ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка заявляет приоритет в соответствии с разделом 119(e) по предварительной заявке США с серийным номером 62/356410, поданной 29 июня 2016 года под названием «Структурные пептидные ингибиторы агрегации альфа-синуклеина», содержание которой включено в данный документ посредством ссылки.
ЗАЯВЛЕНИЕ О ГОСУДАРСТВЕННОЙ ПОДДЕРЖКЕ
Данное изобретение было сделано при государственной поддержке по гранту № AG029430, присужденному Национальным институтом здоровья. Государство обладает определенными правами на данное изобретение.
Перечень последовательностей
Данная заявка включает перечень последовательностей, который был подан в электронном виде в формате с кодировкой ASCII и, таким образом, включен в данный документ посредством ссылки во всей своей полноте. Указанная копия ASCII, созданная 9 августа 2017 года, называется 30435_305-WO-U2_SL.txt и ее размер составляет 18456 байт.
УРОВЕНЬ ТЕХНИКИ
Болезнь Паркинсона (PD), деменция с тельцами Леви (DLB) и множественная системная атрофия (MSA) совместно классифицируются как синуклеинопатии, которые представляют собой класс нейродегенеративных заболеваний, характеризуемых патологическим накоплением белка αсинуклеина (αsyn) в нейронах. Совместно они образуют вторую наиболее распространенную форму нейродегенеративного заболевания. Пресинапатический белок α-синуклеин (αsyn), встречающийся как в растворимых, так и в мембраноассоциированных фракциях головного мозга, накапливается, например, при болезни Паркинсона (PD). Такие агрегаты являются основным компонентом телец Леви, определяющей гистологической характеристикой данного нейродегенеративного заболевания, и, как было показано, сопровождают нейрональное повреждение7. Не ограничиваясь каким-либо определенным механизмом, предполагается, что это и два другие наблюдения указывают на накопление αsyn как молекулярную причину болезни Паркинсона8. Первым является то, что в семьях с наследственными формами PD встречаются мутации αsyn, такие как A53T, и избыток телец Леви9,10,11. Вторым является то, что в семьях с двумя или тремя копиями гена, кодирующего αsyn, развивается PD с ранним началом, предположительно, в связи с тем, что при высоких концентрациях αsyn проникает в амилоид12, 13.
На основании структурных исследований, был предложен ряд различных моделей фибрилл αsyn. Ограниченный протеолиз и исследования на основании NMR указывают на то, что кор фибриллы состоит из остатков 30-100 (Miake H (2002) J Biol Chem 277(21):19213-19219). Исследования кристаллических структур и исследования на основании NMR указывают на различные модели фибрилл αsyn. В одной модели на основании кристаллических структур два мономера на лист образуют удлиненные стерические молнии (Rodriguez JA, et al. (2015) Nature 525(7570):486-490). Во второй модели на основании ssNMR была показана топология типа «греческий ключ» с одним мономером на слой амилоида (Tuttle MD, et al. (2016) Nat Struct Mol Biol 23(5):409-415). В своей совокупности эти исследования указывают на то, что αsyn может образовывать полиморфные фибриллярные архитектуры. Сегмент 68-78 αsyn, называемый NACore, может образовывать кор фибрилл αsyn. NACore находится в домене, состоящем из 35 остатков NAC (неамилоидного β-компонента), обнаруженного в амилоидных отложениях (Rodriguez JA, et al. (2015) Nature 525(7570):486-490). NACore легко агрегирует и агрегаты демонстрируют свойства, такие как паттерн дифракции и цитотоксичность, аналогичные полноразмерному αsyn. Помимо этого, β-синуклеин, гомолог, не содержит остатки 74-84 и не встречается в амилоидных отложениях, и, как было показано ранее, удаление остатков 71-82 не снижает агреграции и токсичности in vitro и в модели на дрозофиле (Giasson et al. (2001) J Biol Chem 276(4):2380-238, и Periquet et al. (2007) J Neurosci 27(12):3338-3346). Кроме этого, модификация в Thr72 предупреждает его способность к агрегации.
Авторы данного изобретения недавно показали, что возможно эффективно останавливать агрегацию связанного с болезнью Альцгеймера тау-белка и полученного из семенной жидкости усилителя инфекции, вызываемой вирусом ВИЧ (SEVI - semen-derived enhancer of HIV virus infection), при использовании коротких аминокислотных ингибиторов, разработанных для специфического «кэппирования» растущих агрегатов (Sievers et al.36; патент США № 875403457). Они также показали, что та же самая стратегия дизайна может привести к разработке проникающего в клетку пептида, который ингибирует образование амилоида p53, восстанавливает функцию мутантного p53 в линиях раковых клеток и снижает опухолевую пролиферацию (заявка на патент США № 2014/03738758). Соответственно имеет место перспективное терапевтическое окно, которое целенаправленно воздействует на агрегацию амилоида αсинуклеина, который представляет собой основной компонент внутриклеточных отложений в форме телец Леви (LB) при болезни Паркинсона и соответствующих нейродегенеративных нарушениях.
Существует необходимость в идентификации агентов, которые предупреждают и/или ингибируют агрегацию и/или цитотоксичность αсинуклеина.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Несмотря на то, что образование αsyn было подробно описано, незначительный прогресс был сделан в дизайне лекарственных препаратов, которые ингибируют агрегацию αsyn или снижают прионоподобное распространение агрегатов αsyn («затравок») из клетки в клетку. Перспективные подходы включают в себя антитела, которые секвестируют агрегаты αsyn, а также низкомолекулярные стабилизаторы, которые связывают мономеры αsyn. (см., напр., Mandler M, et al. (2015) Mol Neurodegener 10(1). doi:10.1186/s13024-015-0008-9; Wrasidlo W, et al. (2016) Brain:aww238). С помощью атомной структуры NACore в качестве матрицы, нами был разработан новый класс ингибиторов, пептидные агенты, которые связывают затравки αsyn и предупреждают их рост и удлинение. Как показано ниже, эти ингибиторы ингибируют образование и затравку фибрилл αсинуклеина в ряде модельных систем, а также распространение агрегатов a-синуклеина.
Данное изобретение, раскрытое в данном документе, имеет ряд вариантов осуществления. В одном варианте осуществления данного изобретения предложена заявленная композиция, содержащая по меньшей мере один ингибиторный пептид, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина. В типичных вариантах осуществления данного изобретения ингибиторный пептид содержит последовательность GAVVWGVTAVKK (SEQ ID NO: 3) или RAVVTGVTAVAE (SEQ ID NO: 4). Необязательно ингибиторный пептид содержит последовательность GAVVWGVTAVKKKKK (SEQ ID NO: 5), GAVVWGVTAVKKGRKKRRQRRRPQ (SEQ ID NO: 6); или YGRKKRRQRRRAVVTGVTAVAE (SEQ ID NO: 7). В определенных вариантах осуществления данного изобретения композиция содержит множество ингибиторных пептидов. В типичном случае ингибиторный(ингибиторные) пептид(пептиды) составляет(составляют) от 6 до 30 аминокислот в длину.
В композициях на основе ингибиторного пептида по данному изобретению по меньшей мере одна из аминокислот в ингибиторном пептиде может содержать не встречающуюся в природе аминокислоту (например, D-аминокислоту или аминокислоту, содержащую фрагмент N-метильной группы); и/или ингибиторный пептид связан с гетерологичным пептидным тэгом. Такие гетерологичные пептидные тэги включают в себя аминокислотные последовательности, которые повышают растворимость пептида; или аминокислотные последовательности, которые облегчают контроль или манипуляцию с пептидом; или аминокислотные последовательности, которые облегчают вхождение пептида в клетку млекопитающего. Необязательно эти пептидные композиции включают фармацевтически приемлемый носитель и эксципиент, стабилизирующий пептид.
В другом варианте осуществления данного изобретения предложен вектор экспрессии, кодирующий ингибиторный пептид, который ингибирует агрегацию αсинуклеина в результате связывания с остатками 68-78 αсинуклеина. В связанном варианте осуществления предложен набор, содержащий пептид, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина, или вектор экспрессии, кодирующий такой пептид. Варианты осуществления данного изобретения также включают в себя способ получения пептида, раскрытого в данном документе, путем его синтеза химическим путем или его получения рекомбинантным путем. В еще одном варианте осуществления данного изобретения предложен комплекс, содержащий α-синуклеин, и пептид, который ингибирует агрегацию αсинуклеина в результате связывания с остатками 68-78 αсинуклеина.
В еще одном варианте осуществления данного изобретения предложен способ снижения или ингибирования агрегации αсинуклеина (SEQ ID NO: 1), включающий приведение в контакт амилоидных фибрилл αсинуклеина с ингибиторным пептидом, раскрытым в данном документе, в количестве, достаточном для снижения или ингибирования агрегации αсинуклеина. Необязательно в этом способе амилоидные фибриллы αсинуклеина включены в вариант осуществления in vivo. Как альтернативный вариант в этом способе амилоидные фибриллы αсинуклеина включены в вариант осуществления in vitro. В связанном варианте осуществления предложен способ модуляции размера или скорости роста амилоидные фибриллы αсинуклеина, включающий приведение в контакт фибриллы с некоторым количеством по меньшей мере одного ингибиторного пептида, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина в среде, в которой ингибиторный пептид контактирует с остатками 68-78 αсинуклеина таким образом, что приведенная в контакт фибрилла αсинуклеина проявляет модулируемый размер или скорость роста.
В еще одном варианте осуществления данного изобретения предложен способ наблюдения наличия или отсутствия амилоидных фибрилл αсинуклеина в биологическом образце, включающий комбинирование биологического образца с пептидом, который связывается с остатками 68-78 αсинуклеина, способствуя связыванию пептида с амилоидными фибриллами α-синуклеина, которые могут присутствовать в биологическом образце, а затем контроля этой комбинации в отношении наличия комплексов, образованных между амилоидными фибриллами αсинуклеина и пептидом; при этом наличие указанных комплексов свидетельствует о наличии амилоидных фибрилл αсинуклеина в биологическом образце. В данном варианте осуществления один из наших ингибиторов может быть связан с визуализирующим агентом, таким как радиоактивная метка, рентгеноконтрастная метка, флуоресцентный краситель, флуоресцентный белок, колориметрическая метка и т.п. (например, для облегчения способа визуализации, такого как МРТ или ПЭТ), и наш ингибитор будет связываться с фибриллами альфа-синуклеина в головном мозге пациента, способствуя визуализации фибрилл для диагностики последующего прогрессирования заболевания.
Другие цели, характеристики и преимущества данного изобретения будут очевидны специалистам в данной области из следующего подробного описания. Однако следует понимать, что подробное описание и конкретные примеры, хотя в них указаны варианты осуществления данного изобретения, приведены лишь с целью иллюстрации, а не ограничения. Многие изменения и модификации в объеме данного изобретения могут быть выполнены без отклонения от его идеи, и данное изобретение включает в себя все такие модификации.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Файл патента или заявки содержит по меньшей мере один графический материал, выполненный в цвете.
Фиг. 1. Дизайн ингибиторов затравки αsyn. (A) Структурная схема ингибиторов агрегации αsyn. Структура NACore состоит из двух самокомплементарных β-листов, образующих стерическую молнию. Были идентифицированы три типа ингибиторов (каштанового, оранжевого цвета и цвета морской волны), которые связывают один или оба конца. (B) Энергии связывания различных ингибиторов, рассчитанные в соответствии с Rossetta, свидетельствуют о том, что S37 и S62 связывают обе поверхности раздела, в то время как S71, как предполагается, связывается только с одной поверхностью раздела. Комплементарность формы всех трех ингибиторов является высокой. (C) Анализ с использованием тиофлавина T в целях измерения агрегации αsyn и влияния ингибиторов. Добавляли 50 мкM αsyn и ингибиторы в 5-кратном молярном избытке.
Фиг. 2. Агрегаты αsyn, образованные в присутствии ингибиторов, не являются компетентными в отношении затравки. (A) Экспериментальная схема анализа затравки в культуре клеток. αsyn агрегировали в присутствии ингибиторов и смесь трансфицировали в клетки HEK293, экспрессирующие меченый YFP αsyn ДТ или A53T αsyn (зеленый). A53T представляет собой родственный вариант альфа-синуклеина, который вызывает ранее начало болезни Паркинсона. При трансфекции эндогенный αsyn образовывал флуоресцентные пятна (красные). (B) NACore не влиял на затравливающую способность фибрилл αsyn. (C,D,E) 50 мкM αsyn, агрегировавшего в 10-, 5-, 2- и 1-кратном избытке S37, S61 и S62, не были компетентными в отношении затравки, измеряемой с помощью подсчета общего количества частиц, образованных на лунку, как в экспрессирующих клетках ДТ, так и в экспрессирующих клетках A53T. (F) 50 мкM αsyn, агрегировавшего в избытке S71, не были компетентными в отношении затравки, измеряемой с помощью подсчета общего количества частиц, образованных на лунку, как в экспрессирующих клетках ДТ, так и в экспрессирующих клетках A53T. Результаты представлены в виде среднего+SD (n=3). Статистическую значимость анализировали с помощью двухфакторного дисперсионного анализа.
Фиг. 3. Ингибиторы предупреждают затравку в культуре клеток. (A) Экспериментальная схема анализа затравки в культуре клеток. 125 нМ рекомбинатных фибрилл αsyn трансфицировали различными количествами ингибиторов и контролировали агрегацию в динамике. (B) NACore не влияет на затравливающую способность. (C, D, E, F) S37, S61, S62 и S71 снижают затравливающую способность αsyn. Все данные представлены в виде частиц в расчете на лунку и нормализованы на частицы, включенные в лунки, обработанные буфером. Результаты представлены в виде среднего+SD (n=3). Статистическую значимость анализировали с помощью двухфакторного дисперсионного анализа.
Фиг. 4. Экстрагированные филаменты из ткани головного мозга, характерной для PD, затравливают агрегацию αsyn in vitro и в культуре клеток. (A) Протокол экстракции нерастворимых в саркозиле белковых филаментов, характерных для PD, из тканей головного мозга. (B,C,D) 2% затравки от 3 различных субъектов индуцируют быструю агрегацию αsyn с 4-10-кратным увеличением флуоресценции ThT. (E,F,G) αsyn, затравленный филаментами от субъектов с PD, индуцирует образование большего количества частиц, чем αsyn в отдельности. Все данные представлены в виде частиц в расчете на лунку и нормализованы на частицы, включенные в лунки, обработанные буфером. Результаты представлены в виде среднего+SD (n=3). Статистическую значимость анализировали с помощью двухфакторного дисперсионного анализа.
Фиг. 5. Ингибиторы предупреждают затравку филаментами, экстрагированными из тканей, характерных для PD. Филаменты от 4 различных субъектов исследовали в отношении агрегации αsyn и контролировали с помощью анализа ThT. S61 и S62 были эффективными в отношении всех затравок, которые были исследованы. S71 был эффективным в отношении затравок B и D.
Фиг. 6. Фибриллы αsyn, образованные в присутствии затравок филаментов, характерных для PD, и ингибиторов, не являются компетентными в отношении затравки. (A) Экспериментальная схема анализа затравки культуры клеток. (B, C, D) Ингибиторы исследовали в отношении инициализации образования агрегатов тремя различными тканями, экстрагированными из тканей головного мозга, характерных для PD. S71 был эффективным в отношении всех трех затравок, в то время как S61 был эффективным в отношении затравок B. Все данные представлены в виде частиц в расчете на лунку и нормализованы на частицы, включенные в лунки, обработанные буфером. Результаты представлены в виде среднего+SD (n=3). Статистическую значимость анализировали с помощью двухфакторного дисперсионного анализа.
Фиг. 7. Ингибиторы снижают инициализацию образования агрегатов фибрилл αsyn, затравленных филаментами PD, в культуре клеток. (A) Экспериментальная схема анализа затравки в культуре клеток. (B,C,D) Фибриллы αsyn, образованные в присутствии двух различных связанных с PD филаментами, трансфицировали в клетки HEK с YFP-αsyn и после этого измеряли флуоресценцию в течение 6 дней. S37 предупреждал затравку при концентрациях 12,5 мкм - 1,25 мкМ. S61 предупреждал затравку с длительным эффектом на 2-е сутки. S71 предупреждал затравку при концентрациях 6,25 мкM. Все данные представлены в виде количества частиц в расчете на лунку и нормализованы на частицы, включенные на 0-е сутки до трансфекции. Результаты представлены в виде среднего+SD (n=3). Статистическую значимость анализировали с помощью двухфакторного дисперсионного анализа.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не указано иное, то предполагается, что все термины в данной области техники, обозначения и другие научные термины или терминология, используемая в данном документе, имеют значения, обычно понимаемые специалистами в области, к которой принадлежит данное изобретение. В некоторых случаях термины с обычно понимаемыми значениями определены в данном документе для ясности и/или для справочного материала, и включение таких определений в данном документе не следует обязательно подразумевать как представление существенной разницы по сравнению с тем, что обычно понимается в данной области техники. Многие из методик и процедур, описанных или упоминаемых в данном документе, широко распространены и обычно применяются с использованием стандартной методологии специалистами в данной области. Все публикации, патенты и заявки на патенты, упомянутые в данном документе, включены в данный документ посредством ссылки во всей своей полноте для всех целей. В описании предпочтительного варианта осуществления можно сделать ссылку на прилагаемые графические материалы, которые образуют его часть, и в котором показан посредством иллюстрации конкретный вариант осуществления, в котором данное изобретение может быть осуществлено на практике. Необходимо понимать, что могут быть применены другие варианты осуществления и могут быть выполнены структурные изменения без отклонения от объема данного изобретения.
αСинуклеин, белок из 140 аминокислот, встречается в амилоидных отложениях в нейронах при патологических состояниях. Причинная связь между образованием амилоида αsyn и прогрессированием заболевания подкрепляется результатами, согласно которым дупликации генов и семейные мутации, которые повышают амилоидную нагрузку, также вызывают PD с ранним началом, и более 90% пациентов со спорадической PD имеют положительное окрашивание в отношении отложений αsyn. В данном изобретении также предложен структурный подход, способствующий остановке агрегации αsyn. Нами была выдвинута гипотеза о том, что атомные структуры NACore и PreNAC сохраняются в затравках амилоида αsyn и рекрутируют эндогенный белок в конформацию молнии. С помощью атомной структуры NACore нами были разработаны ингибиторы, которые препятствуют образованию фибрилл, и была исследована их эффективность in vitro и в культуре клеток. Эта же процедура может быть использована для дизайна ингибиторов на основе PreNAC. Ингибиторы оптимизируют для кэппирования концов фибриллы, предупреждая последующее добавление мономеров. Нами была использована компьютерная программа Rosetta для дизайна пептидных последовательностей, которые эффективно взаимодействуют с сегментом NACore. Функция энергии, используемая Rosetta, принимает во внимание электростатические взаимодействия, водородное связывание, силы Ван-дер-Ваальса, среди других терминов, для определения энергии связывания. После того, как было показано, что конкретный остаток приводил к эффективному связыванию в определенном положении разработанного пептида, он может быть фиксирован и не допущен к последующему дизайну, в то время как оставшуюся часть последовательности совершенствуют дополнительно. Нами был осуществлен этот процесс фиксации и последующего дизайна итерационно до тех пор, пока не была идентифицирована оптимальная совокупность последовательностей. Такой процесс рационального дизайна способствовал компьютерному отбору на порядок больше ингибиторных последовательностей, чем было возможно исследовать экспериментально.
Нами были осуществлены проверки кристаллов пептидных сегментов в домене NAC и прилегающих участках с поиском структурной информации касательно молекулярной основы агрегации и токсичности αsyn. С помощью алгоритма ZipperDB55 в домене NAC были идентифицированы кристаллизуемые сегменты, образующие амилоид. В частности, в центральном сегменте αsyn нами были идентифицированы и концентрированы остатки 68-78, которые обозначаются в данном документе как NACore в связи с их важной ролью как в агрегации, так и в цитотоксичности αsyn. NACore представляет собой образующий фибриллы кор домена NAC (Non Amyloid-β Component), состоящего из 35 остатков (остатки 61-95) полного αсинуклеина, состоящего из 140 остатков. В некоторых исследованиях нами был использован субфрагмент NACore, состоящий из остатков 69-77. В других исследованиях нами была кристаллизовала и определена структура остатков 47-56 αсинуклеина, обозначаемая PreNAC, которая образует другую стерическую молнию. Идентифицированные сегменты синтезировали химическим путем и кристаллизовали, а их трехмерные структуры определяли с помощью микрокристаллографии и микроэлектронной дифракции (MicroED).
В целях обнаружения наиболее эффективных ингибиторов около 100 различных схем исследовали эмпирически с последовательными циклами оптимизации дизайна ингибитора. Например, нами было замечено, что расположение тэга на N- или C-конце может влиять на его эффективность. Кроме того, тип добавляемой модификации также мог влиять на его эффективность. В нашем случае лишь замена Trp в Thr72 была эффективной, в то время как замена Arg не была таковой. Несмотря на то, что компьютерный подход не является достаточно мощным для идентификации одного эффективного дизайна, он может сузить наш поиск кандидатных ингибиторов, которые затем могут быть усовершенствованы посредством рационального дизайна.
Эффективность кэппинг-ингибиторов в предупреждении затравки исследовали с помощью клеточного анализа. В этой системе трансфекция наномолярных количеств затравок αsyn вызывала агрегацию эндогенного белка. Агрегаты проявляли амилоидогенные свойства, а именно связывание с амилоид-специфичными малыми молекулами, точно перенесенными при клеточном делении, и заметной специфичностью (Sanders DW, et al. (2014) Neuron 82(6):1271-1288). Например, фибриллы αsyn могут затравливать только белок αsyn в агрегаты. Примечательно, что в этой системе нами не было замечено острой клеточной смерти при образовании агрегатов, при этом имеет место лишь незначительное замедление клеточной пролиферации. Наши ингибиторы предупреждали образование пятен в этом системе, при этом однократное введение ингибиторов было эффективным в течение 2-3 дней.
Затравка в контексте амилоидного заболевания представляет собой последовательный перенос патологических белковых агрегатов в связанных тканях. Этот процесс способствует прогрессированию и тяжести нейродегенеративных заболеваний. К настоящему времени отсутствуют терапевтические средства, которые специфично целенаправленно воздействуют на затравку, в частности, в связи с отсутствием информации о структурных свойствах патологических затравок. Данная заявка относится, например, к дизайну, синтезу и функциональной характеристике пептидов, которые специфично связываются с агрегатами αсинуклеина (αsyn) и блокируют, ингибируют и/или уменьшают агрегацию αsyn и/или цитотоксичность αsyn. Пептидные ингибиторы специфично «кэппируют» рост агрегатов αсинуклеина. В некоторых вариантах осуществления пептиды сливают с проникающими в клетку пептидами, которые усиливают их доставку в клетки.
Помимо спонтанной агрегации внутриклеточных амилоидных фибрилл αsyn, вторым феноменом, которым способствует прогрессированию заболевания, является прионоподобное распространение агрегатов αsyn (Goedert M (2015). Science 349(6248):1255555-1-1255555-9). Стадирование по Braak показало, что данная патология постепенно распространяется в течение времени через связанные участки головного мозга, а культивирование клеток и животные модели показывают, что незначительные количества агрегатов αsyn может выступать в качестве затравок и индуцировать агрегацию нативного белка (см., например, Braak H, et al. (2003) Neurobiol Aging 24(2):197-211; Braak et al. (2009) Adv Anat Embryol Cell Biol 201:1-119; Masuda-Suzukake M, et al. (2013) Brain 136(4):1128-1138; Desplats P, et al. (2009) Proc Natl Acad Sci U S A 106(31):13010-13015; Luk KC, et al. (2009) Proc Natl Acad Sci 106(47):20051-20056). Несмотря на отличие от классических прионов, которые могут передаваться от человека к человеку, этот феномен «затравки» выглядит движущей силой прогрессирования заболеваний.
Замечено, что экстрагированные у пациентов фибриллы отличаются по затравливающей способности и проявляют штаммоподобные характеристики. In vitro экстрагированные у пациентов с PD фибриллы вызывали существенное повышение агрегации αsyn, а в модели культуры клеток затравленные образцы повышали образование пятен. Примечательно, что в отличие от предыдущих отчетов, в которых описаны полученные от пациентов затравки филаментов αsyn в культуре клеток, в наших анализах нами не было замечено затравки в культуре клеток (Prusiner SB, et al. (2015) Proc Natl Acad Sci 112(38):E5308-E5317; and Woerman AL, et al. (2015) Proc Natl Acad Sci 112(35):E4949-E4958). В предыдущих исследованиях использовали ткани черной субстанции, в то время как нами были использованы ткани лобной и височной области, которые могли отличаться по затравливающей способности. Действительно, как было показано в предыдущих отчетах, фибриллы, экстрагированные из различных областей головного мозга, отличаются по затравливающей способности, аналогично различных штаммам (Prusiner SB, et al. (2015) Proc Natl Acad Sci 112(38):E5308-E5317). Кроме того, различные ингибиторы отличались эффективностью в отношении различных затравок. Например, S61 был эффективным только в отношении затравок A и B, в то время как S71 был эффективным в отношении затравок B и D. Недавно была описана ЯМР-структура фибрилл полноразмерного αsyn, в котором сегмент NACore не был обнаружен в расширенной конформации молнии, несмотря на то, что сегмент 68-78 обнаружен в коре фибриллы (Tuttle MD, et al. (2016) Nat Struct Mol Biol 23(5):409-415). В противном случае может иметь место ситуация, в которой описанные ингибиторы могут связываться с конформацией NACore в ЯМР-струструре. Также возможно, что ЯМР-структура и структура стерической молнии представляют собой различные полимормы. В отсутствие диагностического способа идентификации различных полимформов у субъектов-людей, теоретически мог бы быть пригодным коктейль из различных ингибиторов, целенаправленно воздействующих на различные полиморфы.
Нами была использована комбинация компьютерных способов и рационального дизайна для разработки линии ингибиторов, целенаправленно воздействующих на предупреждение распространения агрегатов αsyn. Наш подход был возможен только благодаря определению атомной структуры кора амилоидных фибрилл αsyn, и такой подход может быть принят для других заболеваний, в которых затравка играет роль в прогрессировании заболеваний. Ингибиторы предупреждают агрегацию αсинуклеина in vitro и в моделях клеточной культуры. Ингибиторы также проявляют эффективность в предупреждении затравки полученными от пациентов фибриллами αсинуклеина как в моделях in vitro, так и в моделях культур клеток. Наши результаты подтверждают, что патологические затравки αsyn содержат стерические молнии, и предлагают терапевтический подход, целенаправленно воздействующий на распространение и прогрессирование, который может быть применим в случае PD и родственных синуклеинопатий. Аналогично наши ингибиторы могут быть применимы для диагностического подхода в случае PD и родственных синуклеинопатий.
Нами была выдвинута гипотеза о том, что мутации, сверхэкспрессия и другие клеточные факторы могут дестабилизировать нативную структуру αсинуклеина, воздействуя на адгезивный сегмент «стерической молнии» в качестве основного строительного блока амилоидных агрегатов 18, 19. В связи с этим, нами были получены с высокоразрешающие изображения амилоидных остовов агрегатов αсинуклеина. Затем нами был использован способ Rosetta 36 в целях дизайна ингибиторов, которые специфично «кэппируют» растущие агрегаты αсинуклеина и, таким образом, нарушают и ингибируют дальнейшую агрегацию αсинуклеина, используя структуру 69-77 αсинуклеина или структуру 68-78 αсинуклеина в качестве матрицы.
Нами была использована атомная структура NACore [68-GAVVTGVTAVA-78] (SEQ ID NO: 46) в качестве матрицы и с помощью компьютерных и структурных подходов разработанные пептидные ингибиторы. Атомная структура NACore демонстрировала самокомплементарные β-листы, образующие стерическую молнию (Sawaya MR, et al. (2007) Nature 447(7143):453-457). В соответствии с компьютерным моделированием Rossetta предполагается, что ингибиторы связывают поверхность раздела стерической молнии и «кэппируют» фибриллы. Нами было идентифицировано 3 кандидатных ингибитора, S37, S61 и S71, которые эффективно связываются с одним или более концами молнии (Фиг. 1). Энергии связывания и комплементарность формы трех ингибиторов также являются благоприятными (Фиг. 1B). Все ингибиторы сохраняют большинство остатков нативной последовательности NACore, однако содержат один или более модифицированных остатков. Rodriquez et al. показали, что сегмент меньшего размера, состоящий из 9 остатков, в NACore [69-AVVTGVTAV-77] (SEQ ID NO: 48) образует агрегаты медленнее, чем NACore, а его структура является аналогичной NACore. Rodriguez et al. (2015) также описывают второй сегмент α-syn, обозначаемый PreNAC, при этом он содержит остатки 47-56 с последовательностью GVVHGVTTV. Разработанные ингибиторы в отношении этой последовательности также могли ингибировать образование фибрилл α-syn.
В целях предупреждения самоагрегации наших разработанных ингибиторов нами был использован более короткий сегмент совместно с одной или более модификациями. S37 имеет мутацию W в Thr72 и дополнительную полилизиновый тэг на C-конце для индукции отталкивания заряд-заряд. Предполагается, что он связывает оба конца фибриллы со стерической молнией. S61 и S62 сохраняют ту же самую ингибиторную последовательность, что и S37, однако вместо полилизинового тэга добавляют тэг TAT для содействия растворимости и предупреждения самоагрегации. S71 имеет метилированный глицин в Gly73, который ослабляет водородное связывание в β-листе, и дополнительный тэг TAT в целях растворимости и проникновения в клетку.
Нами исследована эффективность ингибиторов в анализе агрегации in vitro. Очищенный рекомбинантным путем αsyn агрегировали в присутствии ингибиторов и контролировали с помощью измерения флуоресценции тиофлавина T, красителя, связывающего амилоид. Все три ингибитора предупреждали агрегацию при значительном снижении флуоресценции ThT (Фиг. 1C, 1D, 1E).
Нами была исследована эффективность ингибиторов в предупреждении агрегации в модели культуры клеток. Применительно к этим анализам нами были использованы клетки HEK293T, которые стабильно экспрессируют меченый YFP полноразмерный αsyn ДТ и αsyn A53T (Sanders DW, et al. (2014) Neuron 82(6):1271-1288). В этой модели опосредованная липофектамином трансфекция рекомбинантных фибрилл приводит к агрегации эндогенного меченого YFP белка, который наблюдается в виде флуоресцентных пятен. Помимо этого, со временем эти пятна увеличиваются в размере и количестве. Такая пролиферация агрегатов со временем указывает на феномен «затравки», в соответствии с которым незначительное содержание амилоидных фибрилл индуцирует агрегацию эндогенного белка. Сначала нами был исследован исходный пептидный сегмент, NACore, в целях проверки его влияния на затравку. αSyn агрегировали в присутствии молярного избытка NACore (Фиг. 2B). Смесь трансфицировали в клетки, а образование областей визуализировали и количество пятен подсчитывали в виде частиц на лунку. Как и предполагалось, NACore не вызывал значимого снижения образования пятен ни в какой клеточной линии. Затем нами было агрегировано 50 мкМ αsyn в присутствии 500 мкМ, 250 мкМ, 100 мкМ и 50 мкМ ингибитора, соответствующего 10-, 5-, 2- и 1-кратному избытку. Затем смесь трансфицировали в клетки и агрегацию контролировали в динамике в течение 3 дней с помощью флуоресцентной визуализации (Фиг. 2A). S37 вызывал значимое снижение затравки в течение 2 дней как в клеточных линиях ДТ, так и в клеточных линиях, экспрессирующих A53T. Аналогично S37, S61 (Фиг. 2D) также вызывал снижение образования пятен с максимальной эффективностью при 2-, 5- и 10-кратном избытке в обеих клеточных линиях. Агрегаты, образованные в присутствии S62 (Фиг. 2E), также были некомпетентными в отношении затравки, при этом образовывалось значительно меньшее количество частиц при всех концентрациях ингибиторов. S71 исследовали при эквимолярных и субстехиометрических соотношениях и, было обнаружено, что он снижает затравливающую способность агрегатов (Фиг. 2F). Эти результаты свидетельствуют о том, что ингибиторы предупреждают образование компетентных в отношении затравки агрегатов.
Нами была исследована эффективность ингибиторов в предупреждении затравки в модели культуры клеток (Фиг. 3А). Нами были трансфицированы фибриллы αsyn совместно с различными ингибиторами. NACore, исходный агрегирующий пептид, не влиял на затравливающую способность фибрилл αsyn (Фиг. 3B). S37 вызывал значимое снижение затравки в обеих линиях клеток при концентрациях 12,5 мкM и 6,25 мкМ до 2 дней (Фиг 3C). S61 при концентрациях 2,5 мкM и 1,25 мкм вызывал длительное снижение затравки, продолжающееся до 6 дней в клетках с αsyn ДК и до 2 дней в клетках HEK с αsyn A53T (Фиг. 3D). S62 был эффективным при концентрациях 12,5 мкM и 6,25 мкM в обеих линиях клеток (Фиг. 3E). S71 был эффективным при низких концентрациях, составляющих 1,25 мкM. Интересно, что нами было замечено, что добавление более высоких концентраций этих ингибиторов не предупреждает затравку, свидетельствуя о критическом диапазоне концентраций с максимальной эффективностью. Вместе взятые эти результаты свидетельствуют о том, что ингибиторы могут кэппировать затравки фибрилл и предупреждать их элонгацию.
Нами были экстрагированы нерастворимые белковые агрегаты из замороженных тканей головного мозга, характерных для PD, после вскрытия. Нами были получены ткани от 4 различных субъектов, в том числе черная субстанция и лобные области одного субъекта и височные и лобные области других субъектов. С помощью ранее описанных протоколов (Goedert et al. (1992) Neuron 8(1):159-168), которые включали осаждение ионным детергентом саркозилом, нами были экстрагированы нерастворимые белковые агрегаты (Фиг. 4A). Все образцы устойчиво вызывали затравку агрегации αsyn in vitro и в нашей модели культуры клеток. Добавление in vitro 2% затравок повышало флуоресценцию ThT от 4 до 10 раз (Фиг. 4B, 4C, 4D) совместно с незначительным уменьшением времени задержки. Затравленные образцы затем трансфицировали в клетки HEK. В соответствии с анализом ThT, все затравленные образцы вызывали быстрое образование пятен (Фиг. 4E, 4F, 4G). Таким образом, фибриллы, экстрагированные из тканей головного мозга, характерных для PD, приводят к затравке рекомбинантного белка, а агрегаты, образованные при затравке, индуцируют быстрое образование пятен в культуре клеток.
Нами было исследовано влияние различных ингибиторов в предупреждении агрегации αsyn в присутствии экстрагированных затравок, характерных для PD. S71 и S62 были наиболее эффективными, проявляя эффективность в отношении всех затравок при измерении с помощью флуоресцентного анализа ThT (Фиг. 5). S61 также снижал агрегацию двух различных затравок (Фиг. 5B, 5D), в то время как S37 продемонстрировал крайне ограниченное снижение флуоресценции ThT (Фиг. 5D). Затем нами была исследована затравливающая способность агрегатов αsyn, образованных в присутствии затравок, характерных для PD, и различных ингибиторов (Фиг. 6A). Агрегаты, образованные в присутствии S71 (Фиг. 6B, 6C и 6D), не индуцировали образование пятен как в клетках ДТ, так и в клетках HEK, экспрессирующих A53T. S61 также продемонстрировал эффективность (Фиг. 6C). В соответствии с анализом in vitro, S37 не был эффективным в снижении затравливающей способности агрегатов. Совместно эти результаты свидетельствуют о том, что S62 и S71 могут предупреждать образование компетентных в отношении затравки фибрилл.
Нами была исследована эффективность ингибиторов в предупреждении затравки фибриллами, характерными для PD, в культуре клеток. Агрегаты αsyn, образованные в присутствии фибрилл, характерных для PD, трансфицировали в клетки HEK ДТ совместно с различными ингибиторами (Фиг. 7A). S37 предупреждал образование пятен до 2 дней в случае различных филаментов, характерных для PD, при концентрациях 12,5 мкM - 1,25 мкмM (Фиг. 7B). Аналогично S37, S61 также проявлял эффективность до 2 дней в концентрациях 12,5 мкM - 1,25 мкM, при этом S71 также предупреждал затравку при аналогичных концентрациях. Эти результаты свидетельствуют о том, что ингибиторы устойчиво предупреждают затравку в культуре клеток.
В раскрытии непосредственно выше описано исследование с рядом рабочих вариантов осуществления данного изобретения. В данном изобретении, раскрытом в данном документе, предложены такие ингибиторные пептиды; фармацевтические композиции, содержащие ингибиторные пептиды по данному изобретению и фармацевтически приемлемый носитель; способы применения ингибиторных пептидов для блокирования, ингибирования и/или предупреждения агрегации αсинуклеина и/или цитотоксичности αсинуклеина, включающие приведение в контакт молекулы αсинуклеина (например, мономера, малого агрегата, олигомера или фибриллы) с эффективным количеством пептидного ингибитора по данному изобретению или введение субъекту эффективного количества пептидного ингибитора по данному изобретению; и компьютерные варианты осуществления, такие как способ дизайна и получения ингибиторных пептидов или малых молекул на основании структур, описанных в данном документе.
Преимущества ингибиторных пептидов по данному изобретению включают в себя следующее: (1) синтетические пептиды не являются дорогостоящими; (2) проникновение в клетку и стабильность белка не представляют собой проблему в связи со своим составом и небольшим размером. Кроме того, пептиды могут быть слиты с проникающими в клетку пептидами, которые усиливают их доставку в клетки; (3) они являются неожиданным образом стабильными: они не подвергаются протеолизу и проявляют достаточно длительный период полужизни для того, чтобы функционировать in vivo (например, в организме); (4) пептидные ингибиторы являются специфичными в отношении своих мишеней, и, таким образом, представляют меньше возможностей для побочных эффектов, чем, например, малые молекулы, которые могут связываться со своими мишенями.
Данное изобретение, раскрытое в данном документе, имеет ряд вариантов осуществления. В одном варианте осуществления данного изобретения предложена заявленная композиция, содержащая по меньшей мере один ингибиторный пептид, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина. Как раскрыто в Примерах ниже, рабочие варианты осуществления этих пептидов включают S37, S62 и S71, как показано в Табл. 1 ниже.
В таблице N-me-Gly обозначает глицин с метилированной аминогруппой. Жирным шрифтом представлена ингибиторная последовательность. Нежирным шрифтом показаны тэги, как солюбилизирующие тэги, так и линкеры.
В типичных вариантах осуществления данного изобретения ингибиторный пептид содержит последовательность GAVVWGVTAVKK (SEQ ID NO: 3) или RAVVTGVTAVAE (SEQ ID NO: 4). Необязательно ингибиторный пептид содержит последовательность GAVVWGVTAVKKKKK (SEQ ID NO: 5), GAVVWGVTAVKKGRKKRRQRRRPQ (SEQ ID NO: 6); или YGRKKRRQRRRAVVTGVTAVAE (SEQ ID NO: 7). В определенных вариантах осуществления данного изобретения композиция содержит множество ингибиторных пептидов. В типичном случае ингибиторный(ингибиторные) пептид(пептиды) составляет(составляют) от 6 до 30 аминокислот в длину. Также включены активные варианты любого из этих ингибиторных пептидов. Ингибиторные пептиды, имеющие предыдущие последовательности, в том числе активные варианты, иногда обозначаются в данном документе как «ингибиторные пептиды по данному изобретению».
В композициях на основе ингибиторного пептида по данному изобретению по меньшей мере одна из аминокислот в ингибиторном пептиде содержат не встречающуюся в природе аминокислоту (например, D-аминокислоту или аминокислоту, содержащую фрагмент N-метильной группы); и/или ингибиторный пептид связан с гетерологичным пептидным тэгом. Такие гетерологичные пептидные тэги включают аминокислотные последовательности, которые повышают растворимость пептидов in vivo или in vitro (например, множество аргининовых остатков); или аминокислотные последовательности, которые облегчают контроль или манипуляцию с пептидом in vivo или in vitro (множество лизиновых или гистидиновых аминокислот); или аминокислотные последовательности, которые облегчают вхождение пептидов в клетку млекопитающего (например, последовательность проникающего в клетку пептида). В другом аспекте данного изобретения предложена фармацевтическая композиция, содержащая ингибиторный пептид и фармацевтически приемлемый носитель. Такие фармацевтические композиции иногда обозначаются в данном документе как «фармацевтические композиции по данному изобретению». Необязательно пептидные композиции, раскрытые в данном документе, включают фармацевтически приемлемый носитель и эксципиент, стабилизирующий пептид.
В другом варианте осуществления данного изобретения предложен вектор экспрессии, кодирующий ингибиторный пептид, который ингибирует агрегацию αсинуклеина в результате связывания с остатками 68-78 αсинуклеина. Необязательно вектор экспрессии представляет собой таковой, используемый для доставки полипептидов в клетки млекопитающих, например, лентивирус. В этом контексте другой вариант осуществления данного изобретения представляет способ доставки ДНК, кодирующей ингибиторный пептид, который ингибирует агрегацию αсинуклеина в клетке млекопитающего (например, клетки ex vivo или in vivo) в результате контакта клетки млекопитающего с вектором, который трансдуцирует клетку таким образом, что ДНК экспрессируется в клетке.
В другом варианте осуществления представлен набор, содержащий пептид, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина, или вектор экспрессии, кодирующий такой пептид. Варианты осуществления данного изобретения также включают в себя способ получения пептида, раскрытого в данном документе, путем его синтеза химическим путем или его получения рекомбинантным путем. В еще одном варианте осуществления данного изобретения предложен комплекс, содержащий αсинуклеин, и пептид, который ингибирует агрегацию αсинуклеина в результате связывания с остатками 68-78 αсинуклеина.
В другом варианте осуществления предложен пептид, разработанный на основе структуры PreNAC (остатки 47-56 a-синуклеина: GVVHGVTTVA) для ингибирования образования фибрилл α-синуклеина.
В еще одном варианте осуществления данного изобретения предложен способ снижения или ингибирования агрегации αсинуклеина (SEQ ID NO: 1), включающий приведение в контакт амилоидных фибрилл αсинуклеина с ингибиторным пептидом, раскрытым в данном документе, в количестве, достаточном для снижения или ингибирования агрегации αсинуклеина. Необязательно в этом способе амилоидные фибриллы αсинуклеина включены в вариант осуществления in vivo. Как альтернативный вариант в этом способе амилоидные фибриллы αсинуклеина включены в вариант осуществления in vitro. В связанном варианте осуществления предложен способ модуляции размера или скорости роста амилоидные фибриллы αсинуклеина, включающий приведение в контакт фибриллы с некоторым количеством по меньшей мере одного ингибиторного пептида, который ингибирует агрегацию αсинуклеина (SEQ ID NO: 1) в результате связывания с остатками 68-78 αсинуклеина в среде, в которой ингибиторный пептид контактирует с остатками 68-78 αсинуклеина таким образом, что приведенная в контакт фибрилла αсинуклеина проявляет модулируемый размер или скорость роста.
Пептидные ингибиторы, раскрытые в данном документе, могут быть использованы в качестве терапии для остановки распространения болезни Паркинсона в головном мозге. Как альтернативный вариант, пептидные ингибиторы, раскрытые в данном документе, могут быть использованы в качестве диагностического зонда для распознавания патологических агрегированных затравок белка αсинуклеина при заболеваниях, таких как болезнь Паркинсона, деменция с тельцами Леви и множественная системная атрофия. В этом контексте варианты осуществления данного изобретения включают способы наблюдения агрегированных затравок белка αсинуклеина в биологическом образце, включающие приведение в контакт биологического образца, который связывается с остатками 68-78 αсинуклеина, с последующим наблюдением того, связывается ли пептид с агрегированными затравками белка αсинуклеина, при наличии в этом биологическом образце. В типичном случае в этих способах гетерологичный пептидный тэг связывают с ингибиторным пептидом для того, чтобы облегчить наблюдение пептида. Необязательно в этих способах биологический образец происходит от индивидуума с подозрением на то, что он страдает болезнью Паркинсона, деменцией с тельцами Леви или множественной системной атрофией.
Используемые в данном документе формы существительного единственного числа включают ссылку на формы множественного числа, если контекст четко не определяет иное. Например, в предыдущем случае фармацевтическая композиция может содержать одну или более молекул ингибиторных пептидов по данному изобретению, которые могут быть одинаковыми или разными.
В другом аспекте данного изобретения предложен комплекс, содержащий молекулу αсинуклеина (например, мономер, малый агрегат, олигомер или фибриллу αсинуклеина) и ингибиторный пептид по данному изобретению. Они могут быть связаны, конъюгированы или иным образом ассоциированы друг с другом. αСинуклеин и ингибиторный пепитд могут быть ковалентно связаны или нековалентно связаны.
Другие аспекты данного изобретения включают полинуклеотид, кодирующий ингибиторный пептид по данному изобретению; вектор экспрессии, содержащий полинуклеотид; клетку, трансфицированную полинуклеотидом или вектором экспрессии; и способ получения пептида, включающий экспрессию его в трансфицированной клетке, культивирование клетки и сбор пептида, полученного таким образом.
В другом аспекте данного изобретения предложен способ ингибирования (предупреждения, остановки) агрегации молекулы αсинуклеина (например, мономера, малого агрегата, олигомера или фибриллы αсинуклеина), включающий приведение в контакт молекулы αсинуклеина с эффективным количеством ингибиторного пептида или фармацевтической композиции по данному изобретению. Молекула αсинуклеина может находиться в растворе или в клетке, которая находится в культуре или в субъекте. В одном варианте осуществления приведение в контакт молекулы αсинуклеина, которая представляет собой мономер, олигомер или малый агрегат, предупреждает агрегацию (олигомеризацию, дополнительную олигомеризацию и/или образование фибрилл) молекулы αсинуклеина. В другом варианте осуществления приведение в контакт агрегированной формы αсинуклеина или фибриллы предупреждает дополнительную агрегацию (фибриллирование) агрегированной формы или фибриллы.
В одном варианте осуществления этого способа молекула белка αсинуклеина которую приводят в контакт, находится в субъекте, имеющем заболевание или состояние, которое опосредовано наличием фибриллированного αсинуклеина (иногда обозначаемого в данном документе как опосредованное αсинуклеином заболевание или состояние, или синуклеинопатия), такое как болезнь Паркинсона (PD), деменция с тельцами Леви и множественная системная атрофия. Патология αсинуклеина также встречаются при других родственных нейродегенеративных заболеваниях, таких как, например, как спорадическая, так и семейная болезнь Альцгеймера.
В другом аспекте данного изобретения предложен способ лечения субъекта, имеющего опосредованное αsyn заболевание или состояние, такое как, например, болезнь Паркинсона, деменция с тельцами Леви или множественная системная атрофия, включающий введение субъекту эффективного количества ингибиторного пептида или фармацевтической композиции по данному изобретению. Лечение может приводить к блокаде (предупреждению) или ингибированию агрегации αсинуклеина и/или цитотоксичности αсинуклеина у субъекта, а также распространению патологии (затравке).
В другом аспекте данного изобретения предложен компьютерный способ идентификации пептида, который ингибирует агрегацию αсинуклеина и/или цитотоксичность αсинуклеина, как описано в данном документе. В другом аспекте данного изобретения предложен набор, содержащий ингибиторный пептид по данному изобретению, необязательно упакованный в контейнер. В другом аспекте данного изобретения предложен способ получения ингибиторного пептида по данному изобретению, включающий его синтез химическим путем или его получение рекомбинантным путем.
В еще одном варианте осуществления данного изобретения предложен способ наблюдения наличия или отсутствия амилоидных фибрилл αсинуклеина в биологическом образце, включающий комбинирование биологического образца с пептидом, который связывается с остатками 68-78 αсинуклеина, способствуя связыванию пептида с амилоидными фибриллами αсинуклеина, которые могут присутствовать в биологическом образце, а затем контроля этой комбинации в отношении наличия комплексов, образованных между амилоидными фибриллами αсинуклеина и пептидом; при этом наличие указанных комплексов свидетельствует о наличии амилоидных фибрилл αсинуклеина в биологическом образце. Необязательно в данном способе наличие комплексов между амилоидными фибриллами αсинуклеина и пептидом контролируют с помощью детектируемой метки, которая связана с пептидом (например, гетерологичным пептидным тэгом). В иллюстративных вариантах осуществления данного изобретения пептид содержит последовательность GAVVWGVTAVKK (SEQ ID NO: 3) или RAVVTGVTAVAE (SEQ ID NO: 4). В типичном случае способ осуществляют в биологическом образце, полученном от индивидуума с подозрением на то, что он страдает болезнью Паркинсона. Такие варианты осуществления данного изобретения могут быть использованы, например, в диагностических способах, разработанных для наблюдения наличия или статуса PD, например, для выявления начальных признаков заболевания до наступления клинических симптомов в целях контроля эффективности (или отсутствия эффективности) терапевтического лечения.
Пептидные ингибиторы по данному изобретению связываются специфично (избирательно, предпочтительно) с αсинуклеином, а не с непредусмотренными белками. Белок, с которым пептидный ингибитор связывается, может представлять собой, например, мономер, малый агрегат, олигомер или фибриллу. Например, связывание может быть в 2 раза, в 5 раз, в 10 раз, в 100 раз или 200 раз сильнее, или никакого связывания не может быть выявлено с непредусмотренной мишенью. Для определения специфичности связывания могут быть использованы стандартные способы, такие как, например, анализы конкурентного связывания или другие подходящие аналитические способы.
Также включены активные варианты ингибиторных пептидов, описанных выше. «Активный вариант» представляет собой вариант, который сохраняет по меньшей мере одно из свойств ингибиторных пептидов, описанных в данном документе (например, способность связываться с αсинуклеином и/или блокировать, ингибировать или предупреждать фибриллирование αсинуклеина (агрегацию) и/или цитотоксичность αсинуклеина). Фибриллирование, используемое в данном документе, относится к образованию волокон или фибрилл, таких как амилоидные фибриллы.
Подходящие активные варианты включают в себя соединения-пептидомиметики (любое соединение, содержащее непептидные структурные элементы, которое способно имитировать биохимическое(биохимические) и/или биологическое(биологические) действие(действия) природного имитирующего пептида, в том числе, например, таковые, разработанные для имитации структуры и/или активности связывания (такого как, например, водородные связи и взаимодействия на основе гидрофобной упаковки) пептидов в соответствии со способами, раскрытыми в данном изобретении). Ингибиторные пептиды по данному изобретению, в том числе их активные варианты, иногда обозначаются в данном документе как «пептидные соединения» или «соединения».
В одном варианте осуществления активные варианты ингибиторных пептидов укорачивают на 1-3 (например, 1, 2 или 3) аминокислоты как на N-конце, C-конце, так и на обоих исходного ингибиторного пептида. В другом варианте осуществления активные варианты удлиняют (расширяют) на 1, 2, 3 или 4 аминокислоты на C-терминальном конце исходного ингибиторного пептида, например, с помощью аминокислотных остатков в положении, в котором они встречаются в αсинуклеине.
Включены ряд других типов активных вариантов. В некоторых вариантах осуществления замещают аминокислоты, отличные от таковых, упомянутых выше. Эти аминокислоты могут способствовать защите пептидных ингибиторов от протеолиза или иным способом стабилизировать пептиды и/или способствовать необходимым фармакодинамическим свойствам другими путями. В других вариантах осуществления не встречающиеся в природе аминокислоты способствуют более тесному связыванию ингибитора с мишенью, поскольку боковые цепи оптимизируют водородное связывание и/или неполярные взаимодействия с ней. Помимо этого, не встречающиеся в природе аминокислоты открывают возможность для введения детектируемых маркеров, таких как высокофлуоресцентные маркеры, которые могут быть использованы, например, для измерения значений, таких как константы ингибирования. Также включены пептидомиметики, такие как, например, пептиды, бета-аминокислоты, N-этилированные аминокислоты и низкомолекулярные миметики.
В одном варианте осуществления не встречающиеся в природе аминокислоты замещают на аминокислот в последовательности. Более 100 не встречающихся в природе аминокислот являются коммерчески доступными. Они включают, например, следующие.
В другом варианте осуществления одна или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17) из L-аминокислот замещены D-аминокислотой.
В другом варианте осуществления один или более (например, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16 или 17) N-метилированных остатков включены в пептид.
Ингибиторный пептид по данному изобретению может содержать, например, L-аминокислоты, D-аминокислоты, не встречающиеся в природе аминокислоты или их комбинации.
Активные варианты включают молекулы, содержащие различные тэги на N-конце или C-конце пептида. Например, ингибиторный пептид по данному изобретению может содержать в качестве меток на своем N-конце и/или на своем С-конце: 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более лизиновых остатков; 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более аргининовых остатков; 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более глутаматных остатков; 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 или более аспартатных остатков; комбинации из этих аминокислот; или другие полярные тэги, которые будут очевидны специалисту в данной области техники. Другие активные варианты включают мутации последовательности αсинуклеина, которые повышают аффинность ингибиторных пептидов в отношении αсинуклеина.
В одном варианте осуществления ингибитор представляет собой малую молекулу, которая была разработана с помощью способов, описанных Jiang et al. 63 (которая включена в данный документ посредством ссылки, в частности, в отношении этого способа), с использованием атомной структуры волокна, образующего сегмент αсинуклеина, описанного в данном документе, в качестве основы для дизайна ингибитора. Подходящие малые молекулы, которые могут быть идентифицированы с помощью этого способа Jiang et al. будут очевидны специалисту в данной области техники.
В одном варианте осуществления данного изобретения пептид по данному изобретению модифицируют таким образом, что 1, 2 или 3 из его аминокислот замещают аминокислотой, имеющей не встречающуюся в природе боковую цепь, такой как не встречающимися в природе аминокислотами, рассмотренными выше, или аминокислотой, имеющей боковую цепь, модифицированную с помощью перекрестного сшивания (например, посредством эпсилон-аминогруппы остатка Lys) малой молекулы, которая была разработана Jiang et al. 63. Такие иллюстративные связывающие волокна молекулы представлены ниже. Эти активные варианты не только кэппируют растущие агрегаты αсинуклеина, но также посредством модифицированных боковых цепей могут связываться (скрепляться) со стенками стерической молнии, тем самым, усиливая ингибиторную активность пептида.
Связывающие волокна соединения, разработанные Jiang et al. 63, включают следующие.

В одном варианте осуществления данного изобретения ингибиторный пептид по данному изобретению выделяют или очищают с помощью стандартных методик, таких как способы, описанные в данном документе. Под «выделенным» подразумевается отделенный от компонентов, с которыми он в норме ассоциирован, например, компонентами, присутствующими после того, как пептид синтезирован. Выделенный пептид может представлять собой расщепляемый продукт белка, который содержит пептидную последовательность. «Очищенный» ингибиторный пептид может быть, например, на 90%, 95%, 98% или 99% чистым.
В одном варианте осуществления в целях усиления проницаемости в клетку ингибиторного пептида по данному изобретению пептид сливают с любым из множества проникающих в клетку пептидов (CPP). CPP в типичном случае имеют аминокислотный состав, который либо содержит высокий относительный избыток положительно заряженных аминокислот, таких как лизин или аргинин, либо имеет последовательности, которые содержат чередующийся паттерн полярных/заряженных аминокислот и неполярных гидрофобных аминокислот. Эти два типа структур обозначаются в данном документе как поликатионные или амфипатические соответственно. Третий класс CPP представляют собой гидрофобные пептиды, содержащие только неполярные остатки с низким суммарным зарядом, или имеют гидрофобные аминокислотные группы, которые являются важными для захвата клеткой. Некоторые типичные CPP, которые могут быть слиты с ингибиторным пептидом по данному изобретению, представлены в Табл. 2.
Таблица 2
Название Последовательность
Источник литературы - оригинал или обзор
polyARG nR, где 4 < n < 17 (например, n=5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16) (SEQ ID NO:8)
Wender, P.A., Mitchell, D.J., Pattabiraman, K., Pelkey, E.T., Steinman, L., and Rothbard, J.B. (2000). The design, synthesis, and evaluation of molecules that enable or enhance cellular uptake: peptoid molecular transporters. Proc. Natl. Acad. Sci. U. S. A. 97, 13003-8.
polyLYS nK, где 4 < K < 17 (например, K=5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16)
D-polyARG nR, где 4 < n < 17 (например, n=5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16)
D-polyLYS nK, где 4 < K < 17 (например, K=5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 или 16)
SynB1 RGGRLSYSRRRFSTSTGR (SEQ ID NO:9)
SynB3 RRLSYSRRRF (SEQ ID NO:10)
Пенетратин RQIKIWFQNRRMKWKK (SEQ ID NO:11)
Derossi, D., Joliot, A.H., Chassaing, G., and Prochiantz, A. (1994). The third helix of the Antennapedia homeodomain translocates through biological mem-branes. J. Biol. Chem. 269, 10444-50.
PenArg RQIRIWFQNRRMRWRR (SEQ ID NO:12)
PenLys KQIKIWFQNKKMKWKK (SEQ ID NO:13)
TatP59W GRKKRRQRRRPWQ (SEQ ID NO:14)
Tat (48-60) GRKKRRQRRRPPQ (SEQ ID NO:15)
Vives, E., Brodin, P., and Lebleu, B. (1997). A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus. J. Biol. Chem. 272, 16010-7.
R9-Tat GRRRRRRRRRPPQ (SEQ ID NO:16)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
Tat YGRKKRRQRRR (SEQ ID NO:17)
Vives, E., Brodin, P., and Lebleu, B. (1997). A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus. J. Biol. Chem. 272, 16010-7.
D-Tat GRKKRRQRRRPPQ (SEQ ID NO:18)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
BMVGag (7-25) KMTRAQRRAAARRNRWTAR (SEQ ID NO:19)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
FHVCoat (35-49) RRRRNRTRRNRRRVR (SEQ ID NO:20)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
HTLV-II Rex (4-16) TRRQRTRRARRNR (SEQ ID NO:21)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
P22 N-(14-30) NAKTRRHERRRKLAIER (SEQ ID NO:22)
pVEC LLIILRRRIRKQAHAHSK (SEQ ID NO:23)
Elmquist, A., Lindgren, M., Bartfai, T., and Langel, Ü. (2001). VE-cadherin-derived cell-penetrating peptide, pVEC, with carrier functions. Exp. Cell Res. 269, 237-44.
Транспортан GWTLNSAGYLLGKINLKALAALAKKIL (SEQ ID NO:24)
Pooga, M., Hällbrink, M., Zorko, M., and Langel, Ü. (1998). Cell penetration by transportan. FASEB J. 12, 67-77.
TP10 AGYLLGKINLKALAALAKKIL (SEQ ID NO:25)
Soomets, U., Lindgren, M., Gallet, X., Hällbrink, M., Elmquist, A., Balaspiri, L., Zorko, M., Pooga, M., Brasseur, R., and Langel, Ü. (2000). Dele-tion analogues of transportan. Biochim. Biophys. Acta 1467, 165-76.
PTD-4 PIRRRKKLRRLK (SEQ ID NO:26)
PTD-5 RRQRRTSKLMKR (SEQ ID NO:27)
Pep-1 ac-KETWWETWWTEWSQPKKKRKV-cya (SEQ ID NO:28)
Pep-2 ac-KETWFETWFTEWSQPKKKRKV-cya (SEQ ID NO:29)
Morris, MC, Chaloin, L, Choob, M, Archdeacon, J, Heitz, F and Divita, G (2004). Combination of a new generation of PNAs with a peptide-based carrier enables efficient targeting of cell cycle progression. Gene Ther 11: 757-764.
Pep-3 ac-KWFETWFTEWPKKRK-cya (SEQ ID NO:30)
Morris, MC, Gros, E, Aldrian-Herrada, G, Choob, M, Archdeacon, J, Heitz, F et al. (2007). A non-covalent peptide-based carrier for in vivo delivery of DNA mimics. Nucleic Acids Res 35: e49.
E N(1-22) MDAQTRRRERRAEKQAQWKAAN (SEQ ID NO:31)
B 21 N-(12-29) TAKTRYKARRAELIAERR (SEQ ID NO:32)
U2AF(142-153) SQMTRQARRLYV (SEQ ID NO:33)
PRP6(129-144) TRRNKRNRIQEQLNRK (SEQ ID NO:34)
MAP KLALKLALKLALALKLA (SEQ ID NO:35)
SBP MGLGLHLLVLAAALQGAWSQPKKKRKV (SEQ ID NO:36)
FBP GALFLGWLGAAGSTMGAWSQPKKKRKV (SEQ ID NO:37)
MPG ac-GALFLGFLGAAGSTMGAWSQPKKKRKV-cya (SEQ ID NO:38)
Morris, MC, Vidal, P, Chaloin, L, Heitz, F and Divita, G (1997). A new peptide vector for efficient delivery of oligonucleotides into mammalian cells. Nucleic Acids Res 25: 2730-2736.
MPG(ΔNLS) ac- GALFLGFLGAAGSTMGAWSQPKSKRKV-cya (SEQ ID NO:39)
REV(34-50) TRQARRNRRRRWRERQR (SEQ ID NO:40)
Futaki, S. (2002) Arginine-rich peptides: potential for intracellular delivery of macromolecules and the mystery of the translocation mechanisms. Int. J. Pharm. 245, 1-7.
ACPPs from Jiang et al., PNAS 2004 - lower case indicates D-aa. Символ «__» в некоторых из этих последовательностей указывает положение, в которое может быть включен любой из множества признанных в данной области техники сайтов расщепления протеазой:
EEEEEDDDDK_AXRRRRRRRRRXC (SEQ ID NO:41)
EEEEEDDDDK_ARRRRRRRRRXC (SEQ ID NO:42)
EDDDDK_AXRRRRRRRRRXC (SEQ ID NO:43)
EEDDDDK_ARXRRXRRXRRXRRXC (SEQ ID NO:44)
DDDDDDK_ARRRRRRRRRXC (SEQ ID NO:45)
В другом варианте осуществления CPP представляет собой polyD(1-16).
В целом, рекомендуется, чтобы длина CPP была довольно короткой, например, менее около 30 аминокислот, в целях повышения стабильности и фармакодинамических свойств после того, как молекула проникает в клетку.
В некоторых вариантах осуществления CPP непосредственно присоединяют (сливают) к пептиду по данному изобретению. В других вариантах осуществления желательно отделять сильно заряженный CPP от ингибиторного пептида с помощью линкера, в целях того, чтобы содействовать сохранности ингибитором своей активности. Может быть использован любой из множества линкеров. Размер линкера может варьировать, например, от 1-7 или даже более аминокислот (например, 1, 2, 3, 4, 5, 6 или 7 аминокислот). Например, линкер может представлять собой QVTNVG на N-конце и QKTVEG на C-конце или их усеченный вариант, имеющий 1, 2, 3, 4 или 5 из прилегающих аминокислот, N-концевых в отношении к ингибиторному пептиду.
В вариантах осуществления по данному изобретению ингибиторный пептид является детектируемым образом меченым. Меченые пептиды могут быть использованы, например, для лучшего понимания механизма действия и/или клеточной локализации ингибиторного пептида. Подходящие метки, которые способствуют детекции (например, обеспечивают детектируемый сигнал или могут быть выявлены) являются стандартными и хорошо известны специалистам в данной области техники. Подходящие детектируемые метки включают, например, радиоактивные агенты, флуоресцентные метки и т.п. Способы прикрепления таких меток к белку или анализы для выявления их наличия и/или количества, являются стандартными и хорошо известными.
Ингибиторный пептид по данному изобретению может быть синтезирован (например, химическим путем или с помощью рекомбинантной экспрессии в подходящей клетке-хозяине) посредством любого из множества признанных в данной области техники способов. В целях получения достаточных количеств ингибиторного пептида для применения в способе по данному изобретению, специалист-практик может, например, с помощью стандартных методик, получить нуклеиновую кислоту (например, ДНК), кодирующую пептид, и вставить его в вектор экспрессии, в котором последовательность находится под контролем последовательности контроля экспрессии, такой как промотор или энхансер, которые могут направлять синтез пептида. Например, можно (a) синтезировать ДНК de novo с использованием подходящих линкеров на концах для клонирования ее в вектор; (b) клонировать всю последовательность ДНК в вектор; или (c) начиная с перекрывающихся олигонуклеотидов, соединить их с помощью стандартных способов синтеза генов на основе ПЦР и вставить полученную ДНК в вектор. Подходящие векторы экспрессии (например, плазмидные векторы, в том числе фаги, векторы, искусственные векторы, дрожжевые векторы, эукариторические векторы и т.д.) будут очевидны специалистам в данной области техники, как и способы получения векторов, включающих последовательности, представляющие интерес, экспрессирующие белки, кодируемые нуклеиновой кислотой, а также выделения или очистки экспрессируемых белков.
В другом аспекте данного изобретения предложена фармацевтическая композиция, содержащая один или более ингибиторных пептидов и фармацевтически приемлемый носитель. Компоненты фармацевтической композиции могут быть детектируемым образом мечены, например, с помощью радиоактивной или флуоресцентной метки, например, таковой, которая является подходящей для детекции с помощью позитрон-эмиссионной томографии (ПЭТ) или магнитно-ядерной томографии (МРТ). Например, пептиды по данному изобретению могут быть связаны с детектируемой меткой, выбранной из группы, состоящей из радиоактивной метки, рентгеноконтрастной метки, флуоресцентного красителя, флуоресцентного белка, колориметрической метки и т.п. В некоторых вариантах осуществления ингибиторный пептид присутствует в эффективном количестве для необходимой цели.
«Фармацевтически приемлемый» означает таковой, который является пригодным при получении фармацевтической композиции, которая в целом является безопасной, нетоксичной и ни в биологическом, ни в другом отношении не является нежелательной, а также включает в себя таковое, которое является приемлемым для ветеринарного применения, а также для фармацевтического применения у человека. Например, «фармацевтически приемлемые соли» соединения означают соли, которые являются фармацевтически приемлемыми, как определено в данном документе, и которые обладают необходимой фармакологической активностью исходного соединения.
В другом аспекте данного изобретения предложен полинуклеотид, кодирующий ингибиторный пептид по данному изобретению. В вариантах осуществления данного изобретения полинуклеотид функционально связан с регуляторной контрольной последовательностью (например, промотором или энхансером) для облегчения продуцирования кодируемого белка после введения (например, с помощью трансфекции) в подходящую клетку. Другие варианты осуществления включают клетку, содержащую вектор экспрессии; а также способ получения ингибиторного пептида по данному изобретению, включающий культивирование клетки и сбор пептида, полученного таким образом.
Используемый в заявке термин «около» означает плюс или минус 5% от значения.
В другом аспекте данного изобретения предложен набор для осуществления любого из способов, описанных в данном документе. Набор может содержать подходящее количество ингибиторного пептида по данному изобретению; реагенты для получения пептида; реагенты для анализов в целях измерения его функций или активностей; или т.п. Наборы по данному изобретению могут содержать инструкции для осуществления способа. Другие необязательные элементы набора по данному изобретению включают подходящие буферы, компоненты среды или т.п.; компьютер или машиночитаемый носитель, обеспечивающий структурное представление кристаллической структуры, описанной в данном документе; контейнеры; или упаковочные материалы. Также могут быть включены реагенты для осуществления подходящих контролей. Реагенты набора могут находиться в контейнерах, в которых реагенты являются стабильными, например, в лиофилизированной форме или стабилизированных жидкостях. Реагенты могут находиться также в одноразовой форме, например, в форме для одной реакции для введения субъекту.
Характеристика кандидатных ингибиторных пептидов по данному изобретению может быть осуществлена с помощью любого из ряда стандартных способов. Например, пептиды могут быть проанализированы в отношении способности снижать или ингибировать агрегацию или цитотоксичность αсинуклеина или распространение от клетки к клетке. Анализы можно проводить in vitro или in vivo. Подходящие анализы будут очевидны специалисту в данной области техники; некоторые подходящие анализы описаны в данном документе.
В одном аспекте данного изобретения предложен способ снижения или ингибирования агрегации αсинуклеина, включающий приведение в контакт протофиламентов αсинуклеина с эффективным количеством одного или более ингибиторных пептидов по данному изобретению. Такой способ может быть осуществлен в растворе или в клетке (например, клетках в культуре или в субъекте).
В другом аспекте данного изобретения предложен способ лечения субъекта, имеющего заболевание или состояние, которое опосредовано наличием фибриллированного αсинуклеина (иногда обозначаемого в данном документе как опосредованное αсинуклеином заболевание или состояние), включающий введение субъекту эффективного количества ингибиторного пептида или фармацевтической композиции по данному изобретению. Среди таких заболеваний и состояний присутствуют, например, болезнь Паркинсона (PD), деменция с тельцами Леви или множественная системная атрофия. В другом аспекте данного изобретения предложен способ предупреждения начала таких заболеваний или состояний (например, PD), или лечения субъекта на ранних стадиях таких заболеваний или состояний, или такового, у которого развивается такое заболевание или состояние, в целях предупреждения или ингибирования развития состояния или заболевания.
Ингибиторный пептид или фармацевтическая композиция по данному изобретению иногда обозначается в данном документе как «ингибитор».
«Эффективное количество» ингибитора по данному изобретению представляет собой количество, которое может вызывать измеримое количество необходимого результата, например, ингибрование агрегации или цитотоксичность αсинуклеина; в случае диагностического анализа - количество, которое, может выявлять мишень, представляющую интерес, такую как агрегат αсинуклеина; или в способе лечения - количество, которое может снижать или нормализовать, посредством измеряемого количества, симптом заболевания или состояния, которое подлежит лечению.
«Субъект» может представлять собой любого субъекта (пациента), имеющего агрегированные (фибриллированные) молекулы αсинуклеина, ассоциированные с состоянием или заболеванием, которое можно лечить с помощью способа по данному изобретению. В одном варианте осуществления данного изобретения субъект имеет PD. Типичные субъекты включают позвоночных, таких как млекопитающие, в том числе лабораторные животные, собаки, кошки, отличные от человека приматы и человек.
Ингибиторы по данному изобретению могут быть разработаны в виде фармацевтических композиций в ряде форм, приспособленных к выбранному пути введения, например, перорально, назально, интраперитонеально или парентерально, с помощью внутривенных, внутримышечных, местных или подкожных путей, или с помощью инъекции в ткань.
Подходящие пероральные формы для введения ингибиторов включают в себя пастилки, троше, таблетки, капсулы, шипучие таблетки, перорально распадающиеся таблетки, плавающие таблетки, разработанные для повышения времени задержки в желудке, буккальные пластыри и сублингвальные таблетки.
Ингибиторы по данному изобретению могут быть введены системно, например, перорально, в комбинации с фармацевтически приемлемым носителем, таким как инертный разбавитель или ассимилируемый съедобный носитель, или путем ингаляции или инсуффляции. Они могут быть заключены в покрытые или непокрытые твердой или мягкой оболочкой желатиновые капсулы, могут быть спрессованы в таблетки или могут быть включены непосредственно в пищу рациона пациента. В случае перорального терапевтического введения соединения могут быть комбинированы с одним или более эксципиентами и использованы в форме проглатываемых таблеток, буккальных таблеток, троше, капсул, эликсиров, суспензий, сиропов, облаток и т.п. В случае композиций, подходящих для введения человеку, термин «эксципиент» означает включение, но не ограничиваясь ими, таких компонентов, описанных в Remington: The Science and Practice of Pharmacy, Lippincott Williams & Wilkins, 21st ed. (2006) (далее в данном документе Remington).
Ингибиторы могут быть комбинированы с тонкодисперсным инертным порошковым носителем и ингалированы субъектом или инсуффлированы. Некоторые композиции и препараты должны содержать по меньшей мере 0,1% соединений. Процент композиций и препаратов может, само собой разумеется, варьировать и может для удобства находиться от около 2% до около 60% от веса определенной единичной лекарственной формы.
Таблетки, троше, пилюли, капсулы и т.п. могут также содержать следующее: связывающие вещества, такие как трагакантовая камедь, аравийская камедь, кукурузный крахмал или желатин; эксц, такие как дикальцийфосфат; агент для улучшения распадаемости таблеток, такой как кукурузный крахмал, картофельный крахмал, альгиновая кислота и т.п.; смазывающее вещество, такое как стеарат магния; и подсластитель, такой как сахароза, фруктоза, лактоза или аспартам, или может быть добавлен ароматизатор, такой как масло мяты перечной, винтергреневое масло или вишневый ароматизатор. В случае, если единичная дозированная форма представляет собой капсулу, она может содержать, помимо веществ упомянутого выше типа, жидкий носитель, такой как растительное масло или полиэтиленгликоль. Сироп или эликсир может содержать активное соединение, сахарозу или фруктозу в качестве подсластителя, метил- или пропилпарабены в качестве консервантов, краситель и ароматизатор, такой как вишневый или апельсиновый ароматизатор.
Различные другие вещества могут присутствовать в качестве покрытий или иным образом модифицировать физическую форму твердой единичной лекарственной формы. Например, таблетки, пилюли или капсулы могут быть покрыты желатином, воском, шеллаком или сахаром и т.п. Само собой разумеется, что любое вещество, используемое в приготовлении любой лекарственной формы должно быть фармацевтически приемлемым и по сути нетоксичным в используемых количествах.
Помимо этого, ингибиторы могут быть включены в препараты и изделия с длительным высвобождением. Например, ингибиторы могут быть включены в капсулы с пролонгированным высвобождением, таблетки с пролонгированным высвобождением и пилюли с пролонгированным высвобождением. В некоторых вариантах осуществления композицию вводят с использованием лекарственной формы, выбранной из группы, состоящей из шипучих таблеток, перорально распадающихся таблеток, плавающих таблеток, разработанных для повышения времени задержки в желудке, буккальных пластырей и сублингвальных таблеток.
Ингибиторы могут быть также введены внутривенно или интраперитонеально с помощью инфузии или инъекции. Растворы ингибиторов могут быть приготовлены в воде, необязательно смешанной с нетоксичным поверхностно-активным веществом. Также могут быть приготовлены дисперсии в глицерине, жидких полиэтиленгликолях, триацетине и их смесях и в маслах. При обычных условиях хранения и применения эти препараты могут содержать консервант для предупреждения роста микроорганизмов.
Фармацевтические лекарственные формы, подходящие для инъекции или инфузии, могут включать стерильные водные растворы или стерильные порошки, содержащие соединения, которые приспособлены для экстемпорального приготовления стерильных инъекционных или инфузионных растворов или дисперсий, необязательно инкапсулированных в липосомы. Во всех случаях окончательная лекарственная форма должна быть стерильной, жидкой и стабильной в условиях производства и хранения. Жидкий носитель или основа может представлять собой раствор или жидкую дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.), растительные масла, нетоксичные сложные эфиры глицерина и их подходящие смеси. Подходящая текучесть может поддерживаться, например, за счет образования липосом, с помощью поддержания требуемого размера частиц в случае дисперсий и с использованием поверхностно-активных веществ.
Стерильные инъекционные растворы готовят с помощью включения соединений в требуемом количестве в подходящий растворитель с различными другими компонентами, перечисленными выше, при необходимости, с последующей стерилизацией фильтрацией. В случае стерильных порошков для приготовления стерильных инъекционных растворов предпочтительными способами приготовления являются методики вакуумной сушки и сублимационной сушки, которые приводят к образованию порошка активного компонента совместно с любым дополнительным необходимым компонентом, присутствующим в предварительно стерилизованных фильтрованием растворах.
В случае местного введения соединения могут быть нанесены в чистом виде. Однако в целом будет желательно наносить их на кожу в виде композиций или составов в комбинации с дерматологически приемлемым носителем, который может представлять собой твердое вещество или жидкость.
Пригодные твердые носители включают в себя мелкодисперсные твердые вещества, такие как тальк, глина, микрокристаллическая целлюлоза, кремний, глинозем и т.п. Другие твердые носители включают в себя стандартные нетоксичные полимерные наночастицы или микрочастицы. Пригодные жидкие носители включают в себя воду, спирты или гликоли или смеси вода/спирт/гликоль, в которых соединения могут быть растворены или диспергированы при эффективных уровнях, необязательно с помощью нетоксичных поверхностно-активных веществ. Адъюванты, такие как отдушки и дополнительные антимикробные агенты могут быть добавлены для оптимизации свойств для определенного применения. Полученные жидкие композиции могут быть нанесены из гигроскопических прокладок, используемых для пропитки бандажей и других перевязочных материалов, или распылены на пораженную область с помощью распылителей насосного типа или аэрозольных распылителей.
Пригодные дозы пептидов или фармацевтические композиции по данному изобретению могут быть определены с помощью сравнения их активности in vitro и активности in vivo в животных моделях. Способы экстраполяции эффективных доз у мышей и других животных на человека известны в данной области техники.
Например, концентрация соединений в жидкой композиции, такой как лосьон, может составлять около 0,1-25% по весу, или около 0,5-10% по весу. Концентрация в мягкой или твердой композиции, такой как гель или порошок, может составлять около 0,1-5% по весу, или около 0,5-2,5% по весу.
Эффективные дозы и пути введения агентов по данному изобретению являются стандартными. Точное количество (эффективная доза) агента будет варьировать от субъекта к субъекту в зависимости, например, от вида, возраста, веса и общего и клинического состояния субъекта, тяжести или механизма любого нарушения, подлежащего лечению, определенного используемого агента или основы, способа и режима введения и т.п. Терапевтически эффективная доза может быть определена эмпирически с помощью стандартных процедур, известных специалистам в данной области техники. См., например, The Pharmacological Basis of Therapeutics, Goodman and Gilman, eds., Macmillan Publishing Co., New York. Например, эффективная доза может быть оценена изначально либо в анализах с использованием культур клеток, либо в подходящих животных моделях. Животная модель также может быть использована для определения подходящих диапазонов концентраций и путей введения. Такая информация затем может быть использована для определения пригодных доз и путей введения у человека. Терапевтическая доза также может быть выбрана по аналогии с дозами для сопоставимых терапевтических средств.
Определенный способ введения и схема дозирования будут выбраны лечащим врачом с учетом особенностей случая (например, субъекта, заболевания, соответствующей стадии заболевания и того, является ли лечения профилактическим). Лечение может включать в себя введение одной или более доз в день соединения(соединений) в течение периода времени, состоящего от нескольких дней до месяцев или даже лет.
Однако, как правило, подходящая доза будет находиться в диапазоне от около 0,001 до около 100 мг/кг, например, от около 0,01 до около 100 мг/кг веса тела в день, например, свыше около 0,1 мг на килограмм, или в диапазоне от около 1 до около 10 мг на килограмм веса тела в день. Например, подходящая доза может составлять около 1 мг/кг, 5 мг/кг, 10 мг/кг, 20 мг/кг или 30 мг/кг веса тела в день.
Ингибиторы для удобства вводят в единичной лекарственной форме; например, содержащей от 0,05 до 10000 мг, от 0,5 до 10000 мг, от 5 до 1000 мг или от около 100 мг активного компонента на единичную лекарственную форму. В некоторых вариантах осуществления единичная доза содержит около 0,1 мг, около 0,5 мг, около 1 мг, около 10 мг, около 25 мг, около 50 мг, около 75 мг или около 100 мг активного компонента.
ПРИМЕРЫ
Пример I - Токсический кор αсинуклеина болезни Паркинсона: структура из невидимых кристаллов
Нами были осуществлены проверки кристаллов пептидных сегментов в домене NAC и прилегающих участках с поиском структурной информации касательно молекулярной основы агрегации и токсичности αsyn.
Масштабные кристаллические исследования двух сегментов, NACore, остатков 68GAVVTGVTAVA78 (SEQ ID NO:46) и PreNAC, 47GVVHGVTTVA56 (SEQ ID NO:47), по-видимому, давали некристаллические аморфные агрегаты. Однако при исследовании с помощью электронной микроскопии нами было обнаружено, что агрегаты представляют собой кластеры удлиненных нанокристаллов, составляющих всего 50-300 нм в поперечном разрезе и, таким образом, невидимые с помощью стандартной световой микроскопии. Нами была подтверждена высокоупорядоченная кристалличность NACore на обоих лазерах на свободных электронах SACLA и LCLS. Нами также было обнаружено, что фрагмент, состоящий из 9 остатков, в NACore, который обозначен нами SubNACore, 69AVVTGVTAV77 (SEQ ID NO:48), давал кристаллы в 1000-10000 раз большие по объему, чем нанокристаллы NACore. Таким образом, мы смогли применить синхротронные способы 18,19 в отношении этих более крупных кристаллов для определения структуры амилоид-подобных фибрилл. Несмотря на то, что этот фрагмент, состоящий из 9 остатков, не имел лишь двух остатков по сравнению с NACore, он не был таким же токсичным20, что давало общее представление, описанное ниже, о токсичности αsyn.
Для определения структуры невидимых кристаллов NACore и PreNAC мы обратились к MicroED 21. В MicroED пучок электронов с крайне низкой дозой направляют на нанокристалл в электронном трансмиссионном микроскопе в криогенных условиях с получением паттернов дифракции. Поскольку длина волны, используемая в наших экспериментах, при 200 keV, является очень незначительной (0,025 Å) то сфера Эвальда является по сути плоской, давая паттерны дифракции, которые очень напоминают двумерный разрез через трехмерное обратное пространство. Поскольку кристалл непрерывно вращается в пучке, то собирают ряд таких паттернов дифракции 5. Масштабирование наряду с данными дифракции, полученными от нескольких кристаллов дает полную трехмерную совокупность дифракционных данных. MicroED успешно применяли в отношении хорошо известных структур лизозима белка куриного яйца 6,5, каталазы бычьей печени 22 и Ca2+-АТФазы23. Однако NACore и PreNAC представляют собой первые, ранее не описанные структуры, определенные с помощью MicroED.
В случае NACore и PreNAC нами были собраны паттерны электронной дифракции от нанокристаллов, которые располагаются предпочтительно ориентированными плоскими на поверхности угольной решетки Quantifoil с отверстиями в замороженном гидратированном состоянии. Решетки вначале исследовали в отношении кристаллов подходящего размера, а кандидатные кристаллы исследовали в отношении дифракции. Нами были использованы кристаллы, демонстрирующие выраженную дифракцию в отношении сбора данных с помощью непрерывного вращения в одном направлении около фиксированной оси, при этом получали ряд дифракционных кадров через фиксированные промежутки времени 5. Игольчатые кристаллы в типичном случае превышали длину, необходимую для дифракции; таковые, которые были неизогнутыми и от 100 до 300 нм шириной давали наилучшие паттерны дифракции. Данные от нескольких кристаллов интегрировали, масштабировали и объединяли вместе.
Мультикристаллические совокупности данных NACore и PreNAC синхронизировали по фазе путем молекулярного замещения с помощью атомной модели SubNACore и идеальной бета-цепочечной модели соответственно в качестве зондов. Фазы дифракции рассчитывали исходя из структуры зона SubNACore и структурные факторы NACore давали карту плотности разницы, которая отчетливо показывает положения отсутствующих остатков после последующего очищения, две молекулы воды и несколько атомов водорода. Полные модели NACore и PreNAC очищали по отношению к данным MicroED, при этом получали структуры при разрешении 1,4 Å с допустимыми R-факторами. Факторы рассеяния электронов использовали при расчетах с учетом очищения24.
Структура пептидной цепи NACore представляет собой почти полностью развернутую β-цепь. Эти цепи NACore укладываются в β-листы, как предполагали с помощью сайт-направленного мечения спина 25,26. Листы являются парными, что является обычным в амилоидных спинах, а пары листов образуют типичные протофиламенты со стерическими молниями, ранее наблюдаемые в виде спинов во многих амилоид-подобных фибриллах, образуемых из коротких сегментов образующих фибриллы белков. Необычными свойствами этой стерической молнии было то, что ширина молнии, состоящего из 11 остатков, была длиннее, чем нами наблюдалось ранее, и каждая пара этих листов содержит две молекулы воды, при этом каждая ассоциирована с треониновой боковой цепью в поверхности раздела вместо того, чтобы быть полностью сухими. Кроме того, в наших кристаллах NACore каждый лист образует две прилегающие поверхности раздела: поверхность раздела A с 268 Å2 заглубленной доступной площади поверхности на цепь, является более протяженной и предпочтительно прочной, чем поверхность раздела B (167 Å2), поскольку концевые остатки цепей в противонаправленных листах изгибаются в отношении друг друга. Структура PreNAC показывает пептидную цепь, которая образует β-цепь, изогнутую в остатке глицина 51. Эти цепи упорядочены в пары β-листов, которые подобно структуре NACore, переплетаются с образованием протофиламентов со стерическими молниями (Фиг. 3). Особенно следует отметить, что сегмент PreNAC, состоящий из пяти остатков (51GVTTV55) (SEQ ID NO:49), отличается одним остатком от сегмента NACore, состоящего из пяти остатков (73GVTAV77) (SEQ ID NO:50), а их остовы и идентичные боковые цепи накладываются вплотную с отклонением RMS альфа-углеродного атома, составляющим 1,5 Å. Это означает, что более слабая поверхность раздела B NACore имитирует гипотетическую поверхность раздела между NACore и PreNAC. Ниже мы предполагаем значение возможного контакта между этими двумя сегментами αsyn.
Соответствие структуры NACore фибриллам полноразмерного αsyn определяют на основании сходства их паттернов дифракции. В частности, паттерн дифракции волокон выравненных фибрилл полноразмерного и ацетилированного на N-конце 27 белка αsyn демонстрирует те же самые основные пики, что и дифракция выравненных нанокристаллов NACore. Все три фибриллы демонстрируют выраженное отклонение при 2,4 Å в своих паттернах дифракции. Это отклонение возникает в NACore, поскольку один β-лист стерической молнии перемещается вдоль оси волокна в отношении другого β-листа на 2,4Å, что составляет половину промежутка, составляющего 4,8 Å, между β-цепями, способствуя тому, что два листа плотно переплетаются вместе. Все три фибриллы имеют выраженное отклонение, составляющее 4,6 Å, которое в NACore возникает в результате укладывания β-цепей и чередования между прилегающими β-листами стерической молнии, в то время как общее отклонение при около 8,2 Å, вероятно, возникает в результате расстояния между прилегающими парами β-листов, которые составляют фибриллу αsyn. Это сравнение паттернов дифракции волокон отчетливо свидетельствуют о том, что структура NACore является аналогичной спину наших токсических фибрилл полноразмерного αsyn.
Комбинированные структуры NACore и PreNAC позволяют нам конструировать предположительную модель для большей части упорядоченных сегментов мутантного αsyn А53T, характеризующегося ранним началом болезни. Экспериментальное подтверждение этой модели исходит из соглашения о его имитированной дифракции волокон с измеренными паттернами дифракции αsyn и фибрил N-ацетил-αsyn, а также выравненных нанокристаллов NACore. Выше нами было отмечено, что более слабая поверхность раздела B NACore имитирует гипотетическое взаимодействие PreNAC с NACore. Фактически взаимодействующие боковые цепи в более слабой поверхности раздела В NACore (G73, T75 и V77) являются идентичными боковым цепям (G51, T53, V55), взаимодействующим в гипотетической поверхности раздела PreNAC с NACore.
Идентичность и структура цитотоксического амилоида, образованного αsyn, остается предметом интенсивного исследования 17,28,29,30,31,32. Весомость доказательства в последнее десятилетие отклонило научно обоснованное мнение от полностью сложившихся амилоидных фибрилл, обнаруженных в тельцах Леви в виде токсических фрагментов, к более мелким временным амилоидным олигомерам. Однако недавно количественные аргументы были выдвинуты в пользу фибрилл 33. Не ограничиваясь каким-либо определенным механизмом, предполагается, что наши эксперименты по цитотоксичности NACore в отношении клеток нейробластомы согласуются с точкой зрения о том, что фибриллы являются токсичными: нами обнажено, что NACore, подвергнутый встряхиванию и агрегации в течение 72 часов, демонстрирует многочисленные фибриллы, и является более токсичным, чем свежерастворенный NACore, и сопоставимо токсичный с аналогичным образом агрегированным полноразмерным αsyn. Нами также обнаружена более высокая цитотоксичность NACore по сравнению с SubNACore, который является короче на два остатка. Это согласуется с более быстрым образованием фибрилл NACore по сравнению с SubNACore. Эти наблюдения не исключают образование нефибриллярного олигомерного скопления, присутствующего, но невидимого, в наших агрегированных образцах NACore и αsyn. Само собой разумеется, что NACore является лишь фрагментом полноразмерного αsyn и не имеет большинства мембраносвязывающих мотивов N-конца белка, которые были задействованы в разрушении мембраны 34,35. В то же время ясно, что NACore является минимальным фрагментом, который повторяет все свойства агрегации и токсичности полноразмерного α-syn.
Крайне незначительный размер NACore является типичным для амилоидных кристаллов, а также различных других биологических кристаллов, представляющих интерес. В случае амилоидных кристаллов нашим предположением является то, что небольшой размер является следствием природного изгиба β-листов, которые образуют протофиламенты фибрилл. Кристаллическая решетка сохраняет изгиб, создавая напряжение в этих кристаллах, которое возрастает по мере роста кристаллов. В конечном итоге это напряжение предупреждает добавление β-цепей, ограничивая толщину игольчатых кристаллов. В соответствии с нашим опытом, более длинные сегменты (например, 11 остатков по сравнению с 9 остатками) даже в большей степени ограничивают рост кристаллов; в случае NACore, состоящего из 11 остатков, и PreNAC, состоящего из 10 остатков, напряжение приводит к образованию нанокристаллов, невидимых с помощью оптической микроскопии. Эти кристаллы являются слишком маленькими для крепления и стандартного сбора синхротонных данных, однако идеально подходят для анализа с помощью MicroED. Наши структуры NACore и PreNAC демонстрируют, что MicroED способен определить новые и точные структуры биологического материала при атомном разрешении. Этот результат является основой для применений MicroED в отношении других биологических веществ, представляющих значение, для которых могут быть выращены только нанокристаллы. В нашей конкретной заявке мы смогли изучить расположение атомов кора ключевого домена NAC. Это открывает возможности для структурного дизайна ингибиторов образования амилоида αsyn 36.
Способы
Кристаллизация
Микрокристаллы SubNACore, 69AVVTGVTAV77 (SEQ ID NO:48), выращивали из синтетического пептида, приобретенного в CS Bio. Кристаллы выращивали при комнатной температуре с помощью испарения висячей капли. Лиофилизированный пептид растворяли в воде при концентрации 2,9 мг/мл в 48 мM гидроксида лития. Пептид смешивали при соотношении 2:1 с использованием резервуара, содержащего 0,9 M фосфата аммония и 0,1 M ацетата натрия, pH 4,6.
Нанокристаллы NACore, 68GAVVTGVTAV78 (SEQ ID NO:46), выращивали из синтетического пептида, приобретенного в CS Bio. Десять партий синтезированного пептида (CSBio) при концентрации 1 мг/мл в стерильной воде встряхивали при 37°C на орбитальном смешивающем планшете Torrey Pines при настройке скорости 9 в течение ночи. Нерастворимый материал промывали в 30% (вес/объем) глицерине, затем хранили в воде при комнатной температуре до дифракции. Образец содержат смесь фибрилл и кристаллов.
Нанокристаллы PreNAC, 47GVVHGVTTVA56 (SEQ ID NO:47), выращивали из синтетического пептида, приобретенного в InnoPep. Исследования кристаллизации синтезированного пептида готовили в партии. Пептид взвешивали и растворяли в стерилизованном фильтрацией 50 мM фосфатном буфере, pH 7,0, с 0,1% DMSO при концентрации 5 мг/мл. Этот раствор встряхивали при 37°C на орбитальном смешивающем планшете Torrey Pines при настройке скорости 9 в течение ночи.
Сбор и обработка данных
Данные рентгенодифракции на основе микрокристаллов SubNACore собирали с помощью синхротонного излучения в микрофокусном пучке излучения 24-ID-E Advanced Photon Source, Northeast Collaborative Access Team. Пучок излучения оснащали CCD детектором ADSC Quantum 315. Данные от одного кристалла собирали в 5° клиньях при длине волны 0,9791 Å с использованием диаметра пучка 5 мкм. Мы использовали данные от трех различных срезов вдоль оси иглы. Для сбора данных кристаллы охлаждали криогенным путем (100 K). Данные обрабатывали и сокращали с помощью Denzo/Scalepack из пакета программ HKL37.
Данные рентгенодифракции на основе нанокристаллов NACore собирали с помощью излучения XFEL в инструменте CXI (когерентное рентгеновское излучение) в линейном источнике когерентного света (LCLS)-SLAC. Энергия фотонов рентгеновских импульсов составляла 8,52 keV (1,45 Å). Каждый импульс 40 fs содержал до 6×1011 фотонов в положении образца, принимая во внимание пучок излучения 60%. Диаметр пучка составлял примерно 1 мкм. Нами была использована концентрация из примерно 25 мкл осажденного вещества, суспендированного в 1 мл воды. Образец вводили в пучок XFEL с помощью жидкоструйного инжектора и газодинамической виртуальной насадки38. Ширина микроструи составляла примерно 4 мкм, а скорость потока составляла 40 мкл/мин. Образец вызывал заметное распыление жидкой струи. Данные ХFEL обрабатывали с помощью cctbx.xfel 39,40.
Данные электронной дифракции на основе нанокристаллов NACore и PreNAC собирали с помощью методик MicroED5,6. В типичном случае эти нанокристаллы слипаются вместе. Для разделения скоплений примерно 100 мкл объем нанокристаллов помещали в ультразвуковую баню на 30 минут. Нанокристаллы откладывали на угольную решетку EM с отверстиями Quantifoil в 2-3 мкл капли после соответствующего разведения, которая оптимизировала плотность кристаллов на решетке. Все решетки затем блотировали и стекловали с помощью погружения в жидкий этан с использованием Vitrobot Mark IV (FEI), после чего переносили в жидкий азот для хранения. Замороженные гидратированные решетки затем переносили в крио-ТЭМ с помощью криодержателя Gatan 626. Паттерны дифракции и изображения кристаллов собирали с помощью оснащенного FEG ТЭМ FEI Tecnai F20, функционирующего при 200 кВ и записывающего с помощью фотоаппарата CMOS на нижнем основании TVIPS F416 с размером сенсора 4000 квадратных пикселей, каждый по 15,6 мкм в размере на квадратное измерение. Паттерны дифракции записывали при функционировании детектора в режиме сдвигаемого затвора с группировкой 2×2 пикселя, что приводило к конечному изображению, составляющему 2000 квадратных пикселей на размер. Отдельные кадры изображения снимали при времени экспозиции 3-4 секунды на изображение, с использованием определенной апертуры площади с освещенным размером пятна, составляющим примерно один микрон. Такая геометрия равняется дозе электронов менее 0,1 e−/Å2 на секунду. Во время каждого воздействия кристаллы непрерывно вращали в пучке со скоростью 0,3° в секунду, что соответствовало клину 1,2° на кадр. Данные о дифракции собирали от нескольких кристаллов, при этом каждый находился в разной ориентации по отношению к оси вращения. Каждая из этих совокупностей данных перекрывала клинья обратного пространства, варьируя от 40° до 80°.
Калибровку образца на расстояние детектора осуществляли с помощью поликристаллического золотого стандарта и с помощью ссылки на выраженные отклонения в эксперименте с дифракцией электронов с соответствующими отклонениями в данных XFEL. Калибровку положений x/y 64-модульном детекторе CSPAD осуществляли с помощью cctbx.xfel в результате очистки оптически измеренных положений модулей по отношению к совокупности данных термолизина39.
В целях получения совместимости со стандартными программами обработки рентгеновских данных дифракционные изображения конвертировали из формата tiff или TVIPS в кристаллографический формат SMV. Нами была использована XDS для индексации дифракционных изображений41, а также XSCALE для совместного объединения и масштабирования совокупностей данных, полученных от различных кристаллов. В случае NACore данные от четырех кристаллов объединяли, в то время как в случае PreNAC данные от трех кристаллов объединяли для компоновки конечных совокупностей данных.
Определение структуры
Раствор для молекулярного замещения в случае SubNACore получали с помощью программы Phaser 42. Модель поиска состояла из геометрически идеальной β-цепи, состоящей из девяти аланиновых остатков. Кристаллографические очистки осуществляли с помощью программы REFMAC 43. Раствор для молекулярного замещения в случае NACore получали с помощью программы Phaser 42. Модель поиска состояла из структуры SubNACore, определенной ранее. Кристаллографические очистки осуществляли с помощью программы Phenix 44. Раствор для молекулярного замещения в случае PreNAC получали с помощью программы Phaser 42. Модель поиска состояла из геометрически идеальной β-цепи, состоящей из шести остатков с последовательностью GVTTVA. Кристаллографические очистки осуществляли с помощью программы Phenix 44.
Построение модели для всех сегментов осуществляли с помощью COOT45. Координаты конечных моделей и структурных факторов откладывали в базе данных белковых структур с кодом PDB 4RIK в случае SubNACore, 4RIL - в случае NACore и 4ZNN - в случае PreNAC. Структуры иллюстрировали с помощью Pymol 46.
αСинуклеин
Конструкцию αsyn человека дикого типа характеризовали ранее 47 (pRK172, ампициллин, промотор T7) с помощью последовательности
MDVFMKGLSKAKEGVVAAAEKTKQGVAEAAGKTKEGVLYVGSKTKEGVVHG
VATVAEKTKEQVTNVGGAVVTGVTAVAQKTVEGAGSIAAATGFVKKDQLGKNEEGAPQEGILEDMPVDPDNEAYEMPSEEGYQDYEPEA (SEQ ID NO:1).
Очистка αсинуклеина
Полноразмерный αsyn очищали в соответствии с опубликованными протоколами 31. Конструкцию αsyn трансформировали в золотой стандарт линии клеток экспрессии BL21 E.coli (DE3) (Agilent Technologies, Санта-Клара, Калифорния) в случае экспрессии белка αsyn дикого типа. Одну колонию инкубировали в 100 мл бульона LB Miller (Fisher Scientific, Питтсбург, Пенсильвания) с добавлением 100 мкг/мл ампициллина (Fisher Scientific, Питтсбург, Пенсильвания) и выращивали в течение ночи при 37°C. Один литр бульона LB Miller с добавлением 100 мкг/мл ампициллина в 2 л шейкерных колбах инкубировали с 10 мл ночной культуры и выращивали при 37°C до тех пор, пока культура не достигала OD600 ~0,6-0,8, как измеряли с помощью УФ-ВИС-фотометра BioPhotometer (Eppendorf, Вестбери, Нью-Йорк). IPTG (изопропил-β-D-1-тиогалактопиранозид) добавляли до конечной концентрации 0,5 мМ и выращивали в течение 4-6 часов при 30°C. Клетки собирали с помощью центрифугирования при 5500 x g в течение 10 минут при 4°C. Клеточный осадок замораживали и хранили при -80°C.
Клеточный осадок размораживали на льду и ресуспендировали в буфере для лизиса (100 мM Tris-HCl, pH 8,0, 500 мM NaCl, 1 мM EDTA, pH 8,0), а затем лизировали с помощью ультразвука. Неочищенный клеточный лизат осветляли с помощью центрифугирования при 15000 x g в течение 30 минут при 4°C. Осветленный клеточный лизат кипятили и клеточный остаток удаляли с помощью центрифугирования. Белок в супернатанте осаждали в кислоте при pH 3,5 путем добавления HCl в результате титрования к белковому раствору на льду при помешивании, затем центрифугировали дополнительно при 15000 x g в течение 30 минут при 4°C. Супернатант диализировали против буфера A (20 мM Tris-HCl, pH 8,0). После диализа раствор фильтровали с помощью шприца 0,45 мкм (Корнинг, Нью-Йорк, 14831) перед загрузкой в 20 мл колонку HiPrep Q-HP 16/10 (GE Healthcare, Пискатауэй, Нью-Джерси). Колонку Q-HP промывали с использованием пяти колоночных объемов буфера A, а белок элюировали с помощью линейного градиента до 100% в пяти колоночных объемах буфера B (20 мM Tris-HCl, 1 M NaCl, pH 8,0). Белок элюировали при приблизительно 50-70% буфере B; пиковые фракции объединяли. Объединенные образцы концентрировали примерно в десять раз с помощью центрифужных фильтров Amicon Ultra-15. Примерно 5 мл концентрированного образца загружали в колонку HiPrep 26/60 Sephacryl S-75 HR, уравновешенную буфером для фильтрации (25 мM фосфата натрия, 100 мM NaCl, pH 7,5). Пиковые фракции объединяли из гель-фильтрационной колонки и диализировали против 5 мM Tris-HCl, pH 7,5, концентрировали до 3 мг/мл. Их фильтровали через фильтр с размером пор 0,2 мкм (Корнинг, Нью-Йорк, 14831) и хранили при 4°C.
Рекомбинантно экспрессируемый полноразмерный αsyn с N-концевым ацетилированием получали и очищали следующим образом на основании протокола Der-Sarkissian et al.25
Плазмиду на основе αsyn коэкспрессировали с гетеродимерным белковым ацетилирующим комплексом от S. pombe в целях ацетилирования N-конца (pACYC-DUET, хлорамфеникол, промотор T7)48. Два вектора котрансфицировали в E. coli BL21 (DE3) с помощью среды, содержащей как ампициллин, так и хлорамфеникол. Культуры клеток выращивали в среде TB, содержащей ампициллин и хлорамфеникол, и индуцировали экспрессию αsyn с помощью 0,5 мM IPTG в течение ночи при 25°C. Клетки собирали с помощью центрифугирования, клеточный осадок затем ресуспендировали в буфере для лизиса (100 мM Tris-HCl, pH 8,0, 500 мM NaCl, 1 мM EDTA, pH 8,0, и 1 мM фенилметилсульфонилфторид) и клетки лизировали с помощью гомогенизатора Emulsiflex (Avestin). Лизат кипятили и клеточный остаток удаляли с помощью центрифугирования. Белковую фракцию также удаляли с помощью осаждения при низком значении pH на льду после центрифугирования. Оставшийся супернатант доводили до значения pH с помощью титрования и диализировали против буфера A (20 мM Tris-HCl, pH 8,0, 1 мM DTT, 1 мM EDTA, pH 8,0). Полученный белковый раствор загружали в 5 мл колонку Q-Sepharose FF (GE Healthcare), уравновешенную буфером A, и элюировали против линейного градиента буфера B (1 M NaCl, 20 мM Tris-HCl, pH 8,0, 1 мM DTT, 1 мM EDTA, pH 8,0). Фракции, содержащие αsyn, идентифицировали с помощью SDS-PAGE, собирали, концентрировали и очищали с помощью эксклюзионной хроматографии (Sephacryl S-100 16/60, GE Healthcare) в 20 мM Tris, pH 8,0, 100 мM NaCl, 1 мM DTT, 1 мM EDTA. Чистоту фракций определяли с помощью SDS-PAGE.
Ацетилированный белок характеризовали с помощью LC-MS27,49. Предполагаемая средняя масса: 14460,1 Да в случае альфа-синуклеина и 14502,1 - в случае ацетилированного альфа-синуклеина. Наблюдаемая средняя масса: 14464,0 Да в случае альфа-синуклеина и 14506,0 - в случае ацетилированного альфа-синуклеина. Смещение на 4 Да между наблюдаемыми и предполагаемыми средними массами связан с инструментальной ошибкой.
Синтез ингибиторов
Ингибиторы получали коммерческим путем от Genscript Inc. при более 98% чистоты и солюбилизированными в 100% DMSO при концентрации 10 мM. Солюбилизированные ингибиторы фильтровали с помощью фильтра 0,1 мкM и хранили при -20°C в 20 мкл аликвот до последующего использования.
Анализы с использованием тиофлавина T
Анализы, связанные с образованием фибрилл, осуществляли с использованием концентрации белка, составляющей 50 мкМ, в условиях, идентичных таковым, используемым для агрегации белков в случае анализа затравки, однако в добавлением ThT. Все анализы проводили в черных 96-луночных планшетах с оптическим дном Nunc (Thermo Scientific). Планшеты встряхивали при 600 об./мин. в 3 мм вращательном диаметре в микропланшетом ридере Varioskan (Thermo) при 37°C. Флуоресцентные измерения фиксировали каждые 30 минут с использованием λex=444 нм, λem=482 нм, при времени интеграции 200 мкс.
Анализы агреации
Очищенный αсинуклеин диализировали в 0,1 M сульфате натрия, 25 мМ фосфате натрия и агрегировали посредством встряхивания в шейкерах Torrey Pine при 50 мкM при 37°C со скоростью 9 в течение 5-6 дней.
Экстракция нерастворимых в саркозиле белковых филаментов, характерных для PD из тканей головного мозга человека
Замороженные ткани головного мозга человека получали из UCLA Brain Tumor Translational Resource (BTTR). Нерастворимый в саркозиле белок экстрагировали с помощью ранее описанных протоколов. Вкратце, замороженную ткань гомогенизировали на охлажденном на льду PBS с помощью гомогенизатора Даунса при 200 мг/мл. Гомогенат затем разводили в буфере A (10 мM Tris, рН 7,4, 800 мM NaCl, 1 мM EGTA, 10% сахарозы) до 50 мг/мл в общем объеме 1 мл и центрифугировали при 20700 g в течение 20 мин. при 4°C. Супернатант собирали в ультрацентрифужной пробирке и осадок ресуспендировали в 0,5 мл буфера А с последующим центрифугированием при 20700 g в течение 20 мин. при 4°C. Супернатанты объединяли вместе. 150 мкл 10% саркозила (вес/объем) в воде Millipore затем добавляли и инкубировали в течение 1 ч при комнатной температуре на плоском вращающемся шейкере при 700 об./мин. Затем раствор центрифугировали при 100000 g в течение 1 ч при 4°C с помощью ротора SW 55 (Beckman Coulter). Супернатант снимали и осадок затем промывали с помощью 5 мл буфера A и центрифугировали при 100000 g в течение 20 мин. пи 4°C. Осадок ресуспендировали в 100 µл 50 мM Tris, pH 7,4.
Культура клеток HEK 293T
Клетки HEK 293T, которые стабильно экспрессируют меченый YFP αsyn ДТ и αsyn A53T, представляли собой щедрый подарок от д-ра Марка Даймонда (Marc Diamond). Клетки выращивали в среде Игла в модификации Дульбекко (Gibco) с добавлением 10% фетальной бычьей сыворотки (HyClone), 1% пенициллина/стрептомицина (Gibco) и 1% глютамакса (Gibco) в увлажненном инкубаторе при 37°C, 5% CO2.
Затравка в клетках HEK 293T
10000 клеток в 90 мкл среды высевали в 96-луночные планшет с черными стенками (№ по катал. №3660) и оставляли прикрепляться в течение в течение ночи. αSyn трансфицировали в конечной концентрации мономера 125 нM. Липофектамин 2000 разводили в среде OptiMEM (2,5+17,5 мкл) и инкубировали при комнатной температуре в течение 5 мин. Белковые агрегаты разводили в среде OptiMEM (1:20) и подвергали ультразвуковому воздействию в течение 3 мин. при низком импульсе. Разведенный липофектамин и белковые образцы затем смешивали 1:1 и инкубировали при комнатной температуре в течение 20 мин., после чего к каждой лунке добавляли 10 мкл. Все образцы добавляли в тройных повторностях, а эксперименты повторяли как минимум два раза. В случае котрансфекции фибрилл αsyn ингибиторами фибриллы и ингибиторы разводили в OptiMEM и инкубировали в течение 3 часов с последующим ульльтразвуковым воздействием.
Измерение внутриклеточных пятен в клетках
Образование пятен и рост клеток измеряли в визуализационном клеточном цитометре Celigo, обеспечивая несмещенное измерение. Лунки визуализировали с помощью флуоресцентного канала GFP и конфлюентность измеряли с помощью компьютерной программы для анализа Celigo. Получали изображения всей лунки и частицы подсчитывали с помощью ImageJ для анализа частиц. Во все дни использовали одни и те же настройки для анализа лунок одного планшета. Общее количество частиц в каждой лунке нормализовали в отношении конфлюентности и описывали в виде количества частиц на лунку.
Образование и детекция фибрилл
Очищенный αsyn в 50 мM Tris, 150 мM KCl, pH 7,5, встряхивали при концентрации 500 мкM при 37°C в шейкере Torey Pine. В целях образования фибриллярных образцов SubNACore и NACore лиофилизированные пептиды растворяли до конечной концентрации 500 мкM в 5 мM гидроксида лития, 20 мM фосфата натрия, pH 7,5, и 0,1 M NaCl. Все образцы встряхивали при 37°C в шейкере Torey Pine в течение 72 часов. Свежерастворенные образцы получали с путем разбавления лиофилизированных пептидов перед добавлением к клеткам для анализа.
Измерения мутности использовали для сравнения агрегации NACore и SubNACore. Пептидные образцы растворяли свежими до 1,6 мM в буфере для образца с 5 мM LiOH и 1% DMSO и затем фильтровали с помощью фильтра PVDF (Millipore, 0,1 мкм). Измерения осуществляли с помощью черных 96-луночных планшетов NUNC с 200 мкл образца/лунка (3-4 повторности на образец). Планшет встряхивали при 37°C в 3 мм вращательном диаметре при 300 об./мин. в микропланшетном ридере Varioskan (Thermo). Показатели поглощения считывали каждые 3-15 минут при 340 нм.
Трансмиссионная электронная микроскопия с отрицательным окрашиванием
Образцы для анализа цитотоксичности оценивали в отношении наличия фибрилл с помощью электронной микроскопии. Вкратце, 5 мкл образцов наносили непосредственно на свежепокрытые углеродом решетки электронного микроскопа (Ted Pella, Реддинг, Калифорния). Через 4 мин. после инкубации решетки ополаскивали дважды 5 мл дистиллированной воды и окрашивали 2% уранилацетатом на 1 мин. Образцы исследовали на электронном микроскопе FEI T12.
Дифракция фибрилл
Фибриллы, образованные из очищенного αsyn с наличием и без N-концевого ацетилирования концентрировали с помощью центрифугирования, промывали и ориентировали во время сушки между двумя стеклянными капиллярами. Аналогично нанокристаллы NACore также концентрировали, промывали и оставляли ориентироваться во время сушки между двумя стеклянными капиллярами. Выравненные фибриллы или нанокристаллы помещали на криоиглу для дифракции с помощью 1,54 Å рентгеновских лучей, образуемых с помощью ротационного анодного генератора Rigaku, оснащенного визуализационным планшетом HTC и встроенным в криоохлаждающее устройство X-Stream. Все паттерны собирали на расстоянии 180 мм и анализировали с помощью пакета компьютерных программ Adxv.
Анализы цитотоксичности
Прикрепившиеся клетки PC12 культивировали в составленной на основе ATCC среде RPMI 1640 (ATCC; № по кат. 30-2001) с добавлением 10% лошадиной сыворотки и 5% фетальной бычьей сыворотки и помещали по 10000 на лунку до конечного объема 90 мкл. Все анализы MTT осуществляли с помощью водного нерадиактивного набора для пролиферации клеток Cell Titer 96 (MTT, Promega, № по кат. 4100). Клетки культивировали в 96-луночных планшетах в течение 20 ч при 37°C в 5% CO2 перед добавлением образцов (№ по кат. Costar 3596). 10 мкл образца добавляли к каждой лунке, содержащей 90 мкл среды и инкубировали в течение 24 ч при 37°C в 5% CO2. Затем 15 мкл раствора красителя (Promega, № по кат. 4102) добавляли в каждую лунку с последующей инкубацией в течение 4 ч при 37°C в 5% CO2. За этим следовало добавление 100 мкл смеси для солюбилизации/стоп-раствора (Promega, № по кат. 4101) в каждую лунку. Через 12 ч после инкубации при комнатной температуре поглощение измеряли при 570 нм. Фоновое поглощение фиксировали при 700 нм. Данные нормализовали с использованием клеток, обработанных 1% (вес/объем) SDS до 0% восстановления, а затем клетки обрабатывали буфером для образца до 100% восстановления.
Анализы с использованием лактатдегидрогеназы выполняли с помощью CytoTox-ONE™ Homogeneous Membrane Integrity (Promega, № по кат. G7890), согласно инструкциям производителя. Вкратце, клетки высевали в 96-луночные планшеты для культивирования тканей с черными стенками и прозрачным дном (Fisher, № по кат. 07-200-588) по 10000 клеток на лунку до конечного объема 90 мкл. Клетки инкубировали дополнительно в течение 20 ч при 37°C в 5% CO2 перед добавлением образцов. Затем 10 мкл образца добавляли в каждую лунку, после чего клетки инкубировали еще в течение 24 часов. 100 мкл реагента добавляли в каждую лунку и инкубировали в течение 15 мин. при комнатной температуре. Добавление 50 мкл стоп-раствора останавливало реакцию. Флуоресценцию измеряли с помощью длин волн возбуждения и излучения 560 нм и 590 нм соответственно. Данные нормализовали с помощью клеток, обработанных буфером, в качестве 0% высвобождения, и 0,1% тритоном X-100, в качестве 100% высвобождения.
Атомные координаты структуры NACore: GAVVTGVTAVA (SEQ ID NO:46) представлены ниже в Табл. 3. Атомные координаты PreNAC представлены ниже в Табл. 4.
Атомная структура NACore показывает амилоидную фибриллу, образованную из самокомплементарной пары β-листов, плотно соединенных с образованием стерической молнии. С помощью компьютерной модели Rossetta каждый из ингибиторов разрабатывали в целях связывания конца стерической молнии и, тем самым, «кэппирования» фибрилл, предупреждая добавление других молекул альфа-синуклеина к фибриллам. Нами было разработано 3 кандидатных ингибитора, S37, S61 и S71, которые эффективно связываются с одним или более концами молнии. Рассчитанные энергии связывания и комплементарность форме каждого из трех ингибиторов с фибриллой также являются благоприятными для связывания. Каждый из ингибиторов сохраняет большинство остатков нативной последовательности NACore, однако также содержит один или более модифицированных остатков. Rodriquez et al. показали, что меньший фрагмент, состоящий из 9 остатков, в NACore [69-77] агрегирует медленнее, чем NACore, а его структура является аналогичной NACore. В целях предупреждения самоагрегации наших разработанных ингибиторов нами был использован более короткий сегмент совместно с одной или более модификациями. S37 имеет мутацию W в Thr72 и дополнительную полилизиновый тэг на C-конце для индукции отталкивания заряд-заряд. Предполагается, что он связывает оба конца фибриллы со стерической молнией (т.е., верх и низ). S61 и S62 сохраняют ту же самую ингибиторную последовательность, что и S37, однако вместо полилизинового тэга добавляют тэг TAT для содействия растворимости и предупреждения самоагрегации. S71 имеет метилированный глицин в Gly73, который ослабляет водородное связывание в β-листе, и дополнительный тэг TAT в целях растворимости и проникновения в клетку.
Пример II Компьютерный дизайн
На основании атомной структуры волокна, образующего сегмент αсинуклеина, нами был разработан ряд ингибиторов, которые ингибируют агрегацию. Мы разработали пептидные ингибиторы для специфичного «кэппирования» роста агрегатов αсинуклеина. С помощью алгоритма ZipperDB (Goldschmidt et al.55), в домене NAC были идентифицированы кристаллизуемые сегменты, образующие амилоид. Было отмечено, что этот домен является необходимым и достаточным для агрегации и токсичности αсинуклеина (Bodles et al. 20). Идентифицированные сегменты (сегменты αсинуклеина, состоящие из остатков 69-77 и остатков 68-78) синтезировали химическим путем и кристаллизовали; их трехмерные структуры определяли с помощью микрокристаллографии и микроэлектронной дифракции (MicroED). Затем нами был применен способ Rosetta (Sievers et al. 36) в стадиях дизайна ингибиторов, которые нарушают агрегацию αсинуклеина, с использованием структуры 69-77 αсинуклеина или структуры 68-78 αсинуклеина в виде матрицы. Предполагаемые ингибиторы идентифицировали и отбирали для экспериментальной характеристики.
Пример III - Второй цикл дизайна - рациональный дизайн на основе последовательностей
Во втором цикле дизайна начало осуществляли исходя из нескольких амилоидогенных сегментов альфа-синуклеина: 47-56, 68-78 и 84-94. N-метилирование вводили в двух отдельных остатках в нативной аминокислотной последовательности. В этих N-метильных аминокислотах дополнительная метильная группа замещает аминный водород. Тем самым, это блокирует водородную связь и предупреждает добавление пептидных мономеров на своем растущем конце. Помимо N-метилирования, считается, что дополнительные полярные/заряженные аминокислотные дополнения по сравнению с нативной последовательностью альфа-синуклеина улучшают общую растворимость пептидного ингибитора.
Таблица 3
TITLE ELECTRON DIFFRACTION STRUCTURE OF THE PARKINSON'S DISEASE TOXIC CORE
TITLE 2 OF ALPHA-SYNUCLEIN AMYLOID, GAVVTGVTAVA
COMPND MOL_ID: 1;
COMPND 2 MOLECULE: ALPHA-SYNUCLEIN;
COMPND 3 CHAIN: A;
COMPND 4 SYNONYM: NON-A BETA COMPONENT OF AD AMYLOID, NON-A4 COMPONENT OF
COMPND 5 AMYLOID PRECURSOR, NACP;
COMPND 6 ENGINEERED: YES
SOURCE MOL_ID: 1;
SOURCE 2 SYNTHETIC: YES;
SOURCE 3 ORGANISM_SCIENTIFIC: HOMO SAPIENS;
SOURCE 4 ORGANISM_COMMON: HUMAN;
SOURCE 5 ORGANISM_TAXID: 9606;
SOURCE 6 OTHER_DETAILS: SYNTHETIC PEPTIDE GAVVTGVTAVA CORRESPONDING TO
SOURCE 7 SEGMENT 68-78 OF HUMAN ALPHA-SYNUCLEIN
KEYWDS AMYLOID, ALPHA-SYNUCLEIN, PARKINSON'S DISEASE, TOXIC CORE, NACORE,
KEYWDS 2 LIPID BINDING PROTEIN
EXPDTA ELECTRON CRYSTALLOGRAPHY
AUTHOR J.A.RODRIGUEZ,M.IVANOVA,M.R.SAWAYA,D.CASCIO,F.REYES,D.SHI,L.JOHNSON,
AUTHOR 2 E.GUENTHER,S.SANGWAN,J.HATTNE,B.NANNENGA,A.S.BREWSTER,
AUTHOR 3 M.MESSERSCHMIDT,S.BOUTET,N.K.SAUTER,T.GONEN,D.S.EISENBERG
JRNL AUTH J.A.RODRIGUEZ,M.IVANOVA,M.R.SAWAYA,D.CASCIO,F.REYES,D.SHI,
JRNL AUTH 2 L.JOHNSON,E.GUENTHER,S.SANGWAN,J.HATTNE,B.NANNENGA,
JRNL AUTH 3 A.S.BREWSTER,M.MESSERSCHMIDT,S.BOUTET,N.K.SAUTER,T.GONEN,
JRNL AUTH 4 D.S.EISENBERG
JRNL TITL MICROED STRUCTURE OF THE TOXIC CORE OF ALPHA-SYNUCLEIN
JRNL TITL 2 AMYLOID, THE PROTEIN ASSOCIATED WITH THE DEVELOPMENT OF
JRNL TITL 3 PARKINSONS DISEASE
JRNL REF TO BE PUBLISHED
JRNL REFN
REMARK 2
REMARK 2 RESOLUTION. 1.43 ANGSTROMS.
REMARK 3
REMARK 3 REFINEMENT.
REMARK 3 PROGRAM : BUSTER
REMARK 3 AUTHORS : BRICOGNE,BLANC,BRANDL,FLENSBURG,KELLER,
REMARK 3 : PACIOREK,ROVERSI,SMART,VONRHEIN,WOMACK,
REMARK 3 : MATTHEWS,TEN EYCK,TRONRUD
REMARK 3
REMARK 3 DATA USED IN REFINEMENT.
REMARK 3 RESOLUTION RANGE HIGH (ANGSTROMS) : 1.43
REMARK 3 RESOLUTION RANGE LOW (ANGSTROMS) : 16.43
REMARK 3 DATA CUTOFF (SIGMA(F)) : 0.000
REMARK 3 COMPLETENESS FOR RANGE (%) : 87.9
REMARK 3 NUMBER OF REFLECTIONS : 1073
REMARK 3
REMARK 3 FIT TO DATA USED IN REFINEMENT.
REMARK 3 CROSS-VALIDATION METHOD : THROUGHOUT
REMARK 3 FREE R VALUE TEST SET SELECTION : RANDOM
REMARK 3 R VALUE (WORKING+TEST SET) : 0.251
REMARK 3 R VALUE (WORKING SET) : 0.248
REMARK 3 FREE R VALUE : 0.275
REMARK 3 FREE R VALUE TEST SET SIZE (%) : 11.840
REMARK 3 FREE R VALUE TEST SET COUNT : 127
REMARK 3 ESTIMATED ERROR OF FREE R VALUE : NULL
REMARK 3
REMARK 3 FIT IN THE HIGHEST RESOLUTION BIN.
REMARK 3 TOTAL NUMBER OF BINS USED : 5
REMARK 3 BIN RESOLUTION RANGE HIGH (ANGSTROMS) : 1.43
REMARK 3 BIN RESOLUTION RANGE LOW (ANGSTROMS) : 1.60
REMARK 3 BIN COMPLETENESS (WORKING+TEST) (%) : 87.88
REMARK 3 REFLECTIONS IN BIN (WORKING+TEST SET) : 286
REMARK 3 BIN R VALUE (WORKING+TEST SET) : 0.2642
1
REMARK 3 REFLECTIONS IN BIN (WORKING SET) : 245
REMARK 3 BIN R VALUE (WORKING SET) : 0.2532
REMARK 3 BIN FREE R VALUE : 0.3310
REMARK 3 BIN FREE R VALUE TEST SET SIZE (%) : 14.34
REMARK 3 BIN FREE R VALUE TEST SET COUNT : 41
REMARK 3 ESTIMATED ERROR OF BIN FREE R VALUE : NULL
REMARK 3
REMARK 3 NUMBER OF NON-HYDROGEN ATOMS USED IN REFINEMENT.
REMARK 3 PROTEIN ATOMS : 66
REMARK 3 NUCLEIC ACID ATOMS : 0
REMARK 3 HETEROGEN ATOMS : 0
REMARK 3 SOLVENT ATOMS : 2
REMARK 3
REMARK 3 B VALUES.
REMARK 3 FROM WILSON PLOT (A**2) : 10.33
REMARK 3 MEAN B VALUE (OVERALL, A**2) : 12.75
REMARK 3 OVERALL ANISOTROPIC B VALUE.
REMARK 3 B11 (A**2) : -3.98760
REMARK 3 B22 (A**2) : 3.49080
REMARK 3 B33 (A**2) : 0.49680
REMARK 3 B12 (A**2) : 0.00000
REMARK 3 B13 (A**2) : -1.55090
REMARK 3 B23 (A**2) : 0.00000
REMARK 3
REMARK 3 ESTIMATED COORDINATE ERROR.
REMARK 3 ESD FROM LUZZATI PLOT (A) : 0.305
REMARK 3 DPI (BLOW EQ-10) BASED ON R VALUE (A) : NULL
REMARK 3 DPI (BLOW EQ-9) BASED ON FREE R VALUE (A) : NULL
REMARK 3 DPI (CRUICKSHANK) BASED ON R VALUE (A) : 0.108
REMARK 3 DPI (CRUICKSHANK) BASED ON FREE R VALUE (A) : NULL
REMARK 3
REMARK 3 REFERENCES: BLOW, D. (2002) ACTA CRYST D58, 792-797
REMARK 3 CRUICKSHANK, D.W.J. (1999) ACTA CRYST D55, 583-601
REMARK 3
REMARK 3 CORRELATION COEFFICIENTS.
REMARK 3 CORRELATION COEFFICIENT FO-FC : 0.913
REMARK 3 CORRELATION COEFFICIENT FO-FC FREE : 0.900
REMARK 3
REMARK 3 NUMBER OF GEOMETRIC FUNCTION TERMS DEFINED : 15
REMARK 3 TERM COUNT WEIGHT FUNCTION.
REMARK 3 BOND LENGTHS : 65 ; 2.000 ; HARMONIC
REMARK 3 BOND ANGLES : 90 ; 2.000 ; HARMONIC
REMARK 3 TORSION ANGLES : 16 ; 2.000 ; SINUSOIDAL
REMARK 3 TRIGONAL CARBON PLANES : 1 ; 2.000 ; HARMONIC
REMARK 3 GENERAL PLANES : 10 ; 5.000 ; HARMONIC
REMARK 3 ISOTROPIC THERMAL FACTORS : 65 ; 20.000 ; HARMONIC
REMARK 3 BAD NON-BONDED CONTACTS : NULL ; NULL ; NULL
REMARK 3 IMPROPER TORSIONS : NULL ; NULL ; NULL
REMARK 3 PSEUDOROTATION ANGLES : NULL ; NULL ; NULL
REMARK 3 CHIRAL IMPROPER TORSION : 11 ; 5.000 ; SEMIHARMONIC
REMARK 3 SUM OF OCCUPANCIES : NULL ; NULL ; NULL
REMARK 3 UTILITY DISTANCES : NULL ; NULL ; NULL
REMARK 3 UTILITY ANGLES : NULL ; NULL ; NULL
REMARK 3 UTILITY TORSION : NULL ; NULL ; NULL
REMARK 3 IDEAL-DIST CONTACT TERM : 76 ; 4.000 ; SEMIHARMONIC
REMARK 3
REMARK 3 RMS DEVIATIONS FROM IDEAL VALUES.
REMARK 3 BOND LENGTHS (A) : 0.010
REMARK 3 BOND ANGLES (DEGREES) : 1.65
REMARK 3 PEPTIDE OMEGA TORSION ANGLES (DEGREES) : 4.08
REMARK 3 OTHER TORSION ANGLES (DEGREES) : 6.49
REMARK 3
REMARK 3 TLS DETAILS
REMARK 3 NUMBER OF TLS GROUPS : NULL
2
REMARK 3
REMARK 3 OTHER REFINEMENT REMARKS: NULL
REMARK 4
REMARK 4 4RIL COMPLIES WITH FORMAT V. 3.30, 13-JUL-11
REMARK 100
REMARK 100 THIS ENTRY HAS BEEN PROCESSED BY RCSB ON 09-OCT-14.
REMARK 100 THE RCSB ID CODE IS RCSB087390.
REMARK 240
REMARK 240 EXPERIMENTAL DETAILS
REMARK 240 RECONSTRUCTION METHOD : NULL
REMARK 240 SAMPLE TYPE : NULL
REMARK 240 SPECIMEN TYPE : NULL
REMARK 240 DATA ACQUISITION
REMARK 240 DATE OF DATA COLLECTION : 28-AUG-14
REMARK 240 TEMPERATURE (KELVIN) : 100.0
REMARK 240 PH : NULL
REMARK 240 NUMBER OF CRYSTALS USED : 4
REMARK 240 MICROSCOPE MODEL : TECNAI F20 TEM
REMARK 240 DETECTOR TYPE : TVIPS F416 CMOS CAMERA
REMARK 240 ACCELERATION VOLTAGE (KV) : NULL
REMARK 240 NUMBER OF UNIQUE REFLECTIONS : 1073
REMARK 240 RESOLUTION RANGE HIGH (A) : 1.430
REMARK 240 RESOLUTION RANGE LOW (A) : 16.430
REMARK 240 DATA SCALING SOFTWARE : XSCALE
REMARK 240 COMPLETENESS FOR RANGE (%) : 89.9
REMARK 240 DATA REDUNDANCY : 4.400
REMARK 240 IN THE HIGHEST RESOLUTION SHELL
REMARK 240 HIGHEST RESOLUTION SHELL, RANGE HIGH (A) :1.43
REMARK 240 HIGHEST RESOLUTION SHELL, RANGE LOW (A) :1.60
REMARK 240 COMPLETENESS FOR SHELL (%) : 82.5
REMARK 240 DATA REDUNDANCY IN SHELL : 4.40
REMARK 240 R MERGE FOR SHELL (I) : 0.56500
REMARK 240 METHOD USED TO DETERMINE THE STRUCTURE: MOLECULAR REPLACEMENT
REMARK 240 SOFTWARE USED : PHASER
REMARK 240 STARTING MODEL : 4RIK
REMARK 290
REMARK 290 CRYSTALLOGRAPHIC SYMMETRY
REMARK 290 SYMMETRY OPERATORS FOR SPACE GROUP: C 1 2 1
REMARK 290
REMARK 290 SYMOP SYMMETRY
REMARK 290 NNNMMM OPERATOR
REMARK 290 1555 X,Y,Z
REMARK 290 2555 -X,Y,-Z
REMARK 290 3555 X+1/2,Y+1/2,Z
REMARK 290 4555 -X+1/2,Y+1/2,-Z
REMARK 290
REMARK 290 WHERE NNN -> OPERATOR NUMBER
REMARK 290 MMM -> TRANSLATION VECTOR
REMARK 290
REMARK 290 CRYSTALLOGRAPHIC SYMMETRY TRANSFORMATIONS
REMARK 290 THE FOLLOWING TRANSFORMATIONS OPERATE ON THE ATOM/HETATM
REMARK 290 RECORDS IN THIS ENTRY TO PRODUCE CRYSTALLOGRAPHICALLY
REMARK 290 RELATED MOLECULES.
REMARK 290 SMTRY1 1 1.000000 0.000000 0.000000 0.00000
REMARK 290 SMTRY2 1 0.000000 1.000000 0.000000 0.00000
REMARK 290 SMTRY3 1 0.000000 0.000000 1.000000 0.00000
REMARK 290 SMTRY1 2 -1.000000 0.000000 0.000000 0.00000
REMARK 290 SMTRY2 2 0.000000 1.000000 0.000000 0.00000
REMARK 290 SMTRY3 2 0.000000 0.000000 -1.000000 0.00000
REMARK 290 SMTRY1 3 1.000000 0.000000 0.000000 35.40500
REMARK 290 SMTRY2 3 0.000000 1.000000 0.000000 2.41000
REMARK 290 SMTRY3 3 0.000000 0.000000 1.000000 0.00000
REMARK 290 SMTRY1 4 -1.000000 0.000000 0.000000 35.40500
REMARK 290 SMTRY2 4 0.000000 1.000000 0.000000 2.41000
3
REMARK 290 SMTRY3 4 0.000000 0.000000 -1.000000 0.00000
REMARK 290
REMARK 290 REMARK: NULL
REMARK 300
REMARK 300 BIOMOLECULE: 1, 2
REMARK 300 SEE REMARK 350 FOR THE AUTHOR PROVIDED AND/OR PROGRAM
REMARK 300 GENERATED ASSEMBLY INFORMATION FOR THE STRUCTURE IN
REMARK 300 THIS ENTRY. THE REMARK MAY ALSO PROVIDE INFORMATION ON
REMARK 300 BURIED SURFACE AREA.
REMARK 300 REMARK: THE BIOLOGICAL UNIT IS A PAIR OF BETA-SHEETS. ONE SHEET IS
REMARK 300 COMPOSED OF CHAIN A AND UNIT CELL TRANSLATIONS ALONG THE B
REMARK 300 DIMENSION. THE OTHER SHEET IS COMPOSED OF THE SYMMETRY MATE -X+1/2,
REMARK 300 Y+1/2,-Z, AND UNIT CELL TRANSLATIONS ALONG B.
REMARK 350
REMARK 350 COORDINATES FOR A COMPLETE MULTIMER REPRESENTING THE KNOWN
REMARK 350 BIOLOGICALLY SIGNIFICANT OLIGOMERIZATION STATE OF THE
REMARK 350 MOLECULE CAN BE GENERATED BY APPLYING BIOMT TRANSFORMATIONS
REMARK 350 GIVEN BELOW. BOTH NON-CRYSTALLOGRAPHIC AND
REMARK 350 CRYSTALLOGRAPHIC OPERATIONS ARE GIVEN.
REMARK 350
REMARK 350 BIOMOLECULE: 1
REMARK 350 AUTHOR DETERMINED BIOLOGICAL UNIT: TETRAMERIC
REMARK 350 APPLY THE FOLLOWING TO CHAINS: A
REMARK 350 BIOMT1 1 1.000000 0.000000 0.000000 0.00000
REMARK 350 BIOMT2 1 0.000000 1.000000 0.000000 0.00000
REMARK 350 BIOMT3 1 0.000000 0.000000 1.000000 0.00000
REMARK 350 BIOMT1 2 1.000000 0.000000 0.000000 0.00000
REMARK 350 BIOMT2 2 0.000000 1.000000 0.000000 4.82000
REMARK 350 BIOMT3 2 0.000000 0.000000 1.000000 0.00000
REMARK 350 BIOMT1 3 -1.000000 0.000000 0.000000 35.40500
REMARK 350 BIOMT2 3 0.000000 1.000000 0.000000 2.41000
REMARK 350 BIOMT3 3 0.000000 0.000000 -1.000000 0.00000
REMARK 350 BIOMT1 4 -1.000000 0.000000 0.000000 35.40500
REMARK 350 BIOMT2 4 0.000000 1.000000 0.000000 7.23000
REMARK 350 BIOMT3 4 0.000000 0.000000 -1.000000 0.00000
REMARK 350
REMARK 350 BIOMOLECULE: 2
REMARK 350 AUTHOR DETERMINED BIOLOGICAL UNIT: MONOMERIC
REMARK 350 APPLY THE FOLLOWING TO CHAINS: A
REMARK 350 BIOMT1 1 1.000000 0.000000 0.000000 0.00000
REMARK 350 BIOMT2 1 0.000000 1.000000 0.000000 0.00000
REMARK 350 BIOMT3 1 0.000000 0.000000 1.000000 0.00000
DBREF 4RIL A 1 11 UNP P37840 SYUA_HUMAN 68 78
SEQRES 1 A 11 GLY ALA VAL VAL THR GLY VAL THR ALA VAL ALA
FORMUL 2 HOH *2(H2 O)
CRYST1 70.810 4.820 16.790 90.00 105.68 90.00 C 1 2 1 4
ORIGX1 1.000000 0.000000 0.000000 0.00000
ORIGX2 0.000000 1.000000 0.000000 0.00000
ORIGX3 0.000000 0.000000 1.000000 0.00000
SCALE1 0.014122 0.000000 0.003964 0.00000
SCALE2 0.000000 0.207469 0.000000 0.00000
SCALE3 0.000000 0.000000 0.061861 0.00000
ATOM 1 N GLY A 1 -1.664 2.302 6.349 1.00 23.58 N
ATOM 2 CA GLY A 1 -1.194 2.605 5.007 1.00 23.02 C
ATOM 3 C GLY A 1 0.105 1.901 4.702 1.00 21.89 C
ATOM 4 O GLY A 1 0.121 0.676 4.579 1.00 20.14 O
ATOM 5 N ALA A 2 1.197 2.666 4.558 1.00 16.28 N
ATOM 6 CA ALA A 2 2.509 2.094 4.221 1.00 15.41 C
ATOM 7 C ALA A 2 3.664 2.731 5.005 1.00 12.05 C
ATOM 8 O ALA A 2 3.668 3.942 5.227 1.00 10.77 O
ATOM 9 CB ALA A 2 2.764 2.186 2.712 1.00 15.67 C
ATOM 10 N VAL A 3 4.640 1.905 5.417 1.00 5.98 N
ATOM 11 CA VAL A 3 5.852 2.304 6.129 1.00 3.93 C
ATOM 12 C VAL A 3 7.023 1.700 5.344 1.00 5.93 C
4
ATOM 13 O VAL A 3 7.217 0.485 5.388 1.00 6.66 O
ATOM 14 CB VAL A 3 5.856 1.891 7.629 1.00 7.66 C
ATOM 15 CG1 VAL A 3 7.148 2.337 8.330 1.00 6.70 C
ATOM 16 CG2 VAL A 3 4.661 2.494 8.333 1.00 6.73 C
ATOM 17 N VAL A 4 7.702 2.522 4.533 1.00 3.26 N
ATOM 18 CA VAL A 4 8.782 2.087 3.638 1.00 4.61 C
ATOM 19 C VAL A 4 10.065 2.743 4.088 1.00 4.32 C
ATOM 20 O VAL A 4 10.149 3.973 4.071 1.00 3.00 O
ATOM 21 CB VAL A 4 8.458 2.457 2.171 1.00 8.53 C
ATOM 22 CG1 VAL A 4 9.486 1.874 1.188 1.00 9.58 C
ATOM 23 CG2 VAL A 4 7.041 2.046 1.790 1.00 9.20 C
ATOM 24 N THR A 5 11.066 1.952 4.438 1.00 3.32 N
ATOM 25 CA THR A 5 12.360 2.426 4.896 1.00 3.88 C
ATOM 26 C THR A 5 13.504 1.769 4.092 1.00 5.43 C
ATOM 27 O THR A 5 13.567 0.539 4.039 1.00 6.20 O
ATOM 28 CB THR A 5 12.540 2.051 6.351 1.00 6.84 C
ATOM 29 OG1 THR A 5 11.414 2.503 7.130 1.00 9.13 O
ATOM 30 CG2 THR A 5 13.891 2.524 6.943 1.00 10.98 C
ATOM 31 N GLY A 6 14.417 2.588 3.546 1.00 6.04 N
ATOM 32 CA GLY A 6 15.571 2.100 2.793 1.00 6.46 C
ATOM 33 C GLY A 6 16.874 2.688 3.292 1.00 7.38 C
ATOM 34 O GLY A 6 16.959 3.896 3.501 1.00 6.32 O
ATOM 35 N VAL A 7 17.913 1.864 3.436 1.00 4.11 N
ATOM 36 CA VAL A 7 19.223 2.311 3.884 1.00 5.04 C
ATOM 37 C VAL A 7 20.243 1.730 2.870 1.00 4.68 C
ATOM 38 O VAL A 7 20.244 0.519 2.678 1.00 5.69 O
ATOM 39 CB VAL A 7 19.532 1.802 5.294 1.00 9.15 C
ATOM 40 CG1 VAL A 7 21.007 1.991 5.629 1.00 9.39 C
ATOM 41 CG2 VAL A 7 18.606 2.431 6.367 1.00 9.75 C
ATOM 42 N THR A 8 21.105 2.570 2.279 1.00 3.68 N
ATOM 43 CA THR A 8 22.201 2.169 1.357 1.00 4.35 C
ATOM 44 C THR A 8 23.468 2.783 1.930 1.00 6.01 C
ATOM 45 O THR A 8 23.530 4.005 2.031 1.00 6.48 O
ATOM 46 CB THR A 8 21.874 2.496 -0.083 1.00 12.43 C
ATOM 47 OG1 THR A 8 20.601 1.921 -0.394 1.00 14.76 O
ATOM 48 CG2 THR A 8 22.929 1.954 -1.075 1.00 13.32 C
ATOM 49 N ALA A 9 24.453 1.977 2.339 1.00 7.04 N
ATOM 50 CA ALA A 9 25.628 2.542 2.994 1.00 9.57 C
ATOM 51 C ALA A 9 26.967 1.849 2.849 1.00 11.59 C
ATOM 52 O ALA A 9 27.052 0.628 2.756 1.00 8.18 O
ATOM 53 CB ALA A 9 25.328 2.729 4.478 1.00 10.30 C
ATOM 54 N VAL A 10 28.017 2.646 2.956 1.00 10.58 N
ATOM 55 CA VAL A 10 29.405 2.206 3.043 1.00 12.39 C
ATOM 56 C VAL A 10 29.896 2.880 4.314 1.00 18.51 C
ATOM 57 O VAL A 10 30.171 4.075 4.301 1.00 16.08 O
ATOM 58 CB VAL A 10 30.273 2.561 1.816 1.00 17.83 C
ATOM 59 CG1 VAL A 10 31.744 2.243 2.085 1.00 18.89 C
ATOM 60 CG2 VAL A 10 29.778 1.831 0.567 1.00 17.39 C
ATOM 61 N ALA A 11 29.916 2.136 5.427 1.00 17.81 N
ATOM 62 CA ALA A 11 30.340 2.647 6.726 1.00 23.47 C
ATOM 63 C ALA A 11 31.643 1.988 7.097 1.00 51.68 C
ATOM 64 O ALA A 11 31.622 0.916 7.734 1.00 58.63 O
ATOM 65 CB ALA A 11 29.289 2.350 7.778 1.00 24.49 C
ATOM 66 OXT ALA A 11 32.691 2.485 6.653 1.00 74.50 O
TER 67 ALA A 11
HETATM 68 O HOH A 101 9.830 4.990 7.085 1.00 11.92 O
HETATM 69 O HOH A 102 18.548 3.873 0.260 1.00 20.36 O
MASTER 203 0 0 0 0 0 0 6 68 1 0 1
END
5
Атомные координаты структуры PreNAC: GVVHGVTTVA (SEQ ID NO:47) представлены ниже в Табл. 4.
Таблица 4
HEADER ---- 27-APR-15 xxxx
COMPND ---
REMARK 3
REMARK 3 REFINEMENT.
REMARK 3 PROGRAM : REFMAC 5.8.0073
REMARK 3 AUTHORS : MURSHUDOV,SKUBAK,LEBEDEV,PANNU,
REMARK 3 STEINER,NICHOLLS,WINN,LONG,VAGIN
REMARK 3
REMARK 3 REFINEMENT TARGET : MAXIMUM LIKELIHOOD
REMARK 3
REMARK 3 DATA USED IN REFINEMENT.
REMARK 3 RESOLUTION RANGE HIGH (ANGSTROMS) : 1.41
REMARK 3 RESOLUTION RANGE LOW (ANGSTROMS) : 32.94
REMARK 3 DATA CUTOFF (SIGMA(F)) : ОТСУТСТВУЕТ
REMARK 3 COMPLETENESS FOR RANGE (%) : 86.73
REMARK 3 NUMBER OF REFLECTIONS : 1006
REMARK 3
REMARK 3 FIT TO DATA USED IN REFINEMENT.
REMARK 3 CROSS-VALIDATION METHOD : THROUGHOUT
REMARK 3 FREE R VALUE TEST SET SELECTION : RANDOM
REMARK 3 R VALUE (WORKING+TEST SET) : 0.26104
REMARK 3 R VALUE (WORKING SET) : 0.25243
REMARK 3 FREE R VALUE : 0.33523
REMARK 3 FREE R VALUE TEST SET SIZE (%) : 10.0
REMARK 3 FREE R VALUE TEST SET COUNT : 112
REMARK 3
REMARK 3 FIT IN THE HIGHEST RESOLUTION BIN.
REMARK 3 TOTAL NUMBER OF BINS USED : 20
REMARK 3 BIN RESOLUTION RANGE HIGH : 1.409
REMARK 3 BIN RESOLUTION RANGE LOW : 1.446
REMARK 3 REFLECTION IN BIN (WORKING SET) : 49
REMARK 3 BIN COMPLETENESS (WORKING+TEST) (%) : 51.43
REMARK 3 BIN R VALUE (WORKING SET) : 0.388
REMARK 3 BIN FREE R VALUE SET COUNT : 5
REMARK 3 BIN FREE R VALUE : 0.528
REMARK 3
REMARK 3 NUMBER OF NON-HYDROGEN ATOMS USED IN REFINEMENT.
REMARK 3 ALL ATOMS : 68
REMARK 3
REMARK 3 B VALUES.
REMARK 3 FROM WILSON PLOT (A**2) : NULL
REMARK 3 MEAN B VALUE (OVERALL, A**2) : 19.367
REMARK 3 OVERALL ANISOTROPIC B VALUE.
REMARK 3 B11 (A**2) : -0.23
REMARK 3 B22 (A**2) : -0.23
REMARK 3 B33 (A**2) : 0.49
REMARK 3 B12 (A**2) : 0.00
REMARK 3 B13 (A**2) : -0.30
REMARK 3 B23 (A**2) : -0.00
REMARK 3
REMARK 3 ESTIMATED OVERALL COORDINATE ERROR.
REMARK 3 ESU BASED ON R VALUE (A): 0.118
REMARK 3 ESU BASED ON FREE R VALUE (A): 0.133
REMARK 3 ESU BASED ON MAXIMUM LIKELIHOOD (A): 0.155
REMARK 3 ESU FOR B VALUES BASED ON MAXIMUM LIKELIHOOD (A**2): 4.934
REMARK 3
REMARK 3 CORRELATION COEFFICIENTS.
REMARK 3 CORRELATION COEFFICIENT FO-FC : 0.950
REMARK 3 CORRELATION COEFFICIENT FO-FC FREE : 0.895
REMARK 3
REMARK 3 RMS DEVIATIONS FROM IDEAL VALUES COUNT RMS WEIGHT
REMARK 3 BOND LENGTHS REFINED ATOMS (A): 66 ; 0.017 ; 0.019
REMARK 3 BOND LENGTHS OTHERS (A): 69 ; 0.010 ; 0.020
REMARK 3 BOND ANGLES REFINED ATOMS (DEGREES): 91 ; 1.876 ; 1.905
1
REMARK 3 BOND ANGLES OTHERS (DEGREES): 155 ; 0.648 ; 3.000
REMARK 3 TORSION ANGLES, PERIOD 1 (DEGREES): 9 ; 5.684 ; 5.000
REMARK 3 TORSION ANGLES, PERIOD 2 (DEGREES): 1 ;75.981 ;20.000
REMARK 3 TORSION ANGLES, PERIOD 3 (DEGREES): 7 ; 7.001 ;15.000
REMARK 3 CHIRAL-CENTER RESTRAINTS (A**3): 14 ; 0.098 ; 0.200
REMARK 3 GENERAL PLANES REFINED ATOMS (A): 73 ; 0.005 ; 0.020
REMARK 3 GENERAL PLANES OTHERS (A): 13 ; 0.001 ; 0.020
REMARK 3
REMARK 3 ISOTROPIC THERMAL FACTOR RESTRAINTS. COUNT RMS WEIGHT
REMARK 3 MAIN-CHAIN BOND REFINED ATOMS (A**2): 39 ; 6.545 ; 1.715
REMARK 3 MAIN-CHAIN BOND OTHER ATOMS (A**2): 38 ; 3.948 ; 1.607
REMARK 3 MAIN-CHAIN ANGLE REFINED ATOMS (A**2): 41 ; 8.395 ; 2.554
REMARK 3 MAIN-CHAIN ANGLE OTHER ATOMS (A**2) : 42 ; 8.555 ; 2.605
REMARK 3 SIDE-CHAIN BOND REFINED ATOMS (A**2): 27 ; 5.989 ; 2.165
REMARK 3 SIDE-CHAIN BOND OTHER ATOMS (A**2) : 27 ; 5.243 ; 2.077
REMARK 3 SIDE-CHAIN ANGLE OTHER ATOMS (A**2) : 44 ; 8.091 ; 3.056
REMARK 3 LONG RANGE B REFINED ATOMS (A**2) : 54 ; 9.624 ;14.765
REMARK 3 LONG RANGE B OTHER ATOMS (A**2) : 55 ;11.399 ;15.221
REMARK 3
REMARK 3 NCS RESTRAINTS STATISTICS
REMARK 3 NUMBER OF NCS GROUPS : NULL
REMARK 3
REMARK 3 TWIN DETAILS
REMARK 3 NUMBER OF TWIN DOMAINS : NULL
REMARK 3
REMARK 3
REMARK 3 TLS DETAILS
REMARK 3 NUMBER OF TLS GROUPS : NULL
REMARK 3
REMARK 3
REMARK 3 BULK SOLVENT MODELLING.
REMARK 3 METHOD USED : MASK
REMARK 3 PARAMETERS FOR MASK CALCULATION
REMARK 3 VDW PROBE RADIUS : 1.20
REMARK 3 ION PROBE RADIUS : 0.80
REMARK 3 SHRINKAGE RADIUS : 0.80
REMARK 3
REMARK 3 OTHER REFINEMENT REMARKS:
REMARK 3 HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
REMARK 3 U VALUES : REFINED INDIVIDUALLY
REMARK 3
CRYST1 17.930 4.710 33.030 90.00 94.33 90.00 P 1 21 1
SCALE1 0.055772 0.000000 0.004219 0.00000
SCALE2 -0.000000 0.212314 0.000000 0.00000
SCALE3 0.000000 -0.000000 0.030362 0.00000
ATOM 1 N GLY A 47 -7.911 -0.639 18.278 1.00 20.19 N
ATOM 2 CA GLY A 47 -6.901 0.239 18.839 1.00 16.21 C
ATOM 3 C GLY A 47 -5.632 -0.528 19.111 1.00 16.40 C
ATOM 4 O GLY A 47 -5.600 -1.727 19.011 1.00 22.51 O
ATOM 5 N VAL A 48 -4.568 0.169 19.444 1.00 17.59 N
ATOM 6 CA VAL A 48 -3.313 -0.460 19.729 1.00 15.77 C
ATOM 7 CB VAL A 48 -2.269 -0.088 18.637 1.00 17.32 C
ATOM 8 CG1 VAL A 48 -0.855 -0.462 19.056 1.00 17.64 C
ATOM 9 CG2 VAL A 48 -2.659 -0.731 17.326 1.00 18.74 C
ATOM 10 C VAL A 48 -2.906 0.080 21.062 1.00 13.82 C
ATOM 11 O VAL A 48 -3.040 1.262 21.327 1.00 18.65 O
ATOM 12 N VAL A 49 -2.365 -0.782 21.895 1.00 16.57 N
ATOM 13 CA VAL A 49 -1.779 -0.342 23.136 1.00 16.11 C
ATOM 14 CB VAL A 49 -2.592 -0.887 24.317 1.00 16.48 C
ATOM 15 CG1 VAL A 49 -1.861 -0.622 25.628 1.00 20.39 C
ATOM 16 CG2 VAL A 49 -4.002 -0.324 24.303 1.00 20.73 C
ATOM 17 C VAL A 49 -0.366 -0.882 23.250 1.00 13.58 C
ATOM 18 O VAL A 49 -0.178 -2.043 23.137 1.00 20.25 O
ATOM 19 N HIS A 50 0.614 -0.063 23.557 1.00 16.22 N
2
ATOM 20 CA HIS A 50 2.002 -0.524 23.593 1.00 15.91 C
ATOM 21 CB HIS A 50 2.612 -0.176 22.220 1.00 18.19 C
ATOM 22 CG HIS A 50 4.036 -0.543 22.024 1.00 23.24 C
ATOM 23 ND1 HIS A 50 4.728 -1.390 22.851 1.00 32.41 N
ATOM 24 CE1 HIS A 50 5.964 -1.539 22.399 1.00 29.90 C
ATOM 25 NE2 HIS A 50 6.088 -0.839 21.291 1.00 34.03 N
ATOM 26 CD2 HIS A 50 4.892 -0.215 21.028 1.00 33.27 C
ATOM 27 C HIS A 50 2.680 0.171 24.736 1.00 14.51 C
ATOM 28 O HIS A 50 2.686 1.396 24.776 1.00 21.97 O
ATOM 29 N GLY A 51 3.213 -0.580 25.689 1.00 15.69 N
ATOM 30 CA GLY A 51 3.877 0.021 26.829 1.00 11.67 C
ATOM 31 C GLY A 51 2.957 0.800 27.740 1.00 12.93 C
ATOM 32 O GLY A 51 2.967 2.022 27.786 1.00 20.51 O
ATOM 33 N VAL A 52 2.174 0.081 28.498 1.00 15.10 N
ATOM 34 CA VAL A 52 1.315 0.692 29.514 1.00 14.31 C
ATOM 35 CB VAL A 52 -0.162 0.399 29.278 1.00 14.02 C
ATOM 36 CG1 VAL A 52 -1.002 0.803 30.489 1.00 16.11 C
ATOM 37 CG2 VAL A 52 -0.626 1.146 28.077 1.00 15.58 C
ATOM 38 C VAL A 52 1.778 0.094 30.802 1.00 13.92 C
ATOM 39 O VAL A 52 1.742 -1.133 30.969 1.00 20.14 O
ATOM 40 N THR A 53 2.253 0.954 31.698 1.00 17.96 N
ATOM 41 CA THR A 53 2.886 0.481 32.943 1.00 18.30 C
ATOM 42 CB THR A 53 4.381 0.885 33.010 1.00 20.92 C
ATOM 43 OG1 THR A 53 5.064 0.409 31.840 1.00 29.94 O
ATOM 44 CG2 THR A 53 5.063 0.255 34.175 1.00 22.40 C
ATOM 45 C THR A 53 2.116 1.037 34.132 1.00 14.85 C
ATOM 46 O THR A 53 1.778 2.193 34.103 1.00 21.57 O
ATOM 47 N THR A 54 1.840 0.202 35.151 1.00 15.66 N
ATOM 48 CA THR A 54 1.239 0.607 36.427 1.00 14.08 C
ATOM 49 CB THR A 54 -0.184 0.059 36.602 1.00 18.69 C
ATOM 50 OG1 THR A 54 -1.031 0.491 35.534 1.00 35.75 O
ATOM 51 CG2 THR A 54 -0.768 0.525 37.872 1.00 23.36 C
ATOM 52 C THR A 54 2.061 -0.047 37.500 1.00 12.83 C
ATOM 53 O THR A 54 2.023 -1.258 37.626 1.00 22.40 O
ATOM 54 N VAL A 55 2.821 0.725 38.262 1.00 14.38 N
ATOM 55 CA VAL A 55 3.639 0.203 39.339 1.00 13.58 C
ATOM 56 CB VAL A 55 5.099 0.641 39.138 1.00 15.65 C
ATOM 57 CG1 VAL A 55 5.915 0.347 40.358 1.00 16.16 C
ATOM 58 CG2 VAL A 55 5.696 -0.078 37.939 1.00 17.66 C
ATOM 59 C VAL A 55 3.100 0.756 40.673 1.00 14.57 C
ATOM 60 O VAL A 55 2.916 1.968 40.780 1.00 16.23 O
ATOM 61 N ALA A 56 2.809 -0.130 41.639 1.00 17.51 N
ATOM 62 CA ALA A 56 2.466 0.268 43.022 1.00 26.29 C
ATOM 63 CB ALA A 56 1.113 -0.279 43.465 1.00 21.51 C
ATOM 64 C ALA A 56 3.547 -0.222 43.971 1.00 38.92 C
ATOM 65 O ALA A 56 3.757 -1.425 44.111 1.00 69.25 O
ATOM 66 OXT ALA A 56 4.249 0.549 44.634 1.00 54.03 O
HETATM 67 O HOH B 1 4.715 2.482 29.991 1.00 27.67 O
HETATM 68 O HOH B 2 -1.034 3.065 33.500 1.00 32.66 O
3
ИСТОЧНИКИ ЛИТЕРАТУРЫ
1. Biere, A. L. et al. Parkinson’s disease-associated alpha-synuclein is more fibrillogenic than beta- and gamma-synuclein and cannot cross-seed its homologs. J. Biol. Chem. 275, 34574-34579 (2000).
2. Giasson, B. I., Murray, I. V. J., Trojanowski, J. Q. & Lee, V. M.-Y. A Hydrophobic Stretch of 12 Amino Acid Residues in the Middle of αSynuclein Is Essential for Filament Assembly. J. Biol. Chem. 276, 2380-2386 (2001).
3. Du, H.-N. et al. A peptide motif consisting of glycine, alanine, and valine is required for the fibrillization and cytotoxicity of human alpha-synuclein. Biochemistry (Mosc.) 42, 8870-8878 (2003).
4. Periquet, M., Fulga, T., Myllykangas, L., Schlossmacher, M. G. & Feany, M. B. Aggregated αSynuclein Mediates Dopaminergic Neurotoxicity In Vivo. J. Neurosci. 27, 3338-3346 (2007).
5. Nannenga, B. L., Shi, D., Leslie, A. G. W. & Gonen, T. High-resolution structure determination by continuous-rotation data collection in MicroED. Nat. Methods 11, 927-930 (2014).
6. Shi, D., Nannenga, B. L., Iadanza, M. G. & Gonen, T. Three-dimensional electron crystallography of protein microcrystals. eLife 2, e01345 (2013).
7. Spillantini, M. G. et al. Alpha-synuclein in Lewy bodies. Nature 388, 839-840 (1997).
8. Goedert, M., Spillantini, M. G., Del Tredici, K. & Braak, H. 100 years of Lewy pathology. Nat. Rev. Neurol. 9, 13-24 (2013).
9. Polymeropoulos, M. H. et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 276, 2045-2047 (1997).
10. Krüger, R. et al. Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson’s disease. Nat. Genet. 18, 106-108 (1998).
11. Zarranz, J. J. et al. The new mutation, E46K, of alpha-synuclein causes Parkinson and Lewy body dementia. Ann. Neurol. 55, 164-173 (2004).
12. Ibáñez, P. et al. Causal relation between alpha-synuclein gene duplication and familial Parkinson’s disease. Lancet 364, 1169-1171 (2004).
13. Singleton, A. B. et al. alpha-Synuclein locus triplication causes Parkinson’s disease. Science 302, 841 (2003).
14. Uéda, K. et al. Molecular cloning of cDNA encoding an unrecognized component of amyloid in Alzheimer disease. Proc. Natl. Acad. Sci. U. S. A. 90, 11282-11286 (1993).
15. Han, H., Weinreb, P. H. & Lansbury, P. T. The core Alzheimer’s peptide NAC forms amyloid fibrils which seed and are seeded by beta-amyloid: is NAC a common trigger or target in neurodegenerative disease? Chem. Biol. 2, 163-169 (1995).
16. El-Agnaf, O. M. et al. Aggregates from mutant and wild-type alpha-synuclein proteins and NAC peptide induce apoptotic cell death in human neuroblastoma cells by formation of beta-sheet and amyloid-like filaments. FEBS Lett. 440, 71-75 (1998).
17. Conway, K. A., Harper, J. D. & Lansbury, P. T. Accelerated in vitro fibril formation by a mutant alpha-synuclein linked to early-onset Parkinson disease. Nat. Med. 4, 1318-1320 (1998).
18. Nelson, R. et al. Structure of the cross-beta spine of amyloid-like fibrils. Nature 435, 773-778 (2005).
19. Sawaya, M. R. et al. Atomic structures of amyloid cross-beta spines reveal varied steric zippers. Nature 447, 453-457 (2007).
20. Bodles, A. M., Guthrie, D. J., Greer, B. & Irvine, G. B. Identification of the region of non-Abeta component (NAC) of Alzheimer’s disease amyloid responsible for its aggregation and toxicity. J. Neurochem. 78, 384-395 (2001).
21. Nannenga, B. L. & Gonen, T. Protein structure determination by MicroED. Curr. Opin. Struct. Biol. 27C, 24-31 (2014).
22. Nannenga, B. L. eLife (2014). Structure of catalase determined by MicroED. 2014;3:e03600
23. Yonekura, K., Kato, K., Ogasawara, M., Tomita, M. & Toyoshima, C. Electron crystallography of ultrathin 3D protein crystals: atomic model with charges. Proc. Natl. Acad. Sci. U. S. A. 112, 3368-3373 (2015).
24. Doyle, P. A. & Turner, P. S. Relativistic Hartree-Fock X-ray and electron scattering factors. Acta Crystallogr. Sect. A 24, 390-397 (1968).
25. Der-Sarkissian, A., Jao, C. C., Chen, J. & Langen, R. Structural organization of alpha-synuclein fibrils studied by site-directed spin labeling. J. Biol. Chem. 278, 37530-37535 (2003).
26. Chen, M., Margittai, M., Chen, J. & Langen, R. Investigation of alpha-synuclein fibril structure by site-directed spin labeling. J. Biol. Chem. 282, 24970-24979 (2007).
27. Sarafian, T. A. et al. Impairment of mitochondria in adult mouse brain overexpressing predominantly full-length, N-terminally acetylated human αsynuclein. PloS One 8, e63557 (2013).
28. Caughey, B. & Lansbury, P. T. Protofibrils, pores, fibrils, and neurodegeneration: separating the responsible protein aggregates from the innocent bystanders. Annu. Rev. Neurosci. 26, 267-298 (2003).
29. Danzer, K. M., Schnack, C., Sutcliffe, A., Hengerer, B. & Gillardon, F. Functional protein kinase arrays reveal inhibition of p-21-activated kinase 4 by alpha-synuclein oligomers. J. Neurochem. 103, 2401-2407 (2007).
30. Karpinar, D. P. et al. Pre-fibrillar alpha-synuclein variants with impaired beta-structure increase neurotoxicity in Parkinson’s disease models. EMBO J. 28, 3256-3268 (2009).
31. Winner, B. et al. In vivo demonstration that alpha-synuclein oligomers are toxic. Proc. Natl. Acad. Sci. U. S. A. 108, 4194-4199 (2011).
32. Chen, S. W. et al. Structural characterization of toxic oligomers that are kinetically trapped during αsynuclein fibril formation. Proc. Natl. Acad. Sci. U. S. A. 112, E1994-2003 (2015).
33. Bousset, L. et al. Structural and functional characterization of two alpha-synuclein strains. Nat. Commun. 4, 2575 (2013).
34. Auluck, P. K., Caraveo, G. & Lindquist, S. αSynuclein: membrane interactions and toxicity in Parkinson’s disease. Annu. Rev. Cell Dev. Biol. 26, 211-233 (2010).
35. Lee, J. C., Langen, R., Hummel, P. A., Gray, H. B. & Winkler, J. R. Alpha-synuclein structures from fluorescence energy-transfer kinetics: implications for the role of the protein in Parkinson’s disease. Proc. Natl. Acad. Sci. U. S. A. 101, 16466-16471 (2004).
36. Sievers, S. A. et al. Structure-based design of non-natural amino-acid inhibitors of amyloid fibril formation. Nature 475, 96-100 (2011).
37. Otwinowski, Z. & Minor, W. in Methods in Enzymology (ed. Charles W. Carter, J.) Volume 276, 307-326 (Academic Press, 1997).
38. Weierstall, U., Spence, J. C. H. & Doak, R. B. Injector for scattering measurements on fully solvated biospecies. Rev. Sci. Instrum. 83, 035108 (2012).
39. Hattne, J. et al. Accurate macromolecular structures using minimal measurements from X-ray free-electron lasers. Nat. Methods 11, 545-548 (2014).
40. Sauter, N. K., Hattne, J., Grosse-Kunstleve, R. W. & Echols, N. New Python-based methods for data processing. Acta Crystallogr. D Biol. Crystallogr. 69, 1274-1282 (2013).
41. Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 66, 125-132 (2010).
42. McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658-674 (2007).
43. Murshudov, G. N., Vagin, A. A. & Dodson, E. J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 53, 240-255 (1997).
44. Afonine, P. V. et al. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 68, 352-367 (2012).
45. Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486-501 (2010).
46. Delano, W. The PyMOL Molecular Graphics System. (Schrödinger LLC). at <http://www.pymol.org>
47. Jakes, R., Spillantini, M. G. & Goedert, M. Identification of two distinct synucleins from human brain. FEBS Lett. 345, 27-32 (1994).
48. Johnson, M., Coulton, A. T., Geeves, M. A. & Mulvihill, D. P. Targeted amino-terminal acetylation of recombinant proteins in E. coli. PloS One 5, e15801 (2010).
49. Whitelegge, J. P., Zhang, H., Aguilera, R., Taylor, R. M. & Cramer, W. A. Full subunit coverage liquid chromatography electrospray ionization mass spectrometry (LCMS+) of an oligomeric membrane protein: cytochrome b(6)f complex from spinach and the cyanobacterium Mastigocladus laminosus. Mol. Cell. Proteomics MCP 1, 816-827 (2002).
50. Rao, J. N., Jao, C. C., Hegde, B. G., Langen, R. & Ulmer, T. S. A combinatorial NMR and EPR approach for evaluating the structural ensemble of partially folded proteins. J. Am. Chem. Soc. 132, 8657-8668 (2010).
51. Brunger, A. T. Version 1.2 of the Crystallography and NMR system. Nat. Protoc. 2, 2728-2733 (2007).
52. Fabiola, F., Bertram, R., Korostelev, A. & Chapman, M. S. An improved hydrogen bond potential: impact on medium resolution protein structures. Protein Sci. Publ. Protein Soc. 11, 1415-1423 (2002).
53. Comellas, G. et al. Structured regions of αsynuclein fibrils include the early-onset Parkinson’s disease mutation sites. J. Mol. Biol. 411, 881-895 (2011).
54. Vilar, M. et al. The fold of alpha-synuclein fibrils. Proc. Natl. Acad. Sci. U. S. A. 105, 8637-8642 (2008).
55. Goldschmidt, L., Teng, P. K., Riek, R. & Eisenberg, D. Identifying the amylome, proteins capable of forming amyloid-like fibrils. Proc. Natl. Acad. Sci. 107, 3487-3492 (2010).
56. Read, R. J. Improved Fourier coefficients for maps using phases from partial structures with errors. Acta Crystallogr. A 42, 140-149 (1986).
57. Eisenberg, D.S. et al. Structure-based design of peptide inhibitors of amyloid fibrillation, US Patent No. 8,754,034.
58. Eisenberg, D.S. et al. Structure-based peptide inhibitors of p53 aggregation as a new approach to cancer therapeutics; WO 2014/182961 (2014); US2014/037387.
59. The CCP4 suite: programs for protein crystallography. Acta Crystallogr D Biol Crystallogr 50, 760-3 (1994).
60. Lawrence, M. C. & Colman, P. M. Shape complementarity at protein/protein interfaces. J Mol Biol 234, 946-50 (1993).
61. Kyte J, Doolittle RF (May 1983). "A simple method for displaying the hydropathic character of a protein". J. Mol. Biol. 157 (1): 105-32.
62. Warren L. DeLano "The PyMOL Molecular Graphics System." DeLano Scientific LLC, San Clos, CA, USA. http://www.pymol.org
63. Jiang, L. et al. (2013). Structure-based discovery of fiber-binding compounds that reduce the cytotoxicity of amyloid beta. ELife 2013; 2e00857
Из приведенного выше описания специалист в данной области может легко установить необходимые характеристики данного изобретения, и не отклоняясь от его смысла и области применения, может выполнить изменения и модификации по данному изобретению для адаптации его к различному применению и условиям и применению данного изобретения в его наиболее полном объеме. Предыдущие варианты осуществления следует рассматривать только в качестве иллюстративных и никоим образом не ограничивающих объем данного изобретения никоим образом. Полное раскрытие всех заявок, патентов и публикаций, упоминаемых в данном документе и в фигурах, тем самым включено во всей их полноте посредством ссылки, в частности, в связи с информацией, в отношении которой они упомянуты.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> РЕГЕНТЫ УНИВЕРСИТЕТА КАЛИФОРНИИ (THE REGENTS OF THE UNIVERSITY OF CALIFORNIA)
<120> СТРУКТУРНЫЕ ПЕПТИДНЫЕ ИНГИБИТОРЫ АГРЕГАЦИИ АЛЬФА-СИНУКЛЕИНА
<130> 30435.305-WO-U2
<140> PCT/US2017/040106
<141> 2017-06-29
<150> 62/356,410
<151> 2016-06-29
<160> 56
<170> PatentIn версия 3.5
<210> 1
<211> 140
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Asp Val Phe Met Lys Gly Leu Ser Lys Ala Lys Glu Gly Val Val
1 5 10 15
Ala Ala Ala Glu Lys Thr Lys Gln Gly Val Ala Glu Ala Ala Gly Lys
20 25 30
Thr Lys Glu Gly Val Leu Tyr Val Gly Ser Lys Thr Lys Glu Gly Val
35 40 45
Val His Gly Val Ala Thr Val Ala Glu Lys Thr Lys Glu Gln Val Thr
50 55 60
Asn Val Gly Gly Ala Val Val Thr Gly Val Thr Ala Val Ala Gln Lys
65 70 75 80
Thr Val Glu Gly Ala Gly Ser Ile Ala Ala Ala Thr Gly Phe Val Lys
85 90 95
Lys Asp Gln Leu Gly Lys Asn Glu Glu Gly Ala Pro Gln Glu Gly Ile
100 105 110
Leu Glu Asp Met Pro Val Asp Pro Asp Asn Glu Ala Tyr Glu Met Pro
115 120 125
Ser Glu Glu Gly Tyr Gln Asp Tyr Glu Pro Glu Ala
130 135 140
<210> 2
<211> 9
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 2
Gly Val Val His Gly Val Thr Thr Val
1 5
<210> 3
<211> 12
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 3
Gly Ala Val Val Trp Gly Val Thr Ala Val Lys Lys
1 5 10
<210> 4
<211> 12
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 4
Arg Ala Val Val Thr Gly Val Thr Ala Val Ala Glu
1 5 10
<210> 5
<211> 15
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 5
Gly Ala Val Val Trp Gly Val Thr Ala Val Lys Lys Lys Lys Lys
1 5 10 15
<210> 6
<211> 24
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 6
Gly Ala Val Val Trp Gly Val Thr Ala Val Lys Lys Gly Arg Lys Lys
1 5 10 15
Arg Arg Gln Arg Arg Arg Pro Gln
20
<210> 7
<211> 22
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 7
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Ala Val Val Thr Gly
1 5 10 15
Val Thr Ala Val Ala Glu
20
<210> 8
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MISC_FEATURE
<222> (1)..(16)
<223> Эта последовательность может простираться на 5-16 остатков
<400> 8
Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg Arg
1 5 10 15
<210> 9
<211> 18
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 9
Arg Gly Gly Arg Leu Ser Tyr Ser Arg Arg Arg Phe Ser Thr Ser Thr
1 5 10 15
Gly Arg
<210> 10
<211> 10
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 10
Arg Arg Leu Ser Tyr Ser Arg Arg Arg Phe
1 5 10
<210> 11
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 11
Arg Gln Ile Lys Ile Trp Phe Gln Asn Arg Arg Met Lys Trp Lys Lys
1 5 10 15
<210> 12
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 12
Arg Gln Ile Arg Ile Trp Phe Gln Asn Arg Arg Met Arg Trp Arg Arg
1 5 10 15
<210> 13
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 13
Lys Gln Ile Lys Ile Trp Phe Gln Asn Lys Lys Met Lys Trp Lys Lys
1 5 10 15
<210> 14
<211> 13
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 14
Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Pro Trp Gln
1 5 10
<210> 15
<211> 13
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 15
Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Pro Pro Gln
1 5 10
<210> 16
<211> 13
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 16
Gly Arg Arg Arg Arg Arg Arg Arg Arg Arg Pro Pro Gln
1 5 10
<210> 17
<211> 11
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 17
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg
1 5 10
<210> 18
<211> 13
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 18
Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Pro Pro Gln
1 5 10
<210> 19
<211> 19
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 19
Lys Met Thr Arg Ala Gln Arg Arg Ala Ala Ala Arg Arg Asn Arg Trp
1 5 10 15
Thr Ala Arg
<210> 20
<211> 15
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 20
Arg Arg Arg Arg Asn Arg Thr Arg Arg Asn Arg Arg Arg Val Arg
1 5 10 15
<210> 21
<211> 13
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 21
Thr Arg Arg Gln Arg Thr Arg Arg Ala Arg Arg Asn Arg
1 5 10
<210> 22
<211> 17
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 22
Asn Ala Lys Thr Arg Arg His Glu Arg Arg Arg Lys Leu Ala Ile Glu
1 5 10 15
Arg
<210> 23
<211> 18
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 23
Leu Leu Ile Ile Leu Arg Arg Arg Ile Arg Lys Gln Ala His Ala His
1 5 10 15
Ser Lys
<210> 24
<211> 27
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 24
Gly Trp Thr Leu Asn Ser Ala Gly Tyr Leu Leu Gly Lys Ile Asn Leu
1 5 10 15
Lys Ala Leu Ala Ala Leu Ala Lys Lys Ile Leu
20 25
<210> 25
<211> 21
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 25
Ala Gly Tyr Leu Leu Gly Lys Ile Asn Leu Lys Ala Leu Ala Ala Leu
1 5 10 15
Ala Lys Lys Ile Leu
20
<210> 26
<211> 12
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 26
Pro Ile Arg Arg Arg Lys Lys Leu Arg Arg Leu Lys
1 5 10
<210> 27
<211> 12
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 27
Arg Arg Gln Arg Arg Thr Ser Lys Leu Met Lys Arg
1 5 10
<210> 28
<211> 21
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 28
Lys Glu Thr Trp Trp Glu Thr Trp Trp Thr Glu Trp Ser Gln Pro Lys
1 5 10 15
Lys Lys Arg Lys Val
20
<210> 29
<211> 21
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 29
Lys Glu Thr Trp Phe Glu Thr Trp Phe Thr Glu Trp Ser Gln Pro Lys
1 5 10 15
Lys Lys Arg Lys Val
20
<210> 30
<211> 15
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 30
Lys Trp Phe Glu Thr Trp Phe Thr Glu Trp Pro Lys Lys Arg Lys
1 5 10 15
<210> 31
<211> 22
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 31
Met Asp Ala Gln Thr Arg Arg Arg Glu Arg Arg Ala Glu Lys Gln Ala
1 5 10 15
Gln Trp Lys Ala Ala Asn
20
<210> 32
<211> 18
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 32
Thr Ala Lys Thr Arg Tyr Lys Ala Arg Arg Ala Glu Leu Ile Ala Glu
1 5 10 15
Arg Arg
<210> 33
<211> 12
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 33
Ser Gln Met Thr Arg Gln Ala Arg Arg Leu Tyr Val
1 5 10
<210> 34
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 34
Thr Arg Arg Asn Lys Arg Asn Arg Ile Gln Glu Gln Leu Asn Arg Lys
1 5 10 15
<210> 35
<211> 17
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 35
Lys Leu Ala Leu Lys Leu Ala Leu Lys Leu Ala Leu Ala Leu Lys Leu
1 5 10 15
Ala
<210> 36
<211> 27
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 36
Met Gly Leu Gly Leu His Leu Leu Val Leu Ala Ala Ala Leu Gln Gly
1 5 10 15
Ala Trp Ser Gln Pro Lys Lys Lys Arg Lys Val
20 25
<210> 37
<211> 27
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 37
Gly Ala Leu Phe Leu Gly Trp Leu Gly Ala Ala Gly Ser Thr Met Gly
1 5 10 15
Ala Trp Ser Gln Pro Lys Lys Lys Arg Lys Val
20 25
<210> 38
<211> 27
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 38
Gly Ala Leu Phe Leu Gly Phe Leu Gly Ala Ala Gly Ser Thr Met Gly
1 5 10 15
Ala Trp Ser Gln Pro Lys Lys Lys Arg Lys Val
20 25
<210> 39
<211> 27
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 39
Gly Ala Leu Phe Leu Gly Phe Leu Gly Ala Ala Gly Ser Thr Met Gly
1 5 10 15
Ala Trp Ser Gln Pro Lys Ser Lys Arg Lys Val
20 25
<210> 40
<211> 17
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 40
Thr Arg Gln Ala Arg Arg Asn Arg Arg Arg Arg Trp Arg Glu Arg Gln
1 5 10 15
Arg
<210> 41
<211> 23
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (12)..(12)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (22)..(22)
<223> Любая аминокислота
<400> 41
Glu Glu Glu Glu Glu Asp Asp Asp Asp Lys Ala Xaa Arg Arg Arg Arg
1 5 10 15
Arg Arg Arg Arg Arg Xaa Cys
20
<210> 42
<211> 22
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (21)..(21)
<223> Любая аминокислота
<400> 42
Glu Glu Glu Glu Glu Asp Asp Asp Asp Lys Ala Arg Arg Arg Arg Arg
1 5 10 15
Arg Arg Arg Arg Xaa Cys
20
<210> 43
<211> 19
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (8)..(8)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (18)..(18)
<223> Любая аминокислота
<400> 43
Glu Asp Asp Asp Asp Lys Ala Xaa Arg Arg Arg Arg Arg Arg Arg Arg
1 5 10 15
Arg Xaa Cys
<210> 44
<211> 23
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (10)..(10)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (13)..(13)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (16)..(16)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (19)..(19)
<223> Любая аминокислота
<220>
<221> MOD_RES
<222> (22)..(22)
<223> Любая аминокислота
<400> 44
Glu Glu Asp Asp Asp Asp Lys Ala Arg Xaa Arg Arg Xaa Arg Arg Xaa
1 5 10 15
Arg Arg Xaa Arg Arg Xaa Cys
20
<210> 45
<211> 19
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (18)..(18)
<223> Любая аминокислота
<400> 45
Asp Asp Asp Asp Asp Asp Lys Ala Arg Arg Arg Arg Arg Arg Arg Arg
1 5 10 15
Arg Xaa Cys
<210> 46
<211> 11
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 46
Gly Ala Val Val Thr Gly Val Thr Ala Val Ala
1 5 10
<210> 47
<211> 10
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 47
Gly Val Val His Gly Val Thr Thr Val Ala
1 5 10
<210> 48
<211> 9
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 48
Ala Val Val Thr Gly Val Thr Ala Val
1 5
<210> 49
<211> 5
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 49
Gly Val Thr Thr Val
1 5
<210> 50
<211> 5
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 50
Gly Val Thr Ala Val
1 5
<210> 51
<211> 6
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 51
Gln Val Thr Asn Val Gly
1 5
<210> 52
<211> 6
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 52
Gln Lys Thr Val Glu Gly
1 5
<210> 53
<211> 6
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<400> 53
Gly Val Thr Thr Val Ala
1 5
<210> 54
<211> 22
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MOD_RES
<222> (16)..(16)
<223> nme-Gly
<400> 54
Tyr Gly Arg Lys Lys Arg Arg Gln Arg Arg Arg Ala Val Val Thr Gly
1 5 10 15
Val Thr Ala Val Ala Glu
20
<210> 55
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MISC_FEATURE
<222> (1)..(16)
<223> Эта последовательность может простираться на 5-16 остатков
<400> 55
Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys Lys
1 5 10 15
<210> 56
<211> 16
<212> БЕЛОК
<213> Исскуственная последовательность
<220>
<223> Описание исскуственной последовательности: Синтетический
пептид
<220>
<221> MISC_FEATURE
<222> (6)..(16)
<223> This sequence may encompass 1-16 residues
<400> 56
Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp Asp
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| EGFL6-СПЕЦИФИЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779902C2 |
| ХИМЕРНЫЕ РЕЦЕПТОРЫ АНТИГЕНА ПРОТИВ МЕЗОТЕЛИНА ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2714902C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| КОМПОЗИЦИИ И СПОСОБЫ РЕДАКТИРОВАНИЯ ГЕНОВ | 2019 |
|
RU2804665C2 |
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
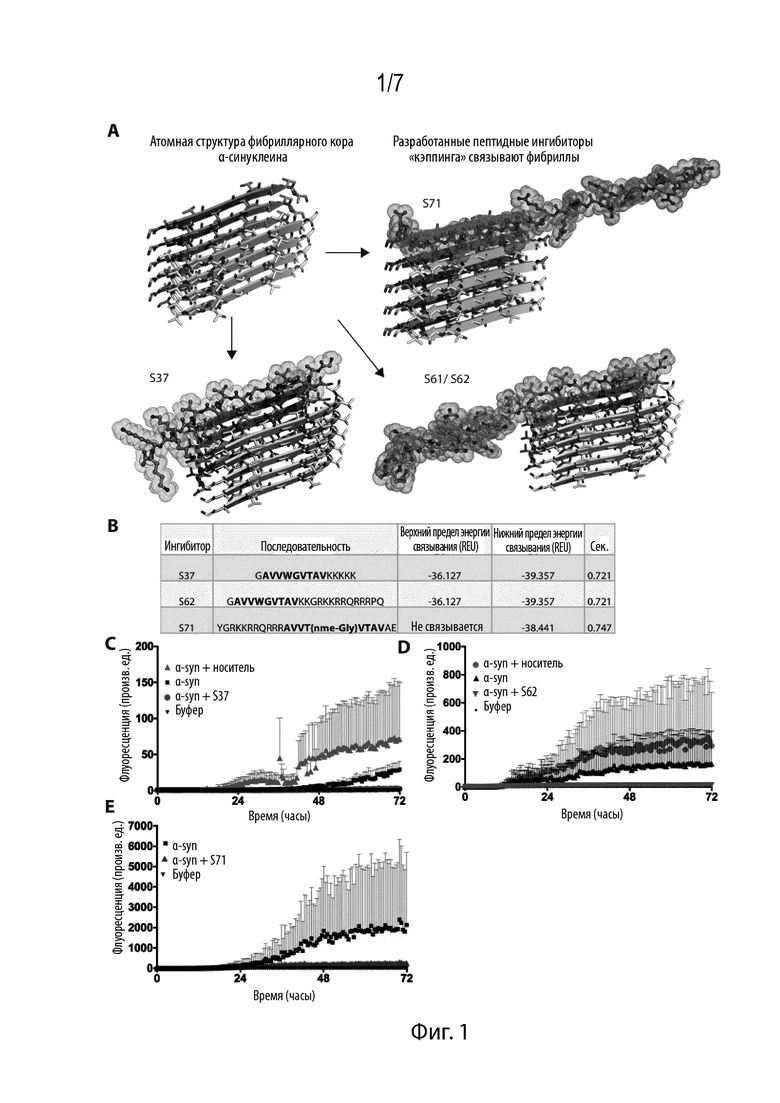
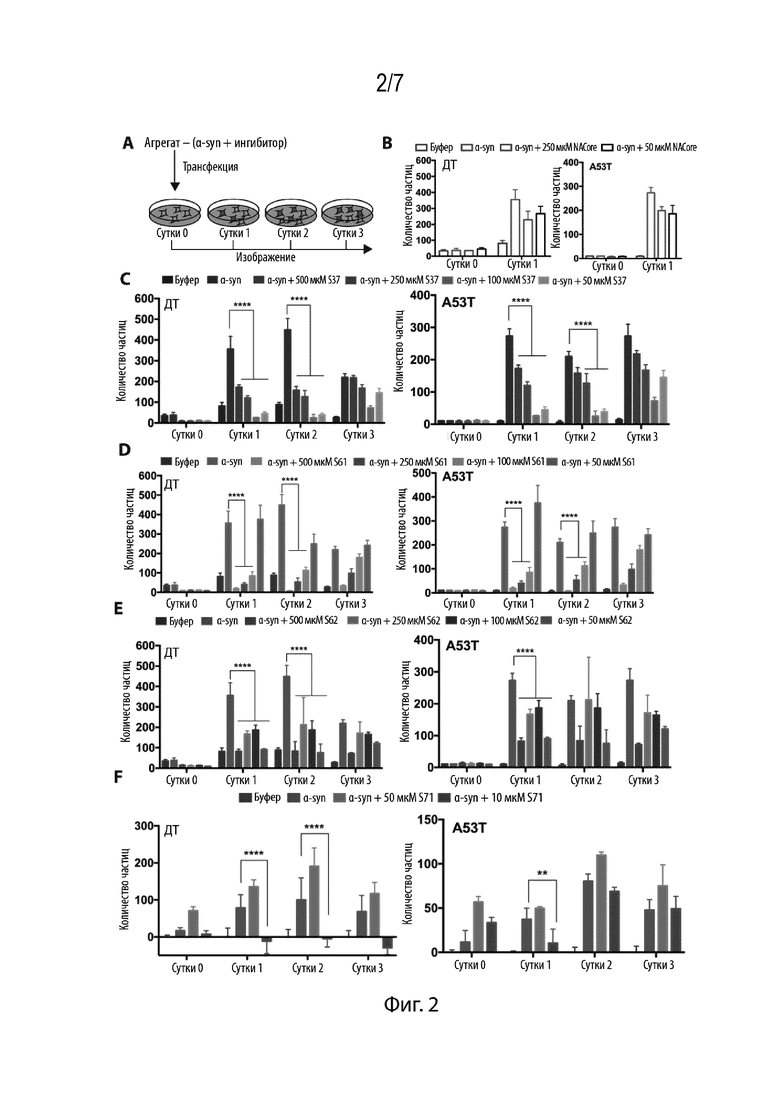

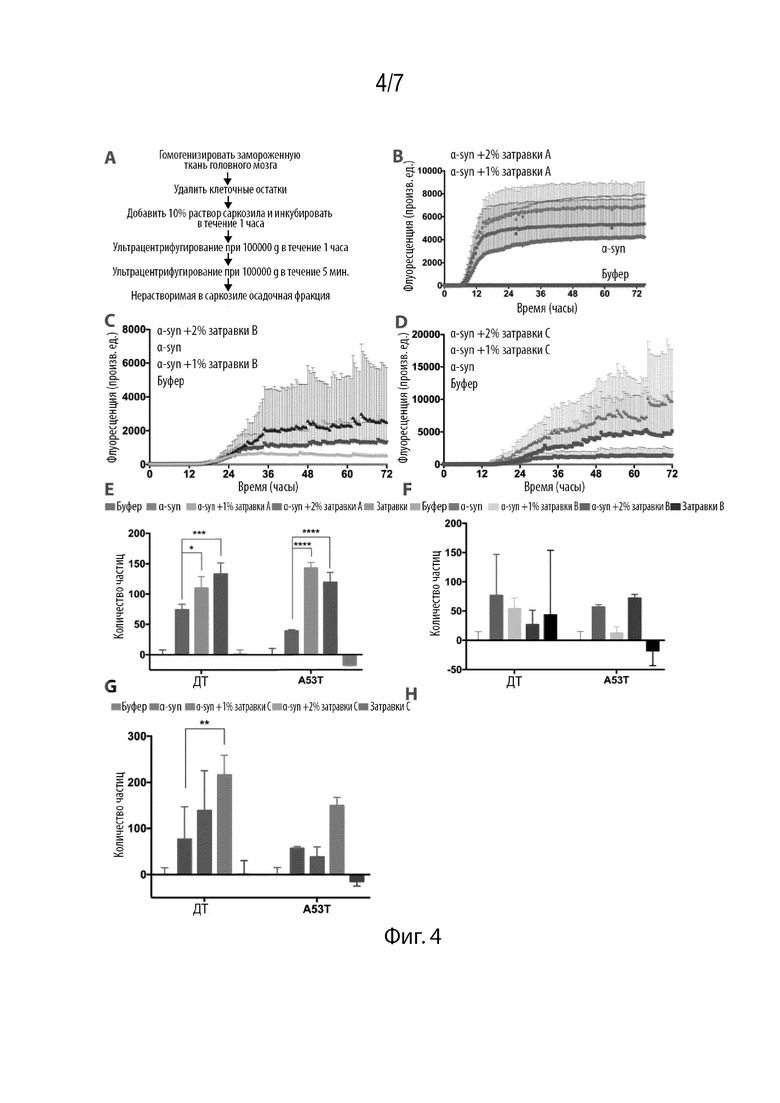
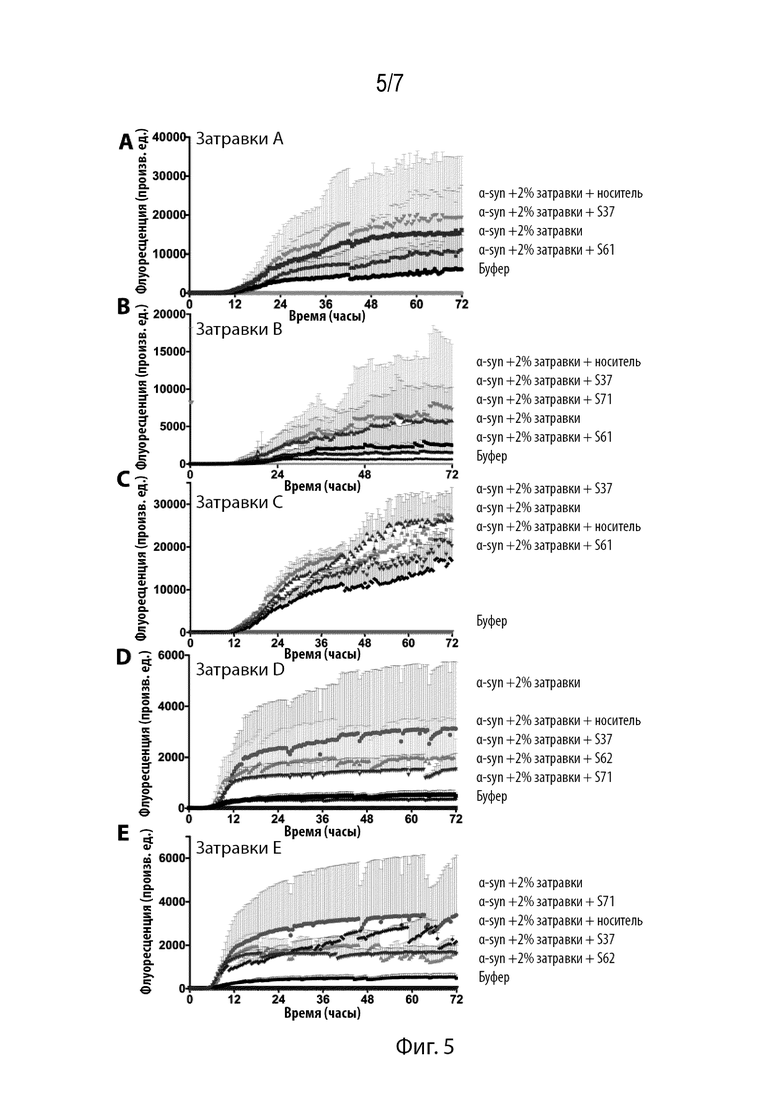

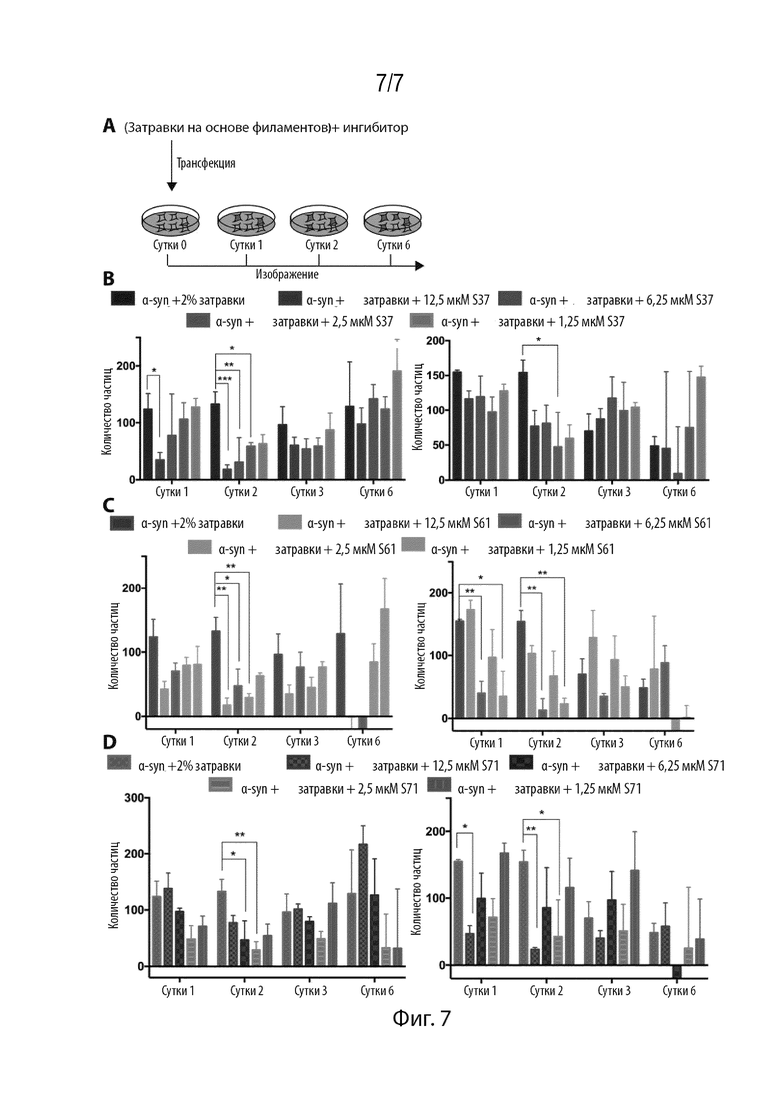
Изобретение относится к области биотехнологии, конкретно к ингибиторным пептидам α–синуклеина, и может быть использовано в медицине для ингибирования агрегации α–синуклеина. Предложен ингибиторный пептид до 30 аминокислот в длину, содержащий GAVVWGVTAVKK или RAVVTGVTAVAE. При этом по меньшей мере одна из аминокислот в ингибиторном пептиде представляет собой не встречающуюся в природе аминокислоту, выбранную из D–аминокислоты или аминокислоты, содержащей фрагмент N–метильной группы. Также ингибиторный пептид содержит гетерологичный пептидный тэг для повышения его растворимости, облегчения его мониторинга и/или вхождения пептида в клетку млекопитающего. Изобретение обеспечивает ингибирование образования, затравки фибрилл и распространения агрегатов aльфа-синуклеина. 2 н. и 2 з.п. ф-лы, 7 ил., 4 табл., 3 пр.
1. Ингибитор агрегации α–синуклеина (SEQ ID NO: 1), состоящий из:
(a) ингибиторного пептида до 30 аминокислот в длину и содержащего последовательность:
GAVVWGVTAVKK (SEQ ID NO: 3); или
RAVVTGVTAVAE (SEQ ID NO: 4); где
по меньшей мере одна из аминокислот в ингибиторном пептиде представляет собой не встречающуюся в природе аминокислоту, где указанная не встречающаяся в природе аминокислота содержит по меньшей мере одну из D–аминокислоты; или аминокислоты, содержащей фрагмент N–метильной группы; и
гетерологичный пептидный тэг, связанный с указанным ингибиторным пептидом, где указанный гетерологичный пептидный тэг содержит:
аминокислотную последовательность, которая повышает растворимость пептида;
аминокислотную последовательность, которая облегчает мониторинг пептида; и/или
аминокислотную последовательность, которая облегчает вхождение пептида в клетку млекопитающего.
2. Ингибитор по п.1, где ингибиторный пептид содержит последовательность:
GAVVWGVTAVKKKKK (SEQ ID NO: 5);
GAVVWGVTAVKKGRKKRRQRRRPQ (SEQ ID NO: 6); или
YGRKKRRQRRRAVVTGVTAVAE (SEQ ID NO: 7).
3. Ингибитор по п.1, где указанный гетерологичный пептидный тэг содержит:
множество аргининовых остатков;
множество лизиновых аминокислотных остатков; и/или
аминокислотную последовательность TAT.
4. Фармацевтическая композиция для ингибирования агрегации α–синуклеина, где указанная фармацевтическая композиция содержит один или более ингибиторов по п.1 и фармацевтически приемлемый носитель.
| BODLES A.M | |||
| et al., Inhibition of fibril formation and toxicity of a fragment of alpha-synuclein by an N 1-methylated peptide analogue, Neurosci | |||
| Lett., 2004, v.359, n.1-2, p.89-93 | |||
| WO 2004009625 A2, 29.01.2004 | |||
| WO 2009027690 A1, 05.03.2009 | |||
| WO 2012068405 A2, 24.05.2012 | |||
| Урологическая грелка | 1929 |
|
SU17611A1 |
| ZHONGLI LUO et al., Fabrication of self-assembling | |||

