Область техники, к которой относится изобретение
Настоящее изобретение относится к области биоинженерии, молекулярной биологии и биотехнологии, в частности к методам тестирования взаимодействий между белковыми молекулами.
Изобретение создано при финансовой поддержке Министерства Науки и Высшего образования Российской Федерации в рамках Соглашения № 075-15-2019-1661 от 31.10.2019.
Уровень техники
Одна из ключевых проблем молекулярной и клеточной биологии связана с исследованием взаимодействий между белками. Среди всех известных способов исследований таких взаимодействий дрожжевая двугибридная система – наиболее быстрый и экономичный метод, на основе которого были проведены первые протеомные исследования с попыткой установить все возможные взаимодействия между дрожжевыми белками и белками дрозофиллы.
Дрожжевая двугибридная система основана на использовании факторов транскрипции, которые характеризуются модульностью строения и состоят из физически и функционально разделимых доменов: ДНК-связывающего (BD – binding domain) и домена, активирующего транскрипцию (AD – activation domain). Физическое разделение доменов BD и AD приводит к инактивации фактора транскрипции: BD отдельно не способен активировать транскрипцию, а AD отдельно не способен связываться со своими специфическими сайтами. Домены BD и AD сливают с исследуемыми белками X («наживка») и Y («добыча») соответственно. Реконструкция фактора становится возможна в случае взаимодействия X с Y, тогда происходит активация транскрипции генов, зависимых от используемого фактора транскрипции (Bartel, Chien et al. 1993). Для создания двугибридной системы чаще всего используют фактор транскрипции Saccharomyces сerevisiae GAL4, разделенный на ДНК-связывающий и активационный домены.
Исходным материалом для анализа являются две плазмиды, в которые в нужной рамке считывания клонируются последовательности кДНК двух белков, исследуемых на предмет взаимодействия. Последовательность первого белка клонируется в единой рамке с BD-доменом в одной плазмиде, а последовательность второго – в единой рамке с AD-доменом GAL4 в другой плазмиде.
Дрожжевые клетки котрансформируют такими плазмидами, белковые конструкции экспрессируются в клетках и направляются с помощью особого сигнала ядерной локализации из цитоплазмы в ядро клетки. BD-домен, соединенный со вторым белком, связывается с промоторами репортерных генов, в которые встроены GAL4-связывающие последовательности ДНК, и если первый и второй белки взаимодействуют, то в результате AD-домен привлекается к регуляторной части репортерного гена и активирует его экспрессию.
В клетках S. cerevisiae тестируемые белки экспрессируются в относительно небольшом количестве с минимального конститутивного ADH1-промотора, в составе вектора также присутствует последовательность терминирующего сигнала ADH1. Экспрессируемый белок направляется в ядро клеток S. cerevisiae с помощью сигнала ядерной локализации, являющегося частью BD, или имеющего гетерологичное происхождение (из вируса SV40). Вектора способны автономно реплицироваться в клетках E. coli и в клетках S. cerevisiae. Вектора содержат ген β-лактамазы, обуславливающий резистентность E. coli к ампициллину. В векторе с активационным доменом GAL4 есть LEU2 ген, с ДНК-связывающим доменом – TRP1 ген, с дополнительным негибридным белком – URA3 ген, позволяющие гетеротрофным клеткам S. cerevisiae расти в условиях, лимитированных по лейцину, триптофану или урацилу, соответственно.
В наиболее распространенном варианте двугибридной системы используются ДНК-связывающий и активационный домены белка GAL4 и дрожжевой штамм, содержащий репортерные гены с GAL4-связывающими сайтами в промоторной области (US5283173A, US5468614A).
Также существуют модификации векторов, используемые в дрожжевой двугибридной системе, в основном направленные на применение других доменов (в отличие от классической с использованием активатора GAL4): например, репрессионные (US5885779A) или домены белков-участников сигнальных путей, связанных с мембранами клеток (US20100075326A1). Кроме этого, существуют модификации векторов, основанные на использовании рекомбинации и упрощающие клонирование кДНК в соответствующие плазмиды, используемые в двугибридной системе (US7323313B2). В патенте RU2529356C нуклеотидные последовательности, кодирующие исследуемые белки, отделены от нуклеотидных последовательностей, кодирующих активационный или ДНК-связывающий домен белка GAL4, при помощи нуклеотидной последовательности, кодирующей пептид, который экспрессируется в виде белкового мостика между исследуемым белком и доменами белка GAL4.
Для исследования взаимодействий белков в трехкомпонентных комплексах, кроме двух гибридных белков, слитых с ДНК-связывающим и активационным доменами GAL4, используют дополнительный негибридный белок. Негибридный белок экспрессируется либо с одного из векторов, кодирующих гибридный белок, либо с третьего дополнительного вектора. В случае четырех- и более компонентных белковых систем каждый дополнительный негибридный белок должен быть клонирован с собственным промотором и терминатором в один из трех используемых векторов, что значительно усложняет схему клонирования (Stynen et al. 2012).
В многокомпонентной дрожжевой двугибридной системе используется дополнительный негибридный белок или белки, необходимые для соединения двух гибридных белков, которые не могут напрямую взаимодействовать друг с другом, либо для стабилизации слабого взаимодействия двух гибридных белков.
Сущность изобретения
Классическая двугибридная дрожжевая система позволяет тестировать двойные или тройные белковые взаимодействия, что значительно снижает возможности применения метода, так как многие белки в живом организме функционируют только в составе многокомпонентных белковых комплексов. Существующие модификации метода для многокомпонентных систем (четыре и более белков) подразумевают клонирование дополнительных белков в отдельной рамке считывания с отдельным промотором и терминатором, что усложняет использование метода.
В настоящем изобретении предлагается новый способ одновременной экспрессии нескольких белков или их доменов в единой рамке считывания с одним промотором и терминатором в дрожжевой двугибридной системе. Вектора, используемые в дрожжевой двугибридной системе, модифицируются таким образом, чтобы кодирующие последовательности исследуемых белков находились в единой рамке считывания и были соединены друг с другом короткими саморасщепляющимися 2А-пептидами. 2А-пептиды представляют собой короткие аминокислотные последовательности, вызывающие терминацию трансляции полипептида без диссоциации рибосомы с мРНК, в результате чего продолжается трансляция следующего полипептида. Механизм их саморасщепления заключается в остановке рибосом на остатке глицина, высвобождении вышележащего белка, связанного с 2A-пептидом, и трансляции нижележащего белка с пролином в качестве первого остатка. С помощью 2А пептидных последовательностей можно добиться одновременной экспрессии нескольких белков. Для подтверждения эффективности выбранного подхода ряд известных взаимодействий, которые не выявляются в обычном варианте метода, протестированы с использованием модифицированных векторов.
Техническим результатом изобретения являются модифицированные плазмиды, содержащие нуклеотидную последовательность, кодирующую саморасщепляющийся пептид, который позволяет при котрансформации в клетки дрожжей одновременно экспрессировать несколько белков и детектировать взаимодействия между белками и их доменами в многокомпонентных белковых комплексах.
Предлагаемое изобретение может быть использовано для детекции взаимодействия между белковыми молекулами в составе многокомпонентных белковых комплексов, которые могут быть использованы в медицинских и исследовательских целях.
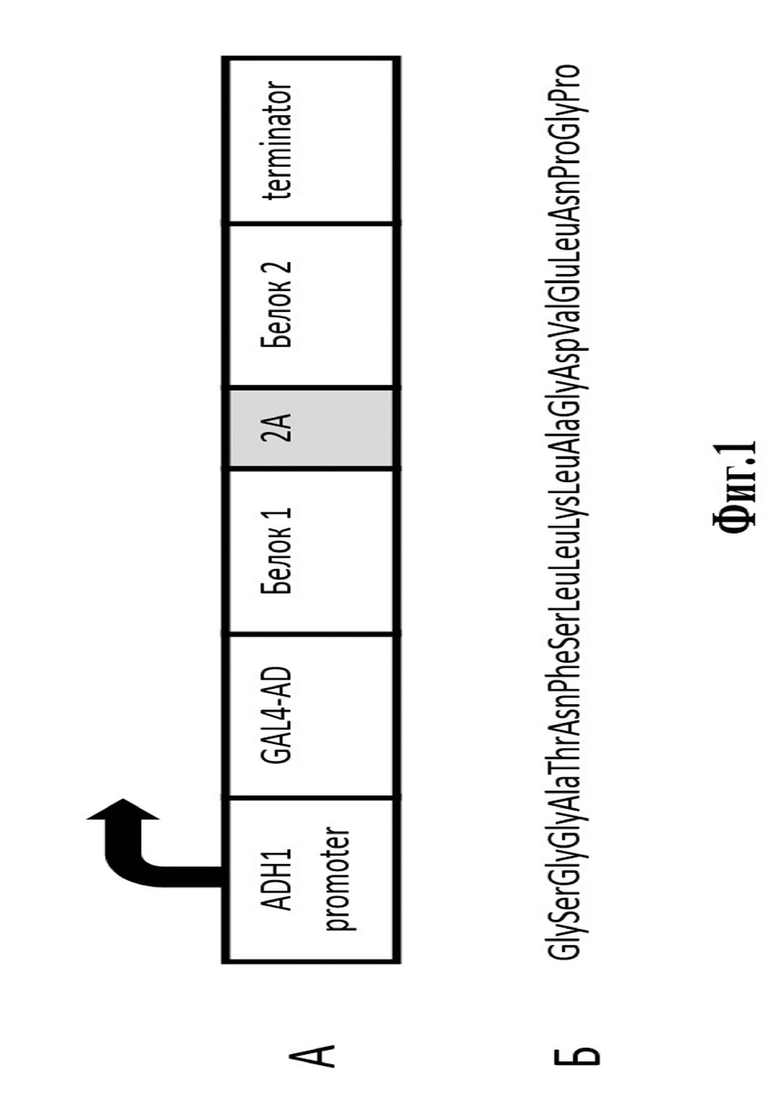
Сущность описываемого изобретения поясняется чертежом (фиг.1).
Краткое описание чертежей
Фигура 1. Схема вектора, предназначенного для экспрессии тестируемых белков, разделенных последовательностью, кодирующей саморасщепляющийся 2А-пептид, и последующего скрининга в дрожжевой двугибридной системе, где
А – вектор, в котором в единой рамке считывания с активационным доменом GAL4 клонированы белки, разделенные последовательностью, кодирующей саморасщепляющийся 2А-пептид;
Б – аминокислотная последовательность саморасщепляющегося 2А-пептида ERBV-1.
Фигура 2. Результаты дрожжевого двугибридного анализа с классическими и модифицированными векторами
Осуществление изобретения
Пример 1. Создание модификаций векторов, предназначенных для дрожжевой двугибридной системы.
Под термином «саморасщепляющийся пептид» следует понимать короткие аминокислотные последовательности, вызывающие терминацию трансляции полипептида без диссоциации рибосомы с мРНК, в результате чего продолжается трансляция следующего полипептида.
В качестве модифицируемого вектора был выбран pGAD424 (Clontech), предназначенный для использования в дрожжевой двугибридной системе и основанный на GAL4-активационном домене.
В качестве саморасщепляющегося пептида был выбран оптимизированный 2A-пептид вируса ринита В лошади (ERBV-1), для которого показатель эффективности саморасщепления в клетках Saccharomyces cerevisiae является одним из самых высоких среди известных последовательностей (более 90%) (Souza-Moreira et al. 2018). Аминокислотная последовательность оптимизированного ф2А-пептида ERBV-1 представлена на Фиг.1Б. (SEQ ID NO: 1).
Осуществление изобретения подразумевает проведение многоэтапного эксперимента.
1. Получение экспрессирующих векторов – двух плазмид, в которых последовательности кДНК исследуемых белков клонированы в единой рамке с ДНК-связывающим и активационным доменом, и третьей дополнительной плазмиды с негибридным белком.
Котрансформация дрожжей тремя типами плазмид и посев трансформантов на чашки Петри с селективной средой, не содержащей селективные аминокислоты (триптофан, лейцин и урацил). В гены биосинтеза этих аминокислот в геноме используемого дрожжевого штамма искусственно внесены мутации, нарушающие образование функционально активных белков. Эти гены без мутаций экспрессируются с трансформированных плазмид. В результате на чашках происходит селекция, если котрансформация прошла успешно, то на чашке через 2-3 суток вырастают отдельные колонии.
Дрожжи S. Cerevisiae штамма Pj69-4A пересеваются в 15 мл пробирку в жидкую среду YPD (2% бактопептон, 1% дрожжевой экстракт, 2% глюкоза) с помощью стерильной петли. Наращиваются в термостатированной качалке при 250-300 об/мин и температуре 30°С до оптической плотности OD600~1. Далее культура разбавляется в 10 раз средой YPDA и выращивается 3 часа при тех же условиях. 1,5 мл полученной суспензии клеток котрансформируется тремя плазмидами. Клетки осаждаются центрифугированием при 4000 g, 10 секунд, удаляется супернатант. К осадку добавляется 1 мл 0,1 М ацетата лития, затем инкубируется 10 минут на качалке при температуре 30°C, центрифугируется при 4000 g, 10 секунд, удаляется супернатант. К осадку клеток добавляется последовательно 270 мкл 50% PEG 3380, растворенного в 0,1 М ацетате лития, 25 мкг денатурированной ДНК лосося и 50 мкл смеси трех плазмид в 0,1 М ацетате лития, по 0,4-0,8 мкг каждой. Смесь ресуспендируется и инкубируется в качалке при 30°С, 30 минут. Затем инкубируется на 42°С 2 минуты. Пробирки охлаждаются во льду, центрифугируются при 4000 g, 10 секунд, удаляется супернатант. К осадку добавляется 150 мкл деионизованной воды (mQ), ресуспендируется и высевается на чашку с селективной средой (0,67% дрожжевая среда без аминокислот, 1,5% агар, 2% глюкоза, 0,030% L-изолейцин, 0,015% L-валин, 0,002% L-аргинин, 0,002% L- хлорид моногидрат гистидина, 0,003% L- хлорид лизина, 0,002% L-метионин, 0,005% L-фенилаланин, 0,02% L-треонин, 0,003% L-тирозин, 0,002% сульфат аденина). Чашки помещаются в термостат 30°С на трое суток, до появления отдельных колоний.
2. Несколько колоний пересеваются истощающим штрихом на чашки Петри с селективной средой (0,67% дрожжевая среда без аминокислот, 1,5% агар, 2% глюкоза, 0,030% L-изолейцин, 0,015% L-валин, 0,002% L-аргинин, 0,003% L- хлорид лизина, 0,002% L-метионин, 0,005% L-фенилаланин, 0,02% L-треонин, 0,003% L-тирозин, 0,002% сульфат аденина). Чашки помещаются в термостат 30°С на несколько суток.
3. Чаще всего, гены биосинтеза гистидина HIS3 и аденина ADE2, а также ген β-галактозидазы LacZ, являются репортерными. Если белки взаимодействуют, то активируются гены ADE2 и HIS3, и, следовательно, колонии вырастают. Если белки не взаимодействуют, то рост колоний не наблюдается. Кроме этого, можно параллельно детектировать уровень активации гена LacZ.
Пример 2. Тестирование модифицированных векторов на предмет детекции взаимодействия между белковыми молекулами
Для проверки эффективности саморасщепляющегося пептида 2А ERBV-1 в дрожжевой дигибридной системе был выбран DUB-модуль SAGA-комплекса Drosophila melanogaster, состоящий из трех белков Eny2, Sgf11 и Nonstop.
На основе модифицированных векторов были созданы плазмиды для тестирования эффективности разработанного подхода. Для этого в вектора для дрожжевой двугибридной системы были клонированы кДНК-последовательности тестируемых белков. В качестве положительного контроля корректности функционирования всех плазмид и эффективности саморасщепления пептида 2А ERBV-1 было использовано взаимодействие белков дрозофилы Eny2, Sgf11 и Nonstop, образующих DUB-модуль SAGA-комплекса. Активационный домен белка GAL4, кДНК Eny2, кДНК саморасщепляющегося пептида 2А ERBV-1 и кДНК Sgf11 находятся в единой рамке трансляции. ДНК-связывающий домен белка GAL4 слит в единой рамке трансляции с кДНК домена UBP Nonstop. В качестве отрицательных контролей были использованы исходные плазмиды. Результаты проверки всех вариантов векторов представлены на Фиг.2, из которых видно, что на селективной среде без гистидина детектируются положительные сигналы только в паре UBPNonstop – Eny2_2A_Sgf11. Для сборки белкового комплекса необходимы три отдельных белка, что возможно только при расщеплении 2A-пептида.
Затем был протестирован белок Ataxin7, который предположительно может связывать белки DUB-модуля с образованием четверного комплекса. ДНК-связывающий домен белка GAL4 слит в единой рамке трансляции с кДНК Ataxin7. Белок Nonstop заклонирован в третьем негибридном векторе Y3H. Из результатов дрожжевого двугибридного анализа следует, что Ataxin7 не взаимодействует ни с одним из белков Nonstop, Eny2 или Sgf11 по отдельности, также Ataxin7 не связывается ни с одной из возможных пар белков DUB-модуля. И только при одновременном присутствии всех трех белков Nonstop, Eny2 и Sgf11, что возможно только при расщеплении 2A-пептида, образуется четверной комплекс с Ataxin7 (Фиг.2).
Список цитированной литературы
Stynen et al. (2012). "Diversity in Genetic In Vivo Methods for Protein-Protein Interaction Studies: from the Yeast Two-Hybrid System to the Mammalian Split-Luciferase System. " Microbiol Mol Biol Rev. 76 (2): 331–382.
Bartel et al. (1993). "Elimination of false positives that arise in using the two-hybrid system." Biotechniques 14 (6): 920-4.
Souza-Moreira et al. (2018). "Screening of 2A peptides for polycistronic gene expression in yeast. " FEMS Yeast Research 18 (5).
<110> Федеральное государственное бюджетное учреждение науки Институт биологии гена Российской академии наук (Institute of Gene Biology Russian Academy of Sciences)
<120> Модификация дрожжевой двугибридной системы, которая позволяет тестировать взаимодействия между белками внутри многокомпонентных белковых комплексов
<160> 1
<210> 1
<211> 23
<212> PRT
<213> ERBV-1
<220>
<223> 2A-пептид вируса ринита В лошади (ERBV-1)
<400> 1
Gly Ser Gly Gly Ala Thr Asn Phe Ser Leu Leu Lys Leu Ala Gly Asp 16
1 5 10 15
Val Glu Leu Asn Pro Gly Pro 23
20

Изобретение относится к дрожжевой клетке, предназначенной для детекции взаимодействия между белками и их доменами в многокомпонентных белковых комплексах. Дрожжевая клетка котрансформирована тремя плазмидами, модифицированными для экспрессии в клетках дрожжей нескольких белков. При этом нуклеотидная последовательность, кодирующая ДНК-связывающий домен белка GAL4, слита с нуклеотидной последовательностью, кодирующей первый белок, и клонирована в одну плазмиду. Нуклеотидная последовательность, кодирующая активационный домен белка GAL4, слита с нуклеотидной последовательностью, кодирующей второй и третий белки и имеющей единую рамку считывания с одним промотором и терминатором, и клонирована во вторую плазмиду, при этом между вторым и третьим белком встроена нуклеотидная последовательность, кодирующая саморасщепляющийся пептид, который позволяет одновременно экспрессировать несколько белков или их доменов в единой рамке считывания с одним промотором и терминатором. Нуклеотидная последовательность, кодирующая негибридный белок, клонирована в третью плазмиду. При этом саморасщепляющийся пептид представляет собой 2A-пептид вируса ринита В лошади, имеющий аминокислотную последовательность GlySerGlyGlyAlaThrAsnPheSerLeuLeuLysLeuAlaGlyAspValGluLeuAsnProGlyPro, представленную в SEQ ID NO: 1. Изобретение может быть использовано для детекции взаимодействия между белковыми молекулами в составе многокомпонентных белковых комплексов, которые могут быть использованы в медицинских и исследовательских целях. 2 ил., 2 пр.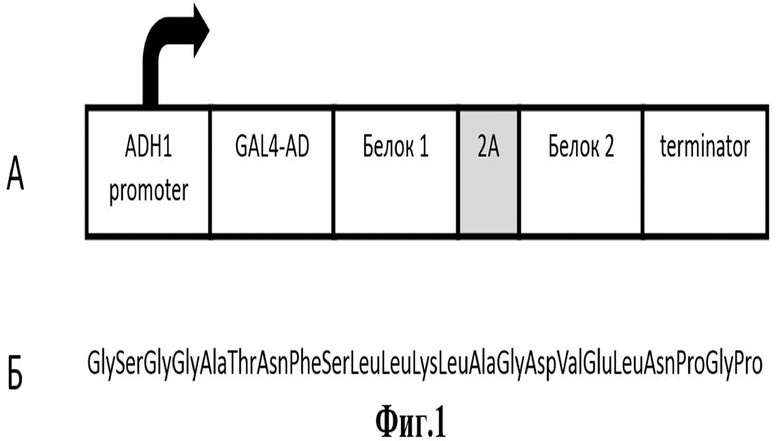
Дрожжевая клетка, предназначенная для детекции взаимодействия между белками и их доменами в многокомпонентных белковых комплексах,
котрансформированная тремя плазмидами, модифицированными для экспрессии в клетках дрожжей нескольких белков,
где нуклеотидная последовательность, кодирующая ДНК-связывающий домен белка GAL4, слита с нуклеотидной последовательностью, кодирующей первый белок, и клонирована в одну плазмиду;
где нуклеотидная последовательность, кодирующая активационный домен белка GAL4, слита с нуклеотидной последовательностью, кодирующей второй и третий белки и имеющей единую рамку считывания с одним промотором и терминатором, и клонирована во вторую плазмиду, при этом между вторым и третьим белком встроена нуклеотидная последовательность, кодирующая саморасщепляющийся пептид, который позволяет одновременно экспрессировать несколько белков или их доменов в единой рамке считывания с одним промотором и терминатором;
и где нуклеотидная последовательность, кодирующая негибридный белок, клонирована в третью плазмиду;
при этом саморасщепляющийся пептид представляет собой 2A-пептид вируса ринита В лошади, имеющий аминокислотную последовательность GlySerGlyGlyAlaThrAsnPheSerLeuLeuLysLeuAlaGlyAspValGluLeuAsnProGlyPro, представленную в SEQ ID NO: 1.
| МОДИФИЦИРОВАННАЯ ДРОЖЖЕВАЯ ДВУГИБРИДНАЯ СИСТЕМА ДЛЯ ЭФФЕКТИВНОГО ИССЛЕДОВАНИЯ ВЗАИМОДЕЙСТВИЯ МЕЖДУ БЕЛКАМИ И ИХ ДОМЕНАМИ. | 2012 |
|
RU2529356C2 |
| ДВУГИБРИДНАЯ СИСТЕМА, ОСНОВАННАЯ НА ОБЕСПЕЧЕНИИ УМОЛЧАНИЯ ГЕНОВ ПУТЕМ ТРАНСКРИПЦИОННОЙ ИНТЕРФЕРЕНЦИИ | 2005 |
|
RU2403287C2 |
| US 20100075326 A1, 25.03.2010 | |||
| SOUZA-MOREIRA Т.М | |||
| ET AL | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

