Область изобретения
Раскрываемая сущность изобретения в целом относится к композициям для защиты от эффектов, возникающих вследствие старения, и их устранения.
Предшествующий уровень техники
Древние предки человека не обладали достаточной энергией, чтобы делать все, что принесло бы им пользу. Ограничения доступности энергии в ходе ранней эволюции человека привели к биологическим компромиссным механизмам, управляющим использованием энергии, что ограничивало использование энергии для защиты от клеточного старения человека и его устранения. Предполагают, что причиной биологического старения человека является клеточное повреждение (Olson С 1987, Holliday R 2004, Kirkwood Т 2005, Gavrilov L 2001). Предполагают, что биологическое старение человека является причиной «болезней старения» у человека (Cutler R 2006).
Частью компромиссных механизмов контроля энергии являются механизмы по типу обратной связи
Механизмы обратной связи в биохимических путях синтеза отводят затраты энергии из тех областей, где она не используется, чтобы повысить эффективность общего использования энергии на уровне всего организма. В процессе эволюции необходимо было сбалансировать использование энергии и питательных веществ и способность к получению этих калорий и питательных веществ из окружающей среды, которая была изменчивой и ограниченной. Отчасти это достигается за счет того, что известно в медицине как принцип «использовать или потерять», который является механизмом повышающей/понижающей регуляции, основанным на потребностях. Примером этого являются системы антиоксидантных ферментов, которые переводятся в режим более низкой активности, если со временем не получают новых окислительных импульсов для поддержания антиоксидантных ферментов в режиме высокой активности.
В клеточную репарацию вовлечены энергетические компромиссы
Энергия, используемая для репарации клеточного повреждения, представляет собой энергию, недоступную для использования для других функций, полезных для функционирования и жизни клеток. Клеточные системы репарации и комплексные иммунные системы человека представляют собой две противостоящие по потребностям в энергии системы, которые конкурируют за использование энергии. В своей книге «Теория старения одноразовой сомы (Disposable Soma Theory of Aging)» Кирквуд и Роуз (Kirkwood and Rose 1991) предположили, что для оптимизации использования энергии биологические системы могут вкладывать большую часть своей энергии в рост и развитие и меньшую - в контроль и репарацию повреждений незародышевых (соматических) клеток. Доступность энергии из пищи влияет на энергетические компромиссы Второй закон термодинамики учит, что состояние энтропии в замкнутой системе может изменяться со временем только в одном направлении. Животным необходимо поедать пищу, поддерживая, таким образом, открытую систему для улучшения, репарации или поддержания своей структуры со временем за счет пищи, которую они поедают, которая приобретает энтропию, становясь веществом фекалий.
В процессе эволюции пища и ее питательные вещества и энергия часто были ограничены и доступны лишь спорадически. Эволюция должна была приспособиться к этому. Во время ограничения калорий энергетические пути метаболизма адаптировались к этим ограничениям. Эти пути метаболизма обладают преимуществами. В недавно проведенном исследовании наблюдали, что фактором, направляющим полезный эффект ограничения калорий, являются ферменты сиртуины. Ферменты сиртуины у человека вовлечены в клеточную репарацию. У человека известно 7 ферментов сиртуинов. Все эти 7 ферментов ситруинов человека используют никотинамиддинуклеотид (НАД) + (Imai S 2000). Никотинамид является конечным продуктом этих реакций сиртуина.
Пример механизма обратной связи в метаболических путях сиртуина состоит в том, что конечный продукт никотинамид способен связываться с ферментами семейства сиртуина и снижать их ферментативные свойства. Механизм обратной связи изменяется при метилировании никотинамида ферментом никотинамид-N-метилтрансферазой (NNMT) человека в клетке с использованием S-5'-аденозил-L-метионина (SAM). Впоследствии вновь метилированный никотинамид неспособен связываться в сайте связывания никотинамида из-за стерического затруднения, связанного с физическим размером вновь присоединенной метильной группы к никотинамиду (Schmeisser K 2013). При данном изменении в результате метилирования ферменты ситруины способны к поддержанию рабочего состояния вместо остановки их активности.
Защита от патогенов возникла за счет энергетических компромиссов
Люди, страдающие заболеванием, особенно хроническим заболеванием, стареют быстрее. Врожденная иммунная система (например, лейкоциты) при выбросе оксидантов (например, Cl-) для уничтожения патогенов создает фоновое повреждение в собственных клетках, которое ведет к более быстрому старению организма. Патогены стали основным убийцей людей, поэтому без энергии для борьбы с этими патогенами индивиды быстро были бы устранены из эволюции. Важным компромиссом в эволюции является выработка компромиссного решения, сколько энергии затрачивать на патогенную атаку, сколько энергии использовать на репарацию повреждения вследствие патогенной атаки и даже сколько энергии использовать для повышения готовности иммунной системы к атаке на иммунитет.
Пример этого компромисса наблюдают в исследовании 684 участников в возрасте старше 100 лет и 536 участников в возрасте от 85 до 99 лет в Японии (Arai Y 2015). Низкие уровни воспаления (4 комплексных показателя переменных иммунитета) были наилучшим прогностическим фактором для людей, которые будут продолжать жить (продолжительности жизни) и сохранять физическое и когнитивное здоровье (продолжительность здоровья). Было обнаружено, что иммунные маркеры (простой показатель сывороточного интерлейкина-6 (ИЛ-6) и фактора некроза опухоли альфа (ФНО-альфа), которые представляют собой два из 4 маркеров Араи (Arai)) являются лучшим прогностическим показателем смертности у 1155 взрослых людей пожилого возраста в 10-летнем исследовании смертности по любой причине после поправки на уже известные переменные, являющиеся причиной смерти (Varadhan R 2014). В проспективном когортном исследовании 1843 человек только один иммунный маркер, сывороточный ИЛ-6, был прогностическим фактором смертности по любой причине, рака, сердечно-сосудистого заболевания и заболевания печени (Lee JK 2012). В этих исследованиях были подтверждены результаты менее обширных предшествующих исследований (Derhovanessian Е 2010, Reuben DB 2002, Taaffe DR 2000).
Биологический клеточный механизм этого компромисса может быть связан с тем, что при высвобождении Keap1 от Nrf2 он становится доступным для захвата субъединицей IKKBeta, ингибируя, таким образом, гены-мишени ядерного фактора каппа-би (NF-kB). Это взаимодействие коррелирует с экспрессией антиоксидантных ферментов под действием Nrf2 и с вводом в действие и выводом из действия иммунной системы под действием NF-kB.
Животные, размножающиеся половым путем, имеют компромиссные механизмы использования энергии, которые отсутствуют у животных, размножающихся бесполым путем
Бесполые животные, подобные морским анемонам, не стареют. У гидры при размножении бесполым путем видимые признаки старения отсутствуют, но эти признаки старения наблюдаются при размножении гидры половым путем (Yoshida К 2006). Гидра имеет 6071 общих генов с человеком (Wenger Y 2013), и по меньшей мере 80% известных генов старения человека также имеется у гидры (Tomczyk S 2014). В исследовании показано, что соматические клетки животных, размножающихся половым путем, стареют быстрее после пубертатного периода и медленнее в случае уменьшения количества половых гормонов. Примером у человека являются евнухи в Индии и Корее, которые не имеют яичек и живут в среднем на 9-13 лет дольше. В экспериментальной модели плоских червей, называемых С. elegans, реакция теплового шока (heat shock response, HSR), существенная для протеостаза и клеточного здоровья, подавляется после наступления половозрелой стадии в соматических (неполовых) клетках зародышевыми (половыми) клетками в результате тройного метилирования в локусах стрессовых генов. Данная конкуренция между интересом к зародышевой линии и соматическим клеткам (Kirkwood TL 2000) определяет скорость старения у половозрелых индивидов (Labbadia J 2015). В исследовании также показаны компромиссы между способностью к деторождению и старением. Примером является применение низкой дозы RU-486, лекарственного средства, вызывающего прерывание беременности, которое приводит к более низкой репродуктивной способности и в среднем более высокой продолжительности жизни (Landis G. 2015). Вынашивание детей и особенно вынашивание детей на позднем этапе жизни (Sun F 2015, Perls ТТ 1997) связано с повышением вероятности более долгой жизни у женщин, хотя причина и влияние до сих пор неизвестны. Время менопаузы также коррелирует со скоростью старения.
Компромиссы использования энергии, наблюдаемые с хронологическим возрастом
В юности у людей имеется избыток или резервуар способности и энергия в избыточном количестве по сравнению с необходимым клеткам и органам в среднем, но с возрастом они уменьшаются. В молодости меньше знаний и мудрости и меньше размер тела, но эволюция восполняет этот недостаток высоким метаболизмом, дающим возможность тратить больше энергии (в частности, на единицу массы тела), поэтому в единицу времени можно прожить больше. Высокий метаболизм коррелирует с более быстрым старением по существу у всех биологических видов, хотя известно, что люди стареют быстрее после пубертатного периода, поэтому это не обязательно является жестким правилом. Теория «Скорости жизни» (Rate of Living) (Pearl R 1928) была усовершенствована, и создана теория «Жизненности» (Livingness) (Sohal R 2012), включающая в себя температуру, гибернацию, продуктивность и метаболический потенциал на основании первоначальных наблюдений потребления кислорода (Rubner М 1908). Индивиды старшего возраста обладают большим опытом, знаниями и мудростью и поэтому способны к поддержанию своей жизни при меньшем потреблении энергии. Эта пониженная выработка энергии может быть по меньшей мере отчасти связана со снижением качества и функции митохондрий, вырабатывающих энергию в течение всей продолжительности жизни.
Дополнительная энергия, используемая у людей для деятельности головного мозга, поступает за счет энергетических компромиссов
У животных известен компромисс между большим размером головного мозга и меньшими запасами жира и меньшим развитием мускулатуры. У людей в процессе эволюции развилось и то, и другое для удовлетворения повышенной потребности в энергии (головной мозг человека использует около 30% энергии организма), необходимой вследствие большего размера (и в 3 раза большей плотности клеток) головного мозга (относительно размера тела). Это указывает на недостаточное обеспечение энергией предков человека в ходе эволюции. Приготовление пищи, чтобы сделать эту энергию более доступной, также способствовало выравниванию этой энергии.
Физические упражнения появились благодаря энергетическим компромиссам Благодаря принципу биологических систем обратной связи «используй или потеряешь», за счет большей тренировки будет продолжаться приток энергии, используемой в повышенном режиме по сравнению с нормальным количеством, к тканям и биологическим системам, таким как мышцы и системы антиоксидантной защиты. Когда эти системы не используются, организм прекращает подачу к ним энергии, чтобы сохранить энергию. Из медицинской науки в течение некоторого времени известно, что долгосрочные физические упражнения полезны для людей, а краткосрочные физические упражнения - вредны.
Механизм этого эффекта видят в том, что вред физических упражнений проистекает вследствие высвобождения окислителей, в том числе окислителей, образующихся в результате выработки энергии митохондриями. Этот импульс окисления вводит в действие механизм защиты и репарации, который, в свою очередь, обеспечивает преимущество для клеток и организма в течение часов дня без физической нагрузки, которое называют окислительным прекондиционированием.
Сон является компромиссом использования энергии
Все животные, имеющие нейроны, спят. За счет сна мы получаем больше времени для репарации клеточного повреждения и, таким образом, продлеваем качество и продолжительность жизни за счет деятельности, которую мы не можем осуществлять в часы сна.
Биологическое старение и хронологическое старение
Показано, что биологическое старение человека меняется в течение хронологического года. В исследовании (Belsky DW 2015) 954 «молодых» людей в третьем и четвертом десятилетии жизни (на момент окончания исследования возраст составлял 38 хронологических лет, и отсутствовали признаки болезней старения), которые все родились в течение одного и того же периода времени, составляющего один год, в одном и том же новозеландском городе, на основании группы из 10 диагностических тестов, выполняемых в три момента времени, определили, что скорость их биологического старения (биологический возраст) изменялась от 1 биологического года на хронологический год до около 3 биологических лет на хронологический год. У 3 из 954 было выявлено даже обратное развитие биологического возраста за период времени. Это количественное варьирование биологического старения человека за хронологический год указывает на то, что скорость биологического старения у людей не является фиксированной величиной, и существует возможность ее изменения.
«Унифицированная теория старения»
За много лет было разработано четыре основных теории старения. Эти четыре общих теории возникли из различных ветвей научного поиска. Четыре основных теории старения состоят в следующем:
Теория старения, основанная на ограничении калорий (МсСау С 1935)
Свободнорадикальная теория старения (в настоящее время называемая редокс-теорией), (Harmon D 1956)
Теория старения, основанная на метилировании в 1967 г. (Vanyushin В 2005), и
Теория старения, основанная на соматических мутациях (Szilard L 1959). Другие теории старения включают:
Теория старения, основанная на скорости жизни (Pearl R 1928, Rubner М 1908, Sohal R 2012)
Теория старения одноразовой сомы (Kirkwood and Rose 1991)
Гипотеза старения, основанная на окислительно-восстановительном (редокс) стрессе (Sohal R 2012)
Инфламмейджинг (старение вследствие окислительного воспалительного процесса) (Franceschi С 2007, 2007, 2014) пара-воспаление (Medzhitov R 2008)
«Гипотеза старения по Мечникову» (Metchnikoff Е 1901)
Между всеми этими девятью теориями старения существуют взаимосвязи и перекрывание, и соединения, композиции, препараты и способы, раскрытые в настоящем документе, дополнительно подтверждают эти теории и фактически обеспечивают их унификацию.
Теория старения, основанная на ограничении калорий (Caloric restriction, CR)
В 1935 г. Клайв Мак-Кей (Clive МсСау) впервые открыл, что ограничение калорий (CR) увеличивает продолжительность жизни у животных. CR представляет собой практику снижения потребляемых калорий, не вызывающего недостаточность питания. Для этого требуется, чтобы организм получал достаточные количества воды, витаминов, минералов и белка, но ограниченно потреблял калории из углеводов и жиров (в количестве, меньшем рекомендуемой нормы потребления (РНП) для человека). CR можно безопасно осуществлять без вреда для здоровья при общих ограничениях потребления калорий в диапазоне, составляющем на 10-40% меньше рекомендуемого РНП. В 1986 г. Ричард Вейнрух (Richard Weindruch) показал, что ограничение калорий до 2/3 нормального количества у мышей приводит к увеличению продолжительности жизни на 40%. К настоящему времени большое количество экспериментов на биологических моделях подтвердило эти результаты. Биологические модели CR также помогли исследователям раскрыть молекулярно-биологические пути, отвечающие за увеличение продолжительности жизни и продолжительности здоровья (Colman RJ 2014). В рандомизированном контролируемом исследовании ограничения калорий у людей (Ravussin Е 2015) показана пригодность и влияние на прогностические факторы продолжительности здоровья и долголетия (продолжительности жизни).
Сиртуины и ограничение калорий
В 1990-е годы группой исследователей митохондрий (МТ), возглавляемой Леонардом Гваренте (Leonard Guarente), обнаружено, что определенный фермент, находящийся в дрожжах, является «питательным сенсором» и, возможно, является молекулярным механизмом, который мог бы объяснить эффекты ограничения калорий (Guarente L 2000). У дрожжей ограничение калорий увеличивало продолжительность жизни дрожжей на 40%. При нокауте этого фермента, называемого сиртуином, продолжительность жизни в ответ на ограничение калорий у дрожжей не была дольше.
Сиртуины, НАД+ и решение о лимитирующей скорость стадии биосинтеза НАД+
Всем ферментам сиртуинам требуется кофактор, называемый никотинамиддинуклеотидом (НАД+) (Imai S 2000). Это соединение встречается в природе, обнаруживается во всех клетках и составляет один из «энергетических потоков» клетки во многом сходно с АТФ. НАД+ является «энергетически обедненной формой» НАД-Н, который представляет собой действующую «форму энергообмена» молекулы. Таким образом, НАД+ является «сигналом» о том, что клетке не хватает энергии, где этот «сигнал» активируется и используется ферментами сиртуинами. Это объясняет, каким образом ограничение калорий, представляющее собой «обедненное энергией состояние», может активировать запуск клеткой клеточных стрессовых метаболических путей, что способствует выживанию. Все 7 сиртуинов, обнаруженных у человека, по-видимому, запускаются клеточным питательным стрессом. НАД+ является пусковым механизмом этого ответа. НАД+ образуется из никотинамидмононуклеотида (NMN), и NMN может быть получен под действием фермента, называемого NAMPT. В клетках, не подвергающихся стрессу, период полураспада НАД+ составляет от 3 до 5 часов (лаборатория Suave А: описано в Canto С 2013). К сожалению, у людей, по видимому, не наблюдается недостатка НАД+, получаемого в организме за счет механизмов регуляции энергопотребления. В 2011 г. было показано, что регуляторной точкой остановки синтеза НАД+ является фермент NAMPT, который преобразует предшественник NMN в соединение NMN. При введении NMN мышам у них образуется НАД+ из NMN за 15 минут. Таким образом, решение «проблемы ограничения синтеза НАД+» состоит в том, чтобы миновать лимитирующую скорость стадию, которой является продукция NMN. Это было продемонстрировано в 2011 г. (Jun Yoshino and Kathryn Mills 2011).
Сиртуины человека 1, 2, 3, 4, 5, 6, 7
Сиртуин 1 (Sirt1)
Сиртуин 1 (Sirt1) локализуется в ядре и в цитоплазме. Он крайне чувствителен к ингибированию H2O2 окисления. Внеклеточные концентрации H2O2, столь низкие, как 1 мкмоль/л, ингибируют Sirt1 посредством окисления критических остатков цистеина в активном центре сиртуина (Jung S-B 2013). В дополнение к этому РНК-связывающий белок HUR связывается с 3'-нетранслируемой областью мРНК, кодирующей Sirt1, что приводит к его стабилизации и повышенным уровням. H2O2 запускают диссоциацию HUR из комплекса HUR-мРНК Sirt1, стимулиря распад мРНК Sirt1, в результате чего количество Sirt1 уменьшается, и этот процесс, по-видимому, претерпевает регуляцию киназой Chk2 (Abdelmohsen K 2007). Было обнаружено, что редокс-фактор-1 (REF-1) химически восстанавливают остатки цистеина Sirt1, стимулирующие его активность (Jung S-B 2013). REF-1, поддерживающий сульфгидрильные (тиольные) группы цистеиновых остатков в Sirt1 в восстановленной форме и защищающий Sirt1 от окисления H2O2, также называют АРЕ1 (апуриновая/апиримидиновая эндонуклеаза)-1, поскольку в отдельном активном центре этого фермента он является ограничивающим скорость ферментом в метаболическом пути эксцизионной репарации оснований млекопитающих. К настоящему времени сиртуин 1 является наиболее изученным сиртуином человека.
Сиртуин 2 (Sirt2)
Sirt2 находится, в основном, в цитоплазме (Yudoh K 2015, Gomes Р 2015). Sirt2 важен в регуляции клеточного цикла (Nie Н 2014). Показано, что он является деацетилазой гистонов (Moscardo А 2015). Показано, что он поддерживает правильное деление и репликацию хромосомы (Kim HS 2011). Описанный механизм этого состоит в деацетилировании под действием Sirt2 АТФ-взаимодействующего белка (ATRIP) в лизине 32 в ответ на репликативный стресс. Мишенью деацетилирования Sirt2 является киназа контрольной точки клеточного цикла BubR1. Посредством деацетилирования 668 Sirt2 стабилизирует BubR1 и поддерживает ее в форме убиквитинирования и деградации. У мышей это приводит к поразительному медианному увеличению продолжительности жизни на 58% (122% у самцов) и максимальному увеличению продолжительности жизни на 21% (North BJ 2014).
Установлена корреляция активности Sirt2 со снижением депрессии (в модельной системе на крысах, в которой депрессию создают посредством стресса), возможно за счет увеличения нейрогенеза (Liu R 2015).
Сиртуин 3 (Sirt3)
Сиртуин 3 локализован во внутренней мембране митохондрий и является важным регулятором клеточного гомеостаза энергии (Nogueiras R 2012). Специфический аллель Sirt3 активировал повышенный уровень активности, и показано, что он необходим для продолжительности жизни более 90 лет у людей (Rose G 2003, Bellizzi D 2005, Halaschek-Wiener J 2009). Sirt 3 обладает доминантной активностью митохондриальной деацетилазы (Lombard DB 2007). Экспрессия Sirt3 в печени возрастает после голодания (Hirschey MD 2010). Экспрессия Sirt3 в мышцах возрастает после физической нагрузки (Hokari F 2010), голодания и ограничения калорий и уменьшается при постоянном употреблении в пищу жиров (Palacios ОМ 2009). В целом все эти исследования указывают на то, что Sirt3 действует как основной пусковой механизм, осуществляющий приспособление к недостатку энергии (Cho Е-Н 2014), поддерживая продукцию АТФ, в том числе изменение направления метаболизма, известное как эффект Варбурга (Guarente L 2014). Sirt3 деацетилируется при остатках лизина 926 и 931, в результате чего активируется митохондриальный гибридный белок ОРА1, повышающий его ГТФазную активность. Около 20% митохондриальных белков могут быть ацетилированы. Считают, что ацетилирование/деацетилирование белков является основным регуляторным механизмом в митохондриях (Kim SC 2006). Показана роль Sirt3 в регуляции биогнеза митохондрий посредством активации комплекса PGC-альфа/ERR-альфа (Giralt А 2012, Hirschey MD 2011, Kong X 2010).
Sirt3-зависимые метаболические пути осуществляют предполагаемую молекулярную связь между бессонницей и нейродегенерацией (Fifel K 2014, Zhang J 2014). Sirt3 опосредует уменьшение окислительного повреждения и предотвращение возрастной потери слуха (Someya S 2010) под действием ОРА1 (Leruez S 2013). Также предполагают участие Sirt3 при болезни Альцгеймера, болезни Хантингтона, болезни Паркинсона, боковом амиотрофическом склерозе (Kincaid В 2013) и неалкогольной жировой болезни печени (Cho Е-Н 2014).
Сиртуин 4 (Sirt4)
Сиртуин 4 локализован в митохондриях. Он представляет собой клеточную липоамидазу (или делипоилазу), удаляющую липоильные модификации из остатков лизина субстратов. Sirt4 делипоилирует и модулирует активность пируватдегидрогеназного комплекса (PHD), которая, в свою очередь, ингибирует продукцию ацетил-КоА (Mathias RA 2014). Он деацетилирует малонил-КоА-декарбоксилазу (MCD) для регуляции катаболизма липидов (Laurent G 2013). Он также осуществляет АДФ-рибозилирование на глутаматдегидрогеназе (GLUDI) (Haigis МС 2006).
Сиртуин 5 (Sirt5)
Сиртуин 5 локализован в митохондриях. Sirt5 десукцинилирует, демалонилирует и деглутарилирует белковые субстраты, такие как карбамоилфосфатсинтаза 1 (CPS1), для регуляции цикла мочевины (Du J 2011, Peng С 2011, Tan М 2014). Деацетилирующая активность Sirt5 является слабой (Du J 2011, Tan М 2014). Предположили, что Sirt5 регулирует продукцию аммиака и индуцированную аммиаком аутофагию и митофагию посредством регулирования метаболизма глутамина (Polletta L 2015).
Сиртуин 6 (Sirt6)
Сиртуин 6 локализован в ядре и представляет собой ассоциированную с хроматином деацетилазу гистонов (Kugel S 2014). Он может деацетилировать лизин 9 гистона Н3 (H3K9), участвуя, таким образом, в регуляции теломерного хроматина и клеточного старения (Michishita Е 2008). Когда он деацетилирует лизин 56 гистона Н3 (H3K56), он уменьшает хроматиновую доступность транскрипционных факторов, таких как NF-kB, Foxo3 и HIF1-альфа, к промоторам их мишеней, в результате чего происходит ингибирование их генов-мишеней (Kugel S 2014). Sirt6 деацетилирует гистон H4K16, который регулирует мейотический аппарат в ооците (Han L 2015). Sirt6 связан с регуляцией продолжительности жизни и продолжительности здоровья (Kanfi Y 2012, Cardus А 2013, Shen J 2013, Liu R 2014, Sharma A 2013). Установлено, что Sirt6 уменьшает атеросклеротические сосудистые заболевания. Экспрессия Sirt6 подавляет клеточное старение и опосредованные NF-kB воспалительные ответы, такие как ФНО-альфа, в коленном суставе человека, которые ведут к развитию остеоартрита (Wu Y 2015). Также предполагают, что повышение активности Sirt6 можно рассматривать в качестве терапии идиопатического фиброза легких (ИФЛ) (Minagawa S 2011).
Сиртуин 7 (Sirt7)
Сиртуин 7 локализован в ядре. Sirt7 функционально связан с его транскрипционной регуляцией. Он осуществляет положительный контроль продукции рибосом посредством прямого взаимодействия с механизмом Poll (Ford Е 2006, Grob А 2009, Chen S 2013). Напротив, Sirt7 осуществляет отрицательную регуляцию транскрипции генов вне повторов рДНК посредством деацетилирования гистона H3K18 (Barber MF 2012). Sirt7 нацелен на ацетилированный лизин в N-концевом хвосте гистона Н3 (H3K18Ac). В каскаде передачи сигнала повреждения ДНК Sirt7 находится после Sirt1 и Sirt6. Рекрутмент Sirt7 в сайтах повреждения ДНК зависит от активности PARP1. Там он может осуществлять деацетилирование H3K18Ac. H3K18Ac влияет на рекрутмент фактора ответа на повреждение 53 ВР1 к двунитевым повреждениям ДНК, приводя к соединению их концов и стабильности генома.
Циклический аденозинмонофосфат (цАМФ)
За открытие цАМФ в качестве второго мессенджера в 1971 г. была присуждена Нобелевская премия. Ограничение калорий повышает уровень цАМФ Уровни цАМФ снижаются с возрастом. В настоящее время установлена корреляция высоких уровней цАМФ с большей продолжительностью жизни. цАМФ осуществляет ряд связанных с метаболизмом процессов передачи сигнала гормонов. НАД+ содержит группировку АМФ. цАМФ взаимодействует с карманом связывания НАД+ сиртуина. Это связывание усиливает гидролиз НАД+ до NAM и 2'-O-ацетил-АДФ-рибозы. Таким образом, цАМФ является стимулятором ферментативной активности сиртуинов (Wang Z 2015), действуя в качестве подкрепления сигнала НАД+ об истощении энергии.
Фосфорилирование
Sirt1 может претерпевать фосфорилирование в высоко консервативном положении остатка серина 434, представляющего собой каталитический сайт сиртуина. Фосфорилирование S434 повышает деацетилазную активность Sirt1 Считают, что фосфорилирование Sirt1 осуществляет протеинкиназа А (PKA) или киназа, следующая по метаболическому пути после PKA. Считают, что эта регуляция путем фосфорилирования регулирует активность сиртуина в более короткой временной рамке (от 5 до 15 минут), чем обычные меры повышения уровней НАД+. Эта более короткая временная рамка обеспечивает возможность для индукции цАМФ/РКА для более краткосрочной утилизации жирных кислот (Gerhart-Hines Z 2011). В дополнение к этому регуляция транскрипционных уровней Sirt1 осуществляется посредством конкуренции сайта связывания с промотором цАМФ-ответного элемента активирующего белка (CREB) и белка, связывающего углеводчувствительный элемент (ChREBP). Сам CREB может претерпевать фосфорилирование, которое приводит к его импорту в ядро, что делает его лучшим конкурентом за сайт промотора на Sirt1 и транскрипцию Sirt1 Эта мРНК достигает максимума за 12-18 часов и возвращается к базальным уровням за 24 часа (Noriega LG 2011), что указывает на желательный отказ от пищи на 12 часов каждый день (Chaix А 2014).
Белок Sirt1 имеет несколько других сайтов фосфорилирования на боковых цепях аминокислоты серина. Ser27 является одним из сайтов, которые претерпевают непрямое фосфорилирование посредством активации JNK2. В случае фосфорилированного состояния сайта Ser27 на Sirt1 белок Sirt1 становится значительно более резистентным к опосредованной протеасомой деградации. Таким образом, увеличивается период полураспада белка Sirt1 от менее 2 часов до более 9 часов. Это составляет очень важную часть поддержания уровней белка Sirt1 внутри клетки (Ford J 2008).
Keap1 функционирует как отрицательный регулятор Nrf2, описанный позже, который активирует антиоксидантные ферменты. Контроль деградации Keap1 в ответ на антиоксиданты осуществляется посредством фосфорилирования тирозина (Kasper JW 2011).
Никотинамидмононуклеотид (NMN) в качестве соединения, препятствующего старению
Только НАД+ активирует все 7 сиртуинов. В 2008 г. было продемонстрировано, что предшественник НАД+ NMN производит эффекты, вызывающие обратное развитие старения у мышей (Ramsey K and Mills K 2008). Впоследствии в 2009 г. было показано, что NMN оказывает мощное воздействие на обратное развитие эффектов индуцированного ожирением сахарного диабета (Imai S 2009), а также возрастного диабета. В 2013 г. было показано, что высокая доза NMN вызывала обратное развитие мышечного старения в течение одной недели введения NMN (Gomes А 2013).
CD38
CD38 является как НАДазой, так и НАДФазой. CD38 может быть внеклеточным (фермент плазматической мембраны II типа) и внутриклеточным. CD38 преобразует НАД+ в никотинамид и cADPR. cADPR является вторым мессенджером, вовлеченным в клеточную функцию. Никотинамид, как отмечено ранее, ингибирует как сиртуиновые ферменты, так и ферменты PARP по механизму обратной связи, как обсуждается в следующем разделе. CD38 обнаружен во многих популяциях клеток. CD38 связан как с системой врожденного иммунитета, так и с системой адаптивного иммунитета. CD38 характеризуется высокой экспрессией в воспалительных клетках, и утрата CD38 связана с нарушенными иммунными ответами. Считают, что CD38 и CD157 обеспечивают восстановление энергии энергетически затратных продуктов, которые иначе были бы израсходованы впустую. В популяции европеоидов известно два аллеля CD38 (Malavasi F 2007). CD157 является вторым членом этого семейства, хотя каталитическая эффективность CD157 в несколько сотен раз ниже, чем CD38 (Hussain AM 1998). CD38 и CD157 могут иметь мономерные или димерные формы. CD38 является основным регулятором Са++, катализирующим образование эндогенного Са++ (Lee S 2015). Высвобождение Са++ может стимулировать продукцию ИЛ-6 (Adebanjo OA 1999, Sun L 2003). В разделе Примеры данного документа было показано снижение ИЛ-6.
Показано, что НАД+ снижается с возрастом. Уровни белка, уровни мРНК и ферментативная активность CD38 с возрастом повышаются (во всех исследованных тканях: печени, белой жировой ткани, селезенке, скелетных мышцах, подвздошной кишке, тощей кишке и почке). Это увеличение количества CD38 требуется для возрастного снижения НАД. Другие белки, которые используют НАД, по-видимому, не являются причиной снижения НАД+ с возрастом; примеры включают PARP1 и Sirt1, которые оба снижаются с возрастом. Между снижением активности CD38 и НАД+ в процессе старения наблюдали высокий коэффициент обратной корреляции. CD38 также способен приводить к деградации предшественника НАД+ никотинамидмононуклеотида (NMN) (Grozio А 2013). Константа скорости ферментативной реакции (Kcat) для NMN+ была в 5 раз выше, чем для НАД+, и характеризуется самой высокой Kcat для любого субстрата CD38 (Sauve АА 1998).
Снижение НАД+ в клетках под действием CD38 приводило к утрате функции митохондрий без изменений уровней Sirt3.
Индукция CD38 происходит посредством следующих механизмов:
i. Окисление связано с активацией CD38 (Zhang AY 2004, Kumasaka S 1999, Wilson HL 2001, Dipp M 2001,Okabe E 2000, Ge Y 2010)
Это противоположно Sirt1, где для сохранения Sirt1 требуется восстановление, а в результате окисления его действие прекращается. Окисление также активирует PARP-1 (Bai Р 2011).
ii. ФНО-альфа является активным индуктором экспрессии CD38 в клетках (Barata Н 2004, Lee CU 2012).
Примечание: Показано, что тройная терапия, описанная в разделе Примеры настоящего документа, вызывает снижение и ФНО-альфа, и ИЛ-6.
a) CD38 имеет рецептор ФНО (Prasad GS 1996);
b) ФНО-альфа также индуцирует двукратную активацию промотора CD38.
Таким образом, ФНО регулирует как уровни (транскрипционную регуляцию)
РНК, так и активность белка. Механизм этой регуляции состоит в том, что ФНО-альфа усиливает связывание с сайтом NF-kB и с некоторыми из сайтов связывания АР-1 (Tirumurugaan K 2008).
На CD38, по-видимому, не влияет конечный продукт механизма обратной связи:
i Никотинамид защищает CD38 от ингибирования синтетическими ингибиторами (Sauve АА 2002, Sauve АА 1998). Никотинамид ингибирует Sirt1 и PARP (другие «пользователи» НАД+). В метилированном состоянии никотинамид не взаимодействует по механизму обратной связи с сиртуинами и PARP и не прекращает действие ферментов, использующих НАД+ (вследствие стерического затруднения).
Ингибирование CD38 происходит за счет следующих механизмов:
ii. Существуют молекулы, отличающиеся от никотинамидмононуклеотида, например, флавоноиды лютеолинидин, куроманин и лютеолин (Kellenberger Е 2011), которые ингибируют CD38, но они, вероятно, также будут ингибировать другие реакции, в которые вовлечены три фермента, образующие НАД+ из NMN, а также другие ферменты, использующие НАД+, такие как SIRT и PARP.
iii. Метилирование гена CD38 может составлять часть этой регуляции (Ferrero Е 1999). В дополнение к влиянию метилирования никотинамида до метил-никотинамида это приводит к изменению механизма обратной связи сиртуина и ферментов PARP. Это метилирование гена (эпигенетика) может объяснять, почему CD38 повышается с возрастом.
iv. Апигенин ингибирует CD38. Он также вводит в действие Nrf2. Апигенин эффективно вызывал обратное развитие гиперметилированного состояния 15 CpG в промоторе Nrf2 дозозависимым образом. Апигенин усиливал ядерную транслокацию Nrf2 и повышал экспрессию мРНК и белка Nrf2 и следующего за Nrf2 гена-мишени NQ01. Апигенин восстанавливал NRF2 из состояния сайленсинга посредством деметилирования CpG (Parededs-Gonzalez X 2014).
v. Восстановление приводит к прекращению действия CD38. Восстановление дисульфида Cys 118-Cys 201 в CD38 приводит к инактивации (Prasad GS 1996). Дисульфид вовлечен в бифункциональную активность в шарнирной области фермента и трехмерную структуру, зависимую от 10 остатков цистеина (Lin Q 2005, Prasad GS 1996). Sirt1, нпротив, поддерживается посредством восстановления.
vi. Выше сайта старта транскрипции CD38 находится потенциальный сайт связывания для транс-взаимодействий с ИЛ-6 (следует отметить, что тройная терапия, описанная в разделе Примеров, приводит к снижению ИЛ-6).
Поли(АДФ-рибоза)полимераза (PARP)
Репарация повреждений ДНК требует большого количества энергии, и для этой цели происходит перераспределение энергии. Количество повреждений ДНК в среднем увеличивается с возрастом отчасти из-за того, что для их репарации энергия не выделяется, даже несмотря на то, что разрыв ДНК мог бы претерпевать репарацию. Поли(АДФ-рибоза)полимеразы (PARP) являются НАД+-зависимыми ферментами, осуществляющими репарацию ДНК (древнюю и эволюционно консервативную биохимическую реакцию; Otto Н 2005), а также ответственными за другие биологические функции. Никотинамид высвобождается в виде конечного продукта этих реакций PARP. Поэтому сиртуиновые ферменты не только конкурируют за доступный НАД+ с PARP и CD38, но сиртуины также претерпевают ингибирование конечным продуктом использования НАД+, который представляет собой никотинамид, образующийся в результате использования сиртуином НАД+, использования PARP НАД+ и использования CD38 НАД+, а также использования НАД+ другими ферментами. Как указано выше, наблюдается снижение НАД+ при старении (Braidy N 2011 and Massudi Н 2012). У людей существует 17 ферментов PARP (Ame J 2004). PARP-1, 2 и 3 (и танкиразы) вовлечены в репарацию ДНК.
Показано, что в репарацию ДНК также вовлечены Sirt1 и Sirt6. Основную часть ДНК-индуцированной активности PARP покрывает PARP-1 (85-90%), тогда как PARP-2 считают ответственной за остальную активность (Szanto М 2012). Регуляция PARP-1 осуществляется посредством а) механизма обратной связи с никотинамидом, b) редокс-баланса; окисление H2O2 активирует PARP-1, с) обратимого метилирования и d) действие PARP-1 прекращается в результате деацетилирования Sirt1. Пролонгированная активация PARP может приводить к обеднению клеточных пулов НАД+, поэтому при избыточном повреждении ДНК происходит значительное падение НАД+. PARP-1 осуществляет высокий каталитический кругооборот НАД+ по сравнению с Sirt1 (Bai Р 2012). Его константа Михаэлиса (Km) (20-60 мкмоль/л - Ame J 1999) в 5 раз ниже, и PARP-1 обладает значительно более сильным Vmax, чем сиртуин-1 (Houtkooper R 2010). Аффинность PARP-2 к НАД+ и его деградация почти аналогичны таковым для Sirt1. PARP-2 связывается с ДНК в проксимальном участке промотора Sirt1. Km большинства сиртуинов к НАД+ находится в диапазоне 100-300 мкмоль/л, и колебания НАД+ описаны как составляющие от 200 до 500 мкмоль/л. Уровни НАД+ обычно колеблются в двухкратном диапазоне (Chen D 2008). Кроме того, колебания НАД+ происходят в соответствии с циркадными ритмами (Ramsey К 2009). Хотя эти количественные определения обладают рядом ограничений, фактором, ограничивающим активность сиртуинов по скорости, является доступность НАД+.
Свободнорадикальная теория старения
В 1956 г. Денемом Хармоном (Denham Harmon), исследующим эффекты рентгеновского излучения, выдвинуто предположение, что причиной старения являются активные формы кислорода, называемые «свободными радикалами», и в настоящее время эта теория называется «свободнорадикальной теорией старения» (Harman D 2009). На основании этих наблюдений за воздействиями рентгеновского излучения на животных д-р Хармон предположил, что подобно тому, как рентгеновское излучение индуцирует образование свободных радикалов, нормальный процесс старения генерирует свободные радикалы и оказывает сходные воздействия на организм. К этому времени источник этих «свободных радикалов» при нормальном процессе старения был неизвестен. Усилия, предпринятые в дальнейшем, позволили подтвердить, что клетки продуцируют свои собственные активные формы кислорода (т.е. свободные радикалы). Свободные радикалы образуются фактически в каждой клетке с рождения до смерти. Многие клеточные биохимические реакции создают активные формы кислорода внутри клетки. Старение связано не с присутствием этих свободных радикалов как таковых, а, вероятнее, с вызванным в клетках повреждающим избытком свободных радикалов в связи с недостаточным захватом свободных радикалов многими ферментами, которые гасят эти активные формы кислорода. Контроль активных форм кислорода (АФК) модифицирован в мышцах старых животных, а высвобождение АФК (супероксида) снижено в старых мышцах (Jackson М. 2011). Эту область исследования свободных радикалов в настоящее время называют окислительно-восстановительной (редокс) биологией (Nathan С 2013), и к настоящему времени появляется все большее количество публикаций, в которых подробно описаны обладающие преимуществом биологические эффекты свободных радикалов, вовлеченных в модулирование метаболических путей передачи клеточных сигналов (Powers and Jackson 2008). В книге «Гипотеза старения вследствие окислительно-восстановительного стресса» (Redox Stress Hypothesis of Aging) концептуально смещена значимость окисления и восстановления для преобразования сигналов и регуляции генов со смещением окислительно-восстановительного состояния клеток с возрастом в сторону окисления, что приводит к переокислению редокс-чувствительных тиолов белков и последующему прерыванию редокс-регулируемых механизмов передачи сигнала. Подтверждения этой теории получены в результате а) результатов наблюдений, что количество окислительных побочных продуктов увеличивается с 25 до 100% от пубертатного до взрослого возраста; b) количество карбонильных групп белков увеличивается с возрастом и уменьшается при ограничении калорий и с) средняя продолжительность жизни пропорциональна количеству карбонильных групп белков (Sohal R 2012).
Чувствительные к окислению тиольные группы белков
Изменение редокс-потенциала чувствительных к окислению тиольных групп белков может обеспечить возможность смены направления между отдельными катаболическими и анаболическими процессами, а также активации метаболических путей выживания. Остатки метионина и цистеина белка особенно чувствительны к окислительной модификации. Метионин является стадией, предшествующей синтезу SAM в метаболическом пути метилирования. Таким образом, метионин связан с метаболическим путем метилирования, и его регуляция осуществляется за счет редокс-баланса. Доля цистеиновых остатков в процентах увеличивается с возрастанием сложности организма, но их распространенность все же значительно меньше, чем частота встречаемости, рассчитанная только на основании использования кодонов. Цистеины, встречающиеся в кластерах, характеризуются высокой консервативностью в эволюции и обычно важны для структуры или функции. На значения pKa для тиольных групп влияет их локальное окружение. Состояния окисления могут находиться в диапазоне от полностью восстановленного тиольного/тиолатного аниона до полностью окисленной сульфоновой кислоты (Cremers СМ 2013). Скорость реакции тиольных групп белка с окислителями, такими как пероксид водорода (Н2О2), охватывает более 7 порядков величины без какой-либо обнаружимой корреляции с кислотностью соответствующего тиола активного центра (Ferrer-Sueta G 2011).
Существуют обратимые и необратимые модификации цистеина. Окисление тиола цистеина (RSH/RS-) под действием АФК, активных форм азота (АФА) или активных форм углерода (АФУ) приводит к образованию сульфеновой кислоты (RSOH), обладающей высокой реакционной способностью, которая может взаимодействовать либо с другим тиолом с образованием дисульфидной связи (RSSR), либо с глутатионом (GSH), в результате чего становится S-глутатионилированной (RSSG). Эти окислительные модификации обратимы, и восстановление катализируется системой Trx и/или Grx. Считают, что дальнейшее окисление сульфеновой кислоты до сульфиновой кислоты (RSO2H) и сульфоновой кислоты (RSO3H), как правило, необратимо in vivo. Многие из белков, регулируемых окислением и восстановлением тиола, действуют как транскрипционные регуляторы (например, OxyR, Yap1p), которые быстро индуцируют экспрессию генов, вовлеченных в механизмы антиоксидантной защиты (Zheng М 1998, Tachibana Т 2009), и других генов, вовлеченных в каскады преобразования сигнала (Gopalakrishna R 2000 and Dinkova-Kostova AT 2005). (см. дополнительные примеры в Дополнении 1 к публикации Cremer СМ 2013).
Примером фермента с тиольными группами в активном центре является глицеральдегидфосфатдегидрогеназа (GAPDH), которая играет критическую роль в гликолизе. Окисление тиолов GAPDH блокирует гликолиз и вносит вклад в образование НАДФ-Н вместо НАД-Н (Shenton D 2003). Другим примером является окисление тиолов в активных центрах, инактивирующее фосфатазную активность SHP1/2, PTEN, Cdc25, усиливая интенсивность передачи сигнала, достигаемой за счет фосфорилирования субстрата, что приводит к активации метаболических путей передачи сигнала NF-kB-индуцирующей киназы/IkB, которая вызывает экспрессию генов, вовлеченных в антиоксидантную защиту (Jung KJ 2009). Третьим примером являются тиольные группы сиртуина в активных центрах сиртуина, очень чувствительные к окислению, которые ингибируют активность сиртуина при окислении. 3 (Cys-67, Cys-268 и Cys-623) из 5 цистеинов сиртуина-1 человека доступны для возможной обратимой модификации тиола посредством редокс-баланса (Autiero I 2008). Cys-67 и Cys-623 находятся в соответствии с посттрансляционной регуляцией этих концевых участков, Cys-268 находится в НАД+-связывающем участке всех членов семейства сиртуинов, в которых каталитический центр высоко консервативен. Результатом связывания НАД+ являются изменения конформации сиртуина, допускающие протекание катализа (Zee R 2010).
Основные компоненты редокс-биологии
Существуют различные типы АФК и АФА (активных форм азота). В совокупности их называют АФКА. Эти формы включают: супероксид, пероксид водорода, гидроксильные радикалы, синглетный кислород, оксид азота, пероксинитрит, гиперхлорит, а также продукты перекисного окисления липидов ПНЖК. Существует различная специфичность АФК. АФК характеризуются типом специфичности, которая является скорее атомной, чем молекулярной. АФК чаще всего обратимо взаимодействуют в клеточной передаче сигнала с серой, которая является одним из наименее распространенных атомов в биологических макромолекулах и, в основном, с боковыми цепями остатков цистеина или метионина в пептидах или белках (Nathan С 2013). Эндогенные ферментативные источники АФК (множественные изоформы обеспечивают большую чувствительность и специфичность в регуляции) включают семь изоформ NADPH-оксидаз (NOX), которые дифференциально экспрессируются (регулируются) в различных клетках и у различных биологических видов, а также в ряде других источников (см. блок 1, стр. 2 Nathan С 2013).
Основными типами антиоксидантных ферментов (многочисленные изоформы обеспечивают более высокую чувствительность и специфичность) (требуется контроль и использование переходных металлов) являются 3 изоформы супероксиддисмутазы (SOD), 5 изоформ глутатионпероксидазы (GPX) и каталаза. Также существует 2 изоформы тиоредоксина (TRX)(c тиоредоксин-редуктазой), при этом рециркуляция тиоредоксина может осуществляться посредством взаимодействия с REF-1 (REF-1 поддерживает тиольные группы сиртуина в восстановленном состоянии), 3 изоформы глутаредоксина (GRX), 6 изоформ пероксиредоксина (PRX) (ответственного за индукцию 90% митохондриального и более цитозольного H2O2 эукариот. Этот процесс может быть приведен в действие и прекращен посредством регуляции по функциональному механизму обратной связи, что дает возможность передачи редокс-сигнала (Sies Н. 2014). Пероксиредоксины составляют филогенетически древнее семейство белков, основной ролью которого является детоксикация H2O2. Они также создают редокс-ритм. Считают, что каталитический цикл гипероксидации пироредоксина и рециркуляции под действием сульфоредоксина может составлять основу транскрипционно-независимо циркадного ритма (Rey, G. 2013). Корреляция уровней НАД+ с биологическими часами характеризуется 2 пиками в сутки, разделенными 12 часами. Считают, что, поскольку клеточное отношение НАД+/НАД-Н более 1 больше, чем клеточное отношение НАДФ+/НАДФ-Н менее 0,01 в цитозоле, это позволяет отделять антиоксидантные и биосинтетические восстанавливающие эквиваленты (НАДФ-Н) в клетке от предназначенных для образования АТФ в митохондриях (НАД-Н). Фосфат НАДФ-Н придает различную субстратную специфичность, но обладает такими же свойствами переноса электронов. Гипероксидация пероксиредоксинов может индуцировать как шаперонную функцию, так и преобразование сигнала.
Малые антиоксидантные молекулы включают: глутатион (GSH), мочевую кислоту, билирубин, аскорбиновую кислоту (витамин С), витамин Е, а также каротиноиды, коэнзим Q10, N-ацетилцистеин (NAC). NAC действует как донор восстановленного тиола и противостоит H2O2, который окисляет тиолы.
Теория старения, основанная на метилировании
Не все гены экспрессируются во всех клетках. Этот контроль «селективной экспрессии генов» 21 800 генов, которые кодируют белки в клетках человека, определяет, станет ли клетка клеткой головного мозга или клеткой сердца. Эту систему регуляции генов называют «эпигенетикой» (Kundaje А 2015). Эпигенетика контролирует скорость старения (Reynolds L 2014). Один из способов, посредством которого осуществляется регуляция генов, является метилирование определенных остатков ДНК, называемых цитозинами. В 1967 г. Борис Ванюшин (Boris Vanyushin) показал, что в процессе старения ДНК утрачивает метилирование (Vanyushin BF 2005). В регуляцию генов вовлечены и другие эпигенетические механизмы помимо метилирования, такие как модификации гистоновых белков, микроРНК и ремоделирование хроматина (соотношение гетерохроматина и эухроматина) (Kundaje А 2015). Кроме того, в процессе старения метилирование некоторых цитозинов ДНК увеличивается, а метилирование других сайтов в процессе старения уменьшается. Понятно, что это метилирование ДНК является формой эпигенетической регуляции генов, коррелирующей со старением. Это было совсем недавно показано Стивеном Хорватом (Steven Horvath), который показал, что можно сконструировать «часы метилирования ДНК» на основании анализа только 353 цистеиновых остатков, и что эти «часы мДНК» (Bocklandt S 2014) имеют 0,96 корреляцию со старением. Важнее, что эти часы показывают время намного лучше, чем любой другой показатель старения, кроме даты рождения. В результате профилирования метилирования ДНК мезенхимных стволовых клеток (МСК), полученных от индивидов в возрасте от 2 до 92 лет было идентифицировано 18 735 гиперметилированных и 45 407 гипометилированных сайтов CpG, ассоциированных со старением. Важнее, что гипометилированные сайты CpG были в значительной степени обогащены маркером активного хроматина H3K4me1 в стволовых и дифференцированных клетках, что позволяет предположить существование независимой от клеточного типа сигнатуры гипометилированной ДНК в процессе старения. Эти результаты указывают на то, что динамика метилирования ДНК в процессе старения зависит от комплексной смеси факторов, включающих последовательность ДНК, клеточный тип и вовлеченный хроматиновый контекст, и что в зависимости от локуса изменения могут претерпевать модуляцию под действием эпигенетических и/или внешних факторов (Fernandez AF 2015). Показано, что ограничение калорий предотвращает возрастные изменения метилирования ДНК у мышей (Chouliaras L 2012). Показано, что два из 7 ферментов сиртуинов (Sirt1 и Sirt6) косвенно влияют на метилирование ДНК посредством их действия на деацетилирование гистонов. Было также показано, что конечный продукт реакции сиртуина никотинамид должен быть метилирован до 1-метилникотинамида, в противном случае конечный продукт никотинамид будет связываться внутри фермента сиртуина и останавливать его ферментативную активность (Schmeisser K 2013). Сиртуин-1 снижает активность NF-kB, который увеличивает триметилирование лизина 36 на гистоне 3 (H3K36me3). Это коррелирует с ускоренным метилированием ДНК. Гены, экспрессия которых претерпевает резкое изменение в процессе старения, отличаются низкими или даже необнаружимыми уровнями H3K36me3 в структуре генов независимо от количества их соответствующей мРНК (Pu М 2015). В клетках человека глобальная утрата триметилирования H3K9 (H3K9me3) воспроизводит ускоренное клеточное старение и изменения в архитектуре гетерохроматина. Эти изменения также коррелируют с дезорганизацией гетерохроматина у людей в возрасте от 7 до 72 лет с увеличением возраста (Zhang W 2015). 30 января 2015 г. возраст крови по метилированию ДНК использовали для прогнозирования смертности по любой причине в пожилом возрасте у людей независимо от состояния здоровья, факторов образа жизни и известных наследственных факторов (Marioni RE 2015). 19 февраля 2015 г. в журнале Nature были опубликованы результаты 111 эпигеномов человека, позволяющие другим исследователям проводить сравнения в будущем и ссылаться на них (Kundaje А 2015).
Метаболический путь метилирования
Метионин особенно чувствителен к окислительной модификации. Метионин является стадией, следующей после синтеза гомоцистеина и предшествующей синтезу SAM в метаболическом пути метилирования. Таким образом, метионин принимает участие в метаболическом пути метилирования, и его регуляция осуществляется за счет редокс-баланса. Цистеин синтезируется из метионина и является основным предшественником сульфида водорода (H2S). Повышенные уровни гомоцистеина связаны с ингибированием образования эндогенного сульфида водорода (H2S) (Tang X 2011). Сульфид водорода (H2S) ослабляет вызванный метионином окислительный стресс (Tyagi N 2009). Гомоцистеин (Нсу) может необратимо подвергаться деградации до сульфида водорода (H2S) посредством метаболического пути транссульфурации, активируемого окислительным стрессом. H2S обладает защитными функциями при гипергомоцистеинемии (Ohashi, R. 2006, Chang L 2008). Жировая ткань является важным органом метаболизма метионина, а также инсулин-чувствительным органом. Увеличение количества H2S в жировой ткани повышает чувствительность к инсулину (Feng X 2009). Высокий уровень H2S в поджелудочной железе подавляет высвобождение инсулина (Wu L 2009). Уровни H2S в крови ниже при диабете 2 типа, чем у здоровых субъектов соответствующего возраста (Jain S 2010). Аспирин является ингибитором арахидоната и может влиять на цикл метионина - гомоцистеина и связан с одноуглеродным метаболизмом и, таким образом, как с метилированием, так и с редокс-балансом (Lupoli R 2015). Терапия H2S донорами H2S Na2S или NaSH ингибирует аспирин дозозависимым образом (Zanardo RC 2006).
Также существует ингибитор метилирования: S-аденозилгомоцистеин (SAH). Когда метионин присутствует в большом количестве, NNMT регулирует только SAH, но не SAM (Ulanovskaya OA 2013).
Радикальные ферменты SAM составляют разнообразное надсемейство белков, использующих химию радикалов (5'-dAdo) для осуществления модификации субстрата. Субстраты этих ферментов отличаются от нуклеофильных субстратов, которые претерпевают метилирование посредством механизма полярности. Известно 4 подкласса этих ферментов (А, В, С, D).
В качестве общего правила, возрастное гипометилирование ДНК является доминантным событием, приводящим к повышенной экспрессии генов, но с возрастом в некоторых промоторных областях ДНК часто происходит гиперметилирование, приводящее к репрессии промотора. Между редокс-балансом и балансом метилирования существует тесная взаимосвязь (Metes-Kosik N 2012).
Взаимосвязь метилирования с редокс-балансом осуществляется путем перехода гомоцистеина в антиоксидант глутатион в окислительных условиях и его перехода в SAM и метилирования в восстанавливающих условиях (Mosharov Е 2000).
Теория старения, основанная на соматических мутациях
Соматические клетки представляют собой клетки, которые при их клонировании могут вырасти до целого животного, которое стареет нормально. Соматические клетки в организмах, размножающихся половым путем, живут, чтобы поддерживать достижение ДНК зародышевых клеток следующего поколения. Известно, что соматические клетки оставляют собственную жизнь во благо организма. Одним из путей достижения этого является программируемая клеточная гибель, также называемая апоптозом, где соматическая клетка погибает организованным образом, который меньше вредит соседним клеткам, чем некротическая гибель клеток. Sirt1 ингибирует апоптоз. Показано, что Sirt2 вовлечен в регуляцию некроптоза, являющегося несколько более организованным вариантом некроза (Narayan N 2012). Некоторые вирусы, такие как вирус осповакцины, имеют антиапоптические гены, поэтому необходимы другие способы гибели клеток. Другим путем является выбраковка соматических клеток, являющихся менее жизнеспособными по сравнению с соседними клетками. При этом специфическом отборе отбираются клетки с более высокой анаболической способностью и более высоким относительным содержанием с-Мус по сравнению с соседними клетками, при этом происходит удаление относительно непригодных клеток с более низким содержанием с-Мус по сравнению с соседними клетками (Merino М 2015). Sirt1 у людей регулирует с-Мус и, следовательно, этот процесс, а также апоптоз. Следует отметить, что изменение концентрации с-Мус происходит в направлении, противоположном направлению изменений уровня ИЛ-6 (Hoffman-Liebermann В 1991). Примечание: в примере, описанном в настоящем документе, концентрации ИЛ-6 в сыворотке крови снижаются. Результаты снижения концентрации mуc у мышей привели к его предложению в качестве терапии против старения (Alic N 2015).
Аутофагия
При аутофагии образуются аминокислоты, сахара, жирные кислоты и нуклеозиды, которые претерпевают рециркуляцию для синтеза макромолекул и выработки энергии, что важно во время голодания и стресса для выживания клетки. НАД+ тесно коррелирует с аутофагией, и НАД+ и его метаболизм может влиять на аутофагию. Механизм контроля аутофагии посредством НАД+ включает метаболические пути, в которые вовлечены: а) НАД+/НАД-Н, b) НАДФ-Н, с) РАRилирование, d) деацетилирование, е) НААДФ и f) cADPR/ADPR. НАД+-зависимое деацетилирование под действием Sirt1 регулирует множественные процессы аутофагии. Катализируемые CD38 метаболиты НАД+ также вовлечены в множественные процессы аутофагии. Sirt1 регулирует аутофагию посредством р53, который играет ключевую роль в восприятии клеточного стресса, включающего повреждение ДНК и окислительный стресс. Также описана связь между р53 и некрозом (Tu Н 2009). Аутофагия представляет собой процесс саморазрушения клеточных компонентов, при котором аутофагосомы с двойной мембраной захватывают органеллы или части цитозоля и сливаются с лизосомами или вакуолями, в результате чего происходит их расщепление под действием резидентных гидролаз. Модификация механизма аутофагии, состоящая в деацетилировании, также требуется для аутофагии, и процесс деацетилирования зависит от НАД-зависимой деацетилазы Sirt1 (Не С 2009).
Связи между различными теориями старения
Различные обсуждаемые в настоящем документе теории старения взаимосвязаны между собой. Например, теория старения, основанная на ограничении калорий/сиртуине, связана с теорией старения, основанной на метилировании, посредством метилирования никотинамида. Никотинамид образуется в результате использования сиртуинами НАД+ и посредством метилирования никотинамида, поскольку метилированный никотинамид не может ингибировать сиртуины по механизму отрицательной обратной связи. PARP и CD-38 также используют НАД+ и образуют никотинамид в виде конечного продукта их реакций, который ингибирует активность PARP и сиртуина по механизму отрицательной обратной связи. Таким образом, метилирование никотинамида может препятствовать действию механизмов отрицательной обратной связи ферментов сиртуина и PARP.
Кроме того, теория старения, основанная на ограничении калорий/сиртуине, связана со свободнорадикальной (в настоящее время окислительно-восстановительной) теорией старения посредством ингибирования NF-kB, являющегося основным компонентом воспаления и иммунной защиты, под действием сиртуинов. Этот процесс приводит к увеличению триметилирования ДНК, усиливая свертывание ДНК, в результате чего снижается смертность по любой причине. Для осуществления активности сиртуина тиольные группы в активном центре сиртуина должны быть восстановлены, что непосредственно связывает две теории.
Кроме того, теория старения, основанная на ограничении калорий/сиртуине, также связана с теорией старения, основанной на соматических мутациях. В отсутствие репарации клеточных повреждений происходит их накопление, что влияет на приспособляемость клеток, и в случае падения функциональности клеток ниже критического уровня индивид умирает. Эту теорию называют теорией старения, основанной на соматических мутациях (Kennedy S 2012 and Szilard L 1959). Sirt1, как и другие сиртуины, влияет на экспрессию Мус. У млекопитающих возможен специфичный отбор клеток с высокой анаболической способностью и удаление относительно непригодных клеток на основании относительной активности Мус (Mareno Е 2014). Эта способность к отбору более приспособленных клеток и удалению непригодных клеток продлевает продолжительность жизни мух на 35% в условиях ограничения калорий (Merino М 2015).
Теория старения, основанная на метилировании, связана со свободнорадикальной (в настоящее время окислительно-восстановительной) теорией старения, что наблюдается при переходе гомоцистеина в метаболическом пути синтеза в S-аденозил-метионин (SAM), необходимый для образования 1-метилникотинамида в приведенном выше примере, под действием антиоксидантной защитной системы, но в условиях окислительного стресса происходит его ответвление в направлении синтеза антиоксиданта глутатиона.
Другие теории старения подкрепляют описанные выше теории старения, как обсуждается в тексте. Как обсуждаемая выше публикация «Теория старения, основанная на скорости жизни» (Rate of Living Theory of Aging) (Pearl R 1928, Rubner A 1908, Sohal R 2012), так и обсуждаемая выше публикация «Теория старения одноразовой сомы» (Disposable Soma Theory of Aging) (Kirkwood and Rose 1991) подкрепляют обсуждения «Ограничение калорий» и «Использование сиртуином НАД+». Как отмечено в данном разделе, «Гипотеза старения вследствие окислительно-восстановительного стресса» (Redox Stress Hypothesis of Aging) (Sohal R 2012) подкрепляет обсуждение свободнорадикальной теории. Также следует отметить, что редокс-баланс как таковой связан с воспалением. Результаты, продемонстрированные в разделе Примеров настоящего документа, где концентрации ИЛ-6 и ФНО-альфа в плазме крови снижаются, коррелируют с «Воспалительной теорией старения» (Inflammaging Theory of Aging, Franceschi С 2007, 2007, 2014), также называемой пара-воспалением (Medzhitov R 2008) и «гипотезой старения Мечникова», которая относится к проницаемости выстилки кишечника для бактерий и их продуктов, приводящей к старению (Metchnikoff Е 1901), а эта проницаемость для бактерий может быть уменьшена за счет голодания посредством метаболического пути, в который вовлечена молекула Crtc, взаимодействующая с CREB, и который связан с обсуждаемыми в настоящем документе метаболическими путями Sirt1, которые активируются голоданием. По-видимому, существует связь между атакой бактерий иммунной системой с ее атакой митохондриальных белков, таких как кардиолипин, которая необходима для активации инфламмосомы N1rp3 (Iyer SS 2013).
Клеточное повреждение является причинным фактором старения, а старение является причинным фактором «болезней старения»
Кроме старения как такового, существуют болезни старения (Goldman DP 2013). При этих болезнях старения старение является причинным фактором заболевания. Болезни старения включают: воспаление, сердечное заболевание (инфаркт миокарда и сердечную недостаточность), инсульт, нейродегенеративное заболевание, такое как болезнь Альцгеймера, сахарный диабет, рак, респираторное заболевание, системное аутоиммунное заболевание и мышечную атрофию.
Поэтому для борьбы с действиями старения необходимы новые композиции и подходы. Раскрытые в настоящем документе композиции и способы направлены на данную потребность.
Сущность изобретения
В соответствии с целями раскрытых материалов и способов, осуществляемых на практике и широко описанных в настоящем документе, раскрываемая сущность изобретения в одном аспекте относится к соединениям, композициям и способам получения и применения соединений и композиций. В конкретных аспектах раскрываемая сущность изобретения относится к композициям для борьбы с одним или более действий старения. В следующих аспектах изобретения раскрыты композиции, содержащие первое соединение, второе соединение и третье соединение, где первое соединение содержит никотинамидадениндинуклеотид (НАД+), предшественник NAD+, такой как никотинамидаденинмононуклеотид (NMN), предшественник или пролекарство NMN, никотинамид рибозид (NR), рибозид никотиновой кислоты (NAR), аденинмононуклеотид никотиновой кислоты (NaMN) (Zhou Т 2002), адениндинуклеотид никотиновой кислоты (NaAD), аналог НАД+, который стимулирует использование НАД+, такой как 1-метилникотинамид (MNM) (Hong S 2015), циклический аденинмонофосфат (цАМФ) (Wang Z 2015), где второе соединение содержит S-5'-аденозил-L-метионин (SAM), предшественник SAM, такой как метионин, бетаин, холин, фолат, витамин В12, и где третье соединение содержит активатор антиоксидантной защиты, такой как активатор эритроидного ядерного фактора 2 (Nrf2) [включая активаторы, усиливающие ядерную транслокацию Nrf2, усиливающие транскрипцию мРНК Nrf2, повышающие экспрессию белка Nrf2 и повышающие экспрессию генов-мишеней, расположенных после Nrf2 по ходу транскрипции, уменьшающих количество ингибиторов Nrf2 (таких как Bach 1, кавеолы, ФНО-бета)], такие как H2O2) генератор H2O2, сульфид водорода (H2S), донор H2S, такой как гидросульфид натрия (NaHS), сульфид натрия (Na2S), и возможно носитель.
Также раскрыты композиции, где первое соединение содержит НАД+, NMN, NR, NaMN, NaAD, NAR, MNM, цАМФ отдельно или в комбинации. Также раскрыты композиции, где первое соединение содержит NMN. Также раскрыты композиции, где первое соединение содержит предшественник или пролекарство NMN, например соединение, повышающее продукцию NMN в организме или метаболизируется до NMN. Также раскрыты композиции, где композиция вызывает снижение суррогатных маркеров старения. Также раскрыты композиции, где суррогатный маркер старения представляет собой IgG к цитомегаловирусу (CMV), С-реактивный белок, фактор некроза опухоли альфа или интерлейкин-6 в сыворотке крови. Также раскрыты композиции, где композиция содержит воду. Также раскрыты композиции, где композиция включена в препарат для инъекций. Также раскрыты композиции, где композиция представляет собой концентрированную форму для растворения в жидкости. Также раскрыты композиции, где композиция принимает форму таблетки или форму аэрозоля. Также раскрыты композиции, где композиция содержит по меньшей мере 1×10-8 моль первого соединения, по меньшей мере 1×10-8 моль второго соединения и по меньшей мере 1×10-9 моль третьего соединения.
В следующих аспектах раскрыты способы уменьшения воспаления у субъекта, включающие введение субъекту композиций, раскрытых в настоящем документе, и возможно носитель. Также раскрыты способы, в которых первое соединение, второе соединение и третье соединение вводят приблизительно в одно и то же время. Также раскрыты способы, в которых первое соединение вводят в пределах 15, 30, 60, 90 или 120 минут от пика биологических часов для НАД+. Также раскрыты способы, в которых композицию вводят субъекту в дозе, составляющей по меньшей мере 1×10-6 моль/кг первого соединения субъекту, 1×10-6 моль/кг второго соединения субъекту и 1×10-7 моль/кг третьего соединения субъекту. Также раскрыты способы, в которых композицию вводят путем инъекции в течение 8-12 дней. Также раскрыты способы, в которых композиция представляет собой аэрозоль, лиофилизат, порошок или эмульсию. Также раскрыты способы, в которых субъект представляет собой человека. Также раскрыты способы, в которых человека лечат в течение по меньшей мере двух месяцев. Также раскрыты способы, в которых композиция представляет собой таблетку, которую принимают перорально по меньшей мере раз в день. Также раскрыты способы, в которых композицию вводят раз в день.
Дополнительные преимущества будут изложены в последующем разделе описания и отчасти будут очевидными на основании описания, либо могут быть изучены путем осуществления на практике описанных ниже аспектов. Описанные ниже преимущества будут реализованы и достигнуты посредством элементов и комбинаций, конкретно указанных в прилагаемой формуле изобретения. Должно быть понятно, что и предшествующее общее описание, и последующее подробное описание имеют лишь иллюстративный и пояснительный, но не ограничивающий характер.
Подробное описание изобретения
Описанные в настоящем документе материалы, соединения, композиции и способы могут быть легче поняты со ссылкой на приведенное ниже подробное описание конкретных аспектов раскрываемой сущности изобретения и включенных в него примеров.
Прежде чем раскрыть и описать настоящие материалы, соединения, композиции и способы, необходимо понять, что описанные ниже аспекты изобретения не ограничены конкретными способами синтеза или конкретными реагентами, которые как таковые могут быть, несомненно, изменены. Также следует понимать, что используемая в настоящем документе терминология предназначена только в целях описания конкретных аспектов и не предназначена для их ограничения.
Также во всем тексте данного описания приведены ссылки на различные публикации. Описания этих публикаций в их полном объеме включены в настоящий документ заявки посредством ссылки в целях более полного описания уровня техники, к которому относится раскрываемая сущность изобретения. Раскрытые ссылки также индивидуально и конкретно включены в настоящий документ посредством ссылки на содержащийся в них материал, который обсуждается в том предложении, к которому относится ссылка.
Исследование ограничения калорий привело к открытию сиртуинов, которые активируются посредством «обедненного энергией» варианта НАД-Н, называемого НАД+. НАД-Н не используется ферментами сиртуинами и является ингибиторным лишь при концентрациях, значительно превышающих прогнозируемые концентрации для клеток. НАД-Н также не используется для образования НАДФ+ цитозольным ферментом NADK, и образованный таким путем НАДФ+ быстро преобразуется в НАДФ-Н (Pollak N 2007). Ограничение калорий вызывает «питательный стресс», приводящий в результате к обеднению запасов энергии в клетках (АТФ, НАД-Н и т.д.). «Обедненными энергией формами» этой запасенной энергии являются цАМФ и НАД+.
НАД+ активирует группу ферментов, называемых сиртуинами, а также PARP. Раскрытые в настоящем документе данные показывают, что в результате обеспечения НАД+ или соединений или композиций, обладающих сходной активностью, уменьшается количество маркеров иммунной системы, что, как показано, связано с действием, препятствующим старению. Эти данные согласуются с повышенной активацией сиртуинов посредством взаимодействия с НАД+ или действующими аналогичным образом молекулами. Однако в настоящем документе также раскрыто, что положительный эффект НАД+ может выравниваться, преимущественно в связи с другими реакциями, протекающими в организме, включая реакции в активном центре самих сиртуинов.
Поэтому посредством раскрытых способов и композиций было дополнительно показано, что в результате добавления дополнительных молекул параллельно с НАД+ или действующими аналогичным образом молекулами полезные эффекты можно продлить путем, например, непрерывного усиленного и поддерживаемого уменьшения количества маркеров воспаления, что связано с действием, препятствующим старению. Эта информация привела к разработке композиций и препаратов, содержащих три категории композиций, или к способам, в которых субъекту вводят три различные категории молекул отдельно, в сочетании или в комбинации.
Раскрыто увеличение продолжительности жизни и продолжительности здоровья путем репарации клеточного повреждения и предотвращения возрастных изменений, которые могут происходить. Представленные в настоящем документе данные показывают, что для уменьшения количества маркеров воспаления следует стремиться к достижению трех широких целей по защите от и репарации повреждений, вызванных старением:
I. НАД+ должен быть доступным для ввода в действие и использования сиртуинами,
II. доноры метила должны быть доступными для метилирования ДНК и других соединений, которые должны претерпевать метилирование, такое как реакция никотинамида до 1-метил-никотинамида под действием фермента никотинамид-N-метилтрансферазы (NNMT), и
III. восстановительный баланс должен быть обеспечен таким образом, чтобы тиольные (серосодержащие) группы в активных центрах таких важных ферментов, как сиртуины, могли поддерживаться в восстановленном состоянии.
В настоящем документе раскрыты соединения, препараты и способы, которые приводят к уменьшению количества связанных со старением маркеров воспаления и согласуются с достижением этих трех целей.
Выполнение этих трех целей возможно, если окисление в форме импульсного низкого уровня H2O2 доступно для ввода в действие прекондиционирования антиоксидантной защиты и системы репарации. В результате ввода этой системы в действие система становится защищенной от понижающей регуляции системы антиоксидантной защиты и репарации энергосберегающими механизмами. Таким образом, в случае провокации системы антиоксидантной защиты окислительной атакой в результате более обширного выброса окислителей эта система способна к защите от окисления, которое привело бы к клеточному повреждению и деструкции клетки.
В одном воплощении изобретения обеспечивают достаточное окисление H2O2, чтобы обеспечить прекондиционирование к сигналу ввода системы антиоксидантной защиты и репарации в действие, но недостаточное для того, чтобы создать окислитеьное повреждение, такое как окисление тиольных групп в активном центре сиртуина, прекращающее активность ферментов сиртуинов. Молекула АРЕ-1/ Ref-1 представляет собой молекулу, которая защищает тиольные группы аминокислот в активном центре сиртуина от окисления под действием H2O2. Это может поддерживать его в активном состоянии. Теоретически предполагают, что такой же или подобный процесс необходим для образования 1-метилникотинамида из никотинамида ферментом никотинамид-N-метилтрансферазой (NNMT) и, следовательно, остановить действие механизма обратной связи, прекращающего действие фермента сиртуина за счет перекрывания поступления никотинамида, который может встраиваться в фермент сиртуин и останавливать его активность.
Для обратного развития старения человека раскрыто полезное решение путем нормализации функции метаболических путей и механизмов эндогенной защиты и репарации у человека. Вследствие установившихся в эволюции молекулярных паттернов для приспособления к недостаточности энергии, эволюционному выбору полового размножения и защите от патогенов эти механизмы в норме функционируют как сохраняющие энергию за счет отведения более полезной энергии и ресурсов от механизмов защиты и репарации. Посредством введения раскрытых соединений, композиций и препаратов эти метаболические пути можно вернуть к нормальному функционированию для повышенной репарации и защиты.
В настоящем документе продемонстрировано, что диетический напиток NMN самостоятельно преобразуется в воде в НАД+ и активирует сиртуины у людей, но эти действия являются кратковременными. В настоящем документе продемонстрировано, что в результате гормезиса/механизма обратной связи полезные эффекты у людей действовали до тех пор, пока эти полезные эффекты не выходили на плато или не развивались в обратном направлении и даже перекрывали первоначальные полезные эффекты в пределах трехмесячной временной рамки. Данное изобретение позволяет решить эту проблему уменьшения полезного эффекта за счет ввода в действие полезных эффектов ферментов сиртуинов, оптимизации их полезных эффектов и поддержания этих полезных эффектов в активном состоянии.
В настоящем документе раскрыты соединения, композиции, препараты и способы, которые вводят в действие, усиливают и в некоторых препаратах поддерживают механизмы защиты и репарации человека, в которые вовлечены ферменты сиртуины. Эти соединения, композиции и препараты содержат одно или более веществ из каждой из трех (3) категорий отдельно или в комбинации, и их можно вводить посредством приема внутрь, инъекции, ингаляции, нанесения на кожу или любыми другими средствами.
При введении раскрытые соединения, композиции и препараты могут осуществлять по меньшей мере один из следующих видов активности:
A) Защищать от дополнительного клеточного повреждения, вызванного процессом старения
B) Осуществлять репарацию клеточного повреждения, вызванного процессом старения
C) Задерживать начало развития болезней старения, где старение является причинным фактором.
Болезни старения включают: воспаление, сердечное заболевание (включая инфаркт миокарда и сердечную недостаточность), инсульт, нейродегенеративное заболевание, такое как болезнь Альцгеймера, сахарный диабет, рак, респираторное заболевание, системное аутоиммунное заболевание (включая артрит) и мышечную атрофию.
D) Способствовать потере массы тела/уменьшению чувства голода
E) Способствовать более продуктивному сну, чтобы просыпаться более отдохнувшим
Соединения, композиции и препараты
Также раскрыты соединения, композиции и препараты, входящие в одну или содержащиеся в одной или более из следующих трех общих категорий:
Категория 1, представляющая собой активаторы системы репарации
Категория 2, представляющая собой доноры метила, и
Категория 3, представляющая собой активаторы антиоксидантной защиты
Раскрыты композиции, содержащие первое соединение, второе соединение и третье соединение, где первое соединение содержит никотинамидадениндинуклеотид (НАД+), предшественник NAD+, такой как никотинамидаденинмононуклеотид (NMN), предшественник или пролекарство NMN, никотинамид рибозид (NR), рибозид никотиновой кислоты (NAR), аденинмононуклеотид никотиновой кислоты (NaMN), адениндинуклеотид никотиновой кислоты (NaAD), аналог НАД+, который стимулирует использование НАД+, такой как 1-метилникотинамид (MNM), циклический аденинмонофосфат (цАМФ), где второе соединение содержит S-5'-аденозил-L-метионин (SAM), предшественник SAM, такой как метионин, бетаин, холин, фолат, витамин В12, и где третье соединение содержит активатор антиоксидантной защиты, такой как активатор эритроидного ядерного фактора 2 (Nrf2) [включая активаторы, усиливающие ядерную транслокацию Nrf2, усиливающие транскрипцию мРНК Nrf2, повышающие экспрессию белка Nrf2 и повышающие экспрессию генов-мишеней, расположенных после Nrf2 по ходу транскрипции, уменьшающих количество ингибиторов Nrf2 (таких как Bach 1, кавеолы, ФНО-бета)], такие как H2O2, генератор H2O2, сульфид водорода (H2S), донор H2S, такой как гидросульфид натрия (NaHS), сульфид натрия (Na2S), и возможно носитель.
Также раскрыты композиции, где первое соединение содержит НАД+, NMN, NR, NaMN, NaAD, NAR, MNM, цАМФ отдельно или в комбинации. Также раскрыты композиции, где первое соединение содержит NMN. Также раскрыты композиции, где первое соединение содержит предшественник или пролекарство NMN, например соединение, повышающее продукцию NMN в организме или метаболизируется до NMN. Также раскрыты композиции, где композиция вызывает снижение суррогатных маркеров старения. Также раскрыты композиции, где суррогатный маркер представляет собой IgG к CMV, С-реактивный белок, фактор некроза опухоли альфа или интерлейкин-6 в сыворотке крови. Также раскрыты композиции, где композиция содержит воду. Также раскрыты композиции, где композиция включена в препарат для инъекций. Также раскрыты композиции, где композиция представляет собой концентрированную форму для растворения в жидкости. Также раскрыты композиции, где композиция принимает форму таблетки или аэрозоля. Также раскрыты композиции, где композиция содержит по меньшей мере 1×10-8 моль первого соединения, по меньшей мере 1×10-8 моль второго соединения и по меньшей мере 1×10-9 моль третьего соединения.
Категория 1, активаторы системы репарации
Ввод в действие и поддержание активности сиртуина обеспечивает полезные эффекты, раскрытые в настоящем документе. Сиртуинам требуется НАД+. Обеспечение активатора системы репарации может вводить в действие сиртуины. Примеры активатора системы репарации включают НАД+, предшественник НАД+, такой как NMN, NR, NaMN, NaAD, NAR, аналог НАД+, который стимулирует использование НАД+, такой как MNM и цАМФ, или любую их комбинацию. Предпочтительным активатором системы репарации является предшественник НАД+NMN (для получения NAD+, который вводится в действие и потребляется ситруинами, что обеспечивает полезный эффект ограничения калорий). У людей НАД+ в характерном случае достигает естественных пиковых концентраций утром и вечером, например, в 8.00 и в 20.00, и поэтому добавление НАД+ или его предшественника, который будет преобразовываться в НАД+, будут осуществлять, например, преимущественно во временной рамке с 7.00 до 8.00 утра и с 19.00 до 20.00 вечера. В определенном аспекте предпочтительно желательны две суточные дозы с интервалом 12 часов, чтобы не прерывать естественный цикл биологических часов. Типичные препараты обеспечивают количество, большее или равное 1,19×10-4 моль/кг массы тела субъекта, NMN, НАД+ или предшественника НАД+, при введении (NMN составляет 334,22 грамма/моль).
Можно также вводить, в характерном случае посредством инъекции, НАД+ или применять никотинамид рибозид (NR), из которого в некоторых клетках организма может образовываться NMN. В характерном случае введение НАД+ и NR менее предпочтительно, поскольку НАД+ недостаточно хорошо абсорбируется посредством пищеварительной системы, а ферменты, которые образуют NMN из NR, обнаруживаются не в каждой клетке организма. Также показано, что при приеме NR перорально большая его часть не достигает мышц.
В конкретном аспекте раскрыто введение NMN (никотинамидмононуклеотида) людям в предпочтительных дозировках приблизительно 0,08 грамма/кг общей массы тела в сутки, разделенных на две равные дозы, которые принимают приблизительно в интервалом 12 часов. В некоторых воплощениях изобретения дозировка может быть скорректирована на абсорбцию. Активатор системы репарации, такой как NMN, предпочтительно вводят посредством воды и напитка. В дополнительных примерах можно также вводить предшественник или пролекарство NMN.
В некоторых воплощениях изобретения активаторы системы репарации вводят для уменьшения количества воспалительных маркеров, связанных со старением. При использовании в настоящем документе активаторы системы репарации представляют собой любое соединение, препарат, молекулу, биологический препарат или вещество, которое активирует ферменты ситрулины. Для этих типов ферментов предпочитают редокс-баланс около или при восстановлении. Примерами таких молекул, которые активируют сиртуин, являются НАД+, предшественник НАД+, такой как NMN, NR, NaMN, NaAD, NAR, аналог НАД+, который стимулирует использование НАД+, такой как MNM и цАМФ.
Раскрыты соединения и композиции, которые будут активировать продукцию NMN. Например, в работе Wang и соавт. обсуждается класс Р7С3 аминопропилкарбазольных химических веществ, соединений и композиций, которые действуют посредством повышения уровней над посредством его NAMPT-опосредованной реутилизации. (Wang et al. 2014)
Категория 2, доноры метила
При добавлении донора метила для метилирования предпочтительно добавление бетаина. Бетаин позволяет избежать необходимости (при использовании холина) в дополнительном количестве НАД+, если он используется, для получения S-5'-аденозил-L-метионина (SAM). SAM может обеспечивать метильную группу для никотинамида, который обладает свойствами старения за счет прекращения работы ферментов сиртуинов. Это метилирование никотинамида происходит посредством N-метилирования под действием N-метилтрансферазы (NNMT) до 1-метилникотинамида. Этот никотинамид с присоединенной метильной группой обеспечивает конкуренцию с доступными молекулами никотинамида, которые могут попадать внутрь фермента сиртуина и снижать реакционную способность фермента сиртуина; таким образом, предотвращение возникновения этого процесса пропорционально концентрации каждого из двух конкурентов. В характерном случае периодичность приема должна быть совмещена с активатором системы репарации, таким как предшественник НАД+ или НАД+.
SAM также обеспечивает метильные группы для снижения гипометилирования, наблюдаемого при старении, и в правильном контексте его можно с пользой применять для борьбы со старением, например: необходимость в метилировании H3K4me3 (Ulanovskaya OA 2013) ДНК наблюдается, в основном, у пожилых людей.
Доноры метила, которые можно использовать в дополнение к бетаину, включают SAM, метионин, холин, фолат и В12. В характерном случае эти альтернативные варианты менее предпочтительны, поскольку при приеме внутрь лишь 2% SAM попадает в организм (McMillan JM 2005); для холина требуется дополнительное количество НАД+ для его преобразования в бетаин, поступление которого в организм недостаточно.
Дозировки бетаина (триметилглицина) могут составлять по меньшей мере 0,03 грамма/кг (3×10-4 моль/кг) общей массы тела субъекта (в расчете на 0,08 грамма (из приведенного выше расчета NMN) в 0,35 раз (для молекулярно-массового отношения бетаин/NMN) =0,03 грамма/кг общей массы тела). Эту дозу можно давать в течение 24 часов и можно делить на две приблизительно равные дозы для приема с интервалом приблизительно 12 часов. Субъект может растворять дозу в воде и употреблять в виде питья. Прием можно совмещать с приемом соединения или композиции категории 1.
В некоторых воплощениях изобретения субъекту вводят доноры метилирования, причем доноры метилирования представляют собой молекулы, вещества, композиции, соединения и препараты, которые повышают метилирование молекул или сами метилируют молекулы. В характерном случае для оптимальной активности доноров метила предпочтителен редокс-баланс вблизи восстановления. Предшественники S-5'-аденозил-L-метионина (SAM) включают метионин, бетаин, холин (предшественник бетаина), фолат, витамин В12 отдельно или в комбинации.
Категория 3, активаторы антиоксидантной защиты
При обеспечении соединения, композиции или препарата категории 3 антиоксидантная защита вводится в действие. В результаты работы ферментов антиоксидантной защиты повышается восстановление тиольных (серосодержащих) групп в активном центре ферментов сиртуинов и других ферментов с подобной регуляцией. Это предотвращает прекращение работы ферментов сиртуинов вследствие окисления тиола.
Пероксид водорода (H2O2)
Одним из путей создания общей восстанавливающей среды является «шок» организма под действием импульсного выброса окислителей, таких как H2O2. Для поддержания образования антиоксидантных ферментов и поддержания их в рабочем состоянии используют прекондиционирование окислителями для осуществления шока системы и поддерживают их путем дополнительных периодических шоковых импульсов окислителей, прежде чем работа антиоксидантных ферментов прекратится вследствие механизмов их регуляции по типу обратной связи, которые прекращают или снижают их работу в отсутствие провокации окислителями. При проведении импульса окислителей для прекондиционирования используют достаточный уровень окислителей, чтобы ввести в действие и поддерживать антиоксидантные ферменты. Предпочтительным выбором окислителя для проведения прекондиционирования является пероксид водорода (H2O2) благодаря его центральной роли в метаболических путях передачи редокс-сигнала и его относительной стабильности для окислителя, а также низкому уровню потенциальных вредных эффектов по сравнению с другими окислителями, с которыми клетка имеет дело в своем жизненом цикле. H2O2 может окислять тиольные группы на белках/ферментах, в результате чего изменяются их ферментативные свойства.
Прекондиционирование низким уровнем окисления H2O2 можно проводить импульсным, контролируемым по времени и контролируемым по дозе образом, чтобы ввести в действие ферменты и процессы, при этом не вызывая избыточное окисление того, что необходимо для введения в действие ферментов антиоксидантной защиты и систем репарации, поскольку избыточное окисление вызывает клеточное повреждение и причиняет вред клетке. Любые низкомолекулярные (неферментные) антиоксиданты следует принимать в другие периоды времени (отличающиеся от периода времени окислительного импульса) так, чтобы не уменьшать этот преходящий эффект окислительного импульса.
Окисление пероксидом водорода (H2O2) и передача редокс-сигнала
Пероксид водорода (H2O2) является повсеместным окислителем, присутствующим у всех аэробных организмов (Marino HS 2014). В настоящее время H2O2 признан молекулой-мессенджером, обеспечивающей чувствительность к передасе редокс-сигнала. H2O2 приводит к окислительной модификации боковых цепей аминокислот в белках - в убывающем порядке реакционной способности и биологичесой обратимости цистеина, метионина, пролина, гистидина и триптофана. Тиольная модификация является ключевым механизмом в обнаружении и восприятии H2O2 в белках. Обнаружено, что пероксид водорода имитирует активность инсулина, вызывает релаксацию легочных артерий, стимулирует пролиферацию клеток и активирует 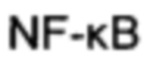 и АР-1. Функциональные последствия передачи сигнала H2O2 затрагивают фундаментальные биологические процессы. С признанием роли окислительных стимулов низкого уровня для изменения заданного значения экспрессии генов для батарей ферментов стало известно такое явление, как гормезис (Helmut Sies 2014). Транскрипционные факторы, на которые влияет H2O2, включают: АР-1, Nrf2, CREB, HSF1, HIF-1, TPSS, , NOTCH, SP1 и SCREB-1, при этом большинство из них вовлечено в регуляцию ответа на клеточное повреждение, клеточной пролиферации (регуляцию клеточного цикла), дифференцировку и апоптоз (Albrecht SC 2011).
и АР-1. Функциональные последствия передачи сигнала H2O2 затрагивают фундаментальные биологические процессы. С признанием роли окислительных стимулов низкого уровня для изменения заданного значения экспрессии генов для батарей ферментов стало известно такое явление, как гормезис (Helmut Sies 2014). Транскрипционные факторы, на которые влияет H2O2, включают: АР-1, Nrf2, CREB, HSF1, HIF-1, TPSS, , NOTCH, SP1 и SCREB-1, при этом большинство из них вовлечено в регуляцию ответа на клеточное повреждение, клеточной пролиферации (регуляцию клеточного цикла), дифференцировку и апоптоз (Albrecht SC 2011).
Ацетилирование белка регулируется H2O2 (Jung S-B 2013). Деацетилирование белков регулируется сиртуинами (Imai, S. 2000). H2O2 приводит к поышению ацетилирования, а сиртуины уменьшают ацетилирование, таким образом, эффекты H2O2 и сиртуинов участвуют в метаболических путях, форсирующих реакцию ацетилирования в противоположных направлениях. Sirt1 характеризуется высокой чувствительностью к H2O2, соответствующей 1 мкмоль внеклеточного H2O2 (Jung S-B 2013). Sirt1 защищен от тиольного окисления (APE1/Ref-1). Эта молекула управляет редокс-состоянием и активностью Sirt1. Она восстанавливает тиольные группы в активном центре Sirt1, а H2O2 окисляет тиолы в активном центре Sirt1. Sirt1 также регулируется редокс-зависимым фосфорилированием (Caito, S. 2010).
Потребность в импульсе сигнальных оксидантов
Низкие уровни H2O2 повышают защиту за счет прекондиционирования и, следовательно, в конечном счете могут защищать от увеличения доли окисленных тиолов в активном центре сиртуина и снижения активности Sirt1 за счет окислительной провокации. Адаптация к H2O2 уменьшает проницаемость плазматических мембран для H2O2. Различные клеточные мембраны характеризуются полным диапазоном проницаемости для H2O2. Аквапорины также регулируют транспорт H2O2 через биологические мембраны (Marinho HS 2014).
Традиционные лекарственные средства, изменяющие уровни H2O2
Наиболее широко назначаемое во всем мире противодиабетическое лекарственное средство метформин повышает уровни пероксида водорода (H2O2); это приводит к повышающей регуляции пероксиредоксина-2 (PRDX-2). Метформин увеличивает продолжительность жизни С. elegans, а удаление гена PRDX-2 устраняет этот эффект. PRDX-2, по-видимому, играет роль в трансляции окислительного стресса в последующий сигнал, способствующий увеличению продолжительности жизни. Воздействие N-ацетилцистеина (NAC) и бутилгидроксианизола (ВНА), которые являются низкомолекулярными антиоксидантами, устраняло положительное влияние метформина на продолжительность жизни (De Haes W 2014). Фармацевтические средства, повышающие концентрацию пероксида водорода в организме, можно также применять для этой категории либо в дополнение к H2O2, либо в качестве замены добавления самого пероксида водорода. Фармацевтические средства, повышающие концентрацию H2O2 в организме, включают метформин (De Haes W 2014) и ацетаминофен (Hinson J 2010).
Фармацевтические средства, повышающие концентрацию H2O2 в организме, необходимо также включать в расчет окислительного импульса, который обеспечивает категория №3. Примером является ацетаминофен (ингредиент препарата Тайленол), который представляет собой известное фармацевтическое средство, повышающее концентрацию H2O2 в организме (Hinson J 2010). N-ацетил-I-цистеин (NAC) представляет собой известное соединение, отвечающее за многие эффекты H2O2 в организме.
Периодичность, продолжительность действия и уровни H2O2
Достаточное окисление, чтобы обеспечить прекондиционирование для сигнала ввода системы антиоксидантной защиты и репарации в действие, является желательным; но оно недостаточно для того, чтобы создать окислитеьное повреждение, такое как окисление тиольных групп в активном центре сиртуина, прекращающее активность ферментов сиртуинов. Этот уровень называют «зоной умеренности». Молекула АРЕ-1/ Ref-1 представляет собой молекулу, которая защищает тиольные группы ферментов сиртуинов, которые должны оставаться активными. Такой же или подобный процесс теоретически предполагают для фермента никотинамид-N-метилтрансферазы (NNMT).
В некоторых воплощениях изобретения можно транзиторно добавлять импульсные низкие уровни пероксида водорода (H2O2) человеку для прекондиционирования систем антиоксидантной защиты и репарации к вводу в действие и поддержанию. В некоторых предпочтительных воплощениях изобретения предпочтительна концентрация 100 мкмоль/л марки, пригодной для пищевых продуктов (коммерческая марка содержит ацетанилид в качестве стабилизатора) H2O2 в 400-500 мл воды на индивидуальную дозу, которую можно принимать отдельно или с соединениями или композициями категории 1 и категории 2. 1 моль H2O2=1+1+16+16= приблизительно 34 грамма. Согласно расчетам 50% H2O2 абсорбируется в кишечнике, поэтому более предпочтительная концентрация для приема составляет приблизительно 200 мкмоль/л (в 500 мл). Например, в некоторых воплощениях изобретения одна капля H2O2 соответствует 0,05 мл. H2O2 марки, пригодной для пищевых продуктов, поставляется в концентрациях 35%. Прием 2 капель 35% H2O2 в 500 мл дистиллированной воды (с каждой дозой/сутки) обеспечивает концентрацию приблизительно 200 мкмоль/л. Распад H2O2 составляет около 10%/год в отсутствие света и загрязняющих веществ в деионизированной/дистиллированной H2O. H2O2 замерзает при -11°С. Поэтому в некоторых воплощениях изобретения прием 4 капель/день или 0,2 мл 35% H2O2/день в 1 литре воды. 35 граммов/100 мл =0,07 граммов/0,2 мл. В некоторых воплощениях изобретения можно применять дозировки в количестве приблизительно 0,0008 граммов H2O2/кг общей массы тела.
Предпочтительным способом применения является прием внутрь H2O2 путем растворения H2O2 в деионизированной/дистиллированной воде и питья. Применение характеризуется такой же предпочтительной периодичностью концентрации дозы, временем приема и длительностью приема, как №1 и №2 при растворении в воде. В некоторых воплощениях изобретения, если H2O2 частично повышается в результате физических упражнений на выносливость, эти упражнения выполняют непосредственно перед приемом или после приема.
Применение метформина (De Haes W 2014) можно осуществлять в жидкой форме препарата Риомет, а также в таблетках. В жидкой форме 5 мл эквивалентно таблетке 500 мг. Он достигает максимальных концентраций в плазме крови за 1-3 часа в форме немедленного высвобождения, а стабильного состояния - в течение от одного до двух дней. В характерном случае его биодоступность в состоянии натощак составляет от 50 до 60%. Эти данные необходимо использовать для расчета соответствующего времени приема и дозы метформина.
Сульфид водорода (H2S)
Помимо использования пероксида водорода (H2O2) для прекондиционирования системы антиоксидантной защиты, как обсуждалось выше, другой путь изменения редокс-потенциала чувствительных к окислению тиолов белков состоит в непосредственном усилении системы антиоксидантной защиты сульфидом водорода (H2S). Воздействие NaSH (донор H2S) (0,025-0,1 миллимоль/литр) дозозависимо противостоит воздействию H2O2. У людей в возрасте от 50 до 80 лет концентрации H2S в плазме крови снижаются (Chen Y 2005), а концентрации H2S в плазме крови у пациентов с сердечно-сосудистым заболеванием (ССЗ) характеризуются статистически значимой обратной корреляцией с тяжестью ССЗ и изменениями в коронарных артериях (Jiang Н 2005). NaSH уменьшает количество АФК и усиливает экспрессию супероксиддисмутазы (SOD), глутатионпероксидазы (GPx) и глутатион-S-трансферазы (GST). В образцах плазмы крови здоровых добровольцев при употреблении обогащенной H2S воды (500 мл/день в течение 2 недель) количество продуктов распада липидов и белков статистически значимо уменьшалось (Benedetti S 2009). 0,1 ммоль NaSH/литр может вызывать повышение Sirt1 зависимым от времени образом (Wu D 2015). Экзогенный H2S обладает защитным эффектом для поддержания циркадного ритма часовых генов посредством изменения отношения НАД+/НАД-Н и усиления белка Sirt1 (Shang Z 2012). H2S также является важным эндогенным ингибитором ключевых элементов острых воспалительных реакций (Zanardo R 2006) за счет понижающей регуляции экспрессии NF-kB или повышающей регуляции экспрессии гемоксигеназы 1 (Jin Н 2008, Kim K 2008, Oh G 2006, Pan L 2011). H2S может активировать АТФ-чувствительные калиевые каналы промежуточной проводимости и малой проводимости посредством S-сульфгидрирования цистеина (Mustafa А 2011, Yang G 2008), что вызывает гиперполяризацию эндотелиальных и гладкомышечных клеток, интернирование которых вызывает вазорелаксацию эндотелия сосудов и снижение артериального давления. H2S обладает прямым ингибиторным эффектом в отношении активности ангиотензинпревращающего фермента (АПФ) (Laggner Н 2007). NaSH повышает экспрессию эндотелиальной NO-синтазы (eNOS) и альфа субъединицы протеинкиназы С-1 (PGC-1Alpha) (Wu D 2015), которые оба играют роль в биогенезе и функции митохондрий (Wu, СС 2013, Lagouge М 2006). H2S осуществляет повышающую регуляцию MAPK (Barr LA 2014, Papapetropoulos А 2009, Yong QC 2008). Предположили, что ограничение калорий может способствовать поддержанию передачи сигнала H2S (Predmore В 2010). Донор H2S медленного высвобождения GYY4237 может зависимым от концентрации образом ингибировать семь различных линий раковых клеток человека (Lee Z 2011). Сульфорафан, также донор H2S, обладает дозозависимыми свойствами против рака предстательной железы (РС-3) (Pei Y 2011).
H2S является газотрансмиттером. Газотрансмиттеры продуцируются эндогенно в низких концентрациях и способны свободно диффундировать через клеточные мембраны, вызывая передачу сигналов на клеточном уровне (Calvert JW 2010). Тремя газотрансмиттерами являются оксид азота (NO), монооксид углерода (СО) и сульфид водорода (H2S).
Сульфид водорода синтезируется из L-цистеина. Эндогенными ферментативными источниками сульфида водорода (H2S) являются цистатион-гамма-лиаза (CSE), цистатионин-бета-синтаза (CBS), цистеинаминотрансфераза (CAT) и 3-меркаптопируватсульфотрансфераза (MST). Для этих ферментов показана некоторая степень продукции H2S в печени, и показано, что H2S регулирует перекисное окисление липидов и активность антиоксидантных ферментов (GPx, T-SOD, Cu/Zn-SOD и Mn-SOD) в печени в результате введения донора H2S NaSH мышам путем инъекции 0,05 ммоль/л NaSH/кг массы тела/сутки, растворенного в 10 мл/кг массы тела физиологического раствора (Wu D 2015). Митохондрии способны использовать H2S в условиях гипоксии и стресса для продукции АТФ (Fu М 2012).
Первоначальные сообщения об антиоксидантной активности H2S относились к тому, что H2S может захватывать супероксид (Geng В 2004), и H2S может осуществлять повышающую регуляцию глутатиона (Kimura Y 2004). Впоследствии получены более детальные сообщения о его активации антиоксидантных ферментов. Показано, что H2S активирует фактор 2, родственный эритроидному ядерному фактору 2 (Nfr2) (Peake BF 2013), который вводит в действие антиоксидантные гены. Ежедневное введение Na2S в течение 7 дней приводило к повышению экспрессии Nrf2 как в цитозольной, так и в ядерной фракциях (Calvert JW 2010). Nrf2, который осуществляет повышающую регуляцию экспрессии генов, регулируемых элементом ответа антиоксидантов, претерпевает повышающую регуляцию под действием H2S (Islam KN 2015). Активация H2S вызывает отделение Nrf2 от его прикрепленного ингибитора, Kelch-подобного ЕСН-ассоциированного белка 1, в цитозоле (Wakabayashi N 2004) с последующим перемещением в ядро и связыванием со специфичной энхансерной последовательностью, известной как элемент антиоксидантного ответа, в промоторной области антиоксидантных генов, включая НО-1 и тиоредоксин 1 (Calvert JW 2009). H2S оказывает влияние на функцию митохондрий (Helmy N 2014, Wang CN 2014), окислительный стресс (Bos ЕМ 2013, Du JT 2013), апоптоз (Yao LL 2010), воспаление (Lo Faro ML 2014), ангиогенез (Szabo С 2013, Coletta С 2012, Wang MJ 2010), сепсис и шок (Kida, F. 2015) и артериальное давление (Polhemus DJ 2014, Ge SN 2014, Yang G 2008).
H2S, как и глутатион, защищает от NO3-. H2S также статистически значимо снижает токсические эффекты HOCl. H2S усиливает антиоксидантные эффекты N-ацетил-1-цистеина (NAC).
Терапевтические эффекты H2S к настоящему времени лучше всего исследованы для заболеваний сердца. Влияние H2S на заболевания сердца включает следующее: макрофаги способны к эндогенной продукции H2S (Zhu XY 2010). Ингибированные NaHS (донор H2S) проатерогенные окисленные липопротеины низкой плотности вызывали образование пенистых клеток в макрофагах (Wang Y 2009). H2S способен к понижающей регуляции АФК в митохондриях, обеспечивая защиту за счет пониженного дыхания (Chen Q 2006). Продукция H2S (10-100 нмоль/л) усиливала митохондриальный транспорт электронов и клеточную биоэнергетику (Modis K 2013), однако в высоких концентрациях H2S токсичен (Hill ВС 1984, Nicholls Р 1982). H2S в рационе питания вызывал уменьшение нежелательного ремоделирования левого желудочка (ЛЖ) при сердечной недостаточности (Kondo K 2013). H2S может осуществлять повышающую регуляцию эндотелиальной синтазы оксида азота, образующей NO (Kondo K 2013), a NO может осуществлять повышающую регуляцию фермента синтеза H2S - цистотионин-γ-лиазы (CSE) (Zhao, W. 2001). У мышей, которым вводили донор H2S, статистически значимо повышалось фосфорилирование, действующее на eNOS, что позволяет предположить активное перекрестное взаимодействие H2S и NO (Kondo K 2013). По-видимому, перекрестное взаимодействие также существует между СО и H2S (Zhange QY 2004, Majid AS 2013). H2S вызывает расширение кровеносных сосудов, приводящее к снижению артериального давления (Cheng Y 2004). H2S в форме Na2S (при введении 10 минутами раньше) предотвращал реперфузионное повреждение (Sodha NR 2008). Экзогенный H2S также приводил к улучшению функции почек (Xu Z 2009).
В условиях in vivo H2S характеризуется крайне коротким периодом полувыведения, который оценивают как составляющий от нескольких секунд до нескольких минут (Wang R 2002, Insko MA 2009). Концентрации H2S в плазме крови находятся в диапазоне от 0,034 до 0,065 ммоль/л (Whiteman М 2009), и в головном мозге они в три раза выше, чем в плазме крови (Hogg Р 2009, Zhao W 2001). Концентрация H2S обратно пропорциональна концентрации O2, и H2S уменьшает потребление O2 клеткой (Olson K 2015). Также описано, что концентрации H2S в крови и плазме крови составляют 0,030 и 0,300 (Olson K 2009). Доноры H2S NaHS и Na2S повышают концентрацию H2S в пределах от нескольких секунд до нескольких минут.
Физиологический диапазон H2S изменяется в широких пределах от 0,005 до 0,300 ммоль/л (Predmore BL 2012). Эндогенные концентрации H2S в головном мозге человека определены как составляющие от 0,05 до 0,16 ммоль/л (Whiteman М 2004); в головном мозге пациентов с болезнью Альгеймера концентрация H2S снижена (Seshadri S 2002, Tang X 2010). Диаллилтрисульфид (DATS) является стабильным донором H2S и проявляет продолжительные эффекты через 30 минут после инъекции. NaHS можно принимать с питьевой водой (Givvimani S 2011). NaHS (донор H2S) в водном растворе высвобождает H2S в питьевую воду в течение 6 недель. При добавлении извне отмечено повышение концентрации H2S в плазме крови (Peake BF 2013, Kondo K 2013). Различие в потреблении воды между группами мышей, которым вводили NaHS, и группами, которым не вводили препарат, отсутствовало. Другие доноры H2S включают GYY 4137 (CAS №106740-09-4), водорастворимый донор H2S, который медленно высвобождает H2S в течение нескольких часов (Li L 2008), и SG 1002 от компании Sulfagenix, Inc. АР97, АР39, АР67 и АР105 также являются донорами H2S медленного высвобождения (Whiteman М 2015, Wallace J 2015, Hancock J 2014). H2S может поступать с пищей, содержащей органические сульфиды, полисульфиды которых могут быть донорами H2S.
В дополнение к приему H2S внутрь в растворенном в воде виде H2S можно вводить путем ингаляции, и в результате ингаляции концентрации H2S в крови повышаются (мышам вводили 40 частей на миллион (ч/млн) за 8 часов в течение 7 дней). Ингаляцию можно также комбинировать с донорами H2S, принимаемыми внутрь, такими как Na2S и NaHS (Kida K 2011 and 2015). Количественное определение H2S в крови и в тканях проведено чувствительными и надежными методами (Wintner Е 2010).
H2S может также запасаться в клетках в форме сульфна серы и транспортироваться и высвобождаться в ответ на физиологический стимул (Ishigami М. 2009).
Активаторы NRF2
Транскрипционный фактор NF-E2 р45-родственный фактор 2 (Nrf2: название гена NFE212) регулирует экспрессию сетей генов, кодирующих белки с различными цитопротективными активностями. Регуляция самого Nrf2 осуществляется главным образом на уровне стабильности белка. Nrf2 является короткоживущим белком, подвергающимся постоянному убиквитинированию и расщеплению протеазами. Известно три системы убиквитинлигазы, которые вносят вклад в деградацию Nrf2: а) Keap-1, адаптерный белок субстрата для куллина-3, b) киназа гликогенсинтазы и с) Е3-убиквитинлигаза Hrd1. Keap-1 также является сенсором для широкого ряда низкомолекулярных активаторов, также называемых индукторами. Когда Nrf2 не претерпевает деградацию и транслоцируется в ядро, он образует димер с низкомолекулярным белком Maf, связывается с элементами антиоксидантного ответа, представляющим собой расположенные выше регуляторные участки его генов-мишеней и инициирует транскрипцию. Nrf2 является основным регулятором клеточного редокс-гомеостаза. (Dinkova-Kostova AT 2015). У человека регуляция более 50 генов осуществляется под действием Nrf2 (Pall ML 2015, Choi B-H 2014). При прямом действии генов воспаления без редокс-механизма Nrf2 также связывается с расположенным выше участком гена ИЛ-6 и при связывании может в значительной степени прерывать рекрутмент РНК-полимеразы II для регуляции транскрипции ИЛ-6 в макрофагах человека.
Регуляция передачи сигнала Nrf2 осуществляется посредством транскрипционных, трансляционных, посттрансляционных и эпигенетических механизмов, а также под действием других белков-партнеров, включающих р62, р21 и IQ мотив-содержащий активирующий ГТФазу белок 1 (Huand Y 2015). Активаторы эритроидного ядерного фактора 2 (Nrf2) включают классы активаторов, обладающих активностями, которые: вызывают ядерную транслокацию Nrf2, повышают транскрипцию мРНК Nrf2, повышают экспрессию белка Nrf2 и повышают нижележащие гены-мишени Nrf2. Существуют также ингибиторы Nrf2 (Bach 1, кавеолы, трансформирующий фактор роста бета-1 (TGF-beta)) (Gegotek А 2015). Метаболический путь Keap1-Nrf2 кооперативно взаимодействует с аутофагией по борьбе с протеотоксичностью (Dodson М 2015).
Keap-1 представляет собой цинковый металлопротеин, локализованный около плазматической мембраны. Он имеет три функциональных домена, по меньшей мере 25 реакционных тиолов, большая часть из которых обнаружена в промежуточном линкерном участке. Keap-1 имеет сайт связывания Nrf2 на каждой субъединице димера, образуя структуру типа «петли и крюка». Keap-1 обладает высокой чувствительностью к окислению, и его различные тиольные группы имеют различные редокс-потенциалы. Три различных цистеиновых остатка создают сенсорную систему (Suzuki Т 2013).
Nrf2 представляет собой 605-аминокислотный транскрипционный фактор, состоящий из шести доменов. N-концевой домен Neh2 представляет собой сайт связывания ингибиторного белка Keap-1. Период полураспада Nrf2 при отделении от Кеар-1 составляет 20 минут (Kasper JW 2011). Keap-1 экспортируется из ядра за 0,5 часа. Активация Nrf2 усиливает активность Sirt1 в клеточной культуре фибробластов мышей (Jodar L 2010).
При высвобождении Keap-1 из Nrf2 он становится доступным для захвата киназным комплексом IKKBeta, что, таким образом, приводит к ингибированию генов-мишеней . Это взаимодействие коррелирует с экспрессией антиоксидантных ферментов под действием Nrf2 и с вводом в действие и выводом из действия иммунной системы под действием NF-kB. Nrf2 и NF-kB конкурируют за CREB-связывающий белок (СВР) (Liu GH 2008). Существует много фитохимических веществ, обладающих способностью к активации Nrf2 посредством взаимодействия с Keap-1 различными путями. Немедленные алкилирующие агенты являются быстро активирующими. «Акцепторы Михаэля», представляющие собой соединения ацетилена, конъюгированные с электроноакцепторной группой, образуют обратимые реакции алкилирования с тиолами-сенсорами Keap-1.
Фенольные соединения, которые, по-видимому, наиболее прямым путем действуют на Nrf2, представляют собой орто- или пара-дигидроксифенолы, которые могут окисляться до хинонов (Kumar Н 2014). Хиноны представляют собой окисленные производные ароматических соединений, и их чаще всего можно легко получить из реакционноспособных ароматических соединений с электронодонорными заместителями, таких как фенолы и катехолы, которые повышают нуклеофильность кольца и вносят вклад в высокий редокс-потенциал, необходимый для разрушения ароматичности. Хиноны являются конъюгированными, но неароматическими. Хиноны представляют собой электрофильные акцепторы Михаэля, стабилизированные конъюгацией. В зависимости от хинона и от центра восстановления восстановление может привести либо к восстановлению ароматичности соединения, либо к разрушению конъюгации. Присоединение конъюгата почти всегда разрушают конъюгацию.
H2O2 и H2S являются активаторами Nrf2 (отдельно указанными выше). Всегда при упоминании активатора Nrf2 он также является активатором системы антиоксидантной защиты, хотя некоторые активируемые Nrf2 соединения можно рассматривать как дополнительные для активации системы антиоксидантной защиты. Активация происходит многочисленными путями, перечисленными выше для поддержания системы Nrf2 в действии. Одной из форм регуляции Nrf2 является обратимое фосфорилирование. Sirt1 и PARP1, как обсуждалось выше, могут также претерпевать обратимое фосфорилирование.
Для активации Nrf2 и ввода в действие системы антиоксидантной защиты необходима корреляция по времени с доступностью НАД+ и доступностью метилирования и синхронизация с пиками НАД+ биологических часов человека. Действие системы Nrf2 действительно необходимо прекращать (пример: около 2 пмоль, когда концентрации НАД+ обычно находятся в нижней точке его суточных биологических часов) так, чтобы организм мог выполнять необходимые ему функции при редокс-балансе, который впоследствии склоняется в сторону окисления.
Соединения категории 3
Активаторы антиоксидантной защиты, такие как активаторы эритроидного ядерного фактора 2 (Nrf2) (включая активности, такие как: ядерная транслокация Nrf2, повышающая транскрипцию мРНК Nrf2, повышенная экспрессия белка Nrf2 и повышение экспрессии нижележащих генов-мишеней Nrf2), H2O2, ROS, RNS, RCS, RSOH,  , O2, H2S, O3, HOCl, HOBr, HOI, ROOH, где R представляет собой алкил, циклоалкил, гетероалкил, гетероциклоалкил, алкенил, гетероалкенил, циклоалкенил или гетероциклоалкенил, генератор H2O2, такой как метформин или ацетаминофен, орто-гидроксифенолы, которые могут окисляться до хинонов (Kumar Н 2014), пара-дигидроксифенолы, которые могут окисляться до хинонов (Kumar Н 2014), хиноны (окисленные производные ароматических соединений), сульфид водорода (H2S), донор сульфида водорода (H2S), такой как NaHS, сульфид натрия (Na2S), диаллилтрифульфид (DATS), GYY4137 (водорастворимый донор H2S (патент №WO 2014018569 А1) (Li L 2008)), SG-1002 (синтетический донор H2S от компании SulfaGENEX) (Kondo K 2013), доноры H2S на основе пеницилламина (Zhao Y 2013), полиорганосульфиды (Tocmo R 2015), 2-меркаптоэтанол, дитиотрейтол, изотиоцианаты, сульфорафан (в брокколи) (Nallasamy Р 2014), глюкорафанин (в брокколи) (Armah CN 2013), куркумин (в куркуме) (Рае Н-O 2007, Не HJ 2012, Balogun Е 2003, Goel А 2007), пирролидон (водорастворимый), теракумин (наночастицы), церумбон (Stefanson AL 2014), аналоги циннамата, содержащие тиокетон-конъюгированную альфа, бета-ненасыщенную группировку (Kumar S 2013), подобные альдегиду коричной кислоты, кверцетин (в луке, яблоках, чае) (Magesh S 2012, Kimura S 2009), изокверцетин (в 2-6 раз лучшая абсорбция), кемпферол (Kang BY 2008), женьшень (Panax ginseng и Panax quinquefolius), карнозиновая кислота, ксантогумол, Dh404, (R)-альфа-липоевая кислота (Flier J 2002, Suh JH 2004, Cao Z 2003), изотиоцианат, бензилизотиоцианат (Sahu RP 2009), неоглюкобрассицин (Stefanson AL 2014), глюкозинолаты (Stefanson AL 2014), гидрофильные окисленные производные ликопина (Stefanson AL 2014), (HNE) 4-гидроксиноненаль (Forman HJ 2008), (15-dPGJ2) 15-деоксидельтапростагландин J2 (Mochizuki M 2005), фалькариндиол (Stefanson AL 2014), гидрокситирозол (Stefanson AL 2014), бета-глюкан ячменя, спермидин (Kwak MK 2003), спермин (Kwak MK 2003), лютеолин (Paredes-Gonzalez X 2015), 4-метилалкилкатехол, 4-винилкатехол, 4-этилкатехол, пирролохинолинхинон (Zhang Q 2012, Liang С 2015), мангафодипир тринатрий (MnDPDP) (контрастный агент, используемый в настоящее время в магнитно-резонансной томографии) (Mosbah IB 2012), N-ацетилцистеин (Wallace J 2015), АТВ-346 от Antibe Therapeutics (Wallace J 2015), NBS-1120 от Колледжа Нью-Йорка (Wallace J 2015), GIC-101 от GI care Pharma (Wallace J 2015), AP39, номер патента WO 2013045951 A1 Эксетерского университета, Alos АР67, АР 97 и АР105, WO 2014018569 A1, сиалор (Wallace J 2015), сульфарлем (Wallace J 2015) и анетола тритион (Wallace J 2015), DHEA (Jeon S 2015), каменноугольная смола (Van den Bogaard EH 2013), чеснок (посредством H2S), β-лапачо (из древесной коры южноамериканского дерева: он осуществляет окисление посредством циклизации клеточного НАД-Н до НАД+), птеростильбен (McCormack D 2013, ресвератрол (Cheng L 2015, Mokni М 2007, Kitada М 2011, апигенин (в петрушке огородной) (Paredes-Gonzalez X 2015 and 2014, Escande С 2013), цинк (Wang F 2015, Sternberg P 2007, Magesh S 2012) и возможно носитель.
, O2, H2S, O3, HOCl, HOBr, HOI, ROOH, где R представляет собой алкил, циклоалкил, гетероалкил, гетероциклоалкил, алкенил, гетероалкенил, циклоалкенил или гетероциклоалкенил, генератор H2O2, такой как метформин или ацетаминофен, орто-гидроксифенолы, которые могут окисляться до хинонов (Kumar Н 2014), пара-дигидроксифенолы, которые могут окисляться до хинонов (Kumar Н 2014), хиноны (окисленные производные ароматических соединений), сульфид водорода (H2S), донор сульфида водорода (H2S), такой как NaHS, сульфид натрия (Na2S), диаллилтрифульфид (DATS), GYY4137 (водорастворимый донор H2S (патент №WO 2014018569 А1) (Li L 2008)), SG-1002 (синтетический донор H2S от компании SulfaGENEX) (Kondo K 2013), доноры H2S на основе пеницилламина (Zhao Y 2013), полиорганосульфиды (Tocmo R 2015), 2-меркаптоэтанол, дитиотрейтол, изотиоцианаты, сульфорафан (в брокколи) (Nallasamy Р 2014), глюкорафанин (в брокколи) (Armah CN 2013), куркумин (в куркуме) (Рае Н-O 2007, Не HJ 2012, Balogun Е 2003, Goel А 2007), пирролидон (водорастворимый), теракумин (наночастицы), церумбон (Stefanson AL 2014), аналоги циннамата, содержащие тиокетон-конъюгированную альфа, бета-ненасыщенную группировку (Kumar S 2013), подобные альдегиду коричной кислоты, кверцетин (в луке, яблоках, чае) (Magesh S 2012, Kimura S 2009), изокверцетин (в 2-6 раз лучшая абсорбция), кемпферол (Kang BY 2008), женьшень (Panax ginseng и Panax quinquefolius), карнозиновая кислота, ксантогумол, Dh404, (R)-альфа-липоевая кислота (Flier J 2002, Suh JH 2004, Cao Z 2003), изотиоцианат, бензилизотиоцианат (Sahu RP 2009), неоглюкобрассицин (Stefanson AL 2014), глюкозинолаты (Stefanson AL 2014), гидрофильные окисленные производные ликопина (Stefanson AL 2014), (HNE) 4-гидроксиноненаль (Forman HJ 2008), (15-dPGJ2) 15-деоксидельтапростагландин J2 (Mochizuki M 2005), фалькариндиол (Stefanson AL 2014), гидрокситирозол (Stefanson AL 2014), бета-глюкан ячменя, спермидин (Kwak MK 2003), спермин (Kwak MK 2003), лютеолин (Paredes-Gonzalez X 2015), 4-метилалкилкатехол, 4-винилкатехол, 4-этилкатехол, пирролохинолинхинон (Zhang Q 2012, Liang С 2015), мангафодипир тринатрий (MnDPDP) (контрастный агент, используемый в настоящее время в магнитно-резонансной томографии) (Mosbah IB 2012), N-ацетилцистеин (Wallace J 2015), АТВ-346 от Antibe Therapeutics (Wallace J 2015), NBS-1120 от Колледжа Нью-Йорка (Wallace J 2015), GIC-101 от GI care Pharma (Wallace J 2015), AP39, номер патента WO 2013045951 A1 Эксетерского университета, Alos АР67, АР 97 и АР105, WO 2014018569 A1, сиалор (Wallace J 2015), сульфарлем (Wallace J 2015) и анетола тритион (Wallace J 2015), DHEA (Jeon S 2015), каменноугольная смола (Van den Bogaard EH 2013), чеснок (посредством H2S), β-лапачо (из древесной коры южноамериканского дерева: он осуществляет окисление посредством циклизации клеточного НАД-Н до НАД+), птеростильбен (McCormack D 2013, ресвератрол (Cheng L 2015, Mokni М 2007, Kitada М 2011, апигенин (в петрушке огородной) (Paredes-Gonzalez X 2015 and 2014, Escande С 2013), цинк (Wang F 2015, Sternberg P 2007, Magesh S 2012) и возможно носитель.
Конкретные композиции
В конкретных примерах раскрытая питательная композиция может содержать никотинамидадениндинуклеотид (НАД+), бетаин и H2O2. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), фолат плюс витамин В12 и H2O2. В конкретных примерах раскрытая питательная композиция может содержать никотинамидадениндинуклеотид (НАД+), метионин и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), метионин и H2O2. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), холин и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), бетаин и NaHS. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), фолат плюс витамин В12 и NaHS. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), метионин и NaHS. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), холин и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), бетаин и Na2S. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), фолат плюс витамин В12 и Na2S. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), метионин и Na2S. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), холин и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), бетаин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), фолат плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), метионин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытая композиция может содержать никотинамидадениндинуклеотид (НАД+), холин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, теростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая питательная композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR) с бетаином и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, фолат плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), фолат плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолат плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), фолат плюс витамин В12 и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин плюс витамин В12 и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин плюс витамин В12 и H2O2.
В конкретных примерах раскрытая питательная композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, метионин и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), метионин и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR) с метионином и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), метионин и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, холин и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), холин и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холин и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), холин и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, S-аденозил-метионин (SAM) и H2O2. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), S-аденозил-метионин (SAM) и H2O2. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионин (SAM) и H2O2. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), S-аденозил-метионин (SAM) и H2O2.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, фолат плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), фолат плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолат плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), фолат плюс витамин В12 и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин плюс витамин В12 и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин плюс витамин В12 и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, метионин и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), метионин и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионин и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), метионин и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, холин и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), холин и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холин и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), холин и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, S-аденозил-метионин (SAM) и NaHS. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), S-аденозил-метионин (SAM) и NaHS. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионин (SAM) и NaHS. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), S-аденозил-метионин (SAM) и NaHS.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, фолат плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), фолат плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолат плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), фолат плюс витамин В12 и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин плюс витамин В12 и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин плюс витамин В12 и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, метионин и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), метионин и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионин и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), метионин и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, холин и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), холин и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холин и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), холин и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, S-аденозил-метионин (SAM) и Na2S. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), S-аденозил-метионин (SAM) и Na2S. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионин (SAM) и Na2S. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), S-аденозил-метионин (SAM) и Na2S.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN) или предшественник или пролекарство NMN, бетаин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), бетаин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN), фолат плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать никотинамида рибозид (NR), фолат плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолат плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), фолат плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN), бетаин плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать никотинамида рибозид (NR), бетаин плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаин плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), бетаин плюс витамин В12 и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN), метионин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), метионин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), метионин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN), холин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать никотинамид рибозид (NR), холин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), холин и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытая композиция может содержать никотинамидмононуклеотид (NMN), S-аденозил-метионин (SAM) и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытая композиция может содержать никотинамида рибозид (NR), S-аденозил-метионин (SAM) и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать одно или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионин (SAM) и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытая композиция может содержать 1-метилникотинамид (MNM) и/или циклический аденозинмонофосфат (цАМФ), S-аденозил-метионин (SAM) и любое одно или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В раскрытых композициях объединенное количество соединений категории 1, 2 и 3 в композиции может составлять по меньшей мере 5 масс. % композиции. Например, активатор системы репарации, донор метила и активатор антиоксидантной защиты могут составлять по меньшей мере 5 масс. % композиции. В другом примере объединенное количество соединений категории 1, 2 и 3 в композиции может составлять по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 55, по меньшей мере 60, по меньшей мере 65, по меньшей мере 70, по меньшей мере 75, по меньшей мере 80, по меньшей мере 85, по меньшей мере 90, по меньшей мере 95 или 100 масс. % композиции, где любая из указанных выше величин может составлять верхнюю или нижнюю конечную точку диапазона.
Система доставки ингредиентов категории 1, 2 и 3
Раскрыты препараты, которые могут быть упакованы в форме порошка или лиофилизированной форме, которую впоследствии добавляют горячую или холодную жидкость для восстановления в виде раствора. Например, раскрытые композиции могут быть смешаны с композициями, такими как композиции в персональных системах напитков, из отдельно упакованных компонентов которых и добавления воды готовят горячий или холодный кофе или чай или горячий шоколад. Раскрытые композиции можно вводить in vivo, либо отдельно, либо в фармацевтически приемлемом носителе. Под «фармацевтически приемлемым» подразумевают материал, не являющийся нежелательным ни с биологической, ни с иной точки зрения, т.е. этот материал можно вводить субъекту наряду с раскрытой в настоящем документе композицией, не вызывая каких-либо нежелательных биологических эффектов. Носитель, естественно, должен быть выбран так, чтобы свести к минимуму какое-либо разложение активного ингредиента и свести к минимуму любые нежелательные побочные эффекты у субъекта, как должно быть хорошо известно специалистам в данной области техники. Эти материалы могут находиться в растворе, суспензии (например, включен в микрочастицы, липосомы или клетки).
Взаимодействие с микробиомом при доставке посредством пищеварительного тракта или кожи
Микробиоту кишечника млекопитающих составляют вплоть до 100 триллионов микробов из более чем 500 родов бактерий из двух основных типов, а именно Bacteroidetes и Firmicutes. Хорошо известный у млекопитающих пробиотик Lactobacillus rhamnosus GG является активным индуктором АФК (Jones R 2014). Передача редокс-сигналов опосредует симбиоз между микробиотой кишечника и кишечником. У мух отмечена корреляция между увеличением продолжительности жизни с усилением образования окислителя H2O2 в икшечнике. H2S защищает выстилку слизистой оболочки желудочно-кишечного тракта от окислительного стресса, а также регулирует различные функции, включающие транспорт текучей среды, воспаление, индуцируемую кислотой секрецию HCO3- (Yonezawa D 2007, Ise F 2011, Wallace J 2009+2010, Fiorucci S 2006, Kasparek M 2008, Takeuchi K 2011+2015). У пожилых людей отмечена корреляция состава микробиоты кишечника с концентрациями ИЛ-6 в плазме крови (Claesson MJ 2012).
Молекула голодания Crtc (CREB-регулируемый транскрипционный коактиватор 1) усиливает иммунитет посредством создания кишечного барьера, менее проницаемого для бактерий. Кишечные бактерии, проходящие через кишечный барьер, вызывают воспаление. Эта молекула Crtc является генетическим переключателем в головном мозге, который контролирует энергетический баланс. Это постоянное сообщение между головным мозгом и ЖК трактом позволяет организму поддерживать пути расходов и запасания энерги. Crtc взаимодействует с CREB (белком, связывающим элемент ответа цАМФ). Партнером Crtc в головном мозге человека является нейропептид Y, заставляющий млекопитающих искать пищу. Активность CREB регулируется энергетическим сенсором Sirt1 и его способностью к деацетилированию CREB (Paz JC 2014). Это связывает уровень НАД+ и чувство голода. Регулируемый глюкозой антагонизм между (еще координированными) CREB и Sirt1 за транскрипцию Hes-1 участвует в метаболической регуляции нейрогенеза. Это важно в связи с тем, что нейрогенез сопровождается старением головного мозга (Bondolfi L 2004), и транскрипционный фактор CREB активируется депривацией питательных веществ, которая коррелирует с ферментативной активностью сиртуина.
Фактор некроза опухоли (ФНО) в кровообращении человека, который появляется как часть процесса старения, нарушает функцию развития воспалительных моноцитов и оказывает вредное действие на антипневмококковый иммунитет. Это действие претерпевает обратное развитие при фармакологическом уменьшении количества ФНО.
Препарат может содержать микроорганизмы, такие как бактерии в микробиоме, которые выделяют любое или все из трех категорий желательных соединений и которые добавляют непосредственно в кишечник. Эти микроорганизмы могут выделять желаемые соединения в желаемом количестве и с желаемой периодичностью. Эти микроорганизмы можно ввести в микробном либо из выбранных микроорганизмов, которые естественно встречаются в микробиоме, либо путем конструирования микроорганизмов, которые естественно встречаются в микробиоме. Эти сконструированные организмы могут быть сконструированы так, чтобы выделять эти соединения в соответствии с введенными микроорганизмами и/или с биологическими часами организма-хозяина. Введенный организм может быть сконструирован так, чтобы выделять желаемое количество соединений или соединений. Для переключения всех видов в кишечнике данного типа с получением желаемого генного типа введенного микроорганизма можно использовать метод генного драйва. В этих введенных видах можно сконструировать блокировку, позволяющую удалить эти сконструированные виды, если в последующее время они нежелательны.
Фармацевтически приемлемые носители
Раскрытые в настоящем документе композиции можно применять в терапии в комбинации с фармацевтически приемлемым носителем.
Приемлемые носители и их препараты описаны в кн. Remington: The Science and Practice of Pharmacy (22nd ed.) ed. L.V. Loyd Jr., CBS Publishers & Distributors Grandville MI USA 2012. В характерном случае в препарате используют соответствующее количество фармацевтически приемлемой соли, чтобы сделать раствор изотоническим. Примеры фармацевтически приемлемого носителя включают без ограничений физиологический раствор, раствор Рингера и раствор декстрозы. рН раствора предпочтительно составляет от около 5 до около 8 и более предпочтительно от около 7 до около 7,5. Дополнительные носители включают препараты пролонгированного высвобождения, такие как полупроницаемые матриксы из твердых гидрофобных полимеров, принимающие форму формованных изделий, например пленок, липосом или микрочастиц. Специалистам в данной области техники будет очевидно, что некоторые носители могут быть более предпочтительными в зависимости от пути введения и от концентрации вводимой композиции.
Фармацевтические носители известны специалистам в данной области техники. Эти носители в наиболее характерном случае должны представлять собой стандартные носители для введения лекарственных средств человеку, включая растворы, такие как стерильная вода, физиологический раствор и буферные растворы при физиологическом рН. Композиции можно вводить внутримышечно или подкожно. Другие соединения будут вводить в соответствии со стандартными методиками, используемыми специалистами в данной области техники.
Фармацевтические композиции могут включать носители, загустители, разбавители, буферы, консерванты, поверхностно-активные вещества (ПАВ) и т.п. в дополнение к выбранной молекуле. Фармацевтические композиции могут также включать один или более активных ингредиентов, таких как противомикробные агенты, противовоспалительные агенты, анестетики и т.п.
Фармацевтическую композицию можно вводить с помощью различных путей в зависимости от того, какое лечение требуется, местное или системное, и от области, подлежащей лечению. Введение можно осуществлять местным путем (включая глазное, вагинальное, ректальное, интраназальное введение), перорально, путем ингаляции или парентерально, например, с помощью капельницы для внутривенного введения, подкожной, интраперитонеальной или внутримышечной инъекции. Раскрытые соединения можно вводить внутривенно, интраперитонеально, внутримышечно, подкожно, с помощью внутриполостного введения или чрескожно.
Препараты для парентерального введения включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая физиологический раствор и забуференные среды. Несущие среды для парентерального введения включают раствор хлорида натрия, раствор Рингера с декстрозой, с декстрозой и хлоридом натрия, растворы Рингера с лактатом или нелетучие масла. Несущие среды для внутривенного введения включают текучую среду и добавки питательных веществ (например, на основе декстрозы Рингера) и т.п. Также могут присутствовать консерванты и другие добавки, такие как, например, противомикробные агенты, хелатирующие агенты и инертные газы и т.п.
Препараты для местного введения могут включать мази, лосьоны, кремы, гели, капли, суппозитории, спреи, жидкости и порошки. Традиционные фармацевтические носители, водные, порошковые или масляные основы, загустители и т.п. могут быть необходимы или желательны.
Композиции для перорального применения включают порошки или гранулы, суспензии или растворы в воде или неводных средах, капсулы, пакетики-саше или таблетки. Могут быть желательны загустители, корригенты, разбавители, эмульгаторы, диспергирующие добавки или связующие вещества.
Некоторые из композиций можно вводить в виде фармацевтически приемлемой кислотно- или основно-аддитивной соли, образованной путем взаимодействия с неорганическими кислотами, такими как хлористоводородная кислота, бромистоводородная кислота, перхлорная кислота, азотная кислота, тиоциановая кислота, серная кислота и фосфорная кислота, и органическими кислотами, такими как муравьиная кислота, уксусная кислота, пропионовая кислота, гликолевая кислота, молочная кислота, пировиноградная кислота, щавелевая кислота, малоновая кислота, янтарная кислота, малеиновая кислота и фумаровая кислота, или путем взаимодействия с неорганическим основанием, таким как гидроксид натрия, гидроксид аммония, гидроксид калия, и органическими основаниями, такими как моно-, ди-, триалкил- и ариламины и замещенные этаноламины.
Различные соединения и композиции категорий 1, 2 и 3 можно принимать в одно и то же или приближенное время, например в пределах 1, 5, 10, 30, 60, 90 или 120 минут.
Дозировки каждого вещества или веществ из категории 1, 2 и 3 являются достаточными, но не избыточными (описанные в молярном отношении к массе тела), и ингредиенты являются такими, чтобы взаимное соотношение этих доз было сбалансированным.
При использовании предпочтительного ингредиента из категории 1, 2 и 3 предпочтительна система доставки в воде. Это поможет выявить правильную периодичность (все 3 предпочтительных ингредиента легко абсорбируются и растворимы в воде). Для некоторых других менее предпочтительных ингредиентов, которые не растворяются в воде или не настолько легко абсорбируются, их доставка приведет в результате к сниженной пользе в отношении импульсной периодичности доставки этих трех категорий ингредиентов.
Раскрыты способы уменьшения воспаления у субъекта, включающие введение субъекту соединений, композиций или препаратов и возможно носителя, как описано в настоящем документе.
Также раскрыты способы, в которых первое соединение, второе соединение и третье соединение вводят приблизительно в одно и то же время.
Также раскрыты способы, в которых первое соединение вводят в пределах 15, 30, 60, 90 или 120 минут от пика биологических часов для НАД+.
Также раскрыты способы, в которых композиции вводят субъекту в дозе, составляющей по меньшей мере 1×10-8 моль первого соединения субъекту, 1×10-8 моль второго соединения субъекту и 1×10-9 моль третьего соединения субъекту.
Также раскрыты способы, в которых композицию вводят путем инъекции в течение 8-12 дней.
Также раскрыты способы, в которых композиция представляет собой аэрозоль, лиофилизат, порошок или эмульсию.
Также раскрыты способы, в которых субъект представляет собой человека.
Также раскрыты способы, в которых человека лечат в течение по меньшей мере двух месяцев.
Также раскрыты способы, в которых композиция представляет собой таблетку, которую принимают перорально по меньшей мере раз в день.
Также раскрыты способы, в которых композицию вводят раз в день.
Раскрытые композиции можно вводить в различных дозировках. Например, соединения категории 1, такие как никотинамидмононуклеотид (NMN), можно вводить в дозировках в сутки, составляющих от 1×10-6 моль/кг до 1×10-2 моль/кг, или от 1×10-5 моль/кг до 1×10-3 моль/кг, или от 1×10-4 моль/кг до 1×10-3 моль/кг, или от 2×10-4 моль/кг до 7×10-4 моль/кг. В некоторых воплощениях изобретения дозировки молекулы категории 1 в сутки могут составлять по меньшей мере 1×10-6 моль/кг, 1×10-5 моль/кг, 1×10-4 моль/кг, 1×10-3 моль/кг или 1×10-2 моль/кг. Дозировки могут также составлять по меньшей мере 2,38 моль/кг в сутки. Такие же дозировки рассмотрены в настоящем документе для других соединений категории 1 НАД+, NR, NaMN, NaAD, NAR, MNM и цАМФ.
Дозировка соединений категории 2, таких как бетаин, может составлять дозировки в сутки от 1×10-6 моль/кг до 1×10-2 моль/кг, или от 1×10-5 моль/кг до 1×10-3 моль/кг, или от 1×10-4 моль/кг до 1×10-3 моль/кг, или от 2×10-4 моль/кг до 7×10-4 моль/кг. В некоторых воплощениях изобретения дозировки соединения категории 2 в сутки могут составлять по меньшей мере 1×10-6 моль/кг, 1×10-5 моль/кг, 1×10-4 моль/кг, 1×10-3 моль/кг или 1×10-2 моль/кг. Дозировки могут также составлять по меньшей мере 5,82×10-4 моль/кг массы тела в сутки.
Дозировки соединений категории 3, таких как H2O2, могут составлять дозировки в сутки от 1×10-7 моль/кг до 1×10-2 моль/кг, или от 1×10-6 моль/кг до 1×10-3 моль/кг, или от 1×10-5 моль/кг до 1×10-4 моль/кг, или от 1×10-5 моль/кг до 7×10-5 моль/кг. В некоторых воплощениях изобретения дозировки соединения категории 3 в сутки могут составлять по меньшей мере 1×10-7 моль/кг, 1×10-6 моль/кг, 1×10-5 моль/кг, 1×10-4 моль/кг или 1×10-3 моль/кг. Дозировки могут также составлять по меньшей мере 2,34×10-5 моль/кг массы тела в сутки.
Дозировки соединений категории 3, таких как NaSH, могут составлять дозировки в сутки от 1×10-8 моль/кг до 1×10-3 моль/кг, или от 1×10-7 моль/кг до 1×10-4 моль/кг, или от 1×10-6 моль/кг до 1×10-5 моль/кг, или от 1×10-6 моль/кг до 7×10-6 моль/кг. В некоторых воплощениях изобретения дозировки соединения категории 3 в сутки могут составлять по меньшей мере 1×10-8 моль/кг, 1×10-7 моль/кг, 1×10-6 моль/кг, 1×10-4 моль/кг или 1×10-3 моль/кг. Дозировки могут также составлять по меньшей мере 3,02×10-6 моль/кг массы тела в сутки.
Конкретные способы
Раскрыты способы нормализации функций биологических путей для защиты от и устранения нарушений, возникающих вследствие старения человека. Эти способы могут привести к уменьшению воспаления у субъекта. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), S-аденозил-метионина (SAM) и H2O2. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), S-аденозил-метионина (SAM) и NaSH. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), S-аденозил-метионина (SAM) и Na2S. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), S-аденозил-метионина (SAM) и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), бетаина и H2O2. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), фолата плюс витамин В12 и H2O2. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), метионина и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), метионина и H2O2. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), холина и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), бетаина и NaHS. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), фолата плюс витамин В12 и NaHS. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), метионина и NaHS. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), холина и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), бетаина и Na2S. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), фолата плюс витамин В12 и Na2S. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), метионина и Na2S. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), холина и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), бетаина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), фолата плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), метионина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидадениндинуклеотида (НАД+), холина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR) с бетаином и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, фолата плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), фолата плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолата плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), фолата плюс витамин В12 и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина плюс витамин В12 и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-мети л никотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина плюс витамин В12 и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, метионина и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), метионина и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR) с метионином и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), метионина и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, холина и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), холина и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холина и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), холина и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, S-аденозил-метионина (SAM) и H2O2. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), S-аденозил-метионина (SAM) и H2O2. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионина (SAM) и H2O2. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), S-аденозил-метионина (SAM) и H2O2.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, фолата плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), фолата плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолата плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), фолата плюс витамин В12 и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина плюс витамин В12 и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина плюс витамин В12 и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, метионина и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), метионина и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионина и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), метионина и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, холина и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), холина и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холина и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), холина и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, S-аденозил-метионина (SAM) и NaHS. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), S-аденозил-метионина (SAM) и NaHS. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионина (SAM) и NaHS. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), S-аденозил-метионина (SAM) и NaHS.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, фолата плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), фолата плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолата плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), фолата плюс витамин В12 и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина плюс витамин В12 и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина плюс витамин В12 и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, метионина и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), метионина и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионина и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), метионина и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, холина и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), холина и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холина и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), холина и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, S-аденозил-метионина (SAM) и Na2S. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), S-аденозил-метионина (SAM) и Na2S. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионина (SAM) и Na2S. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), S-аденозил-метионина (SAM) и Na2S.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, фолата плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), фолата плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), фолата плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), фолата плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, бетаина плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), бетаина плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), бетаина плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метил никотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), бетаина плюс витамин В12 и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, метионина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), метионина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), метионина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), метионина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, холина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), холина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), холина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), холина и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
В конкретных примерах раскрытые способы могут включать введение субъекту никотинамидмононуклеотида (NMN) или предшественника или пролекарства NMN, S-аденозил-метионина (SAM) и любого одного или более из H2S, О3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту никотинамида рибозида (NR), S-аденозил-метионина (SAM) и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту одного или более из никотиновой кислоты аденинмононуклеотида (NaMN), никотиновой кислоты адениндинуклеотида (NaAD) и никотиновой кислоты рибозида (NAR), S-аденозил-метионина (SAM) и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка. В других примерах раскрытые способы могут включать введение субъекту 1-метилникотинамида (MNM) и/или циклического аденозинмонофосфата (цАМФ), S-аденозил-метионина (SAM) и любого одного или более из H2S, O3, метформина, ацетаминофена, сульфорафана, глюкорафанина, куркумина, кверцетина, изокверцетина, женьшеня, (R)-альфа-липоевой кислоты, гидрофильных окисленных производных ликопина, N-ацетилцистеина, DHEA, чеснока, β-лапачо, птеростильбена, ресвератрола, апигенина и цинка.
Суррогатные маркеры старения
Для мониторинга старения можно использовать различные маркеры в качестве суррогатов.
Уровни метилирования ДНК
Уровни метилирования ДНК с возрастом меняются. В исследованиях идентифицированы биомаркеры хронологического возраста, основанные на уровнях метилирования ДНК, называемые «эпигенетическими часами» (Horvath S 2013; на основании 353 динуклеотидных маркеров CpG). Различия между возрастом на основании метилирования ДНК и хронологическим возрастом позволили прийти к выводу, что меры биологического старения, выведенные на основании метилирования ДНК, являются признаками, на основании которых можно прогнозировать смертность независимо от состояния здоровья, факторов образа жизни и известных наследственных факторов (Marioni RE 2015). Эти эпигенетические часы тканеспецифичны, поскольку одни ткани стареют быстрее других. Мозжечок стареет медленнее, чем остальные части тела (Horvath S 2015). На основании этих эпигенетических часов у ВИЧ-1-инфицированных индивидов показано ускоренное старение (Rickabaugh ТМ 2015). Данные по метилированию могут быть собраны на основании циркулирующих Т клеток и моноцитов, что было проведено в когортном исследовании популяции 1264 участников (Reynolds LM 2014).
Повреждение ДНК
В отличие от метилирования, показатель однонитевых и двунитевых повреждений ДНК не был использован для биологических часов, но отмечена его корреляция со старением (Yu Q 2015), поскольку в пожилом возрасте в среднем повреждений больше. Такие компании, как Exogen Biotechnology, имеют возможность проводить исследования на однонитевые и двунитевые повреждения ДНК. НАД+ используется ферментами PARP и сиртуином в репарации ДНК, таким образом, наблюдаемое меньшее количество повреждений ДНК указывает на то, что эти ферментные системы работают.
Маркеры воспаления
Маркеры воспаления можно анализировать на старение, в том числе маркеры, выявленные в исследовании Arai в 2015. Arai были найдены маркеры воспаления, на основании которых можно было прогнозировать, кто продолжит жит (продолжительности жизни), и кто сохранит физическое и когнитивное здоровье (продолжительность здоровья). Используемыми маркерами были IgG к CMV, ИЛ-6, ФНО-альфа и CRP.
Другие маркеры, связанные со старением
В исследовании старения человека, вызванного преждевременным старением вследствие синдрома Вернера, и которое можно было проанализировать, была показана корреляция глобальной утраты H3K9me3 или возникших в результате изменений архитектуры гетерохроматина с биологическим старением (Zhang W 2015).
Ряд соединений в крови коррелирует с возрастом, а также с влиянием возраста, и эти соединения могут быть определены количественно. Примером является ФНО-бета, который ниже у молодых индивидов, чем у пожилых индивидов.
С использованием методики нелинейной регрессии и 13-летнего последующего наблюдения установлена корреляция метаболических показателей со старением.
Длину теломеров лейкоцитов периферической крови можно было измерить и сравнить у 64 637 индивидов известного возраста (Rode L 2015), хотя длина теломеров лишь умеренно коррелирует с возрастом (r равно 0,5), и клеточное старение продолжается независимо от длины теломеров.
Определения
В данном описании и в последующей формуле изобретения будет сделана ссылка на различные термины, для которых определены приведенные ниже значения:
Во всем тексте описания и формулы изобретения этого описания слово «содержать» и другие формы этого слова, такие как «содержащий» и «содержит», означает включение без ограничений и не предназначено для исключения, например, других добавок, компонентов, целых чисел или стадий.
При использовании в описании и в прилагаемой формуле изобретения формы единственного числа включают соответствующие ссылки на множественное число, если контекстом явным образом не продиктовано иное. Так, например, ссылка на «композицию» включает в себя смеси из двух или более таких композиций, ссылка на «соединение» включает в себя смеси из двух или более таких соединений, ссылка на «агент» включает в себя смеси из двух или более таких агентов и т.п.
«Возможный (необязательный)» или «возможно (необязательно)» означает, что описанное впоследствии событие или обстоятельство может либо наступить, либо не наступить, и что описание включает случаи, где это событие или обстоятельство наступает, и случаи, где оно не наступает.
При использовании в настоящем документе «субъект» означает индивида. Так, «субъект» может включать в себя домашних животных (например, кошек, собак и т.д.), скот (например, крупный рогатый скот, лошадей, свиней, овец, коз и т.д.), лабораторных животных (например, мышь, крысу, кролика, морскую свинку и т.д.) и птиц. «Субъект» может также включать в себя млекопитающее, такое как примат или человек.
Список литературы
Abdelmohsen, K., R. Pullmann, Jr., A. Lal, Н.Н. Kim, S. Galban, X. Yang, J.D. Blethrow, M. Walker, J. Shubert, D.A. Gillespie, H. Furneaux and M. Gorospe (2007). "Phosphorylation of HuR by Chk2 regulates SIRT1 expression." Mol Cell 25(4): 543-557.
Adebanjo, О.A., H.K. Anandatheerthavarada, A.P. Koval, B.S. Moonga, G. Biswas, L. Sun, B.R. Sodam, et al. "A New Function for Cd38/Adp-Ribosyl Cyclase in Nuclear Ca2+ Homeostasis. "Nat Cell Biol 1, no. 7 (Nov 1999): 409-14.
Albrecht, S.C, A.G. Barata, J. Grosshans, A.A. Teleman and T.P. Dick (2011). "In vivo mapping of hydrogen peroxide and oxidized glutathione reveals chemical and regional specificity of redox homeostasis." Cell Metab 14(6): 819-829.
Alic N, Partridge L. Мус mouse and anti-ageing therapy. Trends Endocrinol Metab. 2015; 26(4): 163-4.
Ame, J.С, V. Rolli, V. Schreiber, C. Niedergang, F. Apiou, P. Decker, S. Muller, T. Hoger, J. Menissier-de Murcia and G. de Murcia (1999). "PARP-2, A novel mammalian DNA damage-dependent poly(ADP-ribose) polymerase." J Biol Chem 274(25): 17860-17868.
Arai, Y., С.M. Martin-Ruiz, M. Takayama, Y. Abe, T. Takebayashi, S. Koyasu, M. Suematsu, N. Hirose, and T. von Zglinicki. "Inflammation, but Not Telomere Length, Predicts Successful Ageing at Extreme Old Age: A Longitudinal Study of Semi-Supercentenarians." EBioMedicine 2, no. 10 (Oct 2015): 1549-58.
Armah CN, Traka MH, Dainty JR, Defernez M, Janssens A, Leung W, et al. A diet rich in high-glucoraphanin broccoli interacts with genotype to reduce discordance in plasma metabolite profiles by modulating mitochondrial function. Am J Clin Nutr. 2013; 98(3):712-22.
Astrakhantseva IV, Efimov GA, Drutskaya MS, Kruglov AA, Nedospasov SA. Modern anti-cytokine therapy of autoimmune diseases. Biochemistry (Mosc). 2014; 79(12): 1308-21.
Bai P, Canto C, Oudart H, Brunyanszki A, Cen Y, Thomas C, et al. PARP-1 inhibition increases mitochondrial metabolism through SIRT1 activation. Cell Metab. 2011; 13(4):461-8.
Bai, P. and C. Canto (2012). "The role of PARP-1 and PARP-2 enzymes in metabolic regulation and disease." Cell Metab 16(3): 290-295.
Balogun, E., M. Hoque, P. Gong, E. Killeen, C.J. Green, R. Foresti, J. Alam and R. Motterlini (2003). "Curcumin activates the haem oxygenase-1 gene via regulation of Nrf2 and the antioxidant-responsive element." Biochem J 371 (Pt 3): 887-895.
Barata H, Thompson M, Zielinska W, Han YS, Mantilla CB, Prakash YS, et al. The role of cyclic-ADP-ribose-signaling pathway in oxytocin-induced Ca2+ transients in human myometrium cells. Endocrinology. 2004; 145(2):881-9.
Barber MF, Michishita-Kioi E, Xi Y, Tasselli L, Kioi M, Moqtaderi Z, et al. SIRT7 links H3K18 deacetylation to maintenance of oncogenic transformation. Nature. 2012; 487(7405):114-8.
Barr, L.A. and J.W. Calvert (2014). "Discoveries of Hydrogen Sulfide as a Novel Cardiovascular Therapeutic." Circulation Journal 78(9): 2111-2118.
Bellizzi D, Rose G, Cavalcante P, Covello G, Dato S, De Rango F, et al. A novel VNTR enhancer within the SIRT3 gene, a human homologue of SIR2, is associated with survival at oldest ages. Genomics. 2005; 85(2):258-63.
Belsky, D.W., A. Caspi, R. Houts, H.J. Cohen, D.L. Corcoran, A. Danese, H. Harrington, S. Israel, M.E. Levine, J.D. Schaefer, K. Sugden, B. Williams, A.I. Yashin, R. Poulton and Т.E. Moffitt (2015). "Quantification of biological aging in young adults." Proc Natl Acad Sci USA 112(30): E4104-4110.
Ben Mosbah, I., Y. Mouchel, J. Pajaud, C. Ribault, C. Lucas, A. Laurent, K. Boudjema, F. Morel, A. Corlu and P. Compagnon (2012). "Pretreatment with mangafodipir improves liver graft tolerance to ischemia/reperfusion injury in rat." PLoS One 7(11): e50235.
Bettcher BM, Kramer JH. Longitudinal inflammation, cognitive decline, and Alzheimer's disease: a mini-review. Clin Pharmacol Ther. 2014; 96(4):464-9.
Bettcher BM, Watson CL, Walsh CM, Lobach IV, Neuhaus J, Miller JW, et al. Interleukin-6, age, and corpus callosum integrity. PLoS One. 2014; 9(9): e106521.
Bettcher BM, Yaffe K, Boudreau RM, Neuhaus J, Aizenstein H, Ding J, et al. Declines in inflammation predict greater white matter microstructure in older adults. Neurobiol Aging. 2015; 36(2):948-54.
Bocklandt, S., W. Lin, M.E. Sehl, F.J. Sanchez, J.S. Sinsheimer, S. Horvath and E. Vilain (2011). "Epigenetic predictor of age." PLoS One 6(6): e14821.
Bos, E.M., R. Wang, P.M. Snijder, M. Boersema, J. Damman, M. Fu, J. Moser, J.L. Hillebrands, R.J. Ploeg, G. Yang, H.G. Leuvenink and H. van Goor (2013). "Cystathionine gamma-lyase protects against renal ischemia/reperfusion by modulating oxidative stress." J Am Soc Nephrol 24(5): 759-770.
Braidy, N., G.J. Guillemin and R. Grant (2011). "Effects of Kynurenine Pathway Inhibition on NAD Metabolism and Cell Viability in Human Primary Astrocytes and Neurons." Int J Tryptophan Res 4: 29-37.
Braidy, N., G.J. Guillemin, H. Mansour, T. Chan-Ling, A. Poljak and R. Grant (2011). "Age related changes in NAD+ metabolism oxidative stress and Sirt1 activity in wistar rats." PLoS One 6(4): e19194.
Butchart, J., L. Brook, V. Hopkins, J. Teeling, U. Puntener, D. Culliford, R. Sharpies, et al. "Etanercept in Alzheimer Disease: A Randomized, Placebo-Controlled, Double-Blind, Phase 2 Trial." Neurology 84, no. 21 (May 26 2015): 2161-8.
Calvert, J.W., M. Elston, С.K. Nicholson, S. Gundewar, S. Jha, J.W. Elrod, A. Ramachandran and D. J. Lefer (2010). "Genetic and pharmacologic hydrogen sulfide therapy attenuates ischemia-induced heart failure in mice." Circulation 122(1): 11-19.
Calvert, J.W., S. Jha, S. Gundewar, J.W. Elrod, A. Ramachandran, С.B. Pattillo, C.G. Kevil and D.J. Lefer (2009). "Hydrogen sulfide mediates cardioprotection through Nrf2 signaling." Circ Res 105(4): 365-374.
Canto, C, A.A. Sauve and P. Bai (2013). "Crosstalk between poly(ADP-ribose) polymerase and sirtuin enzymes." Mol Aspects Med 34(6): 1168-1201.
Cao Z, Tsang M, Zhao H, Li Y. Induction of endogenous antioxidants and phase 2 enzymes by α-lipoic acid in rat cardiac H9C2 cells: protection against oxidative injury. Biochemical and Biophysical Research Communications. 2003; 310(3):979-85.
Cardus A, Uryga AK, Walters G, Erusalimsky JD. SIRT6 protects human endothelial cells from DNA damage, telomere dysfunction, and senescence. Cardiovasc Res. 2013; 97(3):571-9.
Cesari, M., B.W. Penninx, M. Pahor, F. Lauretani, A.M. Corsi, G. Rhys Williams, J.M. Guralnik, and L. Ferrucci. "Inflammatory Markers and Physical Performance in Older Persons: The Inchianti Study." J Gerontol A Biol Sci Med Sci 59, no. 3 (Mar 2004): 242-8.
Chaix A, Zarrinpar A, Miu P, Panda S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 2014; 20(6):991-1005.
Chen S, Seiler J, Santiago-Reichelt M, Felbel K, Grummt I, Voit R. Repression of RNA polymerase I upon stress is caused by inhibition of RNA-dependent deacetylation of PAF53 by SIRT7. Mol Cell. 2013; 52(3):303-13.
Chen, D., J. Bruno, E. Easlon, S.J. Lin, H.L. Cheng, F.W. Alt and L. Guarente (2008). "Tissue-specific regulation of SIRT1 by calorie restriction." Genes Dev 22(13):1753-1757.
Chen, Q., A.K. Camara, D.F. Stowe, C.L. Hoppel and E.J. Lesnefsky (2007). "Modulation of electron transport protects cardiac mitochondria and decreases myocardial injury during ischemia and reperfusion." Am J Physiol Cell Physiol 292(1): C137-147.
Cheng L. Resveratrol attenuates inflammation and oxidative stress induced by myocardialischemia-reperfusion injury: role of Nrf2/ARE pathway. 2015.
Cheng, Y., J.F. Ndisang, G. Tang, K. Cao and R. Wang (2004). "Hydrogen sulfide-induced relaxation of resistance mesenteric artery beds of rats." Am J Physiol Heart Circ Physiol 287(5): H2316-2323.
Cho EH. SIRT3 as a Regulator of Non-alcoholic Fatty Liver Disease. J Lifestyle Med. 2014; 4(2):80-5.
Chouliaras, L., D.L. van den Hove, G. Kenis, S. Keitel, P.R. Hof, J. van Os, H.W. Steinbusch, C. Schmitz and B.P. Rutten (2012). "Prevention of age-related changes in hippocampal levels of 5-methylcytidine by caloric restriction." Neurobiol Aging 33(8): 1672-1681.
Coletta, C, A. Papapetropoulos, K. Erdelyi, G. Olah, K. Modis, P. Panopoulos, A. Asimakopoulou, D. Gero, I. Sharina, E. Martin and C. Szabo (2012). "Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation." Proc Natl Acad Sci USA 109(23): 9161-9166.
Colman, R.J., Т.M. Beasley, J.W. Kemnitz, S.C. Johnson, R. Weindruch and R.M. Anderson (2014). "Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys." Nat Commun 5: 3557.
Cremers, С.M., and U. Jakob. "Oxidant Sensing by Reversible Disulfide Bond Formation." J Biol Chem 288, no. 37 (Sep 13 2013): 26489-96.
De Haes, W., L. Frooninckx, R. Van Assche, A. Smolders, G. Depuydt, J. Billen, B.P. Braeckman, L. Schoofs and L. Temmerman (2014). "Metformin promotes lifespan through mitohormesis via the peroxiredoxin PRDX-2." Proc Natl Acad Sci USA 111(24): E2501-2509.
Deaglio S, Vaisitti T, Billington R, Bergui L, Omede P, Genazzani AA, et al. CD38/CD19: a lipid raft-dependent signaling complex in human В cells. Blood. 2007; 109(12):5390-8.
Derhovanessian, E., A.B. Maier, R. Beck, G. Jahn, K. Hahnel, P.E. Slagboom, A.J. de Craen, R.G. Westendorp and G. Pawelec (2010). "Hallmark features of immunosenescence are absent in familial longevity." J Immunol 185(8): 4618-4624.
Dinkova-Kostova AT, Abramov AY. The emerging role of Nrf2 in mitochondrial function. Free Radic Biol Med. 2015; 88(Pt B): 179-88.
Dinkova-Kostova, А.Т., K.Т. Liby, K.K. Stephenson, W.D. Holtzclaw, X. Gao, N. Suh, C. Williams, R. Risingsong, T. Honda, G.W. Gribble, M.B. Sporn and P. Talalay (2005). "Extremely potent triterpenoid inducers of the phase 2 response: correlations of protection against oxidant and inflammatory stress." Proc Natl Acad Sci USA 102(12): 4584-4589.
Dipp, M., and A.M. Evans. "Cyclic Adp-Ribose Is the Primary Trigger for Hypoxic Pulmonary Vasoconstriction in the Rat Lung in Situ." Circ Res 89, no. 1 (Jul 6 2001): 77-83.
Dodson M, Redmann M, Rajasekaran NS, Darley-Usmar V, Zhang J. KEAP1-NRF2 signalling and autophagy in protection against oxidative and reductive proteotoxicity. Biochem J. 2015; 469(3):347-55.
Du J, Zhou Y, Su X, Yu JJ, Khan S, Jiang H, et al. Sirt5 is a NAD-dependent protein lysine demalonylase and desuccinylase. Science. 2011; 334(6057):806-9.
Escande, С, V. Nin, N.L. Price, V. Capellini, A.P. Gomes, M.T. Barbosa, L.  , et al. "Flavonoid Apigenin Is an Inhibitor of the Nad+ Ase Cd38: Implications for Cellular Nad+ Metabolism, Protein Acetylation, and Treatment of Metabolic Syndrome." Diabetes 62, no. 4 (Apr 2013): 1084-93.
, et al. "Flavonoid Apigenin Is an Inhibitor of the Nad+ Ase Cd38: Implications for Cellular Nad+ Metabolism, Protein Acetylation, and Treatment of Metabolic Syndrome." Diabetes 62, no. 4 (Apr 2013): 1084-93.
Fernandez, A.F., et al. (2015). "H3K4me1 marks DNA regions hypomethylated during aging in human stem and differentiated cells." Genome Res 25(1): 27-40.
Ferrero, E., and F. Malavasi. "The Metamorphosis of a Molecule: From Soluble Enzyme to the Leukocyte Receptor Cd38." J Leukoc Biol 65, no. 2 (Feb 1999): 151-61.
Fifel K. Sirtuin 3: a molecular pathway linking sleep deprivation to neurological diseases. J Neurosci. 2014; 34(28):9179-81.
Fiorucci S, Distrutti E, Cirino G, Wallace JL. The emerging roles of hydrogen sulfide in the gastrointestinal tract and liver. Gastroenterology. 2006; 131(1):259-71.
Flier J, Van Muiswinkel FL, Jongenelen CA, Drukarch B. The neuroprotective antioxidant alpha-lipoic acid induces detoxication enzymes in cultured astroglial cells. Free Radic Res. 2002; 36(6):695-9.
Ford E, Voit R, Liszt G, Magin C, Grummt I, Guarente L. Mammalian Sir2 homolog SIRT7 is an activator of RNA polymerase I transcription. Genes Dev. 2006; 20(9): 1075-80.
Ford, J., S. Ahmed, S. Allison, M. Jiang and J. Milner (2014). "JNK2-dependent regulation of SIRT1 protein stability." Cell Cycle 7(19): 3091-3097.
Forman, H.J., J.M. Fukuto, T. Miller, H. Zhang, A. Rinna and S. Levy (2008). "The chemistry of cell signaling by reactive oxygen and nitrogen species and 4-hydroxynonenal." Arch Biochem Biophys 477(2): 183-195.
Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014; 69 Suppl 1:S4-9.
Franceschi C, Capri M, Monti D, Giunta S, Olivieri F, Sevini F, et al. Inflammaging and anti-inflammaging: a systemic perspective on aging and longevity emerged from studies in humans. Mech Ageing Dev. 2007; 128(1):92-105.
Franceschi C. Inflammaging as a Major Characteristic of Old People: Can It Be Prevented or Cured? Nutrition Reviews. 2007; 65(12):173-6.
Fu, M., W. Zhang, L. Wu, G. Yang, H. Li and R. Wang (2012). "Hydrogen sulfide (H2S) metabolism in mitochondria and its regulatory role in energy production." Proc Natl Acad Sci USA 109(8): 2943-2948.
Fusco S, Leone L, Barbati SA, Samengo D, Piacentini R, Maulucci G, et al. A CREB-Sirt1-Hes1 Circuitry Mediates Neural Stem Cell Response to Glucose Availability. Cell Rep. 2016; 14(5):1195-205.
Ge Y, Jiang W, Gan L, Wang L, Sun C, Ni P, et al. Mouse embryonic fibroblasts from CD38 knockout mice are resistant to oxidative stresses through inhibition of reactive oxygen species production and Ca(2+) overload. Biochem Biophys Res Commun. 2010; 399(2):167-72.
Ge, S.N., M.M. Zhao, D.D. Wu, Y. Chen, Y. Wang, J.H. Zhu, W.J. Cai, Y.Z. Zhu and Y.C. Zhu (2014). "Hydrogen sulfide targets EGFR Cys797/Cys798 residues to induce Na(+)/K(+)-ATPase endocytosis and inhibition in renal tubular epithelial cells and increase sodium excretion in chronic salt-loaded rats." Antioxid Redox Signal 21(15): 2061-2082.
Gegotek A, Skrzydlewska E. The role of transcription factor Nrf2 in skin cells metabolism. Arch Dermatol Res. 2015; 307(5):385-96.
Geng, В., H. Yan, G.Z. Zhong, C.Y. Zhang, X.B. Chen, H.F. Jiang, C.S. Tang and J.B. Du (2004). "[Hydrogen sulfide: a novel cardiovascular functional regulatory gas factor]." Beijing Da Xue Xue Bao 36(1): 106.
Gerhart-Hines Z, Dominy JE, Jr., Blattler SM, Jedrychowski MP, Banks AS, Lim JH, et al. The cAMP/PKA pathway rapidly activates SIRT1 to promote fatty acid oxidation independently of changes in NAD(+). Mol Cell. 2011; 44(6):851-63.
Giralt A, Villarroya F. SIRT3, a pivotal actor in mitochondrial functions: metabolism, cell death and aging. Biochem J. 2012; 444(1):1-10.
Givvimani, S., C. Munjal, R. Gargoum, U. Sen, N. Tyagi, J.C. Vacek and S.C. Tyagi (2011). "Hydrogen sulfide mitigates transition from compensatory hypertrophy to heart failure." J Appl Physiol (1985) 110(4): 1093-1100.
Goel, A., D.R. Spitz and G.J. Weiner (2012). "Manipulation of cellular redox parameters for improving therapeutic responses in B-cell lymphoma and multiple myeloma." J Cell Biochem 113(2): 419-425.
Gomes P, Outeiro TF, Cavadas C. Emerging Role of Sirtuin 2 in the Regulation of Mammalian Metabolism. Trends Pharmacol Sci. 2015; 36(11):756-68.
Gomes, A.P., N.L. Price, A.J. Ling, J.J. Moslehi, M.K. Montgomery, L. Rajman, J.P. White, J.S. Teodoro, C.D. Wrann, B.P. Hubbard, E.M. Mercken, С.M. Palmeira, R. de Cabo, A.P. Rolo, N. Turner, E.L. Bell and D.A. Sinclair (2013). "Declining NAD(+) induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging." Cell 155(7): 1624-1638.
Goodson NJ, Symmons DP, Scott DG, Bunn D, Lunt M, Silman AJ. Baseline levels of C-reactive protein and prediction of death from cardiovascular disease in patients with inflammatory polyarthritis: a ten-year follow up study of a primary care-based inception cohort. Arthritis Rheum. 2005; 52(8):2293-9.
Grob A, Roussel P, Wright JE, McStay B, Hernandez-Verdun D, Sirri V. Involvement of SIRT7 in resumption of rDNA transcription at the exit from mitosis. J Cell Sci. 2009; 122(Pt 4):489-98.
Grozio A, Sociali G, Sturla L, Caffa I, Soncini D, Salis A, et al. CD73 protein as a source of extracellular precursors for sustained NAD+ biosynthesis in FK866-treated tumor cells. J Biol Chem. 2013; 288(36):25938-49.
Guarente L. The many faces of sirtuins: Sirtuins and the Warburg effect. Nat Med. 2014; 20(1):24-5.
Guarente, L. (2000). "Sir2 links chromatin silencing, metabolism, and aging." Genes Dev 14(9): 1021-1026.
Haigis MC, Mostoslavsky R, Haigis KM, Fahie K, Christodoulou DC, Murphy AJ, et al. SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell. 2006; 126(5):941-54.
Halaschek-Wiener J, Amirabbasi-Beik M, Monfared N, Pieczyk M, Sailer C, Kollar A, et al. Genetic variation in healthy oldest-old. PLoS One. 2009; 4(8):e6641.
Han L, Ge J, Zhang L, Ma R, Hou X, Li B, et al. Sirt6 depletion causes spindle defects and chromosome misalignment during meiosis of mouse oocyte. Sci Rep. 2015; 5:15366.
Harman, D. "Aging: A Theory Based on Free Radical and Radiation Chemistry." J Gerontol 11, no. 3 (Jul 1956): 298-300.
He, C. and D.J. Klionsky (2009). "Regulation mechanisms and signaling pathways of autophagy." Annu Rev Genet 43: 67-93.
He, H.J., G.Y. Wang, Y. Gao, W.H. Ling, Z.W. Yu and T.R. Jin (2012). "Curcumin attenuates Nrf2 signaling defect, oxidative stress in muscle and glucose intolerance in high fat diet-fed mice." World J Diabetes 3(5):94-104.
Hill, В.С, Т.C. Woon, P. Nicholls, J. Peterson, C. Greenwood and A.J. Thomson (1984). "Interactions of sulphide and other ligands with cytochrome с oxidase. An electron-paramagnetic-resonance study." Biochem J 224(2): 591-600.
Hinson, J.A., D.W. Roberts and L.P. James (2010). "Mechanisms of acetaminophen-induced liver necrosis." Handb Exp Pharmacol (196): 369-405.
Hirschey MD, Shimazu T, Goetzman E, Jing E, Schwer B, Lombard DB, et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature. 2010; 464(7285):121-5.
Hirschey MD, Shimazu T, Huang JY, Schwer B, Verdin E. SIRT3 regulates mitochondrial protein acetylation and intermediary metabolism. Cold Spring Harb Symp Quant Biol. 2011; 76:267-77.
Hoffman-Liebermann, В., and D. A. Liebermann. "Interleukin-6- and Leukemia Inhibitory Factor-Induced Terminal Differentiation of Myeloid Leukemia Cells Is Blocked at an Intermediate Stage by Constitutive C-Myc." Mol Cell Biol 11, no. 5 (May 1991): 2375-81.
Hokari F, Kawasaki E, Sakai A, Koshinaka K, Sakuma K, Kawanaka K. Muscle contractile activity regulates Sirt3 protein expression in rat skeletal muscles. J Appl Physiol (1985). 2010; 109(2):332-40.
Holmes C, Butchart J. Systemic inflammation and Alzheimer's disease. Biochem Soc Trans. 2011; 39(4):898-901.
Horvath, S. (2013). "DNA methylation age of human tissues and cell types." Genome Biol 14(10): R115.
Horvath, S. (2015). "Erratum to: DNA methylation age of human tissues and cell types." Genome Biol 16: 96.
Horvath, S., V. Mah, A.T. Lu, J.S. Woo, O.W. Choi, A.J. Jasinska, J.A. Riancho, S. Tung, N.S. Coles, J. Braun, H.V. Vinters and L.S. Coles (2015). "The cerebellum ages slowly according to the epigenetic clock." Aging (Albany NY) 7(5): 294-306.
Huang Y, Li W, Su ZY, Kong AN. The complexity of the Nrf2 pathway: beyond the antioxidant response. J Nutr Biochem. 2015; 26(12):1401-13.
Hussain, A.M., H.C. Lee, and C.F. Chang. "Functional Expression of Secreted Mouse Bst-1 in Yeast." Protein Expr Purif 12, no. 1 (Feb 1998): 133-7.
Imai, S. (2009). "SIRT1 and caloric restriction: an insight into possible trade-offs between robustness and frailty." Curr Opin Clin Nutr Metab Care 12(4): 350-356.
Insko, M.A., T.L. Deckwerth, P. Hill, C.F. Toombs and C. Szabo (2009). "Detection of exhaled hydrogen sulphide gas in rats exposed to intravenous sodium sulphide." Br J Pharmacol 157(6): 944-951.
Ise F, Takasuka H, Hayashi S, Takahashi K, Koyama M, Aihara E, et al. Stimulation of duodenal HCO(3)(-) secretion by hydrogen sulphide in rats: relation to prostaglandins, nitric oxide and sensory neurones. Acta Physiol (Oxf). 2011; 201 (1):117-26.
Iyer SS, He Q, Janczy JR, Elliott EI, Zhong Z, Olivier AK, et al. Mitochondrial cardiolipin is required for Nlrp3 inflammasome activation. Immunity. 2013; 39(2):311-23.
Jackson, M.J. (2011). "Control of reactive oxygen species production in contracting skeletal muscle." Antioxid Redox Signal 15(9): 2477-2486.
Jain, S.K., J. Rains, J. Croad, B. Larson and K. Jones (2009). "Curcumin supplementation lowers TNF-alpha, IL-6, IL-8, and MCP-1 secretion in high glucose-treated cultured monocytes and blood levels of TNF-alpha, IL-6, MCP-1, glucose, and glycosylated hemoglobin in diabetic rats." Antioxid Redox Signal 11(2): 241-249.
Jiang, H.L, H.С. Wu, Z.L. Li, B. Geng and C.S. Tang (2005). "[Changes of the new gaseous transmitter H2S in patients with coronary heart disease]." Di Yi Jun Yi Da Xue Xue Bao 25(8): 951-954.
Jin, H.F., J.B. Du, X.H. Li, Y.F. Wang, Y.F. Liang and C.S. Tang (2006). "Interaction between hydrogen sulfide/cystathionine gamma-lyase and carbon monoxide/heme oxygenase pathways in aortic smooth muscle cells." Acta Pharmacol Sin 27(12): 1561-1566.
Jodar, L., E.M. Mercken, J. Ariza, C. Younts, J.A. Gonzalez-Reyes, F.J. Alcain, I. Buron, R. de Cabo and J. M. Villalba (2011). "Genetic deletion of Nrf2 promotes immortalization and decreases life span of murine embryonic fibroblasts." J Gerontol A Biol Sci Med Sci 66(3): 247-256.
Jung, K.J., A. Dasgupta, K. Huang, S.J. Jeong, C. Pise-Masison, K.V. Gurova and J.N. Brady (2008). "Small-molecule inhibitor which reactivates p53 in human T-cell leukemia virus type 1-transformed cells." J Virol 82(17): 8537-8547.
Jung, S.В., С.S. Kim, Y.R. Kim, A. Naqvi, T. Yamamori, S. Kumar, A. Kumar and K. Irani (2013). "Redox factor-1 activates endothelial SIRTUIN1 through reduction of conserved cysteine sulfhydryls in its deacetylase domain." PLoS One 8(6): e65415.
Kanfi Y, Naiman S, Amir G, Peshti V, Zinman G, Nahum L, et al. The sirtuin SIRT6 regulates lifespan in male mice. Nature. 2012; 483(7388):218-21.
Kang, B.Y., S. Kim, K.H. Lee, Y.S. Lee, I. Hong, M.O. Lee, D. Min, et al. "Transcriptional Profiling in Human Hacat Keratinocytes in Response to Kaempferol and Identification of Potential Transcription Factors for Regulating Differential Gene Expression." Exp Mol Med 40, no. 2 (Apr 30 2008): 208-19.
Kaspar, J.W., S.K. Niture and A.K. Jaiswal (2012). "Antioxidant-induced INrf2 (Keap1) tyrosine 85 phosphorylation controls the nuclear export and degradation of the INrf2-Cul3-Rbx1 complex to allow normal Nrf2 activation and repression." J Cell Sci 125(Pt 4): 1027-1038.
Kasparek MS, Linden DR, Kreis ME, Sarr MG. Gasotransmitters in the gastrointestinal tract. Surgery. 2008; 143(4):455-9.
Kellenberger E, Kuhn I, Schuber F, Muller-Steffner H. Flavonoids as inhibitors of human CD38. Bioorg Med Chem Lett. 2011; 21(13):3939-42.
Kennedy, S.R., L.A. Loeb and A.J. Herr (2012). "Somatic mutations in aging, cancer and neurodegeneration." Mech Ageing Dev 133(4): 118-126.
Kida, K., E. Marutani, R.K. Nguyen and F. Ichinose (2015). "Inhaled hydrogen sulfide prevents neuropathic pain after peripheral nerve injury in mice." Nitric Oxide 46: 87-92.
Kida, K., M. Yamada, K. Tokuda, E. Marutani, M. Kakinohana, M. Kaneki and F. Ichinose (2011). "Inhaled hydrogen sulfide prevents neurodegeneration and movement disorder in a mouse model of Parkinson's disease." Antioxid Redox Signal 15(2): 343-352.
Kim HS, Vassilopoulos A, Wang RH, Lahusen T, Xiao Z, Xu X, et al. SIRT2 maintains genome integrity and suppresses tumorigenesis through regulating APC/C activity. Cancer Cell. 2011; 20(4):487-99.
Kim SC, Sprung R, Chen Y, Xu Y, Ball H, Pei J, et al. Substrate and functional diversity of lysine acetylation revealed by a proteomics survey. Mol Cell. 2006; 23(4):607-18.
Kimura S, Warabi E, Yanagawa T, Ma D, Itoh K, Ishii Y, et al. Essential role of Nrf2 in keratinocyte protection from UVA by quercetin. Biochem Biophys Res Commun. 2009; 387(1):109-14.
Kimura, Y. and H. Kimura (2004). "Hydrogen sulfide protects neurons from oxidative stress." FASEB J 18(10): 1165-1167.
Kincaid B, Bossy-Wetzel E. Forever young: SIRT3 a shield against mitochondrial meltdown, aging, and neurodegeneration. Front Aging Neurosci. 2013; 5:48.
Kirkwood, Т. B. and M. R. Rose (1991). "Evolution of senescence: late survival sacrificed for reproduction." Philos Trans R Soc Lond В Biol Sci 332(1262): 15-24.
Kirkwood, Т. B. (2005). "Understanding the odd science of aging." Cell 120(4):437-447.
Kirkwood, T.L, P. Kapahi and D.P. Shanley (2000). "Evolution, stress, and longevity." J Anat 197 Pt 4: 587-590.
Kitada, M., S. Kume, N. Imaizumi and D. Koya (2011). "Resveratrol improves oxidative stress and protects against diabetic nephropathy through normalization of Mn-SOD dysfunction in AMPK/SIRT1-independent pathway." Diabetes 60(2): 634-643.
Kondo, K., S. Bhushan, A.L. King, S.D. Prabhu, T. Hamid, S. Koenig, T. Murohara, B.L. Predmore, G. Gojon, Sr., G. Gojon, Jr., R. Wang, N. Karusula, С.K. Nicholson, J.W. Calvert and D.J. Lefer (2013). "H(2)S protects against pressure overload-induced heart failure via upregulation of endothelial nitric oxide synthase." Circulation 127(10): 1116-1127.
Kong X, Wang R, Xue Y, Liu X, Zhang H, Chen Y, et al. Sirtuin 3, a new target of PGC-1 alpha, plays an important role in the suppression of ROS and mitochondrial biogenesis. PLoS One. 2010; 5(7): e11707.
Kugel S, Mostoslavsky R. Chromatin and beyond: the multitasking roles for SIRT6. Trends Biochem Sci. 2014; 39(2):72-81.
Kumar, S. and A.K. Pandey (2013). "Chemistry and biological activities of flavonoids: an overview." ScientificWorldJournal 2013: 162750.
Kumasaka, S., et al. (1999). "Novel mechanisms involved in superoxide anion radical-triggered Ca2+ release from cardiac sarcoplasmic reticulum linked to cyclic ADP-ribose stimulation." Antioxid Redox Signal 1(1): 55-69.
Labbadia, J. and R.I. Morimoto (2015). "The biology of proteostasis in aging and disease." Annu Rev Biochem 84: 435-464.
Lagouge, М., C. Argmann, Z. Gerhart-Hines, H. Meziane, C. Lerin, F. Daussin, N. Messadeq, J. Milne, P. Lambert, P. Elliott, B. Geny, M. Laakso, P. Puigserver and J. Auwerx (2006). "Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha." Cell 127(6): 1109-1122.
Landis, G. N., M. P. Salomon, D. Keroles, N. Brookes, T. Sekimura and J. Tower (2015). "The progesterone antagonist mifepristone/RU486 blocks the negative effect on life span caused by mating in female Drosophila." Aging (Albany NY) 7(1): 53-69.
Laurent G, German NJ, Saha AK, de Boer VC, Davies M, Koves TR, et al. SIRT4 coordinates the balance between lipid synthesis and catabolism by repressing malonyl CoA decarboxylase. Mol Cell. 2013; 50(5):686-98.
Lee ВТ, Ahmed FA, Hamm LL, Teran FJ, Chen CS, Liu Y, et al. Association of C-reactive protein, tumor necrosis factor-alpha, and interleukin-6 with chronic kidney disease. BMC Nephrol. 2015; 16:77.
Lee CU, Song EK, Yoo CH, Kwak YK, Han MK. Lipopolysaccharide induces CD38 expression and solubilization in J774 macrophage cells. Mol Cells. 2012; 34(6):573-6.
Lee JK, Bettencourt R, Brenner D, Le ТА, Barrett-Connor E, Loomba R. Association between serum interleukin-6 concentrations and mortality in older adults: the Rancho Bernardo study. PLoS One. 2012; 7(4):e34218.
Lee S, Paudel O, Jiang Y, Yang XR, Sham JS. CD38 mediates angiotensin II-induced intracellular Ca(2+) release in rat pulmonary arterial smooth muscle cells. Am J Respir Cell Mol Biol. 2015; 52(3):332-41.
Lee, Z.W., J. Zhou, C.S. Chen, Y. Zhao, С.H. Tan, L. Li, P.K. Moore and L.W. Deng (2011). "The slow-releasing hydrogen sulfide donor, GYY4137, exhibits novel anticancer effects in vitro and in vivo." PLoS One 6(6): e21077.
Leruez S, Milea D, Defoort-Dhellemmes S, Colin E, Crochet M, Procaccio V, et al. Sensorineural hearing loss in OPA1-linked disorders. Brain. 2013; 136(Pt 7): e236.
Li, L., M. Whiteman, Y.Y. Guan, K.L. Neo, Y. Cheng, S.W. Lee, Y. Zhao, R. Baskar, С.H. Tan and P.K. Moore (2008). "Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137): new insights into the biology of hydrogen sulfide." Circulation 117(18): 2351-2360.
Liu GH, Qu J, Shen X. NF-kappaB/p65 antagonizes Nrf2-ARE pathway by depriving СВР from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta. 2008; 1783(5):713-27.
Liu R, Dang W, Du Y, Zhou Q, Jiao K, Liu Z. SIRT2 is involved in the modulation of depressive behaviors. Sci Rep. 2015; 5:8415.
Liu R, Liu H, Ha Y, Tilton RG, Zhang W. Oxidative stress induces endothelial cell senescence via downregulation of Sirt6. Biomed Res Int. 2014; 2014:902842.
Lombard DB, Alt FW, Cheng HL, Bunkenborg J, Streeper RS, Mostoslavsky R, et al. Mammalian Sir2 homolog SIRT3 regulates global mitochondrial lysine acetylation. Mol Cell Biol. 2007; 27(24):8807-14.
Long AN, Owens K, Schlappal AE, Kristian T, Fishman PS, Schuh RA. Effect of nicotinamide mononucleotide on brain mitochondrial respiratory deficits in an Alzheimer's disease-relevant murine model. BMC Neurol. 2015; 15:19.
Lupoli, R., A. Di Minno, G. Spadarella, M. Franchini, R. Sorrentino, G. Cirino and G. Di Minno (2015). "Methylation reactions, the redox balance and atherothrombosis: the search for a link with hydrogen sulfide." Semin Thromb Hemost 41(4): 423-432.
Magesh, S., Y. Chen and L. Hu (2012). "Small molecule modulators of Keapl-Nrf2-ARE pathway as potential preventive and therapeutic agents." Med Res Rev 32(4): 687-726.
Marinho, H.S., C. Real, L. Cyrne, H. Soares and F. Antunes (2014). "Hydrogen peroxide sensing, signaling and regulation of transcription factors." Redox Biol 2: 535-562.
Marioni, R.E., et al. (2015). "DNA methylation age of blood predicts all-cause mortality in later life." Genome Biol 16: 25.
Martin GR, McKnight GW, Dicay MS, Coffin CS, Ferraz JG, Wallace JL. Hydrogen sulphide synthesis in the rat and mouse gastrointestinal tract. Dig Liver Dis. 2010; 42(2):103-9.
Massudi, H., R. Grant, N. Braidy, J. Guest, B. Farnsworth and G. J. Guillemin (2012). "Age-associated changes in oxidative stress and NAD+ metabolism in human tissue." PLoS One 7(7): e42357.
Mathias RA, Greco TM, Oberstein A, Budayeva HG, Chakrabarti R, Rowland EA, et al. Sirtuin 4 is a lipoamidase regulating pyruvate dehydrogenase complex activity. Cell. 2014; 159(7):1615-25.
McCay C. The Effect of Retarded Growth Upon The Length of Life Span And Upon The Ultimate Body  . 1935.
. 1935.
McCormack D, McFadden D.A review of pterostilbene antioxidant activity and disease modification. Oxid Med Cell Longev. 2013; 2013:575482.
Medzhitov R. Origin and physiological roles of inflammation. Nature. 2008; 454(7203):428-35.
Merino, M.M., C. Rhiner, J.M. Lopez-Gay, D. Buechel, B. Hauert and E. Moreno (2015). "Elimination of unfit cells maintains tissue health and prolongs lifespan." Cell 160(3): 461-476.
Metes-Kosik, N., I. Luptak, P.M. Dibello, D.E. Handy, S.S. Tang, H. Zhi, F. Qin, D.W. Jacobsen, J. Loscalzo and J. Joseph (2012). "Both selenium deficiency and modest selenium supplementation lead to myocardial fibrosis in mice via effects on redox-methylation balance." Mol Nutr Food Res 56(12): 1812-1824.
Michishita E, McCord RA, Berber E, Kioi M, Padilla-Nash H, Damian M, et al. SIRT6 is a histone H3 lysine 9 deacetylase that modulates telomeric chromatin. Nature. 2008; 452(7186):492-6.
Minagawa S, Araya J, Numata T, Nojiri S, Hara H, Yumino Y, et al. Accelerated epithelial cell senescence in IPF and the inhibitory role of SIRT6 in TGF-beta-induced senescence of human bronchial epithelial cells. Am J Physiol Lung Cell Mol Physiol. 2011; 300(3):L391-401.
Mokni M, Elkahoui S, Limam F, Amri M, Aouani E. Effect of resveratrol on antioxidant enzyme activities in the brain of healthy rat. Neurochem Res. 2007; 32(6):981-7.
Mokni, M., S. Hamlaoui, I. Karkouch, M. Amri, L. Marzouki, F. Limam and E. Aouani (2013). "Resveratrol Provides Cardioprotection after Ischemia/reperfusion Injury via Modulation of Antioxidant Enzyme Activities." Iran J Pharm Res 12(4): 867-875.
Moriarty-Craige, S.E., K.N. Ha, P. Sternberg, Jr., M. Lynn, S. Bressler, G. Gensler and D.P. Jones (2007). "Effects of long-term zinc supplementation on plasma thiol metabolites and redox status in patients with age-related macular degeneration." Am J Ophthalmol 143(2): 206-211.
Moscardo A, Valles J, Latorre A, Jover R, Santos MT. The histone deacetylase sirtuin 2 is a new player in the regulation of platelet function. J Thromb Haemost. 2015; 13(7):1335-44.
Mustafa, A.K., G. Sikka, S.K. Gazi, J. Steppan, S.M. Jung, A.K. Bhunia, V.M. Barodka, F.K. Gazi, R.K. Barrow, R. Wang, L. M. Amzel, D.E. Berkowitz and S.H. Snyder (2011). "Hydrogen sulfide as endothelium-derived hyperpolarizing factor sulfhydrates potassium channels." Circ Res 109(11): 1259-1268.
Nallasamy, P., H. Si, P.V. Babu, D. Pan, Y. Fu, E.A. Brooke, H. Shah, W. Zhen, H. Zhu, D. Liu, Y. Li and Z. Jia (2014). "Sulforaphane reduces vascular inflammation in mice and prevents TNF-alpha-induced monocyte adhesion to primary endothelial cells through interfering with the NF-kappaB pathway." J Nutr Biochem 25(8): 824-833.
Nathan, C. and A. Cunningham-Bussel (2013). "Beyond oxidative stress: an immunologist's guide to reactive oxygen species." Nat Rev Immunol 13(5): 349-361.
Neish AS, Jones RM. Redox signaling mediates symbiosis between the gut microbiota and the intestine. Gut Microbes. 2014; 5(2):250-3.
Nie, H., et al. (2014). "SIRT2 plays a key role in both cell cycle regulation and cell survival of BV2 microglia." Int J Physiol Pathophysiol Pharmacol 6(3): 166-171.
Nogueiras R, Habegger KM, Chaudhary N, Finan B, Banks AS, Dietrich MO, et al. Sirtuin 1 and sirtuin 3: physiological modulators of metabolism. Physiol Rev. 2012; 92(3):1479-514.
Noriega LG, Feige JN, Canto C, Yamamoto H, Yu J, Herman MA, et al. CREB and ChREBP oppositely regulate SIRT1 expression in response to energy availability. EMBO Rep. 2011; 12(10):1069-76.
North BJ, Rosenberg MA, Jeganathan KB, Hafner AV, Michan S, Dai J, et al. SIRT2 induces the checkpoint kinase BubR1 to increase lifespan. EMBO J. 2014; 33(13):1438-53.
Okabe, E., et al. (2000). "Calmodulin and cyclic ADP-ribose interaction in Ca2+ signaling related to cardiac sarcoplasmic reticulum: superoxide anion radical-triggered Ca2+ release." Antioxid Redox Signal 2(1): 47-54.
Oliver, A.W., J.C. Ame, S.M. Roe, V. Good, G. de Murcia and L. H. Pearl (2004). "Crystal structure of the catalytic fragment of murine poly(ADP-ribose) polymerase-2." Nucleic Acids Res 32(2): 456-464.
Otto, H., P.A. Reche, F. Bazan, K. Dittmar, F. Haag and F. Koch-Nolte (2005). "In silico characterization of the family of PARP-like poly(ADP-ribosyl)transferases (pARTs)." BMC Genomics 6:139.
Рае, H.O., G.S. Jeong, S.O. Jeong, H.S. Kim, S.A. Kim, Y.C. Kim, S.J. Yoo, H.D. Kim and H.T. Chung (2007). "Roles of heme oxygenase-1 in curcumin-induced growth inhibition in rat smooth muscle cells." Exp Mol Med 39(3): 267-277.
Palacios, О.M., et al. (2009). "Diet and exercise signals regulate SIRT3 and activate AMPK and PGC-1alpha in skeletal muscle." Aging (Albany NY) 1(9): 771-783.
Pall, M.L. and S. Levine (2015). "Nrf2, a master regulator of detoxification and also antioxidant, anti-inflammatory and other cytoprotective mechanisms, is raised by health promoting factors." Sheng Li Xue Bao 67(1): 1-18.
Pan, L.L, X.H. Liu, Q.H. Gong, D. Wu and Y. Z. Zhu (2011). "Hydrogen sulfide attenuated tumor necrosis factor-alpha-induced inflammatory signaling and dysfunction in vascular endothelial cells." PLoS One 6(5): e19766.
Papapetropoulos, A., A. Pyriochou, Z. Altaany, G. Yang, A. Marazioti, Z. Zhou, M.G. Jeschke, L.K. Branski, D.N. Herndon, R. Wang and C. Szabo (2009). "Hydrogen sulfide is an endogenous stimulator of angiogenesis." Proc Natl Acad Sci USA 106(51): 21972-21977.
Paredes-Gonzalez X, Fuentes F, Jeffery S, Saw CL, Shu L, Su ZY, et al. Induction of NRF2-mediated gene expression by dietary phytochemical flavones apigenin and luteolin. Biopharm Drug Dispos. 2015; 36(7):440-51.
Paredes-Gonzalez X, Fuentes F, Su ZY, Kong AN. Apigenin reactivates Nrf2 anti-oxidative stress signaling in mouse skin epidermal JB6 P + cells through epigenetics modifications. AAPS J. 2014; 16(4):727-35.
Paz JC, Park S, Phillips N, Matsumura S, Tsai WW, Kasper L, et al. Combinatorial regulation of a signal-dependent activator by phosphorylation and acetylation. Proc Natl Acad Sci USA. 2014; 111 (48):17116-21.
Реаке, В.F., С.K. Nicholson, J.P. Lambert, R.L. Hood, H. Amin, S. Amin and J.W. Calvert (2013). "Hydrogen sulfide preconditions the db/db diabetic mouse heart against ischemia-reperfusion injury by activating Nrf2 signaling in an Erk-dependent manner." Am J Physiol Heart Circ Physiol 304(9): H1215-1224.
Pearl, R., et al. (1928). "The Form of the Growth Curve of the Canteloup (Cucumis Melo) under Field Conditions." Proc Natl Acad Sci USA 14(12): 895-901.
Peng, C., et al. (2011). "The first identification of lysine malonylation substrates and its regulatory enzyme." Mol Cell Proteomics 10(12): M111 012658.
Perls, Т.Т., et al. (1997). "Middle-aged mothers live longer." Nature 389(6647): 133.
Polhemus, D.J. and D.J. Lefer (2014). "Emergence of hydrogen sulfide as an endogenous gaseous signaling molecule in cardiovascular disease." Circ Res 114(4): 730-737.
Pollak N, Niere M, Ziegler M. NAD kinase levels control the NADPH concentration in human cells. J Biol Chem. 2007; 282(46):33562-71.
Polletta L, Vernucci E, Carnevale I, Arcangeli T, Rotili D, Palmerio S, et al. SIRT5 regulation of ammonia-induced autophagy and mitophagy. Autophagy. 2015; 11(2):253-70.
Powers, S.K. and M.J. Jackson (2008). "Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production." Physiol Rev 88(4): 1243-1276.
Prasad, G.S., et al. (1996). "Crystal structure of Aplysia ADP ribosyl cyclase, a homologue of the bifunctional ectozyme CD38." Nat Struct Biol 3(11): 957-964.
Predmore, B.L., D.J. Lefer and G. Gojon (2012). "Hydrogen sulfide in biochemistry and medicine." Antioxid Redox Signal 17(1): 119-140.
Predmore, B.L., K. Kondo, S. Bhushan, M.A. Zlatopolsky, A.L. King, J.P. Aragon, D.B. Grinsfelder, M.E. Condit and D.J. Lefer (2012). "The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability." Am J Physiol Heart Circ Physiol 302(11): H2410-2418.
Predmore, B.L., M.J. Alendy, K.I. Ahmed, C. Leeuwenburgh and D. Julian (2010). "The hydrogen sulfide signaling system: changes during aging and the benefits of caloric restriction." Age (Dordr) 32(4):467-481.
Ramsey, K.M. and J. Bass (2009). "Obeying the clock yields benefits for metabolism." Proc Natl Acad Sci USA 106(11): 4069-4070.
Ramsey, K.M., J. Yoshino, С.S. Brace, D. Abrassart, Y. Kobayashi, B. Marcheva, H.K. Hong, J.L Chong, E.D. Buhr, C. Lee, J.S. Takahashi, S. Imai and J. Bass (2009). "Orcadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis." Science 324(5927): 651-654.
Ramsey, K.M., K.F. Mills, A. Satoh and S. Imai (2008). "Age-associated loss of Sirt1-mediated enhancement of glucose-stimulated insulin secretion in beta cell-specific Sirt1-overexpressing (BESTO) mice." Aging Cell 7(1): 78-88.
Ravussin, E., L.M. Redman, J. Rochon, S.K. Das, L. Fontana, W.E. Kraus, S. Romashkan, D.A. et al. (2015). "A 2-Year Randomized Controlled Trial of Human Caloric Restriction: Feasibility and Effects on Predictors of Health Span and Longevity." J Gerontol A Biol Sci Med Sci 70(9): 1097-1104.
Reuben, D.В., et al. (2002). "Peripheral blood markers of inflammation predict mortality and functional decline in high-functioning community-dwelling older persons." J Am Geriatr Soc 50(4): 638-644.
Rey, G. and A.B. Reddy (2013). "Protein acetylation links the circadian clock to mitochondrial function." Proc Natl Acad Sci USA 110(9): 3210-3211.
Reynolds, L.M., J.R. Taylor, J. Ding, K. Lohman, C. Johnson, D. Siscovick, G. Burke, W. Post, S. Shea, D.R. Jacobs, Jr., H. Stunnenberg, S.B. Kritchevsky, I. Hoeschele, С.E. McCall, D.M. Herrington, R.P. Tracy and Y. Liu (2014). "Age-related variations in the methylome associated with gene expression in human monocytes and T cells." Nat Commun 5: 5366.
Rickabaugh, Т.M., R.M. Baxter, M. Sehl, J.S. Sinsheimer, P.M. Hultin, L.E. Hultin, A. Quach, O. Martinez-Maza, S. Horvath, E. Vilain and B. D. Jamieson (2015). "Acceleration of age-associated methylation patterns in HIV-1-infected adults." PLoS One 10(3): e0119201.
Ridker, P.M., et al. (2000). "Elevation of tumor necrosis factor-alpha and increased risk of recurrent coronary events after myocardial infarction." Circulation 101(18): 2149-2153.
Roadmap Epigenomics, C., et al. (2015). "Integrative analysis of 111 reference human epigenomes." Nature 518(7539): 317-330.
Rose G, Dato S, Altomare K, Bellizzi D, Garasto S, Greco V, et al. Variability of the SIRT3 gene, human silent information regulator Sir2 homologue, and survivorship in the elderly. Exp Gerontol. 2003; 38(10):1065-70.
Sahu, R.P., S. Batra and S.K. Srivastava (2009). "Activation of ATM/Chk1 by curcumin causes cell cycle arrest and apoptosis in human pancreatic cancer cells." Br J Cancer 100(9): 1425-1433.
Sauve, A.A. and V.L. Schramm (2002). "Mechanism-based inhibitors of CD38: a mammalian cyclic ADP-ribose synthetase." Biochemistry 41(26): 8455-8463.
Sauve, A.A., et al. (1998). "The reaction mechanism for CD38. A single intermediate is responsible for cyclization, hydrolysis, and base-exchange chemistries." Biochemistry 37(38): 13239-13249.
Schmeisser, K., J. Mansfeld, D. Kuhlow, S. Weimer, S. Priebe, I. Heiland, M. Birringer, M. Groth, A. Segref, Y. Kanfi, N.L. Price, S. Schmeisser, S. Schuster, A.F. Pfeiffer, R. Guthke, M. Platzer, Т. Hoppe, H.Y. Cohen, K. Zarse, D.A. Sinclair and M. Ristow (2013). "Role of sirtuins in lifespan regulation is linked to methylation of nicotinamide." Nat Chem Biol 9(11): 693-700.
Seshadri, S., A. Beiser, J. Selhub, P.F. Jacques, I.H. Rosenberg, R.B.  P.W. Wilson and P.A. Wolf (2002). "Plasma homocysteine as a risk factor for dementia and Alzheimer's disease." N Engl J Med 346(7): 476-483.
P.W. Wilson and P.A. Wolf (2002). "Plasma homocysteine as a risk factor for dementia and Alzheimer's disease." N Engl J Med 346(7): 476-483.
Shang, Z., C. Lu, S. Chen, L. Hua and R. Qian (2012). "Effect of H(2)S on the circadian rhythm of mouse hepatocytes." Lipids Health Dis 11: 23.
Sharma A, Diecke S, Zhang WY, Lan F, He C, Mordwinkin NM, et al. The role of SIRT6 protein in aging and reprogramming of human induced pluripotent stem cells. J Biol Chem. 2013; 288(25):18439-47.
Shen J, Ma W, Liu Y. Deacetylase SIRT6 deaccelerates endothelial senescence. Cardiovasc Res. 2013; 97(3):391-2.
Sies, H. (2014). "Role of metabolic H2O2 generation: redox signaling and oxidative stress." J Biol Chem 289(13): 8735-8741.
Sodha, N.R., R.T. Clements, J. Feng, Y. Liu, C. Bianchi, E.M. Horvath, C. Szabo and F. W. Sellke (2008). "The effects of therapeutic sulfide on myocardial apoptosis in response to ischemia-reperfusion injury." Eur J Cardiothorac Surg 33(5): 906-913.
Sohal RS, Orr WC. The redox stress hypothesis of aging. Free Radic Biol Med. 2012; 52(3):539-55.
Someya S, Yu W, Hallows WC, Xu J, Vann JM, Leeuwenburgh C, et al. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell. 2010; 143(5):802-12.
Stefanson, A.L and M. Bakovic (2014). "Dietary regulation of Keap1/Nrf2/ARE pathway: focus on plant-derived compounds and trace minerals." Nutrients 6(9): 3777-3801.
Suh JH, Wang H, Liu RM, Liu J, Hagen TM. (R)-alpha-lipoic acid reverses the age-related loss in GSH redox status in post-mitotic tissues: evidence for increased cysteine requirement for GSH synthesis. Arch Biochem Biophys. 2004; 423(1):126-35.
Sun, L., et al. (2003). "Disordered osteoclast formation and function in a CD38 (ADP-ribosyl cyclase)-deficient mouse establishes an essential role for CD38 in bone resorption." FASEB J 17(3): 369-375.
Suzuki T, Motohashi H, Yamamoto M. Toward clinical application of the Keap1-Nrf2 pathway. Trends Pharmacol Sci. 2013; 34(6):340-6.
Suzuki, Т., et al. (2013). "Regulatory nexus of synthesis and degradation deciphers cellular Nrf2 expression levels." Mol Cell Biol 33(12): 2402-2412.
Szabo, С, C. Coletta, C. Chao, K. Modis, B. Szczesny, A. Papapetropoulos and M.R. Hellmich (2013). "Tumor-derived hydrogen sulfide, produced by cystathionine-beta-synthase, stimulates bioenergetics, cell proliferation, and angiogenesis in colon cancer." Proc Natl Acad Sci USA 110(30): 12474-12479.
Szilard, L. (1959). "On the Nature of the Aging Process." Proc Natl Acad Sci USA 45(1): 30-45.
Taaffe, D.R., et al. (2000). "Cross-sectional and prospective relationships of interleukin-6 and C-reactive protein with physical performance in elderly persons: MacArthur studies of successful aging." J Gerontol A Biol Sci Med Sci 55(12): M709-715.
Tachibana, Т., S. Okazaki, A. Murayama, A. Naganuma, A. Nomoto and S. Kuge (2009). "A major peroxiredoxin-induced activation of Yap1 transcription factor is mediated by reduction-sensitive disulfide bonds and reveals a low level of transcriptional activation." J Biol Chem 284(7): 4464-4472.
Takeuchi K, Kita K, Hayashi S, Aihara E. Regulatory mechanism of duodenal bicarbonate secretion: Roles of endogenous prostaglandins and nitric oxide. Pharmacol Ther. 2011; 130(1):59-70.
Takeuchi, K., et al. (2015). "Muscarinic acetylcholine receptor subtype 4 is essential for cholinergic stimulation of duodenal bicarbonate secretion in mice-relationship to D cell/somatostatin." J Physiol Pharmacol 66(3): 391-401.
Tan M, Peng С, Anderson KA, Chhoy P, Xie Z, Dai L, et al. Lysine glutarylation is a protein posttranslational modification regulated by SIRT5. Cell Metab. 2014; 19(4):605-17.
Tang, X.Q., R.Q. Chen, Y.K. Ren, P.D. Soldato, A. Sparatore, Y.Y. Zhuang, H.R. Fang and C.Y. Wang (2011). "ACS6, a Hydrogen sulfide-donating derivative of sildenafil, inhibits homocysteine-induced apoptosis by preservation of mitochondrial function." Med Gas Res 1(1): 20.
Tirumurugaan KG, Kang BN, Panettieri RA, Foster DN, Walseth TF, Kannan MS. Regulation of the cd38 promoter in human airway smooth muscle cells by TNF-alpha and dexamethasone. Respir Res. 2008; 9:26.
Tocmo, R., D. Liang, Y. Lin and D. Huang (2015). "Chemical and biochemical mechanisms underlying the cardioprotective roles of dietary organopolysulfides." Front Nutr 2: 1.
Tomczyk, S., K. Fischer, S. Austad and B. Galliot (2015).", a powerful model for aging studies." Invertebr Reprod Dev 59(sup1): 11-16.
Tu, H.C, D. Ren, G.X. Wang, D.Y. Chen, T.D. Westergard, H. Kim, S. Sasagawa, J.J. Hsieh and E.H. Cheng (2009). "The p53-cathepsin axis cooperates with ROS to activate programmed necrotic death upon DNA damage." Proc Natl Acad Sci USA 106(4): 1093-1098.
Tyagi, N., K.S. Moshal, U. Sen, T.P. Vacek, M. Kumar, W.M. Hughes, Jr., S. Kundu and S.C. Tyagi (2009). "H2S protects against methionine-induced oxidative stress in brain endothelial cells." Antioxid Redox Signal 11(1): 25-33.
van den Bogaard EH, Bergboer JG, Vonk-Bergers M, van Vlijmen-Willems IM, Hato SV, van der Valk PG, et al. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J Clin Invest. 2013; 123(2):917-27.
Vanyushin, B.F. (2005). "Enzymatic DNA methylation is an epigenetic control for genetic functions of the cell." Biochemistry (Mosc) 70(5): 488-499.
Varadhan R, Yao W, Matteini A, Beamer BA, Xue QL, Yang H, et al. Simple biologically informed inflammatory index of two serum cytokines predicts 10 year all-cause mortality in older adults. J Gerontol A Biol Sci Med Sci. 2014; 69(2):165-73.
Wakabayashi, N., A.T. Dinkova-Kostova, W.D. Holtzclaw, M.I. Kang, A. Kobayashi, M. Yamamoto, T.W. Kensler and P. Talalay (2004). "Protection against electrophile and oxidant stress by induction of the phase 2 response: fate of cysteines of the Keapl sensor modified by inducers." Proc Natl Acad Sci USA 101(7): 2040-2045.
Wallace, J.L. (2010). "Physiological and pathophysiological roles of hydrogen sulfide in the gastrointestinal tract." Antioxid Redox Signal 12(9): 1125-1133.
Wallace, J.L. and R. Wang (2015). "Hydrogen sulfide-based therapeutics: exploiting a unique but ubiquitous gasotransmitter." Nat Rev Drug Discov 14(5): 329-345.
Wang F, Li Y, Cao Y, Li C. Zinc might prevent heat-induced hepatic injury by activating the Nrf2-antioxidant in mice. Biol Trace Elem Res. 2015; 165(1):86-95.
Wang Z, Zhang L, Liang Y, Zhang C, Xu Z, Zhang L, et al. Cyclic AMP Mimics the Anti-ageing Effects of Calorie Restriction by Up-Regulating Sirtuin. Sci Rep. 2015; 5:12012.
Wang, J., S. Zhang, Y. Wang, L. Chen and X.S. Zhang (2009). "Disease-aging network reveals significant roles of aging genes in connecting genetic diseases." PLoS Comput Biol 5(9): e1000521.
Wang, G., et al. (2014) "P7C3 Neuroprotective Chemicals Function by Activating the Rate-Limiting Enzyme in NAD Salvage." Cell 158: 1324-1334.
Wang, R. (2002). "Two's company, three's a crowd: can H2S be the third endogenous gaseous transmitter?" FASEB J 16(13):1792-1798.
Wenger, Y. and B. Galliot (2013). "RNAseq versus genome-predicted transcriptomes: a large population of novel transcripts identified in an Illumina-454 Hydra transcriptome." BMC Genomics 14: 204.
Whiteman, M. and P.K. Moore (2009). "Hydrogen sulfide and the vasculature: a novel vasculoprotective entity and regulator of nitric oxide bioavailability?" J Cell Mol Med 13(3): 488-507.
Wilson HL, Dipp M, Thomas JM, Lad C, Galione A, Evans AM. Adp-ribosyl cyclase and cyclic ADP-ribose hydrolase act as a redox sensor, a primary role for cyclic ADP-ribose in hypoxic pulmonary vasoconstriction. J Biol Chem. 2001; 276(14):11180-8.
Wintner, E.A., T.L. Deckwerth, W. Langston, A. Bengtsson, D. Leviten, P. Hill, M.A. Insko, R. Dumpit, E. VandenEkart, C. F. Toombs and C. Szabo (2010). "A monobromobimane-based assay to measure the pharmacokinetic profile of reactive sulphide species in blood." Br J Pharmacol 160(4): 941-957.
Wu Y, Chen L, Wang Y, Li W, Lin Y, Yu D, et al. Overexpression of Sirtuin 6 suppresses cellular senescence and NF-kappaB mediated inflammatory responses in osteoarthritis development. Sci Rep. 2015; 5:17602.
Wu, С.C. and S.B. Bratton (2013). "Regulation of the intrinsic apoptosis pathway by reactive oxygen species." Antioxid Redox Signal 19(6): 546-558.
Wu, D., Q. Hu, X. Liu, L. Pan, Q. Xiong and Y.Z. Zhu (2015). "Hydrogen sulfide protects against apoptosis under oxidative stress through SIRT1 pathway in H9c2 cardiomyocytes." Nitric Oxide 46: 204-212.
Wu, L. and R. Derynck (2009). "Essential role of TGF-beta signaling in glucose-induced cell hypertrophy." Dev Cell 17(1): 35-48.
Xu, Z., G. Prathapasinghe, N. Wu, S.Y. Hwang, Y.L. Siow and K.О (2009). "Ischemia-reperfusion reduces cystathionine-beta-synthase-mediated hydrogen sulfide generation in the kidney." Am J Physiol Renal Physiol 297(1): F27-35.
Yang, G., L. Wu, B. Jiang, W. Yang, J. Qi, K. Cao, Q. Meng, A.K. Mustafa, W. Mu, S. Zhang, S.H. Snyder and R. Wang (2008). "H2S as a physiologic vasorelaxant: hypertension in mice with deletion of cystathionine gamma-lyase." Science 322(5901): 587-590.
Yao, L.L, X.W. Huang, Y.G. Wang, Y.X. Cao, С.C. Zhang and Y.C. Zhu (2010). "Hydrogen sulfide protects cardiomyocytes from hypoxia/reoxygenation-induced apoptosis by preventing GSK-3beta-dependent opening of mPTP." Am J Physiol Heart Circ Physiol 298(5): H1310-1319.
Yonezawa D, Sekiguchi F, Miyamoto M, Taniguchi E, Honjo M, Masuko T, et al. A protective role of hydrogen sulfide against oxidative stress in rat gastric mucosal epithelium. Toxicology. 2007; 241 (1-2):11-8.
Yong, Q.C, S.W. Lee, C.S. Foo, K.L. Neo, X. Chen and J.S. Bian (2008). "Endogenous hydrogen sulphide mediates the cardioprotection induced by ischemic postconditioning." Am J Physiol Heart Circ Physiol 295(3): H1330-H1340.
Yoshino, J. and S. Imai (2013). "Accurate measurement of nicotinamide adenine dinucleotide (NAD(+)) with high-performance liquid chromatography." Methods Mol Biol 1077: 203-215.
Yu, Q., Y.V. Katlinskaya, C.J. Carbone, B. Zhao, K.V. Katlinski, H. Zheng, M. Guha, N. Li, Q. Chen, T. Yang, C.J. Lengner, R.A. Greenberg, F.B. Johnson and S.Y. Fuchs (2015). "DNA-damage-induced type I interferon promotes senescence and inhibits stem cell function." Cell Rep 11(5): 785-797.
Yudoh K, Karasawa R, Ishikawa J. Age-related Decrease of Sirtuin 2 Protein in Human Peripheral Blood Mononuclear Cells. Curr Aging Sci. 2015; 8(3):256-8.
Zee, R.S., С.B. Yoo, D.R. Pimentel, D.H. Perlman, J.R. Burgoyne, X. Hou, M. E. McComb, С.E. Costello, R.A. Cohen and M.M. Bachschmid (2010). "Redox regulation of sirtuin-1 by S-glutathiolation." Antioxid Redox Signal 13(7): 1023-1032.
Zhang AY, Yi F, Teggatz EG, Zou AP, Li PL. Enhanced production and action of cyclic ADP-ribose during oxidative stress in small bovine coronary arterial smooth muscle. Microvasc Res. 2004; 67(2):159-67.
Zhao, W., J. Zhang, Y. Lu and R. Wang (2001). "The vasorelaxant effect of H(2)S as a novel endogenous gaseous K(ATP) channel opener." EMBO J 20(21):6008-6016.
Zhao, Y., S. Bhushan, C. Yang, H. Otsuka, J.D. Stein, A. Pacheco, B. Peng, N.O. Devarie-Baez, H.C. Aguilar, D.J. Lefer and M. Xian (2013). "Controllable hydrogen sulfide donors and their activity against myocardial ischemia-reperfusion injury." ACS Chem Biol 8(6): 1283-1290.
Zhou T, Kurnasov O, Tomchick DR, Binns DD, Grishin NV, Marquez VE, et al. Structure of human nicotinamide/nicotinic acid mononucleotide adenylyltransferase. Basis for the dual substrate specificity and activation of the oncolytic agent tiazofurin. J Biol Chem. 2002; 277(15):13148-54.
Примеры
Следующие примеры приведены ниже для иллюстрации способов, композиций и результатов в соответствии с раскрываемой сущностью изобретения. Эти примеры не предназначены для включения всех аспектов раскрываемой в настоящем документе сущности изобретения, но вероятнее для иллюстрации репрезентативных способов, композиций и результатов. Эти примеры не предназначены для включения эквивалентов и изменений настоящего изобретения, которые очевидны специалистам в данной области техники.
61-летний мужчина европеоидной расы с массой тела 88 кг в начале лечения получал лечение по отмеченной ниже схеме приема молекул категории 1, категории 2 и категории 3.
Никотинамидмононуклеотид (NMN) (молекулярная масса (MW)=334,22)
Бетаин (триметилглицин) (MW=117,14)
H2O2 (MW=34,01)
NaSH (MW=56,06)
Для введения субъекту растворы различных соединений готовили путем смешивания указанного количества в граммах с 500 мл воды.
Типичные конечные концентрации NMN, принимаемого субъектом, составляли 3,5 граммов в 500 мл H2O, бетаина - 3 грамма в 500 мл H2O, H2O2 - 2 капли раствора 35%-ной концентрации в 500 мл H2O и NaSH - 2 капли при концентрации 66 мкмоль/л на одну каплю в 500 мл H2O).
Количества каждой композиции были определены так, чтобы субъект, полностью выпивающий 500 мл, получил конечную дозировку в 500 мл раствора в виде питья приблизительно 1,19×10-4 моль NMN/кг массы тела на дозу, 2,91×10-4 моль бетаина/кг массы тела на дозу, 1,17×10-5 моль H2O2/кг массы тела на дозу и 1,51×10-6 моль NaSH/кг массы тела на дозу.
При приеме двух аналогичных доз в сутки сумма двух равных ежедневных порций в сутки составляла:
Доза никотинамидмононуклеотида (NMN) -- 2,38×10-4 моль/кг массы тела/сутки
Доза бетаина - 5,82×10-4 моль/кг массы тела/сутки
Доза пероксида водорода (H2O2) -- 2,34×10-5 моль/кг массы тела/сутки
Доза гидросульфида натрия (NaSH) - 3,02×10-6 моль/кг массы тела/сутки
Субъекта взвешивали ежедневно.
Субъект самостоятельно принимал препараты перорально посредством питья раствора приблизительно в 7.00 и в 19.00 ежедневно. Это время было выбрано, поскольку было аппроксимировано к пикам биологических часов НАД+ у субъекта, как определено в работе Ramsey K 2009. Это приводило к эффекту импульсного поступления ингредиентов в организм два раза в день, приблизительно соответствуя по времени биологическим часам субъекта.
Тестирование маркеров было выполнено компанией LabCor Inc. с использованием стандартных протоколов на ежемесячной основе. Время взятия образцов крови находилось в диапазоне от 8:19 до 8:54 утра. Измерения воспалительных маркеров коррелируют с биологическими часами. В LabCor тестировали уровни IgG к CMV, С-реактивного белка, фактора некроза опухоли-альфа и интерлейкина-6 в сыовротке крови.
Раз в месяц в LabCorp у субъекта также получали следующие данные, включающие концентрацию глюкозы в сыворотке крови, концентрацию мочевой кислоты в сыворотке крови, остаточный азот мочевины крови (АМК), сывороточный креатинин, eGRF у не афроамериканцев, отношение АМК/креатинин, сывороточный натрий, сывороточный калий, сывороточный хлорид, общий диоксид углерода, сывороточный кальций, сывороточный фосфор, общий белок сыворотки крови, сывороточный альбумин, сывороточный глобулин, общий глобулин, отношение А/Г, общий билирубин, сывороточную щелочную фосфатазу, лактатдегидрогеназу (ЛДГ), аспартатаминотрансферазу (ACT) (глутамат-оксалоацетат-трансаминазу сыворотки крови (СГОТ)), аланинаминотрансферазу (АЛТ) (сывороточную глутамат-пируватную трансаминазу (СГПТ)), сывороточное железо, общий холестерин, триглицериды, ЛПВП-холестерин, расчет ЛПОНП-холестерина, расчет ЛПНП-холестерина, отношение общий холестерин/ЛПВП-холестерин, оцениваемый риск ССЗ, количество лейкоцитов, количество эритроцитов, концентрацию гемоглобина, гематокрит, средний объем эритроцита (MCV), среднее количество гемоглобина в эритроците (МСН), среднее содержание гемоглобина в эритроцитах (МСНС), ширину распределения эритроцитов по объему (RDW), количество тромбоцитов, количество нейтрофилов, количества лимфоцитов, моноцитов, эозинофилов, базофилов, палочкоядерных клеток, количество нейтрофилов (абсолютное). Количество лимфоцитов (абсолютное), моноцитов (абсолютное), эозинофилов (абсолютное), базофилов (абсолютное), незрелых гранулоцитов, незрелых гранулоцитов (абсолютное), ядросодержащих эритроцитов (NRBC), VAP профиль холестерина, ЛПНП-холестерин, ЛПВП-холестерин, ЛПОНП-холестерин, общий холестерин, триглицериды, не-ЛПВП-холестерин (ЛПНП плюс ЛПОНП), АроВ100 = расчет, ЛПНП-R (фактическое)-С, Lp(a) холестерин, липопротеины промежуточной плотности (ЛППП)-холестерин, ремнантный липопротеин (ЛППП плюс ЛПОНП3), вероятность метаболического синдрома, ЛПВП-2 (наиболее защитные), ЛПВП-3 (меньшая степень защиты), ЛПОНП-3 (малые ремнантные), ЛПНП1 паттерн А, ЛПНП2 паттерн А, ЛПНП3 паттерн В, ЛПНП4 паттерн В, паттерн плотности ЛПНП, толерантность к глюкозе (4 образца крови), глюкоза натощак, глюкоза через 1 час, глюкоза через 2 часа, глюкоза через 3 часа, инсулин натощак, инсулин через 1 час, инсулин через 2 часа, инсулин через 3 часа, кортизол утром, кортизол вечером, ИЛ-1b (сыворотка крови), гемоглобин А1с, фактор ревматоидного артрита, инсулиноподобный фактор роста (IGF)-1, сердечный, опухолевый интерлейкин-8 (сыворотка крови), гомоцистеин (плазма крови), антиядерные антитела прямые, СОЭ по Вестергрену, кортизол (свободный в моче), кортизол, F, мкг, л, Ед., кортизол, F, мкг, 24 ч, Ед., сывороточный иммуноглобулин G, Qn, сывороточный иммуноглобулин A, Qn, сывороточный иммуноглобулин М, Qn, оксЛПНП, IgM к CMV, ферритин и IgG Н. pylori.
В Университете Калифорнии (УКЛА) в г. Сан-Диего измеряли:
a. Спектральная MPT 3 Тесла мышц правой голени до, во время и после нагрузки
b. Спектральная МРТ 3 Тесла печени
c. Структурная МРТ 3 Тесла печени
d. Спектральная МРТ 3 Тесла головного мозга (передняя и задняя)
e. Структурная МРТ 3 Тесла головного мозга
f. Структурная МРТ 3 Тесла правого колена (показатель артрита)
g. 3-Нитротирозин (маркер окислительного/нитративного стресса)
h. Коагулограмма (маркер окислительного стресса)
i. F2-изопростаны (маркер окислительного/нитративного стресса)
j. GSH: GSSH (маркер окислительного/нитративного стресса и защиты от него)
k. Органические кислоты в моче
l. 8-гидроксидеоксигуанозин (8-OHDG) (маркер окислительного/нитративного стресса)
m. Малондиальдегид (маркер окислительного/нитративного стресса)
n. hsCRP (маркер, на которых может оказывать вредное влияние окислительный стресс)
о. Протеомный профиль (маркер окислительного/нитративного стресса)
Были получены ответы на перечень вопросов по медицинскому анамнезу (УКЛА). Исследование жировой массы тела и минералов было проведено в частной клинике. Тредмил-тест (ЭКГ с нагрузкой) был проведен в частной клинике. Были получены образцы биопсии 4 типов ткани (печень (тонкоигольная биопсия), кожа; жировая ткань, мышцы) (хранили при -80 С в УКЛА). Ежедневные тренировки и массу тела записывали в журнал. Также проводили мониторинг глюкозы раз в неделю до и после приема NMN и мониторинг АД до и после приема NMN.
Результаты
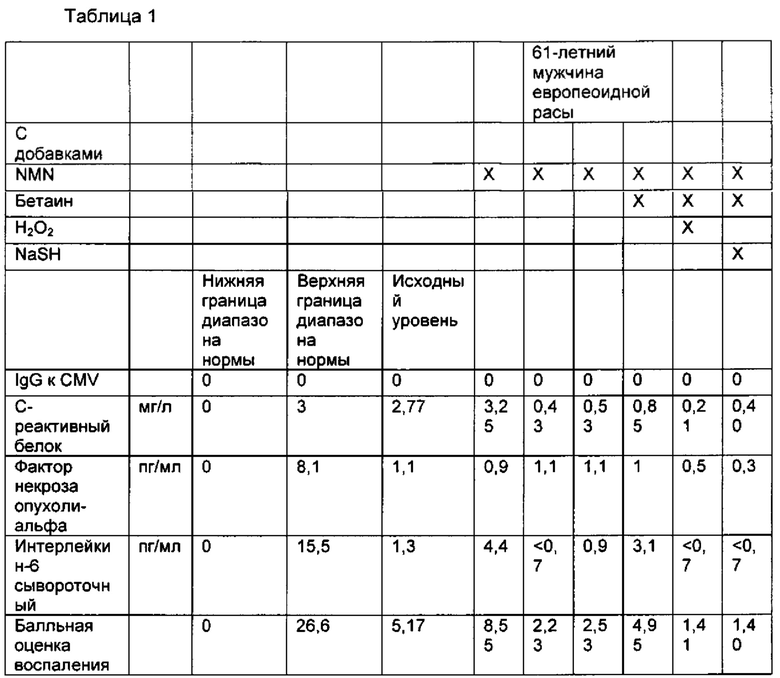
Результаты ежемесячного применения по схеме и исследования субъекта представлены в таблице 1. Как показано в таблице 1, субъект получал препарат по следующей схеме по месяцам, рецептура которого включала монотерапию NMN в течение 3 месяцев, NMN в комбинации с бетаином в течение одного месяца, NMN в комбинации с бетаином и H2O2 в течение одного месяца и NMN в комбинации с бетаином и NaSH в течение одного месяца.
Другие представляющие интерес наблюдения в ходе исследования состоят в том, что субъект был здоров на протяжении всего исследования. На фотографиях видно, что стареющие клетки кожи на руке стали выглядеть более молодыми. Цвет и внешнее строение кожи лица субъекта улучшились в ходе исследования. В ходе исследования субъект значительно потерял массу тела, и его аппетит снизился. В ходе исследования у субъекта исчезла боль в правом колене, связанная с артритом. В ходе исследования у субъекта отмечен значительно более спокойный сон. В ходе исследования у субъекта отмечена повышенная энергия. Обследование глаз субъекта показало улучшение зрения.
Обсуждение
Возраст 61 год коррелирует с возрастом неродственных и родственных 20 семей в исследовании И. Араи (Arai Y) 2015 г., подробно описанном в настоящем документе. Результаты данного исследования в свете исследования И. Араи (Arai Y) 2015 г. показывают, что тройная терапия тремя категориями соединений изменяет прогнозируемый исход неблагополучного старения, идентифицированный И. Араи (Arai Y) в 2015 г., у этого 61-летнего мужчины европеоидной расы с массой тела 88 кг, на прогнозируемый исход благополучного старения. По состоянию этого субъекта на исходном уровне показатели как С-реактивного белка (2,77 мг/л), так и интерлейкина-6 (1,3 пг/мл) были выше уровня как для «неродственных семей» (0,7 мг/л и 1,13 пг/мл) (Arai Y 2015, таблица 1), так и уровня «потомства» (0,7 мг/л и 1,03 пг/мл) (Arai Y. 2015, таблица 1) соответственно. Возраст 61-летнего субъекта мужского пола данного исследования был сходным с возрастом для группы «потомства» и группы «неродственных семей» по Араи. Эти два показателя воспаления в баллах влияют на алгоритм прогнозирования, который на исходном уровне прогнозирует худший исход старения для 61-летнего субъекта по сравнению с группами «потомства» или «неродственных семей» по Араи.
Однако после двух месяцев лечения NMN маркеры этого 61-летнего субъекта достигли лучших уровней по сравнению с группой «потомства» по Араи (С-РБ, 0,43 мг/л и ИЛ-6 менее 0,7 пг/мл). Хотя оба этих маркера незначительно повысились в первый месяц, общий эффект лечения NMN состоит в снижении уровня этих маркеров. У этого 61-летнего мужчины в результате приема NMN на протяжении 3 месяцев сохранялись более низкие или приблизительно сходные уровни по сравнению с группой «потомства» по Араи, но этот эффект, по-видимому, вышел на плато.
При добавлении всех трех категорий ингредиентов все три воспалительных маркера снизились до самого низкого уровня. Уровень ИЛ-6 снизился до необнаружимых уровней, уровень ФНО-альфа снизился почти на 50%, а С-РБ снизился почти в десять раз по сравнению с исходным значением. При применении H2O2 в качестве ингредиента категории 3 в данном примере С-РБ снижался больше, чем при применении NaSH, а при применении NaSH в качестве ингредиента категории 3 ФНО-альфа снижался больше, чем при применении H2O2. В обоих случаях результаты тройной терапии оказались значительно ниже необходимых уровней для предсказания очень благополучного старения. Титры CMV в данном случае не обсуждаются, поскольку у 61-летнего мужчины отсутствовали какие-либо обнаружимые уровни IgG к CMV, что лучше, чем возможность получить измеримое значение.
При сравнении результатов интервенционной терапии этого 61-летнего мужчины в данном эксперименте с результатами, полученными за один или два года ограничения калорий видно, что при данной терапии тремя категориями результаты значительно лучше и могут быть получены значительно легче (Di Francesco А 2015, Ravussin Е 2015).
Корреляции, наблюдаемые другими исследователями между улучшением состояния здоровья человека и снижением ФНО-альфа и ИЛ-6, которые снизились в данном примере;
Другие исследования (подобные Arai Y 2015)
Было обнаружено, что иммунные маркеры (простой показатель сывороточного интерлейкина-6 (ИЛ-6) и фактора некроза опухоли альфа (ФНО-альфа), два из 4 маркеров Араи (Arai)), являются лучшим прогностическим показателем смертности у 1155 взрослых людей пожилого возраста в 10 летнем исследовании смертности по любой причине после поправки на уже известные переменные, являющиеся причиной смерти (Varadhan R 2014). В проспективном когортном исследовании 1843 человек единственный иммунный маркер, сывороточный ИЛ-6, был прогностическим фактором смертности по любой причине, рака, сердечно-сосудистого заболевания и заболевания печени (Lee JK 2012). В этих исследованиях были подтверждены результаты менее обширных предшествующих исследований (Derhovanessian Е 2010, Reuben DB 2002, Taaffe DR 2000).
Возможный механизм действия:
В декабре 2013 г. A. Gomes и соавт. опубликовали исследование, демонстрирующее, что повышение уровней НАД+ предшественником NMN у старых мышей восстанавливается функция митохондрий до уровня молодой мыши. С.Correia-Melo показали, что с возрастом митохондрии управляют клеточным провоспалительным фенотипом, включая секрецию ИЛ-6.
Нарушение иммунитета:
В июле 2014 г. И.В. Астраханцева (I.V. Astrakhantseva и соавт.) сделала доклад, в котором показала преимущество снижения уровней ФНО и ИЛ-6 в качестве эффективных путей контроля симптомов воспаления, таких как деструкция сустава и аутоиммунные заболевания. А. Пушта (A. Puchta) и соавт.выдвинули гипотезу о молекулярном механизме, используя две воспалительные переменные (ФНО и ИЛ-6) для прогнозирования влияния на продолжительность жизни. В этом исследовании показано, как ФНО все в большей степени управляет нарушением иммунитета с возрастом, и что снижение уровней ФНО уменьшает это нарушение.
Заболевание головного мозга:
В сентябре 2014 г. Брайен Беттчер (Brianne Bettcher) и соавт. опубликовали исследование, показывающее, что в пожилом возрасте существует положительная корреляция между повышенными уровнями ИЛ-6 и снижением функции белого вещества в головном мозге. В феврале 2015 г. Брайен Беттчер (Brianne Bettcher) и соавт. опубликовали исследование, показывающее, что уменьшение системного воспаления оказывает положительное влияние на познавательную способность и структуру головного мозга, что может провести к обратному развитию нейродегенеративных процессов.
Заболевание сердца:
В 2000 г. Пол Ридкер (Paul Ridker) и соавт. опубликовали 2 исследования, в которых сделан вывод, что у здоровых на вид людей повышенные уровни ИЛ-6 связаны с повышенным риском инфаркта миокарда в будущем, а ФНО повышает риск рецидивирующих эпизодов болезни коронарных артерий после инфаркта миокарда. В августе 2005 г. Н. Дж. Гудсон (NJ Goodson) и соавт. опубликовали исследование, в котором выявлена связь между повышенными уровнями С-реактивного белка и прогнозом смерти от сердечно-сосудистого заболевания.
Заболевание почек:
В 2015 г. Белинда Ли (Belinda Lee) и соавт. опубликовали исследование, демонстрирующее связь между повышенными уровнями С-РБ, ФНО и ИЛ-6 и хроническим заболеванием почек.
Болезнь Альцгеймера:
Снижение уровней ФНО-альфа и ИЛ-6 уменьшает вероятность развития болезни Альцгеймера и уменьшает отрицательные эффекты болезни Альцгеймера (Butchart J 2015, Holmes С 2011). Добавление NMN в модели болезни Альцгеймера на мышах обладало благоприятным эффектом (Long AN 2015).
Исследование потенциальных преимуществ снижения ФНО-альфа и ИЛ-6 для более эффективного иммунного ответа на вирусы и бактерии:
Мак-Элрой А.К. (McElroy AK), проанализировав кинетику передачи воспалительных сигналов при опасном для жизни заболевании человека, вызванном вирусом лихорадки Эбола, предположил возможную терапевтическую пользу снижения передачи провоспалительных сигналов ИЛ-6 для лечебного вмешательства у этих пациентов. А. Пушта (A. Puchta) предположил возможную терапевтическую пользу снижения ИЛ-6 и ФНО альфа для повышения способности к борьбе против Streptococcus pneumoniae.
Исследование корреляции потенциальных преимуществ снижения ФНО-альфа и ИЛ-6 для лучшей физической работоспособности
Сезари М. (Cesari М) в 2004 г. пришел к выводу, что повышенные уровни ИЛ-6 коррелируют со снижением физической работоспособности у пожилых взрослых людей и являются мишенью для вмешательства. Пузановска-Кузника М. (Puzianowska-Kuznicka М) показала, что ИЛ-6 и С-РБ являются надежными прогностическими факторами физической и когнитивной работоспособности и риска смертности у 3496 индивидов.
Сон:
Ирвин М.P. (Irwin MR) выявил корреляцию нарушений сна с повышенными уровнями С-РБ и ИЛ-6, но не ФНО, в мета-анализе 72 предшествующих исследований сна.
Изобретение относится к композиции для уменьшения воспаления для введения субъекту, содержащей активатор системы репарации, донор метила и активатор антиоксидантной защиты, где активатор системы репарации представляет собой никотинамидмононуклеотид (NMN); донор метила представляет собой бетаин и активатор антиоксидантной защиты выбран из H2O2, NaSH и/или цинка. 3 н. и 27 з.п. ф-лы, 1 табл.
1. Композиция для уменьшения воспаления для введения субъекту, содержащая: активатор системы репарации, донор метила и активатор антиоксидантной защиты, где
активатор системы репарации представляет собой никотинамидмононуклеотид (NMN);
донор метила представляет собой бетаин; и
активатор антиоксидантной защиты выбран из H2O2, NaSH и/или цинка.
2. Композиция по п. 1, где активатор системы репарации, донор метила и активатор антиоксидантной защиты составляют по меньшей мере 5 масс.% композиции.
3. Композиция по п. 1, где активатор системы репарации представляет собой никотинамидмононуклеотид (NMN), никотинамид рибозид (NR) или оба соединения.
4. Композиция по п. 1, где донор метила представляет собой метионин, бетаин или оба соединения.
5. Композиция по п. 1, где активатор антиоксидантной защиты представляет собой H2O2 или NaSH.
6. Композиция по п. 1, где активатор системы репарации, донор метила и активатор антиоксидантной защиты представлены в количестве, достаточном для благоприятного изменения уровня суррогатного маркера старения у человека по сравнению с уровнем суррогатного маркера до введения композиции.
7. Композиция по п. 6, где изменением уровня суррогатного маркера является его снижение.
8. Композиция по п. 7, где суррогатный маркер представляет собой IgG к цитомегаловирусу (CMV), C-реактивный белок, фактор некроза опухоли альфа или
интерлейкин-6.
9. Композиция по п. 6, где изменением уровня суррогатного маркера является его повышение.
10. Композиция по п. 9, где суррогатным маркером является метилирование ДНК.
11. Композиция по п. 1, дополнительно содержащая воду.
12. Композиция по п. 1, содержащая по меньшей мере 1 x 10-8 моль активатора системы репарации, по меньшей мере 1 x 10-8 моль донора метила и по меньшей мере 1 x 10-9 моль активатора антиоксидантной защиты.
13. Композиция по п. 1, содержащая никотинамидмононуклеотид (NMN), бетаин и H2O2.
14. Способ уменьшения воспаления у субъекта, включающий: введение субъекту композиции по п. 1.
15. Способ по п. 14, в котором композицию вводят субъекту в дозе, составляющей по меньшей мере 1 x 10-6 моль/кг активатора системы репарации субъекту, 1 x 10-6 моль/кг донора метила субъекту и 1 x 10-7 моль/кг активатора антиоксидантной защиты субъекту.
16. Способ по п. 14, в котором композицию вводят путем инъекции в течение 8-12 дней.
17. Способ по п. 14, в котором композиция имеет аэрозольную, лиофилизированную, порошковую или эмульсионную форму.
18. Способ по п. 14, в котором субъект представляет собой человека.
19. Способ по п. 18, в котором композицию вводят человеку в течение по меньшей мере двух месяцев.
20. Способ по п. 14, в котором композиция представляет собой таблетку, которую принимают перорально по меньшей мере раз в день.
21. Способ по п. 14, в котором композиция дополнительно содержит воду.
22. Способ по п. 14, в котором композицию вводят субъекту раз в день.
23. Способ по п. 14, в котором композиция содержит никотинамидмононуклеотид (NMN), бетаин и H2O2.
24. Способ уменьшения воспаления у субъекта, включающий: введение субъекту активатора системы репарации, донора метила и активатора антиоксидантной защиты, где
активатор системы репарации представляет собой никотинамидмононуклеотид (NMN);
донор метила представляет собой бетаин; и
активатор антиоксидантной защиты выбран из H2O2, NaSH и/или цинка.
25. Способ по п. 24, где активатор системы репарации, донор метила и активатор антиоксидантной защиты вводят приблизительно в одно и то же время.
26. Способ по п. 24, в котором активатор системы репарации вводят в пределах 15, 30, 60, 90 или 120 минут от пика биологических часов для НАД+.
27. Способ по п. 24, в котором активатор системы репарации, донор метила и активатор антиоксидантной защиты вводят в различные моменты времени.
28. Способ по п. 24, в котором субъект представляет собой человека.
29. Способ по п. 28, в котором активатор системы репарации, донор метила и активатор антиоксидантной защиты вводят человеку в течение по меньшей мере двух месяцев.
30. Способ по п. 24, в котором активатор системы репарации, донор метила и активатор антиоксидантной защиты вводят человеку раз в день.
| US 2009214628 A1, 27.08.2009 | |||
| US 2015072950 A1, 12.03.2015 | |||
| US 2009035275 A1, 05.02.2009 | |||
| Maria Giulia Bacalini et al | |||
| "Present and future of anti-ageing epigenetic diets", Mechanisms of Ageing and Development, 2014, Vol | |||
| Регулятор для ветряного двигателя в ветроэлектрических установках | 1921 |
|
SU136A1 |
| Приспособление для записи звуковых явлений на светочувствительной поверхности | 1919 |
|
SU101A1 |
| СПОСОБ ЛЕЧЕНИЯ И ОЗДОРОВЛЕНИЯ ОРГАНИЗМА ЧЕЛОВЕКА | 2012 |
|
RU2488376C1 |

