ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Данная патентная заявка претендует на приоритет предварительной патентной заявки США № 61/622600, поданной 11 апреля 2012, которая включена в данный документ во всей ее полноте посредством ссылки. Настоящее изобретение было создано при поддержке Правительства с № проекта ZIA BC 011417 Национальным институтом здоровья, Национальным институтом онкологии. Правительство имеет определенные права на данное изобретение.
ВКЛЮЧЕНИЕ ПОСРЕДСТВОМ ССЫЛКИ МАТЕРИАЛА, ПРЕДСТАВЛЕННОГО В ЭЛЕКТРОННОМ ВИДЕ
[0002] Включенный в данный документ в полном объеме посредством ссылки читаемый на компьютере список нуклеотидных/аминокислотных последовательностей, представленный одновременно с данной заявкой и определенный следующим образом: один файл размером 42589 байт ASCII (Text) с названием "712361_ST25.TXT", созданный 14 марта 2013 г.
УРОВЕНЬ ТЕХНИКИ
[0003] Множественная миелома (ММ) представляет собой злокачественное заболевание, которое характеризуется накоплением клональных плазматических клеток (см., например, Palumbo et al., New England J. Med., 364(11): 1046-1060 (2011), и Lonial et al., Clinical Cancer Res., 17(6): 1264-1277 (2011)). Современные подходы к лечению ММ часто вызывают ремиссию, но почти все пациенты в конечном итоге страдают от рецидива и умирают (см., например, Lonial et al., выше, и Rajkumar, Nature Rev. Clinical Oncol., 8(8): 479-491 (2011)). Было показано, что аллогенная трансплантация гемопоэтических стволовых клеток вызывает иммуно-опосредованное удаление клеток миеломы; тем не менее, токсичность этого подхода является высокой, и некоторые пациенты излечиваются (см., например, Lonial et al., выше, и Salit et al., Clin. Lymphoma, Myeloma, and Leukemia, 11(3): 247-252 (2011)). В настоящее время не существует клинически эффективных FDA-утвержденных способов лечения ММ моноклональными антителами или аутологичными Т-клетками (см., например, Richardson et al., British J. Haematology, 154(6): 745-754 (2011), и Yi, Cancer Journal, 15(6): 502-510 (2009)).
[0004] Адоптивный перенос Т-клеток, генетически модифицированных для распознавания связанных со злокачественностью антигенов, выглядит обещающим в качестве нового подхода к лечению рака (см., например, Morgan et al., Science, 314(5796): 126-129 (2006); Brenner et al., Current Opinion in Immunology, 22(2): 251-257 (2010); Rosenberg et al., Nature Reviews Cancer, 8(4): 299-308 (2008), Kershaw et al., Nature Reviews Immunology, 5(12): 928-940 (2005); и Pule et al., Nature Medicine, 14(11): 1264-1270 (2008)). Т-клетки могут быть генетически модифицированы для экспрессии химерных антигенных рецепторов (CAR, chimeric antigen receptor), которые являются гибридными белками, состоящими из фрагмента распознавания антигена и доменов Т-клеточной активации (см., например, Kershaw et al., выше, Eshhar et al., Proc. Natl. Acad. Sci. USA, 90(2): 720-724 (1993), и Sadelain et al., Curr. Opin. Immunol., 21(2): 215-223 (2009)).
[0005] Для злокачественных образований В-клеточных линий был достигнут значительный прогресс в разработке адоптивных Т-клеточных подходов, которые используют анти-CD19-CAR (см., например, Jensen et al., Biology of Blood and Marrow Transplantation, 16: 1245-1256 (2010); Kochenderfer et al., Blood, 116(20): 4099-4102 (2010); Porter et al., The New England Journal of Medicine, 365(8): 725-733 (2011); Savoldo et al., Journal of Clinical Investigation, 121(5): 1822-1826 (2011), Cooper et al., Blood, 101(4): 1637-1644 (2003); Brentjens et al., Nature Medicine, 9(3): 279-286 (2003); и Kalos et al., Science Translational Medicine, 3(95): 95ra73 (2011)). Адоптивно перенесенные Т-клетки, трансдуцированные анти-CD19-CAR, вылечили лейкемию и лимфому у мышей (см., например, Cheadle et al., Journal of Immunology, 184(4): 1885-1896 (2010); Brentjens et al., Clinical Cancer Research, 13(18 Pt 1): 5426-5435 (2007); и Kochenderfer et al., Blood, 116(19): 3875-3886 (2010)). В ранних клинических испытаниях адоптивно перенесенные Т-клетки, трансдуцированные анти-CD19-CAR, удаляли нормальные и злокачественные В-клетки у пациентов с лейкемией и лимфомой (см, например, Kochenderfer et l., Blood, 116(20): 4099-4102 (2010); Porter et al., supra, Brentjens et al., Blood, 118(18): 4817-4828 (2011); и Kochenderfer et al., Blood, December 8, 2011 (электронная публикация перед печатью (2012).). Тем не менее, CD19 только в редких случаях экспрессируется на злокачественных плазматических клетках множественной миеломы (см., например, Gupta et al., Amer. J. Clin. Pathology, 132(5): 728-732 (2009); и Lin et al., Amer. J. Clin. Pathology, 121(4): 482-488 (2004)).
[0006] Таким образом, существует потребность в композициях, которые могут быть использованы в способах лечения множественной миеломы. Это изобретение предусматривает такие композиции и способы.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Данное изобретение относится к выделенной или очищенной нуклеиновокислотной последовательности, кодирующей химерный рецептор антигена (CAR), где CAR включает фрагмент распознавания антигена и фрагмент Т-клеточной активации, и где фрагмент распознавания антигена направлен против антигена созревания В-клеток (ВСМА).
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0008] Фиг. 1А и 1В представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие характер экспрессии ВСМА в различных типах человеческих клеток, определенный с помощью количественной ПЦР. Результаты выражены как число копий кДНК ВСМА на 105 копий кДНК актина.
[0009] Фиг. 2А-2L представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что экспрессия ВСМА на клеточной поверхности была обнаружена на нескольких клеточных линиях миеломы, но не на других типах клеток, как описано в примере 1. Во всех графиках сплошная линия представляет окрашивание анти-ВСМА-антителами, а пунктирная линия представляет окрашивание контрольными антителами, подходящими по изотипу. На всех графиках клетки гейтировали по живым клеткам.
[0010] Фиг. 3А представляет собой схему, на которой изображена нуклеиновокислотная конструкция, кодирующая анти-ВСМА-CAR. В направлении с N-конца к С-концу анти-ВСМА-CAR включает анти-ВСМА scFv, шарнирную и трансмембранную области молекулы CD8α, цитоплазматическую часть молекулы CD28 и цитоплазматическую часть молекулы CD3ζ.
[0011] Фиг. 3B-3D представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что анти-bcma1-CAR, анти-bcma2-CAR и SP6-CAR (описаны в примере 2) экспрессируются на поверхности Т-клеток. Минимальное анти-Fab-окрашивание произошло на нетрансдуцированных (UT, untransduced) клетках. На всех графиках клетки гейтировали по CD3+-лимфоцитам. Цифры на графиках являются процентами клеток в каждом квадранте.
[0012] Фиг. 4A-4C представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что Т-клетки, экспрессирующие анти-ВСМА-CAR, вызывают дегрануляцию Т-клеток ВСМА-специфическим образом, как описано в примере 3. На графиках клетки гейтировали по живым CD3+-лимфоцитам. Цифры на графиках являются процентами клеток в каждом квадранте.
[0013] Фиг. 5A-5D представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что Т-клетки, экспрессирующие анти-ВСМА-CAR, вызывают дегрануляцию Т-клеток ВСМА-специфическим образом, как описано в примере 3. На графиках клетки гейтировали по живым CD3+-лимфоцитам. Цифры на графиках являются процентами клеток в каждом квадранте.
[0014] Фиг. 6А-6С представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что Т-клетки, экспрессирующие анти-ВСМА-CAR, продуцируют цитокины IFNγ, IL-2 и TNF ВСМА-специфическим образом, как описано в примере 3. На графиках клетки гейтировали по живым CD3+-лимфоцитам. Цифры на графиках являются процентами клеток в каждом квадранте.
[0015] Фиг. 7А представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что Т-клетки, экспрессирующие анти-bcma2-CAR, специфически пролиферируют в ответ на ВСМА. Фиг. 6В представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что Т-клетки, экспрессирующие SP6-CAR, не пролиферируют специфически в ответ на ВСМА.
[0016] Фиг. 7С и 7D представляют собой графики, на которых изображены экспериментальные данные, иллюстрирующие, что Т-клетки от донора А, экспрессирующие анти-bcma2-CAR, специфически убивают клеточные линии множественной миеломы H929 (фиг. 6С) и RPMI8226 (фиг. 6D) в четырехчасовом анализе цитотоксичности при различных соотношениях эффектор: клетка-мишень. Т-клетки, трансдуцированные отрицательным контролем SP6-CAR, индуцируют гораздо более низкие уровни цитотоксичности во всех соотношениях эффектор: мишень. Для всех соотношений эффектор: мишень была определена цитотоксичность в двух повторах, и результаты представлены в виде среднего +/- стандартная ошибка среднего.
[0017] Фиг. 8А представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что ВСМА экспрессируется на поверхности первичных клеток множественной миеломы костного мозга от пациента с миеломой 3, как описано в примере 5. На графике клетки гейтировали по CD38высокий CD56+-плазматическим клеткам, которые составляют до 40% клеток костного мозга.
[0018] Фиг. 8В представляет собой график, на котором изображены экспериментальные данные, иллюстрирующие, что аллогенные Т-клетки от донора C, трансдуцированные анти-bcma2-CAR, продуцируют IFNγ после совместного культивирования с необработанными клетками костного мозга от пациента с миеломой 3, как описано в примере 5. Фиг. 7В также иллюстрирует, что Т-клетки от одного и того же аллогенного донора, экспрессирующие анти-bcma2-CAR, продуцируют гораздо меньше IFNγ, когда они культивированы с мононуклеарными клетками периферической крови (РВМС) от пациента с миеломой 3. Кроме того, Т-клетки от донора С, экспрессирующие SP6-CAR, специфически не распознают костный мозг от пациента с миеломой 3.
[0019] Фиг. 8C представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что плазмоцитома, резецированная у пациента с миеломой 1, состоит на 93% из плазматических клеток, и эти первичные плазматические клетки экспрессируют ВСМА, что показано с помощью проточной цитометрии на ВСМА (сплошная линия) и путем окрашивания изотипическим контролем (пунктирная линия). На всех графиках клетки гейтировали по плазматическим клеткам.
[0020] Фиг. 8D представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что Т-клетки от пациента с миеломой 1, экспрессирующие анти-bcma2-CAR, продуцируют IFNγ специфически в ответ на аутологичные клетки плазмоцитомы.
[0021] Фиг. 8E представляет собой график, показывающий экспериментальные данные, иллюстрирующие, что Т-клетки от пациента с миеломой 1, экспрессирующие анти-bcma2-CAR, специфически убивают аутологичные клетки плазмоцитомы при низком соотношении эффектор: мишень. Напротив, Т-клетки от пациента с миеломой 1, экспрессирующие SP6-CAR, демонстрируют низкие уровни цитотоксичности против аутологичных клеток плазмоцитомы. Для всех соотношений эффектор: мишень была определена цитотоксичность в двух повторах, и результаты представлены в виде среднего +/- стандартная ошибка среднего.
[0022] Фиг. 9А представляет собой график, который изображает экспериментальные данные, иллюстрирующие, что Т-клетки, трансдуцированные анти-bcma2-CAR, могут разрушать верифицированные опухоли множественной миеломы у мышей. Фиг. 9В представляет собой график, который изображает выживание мышей с опухолями, получавших Т-клетки, экспрессирующие анти-bcma2-CAR, по сравнению с контрольной группой.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0023] Данное изобретение относится к выделенной или очищенной нуклеиновокислотной последовательности, кодирующей химерный рецептор антигена (CAR), где CAR содержит фрагмент распознавания антигена и фрагмент Т-клеточной активации. Химерный рецептор антигена (CAR) является искусственно созданным гибридным белком или полипептидом, содержащим антигенсвязывающий домен антитела (например, одноцепочечный вариабельный фрагмент (scFv)), связанный с Т-клеточным сигналингом или доменами Т-клеточной активации. CAR имеют возможность перенаправлять Т-клеточную специфичность и реактивность на выбранную мишень MHC-неограниченным образом, используя антигенсвязывающие свойства моноклональных антител. MHC-неограниченное распознавание антигена дает Т-клеткам, экспрессирующим CAR, способность распознавать антиген независимо от процессинга антигена, таким образом минуя главный механизм избегания опухоли. Более того, при экспрессии в Т-клетках CAR преимущественно не димеризуются с альфа- и бета-цепями эндогенного Т-клеточного рецептора (TCR).
[0024] Понятие "нуклеиновокислотной последовательности" охватывает полимеры ДНК или РНК, т.е. полинуклеотиды, которые могут быть одноцепочечными или двуцепочечными, и которые могут содержать неприродные или измененные нуклеотиды. Термины "нуклеиновая кислота" и "полинуклеотид", используемые в данном описании, относятся к полимерной форме нуклеотидов любой длины, рибонуклеотидов (РНК) или дезоксирибонуклеотидов (ДНК). Эти термины относятся к первичной структуре молекулы, и, таким образом, включают дву- и одноцепочечную ДНК, а также дву- и одноцепочечную РНК. Термины включают, в качестве эквивалентов, аналоги РНК или ДНК, изготовленные из нуклеотидных аналогов и модифицированных полинуклеотидов, таких как, но не ограничиваясь ими, метилированные и/или кэппированные полинуклеотиды.
[0025] Под "выделенной" подразумевается удаление нуклеиновой кислоты из ее природной среды. Под "очищенной" подразумевается, что данная нуклеиновая кислота, была ли она выделена из природной среды (в том числе геномная ДНК и мРНК) или синтезирована (в том числе кДНК) и/или амплифицирована в лабораторных условиях, имеет повышенную чистоту, где "чистота" является относительным термином, а не "абсолютной чистотой". Тем не менее, следует понимать, что нуклеиновые кислоты и белки могут быть собраны в состав с разбавителями или адъювантами и по-прежнему для практических целей быть выделены. Например, нуклеиновые кислоты, как правило, смешивают с приемлемым носителем или разбавителем, когда используют их для введения в клетки.
[0026] Нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит фрагмент распознавания антигена, направленный против антигена созревания В-клеток (ВСМА, также известного как CD269). ВСМА является членом суперсемейства рецептора фактора некроза опухоли (см., например, Thompson et al., J. Exp. Medicine, 192(1): 129-135 (2000), и Mackay et al., Annu. Rev. Immunol., 21: 231-264 (2003)). ВСМА связывает фактор В-клеточной активации (BAFF) и лиганд, индуцирующий пролиферацию (APRIL) (см., например, Mackay et al., см. выше, и Kalled et al., Immunological Reviews, 204: 43-54 (2005)). Сообщалось, что среди доброкачественных клеток ВСМА экспрессируется в основном в плазматических клетках и субпопуляции зрелых В-клеток (см., например, Laabi et al., EMBO J., 11(11): 3897-3904 (1992); Laabi et al., Nucleic Acids Res., 22(7): 1147-1154 (1994); Kalled et al., выше; O'Connor et al., J. Exp. Medicine, 199(1): 91-97 (2004); и Ng et al., J. Immunol., 173(2): 807-817 (2004)). Мыши, дефицитные по ВСМА, здоровы и имеют нормальное число В-клеток, но выживание долгоживущих плазматических клеток нарушается (см., например, O'Connor et al, выше; Xu et al., Mol. Cell. Biol., 21(12): 4067-4074 (2001); и Schiemann et al., Science, 293(5537): 2111-2114 (2001)). РНК ВСМА была обнаружена повсеместно в клетках множественной миеломы, а белок ВСМА был обнаружен на поверхности плазматических клеток от пациентов с множественной миеломой несколькими исследователями (см., например, Novak et al., Blood, 103(2): 689-694 (2004); Neri et al., Clinical Cancer Research, 13(19): 5903-5909 (2007); Bellucci et al., Blood, 105(10): 3945-3950 (2005); и Moreaux et al., Blood, 103(8): 3148-3157 (2004)).
[0027] Нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит фрагмент распознавания антигена, содержащий моноклональное антитело, направленное против ВСМА, или его антигенсвязывающую часть. Термин "моноклональные антитела", используемый в данном документе, относится к антителам, которые продуцируются одним клоном В-клеток и связываются с одним и тем же эпитопом. Напротив, "поликлональные антитела" относятся к популяции антител, которые продуцируются различными В-клетками и связываются с различными эпитопами одного и того же антигена. Фрагмент распознавания антигена в CAR, кодируемого нуклеиновокислотной последовательностью согласно изобретению, может быть целым антителом или фрагментом антитела. Целое антитело, как правило, состоит из четырех полипептидов: двух идентичных копий полипептида тяжелой (H) цепи и двух идентичных копий полипептида легкой (L) цепи. Каждая из тяжелых цепей содержит одну N-концевую вариабельную (VH) область и три C-концевых константных (CH1, CH2 и СН3) области, а каждая легкая цепь содержит одну N-концевую вариабельную (VL) область и одну C-концевую константную (CL) область. Вариабельные области каждой пары легкой и тяжелой цепей образуют сайт связывания антигена с антителом. Области VH и VL имеют одинаковую общую структуру, при этом каждая область содержит четыре каркасные области, последовательности которых относительно консервативны. Каркасные области соединены тремя определяющими комплементарность областями (CDR). Три CDR, известные как CDR1, CDR2 и CDR3, образуют "гипервариабельную область" антитела, которая отвечает за связывание антигена.
[0028] Термины "фрагмент антитела", "функциональный фрагмент антитела" и "антигенсвязывающий фрагмент" используются в данном документе взаимозаменяемо для обозначения одного или более чем одного фрагмента или части антитела, которые сохраняют способность специфически связываться с антигеном (см., в целом, Holliger et al., Nat. Biotech., 23(9): 1126-1129 (2005)). Фрагмент распознавания антигена в CAR, кодируемый нуклеиновокислотной последовательностью согласно изобретению, может содержать любой фрагмент ВСМА-связывающего антитела. Желательно, чтобы фрагмент антитела содержал, например, одну или более чем одну CDR, вариабельную область (или ее части), константную область (или ее части) или их комбинации. Примеры фрагментов антител включают, но не ограничиваясь ими, (i) Fab-фрагмент, который представляет собой моновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; (ii) F(ab')2-фрагмент, который представляет собой бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) Fv-фрагмент, состоящий из VL- и VH-доменов одного плеча антитела; (iv) одноцепочечный Fv (scFv), который представляет собой одновалентную молекулу, состоящую из двух доменов Fv-фрагмента (т.е. VL и VH), соединенных синтетическим линкером, который позволяет синтезировать два домена в виде одной полипептидной цепи (см., например, Bird et al., Science, 242: 423-426 (1988); Huston et al., Proc. Natl. Acad. Sci. USA, 85: 5879-5883 (1988); и Osbourn et al., Nat. Biotechnol., 16: 778 (1998)); и (v) двойное антитело, представляющее собой димер полипептидных цепей, где каждая полипептидная цепь содержит VH, соединенный с VL пептидным линкером, который является слишком коротким, чтобы позволить спаривание VH и VL на одной и той же полипептидной цепи, тем самым обеспечивая спаривание комплементарных доменов различных VH-VL полипептидных цепей для образования димерной молекулы, имеющей два функциональных антигенсвязывающих участка. Фрагменты антител известны в данной области и описаны более подробно, например, в публикации патентной заявки США 2009/0093024 A1. В предпочтительном воплощении фрагмент распознавания антигена в CAR, кодируемый нуклеиновокислотной последовательностью согласно изобретению, содержит одноцепочечный анти-ВСМА-Fv (scFv).
[0029] Антигенсвязывающая часть или фрагмент моноклонального антитела может иметь любой размер при условии, что эта часть связывается с ВСМА. В этом отношении антигенсвязывающая часть или фрагмент моноклонального антитела, направленного против ВСМА (также называемого здесь "моноклональным анти-ВСМА-антителом"), желательно, содержит от примерно 5 до 18 аминокислот (например, примерно 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, или в диапазоне, определенном любыми двумя из указанных выше значений).
[0030] В одном воплощении нуклеиновокислотная последовательность согласно изобретению кодирует фрагмент распознавания антигена, который содержит вариабельную область моноклонального анти-ВСМА-антитела. В этом отношении фрагмент распознавания антигена содержит вариабельную область легкой цепи, вариабельную область тяжелой цепи, или и вариабельную область легкой цепи, и вариабельную область тяжелой цепи моноклонального анти-ВСМА-антитела. Предпочтительно, фрагмент распознавание антигена в CAR, кодируемый нуклеиновокислотной последовательностью согласно изобретению, включает вариабельную область легкой цепи и вариабельную область тяжелой цепи моноклонального анти-ВСМА-антитела. Аминокислотные последовательности тяжелой и легкой цепей моноклонального антитела, которое связывается с ВСМА, раскрыты, например, в публикации международной патентной заявки WO 2010/104949.
[0031] В другом воплощении нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который включает сигнальную последовательность. Сигнальная последовательность может быть расположена на амино-конце фрагмента распознавания антигена (например, вариабельная область анти-ВСМА-антитела). Сигнальная последовательность может содержать любую подходящую сигнальную последовательность. В одном воплощении сигнальная последовательность представляет собой последовательность рецептора человеческого гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) или сигнальную последовательность CD8α.
[0032] В другом воплощении CAR содержит шарнирную последовательность. Специалисту в данной области будет понятно, что шарнирная последовательность является короткой последовательностью аминокислот, которая облегчает гибкость антитела (см., например, Woof et al., Nat. Rev. Immunol., 4(2): 89-99 (2004)). Шарнирная последовательность может быть расположена между фрагментом распознавания антигена (например, анти-ВСМА-scFv) и фрагментом Т-клеточной активации. Шарнирная последовательность может быть любой подходящей последовательностью, полученной из любой подходящей молекулы. В одном воплощении, например, шарнирная последовательность получена из молекулы человеческого CD8α или молекулы CD28.
[0033] Нуклеиновокислотная последовательность согласно изобретению кодирует CAR, содержащий фрагмент Т-клеточной активации. Фрагмент Т-клеточной активации может быть любым подходящим фрагментом, полученным из любой подходящей молекулы. В одном воплощении, например, фрагмент Т-клеточной активации содержит трансмембранный домен. Трансмембранный домен может быть любым трансмембранным доменом, полученным из любой молекулы, известной в данной области. Например, трансмембранный домен может быть получен из молекулы CD8α или из молекулы CD28. CD8 является трансмембранным гликопротеином, который выступает в качестве корецептора Т-клеточного рецептора (TCR) и экспрессируется в основном на поверхности цитотоксических Т-клеток. Наиболее распространенная форма CD8 существует в виде димера, состоящего из цепей CD8α и CD8β. CD28 экспрессируется на Т-клетках и обеспечивает костимулирующие сигналы, необходимые для активации Т-клеток. CD28 является рецептором для CD80 (В7.1) и CD86 (B7.2). В предпочтительном воплощении CD8α и CD28 являются человеческими.
[0034] В дополнение к трансмембранному домену фрагмент Т-клеточной активации также содержит внутриклеточный (т.е. цитоплазматический) домен сигналинга Т-клеток. Домен межклеточного сигналинга Т-клеток может быть получен из молекулы CD28, молекулы CD3 дзета (ζ) или их модифицированных версий, из человеческой гамма-цепи Fc-рецептора (FcRγ), молекулы CD27, молекулы OX40, молекулы 4-1BB или других молекул внутриклеточного сигналинга, известных в данной области. Как обсуждалось выше, CD28 представляет собой Т-клеточный маркер, важный для костимуляции Т-клеток. CD3ζ ассоциирует с TCR, чтобы сформировать сигнал, и содержит иммунорецепторные активирующие тирозинсодержащие повторы (ITAM). 4-1ВВ, также известный как CD137, передает мощный костимулирующий сигнал на Т-клетки, усиливая дифференцировку и повышая долгосрочное выживание Т-лимфоцитов. В предпочтительном воплощении CD28, CD3 дзета, 4-1ВВ, OX40 и CD27 являются человеческими.
[0035] Домен Т-клеточной активации CAR, кодируемый нуклеиновокислотной последовательностью согласно изобретению, может содержать любой из вышеупомянутых трансмембранных доменов и любой один или более чем один из вышеупомянутых доменов межклеточного сигналинга Т-клеток в любой комбинации. Например, нуклеиновокислотная последовательность согласно изобретению может кодировать CAR, содержащий трансмембранный домен CD28 и внутриклеточные домены Т-клеточного сигналинга CD28 и CD3 дзета. Альтернативно, например, нуклеиновокислотная последовательность согласно изобретению может кодировать CAR, содержащий трансмембранный домен CD8α и внутриклеточные домены Т-клеточного сигналинга CD28, CD3 дзета, гамма-цепи Fc-рецептора (FcRγ) и/или4-1ВВ.
[0036] В одном воплощении нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит в направлении от 5' к 3' сигнальную последовательность рецептора гранулоцитарно-макрофагального колониестимулирующего фактора (рецептора GM-CSF), анти-ВСМА-scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический домен Т-клеточного сигналинга молекулы человеческого CD28 и домен Т-клеточного сигналинга молекулы человеческого CD3ζ. В другом воплощении нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит в направлении от 5' к 3' сигнальную последовательность человеческого CD8α, анти-ВСМА-scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический домен T-клеточного сигналинга молекулы человеческого CD28 и домен Т-клеточного сигналинга молекулы человеческого CD3ζ. В другом воплощении нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит в направлении от 5' к 3' сигнальную последовательность человеческого CD8α, анти-ВСМА-scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматическую область Т-клеточного сигналинга человеческой молекулы 4-1ВВ и/или цитоплазматическую область Т-клеточного сигналинга молекулы человеческого OX40, и домен Т-клеточного сигналинга молекулы человеческого CD3ζ. Например, нуклеиновокислотная последовательность согласно изобретению содержит или состоит из нуклеиновокислотной последовательности SEQ ID № 1, SEQ ID № 2 или SEQ ID № 3.
[0037] Изобретение также относится к выделенному или очищенному химерному рецептору антигена (CAR), кодируемому нуклеиновокислотной последовательностью согласно изобретению.
[0038] Нуклеиновокислотная последовательность согласно изобретению может кодировать CAR любой длины, т.е. CAR может содержать любое число аминокислот, при условии, что CAR сохраняет свою биологическую активность, например, способность специфически связываться с антигеном, обнаруживать больные клетки у млекопитающих или лечить или предотвращать заболевание у млекопитающего и т.д. Например, CAR может содержать 50 или более (например, 60 или более, 100 или более, или 500 или более аминокислот), но меньше 1000 (например, 900 или менее, 800 или менее, 700 или менее, или 600 или менее) аминокислот. Предпочтительно, CAR имеет от примерно 50 до примерно 700 аминокислот (например, примерно 70, примерно 80, примерно 90, примерно 150, примерно 200, примерно 300, примерно 400, примерно 550, или примерно 650 аминокислот), от примерно 100 до примерно 500 аминокислот (например, примерно 125, примерно 175, примерно 225, примерно 250, примерно 275, примерно 325, примерно 350, примерно 375, примерно 425, примерно 450, или примерно 475 аминокислот), или в диапазоне, определяемом любыми двумя из указанных выше значений.
[0039] В объем данного изобретения включены нуклеиновокислотные последовательности, которые кодируют функциональные части CAR, описанные в данном документе. Термин "функциональная часть", используемый в отношении к CAR, относится к любой части или фрагменту CAR согласно изобретению, где часть или фрагмент сохраняет биологическую активность того CAR, частью которого он является (родительского CAR). Функциональные части охватывают, например, те части CAR, которые сохраняют способность распознавать клетки-мишени или обнаруживать, излечивать или предотвращать заболевание в подобной степени, в той же степени или в большей степени, чем родительский CAR. В отношении нуклеиновокислотной последовательности, кодирующей родительский CAR, нуклеиновокислотная последовательность, кодирующая функциональную часть CAR, может кодировать белок, содержащий, например, примерно 10%, 25%, 30%, 50%, 68%, 80%, 90%, 95% или более родительского CAR.
[0040] Нуклеиновокислотная последовательность согласно изобретению может кодировать функциональную часть CAR, которая содержит дополнительные аминокислоты на амино- или карбокси-конце этой части или на обоих концах, где дополнительные аминокислоты не найдены в аминокислотной последовательности родительского CAR. Желательно, чтобы дополнительные аминокислоты не мешали биологической функции функциональной части, например, распознаванию клетки-мишени, обнаружению рака, лечению или профилактике рака и т.д. Более предпочтительно, чтобы дополнительные аминокислоты повышали биологическую активность CAR по сравнению с биологической активностью родительского CAR.
[0041] Данное изобретение также относится к нуклеиновокислотным последовательностям, кодирующим функциональные варианты вышеупомянутого CAR. Термин "функциональный вариант", используемый в данном документе, относится к CAR, полипептиду или белку, имеющему существенную или значительную идентичность или сходство последовательности с CAR, закодированным нуклеиновокислотной последовательностью согласно изобретению, при этом функциональный вариант сохраняет биологическую активность CAR, вариантом которого она является. Функциональные варианты охватывают, например, те варианты CAR, описанные в данном документе (родительский CAR), которые сохраняют способность распознавать клетки-мишени в подобной степени, в той же степени или в большей степени, чем родительский CAR. В отношении нуклеиновокислотной последовательности, кодирующей родительский CAR, нуклеиновокислотная последовательность, кодирующая функциональный вариант CAR, может быть, например, примерно на 10% идентична, примерно на 25% идентична, примерно на 30% идентична, примерно на 50% идентична, примерно на 65% идентична, примерно на 80% идентична, примерно на 90% идентична, примерно на 95% идентична или примерно на 99% идентична нуклеиновокислотной последовательности, кодирующей родительский CAR.
[0042] Функциональный вариант может, например, включать аминокислотную последовательность CAR, закодированную нуклеиновокислотной последовательностью согласно изобретению, по меньшей мере с одной консервативной аминокислотной заменой. Фраза "консервативная аминокислотная замена" или "консервативная мутация" относится к замене одной аминокислоты другой аминокислотой с общими свойствами. Функциональным способом определить общие свойства у отдельных аминокислот является анализ нормированных частот аминокислотных замен в соответствующих белках гомологичных организмов (Schulz, G. E. and Schirmer, R. H., Principles of Protein Structure, Springer-Verlag, New York (1979)). Согласно таким анализам могут быть определены группы аминокислот, где аминокислоты в пределах группы заменяются преимущественно друг на друга, и поэтому наиболее похожи друг на друга по их влиянию на общую структуру белка (Schulz, G. E. and Schirmer, R. H., см. выше). Примеры консервативных мутаций включают аминокислотные замены аминокислот внутри подгрупп, описанных выше, например, лизина на аргинин и наоборот таким образом, чтобы положительный заряд мог быть сохранен; глутаминовой кислоты на аспарагиновую кислоту и наоборот таким образом, чтобы отрицательный заряд мог быть сохранен; серина на треонин таким образом, чтобы свободный -OH мог быть сохранен; и глутамина на аспарагин таким образом, чтобы свободный -NH2 мог быть сохранен.
[0043] Альтернативно или дополнительно, функциональные варианты могут включать аминокислотную последовательность родительского CAR по меньшей мере с одной неконсервативной аминокислотной заменой. "Неконсервативные мутации" включают замены аминокислот из различных групп, например, лизина на триптофан или фенилаланина на серин и т.д. В этом случае для неконсервативной аминокислотной замены предпочтительно, чтобы она не мешала или не ингибировала биологическую активность функционального варианта. Неконсервативная аминокислотная замена может повысить биологическую активность функционального варианта, так что биологическая активность функционального варианта увеличивается по сравнению с родительским CAR.
[0044] Нуклеиновокислотная последовательность согласно изобретению может кодировать CAR (в том числе его функциональные части и функциональные варианты), который включает синтетические аминокислоты вместо одной или более чем одной природной аминокислоты. Такие синтетические аминокислоты известны в данной области и включают, например, аминоциклогексанкарбоновую кислоту, норлейцин, α-амино-n-декановую кислоту, гомосерин, S-ацетиламинометилцистеин, транс-3- и транс-4-гидроксипролин, 4-аминофенилаланин, 4-нитрофенилаланин, 4-хлорофенилаланин, 4-карбоксифенилаланин, β-фенилсерингидроксифенилаланин, β-фенилглицин, α-нафтилаланин, циклогексилаланин, циклогексилглицин, индолин-2-карбоновую кислоту, 1,2,3,4-тетрагидроизохинолин-3-карбоновую кислоту, аминомалоновую кислоту, моноамид аминомалоновой кислоты, N'-бензил-N'-метиллизин, N',N'-дибензиллизин, 6-гидроксилизин, орнитин, α-аминоциклопентанкарбоновую кислоту, α-аминоциклогексанкарбоновую кислоту, α-аминоциклогептанкарбоновую кислоту, α-(2-амино-2-норборнан)карбоновую кислоту, α,γ-диаминомасляную кислоту, α,β-диаминопропионовую кислоту, гомофенилаланин и α-трет-бутилглицин.
[0045] Нуклеиновокислотная последовательность согласно изобретению может кодировать CAR (в том числе его функциональные части и функциональные варианты), который гликозилирован, амидирован, карбоксилирован, фосфорилирован, этерифицирован, N-ацилирован, циклизован, например, через дисульфидный мостик, или превращен в кислотно-аддитивную соль и/или, возможно, димеризован, полимеризован или конъюгирован.
[0046] В предпочтительном воплощении нуклеиновокислотная последовательность согласно изобретению кодирует CAR, который содержит или состоит из аминокислотной последовательности SEQ ID № 4, SEQ ID № 5, SEQ ID № 6, SEQ ID № 8, SEQ ID № 9, SEQ ID № 10, SEQ ID № 11 или SEQ ID № 12.
[0047] Нуклеиновокислотная последовательность согласно изобретению может быть получена с использованием способов, известных в данной области. Например, нуклеиновокислотные последовательности, полипептиды и белки могут быть получены рекомбинантным путем с использованием стандартных методик рекомбинантной ДНК (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 3rd ed., Cold Spring Harbor Press, Cold Spring Harbor, NY 2001; и Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates and John Wiley & Sons, NY, 1994). Кроме того, синтетически полученная нуклеиновокислотная последовательность, кодирующая CAR, может быть выделена и/или очищена из источника, такого как растение, бактерия, насекомое или млекопитающее, например, крыса, человек и т.д. Способы выделения и очистки хорошо известны в данной области. Альтернативно, нуклеиновокислотные последовательности, описанные в данном документе, могут быть синтезированы коммерческим образом. В этом отношении нуклеиновокислотная последовательность согласно изобретению может быть синтетической, рекомбинантной, выделенной и/или очищенной.
[0048] Данное изобретение также относится к вектору, содержащему нуклеиновокислотную последовательность, кодирующую CAR согласно изобретению. Вектор может быть, например, плазмидой, космидой, вирусным вектором (например, ретровирусным или аденовирусным) или фагом. Подходящие векторы и способы получения векторов хорошо известны в данной области (например, Sambrook et al., см. выше, и Ausubel et al., см. выше).
[0049] В дополнение к нуклеиновокислотной последовательности согласно изобретению, кодирующей CAR, вектор предпочтительно содержит последовательности, контролирующие экспрессию, такие как промоторы, энхансеры, сигналы полиаденилирования, терминаторы транскрипции, внутренние сайты посадки рибосомы (IRES) и т.п., которые обеспечивают экспрессию нуклеиновокислотной последовательности в клетке-хозяине. Иллюстративные последовательности, контролирующие экспрессию, известны в данной области и описаны, например, в Goeddel, Gene Expression Technology: Methods in Enzymology, Vol. 185, Academic Press, San Diego, Calif. (1990).
[0050] Большое число промоторов из множества различных источников, включая конститутивные, индуцибельные и репрессируемые промоторы, хорошо известны в данной области. Типичными источниками промоторов являются, например, вирусы, млекопитающие, насекомые, растения, дрожжи и бактерии, и подходящие промоторы из этих источников легко доступны или могут быть получены синтетическим путем на основании общедоступных последовательностей, например, из депозитариев, таких как АТСС, а также из других коммерческих или личных источников. Промоторы могут быть однонаправленными (т.е. инициировать транскрипцию в одном направлении) или двунаправленными (т.е. инициировать транскрипцию либо в 3'-, либо в 5'-направлении). Неограничивающие примеры промоторов включают, например, систему бактериальной экспрессии Т7, систему бактериальной экспрессии pBAD (araA), промотор цитомегаловируса (CMV), промотор SV40 и промотор RSV. Индуцируемые промоторы включают, например, систему Tet (патенты США 5464758 и 5814618), индуцируемую систему экдизона (No et al., Proc. Natl. Acad. Sci., 93: 3346-3351 (1996)), систему T-REX™ (Invitrogen, Карлсбад, Калифорния), систему LACSWITCH™ (Stratagene, Сан-Диего, Калифорния) и систему тамоксифен-индуцируемой рекомбиназы Cre-ERT (Indra et al., Nuc. Acid. Res., 27: 4324-4327 (1999); Nuc. Acid. Res., 28: e99 (2000); патент США 7112715; и Kramer & Fussenegger, Methods Mol. Biol., 308: 123-144 (2005)).
[0051] Термин "энхансер", используемый в данном документе, относится к последовательности ДНК, которая повышает транскрипцию, например, нуклеиновокислотной последовательности, с которой она функционально связана. Энхансеры могут быть расположены за много тысяч пар нуклеотидов в стороне от кодирующей области нуклеиновокислотной последовательности и могут опосредовать связывание регуляторных факторов, паттерны метилирования ДНК или изменения в структуре ДНК. Большое число энхансеров из множества различных источников хорошо известны в данной области и доступны в виде или внутри клонированных полинуклеотидов (например, из таких депозитариев как АТСС, а также других коммерческих или личных источников). Большое число полинуклеотидов, содержащих промоторы (например, часто используемый промотор CMV), также содержат энхансерные последовательности. Энхансеры могут быть расположены выше, в пределах или ниже кодирующих последовательностей. Термин "энхансеры Ig" относится к энхансерным элементам, полученным из энхансерных областей, картированных в пределах иммуноглобулинового (Ig) локуса (такие энхансеры включают, например, 5'-энхансеры тяжелой цепи (мю), 5'-энхансеры легкой цепи (каппа), интронные энхансеры каппа и мю и 3'-энхансеры (см. в целом Paul W.E. (ed), Fundamental Immunology, 3rd Edition, Raven Press, New York (1993), pages 353-363; и патент США 5885827).
[0052] Вектор также может содержать "ген селективного маркера". Термин "ген селективного маркера", используемый в данном документе, относится к нуклеиновокислотной последовательности, которая позволяет клеткам экспрессировать нуклеиновокислотную последовательность, чтобы быть специфически выбранными в пользу него или против, в присутствии соответствующего селективного агента. Подходящие селективные маркерные гены известны в данной области и описаны, например, в международных патентных заявках WO 1992/08796 и WO 1994/28143; Wigler et al., Proc. Natl. Acad. Sci. USA, 77: 3567 (1980); O'Hare et al., Proc. Natl. Acad. Sci. USA, 78: 1527 (1981); Mulligan & Berg, Proc. Natl. Acad. Sci. USA, 78: 2072 (1981); Colberre-Garapin et al., J. Mol. Biol., 150: 1 (1981); Santerre et al., Gene, 30: 147 (1984); Kent et al., Science, 237: 901-903 (1987); Wigler et al., Cell, 11: 223 (1977); Szybalska & Szybalski, Proc. Natl. Acad. Sci. USA, 48: 2026 (1962); Lowy et al., Cell, 22: 817 (1980); и патентах США 5122464 и 5770359.
[0053] В некоторых воплощениях вектор является "эписомальным экспрессионным вектором" или "эписомой", которая способна к репликации в клетке-хозяине и сохраняется в качестве внехромосомного сегмента ДНК в клетке-хозяине в присутствии соответствующего селективного давления (см., например, Conese et al., Gene Therapy, 11: 1735-1742 (2004)). Типичные коммерчески доступные эписомальные экспрессионные вектора включают, но не ограничиваясь ими, эписомальные плазмиды, которые используют ядерный антиген вируса Эпштейна-Барр 1 (EBNA1) и сайт начала репликации (oriP) вируса Эпштейна-Барр (EBV). Векторы pREP4, рСЕР4, pREP7 и pcDNA3.1 от Invitrogen (Карлсбад, Калифорния) и pBK-CMV от Stratagene (Ла Джолла, Калифорния) являются неограничивающими примерами эписомального вектора, который использует Т-антиген и сайт начала репликация SV40 вместо EBNA1 и oriP.
[0054] Другие подходящие векторы включают интегрирующие экспрессионные векторы, которые могут случайным образом интегрировать в ДНК клетки-хозяина, или могут включать сайт рекомбинации для включения специфической рекомбинации между экспрессионным вектором и хромосомой клетки-хозяина. Такие интегрирующие экспрессионные векторы могут использовать эндогенные последовательности, контролирующие экспрессию хромосом клетки-хозяина, чтобы осуществить экспрессию желаемого белка. Примеры векторов, которые интегрируют сайт-специфическим образом, включают, например, компоненты системы Flp-In от Invitrogen (Карлсбад, Калифорния) (например, pcDNA™5/FRT), или системы Cre-lox, такие, как могут быть найдены в pExchange-6 Core Vectors от Stratagene (Ла Джолла, Калифорния). Примеры векторов, которые случайным образом интегрируют в хромосомы клетки-хозяина, включают, например, pcDNA3.1 (при введении в отсутствие Т-антигена) от Invitrogen (Карлсбад, Калифорния) и pCI или pFN10A (ACT) FLEXI™ от Promega (Мадисон, Висконсин).
[0055] Также могут использоваться вирусные векторы. Типичные вирусные экспрессионные векторы включают, но не ограничиваясь ими, векторы на основе аденовирусов (например, система Per.C6 на основе аденовируса от Crucell, Inc. (Лейден, Нидерланды)), векторы на основе лентивируса (например, pLP1 на основе лентивируса от Life Technologies (Карлсбад, Калифорния)) и ретровирусные векторы (например, pFB-ERV плюс pCFB-EGSH от Stratagene (La Jolla, CA)). В предпочтительном воплощении вирусный вектор представляет собой лентивирусный вектор.
[0056] Вектор, содержащий нуклеиновую кислоту согласно изобретению, кодирующую CAR, может быть введен в клетку-хозяина, которая способна экспрессировать CAR, закодированный таким образом, включая любую подходящую прокариотическую или эукариотическую клетку. Предпочтительными клетками-хозяевами являются те, которые могут быть легко и надежно выращены, имеют достаточно быстрые темпы роста, имеют хорошо охарактеризованные системы экспрессии и могут быть трансформированы или трансфицированы легко и эффективно.
[0057] Используемый в данном документе термин "клетка-хозяин" относится к любому типу клеток, которые могут содержать экспрессионный вектор. Клетка-хозяин может быть эукариотической клеткой, например, клеткой растения, животного, гриба или водоросли, или может быть прокариотической клеткой, например, бактерией или простейшим. Клетка-хозяин может быть культивированной клеткой или первичной клеткой, т.е. выделенной непосредственно из организма, например, человека. Клетка-хозяин может быть прикрепленной клеткой или суспендированной клеткой, т.е. клеткой, которая растет в суспензии. Подходящие клетки-хозяева известны в данной области и включают, например, клетки DH5α E. coli, клетки яичников китайского хомячка, клетки обезьяны VERO, клетки COS, клетки HEK293 и т.п. Для амплификации или репликации рекомбинантного экспрессионного вектора клетка-хозяин может быть прокариотической клеткой, например, клеткой DH5α. Для получения рекомбинантного CAR клетка-хозяин может быть клеткой млекопитающего. Клетка-хозяин предпочтительно является клеткой человека. Клетка-хозяин может быть клеткой любого типа, может происходить из любого типа ткани и может находиться на любой стадии развития. В одном воплощении клетка-хозяин может быть лимфоцитом периферической крови (PBL), мононуклеарной клеткой периферической крови (РВМС) или натуральным киллером (NK). Предпочтительно клетка-хозяин является натуральным киллером (NK). Более предпочтительно, клетка-хозяин представляет собой Т-клетку. Способы выбора подходящих клеток-хозяев млекопитающих и способы трансформации, культивирования, амплификации, скрининга и очистки клеток известны в данной области.
[0058] Данное изобретение относится к выделенной клетки-хозяину, которая экспрессирует нуклеиновокислотную последовательность согласно изобретению, кодирующую CAR, описанный в данном документе. В одном воплощении клетка-хозяин представляет собой Т-клетку. Т-клеток согласно изобретению может быть любой Т-клеткой, такой как культивированная Т-клетка, например, первичная Т-клетка, или Т-клетка из культивированной Т-клеточной линии, или Т-клеткой, полученной от млекопитающего. Если Т-клетка получена от млекопитающего, то она может быть получена из различных источников, включая, но не ограничиваясь ими, кровь, костный мозг, лимфатические узлы, вилочковую железу или другие ткани или жидкости. Т-клетки также могут быть обогащены или очищены. Т-клетка предпочтительно представляет собой Т-клетку человека (например, выделенную из человека). Т-клетка может находиться на любой стадии развития, включая, но не ограничиваясь ими, CD4+/CD8+ дважды положительную Т-клетку, CD4+ Т-хелперную клетку, например, клетку Th1 и Th2, CD8+ Т-клетку (например, цитотоксическую Т-клетку), клетку, инфильтрирующую опухоль, Т-клетку памяти, наивную Т-клетку и т.п. В одном воплощении Т-клетка представляет собой CD8+ Т-клетку или CD4+ Т-клетку. Т-клеточные линии могут быть получены, например, из Американской коллекции типовых культур (АТСС, Манассас, Вирджиния) и Немецкой коллекции микроорганизмов и клеточных культур (DSMZ), и включают, например, клетки Jurkat (АТСС TIB-152), клетки Sup-T1 (АТСС CRL-1942), клетки RPMI 8402 (DSMZ ACC-290), клетки Karpas 45 (DSMZ ACC-545) и их производные.
[0059] В другом воплощении клетка-хозяин представляет собой натуральный киллер (NK). NK-клетки являются типом цитотоксического лимфоцита, который играет определенную роль во врожденной иммунной системе. NK-клетки определяются как большие гранулярные лимфоциты и представляют собой третий вид клеток, дифференцирующихся из общего лимфоидного предшественника, который также дает В- и Т-лимфоциты (см., например, Immunobiology, 5th ed., Janeway et al., eds., Garland Publishing, New York, NY (2001)). NK-клетки дифференцируются и созревают в костном мозге, лимфатических узлах, селезенке, миндалинах и тимусе. После созревания NK-клетки вступают в кровоток в виде больших лимфоцитов с отдельными цитотоксическими гранулами. NK-клетки способны распознавать и убивать аномальные клетки, такие как, например, некоторые опухолевые клетки и инфицированные вирусом клетки, и, по-видимому, играют важную роль во врожденной иммунной защите от внутриклеточных патогенов. Как описано выше в отношении Т-клеток, NK-клетка может быть любой NK-клеткой, такой как культивированная NK-клетка, например, первичная NK-клетка или NK-клетка из культивируемой NK-клеточной линии, или NK-клеткой, полученной от млекопитающего. Если NK-клетка получена от млекопитающего, то она может быть получена из различных источников, включая, но не ограничиваясь ими, кровь, костный мозг, лимфатические узлы, вилочковую железу или другие ткани или жидкости. NK-клетки также могут быть обогащены или очищены. NK-клетка предпочтительно представляет собой NK-клетку человека (например, выделенную из человека). NK-клеточные линии могут быть получены, например, из Американской коллекции типовых культур (АТСС, Манассас, Вирджиния) и включают, например, клетки NK-92 (АТСС CRL-2407), клетки NK92MI (АТСС CRL-2408), а также их производные.
[0060] Нуклеиновокислотная последовательность согласно изобретению, кодирующая CAR, может быть введена в клетку путем "трансфекции", "трансформации" или "трансдукции". Понятия "трансфекции", "трансформации" или "трансдукции", используемые в данном документе, относятся к введению одного или более чем одного экзогенного полинуклеотида в клетку-хозяина с помощью физических или химических способов. Многие методики трансфекции известны в данной области и включают, например, копреципитацию ДНК фосфатом кальция (см., например, Murray E.J. (ed.), Methods in Molecular Biology, Vol. 7, Gene Transfer and Expression Protocols, Humana Press (1991)); DEAE-декстран; электропорацию; опосредованную катионными липосомами трансфекцию; бомбардировку микрочастицами, облегченную частицами вольфрама (Johnston, Nature, 346: 776-777 (1990)); и копреципитацию ДНК фосфатом стронция (Brash et al., Mol. Cell Biol., 7: 2031-2034 (1987)). Фаговые или вирусные векторы могут быть введены в клетки-хозяева после роста инфекционных частиц в подходящих упаковывающих клетках, многие из которых коммерчески доступны.
[0061] Без связи с конкретной теорией или механизмом, полагают, что, вызывая антигенспецифический ответ против ВСМА, CAR, кодируемые нуклеиновокислотной последовательностью согласно изобретению, вызывают одно или более чем одно из следующих явлений: нацеливание и разрушение BCMA-экспрессирующих раковых клеток, уменьшение или устранение раковых клеток, облегчение инфильтрации мест(а) опухоли иммунными клетками и усиление/удлинение противораковых ответов. Таким образом, изобретение предлагает способ разрушения клеток множественной миеломы, который включает контактирование одной или более чем одной из вышеупомянутых выделенных Т-клеток или натуральных киллеров с популяцией клеток множественной миеломы, которые экспрессируют ВСМА, где CAR образуется и связывается с ВСМА на клетках множественной миеломы, и клетки множественной миеломы разрушаются. Как обсуждалось в данном документе, множественная миелома, также известная как миелома плазматических клеток или болезнь Келлера, представляет собой рак плазматических клеток, которые являются одним из видов белых кровяных клеток, в норме отвечающих за продукцию антител (Raab et al., Lancet, 374: 324-329 (2009)). Множественная миелома поражает 1-4 человека на 100000 человек в год. Заболевание чаще встречается у мужчин, и по неизвестным до сих пор причинам в два раза чаще у афроамериканцев, чем у американцев европеоидной расы. Множественная миелома является наиболее редким общегематологическим злокачественным заболеванием (14%) и составляет 1% от всех случаев рака (Raab et al., см. выше). Лечение множественной миеломы обычно включает химиотерапию в высоких дозах с последующей трансплантацией (аллогенной или аутологичной) гемопоэтических стволовых клеток; тем не менее, высокая частота рецидивов является общим признаком пациентов с множественной миеломой, которые прошли такое лечение. Как обсуждалось выше, ВСМА экспрессируется на высоком уровне клетками множественной миеломы (см., например, Novak et al., выше; Neri et al., выше; Bellucci et al., выше; и Moreaux et al., выше).
[0062] Одна или более чем одна выделенная Т-клетка, экспрессирующая нуклеиновокислотную последовательность согласно изобретению, кодирующую анти-ВСМА CAR, описанный в данном документе, может контактировать с популяцией клеток множественной миеломы, которые экспрессируют ВСМА ex vivo, in vivo или in vitro. "Ex vivo" относится к способам, проводимым в или на клетках или ткани в искусственной среде вне организма с минимальным изменением природных условий. Напротив, термин "in vivo" относится к способу, который проводится в живых организмах в их нормальном, неповрежденном состоянии, в то время как способ "in vitro" проводится с использованием компонентов организма, которые были извлечены из своего обычного биологического окружения. Способ согласно изобретению предпочтительно включает компоненты ex vivo и in vivo. В связи с этим, например, выделенные Т-клетки, описанные выше, могут быть культивированы ex vivo в условиях, подходящих для экспрессии нуклеиновокислотной последовательности согласно изобретению, кодирующей анти-ВСМА-CAR, а затем непосредственно перенесены млекопитающему (предпочтительно, человеку), страдающему от множественной миеломы. Такой способ переноса клеток в данной области называют "адоптивным переносом клеток (АСТ)", где полученные иммунные клетки пассивно переносят новому реципиенту, чтобы передать новому хозяину функциональные возможности иммунных клеток, полученных от донора. Способы адоптивного переноса клеток для лечения различных видов рака, в том числе гематологических раков, таких как миелома, известны в данной области и описаны, например, в Gattinoni et al., Nat. Rev. Immunol., 6(5): 383-393 (2006); June, CH, J. Clin. Invest., 117(6): 1466-76 (2007); Rapoport et al., Blood, 117(3): 788-797 (2011); и Barber et al., Gene Therapy, 18: 509-516 (2011)).
[0063] Данное изобретение также предусматривает способ разрушения клеток лимфомы Ходжкина. Лимфома Ходжкина (ранее известная как болезнь Ходжкина) представляет собой рак иммунной системы, который отличается присутствием типа многоядерных клеток, называемых клетками Рида-Штернберга. Два основных типа лимфомы Ходжкина включают классическую лимфому Ходжкина и узловую лимфому Ходжкина с преобладанием лимфоцитов. Лимфому Ходжкина в настоящее время лечат путем лучевой терапии, химиотерапии или трансплантации гемопоэтических стволовых клеток, выбирая способ лечения в зависимости от возраста и пола пациента и стадии, объема и гистологического подтипа заболевания. Экспрессия ВСМА была обнаружена на поверхности клеток лимфомы Ходжкина (см., например, Chiu et al., Blood, 109(2): 729-739 (2007)).
[0064] Когда Т-клетки или NK-клетки вводят млекопитающему, то эти клетки могут быть аллогенными или аутологичными для этого млекопитающего. В "аутологичных" способах введения клетки (например, кроветворные стволовые клетки или лимфоциты) удаляются из организма млекопитающего, сохраняются (и, возможно, модифицируются) и возвращаются обратно тому же млекопитающему. В "аллогенных" способах введения млекопитающее получает клетки (например, кроветворные стволовые клетки или лимфоциты) от генетически подобного, но не идентичного донора. Предпочтительно клетки являются аутологичными млекопитающему.
[0065] Т-клетки или NK-клетки предпочтительно вводятся человеку в форме композиции, такой как фармацевтическая композиция. Альтернативно, нуклеиновокислотная последовательность согласно изобретению, кодирующая CAR, или вектор, содержащий кодирующую CAR нуклеиновокислотную последовательность, могут быть составлены в композицию, такую как фармацевтическая композиция, и введены человеку. Фармацевтическая композиция согласно изобретению может содержать популяцию Т-клеток или NK клеток, которые экспрессируют CAR согласно изобретению. В дополнение к нуклеиновокислотной последовательности согласно изобретению или к клеткам-хозяевам, которые экспрессируют CAR согласно изобретению, фармацевтическая композиция может содержать другие фармацевтически активные агенты или препараты, такие как химиотерапевтические агенты, например, аспарагиназу, бусульфан, карбоплатин, цисплатин, даунорубицин, доксорубицин, фторурацил, гемцитабин, гидроксимочевину, метотрексат, паклитаксел, ритуксимаб, винбластин, винкристин и т.п. В предпочтительном воплощении фармацевтическая композиция содержит выделенную Т-клетку или NK-клетку, экспрессирующую CAR согласно изобретению, более предпочтительно популяцию Т-клеток или NK-клеток, которые экспрессируют CAR согласно изобретению.
[0066] Т-клетки или NK-клетки согласно изобретению могут быть предоставлены в форме соли, например, фармацевтически приемлемой соли. Подходящие фармацевтически приемлемые кислотно-аддитивные соли включают соли, полученные из минеральных кислот, таких как соляная, бромистоводородная, фосфорная, метафосфорная, азотная и серная кислоты, а также из органических кислот, таких как винная, уксусная, лимонная, яблочная, молочная, фумаровая, бензойная, гликолевая, глюконовая, янтарная и арилсульфокислоты, например, p-толуолсульфокислота.
[0067] Выбор носителя будет определяться частично конкретной согласно изобретению нуклеиновокислотной последовательностью, вектором или клетками-хозяевами, экспрессирующими CAR, а также конкретным способом, используемым для введения нуклеиновокислотной последовательности, вектора или клеток-хозяев, экспрессирующих CAR, согласно изобретению. Соответственно, существует множество приемлемых составов фармацевтической композиции данного изобретения. Например, фармацевтическая композиция может содержать консерванты. Подходящие консерванты могут включать, например, метилпарабен, пропилпарабен, бензоат натрия и хлорид бензалкония. Возможно, может быть использована смесь двух или более консервантов. Консервант или его смеси, как правило, присутствуют в количестве от примерно 0,0001% до примерно 2% от общего веса композиции.
[0068] Кроме того, в композиции могут быть использованы буферные агенты. Подходящие буферные агенты включают, например, лимонную кислоту, цитрат натрия, фосфорную кислоту, фосфат калия и различные другие кислоты и соли. Возможно, может быть использована смесь двух или более буферных агентов. Буферный агент или его смеси, как правило, присутствуют в количестве от примерно 0,001% до примерно 4% от общего веса композиции.
[0069] Способы получения вводимой (например, вводимой парентеральным путем) композиции известны специалистам в данной области и описаны более подробно, например, в Remington: The Science and Practice of Pharmacy, Lippincott Williams & Wilkins; 21st ed. (May 1, 2005).
[0070] Композиция, содержащая согласно изобретению нуклеиновокислотную последовательность, кодирующую CAR, или клетки-хозяева, экспрессирующие CAR, может быть составлена как комплекс включения, такой как циклодекстриновый комплекс, или как липосома. Липосомы могут служить для нацеливания клеток-хозяев (например, Т-клеток или NK-клеток) или нуклеиновокислотной последовательности согласно изобретению на конкретную ткань. Липосомы могут быть также использованы для увеличения периода полужизни нуклеиновокислотной последовательности согласно изобретению. Для получения липосом доступны многие способы, такие как описанные, например, в Szoka et al., Ann. Rev. Biophys. Bioeng., 9: 467 (1980), и в патентах США 4235871, 4501728, 4837028 и 5019369.
[0071] Композиция может использовать систему с отсроченным высвобождением, замедленным высвобождением и пролонгированным высвобождением, так что доставка композиции согласно изобретению происходит до и в течение достаточно времени, чтобы вызвать сенсибилизацию места лечения. Многие типы систем доставки с отсроченным высвобождением доступны и известны специалистам в данной области. С помощью таких систем можно избежать повторных введений композиции, тем самым увеличивая удобство для субъекта и врача, и они могут быть особенно подходящими для определенных воплощений композиции данного изобретения.
[0072] Композиция желательно содержит клетки-хозяева, экспрессирующие нуклеиновокислотную последовательность согласно изобретению, кодирующую CAR, или вектор, содержащий нуклеиновокислотную последовательность согласно изобретению, в количестве, которое является эффективным для лечения или предотвращения множественной миеломы или лимфомы Ходжкина. Используемый в данном документе термин "лечение" относится к получению желаемого фармакологического и/или физиологического эффекта. Предпочтительно, эффект является терапевтическим, т.е. эффект частично или полностью излечивает заболевание и/или неблагоприятный симптом, свойственный заболеванию. Для этого способ согласно изобретению включает введение "терапевтически эффективного количества" композиции, содержащей клетки-хозяева, экспрессирующие нуклеиновокислотную последовательность согласно изобретению, кодирующую CAR, или вектор, содержащий нуклеиновокислотную последовательность согласно изобретению. Термин "терапевтически эффективное количество" относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого терапевтического результата. Терапевтически эффективное количество может варьировать в зависимости от таких факторов, как степень заболевания, возраст, пол и вес человека и способность CAR вызывать желаемый ответ у индивидуума. Например, терапевтически эффективное количество CAR согласно изобретению представляет собой количество, которое связывается с ВСМА на клетках множественной миеломы и разрушает их.
[0073] Альтернативно, фармакологический и/или физиологический эффект может быть профилактическим, т.е. эффект полностью или частично предотвращает заболевание или его симптом. В этом отношении способ согласно изобретению включает введение "профилактически эффективного количества" композиции, содержащей клетки-хозяева, экспрессирующие нуклеиновокислотную последовательность согласно изобретению, кодирующую CAR, или вектор, содержащий нуклеиновокислотную последовательность согласно изобретению, млекопитающему, который предрасположен к множественной миеломе или лимфоме Ходжкина. Понятие "профилактически эффективного количества" относится к количеству, эффективному в дозах и в течение периодов времени, необходимых для достижения желаемого профилактического результата (например, предотвращения возникновения заболевания).
[0074] Типичное число клеток-хозяев, вводимых млекопитающему (например, человеку), может находиться, например, в диапазоне от 1 млн до 100 млрд клеток; тем не менее, в объем данного изобретения входят числа ниже или выше этого примерного диапазона. Например, суточная доза клеток-хозяев согласно изобретению может составлять от примерно 1 млн до примерно 50 млрд клеток (например, примерно 5 млн клеток, примерно 25 млн клеток, примерно 500 млн клеток, примерно 1 млрд клеток, примерно 5 млрд клеток, примерно 20 млрд клеток, примерно 30 млрд клеток, примерно 40 млрд клеток, либо диапазон определяется любыми двумя из указанных выше значений), предпочтительно от примерно 10 млн до примерно 100 млрд клеток (например, примерно 20 млн клеток, примерно 30 млн клеток, примерно 40 млн клеток, примерно 60 млн клеток, примерно 70 млн клеток, примерно 80 млн клеток, примерно 90 млн клеток, примерно 10 млрд клеток, примерно 25 млрд клеток, примерно 50 млрд клеток, примерно 75 млрд клеток, примерно 90 млрд клеток, либо диапазон определяется любыми двумя из указанных выше значений), более предпочтительно от примерно 100 млн клеток до примерно 50 млрд клеток (например, примерно 120 миллионов клеток, примерно 250 млн клеток, примерно 350 млн клеток, примерно 450 млн клеток, примерно 650 миллионов клеток, примерно 800 млн клеток, примерно 900 млн клеток, примерно 3 млрд клеток, примерно 30 млрд клеток, примерно 45 млрд клеток, либо диапазон определяется любыми двумя из вышеуказанных значений).
[0075] Терапевтическую или профилактическую эффективность можно контролировать путем периодического обследования пациентов. В случае многократных введений в течение нескольких дней или дольше, в зависимости от состояния лечение повторяют до желательного подавления симптомов заболевания. Тем не менее, другие режимы дозировки также могут быть использованы и входят в объем данного изобретения. Желаемая дозировка может быть доставлена путем однократного болюсного введения композиции, повторных болюсных введений композиции или путем непрерывного инфузионного введения композиции.
[0076] Композицию, содержащую клетки-хозяева, которые экспрессируют кодирующую CAR нуклеиновокислотную последовательность согласно изобретению, или вектор, содержащий кодирующую CAR нуклеиновокислотную последовательность согласно изобретению, можно вводить в организм млекопитающего с помощью стандартных способов введения, в том числе пероральным, внутривенным, внутрибрюшинным, подкожным, легочным, трансдермальным, внутримышечным, интраназальным, трансбуккальным, сублингвальным путем или с помощью суппозиториев. Композиция предпочтительно является подходящей для парентерального введения. Термин "парентеральное", используемый в данном документе, включает внутривенное, внутримышечное, подкожное, ректальное, вагинальное и внутрибрюшинное введение. Более предпочтительно, композицию вводят млекопитающему путем периферийной системной доставки с помощью внутривенной, внутрибрюшинной или подкожной инъекции.
[0077] Композицию, содержащую клетки-хозяева, которые экспрессируют кодирующую CAR нуклеиновокислотную последовательность согласно изобретению, или вектор, содержащий кодирующую CAR нуклеиновокислотную последовательность согласно изобретению, можно вводить с одним или более чем одним дополнительным терапевтическим агентом, который может быть совместно введен млекопитающему. Под "совместном введением" понимается введение одного или более чем одного дополнительного терапевтического агента и композиции, содержащей клетки-хозяева согласно изобретению или вектор согласно изобретению, достаточно близко по времени, так что CAR согласно изобретению может усилить эффект одного или более чем одного дополнительного терапевтического агента, или наоборот. В связи с этим композиция, содержащая клетки-хозяева согласно изобретению или вектор согласно изобретению, может быть введена первой, а один или более чем один дополнительный терапевтический агент может быть введен вторым, или наоборот. Альтернативно, композиция, содержащая клетки-хозяева согласно изобретению или вектор согласно изобретению, и один или более чем один дополнительный терапевтический агент могут быть введены одновременно. Примером терапевтического агента, который может быть введен совместно с композицией, содержащей клетки-хозяева согласно изобретению или вектор согласно изобретению, является IL-2.
[0078] После того как композицию, содержащую клетки-хозяева, экспрессирующие нуклеиновокислотную последовательность согласно изобретению, или вектор, содержащий кодирующую CAR нуклеиновокислотную последовательность согласно изобретению, вводят млекопитающему (например, человеку), биологическая активность CAR может быть измерена любым подходящим способом, известным в данной области. В соответствии со способом согласно изобретению CAR связывается с ВСМА на клетках множественной миеломы, и клетки множественной миеломы разрушаются. Связывание CAR с ВСМА на поверхности клеток множественной миеломы может быть проанализировано с помощью любого подходящего способа, известного в данной области, в том числе, например, ELISA и проточной цитометрии. Способность CAR разрушать клетки множественной миеломы может быть измерена с помощью любого подходящего способа, известного в данной области, такого как анализ цитотоксичности, описанный, например, Kochenderfer et al., J. Immunotherapy, 32(7): 689-702 (2009), и Herman et al. J. Immunological Methods, 285(1): 25-40 (2004). Биологическую активность CAR также можно измерить путем анализа экспрессии некоторых цитокинов, таких как CD107a, IFNγ, IL-2 и TNF.
[0079] Специалист в данной области легко поймет, что нуклеиновокислотная последовательность согласно изобретению, кодирующая CAR, может быть изменена с помощью любого числа способов, например, так, чтобы терапевтическая или профилактическая эффективность CAR увеличилась за счет модификации. Например, CAR может быть конъюгирован прямо или косвенно через линкер с целевой группировкой. Практика конъюгации соединений, например, CAR, с целевыми группировками известна в данной области. См., например, Wadwa et al., J. Drug Targeting 3: 111 (1995) и патент США 5087616.
[0080] Следующие примеры дополнительно иллюстрируют изобретение, но, разумеется, не должны быть истолкованы как каким-либо образом ограничивающие его объем.
Пример 1
[0081] Этот пример демонстрирует характер экспрессии ВСМА на человеческих клетках.
[0082] Количественную полимеразную цепную реакцию (кПЦР, qPCR) проводили на панели образцов кДНК из широкого диапазона нормальных тканей, включенных в панель II Human Major Tissue qPCR (Origine Technologies, Роквилл, Мэриленд), с использованием ВСМА-специфического праймера и набора зондов (Life Technologies, Карлсбад, Калифорния). кДНК из клеток плазмоцитомы, которую резецировали у пациента с прогрессирующей множественной миеломой, анализировали в качестве положительного контроля. РНК экстрагировали из клеток плазмоцитомы с помощью мини-набора RNeasy (Qiagen, Inc., Валенсия, Калифорния), и синтезировали кДНК с помощью стандартных способов. Стандартную кривую для кПЦР ВСМА получали путем разведения плазмиды, кодирующей полноразмерную кДНК ВСМА (Origine Technologies, Роквилл, Мэриленд), в носителе ДНК. кПЦР точно определяла число копий BCMA от 102 до 109 на реакцию. Число копий кДНК β-актина в тех же тканях количественно оценивали с помощью праймера на β-актин Taqman и набора зондов (Life Technologies, Карлсбад, Калифорния). Стандартную кривую β-актина получали путем амплификации серийных разведений плазмиды с β-актином. Все реакции кПЦР проводили на машине Roche LightCycler480 (Roche Applied Sciences, Индианополис, Индиана).
[0083] Результаты анализа кПЦР изображены на фиг. 1А и 1В. 93% клеток из образца плазмоцитомы были плазматическими клетками, что было определено с помощью проточной цитометрии. Экспрессия ВСМА в образце плазмоцитомы была значительно выше, чем экспрессия ВСМА в любой другой ткани. кДНК ВСМА была обнаружена в нескольких гематологических тканях, таких как мононуклеарные клетки периферической крови (РВМС), костный мозг, селезенка, лимфатический узел и миндалина. Низкие уровни кДНК ВСМА были обнаружены в большинстве органов желудочно-кишечного тракта, таких как двенадцатиперстная кишка, прямая кишка и желудок. Экспрессия ВСМА в органах желудочно-кишечного тракта может быть связана с плазматическими клетками и В-клетками, присутствующими в лимфоидной ткани кишечника, например в собственной пластинке и пейеровых бляшках (см., например, Brandtzaeg, Immunological Investigations, 39(4-5): 303-355 (2010)). Низкие уровни кДНК ВСМА также были обнаружены в семенниках и трахее. Низкие уровни кДНК ВСМА, обнаруженные в трахее, могут быть связаны с наличием плазматических клеток в собственной пластинке слизистой оболочки трахеи (см., например, Soutar, Thorax, 31(2):158-166 (1976)).
[0084] Экспрессию ВСМА дополнительно характеризовали с помощью проточной цитометрии на поверхности различных типов клеток (см. фиг. 2А-2L), включая линии клеток множественной миеломы H929, U266 и RPMI8226. Линии клеток множественной миеломы H929, U266 и RPMI8226 все экспрессировали ВСМА на клеточной поверхности. Напротив, клеточная линия саркомы TC71, линия Т-клеточной лейкемии CCRF-CEM и линия клеток почки 293Т-17 не экспрессировали ВСМА на клеточной поверхности. Первичные гемопоэтические CD34+-клетки, первичные эпителиальные клетки малых дыхательных путей, первичные бронхиальные эпителиальные клетки и первичные эпителиальные клетки кишечника не экспрессировали ВСМА на клеточной поверхности.
[0085] Результаты этого примера показывают, что ВСМА экспрессируется на поверхности клеток множественной миеломы и имеет ограниченный характер экспрессии в нормальных тканях.
Пример 2
[0086] Этот пример описывает конструкцию нуклеиновокислотной последовательности согласно изобретению, кодирующей химерные антигенные анти-ВСМА-рецепторы (CAR).
[0087] Последовательности двух мышиных антител против человеческого ВСМА, обозначенных как "C12A3.2" и "C11D5.3", были получены из публикации международной патентной заявки WO 2010/104949 (Kalled et al.). Аминокислотные последовательности вариабельных областей тяжелой цепи и вариабельных областей легкой цепи этих антител использовали для создания одноцепочечных вариабельных фрагментов (scFv), имеющих следующую общую структуру:
вариабельная область легкой цепи - линкер - вариабельная область тяжелой цепи.
[0088] Линкер имеет следующую аминокислотную последовательность: GSTSGSGKPGSGEGSTKG (SEQ ID № 7) (см., например, Cooper et al., Blood, 101(4): 1637-1644 (2003)).
[0089] Были разработаны последовательности ДНК, кодирующие два химерных антигенных рецептора, каждая из которых содержала следующие элементы в направлении от 5' к 3': сигнальную последовательность CD8α, вышеупомянутую анти-ВСМА-scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы CD28 и цитоплазматический фрагмент молекулы CD3ζ. Схема этих нуклеиновокислотных последовательностей, кодирующих CAR, приведена на фиг. 3А. CAR, включающие вариабельные области из C12A3.2 и C11D5.3, были обозначены как анти-bcma1 и анти-bcma2, соответственно.
[0090] На основании описанного выше анти-bcma2-CAR были разработаны последовательности ДНК, кодирующие пять дополнительных химерных антигенных рецепторов, каждый из которых содержал различные сигнальные последовательности и домены Т-клеточной активации. В этом отношении 8ss-анти-bcma2-CAR содержал следующие элементы в направлении от 5' к 3': сигнальную последовательность CD8α, scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы CD28 и цитоплазматический фрагмент молекулы CD3ζ. G-анти-bcma2-CAR содержал следующие элементы в направлении от 5' к 3': сигнальную последовательность рецептора человеческого GM-CSF, scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы CD28 и цитоплазматический фрагмент молекулы CD3ζ. Анти-bcma2-BB-CAR содержал следующие элементы в направлении от 5' к 3': сигнальную последовательность CD8α, scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы 4-1ВВ и цитоплазматический фрагмент молекулы CD3ζ . Анти-bcma2-OX40-CAR содержал следующие элементы в направлении от 5' к 3': сигнальную последовательность CD8α, scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы OX40 (см., например, Latza et al., European Journal of Immunology, 24: 677-683 (1994)), и цитоплазматический фрагмент молекулы CD3ζ. Анти-bcma2-BBOX40 содержал следующие элементы в направлении от 5' к 3': сигнальную последовательность CD8α, scFv, шарнирную и трансмембранную области молекулы человеческого CD8α, цитоплазматический фрагмент молекулы 4-1ВВ, цитоплазматический фрагмент молекулы OX40 и цитоплазматический фрагмент молекулы CD3ζ. Элементы, присутствующие в каждой из семи последовательностей CAR, приведены в таблице 1.
Таблица 1
(аминокислотная)
трансмембранная области
Т-клеточного сигналинга
CD3ζ
CD3ζ
CD3ζ
CD3ζ
CD3ζ
CD3ζ
OX40
CD3ζ
[0091] Последовательности, используемые для CD8α, CD28, CD3ζ, 4-1BB (CD137) и OX40 (CD134), были получены из общедоступной базы данных Национального центра биотехнологической информации (NCBI).
[0092] Нуклеиновокислотные последовательности, кодирующие CAR, были получены с использованием способов, известных в данной области, таких как описанные, например, в Kochenderfer et al., J. Immunology, 32(7): 689-702 (2009) и Zhao et al., J. Immunology, 183(9): 5563-5574 (2009). Нуклеиновокислотную последовательность, кодирующую каждый CAR, подвергали оптимизации по кодонам и синтезировали с использованием технологии GeneArt™ (Life Technologies, Карлсбад, Калифорния) с соответствующими сайтами рестрикции.
[0093] Последовательности, кодирующие анти-bcma1- и анти-bcma2-CAR, лигировали в лентивирусную векторную плазмиду, обозначенную pRRLSIN.cPPT.MSCV.coDMF5.oPRE (см., например, Yang et al., J. Immunotherapy, 33(6): 648-658 (2010)). Часть CoDMF5 этого вектора была заменена нуклеиновокислотными последовательностями, кодирующими CAR, с помощью стандартных способов. Два полученных анти-ВСМА-CAR-вектора были обозначены как pRRLSIN.cPPT.MSCV.anti-bcma1.oPRE и pRRLSIN.cPPT.MSCV.anti-bcma2.oPRE. Также был сконструирован отрицательный контроль CAR, содержащий scFv SP6, который распознает гаптен 2,4,6-тринитрофенил (см., например, Gross et al., Proc. Natl. Acad. Sci. USA, 86(24): 10024-10028 (1989)). Этот CAR был обозначен как SP6. SP6-CAR был клонирован в тот же лентивирусный вектор, что и анти-ВСМА-CAR, и содержал те же сигнальные домены, что и анти-bcma1 и анти-bcma2. Супернатант, содержащий лентивирусы, кодирующие каждый CAR, получали в соответствии с протоколом, описанным в Yang et al., см. выше. В частности, клетки 293T-17 (АТСС CRL-11268) трансфицировали следующими плазмидами: pMDG (кодирующей белок оболочки вируса везикулярного стоматита), pMDLg/pRRE (кодирующей белки ВИЧ Gag и Pol), PRSV-Rev (кодирующей белок RSV Rev), и плазмидами, кодирующими анти-ВСМА-CAR (например, Yang et al., см. выше).
[0094] Последовательности, кодирующие CAR G-анти-bcma2, 8ss-анти-bcma2, анти-bcma2-BB, анти-bcma2-OX40 и анти-bcma2-BBOX40, лигировали в гамма-ретровирусный плазмидный вектор, обозначенный как MSGV (сплайс-gag вектор на основе вируса мышиных стволовых клеток) с использованием стандартных способов, таких как описанные, например, в Hughes et al., Human Gene Therapy, 16: 457-472 (2005). После того как были получены кодирующие CAR гамма-ретровирусные плазмиды, были созданы некомпетентные по репликации ретровирусы с оболочкой RD114 путем временной трансфекции упаковывающих клеток на основе 293, описанных в Kochenderfer et al., J. Immunotherapy, 32(7): 689-702 (2009).
[0095] Некомпетентные по репликации лентивирусы и ретровирусы, кодирующие описанные выше CAR, использовали для трансдукции Т-клеток человека. Для анти-bcma1 и анти-bcma2 Т-клетки культивировали, как описано выше (см., например, Kochenderfer et al., J. Immunotherapy, 32(7): 689-702 (2009)) и стимулировали моноклональным анти-CD3-антителом ОКТ3 (Орто-Biotech, Хоршам, Пенсильвания) в среде AIM V™ (Life Technologies, Карлсбад, Калифорния), содержащей 5% человеческой AB-сыворотки (Valley Biomedical, Винчестер, Вирджиния) и 300 международных единиц (МЕ)/мл интерлейкина-2 (Novartis Diagnostics, Эмеривилл, Калифорния). Через тридцать шесть часов после начала культивирования активированные Т-клетки суспендировали в лентивирусном супернатанте с сульфатом протамина и 300 МЕ/мл IL-2. Клетки центрифугировали в течение 1 часа при 1200g. Затем Т-клетки культивировали в течение трех часов при 37°С. Затем супернатант разводили 1:1 средой RPMI (Mediatech, Inc., Манассас, Вирджиния) + 10% фетальной бычьей сыворотки (Life Technologies, Карлсбад, Калифорния) и IL-2. Т-клетки культивировали в разведенном супернатанте в течение ночи, а затем возвращали в культуру в среде AIM V™ (Life Technologies, Карлсбад, Калифорния) с 5% человеческой AB-сывороткой и IL-2. Т-клетки окрашивали меченными биотином поликлональными козлиными антителами против мышиного F(ab)2 (Jackson ImmunoResearch Laboratories, Inc., Вест Гроув, Пенсильвания) для обнаружения анти-ВСМА-CAR. Высокие уровни экспрессии анти-bcma1-CAR, анти-bcma2-CAR, и SP6-CAR наблюдались на клеточной поверхности трансдуцированных Т-клеток, как показано на фиг. 3B-3D.
[0096] Для CAR G-анти-bcma2, 8ss-анти-bcma2, анти-bcma2-BB, анти-bcma2-OX40 и анти-bcma2-BBOX40 мононуклеарные клетки периферической крови суспендировали в концентрации 1×106 клеток в 1 мл Т-клеточной среды, содержащей 50 нг/мл моноклонального анти-CD3-антитела ОКТ3 (Ortho, Бриджуотер, Нью-Джерси) и 300 МЕ/мл IL-2. Полипептид RETRONECTIN™ (Takara Bio Inc., Шига, Япония), который представляет собой рекомбинантный полипептид из фрагментов фибронектина человека, который связывает вирусы и белки клеточной поверхности, растворяли в концентрации 11 мкг/мл в фосфатно-буферном солевом растворе (PBS), и 2 мл полипептида RETRONECTIN™ в растворе PBS добавляли в каждую лунку 6-луночных планшетов, покрытых неклеточной культурой (BD Biosciences, Франклин Лейкс, Нью-Джерси). Планшеты инкубировали в течение двух часов при комнатной температуре (RT). После инкубации раствор RETRONECTIN™ аспирировали, и в каждую лунку, покрытую RETRONECTIN™, вносили 2 мл блокирующего раствора, состоящего из сбалансированного солевого растворе Хэнкса (HBSS) с 2% бычьим сывороточным альбумином (BSA). Планшеты инкубировали в течение 30 минут при комнатной температуре (RT). Блокирующий раствор аспирировали, и лунки промывали раствором HBSS + 2,5% (4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота) (HEPES). Ретровирусный супернатант быстро размораживали и разводили 1:1 в Т-клеточной среде, и затем в каждую лунку, покрытую RETRONECTIN™, вносили 2 мл разведенного супернатанта. После добавления супернатантов планшеты центрифугировали при 2000g в течение 2 часов при 32°С. Затем супернатант аспирировали из лунок и в каждую лунку добавляли 2×106 Т-клеток, которые были культивированы с ОКТ3-антителом и IL-2 в течение 2 дней. Когда Т-клетки вносили в планшет, покрытый ретровирусом, их суспендировали в концентрации 0,5×106 клеток на 1 мл Т-клеточной среды плюс 300 МЕ/мл IL-2. После того как Т-клетки были добавлены в каждую лунку, планшеты центрифугировали в течение 10 минут при 1000g. Планшеты инкубировали при 37°С в течение ночи. Трансдукцию повторяли на следующий день. После 18-24-часовой инкубации Т-клетки удаляли из планшетов и суспендировали в свежей Т-клеточной среде с 300 МЕ/мл IL-2 в концентрации 0,5×106 клеток на 1 мл и культивировали при 37°С и 5% СО2. Наблюдались высокие уровни экспрессии анти-bcma2-BBOX40, анти-bcma2-BB, и 8ss-анти-bcma2 на поверхности трансдуцированных Т-клеток.
[0097] Результаты этого примера демонстрируют способ получения нуклеиновокислотной последовательности согласно изобретению, кодирующей CAR, и способ экспрессии CAR на поверхности Т-клеток.
Пример 3
[0098] Этот пример описывает серию экспериментов, используемых для определения специфичности CAR согласно изобретению к ВСМА.
Клетки
[0099] NCI-H929, U266 и RPMI8226 все являются ВСМА-положительными клеточными линиями множественной миеломы, которые получены из АТСС (АТСС №№ CRL-9068, TIB-196 и CCL-155, соответственно). А549 (АТСС № CCL-185) представляет собой ВСМА-отрицательную клеточную линию рака легких. TC71 представляет собой ВСМА-отрицательную клеточную линию саркомы. CCRF-CEM представляет собой ВСМА-отрицательную Т-клеточную линию (АТСС № CCL-119). ВСМА-K562 представляет собой клетки К562 (АТСС № CCL-243), которые были трансдуцированы нуклеиновокислотной последовательностью, кодирующей полноразмерный ВСМА. NGFR-K562 представляет собой клетки К562, которые были трансдуцированы геном, кодирующим низкоаффинный рецептор фактора роста нервов (см, например, Kochenderfer et al., J. Immunotherapy., 32(7):689-702 (2009)). Были использованы лимфоциты периферической крови (PBL) от трех пациентов с множественной миеломой (т.е. пациенты с миеломой 1-3), а также PBL от трех других субъектов: донора А, донора В и донора С. Доноры А-С все имели меланому. Первичные CD34+-клетки были получены от трех нормальных здоровых доноров. Образец клеток плазмоцитомы был получен от пациента с миеломой 1, а образец костного мозга был получен от пациента с миеломой 3. Все образцы от людей, упомянутые выше, были получены от пациентов, включенных в IRB-утвержденные клинические испытания в Национальном институте рака. Следующие первичные эпителиальные клетки человека были получены от Lonza, Inc. (Базель, Швейцария): эпителиальные клетки малых дыхательных путей, бронхиальные эпителиальные клетки и эпителиальные клетки кишечника.
ELISA на интерферон-γ и TNF
[0100] ВСМА-положительные или ВСМА-отрицательные клетки объединяли с CAR-трансдуцированными Т-клетками в дубликатах лунок 96-луночного круглодонного планшета (Corning Life Sciences, Лоуэлл, Массачусетс) в среде AIM V™ (Life Technologies, Карлсбад, CA) + 5% человеческой сыворотки. Планшеты инкубировали при 37°С в течение 18-20 часов. После инкубации выполняли анализ ELISA на IFNγ и TNF с использованием стандартных способов (Pierce, Рокфорд, Иллинойс).
[0101] Т-клетки, трансдуцированные анти-bcma1- или анти-bcma2-CAR, продуцировали большие количества IFNγ, когда их культивировали в течение ночи с ВСМА-экспрессирующей клеточной линией ВСМА-K562, но CAR-трансдуцированные Т-клетки продуцировали только фоновые уровни IFNγ, когда их культивировали с отрицательным контролем, клеточной линией NGFR-K562, как указано в таблице 2 (все единицы: пг/мл IFNγ).
Таблица 2
*Эффекторные клетки были Т-клетками от пациента с множественной миеломой (пациента с миеломой 2). Т-клетки трансдуцировали указанным CAR или оставляли нетрансдуцированными.
**Указанные клетки-мишени объединяли с эффекторных клеток для инкубации в течение ночи и проводили ELISA на IFNγ.
[0102] T-клетки, экспрессирующие CAR 8ss-анти-bcma2, анти-bcma2-BB и анти-bcma2-OX40, продуцировали IFNγ специфически в ответ на ВСМА-положительные клетки-мишени, когда Т-клетки и клетки-мишени совместно культивировали в течение ночи, как указано в таблице 3 (все единицы: пг/мл IFNγ).
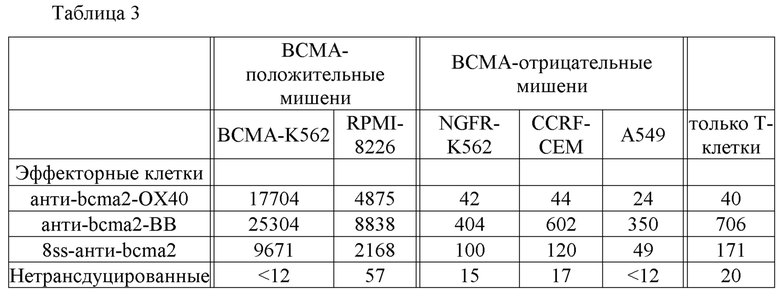
[0103] T-клетки, трансдуцированные анти-ВСМА-CAR, продуцировали большие объемы IFNγ, когда их культивировали в течение ночи с ВСМА-экспрессирующими клеточными линиями множественной миеломы. Напротив, анти-ВСМА-CAR продуцировали значительно более низкие количества IFNγ, когда их культивировали с различными ВСМА-отрицательными клеточными линиями. По сравнению с Т-клетками, трансдуцированными анти-bcma1-CAR, Т-клетки, трансдуцированные анти-bcma2-CAR и его вариантами (т.е. 8ss-анти-bcma2, анти-bcma2-BB и анти-bcma2-OX40), продуцировали больше IFNγ при культивировании с ВСМА-положительными клетками и меньше IFNγ при культивировании с ВСМА-отрицательными клетками.
[0104] Т-клетки, трансдуцированные вариантами анти-bcma2-CAR, продуцировали TNF специфически в ответ на ВСМА-положительные клетки-мишени, когда Т-клетки и клетки-мишени совместно культивировали в течение ночи, как указано в таблице 4 (все единицы: пг/мл фактора некроза опухоли (TNF)).
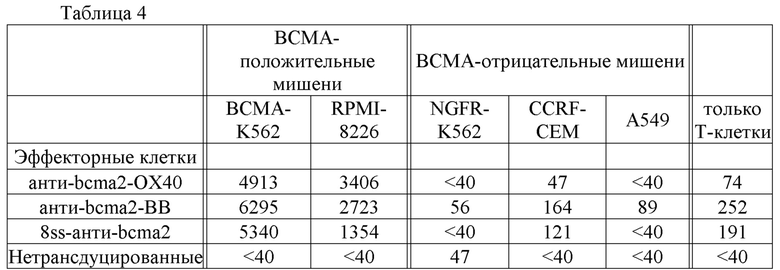
[0105] Так как Т-клетки, трансдуцированные анти-bcma2-CAR и его вариантами, демонстрировали чуть более сильное и более специфическое распознавание ВСМА-экспрессирующих клеток, чем Т-клетки трансдуцированные анти-bcma1-CAR, то в последующих экспериментах были использованы только анти-bcma2-CAR и варианты анти-bcma2-CAR.
Анализ CD107a
[0106] Две популяции Т-клеток были получены в двух отдельных пробирках. Одна пробирка содержала ВСМА-клетки К562, а другая пробирка содержала клетки NGFR-К562. Обе пробирки также содержали Т-клетки, трансдуцированные анти-bcma2-CAR и вариантами анти-bcma2-CAR, 1 мл AIM V™ среды (Life Technologies, Карлсбад, Калифорния) с 5% человеческой сыворотки, титрованную концентрацию анти-CD107a-антитела (eBioscience, Inc., Сан-Диего, Калифорния, клон eBioH4A3) и 1 мкл Golgi Stop (BD Biosciences, Франклин Лейкс, Нью-Джерси). Все пробирки инкубировали при 37°С в течение четырех часов, а затем окрашивали на экспрессию CD3, CD4 и CD8.
[0107] CAR-трансдуцированные Т-клетки от трех различных субъектов имели повышенную регуляцию CD107a специфически в ответ на стимуляцию с ВСМА-экспрессирующими клетками-мишенями (см. фиг. 4А-4С). Это указывает на возникновение ВСМА-специфической дегрануляции Т-клеток, которая является необходимым условием для перфорин-опосредованной цитотоксичности (см., например, Rubio et al., Nature Medicine, 9(11): 1377-1382 (2003)). Кроме того, Т-клетки, экспрессирующие варианты анти-bcma2-CAR (8ss-анти-bcma2, анти-bcma2-ВВ, анти-bcma2-OX40), дегранулировали ВСМА-специфическим образом при стимуляции с клетками-мишенями in vitro, как показано на фиг. 5A-5D.
Анализ внутриклеточного окрашивания цитокинов (intracellular cytokine staining assay, ICCS)
[0108] Популяция клеток ВСМА-К562 и популяция клеток NGFR-К562 были получены в двух отдельных пробирках, как описано выше. Обе пробирки также содержали Т-клетки, трансдуцированные анти-bcma2-CAR, от пациента с миеломой 2, 1 мл AIM V среды (Life Technologies, Карлсбад, Калифорния) с 5% человеческой сывороткой и 1 мкл Golgi Stop (BD Biosciences, Франклин Лейкс, Нью-Джерси). Все пробирки инкубировали при 37°С в течение шести часов. Поверхность клеток окрашивали анти-CD3-, анти-CD4-, и анти-CD8-антителами. Клетки подвергали пермеабилизации и проводили внутриклеточное окрашивание на IFNγ (BD Biosciences, Франклин Лейкс, Нью-Джерси, клон B27), IL-2 (BD Biosciences, Франклин Лейкс, Нью-Джерси, клон MQ1-17H12) и TNF (BD Biosciences, Франклин Лейкс, Нью-Джерси, клон MAb11), следуя инструкциям к набору Cytofix/Cytoperm (BD Biosciences, Франклин Лейкс, Нью-Джерси).
[0109] Большие популяции Т-клеток, трансдуцированных анти-bcma2-CAR, от пациента с миеломой 2 специфически продуцировали цитокины IFNγ, IL-2 и TNF ВСМА-специфическим образом после шестичасовой стимуляции с ВСМА-экспрессирующими клетками-мишенями, как показано на фиг. 6А-6С.
Анализы пролиферации
[0110] Оценивали способность Т-клеток, трансдуцированных анти-bcma2-CAR, к пролиферации при стимуляции с ВСМА-экспрессирующими клетками-мишенями. В частности, 0,5×106 облученных клеток ВСМА-562 или 0,5×106 облученных клеток NGFR-562 культивировали совместно с 1×106 общих Т-клеток, которые были трансдуцированы либо анти-bcma2-CAR, либо SP6-CAR. Т-клетки были помечены карбоксифлуоресцеин диацетат сукцинимидиловым эфиром (CFSE) (Life Technologies, Карлсбад, Калифорния), как описано в Mannering et al., J. Immunological Methods, 283(1-2): 173-183 (2003). Среда, используемая в объединенных культурах, была средой AIM V™ (Life Technologies, Карлсбад, Калифорния) с 5% человеческой AB-сыворотки. IL-2 к среде не добавляли. Через четыре дня после начала живые клетки в каждой объединенной культуре подсчитывали с помощью трипанового синего для исключения мертвых клеток. Затем проводили проточную цитометрию путем окрашивания Т-клеток поликлональными меченными биотином козлиными антителами против человеческого ВСМА (R&D Systems, Миннеаполис), а затем стрептавидином (BD Biosciences, Франклин Лейкс, Нью-Джерси), анти-CD38-антителом (eBioscience, Inc., San Diego, CA) и анти-CD56-антителом (BD Biosciences, Франклин Лейкс, Нью-Джерси). Анализ данных проточной цитометрии выполняли с помощью программного обеспечения FlowJo (Tree Star, Inc., Ашленд, Орегон).
[0111] Т-клетки, которые экспрессировали анти-bcma2-CAR, демонстрировали большее разведение CFSE при культивировании с клетками ВСМА-K562, чем при культивировании с отрицательным контролем, клетками NGFR-К562, как показано на фиг. 7А. Эти результаты указывают на то, что Т-клетки, трансдуцированные анти-bcma2-CAR, специфически пролиферировали при стимуляции с ВСМА-экспрессирующими клетками-мишенями. Напротив, никакой существенной разницы в разведении CFSE не было, когда Т-клетки, экспрессирующие SP6-CAR, культивировали либо с клетками-мишенями ВСМА-K562, либо с клетками-мишенями NGFR-K562 (фиг. 7В), что указывает на отсутствие ВСМА-специфической пролиферации Т-клеток, экспрессирующих SP6-CAR.
[0112] В начале анализа пролиферации 0,8×106 Т-клеток, экспрессирующих анти-bcma2-CAR, культивировали либо с клетками ВСМА-К562, либо с клетками NGFR-К562. После 4 дней культивирования 2,7×106 Т-клеток, экспрессирующих анти-bcma2-CAR, присутствовало в культурах, содержащих клетки ВСМА-К562, в то время как в культурах, содержащих клетки NGFR-К562, присутствовало только 0,6×106 Т-клеток, экспрессирующих анти-bcma2-CAR. Это ВСМА-специфическое увеличение абсолютного числа Т-клеток, экспрессирующих анти-bcma2-CAR, указывает на то, что эти Т-клетки пролиферировали в ответ на ВСМА.
[0113] Результаты этого примера показывают, что Т-клетки, экспрессирующие CAR согласно изобретению, демонстрируют ВСМА-специфическую продукцию цитокинов, дегрануляцию и пролиферацию.
Пример 4
[0114] В этом примере показано, что Т-клетки, экспрессирующие анти-ВСМА-CAR согласно изобретению, могут разрушать клеточные линии множественной миеломы.
[0115] Анализы цитотоксичности выполняли, чтобы определить, могут ли Т-клетки, трансдуцированные анти-bcma2-CAR, описанным в примерах 2 и 3, разрушать ВСМА-экспрессирующие клеточные линии множественной миеломы (ММ). Конкретно, цитотоксичность клеток-мишеней измеряли путем сравнения выживания ВСМА-экспрессирующих клеток-мишеней (например, клеточных линий множественной миеломы H929 и RPMI8226) относительно выживания отрицательного контроля, клеток CCRF-CEM, с использованием анализа, описанного, например, в Kochenderfer et al., J. Immunotherapy, 32(7): 689-702 (2009), и Hermans et al., J. Immunological Methods, 285(1): 25-40 (2004).
[0116] Приблизительно 50000 ВСМА-экспрессирующих клеток-мишеней и 50000 клеток CCRF-CEM объединяли в одних и тех же пробирках с различным числом CAR-трансдуцированных Т-клеток. Клетки отрицательного контроля CCRF-CEM метили флуоресцентным красителем 5-(и-6)-(((4-хлорметил)бензоил)амино)тетраметилродамином (CMTMR) (Life Technologies, Карлсбад, Калифорния), а ВСМА-экспрессирующие клетки-мишени метили CFSE. Во всех экспериментах цитотоксичность эффекторных Т-клеток, трансдуцированных анти-bcma2-CAR, сравнивали с цитотоксичностью эффекторных Т-клеток отрицательного контроля от того же самого субъекта, которые были трансдуцированы SP6-CAR. Объединенные культуры находились в стерильных 5 мл пробирках (BD Biosciences, Франклин Лейкс, Нью-Джерси) в двух экземплярах в следующем соотношении Т-клеток и клеток-мишеней: 20,0:1, 7:1, 2:1 и 0,7:1. Культуры инкубировали в течение четырех часов при 37°С. Сразу же после инкубации добавляли 7-амино-актиномицин D (7AAD BD Biosciences, Франклин Лейкс, Нью-Джерси). Для каждой объединенной культуры Т-клеток/клеток-мишеней определяли процент живых ВСМА-экспрессирующих клеток-мишеней и живых клеток CCRF-CEM отрицательного контроля.
[0117] Для каждой объединенной культуры Т-клеток/клеток-мишеней определяли процент выживаемости ВСМА-экспрессирующих клеток-мишеней по отношению к клеткам CCRF-CEM отрицательного контроля посредством деления процента ВСМА-экспрессирующих клеток на процент клеток CCRF-CEM отрицательного контроля. Скорректированный процент выживания ВСМА-экспрессирующих клеток-мишеней рассчитывали путем деления процента живых ВСМА-экспрессирующих клеток-мишеней в каждой объединенной культуре Т-клеток/клеток-мишеней на отношение процента ВСМА-экспрессирующих клеток-мишеней к проценту клеток CCRF-CEM отрицательного контроля в пробирках, содержащих только ВСМА-экспрессирующие клетки-мишени и клетки CCRF-CEM отрицательного контроля без эффекторных Т-клеток. Эта поправка была необходима для учета различий в исходных количествах клеток и спонтанной гибели клеток-мишеней. Цитотоксичность рассчитывали следующим образом:
% цитотоксичности ВСМА-экспрессирующих клеток-мишеней = 100 - скорректированный % живых ВСМА-экспрессирующих клеток-мишеней
[0118] Результаты анализа цитотоксичности показаны на фиг. 7C и 7D. Т-клетки, трансдуцированные анти-bcma2-CAR, специфически убивали ВСМА-экспрессирующие клеточные линии множественной миеломы H929 и RPMI8226. Напротив, Т-клетки, трансдуцированные SP6-CAR, обладали гораздо более низкими уровнями цитотоксичности против этих клеточных линий.
[0119] Результаты этого примера показывают, что нуклеиновокислотная последовательность согласно изобретению, кодирующая анти-ВСМА-CAR, может быть использована в способе разрушения клеточных линий множественной миеломы.
Пример 5
[0120] В этом примере показано, что Т-клетки, экспрессирующие анти-ВСМА-CAR согласно изобретению, могут разрушать первичные клетки множественной миеломы.
[0121] Первичные клетки множественной миеломы, описанные в примере 2, оценивали на экспрессию ВСМА, а также на ВСМА-специфическую продукцию цитокинов, дегрануляцию и пролиферацию с использованием способов, описанных выше.
[0122] Экспрессия ВСМА на клеточной поверхности была обнаружена в четырех образцах первичной множественной миеломы, а также на первичных клетках множественной миеломы в костном мозге пациента с миеломой 3 (см. фиг. 8А). ВСМА-экспрессирующие плазматические клетки составляли 40% клеток в образце костного мозга от пациента с миеломой 3. Аллогенные Т-клетки от донора C, трансдуцированные анти-bcma2-CAR, продуцировали IFNγ после совместного культивирования с необработанными клетками костного мозга от пациента с миеломой 3, как показано на фиг. 8В. Т-клетки от того же аллогенного донора, трансдуцированные анти-bcma2-CAR, продуцировали гораздо меньше IFNγ, когда их культивировали с мононуклеарными клетками периферической крови (РВМС) от пациента с миеломой 3. Кроме того, SP6-CAR-трансдуцированные Т-клетки от донора С специфически не распознавали костный мозг от пациента с миеломой 3. Ранее сообщалось, что нормальные РВМС не включают клетки, экспрессирующие ВСМА (см., например, Ng et al., J. Immunology, 173(2): 807-817 (2004)). Чтобы подтвердить это наблюдение, РВМС от пациента 3 оценивали на экспрессию ВСМА с помощью проточной цитометрии. РВМС от пациента 3 не включали ВСМА-экспрессирующие клетки, отдельно от небольшой популяции клеток CD56+CD38high, которая составила примерно 0,75% от РВМС. Эта популяция, возможно, состояла из циркулирующих клеток множественной миеломы.
[0123] Плазмоцитома, резецированная у пациента с миеломой 1, состояла на 93% из плазматических клеток, и эти первичные плазматические клетки экспрессировали ВСМА, как показано на фиг. 8C. Т-клетки от пациента с миеломой 2 продуцировали IFNγ при культивировании с аллогенными необработанными клетками плазмоцитомы от пациента с миеломой 1. Т-клетки от пациента с миеломой 2 не продуцировали значительных количеств IFNγ при культивировании с РВМС от пациента с миеломой 1. Т-клетки от пациента с миеломой 2, трансдуцированные SP6-CAR, не продуцировали значительных количеств IFNγ, когда их культивировали либо с клетками плазмоцитомы, либо с РВМС от пациента с миеломой 1. РВМС от пациента с миеломой 1 не экспрессировали ВСМА, что было измерено с помощью проточной цитометрии.
[0124] T-клетки от пациента с миеломой 1, который ранее получил восемь циклов терапии против миеломы, были успешно культивированы и трансдуцированы лентивирусом вектором, кодирующим анти-bcma2-CAR. Через восемь дней после того, как культуры были инициированы, экспрессия анти-bcma2-CAR была обнаружена на 65% Т-клеток. Т-клетки от пациента с миеломой 1, экспрессирующие анти-bcma2-CAR, продуцировали IFNγ специфически в ответ на аутологичные клетки плазмоцитомы (фиг. 8D). Т-клетки от пациента с миеломой 1, экспрессирующие SP6-CAR, не распознавали аутологичные клетки плазмоцитомы. Т-клетки, экспрессирующие анти-bcma2-CAR, и Т-клетки, экспрессирующие SP6-CAR, не распознавали аутологичные РВМС. Т-клетки от пациента с миеломой 1, экспрессирующие анти-bcma2-CAR, также специфически убивали аутологичные клетки плазмоцитомы при низком соотношении с мишенью. Напротив, Т-клетки от пациента с миеломой 1, экспрессирующие SP6-CAR, демонстрировали низкие уровни цитотоксичности против аутологичных клеток плазмоцитомы (фиг. 8E).
[0125] Результаты этого примера показывают, что анти-ВСМА-CAR согласно изобретению может быть использован в способе разрушения первичных клеток множественной миеломы.
Пример 6
[0126] В этом примере показано, что Т-клетки, экспрессирующие анти-ВСМА-CAR согласно изобретению, могут разрушать верифицированные опухоли у мышей.
[0127] Иммунодефицитным мышам NSG (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ, Jackson Laboratory) вводили внутрикожно 8×106 клеток RPMI8226. Опухоли росли в течение 17-19 дней, а затем мыши получали внутривенные инфузии 8×106 человеческих Т-клеток, трансдуцированных либо анти-bcma2-CAR, либо SP6-CAR. Опухоли измеряли циркулем каждые 3 дня. Самую большую длину и длину, перпендикулярную самой большой длине, перемножали, получая размер опухоли (площадь) в мм2. Когда самая большая длина достигала 15 мм, мышей умерщвляли. Исследования на животных были утверждены Комитетом по уходу и использованию животных Национального института рака.
[0128] Результаты этого примера показаны на фиг. 9А и 9В. Примерно на 6-й день мыши, которым вводили анти-bcma2-трансдуцированные Т-клетки, продемонстрировали уменьшение размера опухоли, и опухоли были ликвидированы к 15-му дню. Кроме того, все мыши, которым вводили анти-bcma2-трансдуцированные Т-клетки, дожили до 30 дня после инфузии Т-клеток.
[0129] Результаты этого примера показывают, что анти-BMCA-CAR согласно изобретению может разрушать клетки множественной миеломы in vivo.
[0130] Все ссылки, включая публикации, патентные заявки и патенты, цитируемые в данном документе, включены посредством ссылки в той же степени, как если бы про каждый объект ссылки было отдельно и конкретно указано, что он включен посредством ссылки и изложен в данном документе во всей его полноте.
[0131] Использование терминов в единственном числе и аналогичные объекты ссылки в контексте описания изобретения (особенно в контексте следующей ниже формулы изобретения) должно толковаться как охватывающее единственное число и множественное число, если иное не указано в данном документе или это явно не противоречит контексту. Термины "содержащий", "имеющий" и "включающий" должны быть истолкованы как неограничивающие термины (т.е. означающие "включающий, но не ограниченный этим"), если не указано иное. Указание диапазонов значений в данном документе является только способом быстрой записи с индивидуальной ссылкой на каждое отдельное значение, попадающее в диапазон, если в данном документе не указано иное, и каждое отдельное значение включено в описание, как если бы оно было отдельно приведено в данном документе. Все описанные здесь способы могут быть выполнены в любом подходящем порядке, если это явно не противоречит контексту или не указано иное. Использование в данном документе любых и всех примеров или иллюстративных выражений (например, "такой как") предназначено только для лучшего освещения изобретения и не накладывает ограничений на объем изобретения, если не заявлено иное. Отсутствие выражения в описании следует толковать как указание на любой незаявленный элемент в качестве существенного в практике изобретения.
[0132] В данном документе описаны предпочтительные воплощения данного изобретения, включая наилучший способ, известный авторам изобретения, для осуществления данного изобретения. Вариации этих предпочтительных воплощений могут стать очевидными для специалистов в данной области после прочтения приведенного выше описания. Изобретатели ожидают, что специалисты используют такие вариации в зависимости от обстоятельств, и изобретатели подразумевают, что изобретение будет осуществлено иначе, чем конкретно описано в данном документе. Соответственно, данное изобретение включает все модификации и эквиваленты предмета, изложенного в формуле изобретения, прилагаемой к данному документу, предусмотренные действующим законодательством. Кроме того, любая комбинация описанных выше элементов во всех возможных вариантах охватывается изобретением, если это явно не противоречит контексту или в данном описании не указано иное.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> THE UNITED STATES OF AMERICA, AS REPRESENTED BY THE
SECRETARY, DEPARTMENT OF HEALTH AND HUMAN SERVICES
<120> ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ
В-КЛЕТОК
<130> 712361
<150> US 61/622,600
<151> 2012-04-11
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 1512
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 1
atggccctgc ctgtgacagc tctgctgctg cccctggccc tgctgctcca tgccgccaga 60
cccgacatcg tgctgaccca gagccccccc agcctggcca tgtctctggg caagagagcc 120
accatcagct gccgggccag cgagagcgtg accatcctgg gcagccacct gatctactgg 180
tatcagcaga agcctggcca gccccccacc ctgctgatcc agctggctag caatgtgcag 240
accggcgtgc ccgccagatt cagcggcagc ggcagcagaa ccgacttcac cctgaccatc 300
gaccccgtgg aagaggacga cgtggccgtg tactactgcc tgcagagccg gaccatcccc 360
cggacctttg gcggaggaac aaagctggaa atcaagggca gcaccagcgg ctccggcaag 420
cctggctctg gcgagggcag cacaaaggga cagattcagc tggtgcagag cggccctgag 480
ctgaagaaac ccggcgagac agtgaagatc agctgcaagg cctccggcta caccttccgg 540
cactacagca tgaactgggt gaaacaggcc cctggcaagg gcctgaagtg gatgggccgg 600
atcaacaccg agagcggcgt gcccatctac gccgacgact tcaagggcag attcgccttc 660
agcgtggaaa ccagcgccag caccgcctac ctggtgatca acaacctgaa ggacgaggat 720
accgccagct acttctgcag caacgactac ctgtacagcc tggacttctg gggccagggc 780
accgccctga ccgtgtccag cttcgtgcct gtgttcctgc ccgccaagcc caccaccacc 840
cctgccccta gacctcccac cccagcccca acaatcgcca gccagcctct gtccctgcgg 900
cccgaagcct gtagacctgc tgccggcgga gccgtgcaca ccagaggcct ggatttcgcc 960
tgcgacatct acatctgggc ccctctggcc ggcacctgtg gcgtgctgct gctgagcctg 1020
gtgatcaccc tgtactgcaa ccaccggaac agaagcaagc ggagccggct gctgcacagc 1080
gactacatga acatgacccc aagacggcct ggccccaccc ggaagcacta ccagccttac 1140
gcccctccca gagacttcgc cgcctaccgg tccagagtga agttcagcag atccgccgac 1200
gcccctgcct accagcaggg acagaaccag ctgtacaacg agctgaacct gggcagacgg 1260
gaagagtacg acgtgctgga caagcggaga ggccgggacc ccgagatggg cggaaagccc 1320
agacggaaga acccccagga aggcctgtat aacgaactgc agaaagacaa gatggccgag 1380
gcctacagcg agatcggcat gaagggcgag cggaggcgcg gcaagggcca cgatggcctg 1440
taccagggcc tgagcaccgc caccaaggac acctacgacg ccctgcacat gcaggccctg 1500
ccccccagat ga 1512
<210> 2
<211> 1512
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 2
atggccctgc ctgtgacagc tctgctcctc cctctggccc tgctgctcca tgccgccaga 60
cccgacatcg tgctgaccca gagccccccc agcctggcca tgtctctggg caagagagcc 120
accatcagct gccgggccag cgagagcgtg accatcctgg gcagccacct gatccactgg 180
tatcagcaga agcccggcca gccccccacc ctgctgatcc agctcgccag caatgtgcag 240
accggcgtgc ccgccagatt cagcggcagc ggcagcagaa ccgacttcac cctgaccatc 300
gaccccgtgg aagaggacga cgtggccgtg tactactgcc tgcagagccg gaccatcccc 360
cggacctttg gcggaggcac caaactggaa atcaagggca gcaccagcgg ctccggcaag 420
cctggctctg gcgagggcag cacaaaggga cagattcagc tggtgcagag cggccctgag 480
ctgaagaaac ccggcgagac agtgaagatc agctgcaagg cctccggcta caccttcacc 540
gactacagca tcaactgggt gaaaagagcc cctggcaagg gcctgaagtg gatgggctgg 600
atcaacaccg agacaagaga gcccgcctac gcctacgact tccggggcag attcgccttc 660
agcctggaaa ccagcgccag caccgcctac ctgcagatca acaacctgaa gtacgaggac 720
accgccacct acttttgcgc cctggactac agctacgcca tggactactg gggccagggc 780
accagcgtga ccgtgtccag cttcgtgccc gtgttcctgc ccgccaaacc taccaccacc 840
cctgccccta gacctcccac cccagcccca acaatcgcca gccagcctct gtctctgcgg 900
cccgaagcct gtagacctgc tgccggcgga gccgtgcaca ccagaggcct ggacttcgcc 960
tgcgacatct acatctgggc ccctctggcc ggcacctgtg gcgtgctgct gctgagcctg 1020
gtgatcaccc tgtactgcaa ccaccggaac agaagcaagc ggagccggct gctgcacagc 1080
gactacatga acatgacccc aagacggcct ggccccaccc ggaagcacta ccagccttac 1140
gcccctccca gagacttcgc cgcctaccgg tccagagtga agttcagcag atccgccgac 1200
gcccctgcct accagcaggg acagaaccag ctgtacaacg agctgaacct gggcagacgg 1260
gaagagtacg acgtgctgga caagcggaga ggccgggacc ccgagatggg cggaaagccc 1320
agacggaaga acccccagga aggcctgtat aacgaactgc agaaagacaa gatggccgag 1380
gcctacagcg agatcggcat gaagggcgag cggaggcgcg gcaagggcca cgatggcctg 1440
taccagggcc tgagcaccgc caccaaggac acctacgacg ccctgcacat gcaggccctg 1500
ccccccagat ga 1512
<210> 3
<211> 1512
<212> DNA
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 3
atggctctgc ctgtgacagc tctgctgctg cctctggccc tgctgctgca tgccgccaga 60
cctgatatcg tgctgaccca gagccctccc agcctggcca tgtctctggg caagagagcc 120
accatcagct gcagagccag cgagagcgtg accatcctgg gcagccacct gatctactgg 180
tatcagcaga agcccggcca gccccccaca ctgctgattc agctggcctc caatgtgcag 240
accggcgtgc cagccagatt ttccggcagc ggcagcagaa ccgacttcac cctgaccatc 300
gaccccgtgg aagaggacga cgtggccgtg tactactgcc tgcagagcag aaccatcccc 360
cggacctttg gcggaggcac caagctggaa atcaagggca gcaccagcgg ctccggcaag 420
cctggatctg gcgagggatc taccaaggga cagatccagc tggtgcagag cggccctgag 480
ctgaagaaac ccggcgagac agtgaagatc tcctgcaagg ccagcggcta caccttcacc 540
cactacagca tgaactgggt caagcaggcc cctggcaagg gcctgaagtg gatgggccgg 600
atcaacaccg agacaggcga gcccctgtac gccgacgact ttaagggcag attcgccttc 660
agcctggaaa ccagcgccag caccgcctac ctcgtgatca acaacctgaa gaacgaggac 720
accgccacct ttttctgctc caacgactac ctgtacagct gcgactactg gggccagggc 780
accaccctga cagtgtctag cttcgtgccc gtgttcctgc ccgccaagcc tacaacaacc 840
cctgccccta gacctcccac cccagcccct acaattgcct ctcagcctct gagcctgagg 900
cccgaggctt gtagaccagc tgctggcgga gccgtgcaca ccagaggact ggatttcgcc 960
tgcgacatct acatctgggc ccctctggcc ggcacctgtg gcgtgctgct gctgagcctg 1020
gtgatcaccc tgtactgcaa ccaccggaac agaagcaagc ggagccggct gctgcacagc 1080
gactacatga acatgacccc aagacggcct ggccccaccc ggaagcacta ccagccttac 1140
gcccctccca gagacttcgc cgcctaccgg tccagagtga agttcagcag atccgccgac 1200
gcccctgcct accagcaggg acagaaccag ctgtacaacg agctgaacct gggcagacgg 1260
gaagagtacg acgtgctgga caagcggaga ggccgggacc ccgagatggg cggaaagccc 1320
agacggaaga acccccagga aggcctgtat aacgaactgc agaaagacaa gatggccgag 1380
gcctacagcg agatcggcat gaagggcgag cggaggcgcg gcaagggcca cgatggcctg 1440
taccagggcc tgagcaccgc caccaaggac acctacgacg ccctgcacat gcaggccctg 1500
ccccccagat ga 1512
<210> 4
<211> 503
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 4
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile Tyr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Arg His Tyr Ser Met Asn Trp Val Lys Gln Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Arg Ile Asn Thr Glu Ser Gly Val Pro
195 200 205
Ile Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Val Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Val Ile Asn Asn Leu Lys Asp Glu Asp
225 230 235 240
Thr Ala Ser Tyr Phe Cys Ser Asn Asp Tyr Leu Tyr Ser Leu Asp Phe
245 250 255
Trp Gly Gln Gly Thr Ala Leu Thr Val Ser Ser Phe Val Pro Val Phe
260 265 270
Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro
275 280 285
Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys
290 295 300
Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala
305 310 315 320
Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu
325 330 335
Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg Asn Arg Ser
340 345 350
Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg
355 360 365
Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg
370 375 380
Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
385 390 395 400
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
405 410 415
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
420 425 430
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
435 440 445
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
450 455 460
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
465 470 475 480
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
485 490 495
Met Gln Ala Leu Pro Pro Arg
500
<210> 5
<211> 503
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 5
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro
195 200 205
Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp
225 230 235 240
Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Phe Val Pro Val Phe
260 265 270
Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro
275 280 285
Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys
290 295 300
Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala
305 310 315 320
Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu
325 330 335
Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg Asn Arg Ser
340 345 350
Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg
355 360 365
Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg
370 375 380
Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
385 390 395 400
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
405 410 415
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
420 425 430
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
435 440 445
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
450 455 460
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
465 470 475 480
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
485 490 495
Met Gln Ala Leu Pro Pro Arg
500
<210> 6
<211> 503
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 6
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile Tyr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr His Tyr Ser Met Asn Trp Val Lys Gln Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Arg Ile Asn Thr Glu Thr Gly Glu Pro
195 200 205
Leu Tyr Ala Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Val Ile Asn Asn Leu Lys Asn Glu Asp
225 230 235 240
Thr Ala Thr Phe Phe Cys Ser Asn Asp Tyr Leu Tyr Ser Cys Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Phe Val Pro Val Phe
260 265 270
Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro
275 280 285
Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys
290 295 300
Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala
305 310 315 320
Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu
325 330 335
Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg Asn Arg Ser
340 345 350
Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met Thr Pro Arg
355 360 365
Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala Pro Pro Arg
370 375 380
Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg Ser Ala Asp
385 390 395 400
Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn
405 410 415
Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg
420 425 430
Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly
435 440 445
Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu
450 455 460
Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu
465 470 475 480
Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His
485 490 495
Met Gln Ala Leu Pro Pro Arg
500
<210> 7
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 7
Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly Glu Gly Ser Thr
1 5 10 15
Lys Gly
<210> 8
<211> 507
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 8
Met Leu Leu Leu Val Thr Ser Leu Leu Leu Cys Glu Leu Pro His Pro
1 5 10 15
Ala Phe Leu Leu Ile Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser
20 25 30
Leu Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser
35 40 45
Glu Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val
65 70 75 80
Gln Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp
85 90 95
Phe Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr
100 105 110
Tyr Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr
115 120 125
Lys Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser
130 135 140
Gly Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro
145 150 155 160
Glu Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser
165 170 175
Gly Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro
180 185 190
Gly Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu
195 200 205
Pro Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu
210 215 220
Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu
225 230 235 240
Asp Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp
245 250 255
Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ala Ala Phe
260 265 270
Val Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg
275 280 285
Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg
290 295 300
Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly
305 310 315 320
Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr
325 330 335
Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His
340 345 350
Arg Asn Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn
355 360 365
Met Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr
370 375 380
Ala Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser
385 390 395 400
Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr
405 410 415
Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys
420 425 430
Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn
435 440 445
Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu
450 455 460
Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly
465 470 475 480
His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr
485 490 495
Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505
<210> 9
<211> 506
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 9
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro
195 200 205
Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp
225 230 235 240
Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ala Ala Phe Val
260 265 270
Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro
275 280 285
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
290 295 300
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
305 310 315 320
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
325 330 335
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
340 345 350
Asn Arg Ser Lys Arg Ser Arg Leu Leu His Ser Asp Tyr Met Asn Met
355 360 365
Thr Pro Arg Arg Pro Gly Pro Thr Arg Lys His Tyr Gln Pro Tyr Ala
370 375 380
Pro Pro Arg Asp Phe Ala Ala Tyr Arg Ser Arg Val Lys Phe Ser Arg
385 390 395 400
Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn
405 410 415
Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg
420 425 430
Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro
435 440 445
Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala
450 455 460
Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His
465 470 475 480
Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp
485 490 495
Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505
<210> 10
<211> 512
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 10
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro
195 200 205
Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp
225 230 235 240
Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ala Ala Phe Val
260 265 270
Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro
275 280 285
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
290 295 300
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
305 310 315 320
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
325 330 335
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
340 345 350
Asn Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile
355 360 365
Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp
370 375 380
Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
385 390 395 400
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Gln Gln Gly
405 410 415
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
420 425 430
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
435 440 445
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
450 455 460
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
465 470 475 480
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
485 490 495
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
500 505 510
<210> 11
<211> 502
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 11
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro
195 200 205
Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp
225 230 235 240
Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ala Ala Phe Val
260 265 270
Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro
275 280 285
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
290 295 300
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
305 310 315 320
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
325 330 335
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
340 345 350
Asn Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly
355 360 365
Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His
370 375 380
Ser Thr Leu Ala Lys Ile Arg Val Lys Phe Ser Arg Ser Ala Asp Ala
385 390 395 400
Pro Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu
405 410 415
Gly Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp
420 425 430
Pro Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu
435 440 445
Tyr Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile
450 455 460
Gly Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr
465 470 475 480
Gln Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met
485 490 495
Gln Ala Leu Pro Pro Arg
500
<210> 12
<211> 549
<212> PRT
<213> Artificial Sequence
<220>
<223> Synthetic
<400> 12
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Asp Ile Val Leu Thr Gln Ser Pro Pro Ser Leu
20 25 30
Ala Met Ser Leu Gly Lys Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu
35 40 45
Ser Val Thr Ile Leu Gly Ser His Leu Ile His Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Thr Leu Leu Ile Gln Leu Ala Ser Asn Val Gln
65 70 75 80
Thr Gly Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Arg Thr Asp Phe
85 90 95
Thr Leu Thr Ile Asp Pro Val Glu Glu Asp Asp Val Ala Val Tyr Tyr
100 105 110
Cys Leu Gln Ser Arg Thr Ile Pro Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Ser Thr Ser Gly Ser Gly Lys Pro Gly Ser Gly
130 135 140
Glu Gly Ser Thr Lys Gly Gln Ile Gln Leu Val Gln Ser Gly Pro Glu
145 150 155 160
Leu Lys Lys Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly
165 170 175
Tyr Thr Phe Thr Asp Tyr Ser Ile Asn Trp Val Lys Arg Ala Pro Gly
180 185 190
Lys Gly Leu Lys Trp Met Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro
195 200 205
Ala Tyr Ala Tyr Asp Phe Arg Gly Arg Phe Ala Phe Ser Leu Glu Thr
210 215 220
Ser Ala Ser Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Tyr Glu Asp
225 230 235 240
Thr Ala Thr Tyr Phe Cys Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr
245 250 255
Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ala Ala Phe Val
260 265 270
Pro Val Phe Leu Pro Ala Lys Pro Thr Thr Thr Pro Ala Pro Arg Pro
275 280 285
Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu Ser Leu Arg Pro
290 295 300
Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His Thr Arg Gly Leu
305 310 315 320
Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu Ala Gly Thr Cys
325 330 335
Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr Cys Asn His Arg
340 345 350
Asn Arg Phe Ser Val Val Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile
355 360 365
Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp
370 375 380
Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
385 390 395 400
Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys Pro Pro Gly Gly
405 410 415
Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala Asp Ala His Ser
420 425 430
Thr Leu Ala Lys Ile Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro
435 440 445
Ala Tyr Gln Gln Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly
450 455 460
Arg Arg Glu Glu Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro
465 470 475 480
Glu Met Gly Gly Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr
485 490 495
Asn Glu Leu Gln Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly
500 505 510
Met Lys Gly Glu Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln
515 520 525
Gly Leu Ser Thr Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln
530 535 540
Ala Leu Pro Pro Arg
545
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| CD20 ТЕРАПИЯ, CD22 ТЕРАПИЯ И КОМБИНИРОВАННАЯ ТЕРАПИЯ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) K CD19 | 2016 |
|
RU2752918C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| Т-КЛЕТКИ, МОДИФИЦИРОВАННЫЕ ХИМЕРНЫМ РЕЦЕПТОРОМ АНТИГЕНА, НАЦЕЛЕННЫМ НА CS1, ДЛЯ ЛЕЧЕНИЯ АМИЛОИДОЗА AL | 2018 |
|
RU2774895C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА CLL-1 | 2015 |
|
RU2741120C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
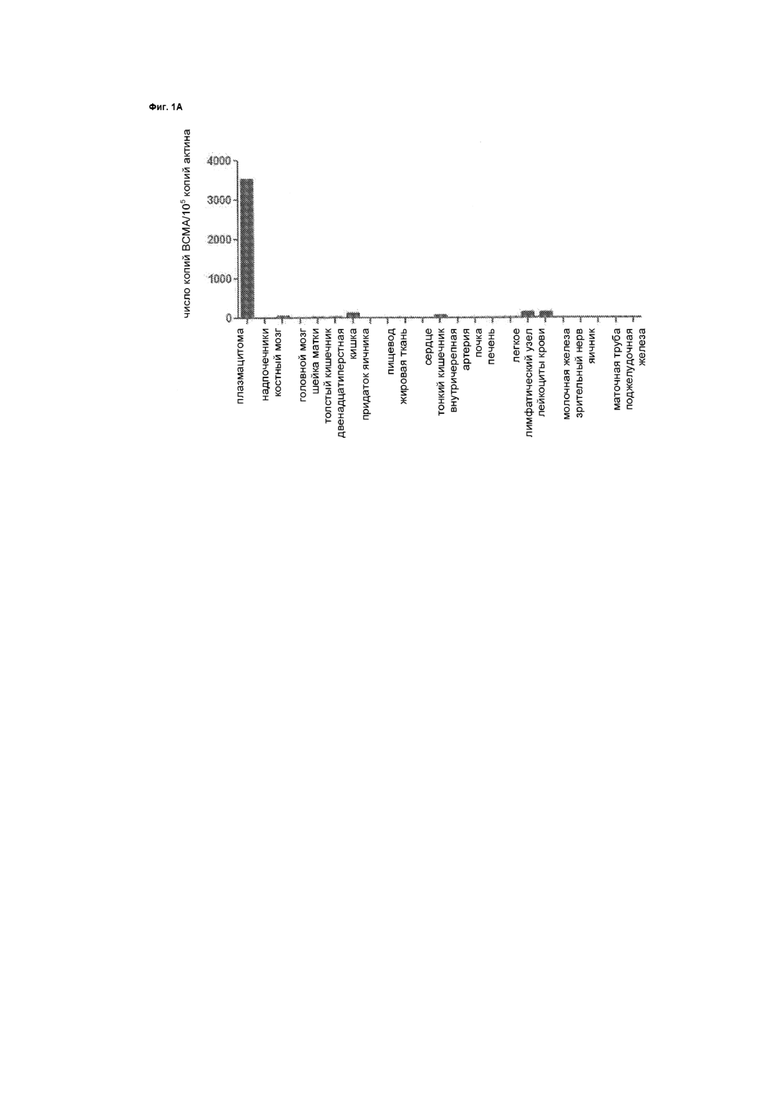
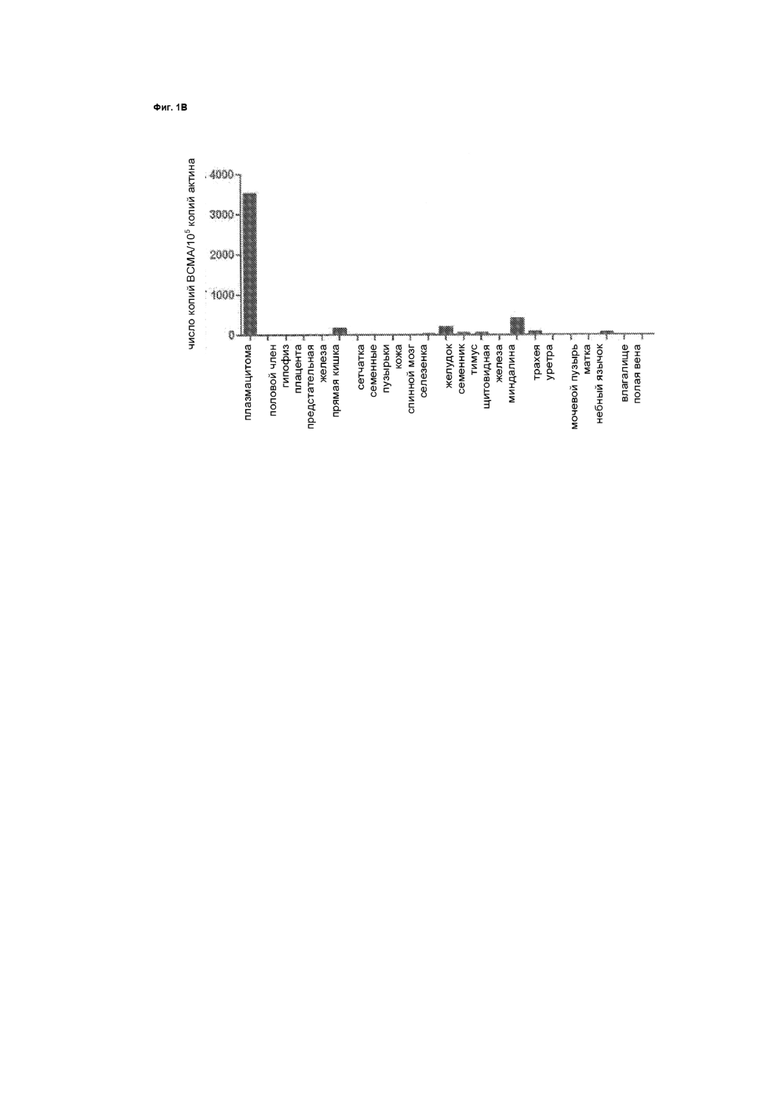

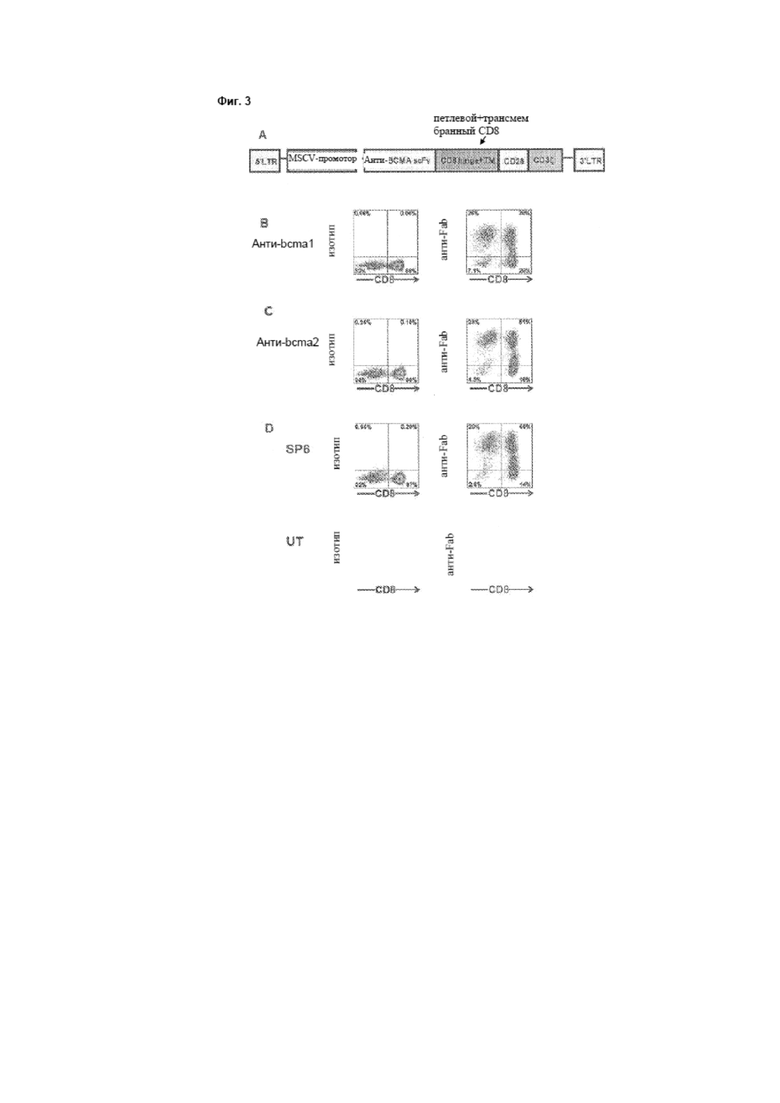

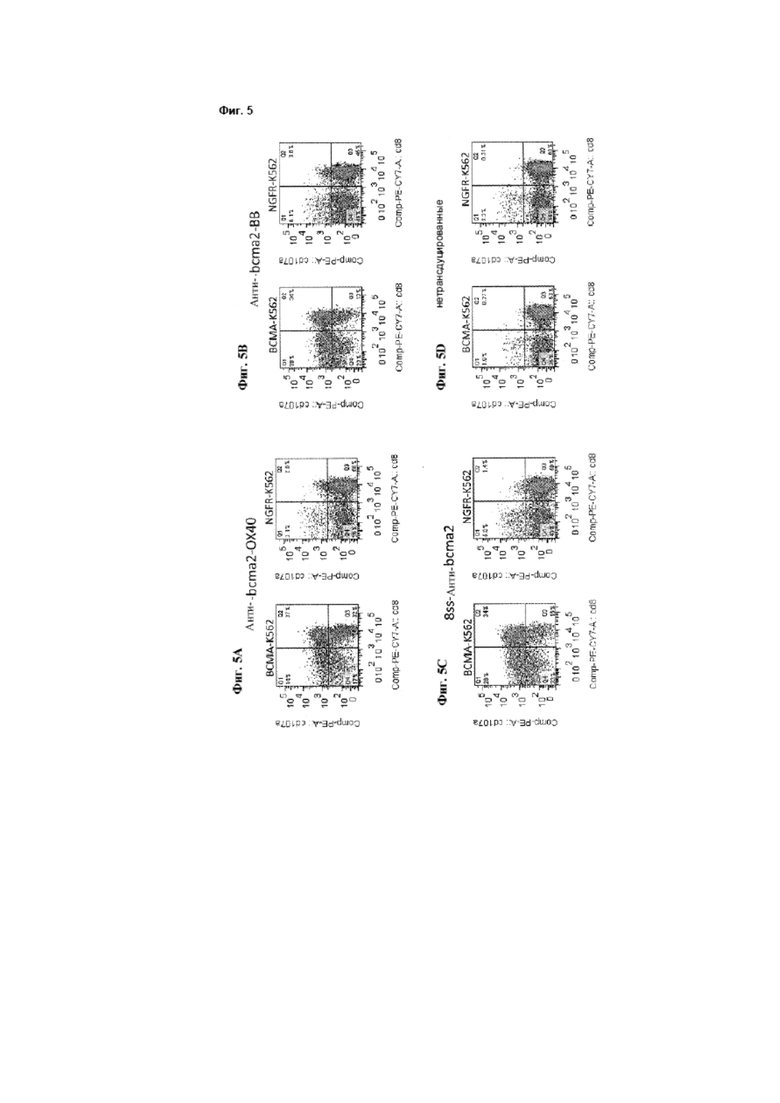

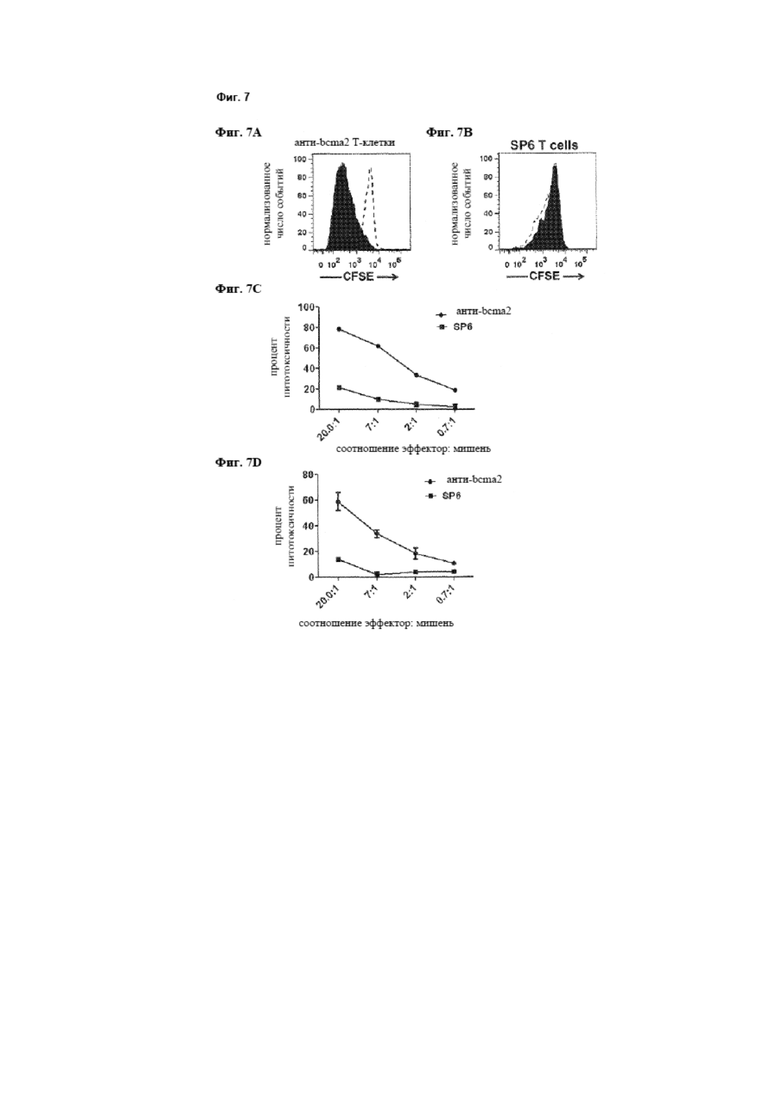
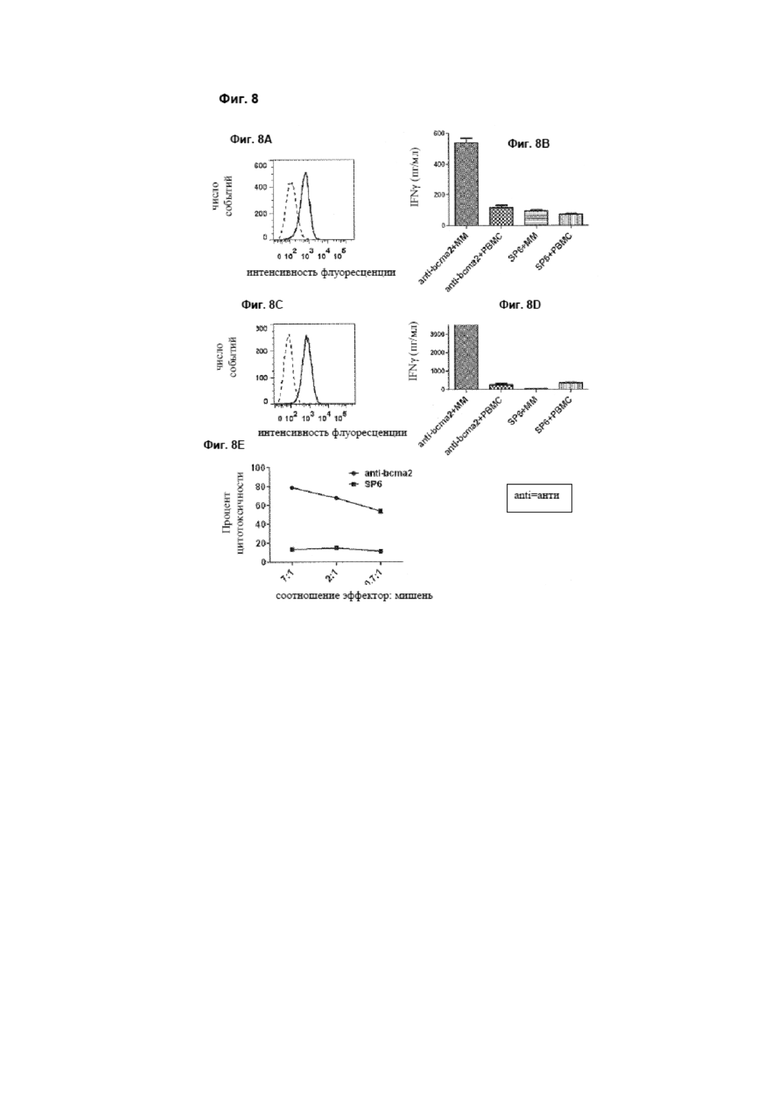
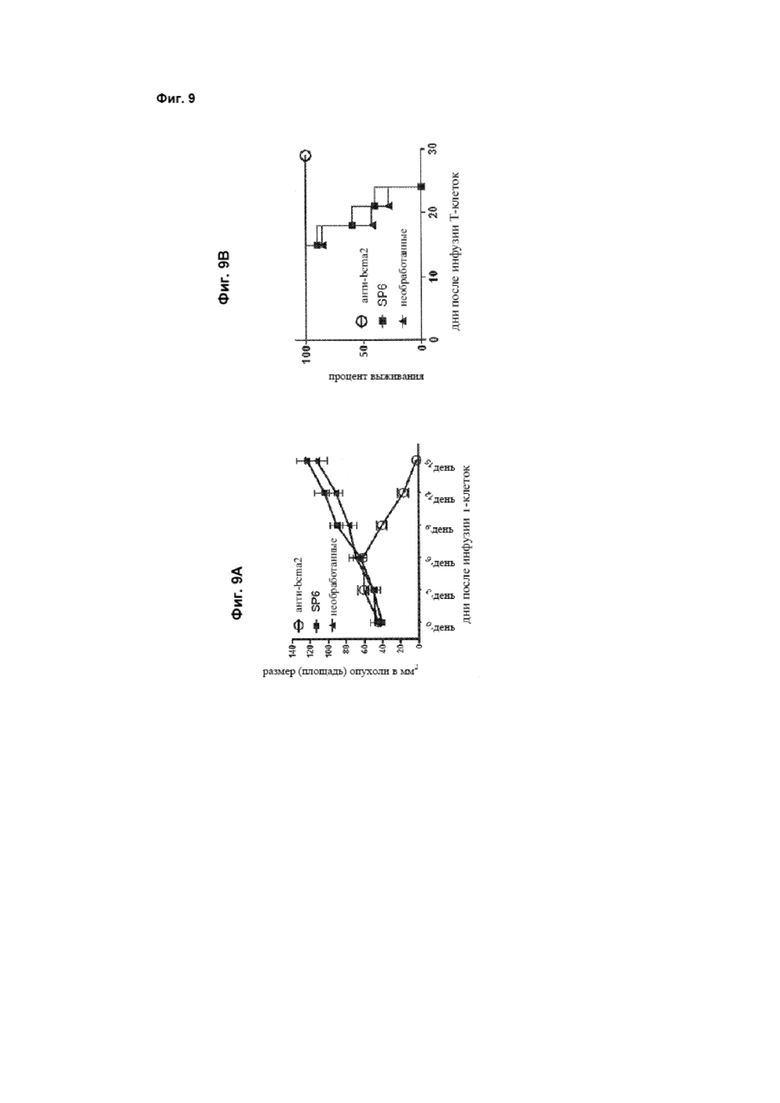
Изобретение относится к области биотехнологии, в частности к химерному рецептору антигена (CAR), направленному против антигена созревания В-клеток (ВСМА), а также к содержащей его композиции. Также раскрыты молекула нуклеиновой кислоты, кодирующая вышеуказанный CAR, а также вектор и клетка, его содержащие. Изобретение эффективно для лечения млекопитающего, страдающего от ВСМА-экспрессирующего рака. 13 н. и 13 з.п. ф-лы, 9 ил., 4 табл., 6 пр.
1. Химерный рецептор антигена (CAR), направленный против антигена созревания В-клеток (ВСМА), включающий:
(a) одноцепочечный вариабельный фрагмент (scFv), направленный против ВСМА, содержащий (i) определяющую комплементарность область (CDR) тяжелой цепи - CDR1, (ii) CDR2 тяжелой цепи, (iii) CDR3 тяжелой цепи, (iv) CDR1 легкой цепи, (v) CDR2 легкой цепи и (vi) CDR3 легкой цепи, имеющий аминокислотную последовательность, выбранную из группы, состоящей из: SEQ ID №4, SEQ ID №5, SEQ ID №6, SEQ ID №8, SEQ ID №9, SEQ ID №10, SEQ ID №11 и SEQ ID №12;
(b) трансмембранный домен CD28 или CD8α; и
(c) один или более внутриклеточных доменов сигналинга Т-клеток, выделенных из белка, выбранного из группы, состоящей из: CD28, CD3ζ, FcRγ, CD27, ОХ40 и белка 4-1ВВ.
2. CAR по п. 1, включающий трансмембранный домен CD28.
3. CAR по п. 1, включающий трансмембранный домен CD8α.
4. CAR по любому из пп. 1-3, включающий внутриклеточные домены сигналинга Т-клеток CD28 и CD3ζ.
5. CAR по любому из пп. 1-3, включающий внутриклеточные домены сигналинга Т-клеток ОХ40 и CD3ζ.
6. CAR по любому из пп. 1-3, включающий внутриклеточные домены сигналинга Т-клеток 4-1ВВ и CD3ζ.
7. CAR по п. 1, включающий трансмембранный домен CD8α и внутриклеточные домены сигналинга Т-клеток 4-1ВВ и CD3ζ.
8. Выделенная молекула нуклеиновой кислоты, кодирующая CAR по любому из пп. 1-7.
9. Экспрессионный вектор, содержащий выделенную молекулу нуклеиновой кислоты по п. 8.
10. Экспрессионный вектор по п. 9, где указанный вектор представляет собой ретровирусный вектор.
11. Экспрессионный вектор по п. 10, где указанный ретровирусный вектор представляет собой лентивирусный вектор.
12. Выделенная клетка для экспрессии CAR по любому из пп. 1-7, содержащая выделенную молекулу нуклеиновой кислоты по п. 8.
13. Выделенная клетка для экспрессии CAR по любому из пп. 1-7, содержащая вектор по любому из пп. 9-11.
14. Выделенная клетка по п. 12 или 13, которая представляет собой Т-клетку.
15. Выделенная клетка по п. 12 или 13, которая является натуральным киллером (NK-клеткой).
16. Выделенная клетка по п. 14, где указанная Т-клетка представляет собой CD8+ Т-клетку.
17. Композиция для лечения млекопитающего, страдающего от ВСМА-экспрессирующего рака, содержащая эффективное количество выделенной клетки по любому из пп. 12-16 и фармацевтически приемлемый носитель или разбавитель.
18. Способ разрушения клеток множественной миеломы, который включает контактирование клеток множественной миеломы, экспрессирующих ВСМА, с одной или более чем одной из выделенных клеток по п. 14, где CAR экспрессируется и связывается с ВСМА на клетках множественной миеломы, и клетки множественной миеломы разрушаются.
19. Способ разрушения клеток множественной миеломы, который включает контактирование клеток множественной миеломы, экспрессирующих ВСМА, с одной или более чем одной из выделенных клеток по п. 15, где CAR экспрессируется и связывается с ВСМА на клетках множественной миеломы, и клетки множественной миеломы разрушаются.
20. Способ по п. 18 или 19, где клетки множественной миеломы являются человеческими.
21. Применение CAR по любому из пп. 1-7 при производстве лекарственного средства для разрушения раковых клеток, экспрессирующих ВСМА.
22. Применение выделенной нуклеиновой кислоты по п. 8 при производстве лекарственного средства для разрушения раковых клеток, экспрессирующих ВСМА.
23. Применение экспрессионного вектора по любому из пп. 9-11 при производстве лекарственного средства для разрушения раковых клеток, экспрессирующих ВСМА.
24. Применение выделенной клетки по любому из пп. 12-16 при производстве лекарственного средства для разрушения раковых клеток, экспрессирующих ВСМА.
25. Применение композиции по п. 17 при производстве лекарственного средства для разрушения раковых клеток, экспрессирующих ВСМА.
26. Применение по любому из пп. 21-25, где раковые клетки представляют собой клетки множественной миеломы или клетки лимфомы Ходжкина.
| WO 2011041093 А1, 07.04.2011 | |||
| RYAN MAUREEN C | |||
| et al., Antibody targeting of B-cell maturation antigen on malignant plasma cells, Mol Cancer Ther, 2007, 6, pp.3009-3018 | |||
| WO 2012031744 A1, 15.03.2012 | |||
| WO 2010104949 A2, 16.09.2010 | |||
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ | 2010 |
|
RU2444570C1 |

