Область техники, к которой относится изобретение
Настоящее изобретение относится к области биоинженерии, молекулярной биологии и биотехнологии, в частности к методам тестирования взаимодействий между белковыми молекулами.
Уровень техники
Одна из ключевых проблем молекулярной и клеточной биологии связана с исследованием взаимодействий между белками. Среди всех известных способов исследований таких взаимодействий дрожжевая двугибридная система - наиболее эффективный и экономичный метод, на основе которого многими фирмами были проведены первые протеомные исследования с попыткой установить все возможные взаимодействия между дрожжевыми и дрозофильными белками. Исходным материалом для анализа являются две плазмиды, в которые в нужной рамке считывания клонируются последовательности к ДНК двух белков (один из них оказывается слит в единой рамке трансляции с ДНК-связывающим доменом фактора транскрипции, а второй - с его доменом, активирующим транскрипцию), исследуемых на предмет взаимодействия. При этом физическое связывание друг с другом исследуемых белков приводит к сближению ДНК-связывающего и активационного доменов и, как следствие, активируется либо репрессируется экспрессия репортерного гена. В наиболее распространенном варианте двугибридной системы используются ДНК-связывающий и активационный домены белка GAL4 и дрожжевой штамм, содержащий репортерные гены с GAL4-связывающими сайтами в промоторной области (US 5283173, US 5468614). Основной недостаток этой системы связан со структурными ограничениями, накладываемыми близко расположенными доменами в химерном белке, в результате которых происходит либо инактивация ДНК-связывающего или активационного домена белка GAL4, либо соответствующих доменов исследуемых белков. В результате более половины всех потенциальных взаимодействий не детектируется, что значительно снижает возможности применения метода.
Функционирование некоторых современных дрожжевых двугибридных систем обеспечивается модификациями, улучшающими детекцию белок-белковых взаимодействий при наличии возможных ложных сигналов путем использования сплит-систем (чаще всего основанные на молекуле убиквитина), которые работают независимо от активации или репрессии транскрипции репортерного гена, на которую могут оказывать сильное влияние тестируемые белковые молекулы (US 6562576, US 6911311, US 20090298089 A1).
Также существуют модификации векторов, используемые в двугибридной системе, в основном направленные на применение других доменов (в отличие от классической с использованием активатора GAL4) в дрожжевой двугибридной системе: например, репрессионные (US 5885779) или домены белков-участников сигнальных путей, связанных с мембранами клеток (US 2010/0075326 А1). Кроме этого существуют модификации векторов, основанные на использовании рекомбинации и используемые в двугибридной системе, которые упрощают клонирование кДНК в соответствующие плазмиды (US 7323313). Наконец, известен вариант применения гибких линкеров между искусственными доменами «цинковые пальцы», использование которых увеличивает аффинность химерных белков к их ДНК-мишеням (US 6479626).
Тем не менее, не существует способов, в которых бы вектора для дрожжевой двугибридной системы модифицировались таким образом, чтобы домены тестируемых белков были бы отделены от ДНК-связывающего и/или активационного доменов гибкими белковыми мостиками, которые бы изолировали домены друг от друга и тем самым нивелировали влияние стерических эффектов.
Сущность изобретения
В настоящем изобретении предлагается новый способ снижения взаимозависимости доменов в дрожжевой двугибридной системе. Вектора, используемые в дрожжевой двугибридной системе, модифицируются таким образом, чтобы исследуемые белки были отделены от ДНК-связывающего или активационного доменов белка GAL4 при помощи нейтрального гибкого белкового мостика. Для подтверждения эффективности выбранного подхода ряд известных взаимодействий, которые не выявляются в обычном варианте метода, протестированы с использованием модифицированных векторов.
Предлагаемое изобретение может быть использовано для детекции взаимодействия между белковыми молекулами, которые могут быть использованы в медицинских и исследовательских целях.
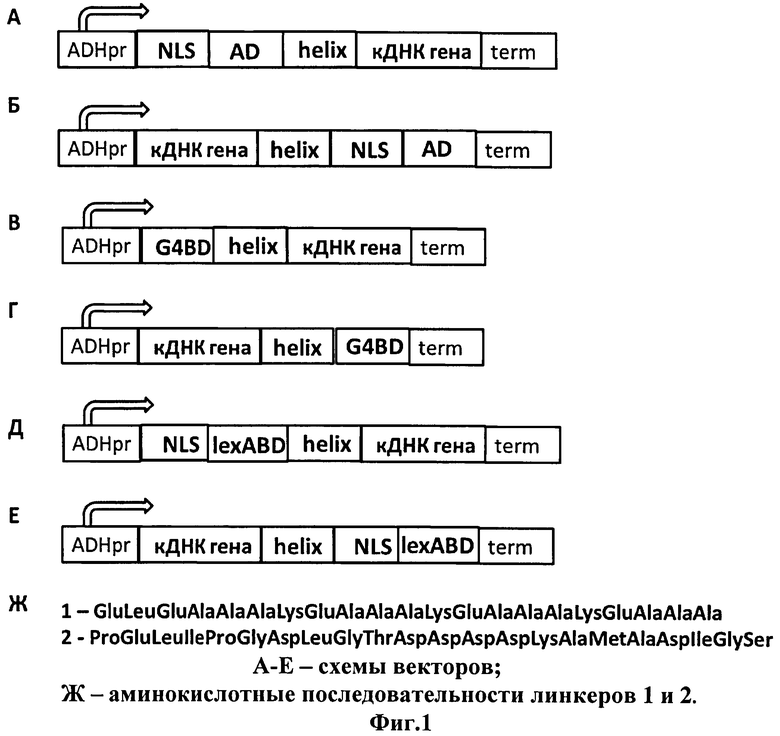
Сущность описываемого изобретения поясняется чертежом (фиг.1), на котором представлены схемы векторов, предназначенные для клонирования в них кДНК-последовательностей тестируемых белков и последующего скрининга в дрожжевой двугибридной системе, где
А - вектора pG4AD_h1 и pG4AD_h2, в которых ниже активационного домена GAL4 клонированы последовательности, кодирующие гибкий белковый мостик 1 или 2;
Б - вектор ph1_G4AD, в котором выше активационного домена GAL4 клонирована последовательность, кодирующая гибкий белковый мостик 1;
В - вектор pG4BD_h1 и pG4BD_h2, в которых ниже ДНК-связывающего домена GAL4 клонированы последовательности, кодирующие гибкий белковый мостик 1 или 2;
Г - вектор ph1_G4BD, в котором выше ДНК-связывающего домена GAL4 клонирована последовательность, кодирующая гибкий белковый мостик 1;
Д - вектор plexABD_h1, в котором ниже ДНК-связывающего домена lexA клонирована последовательность, кодирующая гибкий белковый мостик 1;
Е - вектор ph1_lexABD, в котором выше ДНК-связывающего домена lexA клонирована последовательность, кодирующая гибкий белковый мостик 1;
Ж - аминокислотные последовательности используемых белковых мостиков 1 и 2.
Пример 1. Создание модификаций векторов, предназначенных для дрожжевой двугибридной системы.
Дрожжевая двугибридная система основана на использовании факторов транскрипции, которые характеризуются модульностью строения и состоят из физически и функционально разделимых доменов: ДНК-связывающего (BD - binding domain) и домена, активирующего транскрипцию (AD - activation domain). Физическое разделение BD и AD доменов приводит к инактивации фактора транскрипции. BD и AD домены сливают с исследуемыми белками Х («наживка») и Y («добыча»), соответственно. Реконструкция фактора становится возможна в случае взаимодействия Х с Y, тогда происходит активация транскрипции генов, зависимых от используемого фактора транскрипции (Bartel, Chien et al. 1993). Для создания двугибридной системы чаще всего используют фактор транскрипции S. cerevisiae GAL4, разделенный на ДНК-связывающий и активационный домены. Также в качестве ДНК-связывающего мотива часто используют домен бактериального репрессорного белка lexA.
Исходным материалом для анализа являются две плазмиды, в которые в нужной рамке считывания клонируются последовательности кДНК двух белков, исследуемых на предмет взаимодействия. Последовательность первого белка клонируется в единой рамке с BD-доменом в одной плазмиде, а последовательность второго - в единой рамке с AD-доменом Gal4 в другой плазмиде.
Дрожжевые клетки котрансформируют двумя такими плазмидами, белковые конструкции экспрессируются в клетках и направляются с помощью особого сигнала ядерной локализации из цитоплазмы в ядро клетки. BD-домен, соединенный со вторым белком, связывается с промоторами репортерных генов, в которые встроены либо GAL4-, либо lexA-связывающие последовательности ДНК, и если первый и второй белки взаимодействуют, то в результате AD-домен привлекается к регуляторной части репортерного гена и активирует его экспрессию.
Необходимо отметить, что в клетках S. cerevisiae слитый белок экспрессируется в относительно небольшом количестве с минимального конститутивного ADH1-промотора, в составе вектора также присутствует последовательность терминирующего сигнала ADH1. Слитый белок направляется в ядро клеток S. cerevisiae с помощью сигнала ядерной локализации, являющегося частью BD или имеющего гетерологичное происхождение (из вируса SV40). Вектора способны автономно реплицироваться в клетках Е. coli и в клетках S. cerevisiae. Вектора содержат ген β-лактамазы, обуславливающий резистентность Е. coli к ампициллину. В варианте вектора с активационным доменом GAL4 есть LEU2 ген, а с ДНК-связывающим доменом - TRP1 ген, позволяющие гетеротрофным клеткам S. cerevisiae расти в условиях, лимитированных по лейцину или триптофану, соответственно.
Эксперимент включает несколько этапов.
- Получение экспрессирующих векторов - плазмид, в которых последовательности кДНК исследуемых белков клонированы в единой рамке с ДНК-связывающим или активационным доменом.
- Котрансформация дрожжей двумя типами плазмид и посев трансформантов на чашки Петри с селективной средой, не содержащей селективные аминокислоты (например, в классическом варианте - триптофан и лейцин). В гены биосинтеза этих аминокислот в геноме используемого дрожжевого штамма искусственно внесены мутации, нарушающие образование функционально активных белков. Эти гены без мутаций экспрессируются с трансформированных плазмид. В результате на чашках происходит селекция, если котрансформация прошла успешно, то на чашке через 2-3 суток вырастают отдельные колонии.
- Несколько колоний пересеваются истощающим штрихом на чашки Петри с селективной средой (например, без триптофана, лейцина, гистидина и аденина).
- Чаще всего, гены биосинтеза гистидина и аденина ADE2 и HIS3, а также ген β-галактозидазы LacZ являются репортерными. Если белки взаимодействуют, то активируются гены ADE2 и HIS3 и, следовательно, колонии вырастают. Если белки не взаимодействуют, то рост колоний не наблюдается. Кроме этого можно параллельно детектировать уровень активации гена LacZ.
Основным ограничением в использовании дрожжевой двугибридной системы является частое появление ложноотрицательных и ложноположительных результатов. Есть несколько подходов на пути преодоления данных проблем:
1) использование нескольких генов-репортеров,
2) перемещение в плазмидах доменов белка Gal4/lexA на С-конец химерного белка,
3) проверка не целого белка на взаимодействие, а только его отдельных доменов,
4) использование ингибиторов, которые в определенных концентрациях исключают ложноположительный рост.
Однако, несмотря на все эти подходы, многие взаимодействия не удается детектировать с использованием классического варианта дрожжевой двугибридной системы из-за того, что в последовательности химерного белка домены тестируемого белка, по всей видимости, оказываются жестко связаны с последовательностями, кодирующими ДНК-связывающий/активационный домены. В связи с этим было высказано предположение, согласно которому использование в векторах для дрожжевой двугибридной системы гибких белковых мостиков, отделяющих последовательность тестируемого белка от последовательностей ДНК-связывающего/активационного доменов, позволит стерически изолировать домены белков и добиться детектируемого сигнала взаимодействия белковых молекул.
В качестве модифицируемых векторов были выбраны pGAD424 и pGBT9 (Clontech), предназначенные для использования в дрожжевой двугибридной системе и основанные на GAL4-активаторе. Дополнительно в качестве ДНК-связывающего домена был использован соответствующий мотив lexA, который был клонирован в вектор pGBT9 вместо ДНК-связывающего домена GAL4.
Были разработаны и синтезированы ДНК-последовательности, кодирующие 21- и 22-аминокислотные линкеры, представляющие собой достаточно гибкие α-спирали. Разработанные аминокислотные последовательности представлены на Фиг.1Ж.
ДНК-последовательности линкера были клонированы в вектора для дрожжевой двугибридной системы либо на N- (Фиг.1Б, Г, Е), либо на С-конце (Фиг.1А, В, Д) относительно ДНК-связывающего (Фиг.1В, Г, Д, Е)/активационного (Фиг.1А, Б) доменов. Для клонирования ДНК-последовательности, кодирующей белковый мостик, на N-конец ДНК-связывающего/активационного доменов был использован сайт рестрикции HindIII. Для клонирования ДНК-последовательности, кодирующей белковый мостик, на С-конец ДНК-связывающего/активационного доменов был использован сайт рестрикции EcoRI. Соответствующие рестриктные сайты были заложены в последовательности олигонуклеотидов, используемых для синтеза последовательности ДНК, кодирующей белковый мостик. Последовательности олигонуклеотидов были подобраны таким образом, чтобы открытая рамка считывания клонируемых фрагментов в итоговых конструкциях не была нарушена.
В итоге получено 8 векторов, в которых последовательности изучаемых белков можно клонировать в единой рамке считывания с ДНК-связывающим/активационным доменами либо на N- либо на С-конце, и во всех случаях белковые домены тестируемых белков будут отделены от ДНК-связывающего/активационного доменов 21- или 22-аминокислотным белковым мостиком.
Пример 2. Тестирование модифицированных векторов на предмет детекции взаимодействия между белковыми молекулами.
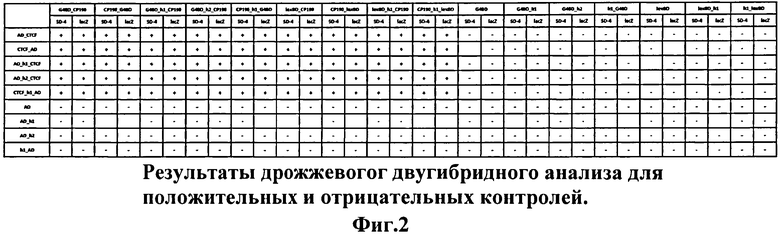
На основе модифицированных векторов были созданы плазмиды для тестирования эффективности разработанного подхода. Для этого как в классические вектора для дрожжевой двугибридной системы (pGAD424 и pGBT9), так и в модифицированные (pG4AD_h1, pG4AD_h2, ph1_G4AD, pG4BD_h1, pG4BD_h2, ph1_G4BD, plexABD_h1, ph1_lexABD) были клонированы кДНК-последовательности тестируемых белков. В качестве положительного контроля корректности функционирования всех плазмид было использовано хорошо изученное взаимодействие белков Дрозофилы CTCF и СР190. В качестве отрицательных контролей были использованы исходные плазмиды. Результаты проверки всех вариантов векторов представлены на Фиг.2, из которой видно, что как на селективной среде без гистидина и аденина, так и в β-галактозидазном тесте детектируются положительные сигналы только в парах dCTCF-CP190.
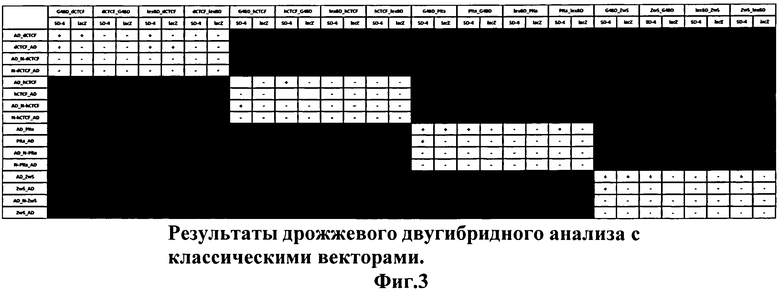
Затем были выбраны белки и отдельные белковые домены, для которых из экспериментов in vitro известно, что они могут формировать гомодимеры, однако в классическом варианте дрожжевой двугибридной системы не удавалось детектировать положительный сигнал. Это полноразмерные белки CTCF Дрозофилы и человека и его N-концевые домены, белки Pita, Zw5 Дрозофилы, и их N-концевые домены, представляющие собой цинковый палец С4-типа (так называемый ZAD-домен). Для всех этих белков в экспериментах in vitro показано, что они могут формировать гомодимеры, и для такой димеризации необходимы N-концевые домены данных белков. На Фиг.3 представлены результаты дрожжевой двугибридной системы, полученные с использованием классических векторов, в которые были клонированы кДНК-последовательности данных белков. Видно, что в случае отдельных доменов во всех экспериментах были получены отрицательные результаты, и только в некоторых вариантах с использованием полноразмерных белков детектируется положительный сигнал.
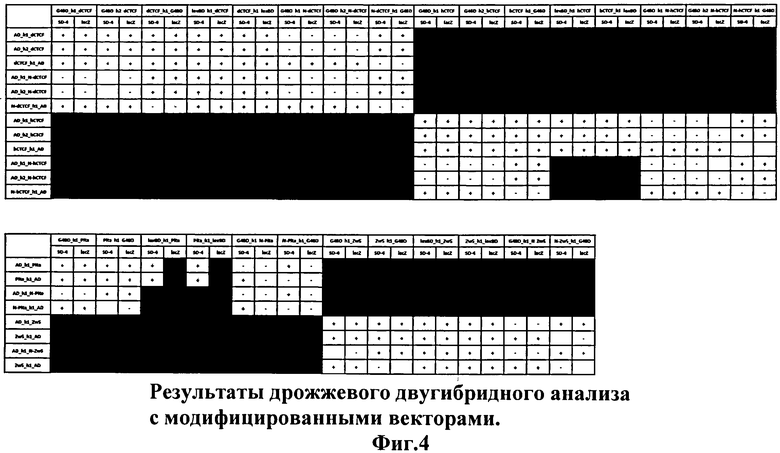
Все изучаемые последовательности были клонированы в модифицированные вектора, и на Фиг.4 представлены результаты данного варианта дрожжевой двугибридной системы. Так, во многих комбинациях, в которых N-конец тестируемых белков оставался свободным и при этом отделялся от ДНК-связывающего/активационного доменов белковым мостиком, удалось детектировать положительные сигналы, которые ранее были недетектируемы. На основании полученных результатов действительно видно, что молекулы изучаемых белков взаимодействуют друг с другом за счет N-концевых доменов.
Список литературы
1. Bartel, Р., С.Т. Chien, et al. (1993). ″Elimination of false positives that arise in using the two-hybrid system.″ Biotechniques 14(6): 920-4.
Изобретение относится к области биоинженерии, молекулярной биологии и биотехнологии. Предложена дрожжевая клетка, предназначенная для детекции взаимодействия между белками и их доменами, котрансформированная двумя плазмидами, модифицированными для экспрессии в клетках дрожжей двух белков, где нуклеотидная последовательность, кодирующая первый белок, клонирована в одну плазмиду, а нуклеотидная последовательность, кодирующая второй белок, - в другую плазмиду, где нуклеотидные последовательности, кодирующие белки, слиты с нуклеотидными последовательностями, кодирующими активационный и ДНК-связывающий домены белка GAL4, при этом нуклеотидные последовательности, кодирующие исследуемые белки, отделены от нуклеотидных последовательностей, кодирующих активационный или ДНК-связывающий домен белка GAL4, при помощи нуклеотидной последовательности, кодирующей пептид, представляющий собой последовательность GluLeuGluAlaAlaAlaLysGluAlaAlaAlaLysGluAlaAlaAlaLysGluAlaAlaAla, который экспрессируется в виде белкового мостика между исследуемым белком и доменами белка GAL4. Изобретение позволяет свести к минимуму взаимовлияние доменов тестируемых белков и ДНК-связывающего/активационного доменов вектора и может быть использовано для детекции взаимодействия между белковыми молекулами в медицинских и исследовательских целях. 4 ил., 2 пр.
Дрожжевая клетка, предназначенная для детекции взаимодействия между белками и их доменами, котрансформированная двумя плазмидами, модифицированными для экспрессии в клетках дрожжей двух белков, где нуклеотидная последовательность, кодирующая первый белок, клонирована в одну плазмиду, а нуклеотидная последовательность, кодирующая второй белок,- в другую плазмиду, где нуклеотидные последовательности, кодирующие белки, слиты с нуклеотидными последовательностями, кодирующими активационный и ДНК-связывающий домены белка GAL4, при этом нуклеотидные последовательности, кодирующие исследуемые белки, отделены от нуклеотидных последовательностей, кодирующих активационный или ДНК-связывающий домен белка GAL4, при помощи нуклеотидной последовательности, кодирующей пептид, представляющий собой последовательность GluLeuGluAlaAlaAlaLysGluAlaAlaAlaLysGluAlaAlaAlaLysGluAlaAlaAla, который экспрессируется в виде белкового мостика между исследуемым белком и доменами белка GAL4.
| US 20070031815 A1, 08.02.2007 | |||
| NAC1-ГЕН РАСТЕНИЯ, КОДИРУЮЩИЙ ФАКТОР ТРАНСКРИПЦИИ, УЧАСТВУЮЩИЙ В РАЗВИТИИ СЕМЯДОЛИ И БОКОВОГО КОРНЯ | 1999 |
|
RU2241748C2 |
| МОДУЛЯТОРЫ НЕЙРОНАЛЬНОЙ РЕГЕНЕРАЦИИ | 2007 |
|
RU2447449C2 |
| ДВУГИБРИДНАЯ СИСТЕМА, ОСНОВАННАЯ НА ОБЕСПЕЧЕНИИ УМОЛЧАНИЯ ГЕНОВ ПУТЕМ ТРАНСКРИПЦИОННОЙ ИНТЕРФЕРЕНЦИИ | 2005 |
|
RU2403287C2 |
| EP 1975620 A2, 01.10.2008 | |||

