессии в эукариотической клетке. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку человека.
[0105] В некоторых вариантах осуществления вышеприведенных композиций содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность tracr содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25.
[0106] В некоторых вариантах осуществления вышеприведенных композиций система CRISPR-Cas (например, первая направляющая РНК, вторая направляющая РНК и Cas-белок) находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты и/или белка клетками.
[0107] В некоторых вариантах осуществления вышеприведенных композиций нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и Cas-белок, экспрессируется в эукариотических клетках. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую направляющую РНК, вторую направляющую РНК и/или Cas-белок, функционально связана с одним или несколькими регуляторными контрольными элементами. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, 7SK или H1. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. В некоторых вариантах осуществления промотор РНК-полимеразы II представляет собой немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40), промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор EF1α, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотор гена родопсина, промотор гена конопсина, промотор гена бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0108] В некоторых вариантах осуществления вышеприведенных композиций нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления вектор представляет собой плазмиду. В некоторых вариантах осуществления вектор находится в комплексе с системой доставки. В некоторых вариантах осуществления вектор находится в комплексе с липидом, катионным липидом, липосомой, поликатионом или средством, которое усиливает поглощение нуклеиновой кислоты клетками.
[0109] В некоторых вариантах осуществления вышеприведенных композиций вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV). В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5.
[0110] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0111] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вектор на основе rHSV получен из rHSV-1 или rHSV-2.
[0112] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV. В некоторых вариантах осуществления ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши. В некоторых вариантах осуществления ITR AAV представляют собой ITR AAV2. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор.
[0113] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид от аденовируса серотипов 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0114] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0115] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
[0116] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид серотипа AAV из клад A-F. В некоторых вариантах осуществления вирусная частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота или AAV мыши, капсид rAAV2/HBoV1. В некоторых вариантах осуществления ITR и капсид вирусной частицы rAAV получены из AAV одного и того же серотипа. В некоторых вариантах осуществления ITR и капсид вирусных частиц rAAV получены из разных серотипов AAV. В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном. В некоторых вариантах осуществления вектор на основе rAAV содержит ITR AAV2.
[0117] Все литературные источники, цитируемые в данном документе, в том числе патентные заявки и публикации, включены с помощью ссылки во всей своей полноте.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0118] ФИГ. 1 представляет собой схематическую диаграмму интронной мутации c.2291+1655 A>G в гене CEP290. Мутация A>G происходит в +5 положении 5'-конца сайта сплайсинга криптического экзона, расположенного в интроне 26. Экзон-интронная структура области от экзона 26 до экзона 27 CEP290 представлена прямоугольниками и линиями, соответственно, вместе со схемой сплайсинга для дикого типа (сплошная линия) и мутантных (пунктирная линия) транскриптов. Мутированный нуклеотид обозначен жирным шрифтом и подчеркиванием, и символизируется закрашенной звездой. Последовательности 5'-конца сайта сплайсинга криптического экзона представлены вместе с их оценками устойчивости сайта сплайсинга, рассчитанными с помощью BDGP: Splice Site Prediction by Neural Network (прогнозирование сайта сплайсинга с использованием нейронной сети) (см. например www.fruitfly.org/seq_tools/splice.html and Reese, M.G. et al. (1997) J. Comput. Biol. 4:311-323). Нуклеотиды экзона показаны заглавными буквами. Размеры экзона и интронов даны в виде примечания под фигурой.
[0119] ФИГ. 2 представляет собой схему, показывающую стратегию делеции интронной области, фланкирующей мутацию c.2991+1655 A>G CEP290. Вышерасположенная sgRNA направляет первое расщепление Cas9, располагающееся выше интронной мутации, и нижерасположенная sgRNA направляет второе расщепление Cas9, расположенное ниже мутации. Обратите внимание, что вышерасположенный целевой локус может располагаться либо выше криптического экзона, либо внутри криптического экзона. Два конца расщепления непосредственно лигируют посредством процесса негомологичного соединения концов (NHEJ) с удалением фрагмента интрона, фланкирующего мутацию c.2991+1655 A>G. Интрон 26 дополнительно удаляют при сплайсинге РНК в ходе процессинга mRNA.
[0120] На ФИГ. 3A-3C показаны уровни экспрессии mRNA (ФИГ. 3A и 3B) и белка (ФИГ. 3C) в различных линиях клеток. На ФИГ. 3A и 3B показаны графики базальных уровней экспрессии mRNA дикого типа (ФИГ. 3A) и мутантной (ФИГ. 3B) в клетках дикого типа (белые столбики), гетерозиготных по хромосоме, несущей мутации c.2991+1655A>G и c.2991+1666C>G (серые столбики), и клетках, гомозиготных по c.2991+1655A>G и c.2991+1666C>G (мутантные клетки; черные столбики), как определено с помощью RT-qPCR. Данные представлены как среднее ± стандартное отклонение образцов из трех независимых трансфекций (n=3). Сравнения проводили с использованием однофакторного дисперсионного анализа ANOVA с последующим ретроспективным анализом с HSD-критерием Тьюки. *=p<0,05, **=p<0,01, ***=p<0,001. ФИГ. 3C представляет собой иммуноблот лизатов, полученных из клеток дикого типа (WT), гетерозиготных клеток (Het) и мутантных клеток (MT). На мембране исследовали CEP290 (верх) и β-актин в качестве контроля нанесения (низ).
[0121] На ФИГ. 4A и 4B показана эффективность целенаправленной делеции с парными sgRNA и SpCas9, определенная с помощью ПЦР (ФИГ. 4A) и секвенирования нового поколения (ФИГ. 4B). На ФИГ. 4A использованы праймеры за пределами ожидаемых областей делеции. Верхние полосы представляют собой продукты ПЦР, амплифицированные из интрона 26 CEP290 дикого типа, тогда как нижние полосы представляют собой продукты ПЦР, амплифицированные из аллеля CEP290 после ожидаемых геномных делеций (отмечено соответственно как "Wt" и "Trunc"). M, ДНК-маркер 1 т.п.н. На ФИГ. 4B показан процент ДНК дикого типа и усеченной в мутантных клетках, трансфицированных парными sgRNA и SpCas9, определенный с помощью секвенирования нового поколения (NGS).
[0122] На ФИГ. 5A-5C показано восстановление экспрессии CEP290 с использованием парных sgRNA и SpCas9. На ФИГ. 5A и 5B показаны уровни экспрессии mRNA дикого типа (ФИГ. 5A) и мутантной (ФИГ. 5B) в клетках дикого типа (белые столбики), гетерозиготных (серые столбики) и мутантных (черные столбики), трансфицированных парными sgRNA и SpCas9, измеренные с помощью RT-qPCR. Данные представлены как среднее ± стандартное отклонение образцов из двух независимых трансфекций (n=2). Сравнения проводили с использованием однофакторного дисперсионного анализа ANOVA с последующим ретроспективным анализом с HSD-критерием Тьюки. *=p<0,05, **=p<0,01. ФИГ. 5C представляет собой иммуноблот лизатов, полученных из мутантных клеток, трансфицированных парными sgRNA и SpCas9. На мембране исследовали CEP290 (верх) и β-актин в качестве контроля нанесения (низ).
[0123] На ФИГ. 6A-6E показана самоограничивающаяся система CRISPR-SpCas9. ФИГ. 6A представляет собой схематическую диаграмму вектора pAAV-SpCas9, используемого в самоограничивающейся системе CRISPR-SpCas9. Последовательность распознавания для нуклеазы SpCas9 (целевая последовательность sgRNA плюс мотив PAM) вводят в сайт вставки 1 (между промотором minCMV и SpCas9) и/или сайт вставки 2 (между SpCas9-NLS и pA SV40). NLS, сигнал внутриядерной локализации. pA SV40, сигнал полиаденилирования вируса обезьян 40. ФИГ. 6B представляет собой иммуноблот лизатов, полученных из мутантных клеток, трансфицированных первой упаковочной плазмидой AAV, экспрессирующей пару sgRNA U1D3, и второй упаковочной плазмидой AAV, экспрессирующей SpCas9. Плазмида SpCas9 содержит последовательность распознавания sgRNA U1 (U1T) и/или последовательность распознавания sgRNA D3 (D3T) в двух сайтах вставки. В качестве контроля в данном эксперименте служили клетки-мутанты, трансфицированные только плазмидой U1D3. В качестве контроля нанесения (низ) на мембране исследовали SpCas9 (верх) и β-актин. На ФИГ. 6C показана целенаправленная делеция после того, как мутантные клетки трансфицировали парой sgRNA U1D3 и самоограничивающимся SpCas9, как определено с помощью ПЦР. Верхние полосы представляют собой продукты ПЦР, амплифицированные из интрона 26 CEP290 дикого типа, тогда как нижние полосы представляют собой продукты ПЦР, амплифицированные из аллеля CEP290 после геномной делеции, направляемой sgRNA U1 и D3. M, ДНК-маркер 1 т.п.н. На ФИГ. 6D и 6E показаны уровни экспрессии mRNA дикого типа (ФИГ. 6D) и мутантной (ФИГ. 6E) в мутантных клетках, трансфицированных парой sgRNA U1D3 и самоограничивающимся SpCas9, измеренные с помощью RT-qPCR. Данные представлены как среднее ± стандартное отклонение образцов из трех независимых трансфекций (n=3). Сравнения проводили с использованием однофакторного дисперсионного анализа ANOVA с последующим ретроспективным анализом с HSD-критерием Тьюки. *=p<0,05, **=p<0,01, ***=p<0,001 по сравнению с клетками, трансфицированными только парой sgRNA U1D3.
[0124] На ФИГ. 7A и 7B показана делеция области в интроне 25 гена Cep290 в сетчатке мыши с помощью системы на основе двух AAV. ФИГ. 7A представляет собой схематическую диаграмму двойных AAV, используемых в субретинальной инъекции. На ФИГ. 7B показана целенаправленная делеция с использованием (1) AAV5-RK-EGFP (контроль) или AAV5-U11D11 sgRNA-пары RK-EGFP и (2) AAV5-SpCas9, как определено с помощью ПЦР. Верхние полосы представляют собой продукты ПЦР, амплифицированные из интрона 25 мышиного Cep290 дикого типа, тогда как нижние полосы представляют собой продукты ПЦР, амплифицированные из аллеля Cep290 после геномной делеции, направляемой sgRNA U11 и D11. M, ДНК-маркер 1 т.п.н.
[0125] На ФИГ. 8A-8C показана целенаправленная делеция с использованием Cas9 S. aureus (SaCas9) и SpCas9, определенная с помощью ПЦР (ФИГ. 8A) и с помощью RT-qPCR (ФИГ. 8B и 8C). Мутантные клетки трансфицировали парами sgRNA, спаренными вместе с SaCas9 либо SpCas9. Обратите внимание, что парные sgRNA и SaCas9 находятся в одной упаковочной плазмиде AAV, тогда как парные sgRNA и SpCas9 находятся в двух отдельных упаковочных плазмидах AAV. На ФИГ. 8A верхние полосы представляют собой продукты ПЦР, амплифицированные из интрона 26 CEP290 дикого типа, тогда как нижние полосы представляют собой продукты ПЦР, амплифицированные из аллеля CEP290 после ожидаемых геномных делеций (отмечено соответственно как "Wt" и "Trunc"). M, ДНК-маркер 1 т.п.н. На ФИГ. 8A и 8B показаны уровни экспрессии mRNA дикого типа (ФИГ. 8А) и мутантной (ФИГ. 8В) в мутантных клетках, трансфицированных sgRNA, спаренными вместе с SaCas9 (белые столбики) либо SpCas9 (серые столбики), как измерено с помощью RT-qPCR. Данные представлены как среднее ± стандартное отклонение образцов из трех независимых трансфекций (n=3). Сравнения проводили с использованием однофакторного дисперсионного анализа ANOVA с последующим ретроспективным анализом с HSD-критерием Тьюки. *=p<0,05, **=p<0,01, ***=p<0,001. #=p<0,05 по сравнению с мутантными клетками, трансфицированными только SaCas9.
ПОДРОБНОЕ ОПИСАНИЕ
[0126] Настоящее изобретение предусматривает композиции, способы и вирусные частицы для редактирования глубоких интронных мутаций. В некоторых вариантах осуществления композиция для лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержит сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В других вариантах осуществления композиция для лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, содержит нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию, и b) нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию.
[0127] В некоторых аспектах настоящее изобретение предусматривает композиции, способы и вирусные частицы для лечения глазных заболеваний. Как описано выше, наиболее частой генетической причиной LCA является глубокая интронная мутация c.2991+1655A>G в интроне 26 гена CEP290, которая создает криптический сайт донора сплайсинга, приводящий к включению аберрантного экзона, содержащего преждевременный стоп-кодон (p.C998X), в mRNA CEP290 (ФИГ. 1). Авторы настоящего изобретения разработали простой и эффективный способ лечения пациентов с LCA, у которых имеется интронная мутация c.2991+1655 A>G в гене CEP290, посредством целенаправленной делеции геномной ДНК в клетках человека с помощью пары единых направляющих РНК (sgRNA), а также системы коротких палиндромных повторов, регулярно расположенных группами (CRISPR), и CRISPR-ассоциированных белков (Cas) (ФИГ. 2). Данный подход эффективно и необратимо удаляет интронную мутацию c.2991+1655 A>G, предотвращая сплайсинг криптического экзона, вставленного в mRNA CEP290, и оставляя тем самым эндогенные генетические регуляторные элементы интактными.
I. Общие методики
[0128] Методики и процедуры, описанные или упоминаемые в данном документе, как правило, широко распространены и часто используются специалистами в данной области техники с использованием традиционной методологии, как например широко используемые методики, описанные в Molecular Cloning: A Laboratory Manual (Sambrook et al., 4th ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 2012); Current Protocols in Molecular Biology (F.M. Ausubel, et al. eds., 2003); серии Methods in Enzymology (Academic Press, Inc.); PCR 2: A Practical Approach (M.J. MacPherson, B.D. Hames and G.R. Taylor eds., 1995); Antibodies, A Laboratory Manual (Harlow and Lane, eds., 1988); Culture of Animal Cells: A Manual of Basic Technique and Specialized Applications (R.I. Freshney, 6th ed., J. Wiley and Sons, 2010); Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., Academic Press, 1998); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, Plenum Press, 1998); Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., J. Wiley and Sons, 1993-8); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds., 1996); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Ausubel et al., eds., J. Wiley and Sons, 2002); Immunobiology (C.A. Janeway et al., 2004); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane, Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995) и Cancer: Principles and Practice of Oncology (V.T. DeVita et al., eds., J.B. Lippincott Company, 2011).
II. Определения
[0129] Используемый в данном документе термин "CRISPR-Cas" относится к двухкомпонентному рибонуклеопротеиновому комплексу направляющей РНК и эндонуклеазы Cas. CRISPR относится к системе коротких палиндромных повторов, регулярно расположенных группами, II типа. Хотя CRISPR была открыта как адаптивная система защиты, которая дает возможность бактериям и археям обнаруживать и выключать чужеродные нуклеиновые кислоты (например, из вирусов или плазмид), она была адаптирована для применения в различных типах клеток, чтобы обеспечить возможность редактирования полинуклеотидов специфическим для последовательности образом (см., например, Jinek, M. et al. (2012) Science 337:816-821 и Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308). В системах II типа направляющая РНК взаимодействует с Cas и направляет нуклеазную активность Cas-фермента на последовательности ДНК-мишени, идентичные направляющим последовательностям в направляющей РНК. Основание направляющей РНК спаривается с противоположной нитью последовательности-мишени. Затем нуклеазная активность Cas создает двухнитевой разрыв в ДНК-мишени. В некоторых вариантах осуществления Cas-белок представляет собой белок Cas9.
[0130] Термин "единая направляющая РНК CRISPR-Cas", используемый в данном документе, (термины "единая направляющая РНК" и "sgRNA" могут использоваться в данном документе взаимозаменяемо) относится с единому виду РНК, способному направлять Cas-опосредованное расщепление ДНК-мишени. В некоторых вариантах осуществления единая направляющая РНК может содержать последовательности, необходимые для нуклеазной активности Cas (например, Cas9), и направляющую последовательность, идентичную ДНК-мишени, представляющей интерес.
[0131] Термины "химерная РНК", "химерная направляющая РНК", "направляющая РНК", "единая направляющая РНК" и "синтетическая направляющая РНК" могут использоваться в данном документе взаимозаменяемо и относятся к полинуклеотидной последовательности, содержащей направляющую последовательность, последовательность tracr и последовательность, сопряженную с tracr. Термин "направляющая последовательность", используемый в данном документе, относится к последовательности длиной приблизительно 20 п.н. в направляющей РНК, которая указывает целевой сайт и может использоваться взаимозаменяемо с терминами "направляющая", или "спейсер", или "протоспейсер". Термин "последовательность, сопряженная с tracr" также можно использовать взаимозаменяемо с термином "прямой повтор(-ы)".
[0132] Термин "направляющая последовательность sgRNA", используемый в данном документе, может относиться к нуклеотидной последовательности sgRNA, которая связывается с противоположными нитями ДНК-мишени и направляет нуклеазную активность Cas (например, Cas9) на этот локус. В некоторых вариантах осуществления направляющая последовательность sgRNA идентична последовательности-мишени. Полная идентичность не является необходимой при условии, что существует достаточное сходство, чтобы вызвать гибридизацию и способствовать формированию комплекса CRISPR. Направляющая последовательность может содержать любой полинуклеотид, такой как полинуклеотиды ДНК или РНК.
[0133] Термин полипептид "Cas", используемый в данном документе, представляет собой полипептид, который функционирует как нуклеаза при образовании комплекса с направляющей РНК, например sgRNA. В некоторых вариантах осуществления полипептид Cas представляет собой полипептид Cas9 (CRISPR-ассоциированный 9, также известный как Csn1). При связывании с направляющей crRNA:tracrRNA или единой направляющей РНК полипептиды Cas (например, Cas9) способны расщеплять ДНК-мишень в последовательности, идентичной направляющей последовательности sgRNA и прилегающей к мотиву PAM. В отличие от других полипептидов, Cas-полипептиды Cas9 характерны для систем CRISPR-Cas II типа (описание Cas-белков других систем CRISPR-Cas см. в Makarova, K.S., et al. (2011) Nat. Rev. Microbiol. 9(6):467-77). Термин "Cas", используемый в данном документе, может относиться к рибонуклеопротеиновому комплексу с sgRNA или полипептидному компоненту комплекса, если не указано иное.
[0134] Термин "CRISPR-РНК (crRNA)", используемый в данном документе, относится к РНК, содержащей направляющую последовательность, используемую системой CRISPR-Cas для направления расщепления последовательности ДНК-мишени. Термин "трансактивирующая crRNA (tracrRNA)", используемый в данном документе, относится к РНК, содержащей последовательность, которая образует структуру, необходимую для эффекторного комплекса CRISPR-Cas, который опосредует расщепление ДНК. В эндогенных системах CRISPR-Cas II типа у бактерий и архей эффекторный комплекс CRISPR-Cas включает в себя Cas-белок (например, белок Cas9) в комплексе с двумя полирибонуклеотидными молекулами: crRNA и tracrRNA. При этом crRNA содержит направляющую последовательность из ~20 нуклеотидов, которая опосредует распознавание мишени, и последовательность, которая образует двойную спираль с tracrRNA. Двойная спираль crRNA:tracrRNA связывает Cas-белок и необходима для работы эффекторного комплекса CRISPR-Cas. В некоторых вариантах осуществления функции crRNA и tracrRNA могут быть выполнены одной РНК (единой направляющей РНК или sgRNA), которая содержит как последовательность, которая опосредует распознавание мишени, так и последовательность, которая образует структуру, требуемую для эффекторного комплекса CRISPR-Cas.
[0135] Термин "глубокая интронная мутация", используемый в данном документе, относится к мутации внутри интронной последовательности в области вне последовательностей акцептора сплайсинга и донора сплайсинга дикого типа. В некоторых случаях глубокая интронная мутация может приводить к изменению сплайсинга ассоциированного гена, например включению интронной последовательности в зрелую mRNA. В неограничивающих примерах глубокая интронная мутация находится более чем на приблизительно 100 п.н. ниже (т. e. 3'-конец) экзона, более чем на приблизительно 100 п.н. выше (т. e. 5'-конец) экзона или более чем на приблизительно 100 п.н. ниже первого экзона и более чем на приблизительно 100 п.н. выше второго экзона.
[0136] Термин "врожденный амавроз Лебера (LCA)", используемый в данном документе, относится к группе нарушений с ранним началом, характеризующихся потерей зрения, дисфункцией сетчатки и нистагмом. В LCA вовлечены различные мутации, но обычно LCA наследуется как аутосомно-рецессивное нарушение. Более подробное описание, а также иллюстративные гены и локусы для заболевания LCA, см. например в OMIM Entry 204000.
[0137] Термин "CEP290", используемый в данном документе, относится к генам, кодирующим центросомальный белок, вовлеченный в цилиогенез, также известный как MKS4, CT87, POC3, rd16, BBS14, LCA10, JBTS5, NPHP6, SLSN6 и 3H11Ag. Мутации в CEP290 вовлечены в LCA. Одним из примеров такой мутации является мутация c.2991+1655A>G, которая вводит криптический сайт донора сплайсинга, приводя к включению аберрантного экзона с преждевременным стоп-кодоном. Иллюстративные последовательности генов и белков человека см., например, соответственно в NCBI Gene ID № 80184 и UniProt ID № O15078. Другие примеры генов CEP290 включают без ограничения CEP290 мыши (например, NCBI Gene ID № 216274), CEP290 крысы (например, NCBI Gene ID № 314787), CEP290 макака-резуса (например, NCBI Gene ID № 708286), CEP290 данио-рерио (например, NCBI Gene ID № 560588), CEP290 собаки (например, NCBI Gene ID № 482591), CEP290 шимпанзе (например, NCBI Gene ID № 452113), CEP290 кошки (например, NCBI Gene ID № 100113471), CEP290 курицы (например, NCBI Gene ID № 417887) и CEP290 коровы (например, NCBI Gene ID № 282707). В некоторых вариантах осуществления ген CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0138] Термин "вектор", используемый в данном документе, относится к рекомбинантной плазмиде или вирусу, содержащим нуклеиновую кислоту, которую необходимо доставить в клетку-хозяина либо in vitro, либо in vivo.
[0139] Термины "полинуклеотид" или "нуклеиновая кислота", используемые в данном документе, относятся к полимерной форме нуклеотидов любой длины, либо рибонуклеотидов, либо дезоксирибонуклеотидов. Таким образом, данный термин включает без ограничения одно-, двух- или многонитевые ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК или полимер, содержащий пуриновые и пиримидиновые основания или другие природные, химически или биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания. Остов полинуклеотида может содержать сахара и фосфатные группы (которые обычно могут обнаруживаться в РНК или ДНК) или модифицированные либо замещенные сахарные или фосфатные группы. Как альтернатива, остов полинуклеотида может включать в себя полимер из синтетических субъединиц, таких как фосфорамидаты, и таким образом может представлять собой олигодезоксинуклеозидный фосфорамидат (P-NH2) или смешанный олигомер фосфорамидата-сложного фосфодиэфира. Кроме того, двухнитевой полинуклеотид можно получить из однонитевого полинуклеотидного продукта химического синтеза либо путем синтеза комплементарной нити и гибридизации нитей в соответствующих условиях, либо путем синтеза комплементарной нити de novo с использованием ДНК-полимеразы с соответствующим праймером.
[0140] Термины "полипептид" и "белок" используются взаимозаменяемо для обозначения полимера из аминокислотных остатков и не ограничены минимальной длиной. Такие полимеры из аминокислотных остатков могут содержать природные или неприродные аминокислотные остатки, и включают без ограничения пептиды, олигопептиды, димеры, тримеры и мультимеры из аминокислотных остатков. Данным определением охватываются как полноразмерные белки, так и их фрагменты. Термины включают также постэкспрессионные модификации полипептида, например, гликозилирование, сиалирование, ацетилирование, фосфорилирование и т. п. Кроме того, применительно к целям настоящего изобретения термин "полипептид" относится к белку, нативная последовательность которого содержит модификации, такие как делеции, добавления и замены (как правило, консервативные по своей природе), при условии, что белок сохраняет требуемую активность. Эти модификации могут быть преднамеренными, как например полученными с помощью сайт-направленного мутагенеза, или могут быть случайными, как например возникшими вследствие мутаций у хозяев, которые продуцируют белки, или ошибок, обусловленных ПЦР-амплификацией.
[0141] Термин "рекомбинантный вирусный вектор" относится к рекомбинантному полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. е. последовательность нуклеиновой кислоты, не происходящую от вируса). В случае рекомбинантных векторов на основе AAV, рекомбинантная нуклеиновая кислота фланкирована по меньшей мере одной последовательностью инвертированного концевого повтора (ITR). В некоторых вариантах осуществления рекомбинантная нуклеиновая кислота фланкирована двумя ITR.
[0142] Термин "рекомбинантный вектор на основе AAV (вектор на основе rAAV)" относится к полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. e. последовательностей нуклеиновых кислот, не происходящих из AAV), которые фланкированы по меньшей мере одной последовательностью инвертированного концевого повтора (ITR). Такие векторы на основе rAAV могут реплицироваться и упаковываться в инфекционные вирусные частицы, если они присутствуют в клетке-хозяине, которая была инфицирована подходящим вирусом-помощником (или которая экспрессирует подходящие хелперные функциональные элементы) и которая экспрессирует продукты генов rep и cap AAV (т. e. белки Rep и Cap AAV). Если вектор на основе rAAV встроен в более крупный полинуклеотид (например, в хромосому или в другой вектор, такой как плазмида, применяемая для клонирования или трансфекции), то вектор на основе rAAV можно называть "провектором", который может быть "восстановлен" с помощью репликации и заключения в капсид в присутствии упаковывающих функциональных элементов и подходящих функциональных элементов помощника AAV. Вектор на основе rAAV может находиться в любой из множества форм, включающих без ограничения плазмиды, линейные искусственные хромосомы, образующие комплексы с липидами, инкапсулированные в липосомы и заключенные в капсид вирусной частицы, например частицы AAV. Вектор на основе rAAV может быть упакован в капсид вируса AAV с получением "частицы рекомбинантного аденоассоциированного вируса (частица rAAV)".
[0143] Термины "вирус rAAV" или "вирусная частица rAAV" относятся к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и заключенного в капсид генома вектора на основе rAAV.
[0144] Термин "рекомбинантный аденовирусный вектор" относится к полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. е. последовательность нуклеиновой кислоты, не происходящую из аденовируса), которые фланкированы по меньшей мере одной последовательностью инвертированного концевого повтора (ITR) аденовируса. В некоторых вариантах осуществления рекомбинантная нуклеиновая кислота фланкирована двумя последовательностями инвертированных концевых повторов (ITR). Такие векторы на основе рекомбинантных вирусов могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они находятся в клетке-хозяине, которая экспрессирует жизненно важные гены аденовируса, которые были подвержены делеции в геноме рекомбинантного вируса (например, гены E1, гены E2, гены E4 и т. д.). Если рекомбинантный вирусный вектор включен в более крупный полинуклеотид (например, в хромосому или в другой вектор, такой как плазмида, используемая для клонирования или трансфекции), то рекомбинантный вирусный вектор можно обозначить как "провектор", который может быть "восстановлен" посредством репликации и заключения в капсид в присутствии упаковывающих функциональных элементов аденовируса. Рекомбинантный вирусный вектор может находиться в любой из множества форм, в том числе без ограничения в форме плазмид, линейных искусственных хромосом, образующих комплексы с липидами, инкапсулированными в липосомах и заключенными в капсид в вирусной частице, например аденовирусной частице. Рекомбинантный вирусный вектор может быть упакован в вирусный капсид аденовируса с получением "рекомбинантной аденовирусной частицы".
[0145] Термин "вектор на основе рекомбинантного лентивируса" относится к полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. е. последовательность нуклеиновой кислоты, не происходящую из лентивируса), которые фланкированы по меньшей мере одной последовательностью концевого повтора лентивируса (LTR). В некоторых вариантах осуществления рекомбинантная нуклеиновая кислота фланкирована двумя последовательностями концевых повторов лентивируса (LTR). Такие векторы на основе рекомбинантных вирусов могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они находятся в клетке-хозяине, которая была инфицирована подходящими хелперными функциональными элементами. Вектор на основе рекомбинантного лентивируса может быть упакован в капсид лентивируса с получением "рекомбинантной лентивирусной частицы".
[0146] Термин "вектор на основе рекомбинантного вируса простого герпеса (вектор на основе рекомбинантного HSV)" относится к полинуклеотидному вектору, содержащему одну или несколько гетерологичных последовательностей (т. е. последовательность нуклеиновой кислоты, не происходящую из HSV), которые фланкированы последовательностями концевых повторов HSV. Такие векторы на основе рекомбинантных вирусов могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они находятся в клетке-хозяине, которая была инфицирована подходящими хелперными функциональными элементами. Если рекомбинантный вирусный вектор включен в более крупный полинуклеотид (например, в хромосому или в другой вектор, такой как плазмида, используемая для клонирования или трансфекции), то рекомбинантный вирусный вектор можно обозначить как "провектор", который может быть "восстановлен" посредством репликации и инкапсидирования в присутствии упаковывающих функциональных элементов HSV. Рекомбинантный вирусный вектор может находиться в любой из множества форм, включающих без ограничения плазмиды, линейные искусственные хромосомы, образующие комплексы с липидами, инкапсулированные в липосомы и заключенные в капсид вирусной частицы, например частицы HSV. Рекомбинантный вирусный вектор может быть упакован в капсид HSV с получением "частицы рекомбинантного вируса простого герпеса".
[0147] Термин "гетерологичный" означает полученный из объекта, генотипически отличающегося от остальной части объекта, с которым его сравнивают или в который его вводят или встраивают. Например, полинуклеотид, введенный с помощью методик генной инженерии в клетки другого типа, является гетерологичным полинуклеотидом (и при экспрессии может кодировать гетерологичный полипептид). Аналогично, клеточная последовательность (например, ген или его часть), встроенная в вирусный вектор, является гетерологичной нуклеотидной последовательностью по отношению к вектору.
[0148] Термин "трансген" относится к полинуклеотиду, вводимому в клетку и способному к транскрипции в РНК, и необязательно к трансляции и/или экспрессии в соответствующих условиях. В некоторых аспектах он придает требуемое свойство клетке, в которую он был введен, или иным образом приводит к требуемому терапевтическому или диагностическому эффекту. В другом аспекте он может транскрибироваться в молекулу, которая опосредует РНК-интерференцию, такую как miРНК, siРНК или shРНКA.
[0149] Термины "геномные частицы (gp)", "геномные эквиваленты" или "копии генома", используемые в отношении вирусного титра, относятся к числу вирионов, содержащих ДНК-геном рекомбинантного AAV, вне зависимости от инфекционности или функциональности. Число геномных частиц в конкретном векторном препарате можно определять с помощью процедур, таких как описанные в примерах данного документа или, например, в Clark et al. (1999) Hum. Gene Ther., 10:1031-1039; Veldwijk et al. (2002) Mol. Ther., 6:272-278.
[0150] Термин "геном вектора (vg)", используемый в данном документе, может относиться к одному или нескольким полинуклеотидам, содержащим набор полинуклеотидных последовательностей вектора, например вирусного вектора. Геном вектора может быть заключен в капсид вирусной частицы. В зависимости от конкретного вирусного вектора, геном вектора может содержать однонитевую ДНК, двухнитевую ДНК, или однонитевую РНК, или двухнитевую РНК. Геном вектора может содержать эндогенные последовательности, ассоциированные с конкретным вирусным вектором, и/или любые гетерологичные последовательности, встроенные в конкретный вирусный вектор посредством рекомбинантных методик. Например, геном рекомбинантного вектора на основе AAV может содержать по меньшей мере одну последовательность ITR, фланкирующую промотор, "лишний" фрагмент, последовательность, представляющую интерес (например, средство для RNAi), и последовательность полиаденилирования. Полный геном вектора может содержать полный набор полинуклеотидных последовательностей вектора. В некоторых вариантах осуществления титр нуклеиновых кислот вирусного вектора можно измерять в единицах vg/мл. Способы, подходящие для измерения данного титра, известны из уровня техники (например, количественная ПЦР).
[0151] Термины "инфекционная единица (iu)", "инфекционная частица" или "единица репликации", используемые в отношении вирусного титра, относятся к числу инфекционных и репликационно компетентных частиц вектора на основе рекомбинантного AAV, как измерено с помощью анализа центров инфекции, также известного как анализ центров репликации, описанного например в McLaughlin et al. (1988) J. Virol., 62:1963-1973.
[0152] Термин "трансдуцирующая единица (tu)", используемый в отношении вирусного титра, относится к числу инфекционных частиц вектора на основе рекомбинантного AAV, которые приводят к получению функционального трансгенного продукта, измеряемому в функциональных анализах, таких как описанные в примерах данного документа или, например, в Xiao et al. (1997) Exp. Neurobiol., 144:113-124; или в Fisher et al. (1996) J. Virol., 70:520-532 (анализ LFU).
[0153] Последовательность "инвертированного концевого повтора" или "ITR" является термином, широко распространенным в уровне техники, и относится к относительно коротким последовательностям, встречающимся на концах вирусных геномов, которые имеют противоположную ориентацию.
[0154] Последовательность "инвертированного концевого повтора (ITR) AAV", термин, хорошо известный из уровня техники, означает последовательность из примерно 145 нуклеотидов, которая присутствует на обоих концах нативного однонитевого генома AAV. Крайние 125 нуклеотидов ITR могут присутствовать в любой из двух альтернативных ориентаций, что обуславливает гетерогенность между различными геномами AAV и между двумя концами одного генома AAV. Крайние 125 нуклеотидов также содержат несколько более коротких областей самокомплементарности (обозначаемых как A-, A'-, B-, B'-, C-, C'- и D-области), которые обеспечивают возможность образования внутринитевого спаривания оснований в пределах данной части ITR.
[0155] "Последовательность концевого разрешения" или "trs" представляет собой последовательность в D-области ITR AAV, которая отщепляется белками rep AAV в ходе репликации вирусной ДНК. Мутантная последовательность концевого разрешения устойчива к отщеплению белками rep AAV.
[0156] Термин "вирус-помощник" для AAV относится к вирусу, который обеспечивает возможность репликации и упаковки AAV (который является дефектным парвовирусом) в клетке-хозяине. Было идентифицировано множество таких вирусов-помощников, в том числе аденовирусы, герпесвирусы и поксвирусы, такие как вирус осповакцины. Аденовирусы охватывают множество различных подгрупп, однако наиболее широко применяется аденовирус 5 типа подгруппы C (Ad5). Многочисленные аденовирусы человека, млекопитающих, отличных от человека, и птиц, известны и доступны из депозитариев, таких как ATCC. Вирусы семейства герпесвирусов, которые также доступны из депозитариев, таких как ATCC, включают, например, вирусы простого герпеса (HSV), вирусы Эпштейна-Барр (EBV), цитомегаловирусы (CMV) и вирусы псевдобешенства (PRV).
[0157] Термин "процентная (%) идентичность последовательностей" относительно эталонной полипептидной последовательности или последовательности нуклеиновой кислоты определяется как процентная доля аминокислотных остатков или нуклеотидов в последовательности-кандидате, которые идентичны аминокислотным остаткам или нуклеотидам в эталонной полипептидной последовательности или последовательности нуклеиновой кислоты после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимальной процентной идентичности последовательностей, и не учитывая любые консервативные замены как часть идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей или последовательностей нуклеиновых кислот может быть достигнуто различными способами, которые находятся в пределах компетенции специалиста в данной области техники, например с помощью общедоступных компьютерных программ, например описанных в Current Protocols in Molecular Biology (Ausubel et al., eds., 1987), прилож. 30, раздел 7.7.18, таблица 7.7.1, и в том числе программного обеспечения BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Примером программы для выравнивания является ALIGN Plus (Scientific and Educational Software, Пенсильвания). Специалисты в данной области техники способны определить соответствующие параметры для оценки выравнивания, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по полной длине сравниваемых последовательностей. Для целей данного документа % идентичность аминокислотной последовательности, свойственная данной аминокислотной последовательности А, в отношении данной аминокислотной последовательности В, с ней или в сравнении с ней (что в качестве альтернативы можно перефразировать как данная аминокислотная последовательность A, которая характеризуется или обладает определенной % идентичностью аминокислотной последовательности в отношении данной аминокислотной последовательности В, с ней или в сравнении с ней), рассчитывается следующим образом: 100 умножить на частное от X/Y, где X представляет собой число аминокислотных остатков, учитываемых в качестве идентичных совпадений программой для выравнивания последовательностей в данном программном выравнивании A и B, и где Y представляет собой общее число аминокислотных остатков в B. Следует принимать во внимание, что если длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, то % идентичность аминокислотной последовательности A в отношении B не будет равна % идентичности аминокислотной последовательности B в отношении A. Для целей данного документа % идентичность последовательности нуклеиновой кислоты, свойственная данной последовательности нуклеиновой кислоты C, в отношении данной последовательности нуклеиновой кислоты D, с ней или в сравнении с ней (что в качестве альтернативы можно перефразировать как данная последовательность нуклеиновой кислоты C, которая характеризуется или обладает определенной % идентичностью последовательности нуклеиновой кислоты в отношении данной последовательности нуклеиновой кислоты D, с ней или в сравнении с ней), рассчитывается следующим образом: 100 умножить на частное от W/Z, где W представляет собой число нуклеотидов, учитываемых в качестве идентичных совпадений программой для выравнивания последовательностей в данном программном выравнивании C и D, и где Z представляет собой общее число нуклеотидов в D. Следует принимать во внимание, что если длина последовательности нуклеиновой кислоты C не равна длине последовательности нуклеиновой кислоты D, то % идентичность последовательности нуклеиновой кислоты C в отношении D не будет равна % идентичности последовательности нуклеиновой кислоты D в отношении C.
[0158] Термин "выделенная" молекула (например, нуклеиновая кислота или белок) или клетка означает, что она была идентифицирована и отделена и/или извлечена из компонента своего природного окружения.
[0159] "Эффективное количество" представляет собой количество, достаточное для достижения благоприятных или требуемых результатов, в том числе клинических результатов (например, уменьшения интенсивности симптомов, достижения клинических конечных точек и т. п.). Эффективное количество можно вводить за одно или несколько введений. Применительно к болезненному состоянию, эффективным количеством является количество, достаточное для уменьшения интенсивности, стабилизации или задержки развития заболевания.
[0160] "Индивидуум" или "субъект" представляет собой млекопитающее. Млекопитающие включают без ограничения одомашненных животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и отличных от человека приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых вариантах осуществления индивидуум или субъект является человеком.
[0161] Термин "лечение", используемый в данном документе, представляет собой подход для получения благоприятных или требуемых клинических результатов. Применительно к целям настоящего изобретения благоприятные или требуемые клинические результаты включают без ограничения облегчение симптомов, уменьшение степени тяжести заболевания, стабилизацию (например, отсутствие ухудшения) болезненного состояния, предупреждение распространения (например, метастазирования) заболевания, задержку или замедление прогрессирования заболевания, уменьшение интенсивности или временное ослабление проявлений болезненного состояния и ремиссию (как частичную, так и полную), как выявляемые, так и невыявляемые. Термин "лечение" может означать также продление выживаемости по сравнению с ожидаемой выживаемостью в случае неполучения лечения.
[0162] Термин "профилактическое лечение", используемый в данном документе, относится к лечению, при котором у индивидуума известно или подозревается наличие или имеется риск наличия нарушения, однако не проявляются симптомы или проявляются минимальные симптомы нарушения. Индивидуума, подвергаемого профилактическому лечению, можно подвергать лечению до начала проявления симптомов.
[0163] Термин "терапевтическое" средство (например, терапевтические полипептид, нуклеиновая кислота или трансген), используемый в данном документе, представляет собой средство, обеспечивающее благоприятный или требуемый клинический результат, как например иллюстративные клинические результаты, описанные выше. Таким образом, терапевтическое средство можно применять в лечении, как описано выше.
[0164] Термин "центральная зона сетчатки", используемый в данном документе, относится к наружной части желтого пятна, и/или к внутренней части желтого пятна, и/или к центральной ямке. Термин "типы клеток центральной зоны сетчатки", используемый в данном документе, относится к типам клеток центральной зоны сетчатки, например таким как RPE и фоторецепторные клетки.
[0165] Термин "желтое пятно" относится к области центральной зоны сетчатки у приматов, которая содержит фоторецепторные клетки, в частности палочки и колбочки, в более высокой относительной концентрации по сравнению с периферической зоной сетчатки. Термин "наружная часть желтого пятна", используемый в данном документе, также может упоминаться как "периферическая часть желтого пятна". Термин "внутренняя часть желтого пятна", используемый в данном документе, также может упоминаться как "центральная часть желтого пятна".
[0166] Термин "центральная ямка" относится к небольшой области в центральной зоне сетчатки приматов, диаметр которой примерно равен или составляет менее 0,5 мм, которая содержит фоторецепторные клетки, в частности колбочки, в более высокой относительной концентрации по сравнению с периферической зоной сетчатки и желтым пятном.
[0167] Термин "субретинальное пространство", используемый в данном документе, относится к местоположению в сетчатке между фоторецепторными клетками и клетками пигментного эпителия сетчатки. Субретинальное пространство может являться предполагаемым пространством, как например до какой-либо субретинальной инъекции жидкости. Субретинальное пространство может также содержать жидкость, введенную с помощью инъекции в предполагаемое пространство. В данном случае жидкость находится "в контакте с субретинальным пространством". Клетки, которые находятся "в контакте с субретинальным пространством", предусматривают клетки, которые ограничивают субретинальное пространство, такие как RPE и фоторецепторные клетки.
[0168] Термин "пузырек", используемый в данном документе, относится к жидкому пространству в пределах субретинального пространства глаза. Пузырек по настоящему изобретению можно создать с помощью однократной инъекции жидкости в одно пространство, с помощью нескольких инъекций одной или нескольких жидкостей в одно и то же пространство или с помощью нескольких инъекций в несколько пространств, которые при перемещении создают общее жидкое пространство, применимое для достижения терапевтического эффекта на требуемой части субретинального пространства.
[0169] Ссылка на "приблизительно" в отношении значения или параметра в данном документе включает (и описывает) варианты осуществления, которые направлены на данное значение или параметр per se. Например, описание, относящееся к "приблизительно X", включает описание "X".
[0170] Формы единственного числа, используемые в данном документе, включают ссылки на множественное число, если не указано иное.
[0171] Понятно, что аспекты и варианты осуществления настоящего изобретения, описанные в данном документе, включают аспекты и варианты осуществления с терминами "содержащий", "состоящий из" и/или "состоящий практически из".
III. CRISPR-Cas
[0172] Определенные аспекты настоящего раскрытия относятся к сконструированным не встречающимся в природе системам на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) -CRISPR-ассоциированных генов (Cas) (CRISPR-Cas). Эти системы могут быть использованы, inter alia, для лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, например глазного заболевания или нарушения, связанных с глубокой интронной мутацией. В некоторых вариантах осуществления системы CRISPR-Cas включают первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию; и нуклеотидную последовательность, кодирующую Cas-белок, где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию.
[0173] Как описано выше, системы CRISPR-Cas первоначально были обнаружены как адаптивная защита от чужеродных нуклеиновых кислот у бактерий и архей. Фактически, локусы CRISPR были идентифицированы у более чем 40 прокариот (см., например, Jansen, R. et al. (2002) Mol. Microbiol. 43:1565-1575 и Mojica, F.J. et al. (2005) J. Mol. Evol. 60:174-182), включая без ограничения Aeropyrum, Pyrobaculum, Sulfolobus, Archaeoglobus, Halocarcula, Methanobacteriumn, Methanococcus, Methanosarcina, Methanopyrus, Pyrococcus, Picrophilus, Thernioplasnia, Corynebacterium, Mycobacterium, Streptomyces, Aquifrx, Porphvromonas, Chlorobium, Thermus, Bacillus, Listeria, Staphylococcus, Clostridium, Thermoanaerobacter, Mycoplasma, Fusobacterium, Azarcus, Chromobacterium, Neisseria, Nitrosomonas, Desulfovibrio, Geobacter, Myrococcus, Campylobacter, Wolinella, Acinetobacter, Erwinia, Escherichia, Legionella, Methylococcus, Pasteurella, Photobacterium, Salmonella, Xanthomonas, Yersinia, Treponema и Thermotoga.
[0174] У бактерий Cas-белки (например, Cas9) связываются с двумя различными направляющими РНК: CRISPR-РНК (crRNA) и трансактивирующей crRNA (tracrRNA). Основания рибонуклеотидов crRNA и tracrRNA спариваются и образуют структуру, необходимую для Cas-опосредованного расщепления ДНК-мишени. Однако недавно было продемонстрировано, что можно сконструировать единую направляющую РНК (sgRNA) для образования структуры crRNA:tracrRNA и направления Cas-опосредованного расщепления ДНК-мишени (Jinek, M., et al. (2012) Science 337(6096):816-21). Поскольку специфичность нуклеазной активности Cas определяется направляющей РНК, то система CRISPR-Cas изучалась как инструмент для направления двухнитевых разрывов ДНК в гетерологичных клетках, что дает возможность осуществлять индивидуально настраиваемое редактирование генома (Mali, P., et al. (2013) Science 339(6121):823-6). Дополнительные описания иллюстративных систем CRISPR-Cas и способов применения, связанных с ними, можно обнаружить, inter alia, в патенте США № 8697359. В некоторых варианта осуществления направляющая РНК, описываемая в данном документе (например, первая или вторая направляющая РНК), включает в себя единую направляющую РНК (sgRNA), содержащую CRISPR-РНК (crRNA) и транс-активирующую РНК crRNA (tracrRNA). В некоторых варианта осуществления направляющая РНК, описываемая в данном документе (например, первая или вторая направляющая РНК), слита с последовательностью трансактивирующей cr (tracr). В некоторых вариантах осуществления последовательность трансактивирующей cr (tracr) содержит последовательность SEQ ID NO: 25.
[0175] Таким образом, описанные в данном документе системы CRISPR-Cas могут отличаться от встречающейся в природе системы CRISPR-Cas по многим причинам. Например, inter alia, системы CRISPR-Cas по настоящему раскрытию могут включать в себя одну или несколько направляющих РНК, которые гибридизуются с неприродной последовательностью (например, эукариотическим интроном). Встречающиеся в природе системы CRISPR-Cas распознают последовательность, которая обычно воздействует на бактерии и археи, например последовательность плазмиды или фага. Кроме того, многие из систем CRISPR-Cas по настоящему раскрытию включают в себя единую направляющую РНК, тогда как встречающиеся в природе системы CRISPR-Cas обычно включают отдельные CRISPR-РНК (crRNA) и трансактивирующие crRNA (tracrRNA).
[0176] В некоторых вариантах осуществления описанная в данном документе система CRISPR-Cas может быть самоограничивающейся. Например, как описано ниже, система CRISPR-Cas может включать в себя одну или несколько направляющих РНК, которые гибридизуются с последовательностью-мишенью(-ями) в самой системе, например последовательностью(-ями), расщепление которой влияет на уровень экспрессии компонентов системы, таких как Cas-белок. Не желая быть связанным теорией, считают, что поскольку система CRISPR-Cas не обязательно должна постоянно экспрессироваться в клетке-хозяине, конструирование системы как "самоограничивающейся" (например, характеризующейся уменьшенной длительностью существования и/или экспрессией) может быть выгодным, например, для уменьшения побочных эффектов, снижения вероятности нежелательных иммунных ответов и/или проблем безопасности и т. д.
[0177] В самоограничивающейся системе CRISPR-Cas комплекс CRISPR-Cas нацелен на один или несколько сайтов в векторе, используемом для экспрессии одного или нескольких компонентов самого комплекса. Таким образом, при экспрессии направляющей(-их) РНК и Cas-белка система CRISPR-Cas нацелена на локус, представляющий интерес (например, сайт мутации, описываемый в данном документе), а также на одну или несколько мишеней в векторе Cas, что в конечном итоге приводит к расщеплению вектора Cas и уменьшению или устранению экспрессии Cas-белка (после расщепления в локусе, представляющем интерес). В примере 3 infra показано, что данная самоограничивающаяся система CRISPR-Cas характеризуется уменьшением времени существования Cas, сохраняя при этом эффективное расщепление последовательности-мишени(-ей), представляющей интерес (например, вырезание глубокой интронной мутации).
[0178] Определенные аспекты настоящего раскрытия относятся к способам лечения заболевания или нарушения, связанных с мутацией в гене индивидуума, с использованием самоограничивающейся системы CRISPR-Cas. Например, мутация может быть нежелательной последовательностью (например, глубокой интронной мутацией), которую вырезают из гена индивидуума с помощью системы CRISPR-Cas. В других вариантах осуществления мутация может представлять собой миссенс, точковую или другую мутацию, которую корректируют с помощью системы CRISPR-Cas (например, восстановление гомологичной ДНК в расщепленной последовательности ДНК, в частности если включена гомологичная матрица). В некоторых вариантах осуществления система CRISPR-Cas находится в композиции. В некоторых вариантах осуществления композицию вводят индивидууму в терапевтически эффективном количестве.
[0179] В некоторых вариантах осуществления композиция включает a) нуклеиновую кислоту, кодирующую систему CRISPR-Cas, которая содержит первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию, представляющую интерес (включая без ограничения глубокую интронную мутацию, описанную в данном документе); и b) кассету экспрессии Cas. В некоторых вариантах осуществления кассета экспрессии Cas включает в себя нуклеотидную последовательность, кодирующую Cas-белок и целевой сайт направляющей РНК. Первая или вторая направляющая РНК гибридизуется с целевым сайтом направляющей РНК, что дает возможность системе CRISPR-Cas катализировать расщепление в целевом сайте направляющей РНК. Целевой сайт направляющей РНК может также включать прилегающий к протоспейсеру мотив (PAM), специфичный для Cas-белка, примыкающий к последовательности, которая гибридизуется с направляющей РНК. При экспрессии Cas-белка Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию. При экспрессии Cas-белка Cas-белок также расщепляет кассету экспрессии Cas по целевому сайту направляющей РНК, уменьшая тем самым экспрессию Cas-белка. Как таковая, экспрессия Cas первоначально возрастает после введения кассеты экспрессии Cas в клетку, однако по мере накопления Cas-белка в клетке Cas-белок расщепляет кассету экспрессии Cas. По мере того как все большая часть кассет экспрессии Cas прерывается Cas-белком, экспрессия дополнительного Cas-белка уменьшается (т. е. Cas ограничивает экспрессию своей собственной кассеты экспрессии и может считаться самоограничивающейся кассетой экспрессии Cas). В некоторых вариантах осуществления экспрессия Cas-белка характеризуется начальным увеличением с последующим снижением экспрессии после расщепления в целевом сайте направляющей РНК. Как описано в данном документе, снижение экспрессии Cas-белка может относиться к уменьшению количества и/или длительности существования Cas-белка. В некоторых вариантах осуществления экспрессию Cas-белка можно уменьшить по сравнению с экспрессией до расщепления кассеты экспрессии Cas (например, по сравнению с исходной экспрессией Cas). В некоторых вариантах осуществления экспрессию Cas-белка можно уменьшить по сравнению с экспрессией при применении кассеты экспрессии Cas, лишенной целевого сайта направляющей РНК. В некоторых вариантах осуществления композицию, содержащую самоограничивающуюся кассету экспрессии Cas, можно использовать для расщепления нуклеиновой кислоты-мишени. В некоторых вариантах осуществления композицию можно использовать для расщепления нуклеиновой кислоты-мишени in vitro или in vivo. В некоторых вариантах осуществления композицию можно использовать для расщепления нуклеиновой кислоты-мишени, содержащей мутацию (например, глубокую интронную мутацию). Например, самоограничивающуюся кассету экспрессии Cas используют для лечения заболевания или нарушения, связанных с мутацией в нуклеиновой кислоте.
[0180] В некоторых вариантах осуществления кассета экспрессии Cas дополнительно включает в себя целевой сайт второй направляющей РНК, где первая направляющая РНК или вторая направляющая РНК гибридизуются с целевым сайтом второй направляющей РНК, и где целевой сайт второй направляющей РНК примыкает к прилегающему к протоспейсеру мотиву (PAM), специфичному для Cas-белка. При экспрессии Cas-белка Cas-белок расщепляет последовательности ДНК-мишени, фланкирующие мутацию, вырезая таким образом часть ДНК-мишени, содержащую мутацию. Далее при экспрессии Cas-белка Cas-белок расщепляет кассету экспрессии Cas в обоих целевых сайтах направляющей РНК, уменьшая тем самым экспрессию Cas-белка. Как проиллюстрировано ниже, одна направляющая РНК может гибридизоваться с целевыми сайтами двоих направляющих РНК; при этом первая направляющая РНК может гибридизоваться с целевым сайтом первой направляющей РНК и вторая направляющая РНК может гибридизоваться с целевым сайтом второй направляющей РНК; или вторая направляющая РНК может гибридизоваться с целевым сайтом первой направляющей РНК и первая направляющая РНК может гибридизоваться с сайтом второй направляющей РНК.
[0181] В некоторых вариантах осуществления кассета экспрессии Cas может включать один или несколько промоторов, энхансеров, интронов, последовательностей полиаденилирования (polyA), терминаторов, регуляторных элементов, присутствующих в 5'- или 3'-нетранслируемых областях, и так далее, полезных для направления/активации экспрессии Cas-белка. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, может быть функционально связана с промотором. В некоторых вариантах осуществления целевой сайт направляющей РНК может находиться между промотором и нуклеотидной последовательностью, кодирующей Cas-белок, что приводит к уменьшению экспрессии Cas-белка при расщеплении. В некоторых вариантах осуществления экспрессия Cas-белка уменьшается при расщеплении функциональной связи между промотором и нуклеотидной последовательностью, кодирующей Cas-белок. В некоторых вариантах осуществления нуклеотидная последовательность, кодирующая Cas-белок, может быть функционально связана с последовательностью polyA. В некоторых вариантах осуществления целевой сайт направляющей РНК может находиться между последовательностью polyA и нуклеотидной последовательностью, кодирующей Cas-белок, что приводит к уменьшению экспрессии Cas-белка при расщеплении. В некоторых вариантах осуществления экспрессия Cas-белка уменьшается при расщеплении функциональной связи между нуклеотидной последовательностью, кодирующей Cas-белок, и последовательностью полиаденилирования. В некоторых вариантах осуществления целевой сайт первой направляющей РНК может находиться между промотором и нуклеотидной последовательностью, кодирующей Cas-белок, а целевой сайт второй направляющей РНК может находиться между последовательностью polyA и нуклеотидной последовательностью, кодирующей Cas-белок, что приводит к уменьшению экспрессии Cas-белка при расщеплении. В некоторых вариантах осуществления Cas-белок может быть слит в рамке считывания с одним или несколькими NLS, и целевой сайт направляющей РНК может находиться между последовательностью, кодирующей один или несколько NLS, и последовательностью polyA (в частности, если NLS слиты с С-концом Cas-белка), что приводит к уменьшению экспрессии слитого с NLS Cas-белка при расщеплении.
[0182] Как описано в данном документе, система CRISPR-Cas, такая как самоограничивающаяся система CRISPR-Cas, может быть закодирована на одном или нескольких векторах, таких как любой из векторов или вирусных векторов/частиц, описанных в данном документе. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая первую и вторую направляющие РНК, может находиться в том же векторе, что и кассета экспрессии Cas. В других вариантах осуществления нуклеиновая кислота, кодирующая первую и вторую направляющие РНК, может быть другим вектором, чем кассета экспрессии Cas. Например, первая и вторая направляющие РНК могут быть закодированы первым вектором на основе rAAV, и кассета экспрессии Cas может быть закодирована вторым вектором на основе rAAV. В некоторых вариантах осуществления клетка-мишень может быть трансфицирована обоими векторами, что приводит к экспрессии самоограничивающейся системы CRISPR-Cas.
[0183] В некоторых вариантах осуществления Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим мутацию (например, глубокую интронную мутацию), вырезая таким образом часть ДНК-мишени, содержащую мутацию. Например, процесс репарации ДНК, такой как негомологичное соединение концов (NHEJ), может восстанавливать расщепленную последовательность ДНК путем соединения расщепленных концов, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию. В других примерах гомологичная репарация ДНК может восстанавливать расщепленную последовательность ДНК, особенно если включена гомологичная матрица. Как описано и проиллюстрировано в данном документе, использование двух направляющих РНК, которые фланкируют целевую последовательность ДНК (например, последовательность, несущую глубокую интронную мутацию), дает возможность вырезать часть последовательности ДНК-мишени, например последовательность, несущую глубокую интронную мутацию (см. также Brandl, C. et al. (2014) FEBS Open Bio. 5:26-35; Zheng, Q. et al. (2014) Biotechniques 57:115-124 для описания иллюстративных стратегий делеции генов с использованием систем CRISPR-Cas). В некоторых вариантах осуществления вырезанная часть последовательности ДНК-мишени содержит интронную ДНК. В некоторых вариантах осуществления вырезанная часть последовательности ДНК-мишени состоит только из интронной ДНК.
[0184] В некоторых вариантах осуществления первая и/или вторая направляющие РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих мутацию (например, глубокую интронную мутацию). Не желая быть связанными теорией, считают, что первая и/или вторая направляющие РНК могут гибридизоваться с противоположными нитями последовательностей ДНК-мишени, расположенных внутри интрона на любом расстоянии от глубокой интронной мутации. В некоторых вариантах осуществления первая и/или вторая направляющие РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени на расстоянии от 1 пары оснований до 10000 пар оснований от глубокой интронной мутации. В некоторых вариантах осуществления первая и/или вторая направляющие РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, расположенных менее чем на приблизительно любом из следующих расстояний от глубокой интронной мутации (в нуклеотидах): 10000; 9500; 9000; 8500; 8000; 7500; 7000; 6500; 6000; 5500; 5000; 4500; 4000; 3500; 3000; 2500; 2000; 1500; 1000; 950; 900; 850; 800; 750; 700; 650; 600; 550; 500; 450; 400; 350; 300; 250; 200; 150; 100; 95; 90; 85; 80; 75, 70, 65, 60, 55, 50, 45, 40, 35, 30, 25, 20, 15, 10, 9, 8, 7, 6, 5, 4; 3; 2 или любое значение между ними. В некоторых вариантах осуществления первая и/или вторая направляющие РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, расположенных более чем на приблизительно любом из следующих расстояний от глубокой интронной мутации (в нуклеотидах): 1; 2; 3; 4; 5; 6; 7; 8; 9; 10; 15; 20; 25; 30; 35; 40; 45; 50; 55; 60; 65; 70; 75; 80; 85; 90; 95; 100; 150; 200; 250; 300; 350; 400; 450; 500; 550; 600; 650; 700; 750; 800, 850, 900, 950, 1000, 1500, 2000, 2500, 3000, 3500, 4000, 4500, 5000, 5500, 6000, 6500, 7000, 7500, 8000, 8500; 9000; 9500 или любое значение между ними. То есть первая и/или вторая направляющие РНК могут гибридизоваться с противоположными нитями последовательностей ДНК-мишени, расположенных на расстоянии от глубокой интронной мутации, которое может быть любым из диапазона расстояний (в нуклеотидах), имеющего верхний предел 10000; 9500; 9000; 8500; 8000; 7500; 7000; 6500; 6000; 5500; 5000; 4500; 4000; 3500; 3000; 2500; 2000; 1500; 1000; 950; 900; 850; 800; 750; 700; 650; 600; 550; 500; 450; 400; 350; 300; 250; 200; 150; 100; 95; 90; 85; 80; 75, 70, 65, 60, 55, 50, 45, 40, 35, 30, 25, 20, 15, 10, 9, 8, 7, 6, 5, 4; 3; 2 или любое значение между ними, и независимо выбранный нижний предел из 1; 2; 3; 4; 5; 6; 7; 8; 9; 10; 15; 20; 25; 30; 35; 40; 45; 50; 55; 60; 65; 70; 75; 80; 85; 90; 95; 100; 150; 200; 250; 300; 350; 400; 450; 500; 550; 600; 650; 700; 750; 800; 850; 900; 950; 1000; 1500; 2000; 2500; 3000; 3500; 4000; 4500; 5000; 5500; 6000; 6500; 7000; 7500; 8000; 8500; 9000; 9500 или любого значения между ними, причем нижний предел меньше верхнего предела.
[0185] Неограничивающие примеры Cas-белков включают Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (также известный как Csn1 и Csx12), Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, их гомологи или их модифицированные версии. Эти ферменты общеизвестны из уровня техники.
[0186] В некоторых вариантах осуществления Cas-белок (например, фермент CRISPR) представляет собой белок Cas9. В некоторых вариантах осуществления немодифицированный фермент CRISPR характеризуется расщепляющей активностью ДНК, такой как Cas9. Иллюстративные белки Cas9 включают без ограничения Cas9 S. pyogenes (см., например, номер доступа базы данных SwissProt Q99ZW2), Cas9 S. aureus (см., например, номер доступа GenBank CCK74173), Cas9 S. thermophilus (см., например, номер доступа базы данных SwissProt G3ECR1), Cas9 N. meningitidis (см., например, номер доступа UniProt C9X1G5) и Cas9 T. denticola (см., например, номер доступа GenBank EMB41078). В некоторых вариантах осуществления Cas9 происходит из S. pyogenes или S. pneumoniae. В некоторых вариантах осуществления фермент CRISPR направляет расщепление одной или обеих нитей в месте расположения последовательности-мишени, как например в пределах последовательности-мишени и/или в пределах комплемента к последовательности-мишени. В некоторых вариантах осуществления фермент CRISPR направляет расщепление одной или обеих нитей в пределах приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 50, 100, 200, 500 или более пар оснований от первого или последнего нуклеотида последовательности-мишени.
[0187] В некоторых вариантах осуществления кодирующая последовательность фермента, кодирующая Cas-белок по настоящему раскрытию, является кодон-оптимизированной для экспрессии в конкретной клетке, такой как эукариотическая клетка. Эукариотические клетки могут быть клетками-организмами или полученными из определенного организма, такого как млекопитающее, включая без ограничения человека, мышь, крысу, кролика, собаку или примата, отличного от человека. В целом, оптимизация кодонов относится к процессу модификации последовательности нуклеиновой кислоты для улучшения экспрессии в клетках-хозяевах, представляющих интерес, путем замены по меньшей мере одного кодона нативной последовательности кодонами, которые более часто или чаще всего используются в генах данной клетки-хозяина, при сохранении нативной аминокислотной последовательности. Различные виды демонстрируют специфическую склонность к определенным кодонам для конкретной аминокислоты. Таблицы использования кодонов легко доступны, например, в "Codon Usage Database", и эти таблицы можно адаптировать несколькими способами (см. например Nakamura, Y. et al. (2000) Nucleic Acids Res. 28:292). Также доступны компьютерные алгоритмы оптимизации кодонов определенной последовательности для экспрессии в конкретной клетке-хозяине, такие как Gene Forge (Aptagen, Джакобус, Пенсильвания).
[0188] В некоторых вариантах осуществления Cas-белок представляет собой белок слияния, содержащий один или несколько гетерологичных белковых доменов (например, приблизительно или более чем приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более доменов в дополнение к ферменту CRISPR). Белок слияния Cas может содержать любую дополнительную белковую последовательность, и необязательно линкерную последовательность между любыми двумя доменами. Примеры белковых доменов, которые можно сливать с Cas-белком, включают без ограничения метки эпитопа, последовательности репортерного гена и белковые домены, характеризующиеся одной или несколькими из следующих видов активности: метилазная активность, деметилазная активность, активность активации транскрипции, активность репрессии транскрипции, активность фактора освобождения транскрипции, активность модификации гистонов, расщепляющую активность РНК и активность связывания нуклеиновой кислоты. Примеры меток эпитопа включают без ограничения гистидиновые (His) метки, V5-метки, FLAG-метки, метки гемагглютинина (HA) вируса гриппа, Myc-метки, VSV-G-метки и метки тиоредоксина (Trx). Примеры репортерных генов включают без ограничения глутатион-5-трансферазу (GST), пероксидазу хрена (HRP), хлорамфениколацетилтрансферазу (CAT), бета-галактозидазу, бета-глюкуронидазу, люциферазу и флуоресцентные белки (например, GFP, CFP, YFP, BFP и т. д.). Cas-белок можно сливать с последовательностью гена, кодирующей белок или фрагмент белка, которые связывают молекулы ДНК или связывают другие клеточные молекулы, включая без ограничения мальтозосвязывающий белок (MBP), S-метку, слитые молекулы на основе ДНК-связывающего (DBD) домена Lex A, слитые молекулы на основе ДНК-связывающего домена GAL4A и слитые молекулы на основе белка BP16 вируса простого герпеса (HSV). В некоторых вариантах осуществления Cas-белок модифицирован для улучшения функции.
[0189] В некоторых вариантах осуществления Cas-белок представляет собой мутантный Cas-белок. В некоторых вариантах осуществления Cas-белок (например, белок Cas9) представляет собой мутанта-никазу (Ran et al., 2013 Cell 156(6):1380-9). В некоторых вариантах осуществления мутанта-никазу Cas9 используют с парами направляющих РНК для введения целенаправленных двухнитевых разрывов со сниженным нецелевым расщеплением ДНК.
[0190] В некоторых вариантах осуществления Cas-белок представляет собой вариант Cas-белка высокой точности воспроизведения, несущий аминокислотные изменения и демонстрирующий устойчивую целевую активность, но незначительное нецелевое расщепление (Slaymaker, I.M. et al. (2016) Science 351(6268):84-88; Kleinstiver, B.P. et al. (2016) Nature 529:490-495).
[0191] В некоторых вариантах осуществления система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS). Например, Cas-белок (например, белок Cas9) может содержать один или несколько NLS. Иллюстративные плазмиды, включающие в себя Cas9 с NLS, можно найти в Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308. Из уровня техники известно множество NLS, подходящих для ряда клеток-хозяев. Например, без ограничения NLS может представлять собой NLS SV40 (например, как описано в Mali, P., et al. (2013) Science 339(6121):823-6), однокомпонентный NLS большого Т-антигена SV40, NLS нуклеоплазмина и NLS hnRNP A1. Иллюстративные последовательности NLS, которые можно использовать, включают в себя без ограничения PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27) (см. например Jinek, M. et al. (2013) eLife 2:e00471).
[0192] Образование комплекса CRISPR (включающего в себя направляющую последовательность, гибридизованную с противоположной нитью последовательности-мишени и образующую комплекс с одним или несколькими Cas-белками) обычно приводит к расщеплению одной или обеих нитей внутри или вблизи (например, в пределах 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 или более пар оснований) последовательности-мишени в эндогенной системе CRISPR. Также считается, что последовательность tracr, которая может содержать всю или часть последовательности tracr дикого типа, может составлять часть комплекса CRISPR, например путем гибридизации вдоль по меньшей мере части последовательности tracr со всей или частью сопряженной с tracr последовательности, например сопряженной с tracr последовательности, функционально связанной с направляющей последовательностью.
[0193] В целом, сопряженная с tracr последовательность может включать любую последовательность, которая имеет достаточную комплементарность с последовательностью tracr, чтобы стимулировать вырезание направляющей последовательности, фланкированной сопряженными с tracr последовательностями в клетке, содержащей соответствующую последовательность tracr; образование комплекса CRISPR, включающего сопряженную с tracr последовательность, гибридизованную с последовательностью tracr в последовательности-мишени; или оба. В некоторых вариантах осуществления последовательность tracr имеет достаточную комплементарность для гибридизации с сопряженной с tracr последовательностью и участия в формировании комплекса CRISPR. Как описано ниже в отношении последовательностей-мишеней, считается, что необходима комплементарность, достаточная по меньшей мере для функциональности (т. e. полная комплементарность между tracr и сопряженной с tracr последовательностями не требуется).
[0194] Как правило, степень комплементарности относится к оптимальному выравниванию сопряженной с tracr последовательности и последовательности tracr по длине более короткой из двух последовательностей. Оптимальное выравнивание можно определить с помощью любого подходящего алгоритма выравнивания (например, как описано в данном документе) и можно дополнительно учитывать вторичные структуры, такие как самокомплементарность, в последовательности tracr или в сопряженной с tracr последовательности. В некоторых вариантах осуществления последовательность tracr составляет приблизительно или более чем приблизительно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 40, 50 или более нуклеотидов в длину. В некоторых вариантах осуществления последовательность tracr характеризуется по меньшей мере 50%, 60%, 70%, 80%, 90%, 95% или 99% комплементарностью последовательности по длине с сопряженной с tracr последовательностью при оптимальном выравнивании (например, как определено с помощью любого из иллюстративных способов выравнивания, описанных в данном документе).
[0195] Не желая быть связанными теорией, считают, что любая требуемая последовательность ДНК-мишени, представляющая интерес, может быть нацелена с помощью направляющей последовательности sgRNA, и что единственным требованием для последовательности ДНК-мишени является наличие прилегающего к протоспейсеру мотива (PAM), примыкающего к последовательности-мишени sgRNA (Mali, P., et al. (2013) Science 339(6121):823-6). Известно, что различные комплексы Cas имеют разные мотивы PAM. Например, Cas9 из Streptococcus pyogenes имеет GG-динуклеотидный мотив PAM. Для дополнительных примеров, мотив PAM из Cas9 S. aureus представляет собой GRRT, в котором R представляет собой пурин (A или G), мотив PAM из Cas9 N. meningitidis представляет собой GATT, мотив PAM из Cas9 S. thermophilus представляет собой AGAA и мотив PAM из Cas9 T. denticola представляет собой AAAAC.
[0196] В целом, направляющая последовательность может представлять собой любую полинуклеотидную последовательность, имеющую достаточное сходство с полинуклеотидной последовательностью-мишенью для гибридизации с противоположной нитью последовательности-мишени и направления сайт-специфичного связывания комплекса CRISPR с последовательностью-мишенью. В некоторых вариантах осуществления степень идентичности между направляющей последовательностью и соответствующей ей последовательностью-мишенью составляет приблизительно или более чем приблизительно 50%, 60%, 75%, 80%, 85%, 90%, 95%, 97,5%, 99% или более при оптимальном выравнивании с использованием подходящего алгоритма выравнивания. Оптимальное выравнивание можно определить с использованием любого подходящего алгоритма для выравнивания последовательностей; неограничивающие примеры включают алгоритм Смита-Уотермана, алгоритм Нидлмана-Вунша, алгоритмы, основанные на преобразовании Барроуза-Уилера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAT, Novoalign (Novocraft Technologies), ELAND (Illumina, Сан Диего, Калифорния), SOAP (доступно на soap.genomics.org.cn), Maq (доступно на maq.sourceforge.net) и тому подобное.
[0197] В некоторых вариантах осуществления направляющая последовательность составляет приблизительно или более чем приблизительно любое значение из 5, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 75 или более нуклеотидов в длину. В некоторых вариантах осуществления направляющая последовательность составляет менее чем приблизительно любое значение из 75, 50, 45, 40, 35, 30, 25, 20, 15, 12 или менее нуклеотидов в длину. Анализы для определения способности направляющей последовательности направлять сайт-специфичное связывание комплекса CRISPR с последовательностью-мишенью известны из уровня техники. Например, компоненты системы CRISPR, достаточные для образования комплекса CRISPR, включая направляющую последовательность, подлежащую тестированию, могут быть доставлены в клетку-хозяина, имеющую соответствующую последовательность-мишень, например путем трансфекции векторами, кодирующими компоненты последовательности CRISPR, после чего следует оценка предпочтительного расщепления в последовательности-мишени, например как описано в патенте США № 8697359.
[0198] В некоторых вариантах осуществления направляющая РНК по настоящему раскрытию (например, первая направляющая РНК) содержит последовательности под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9) или SEQ ID NO:47 (для SaCas9). В некоторых вариантах осуществления направляющая РНК по настоящему раскрытию (например, первая направляющая РНК) содержит последовательность под SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9). В некоторых вариантах осуществления направляющая РНК содержит 1, 2, 3, 4 или 5 замен, делеций или вставок последовательностей под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9), SEQ ID NO:47 (для SaCas9), SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9), сохраняя при этом свою функцию в качестве направляющей РНК для Cas-расщепления гена CEP290. В некоторых вариантах осуществления направляющая РНК представляет собой вариант направляющей РНК под SEQ ID NO:41 (для SpCas9), SEQ ID NO:45 (для SaCas9), SEQ ID NO:46 (для SaCas9), SEQ ID NO:47 (для SaCas9), SEQ ID NO:19 (для SpCas9), SEQ ID NO:50 (для SaCas9), SEQ ID NO:51 (для SaCas9) или SEQ ID NO:52 (для SaCas9) с повышенной функцией в качестве направляющей РНК для Cas-расщепления гена CEP290.
[0199] В некоторых вариантах осуществления направляющая РНК по настоящему раскрытию (например, вторая направляющая РНК) содержит последовательности под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9) или SEQ ID NO:49 (для SaCas9). В некоторых вариантах осуществления направляющая РНК по настоящему раскрытию (например, вторая направляющая РНК) содержит последовательности под SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9). В некоторых вариантах осуществления направляющая РНК содержит 1, 2, 3, 4 или 5 замен, делеций или вставок последовательностей под SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:17 или SEQ ID NO:18, сохраняя при этом свою функцию в качестве направляющей РНК для Cas-расщепления гена CEP290. В некоторых вариантах осуществления направляющая РНК представляет собой вариант направляющей РНК под SEQ ID NO:42 (для SpCas9), SEQ ID NO:43 (для SpCas9), SEQ ID NO:44 (для SpCas9), SEQ ID NO:48 (для SaCas9), SEQ ID NO:49 (для SaCas9), SEQ ID NO:20 (для SpCas9), SEQ ID NO:21 (для SpCas9), SEQ ID NO:22 (для SpCas9), SEQ ID NO:53 (для SaCas9) или SEQ ID NO:54 (для SaCas9) с повышенной функцией в качестве направляющей РНК для Cas-расщепления гена CEP290.
[0200] В некоторых вариантах осуществления последовательность tracr и последовательность, сопряженная с tracr, содержатся в одном транскрипте, так что гибридизация между обеими создает транскрипт, имеющий вторичную структуру (например, шпильку). В некоторых вариантах осуществления последовательности, образующие петлю, для применения в структурах шпильки имеют четыре нуклеотида в длину. В некоторых вариантах осуществления последовательности, образующие петлю, имеют последовательность GAAA. Однако можно использовать более длинные или более короткие последовательности петли, а можно также альтернативные последовательности. Примеры других последовательностей, образующих петлю, включают без ограничения CAAA и AAAG. В некоторых вариантах осуществления транскрипт или транскрибированная полинуклеотидная последовательность имеет по меньшей мере две или более шпильки, например две, три, четыре или пять шпилек. В некоторых вариантах осуществления единый транскрипт дополнительно включает в себя последовательность терминации транскрипции, например последовательность polyT, такую как шесть нуклеотидов Т.
IV. Глубокие интронные мутации
[0201] Определенные аспекты настоящего раскрытия относятся к глубоким интронным мутациям. Как описано выше, глубокая интронная мутация может приводить к аберрантному включению интронной последовательности в зрелую (например, сплайсированную) mRNA. Например, глубокая интронная мутация может вводить в ген сайт донора сплайсинга, сайт акцептора сплайсинга или сайт энхансера сплайсинга. В результате интронная последовательность может быть включена в виде криптического экзона. Как правило, это дает в результате мутированный полипептид, особенно если криптический экзон включает мутацию сдвига рамки или преждевременный стоп-кодон.
[0202] Как описано выше, глубокая интронная мутация относится к мутации вне последовательностей акцептора сплайсинга и донора сплайсинга дикого типа, т. e. в отличие от мутации в эндогенном акцепторе сплайсинга или доноре сплайсинга. Глубокая интронная мутация обычно происходит на некотором расстоянии от эндогенных сайтов акцептора сплайсинга/донора сплайсинга.
[0203] В некоторых вариантах осуществления глубокая интронная мутация расположена на расстоянии по меньшей мере приблизительно 100 нуклеотидов от 5'-конца сайта донора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на расстоянии по меньшей мере приблизительно 100 нуклеотидов от 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация расположена на расстоянии по меньшей мере приблизительно 100 нуклеотидов от 5'-конца сайта донора сплайсинга и по меньшей мере приблизительно 100 нуклеотидов от 3'-конца сайта акцептора сплайсинга гена. В некоторых вариантах осуществления глубокая интронная мутация происходит на расстоянии более чем приблизительно 100 нуклеотидов, более чем приблизительно 150 нуклеотидов, более чем приблизительно 200 нуклеотидов, более чем приблизительно 250 нуклеотидов, более чем приблизительно 300 нуклеотидов, более чем приблизительно 350 нуклеотидов, более чем приблизительно 400 нуклеотидов, более чем приблизительно 450 нуклеотидов, более чем приблизительно 500 нуклеотидов, более чем приблизительно 550 нуклеотидов, более чем приблизительно 600 нуклеотидов, более чем приблизительно 650 нуклеотидов, более чем приблизительно 700 нуклеотидов, более чем приблизительно 750 нуклеотидов, более чем приблизительно 800 нуклеотидов, более чем приблизительно 850 нуклеотидов, более чем приблизительно 900 нуклеотидов, более чем приблизительно 950 нуклеотидов, более чем приблизительно 1000 нуклеотидов, более чем приблизительно 1500 нуклеотидов, более чем приблизительно 2000 нуклеотидов, более чем приблизительно 2500 нуклеотидов, более чем приблизительно 3000 нуклеотидов, более чем приблизительно 3500 нуклеотидов, более чем приблизительно 4000 нуклеотидов, более чем приблизительно 4500 нуклеотидов, более чем приблизительно 5000 нуклеотидов, более чем приблизительно 5500 нуклеотидов, более чем приблизительно 6000 нуклеотидов, более чем приблизительно 6500 нуклеотидов, более чем приблизительно 7000 нуклеотидов, более чем приблизительно 7500 нуклеотидов, более чем приблизительно 8000 нуклеотидов, более чем приблизительно 8500 нуклеотидов, более чем приблизительно 9000 нуклеотидов, более чем приблизительно 9500 нуклеотидов, более чем приблизительно 10000 нуклеотидов, более чем приблизительно 15000 нуклеотидов, более чем приблизительно 20000 нуклеотидов, более чем приблизительно 25000 нуклеотидов, более чем приблизительно 30000 нуклеотидов, более чем приблизительно 35000 нуклеотидов, более чем приблизительно 40000 нуклеотидов, более чем приблизительно 45000 нуклеотидов, более чем приблизительно 50000 нуклеотидов, более чем приблизительно 55000 нуклеотидов, более чем приблизительно 60000 нуклеотидов, более чем приблизительно 65000 нуклеотидов, более чем приблизительно 70000 нуклеотидов, более чем приблизительно 75000 нуклеотидов, более чем приблизительно 80000 нуклеотидов или более чем приблизительно 85000 нуклеотидов от эндогенного сайта акцептора сплайсинга и/или донора сплайсинга (например, 5'-конца сайта донора сплайсинга и/или 3'-конца сайта акцептора сплайсинга).
[0204] В некоторых вариантах осуществления глубокая интронная мутация происходит на расстоянии от эндогенного сайта акцептора сплайсинга и/или донора сплайсинга (например, 5'-конца сайта донора сплайсинга или 3'-конца акцептора сплайсинга), которое составляет менее чем приблизительно любое значение из следующих расстояний (в нуклеотидах): 85000; 80000; 75000; 70000; 65000; 60000; 55000; 50000; 45000; 40000; 35000; 30000; 25000; 20000; 15000; 10000; 9500; 9000; 8500; 8000; 7500; 7000; 6500; 6000; 5500; 5000; 4500; 4000; 3500; 3000; 2500; 2000; 1500; 1000; 950; 900; 850; 800; 750; 700; 650; 600; 550; 500; 450; 400; 350; 300; 250; 200 или 150. В некоторых вариантах осуществления глубокая интронная мутация происходит на расстоянии от эндогенного сайта акцептора сплайсинга и/или донора сплайсинга, которое составляет более чем приблизительно любое значение из следующих расстояний (в нуклеотидах): 100; 150; 200; 250; 300; 350; 400; 450; 500; 550; 600; 650; 700; 750; 800; 850; 900; 950; 1000; 1500; 2000; 2500; 3000; 3500; 4000; 4500; 5000; 5500; 6000; 6500; 7000; 7500; 8000; 8500; 9000; 9500; 10000; 15000; 20000; 25000; 30000; 35000; 40000; 45000; 50000; 55000; 60000; 65000; 70000; 75000 или 80000. То есть расстояние от глубокой интронной мутации до эндогенного сайта акцептора сплайсинга и/или донора сплайсинга (например, 5'-конца сайта донора сплайсинга) может быть любым из диапазона расстояний (в нуклеотидах), имеющего верхний предел 85000; 80000; 75000; 70000; 65000; 60000; 55000; 50000; 45000; 40000; 35000; 30000; 25000; 20000; 15000; 10000; 9500; 9000; 8500; 8000; 7500; 7000; 6500; 6000; 5500; 5000; 4500; 4000; 3500; 3000; 2500; 2000; 1500; 1000; 950; 900; 850; 800; 750; 700; 650; 600; 550; 500; 450; 400; 350; 300; 250; 200 или 150, и независимо выбранный нижний предел 100; 150; 200; 250; 300; 350; 400; 450; 500; 550; 600; 650; 700; 750; 800; 850; 900; 950; 1000; 1500; 2000; 2500; 3000; 3500; 4000; 4500; 5000; 5500; 6000; 6500; 7000; 7500; 8000; 8500; 9000; 9500; 10000; 15000; 20000; 25000; 30000; 35000; 40000; 45000; 50000; 55000; 60000; 65000; 70000; 75000 или 80000 или любого значения между ними, причем нижний предел меньше верхнего предела.
[0205] Определенные аспекты настоящего раскрытия относятся к способам, наборам, композициям и вирусным частицам, которые можно использовать, inter alia, для лечения заболевания или нарушения, связанных с глубокой интронной мутацией, например в гене индивидуума. Из уровня техники известно множество глубоких интронных мутаций. Иллюстративные глубокие интронные мутации, ассоциированные с заболеванием, приведены в таблице 1 ниже (обратите внимание, что конкретные мутации, гены и размеры интронов, приведенные в таблице 1, относятся к последовательности ДНК человека в качестве эталона).
Таблица 1. Иллюстративные заболевания, связанные с глубокими интронными мутациями
[0206] В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляют собой афибриногенемию, синдром Альпорта, амиотрофический латеральный склероз, атаксию-телеангиэктазию, аутосомно-рецессивное поликистозное заболевание почек, синдром Барта, бета-талассемию, врожденную афибриногенемию, синдром врожденной катаракты, лицевого дисморфизма и невропатии, врожденное нарушение гликозилирования Ia типа, врожденное нарушение гликозилирования II типа, кистозный фиброз, дефицит дигидроптеридинредуктазы, заболевание Фабри, семейное тромбоцитарное нарушение с предрасположенностью к острому миелогенному лейкозу, анемию Фанкони, синдром Гительмана, нечувствительность к гормону роста, атаксию Фридрейха, гемофилию A, наследственную мегалобластную анемию 1, синдром Германски-Пудлака, гомоцистинурию, заболевание мочи с запахом кленового сиропа, синдром Марфана, дефицит метионинсинтазы, метилмалоновую ацидемию, дефицит митохондриального трифункционального белка, мукополисахаридоз II типа, доброкачественную врожденную миопатию, мышечную дистрофию, нейрофиброматоз I типа, заболевание Ниманна-Пика типа С, глазной альбинизм I типа, дефицит орнитин-дельта-аминотрансферазы, предрасположенность к системной красной волчанке, пропионовую ацидемию, рабдоидные опухоли, синдром Шварца-Джампеля, синдром Стиклера, системную красную волчанку, туберозный склероз, синдром Вернера, Х-сцепленную гипериммуноглобулинемию М или Х-сцепленную гипофосфатемию. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 1.
[0207] В некоторых вариантах осуществления заболевание или нарушение, связанные с глубокой интронной мутацией, представляет собой глазное заболевание. Используемый в данном документе термин "глазное заболевание" используется в самом широком смысле и может относиться к заболеванию, возникающему во время или в результате патологического состояния любой структуры глаза, включая без ограничения роговицу, радужную оболочку, хрусталик, сетчатку, зрительный нерв, водянистую влагу, конъюнктиву, одну или несколько глазных мышц, склеру, стекловидное тело, желтое пятно, центральную ямку, цилиарное тело, одну или несколько связок или цилиарных связок, зрачок, переднюю камеру и/или заднюю камеру. Иллюстративные глубокие интронные мутации, ассоциированные с глазным заболеванием, приведены в таблице 2 ниже (обратите внимание, что конкретные мутации, гены и размеры интронов, приведенные в таблице 2, относятся к последовательности ДНК человека в качестве эталона).
Таблица 2. Иллюстративные глазные заболевания, связанные с глубокими интронными мутациями
[0208] В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера, атрофию зрительного нерва, пигментный ретинит, ретинобластому, заболевание Штаргардта, синдром Ашера или Х-сцепленный пигментный ретинит. В некоторых вариантах осуществления глубокая интронная мутация представляет собой глубокую интронную мутацию, представленную в таблице 2.
[0209] В некоторых вариантах осуществления глазное заболевание представляет собой врожденный амавроз Лебера. Врожденный амавроз Лебера (LCA) относится к группе глазных нарушений, характеризующихся такими симптомами, как тяжелая потеря зрения, нистагм, дисфункция сетчатки, светобоязнь, окулодигитальные признаки (например, трение, жжение и давление глаза) и кератоконус. LCA обычно проявляется при рождении в виде глубокой потери зрения и нистагма. Хотя случаи LCA обычно наследуются как аутосомно-рецессивное нарушение, вовлечены мутации в различных генетических локусах. Например, в таблице 3 перечислены некоторые из локусов, вовлеченных в формы LCA.
Таблица 3. Локусы заболевания LCA.
[0210] Наиболее частый тип LCA вызван мутацией в CEP290, который кодирует центросомальный белок, играющий важную роль в развитии центросомы и цилий (см.например NCBI Gene ID № 80184 и UniProt ID № O15078 для иллюстративных последовательностей соответственно гена и белка человека). CEP290 имеет принципиальное значение для образования первичной цилии на клеточной мембране, которая играет важную роль в фоторецепторах в задней части сетчатки и в почках, головном мозге и многих других органах тела. CEP290 также известен как MKS4, CT87, POC3, rd16, BBS14, LCA10, JBTS5, NPHP6, SLSN6 и 3H11Ag. В некоторых вариантах изобретения каузальной мутацией представляет собой мутацию c.2991+1655A>G, которая вводит криптический сайт донора сплайсинга, приводя к включению аберрантного экзона с преждевременным стоп-кодоном. В некоторых вариантах осуществления последовательности первой направляющей РНК и второй направляющей РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию гена центросомального белка размером 290 кДа (CEP290). В определенных вариантах осуществления глубокая интронная мутация представляет собой мутацию c.2991+1655A>G. В некоторых вариантах осуществления CEP290 представляет собой CEP290 человека (см. например последовательность CEP290 человека в соответствии с эталонной последовательностью NCBI NG_008417). В некоторых вариантах осуществления CEP290 содержит глубокую интронную мутацию в последовательности, изложенной под SEQ ID NO:23.
[0211] Определенные аспекты настоящего раскрытия дополнительно относятся к способам получения in vitro модели глазного заболевания, связанного с глубокой интронной мутацией в гене. В некоторых вариантах осуществления способы предусматривают введение в эукариотические клетки нуклеиновой кислоты, кодирующей систему CRISPR-Cas, где система CRISPR-Cas содержит i) единую направляющую РНК для нацеливания на последовательности ДНК интрона в гене, ii) нуклеотидную последовательность, кодирующую Cas-белок, iii) однонитевой олигонуклеотид, содержащий матрицу для репарации, направляемой гомологией (HDR), содержащую гомологичные "плечи", фланкирующие требуемую интронную мутацию, и прилегающий к протоспейсеру мотив (PAM); и выделение клеток, которые содержат мутацию, включенную в ген. Иллюстративные способы, связанные с этим подходом, проиллюстрированы в примерах ниже. Как описано выше, CRISPR-Cas-опосредованное расщепление ДНК можно восстановить с помощью NHEJ, что приводит, например, к вырезанию последовательности ДНК-мишени. Однако CRISPR-Cas-опосредованное расщепление ДНК также можно восстановить с помощью репарации, направляемой гомологией (HDR), что приводит, например, к введению последовательности, присутствующей в матрице HDR (например, введенная глубокая интронная мутация). См., например, Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308. Эти способы могут включать в себя любые признаки, аспекты или элементы, описанные в данном документе в отношении способов лечения, вирусных частиц, композиций и наборов.
[0212] Матрица для репарации, направляемой гомологией (HDR), может принимать различные формы, такие как двухнитевой полинуклеотид ДНК или однонитевой олигонуклеотид ДНК (ssODN). Матрица HDR может включать одно или несколько гомологичных "плеч", фланкирующих требуемую интронную мутацию. Эти гомологичные "плечи" могут располагаться в смысловом или в антисмысловом направлении относительно локуса-мишени. В некоторых вариантах осуществления одно или несколько гомологичных "плеч" могут располагаться на расстоянии по меньшей мере приблизительно 40 пар оснований, по меньшей мере приблизительно 50 пар оснований, по меньшей мере приблизительно 60 пар оснований, по меньшей мере приблизительно 70 пар оснований, по меньшей мере приблизительно 80 пар оснований, по меньшей мере приблизительно 90 пар оснований или по меньшей мере приблизительно 100 пар оснований от локуса-мишени.
[0213] Матрица HDR может также включать в себя прилегающий к протоспейсеру мотив (PAM). В некоторых вариантах осуществления PAM содержит мутацию, позволяющую избежать расщепления однонитевого олигонуклеотида экспрессированным Cas-белком в клетках. Например, PAM может быть мутирован таким образом, что конкретный экспрессированный Cas-белок не расщепляет матрицу HDR, или мутирован таким образом, что идентифицирующая последовательность (например, уникальный сайт рестрикции, как показано в примерах ниже) вводится после CRISPR-Cas-опосредованного редактирования геномного локуса.
[0214] Для модели in vitro можно использовать любую подходящую эукариотическую клетку. В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку млекопитающего или человека. В некоторых вариантах осуществления эукариотическая клетка представляет собой линию клеток, такую как линия клеток человека (например, HeLa, A549, 293 и т. д.), линия клеток млекопитающего, линия клеток позвоночного или линия клеток насекомого (например, Sf9 или S2). В некоторых вариантах осуществления эукариотическая клетка представляет собой клетку сетчатки (например, клетку WERI), такую как фоторецепторная клетка.
V. Способы доставки
[0215] Определенные аспекты настоящего раскрытия относятся к лечению заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума. В некоторых аспектах настоящее изобретение предусматривает способы лечения заболевания или нарушения, связанных с глубокой интронной мутацией в гене индивидуума, включающие введение индивидууму терапевтически эффективного количества композиции по настоящему раскрытию, например композиции, содержащей нуклеиновую кислоту, которая кодирует сконструированную не встречающуюся в природе систему CRISPR-Cas по настоящему раскрытию. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая сконструированную не встречающуюся в природе систему CRISPR-Cas, может представлять собой ДНК или РНК. В некоторых вариантах осуществления Cas-белок доставляют в виде белка.
Векторы
[0216] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы. В некоторых вариантах осуществления один или несколько векторов, управляющих экспрессией одного или нескольких элементов системы CRISPR, вводят в клетку, так что экспрессия элементов системы CRISPR направляет образование комплекса CRISPR в одном или нескольких сайтах-мишенях. Например, фермент Cas, направляющая последовательность, связанная с сопряженной с tracr последовательностью, и последовательность tracr могут быть функционально связаны с отдельными регуляторными элементами на отдельных векторах. Как альтернатива, два или более из вышеописанных элементов могут экспрессироваться из одних и тех же или разных регуляторных элементов и/или могут быть объединены в одном векторе, необязательно с одним или несколькими дополнительными векторами, обеспечивающими любые компоненты системы CRISPR, не включенные в первый вектор. Элементы системы CRISPR, которые объединены в вектор, могут располагаться в любой подходящей ориентации. Например, кодирующая последовательность одного элемента может быть расположена на той же или противоположной нити кодирующей последовательности второго элемента и/или ориентирована в том же или противоположном направлении.
[0217] В некоторых вариантах осуществления один промотор управляет экспрессией транскрипта, кодирующего Cas-белок и одно или несколько из направляющей последовательности, сопряженной с tracr последовательности (необязательно функционально связанной с направляющей последовательностью) и последовательности tracr, внедренных в одну или несколько интронных последовательностей (например, каждая в другом интроне, две или более по меньшей мере в одном интроне или все в одном интроне). В некоторых вариантах осуществления Cas-белок, направляющая последовательность, сопряженная с tracr последовательность и последовательность tracr функционально связаны с одним и тем же промотором и экспрессируются с одного и того же промотора. В некоторых вариантах осуществления первая направляющая РНК, вторая направляющая РНК и/или нуклеиновая кислота, кодирующая Cas-белок, функционально связаны с одним или несколькими регуляторными контрольными элементами и/или промоторами.
[0218] В некоторых вариантах осуществления вектор представляет собой плазмиду.
[0219] В некоторых вариантах осуществления первая направляющая РНК, вторая направляющая РНК и Cas-белок экспрессируются в эукариотических клетках. В некоторых вариантах осуществления первая направляющая РНК, вторая направляющая РНК и Cas-белок функционально связаны с одним или несколькими промоторами, которые обеспечивают экспрессию в эукариотической клетке. Из уровня техники известно множество промоторов, которые экспрессируются в эукариотических клетках. Иллюстративные промоторы предоставлены без ограничения ниже.
[0220] В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III. Промоторы РНК-полимеразы III могут включать в себя полноразмерный промотор или его фрагмент, достаточный для осуществления транскрипции РНК-полимеразой III. Для более подробного описания типов промоторов РНК-полимеразы III, структурных особенностей и взаимодействий с РНК-полимеразой III, а также подходящих промоторов РНК-полимеразы III, см. Schramm, L. and Hernandez, N. (2002) Genes Dev. 16:2593-620. Можно использовать любой подходящий промотор РНК-полимеразы III, известный из уровня техники, включая без ограничения промоторы для tRNA, РНК 5S, snRNA U6, H1, 7SK, РНказы P, РНК-компонента частицы распознавания сигнала и snoRNA (см., например, Ma, H. et al. (2014) Mol. Ther. Nucleic Acids 3:e161). В некоторых вариантах осуществления промотор РНК-полимеразы III представляет собой промотор U6, H1 или 7SK. В некоторых вариантах осуществления первая направляющая РНК и/или вторая направляющая РНК функционально связаны с терминатором РНК-полимеразы III. Примеры терминаторов РНК-полимеразы III могут включать без ограничения цепочку уридиновых нуклеотидов длиной по меньшей мере 5-6 оснований (для получения дополнительной информации о терминаторах РНК-полимеразы III см. Marck, C., et al. (2006) Nucleic Acids Res 34(6):1816-35).
[0221] В некоторых вариантах осуществления нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II. Промоторы РНК-полимеразы II могут включать в себя полноразмерный промотор или его фрагмент, достаточный для осуществления транскрипции РНК-полимеразой II. Можно использовать любой подходящий промотор РНК-полимеразы II, известный из уровня техники, включая без ограничения немедленно-ранний промотор цитомегаловируса (CMV), минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотор гена фосфоглицераткиназы-1 (PGK), промотор вируса обезьян 40 (SV40) и промотор CK6, промотор гена транстиретина (TTR), промотор TK, тетрациклин-чувствительный промотор (TRE), промотор HBV, промотор hAAT, промотор LSP, химерные печень-специфичные промоторы (LSP), промотор E2F, промотор гена теломеразы (hTERT); составной промотор энхансер цитомегаловируса/промотор гена бета-актина курицы/гена β-глобина кролика (промотор CAG; Niwa et al., Gene, 1991, 108(2):193-9) и промотор фактора элонгации 1-альфа (EF1-альфа) (Kim et al., Gene, 1990, 91(2):217-23 и Guo et al., Gene Ther., 1996, 3(9):802-10). В некоторых вариантах осуществления промотор предусматривает промотор β-глюкуронидазы человека или промотор из энхансера цитомегаловируса, соединенного с промотором β-актина курицы (CBA). Промотор может представлять собой конститутивный, индуцируемый или репрессируемый промотор. В некоторых вариантах осуществления промотор способен обеспечивать экспрессию гетерологичной нуклеиновой кислоты в клетке глаза. В некоторых вариантах осуществления промотор способен обеспечивать экспрессию гетерологичной нуклеиновой кислоты в фоторецепторных клетках или RPE. В вариантах осуществления промотор представляет собой промотор родопсинкиназы (RK); например промотор RK человека. В некоторых вариантах осуществления промотор представляет собой промотор опсина; например промотор опсина человека или промотор опсина мыши. В некоторых вариантах осуществления промотор представляет собой промотор родопсина, промотор конопсина, промотор бета-фосфодиэстеразы (PDE), промотор гена, связанного с пигментным ретинитом (RP1), или промотор гена межфоторецепторного ретинол-связывающего белка (IRBP).
[0222] Индуцируемые промоторы обеспечивают возможность регуляции экспрессии генов, и они могут регулироваться экзогенно вводимыми соединениями, факторами окружающей среды, такими как температура, или наличием конкретного физиологического состояния, например, острой фазы, определенного состояния дифференцировки клетки, или функционируют только в размножающихся клетках. Индуцируемые промоторы и индуцируемые системы доступны из ряда коммерческих источников, включающих без ограничения Invitrogen, Clontech и Ariad. Многие другие системы были описаны и могут быть без труда выбраны специалистом в данной области техники. Примеры индуцируемых промоторов, регулируемых экзогенно предоставляемыми соединениями, включают индуцируемый цинком промотор металлотионеина овцы (MT), индуцируемый дексаметазоном (Dex) промотор вируса опухоли молочной железы мышей (MMTV), промоторную систему полимеразы T7 (WO 98/10088); промотор, индуцируемый экдизоном насекомых (No et al., Proc. Natl. Acad. Sci. USA, 93:3346-3351 (1996)), систему с тетрациклин-зависимой репрессией (Gossen et al, Proc. Natl. Acad. Sci. USA, 89:5547-5551 (1992)), систему с тетрациклин-зависимой индукцией (Gossen et al, Science, 268:1766-1769 (1995), см. также Harvey et al, Curr. Opin. Chem. Biol., 2:512-518 (1998)), систему с RU486-зависимой репрессией (Wang et al, Nat. Biotech., 15:239-243 (1997) и Wang et al, Gene Ther., 4:432-441 (1997)) и систему с рапамицин-зависимой репрессией (Magari et al, J. Clin. Invest., 100:2865-2872 (1997)). Несколько других типов индуцируемых промоторов, которые могут быть применимыми в данном контексте, представляют собой промоторы, регулируемые конкретным физиологическим состоянием, например, температурой, острой фазой, определенным состоянием дифференцировки клетки, или функционирующие только в размножающихся клетках.
[0223] В другом варианте осуществления для трансгена будет применяться нативный промотор или его фрагмент. Нативный промотор можно применять в тех случаях, когда является желательным, чтобы экспрессия трансгена имитировала экспрессию нативного гена. Нативный промотор можно применять в тех случаях, когда экспрессия трансгена должна регулироваться во времени, или в зависимости от стадии развития, или тканеспецифичным образом, или в ответ на конкретные транскрипционные стимулы. В дополнительном варианте осуществления для имитации экспрессии нативного гена также можно применять другие нативные элементы, осуществляющие контроль экспрессии, такие как энхансерные элементы, сайты полиаденилирования или консенсусные последовательности Козак.
[0224] В некоторых вариантах осуществления регуляторные последовательности придают способности к тканеспецифичной экспрессии генов. В некоторых случаях тканеспецифичные регуляторные последовательности связывают тканеспецифичные факторы транскрипции, которые индуцируют транскрипцию тканеспецифичным образом. Эти тканеспецифичные регуляторные последовательности (например, промоторы, энхансеры и т. д.) хорошо известны из уровня техники. Иллюстративные тканеспецифичные регуляторные последовательности включают без ограничения следующие тканеспецифичные промоторы: промоторы генов нейронов, такие как промотор гена нейрон-специфичной енолазы (NSE) (Andersen et al., Cell. Mol. Neurobiol., 13:503-15 (1993)), промотор гена легкой цепи нейрофиламента (Piccioli et al., Proc. Natl. Acad. Sci. USA, 88:5611-5 (1991)) и промотор нейрон-специфичного гена vgf (Piccioli et al., Neuron, 15:373-84 (1995)). В некоторых вариантах осуществления тканеспецифичный промотор представляет собой промотор гена, выбранного из гена ядерного белка нейронов (NeuN), гена глиофибриллярного кислого белка (GFAP), гена, обусловливающего аденоматозный полипоз толстой кишки (APC), и гена адаптерной молекулы 1, связывающей ионизированный кальций (Iba-1). Другие подходящие тканеспецифичные промоторы будут очевидны специалисту в данной области. В некоторых вариантах осуществления промотор представляет собой промотор бета-актина курицы.
[0225] В настоящем изобретении рассматривается применение рекомбинантного вирусного генома для введения одной или нескольких последовательностей нуклеиновых кислот (например, направляющей РНК и/или нуклеотидной последовательности, кодирующей Cas-белок), для упаковки в вирусную частицу, например нижеописанную вирусную частицу. Рекомбинантный вирусный геном может содержать любой элемент для осуществления экспрессии направляющей РНК и/или нуклеотидной последовательности, кодирующей Cas-белок, например, промотор, ITR, элемент связывания рибосомы, терминатор, энхансер, селективный маркер, интрон, сигнал polyA и/или точку начала репликации. Иллюстративные элементы вирусного генома и способы доставки для ряда вирусных частиц описаны более подробно ниже.
Невирусные системы доставки
[0226] Общепринятые невирусные способы переноса генов также можно использовать для введения нуклеиновых кислот в клетки или ткани-мишени. Невирусные векторные системы доставки включают в себя ДНК-плазмиды, РНК (например, направляющую РНК и/или нуклеотидную последовательность, кодирующую Cas-белок), голую нуклеиновую кислоту (например, ДНК или РНК) и нуклеиновую кислоту (например, ДНК или РНК) в комплексе с системой доставки. Например, вектор может находиться в комплексе с липидом (например, катионным или нейтральным липидом), липосомой, поликатионом, наночастицей или средством, которое усиливает клеточный захват нуклеиновой кислоты. Вектор может находиться в комплексе со средством, подходящим для любого из способов доставки, описанных в данном документе.
[0227] Способы невирусной доставки нуклеиновых кислот включают липофекцию, нуклеофекцию, микроинъекцию, биолистику, наночастицы (см., например, Jin, S. et al. (2009) Methods Mol. Biol. 544:547-557), виросомы, липосомы, иммунолипосомы, конъюгаты поликатион/липид:нуклеиновая кислота, голую ДНК, искусственные вирионы и средство усиления захвата ДНК. Липофекция описана, например, в патентах США №№ 5049386, 4946787 и 4897355); а реагенты для липофекции продаются на коммерческой основе (например, Lipofectamine®, Transfectam™ и Lipofectin™). Катионные и нейтральные липиды, которые подходят для эффективной рецептор-распознающей липофекции полинуклеотидов, включают в себя те, которые принадлежат Felgner, WO 91/17424; WO 91/16024.
[0228] Получение комплексов липид:нуклеиновая кислота, включая нацеленные липосомы, такие как иммунолипидные комплексы, хорошо известно специалисту в данной области техники (см., например, Crystal, Science 270:404-410 (1995); Blaese et al., Cancer Gene Ther. 2:291-297 (1995): Behr et al., Bioconjugate Chem. 5:382-389 (1994); Remy et al., Bioconjugate Chem. 5:647-654 (1994); Gao et al., Gene Therapy 2:710-722 (1995); Ahmad et al., Cancer Res. 52:4817-4820 (1992); патенты США №№ 4186183, 4217344, 4235871, 4261975, 4485054, 4501728, 4774085, 4837028 и 4946787).
[0229] Другие соединения, которые могут находиться в комплексе с вектором по настоящему раскрытию, включают без ограничения катионный пептид (например, поли-L-лизин), соль (например, фосфат кальция), декстран DEAE, дендример (например, полиамидоамин или PAMAM), полиэтиленгликоль, полиэтиленимин (PEI) и их конъюгаты, и т. п. Для более подробного обсуждения таких средств см., например, Luo, D. and Saltzman, W.M. (2000) Nature Biotechnology 18:33-37.'
[0230] В некоторых вариантах осуществления нуклеиновая кислота находится в фармацевтическом составе. В некоторых вариантах осуществления фармацевтический состав дополнительно включает фармацевтически приемлемый носитель. Такие носители хорошо известны из уровня техники (см., например, Remington's Pharmaceutical Sciences, 15th Edition, стр. 1035-1038 и 1570-1580). В некоторых вариантах осуществления фармацевтические композиции, содержащие нуклеиновую кислоту, описанную в данном документе, и фармацевтически приемлемый носитель, пригодны для глазной инъекции. Такими фармацевтически приемлемыми носителями могут быть стерильные жидкости, такие как вода и масло, в том числе стерильные жидкости на основе нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло и т. п. В качестве жидких носителей, в частности для инъекционных растворов, также могут использоваться солевые растворы и водные растворы декстрозы, полиэтиленгликоля (PEG) и глицерина. Кроме того, фармацевтическая композиция может содержать дополнительные ингредиенты, например, консерванты, буферы, средства, регулирующие тоничность, антиоксиданты и стабилизаторы, неионные увлажняющие и осветляющие средства, загустители и т. п. Фармацевтические композиции, описанные в данном документе, могут быть упакованы в единичные дозированные формы или многодозовые формы. Как правило, композиции составляют в виде стерильного или практически изотонического раствора.
Вирусные частицы
[0231] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV).
Частицы rAAV
[0232] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного AAV (rAAV). В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной или несколькими последовательностями инвертированных концевых повторов (ITR) AAV. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу AAV, которая содержит нуклеиновую кислоту, содержащую гетерологичный трансген, фланкированный одним или двумя ITR. В некоторых вариантах осуществления нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована двумя ITR AAV.
[0233] В некоторых вариантах осуществления нуклеиновая кислота содержит одну или две направляющих РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию, функционально связанные компоненты в направлении транскрипции, контролирующие последовательности, в том числе последовательности инициации и терминации, с образованием таким образом кассеты экспрессии. Кассета экспрессии фланкирована на 5'- и 3'-конце по меньшей мере одной функциональной последовательностью ITR AAV. Под термином "функциональные последовательности ITR AAV" подразумевается, что последовательности ITR функционируют надлежащим образом в отношении восстановления, репликации и упаковки вириона AAV. См. Davidson et al., PNAS, 2000, 97(7)3428-32; Passini et al., J. Virol., 2003, 77(12):7034-40; и Pechan et al., Gene Ther., 2009, 16:10-16, все из которых включены в данный документ с помощью ссылки во всей своей полноте. Для осуществления на практике некоторых аспектов настоящего изобретения рекомбинантные векторы содержат по меньшей мере все последовательности AAV, необходимые для заключения в капсид, и физические структуры для инфицирования с помощью rAAV. ITR AAV для использования в векторах по настоящему изобретению не требуют наличия нуклеотидной последовательности дикого типа (например, как описано у Kotin, Hum. Gene Ther., 1994, 5:793-801), и они могут быть изменены с помощью вставки, делеции или замены нуклеотидов, или ITR AAV могут происходить из любого из нескольких серотипов AAV. На сегодняшний день известно более 40 серотипов AAV, и продолжают идентифицировать новые серотипы и варианты существующих серотипов. См. Gao et al., PNAS, 2002, 99(18): 11854-6; Gao et al., PNAS, 2003, 100(10):6081-6; и Bossis et al., J. Virol., 2003, 77(12):6799-810.
[0234] Применение AAV любого серотипа рассматривается в пределах объема настоящего изобретения. В некоторых вариантах осуществления вектор на основе rAAV представляет собой вектор, полученный из серотипа AAV, содержащий без ограничения ITR AAV, которые представляют собой ITR AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или ITR AAV мыши и т. п. В некоторых вариантах осуществления нуклеиновая кислота в AAV содержит ITR из ITR AAV, который представляет собой AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV коз, AAV крупного рогатого скота или AAV мышей и т. п. В определенных вариантах осуществления ITR AAV представляют собой ITR AAV2.
[0235] В некоторых вариантах осуществления вектор может включать в себя спейсерную последовательность нуклеиновой кислоты. В некоторых вариантах осуществления спейсерная последовательность нуклеиновой кислоты может кодировать зеленый флуоресцентный белок. В некоторых вариантах осуществления спейсерная последовательность нуклеиновой кислоты может располагаться между промотором и одним или более из одной или двух направляющих РНК по настоящему раскрытию и/или нуклеотидной последовательности, кодирующей Cas-белок по настоящему раскрытию.
[0236] В некоторых аспектах настоящее изобретение предусматривает вирусные частицы, содержащие рекомбинантный самокомплементарный геном. В некоторых вариантах осуществления вектор представляет собой самокомплементарный вектор. Вирусные частицы AAV с самокомплементарными геномами и способы применения самокомплементарных геномов AAV описаны в патентах США №№ 6596535; 7125717; 7765583; 7785888; 7790154; 7846729; 8093054 и 8361457; а также Wang Z., et al., (2003) Gene Ther 10:2105-2111, каждый из которых включен в данный документ с помощью ссылки во всей своей полноте. Благодаря своим частично комплементарным последовательностям (например, комплементарным кодирующим и некодирующим цепям трансгена), rAAV, содержащий самокомплементарный геном, будет быстро образовывать двухнитевую молекулу ДНК. В некоторых вариантах осуществления настоящее изобретение предусматривает вирусную частицу AAV, содержащую геном AAV, где геном rAAV содержит первую гетерологичную полинуклеотидную последовательность (например, одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию) и вторую гетерологичную полинуклеотидную последовательность (например, некодирующую или антисмысловую нить одной или двух направляющих РНК по настоящему раскрытию и/или нуклеотидной последовательности, кодирующей Cas-белок по настоящему раскрытию), где первая гетерологичная полинуклеотидная последовательность может образовывать внутринитевые пары оснований со второй полинуклеотидной последовательностью вдоль большей части или всей ее длины.
[0237] В некоторых вариантах осуществления первая гетерологичная полинуклеотидная последовательность и вторая гетерологичная полинуклеотидная последовательность соединены с помощью последовательности, которая облегчает внутринитевое спаривание оснований, например шпилечной структуры ДНК. Шпилечные структуры известны из уровня техники, например в молекулах siRNA. В некоторых вариантах осуществления первая гетерологичная полинуклеотидная последовательность и вторая гетерологичная полинуклеотидная последовательность связаны с помощью мутированного ITR (например, правого ITR). Мутантный ITR характеризуется делецией D-области, содержащей последовательность концевого разрешения. В результате, при репликации вирусного генома AAV белки rep не будут расщеплять вирусный геном по мутантному ITR, и в связи с этим в вирусный капсид будет упакован рекомбинантный вирусный геном, содержащий следующее в порядке от 5'- к 3'-концу: ITR AAV, первую гетерологичную полинуклеотидную последовательность, в том числе регуляторные последовательности, мутантный ITR AAV, второй гетерологичный полинуклеотид в обратной ориентации по отношению к первому гетерологичному полинуклеотиду и третий ITR AAV.
[0238] В некоторых вариантах осуществления первая гетерологичная последовательность нуклеиновой кислоты и вторая гетерологичная последовательность нуклеиновой кислоты соединены с помощью мутантного ITR (например, правого ITR). В некоторых вариантах осуществления ITR содержит полинуклеотидную последовательность 5'-CACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCACGCCCGG GCTTTGCCCGGGCG-3' (SEQ ID NO:24). Мутантный ITR характеризуется делецией D-области, содержащей последовательность концевого разрешения. В результате, при репликации вирусного генома AAV белки rep не будут расщеплять вирусный геном по мутантному ITR, и в связи с этим в вирусный капсид будет упакован рекомбинантный вирусный геном, содержащий следующее в порядке от 5'- к 3'-концу: ITR AAV, первую гетерологичную полинуклеотидную последовательность, в том числе регуляторные последовательности, мутантный ITR AAV, второй гетерологичный полинуклеотид в обратной ориентации по отношению к первому гетерологичному полинуклеотиду и третий ITR AAV.
[0239] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную вирусную частицу AAV, содержащую вектор на основе рекомбинантного AAV. Для оптимизации трансдукции конкретных клеток-мишеней или для нацеливания на определенные типы клеток в пределах конкретной ткани-мишени (например, ткани глаза) применяют различные серотипы AAV. Частица rAAV может содержать белки вируса и нуклеиновые кислоты вируса одного и того же серотипа или смешанного серотипа. Например, в некоторых вариантах осуществления частица rAAV может содержать капсидные белки AAV2 по настоящему изобретению и по меньшей мере один ITR AAV2, или она может содержать капсидные белки AAV2 и по меньшей мере один ITR AAV1. В данном документе для получения частицы rAAV предусматривается любая комбинация серотипов AAV, как если бы каждая комбинация была описана в данном документе в явной форме. В некоторых вариантах осуществления настоящее изобретение предусматривает частицы rAAV, содержащие капсид AAV2 по настоящему изобретению. В некоторых вариантах осуществления настоящее изобретение предусматривает частицы rAAV, содержащие капсид AAVrh8R по настоящему изобретению.
[0240] В некоторых вариантах осуществления частицы rAAV содержат капсид AAV1, капсид AAV2, капсид AAV3, капсид AAV4, капсид AAV5, капсид AAV6 (например, капсид AAV6 дикого типа или вариант капсида AAV6, такой как ShH10, как описано в публикации заявки № 2012/0164106, опубликованной до выдачи патента США), капсид AAV7, капсид AAV8, капсид AAVrh8, капсид AAVrh8R, капсид AAV9 (например, капсид AAV9 дикого типа или модифицированный капсид AAV9, как описано в публикации заявки № 2013/0323226, опубликованной до выдачи патента США), капсид AAV10, капсид AAVrh10, капсид AAV11, капсид AAV12, капсид с мутацией по тирозину, капсид с мутацией, влияющей на связывание с гепарином, капсид AAV2R471A, капсид AAVAAV2/2-7m8, капсид AAV DJ (например, капсид AAV-DJ/8, капсид AAV-DJ/9 или любые другие капсиды, описанные в публикации заявки 2012/0066783, опубликованной до выдачи патента США), капсид AAV2 N587A, капсид AAV2 E548A, капсид AAV2 N708A, капсид AAV V708K, капсид AAV козы, химерный капсид AAV1/AAV2, капсид AAV крупного рогатого скота, капсид AAV мыши, капсид rAAV2/HBoV1 или капсид AAV, описанный в патенте США № 8283151 или в Международной публикации № WO/2003/042397. В некоторых вариантах осуществления мутантный капсидный белок сохраняет способность формировать капсид AAV. В некоторых вариантах осуществления частица rAAV содержит мутантный по тирозину капсид AAV5 (Zhong L. et al., (2008) Proc Natl Acad Sci U S A 105(22):7827-7832. В дополнительных вариантах осуществления частица rAAV содержит капсидные белки серотипа AAV из клад A-F (Gao, et al., J. Virol. 2004, 78(12):6381). В некоторых вариантах осуществления частица rAAV содержит капсидный белок AAV1 или его мутант. В других вариантах осуществления частица rAAV содержит капсидный белок AAV2 или его мутант. В некоторых вариантах осуществления серотип AAV представляет собой AAV1, AAV2, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10 или AAVrh10. В некоторых вариантах осуществления частица rAAV содержит капсид AAV серотипа 1 (AAV1). В некоторых вариантах осуществления частица rAAV содержит капсид AAV серотипа 2 (AAV2). В некоторых вариантах осуществления рекомбинантная вирусная частица AAV содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10. В некоторых вариантах осуществления капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10 содержит мутацию, затрагивающую тирозин, или мутацию гепаран-связывающего сайта, например, как описано ниже.
[0241] Известно, что капсид AAV (например, AAV2, AAVrh8R и т. д.) содержит три капсидных белка: VP1, VP2 и VP3. Эти белки содержат значительные количества перекрывающихся аминокислотных последовательностей и уникальные N-концевые последовательности. Капсид AAV2 содержит 60 субъединиц, расположенных по принципу икосаэдрической симметрии (Xie, Q., et al. (2002) Proc. Natl. Acad. Sci. 99(16):10405-10). Было обнаружено, что VP1, VP2 и VP3 присутствуют в соотношении 1:1:10.
[0242] Известно, что связывание между капсидными белками AAV2 и HSPG происходит посредством электростатических взаимодействий между основными остатками капсидных белков AAV2 и отрицательно заряженными остатками гликозаминогликана (Opie, SR et al., (2003) J. Virol. 77:6995-7006; Kern, A et al., (2003) J. Virol. 77:11072-11081). Конкретные остатки капсида, вовлеченные в эти взаимодействия, включают в себя R484, R487, K532, R585 и R588. Было показано, что мутации в этих остатках ослабляют связывание AAV2 с клетками HeLa и самим гепараном (Opie, SR et al., (2003) J. Virol. 77:6995-7006; Kern, A et al., (2003) J. Virol. 77:11072-11081; WO 2004/027019 A2, патент США № 7629322). Дополнительно, не желая ограничиваться какой-либо теорией, полагают, что посредством аминокислотной замены(-мен) одного или нескольких остатков, соответствующих аминокислотам 484, 487, 532, 585 или 588, нумерация которых приведена согласно нумерации VP1 AAV2, можно модулировать трансдукционные свойства типов капсидов AAV, которые не связываются с HSPG, или можно модулировать трансдукционные свойства типов капсидов AAV независимо от их способности к связыванию с HSPG.
[0243] В некоторых вариантах осуществления частицы rAAV несут мутацию в капсидном белке в остатке, который взаимодействует с HSPG, или в одном или нескольких остатках, соответствующих аминокислотам 484, 487, 532, 585 или 588, нумерация которых приведена согласно нумерации VP1 AAV2. Соответственно, в некоторых вариантах осуществления после доставки гетерологичная нуклеиновая кислота, кодируемая вектором на основе rAAV, экспрессируется при повышенном уровне экспрессии по сравнению с уровнем экспрессии гетерологичной нуклеиновой кислоты частицы rAAV, содержащей капсид rAAV, который содержит эталонный капсидный белок rAAV (например, капсидный белок rAAV дикого типа). В некоторых вариантах осуществления уровень экспрессии нуклеиновой кислоты повышен по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 25%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 75% или по меньшей мере на приблизительно 100%. В некоторых вариантах осуществления после доставки частица rAAV вызывает сниженное нейровоспаление по сравнению с частицей rAAV, содержащей эталонный капсидный белок rAAV (например, капсидный белок rAAV дикого типа). В некоторых вариантах осуществления нейровоспаление снижено по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 25%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 75% или по меньшей мере на приблизительно 100%. Подходящий эталонный капсидный белок rAAV может включать в себя любой капсидный белок, у которого отсутствуют одна или несколько аминокислотных замен в одном или нескольких положениях, в которых происходит взаимодействие с гепарансульфатсодержащим протеогликаном (эталонный капсид может, таким образом, содержать одну или несколько "фоновых" замен, которые не изменяют связывание с HSPG).
[0244] В некоторых вариантах осуществления частица rAAV содержит a) капсид rAAV, содержащий капсидные белки rAAV, которые содержат одну или несколько аминокислотных замен в одном или нескольких положениях, в которых происходит взаимодействие с гепарансульфатсодержащим протеогликаном, и b) вектор на основе rAAV, содержащий гетерологичную нуклеиновую кислоту и по меньшей мере один инвертированный концевой повтор AAV.
[0245] В некоторых вариантах осуществления одна или несколько аминокислотных замен ослабляют связывание частицы rAAV с гепарансульфатсодержащим протеогликаном приблизительно по меньшей мере на 10%, приблизительно по меньшей мере на 25%, приблизительно по меньшей мере на 50%, приблизительно по меньшей мере на 75% или приблизительно по меньшей мере на 100%. В некоторых вариантах осуществления одна или несколько аминокислотных замен ослабляют связывание частицы rAAV с гепарансульфатсодержащим протеогликаном приблизительно по меньшей мере на 10%, приблизительно по меньшей мере на 15%, приблизительно по меньшей мере на 20%, приблизительно по меньшей мере на 25%, приблизительно по меньшей мере на 30%, приблизительно по меньшей мере на 35%, приблизительно по меньшей мере на 40%, приблизительно по меньшей мере на 45%, приблизительно по меньшей мере на 50%, приблизительно по меньшей мере на 55%, приблизительно по меньшей мере на 60%, приблизительно по меньшей мере на 65%, приблизительно по меньшей мере на 70%, приблизительно по меньшей мере на 75%, приблизительно по меньшей мере на 80%, приблизительно по меньшей мере на 85%, приблизительно по меньшей мере на 90%, приблизительно по меньшей мере на 95% или приблизительно по меньшей мере на 100% (по сравнению со связыванием частицы rAAV, содержащей капсид дикого типа). В некоторых вариантах осуществления одна или несколько аминокислотных замен ослабляют связывание частицы rAAV с гепарансульфатсодержащим протеогликаном на любое из значений от приблизительно 10% до приблизительно 100%, от приблизительно 20% до приблизительно 100%, от приблизительно 30% до приблизительно 100%, от приблизительно 40% до приблизительно 100%, от приблизительно 50% до приблизительно 100%, от приблизительно 60% до приблизительно 100%, от приблизительно 70% до приблизительно 100%, от приблизительно 80% до приблизительно 100%, от приблизительно 90% до приблизительно 100%, от приблизительно 10% до приблизительно 90%, от приблизительно 20% до приблизительно 90%, от приблизительно 30% до приблизительно 90%, от приблизительно 40% до приблизительно 90%, от приблизительно 50% до приблизительно 90%, от приблизительно 60% до приблизительно 90%, от приблизительно 70% до приблизительно 90%, от приблизительно 80% до приблизительно 90%, от приблизительно 10% до приблизительно 80%, от приблизительно 20% до приблизительно 80%, от приблизительно 30% до приблизительно 80%, от приблизительно 40% до приблизительно 80%, от приблизительно 50% до приблизительно 80%, от приблизительно 60% до приблизительно 80%, от приблизительно 70% до приблизительно 80%, от приблизительно 10% до приблизительно 70%, от приблизительно 20% до приблизительно 70%, от приблизительно 30% до приблизительно 70%, от приблизительно 40% до приблизительно 70%, от приблизительно 50% до приблизительно 70%, от приблизительно 60% до приблизительно 70%, от приблизительно 10% до приблизительно 60%, от приблизительно 20% до приблизительно 60%, от приблизительно 30% до приблизительно 60%, от приблизительно 40% до приблизительно 60%, от приблизительно 50% до приблизительно 60%, от приблизительно 10% до приблизительно 50%, от приблизительно 20% до приблизительно 50%, от приблизительно 30% до приблизительно 50%, от приблизительно 40% до приблизительно 50%, от приблизительно 10% до приблизительно 40%, от приблизительно 20% до приблизительно 40%, от приблизительно 30% до приблизительно 40%, от приблизительно 10% до приблизительно 30%, от приблизительно 20% до приблизительно 30% или от приблизительно 10% до приблизительно 20% (по сравнению со связыванием частицы rAAV, содержащей капсид дикого типа). В некоторых вариантах осуществления одна или несколько аминокислотных замен приводят к невыявляемому связыванию частицы rAAV с гепарансульфатсодержащим протеогликаном по сравнению со связыванием частицы rAAV дикого типа. Способы измерения связывания частиц AAV с HSPG; например связывания с хроматографической средой на основе гепарансульфата или связывания с клеткой, о которой известно, что она экспрессирует HSPG на своей поверхности, известны из уровня техники. Например, см. Opie, SR et al., (2003) J. Virol. 77:6995-7006 и Kern, A et al., (2003) J. Virol. 77:11072-11081.
[0246] В некоторых вариантах осуществления частицы rAAV содержат одну или несколько аминокислотных замен в капсидных белках, которые ослабляют или устраняют связывание частицы rAAV с гепарансульфатсодержащим протеогликаном, и/или где одна или несколько аминокислотных замен находятся в положениях 484, 487, 532, 585 или 588, нумерация которых приведена согласно нумерации VP1 AAV2. Термин "нумерация приведена согласно VP1 AAV2", используемый в данном документе, относится к аминокислоте упомянутого капсидного белка, соответствующей упомянутой аминокислоте VP1 AAV2. Например, если одна или несколько аминокислотных замен находятся в положениях 347, 350, 390, 395, 448, 451, 484, 487, 527, 532, 585 и/или 588, нумерация которых приведена согласно VP1 AAV2, то одна или несколько аминокислотных замен произведены в отношении аминокислоты(-лот) упомянутого капсидного белка, соответствующих аминокислотам 347, 350, 390, 395, 448, 451, 484, 487, 527, 532, 585 и/или 588 VP1 AAV2. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся в положениях 484, 487, 532, 585 или 588 VP1 AAV2. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся в положениях 484, 487, 532, 585 или 588 VP1 AAV3, нумерация которых приведена согласно VP1 AAV2. В некоторых вариантах осуществления одна или несколько аминокислотных замен находятся в положениях 485, 488, 528, 533, 586 или 589, нумерация которых приведена согласно нумерации VP1 AAVrh8R. В некоторых вариантах осуществления одна или несколько аминокислот в положении(-ях), соответствующих аминокислотам 585 и/или 588 (нумерация которых приведена согласно VP1 AAV2), замещены аргининовыми остатками (например, S586 и/или T589 для AAV1 или AAV6; S586 и/или A589 для AAV9; A586 и/или T589 для AAVrh8R; Q588 и/или T591 для AAV8 и Q588 и/или A591 для AAVrh10). В других вариантах осуществления одна или несколько аминокислот (например, аргинин или лизин) в положении(-ях), соответствующих аминокислотам 484, 487, 527 и/или 532 (нумерация которых приведена согласно VP1 AAV2), замещены аминокислотами, не являющимися положительно заряженными, такими как аланин (например, R485, R488, K528 и/или K533 для AAV1 или AAV6; R485, R488, K528 и/или R533 для AAV9 или AAVrh8R и R487, R490, K530 и/или R535 для AAV8 или AAVrh10).
Другие вирусные частицы
[0247] В некоторых вариантах осуществления вектор представляет собой рекомбинантный аденовирусный вектор. В некоторых вариантах осуществления вирусная частица представляет собой аденовирусную частицу. В некоторых вариантах осуществления аденовирусная частица представляет собой рекомбинантную аденовирусную частицу, например полинуклеотидный вектор, содержащий между двумя ITR одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. В некоторых вариантах осуществления в аденовирусной частице отсутствует или она содержит дефектную копию одного или нескольких генов E1, которые делают аденовирус дефектным по репликации. Аденовирусы содержат линейний двухнитевой ДНК-геном внутри крупного (~950Å), не имеющего оболочки икосаэдрического капсида. Аденовирусы имеют крупный геном, который может включать в себя более 30 т.п.о. гетерологичной последовательности (например, вместо области E1 и/или E3), что делает их исключительно подходящими для применения с более крупными гетерологичными генами. Известно также, что они инфицируют делящиеся и неделящиеся клетки, и обычно не интегрируются в геном хозяина (хотя гибридные варианты могут обладать такой способностью). В некоторых вариантах осуществления аденовирусный вектор может представлять собой аденовирусный вектор первого поколения с гетерологичной последовательностью вместо E1. В некоторых вариантах осуществления аденовирусный вектор может представлять собой аденовирусный вектор второго поколения с дополнительными мутациями или делециями в E2A, E2B и/или E4. В некоторых вариантах осуществления аденовирусный вектор может представлять собой аденовирусный вектор третьего поколения или "выпотрошенный" аденовирусный вектор, в котором отсутствуют все кодирующие вирус гены с сохранением только ITR и сигнала упаковки, и для репликации и упаковки которого требуется наличие аденовируса-помощника в транс-положении. Аденовирусные частицы исследовали в отношении возможности применения в качестве векторов для транзиентной трансфекции клеток млекопитающих, а также в качестве векторов для генной терапии. Для дополнительного описания см., например, Danthinne, X. and Imperiale, M.J. (2000) Gene Ther. 7:1707-14 и Tatsis, N. and Ertl, H.C. (2004) Mol. Ther. 10:616-29.
[0248] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу, содержащую нуклеиновую кислоту, которая содержит одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. Использование аденовируса любого серотипа рассматривается в пределах объема настоящего изобретения. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор представляет собой вектор, полученный из аденовируса серотипов, включая без ограничения аденовирус серотипа 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантный аденовирусный вектор получен из аденовируса серотипа 2 или варианта аденовируса серотипа 5.
[0249] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную аденовирусную частицу с заключенным в капсид рекомбинантным аденовирусным вектором. В некоторых вариантах осуществления рекомбинантные вирусные частицы содержат аденовирусную частицу в комбинации с одним или несколькими капсидными белками стороннего вируса. Такие комбинации могут быть обозначены как псевдотипированные рекомбинантные аденовирусные частицы. В некоторых вариантах осуществления капсидные белки стороннего вируса, применяемые в создании псевдотипированных рекомбинантных аденовирусных частиц, получены из стороннего вируса или аденовируса другого серотипа. В некоторых вариантах осуществления капсидные белки стороннего вируса получены, в том числе без ограничения, из реовируса типа 3. Примеры комбинаций векторов и капсидных белков, применяемых в создании псевдотипированных аденовирусных частиц, можно найти в следующих литературных источниках (Tatsis, N. et al. (2004) Mol. Ther. 10(4):616-629 и Ahi, Y. et al. (2011) Curr. Gene Ther. 11(4):307-320). Для оптимизации трансдукции конкретных клеток-мишеней или для нацеливания специфических типов клеток в пределах определенной ткани-мишени (например, ткани, пораженной заболеванием) можно применять аденовирусы различных серотипов. Ткани или клетки, на которые целенаправленно воздействуют аденовирусы конкретных серотипов, включают без ограничения ткани или клетки легкого (например, HuAd3), селезенки и печени (например, HuAd37), гладких мышц, синовиоциты, дендритные клетки, клетки сердечно-сосудистой системы, линии клеток опухолей (например, HuAd11) и дендритные клетки (например, HuAd5, псевдотипированный с ретровирусом типа 3, HuAd30 или HuAd35). Для дополнительного описания см. Ahi, Y. et al. (2011) Curr. Gene Ther. 11(4):307-320, Kay, M. et al. (2001) Nat. Med. 7(1):33-40, and Tatsis, N. et al. (2004) Mol. Ther. 10(4):616-629. В некоторых вариантах осуществления рекомбинантная аденовирусная частица может содержать капсид аденовируса серотипа, включая без ограничения 2, 1, 5, 6, 19, 3, 11, 7, 14, 16, 21, 12, 18, 31, 8, 9, 10, 13, 15, 17, 19, 20, 22, 23, 24-30, 37, 40, 41, AdHu2, AdHu 3, AdHu4, AdHu24, AdHu26, AdHu34, AdHu35, AdHu36, AdHu37, AdHu41, AdHu48, AdHu49, AdHu50, AdC6, AdC7, AdC69, Ad типа 3 крупного рогатого скота, Ad типа 2 собаки, Ad овцы или Ad типа 3 свиньи. В некоторых вариантах осуществления рекомбинантная аденовирусная частица содержит капсид аденовируса серотипа 2 или вариант капсида аденовируса серотипа 5.
[0250] В некоторых вариантах осуществления вектор представляет собой вектор на основе рекомбинантного лентивируса. В некоторых вариантах осуществления вирусная частица представляет собой лентивирусную частицу. В некоторых вариантах осуществления лентивирусная частица представляет собой рекомбинантную лентивирусную частицу, например полинуклеотидный вектор, содержащий между двумя LTR одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. Лентивирусы представляют собой ретровирусы с положительно-смысловой ssRNA с геномом размером примерно 10 т.п.о. Известно, что лентивирусы интегрируются в геном делящихся и неделящихся клеток. Лентивирусные частицы можно получить, например, путем трансфекции множеством плазмид (обычно геном лентивируса и гены, необходимые для репликации и/или упаковки, разделены для предотвращения репликации вируса) в пакующую линию клеток, в которых модифицированный геном лентивируса пакуется в лентивирусные частицы. В некоторых вариантах осуществления лентивирусная частица может относиться к вектору первого поколения, у которого отсутствует ген белка оболочки. В некоторых вариантах осуществления лентивирусная частица может относиться к вектору второго поколения, у которого отсутствуют все гены, за исключением областей gag/pol и tat/rev. В некоторых вариантах осуществления лентивирусная частица может относиться к вектору третьего поколения, который содержит только эндогенные гены rev, gag и pol и имеет химерный LTR для трансдукции без гена tat (см. Dull, T. et al. (1998) J. Virol. 72:8463-71). Для дополнительного описания см. Durand, S. and Cimarelli, A. (2011) Viruses 3:132-59.
[0251] В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу, содержащую нуклеиновую кислоту, которая содержит одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. Применение любого лентивирусного вектора рассматривается в пределах объема настоящего изобретения. В некоторых вариантах осуществления лентивирусный вектор получен из лентивируса, в том числе без ограничения вируса иммунодефицита человека 1 (HIV-1), вируса иммунодефицита человека 2 (HIV-2), вируса иммунодефицита обезьян (SIV), вируса иммунодефицита кошек (FIV), вируса инфекционной анемии лошадей (EIAV), вируса иммунодефицита крупного рогатого скота (BIV), вируса заболевания Джембрана (JDV), вируса Висна (VV) и вируса артрита-энцефалита коз (CAEV).
[0252] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную лентивирусную частицу с заключенным в капсид вектором на основе рекомбинантного лентивируса. В некоторых вариантах осуществления рекомбинантные вирусные частицы содержат лентивирусный вектор в комбинации с одним или несколькими капсидными белками стороннего вируса. Такие комбинации могут быть обозначены как псевдотипированные рекомбинантные лентивирусные частицы. В некоторых вариантах осуществления капсидные белки стороннего вируса, применяемые в создании псевдотипированных рекомбинантных лентивирусных частиц, получены из стороннего вируса. В некоторых вариантах осуществления капсидный белок стороннего вируса, применяемый в создании псевдотипированных рекомбинантных лентивирусных частиц, представляет собой гликопротеин вируса везикулярного стоматита (VSV-GP). VSV-GP взаимодействует с универсальным клеточным рецептором, обеспечивая широкий тканевый тропизм для псевдотипированных рекомбинантных лентивирусных частиц. Кроме того, считается, что VSV-GP обеспечивает более высокую стабильность псевдотипированных рекомбинантных лентивирусных частиц. В других вариантах осуществления капсидные белки стороннего вируса получены, в том числе без ограничения, из вируса Чандипура, вируса бешенства, вируса Мокола, вируса лимфатического хориоменингита (LCMV), вируса Росс-ривер (RRV), вируса Синдбис, вируса леса Семлики (SFV), вируса венесуэльского энцефалита лошадей, эболавируса Рестон, эболавируса Заир, вируса Марбург, вируса Ласса, вируса лейкоза птиц (ALV), ретровируса овец Jaagsiekte (JSRV), вируса лейкоза мышей Молони (MLV), вируса лейкоза гиббонов (GALV), эндогенного ретровируса кошек (RD114), Т-лимфотропного вируса человека 1 (HTLV-1), пенящегося вируса человека, вируса Меди-Висна (MVV), SARS-CoV, вируса Сендай, респираторно-синцитиального вируса (RSV), вируса парагриппа человека типа 3, вируса гепатита С (HCV), вируса гриппа, вируса чумы домашней птицы (FPV) или вируса множественного ядерного полиэдроза совки калифорнийской люцерновой (AcMNPV).
[0253] В некоторых вариантах осуществления вектор на основе рекомбинантного лентивируса получен из лентивируса, псевдотипированного вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами. Примеры комбинаций векторов и капсидных белков, применяемых в создании псевдотипированных лентивирусных частиц, можно найти, например, в Cronin, J. et al. (2005). Curr. Gene Ther. 5(4):387-398. Для оптимизации трансдукции определенных клеток-мишеней или для нацеливания специфичных типов клеток в пределах определенной ткани-мишени (например, пораженной заболеванием ткани) можно использовать различные псевдотипированные рекомбинантные лентивирусные частицы. Например, ткани, на которые целенаправленно воздействуют конкретными псевдотипированными рекомбинантными лентивирусными частицами, включают без ограничения ткань печени (например, псевдотипированная F-белком VSV-G, LCMV, RRV или SeV), ткань легкого (например, псевдотипированная белком вирусов Эбола, Марбург, F-белком и HN-белком SeV или белком JSRV), островковые клетки поджелудочной железы (например, псевдотипированные белком LCMV), ткань центральной нервной системы (например, псевдотипированная белком VSV-G, LCMV, вируса бешенства или Мокола), ткань сетчатки (например, псевдотипированная белком VSV-G или вируса Мокола), моноциты или мышцы (например, псевдотипированные белком вируса Мокола или Эбола), ткань гематопоэтической системы (например, псевдотипированная белком RD114 или GALV) или раковые клетки (например, псевдотипированные белком GALV или LCMV). Для дополнительного описания см. Cronin, J. et al. (2005). Curr. Gene Ther. 5(4):387-398 and Kay, M. et al. (2001) Nat. Med. 7(1):33-40. В некоторых вариантах осуществления рекомбинантная лентивирусная частица содержит капсид, псевдотипированный вирусом везикулярного стоматита (VSV), вирусом лимфатического хориоменингита (LCMV), вирусом Росс-ривер (RRV), вирусом Эбола, вирусом Марбург, вирусом Мокола, вирусом бешенства, RD114 или их вариантами.
[0254] В некоторых вариантах осуществления вектор представляет собой вектор на основе rHSV. В некоторых вариантах осуществления вирусная частица представляет собой частицу вируса простого герпеса (HSV). В некоторых вариантах осуществления частица HSV представляет собой частицу rHSV, например полинуклеотидный вектор, содержащий между двумя TR одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. HSV представляет собой вирус с двухнитевой ДНК, покрытой оболочкой, с геномом размером примерно 152 т.п.о. Преимущественно, примерно половина его генов является несущественной и может быть удалена, чтобы вместить гетерологичную последовательность. Частицы HSV инфицируют неделящиеся клетки. Кроме того, они природным путем обеспечивают переход в неактивное состояние в нейронах, перемещаются с помощь ретроградного транспорта и могут переноситься через синапсы, что делает их преимущественными для трансфекции нейронов и/или для подходов на основе генной терапии, задействующих нервную систему. В некоторых вариантах осуществления частица HSV может быть дефектной по репликации или репликационно компетентной (например, компетентной в отношении одного цикла репликации посредством инактивации одного или нескольких "поздних" генов). Для дополнительного описания см. Manservigi, R. et al. (2010) Open Virol. J. 4:123-56.
[0255] В некоторых вариантах осуществления вирусная частица представляет собой частицу rHSV, содержащую нуклеиновую кислоту, которая содержит одну или две направляющие РНК по настоящему раскрытию и/или нуклеотидную последовательность, кодирующую Cas-белок по настоящему раскрытию. Применение любого вектора HSV рассматривается в пределах объема настоящего изобретения. В некоторых вариантах осуществления вектор HSV получен из серотипа HSV, в том числе без ограничения HSV-1 и HSV-2.
[0256] В некоторых вариантах осуществления вектор заключен в капсид вирусной частицы. В некоторых вариантах осуществления вирусная частица представляет собой рекомбинантную частицу HSV с заключенным в капсид вектором на основе рекомбинантного HSV. В некоторых вариантах осуществления рекомбинантные вирусные частицы содержат вектор HSV в комбинации с одним или несколькими капсидными белками стороннего вируса. Такие комбинации могут быть обозначены как псевдотипированные частицы rHSV. В некоторых вариантах осуществления капсидные белки стороннего вируса, применяемые в создании псевдотипированных частиц rHSV, получены из стороннего вируса или HSV другого серотипа. В некоторых вариантах осуществления капсидный белок стороннего вируса, применяемый в создании псевдотипированной частицы rHSV, представляет собой гликопротеин вируса везикулярного стоматита (VSV-GP). VSV-GP взаимодействует с универсальным клеточным рецептором, обеспечивая широкий тканевый тропизм для псевдотипированных частиц rHSV. Кроме того, считается, что VSV-GP обеспечивает более высокую стабильность псевдотипированных частиц rHSV. В других вариантах осуществления капсидный белок стороннего вируса может происходить из HSV другого серотипа. Например, вектор HSV-1 может содержать один или несколько капсидных белков HSV-2. Для оптимизации трансдукции определенных клеток-мишеней или для нацеливания специфичных типов клеток в пределах определенной ткани-мишени (например, пораженной заболеванием ткани) можно использовать HSV различных серотипов. Ткани или клетки, на которые целенаправленно воздействуют аденовирусы конкретных серотипов, включают без ограничения ткани и клетки центральной нервной системы и нейроны (например, HSV-1). Для дополнительного описания см. Manservigi, R. et al. (2010) Open Virol J 4:123-156, Kay, M. et al. (2001) Nat. Med. 7(1):33-40, и Meignier, B. et al. (1987) J. Infect. Dis. 155(5):921-930. В некоторых вариантах осуществления рекомбинантная частица HSV представляет собой частицу rHSV-1 или вирусную частицу rHSV-2.
Получение вирусных частиц
[0257] Из уровня техники известно множество способов получения частиц, представляющих собой аденовирусные векторы. Например, в случае "выпотрошенного" аденовирусного вектора, геном аденовирусного вектора и геном аденовируса-помощника можно ввести путем трансфекции в пакующую линию клеток (например, линию клеток 293). В некоторых вариантах осуществления геном аденовируса-помощника может содержать сайты рекомбинации, фланкирующие его сигнал упаковки, и оба генома можно ввести путем трансфекции в пакующую линию клеток, которые экспрессируют рекомбиназу (например, можно применять систему Cre/loxP), вследствие чего аденовирусный вектор, представляющий интерес, упаковывается более эффективно, чем аденовирус-помощник (см., например, Alba, R. et al. (2005) Gene Ther. 12 Suppl 1:S18-27). Аденовирусные векторы можно собирать и очищать с использованием стандартных способов, таких как описанные в данном документе.
[0258] Из уровня техники известно множество способов получения частиц, представляющих собой лентивирусные векторы. Например, в случае лентивирусного вектора третьего поколения, вектор, содержащий лентивирусный геном, представляющий интерес, с генами gag и pol, вместе с вектором, содержащим ген rev, можно ввести путем совместной трансфекции в пакующую линию клеток (например, линию клеток 293). Лентивирусный геном, представляющий интерес, также содержит химерный LTR, который способствует осуществлению транскрипции в отсутствие Tat (см. Dull, T. et al. (1998) J. Virol. 72:8463-71). Лентивирусные векторы можно собирать и очищать с использованием способов (например, Segura MM, et al., (2013) Expert Opin Biol Ther. 13(7):987-1011), описанных в данном документе.
[0259] Из уровня техники известно множество способов получения частиц HSV. Векторы HSV можно собирать и очищать с использованием стандартных способов, таких как описанные в данном документе. Например, в случае вектора на основе дефектного по репликации HSV, геном HSV, представляющий интерес, в котором отсутствуют все немедленно-ранние (IE) гены, можно вводить путем трансфекции в дополняющую линию клеток, которые предоставляют гены, необходимые для продуцирования вирусов, как например ICP4, ICP27 и ICP0 (см., например, Samaniego, L.A. et al. (1998) J. Virol. 72:3307-20). Векторы на основе HSV можно собирать и очищать с использованием описанных способов (например, Goins, WF et al., (2014) Herpes Simplex Virus Methods in Molecular Biology 1144:63-79).
[0260] Из уровня техники известны многочисленные способы получения векторов на основе rAAV, в том числе трансфекция, получение стабильных линий клеток и применение систем продуцирования на основе инфекционных гибридных вирусов, которые включают в себя гибриды аденовирус-AAV, гибриды герпесвирус-AAV (Conway, JE et al., (1997) J. Virology 71(11):8780-8789) и гибриды бакуловирус-AAV. Для получения вирусных частиц rAAV, всем культурам для продуцирования rAAV требуется: 1) подходящие клетки-хозяева, 2) подходящий функциональный элемент вируса-помощника, 3) гены rep и cap AAV и продукты генов; 4) нуклеиновая кислота (такая как терапевтическая нуклеиновая кислота), фланкированная по меньшей мере одной последовательностью ITR AAV (например, увеличенный в размере геном вектора на основе rAAV); а также 5) подходящая среда и компоненты среды для способствования получению rAAV. В некоторых вариантах осуществления подходящая клетка-хозяин представляет собой клетку-хозяина, полученную от примата. В некоторых вариантах осуществления подходящая клетка-хозяин представляет собой линии клеток, полученные от человека, такие как клетки HeLa, A549, 293 или Perc.6. В некоторых вариантах осуществления подходящий функциональный элемент вируса-помощника обеспечивается аденовирусом дикого типа или мутантным аденовирусом (таким как термочувствительный аденовирус), вирусом герпеса (HSV), бакуловирусом или плазмидной конструкцией, обеспечивающей функциональные элементы помощника. В некоторых вариантах осуществления продукты генов rep и cap AAV могут происходить из AAV любого серотипа. В целом, но не обязательно, продукт гена rep AAV относится к тому же серотипу, что и ITR из генома вектора на основе rAAV, при условии, что продукты гена rep могут обеспечивать возможность репликации и упаковки генома rAAV. Для получения векторов на основе rAAV можно применять подходящие среды, известные из уровня техники. Эти среды включают без ограничения среды, производимые Hyclone Laboratories и JRH, в том числе модифицированную среду Игла (MEM), среду Игла, модифицированную по Дульбекко (DMEM), изготавливаемые по заказу составы, такие как описанные в патенте США № 6566118, и среду Sf-900 II SFM, описанную в патенте США № 6723551, каждый из которых включен в данный документ с помощью ссылки во всей своей полноте, в особенности в отношении изготавливаемых по заказу составов сред для применения в производстве рекомбинантных векторов на основе AAV. В некоторых вариантах осуществления функциональные элементы помощника AAV представлены аденовирусом или HSV. В некоторых вариантах осуществления функциональные элементы помощника AAV представлены бакуловирусом, и клетка-хозяин представляет собой клетку насекомого (например, клетки Spodoptera frugiperda (Sf9)). Примеры функциональных элементов аденовируса-помощника для репликации AAV включают функциональные элементы E1A, функциональные элементы E1B, функциональные элементы E2A, функциональные элементы VA и функциональные элементы E4orf6. Бакуловирусы, доступные из депозитариев, включают вирус ядерного полиэдроза Autographa californica.
[0261] Частицы rAAV можно получать с помощью способов, известных из уровня техники. См., например, патенты США №№ 6566118; 6989264 и 6995006. При практическом использовании настоящего изобретения клетки-хозяева для продукции частиц rAAV включают в себя клетки млекопитающих, клетки насекомых, растительные клетки, микроорганизмы и дрожжи. Клетки-хозяева также могут представлять собой упаковывающие клетки, в которых гены rep и cap AAV стабильно сохраняются в клетке-хозяине или клетках-продуцентах, в которых стабильно сохраняется геном вектора AAV. Иллюстративные упаковывающие клетки и клетки-продуценты получены из клеток 293, A549 или HeLa. Векторы AAV очищают и помещают в составы с помощью стандартных методик, известных из уровня техники.
[0262] В некоторых вариантах осуществления частицы rAAV можно получать с помощью способа тройной трансфекции, такого как иллюстративный способ тройной трансфекции, представленный ниже. Вкратце, плазмиду, содержащую ген rep и ген капсидного белка, вместе с аденовирусной плазмидой-помощником, можно ввести путем трансфекции (например, с использованием способа с фосфатом кальция) в линию клеток (например, клетки HEK-293), и вирус можно собирать и необязательно очищать.
[0263] В некоторых вариантах осуществления частицы rAAV можно получить с помощью способа с линией клеток-продуцентов (см., например, Martin et al., (2013) Human Gene Therapy Methods 24:253-269). Вкратце, линию клеток (например, линию клеток HeLa) можно стабильно трансфицировать плазмидой, содержащей ген rep, ген капсидного белка и последовательность промотор-трансген. Линии клеток можно подвергать скринингу для выбора лидерного клона для производства rAAV, который можно затем размножать в производственном биореакторе и инфицировать аденовирусом (например, аденовирусом дикого типа) в качестве помощника для начала продуцирования rAAV. Затем вирус можно собрать, аденовирус можно инактивировать (например, под действием тепла) и/или удалить, а частицы rAAV можно очистить. В некоторых вариантах осуществления линия клеток-продуцентов получена из клеток HeLa, 293, A549 или Perc.6. В некоторых вариантах осуществления линия клеток-продуцентов адаптирована для роста в суспензии. В некоторых вариантах осуществления функциональные элементы помощника AAV представлены аденовирусом, HSV или бакуловирусом.
[0264] В некоторых вариантах осуществления частицы rAAV собирают спустя от приблизительно 48 часов до приблизительно 96 часов после обеспечения функциональных элементов помощника. Например, в некоторых вариантах осуществления частицы rAAV собирают приблизительно через 48 часов, приблизительно 60 часов, приблизительно 72 часа, приблизительно 84 часа или приблизительно 96 часов после обеспечения функциональных элементов помощника. В некоторых вариантах осуществления частицы rAAV собирают спустя от приблизительно 48 часов до приблизительно 96 часов, от приблизительно 48 часов до приблизительно 84 часа, от приблизительно 48 часов до приблизительно 72 часа, от приблизительно 48 часов до приблизительно 60 часов, от приблизительно 60 часов до приблизительно 96 часов, от приблизительно 60 часов до приблизительно 84 часа, от приблизительно 60 часов до приблизительно 72 часа, от приблизительно 72 часа до приблизительно 96 часов, от приблизительно 72 часа до приблизительно 84 часа или от приблизительно 84 часа до приблизительно 96 часов после обеспечения функциональных элементов помощника.
[0265] Подходящие культуральные среды для продуцирования rAAV по настоящему изобретению могут быть дополнены сывороткой или рекомбинантными белками, полученными из сыворотки, на уровне 0,5% - 20% (об./об. или вес/об.). Как альтернатива, как известно из уровня техники, векторы на основе rAAV можно получать в бессывороточных условиях, которые также могут называться средой, не содержащей продуктов животного происхождения. Специалист в данной области техники может понять, что коммерческие или оригинальные среды, разработанные для способствования продуцированию векторов на основе rAAV, также могут быть дополнены одним или несколькими компонентами для культур клеток, известными из уровня техники, включающими без ограничения глюкозу, витамины, аминокислоты и/или факторы роста, в целях повышения титра rAAV в культурах для продуцирования.
[0266] Культуры для продуцирования rAAV можно выращивать при различных условиях (в широком диапазоне температур, в течение различных промежутков времени и т. п.), подходящих для конкретной используемой клетки-хозяина. Как известно из уровня техники, культуры для продуцирования rAAV включают культуры, зависимые от прикрепления, которые можно культивировать в подходящих сосудах для культур, зависимых от прикрепления, как например, в роллерных флаконах, на фильтрах из полых волокон, на микроносителях и в биореакторах со слоем носителя или псевдоожиженным слоем. Культуры для получения векторов на основе rAAV также могут включать клетки-хозяева, приспособленные к суспензионному росту, такие как клетки HeLa, 293 и SF-9, которые можно культивировать разнообразными способами, в том числе, например, во флаконах с перемешиванием, биореакторах с механическим перемешиванием и одноразовых системах, таких как резервуарная система Wave.
[0267] Частицы, представляющие собой вектор на основе rAAV по настоящему изобретению, можно собирать из культур для производства rAAV посредством лизиса клеток-хозяев в культурах для продуцирования или посредством сбора отработанной среды из культуры для продуцирования, при условии, что клетки культивируют при условиях, которые, как известно из уровня техники, вызывают высвобождение частиц rAAV в среду из интактных клеток, как более подробно описано в патенте США № 6566118. Подходящие способы лизиса клеток также известны из уровня техники и включают, например, несколько циклов замораживания/размораживания, обработку ультразвуком, микрофлюидизацию и обработку химическими веществами, такими как детергенты и/или протеазы.
[0268] В дополнительных вариантах осуществления вирусные частицы очищают. Термин "очищенный", используемый в данном документе, подразумевает препарат из вирусных частиц, в котором отсутствуют по меньшей мере некоторые из других компонентов, которые также могут присутствовать в среде, где вирусные частицы находятся в естественном состоянии или из которой они изначально получены. Таким образом, например, выделенные вирусные частицы можно получать с использованием методики очистки для обогащения ними исходной смеси, такой как лизат культуры или надосадочная жидкость культуры для продуцирования. Обогащение можно измерять с помощью множества способов, таких как, например, по доле устойчивых к ДНКазе частиц (DRP) или копий генома (gc), присутствующих в растворе, или по инфекционности, или его можно измерять по отношению ко второму, потенциально мешающему веществу, присутствующему в исходной смеси, такому как контаминанты, в том числе контаминанты из культуры для продуцирования или контаминанты в ходе продуцирования, в том числе вирус-помощник, компоненты среды и т. п.
[0269] В некоторых вариантах осуществления сбор культуры для продуцирования вирусов очищают от примесей для удаления дебриса клеток-хозяев. В некоторых вариантах осуществления сбор культуры для продуцирования очищают от примесей посредством фильтрации через серию пористых фильтров, включающих, например, модульный фильтр Millipore Millistak+ HC марки DOHC, модульный фильтр Millipore Millistak+ HC марки A1HC и 0,2 мкм фильтр Opticap XL1O с гидрофильной мембраной фильтра Millipore Express SHC. Очистку от примесей также можно осуществлять с помощью ряда других стандартных методик, известных из уровня техники, таких как центрифугирование или фильтрация через любой фильтр из ацетата целлюлозы с размером пор 0,2 мкм или больше, известные из уровня техники.
[0270] В некоторых вариантах осуществления сбор культуры для продуцирования вирусов дополнительно обрабатывают с помощью Benzonase® с целью расщепления любой высокомолекулярной ДНК, присутствующей в культуре для продуцирования. В некоторых вариантах осуществления расщепление с помощью Benzonase® осуществляют в стандартных условиях, известных из уровня техники, в том числе, например, при конечной концентрации 1-2,5 единиц Benzonase®/мл, при температуре, варьирующей в диапазоне от температуры окружающей среды до 37°C, в течение периода времени от 30 минут до нескольких часов.
[0271] Частицы rAAV можно выделять или очищать с использованием одной или нескольких из следующих стадий очистки: равновесного центрифугирования; проточной анионообменной фильтрации; тангенциальной поточной фильтрации (TFF) для концентрирования частиц rAAV; захвата rAAV с помощью хроматографии на апатите; термоинактивации вируса-помощника; захвата rAAV с помощью хроматографии с гидрофобными взаимодействиями; замены буфера с помощью эксклюзионной хроматографии (SEC); нанофильтрации и захвата rAAV с помощью анионообменной хроматографии, катионообменной хроматографии или аффинной хроматографии. Эти стадии можно применять в отдельности, в различных комбинациях или в различном порядке. Способы очистки частиц rAAV приведены, например, в Xiao et al., (1998) Journal of Virology 72:2224-2232; патентах США №№ 6989264 и 8137948; а также WO 2010/148143. Способы очистки аденовирусных частиц находятся, например, в Bo, H et al., (2014) Eur. J. Pharm. Sci. 67C:119-125. Способы очистки лентивирусных частиц находятся, например, в Segura MM, et al., (2013) Expert Opin Biol Ther. 13(7):987-1011. Способы очистки частиц HSV находятся, например, в Goins, WF et al., (2014) Herpes Simplex Virus Methods in Molecular Biology 1144:63-79.
[0272] В некоторых вариантах осуществления вирусная частица находится в фармацевтическом составе. В некоторых вариантах осуществления фармацевтический состав дополнительно включает фармацевтически приемлемый носитель. Такие носители хорошо известны из уровня техники (см., например, Remington's Pharmaceutical Sciences, 15th Edition, стр. 1035-1038 и 1570-1580). В некоторых вариантах осуществления фармацевтические композиции, содержащие вирусную частицу, описанную в данном документе, и фармацевтически приемлемый носитель, пригодны для глазной инъекции. Такими фармацевтически приемлемыми носителями могут быть стерильные жидкости, такие как вода и масло, в том числе стерильные жидкости на основе нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло и т. п. В качестве жидких носителей, в частности для инъекционных растворов, также могут использоваться солевые растворы и водные растворы декстрозы, полиэтиленгликоля (PEG) и глицерина. Кроме того, фармацевтическая композиция может содержать дополнительные ингредиенты, например, консерванты, буферы, средства, регулирующие тоничность, антиоксиданты и стабилизаторы, неионные увлажняющие и осветляющие средства, загустители и т. п. Фармацевтические композиции, описанные в данном документе, могут быть упакованы в единичные дозированные формы или многодозовые формы. Как правило, композиции составляют в виде стерильного или практически изотонического раствора.
VI. Способы лечения
[0273] Некоторые аспекты настоящего раскрытия предусматривают введение индивидууму терапевтически эффективного количества композиции, содержащей нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему CRISPR-Cas по настоящему изобретению, например как описано выше.
[0274] В некоторых вариантах осуществления аспекты настоящего раскрытия предусматривают введение индивидууму терапевтически эффективного количества композиции, содержащей Cas-белок по настоящему раскрытию и одну или несколько нуклеиновых кислот, содержащих первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующими мутацию, в том числе глубокую интронную мутацию, например как описано выше. То есть для любого из способов, композиций и наборов по настоящему раскрытию Cas-белок можно доставить в виде нуклеотидной последовательности, кодирующей Cas-белок, или в виде полипептида. В качестве неограничивающего примера Cas-белок можно вводить с одной или несколькими направляющими РНК (например, sgRNA) с использованием опосредованной катионным липидом доставки (см., например, Zuris, J.A. et al. Nat Biotechnol. 33:73-80). В некоторых вариантах осуществления Cas-белок можно вводить в комплексе с одной или несколькими направляющими РНК, например как в эффекторном комплексе CRISPR-Cas. Понятно, что любой из способов доставки и/или введения, описанных ниже, можно использовать для доставки Cas-белка, вводимого с одной или несколькими направляющими РНК. В некоторых вариантах осуществления Cas-белок экспрессируется из самоограничивающейся кассеты экспрессии, как описано выше.
[0275] В некоторых вариантах осуществления композицию, содержащую нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему CRISPR-Cas по настоящему раскрытию (или композицию, содержащую Cas-белок по настоящему раскрытию и одну или несколько направляющих РНК по настоящему раскрытию), вводят внутривенно, внутримышечно, подкожно, местно, перорально, трансдермально, внутрибрюшинно, внутриорбитально, путем имплантации, путем ингаляции, интратекально, интравентрикулярно или интраназально.
[0276] В некоторых вариантах осуществления композицию, содержащую нуклеиновую кислоту, кодирующую сконструированную не встречающуюся в природе систему CRISPR-Cas по настоящему раскрытию (или композицию, содержащую Cas-белок по настоящему раскрытию и одну или несколько направляющих РНК по настоящему раскрытию), вводят субретинально или интравитреально. Протоколы генной терапии для заболеваний сетчатки, таких как глазные заболевания, связанные с глубокой интронной мутацией, требуют локализованной доставки нуклеиновой кислоты в клетки сетчатки. Клетки, которые будут мишенями для лечения при данных заболеваниях, представляют собой фоторецепторные клетки сетчатки или клетки RPE, лежащие под нейросенсорной частью сетчатки. Доставка нуклеиновых кислот в эти клетки требует инъекции в субретинальное пространство между сетчаткой и RPE.
[0277] В некоторых аспектах настоящее изобретение предусматривает композиции, содержащие любые из нуклеиновых кислот, описанных в данном документе, необязательно в фармацевтически приемлемом вспомогательном веществе. Как хорошо известно из уровня техники, фармацевтически приемлемые вспомогательные вещества представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества, и могут поставляться в виде жидких растворов или суспензий, в виде эмульсий или в виде твердых форм, подходящих для растворения или суспендирования в жидкости перед применением. Например, вспомогательное вещество может придавать форму или консистенцию или выступать в качестве разбавителя. Подходящие вспомогательные вещества включают без ограничения стабилизирующие средства, смачивающие и эмульгирующие средства, соли для изменения осмолярности, инкапсулирующие средства, буферные вещества для поддержания pH и буферы. Такие наполнители включают любое фармацевтическое средство, подходящее для прямой доставки в глаз, которое можно вводить без излишней токсичности. Фармацевтически приемлемые вспомогательные вещества включают без ограничения сорбит, любое из различных соединений TWEEN и жидкости, такие как вода, солевой раствор, глицерин и этанол. В них могут быть включены фармацевтически приемлемые соли, например соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и т. п.; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и т. п. Всестороннее обсуждение фармацевтически приемлемых вспомогательных веществ доступно в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub. Co., N.J. 1991).
[0278] Как правило, эти композиции составляют для введения путем субретинальной инъекции. Соответственно, эти композиции можно объединять с фармацевтически приемлемыми основами, такими как физиологический раствор, сбалансированный солевой раствор Рингера (pH 7,4) и т. п. Хотя это и не требуется, композиции необязательно могут поставляться в виде стандартной лекарственной формы, подходящей для введения точного количества.
Способы субретинальной доставки
[0279] Способы субретинальной доставки известны из уровня техники. Например, см. WO 2009/105690, включенную в данный документ с помощью ссылки. Вкратце, общий способ доставки композиции (например, нуклеиновой кислоты, кодирующей сконструированную не встречающуюся в природе систему CRISPR-Cas по настоящему раскрытию, которую можно доставить посредством вирусной или невирусной доставки, как описано выше) в субретинальную область желтого пятна и центральной ямки можно проиллюстрировать следующим кратким описанием. Данный пример направлен лишь на иллюстрацию определенных признаков способа и никоим образом не подразумевается как ограничивающий.
[0280] Как правило, вектор можно доставить в форме композиции, вводимой путем внутриглазной инъекции (субретинально), при непосредственном наблюдении с помощью операционного микроскопа. Данная процедура может включать удаление стекловидного тела с последующей инъекцией суспензии вектора в субретинальное пространство с помощью тонкой канюли через один или несколько небольших разрезов сетчатки.
[0281] Вкратце, инфузионную канюлю можно закреплять на месте швом с целью сохранения нормального объема глазного яблока путем инфузии (например, физиологического раствора) на протяжении всей операции. Удаление стекловидного тела осуществляют с помощью канюли с подходящим размером канала (например, 20-27 калибра), где объем удаляемого гелеобразного стекловидного тела замещают путем инфузии физиологического раствора или другого изотонического раствора из инфузионной канюли. Преимущественно осуществляют удаление стекловидного тела, поскольку (1) удаление его коркового слоя (задней гиалоидной мембраны) облегчает проникновение канюли в сетчатку; (2) его удаление и замещение жидкостью (например, физиологическим раствором) создает пространство для обеспечения внутриглазной инъекции вектора, и (3) его контролируемое удаление снижает вероятность разрывов сетчатки и незапланированного отслоения сетчатки.
[0282] В некоторых вариантах осуществления композицию на основе rAAV вводят путем прямой инъекции в субретинальное пространство за пределами центральной зоны сетчатки при использовании канюли с подходящим размером канала (например, 27-45 калибра), создавая таким образом пузырек в субретинальном пространстве. В других вариантах осуществления субретинальной инъекции композиции на основе вектора предшествует субретинальная инъекция небольшого объема (например, от приблизительно 0,1 до приблизительно 0,5 мл) подходящей жидкости (такой как физиологический раствор или раствор Рингера) в субретинальное пространство за пределами центральной зоны сетчатки. Посредством этой первоначальной инъекции в субретинальное пространство в субретинальном пространстве формируется первоначальный пузырек жидкости, что вызывает локальное отслоение сетчатки в местоположении первоначального пузырька. Этот первоначальный пузырек жидкости может облегчать целенаправленную доставку композиции на основе вектора в субретинальное пространство (с помощью определения плоскости инъекции перед доставкой вектора) и обеспечивать сведение к минимуму возможности введения вектора в сосудистую оболочку глаза и вероятности инъекции или обратного тока в полость стекловидного тела. В некоторых вариантах осуществления в этот первоначальный пузырек жидкости можно дополнительно вводить с жидкостями, содержащими одну или несколько композиций на основе rAAV и/или одно или несколько дополнительных терапевтических средств, путем введения этих жидкостей непосредственно в первоначальный пузырек жидкости с помощью тех же или дополнительных канюль с узким просветом.
[0283] Внутриглазное введение композиций на основе вектора и/или первоначального небольшого объема жидкости можно осуществлять с помощью канюли с узким просветом (например, 27-45 калибра), прикрепленной к шприцу. В некоторых вариантах осуществления поршень этого шприца может управляться механическим устройством, как например с помощью нажатия на ножную педаль. Посредством проведения склеротомии канюлю с узким просветом продвигают через полость стекловидного тела в участок сетчатки, предварительно определенный для каждого субъекта в соответствии с зоной сетчатки, подлежащей целенаправленному воздействию (но за пределами центральной зоны сетчатки). При непосредственной визуализации суспензию вектора вводят путем механической инъекции под нейросенсорную часть сетчатки, что вызывает локальное отслоение сетчатки с самозатягивающимся нерасширяющимся рассечением сетчатки. Как отмечено выше, композицию на основе rAAV можно вводить либо с помощью прямой инъекции в субретинальное пространство, создавая пузырек за пределами центральной зоны сетчатки, либо вектор можно вводить с помощью инъекции в первоначальный пузырек за пределами центральной зоны сетчатки, вызывая его расширение (и расширение зоны отслоения сетчатки). В некоторых вариантах осуществления за инъекцией композиции на основе вектора следует инъекция другой жидкости в пузырек.
[0284] Не желая ограничиваться какой-либо теорией, полагают, что скорость и местоположение субретинальной инъекции(-ий) могут обусловливать возникновение локальных сил сдвига, которые могут повреждать желтое пятно, центральную ямку и/или нижележащие клетки RPE. Субретинальные инъекции можно осуществлять со скоростью, при которой силы сдвига сводятся к минимуму или исключаются. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции в течение приблизительно 15-17 минут. В некоторых вариантах осуществления вектор вводят с помощью инъекции в течение приблизительно 17-20 минут. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции в течение приблизительно 20-22 минут. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости от приблизительно 35 до приблизительно 65 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 35 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 40 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 45 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 50 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 55 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 60 мкл/мин. В некоторых вариантах осуществления композицию на основе вектора вводят с помощью инъекции при скорости приблизительно 65 мкл/мин. Специалисту в данной области техники будет понятно, что скорость и продолжительность инъекции в пузырек может определяться, например, объемом композиции на основе rAAV или размером пузырька, необходимыми для создания достаточного отслоения сетчатки для получения доступа к клеткам центральной зоны сетчатки, размером канюли, применяемой для доставки композиции на основе rAAV, и возможностью безопасного сохранения положения канюли по настоящему изобретению.
[0285] В некоторых вариантах осуществления настоящего изобретения объем композиции, вводимой с помощью инъекции в субретинальное пространство сетчатки, превышает приблизительно любое значение из 1 мкл, 2 мкл, 3 мкл, 4 мкл, 5 мкл, 6 мкл, 7 мкл, 8 мкл, 9 мкл, 10 мкл, 15 мкл, 20 мкл, 25 мкл, 50 мкл, 75 мкл, 100 мкл, 200 мкл, 300 мкл, 400 мкл, 500 мкл, 600 мкл, 700 мкл, 800 мкл, 900 мкл или 1 мл или любое количество между ними.
[0286] В некоторых вариантах осуществления способы предусматривают введение в глаз (например, при субретинальном и/или интравитреальном введении) эффективного количества рекомбинантных вирусных частиц, содержащих вектор по настоящему раскрытию. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 1012, 6 × 1012, 7 × 1012, 8 × 1012, 9 × 1012, 10 × 1012, 11 × 1012, 15 × 1012, 20 × 1012, 25 × 1012, 30 × 1012 или 50 × 1012 копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое значение из от 5 × 1012 до 6 × 1012, от 6 × 1012 до 7 × 1012, от 7 × 1012 до 8 × 1012, от 8 × 1012 до 9 × 1012, от 9 × 1012 до 10 × 1012, от 10 × 1012 до 11 × 1012, от 11 × 1012 до 15 × 1012, от 15 × 1012 до 20 × 1012, от 20 × 1012 до 25 × 1012, от 25 × 1012 до 30 × 1012, от 30 × 1012 до 50 × 1012 или от 50 × 1012 до 100 × 1012 копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 1012 до 10 × 1012, от 10 × 1012 до 25 × 1012 или от 25 × 1012 до 50 × 1012 копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 109, 6 × 109, 7 × 109, 8 × 109, 9 × 109, 10 × 109, 11 × 109, 15 × 109, 20 × 109, 25 × 109, 30 × 109 или 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 109 до 6 × 109, от 6 × 109 до 7 × 109, от 7 × 109 до 8 × 109, от 8 × 109 до 9 × 109, от 9 × 109 до 10 × 109, от 10 × 109 до 11 × 109, от 11 × 109 до 15 × 109, от 15 × 109 до 20 × 109, от 20 × 109 до 25 × 109, от 25 × 109 до 30 × 109, от 30 × 109 до 50 × 109 или от 50 × 109 до 100 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 109 до 10 × 109, от 10 × 109 до 15 × 109, от 15 × 109 до 25 × 109 или от 25 × 109 до 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 1010, 6 × 1010, 7 × 1010, 8 × 1010, 9 × 1010, 10 × 1010, 11 × 1010, 15 × 1010, 20 × 1010, 25 × 1010, 30 × 1010, 40 × 1010 или 50 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое из значений от 5 × 1010 до 6 × 1010, от 6 × 1010 до 7 × 1010, от 7 × 1010 до 8 × 1010, от 8 × 1010 до 9 × 1010, от 9 × 1010 до 10 × 1010, от 10 × 1010 до 11 × 1010, от 11 × 1010 до 15 × 1010, от 15 × 1010 до 20 × 1010, от 20 × 1010 до 25 × 1010, от 25 × 1010 до 30 × 1010, от 30 × 1010 до 40 × 1010, от 40 × 1010 до 50 × 1010 или от 50 × 1010 до 100 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое из значений от 5 × 1010 до 10 × 1010, от 10 × 1010 до 15 × 1010, от 15 × 1010 до 25 × 1010 или от 25 × 1010 до 50 × 1010 инфекционных единиц/мл.
[0287] В некоторых вариантах осуществления способы предусматривают введение в глаз (например, при субретинальном и/или интравитреальном введении) индивидууму (например, человеку) эффективного количества рекомбинантных вирусных частиц, содержащих вектор по настоящему раскрытию. В некоторых вариантах осуществления доза вирусных частиц, вводимых индивидууму, составляет по меньшей мере любое значение от приблизительно 1 × 108 до приблизительно 1 × 1013 копий генома/кг веса тела. В некоторых вариантах осуществления доза вирусных частиц, вводимых индивидууму, составляет любое из значений от приблизительно 1 × 108 до приблизительно 1 × 1013 копий генома/кг веса тела.
[0288] Можно создать один или несколько (например, 2, 3 или больше) пузырьков. Как правило, общий объем пузырька или пузырьков, создаваемых с помощью способов и систем по настоящему изобретению, не может превышать объем жидкости в глазу, например приблизительно 4 мл у типичного субъекта-человека. Общий объем каждого отдельного пузырька может составлять по меньшей мере приблизительно 0,3 мл или по меньшей мере приблизительно 0,5 мл для обеспечения отслоения сетчатки достаточного размера с целью обнажения типов клеток центральной зоны сетчатки и создания пузырька с достаточной способностью к оседанию для оптимальных манипуляций. Специалисту в данной области техники будет понятно, что при создании пузырька в соответствии со способами и системами по настоящему изобретению должно поддерживаться соответствующее внутриглазное давление, во избежание повреждения структур глаза. Размер каждого отдельного пузырька может составлять, например, от приблизительно 0,5 до приблизительно 1,2 мл, от приблизительно 0,8 до приблизительно 1,2 мл, от приблизительно 0,9 до приблизительно 1,2 мл, от приблизительно 0,9 до приблизительно 1,0 мл, от приблизительно 1,0 до приблизительно 2,0 мл, от приблизительно 1,0 до приблизительно 3,0 мл. Таким образом, в одном примере для инъекции в общей сложности 3 мл суспензии композиции на основе вектора можно сформировать 3 пузырька приблизительно по 1 мл каждый. Общий объем всех пузырьков в комбинации может составлять, например, от приблизительно 0,5 до приблизительно 3,0 мл, от приблизительно 0,8 до приблизительно 3,0 мл, от приблизительно 0,9 до приблизительно 3,0 мл, от приблизительно 1,0 до приблизительно 3,0 мл, от приблизительно 0,5 до приблизительно 1,5 мл, от приблизительно 0,5 до приблизительно 1,2 мл, от приблизительно 0,9 до приблизительно 3,0 мл, от приблизительно 0,9 до приблизительно 2,0 мл, от приблизительно 0,9 до приблизительно 1,0 мл.
[0289] В целях безопасной и эффективной трансдукции в целевых зонах сетчатки (например, в центральной зоне сетчатки) за пределами границ исходного местоположения пузырька пузырек можно подвергнуть манипуляциям для перемещения пузырька в целевую зону для трансдукции. Манипуляции с пузырьком могут происходить благодаря способности пузырька к оседанию, создаваемой объемом пузырька, с помощью перемещения глаза, содержащего пузырек, перемещения головы человека с глазом или глазами, содержащими один или несколько пузырьков, и/или с помощью обмена жидкость-воздух. Это особенно относится к центральной зоне сетчатки, поскольку данная зона обычно является устойчивой к отслоению в результате субретинальной инъекции. В некоторых вариантах осуществления для перемещения пузырька используют обмен жидкость-воздух; жидкость из инфузионной канюли временно замещают воздухом, например воздухом, выдуваемым на поверхность сетчатки. Поскольку объем воздуха вытесняет жидкость полости стекловидного тела с поверхности сетчатки, то жидкость из полости стекловидного тела может вытекать из канюли. Временное отсутствие давления жидкости из полости стекловидного тела вызывает движение и перемещение пузырька под действием силы тяжести в нижерасположенную часть глаза. С помощью соответствующего размещения глазного яблока, с пузырьком с субретинально введенной композицией на основе вектора производят манипуляции с целью охвата прилегающих зон (например, желтого пятна и/или центральной ямки). В некоторых случаях масса пузырька является достаточной, чтобы вызвать его перемещение под действием силы тяжести даже без применения обмена жидкость-воздух. Движение пузырька в требуемое местоположение можно дополнительно облегчить путем изменения положения головы субъекта для того, чтобы дать возможность пузырьку переместиться под действием силы тяжести в требуемое местоположение в глазу. После достижения требуемой конфигурации пузырька жидкость возвращают в полость стекловидного тела. Жидкость представляет собой подходящую жидкость, например свежеприготовленный физиологический раствор. Как правило, субретинально введенную композицию на основе вектора можно оставить in situ без ретинопексии в отношении рассечения сетчатки и без внутриглазной тампонады, и сетчатка самопроизвольно повторно прикрепляется в течение приблизительно 48 часов.
[0290] Благодаря безопасной и эффективной трансдукции клеток глаза (например, RPE и/или фоторецепторных клеток, например желтого пятна и/или центральной ямки) вектором по настоящему раскрытию, способы по настоящему изобретению можно применять для лечения индивидуума, например человека, у которого имеется глазное нарушение, связанное с глубокой интронной мутацией, где трансдуцированные клетки продуцируют систему CRISPR-Cas в количестве, достаточном для лечения глазного нарушения.
[0291] Эффективное количество вектора (в некоторых вариантах осуществления в форме вирусных частиц) вводят в зависимости от целей лечения. Например, если при низкой процентной доле трансдукции можно достичь требуемого терапевтического эффекта, то целью лечения, как правило, является соответствие данному уровню трансдукции или его превышение. В некоторых случаях этого уровня трансдукции можно достичь путем трансдукции лишь приблизительно 1-5% клеток-мишеней, в некоторых вариантах осуществления по меньшей мере приблизительно 20% клеток требуемого типа ткани, в некоторых вариантах осуществления по меньшей мере приблизительно 50%, в некоторых вариантах осуществления по меньшей мере приблизительно 80%, в некоторых вариантах осуществления по меньшей мере приблизительно 95%, в некоторых вариантах осуществления по меньшей мере приблизительно 99% клеток требуемого типа ткани. В качестве руководства, число частиц, вводимых в расчете на одну инъекцию, составляет как правило от приблизительно 1×106 до приблизительно 1×1014 частиц, от приблизительно 1×107 до 1×1013 частиц, от приблизительно 1×109 до 1×1012 частиц или приблизительно 1×1011 частиц. Композицию на основе вектора можно вводить с помощью одной или нескольких субретинальных инъекций, производимых в ходе одной и той же процедуры или разделенных несколькими днями, неделями, месяцами или годами. В некоторых вариантах осуществления для лечения человека можно применять несколько векторов.
[0292] В некоторых вариантах осуществления введение в сетчатку эффективного количества вектора или нуклеиновой кислоты по настоящему раскрытию трансдуцирует фоторецепторные клетки в участке введения или рядом с ним. В некоторых вариантах осуществления показатель трансдукции фоторецепторных клеток превышает приблизительно любое значение из 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75% или 100%. В некоторых вариантах осуществления от приблизительно 5% до приблизительно 100%, от приблизительно 10% до приблизительно 50%, от приблизительно 10% до приблизительно 30%, от приблизительно 25% до приблизительно 75%, от приблизительно 25% до приблизительно 50% или от приблизительно 30% до приблизительно 50% фоторецепторных клеток являются трансдуцированными. Способы идентификации трансдуцированных фоторецепторных клеток известны из уровня техники; например, для выявления трансдукции можно применять иммуногистохимический анализ или маркер, такой как усиленный зеленый флуоресцентный белок.
[0293] В некоторых вариантах осуществления настоящего изобретения способы предусматривают введение в субретинальную область (например, в субретинальное пространство) млекопитающего эффективного количества вектора или нуклеиновой кислоты по настоящему раскрытию для лечения индивидуума с глазным нарушением, например человека с глазным нарушением, связанным с глубокой интронной мутацией. В некоторых вариантах осуществления композицию вводят с помощью инъекции в одно или несколько местоположений в субретинальной области для обеспечения экспрессии нуклеиновой кислоты в фоторецепторных клетках. В некоторых вариантах осуществления композицию вводят с помощью инъекции в любое из одного, двух, трех, четырех, пяти, шести, семи, восьми, девяти, десяти или более десяти местоположений в субретинальной области.
[0294] В некоторых вариантах осуществления композицию вводят в более чем одно местоположение одновременно или последовательно. В некоторых вариантах осуществления многократные инъекции проводят с интервалом, составляющим не более чем один час, два часа, три часа, четыре часа, пять часов, шесть часов, девять часов, двенадцать часов или 24 часа. В некоторых вариантах осуществления многократные инъекции проводят по отдельности с интервалом один день, два дня, три дня, четыре дня, пять дней, шесть дней, семь дней, десять дней, 15 дней, 20 дней, 25 дней или 30 дней. В некоторых вариантах осуществления многократные инъекции проводят по отдельности с интервалом один месяц, два месяца, три месяца, четыре месяца, пять месяцев, шесть месяцев, восемь месяцев, десять месяцев или одиннадцать месяцев. В некоторых вариантах осуществления многократные инъекции проводят по отдельности с интервалом один год, два года, три года, четыре года, пять лет, шесть лет, восемь лет, десять лет, 15 лет, 20 лет, 25 лет, 30 лет, 35 лет, 40 лет, 45 лет, 50 лет, 55 лет или 60 лет.
Способы интравитреальной инъекции
[0295] Общий способ интравитреальной инъекции можно проиллюстрировать следующим кратким обзором. Данный пример направлен лишь на иллюстрацию определенных признаков способа и никоим образом не подразумевается как ограничивающий. Процедуры интравитреальной инъекции известны из уровня техники (см., например, Peyman, G.A., et al. (2009) Retina 29(7):875-912 и Fagan, X.J. and Al-Qureshi, S. (2013) Clin. Experiment. Ophthalmol. 41(5):500-7).
[0296] Вкратце, субъекта можно подготовить к процедуре интравитреальной инъекции с помощью расширения зрачков, стерилизации глаза и введения анестетика. Для расширения зрачков можно применять любое подходящее мидриатическое средство, известное из уровня техники. Перед лечением можно подтвердить надлежащее расширение зрачков. Стерилизацию можно осуществлять с помощью применения стерилизующей обработки глаза, например, йодидсодержащего раствора, такого как повидон-йод (BETADINE®). Аналогичный раствор также можно применять для очистки века, ресниц и любых других соседних тканей (например, кожи). Можно применять любой подходящий анестетик, такой как лидокаин или пропаракаин, в любой подходящей концентрации. Анестетик можно вводить с помощью любого способа, известного из уровня техники, в том числе без ограничения в составе капель, гелей или желе местного действия, и с помощью субконъюнктивального применения анестетика.
[0297] Перед инъекцией, для удаления ресниц от зоны, можно применять стерилизованный векорасширитель. Место инъекции можно отметить шприцем. Место инъекции можно выбрать в зависимости от хрусталиков пациента. Например, место инъекции может находиться на расстоянии 3-3,5 мм от лимба роговицы у пациентов с псевдофакичным или афакичным глазом и на расстоянии 3,5-4 мм от лимба роговицы у пациентов с факичным глазом. Пациент может смотреть в сторону, противоположную месту инъекции.
[0298] В некоторых вариантах осуществления способы предусматривают введение в глаз (например, при субретинальном и/или интравитреальном введении) эффективного количества рекомбинантных вирусных частиц, содержащих вектор по настоящему раскрытию. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 1012, 6 × 1012, 7 × 1012, 8 × 1012, 9 × 1012, 10 × 1012, 11 × 1012, 15 × 1012, 20 × 1012, 25 × 1012, 30 × 1012 или 50 × 1012 копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 1012 до 6 × 1012, от 6 × 1012 до 7 × 1012, от 7 × 1012 до 8 × 1012, от 8 × 1012 до 9 × 1012, от 9 × 1012 до 10 × 1012, от 10 × 1012 до 11 × 1012, от 11 × 1012 до 15 × 1012, от 15 × 1012 до 20 × 1012, от 20 × 1012 до 25 × 1012, от 25 × 1012 до 30 × 1012, от 30 × 1012 до 50 × 1012 или от 50 × 1012 до 100 × 1012 копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 1012 до 10 × 1012, от 10 × 1012 до 25 × 1012 или от 25 × 1012 до 50 × 1012копий генома/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 109, 6 × 109, 7 × 109, 8 × 109, 9 × 109, 10 × 109, 11 × 109, 15 × 109, 20 × 109, 25 × 109, 30 × 109 или 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 109 до 6 × 109, от 6 × 109 до 7 × 109, от 7 × 109 до 8 × 109, от 8 × 109 до 9 × 109, от 9 × 109 до 10 × 109, от 10 × 109 до 11 × 109, от 11 × 109 до 15 × 109, от 15 × 109 до 20 × 109, от 20 × 109 до 25 × 109, от 25 × 109 до 30 × 109, от 30 × 109 до 50 × 109 или от 50 × 109 до 100 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет приблизительно любое из значений от 5 × 109 до 10 × 109, от 10 × 109 до 15 × 109, от 15 × 109 до 25 × 109 или от 25 × 109 до 50 × 109 трансдуцирующих единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое значение из 5 × 1010, 6 × 1010, 7 × 1010, 8 × 1010, 9 × 1010, 10 × 1010, 11 × 1010, 15 × 1010, 20 × 1010, 25 × 1010, 30 × 1010, 40 × 1010 или 50 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое из значений от 5 × 1010 до 6 × 1010, от 6 × 1010 до 7 × 1010, от 7 × 1010 до 8 × 1010, от 8 × 1010 до 9 × 1010, от 9 × 1010 до 10 × 1010, от 10 × 1010 до 11 × 1010, от 11 × 1010 до 15 × 1010, от 15 × 1010 до 20 × 1010, от 20 × 1010 до 25 × 1010, от 25 × 1010 до 30 × 1010, от 30 × 1010 до 40 × 1010, от 40 × 1010 до 50 × 1010 или от 50 × 1010 до 100 × 1010 инфекционных единиц/мл. В некоторых вариантах осуществления вирусный титр в композиции составляет по меньшей мере приблизительно любое из значений от 5 × 1010 до 10 × 1010, от 10 × 1010 до 15 × 1010, от 15 × 1010 до 25 × 1010 или от 25 × 1010 до 50 × 1010 инфекционных единиц/мл.
[0299] В некоторых вариантах осуществления способы предусматривают введение в глаз (например, при субретинальном и/или интравитреальном введении) индивидууму (например, человеку) эффективного количества рекомбинантных вирусных частиц, содержащих вектор по настоящему раскрытию. В некоторых вариантах осуществления доза вирусных частиц, вводимых индивидууму, составляет по меньшей мере любое значение от приблизительно 1 × 108 до приблизительно 1 × 1013 копий генома/кг веса тела. В некоторых вариантах осуществления доза вирусных частиц, вводимых индивидууму, составляет любое из значений от приблизительно 1 × 108 до приблизительно 1 × 1013 копий генома/кг веса тела.
[0300] Во время инъекции игла может быть вставлена перпендикулярно склере и направлена в центр глаза. Игла может быть вставлена таким образом, чтобы кончик заканчивался в стекловидном теле, а не в субретинальном пространстве. Для инъекции можно применять любой подходящий объем, известный из уровня техники. После инъекции глаз можно обработать стерилизующим средством, таким как антибиотик. Также глаз можно промыть с целью удаления избытка стерилизующего средства.
Структура сетчатки и способы определения эффективности доставки нуклеиновой кислоты
[0301] Известно, что сетчатка состоит из несколько слоев. Слои клеток сетчатки могут включать внутреннюю пограничную мембрану, слой нервных волокон, слой ганглионарных клеток, внутренний сплетениевидный слой, внутренний ядерный слой, внешний сплетениевидный слой, внешний ядерный слой, внешнюю пограничную мембрану, фоторецепторный слой и слои пигментного эпителия сетчатки. Наиболее близко расположенным к стекловидному телу слоем является внутренняя пограничная мембрана. Этот слой может содержать клетки Мюллера, класс глиальных клеток. Слой нервных волокон может содержать аксоны ганглионарных клеток, которые образуют зрительный нерв. Слой ганглионарных клеток может содержать ганглионарные клетки и амакриновые клетки. Внутренний сплетениевидный слой может содержать синапсы между дендритами ганглионарных и амакриновых клеток и аксонами биполярных клеток. Внутренний ядерный слой может содержать клеточные ядра амакриновых, биполярных и горизонтальных клеток. Внешний сплетениевидный слой может содержать синапсы между дендритами горизонтальных клеток и отростками фоторецепторных клеток. Внешний ядерный слой может содержать тела фоторецепторных клеток. Внешняя или наружная пограничная мембрана может содержать межклеточные соединения, такие как адгезионные контакты и десмосомы, между апикальными дендритами клеток Мюллера, а также между этими выростами и внутренними сегментами фоторецепторных клеток. Фоторецепторный слой, также известный как слой палочек и колбочек и мембрана Джейкоба, может содержать фоторецепторные клетки, включающие в себя палочки и колбочки. Слоем сетчатки, наиболее удаленным от стекловидного тела, является пигментный эпителий сетчатки (RPE), который может содержать слой гексагональных эпителиальных клеток, содержащих пигментные гранулы.
[0302] Также известно, что сетчатка содержит много различных типов клеток. Нейроны сетчатки могут включать в себя фоторецепторные клетки, биполярные клетки, ганглионарные клетки, амакриновые клетки и горизонтальные клетки. Фоторецепторные клетки чувствительны к свету. Они могут воспринимать свет и отвечать путем передачи сигналов в зрительный нерв через биполярные клетки и ганглионарные клетки. Фоторецепторные клетки могут включать в себя палочковые клетки, которые как правило воспринимают свет в условиях тусклого освещения, и колбочковые клетки, которые как правило воспринимают цвет и более яркий свет. Биполярные клетки могут получать входные сигналы от фоторецепторных клеток и передавать их по синапсам к амакриновым или ганглионарным клеткам. Ганглионарные клетки могут получать информацию от амакриновых клеток или горизонтальных клеток, и их аксоны образуют зрительный нерв. Горизонтальные клетки могут интегрировать входные сигналы от нескольких фоторецепторов и содействовать адаптации к уровням освещения. Амакриновые клетки представляют собой вставочные нейроны, которые способствуют регуляции активности биполярных клеток и передают входные сигналы в ганглионарные клетки. Глиальные клетки сетчатки могут включать в себя клетки Мюллера, астроглии и микроглии.
[0303] Эффективность доставки нуклеиновой кислоты с помощью субретинальной или интравитреальной инъекции можно контролировать по нескольким критериям, описанным в данном документе. В некоторых вариантах осуществления эффективность анализируют путем выявления делеции глубокой интронной мутации и/или фланкирующей последовательности в образце, включающем в себя одну или несколько клеток, к которым была доставлена нуклеиновая кислота(-ы). Делеции можно выявить любыми способами, известными из уровня техники, включая без ограничения Саузерн-блоттинг, ПЦР, qPCR, секвенирование ДНК (например, секвенирование методом Сэнгера и секвенирование нового поколения), гибридизация in situ, микрочипы ДНК и анализ с нуклеазой Surveyor (см., например, Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308). Иллюстративные способы продемонстрированы в примерах ниже.
[0304] В некоторых вариантах осуществления эффективность оценивают функционально. Например, после лечения субъекта с применением способов по настоящему изобретению у субъекта можно оценивать, например, улучшение, и/или стабилизацию, и/или задержку прогрессирования одного или нескольких признаков или симптомов болезненного состояния по одному или нескольким клиническим параметрам. Примеры таких тестов известны из уровня техники и включают объективные, а также субъективные (например, сообщаемые субъектами) показатели. Например, для измерения эффективности лечения в отношении зрительной функции субъекта можно оценивать одно или несколько из следующего: субъективное качество зрения или улучшенная функция центрального зрения субъекта (например, по улучшению способности субъекта к свободному чтению и узнаванию лиц), визуальная подвижность субъекта (например, по уменьшению времени, необходимого для прохождения лабиринта), острота зрения (например, по улучшению количественного показателя субъекта по LogMAR), показатели микропериметрии (например, по улучшению количественного показателя субъекта в dB), показатели периметрии глаза, адаптированного к темноте (например, по улучшению количественного показателя субъекта в dB), показатели тонкого матричного картирования (например, по улучшению количественного показателя субъекта в dB), показатели периметрии по Гольдману (например, по уменьшению размера зоны скотомы (т. е. зон слепоты) и улучшению способности к различению более мелких целей), показатели чувствительности к мельканиям (например, по улучшению в герцах), аутофлуоресценции и электрофизиологических показателей (например, по улучшению ERG). В некоторых вариантах осуществления зрительную функцию измеряют по визуальной подвижности субъекта. В некоторых вариантах осуществления зрительную функцию измеряют по остроте зрения субъекта. В некоторых вариантах осуществления зрительную функцию измеряют с помощью микропериметрии. В некоторых вариантах осуществления зрительную функцию измеряют с помощью периметрии глаза, адаптированного к темноте. В некоторых вариантах осуществления зрительную функцию измеряют с помощью ERG. В некоторых вариантах осуществления зрительную функцию измеряют по субъективному качеству зрения субъекта.
[0305] В случае заболеваний, приводящих к прогрессирующей дегенерации зрительной функции, лечение субъекта в раннем возрасте может не только приводить к замедлению или остановке прогрессирования заболевания, но также может уменьшать интенсивность утраты зрительной функции вследствие приобретенной амблиопии или предупреждать ее. Амблиопия может быть двух типов. В исследованиях на приматах, отличных от человека, и котятах, которых выдерживали в полной темноте от рождения вплоть до возраста нескольких месяцев, у животных даже при последующем воздействии света была функциональная необратимая слепота, несмотря на наличие посылаемых сетчаткой функциональных сигналов. Эта слепота имеет место по причине остановки в развитии нейронных связей и "обучении" коры головного мозга от рождения вследствие блокирования воздействия раздражителя. Неизвестно, можно ли будет когда-либо восстановить данную функцию. В случае заболеваний по типу дегенерации сетчатки нормальная нейронная сеть зрительной коры головного мозга была изначально "обученной" или соответствовала уровню развития до момента, в который дегенерация вызывала значительную дисфункцию. Утрата зрительного раздражителя вызывает "приобретенную" или "обученную" дисфункцию ("приобретенную амблиопию") в том, что касается передачи сигналов в дисфункциональном глазу, что приводит к неспособности головного мозга к интерпретации сигналов или к "использованию" этого глаза. В этих случаях "приобретенной амблиопии" неизвестно, может ли улучшение передачи сигналов от сетчатки в результате генной терапии амблиопичного глаза когда-либо привести к приобретению более нормальной функции, в дополнение к замедлению прогрессирования или стабилизации болезненного состояния. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 30 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 20 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 18 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 15 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 14 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 13 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 12 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 10 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 8 лет. В некоторых вариантах осуществления человек, подвергаемый лечению, имеет возраст менее 6 лет.
[0306] При некоторых глазных нарушениях имеют место феномены "клеток-нянь", при которых улучшение функции одного типа клеток приводит к улучшению функции другого. Например, трансдукция RPE центральной зоны сетчатки, опосредованная нуклеиновой кислотой по настоящему изобретению, может в таком случае улучшать функцию палочек и, в свою очередь, улучшение функции палочек приводит к улучшению функции колбочек. Соответственно, лечение в отношении одного типа клеток может приводить к улучшению функции другого.
[0307] Выбор конкретного вектора и композиции зависит от ряда различных факторов, включающих без ограничения индивидуальный анамнез человека и характерные особенности состояния и индивидуума, подвергаемых лечению. Оценка этих характерных особенностей и разработка соответствующего режима терапии в конечном счете является ответственностью лечащего врача.
[0308] В некоторых вариантах осуществления у человека, подлежащего лечению, имеется генетическое глазное нарушение (например, ассоциированное с глубокой интронной мутацией), но еще не проявляются клинические признаки или симптомы. В некоторых вариантах осуществления у человека, подлежащего лечению, имеется глазное нарушение (например, ассоциированное с глубокой интронной мутацией). В некоторых вариантах осуществления у человека, подлежащего лечению, проявляются один или несколько признаков или симптомов глазного нарушения (например, ассоциированного с глубокой интронной мутацией). В некоторых вариантах осуществления глубокая интронная мутация была идентифицирована у человека, подлежащего лечению.
[0309] Композиции по настоящему изобретению можно применять либо в отдельности, либо в комбинации с одним или несколькими дополнительными терапевтическими средствами для лечения глазных нарушений. Интервал между последовательными введениями может измеряться сроками, составляющими по меньшей мере (или, в качестве альтернативы, менее чем) минуты, часы или дни.
[0310] В некоторых вариантах осуществления в субретинальную область или стекловидное тело можно вводить одно или несколько дополнительных терапевтических средств (например, посредством интравитреального введения). Неограничивающие примеры дополнительных терапевтических средств включают полипептидные нейротрофические факторы (например, GDNF, CNTF, BDNF, FGF2, PEDF, EPO), полипептидные антиангиогенные факторы (например, sFlt, ангиостатин, эндостатин), антиангиогенные нуклеиновые кислоты (например, siRNA, miRNA, рибозим), например антиангиогенные нуклеиновые кислоты, направленные против VEGF, антиангиогенные морфолиновые олигонуклеотиды, например антиангиогенные морфолиновые олигонуклеотиды, направленные против VEGF, антиангиогенные антитела и/или фрагменты антител (например, Fab-фрагменты), например антиангиогенные антитела и/или фрагменты антител, направленные против VEGF.
VII. Наборы
[0311] Композиции, нуклеиновые кислоты и вирусные частицы, описанные в данном документе, могут содержаться в наборе, предназначенном для применения в одном из способов по настоящему изобретению, описанных в данном документе.
[0312] Композиции, нуклеиновые кислоты и вирусные частицы по настоящему изобретению могут быть дополнительно упакованы в наборы, где наборы могут дополнительно содержать инструкции по применению. В некоторых вариантах осуществления инструкции по применению включают инструкции в соответствии с одним из способов, описанных в данном документе.
[0313] В некоторых вариантах осуществления наборы дополнительно содержат буферы и/или фармацевтически приемлемые вспомогательные вещества. Как хорошо известно из уровня техники, фармацевтически приемлемые вспомогательные вещества представляют собой относительно инертные вещества, которые облегчают введение фармакологически эффективного вещества, и могут поставляться в виде жидких растворов или суспензий, в виде эмульсий или в виде твердых форм, подходящих для растворения или суспендирования в жидкости перед применением. Например, вспомогательное вещество может придавать форму или консистенцию или выступать в качестве разбавителя. Подходящие вспомогательные вещества включают без ограничения стабилизирующие средства, смачивающие и эмульгирующие средства, соли для изменения осмолярности, инкапсулирующие средства, буферные вещества для поддержания pH и буферы. Такие наполнители включают любое фармацевтическое средство, подходящее для прямой доставки в глаз, которое можно вводить без излишней токсичности. Фармацевтически приемлемые вспомогательные вещества включают без ограничения сорбит, любое из различных соединений TWEEN и жидкости, такие как вода, солевой раствор, глицерин и этанол. В них могут быть включены фармацевтически приемлемые соли, например соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и т. п.; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и т. п. Всестороннее обсуждение фармацевтически приемлемых вспомогательных веществ доступно в REMINGTON'S PHARMACEUTICAL SCIENCES (Mack Pub. Co., N.J. 1991).
[0314] В некоторых вариантах осуществления фармацевтически приемлемые вспомогательные вещества могут включать фармацевтически приемлемые носители. Такими фармацевтически приемлемыми носителями могут быть стерильные жидкости, такие как вода и масло, в том числе стерильные жидкости на основе нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло и т. п. В качестве жидких носителей, в частности для инъекционных растворов, также могут использоваться солевые растворы и водные растворы декстрозы, полиэтиленгликоля (PEG) и глицерина. Также можно применять дополнительные ингредиенты, например, консерванты, буферы, средства, регулирующие тоничность, антиоксиданты и стабилизаторы, неионные увлажняющие или осветляющие средства, средства, повышающие вязкость, и т. п. Наборы, описанные в данном документе, могут быть упакованы в единичные дозированные формы или в многодозовые формы. Как правило, содержание наборов составляют в виде стерильного и практически изотонического раствора.
ПРИМЕРЫ
[0315] Настоящее изобретение будет более понятным со ссылкой на следующие примеры. Тем не менее, их не следует истолковывать как ограничивающие объем настоящего изобретения. Понятно, что описанные в данном документе примеры и варианты осуществления служат только в качестве иллюстрации, и что различные модификации или изменения с их учетом будут понятны специалистам в данной области техники, и они должны быть включены в сущность и содержание данной заявки, а также объем прилагаемой формулы изобретения.
Пример 1. Создание in vitro модели врожденного амавроза Лебера с применением технологии Crispr-Cas9
Способы
Плазмиды
[0316] Плазмиду pSpCas9, которая экспрессирует SpCas9, заказали в Sigma (номер по каталогу: CAS9P-1EA). Сайт рестрикции BbsI в polyA BGH удаляли с использованием набора сайт-направленного мутагенеза QuikChange Lightning (Stratagene) и пары праймеров для мутагенеза (SEQ ID NO: 28-29) в соответствии с протоколом изготовителя. Кассету промотор U6-BbsI:BbsI-каркас sgRNA-терминатор U6 (SEQ ID NO: 30) синтезировали с помощью GeneArt (Life Technologies) и вставляли в сайты рестрикции PciI и NruI нулевой плазмиды pSpCas9-BbsI для создания плазмиды pSpCas9(BB). Затем олигонуклеотиды sgRNA (SEQ ID NO: 1-2) субклонировали в два сайта рестрикции BbsI плазмиды pSpCas9(BB) в соответствии с протоколом, описанным ранее (Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308).
Нуклеофекция
[0317] Совместно трансфицировали 2,5 микрограмма ДНК плазмиды pSpCas9(BB)-U6-sgRNA и 5 микролитров ssODN (10 микромолей) (SEQ ID NO: 3) в 1×106 клеток HEK 293FT с использованием набора линии клеток Amaxa SF cell line 4D-Nucleofector X kit L (Lonza) и программы CM-130 в системе 4D-Nucleofector System (Lonza) в соответствии с протоколом изготовителя.
Скрининг
[0318] Для идентификации клонов, несущих мутацию c.2991+1655A>G CEP290, клетки разделяли на отдельные клетки через 48 ч. после совместной трансфекции и серийно разбавляли до конечной концентрации 0,5 клеток на 100 микролитров с целью снижения вероятности наличия нескольких клеток на лунку. Высевали 100 микролитров разбавленных клеток в каждую лунку девяти 96-луночных планшетов. Клетки размножали в инкубаторе при 5% CO2, 37°C в течение 2 недель.
[0319] 235 одноклеточных клонов идентифицировали и подвергли скринингу в отношении мутации c.2991+1655A>G CEP290. Геномную ДНК экстрагировали с использованием раствора для экстракции ДНК QuickExtract (Epicenter) и амплифицировали с помощью GoTaq Hot Start Green Master Mix (Promega) и праймеров ПЦР, фланкирующих интронную мутацию (SEQ ID NO: 4-5). Амплификацию продуктов ПЦР выполняли со следующими параметрами циклов: 1 цикл при 95°С в течение 2 мин.; 35 циклов 95°C в течение 30 с, 60°C в течение 30 с и 72°C в течение 3 мин.; 1 цикл при 72°С в течение 15 мин. Продукты ПЦР подвергали расщеплению с помощью SnaBI и секвенированию методом Сэнгера с помощью секвенирующего праймера (SEQ ID NO: 6).
RT-qPCR
[0320] Из клеток WT, Het и MT экстрагировали mRNA с использованием набора RNeasy Plus Mini Kit (Qiagen) в соответствии с протоколом производителя. 1 мкг общей РНК использовали для синтеза кДНК с помощью набора для синтеза кДНК iScript (Bio-Rad) в соответствии с протоколом производителя. кДНК подвергали ПЦР-амплификации в реальном времени в буфере, содержащем Fast Plus EvaGreen qPCR Master Mix с низким ROX (Biotium) и праймеры, которые специфически выявляют mRNA CEP290 дикого типа (SEQ ID NO: 7-8) и мутантную mRNA CEP290 (SEQ ID NO: 9-10), соответственно, на системе для ПЦР в реальном времени ABI Prism 7500 (Applied Biosystems). При этом использовали следующие условия: 1 цикл при 50°С в течение 2 мин.; 1 цикл при 95°С в течение 10 мин.; 40 циклов при 95°C в течение 15 с и 60°C в течение 60 с. Специфичность продуктов амплификации определяли по анализу кривой плавления, проводимому в конце каждого прогона, с использованием цикла при 95°С в течение 15 с, 60°С в течение 60 с, 95°С в течение 15 с и 60°С в течение 15 с. Данные анализировали с использованием программного обеспечения SDS 2.3 (Applied Biosystems). Уровни экспрессии CEP290 нормализовали к уровням экспрессии mRNA PPIA (для последовательностей праймеров см. SEQ ID NO: 31-32).
Вестерн-блот-анализ
[0321] Клетки лизировали в буфере для лизиса RIPA (Cell Signaling Technology), дополненном 1 милимоль/литр фенилметилсульфонилфторида (PMSF; Cell Signaling Technology) и 1X смесью ингибитора протеазы (Cell Signaling Technology) на льду. Затем клетки соскабливали, собирали в пробирки Эппендорф и лизаты осветляли центрифугированием при 13000 об/мин в течение 6 мин. при 4oC. Образцы готовили с помощью добавления буфера для образцов 4X LDS NuPage и 10X восстановителя NuPage (оба от Life Technologies), нагревания при 70°C в течение 10 мин. и центрифугирования при 13000 об/мин в течение 1 мин. Образцы белков загружали на 3-8% трис-ацетатный гель NuPAGE вместе с предварительно окрашенным эталоном белка HiMark (оба от Life Technologies). Образцы разделяли с помощью гель-электрофореза при 180 вольт в течение 1 часа. В качестве подвижного буфера использовали подвижный трис-ацетатный буфер SDS (Life Technologies). Для переноса, мембраны из поливинилидинфторида (PVDF) кратковременно обрабатывали в метаноле и промывали водой, чтобы сделать гидрофильными. "Сэндвич" для переноса получали путем переслаивания мембраны PVDF и геля между фильтровальной бумагой и губчатым материалом в модуле для блота XCell II XCell II (Life Technologies). В качестве буфера для переноса использовали 20X буфер для переноса NuPage (Life Technologies) с 20% метанолом. Перенос проводили при 30 вольт в течение 2 часов в мини-ячейках XCell SureLock (Life Technologies). После переноса мембраны PVDF блокировали в буфере TBST Pierce (трис-забуференный физиологический раствор с детергентом Tween 20, Thermo Fisher Scientific), содержащем 1% обезжиренного сухого молока, встряхивая при комнатной температуре в течение 1 часа. Затем блоты инкубировали в растворе первичного антитела, приготовленном в блокирующем растворе, с покачиванием в течение ночи при 4oC. В качестве первичных антител использовали кроличье поликлональное антитело к CEP290 (любезный подарок от профессора Хеманта Кханны из Университета Массачусетса в Вустере), моноклональное антитело к Cas9 мыши (клон 7A9, Millipore), конъюгированное с HRP моноклональное антитело к β-актина кролика (клон 13E5, Cell Signaling Technology). Несвязанные первичные антитела отмывали 3 раза с TBST, в течение 10 мин. каждый. Затем к мембранам добавляли вторичные антитела (антитело к IgG кролика или мыши; Cell Signaling Technology, конъюгированное с Alexa Fluor 647) в блокирующем растворе и оставляли на шейкере в течение 1 часа при комнатной температуре. Мембраны промывали 3 раза с TBST, в течение 10 минут каждый, для уменьшения неспецифичного фона. Мембраны обрабатывали в течение 4 мин. с использованием субстрата для вестерн-блоттинга с усиленной хемилюминесценцией (ECL) Pierce (Thermo Fisher Scientific). Полосы белка в блотах окончательно визуализировали при экспонировании пленки в течение различных временных интервалов в обрабатывающем устройстве Kodak X-OMAT 2000. Для вспомогательного зондирования блота с антителом к β-актина, мембраны сначала снимали с помощью инкубации при 37°С в течение 30 мин. в буфере для снятия вестерн-блота Restore (Thermo Fisher Scientific), а затем подвергали вспомогательному зондированию с антителом против β-актина. Данные блоттинга, показанные в этой работе, представляют по меньшей мере три независимых эксперимента.
Результаты
[0322] Во-первых, технологию редактирования генома CRISPR-Cas9 использовали для создания клеточной модели, несущей интронную мутацию сплайсинга c.2991+1655 A>G в CEP290. Эта клеточная модель является ценным инструментом оценки терапевтических средств для лечения пациентов с LCA, несущих мутацию c.2991+1655 A>G в CEP290.
[0323] Редактирование генома с помощью бактериальной системы CRISPR-Cas9 II типа инициируется внесением двухнитевого разрыва (DSB) в намеченный геномный локус, определяемый направляющей последовательностью sgRNA и прилегающим к протоспейсеру мотивом (PAM), с последующим восстановлением DSB посредством либо гомологичной репарации (HDR), либо негомологичного соединения концов (NHEJ) (Jinek, M. et al. (2012) Science 337:816-821; Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308). При наличии матрицы HDR, систему CRISPR-Cas9 можно использовать для создания точных и определенных модификаций в намеченном локусе с помощью процесса HDR.
[0324] Для получения намеченной замены в геномной ДНК плазмиду, которая экспрессирует как sgРНКA, так и Cas9 S. pyogenes (SpCas9), вместе с линейной матрицей HDR вводили в клетки 293FT с помощью нуклеофекции. Матрица HDR представляет собой однонитевой олигонуклеотид ДНК (ssODN, SEQ ID NO: 3), который содержит гомологичные "плечи" 75 п.о., фланкирующие мутацию c.2991+1655A>G вместе с мутантным PAM (c.2991+1666C>G). Мутантный PAM будет предотвращать деградацию донорного ssODN посредством Cas9 в клетках и в то же время вводить уникальный сайт рестрикции SnaBI в интрон 26 CEP290.
[0325] Для получения клеток, несущих мутацию c.2991+1655A>G CEP290, 235 одноклеточных клонов выделяли из совместно трансфицированных клеток sgRNA/SpCas9 и ssODN и подвергали скринингу. Геномную ДНК экстрагировали и амплифицировали с помощью ПЦР-праймеров, фланкирующих интронную мутацию (SEQ ID NO: 4-5). Продукты ПЦР подвергали расщеплению посредством SnaBI и секвенированию методом Сэнгера с секвенирующим праймером (SEQ ID NO: 6).
[0326] Среди 235 одноклеточных клонов один клон содержал мутации c.2991+1655A>G и c.2991+1666C>G на обоих аллелях CEP290 (в дальнейшем называемые "мутантными клетками" или "MT-клетками"). Другой клон содержал две мутации на одном аллеле CEP290 и эндогенную ДНК CEP290 дикого типа на другом аллеле (в дальнейшем называемые "гетерозиготными клетками" или "Het-клетками"). Клетки, которые содержали два аллеля эндогенного CEP290 дикого типа, в дальнейшем называются "клетками дикого типа" или "клетками WT".
[0327] Уровни экспрессии mRNA дикого типа и мутантного CEP290 в клетках дикого типа, гетерозиготных и мутантных, измеряли методом количественной ПЦР с обратной транскрипцией (RT-qPCR) с использованием праймеров, которые специфически определяют mRNA CEP290 дикого типа (SEQ ID NO: 7-8) и мутантную mRNA CEP290 (SEQ ID NO: 9-10), соответственно. Результаты нормализовали к уровням экспрессии mRNA PPIA (SEQ ID NO: 31-32).
[0328] По сравнению с клетками дикого типа уровни mRNA CEP290 дикого типа были снижены соответственно на 27% и 48% в гетерозиготных и мутантных клетках (ФИГ. 3А). Как и ожидалось, клетки дикого типа не экспрессировали mRNA мутантного CEP290, тогда как ее уровни были на 24% выше в мутантных клетках, чем в гетерозиготных клетках (ФИГ. 3В). По сравнению с гетерозиготными клетками мутантные клетки экспрессировали значительно более низкие уровни mRNA CEP290 дикого типа и значительно более высокие уровни mRNA мутантного CEP290. Следовательно, данная клеточная модель повторяла фенотип пациентов с LCA, несущих мутацию c.2991+1655 A>G, поскольку эти пациенты экспрессировали на ~50% сниженный уровень mRNA CEP290 дикого типа (denHollander,A.I.etal.(2006)Am.J.Hum.Genet.79:556-561).
[0329] Уровни белка CEP290 в дикого типа, гетерозиготных и мутантных клетках исследовали с помощью вестерн-блот-анализа. По сравнению с клетками дикого типа уровни белка CEP290 в гетерозиготных и мутантных клетках были значительно снижены (ФИГ. 3C). Мутантные клетки экспрессировали более низкие уровни белка CEP290 по сравнению с гетерозиготными клетками (ФИГ. 3C). Следовательно, данные иммуноблоттинга соответствуют данным RT-qPCR.
Пример 2. Целенаправленная делеция мутации c.2991+1655 A>G
Способы
Плазмиды
[0330] Вектор экспрессии "все-в-одном" конструировали с использованием способа клонирования Golden Gate, как описано ранее (Sakuma, T. et al. (2014) Sci. Rep. 4:5400). Вкратце, фрагмент ДНК, который содержит промотор U6-BbsI:BbsI-каркас sgRNA-терминатор U6-промотор CMV (SEQ ID NO: 33) синтезировали с помощью GeneArt (Life Technologies) и вставляли в сайты рестрикции PciI и NruI нулевой плазмиды pSpCas9-BbsI, описанной выше, для создания плазмиды pSpCas9(BBU), которую использовали для субклонирования вышерасположенных направляющих последовательностей sgRNA. Для конструирования плазмиды pSpCas9(BBD), которую использовали для субклонирования нижерасположенных направляющих последовательностей sgRNA, осуществляли реакцию ПЦР с использованием ДНК-полимеразы высокой точности Phusion (New England BIoLabs Inc.) с ДНК плазмиды pSpCas9(BB) в качестве матрицы ДНК и парой ПЦР-праймеров (SEQ ID NO: 34-35). Параметры проведения циклов: 1 цикл при 98°С в течение 1 мин.; 35 циклов 98°C в течение 20 с и 72°C в течение 30 с; 1 цикл при 72°С в течение 5 мин. ПЦР-продукт вводили в сайты PciI и KpnI плазмиды pSpCas9(BBU). В этой плазмиде pSpCas9(BBD) два сайта BsaI фланкировали sgRNA под управлением промотора U6. Олигонуклеотиды sgRNA U1 (SEQ ID NO: 11-12) отжигали и затем субклонировали в два сайта рестрикции BbsI плазмиды pSpCas9(BBU), и олигонуклеотиды sgRNA D1, D2 и D3 (SEQ ID NO: 13-18) отжигали и затем субклонировали в два сайта рестрикции BbsI плазмиды pSpCas9(BBD) в соответствии с протоколом, описанным ранее (Ran, F.A. et al. (2013) Nat. Protoc. 8:2281-2308). Полученные в результате sgRNA D1-D3 вместе с промотором U6 дополнительно вырезали из плазмид pSpCas9(BBD) с использованием рестриктазы BsaI и затем субклонировали в два сайта BsaI в плазмиде pSpCas9(BBU)-U1 sgRNA. Полученные плазмиды pSpCas9(BBUD) экспрессировали две sgRNA под управлением промотора U6 и один SpCas9 под управлением промотора CMV. Олигонуклеотиды для двух контрольных sgRNA (SEQ ID NO: 36-39) субклонировали в сайты рестрикции BbsI такого же вектора экспрессии "все-в-одном" с использованием способа, описанного выше, в качестве контрольной плазмиды.
Трансфекция и ПЦР-анализ
[0331] Плазмиды с парными sgRNA-SpCas9 трансфицировали в клетки дикого типа, гетерозиготные и мутантные с использованием реагента трансфекции Lipofectamine 3000 (Life Technologies) согласно протоколу производителя.
[0332] Через 48 ч. после трансфекции геномную ДНК экстрагировали из клеток с использованием раствора для экстракции ДНК QuickExtract (Epicenter) и амплифицировали с помощью GoTaq Hot Start Green Master Mix (Promega) и праймеров ПЦР, фланкирующих интронную мутацию (SEQ ID NO: 4-5). Амплификацию продуктов ПЦР выполняли со следующими параметрами циклов: 1 цикл при 95°С в течение 2 мин.; 35 циклов при 95°C в течение 30 с, 60°C в течение 30 с и 72°C в течение 3 мин.; 1 цикл при 72°С в течение 15 мин. Продукты ПЦР подвергали электрофорезу в агарозном геле.
Секвенирование нового поколения (NGS)
[0333] Продукты ПЦР, полученные выше, отправляли в ACGT (Уилинг, Иллинойс) для секвенирования NGS. ДНК образцов фрагментировали до среднего размера целевого фрагмента 350 п.о. С помощью ультразвуковой обработки и использовали для построения библиотеки секвенирования с использованием набора для подготовки образцов ДНК без ПЦР Illumina TruSeq. Библиотеку количественно оценивали посредством Qubit и 2100 Bioanalyzer и загружали на платформу Illumina для генерации прочтений PE150. Генерировали приблизительно 150 тыс. прочтений (±20%) на образец. Необработанные данные Illumina демультиплексировали и конвертировали в формат.fastq и отфильтровывали прочтения низкого качества (Q<20) и короткие (N<50). Отфильтрованные прочтения выравнивали с эталонными последовательностями (ДНК дикого типа и усеченная ДНК) с использованием Bowtie2. Для количественной оценки, число прочтений, выровненных по последовательности 40 п.о., фланкирующей сайт расщепления sgRNA U1 (20 п.о. до и 20 п.о. после сайта расщепления), которые являются уникальными либо для ДНК дикого типа, либо для усеченной ДНК, использовали для вычисления процента ДНК дикого типа и усеченной в каждом образце.
Результаты
[0334] Затем вышеописанную модель использовали для тестирования подхода к созданию целенаправленной делеции мутации c.2991+1655 A>G.
[0335] Одну вышерасположенную sgRNA (U1, SEQ ID NO: 19), которая нацеливает локус в криптическом экзоне, экспрессировали с каждой из трех нижерасположенных sgRNA (D1, D2, D3; SEQ ID NO: 20-22) в том же векторе, что и SpCas9. Пары выше и ниже расположенных sgRNA фланкировали мутацию c.2991+1655A>G и, как прогнозировалось, генерировали геномные делеции размером соответственно 283 п.о., 187 п.о. и 231 п.о., поскольку предыдущие исследования показали, что репарация индуцированной парными sgRNA геномной делеции в основном достигается точным соединением концов (Brandl, C. et al. (2014) FEBS Open Bio. 5:26-35; Zheng, Q. et al. (2014) Biotechniques 57:115-124). Две контрольные sgRNA субклонировали в ту же плазмиду, что и контроль.
[0336] Плазмиды с парными sgRNA-SpCas9 трансфицировали в клетки дикого типа, гетерозиготные и мутантные, как описано выше, с последующим ПЦР-анализом с использованием праймеров, фланкирующих области делеции (SEQ ID NO: 4-5). Геномные фрагменты дикого типа и усеченные разделяли с помощью гель-электрофореза. Для всех трех тестируемых вышерасположенных/нижерасположенных пар sgRNA обнаружили продукты ПЦР, которые указывали на наличие ожидаемых геномных делеций, которые отсутствовали в клетках, трансфицированных контрольными sgRNA (ФИГ. 4A). Эти результаты показывают, что парные sgRNA и SpCas9 способны удалять мутацию c.2991+1655A>G. Анализ методом секвенирования нового поколения (NGS) 4 образцов ПЦР, полученных из мутантных клеток, дополнительно показал, что 60,7%, 65,9% и 72,4% NGS-прочтений были выровнены с усеченной ДНК соответственно в U1D1-, U1D2- и U1D3-трансфицированных клетках. Следовательно, все три пары sgRNA и SpCas9 высокоэффективны при удалении мутации c.2991+1655A>G.
[0337] Для подтверждения того, что парные sgRNA способны восстанавливать уровни экспрессии CEP290 дикого типа в гетерозиготных и мутантных клетках, клетки дикого типа, гетерозиготные и мутантные трансфицировали плазмидами, которые экспрессировали парные sgRNA и SpCas9, с последующим анализом RT-qPCR для mRNA CEP290, как описано выше. Результаты нормализовали к уровням экспрессии mRNA PPIA.
[0338] По сравнению с контрольными sgRNA, пары sgRNA, особенно пара U1D3, заметно восстанавливали уровни mRNA CEP290 дикого типа (ФИГ. 5A) и снижали уровни mRNA мутантного CEP290 (ФИГ. 5B) в гетерозиготных и мутантных клетках. Важно отметить, что ни одна из трех пар sgRNA значительно не изменяла уровни mRNA CEP290 дикого типа (ФИГ. 5A), что указывало на то, что целенаправленные геномные делеции не мешали нормальному сплайсингу транскриптов CEP290. Пара sgRNA U1D3 значительно восстанавливала уровни mRNA CEP290 дикого типа в гетерозиготных и мутантных клетках до уровней, сравнимых с уровнями в клетках дикого типа (ФИГ. 5A). Пара sgRNA U1D2 также значительно повышала уровни mRNA CEP290 дикого типа в гетерозиготных клетках (ФИГ. 5A). Все три пары sgRNA значительно снижали уровни mRNA мутантного CEP290 в гетерозиготных клетках (ФИГ. 5B). Пары sgRNA U1D2 и U1D3 значительно снижали уровни mRNA мутантного CEP290 в MT-клетках (ФИГ. 5B).
[0339] Для подтверждения того, что парные sgRNA были способны восстанавливать экспрессию белка CEP290 дикого типа в мутантных клетках, мутантные клетки трансфицировали парными sgRNA и SpCas9 и затем подвергали вестерн-блот-анализу, как описано выше. По сравнению с контрольными sgRNA все три пары sgRNA восстанавливали экспрессию белка CEP290 (ФИГ. 5C).
[0340] В совокупности эти данные показывают, что вышерасположенные/нижерасположенные пары sgRNA, особенно пара U1D3, являются высокоэффективными в предотвращении сплайсинга мутантного криптического экзона и восстановлении экспрессии CEP290 дикого типа. Эти результаты показывают, что пара направляющих РНК, связанная с Cas9, может необратимо удалять область интрона, фланкирующую мутацию c.2991+1655 A>G CEP290, для предотвращения сплайсинга криптического экзона, введенного в mRNA CEP290.
Пример 3. Разработка самоограничивающейся системы Crispr-Cas9 для ограничения времени существования SpCas9
Способы
Плазмиды
[0341] Для самоограничивающейся системы Crispr-Cas9 сконструировали упаковочную плазмиду AAV pAAV-minCMV-SpCas9-NLS-SV40 pA. Вкратце, фрагмент ДНК, который содержал минимальный промоторный фрагмент, полученный из промотора CMV (промотор minCMV) (SEQ ID NO: 56), синтезировали с помощью GeneArt (Life Technologies) и вставляли в сайты рестрикции MluI и ApoI плазмиды pSpCas9 от Sigma, описанной выше, с получением плазмиды pminCMV-SpCas9-NLS-BGH pA. Затем фрагмент ДНК, который содержал ранний сигнал poly(A) SV40 (SV40 pA) (SEQ ID NO: 57), синтезировали с помощью GeneArt (Life Technologies) и вставляли в сайты рестрикции XhoI и BbsI вышеприведенной плазмиды с получением плазмиды pminCMV-SpCas9-NLS-SV40 pA. Наконец, фрагмент minCMV-SpCas9-NLS-SV40 pA субклонировали в упаковочную плазмиду AAV с получением pAAV-minCMV-SpCas9-NLS-SV40 pA.
[0342] Для конструирования плазмиды с самоограничивающимся pSpCas9 последовательности распознавания для нуклеазы SpCas9 (направляющие последовательности sgRNA U1 и D3 плюс соответствующие мотивы PAM; SEQ ID NO: 58-59) субклонировали в сайт вставки 1 (между промотором minCMV и SpCas9) и/или сайт вставки 2 (между сигналом внутриядерной локализации (NLS) SpCas9 и сигналом poly(A) SV40).
[0343] Второй упаковочной плазмидой AAV для самоограничивающейся системы Crispr-Cas9 являлась pAAV-U6-U1 sgRNA-U6-D3 sgRNA-BGH pA. Для конструирования этой плазмиды фрагмент U6-U1 sgRNA-U6-D3 sgRNA вырезали из плазмиды pSpCas9(BBUD)-U1D3 с использованием рестриктаз PciI и KpnI и затем субклонировали в сайты EcoRV и KpnI упаковочной плазмиды AAV с получением плазмиды pAAV-U6-U1 sgRNA-U6-D3 sgRNA. С целью титрования AAV фрагмент BGH pA (SEQ ID NO: 60) клонировали в сайты SpeI и HindIII этой плазмиды.
Результаты
[0344] CRISPR-Cas9 является мощным инструментом для редактирования генома, однако неясно, как будет переноситься экспрессия бактериального белка Cas9 у пациентов. CRISPR-Cas9 работает быстро (от нескольких часов до нескольких дней), и ее длительное присутствие в клетках не требуется. Нежелательные иммунные ответы и потенциальные проблемы безопасности могут быть вызваны длительной экспрессией экзогенного белка. Например, экспрессия экзогенного маркерного белка, зеленого флуоресцентного белка (GFP) может вызывать значительные нежелательные иммунологические ответы (Stripecke R et al. (1999) Gene Ther. 6:1305-1312). Кроме того, в недавнем докладе было высказано предположение о том, что имеет место индукция гуморального иммунитета против Cas9 и потенциальное присутствие специфичного к Cas9 клеточного иммунного ответа после доставки CRISPR-Cas9 in vivo (Wang, D. et al. (2015) Hum. Gene Ther. 26:432-442). Следовательно, подход "попасть и уйти", при котором воздействие белка Cas9 ограничено, может быть полезным при доставке CRISPR-Cas9 in vivo. Такой подход "попасть и уйти" также может уменьшать побочные эффекты, поскольку это уменьшит время взаимодействия между Cas9 и непредусмотренными мишенями.
[0345] С этой целью авторы разработали самоограничивающуюся систему Crispr-Cas9 для ограничения времени существования SpCas9, включив сайт(-ы) распознавания для sgRNA в сам вектор SpCas9, так что вектор будет разрезан и разрушен вскоре после начала экспрессии SpCas9. Самоограничивающаяся система Crispr-Cas9 содержит два вектора: вектор pAAV-U6-U1 sgRNA-U6-D3 sgRNA, который экспрессирует пару sgRNA U1 и D3, и вектор pAAV-minCMV-SpCas9-NLS-SV40 pA, который экспрессирует SpCas9 под управлением промотора minCMV. Последовательности распознавания для sgRNA U1 и/или D3 (направляющие последовательности плюс соответствующие мотивы PAM) включены в один или два из двух сайтов вставки во втором векторе. Сайт вставки 1 расположен между промотором minCMV и кодирующей последовательностью SpCas9, а сайт вставки 2 расположен между сигналом внутриядерной локализации (NLS) и сигналом poly(A) SV40 (ФИГ. 6A). Таким образом, в этой самоограничивающейся системе Crispr-Cas9 sgRNA U1 и D3 будут направлять белок SpCas9 как для целенаправленной геномной делеции, так и для расщепления самоограничивающегося вектора SpCas9, что лишит вектор возможности создания избыточного количества белка SpCas9.
[0346] Для тестирования самоограничивающейся системы Crispr-Cas9 in vitro, мутантные клетки трансфицировали двумя вышеописанными векторами с последующим вестерн-блот-анализом (ФИГ. 6В), ПЦР геномной ДНК (ФИГ. 6С) и анализом RT-qPCR mRNA CEP290 (ФИГ. 6D и 6E), как описано выше.
[0347] Когда одиночную последовательность распознавания sgRNA (U1T или D3T; SEQ ID NO: 58-59) вводили в самоограничивающийся вектор SpCas9, уровни белка SpCas9 значительно снижались по сравнению с контрольным вектором SpCas9, который не содержал последовательность распознавания sgRNA(ФИГ. 6B). В случае если в самоограничивающийся вектор SpCas9 вводили два U1T, два D3T или D3T и U1T в комбинации, уровень белка SpCas9 практически исчезал(ФИГ. 6B). Следовательно, самоограничивающаяся система Crispr-Cas9 способна эффективно ограничивать экспрессию SpCas9.
[0348] Для подтверждения того, что самоограничивающаяся система Crispr-Cas9 все еще способна индуцировать целенаправленную геномную делецию и удалять мутацию LCA10 c.2991+1655A>G, авторы провели ПЦР-анализ геномной ДНК в мутантных клетках, трансфицированных двойными векторами (ФИГ. 6C). Полоса ПЦР, соответствующая усеченной ДНК, присутствовала как в клетках, трансфицированных контрольным вектором SpCas9, так и в клетках, трансфицированных самоограничивающимися векторами SpCas9, что указывало на то, что самоограничивающаяся система Crispr-Cas9 все еще была способна удалять интронную мутацию LCA10, несмотря на короткое время существования SpCas9. Следует отметить, что геномная делеция, показанная на ФИГ. 6С, не была такой надежной, как делеция, показанная на ФИГ. 4A. Это может быть связано с различными условиями эксперимента в этих двух исследованиях. Во-первых, на ФИГ. 4А авторы использовали вектор "все-в-одном", а на ФИГ. 6C использовали двойные векторы. Во-вторых, экспрессия SpCas9 на ФИГ. 4А находилась под управлением промотора CMV, а на ФИГ. 6C находилась под управлением промотора minCMV.
[0349] Затем авторы исследовали уровни mRNA дикого типа и мутантного CEP290 в мутантных клетках, трансфицированных двойными векторами. По сравнению с одной sgRNA U1D3 (контроль), контрольный вектор SpCas9 значительно восстанавливал уровни mRNA CEP290 дикого типа в мутантных клетках (ФИГ. 6D). Аналогично, вектор SpCas9, который содержал последовательность распознавания sgRNA D3 в любом сайте вставки, значительно увеличивал уровни mRNA CEP290 дикого типа (ФИГ. 6D). Другие самоограничивающиеся векторы SpCas9 также увеличивали уровни mRNA CEP290 дикого типа, хотя по сравнению с контролем различия не были статистически значимыми (ФИГ. 6D). Кроме того, все самоограничивающиеся векторы SpCas9 и контрольный вектор SpCas9 значительно уменьшали уровни mRNA мутантного CEP290 (ФИГ. 6Е).
[0350] В совокупности эти данные демонстрируют, что самоограничивающаяся система Crispr-Cas9 способна ограничивать время существования SpCas9, удалять мутацию LCA10 c.2991+1655A>G, восстанавливать экспрессию CEP290 дикого типа и уменьшать экспрессию мутантного CEP290.
Пример 4. Целенаправленная делеция интронной области гена Cep290 мыши в сетчатке мыши
Способы
Плазмиды
[0351] Авторы использовали систему на основе двух AAV для целенаправленной геномной делеции в сетчатке мыши. Первый AAV получали с помощью упаковочной плазмиды AAV pAAV-minCMV-SpCas9-NLS-SV40 pA, как описано выше. Второй AAV получали с помощью упаковочной плазмиды AAV pAAV-U6-U11 sgRNA-U6-D11 sgRNA-RK-EGFP-BGH pA. Для конструирования этой плазмиды направляющую последовательность sgRNA U11 (SEQ ID NO: 61) и направляющую последовательность sgRNA D11 (SEQ ID NO: 62) субклонировали в плазмиду pSpCas9(BBUD), как описано выше, с получением плазмиды pSpCas9(BBUD)-U11D11. Фрагмент промотор RK-EGFP-BGH pA (SEQ ID NO: 63) вводили в сайты KpnI и XhoI этой плазмиды, и затем полную кассету U6-U11 sgRNA-U6-D11 sgRNA-RK-EGFP-BGH pA вырезали из этой плазмиды с помощью рестриктаз PciI и PmeI и субклонировали в сайты BamHI и SphI упаковочной плазмиды AAV, описанной выше, с получением конечной плазмиды. Также создавали контрольную плазмиду pAAV-RK-EGFP-BGH pA.
Продуцирование AAV
[0352] Рекомбинантные векторы на основе AAV получали с помощью тройной трансфекции клеток эмбриональной карциномы почки человека 293 (HEK-293), как описано ранее (Xiao et al. (1998) J. Virol. 72:2224-2232). Вкратце, использовали упаковочную плазмиду AAV, плазмиду, содержащую ген rep из серотипа 2 и ген капсидного белка из серотипа 5, и аденовирусную плазмиду-помощника (Stratagene). Вирус собирали через 72 часа после трансфекции, очищали с помощью аффинной хроматографии в сефарозе AVB (GE Healthcare). Титры копий генома (GC) векторов на основе AAV определяли с помощью количественного ПЦР-анализа на основе TaqMan (Applied Biosystems), как описано ранее (Gerits, A. et al. (2015) Neurophotonics. 2:031209).
Животные
[0353] Самок мышей C57BL/6J возрастом 8-10 недель приобретали у Jackson Laboratories (Бар-Харбор, Мэн) и выдерживали в виварии Sanofi. Животным предоставляли свободный доступ к пище и воде в течение всего периода проведения исследования. Все процедуры для животных проводили согласно Закону о благополучии животных, Руководству по уходу и использованию лабораторных животных, Управлению по благополучию лабораторных животных и в соответствии с Заявлением ARVO об использовании животных в офтальмологических исследованиях и исследованиях зрения.
Субретинальная инъекция AAV
[0354] Мышей седатировали с помощью 3,5% изофлурана, вводимого с кислородом, доставляемом животному через дыхательный контур при скорости 800 мл/мин. У мышей индуцировали мидриаз и циклоплегию с помощью местного применения тропикамида (Alcon, Форт-Уэрт, Техас). Экспериментальный разрез выполняли в роговице, и иглу с тупым наконечником 33 калибра направляли через разрез и продвигали назад между радужкой и капсулой хрусталика до тех пор, пока наконечник не проникал в заднюю нейросенсорную сетчатку. Доставляли 1 × 109 частиц вирусного генома AAV5-minCMV-SpCas9-NLS-SV40 pA (AAV5-SpCas9 на ФИГ. 7A) вместе с 1×109 частиц вирусного генома AAV5-U6-U11 sgRNA-U6-D11 sgRNA-RK-EGFP-BGH pA (AAV5-U11D11-RK-EGFP на ФИГ. 7A) или контрольной AAV5-U6-RK-EGFP-BGH pA (AAV5-RK-EGFP на ФИГ. 7B) в OS глаза каждой мыши в объеме одного микролитра в течение одной секунды. Иглу удерживали в рабочем положении в течение примерно пяти секунд перед извлечением. Животному давали возможность восстановиться после анестезии перед возвращения его в клетку.
[0355] Экспрессию EGFP в сетчатке оценивали у живых животных с использованием микроскопа для визуализации сетчатки Micron IV (Phoenix Research Labs) через две и четыре недели после инъекции, и при этом мышей, не имеющих экспрессии EGFP, исключали из исследования. Всех животных подвергали эвтаназии через четыре недели после инъекции.
Препарирование сетчатки
[0356] Глаза мыши энуклеировали и помещали в фосфатно-солевой буфер (PBS). Сетчатку выделяли ножницами для микродиссекции под препаровальной лупой. Слой пигментного эпителия сетчатки (RPE) тщательно удаляли.
Экспрессия геномной ДНК и ПЦР-анализ
[0357] Сетчатку гомогенизировали в растворе для экстракции ДНК QuickExtract (Epicenter) с пестиками, приводимыми в движение беспроводным мотором (VWR). Геномную ДНК экстрагировали в соответствии с инструкцией производителя (Epicenter), разбавляли до 10 нанограмм/микролитр и затем амплифицировали с помощью GoTaq Hot Start Green Master Mix (Promega) и праймеров ПЦР, фланкирующих удаляемую область (SEQ ID NO: 64-65). Амплификацию продуктов ПЦР выполняли со следующими параметрами циклов: 1 цикл при 95°С в течение 2 мин.; 35 циклов при 95°C в течение 30 с, 62°C в течение 30 с и 72°C в течение 2 мин.; 1 цикл при 72°С в течение 12 мин. Продукты ПЦР подвергали электрофорезу в агарозном геле.
Результаты
[0358] Исследование на клеточной модели LCA10 подтвердило эффективность подхода авторов с парными sgRNA и SpCas9 для удаления мутации LCA10 c.2991+1655A>G in vitro. Затем авторы оценили свой подход CRISPR-Cas9 в условиях in vivo. Поскольку в настоящее время нет модели животных, которая экспрессирует криптический экзон, вызванный мутацией сплайсинга интрона LCA10, авторы таким образом испытывали, могла бы пара sgRNA вместе с SpCas9 индуцировать целенаправленную геномную делецию в интроне 25 гена Cep290 мыши, который гомологичен интрону 26 гена CEP290 человека, где расположена мутация c.2991+1655A>G.
[00100] Авторы решили использовать векторы на основе AAV для доставки гена in vivo из-за их низкой иммуногенности и диапазона серотипов, обеспечивающих возможность преимущественного заражения определенных типов клеток. Клетками сетчатки, наиболее подверженными мутациям CEP290, являются фоторецепторы, причем предыдущие исследования показали, что AAV серотипа 5 (AAV5) мог эффективно трансдуцировать фоторецепторы с помощью субретинальной инъекции (Boye, S.E. et al. (2012) Hum. Gene Ther. 23:1101-1115). Поэтому авторы решили использовать AAV5 для доставки SpCas9 и парных sgRNA к фоторецепторам.
[0359] Из-за низкой упаковочной способности вектора на основе AAV (~4,8 т.п.н.) и длины кодирующей последовательности SpCas9, измеряемой при 4,1 т.п.н. (см. SEQ ID NO: 40 для аминокислотной последовательности), сложно упаковать один компонент SpCas9 и два компонента U6-sgRNA в один вектор на основе AAV. Поэтому авторы использовали систему на основе двух AAV для своего валидационного исследования in vivo (ФИГ. 7A). Первый вектор на основе AAV представлял собой AAV5-U11D11-RK-EGFP, который экспрессировал вышерасположенную sgRNA U11 (SEQ ID NO: 61) и нижерасположенную sgRNA D11 (SEQ ID NO: 62), а также репортерный ген EGFP под управлением промотора родопсинкиназы (RK). Предполагается, что пара sgRNA U11 и D11 генерировала геномную делецию 557 п.о. в интроне 25 гена Cep290 мыши. Также изготовили контрольный AAV AAV5-RK-EGFP. Второй вектор на основе AAV представлял собой AAV5-SpCas9, который экспрессировал SpCas9 под управлением промотора minCMV.
[0360] Совместно вводили AAV5-U11D11-RK-EGFP (или AAV5-RK-EGFP) и AAV5-SpCas9 (1 × 109 частиц вирусного генома каждый) с помощью инъекции в субретинальное пространство взрослых мышей C57BL/6J. Экспрессию EGFP в сетчатке оценивали у живых животных, и мышей, лишенных экспрессии EGFP, исключали из исследования. Через четыре недели после инъекции животных подвергали эвтаназии и геномную ДНК экстрагировали из сетчатки с последующим ПЦР-анализом с использованием праймеров, фланкирующих области делеции (SEQ ID NO: 64-65). Геномные фрагменты дикого типа и усеченные разделяли с помощью гель-электрофореза. У животных, которым вводили AAV5-U11D11-RK-EGFP и AAV5-SpCas9, обнаружили продукт ПЦР, который указывал на наличие ожидаемой геномной делеции с парой sgRNA U11 и D11, которая отсутствовала у животных, которым вводили AAV5-RK-EGFP и AAV5-SpCas9 (ФИГ. 7В). Эти результаты показывают, что данная система на основе двух AAV способна индуцировать целенаправленную геномную делецию in vivo.
Пример 5. Использование пары вышерасположенная/нижерасположенная sgRNA и SaCas9 для удаления мутации LCA10 c.2991+1655A>G
Способы
Плазмиды
[0361] Для конструирования плазмиды SaCas9 каркасный фрагмент minCMV-SaCas9-NLS-FLAG-BGH pA-U6-BsaI:BsaI-sgRNA (SEQ ID NO: 66) синтезировали в GenScript и субклонировали в сайты рестрикции PciI и BbsI плазмиды pSpCas9 от Sigma для замещения кассеты CMV-SpCas9-BGH pA и создания плазмиды pminCMV-SaCas9-BGH pA-U6. Пять направляющих последовательностей sgRNA (aU1, aU2, aU3, aD1, aD2; SEQ ID NO: 45-49) идентифицировали с использованием онлайн-инструмента разработки Benchling для редактирования генома и затем субклонировали в два сайта рестрикции BsaI вышеуказанной плазмиды. Для образования пары вышерасположенных sgRNA (aU1, aU2, aU3) и нижерасположенных sgRNA (aD1, aD2), кассету sgRNA U6-aD1 и кассету sgRNA U6-aD2 вырезали из их плазмид с использованием рестриктаз KpnI и NotI и затем субклонировали в сайт NotI плазмиды, которая экспрессировала sgRNA aU1, aU2 или aU3. В результате авторы создали шесть плазмид, которые экспрессировали шесть различных пар sgRNA (aU1aD1, aU1aD2, aU2aD1, aU2aD2, aU4aD1, aU4aD2). Наконец, фрагменты minCMV-SaCas9-BGH pA-U6-парные sgRNA в этих шести плазмидах субклонировали в упаковочную плазмиду AAV с получением плазмид pAAV-minCMV-SaCas9-BGH pA-U6-парные sgRNA. Также создавали контрольную плазмиду pAAV-minCMV-SaCas9-BGH pA, которая не экспрессировала sgRNA.
Результаты
[0362] В результате вышеприведенных исследований авторы продемонстрировали, что пара из вышерасположенной/нижерасположенной sgRNA может эффективно направлять SpCas9 для удаления мутации LCA10 c.2991+1655A>G и восстановления экспрессии CEP290 дикого типа. Длина кодирующей последовательности SpCas9 (~4,1 т.п.н.) затрудняет упаковку одного компонента minCMV-SpCas9 и двух компонентов U6-sgRNA в одном векторе на основе AAV. Ran et al. (Ran, F.A. et al. (2015) Nature 520:186-191) ранее обнаружили более короткий фермент Cas9 - Cas9 S. aureus (SaCas9; 1053 аминокислоты; SEQ ID NO: 55), который также демонстрировал расщепляющую активность в клетках млекопитающих. Меньший размер SaCas9 давал возможность упаковать один компонент minCMV-SaCas9 и два компонента U6-sgRNA в один вектор на основе AAV. Чтобы проверить, могут ли парные sgRNA и SaCas9 также удалять интронную мутацию LCA10, авторы разработали три вышерасположенные направляющие последовательности sgRNA (aU1, aU2, aU3, SEQ ID NO: 45-47) и две нижерасположенные направляющие последовательности sgRNA (aD1, aD2, SEQ ID NO: 48-49) специально для SaCas9, используя онлайн-инструмент разработки Benchling для редактирования генома. Вышерасположенные/нижерасположенные sgRNA спаривали и субклонировали в упаковочную плазмиду AAV. Те же плазмиды также экспрессировали оптимизированный человеческими кодонами SaCas9 под управлением промотора miCMV (SEQ ID NO: 66).
[0363] Плазмиды с парными sgRNA-SaCas9 трансфицировали в мутантные клетки, как описано выше. Чтобы сравнить SaCas9 с SpCas9, мутантные клетки также трансфицировали двойными плазмидами SpCas9 AAV (pAAV-minCMV-SpCas9+пара sgRNA pAAV-U1D3) или только отдельными плазмидами. ПЦР-анализ проводили с использованием праймеров, фланкирующих области делеции (SEQ ID NO: 4-5). Геномные фрагменты дикого типа и усеченные разделяли с помощью гель-электрофореза. Для всех тестируемых вышерасположенных/нижерасположенных пар sgRNA обнаружили продукты ПЦР, которые указывали на наличие ожидаемых геномных делеций, которые отсутствовали в клетках, трансфицированных контрольными плазмидами (ФИГ. 8A). Эти результаты демонстрируют, что как SaCas9, так и SpCas9 способны работать с парными sgRNA для удаления интронной мутации сплайсинга LCA10.
[0364] Затем авторы исследовали уровни mRNA дикого типа и мутантного CEP290 в мутантных клетках, трансфицированных вышеуказанными упаковочными плазмидами AAV. Удивительно, однако ни одна из пар вышерасположенной/нижерасположенной sgRNA значительно не увеличивала уровни mRNA CEP290 дикого типа по сравнению с плазмидой одного SaCas9, хотя пара aU2aD1 и пара aU2aD2 продемонстрировали тенденцию к увеличению (ФИГ. 8B). В отличие от этого, двойные AAV-плазмиды для SpCas9 значительно восстанавливали уровни mRNA CEP290 дикого типа по сравнению с отдельными плазмидами (ФИГ. 8B). Все плазмиды с парными sgRNA-SaCas9 значительно уменьшали уровни mRNA мутантного CEP290 по сравнению с плазмидой одного SaCas9 (ФИГ. 8C). Аналогично, двойные AAV-плазмиды для SpCas9 значительно уменьшали уровни mRNA мутантного CEP290 по сравнению с отдельными плазмидами (ФИГ. 8C). Хотя в настоящем исследовании SaCas9 не так эффективен, как SpCas9 в отношении восстановления уровней mRNA CEP290 дикого типа в мутантных клетках, авторы не могут исключить возможность того, что действенные пары sgRNA могут направлять SaCas9 для эффективного удаления интронной мутации сплайсинга LCA10 и значительного восстановления уровней mRNA CEP290 дикого типа в мутантных клетках.
ПОСЛЕДОВАТЕЛЬНОСТИ
Все последовательности нуклеиновых кислот представлены в направлении от 5'- к 3'-концу, если не указано иное.
Все аминокислотные последовательности представлены в направлении от N-конца к C-концу, если не указано иное.
Олигонуклеотид верхней нити для sgRNA, используемой в гомологичной репарации (HDR) (SEQ ID NO: 1)
caccgAAGACACTGCCAATAGGGAT
Олигонуклеотид нижней нити для sgRNA, используемой в гомологичной репарации (HDR) (SEQ ID NO: 2)
aaacATCCCTATTGGCAGTGTCTTc
Матрица HDR (SEQ ID NO: 3)
CCACCCGCCTCGGCCTCCTAAAGTGCTGGGATTACAGATGTGAGCCACCGCACCTGGCCCCAGTTGTAATTGTGAGTATCTCATACGTATCCCTATTGGCAGTGTCTTAGTTTTATTTTTTATTATCTTTATTGTGGCAGCCATTATTCCTGTCTCTA
F-праймер нуклеиновой кислоты для интрона 26 гена CEP290 (SEQ ID NO: 4)
GGTCCCTGGCTTTTGTTCCT
R-праймер нуклеиновой кислоты для интрона 26 гена CEP290 (SEQ ID NO: 5)
CAGGAGGCTGAGGGTGTTTT
Секвенирующий праймер для интрона 26 гена CEP290 (SEQ ID NO: 6)
AGTAGAGATGGGGTTTCACC
F-праймер нуклеиновой кислоты для гена CEP290 дикого типа (SEQ ID NO: 7)
TGACTGCTAAGTACAGGGACATCTTG
R-праймер нуклеиновой кислоты для гена CEP290 дикого типа (SEQ ID NO: 8)
AGGAGATGTTTTCACACTCCAGGT
F-праймер нуклеиновой кислоты для мутантного гена CEP290 (SEQ ID NO: 9)
CTGGCCCCAGTTGTAATTTGTGA
R-праймер нуклеиновой кислоты для мутантного гена CEP290 (SEQ ID NO: 10)
CTGTTCCCAGGCTTGTTCAATAGT
Олигонуклеотид верхней нити для sgRNA U1 (SEQ ID NO: 11)
caccGGCGGGTGGATCACGAGTTC
Олигонуклеотид нижней нити для sgRNA U1 (SEQ ID NO: 12)
aaacGAACTCGTGATCCACCCGCC
Олигонуклеотид верхней нити для sgRNA D1 (SEQ ID NO: 13)
caccgAAAGCTACCGGTTACCTGAA
Олигонуклеотид нижней нити для sgRNA D1 (SEQ ID NO: 14)
aaacTTCAGGTAACCGGTAGCTTTc
Олигонуклеотид верхней нити для sgRNA D2 (SEQ ID NO: 15)
caccgTCATTCTTGTGGCAGTAAGG
Олигонуклеотид нижней нити для sgRNA D2 (SEQ ID NO: 16)
aaacCCTTACTGCCACAAGAATGAc
Олигонуклеотид верхней нити для sgRNA D3 (SEQ ID NO: 17)
caccGGAGTCACATGGGAGTCACA
Олигонуклеотид нижней нити для sgRNA D3 (SEQ ID NO: 18)
aaacTGTGACTCCCATGTGACTCC
Последовательность sgRNA U1 (SEQ ID NO: 19)
GGCGGGTGGATCACGAGTTCGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT
Последовательность sgRNA D1 (SEQ ID NO: 20)
GAAAGCTACCGGTTACCTGAAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT
Последовательность sgRNA D2 (SEQ ID NO: 21)
GTCATTCTTGTGGCAGTAAGGGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT
Последовательность sgRNA D3 (SEQ ID NO: 22)
GGAGTCACATGGGAGTCACAGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTT
Последовательность интрона 26 CEP290 (примечание: мутация c.2991+1655A>G представляет собой 1655-ый нуклеотид в данной последовательности и выделена жирным шрифтом, а также подчеркнута) (SEQ ID NO: 23)
GTAAGTTTGTGTGATTCTTGAACCTTGTGAAATTAGCCATTTTTCTTCAATATTTTTGTGTTTGGGGGGATTTGGCAGATTTTAATTAAAGTTTGCCTGCATTTATATAAATTTAACAGAGATATAATTATCCATATTATTCATTCAGTTTAGTTATAAATATTTTGTTCCCACATAACACACACACACACACACAATATATTATCTATTTATAGTGGCTGAATGACTTCTGAATGATTATCTAGATCATTCTCCTTAGGTCACTTGCATGATTTAGCTGAATCAAACCTCTTTTAACCAGACATCTAAGAGAAAAAGGAGCATGAAACAGGTAGAATATTGTAATCAAAGGAGGGAAGCACTCATTAAGTGCCCATCCCTTTCTCTTACCCCTGTACCCAGAACAAACTATTCTCCCATGGTCCCTGGCTTTTGTTCCTTGGAATGGATGTAGCCAACAGTAGCTGAAATATTAAGGGCTCTTCCTGGACCATGGATGCACTCTGTAAATTCTCATCATTTTTTATTGTAGAATAAATGTAGAATTTTAATGTAGAATAAATTTATTTAATGTAGAATAAAAAATAAAAAAACTAGAGTAGAATATCATAAGTTACAATCTGTGAATATGGACCAGACCCTTTGTAGTTATCTTACAGCCACTTGAACTCTATACCTTTTACTGAGGACAGAACAAGCTCCTGATTTGTTCATCTTCCTCATCAGAAATAGAGGCTTATGGATTTTGGATTATTCTTATCTAAGATCCTTTCACAGGAGTAGAATAAGATCTAATTCTATTAGCTCAAAAGCTTTTGCTGGCTCATAGAGACACATTCAGTAAATGAAAACGTTGTTCTGAGTAGCTTTCAGGATTCCTACTAAATTATGAGTCATGTTTATCAATATTATTTAGAAGTAATCATAATCAGTTTGCTTTCTGCTGCTTTTGCCAAAGAGAGGTGATTATGTTACTTTTTATAGAAAATTATGCCTATTTAGTGTGGTGATAATTTATTTTTTTCCATTCTCCATGTCCTCTGTCCTATCCTCTCCAGCATTAGAAAGTCCTAGGCAAGAGACATCTTGTGGATAATGTATCAATGAGTGATGTTTAACGTTATCATTTTCCCAAAGAGTATTTTTCATCTTTCCTAAAGATTTTTTTTTTTTTTTTTTGAGATGGAGTTTCATTCTGTCACCCAGGCTGAGTGCAGTGGCACGATCTCGGCTTAACGCTTACTGCATCCTCTGCCTCCCAGATTCAAGCAGTTCTCCTGCCTCAGCCTCTGAGTAGCTGGGATTACAGGTGTGCACCACCACACCAGCTAATTTTTTTTTTTTTTTTTTTTTTTTTGAGGCAGAGTCTCGCTCTGTCACCCAGGCTGGAGTGCAGTGGCGCCATCTTGGCTCACTGCAAGCTCCACCTCCCGGGTTCAGGCCGTTCTCCTGCCTCAGCCTCCTGAGTAGCTGGTACCACAGGCACCCACCATCATGCCCGGCTAATTTTTTGTATTTTTAGTAGAGATGGGGTTTCACCTTGTTAGCCAGGATGGTGTCGATCTCCTGAACTCGTGATCCACCCGCCTCGGCCTCCTAAAGTGCTGGGATTACAGATGTGAGCCACCGCACCTGGCCCCAGTTGTAATTGTGAGTATCTCATACCTATCCCTATTGGCAGTGTCTTAGTTTTATTTTTTATTATCTTTATTGTGGCAGCCATTATTCCTGTCTCTATCTCCAGTCTTACATCCTCCTTACTGCCACAAGAATGATCATTCTAAACATGAATCCTACCCTGTGACTCCCATGTGACTCCCCGCCTTAAAAACTGTCAAAAGCTACCGGTTACCTGAAGGGTAAAAGTCAAGTCCCCTACTTACCTCATGTCATCTAGAGCAAGAGATGAACTAGCTGAGTTTTCTGACCACAGTGTTCTTTCTTATGTATGTTCTTTTGTACGTGCTCTTTTCTATATATAGGGAACCATTTCTCTCTTCCAGTTGTTTTGCTCAGTGAATTTCTATTCCTGTTTCAAAACTTGTTCAGGCATTACCTTTTTTTTCTTAAGCATACTTTTTTTAATGGAACAAAGTCACTCCTGTCTACACTAGTTCTGCATCTTATACATAGGTTTTGTACATAGTACATATTTATATCACATCAAATTATATGTGTTTACATATCTGTCTTCCTTAATGGAATATAAGTCTTTTGATATAAGGAACTATTTAATTTGTTTCTGTGTGTTGAGTATCTCCTGTTTGGCACAGAGTTCAAGCTAATACATGAGAGTGATTAGTGGTGGAGAGCCACAGTGCATGTGGTGTCAAATATGGTGCTTAGGAAATTATTGTTGCTTTTTGAGAGGTAAAGGTTCATGAGACTAGAGGTCACGAAAATCAGATTTCATGTGTGAAGAATGGAATAGATAATAAGGAAATACAAAAACTGGATGGGTAATAAAGCAAAAGAAAAACTTGAAATTTGATAGTAGAAGAAAAAAGAAATAGATGTAGATTGAGGTAGAATCAAGAAGAGGATTCTTTTTTTGTTGTTTTTTTTTTTGAAACAGAGTCTCACTGTGTTGCCCAGGCTGGAGTGCAGTGGAGTGATCTTGGCTTACTGCAACCTCTGCCTCCCAGGTTCAAGCGATTCTTCTGCTTCAGTCTCCCGAGTAGCTGGAATTACAGGTGCCCACCAGCACGGCCGGCTAATTTAGTAGAGACAGGGTTTTGCCATGTTGGCCGGGCTGGTCTCAAACTTTGGATCTCAGGTAATCCGCCAGCCTCAACTTCCCAAAGTGCTGGGATTACAGGCATGAGCCACTGTGCCCAGCCTGTTTTTTTTTTTTTAAAGGAGACCAGTGAAGTTTCAGGAGGAGGGAAAGAAAATTTAGAGTTACTAGGGAGAGAGTGATGAAGATAAGAGATGAAAGTGGTAATAAGGGAAATAGCAAAATATCAGGGTAGGTGGGAGAAAAAGAGATTTGTAACAAACAATAGGATTATCCTGTGAAAAAGGATGAAAGGAAGAAAAAAATGGATAGAAAGATATTTAAAACACCCTCAGCCTCCTGTTTTCCCTCCTGTGTATTCATAGTATATAAAACTATAATTATGTACTTTACTTAAAAAATATATTATTATTACCTTATCGTGCTTATTTAATCATAGCATGTCCTCTTTTTAGTCTCATTACCCTGTTTGTATTATTCTTCATAACACTTAATACCTGACATTGTATTATATATTGGCTTATTTTCCAGGTACTCCACTCAAATATAAGTTCTAGGATATAATTTATTTATCACTGAAATCCATTGCTTAGAGTACCTGGCATGTAGTAAATAGGCATTCTGTTTTTTCAAATAAAAAATAAAGGAACTTAAGATATATATTTATGTTATATCGCCAGCCTTTTTCCTCACAGCTCTATTCTGTTGTACAGAATTACCTACTTTACAATTCCTGTGTTTCAAGGGGATCTCAAATTTAACGTGTCCACAATGAACTCCTGATTTCTGTTTCTCTCCTAGTCATTCTTATTTCAATATATGTTCAGTTACCTAACCAGCTAGTCAAGGCAGATACTTTAGAGTTATTCTGTAGTCATTCTTTTTCCCTACCATTTTTGTTTTCCAAATGTAATTTATGTGTGTCTTCTTCATCCTCGCAGCTCTAACCCTTGTCCAAACCAGCATCATCACTCATCTGGAGTTCCACAATGTCTTTCTGGCTAGTTTCCCTGATTTCTCTATTGACCCCTTTATTCTCCACAGTGCAGCCAGAATGATTGTTTAAAACTTCCTCCTTAAAATCTTTAAATTGTTTTCTTTTATACGTTAAGTTAAATTCCAGTTCCTTGTCTTGGCATGCCATGCCCTGCCTGGTGTGGCCCCTGATGGTCTCTCCAACTTCATGTTTTACTACTATTGACTCTTATTTTTGCTTACTCTGCTTGGGTGCTCCAGTCCTCCAAATCATTTCCTGCTCCAATCATTTCAATCATTTTTTCCTCTCAGATCTTATAGTATTCCAAATGCTTTCTTCCTTTGGAGCATCTGGGTTTACTAATAAATACTTCGTACCTCACAGTTCAGCTTAAATATCAATTATTTGGTGGTTAAGACATCCTTCAACCGCTCTATCTAAATGTTCCTTTCTATTATTCACTGGCTCAGTACTCTGTTTTTATTTTCTTTCTAAATGTCAACTTTTTTTTTTTTGAGTCAGGGTCTCACTGTTGCCCAGGCTCGAGTGCAGTTGCACAATCATAGCTCATTGCAGCCTTGCCCTCCTGGGATCAAGTAATTCTCCCACCTCAGCCTCCAAAATAGCTGGGATTACAGGTATGCATCACCATGCTCAGCTAATTTTTTGTGTTTTTTTGTAGAGATGAGGTCTCACTTTGTTGCCCAGGCTGGTCTCAAACTCCTGGACTCAAGTGATTCTCCCACCTCAGCCTCCCAAAGTGCTGGGGTTACAGGTGTGAGCCACTGCACCTGGTCGATACTGACTTTTTTTTTTTTTTGAGATGGAGTTTTGCTCTGTTGCCCAGGCTAGAGCGCAGTGGTGTGATCTCAGCTCACTGCAACCTCCACCTCCCAGGTTAAAGGGATTCTTCTGCCTCAGTCTCCTGAGTAGCTGGGATTACAGGCAAGTGCCATCATGACTGGCTAATTTTTGTATTTTTAGCACTATGTTTAGTACTGTGTTGGCCAGGCTTGTCTCGAACTCCTGACCTCAAGTGATCCACCCACCTCAGCCTCCCAAAGTGCTGGGATTACAGGTGTGAGCCACCGTAATCGGCCAACATTGACATTTTTAGTAGACTTTTTGTTTGTTTACTTGCTTATTATCTGCTGCCTTCCACACTCTGGCGAAATCCTGCCACCCACCCACACACACATAGGCACTGAATGGGCAGAACTCTGAAGGCCAGAATTTTATATTTCTTTTCACTGTAAACATCATCATCTGTCACTGATGGCACACTAGGATGCTCAGCAACTGTGTGCATGAAGGAAGTAAGCACTAGTTTGTGAAGGCTGCAAAACTCTTGAGTATTCTAAGAGTTTTGGCCAAAATGAATGTACAGCTTTAGTGGCAGAAGCTAATACTCAGAAATTGAGGCCGTATATTGGATAACACAGGATTTGGATGATTATTTTAAAATAATATTTTACATTGTATATATGTGTGTGTGTGTGTGTGTGTGTGTGTGTATGTGTGTGTGTGTGTATATATATATGTATGTATGTGTATTAGTCCGTTCTCATGCTGCTATGAAGAAATACCTGAGACTGGGTAATTTATAAAGGAAAGAGGTTTAATTGACTCACAGTTCCACAGAGCTGGGGAGGCCTCAGAAAACTTAACAGTTATGGCAGAAGGGGAAGCAAACACATTTTTCTTCACATGGTGGCCGGAATTAGAAGAATGTGAGCCGAGCAAAGGGGAAAGCCCCTTATAAAACCATCAGACATCGTGAGAACTTACTATTATGAGAATAGCGTGGGGGAAACCACCCCCACGATTCAATTACCTCCCACCAAATCCCTCCCATGACATATGAGGATTATGGGAACTATGATTCAAGATGAGATTTGGGTAGGGACACAGCCAAACCATATCAGTATGTATATGTATACAAGTATTATATATATATGTATGTGTTTGTATGCATACATGTATTATATATGGAGGAAATTCTAATTTTGTAAAAAACTGGATTGTGAGTTTTAAGGAGATGTTATATAAAGTTAAGACAATGTCATTTTGTGGTATTGGTCTGAATTACAATGTAGTTTCTTAGTGATATTTTTCCTTTATTCAG
Мутантная последовательность ITR (SEQ ID NO: 24)
CACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCACGCCCGGGCTTTGCCCGGGCG
Последовательность tracr (SEQ ID NO: 25)
TAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGC
Последовательности NLS
PKKKRKV (SEQ ID NO: 26)
PKKKRKVEDPKKKRKVD (SEQ ID NO: 27)
Прямой праймер нуклеиновой кислоты для мутации сайта рестрикции BbsI (SEQ ID NO: 28)
GGGAGGATTGGGAAGAGAATAGCAGGCATGCTG
Обратный праймер нуклеиновой кислоты для мутации сайта рестрикции BbsI (SEQ ID NO: 29)
CAGCATGCCTGCTATTCTCTTCCCAATCCTCCC
Кассета промотор U6-BbsI:BbsI-каркас sgРНК-терминатор U6 для конструирования pSpCas9(BB) с подчеркнутой последовательностью промотора U6 (SEQ ID NO: 30)
CACATGTGAGGGCCTATTTCCCATGATTCCTTCATATTTGCATATACGATACAAGGCTGTTAGAGAGATAATTGGAATTAATTTGACTGTAAACACAAAGATATTAGTACAAAATACGTGACGTAGAAAGTAATAATTTCTTGGGTAGTTTGCAGTTTTAAAATTATGTTTTAAAATGGACTATCATATGCTTACCGTAACTTGAAAGTATTTCGATTTCTTGGCTTTATATATCTTGTGGAAAGGACGAAACACCGGGTCTTCGAGAAGACCTGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTTGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTTTTAGCGCGTGCGCCAATTCTGCAGACAAATGGCTCTAGAGGTACCC
F-праймер нуклеиновой кислоты для PPIA (SEQ ID NO: 31)
TTCATCTGCACTGCCAAGAC
R-праймер нуклеиновой кислоты для PPIA (SEQ ID NO: 32)
TCGAGTTGTCCACAGTCAGC
Промотор U6-BbsI:BbsI-каркас sgRNA-терминатор U6-промотор CMV для конструирования pSpCas9(BBU) с подчеркнутой последовательностью промотора U6 и промотора CMV (SEQ ID NO: 33)
CTCACATGTGAGGGCCTATTTCCCATGATTCCTTCATATTTGCATATACGATACAAGGCTGTTAGAGAGATAATTGGAATTAATTTGACTGTAAACACAAAGATATTAGTACAAAATACGTGACGTAGAAAGTAATAATTTCTTGGGTAGTTTGCAGTTTTAAAATTATGTTTTAAAATGGACTATCATATGCTTACCGTAACTTGAAAGTATTTCGATTTCTTGGCTTTATATATCTTGTGGAAAGGACGAAACACCGGGTCTTCGAGAAGACCTGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTTGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTTTTAGCGCGTGCGCCAATTCTGCAGACAAATGGCTCTAGAGACCGGCGCCGCTACAGGCTTTCCACCGGTGGTCTCTTCTAGAGGTACCCGTTACATCTAGTTATTAATAGTAATCAATTACGGGGTCATTAGTTCATAGCCCATATATGGAGTTCCGCGTTACATAACTTACGGTAAATGGCCCGCCTGGCTGACCGCCCAACGACCCCCGCCCATTGACGTCAATAATGACGTATGTTCCCATAGTAACGCCAATAGGGACTTTCCATTGACGTCAATGGGTGGAGTATTTACGGTAAACTGCCCACTTGGCAGTACATCAAGTGTATCATATGCCAAGTACGCCCCCTATTGACGTCAATGACGGTAAATGGCCCGCCTGGCATTATGCCCAGTACATGACCTTATGGGACTTTCCTACTTGGCAGTACATCTACGTATTAGTCATCGCTATTACCATGGTGATGCGGTTTTGGCAGTACATCAATGGGCGTGGATAGCGGTTTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCTCTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGAACCAAGCTGGCTAGCCGC
Прямой праймер нуклеиновой кислоты для конструирования pSpCas9(BBD) (SEQ ID NO: 34)
ATAACATGTGGTCTCACTCTAGAGGCATGTGAGGGCCTATTTCCC
Обратный праймер нуклеиновой кислоты для конструирования pSpCas9(BBD) (SEQ ID NO: 35)
TATGGTACCGGTCTCATAGAGCCATTTGTCTGCAGA
Олигонуклеотид верхней нити для контрольной sgRNA 1 (SEQ ID NO: 36)
caccGCACTACCAGAGCTAACTCA
Олигонуклеотид нижней нити для контрольной sgRNA 1 (SEQ ID NO: 37)
aaacTGAGTTAGCTCTGGTAGTGC
Олигонуклеотид верхней нити для контрольной sgRNA 2 (SEQ ID NO: 38)
caccgTGCGAATACGCCACGCGAT
Олигонуклеотид нижней нити для контрольной sgRNA 2 (SEQ ID NO: 39)
aaacATCGCGTGGCGTATTCGCAc
Аминокислотная последовательность Cas9 S. pyogenes (SEQ ID NO: 40)
MDKKYSIGLDIGTNSVGWAVITDEYKVPSKKFKVLGNTDRHSIKKNLIGALLFDSGETAEATRLKRTARRRYTRRKNRICYLQEIFSNEMAKVDDSFFHRLEESFLVEEDKKHERHPIFGNIVDEVAYHEKYPTIYHLRKKLVDSTDKADLRLIYLALAHMIKFRGHFLIEGDLNPDNSDVDKLFIQLVQTYNQLFEENPINASGVDAKAILSARLSKSRRLENLIAQLPGEKKNGLFGNLIALSLGLTPNFKSNFDLAEDAKLQLSKDTYDDDLDNLLAQIGDQYADLFLAAKNLSDAILLSDILRVNTEITKAPLSASMIKRYDEHHQDLTLLKALVRQQLPEKYKEIFFDQSKNGYAGYIDGGASQEEFYKFIKPILEKMDGTEELLVKLNREDLLRKQRTFDNGSIPHQIHLGELHAILRRQEDFYPFLKDNREKIEKILTFRIPYYVGPLARGNSRFAWMTRKSEETITPWNFEEVVDKGASAQSFIERMTNFDKNLPNEKVLPKHSLLYEYFTVYNELTKVKYVTEGMRKPAFLSGEQKKAIVDLLFKTNRKVTVKQLKEDYFKKIECFDSVEISGVEDRFNASLGTYHDLLKIIKDKDFLDNEENEDILEDIVLTLTLFEDREMIEERLKTYAHLFDDKVMKQLKRRRYTGWGRLSRKLINGIRDKQSGKTILDFLKSDGFANRNFMQLIHDDSLTFKEDIQKAQVSGQGDSLHEHIANLAGSPAIKKGILQTVKVVDELVKVMGRHKPENIVIEMARENQTTQKGQKNSRERMKRIEEGIKELGSQILKEHPVENTQLQNEKLYLYYLQNGRDMYVDQELDINRLSDYDVDHIVPQSFLKDDSIDNKVLTRSDKNRGKSDNVPSEEVVKKMKNYWRQLLNAKLITQRKFDNLTKAERGGLSELDKAGFIKRQLVETRQITKHVAQILDSRMNTKYDENDKLIREVKVITLKSKLVSDFRKDFQFYKVREINNYHHAHDAYLNAVVGTALIKKYPKLESEFVYGDYKVYDVRKMIAKSEQEIGKATAKYFFYSNIMNFFKTEITLANGEIRKRPLIETNGETGEIVWDKGRDFATVRKVLSMPQVNIVKKTEVQTGGFSKESILPKRNSDKLIARKKDWDPKKYGGFDSPTVAYSVLVVAKVEKGKSKKLKSVKELLGITIMERSSFEKNPIDFLEAKGYKEVKKDLIIKLPKYSLFELENGRKRMLASAGELQKGNELALPSKYVNFLYLASHYEKLKGSPEDNEQKQLFVEQHKHYLDEIIEQISEFSKRVILADANLDKVLSAYNKHRDKPIREQAENIIHLFTLTNLGAPAAFKYFDTTIDRKRYTSTKEVLDATLIHQSITGLYETRIDLSQLGGD
Направляющая/протоспейсерная последовательность sgRNA U1 (SEQ ID NO: 41)
GGCGGGTGGATCACGAGTTC
Направляющая/протоспейсерная последовательность sgRNA D1 (SEQ ID NO: 42)
AAAGCTACCGGTTACCTGAA
Направляющая/протоспейсерная последовательность sgRNA D2 (SEQ ID NO: 43)
TCATTCTTGTGGCAGTAAGG
Направляющая/протоспейсерная последовательность sgRNA D3 (SEQ ID NO: 44)
GGAGTCACATGGGAGTCACA
Направляющая/протоспейсерная последовательность sgRNA aU1 (SEQ ID NO: 45)
TTTAACGTTATCATTTTCCCA
Направляющая/протоспейсерная последовательность sgRNA aU2 (SEQ ID NO: 46)
AGTTTCATTCTGTCACCCAGG
Направляющая/протоспейсерная последовательность sgRNA aU3 (SEQ ID NO: 47)
AAAAATTAGCCGGGCATGATG
Направляющая/протоспейсерная последовательность sgRNA aD1 (SEQ ID NO: 48)
TGTAAGACTGGAGATAGAGAC
Направляющая/протоспейсерная последовательность sgRNA aD2 (SEQ ID NO: 49)
CTTTTGACAGTTTTTAAGGCG
Последовательность sgRNA aU1 (SEQ ID NO: 50)
GTTTAACGTTATCATTTTCCCAGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTT
Последовательность sgRNA aU2 (SEQ ID NO: 51)
GAGTTTCATTCTGTCACCCAGGGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTT
Последовательность sgRNA aU3 (SEQ ID NO: 52)
GAAAAATTAGCCGGGCATGATGGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTT
Последовательность sgRNA aD1 (SEQ ID NO: 53)
GTGTAAGACTGGAGATAGAGACGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTT
Последовательность sgRNA aD2 (SEQ ID NO: 54)
GCTTTTGACAGTTTTTAAGGCGGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTT
Аминокислотная последовательность Cas9 S. aureus (SEQ ID NO: 55)
MKRNYILGLDIGITSVGYGIIDYETRDVIDAGVRLFKEANVENNEGRRSKRGARRLKRRRRHRIQRVKKLLFDYNLLTDHSELSGINPYEARVKGLSQKLSEEEFSAALLHLAKRRGVHNVNEVEEDTGNELSTKEQISRNSKALEEKYVAELQLERLKKDGEVRGSINRFKTSDYVKEAKQLLKVQKAYHQLDQSFIDTYIDLLETRRTYYEGPGEGSPFGWKDIKEWYEMLMGHCTYFPEELRSVKYAYNADLYNALNDLNNLVITRDENEKLEYYEKFQIIENVFKQKKKPTLKQIAKEILVNEEDIKGYRVTSTGKPEFTNLKVYHDIKDITARKEIIENAELLDQIAKILTIYQSSEDIQEELTNLNSELTQEEIEQISNLKGYTGTHNLSLKAINLILDELWHTNDNQIAIFNRLKLVPKKVDLSQQKEIPTTLVDDFILSPVVKRSFIQSIKVINAIIKKYGLPNDIIIELAREKNSKDAQKMINEMQKRNRQTNERIEEIIRTTGKENAKYLIEKIKLHDMQEGKCLYSLEAIPLEDLLNNPFNYEVDHIIPRSVSFDNSFNNKVLVKQEENSKKGNRTPFQYLSSSDSKISYETFKKHILNLAKGKGRISKTKKEYLLEERDINRFSVQKDFINRNLVDTRYATRGLMNLLRSYFRVNNLDVKVKSINGGFTSFLRRKWKFKKERNKGYKHHAEDALIIANADFIFKEWKKLDKAKKVMENQMFEEKQAESMPEIETEQEYKEIFITPHQIKHIKDFKDYKYSHRVDKKPNRELINDTLYSTRKDDKGNTLIVNNLNGLYDKDNDKLKKLINKSPEKLLMYHHDPQTYQKLKLIMEQYGDEKNPLYKYYEETGNYLTKYSKKDNGPVIKKIKYYGNKLNAHLDITDDYPNSRNKVVKLSLKPYRFDVYLDNGVYKFVTVKNLDVIKKENYYEVNSKCYEEAKKLKKISNQAEFIASFYNNDLIKINGELYRVIGVNNDLLNRIEVNMIDITYREYLENMNDKRPPRIIKTIASKTQSIKKYSTDILGNLYEVKSKKHPQIIKKG
Последовательность промотор minCMV-SpCas9 для конструирования pAAV-minCMV-SpCas9-NLS-SV40 pA с подчеркнутым промотором minCMV (SEQ ID NO: 56)
TATACGCGTGTTGACACTAGTTCGCGAAATATTGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCGTTTAGTGAACCGTCAGATCGCCGCCACCATGGACAAGAAGTACAGCATCGGCCTGGACATCGGCACCAACTCTGTGGGCTGGGCCGTGATCACCGACGAGTACAAGGTGCCCAGCAAGAAATTCAAG
Последовательность pA SV40 для конструирования pAAV-minCMV-SpCas9-NLS-SV40 pA с подчеркнутым ранним сигналом poly(A) SV40 (SEQ ID NO: 57)
TGACTCGAGAACTTGTTTATTGCAGCTTATAATGGTTACAAATAAAGCAATAGCATCACAAATTTCACAAATAAAGCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTCCAAACTCATCAATGTATCTTATCATGTCTGCAATATTTCGCGAGAAGACAATAGCAGG
Последовательность распознавания sgRNA U1 (U1T; направляющая последовательность sgRNA U1+PAM) (SEQ ID NO: 58)
GGCGGGTGGATCACGAGTTCAGG
Последовательность распознавания sgRNA D3 (D3T; направляющая последовательность sgRNA D3+PAM) (SEQ ID NO: 59)
GGAGTCACATGGGAGTCACAGGG
pA BGH-содержащий фрагмент с подчеркнутым pA BGH (SEQ ID NO: 60)
CTAGTGGCGGCCGCTCGAGCATGCATCTAGAGGGCCCTATTCTATAGTGTCACCTAAATGCTAGAGCTCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGAGCTAGAGTCGACCGGACCGCTGCAGGCATGCA
Направляющая/протоспейсерная последовательность sgRNA U11 (SEQ ID NO: 61)
GCATAAGGACTAAAGACCTA
Направляющая/протоспейсерная последовательность sgRNA D11 (SEQ ID NO: 62)
GGTAGTGGTTGAACTCACAA
Фрагмент промотор RK-химерный интрон-EGFP-BGH pA (SEQ ID NO: 63)
GGGCCCCAGAAGCCTGGTGGTTGTTTGTCCTTCTCAGGGGAAAAGTGAGGCGGCCCCTTGGAGGAAGGGGCCGGGCAGAATGATCTAATCGGATTCCAAGCAGCTCAGGGGATTGTCTTTTTCTAGCACCTTCTTGCCACTCCTAAGCGTCCTCCGTGACCCCGGCTGGGATTTAGCCTGGTGCTGTGTCAGCCCCGGTCTCCCAGGGGCTTCCCAGTGGTCCCCAGGAACCCTCGACAGGGCCCGGTCTCTCTCGTCCAGCAAGGGCAGGGACGGGCCACAGGCCAAGGGCGGAGTCGCTGCGACGCTGCCTTCGCCCCGTGCCCCGCTCCGCCGCCGCCTCGCGCCGCCCGCCCCGGCTCTGACTGACCGCGTTACTCCCACAGGTGAGCGGGCGGGACGGCCCTTCTCCTCCGGGCTGTAATTAGCGCTTGGTTTAATGACGGCTTGTTTCTTTTCTGTGGCTGCGTGAAAGCCTTGAGGGGCTCCGGGAGGGCCCTTTGTGCGGGGGGAGCGGCTCGGGGGGTGCGTGCGTGTGTGTGTGCGTGGGGAGCGCCGCGTGCGGCTCCGCGCTGCCCGGCGGCTGTGAGCGCTGCGGGCGCGGCGCGGGGCTTTGTGCGCTCCGCAGTGTGCGCGAGGGGAGCGCGGCCGGGGGCGGTGCCCCGCGGTGCGGGGGGGGCTGCGAGGGGAACAAAGGCTGCGTGCGGGGTGTGTGCGTGGGGGGGTGAGCAGGGGGTGTGGGCGCGTCGGTCGGGCTGCAACCCCCCCTGCACCCCCCTCCCCGAGTTGCTGAGCACGGCCCGGCTTCGGGTGCGGGGCTCCGTACGGGGCGTGGCGCGGGGCTCGCCGTGCCGGGCGGGGGGTGGCGGCAGGTGGGGGTGCCGGGCGGGGCGGGGCCGCCTCGGGCCGGGGAGGGCTCGGGGGAGGGGCGCGGCGGCCCCCGGAGCGCCGGCGGCTGTCGAGGCGCGGCGAGCCGCAGCCATTGCCTTTTATGGTAATCGTGCGAGAGGGCGCAGGGACTTCCTTTGTCCCAAATCTGTGCGGAGCCGAAATCTGGGAGGCGCCGCCGCACCCCCTCTAGCGGGCGCGGGGCGAAGCGGTGCGGCGCCGGCAGGAAGGAAATGGGCGGGGAGGGCCTTCGTGCGTCGCCGCGCCGCCGTCCCCTTCTCCCTCTCCAGCCTCGGGGCTGTCCGCGGGGGGACGGCTGCCTTCGGGGGGGACGGGGCAGGGCGGGGTTCGGCTTCTGGCGTGTGACCGGCGGCTCTAGAGCCTCTGCTAACCATGTTCATGCCTTCTTCTTTTTCCTACAGCTCCTGGGCAACGTGCTGGTTATTGTGCTGTCTCATCATTTTGGCAAAGAATTCTTCGAAAGATCTGCTAGCTTAATTAACCCGGTCGCCACCATGGTGAGCAAGGGCGAGGAGCTGTTCACCGGGGTGGTGCCCATCCTGGTCGAGCTGGACGGCGACGTAAACGGCCACAAGTTCAGCGTGTCCGGCGAGGGCGAGGGCGATGCCACCTACGGCAAGCTGACCCTGAAGTTCATCTGCACCACCGGCAAGCTGCCCGTGCCCTGGCCCACCCTCGTGACCACCCTGACCTACGGCGTGCAGTGCTTCAGCCGCTACCCCGACCACATGAAGCAGCACGACTTCTTCAAGTCCGCCATGCCCGAAGGCTACGTCCAGGAGCGCACCATCTTCTTCAAGGACGACGGCAACTACAAGACCCGCGCCGAGGTGAAGTTCGAGGGCGACACCCTGGTGAACCGCATCGAGCTGAAGGGCATCGACTTCAAGGAGGACGGCAACATCCTGGGGCACAAGCTGGAGTACAACTACAACAGCCACAACGTCTATATCATGGCCGACAAGCAGAAGAACGGCATCAAGGTGAACTTCAAGATCCGCCACAACATCGAGGACGGCAGCGTGCAGCTCGCCGACCACTACCAGCAGAACACCCCCATCGGCGACGGCCCCGTGCTGCTGCCCGACAACCACTACCTGAGCACCCAGTCCGCCCTGAGCAAAGACCCCAACGAGAAGCGCGATCACATGGTCCTGCTGGAGTTCGTGACCGCCGCCGGGATCACTCTCGGCATGGACGAGCTGTACAAGTAAAGCGGCCAAATCGTACGCCTAGGTGATCAAGATCTGCTAGCTTAATTAACCCGGGACTAGTGGCGGCCGCTCGAGCATGCATCTAGAGGGCCCTATTCTATAGTGTCACCTAAATGCTAGAGCTCGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGA
F-праймер нуклеиновой кислоты для интрона 25 гена Cep290 мыши (SEQ ID NO: 64)
CCCCTCGCCTGTACTGAAAG
R-праймер нуклеиновой кислоты для интрона 25 гена Cep290 мыши (SEQ ID NO: 65)
GCACATCATCTGAGGCAGGT
Каркасный фрагмент minCMV-SaCas9-NLS-FLAG-BGH pA-U6-BsaI:BsaI-sgRNA с подчеркнутой последовательностью minCMV и SaCas9 (SEQ ID NO: 66)
ATGAATTCTCTAGACAATTGGACTCACGGGGATTTCCAAGTCTCCACCCCATTGACGTCAATGGGAGTTTGTTTTGGCACCAAAATCAACGGGACTTTCCAAAATGTCGTAACAACTCCGCCCCATTGACGCAAATGGGCGGTAGGCGTGTACGGTGGGAGGTCTATATAAGCAGAGCTCGTTTAGTGAACCGTCAGATCACGCGTGCCACCATGAAGCGGAACTACATCCTGGGCCTGGACATCGGCATCACCAGCGTGGGCTACGGCATCATCGACTACGAGACACGGGACGTGATCGATGCCGGCGTGCGGCTGTTCAAAGAGGCCAACGTGGAAAACAACGAGGGCAGGCGGAGCAAGAGAGGCGCCAGAAGGCTGAAGCGGCGGAGGCGGCATAGAATCCAGAGAGTGAAGAAGCTGCTGTTCGACTACAACCTGCTGACCGACCACAGCGAGCTGAGCGGCATCAACCCCTACGAGGCCAGAGTGAAGGGCCTGAGCCAGAAGCTGAGCGAGGAAGAGTTCTCTGCCGCCCTGCTGCACCTGGCCAAGAGAAGAGGCGTGCACAACGTGAACGAGGTGGAAGAGGACACCGGCAACGAGCTGTCCACCAAAGAGCAGATCAGCCGGAACAGCAAGGCCCTGGAAGAGAAATACGTGGCCGAACTGCAGCTGGAACGGCTGAAGAAAGACGGCGAAGTGCGGGGCAGCATCAACAGATTCAAGACCAGCGACTACGTGAAAGAAGCCAAACAGCTGCTGAAGGTGCAGAAGGCCTACCACCAGCTGGACCAGAGCTTCATCGACACCTACATCGACCTGCTGGAAACCCGGCGGACCTACTATGAGGGACCTGGCGAGGGCAGCCCCTTCGGCTGGAAGGACATCAAAGAATGGTACGAGATGCTGATGGGCCACTGCACCTACTTCCCCGAGGAACTGCGGAGCGTGAAGTACGCCTACAACGCCGACCTGTACAACGCCCTGAACGACCTGAACAATCTCGTGATCACCAGGGACGAGAACGAGAAGCTGGAATATTACGAGAAGTTCCAGATCATCGAGAACGTGTTCAAGCAGAAGAAGAAGCCCACCCTGAAGCAGATCGCCAAAGAAATCCTCGTGAACGAAGAGGATATTAAGGGCTACAGAGTGACCAGCACCGGCAAGCCCGAGTTCACCAACCTGAAGGTGTACCACGACATCAAGGACATTACCGCCCGGAAAGAGATTATTGAGAACGCCGAGCTGCTGGATCAGATTGCCAAGATCCTGACCATCTACCAGAGCAGCGAGGACATCCAGGAAGAACTGACCAATCTGAACTCCGAGCTGACCCAGGAAGAGATCGAGCAGATCTCTAATCTGAAGGGCTATACCGGCACCCACAACCTGAGCCTGAAGGCCATCAACCTGATCCTGGACGAGCTGTGGCACACCAACGACAACCAGATCGCTATCTTCAACCGGCTGAAGCTGGTGCCCAAGAAGGTGGACCTGTCCCAGCAGAAAGAGATCCCCACCACCCTGGTGGACGACTTCATCCTGAGCCCCGTCGTGAAGAGAAGCTTCATCCAGAGCATCAAAGTGATCAACGCCATCATCAAGAAGTACGGCCTGCCCAACGACATCATTATCGAGCTGGCCCGCGAGAAGAACTCCAAGGACGCCCAGAAAATGATCAACGAGATGCAGAAGCGGAACCGGCAGACCAACGAGCGGATCGAGGAAATCATCCGGACCACCGGCAAAGAGAACGCCAAGTACCTGATCGAGAAGATCAAGCTGCACGACATGCAGGAAGGCAAGTGCCTGTACAGCCTGGAAGCCATCCCTCTGGAAGATCTGCTGAACAACCCCTTCAACTATGAGGTGGACCACATCATCCCCAGAAGCGTGTCCTTCGACAACAGCTTCAACAACAAGGTGCTCGTGAAGCAGGAAGAAAACAGCAAGAAGGGCAACCGGACCCCATTCCAGTACCTGAGCAGCAGCGACAGCAAGATCAGCTACGAAACCTTCAAGAAGCACATCCTGAATCTGGCCAAGGGCAAGGGCAGAATCAGCAAGACCAAGAAAGAGTATCTGCTGGAAGAACGGGACATCAACAGGTTCTCCGTGCAGAAAGACTTCATCAACCGGAACCTGGTGGATACCAGATACGCCACCAGAGGCCTGATGAACCTGCTGCGGAGCTACTTCAGAGTGAACAACCTGGACGTGAAAGTGAAGTCCATCAATGGCGGCTTCACCAGCTTTCTGCGGCGGAAGTGGAAGTTTAAGAAAGAGCGGAACAAGGGGTACAAGCACCACGCCGAGGACGCCCTGATCATTGCCAACGCCGATTTCATCTTCAAAGAGTGGAAGAAACTGGACAAGGCCAAAAAAGTGATGGAAAACCAGATGTTCGAGGAAAAGCAGGCCGAGAGCATGCCCGAGATCGAAACCGAGCAGGAGTACAAAGAGATCTTCATCACCCCCCACCAGATCAAGCACATTAAGGACTTCAAGGACTACAAGTACAGCCACCGGGTGGACAAGAAGCCTAATAGAGAGCTGATTAACGACACCCTGTACTCCACCCGGAAGGACGACAAGGGCAACACCCTGATCGTGAACAATCTGAACGGCCTGTACGACAAGGACAATGACAAGCTGAAAAAGCTGATCAACAAGAGCCCCGAAAAGCTGCTGATGTACCACCACGACCCCCAGACCTACCAGAAACTGAAGCTGATTATGGAACAGTACGGCGACGAGAAGAATCCCCTGTACAAGTACTACGAGGAAACCGGGAACTACCTGACCAAGTACTCCAAAAAGGACAACGGCCCCGTGATCAAGAAGATTAAGTATTACGGCAACAAACTGAACGCCCATCTGGACATCACCGACGACTACCCCAACAGCAGAAACAAGGTCGTGAAGCTGTCCCTGAAGCCCTACAGATTCGACGTGTACCTGGACAATGGCGTGTACAAGTTCGTGACCGTGAAGAATCTGGATGTGATCAAAAAAGAAAACTACTACGAAGTGAATAGCAAGTGCTATGAGGAAGCTAAGAAGCTGAAGAAGATCAGCAACCAGGCCGAGTTTATCGCCTCCTTCTACAACAACGATCTGATCAAGATCAACGGCGAGCTGTATAGAGTGATCGGCGTGAACAACGACCTGCTGAACCGGATCGAAGTGAACATGATCGACATCACCTACCGCGAGTACCTGGAAAACATGAACGACAAGAGGCCCCCCAGGATCATTAAGACAATCGCCTCCAAGACCCAGAGCATTAAGAAGTACAGCACAGACATTCTGGGCAACCTGTATGAAGTGAAATCTAAGAAGCACCCTCAGATCATCAAAAAGGGCGGATCCCCCAAGAAAAAGCGCAAAGTGGACTACAAAGACGATGACGACAAGTGAGCTAGCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGAGAATAGCAGGCATGCTGGTACCTGAGGGCCTATTTCCCATGATTCCTTCATATTTGCATATACGATACAAGGCTGTTAGAGAGATAATTGGAATTAATTTGACTGTAAACACAAAGATATTAGTACAAAATACGTGACGTAGAAAGTAATAATTTCTTGGGTAGTTTGCAGTTTTAAAATTATGTTTTAAAATGGACTATCATATGCTTACCGTAACTTGAAAGTATTTCGATTTCTTGGCTTTATATATCTTGTGGAAAGGACGAAACACCGGAGACCACGGCAGGTCTCAGTTTTAGTACTCTGGAAACAGAATCTACTAAAACAAGGCAAAATGCCGTGTTTATCTCGTCAACTTGTTGGCGAGATTTTTGCGGCCGCGTCGACAT
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> RUAN, Guoxiang
SCARIA, Abraham
<120> ГЕННОЕ РЕДАКТИРОВАНИЕ ГЛУБОКИХ ИНТРОННЫХ МУТАЦИЙ
<130> 15979013440
<140> Not Yet Assigned
<141> Concurrently Herewith
<150> 62/162,720
<151> 2015-05-16
<160> 66
<170> FastSEQ для Windows версии 4.0
<210> 1
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
caccgaagac actgccaata gggat 25
<210> 2
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
aaacatccct attggcagtg tcttc 25
<210> 3
<211> 158
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 3
ccacccgcct cggcctccta aagtgctggg attacagatg tgagccaccg cacctggccc 60
cagttgtaat tgtgagtatc tcatacgtat ccctattggc agtgtcttag ttttattttt 120
tattatcttt attgtggcag ccattattcc tgtctcta 158
<210> 4
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 4
ggtccctggc ttttgttcct 20
<210> 5
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 5
caggaggctg agggtgtttt 20
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
agtagagatg gggtttcacc 20
<210> 7
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 7
tgactgctaa gtacagggac atcttg 26
<210> 8
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 8
aggagatgtt ttcacactcc aggt 24
<210> 9
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 9
ctggccccag ttgtaatttg tga 23
<210> 10
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 10
ctgttcccag gcttgttcaa tagt 24
<210> 11
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 11
caccggcggg tggatcacga gttc 24
<210> 12
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 12
aaacgaactc gtgatccacc cgcc 24
<210> 13
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 13
caccgaaagc taccggttac ctgaa 25
<210> 14
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 14
aaacttcagg taaccggtag ctttc 25
<210> 15
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 15
caccgtcatt cttgtggcag taagg 25
<210> 16
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 16
aaacccttac tgccacaaga atgac 25
<210> 17
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 17
caccggagtc acatgggagt caca 24
<210> 18
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 18
aaactgtgac tcccatgtga ctcc 24
<210> 19
<211> 102
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 19
ggcgggtgga tcacgagttc gttttagagc tagaaatagc aagttaaaat aaggctagtc 60
cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt tt 102
<210> 20
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 20
gaaagctacc ggttacctga agttttagag ctagaaatag caagttaaaa taaggctagt 60
ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt ttt 103
<210> 21
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 21
gtcattcttg tggcagtaag ggttttagag ctagaaatag caagttaaaa taaggctagt 60
ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt ttt 103
<210> 22
<211> 102
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 22
ggagtcacat gggagtcaca gttttagagc tagaaatagc aagttaaaat aaggctagtc 60
cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt tt 102
<210> 23
<211> 5838
<212> ДНК
<213> Homo sapiens
<400> 23
gtaagtttgt gtgattcttg aaccttgtga aattagccat ttttcttcaa tatttttgtg 60
tttgggggga tttggcagat tttaattaaa gtttgcctgc atttatataa atttaacaga 120
gatataatta tccatattat tcattcagtt tagttataaa tattttgttc ccacataaca 180
cacacacaca cacacaatat attatctatt tatagtggct gaatgacttc tgaatgatta 240
tctagatcat tctccttagg tcacttgcat gatttagctg aatcaaacct cttttaacca 300
gacatctaag agaaaaagga gcatgaaaca ggtagaatat tgtaatcaaa ggagggaagc 360
actcattaag tgcccatccc tttctcttac ccctgtaccc agaacaaact attctcccat 420
ggtccctggc ttttgttcct tggaatggat gtagccaaca gtagctgaaa tattaagggc 480
tcttcctgga ccatggatgc actctgtaaa ttctcatcat tttttattgt agaataaatg 540
tagaatttta atgtagaata aatttattta atgtagaata aaaaataaaa aaactagagt 600
agaatatcat aagttacaat ctgtgaatat ggaccagacc ctttgtagtt atcttacagc 660
cacttgaact ctataccttt tactgaggac agaacaagct cctgatttgt tcatcttcct 720
catcagaaat agaggcttat ggattttgga ttattcttat ctaagatcct ttcacaggag 780
tagaataaga tctaattcta ttagctcaaa agcttttgct ggctcataga gacacattca 840
gtaaatgaaa acgttgttct gagtagcttt caggattcct actaaattat gagtcatgtt 900
tatcaatatt atttagaagt aatcataatc agtttgcttt ctgctgcttt tgccaaagag 960
aggtgattat gttacttttt atagaaaatt atgcctattt agtgtggtga taatttattt 1020
ttttccattc tccatgtcct ctgtcctatc ctctccagca ttagaaagtc ctaggcaaga 1080
gacatcttgt ggataatgta tcaatgagtg atgtttaacg ttatcatttt cccaaagagt 1140
atttttcatc tttcctaaag attttttttt tttttttttg agatggagtt tcattctgtc 1200
acccaggctg agtgcagtgg cacgatctcg gcttaacgct tactgcatcc tctgcctccc 1260
agattcaagc agttctcctg cctcagcctc tgagtagctg ggattacagg tgtgcaccac 1320
cacaccagct aatttttttt tttttttttt tttttttgag gcagagtctc gctctgtcac 1380
ccaggctgga gtgcagtggc gccatcttgg ctcactgcaa gctccacctc ccgggttcag 1440
gccgttctcc tgcctcagcc tcctgagtag ctggtaccac aggcacccac catcatgccc 1500
ggctaatttt ttgtattttt agtagagatg gggtttcacc ttgttagcca ggatggtgtc 1560
gatctcctga actcgtgatc cacccgcctc ggcctcctaa agtgctggga ttacagatgt 1620
gagccaccgc acctggcccc agttgtaatt gtgagtatct catacctatc cctattggca 1680
gtgtcttagt tttatttttt attatcttta ttgtggcagc cattattcct gtctctatct 1740
ccagtcttac atcctcctta ctgccacaag aatgatcatt ctaaacatga atcctaccct 1800
gtgactccca tgtgactccc cgccttaaaa actgtcaaaa gctaccggtt acctgaaggg 1860
taaaagtcaa gtcccctact tacctcatgt catctagagc aagagatgaa ctagctgagt 1920
tttctgacca cagtgttctt tcttatgtat gttcttttgt acgtgctctt ttctatatat 1980
agggaaccat ttctctcttc cagttgtttt gctcagtgaa tttctattcc tgtttcaaaa 2040
cttgttcagg cattaccttt tttttcttaa gcatactttt tttaatggaa caaagtcact 2100
cctgtctaca ctagttctgc atcttataca taggttttgt acatagtaca tatttatatc 2160
acatcaaatt atatgtgttt acatatctgt cttccttaat ggaatataag tcttttgata 2220
taaggaacta tttaatttgt ttctgtgtgt tgagtatctc ctgtttggca cagagttcaa 2280
gctaatacat gagagtgatt agtggtggag agccacagtg catgtggtgt caaatatggt 2340
gcttaggaaa ttattgttgc tttttgagag gtaaaggttc atgagactag aggtcacgaa 2400
aatcagattt catgtgtgaa gaatggaata gataataagg aaatacaaaa actggatggg 2460
taataaagca aaagaaaaac ttgaaatttg atagtagaag aaaaaagaaa tagatgtaga 2520
ttgaggtaga atcaagaaga ggattctttt tttgttgttt ttttttttga aacagagtct 2580
cactgtgttg cccaggctgg agtgcagtgg agtgatcttg gcttactgca acctctgcct 2640
cccaggttca agcgattctt ctgcttcagt ctcccgagta gctggaatta caggtgccca 2700
ccagcacggc cggctaattt agtagagaca gggttttgcc atgttggccg ggctggtctc 2760
aaactttgga tctcaggtaa tccgccagcc tcaacttccc aaagtgctgg gattacaggc 2820
atgagccact gtgcccagcc tgtttttttt tttttaaagg agaccagtga agtttcagga 2880
ggagggaaag aaaatttaga gttactaggg agagagtgat gaagataaga gatgaaagtg 2940
gtaataaggg aaatagcaaa atatcagggt aggtgggaga aaaagagatt tgtaacaaac 3000
aataggatta tcctgtgaaa aaggatgaaa ggaagaaaaa aatggataga aagatattta 3060
aaacaccctc agcctcctgt tttccctcct gtgtattcat agtatataaa actataatta 3120
tgtactttac ttaaaaaata tattattatt accttatcgt gcttatttaa tcatagcatg 3180
tcctcttttt agtctcatta ccctgtttgt attattcttc ataacactta atacctgaca 3240
ttgtattata tattggctta ttttccaggt actccactca aatataagtt ctaggatata 3300
atttatttat cactgaaatc cattgcttag agtacctggc atgtagtaaa taggcattct 3360
gttttttcaa ataaaaaata aaggaactta agatatatat ttatgttata tcgccagcct 3420
ttttcctcac agctctattc tgttgtacag aattacctac tttacaattc ctgtgtttca 3480
aggggatctc aaatttaacg tgtccacaat gaactcctga tttctgtttc tctcctagtc 3540
attcttattt caatatatgt tcagttacct aaccagctag tcaaggcaga tactttagag 3600
ttattctgta gtcattcttt ttccctacca tttttgtttt ccaaatgtaa tttatgtgtg 3660
tcttcttcat cctcgcagct ctaacccttg tccaaaccag catcatcact catctggagt 3720
tccacaatgt ctttctggct agtttccctg atttctctat tgaccccttt attctccaca 3780
gtgcagccag aatgattgtt taaaacttcc tccttaaaat ctttaaattg ttttctttta 3840
tacgttaagt taaattccag ttccttgtct tggcatgcca tgccctgcct ggtgtggccc 3900
ctgatggtct ctccaacttc atgttttact actattgact cttatttttg cttactctgc 3960
ttgggtgctc cagtcctcca aatcatttcc tgctccaatc atttcaatca ttttttcctc 4020
tcagatctta tagtattcca aatgctttct tcctttggag catctgggtt tactaataaa 4080
tacttcgtac ctcacagttc agcttaaata tcaattattt ggtggttaag acatccttca 4140
accgctctat ctaaatgttc ctttctatta ttcactggct cagtactctg tttttatttt 4200
ctttctaaat gtcaactttt ttttttttga gtcagggtct cactgttgcc caggctcgag 4260
tgcagttgca caatcatagc tcattgcagc cttgccctcc tgggatcaag taattctccc 4320
acctcagcct ccaaaatagc tgggattaca ggtatgcatc accatgctca gctaattttt 4380
tgtgtttttt tgtagagatg aggtctcact ttgttgccca ggctggtctc aaactcctgg 4440
actcaagtga ttctcccacc tcagcctccc aaagtgctgg ggttacaggt gtgagccact 4500
gcacctggtc gatactgact tttttttttt tttgagatgg agttttgctc tgttgcccag 4560
gctagagcgc agtggtgtga tctcagctca ctgcaacctc cacctcccag gttaaaggga 4620
ttcttctgcc tcagtctcct gagtagctgg gattacaggc aagtgccatc atgactggct 4680
aatttttgta tttttagcac tatgtttagt actgtgttgg ccaggcttgt ctcgaactcc 4740
tgacctcaag tgatccaccc acctcagcct cccaaagtgc tgggattaca ggtgtgagcc 4800
accgtaatcg gccaacattg acatttttag tagacttttt gtttgtttac ttgcttatta 4860
tctgctgcct tccacactct ggcgaaatcc tgccacccac ccacacacac ataggcactg 4920
aatgggcaga actctgaagg ccagaatttt atatttcttt tcactgtaaa catcatcatc 4980
tgtcactgat ggcacactag gatgctcagc aactgtgtgc atgaaggaag taagcactag 5040
tttgtgaagg ctgcaaaact cttgagtatt ctaagagttt tggccaaaat gaatgtacag 5100
ctttagtggc agaagctaat actcagaaat tgaggccgta tattggataa cacaggattt 5160
ggatgattat tttaaaataa tattttacat tgtatatatg tgtgtgtgtg tgtgtgtgtg 5220
tgtgtgtatg tgtgtgtgtg tgtatatata tatgtatgta tgtgtattag tccgttctca 5280
tgctgctatg aagaaatacc tgagactggg taatttataa aggaaagagg tttaattgac 5340
tcacagttcc acagagctgg ggaggcctca gaaaacttaa cagttatggc agaaggggaa 5400
gcaaacacat ttttcttcac atggtggccg gaattagaag aatgtgagcc gagcaaaggg 5460
gaaagcccct tataaaacca tcagacatcg tgagaactta ctattatgag aatagcgtgg 5520
gggaaaccac ccccacgatt caattacctc ccaccaaatc cctcccatga catatgagga 5580
ttatgggaac tatgattcaa gatgagattt gggtagggac acagccaaac catatcagta 5640
tgtatatgta tacaagtatt atatatatat gtatgtgttt gtatgcatac atgtattata 5700
tatggaggaa attctaattt tgtaaaaaac tggattgtga gttttaagga gatgttatat 5760
aaagttaaga caatgtcatt ttgtggtatt ggtctgaatt acaatgtagt ttcttagtga 5820
tatttttcct ttattcag 5838
<210> 24
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 24
cactccctct ctgcgcgctc gctcgctcac tgaggccggg cgaccaaagg tcgcccacgc 60
ccgggctttg cccgggcg 78
<210> 25
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 25
tagcaagtta aaataaggct agtccgttat caacttgaaa aagtggcacc gagtcggtgc 60
<210> 26
<211> 7
<212> БЕЛОК
<213> Вакуолизирующий вирус обезьян 40
<400> 26
Pro Lys Lys Lys Arg Lys Val
1 5
<210> 27
<211> 17
<212> БЕЛОК
<213> Вакуолизирующий вирус обезьян 40
<400> 27
Pro Lys Lys Lys Arg Lys Val Glu Asp Pro Lys Lys Lys Arg Lys Val
1 5 10 15
Asp
<210> 28
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 28
gggaggattg ggaagagaat agcaggcatg ctg 33
<210> 29
<211> 33
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 29
cagcatgcct gctattctct tcccaatcct ccc 33
<210> 30
<211> 447
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 30
cacatgtgag ggcctatttc ccatgattcc ttcatatttg catatacgat acaaggctgt 60
tagagagata attggaatta atttgactgt aaacacaaag atattagtac aaaatacgtg 120
acgtagaaag taataatttc ttgggtagtt tgcagtttta aaattatgtt ttaaaatgga 180
ctatcatatg cttaccgtaa cttgaaagta tttcgatttc ttggctttat atatcttgtg 240
gaaaggacga aacaccgggt cttcgagaag acctgtttta gagctagaaa tagcaagtta 300
aaataaggct agtccgttat caacttgaaa aagtggcacc gagtcggtgc ttttttgttt 360
tagagctaga aatagcaagt taaaataagg ctagtccgtt tttagcgcgt gcgccaattc 420
tgcagacaaa tggctctaga ggtaccc 447
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 31
ttcatctgca ctgccaagac 20
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 32
tcgagttgtc cacagtcagc 20
<210> 33
<211> 1153
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 33
ctcacatgtg agggcctatt tcccatgatt ccttcatatt tgcatatacg atacaaggct 60
gttagagaga taattggaat taatttgact gtaaacacaa agatattagt acaaaatacg 120
tgacgtagaa agtaataatt tcttgggtag tttgcagttt taaaattatg ttttaaaatg 180
gactatcata tgcttaccgt aacttgaaag tatttcgatt tcttggcttt atatatcttg 240
tggaaaggac gaaacaccgg gtcttcgaga agacctgttt tagagctaga aatagcaagt 300
taaaataagg ctagtccgtt atcaacttga aaaagtggca ccgagtcggt gcttttttgt 360
tttagagcta gaaatagcaa gttaaaataa ggctagtccg tttttagcgc gtgcgccaat 420
tctgcagaca aatggctcta gagaccggcg ccgctacagg ctttccaccg gtggtctctt 480
ctagaggtac ccgttacatc tagttattaa tagtaatcaa ttacggggtc attagttcat 540
agcccatata tggagttccg cgttacataa cttacggtaa atggcccgcc tggctgaccg 600
cccaacgacc cccgcccatt gacgtcaata atgacgtatg ttcccatagt aacgccaata 660
gggactttcc attgacgtca atgggtggag tatttacggt aaactgccca cttggcagta 720
catcaagtgt atcatatgcc aagtacgccc cctattgacg tcaatgacgg taaatggccc 780
gcctggcatt atgcccagta catgacctta tgggactttc ctacttggca gtacatctac 840
gtattagtca tcgctattac catggtgatg cggttttggc agtacatcaa tgggcgtgga 900
tagcggtttg actcacgggg atttccaagt ctccacccca ttgacgtcaa tgggagtttg 960
ttttggcacc aaaatcaacg ggactttcca aaatgtcgta acaactccgc cccattgacg 1020
caaatgggcg gtaggcgtgt acggtgggag gtctatataa gcagagctct ctggctaact 1080
agagaaccca ctgcttactg gcttatcgaa attaatacga ctcactatag ggagaaccaa 1140
gctggctagc cgc 1153
<210> 34
<211> 45
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 34
ataacatgtg gtctcactct agaggcatgt gagggcctat ttccc 45
<210> 35
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 35
tatggtaccg gtctcataga gccatttgtc tgcaga 36
<210> 36
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 36
caccgcacta ccagagctaa ctca 24
<210> 37
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 37
aaactgagtt agctctggta gtgc 24
<210> 38
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 38
caccgtgcga atacgccacg cgat 24
<210> 39
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 39
aaacatcgcg tggcgtattc gcac 24
<210> 40
<211> 1368
<212> БЕЛОК
<213> Streptococcus pyogenes
<400> 40
Met Asp Lys Lys Tyr Ser Ile Gly Leu Asp Ile Gly Thr Asn Ser Val
1 5 10 15
Gly Trp Ala Val Ile Thr Asp Glu Tyr Lys Val Pro Ser Lys Lys Phe
20 25 30
Lys Val Leu Gly Asn Thr Asp Arg His Ser Ile Lys Lys Asn Leu Ile
35 40 45
Gly Ala Leu Leu Phe Asp Ser Gly Glu Thr Ala Glu Ala Thr Arg Leu
50 55 60
Lys Arg Thr Ala Arg Arg Arg Tyr Thr Arg Arg Lys Asn Arg Ile Cys
65 70 75 80
Tyr Leu Gln Glu Ile Phe Ser Asn Glu Met Ala Lys Val Asp Asp Ser
85 90 95
Phe Phe His Arg Leu Glu Glu Ser Phe Leu Val Glu Glu Asp Lys Lys
100 105 110
His Glu Arg His Pro Ile Phe Gly Asn Ile Val Asp Glu Val Ala Tyr
115 120 125
His Glu Lys Tyr Pro Thr Ile Tyr His Leu Arg Lys Lys Leu Val Asp
130 135 140
Ser Thr Asp Lys Ala Asp Leu Arg Leu Ile Tyr Leu Ala Leu Ala His
145 150 155 160
Met Ile Lys Phe Arg Gly His Phe Leu Ile Glu Gly Asp Leu Asn Pro
165 170 175
Asp Asn Ser Asp Val Asp Lys Leu Phe Ile Gln Leu Val Gln Thr Tyr
180 185 190
Asn Gln Leu Phe Glu Glu Asn Pro Ile Asn Ala Ser Gly Val Asp Ala
195 200 205
Lys Ala Ile Leu Ser Ala Arg Leu Ser Lys Ser Arg Arg Leu Glu Asn
210 215 220
Leu Ile Ala Gln Leu Pro Gly Glu Lys Lys Asn Gly Leu Phe Gly Asn
225 230 235 240
Leu Ile Ala Leu Ser Leu Gly Leu Thr Pro Asn Phe Lys Ser Asn Phe
245 250 255
Asp Leu Ala Glu Asp Ala Lys Leu Gln Leu Ser Lys Asp Thr Tyr Asp
260 265 270
Asp Asp Leu Asp Asn Leu Leu Ala Gln Ile Gly Asp Gln Tyr Ala Asp
275 280 285
Leu Phe Leu Ala Ala Lys Asn Leu Ser Asp Ala Ile Leu Leu Ser Asp
290 295 300
Ile Leu Arg Val Asn Thr Glu Ile Thr Lys Ala Pro Leu Ser Ala Ser
305 310 315 320
Met Ile Lys Arg Tyr Asp Glu His His Gln Asp Leu Thr Leu Leu Lys
325 330 335
Ala Leu Val Arg Gln Gln Leu Pro Glu Lys Tyr Lys Glu Ile Phe Phe
340 345 350
Asp Gln Ser Lys Asn Gly Tyr Ala Gly Tyr Ile Asp Gly Gly Ala Ser
355 360 365
Gln Glu Glu Phe Tyr Lys Phe Ile Lys Pro Ile Leu Glu Lys Met Asp
370 375 380
Gly Thr Glu Glu Leu Leu Val Lys Leu Asn Arg Glu Asp Leu Leu Arg
385 390 395 400
Lys Gln Arg Thr Phe Asp Asn Gly Ser Ile Pro His Gln Ile His Leu
405 410 415
Gly Glu Leu His Ala Ile Leu Arg Arg Gln Glu Asp Phe Tyr Pro Phe
420 425 430
Leu Lys Asp Asn Arg Glu Lys Ile Glu Lys Ile Leu Thr Phe Arg Ile
435 440 445
Pro Tyr Tyr Val Gly Pro Leu Ala Arg Gly Asn Ser Arg Phe Ala Trp
450 455 460
Met Thr Arg Lys Ser Glu Glu Thr Ile Thr Pro Trp Asn Phe Glu Glu
465 470 475 480
Val Val Asp Lys Gly Ala Ser Ala Gln Ser Phe Ile Glu Arg Met Thr
485 490 495
Asn Phe Asp Lys Asn Leu Pro Asn Glu Lys Val Leu Pro Lys His Ser
500 505 510
Leu Leu Tyr Glu Tyr Phe Thr Val Tyr Asn Glu Leu Thr Lys Val Lys
515 520 525
Tyr Val Thr Glu Gly Met Arg Lys Pro Ala Phe Leu Ser Gly Glu Gln
530 535 540
Lys Lys Ala Ile Val Asp Leu Leu Phe Lys Thr Asn Arg Lys Val Thr
545 550 555 560
Val Lys Gln Leu Lys Glu Asp Tyr Phe Lys Lys Ile Glu Cys Phe Asp
565 570 575
Ser Val Glu Ile Ser Gly Val Glu Asp Arg Phe Asn Ala Ser Leu Gly
580 585 590
Thr Tyr His Asp Leu Leu Lys Ile Ile Lys Asp Lys Asp Phe Leu Asp
595 600 605
Asn Glu Glu Asn Glu Asp Ile Leu Glu Asp Ile Val Leu Thr Leu Thr
610 615 620
Leu Phe Glu Asp Arg Glu Met Ile Glu Glu Arg Leu Lys Thr Tyr Ala
625 630 635 640
His Leu Phe Asp Asp Lys Val Met Lys Gln Leu Lys Arg Arg Arg Tyr
645 650 655
Thr Gly Trp Gly Arg Leu Ser Arg Lys Leu Ile Asn Gly Ile Arg Asp
660 665 670
Lys Gln Ser Gly Lys Thr Ile Leu Asp Phe Leu Lys Ser Asp Gly Phe
675 680 685
Ala Asn Arg Asn Phe Met Gln Leu Ile His Asp Asp Ser Leu Thr Phe
690 695 700
Lys Glu Asp Ile Gln Lys Ala Gln Val Ser Gly Gln Gly Asp Ser Leu
705 710 715 720
His Glu His Ile Ala Asn Leu Ala Gly Ser Pro Ala Ile Lys Lys Gly
725 730 735
Ile Leu Gln Thr Val Lys Val Val Asp Glu Leu Val Lys Val Met Gly
740 745 750
Arg His Lys Pro Glu Asn Ile Val Ile Glu Met Ala Arg Glu Asn Gln
755 760 765
Thr Thr Gln Lys Gly Gln Lys Asn Ser Arg Glu Arg Met Lys Arg Ile
770 775 780
Glu Glu Gly Ile Lys Glu Leu Gly Ser Gln Ile Leu Lys Glu His Pro
785 790 795 800
Val Glu Asn Thr Gln Leu Gln Asn Glu Lys Leu Tyr Leu Tyr Tyr Leu
805 810 815
Gln Asn Gly Arg Asp Met Tyr Val Asp Gln Glu Leu Asp Ile Asn Arg
820 825 830
Leu Ser Asp Tyr Asp Val Asp His Ile Val Pro Gln Ser Phe Leu Lys
835 840 845
Asp Asp Ser Ile Asp Asn Lys Val Leu Thr Arg Ser Asp Lys Asn Arg
850 855 860
Gly Lys Ser Asp Asn Val Pro Ser Glu Glu Val Val Lys Lys Met Lys
865 870 875 880
Asn Tyr Trp Arg Gln Leu Leu Asn Ala Lys Leu Ile Thr Gln Arg Lys
885 890 895
Phe Asp Asn Leu Thr Lys Ala Glu Arg Gly Gly Leu Ser Glu Leu Asp
900 905 910
Lys Ala Gly Phe Ile Lys Arg Gln Leu Val Glu Thr Arg Gln Ile Thr
915 920 925
Lys His Val Ala Gln Ile Leu Asp Ser Arg Met Asn Thr Lys Tyr Asp
930 935 940
Glu Asn Asp Lys Leu Ile Arg Glu Val Lys Val Ile Thr Leu Lys Ser
945 950 955 960
Lys Leu Val Ser Asp Phe Arg Lys Asp Phe Gln Phe Tyr Lys Val Arg
965 970 975
Glu Ile Asn Asn Tyr His His Ala His Asp Ala Tyr Leu Asn Ala Val
980 985 990
Val Gly Thr Ala Leu Ile Lys Lys Tyr Pro Lys Leu Glu Ser Glu Phe
995 1000 1005
Val Tyr Gly Asp Tyr Lys Val Tyr Asp Val Arg Lys Met Ile Ala Lys
1010 1015 1020
Ser Glu Gln Glu Ile Gly Lys Ala Thr Ala Lys Tyr Phe Phe Tyr Ser
1025 1030 1035 1040
Asn Ile Met Asn Phe Phe Lys Thr Glu Ile Thr Leu Ala Asn Gly Glu
1045 1050 1055
Ile Arg Lys Arg Pro Leu Ile Glu Thr Asn Gly Glu Thr Gly Glu Ile
1060 1065 1070
Val Trp Asp Lys Gly Arg Asp Phe Ala Thr Val Arg Lys Val Leu Ser
1075 1080 1085
Met Pro Gln Val Asn Ile Val Lys Lys Thr Glu Val Gln Thr Gly Gly
1090 1095 1100
Phe Ser Lys Glu Ser Ile Leu Pro Lys Arg Asn Ser Asp Lys Leu Ile
1105 1110 1115 1120
Ala Arg Lys Lys Asp Trp Asp Pro Lys Lys Tyr Gly Gly Phe Asp Ser
1125 1130 1135
Pro Thr Val Ala Tyr Ser Val Leu Val Val Ala Lys Val Glu Lys Gly
1140 1145 1150
Lys Ser Lys Lys Leu Lys Ser Val Lys Glu Leu Leu Gly Ile Thr Ile
1155 1160 1165
Met Glu Arg Ser Ser Phe Glu Lys Asn Pro Ile Asp Phe Leu Glu Ala
1170 1175 1180
Lys Gly Tyr Lys Glu Val Lys Lys Asp Leu Ile Ile Lys Leu Pro Lys
1185 1190 1195 1200
Tyr Ser Leu Phe Glu Leu Glu Asn Gly Arg Lys Arg Met Leu Ala Ser
1205 1210 1215
Ala Gly Glu Leu Gln Lys Gly Asn Glu Leu Ala Leu Pro Ser Lys Tyr
1220 1225 1230
Val Asn Phe Leu Tyr Leu Ala Ser His Tyr Glu Lys Leu Lys Gly Ser
1235 1240 1245
Pro Glu Asp Asn Glu Gln Lys Gln Leu Phe Val Glu Gln His Lys His
1250 1255 1260
Tyr Leu Asp Glu Ile Ile Glu Gln Ile Ser Glu Phe Ser Lys Arg Val
1265 1270 1275 1280
Ile Leu Ala Asp Ala Asn Leu Asp Lys Val Leu Ser Ala Tyr Asn Lys
1285 1290 1295
His Arg Asp Lys Pro Ile Arg Glu Gln Ala Glu Asn Ile Ile His Leu
1300 1305 1310
Phe Thr Leu Thr Asn Leu Gly Ala Pro Ala Ala Phe Lys Tyr Phe Asp
1315 1320 1325
Thr Thr Ile Asp Arg Lys Arg Tyr Thr Ser Thr Lys Glu Val Leu Asp
1330 1335 1340
Ala Thr Leu Ile His Gln Ser Ile Thr Gly Leu Tyr Glu Thr Arg Ile
1345 1350 1355 1360
Asp Leu Ser Gln Leu Gly Gly Asp
1365
<210> 41
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 41
ggcgggtgga tcacgagttc 20
<210> 42
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 42
aaagctaccg gttacctgaa 20
<210> 43
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 43
tcattcttgt ggcagtaagg 20
<210> 44
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 44
ggagtcacat gggagtcaca 20
<210> 45
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 45
tttaacgtta tcattttccc a 21
<210> 46
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 46
agtttcattc tgtcacccag g 21
<210> 47
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 47
aaaaattagc cgggcatgat g 21
<210> 48
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 48
tgtaagactg gagatagaga c 21
<210> 49
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 49
cttttgacag tttttaaggc g 21
<210> 50
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 50
gtttaacgtt atcattttcc cagttttagt actctggaaa cagaatctac taaaacaagg 60
caaaatgccg tgtttatctc gtcaacttgt tggcgagatt ttt 103
<210> 51
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 51
gagtttcatt ctgtcaccca gggttttagt actctggaaa cagaatctac taaaacaagg 60
caaaatgccg tgtttatctc gtcaacttgt tggcgagatt ttt 103
<210> 52
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 52
gaaaaattag ccgggcatga tggttttagt actctggaaa cagaatctac taaaacaagg 60
caaaatgccg tgtttatctc gtcaacttgt tggcgagatt ttt 103
<210> 53
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 53
gtgtaagact ggagatagag acgttttagt actctggaaa cagaatctac taaaacaagg 60
caaaatgccg tgtttatctc gtcaacttgt tggcgagatt ttt 103
<210> 54
<211> 103
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 54
gcttttgaca gtttttaagg cggttttagt actctggaaa cagaatctac taaaacaagg 60
caaaatgccg tgtttatctc gtcaacttgt tggcgagatt ttt 103
<210> 55
<211> 1053
<212> БЕЛОК
<213> Staphylococcus aureus
<400> 55
Met Lys Arg Asn Tyr Ile Leu Gly Leu Asp Ile Gly Ile Thr Ser Val
1 5 10 15
Gly Tyr Gly Ile Ile Asp Tyr Glu Thr Arg Asp Val Ile Asp Ala Gly
20 25 30
Val Arg Leu Phe Lys Glu Ala Asn Val Glu Asn Asn Glu Gly Arg Arg
35 40 45
Ser Lys Arg Gly Ala Arg Arg Leu Lys Arg Arg Arg Arg His Arg Ile
50 55 60
Gln Arg Val Lys Lys Leu Leu Phe Asp Tyr Asn Leu Leu Thr Asp His
65 70 75 80
Ser Glu Leu Ser Gly Ile Asn Pro Tyr Glu Ala Arg Val Lys Gly Leu
85 90 95
Ser Gln Lys Leu Ser Glu Glu Glu Phe Ser Ala Ala Leu Leu His Leu
100 105 110
Ala Lys Arg Arg Gly Val His Asn Val Asn Glu Val Glu Glu Asp Thr
115 120 125
Gly Asn Glu Leu Ser Thr Lys Glu Gln Ile Ser Arg Asn Ser Lys Ala
130 135 140
Leu Glu Glu Lys Tyr Val Ala Glu Leu Gln Leu Glu Arg Leu Lys Lys
145 150 155 160
Asp Gly Glu Val Arg Gly Ser Ile Asn Arg Phe Lys Thr Ser Asp Tyr
165 170 175
Val Lys Glu Ala Lys Gln Leu Leu Lys Val Gln Lys Ala Tyr His Gln
180 185 190
Leu Asp Gln Ser Phe Ile Asp Thr Tyr Ile Asp Leu Leu Glu Thr Arg
195 200 205
Arg Thr Tyr Tyr Glu Gly Pro Gly Glu Gly Ser Pro Phe Gly Trp Lys
210 215 220
Asp Ile Lys Glu Trp Tyr Glu Met Leu Met Gly His Cys Thr Tyr Phe
225 230 235 240
Pro Glu Glu Leu Arg Ser Val Lys Tyr Ala Tyr Asn Ala Asp Leu Tyr
245 250 255
Asn Ala Leu Asn Asp Leu Asn Asn Leu Val Ile Thr Arg Asp Glu Asn
260 265 270
Glu Lys Leu Glu Tyr Tyr Glu Lys Phe Gln Ile Ile Glu Asn Val Phe
275 280 285
Lys Gln Lys Lys Lys Pro Thr Leu Lys Gln Ile Ala Lys Glu Ile Leu
290 295 300
Val Asn Glu Glu Asp Ile Lys Gly Tyr Arg Val Thr Ser Thr Gly Lys
305 310 315 320
Pro Glu Phe Thr Asn Leu Lys Val Tyr His Asp Ile Lys Asp Ile Thr
325 330 335
Ala Arg Lys Glu Ile Ile Glu Asn Ala Glu Leu Leu Asp Gln Ile Ala
340 345 350
Lys Ile Leu Thr Ile Tyr Gln Ser Ser Glu Asp Ile Gln Glu Glu Leu
355 360 365
Thr Asn Leu Asn Ser Glu Leu Thr Gln Glu Glu Ile Glu Gln Ile Ser
370 375 380
Asn Leu Lys Gly Tyr Thr Gly Thr His Asn Leu Ser Leu Lys Ala Ile
385 390 395 400
Asn Leu Ile Leu Asp Glu Leu Trp His Thr Asn Asp Asn Gln Ile Ala
405 410 415
Ile Phe Asn Arg Leu Lys Leu Val Pro Lys Lys Val Asp Leu Ser Gln
420 425 430
Gln Lys Glu Ile Pro Thr Thr Leu Val Asp Asp Phe Ile Leu Ser Pro
435 440 445
Val Val Lys Arg Ser Phe Ile Gln Ser Ile Lys Val Ile Asn Ala Ile
450 455 460
Ile Lys Lys Tyr Gly Leu Pro Asn Asp Ile Ile Ile Glu Leu Ala Arg
465 470 475 480
Glu Lys Asn Ser Lys Asp Ala Gln Lys Met Ile Asn Glu Met Gln Lys
485 490 495
Arg Asn Arg Gln Thr Asn Glu Arg Ile Glu Glu Ile Ile Arg Thr Thr
500 505 510
Gly Lys Glu Asn Ala Lys Tyr Leu Ile Glu Lys Ile Lys Leu His Asp
515 520 525
Met Gln Glu Gly Lys Cys Leu Tyr Ser Leu Glu Ala Ile Pro Leu Glu
530 535 540
Asp Leu Leu Asn Asn Pro Phe Asn Tyr Glu Val Asp His Ile Ile Pro
545 550 555 560
Arg Ser Val Ser Phe Asp Asn Ser Phe Asn Asn Lys Val Leu Val Lys
565 570 575
Gln Glu Glu Asn Ser Lys Lys Gly Asn Arg Thr Pro Phe Gln Tyr Leu
580 585 590
Ser Ser Ser Asp Ser Lys Ile Ser Tyr Glu Thr Phe Lys Lys His Ile
595 600 605
Leu Asn Leu Ala Lys Gly Lys Gly Arg Ile Ser Lys Thr Lys Lys Glu
610 615 620
Tyr Leu Leu Glu Glu Arg Asp Ile Asn Arg Phe Ser Val Gln Lys Asp
625 630 635 640
Phe Ile Asn Arg Asn Leu Val Asp Thr Arg Tyr Ala Thr Arg Gly Leu
645 650 655
Met Asn Leu Leu Arg Ser Tyr Phe Arg Val Asn Asn Leu Asp Val Lys
660 665 670
Val Lys Ser Ile Asn Gly Gly Phe Thr Ser Phe Leu Arg Arg Lys Trp
675 680 685
Lys Phe Lys Lys Glu Arg Asn Lys Gly Tyr Lys His His Ala Glu Asp
690 695 700
Ala Leu Ile Ile Ala Asn Ala Asp Phe Ile Phe Lys Glu Trp Lys Lys
705 710 715 720
Leu Asp Lys Ala Lys Lys Val Met Glu Asn Gln Met Phe Glu Glu Lys
725 730 735
Gln Ala Glu Ser Met Pro Glu Ile Glu Thr Glu Gln Glu Tyr Lys Glu
740 745 750
Ile Phe Ile Thr Pro His Gln Ile Lys His Ile Lys Asp Phe Lys Asp
755 760 765
Tyr Lys Tyr Ser His Arg Val Asp Lys Lys Pro Asn Arg Glu Leu Ile
770 775 780
Asn Asp Thr Leu Tyr Ser Thr Arg Lys Asp Asp Lys Gly Asn Thr Leu
785 790 795 800
Ile Val Asn Asn Leu Asn Gly Leu Tyr Asp Lys Asp Asn Asp Lys Leu
805 810 815
Lys Lys Leu Ile Asn Lys Ser Pro Glu Lys Leu Leu Met Tyr His His
820 825 830
Asp Pro Gln Thr Tyr Gln Lys Leu Lys Leu Ile Met Glu Gln Tyr Gly
835 840 845
Asp Glu Lys Asn Pro Leu Tyr Lys Tyr Tyr Glu Glu Thr Gly Asn Tyr
850 855 860
Leu Thr Lys Tyr Ser Lys Lys Asp Asn Gly Pro Val Ile Lys Lys Ile
865 870 875 880
Lys Tyr Tyr Gly Asn Lys Leu Asn Ala His Leu Asp Ile Thr Asp Asp
885 890 895
Tyr Pro Asn Ser Arg Asn Lys Val Val Lys Leu Ser Leu Lys Pro Tyr
900 905 910
Arg Phe Asp Val Tyr Leu Asp Asn Gly Val Tyr Lys Phe Val Thr Val
915 920 925
Lys Asn Leu Asp Val Ile Lys Lys Glu Asn Tyr Tyr Glu Val Asn Ser
930 935 940
Lys Cys Tyr Glu Glu Ala Lys Lys Leu Lys Lys Ile Ser Asn Gln Ala
945 950 955 960
Glu Phe Ile Ala Ser Phe Tyr Asn Asn Asp Leu Ile Lys Ile Asn Gly
965 970 975
Glu Leu Tyr Arg Val Ile Gly Val Asn Asn Asp Leu Leu Asn Arg Ile
980 985 990
Glu Val Asn Met Ile Asp Ile Thr Tyr Arg Glu Tyr Leu Glu Asn Met
995 1000 1005
Asn Asp Lys Arg Pro Pro Arg Ile Ile Lys Thr Ile Ala Ser Lys Thr
1010 1015 1020
Gln Ser Ile Lys Lys Tyr Ser Thr Asp Ile Leu Gly Asn Leu Tyr Glu
1025 1030 1035 1040
Val Lys Ser Lys Lys His Pro Gln Ile Ile Lys Lys Gly
1045 1050
<210> 56
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 56
tatacgcgtg ttgacactag ttcgcgaaat attgactcac ggggatttcc aagtctccac 60
cccattgacg tcaatgggag tttgttttgg caccaaaatc aacgggactt tccaaaatgt 120
cgtaacaact ccgccccatt gacgcaaatg ggcggtaggc gtgtacggtg ggaggtctat 180
ataagcagag ctcgtttagt gaaccgtcag atcgccgcca ccatggacaa gaagtacagc 240
atcggcctgg acatcggcac caactctgtg ggctgggccg tgatcaccga cgagtacaag 300
gtgcccagca agaaattcaa g 321
<210> 57
<211> 168
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 57
tgactcgaga acttgtttat tgcagcttat aatggttaca aataaagcaa tagcatcaca 60
aatttcacaa ataaagcatt tttttcactg cattctagtt gtggtttgtc caaactcatc 120
aatgtatctt atcatgtctg caatatttcg cgagaagaca atagcagg 168
<210> 58
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 58
ggcgggtgga tcacgagttc agg 23
<210> 59
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 59
ggagtcacat gggagtcaca ggg 23
<210> 60
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 60
ctagtggcgg ccgctcgagc atgcatctag agggccctat tctatagtgt cacctaaatg 60
ctagagctcg ctgatcagcc tcgactgtgc cttctagttg ccagccatct gttgtttgcc 120
cctcccccgt gccttccttg accctggaag gtgccactcc cactgtcctt tcctaataaa 180
atgaggaaat tgcatcgcat tgtctgagta ggtgtcattc tattctgggg ggtggggtgg 240
ggcaggacag caagggggag gattgggaag acaatagcag gcatgctggg gagctagagt 300
cgaccggacc gctgcaggca tgca 324
<210> 61
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 61
gcataaggac taaagaccta 20
<210> 62
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 62
ggtagtggtt gaactcacaa 20
<210> 63
<211> 2471
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 63
gggccccaga agcctggtgg ttgtttgtcc ttctcagggg aaaagtgagg cggccccttg 60
gaggaagggg ccgggcagaa tgatctaatc ggattccaag cagctcaggg gattgtcttt 120
ttctagcacc ttcttgccac tcctaagcgt cctccgtgac cccggctggg atttagcctg 180
gtgctgtgtc agccccggtc tcccaggggc ttcccagtgg tccccaggaa ccctcgacag 240
ggcccggtct ctctcgtcca gcaagggcag ggacgggcca caggccaagg gcggagtcgc 300
tgcgacgctg ccttcgcccc gtgccccgct ccgccgccgc ctcgcgccgc ccgccccggc 360
tctgactgac cgcgttactc ccacaggtga gcgggcggga cggcccttct cctccgggct 420
gtaattagcg cttggtttaa tgacggcttg tttcttttct gtggctgcgt gaaagccttg 480
aggggctccg ggagggccct ttgtgcgggg ggagcggctc ggggggtgcg tgcgtgtgtg 540
tgtgcgtggg gagcgccgcg tgcggctccg cgctgcccgg cggctgtgag cgctgcgggc 600
gcggcgcggg gctttgtgcg ctccgcagtg tgcgcgaggg gagcgcggcc gggggcggtg 660
ccccgcggtg cggggggggc tgcgagggga acaaaggctg cgtgcggggt gtgtgcgtgg 720
gggggtgagc agggggtgtg ggcgcgtcgg tcgggctgca accccccctg cacccccctc 780
cccgagttgc tgagcacggc ccggcttcgg gtgcggggct ccgtacgggg cgtggcgcgg 840
ggctcgccgt gccgggcggg gggtggcggc aggtgggggt gccgggcggg gcggggccgc 900
ctcgggccgg ggagggctcg ggggaggggc gcggcggccc ccggagcgcc ggcggctgtc 960
gaggcgcggc gagccgcagc cattgccttt tatggtaatc gtgcgagagg gcgcagggac 1020
ttcctttgtc ccaaatctgt gcggagccga aatctgggag gcgccgccgc accccctcta 1080
gcgggcgcgg ggcgaagcgg tgcggcgccg gcaggaagga aatgggcggg gagggccttc 1140
gtgcgtcgcc gcgccgccgt ccccttctcc ctctccagcc tcggggctgt ccgcgggggg 1200
acggctgcct tcggggggga cggggcaggg cggggttcgg cttctggcgt gtgaccggcg 1260
gctctagagc ctctgctaac catgttcatg ccttcttctt tttcctacag ctcctgggca 1320
acgtgctggt tattgtgctg tctcatcatt ttggcaaaga attcttcgaa agatctgcta 1380
gcttaattaa cccggtcgcc accatggtga gcaagggcga ggagctgttc accggggtgg 1440
tgcccatcct ggtcgagctg gacggcgacg taaacggcca caagttcagc gtgtccggcg 1500
agggcgaggg cgatgccacc tacggcaagc tgaccctgaa gttcatctgc accaccggca 1560
agctgcccgt gccctggccc accctcgtga ccaccctgac ctacggcgtg cagtgcttca 1620
gccgctaccc cgaccacatg aagcagcacg acttcttcaa gtccgccatg cccgaaggct 1680
acgtccagga gcgcaccatc ttcttcaagg acgacggcaa ctacaagacc cgcgccgagg 1740
tgaagttcga gggcgacacc ctggtgaacc gcatcgagct gaagggcatc gacttcaagg 1800
aggacggcaa catcctgggg cacaagctgg agtacaacta caacagccac aacgtctata 1860
tcatggccga caagcagaag aacggcatca aggtgaactt caagatccgc cacaacatcg 1920
aggacggcag cgtgcagctc gccgaccact accagcagaa cacccccatc ggcgacggcc 1980
ccgtgctgct gcccgacaac cactacctga gcacccagtc cgccctgagc aaagacccca 2040
acgagaagcg cgatcacatg gtcctgctgg agttcgtgac cgccgccggg atcactctcg 2100
gcatggacga gctgtacaag taaagcggcc aaatcgtacg cctaggtgat caagatctgc 2160
tagcttaatt aacccgggac tagtggcggc cgctcgagca tgcatctaga gggccctatt 2220
ctatagtgtc acctaaatgc tagagctcgc tgatcagcct cgactgtgcc ttctagttgc 2280
cagccatctg ttgtttgccc ctcccccgtg ccttccttga ccctggaagg tgccactccc 2340
actgtccttt cctaataaaa tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct 2400
attctggggg gtggggtggg gcaggacagc aagggggagg attgggaaga caatagcagg 2460
catgctgggg a 2471
<210> 64
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 64
cccctcgcct gtactgaaag 20
<210> 65
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 65
gcacatcatc tgaggcaggt 20
<210> 66
<211> 4009
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 66
atgaattctc tagacaattg gactcacggg gatttccaag tctccacccc attgacgtca 60
atgggagttt gttttggcac caaaatcaac gggactttcc aaaatgtcgt aacaactccg 120
ccccattgac gcaaatgggc ggtaggcgtg tacggtggga ggtctatata agcagagctc 180
gtttagtgaa ccgtcagatc acgcgtgcca ccatgaagcg gaactacatc ctgggcctgg 240
acatcggcat caccagcgtg ggctacggca tcatcgacta cgagacacgg gacgtgatcg 300
atgccggcgt gcggctgttc aaagaggcca acgtggaaaa caacgagggc aggcggagca 360
agagaggcgc cagaaggctg aagcggcgga ggcggcatag aatccagaga gtgaagaagc 420
tgctgttcga ctacaacctg ctgaccgacc acagcgagct gagcggcatc aacccctacg 480
aggccagagt gaagggcctg agccagaagc tgagcgagga agagttctct gccgccctgc 540
tgcacctggc caagagaaga ggcgtgcaca acgtgaacga ggtggaagag gacaccggca 600
acgagctgtc caccaaagag cagatcagcc ggaacagcaa ggccctggaa gagaaatacg 660
tggccgaact gcagctggaa cggctgaaga aagacggcga agtgcggggc agcatcaaca 720
gattcaagac cagcgactac gtgaaagaag ccaaacagct gctgaaggtg cagaaggcct 780
accaccagct ggaccagagc ttcatcgaca cctacatcga cctgctggaa acccggcgga 840
cctactatga gggacctggc gagggcagcc ccttcggctg gaaggacatc aaagaatggt 900
acgagatgct gatgggccac tgcacctact tccccgagga actgcggagc gtgaagtacg 960
cctacaacgc cgacctgtac aacgccctga acgacctgaa caatctcgtg atcaccaggg 1020
acgagaacga gaagctggaa tattacgaga agttccagat catcgagaac gtgttcaagc 1080
agaagaagaa gcccaccctg aagcagatcg ccaaagaaat cctcgtgaac gaagaggata 1140
ttaagggcta cagagtgacc agcaccggca agcccgagtt caccaacctg aaggtgtacc 1200
acgacatcaa ggacattacc gcccggaaag agattattga gaacgccgag ctgctggatc 1260
agattgccaa gatcctgacc atctaccaga gcagcgagga catccaggaa gaactgacca 1320
atctgaactc cgagctgacc caggaagaga tcgagcagat ctctaatctg aagggctata 1380
ccggcaccca caacctgagc ctgaaggcca tcaacctgat cctggacgag ctgtggcaca 1440
ccaacgacaa ccagatcgct atcttcaacc ggctgaagct ggtgcccaag aaggtggacc 1500
tgtcccagca gaaagagatc cccaccaccc tggtggacga cttcatcctg agccccgtcg 1560
tgaagagaag cttcatccag agcatcaaag tgatcaacgc catcatcaag aagtacggcc 1620
tgcccaacga catcattatc gagctggccc gcgagaagaa ctccaaggac gcccagaaaa 1680
tgatcaacga gatgcagaag cggaaccggc agaccaacga gcggatcgag gaaatcatcc 1740
ggaccaccgg caaagagaac gccaagtacc tgatcgagaa gatcaagctg cacgacatgc 1800
aggaaggcaa gtgcctgtac agcctggaag ccatccctct ggaagatctg ctgaacaacc 1860
ccttcaacta tgaggtggac cacatcatcc ccagaagcgt gtccttcgac aacagcttca 1920
acaacaaggt gctcgtgaag caggaagaaa acagcaagaa gggcaaccgg accccattcc 1980
agtacctgag cagcagcgac agcaagatca gctacgaaac cttcaagaag cacatcctga 2040
atctggccaa gggcaagggc agaatcagca agaccaagaa agagtatctg ctggaagaac 2100
gggacatcaa caggttctcc gtgcagaaag acttcatcaa ccggaacctg gtggatacca 2160
gatacgccac cagaggcctg atgaacctgc tgcggagcta cttcagagtg aacaacctgg 2220
acgtgaaagt gaagtccatc aatggcggct tcaccagctt tctgcggcgg aagtggaagt 2280
ttaagaaaga gcggaacaag gggtacaagc accacgccga ggacgccctg atcattgcca 2340
acgccgattt catcttcaaa gagtggaaga aactggacaa ggccaaaaaa gtgatggaaa 2400
accagatgtt cgaggaaaag caggccgaga gcatgcccga gatcgaaacc gagcaggagt 2460
acaaagagat cttcatcacc ccccaccaga tcaagcacat taaggacttc aaggactaca 2520
agtacagcca ccgggtggac aagaagccta atagagagct gattaacgac accctgtact 2580
ccacccggaa ggacgacaag ggcaacaccc tgatcgtgaa caatctgaac ggcctgtacg 2640
acaaggacaa tgacaagctg aaaaagctga tcaacaagag ccccgaaaag ctgctgatgt 2700
accaccacga cccccagacc taccagaaac tgaagctgat tatggaacag tacggcgacg 2760
agaagaatcc cctgtacaag tactacgagg aaaccgggaa ctacctgacc aagtactcca 2820
aaaaggacaa cggccccgtg atcaagaaga ttaagtatta cggcaacaaa ctgaacgccc 2880
atctggacat caccgacgac taccccaaca gcagaaacaa ggtcgtgaag ctgtccctga 2940
agccctacag attcgacgtg tacctggaca atggcgtgta caagttcgtg accgtgaaga 3000
atctggatgt gatcaaaaaa gaaaactact acgaagtgaa tagcaagtgc tatgaggaag 3060
ctaagaagct gaagaagatc agcaaccagg ccgagtttat cgcctccttc tacaacaacg 3120
atctgatcaa gatcaacggc gagctgtata gagtgatcgg cgtgaacaac gacctgctga 3180
accggatcga agtgaacatg atcgacatca cctaccgcga gtacctggaa aacatgaacg 3240
acaagaggcc ccccaggatc attaagacaa tcgcctccaa gacccagagc attaagaagt 3300
acagcacaga cattctgggc aacctgtatg aagtgaaatc taagaagcac cctcagatca 3360
tcaaaaaggg cggatccccc aagaaaaagc gcaaagtgga ctacaaagac gatgacgaca 3420
agtgagctag cgactgtgcc ttctagttgc cagccatctg ttgtttgccc ctcccccgtg 3480
ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa tgaggaaatt 3540
gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg gcaggacagc 3600
aagggggagg attgggaaga gaatagcagg catgctggta cctgagggcc tatttcccat 3660
gattccttca tatttgcata tacgatacaa ggctgttaga gagataattg gaattaattt 3720
gactgtaaac acaaagatat tagtacaaaa tacgtgacgt agaaagtaat aatttcttgg 3780
gtagtttgca gttttaaaat tatgttttaa aatggactat catatgctta ccgtaacttg 3840
aaagtatttc gatttcttgg ctttatatat cttgtggaaa ggacgaaaca ccggagacca 3900
cggcaggtct cagttttagt actctggaaa cagaatctac taaaacaagg caaaatgccg 3960
tgtttatctc gtcaacttgt tggcgagatt tttgcggccg cgtcgacat 4009
<---
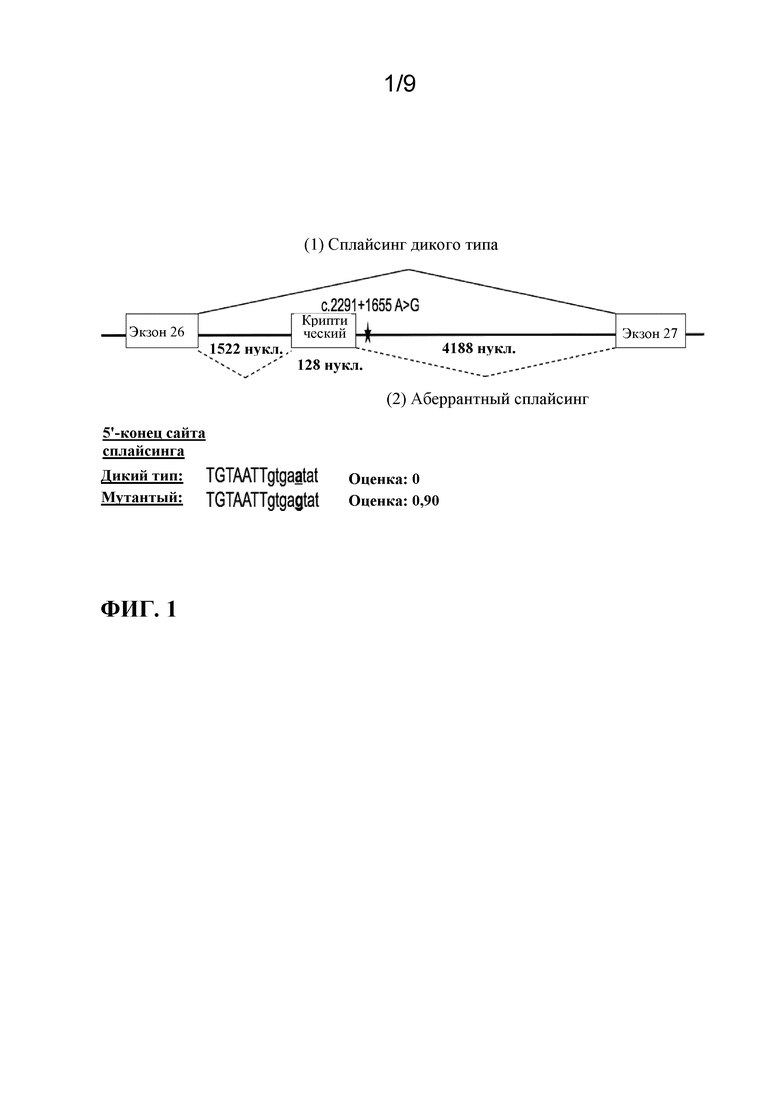
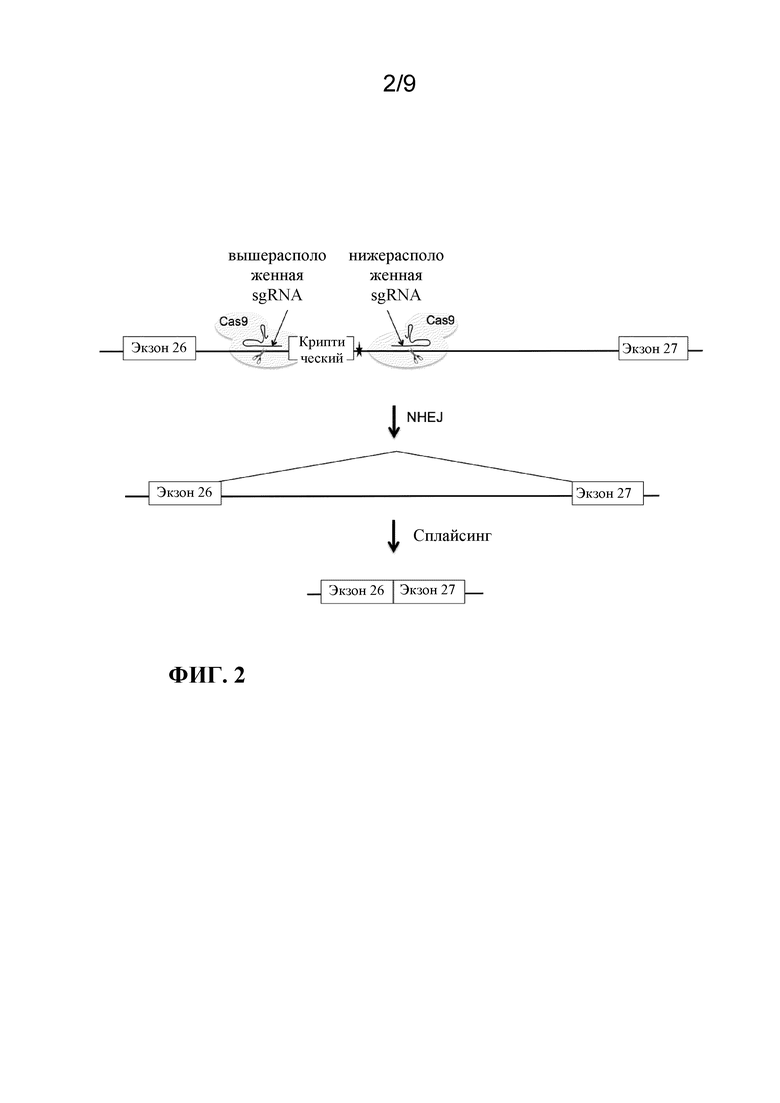
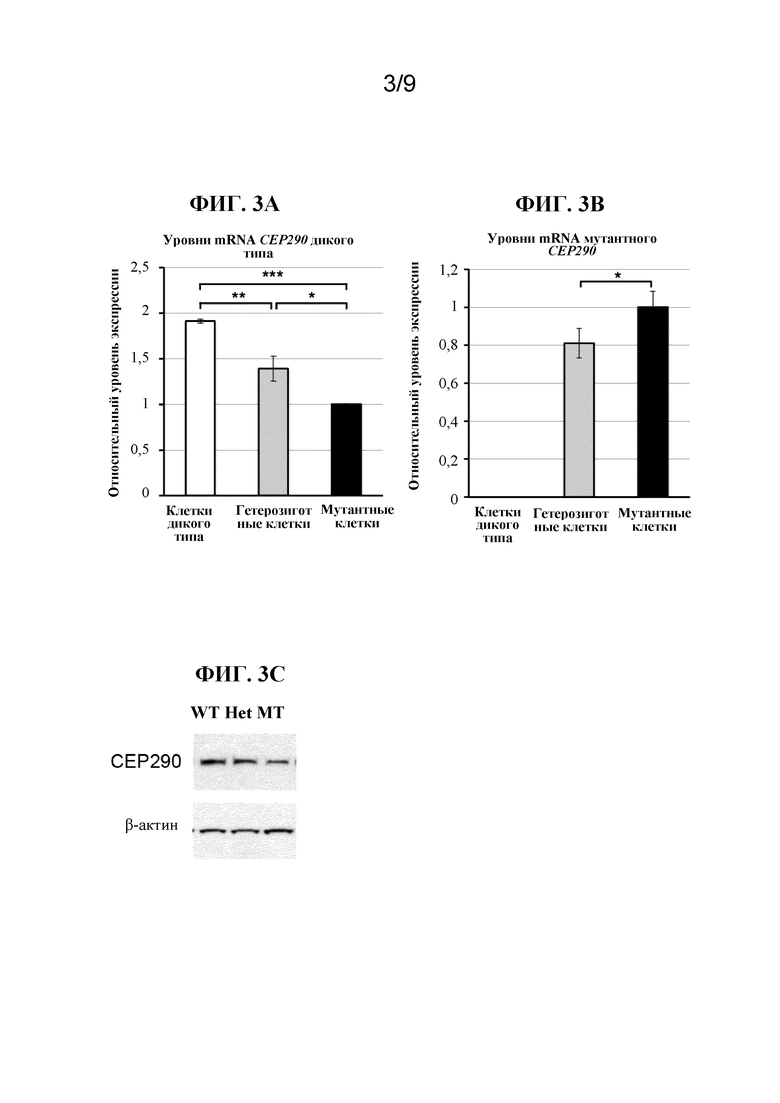
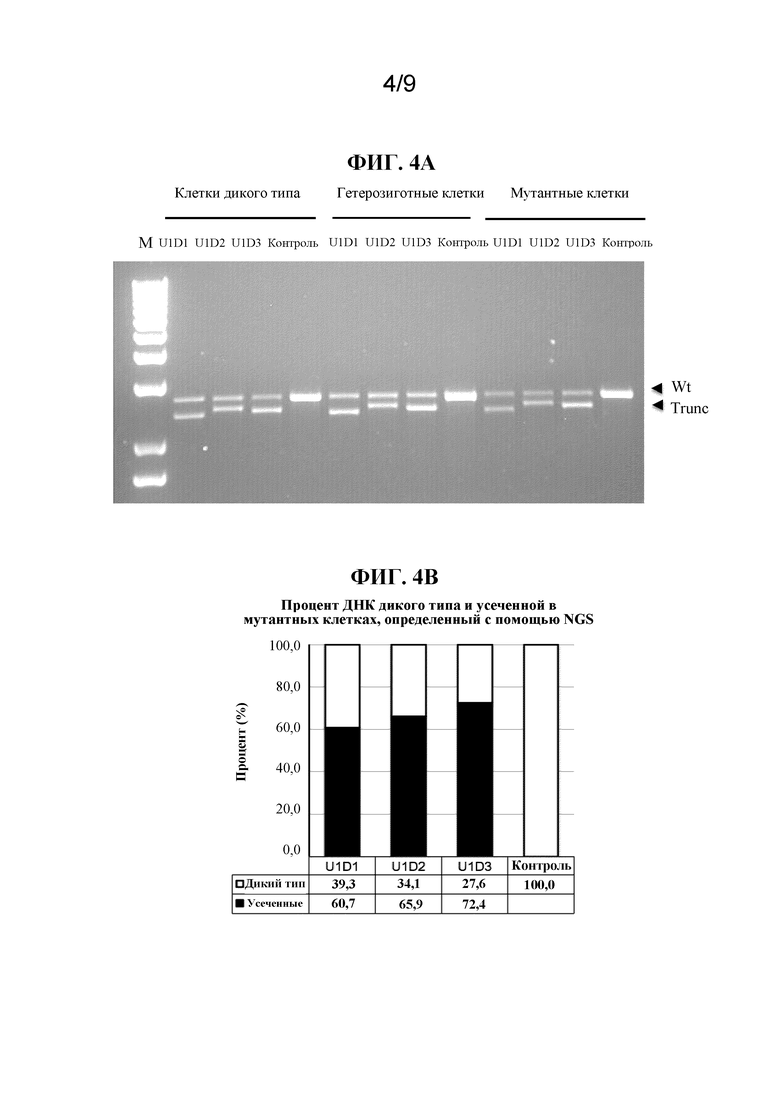
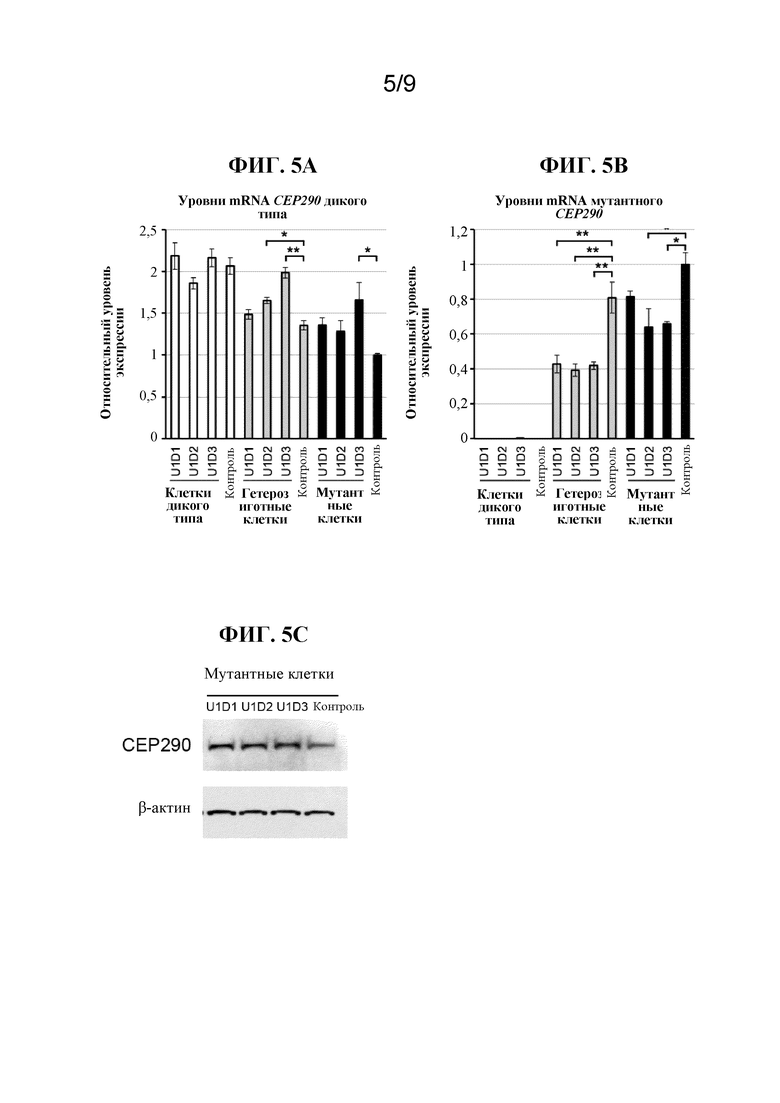
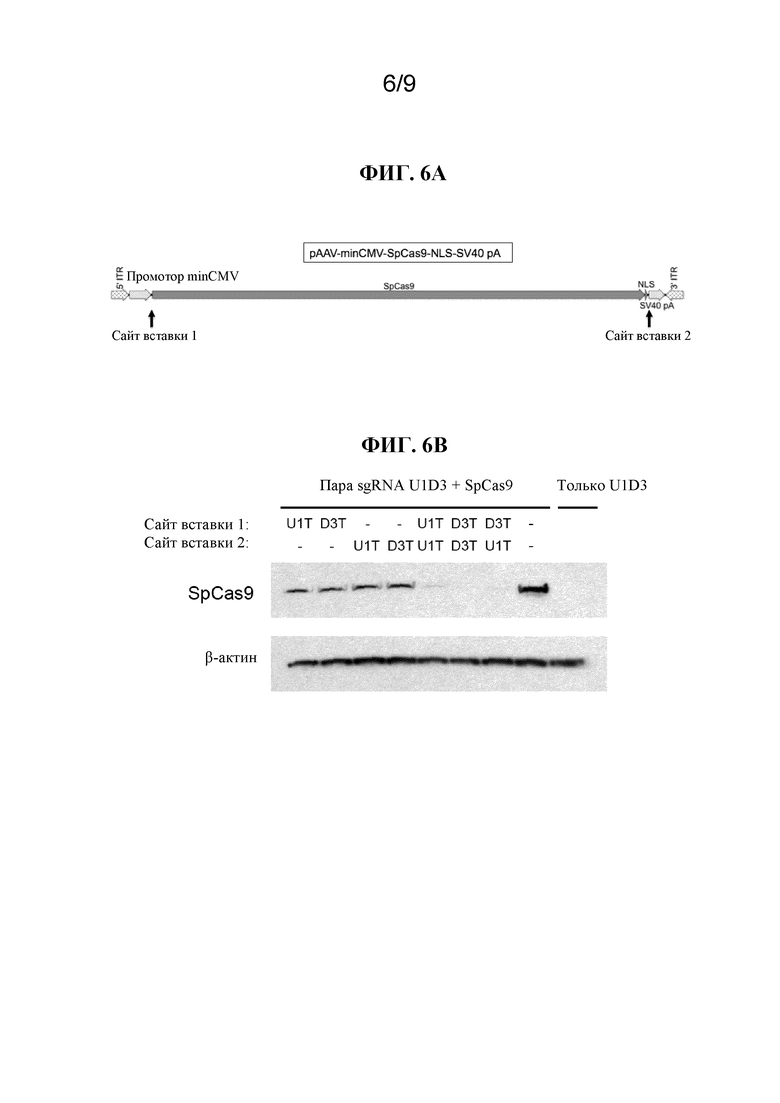
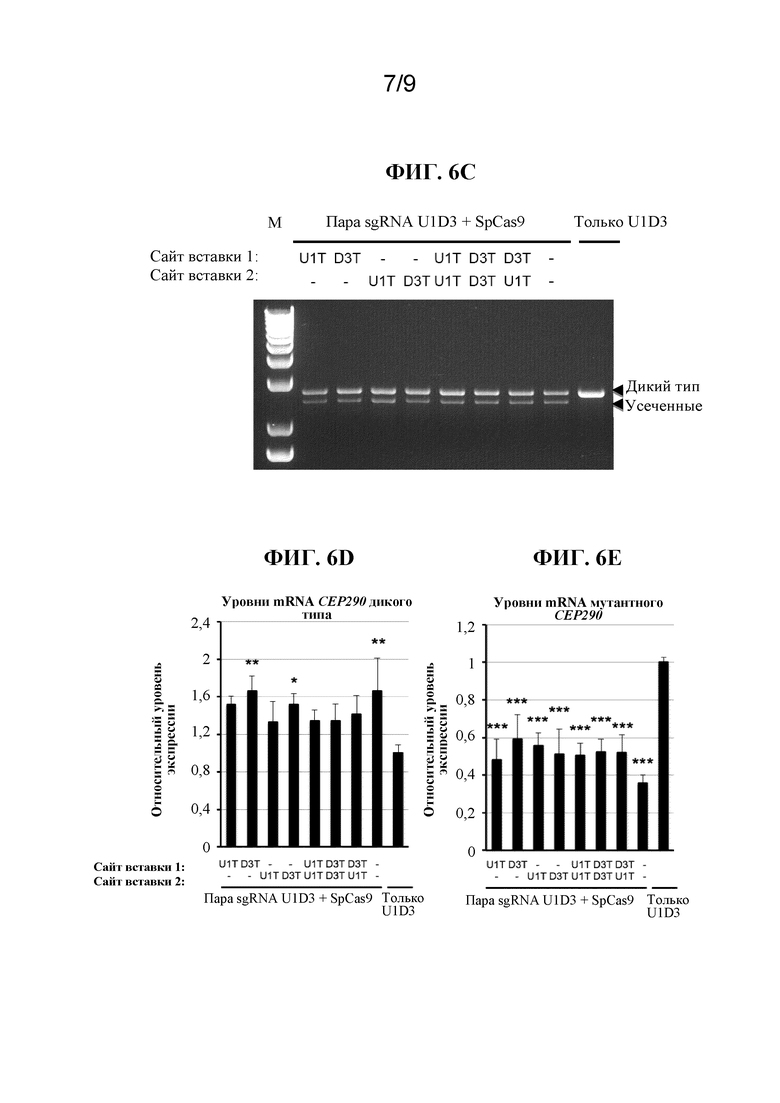
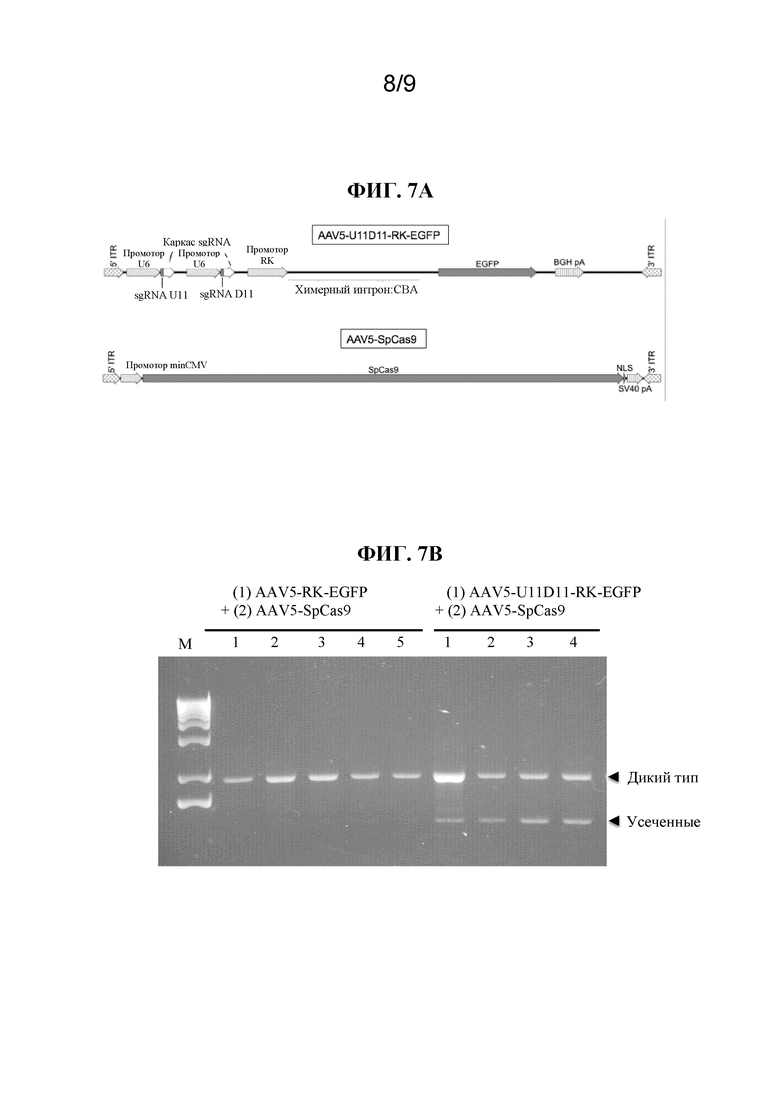
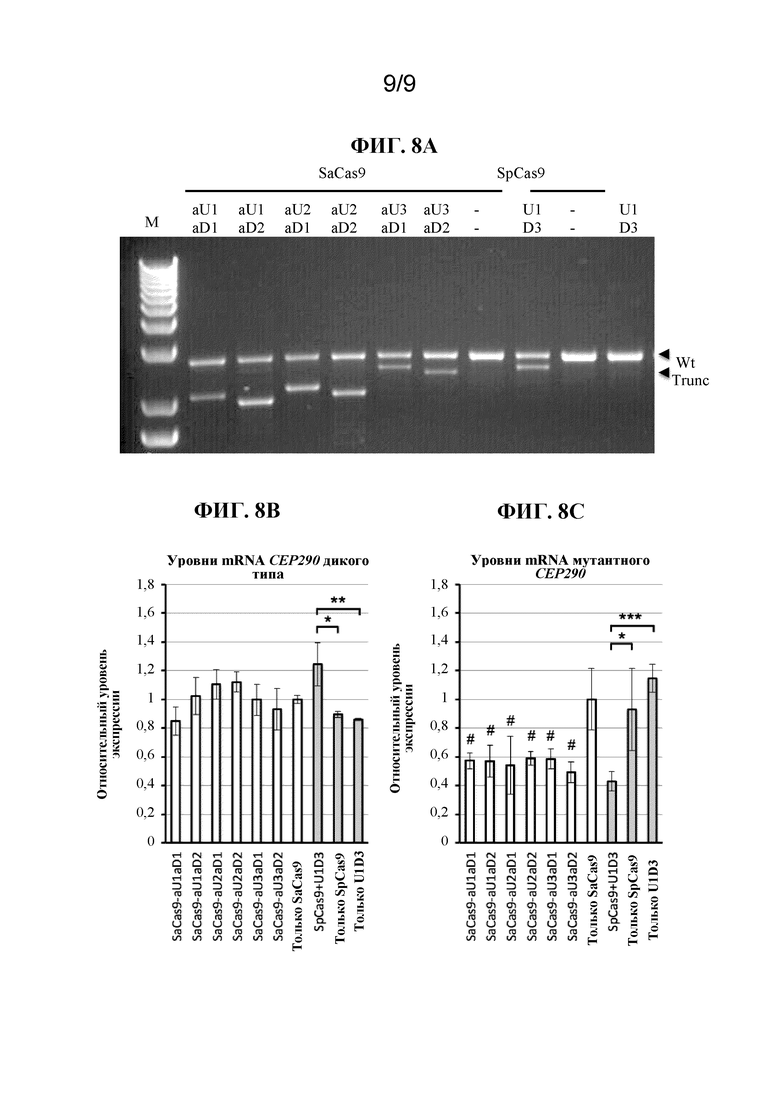
Изобретение относится к области биотехнологии, в частности к композиции для лечения врожденного амавроза Лебера, связанного с глубокой интронной мутацией в гене CEP290 индивидуума на основе системы CRISPR-Cas, где Cas-белок расщепляет кассету экспрессии Cas, тем самым снижая экспрессию Cas-белка, а также к способу лечения врожденного амавроза Лебера с ее использованием. Также представлены: набор, содержащий вышеуказанную композицию, и вирусная частица, содержащая вышеуказанную систему CRISPR-Cas. Изобретение обеспечивает благоприятный клинический результат при лечении врожденного амавроза Лебера. 4 н. и 9 з.п. ф-лы, 8 ил., 3 табл., 5 пр.
1. Композиция для лечения врожденного амавроза Лебера, связанного с глубокой интронной мутацией в гене CEP290 индивидуума, содержащая:
(i) одну или более нуклеиновую кислоту (нуклеиновых кислот), кодирующую сконструированную, не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) – CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую
a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с нитью ДНК, противоположной ДНК-мишени, где ДНК-мишень содержит глубокую интронную мутацию в гене CEP290 и последовательности, фланкирующие глубокую интронную мутацию, где первая направляющая и вторая направляющая гибридизуются с сайтами, противоположными фланкирующим последовательностям, и
b) кассету экспрессии Cas, содержащую нуклеотидную последовательность, кодирующую Cas-белок, и целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК способна гибридизоваться с целевым сайтом первой направляющей РНК; или
(ii) вирусную частицу, содержащую вирусный вектор, где вирусный вектор кодирует одну или более нуклеиновую кислоту (нуклеиновых кислот), кодирующую сконструированную, не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) – CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую
a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с нитью ДНК, противоположной ДНК-мишени, где ДНК-мишень содержит глубокую интронную мутацию в гене CEP290 и последовательности, фланкирующие глубокую интронную мутацию, где первая направляющая и вторая направляющая гибридизуются с сайтами, противоположными фланкирующим последовательностям, при этом противоположные нити последовательностей ДНК-мишени фланкируют глубокую интронную мутацию, и
b) кассету экспрессии Cas, содержащую нуклеотидную последовательность, кодирующую Cas-белок, и целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК способна гибридизоваться с целевым сайтом первой направляющей РНК;
где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию; где Cas-белок расщепляет кассету экспрессии Cas в первом целевом сайте направляющей РНК, тем самым снижая экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas; где ДНК-мишень не является ДНК эмбриональной линии человека.
2. Композиция по п. 1, где глубокая интронная мутация
(i) представляет собой глубокую интронную мутацию последовательности, представленной SEQ ID NO:23;
(ii) представляет собой мутацию c.2991+1655A>G;
(iii) расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов ниже 5'-конца сайта донора сплайсинга нуклеиновой кислоты;
(iv) расположена на приблизительно 1-10000 нуклеотидов, приблизительно 1-1000 нуклеотидов или приблизительно 100-1000 нуклеотидов выше 3'-конца сайта акцептора сплайсинга нуклеиновой кислоты; или
(v) вводит сайт донора сплайсинга или сайт акцептора сплайсинга в нуклеиновую кислоту.
3. Композиция по п. 1 или 2, где
(i) направляющие последовательности, первая направляющая РНК и вторая направляющая РНК, гибридизуются с противоположными нитями последовательностей ДНК-мишени, фланкирующих глубокую интронную мутацию последовательности, изложенной под SEQ ID NO:23;
(ii) первая направляющая РНК кодируется ДНК, содержащей последовательности под SEQ ID NO:41, SEQ ID NO:45, SEQ ID NO:46, SEQ ID NO:47; SEQ ID NO:19, SEQ ID NO:50, SEQ ID NO:51 или SEQ ID NO:52;
(iii) вторая направляющая РНК кодируется ДНК, содержащей последовательности под SEQ ID NO:42, SEQ ID NO:43, SEQ ID NO:44, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:20, SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:53 или SEQ ID NO:54;
(iv) содержащиеся первая направляющая РНК и/или вторая направляющая РНК слиты с последовательностью трансактивирующей cr (tracr), при этом последовательность tracr необязательно содержит нуклеотидную последовательность, кодируемую SEQ ID NO:25;
(v) первая направляющая РНК, вторая направляющая РНК и Cas-белок экспрессируются в эукариотических клетках;
(vi) первая направляющая РНК, вторая направляющая РНК и/или нуклеиновая кислота, кодирующая Cas-белок, функционально связаны с одним или несколькими регуляторными контрольными элементами;
(vii) первая направляющая РНК и/или вторая направляющая РНК функционально связаны с промотором РНК-полимеразы III, необязательно промотором U6, 7SK или H1; и/или
(viii) нуклеиновая кислота, кодирующая Cas-белок, функционально связана с промотором РНК-полимеразы II, необязательно немедленно-ранним промотором цитомегаловируса (CMV), минимальным фрагментом промотора, полученным из промотора CMV (промотор minCMV), LTR RSV, LTR MoMLV, промотором гена фосфоглицераткиназы-1 (PGK), промотором вируса обезьян 40 (SV40), промотором CK6, промотором гена транстиретина (TTR), промотором TK, тетрациклин-чувствительным промотором (TRE), промотором HBV, промотором hAAT, LSP-промотором, химерными печень-специфичными промоторами (LSP), промотором E2F, промотором EF1α, промотором гена теломеразы (hTERT); составным промотором энхансер цитомегаловируса/промотор гена бета-актина курицы/интрон гена β-глобина кролика (CAG), промотором гена родопсина, промотором гена конопсина, промотором гена бета-фосфодиэстеразы (PDE), промотором гена, связанного с пигментным ретинитом (RP1), или промотором гена межфоторецепторного ретинол-связывающего белка (IRBP).
4. Композиция по любому из пп. 1-3, где Cas-белок представляет собой белок Cas9, необязательно
(i) белок Cas9 Streptococcus pyogenes, белок Cas9 Staphylococcus aureus, белок Cas9 Streptococcus thermophilus, белок Cas9 Neisseria meningitidis или белок Cas9 Treponema denticola; и/или
(ii) кодон-оптимизированный для экспрессии в эукариотической клетке, необязательно клетке млекопитающего или клетке человека.
5. Композиция по любому из пп. 1-4, где система CRISPR-Cas дополнительно содержит один или несколько сигналов внутриядерной локализации (NLS), при этом необязательно
(i) Cas-белок содержит один или несколько NLS;
(ii) NLS представляет собой С-концевую последовательность в большом T-антигене SV40,
(iii) NLS содержит последовательность PKKKRKV (SEQ ID NO:26) или PKKKRKVEDPKKKRKVD (SEQ ID NO:27).
6. Композиция по любому из пп. 1-5, где нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, расположена на одном и том же или на разных векторах системы, где вектор необязательно представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV).
7. Композиция по п. 6, где вектор представляет собой вектор на основе рекомбинантного AAV (rAAV) и где нуклеиновая кислота, кодирующая одно или несколько из первой направляющей РНК, второй направляющей РНК или Cas-белка, фланкирована одной, двумя или более последовательностями инвертированных концевых повторов (ITR) AAV.
8. Композиция по п. 7, где ITR AAV представляют собой ITR с серотипами капсида AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV DJ, AAV козы, AAV крупного рогатого скота или AAV мыши и частица AAV содержит капсид серотипов AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAVrh8, AAVrh8R, AAV9, AAV10, AAVrh10, AAV11, AAV12, AAV2R471A, AAV2/2-7m8, AAV DJ, AAV2 N587A, AAV2 E548A, AAV2 N708A, AAV V708K, AAV козы, химерный AAV1/AAV2, AAV крупного рогатого скота, AAV мыши, капсид rAAV2/HBoV1 или содержит капсид AAV1, AAV2, AAV8, AAVrh8R, AAV9 и/или AAVrh10, содержащий мутацию, затрагивающую тирозин, или мутацию, влияющую на связывание с гепараном; где вектор необязательно представляет собой самокомплементарный вектор.
9. Способ лечения врожденного амавроза Лебера, связанного с глубокой интронной мутацией в гене CEP290 индивидуума, предусматривающий введение индивидууму терапевтически эффективного количества композиции по любому из пп. 1-8.
10. Способ по п. 9, где глубокая интронная мутация представляет собой
(i) глубокую интронную мутацию последовательности, представленной SEQ ID NO:23; или
(ii) мутацию c.2991+1655A>G.
11. Способ по п. 9 или 10, где композицию вводят в глаз индивидуума; при этом введение необязательно является субретинальным или интравитреальным.
12. Набор для лечения врожденного амавроза Лебера, связанного с глубокой интронной мутацией в гене CEP290 индивидуума, содержащий композицию по любому из пп. 1-8.
13. Вирусная частица для лечения врожденного амавроза Лебера, связанного с глубокой интронной мутацией в гене CEP290 индивидуума, содержащая вирусный вектор, где вирусный вектор содержит одну или более нуклеиновую кислоту (нуклеиновых кислот), кодирующую сконструированную, не встречающуюся в природе систему на основе коротких палиндромных повторов, регулярно расположенных группами (CRISPR) – CRISPR-ассоциированных генов (Cas) (CRISPR-Cas), содержащую
a) первую направляющую РНК и вторую направляющую РНК, где первая направляющая РНК и вторая направляющая РНК гибридизуются с нитью ДНК, противоположной ДНК-мишени, где ДНК-мишень содержит глубокую интронную мутацию и последовательности, фланкирующие глубокую интронную мутацию, где первая направляющая и вторая направляющая гибридизуются с сайтами, противоположными фланкирующим последовательностям, и
b) кассету экспрессии Cas, содержащую нуклеотидную последовательность, кодирующую Cas-белок, и целевой сайт первой направляющей РНК, где первая направляющая РНК или вторая направляющая РНК способна гибридизоваться с целевым сайтом первой направляющей РНК,
где Cas-белок расщепляет молекулу ДНК-мишени по сайтам, фланкирующим глубокую интронную мутацию, вырезая таким образом часть ДНК-мишени, содержащую глубокую интронную мутацию; где Cas-белок расщепляет кассету экспрессии Cas в первом целевом сайте направляющей РНК, тем самым снижая экспрессию Cas-белка по сравнению с экспрессией Cas-белка до расщепления кассеты экспрессии Cas; и где вирусный вектор представляет собой вектор на основе рекомбинантного аденоассоциированного вируса (rAAV), рекомбинантный аденовирусный вектор, вектор на основе рекомбинантного лентивируса или вектор на основе рекомбинантного вируса простого герпеса (HSV); где ДНК-мишень не является ДНК эмбриональной линии человека.
| MAEDER M.L | |||
| et al., 687 | |||
| РЕЛЬСОВАЯ ПЕДАЛЬ | 1920 |
|
SU290A1 |
| WO2015070083 A1, 14.05.2015 | |||
| OUSTEROUT D.G | |||
| et al., Multiplex CRISPR/Cas9-based genome editing for correction of dystrophin mutations that cause | |||

