Настоящее изобретение относится к новой двугибридной системе, основанной на обеспечении умолчания генов путем транскрипционной интерференции. Оно также относится к применению этой системы для идентификации биологически активных соединений, оказывающих влияние на взаимодействие белок-белок.
Двугибридная система (Y2H: «Yeast Two hybrid system») была использована экстенсивным образом в случае дрожжей для идентификации взаимодействий белок-белок (см., в частности, Chien C.T. и др., 1991; Fields S. и O. Song, 1989; Cagney G. и др., 2000; Causier B. и B. Davis, 2002; Coates P.J. и P.A. Hall, 2003).
Двугибридная технология основана на том факте, что ДНК-связывающий домен и домен активации транскрипции, содержащиеся в разных химерных белках, могут активировать транскрипцию гена, когда эти два домена являются достаточно близкими друг к другу. Обычно используемый вариант двугибридной системы у дрожжей является полезным в отношении свойств белка GAL4p дрожжей Saccharomyces cerevisiae (Fields. S и O. Song, 1989).
Белок GAL4p, кроме того, активирует транскрипцию гена GAL1, необходимого для использования галактозы дрожжами, и образован двумя доменами: N-концевой домен, который специфически связывает последовательность ДНК (UASG, активируемая, расположенная выше хода транскрипции последовательность генов дрожжей GAL), и С-концевой домен, содержащий кислотную область, позволяющую осуществляться транскрипционной активации.
В этой первой двугибридной системе принимают участие три активных компонента: два гибридных белка и конструкция, в которой ген-репортер находится под контролем промотора гена GAL1 (Pgal1). Первый гибридный белок с ДНК-связывающим доменом (DBD, ДНК-связывающий домен) белка GAL4p экспрессируется дрожжами S.cerevisiae, несущими вышеуказанную конструкцию. Второй гибридный белок с доменом активации транскрипции (AD, активация транскрипции) GAL4p соэкспрессируется в тех же самых дрожжах. В случае взаимодействия двух гибридных белков сближение домена активации транскрипции GAL4-AD и ДНК-связывающего домена позволяет осуществляться реконструкции функционального фактора транскрипции GAL4p, что, в свою очередь, позволяет индуцировать экспрессию гена-репортера, находящегося под контролем Pgal1.
Выбор в качестве гена-репортера гена, существенного для жизнеспособности дрожжей (как, например, ген HIS3), позволяет легко осуществлять скрининг штаммов, которыми экспрессируется ген-репортер и, следовательно, в случае которых происходит взаимодействие между двумя гибридными белками. Штаммы, в случае которых такое взаимодействие не имеет места, не могут активировать Pgal1 и не могут расти, если геном-репортером является ген, необходимый для жизнеспособности дрожжей.
Недавно были получены так называемые обратные двугибридные системы (rY2H: «reverse Yeast Two hybrid system»). В этих системах активация гена-репортера индуцируется в отсутствие взаимодействия белок-белок, что является пригодным, в частности, для идентификации мутантных белков, утративших способность взаимодействовать со своим партнером, а также для идентификации молекул, специфически ингибирующих взаимодействие белок-белок.
Так, Young и др. разработали высокоэффективный тест скрининга, основанный на обратной двугибридной системе, для идентификации модуляторов кальциевых каналов (Young K. и др., 1998). В этой системе rY2H используются свойства гена CYH2. Ген CYH2 кодирует рибосомный белок L29, компонент рибосомной субъединицы 60S, который придает клеткам чувствительность к циклогексимиду (Kaufer N.F. и др., 1983). Репортерную кассету Pgal1-CYH2 вводят в резистентный к циклогексимиду штамм дрожжей, обладающий эндогенным мутированным геном CYH2. При доминировании аллеля CYH2 дикого типа рост клетки-репортера ингибируется на среде, содержащей циклогексимид, когда происходит взаимодействие между двумя белками (и, следовательно, когда репортерная кассета является транскриптом). Прерывание взаимодействия уменьшает токсический эффект циклогексимида и позволяет клетке-репортеру расти на среде, содержащей циклогексимид (Leanna C.A. и M. Hannink, 1996).
Однако проблемы, связанные со стабильностью мРНК и белка, а также различиями на уровне активности белков CYH2p дикого типа и мутированных делают эту систему сложной для использования и довольно малочувствительной для детектирования прекращения взаимодействий белок-белок (обратное Y2H взаимодействие).
Эта система, кроме того, обладает тем недостатком, что ее относительно трудно использовать в целях достижения максимальной чувствительности, так как ее ответ по отношению к обратным Y2H взаимодействиям зависит от нескольких молекул. Так, конкуренция между CYH2p дикого типа и мутантным CYH2p на уровне включения в рибосомы влияет на чувствительность репортера. Кроме того, флуктуации на уровне захвата циклогексимида и его реактивности в отношении рибосом с высокой степенью вероятности влияют на ответную способность системы.
Также описана другая обратная двугибридная система, основанная на использовании гена-репортера URA3 в комбинации с его протоксическим субстратом: 5-фтороротовая кислота (5-FOA) (Huang J. и S.L. Schraiber, 1997; Vidal M. и др., 1996; WO-96/32503). В этой системе индукция экспрессии белка URA3p ингибирует клеточный рост, так как 5-FOA тогда превращается в свой токсический аналог: 5-фтор-UTP. Инактивация URA3, напротив, способствует клеточному росту, так как 5-FOA тогда более не метаболизируется и клетки используют введенный в среду урацил. Эту систему использовали для идентификации мутантных белков, утративших свою способность взаимодействовать с их обычным партнером (Burke T.W. и др., 2001; Daros J.A. и др., 1999; Puthalakath H. и др., 1999).
Конфигурация и механизм этой системы подобны таковым обратной двугибридной системы, описанной выше (Young K. и др., 1998). Следовательно, эта система обладает вышеуказанными недостатками. Кроме того, система, основанная на использовании 5-FOA, обладает тем недостатком, что генерирует позитивные имитации, связанные с самой же системой. В самом деле, клеточный рост связан с инактивацией URA3 дикого типа. Следовательно, соединения, ингибирующие белок URA3p, способствуют клеточному росту. Эти соединения, таким образом, генерируют ложный положительный сигнал во время использования системы для идентификации молекул, специфически ингибирующих взаимодействие белок-белок.
Эта система также обладает тем недостатком, что в ней используется 5-FOA, которая представляет собой относительно дорогостоящий химический реагент.
Наконец, также описана промежуточная система, включающая каскад из двух генов-репортеров и так называемый «расщепленный гибрид» (WO-95/26400 и WO-98/13502). Эта система rY2H была использована для идентификации мутаций в белке CREB, которые прерывают его ассоциацию с СВР (Crispino J.D. и др., 1999). В этих системах двугибридное взаимодействие активирует экспрессию первого белка-репортера, который, в свою очередь, контролирует экспрессию второго гена-репортера, используемого для селекции роста. Эта система, однако, является сложной для использования и довольно малочувствительна для детектирования обратных Y2H взаимодействий.
Вышеописанные системы, следовательно, зависят от активности нескольких молекул (два гена-репортера, ген-репортер и токсическое вещество, ген-репортер дикого типа и мутированный ген-репортер). Это повышает сложность их осуществления, в частности, когда стремятся к максимальной чувствительности. Желательно иметь систему, одновременно более простую, более экономичную, обладающую очень хорошей чувствительностью.
Общее описание изобретения
Настоящее изобретение относится к новой двугибридной системе, основанной на обеспечении умолчания гена путем транскрипционной интерференции. Двугибридная система согласно настоящему изобретению является простой, не требует добавления в среду токсических веществ и может быть использована без необходимости интеграции в геном.
Первый объект изобретения относится к клетке, включающей интерференционную конструкцию ДНК, причем вышеуказанная конструкция включает:
- ген-репортер, находящийся под контролем первого промотора;
- один или несколько индуцируемый(ых) промотор(ов), так называемый(ых) промотор(ов) интерференции, выбираемый(ых) и фиксированный(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера;
причем вышеуказанная клетка, кроме того, экспрессирует:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированный с белком Y, способным взаимодействовать с белком-партнером Х;
- второй химерный белок (Х-DBD), образованный первым, ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Х-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции.
Согласно одному варианту осуществления объектом настоящего изобретения является клетка, такая как описанная выше, промотор которой, регулирующий экспрессию гена-репортера, представляет собой индуцируемый промотор, белок Y которого способен взаимодействовать с двумя белками-партнерами Х и Z, причем клетка, кроме того, экспрессирует третий химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
Согласно предпочтительному варианту осуществления, объектом настоящего изобретения является клетка, такая как указанная выше, индуцируемый, регулирующий экспрессию гена-репортера промотор которой включает последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Z-DBD).
Объектом изобретения также является клетка, такая как указанная выше, промотор которой, регулирующий экспрессию гена-репортера, представляет собой конститутивный промотор.
Изобретение также относится к клетке, такой как описанная выше, отличающейся тем, что она представляет собой клетку-хозяин, трансформированную или трансфицированную с помощью интерференционной конструкции ДНК и конструкций ДНК, кодирующих химерные белки, причем совокупность этих конструкций несут один или несколько неинтегративных векторов.
Другой аспект изобретения относится к клетке, такой как указанная выше, отличающейся тем, что она представляет собой клетку-хозяин, трансформированную или трансфицированную конструкциями ДНК, кодирующими химерные белки, причем эти конструкции несут один или несколько неинтегративных векторов, и тем, что интерференционная конструкция ДНК интегрирована в геном клетки.
Изобретение относится также к клетке, такой как указанная выше, в случае которой интерференционная конструкция ДНК и конструкции ДНК, кодирующие химерные белки, интегрированы в геном клетки.
Согласно предпочтительному варианту осуществления настоящее изобретение относится к клетке, такой как описанная выше, в случае которой интерференционная конструкция ДНК интегрирована в локус, лишенный пертурбированных геномных активностей транскрипции.
Изобретение также относится к клетке, такой как указанная выше, отличающейся тем, что ее выбирают из группы, состоящей из клеток млекопитающих, насекомых, растений и дрожжей.
Объектом изобретения также является клетка, такая как описанная выше, отличающаяся тем, что она представляет собой дрожжевую клетку.
Согласно предпочтительному варианту осуществления изобретение относится к клетке, такой как указанная выше, отличающейся тем, что она является клеткой дрожжей вида Saccharomyces cerevisiaе, Schizosaccharomyces pombe, Kluyveromyces lactis, Pichia pastoris, Saccharomyces carlsbergensis или Candida albicans.
Изобретение относится также к клетке, такой как указанная выше, по меньшей мере с одним промотором интерференции, расположенным ниже по ходу транскрипции гена-репортера и первого промотора и в ориентации, противоположной этому последнему (DI).
Изобретение относится, кроме того, к клетке, такой как описанная выше, по меньшей мере с одним промотором интерференции, расположенным ниже по ходу транскрипции гена-репортера и первого промотора и в той же ориентации, что и таковая этого последнего (nDI).
Изобретение также относится к клетке, такой как указанная выше, по меньшей мере с одним промотором интенференции, расположенным выше хода транскрипции гена-репортера и первого промотора и в той же самой ориентации, что и таковая этого последнего (UI).
Другой аспект изобретения относится к клетке, такой как описанная выше, по меньшей мере с одним промотором интерференции, фиксированным с той и другой стороны первым промотором и геном-репортером, промотором или промоторами интерференции, фиксированным(ми) ниже по ходу транскрипции первого промотора и гена-репортера, обладающим(ми) ориентацией, конвергентной по отношению к первому промотору, и промотором или промоторами интерференции, фиксированным(ми) выше хода транскрипции первого промотора и гена-репортера, обладающим(ми) ориентацией, идентичной таковой первого промотора (UDI).
Изобретение относится, кроме того, к клетке, такой как описанная выше, по меньшей мере с одним промотором интерференции, фиксированным с той и другой стороны первым промотором и геном-репортером, промотором или промоторами интерференции, фиксированным(ми) ниже по ходу транскрипции первого промотора и гена-репортера, обладающим(ми) ориентацией, тандемной по отношению к первому промотору, и промотором или промоторами интерференции, фиксированным(ми) выше хода транскрипции первого промотора и гена-репортера, обладающим(ми) ориентацией, идентичной таковой первого промотора (nUDI).
Согласно предпочтительному варианту осуществления изобретение относится к клетке, такой как указанная выше и геном-репортером которой является ген, необходимый для выживания клетки.
Изобретение также относится к клетке, такой как описанная выше и геном-репортером которой является ген, необходимый для первичного метаболизма, клеточного деления, синтеза белков, синтеза ДНК или синтеза РНК.
Объектом изобретения также является клетка, такая как указанная выше, ген-репортер которой сам по себе не является необходимым для выживания клетки, но является необходимым для выживания клетки, когда ее транскрипция ингибирована, за счет ассоциации с одним или несколькими генами-репортерами одного и того же типа, экспрессия которых контролируется или нет системой транскрипционной интерференции.
Согласно предпочтительному варианту осуществления изобретение относится к клетке, такой как описанная выше, индуцируемый промотор или индуцируемые промоторы интерференции которой включают последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Х-DBD).
Изобретение относится также к клетке, такой как указанная выше, индуцируемый промотор или индуцируемые промоторы интерференции которой включают последовательность, способную взаимодействовать с белком, имеющим ДНК-связывающий домен (DBD), выбираемый из группы, состоящей из: GAL4 UAS, LexAop, cIop и TetRop, и ДНК-связывающим доменом химерного белка Х-DBD которой является соответствующий DBD (соответственно, GAL4, LexA, cI и TetR).
Другой аспект изобретения относится к клетке, такой как описанная выше, домен активации транскрипции (AD) химерного белка Y-AD которой выбирают из группы, состоящей из доменов активации транскрипции следующих белков: В42, VP16 и GAL4p.
Согласно предпочтительному варианту осуществления объектом изобретения является клетка, такая как указанная выше, интерференционная конструкция ДНК которой фланкирована на своих концах одним или несколькими, одно- или двудирекционными, терминаторами транскрипции.
Другой аспект изобретения относится к способу идентификации соединения, ингибирующего взаимодействие первого белка Х со вторым белком Y, включающему следующие стадии:
а) культивирование клеток, таких как указанные выше;
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
с) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка Х с белком-партнером Y, экспрессируемых культивированными клетками.
Изобретение относится, кроме того, к применению клеток, таких как указанные выше, для идентификации соединений, ингибирующих взаимодействие белок-белок.
Объектом изобретения является также применение клеток, таких как указанные выше, для скрининга банков кДНК или банков пептидов с целью идентификации пептидов или белковых факторов, специфически ликвидирующих взаимодействие белок-белок.
Другой аспект изобретения относится к набору для получения двугибридной системы, включающему:
- первую конструкцию ДНК, включающую:
- ген-репортер, находящийся под контролем первого промотора;
- один или несколько индуцируемых промоторов, выбираемый(ых) и фиксированный(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера;
- вторую конструкцию ДНК, кодирующую:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с первым партнером Y, способным взаимодействовать с белком-партнером Х;
- третью конструкцию ДНК, кодирующую:
- второй химерный белок (Х-DBD), образованный первым ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции, когда два химерных белка экспрессируются в клетке-хозяине.
Изобретение относится также к набору, такому как указанный выше, отличающемуся тем, что промотором, регулирующим экспрессию гена-репортера, является индуцируемый промотор; белок Y способен взаимодействовать с двумя белками-партнерами Х и Z; и вышеуказанный набор включает четвертую конструкцию ДНК, кодирующую третий химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
Следующий аспект изобретения относится к способу идентификации соединения, ингибирующего взаимодействие первого белка Х со вторым белком Y, но не ингибирующего или менее ингибирующего взаимодействие между белком Y и третьим белком Z, включающему следующие стадии:
а) культивирование клеток, таких как указанные выше;
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
с) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка Х с белком-партнером Y, но что этот продукт не ингибирует или менее ингибирует взаимодействие между белком Y и белком Z.
Изобретение относится, наконец, к вектору интеграции дрожжей, включающему два фрагмента, гомологичных областям, расположенным выше хода транскрипции и ниже по ходу транскрипции открытой рамки считывания гена URA3 S. cerevisiae, и способствующих интеграции путем гомологичной рекомбинации на уровне локуса URA3 последовательности, встраиваемой между этими двумя фрагментами.
Подробное описание изобретения
Настоящее изобретение относится к новой двугибридной системе, основанной на обеспечении умолчания гена путем транскрипционной интерференции. Двугибридная система согласно настоящему изобретению является простой, не требует добавления в среду токсических веществ и может быть использована без необходимости интеграции в геном.
Транскрипционную интерференцию определяют как пертурбацию активности первого промотора, когда второй промотор активирован. Следовательно, активность транскрипции одного промотора уменьшает транскрипцию, инициированную на уровне другого промотора. Транскрипционная интерференция описана, например, в случае дрожжей (Greger I.H. и N.J. Proudfoot, 1998; Springer C.O. и др., 1997; Peterson J.A. и A.M. Myers, 1993; Puig S. и др., 1999; Martens и др., 2004).
Первый объект изобретения относится к клеткам, несущим одну или несколько конструкций, позволяющих осуществляться экспрессии гена-репортера в отсутствие особого взаимодействия белок-белок, тогда как, когда это взаимодействие имеет место, констатируют поддающееся обнаружению уменьшение экспрессии вышеуказанного гена-репортера.
Более конкретно, клетки согласно настоящему изобретению включают одну или несколько конструкций ДНК, так называемых интерференционных конструкций, причем вышеуказанная конструкция или вышеуказанные конструкции включают:
- ген-репортер, находящийся под контролем первого промотора;
- один или несколько индуцируемый(ых) промотор(ов), так называемый(ых) промотор(ов) интерференции, выбираемый(ых) и фиксированный(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера;
причем вышеуказанная клетка, кроме того, экспрессирует:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с белком Y, способным взаимодействовать с белком-партнером Х;
- второй химерный белок (Х-DBD), образованный первым, ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Х-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции.
Согласно первому варианту осуществления клетки согласно настоящему изобретению представляют собой клетки-хозяева, трансформированные или трансфицированные с помощью конструкции ДНК, включающей ген-репортер (интерференционная конструкция), и они также трансформированы или трансфицированы двумя конструкциями ДНК, кодирующими каждый из двух химерных белков. Три конструкции может нести один и тот же вектор ДНК или они могут находиться на двух или трех различных векторах. Все комбинации предусматриваемы: три конструкции на одном и том же векторе; три конструкции на трех разных векторах; две конструкции, кодирующие два химерных белка, на одном и том же векторе, тогда как третью конструкцию несет второй вектор; конструкция, кодирующая один из двух химерных белков, и интерференционная конструкция на одном и том же векторе, тогда как конструкцию, кодирующую второй химерный белок, несет второй вектор. Согласно этому первому варианту осуществления векторы представляют собой неинтегративные векторы.
Согласно второму варианту осуществления изобретения интерференционная конструкция интегрирована в геном клеток и эти последние трансформированы или трансфицированы двумя конструкциями ДНК, кодирующими каждый из двух химерных белков. Конструкции ДНК, кодирующие химерные белки, могут находиться на одном и том же векторе ДНК или находиться на двух различных векторах, причем этот вектор или эти векторы являются неинтегративными векторами.
Согласно третьему варианту осуществления изобретения интерференционная конструкция, а также конструкции ДНК, кодирующие каждый из двух химерных белков, интегрированы в геном клеток. Также возможно, что одна из двух конструкций, кодирующих один из двух химерных белков, интегрирована в геном, а вторая конструкция находится на неинтегративном векторе. Наконец, согласно последнему варианту осуществления интерференционная конструкция и одна из двух конструкций, кодирующих один из двух химерных белков, находятся на двух неинтегративных векторах, тогда как конструкция, кодирующая второй химерный белок, интегрирована в геном клеток.
Ген-репортер обычно определяют как ген, кодирующий легко поддающийся обнаружению продукт. Согласно предпочтительному варианту осуществления геном-репортером, используемым в рамках настоящего изобретения, является ген, экспрессия которого необходима для выживания клетки. Она может представлять собой, например, ген, необходимый для первичного метаболизма, клеточного деления, синтеза белков (как рибосомы), синтеза ДНК или синтеза РНК и т.д.
В качестве примера этого типа гена можно назвать все гены, описанные как необходимые для выживания дрожжей, то есть примерно 1/6 часть генов, образующих их геном (Winzeler E.F. и др., 1999).
Также он может представлять собой ген, принимающий активное участие в первичном метаболизме и необходимый для этого последнего, или ген, необходимый для клеточного деления. Более конкретно, он представляет собой ген, кодирующий фермент, участвующий в биосинтезе необходимого метаболита, который может быть добавлен в культуральную среду или удален из нее.
В качестве примера этого типа гена можно назвать гены HISS3, URA3, LEU2, LYS2, TRP1, ADE2, MET15 и ARG4, широко используемые в случае дрожжей S.cerevisiae, так как легко комплементабельны по отношению к соответствующей аминокислоте или соответствующему основанию нуклеиновой кислоты в культуральной среде дрожжей. Могут быть использованы другие маркеры, основанные на метаболизме, в качестве примера можно назвать ген HIS5 (гены дрожжей S.pombe), гены URA3 и LEU2 (гены дрожжей K.lactis), URA3 (ген дрожжей C.albicans), LEU2 (ген дрожжей A.gossypii).
Также могут быть использованы гены, принимающие активное участие в первичном метаболизме и не являющиеся необходимыми для этого последнего, или гены, не являющиеся необходимыми для клеточного деления, когда они используются индивидуально. Напротив, если их транскрипция ингибирована за счет ассоциации с одним или несколькими генами-репортерами одного и того же типа, ассоциация эффектов может приводить к ингибированию необходимой для выживания дрожжей функции. Может быть, например, использована совокупность генов, «ассоциированные» роли которых приводят к функции, необходимой для первичного метаболизма, клеточного деления, синтеза белков (как рибосомы), синтеза ДНК или синтеза РНК и т.д.
В качестве примера этого типа гена можно назвать все гены, описанные как не являющиеся необходимыми для выживания дрожжей (то есть примерно 5/6 числа генов, которые образуют их геном (Winzler E.A. и др., 1999)), которые за счет ассоциации с одним или несколькими генами одного и того же типа становятся тогда необходимыми.
Однако в рамках настоящего изобретения могут быть также использованы другие гены. В качестве примера можно назвать ген, кодирующий фермент САТ (хлорамфениколацетилтрансфераза), ген luc, кодирующий люциферазу, гены, кодирующие флуоресцирующий белок, как GFP («Green Fluorescent Protein»), CFP («Сyan Fluorescent Protein»), YFP («Yellow Fluorescent Protein») или RFP («Red Fluorescent Protein»), бета-галактозидазу, бета-лактамазу, бета-глюкуронидазу.
Также может быть использован маркер, придающий резистентность к токсичному продукту: ген kan транспозона Tn903 (придающий резистентность к генетицину (G418)), ген ble транспозона Tn5 (придающий резистентность к флеомицину), ген CYHr (придающий резистентность к циклогексимиду), pat (ген S.viridochromogenes, придающий резистентность к биалафосу), nat1 (ген S.noursei, придающий резистентность к нурзеотрицину), hph (ген E.coli, придающий резистентность к гигромицину В). Все эти гены являются функциональными, особенно у дрожжей (Gutherie и Fink, 2002).
Ген-репортер находится под контролем первого промотора. Согласно предпочтительному варианту осуществления, этот первый промотор представляет собой конститутивный промотор. Конститутивным промотором, как его обычно называют, является промотор, способствующий экспрессии, относительно независимой от окружающих условий.
В качестве примера в случае дрожжей ген-репортер может быть экспрессирован под контролем мутантного конститутивного промотора ADH1(700) (мутированный промотор гена ADH1 (Padh1)) (Ruohonen и др., 1995). Однако в рамках настоящего изобретения могут быть использованы другие конститутивные промоторы дрожжей. В качестве примера можно назвать промотор TPI (Alber и Kawasaki, 1982), TEF1, TDH3, KEX2 (Nacken и др., 1996) и АСТ1 (Ernst, 1986).
В том, что касается клеток млекопитающих, в качестве примера можно назвать промотор CMV (предранний энхансер/промотор человеческого цитомегаловируса (CMV)) (Foecking и Hofstetter, 1986) (Kronman и др., 1992), промотор EF-1α (Mizushima и Nagata, 1990), промотор SV40 (ранний энхансер/промотор вируса обезьян 40) (Das и др., 1985), промотор UB (промотор человеческого гена убикитина С (hUbC)) (Nenoi и др., 1996; Schorpp и др., 1996), промотор RSV LTR (длинный концевой повтор на концах ДНК-копии генома вируса саркомы Рауса) (Yamamoto и др., 1980).
Промотор или промоторы, так называемый(ые) «промотор(ы) интерференции» представляют собой индуцируемые промоторы. Этот промотор или эти промоторы выбирают и фиксируют таким образом, что их активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера.
Промоторы интерференции, которые могут быть использованы в рамках настоящего изобретения, представляют собой индуцируемые промоторы, активация которых индуцируется фактором транскрипции, взаимодействующим через ДНК-связывающий домен (DBD) с последовательностью вышеуказанного промотора (последовательность ДНК, связанная с помощью DBD), причем вышеуказанный фактор транскрипции способен инициировать транскрипцию, управляемую промотором. Так, промоторами интерференции, которые могут быть использованы в рамках настоящего изобретения, являются все промоторы, пригодные для получения двугибридной системы в случае рассматриваемого организма.
Предпочтительно промоторы интерференции согласно настоящему изобретению включают последовательность, способную взаимодействовать с белком, обладающим ДНК-связывающим доменом (DBD) (последовательность ДНК, связанная с помощью DBD (UAS)). Более конкретно, индуцируемый промотор или индуцируемые промоторы интерференции включает(ют) последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Х-DBD).
Речь может идти, например, о промоторах Pgal1, Pgal10, Pgal5, Pgal80, Pmel1, Pgal2, Pgal7, Pgal3, Pgcyl, Plth1, Ppcl10 и Pfur4 (Ren и др., 2000; Svetlov и Cooher, 1995), индуцируемых фактором транскрипции GAL4p (в клетке дикого типа фактор транскрипции GAL4p принимает участие, в частности, в экспрессии генов, индуцируемых галактозой). Предпочтительно используемым индуцируемым промотором является промотор Pgal1.
Однако согласно другим вариантам осуществления изобретения в рамки настоящего изобретения также входит использование других систем индуцируемой экспрессии. В таблице 1 представлено некоторое количество индуцируемых систем, уже использованных в двугибридных системах. Индуцируемые промоторы этих систем, описанных в литературе, представляют собой также промоторы интерференции в смысле настоящего изобретения. С целью получения функциональной двугибридной системы предусматриваемыми являются все возможные комбинации между DBDs и Ads, представленные в таблице 1.
Индуцируемый промотор или индуцируемые промоторы интерференции фиксированы таким образом, что их активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера.
Промотор или промоторы интерференции также могут быть фиксированы ниже по ходу транскрипции первого промотора и гена-репортера, тогда речь идет об интерференции в прямом направлении, так как активация промотора интерференции, расположенного ниже по ходу транскрипции, интерферирует с активностью первого промотора, расположенного выше хода транскрипции.
В рамках интерференции в прямом направлении промотор интерференции, однако, может иметь две возможные ориентации. Первая ориентация соответствует ориентации, конвергентной по отношению к первому промотору; эта конфигурация была названа как интерференция в прямом направлении (или DI, «Downstream Interference»). Промотор интерференции также может иметь ориентацию, тандемную по отношению в первому промотору; эта вторая конфигурация была названа как несмысловая интерференция в прямом направлении (или nDI, «non-sense Downstream Interfere-nce»), которая показывает также ингибирование экспрессии гена-репортера (см. относящуюся к примеру часть).
Промотор или промоторы интерференции также могут быть фиксированы выше хода транскрипции первого промотора и гена-репортера; тогда речь идет об интерференции против хода транскрипции, так как активация промотора интерференции, расположенного выше хода транскрипции, интерферирует с активностью первого промотора, расположенного ниже по ходу транскрипции. Промотор интерференции тогда имеет ориентацию, идентичную таковой первого промотора (оба промотора, следовательно, имеют тандемную конфигурацию); эта конфигурация была названа как интерференция против хода транскрипции (или UI, «Upstream Interference»).
Другой возможной конфигурацией является комбинация двух вышеуказанных конфигураций (система UI и DI). В этом случае по меньшей мере один индуцируемый промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером. Речь идет тогда об интерференции в прямом-обратном направлении, так как активация промотора(ов) интерференции, расположенного(ных) выше хода транскрипции, интерферирует с активностью первого промотора, расположенного ниже по ходу транскрипции, и активация промотора или промоторов интерференции, расположенного(ных) ниже по ходу транскрипции, интерферирует с активностью первого промотора, расположенного выше хода транскрипции.
Промотор или промоторы интерференции, расположенный(ые) ниже по ходу транскрипции первого промотора и гена-репортера, имеют ориентацию, конвергентную по отношению к первому промотору (см. система DI выше). Промотор или промоторы интерференции, фиксированный(ые) выше хода транскрипции первого промотора и гена-репортера, обладает(ют) ориентацией, идентичной таковой первого промотора (оба промотора, следовательно, имеют тандемную конфигурацию) (см. система UI выше).
Эта конфигурация была названа интерференцией в прямом-обратном направлении (или UDI, «Upstream-Downstream Interfere-nce».
Другой возможной конфигурацией является комбинация двух вышеуказанных конфигураций (система UI и nDI). В этом случае по меньшей мере один индуцируемый промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером. Речь идет тогда об интерференции в прямом-обратном направлении, так как активация промотора или промоторов интерференции, расположенного(ных) выше хода транскрипции, интерферирует с активностью первого промотора, расположенного ниже по ходу транскрипции, и активация промотора или промоторов интерференции, расположенного(ных) ниже по ходу транскрипции, интерферирует с активностью первого промотора, расположенного выше хода транскрипции.
Промотор или промоторы интерференций, расположенные ниже по ходу транскрипции первого промотора и гена-репортера, имеют ориентацию, тандемную по отношению к первому промотору (см. система nDI выше). Промотор или промоторы интерференции, фиксированный(ые) выше хода транскрипции первого промотора и гена-репортера, имеет(имеют) ориентацию, идентичную таковой первого промотора (см. система UI выше), следовательно, три промотора имеют тандемную конфигурацию.
Эта конфигурация была названа как несмысловая в прямом-обратном направлении интерференция (или nUDI, «non-sense Upstream-Downstream Interference»).
В случае конструкции типа UI активация промотора интерференции приводит к снижению активности первого промотора вследствие транскрипционной интерференции. Активность промотора интерференции, расположенного выше хода транскрипции, однако, может еще способствовать продуцированию смыслового транскрипта гена-репортера.
С целью предотвращения трансляции этого функционального смыслового транскрипта короткую открытую рамку считывания, сопровождаемую одним или несколькими (например, 2 или 3) стоп-кодонами, предпочтительно встраивают между промотором интерференции и первым промотором. Транскрипционная интерференция в системе UI тогда продуцирует бисцитронную мРНК, из которой трансдуцируется только первая короткая открытая рамка считывания.
Это также имеет место в случае конфигурации типа UDI и nUDI. В самом деле, в случае конструкции типа UDI или nUDI активация промотора или промоторов интерференции, расположенного(ных) выше хода транскрипции первого промотора, приводит к снижению активности первого промотора вследствие транскрипционной интерференции. Активность промотора интерференции, расположенного выше хода транскрипции, однако, позволяет еще продуцировать смысловой транскрипт гена-репортера.
С целью предотвращения трансляции этого функционального смыслового транскрипта короткую открытую рамку считывания, сопровождаемую одним или несколькими стоп-кодонами, предпочтительно встраивают между промотором интерференции и первым промотором. Транскрипционная интерференция в системе UDI или nUDI тогда продуцирует бисцитронную мРНК, из которой трансдуцируется только первая короткая открытая рамка считывания.
Предпочтительно вышеописанные конструкции интерференций фланкированы на своих концах двудирекционными терминаторами интерференции. Для фланкирования конструкций согласно изобретению может быть использован любой функциональный и двудирекционный терминатор транскрипции. В качестве примера может быть использован терминатор транскрипции гена CYCI (Tcycl) (описан Osborne B.I. и L. Guarente, 1989), а также терминатор гена ADHI (Tadh1) (описан Iringer S. и др., 1991).
Двудирекционные терминаторы транскрипции блокируют транскрипцию в двух направлениях. Это предотвращает распространение транскрипции, регулируемой первым промотором или промотором или промоторами интерференции, сверх конструкции и также защищает систему от возможного влияния активностей транскрипции, расположенных вне конструкции.
С той же самой целью также можно комбинировать однодирекционные терминаторы в виде конфигурации, которая блокирует транскрипцию в двух направлениях (комбинируя два однодирекционных терминатора, блокирующих транскрипцию, в дивергентной или конвергентной ориентации, чтобы получить двудирекционный терминатор). В качестве примера однодирекционных терминаторов можно назвать следующие терминаторы: Tpgk1 (Picard и др., 1990), Ttef (A.gossypii) (Steiner и Philippsen, 1994), This3 (S.kluyveri) (Weinstock и Strathem, 1993).
Промотор интерференции согласно настоящему изобретению представляет собой индуцируемый промотор, и его выбирают и фиксируют таким образом, что его активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера.
Индуцируемый промотор активируется обычным образом. В рамках настоящего изобретения он активируется фактором транскрипции, когда он является активным. Согласно настоящему изобретению фактор транкрипции, активирующий индуцируемый промотор, образован совокупностью двух химерных белков (X-DBD и Y-AD), реконструирующих активный фактор транскрипции, способный активировать индуцируемый промотор, когда имеет место взаимодействие между двумя химерными белками X-DBD и Y-AD.
Как было уточнено выше, клетки согласно настоящему изобретению экспрессируют два химерных белка:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с первым партнером Y, способным взаимодействовать с белком-партнером Х;
- второй химерный белок (Х-DBD), образованный первым ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции.
ДНК-связывающий домен (DBD) второго химерного белка X-DBD, следовательно, выбирают в зависимости от индуцируемого промотора, чтобы связывать ДНК на уровне вышеуказанного промотора. Предпочтительно ДНК-связывающий домен (DBD) этого химерного белка выбирают так, чтобы происходило взаимодействие с последовательностью, имеющейся в вышеуказанном промоторе (последовательность ДНК, связанной с помощью DBD (UAS) (активирующая против хода транскрипции последовательность).
Домен активации транскрипции первого химерного белка Y-AD позволяет активировать транскрипцию индуцируемого промотора в клетке. Так, промотор или промоторы интерференций (индуцируемые промоторы) индуктируются функциональным фактором транскрипции, образованным совокупностью двух химерных белков Y-AD и X-DBD (взаимодействующих между собой), причем этот функциональный фактор транскрипции взаимодействует через свой ДНК-связывающий домен (DBD) с промоторной последовательностью и способен инициировать транскрипцию, управляемую промотором через его домен активации транскрипции (AD).
Это представляет собой тот же принцип двугибридной технологии, и специалист в данной области сам способен выбирать многочисленные пары промотор/фактор транскрипции, позволяющие осуществлять настоящее изобретение.
В обычно используемом варианте двугибридной системы в случае дрожжей оказываются полезными свойства белка GAL4p дрожжей Saccharomyces cerevisiae (Fields S. и O. Song, 1989) и промотора Pgal1. Однако в рамках настоящего изобретения могут быть использованы другие описанные двугибридные системы, в частности таковые, указанные в таблице 1.
Белки-партнеры Х и Y, гибридизированные с ДНК-связывающим доменом (DBD) и доменом активации транскрипции (AD), соответственно, могут быть определены как любая пара белков, которые, гибридизированные с DBD и AD соответственно, взаимодействуют в клетке (даже если это не происходит в их исходном организме), приводя к образованию функционального фактора транскрипции (Х-DBD/Y-AD), активирующего индуцируемый промотор или индуцируемые промоторы, такой(такие), как описанные выше. Речь может идти о полных белках или фрагментах этих белков. Так, в рамках настоящего изобретения могут быть использованы химерные белки, генерирующие двугибридный сигнал. То есть химерные белки, два белка-партнера (или фрагменты белков-партнеров) которых взаимодействуют друг с другом с достаточной аффинностью в используемой клетке, образуя функциональный фактор транскрипции, активирующий индуцируемый промотор или индуцируемые промоторы, причем вышеуказанный фактор транскрипции связывает ДНК через домен связывания в первом химерном белке и активирует промотор или промоторы через домен активации транскрипции (AD) второго химерного белка.
Клетки согласно изобретению могут нести конструкцию ДНК и конструкции, кодирующие химерные белки, либо интегрированным образом, либо на неинтегрированном векторе. Установлено, что наилучших результатов достигают при использовании интегрированной интерференционной конструкции ДНК.
Предпочтительно интерференционная конструкция ДНК интегрирована в геном клетки-хозяина. Интерференционные конструкции преимущественно нацелены на локус, лишенный потенциально пертурбируемых геномных активностей транскрипции, с целью защиты систем интерференции от возможного влияния транскрипционных активностей, инициированных вне репортерной кассеты.
Для этого конструируют новый вектор интеграции дрожжей pRB2. Этот вектор интеграции нацеливает интерференционные конструкции на локус, лишенный потенциально пертурбируемых геномных активностей транскрипции (см. фиг.3).
Также показано, что не интегрированные в вектор системы интерференции согласно настоящему изобретению являются генетически стабильными, что приводит к отсутствию необходимости конструкции штаммов, обладающих интегрированными репортерными кассетами. Эта характеристика представляет большой интерес для скрининга лекарственного средства, так как, таким образом, можно легко оценивать многочисленные штаммы, обладающие разными мутациями, в отношении повышения проницаемости молекул маленького размера.
Клетки, используемые для осуществления двугибридной системы, являющейся объектом настоящего изобретения, представляют собой в целом клетку, способную экспрессировать химерные белки, необходимые для исследуемой двугибридной системы, описанной выше, и которые могут быть трансформированы или трансфицированы с помощью интерференционной конструкции ДНК, описанной выше. Эти клетки-хозяева являются эукариотическими или прокариотическими клетками-хозяевами, причем речь может идти о клетках млекопитающих, насекомых, растений или, более предпочтительно, дрожжей.
Согласно предпочтительному варианту осуществления клетки согласно настоящему изобретению выбирают из царства Fungi. Предпочтительно организмы царства Fungi выбирают из типа Ascomycetes, более предпочтительно их выбирают из подтипа Saccharomecotina, еще более предпочтительно их выбирают из класса Saccharomycetes или Schizosaccharomycetes, еще более предпочтительно их выбирают из отряда Saccharomycetales или Schizosaccharomycetales, еще более предпочтительно их выбирают из семейства Saccharomycetaceae или Schizosaccharomycetaceae, еще более предпочтительно их выбирают из рода Saccharomyces или Schizosaccharomyces, в высшей степени предпочтительно организмы царства Fungi согласно изобретению относятся к виду Saccharomyces cerevisiae или Schizosaccharomyces pombe. Дрожжи видов Kluyveromyces lactis, Pichia pastoris, Saccharomyces carlsbergensis или Candida albicans также являются клетками, входящими в рамки настоящего изобретения.
В противоположность системам, описанным вплоть до настоящего времени, двугибридные системы согласно настоящему изобретению не требуют добавки токсичных или нетоксичных веществ. Это является преимуществом, так как эти вещества, вероятно, оказывают дополнительные воздействия на физиологию клетки и могут интерферировать с детектированием некоторых обратных Y2H взаимодействий. Использование токсичного или нетоксичного вещества, однако, остается возможным. Например, может представлять интерес использование вещества, ингибирущего вырабатываемый геном-репортером продукт в случае использования гена-репортера, необходимого для выживания, например, гена, необходимого для первичного метаболизма, клеточного деления, синтеза белков (как рибосомы), синтеза ДНК или синтеза РНК и т.д. В самом деле, если ген-репортер еще в достаточной степени экспрессируется несмотря на транскрипционную интерференцию и если стремятся к полному отсутствию клеточного роста, может быть использовано вещество, ингибирующее вырабатываемый таким геном-репортером продукт, для повышения чувствительности двугибридной системы. Так, например, если геном-репортером является ген HIS3, клеточный рост интерференционных штаммов дрожжей (например, S.cerevisiae) может быть оценен при использовании среды, лишенной гистидина. В культуральную среду, в случае необходимости, может быть добавлен конкурентный ингибитор белка HIS3p (3-амино-1,2,4-триазол (3-АТ)) с целью обнаружения различий в уровне активности гена-репортера HIS3. Это может способствовать достижению гораздо большей чувствительности и калибровке системы (см. пример 6).
Если, однако, ген-репортер еще в достаточной степени экспрессируется несмотря на транскрипционную интерференцию и стремится к полному отсутствию клеточного роста, существует возможность использования в виде ассоциации других генов-репортеров, контролируемых или нет системами интерференции, описанными выше (UI, DI, nDI, UDI и nUDI), следствием чего является повышение чувствительности двугибридной системы. Даже частичное ингибирование экспрессии этих генов-репортеров приводит за счет добавления воздействий к полному ингибированию роста. Так, например, если геном-репортером является ген HIS3 и одним из других генов является ген URA3, причем оба контролируются системой интерференции, рост интерференционных дрожжей (например, S.cerevisiae) может быть оценен при использовании среды, лишенной гистидина и урацила. Тогда как в случае среды, лишенной только гистидина или урацила, может еще происходить рост дрожжей, обладающих частичной транскрипционной интерференцией генов-репортеров, на среде, лишенной одновременно урацила и гистидина, рост может быть полностью прекращен.
Двугибридная система согласно настоящему изобретению может иметь многочисленные применения. Так, она особенно применима для скрининга молекул, обладающих ингибирующей активностью в отношении взаимодействия белок-белок.
Идентификация взаимодействий белок-белок, принимающих участие в патологиях, представляет собой большой интерес, так как эти взаимодействия представляют собой потенциальные мишени для разработки новых лекарственных средств. Как только взаимодействие белок-белок, принимающее участие в заболевании, идентифицировано, часто представляет большой интерес иметь синтетические или природные молекулы, специфически приводящие к прекращению этого взаимодействия белок-белок, чтобы, с одной стороны, признать действительным существенность этого взаимодействия в комплексных биологических системах и располагать молекулами-кандидатами для разработки лекарственного средства (см. Vidal M. и H. Endoh, 1999).
Двугибридные системы согласно настоящему изобретению представляют собой простые генетические системы для скрининга ингибитора данного взаимодействия белок-белок. Так, в двугибридных системах, согласно настоящему изобретению молекулы, вызывающие прекращение взаимодействия белок-белок, провоцируют экспрессию гена-репортера, что позволяет осуществлять их идентификацию.
Может быть существенным выбор гена-репортера, экспрессия которого является необходимой для выживания клетки. Так, способствуя возобновлению экспрессии этого необходимого гена, молекулы, ингибирующие взаимодействие белок-белок, следовательно, могут быть легко идентифицированы по клеточному росту.
Так, скрининг искомого ингибитора взаимодействия белок-белок может быть осуществлен, например, путем диффузионных анализов на агаре с высокой производительностью (Young K. и др., 1998), которые позволяют оценивать взаимодействия по отношению к градиенту концентрации каждого ингибитора-кандидата.
Таким образом, объектом настоящего изобретения является способ идентификации соединения, ингибирующего взаимодействие первого белка Х со вторым белком Y, включающий следующие стадии:
а) культивирование клеток, таких как описанные выше;
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
с) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка Х с белком-партнером Y, экспрессируемых культивированными клетками.
Другим применением системы интерференции согласно настоящему изобретению является скрининг банков кДНК и банков пептидов для идентификации пептидов или белковых факторов, которые специфически вызывают прекращение исследуемого взаимодействия белок-белок (Zutshi R. и др., 1998).
Как только идентифицировано и установлено надлежащее взаимодействие белок-белок на уровне заболевания, представляет интерес охарактеризовывания структуры и регуляции обнаруженного взаимодействия. Идентификация мутаций на уровне каждого партнера пары взаимодействующих белков, которые прерывают взаимодействие, пригодна не только для исследования структурных компонентов взаимодействия, но и также в качестве возможности получения генетических инструментов, как трансдоминантные отрицательные мутанты, для характеристики функции in vivo (Serebriiskii I.G. и др., 2001). Это является особенно важным для белков, которые имеют многочисленных партнеров взаимодействия. Системы интерференции согласно настоящему изобретению могут быть использованы в скрининге путем мутагенеза для идентификации прерывающих взаимодействие мутаций. Банки взаимодействующих партнеров, подвергнутых мутагенезу, должны быть созданы и могут быть подвергнуты скринингу в высокопроизводительном масштабе в отношении репарации клеточного роста (Gutherie C. и G.R. Fink (ред.), 2002). С помощью этого подхода можно быстро идентифицировать совокупность мутантных белков, дефицитных на уровне взаимодействия, в частности мутантов, обладающих доминирующими отрицательными свойствами. Совокупность мутантов также может способствовать определению поверхностей белка, принимающих участие во взаимодействии белок-белок, и оказывать помощь биологу, специалисту в области структуры, в отношении улучшения структуры ингибиторов-кандидатов во время исследований зависимости структура-аффинность.
Объектом настоящего изобретения также является набор для получения двугибридной системы, такой как описанная выше, включающий:
- интерференционную конструкцию ДНК, такую как указанная выше;
- вторую конструкцию ДНК, кодирующую:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с первым партнером-белком Y, способным взаимодействовать с белком-партнером Х;
- третью конструкцию ДНК, кодирующую:
- второй химерный белок (Х-DBD), образованный первым ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции, когда два химерных белка экспрессируются в клетке-хозяине.
Три конструкции ДНК набора, описанного выше, могут находиться на одной и той же молекуле ДНК или на двух или трех разных молекулах ДНК. Три конструкции также может нести один и тот же вектор ДНК, или они могут находиться на двух или трех разных векторах. Две конструкции, кодирующие два химерных белка, например, могут находиться на одном и том же векторе, тогда как третью конструкцию несет второй вектор.
Согласно настоящему изобретению, таким образом, получают совокупность простых генетических систем для идентификации мутаций и молекул, которые индуцируют прерывание взаимодействий белок-белок. Настоящее изобретение представляет собой инструмент, пригодный для охарактеризовывания взаимодействий белок-белок и также для разработки новых лекарственных средств.
Настоящее изобретение также может оказаться очень полезным в случае взаимодействия белка Y с двумя другими белками X и Z и когда желательно идентифицировать молекулы, ингибирующие взаимодействие между Y и Х, но не ингибирующие взаимодействие между Y и Z. Клетки, позволяющие осуществляться такой селекции, несут, например, генетическую конструкцию, представленную на фиг.7, и экспрессируют, кроме того, по меньшей мере три химерных белка, таких как представленные на фиг.7. Эти клетки позволяют идентифицировать специфические ингибиторы взаимодействия между белком Х и белком Y, если белок Y взаимодействует также с другим белком Z.
Объектом изобретения, следовательно, также являются клетки, включающие конструкцию ДНК, так называемую интерференционную конструкцию, причем вышеуказанная конструкция включает:
- ген-репортер, находящийся под контролем первого индуцируемого промотора;
- один или несколько индуцируемый(ых) промотор(ов), так называемый(ых) промотор(ов) интерференции, выбираемый(ых) и фиксированных(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого индуцируемого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера, когда индуцирована экспрессия этого последнего;
причем вышеуказанная клетка, кроме того, экспрессирует:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированный с белком Y, способным взаимодействовать с двумя белками-партнерами Х и Z;
- второй химерный белок (Х-DBD), образованный первым, ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Х-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции;
- третий химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
Согласно первому варианту осуществления клетки согласно настоящему изобретению представляют собой клетки-хозяева, трансформированные или трансфицированные с помощью конструкции ДНК, включающей ген-репортер, (интерференционная конструкция), и они также трансформированы или трансфицированы тремя конструкциями ДНК, кодирующими каждый из трех химерных белков. Четыре конструкции может нести один и тот же вектор ДНК, или они могут находиться на двух или трех различных векторах. Все комбинации возможны: четыре конструкции на одном и том же векторе; четыре конструкции на четырех разных векторах; три конструкции, кодирующие три химерных белка, на одном и том же векторе, тогда как четвертую конструкцию (интерференционная конструкция) несет второй вектор; конструкция, кодирующая один из трех химерных белков, и интерференционная конструкция на одном и том же векторе, тогда как конструкцию, кодирующую два других химерных белка, несет второй вектор, и т.д. Согласно этому первому варианту осуществления векторы представляют собой неинтегративные векторы.
Согласно второму варианту осуществления интерференционная конструкция интегрирована в геном клеток и эти последние трансформированы или трансфицированы тремя конструкциями ДНК, кодирующими каждый из трех химерных белков. Конструкции ДНК, кодирующие химерные белки, могут находиться на одном и том же векторе ДНК или находиться на двух различных векторах, причем этот вектор или эти векторы являются неинтегративными векторами.
Согласно третьему варианту осуществления изобретения интерференционная конструкция, а также конструкции ДНК, кодирующие химерные белки, интегрированы в геном клеток. Также возможно, что только одна или две из трех конструкций, кодирующих один или два из трех химерных белков, интегрирована(ны) в геном и что конструкцию или конструкции несет неинтегративный вектор, тогда как интерференционная конструкция интегрирована в геном. Наконец, согласно последнему варианту осуществления интерференционную конструкцию и одну или две из конструкций, кодирующих один или два из трех химерных белков, несут неинтегративные векторы, тогда как конструкция, кодирующая третий химерный белок, интегрирована в геном клеток.
Геном-репортером является такой, как указанный выше. Ген-репортер находится под контролем индуцируемого промотора. Предпочтительно индуцируемый промотор, регулирующий экспрессию гена-репортера, включает последовательность, способную взаимодействовать с белком, обладающим ДНК-связывающим доменом (DBD) (последовательность ДНК, связанная с помощью DBD (UAS)). Более конкретно, индуцируемый промотор, регулирующий экспрессию гена-репортера, включает последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Z-DBD).
Промотор или промоторы, так называемые «промоторы интерференции», также являются индуцируемыми промоторами. Этот или эти последние выбирают и фиксируют таким образом, что их активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера, когда экспрессия этого последнего индуктирована.
Промоторы интерференции, которые могут быть использованы, представляют собой индуцируемые промоторы, такие как указанные выше. Предпочтительно промоторы интерференции, которые могут быть использованы, включают последовательность, способную взаимодействовать с белком, обладающим ДНК-связывающим доменом (DBD) (последовательность ДНК, связанная с помощью DBD (UAS)). Более конкретно, индуцируемые промоторы интерференции включают последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Х-DBD).
Промотор или промоторы интерференции могут быть фиксированы, как описано выше. Так, они могут быть фиксированы ниже по ходу транскрипции первого промотора и гена-репортера, тогда речь идет об интерференции в прямом направлении. Промотор или промоторы интерференции также могут быть фиксированы выше хода транскрипции первого промотора и гена-репортера, тогда речь идет об интерференции против хода транскрипции.
В рамках интерференции в прямом направлении промотор интерференции, однако, может иметь две возможные ориентации. Первая ориентация соответствует ориентации, конвергентной по отношению к первому промотору; эта конфигурация была названа как интерференция в прямом направлении (или DI, «Downstream Interference»). Промотор интерференции также может иметь ориентацию, тандемную по отношению в первому промотору; эта вторaя конфигурация была названа как несмысловая интерференция в прямом направлении (или nDI, «non-sense Downstream Interference»).
Промотор или промоторы интерференции также могут быть фиксированы выше хода транскрипции первого промотора и гена-репортера. Промотор интерференции тогда имеет ориентацию, идентичную таковой первого промотора (два промотора, следовательно, имеют тандемную конфигурацию); эта конфигурация была названа как интерференция против хода транскрипции (или UI, «Upstream Interference»).
Другой возможной конфигурацией является комбинация двух вышеуказанных конфигураций (система UI и DI). В этом случае по меньшей мере один (индуцируемый) промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером. Речь идет тогда об интерференции в прямом-обратном направлении, так как активация промотора или промоторов интерференции, расположенного(ных) выше хода транскрипции, интерферирует с активностью первого промотора, расположенного ниже по ходу транскрипции, и активация промотора или промоторов интерференции, расположенного(ных) ниже по ходу транскрипции, интерферирует с активностью первого промотора, расположенного выше хода транскрипции.
Промотор или промоторы интерференции, расположенный(ые) ниже по ходу транскрипции первого промотора и гена-репортера, имеют ориентацию, конвергентную по отношению к первому промотору (см. система DI выше). Промотор или промоторы интерференции, фиксированные(ые) выше хода транскрипции первого промотора и гена-репортера, обладает(ют) ориентацией, идентичной таковой первого промотора (два промотора, следовательно, имеют тандемную конфигурацию) (см. система UI выше).
Эта конфигурация была названа интерференцией в прямом-обратном направлении (или UDI, «Upstream-Downstream Interference»).
Другой возможной конфигурацией является комбинация двух вышеуказанных конфигураций (система UI и nDI). В этом случае по меньшей мере один индуцируемый промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером. Речь идет тогда об интерференции в обратном-прямом направлении, так как активация промотора или промоторов интерференции, расположенного(ных) выше хода транскрипции, интерферирует с активностью первого промотора, расположенного ниже по ходу транскрипции, и активация промотора или промоторов интерференции, расположенного(ных) ниже по ходу транскрипции, интерферирует(ют) с активностью первого промотора, расположенного выше хода транскрипции.
Промотор или промоторы интерференции, расположенный(ые) ниже по ходу транскрипции первого промотора и гена-репортера, имеют ориентацию, тандемную по отношению к первому промотору (см. система nDI выше). Промотор или промоторы интерференции, фиксированный(ые) выше хода транскрипции первого промотора и гена-репортера, имеет(имеют) ориентацию, идентичную таковой первого промотора (см. система UI выше), следовательно, три промотора имеют тандемную конфигурацию.
Эта конфигурация была названа как несмысловая интерференция в прямом-обратном направлении (или nUDI, «non-sense Upstream-Downstream Interference»).
В случае конструкции типа UI, UDI nUDI короткая открытая рамка считывания, сопровождаемая одним или несколькими (например, 2 или 3) стоп-кодонами, предпочтительно встраивается между промотором интерференции и первым промотором, как описывается выше.
Предпочтительно интерференционные конструкции фланкированы на обоих концах двудирекционными терминаторами транскрипции, такими как описанные выше.
Промотором интерференции является индуцируемый промотор, который активируется обычным образом. В рамках настоящего изобретения он активируется фактором транскрипции, когда тот является активным. Согласно настоящему изобретению фактор транскрипции, активирующий индуцируемый промотор, образован совокупностью двух химерных белков (X-DBD и Y-AD), реконструирующих активный фактор транскрипции, способный активировать индуцируемый промотор, когда имеет место взаимодействие между двумя химерными белками X-DBD и Y-AD.
Как было уточнено выше, согласно этому конкретному варианту осуществления настоящего изобретения клетки экспрессируют три химерных белка. Так, они экспрессируют:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с первым белком-партнером Y, способным взаимодействовать с двумя белками-партнерами Х и Z;
- второй химерный белок (Х-DBD), образованный первым ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции;
- третий экспрессируемый химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
ДНК-связывающий домен (DBD) второго химерного белка выбирают в зависимости от промотора интерференции, чтобы связывать ДНК на уровне вышеуказанного промотора. Предпочтительно ДНК-связывающий домен (DBD) второго химерного белка выбирают так, чтобы происходило взаимодействие с последовательностью, имеющейся в вышеуказанном промоторе (последовательность ДНК, связанная с помощью DBD (UAS)) (активирующая против хода транскрипции последовательность). Индуцируемый промотор или индуцируемые промоторы интерференции включают последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Х-DBD).
Домен активации транскрипции первого химерного белка (домен AD Y-AD) позволяет активировать транскрипцию промотора интерференции в клетке. Так, промотор или промоторы интерференции (индуцируемые промоторы) индуктируются функциональным фактором транскрипции, образованным двумя химерными белками X-DBD и Y-AD, взаимодействующими между собой. Этот функциональный фактор транскрипции взаимодействует через свой ДНК-связывающий домен (DBD) с последовательностью промотора интерференции и способен инициировать транскрипцию, управляемую этим промотором через его домен активации транскрипции (AD).
Это представляет собой тот же принцип двугибридной технологии, и специалист в данной области сам способен выбирать многочисленные пары промотор/фактор транскрипции, позволяющие осуществлять настоящее изобретение, как уточнено выше.
ДНК-связывающий домен (DBD) третьего химерного белка (Z-DBD) выбирают в зависимости от индуцируемого промотора, регулирующего экспрессию гена-репортера, так чтобы связывать ДНК на уровне вышеуказанного промотора. Предпочтительно ДНК-связывающий домен (DBD) химерного белка Z-DBD выбирают так, чтобы происходило взаимодействие с последовательностью, имеющейся в промоторе, регулирующим экспрессию гена-репортера (последовательность ДНК, связанная с помощью DBD (UAS; активирующая против хода транскрипции последовательность)).
Индуцируемый промотор, регулирующий экспрессию гена-репортера, и промотор или промоторы интерференции представляют собой разные промоторы. В частности, индуцируемый промотор, регулирующий экспрессию гена-репортера, выбирают таким образом, чтобы не связывать химерный белок Х-DBD (этот промотор, следовательно, не включает последовательность ДНК, связанную с помощью DBD (UAS; активирующая против хода транскрипции последовательность) X-DBD). Кроме того, индуцируемый промотор или индуцируемые промоторы интерференции выбирают так, что они не связывают химерный белок Z-DBD (этот промотор, следовательно, не включает последовательность ДНК, связанную с помощью DBD (UAS; активирующая против хода транскрипции последовательность) Z-DBD).
Домен активации транскрипции первого химерного белка (домен AD Y-AD) позволяет активировать транскрипцию промотора, регулирующего экспрессию гена-репортера в клетке. Таким образом, промотор, регулирующий экспрессию гена-репортера, индуцируется функциональным фактором транскрипции, образованным двумя химерными белками Z-DBD и Y-AD, взаимодействующими друг с другом. Этот функциональный фактор транскрипции взаимодействует через свой ДНК-связывающий домен (DBD) c последовательностью промотора, регулирующего экспрессию гена-репортера, и способен инициировать транскрипцию, управляемую этим промотором через его домен активации транскрипции (AD).
Это представляет собой тот же принцип двугибридной технологии, и специалист в данной области сам способен выбирать многочисленные пары промотор/фактор транскрипции, позволяющие осуществлять настоящее изобретение, как уточнено выше.
Так, в случае взаимодействия между белками Y и Х, промотор интерференции активируется, что приводит к интерференции в транскрипции гена-репортера. Если взаимодействие между Y и Х пертурбировано (например, за счет воздействия химического ингибитора, ингибирующего взаимодействие между Y и Х), промотор интерференции будет менее активен или неактивен, приводя, таким образом, к повышенной экспрессии гена-репортера.
Белки-партнеры Х, Y и Z, гибридизированные с ДНК-связывающим доменом (DBD) и доменом активации транскрипции (AD), могут быть определены как любая группа белков, в случае которых существует взаимодействие между Y и X и между Z и Y и которые, когда Y гибридизирован с AD и Z и Х гибридизированы с разными DBD, взаимодействуют в клетке (даже если это не происходит в их исходных организмах), приводя к конструкции двух функциональных факторов транскрипции (X-DBD/Y-AD и Z-DBD/Y-AD соответственно), активирующих индуцируемые промоторы, такие как описанные выше. Они могут представлять собой полные белки или фрагменты этих последних. Таким образом, в рамках настоящего изобретения могут быть использованы химерные белки, генерирующие двугибридный сигнал. То есть химерные белки, два белка-партнера (или фрагменты белков-партнеров) которых взаимодействуют между собой с достаточной аффинностью в используемой клетке, образуя функциональный фактор транскрипции, активирующий индуцируемый промотор или индуцируемые промоторы, причем вышеуказанный фактор транскрипции связывает ДНК через связывающий домен в первом химерном белке и активирует промотор или промоторы с помощью домена активации транскрипции (AD) второго химерного белка.
Когда интерференционная конструкция ДНК интегрирована в геном клетки-хозяина, эта конструкция преимущественно нацелена на локус, лишенный потенциально пертурбируемых геномных активностей транскрипции, как описано выше.
Клетки, используемые в рамках этого варианта осуществления изобретения (три химерных белка), представляют собой вышеописанные клетки.
Как было указано выше, двугибридная система согласно изобретению может иметь многочисленные применения. Так, она особенно применима для скрининга молекул, обладающих ингибирующей активностью в отношении взаимодействия белок-белок. В рамках варианта осуществления изобретения с тремя химерными белками клетки, описанные выше, представляют в особенности интерес в том, что касается идентификации молекул, ингибирующих взаимодействие между белками Y и Х, однако, не ингибирующих взаимодействие между Y и Z. Таким образом, в этих клетках молекулы, прекращающие взаимодействие белок-белок между Y и Х, провоцируют экспрессию гена-репортера, что позволяет осуществлять их идентификацию.
Объектом настоящего изобретения, следовательно, также является способ идентификации соединения, ингибирующего взаимодействие первого белка Х со вторым белком Y, однако не ингибирующего или менее ингибирующего взаимодействие между белком Y и третьим белком Z, включающий следующие стадии:
а) культивирование клеток, таких как указанные выше (вариант осуществления с тремя химерными белками);
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
с) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка Х с белком-партнером Y, но что этот продукт не ингибирует или менее ингибирует взаимодействие между белком Y и белком Z.
Другим применением системы интерференции с тремя химерными белками является скрининг банков кДНК и банков пептидов для идентификации пептидов или белковых факторов, которые специфически вызывают прекращение взаимодействия между исследуемым белком Х и исследуемым белком Y (Zutshi R. и др., 1998), но которые не затрагивают взаимодействия между белком Х и белком Z.
Другим применением является идентификация мутаций в белке Y, которые прерывают взаимодействие между белком Y и белком Х, но которые не затрагивают взаимодействия между белком Y и белком Z. Это является пригодным не только для исследования структурных компонентов конкретного взаимодействия, но и также в качестве возможности получения генетических инструментов, как трансдоминантные отрицательные мутанты, для характеристики функции in vivo (Serebriiskii I.G. и др., 2001) (см. выше).
Объектом настоящего изобретения также является набор для получения двугибридной системы, такой как описанная выше, включающий:
- конструкцию ДНК, такую как указанная выше;
- вторую конструкцию ДНК, кодирующую:
- первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), гибридизированным с первым белком-партнером Y, способным взаимодействовать с двумя белками-партнерами Х и Z;
- третью конструкцию ДНК, кодирующую:
- второй химерный белок (Х-DBD), образованный первым ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Х, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции;
- четвертую конструкцию ДНК, кодирующую:
- третий химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
Четыре конструкции ДНК набора, описанного выше, могут находиться на одной и той же молекуле ДНК, или на двух, или трех, или четырех разных молекулах ДНК. Четыре конструкции, таким образом, может нести один и тот же вектор ДНК, или они могут находиться на двух, трех или четырех разных векторах. Три конструкции, кодирующие три химерных белка, могут находиться, например, на одном и том же векторе, тогда как четвертую конструкцию несет второй вектор.
Настоящее изобретения представляет собой инструмент, пригодный для характеристики взаимодействий белок-белок и, однако, также для разработки новых лекарственных средств.
Описание фигур
Фигура 1:
Системы rY2H, основанные на транскрипционной интерференции
Схематическое представление, иллюстрирующее применение транскрипционной интерференции для детектирования обратных Y2H взаимодействий. В отсутствие взаимодействия Y2H ген-репортер HIS3 обычным образом экспрессируется и клетки могут расти на среде без гистидина (фиг.1А). Наличие взаимодействия Y2H уменьшает уровень транскрипции HIS3, что провоцирует уменьшение клеточного роста ауксотрофного к гистидину репортерного штамма (фиг.1В). Открытая рамка считывания HIS3 символизирована незакрашенным прямоугольником, причем маленькая черная стрелка обозначает положение исходного кодона. Расположенная выше серая стрелка открытой рамки считывания HIS3 символизирует транскрипцию, инициируемую на уровне Padh1, тогда как черная стрелка означает транскрипцию Pgl1. Толщина стрелок обозначает активность транскрипции промоторов. Стрелки указываются в направлении транскрипции. Взаимодействующие белки Х и Y, используемые для GAL4-DBD (X-DBD) и GAL4-AD (Y-AD), представлены в овальной форме. Padh1: промотор ADH1; Pgal1: промотор GALI; HIS3: открытая рамка считывания HIS3; DI: интерференция в прямом направлении; UI: интерференция против хода транскрипции. Фигура не масштабирована.
Фигура 2:
Конструкции транскрипционной интерференции
Используют пять различных конструкций для получения систем rY2H, основанных на транскрипционной интерференции; пять конструкций представлены в масштабе (фиг.2А-2Е). Указаны сайты рестрикции SalI и SacI и их положения на соответствующих векторах pBluescript. Указаны названия типичных плазмид (см. пример 4 и таблицу 2). Аббревиатуры: Padh1: промотор ADH1; Pgal1: промотор GALI; HIS3: открытая рамка считывания HIS3; Tadh1: терминатор ADHI; Tcyc1: терминатор CYC1; ORF: короткая открытая рамка считывания, сопровождаемая двумя стоп-кодонами; DI: интерференция в прямом направлении (2А); UI: интерференция против хода транскрипции (2В); UDI: интерференция в прямом-обратном направлении (2С); nDI: несмысловая интерференция в прямом направлении (2D); nUI: несмысловая интерференция против хода транскрипции (2Е).
Фигура 3:
Геномная интеграция интерференционных конструкций
Пример, иллюстрирующий геномную интеграцию конструкций транскрипционной интерференции. Вектор интеграции pRB23 сконструирован путем встраивания фрагмента SalI-SacI pRB17 в pRB2. Затем pRB23 использовали для трансформации штамма-мишени уСМ15, куда он интегрируется на уровне локуса URA3 путем гомологичной рекомбинации. Гомологичные последовательности вектора в локусе URA3 (выше/ниже URA3) представлены в заштрихованном виде. Представлены открытые рамки считывания, примыкающие к гену URA3 (GEA2, TIM9 и RPR), и представлена примерная делеция ura3DO (Brachmann C.B. и др., 1998). Аббревиатуры: Padh1: промотор ADH1; Pgal1: промотор GALI; HIS3: открытая рамка считывания HIS3; Tadh1: терминатор ADHI; Tcyc1: терминатор CYC1; 3НА: кодируемые последовательности tag на pBR2; Ptef: промотор TEF; G418r: открытая рамка считывания, кодирующая резистентность к G418; ampR: ген резистентности к ампициллину.
Фигура 4
Функциональная оценка интегрированных систем интерференции
Штаммы, несущие интегрированные копии систем интерференции (уRB48 (DI), yRB49 (UI), yRB50 (nUI), yRB83 (UDI) и yRB82 (nDI)) трансформировали с помощью серий плазмид pCL1/pFL39 (+) и pTAL20/pDBT (-) соответственно и клеточный рост трансформантов исследовали при использовании среды с гистидином (фиг.4А) или без него (фиг.4В-4Е) по капельно-чашечному методу оценок. Готовят серии 10-кратных разведений для каждого трансформанта. В культуральную среду, в случае необходимости, добавляют конкурентный ингибитор белка HIS3p (3-амино-1,2,4-триазол (3-АТ)) в возрастающих концентрациях, как указано (от 0 мМ до 22 мМ) (фиг.4В-4Е). Аббревиатуры: DI: штамм интерференции в прямом направлении; UI: штамм интерференции против хода транскрипции; nDI: штамм несмысловой интерференции в прямом направлении; nUI: штамм несмысловой интерференции против хода транскрипции; UDI: штамм интерференции в прямом-обратном направлении.
Фигура 5
Функциональная оценка систем интерференции, базирующихся на плазмидах (не интегрированных в геном)
Штамм уСМ15 котрансформировали, независимо, с помощью вариантов, базирующихся на плазмидах систем интерференции (pRB20 (DI), pRB21 (UI) или pRB28 (UDI)), и серий плазмид pCL1/pFL39 (+) и pTAL20/pDBT (-) соответственно. Клеточный рост каждого из этих тройных трансформантов исследовали при использовании чашек с агаровой средой с гистидином (фиг.5А) и без него (фиг.5В-5D) и, в случае чашек без гистидина, без или с возрастающими концентрациями 3-АТ (от 0 мМ до 16 мМ) (фиг.5В-5D). Готовят серии десятикратного разведения и оценивают два независимых клона на каждый трансформированный штамм. Аббревиатуры: DI: плазмида интерференции в прямом направлении; UI: плазмида интерференции против хода транскрипции; UDI: плазмида интерференции в прямом-обратном направлении.
Фигура 6
Транскрипционная интерференция, индуцированная взаимодействиями Y2H
Фигура 6А и фигура 6F:
Взаимодействия белок-белок, идентифицированные выше, между белками Fe65 и Арр и р53 и mdm2, в достаточной степени индуцируют конструкцию транскрипционной интерференции для ингибирования клеточного роста дрожжей.
Во-первых, контролировали активность конструкций (фиг.6А и 6F). Для этого репортерный штамм Y187 трансформировали с помощью следующих пар плазмид: pRB34+pRB35 (Fe65/App), pRB34+pDBT (Fe65/DBD) и pTAL20+pRB35 (AD/App) (фиг.6А) и pDBT-mdm2+pVP16-p53Nt (p53/mdm2), pVP16-p53Nt+pDBT (p53/DBD) и pTAL20+pDBT-mdm2 (AD/mdm2) (фиг.6F). Два клона из каждого из этих трансформантов культивировали на минимальной среде с урацилом и без него (фиг.6А и 6F).
Штаммы yRB48 (система DI) и yRB49 (система UI) затем сотрансформировали, каждый, с помощью пары плазмид pRB34 (Fe65) и pRB35 (App), позволяющих осуществляться экспрессии двух гибридных белков AD-Fe65 и DBD-AppCt. Штаммы yRB48 (система DI) и yRB49 (система UI) также котрансформировали, каждый, с помощью пар плазмид pRB34+pDBT (Fe65/DBD) и pTAL20+pRB35 (AD/App) в качестве контролей.
Клеточный рост двух клонов из каждого из этих двойных трансформантов исследовали при использовании чашек с агаровой средой с гистидином и без него и, в случае чашек без гистидина, без или с возрастающими концентрациями 3-АТ (фиг.6В-6Е).
Штамм yRB49 (система UI) также котрансформировали с помощью пары плазмид pDBT-mdm2+pVP16-p53Nt (p53/mdm2), позволяющих осуществляться экспрессии двух гибридных белков DBD-mdm2 и VP16-p53(1-55). Штамм yRB49 (система UI) также котрансформировали с помощью пары плазмид pDBT-mdm2+pTAL20 (AD/mdm2) в качестве контролей. Два клона, котрансформированных двумя плазмидами, отбирали для каждой из этих двух пар плазмид.
Клеточный рост каждого из этих клонов этих двойных трансформантов исследовали при использовании чашек с агаровой средой с гистидином и без него и, в случае чашек без гистидина, без или с возрастающими концентрациями 3-АТ, как указано на фигуре.
Фигура 7
Системы rY2H, основанные на транскрипционной интерференции согласно настоящему изобретению, позволяющие осуществлять идентификацию молекул, ингибирующих взаимодействие между двумя белками Y и Х, однако не ингибирующих взаимодействие между белком Y и третьим белком Z
Схематическое представление, иллюстрирующее использование транскрипционной интерференции для детектирования обратных Y2H взаимодействий, позволяющих осуществлять идентификацию молекул, ингибирующих взаимодействие между двумя белками Y и Х, однако не ингибирующих взаимодействие между белком Y и третьим белком Z. В отсутствие Y2H взаимодействия между белком Х и белком Y ген-репортер HIS3 экспрессируется обычным образом и клетки могут расти на среде без гистидина (фиг.7А). Это происходит, когда присутствует ингибитор взаимодействия белка Х с белком-партнером Y. Наличие Y2H взаимодействия между X-DBD и Y-AD снижает уровень транскрипции HIS3, что провоцирует уменьшение клеточного роста штамма-репортера, ауксотрофного к гистидину (фиг.7В). Открытая рамка считывания HIS3 символизирована незакрашенным прямоугольником, причем маленькая черная стрелка указывает положение исходного кодона. Серая стрелка выше открытой рамки считывания HIS3 символизирует транскрипцию, инициируемую на уровне Plex, тогда как черная стрелка означает транскрипцию Pgal1. Толщина стрелок символизирует активность транскрипции промоторов. Стрелки указываются в направлении транскрипции. Используемые взаимодействующие белки Х/Y и Z/Y представлены в овальной форме. Plex: промотор LexA; Pgal1: промотор GALI; HIS3: открытая рамка считывания HIS3; фигура не масштабирована.
Фигура 8
Определение экспериментальных условий для скрининга ингибиторов взаимодействия Fe65/App
Фигура 8А:
Определение количества клеток
Штамм, содержащий систему транскрипционной интерференции UI (yRB49), котрансформировали c помощью плазмид pRB34и pRB35 (взаимодействие Fe65/App). Культуру трансформированного штамма разводили до достижения оптической плотности (DO)=0,1 и 0,01 и по 10 мл каждого разведения наносили тонким слоем на среду, дополненную гистидином (+HIS). Чашки инкубировали в течение 48 часов при температуре 30°С.
Фигура 8В:
Определение концентрации 3-АТ
Штамм, содержащий систему транскрипционной интерференции UI (yRB49), котрансформированный c помощью плазмид pRB34 и pRB35 (взаимодействие Fe65/App) или с помощью контрольных плазмид pTAL20 и pDBT (AD/DBD), наносили тонким слоем на среду с гистидином и без 3-АТ, без гистидина и без 3-АТ и с гистидином и с 0,5 мМ или 1 мМ 3-АТ. В каждую чашку затем вносили 1 мкл 100% ДМСО, 1 мкл раствора молекулы «х» в концентрации 10 мМ в ДМСО и 1 мкл раствора молекулы «у» в концентрации 10 мМ в ДМСО. Чашки инкубировали в течение 48 часов при температуре 30°С.
Фигура 8С:
Репарация роста с помощью гистидина
Штамм, содержащий систему транскрипционной интерференции UI (yRB49) и котрансформированный с помощью плазмид pRB34 и pRB35 (взаимодействие Fe65/App), наносили на среды без гистидина и содержащие 1 мМ 3-АТ. В центр каждой чашки вносили 1 мкл урацила (U), аденина (А), лизина (К) или гистидина (Н) в концентрациях 0,125%, 0,25%, 0,5% или 1%. Чашки инкубировали в течение 48 часов при температуре 30°С.
Фигура 9
Детектирование ингибиторов взаимодействия Fe65/App путем репарации роста
Штамм, содержащий систему транскрипционной интерференции UI (yRB49) и котрансформированный с помощью плазмид pRB34 и pRB35 (взаимодействие Fe65/App), наносили на среду без гистидина и содержащую 1 мМ 3-АТ. В эту чашку вносили 1 мкл 10 мМ раствора молекулы 1, предполагаемого ингибитора взаимодействия Fe65/App, 1 мкл 10 мМ раствора молекулы 2, предполагаемого ингибитора взаимодействия Fe65/App, и 1 мкл 10 мМ раствора молекулы 3, неактивной в отношении взаимодействия Fe65/App. Чашки инкубировали в течение 48 часов при температуре 30°С.
Фигура 10
Сравнение взаимодействий Fe65/App и p53/MDM2 в отношении активации системы интерференции
Штамм, содержащий систему транскрипционной интерференции UI (yRB49), котрансформировали при использовании пар плазмид pRB34 и pRB35 (взаимодействие Fe65/App), pDBT-mdm2 и pVP16-p53Nt (взаимодействие mdm2/p53) или pTAL20 и pDBT (контроль). В случае каждой трансформации культивировали два независимых клона. После инкубации в течение ночи при температуре 30°С все культуры приводили к одному и тому же значению оптической плотности. Осуществляли серии двудесятикратного разведения для каждой культуры и каплю каждого разведения наносили на среду без гистидина и дополненную различными концентрациями 3-АТ (от 1 мМ до 32 мМ). Чашки инкубировали в течение 48 часов при температуре 30°С.
Фигура 11
Подтверждение специфичности ингибиторов взаимодействия Fe65/App
Штамм, содержащий систему транскрипционной интерференции UI (yRB49), котрансформировали при использовании пар плазмид pRB34 и pRB35 (взаимодействие Fe65/App) или pDBT-mdm2 и pVP16-p53Nt (взаимодействие mdm2/p53). Штамм, содержащий плазмиды pRB34 и pRB35 (взаимодействие Fe65/App), вносили в 5 чашек со средой без гистидина, дополненной 1 мМ 3-АТ, (представлено вверху); штамм, содержащий плазмиды pDBT-mdm2 и pVP16-p53Nt (взаимодействие mdm2/p53), точно также вносили в 5 чашек со средой без гистидина, дополненной 20 мМ 3-АТ, (представлено внизу).
Для каждого взаимодействия реализовали контрольную чашку, внося 1 мкл гистидина в концентрации 0,125% масса/объем в центр этой чашки. В каждую из 4 других чашек вносили другую молекулу, А, В, С или D, согласно следующей схеме: в верхний левый угол помещали 1 мкл продукта в концентрации 0,1 мМ; в верхний правый угол помещали 1 мкл того же самого продукта в концентрации 1 мМ; в центр помещали 1 мкл этого продукта в концентрации 10 мМ.
Чашки инкубировали в течение 48 часов при температуре 30°С.
Настоящее изобретение поясняется с помощью нижеследующих примеров, которые нужно рассматривать как иллюстративные и не ограничивающие объема охраны изобретения.
Способы молекулярной биологии, используемые в нижеследующих примерах, соответствуют стандартным протоколам молекулярной биологии и описаны Sambrook J. и др., 1989. Последовательности ДНК амплифицировали, используя систему полимеразной цепной реакции (РСR) с высокой точностью воспроизведения (Expand High Fidelity PCR System, Roche Diagnostics, Penzberg, Германия).
Пример 1
Конструкция плазмиды pRBV
Фрагмент из 761 пары оснований, соответствующий полному конститутивному промотору гена ADH1, генерировали путем PCR, используя в качестве праймеров следующие олигонуклеотиды:
RB43 5'-AAAATCTAGAGGCGCCATATCCTTTTGTTGTTTCCGG-3' (идентифицированная последовательность № 1) и
RB44 5'-AAAAACGCGTGGCGCCCATCTTTCAGGAGGCTTGC-3' (идентифицированная последовательность № 2) (сайты рестрикции XbaI и MluI подчеркнуты), и используя в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae (доступный, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под номером 28383).
Два однонитевых фосфорилированных олигонуклеотида: RB75 с последовательностью 5'-CGCGTGGGGGGTTAATTAAAAAAAAGC-3' (идентифицированная последовательность № 3) и RB76 с последовательностью 5'-GGCCGCTTTTTTTTAATTAACCCCCCA-3' (идентифицированная последовательность № 4) гибридизировали с целью получения двунитевого связывающего плеча (линкер), имеющего по краям совместимые липкие концы MluI и NotI соответственно. Для этого два однонитевых олигонуклеотида смешивали в эквимолярных концентрациях в воде. Смесь нагревали при температуре 90°С в течение 5 минут в нагревательном блоке, затем блок оставляли охлаждаться до комнатной температуры (примерно 2 часа).
С целью конструирования вектора pRB3 фрагмент из 761 пары оснований, полученный путем PCR, (фрагмент Padh1), сначала рестриктировали ферментами рестрикции XbaI и MluI. Таким образом рестриктированный фрагмент затем встраивали с линкером в вектор pBluescript KS+ (коммерчески доступный, выпускаемый фирмой Stratagene (LaJolla, Калифорния, США)), предварительно рестриктированный с помощью XbaI-NotI. Таким образом полученная плазмида была названа pRB3.
Фрагмент из 660 пар оснований, соответствующий открытой рамке считывания гена HIS3, затем амплифицировали путем PCR, используя в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae и в качестве праймеров следующие олигонуклеотиды:
RB45 5'-AAAAACGCGTACAGAGCAGAAAGCCCTAG-3' (идентифицированная последовательность №5) и RB46 5'-AAAAAAGCGGCCGCGGCGCGCCTTAATTAACTACATAAGAACACCTTTGGTG-3' (идентифицированная последовательность № 6) (сайты MluI, NottI и AscI подчеркнуты). Этот фрагмент рестриктировали с помощью ферментов рестрикции MluI и NotI и клонировали в вектор pRB3, предварительно линеаризированный с помощью ферментов рестрикции MluI и NotI. Таким образом полученный вектор был назван pRB4.
Фрагмент PCR из 23 пар оснований, кодирующий терминатор гена ADHI (Tadh1), затем амплифицировали путем PCR, используя в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae и в качестве праймеров следующие олигонуклеотиды: RB51 5'-AAAAGGCGCGCCTAATTCCGGGCGAATTTCT-3' (идентифицированная последовательность № 7) и RB51 5'-AAAAGAGCTCTGCATGCCGGTAGAGGTG-3' (идентифицированная последовательность № 8) (сайты AscI SacI подчеркнуты). Полученный фрагмент рестриктировали с помощью ферментов рестрикции AscI и SacI и встраивали в вектор pRB4, предварительно рестриктированный ферментами AscI и SacI. Таким образом полученная плазмида была названа pRB5.
Фрагмент из 88 пар оснований, кодирующий терминатор гена CYC1 (Tcyc1), получали путем гибридизации двух однонитевых фосфорилированных олигонуклеотидов: RB98 с последовательностью: 5'-CGATCGCGTTTGTACAGAAAAAAAAGAAAAATTTGAAATATAAATAA CGTTCTTAATACTAACATAACTATTAAAAAAAATAAATAGGGACCG-3' (идентифицированная последовательность № 9) и RB99 с последовательностью: 5'-AATTCGGTCCCTATTTATTTTTTTTAATAGTTATGTTAGTATTAAGAAC GTTATTTATATTTCAAATTTTTCTTTTTTTTCTGTACAAACGCGAT-31 (идентифицированная последовательность № 10). Для этого два однонитевых олигонуклеотида смешивали в эквимолярных концентрациях в воде. Смесь нагревали при температуре 90°С в течение 5 минут в нагревательном блоке, затем блок оставляли охлаждаться до комнатной температуры (примерно 2 часа). Таким образом полученный двунитевой фрагмент имеет по своим краям совместимые липкие концы EcoRI и ClaI соответственно. Это позволяет осуществлять встраивание этого двунитевого фрагмента (соответствующего терминатору Tcyc1) в вышеописанный вектор pRB5, предварительно рестриктированный ферментами EcoRI и ClaI. Таким образом полученный вектор был назван pRBV.
Последовательность этой конструкции подтверждена секвенированием при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
Пример 2
Конструкция систем транскрипционной интерференции
2.1: Конструкция вектора интерференции в прямом направлении pRB17 и вектора несмысловой интерференции в прямом направлении pRB16:
Вектор интерференции в прямом направлении PRB17 (называемый также DI согласно «Dowstream Interference») и вектор несмысловой интерференции в прямом направлении PRB16 (называемый также nDI согласно «non-sense Dowstream Interference») получали путем встраивания фрагмента PCR из 549 пар оснований, соответствующего промотору Pgal1, в вектор pRBV (см. пример 1).
Этот фрагмент PCR получали, используя в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae и в качестве праймеров следующие олигонуклеотиды: RB49 5'-AAAAGGCGCGCCGTAAAGAGCCCCATTATCTTAG-3' (идентифицированная последовательность №11) и RB50 5'-AAAAGGCGCGCCTTTGAGATCCGGGTTTTTTCT-3' (идентифицированная последовательность №12) (сайты рестрикции AscI подчеркнуты). Этот фрагмент PCR затем рестриктировали ферментом AscI и встраивали в вектор pRBV, линеаризированный с помощью фермента AscI. Фрагмент может встраиваться в двух направлениях, что позволяет получать две конструкции: вектор интерференции в прямом направлении pRB17 (называемый также DI и в котором Padh1 и Pgal1 ориентированы в противоположных направлениях) (см. фиг.2А) и вектор несмысловой интерференции в прямом направлении pRB16 (называемый также nDI и в котором Padh1 и Pgal1 имеют одну и ту же ориентацию) (см. фиг.2D).
Последовательность этих конструкций подтверждена секвенированием при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
Используемый однодирекционный промотор Pgal1 происходит от дивергентного двудирекционного промотора GAL1-10 (описан Johnston M. и R.W. Davis, 1984) и содержит четыре консенсусных сайта «GAL1-responsive DNA enhancer elements», а также основной промотор гена GAL1, но не включает фрагмент ТАТА промотора гена GAL10.
Промотор GAL1 регулируется фактором транскрипции GAL4p, в котором имеется двугибридная пара (Y2H) (отдельные домены AD и DBD). Этот промотор хорошо охарактеризован и с успехом используется в случае применений, где необходима индуцируемая экспрессия белка, как в случае очистки белков, исследования функции необходимого гена и т.д.
2.2. Конструкция вектора интерференции в обратном направлении pRB18 и вектора несмысловой интерференции в обратном направлении pRB19:
С целью конструирования вектора интерференции в обратном направлении pRB18 (называемого также как UI согласно «Upstream Interference») и вектора несмысловой интерференции в обратном направлении pRB19 (называемого также nUI согласно «non-sense Upstream Interference») получали короткую открытую рамку считывания, кодирующую аминокислоты карбокси-конца белка GST (C-ter GST) путем гибридизации двух фосфорилированных однонитевых олигонуклеотидов: RB81 с последовательностью
5'-CCGGGGCGGCCGCAACGTTTGGTGGTGGCGACCATCCTCCAAAATCGGATCTGGTTCCGCGTTGAGTAGCTGAATAAGTGAATAGGCGGy CCGCT-3' (идентифицированная последовательность № 13) и RB82 с последовательностью
5'-CTAGAGCGGCCGCCTATTCACTTATTCAGCTACTCAACGCGGAACCAGATCCGATTTTGGAGGATGGTCGCCACCACCAAACGTTGCGGCCGCC-3' (идентифицированная последовательность № 14) (сайт № l1 подчеркнут).
Для этого два однонитевых олигонуклеотида смешивали в эквимолярных концентрациях в воде. Смесь нагревали при температуре 90°С в течение 5 минут в нагревательном блоке, затем блок оставляли охлаждаться до комнатной температуры (примерно 2 часа). Таким образом полученный двунитевой фрагмент имеет по своим краям совместимые липкие концы XmaI и XbaI соответственно.
Параллельно фрагмент из 552 пар оснований, соответствующий промотору гена GALI (речь идет о том же самом промоторе, что и таковой, используемый для конструкции вектора интеграции в прямом направлении pRB17, однако фрагмент является на 3 пары оснований длиннее, чем таковой, используемый для конструкции в прямом направлении, так как он содержит стартовый кодон ATG для инициации трансляции короткой открытой рамки считывания), затем амплифицировали путем PCR. Этот фрагмент получали, используя в качестве матрицы геномную ДНК штамма FL100 S.cereviciae и в качестве праймеров следующие олигонуклеотиды: RB55 5'-AAAACCCGGGGTAAAGAGCCCCATTATCTTAG-3' (идентифицированная последовательность № 15) и RB56 5'-AAAACCCGGGCATTTTGAGATCCGGGTTTTTTC-3' (идентифицированная последовательность № 16) (сайты XmaI подчеркнуты). Этот фрагмент затем рестриктировали с помощью фермента XmaI.
Затем осуществляли лигирование трех фрагментов (два фрагмента, полученные выше,+вектор pRBV). Это лигирование состоит во встраивании фрагмента Pgal1, рестриктированного с помощью фермента XmaI, и короткой открытой рамки считывания С-ter GST (совместимые липкие концы XmaI и XbaI) в вектор pRBV (предварительно рестриктированный с помощью XmaI-XbaI). Фрагмент Pgal1 может встраиваться в двух направлениях. Следовательно, получали две конструкции: вектор интерференции в обратном направлении pRB18 (называемый также UI и в котором Padh1 и Pgal1 находятся в одной и той же ориентации) (см. фиг.2В) и вектор несмысловой интерференции в обратном направлении pRB19 (называемый также nUI и в котором Padh1 и Pgal1 ориентированы в противоположных направлениях) (см. фиг.2Е).
Последовательность этих конструкций подтверждена секвенированием при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
2.3: Конструкция вектора интерференции в прямом-обратном направлении pRB27
Плазмиду интерференции в прямом-обратном направлении pRB27 (называемую также UDI согласно «Upstream-Downstream Interference») конструировали путем встраивания такого же фрагмента PCR из 549 пар оснований, соответствующего промотору Pgal1, как таковой, используемый для конструкции pRB16 и pRB17 (см. пример 2: этот фрагмент PCR получали, используя в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae и в качестве праймеров олигонуклеотиды RB49 и RB50) в вектор pRB18. Для этого фрагмент PCR рестриктировали с помощью фермента AscI и встраивали в вектор pRB18, линеаризированный путем рестрикции с помощью фермента AscI. Более конкретно, отбирали (по профилю рестрикции) фрагмент, который может встраиваться в двух направлениях, плазмиду со вставкой в желательной ориентации (в которой два промотора Pgal1 ориентированы в противоположных направлениях) (см. фиг.2С).
Таким образом полученный вектор был назван pRB27. Последовательность этой конструкции подтверждена секвенированием при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
2.4: Принцип конструкции систем транскрипционной интерференции
Заявитель представил два типа систем rY2H (обратная двугибридная), которые оба основаны на обеспечении умолчания гена путем транскрипционной интерференции. В первой конфигурации Pgal1 расположен ниже по ходу транскрипции гена HIS3 и Padh1 и Pgal1 обладают конвергентной ориентацией. Эта конфигурация была названа интерференцией в прямом направлении (DI), так как активация в прямом направлении Pgal1 интерферирует с транскрипцией гена HIS3. Во второй конфигурации Pgal1 расположен выше хода транскрипции Padh1 и два промотора обладают тандемной ориентацией. Следовательно, эта конфигурация была названа интерференцией против хода транскрипции (UI), так как активация Pgal1 против хода транскрипции интерферирует с активностью промотора Padh1.
На основе конфигураций UI и DI были созданы конструкции транскрипционной интерференции rY2H (см. фиг.2а-2Е).
Активация Pgal1 в системе UI приводит к снижению активности Padh1 вследствие транскрипционной интерференции. Активность Pgal1, однако, может еще позволять осуществляться продуцированию смыслового транскрипта гена HIS3. С целью предотвращения трансляции этого смыслового транскрипта в случае функциональной IGP-дегидратазы короткую открытую рамку считывания из 22 аминокислот, сопровождаемую двумя стоп-кодонами, встраивали между Pgal1 и Padh1. Следовательно, за счет транскрипционной интерференции в системе UI продуцируется бисцитронная мРНК, начиная с которой выражается первая короткая открытая рамка считывания.
Третьей конструкцией является комбинация систем UI и DI. В этой системе, называемой системой интерференции в прямом-обратном направлении (UDI), Pgal1 встроен выше хода транскрипции и в тандемной ориентации по отношению к Padh1-HIS3 и дополнительная копия Pgal1 расположена ниже по ходу транскрипции открытой рамки считывания HIS3 и это в конвергентной ориентации по отношению к Padh1 (см. фиг.2С).
С целью контролирования специфичности транскрипционной интерференции, индуцируемой Pgal1, конструировали систему несмысловой интерференции против хода транскрипции (nUI) и несмысловой интерференции в прямом направлении (nDI). В этих двух контрольных конструкциях ориентация Pgal1 является обратной по отношению к конструкциям UI и DI соответственно. Никакой транскрипционной интерференции не должно наблюдаться во время использования этих контрольных конструкций, даже в присутствии имеющегося взаимодействия Y2H.
Пять вышеописанных конструкций (пять плазмидных вставок) фланкированы на своих концах двудирекционными терминаторами транскрипции (см. пример 1). Таким образом, с одной стороны, находится фрагмент, кодирующий терминатор транскрипции гена CYC1 (Tcyc1) (описан Osborne B.I. и L. Guarente, 1989), и, с другой стороны, находится фрагмент, кодирующий терминатор гена ADH1 (Tadh1) (описан Irniger S. и др., 1991). Эти два элемента блокируют транскрипцию двудирекционным образом. Это предотвращает распространение транскрипции, управляемой с помощью Pgal1 или Padh1, за сайты рестрикции SalI/SacI и защищает также системы rY2H от возможного влияния активностей транскрипции вне кассеты гена-репортера. Конструкции интерференции сначала получали на бактериальных векторах и сайты рестрикции SalI и SacI использовали для субклонирования в векторы репликации и интеграции дрожжей (см. ниже).
Пример 3
Конструкция нового вектора интеграции pRB2
Фрагмент PCR из 871 пары оснований, соответствующий области от +29 до +90, расположенной ниже по ходу транскрипции стоп-кодона ТАА гена URA3, получали путем PCR, используя в качестве праймеров следующие олигонуклеотиды:
RB83 5'-AAAAAAGAGCTCTACTAAACTC ACAAATTAGAGC-3' (идентифицированная последовательность № 17) и RB84 5'-AAAAAAGAATTCGCGGCCGCAAATATACTGGGGAACCAGTC-3' (идентифицированная последовательность № 18) (сайты EcoRI и SacI подчеркнуты) и в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae.
Продукт PCR рестриктировали с помощью EcoRI-SacI и клонировали на уровне сайтов EcoRI-SacI вектора pFA6a-kanMX-PGAL1-3HA (описано в работах Longtine M.S. и др., 1998, и Wach A. и др., 1997, и техническое содержание которых, в том что касается конструкции этого вектора, включено в настоящее описание путем ссылки). Таким образом полученный вектор был назван pRB12.
Параллельно, фрагмент из 769 пар оснований, соответствующий области от -223 до -992, расположенной выше хода транскрипции ATG гена URA3, генерировали путем PCR, используя в качестве праймеров следующие олигонуклеотиды:
RB85 5'-AAAAAACGTACGGCGGCCGCGATAAGGAGAATCCATACAAG-3' (идентифицированная последовательность № 19) и RB86 5'-AAAAAACGTACGTTTATGGACCCTGAAACCAC-3' (идентифицированная последовательность № 20) (сайты BsiWI подчеркнуты) и в качестве матрицы геномную ДНК штамма FL100 S.cerevisiae. Продукт PCR рестриктировали ферментом BsiWI и клонировали на уровне единственного сайта ВsiWI pRB12. Фрагмент, который может встраиваться в двух направлениях, плазмиду со вставкой в желательной ориентации (в которой два промотора Pgal1 ориентированы в противоположных направлениях) отбирали путем анализа профиля рестрикции.
Последовательность этой конструкции подтверждена путем секвенирования при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, CША).
Вектор pRB2 содержит два фрагмента, гомологичных областям, расположенным выше хода транскрипции и ниже по ходу транскрипции, соответственно, открытой рамки считывания гена URA3 S.cerevisiae. Последовательность, встроенная между двумя фрагментами, нацеливает интеграцию на уровне локуса URA3, причем его геномная интеграция вызывает делецию 1104 пар оснований, что удаляет открытую рамку считывания URA3 (см. фиг.3).
Эта делеция URA3 является близкой к делеции, известной в случае модели ura3D0(3), причем различие по отношению к ura3D0 состоит в том, что увеличивается на 22 пары оснований по меньшей мере область ниже по ходу транскрипции URA3 во время интеграции pRB2.
Ген GEA2 расположен выше хода транскрипции URA3, и ранее было показано, что уровень транскрипции и размер транскриптов гена GEA2 не затрагиваются делецией ura3D0 (Brachmann C.B. и др., 1998).
Ген TIM9 локализован ниже по ходу транскрипции URA3, и он представляет собой ген, необходимый в случае дрожжей S.cerevisiae. Его не соотносят с небольшой степенью выживания штаммов ura3D0, оказывается, что TIM9 остается полностью функциональным в штаммах ura3D0. Следовательно, оказывается, что делеция ura3D0 не затрагивает транскрипцию примыкающих генов.
Таким образом, системы интерференции, интегрированные в pRB2, вероятно, не будут ни затрагивать, ни затрагиваться транскрипцией GEA2 и TIM9. Преимущество векторов pRB2 состоит в том, что их можно также использовать в мутантных штаммах URA3, как ura3-52 и ura3D0.
Пример 4
Субклонирование различных вставок векторов интерференции в векторы интеграции и репликации дрожжей pRB2
Различные системы интерференций и контролей (DI, nDI, UI, nUI UDI), описанные в примерах 2 (см. фиг.2А-2Е), на векторах pBluescript субклонировали в форме фрагментов SalI-SacI в вектор интеграции pRB2. Для этого различные плазмиды pRB16, pRB17, pRB18, pRB19 и pRB27 рестриктировали ферментами SalI-SacI и вставку субклонировали в вектор интеграции pRB2, предварительно рестриктированный ферментами SalI-SacI. Полученные различные конструкции были названы pRB31, pRB23, pRB24, pRB25 и pRB29 соответственно (см. таблицу 2).
После интеграции систем интерференции в геном ген-репортер HIS3 является тандемным по отношению к генам GEA2 и TIM9.
Аббревиатуры: nDI: несмысловая интерференция в прямом направлении; DI: интерференция в прямом направлении; UI: интерференция против хода транкрипции; nUI: несмысловая интерференция против хода транскрипции; UDI: интерференция в прямом-обратном направлении.
Пример 5
Конструкция штаммов транскрипционной интерференции с геномной интеграцией векторов интерференции
5.1: Геномная интеграция векторов интерференции
Используемыми культуральными средами являются такие, как описанные Gutherie C. и G.R. Fink, 2002. В случае всех экспериментов клетки выращивали на определенной минимальной среде YNB (Difco Laboratories, Детройт, США) +2% глюкозы (Sigma-Aldrich, Лион, Франция) при температуре 30°С. Для пополнения маркеров ауксотрофии добавляют незаменимые аминокислоты (Sigma-Aldrich, Лион, Франция).
Штамм yCM15 (MAT a Δgal4 Δgal80 ura3-52 lys2-801 his3Δ200 trp1-Δ63 leu2 ade2-101) использовали для конструкций штаммов с геномной интеграцией. Штамм уСМ15 конструировали, исходя из штамма PCY2 (описан Chevray P.M. и Nathans D., 1992), в котором удалена репортерная кассета.
Штамм PCY2 (MAT a Δgal4 Δgal80 URA3::GAL1-lacZ lys2-801 ade2-101 trp1-Δ63 his3-Δ200 leu2) получали путем гибридизации штамма YPH499 (MAT a ura3-52 lys2-801 ade2-101 trp1-Δ63 his3-Δ200 leu2-Δ1: ATCC 76625, 204679; (Sikorski и Hieter, 1989)) (доступный, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под номером 204679) со штаммом GGY1::171 (MAT a Δgal4 Δgal80 leu2 his3 URA3::GAL1-lacZ) (Gill Ptashne, 1987).
Штамм GGY1::171 конструировали путем интеграции кассеты URA3::GAL1-lacZ (Yocum и др., 1984), которая несет ген-репортер lacZ под контролем промотора гена GAL1, активированного с помощью UASG, в штамм GGY1 (MAT a Δgal4 Δgal80 leu2 his3 (Gill и Ptashne, 1987)). Штамм GGY1 также получали в результате гибридизации штамма DBY745 (MAT a Δgal4 Δgal80 leu2-3,112 ade1-100 ura3-52; (Graham и Chambers, 1996; Ferreiro и др., 2004; Nagarajan и Stroms, 1997; Dormer и др., 2000) со штаммом YM709 (Pearlberg J., 1995).
Вместо штамма PCY2 может быть использован любой штамм дрожжей S.cerevisiae, имеющий генотип (gal4-gal80-trp1-his3-leu2-ura3-). Например, вместо штамма PCY2 может быть использован штамм Y187 (описанный Harper J.W. и др., 1993, и доступный, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под регистрационным номером 96399).
В таблице 3 представлены другие примеры штаммов, которые могут заменять штамм PCY2. Представлены их названия, краткое описание этих штаммов и их регистрационные номера в Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США), организм, из которого эти штаммы доступны.
С целью конструирования штамма уСМ15 106-107 клеток клона штамма PCY2 суспендировали в стерильной воде и вносили в чашку с агаровой средой YNB +2% глюкозы, дополненной аминокислотами lys +ade+his+ura+trp+leu. Чашка с агаровой средой содержит также 5-фтороротовую кислоту (5-FOA) в концентрации 1 мг/мл; 5-FOA токсична для клеток, экспрессирующих ген URA3. Соответственно, из этих чашек, содержащих 5-FOA, отбирали мутанты штамма PCY2, которые утратили репортерную кассету URA3::GAL1-lacZ (Boеke и др., 1987). Отсутствие репортерной кассеты в полученных мутантах подтверждали, тестируя активность репортера lacZ. Мутанты затем трансформировали с помощью плазмиды pCL1 (Fields S. и O. Song, 1989), кодирующей фактор транскрипции GAL4p, который является очень сильным активатором промотора GAL1, и измеряли активность lacZ (Breeden L. и K. Nasmyth, 1985). Отбирали резистентный к 5-FOA и без активности lacZ мутант, и он был назван уСМ15.
Векторы интеграции pRB31, pRB23, pRB24, pRB25 и pRB29 линеаризировали путем рестрикции с помощью фермента DrdI. Штамм уСМ15 трансформировали, независимо, с помощью 1 мкг линеаризованной ДНК каждого из векторов интеграции, используя метод трансформации с ацетатом лития (такой, как описанный Gietz R.D. и A. Sugino, 1988), и клетки культивировали на среде, лишенной гистидина (YNB (Difco Laboratories, Детройт, США) +2% глюкозы (Sigma-Aldrich, Лион, Франция), дополненной аминокислотами lys+аde+trp+leu и урацилом (Sigma-Aldrich, Лион, Франция). Выделяли ауксотрофные к гистидину клоны и интеграцию репортерных кассет на уровне локуса URA3 подтверждали путем PCR колонии (описано Gutherie C. и др., 2002), используя внутренние и внешние праймеры, специфические для каждого связывания (см. таблицу 4).
Последовательности используемых праймеров являются следующими:
RB38 : 5'-GGGGTAATTAATCAGCGAAGCGATG-'3 (последовательность № 21)
RB61 : 5'-CGATGTATGGGTTTGGTTG-'3 (последовательность № 22)
RB66 : 5'-TCTTGCGAGATGATCCCGC-'3 (последовательность № 23)
RB68 : 5'-TAAGCGTATTACTGAAAGTTCC-'3 (последовательность № 24)
RB103 : 5'-GCAATGAAAGAGCAGAGCGAGAG-'3 (последовательность № 25)
RB109 : 5'-GTTATCAGATATTATCAGGTGGGAA-'3 (последовательность № 26)
Штаммы, полученные путем независимой трансформации штамма уСМ15 с помощью каждого из векторов интеграции pRB31, pRB23, pRB24, pRB25, pRB29 и интеграция в которые каждой из репортерных кассет на уровне локуса URA3 доказана, были названы yRB82, yRB48, yRB49, yRB50, yRB83 соответственно (см. таблицу 2).
5.2: Активация интегрированных интерференционных конструкций
С целью оценки активности систем интерференции репортерные штаммы, полученные согласно примеру 5.1, трансформировали с помощью двух пар плазмид. Первую пару используют для исследования ответа системы интерференции на сильную активацию Pgal1 вследствие экспрессии фактора транскрипции GAL4p полной длины (Fields S. и O. Song, 1989). Вторая пара плазмид позволяет происходить отдельно экспрессии домена активации (DI) и ДНК-связывающего домена (DBD) GAL4p и служит отрицательным контролем, так как различные домены индивидуально не могут активировать Pgal1.
Так, штаммы, полученные согласно примеру 5.1 (штаммы yRB82, yRB48, yRB49, yRB50, yRB83), котрансформировали с помощью центромерной плазмиды pCL1, кодирующей фактор транскрипции GAL4p (описана Fields S. и O. Song, 1989), и плазмиды без центромерной вставки pFL39 (описана Bonneaud N. и др., 1991). Эти плазмиды, реплицирующиеся в дрожжах, несут ген LEU2 или TRP1 в качестве селекционного маркера.
Трансформантов селекционировали на среде YNB (Difco Laboratories, Детройт, США) +2% глюкозы (Sigma-Aldrich, Лион, Франция), дополненной аминокислотами lys+ade+his и урацилом (Sigma-Aldrich, Лион, Франция).
Полученные трансформанты были названы соответственно nDI+, DI+, UI+, nUI+ и UDI+.
Отрицательные контроли получали путем котрансформации штаммов, описанных в примере 5.1 (штаммы yRB82, yRB48, yRB49, yRB50, yRB83), с помощью векторов pTAL20 и pDBT. Векторы pTAL20 и pDBT позволяют осуществляться экспрессии домена активации (AD) GAL4p и ДНК-связывающего домена (DBD) GAL4p (эти векторы описаны Navarro P. и др., 1997).
Таким образом полученные трансформанты были названы соответственно nDI-, DI-, UI-, nUI- и UDI-.
Пример 6
Исследование роста дрожжей с интегрированной в их геном системой транскрипционной интерференции
Клеточный рост трансформантов исследовали капельно-чашечным методом оценок. Для этого клетки помещают в чашки с агаровой средой, содержащей гистидин, и инкубируют их в течение ночи при температуре 30°С. Затем клетки суспендируют в воде с получением приблизительно 106 клеток на мл и приготовляют серии десятикратных разведений. Отбирают пипеткой капли по 5 мкл из каждого разведения и помещают их в предварительно высушенные чашки с желательной агаровой средой. Чашки инкубируют при температуре 30°С.
Клеточный рост трансформантов интерференции, таким образом, оценивали при использовании среды, лишенной гистидина. В случае необходимости в культуральную среду добавляли конкурентный ингибитор белка HIS3p (3-амино-1,2,4-триазол (3-АТ)) в возрастающих концентрациях с целью обнаружения различий на уровне активности гена-репортера HIS3 между разными штаммами интерференции (фиг.4). Среды, содержащие ингибитор, приготовляли из исходного раствора с 1 М концентрацией конкурентного ингибитора 3-амино-1,2,4-триазола (3-АТ; Sigma) в воде качества Millipore. Перед добавлением 3-АТ в желательных концентрациях агар охлаждали вплоть до температуры 60°С. Различные тестируемые культуральные среды представляют собой минимальную среду (YNBG (Difco Laboratories, Детройт, США) +2% глюкозы (Sigma-Aldrich, Лион, Франция), дополненную аминокислотами lys+ade и урацилом (Sigma-Aldrich, Лион, Франция), без 3-АТ или с 3-АТ в концентрациях 8 мМ, 16 мМ, 22 мМ, 32 мМ. Также реализовали положительный контроль (культуральная среда с гистидином и без 3-АТ). Чашки инкубировали при температуре 30°С в течение 48-72 часов. Часть результатов представлена на фиг.4А-4Е.
В присутствии GAL4p (штаммы +) наблюдают более сильное снижение клеточного роста в случае трансформантов UI+. Клетки UI, экспрессирующие GAL4p, не могут расти в отсутствие гистидина, тогда как клетки, несущие контроль AD/DBD (трансформанты UI-), нормально растут при концентрациях 3-АТ выше 32 мМ. Ни GAL4p, ни контроль AD/DBD не ингибируют клеточного роста контрольных трансформантов nUI+ и nUI-, что указывает на то, что ингибирование роста клеток
UI+, экспрессирующих GAL4p, возникает вследствие активации транскрипции Pgal1, направленной к Padh1.
В клетках DI+, экспрессия GAL4p не затрагивает клеточного роста, когда отсутствует ингибитор 3-АТ (см. фиг.4В), однако рост полностью ингибируется, когда присутствует 3-АТ в концентрации 8 мМ или в более высоких концентрациях (см. фиг.4С-4Е). Напротив, клеточный рост трансформантов с контролем DBD/AD (трансформанты DI-) не затрагивается даже при концентрации 3-АТ, составляющей 32 мМ (см. фиг.4Е). Следовательно, транскрипция, управляемая Pgal1 антисмысловой нити гена HIS3, значительно повышает уровень ауксотрофии к гистидину клеток DI+.
Неожиданным образом, GAL4p также ингибирует клеточный рост в случае трансформантов nDI+ (рост ингибируется, когда 3-АТ присутствует в концентрации 8 мМ или в более высоких концентрациях (см. фиг.4С-4Е)). В случае этих клеток управляемая с помощью Pgal1 транскрипция направлена к Tadh1 (см. фиг.2D), причем оказалось неожиданным, что она интерферирует с экспрессией HIS3. Ингибирование роста возникает вследствие специфической активации Pgal1, так как не наблюдают ингибирования роста, когда в клетках nDI- присутствует контроль AD/DBD (см. фиг.4В-4Е).
В присутствии GAL4p трансформанты UDI+ обнаруживают уровень ингибирования роста, подобный таковому клеток DI+. Контроль DBD/AD не затрагивает роста клеток UDI- (см. фиг.4В-4Е).
Авторы делают вывод, что в случае штаммов UI+, DI+ и UDI+ активация, вызываемая GAL4p промотора Pgal1, приводит к значительному ингибированию клеточного роста и что это ингибирование возникает вследствие пониженной активности репортера HIS3. Ингибирование роста слегка сильнее в случае клеток UI, чем клеток DI и UDI.
Индуцированное GAL4p снижение активности HIS3, следовательно, значительно замедляет клеточный рост; однако полностью он не ингибируется. Тогда как в условиях интерференции не обнаруживают колоний на чашках в течение первых 6-7 дней инкубации, маленькие колонии становятся видимыми спустя более длительные времена инкубации.
Авторы обнаружили, что интерференция является более сильной в случае системы UI, чем систем DI, UDI или nDI, что может проистекать от молекулярного механизма, генерирующего транскрипционную интерференцию. Подробные исследования интерференции против хода транскрипции показали, что активация промотора, расположенного выше хода транскрипции, снижает активность промотора, находящегося ниже по ходу транскрипции, за счет удаления факторов активации из их сайтов связывания на уровне промотора, расположенного ниже по ходу транскрипции (Greger I.H. и др., 2000; Greger I.H. и др., 1998; Valerius O.C. и др., 2002; Gallen B.P. и др., 2004). Если генерирующий интерференцию промотор расположен более близко по отношению к промотору гена-репортера в конструкции UI, то интерференция может быть более эффективной в конструкции UI, чем в DI.
Также возможно, что интерференция в прямом направлении происходит по другому молекулярному механизму. В системе DI РНК-полимеразы, транскрибирующие конвергентным образом, могут вступать в коллизию, и это может приводить к прекращению транскрипции и обеспечению умолчания гена HIS3. Фронтальная коллизия РНК-полимераз может быть менее эффективным способом обеспечения умолчания HIS3, чем удаление факторов активации транскрипции Padh1. Кроме того, возможно, что в системе DI активация Padh1 инактивирует Pgal1 за счет транскрипционной интерференции, что также может объяснять тот факт, почему система UDI не превосходит ни систему UI, ни систему DI.
Неожиданно заявитель обнаружил, что транскрипционная интерференция в прямом направлении существует также тогда, когда Pgal1 встроен тандемно и ниже по ходу транскрипции по отношению к репортерной кассете HIS3 (конструкция nDI). Уровень транскрипции гена зависит от потенции промотора, а также от качества сигнала прекращения транскрипции (Wahle E. и U. Ruegsegger, 1999). Если Pgal1 встроен между открытой рамкой считывания HIS3 и его терминатором транскрипции Tadh1, активация Pgal1 может интерферировать с соответствующей блокировкой транскрипта HIS3 в конструкции nDI, что, таким образом, уменьшает активность HIS3 в клетках.
Пример 7
Субклонирование различных вставок векторов интерференции в вектор pLac33
С целью тестирования, являются ли системы транскрипционной интерференции функциональными, когда они находятся на плазмиде и не интегрированы в геном, различные системы интерференций (DI, UI и UDI), описанные в примере 2 (см. фиг.2А-2Е), на векторах pBluescript субклонировали в виде фрагментов SalI-SacI в вектор интеграции pLac33. Этот вектор содержит ген URA3 дрожжей в качестве селекционного маркера и описан Gietz R.D. и A. Sugino, 1988. Для этого различные плазмиды pRB27, pRB17, pRB18 рестриктировали ферментами SalI-SacI и вставку субклонировали в репликативный вектор pLac33, предварительно рестриктированный ферментами SalI-SacI. Полученные различные конструкции были названы pRB28, pRB20, pRB21 соответственно.
Пример 8
Исследование роста дрожжей с неинтегрированной в их геном системой транскрипционной интерференции
Также исследовали, являются ли конструкции транскрипционной интерференции UI, DI и UDI также функциональными, когда они находятся на плазмиде и не интегрированы в геном. Для этого векторы, происходящие от плазмиды pLac33, несущие различные конструкции транскрипционной интерференции и описанные в примере 7, использовали для трансформации, независимо, штамма уСМ15.
Штамм уСМ15 независимо котрансформировали с помощью 0,3 мкг ДНК каждого из векторов pRB28, pRB20 или pRB21 (см. пример 7), ассоциированных с векторами pCL1 и pFL39. Штамм уСМ15, следовательно, трансформировали независимо с помощью трех групп из трех плазмид (pRB28+pCL1+pFL39, или pRB20+pCL1+pFL39, или pRB21+pCL1+pFL39). Центромерная плазмида pCL1 кодирует фактор транскрипции GAL4p (описывается Fields S. и O. Song, 1989), и плазмидой pFL39 является плазмида без центромерной вставки (описывается Bonneaud N. и др., 1991). Эти плазмиды репликации дрожжей несут ген LEU2 или TRP1 в качестве селекционного маркера.
Штамм уСМ15 также котрансформировали независимо с помощью 0,3 мкг ДНК каждого из векторов pRB28, pRB20 или pRB21 (см. пример 7), ассоциированных с векторами pTAL20 и pDBT (способствующими протеканию экспрессии домена активации (AD) GAL4p и ДНК-связывающего домена (DBD) GAL4p (эти векторы описаны Navarro P и др., 1997)). Штамм уСМ15, таким образом, трансформировали независимо с помощью трех групп из трех плазмид (pRB28+pTAl20+pDBT, или pRB20+pTAL20+pDBT, или pRB21+pTAL20+pDBT).
Клеточный рост каждого из этих тройных трансформантов исследовали при использовании чашек с агаровой средой с гистидином или без него и в случае чашек без гистидина без или с возрастающими концентрациями 3-АТ, как описывается в примере 6.
Результаты, полученные при использовании интерференционных конструкций, основанных на неинтегрированных плазмидах, подобны таковым, полученным в случае интегрированных интерференционных конструкций (см. фиг.5А-5D). Так, когда среда лишена гистидина и 3-АТ, клетки UI, экспрессирующие GAL4p (тройные трансформанты pRB21+pСL1+pFL39), показывают очень сильно сниженный рост по сравнению с клетками UI, не экспрессирующими активный GAL4p (тройные трансформанты pRB21+pTAL20+pDBT). Уровень ингибирования роста UI, однако, не является столь сильным, как таковой, наблюдаемый в случае интегрированного варианта конструкции UI (см. пример 6).
Клетки DI (тройные трансформанты pRB20+pСL1+pFL39 (экспрессирующие активный GAL4p) или pRB20+pTAL20+pDBT (не экспрессирующие активный GAL4p)) и клетки UDI (тройные трансформанты pRB28+pСL1+pFL39 (экспрессирующие активный GAL4p) или pRB28+pTAL20+pDBT (не экспрессирующие активный GAL4p)) растут почти нормально, экспрессируют ли они или нет активный GAL4p.
Когда среда содержит 3-АТ в концентрации 8 мМ или в более высокой концентрации, клетки, экспрессирующие GAL4p и трансформированные конструкцией UI, DI или UDI, более совсем не растут (см. фиг.5С и 5D). Этот результат сильно отличается от результатов, полученных в случае клеток, служащих отрицательным контролем (не экспрессирующие активного GAL4p (AD/DBD)) и трансформированных плазмидами интерференции, клеточный рост которых не затронут (см. фиг.5С и 5D).
Заявитель делает вывод, что варианты, основанные на плазмидах UI, DI и UDI, являются функциональными, однако несколько менее чувствительными к активации Pgal1, чем сответствующие интегрированные варианты интерференционных конструкций.
Пример 9
Независимость систем транскрипционной интерференции по отношению к «генетическому фону» штамма-хозяина
С целью исследования, зависит ли активность систем транскрипционной интерференции от «генетического фона» штамма-хозяина, также оценивали конструкции, основанные на плазмидах (таких, как описанные в примере 7), в случае второго штамма дрожжей: штамм yRB31. Штамм yRB31 (MAT a ura3-52 his3-Δ200 ade2-101 trp1-901 leu2-3,112 met-Δgal4 Δgal80) конструировали путем удаления кассеты URA3::GAL1(UAS)-GAL1(TATA)-lacZ из штамма Y187 (штамм Y187 описан Harper J.W. и др., 1993, и является доступным, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под регистрационным номером 96399).
С целью конструирования штамма yRB31 106-107 клеток клона штамма Y187 суспендировали в стерильной воде и вносили в виде тонкого слоя в чашку с агаровой средой YNB+2% глюкозы, дополненной аминокислотами met+ade+his+ura+trp+leu. Чашка с агаровой средой содержит также 5-фтороротовую кислоту ( 5-FOA) в концентрации 1 мг/мл,; 5-FOA токсична для клеток, экспрессирующих ген URA3. Соответственно, из этих чашек, содержащих 5-FOA, отбирали мутанты штамма Y187, которые утратили репортерную кассету URA3::GAL1(UAS)-GAL1(TATA)-lacZ (Boeke и др., 1987). Отсутствие репортерной кассеты в полученных мутантах подтверждено путем тестирования активности репортера lacZ. Мутантов трансформировали с помощью плазмиды pCL1 (Fields S. и O. Song, 1989), кодирующей фактор транскрипции GAL4p, который является очень сильным активатором промотора GAL1, и определяли активность lacZ (Breeden L. и K. Nasmyth, 1985). Был отобран мутант, резистентный к 5-FOA и без активности lacZ, и назван yRB31.
Штамм yRB31 трансформировали таким же образом, как описывается в примере 8 в случае штамма yCM15. Полученные трансформанты исследовали в отношении того, что касается их клеточного роста, таким же образом, как описывается в примере 8. Не было обнаружено различия между штаммами уСМ15 и yRB31 в том, что касается активности плазмид интерференции.
Пример 10
Индукция экспрессии систем интерференции UI и DI путем взаимодействия белок-белок и воздействие на ингибирование клеточного роста
10.1: Индукция экспрессии систем интерференции путем идентифицированного выше взаимодействия белка Fe65 с белком АРР
Исследовали, способно ли идентифицированное выше взаимодействие белок-белок между белками Fe65 и АРР (McLoughlin D.M. и Miller C.C., 1996; Fiore F. и др., 1995; Zambrano N. и др., 1997) индуцировать в достаточной степени конструкцию транскрипционной интерференции для ингибирования клеточного роста дрожжей.
Использовали интегрированные варианты конструкций UI и DI. Гибридные белки AD-Fe65 и DBD-AppCt экспрессировали под контролем сильно укороченного активного промотора ADH1 (промотор, описанный Ruohonen L. и др., 1995).
Плазмида pRB34 позволяет осуществляться экспрессии гибридного белка AD-Fe65 под контролем сильно укороченного активного промотора ADH1.
Конструирование плазмиды pRB34 осуществляли в несколько стадий. На первой стадии часть рамки считывания гена Fe65 клонировали в вектор pGAD424 (BD Biosciences Clontech (Palo Alto, Калифорния, США)). Фрагмент PCR из 1694 пар оснований, соответствующий аминокислотам 152-708 гена Fe65 (номер доступа в банке генов: ВС010854), генерировали путем PCR, используя в качестве праймеров следующие олигонуклеотиды:
RB250 5'-AGATCGAATTCAAGGCGGCCGGGGAGGCCGAGG-3' (идентифицированная последовательность № 27) и RB251 5'-GCAGGTCGACTCATGGGGTATGGGCCCCCAGC-3' (идентифицированная последовательность № 28) (сайты EcoRI и SalI подчеркнуты) и в качестве матрицы банк плазмидной ДНК коммерчески доступной кДНК головного мозга человека (BD Biosciences Clontech (Palo Alto, Калифорния, США)). Продукт PCR рестриктировали ферментами EcoRI-SalI и клонировали на уровне сайтов EcoRI-SalI вектора pGAD424. Таким образом полученный вектор был назван pRBSGl1. Последовательность этой конструкции подтверждена путем секвенирования при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
На второй стадии осуществляли частичную рестрикцию плазмиды pRBSG11 с помощью фермента SphI и фрагмент, кодирующий слегка укороченный активный промотор ADH1, гибридный белок AD-Fe65 и терминатор ADHI встраивали в сайт SphI центромерной плазмиды pFL36 (описана Bonneaud N. и др., 1991); это позволяет получить плазмиду pRBIM54. Получение пригодной конструкции подтверждено путем рестрикционного анализа. Фрагмент может быть встроен в двух направлениях, и отбирали клон, обладающий желательной ориентацией, в котором промотор ADHI близок к единственному сайту SacI плазмиды pFL36. На третьей стадии фрагмент, кодирующий сильно укороченный активный промотор ADH1, амплифицировали путем PCR, исходя из геномной ДНК штамма FL100, при использовании праймеров RB136 (5'-TTGTAAAACGACGGCCAGTGAATTCCGTACGATATCCTTTTGTTGTTTCCGGGTG-3' (идентифицированная последовательность № 29) и RB137 (5'-AGTTGATTGTATGCTTGGTATAGC-3' (идентифицированная последовательность № 30)). Используя метод клонирования путем гомологичной рекомбинации (GAP-репарация) (DeMarini и др., 2001; Orr-WEAVER и др., 1983), использовали фрагмент PCR для замены слегка укороченного активного промотора ADH1 сильно укороченным активным промотором ADH1 в плазмиде pRBIM54, предварительно рестриктированной ферментом SacI. Гомологичную рекомбинацию осуществляли в штамме уСМ15. Конструкцию новой плазмиды подтверждали путем рестрикционного анализа, и она была названа pRB34. Последовательность этой плазмиды представлена в виде идентифицированной последовательности № 31.
Плазмида pRB35 позволяет осуществляться экспрессии гибридного белка DBD-AppCt под контролем сильно укороченного активного промотора ADFH1. Конструирование плазмиды pRB35 осуществляли в несколько стадий.
На первой стадии карбокси-концевую часть рамки считывания гена HSAFPA4 (App, номер доступа в EMBL: Y00264) клонировали в вектор pGBT9 (BD Biosciences Clontech (Palo Alto, Калифорния, США)). Фрагмент из 143 пар оснований, кодирующий аминокислоты 650-695 гена HSAFPA4, получали путем гибридизации двух однонитевых фосфорилированных олигонуклеотидов: RB252 с последовательностью 5'-AATTCAAGAAACAGTACACATCCATTCATCATGGTGTGGTGGAGGTTGACG CCGCTGTCACCCCAGAGGAGCGCCACCTGTCCAAGATGCAGCAGAACGGCT ACGAAAATCCAACCTACAAGTTCTTTGAGCAGATGCAGAACTAGG-3' (идентифицированная последовательность № 32) и RB253 с последовательностью 5'-TCGACCTAGTTCTGCATCTGCTCAAAGAACTTGTAGGTTGGAmTCGTA GCCGTTCTGCTGCATCTTGGACAGGTGGCGCTCCTCTGGGGTGACAGCGGC GTCAACCTCCACCACACCATGATGAATGGATGTGTACTGTTTCTTG -3' (идентифицированная последовательность № 33). Для этого два однонитевых олигонуклеотида смешивали в эквимолярных концентрациях в воде. Смесь нагревали при температуре 90°С в течение 5 минут в нагревательном блоке, затем блок оставляли охлаждаться до комнатной температуры (примерно 2 часа). Таким образом полученный двунитевой фрагмент обладает по своим краям совместимыми липкими концами EcoRI и SalI соответственно. Это позволяет осуществлять встраивание этого двунитевого фрагмента (соответствующего карбокси-концевой части HSAFPA4) в вектор pGBT9 (BD Biosciences Clontech). Таким образом полученный вектор позволяет осуществляться экспрессии гибридного белка между GAL4DBD и аминокислотами 650-695 Арр (DBD-AppCt), и он был назван pSG14. Последовательность этой конструкции подтверждена путем секвенирования при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
На второй стадии плазмиду pSG14 частично рестриктировали с помощью фермента SphI. Затем фрагмент, кодирующий слегка укороченный активный промотор ADH1, гибридный белок DBD-AppCt и терминатор ADHI встраивали в вектор pFL39 (описан Bonneaud N. и др., 1991), предварительно рестриктированный ферментом SphI. Полученная в результате плазмида была названа pIM55. Получение пригодной конструкции подтверждено путем рестрикционного анализа. Фрагмент может быть встроен в двух направлениях, и отбирали клон, обладающий желательной ориентацией, в котором промотор ADHI близок к единственному сайту SacI плазмиды pFL36.
Затем получали плазмиду pRB35 путем замены слегка укороченного активного промотора ADHI pIM55 сильно укороченным активным промотором ADH1. Такой же фрагмент PCR, как и таковой, используемый для конструкции pRB34, встраивали в pIM55, предварительно рестриктированную ферментом SphI, используя такой же способ GAP-репарации. Плазмидная конструкция подтверждена рестрикционным анализом и она была названа pRB35. Последовательность этой плазмиды представлена в виде идентифицированной последовательности № 34.
Во-первых, была подтверждена активность конструкций. Для этого репортерный штамм Y187 (штамм Y187 имеет ген URA3, находящийся под контролем промотора Pgal1, описан Harper J.W. и др., 1993, и доступен, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под регистрационным номером 96399) трансформировали тремя парами плазмид. Эти пары представляют собой следующие: pRB34+pRB35, pRB34+pDBT и pTAL20+pRB35. Отбирали два клона, котрансформированных с помощью каждой из этих пар плазмид. Исследовали их способность расти на минимальной среде с урацилом или без него. Показано, что шесть соответствующих клонов способны расти на минимальной среде в присутствии урацила (см. фиг.6А). Однако в отсутствие урацила способны расти только два клона, котрансформированных с помощью плазмид pRB34+pRB35 (см. фиг.6А). Это указывает на то, что ген URA3 активен в случае штамма Y187 и это благодаря взаимодействию между AD-Fe65 и DBD-AppCt, реконструирующему активный фактор транскрипции GAL4p, способный активировать промотор Pgal1. Следовательно, доказана активность используемых конструкций.
Штаммы yRB48 (система DI) и yRB49 (система UI) затем котрансформировали, каждый, с помощью пары плазмид pRB34 и pRB35, позволяющих осуществляться экспрессии двух гибридных белков AD-Fe65 и DBD-AppCt. Штаммы yRB48 (система DI) и yRB49 (система UI) также котрансформировали, каждый, с помощью пар плазмид pRB34+pDBT и pTAL20+pRB35 в качестве контролей. Отбирали два клона, котрансформированных с помощью каждой из этих пар плазмид. Клеточный рост каждого из двух клонов этих двойных трансформантов исследовали при использовании чашек с агаровой средой с гистидином или без него и в случае чашек без гистидина без или с возрастающими концентрациями 3-АТ, как описано в примере 6.
Как показано на фиг.6В-6E, взаимодействие между гибридными белками AD-Fe65 и DBD-AppCt ингибирует клеточный рост штаммов, несущих интерференционные конструкции UI и DI, тогда как клетки, экспрессирующие гибридный белок AD-65 и домен DBD или гибридный белок DBD-AppCt и домен AD, растут нормально. В этом эксперименте различия на уровне клеточного роста обнаруживали тогда, когда добавляли 3-АТ в более высоких концентрациях, составляющих 15 мМ.
Может быть сделан вывод, что транскрипционная интерференция, индуктированная двугибридным взаимодействием, может быть легко определена с помощью систем rY2H и DI.
10.2: Индукция экспрессии систем интерференции за счет идентифицированного выше взаимодействия белка р53 с белком mdm2
Также исследовали, способно ли другое, идентифицированное выше, взаимодействие белок-белок между белками р53 и mdm2 (Oliver J.D. и др., 1993; Chen J. и др., 1993; Momand J. и др., 1992) индуцировать в достаточной степени конструкцию транскрипционной интерференции для ингибирования клеточного роста дрожжей. Для этого использовали интегрированный вариант конструкции UI. Гибридные белки DBD-mdm2 и VP16-p53(1-55) экспрессировали под контролем полного промотора гена ADHI, исходя из плазмид 2μ.
С целью конструирования вектора pVP16-p53Nt генерировали открытую рамку считывания, кодирующую 55 аминокислот амино-конца человеческого белка р53 (регистрационный номер в банке генов: ВС003596), путем гибридизации двух однонитевых фосфорилированных олигонуклеотидов: RB200
5'-GGCCGCAGTGAACCATTGTTCAATATCGTCCGGGGACAGCATCAAATCATCCATTGCTTGGGACGGCAAGGGGGACAGAACGTTGTTTTCAGGAAGTAGTTTCCATAGGTCTGAAAATGTTTCCTGACTCAGAGGGGGCTCGACGCTAGGATCTGACTGCGGCTCCTCCATCTG-'3 (идентифицированная последовательность № 35) и RB201
5'-GATCCAGATGGAGGAGCCGCAGTCAGATCCTAGCGTCGAGCCCCCTCTGAGTCAGGAAACATTTTCAGACCTATGGAAACTACTTCCTGAAAACAACGTTCTGTCCCCCTTGCCGTCCCAAGCAATGGATGATTTGATGCTGTCCCCGGACGATATTGAACAATGGTTCACTGC-3' (идентифицированная последовательность № 36) (сайты NotI и BamH1 подчеркнуты). Для этого два однонитевых комплиментарных олигонуклеотида смешивали в эквимолярных концентрациях в воде. Смесь нагревали при температуре 90°С в течение 5 минут в нагревательном блоке, затем блок оставляли охлаждаться до комнатной температуры (примерно 2 часа). Таким образом полученный двунитевой фрагмент имеет на краях совместимые липкие концы NotI и BamH1 соответственно.
Затем осуществляли лигирование фрагмента с вектором pVP16 (Vojtek и Hollenberg, 1995; Vojtek и др., 1993), предварительно рестриктированным с помощью ферментов NotI и BamH1. Последовательность этой конструкции подтверждена путем секвенирования при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
С целью конструирования вектора pDTB-mdm2 ген, кодирующий человеческий целый белок Mdm2 (регистрационный номер в банке генов: NM_002392), амплифицировали путем PCR, используя вектор pCR3-mdm2fl (Sigalas и др., 1996) в качестве матрицы и следующие олигонуклеотиды:
RB202 (5'-CCCGGGAATTCAGATCCATATGTGCAATACCAACATGTCTGTAC-3') (идентифицированная последовательность № 37) и RB203 (5'-ACTTAGAGCTCTAGGGGAAATAAGTTAGCACAATC-3') (идентифицированная последовательность № 38) (сайты EcoRI и SacI подчеркнуты). Продукт PCR рестриктировали с помощью ферментов EcoRI и SacI и клонировали на уровне сайтов EcoRI и SacI в вектор pDBT (Navarro и др., 1997). Последовательность этой конструкции подтверждена путем секвенирования при использовании набора «Big Dye Terminator» (Perkin-Elmer, Wellesley, Massachusetts, США).
Подтверждена активность полученных конструкций. Для этого репортерный штамм Y187 (штамм Y187 имеет ген URA3, находящийся под контролем Pgal1, описан Harper J.W. и др., 1993, и доступен, в частности, из Американской коллекции типовых культур (АТСС) (Manassas, Virginie, США) под регистрационным номером 96399) трансформировали с помощью трех пар плазмид. Эти пары являются следующими: pDBT-mdm2+pVP16-p53Nt, pVP16-p53Nt+pDBT и pTAL20+pDBT-mdm2. Два клона, котрансформированных с помощью двух плазмид, отбирали в случае каждой из этих пар плазмид. Исследовали их способность расти на минимальной среде с урацилом и без него. Показано, что шесть соответствующих клонов способны расти на минимальной среде в присутствии урацила (см. фиг.6F). Однако в отсутствие урацила способны расти только два клона, котрансформированных с помощью плазмид pDBT-mdm2+pVP16-p53Nt. Это указывает на то, что ген URA3 активирован в штамме Y187 и это благодаря взаимодействию между pDBT-mdm2 и pVP16-p53(1-55), реконструирующему активный фактор транскрипции, способный активировать промотор Pgal1. Следовательно, доказана активность используемых конструкций.
Штамм yRB49 (система UI) затем котрансформировали с помощью пары плазмид pDBT-mdm2+pVP16-p53Nt, позволяющих осуществляться экспрессии двух гибридных белков pDBT-mdm2 и pVP16-p53(1-55). Штамм yRB49 (система UI) также котрансформировали с помощью пары плазмид pDBT-mdm2+pTAL20 в качестве контролей. Два клона, котрансформированных с помощью двух плазмид, отбирали в случае каждой из этих двух пар плазмид. Клеточный рост каждого из двух клонов этих двойных трансформантов исследовали при использовании чашек с агаровой средой с гистидином и без него и в случае чашек без гистидина без или с возрастающими концентрациями 3-АТ, как описано в примере 6. Как показано на фигуре 6G, взаимодействие между гибридными белками pDBT-mdm2 и pVP16-p53(1-55) ингибирует клеточный рост штаммов, несущих интерференционные конструкции UI, тогда как клетки, экспрессирующие гибридный белок DBD-mdm2 и домен AD, растут нормально.
В этом эксперименте различия на уровне клеточного роста определяли в присутствии концентрации 3-АТ, равной или выше 5 мМ.
Может быть сделан вывод, что транскрипционная интерференция, индуктированная двугибридным взаимодействием, может быть легко определена при использовании систем rY2H UI.
Пример 11
Применение системы транскрипционной интерференции rY2H UI для скрининга ингибиторов взаимодействий белок-белок
11.1: Калибровка теста репарации роста дрожжей на твердых средах
Штамм дрожжей, содержащий систему транскрипционной интерференции rY2H UI, интегрированную в его геном (yRB49), использовали для выделения ингибиторов взаимодействий белок-белок с помощью теста репарации роста.
Для осуществления этого теста штамм дрожжей, содержащий систему транскрипционной интерференции rY2H UI, связанную со специфическим взаимодействием белок-белок, наносили тонким слоем на твердую среду, содержащую 3-АТ. Капли растворов химических соединений вносили в чашку, в которую произведен посев. Диффузия химических соединений в агар создает градиент концентрации вокруг зоны нанесения. Продукты, способные ингибировать взаимодействие белок-белок, позволяют осуществляться росту клеток, что выражается в образовании ореола репарации роста, окружающего зону нанесения.
С целью определения условий для разработки такого теста осуществляли серию предварительных экспериментов. Так, стремились к оценке, во-первых, количества клеток, помещаемых в чашки Петри, и, во-вторых, минимальной концентрации 3-АТ в культуральной среде, позволяющей ингибировать рост клеток.
Сначала дрожжи, содержащие интегрированный вариант системы UI (yRB49), котрансформировали плазмидами pRB34 и pRB35 с целью получения системы транскрипционной интерференции в зависимости от взаимодействующей пары белок-белок Fe65/App. Эти дрожжи затем культивировали на жидкой среде, содержащей гистидин. Как только оптическая плотность (DO) культуры достигала значения 3 (3·107 клеток на мл), клетки подвергали центрифугированию в течение 10 минут со скоростью 3000 оборотов в минуту, промывали два раза стерильной водой и снова растворяли при достижении DO=0,1 или 0,01. 10 мл этой суспензии равномерно наносили тонким слоем на всю поверхность твердой культуральной среды, предварительно помещенной в квадратные чашки Петри со стороной 120 мм. Спустя одну минуту жидкость удаляли путем аспирации. Чашки высушивали в течение 15 минут в вытяжном шкафу с ламинарным потоком, затем инкубировали при температуре 30°С в течение 48 часов.
На фиг.8А показано, что оптимального обсеменения чашек достигали при наиболее низшем разведении, или DO=0,01 (правая чашка).
С целью определения минимальной концентрации 3-АТ, способной ингибировать рост дрожжей, штамм yRB49 трансформировали, как ранее, с помощью плазмид pRB34 и pRB35 (Fe65/App) или с помощью контрольных плазмид pTAL20/pDBT (не экспрессирующих взаимодействие белок-белок). Два трансформированных штамма наносили тонким слоем на среду, либо содержащую гистидин, либо без гистидина и дополненную с помощью 0 мМ, 0,5 мМ или 1 мМ 3-АТ. Как показано на фиг.8В, штамм, «несущий» взаимодействие Fe65/App, имеет очень замедленный рост на среде, содержащей 1 мМ 3-АТ, по сравнению со штаммом, содержащим контрольные плазмиды, рост которого не затронут в тех же условиях.
В том же самом эксперименте с целью быстрого тестирования токсичности молекул в растворе в случае этого теста репарации роста были выбраны, по случаю, две молекулы из библиотеки 10000 соединений. После растворения при концентрации 10 мМ в ДМСО 1 мкл каждого раствора помещали в каждую чашку, а также 1 мкл ДМСО. Ни одна из этих молекул неспособна вызывать репарацию роста дрожжей. Однако они продуцируют диск ингибирования роста вокруг зоны нанесения молекул, связанный с токсическими эффектами этих молекул в сильной концентрации (фиг.8В).
Добавление гистидина в среду корректирует неспособность штамма к синтезу гистидина, высвобождая систему транскрипционной интерференции. С целью подтверждения того, что описанные выше экспериментальные условия позволяют детектировать репарацию роста дрожжей, штамм yRB49, котрансформированный с помощью пары плазмид pRB34 и pRB35 (взаимодействие Fe65/App), наносили тонким слоем на предыдущую среду без гистидина и дополненную 1 мМ 3-АТ, затем в центр каждой чашки вносили возрастающие количества урацила, аденина, лейцина или гистидина.
Как показано на фиг.8С, только гистидин способен вызывать репарацию роста дрожжей вокруг места нанесения и независимо от его концентрации.
11.2: Детекция репарации роста дрожжей, индуктированной ингибитором взаимодействия белок-белок
Экспериментальные условия, указанные выше, использовали в большом масштабе для скрининга коллекции из 10000 молекул. В этих условиях каждую молекулу коллекции помещали, примерно по 0,2 мкл 10 мМ раствора в ДМСО, благодаря 96-игольчатому репликатору на твердую среду, лишенную гистидина и содержащую 1 мМ 3-АТ. Робототехническое устройство, используемое для этого скрининга, описано ранее Beydon и др. (2000). Так, были выделены две молекулы, которые способны вызывать репарацию роста дрожжей: в случае каждого из этих продуктов интенсивный белый ореол репарации роста легко различим вокруг зоны ингибирования роста, связанной с токсическим воздействием этих молекул в сильных концентрациях, тогда как в случае контрольного продукта 3 никакого эффекта не обнаружено, как и ожидалось (фиг.9).
Для подтверждения специфичности воздействия выделенных согласно этому тесту молекул воздействие трех других молекул, потенциально ингибирующих взаимодействие Fe65/App (называемых А, В и С на фиг.11), тестировали в случае другого типа взаимодействия белок-белок, используя эту систему интерференции, взаимодействия р53/mdm2 (см. пример 10.2).
Сначала тестировали путем капельного теста относительный ответ системы интерференции UI в зависимости от взаимодействий Fe65/App и p53/mdm2. Таким образом, наблюдали, что комплекс р53/mdm2 ингибирует рост дрожжей на среде без гистидина, но при более сильной концентрации 3-АТ, чем таковая, используемая в случае штамма, включающего взаимодействие Fe65/App (фиг.10). Это наводит на мысль, что взаимодействие р53/mdm2 может быть более слабым, чем взаимодействие Арр/Fe65, или же что интерактанты менее хорошо экспрессируются дрожжами.
Затем трансформировали штамм yRB49 с помощью пары плазмид pRB34 и pRB35, приводящей к экспрессии взаимодействия Fe65/Apр, или пары pDBN-mdm2 и pVP16-p53Nt, приводящей к экспрессии взаимодействия р53/mdm2.
Два штамма Fe65/App и p53/mdm2, каждый, вносили в виде тонкого слоя в чашки со средой, лишенной гистидина и дополненной, соответственно, с помощью 1 мМ и 20 мМ 3-АТ. В случае каждой из трех потенциально ингибирующих молекул помещали три капли по 1 мкл растворов с концентрацией 0,1 мМ, 1 мМ и 10 мМ в ДМСО на чашку. Кроме того, 1 мкл гистидина (0,125% мас./об.) помещали в центр другой чашки в качестве положительного контроля репарации роста. В качестве отрицательного контроля использовали продукт, выбранный по случаю из коллекции и используемый в тех же самых концентрациях, что и таковые трех тестируемых молекул.
Результаты, представленные на фиг.11, показывают, что продукты А и В являются активными, поскольку они индуцируют ореол репарации роста штамма Fe65/App, размер которого зависит от используемой концентрации. Такой ореол в зависимости от концентрации в случае штамма р53/mdm2 не наблюдается, что указывает на специфичность ингибирующего воздействия этих двух молекул. Напротив, продукт С индуцирует рост двух штаммов, Fe65/App и p53/mdm2, и, следовательно, может быть классифицирован как активный, но не специфический. Как ожидалось, продукт D не оказывает воздействия на два тестируемых штамма; он рассматривается как неактивный.
Тест на репарацию роста, следовательно, позволяет выделять из библиотеки в несколько тысяч молекул молекулы, которые являются специфическими ингибиторами взаимодействия белок-белок.
Все указанные публикации и патенты включены в настоящую заявку путем ссылки.
Библиография:
























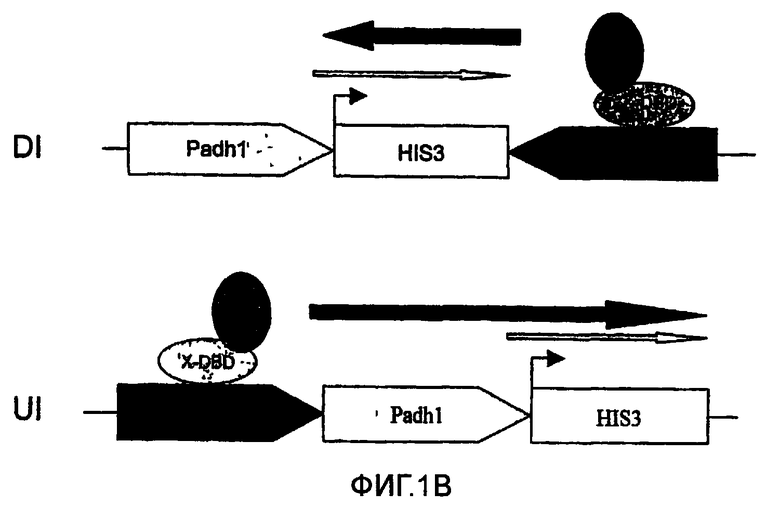
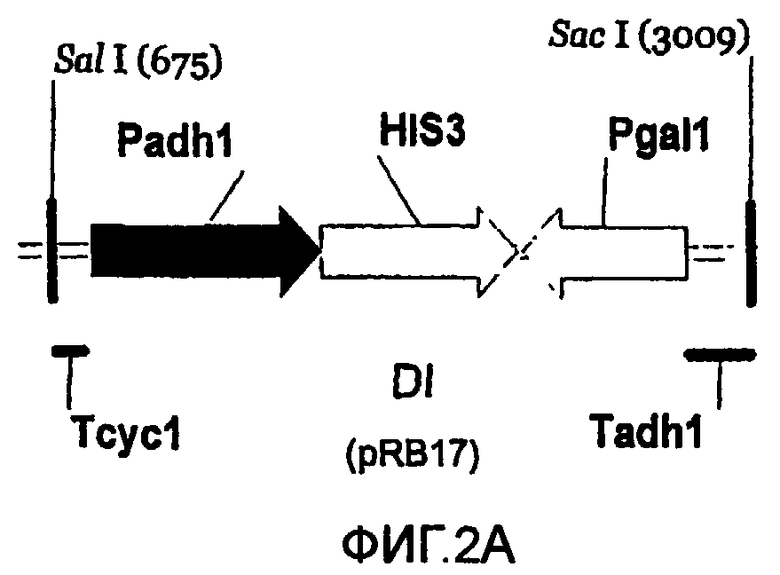
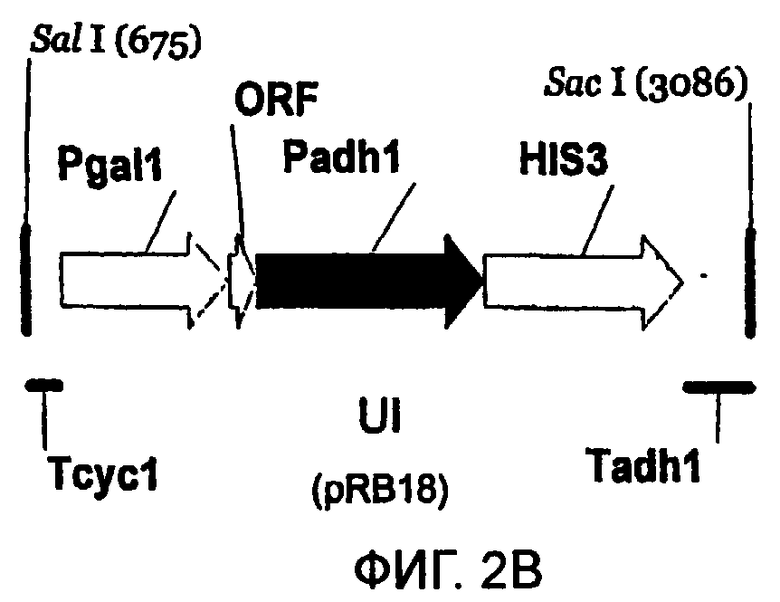
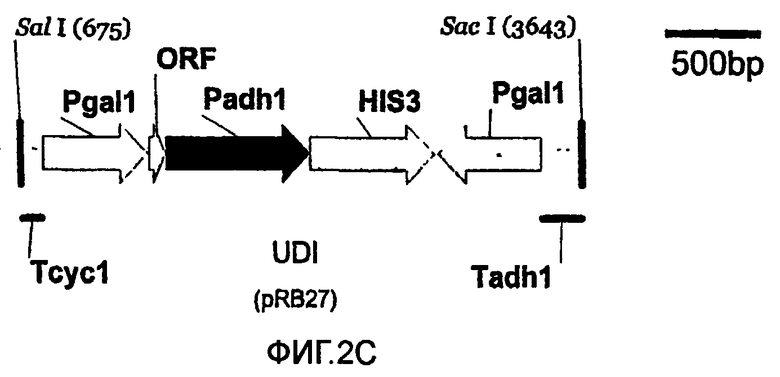


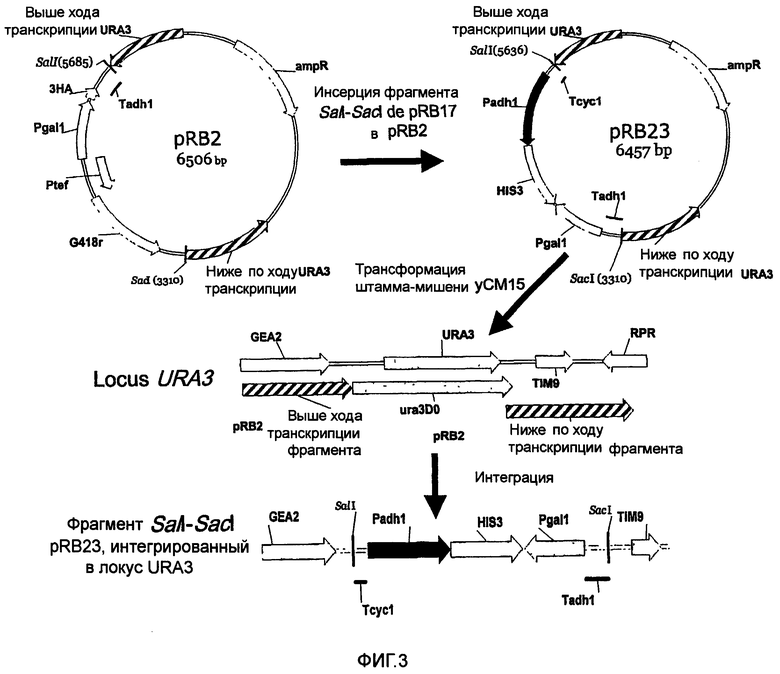
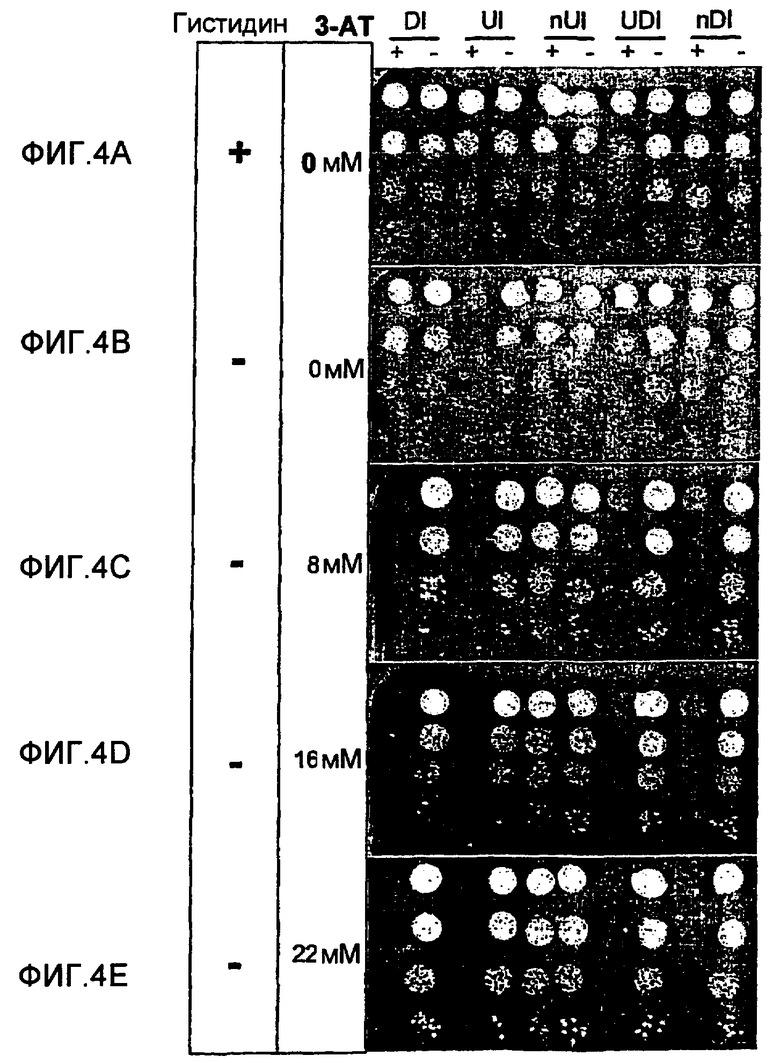
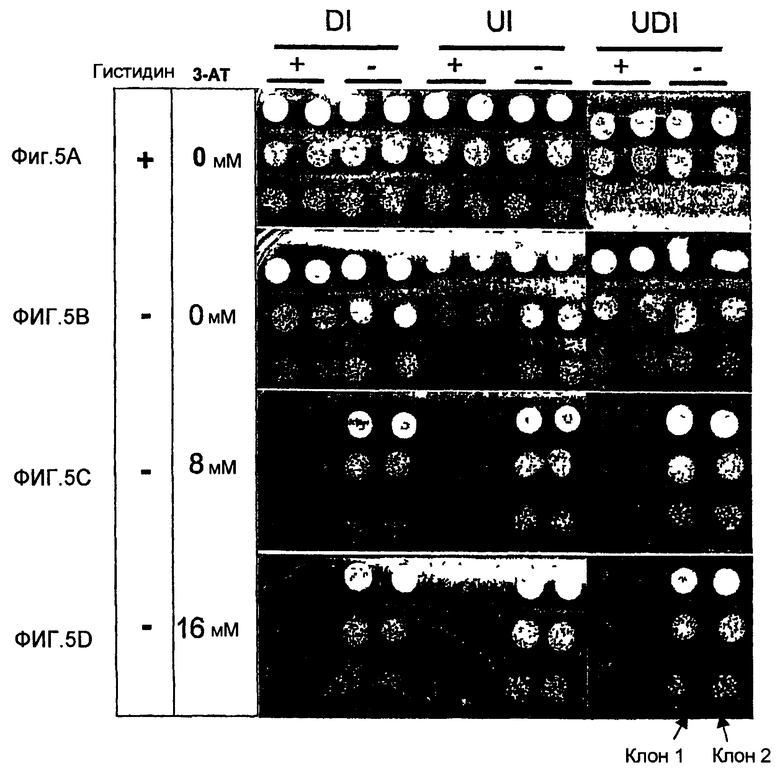


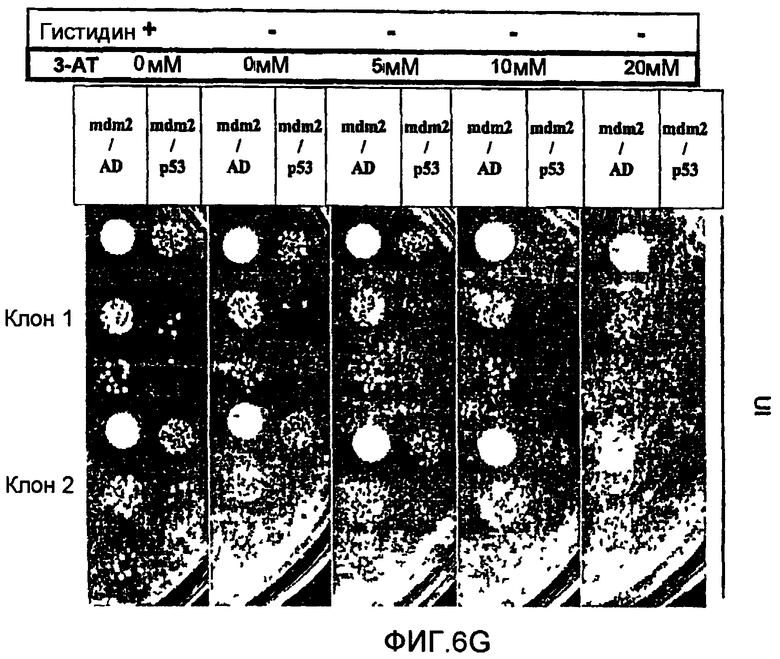
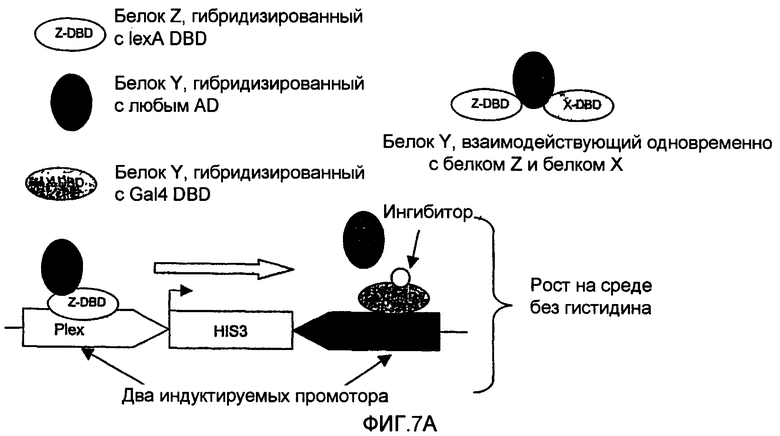
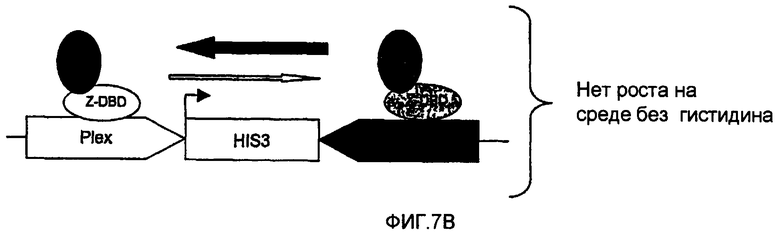

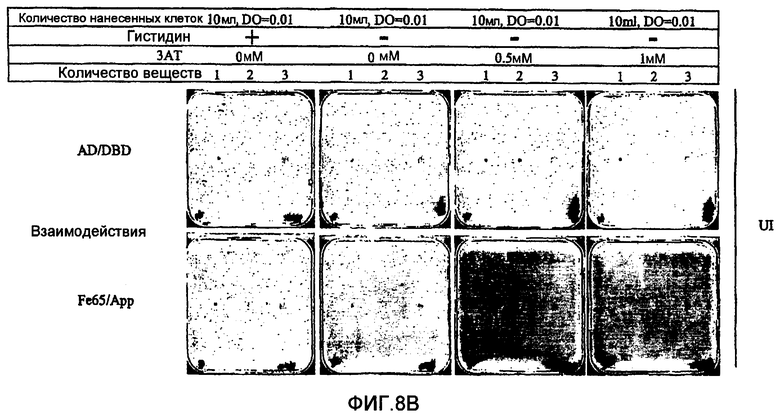
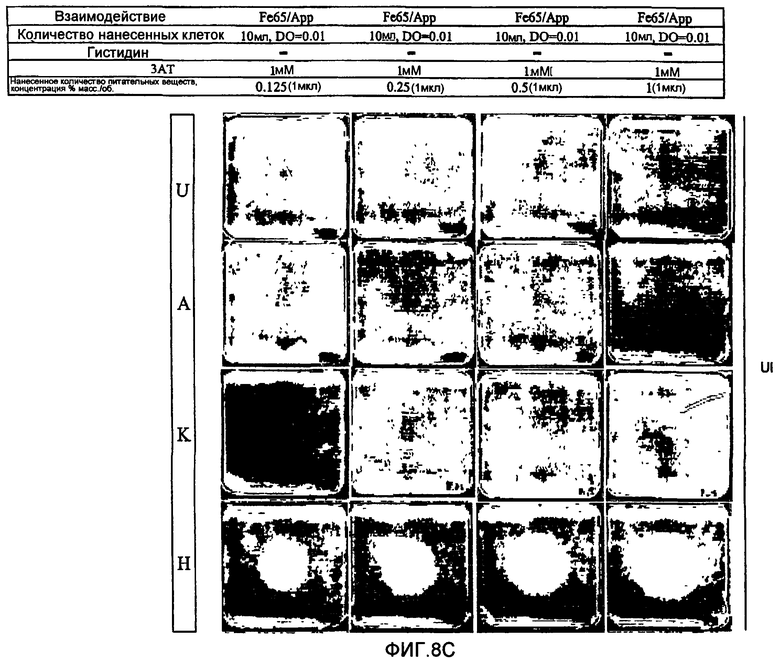
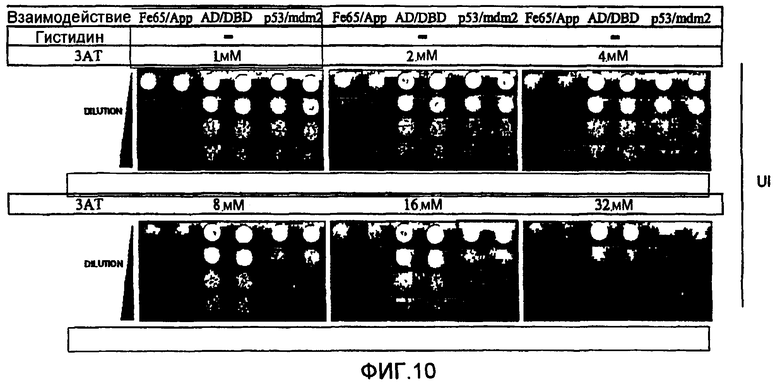
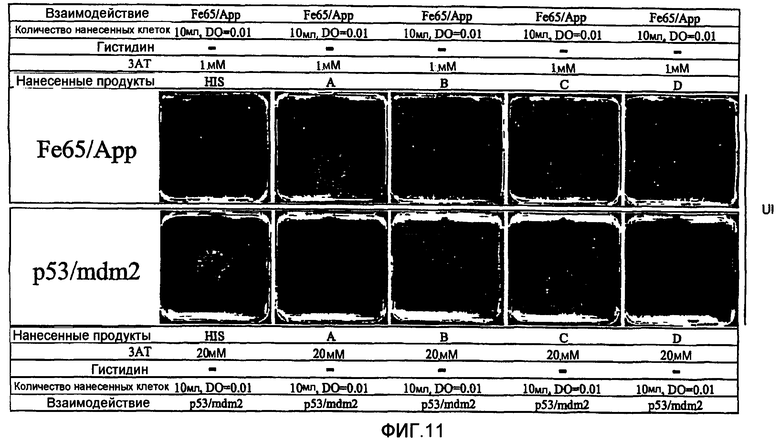
Изобретение относится к биотехнологии. Описана дрожжевая клетка, включающая интерференционную конструкцию ДНК. Конструкция включает: ген-репортер, находящийся под контролем первого промотора; один или несколько индуцируемый(ых) промотор(ов), так называемый(ых) промотор(ов) интерференции, выбираемый(ых) и фиксированный(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера. При этом клетка экспрессирует: первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), слитый с белком Y, способным взаимодействовать по меньшей мере с белком-партнером X; второй химерный белок (X-DBD), образованный первым, ДНК-связывающим доменом (DBD), слитым со вторым доменом, образованным белком X, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции. Преложен способ идентификации соединения, ингибирующего взаимодействие первого белка X со вторым белком Y. Изобретение позволяет осуществлять детектирование прерывания взаимодействия белок-белок, а также осуществлять идентификацию дефицитных аллелей на уровне взаимодействия белков, принимающих участие во взаимодействиях белок-белок. 5 н. и 22 з.п. ф-лы, 32 ил., 4 табл.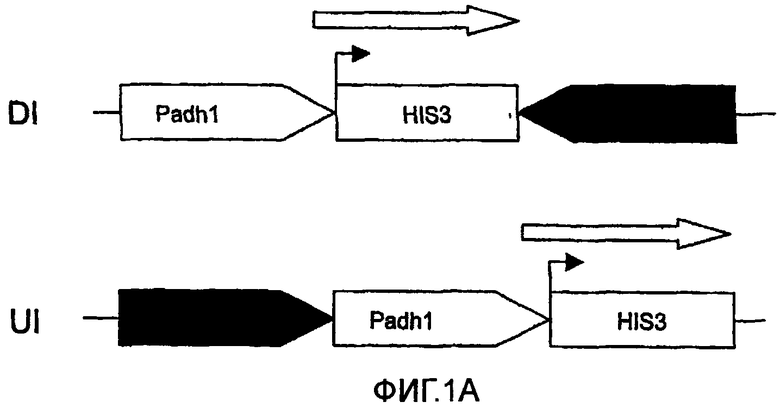
1. Дрожжевая клетка, включающая интерференционную конструкцию ДНК, причем вышеуказанная конструкция включает:
ген-репортер, находящийся под контролем первого промотора;
один или несколько индуцируемый(ых) промотор(ов), так называемый(ых) промотор(ов) интерференции, выбираемый(ых) и фиксированный(ых) таким образом, что его(их) активация вызывает транскрипционную интерференцию первого промотора, приводящую к поддающемуся обнаружению уменьшению экспрессии гена-репортера;
причем вышеуказанная клетка, кроме того, экспрессирует:
первый химерный белок (Y-AD), образованный доменом активации транскрипции (AD), слитый с белком Y, способным взаимодействовать по меньшей мере с белком-партнером X;
второй химерный белок (X-DBD), образованный первым ДНК-связывающим доменом (DBD), слитым со вторым доменом, образованным белком X, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков X-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего промотор или промоторы интерференции.
2. Клетка по п.1, отличающаяся тем, что промотор, регулирующий экспрессию гена-репортера, представляет собой индуцируемый промотор, белок Y способен взаимодействовать с двумя белками-партнерами X и Z и клетка экспрессирует третий химерный белок (Z-DBD), образованный ДНК-связывающим доменом (DBD), гибридизированным со вторым доменом, образованным белком Z, способным взаимодействовать с белком Y, причем взаимодействие двух химерных белков Z-DBD и Y-AD приводит к конструкции функционального фактора транскрипции, активирующего экспрессию гена-репортера.
3. Клетка по п.2, отличающаяся тем, что индуцируемый, регулирующий экспрессию гена-репортера промотор включает последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (Z-DBD).
4. Клетка по п.1, отличающаяся тем, что промотор, регулирующий экспрессию гена-репортера, представляет собой конститутивный промотор.
5. Клетка по п.1, отличающаяся тем, что она представляет собой клетку-хозяина, трансформированную или трансфицированную с помощью интерференционной конструкции ДНК и конструкций ДНК, кодирующих химерные белки, причем совокупность этих конструкций несут один или несколько неинтегративных векторов.
6. Клетка по п.1, отличающаяся тем, что она предтавляет собой клетку-хозяина, трансформированную или трансфицированную конструкциями ДНК, кодирующими химерные белки, причем эти конструкции несут один или несколько неинтегративных векторов, и интерференционная конструкция ДНК интегрирована в геном клетки.
7. Клетка по п.1, отличающаяся тем, что интерференционная конструкция ДНК и конструкции ДНК, кодирующие химерные белки, интегрированы в геном клетки.
8. Клетка по п.6, отличающаяся тем, что интерференционная конструкция ДНК интегрирована в локус, лишенный пертурбированных геномных активностей транскрипции.
9. Клетка по п.1, отличающаяся тем, что ее выбирают из группы, состоящей из клеток млекопитающих, насекомых, растений и дрожжей.
10. Клетка по п.9, отличающаяся тем, что она представляет собой дрожжевую клетку.
11. Клетка по п.10, отличающаяся тем, что она происходит от дрожжей вида Saccharomyces cerevisiae, Schizosaccharomyces pombe, Kluyveromyces lactis, Pichia pastoris, Saccharomyces carlsbergensis или Candida albicans.
12. Клетка по п.1, отличающаяся тем, что по меньшей мере один промотор интерференции фиксирован ниже по ходу транскрипции гена-репортера и первого промотора и в ориентации, противоположной этому последнему (DI).
13. Клетка по п.1, отличающаяся тем, что по меньшей мере один промотор интерференции фиксирован ниже по ходу транскрипции гена-репортера и первого промотора и в той же ориентации, что и таковая этого последнего (nDI).
14. Клетка по п.1, отличающаяся тем, что по меньшей мере один промотор интенференции фиксирован выше хода транскрипции гена-репортера и первого промотора и в той же самой ориентации, что и таковая этого последнего (UI).
15. Клетка по п.1, отличающаяся тем, что по меньшей мере один промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером, причем промотор или промоторы интерференции, расположенные ниже по ходу транскрипции первого промотора и гена-репортера, обладают ориентацией, конвергентной по отношению к первому промотору, и промотор или промоторы интерференции, фиксированные выше хода транскрипции первого промотора и гена-репортера, обладают ориентацией, идентичной таковой первого промотора (UDI).
16. Клетка по п.1, отличающаяся тем, что по меньшей мере один промотор интерференции фиксирован с той и другой стороны первым промотором и геном-репортером, причем промотор или промоторы интенференции, расположенные ниже по ходу транскрипции первого промотора и гена-репортера, обладают ориентацией, тандемной по отношению к первому промотору, и промотор или промоторы интерференции, фиксированные выше хода транскрипции первого промотора и гена-репортера, обладают ориентацией, идентичной таковой первого промотора (nUDI).
17. Клетка по п.1, отличающаяся тем, что геном-репортером является ген, необходимый для выживания клетки.
18. Клетка по п.17, отличающаяся тем, что геном-репортером является ген, необходимый для первичного метаболизма, клеточного деления, синтеза белков, синтеза ДНК или синтеза РНК.
19. Клетка по п.1, отличающаяся тем, что ген-репортер сам по себе не является необходимым для выживания клетки, но является необходимым для выживания клетки, когда ее транскрипция ингибирована, за счет ассоциации с одним или несколькими генами-репортерами одного и того же типа, экспрессия которых контролируется или нет системой транскрипционной интерференции.
20. Клетка по п.1, отличающаяся тем, что индуцируемый промотор или индуцируемые промоторы интерференции включают последовательность, способную взаимодействовать с ДНК-связывающим доменом (DBD) химерного белка (X-DBD).
21. Клетка по п.1, отличающаяся тем, что индуцируемый промотор или индуцируемые промоторы интерференции включают последовательность, способную взаимодействовать с белком, обладающим ДНК-связывающим доменом (DBD), выбираемым из группы, состоящей из GAL4 UAS, LexAop, clop и TetRop, и ДНК-связывающим доменом химерного белка X-DBD является соответствующий DBD (соответственно, GAL4, LexA, cl и TetR).
22. Клетка по п.1, отличающаяся тем, что домен активации транскрипции (AD) химерного белка Y-AD выбирают из группы, состоящей из доменов активации транскрипции следующих белков: B42, VP16 и GAL4p.
23. Клетка по п.1, отличающаяся тем, что интерференционная конструкция ДНК фланкирована на своих концах одним или несколькими одно- или двудирекционными терминаторами транскрипции.
24. Способ идентификации соединения, ингибирующего взаимодействие первого белка X со вторым белком Y, включающий следующие стадии:
a) культивирование клеток, таких, как указанные по любому из пп.1-23;
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
c) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка X с белком-партнером Y, экспрессируемых культивированными клетками.
25. Применение клеток по любому из пп.1-23 для идентификации соединений, ингибирующих взаимодействие белок-белок.
26. Применение клеток по любому из пп.1-23 для скрининга банков кДНК или банков пептидов с целью идентификации пептидов или белковых факторов, специфически ликвидирующих взаимодействие белок-белок.
27. Способ идентификации соединения, ингибирующего взаимодействие первого белка X со вторым белком Y, но не ингибирующего или менее ингибирующего взаимодействие между белком Y и третьим белком Z, включающий следующие стадии:
a) культивирование клеток по п.2 или 3;
b) инкубация вышеуказанных клеток в присутствии тестируемого соединения;
c) сравнение экспрессии гена-репортера в присутствии и в отсутствие вышеуказанного соединения, причем увеличение экспрессии гена-репортера является указанием на то, что тестируемое соединение является ингибитором взаимодействия белка X с белком-партнером Y, но что этот продукт не ингибирует или менее ингибирует взаимодействие между белком Y и белком Z.
| Мартынюк А | |||
| Изучение белок-белковых взаимодействий | |||
| Метод двойного гибрида | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

