Изобретение относится к биотехнологии, генетической и белковой инженерии, а именно, к получению генетической конструкции, обеспечивающей нокаут гена MYC с целью снижения экспрессии транскрипционного фактора c-Myc в клетках человека, которые могут быть использованы для исследования механизмов канцерогенеза, метастазирования и регенерации.
Получение генетических конструкций с целью создания модельных клеточных линий для исследования процессов дедифференцировки клеток крайне востребовано в современной биомедицине, в частности в онкологии и регенеративной медицине.
Ген MYC расположен в локусе 8q24.21 хромосомы 8 человека, содержит 3 экзона, 4 промотора P0, P1, P2, P3, два альтернативных старт-кодона (CTG и ATG), в связи с этим существуют две изоформы мРНК MYC и две изоформы белка - c-Myc с молекулярной массой 64 кДа и минорный белок с молекулярной 67 кДа [1].
Белок Myc является плейотропным фактором транскрипции, регулирующим экспрессию множества генов, влияющих на ключевые процессы, протекающие в клетке такие как клеточный цикл, апоптоз, ответ на повреждение ДНК, гемопоэз, дифференцировка, метаболизм [2, 3]. Белок Myc человека содержит несколько высококонсервативных областей, которые функционально важны и организованы одинаково среди трех паралогов: с-MYC, n-MYC и l-MYC.
MYC является онкогеном, ассоциированным с появлением и развитием различных злокачественных опухолей. Белок c-Myc сверхэкспрессирован в более чем 70% злокачественных новообразований человека, включая лимфомы, лейкемию, карциномы, нейробластомы, меланомы, опухоли молочной железы, яичников, предстательной железы и печени. Большинство злокачественных новообразований, демонстрирующих высокую экспрессию c-Myc, являются высокоагрессивными и/или плохо поддаются лечению [3, 4].
Ген MYC представляет собой одну из наиболее перспективных мишеней для лечения рака. Продемонстрировано, что временное ингибирование MYC in vivo приводит к устойчивой регрессии опухоли, способствуя остановке пролиферации, апоптозу и клеточному старению в раковых клетках. Однако важные функциональные домены с-Myc по своей природе не упорядочены и не имеют сайта ферментативной активности, что является препятствием для дизайна лекарств, которые можно было бы нацеливать на белок с целью понижения экспрессии MYC и предотвращения развития опухоли [3]. Одним из способов решения данной проблемы является поиск ингибиторов, оказывающих свое влияние не на непосредственно белок Myc, а на его гетеродимеризацию с белком Max - обязательным партнером Myc, без которого невозможно связывание димера Myc-Max с промоторами генов. Примерами таких ингибиторов являются следующие химические соединения: SaJM589 [4], MYCMI-6 [5], KSI-3716 [6].
Еще одним подходом к подавлению действия белка Myc является поиск ингибиторов, которые конкурируют с белком Myc за связывание с промоторами его генов-мишеней, предотвращая их транскрипцию, например, Omomyc - небольшой белок, состоящий из 90 аминокислот, прошедший клиническую фазу исследований у пациентов с раком легкого, толстой кишки и молочной железы [3].
Альтеративно предлагают подходы снижения экспрессии с-Myc с использованием shRNA к гену POU5F1, кодирующего белок Oct4 и являющегося энхансером MYC; или в результате нокаута гена MAPKAPK2, кодирующего киназу Mk2, участвующую в фосфорилировании и последующей активации Oct4 [7].
Для проведения исследований направленных на разработку подходов к снижению экспрессии c-Myc требуется создание модельных клеточных линий, различающихся уровнем экспрессии гена MYC. Подобные клеточные линии высоко востребованы как для проведения исследований in vitro, так и для исследований in vivo на моделях ксенографтов опухолей человека. Например, компания ATCC предлагает исследовательскую панель, предназначенную для проведения фундаментальных исследований и разработки противораковых препаратов, состоящую из 9 типов культур клеток, которые содержат различные мутации MYC или разное число копий гена MYC - Genetic Alteration Cell Panel (ATCC). В панель включены линии клеток В-лимфом CRL-1647 ST486, CRL-1648 CA46 и HTB-62 P3HR-1, рака желудка CRL-5974 SNU-16, рака легких HTB-175 NCI-H82, CRL-2081 MSTO-211H и HTB-171 NCI-H446, лейкемии CCL-240 HL-60 и плазмацитомы CRL-9068 NCI-H929.
Для нокаута гена MYC может быть использована система редактирования генома CRISPR/Cas. Система CRISPR/Cas была впервые описана как адаптивная иммунная система у бактерий и архей, а теперь она также является биотехнологическим инструментом для редактирования генома живых клеток [8, 9]. Одним из вариантов систем CRISPR/Cas является система CRISPR/Cas9, функционирующая за счет РНК (gRNA), которая формирует комплекс gRNA-Cas9 и направляет нуклеазу Cas9, выделенную, например, из Streptococcus pyogenes (SpyCas9), к заданным участкам генома. Комплекс gRNA-Cas9 распознает целевую последовательность, связывается с ней за счет комплементарных взаимодействий и катализирует разрезание двухцепочечной ДНК.
Разрыв активирует репарацию посредством негомологичного соединения концов (NHEJ) или гомологичной репарации (HDR). В подавляющем большинстве случаев активируется NHEJ, что приводит к вставкам и/или делециям, которые нарушают последовательность целевого гена, нокаутируя его. Нацеливание gRNA-Cas9 к ДНК осуществляется за счет наличия в gRNA последовательности, комплементарной последовательности гена-мишени. Целевая последовательность в геноме обычно имеет длину порядка 20 п.н. и должна примыкать к PAM-последовательности 5'-NGG-3', которая необходима для распознавания нуклеазой целевого сайта.
Технической задачей изобретения является создание рекомбинантных плазмид, обеспечивающих нокаут гена MYC в клетках человека и наиболее приемлемого и экономически выгодного способа получения линии клеток человека с пониженной экспрессией фактора транскрипции c-Myc для научных исследований и разработок.
Известен лентивирусный вектор Lenti-sh1368 knockdown c-myc, направляющий экспрессию интерферирующей shRNA к гену c-Myc для ингибирования экспрессии c-Myc в клетках млекопитающих [10]. Известна плазмида pX459-puro-hMYC для экспрессии в клетках млекопитающих направляющей РНК, комплементарной последовательности 5’-TGCTGCCAAGAGGGTCAAGT-3’ гена MYC человека [11]. Наиболее близким к заявляемому техническому решению по технической сущности и достигаемому техническому результату является лентивирусный плазмидный вектор pXPR_BRD003-sgMYC, обеспечивающий экспрессию направляющей РНК к гену MYC [12]; в оригинальной работе вектор был использован как положительный контроль работоспособности метода нокаута гена WRN-хеликазы. Данный лентивирусный плазмидный вектор обеспечивал экспрессию направляющей РНК к последовательности 5’-GACAACGTCTTGGAGCGCCAG-3’, расположенной в кодирующей области гена MYC (2 экзон); нокаут гена MYC достигался за счет внесения двухцепочечного разреза в ДНК гена MYC нуклеазой Cas9, формирующей комплекс с данной направляющей РНК.
Известно, что использование системы CRISPR/Cas9 сопряжено с риском внесения нецелевых разрывов в геномную ДНК. Для повышения специфичности редактирования может быть использована мутантная форма нуклеазы Cas9 - Cas9D10A, или никаза, которая вносит разрез только в одну из цепей двухцепочечной ДНК [13]. Для нокаута гена с использованием Cas9D10A подбирают пару направляющих РНК, комплементарных верхней и нижней цепи ДНК, обеспечивающих направление никазы к участкам заданного гена (участки должны быть ограниченны PAM-последовательностью 5’-NGG-3’) таким образом, чтобы в результате гидролиза никазой в каждой из цепей ДНК образовывалось по одному разрезу, расположенному на расстоянии 40-70 п.н. друг от друга. Векторы, позволяющие выполнять нокаут гена MYC по данному способу, в литературе не описаны.
Новый технический результат достигают получением рекомбинантных плазмидных ДНК PX458D10A_Myc, обеспечивающих нокаут гена MYC в клетках человека за счет способности экспрессировать РНК, содержащую участок длиной 20 нуклеотидов, комплементарный ДНК второго экзона гена MYC человека, и способную формировать комплекс c Cas9D10A, вносящих разрез в молекулу ДНК для обеспечения нокаута гена, представляющих собой плазмидные векторы PX458D10A гидролизованные по сайту BbsI (BpiI), содержащие сайт инициации репликации плазмиды pBR322, ген кодирующий нуклеазу Cas9D10A, слитую с сигналом ядерной локализации вируса SV40 (NLS), и транскрибируемый в клетках млекопитающих в виде единого транскрипта, разделенного T2A последовательностью пептида 2А капсида вируса Thosea asigna, с последовательностью, кодирующей зеленый флуоресцентный белок EGFP, промотор U6 РНК полимеразы III человека, фрагмент размером 20 п.н., комплементарный кодирующей последовательности гена MYC человека, экспрессирующийся в составе молекулы направляющей РНК из Streptococcus pyogenes системы CRISPR/Cas9 и формирующей комплекс с Cas9D10A, способной вносить разрез в одну из цепей ДНК, генетический маркер bla ген β-лактамазы, определяющий устойчивость к ампициллину при трансформации плазмидой клеток Escherichia coli.
Также, новым техническим результатом является способ получения моноклональной клеточной линии PX458D10A_Myc, предусматривающий трансформацию клеток E.coli Stable3, парой сконструированных плазмид PX458D10A_Myc, выращивание трансформированных клеток в среде с ампициллином, выделение рекомбинантной плазмидной ДНК, трансфекцию клеток человека плазмидной ДНК и получение клона единичной клетки с пониженной экспрессией транскрипционного фактора c-Myc.
Сущность изобретения заключается в том, что конструируют пару плазмид PX458D10A_Myc путем встраивания в плазмидный вектор PX458D10A фрагмента ДНК, кодирующего РНК комплементарную нижней или верхней цепи ДНК второго экзона гена MYC методом Golden Gate, трансформируют лигазной смесью клетки Escherichia coli Stable3, обеспечивающие амплификацию соответствующей рекомбинантной плазмиды, выделяют рекомбинантную плазмидную ДНК; трансфицируют клетки человека парой плазмидных ДНК, кодирующих направляющие РНК к верхней и нижней цепи ДНК, получают клон единичной клетки с пониженной экспрессией транскрипционного фактора c-Myc.
Способ осуществляют следующим образом:
генно-инженерными методами получают пары плазмид PX458D10A_Myc, содержащих фрагмент ДНК, комплементарный смысловой (верхней, top) или матричной (нижней, bottom) цепям ДНК кодирующей области гена MYC и полученный методом отжига двух синтетических олигонуклеотидных последовательностей.
Клетки E.coli Stable3 трансформируют сконструированными плазмидами PX458D10A_Myc_t и PX458D10A_Myc_b и выращивают в течение ночи в среде с добавлением ампициллина. Далее клетки используют для выделения плазмидной ДНК. Далее парами плазмидных ДНК, содержащих участники, кодирующие РНК, комплементарные верхней и нижней цепям ДНК - PX458D10A_Myc_top и PX458D10A_Myc_bottom - трансфицируют клетки человека. После трансфекции клетки выращивают в полной питательной среде 24 часа, снимают с поверхности пластика, разводят до концентрации 10 клеток в 1 мл и переносят по 100 мкл в лунки культурального планшета. На следующий день методом микроскопии отмечают те лунки планшета, в которых находится 1 клетка, и культивируют планшеты до достижения 80% конфлуентности. Далее клетки моноклона последовательно переносят в культуральные емкости большего объема.
Исходным генетическим материалом для конструирования рекомбинантных плазмид PX458D10A_Myc являются:
а) плазмидный вектор PX458D10A, обеспечивающий встройку фрагмента ДНК, комплементарного кодирующей области гена-мишени и его экспрессию в составе молекулы РНК, формирующей комплекс с никазой Cas9D10A, способной вносить разрез в одну из цепей ДНК, и ген Cas9D10A;
б) фрагмент ДНК, комплементарный кодирующей области гена MYC человека, который получают в результате отжига синтетических олигонуклеотидов.
Полученные в результате плазмиды серии PX458D10A_Myc используют попарно (фиг. 1) и характеризуются следующими признаками:
- имеют молекулярную массу 5,7 МДа и размер 9292 п.н.;
- кодирует молекулу РНК, содержащую фрагмент комплементарный верхней или нижней цепи ДНК кодирующей области гена MYC, обеспечивающую сборку комплекса с никазой Cas9D10A и его позиционирование на экзоне 2 гена MYC человека;
- состоит из следующих элементов:
а) фрагмента ДНК, размером 20 п.н., комплементарного верхней или нижней цепи кодирующей области гена MYC, расположенной в геноме человека на хромосоме 8;
б) плазмиды PX458D10A, обеспечивающей в клетках млекопитающих экспрессию РНК и никазы Cas9D10A, которые формируют комплекс вносящий разрез в одну из цепей ДНК последовательности экзона 2 гена MYC человека;
- содержит:
а) сайт инициации репликации плазмиды pBR322;
б) генетический маркер bla ген β-лактамазы, определяющий устойчивость к ампициллину при трансформации клеток Escherichia coli;
в) ген, кодирующий молекулу направляющей РНК из Streptococcus pyogenes системы CRISPR/Cas9 под промотором U6 полимеразы III человека;
г) ген, кодирующий нуклеазу Cas9D10A из Streptococcus pyogenes системы CRISPR/Cas9, слитый с последовательностью, кодирующей сигнал ядерной локализации вируса SV40, под промотором гена бета-актина цыпленка;
Таким образом, получены две пары плазмидных ДНК PX458D10A_Myc, направляющих продукцию в клетках человека никазы Cas9D10A и пары РНК, формирующих комплекс с никазой Cas9D10A и обеспечивающих разрезание двухцепочечной ДНК в экзоне 2 гена MYC человека, описан способ получения и получена моноклональная линия клеток рака молочной железы человека MDA-MB231_Myc с пониженной экспрессией транскрипционного фактора c-Myc.
Изобретение иллюстрируется следующими чертежами:
фиг. 1. Общая схема структурной организации плазмиды PX458D10A_Myc, содержащей фрагмент ДНК, размером 20 п.н., комплементарный кодирующей области гена MYC, экспрессирующийся в составе РНК, которая формирует комплекс с никазой Cas9D10A и обеспечивает связывание с участком ДНК в экзоне 2 гена MYC человека, промотор U6 полимеразы III человека, сайт инициации репликации плазмиды pBR322, генетический маркер bla ген β-лактамазы, определяющий устойчивость к ампициллину при трансформации Escherichia coli, ген, кодирующий мутантную форму нуклеазы Cas9D10A из Streptococcus pyogenes системы CRISPR/Cas9, слитый с последовательностью, кодирующей сигнал ядерной локализации вируса SV40, под промотором гена бета-актина цыпленка;
фиг. 2. Рестрикционный анализ рекомбинантных плазмид PX458D10A_Myc с использованием эндонуклеазы BbsI. К - плазмида PX458D10A, M12 - маркер молекулярного веса M12 (СибЭнзим, Россия) (10000, 8000, 6000, 5000, 4000, 3000, 2500, 2000, 1500, 1000, 750, 500 п.о.), шифр плазмид, выделенных из отобранных для скининга клонов, указан сверху. Примечание. * отмечены образцы плазмид после обработки эндонуклеазой, плазмиды не обработанные эндонуклеазой без пометок;
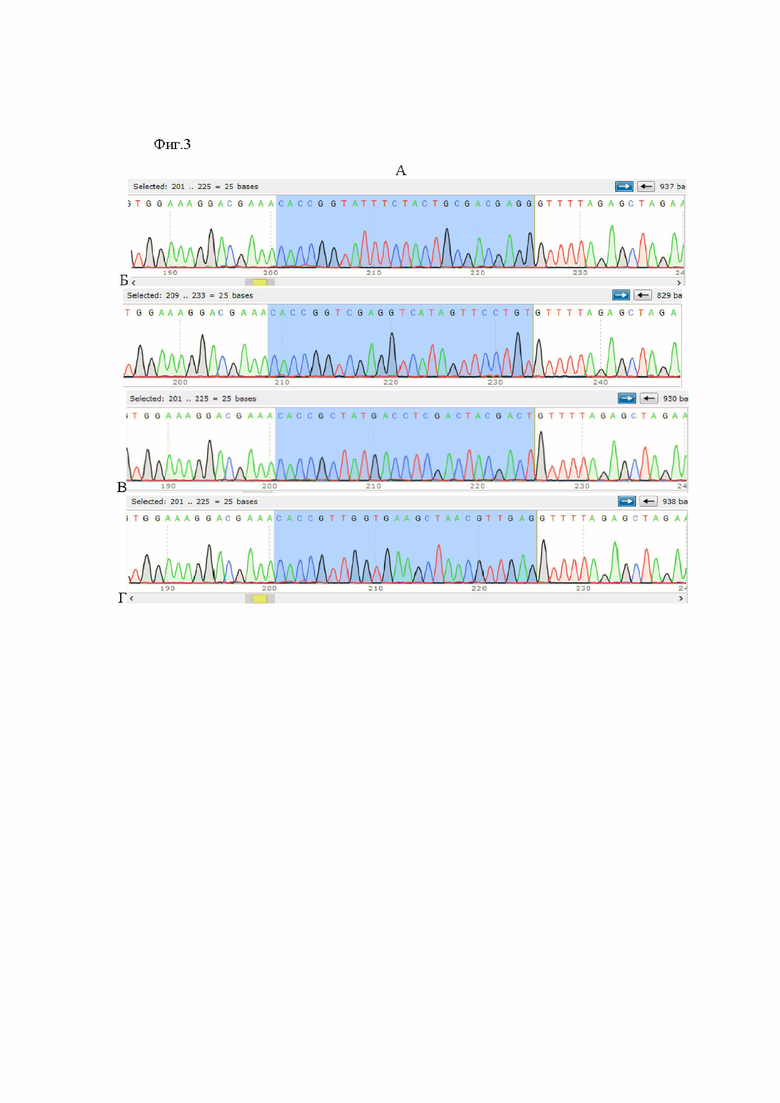
фиг. 3. Фрагменты хроматограмм, полученных после секвенирования по Сенгеру плазмидной ДНК, выделенной из рекомбинантных клонов (А) PX458D10A_MycGt, (Б) PX458D10A_MycGb, (В) PX458D10A_MycPt, (Г) PX458D10A_MycPb;
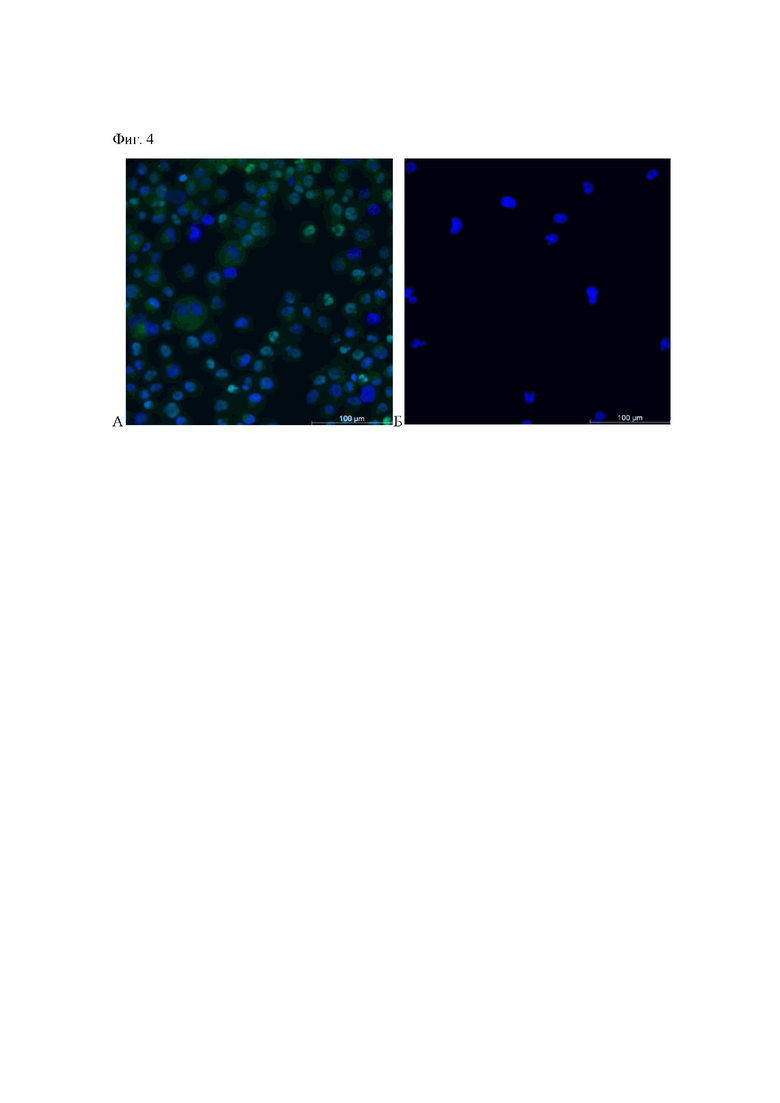
фиг. 4. Анализ уровня экспрессии транскрипционного фактора c-Myc в (A) клетках исходной линии MDA-MB231 и (Б) клетках моноклональной линии MDA-MB231_Myc. Клетки окрашены Anti-c-Myc антителами (зеленый сигнал), ядра окрашены DAPI (синий сигнал).
Пример 1. Получение рекомбинантной плазмиды PX458D10A_Myc
Для получения фрагмента, комплементарного участку экзона 2 гена MYC человека, используют 4 пары олигонуклеотидов длинной 25 н.о.
MYC_G1_t_f 5’-CACCGGTATTTCTACTGCGACGAGG-3’
MYC_G1_t_r 5’-AAACCCTCGTCGCAGTAGAAATACC-3’
MYC_G2_b_f 5’-CACCGGTCGAGGTCATAGTTCCTGT-3’
MYC_G2_b_r 5’-AAACACAGGAACTATGACCTCGACC-3’
MYC_P1 _t_f 5’-CACCGCTATGACCTCGACTACGACT-3’
MYC_P1 _t_r 5’-AAACAGTCGTAGTCGAGGTCATAGC-3’
MYC_P2_b_f 5’-CACCGTTGGTGAAGCTAACGTTGAG-3’
MYC_P2_b_r 5’-AAACCTCAACGTTAGCTTCACCAAC-3’
Готовят реакционную смесь, содержащую по 10 мкМ каждого праймера из пары, Т4 лигазный буфер (Евроген, Россия), 10 е.а. Т4 полинуклеотидкиназы (NEB, Великобритания). Реакцию проводят в термоциклере (Applied Biosystems, США) при 37°С в течение 30 минут, далее при 95°С в течение 5 минут и охлаждают смесь с скоростью 5°С/мин до 25°С. Реакционную смесь разводят в десять раз водой, отбирают 1 мкл и смешивают в Quick ligation буфер (Евроген) с добавлением 0,25 мг/мл бычьего сывороточного альбумина (Sigma), 10 е.а. эндонуклеазы рестрикции BpiI (BbsI), (Thermo Scientific), 375 е.а. Т7 лигазы (NEB), 25 нг плазмидной ДНК PX458D10A. Смесь инкубируют в термоциклере (Applied Biosystems, США) в течение 15 циклов, включающих 2 шага: 37°С 5 мин, 20°С 5 мин. Полученную реакционную смесь используют для трансформации компетентных клеток Escherichia coli Stbl3. Трансформированные клетки культивируют на чашках Петри на агаризованной LB-среде с добавлением ампициллина до конечной концентрации 50 мкг/мл в течение ночи при 30°С. Для выделения рекомбинантных плазмид отбирают по 3 колонии, выросшие на селективной среде, переносят в 5 мл LB-среды (с добавлением ампициллина 50 мкг/мл) и инкубируют в течение ночи при 30°С при постоянном перемешивании при 180 rpm. Выделение плазмидной ДНК проводят набором Plasmid Miniprep (Евроген, Россия) согласно инструкции производителя.
Выполняют рестрикционный анализ выделенных плазмид с использованием эндонуклеазы рестрикции BsuI (СибЭнзим, Россия), реакцию проводят в течение 1 часа при 37°С. Фрагменты ДНК, полученные после расщепления эндонуклеазой, разделяют методом электрофореза в 1% агарозном геле с добавлением интеркалирующего красителя бромистого этидия. Образцы плазмидной ДНК после рестрикции визуализируют с использованием гель-документирующей системы G-box (Syngen, Великобритания). В результате успешного клонирования фрагмента, кодирующего направляющую РНК, в плазмиду PX458D10A сайты узнавания для эндонуклеазы рестрикции BbsI будут удалены и на электрофореграмме будут выявляться полосы, соответствующие линейной, кольцевой и суперскрученной формам плазмиды (фиг. 2).
Соответствие последовательности фрагмента, клонированного в плазмидный вектор PX458D10A, ожидаемой подтверждают методом секвенирования по Сэнгеру с использованием праймера CRISPR-Seq_F 5’-GAGGGCCTATTTCCCATGATTCCTTCATAT-3’ (фиг. 3).
Пример 2. Получение моноклональной линии клеток человека с нокаутом гена MYC
Клетки линии аденокациномы молочной железы человека MDA-MB231 (ECACC) культивируют в полной питательной среде DMEM/F12 (Gibco, Великобритания) с добавлением 10% FBS (Gibco, Великобритания) и 1/100 GlutaMAX 100× (Gibco, Великобритания) в СО2-инкубаторе MCO-5M (Sanyo, Япония) при 5% СО2, 37°С. До начала исследования in vitro клетки культивируют не менее чем до 3 пассажа. Для снятия клеток с пластика при пересеве используют TrypLE (Thermo Fisher Scientific, Дания). Количество живых клеток определяют методом окрашивания с Trypan Blue stain 0,4% (Invitrogen, США) на автоматическом счетчике Countess II FL (Thermo Fisher Scientific, Сингапур).
Клетки MDA-MB231 рассаживают в лунки 6-луночного планшета. На следующий день среду удаляют и приливают 2 мл полной питательной среды DMEM/F12. Готовят смесь для трансфекции: для этого 2 мкл Metafectene (Biontex, Германия) переносят в 45 мкл 1× фосфатно-солевого буфера (ФСБ). Вносят 50 мкл ФСБ содержащего 2 мкг плазмид PX458D10A_Myc_Gt и PX458D10A_Myc_Gb (или PX458D10A_Myc_Pt и PX458D10A_Myc_Pb) в массовом соотношении 1: 1. Смесь мягко перемешивают пипетированием, инкубируют 15 мин при комнатной температуре и по каплям вносят в соответствующие лунки планшета с клетками MDA-MB231.
Через 24 часа после трансфекции клетки снимают с поверхности пластика, готовят суспензию клеток в полной питательной среде DMEM/F12 методом последовательных разведений до концентрации 10 клеток/мл и разносят по 100 мкл в лунки 96-луночного планшета. Планшеты инкубируют в СО2 инкубаторе при 37°С. Через 24 часа после рассадки клеток отмечают лунки, в которых обнаруживается только 1 клетка, экспрессирующая EGFP, при анализе с использованием инвертированного флуоресцентного микроскопа Leica DMi8 (Leica Microsystems GmbH, Германия). Клетки культивируют в стандартных условиях, замену среды проводят по мере необходимости, но не реже двух раза в неделю. После достижения 70-80% конфлюентности клетки моноклональной линии MDA-MB231_Myc в отмеченных лунках последовательно пересаживают в 24-, 6-луночные планшеты, и далее в культуральный флакон Т25.
Экспрессию гена MYC в клетках моноклональной линии MDA-MB231_Myc оценивают методом иммунофлуоресцентного анализа, для окрашивания используют антитела Anti-c-Myc (1:100, Recombinant Anti-c-Myc rabbit monoclonal [Y69] antibody ab32072, Abcam, Великобритания) (фиг. 4). Отсутствие окрашивания клеток антителами указывает на нокаут в них гена MYC.
Список использованных источников
1. Carabet, L.A., P.S. Rennie, and A. Cherkasov, Therapeutic Inhibition of Myc in Cancer. Structural Bases and Computer-Aided Drug Discovery Approaches. Int J Mol Sci., 2018 20: p.120.
2. Ahmadi, S.E., et al., MYC: a multipurpose oncogene with prognostic and therapeutic implications in blood malignancies. J Hematol Oncol., 2021. 14: p.121.
3. Llombart, V. and M.R. Mansoura, eBioMedicine, 2022. 75: p.103756.
4. Choi, S.H., et al., Targeted Disruption of Myc-Max Oncoprotein Complex by a Small Molecule. ACS Chem Biol., 2017. 12: p.2715-2719.
5. Castell, A., et al., A selective high affinity MYC-binding compound inhibits MYC:MAX interaction and MYC-dependent tumor cell proliferation. Sci Rep., 2018. 8: p.10064.
6. Jeong, K.C., et al., Intravesical instillation of c-MYC inhibitor KSI-3716 suppresses orthotopic bladder tumor growth. J Urol., 2014 191: p.510-518.
7. Wei, S.J., et al., MYC transcription activation mediated by OCT4 as a mechanism of resistance to 13-cisRA-mediated differentiation in neuroblastoma. Cell Death Dis., 2020 11: p.368.
8. Ma, Y., L. Zhang, and X. Huang, Genome modification by CRISPR/Cas9. FEBS J., 2014. 281: p.5186-5193.
9. Gasiunas, G., et al., Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci U S A, 2012. 109: p.E2579-E2586.
10. Lin, C.H., et al., Myc-regulated microRNAs attenuate embryonic stem cell differentiation. EMBO J., 2009 28: p.3157-3170.
11. RRID:Addgene_185376), p.-p.-h.w.a.g.f.A.N.A.p.h.n.t.n.a.
12. Chan, E.M., et al., RN helicase is a synthetic lethal target in microsatellite unstable cancers. Nature, 2019 568: p.551-556.
13. Mali, P., et al., CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nat Biotechnol., 2013. 31: p.833-838.


Изобретение относится к области биотехнологии, в частности к рекомбинантной плазмидной ДНК PX458D10A_Myc. Плазмида обеспечивает нокаут гена MYC в линиях клеток рака молочной железы человека MDA-MB231_Myc, имеет молекулярную массу 5,7 МДа и представляет собой плазмидный вектор PX458D10A, гидролизованный по сайту BpiI, содержащий сайт инициации репликации плазмиды pBR322, фрагмент размером 20 п.н., комплементарный ДНК гена MYC человека, имеющий последовательность SEQ ID NO 1-4. Кроме того, настоящее изобретение также относится к способу получения линии клеток рака молочной железы человека - линии MDA-MB231 с подавленной экспрессией гена MYC, включающему трансфекцию клеток человека плазмидной ДНК PX458D10A_Myc. Настоящее изобретение обеспечивает получение рекомбинантных плазмид, обеспечивающих нокаут гена MYC в клетках рака молочной железы человека линии MDA-MB231, и наиболее приемлемый и экономически выгодный способ получения линии клеток человека с пониженной экспрессией фактора транскрипции c-Myc для научных исследований и разработок. 2 н.п. ф-лы, 4 ил., 2 пр.
1. Рекомбинантная плазмидная ДНК PX458D10A_Myc, обеспечивающая нокаут гена MYC в линиях клеток рака молочной железы человека MDA-MB231_Myc, имеющая молекулярную массу 5,7 МДа и представляющая собой плазмидный вектор PX458D10A, гидролизованный по сайту BpiI, содержащий сайт инициации репликации плазмиды pBR322, фрагмент размером 20 п.н., комплементарный ДНК гена MYC человека, имеющий последовательность SEQ ID NO 1-4, который экспрессируется под контролем U6 промотора полимеразы III человека в составе направляющей РНК из Streptococcus pyogenes системы CRISPR/Cas9, способной формировать вторичные структуры, обеспечивающие взаимодействие РНК с нуклеазой Cas9, а также с ее мутантной формой - никазой Cas9D10A, ген, кодирующий никазу Cas9D10A, генетический маркер: bla ген β-лактамазы, определяющий устойчивость к ампициллину.
2. Способ получения линии клеток рака молочной железы человека - линии MDA-MB231 с подавленной экспрессией гена MYC, заключающийся в том, что проводят трансфекцию клеток человека плазмидной ДНК PX458D10A_Myc по п.1, получают клон единичной клетки с подавленной экспрессией гена MYC методом предельных разведений.
| LOFTINEJAD P | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Прибор для определения всасывающей силы почвы | 1921 |
|
SU138A1 |
| PIRITY M | |||
| et al | |||
| Lessons learned from Myc/Max/Mad knockout mice, Curr Top Microbiol Immunol, 2006, vol | |||
| Дровопильное устройство | 1921 |
|
SU302A1 |
| YOSHIDA G.J | |||
| Emerging roles of Myc in stem | |||

