Область техники, к которой относится изобретение
Изобретение относится к области генной инженерии и биотехнологии и представляет способ внесения точечных нуклеотидных замен в бактериальную хромосому без сопутствующей интеграции детерминант антибиотикоустойчивости.
Уровень техники
До начала 2000-х годов для направленной модификации хромосомы Е. coli применялся трудоемкий метод с использованием протяженных гомологичных фрагментов длиной порядка тысячи пар нуклеотидов. Промежуточный этап включал в себя сборку плазмиды, несущей селективный маркер, фланкированный фрагментами для рекомбинации. Для введения линеаризованной плазмиды использовались специальные штаммы, дефектные по экзонуклеазам, но сохранившим способность к рекомбинации - recBC sbcB (Marinus MG, et al. Insertion mutations in the dam gene of Escherichia coli K-12. Mol Gen Genet. 1983; 192(1-2):288-289.; Jasin M, Schimmel P. Deletion of an essential gene in Escherichia coli by site-specific recombination with linear DNA fragments. J Bacteriol. 1984; 159(2):783-786.; Winans SC, et al. Site-directed insertion and deletion mutagenesis with cloned fragments in Escherichia coli. J Bacteriol. 1985; 161(3):1219-1221.) или recD (Russell CB, et al. Chromosomal transformation of Escherichia coli recD strains with linearized plasmids. J Bacteriol. 1989; 171(5):2609-2613).
Внедрение в практику высокоэффективных фаговых рекомбиназ (Murphy KC. Use of bacteriophage lambda recombination functions to promote gene replacement in Escherichia coli. J Bacteriol. 1998; 180(8):2063-2071.; Zhang Y, et al. A new logic for DNA engineering using recombination in Escherichia coli. Nat Genet. 1998; 20(2):123-128) и внесение ряда последовательных улучшений, таких как конструирование хелперных плазмид, обеспечивающих индуцируемую экспрессию рекомбиназ (Datsenko KA, Wanner BL. One step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA. 2000; 97(12):6640-6645), и создание специальных штаммов с инактивированной системой репарации для повышения количества рекомбинантных клонов (Yu D, et al. An efficient recombination system for chromosome engineering in Escherichia coli. Proc Natl Acad Sci USA. 2000; 97(11):5978-5983), открыло новую страницу в генетической инженерии, позволив модифицировать хромосому с использованием ПЦР-ампликонов, несущих короткие (35-50 п.н.) нуклеотидные области на концах, гомологичных модифицируемой области хромосомы. Новый подход, получивший название рекомбиниринга, от терминов "рекомбинация" и "инженерия", позволяет вводить как точечные модификации, так и протяженные делеции и инсерции (Sawitzke JA, et al. Recombineering: highly efficient in vivo genetic engineering using single-strand oligos. Methods Enzymol. 2013; 533:157-177), вводя последовательности изменяемого локуса в свешивающиеся концы праймеров для амплификации, или используя синтетические олигонуклеотиды длиной 70-100 оснований.
Для Е. coli описано использование двух фаговых систем рекомбинации - Red и RecET. Первая система (система Red) была найдена в бактериофаге λ. (Stahl FW, et al. Break-join recombination in phage lambda. Genetics. 1990; 125(3):463-474). Она включает в себя три белка: Ехо - 5'-3' нуклеаза, деградирующая одну цепь двуцепочечной ДНК (Mosberg JA, et al. Lambda red recombineering in Escherichia coli occurs through a fully single-stranded intermediate. Genetics. 2010; 186(3):791-799), Bet - основной компонент рекомбинации, который обеспечивает спаривание с комплементарной последовательностью в хромосоме, связываясь с одноцепочечной ДНК и защищающий от деструкции хозяйскими экзонуклеазами (Murphy KC, Marinus MG. RecA-independent single-stranded DNA oligonucleotide-mediated mutagenesis. F1000 Biol Rep. 2010 Jul 22; 2:56), и Gam - ингибитор хозяйских нуклеаз в E. coli, что обеспечивает возможность рекомбинации практически в любых штаммах, несмотря на наличие системы репарации. Bet в настоящее время включен в широкий класс SSAP-белков - белков гибридизации одноцепочечных ДНК (от английского «single strand annealing proteins»).
Вторая система RecET дефектного Rac профага, локализованного в хромосоме Е. coli, состоит из двух генов recE и recT, кодирующих белки экзонуклеазы и SSAP, функционально аналогичных белкам Ехо и Beta системы Red, несмотря на различия в их структуре (Zhang Y, et al. A new logic for DNA engineering using recombination in Escherichia coli. Nat Genet. 1998; 20(2):123-128). Обе эти системы стали доступны исследователям в виде индуцируемых оперонов, локализованных в хромосоме (Yu D, et al. An efficient recombination system for chromosome engineering in Escherichia coli. Proc Natl Acad Sci USA. 2000; 97(11):5978-5983), или в виде хелперных плазмид (Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA. 2000; 97(12):6640-6645), что более удобно для практического применения.
В большинстве случаев для получения модификаций предпочтительно использование маркеров антибиотикоустойчивости. Фланкирование генов сайтами специфичной фаговой рекомбинации, такими как FRT, LoxP, attL и attR фага λ, позволяет в дальнейшем «вырезать» маркер индукцией фаговых рекомбиназ, для проведения последующих манипуляций с геномом. Немаловажно, что в биотехнологическом производстве предпочтительно использование чувствительных к антибактериальным препаратам штаммов-продуцентов для предотвращения неконтролируемого распространения генов лекарственной резистентности в природе.
Для прецизионной модификации генома, например внесения точечных замен в ген, использование маркера устойчивости, фланкированного сайтами специфичной рекомбинации, невозможно, так как после эксцизии остается либо один исходный сайт (loxP, FRT) в случае симметричной рекомбинации (Zhang Y, et al. A new logic for DNA engineering using recombination in Escherichia coli. Nat Genet. 1998; 20(2):123-128; Datsenko KA, Wanner BL. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA. 2000; 97(12):6640-6645), либо гибридный attB в случае использования системы интеграции фага λ. Для решения этой задачи можно использовать двустадийную модификацию, включающую в себя полное удаление одной части гена от модифицируемого сайта до межгенной области с последующей инсерцией этого же фрагмента с уже внесенной мутацией. Таким образом можно перенести специфический сайт вырезания в межгенную область, где он не будет оказывать сильного воздействия на фенотип.
Альтернативным, одностадийным, подходом модификации генома является электропорация одноцепочечным олигонукдеотидом длиной в 70-100 пар оснований, содержащим необходимую замену (Ellis НМ, et al. High efficiency mutagenesis, repair, and engineering of chromosomal DNA using single-stranded oligonucleotides. Proc Natl Acad Sci USA. 2001; 98(12):6742-6746). В этом случае для осуществления рекомбинации функция экзонуклеазы избыточна, и используется либо Bet, в опциональном сочетании с ингибитором хозяйских нуклеаз Gam, либо один RecT. Высев клеток после электропорации осуществляется на неселективную среду, а отбор колоний проводится с помощью аллель-специфичной ПЦР. В данном случае эффективности протекания электропорации и рекомбинации становятся критически важными параметрами, напрямую влияющими на необходимое для проверки число колоний при поиске клона с желаемой модификацией.
Основным условием высокой частоты рекомбинационных событий является учет направления движения репликационной вилки: олигонуклеотид должен быть выбран так, чтобы он мог встроиться в запаздывающую цепь. Разница в эффективностях рекомбинации с комплементарным ему олигонуклеотидом может достигать четырех порядков (van Kessel JC, Hatfull GF. Efficient point mutagenesis in mycobacteria using single-stranded DNA recombineering: characterization of antimycobacterial drug targets. Mol Microbiol. 2008; 67(5): 1094-1107).
Известно, что система метил-зависимой репарации неспаренных оснований (MMR) в Е. coli снижает эффективность рекомбинации. В этом случае для улучшения рекомбинации используется либо специальный дизайн олигонуклеотидов (например, олигонуклеотид содержит одновременно пять замен), либо специальный штамм, дефицитный по MMR системе (Sawitzke JA, et al. Recombineering: highly efficient in vivo genetic engineering using single-strand oligos. Methods Enzymol. 2013; 533:157-177).
Другими вариантами повышения эффективности рекомбинации являются использование модифицированных нуклеотидов при синтезе рекомбинирующего олигонуклеотида для предотвращения его нуклеазной деградации в клетке, оптимизация условий электропорации для модифицируемого штамма бактерий (Tu Q, et al. Room temperature electrocompetent bacterial cells improve DNA transformation and recombineering efficiency. Sci Rep. 2016; 6:24648).
Таким образом, при учете всех вышеописанных аспектов, влияющих на рекомбинацию, в некоторых случаях до 50% неселектированных колоний Е. coli может нести необходимую замену (Sawitzke JA, et al. Recombineering: highly efficient in vivo genetic engineering using single-strand oligos. Methods Enzymol. 2013; 533:157-177).
Тем не менее, для гарантированного результата при отборе колоний, несущих необходимую замену, предпочтительней использование двустадийной модификации, с использованием селективного и контрселективного маркера в составе одной генной кассеты. Классическим примером является кассета cat-sacB, несущая ген хлорамфеникол-ацетилтрансферазы, кодирующий устойчивость к хлорамфениколу и ген левансукразы, который приводит к чувствительности Е. coli к сахарозе (Gay Р, et al. Positive selection procedure for entrapment of insertion sequence elements in gram-negative bacteria. J Bacteriol. 1985; 164(2):918-921).
На первом этапе в модифицируемый сайт вводится двойной маркер по гомологичным фланкирующим коротким последовательностям, с отбором рекомбинантных клонов на антибиотике, в данном случае - на хлорамфениколе. На втором этапе, проводится электропорация олигонуклеотидом, содержащим необходимую замену и те же самые гомологичные последовательности. Контрселекция на сахарозе отбирает клетки, утратившие гибридную кассету за счет рекомбинации олигонуклеотида, что позволяет достигнуть почти 100% эффективности отбора нужных клонов.
Альтернативный подход заключается в использовании системы CRISPR для индуцированного внесения двуцепочечного разрыва в хромосому и стимулирования рекомбинации. Эффективность внесения точечных замен с использованием такой системы в Е. coli достигает 65%, т.е. 65% выросших клонов несут желаемую замену (Jiang W, et al. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat Biotechnol. 2013; 31(3):233-239). Данный подход был расширен для создания протяженных делеций и инсерций с эффективностью 1%-47% (Pyne ME, et al. Coupling the CRISPR/Cas9 System with Lambda Red Recombineering Enables Simplified Chromosomal Gene Replacement in Escherichia coli. Appl Environ Microbiol. 2015 Aug; 81(15):5103-14). Предполагается, что данный метод является одностадийным, но до момента проведения собственно электропорации олигонуклеотидом необходимо создать специфичную хелперную плазмиду, несущую gRNA для CRISPR системы. Более того, далеко не во всех случаях данная система применима за счет ограничений специфичности CRISPR системы - необходимости в наличии консервативного сайта РАМ в модифицируемом локусе.
Гомологичные фаговые системы рекомбинации обнаружены во многих организмах (Datta S, et al. Identification and analysis of recombineering functions from Gram-negative and Gram-positive bacteria and their phages. Proc Natl Acad Sci USA. 2008; 105(5): 1626-1631), и аналогичные системы модификации хромосомы широко применяются в самых разных микроорганизмах (Ranallo RT, et al. Developing live Shigella vaccines using lambda Red recombineering. FEMS Immunol Med Microbiol. 2006 Aug; 47(3):462-9; van Kessel JC, Hatfull GF. Recombineering in Mycobacterium tuberculosis. Nat Methods. 2007; 4(2): 147-152; Gerlach RG, et al. Rapid oligonucleotide-based recombineering of the chromosome of Salmonella enterica. Appl Environ Microbiol. 2009 Mar; 75(6): 1575-80; Swingle B, et al. Recombineering using RecTE from Pseudomonas syringae. Appl Environ Microbiol. 2010 Aug; 76(15):4960-8; Bryan A, Swanson MS. Oligonucleotides stimulate genomic alterations of Legionella pneumophila. Mol Microbiol. 2011 Apr; 80(1):231-47).
Для Mycobacteria в микобактериофаге Che9C были найдены гомологи RecE и RecT, и продемонстрирована возможность аллельной замены как для М. tuberculosis, так и для широко-используемого модельного организма М. smegmatis (van Kessel JC, Hatfull GF. Recombineering in Mycobacterium tuberculosis. Nat Methods. 2007; 4(2): 147-152; van Kessel JC, Hatfull GF. Efficient point mutagenesis in mycobacteria using single-stranded DNA recombineering: characterization of antimycobacterial drug targets. Mol Microbiol. 2008; 67(5): 1094-1107). Отсутствие MMR системы в Mycobacteria позволяет использовать RecT рекомбинацию для распространенных лабораторных, и даже клинических штаммов, без необходимости создания специальных дефектных штаммов, как в случае Е. coli, обладающих к тому же измененной физиологией и сниженной адаптивностью.
Однако эффективная двухстадийная схема с селекцией и контрселекцией в Mycobacteria нереализуема за счет высокой частоты «запрещенной» (illegitimate), или негомологичной, рекомбинации (Kalpana GV, et al. Insertional mutagenesis and illegitimate recombination in mycobacteria. Proc Natl Acad Sci USA. 1991; 88(12):5433-5437; Muttucumaru DG, Parish T. The molecular biology of recombination in Mycobacteria: what do we know and how can we use it? Curr Issues Mol Biol. 2004; 6(2):145-157.; Khattak FA, et al. Illegitimate recombination: an efficient method for random mutagenesis in Mycobacterium avium subsp. hominissuis. BMC Microbiol. 2012 Sep 11; 12:204). Для встраивания в геном протяженных фрагментов, несущих гены антибиотикорезистентности, необходимо наличие достаточно длинных областей гомологии (более 120 п.н.), аналогично ранним методам модификации генома в Escherichia до открытия фаговых рекомбиназ (Murphy KC, et al. Mycobacterial recombineering. Methods Mol Biol. 2015; 1285:177-199).
Для M. tuberculosis и M. smegmatis при использовании SSAP-белка RecT фага Che9C (плазмида pKM402) заявляется эффективность в 10-3 мутантов на исходное количество клеток, при рекомбинации одноцепочечным олигонуклеотидом (Murphy KC, et al. Mycobacterial recombineering. Methods Mol Biol. 2015; 1285:177-199). Так как зачастую замены не обладают выраженным фенотипом, или вносятся для проверки фенотипа, то после электропорации и «оживления», клетки высеваются на неселективную среду. Вследствие чего встает задача отбора мутантных клонов, решаемая с помощью аллель-специфичной ПЦР. Соответственно, при заявляемой эффективности количество клеток, которое необходимо проверить для идентификации штамма с нужной заменой, составляет 2-3 тысячи клонов.
Для увеличения относительного выхода рекомендован метод одновременной электропорации двумя фрагментами, один из которых несет замену в 43 кодоне гена rplC, приводящую к устойчивости штамма к стрептомицину. Тогда после «оживления» клетки высеваются на среду со стрептомицином. Частота второго рекомбинантного события в этих клонах значительно выше и оценивается в 10-2 (Murphy KC, et al. Mycobacterial recombineering. Methods Mol Biol. 2015; 1285:177-199; Ioerger TR, et al. Identification of new drug targets and resistance mechanisms in Mycobacterium tuberculosis. PLoS One. 2013 Sep 23; 8(9):e75245). Вполне очевидно, что при таком подходе (устойчивость к стрептомицину) отбирается компетентная и рекомбиногенная субпопуляция клеток. Очевидными недостатками этого подхода являются невозможность внесения последующих замен на фоне выработанной устойчивости к стрептомицину, невозможность изучения устойчивости к самому стрептомицину, возможная интерференция с другими антибактериальными препаратами, действующими на рибосому, неизвестное эпистатическое влияние замены в rpsL.
Ключевые принципы метода внесения мутаций в бактериальную хромосому с использованием коротких фрагментов защищены рядом патентов. Использование рекомбиназы Bet Red-системы фага Лямбда описано в Патенте WO 2002014495 B1 (Donald L. Court, Daiguan Yu, E-Chiang Lee, Hilary M. Ellis, Nancy A. Jenkins, Neal G. Copeland, Enhanced homologous recombination mediated by lambda recombination proteins, приоритет от 14 августа 2000 г.). Полная Red-система рекомбинации Bet, Gam и Ехо защищена Патентом US 8067239 В2 (Kenan С. Murphy, Reagents for recombinogenic engineering and use thereof, приоритет от 26 февраля 2003 г.). Схема, заключающаяся в использовании хелперных плазмид или хелперного бактериофага, обеспечивающего рекомбинацию в различных штаммах защищена патентами US 7674621 B2, US 8541229 B2, US 8859277 B2, US 9371532 B2 Court DL et al. (Donald L. Court, Simanti Datta, Nina Costantino, Plasmids and phages for homologous recombination and methods of use, приоритет от 21 мая 2004 г.).
Таким образом все описанные методы введения мутаций в бактериальную хромосому позволяют получить рекомбинантные клоны с эффективностью порядка 10-4-10-3 на общее количество клеток и требуют проверки тысяч колоний для отбора нужных мутантов. Поэтому существует необходимость в способе введения мутаций в хромосому, так, чтобы получать рекомбинантные клоны с высокой эффективностью.
Раскрытие сущности изобретения
Данное изобретение основано на открытии повышения частоты отбора рекомбинантов за счет введения в клетку экстрахромосомной коселектирующей ДНК.
Данное изобретение касается способа внесения безмаркерных точечных модификаций в хромосому микобактерий заключающегося в указанных последовательных стадиях:
- введения в модифицируемый штамм микобактерий хелперной плазмиды, обеспечивающей индуцируемую экспрессию белков рекомбинации;
- индукции экспрессии белков рекомбинации в модифицируемом штамме микобактерий;
- одновременной электропорации модифицируемого штамма микобактерий: одноцепочечным олигонуклеотидом, содержащим желаемую замену и участки гомологии вокруг указанной желаемой замены, и коселективной молекулой ДНК, неспособной к встраиванию в хромосому модифицируемого штамма микобактерий и несущей ген(ы) устойчивости к антибактериальному препарату, отличному от препарата, используемого для поддержания хелперной плазмиды;
- обеспечения рекомбинации и экспрессии гена(ов) устойчивости коселективной молекулы ДНК на среде без селекции;
- отбор колоний модифицированного штамма микобактерий, устойчивых к указанному антибактериальному препарату, чей фенотип обеспечен маркером резистентности коселективной ДНК, путем высева культуры модифицируемого штамма микобактерий на селективную среду, содержащую антибактериальный препарат, отличный от препарата, используемого для поддержания хелперной плазмиды;
- излечивания модифицированного штамма микобактерий от хелперной плазмиды и коселективной молекулы ДНК.
Бактерии
Данное изобретение касается способа осуществления рекомбинации и отбора рекомбинантных клеток в бактериях рода Mycobacterium. В одном из вариантов осуществления способа бактерия относится к виду Mycolicibacterium smegmatis.
Белки рекомбинации
Исторически первой системой описанной и примененной для рекомбинации в М. smegmatis и М. tuberculosis стала система RecET бактериофага Che9c, открытого в 2002 году. Для микобактерий описаны десятки тысяч бактериофагов, и некоторые также содержат белки гомологи RecET и Lambda beta и ехо. Сравнимый уровень рекомбинации был продемонстрирован для RecT гомолога микобактерофага Halo (van Kessel JC et al., Recombineering mycobacteria and their phages. Nat Rev Microbiol. 2008 Nov; 6(11):851-7).
Фаговые белки рекомбинации обладают нестрогой видоспецифичностью, так Red beta Е. coli не обеспечивает эффективную рекомбинацию в М. smegmatis. Для RecT из Rac профага тем не менее зарегистрировано успешное получение мутантов по двум различным мишеням (van Kessel JC et al., Recombineering mycobacteria and their phages. Nat Rev Microbiol. 2008 Nov; 6(11):851-7).
В одном из вариантов осуществления изобретения используется плазмида pKD402, в которой экспрессия гена gp60 (гомолог recT) фага Che9c находится под контролем индуцируемого tetAR промотора. Индукция осуществляется добавлением ангидротетрациклина в среду с последующей инкубацией при физиологичной (при +37°С) температуре в течение 3 часов.
В результате электропорации в штамм с индуцированной экспрессией рекомбиназы одновременно вводится одноцепочечный олигонуклеотид, содержащий желаемую замену, и коселективную плазмиду.
Одноцепочечный олигонуклеотид
Одноцепочечный олигонуклеотид для внесения точечных замен конструируется на основе последовательности генома модифицируемого штамма. Он содержит последовательность с желаемой заменой, или заменами, длиной от 1 до 20 нуклеотидов, фланкированных гомологичными плечами длиной более 30 нуклеотидов.
Под термином «гомологичная» подразумевается, что указанные последовательности идентичны или сходны с последовательностью геномной мишени, чтобы обеспечивать достаточную эффективность гомологичной рекомбинации с помощью используемых фаговых рекомбиназ. В варианте использования RecT для осуществления рекомбинации применяются плечи длиной от 30 нуклеотидов.
Важным аспектом осуществления метода внесения точечных мутаций является выбор цепи из двух комплементарных друг другу вариантов для использования в качестве основы для написания последовательности олигонуклеотида. Условием высокой частоты рекомбинационных событий является учет направления движения репликационной вилки: олигонуклеотид должен быть выбран так, чтобы он мог встроиться в запаздывающую цепь при синтезе.
В одном из вариантов исполнения одноцепоченого олигуонуклеотида замена является точечной, т.е. производится замена только одного нуклеотида, а фланкированные последовательности длиной около 30 нуклеотидов полностью идентичны последовательности генома дикого типа.
Конструкция коселективной ДНК
Конструкция коселективной ДНК включает в себя по меньшей мере следующие основные компоненты: селективный маркер, или ген(ы) устойчивости к антибактериальному препарату, отличному от препарата, используемого для поддержания хелперной плазмиды и плазмидный ориджин репликации. Важным свойством коселективной ДНК является ее неспособность к встраиванию в хромосому модифицируемого штамма микобактерий.
Селективные маркеры
Селективные маркеры используются как для поддержания хелперной плазмиды, так и для поддержания коселективной ДНК. В данном описании термин «селективный маркер» используется для обозначения позитивно-селектируемого признака, обеспечивающего отбор клеток в присутствии агента. В большинстве случаев при создании рекомбинантных бактериальных клеток в роли таких маркеров используются гены устойчивости к антибиотикам.
Микобактерий устойчивы к ряду антибактериальных препаратов, в частности бета-лактамным антибиотикам, поэтому спектр маркеров, пригодных для селекции более узок по сравнению, например, с Е. coli. В варианте осуществрения рекомбинации в М. smegmatis применяются гены устойчивости к канамицину, гигромицину, апрамицину, стрептомицину, тетрациклину, гентамицину и сульфонамидам (Goude R, et al. Electroporation of mycobacteria. Methods Mol Biol. 2015; 1285:117-30).
В варианте реализации метода с использованием хелперной плазмиды pKM402, при создании коселективной ДНК исключаются также гены устойчивости к канамицину и хлорамфениколу, так как они присутствуют в составе pKM402.
Плазмидные ориджины репликации микобактерий
Подавляющее большинство плазмидных конструкций, используемых в микобактериях, содержат ориджин репликации pAL5000 (Ranes MG et al. Functional analysis of pAL5000, aplasmid from Mycobacterium fortuitum: construction of a "mini" mycobacterium-Escherichia coli shuttle vector. J Bacteriol. 1990 May; 172(5):2793-7). Тем не менее, существуют и описаны и другие ориджины репликации, пригодные для создания плазмидных конструкций (Lee et al. The development of a novel Mycobacterium-Escherichia coli shuttle vector system using pMyong2, a linear plasmid from Mycobacterium yongonense DSM 45126T. PLoS One. 2015 Mar 30; 10(3):e0122897).
В одном из вариантов осуществления изобретения коселективная ДНК может нести также ориджин репликации Е. coli для облегчения наработки необходимого для электропорации количества ДНК в стандартном объекте генетической инженерии.
После электропорации одновременно двумя молекулами ДНК необходимо обеспечить прохождение рекомбинации, а также экспрессию гена устойчивости коселективной молекулы ДНК. Для этого к суспензии клеток добавляется свежая жидкая среда без антибиотиков и проводится инкубация при физиологичной (+37°С) температуре. Время инкубации обуславливается временем нормального деления клеток, необходимого для осуществления рекомбинации. Также учитывается то, что после отмывки водным раствором глицерина и проведения электропорации, клетки находятся некоторое время в неделящемся состоянии, обусловленном шоком. В варианте осуществления изобретения на примере М. smegmatis время инкубации составляет 6 часов и более.
Отбор колоний
Отбор колоний модифицированного штамма микобактерий проводится путем высева культуры модифицируемого штамма микобактерий на агаровые чашки с селективной средой, содержащей препарат, устойчивость к которому обусловлена селективным маркером коселективной плазмиды. Полученные устойчивые колонии могут быть проверены на наличие желаемой мутации аллель-специфичной ПЦР, либо секвенированием. Эффективность выхода рекомбинантов оценивается как отношение мутантных колоний к общему числу проверенных устойчивых колоний. В одном из вариантов реализации способа селективным препаратом является гигромицин.
Контрселективные маркеры
В одном из вариантов осуществления способа коселективная плазмида несет маркер контрселекции, обеспечивающий отбор колоний, утерявших названную плазмиду при культивации на специальных средах. В результате возможно отобрать штамм колоний, несущих только желаемые модификации в геноме, не обладающих дополнительной внехромосомной ДНК и маркерами антибиотикорезистентности.
Примерами используемых маркеров контрселекции являются гены sacB, rpsL, tetAR, pheS, thyA, lacY, gata-1, ccdB, I-SceI (Reyrat JM et al. Counterselectable markers: untapped tools for bacterial genetics and pathogenesis. Infect Immun. 1998 Sep; 66(9):4011-7), galK (Barkan D, Stallings CL, Glickman MS. An improved counterselectable marker system for mycobacterial recombination using galK and 2-deoxy-galactose. Gene. 2011 Jan 1; 470(l-2):31-6).
В одном из вариантов в качестве контрселективного маркера используется ген левансукразы sacB, а среда для культивации содержит сахарозу (Gay Р, et al. Positive selection procedure for entrapment of insertion sequence elements in gram-negative bacteria. J Bacteriol. 1985; 164(2):918-921). В этом случае образуются токсичные для бактерий леваны - полимеры фруктозы. Токсичность леванов и возможность контрселекции с помощью sacB были также продемонстрированы и для микобактерий (Pelicic V, Reyrat JM, Gicquel В. Expression of the Bacillus subtilis sacB gene confers sucrose sensitivity on mycobacteria. J Bacteriol. 1996 Feb; 178(4):1197-9).
Краткое описание фигур и таблиц
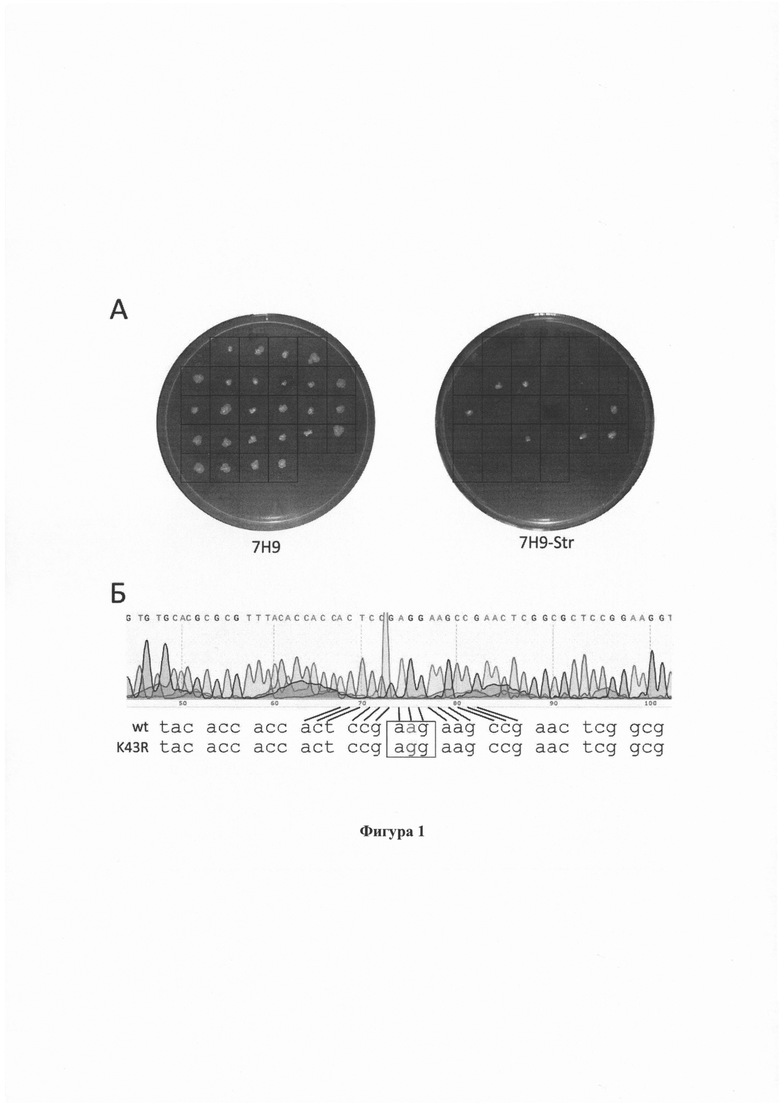
Фигура 1. Оценка эффективности отбора рекомбинантов и анализ первичной последовательности генома в локусе rplC. (А) Параллельный перенос выросших HygR колоний на неселективную среду и среду со стрептомицином. (Б) Хроматограмма реакции секвенирования геномной ДНК в окрестности 43 кодона гена rpsL для одной из колоний, давшей рост на среде со стрептомицином, и выравнивание первичной последовательности данного локуса для штамма дикого типа и штамма несущего замену K43R.
Фигура 2. Оценка эффективности отбора рекомбинантов и анализ первичной последовательности генома в локусе gyrA. (А) Параллельный перенос выросших HygR колоний на неселективную среду и среду с ципрофлоксацином. (Б) Хроматограмма реакции секвенирования геномной ДНК в окрестности 95 кодона гена gyrA для одной из колоний, давшей рост на среде с ципрофлоксацином, и выравнивание первичной последовательности данного локуса для штамма дикого типа и штамма, несущего замену D95G.
Осуществление изобретения
Для осуществления изобретения необходимо было создать специальную коселективную ДНК, обладающую следующими характеристиками: обеспечивать репликацию в М. smegmatis; нести ген устойчивости, отличный от гена устойчивости использованного на хелперной плазмиде; нести контрселективный маркер, позволяющий отбирать колонии, утерявшие коселективную плазмиду.
Первичный анализ эффективности введения точечных замен олигонуклеотидом был осуществлен на модели с устойчивостью к стрептомицину. Устойчивость бактерий к стрептомицину обусловлена модификацией мишени препарата - рибосомы (Ito Т, Kosugi Н, Higo K, Osawa S. Ribosomal proteins from streptomycin-resistant and dependent mutants, and revertants from streptomycin-dependence to independence in Bacillus subtilis. Mol Gen Genet. 1975 Sep 8; 139(4):293-301). Для M. tuberculosis как клинические резистентные, так и селектированные in vitro штаммы, в основном, несут мутации в гене рибосомального белка S12 и гене 16S рРНК. При этом, так как М. smegmatis имеет два рибосомальных оперона в отличие от М. tuberculosis, введение замены в одну копию гена 16S не приводит к формированию устойчивости (Springer В, Kidan YG, Prammananan Т, Ellrott K, Böttger ЕС, Sander P. Mechanisms of streptomycin resistance: selection of mutations in the 16S rRNA gene conferring resistance. Antimicrob Agents Chemother. 2001 Oct; 45(10):2877-84). Замены же в 43 ко доне гена rpsL, кодирующего рибосомальный белок S12, драматически повышают МИК стрептомицина, а частота получения спонтанных мутантов равна 10-8 (в ко доне гена rpsL), что позволяет использовать данную модель для оценки эффективности рекомбинации при использовании различных подходов.
Эксперимент проводили для оценки эффективности внесения точечных замен рекомбинацией синтетического олигонуклеотида. Для осуществления рекомбинации был синтезирован олигонуклеотид с заменой А на G относительно последовательности дикого типа и гомологичными плечами длиной 34 и 35 нуклеотидов (SEQ ID NO: 3). Так как ген rpsL локализован в области 1,5 Мб между OriC (0 Мб) и OriT (3,4 Мб), и репликационная вилка перемещается по направлению гена, в качестве исходной последовательности использовалась минус-цепь последовательности гена. При рекомбинации должна была происходить замена 43 кодона гена AAG (Lys) на AGG (Arg). Исследовали три варианта рекомбинации: электропорация олигонуклеотида в штамм дикого типа, в штамм с введенной хелперной плазмидой, экспрессирующей белок RecT, с индукцией и без индукции.
Аналогичный подход с использованием модели устойчивости к стрептомицину был использован при проверке эффективности отбора колоний с использованием коселективной ДНК.
Для независимого подтверждения высокой эффективности введения точечных замен олигонуклеотидом с помощью коселективного маркера была выбрана модель устойчивости к фторхинолонам. Основным механизмом резистентности микобактерий являются замены в GyrA субъединице ДНК-гиразы. Наиболее часто встречающейся заменой, обнаруживаемой в устойчивых клинических изолятах возбудителя туберкулеза, является замена D94G. Дикий штамм М. smegmatis обладает низким уровнем устойчивости к антибиотикам, с минимальной ингибирующей концентрацией (МИК) ципрофлоксацина равной 0,25 мкг/мл. Мутация в гене GyrA, приводящая к аминокислотной замене D95G (соответсвует D94G М. tuberculosis), повышает МИК ципрофлоксацина до 5 мкг/мл (Lu Т et al. Enhancement of fluoroquinolone activity by C-8 halogen and methoxy moieties: action against a gyrase resistance mutant of Mycobacterium smegmatis and a gyrase-topoisomerase IV double mutant of Staphylococcus aureus. Antimicrob Agents Chemother. 2001; 45(10):2703-2709).
Если не указано иное, все технические и научные термины, использованные в данном документе, имеют то же значение, которое обычно подразумевается средним специалистом в данной области, к которой относится это изобретение.
Далее изобретение будет проиллюстрировано примерами, которые предназначены для обеспечения лучшего понимания сущности заявленного изобретения, но не должны рассматриваться как ограничивающие данное изобретение.
Пример 1. Конструирование коселективной плазмиды pMyCA-sacB.
Пример служит для иллюстрации одного из вариантов исполнения коселективной молекулы ДНК в виде кольцевой плазмиды, содержащей два ориджина репликации для Escherichia и Mycobacteria, ген устойчивости к гигромицину и контрселективный маркер sacB.
В качестве донора векторной части использовали плазмиду pMyCA-mCherry (Magaña Vergara С, et al. A versatile vector for mycobacterial protein production with a functional minimized acetamidase regulon. Protein Sci. 2017 Nov; 26(11):2302-2311, Addgene plasmid # 84272). Препарат плазмидной ДНК нарабатывали по стандартной методике, включающей Са-зависимую трансформацию, отбор колоний, наработку бактериальной массы на селективной среде с антибиотиком гигромицином и выделение плазмидной ДНК методом щелочного лизиса. Препарат подвергали двум последовательным реакциям рестрикции ферментами BstAUI и XbaI (НПО "СибЭнзим", Новосибирск, Россия). В результате расщепления образовывалось три фрагмента длиной около 4125, 783 и 1708 п.н., как и было предсказано исходя из первичной последовательности плазмиды. Векторную часть, содержащую два ориджина репликации и маркер устойчивости к гигромицину, длиной 4125 п.н. выделяли из геля набором Gel Extaction Kit (QIAGEN, Германия).
В качестве источника маркера sacB использовали плазмиду pKM461 (Murphy KC, et al. ORBIT: aNew Paradigm for Genetic Engineering of Mycobacterial Chromosomes. mBio. 2018 Dec 11; 9(6):e01467-18, Addgene plasmid # 108320). Фрагмент ДНК амплифицировали с помощью праймеров SEQ ID NO: 1 и SEQ ID NO: 2, содержащих сайты узнавания рестриктазами XbaI и PspLI в составе свешивающихся концов. После проведения ПЦР фрагмент ДНК очищали из геля и последовательно обрабатывали рестриктазами XbaI и PspLI (НПО "СибЭнзим", Новосибирск, Россия) и очищали переосаждением в этаноле.
Лигирование векторного фрагмента и вставки проводили Т4 ДНК лигазой (НПО "СибЭнзим", Новосибирск, Россия) в течении 40 минут при +16°С. Са-зависимая трансформация лигазной смеси, пересев колоний, наработка и выделение плазмидной ДНК проводили стандартными методами (Green MR, Sambrook J. Molecular Cloning: A Laboratory Manual (Fourth Edition). 2012. Cold Spring Harbor Laboratory Press).
Проверку контрселективных свойств маркера sacB проводили путем посева колоний, содержащих плазмиду, на чашки с сахарозой и без антибиотика, с последующим переносом выросших колоний на чашки с гигромицином и контрольную чашку с неселективной средой, тем самым подтверждали отсутствие исходной плазмиды в колониях.
Отобраннные функциональные клоны коселективной плазмиды были наработаны в количестве, достаточном для дальнейшего использования в экспериментах с рекомбинацией.
Пример 2. Оценка эффективности рекомбинации олигонуклеотида.
Эксперимент проводили для оценки эффективности внесения точечных замен рекомбинацией синтетического олигонуклеотида. Исследовали три варианта рекомбинации: электропорация олигонуклеотида в штамм дикого типа, в штамм с введенной хелперной плазмидой, экспрессирующей белок RecT, с индукцией и без индукции.
В качестве штамма-реципиента использовали штамм М. smegmatis МС2 155 - основной лабораторный штамм дикого типа, с известной геномной последовательностью. Рекомбинация проводили в соответствии с опубликованным протоколом (Murphy KC, et al. Mycobacterial recombineering. Methods Mol Biol. 2015; 1285:177-199). Хелперная плазмида pKM402, использованная в этом протоколе, была получена из международного банка Addgene (Addgene plasmid #107770). Для рекомбинации использовали контрольный олигонуклеотид из этого протокола, обеспечивающий замену K43R в рибосомальном белке RpsL, приводящую к фенотипической устойчивости М. smegmatis к стрептомицину (SEQ ID NO: 3). Олигонуклеотид содержит в середине нуклеотидную замену: А на G, относительно последовательности дикого типа, приводящую к замене ко дона AAG (Lys) на AGG (Arg). При общей длине олигонуклеотида в 70 нуклеотидов, плечи для гомологичной рекомбинации составляют 34 и 35 нуклеотидов.
Введение pKM402 в М. smegmatis МС2 155 проводили методом электропорации (метод Мерфи) (Goude R, et al. Electroporation of mycobacteria. Methods Mol Biol. 2015; 1285:117-30). Трансформанты высевали и в дальнейшем поддерживали на чашках с агаризованной средой Middlebrook 7H9-OADC (M197 и FD018, Himedia, Индия) с 20 мкг/мл канамицина (11860-038, Gibco, Великобритания).
Для рекомбинации выращивали ночную культуру микобактерий, посеянную со свежей чашки (1-2 нед.), из расчета 4 мл культуры на образец. Для культивирования использовали жидкую среду Middlebrook 7H9-ADC M198 и FD019, Himedia, Индия) с добавлением 0,05% Tween-80 (Р8074, SIGMA, США). Для штамма, содержащего хелперную pKM402 в среду также добавляли канамицин до концентрации 20 мкг/мл. Культуры доращивали до оптической плотности 0,5-0,8, затем проводили индукцию экспрессии рекомбиназы добавлением 300 мкг/мл ангидротетрациклина (94664, SIGMA, США) и культивировали в течение 3 часов с постоянной аэрацией. Оценку количества клеток проводили серийными разведениями культуры, высевом на чашки с агаризованной средой и последующим подсчетом выросших колоний. В дальнейшем использовали калибровочную кривую зависимости числа колониеобразующих единиц от оптической плотности OD600.
После тройной отмывки ледяным 10% глицерином и концентрировании исходных 4 мл культуры до 400 мкл, к клеткам добавляли олигонуклеотид (SEQ ID NO: 3) для рекомбинации, растворенный в воде, в количестве 500-1000 нг в 1-2 мкл. Электропорацию проводили на приборе BIO-RAD GenePulser, в 2 мм кюветах, поставляемых с прибором, использовался профиль Ес2 (2.5 kV, 1,000 Ω, 25 μF), как рекомендовано в протоколе рекомбинации.
Для обеспечения восстановления физиологических функций клеток после электропорации («оживление» клеток), необходимого также для прохождения рекомбинации, репликации и деления, клетки переносили в 15 мл пробирки, с добавлением 2 мл свежей среды Middlebrook 7H9-OADC-Tween. Затем проводили инкубацию в течение ночи при +37°С на роллерном шейкере со скоростью вращения около 1 об. / 5 с.
После ночной инкубации клетки высевали на чашки, содержащие 30 мкг/мл стрептомицина, и проводили инкубацию в течение 72 часов при +37°С. Отношение подсчитанного количества StrR колоний на число КОЕ в культуре, использованной для электропорации, принималось за эффективность протокола рекомбинации.
Так, эффективность рекомбинации олигонуклеотида (SEQ ID NO: 3) в штамме дикого типа, не экспрессирующего дополнительные рекомбиназы, М. smegmatis МС2 155 составила 4,2*10-9.
Эффективность рекомбинации в штамме М. smegmatis МС2 155::pKM402, составила в среднем 2,9*10-5 (диапазон от 1,2*10-6 до 9,9*10-5 по данным 9 независимых экспериментов). Контрольный эксперимент по рекомбинации в штамм с хелперной pKM402 без индукции продемонстрировал промежуточную эффективность в 6,3*10-7. Очевидно, что использованная в pKM402 транскрипционная система гена recT на основе тетрациклинового репрессора обладает ненулевым уровнем экспрессии в отсутствии индуктора - ангидротетрациклина.
В итоге, даже максимально достигнутая в наших условиях эффективность получения рекомбинантов в 10-4 требует проверки десятков тысяч колоний для поиска желаемых мутантов аллель-специфичной ПЦР.
Пример 3. Внесение замены rpsL K43R в М. smegmatis МС2 155 с использованием коселективной ДНК.
Использованный в Примере 2 метод внесения точечных замен рекомбинацией олигонуклеотидом по Мёрфи не обеспечивал приемлемой эффективности получения рекомбинантов без селекции. В данном и последующем примерах описано получение различных мутаций в геноме М. smegmatis с помощью использования коселективной ДНК.
В качестве коселективной плазмиды использовали плазмиду pMycA-sacB из Примера 1. Протокол рекомбинации отличался от вышеописанного в двух пунктах. Во-первых, клетки трансформировали сразу обеими ДНК - синтетическим олигонуклеотидом для рекомбинации и коселективной плазмидой. Во-вторых, после «оживления» в течение ночи, клетки высевались на селективную среду с гигромицином - соответствующий маркер устойчивости к нему содержится на коселективной плазмиде. Таким образом выявляли компетентную субпопуляцию клеток, способных к поглощению чужеродной ДНК, что повышает вероятность обнаружения клонов, в которых произошла рекомбинация с помощью олигонуклеотида.
В данном примере использовали тот же самый олигонуклеотид (SEQ ID NO: 3) для внесения замены в RpsL, что и в Примере 2 в количестве 500 или 1000 нг.Одновременно к клеткам добавлялось 50 нг коселективной плазмиды. Устойчивые к гигромицину колонии переносили как на чашку с неселективной средой, так и на чашку, содержащую стрептомицин (Фигура 1.). Стрептомицин-устойчивые колонии подсчитывали, и их отношение к полному числу перенесенных колоний соответствовало относительной эффективности получения рекомбинантов.
Для подтверждения получения желаемой замены часть колонии снимали стерильной петлей с чашки с неселективной средой и ресуспендировали в лизирующем буфере, содержащем 10 мМ Трис (Am-O497, VWR Life Science AMRESCO, США), 1 mM ЭДТА (Am-O105B, VWR Life Science AMRESCO, США) и 10 мг/мл Triton Х-100 (O694, VWR Life Science AMRESCO, США). Геномную ДНК выделяли по следующему протоколу: прогрев 30 мин при температуре +95°С, инкубация на льду 10 мин, центрифугирование в течение 10 мин при 12000 об/мин при комнатной температуре. Один микролитр надосадочной жидкости использовали для ПЦР амплификации с помощью праймеров SEQ ID NO: 4 и SEQ ID NO: 5. Секвенирование ДНК проводили с помощью набора реактивов ABI PRISM® BigDye™ Terminator v. 3.1 с последующим анализом продуктов реакции на автоматическом секвенаторе Applied Biosystems 3730 DNA Analyzer (Фигура 1Б).
Эффективность получения StrR колоний на фоне предселекции на гигромицине зависела от количества олигонуклеотида и составила в среднем 1,1*10-1 и 2,5*10-1 для 500 нг и 1000 нг, соответственно. Общий диапазон эффективностей заключался в границах от 2,8*10-2 до 3,3*10-1 по результатам 10 экспериментов. Таким образом, от 3 до 33% отобранных на чашках с гигромицином колоний были устойчивы к стрептомицину и содержали желаемую замену в гене rpsL. Соответственно, при отборе рекомбинантных клонов без селекции, полученных описанным методом, необходимо проверять всего несколько десятков колоний. На Фигуре 1. приведены фотографии культуральных чашек с пересеянными колониями в одном из экспериментов: так из 26 проверенных колоний (среда 7Н9), 7 обладали резистентным фенотипом (7H9-Str) и несли желаемую нуклеотидную замену в геноме.
Пример 4. Внесение замены gyrA D95G в М. smegmatis МС2 155.
Схема эксперимента была аналогична использованной в Примере 3. Для осуществления рекомбинации был синтезирован олигонуклеотид с заменой Т на С относительно последовательности дикого типа и гомологичными плечами длиной 37 нуклеотидов (SEQ ID NO: 6). Так как ген GyrA расположен рядом с ориджином репликации, и репликационная вилка перемещается по направлению гена, в качестве исходной последовательности использовалась минус-цепь последовательности гена. При рекомбинации должна была происходить замена кодона GAC на GGC.
Для подсчета и анализа колоний после отбора на гигромицине использовали чашки с агаризованной средой Middlebrook 7H9-OADC и 7H9-OADC с добавлением 3 мкг/мл ципрофлоксацина (17850-5G-F, SIGMA, USA). При анализе первичной последовательности генома полученных клонов также использовали колонии, выросшие на неселективной среде и не подвергавшиеся действию ципрофлоксацина. В качестве праймеров для ПЦР использовали олигонуклеотиды SEQ ID NO: 7 и SEQ ID NO: 8.
На Фигуре 2А приведены фотографии чашек, с перенесенными с гигромициновой среды колониями. Всего было перепечатано 144 колонии, из которых две также дали рост на среде с ципрофлоксацином. Секвенирование фрагмента гена подтвердило наличие желаемой мутации в геноме обеих полученных штаммов (Фигура 2Б).
Эффективность, или относительный выход рекомбинантов, составил 1.4*10-2, что несколько ниже средней эффективности, полученной в Примере 3, но, тем не менее, превышает на 2 порядка эффективность стандартного метода без использования коселективной плазмиды.
Пример 5. Излечивание модифицированного штамма микобактерий от хелперной плазмиды и коселективной молекулы ДНК.
Данный пример демонстрирует излечивание рекомбинантных колоний, полученных в Примере 3 и Примере 4, от хелперной и коселективной плазмид.
Индивидуальные колонии после рекомбинации и проверки наличия мутации помещали в жидкую среду 7H9-ADC-Tween и подращивали при +37°С с перемешиванием до логарифмической фазы (OD600 ~0,5). Полученные культуры два раза последовательно разводили в 10 раз свежей средой, и по 50 мкл исходной и полученных суспензий клеток наносились на агаровую среду 7H9-OADC с добавлением 3% сахарозы (S0389, SIGMA, США). Индивидуальные, выросшие на сахарозе, колонии перепечатывались параллельно на чашки без селекции (среда 7H9-OADC), на чашки с гигромицином (среда 7H9-OADC+100 мкг/мл гигромицина) и на чашки с канамицином (среда 7H9-OADC + 50 мкг/мл канамицина). Чувствительные к гигромицину и канамицину колонии подвергали дополнительной проверке на отсутствие коселективной плазмиды. Для этого выделяли тотальную ДНК, как описано в Примере 3, и проводили ПЦР амплификацию с помощью праймеров SEQ ID NO: 1 и SEQ ID NO: 2, использованных при конструировании коселективной ДНК, как описано в Примере 1. Отсутствие полосы на электрофорезе свидетельствовало о потере коселективной плазмиды. В качестве контроля использовалась ДНК штамма после рекомбинации, не подвергавшегося действию сахарозы, и сохранившего устойчивость к гигромицину.
Таким образом, двумя независимыми свидетельствами утери коселективной плазмиды являлись потеря устойчивости к гигромицину и отсутствие гена sacB в рекомбинантном штамме. Излечивание от хелперной плазмиды подтверждалось утратой маркера устойчивости к канамицину. Дополнительных тестов в этом случае не было необходимости проводить, так как одинаковые репликоны двух плазмид являются несовместимыми, и утрата хелперной плазмиды происходит еще на этапе электропорации и селекции штаммов, несущих коселективную плазмиду, на среде с гигромицином.
--->
<210> 1
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 1
ccgccctcta gacgcccatg caacagaaac ta (SEQ ID NO: 1)
<210> 2
<211> 32
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 2
tgcagtcgta cggcgaaaaa ggcatccgtc ag (SEQ ID NO: 2)
<210> 3
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид для рекомбинации
<400> 3
cgcgcgcgac cttccggagc gccgagttcg gcttcctcgg agtggtggtgtaaacgcgcg tgcacacgcc
(SEQ ID NO: 3)
<210> 4
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 4
aagatcgcca aggtgaagac c (SEQ ID NO: 4)
<210> 5
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 5
ctctcccgtg acacccttct (SEQ ID NO: 5)
<210> 6
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> олигонуклеотид для рекомбинации
<400> 6
gcgcaacgac cacggctggg ccatgcggac cagggtgccg tagatcgagg cgtcgccgtg cggatggtag ttacc
(SEQ ID NO: 6)
<210> 7
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 7
gatgtacgac tcgggcttcc (SEQ ID NO: 7)
<210> 8
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер
<400> 8
gagtcgcgct tcggtgtaa (SEQ ID NO: 8)
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕСТ-СИСТЕМА Mycobacterium smegmatis aphVIII+ ДЛЯ СКРИНИНГА ИНГИБИТОРОВ СЕРИН-ТРЕОНИНОВЫХ ПРОТЕИНКИНАЗ ЭУКАРИОТИЧЕСКОГО ТИПА | 2014 |
|
RU2566998C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНЫХ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA, И СПОСОБ ПРОДУКЦИИ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ PANTOEA | 2006 |
|
RU2418069C2 |
| Способ репликации человеческого митохондриального генома в клетках дрожжей Yarrowia lipolytica | 2016 |
|
RU2660715C2 |
| Плазмида для редактирования генома бактерий рода Bacillus и способ внесения модификаций в геном бактерий рода Bacillus | 2022 |
|
RU2815835C1 |
| КОЛЬЦЕВАЯ МОЛЕКУЛА ДНК, ХАРАКТЕРИЗУЮЩАЯСЯ ЗАВИСИМОЙ ОТ УСЛОВИЙ ТОЧКОЙ НАЧАЛА РЕПЛИКАЦИИ, СПОСОБ ИХ ПОЛУЧЕНИЯ И ИХ ПРИМЕНЕНИЕ В ГЕННОЙ ТЕРАПИИ | 2003 |
|
RU2307870C2 |
| Средство редактирования генома на основе белка LigD из бактерии Pseudomonas putida и Cas9 комплекса | 2022 |
|
RU2797049C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI-ПРОДУЦЕНТ L-ТРЕОНИНА | 2013 |
|
RU2546237C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2007 |
|
RU2364628C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2012 |
|
RU2528056C2 |
| СПОСОБ ХРОМОСОМНОЙ ИНТЕГРАЦИИ И ЗАМЕНЫ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК В CLOSTRIDIA | 2006 |
|
RU2464317C2 |

Изобретение относится к генной инженерии. Описан способ внесения точечных замен в бактериальную хромосому без сопутствующей интеграции детерминант антибиотикоустойчивости. Способ основан на широко применяемом методе фаговой рекомбинации для внесения мутаций в бактериальную хромосому с использованием коротких фрагментов. Эффективность получения рекомбинантных клонов в Mycobacteria составляет порядка 10-4-10-3 на общее количество клеток. Соответственно, отбор нужных мутантов требует проверки тысяч колоний. Предложенные ранее способы повышения эффективности отбора клонов включают в себя либо использование маркеров антибиотикоустойчивости, либо подразумевают создание сложных генетических конструкций персонально для каждой вводимой замены. Повышение относительного выхода рекомбинантных колоний достигается за счет трансформации рекомбинирующего олигонуклеотида совместно с коселективной ДНК, неспособной к встраиванию в хромосому. Отбор колоний по фенотипической устойчивости, обеспечиваемой маркером резистентности коселективной ДНК, позволяет отобрать компетентную субпопуляцию клеток и повысить эффективность метода до 10-2-10-1 рекомбинантов. Утрата внехромосомного маркера резистентности обеспечивается либо серийными пассажами культуры на среду без антибиотика, либо использованием контрселективных маркеров. 8 з.п. ф-лы, 2 ил., 5 пр.
1. Способ внесения немаркированных точечных замен в хромосому микобактерий, включающий стадии:
- введения в модифицируемый штамм микобактерий хелперной плазмиды, обеспечивающей индуцируемую экспрессию белков рекомбинации;
- индукции экспрессии белков рекомбинации в модифицируемом штамме микобактерий;
- одновременной электропорации модифицируемого штамма микобактерий одноцепочечным олигонуклеотидом, содержащим желаемую замену и участки гомологии вокруг указанной желаемой замены, и коселективной молекулой ДНК, неспособной к встраиванию в хромосому модифицируемого штамма микобактерий и несущей ген(ы) устойчивости к антибактериальному препарату, отличному от препарата, используемого для поддержания хелперной плазмиды;
- обеспечения рекомбинации и экспрессии гена(ов) устойчивости коселективной молекулы ДНК на среде без селекции;
- отбора колоний модифицированного штамма микобактерий, устойчивых к указанному антибактериальному препарату, чей фенотип обеспечен маркером резистентности коселективной ДНК, путем высева культуры модифицируемого штамма микобактерий на селективную среду, содержащую антибактериальный препарат, отличный от препарата, используемого для поддержания хелперной плазмиды;
- излечивания модифицированного штамма микобактерий от хелперной плазмиды и коселективной молекулы ДНК.
2. Способ по п. 1, в котором в качестве модифицируемого штамма используют бактериальные штаммы, принадлежащие к видам Mycobacterium tuberculosis и Mycolicibacterium smegmatis.
3. Способ по п. 1, в котором указанная хелперная плазмида обеспечивает экспрессию белка рекомбинации RecT.
4. Способ по п. 3, в котором указанной хелперной плазмидой является плазмида pKM402.
5. Способ по п. 1, в котором указанный одноцепочечный олигонуклеотид содержит желаемую замену и участки гомологии длиной 24-50 нуклеотидов с каждой стороны от указанной замены.
6. Способ по п. 1, в котором указанная коселективная молекула ДНК содержит ориджин репликации, обеспечивающий поддержание плазмиды в Mycobacteria.
7. Способ по п. 6, в котором в качестве коселективной молекулы ДНК используют кольцевую плазмидную ДНК.
8. Способ по п. 7, в котором коселективная плазмидная ДНК содержит ген устойчивости к гигромицину.
9. Способ по п. 1, при котором отбор колоний после электропорации проводят на селективной среде с гигромицином.
| Zhang Y, et al | |||
| A new logic for DNA engineering using recombination in Escherichia coli | |||
| Nat Genet | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Van Kessel JC, Hatfull GF | |||
| Recombineering in Mycobacterium tuberculosis | |||
| Nat Methods | |||
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| RU 2015134995 A, 28.03.2019. | |||

