Предшествующий уровень техники настоящего изобретения
Область техники настоящего изобретения
[0001] Настоящее изобретение относится к антителу. Более конкретно, настоящее изобретение относится к антителу для лечения рака.
Описание предшествующего уровня техники
[0002] Два основных типа лимфоцитов у людей представляют собой Т (происходящие из тимуса) и В (происходящие из костного мозга). Эти клетки происходят из гемопоэтических стволовых клеток в костном мозге и печени плода, которые участвуют в лимфоидном пути развития. Потомство этих стволовых клеток следует по разным путям, созревая в B- или Т-лимфоциты. Развитие В-лимфоцитов человека происходит исключительно в костном мозге. С другой стороны, Т-клетки развиваются из незрелых предшественников, которые покидают костный мозг и проходят через кровоток к тимусу, где они пролиферируют и дифференцируются в зрелые Т-лимфоциты.
[0003] Т-клетки
[0004] Т-клетки представляют собой наиболее распространенные (приблизительно 75% лимфоцитов крови) и сильные иммунные клетки-киллеры. Роль эффекторных Т-клеток в противоопухолевом иммунном ответе полностью подтверждается исследованиями in vitro и наблюдением того, что высокая инфильтрация CD8+ Т-клеток в нескольких типах опухолей коррелирует с благоприятным клиническим прогнозом (Fridman et al., 2012). Активация эффекторных наивных Т-клеток требует по меньшей мере трех дополнительных сигналов: (i) взаимодействие TCR-CD3/Ag-MHC с помощью корецепторов (CD4 или CD8), (ii) связывание костимулирующих молекул, таких как CD80 или CD86, с CD28, CD40/CD40L, и (iii) вспомогательные молекулы, такие как цитокины.
[0005] Костимуляция или предоставление двух различных сигналов Т-клеткам представляет собой широко распространенную модель активации лимфоцитов покоящихся Т-лимфоцитов антигенпрезентирующими клетками (АРС) (Lafferty and Cunningham, 1975). Эта модель дополнительно обеспечивает различение ауто от не-ауто и иммунную толерантность (Bretscher and Cohn, 1970; Bretscher, 1999; Jenkins and Schwartz, 1987). Первичный сигнал, или антигенспецифический сигнал, передается через Т-клеточный рецептор (TCR) после распознавания чужеродного антигенного пептида, представленного в контексте основного комплекса гистосовместимости (МНС). Второй или костимулирующий сигнал доставляется к Т-клеткам костимулирующими молекулами, экспрессируемыми на антигенпрезентирующих клетках (АРС), и индуцирует Т-клетки активировать клональную экспансию, секрецию цитокинов и эффекторную функцию (Lenschow et al., 1996). В отсутствие костимуляции Т-клетки могут стать невосприимчивыми к стимуляции антигенами, не вызывать эффективный иммунный ответ и, кроме того, могут привести к истощению или толерантности к чужеродным антигенам.
[0006] Белок иммунной контрольной точки: PD-L1 и ОХ40
[0007] Иммунные контрольные точки относятся к группе ингибирующих и стимулирующих путей, в основном инициируемых взаимодействием лиганд-рецептор, связывающих иммунную систему, в частности, опосредованный Т-клетками иммунитет, для поддержания собственной толерантности и модулирования продолжительности и амплитуды физиологических ответов в периферических тканях для того, чтобы минимизировать повреждения коллатеральных тканей в норме (Pardoll, 2012). Опухолевые клетки выбирают определенные пути контрольных точек в качестве основного механизма иммунного сопротивления. Например, лиганд белка запрограммированной гибели клеток 1, PD-L1, как правило, активируется на поверхности опухолевых клеток рака человека. Взаимодействие PD-L1 с его рецептором, PD-1, экспрессируемым на инфильтрирующих опухоль лимфоцитах (TIL), в частности на Т-клетках, ингибирует опосредованный локальными Т-клетками ответ, избегая иммунного надзора (Liang et al. al., 2006; Sznol and Chen, 2013). Таким образом, ингибирование иммуносупрессивных сигналов на раковых клетках или прямая агонистическая стимуляция Т-клеток приводит к сильному устойчивому противоопухолевому иммунному ответу и/или индуцирует его. Недавние клинические исследования убедительно свидетельствуют о том, что блокирование белков иммунных контрольных точек через антитела или модулирование растворимыми лигандами или рецепторами является наиболее перспективным подходом к активации терапевтического противоопухолевого иммунитета (Topalian et al., 2014). В настоящее время антитела к PD-1 и к CTLA-4 (цитотоксический Т-лимфоцит-ассоциированный антиген-4) были одобрены FDA для лечения таких заболеваний, как меланомы.
[0008] Другой молекулой-стимулятором является рецептор ОХ40 (CD134), представитель суперсемейства TNFR, который является мембраносвязанным и экспрессируется преимущественно на активированных CD4+ Т-клетках (Paterson et al., 1987). Передача сигналов через рецептор ОХ40 (далее «ОХ40») является костимулирующим для эффекторных Т-клеток и вызывает пролиферацию Т-клеток (Watts, 2005; Weinberg et al., 1994). Исследования OX40 предполагают, что его основная роль заключается в определении количества эффекторных Т-клеток, которые накапливаются в первичных иммунных ответах и, в конечном итоге, определяют количество Т-клеток памяти, которые впоследствии развиваются и выживают (Croft, 2003). Ряд исследований in vitro показали, что ОХ40 обеспечивает костимулирующий сигнал, приводящий к усилению пролиферации Т-клеток и производства цитокинов.
[0009] Биспецифическое/бифункциональное антитело
[0010] Идея использования биспецифических антител для эффективного перенацеливания эффекторных иммунных клеток на опухолевые клетки появилась в 1980-х годах (Karpovsky et al., 1984; Perez et al., 1985; Staerz et al., 1985). Биспецифические каркасы, как правило, классифицируются на две основные группы с различными фармакокинетическими свойствами, основанными на отсутствии или присутствии фрагмента Fc, IgG-подобных молекул и небольших рекомбинантных биспецифических форматов, большинство из которых происходит из одноцепочечного вариабельного фрагмента (scFv). Благодаря своему компактному размеру фрагменты антител, как правило, проникают в опухоли более эффективно, чем IgG-подобные молекулы, но это преимущество снижается за счет короткого периода полужизни в сыворотке (несколько часов), ограничивающего общее поглощение и время пребывания в опухоли (Goldenberg et al., 2007). В отличие от этого, наличие фрагмента Fc, который связывается с Fc-рецепторами новорожденных, обеспечивает длительное время полужизни в сыворотке (>10 дней) для IgG-подобных форматов, способствуя поглощению и удержанию в опухоли, но ограничивая проникновение в опухоль.
[0011] Недавние исследования выявили терапевтическую эффективность иммунотерапии, класса лечения рака, в котором для разрушения раковых клеток используется собственная иммунная система пациента. Присутствие внутри опухоли семейства негативных регуляторных молекул, известных под общим названием «ингибиторы контрольной точки», может ингибировать функцию Т-клеток для подавления противоопухолевого иммунитета. Ингибиторы контрольной точки, такие как CTLA-4 и PD-1, ослабляют пролиферацию Т-клеток и выработку цитокинов. Целевая блокада CTLA-4 или PD-1 с помощью моноклональных антител-антагонистов (mAb) освобождает «тормоза» Т-клеток для усиления противоопухолевого иммунитета. Для получения оптимальных «уничтожающих» ответов CD8 Т-клеток также требуется активация рецептора Т-клеток плюс костимуляция, которая может быть обеспечена путем лигирования представителей семейства рецепторов фактора некроза опухолей, включая в себя ОХ40 (CD134) и 4-1ВВ (CD137). OX40 представляет особый интерес, так как лечение активирующим (агонистическим) mAb к ОХ40 увеличивает дифференцировку и цитолитическую функцию Т-клеток, ведущую к усилению противоопухолевого иммунитета против различных опухолей. При использовании в качестве отдельных средств эти лекарственные средства могут вызывать сильные клинические и иммунологические ответы у пациентов с метастатическим заболеванием (Linch et al., 2015).
Краткое раскрытие настоящего изобретения
[0012] Настоящее раскрытие предназначено для исследования биспецифического антитела с иммуномодулирующим действием с целью лечения пациента с раковыми заболеваниями, такими как рак простаты, рак легких, NSCLC, меланома, лимфома, рак молочной железы, рак головы и шеи, RCC или рак яичников.
[0013] Настоящее изобретение относится к антителу или его антигенсвязывающей части, связывающейся с ОХ40 (CD134), содержащей: вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO. 6, SEQ ID NO. 8, аминокислот 128-246 из SEQ ID NO. 10 и аминокислот 124-241 из SEQ ID NO. 13, и вариабельную область легкой цепи, содержащей аминокислоту последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-108 из SEQ ID NO. 5, 1-108 из SEQ ID NO. 7, 1-112 из SEQ ID NO. 10 и 1-108 из SEQ ID NO. 13. [0014] Согласно одному варианту осуществления антитело или его антигенсвязывающая часть представляет собой последовательность одноцепочечного вариабельного фрагмента (scFv), выбранную из группы, состоящей из SEQ ID NO. 10, 11, 12 и 13.
[0015] Согласно одному варианту осуществления антитело или его антигенсвязывающая часть представляет собой биспецифическое антитело.
[0016] Согласно одному варианту осуществления биспецифическое антитело представляет собой сайт связывания белка иммунной контрольной точки.
[0017] Согласно одному варианту осуществления сайт связывания белка иммунной контрольной точки включает в себя сайт связывания лиганда белка запрограммированной гибели клеток 1 (PD-L1), сайт связывания PD-1, сайт связывания рецептора эпидермального фактора роста (EGFR), сайт связывания рецептора эпидермального фактора роста человека 2 (HER2), сайт связывания цитотоксического Т-лимфоцит-ассоциированного антигена 4 (CTLA-4) или сайт связывания гена активации лимфоцитов 3 (LAG3).
[0018] В настоящем изобретении также предусмотрено антитело или его антигенсвязывающая часть, связывающаяся с PD-L1, содержащая вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO. 2 и SEQ ID NO. 4, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-111 из SEQ ID NO. 1 и 1-110 из SEQ ID NO. 3.
[0019] В настоящем изобретении также предусмотрено биспецифическое антитело, содержащее по меньшей мере одну полипептидную цепь, причем полипептидная цепь содержит сайт связывания ОХ40 и сайт связывания PD-L1. Сайт связывания ОХ40 содержит вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO. 6, SEQ ID NO. 8, аминокислот 128-246 из SEQ ID NO. 10 и аминокислот 124-241 из SEQ ID NO. 13, и вариабельную область легкой цепи, содержащую аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислоты 1-108 из SEQ ID NO. 5, 1-108 из SEQ ID NO. 7, 1-112 из SEQ ID NO. 10 и 1-108 из SEQ ID NO. 13. Сайт связывания PD-L1 содержит вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO. 2 и SEQ ID NO. 4, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, которая характеризуется гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-111 из SEQ ID NO. 1 и 1-110 из SEQ ID NO. 3.
[0020] Согласно одному варианту осуществления полипептидная цепь дополнительно содержит Fc-домен, Fab-фрагмент и scFv. Fab-фрагмент связан с N-концом Fc-домена, и Fab-фрагмент содержит сайт связывания PD-L1. scFv соединен с С-концом Fc-домена, и scFv содержит сайт связывания ОХ40.
[0021] Согласно одному варианту осуществления полипептидная цепь дополнительно содержит линкер между доменом Fc и scFv.
[0022] Согласно одному варианту осуществления scFv содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислот 455-707 из SEQ ID NO. 18, 455-708 из SEQ ID NO. 19, 455-701 из SEQ ID NO. 20, 455-706 из SEQ ID NO. 21, 455-706 из SEQ ID NO. 22, 455-706 из SEQ ID NO. 23, 455-706 из SEQ ID NO. 24, 455-706 из SEQ ID NO. 25, 455-706 из SEQ ID NO. 26, 455-706 из SEQ ID NO. 27, 455-706 из SEQ ID NO. 28 и 455-706 из SEQ ID NO. 29.
[0023] Согласно одному варианту осуществления биспецифическое антитело содержит одну пару полипептидных цепей.
[0024] Согласно одному варианту осуществления биспецифическое антитело представляет собой антитело IgG, IgE, IgM, IgD, IgA или IgY.
[0025] Согласно одному варианту осуществления биспецифическое антитело представляет собой антитело IgG.
[0026] Согласно одному варианту осуществления антитело IgG представляет собой антитело IgG1, IgG2, IgG3 или IgG4.
[0027] В настоящем раскрытии также предусмотрен конъюгат антитело-лекарственное средство, содержащий терапевтическое средство и антитело или антигенсвязывающую часть, связывающую PD-L1 и/или ОХ40, причем терапевтическое средство ковалентно конъюгировано с антителом или антигенсвязывающей частью с помощью линкера.
[0028] Согласно одному варианту осуществления антитело или его антигенсвязывающая часть выбраны из вышеуказанного антитела или антигенсвязывающей части.
[0029] В настоящем раскрытии также предусмотрена фармацевтическая композиция, содержащая антитело, его антигенсвязывающую часть или биспецифическое антитело, как указано выше, и по меньшей мере один фармацевтически приемлемый носитель.
[0030] В настоящем раскрытии также предусмотрен способ лечения рака, предусматривающий введение нуждающемуся в этом субъекту эффективного количества антитела, его антигенсвязывающей части или биспецифического антитела, как указано выше.
[0031] Согласно одному варианту осуществления рак выбран из группы, состоящей из рака предстательной железы, рака легкого, немелкоклеточного рака легкого (NSCLC), меланомы, лимфомы, рака молочной железы, рака головы и шеи, почечно-клеточной карциномы (RCC) и рака яичников.
[0032] В настоящем раскрытии также предусмотрена молекула нуклеиновой кислоты, кодирующая антитело, его антигенсвязывающую часть или биспецифическое антитело, как указано выше.
[0033] Следует понимать, что как последующее общее описание, так и последующее подробное описание приведены в качестве примеров и предназначены для обеспечения дополнительного объяснения настоящего изобретения, как заявлено.
Краткое описание графических материалов
[0034] Настоящее изобретение может быть более полно понято при прочтении следующего подробного описания варианта осуществления со ссылкой на прилагаемые графические материалы следующим образом:
[0035] На фиг.1 показаны иммунные контрольные точки, модулирующие опосредованный Т-клеточный иммунитет.Антитело, либо агонистическое, либо антагонистическое в отношении контрольных точек, такое как к ICOS, к CD28, к ОХ40 и к CD27, или к PD-1, к CTLA4, к LAG3, к BTLA можно использовать для конструирования бифункционального слитого белка в зависимости от применения.
[0036] На фиг. 2А и 2В показан скрининг клона фага прямым ИФА для экспрессированных клетками НЕК293 PD-L1.
[0037] На фиг. 3А и 3 В показан скрининг клона фага прямым ИФА на основе клеток с экспрессированными клетками НЕК293 ОХ40.
[0038] На фиг. 4 показаны очищенные прототипы антител, специфические для PD-L1, с помощью ДСН-ПААГ с невосстанавливающим реагентом для выявления целостности и чистоты.
[0039] На фиг. 5 показаны очищенные прототипы антител, специфические для ОХ40, с помощью ДСН-ПААГ с невосстанавливающим или восстанавливающим реагентом для выявления целостности и чистоты.
[0040] На фиг. 6 показаны примеры активности прямого связывания лиганда очищенных прототипов антител к белкам иммунной контрольной точки и к PD-L1 против PD-L1. Лунки, предварительно покрытые лигандом, сначала инкубировали с различными концентрациями прототипов антител, как указано. Связанные белки затем обнаруживали с помощью конъюгированного с HRP козьего специфического антитела к Fab IgG человека, и показания OD450 наносили на график.
[0041] На фиг. 7 показаны примеры активности прямого связывания лиганда очищенных прототипов антител к белкам иммунной контрольной точки и к ОХ40 против ОХ40. Сначала лунки с предварительно нанесенным лигандом инкубировали с различными концентрациями прототипов антител, как указано. Белки затем обнаруживали с помощью конъюгированного с HRP козьего специфического антитела к Fab IgG человека, и показания OD450 наносили на график.
[0042] На фиг. 8 показан проточный анализ с использованием экспрессирующих PD-L1 клеток 293. Экспрессирующие PD-L1 клетки НЕК293 сначала инкубировали с очищенными прототипами антител, а связанные антитела обнаруживали с помощью конъюгированного с А1еха-488 козьего антитела к человеческому IgG (H+L) с последующим анализом посредством клеточного сортера с активацией флуоресценции (FACS).
[0043] На фиг. 9 показан проточный анализ с использованием экспрессирующих ОХ40 клеток 293. Экспрессирующие ОХ40 клетки НЕК293 сначала инкубировали с очищенными прототипами антител к ОХ40, и связанные антитела обнаруживали с помощью конъюгированного с А1еха-488 козьего антитела к человеческому IgG (H+L) с последующим анализом FACS. NS: нет окрашивания.
[0044] На фиг. 10 показана блокировка взаимодействия PD-1/PD-L1 с очищенными антителами к PD-L1. Очищенные антитела, как указано, применяли к биотинилированным PD-L1-Fc и рекомбинантным PD-1/His (hPD-1/His) человека для оценки активности ингибирования взаимодействия PD-1/PD-L1. Связывание рекомбинантного PD-L1-Fc и hPD-1/His обнаруживали с помощью стрептавидина-HRP и анализировали с помощью ИФА.
[0045] На фиг. 11А и 11 В показано, что прототипы антител к PD-L1 в дозе 1 или 10 мкг/мл стимулируют пролиферацию Т-клеток и индуцируют производство IL-2 и/или IFN-γ в анализе с реакцией смешанных лимфоцитов (MLR) через 3 дня (фиг. 11А) или 5 дней (фиг. 11В) после воздействия антителом.
[0046] На фиг. 12А показана способность антитела к ОХ40 приводить к усилению активации CD3+ Т-клеток с помощью дозового ответа, а также эталонного антитела. На фиг. 12 В показана концентрация человеческого IL-2 и IFN-α, присутствующего в среде для культивирования клеток после 3-х дней стимуляции Т-клеток человека связанными с планшетами антителами к CD3 и несколькими концентрациями прототипов антител к ОХ40.
[0047] На фиг. 13А и 13В показана концентрация человеческого IL-2 (фиг. 13А) и IFN-γ (фиг. 13В), присутствующих в средах для культивирования клеток после 3-х дней стимуляции Т-клеток человека с помощью связанного с планшетом антитела к CD3 и нескольких концентраций ОХ40-специфических прототипов антител.
[0048] На фиг. 14 показана структура Fc тяжелой цепи антитела, слитого с ОХ40-специфическим scFv-доменом.
[0049] На фиг. 15 показаны примеры анализа ПААГ-геля слитых белков антител к иммунным контрольным точкам с ОХ40 человека. Показано, что очищенные слитые белки, слитые белки анти-PD-L1-ОХ40 scFv характеризуются молекулярной массой приблизительно 220 кДа (невосстанавливающий), и слияние тяжелой цепи составляет приблизительно 85 кДа, а легкая цепь составляет приблизительно 25 кДа (восстанавливающий) при слиянии обоих антител.
[0050] На фиг. 16А и 16В показаны синергические биспецифические антитела, стимулирующие активацию Т-клеток для производства IL-2 и IFN-γ в анализе с реакцией смешанных лимфоцитов (MLR) через 3 дня (фиг. 16А) или 5 дней (фиг. 16В) с воздействием моно-, комбинированным или анти-PD-L1-ОХ40 scFv биспецифическими антителами.
[0051] На фиг. 17А-17Е соответственно показаны агрегация и определение чистоты биспецифических антител, анти-PD-L1-ОХ40 Ab и анти-PD-L1-ОХ40 Ab от V1 до V4, с 5 различными линкерами в ОХ40 scFv.
[0052] На фиг. 18 показаны варианты последовательности среди scFv клона В17 ОХ40 анти-PD-L1-ОХ40 Ab от V4 до V12 (SEQ ID NO. 30-38).
[0053] На фиг. 19 показаны примеры анализа ПААГ-геля слитых белков антител к иммунным контрольным точкам с ОХ40 человека. Показано, что очищенные слитые белки, слитые белки анти-PD-L1-OX40 Ab-V5, характеризуются молекулярной массой приблизительно 220 кДа (невосстанавливающий), и слияние тяжелой цепи составляет приблизительно 80 кДа, а легкая цепь составляет приблизительно 30 кДа (восстанавливающий) в обоих слияниях антител.
[0054] На фиг. 20 показана блок-схема, иллюстрирующая способ ИФА для оценки активности связывания вариантов биспецифических антител.
[0055] На фиг. 21 показана активность связывания человеческого PD-L1 с вариантами биспецифического антитела и его ЕС50.
[0056] На фиг. 22 показана активность связывания человеческого ОХ40 с вариантами биспецифического антитела и его ЕС50.
[0057] На фиг. 23 показана стабильность в сыворотке ex vivo варианта биспецифического антитела, анти-PD-L1-ОХ40 Ab-V5.
[0058] На фиг. 24А и 24В показано производство IL-2 в течение 3 дней (фиг. 24А) и производство IFN-γ в течение 5 дней (фиг. 24В) после модуляции Т-клетки воздействием моно-, комбинированными или биспецифическими антителами анти-PD-L1-ОХ40 Ab-V5.
[0059] На фиг. 25 представлен график, показывающий влияние воздействия биспецифическими антителами анти-PD-L1-ОХ40 Ab-V5 и воздействия моноклональными антителами на рост опухоли РС-3 у мышей Fox Chase SCID® Beige.
Подробное описание настоящего изобретения
[0060] Теперь будет сделана подробная ссылка на настоящие варианты осуществления настоящего изобретения, примеры которых проиллюстрированы на прилагаемых графических материалах. Везде, где возможно, используются одни и те же идентификаторы на графических материалах и в описании для ссылки на те же самые или подобные части.
[0061] Настоящее изобретение описывает экспрессию, очистку и характеристику бифункциональных белков с выделенным функциональным агонистическим scFv ОХ40, слитым с С-концом Fc-домена антител к белкам иммунной контрольной точки. Эти белки взаимодействуют с их соответствующей мишенью контрольной точки и должны передавать ингибирующий или стимулирующий сигнал для модуляции иммунитета, связанного с Т-клетками. Компоненты слитых белков Fc в настоящем изобретении имеют все человеческое происхождение и, таким образом, как ожидается, будут неиммуногенными и могут использоваться в качестве терапевтических средств для человека.
[0062] Биспецифические молекулы, такие как биспецифические антитела (BsAb), обеспечивают средство одновременного нацеливания нескольких эпитопов на одну и ту же молекулярную мишень или разные мишени с помощью одного терапевтического средства. Как противораковые терапевтические средства, они обладают потенциалом для придания новых или более сильных активностей, снижения стоимости товаров и облегчения разработки новых схем лечения в отличие от смеси двух mAb (Chames and Baty, 2009; Hollander, 2009; Thakur and Lum, 2010). Недавно, катумаксомаб, трифункциональное биспецифическое антитело, нацеленное на адгезивную молекулу эпителиальных клеток человека (ЕрСАМ) и CD3, продемонстрировало явное клиническое преимущество у пациентов с карциноматозом брюшины среди форм рака эпителия (Heiss et al., 2010), а биспецифическое антитело активатор Т-клеток (BiTE) с двойной специфичностью к CD19 и CD3 также продемонстрировал обнадеживающую клиническую активность у пациентов с экспрессирующими CD19 гематологическими злокачественными новообразованиями (Bargou et al., 2008). Несмотря на сильный интерес к развитию биспецифических молекул в качестве терапевтических средств для лечения рака, технические проблемы в производстве стабильных и активных биспецифических молекул в прошлом затрудняли клиническую оценку большинства биспецифических форматов. Многие форматы сконструированных антител, включая в себя IgG-подобные биспецифические антитела, характеризовались нарушенной стабильностью или растворимостью (Bargou et al., 2008; Demarest and Glaser, 2008; Lu et al., 2005). Было предпринято несколько стратегий для повышения качества продукта и стабильности биспецифических молекул in vivo, включая в себя пегилирование, конъюгацию с человеческим сывороточным альбумином и конструирование Fc (Muller et al., 2007; Ridgway et al., 1996). Биспецифические одноцепочечные антитела общей формы, описанные выше, характеризуются тем преимуществом, что нуклеотидная последовательность, кодирующая четыре V-домена, два линкера и один спейсер, может быть включена в подходящий организм-хозяин экспрессии под контролем одного промотора. Это увеличивает как гибкость, с которой эти конструкции могут быть разработаны, так и степень контроля экспериментатора во время их производства. Кроме того, Fc IgG представляет собой еще один привлекательный каркас для разработки новых терапевтических средств, поскольку он содержит все функции антител, кроме способности к связыванию. Конструирование Fc важно для повышения эффективности биспецифических антител. Таким образом, конформация на основе IgG используется в настоящем изобретении для двух независимых мишеней на иммунных клетках или клетках-мишенях в иммунотерапии.
[0063] Нацеливание на белки иммунных контрольных точек представляет собой многообещающий подход для активации противоопухолевого иммунитета. Белки иммунных контрольных точек, такие как PD-1, PD-L1, CTLA-4, LAG3 и т.д., в настоящее время оцениваются клинически (фиг. 1). По предварительным данным с блокаторами белков иммунных контрольных точек было показано, что они способны усиливать противоопухолевый иммунитет с потенциалом вызывать длительные клинические ответы. Однако, несмотря на замечательную клиническую эффективность этих средств при ряде злокачественных новообразований, стало ясно, что они не являются достаточно активными для многих пациентов. Многочисленные дополнительные иммуномодулирующие пути, а также ингибирующие факторы, экспрессируемые или секретируемые миелоидными и стромальными клетками в опухолевом микроокружении, являются потенциальными мишенями для синергизма с иммунной контрольной точкой. Таким образом, комбинируемая противоопухолевая или биспецифическая терапия антителами необходима для достижения полной ремиссии и лечения больных раком.
[0064] В настоящем изобретении описано конструирование, экспрессия и характеристика Fc антитела к белку иммунной контрольной точки, слитого со специфическим к другому белку иммунной контрольной точки белком scFv. scFv ОХ40, расположенный на С-конце в слитых конструкциях, должен позволить расширить мощность слитых белков за пределы подхода к активации ОХ40, если партнер слияния представляет собой потенцирующее средство иммунной системы, такое как, например, антитело к EGFR, к HER2 или к CTLA-4.
[0065] Получение антител из библиотеки OmniMab
[0066] Для получения терапевтических антител против PD-L1 или ОХ40 проводили отбор с помощью фагемидной библиотеки OmniMab. Фагемидную библиотеку производит АР Biosciences Inc. (APBio Inc.) из коллекции В-клеток более ста здоровых доноров. Фаги для 1-го раунда пэннинга были подготовлены Hyperphage (M13K07ΔρIII, Progen, Хайдельберг, Германия). Твердофазный пэннинг и пэннинг клеток против PD-L1 или ОХ40 применяли для отбора специфических связующих PD-L1 или ОХ40 и выделения из OmniMab. Твердофазный пэннинг проводили с использованием рекомбинантного человеческого PD-L1-Fc или OX40-Fc (APBio Inc.) в первом раунде отбора, а затем клетки НЕК293, экспрессирующие PD-L1 или ОХ40, использовали для двух и трех раундов обогащения. После трех раундов отбора специфические связующие для PD-L1 или ОХ40 подвергали скринингу и выделяли прямым анализом ИФА или ИФА на основе клеток с соответствующим рекомбинантным белком (фиг. 2А, 2В, 3А и 3В). Предварительно покрытые PD-L1-Fc рекомбинантные белки или экспрессирующие ОХ40 клетки 293 подвергали блоттингу с супернатантом, содержащим обнаруженные фаги в течение 1 часа и трижды промывали PBS, содержащим 0,1% Твин-20. Связанные фаги обнаруживали с помощью конъюгированного с HRP антитела к М13 (Roche) и субстрат ТМВ использовали для развития сигнала. Регистрировали показания OD450. Положительные связующие вещества выделяли и отправляли для секвенирования для подтверждения последовательности и разнообразия тяжелой цепи и легкой цепи. Вариабельную область тяжелой цепи и легкой цепи, специфическую для PD-L1 или ОХ40, описывали от SEQ ID NO. 1 до SEQ ID NO. 8: SEQ ID NO. 1 представляет собой легкую цепь клона 6 PD-L1, SEQ ID NO. 2 представляет собой вариабельную область тяжелой цепи клона 6 PD-L1, SEQ ID NO. 3 представляет собой легкую цепь клона 32PD-L1, SEQ ID NO. 4 представляет собой вариабельную область тяжелой цепи клона 32 PD-L1, SEQ ID NO. 5 представляет собой легкую цепь клона В17 ОХ40, SEQ ID NO. 6 представляет собой вариабельную область тяжелой цепи клона В17 ОХ40, SEQ ID NO. 7 представляет собой легкую цепь клона В19 ОХ40, SEQ ID NO. 8 представляет собой вариабельную область тяжелой цепи клона В19 ОХ40. Как показано на фиг. 2А, 2В, 3А и 3 В было выделено несколько клонов, и известно, что они специфично распознаются соответствующим антигеном по сравнению с отрицательным контролем.
[0067] Субклонирование и экспрессия/очистка выбранного специфического связующего для PD-L1 или ОХ40 в формате IgG
[0068] Для облегчения быстрого скрининга специфического связующего с функциональностью активации Т-клеток тяжелые цепи и легкие цепи положительных связующих против PD-L1 или ОХ40 с помощью анализа ИФА затем амплифицировали, расщепляли и субклонировали в экспрессирующий вектор APGio-специализированных IgG, несущий константную область IgG4 (SEQ ID NO. 9). После подтверждения последовательности плазмиды затем готовили и трансфицировали в клетки НЕК293 для экспрессии антител с помощью реагента для трансфекции 293 фектина (Invitrogen). После 4-дневного культивирования антитело, секретированное в бессывороточную среду, аффинно очищают от культурального супернатанта хроматографией на основе связывания с белком G. Затем очищенное антитело концентрируют с последующим диализом в буфере PBS. Конечную концентрацию диализированного белка определяют с помощью спектрофотометра NanoDrop2000, а чистоту и целостность определяют с помощью ДСН-ПААГ с восстанавливающим реагентом или без него, как показано на фиг. 4 и 5. Целостность различных очищенных прототипов антител, специфических к PD-L1 или ОХ40, является нормальной в клетках НЕК293, также как у эталонного антитела, MPDL3280A для PD-L1 или GSK3174998 для ОХ40.
[0069] Согласно одному варианту осуществления в настоящем раскрытии предусмотрено антитело или его антигенсвязывающая часть, связывающаяся с ОХ40 (CD134), содержащая вариабельную область тяжелой цепи и вариабельную область легкой цепи. Вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO. 6, SEQ ID NO. 8, аминокислот 128-246 из SEQ ID NO. 10 и аминокислот 124-241 SEQ ID NO. 13. В некоторых примерах вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 85%, 90% или 95% по отношению к аминокислотной последовательности, как указано выше. Вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-108 из SEQ ID NO. 5, 1-108 из SEQ ID NO. 7, 1-112 из SEQ ID NO. 10 и 1-108 из SEQ ID NO. 13. В некоторых примерах вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 85%, 90% или 95% по отношению к аминокислотной последовательности, как указано выше.
[0070] Согласно одному варианту осуществления в настоящем раскрытии предусмотрено антитело или его антигенсвязывающая часть, связывающаяся с PD-L1, содержащая вариабельный домен тяжелой цепи и вариабельный домен легкой цепи. Вариабельный домен тяжелой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 2 и SEQ ID NO. 4. В некоторых примерах вариабельная область тяжелой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 85%, 90% или 95% по отношению к аминокислотной последовательностью, как указано выше. Вариабельный домен легкой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 80% по отношению к аминокислотной последовательности, выбранной из группы, состоящей из аминокислот 1-111 из SEQ ID NO. 1 и 1-110 из SEQ ID NO. 3. В некоторых примерах вариабельная область легкой цепи содержит аминокислотную последовательность, характеризующуюся гомологией последовательности, составляющей по меньшей мере приблизительно 85%, 90% или 95% по отношению к аминокислотной последовательности, как указано выше.
[0071] Определение активности связывания для специфических к PD-L1, ОХ40 прототипов IgG посредством прямого анализа ИФА
[0072] Очищенные прототипы антител против PD-L1 или ОХ40 (прототипы антител к PD-L1 или прототипы антител к ОХ40) затем применяли для характеристики связывания анализом ИФА на человеческом PD-L1-Fc или OX40-Fc в постановке с прямым покрытием. На фиг. 6 и 7 показан результат связывания ИФА для антител к PD-L1 и к ОХ40, соответственно. Для антител, специфических для PD-L1, большинство прототипов продемонстрировали сходную или лучшую активность связывания по сравнению с эталонным антителом (Ref Ab, MPDL3280A, Roche).
[0073] Очищенную химеру Fc IgG1 PD-L1 или ОХ40 человека (PD-L1-Fc или ОХ40-Fc, APBio) подвергали диализу в фосфатно-солевом буфере (PBS), доводили до 1 мг/мл и затем разбавляли PBS до конечной концентрации 1 мкг/мл. 96-луночные планшеты Nunc-Immuno Maxisorp покрывали 0,1 мл на лунку рекомбинантной химерой PD-L1-Fc или OX40-Fc, оставляя пустые лунки для контроля неспецифического связывания и инкубировали при температуре 4°С в течение ночи. Раствор химер PD-L1-Fc или OX40-Fc удаляли и планшеты трижды промывали промывочным буфером объемом 0,4 мл (0,1% твин-20 в PBS). Добавляли блокирующий буфер объемом 0,4 мл (5% сухое молоко с низким содержанием жира в PBS) во все лунки и инкубировали при комнатной температуре в течение 1 часа при перемешивании. Блокирующий буфер удаляли и планшеты трижды промывали 0,4 мл промывочного буфера. Готовили серийные разведения исследуемых антител к PD-L1 или ОХ40 в PBS и 0,1 мл разведенного антитела добавляли на лунку. Планшеты инкубировали 1 час при комнатной температуре. Раствор антитела удаляли и планшеты промывали три раза 0,4 мл буфера для промывания на лунку. Меченное пероксидазой хрена F(ab')2 козьего антитела к F(ab')2 IgG человека (Jackson Immunoresearch #109-036-097) разводили 1:2000 PBS и добавляли 0,1 мл на лунку. Планшеты инкубировали 1 час при комнатной температуре и промывали 0,4 мл на лунку буфером для промывания. Добавляли 0,1 мл реагента ТМВ (Invitrogen) и инкубировали в течение от 1 до 5 минут при комнатной температуре. Реакцию останавливали путем добавления 0,05 мл 1 н. HCl, и абсорбцию считывали при 450 нм на спектрометре Bio-Tek. Рассчитанное ЕС50 для прототипов антител к PD-L1 к PD-L1 показало, что большинство прототипов обладают хорошей связывающей активностью, также как MPDL3280A (Ref Ab) посредством прямого анализа ИФА (фиг. 6). Напротив, большинство прототипов антител к ОХ40 показало гораздо более низкую активность связывания по сравнению с эталонным антителом (Ref Ab, GSK3174998) (фиг. 7).
[0074] Определение связывающей активности для специфических к PD-L1 и ОХ40 прототипов IgG с помощью FACS
[0075] Очищенные прототипы антител (прототипы антител к PD-L1 или прототипы антител к ОХ40) также применяли для проточной цитометрии для определения и сравнения активности связывания с экспрессированными клетками НЕК293 PD-L1 или ОХ40. На фиг. 8 и 9 показаны активности связывания соответствующих прототипов антитела, как показано посредством FACS, со стабильно экспрессированными клетками НЕК293 PD-L1 или ОХ40.
[0076] Анализ FACS клеток 293 со стабильной экспрессией PD-L1, окрашенных прототипами антител к PD-L1, приводит к исследованию активности связывания PD-L1, клетки со стабильной экспрессией инкубировали с 1 мкг/мл очищенных прототипов антитела к PD-L1, эталонным антителом (Ref Ab MPDL3280A) или изотипическим антителом в качестве отрицательного контроля на льду в течение 1 ч. Клетки трижды промывали 1-кратным PBS, а затем инкубировали с конъюгированным с Alexa-488 козьим антителом к человеческому IgG (H+L) (Invitrogen Inc.) на льду в течение дополнительного 1 ч. После окрашивания клетки промывали три раза 1-кратным PBS, ресуспендировали в 1-кратном PBS/2% FBS перед анализом в FACS Calibur (BD Biosciences, Inc.) и FlowJo (TreeStar, LLC). По тому же сценарию активность связывания прототипов антител к ОХ40 для стабильно экспрессированных клетками НЕК293 ОХ40 на фиг. 9, также выполняли с аналогичной стратегией и анализировали, как описано выше. Как показано на фиг. 8, большинство прототипов антител к PD-L1 обладают хорошей активностью связывания, также как эталонные антитела. Это указывает на то, что клоны фага, выбранные из библиотеки OmniMab, действительно распознают нативный PD-L1 в клетках.
[0077] Это явление также наблюдается для прототипов антител к ОХ40, как показано на фиг. 9. Анализ FACS стабильно экспрессирующих ОХ40 клеток 293 клона 2D5, окрашенных очищенными прототипами антител к ОХ40 для исследования активности связывания ОХ40, проводили следующим образом: стабильно экспрессирующие клетки инкубировали с 2 мкг/мл эталонных антител к ОХ40 (ОХ40 ref.) или эталонных антител к CD137 (CD137 ref.) в качестве контрольного антитела на льду в течение 1 ч. Клетки трижды промывали 1-кратным PBS и затем инкубировали с конъюгированным с Alexa-488 козьим антителом к IgG человека (H+L) (Invitrogen Inc.) на льду в течение дополнительного 1 часа. После окрашивания клетки трижды промывали 1-кратным PBS, ресуспендировали в 1-кратном PBS/2%) FBS перед анализом с помощью FACS Calibur (BD Biosciences, Inc.) и FlowJo (TreeStar, LLC).
[0078] Конкурентное связывание лиганда (анализ ИФА)
[0079] Было показано, что анализ селективности и аффинности связывания прототипов антител использовали для оценки прототипов антител к PD-L1 по настоящему изобретению на их способность блокировать связывание PD-L1 с PD-1. [0080] Антитела исследовали на их способность блокировать связывание химеры PD-L1-Fc человека (PD-L1-Fc) с рекомбинантным человеческим PD-1/His (hPD-1/His) с помощью ИФА. Очищенный рекомбинантный hPD-1/His (APBio) диализовали до 1 мг/мл в PBS и затем конъюгировали с биотином (Abcam). 96-луночный планшет Nunc Maxisorp покрывали 250 нг hPD-1/His на лунку в PBS в течение ночи. Раствор hPD-1/His удаляли и планшеты трижды промывали 0,4 мл промывочного буфера (0,1% Твин-20 в PBS). 0,4 мл блокирующего буфера (5% обезжиренного сухого молока в PBS) добавляли во все лунки и инкубировали при комнатной температуре в течение 1 часа с перемешиванием. Во время стадии блокирования маточный раствор антител разбавляли в диапазоне от 200 нМ до 0 нМ в PBS с 2-кратным серийным разведением. Очищенную рекомбинантную биотинилированную химеру PD-L1-Fc разбавляли до 4 мкг/мл в PBS. Покрытые PD-1/His планшеты трижды промывали 0,2 мл промывочного буфера (0,1% Твин-20 в PBS). Разбавления 60 мкл антител (прототипы антител к PD-L1 или эталонное антитело MPDL3280A) добавляли отдельно с 60 мкл химеры биотинилированный-PD-L1-Fc и инкубировали при комнатной температуре в течение 1 ч. Планшеты промывали, как описано ранее. Стрептавидин-HRP разводили 1:2000 в PBS, добавляли 100 мкл полученного раствора в лунки промытых планшетов и сушили при комнатной температуре в течение 1 часа. Планшеты промывали, как описано ранее, в каждую лунку добавляли 100 мкл раствора субстрата ТМВ и перемешивали в течение 10 минут. Реакцию останавливали с помощью 50 мкл 1 н. HCl и оптическую плотность при 450 нм считывали с использованием ридера Bio-Tek и представили на фиг. 10. Показано, что частичные прототипы антител ингибируют взаимодействие между PD-PD-L1 посредством конкурентного анализа ИФА. Большинство прототипов антител выявили аналогичную блокирующую активность по сравнению с эталонным антителом (Ref Ab MPDL3280A).
[0081] Усиленная стимуляция активации Т-клеток путем ингибирования взаимодействия лиганда PD-1:PD-L1 для антитела к PD-L1
[0082] Сигнальный путь PD-1 ингибирует умеренные костимулирующие сигналы TCR/CD28, с производством цитокина, сначала снижающимся без снижения пролиферации Т-клеток. Поскольку костимулирующие сигналы TCR/CD28 ослабевают, доминирует путь PD-1 с большим уменьшением производства цитокинов, сопровождаемым уменьшением пролиферации. Соответственно, чтобы подтвердить, что ингибирование PD-1 посредством ингибирования взаимодействия с PD-L1 антителами человека по настоящему изобретению усиливает активацию Т-клеток, выполняются реакции смешанных лимфоцитов (MLR).
[0083] Моноциты из цельной крови человека обогащали с помощью коктейля по обогащению моноцитов человека RosetteSep™ (кат.№15068) и культивировали в среде для дифференцировки, RPMI 1640 с 10% FBS, 100 нг/мл (1000 Ед/мл) GM-CSF, 100 нг/мл (500 Ед/мл) в течение 6 дней. Дифференцированные дендритные клетки (DC) из моноцитов проверяли с помощью DC-SIGN-PE, конъюгированного с FITC антитела к CD14, конъюгированного с РЕ антитела к CD83 или конъюгированного с FITC антитела к CD80 для подтверждения дифференциации и использовали в качестве АРС в MLR.
[0084] Аллогенные CD4+ Т-клетки из цельной крови человека выделяли с помощью коктейля для обогащения CD4+ Т-клеток человека RosetteSep™ (кат.№15062). Чистоту CD4+ Т-клеток проверяли с помощью конъюгированного с АРС антитела к CD4, чтобы убедиться в чистоте выше 95% и затем метили 1 мкМ CFSE (набор для пролиферации клеток CFSE CellTraceTM, Life Technologies, кат. №С34554) для анализа пролиферации Т-клеток. Меченые CD4+Т-клетки использовали для совместного культивирования с незрелыми DC с различными прототипами антител, как указано в течение 3 и 5 дней, чтобы увидеть, могут ли прототипы антитела восстанавливать активацию Т-клеток путем блокирования взаимодействия между PD-1 и PD-L1. После 3-й 5-дневной инкубации супернатант собирали для определения цитокина, такого как количественное определение IL-2 и IFN-γ с помощью ИФА. Добавление прототипов антител к PD-L1 (клоны 6, 32, 28, 51, 64, 27 и 37) к культурам незрелых дендритных клеток плюс аллогенных Т-клеток, как ожидается, приведет к увеличение пролиферации Т-клеток и производству цитокинов, по сравнению с культурами, обработанными изотипом IgG (iso#1, #2), и они показаны на фиг. 11А и 11В. Производство IL-2 и IFN-γ значительно увеличивается в MLR по сравнению с воздействием изотипическим антителом через 3 дня (фиг. 11А) или 5 дней (фиг. 11В) воздействия антителом, особенно для клона 6 антитела к PD-L1. Инкремент цитокинов остается очевидным после 5-дневного воздействия антителами и аналогичен эталонному антителу (ref), MPDL3280A. Это указывает на то, что клон 6 антитела к PD-L1 должен быть одним из потенциальных прототипов для композита биспецифического антитела.
[0085] Анализ агонистической активности антитела к ОХ40
[0086] Чтобы активировать костимуляцию ОХ40 пролиферации Т-клеток и производства цитокинов, очищенные прототипы антител функционально подвергали скринингу на их способность усиливать производство, пролиферацию цитокинов и индуцировать пролиферацию в CD3+ Т-клетках человека. Прототипы антитела к CD3 (ОКТЗ, BioLegend, кат. №317304) и антитела к ОХ40 (клоны В6, В70, В120, А4, В17, В19 и В30), эталонное антитело (GSK3174998) или изотопические антитела (iso#1, #2) использовали для покрытия в 96-луночном планшете Maxisorp.Тем временем из человеческой крови от здоровых взрослых добровольцев выделяли наивные человеческие CD3+ Т-клетки с использованием коммерчески доступного коктейля для обогащения Т-клеток человека Rosette Sep™ (STEMCELL, кат. №15061), как описание изготовителем. Выделенные CD3+ Т-клетки затем метили с помощью CFSE (набор для пролиферации клеток CFSE CellTrace™, Life Technologies, кат. №C34554) и высевали в количестве 1×106 клеток/мл в предварительно покрытую антителом лунку, содержащую среду RPMI 1640, 10% эмбриональную бычью сыворотку и 2,5 мМ L-глутамин, для определения пролиферации клеток и производства цитокинов. После 3-дневного культивирования клетки собирали для анализа пролиферации с помощью проточной цитометрии, а затем среду анализировали на производство IL-2 и IFN-γ путем количественного анализа ИФА.
[0087] Скрининг прототипов антител к ОХ40 с агонистической активностью в активации Т-клеток показан на фиг. 12А. Все прототипы антител к ОХ40 показали способность усиливать активацию CD3+ Т-клеток с помощью дозового ответа, также как и эталонное антитело. Воздействие антителом в более высоких дозах показало явно более высокую активность активации Т-клеток. Между тем, производство цитокинов (фиг. 12В), таких как IL-2 и IFN-γ, также выявило сходный ответ на активацию Т-клеток, особенно для клона В17 прототипа антитела к ОХ40. Цитокин обладает высокой индукцией после 3-дневного воздействия антителом к ОХ40 В17. Усиление намного выше, чем при воздействии эталонным антителом, этот предполагаемый клон В17 должен быть одним из кандидатов на конструирование биспецифических антител.
[0088] Как показывают данные, показанные на фиг. 13А и 13В, оба прототипа антитела к ОХ40, клоны В17 и В19, показали лучшую агонистическую активность в анализе после 3-дневного воздействия прототипами антитела к ОХ40 (В17 или В19). Как производство IL-2, так и производство IFN-γ демонстрирует очевидное улучшение при воздействии антителами и выявляет зависимую от дозы корреляцию. Более высокое производство цитокинов регистрировали при воздействии антителами в более высоких дозах.
[0089] Для оценки агонистической активности прототипов антител к ОХ40, В17 и B19, также определяли ЕС50, а также анализ агонистической активности и производство цитокинов регистрировали для сравнения.
[0090] Конструирование, экспрессия и очистка антитела анти-PD-L1-ОХ40 scFv
[0091] Поскольку биспецифическое антитело разрабатывали на основе IgG, слитого с форматом scFv, структура Fc антитела к иммунной контрольной точки слита на конце с scFv ОХ40. Антитело может представлять собой ингибирующие антитела к иммунным контрольным точкам, такие как к PD-L1, к PD-1, к CTLA4, к LAG3 и т.д., или стимулирующие антитела, такие как к CD28, к CD137, к CD27, к ICOS и др. Линкер расположен между Fc антитела и scFv ОХ40 для получения биспецифического антитела, как изображено на фиг. 14.
[0092] Согласно некоторым вариантам осуществления клон 6 прототипа антитела к PD-L1 назначается представлять собой форму IgG, с другой стороны, прототип антитела к ОХ40 трансформируется как формат scFv для слияния на С-конце области Fc в клоне 6 прототипа антитела к PD-L1. Трансформация из антитела в формат scFv может привести к снижению активности или специфичности связывания, поэтому несколько прототипов антитела к ОХ40 использовали для трансформации scFv. Конструирование Fc бифункционального антитела к PD-L1, слитого с полноразмерным scFv ОХ40 (SEQ ID NO. 10 в качестве клона А4, SEQ ID NO. 11 в качестве клона В17, SEQ ID NO. 12 в качестве клона В19 или SEQ ID NO. 13 в качестве клона В120). Короткий гибкий пептидный линкер, (GGGGS)2 (SEQ ID NO. 14) помещали, например, между С-концом области Fc тяжелой цепи антитела к PD-L1 и N-концевым модулем scFv ОХ40, чтобы обеспечить правильность сворачивания и минимизацию стерических помех. Кодирующие последовательности антител анти-PL-L1-ОХ40 scFv были показаны в SEQ ID N0. 16 (тяжелая цепь клона 6 к PD-L1 - scFv клона В17 к ОХ40) и NO. 17 (тяжелая цепь клона 6 к PD-L1 - scFv клона В19 к ОХ40). Сконструированные слитые белки Fc антитела были выведены сигнальным пептидом (SEQ ID NO. 15) и экспрессированы клетками млекопитающих и очищены от супернатанта трансфицированных клеточных культур с помощью одностадийной хроматографии на основе связывания с белком G. Как показано на фиг. 15, чистота более чем 90% может быть получена в одностадийном процессе очистки и показывает, что очищенные слитые белки характеризуются правильной молекулярной массой (ММ=220 кДа).
[0093] Усиленная стимуляция активации Т-клеток для прототипов биспецифических антител анти-PD-L1-ОХ40 scFv в MLR
[0094] Для определения синергического взаимодействия биспецифического антитела в усилении активации Т-клеток посредством ингибирования взаимодействия между PD-1 и PD-L1 и агонистической активации передачи сигналов ОХ40, прототипы биспецифических антител, анти-PD-L1-OX40 scFv, вносили в MLR, как описано выше. Производство IL-2 и IFN-γ затем регистрировали через 3 или 5 дней после воздействия антителом. Моно-, комбинированное или биспецифическое антитело применяли в виде равного количества или равного моля для сравнения синергетического эффекта в усилении активации Т-клеток, а изотипический IgG использовали в качестве отрицательного контроля. Как показывают данные, показанные на фиг. 16А и 16В, одни только прототипы антител к PD-L1 показали значительную индукцию IL-2 после 3-дневного воздействия, также как и эталонное антитело, MPDL3280A, напротив, прототипы антител к ОХ40 не способны явно увеличивать положительную регуляцию производства цитокинов, как через 3, так и 5 дней воздействия антителом. Он соответствует эталонному антителу GSK3174998. Однако комбинация антител к ОХ40 и антител к PD-L1 показала значительную активацию производства цитокинов через 3 и 5 дней воздействия антителом. Синергетический эффект также наблюдается при воздействии прототипами биспецифических антител, и увеличение производства цитокинов также аналогично, как и при комбинированной терапии. Это указывает на то, что прототипы биспецифических антител анти-PDF-L1-OX40 scFv также функционируют как комбинированное воздействие антител без потери какой-либо связывающей активности в трансформации scFv.
[0095] Определение агрегации и чистоты биспецифического антитела
[0096] Поскольку очищенное антитело анти-PD-L1-клон 6-ОХ40 клон В17 scFv обнаружило более низкую чистоту (74,07%) с помощью анализа эксклюзионной ВЭЖХ после очистки хроматографией на одной колонке на основе связывания с белком А, следовательно, было создано несколько вариантов антител для улучшения чистоты и снижения агрегации для биспецифического антитела в настоящем изобретении. Описанные выше линкеры использовали для замены линкера в scFv В17 ОХ40 в биспецифическом антителе, анти-PD-L1-ОХ40 Ab (SEQ ID NO. 16), и производили как анти-PD-L1-ОХ40 Ab-V1-V4 (SEQ ID NO. 18 - SEQ ID NO. 21) в клетках СНО. Затем эти варианты очищали и анализировали на колонке XBridge Protein ВЕН SEC-HPLC (Waters, кат.№186007640). Данные обобщали, как показано ниже в таблице 1, один из вариантов биспецифических антител, анти-PD-L1-OX40 Ab-V4, выявил значительное улучшение чистоты антител. Чистота повышена с 74,07 до 92,27%. Поэтому анти-PD-L1-ОХ40 Ab-V4 использовали для дальнейшего улучшения чистоты антител.
[0097] Для характеристики распределения по размерам биспецифических антител образцы загружали на колонку XBridge Protein ВЕН SEC-HPLC (Waters, кат.№186007640) с использованием модуля разделения Waters Alliance 2695. Пик белка обнаруживали при 280 нм с использованием детектора Water 2996 PDA. Подвижной фазой был изократический фосфат натрия в концентрации 25 мМ (Sigma, кат.№04272 и кат.№04269) с NaCl в концентрации 200 мМ (AMRESCO, кат.№0241), рН 6,8, при скорости потока 0,4 мл/мин. Процент пиков определяли по участкам площади пиков, как показано на фиг. 17А-17Е.
[0098] Анти-PD-L1-OX40 Ab-V4 выявилозначительное улучшение чистоты (фиг. 17Е). Биспецифическое антитело дополнительно конструировали в фрагмент scFv В17 ОХ40, чтобы снова улучшить чистоту. Несколько остатков в scFv В17 ОХ40, показанных на фиг. 18, заменяли на отличающиеся аминокислоты, и варианты тяжелой цепи объединяли в пару с легкой цепью клона 6 к PD-L1 для получения нескольких вариантов биспецифического антитела, от анти-PD-L1-ОХ40 Ab-V5 до V12 (SEQ ID NO. 22 - SEQ ID NO. 29), а затем экспрессировали и очищали, как указано выше. Чистота вариантов биспецифических антител обобщена, как показано в таблице 2, анти-PD-L1-OX40 scFv-V5 выявило наилучшую чистоту в этих вариантах антител. Чистота повышена до 96,46%. Это показало превосходную чистоту сконструированного биспецифического антитела, а также выявило хорошую способность к развитию этого биспецифического антитела в будущем. Как показано на фиг. 19, целостность анти-PD-L1-ОХ40 Ab-V5 также анализировали с помощью ДСН-ПААГ и показали хорошую целостность в восстанавливающих и невосстанавливающих условиях.

[0099] Между тем, сконструированные варианты биспецифических антител также применяли для оценки активности связывания для PD-L1 и ОХ40 человека с помощью прямого анализа ИФА, как показано на фиг. 20. Все варианты биспецифических антител, как указано, демонстрировали одинаковую активность связывания для PD-L1 человека, (фиг. 21), эта активность связывания аналогична антителу анти-PD-L1 6. Это явление также наблюдалось в анализе связывания ОХ40 человека (фиг. 22). Только анти-PD-L1-ОХ40 Ab-V10 показало более слабую активность связывания для человеческого ОХ40 по сравнению с другими вариантами. Было указано, что конструирование scFv ОХ40 не влияет на активность связывания ОХ40. Активность связывания сохраняется либо для PD-L1, либо для ОХ40. Поскольку анти-PD-L1-ОХ40 Ab-V5 показало превосходную чистоту антител и активность связывания для PD-L1 и ОХ40, поэтому анти-PD-L1-ОХ40 Ab-V5 было выбрано для анализа стабильности в сыворотке.
[00100] Стабильность в сыворотке ex vivo
[00101] Стабильность оценивали в сыворотке человека (BioreclamationIVT, кат.№HMSRM), а также в сыворотке соответствующих доклинических видов: макаки-резуса (BioreclamationlVT, кат.№RHSSRM) и мыши CD1 (BioreclamationlVT, кат.№MSESRM). Образцы добавляли в сыворотку разных видов до конечной концентрации 15 мкг/мл и инкубировали на водяной бане при температуре 37°С. Образцы сыворотки собирали после инкубационного периода 0, 1, 2, 3, 7, 10 и 14 дней и хранили в замороженном виде при температуре -80°С до анализа.
[00102] Количественный сэндвич-анализ ИФА
[00103] OX40-FC покрывали в планшете для ИФА (NUNC, кат.№442404) 100 мкл в концентрации 1 мкг/мл в PBS и оставляли на ночь при температуре 4°С. Промывочный буфер готовили в виде PBS с 0,1% Твин-20 (Sigma, кат. №Р2287-500 мл) и блокирующий буфер готовили в виде 1% BSA (UniRegion, кат. №UR-BSA001-100G) в промывочном буфере. Образцы сыворотки готовили с 10-кратным разведением с 3-кратным серийным разведением в блокирующем буфере, а стандарты готовили в концентрации 10 нМ с 3-кратным серийным разведением в блокирующем буфере. Биотинилированный PD-Ll-Fc метили набором для быстрого конъюгирования с биотином (abcam, кат.№ ab201796) с использованием стандартного протокола и готовили в концентрации 30 нМ в блокирующем буфере. Стрептавидин-HRP (abcam, кат.№ ab7403) готовили в концентрации 1 мкг/мл в блокирующем буфере. Все образцы добавляли в каждую лунку по 100 мкл после того, как планшеты 3 раза промывали промывочным буфером и инкубировали в течение 1 часа при температуре окружающей среды. Развитие ТМВ осуществляли с 100 мкл раствора ТМВ (Invitrogen, кат.№00-2023) в течение 2 минут и остановкой с использованием 100 мкл раствора 1н. НС1 (Merck, кат.№1.00317.1000). Поглощение при длине волны 450 мкм считывали с помощью устройства для считывания ИФА (Biotek, Powerwave XS).
[00104] Анти-PD-L1-ОХ40 Ab-V5 был выбран для анализа стабильности в сыворотке ex vivo из-за его превосходной чистоты и активности связывания для PD-L1 и ОХ40. Очищенные биспецифические антитела смешивали с сывороткой разных видов, таких как человек, мышь или обезьяна. После нескольких дней культивирования образцы отбирали и анализировали с помощью сэндвич-анализа ИФА для определения количества антител. Как показано на фиг. 23, анти-PD-L1-ОХ40 Ab-V5 по-прежнему демонстрировало хорошую стабильность в сыворотке после 14 дней культивирования при температуре 37°С. Концентрация антитела все еще выше 70% у человека, мыши или обезьяны. Указано, что антитело также характеризуется хорошей стабильностью в сыворотке.
[00105] Для измерения способности анти-PD-L1-ОХ40 Ab-V5 модулировать реактивность Т-клеток, очищенные Т-клетки будут культивировать с аллогенными дендритными клетками, полученными посредством культивирования моноцитов в GM-CSF и IL-4 в течение нескольких дней. Устанавливали параллельные планшеты для сбора супернатантов в день 3 и день 5 для измерения IL-2 и IFN-γ, соответственно, с использованием коммерческого набора ИФА. Как показывают данные, показанные на фиг. 24А и 24В, производство IL-2 и IFN-γ сильно повышено при воздействии биспецифическими антителами (V5), а также при комбинированном воздействии после 3 или 5 дней воздействия антителами. Кроме того, усиление явно превосходит лечение только Ab к PD-L1 или Ab к ОХ40. Это подразумевало, что сконструированное биспецифическое антитело, V5, все еще обладает агонистической активностью, также как комбинированное лечение без потери функциональности и может быть разработано в качестве терапевтического антитела для различных солидных опухолей или рака в будущем.
[00106] Противоопухолевая активность биспецифического антитела (модель in vivo)
[00107] Отсутствие перекрестной реактивности PD-L1 и ОХ40 у грызунов в биспецифических антителах препятствовало использованию стандартных моделей мышиной сингенной или человеческой ксенотрансплантатной опухоли для оценки эффективности антител против опухолей человека. Соответственно, новую модель ксенотрансплантата опухоли мыши huPBL-SCID-Bg получали с использованием мыши SCID-Bg (CB.17/Icr.Cg PkrdcscidLystbg/CrI), которая содержит мутацию beige (Bg) с отсутствием мышиных Т- и В-лимфоцитов и функциональных NK-клеток. Эффективность биспецифических антител против опухолей человека оценивали с использованием этой модели, как описано ниже.
[00108] Простату человека РС-3 получали из Американской коллекции типовых культур и культивировали в RPMI-1640 (Invitrogen) с L-глутамином, пируватом натрия, пенициллином/стрептомицином и 10% инактивированной нагреванием фетальной бычьей сывороткой (FBS, Gibco, кат. №10437). Клетки выращивали до слияния в колбах Falcon Т-150. Затем клетки трипсинизировали (трипсин 0,25%-EDTA; Invitrogen), и рост масштабировали до достаточного количества клеток для инокуляции. Лимфоциты периферической крови (РВМС) выделяли из гепаринизированной крови с использованием Lymphoprep™ в соответствии с протоколом производителей (STEMCELL Technologies Inc.). Подсчитанные клеточные суспензии объединяли таким образом, чтобы каждая мышь получала инъекцию 0,75×106 РВМС и 3×106 опухолевых клеток в одном болюсе инъекции 0,1 мл в PBS. Для облегчения роста опухолевых клеток у мыши, другой объем 0,1 мл матригеля затем смешивали с объединенной клеточной суспензией и затем немедленно инъецировали подготовленным мышам.
[00109] Каждой мыши подкожно инъецировали 0,2 мл объема объединенной клеточной суспензии в правый бок животного. После 14-дневной инокуляции образуется солидная опухоль, достигающая объема приблизительно 250-300 мм3, и биспецифическое антитело (3 мг/кг анти-PD-L1-ОХ40 Ab-V5), эталонное антитело PD-L1 (Ref Ab, MPDL3280A) или контрольное антитело (изотипическое) вводят дважды в неделю в течение трех недель посредством внутрибрюшинной инъекцией (i.p.). Измерения опухоли делают посредством штангенциркуля Preisser два раза в неделю, а также регистрируют введение исследуемого образца в течение продолжительности экспериментов и массу тела. Объем опухоли рассчитывали с использованием следующего расчета: длина × ширина2 × 0,44 = объем (мм3) и наносили на график на фиг. 25 Мышей удаляли из исследования до прекращения эксперимента в том случае, если объем опухоли достигал 2000 мм3, или животное теряло 20% массы тела. Аналогичные результаты наблюдали, когда опухоли измеряли на 7-й день после инокуляции, и животных рандомизировали в соответствии с объемом опухоли. Для исследования на животных каждая группа содержала 6 мышей. Как показывают данные, представленные на фиг. 25, биспецифическое антитело демонстрировало значительную противоопухолевую эффективность в мышиной модели ксенотрансплантата РС-3. Размер опухоли уменьшался через 18 дней после инокуляции опухоли, а также эталонного антитела к PD-L1 и продолжал уменьшаться ниже 100 мм3. Мышиная модель ксенотрансплантата РС-3 предварительно демонстрировала противоопухолевое действие биспецифического антитела и выявляла его потенциал в качестве терапевтического потенциального лекарственного средства в будущем.
[00110] В совокупности эти результаты показали, что биспецифическое антитело поддерживает свою блокировку иммунной контрольной точки в передаче сигналов PD-1/PD-L1 и агонистическую активность для передачи сигналов ОХ40. Продолжаются исследования для дальнейшего изучения биологической активности этих белков с использованием подходящей модели на животных, такой как опухоль РС-3 в гуманизированной модели NOD.Cg-PrkdcscidIl22rgtmIwjI/SzJ (NSG).
[00111] Fc-область в настоящем изобретении может быть из любых изотипов иммуноглобулина, подклассов, аллотипов или сконструированных мутантов, таких как фрагмент(ы) Fc по типу «выступы во впадинах».
[00112] ПРИМЕРЫ
[00113] В приведенном ниже примере описано получение моноклональных антител, подходящих для терапевтических целей, нацеленных на PD-L1 и ОХ40 человека. Композитные человеческие антитела против PD-L1 и против ОХ40 человека получали из клона 6 антитела к PD-L1 и клона В17 антитела к ОХ40, соответственно. Сегменты последовательности V-области человека получали из баз данных последовательностей неродственных антител человека (зародышевой и не зародышевой линии).
[00114] Пример 1. Получение антител IgG, которые связываются с PD-L1 и ОХ40.
[00115] Некоторые антитела, предоставленные в соответствии с настоящим изобретением, первоначально получали из Fab, связывающихся с PD-L1 или ОХ40 человека. Fab отбирали из библиотеки фагового дисплея, библиотеки фагемид OmniMab, после чередования пэннинга на соответствующих слитых белках Fc (PD-Ll-Fc или ОХ40-Fc) и клетках, экспрессирующих соответствующий белок человека (PD-L1 или ОХ40). После прямого ИФА или скрининга ИФА на основе клеток, затем секвенировали положительные клоны для тяжелой цепи и легкой цепи. Эти Fab включали в себя те, которые обозначены как «OM-PD-Ll-6», «OM-PDLl-32» и т.д. для PD-L1; «ОМ-ОХ40-А4», «ОМ-ОХ40-В17» и «ОМ-ОХ40-В19» и т.д. для антител ОХ40. PD-L1-клон 3, PD-L1-клон 6 и PD-L1-клон 32 антител к PD-L1, раскрытые в настоящей заявке, получали из «OM-PD-L1-6» и «OM-PDL1-32». Между тем, ОХ40-А4, ОХ40-В17 и ОХ40-В19 антител ОХ40, раскрытые в настоящей заявке, получали из «ОМ-ОХ40-А4»,« ОМ-ОХ40-В17» и «ОМ-ОХ40-В19» в клетке НЕК293 или клетках CHO-S. И биспецифическое антитело, нацеленное на PD-L1 и ОХ40 одновременно, разрабатывали как антитело анти-PD-L1 6-ОХ40 scFv В17 и антитело анти-PD-L1 6-ОХ40 scFv В19. Аминокислотная последовательность вариабельной области легкой цепи и вариабельной области тяжелой цепи данного Fab идентичны аминокислотной последовательности вариабельной области легкой цепи и вариабельной области тяжелой цепи, соответственно.
[00116] Пример 2. Связывание in vitro анти-PD-L1-ОХ40 scFv с соответствующей мишенью.
[00117] Биспецифическое антитело к PD-L1-OX40 конструировали, как показано на фиг. 13, и экспрессировали в клетках НЕК293 или клетке CHO-S. Среду, содержащую биспецифическое антитело, очищали аффинно из супернатанта культуры хроматографией на основе связывания с белком G. Очищенное антитело затем концентрируют с последующим диализом в буфере PBS и анализируют с помощью ДСН-ПААГ, как показано на фиг. 14. Для исследования прямого связывания очищенных слитых белков с PD-L1 или ОХ40 с помощью ИФА, 100 нг/лунку рекомбинантного PD-L1 или ОХ40 использовали для покрытия в 96-луночном планшете для ИФА. Затем в каждую лунку добавляли различные концентрации очищенного анти-PD-L1-ОХ40 scFv и инкубировали в течение 1 ч. После промывания разведение 1:5000 конъюгата HRP к Fab (Jackson Immunochemicals) добавляли в каждую лунку и инкубировали в течение еще одного часа. После заключительного промывания добавляли субстрат ТМВ (Invitrogen Inc.) и измеряли оптическую плотность при 450 нм. Данные анализировали путем нанесения точек на сигмоидальную кривую с использованием GraphPad Prism 5 и вычисляли ЕС50.
[00118] Пример 3. Антигенсвязывающая специфичность анти-PD-L1-ОХ40 scFv с помощью анализа FACS
[00119] Для исследования специфичности связывания антитела анти-PD-L1-ОХ40 scFv стабильно экспрессирующие PD-L1 клетки 293, стимулированные IFNy А549 или WiDr, окрашивали 1 мкг/мл антитела анти-PD-L1-ОХ40 scFv в течение 1 ч на льду, перед тем как промыть три раза 1-кратным PBS. Связанные слитые белки антител обнаруживали с использованием конъюгированного с А1еха-488 козьего антитела к IgG человека (H+L) с последующим анализом FACS. Изотипическое антитело использовали в качестве отрицательного контроля для исследования. Результаты показали, что анти- PD-L1-OX40 scFv сохраняет свою антигенсвязывающую специфичность по сравнению с одним анти-PD-L1. Специфичность связывания антитела анти-PD-L1-ОХ40 scFv также исследовали с использованием стабильно экспрессирующих ОХ40 клеток 293.
[00120] Пример 4. Иммуномодулирующее действие бифункциональных белков in vitro.
[00121] Для измерения способности антител анти-PD-L1-ОХ40 scFv модулировать реактивность Т-клеток, очищенные Т-клетки будут культивироваться с аллогенными дендритными клетками, полученными путем культивирования моноцитов в GM-CSF и IL-4 в течение нескольких дней. Параллельные планшеты были установлены для сбора супернатантов в день 3 и день 5 для измерения IL-2 и IFN-γ, соответственно, с использованием коммерческого набора ИФА. Гуманизированное анти-PD-L1 компании Genentech/Roche, MPDL3280A, будет производиться собственными силами и использоваться в качестве положительного контроля. Как показывают данные, представленные на фиг. 16А и 16В, производство IL-2 и IFN-γ сильно повышено при воздействии биспецифическими антителами, а также при комбинированном воздействии после 3- или 5-дневного лечения антителами. Особенно биспецифические антитела, составленные из клона 6 антитела к PD-L1 и клона В17 антитела к ОХ40 (анти-PD-L1-ОХ40 scFv В17 Ab), или комбинация (анти-PD-Ll клон 6 Ab плюс анти-ОХ40 клон В17 Ab) показали усиление активации Т-клеток выше, чем комбинация эталонных антител к PD-L1 и ОХ40 (PD-L1 Ref Ab плюс ОХ40 Ref Ab). Это указывает на то, что антитело В17 к ОХ40 может обладать специальным связыванием эпитопа, что приводит к лучшей агонистической активности по сравнению с эталонным антителом к ОХ40, GSK3174998.
[00122] Пример 5 Экспансия лейкоцитов человека, индуцированная биспецифическими антителами in vivo
[00123] Отсутствие обнаруживаемой перекрестной реактивности антител к PD-L1 или ОХ40 с мышиными PD-L1 или ОХ40 и потребность в наличии иммунных клеток человека требовали разработки моделей для функциональной оценки биспецифических антител in vivo. Мыши с генетическим фоном NOD, несущие тяжелую комбинированную иммунодефицитную (SCID) мутацию и дефицит в общей гамма-цепи рецептора IL-2 (обычно называемый NSG), способны поддерживать приживление большого количества лейкоцитов периферической крови человека (huPBL) и поддерживать приживление в течение по меньшей мере 30 дней (King et al., 2008). Эту мышиную модель, также известную как модель huPBL-NSG, использовали для оценки функционального эффекта системного введения антител in vivo на иммунные клетки человека.
[00124] В частности, 6 миллионов свежевыделенных человеческих РВМС адоптивно переносили посредством внутривенной инъекции мышам huPBL-NSG. Через девять дней после инъекций РВМС животным вводили однократную дозу 1 мг/кг моноантитела, биспецифического антитела или изотипического контрольного антитела IgG4 посредством внутрибрюшинной инъекции. На 24-28 день после приживления РВМС РВМС окрашивали антителами к CD45 человека и мыши, оцениваемыми с помощью проточной цитометрии. Для определения гейта лимфоцитов использовали профили прямого и бокового рассеяния. Биспецифические антитела были способны усиливать экспансию лейкоцитов человека, о чем свидетельствует увеличение доли CD45+ клеток человека в периферической крови привитых мышей. Для каждой группы n ≥ 6 мышей.
[00125] Пример 6. Пнгибирование роста опухолевых клеток РС-3 или А498 в huPBL-NSG антителом анти-PD-L1-ОХ40 scFv.
[00126] Положительную в отношении PD-L1 клеточную линию рака простаты человека, РС-3 (АТСС# CRL-1435) или клеточную линию рака почки, А498 (АТСС® НТВ-44™) можно использовать для создания моделей ксенотрансплантатов у мышей huPBL-NSG. Для формирования опухоли 3×106 клеток РС-3 (или клеток А498) /мышь будут инъецированы подкожно мышам huPBL-NSG, как описано выше. Чтобы оценить ингибирующее влияние на рост опухоли, различные концентрации антитела анти-PD-L1-ОХ40 scFv, эталонного антитела или изотипического антитела в дозе от 0,1-3 мг/кг вводят мышам внутривенно два раза в неделю в течение 4 недель после 14 дней имплантации опухолевых клеток. Рост опухоли будет измеряться два раза в неделю до 5 недель, как описано на мышиной модели Fox Chase SCLD®Beige.
[00127] Пример 7. Фармакокинетическая оценка анти-PD-L1-ОХ40 scFv у мышей и обезьян
[00128] 10-40 мг/кг бифункциональных белков, анти-PD-L1-ОХ40 scFv, будут вводить мышам или обезьянам посредством подкожной инъекции или внутривенной инъекции. Образцы сыворотки будут отбираться в различные моменты времени после инъекции до 15 дней. Концентрации слитого белка Fc в образцах сыворотки будут определять с помощью сэндвич-анализа ИФА.
[00129] Хотя настоящее изобретение было описано довольно подробно со ссылкой на определенные его варианты осуществления, возможны другие варианты осуществления. Таким образом, сущность и объем прилагаемой формулы изобретения не должны быть ограничены описанием вариантов осуществления, содержащихся в настоящем документе.
[00130] Специалистам в настоящей области техники будет очевидно, что в структуру настоящего изобретения могут быть внесены различные модификации и изменения, не выходя за пределы объема или сущности настоящего изобретения. Исходя из вышеизложенного предполагается, что настоящее изобретение охватывает модификации и варианты этого изобретения при условии, что они попадают в объем следующей формулы изобретения.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
--->
<110> ЭйПи БАЙОСАЙЕНСИЗ, ИНК.
<120> МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ
С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА
<130> 9022-1808584
<140> PCT/US2018/067868
<141> 2018-12-28
<150> US 62/611,543
<151> 2017-12-29
<160> 43
<170> PatentIn версия 3.5
<210> 1
<211> 217
<212> PRT
<213> Homo sapiens
<400> 1
Gln Ser Val Leu Thr Gln Pro Pro Ser Ala Ser Gly Thr Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Asn Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Ser Asn Asn Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Ser Gly Leu Gln
65 70 75 80
Ser Glu Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Trp Asp Asp Ser Leu
85 90 95
Asn Ala Trp Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 2
<211> 118
<212> PRT
<213> Homo sapiens
<400> 2
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 3
<211> 216
<212> PRT
<213> Homo sapiens
<400> 3
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Asn Asp Val Gly Ser Tyr
20 25 30
Asn Ser Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Val Ile Tyr Glu Val Ala Asn Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Leu Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 4
<211> 121
<212> PRT
<213> Homo sapiens
<400> 4
Gln Val Gln Leu Val Gln Ser Gly Ala Lys Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Ile Pro Ile Leu Gly Ile Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Val Pro Gly Tyr Ser Tyr Gly Pro Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 5
<211> 214
<212> PRT
<213> Homo sapiens
<400> 5
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe
35 40 45
Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His
85 90 95
Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 6
<211> 126
<212> PRT
<213> Homo sapiens
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr
20 25 30
Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala Leu Asp Ala
100 105 110
Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120 125
<210> 7
<211> 214
<212> PRT
<213> Homo sapiens
<400> 7
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe
35 40 45
Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His
85 90 95
Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Ala Glu Cys Ser
210
<210> 8
<211> 118
<212> PRT
<213> Homo sapiens
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Ser Ser Ser Arg Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gln Asn Glu Asp Val Ala Phe Gly Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser
115
<210> 9
<211> 327
<212> PRT
<213> Homo sapiens
<400> 9
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 10
<211> 246
<212> PRT
<213> Искусственная последовательность
<220>
<223> OX40 scFv (клон A4)
<400> 10
Gln Ser Ala Leu Thr Gln Pro Pro Ser Ala Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Val Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Glu Val Ser Gln Arg Pro Ser Gly Val Pro Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Val Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Ala Gly Ser
85 90 95
Asn Asn Leu Gly Val Phe Gly Gly Gly Thr Lys Leu Thr Val Ser Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Glu
145 150 155 160
Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
165 170 175
Tyr Ile Ser Ser Ser Gly Ser Thr Ile Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
210 215 220
Arg Gly Gly Val Gly Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met
225 230 235 240
Val Thr Val Ser Ser Ala
245
<210> 11
<211> 250
<212> PRT
<213> Искусственная последовательность
<220>
<223> OX40 scFv (клон B17)
<400> 11
Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys Thr
1 5 10 15
Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser
20 25 30
Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr
35 40 45
Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn
50 55 60
Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp
65 70 75 80
Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val
85 90 95
Ile Phe Gly Gly Gly Thr Lys Leu Thr Val Ser Ser Gly Gly Gly Gly
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val
115 120 125
Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
130 135 140
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
145 150 155 160
Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Leu Ile Ser Trp
165 170 175
Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
180 185 190
Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser
195 200 205
Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Ser Ile Tyr Asp
210 215 220
Pro Pro Pro Gly Ser Arg Ser Ala Leu Asp Ala Phe Asp Ile Trp Gly
225 230 235 240
Gln Gly Thr Met Val Thr Val Ser Ser Ala
245 250
<210> 12
<211> 243
<212> PRT
<213> Искусственная последовательность
<220>
<223> OX40 scFv (клон B19)
<400> 12
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys
1 5 10 15
Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe
35 40 45
Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His
85 90 95
Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val Ser Ser Gly Gly Gly
100 105 110
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
115 120 125
Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly Ser Leu Arg Leu
130 135 140
Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ser Met Asn Trp
145 150 155 160
Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser
165 170 175
Ser Ser Ser Arg Tyr Ile Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe
180 185 190
Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn
195 200 205
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gln Asn
210 215 220
Glu Asp Val Ala Phe Gly Ile Trp Gly Gln Gly Thr Met Val Thr Val
225 230 235 240
Ser Ser Ala
<210> 13
<211> 241
<212> PRT
<213> Искусственная последовательность
<220>
<223> OX40 scFv (клон B120)
<400> 13
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Thr Leu Gly Asp Lys Tyr Ala
20 25 30
Ser Trp Tyr Gln Gln Lys Pro Gly Arg Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Ser Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Trp Asp Ser Ser Thr Ser Tyr
85 90 95
Val Phe Gly Thr Gly Thr Lys Val Thr Val Ser Ser Gly Gly Gly Gly
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln
115 120 125
Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu Thr Leu Ser Leu Thr
130 135 140
Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr Tyr Trp Ser Trp Ile
145 150 155 160
Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile Gly Glu Ile Asn His
165 170 175
Ser Gly Ser Thr Asn Tyr Asn Pro Ser Leu Lys Ser Arg Val Thr Ile
180 185 190
Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu Lys Leu Ser Ser Val
195 200 205
Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Arg Trp Gly
210 215 220
Gly Ser Phe Ile Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
225 230 235 240
Ala
<210> 14
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> пептидный линкер
<400> 14
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5
<210> 15
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> сигнальный пептид
<400> 15
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly
20
<210> 16
<211> 704
<212> PRT
<213> Искусственная последовательность
<220>
<223> Антитело анти-PD-L1-клон 6 тяжелая цепь-OX40 клон B17 scFv
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
565 570 575
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln Pro Gly
580 585 590
Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp
595 600 605
Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
610 615 620
Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser
625 630 635 640
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu
645 650 655
Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr
660 665 670
Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala Leu Asp
675 680 685
Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala
690 695 700
<210> 17
<211> 696
<212> PRT
<213> Искусственная последовательность
<220>
<223> Антитело анти-PD-L1-клон 6 тяжелая цепь-OX40 клон B19 scFv
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
565 570 575
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly
580 585 590
Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
595 600 605
Tyr Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
610 615 620
Val Ser Ser Ile Ser Ser Ser Ser Arg Tyr Ile Tyr Tyr Ala Asp Ser
625 630 635 640
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu
645 650 655
Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr
660 665 670
Cys Ala Arg Gln Asn Glu Asp Val Ala Phe Gly Ile Trp Gly Gln Gly
675 680 685
Thr Met Val Thr Val Ser Ser Ala
690 695
<210> 18
<211> 707
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V1
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser
565 570 575
Arg Ser Ser Leu Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val
580 585 590
Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr
595 600 605
Phe Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly
610 615 620
Leu Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr
625 630 635 640
Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys
645 650 655
Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala
660 665 670
Leu Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser
675 680 685
Ala Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val
690 695 700
Ser Ser Ala
705
<210> 19
<211> 708
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V2
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Gly Lys Gly Ser Gly Gly Lys Gly Thr Gly Gly Lys Gly Ser
565 570 575
Gly Gly Lys Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Val Val
580 585 590
Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe
595 600 605
Thr Phe Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys
610 615 620
Gly Leu Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr
625 630 635 640
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser
645 650 655
Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr
660 665 670
Ala Leu Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg
675 680 685
Ser Ala Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr
690 695 700
Val Ser Ser Ala
705
<210> 20
<211> 701
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V3
<400> 20
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Ala Ser Ala Pro Thr Leu Phe Pro Leu Val Ser Glu Val
565 570 575
Gln Leu Val Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu
580 585 590
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met
595 600 605
His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Leu
610 615 620
Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
625 630 635 640
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Ser Leu Tyr Leu Gln
645 650 655
Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Ser
660 665 670
Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala Leu Asp Ala Phe Asp
675 680 685
Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser Ala
690 695 700
<210> 21
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V4
<400> 21
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Lys Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 22
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V5
<400> 22
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Gln Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 23
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V6
<400> 23
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Glu Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 24
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V7
<400> 24
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Gln Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 25
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V8
<400> 25
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Ala Pro Gly Lys Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Pro Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Ile Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 26
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V9
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Ala Pro Gly Lys Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Pro Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 27
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V10
<400> 27
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Ala Pro Gly Gln Thr Ala Arg Ile Thr Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Pro Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 28
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V11
<400> 28
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Glu Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 29
<211> 706
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V12
<400> 29
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Arg Arg Tyr
20 25 30
Ser Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Val Phe Gly Ala Ala Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Glu Phe Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Ser Gly Asp Ser Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr
100 105 110
Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly
435 440 445
Ser Gly Gly Gly Gly Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser
450 455 460
Val Thr Pro Gly Gln Thr Ala Arg Ile Ala Cys Gly Gly Asn Asn Leu
465 470 475 480
Glu Ser Lys Ile Val Ser Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
485 490 495
Val Leu Val Ile Phe Tyr Asp Thr Asp Arg Pro Ser Gly Ile Ser Glu
500 505 510
Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser
515 520 525
Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp
530 535 540
Ser Asn Ser Asp His Val Ile Phe Gly Gly Gly Thr Lys Leu Thr Val
545 550 555 560
Leu Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser
565 570 575
Thr Lys Gly Glu Val Gln Leu Val Glu Ser Gly Gly Val Val Val Gln
580 585 590
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
595 600 605
Asp Asp Tyr Thr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
610 615 620
Glu Trp Val Ser Leu Ile Ser Trp Asp Gly Gly Ser Thr Tyr Tyr Ala
625 630 635 640
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
645 650 655
Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Thr Glu Asp Thr Ala Leu
660 665 670
Tyr Tyr Cys Ala Ser Ile Tyr Asp Pro Pro Pro Gly Ser Arg Ser Ala
675 680 685
Leu Asp Ala Phe Asp Ile Trp Gly Gln Gly Thr Met Val Thr Val Ser
690 695 700
Ser Ala
705
<210> 30
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V4
<400> 30
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Lys Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Ile Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 31
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V5
<400> 31
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Glu
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Gln Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Ile Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 32
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V6
<400> 32
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Glu Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Val Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 33
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V7
<400> 33
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Gln Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Val Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 34
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V8
<400> 34
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Arg
20 25 30
Ile Thr Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Ile Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Ile Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 35
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V9
<400> 35
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Arg
20 25 30
Ile Thr Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Val Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 36
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V10
<400> 36
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Gln Thr Ala Arg
20 25 30
Ile Thr Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Val Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 37
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V11
<400> 37
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Glu Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Ile Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 38
<211> 176
<212> PRT
<213> Искусственная последовательность
<220>
<223> Анти-PD-L1-OX40 Ab-V12
<400> 38
Leu Ser Leu Gly Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Tyr Val
1 5 10 15
Leu Thr Gln Pro Pro Ser Val Ser Val Thr Pro Gly Gln Thr Ala Arg
20 25 30
Ile Ala Cys Gly Gly Asn Asn Leu Glu Ser Lys Ile Val Ser Trp Tyr
35 40 45
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Phe Tyr Asp Thr
50 55 60
Asp Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser Gly Ser Asn Ser Gly
65 70 75 80
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
85 90 95
Asp Tyr Tyr Cys Gln Val Trp Asp Ser Asn Ser Asp His Val Ile Phe
100 105 110
Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Ser Thr Ser Gly Ser Gly
115 120 125
Lys Pro Gly Ser Gly Glu Gly Ser Thr Lys Gly Glu Val Gln Leu Val
130 135 140
Glu Ser Gly Gly Val Val Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
145 150 155 160
Cys Ala Ala Ser Gly Phe Thr Phe Asp Asp Tyr Thr Met His Trp Val
165 170 175
<210> 39
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкер
<400> 39
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 40
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкер
<400> 40
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
1 5 10 15
Ser Ser Leu
<210> 41
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкер
<400> 41
Gly Gly Lys Gly Ser Gly Gly Lys Gly Thr Gly Gly Lys Gly Ser Gly
1 5 10 15
Gly Lys Gly Ser
20
<210> 42
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкер
<400> 42
Gly Ser Ala Ser Ala Pro Thr Leu Phe Pro Leu Val Ser
1 5 10
<210> 43
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> линкер
<400> 43
Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr
1 5 10 15
Lys Gly
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ КОСТИМУЛЯТОРНОГО TNF-РЕЦЕПТОРА | 2016 |
|
RU2761115C1 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2766199C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| СВЯЗЫВАЮЩИЕСЯ С CD38 И PD-L1 МОЛЕКУЛЫ | 2017 |
|
RU2764201C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2787783C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ НА ОСНОВЕ АКТИВИРУЮЩИХ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИХ АНТИГЕНСВЯЗЫВАЮЩИХ МОЛЕКУЛ ПРОТИВ CD3 И ФОЛАТНОГО РЕЦЕПТОРА 1 (FOLR1) И АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХСЯ С ОСЬЮ PD-1 | 2015 |
|
RU2753902C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |



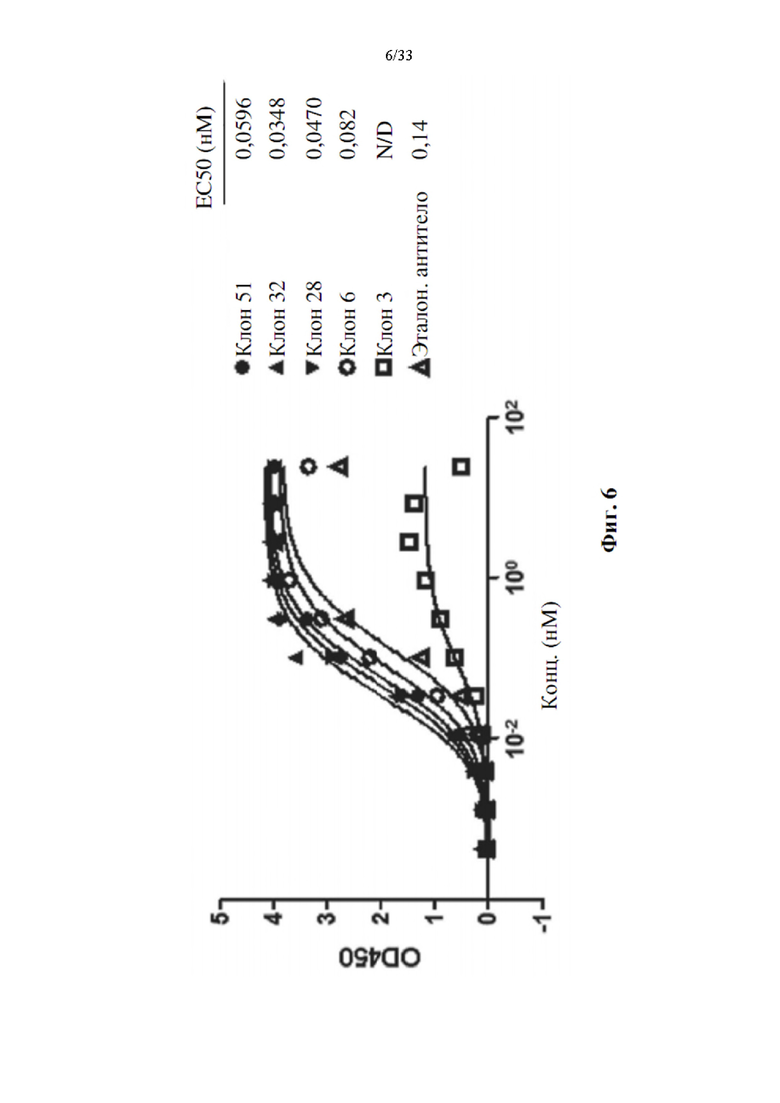
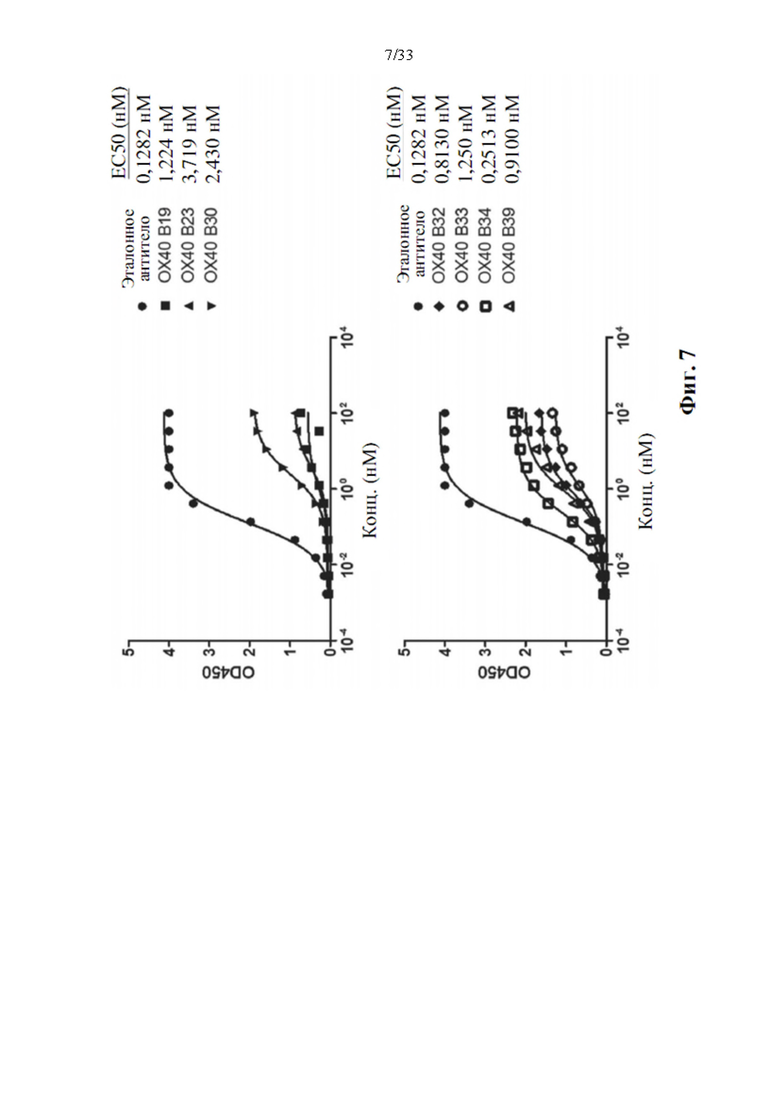

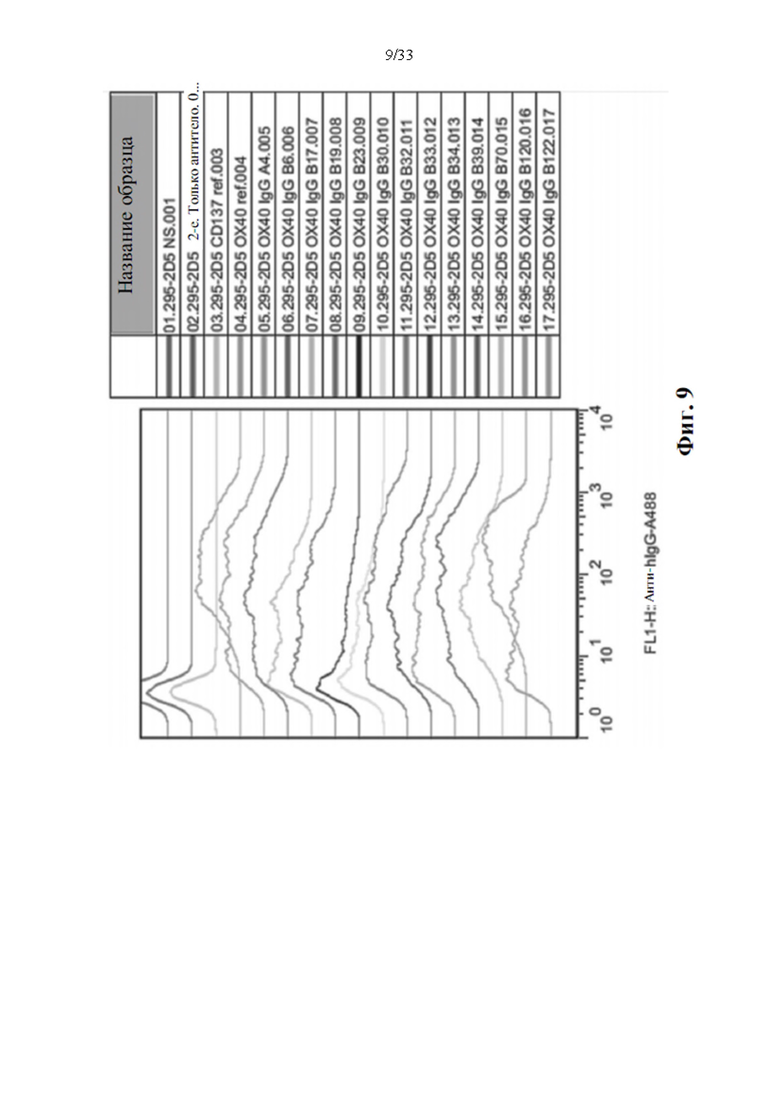

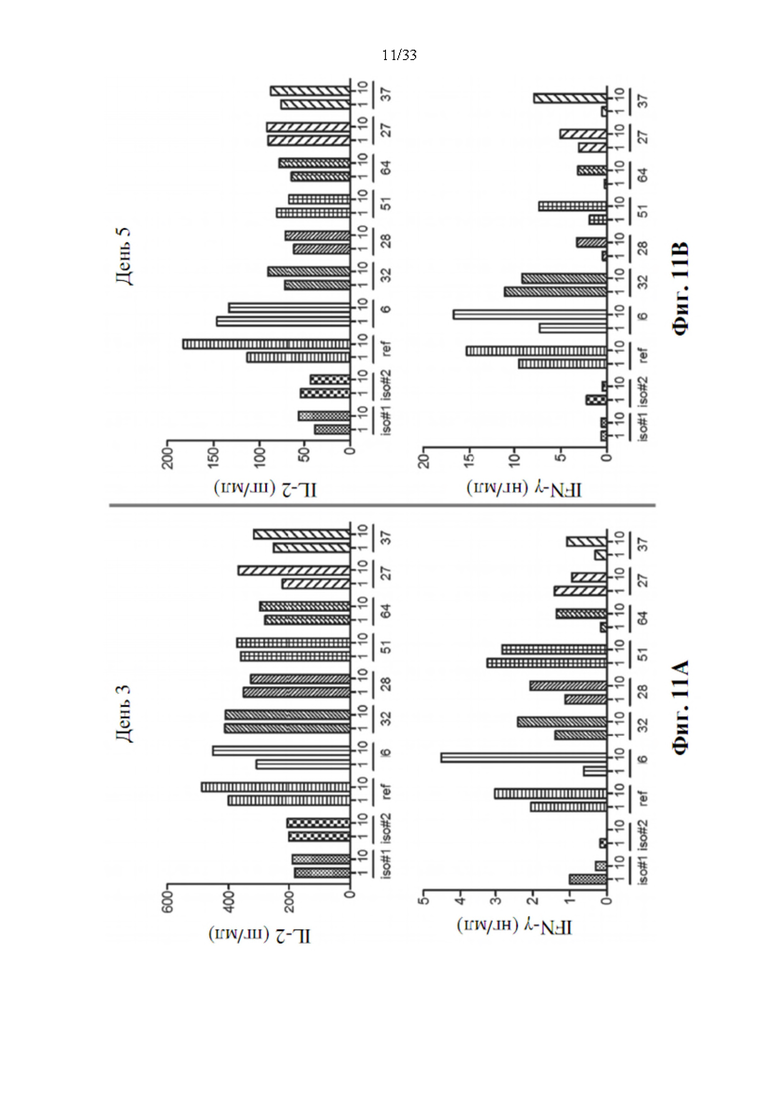
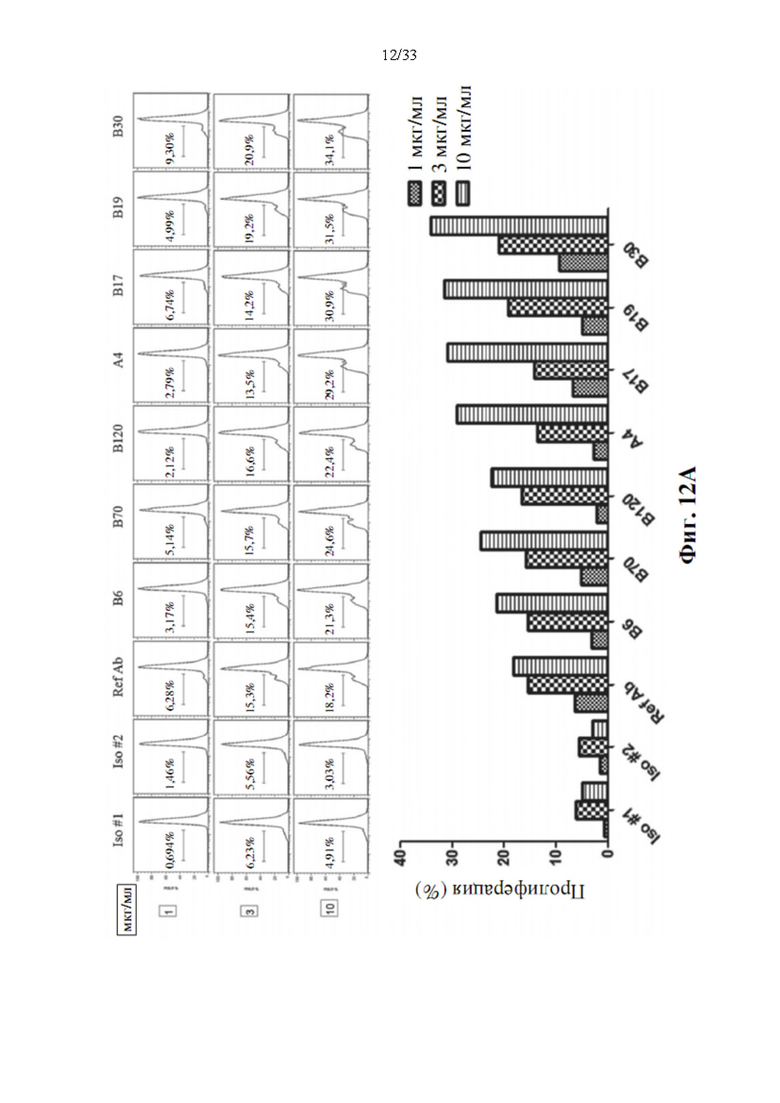
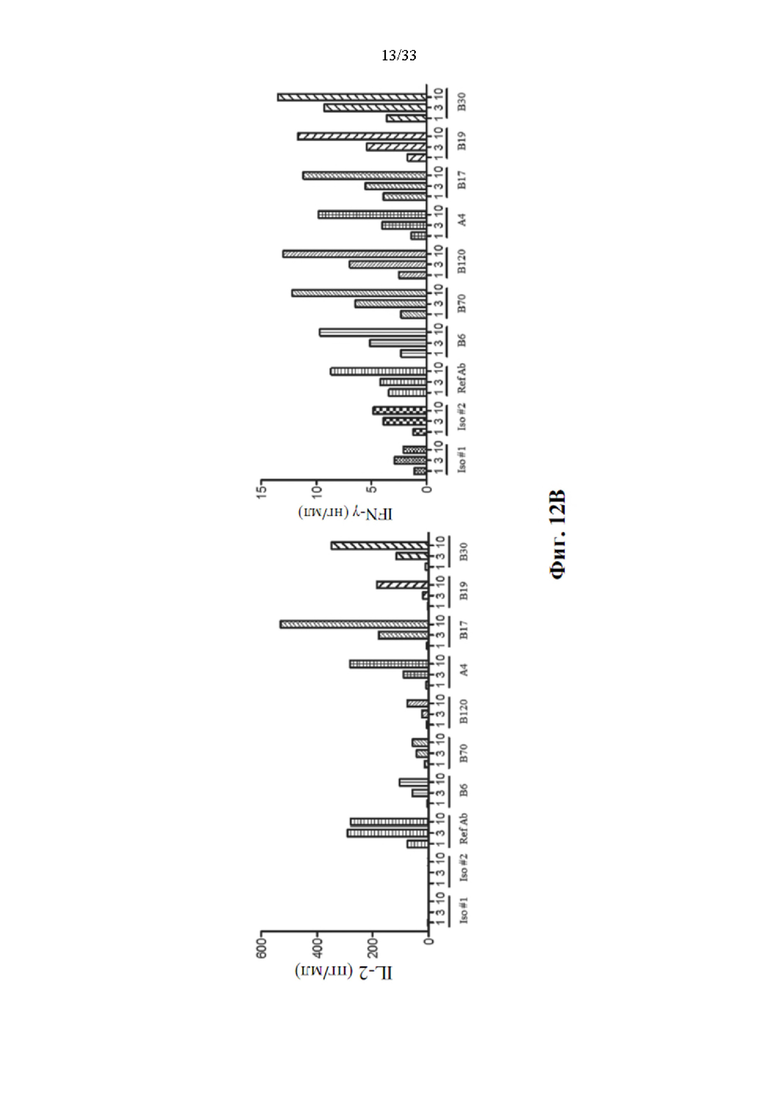
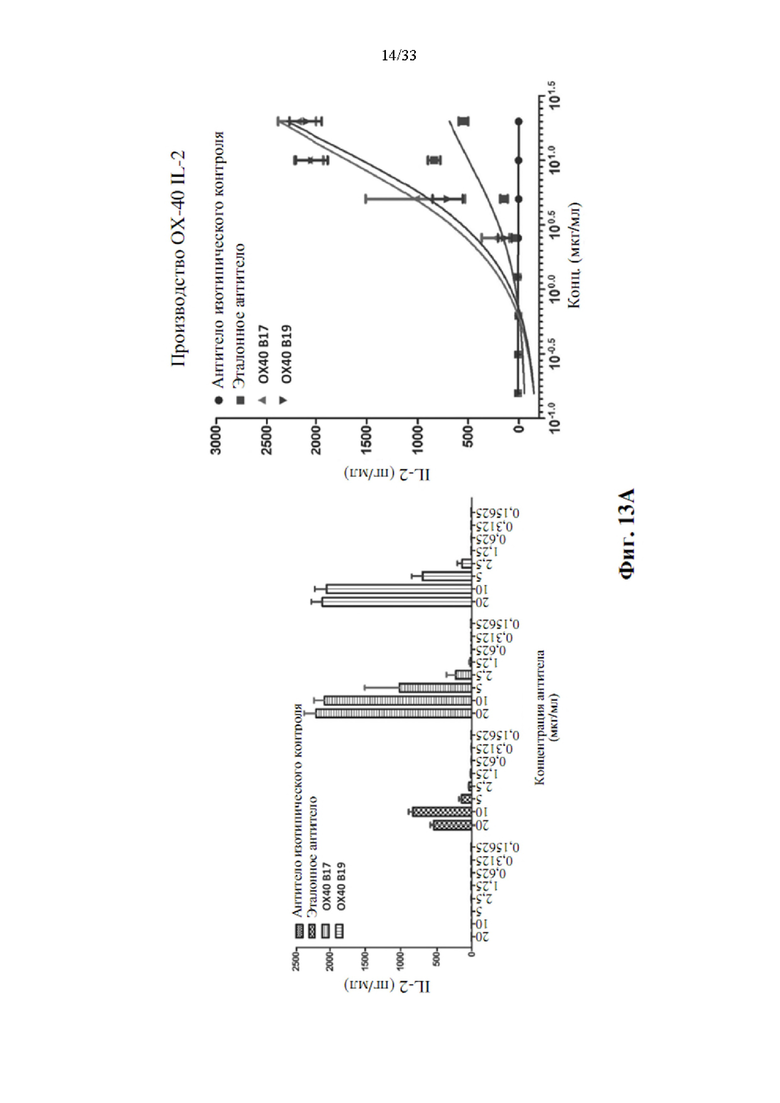
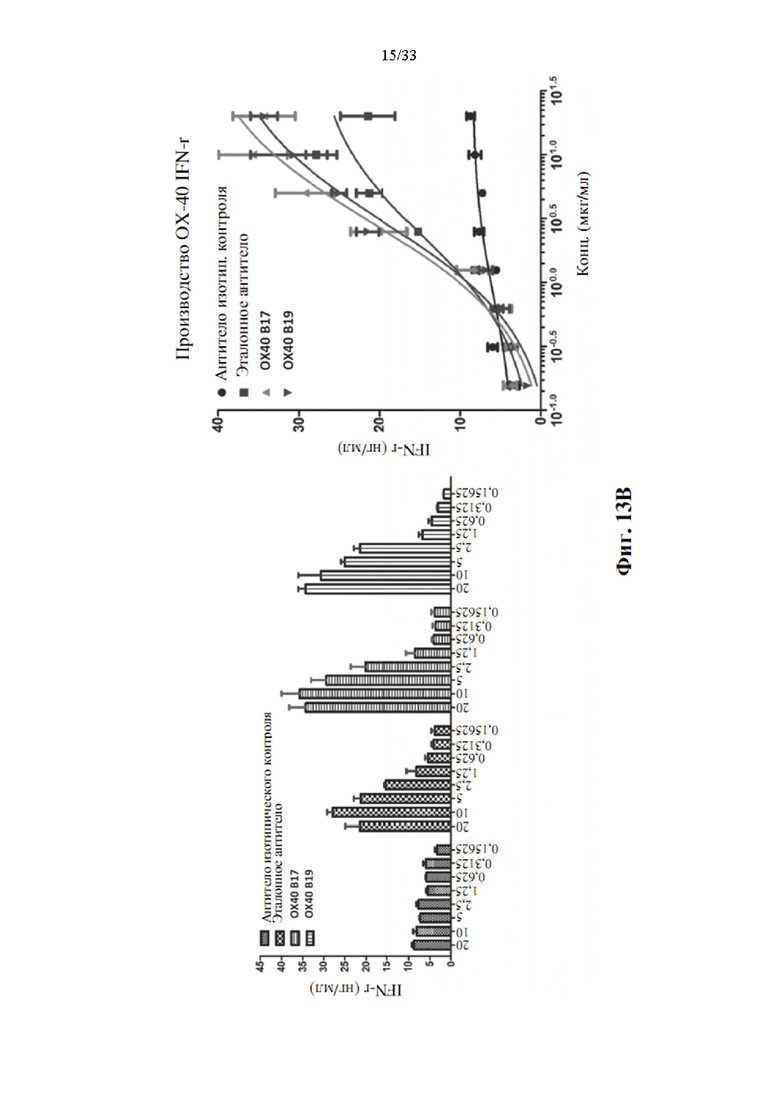
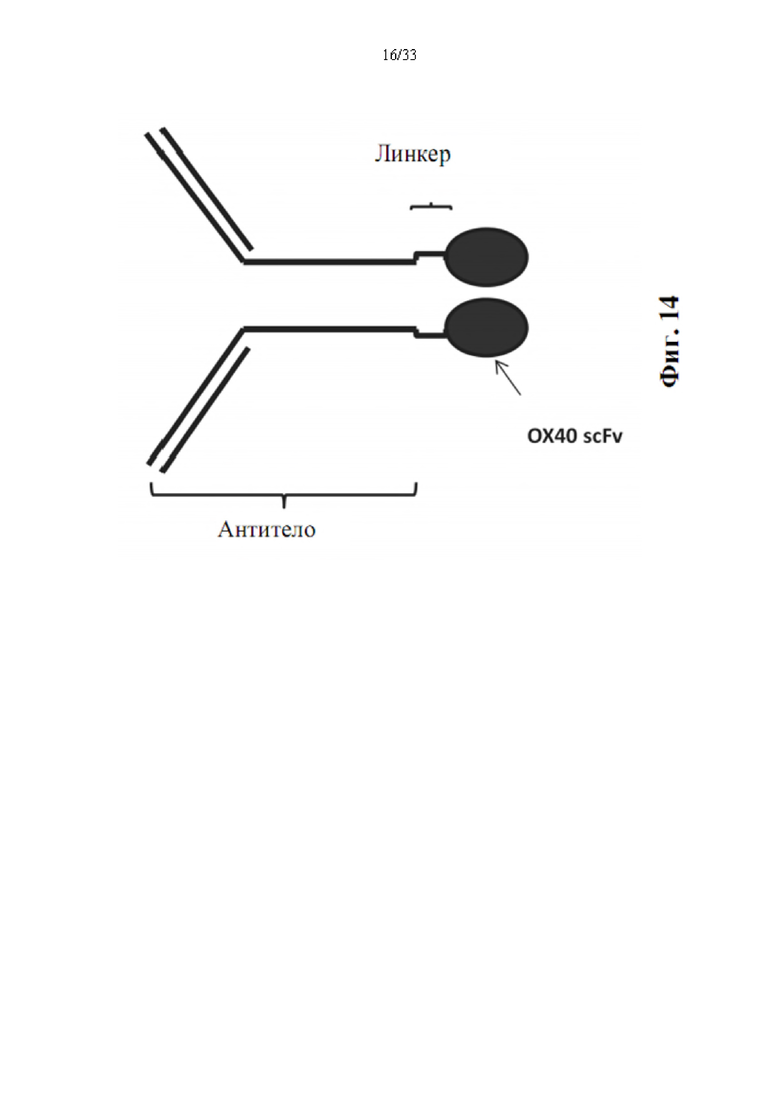
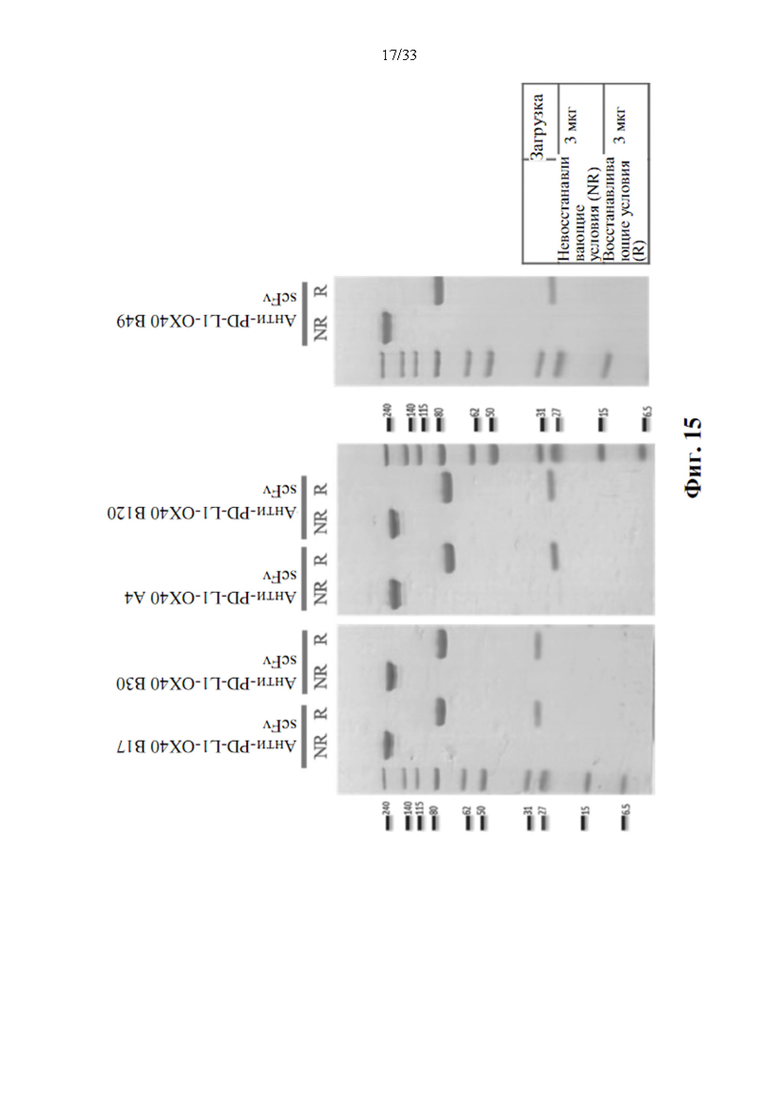
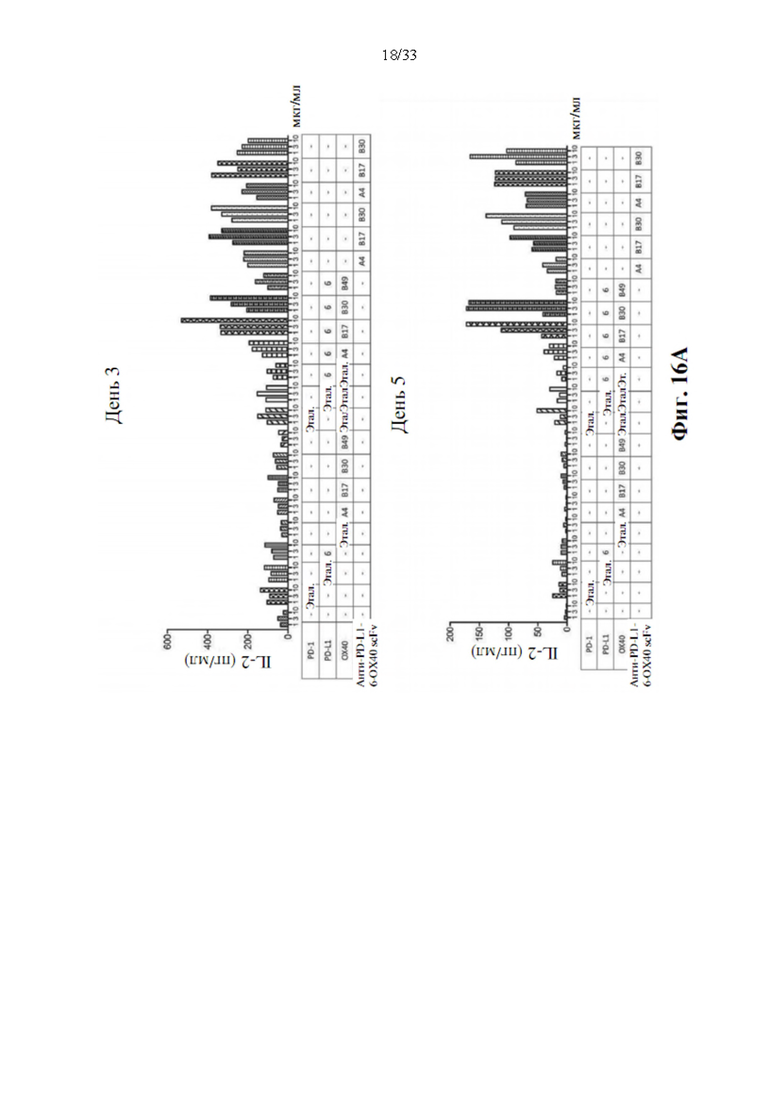
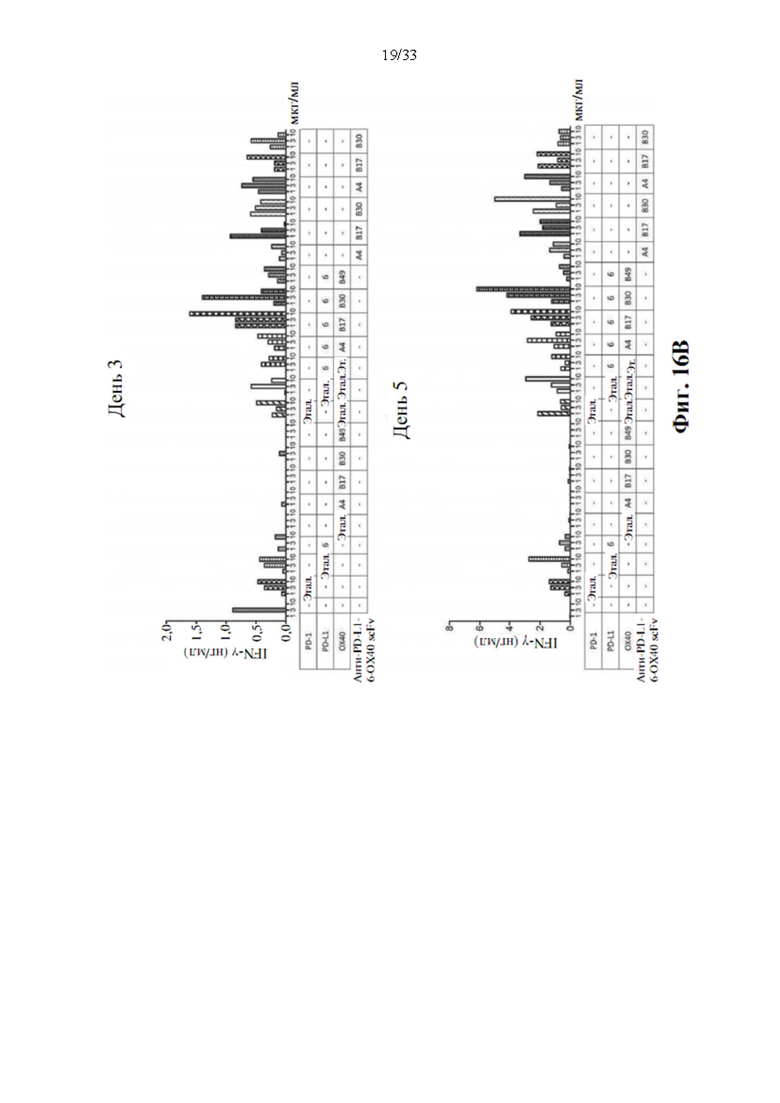
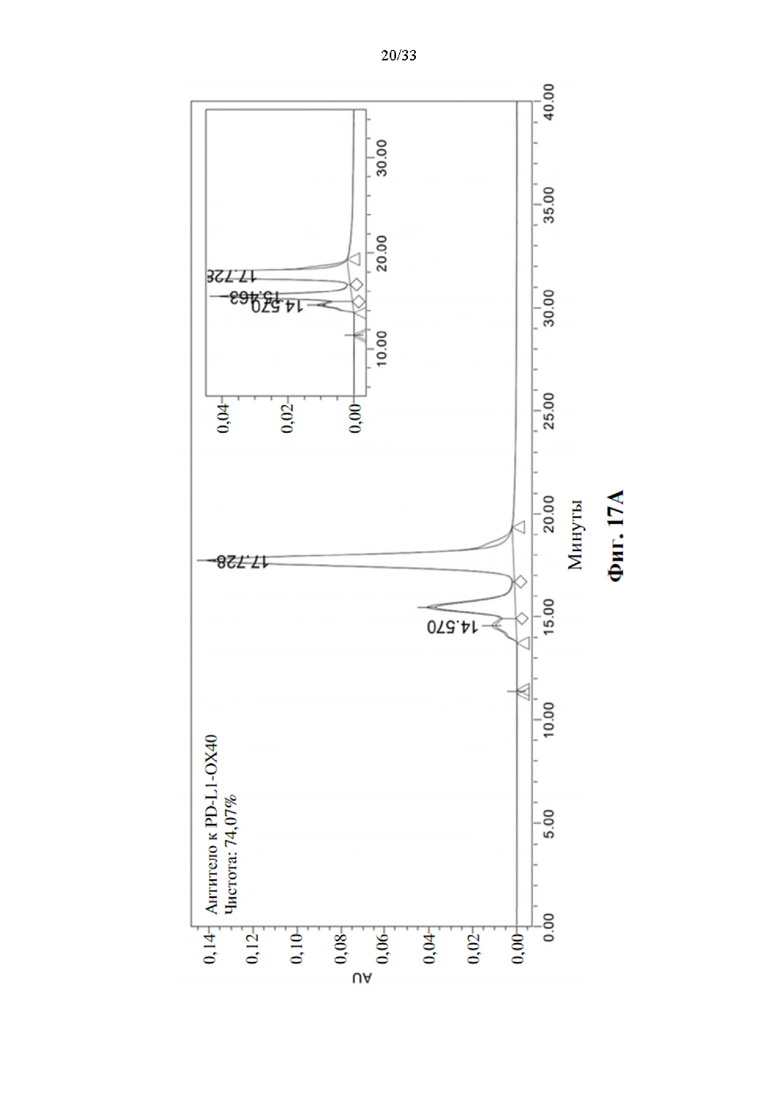
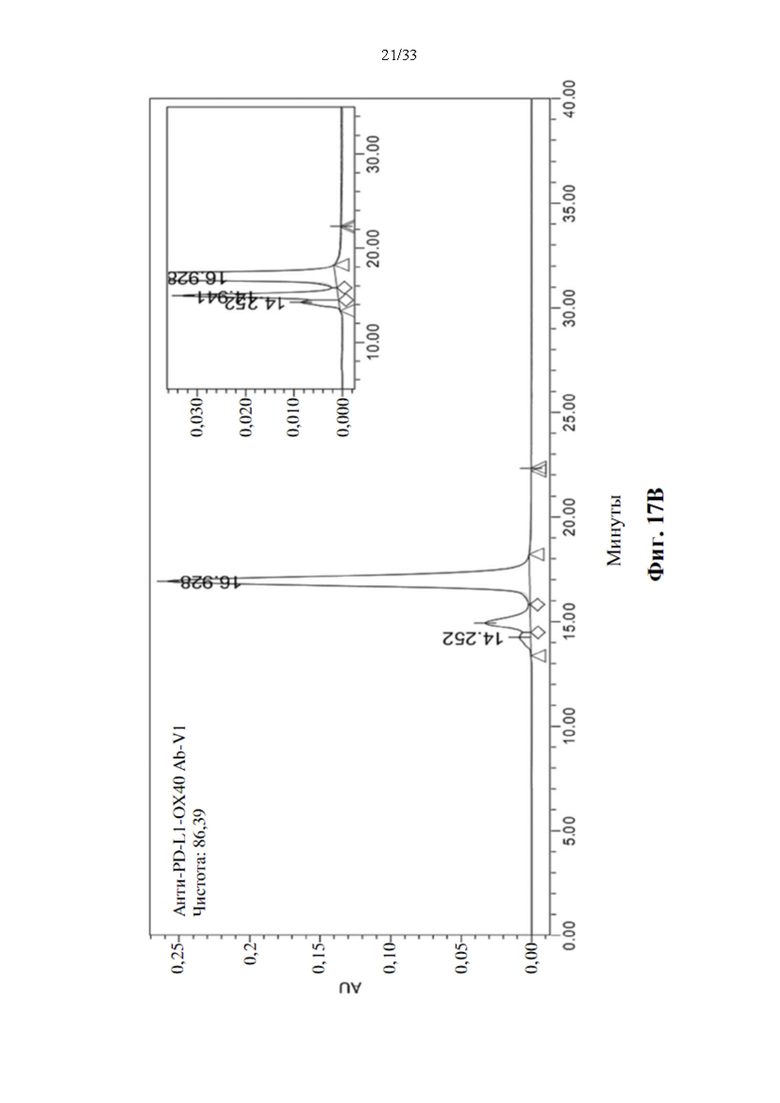
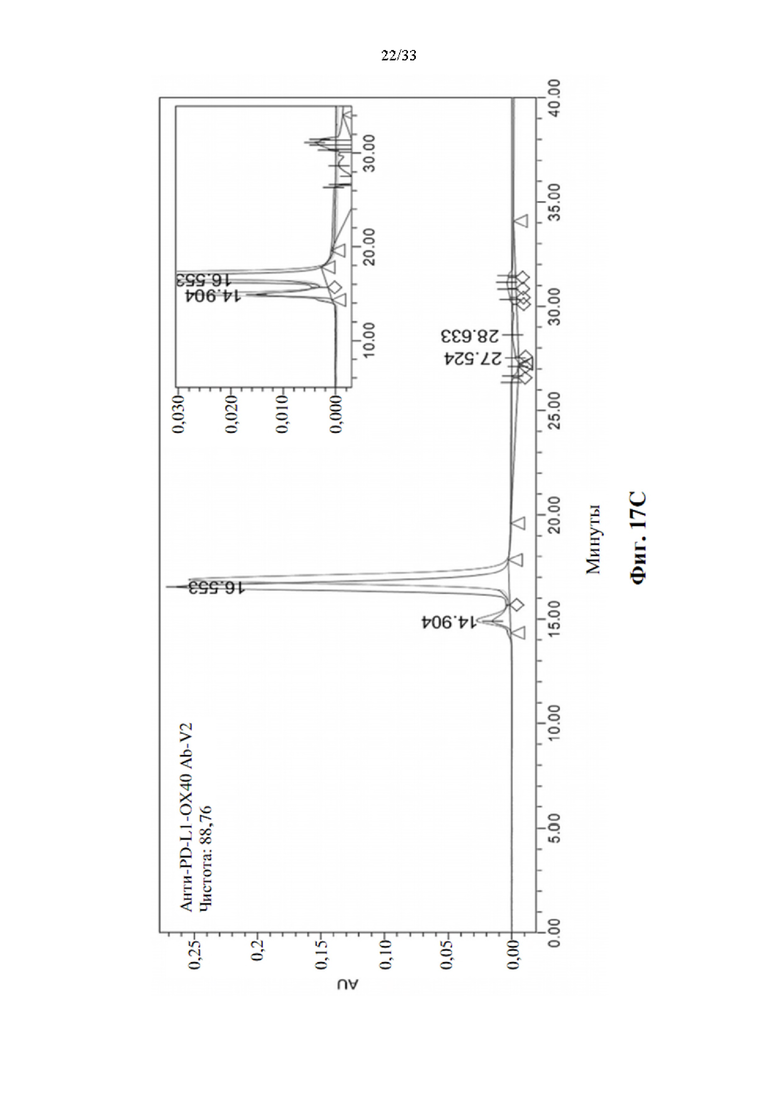
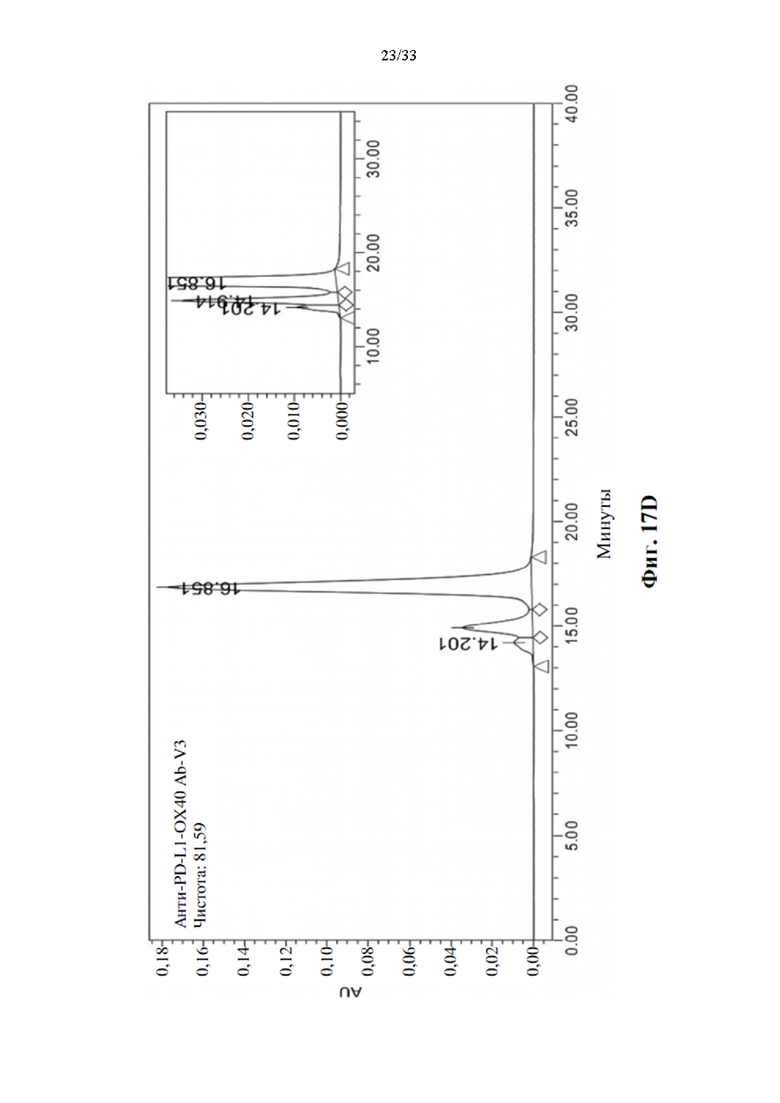


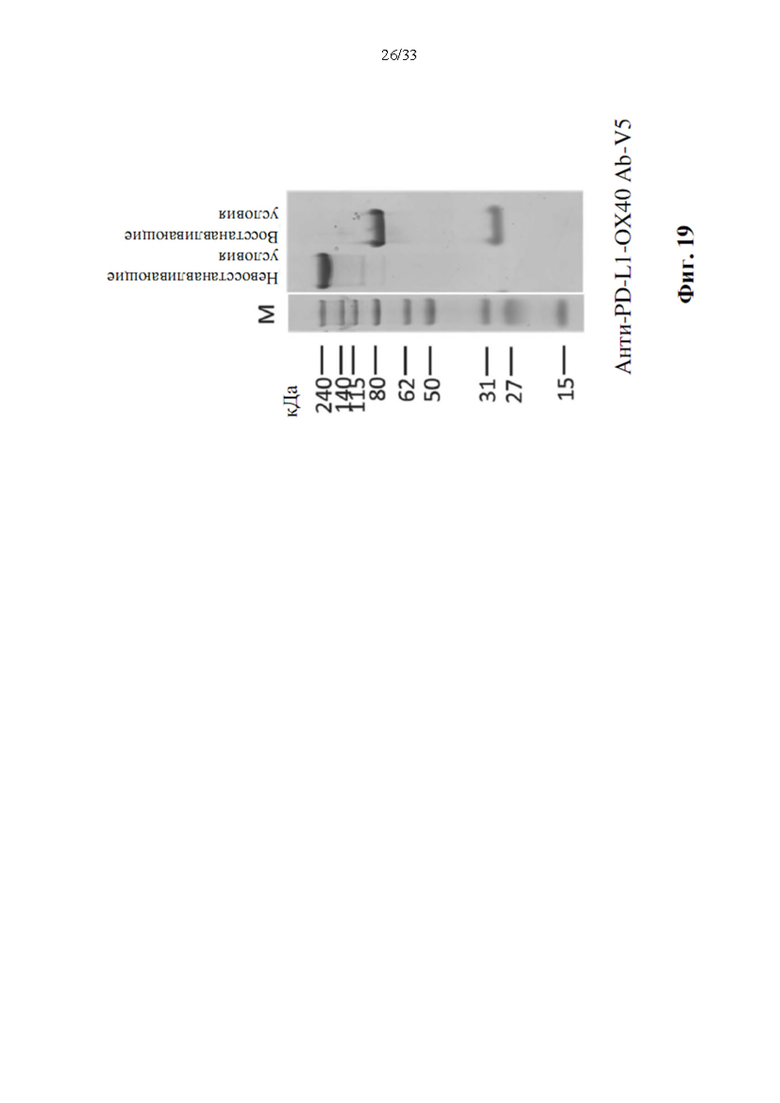

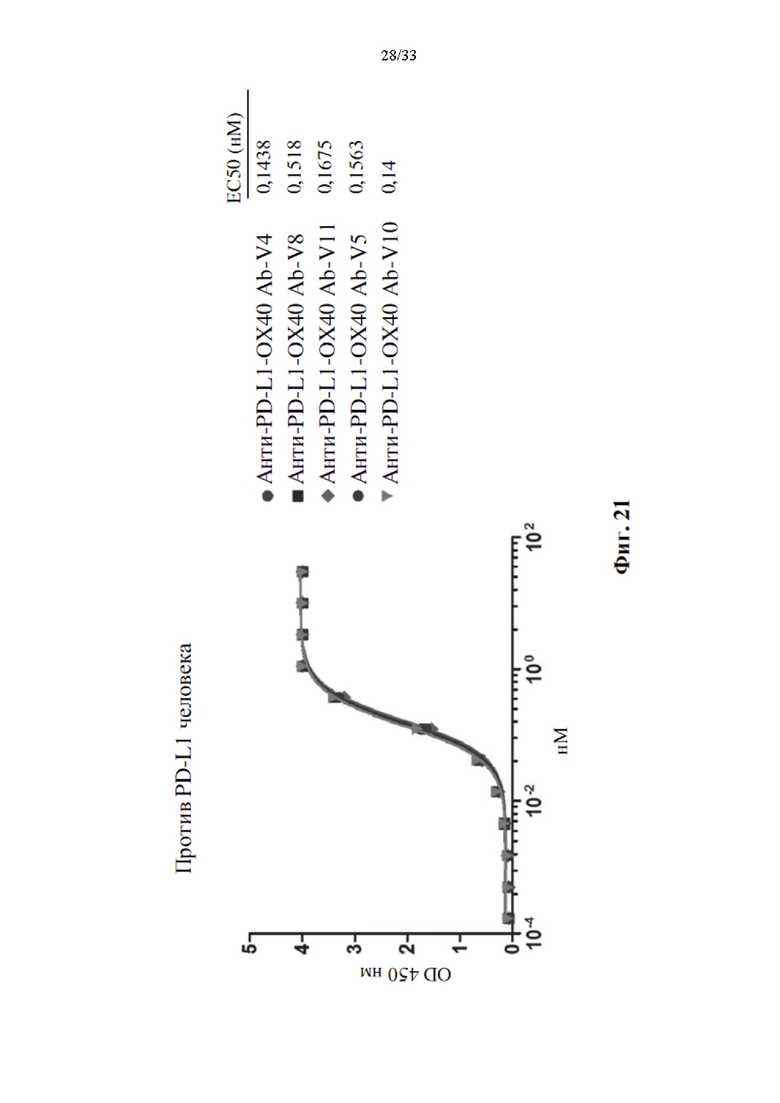
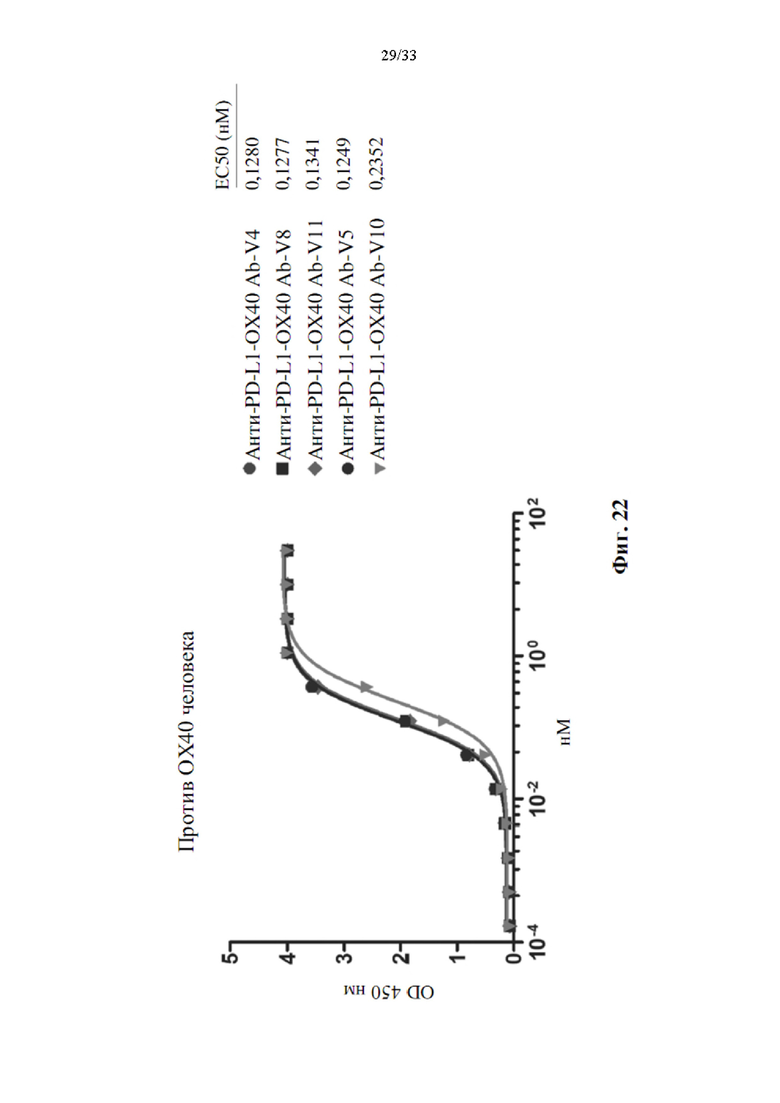
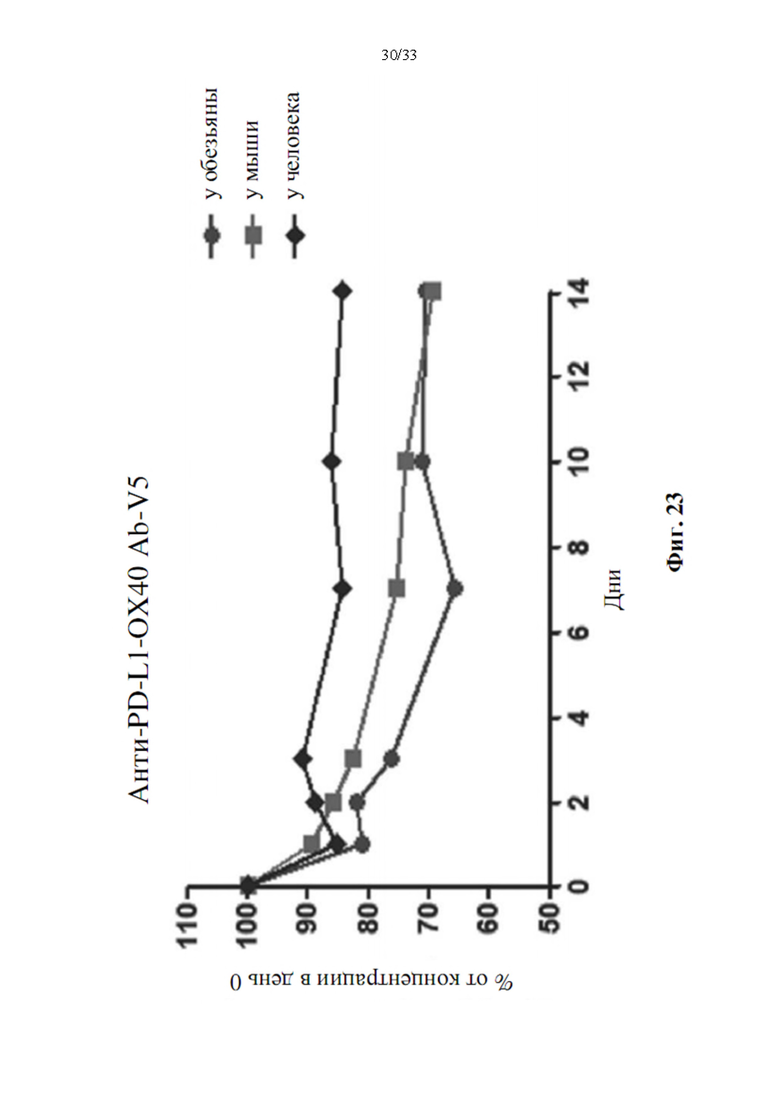
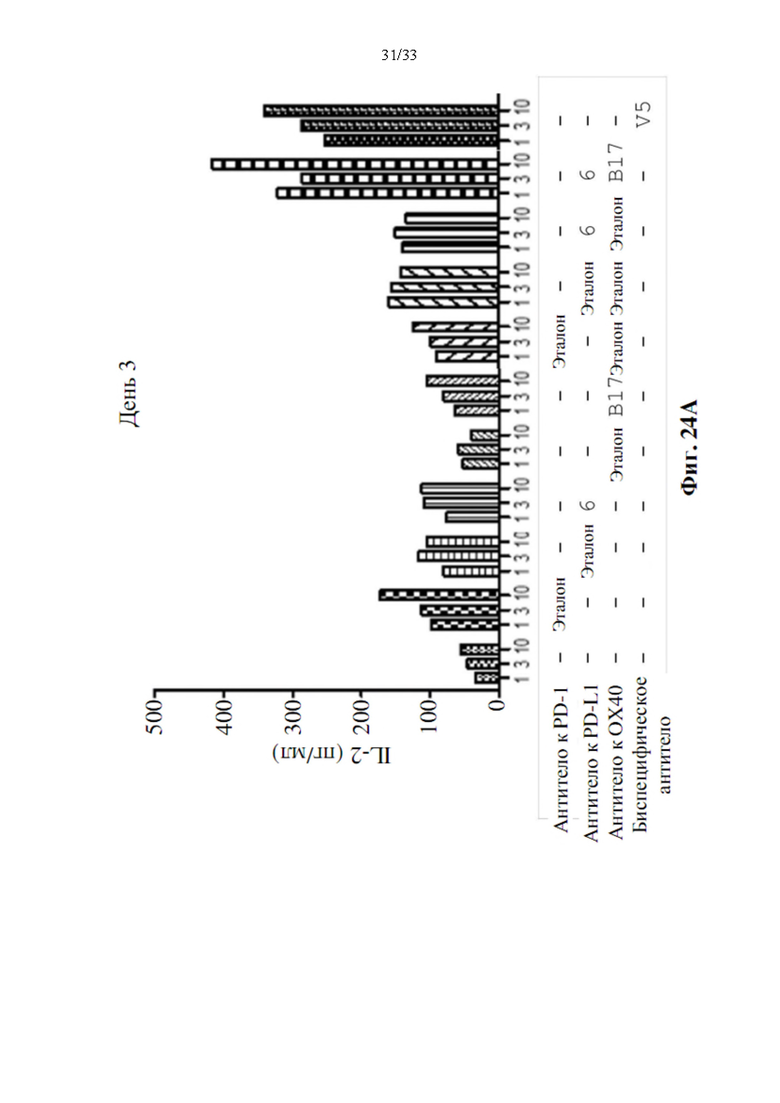
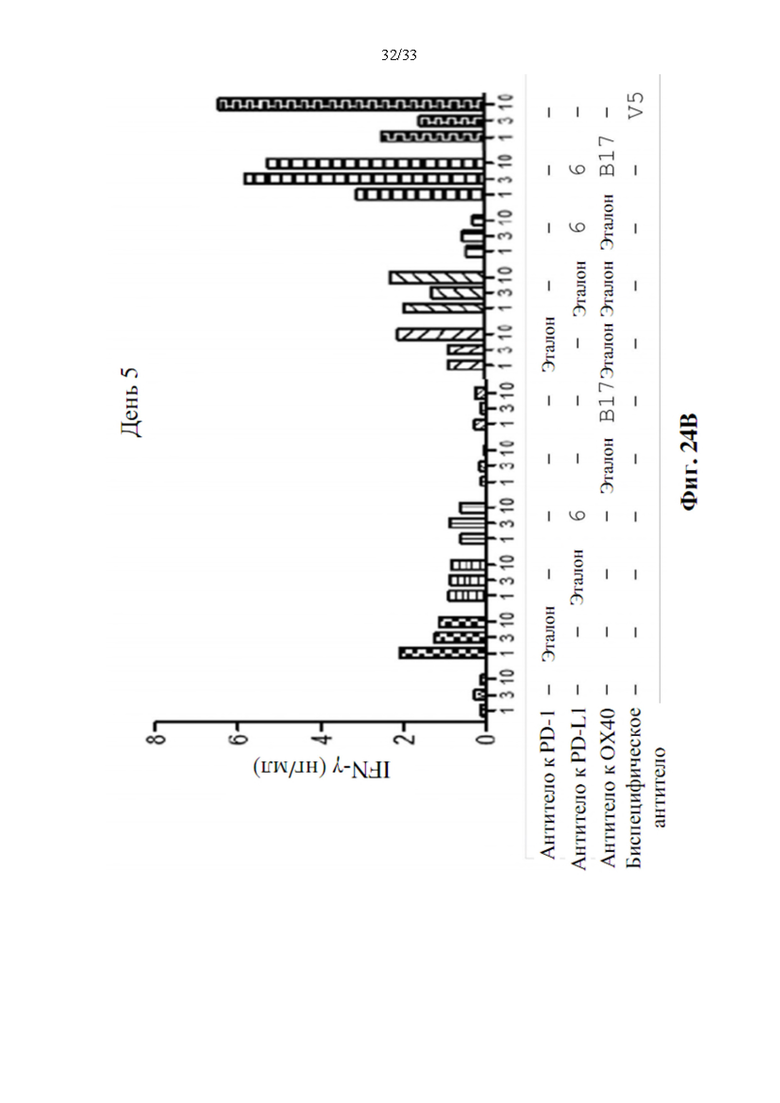
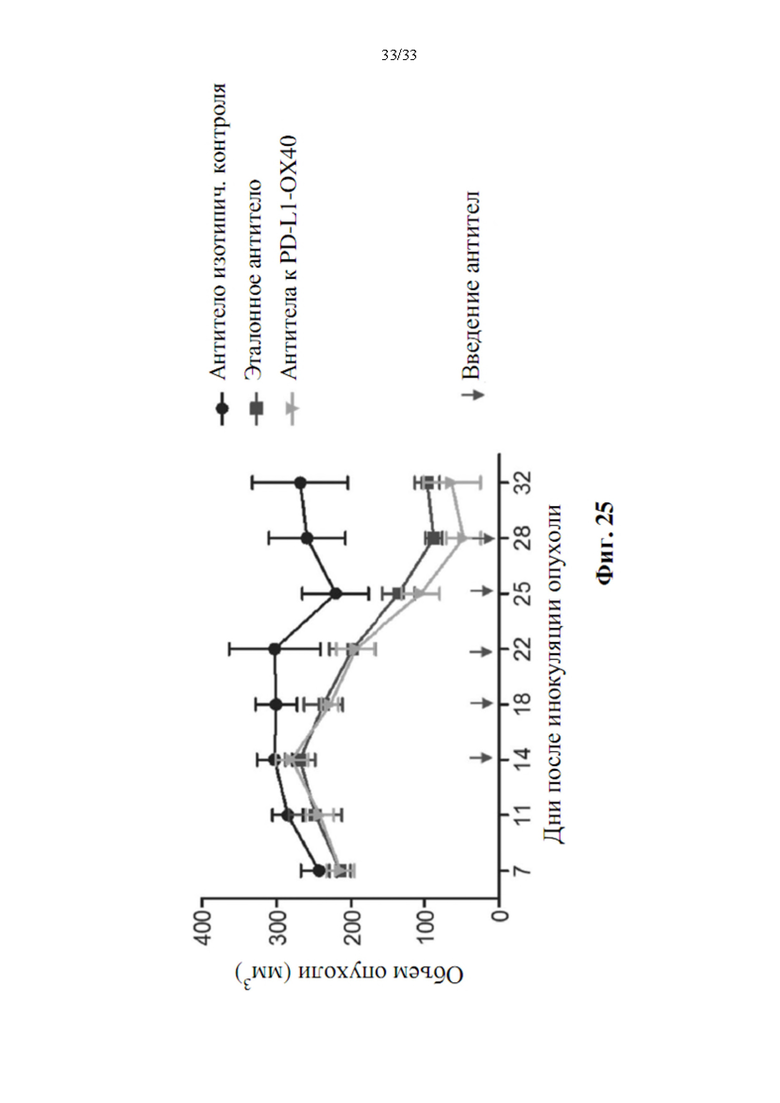
Изобретение относится к области биотехнологии. Предложено антитело или его антигенсвязывающая часть, связывающееся с OX40 (CD134). Также предложено биспецифическое антитело, связывающееся с OX40 и лигандом белка 1 запрограммированной клеточной гибели (PD-L1). Также раскрыты нуклеиновые кислоты, кодирующие указанные антитела. Кроме того, предложено применение указанных антител, в том числе, в составе фармацевтической композиции, для лечения рака, выбранного из рака предстательной железы, рака легкого, немелкоклеточного рака легкого, меланомы, лимфомы, рака молочной железы, рака головы и шеи, почечно-клеточной карциномы и рака яичников. Изобретение обеспечивает улучшенную агонистическую активность антител в активации Т-клеток. 5 н. и 9 з.п. ф-лы, 36 ил., 2 табл., 7 пр.
1. Антитело или его антигенсвязывающая часть, связывающееся с OX40 (CD134), содержащее по меньшей мере одну полипептидную цепь, которая содержит сайт связывания OX40 (CD134), содержащий:
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 6 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 5;
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 8 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 7;
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 128-246 из SEQ ID NO. 10 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-112 из SEQ ID NO. 10; или
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 124-241 из SEQ ID NO. 13 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 13,
где по меньшей мере 90% идентичность по отношению к аминокислотной последовательности исключает определяющие комплементарность области (CDR).
2. Антитело или его антигенсвязывающая часть по п. 1, причем сайт связывания OX40 представляет собой последовательность одноцепочечного вариабельного фрагмента (scFv), выбранную из группы, состоящей из SEQ ID NO. 10, 11, 12 и 13.
3. Биспецифическое антитело, связывающееся с OX40 и лигандом белка 1 запрограммированной клеточной гибели (PD-L1), содержащее полипептидную цепь, причем полипептидная цепь содержит сайт связывания OX40 и сайт связывания PD-L1, где сайт связывания OX40 содержит:
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 6 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 5;
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 8 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 7;
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 128-246 из SEQ ID NO. 10 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-112 из SEQ ID NO. 10; или
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 124-241 из SEQ ID NO. 13 и вариабельную область легкой цепи, содержащую аминокислотную последовательность, характеризующуюся по меньшей мере приблизительно 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-108 из SEQ ID NO. 13,
где сайт связывания PD-L1 содержит:
вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 2 и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью по отношению к аминокислотной последовательности, состоящей из аминокислот 1-111 из SEQ ID NO. 1, и
вариабельный домен тяжелой цепи, содержащий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности SEQ ID NO. 4, и вариабельный домен легкой цепи, содержащий аминокислотную последовательность, характеризующуюся по меньшей мере 90% идентичностью к аминокислотной последовательности, составляющей по меньшей мере приблизительно 90% по отношению к аминокислотной последовательности аминокислот 1-110 из SEQ ID NO. 3,
где по меньшей мере 90% идентичность по отношению к аминокислотной последовательности исключает определяющие комплементарность области (CDR).
4. Биспецифическое антитело по п. 3, в котором полипептидная цепь дополнительно содержит:
домен Fc;
фрагмент Fab, связанный с N-концом домена Fc, и фрагмент Fab, содержащий сайт связывания PD-L1, и
scFv, связанный с C-концом домена Fc, и scFv, содержащий сайт связывания OX40.
5. Биспецифическое антитело по п. 4, в котором полипептидная цепь дополнительно содержит линкер между доменом Fc и scFv.
6. Биспецифическое антитело по п. 5, в котором аминокислотная последовательность линкера представляет собой SEQ ID NO. 14.
7. Биспецифическое антитело по п. 4, в котором scFv содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислот 455-707 из SEQ ID NO. 18, 455-708 из SEQ ID NO. 19, 455-701 из SEQ. ID NO. 20, 455-706 из SEQ ID NO. 21, 455-706 из SEQ ID NO. 22, 455-706 из SEQ ID NO. 23, 455-706 из SEQ ID NO. 24, 455-706 из SEQ ID NO. 25, 455-706 из SEQ ID NO. 26, 455-706 из SEQ ID NO. 27, 455-706 из SEQ ID NO. 28 и 455-706 из SEQ ID NO. 29.
8. Биспецифическое антитело по п. 3, причем биспецифическое антитело содержит одну пару полипептидных цепей.
9. Биспецифическое антитело по п. 8, причем биспецифическое антитело представляет собой антитело IgG, IgE, IgM, IgD, IgA или IgY.
10. Биспецифическое антитело по п. 9, причем биспецифическое антитело представляет собой антитело IgG.
11. Биспецифическое антитело по п. 10, причем антитело IgG представляет собой антитело IgG1, IgG2, IgG3 или IgG4.
12. Фармацевтическая композиция для лечения рака, содержащая антитело или его антигенсвязывающую часть по п. 1 или 2, или биспецифическое антитело по любому из пп. 3-11 и по меньшей мере один фармацевтически приемлемый носитель, причем рак выбирают из группы, состоящей из рака предстательной железы, рака легкого, немелкоклеточного рака легкого (NSCLC), меланомы, лимфомы, рака молочной железы, рака головы и шеи, почечно-клеточной карциномы (RCC) и рака яичников.
13. Применение антитела или его антигенсвязывающей части по п. 1 или 2, или биспецифическое антитело по любому из пп. 3-11 при производстве лекарственного средства для лечения рака, где рак выбирают из группы, состоящей из рака предстательной железы, рака легкого, немелкоклеточного рака легкого (NSCLC), меланомы, лимфомы, рака молочной железы, рака головы и шеи, почечно-клеточной карциномы (RCC) и рака яичников.
14. Молекула нуклеиновой кислоты, кодирующая антитело или его антигенсвязывающую часть по п. 1 или 2, или кодирующая биспецифическое антитело по любому из пп. 3-11.
| WO 2017172981 A2, 05.10.2017 | |||
| US 2017281765 A1, 05.10.2017 | |||
| US 2017198051 A1, 13.07.2017 | |||
| HUSENI M | |||
| et al., Anti-tumor efficacy and biomarker evaluation of agonistic anti-OX40 antibodies in preclinical models, Journal for ImmunoTherapy of Cancer, 2014, 2(Suppl 3):P105 | |||
| BULLIARD Y | |||
| et al, OX40 engagement depletes intratumoral Tregs via activating |

