Перекрестная ссылка на родственные заявки
[0001] Настоящая заявка испрашивает приоритет и преимущество по предварительной заявке на патент США № 62/332788, поданной 6 мая 2016 г., раскрытие которой включено в данный документ посредством ссылки во всей своей полноте.
Перечень последовательностей
[0002] Настоящая заявка подается вместе с перечнем последовательностей в электронном формате. Перечень последовательностей представлен в виде файла под названием IOBS_100_ST25.txt, созданного 2 мая 2017 г., размер которого составляет 244 кБ. Информация из перечня последовательностей в электронном формате включена в данный документ посредством ссылки во всей своей полноте.
Предпосылки создания изобретения
[0003] Рак по-прежнему остается основной глобальной проблемой здравоохранения. Несмотря на прогресс в лечении рака, по-прежнему существует неудовлетворенная медицинская потребность в более эффективных и менее токсичных видах терапии, особенно для пациентов с заболеванием на поздней стадии или с формами рака, которые являются невосприимчивыми к существующим терапевтическим препаратам.
[0004] Роль иммунной системы, в частности, опосредованной T-клетками цитотоксичности, в контроле опухолей хорошо известна. Появляется все больше доказательств того, что Т-клетки контролируют рост опухоли и выживаемость у пациентов, у которых имеется рак, как на ранних, так и на поздних стадиях заболевания. Однако опухолеспецифичные T-клеточные ответы у пациентов, у которых имеется рак, трудно формировать и поддерживать. Продолжающееся продвижение и успехи в отношении видов иммунотерапии рака, с помощью которых стимулируются или усиливаются врожденные иммунные ответы против рака, делают такую терапию привлекательным вариантом лечения по сравнению с видами терапии, в которых применяются неспецифические химиотерапевтические средства и/или облучение.
[0005] Несколько молекулярных мишеней были идентифицированы в отношении их потенциальной полезности в качестве иммуноонкологических (IO) терапевтических препаратов против рака. Некоторые молекулярные мишени, которые исследуются в отношении их терапевтического потенциала в области иммуноонкологической терапии, включают цитотоксический Т-лимфоцитарный антиген-4 (CTLA-4 или CD152), лиганд-1 запрограммированной смерти клетки (PD-L1, или B7-H1, или CD274), белок-1 запрограммированной смерти (PD-1), OX40 (CD134 или TNFRSF4) и T-клеточный ингибиторный рецептор, белок 3, содержащий домены Т-клеточного иммуноглобулина и муцина (TIM3). Хотя некоторые из этих мишеней были успешно использованы в терапевтических целях (например, PD-1 и CTLA-4), многие пациенты не проявили ответа на терапевтические препараты, которые были разработаны. И хотя может быть рассмотрена терапевтическая схема, которая предусматривает более высокие дозы и/или комбинацию видов иммунотерапии, такие способы терапии могут быть связаны с повышенным риском побочных эффектов, которые проявляют тенденцию к увеличению с более высокими дозами и кумулятивным воздействием и, как кажется, являются добавочными при применении с видами комбинированной иммунотерапии. Некоторые распространенные побочные эффекты включают гипофизит, тиреоидит, недостаточность надпочечников, энтероколит, дерматит, пневмонию, гепатит, панкреатит, моторные и сенсорные невропатии и артрит. Кроме того, поскольку иммунотерапия обычно связана с высокими затратами, терапия, которая включает комбинацию иммунотерапевтических средств, может быть дорогостоящей для пациентов.
[0006] Таким образом, по-прежнему необходимо продолжать выявлять мишени-кандидаты для IO терапевтических препаратов, разрабатывать новые терапевтические препараты для существующих мишеней и разрабатывать терапевтические стратегии, которые лишены недостатков применяемых в настоящее время видов иммунотерапии, включающих отсутствие ответа пациента и повышенный риск побочных эффектов, связанных с комбинированным лечением. IO терапевтические препараты (например, связывающие белки), которые являются биспецифическими в отношении комбинации молекул-мишеней, особенно те, которые проявляют более высокую аффинность связывания в отношении молекул-мишеней по сравнению с аффинностью связывания комбинации отдельных моноспецифических связывающих белков, представляют собой класс особенно требуемых молекул для терапевтического потенциала.
Краткое описание изобретения
[0007] Настоящее изобретение относится к биспецифическим молекулам или белкам, которые связывают два эпитопа (например, первый и второй эпитопы), и которые являются бивалентными в отношении связывания с каждым из первого и второго эпитопов. Настоящее изобретение также относится к способам индукции иммунного ответа у субъекта, а также к способам лечения или предупреждения рака у субъекта (например, субъекта-человека) путем введения субъекту белков, молекул нуклеиновых кислот и/или композиций.
[0008] В одном аспекте настоящее изобретение предусматривает белок, содержащий: первый связывающий домен (BD1), который связывается с первым эпитопом, второй связывающий домен (BD2), который связывается со вторым эпитопом, и Fc-область, содержащую CH2- и CH3-домены; где Fc-область содержит BD2 в гидрофильной петле в CH2-домене, CH3-домене или на границе CH2- и CH3-доменов; и где белок является бивалентным в отношении связывания с каждым из первого и второго эпитопов.
[0009] В другом аспекте настоящее изобретение предусматривает композицию, содержащую белок или антитело в соответствии с любым из аспектов в данном документе и фармацевтически приемлемый носитель.
[0010] В другом аспекте настоящее изобретение предусматривает способ лечения или предупреждения рака у субъекта, способ, включающий введение субъекту (например, субъекту-человеку) белка или антитела в соответствии с любым из аспектов в данном документе. В различных вариантах осуществления рак представляет собой один или несколько из рака яичников, рака молочной железы, колоректального рака, рака предстательной железы, рака шейки матки, рака матки, рака яичек, рака мочевого пузыря, рака головы и шеи, меланомы, рака поджелудочной железы, почечно-клеточной карциномы и рака легких.
[0011] В другом аспекте настоящее изобретение предусматривает способ индукции иммунного ответа у субъекта, при этом способ включает введение субъекту (например, субъекту-человеку) белка или антитела в соответствии с любым из аспектов данного документа.
[0012] В другом аспекте настоящее изобретение предусматривает молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую белок или антитело в соответствии с любым из аспектов данного документа.
[0013] В другом аспекте настоящее изобретение предусматривает вектор, содержащий молекулу нуклеиновой кислоты в соответствии с любым из аспектов данного документа.
[0014] В другом аспекте настоящее изобретение предусматривает клетку-хозяина, содержащую вектор в соответствии с любым из аспектов данного документа.
[0015] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:1, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:2.
[0016] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:3, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:4.
[0017] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:5, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:6.
[0018] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и CTLA-4, содержащий первую тяжелую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 9, первую легкую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 7, вторую тяжелую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 12, и вторую легкую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 4.
[0019] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-L1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:14, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:15.
[0020] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-L1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:16, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:17.
[0021] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-L1 и CTLA-4, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:18, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:19.
[0022] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и TIM3, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:22, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:23.
[0023] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и TIM3, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:24 или SEQ ID NO:91, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:23 или SEQ ID NO: 92.
[0024] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с PD-1 и TIM3, содержащий первую тяжелую цепь, имеющую аминокислотную последовательность под SEQ ID NO:9, первую легкую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 7, вторую тяжелую цепь, имеющую аминокислотную последовательность под SEQ ID NO:27 или SEQ ID NO: 30, и вторую легкую цепь, имеющую аминокислотную последовательность под SEQ ID NO: 26 или SEQ ID NO: 28.
[0025] В одном аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с OX40 и PD-L1, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:34, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:32.
[0026] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с OX40 и PD-L1, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:35, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:32.
[0027] В другом аспекте настоящее изобретение предусматривает биспецифический связывающий белок, который связывается с OX40 и PD-L1, содержащий первый пептид, который содержит аминокислотную последовательность под SEQ ID NO:36 или SEQ ID NO:94, и второй пептид, который содержит аминокислотную последовательность под SEQ ID NO:32 или SEQ ID NO:93.
[0028] В одном аспекте настоящее изобретение предусматривает антитело или его антигенсвязывающий фрагмент, которые связываются с TIM3, содержащие тяжелую цепь, которое содержит CDR1, CDR2 и CDR3, и легкую цепь, которое содержит CDR1, CDR2 и CDR3, где CDR1 тяжелой цепи содержит SEQ ID NO:88, CDR2 тяжелой цепи содержит SEQ ID NO:80, CDR3 тяжелой цепи содержит SEQ ID NO:81, и CDR1 легкой цепи содержит SEQ ID NO:82, CDR2 легкой цепи содержит SEQ ID NO:83, CDR3 легкой цепи содержит SEQ ID NO:84.
[0029] В других аспектах настоящее изобретение предусматривает композицию, содержащую биспецифический связывающий белок и фармацевтически приемлемый носитель; молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, кодирующую биспецифический связывающий белок; способы лечения или предупреждения рака у субъекта путем введения биспецифического связывающего белка; и способы усиления иммунного ответа у субъекта путем введения биспецифического связывающего белка.
[0030] В различных вариантах осуществления любого из аспектов, описанных в данном документе, Fc-область содержит BD2 в гидрофильной петле в аминокислотной последовательности в CH2-домене, в CH3-домене или на границе CH2- и CH3-доменов.
[0031] В различных вариантах осуществления любого из аспектов, описанных в данном документе, гидрофильная петля содержит аминокислотную последовательность из CH2-домена. В конкретных вариантах осуществления гидрофильная петля содержит аминокислотную последовательность ISRTP (SEQ ID NO: 39).
[0032] В различных вариантах осуществления любого из аспектов, описанных в данном документе, гидрофильная петля содержит аминокислотную последовательность из CH3-домена. В конкретных вариантах осуществления гидрофильная петля содержит аминокислотную последовательность SNG.
[0033] В различных вариантах осуществления любого из аспектов, описанных в данном документе, гидрофильная петля содержит аминокислотную последовательность из границы CH2-домена и CH3-домена. В конкретных вариантах осуществления гидрофильная петля в белке по п. 7 содержит аминокислотную последовательность AKGQP (SEQ ID NO: 40).
[0034] В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 представляет собой одноцепочечный вариабельный фрагмент (scFv) или содержит его.
[0035] В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD1 представляет собой или содержит связывающий домен, который представляет собой один или несколько из Fab-домена, scFv, однодоменного антитела и вариабельного домена антитела. В конкретных вариантах осуществления BD1 содержит Fab-домен.
[0036] В различных вариантах осуществления любого из аспектов, описанных в данном документе, Fab-домен связан с Fc-областью посредством шарнирной области антитела. В определенных вариантах осуществления Fc-область представляет собой или включает домен, который представляет собой одну или несколько Fc-областей из IgG1, IgG2, IgG3, IgG4, IgA, IgM, IgE или IgD. В конкретных вариантах осуществления Fc-область содержит вариантную Fc-область. В некоторых вариантах осуществления Fc-область является негликозилированной, дегликозилированной и/или нефукозилированной или характеризуется сниженным фукозилированием.
[0037] В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок дополнительно содержит белковый линкер L1 между BD2 и Fc-областью. В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок дополнительно содержит первый белковый линкер L1 и второй белковый линкер L2 между BD2 и Fc-областью. В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 связан с Fc-областью посредством белкового линкера L1. В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 связан с Fc-областью посредством двух белковых линкеров, L1 и L2. В определенных вариантах осуществления L1 и L2 независимо выбраны из (G4S)2 (SEQ ID NO:41), (G4S)3 (SEQ ID NO:42) и (G4S)4 (SEQ ID NO:43).
[0038] В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок содержит химерную тяжелую цепь, содержащую следующие полипептидные домены от N-конца к С-концу: VH1-CH1-CH2(N-конец)-BD2-CH2(C-конец)-CH3; и BD1 содержит Fab-домен; где VH1 содержит вариабельный домен тяжелой цепи Fab-домена, и CH1 содержит константный домен 1 тяжелой цепи Fab.
[0039] В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок содержит химерную тяжелую цепь, содержащую следующие полипептидные домены от N-конца к С-концу: VH1-CH1-CH2-BD2-CH3; и BD1 содержит Fab-домен; где VH1 содержит вариабельный домен тяжелой цепи Fab-домена и содержит константный домен 1 тяжелой цепи Fab.
[0040] В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок содержит химерную тяжелую цепь, содержащую следующие полипептидные домены от N-конца к С-концу: VH1-CH1-CH2-CH3(N-конец)-BD2-CH3(C-конец); и BD1 содержит Fab-домен; где VH1 содержит вариабельный домен тяжелой цепи Fab-домена, и CH1 содержит константный домен 1 тяжелой цепи Fab.
[0041] В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 представляет собой или содержит scFv. В конкретных вариантах осуществления scFv содержит от N-конца к C-концу: VH2-полипептидный линкер-VL2 или VL2-полипептидный линкер-VH2; где VH2 содержит вариабельный домен тяжелой цепи scFv, и VL2 содержит вариабельный домен легкой цепи scFv.
[0042] В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок дополнительно содержит белковый линкер L1 между BD2 и Fc-областью. В различных вариантах осуществления любого из аспектов, описанных в данном документе, белок дополнительно содержит первый белковый линкер L1 и второй белковый линкер L2 между BD2 и Fc-областью.
[0043] В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 связан посредством линкера (L1) с CH2-доменом, CH2-доменом или границей CH2- и CH3-доменов Fc-области.
[0044] В различных вариантах осуществления любого из аспектов, описанных в данном документе, BD2 связан посредством двух белковых линкеров, L1 и L2, с CH2-доменом, CH3-доменом или границей CH2- и CH3-доменов Fc-области. В различных вариантах осуществления L1 и L2 независимо выбраны из белковых линкеров, имеющих длину 1-25 аминокислот. В конкретных вариантах осуществления L1 и L2 независимо выбраны из (G4S)2 (SEQ ID NO:41), (G4S)3 (SEQ ID NO:42) и (G4S)4 (SEQ ID NO:43).
[0045] В различных вариантах осуществления любого из аспектов, описанных в данном документе, первый и второй эпитопы являются разными. В различных вариантах осуществления любого из аспектов, описанных в данном документе, первый и второй эпитопы являются одинаковыми.
Краткое описание графических материалов
[0046] С целью иллюстрации настоящего изобретения в графических материалах изображены определенные аспекты настоящего изобретения. Тем не менее, настоящее изобретение не ограничено точными схемами и средствами из аспектов, изображенных на графических материалах.
[0047] На фигурах 1A-1F представлено общее схематическое изображение определенных иллюстративных белков, описанных в данном документе. CH2- и CH3-области проиллюстрированы на фигурах 1A-1C с использованием PyMOL, а области гидрофильных петель на поверхности определены в виде сфер. На фигуре 1A изображена петля в CH2-области; на фигуре 1B изображена петля на границе CH2-CH3; и на фигуре 1C изображена петля в CH3-области. Иллюстративные конструкции, которые включают репрезентативные BD1- и BD2-домены в качестве Fab- и scFv-доменов соответственно, проиллюстрированы на фигурах 1D-1F. На фигуре 1D изображен BD1, присоединенный в шарнирной области, и BD2, присоединенный в гидрофильных петлях в CH2-области. На фигуре 1E изображен BD1, присоединенный в шарнирной области, и BD2, присоединенный в гидрофильных петлях на границе CH2-CH3. На фигуре 1F изображен BD1, присоединенный в шарнирной области, и BD2, присоединенный в гидрофильных петлях в CH3-области.
[0048] На фигурах 2A-2C представлен расширенный обзор последовательностей гидрофильных петель в CH2, на границе CH2-CH3 и в CH3, как описано в данном документе. Примеры конструкций, содержащих BD2 (scFv), представлены на каждой из фигур 2A-2C. На фигуре 2A проиллюстрирована типичная последовательность петли ISRTP (SEQ ID NO:39), идентифицированная в петле CH2 выше границы CH2-CH3. На фигуре 2B проиллюстрирована типичная последовательность петли AKGQP (SEQ ID NO:40) на границе CH2-CH3. На фигуре 2C проиллюстрирована типичная последовательность петли SNG в CH3-области ниже границы CH2-CH3.
[0049] На фигуре 3 продемонстрировано одновременное связывание конструкций BiS2, BiS3 и BiS5, которые нацеливаются на PD-1 и CTLA-4. Кривая A9 показывает связывание BiS2 с PD-1/CTLA-4, кривая B9 показывает связывание Bis3 с PD-1/CTLA-4, и кривая C9 показывает связывание Bis5 с PD-1/CTLA-4.
[0050] На фигуре 4 показано схематическое изображение предлагаемого механизма для блокады PD-1/CTLA-4.
[0051] На фигуре 5 показаны результаты анализа связывания с помощью Octet, которые демонстрируют одновременное связывание конструкции DuetMab, которая нацеливается на PD-1 и CTLA-4.
[0052] На фигурах 6A-D показано, что биспецифические связывающие PD-1/CTLA-4 белки ингибируют пути PD-1 и CTLA-4 в анализах с репортерным геном. На фигуре 6A изображена активация Т-клеток посредством блокады PD-1. На фигуре 6B изображена активация Т-клеток посредством блокады CTLA-4. На фигуре 6C показаны результаты анализа PD-1 с репортерным геном. На фигуре 6D показаны результаты анализа CTLA-4 с репортерным геном.
[0053] На фигуре 7 показаны результаты анализа SEB, которые показывают, что DuetMab и BiS5Ab к PD-1/CTLA-4 характеризуются эквивалентной активностью в анализе с применением стафилококкового энтеротоксина B (SEB).
[0054] На фигурах 8A-B показана активность DuetMab к PD-1/CTLA-4 в анализах SEB по сравнению с контролями, представляющими собой изотипические и исходные mAb.
[0055] На фигурах 9A-B показана активность BiS5Ab к PD-1/CTLA-4 по сравнению с DuetMab к PD-1/CTLA-4 в анализах SEB.
[0056] На фигурах 10A-C показано, что DuetMab и BiS5Ab к PD-1/CTLA-4 характеризуются эквивалентной активностью в анализе с использованием реакции смешанных лимфоцитов (MLR). На фигуре 10A представлено схематическое изображение анализа (n=4 донора; 2 независимых эксперимента).
[0057] На фигурах 11A-D показана активность DuetMab к PD-1/CTLA-4 в анализах MLR по сравнению с изотипическими контролями (n=2 донора; 1 эксперимент).
[0058] На фигурах 12A-D показана активность DuetMab к PD-1/CTLA-4 в анализах MLR по сравнению с исходными контролями mAb (n=2 донора; 1 эксперимент).
[0059] фигурах 13A-D показана активность DuetMab к PD-1/CTLA-4 в анализах MLR по сравнению с конкурентными антителами (n=2 донора; 1 эксперимент).
[0060] На фигуре 14 показана схема исследования для изучения фармакокинетики/фармакодинамики (PK/PD) с применением однократной дозы у яванских макаков.
[0061] На фигурах 15A-B показано, что DuetMab к PD-1/CTLA-4 продемонстрировал явно выраженную фармакодинамику (PD) у яванских макаков.
[0062] На фигурах 16A-B показан Т-клеточно-зависимый иммунный ответ (TDAR) у яванских макаков, получавших дозу DuetMab к PD-1/CTLA-4 (MEDI5752) и BiS5Ab к PD-1/CTLA-4 (MEDI8500).
[0063] На фигуре 17 показана модельная система для изучения биспецифических молекул, связывающих PD-1/CTLA-4, в которой стабильные клетки СНО экспрессируют разные уровни PD-1 и/или CTLA-4 человека.
[0064] На фигурах 18A-C показано, что DuetMabPD-1/CTLA-4 одновременно связывает PD-1 и CTLA-4 на поверхности одной и той же клетки.
[0065] На фигурах 19A-C показан эксперимент по определению того, различается ли кооперативное связывание от комбинации антител к PD-1 и антител к CTLA-4 в насыщении CTLA-4 на клетках, экспрессирующих избыток PD-1.
[0066] На фигурах 20A-D показано, что исходные моноклональные антитела к PD-1 и CTLA-4 связывают и занимают свой рецептор-мишень без заметного эффекта в отношении рецептора, который не является мишенью.
[0067] На фигурах 21A-D показано, что DuetMab к PD-1/CTLA-4 обеспечивает насыщение CTLA-4 на клетках CHO, экспрессирующих избыток PD-1, при концентрациях ~250 раз ниже по сравнению с комбинацией моноклональных антител.
[0068] На фигурах 22A-F показано, что DuetMab к PD-1/CTLA-4 обеспечивает насыщение CTLA-4 на клетках CHO, экспрессирующих избыток PD-1, при концентрациях ~500 раз ниже по сравнению с клетками, экспрессирующими только CTLA-4.
[0069] На фигурах 23A-B показано, что DuetMab к PD-1/CTLA-4 преимущественно связывается с PD-1 и CTLA-4 в цис-положении на поверхности одной и той же клетки. Treme на фигуре 23B представляет собой mAb к CTLA-4.
[0070] На фигурах 24A-D показано связывание и интернализация DuetMab к PD-1/CTLA-4 и исходных моноклональных антител культивируемыми Т-клетками. DuetMab к PD-1/CTLA-4 обладает свойствами интернализации тремелимумаба.
[0071] На фигуре 25A показано схематическое изображение анализа интернализации. На фигуре 25B показано, что DuetMab к PD-1/CTLA-4 приобретает свойства интернализации тремелимумаба в стабильных клетках СНО, экспрессирующих 10-кратный избыток PD-1.
[0072] На фигуре 26 продемонстрировано конкурентное связывание конструкций BiS2, BiS3 и BiS5, которые нацеливаются на PD-L1 и CTLA-4. Кривая A11 показывает связывание Bis2 с PD-L1/CTLA-4, кривая B11 показывает связывание Bis3 с PD-L1/CTLA-4, и кривая C11 показывает связывание Bis5 с PD-L1/CTLA-4.
[0073] На фигуре 27A продемонстрировано одновременное связывание конструкции BiS3, которая нацеливается на PD-1 и TIM3 (клон 62 дикого типа). На фигуре 27B продемонстрировано одновременное связывание конструкции DuetMab, которая нацеливается на PD-1 и TIM3 (клон 62 дикого типа).
[0074]
На фигурах 28A-28C представлен краткий обзор активности по уничтожению клеток моноспецифических к TIM3 и биспецифических к PD-1_TIM3PD-1 конструкций в анализе уничтожения клеток. На фигуре 28A показаны изображения, полученные с помощью метода светового поля, совместно культивированных лунок через 18 ч. Комбинация антитело к TIM-3+антитело к PD1 или биспецифические форматы, связывающие TIM-3/PD-1, усиливают гибель опухолевых клеток и повышают активацию Т-клеток, как оценено с помощью снижения количества адгезивных клеток и улучшенного бластирования (агглютинации) Т-клеток. На фигуре 28B показана оценка жизнеспособности по поглощению красителя опухолевыми клетками через 18 ч. совместного культивирования с CD8+ T-клетками, специфичными к меланоме. На фигуре 28C показана секреция IFNγ через 18 ч. совместного культивирования. Биспецифические конструкции, как правило, демонстрируют лучшую активность по уничтожению, причем формат DuetMab демонстрирует самую сильную активность по уничтожению.
[0075] На фигуре 29 продемонстрировано одновременное связывание конструкции DuetMab к PD-1/TIM3, содержащей последовательность плеча TIM3, которая представляет собой вариант клона 62 с созревшей аффинностью.
[0076] На фигуре 30 показано связывание биспецифических антител к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, с клетками CHO, сверхэкспрессирующими TIM3 человека или PD1 человека. Данные по экспрессии PD-1 и TIM3 показаны на вставке.
[0077] На фигуре 31 показаны данные по экспрессии TIM3 и PD-1 на активированном Т-клеточном клоне (DMF4).
[0078] На фигурах 32A-C изображены результаты анализа вторичного иммунного ответа антигена к CMV, показывающие, что биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, демонстрируют повышенную активность по сравнению с обработкой изотипическими антителами (3 донора (1-2 повторности на обработку/на донора), 1 эксперимент).
[0079] На фигурах 33A-D показано, что биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, увеличивали выработку интерферона (IFNγ) при концентрациях выше 8 нМ в анализе реакции смешанных лимфоцитов (MLR) (2-4 лунки для реплицирования на обработку/1 донорную пару/1 из 2 независимых экспериментов).
[0080] На фигурах 34A-C показаны результаты анализа PD-1 с репортерным геном (двухклеточная система) с использованием биспецифических антител к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab. Все биспецифические форматы демонстрируют активность, аналогичную активности исходного IgG1 LO115 (компиляция из 3-5 независимых экспериментов/3 биологических повторностей на обработку).
[0081] На фигуре 35 показаны результаты анализа биспецифических молекул BiS2 и BiS3, связывающих OX40/PD-L1, с применением Octet.
[0082] На фигурах 36A-B показаны результаты анализа SEB с использованием биспецифических молекул BiS2 и BiS3, связывающих OX40/PD-L1.
[0083] На фигуре 37 показаны результаты анализа PD-L1 с репортерным геном с использованием биспецифических молекул BiS2 и BiS3, связывающих OX40/PD-L1.
[0084] На фигуре 38 показаны результаты анализа вторичного иммунного ответа на CMV Ag с использованием биспецифических молекул BiS2 и BiS3, связывающих OX40/PD-L1.
[0085] На фигуре 39 показаны результаты анализа связывания с помощью Octet, которые демонстрируют одновременное связывание конструкции BiS5 к OX40(SLR)/PD-L1, которая нацеливается на PD-L1 и OX40.
[0086] На фигурах 40A-F показано связывание биспецифических конструкций, которые нацеливаются на PD-L1 и OX40, с клетками CHO, экспрессирующими X40 и PD-L1/B7H1 человека и яванского макака.
[0087] На фигуре 41 показано связывание биспецифических конструкций, которые нацеливаются на PD-L1 и OX40, с репортерными клетками Jurkat OX40, клетками NCI H358, CHOK1 B7H1(PD-L1)/OKT3 и клетками HEK CD32a, измеренное с помощью проточной цитометрии (HyperCyt).
[0088] На фигурах 42A-B показаны результаты анализа PD-L1 с репортерным геном с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0089] На фигурах 43A-B показаны результаты анализа OX40 с репортерным геном в клетках HEK CD32a с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0090] На фигурах 44A-B показаны результаты анализа OX40 с репортерным геном в сверхэкспрессирующих PD-L1 клетках CHOK с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0091] На фигурах 45 показан опосредованный PD-L1 агонизм OX40 с помощью опухолевых клеток с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0092] На фигурах 46A-D показаны результаты, отмечающие отсутствие агонизма, которое обнаружено с клетками NCI H358 PD-L1 KO с использованием биспецифических молекул, связывающих OX40/PD-L1, в контролях для анализа OX40/опухолевых клеток.
[0093] На фигурах 47A-D показаны результаты анализа SEB с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0094] На фигуре 48A показано схематическое изображение эксперимента по анализу супрессии Treg для тестирования биспецифических молекул, связывающих OX40/PD-L1. На фигурах 48B-C показана супрессия Treg на основе связывания биспецифической молекулы.
[0095] На фигуре 49 показаны результаты анализа супрессии Treg с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0096] На фигуре 50 показаны результаты анализа супрессии Treg с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0097] На фигурах 51A-B показана схема эксперимента с помощью анализа реакции смешанных лейкоцитов (MLR) для тестирования биспецифических молекул, связывающих OX40/PD-L1.
[0098] На фигурах 52A-E показаны результаты MLR-анализа с использованием биспецифических молекул, связывающих OX40/PD-L1.
[0099] На фигуре 53 показано, что биспецифические молекулы BiS2 и BiS5, связывающие OX40/PD-L1, опосредуют антителозависимую клеточно-опосредованную цитотоксичность (ADCC) природных клеток-киллеров (NK) в отношении клеток СНО, экспрессирующих PD-L1 или OX40.
[00100] На фигуре 54 показано, что биспецифические молекулы BiS2 и BiS5, связывающие OX40/PD-L1, опосредуют антителозависимую клеточно-опосредованную цитотоксичность (ADCC) NK-клеток в отношении клеток СНО, экспрессирующих PD-L1 или OX40.
[00101] На фигуре 55 показано, что биспецифические молекулы BiS2 и BiS5, связывающие OX40/PD-L1, увеличивают мобилизацию CD107a NK-клеток в отношении клеток СНО, экспрессирующих PD-L1 или OX40, при антителозависимой клеточно-опосредованной цитотоксичности (ADCC).
[00102] На фигуре 56 показано, что биспецифические молекулы BiS2 и BiS5, связывающие OX40/PD-L1, опосредуют антителозависимую клеточно-опосредованную цитотоксичность (ADCC) NK-клеток в отношении активированных аллогенных Т-клеток.
[00103] На фигурах 57A-B показано, что BiS5 к OX40/PD-L1 увеличивает мобилизацию CD107a NK-клеток от двух разных доноров в отношении активированных аллогенных Т-клеток при антителозависимой клеточно-опосредованной цитотоксичности (ADCC).
[00104] На фигуре 58 показана схема исследования для сравнения PK/PD биспецифических молекул, связывающих OX40/PD-L1.
[00105] На фигуре 59 показано сравнение профилей концентрации-времени в сыворотке крови для биспецифических молекул, связывающих PD-L1/OX40, у яванских макаков.
[00106] На фигуре 60 показано снижение содержания растворимого PD-L1 в сыворотке крови посредством биспецифических молекул, связывающих PD-L1/OX40.
[00107] На фигурах 61A-F представлен краткий обзор фармакодинамических данных для биспецифических молекул, связывающих PD-L1/OX40. Исходный уровень определен как среднее значение предварительной дозы в день 5 и день 0.
[00108] На фигуре 62 продемонстрировано схематическое изображение моноклонального антитела (mAb) IgG4P BiS2 к PD-1/OX40.
[00109] На фигуре 63 изображен потенциальный механизм действия mAb BiS2 к PD-1/OX40.
[00110] На фигуре 64 изображена активность одновременного связывания для двух разных партий mAb BiS2 к PD-1(LO115)/OX40 с PD1-His и OX40-Fc человека.
[00111] На фигуре 65A продемонстрировано схематическое изображение анализа OX40 с репортерным геном. На фигуре 65B показаны результаты анализа OX40 с репортерным геном с использованием mAb к PD1 LO115, mAb к OX40, контрольного mAb и mAb BiS2 к PD1(LO115)/OX40.
[00112] На фигуре 66A представлено схематическое изображение анализа PD-1/PD-L1 с репортерным геном. На фигуре 66B показаны результаты анализа PD1/PD-L1 с репортерным геном с использованием mAb к PD1 LO115, mAb к OX40, контрольного mAb и mAb BiS2 к PD1(LO115)/OX40.
[00113] На фигуре 67 показаны результаты анализа SEB с использованием варианта BiS2 биспецифической молекулы, связывающей PD-1(LO115)/OX40, и контролей.
[00114] На фигуре 68 показаны результаты анализа вторичного иммунного ответа на антиген CMV с использованием варианта BiS2 биспецифической молекулы, связывающей PD-1(LO115)/OX40, и контролей.
[00115] На фигуре 69 показаны результаты анализа вторичного иммунного ответа на антиген CMV с использованием вариантов BiS2 и BiS3 биспецифических молекул, связывающих PD-1(AMP514)/OX40, и контролей.
[00116] На фигуре 70 показаны профили концентрации-времени mAb BiS2 к PD1(LO115)/OX40 в сыворотке крови после однократной IV дозы у яванских макаков. Данные представляют среднее ± стандартное отклонение для 3 самцов/группа. LLOQ (5 нг/мл) показан пунктирной линией. PKC=фармакокинетическая концентрация; LLOQ=нижний предел количественного определения.
[00117] На фигуре 71 показано процентное содержание Ki67-положительных CD4+ и CD8+ Т-клеток памяти после однократного IV введения mAb BiS2 к PD-1(LO115)/OX40 у яванских макаков. Данные представляют среднее ± стандартное отклонение 3 самца/группа. На левой панели A представлены CD4+ Т-клетки памяти, а на правой панели показаны CD8+ Т-клетки памяти. IV=внутривенный.
[00118] На фигуре 72 показана типичная стандартная кривая количественного определения PD-1/OX40 в сыворотке крови яванских макаков.
[00119] На фигурах 73A-73E представлены иллюстративные термограммы DSC биспецифического связывающего белка ("BiS5"), раскрытого в данном документе, относительно другого формата BiS ("BiS4") при различных значениях рН. На фигуре 73A проиллюстрирован эффект pH в отношении термической стабильности BiS4. На фигуре 73B проиллюстрирован эффект pH в отношении термической стабильности BiS5. На фигуре 73C изображена типичная криволинейная термограмма DSC для BiS4 с Tonset, Tm1, Tm2 и Tm3. На фигуре 73D изображены типичные криволинейные термограммы DSC для BiS5 с Tonset, Tm1, Tm2 и Tm3. На фигуре 73E изображен график, представляющий эффект рН в отношении Tonset, Tm1, Tm2 и Tm3 для форматов BiS4 и BiS5.
[00120] На фигурах 74A и 74B изображен анализ HP-SEC образцов при pH 7,5 до и после хранения при 40°C в течение 4 недель. На фигуре 74A представлено наложение графиков хроматограмм SEC для BiS4 и BiS5 до и после термического стресса, при этом сплошные линии соответствуют образцам BiS4 и BiS5 в момент времени ноль (без стресса), а пунктирные линии соответствуют образцам BiS4 и BiS5, инкубированным при 40°C в течение 4 недель (при стрессе). На фигуре 74B представлена гистограмма, показывающая эффект pH 7,5 в отношении различных разновидностей (мономер, фрагменты и агрегаты) BiS4 и BiS5, измеренный в нулевой день и через 4 недели при 40°C.
[00121] На фигурах 75A-75C представлены кинетические графики, показывающие эффект pH 7,5 в отношении стабильности при ускоренном и кратковременном хранении при 40°C для BiS4 (кривая с треугольниками) и BiS5 (кривая с кружками). На фигуре 75A показано процентное содержание оставшегося мономера, измеренное с помощью HP-SEC, за период времени, составляющий 4 недели. На фигуре 75B показано процентное содержание фрагментированных белков, измеренное с помощью HP-SEC, за период времени, составляющий 4 недели. На фигуре 75C показано процентное содержание агрегированных белков, измеренное с помощью HP-SEC, за период времени, составляющий 4 недели. Представленные данные получены помощью анализа в одиночных флаконах.
[00122] На фигурах 76A-76C изображены графики профиля скорости изменения параметров в зависимости от рН для BiS4 (треугольники) и BiS5 (кружки). На фигуре 76A показан эффект различных условий рН на скорость потери мономеров при 40°C. На фигуре 76B показан эффект различных условий рН на скорость фрагментации при 40°C. На фигуре 76C показан эффект различных условий рН на скорость агрегации при 40°C.
[00123] На фигурах 77A и 77B изображен дополнительный анализ фрагментации BiS4 и BiS5. На фигуре 77A показано, что при pH 7,5 и 40°C (в момент времени=0) ни одна из молекул не проявляет заметной фрагментации. На фигуре 77B показано, что при тех же условиях, что и на фигуре 77A, но после хранения в течение 2 недель при 40°C, наблюдается заметная фрагментация для BiS4 и минимальная фрагментация для BiS5
[00124] На фигуре 78 изображен анализ фрагментации (левая панель) и агрегации (правая панель) BiS4 и BiS5 в зависимости от рН. Оба формата характеризуются уменьшенной фрагментацией и агрегацией при более низком значении рН (5,5), тогда как BiS5 обладает превосходными характеристиками при обоих значениях рН как для фрагментации, так и для агрегации.
[00125] На фигуре 79 изображены термограммы DSC для нескольких конструкций биспецифического связывающего белка BiS5, раскрытого в данном документе: (левая панель) A, B, C и D, а также (правая панель) E, F, G и H. Конструкции A и E содержат scFv в IS-RTP; B и F содержат scFv в AK-GQP; C и G содержат scFv в S-NG; и D и H содержат scFv в SN-G. У конструкций A, B, C и D scFv представляет собой 2F4 (IgG представляет собой LC10), тогда как у E, F, G и H scFv представляет собой LC10 (IgG представляет собой 2F4). Различные значения TM связаны со следующими доменами: TM1=CH2/scFv; TM2=Fab; TM3=CH3.
[00126] На фигуре 80 изображен типичный набор данных для связывания FcRn с биспецифическим связывающим белком, раскрытым в данном документе (конструкции D и H). Расположение scFv в домене CH2-CH3 (т. е внутри петли ISTRP) может характеризоваться эффектом на активность связывания FcRn.
[00127] На фигуре 81 изображен типичный набор данных для связывания FcγR с биспецифическим связывающим белком, раскрытым в данном документе (конструкции E, G и H с FcγRIIIa-158V). На вставке отражены те же данные, что и на основной фигуре, перенормированные при инъекции FcγRIIIa-158V. Все конструкции были способны связывать FcγR с наблюдаемой разницей в значениях аффинности, основанной на местоположении доменов scFv.
[00128] На фигуре 82 показано, что интактная петля ISRTP важна для связывания FcRn иллюстративных биспецифических связывающих конструкций (например, A и E). Введение петли N3 не компенсирует нарушения петли ISRTP (BiS5E+N3). Введение scFv в петлю N3, вставленную в Fc IgG1, делает IgG неспособным связывать FcRn. Время на оси х измеряется в секундах.
[00129] На фигуре 83 изображено общее схематическое изображение структурного формата каждой из конструкций BiS1, BiS2, BiS3, BiS4 и BiS5. Обозначения "k1" и "k2" указывают на паттерны фрагментации, применяемые в кинетическом анализе, обсуждаемом в примере 3.
[00130] На фигуре 84 изображена скорость фрагментации каждого из BiS1, BiS2, BiS3, BiS4 и BiS5 в зависимости от pH.
[00131] На фигуре 85 изображена скорость агрегации каждого из BiS1, BiS2, BiS3, BiS4 и BiS5 в зависимости от pH.
[00132] На фигуре 86 изображена скорость потери мономеров каждого из BiS1, BiS2, BiS3, BiS4 и BiS5 в зависимости от pH.
[00133] На фигуре 87 представлено изображение паттерна фрагментации и соответствие пикам на хроматограммах HPSEC для каждого из BiS1, BiS2, BiS3, BiS4 и BiS5.
[00134] На фигуре 88 изображен типичный анализ паттерна фрагментации в восстанавливающих условиях.
[00135] На фигуре 89 изображена структурная схема BiS5 в восстанавливающих и невосстанавливающих условиях.
[00136] На фигуре 90 изображен формат анализа SEB.
Подробное описание изобретения
[00137] Прежде чем продолжить описание настоящего изобретения более подробно, следует понимать, что настоящее изобретение не ограничивается конкретными композициями или стадиями способа, поскольку они могут варьироваться. Следует отметить, что используемые в настоящем описании и прилагаемой формуле изобретения формы единственного числа включают определяемые объекты во множественном числе, если из контекста явно не следует иное.
[00138] Если не определено иное, то все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается средним специалистом в области техники, к которой относится настоящее изобретение. Например, the Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; the Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press и Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press, обеспечивают специалиста общим словарем многих терминов, используемых в настоящем изобретении.
[00139] Аминокислоты могут называться в данном документе либо по их общеизвестным трехбуквенным символам, либо по однобуквенным символам, рекомендованным комиссией по биохимической номенклатуре IUPAC-IUB. Аналогично, нуклеотиды могут называться по их общепринятым однобуквенным кодам.
[00140] При нумерации аминокислот в вариабельном домене определяющие комплементарность области (CDR) и каркасные области (FR) антитела следуют, если не указано иное, определению согласно Kabat, как изложено в документе Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Бетезда, Мэриленд. (1991). При использовании данной системы нумерации фактическая линейная аминокислотная последовательность может содержать меньшее количество аминокислот или дополнительные аминокислоты, соответствующие укорачиванию FR или CDR вариабельного домена или вставке в них. Например, вариабельный домен тяжелой цепи может включать в себя вставку одной аминокислоты (остаток 52a согласно Kabat) после остатка 52 в H2 и вставленные остатки (например, остатки 82a, 82b и 82c и т. д., согласно Kabat) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Kabat можно определить для данного антитела путем выравнивания последовательности антитела в областях гомологии со "стандартной" последовательностью с нумерацией по Kabat. Максимальное выравнивание остатков каркасных участков зачастую требует вставки "спейсерных" остатков в систему нумерации, чтобы использовать ее для Fv-области. Кроме того, идентичность определенных отдельных остатков в любом данном участке с нумерацией согласно Kabat может варьироваться от цепи антитела к цепи антитела из-за межвидовой или аллельной дивергенции.
[00141] Термины "антитело" и "антитела", используемые в данном документе, также известные как иммуноглобулины, охватывают моноклональные антитела (в том числе полноразмерные моноклональные антитела), поликлональные антитела, мультиспецифические антитела, образованные по меньшей мере из двух разных эпитопсвязывающих фрагментов (например, мультиспецифические антитела, например, PCT-публикация WO 2009/018386, PCT-заявка № PCT/US2012/045229, включенные в данный документ посредством ссылки во всей их полноте), biMab, антитела человека, гуманизированные антитела, верблюжьи антитела, одноцепочечные Fv (scFv), одноцепочечные антитела, однодоменные антитела, доменные антитела, Fab-фрагменты, фрагменты F(аb')2, фрагменты антител, которые проявляют требуемую биологическую активность (например антигенсвязывающая часть), дисульфидно-связанные Fv (dsFv) и антиидиотипические (anti-Id) антитела (в том числе, например, антитела anti-Id к антителам по настоящему изобретению), интратела и эпитопсвязывающие фрагменты любого из вышеупомянутых. В частности, антитела включают в себя молекулы иммуноглобулинов и иммунологически активные фрагменты молекул иммуноглобулинов, т. е. молекулы, которые содержат по меньшей мере один антигенсвязывающий участок. Антитела также включают в себя слитые пептиды с антителами или их частями, как например белок, слитый с Fc-доменом. Молекулы иммуноглобулинов могут быть любого изотипа (например, IgG, IgE, IgM, IgD, IgA и IgY), субизотипа (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или аллотипа (например, Gm, например, G1m(f, z или x), G2m(n), G3m(g, b или c), Am, Em, и Km(1, 2 или 3)). Антитела могут быть получены от любого млекопитающего, в том числе без ограничения людей, обезьян, свиней, лошадей, кроликов, собак, котов, мышей и т. д., или других животных, таких как птицы (например, куры).
[00142] CTLA-4
[00143] Цитотоксический Т-лимфоцит-ассоциированный белок 4 (CTLA-4) экспрессируется на активированных Т-клетках и служит в качестве коингибитора для удерживания Т-клеточных ответов под контролем после CD28-опосредованной активации Т-клеток. Полагают, что CTLA-4 регулирует амплитуду ранней активации не подвергнутых влиянию Т-клеток и Т-клеток памяти после вовлечения TCR и является частью центрального ингибирующего пути, который оказывает влияние как на противоопухолевый иммунитет, так и на аутоиммунитет. CTLA-4 экспрессируется исключительно на Т-клетках, и экспрессия его лигандов CD80 (B7.1) и CD86 (B7.2) в значительной степени ограничивается антигенпрезентирующими клетками, Т-клетками и другими клетками, опосредующими иммунитет. Сообщалось о том, что антагонистические антитела к CTLA-4, которые блокируют сигнальный путь CTLA-4, усиливают активацию Т-клеток. Одно такое антитело, ипилимумаб, было одобрено FDA в 2011 году для лечения метастатической меланомы. Было предложено использование антител к CTLA-4 для лечения инфекций и опухолей и повышения модуляции адаптивного иммунного ответа (см., патенты США №№ 6682736; 7109003; 7132281; 7411057; 7824679; 8143379 7807797; 8491895; 8883984 и публикацию патента США № 20150104409, включенные в данный документ посредством ссылки во всей их полноте).
[00144] PD-L1
[00145] Лиганд 1 запрограммированной смерти клетки (PD-L1) также является частью комплексной системы рецепторов и лигандов, которые вовлечены в осуществление контроля активации T-клеток. В нормальной ткани PD-L1 экспрессируется на T-клетках, B-клетках, дендритных клетках, макрофагах, мезенхимальных стволовых клетках, тучных клетках, происходящих из костного мозга, а также на различных клетках, не являющихся гемопоэтическими. Его нормальной функцией является регуляция баланса между активацией и толерантностью T-клеток посредством взаимодействия с его двумя рецепторами: белком 1 программируемой смерти клеток (также известным как PD-1 или CD279) и CD80 (также известным как B7-1 или B7.1). PD-L1 также экспрессируется опухолями и действует во множестве участков, помогая опухолям избегать выявления и устранения иммунной системой хозяина. PD-L1 с высокой частотой экспрессируется при широком спектре форм рака. При некоторых формах рака экспрессия PD-L1 была ассоциирована с уменьшенной выживаемостью и неблагоприятным прогнозом. Антитела, которые блокируют взаимодействие между PD-L1 и его рецепторами, способны к ослаблению PD-L1-зависимых иммуносупрессивных эффектов и к усилению цитотоксической активности противоопухолевых T-клеток in vitro. Дурвалумаб представляет собой моноклональное антитело человека, направленное против PD-L1 человека, которое способно блокировать связывание PD-L1 с обоими рецепторами PD-1 и CD80. Было предложено использование антител к PD-L1 для лечения инфекций и опухолей и усиления адаптивного иммунного ответа (см., патенты США №№ 8779108 и 9493565, включенные в данный документ посредством ссылки во всей их полноте).
[00146] PD-1
[00147] Белок-1 запрограммированной смерти клетки ("PD-1") представляет собой представителя мембранного белка I типа размером примерно 31 кДа из обширного семейства CD28/CTLA-4 регуляторов T-клеток (см. Ishida, Y. et al. (1992) Induced Expression Of PD-1, A Novel Member Of The Immunoglobulin Gene Superfamily, Upon Programmed Cell Death," EMBO J. 11:3887-3895.
[00148] PD-1 экспрессируется на активированных Т-клетках, В-клетках и моноцитах (Agata, Y. et al. (1996) "Expression of the PD-1 Antigen on the Surface of Stimulated Mouse T and B Lymphocytes," Int. Immunol. 8(5):765-772; Martin-Orozco, N. et al. (2007) "Inhibitory Costimulation and Anti-Tumor Immunity," Semin. Cancer Biol. 17(4):288-298). PD-1 представляет собой рецептор, ответственный за понижающую регуляцию иммунной системы после активации с помощью связывания PDL-1 или PDL-2 (Martin-Orozco, N. et al. (2007) "Inhibitory Costimulation and Anti-Tumor Immunity," Semin. Cancer Biol. 17(4):288-298) и который функционирует как индуктор гибели клеток (Ishida, Y. et al. (1992) "Induced Expression of PD-1, A Novel Member of The Immunoglobulin Gene Superfamily, Upon Programmed Cell Death," EMBO J. 11: 3887- 3895; Subudhi, S.K. et al. (2005) "The Balance of Immune Responses: Costimulation Verse Coinhibition," J. Molec. Med. 83: 193-202). Этот процесс используется во многих опухолях посредством сверхэкспрессии PD-L1, что приводит к ослабленному иммунному ответу.
[00149] PD-1 является хорошо подтвержденной мишенью для иммуноопосредованной терапии в онкологии, с положительными результатами клинических испытаний при лечении, среди прочего, меланомы и форм немелкоклеточного рака легкого (NSCLC). Антагонистическое ингибирование взаимодействия PD-1/PD-L-1 усиливает активацию Т-клеток, увеличивая распознавание и элиминацию опухолевых клеток иммунной системой хозяина. Было предложено использование антител к PD-L1 для лечения инфекций и опухолей и усиления адаптивного иммунного ответа (см. патенты США №№ 7521051; 7563869; 7595048).
[00150] OX40
[00151] OX40 (CD134; TNFRSF4) представляет собой рецептор фактора некроза опухолей, встречающийся преимущественно на активированных CD4+ и CD8+ T-клетках, регуляторных T-клетках (Treg) и природных клетках-киллерах (NK) (Croft et al., 2009, Immunol Rev. 229:173-91). OX40 содержит один известный эндогенный лиганд, лиганд OX40 (OX40L; CD152; TNFSF4), который существует в тримерной форме и может вызывать кластеризацию OX40, что приводит к эффективным явлениям клеточной передачи сигнала в Т-клетках. (там же). Передача сигналов с помощью OX40 на активированных CD4+ и CD8+ T-клетках приводит к повышенной выработке цитокинов, высвобождению гранзимов и перфоринов и экспансии популяций эффекторных Т-клеток и Т-клеток памяти (Jensen et al., 2010, Semin Oncol. 37:524-32). Кроме того, передача сигнала от OX40 на клетках Treg ингибирует размножение Treg, останавливает индукцию Treg и блокирует супрессорную функцию Treg (Voo et al., 2013, J Immunol. 191:3641-50; Vu et al., 2007, Blood. 110:2501-10).
[00152] В иммуногистохимических исследованиях и ранних анализах с применением проточной цитометрии было показано, что OX40 экспрессируется на T-клетках, инфильтрирующих широкий спектр форм рака человека (Baruah et al., 2011, Immunobiology 217:668-675; Curti et al, 2013, Cancer Res. 73:7189-98; Ladanyi et al, 2004, Clin Cancer Res. 10:521-30; Petty et al, 2002, Am J Surg. 183:512-8; Ramstad et al, 2000, Am J Surg. 179:400-6; Sarff et al, 2008, Am J Surg. 195:621-5; обсуждение 625; Vetto et al, 1997, Am J Surg. 174:258-65). Без привязки к какой-либо конкретной теории, экспрессия OX40 на лимфоцитах, инфильтрирующих опухоли, коррелирует с более длительной выживаемостью при некоторых формах рака человека, что указывает на то, что сигналы от OX40 могут играть роль в формировании противоопухолевого иммунного ответа (Ladanyi et al., 2004, Clin Cancer Res. 10:521-30; Petty et al., 2002, Am J Surg. 183:512-8).
[00153] В ряде неклинических мышиных моделей опухолей агонисты OX40, в том числе антитела и слитые белки, представляющие собой лиганды OX40, успешно применялись с обнадеживающими результатами (Kjaergaard et al., 2000, Cancer Res. 60:5514-21; Ndhlovu et al., 2001, J Immunol. 167:2991-9; Weinberg et al., 2000, J Immunol. 164:2160-9). Совместная стимуляция Т-клеток посредством OX40 содействовала противоопухолевой активности, которая в некоторых случаях была длительной, обеспечивая продолжительную защиту в отношении последующего введения опухоли (Weinberg et al., 2000, J Immunol. 164:2160-9). Было показано, что подавление Treg-клеток и совместная стимуляция эффекторных T-клеток необходимы для подавления роста опухоли агонистами OX40 (Piconese et al., 2008, J Exp Med. 205:825-39). Было изучено множество стратегий и технологий усиления противоопухолевого эффекта терапии агонистами OX40 посредством комбинаций с вакцинами, химиотерапией, лучевой терапией и иммунотерапией (Jensen et al., 2010, Semin Oncol. 37:524-32; Melero et al., 2013, Clin Cancer Res. 19:997-1008). Было предложено использование антител к OX40 для лечения инфекций и опухолей и повышающей модуляции адаптивного иммунного ответа (см. публикацию патента США № 2016/0137740, включенную в данный документ посредством ссылки во всей ее полноте).
[00154] TIM3
[00155] T-клеточный ингибиторный рецептор Tim-3 (белок 3, содержащий домены Т-клеточного иммуноглобулина и муцина) играет роль в осуществлении регуляции противоопухолевого иммунитета, поскольку он экспрессируется на продуцирующих IFN-гамма CD4+ Т-хелперах 1 (Th1) и CD8+ T-цитотоксических 1 (Tc1) T-клетках. Первоначально он был идентифицирован как T-клеточный ингибиторный рецептор, действующий как рецептор иммунной контрольной точки, который функционирует специфически для ограничения продолжительности и величины Т-клеточных ответов Th1 и Tc1. Дальнейшее исследование показало, что путь Tim-3 может взаимодействовать с путем PD-1, чтобы способствовать развитию тяжелого дисфункционального фенотипа в CD8+ Т-клетках при раке. Он также был экспрессирован на регуляторных Т-клетках (Treg) при определенных формах рака. Принимая во внимание участие пути TIM3 в популяциях ключевых иммунных клеток, которые являются иммуносупрессироваными при некоторых формах рака, он представляет собой привлекательного кандидата для иммуноонкологической терапии. См. Anderson, A.C., Cancer Immunol Res., (2014) 2:393-398; и Ferris, R.L., et al., J Immunol. (2014) 193:1525-1530.
A. Биспецифические связывающие белки
[00156] Добавление нескольких участков связывания к молекуле, характеризующейся специфичностью одного связывающего домена, может значительно улучшить возможности (например, терапевтические, диагностические и т.д.) молекулы. Например, биспецифическое антитело может связываться с более чем одной областью одной и той же биомолекулы-мишени, обеспечивая большую специфичность, чем у моноспецифического полипептида, который связывается только с одним эпитопом на мишени. В качестве альтернативы биспецифическое антитело может связываться с несколькими биомолекулами-мишенями, такими как мишени, которые присутствуют в комплексе, или мишени, для которых требуется секвестрация и/или кластеризация. В третьем варианте одно и то же биспецифическое антитело может выполнять различные функции в любой момент времени в зависимости от локализации и/или экспрессии его молекул-мишеней.
[00157] В данном документе описаны новые связывающие белки. Одна такая конфигурация данных новых связывающих белков называется "DuetMab". DuetMab характеризуется следующей основной структурой: Fc-область, содержащая модифицированную тяжелую цепь, где CH1-область модифицированной тяжелой цепи содержит замену нативного цистеина на аминокислоту, отличную от цистеина, и замену нативной аминокислоты, отличной от цистеина, на аминокислоту цистеин; модифицированную соответствующую легкую цепь, где CL-область модифицированной легкой цепи также содержит замену нативного цистеина на аминокислоту, отличную от цистеина, и замену нативной аминокислоты, отличной от цистеина, на аминокислоту цистеин; вторая Fc-область, содержащая вторую тяжелую цепь; и вторую соответствующую модифицированную легкую цепь, где модифицированная тяжелая цепь непосредственно связана с соответствующей модифицированной легкой цепью и на отдельном связывающем мишень плече вторая тяжелая цепь непосредственно связана со второй соответствующей легкой цепью, и где замененный цистеин модифицированной тяжелой цепи, полученный в результате замены нативной аминокислоты, отличной от цистеина, на аминокислоту цистеин и замененный цистеин соответствующей модифицированной легкой цепи, полученный в результате замены нативной аминокислоты, отличной от цистеина, на аминокислоту цистеин, могут образовывать дисульфидную связь. Раскрытие, относящееся к DuetMab, можно найти, например, в патенте США № 9527927, включенном в данный документ посредством ссылки во всей его полноте.
[00158] Дополнительные иллюстративные конфигурации данных новых связывающих белков называются "BiSAb". В данном документе представлены схематические изображения иллюстративных BiSAb, а также конкретные примеры определенных BiSAb. В более общем случае BiSAb представляет собой полипептид, содержащий две связывающие единицы, каждая из которых связывается с эпитопом (например, связывающая единица 1 связывается с первым эпитопом, и связывающая единица 2 связывается со вторым эпитопом). Исходный BiSAb является бивалентным в отношении связывания с каждым из двух эпитопов (например, полипептид содержит две связывающие единицы 1 ("BD1" или "BU1") и две связывающие единицы 2 ("BD2" или "BU2")). Таким образом, если связывающие единицы 1 и 2 связывают разные эпитопы, то BiSAb характеризуется мультиспецифичностью обычного биспецифического антитела и бивалентностью обычной молекулы антитела. В вариантах осуществления, в которых связывающие единицы 1 и 2 связывают один и тот же эпитоп, то BiSAb характеризуется моноспецифичностью обычного антитела, однако является четырехвалентным. В дополнение к связывающим единицам BiSAb также содержат линкерные полипептиды и Fc-часть. Настоящее изобретение относится к широкому набору биспецифических связывающих белков, таких как BiSAb и белки, содержащие ядро BiSAb, которые нацеливаются на молекулы, модулирующие иммунный ответ. Как правило, платформы на основе новых связывающих белков и иллюстративные биспецифические связывающие белки (BiSAb), описанные в данном документе, содержат связывающие единицы/домены, линкерные полипептиды и Fc-часть. Настоящее изобретение также предусматривает молекулы нуклеиновой кислоты, кодирующие такие BiSAb, а также векторы и клетки-хозяева, которые содержат такие нуклеиновые кислоты и которые могут быть использованы в способах получения и применения таких BiSAb. BiSAb, связывающие белки, содержащие ядро BiSAb, и различные части BiSAb более подробно описаны в данном документе.
[00159] В некоторых аспектах BiSAb может содержать две пары тяжелая-легкая цепь, полученные из специфического связывающего белка (т.е. антитела), где каждая из тяжелых и легких цепей содержит вариабельную область (например, VL и VH), которые вместе образуют первую связывающую единицу, и где каждая из тяжелых цепей дополнительно содержит вторую связывающую единицу (например, домен scFv, присоединенный к Fc или Fab). Если первая и вторая связывающие единицы связывают разные эпитопы, то каждая пара тяжелая-легкая цепь является биспецифической, а две пары вместе являются бивалентными для каждого эпитопа. Если первая и вторая связывающие единицы связывают один и тот же эпитоп, то каждая пара тяжелая-легкая цепь является моноспецифической, а две пары вместе являются четырехвалентными для эпитопа. В некоторых аспектах две пары тяжелая-легкая цепь являются одинаковыми. В некоторых аспектах две пары тяжелая-легкая цепь не одинаковы.
[00160] В конкретных вариантах осуществления домены BiSAb могут быть основаны на известных доменах иммуноглобулинов. Молекулы иммуноглобулинов, такие как моноклональные антитела (mAb), широко применяются в качестве диагностических и терапевтических средств, и способы разработки связывающих фрагментов mAb хорошо известны из уровня техники. Моноклональные антитела, как и все молекулы иммуноглобулинов, состоят из пептидных субъединиц тяжелой цепи и легкой цепи, каждая из которых содержит вариабельный и константный домены, которые придают специфичность связывания (вариабельный домен) и изотип (константный домен).
[00161] Раскрытые в данном документе BiSAb могут характеризоваться общей структурой, сходной с обычным антителом, однако отличаются наличием дополнительной связывающей единицы, которая присоединена в участке, расположенном в пределах Fab-домена, присоединена в участке вне Fab-домена и в пределах шарнирной или Fc-областей (например, в пределах CH2-, CH3- или CH4-областей или на границе таких областей, как, например, на границе CH2-CH3). Таким образом, в отличие от обычных антител, которые являются бивалентными в отношении связывания с одним эпитопом, BiSAb являются бивалентными в отношении связывания с двумя эпитопами. Однако, как описано в данном документе, BiSAb все еще могут сохранять многочисленные требуемые свойства обычных антител, такие как способность связывать FcRn и способность связывать рецепторы C1q и Fcγ (например, характерную способность опосредовать антителозависимую и комплементзависимую цитотоксичность).
[00162] Описанные в данном документе связывающие домены могут содержать антигенсвязывающие фрагменты, которые содержат только части молекулы mAb, такие как фрагменты Fab, F(ab')2, Fab', scFv, di-scFv, sdAb, поскольку эти фрагменты нашли применение в качестве диагностических или терапевтических средств. Кроме того, специфические остатки в вариабельных доменах могут быть изменены для улучшения специфичности связывания и/или стабильности антител и фрагментов антител. Другие остатки, не вовлеченные непосредственно в связывание антигена, могут быть заменены с целью "гуманизации" областей антител, не являющихся человеческими, и снижения иммуногенности mAb.
[00163] Хотя BiSAb отличаются от обычных антител, например, они являются бивалентными в отношении связывания с двумя разными эпитопами (или четырехвалентными в отношении связывания с одним эпитопом), многие из частей BiSAb аналогичны частям обычных антител или получены из них. В описанных в данном документе BiSAb могут применяются любые домены и/или фрагменты mAb, известные из уровня техники. В частности, BiSAb может содержать фрагменты Fab и/или scFv или их варианты. Иллюстративные неограничивающие варианты scFv включают без ограничения тандемные di-scFv, тандемные три-scFv, диатела и три(a)тела.
[00164] Настоящее изобретение в целом относится к новым связывающим белкам, иллюстративным примером которых являются BiSAb. Дополнительными примерами являются связывающие белки, содержащие ядро BiSAb, а также одну или несколько дополнительных связывающих единиц, и/или связывающие белки, содержащие увеличенное ядро BiSAb. Следует понимать, что всякий раз, когда в данном документе описаны BiSAb или признаки BiSAb, такое описание обычно относится к новым связывающим белкам по настоящему изобретению независимо от того, содержат ли такие связывающие белки две связывающие единицы или более двух связывающих единиц. Соответственно, термин BiSAb является иллюстративным для связывающих белков, описанных в данном документе, и, если позволяет контекст, любая такая ссылка на BiSAb также может использоваться для описания связывающих белков, содержащих ядро BiSAb.
[00165] Новая структурная платформа на основе BiSAb.
[00166] В одном аспекте настоящее изобретение предусматривает связывающие белки BiSAb, которые характеризуются структурной платформой, содержащей домены, которые в целом проиллюстрированы с помощью схематических изображений на фигурах 1A-1F. Данные изображения являются иллюстративными, и, таким образом, вставка между дополнительными остатками также находится в пределах объема раскрытых связывающих белков. На фигурах 1A-1C изображена Fc-область антитела на границе CH2-CH3 IgG1, которая смоделирована с использованием PyMOL и иллюстрирует несколько служащих в качестве примера BiSAb по настоящему изобретению. Были идентифицированы три гидрофильные петли на границе CH2-CH3 или вблизи нее, которые, вероятно, были способны препятствовать вставке второго связывающего фрагмента (например, scFv) без ущерба для структурной целостности или стабильности IgG или второго связывающего фрагмента. На фигуре 1A представлено схематическое изображение одной из таких типичных петель ISRTP (SEQ ID NO:39), идентифицированной в CH2-области около границы CH2-CH3. На фигуре 1D также показана типичная конструкция IS-scFv-RTP, в которой scFv вставлен между S и R петли ISRTP. На фигуре 1B представлено схематическое изображение типичной петли AKGQP (SEQ ID NO:40), идентифицированной на границе CH2-CH3. На фигуре 1E показана типичная конструкция AK-scfv-GQP, где scFv вставлен между K и G петли AKGQP. На фигуре 1C представлено схематическое изображение типичной петли SNG, идентифицированной в CH3-области ниже границы CH2-CH3. На фигуре 1F также показана типичная конструкция S-scfv-NG, где scFv вставлен между S и N петли SNG. В примерах в данном документе представлена иллюстрация конструкций, ориентированных в виде SN-scFv-G, где scFv вставлен между N и G петли SNG.
[00167] Таким образом, один аспект данного изобретения относится к BiSAb, который содержит две идентичные пары тяжелая-легкая цепь, где каждая пара тяжелая-легкая цепь является биспецифической, а две идентичные пары вместе являются бивалентными по отношению к каждому эпитопу. Каждая пара тяжелая-легкая цепь содержит связывающий домен (BD), который может содержать Fab-домен, который связывает первый эпитоп (связывающая единица 1), второй связывающий домен (BD2), который связывает второй эпитоп (или связывающая единица 2, которая может представлять собой, например, scFv) и Fc-область. В некоторых вариантах осуществления второй связывающий домен может быть связан с Fab-доменом. В некоторых вариантах осуществления Fc-область BiSAb может быть связана со вторым связывающим доменом (BD2), который связывает второй эпитоп (связывающая единица 2, изображенная как scFv на фигурах 1D-1F).
[00168] В некоторых вариантах осуществления настоящее изобретение предусматривает BiSAb, характеризующийся общей структурой платформы, которая содержит две химерные тяжелые цепи, каждая из которых содержит вариабельную область тяжелой цепи (VH1), константную область тяжелой цепи (CH1), шарнирную область или область полипептидного линкера, Fc-область, содержащую CH2-домен и CH3-домен, где второй связывающий домен (BD2), необязательно фланкированный на одной или обеих сторонах полипептидным линкером (L1 и/или L2), связан с гидрофильными петлями в Fc-области в последовательности (i) CH2-области, (ii) границы CH2- и CH3-областей или (iii) CH3-области. BiSAb по данному аспекту настоящего изобретения также содержит две легкие цепи обычного антитела, каждая из которых содержит вариабельную область легкой цепи (VL1) и константную область легкой цепи (CL), которые образуют часть первого связывающего домена (BD1). Связывающий домен (BD2) конкретного BiSAb, проиллюстрированного на фигурах 1D-1F, представляет собой scFv.
[00169] На фигурах 1D-1F представлено применимое схематическое изображение BiSAb, которое также может называться в данном документе "ядро" BiSAb. Полипептидная цепь, показанная на фигуре 1D, содержит тяжелую цепь, которая содержит: VH1-домен, CH1-домен, шарнир/линкер, частичный N-концевой CH2-домен, необязательный линкер (называемый в данном документе L1 или первым полипептидным линкером), связывающая единица 2 (такая как VL2 и VH2 scFv), еще один необязательный линкер (например, L2 или второй полипептидный линкер), оставшийся C-концевой CH2-домен и CH3-домен. Поскольку эта тяжелая цепь может содержать BD2, который содержит альтернативные связывающие белки и/или области традиционных легких цепей, в данном документе она называется химерной тяжелой цепью. BiSAb содержит две такие химерные тяжелые цепи, и они могут быть одинаковыми или разными. Следует отметить, что вариабельный домен тяжелой цепи (VH) для связывающей единицы 1 называется VH1. В определенных аспектах это вариабельный домен тяжелой цепи Fab, который связывается с первым эпитопом. Аналогичным образом, вариабельный домен легкой цепи (VL) для связывающей единицы 1 называется VL1. В определенных аспектах это вариабельный домен легкой цепи Fab, который связывается с первым эпитопом. Напротив, домены связывающей единицы два обозначены числом "2", например, как VH2 и VL2 для аспектов, в которых связывающая единица 2 представляет собой scFv, который связывается со вторым эпитопом.
[00170] Аналогичным образом, полипептидная цепь, показанная на фигуре 1E, содержит тяжелую цепь, которая содержит: VH1-домен, CH1-домен, шарнир/линкер, CH2-домен, необязательный линкер (называемый в данном документе L1 или первым полипептидным линкером), связывающую единицу 2 (такую как VL2 и VH2 scFv), еще один необязательный линкер (например, L2 или второй полипептидный линкер) и CH3-домен. В данном варианте осуществления BiSAb содержит второй связывающий домен, проиллюстрированный в виде scFv, связанного с Fc в последовательности на границе CH2- и CH3-областей.
[00171] Полипептидная цепь, показанная на фигуре 1F, содержит тяжелую цепь, которая содержит: VH1-домен, CH1-домен, шарнир/линкер, CH2-домен, частичный CH3-домен, необязательный линкер (называемый в данном документе L1 или первым полипептидным линкером), связывающую единицу 2 (такую как VL2 и VH2 scFv), еще один необязательный линкер (например, L2 или второй полипептидный линкер), CH3-домен.
[00172] В данных вариантах осуществления BiSAb обычно содержат типичную или модифицированную шарнирную область антитела в последовательностях химерных тяжелых цепей. Неограничивающие примеры аминокислотных последовательностей, которые содержат шарнирную область, включают: EPKSCDKTHTCPPCP (SEQ ID NO:44); EPKSCDKT (SEQ ID NO:45); EPKSCGKT (SEQ ID NO:46); EPKSC (SEQ ID NO:47).
[00173] После описания общего формата для аспектов, относящихся к конкретной структурной платформе для определенных молекул BiSAb, раскрытых в данном документе, различные части и иллюстративные функциональные свойства раскрытых BiSAb более подробно описаны ниже. В других вариантах осуществления в настоящем изобретении рассмотрены и представлены другие связывающие белки BiSAb, которые предусматривают альтернативные структурные форматы и схемы, которые кратко описаны в данном документе, а также в других раскрытиях (см., например, публикацию США № 2009/0155275 и патент США № 9580509), каждый из которых включен в данный документ посредством ссылки.
1. Связывающие единицы
[00174] BiSAb по настоящему изобретению содержат по меньшей мере две связывающие единицы или связывающих домена (связывающая единица/домен 1 и связывающая единица/домен 2). В определенных аспектах каждое связывающая единица связывается с другим эпитопом, либо с разными эпитопами, расположенными на одной и той же молекуле-мишени, либо с эпитопами на разных мишенях. Поскольку каждая связывающая единица BiSAb присутствует в виде пары (имеются две связывающие единицы 1 и две связывающие единицы 2), BiSAb проявляют бивалентное связывание с каждым эпитопом. Из приведенных в данном документе идей будет понятно, что когда каждая связывающая единица связывает один и тот же эпитоп, BiSAb будет проявлять четырехвалентное связывание с эпитопом.
[00175] В определенных аспектах первая связывающая единица представляет собой Fab-фрагмент, например, Fab-фрагмент обычного моноклонального антитела или антигенсвязывающий фрагмент, полученный рекомбинантным путем, содержащий вариабельный домен легкой цепи (VL1), константный домен легкой цепи (CL), вариабельный домен тяжелой цепи (VH1) и часть константного домена тяжелой цепи (CH1). Необязательно легкая и тяжелая цепи Fab могут быть связаны между собой посредством одной или нескольких дисульфидных связей, как, например, посредством подходящей шарнирной области антитела. Fab связывается с первым эпитопом.
[00176] В определенных аспектах Fab получен из последовательности обычного моноклонального антитела, такого как обычное мышиное, гуманизированное или человеческое антитело, или создан на основе ее. В определенных аспектах BiSAb, содержащий Fab, полученный из последовательности обычного моноклонального антитела или созданный на основе ее, сохраняет одну или несколько функциональных форм активности обычного антитела (например, сохраняет по меньшей мере 80% или больше (80%, 85%, 90%, 95%, 97%, 98%, 99% или 100%) функциональной активности). Например, в определенных аспектах BiSAb, содержащий такой Fab, сохраняет одно или несколько из аффинности по отношению к антигену, ингибирующей активности, активности модуляции иммунной системы, активации или индукции иммунного ответа и/или активности в отношении уничтожения клеток (например, раковых клеток) обычного антитела.
[00177] В определенных аспектах BiSAb по настоящему изобретению содержат связывающую единицу 2, и при этом связывающая единица 2 содержит связывающий домен, который связывает второй эпитоп. Связывающая единица 2 (или связывающий домен 2 (BD2)) может быть связана с BiSAb с помощью любой подходящей стратегии. Используемый в данном документе термин BD2, который "связан" с BiSAb (например, в пределах Fc-области в некоторых вариантах осуществления, в пределах Fab-области в других вариантах осуществления), означает, что две молекулы взаимодействуют между собой таким образом, что BD2 сохраняет ориентацию для связывания мишени и связь с Fc-частью или с Fab-частью структуры BiSAb. Примеры таких взаимодействий включают образование ковалентных связей посредством аминокислотных линкеров, образование ковалентных связей с помощью рекомбинантной экспрессии BD2 в пределах Fab-области, в пределах шарнирной области или в пределах Fc-области в CH2, CH3, или на границе CH2 и CH3, или в CH4-области, а также нековалентные взаимодействия, такие как вандерваальсовы взаимодействия и взаимодействия посредством водородных связей в тех же самых областях. Неограничивающие примеры связывающих доменов (или "BD", или "связывающих единиц"), находящихся в пределах объема настоящего изобретения, включают вариабельные области антител, фрагменты антител, scFv, одноцепочечные диатела или другие связывающие домены, известные из уровня техники. Связывающие домены также включают биспецифические одноцепочечные диатела или одноцепочечные диатела, предназначенные для связывания двух различных эпитопов. В одном аспекте связывающие эпитопы домены, применимые при конструировании мультиспецифических связывающих эпитопы доменов по настоящему изобретению, приведены в качестве примера в US 2010/0298541 и US 2013/0079280, которые включены в данный документ посредством ссылки для всех целей.
[00178] В определенных аспектах BiSAb может содержать связывающий домен, который содержит scFv. Таким образом, в определенных аспектах связывающая единица 2 содержит scFv. Следует понимать, что scFv охватывает полипептидную цепь, содержащую вариабельный домен тяжелой цепи (VH), связанный с вариабельным доменом легкой цепи (VL) посредством гибкого полипептидного линкера. На фигурах 1D-1F показано схематическое изображение иллюстративного BiSAb, где BD (в данном документе обозначен как связывающая единица 2) представляет собой scFv, содержащий домены, как описано в данном документе, которые могут быть обозначены как VL2 и VH2. В некоторых аспектах полипептидный линкер между VH2 и VL2 содержит сайт расщепления протеазами. VH- и VL-домены scFv могут быть получены из одного и того же или из разных антител. В некоторых аспектах VH или VL scFv могут содержать одну или несколько CDR, которые связываются с представляющей интерес мишенью, тогда как остальная часть VH- или VL-домена получена из другого антитела или является синтетической. В некоторых аспектах scFv содержит по меньшей мере одну CDR антитела, например, антитела, которое, как известно из уровня техники, связывается с представляющей интерес мишенью. В некоторых аспектах scFv содержит по меньшей мере две CDR данного антитела. В некоторых аспектах scFv содержит по меньшей мере три CDR данного антитела. В некоторых аспектах scFv содержит по меньшей мере четыре CDR данного антитела. В некоторых аспектах scFv содержит по меньшей мере пять CDR данного антитела. В некоторых аспектах scFv содержит по меньшей мере шесть CDR данного антитела.
[00179] В определенных аспектах BD может содержать лигандсвязывающий домен рецептора или рецепторсвязывающий домен лиганда. В некоторых аспектах BD содержит последовательность, которая характеризуется аффинностью связывания по отношению к одному или нескольким эпитопам на мишени, выбранной из группы, состоящей из CTLA-4, PD-1, PD-L1, OX40 и TIM3, как описано выше. В некоторых вариантах осуществления связывающий домен проявляет активность специфического связывания в отношении мишени, выбранной из группы, состоящей из CTLA-4, PD-1, PD-L1, OX40 и TIM3. Раскрытые в данном документе BiSAb могут содержать любую комбинацию связывающих доменов, которые характеризуется аффинностью связывания или активностью специфического связывания по отношению к молекулярным мишеням, раскрытым данном документе. Например, раскрытые в данном документе BiSAb могут содержать комбинацию связывающих доменов, которые обеспечивают биспецифическое связывание с мишенями, в том числе: CTLA-4 и PD-1; CTLA-4 и PD-L1; CTLA-4 и TIM3; PD-1 и PD-L1; PD-L1 и OX40; PD-1 и TIM3; PD-L1 и TIM3; а также TIM3. BiSAb, которые содержат связывающие домены, которые связывают конкретные комбинации мишеней, проиллюстрированы в примерах и включают в себя неограничивающие комбинации PD-1/CTLA-4; PD-L1/CTLA-4; PD-1/OX40; PD-L1/OX40 и PD-1/TIM3.
[00180] В некоторых дополнительных вариантах осуществления BiSAb проявляют активность связывания (например, аффинность связывания и/или специфичность связывания) по отношению к по меньшей мере одной из молекул-мишеней, которая является больше, чем активность связывания исходной моноспецифической связывающей последовательности, используемой для получения BiSAb. В аналогичных вариантах осуществления BiSAb могут проявлять активность связывания (например, аффинность связывания и/или специфичность связывания) в отношении обеих молекул-мишеней, которая является больше, чем активность связывания обеих исходных моноспецифических связывающих последовательностей, используемых для получения BiSAb. В еще дополнительном варианте осуществления BiSAb могут проявлять активность связывания (например, аффинность связывания и/или специфичность связывания) в отношении обеих молекул-мишеней, которая является больше, чем активность связывания комбинации исходных моноспецифических связывающих последовательностей, используемых для получения BiSAb. Улучшение связывающих свойств BiSAb относительно исходных моноспецифических связывающих последовательностей либо отдельно, либо в комбинации обеспечивает неожиданные преимущества по сравнению с использованием моноспецифических терапевтических средств, которые нацеливаются на те же молекулы, даже если они используются в комбинации.
[00181] В некоторых вариантах осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, который связывается с мишенью, выбранной из группы, состоящей из CTLA-4, PD-1, PD-L1, OX40 и TIM3. В таких вариантах осуществления антитело или его антигенсвязывающий фрагмент могут содержать последовательность тяжелой цепи и последовательность легкой цепи или часть последовательности тяжелой цепи и последовательности легкой цепи, которая содержит последовательности CDR1, CDR2 и CDR3 последовательностей тяжелой и легкой цепей. В других вариантах осуществления антитело или его антигенсвязывающий фрагмент могут содержать последовательность вариабельной области тяжелой цепи (HCv) и последовательность вариабельной области легкой цепи (LCv) или часть HCv и LCv, которая содержит последовательности CDR1, CDR2 и CDR3 последовательностей тяжелой и легкой цепей. В еще других вариантах осуществления антитело или его антигенсвязывающий фрагмент могут содержать последовательности CDR1, CDR2 и CDR3 последовательностей тяжелой и легкой цепей. В некоторых вариантах осуществления антитело может представлять собой химерное, гуманизированное или человеческое антитело. В некоторых вариантах осуществления антитело может представлять собой поликлональное или моноклональное антитело. В дополнительных вариантах осуществления антитело представляет собой моноклональное антитело.
[00182] В некоторых вариантах осуществления домены, которые содержат всю антигенсвязывающую область таких "исходных" антител или ее часть, как обсуждалось выше, могут быть использованы для получения биспецифических связывающих белков (BiSAb), которые раскрыты в данном документе. Неограничивающие варианты осуществления, которые проиллюстрированы в примерах, предоставляют описание, относящееся к тому, как последовательности антител могут быть идентифицированы и объединены для получения BiSAb, который проявляет биспецифическое связывание с комбинацией молекулярных мишеней.
[00183] Для улучшения стабильности BiSAb, содержащего scFv-молекулу, можно применять несколько методик отдельно или в комбинации. Одна потенциальная методика, которая может применяться отдельно или в комбинации с одной или несколькими из других описанных в данном документе методик, представляет собой конструирование длины и/или состава линкера, соединяющего scFv-домены для стабилизации scFv-части.
[00184] Другая потенциальная методика, которая может применяться для введения по меньшей мере двух аминокислотных замен (также называемых модификациями или мутациями) в VH- и/или VL-домены scFv с целью облегчения образования дисульфидных связей (см., например, Brinkmann et al., 1993, PNAS, 90:7538-42; Zhu et al., 1997, Prot. Sci. 6:781-8; Reiter et al., 1994, Biochem. 33:5451-9; Reiter et al., 1996, Nature 14: 1239-45; Luo et al., 1995, J. Biochem. 118:825-31; Young et al., 1995, FEBS Let. 377:135-9; Glockshuber et al., 1990, Biochem. 29:1362-7). Данный способ может применяться отдельно или в комбинация с одной или несколькими другими методиками, описанными в данном документе.
[00185] В определенных аспектах одна или несколько мутаций могут быть введены в каждый из VH- и VL-доменов scFv для облегчения образования межцепочечных дисульфидных связей между VH- и VL-доменами после экспрессии BiSAb, содержащего scFv. В другом аспекте две мутации вводят в один и тот же домен цепи. В определенном аспекте две мутации вводят в разные цепи. В определенных аспектах вводятся множественные комплементарные мутации для облегчения образования множественных дисульфидных связей или других стабилизирующих взаимодействий. В определенных аспектах вводится цистеин для облегчения образования дисульфидных связей. Иллюстративные аминокислоты, которые могут быть подвергнуты мутации с заменой на цистеин, включают аминокислоты 43, 44, 45, 46, 47, 103, 104, 105 и 106 VH2 и аминокислоты 42, 43, 44, 45, 46, 98, 99, 100 и 101 VL2. Вышеизложенная нумерация основана на нумерации согласно Kabat, определяющей положение только по отношению к VH2 и VL2 scFv (а не по отношению к положению аминокислоты в полноразмерной последовательности BiSAb или SEQ ID NO, приведенных в данном документе). Иллюстративные комбинации положений аминокислот, которые могут быть подвергнуты мутации с заменой на остатки цистеина, включают: VH44-VL100, VH105-VL43, VH105-VL42, VH44-VL101, VH106-VL43, VH104-VL43, VH44-VL99, VH45-VL98, VH46-VL98, VH103-VL43, VH103-VL44 и VH103-VL45. В некоторых аспектах аминокислота 44 VH и аминокислота 100 VL подвергнуты мутации с заменой на цистеины.
[00186] Другой способ, который может применяться отдельно или в комбинации с одним или несколькими другими способами, описанными в данном документе, заключается в выборе порядка доменов scFv. В определенных аспектах ориентация VH-домена по отношению к VL-домену оптимизирована в целях стабильности. В определенных аспектах scFv находится в ориентации VH-линкер-VL. В определенных аспектах scFv находится в ориентации VL-линкер-VH. В вариантах осуществления, относящихся к новому формату BiSAb, раскрытому в данном документе, ориентация доменов в scFv может определять, как scFv связывается с Fc-частью BiSAb. Вместе с тем это подробно описано ниже в контексте полипептидных линкеров. Однако вкратце, учитывая что BD (например, scFv) связан с CH2, CH3 или с границей CH2 и CH3 с помощью необязательных полипептидных линкеров (L1) и (L2), порядок доменов определяет, какая часть scFv связана с L1 и какая часть scFv связана с L2.
[00187] Дополнительный способ, который может применяться отдельно или в комбинации с другими способами, описанными в данном документе, заключается во введении одной или нескольких стабилизирующих мутаций путем мутирования одного или нескольких поверхностных остатков scFv. В некоторых аспектах один, два, три, четыре, пять, шесть или более шести остатков подвергнуты мутации в одном или обоих из VH- и/или VL-доменов scFv. В определенных аспектах изменения осуществлены только в VH-домене scFv. В определенных аспектах изменения осуществлены только в VL-домене scFv. В определенных аспектах изменения осуществлены в обоих из VH- и VL-доменов scFv. В каждом домене может быть осуществлено одинаковое количество изменений или в каждом домене может быть осуществлено разное количество изменений. В определенных аспектах одно или несколько изменений представляют собой консервативную аминокислотную замену в остатке, присутствующем в немодифицированном исходном scFv. В других аспектах одно или несколько изменений представляет собой неконсервативную аминокислотную замену в остатке, присутствующем в немодифицированном исходном scFv. При проведении множественных замен в одном из VH- или VL-доменов scFv или в обоих из VH- или VL-доменов scFv каждая замена независимо представляет собой консервативную или неконсервативную замену. В определенных аспектах все замены представляют собой консервативные замены. В определенных аспектах все замены являются неконсервативными. В определенных аспектах по меньшей мере одна из замен является консервативной. В определенных аспектах по меньшей мере одна из замен является неконсервативной.
[00188] Еще другой способ, который может применяться отдельно или в комбинации с другими способами, заключается во введении одной или нескольких аминокислотных замен путем мутирования одного или нескольких остатков, присутствующих в VH- и/или VL-домене scFv, с целью сопоставления наиболее часто встречающегося остатка в указанном конкретном положении консенсусной последовательности VH- и/или VL-домена известных антител. В определенных аспектах замены вводят в одно, два, три, четыре, пять, шесть или более шести положений в одном или обоих из VH- и/или VL-доменов scFv. В каждом домене может быть осуществлено одинаковое количество изменений или в каждом домене может быть осуществлено разное количество изменений. В определенных аспектах одно или несколько изменений в последовательности, соответствующих таковым в данной консенсусной последовательности, представляют собой консервативную аминокислотную замену в остатке, присутствующем в немодифицированной последовательности VH и/или VL. В других аспектах одно или несколько изменений представляют собой неконсервативную аминокислотную замену в остатке, присутствующем в немодифицированной последовательности VH и/или VL. При осуществлении множественных замен либо в одном, либо в обоих из VH- или VL-доменов scFv каждая замена независимо представляет собой консервативную или неконсервативную замену. В определенных аспектах все замены представляют собой консервативные замены. В определенных аспектах все замены представляют собой неконсервативные замены. В определенных аспектах по меньшей мере одна из замен является консервативной. В определенных аспектах по меньшей мере одна из замен является неконсервативной.
[00189] Следует отметить, что для модификации Fab-части может применяться любая из модификаций, описанных в качестве применимых для модификации или стабилизации scFv-части. Например, вариабельные домены Fab-части BiSAb могут быть модифицированы с целью улучшения стабильности, связывания с антигеном и т. п. Более того, для снижения иммуногенности можно модифицировать либо Fab-часть, либо scFv-часть.
[00190] В определенных аспектах связывающая единица 2 (BD) представляет собой scFv, например, scFv, полученный из обычного моноклонального антитела, содержащего вариабельный домен легкой цепи (VL2) и вариабельный домен тяжелой цепи (VH2), соединенные между собой гибким линкером, таким как глицин-сериновый линкер. Необязательно вариабельный домен легкой и вариабельнай домен тяжелой цепи scFv могут быть дополнительно связаны между собой с помощью одной или нескольких дисульфидных связей и, как описано выше, могут содержать одну или несколько мутаций или вариаций. ScFv связывается со вторым эпитопом. В определенных аспектах второй эпитоп отличается от первого эпитопа, связываемого помощью связывающей единицы 1. В других аспектах второй эпитоп является таким же, как и первый эпитоп, связываемый с помощью связывающей единицы 1. В определенных аспектах scFv получен из последовательности обычного моноклонального антитела, такого как обычное мышиное, гуманизированное или человеческое антитело, или создан на ее основе. В определенных аспектах BiSAb, содержащий scFv, полученный из последовательности обычного моноклонального антитела, или созданный на ее основе, сохраняет одну или несколько функциональных активностей обычного антитела (например, сохраняет по меньшей мере 80% или больше (80%, 85%, 90%, 95%, 97%, 98%, 99% или 100%) функциональной активности). Например, в определенных аспектах BiSAb, содержащий такой scFv, сохраняет одно или несколько из аффинности по отношению к антигену, ингибирующей активности или активности в отношении уничтожения клеток обычного антитела.
[00191] В определенных аспектах BiSAb содержит любую из связывающих единиц 1 и связывающих единиц 2, описанных в данном документе, в том числе любую комбинацию связывающей единицы 1 и связывающей единицы 2. Например, в определенных аспектах настоящее изобретение предусматривает полипептид, содержащий Fab, который связывается с конкретной мишенью (например, который связывается с эпитопом на конкретной мишени), такой как Fab, содержащий определенную аминокислотную последовательность или кодируемый конкретной нуклеотидной последовательностью, и/или scFv, который связывается с конкретной мишенью (например, который связывается с эпитопом на конкретной мишени), такой как scFv, содержащий определенную аминокислотную последовательность или кодируемый определенной нуклеотидной последовательностью.
[0192] Как подробно описано выше, связывающая единица 1 и связывающая единица 2 могут быть связаны с BiSAb посредством ковалентной связи с помощью линкерного полипептида 1 (L1, L2). Как правило, связь осуществляется посредством химерной тяжелой цепи BiSAb таким образом, что соединение осуществляется посредством CH2-домена тяжелой цепи, CH3-домена тяжелой цепи или на границе CH2-домена и CH3-домена тяжелой цепи или, в некоторых вариантах осуществления, в пределах шарнирной области или Fab-домена. L1 и L2 могут различаться по длине и последовательности независимо друг от друга, а иллюстративные конфигурации описаны в данном документе. В настоящем изобретении рассматриваются BiSAb, содержащие любую комбинацию связывающих единиц и линкерных полипептидов, в том числе любую комбинацию специфических связывающих единиц, которые связывают требуемую(-ые) мишень(-и), и специфических полипептидных линкеров L1 и L2, описанных в данном документе.
2. Fc-область
[0193] Используемый в данном документе термин "Fc-область" охватывает домены, полученные из константной области иммуноглобулина, предпочтительно иммуноглобулина человека, в том числе фрагмента, аналога, варианта, мутанта или производного константной области. Подходящие иммуноглобулины включают IgG1, IgG2, IgG3, IgG4 и другие классы, такие как IgA, IgD, IgE и IgM. Fc-область может представлять собой нативную последовательность Fc-области или измененной Fc-области. Fc-область иммуноглобулина, как правило, содержит два константных домена, CH2-домен и CH3-домен, и необязательно содержит CH4-домен. BiSAb по настоящему изобретению содержат Fc-область, которая содержит CH2-домен и CH3-домен.
a. Измененные Fc-области
[0194] Измененные Fc-области (также называемые в данном документе "вариантными Fc-областями") могут использоваться для изменения эффекторной функции и/или времени полужизни BiSAb по настоящему изобретению. Одно или несколько изменений могут быть осуществлены в Fc-области с целью изменения функциональных и/или фармакокинетических свойств молекул. Такие изменения могут приводить к снижению или повышению связывания Clq и комплементзависимой цитотоксичности (CDC) или связывания FcγR для IgG и антителозависимой клеточной цитотоксичности (ADCC) или антителозависимого клеточноопосредованного фагоцитоза (ADCP). Настоящее изобретение охватывает BiSAb, в которых были осуществлены изменения для тонкой регулировки эффекторной функции либо путем усиления или ослабления функции, либо обеспечения требуемой эффекторной функции. Соответственно, в одном аспекте настоящего изобретения BiSAb содержат вариантную Fc-область (т. е. Fc-области, которые были изменены, как описано ниже). BiSAb, содержащие вариантную Fc-область, также называются в данном документе "BiSAb с вариантным Fc". Используемый в данном документе термин "нативный" относится к немодифицированной исходной последовательности, и BiSAb, содержащий нативную Fc-область, называется в данном документе "BiSAb с нативным Fc". BiSAb с вариантным Fc могут быть получены с помощью многочисленных способов, хорошо известных специалисту в данной области техники. Неограничивающие примеры включают выделение областей, кодирующих антитело (например, из гибридомы), и осуществление одной или нескольких требуемых замен в Fc-области. Как качестве альтернативы антигенсвязывающая часть (например, вариабельные области) BiSAb может быть субклонирована в вектор, кодирующий вариантную Fc-область. В одном аспекте вариантная Fc-область проявляет сходный уровень индукции эффекторной функции по сравнению с нативной Fc-областью. В другом аспекте вариантная Fc-область проявляет более высокую индукцию эффекторной функции по сравнению с нативным Fc. В другом аспекте вариантная Fc-область проявляет более низкую индукцию эффекторной функции по сравнению с нативным Fc. Некоторые конкретные аспекты вариантных Fc-областей подробно описаны ниже. Способы измерения эффекторной функции хорошо известны из уровня техники.
[0195] Как правило, эффекторную функцию модифицируют посредством изменений в Fc-области, в том числе без ограничения аминокислотных замен, добавлений аминокислот, делеций аминокислот и изменений по типу посттрансляционных модификаций в аминокислотах Fc (например, гликозилирования). Способы, описанные ниже, могут применяться для точной регулировки эффекторной функции BiSAb по настоящему изобретению, соотношения связывающих свойств Fc-области для FcR (например, аффинности и специфичности), что приводит к получению BiSAb с требуемыми свойствами.
[0196] Следует понимать, что Fc-область, как используется в данном документе, включает в себя полипептиды, содержащие константную область молекулы антитела, за исключением первого домена константной области иммуноглобулина. Таким образом, Fc относится к последним двум доменам константной области иммуноглобулинов IgA, IgD и IgG и последним трем доменам константной области иммуноглобулинов IgE и IgM и необязательно ко всему гибкому шарнирному участку на N-конце этих доменов или ее части. В случае IgA и IgM Fc может содержать J-цепь. В случае IgG Fc содержит домены иммуноглобулина C-гамма-2 и C-гамма-3 (Cγ2 и Cγ3) и необязательно часть нижнего шарнирного участка между C-гамма-1 (Cγ1) и C-гамма-2 (Cγ2). Хотя границы Fc-области могут варьироваться, используемая в данном документе Fc-область тяжелой цепи IgG человека содержит остатки A231 на своем карбоксильном конце, где нумерация приведена согласно EU-индексу, как изложено у Kabat. Fc может относиться к данной выделенной области или к данной области в контексте антитела, фрагмента антитела или Fc-слитого белка. Наблюдались полиморфизмы в ряде различных положений Fc, в том числе без ограничения в положениях 270, 272, 312, 315, 356 и 358 IgG1, пронумерованных в соответствии с EU-индексом, и, следовательно, могут существовать незначительные различия между представленной последовательностью и последовательностями из предыдущего уровня техники.
[0197] В одном аспекте настоящее изобретение охватывает BiSAb с вариантным Fc, которые характеризуются измененными связывающими свойствами в отношении лиганда Fc (например, рецептора Fc, C1q) по сравнению с BiSAb с нативным Fc. Примеры связывающих свойств включают без ограничения специфичность связывания, равновесную константу диссоциации (Kd), скорости диссоциации и ассоциации (соответственно koff и kon), аффинность и/или авидность связывания. Из уровня техники известно, что равновесная константа диссоциации (Kd) определяется как koff/kon. В некоторых аспектах BiSAb, содержащий вариантную Fc-область, с низким значением Kd, может являться более целесообразным, чем BiSAb с высоким значением Kd. Однако в некоторых случаях значение kon или koff может быть более значимым, чем значение Kd. Специалист в данной области техники способен определить, какой кинетический параметр является наиболее важным для данного применения. Например, модификация, которая снижает связывание с одним или несколькими положительными регуляторами (например, FcγRIIIA) и/или улучшает связывание с ингибирующим Fc-рецептором (например, FcγRIIB), была бы подходящей для снижения активности ADCC. Соответственно, соотношение аффинностей связывания (например, соотношение равновесных констант диссоциации (Kd)) различных рецепторов может указывать, является ли активность ADCC у BiSAb с вариантным Fc по настоящему изобретению повышенной или сниженной. Кроме того, модификация, которая снижает связывание с C1q, была бы подходящей для снижения или устранения активности CDC.
[0198] В одном аспекте BiSAb с вариантным Fc демонстрируют измененную аффинность связывания в отношении одного или нескольких Fc-рецепторов, в том числе без ограничения FcRn, FcγRI (CD64), в том числе изоформ FcγRIA, FcγRIB и FcγRIC; FcγRII (CD32, в том числе изоформ FcγRIIA, FcγRIIB и FcγRIIC) и FcγRIII (CD16, в том числе изоформ FcγRIIIA и FcγRIIIB) по сравнению с BiSAb с нативным Fc.
[0199] В определенных аспектах BiSAb с вариантным Fc характеризуется повышенной аффинностью в отношении Fc-лиганда. В других аспектах BiSAb с вариантным Fc характеризуется сниженной аффинностью в отношении Fc-лиганда по сравнению с BiSAb с нативным Fc.
[0200] В конкретном аспекте BiSAb с вариантным Fc характеризуется повышенным связыванием с Fc-рецептором FcγRIIIA. В другом конкретном аспекте BiSAb с вариантным Fc характеризуется повышенным связыванием с Fc-рецептором FcγRIIB. В дополнительном конкретном аспекте BiSAb с вариантным Fc характеризуется повышенным связыванием с Fc-рецепторами как FcγRIIIA, так и FcγRIIB. В определенных аспектах BiSAb с вариантным Fc, которые характеризуются повышенным связыванием с FcγRIIIA, не характеризуются при этом одновременно повышением связывания с рецептором FcγRIIB по сравнению с BiSAb с нативным Fc. В конкретном аспекте BiSAb с вариантным Fc характеризуется сниженным связыванием с Fc-рецептором FcγRIIIA. В дополнительном конкретном аспекте BiSAb с вариантным Fc характеризуется сниженным связыванием с Fc-рецептором FcγRIIB. В другом конкретном аспекте BiSAb с вариантным Fc характеризуется повышенным связыванием с Fc-рецептором FcRn. В еще одном конкретном аспекте BiSAb с вариантным Fc, проявляющий измененную аффинность в отношении FcγRIIIA и/или FcγRIIB, характеризуется повышенным связыванием с Fc-рецептором FcRn. В еще одном конкретном аспекте BiSAb с вариантным Fc, проявляющий измененную аффинность в отношении FcγRIIIA и/или FcγRIIB, характеризуется измененным связыванием с C1q по сравнению с BiSAb с нативным Fc.
[0201] В другом аспекте BiSAb с вариантным Fc проявляют повышенные или пониженные значения аффинности в отношении C1q по сравнению с BiSAb с нативным Fc. В еще другом конкретном аспекте BiSAb с вариантным Fc, проявляющий измененную аффинность в отношении C1q, характеризуется повышенным связыванием с Fc-рецептором FcRn. В еще одном конкретном аспекте BiSAb с вариантным Fc, проявляющий измененную аффинность в отношении C1q, характеризуется измененным связыванием с FcγRIIIA и/или FcγRIIB по сравнению с BiSAb с нативным Fc.
[0202] Известно, что антитела способны направлять атаку и разрушение антигена-мишени посредством многочисленных процессов, в совокупности известных из уровня техники как эффекторные функции антитела. Один из этих процессов, известный как "антителозависимая клеточно-опосредованная цитотоксичность", или "ADCC", относится к форме цитотоксичности, при которой секретируемый Ig, связанный с Fc-гамма-рецепторами (FcγR), присутствующими на определенных цитотоксических клетках (например, природных клетках-киллерах (NK), нейтрофилах и макрофагах), дает возможность этим цитотоксическим эффекторным клеткам специфически связываться с несущей антиген клеткой-мишенью и впоследствии уничтожать клетку-мишень с помощью цитотоксинов. Специфические высокоаффинные антитела IgG, направленные на поверхность клеток-мишеней, "вооружают" цитотоксические клетки и являются необходимыми для такого уничтожения. Лизис клетки-мишени является внеклеточным, требует непосредственного межклеточного контакта и не вовлекает систему комплемента. Другим процессом, охватываемым термином "эффекторная функция", является комплементзависимая цитотоксичность (далее в данном документе называемая "CDC"), которая относится к биохимическому явлению опосредованного антителами разрушения клеток-мишеней посредством системы комплемента. Система комплемента представляет собой сложную систему белков, обнаруживаемых в нормальной плазме крови, которая в сочетании с антителами разрушает патогенные бактерии и другие чужеродные клетки. Еще одним процессом, охватываемым термином "эффекторная функция", является антителозависимый клеточноопосредованный фагоцитоз (ADCP), который относится к клеточноопосредованной реакции, где неспецифические цитотоксические клетки, которые экспрессируют один или несколько эффекторных лигандов, распознают связанное антитело на клетке-мишени и впоследствии вызывают фагоцитоз клетки-мишени.
[0203] Предполагается, что характеристики BiSAb с вариантным Fc получают с помощью функциональных in vitro анализов для определения одной или нескольких FcγR-опосредованных эффекторных клеточных функций. В определенных аспектах BiSAb с вариантным Fc характеризуются связывающими свойствами и эффекторными клеточными функциями в моделях in vivo (таких как описанные и раскрытые в данном документе), сходными с таковыми в анализах на основе in vitro. Однако настоящее изобретение не исключает BiSAb с вариантным Fc, которые не проявляют требуемого фенотипа в анализах на основе in vitro, однако проявляют требуемый фенотип in vivo.
[0204] Время полужизни в сыворотке крови белков, содержащих Fc-области, может быть увеличено путем повышения аффинности связывания Fc-области с FcRn. Термин "время полужизни антитела", используемый в данном документе, означает фармакокинетическое свойство антитела, которое представляет собой показатель среднего времени существования молекул антител после их введения. Время полужизни антитела можно выражать как время, требуемое для выведения 50 процентов известного количества иммуноглобулина из организма пациента (или другого млекопитающего) или его конкретного компартмента, например, измеренное в сыворотке крови, т. е. время полужизни в кровотоке или в других тканях. Время полужизни может варьировать среди разных иммуноглобулинов или классов иммуноглобулинов. Как правило, увеличение времени полужизни антитела (или BiSAb) приводит в результате к увеличению среднего времени удержания (MRT) в кровотоке введенного BiSAb.
[0205] Увеличение времени полужизни дает возможность уменьшить количество лекарственного средства, принимаемого пациентом, а также снизить частоту введения. Для увеличения времени полужизни BiSAb в сыворотке крови в BiSAb (в особенности, во фрагмент антитела) может вводиться эпитоп, связывающийся с рецептором реутилизации, как описано, например, в патенте США № 5739277. Используемый в данном документе термин "эпитоп, связывающийся с рецептором реутилизации" относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), который отвечает за увеличение in vivo времени полужизни в сыворотке крови молекулы IgG. В качестве альтернативы BiSAb по настоящему изобретению с увеличенными показателями времени полужизни могут быть получены с помощью модификации аминокислотных остатков, идентифицированных как вовлеченные во взаимодействие между Fc и рецептором FcRn (см., например, патенты США №№ 6821505 и 7083784). Кроме того, время полужизни BiSAb по настоящему изобретению может быть увеличено путем конъюгирования с PEG или альбумином с помощью методик, широко используемых в данной области техники.
[0206] Предполагается, что на активность Fc может оказывать влияние либо вставка дополнительных связывающих доменов в Fc-область, как описано в данном документе, и/или последующее связывание антигеном. Например, связывающий антиген может повышать или понижать аффинность и активность связывания в отношении FcgR, C1q и FcRn. Это создало бы антигензависимый механизм переключения для модуляции различных антителозависимых процессов. В одном аспекте связывание антигена может уменьшать взаимодействие с FcRn, давая возможность свободному BiSAb взаимодействовать с FcRn и характеризоваться нормальным временем полужизни, однако допускает быстрое выведение/клеточную интернализацию комплексов BiSAb-Ag. Далее, это могло бы дать возможность взаимодействиям, опосредованным BD2-антигеном, оказывать эффект на выведение антигенов, связанных BD1. В дополнительном аспекте BiSAb могло бы содержать Fc-область, непосредственно вставленную в BD2 (Fc-BD2).
[0207] В одном аспекте настоящее изобретение предусматривает варианты Fc, где Fc-область содержит модификацию (например, аминокислотные замены, аминокислотные вставки, делеции аминокислот) в одном или нескольких положениях, выбранных из группы, состоящей из 221, 225, 228, 234, 235, 236, 237, 238, 239, 240, 241, 243, 244, 245, 247, 250, 251, 252, 254, 255, 256, 257, 262, 263, 264, 265, 266, 267, 268, 269, 279, 280, 284, 292, 296, 297, 298, 299, 305, 308, 313, 316, 318, 320, 322, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 339, 341, 343, 370, 373, 378, 392, 416, 419, 421, 428, 433, 434, 435, 436, 440 и 443 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. Необязательно Fc-область может содержать модификацию в дополнительных и/или альтернативных положениях, известных специалисту в данной области техники (см., например, патенты США 5624821; 6277375; 6737056; 7083784; 7317091; 7217797; 7276585; 7355008). Дополнительные применимые положения и конкретные замены аминокислот приведены в качестве примеров в таблицах 2 и 6-10 из US 6737556; в таблицах, представленных на фигуре 41 из US 2006/024298; в таблицах, представленных на фигурах 5, 12 и 15 из US 2006/235208; в таблицах, представленных на фигурах 8, 9 и 10 из US 2006/0173170, и в таблицах, представленных на фигурах 8-10, 13 и 14 из WO 09/058492.
[0208] В конкретном аспекте настоящее изобретение предусматривает вариант Fc, где Fc-область содержит по меньшей мере одну замену, выбранную из группы, состоящей из 221K, 221Y, 225E, 225K, 225W, 228P, 234D, 234E, 234N, 234Q, 234T, 234H, 234Y, 234I, 234V, 234F, 235A, 235D, 235R, 235W, 235P, 235S, 235N, 235Q, 235T, 235H, 235Y, 235I, 235V, 235E, 235F, 236E, 237L, 237M, 237P, 239D, 239E, 239N, 239Q, 239F, 239T, 239H, 239Y, 240I, 240A, 240T, 240M, 241W, 241 L, 241Y, 241E, 241 R. 243W, 243L 243Y, 243R, 243Q, 244H, 245A, 247L, 247V, 247G, 250E, 250Q, 251F, 252L, 252Y, 254S, 254T, 255L, 256E, 256F, 256M, 257C, 257M, 257N, 262I, 262A, 262T, 262E, 263I, 263A, 263T, 263M, 264L, 264I, 264W, 264T, 264R, 264F, 264M, 264Y, 264E, 265A, 265G, 265N, 265Q, 265Y, 265F, 265V, 265I, 265L, 265H, 265T, 266I, 266A, 266T, 266M, 267Q, 267L, 268E, 269H, 269Y, 269F, 269R, 270E, 280A, 284M, 292P, 292L, 296E, 296Q, 296D, 296N, 296S, 296T, 296L, 296I, 296H, 296G, 297S, 297D, 297E, 298A, 298H, 298I, 298T, 298F, 299I, 299L, 299A, 299S, 299V, 299H, 299F, 299E, 305I, 308F313F, 316D, 318A, 318S, 320A, 320S, 322A, 322S, 325Q, 325L, 325I, 325D, 325E, 325A, 325T, 325V, 325H, 326A, 326D, 326E, 326G, 326M, 326V, 327G, 327W, 327N, 327L, 328S, 328M, 328D, 328E, 328N, 328Q, 328F, 328I, 328V, 328T, 328H, 328A, 329F, 329H, 329Q, 330K, 330G, 330T, 330C, 330L, 330Y, 330V, 330I, 330F, 330R, 330H, 331G, 331A, 331L, 331M, 331F, 331W, 331K, 331Q, 331E, 331S, 331V, 331I, 331C, 331Y, 331H, 331R, 331N, 331D, 331T, 332D, 332S, 332W, 332F, 332E, 332N, 332Q, 332T, 332H, 332Y, 332A, 333A, 333D, 333G, 333Q, 333S, 333V, 334A, 334E, 334H, 334L, 334M, 334Q, 334V, 334Y, 339T, 370E, 370N, 378D, 392T, 396L, 416G, 419H, 421K, 428L, 428F, 433K, 433L, 434A, 424F, 434W, 434Y, 436H, 440Y и 443W согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. Необязательно Fc-область может содержать дополнительные и/или альтернативные замены аминокислот, известные специалисту в данной области, в том числе без ограничения приведенные в качестве примеров в таблицах 2 и 6-10 из US 6737056; в таблицах, представленных на фигуре 41 из US 2006/024298; в таблицах, представленных на фигурах 5, 12 и 15 из US 2006/235208; в таблицах, представленных на фигурах 8, 9 и 10 из US 2006/0173170, и таблицах, представленных на фигурах 8, 9 и 10 из US20090041770, все из которых включены в данный документ посредством ссылки.
[0209] В конкретном аспекте настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит по меньшей мере одну модификацию (например, аминокислотные замены, аминокислотные вставки, делеции аминокислот) в одном или нескольких положениях, выбранных из группы, состоящей из 228, 234, 235 и 331 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В одном аспекте модификация представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из 228P, 234F, 235E, 235F, 235Y и 331S согласно нумерации в соответствии с EU-индексом, как изложено у Kabat.
[0210] В другом конкретном аспекте настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область представляет собой Fc-область IgG4 и содержит по меньшей мере одну модификацию в одном или нескольких положениях, выбранных из группы, состоящей из 228 и 235 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В еще одном конкретном аспекте Fc-область представляет собой Fc-область IgG4, а не встречающиеся в природе аминокислоты выбраны из группы, состоящей из 228P, 235E и 235Y согласно нумерации в соответствии с EU-индексом, как изложено у Kabat.
[0211] В другом конкретном аспекте настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит по меньшей мере одну не встречающуюся в природе аминокислоту в одном или нескольких положениях, выбранных из группы, состоящей из 239, 330 и 332 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В одном аспекте модификация представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из 239D, 330L, 330Y и 332E согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. См. патент США № 7317091, включенный в данный документ посредством ссылки во всей своей полноте.
[0212] В конкретном аспекте настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит по меньшей мере одну не встречающуюся в природе аминокислоту в одном или нескольких положениях, выбранных из группы, состоящей из 252, 254 и 256 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В одном аспекте модификация представляет собой по меньшей мере одну замену, выбранную из группы, состоящей из 252Y, 254T и 256E согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. См. патент США № 7083784, включенный в данный документ посредством ссылки во всей своей полноте.
[0213] В определенных аспектах настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит не встречающуюся в природе аминокислоту в положении 428 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В одном аспекте модификация в положении 428 выбрана из группы, состоящей из 428T, 428L, 428F и 428S согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. См. патент США № 7670600, включенный в данный документ посредством ссылки во всей своей полноте. В другом аспекте BiSAb с вариантным Fc может дополнительно содержать не встречающуюся в природе аминокислоту в положении 434 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В одном аспекте модификация в положении 434 выбрана из группы, состоящей из 434A, 434S и 434F согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В других аспектах настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит не встречающуюся в природе аминокислоту в положениях 428 и 434 согласно нумерации в соответствии с EU-индексом, как изложено у Kabat. В конкретном аспекте Fc-область содержит 428L, 434S. См. патент США № 8088376.
[0214] В определенных аспектах эффекторные функции, проявляемые антителами IgG, в значительной степени зависят от углеводного фрагмента, соединенного с Fc-областью белка (Claudia Ferrara et al., 2006, Biotechnology and Bioengineering 93:851-861). Таким образом, гликозилирование Fc-области может быть модифицировано с целью усиления или ослабления эффекторной функции (см., например, Umana et al., 1999, Nat. Biotechnol 17:176-180; Davies et al., 2001, Biotechnol Bioeng 74:288-294; Shields et al, 2002, J Biol Chem 277:26733-26740; Shinkawa et al., 2003, J Biol Chem 278:3466-3473; патенты США №№ 6602684; 6946292; 7064191; 7214775;7393683; 7425446; 7504256; технологию POTELLIGENT™ (Biowa, Inc. Принстон, Нью-Джерси); инженерную технологию гликозилирования GLYCOMAB™ (GLYCART biotechnology AG, Цюрих, Швейцария)). Соответственно, в одном аспекте Fc-области BiSAb по настоящему изобретению предусматривают измененное гликозилирование аминокислотных остатков. В другом аспекте измененное гликозилирование аминокислотных остатков приводит к ослаблению эффекторной функции. В другом аспекте измененное гликозилирование аминокислотных остатков приводит к усилению эффекторной функции. В конкретном аспекте Fc-область характеризуется пониженным фукозилированием. В другом аспекте Fc-область является нефукозилированной (см., например, публикацию заявки на патент США № 2005/0226867). В одном аспекте данные BiSAb с усиленной эффекторной функцией, в частности, ADCC, получены в клетках-хозяевах (например, клетках CHO, Lemna minor), сконструированных для выработки полипептида с высокой степенью дефукозилирования, характеризующегося повышенной более чем в 100 раз ADCC по сравнению с полипептидом, вырабатываемым исходными клетками (Mori et al., 2004, Biotechnol Bioeng 88:901-908; Cox et al., 2006, Nat Biotechnol., 24:1591-7).
[0215] Добавление сиаловой кислоты к олигосахаридам на молекулах IgG может повышать их противовоспалительную активность и изменять их цитотоксичность (Keneko et al., Science, 2006, 313:670-673; Scallon et al., Mol. Immuno. 2007 Mar; 44(7):1524-34). Вышеупомянутые исследования показывают, что молекулы IgG с повышенным сиалированием характеризуются противовоспалительными свойствами, тогда как молекулы IgG с пониженным сиалированием характеризуются усиленными иммуностимулирующими свойствами (например, повышенной активностью ADCC). Следовательно, BiSAb может быть модифицирован с помощью подходящего профиля сиалирования для конкретного применения (публикация США № 2009/0004179 и Международная публикация № WO 2007/005786).
[0216] В одном аспекте Fc-области BiSAb по настоящему изобретению предусматривают профиль измененного сиалирования по сравнению с нативной Fc-областью. В одном аспекте Fc-области BiSAb по настоящему изобретению предусматривают профиль повышенного сиалирования по сравнению с нативной Fc-областью. В другом аспекте Fc-области BiSAb по настоящему изобретению предусматривают профиль пониженного сиалирования по сравнению с нативной Fc-областью.
[0217] В одном аспекте варианты Fc по настоящему изобретению можно объединять с другими известными вариантами Fc, такими как раскрытые в Ghetie et al., 1997, Nat Biotech. 15:637-40; Duncan et al, 1988, Nature 332:563-564; Lund et al., 1991, J. Immunol 147:2657-2662; Lund et al, 1992, Mol Immunol 29:53-59; Alegre et al, 1994, Transplantation 57:1537-1543; Hutchins et al., 1995, Proc Natl. Acad Sci U S A 92:11980-11984; Jefferis et al, 1995, Immunol Lett. 44:111-117; Lund et al., 1995, Faseb J 9:115-119; Jefferis et al, 1996, Immunol Lett 54:101-104; Lund et al, 1996, J Immunol 157:4963-4969; Armour et al., 1999, Eur J Immunol 29:2613-2624; Idusogie et al, 2000, J Immunol 164:4178-4184; Reddy et al, 2000, J Immunol 164:1925-1933; Xu et al., 2000, Cell Immunol 200:16-26; Idusogie et al, 2001, J Immunol 166:2571-2575; Shields et al., 2001, J Biol Chem 276:6591-6604; Jefferis et al, 2002, Immunol Lett 82:57-65; Presta et al., 2002, Biochem Soc Trans 30:487-490); патентах США №№ 5624821; 5885573; 5677425; 6165745; 6277375; 5869046; 6121022; 5624821; 5648260; 6528624; 6194551; 6737056; 7122637; 7183387; 7332581; 7335742; 7371826; 6821505; 6180377; 7317091; 7355008. Другие модификации, и/или замены, и/или добавления, и/или делеции в Fc-домене будут полностью очевидны специалисту в данной области техники.
[0218] Примечательно, что полипептиды, представленные в формате BiSAb, содержащие нативный Fc, сохраняют способность связывать FcRn и C1q и опосредовать ADCC, как показано в примерах. Таким образом, в определенных аспектах BiSAb сохраняет способность связывать FcRn, и/или C1q, и/или один или несколько Fc-гамма-рецепторов (FcγR). Например, в некоторых аспектах BiSAb сохраняет по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% способности связывать FcRn, и/или C1q, и/или один или несколько FcγR по сравнению с обычным антителом, которое связывается с одним из эпитопов, с которым связывается BiSAb. В определенных аспектах BiSAb получают из связывающих доменов одного или двух обычных антител, и сравнение активности производится с одним или обоими из этих обычных антител.
[0219] Измененные Fc-области также могут использоваться для получения гетеродимеров тяжелой цепи, в результате чего BiSAb содержат две различные пары тяжелая-легкая цепь. Для облегчения образования гетеродимеров граница между парами Fc-областей сконструирована для максимизации процентной доли гетеродимеров, которые извлекаются из культуры рекомбинантных клеток. В определенных аспектах граница содержит по меньшей мере часть CH3-домена. В данном способе "ответвление" получают путем замены одной или нескольких небольших боковых цепей аминокислот в участке границы первой молекулы антитела на более крупные боковые цепи (например, тирозина или триптофана). Компенсационные "полости", имеющие одинаковый или сходный размер с большой(-ыми) боковой(-ыми) цепью(-ями) образуются в участке границы второй молекулы антитела путем замены больших боковых цепей аминокислот на меньшие цепи (например, аланина или треонина). Это обеспечивает механизм увеличения выхода гетеродимера по сравнению с другими нежелательными конечными продуктами, такими как гомодимеры. Модификации CH3 включают, например, Y407V/T366S/L368A на одной тяжелой цепи и T366W на другой тяжелой цепи; S354C/T366W на одной тяжелой цепи и Y349C/Y407V/T366S/L368A на другой тяжелой цепи. Дополнительные модификации, приводящие в результате к образованию ответвления на одной цепи и полости на другой, описаны в U.S. 7183076; US 2014/0348839 и Merchant et al., 1998, Nat. Biotech 16:677-681. Некоторые неограничивающие примеры модификаций, которые могут в результате приводить к образованию схемы ответвление-полость, представлены в таблице 1а. Другие модификации, которые могут применяться для получения гетеродимеров, включают без ограничения таковые, которые изменяют полярность заряда по участку границы Fc-димера таким образом, что совместная экспрессия электростатически соответствующих Fc-областей приводит к гетеродимеризации. Модификации, которые изменяют полярность заряда, включают без ограничения таковые, которые представлены в таблице 1b ниже (см. также US 2009/0182127; Gunasekaran et al., 2010, JBC 285:19637-46). Кроме того, Davis et al. (2010, Prot. Eng. Design & Selection 23:195-202) описывают гетеродимерную Fc-платформу с использованием CH3-областей с доменом, сконструированным посредством обмена нитей (SEED), которые являются производными CH3-доменов IgG и IgA человека (также см. WO 2007/110205).
Таблица 1a. Модификации CH3 для гетеродимеризации (ответвление-полость).
Таблица 1b. Модификации CH3 для гетеродимеризации
[0220] Специалисту в данной области техники будет понятно, что в некоторых аспектах Fc-слитый белок может образовывать димеры по причине гомодимерной природы молекул, содержащих Fc-область. В некоторых аспектах Fc-области связывающего белка (например, BiSAb) могут быть выборочно сконструированы с мутациями, чтобы способствовать и/или поддерживать гетеродимеризацию (например, химерные мутации, комплементарные мутации, мутации по типу "замок на причале", мутации по типу "выступ во впадине", мутации сконструированных доменов посредством обмена нитей (SEED) и т. д., см., например, патент США № 7183076, Merchant et al. (1998) Nat. Biotech 16:677-681; Ridgway et al. (1996) Protein Engineering 9:617-621; Davis et al. (2010) Prot. Eng. Design & Selection 23: 195-202; WO 2007/110205; WO 2007/147901; Gunasekaran et al. (2010) JBC 285: 19637-46, все включены в данный документ посредством ссылки). Соответственно, связывающий белок может быть сконструирован с образованием гетеродимера, содержащего, например, первый связывающий белок, связывающий домен или BiSAb, слитые с первой Fc-областью или ее фрагментом, и второй (т. е. отличающийся) связывающий белок, связывающий домен или BiSAb, слитые со второй Fc-областью или фрагментом, где первая и вторая Fc-области или их фрагменты были сконструированы для гетеродимеризации.
3. Гликозилирование
[0221] Помимо способности гликозилирования изменять эффекторную функцию полипептидов, модификация гликозилирования в вариабельной области может изменять аффинность антитела (или BiSAb) к антигену-мишени. В одном аспекте гликозилирование в вариабельной области BiSAb по настоящему изобретению является модифицированным. Например, может быть получен негликозилированный BiSAb (т. е. BiSAb с отсутствием гликозилирования). Гликозилирование может быть изменено, например, для повышения аффинности BiSAb к антигену-мишени. Такие углеводные модификации можно осуществить с помощью, например, изменения одного или нескольких сайтов гликозилирования в пределах последовательности BiSAb. Например, могут быть осуществлены одна или несколько аминокислотных замен, что приводит к удалению одного или нескольких сайтов гликозилирования из каркаса вариабельной области с устранением таким образом гликозилирования в данном сайте. Такое гликозилирование может повышать аффинность BiSAb к антигену. Такой подход более подробно описан в патентах США №№ 5714350 и 6350861. Также могут быть получены одна или несколько аминокислотных замен, что приводит к удалению сайта гликозилирования, присутствующего в Fc-области (например, аспарагина 297 из IgG). Кроме того, негликозилированные BiSAb могут быть получены в бактериальных клетках, которые не имеют необходимого аппарата гликозилирования.
4. Полипептидные линкеры
[0222] Линкеры могут применяться для присоединения доменов/областей химерной тяжелой цепи BiSAb к смежной молекуле. Как описано в данном документе, BiSAb может содержать один, два или больше линкерных полипептидов (например, L1 и L2). Кроме того, BiSAb может содержать дополнительные линкеры, такие как гибкий линкер, соединяющий между собой вариабельные области тяжелых и легких цепей scFv. Кроме того, BiSAb может содержать дополнительные линкеры, такие как гибкий линкер, соединяющий между собой вариабельные области тяжелых и легких цепей scFv, и другие линкеры, которые соединяют другие связывающие единицы со структурой ядра BiSAb.
[0223] Иллюстративным неограничивающим примером линкера является полипептидная цепь, содержащая по меньшей мере 4 остатка. Части таких линкеров могут быть гибкими, гидрофильными и характеризоваться своей собственной незначительной вторичной структурой или не иметь ее (части линкеров или части гибких линкеров). Линкеры из по меньшей мере 4 аминокислот могут применяться для соединения доменов и/или областей, которые расположены рядом друг с другом после сборки молекулы. Также могут применяться более длинные или более короткие линкеры. Таким образом, длина линкеров может составлять приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 25, 30, 35, 40, 45 или примерно 50 остатков. В случае, когда для соединения между собой частей молекулы используют несколько линкеров, линкеры могут быть одинаковыми или разными (например, иметь одинаковую или разную длину и/или аминокислотную последовательность).
[0224] Линкеры могут представлять собой расщепляемые линкеры, которые содержат по меньшей мере одну связь, которая может избирательно расщепляться реагентом для расщепления. Расщепляемые линкеры могут применяться для облегчения удаления всей линкерной последовательности или ее части. Линкеры могут быть сконструированы, чтобы содержать сайты расщепления протеазой таким образом, что расщепление происходит в средней части линкера или по меньшей мере на одном конце линкера. Например, сайты тромбина могут быть сконструированы на каждом из двух фланкирующих концов линкера. В зависимости от типа используемого линкера, расщепление может также быть опосредовано средствами, такими как TCEP, TFA и DTT. Линкеры могут быть сконструированы таким образом, чтобы реагенты для расщепления удаляли все остатки из линкера из продукта расщепления. Другие иллюстративные неограничивающие линкеры включают линкеры пролекарств, связи в которых могут избирательно расщепляться в условиях in vivo, например, в присутствии эндогенных ферментов или других эндогенных факторов или просто в водных жидкостях, присутствующих в организме или в клетках организма. Если BiSAb содержат более одного полипептидного линкера, каждый из линкеров может быть отличающимся или по меньшей мере один из линкеров может отличаться от других. В некоторых аспектах BiSAb содержит расщепляемый линкер. В конкретном аспекте BiSAb содержит scFv, где scFv содержит расщепляемый линкер между VH2 и VL2.
[0225] Линкер(линкеры) облегчают образование требуемой структуры. Линкеры могут содержать остатки (Gly-Ser)n, причем некоторые остатки Glu или Lys распределены по всей длине для увеличения растворимости. Как альтернатива или дополнительно, линкеры могут не содержать каких-либо остатков серина, причем такие линкеры могут быть предпочтительными, если линкер подвергается O-связанному гликозилированию. В некоторых аспектах линкеры могут содержать остатки цистеина, например, если димеризация линкеров используется для приведения доменов BiSAb в их правильно свернутую конфигурацию. В некоторых аспектах BiSAb содержит по меньшей мере два полипептидных линкера, которые соединяют домены полипептида. В других аспектах BiSAb содержит по меньшей мере три полипептидных линкера. В других аспектах BiSAb содержит четыре или больше полипептидных линкеров.
[0226] В некоторых аспектах полипептидный линкер содержит часть Fc-фрагмента. Например, в некоторых аспектах полипептидный линкер может содержать часть шарнирного домена иммуноглобулина из антитела IgG1, IgG2, IgG3 и/или IgG4. В некоторых аспектах полипептидный линкер содержит часть мутированного шарнирного домена иммуноглобулина из IgG1, IgG2, IgG3 и/или IgG4. В некоторых аспектах полипептидный линкер содержит по меньшей мере 5, 7, 8 или 15 аминокислотных остатков шарнирных области/домена иммуноглобулина из антитела IgG1, IgG2, IgG3 и/или IgG4. В некоторых аспектах полипептидный линкер содержит по меньшей мере 5, 7, 8 или 15 аминокислотных остатков модифицированных шарнирных области/домена иммуноглобулина из антитела IgG1, IgG2, IgG3 и/или IgG4.
[0227] Полипептидный линкер может содержать всю шарнирную область, которая в природных условиях содержит три цистеина, или ее часть. В определенных аспектах выбранная шарнирная область усечена или иным образом изменена или подвергнута замене относительно полной и/или встречающейся в природе шарнирной области таким образом, что остаются только один или два остатка цистеина. Аналогичным образом в некоторых других аспектах полипептидный линкер может содержать мутированную или иным образом измененную часть шарнирной области, в которой количество остатков цистеина уменьшено путем замены или делеции аминокислот, например, мутированную или иным образом измененную шарнирную область, содержащую ноль, один или два остатка цистеина, как описано в данном документе.
[0228] Таким образом, мутированный или иным образом измененный шарнирный домен может быть получен или сконструирован из (или с использованием) шарнирного домена иммуноглобулина дикого типа, который содержит один или несколько остатков цистеина. В определенных аспектах мутированная или иным образом измененная часть шарнирной области может содержать ноль или только один остаток цистеина, где мутированная или иным образом измененная шарнирная область представляет собой шарнирную область иммуноглобулина дикого типа, которая содержит соответственно один или несколько или два или более остатков цистеина, или была получена из нее. В мутированной или иным образом измененной части шарнирной области остатки цистеина из шарнирной области иммуноглобулина дикого типа предпочтительно удалены или заменены аминокислотами, которые неспособны образовывать дисульфидную связь. В некоторых аспектах мутированная или иным образом измененная часть шарнирной области представляет собой шарнирную область IgG дикого типа человека, который может включать любой из четырех подклассов изотипа IgG человека, IgG1, IgG2, IgG3 или IgG4, или была получена из нее.
[0229] В некоторых аспектах полипептидный линкер содержит часть шарнирной области, содержащую остаток цистеина, который образует дисульфидную связь с легкой цепью иммуноглобулина (остаток 220 согласно EU). В некоторых аспектах полипептидный линкер содержит измененную часть шарнирной области, содержащую аминокислотную замену в остатке C220 согласно EU. В некоторых аспектах полипептидный линкер содержит аминокислотную замену C220V.
[0230] В некоторых аспектах полипептидный линкер содержит аминокислотную замену, которая предотвращает шарнир-зависимое спонтанное саморасщепление. В некоторых аспектах полипептидный линкер содержит аминокислотную замену в положении D221 согласно EU. В некоторых аспектах полипептидный линкер содержит аминокислотную замену D221G. В некоторых аспектах полипептидный линкер не содержит аминокислоту D221.
[0231] Как обсуждалось выше, некоторые варианты осуществления предусматривают один или несколько полипептидных линкеров, которые содержат линкер gly-ser или состоят из него. Используемый в данном документе термин "линкер gly-ser" относится к пептиду, который состоит из остатков глицина и серина. Иллюстративный линкер gly-ser содержит аминокислотную последовательность формулы (Gly4Ser)n, где n представляет собой целое положительное число (например, 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10) включительно. Некоторые предпочтительные и неограничивающие примеры линкера gly-ser включают (Gly4Ser)2 (SEQ ID NO:41) и (Gly4Ser)4 (SEQ ID NO:42), а также (Gly4Ser)3 (SEQ ID NO:43). В еще других аспектах два или более линкеров gly-ser введены последовательно в полипептидный линкер. В некоторых аспектах полипептидный линкер содержит по меньшей мере часть шарнирной области (например, полученной из молекулы IgG1, IgG2, IgG3 или IgG4) и ряд аминокислотных остатков gly-ser (например, линкер gly-ser, такой как (Gly4Ser)n, где n равняется 2, 3 или 4).
[0232] В некоторых аспектах линкеры (например, L1, и/или L2, и/или L3 и т.д.) содержат как шарнирную часть, так и линкерную часть, такую как линкерная часть, содержащая линкер gly-ser. В других аспектах L1 и/или L2 содержат только шарнирную часть или только линкерную часть, такую как линкер gly-ser. В других аспектах L1 и L2 содержат часть линкера gly-ser. В некоторых аспектах линкер gly-ser в BiSAb характеризуется одинаковой длиной, тогда как в других аспектах части линкера gly-ser в BiSAb (например, L1 и L2) характеризуются разными значениями длины. В случае если BiSAb содержит scFv, тяжелая и легкая цепи scFv могут быть соединены с BiSAb (например, BD1, Fab, Fc и т. д.) гибким линкером. Этот гибкий линкер, как правило, не содержит шарнирной части, а скорее представляет собой линкер gly-ser или другой гибкий линкер. Длину и аминокислотную последовательность гибкого линкера, соединяющего между собой домены scFv, можно легко выбрать и оптимизировать (например, (Gly4Ser)n (SEQ ID NO:48), где n равняется 2, 3 или 4 или больше).
[0233] Независимо от полипептидного линкера, используемого для соединения между собой различных связывающих единиц и доменов (например, связывающих доменов/единиц между собой (например, Fab-scFv) или связывающих домена/единицы с Fc (например, scFv посредством L1 и L2), BiSAb может необязательно содержать дополнительные полипептидные линкеры. Значения длины и последовательность таких дополнительных полипептидных линкеров выбирают независимо. Например, BiSAb может дополнительно содержать гибкий полипептидный линкер, соединяющий между собой вариабельные области тяжелых и легких цепей scFv. Данный гибкий полипептидный линкер может содержать линкер gly-ser. Как правило, этот линкер не содержит шарнирной части.
[0234] В данном документе предполагается, что изменение длины линкеров, фланкирующих BD2, может влиять на ориентацию антигенсвязывающего сайта BD2 и пространственное распределение относительно остальной части молекулы BiSAb. Например, короткий N-концевой линкер и длинный C-концевой линкер могут приводить к ориентации, где связывающий сайт согласован в одном направлении, в то время как длинный N-концевой и короткий C-концевой линкеры могут придавать противоположную конформационную ориентацию. Соответственно, длина линкера может быть модулирована с целью ориентации антигенсвязывающего сайта BD2, и она обладает важными параметрами воздействия на появление или исключение стерических эффектов между BD1 и BD2 и/или BD2 и другими объектами, которые связывают молекулу антитела в Fc или других доменах.
5. Конкретная конфигурация BiSAb
[0235] Как обсуждалось выше, один аспект настоящего изобретения относится к структурной схеме (платформе) BiSAb, которая предусматривает две пары тяжелая-легкая цепь (проиллюстрировано на фигурах 1A-1F). В некоторых вариантах осуществления данного аспекта полипептидная последовательность химерной тяжелой цепи BiSAb может содержать полипептидную последовательность, содержащую вариабельный домен тяжелой цепи (VH1) антитела, полипептидную последовательность, содержащую константный домен 1 тяжелой цепи (CH1) антитела, часть Fc-домена, полипептидную последовательность, содержащую первый полипептидный линкер (L1), полипептидную последовательность, содержащую связывающий домен (BD2), полипептидную последовательность, содержащую второй полипептидный линкер (L2), и полипептидную последовательность, содержащую оставшуюся часть Fc-домена. В некоторых аспектах Fc-домен содержит CH2-домен и CH3-домен. Таким образом, некоторые варианты осуществления предусматривают химерную тяжелую цепь BiSAb, которая может содержать полипептидные последовательности в следующей ориентации от N-конца к C-концу: VH1-CH1-CH2(N-конец)-L1-BD2-L2-CH2(C-конец)-CH3; VH1-CH1-CH2-L1-BD2-L2-CH3 и VH1-CH1-CH2-CH3(N-конец)-L1-BD2-L2-CH3(C-конец). Полипептидная последовательность легкой цепи BiSAb может содержать вариабельный домен легкой цепи (VL1) и константный домен легкой цепи (CL). Таким образом, легкая цепь BiSAb может содержать полипептидную последовательность в следующей ориентации от N-конца к C-концу: VL1-CL. Следует отметить, что VH1, VL1 и CL используются для обозначения частей "связывающей единицы 1" (BD1), которая связывает первый эпитоп. BD2 используется для обозначения частей "связывающей единицы 2", которая связывает второй эпитоп.
[0236] В аспектах, где связывающий домен представляет собой scFv, химерная тяжелая цепь BiSAb может содержать полипептидную последовательность, содержащую вариабельный домен тяжелой цепи (VH1) антитела, полипептидную последовательность, содержащую константный домен 1 тяжелой цепи (CH1) антитела, полипептидную последовательность, содержащую первый полипептидный линкер (L1), полипептидную последовательность, содержащую вариабельный домен легкой цепи (VL2) антитела, полипептидную последовательность, содержащую гибкий линкер, полипептидную последовательность, содержащую вариабельный домен тяжелой цепи (VH2) антитела, полипептидную последовательность, содержащую второй полипептидный линкер (L2), и полипептидную последовательность, содержащую Fc-домен антитела. Таким образом, химерная тяжелая цепь BiSAb, содержащая scFv в качестве BD2, может содержать полипептидные последовательности в следующей ориентации от N-конца к C-концу: VH1-CH1-CH2(N-конец)-L1-VL2-L3-VH2-L2-CH2(C-конец)-CH3; VH1-CH1-CH2-L1-VL2-L3-VH2-L2-CH3; VH1-CH1-CH2-CH3(N-конец)-L1-VL2-L3-VH2-L2-CH3(C-конец); VH1-CH1-CH2(N-конец)-L1-VH2-L3-VL2-L2-CH2(C-конец)-CH3; VH1-CH1-CH2-L1-VH2-L3-VL2-L2-CH3 и VH1-CH1-CH2-CH3(N-конец)-L1-VH2-L3-VL2-L2-CH3(C-конец).
[0237] Химерная тяжелая цепь представляет собой полипептидную цепь, содержащую аминокислотную последовательность (например, аминокислотную последовательность каждого из полипептидных доменов). Химерная тяжелая цепь представляет собой полипептидную цепь, содержащую аминокислотную последовательность (например, аминокислотную последовательность каждого из полипептидных доменов). Следует отметить, что VH1, VL1 и CL используются для обозначения частей связывающей единицы 1, при этом VH1 и VL1 обозначают ту часть, которая связывает первый эпитоп. VH2 и VL2 используются для обозначения частей связывающей единицы 2, которая связывает второй эпитоп. В определенных аспектах дополнительные связывающие домены scFv присутствуют на N-концевых и/или С-концевых участках полипептидов, которые составляют ядро BiSAb (где ядро BiSAb дополнительно содержит связывающую единицу (BD) 3, и/или 4, и/или 5). В определенных аспектах в ядре BiSAb присутствует больше одного связывающего домена scFv. Каждый дополнительный scFv содержит вариабельную область тяжелой цепи антитела, обозначенную как VH3, VH4, VH5, и соответствующую вариабельную область легкой цепи антитела, обозначенную как VL3, VL4, VL5.
6. Метки, конъюгаты и фрагменты
[00238] В определенных признаках лекарственные средства и другие молекулы могут быть нацелены на BiSAb посредством сайт-специфической конъюгации. Например, BiSAb могут содержать сконструированные с цистеином домены (содержащие цистеин(цистеины), сконструированный(-ые) в связывающей единице и/или Fc-домене), что приводит к образованию свободных тиольных групп для реакций конъюгации. В определенных аспектах BiSAb сконструировано с включением специфических сайтов конъюгации. В некоторых аспектах настоящее изобретение предусматривает BiSAb с вариантным Fc, где Fc-область содержит аминокислотную замену в одном или нескольких положениях из 239, 282, 289, 297, 312, 324, 330, 335, 337, 339, 356, 359, 361, 383, 384, 398, 400, 440, 422 и 442 согласно нумерации в соответствии с EU-индексом. В некоторых аспектах Fc-область содержит замены в одной или нескольких из следующих групп положений: a) 289 и 440; b) 330 и 440; c) 339 и 440; d) 359 и 440; e) 289 и 359; f) 330 и 359; g) 339 и 359; h) 289 и 339; i) 330 и 339; j) 289 и 330; k) 339 и 442; l) 289, 339 и 442; m) 289, 330 и 339; n) 330, 339 и 442 и o) 289, 330 и 442. В других аспектах настоящее изобретение предусматривает BiSAb, где CH1-домен Fab-плеча содержит замену в одном или нескольких положениях из 131, 132, 134, 135, 136 и 139 согласно нумерации в соответствии с EU-индексом. В одном аспекте замена предусматривает замену на аминокислоту, выбранную из цистеина, лизина, тирозина, гистидина, селеноцистеина и селенометионина. В конкретном аспекте замена представляет собой замену на цистеин. Способы получения стабильных сконструированных с цистеином антител описаны в патентах США №№ 7855275, 2011/0033378 и 2012/0213705, содержание которых включено в данный документ посредством ссылки во всей их полноте.
7. Иллюстративные мишени
[00239] Хотя аспекты и варианты осуществления, относящиеся к разным платформам DuetMab и BiSAb, описанным в данном документе, могут быть получены для связывания с любой требуемой мишенью или мишенями, описанные в данном документе BiSAb предпочтительно нацеливаются на конкретные пары молекул-мишеней (например, связывающая единица 1 связывает одну из мишеней, а связывающая единица 2 связывает другую мишень). Как обсуждалось выше и как приведено в качестве примера в иллюстративных примерах ниже, антитела, DuetMab и BiSAb, раскрытые в данном документе, нацелены на молекулу, которая модулирует иммунный ответ у субъекта-реципиента или в иммунных клетках в культуре. В некоторых вариантах осуществления связывающий домен проявляет активность специфического связывания в отношении мишени, выбранной из группы, состоящей из CTLA-4, PD-1, PD-L1, OX40 и TIM3. DuetMab и BiSAb могут содержать комбинацию различных связывающих доменов в различных вариантах порядка и ориентации, где домены характеризуются аффинностью связывания в отношении мишеней, раскрытых в данном документе, или специфически связываются с ними. Например, раскрытые в данном документе DuetMab и BiSAb могут содержать комбинацию связывающих доменов, которые обеспечивают биспецифическое связывание с мишенями, в том числе CTLA-4 и PD-1; CTLA-4 и PD-L1; и CTLA-4; CTLA-4 и TIM3; PD-1 и PD-L1; PD-L1 и OX40; PD-1 и TIM3; PD-L1 и TIM3. DuetMab и BiSAb, которые содержат связывающие домены, которые связывают конкретные комбинации мишеней, проиллюстрированы в примерах и включают неограничивающие комбинации PD-1/CTLA-4; PD-L1/CTLA-4; PD-1/TIM3 и PD-L1/OX40. В определенных вариантах осуществления BiSAb характеризуются улучшенными связывающими свойствами по сравнению со связывающими свойствами комбинации отдельных моноспецифических связывающих белков, которые используются для получения BiSAb.
[00240] В определенных аспектах DuetMab или BiSAb по настоящему изобретению связывают два разных эпитопа на одной и той же мишени (например, связывающая единица 1 связывает первый эпитоп на мишени, а связывающая единица 2 связывает второй эпитоп на той же мишени).
[00241] В некоторых аспектах мультимерная природа DuetMab или BiSAb по настоящему изобретению дает возможность нацеливать метки или терапевтические препараты на конкретный тип клеток или молекулярную мишень. Например, один функциональный домен в DuetMab или BiSAb может связываться с мишенью на поверхности клетки, тогда как другой функциональный домен в том же DuetMab или BiSAb связывается с гаптеном или средством для мечения, применимыми для обнаружения. Аналогичным образом, один функциональный домен может связываться с клеточной мишенью, тогда как второй функциональный домен связывается с токсином. Поскольку обе реакции связывания опосредованы одной молекулой, то токсин может быть расположен вблизи клеточной мишени, где он оказывает влияние на цитотоксическую функцию.
B. Молекулы нуклеиновой кислоты, кодирующие BiSAb
[0242] Настоящее изобретение предусматривает молекулы нуклеиновой кислоты, которые кодируют BiSAb. Один аспект настоящего изобретения предусматривает молекулы нуклеиновой кислоты, кодирующие любой из BiSAb по настоящему изобретению. Молекула нуклеиновой кислоты может кодировать тяжелую цепь и/или легкую цепь любой из молекул BiSAb, которые раскрыты в данном документе, а также любой из отдельных связывающих доменов (например, scFv), которые раскрыты в данном документе. Специалисту в данной области техники будет понятно, что такие полинуклеотидные молекулы могут различаться в нуклеотидной последовательности, принимая во внимание вырожденность кодонов данной нуклеиновой кислоты, а также частоту кодонов для конкретных организмов, как обычно известно из уровня техники.
C. Векторы и клетки-хозяева для получения BiSAb и последующей очистки
[00243] Настоящее изобретение относится к способам получения BiSAb. В некоторых аспектах молекулы рекомбинантной нуклеиновой кислоты, которые кодируют полностью или часть BiSAb, раскрытых в данном документе, могут быть функционально связаны с одной или несколькими регуляторными нуклеотидными последовательностями в конструкции для экспрессии. Последовательности нуклеиновой кислоты, кодирующие легкие и химерные тяжелые цепи BiSAb, могут быть клонированы в один и тот же вектор экспрессии в любой ориентации (например, легкая цепь перед тяжелой цепью или наоборот) или могут быть клонированы в два разных вектора. Если экспрессия осуществляется с использованием одного вектора, то два кодирующих гена могут иметь свои собственные генетические элементы (например, промотор, RBS, лидер, стоп, полиА и т. д.), или они могут быть клонированы с одним единственным набором генетических элементов, но связаны с цистронным элементом. Как правило, регуляторные нуклеотидные последовательности будут являться подходящими для клетки-хозяина, используемой для экспрессии. Множество типов соответсвующих векторов экспрессии и подходящих регуляторных последовательностей являются известными из уровня техники для множества клеток-хозяев. Как правило, указанные одна или несколько регуляторных нуклеотидных последовательностей могут включать без ограничения промоторные последовательности, лидерные или сигнальные последовательности, сайты связывания рибосомы, последовательности инициации и терминации транскрипции, последовательности инициации и терминации трансляции и последовательности энхансеров и активаторов. В настоящем изобретении предусмотрены конститутивные или индуцируемые промоторы, известные из уровня техники. Промоторы могут представлять собой либо промоторы, встречающиеся в природе, либо гибридные промоторы, которые объединяют элементы из более чем одного промотора. Конструкция для экспрессии может присутствовать в клетке на эписоме, такой как плазмида, или конструкция для экспрессии может быть вставлена в хромосому.
[00244] В определенных аспектах вектор экспрессии содержит ген селектируемого маркера для обеспечения возможности отбора трансформированных клеток-хозяев. Гены селектируемых маркеров хорошо известным из уровня техники и будут варьироваться в зависимости от используемой клетки-хозяина. В определенных аспектах данное изобретение относится к вектору экспрессии, содержащему нуклеотидную последовательность, кодирующую полипептид и функционально связанную с по меньшей мере одной регуляторной последовательностью. Регуляторные последовательности являются принятыми в данной области техники и выбраны для управления экспрессией кодируемого полипептида. Соответственно, термин "регуляторная последовательность" включает промоторы, энхансеры и другие элементы контроля экспрессии. Иллюстративные неограничивающие регуляторные последовательности описаны в Goeddel; Gene Expression Technology: Methods in Enzymology, Academic Press, San Diego, CA (1990). Следует понимать, что конструирование вектора экспрессии может зависеть от таких факторов, как выбор клетки-хозяина, подлежащей трансформации, и/или тип белка, экспрессия которого требуется. Более того, следует также рассматривать количество копий конкретного вектора, способность контролировать данное количество копий и экспрессию любого другого белка, кодируемого вектором, такого как маркеры устойчивости к антибиотикам.
[00245] Настоящее изобретение дополнительно относится к способам получения BiSAb по настоящему изобретению. Например, клетка-хозяин, трансфицированная с помощью одного или нескольких векторов экспрессии, кодирующих BiSAb (например, одного вектора, кодирующего химерную тяжелую и легкую цепь, или двух векторов - одного, кодирующего химерную тяжелую цепь, и одного, кодирующего легкую цепь), может быть культивирована в подходящих условиях, чтобы обеспечивать осуществление экспрессии полипептида. BiSAb может быть секретирован и выделен из смеси клеток и среды, содержащей полипептид. Как альтернатива, BiSAb может оставаться в цитоплазме или в мембранной фракции, а клетки могут быть собраны, лизированы, а белок выделен. Культура клеток включает клетки-хозяева, среды и другие побочные продукты. Подходящие среды для культуры клеток хорошо известны из уровня техники. BiSAb могут быть выделены из среды для культивирования клеток, клеток-хозяев или из того и другого с применением методик, известных из уровня техники для очистки белков, в том числе ионообменной хроматографии, гель-фильтрационной хроматографии, ультрафильтрации, электрофореза и иммуноаффинной очистки. В определенных аспектах BiSAb получают в виде слитого белка, содержащего домен, который облегчает его очистку.
[00246] Рекомбинантная нуклеиновая кислота может быть получена путем лигирования клонированного гена или его части в вектор, подходящий для экспрессии либо в прокариотических клетках, либо в эукариотических клетках (дрожжей, птиц, насекомых или млекопитающих), либо и в тех и других. Средства экспрессии для получения рекомбинантного полипептида предусматривают плазмиды и другие векторы. К примеру, подходящие векторы включают плазмиды следующих типов: pBR322-производные плазмиды, pEMBL-производные плазмиды, pEX-производные плазмиды, pBTac-производные плазмиды и pUC-производные плазмиды для экспрессии в прокариотических клетках, таких как E. coli. В определенных аспектах векторы экспрессии для млекопитающих содержат как прокариотические последовательности для облегчения размножения вектора в бактериях, так и одну или несколько эукариотических транскрипционных единиц, которые экспрессируются в эукариотических клетках. Векторы-производные pcDNAI/amp, pcDNAI/neo, pRc/CMV, pSV2gpt, pSV2neo, pSV2-dhfr, pTk2, pRSVneo, пМSG, pSVT7, pko-neo и pHyg представляют собой примеры векторов экспрессии для млекопитающих, подходящих для трансфекции эукариотических клеток. Некоторые из этих векторов модифицируют последовательностями из бактериальных плазмид, таких как pBR322, для облегчения репликации и отбора по устойчивости к лекарственному средству как у прокариотических, так и у эукариотических клеток. В качестве альтернативы производные вирусов, таких как вирус папилломы крупного рогатого скота (BPV-1) или вирус Эпштейна-Барра (pHEBo, pREP-производный и p205), могут использоваться для временной экспрессии белков в эукариотических клетках. Различные способы, применяемые в получении плазмид и в трансформации организмов-хозяев, известны из уровня техники. В отношении других подходящих систем экспрессии как для прокариотических, так и для эукариотических клеток, а также общих процедур рекомбинации см. Molecular Cloning A Laboratory Manual, 2nd Ed., ed. by Sambrook, Fritsch and Maniatis (Cold Spring Harbor Laboratory Press, 1989), главы 16 и 17. В некоторых случаях может потребоваться экспрессировать рекомбинантный полипептид с использованием бакуловирусной системы экспрессии. Примеры подобных бакуловирусных систем экспрессии включают pVL-производные векторы (такие как pVL1392, pVL1393 и pVL941), pAcUW-производные векторы (такие как pAcUW1), и pBlueBac-производные векторы (такие как pBlueBac III, содержащие ген ß-галактозидазы).
[00247] Непосредственно после получения молекулы она может быть очищена любым способом, известным из уровня техники для очистки белка, молекулы иммуноглобулина или других мультимерных молекул, с использованием методик, таких как, например, хроматография (например, ионообменная, аффинная, в частности, по аффинности к специфическим антигенам, белку A или белку G, и колоночная хроматография размеров), центрифугирование, дифференциальная растворимость, или с помощью любой другой стандартной методики очистки белков. Далее, раскрытые в данном документе молекулы могут быть слиты с гетерологичными полипептидными последовательностями (например, аффинными метками), которые обычно используются для облегчения очистки.
[00248] Независимо от того, как получают и очищают BiSAb, для подтверждения функциональной активности связывания BiSAb могут быть проведены анализы связывания, например, анализы с помощью двойного ELISA (до и/или после очистки). Как правило, такие анализы связывания известны из уровня техники.
D. Фармацевтические составы
[00249] В определенных аспектах настоящее изобретение предусматривает фармацевтические композиции. Такие фармацевтические композиции могут представлять собой композиции, содержащие молекулу нуклеиновой кислоты, которая кодирует BiSAb. Такие фармацевтические композиции могут также представлять собой композиции, содержащие DuetMab, BiSAb, комбинацию DuetMab или комбинацию BiSAb и фармацевтически приемлемое вспомогательное вещество. В определенных аспектах фармацевтические композиции по настоящему изобретению применяются в качестве лекарственного препарата.
E. Пути применения
[00250] Как обсуждалось в данном документе, DuetMab и BiSAb могут использоваться для связывания мишеней, ассоциированных с раковыми и клеточнопролиферативными заболеваниями или расстройствами, которые могут давать ответную реакцию на иммунотерапию, например, путем подавления иммуносупрессорной активности и/или путем индуцирования иммунного ответа, который связан с молекулой(молекулами)-мишенью(мишенями). Например, аберрантная передача сигналов и/или подавленный иммунный ответ могут способствовать нежелательной клеточной пролиферации и раку. Соответственно, DuetMab, BiSAb и антитела, раскрытые в данном документе, могут применяться для лечения нежелательной клеточной пролиферации и/или рака, ассоциированных с подавленным, сниженным или недостаточным иммунным ответом, нацеленным против рака. В частности, кривая опухолевого роста для опухоли и/или объема опухоли может быть снижена путем введения DuetMab BiSAb, которые индуцируют и/или стимулируют иммунный ответ у субъекта, такого как, например, пациент-человек, страдающий от рака.
[00251] Таким образом, настоящее изобретение также относится к различным способам, которые предусматривают введение связывающих белков, раскрытых в данном документе, нуждающемуся в этом субъекту. В одном аспекте настоящее изобретение относится к способу индуцирования иммунного ответа у субъекта, имеющего рак или подверженного риску его развития, предусматривающему введение субъекту связывающего белка, раскрытого в данном документе. В некоторых вариантах осуществления с помощью способа активируют иммунный ответ против рака у субъекта. В некоторых вариантах осуществления с помощью способа усиливают иммунный ответ против рака у субъекта. В некоторых вариантах осуществления с помощью способа активируют путь иммунного ответа, который подавлен у субъекта, причем активация усиливает иммунный ответ, который нацелен на рак у субъекта. В некоторых вариантах осуществления с помощью способа улучшают путь иммунного ответа, который нацелен на рак у субъекта.
[00252] В другом аспекте настоящее изобретение относится к способу лечения рака у нуждающегося в этом субъекта, включающему введение субъекту связывающего белка, раскрытого в данном документе. В одном варианте осуществления способ лечения рака предусматривает прекращение или замедление роста рака у субъекта. В одном варианте осуществления способ лечения рака предусматривает прекращение или замедление метастазирования рака в другие области у субъекта. В одном варианте осуществления способ лечения рака предусматривает уничтожение раковых клеток у субъекта. В одном варианте осуществления способ лечения рака предусматривает приостановление пролиферации и/или распространения раковых клеток у субъекта.
[00253] В различных вариантах осуществления вышеуказанных аспектов способы относятся к лечению у субъекта опухолевого заболевания и/или ракового заболевания. В вариантах осуществления рак выбран из группы форм рака, которые восприимчивы к иммунному ответу, индуцированному у субъекта. В некоторых вариантах осуществления рак представляет собой один или несколько из рака яичников, рака молочной железы, колоректального рака, рака предстательной железы, рака шейки матки, рака матки, рака яичек, рака мочевого пузыря, рака головы и шеи, меланомы, рака поджелудочной железы, почечно-клеточной карциномы или рака легких. В некоторых вариантах осуществления рак выбран из форм рака пищеварительной системы или желудочно-кишечного тракта (например, рака анального отверстия; рака желчного протока; рака внепеченочного желчного протока; рака аппендикса; карциноидной опухоли, желудочно-кишечного рака; колоректального рака; колоректального рака, в том числе колоректального рака у детей; рака пищевода, в том числе рака у детей; рака желчного пузыря; рака желудка (желудочного рака), в том числе рака желудка у детей; гепатоцеллюлярного рака (например, гепатоцеллюлярной карциномы), в том числе (первичного) гепатоцеллюлярного рака у взрослых и гепатоцеллюлярного рака у детей; рака поджелудочной железы, в том числе рака поджелудочной железы у детей; саркомы, рабдомиосаркомы; рака островковых клеток поджелудочной железы; рака прямой кишки и рака тонкого кишечника); рака легкого (например, немелкоклеточного рака легкого (NSCLC) и мелкоклеточного рака легкого (SCLC)); рака головы и шеи (например, рака губы и ротовой полости; рака ротовой полости, в том числе рака ротовой полости у детей; гипофарингеального рака; рака гортани, в том числе рака гортани у детей; метастатического плоскоклеточного рака шеи со скрытой первичной опухолью; рака рта; рака носовой полости и околоносовых пазух; рака носоглотки, в том числе рака носоглотки у детей; рака ротоглотки; рака паращитовидной железы; рака глотки; рака слюнных желез, в том числе рака слюнных желез у детей; рака горла и рака щитовидной железы); рака яичника и молочной железы.
[00254] В вышеприведенных способах количество связывающего белка, которое вводится субъекту, является эффективным для индуцирования иммунного ответа, усиления иммунного ответа, прекращения или замедления роста рака, прекращения или замедления метастазирования рака, уничтожения раковых клеток и/или замедления или прекращения пролиферации и/или распространения раковых клеток у субъекта.
[00255] В вариантах осуществления вышеуказанных способов связывающий белок содержит DuetmAb или BiSAb, раскрытые в данном документе. В некоторых вариантах осуществления вышеуказанных способов связывающий белок содержит антитело или его антигенсвязывающий фрагмент, раскрытые в данном документе.
[00256] Используемый в данном документе термин "субъект" включает человека и животных, отличных от человека, в частности, млекопитающих. Примеры субъектов включают субъектов-людей, например, пациента-человека, у которого имеется расстройство, например, расстройство, описанное в данном документе, такое как рак, или здорового субъекта. Термин "животное, отличное от человека", включает всех позвоночных, например, не относящихся к млекопитающим (таких как куры, амфибии, рептилии), и млекопитающих, таких как приматы, отличные от человека, домашние и/или сельскохозяйственные животные (такие как овцы, собаки, кошки, коровы, свиньи и т.д.) и грызуны (такие как мыши, крысы, хомяки, морские свинки и т.д.). В конкретных вариантах осуществления субъект представляет собой пациента-человека.
[00257] Термины "лечение" или "лечить" относятся как к терапевтическому лечению, так и к профилактическим или превентивным мерам. К субъектам, нуждающимся в лечении, относят тех, у которых уже есть расстройство, а также тех, которые предрасположены к расстройству, или тех, у которых требуется предупредить развитие расстройства. В случае использования со ссылкой на заболевание или субъекта, нуждающегося в лечении, термины соответственно включают без ограничения приостановление или замедление прогрессирования заболевания, ремиссию заболевания, профилактику симптомов, уменьшение тяжести заболевания и/или симптома или уменьшение продолжительности заболевания по сравнению с субъектом, не получавшим лечение. В вариантах осуществления с помощью способов лечения можно ослаблять один или несколько клинических признаков конкретного заболевания, лечение которого осуществляют.
[00258] Приведенные ниже примеры приведены для иллюстрации конкретных аспектов и вариантов осуществления настоящего изобретения и не должны интерпретироваться как ограничивающие объем описания или объект изобретения, заявляемый в прилагаемой формуле изобретения.
Примеры
Материалы и способы
[00259] Анализ модуляции иммунного ответа
[00260] Анализ вторичного иммунного ответа на антиген цитомегаловируса (CMV) использовали для оценки потенциального иммунного ответа, индуцируемого некоторыми из описанных в данном документе иммунотерапевтических молекул. Реагенты для анализа включают:
- замороженные мононуклеарные клетки периферической крови (РВМС), реагирующие с CMV;
- среду AIM V® (Life Technologies, № по кат. 12055-091);
- фосфатно-солевой буферный раствор (PBS, Life technologies, № по кат. 20012-043);
- PepTivator®, пептидный пул CMVpp65, (Miltenyi Biotec, № по кат. 130-093-438, 50 мкг/мл);
- овальбумин, (Thermo scientific № по кат. 77120, 1 мг/мл);
- Costar, 96-луночный планшет, не обработанный TC (Corning, № по кат. 3788); и
- иммунотерапевтическую молекулу.
[00261] Общий протокол анализа
[00262] За день до проведения анализа замороженные РВМС оттаивали в теплой среде AIM V. Клетки промывали дважды в 96-луночном планшете Costar с круглым дном лунок. Концентрацию клеток доводили до концентрации, составляющей 1×106 клеток/мл.
[00263] Аликвоты (100 мкл) клеток распределяли по отдельным лункам, оставляя внешние колонки и ряды планшета пустыми. Клетки оставляли в состоянии покоя в течение ночи.
[00264] На следующий день в лунки добавляли 100 мкл среды AIMV, содержащей 2X пептидный пул CMV PepTivator (конечная концентрация 0,1 мкг/мл - 0,05 мкг/мл) и 2X иммунотерапевтическую молекулу.
[00265] Через 72 часа 25 мкл надосадочной жидкости из каждой лунки переносили на предварительно блокированные и промытые планшеты MSD (антитело к IFN-гамма человека). После добавления стандартов планшеты инкубировали в течение 2 часов при комнатной температуре. После инкубационного периода планшеты MSD промывали три раза. После промывания добавляли 25 мкл антитела для выявления SULFO-TAG и обеспечивали протекание реакции в течение 1 часа при комнатной температуре. Планшеты снова промывали и добавляли 150 мкл 2X буфера для считывания MSD до того, как были проделаны считывания.
[00266] Протокол анализа стафилококкового энтеротоксина A/B (SEA/SEB)
[00267] Реагенты, используемые в протоколе анализа либо SEB, либо SEA для определения эффекта DuetMab или BiSAb в отношении иммунного ответа IL-2, включают:
- конические лейкоцитарные фильтры (код NC24 NHSBT; из Addenbrookes Hospital);
- 50 мл пробирки Falcon (BD 352070);
- Ficoll-Paque PLUS (GE Healthcare 17-1440-02);
- антитело к CD3 (клон OKT3; 1 мг/мл; eBioscience; № по кат.:16-0037-85);
- раствор хлорида аммония (Stemcell Technologies 07850);
- исходные растворы стафилококкового энтеротоксина A (SEA; Sigma, S-9399) или стафилококкового энтеротоксина B (SEB; Sigma, S-4881) при 1 мг/мл, хранили при -20°C;
- культуральные среды (все от Life Technologies): RPMI1640 с глутамаксом (61870) с добавлением 10% об./об. инактивированной нагреванием FCS (90005M) и 100 ЕД/мл пенициллина+100 мкг/мл стрептомицина (15140-122);
- планшет с лунками с V-образным дном (Greiner BioOne 651201);
- 96-луночные планшеты с плоскодонными лунками (Corning Costar 7107).
[00268] Реагенты для ELISA DELFIA IL-2 включают:
- планшеты ELISA Maxisorp FLUONUNC (№ по кат. 437958);
- стрептавидин, меченый европием, SA-Eu (Perkin-Elmer 1244-360);
- аналитический буфер DELFIA® (Perkin-Elmer, № 4002-0010);
- усиливающий раствор DELFIA® (Perkin-Elmer 4001-0010); при к.т. перед использованием;
- разбавитель для анализа: промывочный буфер DELFIA (0,05% Tween-20, 20 мМ Трис, 150 мМ NaCl, pH 7,2-7,4), дополненный 0,1% BSA, стерилизованный фильтрацией;
- сухое молоко (Marvel; Premier Foods);
- разбавитель образца (RPMI1640+10% FCS+1% пенициллин/стрептомицин, как указано выше);
- PBS (ThermoFisher 14190235);
- PBS-Tween (0,01% Tween-20 в PBS);
- набор для ELISA IL-2 человека (Duoset DY202, R&D Systems);
- устройство для промывки планшетов Biotek (EL406) с автоматическим загрузчиком планшетов (Biostack).
[00269] Общий протокол анализа
[00270] РВМС выделяли из конических лейкоцитарных фильтров крови человека (Служба крови и трансплантации NHS, код NC24) с использованием центрифугирования с градиентом плотности (Ficoll-Paque PLUS; GE Healthcare), затем эритроциты лизировали в растворе хлорида аммония (Stemcell Technologies). Антителом к CD3 человека (клон OKT3 при 0,5 мкг/мл в PBS, eBioscience) покрывали 96-луночные планшеты с плоскодонными лунками (Corning Costar 7107) в течение 2 ч при 37°C. Затем в каждую лунку добавляли по 0,2×106 клеток РВМС в культуральных средах (RPMI1640-глутамакс, дополненная 10% об./об. инактивированной бычьей сывороткой и 100 ЕД/100 мкг на мл стрептомицина/пенициллина (соответственно) (Life Technologies). PBMC дополнительно стимулировали добавлением стафилококкового энтеротоксина A или B (SEB; Sigma Aldrich) в диапазоне 0,0088-0,1 мкг/мл, а кандидатные DuetMab или BiSAb добавляли до конечных тестируемых концентраций. После 3-дневного культивирования при 37°C и 5% CO2 надосадочные жидкости удаляли из клеток и секрецию IL-2 определяли с использованием коммерческого ELISA в соответствии с инструкциями производителя (R&D Systems Duoset, код продукта DY202). См. фигуру 90.
[00271] Протокол анализа реакции смешанных лейкоцитов (MLR) (свежая кровь)
[00272] MLR-анализ на основе клеток также использовали для получения in vitro корреляции функции Т-клеток в ответ на раскрытые в данном документе DuetMab и BiSAb. Реагенты, используемые при проведении MLR-анализа на свежих образцах крови, включают:
- 8 мл CPT-пробирки с гепарином;
- среду AIM-V (без сыворотки) Gibco № 12055-091, без добавок;
- 50 мл конические пробирки;
- 2 мл флаконы для криоконсервации;
- лизирующий буфер ACK (Gibco № A10492-01);
- 96-луночные обработанные планшеты для культуры тканей BD Falcon с U-образным дном, № 3077;
- PHA (Roche), 1 мг/мл (конечная концентрация 10 мкг/мл), в качестве положительного контроля.
[00273] Общий протокол анализа
[00274] PBMC получали из образцов крови, втянутых в пробирки CPT с гепарином. Пробирки центрифугировали в течение 20 мин. при 2700 об/мин без торможения при 25°C. Верхний слой из сыворотки отсасывают. Оставшийся материал аккуратно пипетировали и все, что находилось над пробкой пробирки CPT, собирали и помещали в 50 мл конические пробирки. К клеткам добавляли среду AIM-V для промывки клеток (3 раза при 1500 об/мин с торможением при 25°C в течение 5 минут). Какие-либо оставшиеся эритроциты лизировали с использованием лизирующего буфера для эритроцитов (например, приблизительно 5 мин с приблизительно 3 мл буфера). Оставшиеся клетки дважды промывали средой AIM-V (1500 об/мин с торможением при 25°С в течение 5 минут). При необходимости осадки объединяли в одной пробирке и ресуспендировали в среде AIM-V и осуществляли подсчет клеток.
[00275] Для проведения анализа MLR клетки помещали в 96-луночные планшеты с 200000 клеток на донора на лунку в среде AIM-V, 50 мкл на донора (всего 400000/100 мкл). Добавляли кандидатные молекулы (4х) по 50 мкл на лунку, разбавленные в бессывороточной среде AIM-V. Через 72 ч планшеты визуализировали, а 30 мкл надосадочной жидкости удаляли для анализа цитокинов человека TH1/TH2 (MSD).
[00276] 10-плексный протокол MSD TH1/TH2 человека
[00277] Этот анализ использовали для определения количеств цитокинов, присутствующих в культуральных надосадочных жидкостях, в ответ на введение раскрытых в данном документе DuetMab и BiSAb. Для проведения анализа блокирующее средство получали путем растворения 200 мг блокатора В в 20 мл PBS на планшет. В каждую лунку добавляли по 150 мкл растворенного блокатора. Планшет герметизировали и встряхивали в течение 2 ч при комнатной температуре или в течение ночи при 4°C. Лунки промывали 3x буфером PBST. Калибратор получали путем разбавления 10 мкл смеси замороженного калибратора в 1 мл разбавителя и дополнительно 4-кратно последовательно разбавляли. Для разделения лунок добавляли 25 мкл калибратора (стандарта) и 25 мкл образцов. Лунки инкубировали в течение 2 ч при комнатной температуре со встряхиванием. После инкубации лунки промывали 3x с помощью PBST.
[00278] Антитело для выявления получали и разбавляли до необходимой концентрации и добавляли в каждую лунку. После 2 ч инкубации при комнатной температуре со встряхиванием лунки промывали (3x) в PBST. Перед считыванием на оборудовании для MSD в каждую лунку добавляли буфер для считывания.
[00279] Протокол анализа специфического уничтожения опухоли
[00280] Линия CD8+ T-клеток человека (JR6C12) с реакционной способностью к пептиду gp100209-217 человека любезно предоставлена доктором Стивеном Розенбергом (Национальный институт рака, Бетезда, Мэриленд). Клетки JR6C12 совместно культивировали с линией клеток меланомы человека (Mel624), меченных CFSE (набор CellTrace CFSE для анализа пролиферации клеток, ThermoFisher), в течение 18 часов при 37°C при соотношении 1:1 (20000 JR6C12+20000 Mel624) в 96-луночном планшете с плоскодонными лунками. Кандидатные молекулы добавляли в момент времени 0 совместного культивирования при концентрации 69 нМ. Через 18 ч лунки визуализировали с помощью микроскопии по методу светлого поля. Надосадочные жидкости собирали для анализа MSD, а адгезивные клетки трипсинизировали и промывали (2x) в PBS перед окрашиванием для выявления жизнеспособности (набор Zombie UV Fixable Viability, Biolegend). Устойчивое поглощение красителя клетками, меченными CFSE, оценивали с помощью проточной цитометрии на LSRFortessa (BD).
Пример 1. Идентификация кандидатных участков расположения Fc для присоединения связывающего домена
[00281] С использованием системы для молекулярной визуализации PyMOL с открытым исходным кодом структуру антител исследовали в CH2- и CH3-областях, а также на границе CH2-CH3 или вблизи нее для того, чтобы идентифицировать кандидатные области, такие как гидрофильные петли, для присоединения связывающего домена. Такие области можно было бы приспособить для вставки второго связывающего домена (например, scFv) без ущерба для структурной целостности или стабильности IgG или самого второго связывающего домена. В результате анализа идентифицировали три области (представленные в виде сфер на фигурах 1A-1C). На фигурах 1D, 1E и 1F изображены варианты осуществления связывающих белков, на которых показано прикрепление второго связывающего домена (с scFv в целях иллюстрации) в каждой из петель, обозначенных на фигурах 1A, 1B и 1C соответственно.
[00282] На фигуре 2A представлено более подробное схематическое изображение аминокислотной последовательности одной из идентифицированных типичных петель, определенной в CH2-области около границы CH2-CH3 и содержащей последовательность ISRTP (SEQ ID NO:39). Связывающий домен может быть вставлен в данной аминокислотной последовательности для получения любого количества типичных конструкций, таких как, например, вставка scFv-доменов, как проиллюстрировано в примерах (например, I-scFv-SRTP, IS-scFv-RTP, ISR-scFv-TP или ISRT-scFv-P scFV-ISRTP и ISRTP-scFV), где "-scFv-" определяет точку в нативной последовательности петли, с которой может быть связан связывающий домен. На фигуре 2B представлено аналогичное схематическое изображение, которое представляет петлю, идентифицированную на границе CH2-CH3 и содержащую аминокислотную последовательность AKGQP (SEQ ID NO:40). Типичные конструкции, описанные в данном документе, могут содержать связывающий домен (например, такой как scFv-домен), присоединенный к этой последовательности петли, как описано в данном документе, в том числе A-scFv-KGQP, AK-scFv-GQP, AKG-scFv-QP, AKGQ-scFv-P, scFV-AKGQ, AKGQ-scFV, где "-scFv-" определяет точку в нативной последовательности петли, с которой может быть связан связывающий домен. На фигуре 2C представлено схематическое изображение типичной петли, идентифицированной ниже границы CH2-CH3 в CH3-области и содержащей аминокислотную последовательность SNG. Типичные конструкции для данной последовательности петли, обсуждаемые в отношении иллюстративных вариантов осуществления других двух вышеуказанных областей петель, включают scFV-SNG, S-scFv-NG, SN-scFv-G и SNG-scFV.
Пример 2. Получение и определение характеристик серии исходных антител и биспецифических связывающих белков, в том числе комбинации связывающих единиц
[00283] Разрабатывали и определяли характеристики серии моноклональных антител. С использованием комбинаций антигенсвязывающих последовательностей (например, CDR, HCv, LCv, HC, LC), полученных из этих "исходных" антител, получали серию биспецифических связывающих белков, и было показано, что они характеризуются биспецифической связывающей активностью в отношении комбинированных антигенов-мишеней. Биспецифические связывающие белки разрабатывали таким образом, чтобы они имели мотив конкретной структурной платформы, который раскрыт в данном документе (т.е. "BiS5").
[00284] Последовательности исходных антител описаны в следующих таблицах.
Таблица 2a. Последовательности исходных антител
PD-1
CTLA-4
мишень
PD-L1
мишень
HCv
LC
LCv
(MEDI4736) HCv
(MEDI4736)
LC
(MEDI4736) LCv
TIM3
мишень
OX40
мишень
Таблица 2b. Последовательности антигенов
человека
человека
человека
человека
человека
Пример 2(a). Биспецифические связывающие белки для связывания PD-1/CTLA-4
[00285] Не ограничиваясь теорией, существует сильное клиническое и доклиническое обоснование комбинации блокады PD-1 и CTLA-4. Таким образом, было бы желательно максимизировать соотношение риск/польза для комбинации PD-1 и CTLA-4 (фигура 4).
[00286] Следующие биспецифические связывающие белки, которые связывают PD-1 и CTLA-4, создавали с использованием исходных последовательностей, указанных выше в таблице 2. Белки, обозначенные как Bis2, Bis3 и Bis5, получали с указанными ниже последовательностями и оценивали в отношении одновременной антигенсвязывающей активности с использованием анализа связывания с помощью Octet, как обсуждается ниже.
Таблица 3. Конструкции BiS для связывания PD-1/CTLA-4
Bis2
для связывания PD-1/CTLA-4
Bis2
для связывания PD-1/CTLA-4
Bis3
для связывания PD-1/CTLA-4
Bis3
для связывания PD-1/CTLA-4
Bis5
для связывания PD-1/CTLA-4
Bis5
для связывания PD-1/CTLA-4
[00287] Анализ связывания с помощью Octet (BiS2, BiS3 и BiS5)
[00288] Для оценки связывания раскрытых в данном документе биспецифических связывающих молекул применяли Octet QK, снабженный наконечниками биосенсора Ni-NTA, и 10X буфер для кинетического анализа (ForteBio, Менло-Парк, Калифорния). Для данной конкретной серии биспецифических связывающих белков His-меченый PD-L1-Fc, his-меченый PD-1-Fc и CTLA-4-Fc (рекомбинантные белки человека) приобретали у R&D Systems (Миннеаполис, Миннесота). Все анализы связывания осуществляли при 25°C.
[00289] Перед анализом планшеты для образцов перемешивали при 1000 об/мин. Наконечники биосенсора Ni-NTA предварительно смачивали в течение 5 мин в 1X кинетическом буфере. 1X кинетический буфер также служил в качестве подвижного буфера для определения исходного уровня и в качестве буфера для разбавления антигенов и биспецифических антител. Наконечники биосенсора Ni-NTA погружали в 100 нМ his-меченый PD-L1-Fc (см. (b) ниже) или his-меченый PD-1-Fc для захвата антигена в течение приблизительно 1 мин. Каждый из наконечников биосенсора, покрытых антигеном, погружали в раствор 10 мкг/мл биспецифических антител в течение ~5 минут, а затем перемещали в колонку с лунками, содержащими 100 нМ антигена CTLA-4-Fc, на 2 минуты. Результаты связывания показаны на фигуре 3.
[00290] Биспецифический связывающий белок в формате DuetMab, который связывает PD-1 и CTLA-4, получали с использованием исходных последовательностей, указанных выше в таблице 2. DuetMab, связывающий PD-1/CTLA-4, получали с последовательностями в таблице 4 ниже и оценивали, как обсуждается ниже, в том числе при сравнении с Bis5, связывающим PD-1/CTLA-4.
Таблица 4. Конструкции DuetMab для связывания PD-1/CTLA-4
DuetMab
для связывания PD-1(LO115)
Аминокислотная последовательность
DuetMab
для связывания PD-1(LO115)
Последовательность нуклеиновой кислоты
DuetMab
для связывания PD-1(LO115)
Аминокислотная последовательность
DuetMab
для связывания PD-1(LO115)
Последовательность нуклеиновой кислоты
DuetMab
для связывания CTLA-4
Аминокислотная последовательность
DuetMab
для связывания CTLA-4
Последовательность нуклеиновой кислоты
DuetMab
для связывания CTLA-4
Аминокислотная последовательность
DuetMab
CTLA-4
Последовательность нуклеиновой кислоты
[00291] Анализ связывания с помощью Octet (DuetMab)
[00292] Исследования одновременного связывания с двумя различными антигенами проводили с помощью анализа Octet. Биотинилированный PD-1 человека загружали на стрептавидиновые сенсоры с последующим последовательными взаимодействиями сначала с DuetMab, связывающим PD-1/CTLA-4, а затем с растворимым антигеном CTLA-4. Стрептавидиновые (SA) биосенсоры (ForteBio) использовали для захвата биотинилированного PD-1 человека при 5 мкг/мл в PBS, рН 7,2, 3 мг/мл BSA, 0,05% (об./об.) Tween-20 (аналитический буфер). После стадии промывки загруженные биосенсоры подвергали последовательным взаимодействиям по типу ассоциации и диссоциации сначала с лунками с образцами, содержащими биспецифическое антитело DuetMab к PD-1/CTLA-4 при 133 нМ, а затем с лунками, содержащими антиген CTLA-4 человека при 200 нМ. Результаты связывания показаны на фигуре 5.
[00293] Свойственную биспецифическому антителу DuetMab к PD-1/CTLA-4 кинетику также оценивали посредством BiaCore. Эксперименты по связыванию проводили с использованием прибора BIAcore T200 (BIAcore). Для захвата антитела антитело мыши к Fab IgG человека иммобилизовали на чипе CM5 до достижения целевого ответа, составляющего 2000 RU. Пропускали 100 нМ DuetMab или mAb при 20 мкл/мин. в течение 5 мин., чтобы получить приблизительно 100 единиц ответа захваченного антитела. Затем антиген вводили последовательно со скоростью потока 50 мкл/мин. в течение 5 мин. Кинетические параметры (kon и koff) и константу диссоциации (KD) рассчитывали с помощью нелинейной подгонки с использованием программного обеспечения BIAevaluation 4.1. Результаты связывания показаны в таблице 5.
Таблица 5. Данные BiaCore для PD-1/CTLA-4
4 с-1)
[00294] Анализы с использованием репортерного гена
[00295] Результаты анализов с использованием репортерного гена показывают, что биспецифические связывающие белки для связывания PD-1/CTLA-4 ингибировали пути PD-1 и CTLA-4 (фигуры 6A-D). Связывающий белок BiS5 сохранял активность в отношении PD-1 по сравнению с исходным белком, но характеризовался ~3-кратной сниженной эффективностью по сравнению с IgG к CTLA-4. Антитело DuetMab характеризовалось ~9-кратным снижением эффективности в отношении PD-1 и ~16-кратным снижением в отношении CTLA-4 (по сравнению с IgG4P). В уровне техники существует потребность в молекуле, которая характеризуется сниженным нацеливанием на CTLA-4, однако сохраняет функциональную активность (например, как показано ниже в анализе SEB). Таким образом, биспецифические связывающие белки для связывания PD-1/CTLA-4 характеризуются потенциалом для обеспечения преимущества безопасности пациентам.
[00296] Анализ с использованием стафилококкового энтеротоксина B (SEB)
[00297] Результаты анализа SEB показывают, что DuetMab и BiS5Ab характеризовались эквивалентной активностью в анализах SEB с первичными иммунными клетками (фигура 7), и при этом DuetMab демонстрировало большую активность по сравнению с контрольными плечами DummyDuet (фигура 8A). DuetMab демонстрировало активность, примерно эквивалентную комбинации исходных молекул, и большую активность по сравнению с антителом MEDI1123 (тремелимумаб) к LO115 или CTLA-4 (фигура 8B). Наконец, BiS5 и DuetMab демонстрировали большую активность по сравнению с комбинацией новых изотипических контролей (фигуры 9A-B). Данные были получены от четырех доноров в двух независимых экспериментах, и для этих конкретных анализов требовалось использование IFNγ.
[00298] Анализ реакции смешанных лейкоцитов (MLR)
[00299] Анализы MLR проводили для тестирования биспецифических молекул, связывающих PD-1/CTLA-4. DuetMab и BiS5Ab к PD-1/CTLA-4 характеризовались эквивалентной активностью в анализе реакции смешанных лимфоцитов (MLR) (фигуры 10A-C). DuetMab к PD-1/CTLA-4 характеризовалось большей активностью по сравнению с комбинацией контрольных плечей DummyDuet/изотипа. (Фигуры 11A-D). DuetMab к PD-1/CTLA-4 характеризовалось приблизительно эквивалентной активностью по сравнению с комбинацией контролей исходных антител (фигуры 12A-D). Наконец, DuetMab к PD-1/CTLA-4 характеризовалось приблизительно эквивалентной активностью по сравнению с конкурирующей комбинацией к PD-1/CTLA-4 и характеризовался большей активностью, чем антитело к PD-1 отдельно (например, пембролизумаб и ниволумаб) (фигуры 13A-D).
[00300] Фармакокинетические и фармакодинамические (PK/PD) исследования
[00301] Проводили исследование для изучения фармакокинетики/фармакодинамики (PK/PD) однократной дозы у яванских макаков. Схема исследования показана на фигуре 14. DuetMab продемонстрировало четко выраженную фармакодинамику (PD) у яванских макаков, и для обеих молекул наблюдались устойчивые PD-ответы (фигуры 15A-B). Таким образом подтверждали жизнеспособность биспецифического связывающего белка для связывания PD-1/CTLA-4 в условиях in vivo.
[00302] Т-клеточно-зависимый иммунный ответ (TDAR)
[00303] Яванским макакам вводили внутривенно (в подкожную или подкожную латеральную вену передней лапы) указанную дозу (0,5, 5, 50 мг/кг) биспецифических молекул DuetMab или BiS5. Белок гемоцианин лимфы улитки (KLH) восстанавливали соответствующим количеством стерильной воды для инъекций в стерильных условиях. Раствор KLH при низкой дозе вводили подкожно в спину каждого животного в двух случаях (день 1 и день 29). Для дальнейшего анализа у всех животных получали образцы крови. Осуществляли оценку титров антител IgM и IgG, специфичных к KLH. Антитела к KLH в сыворотке крови обезьян обнаруживали с использованием ELISA.
[00304] Т-клеточно-зависимый иммунный ответ (TDAR) наблюдали у яванских макаков, получавших дозу DuetMab к PD-1/CTLA-4 (фигура 16A) и BiS5Ab к PD-1/CTLA-4 (фигура 16B).
[00305] Клетки CHO, экспрессирующие разные уровни PD-1 и/или CTLA-4 человека
[00306] Модельную систему разрабатывали для изучения биспецифических молекул, связывающих PD-1/CTLA-4, с использованием стабильных клеток СНО, экспрессирующих разные уровни PD-1 и/или CTLA-4 человека (фигура 17). Свободные антигенсвязывающие плечи на связанных с клетками DuetMab выявляли с помощью проточной цитометрии с использованием флуоресцентно-меченых растворимых белков PD-1 и CTLA-4. Результаты этого анализа демонстрируют, что DuetMab к PD-1/CTLA-4 одновременно связывает PD-1 и CTLA-4 на поверхности одной и той же клетки (фигуры 18A-C).
[00307] CTLA-4 непрерывно подвергается эндоцитозу в ямках, окаймленных клатрином, в результате чего в любой данный момент времени на поверхности клетки экспрессируется только небольшая часть рецептора. Рециркуляция CTLA-4 на клеточной поверхности происходит быстро, причем более 80% поверхностного CTLA-4 интернализуется в течение 5 минут. Таким образом, проводили эксперимент для определения того, различается ли кооперативное связывание DuetMab к PD-1/CTLA-4 по сравнению с комбинацией антител к PD-1 и антител к CTLA-4 в насыщении CTLA-4 на клетках, экспрессирующих избыток PD-1 (фигуры 19A-C). Занятость рецепторов для каждого антигена-мишени определяли независимо с использованием меченых mAb к PD-1 и к CTLA-4.
[00308] Обнаружили, что исходные моноклональные антитела связывались и занимали свой рецептор-мишень без заметного эффекта в отношении рецептора, не являющегося мишенью (фигуры 20A-D). Насыщение CTLA-4 на клетках CHO, экспрессирующих избыток PD-1, с помощью DuetMab к PD-1/CTLA-4 происходило при ~250 раз более низких концентрациях по сравнению с комбинацией моноклональных антител (фигуры 21A-D). Насыщение CTLA-4 на клетках CHO, экспрессирующих избыток PD-1, с помощью DuetMab к PD-1/CTLA-4 происходило при ~500 раз более низких концентрациях по сравнению с клетками, экспрессирующими только CTLA-4 (фигуры 22A-F). DuetMab к PD-1/CTLA-4 связываются с PD-1 и CTLA-4 преимущественно в цис-положении на поверхности одной и той же клетки, как определено путем количественного определения образования дуплетов во всей предварительно смешанной популяции CHO (фигуры 23A-B). Однако DuetMab к PD-1/CTLA-4 также может связываться в транс-положении с клетками, экспрессирующими только один из белков. DuetMab к PD-1/CTLA-4 приобрел свойства интернализации исходного антитела к CTLA-4, тремелимумаба (фигуры 24A-D). Не ограничиваясь теорией, эффект, проявляемый данной молекулой, характеризуется потенциалом индукции понижающей регуляции PD-1. Свойства интернализации DuetMab к PD-1/CTLA-4 также наблюдались в стабильных клетках СНО, экспрессирующих 10-кратный избыток PD-1 (фигура 25B).
Пример 2(b). Биспецифические связывающие белки для связывания PD-L1/CTLA-4
[00309] Следующие биспецифические связывающие белки, которые связывают PD-L1 и CTLA-4, получали с использованием исходных последовательностей, указанных выше в таблице 2. Белки, указанные как Bis2, Bis3 и Bis5, получали с последовательностями из таблицы 6 ниже, и последовательности, указанные ниже, оценивали в отношении одновременной антигенсвязывающей активности с использованием анализа связывания с помощью Octet, как описано выше в разделе 2(a) (фигура 26).
Таблица 6. Конструкции BiS для связывания PD-L1/CTLA-4
Bis2
для связывания PD-L1/CTLA-4
Bis2
для связывания PD-L1/CTLA-4
Bis3
для связывания PD-L1/CTLA-4
Bis3
для связывания PD-L1/CTLA-4
Bis5
для связывания PD-L1/CTLA-4
Bis5
для связывания PD-L1/CTLA-4
Пример 2(c). Биспецифические связывающие белки для связывания PD-1/TIM3
[00310] Следующие биспецифические связывающие белки (таблица 7), которые связывают PD-1 и TIM3, получали с использованием исходных последовательностей, указанных выше в таблице 2. Белки, обозначенные как Bis3, Bis5 и DuetMab, получали с указанными ниже последовательностями и оценивали для исследований одновременного связывания с использованием анализа с помощью Octet. Вкратце, стрептавидиновые (SA) биосенсоры (ForteBio) использовали для захвата биотинилированного домена TIM3-IgV человека при 2 мкг/мл в PBS, рН 7,2, 3 мг/мл BSA, 0,05% (об./об.) Tween-20 (аналитический буфер). После стадии промывки загруженные биосенсоры подвергали последовательным взаимодействиям по типу ассоциации и диссоциации сначала с лунками с образцами, содержащими биспецифические антитела при 200 нМ, а затем с лунками, содержащими антиген PD-1 при 200 нМ. Биотинилированный домен TIM3-IgV человека загружали на стрептавидиновые сенсоры с последующими последовательными взаимодействиями сначала с биспецифическими молекулами, а затем с антигеном PD-1. Результаты связывания показаны на фигурах 27A-27B.
Таблица 7. Конструкции BiS для связывания PD-1/TIM3
(SEQ ID NO:21)
Тяжелая цепь антитела к PD-1+scFv антитела к TIM3 (WT №62)
LC антитела к PD-1
Тяжелая цепь антитела к PD-1+scFv антитела к TIM3 (WT №62)
(SEQ ID NO:24)
LC антитела к PD-1
Тяжелая цепь антитела к PD-1
LC
антитела к TIM3
HC антитела к TIM3
Выступ
[00311] Анализ активности в отношении опухолеспецифеского уничтожения
[00312] Анализ уничтожения клеток клона Розенберга и меланомы
[00313] Общую активность биспецифических молекул, связывающих TIM3/PD-1, и исходного антитела к TIM3 в отношении уничтожения клеток тестировали с использованием клеток клона Розенберга JR6C12 и линии клеток меланомы Mel324.
[00314] Общий протокол анализа
[00315] Клетки JR6C12 функционировали как эффекторы и представляли собой линию CD8+ T-клеток человека, размноженную из меланомы пациента и специфичную в отношении антигена gp100 меланомы. Для оценки терапевтического потенциала опухолевые клетки Mel624 флуоресцентно метили и добавляли эффекторы (JR6C12) и кандидатное антитело, которое связывает TIM3 и/или PD-1. Клетки совместно культивировали в течение 16 часов. Несколько панелей на фигуре 28A дают визуальное представление о том, что добавление TIM3 62 либо в комбинации с антителом к PD1, либо в виде биспецифических молекул, связывающих PD-1/TIM3 (как описано в таблице 7), усиливает активацию Т-клеток и уничтожение опухоли.
[00316] Кроме того, как показано на фигурах 28B-28C, биспецифические молекулы, связывающие PD-1/TIM3, демонстрировали наибольшую эффективность в отношении уничтожения опухоли по сравнению с монотерапией с помощью антитела к TIM3, антитела к PD-1 или изотипического контроля, оцененную с помощью (b) показателя жизнеспособности опухолевых клеток по поглощению красителя и (c) секреции IFN-гамма.
[00317] Кроме клона 62, получали другой биспецифический связывающий белок, который связывает PD-1 и TIM3 в формате DuetMab, с использованием исходных последовательностей, указанных выше в таблице 2. DuetMab к PD-1/TIM3 получали с последовательностями из таблицы 8 ниже. Последовательность плеча для связывания TIM3 получали из O13-1, который представляет собой вариант клона 62 с созревшей аффинностью, и последовательность плеча антитела к PD-1 получали из LO115, которое идентично плечу антитела к PD-1, используемому для вышеописанного биспецифического антитела DuetMab к PD-1/CTLA-4. Биспецифическое антитело к PD-1 (LO115)/TIM3 (O13-1) оценивали, как описано ниже, в том числе при сравнении с BiS3 и BiS5, связывающими PD-1/TIM3.
Таблица 8. Конструкция DuetMab для связывания PD-1/TIM3
DuetMab
для связывания PD-1 (LO115)
Аминокислотная последовательность
LC
антитела к PD-1 (LO115)
Последовательность нуклеиновой кислоты
HC
антитела к PD-1 (LO115)
Аминокислотная последовательность
DuetMab
для связывания PD-1 (LO115)
Последовательность нуклеиновой кислоты
DuetMab
для связывания TIM3 (O13-1)
Аминокислотная последовательность
DuetMab
для связывания TIM3 (O13-1)
Последовательность нуклеиновой кислоты
DuetMab
для связывания TIM3 (O13-1)
Аминокислотная последовательность
Аминокислотная последовательность
(SEQ ID NO:89)
Аминокислотная последовательность
Аминокислотная последовательность
Аминокислотная последовательность
DuetMab
для связывания TIM3 (O13-1)
Последовательность нуклеиновой кислоты
[00318] Анализ связывания с помощью Octet (DuetMab, плечо для связывания TIM3, вариант с созревшей аффинностью)
[00319] Исследования одновременного связывания с двумя различными антигенами, PD-1 и TIM3, проводили с помощью анализа Octet. Биотинилированный TIM3 человека загружали на стрептавидиновые сенсоры с последующим последовательными взаимодействиями сначала с DuetMab к PD-1/TIM3, а затем с растворимым антигеном PD-1. Стрептавидиновые (SA) биосенсоры (ForteBio) использовали для захвата биотинилированного TIM3 человека при 5 мкг/мл в PBS, рН 7,2, 3 мг/мл BSA, 0,05% (об./об.) Tween-20 (аналитический буфер). После стадии промывки загруженные биосенсоры подвергали последовательным взаимодействиям по типу ассоциации и диссоциации сначала с лунками с образцами, содержащими биспецифическое антитело DuetMab к PD-1/CTLA-4, содержащее плечо к TIM3 (O13-1), которое представляет собой вариант с созревшей аффинностью клона 62 антитела к TIM3, загружали при 200 нМ, а затем с лунками, содержащими антиген CTLA-4 человека при 200 нМ. Результаты связывания показаны на фигуре 29.
[00320] Свойственную биспецифическому антителу DuetMab к PD-1/TIM3 кинетику также оценивали посредством BiaCore. Эксперименты по связыванию проводили с использованием прибора BIAcore T200 (BIAcore). Для захвата антитела антитело мыши к Fab IgG человека иммобилизовали на чипе CM5 до достижения целевого ответа, составляющего 2000 RU. Пропускали 100 нМ DuetMab или mAb при 20 мкл/мин в течение 5 мин, чтобы получить приблизительно 100 единиц ответа захваченного антитела. Затем антиген вводили последовательно со скоростью потока 50 мкл/мин в течение 5 мин. Кинетические параметры (kon и koff) и константу диссоциации (KD) рассчитывали с помощью нелинейной подгонки с использованием программного обеспечения BIAevaluation 4.1. Результаты связывания показаны в таблице 9.
Таблица 9. Данные BiaCore для PD-1/TIM3
(M-1с-1)
(с-1)
06
06
06
[00321] Биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, связанные с клетками CHO, сверхэкспрессирующими TIM3 человека или PD-1 человека (фигура 30 и таблица 25), экспрессия PD-1 и TIM3 (DMF4) показаны на фигуре 31.
Таблица 25
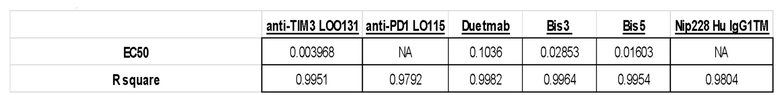
[00322] Анализ вторичного иммунного ответа на Ag CMV
[00323] Биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, улучшали пролиферацию CD8+ T-клеток в анализе вторичного иммунного ответа на антиген CMV по сравнению с обработкой изотипическими антителами (фигуры 32A-C).
[00324] Анализ реакции смешанных лейкоцитов (MLR)
[00325] Биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, увеличивали секрецию интерферона (IFNγ) в анализе реакции смешанных лимфоцитов (MLR) с активностью, обладающей тенденцией к превышению по сравнению с моно- и комбинированной терапией (фигуры 33A-D).
Биспецифические антитела к PD-1/TIM3, в том числе BiS3, BiS5 и DuetMab, демонстрировали аналогичную активность по сравнению с исходным IgG1 к LO115 в линии репортерных клеток Jurkat NFκB, которые преимущественно экспрессирует PD-1 (87% положительных по только PD-1) (фигуры 34A-C).
[00326] Таким образом, для связывания PD-1/TIM-3 получали три биспецифических формата (DuetMab, Bis3 и Bis5). Все биспецифические форматы демонстрируют функциональность in vitro, эквивалентную таковой антитела к PD-1, или лучше, что дает основания предполагать, что эти молекулы могут обеспечивать превосходное преимущество современным стратегиям иммуноонкологии.
Пример 2(d). Биспецифические связывающие белки для связывания OX40/PD-L1
[00327] Следующие биспецифические связывающие белки, которые связывают PD-L1 и OX40, получали с использованием исходных последовательностей, указанных выше в таблице 2. Белки, обозначенные как Bis2, Bis3 и Bis5, получали с помощью последовательностей из таблицы 10 ниже и оценивали в отношении активности одновременного связывания антигенов с использованием анализа связывания с помощью Octet, как обсуждается ниже.
Таблица 10. Конструкции BiS для связывания OX40/PD-L1
PD-L1 - HC OX40-4P
HC OX40-4P - PD-L1
HC OX40-4P - PD-L1
[00328] Анализ связывания с помощью Octet
[00329] Для оценки связывания раскрытых в данном документе биспецифических связывающих молекул применяли Octet QK, снабженный наконечниками биосенсора Ni-NTA, и 10X буфер для кинетического анализа (ForteBio, Менло-Парк, Калифорния). Для данной конкретной серии биспецифических связывающих белков His-меченый PD-L1-Fc, his-меченый PD-1-Fc и hOX40-Fc (рекомбинантные белки человека) приобретали у R&D Systems (Миннеаполис, Миннесота). Все анализы связывания осуществляли при 25°C.
[00330] Перед анализом планшеты для образцов перемешивали при 1000 об/мин. Наконечники биосенсора Ni-NTA предварительно смачивали в течение 5 мин. в 1X кинетическом буфере. 1X кинетический буфер также служил в качестве подвижного буфера для определения исходного уровня и в качестве буфера для разбавления антигенов и биспецифических антител. Наконечники биосенсора Ni-NTA погружали в 100 нМ his-меченый PD-L1-Fc (см. (b) ниже) или his-меченый PD-1-Fc для захвата антигена в течение приблизительно 1 мин. Каждый из наконечников биосенсора, покрытых антигеном, погружали в раствор 10 мкг/мл биспецифических антител в течение ~5 минут, а затем перемещали в колонку с лунками, содержащими 100 нМ антигена hOX40-Fc, на 2 минуты. Результаты связывания показывают, что молекулы BiS2/BiS3 к OX40Ab/PD-L1 связываются с PD-L1-His и hOX40-Fc, и что BiS2 к OX40Ab/PD-L1 связывается с большей аффинностью, чем BiS3 к OX40Ab/PD-L1. В качестве контроля использовали BiS2 к PD-1/OX40 (фигура 35).
[00331] Анализ с использованием стафилококкового энтеротоксина B (SEB)
[00332] В анализе SEB с использованием протокола, описанного выше, продемонстрировано, что биспецифическая молекула, связывающая OX40/PD-L1, была активной в форматах как BiS2, так и BiS3 (фигуры 36A-B).
[00333] Анализ PD-L1 с репортерным геном
[00334] Материалы
- Линии клеток и условия культивирования:
- клетки Jurkat с репортерным геном люциферазы под контролем NFAT, клон 2, экспрессирующие PD-1 человека;
- клетки CHO scFv OKT3 (UBC), экспрессирующие PD-L1 (все клетки поддерживали в среде RPMI 1640 плюс 10% FBS и 1X антибиотиками пеницил./стрепток. (полная среда RPMI) при 37°C в инкубаторе с повышенной влажностью для культивирования тканей).
- RPMI-1640, LifeTechnologies, № по кат. A1049101;
- инактивированная нагреванием сыворотка крови новорожденного теленка (FBS), LifeTechnologies, № по кат. 26010074;
- полная среда RPMI: RPMI-1640 плюс 10% FBS;
- 100X пенициллин/стрептомицин, LifeTechnologies, № по кат. 15140-122;
- 96-луночные культуральные планшеты с плоскодонными лунками, обработанные TC, Costar 3903, VWR, № по кат. 29444-010;
- система SteadyGlo для анализа люциферазы, Promega, № по кат. E2510;
- тестируемые антитела;
- планшет-ридер EnVision Multilabel, Perkin Elmer.
[00335] Способы
[00336] Для 2-клеточного анализа биологической активности для нейтрализации ингибирования PD-L1 клетки CHO scFv OKT3, экспрессирующие PD-L1, трипсинизировали, нейтрализовали с помощью теплой полной среды RPMI и собирали в 50 мл коническую пробирку. Клетки осаждали при 380g в течение 5 мин при к.т., а затем суспендировали в свежей полной среде RPMI и подсчитывали на устройстве для подсчета клеток Vi-Cell. Клетки CHO scFv OKT3, экспрессирующие PD-L1, доводили до 0,4e6/мл и высевали 25 мкл (10000 клеток) на лунку, как показано на схеме для планшета. Клеткам обеспечивали прилипание к планшетам в течение 3 часов. После этого 50 мкл RPMI, содержащей тестовые реагенты (2X конечной конц.), разделяли на аликвоты для клеток СНО и инкубировали еще 1 час. Эта инкубация обеспечивает время для связывания тестового реагента с PD-L1 на поверхности клеток CHO. Через 1 час клетки Jurkat с репортерным геном люциферазы под контролем NFAT, экспрессирующие PD-1, собирали в 50 мл коническую пробирку, осаждали при 380 g в течение 5 мин при к.т. и повторно суспендировали в свежих теплых полных средах RPMI. Клетки доводили до 1,2e6/мл и 25 мкл (30000) клеток помещали в лунки с клетками CHO scFv OKT3, экспрессирующими PD-L1, и тестовыми препаратами.
[00337] Клетки и тестовый реагент дополнительно инкубировали в течение 18 часов для активации репортерных клеток Jurkat PD-1. После этого получали реагент для люциферазы SteadyGlo и разделяли на аликвоты по 100 мкл в каждую лунку. Полного лизиса достигали путем осторожного встряхивания при к.т. (200 об/мин на орбитальном встряхивателе) в течение 15 мин. После лизиса активность люциферазы измеряли на планшет-ридере Envision Multilabel с использованием протокола измерения люминесценции US96. RLU люциферазы наносили на график относительно log [тестовый реагент] с помощью программного обеспечения Graphpad Prism и значения EC50 для антагонизма PD-L1 определяли с использованием нелинейного регрессионного анализа, 4-параметрической аппроксимации сигмоидальных кривых зависимости доза-ответ.
[00338] Результаты
[00339] BiS2/3 к OX40/PD-L1 тестировали при сравнении с исходными PD-L1/PD-1 и контролями NIP228(G4P) с использованием пятиточечного титрования дозы с начальной точкой при 100 нМ (PD-L1). Показано, что оба BisAb к OX40-PD-L1 являлись активными и характеризовались более сильным агонизмом, чем исходный PD-L1(4736) (фигура 37). Форматы BiS2 и BiS3 характеризовались сходным образом.
[00340] Анализ вторичного иммунного ответа на Ag CMV
[00341] В анализе вторичного иммунного ответа на Ag CMV (с применением протокола, описанного выше) молекулы BiS2 и BiS3 демонстрировали равную активность при сравнении с комбинацией (фигура 38).
[00342] Все анализы связывания и иммунного ответа, рассмотренные выше, предоставляют иллюстративные данные о том, что раскрытые в данном документе биспецифические связывающие молекулы проявляют специфическое связывание с обеими молекулами-мишенями, в некоторых случаях более сильную связывающую активность, чем комбинация отдельных моноспецифических исходных связывающих молекул (антител), и могут индуцировать или усиливать иммунный ответ. Кроме того, показано, что молекулы обладают активностью уничтожения клеток в отношении линии раковых клеток. Таким образом, данные показывают, что эти молекулы и структура(структуры) биспецифических платформ представляют собой превосходных кандидатов для иммуноонкологических терапевтических препаратов.
[00343] Анализ связывания с помощью Octet (BiS5 к OX40(SLR)/PD-L1)
[00344] Для оценки связывания раскрытых в данном документе биспецифических связывающих молекул использовали Octet QK, снабженный наконечниками биосенсора Ni-NTA, и 10X буфер для кинетического анализа (ForteBio, Менло-Парк, Калифорния). Для данной конкретной серии биспецифических связывающих белков His-меченый PD-L1-Fc, his-меченый PD-1-Fc и hOX40-Fc (рекомбинантные белки человека) приобретали у R&D Systems (Миннеаполис, Миннесота). Все анализы связывания проводили при 25°С. Результаты связывания показывают, что молекулы BiS5 к OX40Ab/PD-L1 связываются как с PD-L1-His, так и с hOX40-Fc (фигура 39).
[00345] BiS5 к PD-L1/OX40 связываются с клетками СНО, экспрессирующими OX40 и PD-L1/B7H1 человека или яванских макаков (фигуры 40A-F). Связывание конструкций BiS5 к PD-L1/OX40 также измеряли с помощью проточной цитометрии (HyperCyt) (фигура 42). IgG4P к OX40 и биспецифические молекулы, связывающие OX40/PD-L1, связывались с репортерными клетками Jurkat OX40. IgG к PD-L1 и биспецифические молекулы, связывающие OX40/PD-L1, связывались с клетками NCI H358 и CHOK1 B7H1(PD-L1)/OKT3. Все IgG и биспецифические молекулы связывались с клетками HEK CD32a.
[00346] Анализ PD-L1 и OX40 с репортерным геном
[00347] В анализе PD-L1 с репортерным геном (с использованием протокола, описанного выше) все содержащие scFv к PDF-L1 биспецифические молекулы и IgG положительного контроля проявляли активность (фигуры 42A-B). Контроли с одним плечом для связывания OX40 и изотипические контроли не проявляли какой-либо активности в данном анализе. Значения EC50 и углы наклона согласуются со значениями, полученными в предыдущих анализах для контролей исходных антител к PD-L1 и конструкций Bis2, Bis3 и Bis5, связывающих PD-L1.
[00348] В анализе OX40 с репортерным геном с использованием клеток HEK CD32a биспецифические конструкции характеризовались равной активностью при сравнении друг с другом, и при этом наблюдался Fc-опосредованный агонизм (фигуры 43A-B). IgG1 N434A Bis5 к OX40/PD-L1 характеризовалось эквивалентной активностью EC50 при сравнении с IgG4P к OX40 и MEDI0562 (IgG1 к OX40).
[00349] В анализе OX40 с репортерным геном с использованием сверхэкспрессирующих PD-L1 клеток CHOK1 биспецифические молекулы, связывающие OX40/PD-L1, характеризовались равным агонизмом (фигуры 44A-B). IgG1 N434A Bis5 к OX40/PD-L1 характеризовался эквивалентной активностью EC50 при сравнении с другими Fc-вариантами тестируемого биспецифического Mab Bis5 к OX40/PD-L1. Какого-либо агонизма с IgG к OX40 или IgG к PD-L1 не обнаружили. Таким образом, продемонстрировали опосредованный PD-L1 агонизм OX40.
[00350] Выявили опосредованный PD-L1 агонизм OX40 с опухолевыми клетками с использованием биспецифических молекул, связывающих OX40/PD-L1 (фигуры 45A-B). Биспецифические молекулы, связывающие OX40/PD-L1, характеризовались равным агонизмом в данном анализе - колоколообразные кривые. Не наблюдался какой-либо агонизм с IgG к OX40, таким образом демонстрировалось преимущество использования биспецифических молекул по сравнению с комбинацией IgG к OX40 плюс IgG к PD-L1. Не наблюдалось какого-либо агонизма с клетками NCI H358 PD-L1 KO (фигуры 46A-D), что указывало на то, что агонизм NCI H358, наблюдаемый с клетками, является PD-L1-специфическим.
[00351] Анализ с использованием стафилококкового энтеротоксина B (SEB)
[00352] В анализе SEB биспецифические молекулы, связывающие OX40/PD-L1, характеризовались большей активностью, чем комбинация отдельных антител к OX40 и PD-L1 (фигуры 47A-D). В частности, конструкция G4P характеризовалась большей активностью, чем конструкция G1. Дикий тип, вариант, содержащий YTE, и вариант N434A характеризовались эквивалентной активностью.
[00353] Анализ супрессии Treg
[00354] Проводили анализ супрессии Treg для тестирования биспецифических молекул, связывающих OX40/PD-L1 (фигуры 4A-D). Биспецифические молекулы, связывающие OX40/PD-L1, были активны на Teff CD4+ только в присутствии PD-L1 (фигуры 49 и 50). Не ограничиваясь теорией, это указывало на перекрестное связывание OX40 в транс-положении. Биспецифические молекулы, связывающие OX40/PD-L1, подавляли ингибирующие эффекты Treg, но только в случае перекрестного связывания с помощью связывания с PD-L1, иммобилизованным на планшете.
[00355] Анализ реакции смешанных лейкоцитов (MLR)
[00356] Анализ MLR проводили для тестирования биспецифических молекул, связывающих OX40/PD-L1 (фигуры 51A-B). Биспецифические молекулы, связывающие OX40/PD-L1, характеризовались большей активностью, чем комбинация отдельных антител к OX40 и PD-L1 (фигуры 52A-E).
[00357] Анализ антителозависимой клеточноопосредованной цитотоксичности (ADCC)
[00358] Анализ ADCC проводили для тестирования биспецифических молекул, связывающих OX40/PD-L1. Анализ ADCC с использованием свежевыделенных NK-клеток в качестве эффекторных клеток и клеток CHOK1, сверхэкспрессирующих PD-L1 B7H1 и CHOK1, сверхэкспрессирующих OX40, соответственно, в качестве клеток-мишеней при соотношении эффектора и мишени (E:T), составляющем 20:1. Лизис клеток-мишеней анализировали с использованием высвобождения европия из меченых клеток-мишеней через 5 часов. В анализе ADCC BiS2 и BiS5 к OX40/PD-L1 опосредовали ADCC против клеток CHO, экспрессирующих PD-L1 или OX40 (фигуры 53 и 54).
[00359] Анализ мобилизации CD107a проводили с использованием свежевыделенных NK-клеток в качестве эффекторных клеток и клеток K1 CHO, сверхэкспрессирующих PD-L1 и OX40, в качестве клеток-мишеней при соотношении E:T, составляющем 10:1. Мобилизацию CD107a к клеточной поверхности NK-клеток анализировали с помощью проточной цитометрии через 4 часа. Биспецифические молекулы BiS2 и BiS5 к OX40/PD-L1 увеличивали мобилизацию CD107a NK-клеток против клеток СНО, экспрессирующих PD-L1 или OX40 в анализах антителозависимой клеточноопосредованной цитотоксичности (ADCC) (фигура 55). Биспецифические молекулы BiS2 и BiS5 к OX40/PD-L1 увеличивали мобилизацию CD107a NK-клеток против активированных аллогенных Т-клеток, которые характеризовались повышенной регуляцией OX40 и PD-L1 (фигура 56). BiS5 к OX40/PD-L1 увеличивало мобилизацию CD107a NK-клеток от двух разных доноров против активированных аллогенных Т-клеток (фигуры 57A-B).
[00360] Фармакокинетические и фармакодинамические (PK/PD) исследования
[00361] Разрабатывали исследование для сравнения PK/PD биспецифических молекул, связывающих OX40/PD-L1 (фигура 58). Профили концентрации-времени в сыворотке крови для биспецифических молекул, связывающих PD-L1/OX40, сравнивали у яванских макаков (фигура 59 и таблица 11). Среднее значение T1/2 молекулы IgG1 N434A Bis5 к OX40/PD-L1 составляло больше, чем молекулы Bis5 WT; скорость выведения молекулы IgG1 N434A Bis5 к OX40/PD-L1 составляла меньше по сравнению с молекулой Bis5 WT. Обе молекулы сходным образом уменьшали содержание растворимого PD-L1 в сыворотке крови и вызывали значительное увеличение процентного содержания Ki67+ клеток для общего количества CD4+ T-клеток памяти, общего количества CD8+ Т-клеток памяти и NK-клеток.
Таблица 11. Фармакокинетические параметры BiS5-OX40/PD-L1-G1
мл)
кг)
[00362] Биспецифические молекулы, связывающие OX40/PD-L1, снижали значения концентрации растворимого PD-L1 в сыворотке крови ниже значения LLOQ анализа (фигура 60). Мутация N434A улучшала фармакокинетику BiS5-OX40/PD-L1-G1. В частности, значение CL уменьшилось примерно наполовину; произошло соответствующее двукратное увеличение T1/2 и AUCinf; а на Cmax и Vss влияния не оказывалось. Это соответствовало эффектам этой мутации, о которых сообщалось ранее, на PK моноклональных антител. Таким образом, достигнут прогресс в отношении mAb-подобной PK для IO BisAb BiS5-OX40/PD-L1-G1. Через 2 недели значения концентрации BiSAb BiS5-OX40/PD-L1-G1 в сыворотке крови были ниже предела количественного определения (BLOQ) и, возможно, были связаны с ADA.
[00363] Показатели существенного и статистически значимого увеличения наблюдались в отношении пролиферации общего количества клеток памяти CD4, общего количества клеток памяти CD8 и NK-клеток (процентное содержание Ki67+ клеток) при сравнении групп Bis5, связывающих PD-L1 и OX40, относительно группы контрольного (антитело к PcrV-Psl) Ab (фигуры 61A-F). Тенденция к существенным различиям наблюдалась между группами Bis5 к PD-1/LO115 и к PD-L1/OX40 в пролиферации (Ki67+) общего количества клеток памяти CD4, общего количества клеток памяти CD8 и NK-клеток. Не было каких-либо статистически значимых различий в пролиферации между версией N434A Bis5 к PD-L1/OX40 (увеличенное время полужизни) и версией G1. Версии N434A и IgG1 Bis5 к PD-L1/OX40 представляют собой биологически активные биспецифические молекулы.
Пример 2(e). Биспецифические связывающие белки для связывания OX40/PD-1
[00364] Следующие биспецифические связывающие белки, которые связывают PD-1 и OX40, получали с использованием исходных последовательностей, указанных выше в таблице 2. Белки, обозначенные как Bis2 и Bis3, получали с помощью последовательностей из таблицы 24 ниже и оценивали в отношении активности одновременного связывания антигенов с использованием анализа связывания с помощью Octet, как обсуждается ниже.
Таблица 24. Конструкции BiS для связывания OX40/PD-1
PD-1-HC OX40-4P
HC OX40-4P -PD-1
[00365] Моноклональное антитело (mAb) BiS2 к PD-1/OX40 представляет собой биспецифическое антитело (фигура 62, белки, связывающие PD-1, изображены серым цветом, и белки, связывающие OX40, изображены светло-серым цветом), сконструированное для одновременного связывания с PD-1 человека и яванского макака и с OX40 человека и яванского макака. Не ограничиваясь теорией, предлагаемый механизм действия предусматривает двойные сигнальные эффекты в отношении Т-клеток после связывания в цис-положении как с OX40, так и с PD-1, агонизм Т-клеточного совместно стимулирующего поверхностного рецептора OX40 и блокаду иммуносупрессорного PD-1 (фигура 63).
[00366] Анализ связывания с помощью Octet
[00368] Активность одновременного связывания с PD1-His и OX40-Fc человека показывали для двух разных партий mAb BiS2 к PD-1(LO115)/OX40 (фигура 64).
[00368] Анализ OX40 с репортерным геном
[00370] mAb BiS2 к PD-1(LO115)/OX40 демонстрировало активность, сравнимую с другими агонистами OX40 (фигуры 65A-B). Белки хранили при 4°C и немедленно использовали, замораживали/оттаивали три раза, хранили при 4°C в течение 7 дней и хранили при 40°C в течение 7 дней. Активность регистрировали в виде относительных световых единиц относительно концентрации mAb. Значение EC50 составляло ~2 нМ для mAb BiS2 к PD1(LO115)/OX40 при 4°C в день 0.
[00370] Анализ PD-1/PD-L1 с репортерным геном
[00371] mAb PD-1(LO115)/OX40 BiS2 продемонстрировало активность, сравнимую с другими агонистами OX40 (фигуры 66A-B). Белки хранили при 4°C и немедленно использовали, замораживали/оттаивали три раза, хранили при 4°C в течение 7 дней и хранили при 40°C в течение 7 дней. Активность регистрировали в виде относительных световых единиц относительно концентрации mAb. Значение EC50 составляло ~1 нМ для mAb BiS2 к PD1(LO115)/OX40 при 4°C в день 0. Проводили две серии первичных анализов эффективности in vitro у человека: анализ вторичного иммунного ответа Т-клеток на антиген и совместной стимуляции Т-клеток с использованием стафилококкового энтеротоксина В (SEB).
[00372] Анализ с использованием стафилококкового энтеротоксина B (SEB)
[00373] В анализе SEB mAb BiS2 к PD-1(LO115)/OX40 вызывало увеличение уровней IL-2, обнаруженного в надосадочной жидкости клеток через 3 дня в культуре (фигура 67). Таким образом, mAb BiS2 к PD-1/OX40 может одновременно связываться со своими антигенами-мишенями у человека и может совместно стимулировать Т-клетки in vitro.
[00374] В анализе вторичного иммунного ответа на антиген mAb BiS2 к PD-1/OX40 приводил к увеличению уровней содержания интерферона (IFN)-гамма при сравнении с исходными mAb и комбинацией исходных mAb (фигуры 68 и 69).
[00375] Анализ вторичного иммунного ответа на Ag CMV
[00376] Результаты анализа вторичного иммунного ответа на Ag CMV (с использованием протокола, описанного выше) показали, что молекулы BiS2 и BiS3 не демонстрировали равную активность при сравнении с комбинацией (фигура 70). Данные показывают, что mAb IgG4P BiS2 к PD-1/OX40 является активным in vitro и in vivo. BiS3 к PD-1/OX40, который отличается по структуре от BiS2 к PD1/OX40, не обладал выявляемой активностью. Таким образом, BiS3 к PD-1/OX40 (не активный) отличается от BiS2 (активного).
[00377] Фармакокинетические и фармакодинамические (PK/PD) исследования
[00378] Яванский макак считался фармакологически значимым неклиническим видом для тестирования функциональной активности mAb BiS2 к PD-1/OX40. Фармакокинетику (PK) и фармакодинамику (PD) mAb BiS2 к PDS-1/OX40 оценивали в исследовании, не соответствующем требованиям GLP (Надлежащей лабораторной практики), на яванских макаках. PK и PD mAb BiS2 к PD-1(LO115)/OX40 (процент Ki67-положительных CD4+ и CD8+ клеток в общем содержании Т-клеток памяти) оценивали у яванских макаков (n=3; самцы) после однократной внутривенной (IV) дозы в диапазоне доз от 0,1 мг/кг до 30 мг/кг. PBMC собирали перед введением дозы и в дни 1, 8, 11 и 15 после введения дозы, криоконсервировали и оттаивали перед проведением анализа с помощью проточной цитометрии. Таким образом, mAb BiS2 к PD1 (LO115)/OX40 характеризовалось примерно линейной PK с коротким временем полужизни, составляющим 0,6-1,7 дня (фигура 70, таблица 12).
[00379] Таблица 12. Средние фармакокинетические параметры mAb BiS2 к PD1 (LO115)/OX40.
мл)
мл)
мл)
кг)
кг)
Значения представлены как среднее (стандартное отклонение). AUClast=площадь под кривой времени-концентрации до последней измеряемой концентрации; AUCINF=площадь под кривой времени-концентрации до бесконечного времени; Cmax=максимальная наблюдаемая концентрация; CL: системное выведение; T1/2=время полужизни; Vss: объем распределения в конечной фазе; Vss: объем распределения в равновесном состоянии.
[00380] Средние максимальные концентрации (Cmax) увеличивались примерно пропорционально от дозы, составляя от 2,0 мкг/мл при 0,1 мг/кг до 607 мкг/мл при 30 мг/кг. AUC∞ увеличивались примерно пропорционально дозе, составляя от 1,7 мкг•день/мл при 0,1 мг/кг до 577 мкг•день/мл при 30 мг/кг. Средний уровень выведения из сыворотки крови варьировался от 41,8 мл/день/кг до 60,2 мл/день/кг. Объем распределения в равновесном состоянии составлял от 43,2 мл/кг до 85,6 мл/кг. Результаты PD (фигура 71) показали дозозависимое увеличение общей пролиферации CD4+ Т-клеток памяти (Ki67) и увеличение общей пролиферации CD8+ Т-клеток памяти (Ki67). Показана типичная стандартная кривая количественного определения PD-1/OX40 в сыворотке крови яванских макаков (фигура 72).
Пример 3. Физическая и химическая стабильность конструкций BiSAb
[00381] Проводили серию экспериментов с целью определения и оценки физической и химической стабильности конструкций BiSAb, описанных в данном документе, по сравнению с другими структурными стратегиями и платформами биспецифических связывающих белков. В частности, в серии исследований стабильности, обсуждаемых ниже, идентифицировали и анализировали влияние различных диапазонов рН на стабильность BiSAb (например, гидролиз, фрагментацию, агрегацию, термическую стабильность). Как показывают данные, из различных иллюстративных вариантов осуществления разных форматов BiSAb раскрытое в данном документе BiSAb (обозначенное как "BiS5" в нижеприведенных исследованиях и в формате D/H, как показано в таблице 13) демонстрировало неожиданную и удивительную физическую и химическую стабильность по сравнению со всеми другими структурными мотивами BiSAb.
Таблица 13. Исследование стабильности конструкций BiS5
IgG: LC10
scFv: 2F4
SN-(scFv)-G
Аминокислотная последовательность
V-области подчеркнуты, линкеры выделены курсивом. Последовательность выглядит следующим образом: LC10VH, CH1, шарнир, CH2, CH3 (N-конец), L1-2F4VL-линкер-2F4VH-L2, CH3 (C-конец)
IgG: 2F4
scFv: LC10
SN-(scFv)-G
Аминокислотная последовательность
V-области подчеркнуты, линкеры выделены курсивом. Последовательность выглядит следующим образом: 2F4VH, CH1, шарнир, CH2, CH3 (N-конец), L1-LC10VL-линкер-LC10VH-L2, CH3 (C-конец)
Пример 3.1
[00382] Дальнейшее сравнение проводили между раскрытым в данном документе форматом BiS ("BiS5") и другим форматом BiS, который содержит два связывающих домена (домена scFv), связанных в шарнирной области (т.е. между Fc- и Fab-областями), обозначенным как "BiS4". Белки BiS4 и BiS5 экспрессировали в клетках яичников китайского хомячка (CHO) и очищали обычными способами хроматографии. Как отмечалось выше, два данных формата имеют идентичные последовательности Fab и scFv, причем основное различие между ними заключается в расположении scFv-домена (что касается BiS4, то scFv расположен в шарнирной области, что касается этого конкретного BiS5, scFv расположен в петле SNG в CH3-домене, как обсуждалось в данном документе). Очищенные молекулы BiS подавали в PBS-буфере и определяли концентрацию белка с использованием NanoDrop ND-1000 (Thermo Scientific, Уилмингтон, Делавэр) с использованием коэффициента экстинкции, составляющего 1,54 M-1 см-1.
Исследование pH-скрининга и кратковременной стабильности
[00383] Для исследований, связанных с рН-скринингом, антитела BiS4 и BiS5 концентрировали до ~12 мг/мл и подвергали диализу относительно 6 различных условий рН, 20 мМ сукцината натрия (рН 5,0), гистидина/гистидина HCl (рН 5,5, 6,0 и 6,5) и фосфата натрия (рН 7,0 и 7,5), все содержащие 240 мМ сахарозы. Диализ осуществляли с использованием кассет для диализа Slide-A-Lyzer (отсечение молекулярной массы 10 кДа (MWCO), Thermo-Fisher, Рокфорд, Иллинойс). После завершения диализа добавляли 0,02% полисорбата 80 и конечную концентрацию белка доводили до ~10 мг/мл. Составы BiS4 и BiS5 стерилизовали с использованием 0,22 мкм фильтра (Millipore, Биллирика, Массачусетс) в предварительно дезинфицированном вытяжном шкафу с ламинарным потоком. Аликвоты по одному миллилитру распределяли в 3 мл флаконы из боросиликатного стекла типа I (West Pharmaceutical Services, Экстон, Пенсильвания). Образцы хранили при 40°С и анализировали с помощью SEC в момент времени ноль и после хранения в течение 1, 2, 3 и 4 недель.
Дифференциальная сканирующая калориметрия (DSC)
[00384] Термограммы дифференциальной сканирующей калориметрии для образцов в момент времени ноль получали с использованием VP-Capillary DSC, подключенного к автоматическому пробоотборнику с регулируемой температурой (Malvern Instruments Ltd., Вестбороу, Массачусетс). Для получения термограмм использовали концентрацию белка 1 мг/мл вместе со скоростью сканирования 90°С/ч. в диапазоне температур 20°C-100°C. Термограммы BiS4 и BiS5, полученные в различных условиях рН в диапазоне от 5,0 до 7,5, регулировали с учетом буфера и корректировали относительно исходного уровня. Анализ данных выполняли с использованием плагина DSC для пакета программного обеспечения Origin 7 SR4. Экспериментальные результаты согласовывали с моделью со многими состояниями с тремя переходами для расчета значений температуры плавления (Tm). Точка, в которой значение теплоемкости (Cp) для первого теплового перехода достигало 500 кал⋅моль-1⋅°C-1, рассматривалась в качестве температуры начала разложения (Tonset).
Высокоэффективная эксклюзионная хроматография (HP-SEC)
[00385] Для отделения структур агрегатов и фрагментов от мономера на основании размера образцы из анализа стабильности анализировали с использованием системы для высокоэффективной жидкостной хроматографии Agilent с детектором на фотодиодной матрице, способным регистрировать спектры поглощения УФ-излучения 200-400 нм с 7,8 × 30 см2, 5 мкм, 250Å, Tosho TSKgel G3000SWxl (TOSOH Biosciences, Кинг оф Пруссия, Пенсильвания) и соответствующей предколонки. Для разделения структур использовали подвижную фазу, содержащую 0,1 М безводного двухосновного фосфата натрия, 0,1 М сульфата натрия, 0,01% азида натрия, рН 6,8, и скорость потока 1 мл⋅мин.-1. Количество вводимого белка составляло приблизительно 250 мкг. Отделение BiS4 и BiS5 контролировали с использованием спектра поглощения 280 нм. Количественно определяли площади пиков растворимых агрегатов (мультимера и димера), мономера и фрагментов. Затем рассчитывали процентное содержание каждой из структур и наносили на график относительно времени инкубации для получения кинетических графиков. Кривые профиля рН для скорости потери мономеров, фрагментации и агрегации в месяц получали путем вычисления наклона каждого кинетического графика.
Термическая стабильность BiS4 и BiS5
[00386] Эффект рН в отношении термической стабильности BiS4 и BiS5 оценивали посредством анализа термограмм, полученных с использованием капиллярной DSC, и полученных при шести различных условиях рН. На фигурах 74A и 74B показаны наложения термограмм DSC для BiS4 и BiS5 соответственно при pH от 5,0 до 7,5. Как показано на фигурах 74C и 74D, на каждой термограмме представлены три события теплового развертывания с температурами перехода Tm1, Tm2 и Tm3. Первый переход (Tm1) вероятно связан с одновременным развертыванием CH2- и scFv-доменов, тогда как второй (Tm2) и третий (Tm3) переходы связаны с развертыванием CH3- и Fab-доменов. Для обоих форматов наблюдали повышение Tonset, Tm1, Tm2 и Tm3 с увеличением рН до 6,5 (фигуры 73A, 73B, 73E и таблица 14 ниже). Для BiS4 и BiS5 различий в Tonset, Tm2 и Tm3 не наблюдали при всех условиях pH (фигура 73E и таблица 15), что указывает на то, что присутствие scFv либо в шарнирной области, либо в CH3-домене не влияет на термическую стабильность CH3 и Fab. Интересно отметить, что небольшое повышение Tm1 наблюдали для BiS5 при всех условиях рН, что указывает на увеличение термической стабильности или scFv, или CH2, или обоих из них, если scFv расположен в CH3-домене.
Таблица 14. Эффект pH на температуру начала термического разложения (Tonset) и температур термического плавления (Tm1, Tm2 и Tm3) для BiS4 и BiS5, как измерено с помощью капиллярной DSC.
Физическая и химическая стабильность BiS4 и BiS5
[00387] Физическую и химическую стабильность форматов BiS4 и BiS5 при различных значениях рН (в диапазоне от 5,0 до 7,5) оценивали при 40°С в течение периода до 4 недель. Хроматограммы HP-SEC "в момент времени ноль" использовали для сравнения общей площади, содержания мономеров, агрегатов и фрагментов, полученных из хроматограмм HP-SEC для других временных точек. Типичные хроматограммы BiS4 и BiS5 при рН 7,5 в момент времени ноль по сравнению с 4 неделями показаны на фигуре 74A. Все образцы содержат преимущественно мономер с низкими уровнями содержания растворимых агрегатов и с фрагментами или без них. В момент времени ноль (сплошные линии) большая часть образца представляет собой мономер без каких-либо других заметных различий, за исключением небольшой разницы в высоте пиков между двумя образцами, вероятно, по причине небольшой разницы в концентрации (фигура 74A). Пунктирными линиями показано наложение хроматограмм HP-SEC обоих форматов в тех же условиях pH после хранения в течение 4 недель при 40°C. В условиях ускоренного температурного стресса оба формата демонстрируют дополнительный пик, ранний элюируемый пик (мультимерные структуры), снижение содержания мономера и повышенные уровни содержания фрагментов (фигура 74A). Потеря мономера из-за фрагментации была более заметной у BiS4 по сравнению с BiS5, что указывает на то, что BiS5 является более химически стабильным. Исходя из их структуры, возможных участков фрагментации и времени удерживания, авторы изобретения предполагают, что пик малого фрагмента (к.т. ~10,8 мин) представляет собой Fab, а пик большого фрагмента (к.т. ~9,8 мин) и пик в виде плеча (к.т. ~8,7 мин) представляют собой Fab с scFv и его соответствующий фрагмент с более высокой молекулярной массой (HMWF) с Fab, scFv и Fc соответственно.
[00388] Чтобы лучше оценить эффект расположения scFv на физическую и химическую стабильность BiS4 и BiS5, процент от общей площади для каждой структуры наносили на гистограмму для момента времени ноль и 4 недели при 40°C для pH 7,5 (фигура 74B). Как показано на фигуре 74B, в момент времени ноль чистота мономеров у BiS4 и BiS5 является одинаковой. Образцы, инкубированные при 40°С в течение периода до 4 недель, продемонстрировали значительные различия в типе и диапазоне образованных фрагментов. В отношении образцов BiS4 образовывались пик в виде плеча (к.т. ~8,7 мин), пик большого фрагмента (к.т. ~9,8 мин) и пик малого фрагмента (к.т. ~10,8 мин), составляющие 11,8%, 7,2% и 3,5% соответственно (фигура 74B). Неожиданно образец BiS5 продемонстрировал лишь 1,4% для малого фрагмента (к.т. ~10,8 мин), вероятно из-за связывания scFv по обеим сторонам домена с Fc (фигура 89).
[00389] На фигурах 75A-75C показана кинетика агрегации, фрагментации и потери мономера для BiS4 и BiS5 в случае образцов, инкубированных при 40°С при pH 7,5. Образцы BiS4 демонстрировали более высокую скорость потери мономера по сравнению с BiS5 (фигура 75A). Скорость потери мономера для BiS4 и BiS5 при рН 7,5 составляла 27,4%/месяц и 4,5%/месяц соответственно (фигура 75A). Для BiS4 большая часть потери мономера обусловлена фрагментацией, которая составляла 23,9%/месяц, и в меньшей степени по причине агрегации, которая составляла 3,5%/месяц (фигуры 75B и 75C). Интересно отметить, что для BiS5 скорость агрегации, по-видимому, была несколько выше (2,8%/месяц) по сравнению со скоростью фрагментации (1,7%/месяц) (фигуры 75B-75C).
[00390] Дополнительный анализ эффекта рН на физическую и химическую стабильность форматов BiS4 и BiS5, скорость потери мономера, фрагментации и агрегации в месяц проводили путем нанесения данных значений на график в зависимости от 6 условий pH (фигуры 76A-76C). При всех шести условиях рН в диапазоне рН 5,0-7,5 скорость потери мономера была ниже для формата BiS5 по сравнению с BiS4 (фигура 76A), что дает основания предполагать, что раскрытый в данном документе формат BiS5 характеризуется неожиданно превосходной физической и химической стабильностью по отношению к другим форматам биспецифических белков. Для BiS4 по большей части разрушение мономера обусловливалось фрагментацией даже при более низких условиях pH (фигура 76B). BiS5 демонстрировал более низкие скорости фрагментации по сравнению с BiS4 при всех тестируемых условиях pH. Неожиданно то, что значения скорости фрагментации в BiS5, по-видимому, находились в плоскости и являлись более низкими в более широком диапазоне рН по сравнению с BiS4. Не ограничиваясь какой-либо конкретной теорией, возможно более низкие значения скорости фрагментации, наблюдаемые для BiS5, могут обусловливаться линкерами G4S на любом из концов scFv, соединяющими его с Fc. Фрагментация на одном из линкеров G4S, соединяющих с Fc, может не высвобождать scFv, поскольку он может быть все еще соединен с Fc посредством другого линкера G4S. Для BiS4 и BiS5 значения скорости агрегации, по-видимому, были сходными при всех тестируемых условиях pH (фигура 76C), что дает основания предполагать, что расположение scFv оказывает минимальный эффект на кинетику агрегации, что также подтверждается наблюдаемым отсутствием изменений в Tonset между двумя форматами при всех условиях рН, как измерено с использованием капиллярной DSC (фигура 73E и таблица 14, рассмотренные выше). При рН 7,5 и 40°С (в момент времени=0) ни одна из молекул не проявляла заметной фрагментации (фигура 77A), но при тех же условиях после хранения в течение 2 недель при 40°С наблюдалась заметная фрагментация для BiS4 и минимальная фрагментация для BiS5 (фигура 77B). Как BiS4, так и BiS5 характеризуются сниженной фрагментацией и агрегацией при более низком значении рН (5,5), тогда как BiS5 обладает превосходными характеристиками как фрагментации, так и агрегации при обоих значениях рН (фигура 78). Данная серия экспериментов демонстрирует, что раскрытый в данном документе BiS5 обладает превосходной химической стабильностью и сходной физической стабильностью по сравнению с таковыми BiS4.
Пример 3.2
[00391] Дополнительные исследования проводили для оценки физической и химической стабильности различных вариантов осуществления биспецифических связывающих белков, которые раскрыты в данном документе и обозначены как конструкции A-H (см., например, таблицу 13 и соответствующие примеры выше). Данные конструкции анализировали с использованием DSC, анализов стабильности при ускоренном хранении и связывания FcRn и FcgR, как следует ниже.
Анализ с использованием дифференциальной сканирующой калориметрии
[00392] Эксперименты DSC для этого набора данных проводили с использованием сканирующего микрокалориметра Microcal VP-DSC (Microcal). Все растворы и образцы, используемые для DSC, фильтровали с использованием 0,22 мкм фильтра и дегазировали перед загрузкой в калориметр. Антитела, используемые для исследований DSC, содержали >98% мономера, как определено с помощью аналитической SEC. Перед анализом DSC все образцы подвергали полному диализу (по меньшей мере 3 буферных обмена) в 25 мМ гистидина-HCl (pH 6,0). Буфер из этого диализа использовали в качестве эталонного буфера для последующих экспериментов DSC. Перед измерением образца вычитали измерения исходного уровня (буфер относительно буфера) из измерения образца. Подвергнутые диализу образцы (при концентрации 1 мг/мл) добавляли в лунку с образцом и проводили измерения с помощью DSC со скоростью сканирования 1°С/мин. Анализ данных и деконволюцию выполняли с использованием программного обеспечения Origin™ DSC, предоставленного Microcal. Анализ деконволюции осуществляли с использованием модели для белков, не являющихся бистабильными, и при этом наилучшие соответствия получали с использованием 100 итерационных циклов. Tm,onset определяют как определяющую качество температуру, при которой термограмма выглядит как имеющая ненулевой наклон, Tm определяют как температуру, при которой развернута половина молекул в наборе, и ее рассчитывают как значение температуры, соответствующее каждому максимуму пиков на термограмме.
[00393] Результаты для разных конструкций представлены на фигуре 79. Как правило, конструкции A, C, D, которые содержат 2F4 в качестве scFv, характеризуются более низкой TM1 по сравнению с конструкциями E, G, H, которые содержат LC10 в качестве scFv. Не ограничиваясь теорией, разница в значении TM1 может быть обусловлена лучшей термической стабильностью scFv-домена LC10 по своей природе по сравнению с вариабельным доменом 2F4. Данные дают основания предполагать, что конструкции A-D, содержащие 2F4 в качестве scFv, будут менее термически стабильными, чем конструкции E-H с LC10 в качестве scFv.
Анализ стабильности при ускоренном хранении
[00394] Концентрации конструкций нормализовали до 1 мг/мл. По 1 мл каждой биспецифической конструкции или IgG контроля разделяли на аликвоты в 1,5 мл пробирках Эппендорфа. Образцы инкубировали в статическом инкубаторе в течение 2 недель при 45°. Образцы анализировали через 3, 7 и 14 дней и оценивали на стабильность. В каждой временной точке проводили визуальный осмотр для регистрации любой повышенной мутности или осаждения. Образцы фильтровали с использованием 0,2 мкм центрифужной колонки и раззделяли на аликвоты по 120 мкл образца во флакон для HPLC, убедившись, что на дне флакона не осталось пузырьков воздуха. Затем образцы тестировали на устройстве Agilent серии 1100 HPLC-SEC, чтобы проверить агрегацию и разрушение, с использованием колонки TSK-GEL G3000SWXL (300×7,8 мм) Tosoh Bioscience с 0,1 М фосфатом натрия, 0,1 М сульфатом натрия, рН 6,8, в качестве подвижного буфера. Вводили 60 мкл образца и прогоняли при скорости потока 1 мл/мин. Регистрировали и использовали для аналитического анализа SEC время удерживания мономера (мин.), общую площадь пика, % мономера, % агрегата, % фрагмента, % потери мономера. Результаты подытожены в таблице 15.
Таблица 15. Исследования стабильности при ускоренном хранении
0-й день
0-й день
0-й день
7-й день
0-й день
7-й день
[00395] Как описано в данном документе, расположение scFv-домена в вышеуказанных конструкциях выглядит следующим образом (где "-" обозначает scFv): A и E находятся в IS-RTP; B и F находятся в AK-GQP; C и G находятся в S-NG и D и H находятся в SN-G. Различные значения TM ассоциированы со следующими доменами: TM1=CH2/scFv; TM2=Fab; TM3=CH3. Данные показывают тенденцию того, что конструкции A и C с scFv 2F4, вставленным в петли ISRTP (A) и SNG (C), более склонны к агрегации, чем конструкции E и G, которые имеют scFv LC10, вставленный в тех же местах. Данное наблюдение дает основание предполагать, что идентичность последовательностей и поведение scFv-домена могут иметь влияние на стабильность конструкций биспецифических связывающих белков. Кроме того, из вышесказанного можно было бы предсказать, что конструкция D, которая содержит scFv 2F4, будет характеризоваться более низкой стабильностью, сходной с таковой A и C; однако, по-видимому, вставка scFv 2F4 в петлю SNG стабилизирует молекулу и уменьшает тенденцию к образованию агрегатов. В совокупности в данном исследовании стабильности при ускоренном хранении показано, что последовательность и расположение scFv в Fc-области могут играть значимую роль в стабильности конструкции BiSAb.
Анализ связывания FcRn и FcγR
[00396] Эксперименты по связыванию проводили с использованием прибора BIAcore 3000 (BIAcore). Для захвата антитела 1000 RU антигена IsdH (Fab) иммобилизовывали на чипе CM5. Для захвата антитела 100 нМ конструкций BiSAb или контролей mAb пропускали при 20 мкл/мин в течение 5 мин. Пропускали 5 мкМ huFcRn или FcγR I, IIa, IIb, IIIa-158V или IIIA158F при 5 мкл/мин в течение 20 мин Связывание FcRn осуществляли в PBS+5 мкМ EDTA при рН 6,0, а связывание FcRn осуществляли в PBS+5 мкМ EDTA при рН 7,4.
[00397] Конструкции A, C, D, E, G и H оценивали в отношении связывания FcRn. Репрезентативные данные связывания с FcRn каждой из биспецифических конструкций E и H и IgG 2F4 показаны на фигуре 80. Конструкции, содержащие scFv ниже границы CH2-CH3, кажется, сохраняют связывание с FcRn (например, конструкции D и H). Конструкции, содержащие scFv, расположенные в пределах петли ISRTP выше границы CH2-CH3, по-видимому, характеризуются устранением выявляемого связывания FcRn (например, конструкции A и E). Петля ISRTP находится в области мутаций YTE в Fc (M252Y/S254T/T256E), известных как удлиняющие время полужизни, которые, как известно, важны для связывания FcRn.
[00398] Конструкции A, C, D, E, G и H тестировали в отношении связывания с FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa-158F и FcγRIIIa-158V. Репрезентативные данные показаны на фигуре 81 для конструкций E, G и H, связывающихся с FcγRIIIa-158V. Все тестируемые конструкции сохраняли связывание с FcγR, хотя и с различными значениями аффинности (фигура 81, вставка). В таблице 16 показаны наблюдаемые закономерности связывания различных конструкций с FcγRs.
Таблица 16. Закономерности связывания FcγR
[00399] Различия, наблюдаемые при связывании FcγR с конструкциями, содержащими scFv, вставленный в петлю ISRTP выше границы CH2-CH3 (A и E), последовательно показывают уменьшенное связывание FcγR при сравнении с другими конструкциями, содержащими scFv, вставленный в петлю SNG ниже границы CH2-CH3 (C, D, G и H).
[00400] Попытки оценить, можно ли улучшить или восстановить связывание FcRn в конструкциях A и E, предпринимали путем введения петли, продлевающей время полужизни (N3), в Fc-область. На фигуре 82 представлены репрезентативные данные и показано, что в случае конструкций E ни E BiS5Ab, ни конструкция E с введенной петлей N3 (E BiS5Ab+N3) не способны связывать FcRn. Более того, вставка scFv LC10 в петлю N3 (scFv N3) и сохранение без изменений петли ISRTP также уменьшали связывание FcRn ниже уровней обнаружения. Эти данные показывают, что по меньшей мере для конструкции E, если только не для каждой конструкции, раскрытой в данном документе, как петля ISRTP, так и петля N3, в случае их наличия, должны быть без изменений и немодифицированными с целью сохранения связывания FcRn.
Пример 3.3
[00401] В дополнение к сравнению между BiS4 и биспецифическими связывающими конструкциями, раскрытыми в данном документе (BiS5), проводили исследование для оценки трех других платформ структурных мотивов BiS, обозначенных как BiS1, BiS2 и BiS3 (см. фигуру 83). Как будет понятно со ссылкой на фигуру 83, эти платформы различаются в зависимости от расположения одного из связывающих доменов (проиллюстрированного как scFv-домен). Из пяти мотивов только BiS4 и BiS5 содержат два линкерных фрагмента в качестве точек присоединения к белку большего размера, причем другие (BiS1, BiS2 и BiS3) присоединяются с помощью одного линкера.
[00402] Вкратце, типичные молекулы каждой конструкции анализировали на стабильность с использованием методик, рассмотренных выше в примерах 3.1 и 3.2. Образцы каждой конструкции добавляли к буферам с рН 5,0, 5,5, 6,0, 6,5, 7,0 и 7,5 и хранили при 40°С в течение периода, составляющего два месяца. Затем образцы анализировали на скорость фрагментации (фигура 84), скорость агрегации (фигура 85) и скорость потери мономера (фигура 86) с использованием HP-SEC. В данных условиях анализ показал, что раскрытый в данном документе формат биспецифического связывающего белка ("BiS5" и в формате D/H, как указано выше в таблице 13) обладал превосходной физической и химической стабильностью по сравнению со всеми другими форматами при всех условиях рН.
[00403] Данные SEC также использовали для сопоставления различных пиков с соответствующими фрагментами молекул BiS (фигура 87). Сопоставление основывали на предположениях, которые включали то, что фрагментация происходит в шарнирной и линкерной областях молекул, включали размер фрагмента, теоретическую фрагментацию и то, как ожидаемые структуры фрагментов выравниваются по отношению к структурам фрагментов, наблюдаемым в других форматах. Несмотря на то, что в каждом формате имелось хорошее разрешение между фрагментами с более низкой молекулярной массой (LMWF), также имело место плохое разрешение между мономером и фрагментами с более высокой молекулярной массой (HMWF), или оно вообще отсутствовало, во всех форматах. На основании информации, приведенной в таблице 17, сделали вывод о том, что методика HP-SEC в недостаточной степени оценивает фрагментацию в форматах BiS в большей мере, чем для моноклональных антител. Разрабатывали альтернативный анализ, как обсуждается ниже.
Таблица 17. Анализ фрагментации - недостаточная оценка скорости фрагментации форматов BiS с помощью SEC.
(кДа)
1Fab+2scFv+Fc2
150
7,9
x (k1)
*Время удерживания.
1 Потеря мономера включают потерю только по причине фрагментации и не включает агрегацию.
2 Скорости могут быть недостаточно оцениваться по причине совместного элюирования HMWF с мономером.
3 Скорости для пика в виде плеча могут варьироваться из-за спадающей интеграции.
[00404] Разрабатывали альтернативный анализ с целью расчета скоростей фрагментации HMWF с использованием молярного коэффициента экстинкции на основе предположений о том, что (i) во время разрушения, если обнаружен небольшой фрагмент, то также должен присутствовать соответствующий большой фрагмент; (ii) вторичная фрагментация (фрагменты фрагментов) во время исследования стабильности существенно не происходит; и (iii) фрагментация происходит в линкерной области и/или в шарнирной области. Коэффициенты фрагментации определяли на основе следующих соотношений:
Мономер = 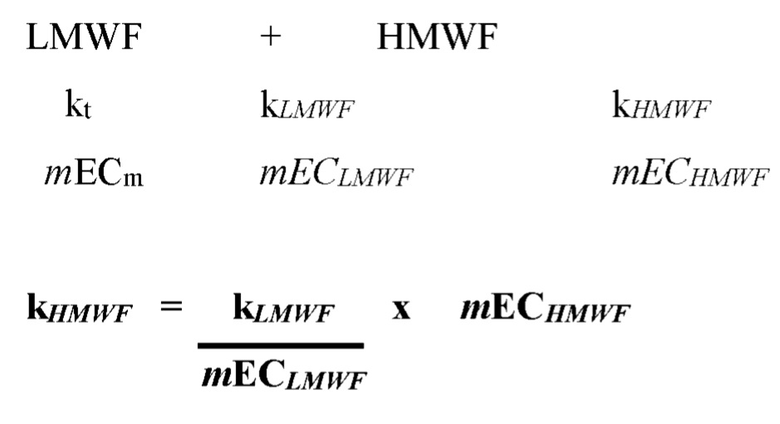
Таблица 18. Анализ предполагаемой фрагментации
(кДа)
1Fab+2scFv+Fc2
150
7,9
1,6 (k1)
1 Потеря мономера включают потерю только по причине фрагментации и не включает агрегацию.
2 Скорости могут быть недостаточно оцениваться по причине совместного элюирования HMWF с мономером.
3 Скорости для пика в виде плеча могут варьироваться из-за интеграции не полностью разрешенных пиков.
[00405] Далее проводили анализ скоростей фрагментации с конструкциями в восстанавливающих условиях, чтобы определить, оказало ли образование дисульфидных связей эффект в отношении фрагментации и стабильности. Репрезентативные данные этого анализа представлены на (фигуре 86). В восстанавливающих условиях более высокие скорости фрагментации наблюдались для всех форматов BiS, за исключением BiS1 (таблица 19). Сделали вывод о том, что более высокие скорости фрагментации в восстанавливающих условиях подтверждают, что фрагмент scFv в конструкции BiS5 связан с CH3-областью (фигура 89).
Таблица 19. Анализ фрагментации - восстанавливающие условия
(кДа)
(SEC/Red-GXII)
Fc2
1Fab+2scFv+
Fc2
150
7,9
x (k1)
Fc3
Таблица 20. Обзор анализов фрагментации
[00406] Дополнительно исследовали характеристики стабилизирующих дисульфидных связей, раскрытых выше. Результаты показаны в таблицах 23 и 24 ниже. Биспецифические антитела, соответствующие двум разным специфичностям, получали в форматах BiS4 и BiS5 (scFv вставлен в петлю SN-G) со стабилизирующей дисульфидной связью в scFv и без нее. Исследование стабильности при ускоренном хранении показало, что конструкции BiS4 без стабилизирующей дисульфидной связи характеризовались значительными потерями мономера из-за разрушения, что предотвращалось путем введения стабилизирующей дисульфидной связи VL-VH. Данные результаты показывают, что удаление стабилизирующей дисульфидной связи в scFv в конструкции BiS5 не оказало существенного эффекта в отношении ее стабильности.
Таблица 21
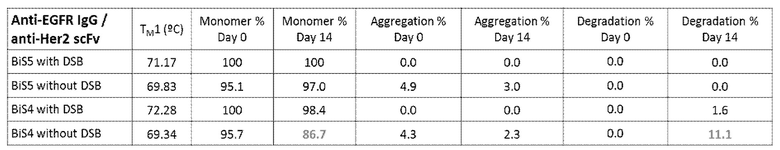
Таблица 22
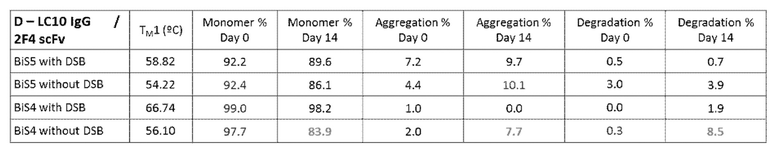
[00407] Исходя из всех вышеперечисленных данных, по-видимому, раскрытый в данном документе формат биспецифического связывающего белка является наиболее стабильным из всех тестируемых форматов. Более того, BiSAb5 представляется наиболее стабильным при более низких тестируемых значениях pH, (например, 5,0, 5,5 и 6,0) с точки зрения минимизации как фрагментации, так и агрегации. Таким образом, неожиданные и удивительные характеристики стабильности BisAb, раскрытых в данном документе, обеспечивают дополнительное преимущество по сравнению с другими структурными платформами и форматами, которые используются в разработке биспецифических связывающих молекул.
Включение посредством ссылки
[00408] Все публикации и патенты, упомянутые в данном документе, настоящим включены в данный документ посредством ссылки в полном их объеме, как если бы каждая отдельная публикация или патент были конкретно и индивидуально указаны как включенные посредством ссылки.
[00409] Хотя обсуждались конкретные аспекты объекта настоящего изобретения, - вышеуказанное описание носит иллюстративный, а не ограничительный характер. Многие варианты настоящего изобретения будут очевидны специалистам в данной области техники после рассмотрения настоящего описания и приведенной ниже формулы изобретения. Полный объем настоящего изобретения должен определяться со ссылкой на пункты формулы изобретения вместе с полным объемом их эквивалентов, и описанием вместе с такими вариантами.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MedImmune, LLC
<120> БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И ПУТИ ИХ ПРИМЕНЕНИЯ
<130> IOBS-100
<150> 62/332,788
<151> 2016-05-06
<160> 97
<170> PatentIn version 3.5
<210> 1
<211> 711
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 1
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
[001] 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly
145 150 155 160
Met His Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala
165 170 175
Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
210 215 220
Arg Asp Pro Arg Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp
225 230 235 240
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly
245 250 255
Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly
260 265 270
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
275 280 285
Phe Thr Phe Ser Asp Tyr Gly Met His Trp Val Arg Gln Ala Pro Gly
290 295 300
Lys Gly Leu Glu Trp Val Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile
305 310 315 320
Tyr Ser Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
325 330 335
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
340 345 350
Thr Ala Val Tyr Tyr Cys Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu
355 360 365
Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
370 375 380
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
385 390 395 400
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
405 410 415
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
420 425 430
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
435 440 445
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
450 455 460
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
465 470 475 480
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
485 490 495
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
500 505 510
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
515 520 525
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
530 535 540
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
545 550 555 560
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
565 570 575
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
580 585 590
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
595 600 605
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
610 615 620
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
625 630 635 640
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
645 650 655
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
660 665 670
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
675 680 685
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
690 695 700
Leu Ser Leu Ser Leu Gly Lys
705 710
<210> 2
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 2
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 3
<211> 711
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 3
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
450 455 460
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
465 470 475 480
Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr Gln
485 490 495
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser
500 505 510
Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
515 520 525
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
530 535 540
Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Cys Gly
545 550 555 560
Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
565 570 575
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Glu
580 585 590
Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
595 600 605
Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg
610 615 620
Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile Trp Tyr Asp
625 630 635 640
Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
645 650 655
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
660 665 670
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly
675 680 685
Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly
690 695 700
Thr Thr Val Thr Val Ser Ser
705 710
<210> 4
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 4
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Cys Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Val Ser
210
<210> 5
<211> 721
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met
385 390 395 400
Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr
405 410 415
Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr
420 425 430
Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser
435 440 445
Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
450 455 460
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
465 470 475 480
Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Cys
485 490 495
Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly
500 505 510
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val
515 520 525
Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser
530 535 540
Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val
545 550 555 560
Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile Trp Tyr
565 570 575
Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
580 585 590
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
595 600 605
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg
610 615 620
Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln
625 630 635 640
Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
645 650 655
Gly Ser Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
660 665 670
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
675 680 685
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
690 695 700
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
705 710 715 720
Lys
<210> 6
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 6
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 7
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 7
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 8
<211> 610
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 8
cgagagagcc accctgagct gctccgcctc ctccaagcac accaacctgt actggtcccg 60
gcacatgtac tggtatcagc agaagcccgg ccaggcccct cggctgctga tctacctgac 120
ctctaaccgg gccaccggca tccctgccag attctccggc tctggctccg gcaccgactt 180
caccctgacc atctccagcc tggaacccga ggacttcgcc gtgtactact gccagcagtg 240
gtcctccaac cccttcacct tcggccaggg caccaagctg gaaatcaagc gtacggtggc 300
tgcaccatct gtcttcatct tcccgccatc tgatgagcag ttgaaatctg gaactgcctc 360
tgttgtgtgc ctgctgaata acttctatcc cagagaggcc aaagtacagt ggaaggtgga 420
taacgccctc caatcgggta actcccagga gagtgtcaca gagcaggaca gcaaggacag 480
cacctacagc ctcagcagca ccctgacgct gagcaaagca gactacgaga aacacaaagt 540
ctacgcctgc gaagtcaccc atcagggcct gagctcgccc gtcacaaaga gcttcaacag 600
gggagagtgt 610
<210> 9
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 10
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полинуклеотид
<400> 10
gaggtgcagc tggtggaatc cggcggagga ctggtgcagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt cacattctcc gactacggca tgcactgggt ccgacaggcc 120
cctggaaagg gcctggaatg ggtggcctac atctcctccg gctcctacac catctactcc 180
gccgactccg tgaagggccg gttcaccatc tcccgggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgcg ggccgaggac acagccgtgt actactgtgc cagacgggcc 300
cctaactcct tctacgagta ctacttcgac tactggggcc agggcaccac cgtgaccgtg 360
tcctctgcta gcaccaaagg tccgagcgtt tttccgctgg caccgagcag caaaagcacc 420
tctggtggca ccgcagcact gggttgtctg gtgaaagatt attttccgga accggttacc 480
gtttcttgga atagcggtgc actgaccagc ggtgttcata cctttccggc agttctgcag 540
agcagcggtc tgtatagcct gtctagcgtt gttaccgttc cgagcagcag cctgggcacc 600
cagacctata tttgcaatgt gaatcataaa ccgagcaata caaaagttga taaacgcgtt 660
gaaccgaaaa gctgtgacaa aactcacacg tgcccaccgt gcccagcacc tgagttcgag 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 780
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagccagcat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtct gcaccctgcc cccatcccgg 1080
gaggagatga ccaagaacca ggtcagcctg agctgcgcgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctcgttagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcctctc cctgtctccg ggtaaa 1356
<210> 11
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 11
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattaac agctatttag attggtatca gcagaaacca 120
gggaaagccc ctaaactcct gatctatgct gcatccagtt tgcaaagtgg ggtcccatca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag tattacagta ctccattcac tttcggccct 300
gggaccaaag tggaaatcaa aggtcagccc aaggcggccc cctcggtcac tctgttcccg 360
ccctgctctg aggagcttca agccaacaag gccacactgg tgtgtctcat aagtgacttc 420
tacccgggag ccgtgacagt ggcctggaag gcagatagca gccccgtcaa ggcgggagtg 480
gagaccacca caccctccaa acaaagcaac aacaagtacg cggccagcag ctacctgagc 540
ctgacgcctg agcagtggaa gtcccacaga agctacagct gccaggtcac gcatgaaggg 600
agcaccgtgg agaagacagt ggcccctaca gaagtgtca 639
<210> 12
<211> 455
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 12
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Arg Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Cys Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
210 215 220
Pro Lys Ser Val Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Phe Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 13
<211> 1365
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 13
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cgtctggatt caccttcagt agctatggca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatggtatg atggaagtaa taaatactat 180
gcagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggctgtgt attactgtgc gagagatccg 300
aggggagcta ccctttacta ctactactac ggtatggacg tctggggcca agggaccacg 360
gtcaccgtct cctcagcgtc gaccaaaggt ccgagcgtgt gcccgctggc accgagcagc 420
aaaagcacct ctggtggcac cgcagcactg ggttgtctgg tgaaagatta ttttccggaa 480
ccggttaccg tttcttggaa tagcggtgca ctgaccagcg gtgttcatac ctttccggca 540
gtcctgcaga gcagcggtct gtatagcctg tctagcgttg ttaccgttcc gagcagcagc 600
ctgggcaccc agacctatat ttgcaatgtg aatcataaac cgagcaatac caaagttgat 660
aaacgcgttg aaccgaaaag cgtggacaaa actcacacgt gcccaccgtg cccagcacct 720
gagttcgagg ggggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 780
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 840
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 900
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 960
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agccagcatc 1020
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtcta caccctgccc 1080
ccatgccggg aggagatgac caagaaccag gtcagcctgt ggtgcctggt caaaggcttc 1140
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 1200
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctatagcaa gctcaccgtg 1260
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 1320
cacaaccact acacgcagaa gagcttaagc ctgtctccgg gtaaa 1365
<210> 14
<211> 710
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
115 120 125
Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser
130 135 140
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly
145 150 155 160
Met His Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala
165 170 175
Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys
180 185 190
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
195 200 205
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
210 215 220
Arg Asp Pro Arg Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp
225 230 235 240
Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly
245 250 255
Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly
260 265 270
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
275 280 285
Phe Thr Phe Ser Arg Tyr Trp Met Ser Trp Val Arg Gln Ala Pro Gly
290 295 300
Lys Gly Leu Glu Trp Val Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys
305 310 315 320
Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn
325 330 335
Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp
340 345 350
Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu
355 360 365
Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala
370 375 380
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser
385 390 395 400
Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
405 410 415
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
420 425 430
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
435 440 445
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr
450 455 460
Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg
465 470 475 480
Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
485 490 495
Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
500 505 510
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
515 520 525
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
530 535 540
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
545 550 555 560
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
565 570 575
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
580 585 590
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
595 600 605
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
610 615 620
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
625 630 635 640
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
645 650 655
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
660 665 670
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
675 680 685
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
690 695 700
Ser Leu Ser Leu Gly Lys
705 710
<210> 15
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 15
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 16
<211> 710
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln
450 455 460
Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr
465 470 475 480
Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr Gln Gln
485 490 495
Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu
500 505 510
Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
515 520 525
Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr
530 535 540
Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Cys Gly Thr
545 550 555 560
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
565 570 575
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Glu Ser
580 585 590
Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala
595 600 605
Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg Gln
610 615 620
Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile Trp Tyr Asp Gly
625 630 635 640
Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser
645 650 655
Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg
660 665 670
Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly Ala
675 680 685
Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr
690 695 700
Thr Val Thr Val Ser Ser
705 710
<210> 17
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 17
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 18
<211> 720
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
385 390 395 400
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
405 410 415
Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr Leu Asp Trp Tyr Gln
420 425 430
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser
435 440 445
Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
450 455 460
Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
465 470 475 480
Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe Thr Phe Gly Cys Gly
485 490 495
Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
500 505 510
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Glu
515 520 525
Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
530 535 540
Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg
545 550 555 560
Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Val Ile Trp Tyr Asp
565 570 575
Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
580 585 590
Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu
595 600 605
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly
610 615 620
Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly
625 630 635 640
Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
645 650 655
Ser Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
660 665 670
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
675 680 685
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
690 695 700
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
705 710 715 720
<210> 19
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 19
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 20
<211> 250
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 20
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr
130 135 140
Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala
145 150 155 160
Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp
165 170 175
Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp
180 185 190
Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser Asn Ser
195 200 205
Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly Asp Glu
210 215 220
Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His Trp Leu
225 230 235 240
Phe Gly Cys Gly Thr Lys Leu Thr Val Leu
245 250
<210> 21
<211> 709
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Leu
450 455 460
Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
465 470 475 480
Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp Val
485 490 495
Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser Ala Ile Ser Gly
500 505 510
Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
515 520 525
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
530 535 540
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Tyr
545 550 555 560
Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp Gly Arg Gly Thr Leu
565 570 575
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Leu Thr Gln Pro
595 600 605
Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Ser Ile Ser Cys Gly
610 615 620
Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp Tyr Gln Gln Lys Pro
625 630 635 640
Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser Asp Arg Pro Ser
645 650 655
Gly Ile Pro Gln Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
660 665 670
Leu Thr Ile His Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
675 680 685
Gln Val Leu Asp Arg Arg Ser Asp His Trp Leu Phe Gly Cys Gly Thr
690 695 700
Lys Leu Thr Val Leu
705
<210> 22
<211> 719
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
385 390 395 400
Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu
405 410 415
Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp
420 425 430
Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser Ala Ile Ser
435 440 445
Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe
450 455 460
Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn
465 470 475 480
Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser
485 490 495
Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp Gly Arg Gly Thr
500 505 510
Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
515 520 525
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Leu Thr Gln
530 535 540
Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Ser Ile Ser Cys
545 550 555 560
Gly Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp Tyr Gln Gln Lys
565 570 575
Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser Asp Arg Pro
580 585 590
Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala
595 600 605
Thr Leu Thr Ile His Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr
610 615 620
Cys Gln Val Leu Asp Arg Arg Ser Asp His Trp Leu Phe Gly Cys Gly
625 630 635 640
Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
645 650 655
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
660 665 670
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
675 680 685
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
690 695 700
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
705 710 715
<210> 23
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 23
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 24
<211> 709
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Leu
450 455 460
Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser
465 470 475 480
Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser Trp Val
485 490 495
Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser Ala Ile Ser Gly
500 505 510
Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
515 520 525
Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
530 535 540
Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Gly Ser Tyr
545 550 555 560
Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp Gly Arg Gly Thr Leu
565 570 575
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
580 585 590
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Thr Val Leu Thr Gln Pro
595 600 605
Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Ser Ile Ser Cys Gly
610 615 620
Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp Tyr Gln Gln Lys Pro
625 630 635 640
Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser Asp Arg Pro Ser
645 650 655
Gly Ile Pro Gln Arg Phe Ser Gly Ser Asn Ser Gly Asn Thr Ala Thr
660 665 670
Leu Thr Ile His Arg Val Glu Ala Gly Asp Glu Ala Asp Tyr Tyr Cys
675 680 685
Gln Val Leu Asp Arg Arg Ser Asp His Trp Leu Phe Gly Cys Gly Thr
690 695 700
Lys Leu Thr Val Leu
705
<210> 25
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 26
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 26
Gln Thr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Cys Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Val Ser
210
<210> 27
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 27
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Cys Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Val Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly
450
<210> 28
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 28
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Phe Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Cys Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Val Ser
210
<210> 29
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 29
agctacgtgc tgacgcagcc gccgtcagtg tcagtggccc caggaaagac ggccaggatt 60
acctgtgggg gagacaacat tggaggtaaa agtgttcact ggtaccagca gaagccaggc 120
caggcccctg tgttggtcat ctattatgat agtgaccggc cctcaggcat ccctgagcga 180
ttctctggct ccaactctgg gaacacggcc accctgacca tcagcagggt cgaagccggg 240
gatgaggccg attattactg tcaggtgttg gatcgtcgta gtgatcattt cctgttcggc 300
ggagggacca agctgaccgt cctaggtcag cccaaggcgg cgccctcggt cactctgttc 360
ccgccctgct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac 420
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga 480
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg 540
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa 600
gggagcaccg tggagaagac agtggcccct acagaagtgt ca 642
<210> 30
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 30
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Cys Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Val Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 31
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 31
gaggtgcagc tgttggagtc tgggggaggc ttggtacagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggatt cacctttagc agctatgcca tgagctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtctcagct attagtggta gtggtggtag cacatactac 180
gcagactccg tgaagggccg gttcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acagcctgag agccgaggac acggccgtgt attactgtgc gagagggtcc 300
tatggtacct actacggaaa ctactttgaa tactggggcc agggcaccct ggtcaccgtc 360
tcgagtgcgt cgaccaaagg tccgagcgtg tgcccgctgg caccgagcag caaaagcacc 420
tctggtggca ccgcagcact gggttgtctg gtgaaagatt attttccgga accggttacc 480
gtttcttgga atagcggtgc actgaccagc ggtgttcata cctttccggc agtcctgcag 540
agcagcggtc tgtatagcct gtctagcgtt gttaccgttc cgagcagcag cctgggcacc 600
cagacctata tttgcaatgt gaatcataaa ccgagcaata ccaaagttga taaacgcgtt 660
gaaccgaaaa gcgtggacaa aactcacacg tgcccaccgt gcccagcacc tgagttcgag 720
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 780
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 840
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 900
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 960
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagccagcat cgagaaaacc 1020
atctccaaag ccaaagggca gccccgagaa ccacaggtct acaccctgcc cccatgccgg 1080
gaggagatga ccaagaacca ggtcagcctg tggtgcctgg tcaaaggctt ctatcccagc 1140
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1200
cccgtgctgg actccgacgg ctccttcttc ctctatagca agctcaccgt ggacaagagc 1260
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1320
tacacgcaga agagcttaag cctgtctccg ggtaaa 1356
<210> 32
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 32
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Lys Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 33
<211> 215
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 33
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 34
<211> 707
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 34
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly
100 105 110
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
130 135 140
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
145 150 155 160
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val
165 170 175
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
180 185 190
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
195 200 205
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
210 215 220
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
225 230 235 240
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
245 250 255
Gly Gly Ser Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
260 265 270
Pro Ser Gln Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe
275 280 285
Ser Ser Gly Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu
290 295 300
Glu Tyr Ile Gly Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro
305 310 315 320
Ser Leu Lys Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln
325 330 335
Tyr Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr
340 345 350
Tyr Cys Ala Arg Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp
355 360 365
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
370 375 380
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
385 390 395 400
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
405 410 415
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
420 425 430
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
435 440 445
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
450 455 460
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
465 470 475 480
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
485 490 495
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
500 505 510
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
515 520 525
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
530 535 540
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
545 550 555 560
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
565 570 575
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
580 585 590
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
595 600 605
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
610 615 620
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
625 630 635 640
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
645 650 655
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
660 665 670
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
675 680 685
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
690 695 700
Leu Gly Lys
705
<210> 35
<211> 707
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 35
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln
450 455 460
Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser
465 470 475 480
Cys Arg Ala Ser Gln Arg Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln
485 490 495
Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Ser
500 505 510
Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
515 520 525
Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val
530 535 540
Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro Trp Thr Phe Gly Cys Gly
545 550 555 560
Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
565 570 575
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
580 585 590
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
595 600 605
Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr Trp Met Ser Trp Val Arg
610 615 620
Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Asn Ile Lys Gln Asp
625 630 635 640
Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile
645 650 655
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
660 665 670
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Gly Trp
675 680 685
Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
690 695 700
Val Ser Ser
705
<210> 36
<211> 707
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 36
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr Gln
450 455 460
Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu Ser
465 470 475 480
Cys Arg Ala Ser Gln Arg Val Ser Ser Ser Tyr Leu Ala Trp Tyr Gln
485 490 495
Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser Ser
500 505 510
Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr
515 520 525
Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala Val
530 535 540
Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro Trp Thr Phe Gly Cys Gly
545 550 555 560
Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
565 570 575
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
580 585 590
Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys
595 600 605
Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr Trp Met Ser Trp Val Arg
610 615 620
Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Asn Ile Lys Gln Asp
625 630 635 640
Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile
645 650 655
Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu
660 665 670
Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Glu Gly Gly Trp
675 680 685
Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
690 695 700
Val Ser Ser
705
<210> 37
<211> 718
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 37
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Asp Met His Trp Val Arg Gln Ala Thr Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Gly Ile Gly Thr Ala Gly Asp Thr Tyr Tyr Pro Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Glu Asn Ala Lys Asn Ser Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Gly Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Arg Tyr Ser Pro Thr Gly His Tyr Tyr Gly Met Asp Val Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
385 390 395 400
Ile Val Met Thr Gln Ser Pro Ala Thr Leu Ser Val Ser Pro Gly Glu
405 410 415
Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu
420 425 430
Gly Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr
435 440 445
Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly Ser
450 455 460
Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Ser Glu
465 470 475 480
Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gln Asn Trp Pro Leu Leu
485 490 495
Thr Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser
500 505 510
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
515 520 525
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
530 535 540
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Tyr Met
545 550 555 560
Met Gln Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser
565 570 575
Ser Ile Trp Pro Ser Gly Gly Lys Thr Tyr Tyr Ala Asp Ser Val Lys
580 585 590
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
595 600 605
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
610 615 620
Arg Val Arg Arg Gly Gly Ala Thr Asp Tyr Trp Gly Gln Gly Thr Leu
625 630 635 640
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
645 650 655
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
660 665 670
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
675 680 685
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
690 695 700
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
705 710 715
<210> 38
<211> 716
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 38
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Met Met Gln Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Trp Pro Ser Gly Gly Lys Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Arg Arg Gly Gly Ala Thr Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
385 390 395 400
Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
405 410 415
Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp Leu Ala Trp Tyr Gln
420 425 430
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Lys Ala Ser Ser
435 440 445
Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
450 455 460
Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Asp Asp Phe Ala Thr
465 470 475 480
Tyr Tyr Cys Lys Gln Tyr Ala Asp Tyr Trp Thr Phe Gly Cys Gly Thr
485 490 495
Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
500 505 510
Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser
515 520 525
Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala
530 535 540
Ala Ser Gly Phe Thr Phe Ser Ser His Asp Met His Trp Val Arg Gln
545 550 555 560
Ala Thr Gly Lys Cys Leu Glu Trp Val Ser Gly Ile Gly Thr Ala Gly
565 570 575
Asp Thr Tyr Tyr Pro Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg
580 585 590
Glu Asn Ala Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala
595 600 605
Gly Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Arg Tyr Ser Pro Thr
610 615 620
Gly His Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr
625 630 635 640
Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gln Pro
645 650 655
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
660 665 670
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
675 680 685
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
690 695 700
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
705 710 715
<210> 39
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 39
Ile Ser Arg Thr Pro
1. 5
<210> 40
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 40
Ala Lys Gly Gln Pro
1. 5
<210> 41
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 41
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10
<210> 42
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 42
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1. 5 10 15
<210> 43
<211> 20
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 43
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1. 5 10 15
Gly Gly Gly Ser
20
<210> 44
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 44
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1. 5 10 15
<210> 45
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 45
Glu Pro Lys Ser Cys Asp Lys Thr
1. 5
<210> 46
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 46
Glu Pro Lys Ser Cys Gly Lys Thr
1. 5
<210> 47
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 47
Glu Pro Lys Ser Cys
1. 5
<210> 48
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 48
Gly Gly Gly Gly Ser
1. 5
<210> 49
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 49
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 50
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 50
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys
<210> 51
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 52
<211> 125
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 52
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Pro Arg Gly Ala Thr Leu Tyr Tyr Tyr Tyr Tyr Gly Met
100 105 110
Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 53
<211> 167
<212> Белок
<213> Искусственная последовательность
<220>
<223> CTLA-4 HC
<400> 53
Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser
1. 5 10 15
Gly Phe Thr Phe Ser Ser Tyr Gly Met His Trp Val Arg Gln Ala Pro
20 25 30
Gly Lys Gly Leu Glu Trp Val Ala Val Ile Trp Tyr Asp Gly Ser Asn
35 40 45
Lys Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp
50 55 60
Asn Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu
65 70 75 80
Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Pro Arg Gly Ala Thr Leu
85 90 95
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His
165
<210> 54
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys
100 105
<210> 55
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 55
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asn Ser Tyr
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 56
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Thr Ala Ser Gly Tyr Thr Phe Pro Asp Tyr
20 25 30
Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Asp Pro Asn Tyr Gly Gly Thr Thr Tyr Asn Ala Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Arg Ser Lys Ser Ile Ala Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Leu Thr Asp Trp Gly Gln Gly Thr Met Val Thr Val
100 105 110
Ser Ser
<210> 57
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 57
Gln Ile Gln Leu Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
Tyr Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Tyr
35 40 45
Ala Thr Phe Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Asn Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 58
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 58
Gln Ile Gln Leu Thr Gln Ser Pro Ser Ile Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
Tyr Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Pro Leu Ile Tyr
35 40 45
Ala Thr Phe Asn Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Asn Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 59
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 60
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 60
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 61
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 61
Gln Thr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 62
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 62
Gln Thr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 63
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 63
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys
<210> 64
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 64
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 65
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 65
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 66
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 66
Ser Tyr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Arg Ile Thr Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 67
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 67
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys
<210> 68
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 68
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 69
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 69
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 70
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 70
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Lys Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 71
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 71
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 72
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 72
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Lys Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 73
<211> 288
<212> Белок
<213> Homo sapiens
<400> 73
Met Gln Ile Pro Gln Ala Pro Trp Pro Val Val Trp Ala Val Leu Gln
1. 5 10 15
Leu Gly Trp Arg Pro Gly Trp Phe Leu Asp Ser Pro Asp Arg Pro Trp
20 25 30
Asn Pro Pro Thr Phe Ser Pro Ala Leu Leu Val Val Thr Glu Gly Asp
35 40 45
Asn Ala Thr Phe Thr Cys Ser Phe Ser Asn Thr Ser Glu Ser Phe Val
50 55 60
Leu Asn Trp Tyr Arg Met Ser Pro Ser Asn Gln Thr Asp Lys Leu Ala
65 70 75 80
Ala Phe Pro Glu Asp Arg Ser Gln Pro Gly Gln Asp Cys Arg Phe Arg
85 90 95
Val Thr Gln Leu Pro Asn Gly Arg Asp Phe His Met Ser Val Val Arg
100 105 110
Ala Arg Arg Asn Asp Ser Gly Thr Tyr Leu Cys Gly Ala Ile Ser Leu
115 120 125
Ala Pro Lys Ala Gln Ile Lys Glu Ser Leu Arg Ala Glu Leu Arg Val
130 135 140
Thr Glu Arg Arg Ala Glu Val Pro Thr Ala His Pro Ser Pro Ser Pro
145 150 155 160
Arg Pro Ala Gly Gln Phe Gln Thr Leu Val Val Gly Val Val Gly Gly
165 170 175
Leu Leu Gly Ser Leu Val Leu Leu Val Trp Val Leu Ala Val Ile Cys
180 185 190
Ser Arg Ala Ala Arg Gly Thr Ile Gly Ala Arg Arg Thr Gly Gln Pro
195 200 205
Leu Lys Glu Asp Pro Ser Ala Val Pro Val Phe Ser Val Asp Tyr Gly
210 215 220
Glu Leu Asp Phe Gln Trp Arg Glu Lys Thr Pro Glu Pro Pro Val Pro
225 230 235 240
Cys Val Pro Glu Gln Thr Glu Tyr Ala Thr Ile Val Phe Pro Ser Gly
245 250 255
Met Gly Thr Ser Ser Pro Ala Arg Arg Gly Ser Ala Asp Gly Pro Arg
260 265 270
Ser Ala Gln Pro Leu Arg Pro Glu Asp Gly His Cys Ser Trp Pro Leu
275 280 285
<210> 74
<211> 176
<212> Белок
<213> Homo sapiens
<400> 74
Met Arg Ile Phe Ala Val Phe Ile Phe Met Thr Tyr Trp His Leu Leu
1. 5 10 15
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
20 25 30
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
35 40 45
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
50 55 60
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
65 70 75 80
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
85 90 95
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
100 105 110
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr His Leu Val
115 120 125
Ile Leu Gly Ala Ile Leu Leu Cys Leu Gly Val Ala Leu Thr Phe Ile
130 135 140
Phe Arg Leu Arg Lys Gly Arg Met Met Asp Val Lys Lys Cys Gly Ile
145 150 155 160
Gln Asp Thr Asn Ser Lys Lys Gln Ser Asp Thr His Leu Glu Glu Thr
165 170 175
<210> 75
<211> 223
<212> Белок
<213> Homo sapiens
<400> 75
Met Ala Cys Leu Gly Phe Gln Arg His Lys Ala Gln Leu Asn Leu Ala
1. 5 10 15
Thr Arg Thr Trp Pro Cys Thr Leu Leu Phe Phe Leu Leu Phe Ile Pro
20 25 30
Val Phe Cys Lys Ala Met His Val Ala Gln Pro Ala Val Val Leu Ala
35 40 45
Ser Ser Arg Gly Ile Ala Ser Phe Val Cys Glu Tyr Ala Ser Pro Gly
50 55 60
Lys Ala Thr Glu Val Arg Val Thr Val Leu Arg Gln Ala Asp Ser Gln
65 70 75 80
Val Thr Glu Val Cys Ala Ala Thr Tyr Met Met Gly Asn Glu Leu Thr
85 90 95
Phe Leu Asp Asp Ser Ile Cys Thr Gly Thr Ser Ser Gly Asn Gln Val
100 105 110
Asn Leu Thr Ile Gln Gly Leu Arg Ala Met Asp Thr Gly Leu Tyr Ile
115 120 125
Cys Lys Val Glu Leu Met Tyr Pro Pro Pro Tyr Tyr Leu Gly Ile Gly
130 135 140
Asn Gly Thr Gln Ile Tyr Val Ile Asp Pro Glu Pro Cys Pro Asp Ser
145 150 155 160
Asp Phe Leu Leu Trp Ile Leu Ala Ala Val Ser Ser Gly Leu Phe Phe
165 170 175
Tyr Ser Phe Leu Leu Thr Ala Val Ser Leu Ser Lys Met Leu Lys Lys
180 185 190
Arg Ser Pro Leu Thr Thr Gly Val Tyr Val Lys Met Pro Pro Thr Glu
195 200 205
Pro Glu Cys Glu Lys Gln Phe Gln Pro Tyr Phe Ile Pro Ile Asn
210 215 220
<210> 76
<211> 301
<212> Белок
<213> Homo sapiens
<400> 76
Met Phe Ser His Leu Pro Phe Asp Cys Val Leu Leu Leu Leu Leu Leu
1. 5 10 15
Leu Leu Thr Arg Ser Ser Glu Val Glu Tyr Arg Ala Glu Val Gly Gln
20 25 30
Asn Ala Tyr Leu Pro Cys Phe Tyr Thr Pro Ala Ala Pro Gly Asn Leu
35 40 45
Val Pro Val Cys Trp Gly Lys Gly Ala Cys Pro Val Phe Glu Cys Gly
50 55 60
Asn Val Val Leu Arg Thr Asp Glu Arg Asp Val Asn Tyr Trp Thr Ser
65 70 75 80
Arg Tyr Trp Leu Asn Gly Asp Phe Arg Lys Gly Asp Val Ser Leu Thr
85 90 95
Ile Glu Asn Val Thr Leu Ala Asp Ser Gly Ile Tyr Cys Cys Arg Ile
100 105 110
Gln Ile Pro Gly Ile Met Asn Asp Glu Lys Phe Asn Leu Lys Leu Val
115 120 125
Ile Lys Pro Ala Lys Val Thr Pro Ala Pro Thr Arg Gln Arg Asp Phe
130 135 140
Thr Ala Ala Phe Pro Arg Met Leu Thr Thr Arg Gly His Gly Pro Ala
145 150 155 160
Glu Thr Gln Thr Leu Gly Ser Leu Pro Asp Ile Asn Leu Thr Gln Ile
165 170 175
Ser Thr Leu Ala Asn Glu Leu Arg Asp Ser Arg Leu Ala Asn Asp Leu
180 185 190
Arg Asp Ser Gly Ala Thr Ile Arg Ile Gly Ile Tyr Ile Gly Ala Gly
195 200 205
Ile Cys Ala Gly Leu Ala Leu Ala Leu Ile Phe Gly Ala Leu Ile Phe
210 215 220
Lys Trp Tyr Ser His Ser Lys Glu Lys Ile Gln Asn Leu Ser Leu Ile
225 230 235 240
Ser Leu Ala Asn Leu Pro Pro Ser Gly Leu Ala Asn Ala Val Ala Glu
245 250 255
Gly Ile Arg Ser Glu Glu Asn Ile Tyr Thr Ile Glu Glu Asn Val Tyr
260 265 270
Glu Val Glu Glu Pro Asn Glu Tyr Tyr Cys Tyr Val Ser Ser Arg Gln
275 280 285
Gln Pro Ser Gln Pro Leu Gly Cys Arg Phe Ala Met Pro
290 295 300
<210> 77
<211> 278
<212> Белок
<213> Homo sapiens
<400> 77
Met Gly Ser Pro Gly Met Val Leu Gly Leu Leu Val Gln Ile Trp Ala
1. 5 10 15
Leu Gln Glu Ala Ser Ser Leu Ser Val Gln Gln Gly Pro Asn Leu Leu
20 25 30
Gln Val Arg Gln Gly Ser Gln Ala Thr Leu Val Cys Gln Val Asp Gln
35 40 45
Ala Thr Ala Trp Glu Arg Leu Arg Val Lys Trp Thr Lys Asp Gly Ala
50 55 60
Ile Leu Cys Gln Pro Tyr Ile Thr Asn Gly Ser Leu Ser Leu Gly Val
65 70 75 80
Cys Gly Pro Gln Gly Arg Leu Ser Trp Gln Ala Pro Ser His Leu Thr
85 90 95
Leu Gln Leu Asp Pro Val Ser Leu Asn His Ser Gly Ala Tyr Val Cys
100 105 110
Trp Ala Ala Val Glu Ile Pro Glu Leu Glu Glu Ala Glu Gly Asn Ile
115 120 125
Thr Arg Leu Phe Val Asp Pro Asp Asp Pro Thr Gln Asn Arg Asn Arg
130 135 140
Ile Ala Ser Phe Pro Gly Phe Leu Phe Val Leu Leu Gly Val Gly Ser
145 150 155 160
Met Gly Val Ala Ala Ile Val Trp Gly Ala Trp Phe Trp Gly Arg Arg
165 170 175
Ser Cys Gln Gln Arg Asp Ser Gly Asn Ala Phe Tyr Ser Asn Val Leu
180 185 190
Tyr Arg Pro Arg Gly Ala Pro Lys Lys Ser Glu Asp Cys Ser Gly Glu
195 200 205
Gly Lys Asp Gln Arg Gly Gln Ser Ile Tyr Ser Thr Ser Phe Pro Gln
210 215 220
Pro Ala Pro Arg Gln Pro His Leu Ala Ser Arg Pro Cys Pro Ser Pro
225 230 235 240
Arg Pro Cys Pro Ser Pro Arg Pro Gly His Pro Val Ser Met Val Arg
245 250 255
Val Ser Pro Arg Pro Ser Pro Thr Gln Gln Pro Arg Pro Lys Gly Phe
260 265 270
Pro Lys Val Gly Glu Glu
275
<210> 78
<211> 277
<212> Белок
<213> Homo sapiens
<400> 78
Met Cys Val Gly Ala Arg Arg Leu Gly Arg Gly Pro Cys Ala Ala Leu
1. 5 10 15
Leu Leu Leu Gly Leu Gly Leu Ser Thr Val Thr Gly Leu His Cys Val
20 25 30
Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His Glu Cys Arg Pro
35 40 45
Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln Asn Thr Val Cys
50 55 60
Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val Ser Ser Lys Pro
65 70 75 80
Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly Ser Glu Arg Lys
85 90 95
Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg Cys Arg Ala Gly
100 105 110
Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp Cys Ala Pro Cys
115 120 125
Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala Cys Lys Pro Trp
130 135 140
Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln Pro Ala Ser Asn
145 150 155 160
Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro Ala Thr Gln Pro
165 170 175
Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr Val Gln Pro Thr
180 185 190
Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr Arg Pro Val Glu
195 200 205
Val Pro Gly Gly Arg Ala Val Ala Ala Ile Leu Gly Leu Gly Leu Val
210 215 220
Leu Gly Leu Leu Gly Pro Leu Ala Ile Leu Leu Ala Leu Tyr Leu Leu
225 230 235 240
Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly Gly
245 250 255
Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His Ser
260 265 270
Thr Leu Ala Lys Ile
275
<210> 79
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 79
Gly Phe Thr Phe Ser Ser Tyr Ala Met Ser
1. 5 10
<210> 80
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 80
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1. 5 10 15
Gly
<210> 81
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 81
Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr
1. 5 10
<210> 82
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 82
Gly Gly Asp Asn Ile Gly Gly Lys Ser Val His
1. 5 10
<210> 83
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 83
Tyr Asp Ser Asp Arg Pro Ser
1. 5
<210> 84
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 84
Gln Val Leu Asp Arg Arg Ser Asp His
1. 5
<210> 85
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 85
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 86
<211> 108
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 86
Gln Thr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 87
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 87
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp
100 105 110
Gly Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys
<210> 88
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический полипептид
<400> 88
Gln Thr Val Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys
1. 5 10 15
Thr Ala Ser Ile Ser Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Tyr Asp Ser Asp Arg Pro Ser Gly Ile Pro Gln Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile His Arg Val Glu Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His
85 90 95
Trp Leu Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 89
<211> 712
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS3 PD-1(LO115)/TIM3(O13-1) HC
<400> 89
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
450 455 460
Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
465 470 475 480
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala Met
485 490 495
Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser Ala
500 505 510
Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys Gly
515 520 525
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu Gln
530 535 540
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
545 550 555 560
Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp Gly Gln
565 570 575
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
580 585 590
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr Val Leu
595 600 605
Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Arg Ile
610 615 620
Thr Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp Tyr Gln
625 630 635 640
Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser Asp
645 650 655
Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly Asn
660 665 670
Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala Asp
675 680 685
Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His Phe Leu Phe Gly
690 695 700
Cys Gly Thr Lys Leu Thr Val Leu
705 710
<210> 90
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS3 PD-1(LO115)/TIM3(O13-1) LC
<400> 90
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 91
<211> 722
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS5 PD-1(LO115)/TIM3(O13-1) HC
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Phe Glu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Ser
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
385 390 395 400
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
405 410 415
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala
420 425 430
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ser
435 440 445
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
450 455 460
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
465 470 475 480
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
485 490 495
Arg Gly Ser Tyr Gly Thr Tyr Tyr Gly Asn Tyr Phe Glu Tyr Trp Gly
500 505 510
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
515 520 525
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ser Tyr Val
530 535 540
Leu Thr Gln Pro Pro Ser Val Ser Val Ala Pro Gly Lys Thr Ala Arg
545 550 555 560
Ile Thr Cys Gly Gly Asp Asn Ile Gly Gly Lys Ser Val His Trp Tyr
565 570 575
Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr Tyr Asp Ser
580 585 590
Asp Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser Asn Ser Gly
595 600 605
Asn Thr Ala Thr Leu Thr Ile Ser Arg Val Glu Ala Gly Asp Glu Ala
610 615 620
Asp Tyr Tyr Cys Gln Val Leu Asp Arg Arg Ser Asp His Phe Leu Phe
625 630 635 640
Gly Cys Gly Thr Lys Leu Thr Val Leu Gly Gly Gly Gly Ser Gly Gly
645 650 655
Gly Gly Ser Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
660 665 670
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
675 680 685
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
690 695 700
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
705 710 715 720
Gly Lys
<210> 92
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS5 PD-1(LO115)/TIM3(O13-1) LC
<400> 92
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 93
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> OX40SLR LCv kappa
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Lys Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 94
<211> 720
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS5-OX40SLR HC-G1-N434A-PD-L1
<400> 94
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly
20 25 30
Tyr Trp Asn Trp Ile Arg Lys His Pro Gly Lys Gly Leu Glu Tyr Ile
35 40 45
Gly Tyr Ile Ser Tyr Asn Ala Ile Thr Tyr His Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Tyr Lys Tyr Asp Tyr Glu Gly Gly His Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val
385 390 395 400
Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser Leu
405 410 415
Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr Trp Met
420 425 430
Ser Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Val Ala Asn
435 440 445
Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys Gly
450 455 460
Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr Leu Gln
465 470 475 480
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
485 490 495
Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly Gln Gly
500 505 510
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
515 520 525
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Ile Val Leu Thr
530 535 540
Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu Arg Ala Thr Leu
545 550 555 560
Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser Tyr Leu Ala Trp Tyr
565 570 575
Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile Tyr Asp Ala Ser
580 585 590
Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly
595 600 605
Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro Glu Asp Phe Ala
610 615 620
Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro Trp Thr Phe Gly Cys
625 630 635 640
Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly
645 650 655
Ser Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
660 665 670
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
675 680 685
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
690 695 700
Leu His Ala His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
705 710 715 720
<210> 95
<211> 707
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS2-PD-1- OX40 HC -4P
<400> 95
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Lys Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp
85 90 95
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser
100 105 110
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
115 120 125
Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln Thr
130 135 140
Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly Tyr
145 150 155 160
Trp Asn Trp Ile Arg Lys His Pro Gly Lys Cys Leu Glu Tyr Ile Gly
165 170 175
Tyr Ile Ser Tyr Asn Gly Ile Thr Tyr His Asn Pro Ser Leu Lys Ser
180 185 190
Arg Ile Thr Ile Asn Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu Gln
195 200 205
Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
210 215 220
Tyr Lys Tyr Asp Tyr Asp Gly Gly His Ala Met Asp Tyr Trp Gly Gln
225 230 235 240
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
245 250 255
Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro
260 265 270
Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser
275 280 285
Asp Tyr Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
290 295 300
Trp Val Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp
305 310 315 320
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
325 330 335
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
340 345 350
Tyr Cys Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp
355 360 365
Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys
370 375 380
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
385 390 395 400
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
405 410 415
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
420 425 430
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
435 440 445
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
450 455 460
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
465 470 475 480
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
485 490 495
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
500 505 510
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
515 520 525
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
530 535 540
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
545 550 555 560
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
565 570 575
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
580 585 590
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
595 600 605
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
610 615 620
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
625 630 635 640
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
645 650 655
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
660 665 670
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
675 680 685
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
690 695 700
Leu Gly Lys
705
<210> 96
<211> 707
<212> Белок
<213> Искусственная последовательность
<220>
<223> BiS3-OX40 HC-4P-PD-1
<400> 96
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Ser Gly Ser Tyr Thr Ile Tyr Ser Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Ala Pro Asn Ser Phe Tyr Glu Tyr Tyr Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
210 215 220
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
260 265 270
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
435 440 445
Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr
450 455 460
Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile
465 470 475 480
Thr Cys Arg Ala Ser Gln Asp Ile Ser Asn Tyr Leu Asn Trp Tyr Gln
485 490 495
Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Ser Lys
500 505 510
Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr
515 520 525
Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr
530 535 540
Tyr Tyr Cys Gln Gln Gly Ser Ala Leu Pro Trp Thr Phe Gly Cys Gly
545 550 555 560
Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
565 570 575
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Glu
580 585 590
Ser Gly Pro Gly Leu Val Lys Pro Ser Gln Thr Leu Ser Leu Thr Cys
595 600 605
Ala Val Tyr Gly Gly Ser Phe Ser Ser Gly Tyr Trp Asn Trp Ile Arg
610 615 620
Lys His Pro Gly Lys Cys Leu Glu Tyr Ile Gly Tyr Ile Ser Tyr Asn
625 630 635 640
Gly Ile Thr Tyr His Asn Pro Ser Leu Lys Ser Arg Ile Thr Ile Asn
645 650 655
Arg Asp Thr Ser Lys Asn Gln Tyr Ser Leu Gln Leu Asn Ser Val Thr
660 665 670
Pro Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Tyr Lys Tyr Asp Tyr
675 680 685
Asp Gly Gly His Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr
690 695 700
Val Ser Ser
705
<210> 97
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> PD1 LCv kappa
<400> 97
Gln Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Ser Ala Ser Ser Lys His Thr Asn Leu
20 25 30
Tyr Trp Ser Arg His Met Tyr Trp Tyr Gln Gln Lys Pro Gly Gln Ala
35 40 45
Pro Arg Leu Leu Ile Tyr Leu Thr Ser Asn Arg Ala Thr Gly Ile Pro
50 55 60
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Trp
85 90 95
Ser Ser Asn Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2793167C2 |
| КОМБИНАЦИЯ | 2015 |
|
RU2744841C2 |
| БЕЛКИ УПРАВЛЕНИЯ, НАВИГАЦИИ И КОНТРОЛЯ И СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2019 |
|
RU2811457C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2811477C2 |
| ИММУНОГЛОБУЛИН С ТАНДЕМНЫМ РАСПОЛОЖЕНИЕМ FAB-ФРАГМЕНТОВ И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2769133C2 |
| МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2837770C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2815683C2 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2727836C2 |

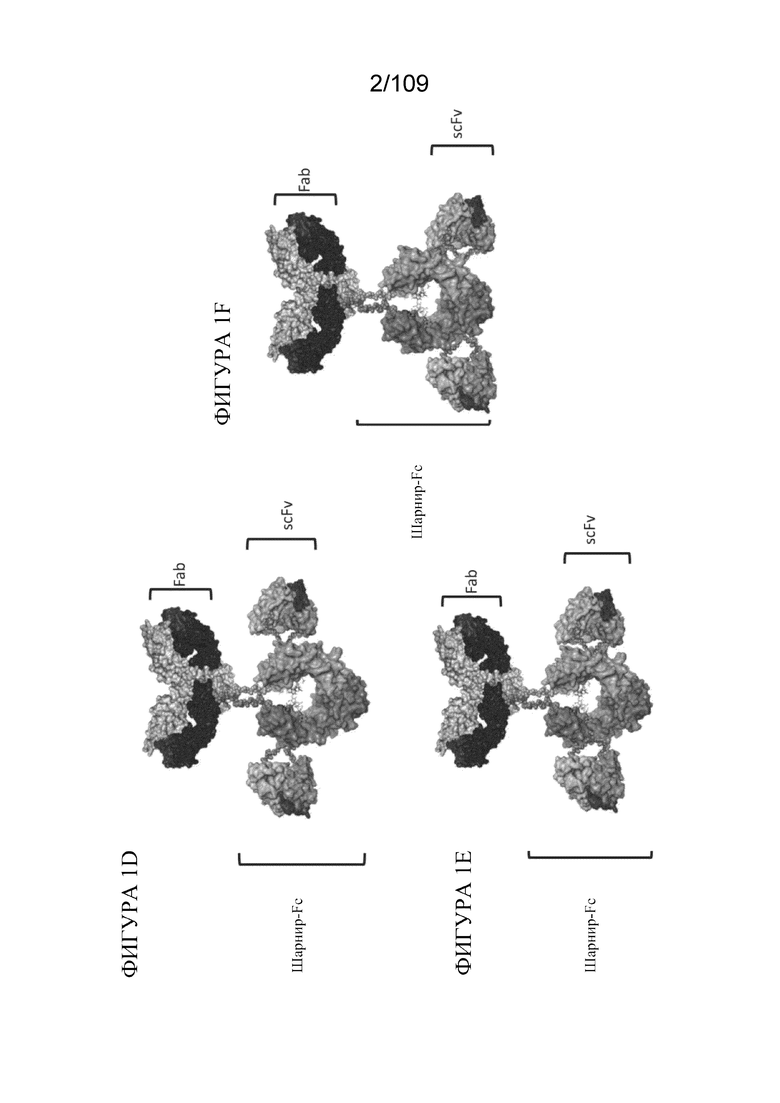
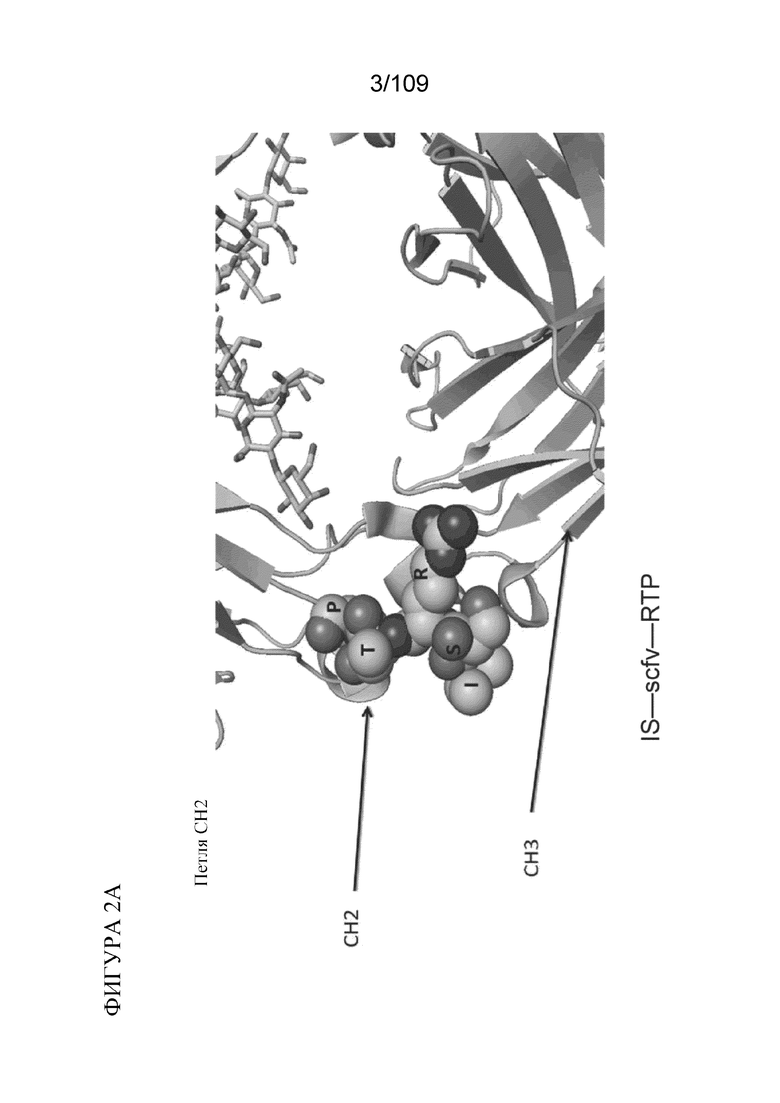
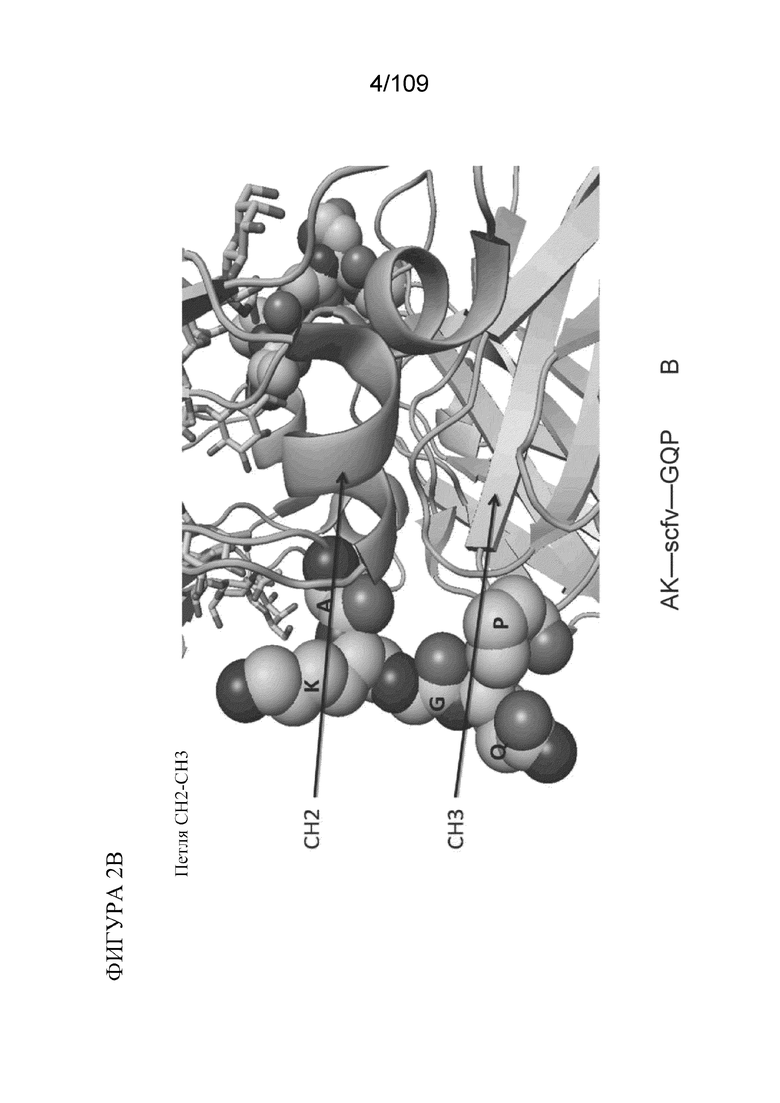
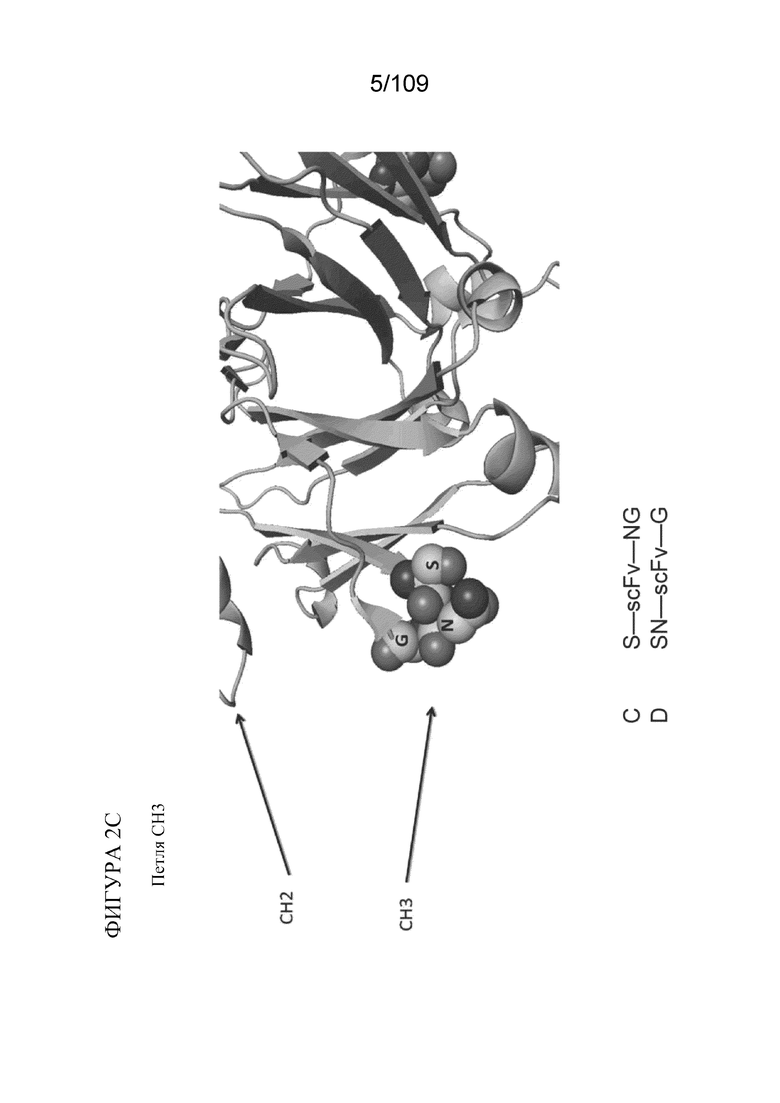
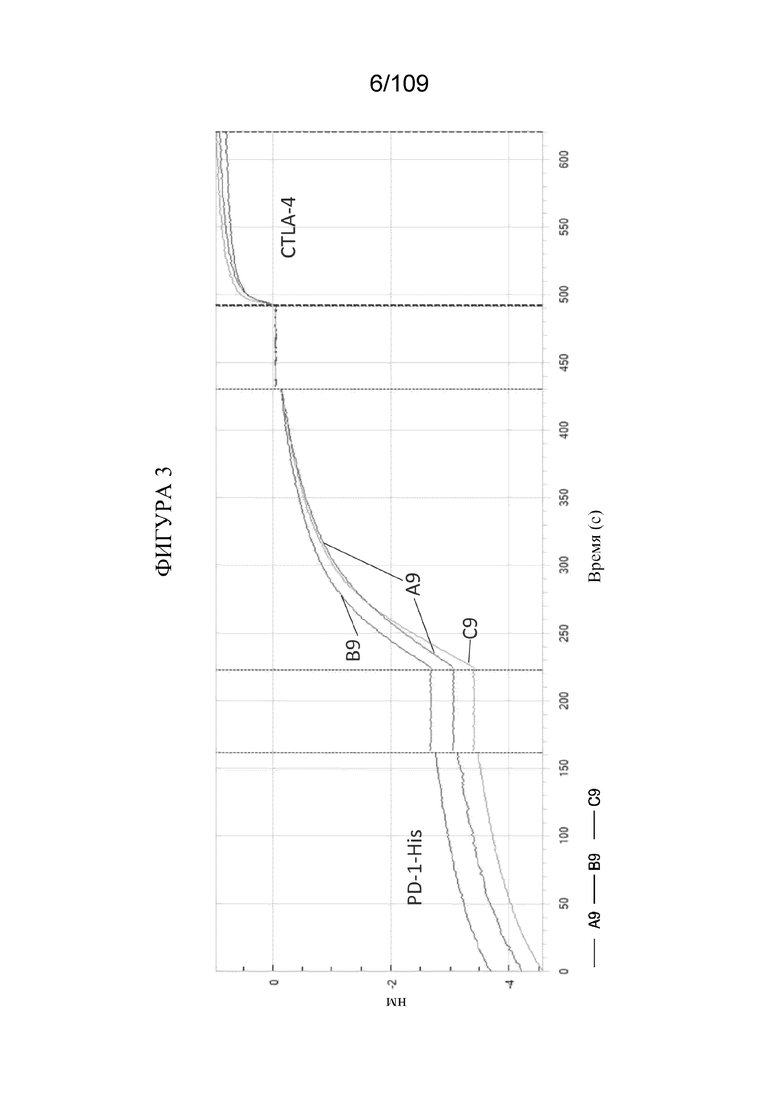

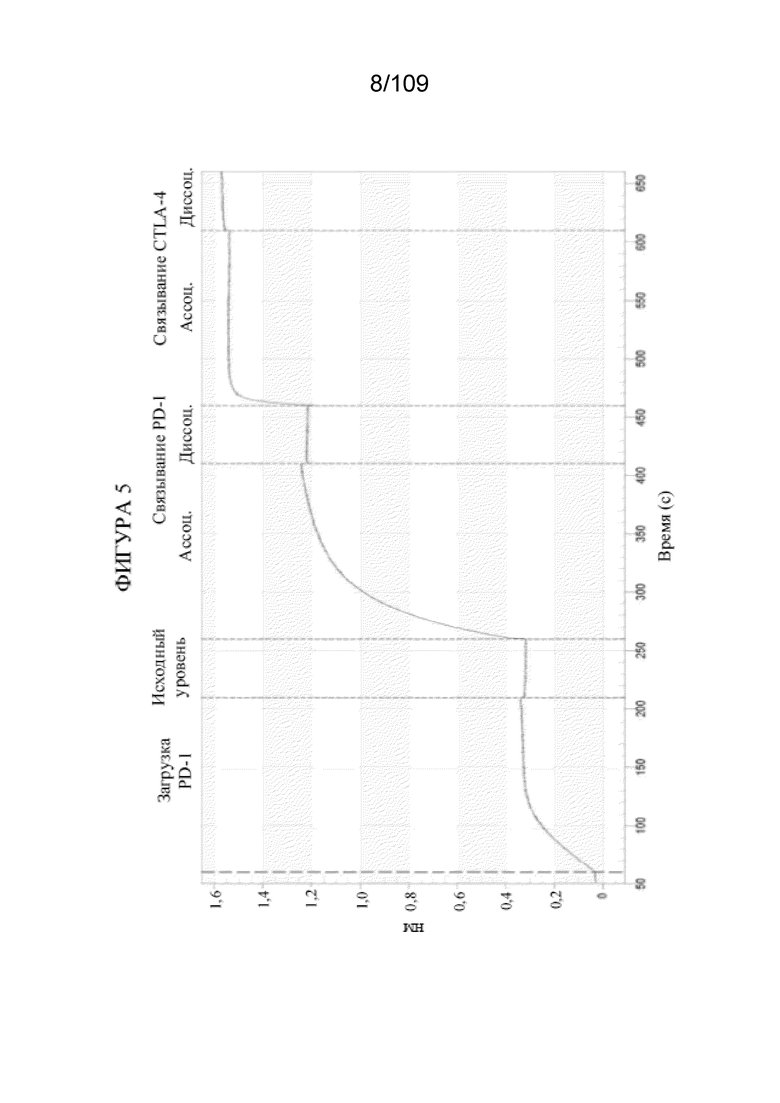
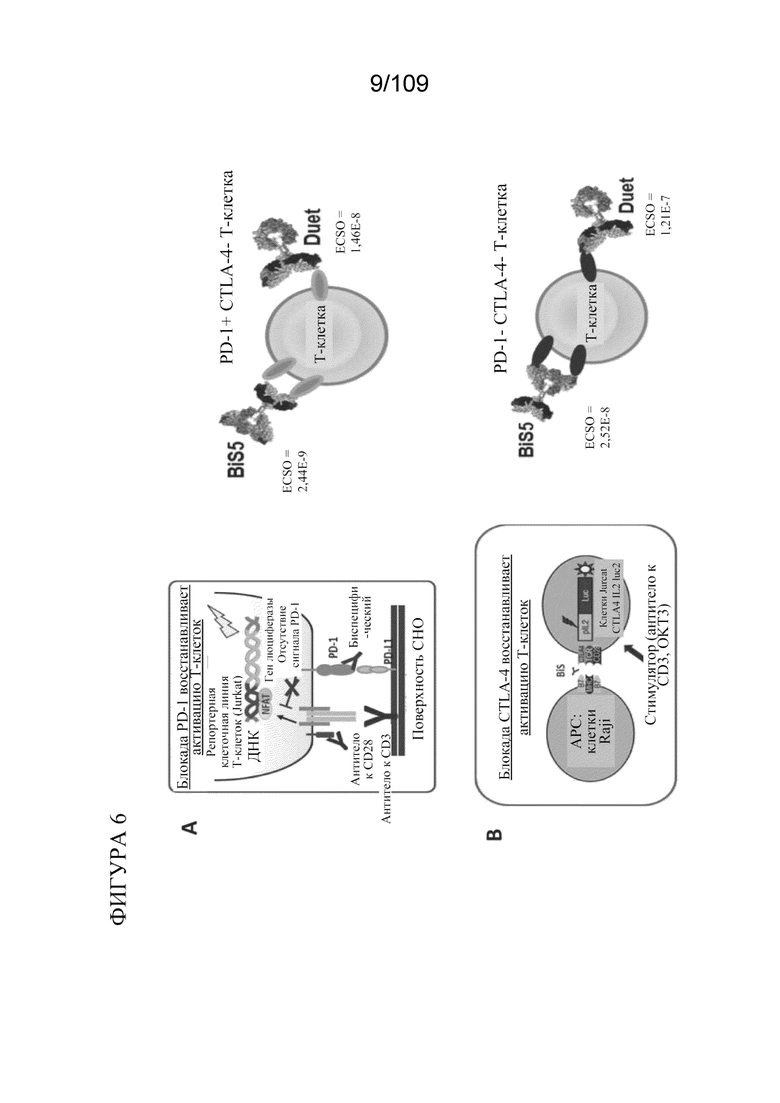

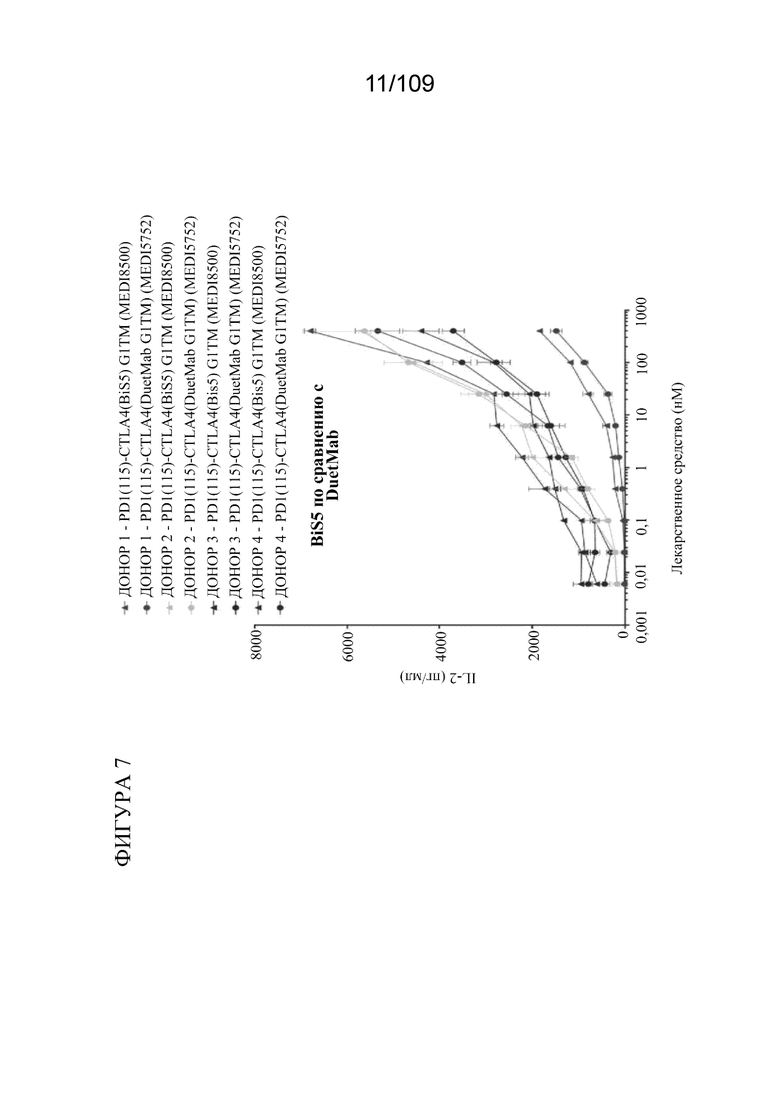
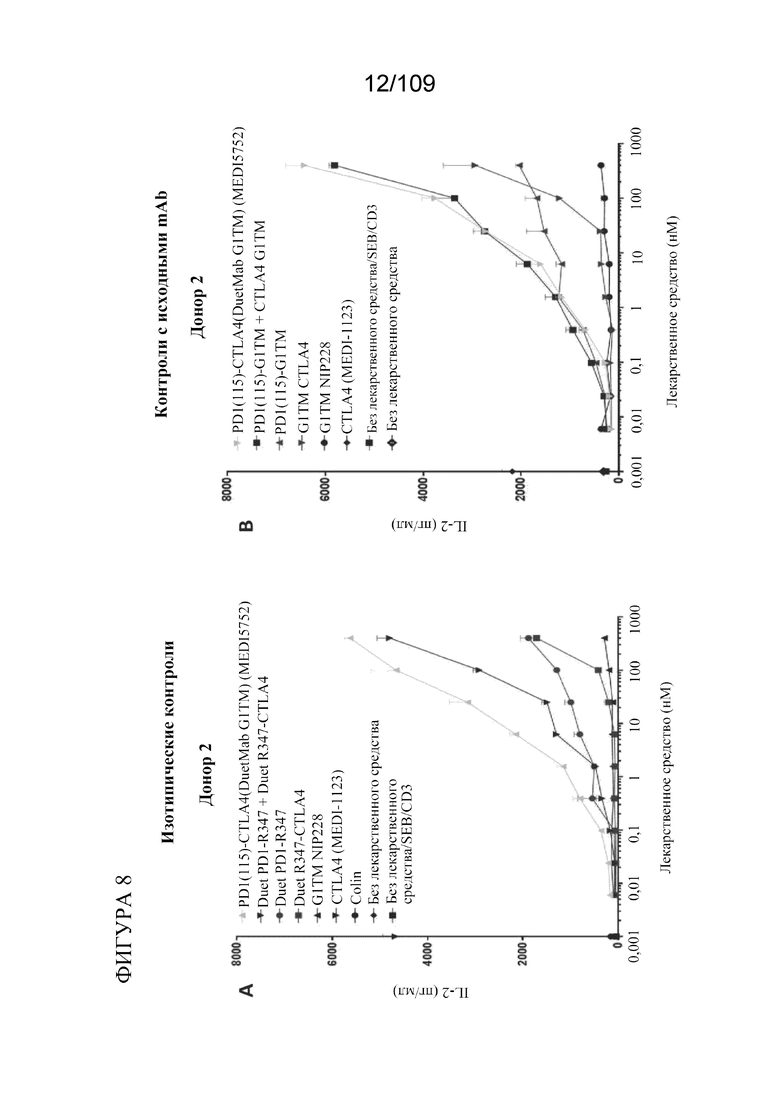
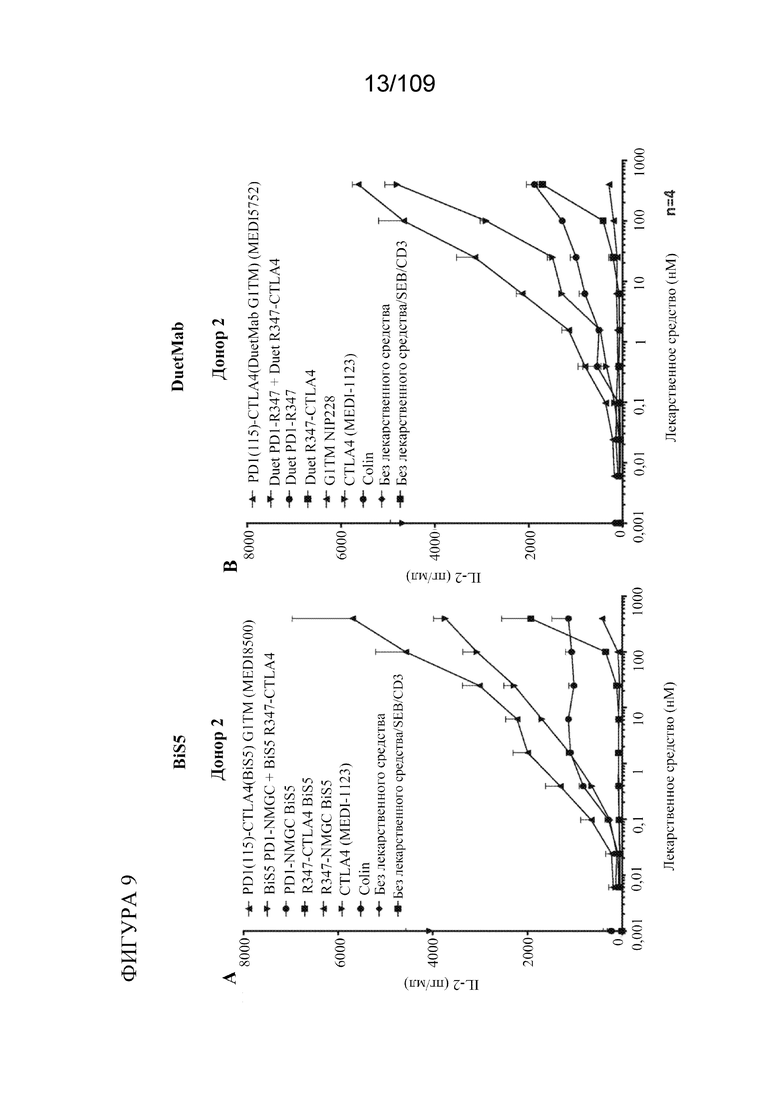
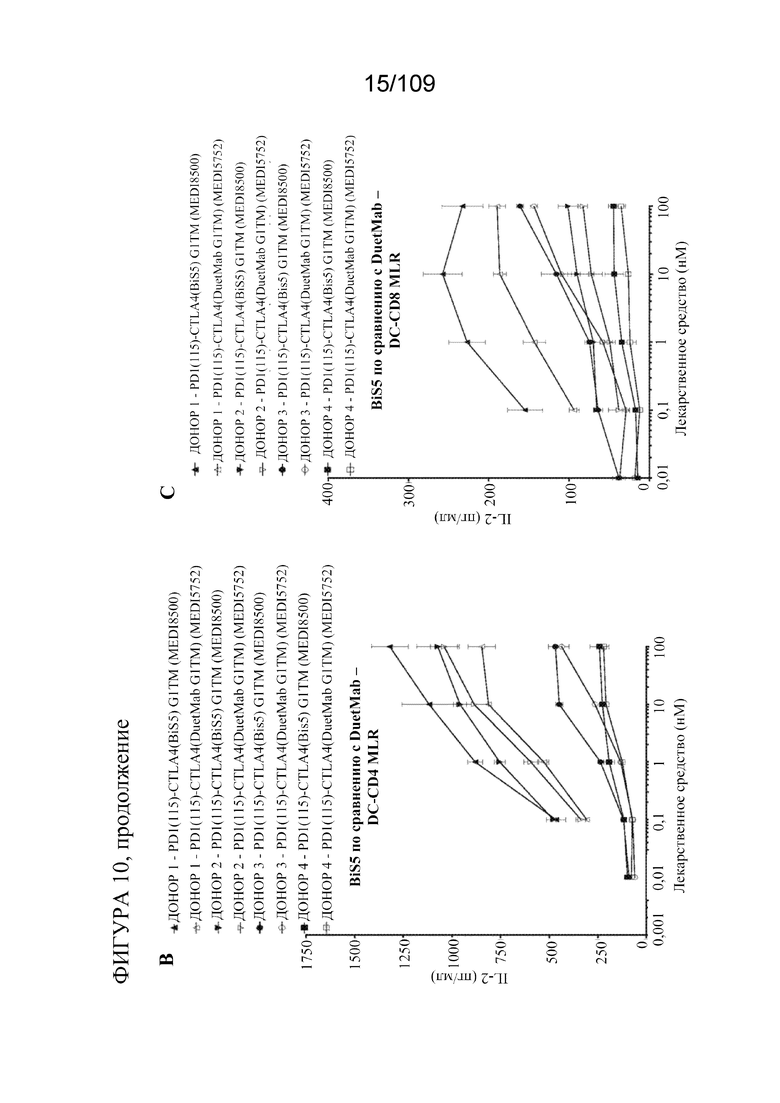
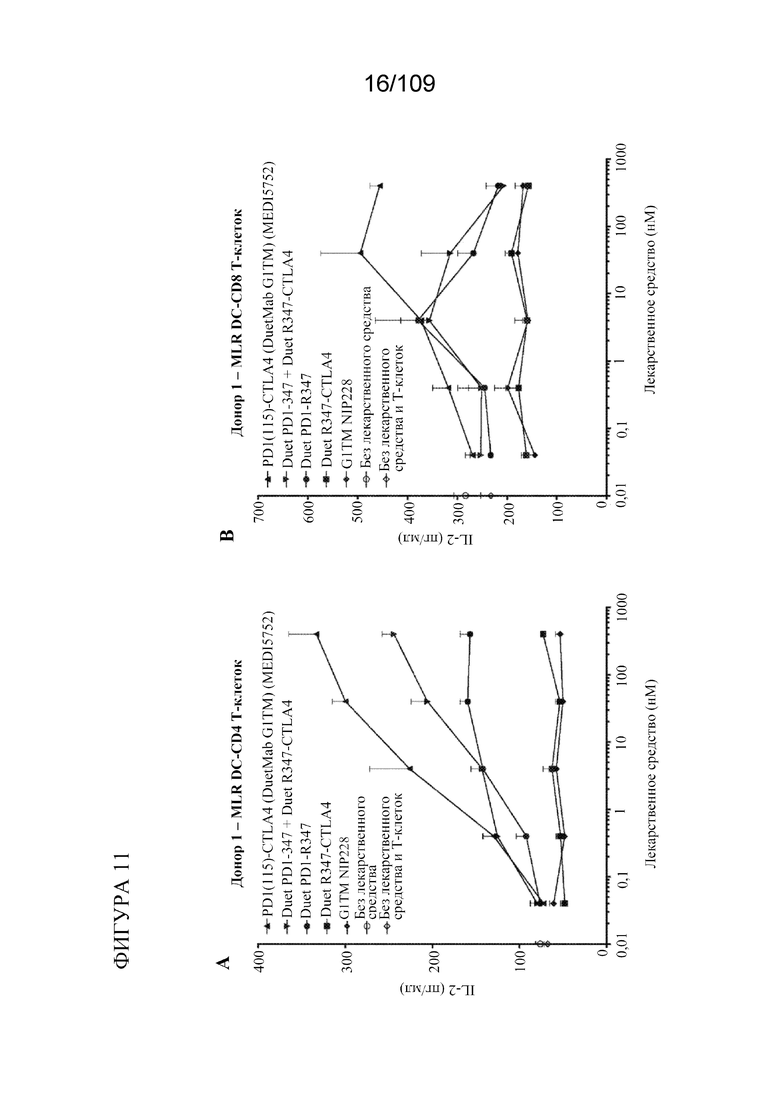
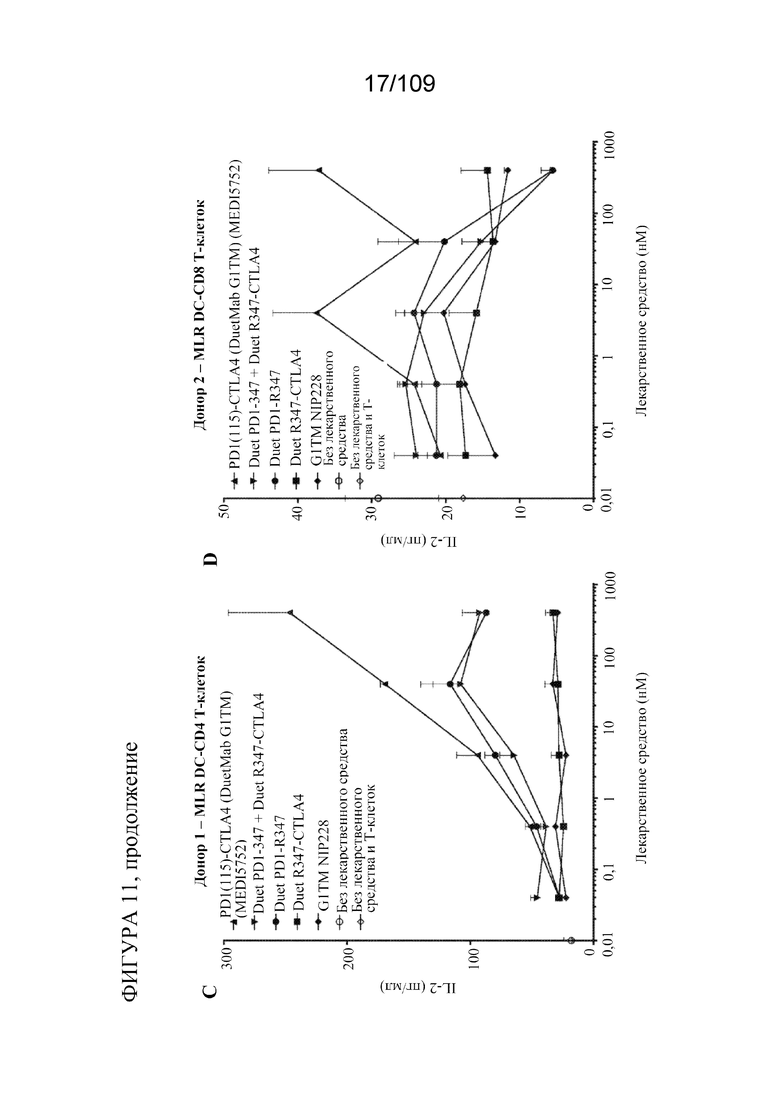
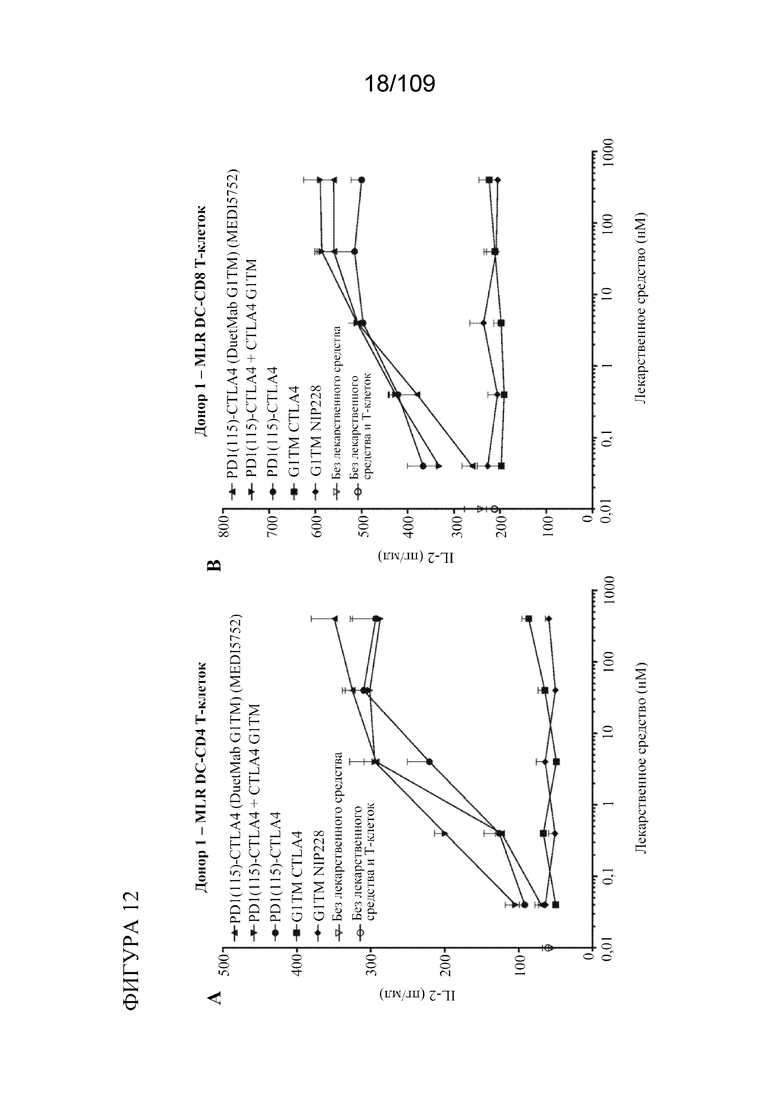
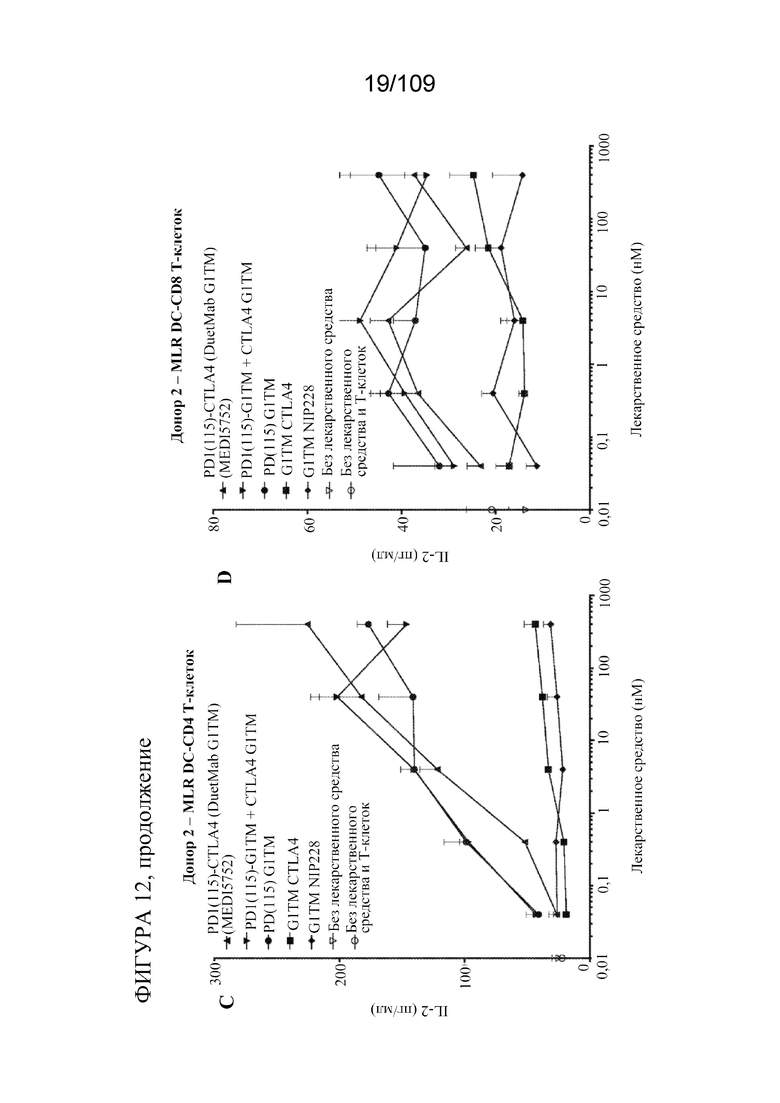
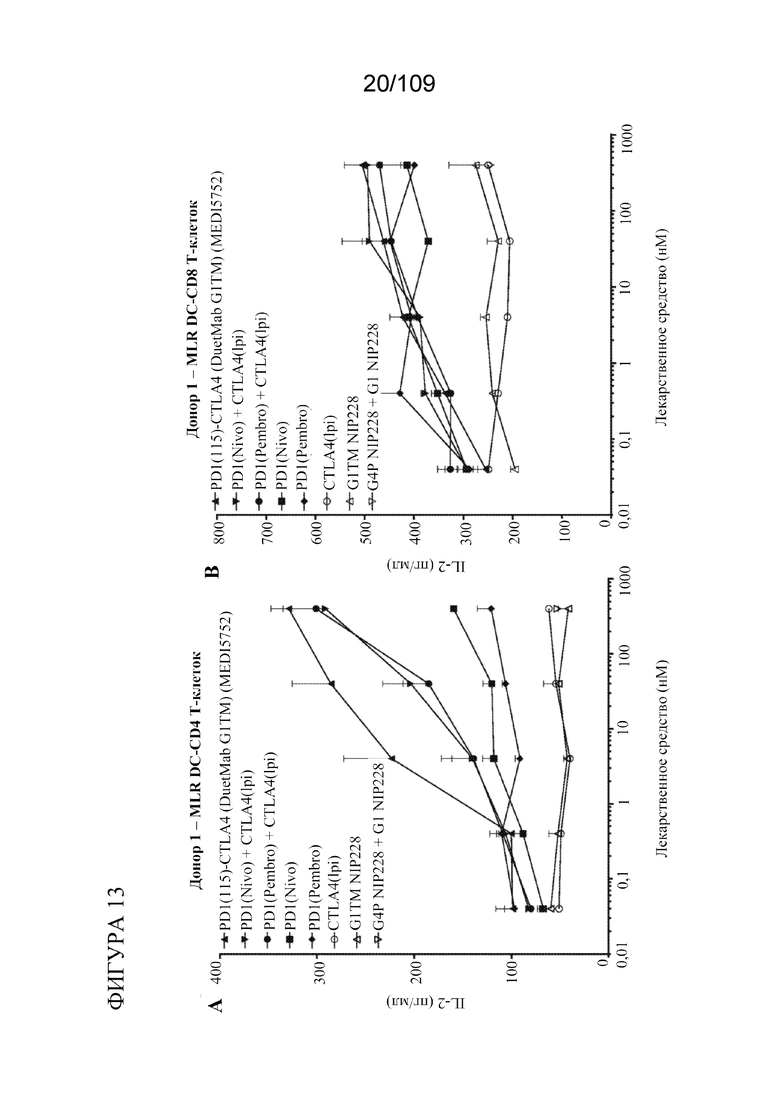
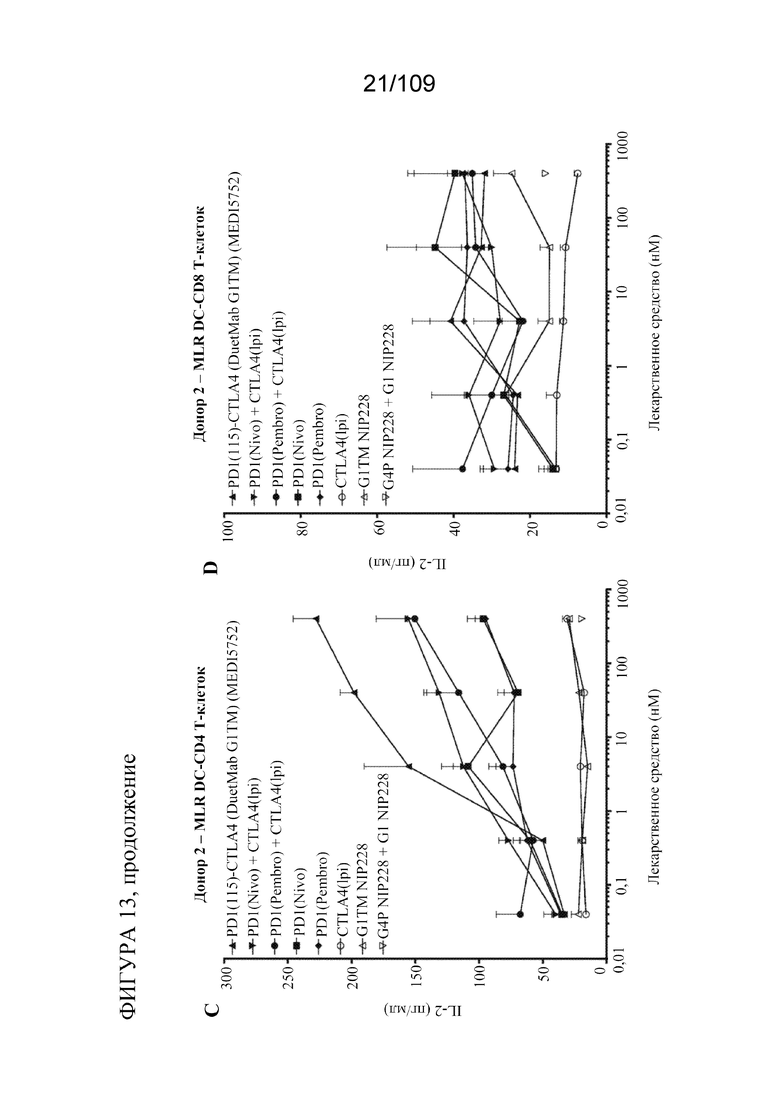
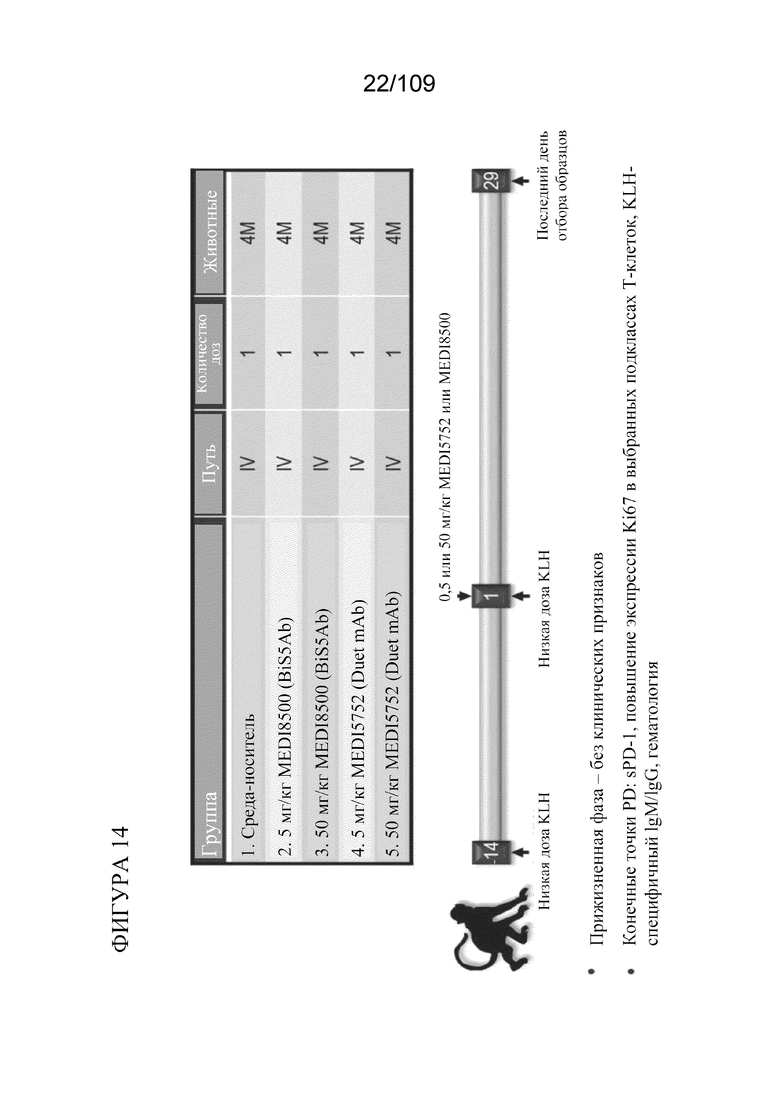
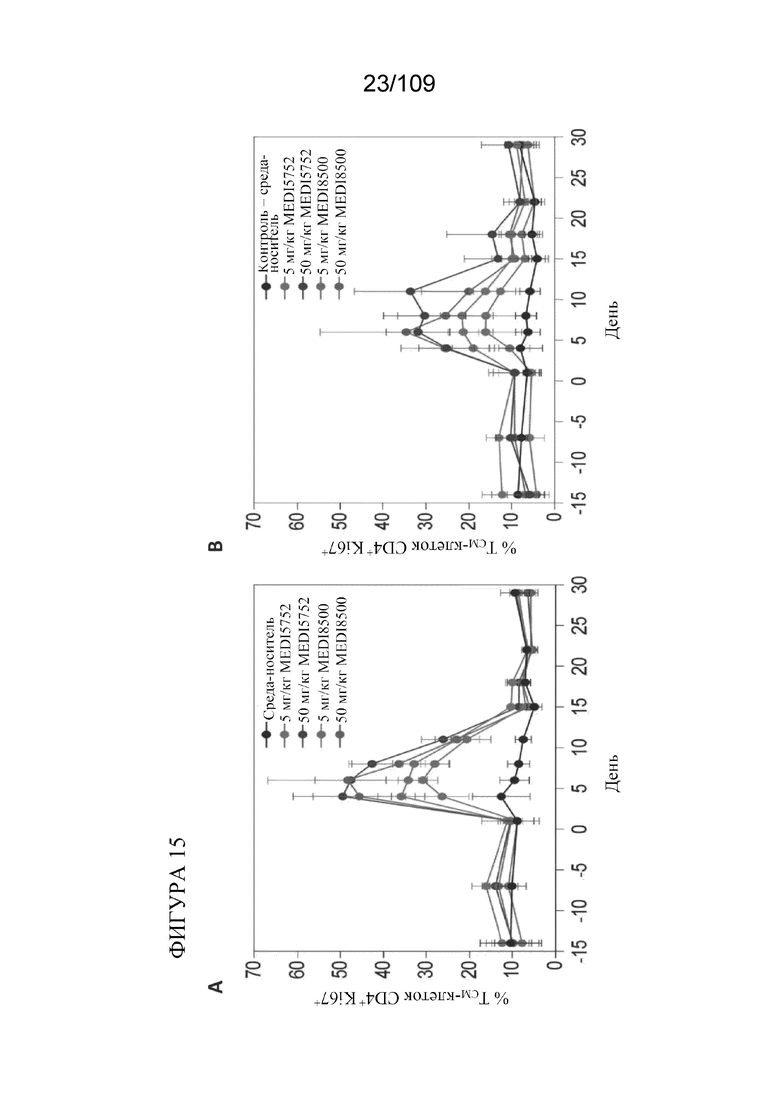
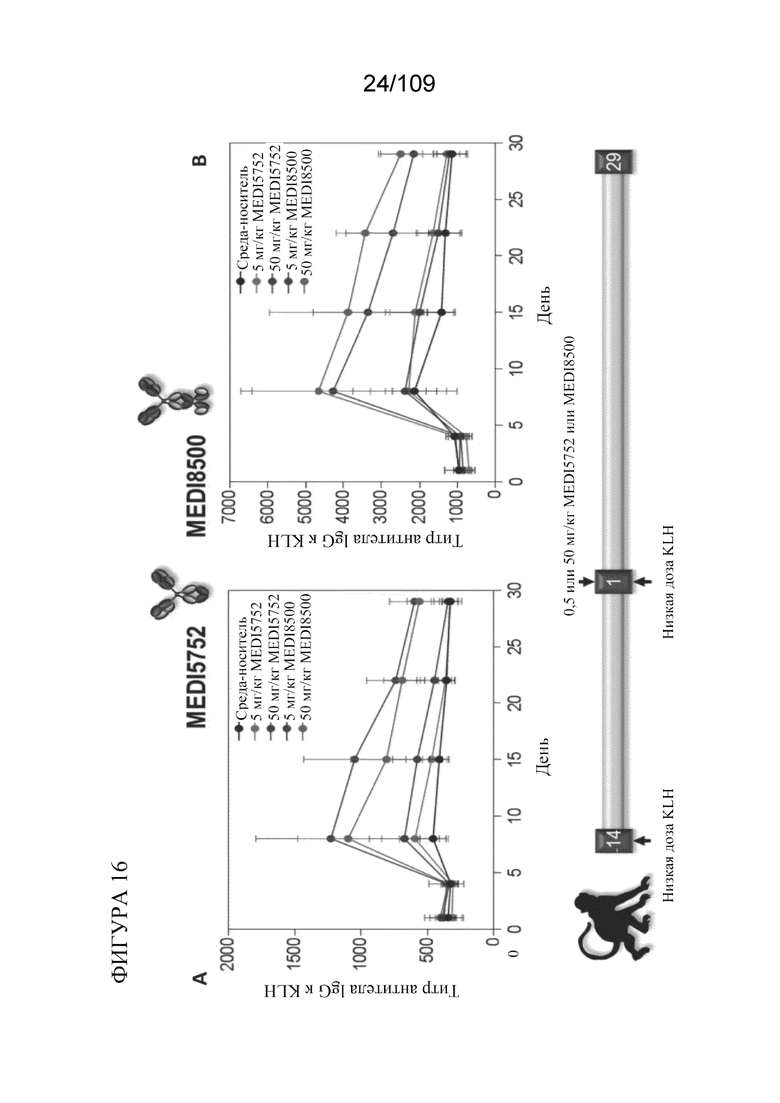
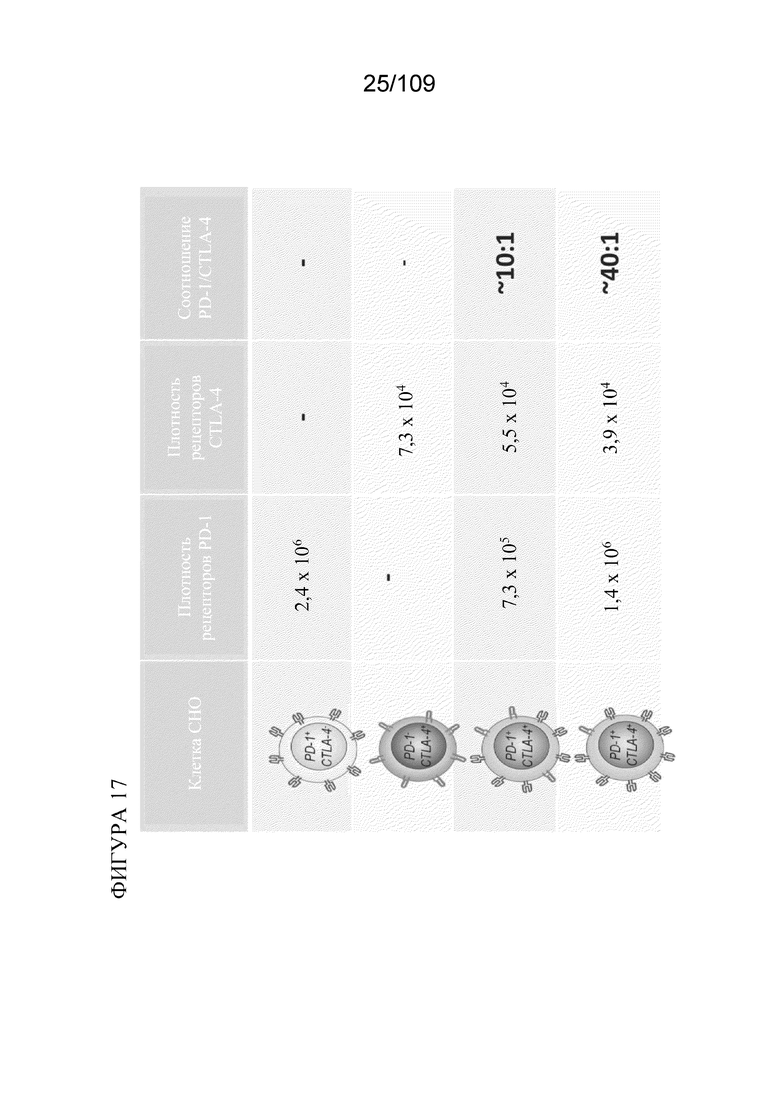
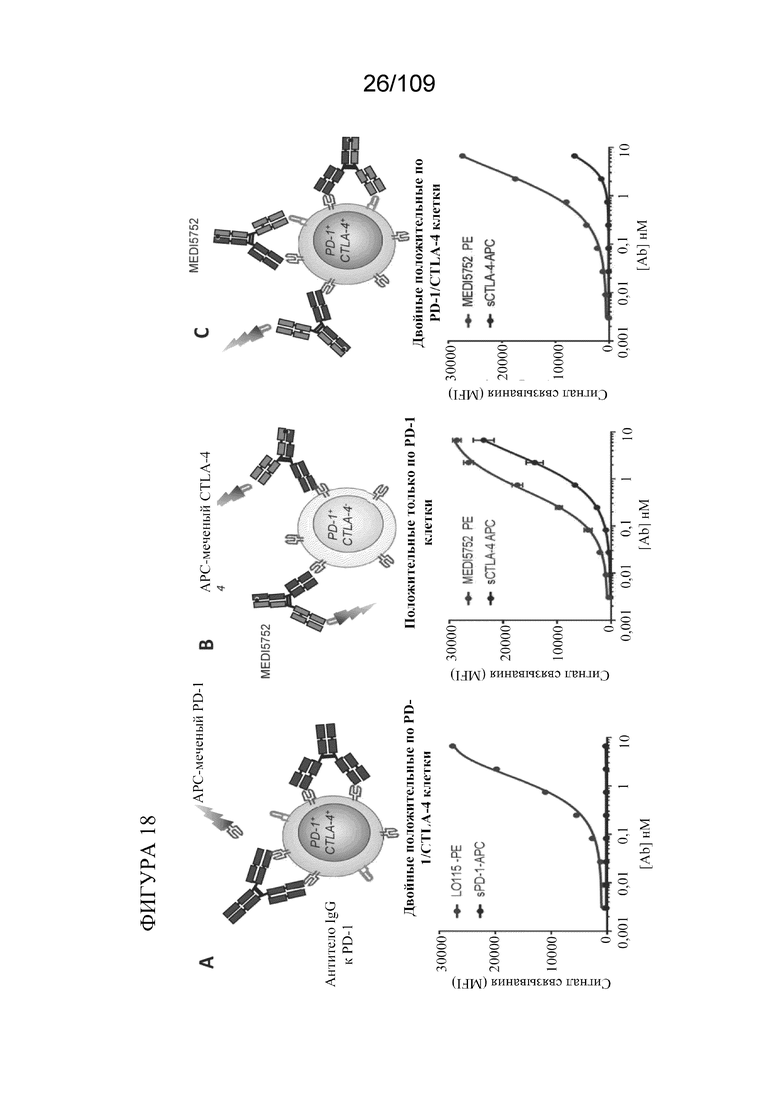
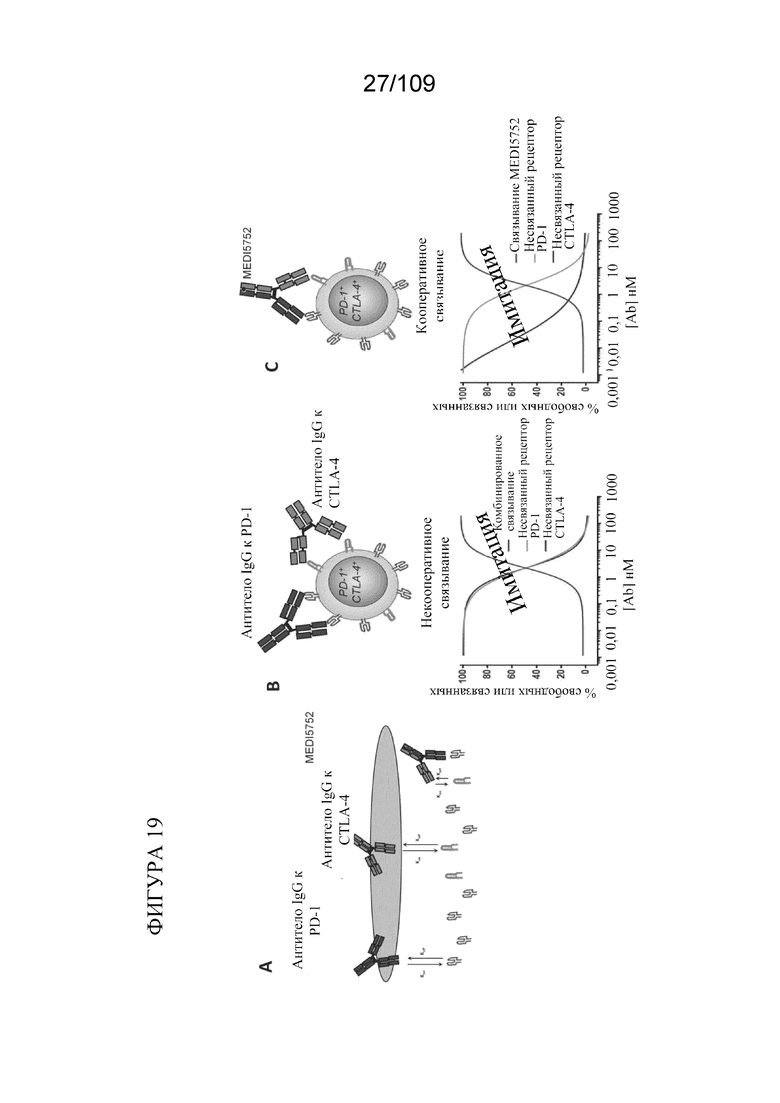
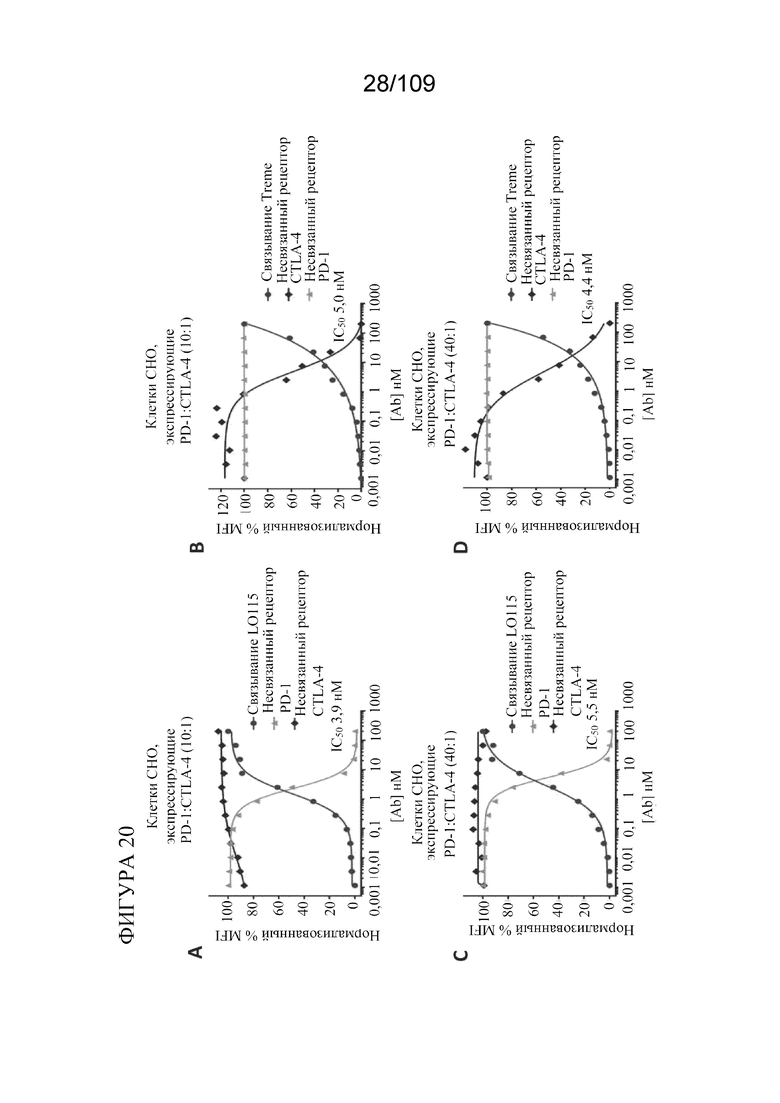
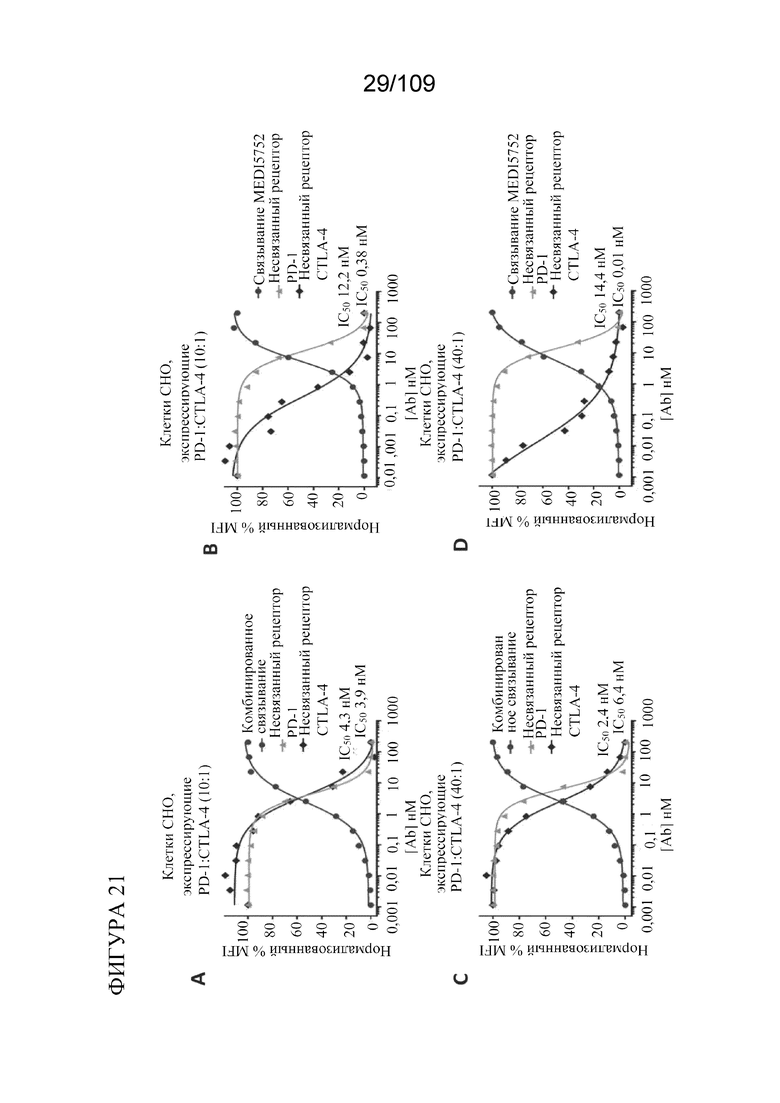
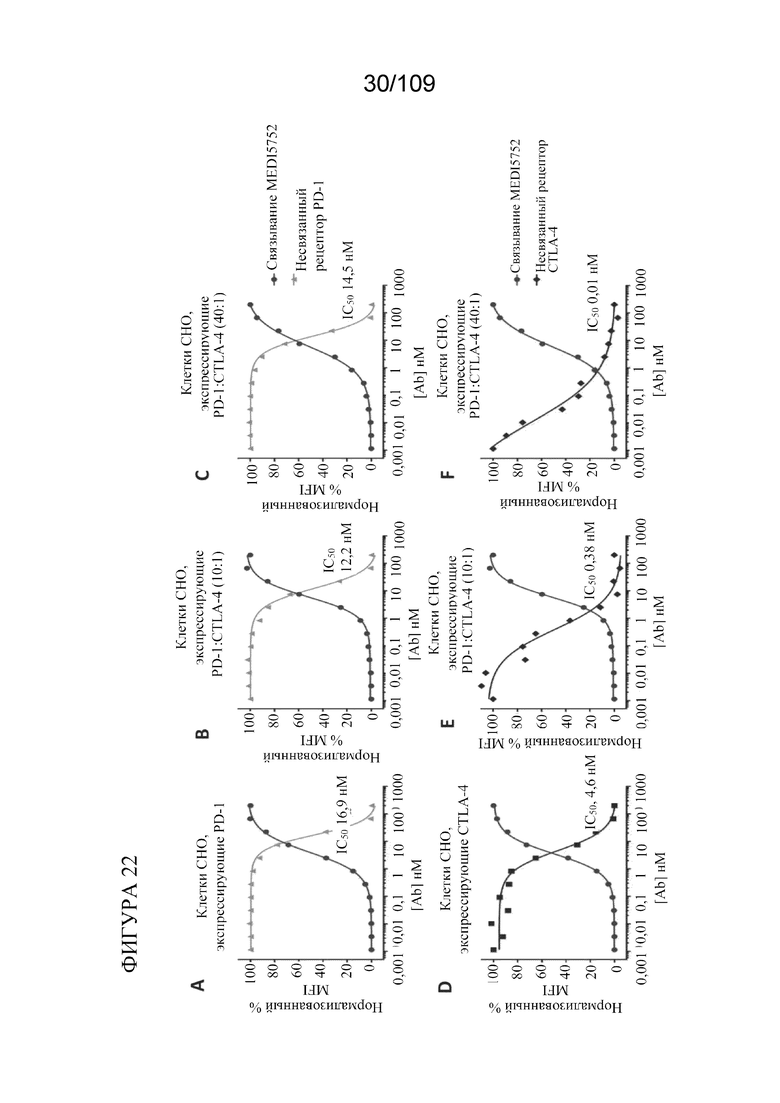
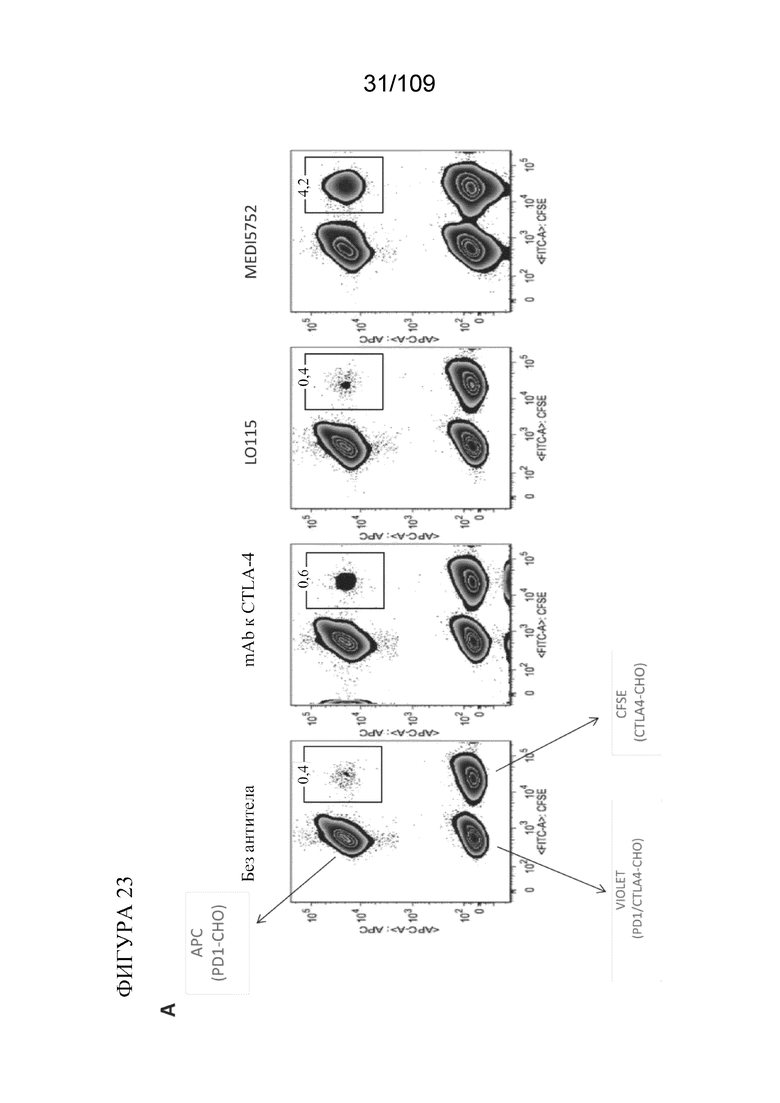
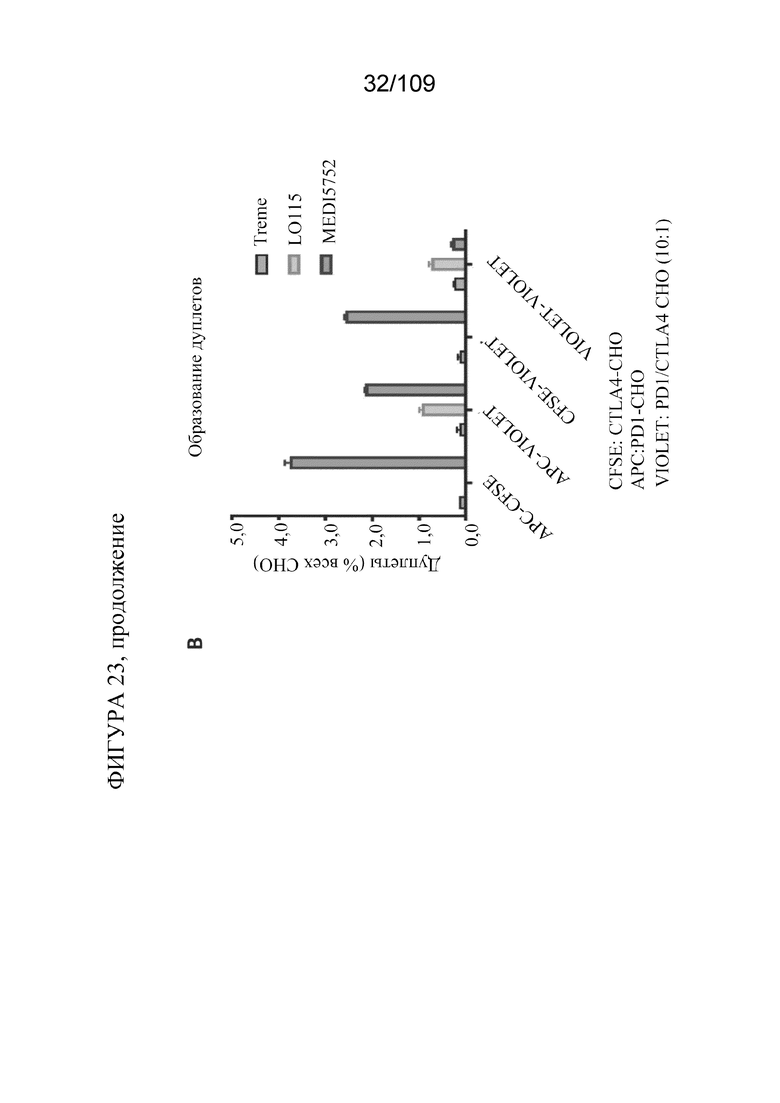
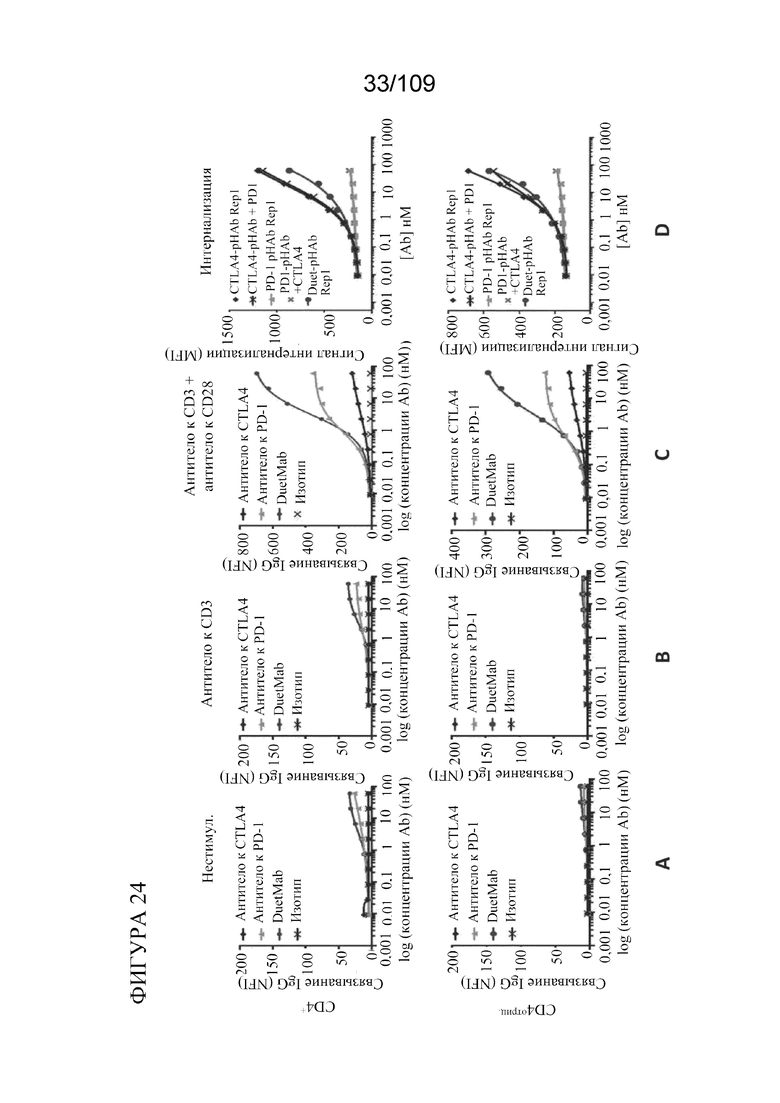
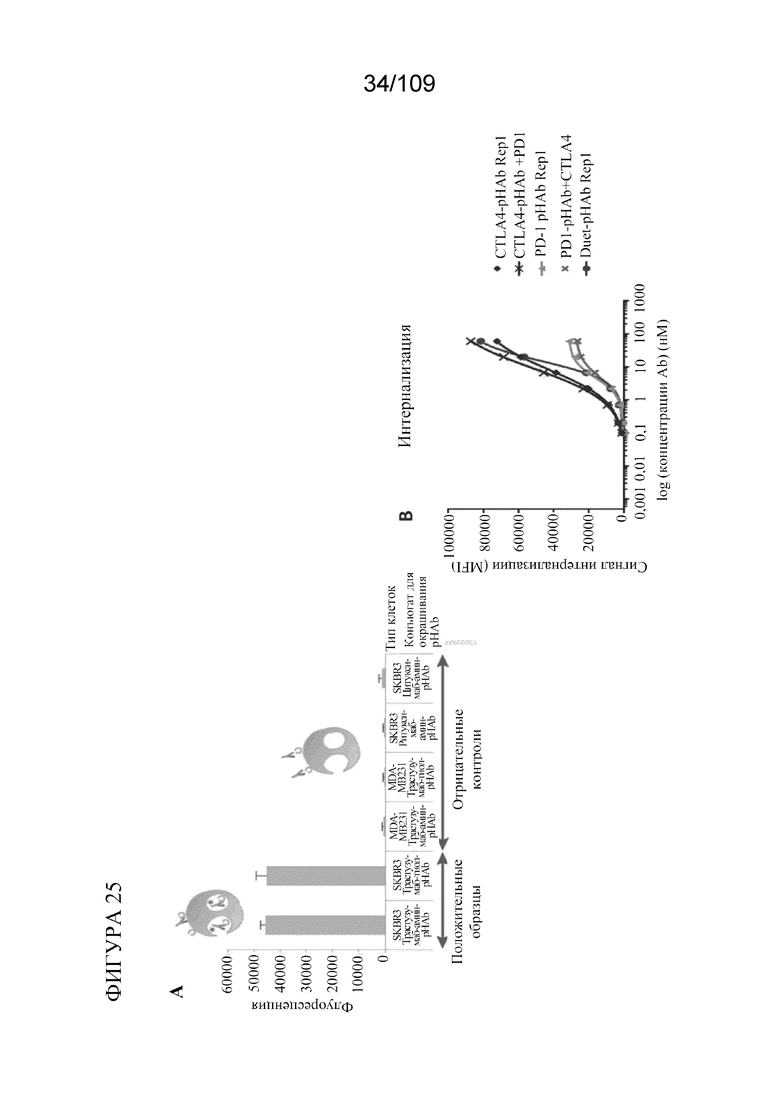
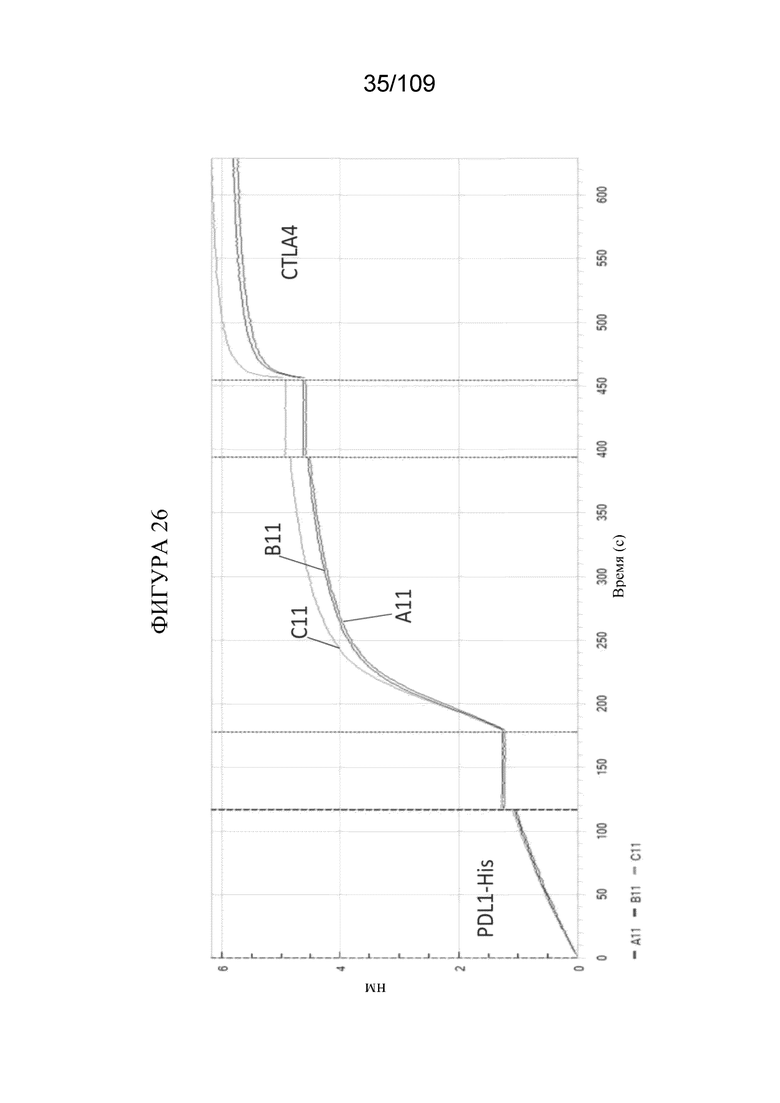
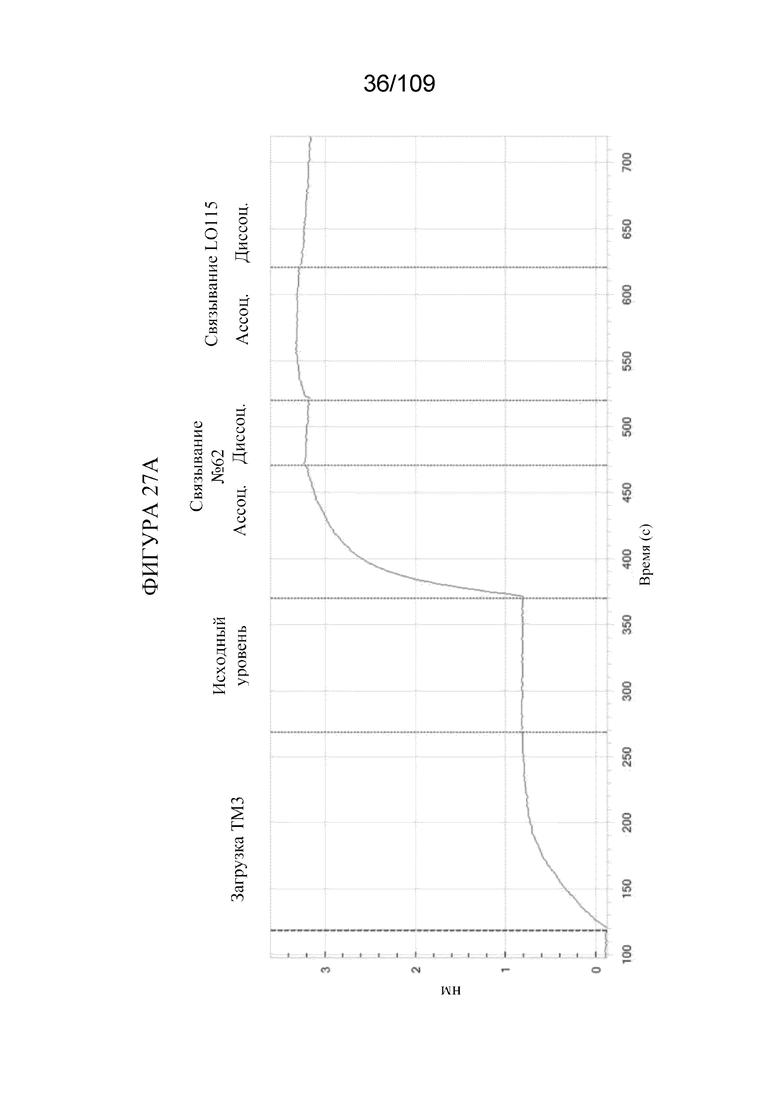
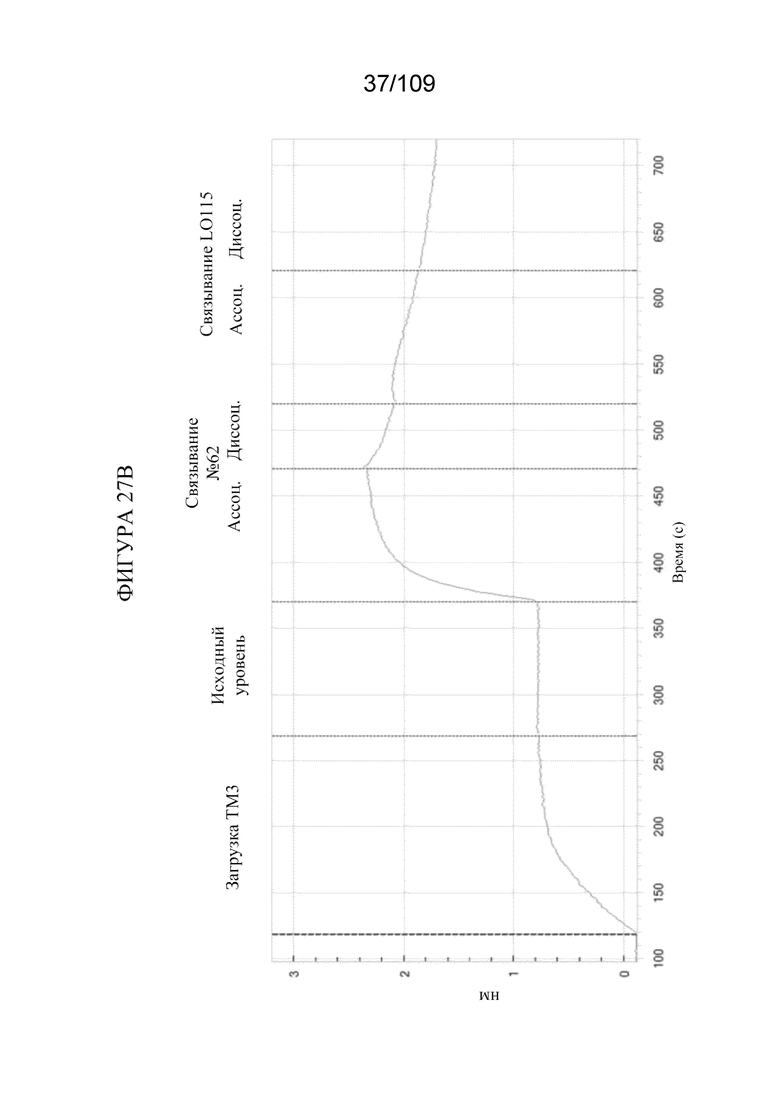
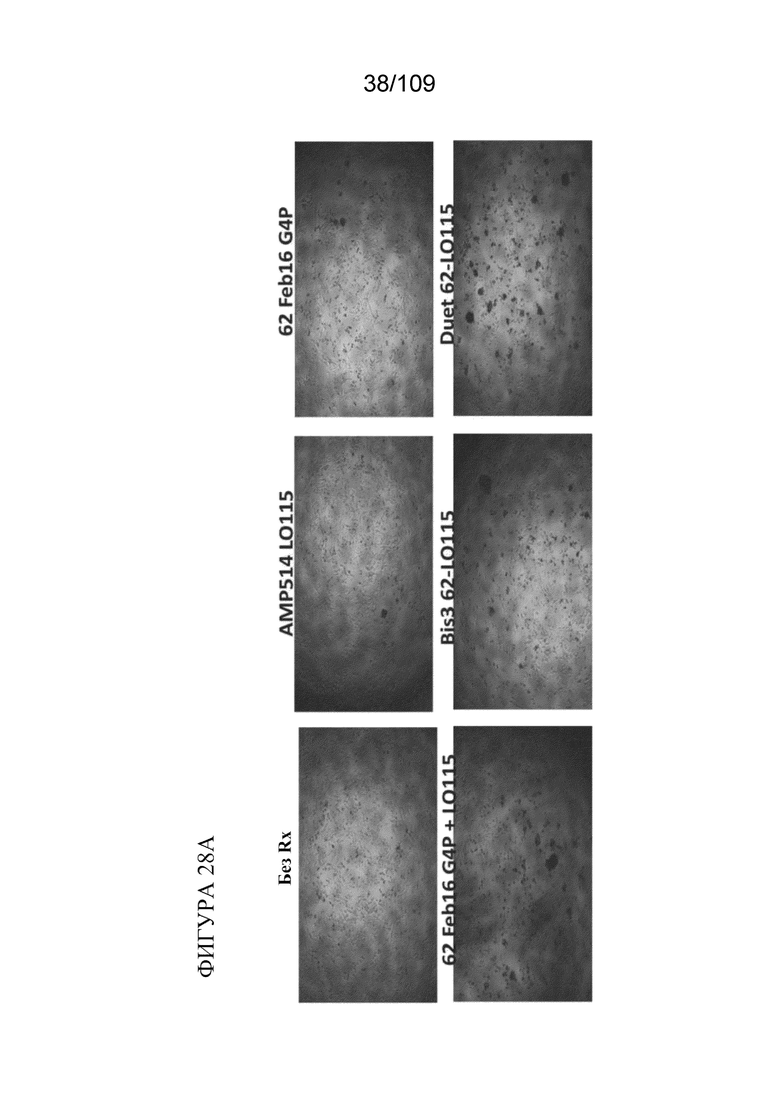
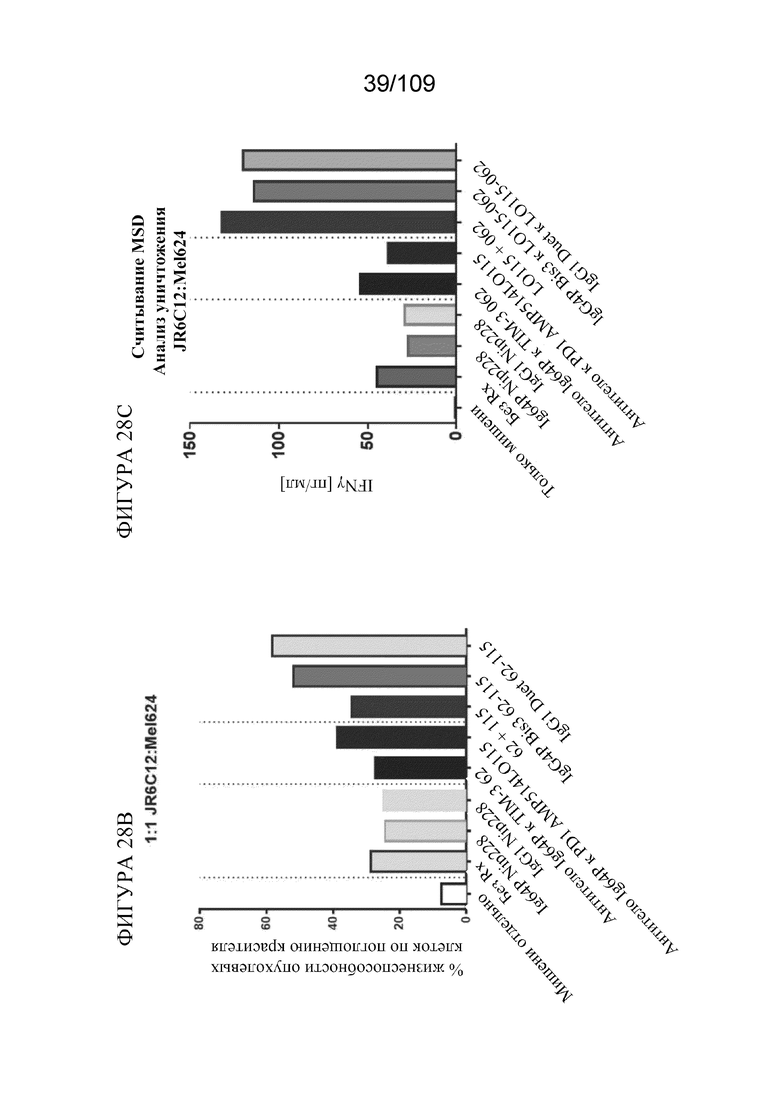
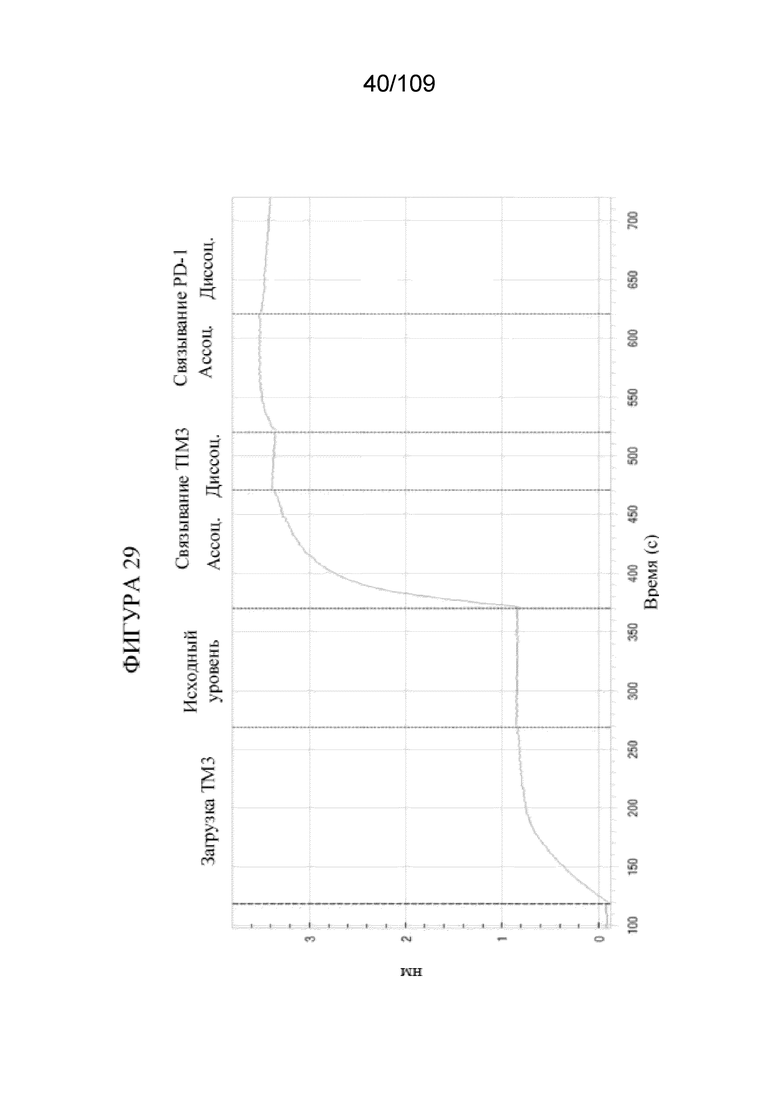
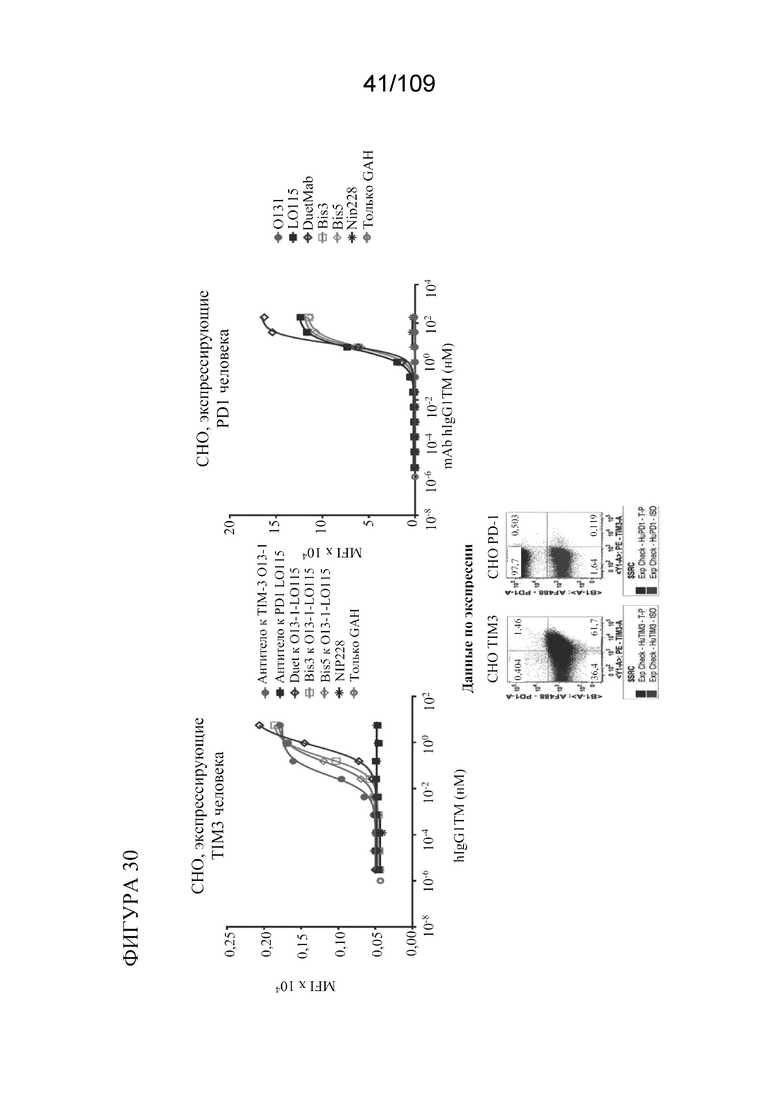
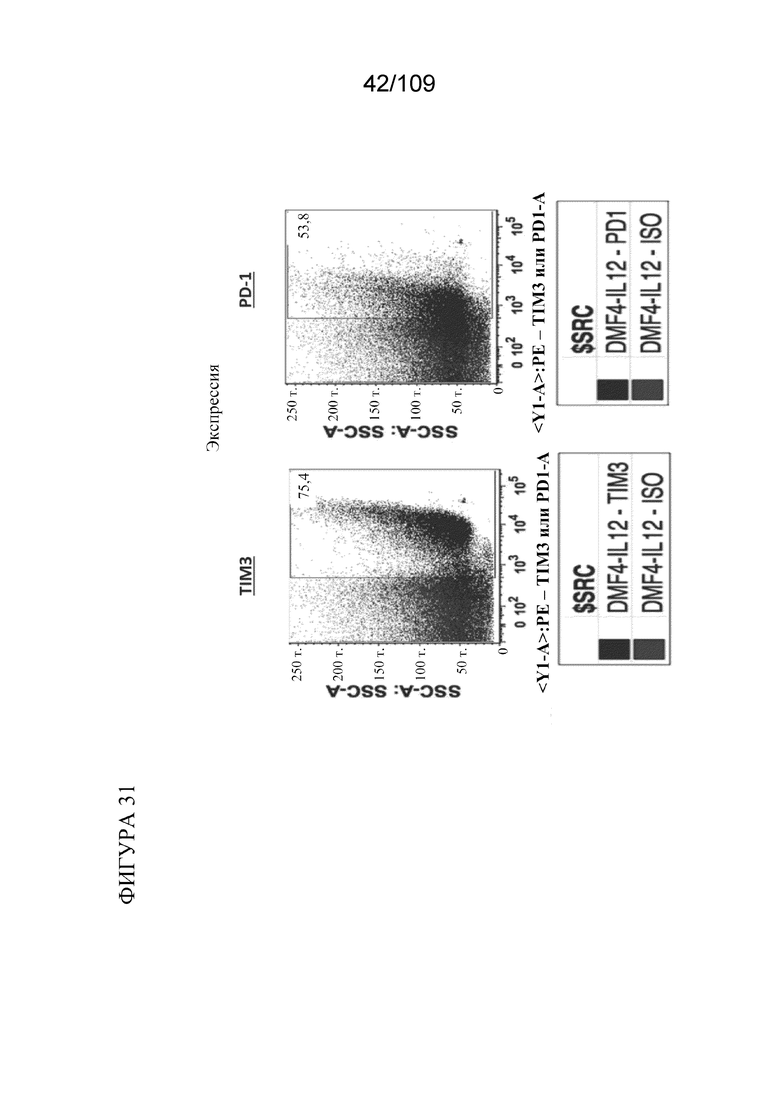
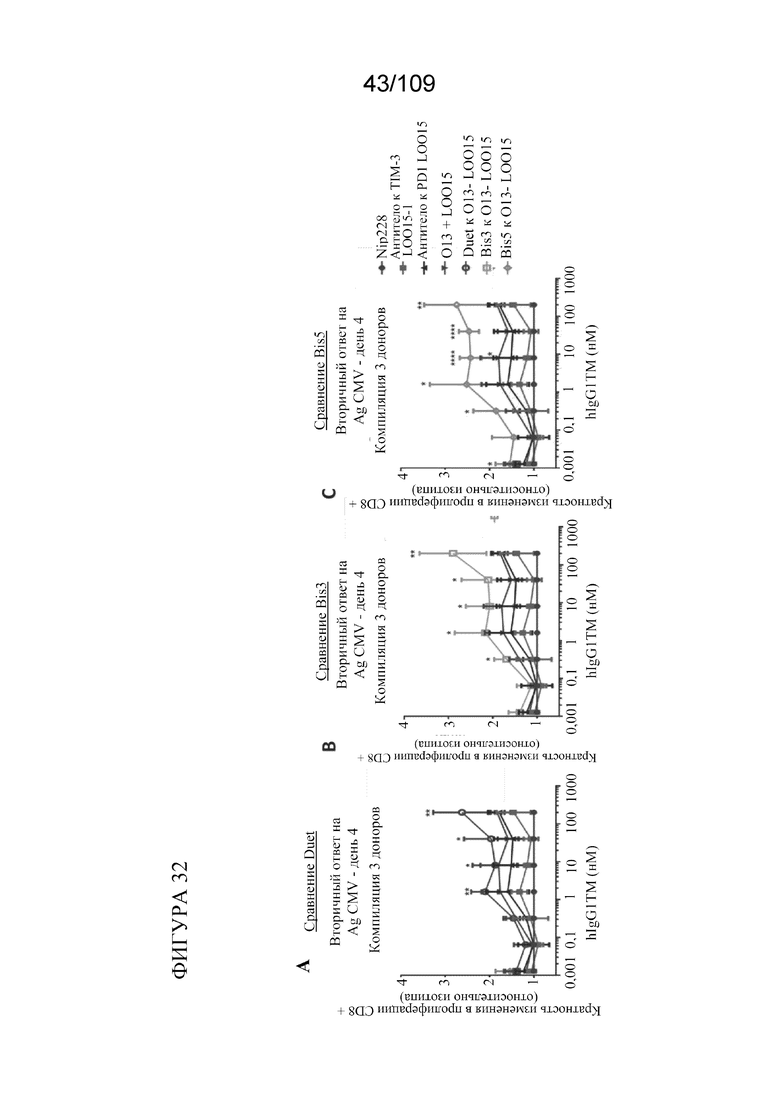
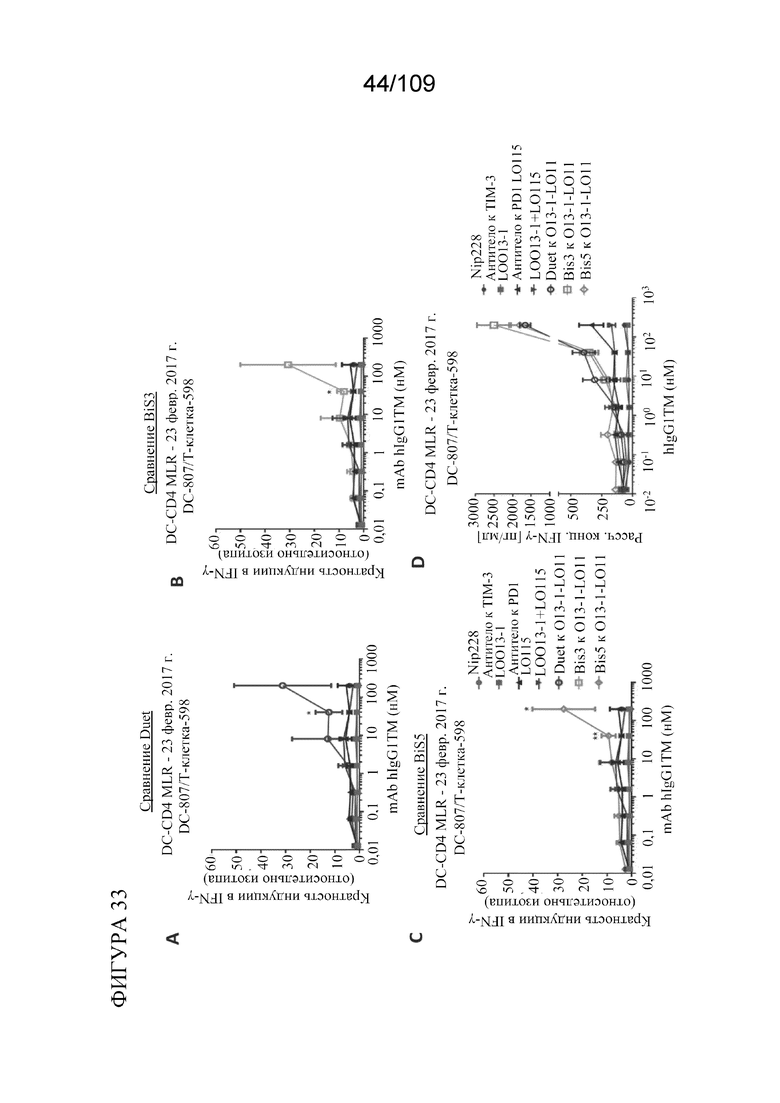
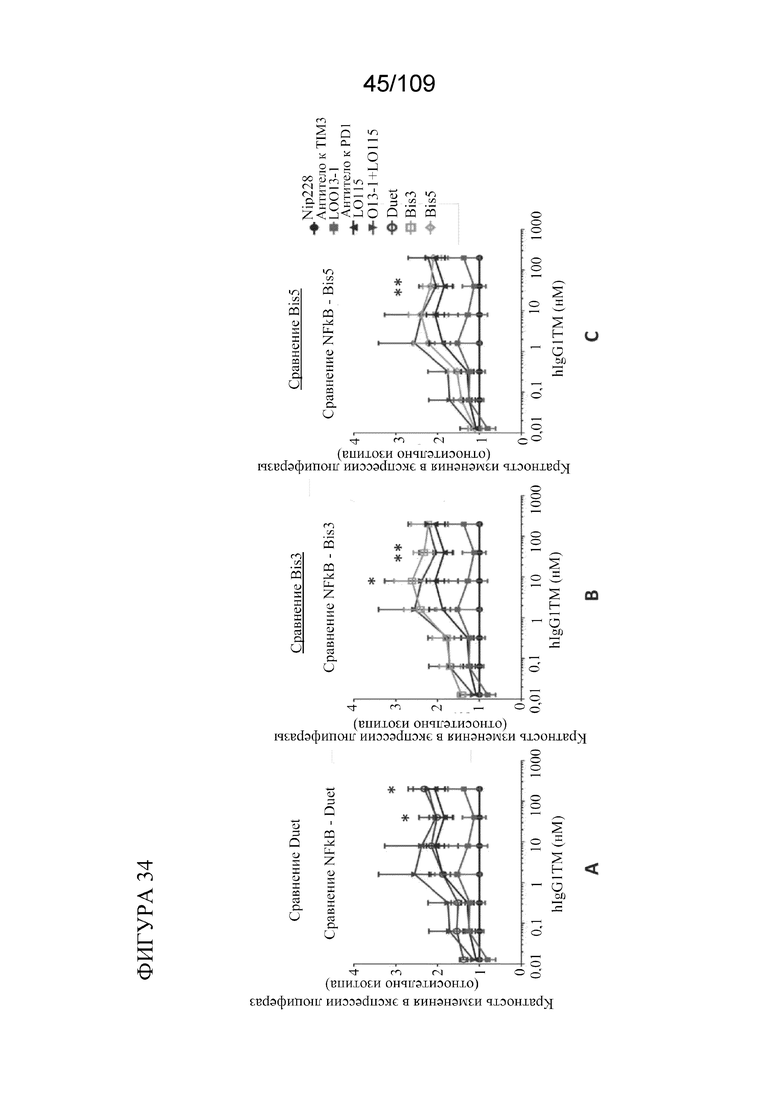
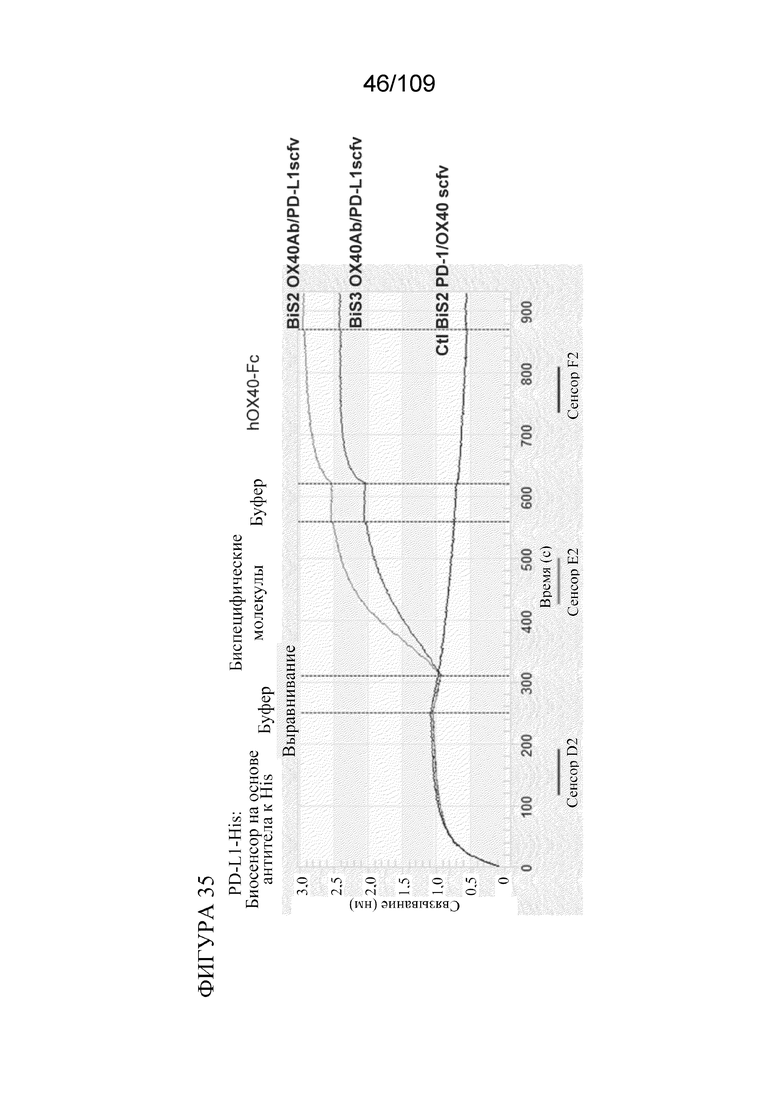
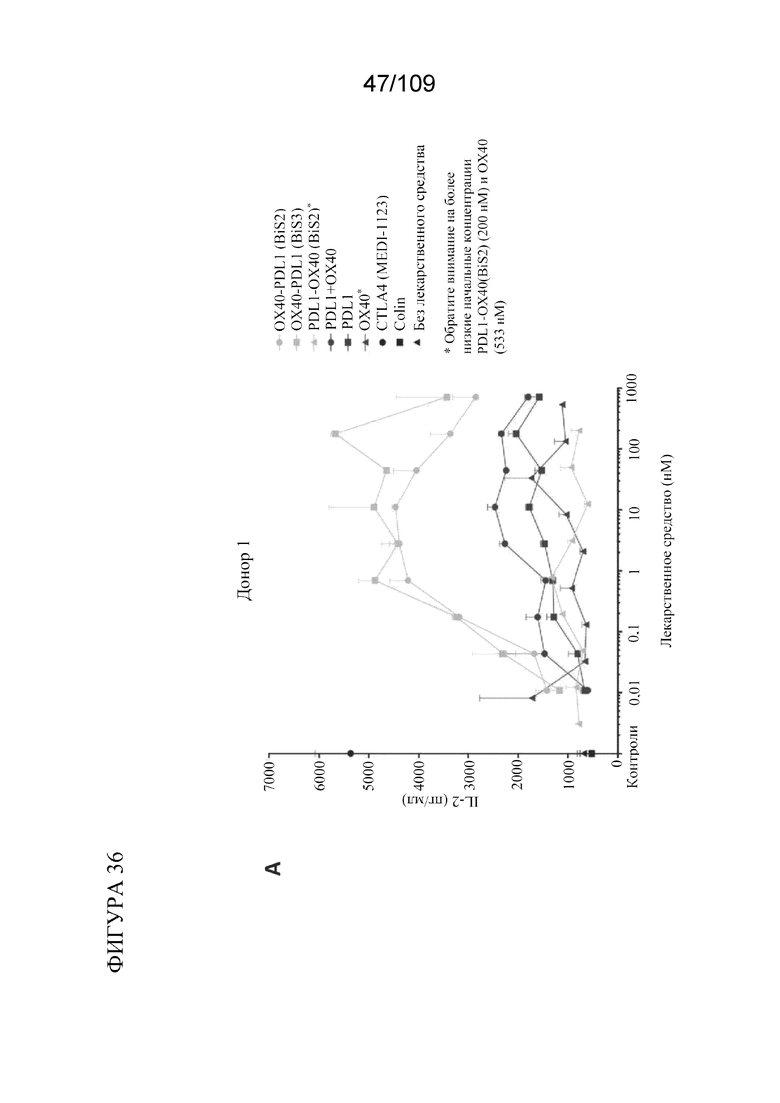
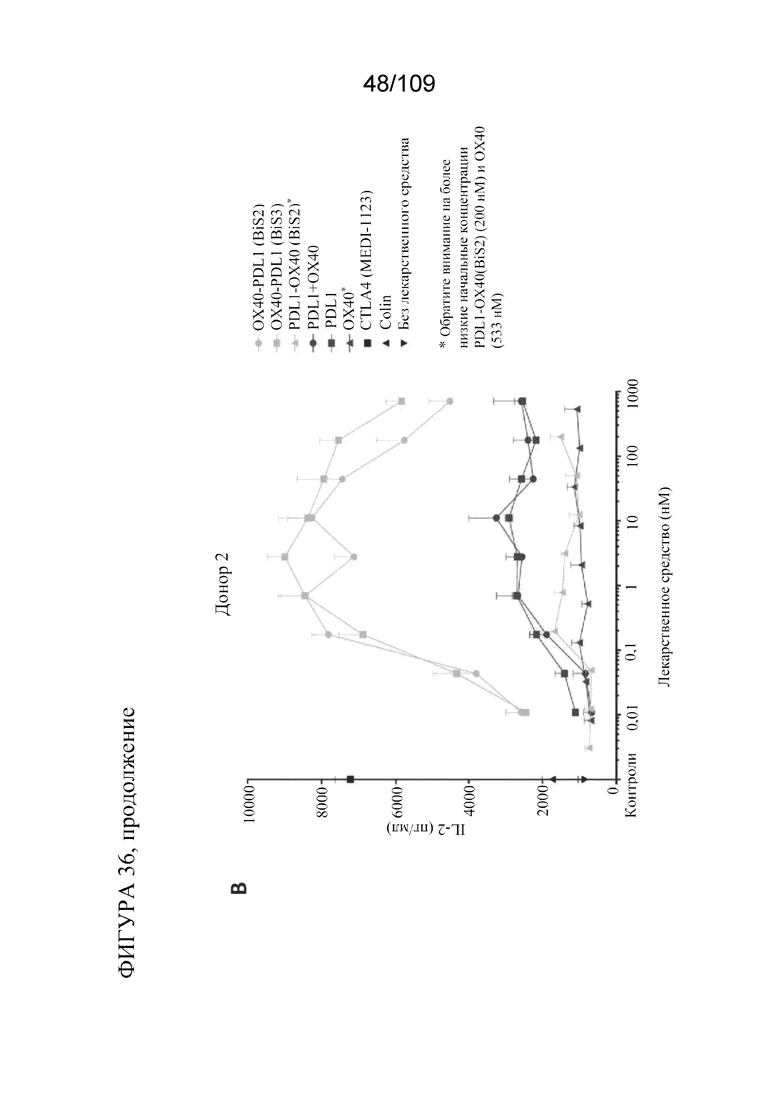
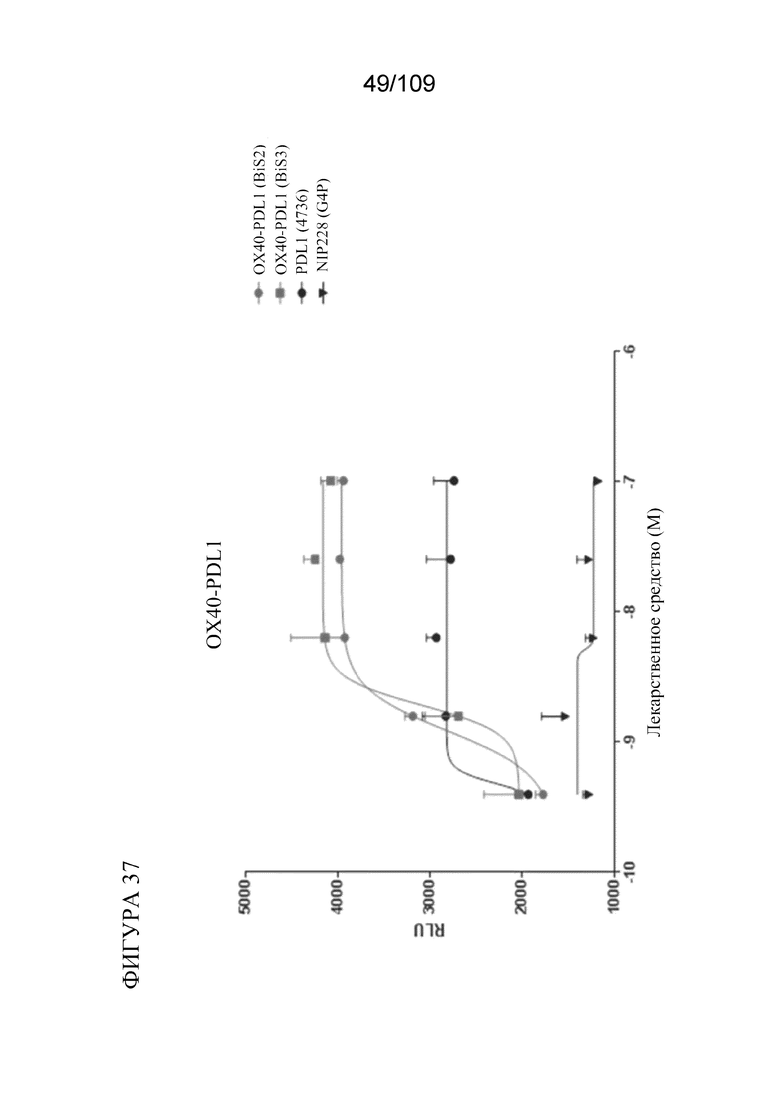
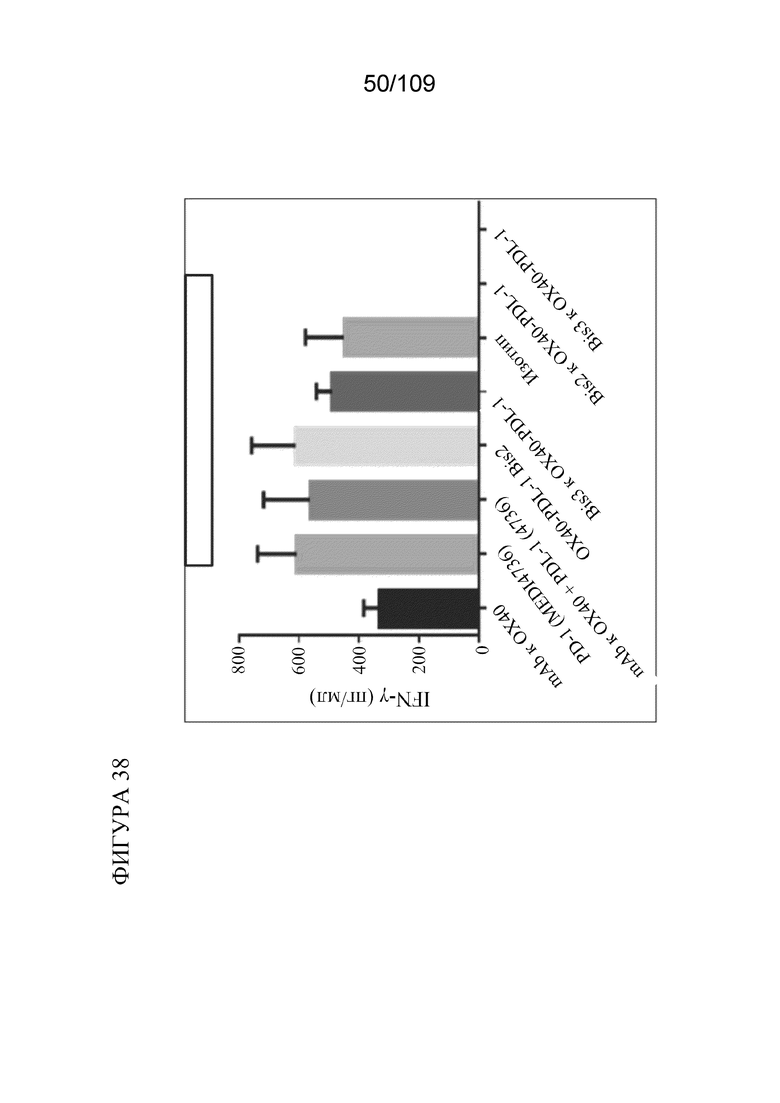
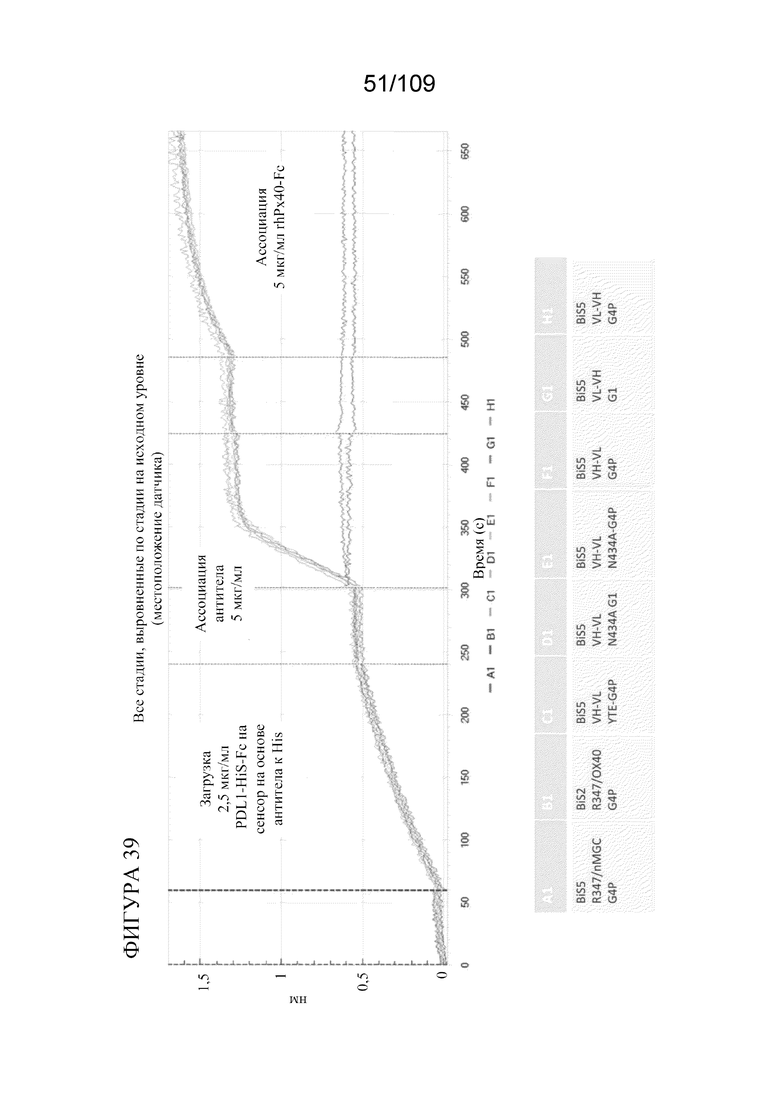
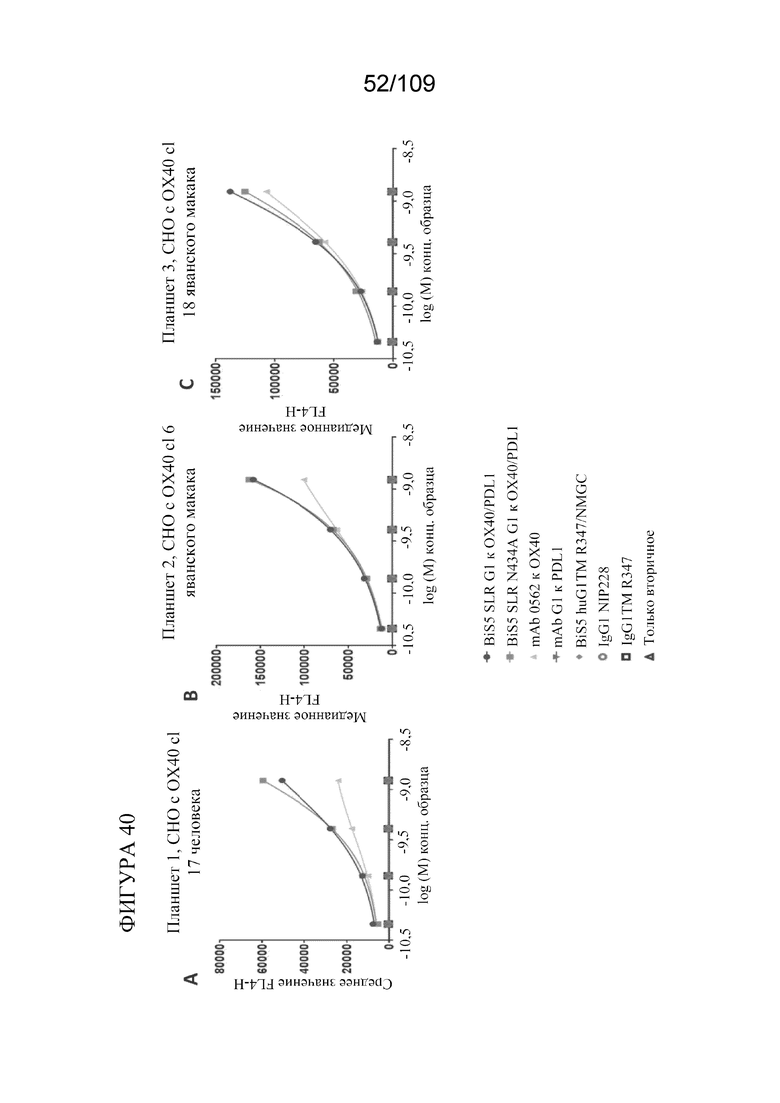
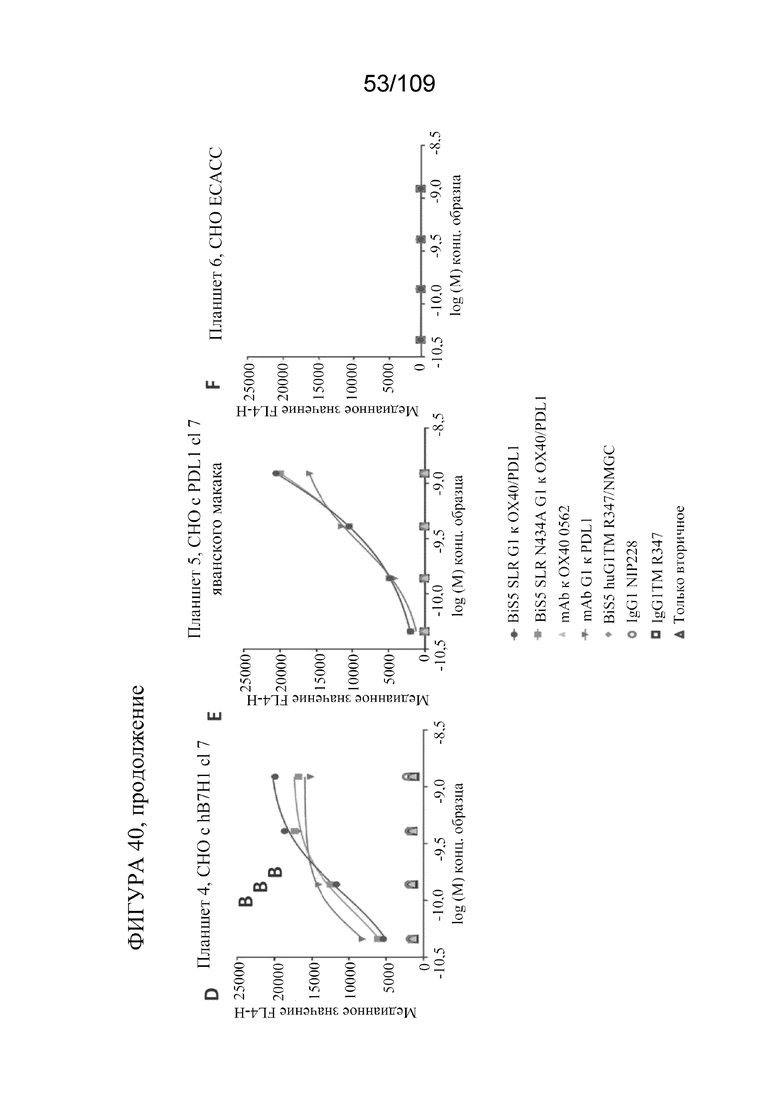
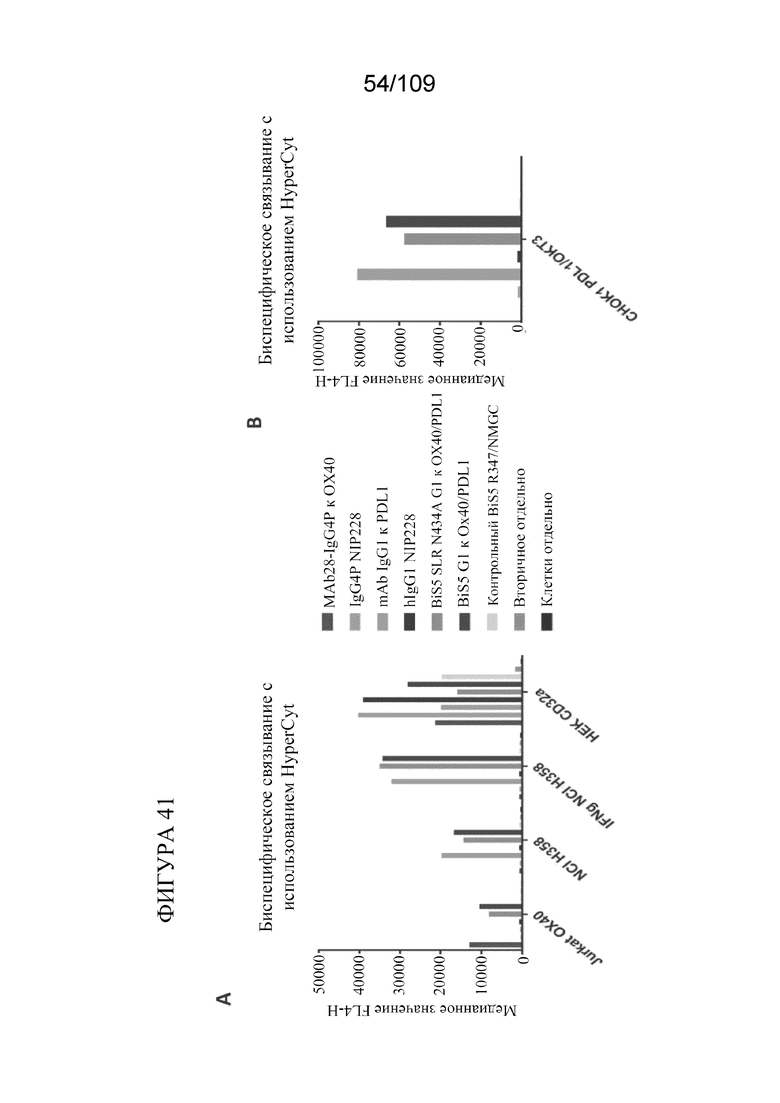
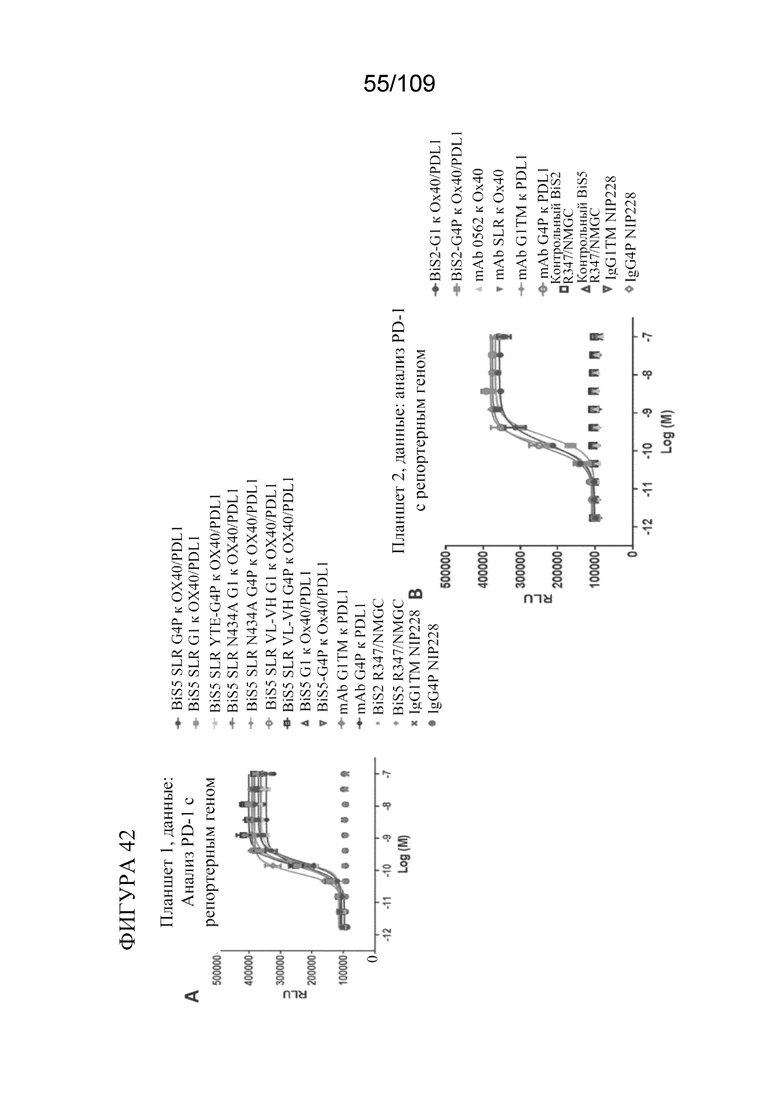
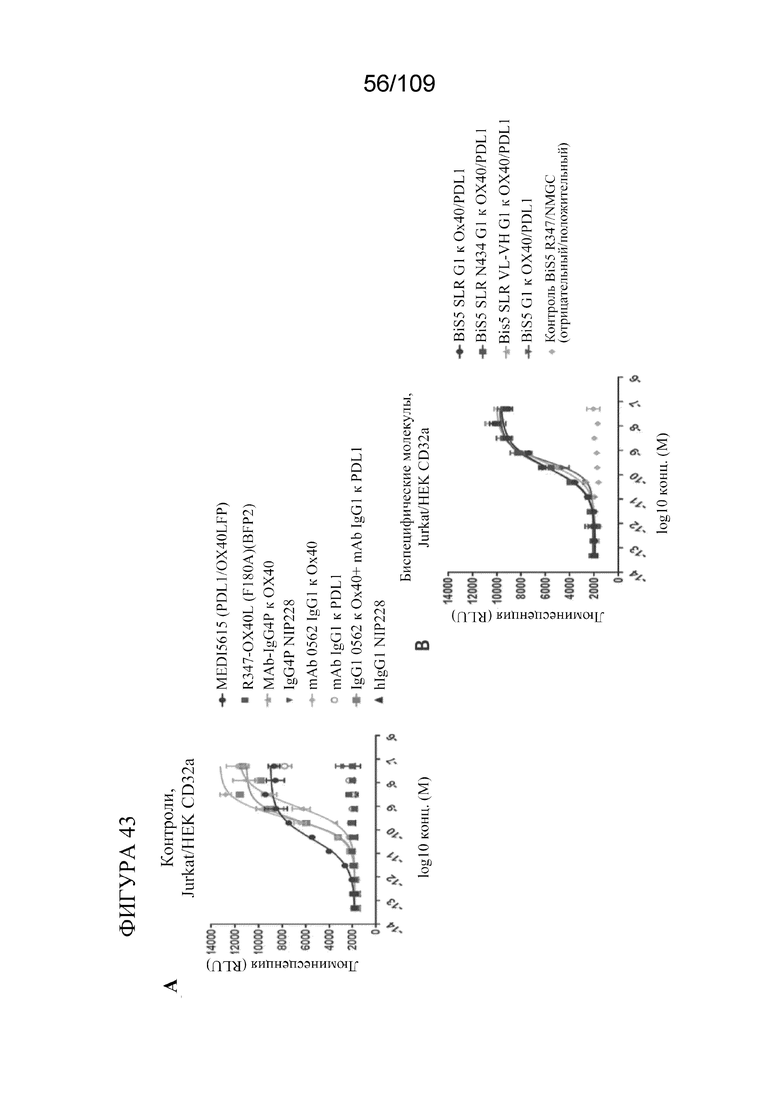
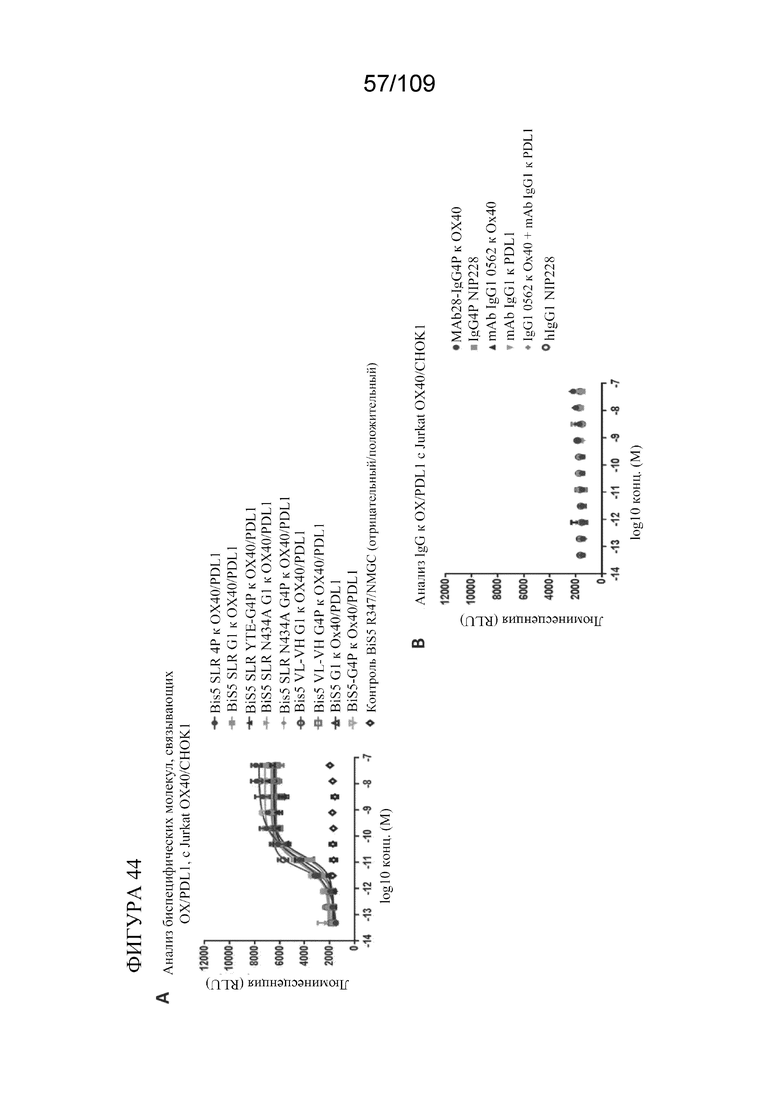
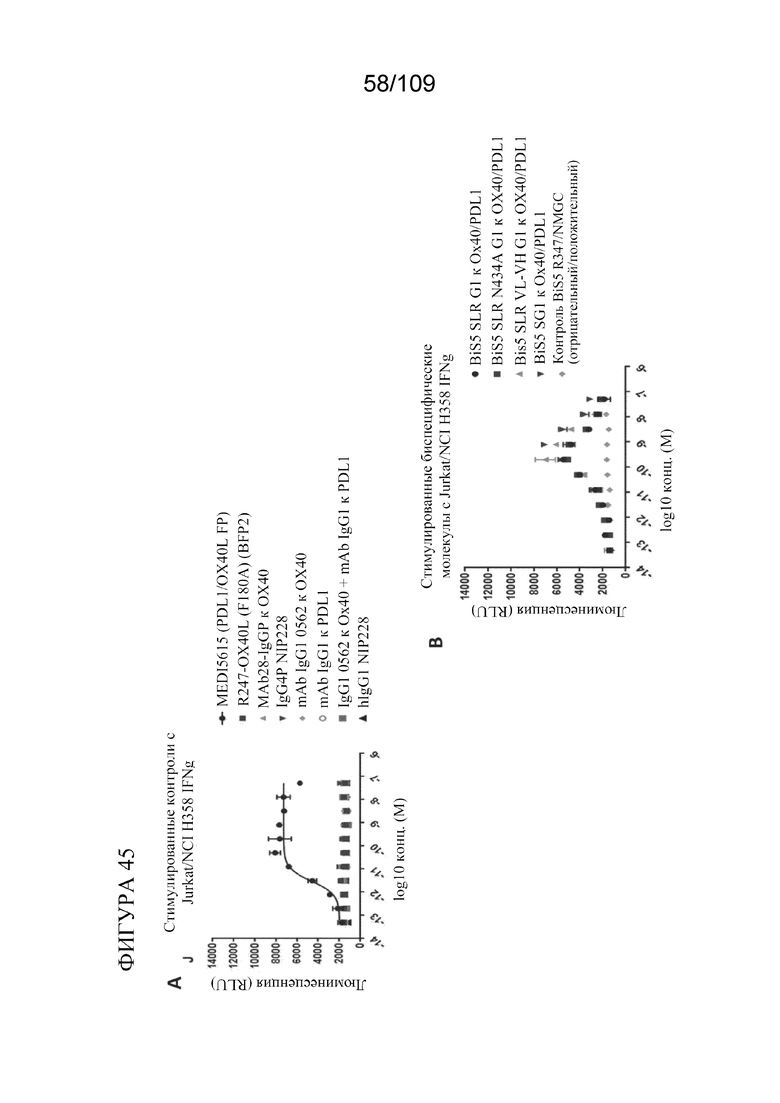
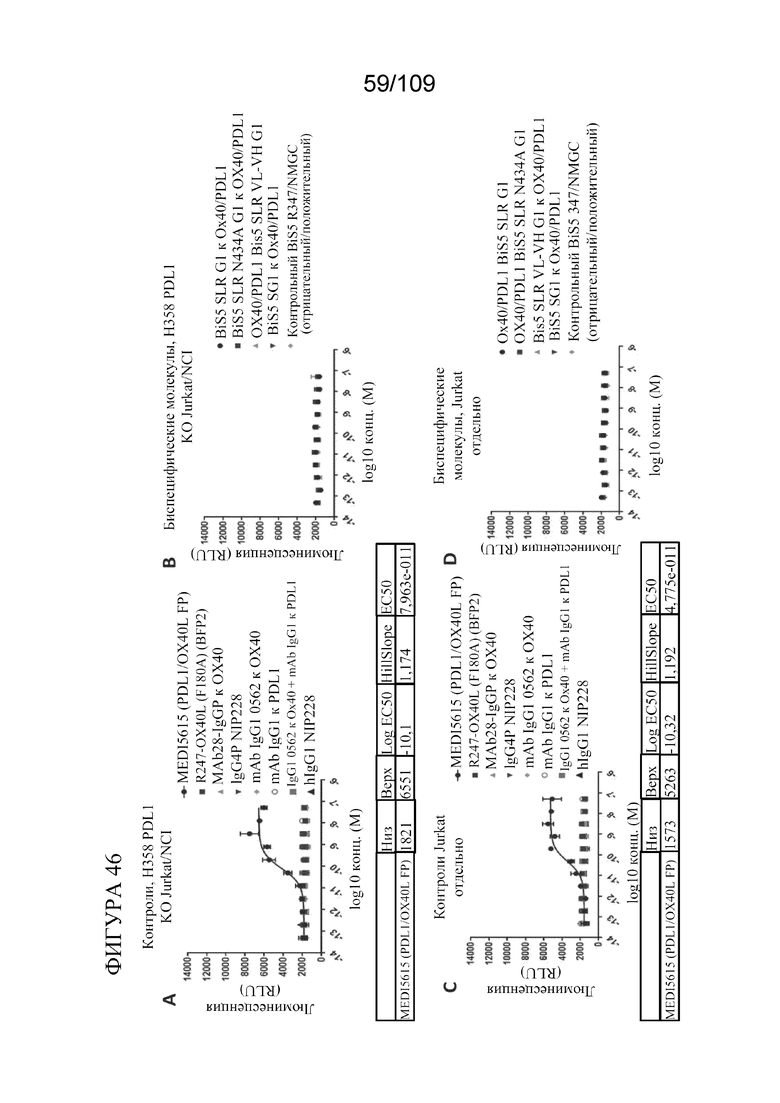
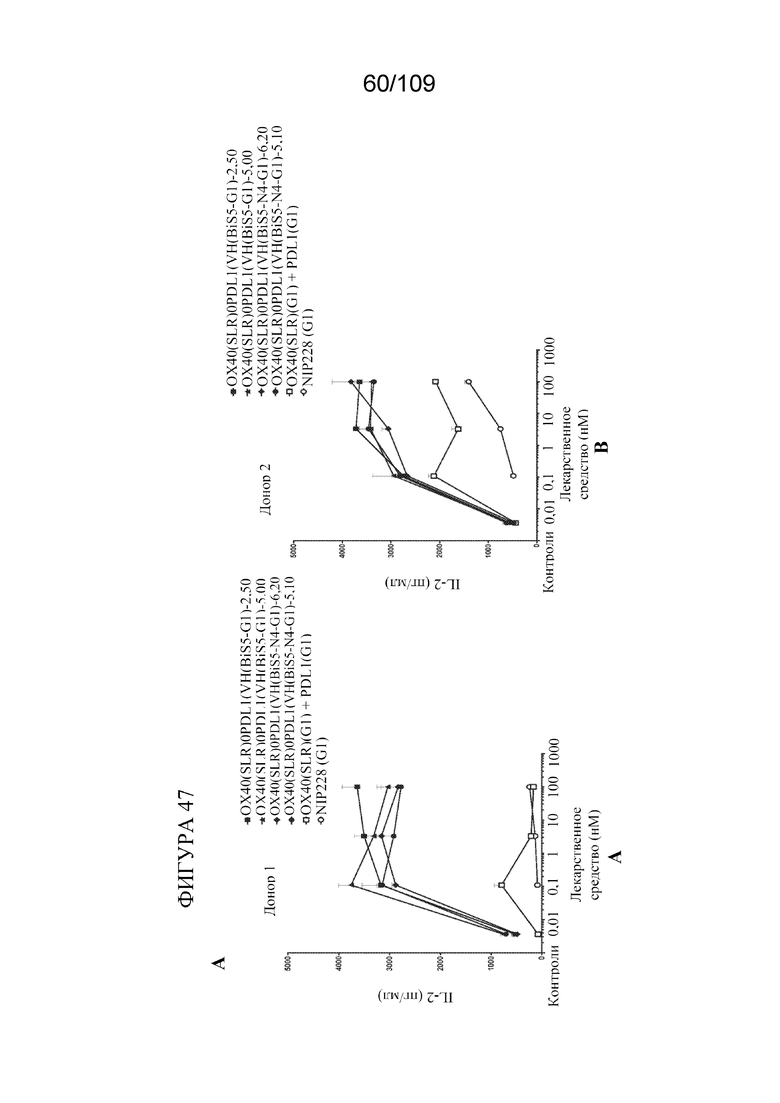
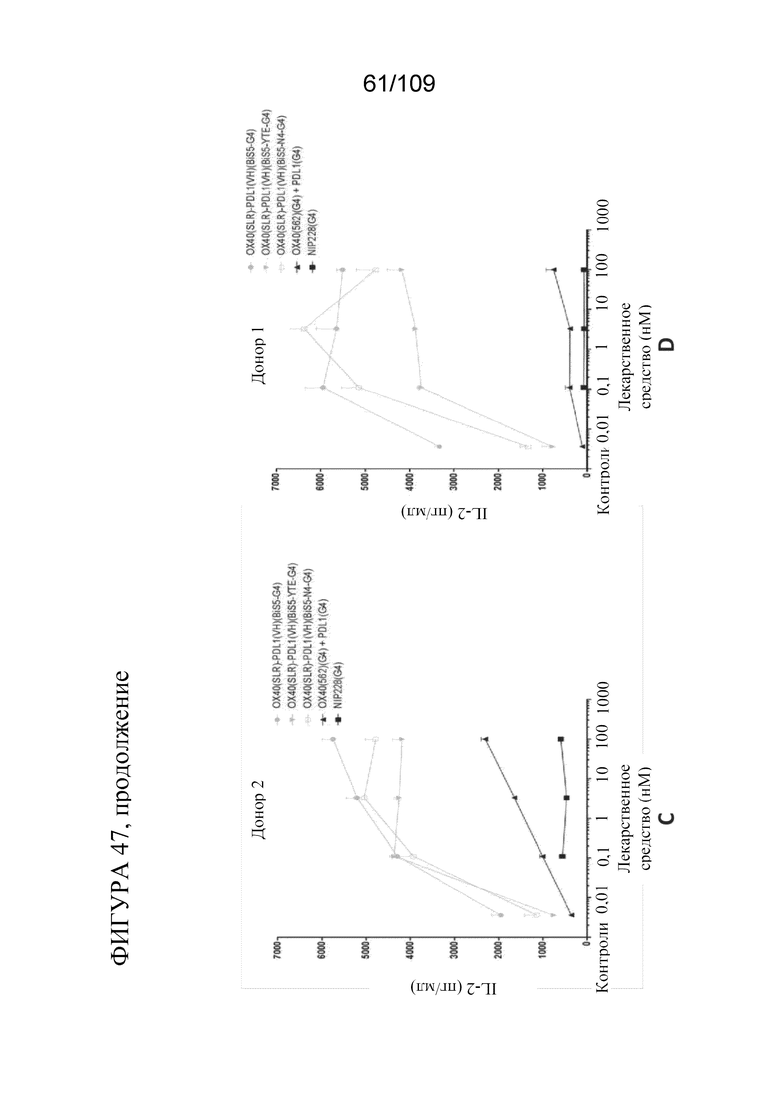
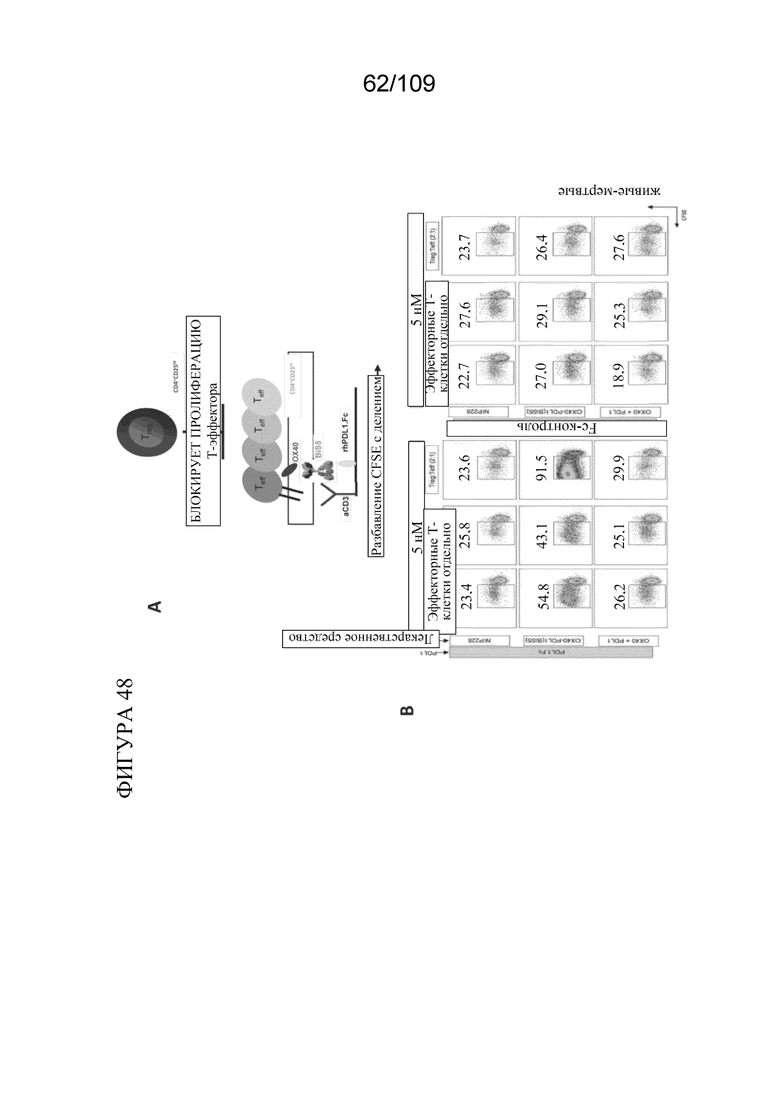
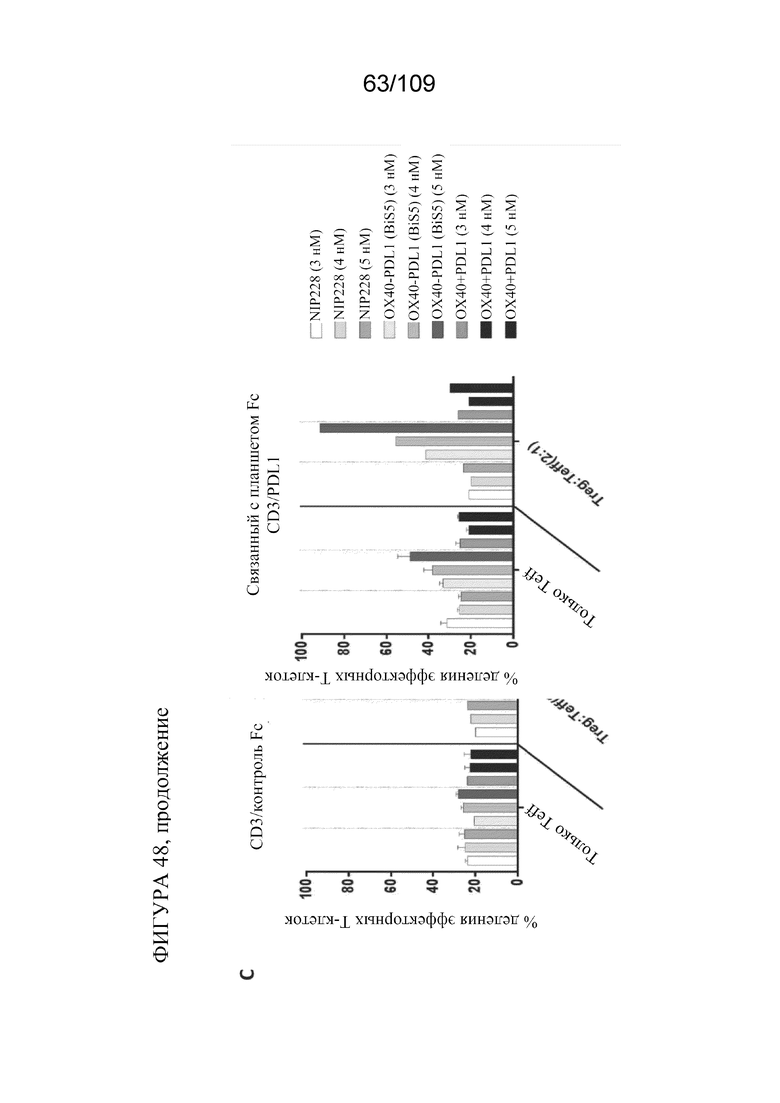
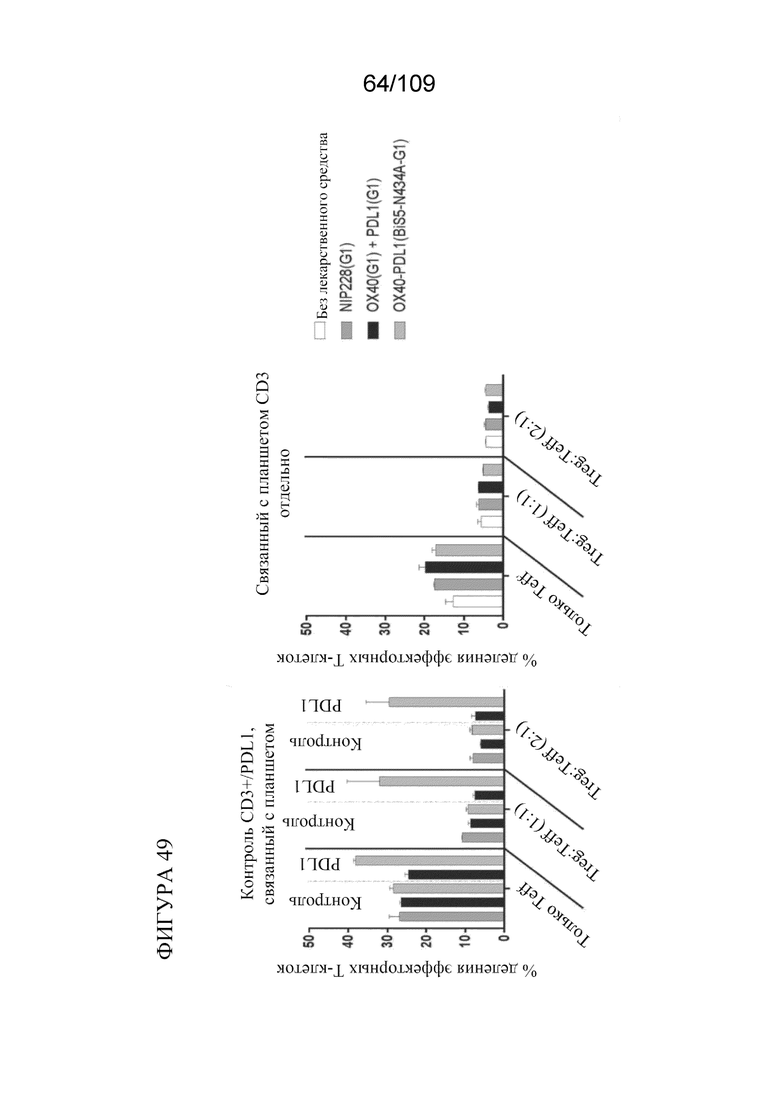
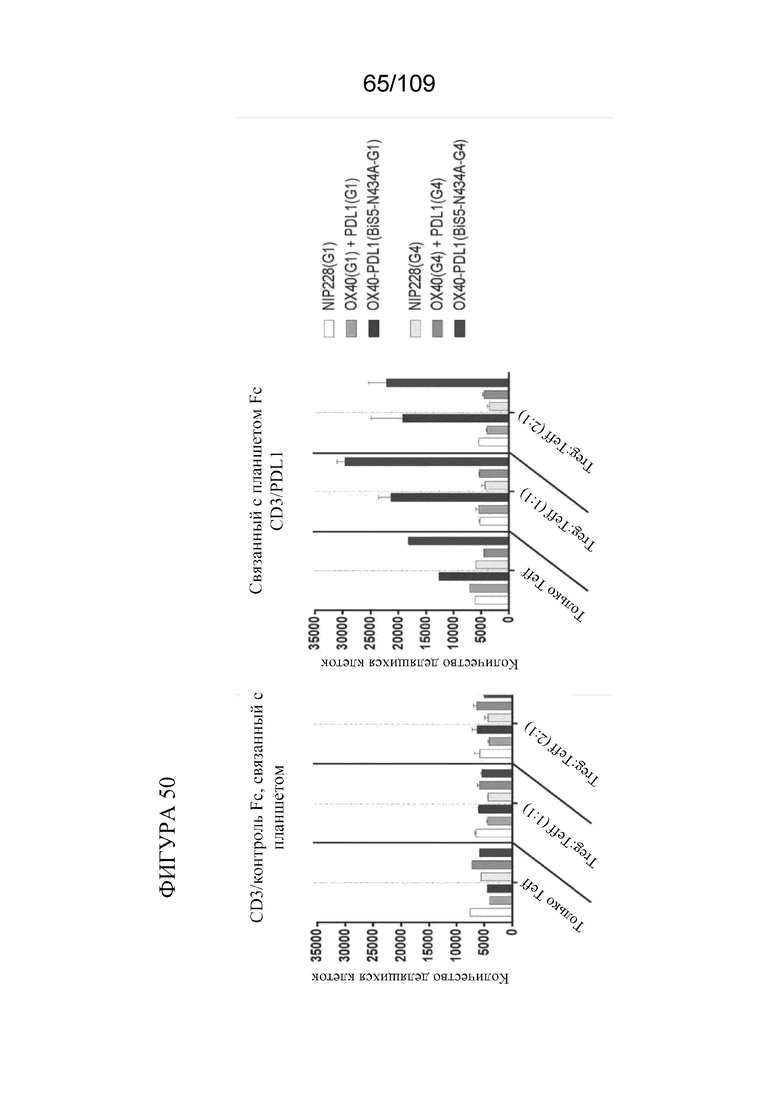
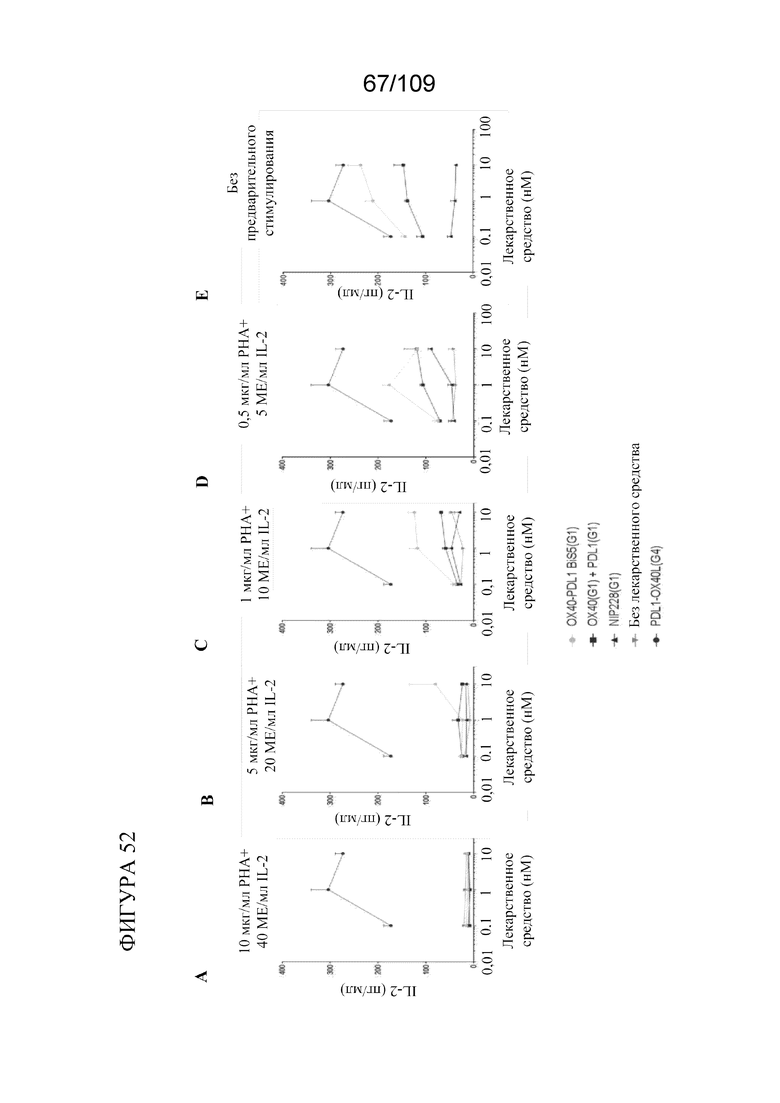
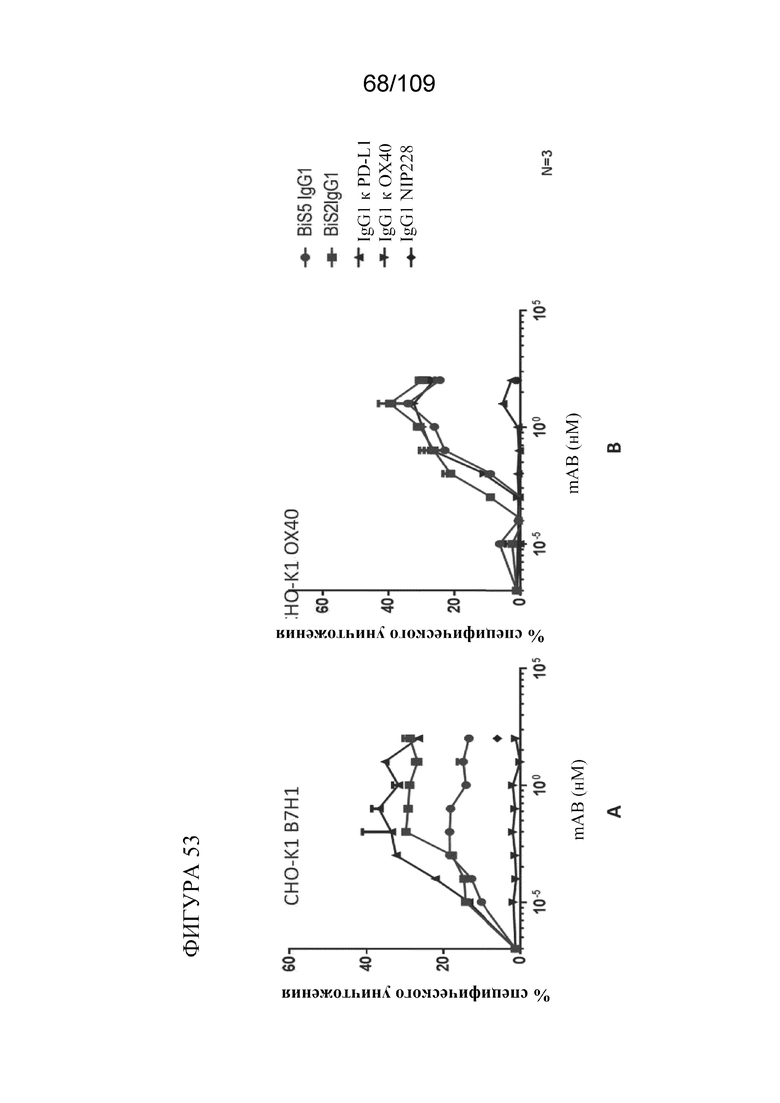
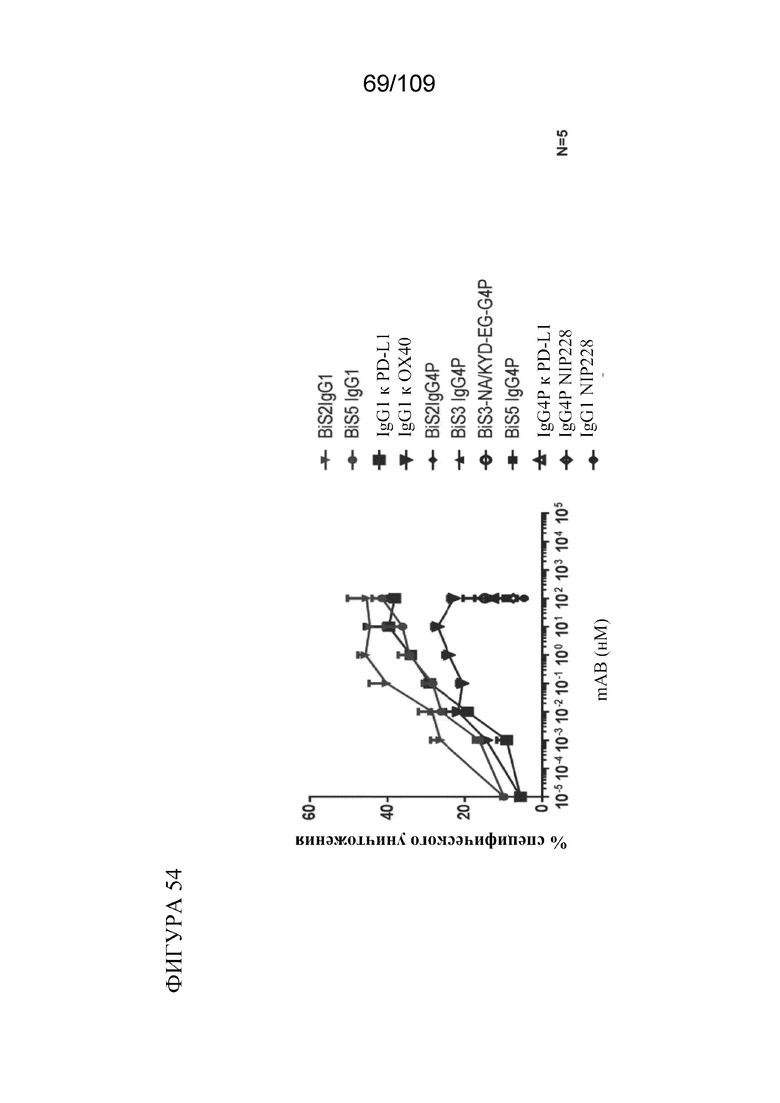
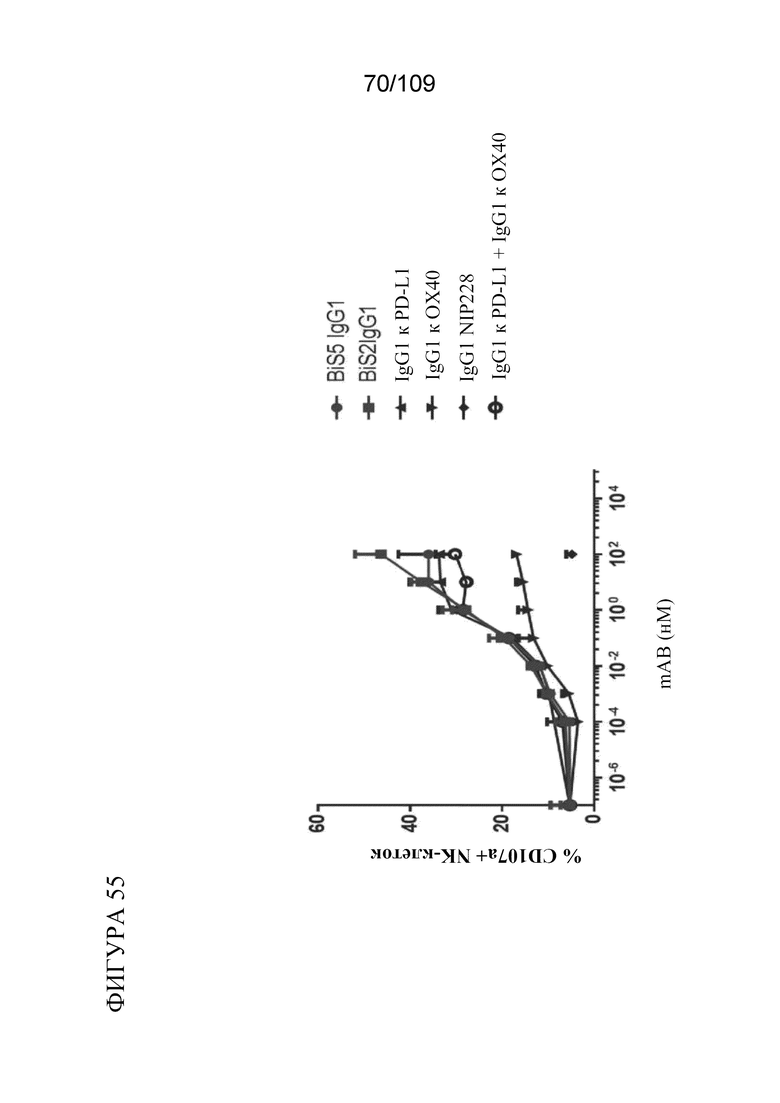
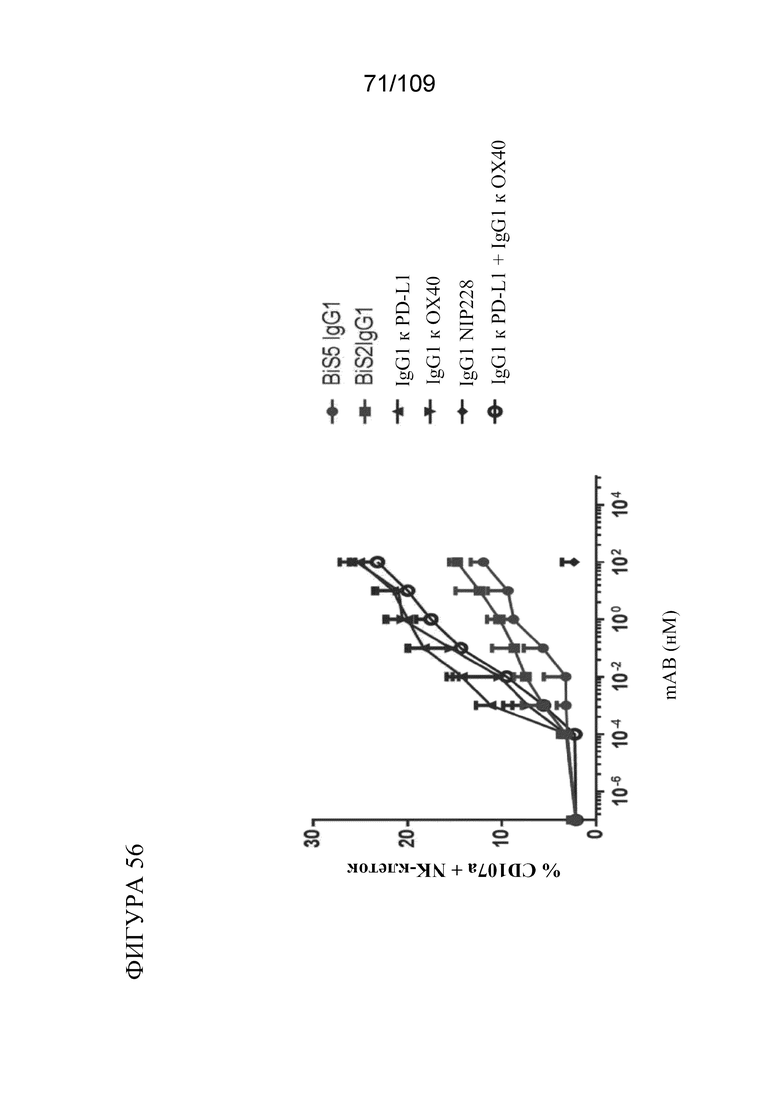
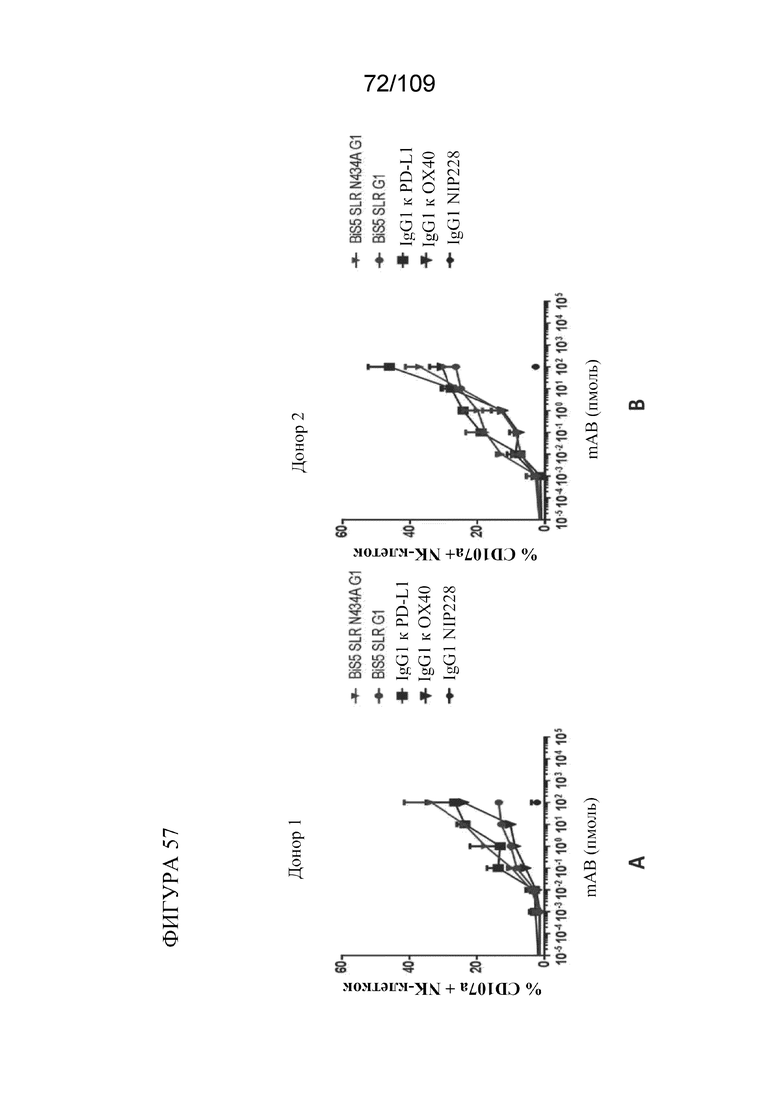

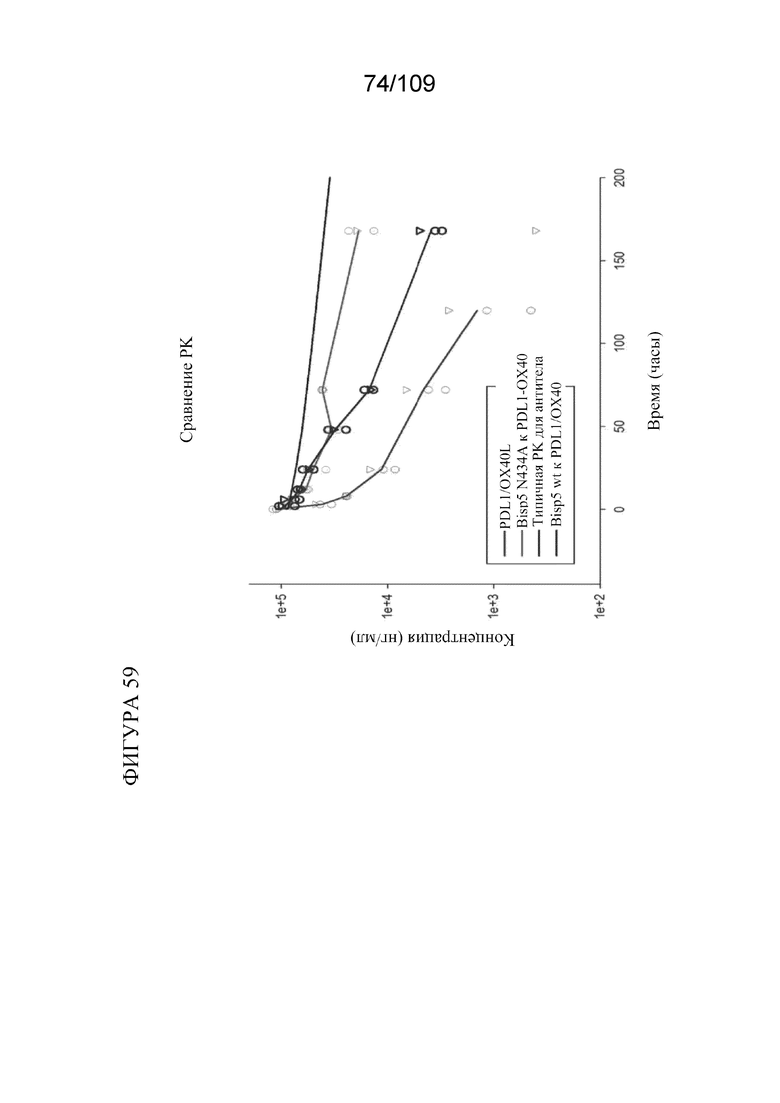
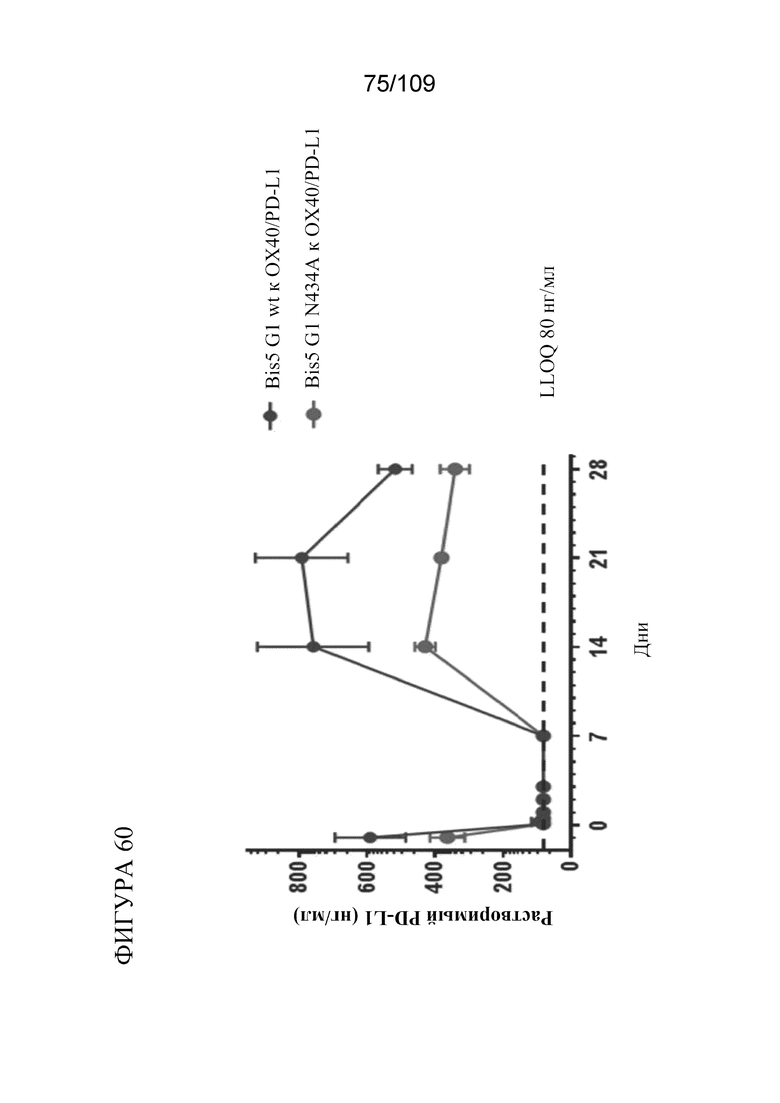
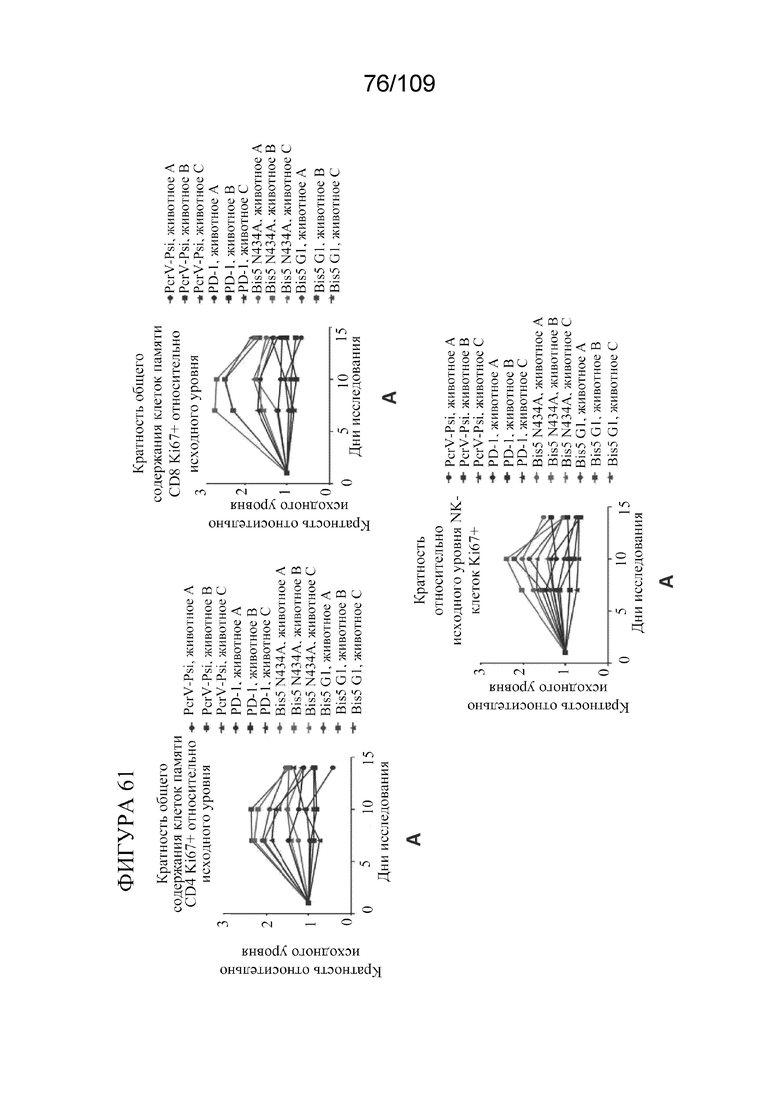
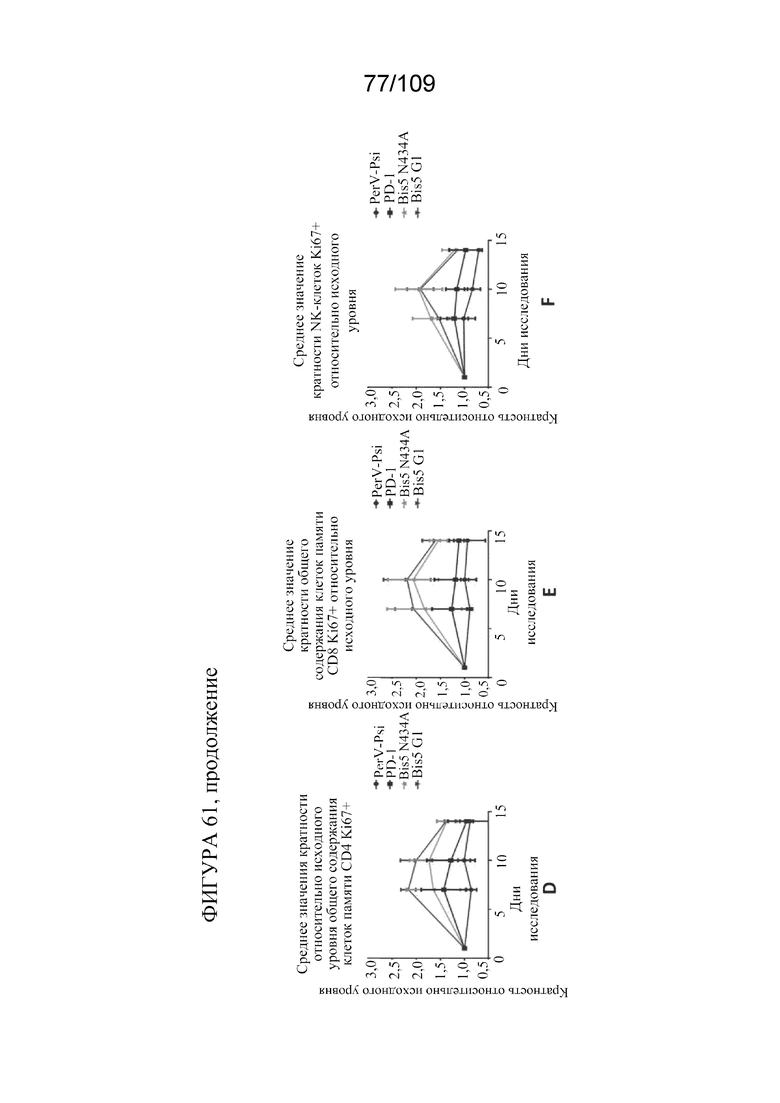

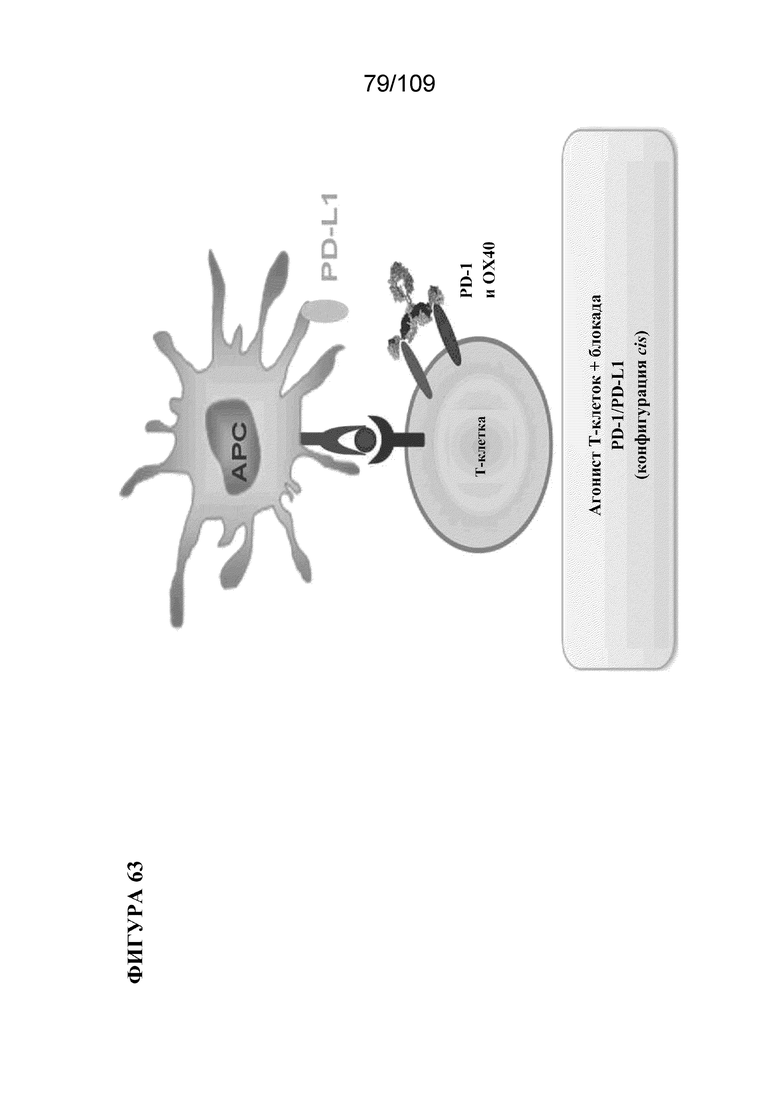
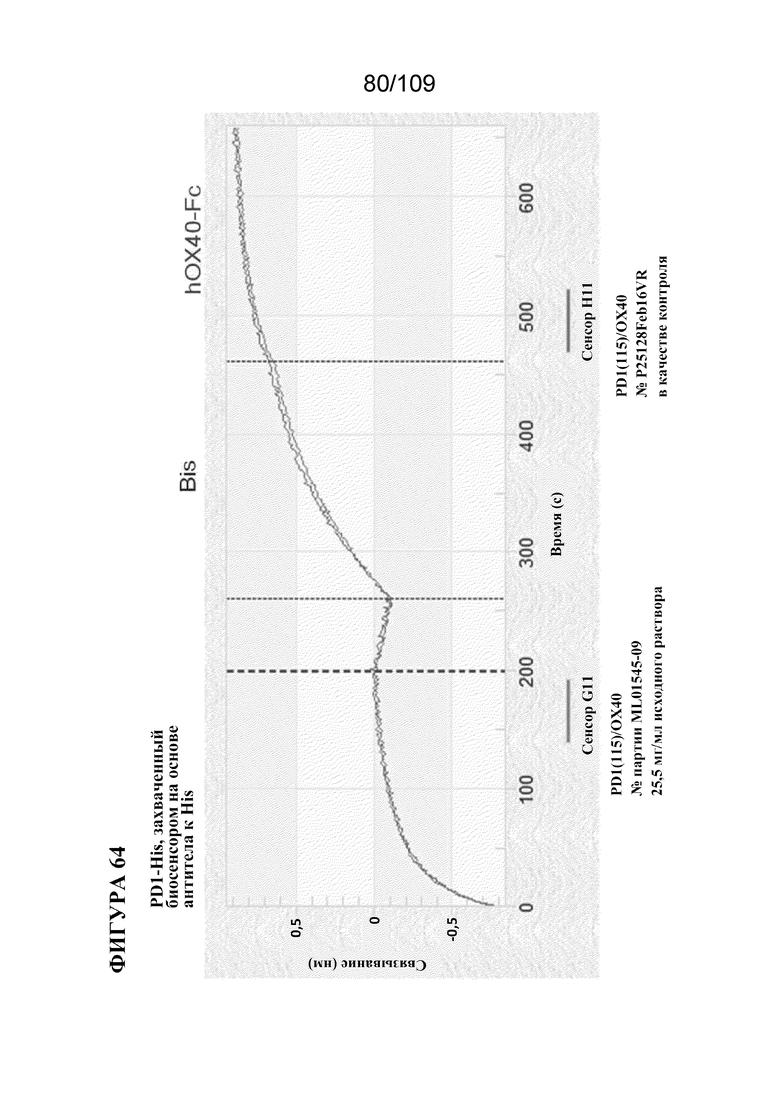
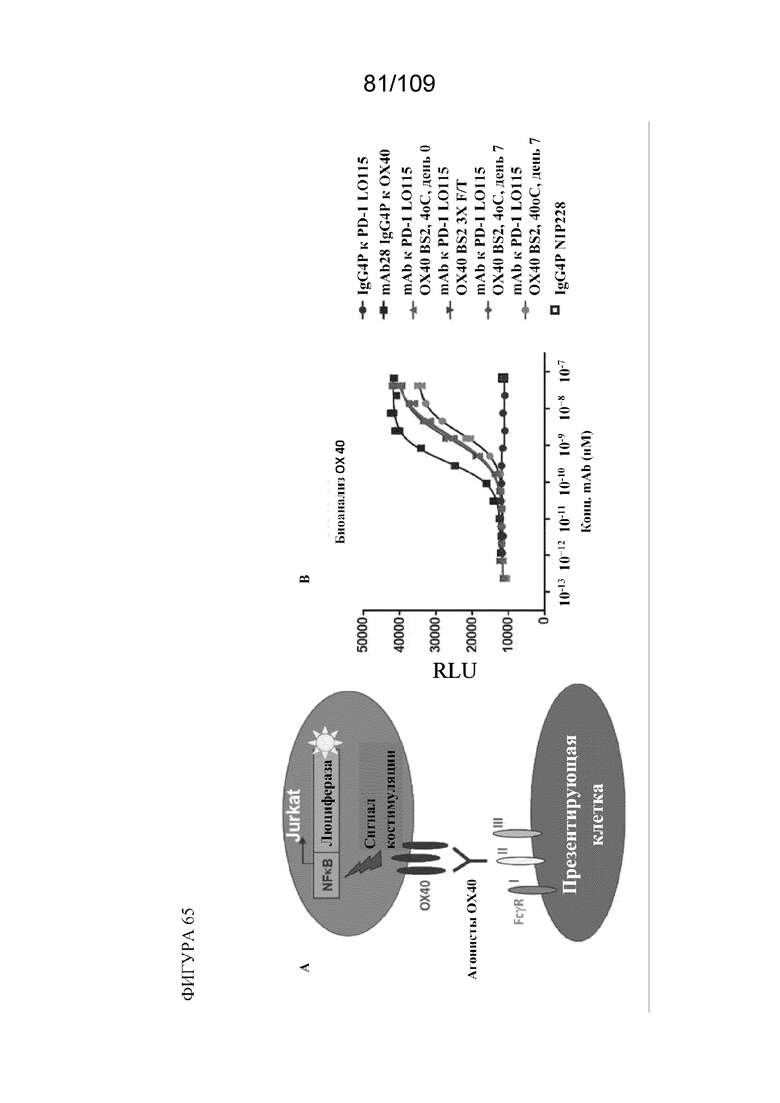
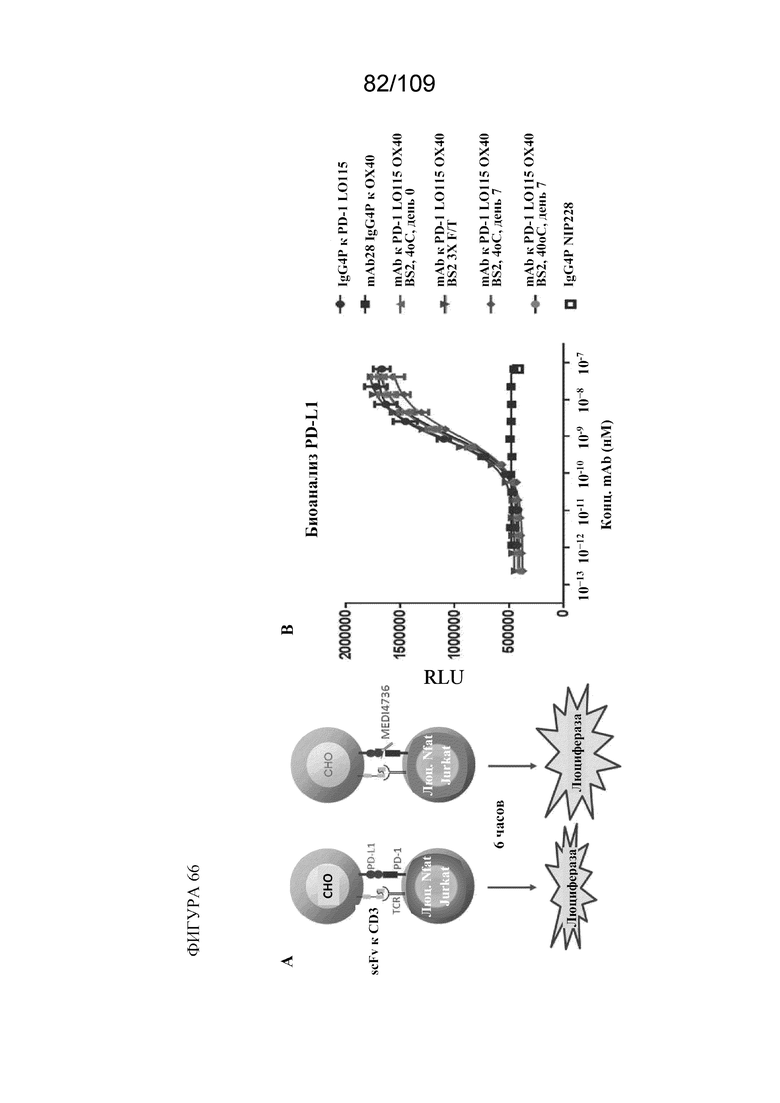
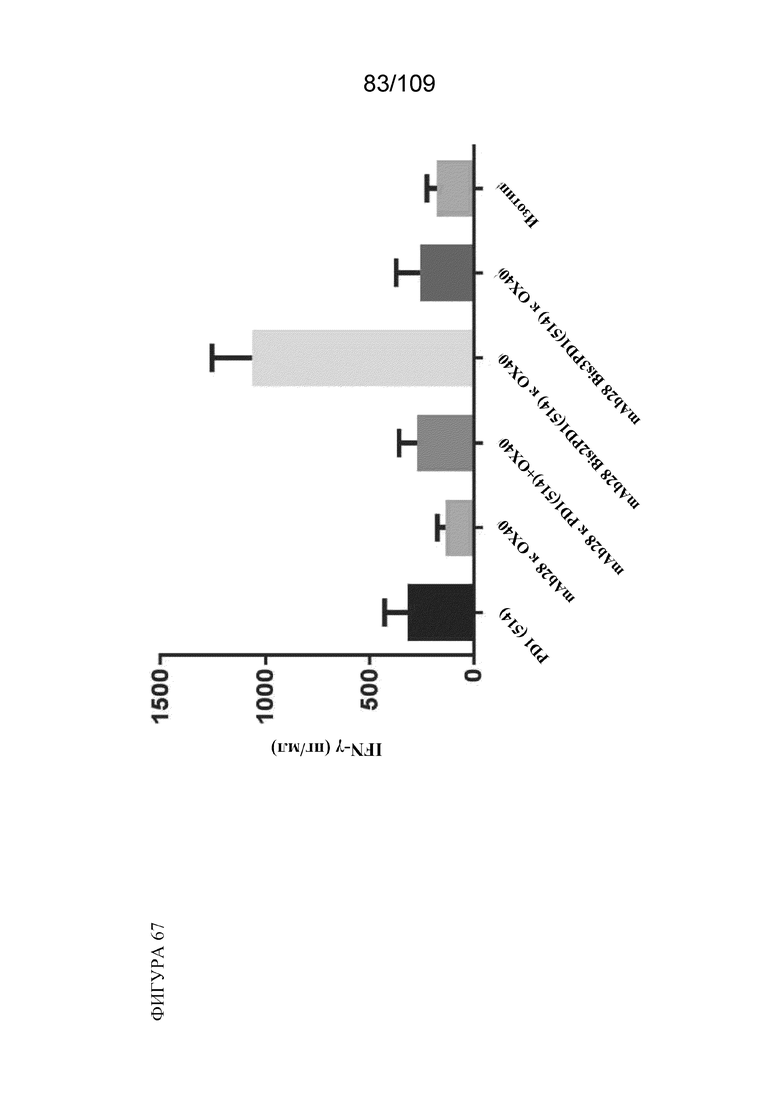
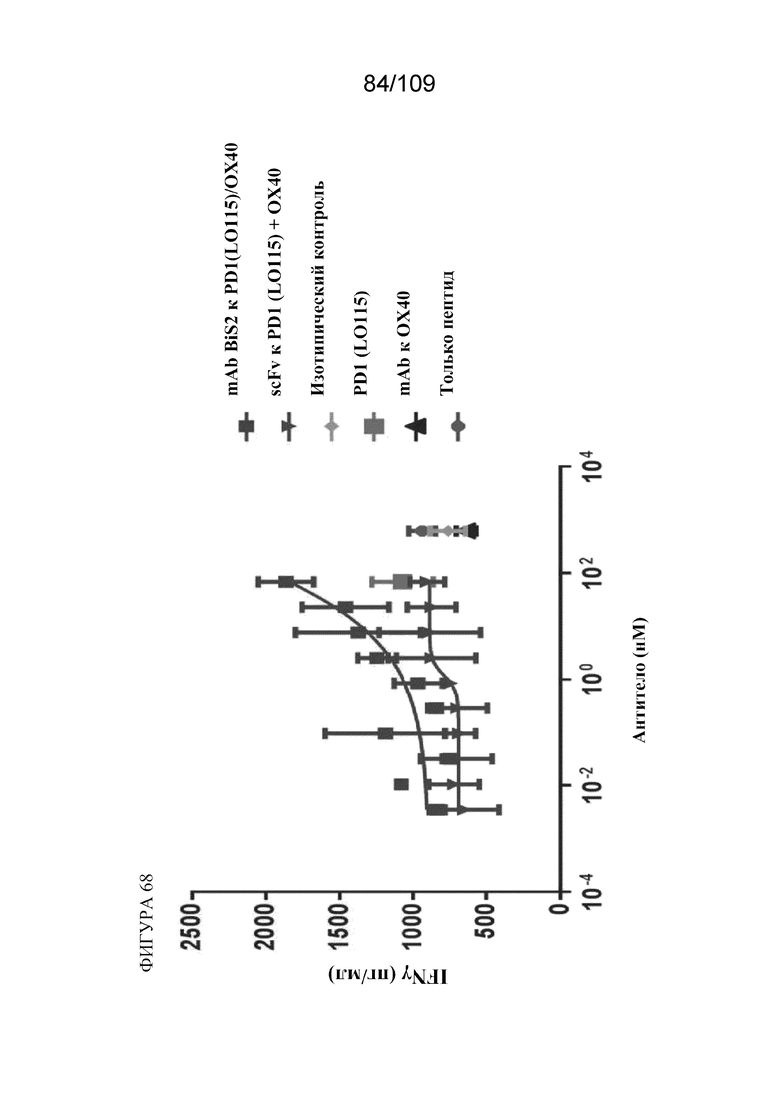
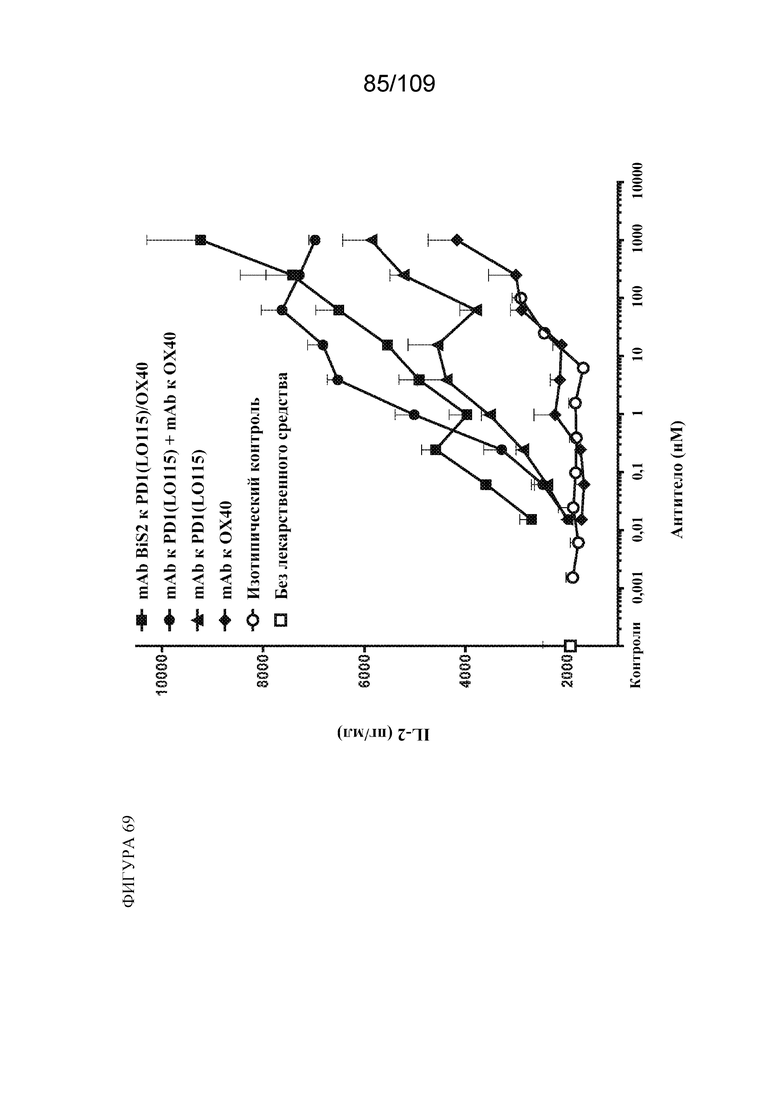
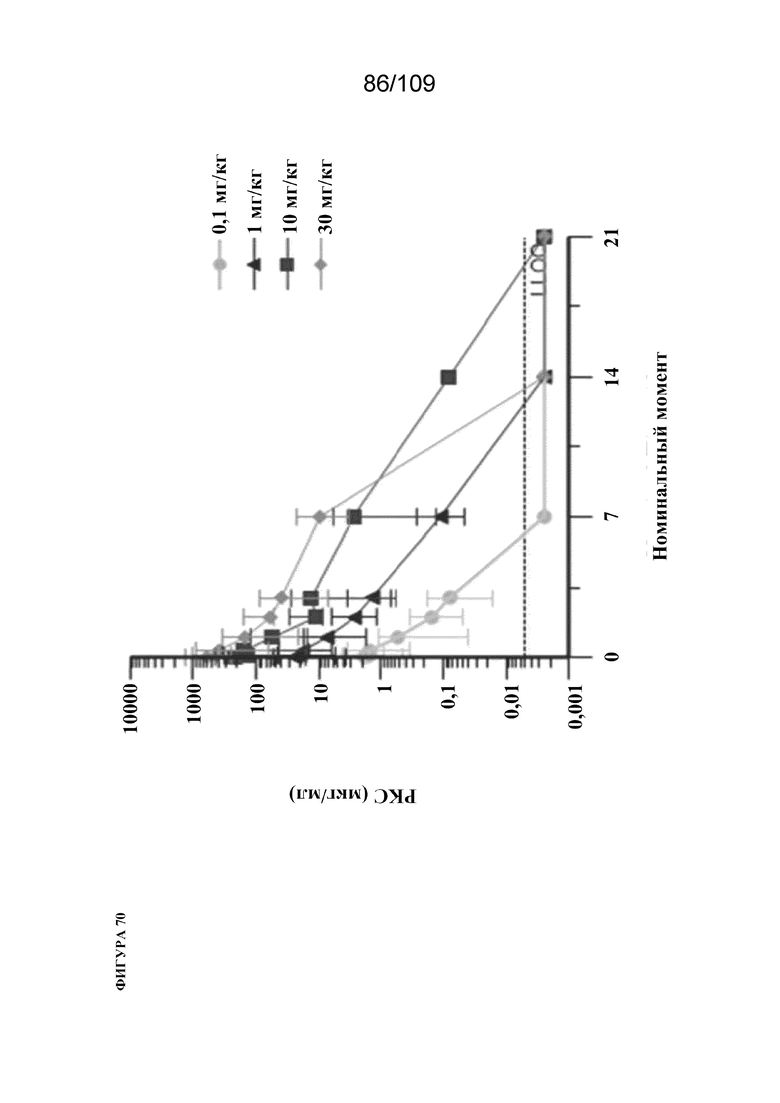
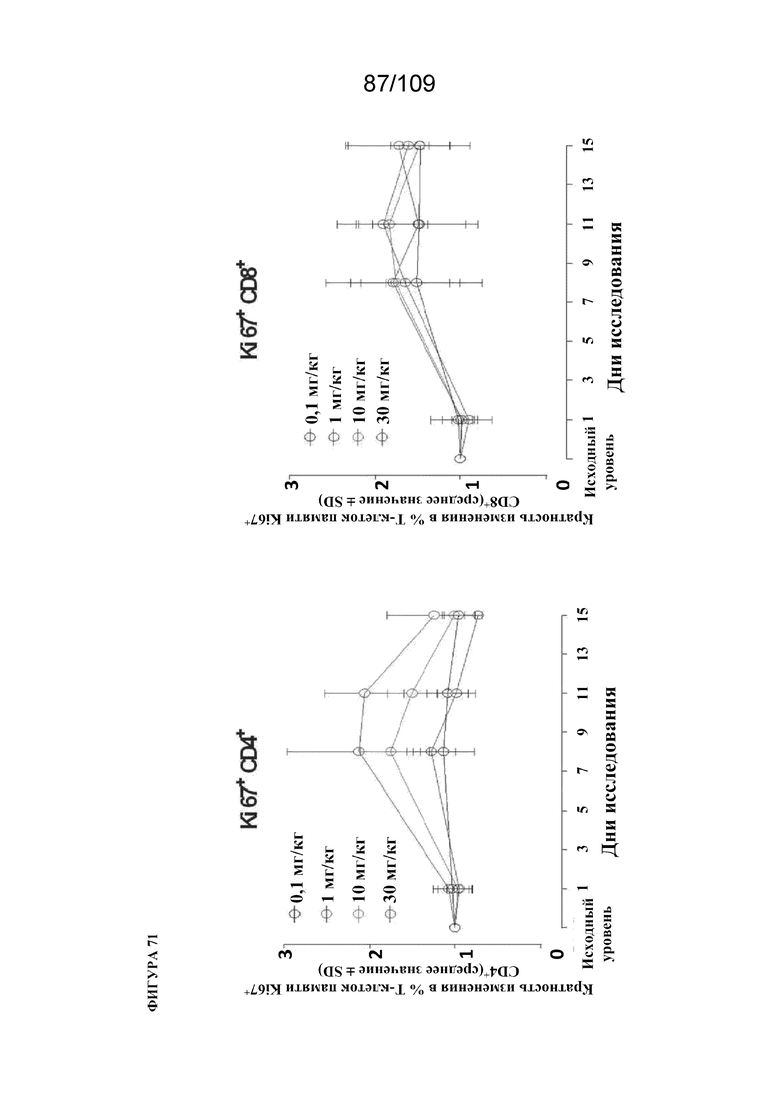
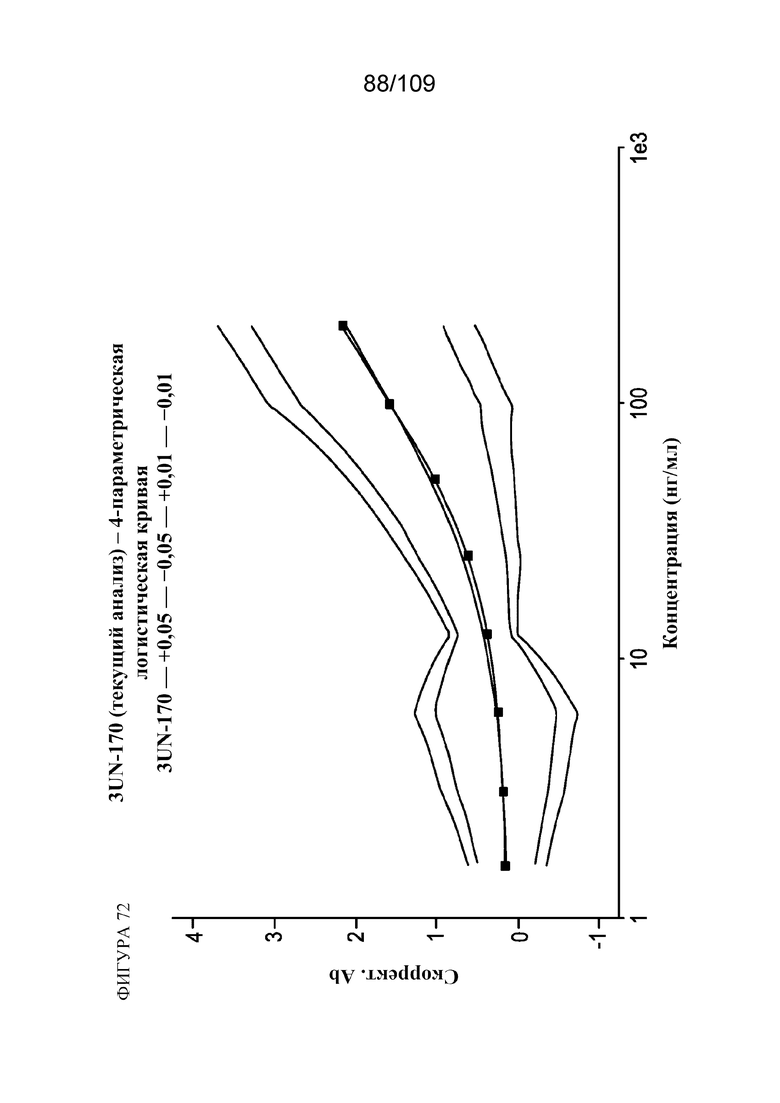
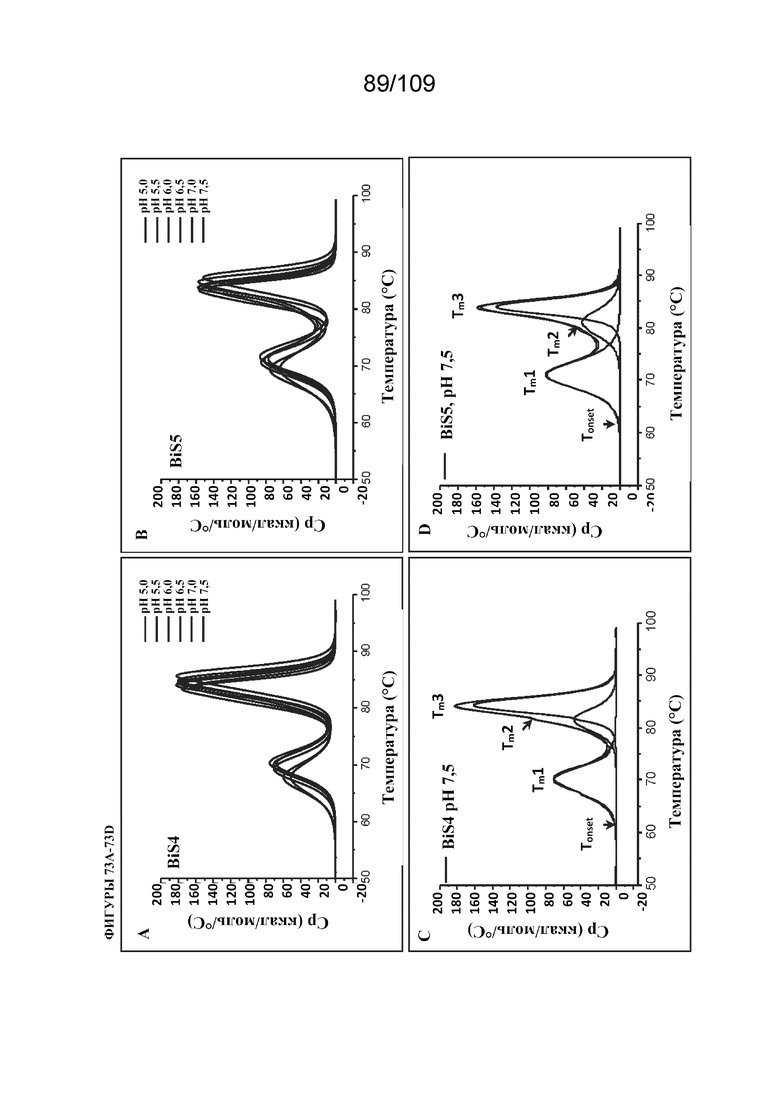
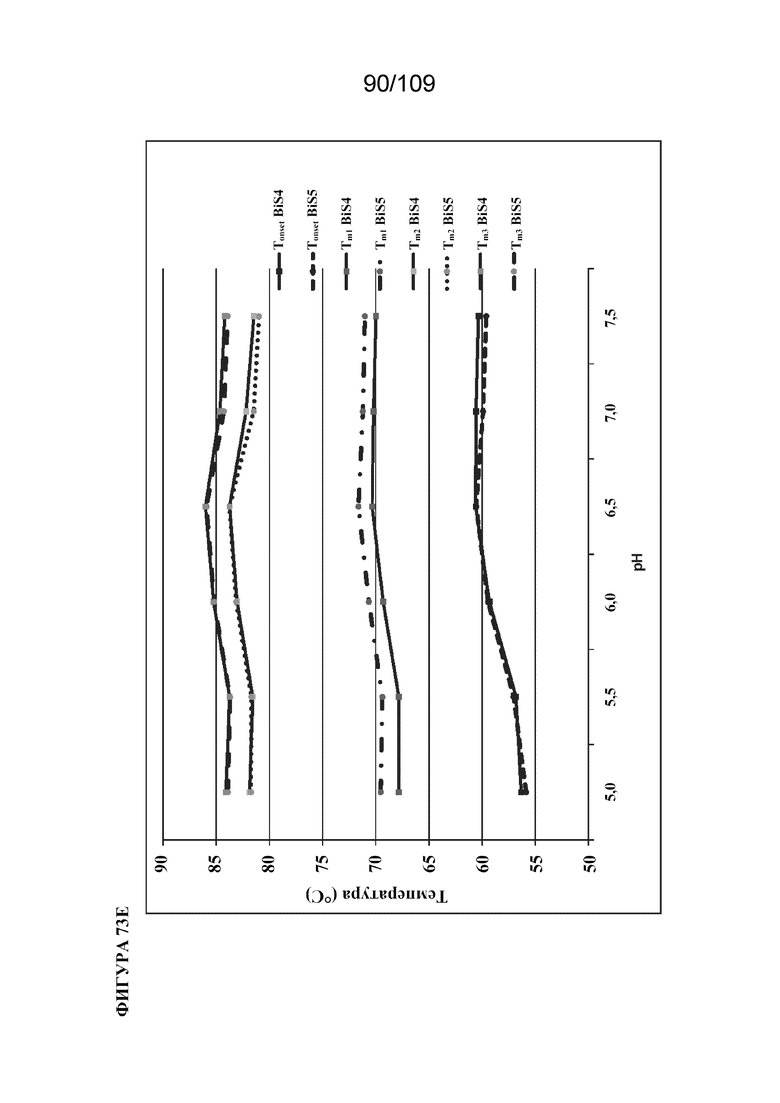
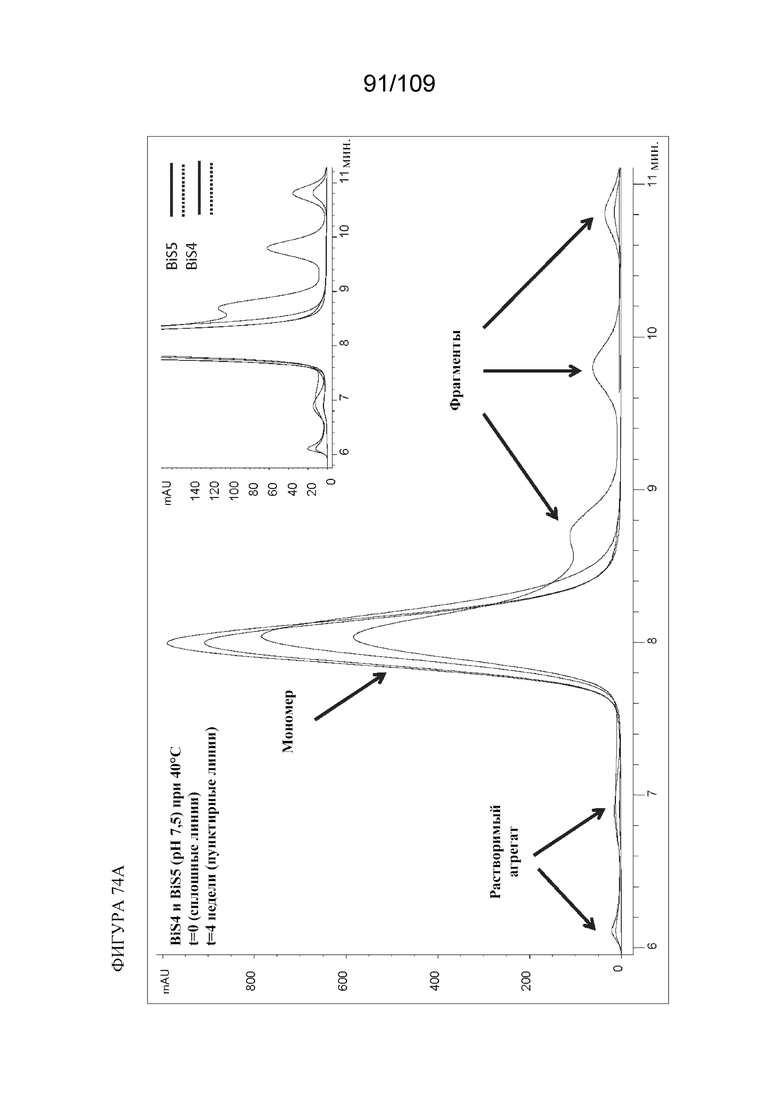
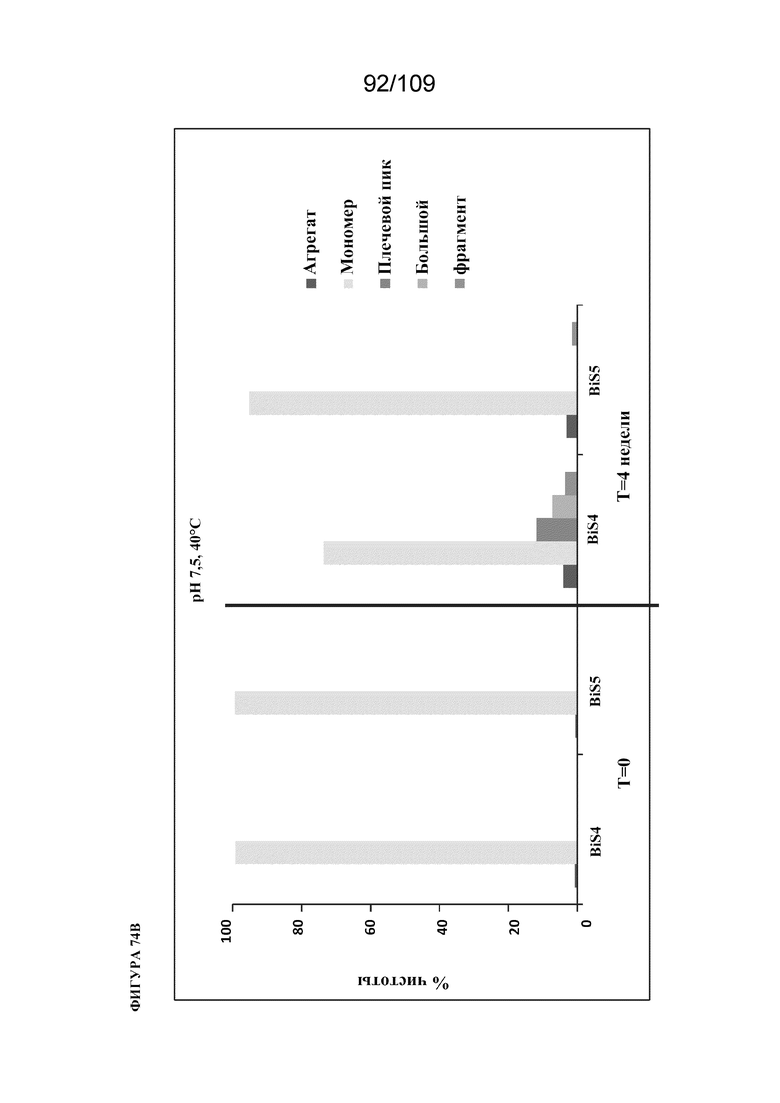
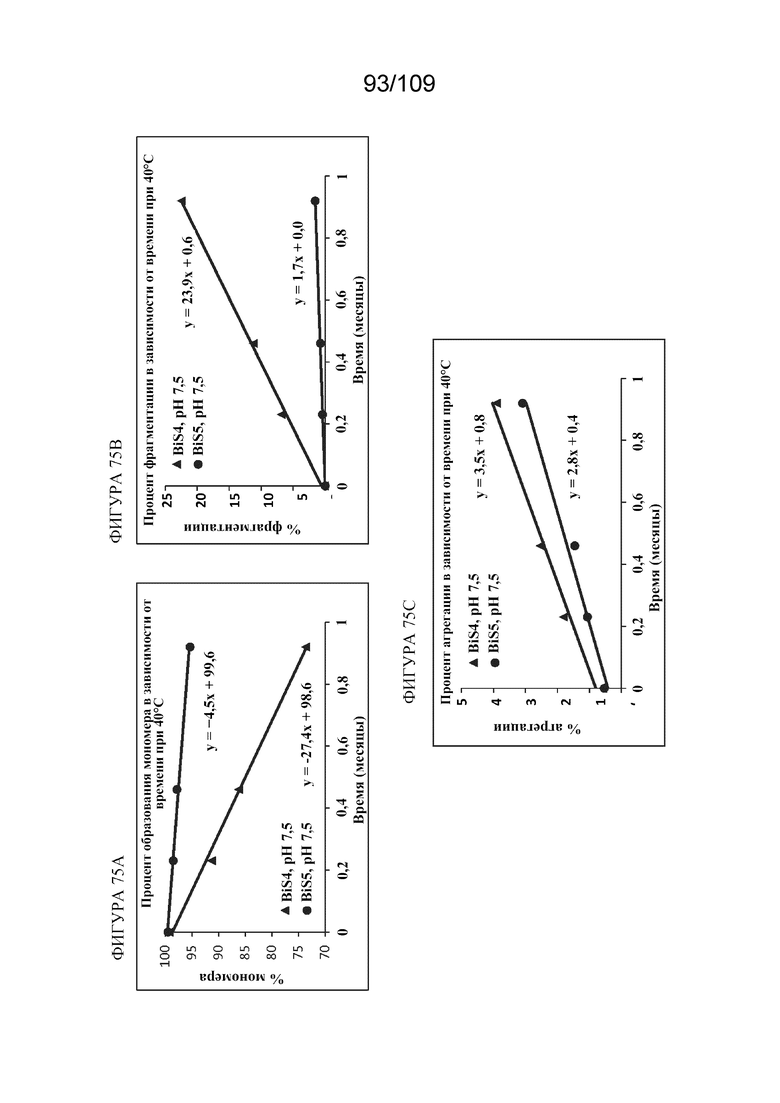
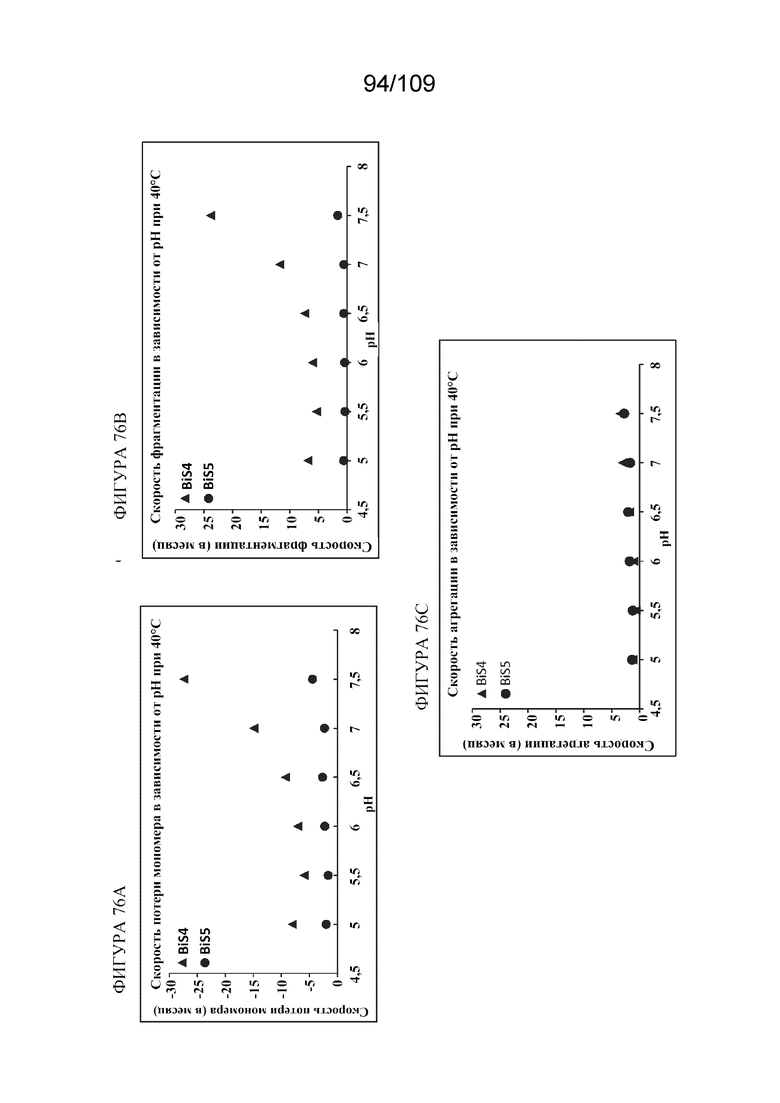
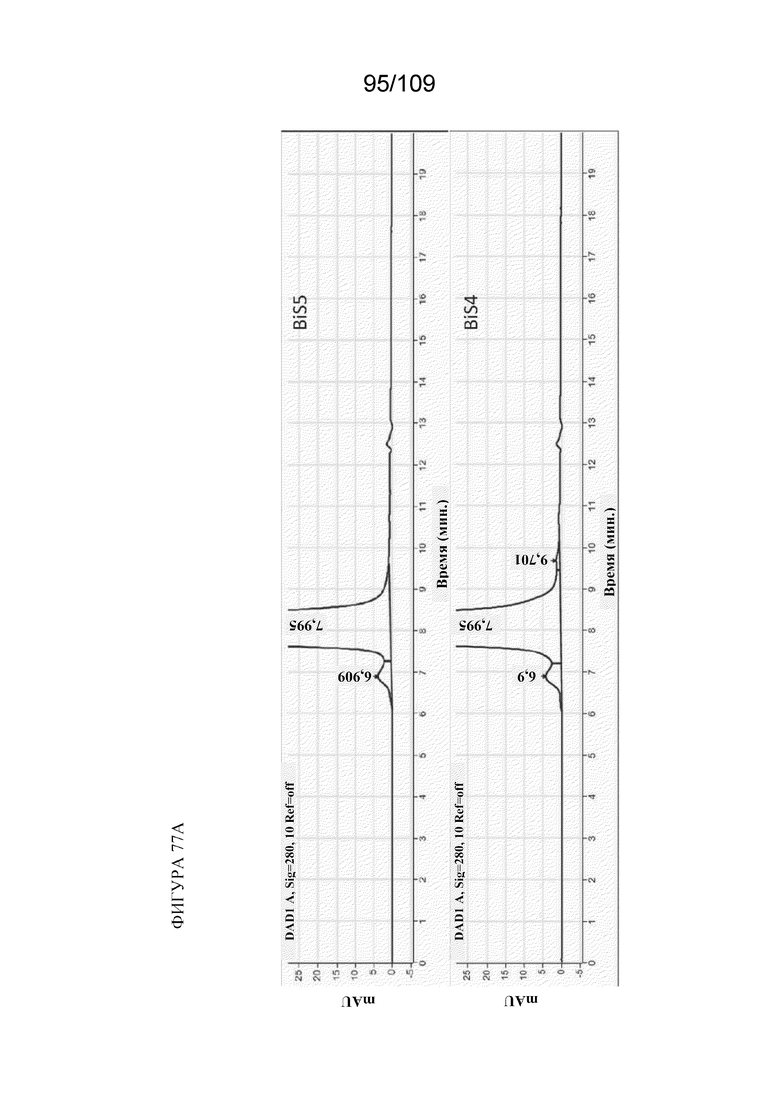

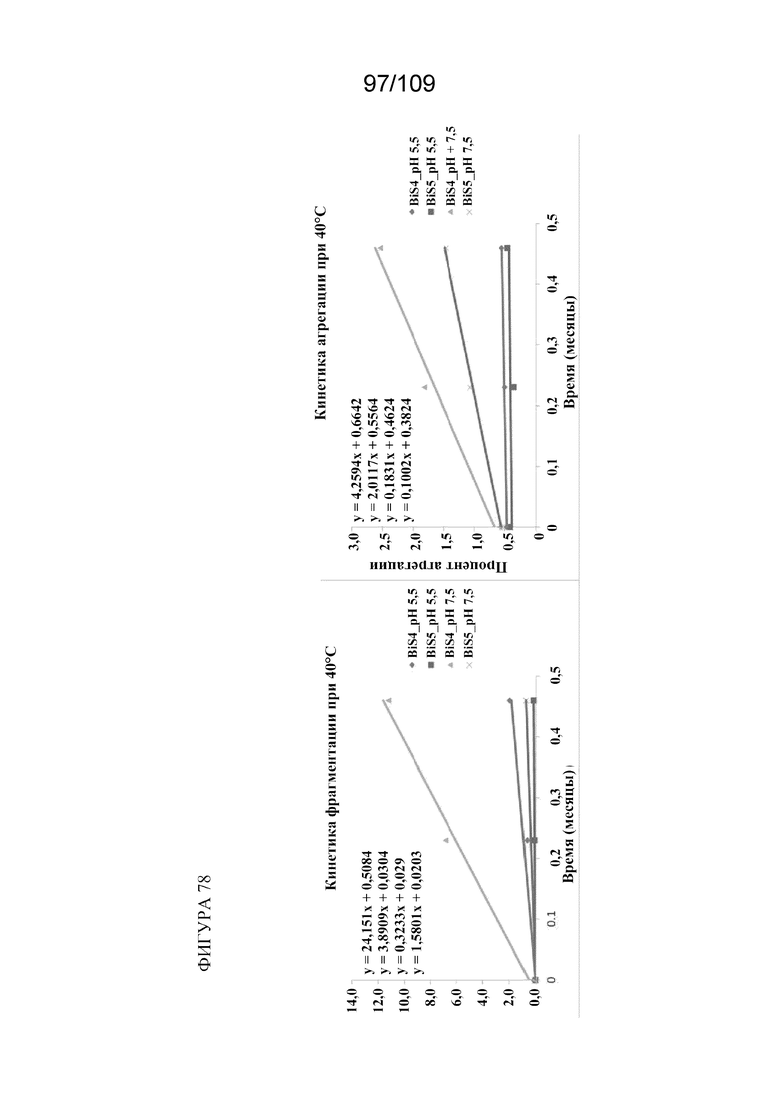
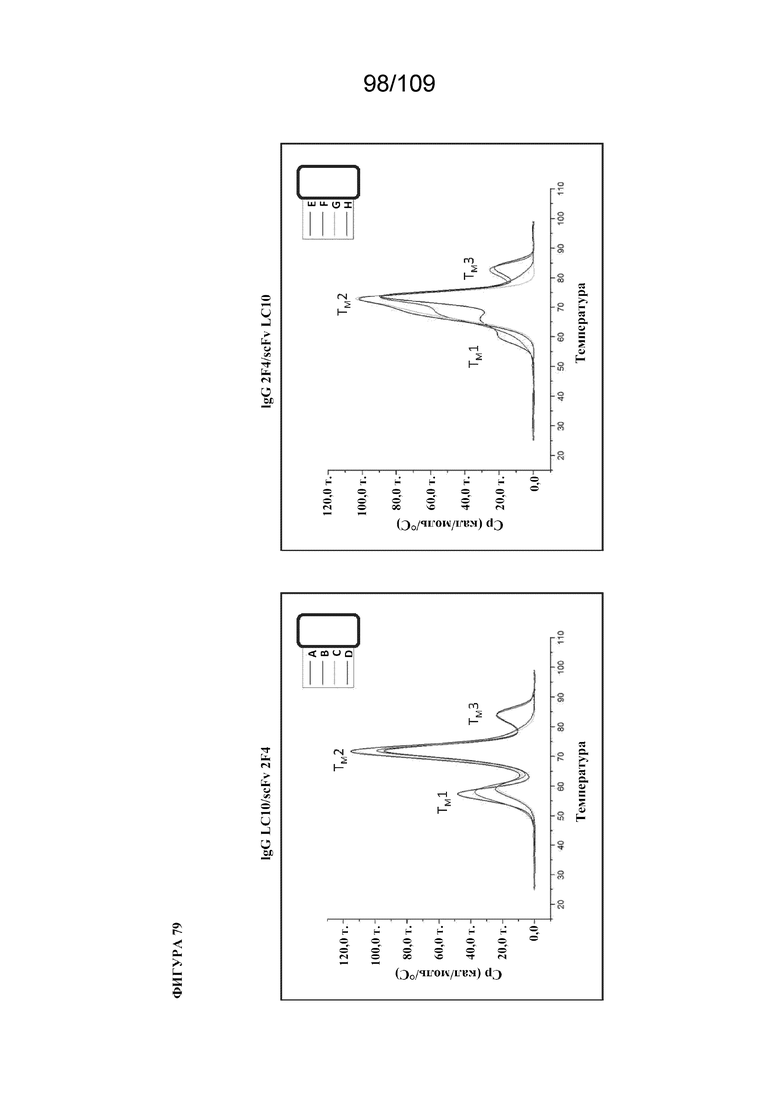
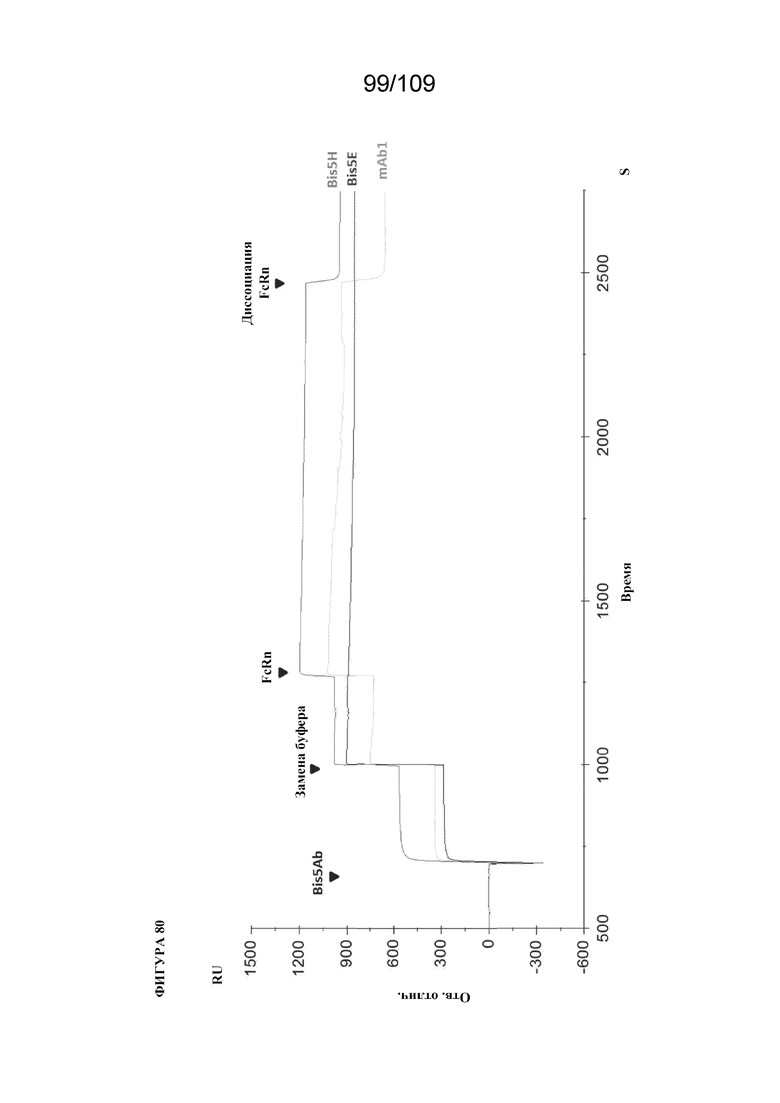
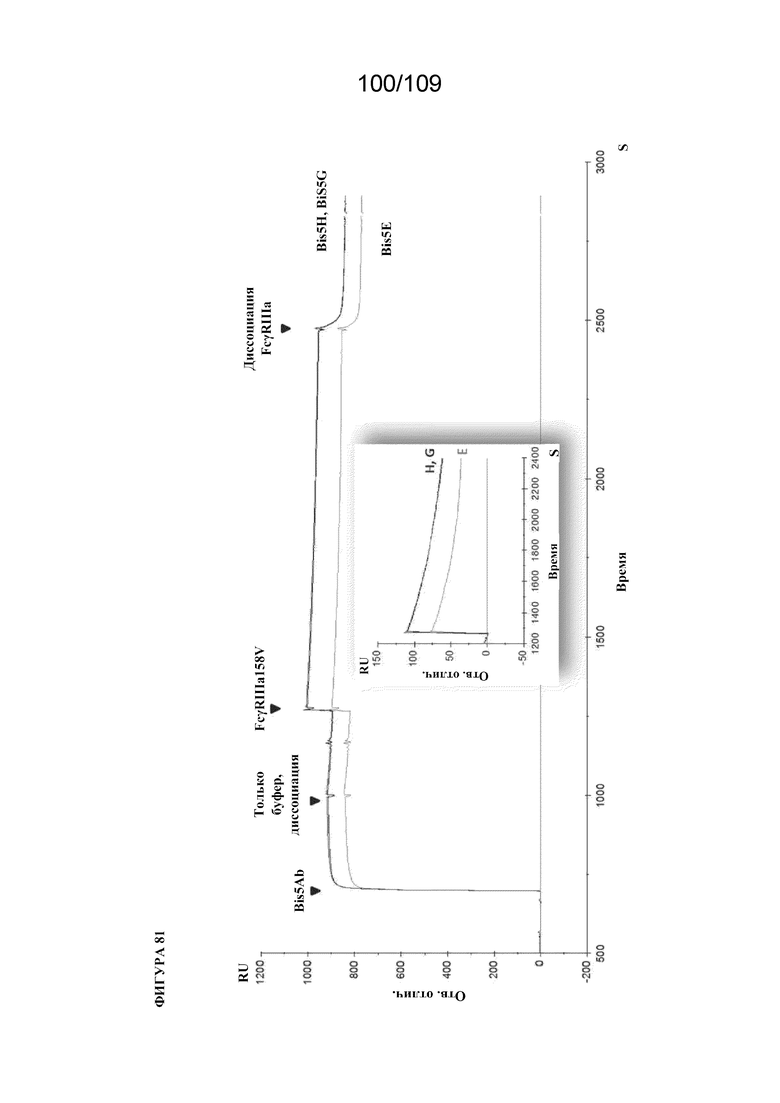
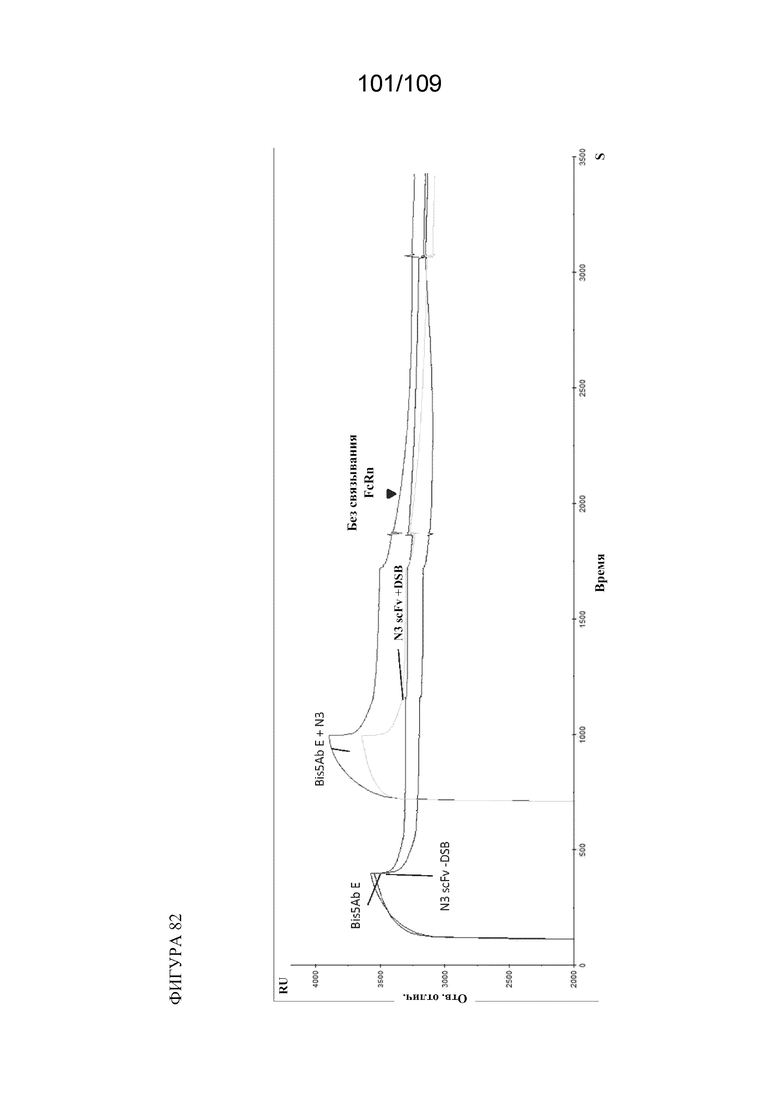
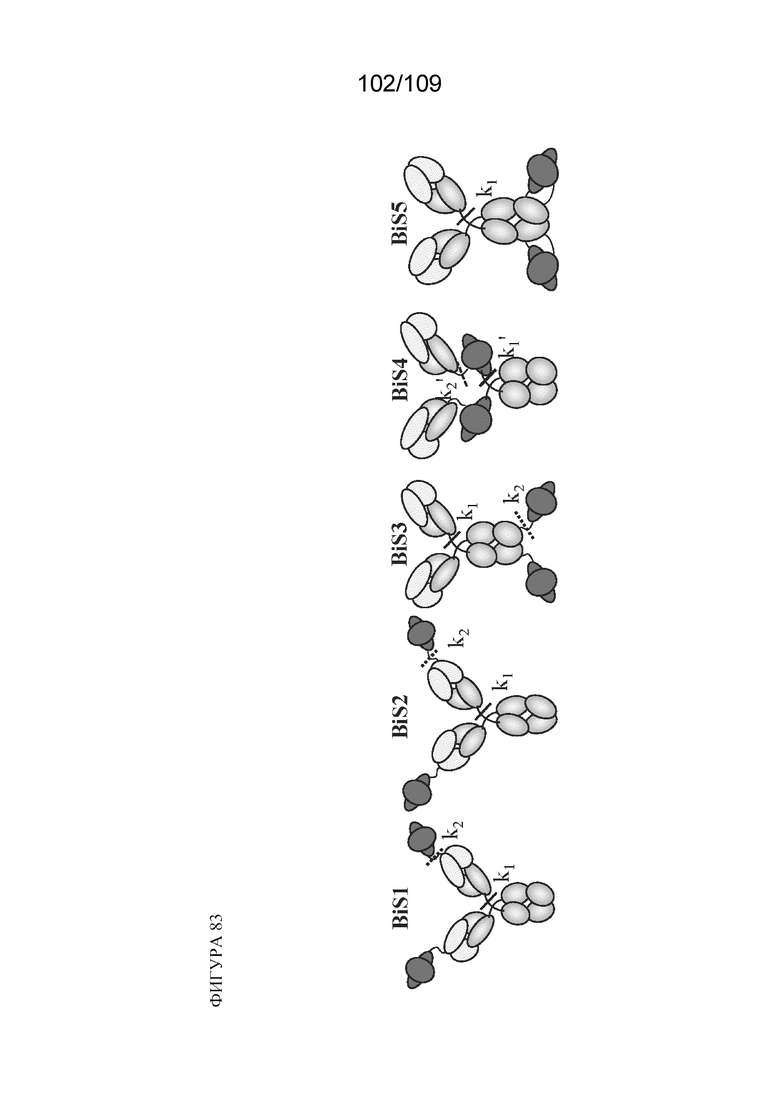
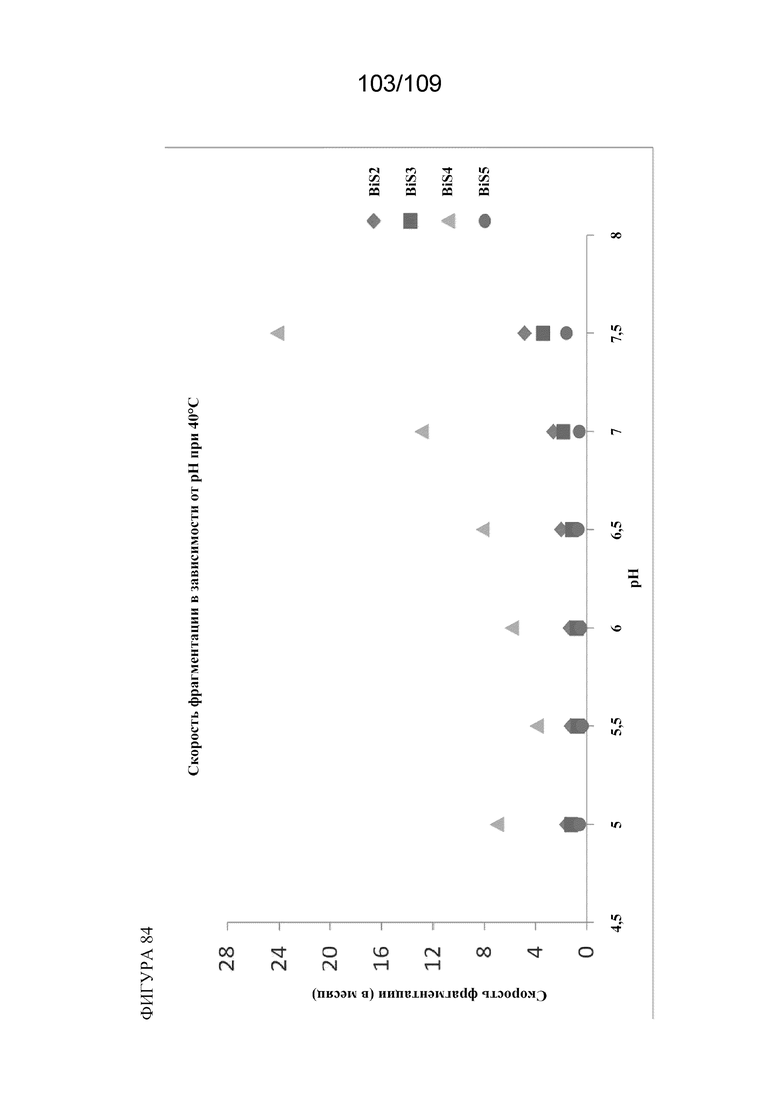
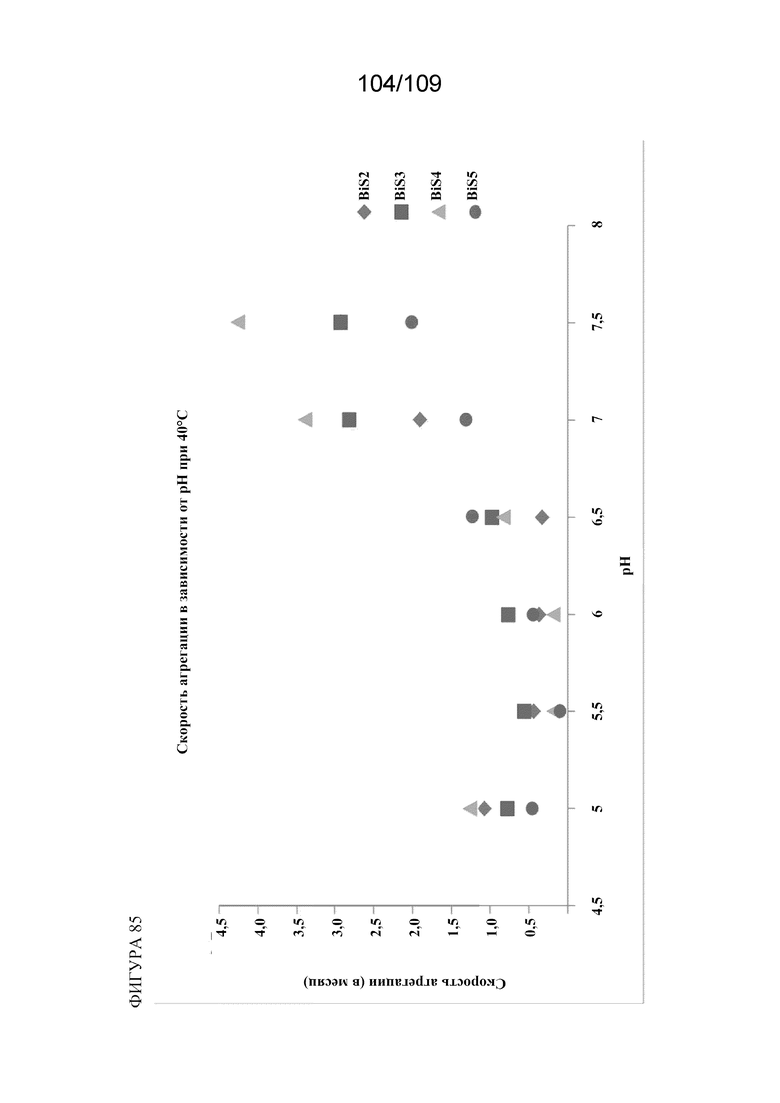
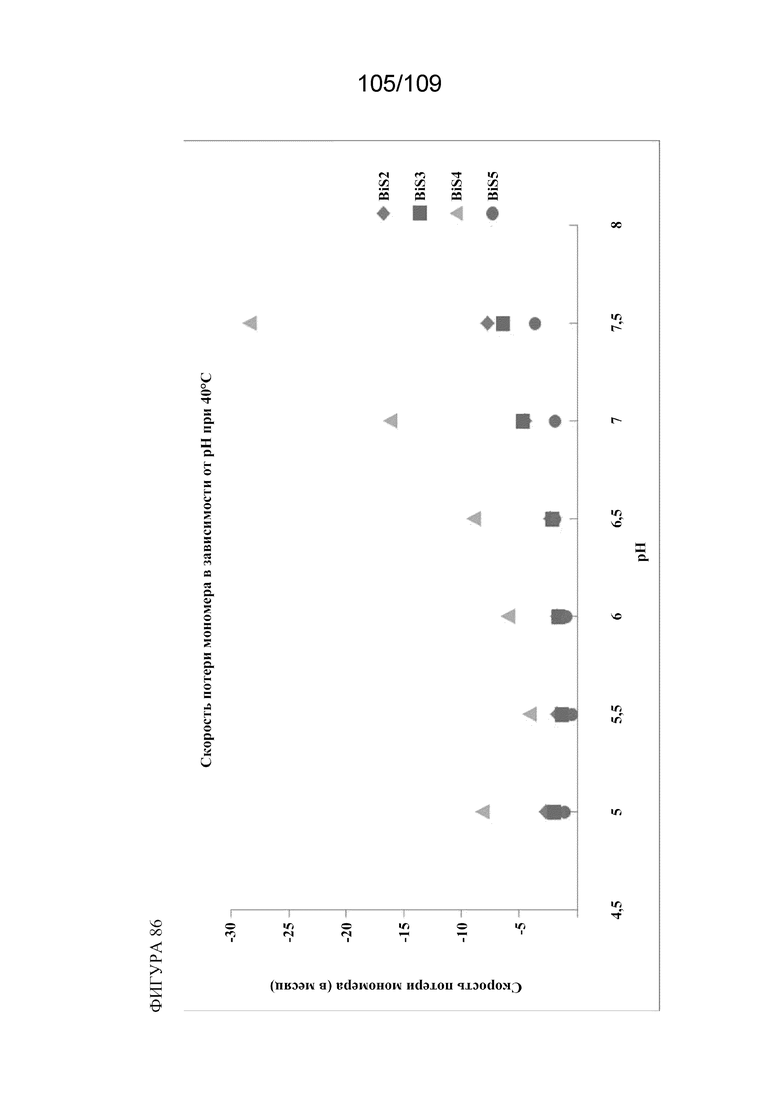
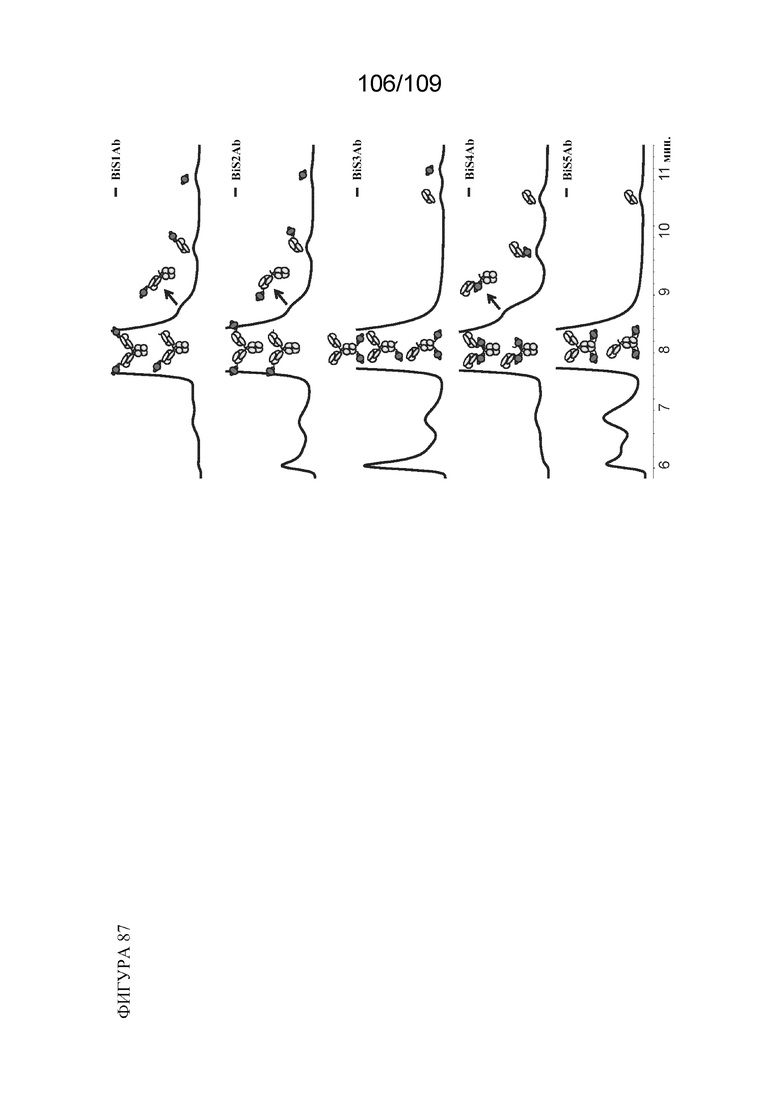
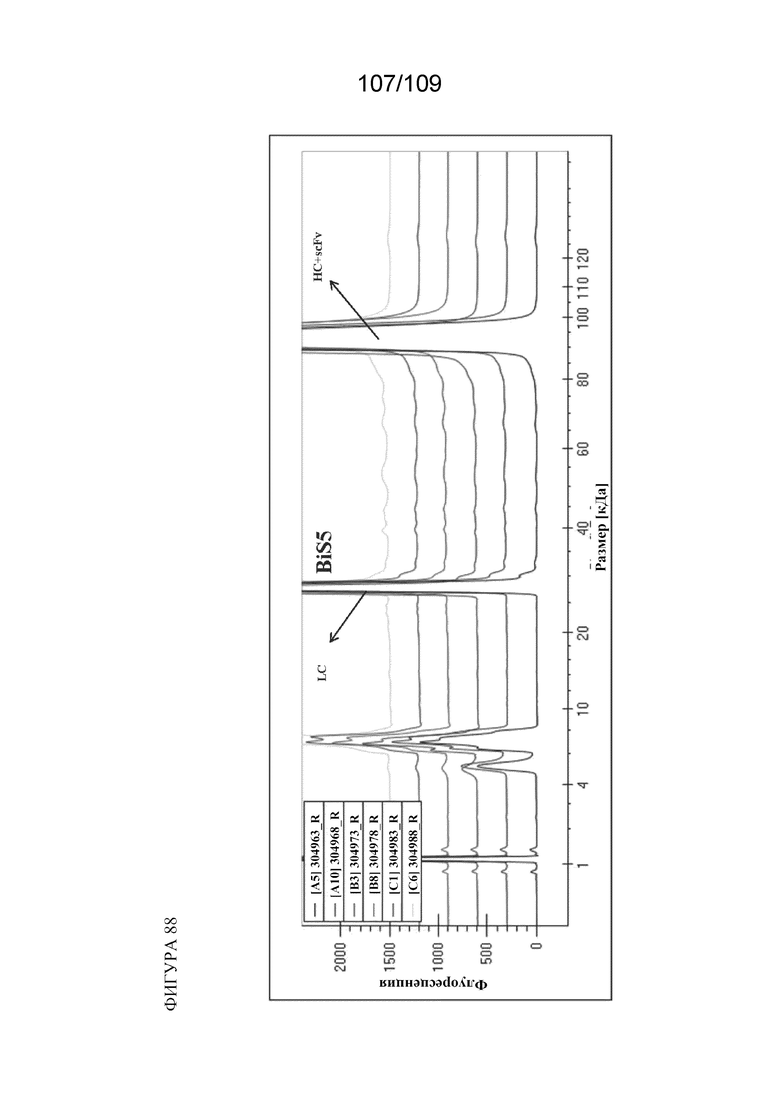
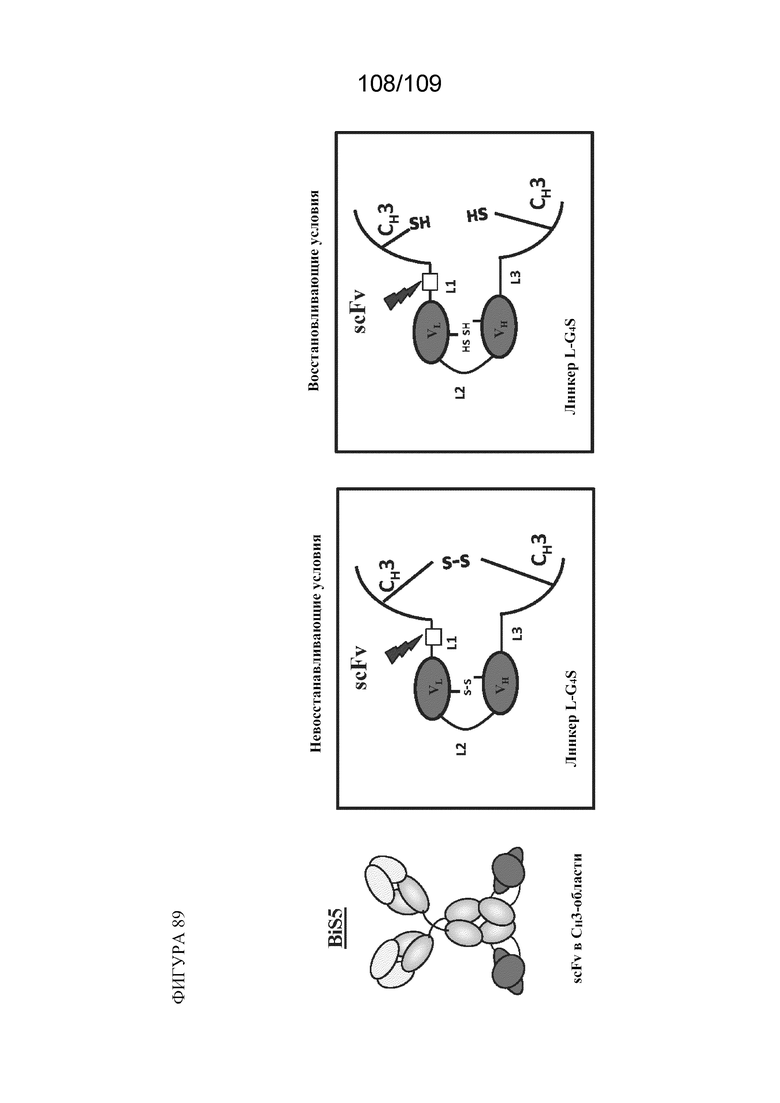
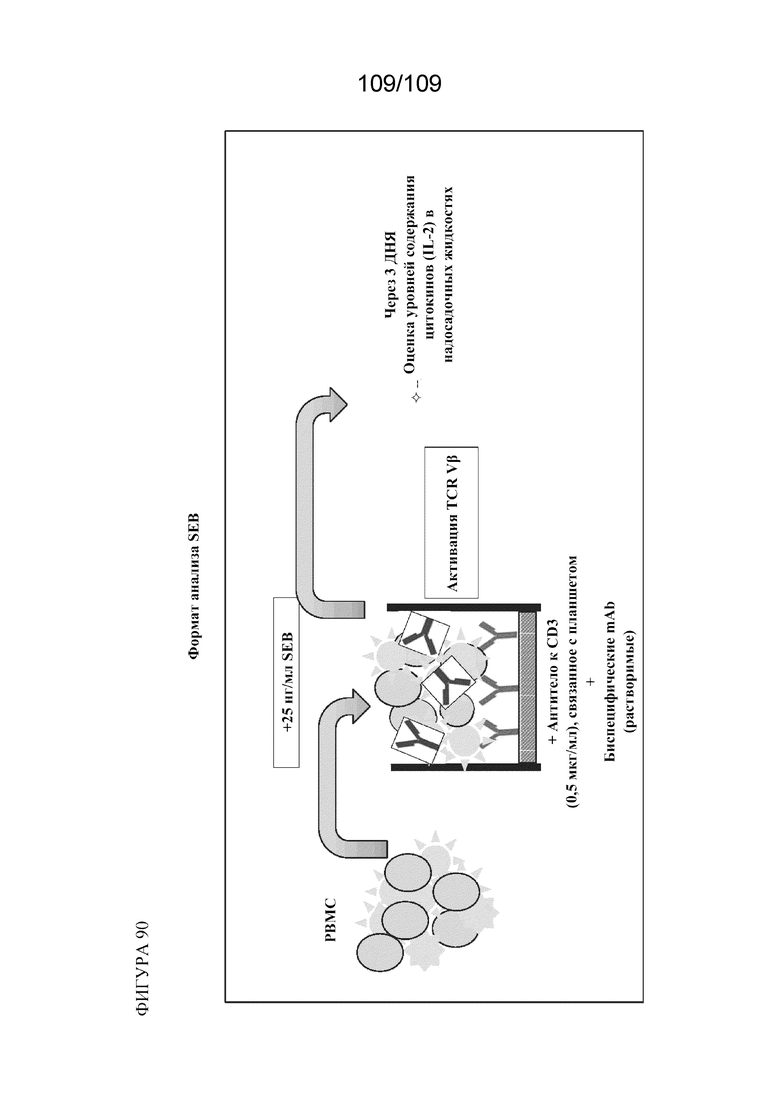
Группа изобретений относится к биотехнологии и иммуноонкологии, в частности к биспецифическому связывающему белку, который связывается с PD-1 и CTLA-4. Белок содержит первую тяжелую цепь, включающую CDR1 - GFTFSDYGMH, CDR2 - YISSGSYTIYSADSVKG, CDR3 - RAPNSFYEYYFDY; первую легкую цепь, включающую CDR1 - SASSKHTNLYWSRHMYWY, CDR2 - LTSNRAT, CDR3 - QQWSSNP; вторую тяжелую цепь, включающую CDR1 - GFTFSSYGMH, CDR2 - VIWYDGSNKYYADSVKG, CDR3 - DPRGATLYYYYYGMDV; и вторую легкую цепь, включающую CDR1 - RASQSINSYLD, CDR2 - AASSLQS, CDR3 - QQYYSTP. Также предложены композиция, содержащая такой белок, молекула нуклеиновой кислоты, кодирующая белок, клетка-хозяин для его экспрессии, способ лечения или предупреждения рака у субъекта и способ усиления иммунного ответа у субъекта, имеющего рак или подверженного риску его развития. Изобретение позволяет получать биспецифические связывающие белки с улучшенными связывающими свойствами относительно исходных моноспецифических связывающих белков, которые нацеливаются на те же молекулы, даже если они используются в комбинации. 7 н. и 5 з.п. ф-лы, 90 ил., 22 табл., 3 пр.
1. Биспецифический связывающий белок, который связывается с PD-1 и CTLA-4, содержащий
первую тяжелую цепь, которая содержит определяющие комплементарность области (CDR) CDR1, CDR2 и CDR3, где CDR1 первой тяжелой цепи содержит аминокислотную последовательность GFTFSDYGMH, CDR2 первой тяжелой цепи содержит аминокислотную последовательность YISSGSYTIYSADSVKG, CDR3 первой тяжелой цепи содержит аминокислотную последовательность RAPNSFYEYYFDY;
первую легкую цепь, которая содержит определяющие комплементарность области (CDR) CDR1, CDR2 и CDR3, где CDR1 первой легкой цепи содержит аминокислотную последовательность SASSKHTNLYWSRHMYWY, CDR2 первой легкой цепи содержит аминокислотную последовательность LTSNRAT, CDR3 первой легкой цепи содержит аминокислотную последовательность QQWSSNP;
вторую тяжелую цепь, которая содержит определяющие комплементарность области (CDR) CDR1, CDR2 и CDR3, где CDR1 второй тяжелой цепи содержит аминокислотную последовательность GFTFSSYGMH, CDR2 второй тяжелой цепи содержит аминокислотную последовательность VIWYDGSNKYYADSVKG, CDR3 второй тяжелой цепи содержит аминокислотную последовательность DPRGATLYYYYYGMDV; и
вторую легкую цепь, которая содержит определяющие комплементарность области (CDR) CDR1, CDR2 и CDR3, где CDR1 второй легкой цепи содержит аминокислотную последовательность RASQSINSYLD, CDR2 второй легкой цепи содержит аминокислотную последовательность AASSLQS, CDR3 второй легкой цепи содержит аминокислотную последовательность QQYYSTP.
2. Белок по п. 1, содержащий Fc-область, где указанная Fc-область является негликозилированной.
3. Белок по п. 1, содержащий Fc-область, где указанная Fc-область является дегликозилированной.
4. Белок по п. 1, содержащий Fc-область, где указанная Fc-область характеризуется сниженным фукозилированием или является нефукозилированной.
5. Композиция для лечения или предотвращения рака, содержащая белок по п. 1 и фармацевтически приемлемый носитель.
6. Молекула нуклеиновой кислоты, кодирующая белок по п. 1.
7. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 6.
8. Выделенная клетка-хозяин для экспрессии биспецифического связывающего белка, который связывается с PD-1 и CTLA-4, по п. 1, содержащая вектор по п. 7.
9. Способ лечения или предупреждения рака у субъекта, где указанный способ включает введение субъекту белка по п. 1.
10. Способ по п. 9, где рак представляет собой один или более из рака яичника, рака молочной железы, колоректального рака, рака предстательной железы, рака шейки матки, рака матки, рака яичка, рака мочевого пузыря, рака головы и шеи, меланомы, рака поджелудочной железы, почечно-клеточной карциномы и рака легких.
11. Способ усиления иммунного ответа у субъекта, имеющего рак или подверженного риску его развития, где указанный способ включает введение субъекту белка по п. 1.
12. Молекула нуклеиновой кислоты по п. 6, где указанная нуклеотидная последовательность, кодирующая белок по п. 1, содержит нуклеотидную последовательность, кодирующую первую тяжелую цепь, содержащую нуклеотидную последовательность под SEQ ID NO:10; нуклеотидную последовательность, кодирующую первую легкую цепь, содержащую нуклеотидную последовательность под SEQ ID NO:8; нуклеотидную последовательность, кодирующую вторую тяжелую цепь, содержащую нуклеотидную последовательность под SEQ ID NO:13; и нуклеотидную последовательность, кодирующую вторую легкую цепь, содержащую нуклеотидную последовательность под SEQ ID NO:11.
| CN 104974253 A, 14.10.2015 | |||
| WO 2013164694 A1, 07.11.2013 | |||
| CN 104987421 A, 21.10.2015 | |||
| US 2014348839 A1, 27.11.2014 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА К БЕЛКУ ПРОГРАММИРУЕМОЙ СМЕРТИ 1 (PD-1) И СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТИ-PD-1-АНТИТЕЛ САМОСТОЯТЕЛЬНО ИЛИ В КОМБИНАЦИИ С ДРУГИМИ ИММУНОТЕРАПЕВТИЧЕСКИМИ СРЕДСТВАМИ | 2006 |
|
RU2494107C2 |
| ОДНОЦЕПОЧЕЧНЫЕ МУЛЬТИВАЛЕНТНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ЭФФЕКТОРНОЙ ФУНКЦИЕЙ | 2007 |
|
RU2487888C2 |

