Область техники, к которой относится изобретение
[0001]
Настоящее изобретение относится к антителу против CD147, обладающему высокой противоопухолевой эффективностью, к способу получения антитела против CD147 и к противоопухолевому средству, включающему антитело против CD147.
Уровень техники
[0002]
Достижения в области создания методов и средств лечения рака показали, что раковые заболевания, ранее считавшиеся неизлечимыми, поддаются лечению, и даже могут быть полностью излечимы. Антитело против CTLA4 и антитело против PD1, на основе которых были разработаны лекарственные препараты, проявляющие чрезвычайно высокую стабильность и специфичность, характеризуются высокой частотой объективных ответов при лечении меланомы и некоторых типов солидного рака, а, в некоторых случаях, и достижением излечения, в результате активации иммуноцитов, в том числе Т-клеток, что является обнадеживающей новостью для больных раком. Были предприняты попытки лечения наиболее трудноизлечимых типов солидного рака с помощью этих лекарственных средств. Однако большинство видов рака поджелудочной железы и печени и другие подобные типы рака не восприимчивы к воздействию этих лекарственных средств и характеризуются высокой частотой рецидивов даже при лечении путем хирургического удаления или при лечении на основе применения традиционных противораковых средств. Поэтому, непрестанно проводятся поиски методов лечения и лекарственных средств, которые позволяли бы достигать полного излечения рака.
[0003]
CD147 представляет собой однопроходный трансмембранный белок, имеющий от 2 до 3 иммуноглобулиноподобных доменов, и известно, что он активирует связанные с нисходящим сигнальным путем молекулы, такие как молекулы семейства FAK, MEK, Erk, JAK/STAT, AKT и MAPK, в результате взаимодействий между множеством белков CD147 или в результате взаимодействий с молекулами поверхности внеклеточной или клеточной мембраны, такими как CD44, молекулы семейства интегринов, CD98, VEGFR, CypA/B и MCT1/3/4, которые участвуют в росте, инвазии и воспалении, вследствие чего ускоряется продуцирование протеаз, таких как MMP, и рост рака, метастазирование и инвазия. Кроме того, сообщалось, что высокий уровень экспрессии CD147 обуславливает непродолжительный период выживаемости и неблагоприятный прогноз. Поэтому, CD147 рассматривают в качестве одной из молекул-мишеней при лечении рака.
[0004]
Известно, что антитела против CD147, такие как ABX-CBL и ликартин, использовали в клинике для введения людям. ABX-CBL ингибирует связывание между CD147 и циклофилином А и не только подавляет активность Т-клеток, но также обладает комплементзависимым цитотоксическим действием в отношении нормальных клеток, в том числе CD147-позитивных Т-клеток, в крови. Клинические испытания, проведенные для реакции "трансплантат против хозяина" (GVHD) в качестве заболевания-мишени, показали недостаточную эффективность ABX-CBL и возникновение сильных мышечных болей, и, поэтому, это антитело не было одобрено к применению в качестве лекарственного средства (патентный документ 1, непатентный документ 1).
[0005]
Ликартин является биофармацевтическим препаратом, получаемым путем добавления радиоактивного изотопа йода 131I к Fab'2-фрагменту антитела HAb18, и он был одобрен к применению в качестве лекарственного средства в Китае, причем для лечения рака печени (непатентный документ 2, непатентный документ 3). Ликартин не содержит Fc-фрагмента антитела для активации иммуноцитов и комплементов, и не было сообщений о том, что он обладает иммунообусловленной токсичностью, а также о том, что в клинических условиях достигалось полное излечение рака печени.
[0006]
Что касается других антител против CD147, то известны моноклональное антитело против CD147, блокирующее биологическую активность, связанную с CD147, такую как васкуляризация или продукция VEGF-продуцирующей матриксной металлопротеиназы (патентный документ 2), моноклональное антитело против CD147, ингибирующее активацию Т-клеток (непатентный документ 4), и антитело, специфически связывающееся с молекулой CD147, характеризующееся наличием опосредованной антителами клеточной цитотоксичности (ADCC) и комплементзависимой цитотоксичности (CDC) (патентный документ 3). Однако, неизвестно антитело против CD147, у которого бы отсутствовали эффекторные функции, но которое обладало бы эффективным противоопухолевым действием. Кроме того, неизвестна взаимозависимость между активацией системы клеточной сигнальной трансдукции через CD147 и противоопухолевым действием.
Список цитируемых в изобретении документов
Патентные документы
[0007]
Патентный документ 1: WO1999/045031
Патентный документ 2: WO2010/036460
Патентный документ 3: WO2017/061602
Непатентные документы
[0008]
Непатентный документ 1: Deeg H, et al., J. Blood., 2052-2058, 2001
Непатентный документ 2: Chen Z, et al., Int. J. Radiation Oncology Biol. Phys., 435-444, 2006
Непатентный документ 3: Xu J, et al., Hepatology, 269-276, 2007
Непатентный документ 4: Koch C, et al., International Immunology, 777-786, 1999
Сущность изобретения
Задача, решаемая изобретением
[0009]
Задачей настоящего изобретения является создание нового антитела против CD147, обладающего новым фармакологическим действием, высокой безопасностью и высокой противоопухолевой активностью, разработка фармацевтического препарата, включающего антитело, и разработка способа лечения опухолей с применением антитела или его фармацевтического препарата и другие подобные задачи.
Решение задачи
[0010]
В результате интенсивных исследований, направленных на решение вышеуказанной задачи, авторы настоящего изобретения впервые обнаружили, что активация связанных с сигнальным путем молекул посредством CD147 ассоциируется с противоопухолевой активностью. Авторы настоящего изобретения успешно получили CD147-специфическое антитело, активирующее CD147 и проявляющее высокую противоопухолевую активность. Антитело по настоящему изобретению способно проявлять высокую противоопухолевую активность, не связанную с эффекторными функциями. Сообщалось об антителах, проявляющих противоопухолевую активность, связанную с эффекторной функции. Однако антитело по настоящему изобретению характеризуется способностью не воздействовать на Т-клетки и мононуклеарные клетки периферической крови (РВМС) и проявлять высокую противоопухолевую активность, не связанную с эффекторными функциями, и, поэтому, оно является превосходным антителом, которое, как ожидается, будет безопасным в качестве фармацевтического препарата. Антитело по настоящему изобретению проявляет активность в клетках рака печени, которая является значительно более высокой, чем активность сорафениба, используемого в качестве одного из стандартных лекарственных средств для лечения рака печени. Антитело по настоящему изобретению проявляет активность в клетках рака поджелудочной железы, которая является значительно более высокой, чем активность гемцитабина, используемого в качестве одного из стандартных лекарственных средств для лечения рака поджелудочной железы. Антитело по настоящему изобретению проявляет активность в клетках хронического миелолейкоза, которая является значительно более высокой, чем у иматиниба, используемого в качестве одного из стандартных лекарственных средств для лечения хронического миелолейкоза. Авторы настоящего изобретения выяснили, что антитело против CD147 по настоящему изобретению активирует систему сигнальной трансдукции p38MAPK и SMAD в раковых клетках. Авторы настоящего изобретения выяснили, что антитело против CD147 по настоящему изобретению проявляет превосходную противоопухолевую активность в SMAD4-позитивных клетках.
[0011]
Настоящее изобретение включает в себя следующие аспекты.
[1]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое конкурирует, по меньшей мере, с одним антителом, выбранным из группы, состоящей из следующих антител по пунктам (A)-(F), за связывание с CD147, и которое активирует сигнальную трансдукцию посредством CD147:
(A) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:71, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:69;
(B) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:51, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:49;
(C) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:61, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:59,
(D) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:81, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:79;
(E) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:10, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:8; и
(F) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:20, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:18.
[2]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое связывается с эпитопом, с котором связывается, по меньшей мере, одно антитело, выбранное из группы, состоящей из следующих антител по пунктам (A)-(F), и которое активирует сигнальную трансдукцию посредством CD147:
(A) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:71, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:69;
(B) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:51, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:49;
(C) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:61, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:59;
(D) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:81, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:79;
(E) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:10, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:8; и
(F) антитело, включающее вариабельную область тяжелой цепи, состоящую из аминокислотной последовательности SEQ ID NO:20, и вариабельную область легкой цепи, состоящую из аминокислотной последовательности SEQ ID NO:18.
[3]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по пункту [1] или [2], которое характеризуется пониженной опосредованной антителами клеточной цитотоксичностью (ADCC) или отсутствием ADCC.
[4]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по любому из пунктов [1]-[3], которое характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или отсутствием CDC.
[5]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по любому из пунктов [1]-[4], которое характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или отсутствием ADCP.
[0012]
[6]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[5], которое связывается с эпитопом, включающим остатки аргинина (Arg) в положении 106 до остатков глицина (Gly) в положении 165 в последовательности SEQ ID NO:3.
[7]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[6], которое связывается с эпитопом, включающим остатки аргинина (Arg) в положении 106, лизина (Lys) в положении 108, аланина (Ala) в положении 109, валина (Val) в положении 110, лизина (Lys) в положении 127, серина (Ser) в положении 128, глутаминовой кислоты (Glu) в положении 129, серина (Ser) в положении 130, валина (Val) в положении 131, пролина (Pro) в положении 132, пролина (Pro) в положении 133, валина (Val) в положении 134, глутамина (Gln) в положении 164 и глицина (Gly) в положении 165 в аминокислотной последовательности, представленной в последовательности SEQ ID NO:3.
[8]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[7], где:
в последовательности тяжелой цепи, которая включает вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, CDRH1 состоит из аминокислотной последовательности SEQ ID NO:75, CDRH2 состоит из аминокислотной последовательности SEQ ID NO:76, и CDRH3 состоит из аминокислотной последовательности SEQ ID NO:77; и
в последовательности легкой цепи, которая включает вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, CDRL1 состоит из аминокислотной последовательности SEQ ID NO:72, CDRL2 состоит из аминокислотной последовательности SEQ ID NO:73, и CDRL3 состоит из аминокислотной последовательности SEQ ID NO:74.
[9]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[5], которое связывается с эпитопом, включающим аминокислотную последовательность, представленную в последовательности SEQ ID NO:143, или аминокислотную последовательность, в которой одна или несколько аминокислот удалены, замещены или добавлены в последовательность SEQ ID NO:143.
[10]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[5] или [9], где:
в последовательности тяжелой цепи, которая включает вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, CDRH1 состоит из аминокислотной последовательности SEQ ID NO:55, CDRH2 состоит из аминокислотной последовательности SEQ ID NO:56, и CDRH3 состоит из аминокислотной последовательности SEQ ID NO:57; и
в последовательности легкой цепи, которая включает вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, CDRL1 состоит из аминокислотной последовательности SEQ ID NO:52, CDRL2 состоит из аминокислотной последовательности SEQ ID NO:53, и CDRL3 состоит из аминокислотной последовательности SEQ ID NO:54.
[0013]
[11]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[5] или [9], где:
в последовательности тяжелой цепи, которая включает вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, CDRH1 состоит из аминокислотной последовательности SEQ ID NO:65, CDRH2 состоит из аминокислотной последовательности SEQ ID NO:66, и CDRH3 состоит из аминокислотной последовательности SEQ ID NO:67; и
в последовательности легкой цепи, которая включает вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, CDRL1 состоит из аминокислотной последовательности SEQ ID NO:62, CDRL2 состоит из аминокислотной последовательности SEQ ID NO:63, и CDRL3 состоит из аминокислотной последовательности SEQ ID NO:64.
[12]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[5] или [9], где:
в последовательности тяжелой цепи, которая включает вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, CDRH1 состоит из аминокислотной последовательности SEQ ID NO:85, CDRH2 состоит из аминокислотной последовательности SEQ ID NO:86, и CDRH3 состоит из аминокислотной последовательности SEQ ID NO:87; и
в последовательности легкой цепи, которая включает вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, CDRL1 состоит из аминокислотной последовательности SEQ ID NO:82, CDRL2 состоит из аминокислотной последовательности SEQ ID NO:83, и CDRL3 состоит из аминокислотной последовательности SEQ ID NO:84.
[13]
Антигенсвязывающий фрагмент антитела по любому из пунктов [1]-[12], который выбирают из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
[14]
Антитело по любому из пунктов [1]-[12], которое представляет собой scFv.
[15]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[12], которое представляет собой химерное антитело.
[0014]
[16]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[12], которое является гуманизированным.
[17]
Антитело по любому из пунктов [1]-[16], где тяжелая цепь включает константную область тяжелой цепи человеческого иммуноглобулина G1, константную область тяжелой цепи человеческого иммуноглобулина G2 или константную область тяжелой цепи человеческого иммуноглобулина G4, и легкая цепь включает константную область легкой цепи человеческого иммуноглобулина κ.
[18]
Антитело по пункту [17], где тяжелая цепь включает константную область тяжелой цепи человеческого иммуноглобулина G4.
[19]
Антитело по пункту [18], где в константной области тяжелой цепи человеческого иммуноглобулина G4, серин (Ser) в положении 228, указанном в соответствии с системой нумерации аминокислот в белке, заменен на пролин (Pro).
[20]
Антитело по пункту [18], где в константной области тяжелой цепи человеческого иммуноглобулина G4, фенилаланин (Phe) в положении 234, указанном в соответствии с системой нумерации аминокислот в белке, заменен на аланин (Ala), и лейцин (Leu) в положении 235, указанном в соответствии с системой нумерации аминокислот в белке, заменен на аланин (Ala).
[0015]
[21]
Антитело по пункту [18], где в константной области тяжелой цепи человеческого иммуноглобулина G4, серин (Ser) в положении 228, указанном в соответствии с системой нумерации аминокислот в белке, заменен на пролин (Pro), фенилаланин (Phe) в положении 234, указанном в соответствии с системой нумерации аминокислот в белке, заменен на аланин (Ala), и лейцин (Leu) в положении 235, указанном в соответствии с системой нумерации аминокислот в белке, заменен на аланин (Ala).
[22]
Антитело по пункту [17], где тяжелая цепь включает константную область тяжелой цепи человеческого иммуноглобулина G2.
[23]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое включает следующие вариабельные области по пунктам (c) и (d), и активирует сигнальную трансдукцию посредством CD147:
(c) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (c1)-(c4):
(c1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135;
(c2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147;
(c3) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% или более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (c1) или (c2); и
(c4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (c1)-(c3); и
(d) вариабельную область легкой цепи, выбранной из группы, состоящей из следующих последовательностей по пунктам (d1)-(d5):
(d1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137;
(d2) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149;
(d3) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151;
(d4) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (d1) и (d3); и
(d5) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (d1)-(d4).
[24]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137.
[0016]
[25]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:135; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:137.
[26]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149.
[27]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:149.
[28]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151.
[29]
Антитело или его антигенсвязывающий фрагмент по пункту [23], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:151.
[30]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое включает следующие области по пунктам (a) и (b), и активирует сигнальную трансдукцию посредством CD147:
(a) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (a1)-(a4):
(a1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123;
(a2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125;
(a3) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (a1) или (a2); и
(a4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (a1)-(a3); и
(b) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (b1)-(b3):
(b1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127;
(b2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (b1); и
(b3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (b1) или (b2).
[0017]
[31]
Антитело или его антигенсвязывающий фрагмент по пункту [30], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123, или вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127.
[32]
Антитело или его антигенсвязывающий фрагмент по пункту [30], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-466 в аминокислотной последовательности SEQ ID NO:123, или тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-467 в аминокислотной последовательности SEQ ID NO:125; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:127.
[33]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое включает следующие области по пунктам (e) и (f), и активирует сигнальную трансдукцию посредством CD147:
(e) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (e1)-(e4):
(e1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129;
(e2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131;
(e3) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (e1) или (e2); и
(e4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (e1)-(e3); и
(f) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (f1)-(f3):
(f1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133;
(f2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (f1); и
(f3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (f1) или (f2).
[34]
Антитело или его антигенсвязывающий фрагмент по пункту [33], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129, или вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133.
[35]
Антитело или его антигенсвязывающий фрагмент по пункту [33], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:129, или тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:131; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:133.
[0018]
[36]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент, которое включает следующие области по пунктам (g) и (h), и активирует сигнальную трансдукцию посредством CD147:
(g) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (g1)-(g3):
(g1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139;
(g2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (g1); и
(g3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (g1) или (g2); и
(h) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих последовательностей по пунктам (h1)-(h3):
(h1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141;
(h2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (h1); и
(h3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (h1) или (h2).
[37]
Антитело или его антигенсвязывающий фрагмент по пункту [36], которое включает:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141.
[38]
Антитело или его антигенсвязывающий фрагмент по пункту [36], которое включает:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:139; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:141.
[39]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по любому из пунктов [23]-[38], которое характеризуется опосредованной антителами клеточной цитотоксичностью (ADCC) или отсутствием ADCC.
[40]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по любому из пунктов [23]-[39], которое характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или отсутствием CDC.
[0019]
[41]
Антитело против человеческого CD147 или его антигенсвязывающий фрагмент по любому из пунктов [23]-[40], которое характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или отсутствием ADCP.
[42]
Фармацевтическая композиция, включающая, по меньшей мере, одно из антител или один из их антигенсвязывающих фрагментов по любому из пунктов [1]-[41].
[43]
Фармацевтическая композиция по пункту [42], которая представляет собой противоопухолевое средство.
[44]
Фармацевтическая композиция по пункту [43], где опухолью является опухоль, экспрессирующая CD147.
[45]
Фармацевтическая композиция по пункту [43] или [44], где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, лимфому, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль (GIST), лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
[0020]
[46]
Фармацевтическая композиция по любому из пунктов [43]-[45], где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
[47]
Фармацевтическая композиция по любому из пунктов [43]-[46], где опухоль представляет собой SMAD4-положительную опухоль или опухоль, имеющую пониженный уровень экспрессии или характеризующуюся отсутствием экспрессии KLF5.
[48]
Фармацевтическая композиция по любому из пунктов [42]-[47], дополнительно включающая еще одно противоопухолевое средство.
[49]
Способ лечения опухоли, включающий введение пациенту антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41] или фармацевтической композиции по любому из пунктов [42]-[48].
[50]
Способ по пункту [49], где опухоль представляет собой опухоль, экспрессирующую CD147.
[0021]
[51]
Способ по любому из пунктов [49] или [50], где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, лимфому, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль (GIST), лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
[52]
Способ по любому из пунктов [49]-[51], где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
[53]
Способ по любому из пунктов [49]-[52], где опухоль представляет собой SMAD4-положительную опухоль или опухоль, имеющую пониженный уровень экспрессии или характеризующуюся отсутствием экспрессии KLF5.
[54]
Способ по любому из пунктов [49]-[53], где антитело или его антигенсвязывающий фрагмент, или фармацевтическую композицию вводят в комбинации с еще одним противоопухолевым средством.
[55]
Полинуклеотид, который кодирует антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[41].
[0022]
[56]
Полинуклеотид по пункту [55], который включает полинуклеотид, выбранный из группы, состоящей из следующих полинуклеотидов по пунктам (j1)-(j3):
(j1) полинуклеотида, кодирующего CDRH1, состоящий из аминокислотной последовательности SEQ ID NO:75, CDRH2, состоящий из аминокислотной последовательности SEQ ID NO:76, и CDRH3, состоящий из аминокислотной последовательности SEQ ID NO:77, и полинуклеотид, кодирующего CDRL1, состоящий из аминокислотной последовательности SEQ ID NO:72, CDRL2, состоящий из аминокислотной последовательности SEQ ID NO:73, и CDRL3, состоящий из аминокислотной последовательности SEQ ID NO:74;
(j2) полинуклеотида, имеющего идентичность, по меньшей мере, 95% с последовательностью нуклеотида по пункту (j1); и
(j3) полинуклеотида, в котором от одного до нескольких нуклеотидов заменены, удалены или добавлены в полинуклеотиде по пунктам (j1) или (j2).
[57]
Полинуклеотид по пункту [55], который включает полинуклеотид, выбранный из группы, состоящей из следующих полинуклеотидов по пунктам (i1)-(i3):
(i1) полинуклеотида, кодирующего CDRH1, состоящий из аминокислотной последовательности SEQ ID NO:55, CDRH2, состоящий из аминокислотной последовательности SEQ ID NO:56, и CDRH3, состоящий из аминокислотной последовательности SEQ ID NO:57, и полинуклеотида, кодирующего CDRL1, состоящий из аминокислотной последовательности SEQ ID NO:52, CDRL2, состоящий из аминокислотной последовательности SEQ ID NO:53, и CDRL3, состоящий из аминокислотной последовательности SEQ ID NO:54;
(i2) полинуклеотида, имеющего идентичность, по меньшей мере, 95% с последовательностью нуклеотида по пункту (i1); и
(i3) полинуклеотида, в котором от одного до нескольких нуклеотидов заменены, удалены или добавлены в полинуклеотиде по пунктам (i1) или (i2).
[58]
Полинуклеотид по пункту [55], который включает полинуклеотид выбранный из группы, состоящей из следующих полинуклеотидов по пунктам (k1)-(k3):
(k1) полинуклеотида, кодирующего CDRH1, состоящий из аминокислотной последовательности SEQ ID NO:65, CDRH2, состоящий из аминокислотной последовательности SEQ ID NO:66, и CDRH3, состоящий из аминокислотной последовательности SEQ ID NO:67, и полинуклеотида, кодирующего CDRL1, состоящий из аминокислотной последовательности SEQ ID NO:62, CDRL2, состоящий из аминокислотной последовательности SEQ ID NO:63, и CDRL3, состоящий из аминокислотной последовательности SEQ ID NO:64;
(k2) полинуклеотида, имеющего идентичность, по меньшей мере, 95% с последовательностью нуклеотида по пункту (k1); и
(k3) полинуклеотида, в котором от одного до нескольких нуклеотидов заменены, удалены или добавлены в полинуклеотиде по пунктам (k1) или (k2).
[59]
Полинуклеотид по пункту [55], который включает полинуклеотид выбранный из группы, состоящей из следующих полинуклеотидов по пунктам (m1)-(m3):
(m1) полинуклеотида, кодирующего CDRH1, состоящий из аминокислотной последовательности SEQ ID NO:85, CDRH2, состоящий из аминокислотной последовательности SEQ ID NO:86, и CDRH3, состоящий из аминокислотной последовательности SEQ ID NO:87, и полинуклеотида, кодирующего CDRL1, состоящий из аминокислотной последовательности SEQ ID NO:82, CDRL2, состоящий из аминокислотной последовательности SEQ ID NO:83, и CDRL3 и состоящий из аминокислотной последовательности SEQ ID NO:84;
(m2) полинуклеотида, имеющего идентичность, по меньшей мере, 95% с последовательностью нуклеотида по пункту (m1); и
(m3) полинуклеотида, в котором от одного до нескольких нуклеотидов заменены, удалены или добавлены в полинуклеотиде по пунктам (m1) или (m2).
[60]
Экспрессирующий вектор, включающий полинуклеотид по любому из пунктов [55]-[59].
[0023]
[61]
Клетка-хозяин, трансформированная с помощью экспрессирующего вектора по пункту [60].
[62]
Способ получения антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41], включающий стадию культивирования клетки-хозяина по пункту [61] и собирание целевого антитела или его антигенсвязывающего фрагмента из жидкости над осадком культуры клеток.
[63]
Антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[41], где активация сигнальной трансдукции посредством CD147 представляет собой активацию p38 и/или активацию SMAD4.
[64]
Антитело или его антигенсвязывающий фрагмент по пункту [63], где активация p38MAPK и/или активация SMAD4 представляет собой повышение уровня экспрессии p38MAPK, фосфорилирование p38MAPK, фосфорилирование HSP27, повышение уровня экспрессии CXCL8, повышение уровня экспрессии rhoB, снижение KLF5 мРНК или снижение уровня экспрессии белка KLF5.
[65]
Способ лечения опухоли, включающий введение антитела или его антигенсвязывающего фрагмента по пункту [63] или [64].
[0024]
[66]
Способ прогнозирования восприимчивости к противораковому лечению, включающий:
выявление экспрессии SMAD4 или экспрессии KLF5 в биологическом образце, полученном от пациента с раковым заболеванием; и
определение восприимчивости пациента к противораковому лечению с помощью антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41] или фармацевтической композиции по любому из пунктов [42]-[48] в качестве положительной в случае, когда у пациента выявляют экспрессию SMAD4, или, когда у пациента выявляют снижение уровня экспрессии KLF5 или отсутствие экспрессии.
[67]
Способ отбора пациентов для лечения рака, включающий:
выявление экспрессии SMAD4 или экспрессии KLF5 в биологическом образце, полученном от пациента с раковым заболеванием; и
отбор пациента в качестве пациента для противоракового лечения с помощью антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41] или фармацевтической композиции по любому из пунктов [42]-[48], в случае, когда у пациента выявляют экспрессию SMAD4, или когда у пациента выявляют снижение уровня экспрессии KLF5 или отсутствие экспрессии.
[68]
Способ лечения злокачественного новообразования, включающий:
использование биологического образца, полученного от пациента с раковым заболеванием, для выявления присутствия или отсутствия экспрессии SMAD4 или выявления экспрессии KLF5 в биологическом образце; и
введение антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41] или фармацевтической композиции по любому из пунктов [42]-[48] пациенту, у которого выявляется экспрессия SMAD4, или пациенту, у которого выявляется снижение уровня экспрессии KLF5 или отсутствие экспрессии KLF5.
[69]
Набор для определения восприимчивости к противораковому лечению с помощью антитела или его антигенсвязывающего фрагмента по любому из пунктов [1]-[41] или фармацевтической композиции по любому из пунктов [42]-[48], где набор включает, по меньшей мере, одно средство для выявления экспрессии SMAD4 или экспрессии KLF5 в биологическом образце, полученном от пациента с раковым заболеванием.
[70]
Комплекс антитело-лекарственное средство, включающий антитело или его антигенсвязывающий фрагмент по любому из пунктов [1]-[41], конъюгированное с другим лекарственным средством.
[0025]
[71]
Биспецифическое антитело, включающее: антигенсвязывающий фрагмент антитела по любому из пунктов [1]-[41]; и антигенсвязывающий фрагмент, который связывается с антигеном, не являющимся CD147.
Полезные эффекты изобретения
[0026]
Антитело по настоящему изобретению представляет собой антитело, которое специфически распознает CD147 и которое характеризуется активацией молекул, связанных с передачей сигнала посредством CD147, и наличием высокой противоопухолевой активностью. CD147 экспрессируется не только в опухолевых клетках, но и в клетках крови. Однако, антитело по настоящему изобретению не воздействует на Т-клетки или мононуклеарные клетки периферической крови (РВМС) и не зависит от эффекторных функций, и, поэтому, имеет преимущество, связанное с решением меньшего количества проблем безопасности при его разработке в качестве противоопухолевого средства. Антитело по настоящему изобретению проявляет чрезвычайно высокую противоопухолевую активность. Антитело по настоящему изобретению проявляет активность в клетках рака печени, которая является значительно более высокой, чем активность сорафениба, используемого в качестве одного из стандартных лекарственных средств для лечения рака печени. Антитело по настоящему изобретению проявляет активность в клетках рака поджелудочной железы, которая является значительно более высокой, чем активность гемцитабина, используемого в качестве одного из стандартных лекарственных средств для лечения рака поджелудочной железы. Антитело по настоящему изобретению проявляет активность в клетках хронического миелолейкоза, которая является значительно более высокой, чем у иматиниба, используемого в качестве одного из стандартных лекарственных средств для лечения хронического миелолейкоза.
Краткое описание чертежей
[0027]
На фигурах 1(a)-(c) представлены данные по изменению объема опухоли в ксенотрансплантатной модели PANC-1 (рака поджелудочной железы).
На фигуре 2-1 представлены данные по активности связывания с человеческим CD147 или обезьяньим CD147.
На фигуре 2-2 представлены данные по активности связывания с человеческим CD147 или обезьяньим CD147.
На фигуре 2-3 представлены данные по активности связывания с человеческим CD147 или обезьяньим CD147.
На фигуре 3 приведено сравнение аминокислотных последовательностей между человеческим CD147 и CD147 яванского макака и положения от mu1 до mu9 в человеческом CD147 и в CD147 яванского макака.
На фигуре 4-1 приведена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи LN22R8 и аминокислотные последовательности CDRL1, CDRL2, и CDRL3 легкой цепи LN22R8.
На фигуре 4-2 приведена нуклеотидная последовательность и аминокислотная последовательность вариабельной области тяжелой цепи LN22R8 и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи LN22R8.
На фигуре 5-1 приведена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи 2P10F2 и аминокислотные последовательности CDRL1, CDRL2 и CDRL3 легкой цепи 2P10F2.
На фигуре 5-2 приведена нуклеотидная последовательность и аминокислотная последовательность вариабельной области тяжелой цепи 2P10F2 и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи 2P10F2.
На фигуре 6 представлены данные по опосредованной антителами клеточной цитотоксичности (ADCC) для человеческих химерных антител.
На фигуре 7 представлены данные по комплементзависимой цитотоксичности (CDC) для человеческих химерных антител.
На фигуре 8 представлены данные по антителозависимому клеточному фагоцитозу (ADCP) для человеческих химерных антител.
На фигурах 9-1(a)-(d) представлены данные по противоопухолевой активности антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2 иммунодефицитным мышам линии NOD-scid.
На фигурах 9-2(e)-(g) представлены данные по противоопухолевой активности антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2 иммунодефицитным мышам линии NOD-scid.
На фигуре 10 представлены данные по противоопухолевой активности LN22R8chIgG4P в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2 мышам с тяжелым иммунодефицитом линии NOG.
[0028]
На фигурах 11(a)-(d) представлены данные по противоопухолевой активности r#84, r#101, r#110 и r#131 в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2 иммунодефицитным мышам линии NOD-scid.
На фигуре 12 представлены данные конкурентного анализа 2P10F2 в сравнении с r#84, r#101, r#110 и r#131.
На фигурах 13-1(a)-(d) представлены данные по противоопухолевой активности химерных антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2.
На фигурах 13-2(e)-(h) представлены данные по противоопухолевой активности химерных антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2.
На фигурах 13-3 (i)-(l) представлены данные по противоопухолевой активности химерных антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2.
На фигуре 13-4 (m) и (n) представлены данные по противоопухолевой активности химерных антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2.
На фигуре 14(a) приведена конструкция вариабельной области тяжелой цепи #84H1h гуманизированного антитела, и на фигуре 14(b) приведена конструкция вариабельной области легкой цепи #84L2h гуманизированного антитела.
На фигуре 15(a) приведена конструкция вариабельной области тяжелой цепи #101H1h гуманизированного антитела, и на фигуре 15(b) приведена конструкция вариабельной области легкой цепи #101L2h гуманизированного антитела.
На фигуре 16(a) приведена конструкция вариабельной области тяжелой цепи #110H1h гуманизированного антитела, на фигуре 16(b) приведена конструкция вариабельной области тяжелой цепи #110H13h гуманизированного антитела, на фигуре 16(c) приведена конструкция вариабельной области легкой цепи #110L4h гуманизированного антитела, на фигуре 16(d) приведена конструкция вариабельной области легкой цепи #110L2h гуманизированного антитела, и на фигуре 16(e) приведена конструкция вариабельной области легкой цепи #110L12h гуманизированного антитела.
На фигуре 17(a) приведена конструкция вариабельной области тяжелой цепи #131H2h гуманизированного антитела, и на фигуре 17(b) приведена конструкция вариабельной области легкой цепи #131L2h гуманизированного антитела.
На фигурах 18(a)-(d) представлены данные по противоопухолевой активности гуманизированных антител в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток MIA PaCa-2.
На фигуре 19 представлены данные по фосфорилированию p38MAPK при воздействии человеческого химерного антитела против человеческого CD147.
На фигурах 20 (a) и (b) представлены данные по фосфорилированию HSP27 при воздействии человеческого химерного антитела против человеческого CD147.
[0029]
На фигурах 21(a)-(c) представлены данные по фосфорилированию p38MAPK при воздействии человеческого химерного антитела против человеческого CD147 в опухолях.
На фигурах 22(a)-(c) представлены данные по экспрессии ipo8, cxcl8 и rhoB при воздействии человеческих химерных антител против человеческого CD147.
На фигуре 23 (a)-(c) представлены данные по экспрессии ipo8, cxcl8 и rhoB при воздействии крысиного антитела и его человеческого химерного антитела против человеческого CD147.
На фигуре 24 приведены результаты оценки противоопухолевой активности антител против CD147 и гемцитабина с использованием клеток BxPC-3 SMAD4-положительного рака поджелудочной железы.
На фигурах 25(a)-(c) приведены результаты оценки противоопухолевой активности человеческих химерных антител против CD147 с использованием клеток BxPC-3, стабильно экспрессирующих SMAD4.
На фигурах 26(a) и (b) представлены данные по фосфорилированию p38MAPK при воздействии человеческого химерного антитела против человеческого CD147 в модели рака поджелудочной железы, вызываемого путем подкожной имплантации клеток BxPC-3, экспрессирующих SMAD4.
На фигуре 27 представлены данные по противоопухолевой активности человеческого химерного антитела против CD147 в модели резистентного к гемцитабину рака поджелудочной железы.
На фигуре 28(a) представлено подтверждение экспрессии CD147 в клетках Hep G2 с использованием метода проточной цитометрии. На фигуре 28(b) представлены данные по активации p38MAPK при воздействии гуманизированных антител против CD147 в клетках рака печени.
На фигурах 29(a)-(d) приведены результаты сравнения противоопухолевой активности антител против CD147 с сорафенибом при раке печени.
На фигуре 30 показано CD147-APC связывание в CD3- и CD4-положительных клетках и CD3- и CD8-положительных клетках.
[0030]
На фигуре 31 приведены результаты оценки влияния антител против человеческого CD147 на рост мононуклеарных клеток периферической крови человека.
На фигуре 32 приведены результаты оценки влияния антител против человеческого CD147 на продукцию цитокинов лимфоцитами периферической крови человека.
На фигуре 33-1 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи крысиного_CD147_#84, и аминокислотные последовательности CDRL1, CDRL2 и CDRL3 легкой цепи крысиного_CD147_#84.
На фигуре 33-2 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области тяжелой цепи крысиного_CD147_#84, и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи крысиного_CD147_#84.
На фигуре 34-1 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи крысиного_CD147_#101, и аминокислотные последовательности CDRL1, CDRL2 и CDRL3 легкой цепи крысиного_CD147_#101.
На фигуре 34-2 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области тяжелой цепи крысиного_CD147_#101, и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи крысиного_CD147_#101.
На фигуре 35-1 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи крысиного_CD147_#110, и аминокислотные последовательности CDRL1, CDRL2 и CDRL3 легкой цепи крысиного_CD147_#110.
На фигуре 35-2 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельная область тяжелой цепи крысиного_CD147_#110, и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи крысиного_CD147_#110.
На фигуре 36-1 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области легкой цепи крысиного_CD147_#131, и аминокислотные последовательности CDRL1, CDRL2 и CDRL3 легкой цепи крысиного_CD147_#131.
На фигуре 36-2 представлена нуклеотидная последовательность и аминокислотная последовательность вариабельной области тяжелой цепи крысиного_CD147_#131, и аминокислотные последовательности CDRH1, CDRH2 и CDRH3 тяжелой цепи крысиного_CD147_#131.
На фигуре 37-1 представлена аминокислотная последовательность и нуклеотидная последовательность #84H1hIgG2.
На фигуре 37-2 представлена аминокислотная последовательность и нуклеотидная последовательность #84H1hIgG4P.
На фигуре 37-3 представлена аминокислотная последовательность и нуклеотидная последовательность #84L2h.
На фигуре 38-1 представлена аминокислотная последовательность и нуклеотидная последовательность #101H1hIgG2.
На фигуре 38-2 представлена аминокислотная последовательность и нуклеотидная последовательность #101H1hIgG4P.
На фигуре 38-3 представлена аминокислотная последовательность и нуклеотидная последовательность #101L2h.
На фигуре 39-1 представлена аминокислотная последовательность и нуклеотидная последовательность #110H1hIgG4P.
На фигуре 39-2 представлена аминокислотная последовательность и нуклеотидная последовательность #110H13hIgG4P.
На фигуре 39-3 представлена аминокислотная последовательность и нуклеотидная последовательность #110L4h.
На фигуре 39-4 представлена аминокислотная последовательность и нуклеотидная последовательность #110L2h.
На фигуре 39-5 представлена аминокислотная последовательность и нуклеотидная последовательность #110L12h.
На фигуре 40-1 представлена аминокислотная последовательность и нуклеотидная последовательность #131H2hIgG2.
На фигуре 40-2 представлена аминокислотная последовательность и нуклеотидная последовательность #131L2h.
[0031]
На фигуре 41 изображена ленточная схема, показывающая два комплекса, содержащихся в асимметричном элементе. CD147 изображен черным цветом, а тяжелая цепь (H CHAIN) и легкая цепь (L CHAIN) антитела изображены серым цветом.
На фигуре 42 изображена поверхность взаимодействия между CD147 и антителом. Аминокислоты в CD147 вблизи антитела представлены в форме веточек и обозначены буквами. Другие части CD147 представлены в форме черных ленточек. При этом, аминокислоты антитела вблизи CD147 представлены в форме тонких линий, а другие части антитела представлены в форме серых ленточек.
На фигуре 43 представлены данные по противоопухолевой активности химерных и гуманизированных антител против CD147 в модели рака желудка.
На фигуре 44 представлены данные по противоопухолевой активности гуманизированного антитела против CD147 в модели хронического миелолейкоза (CML).
На фигуре 45 представлены данные по противоопухолевой активности гуманизированных антител против CD147 в модели рака толстой кишки.
На фигуре 46 представлены данные по противоопухолевой активности гуманизированных антител против CD147 в модели рака почки.
На фигуре 47 представлены данные по противоопухолевой активности гуманизированного антитела против CD147 в модели острого миелолейкоза (AML).
На фигуре 48 представлены данные по противоопухолевой активности гуманизированных антител против CD147 в модели рака поджелудочной железы.
На фигуре 49 приведены результаты конкурентного твердофазного иммуноферментного анализа (ELISA).
На фигуре 50 (a) представлены данные по противоопухолевой активности гуманизированного антитела против CD147 при использовании клеток MIA PaCa-2, и на фигуре 50 (b) для сравнения представлены данные по противоопухолевой активности гуманизированного антитела против CD147 при использовании клеток MIA PaCa-2, экспрессирующих KLF5.
Описания вариантов осуществления изобретения
[0032]
Предпочтительные варианты осуществления настоящего изобретения описываются ниже со ссылкой на чертежи. Описываемые далее варианты осуществления иллюстрируют примеры типичных вариантов осуществления изобретения, и не следует считать, что объем изобретения ограничивается этими вариантами осуществления.
[0033]
Определение
Используемые в изобретении термины "рак (раковые заболевания)" и "опухоль (опухоли)" применяют как синонимы, и каждый термин используют для обозначения солидного рака, несолидного рака или и того и другого, за исключением тех случаев, когда для значения этого термина вводят ограничение.
[0034]
Примеры используемого в изобретении термина "ген (гены)" включают не только ДНК, но также ее мРНК, и кДНК и ее кРНК.
[0035]
Используемый в изобретении термин "полинуклеотид (полинуклеотиды)" или "нуклеотид (нуклеотиды)" применяется в качестве синонима "нуклеиновой кислоты", и его примеры включают ДНК, РНК, зонды, олигонуклеотиды и праймеры.
[0036]
Термины "полипептид (полипептиды)" и "белок (белки)" используются в изобретении взаимозаменяемо.
[0037]
Примеры используемого в изобретении термина "клетки" включают клетки индивидуального животного и культивируемые клетки.
[0038]
Используемый в изобретении термин "CD147" применяют как синоним "белка CD147".
[0039]
Используемый в изобретении термин "функциональный фрагмент антитела", называемый также "антигенсвязывающим фрагментом антитела", относится к отдельному фрагменту антитела, обладающему активностью связывания с антигеном. Его примеры включают Fab, F(ab')2, Fv, scFv, диатело, линейное антитело и полиспецифическое антитело, образованное из фрагментов антител. Примеры антигенсвязывающего фрагмента антитела также включают Fab', который представляет собой моновалентный фрагмент вариабельной области антитела и который получают путем обработки с помощью F(ab')2 в восстановительных условиях. Однако, примеры антигенсвязывающего фрагмента не ограничиваются этими молекулами, могут применяться и другие молекулы при условии, что они способны связываться с антигеном. Примеры этих антигенсвязывающих фрагментов включают не только фрагменты, полученные путем обработки молекул белка полноразмерного антитела с помощью соответствующих ферментов, но также белки, продуцированные соответствующими клетками-хозяина с использованием генов антитела, модифицированных методами генной инженерии.
[0040]
Используемый в изобретении термин "эффекторная активность" относится или к одному типу или к нескольким типам опосредованной антителами клеточной цитотоксичности (далее обозначаемой как "ADCC"), комплементзависимой цитотоксичности (далее обозначаемой как "CDC") и антителозависимого клеточного фагоцитоза (далее обозначаемого как "ADCP").
[0041]
Используемый в изобретении термин "эффекторные функции" относится к проявлению одной или более "эффекторной активности".
[0042]
Опосредованной антителами клеточная цитотоксичность (ADCC) может быть оценена методом анализа, основанном на высвобождении 51Cr, который позволяет измерять гибель клеток, вызванную контактированием иммуноцитов, обладающих эффекторной активностью, антитела и клеток-мишеней, меченных изотопом 51Cr. ADCC антитела против человеческого CD147 по настоящему изобретению измеряют следующим образом. При определении ADCC антитела против человеческого CD147, в качестве эффекторных клеток используют мононуклеарные клетки периферической крови человека (PBMC), а в качестве клеток-мишеней при оценке ADCC используют клеточную линию CD147-позитивного рака человека (например, линию клеток MIA PaCa-2 рака поджелудочной железы). Раковые клетки, меченные радиоизотопом 51Cr, и антитело, подлежащее оценке, подвергают обработке при концентрации 0,5 или 5 мкг/мл при 4°C в течение 30 минут, добавляют PBMC, выделенные из периферической крови человека, в количестве в 20 раз большем, чем раковых клеток, а затем клетки культивируют в течение 4 часов при 37°С в присутствии 5% СО2. Суммарную величину высвобождения получают путем измерения 51Cr, высвобождающегося в надосадочную жидкость, с использованием прибора TopCount NXT v2,53. Измеренное величину 51Cr, высвобождаемого из меченных 51Cr раковых клеток, обработанных неионным поверхностно-активным веществом Triton-100, обозначают как "максимальную величину высвобождения", а измеренную величину 51Cr, высвобождаемого из меченных 51Cr раковых клеток, обработанных антителом в отсутствие РВМС, обозначают как "величину самопроизвольного высвобождения". Используя эти величины, рассчитывают % специфического высвобождения по следующей формуле. В качестве отрицательного контрольного образца используют человеческий IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Cat. 009-000-003). Измерение выполняют три раза, и затем рассчитывают среднее величину и стандартное отклонение.
[0043]
% Специфического высвобождения = (суммарная величина высвобождения-величина самопроизвольного высвобождения) / максимальная величина высвобождения
Комплементзависимая цитотоксичность (CDC) может быть определена путем оценки гибели клеток, вызванной контактированием комплемента, содержащегося в крови, антитела и клеток-мишеней. CDC антитела против человеческого CD147 по настоящему изобретению измеряют следующим образом. Комплементзависимую цитотоксичность (CDC) антитела против CD147 человека оценивают с использованием линии клеток MIA PaCa-2 поджелудочной железы человека в качестве клеток-мишеней. В качестве комплемента используют выпускаемый промышленностью кроличий комплемент (Low Tox-M Rabbit Complement, CEDARLANE LABORATORIES LIMITED, Cat. CL3051). В качестве CDC-отрицательного контрольного антитела используют человеческий IgG (hIgG, ChromPureHuman IgG, Jackson ImmunoResearch Laboratories, Cat. 009-000-003). Клетки-мишени обрабатывают антителом, подлежащим оценке, и антителом отрицательного контроля при концентрациях 0, 0,1, 1 или 10 мкг/мл в течение 1 часа при 4°С, добавляют кроличий комплемент в таком количестве, чтобы его конечная концентрация составляла 7,5%, и полученную смесь нагревают при 37°С в присутствии 5% СО2 в течение 3 часов, а затем измеряют внутриклеточный АТФ, содержащийся в живых клетках, с использованием люминесцентного метода анализа жизнеспособности клеток CellTiter-Glo Lumimescent Cell Viability Assay (Promega, Cat. G7572). Люминесцентные сигналы, полученные с помощью люминесцентного метода анализа жизнеспособности клеток CellTiter-Glo Lumimescent Cell Viability Assay, измеряют с использованием мультимодального ридера EnVision 2104 Multilabel Reader (Perkin Elmer). Измерение выполняют три раза, и рассчитывают среднюю величину и стандартное отклонение. Люминесцентные сигналы, полученные от не подвергавшихся обработке клеток, обозначают как 100%, а пониженные люминесцентные сигналы, которые зависят от антитела и комплемента, обозначают как CDC.
[0044]
Антителозависимый клеточный фагоцитоз (ADCP) может быть оценен путем измерения фагоцитоза, вызванного контактированием иммуноцитов, способных к фагоцитозу, антитела и клеток-мишеней, методом двойной флуоресцентной маркировки.
[0045]
ADCP антитела против человеческого CD147 по настоящему изобретению измеряют следующим образом. Сообщалось, что человеческое антитело IgG проявляет цитотоксическую активность против раковых клеток путем индуцирования антителозависимого клеточного фагоцитоза (ADCP) моноцитами и макрофагами через взаимодействие с Fcγ-рецептором мыши (Overdijk et al., Journal of Immunology, 1-9, 2012 ). ADCP человеческого химерного антитела по настоящему изобретению оценивают с использованием RAW264.7, (ATCC, TIB-71) в качестве эффекторных клеток, и линии клеток поджелудочной железы человека PANC-1 или MIA PaCa-2, в качестве клеток-мишеней для ADCP. После 1 часа обработки клеток-мишеней для ADCP, меченных с использованием мини-набора для общей маркировки клеточных мембран PKH67 Green Fluorescent Cell Linker Mini Kit for General Cell Membrane Labeling (SIGMA, Cat. MINI67-1KIT), с помощью подлежащего оценке антитела с концентрацией 20 мкг/мл при 4°C, добавляют клетки RAW264.7, меченные с использованием набора PKH26 Red Fluorescent Cell Linker Kit for General Cell Membrane Labeling (SIGMA, Cat. PKH26GL-1KT), в количестве в 5 раз большим, чем количество клеток-мишеней для ADCP, затем нагревают в течение 3 часов при 37°C в присутствии 5% CO2. Используя проточный цитометр (BD (Becton, Dickinson and Company), CantoII), измеряют долю PKH26-позитивных клеток, которые относят к положительному статусу сигнала PKH67 вследствие фагоцитоза. В качестве негативного контрольного образца, проводят аналогичное измерение для образца, обработанного человеческим IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Cat. 009-000-003). Измерение выполняют три раза, и затем рассчитывают среднее величину и стандартное отклонение.
[0046]
Используемое в изобретении выражение "практически отсутствует эффекторная активность" или "характеризуется пониженной эффекторной активностью или отсутствием эффекторной активности" означает, что антитело не проявляет, по меньшей мере, один из видов ADCC, CDC или ADCP, или что уровни этих видов токсичности являются настолько низкими, что эффекторные функции практически не проявляются. Выражение "практически отсутствует эффекторная активность" или "характеризуется пониженной эффекторной активностью или отсутствием эффекторной активности" означает, например, что эффекторная активность подлежащего оценке антитела находится на том же уровне, что и эффекторная активность отрицательного контроля, определяемая одним из упомянутых выше методов оценки эффекторной активности.
[0047]
Используемое в изобретении выражение "характеризуется пониженной опосредованной антителами клеточной цитотоксичностью (ADCC) или отсутствием ADCC" означает, что подлежащее оценке антитело не проявляет ADCC, или ADCC находится на низком уровне, так что она не проявляется в достаточной степени. Выражение "характеризуется пониженной опосредованной антителами клеточной цитотоксичностью (ADCC) или отсутствием ADCC" означает, например, что цитотоксичность подлежащего оценке антитела находится на том же уровне, что и цитотоксичность отрицательного контроля, определяемая упомянутым выше методом оценки токсичности.
[0048]
Выражение "характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или отсутствием CDC" означает, что подлежащее оценке антитело не проявляет CDC или CDC находится на низком уровне, так что она не проявляется в достаточной степени. Выражение "характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или отсутствием CDC" означает, например, что цитотоксичность подлежащего оценке антитела находится на том же уровне, что и цитотоксичность отрицательного контроля, определяемая упомянутым выше методом оценки токсичности.
[0049]
Выражение "характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или отсутствием ADCP" означает, что подлежащее оценке антитело не проявляет ADCP или что ADCP находится на низком уровне, так что он не проявляется в достаточной степени. Выражение "характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или отсутствием ADCP" означает, например, что ADCP подлежащего оценке антитела находится на том же уровне, что и ADCP отрицательного контроля, определяемый упомянутым выше методом оценки ADCP.
[0050]
Выражение "характеризуется пониженной опосредованной антителами клеточной цитотоксичностью (ADCC) или отсутствием ADCC", "характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или отсутствием CDC" или "характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или отсутствием ADCP" означает, например, что токсичность каждого подлежащего оценке антитела находится на том же уровне, что и токсичность отрицательного контроля, определяемая упомянутыми выше методами оценки токсичности.
[0051]
Используемое в изобретении выражение "активирует сигнальную трансдукцию посредством CD147", "активация связанных с сигнальным путем молекул посредством CD147", "активация CD147" или "активирует CD147", относится к активации одной или нескольких систем клеточной сигнальной трансдукции посредством CD147 и означает, что активируется, по меньшей мере, одна из связанных с сигнальным путем молекул, расположенных по потоку ниже CD147. Активация сигнальной трансдукции посредством CD147 означает, что ускоряется или уменьшается экспрессия гена, нисходящего относительно сигнала CD147, ускоряется или уменьшается экспрессия белка или ускоряется или уменьшается фосфорилирование белка. Примеры связанных с сигнальным путем молекул, которые расположены ниже CD147, включают FAK, MEK, Erk, JAK/STAT, AKT или митоген-активируемую протеинкиназу (MAPK) или активацию сигнальных молекул, расположенных ниже CD147. Примеры MAPK включают ERK1/2, JNK или p38MAPK, и, более предпочтительно, p38MAPK. Примеры сигнальных молекул, расположенных ниже по потоку от MAPK, включают HSP27, cxcl8 или SMAD (например, SMAD2, SMAD3 и/или SMAD4). Примеры "активации CD147" включают повышение уровня экспрессии p38MAPK мРНК, повышение уровня экспрессии белка p38MAPK, фосфорилирование p38MAPK, фосфорилирование HSP27 (например, фосфорилирование Ser82 HSP27 или фосфорилирование Ser15 HSP27), повышение уровня экспрессии cxcl8 мРНК, повышение уровня экспрессии белка cxcl8, повышение уровня экспрессии rhoB-мРНК или повышение уровня экспрессии белка rhoB через активацию сигнала SMAD, или снижение мРНК снижение уровня экспрессии KLF5 или белка KLF5.
[0052]
Используемый в изобретении термин "эпитоп" относится к неполному пептиду или к неполной трехмерной структуре CD147, с которыми связывается конкретное антитело против CD147. Вышеупомянутый эпитоп, который является неполным пептидом CD147, может быть определен методом, хорошо известным специалистам в данной области, таким как метод иммуноанализа. Сначала готовят различные неполные структуры антигенов. При получении неполных структур могут быть использованы известные методы синтеза олигонуклеотидов. Например, ряд полипептидов получают методами рекомбинации генов, известными специалистам в данной области техники, следующим образом: последовательно сокращают длины полипептидов от С-конца или N-конца CD147, исследуют реакционную способность антитела к этим полипептиды, приблизительно определяют сайты узнавания, синтезируют пептиды, которые еще короче, чем полипептиды, и затем исследуют реакционную способность по отношению к этим пептидам, и таким образом может быть определен эпитоп. Кроме того, когда эпитоп антитела, который связывается с мембранным белком, состоящим из множества внеклеточных доменов, представляет собой трехмерную структуру, состоящую из множества доменов, может быть модифицирована аминокислотная последовательность конкретного внеклеточного домена, для того чтобы модифицировать трехмерную структуру, в результате чего можно определить конкретный домен, с которым связывается антитело. Эпитоп, который представляет собой неполную трехмерную структуру антигена, с которой связывается специфическое антитело, также может быть определен путем установления аминокислотных остатков антигена, расположенных рядом с упомянутым выше антителом, с помощью рентгеноструктурного анализа.
[0053]
Если второе антитело связывается с неполным пептидом или с неполной трехмерной структурой, с которыми связывается первое антитело, то может быть установлено, что первое антитело и второе антитело имеют общий эпитоп. Путем подтверждения того, что второе антитело перекрестно конкурирует с первым антителом за связывание с антигеном (в частности, второе антитело ингибирует связывание первого антитела с антигеном), может быть установлено, что первое антитело и второе антитело связываются с одним и тем же эпитопом, даже если не определены конкретная последовательность или структура эпитопа. В случае, когда первое антитело и второе антитело связываются с одним и тем же эпитопом, и первое антитело обладает особым эффектом, таким как противоопухолевая эффективность, то можно ожидать, что второе антитело имеет аналогичную активность.
[0054]
Известно, что каждая из тяжелых цепей и легких цепей молекулы антитела имеет 3 определяющие комплементарность области (CDR). Определяющие комплементарность области также называют гипервариабельными областями (гипервариабельными доменами), и они расположены в тяжелой цепи и в вариабельных областях легких цепей антитела, и являются сайтами, где вариабельность первичной структуры особенно высока, и расположены отдельно в первичной структуре полипептидных цепей тяжелых цепей и легких цепей. Что касается используемого в изобретении термина "определяющие комплементарность области антитела», то определяющие комплементарность области тяжелой цепи обозначаются как CDRH1, CDRH2 и CDRH3 с амино-концевой стороны тяжелой цепи аминокислотной последовательности, а определяющие комплементарность области легкой цепи обозначаются как CDRL1, CDRL2 и CDRL3 с амино-концевой стороны легкой цепи аминокислотной последовательности. Эти сайты находятся близко друг от друга на трехмерной структуре и определяют специфичность к антигену, с которым связывается антитело.
[0055]
Термин "несколько", используемый в изобретении в выражениях "от 1 до некоторой величины" и "1 или несколько" обозначает от 2 до 10, предпочтительно, 10 или менее, более предпочтительно, 5 или 6 или менее, еще более предпочтительно, 2 или 3.
[0056]
(CD147)
CD147 представляет собой однопроходный трансмембранный белок, имеющий от 2 до 3 иммуноглобулиноподобных доменов, и известно, что он активирует связанные с нисходящим сигнальным путем молекулы, такие как молекулы семейства FAK, MEK, Erk, JAK/STAT, AKT и MAPK, в результате взаимодействий между множеством белков CD147 или в результате взаимодействий с молекулами поверхности внеклеточной или клеточной мембраны, которые участвуют в росте, инвазии и воспалении, такими как CD44, молекулы семейства интегринов, CD98, VEGFR, CypA/B и MCT1/3/4, вследствие чего ускоряется продуцирование протеаз, таких как MMP, и рост рака, метастазирование и инвазия.
[0057]
Известны три варианта человеческого CD147. Вариант 1 экспрессируется специфично в сетчатке глаза и представляет собой однопроходный трансмембранный белок, имеющий 3 иммуноглобулиноподобных домена (эти домены могут обозначаться в изобретении как D0, D1 и D2, соответственно). Вариант 2 экспрессируется в T-клетках или различных нормальных клетках и представляет собой однопроходный трансмембранный белок, имеющий 2 иммуноглобулиноподобных домена (D1, D2), и сообщалось об его повышенной экспрессии в различных раковых тканях. Вариант 3 представляет собой однопроходный трансмембранный белок, имеющий 1 иммуноглобулиноподобный домен.
[0058]
Аминокислотная последовательность и нуклеотидная последовательность варианта 1 человеческого CD147 доступны в базе данных GenBank по номеру доступа Accession Nos. NP_001719.2 и NM_001728.3, и аминокислотная последовательность представлена в изобретении последовательностью SEQ ID NO:1, и нуклеотидная последовательность представлена в изобретении последовательностью SEQ ID NO:2. Три иммуноглобулиноподобных домена варианта 1 имеют аминокислоты в диапазоне номеров 22-138 (D0), аминокислоты в диапазоне номеров 140-218 (D1) и аминокислоты в диапазоне номеров 223-323 (D2), соответственно, в последовательности SEQ ID NO:1 (Redzic, J., J. Mol. Biol., 2011, 68-82)(Grass et al., Biosol. Rep, 2016, 1-16). Трансмембранная область варианта 1 имеют аминокислоты в диапазоне номеров 324-344 в последовательности SEQ ID NO:1.
[0059]
Аминокислотная последовательность и нуклеотидная последовательность варианта 2 человеческого CD147 доступны в базе данных GenBank по номеру доступа Accession Nos. NP_940991.1 и NM_198589.2, и аминокислотная последовательность представлена в изобретении последовательностью SEQ ID NO:3, и нуклеотидная последовательность представлена в изобретении последовательностью SEQ ID NO:4. Два иммуноглобулиноподобных домена (D1, D2) варианта 2 имеют аминокислоты в диапазоне номеров 24-102 (D1) и аминокислоты в диапазоне номеров 107-207 (D2), соответственно, в последовательности SEQ ID NO:3. The трансмембранная область варианта 2 имеет аминокислоты в диапазоне номеров 208-228 в последовательности SEQ ID NO:3 (Grass et al., Biosol. Rep, 2016, 1-16).
[0060]
Аминокислотная последовательность и нуклеотидная последовательность варианта 3 человеческого CD147 доступны в базе данных GenBank по номеру доступа Accession Nos. NP_940992.1 и NM_198590.2. Человеческий ген CD147 также доступен из коммерческого источника.
[0061]
Аминокислотная последовательность и нуклеотидная последовательность CD147 яванского макака (обозначаемая в изобретении также как обезьяний CD147) доступны в базе данных GenBank по номеру доступа Accession Nos. XP_005587354.1 и XM_005587297.1. Ген обезьяньего CD147 доступен также из коммерческого источника. Аминокислотная последовательность и нуклеотидная последовательность мышиного CD147 доступны в базе данных GenBank по номеру доступа Accession Nos. NP_001070652.1 и NM_001077184.1. Ген мышиного CD147 доступен также из коммерческого источника.
[0062]
Используемый в настоящем изобретении CD147 может быть получен путем синтеза CD147 in vitro или путем использования созданных методами генной инженерии клеток-хозяина для продуцирования CD147. В частности, CD147 кДНК вводят в вектор, способный его экспрессировать, и затем синтезируют CD147 в растворе, содержащем фермент, субстрат и энергетические вещества, требующиеся для транскрипции и трансляции, или клетки-хозяина других прокариотов или эукариотов трансформируют для экспрессии CD147, в результате чего может быть получен белок.
[0063]
кДНК CD147 может быть получена полимеразной цепной реакцией (далее обозначаемой в изобретении "PCR"), а именно, например, методом PCR с использованием библиотеки кДНК, экспрессирующей кДНК CD147, в качестве матрицы, и праймеров для специфической амплификации кДНК CD147 (Saiki, R. K., et al., Science, (1988)239, 487-49).
[0064]
Примеры кДНК CD147 включают полинуклеотид, гибридизирующийся в жестких условиях в полинуклеотид, состоящий из нуклеотидной последовательности, комплементарной к нуклеотидной последовательности, кодирующей человеческий, обезьяний или мышиный CD147, и кодирующий белок, имеющий биологическую активность, эквивалентную активности CD147. Кроме того, примеры кДНК CD147 включают полинуклеотид, который является сплайсированным вариантом, транскрибированным из человеческого, обезьяньего или мышиного генного локуса CD147, или полинуклеотид, гибридизирующийся в него в жестких условиях и кодирующий белок, имеющий биологическую активность, эквивалентную активности CD147.
[0065]
Примеры CD147 дополнительно включают белок, состоящий из аминокислотной последовательности человеческого, обезьяньего или мышиного CD147, или из аминокислотной последовательности, в которой одна или несколько аминокислот заменены, удалены или добавлены в аминокислотной последовательности человеческого, обезьяньего или мышиного CD147, из которого удалена сигнальная последовательность; и имеющий биологическую активность, эквивалентную активности CD147. Кроме того, примеры CD147 включают белок, состоящий из аминокислотной последовательности, кодированной сплайсированным вариантом, транскрибированным из человеческого, обезьяньего или мышиного генного локуса CD147, или из аминокислотной последовательности, в которой заменены, удалены или добавлены одна или несколько аминокислот; и имеющий биологическую активность, эквивалентную активности CD147.
[0066]
(Получение антитела против CD147)
Антитело против CD147 по настоящему изобретению может быть получено следующим образом. А именно, низшее животное подвергают иммунизации представляющим интерес антигеном, после выработки иммунитета у животного собирали лимфу, лимфатическую ткань, образец гемацита или клетки спинного мозга, и затем продуцирующие антитело клетки, которые продуцируют антитело против CD147, и миеломные клетки подвергают слиянию известным методом (например, как описано в публикации Kohler and Milstein, Nature(1975)256, p.495-497, Kennet, R. ed., Monoclonal Antibodies, p.365-367, Plenum Press, N. Y. (1980)) с образованием гибридом, и в результате может быть получено моноклональное тело. Конкретные примеры такого метода описаны в патентных документах WO2009/48072 (зарегистрирован 16 апреля 2009 года) и WO2010/117011 (зарегистрирован 14 октября 2010 года). Примеры полученного таким образом моноклонального антитела могут включать LN22R8, 2P10F2, rat_CD147_#84, rat_CD147_#101, rat_CD147_#110 и rat_CD147_#131. Однако, методы получения моноклонального антитела подпадают под категорию уже известных методов, и их примеры не ограничиваются перечисленными выше конкретными примерами.
[0067]
Примеры антитела по настоящему изобретению включают, помимо упомянутого выше моноклонального антитела против CD147, ген рекомбинантного антитела, который искусственно модифицирован для снижения гетерологичных антигенных свойств в отношении людей, например, химерное антитело, гуманизированное антитело и человеческое антитело. Эти антитела могут быть получены известными методами.
[0068]
Примеры химерного антитела могут включать антитела, в которых вариабельные области и константные области антитела являются гетерологичными друг другу, например, химерное антитело, в котором вариабельная область мышиного или крысиного антитела объединена с константной областью человеческого антитела (смотрите публикацию Proc. Natl. Acad. Sci. USA., 81, 6851-6855, (1984)). Примером химерного антитела, произведенного из LN22R8, является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-471 последовательности SEQ ID NO:33, из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-467 последовательности SEQ ID NO:35, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-468 последовательности SEQ ID NO:37, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:31 в перечне последовательностей. Примером 2P10F2-произведенного химерного антитела является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-466 последовательности SEQ ID NO:43, тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-462 последовательности SEQ ID NO:45, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-463 последовательности SEQ ID NO:47, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:41, в перечне последовательностей.
[0069]
Примером химерного антитела, произведенного из rat_CD147_#84, является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-470 последовательности SEQ ID NO:92, тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-466 последовательности SEQ ID NO:94, и тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-467 последовательности SEQ ID NO:96, тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-470 последовательности SEQ ID NO:98, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-467 последовательности SEQ ID NO:100, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:90, в перечне последовательностей.
[0070]
Примером химерного антитела, произведенного из rat_CD147_#101, является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-463 последовательности SEQ ID NO:104, тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-464 последовательности SEQ ID NO:106, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-464 последовательности SEQ ID NO:108, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:102, в перечне последовательностей.
[0071]
Примером химерного антитела, произведенного из rat_CD147_#110, является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-462 последовательности SEQ ID NO:112, тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-463 последовательности SEQ ID NO:114, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-463 последовательности SEQ ID NO:116, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:110, в перечне последовательностей.
[0072]
Примером химерного антитела, произведенного из rat_CD147_#131, является антитело, состоящее из тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-464 последовательности SEQ ID NO:120, или тяжелой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 20-465 последовательности SEQ ID NO:122, и легкой цепи, имеющей аминокислотную последовательность, состоящую из аминокислотных остатков в положениях 21-234 последовательности SEQ ID NO:118, в перечне последовательностей.
[0073]
Примеры гуманизированного антитела могут включать антитело, в котором только CDRs введены в человеческое антитело (смотрите публикацию Nature(1986)321, p.522-525), антитело, в котором аминокислотные остатки неполного каркаса в дополнении к CDR последовательности также введены в человеческое антитело методом трансплантации CDR (патентный документ International Publication WO90/07861).
[0074]
Примеры антитела по настоящему изобретению включают любое гуманизированное антитело, произведенное из rat_CD147_#84 антитела, при условии, что оно сохраняет все 6 типов CDR последовательности rat_CD147_#84, имеет активность связывания с CD147, и является антителом, активирующим CD147. Следует отметить, что вариабельная область тяжелой цепи rat_CD147_#84 антитела включает CDRH1, состоящую из аминокислотной последовательности SEQ ID NO:55, CDRH2, состоящую из аминокислотной последовательности SEQ ID NO:56, и CDRH3, состоящую из аминокислотной последовательности SEQ ID NO:57. Вариабельная область легкой цепи rat_CD147_#84 антитела включает CDRL1, состоящую из аминокислотной последовательности SEQ ID NO:52, CDRL2, состоящую из аминокислотной последовательности SEQ ID NO:53, и CDRL3, состоящую из аминокислотной последовательности SEQ ID NO:54. Аминокислотная последовательность и нуклеотидная последовательность вариабельной области легкой цепи или вариабельной области тяжелой цепи rat_CD147_#84 антитела и аминокислотные последовательности CDRs также представлены на фигуре 33-1 и фигуре 33-2.
[0075]
Примеры антитела по настоящему изобретению включают гуманизированное антитело, произведенное из rat_CD147_#101 антитела, при условии, что оно сохраняет все 6 типов CDR последовательности rat_CD147_#101, имеет активность связывания с CD147, и является антителом, активирующим CD147. Следует отметить, что вариабельная область тяжелой цепи rat_CD147_#101 антитела включает CDRH1, состоящую из аминокислотной последовательности SEQ ID NO:65, CDRH2, состоящую из аминокислотной последовательности SEQ ID NO:66, и CDRH3, состоящую из аминокислотной последовательности SEQ ID NO:67. Вариабельная область легкой цепи rat_CD147_#101 антитела включает CDRL1, состоящую из аминокислотной последовательности SEQ ID NO:62, CDRL2, состоящую из аминокислотной последовательности SEQ ID NO:63, и CDRL3, состоящую из аминокислотной последовательности SEQ ID NO:64. Аминокислотная последовательность и нуклеотидная последовательность вариабельной области легкой цепи или вариабельной области тяжелой цепи rat_CD147_#101 антитела и аминокислотные последовательности CDRs также представлены на фигуре 34-1 и фигуре 34-2.
[0076]
Примеры антитела по настоящему изобретению включает гуманизированное антитело, произведенное из rat_CD147_#110 антитела, при условии, что оно сохраняет все 6 типов CDR последовательности rat_CD147_#110, имеет активность связывания с CD147, и является антителом, активирующим CD147. Следует отметить, что вариабельная область тяжелой цепи rat_CD147_#110 антитела включает CDRH1, состоящую из аминокислотной последовательности SEQ ID NO:75, CDRH2, состоящую из аминокислотной последовательности SEQ ID NO:76, и CDRH3, состоящую из аминокислотной последовательности SEQ ID NO:77. Вариабельная область легкой цепи rat_CD147_#110 антитела включает CDRL1, состоящую из аминокислотной последовательности SEQ ID NO:72, CDRL2, состоящую из аминокислотной последовательности SEQ ID NO:73, и CDRL3, состоящую из аминокислотной последовательности SEQ ID NO:74. Аминокислотная последовательность и нуклеотидная последовательность вариабельной области легкой цепи или вариабельной области тяжелой цепи rat_CD147_#110 антитела и аминокислотные последовательности CDRs также представлены на фигуре 35-1 и фигуре 35-2.
[0077]
Примеры антитела по настоящему изобретению включают гуманизированное антитело, произведенное из rat_CD147_#131 антитела, при условии, что оно сохраняет все 6 типов CDR последовательности rat_CD147_#131, имеет активность связывания с CD147, и является антителом, активирующим CD147. Следует отметить, что вариабельная область тяжелой цепи rat_CD147_#131 антитела включает CDRH1, состоящую из аминокислотной последовательности SEQ ID NO:85, CDRH2, состоящую из аминокислотной последовательности SEQ ID NO:86, и CDRH3, состоящую из аминокислотной последовательности SEQ ID NO:87. Вариабельная область легкой цепи rat_CD147_#131 антитела включает CDRL1, состоящую из аминокислотной последовательности SEQ ID NO:82, CDRL2, состоящую из аминокислотной последовательности SEQ ID NO:83, и CDRL3, состоящую из аминокислотной последовательности SEQ ID NO:84. Аминокислотная последовательность и нуклеотидная последовательность вариабельной области легкой цепи или вариабельной области тяжелой цепи rat_CD147_#131 антитела и аминокислотные последовательности CDRs также представлены на фигуре 36-1 и фигуре 36-2.
[0078]
Примеры антитела по настоящему изобретению дополнительно включают CDR-модифицированное гуманизированное антитело, в котором от 1 до 3 аминокислотных остатков в каждой CDR заменены на другие аминокислотные остатки, при условии, что это антитело имеет активность связывания с CD147, и является антителом, активирующим CD147.
Примеры гуманизированного антитела, произведенного из rat_CD147_#84 антитела, включают гуманизированное антитело против CD147 или антигенсвязывающий фрагмент антитела, имеющее следующие области (a) и (b):
(a) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих областей (a1)-(a4):
(a1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123;
(a2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125;
(a3) аминокислотной последовательности, имеющая идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (a1) или (a2); и
(a4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (a1)-(a3); и
(b) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих последовательностей (b1)-(b3):
(b1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127;
(b2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (b1); и
(b3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (b1) и (b2).
[0079]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#84 антитела, включают антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127, или антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127.
[0080]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#84 антитела, включают антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-467 в аминокислотной последовательности SEQ ID NO:125, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:127, или антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-466 в аминокислотной последовательности SEQ ID NO:123, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:127.
[0081]
Примеры гуманизированного антитела, произведенного из rat_CD147_#101 антитела, включают гуманизированное антитело против CD147 или антигенсвязывающий фрагмент антитела, имеющие следующие области (e) и (f):
(e) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих областей (e1)-(e4):
(e1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129;
(e2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131;
(e3) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (e1) или (e2); и
(e4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (e1) -- (e3); и
(f) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих областей (f1)-(f3):
(f1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133;
(f2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (f1); и
(f3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (f1) или (f2).
[0082]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#101 антитела, включают антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133, или антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133.
[0083]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#101 антитела, включают антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:129, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:133, или антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:131, и легкую цепь, состоящая из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:133.
[0084]
Примеры гуманизированного антитела, произведенного из rat_CD147_#110 антитела, включают гуманизированное антитело против CD147 или антигенсвязывающий фрагмент антитела, имеющее следующие области (c)-(d):
(c) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих областей (c1)-(c4):
(c1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135;
(c2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147;
(c3) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (c1) или (c2); и
(c4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (c1)-(c3),
(d) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих областей (d1)-(d5):
(d1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137;
(d2) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149;
(d3) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151;
(d4) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (d1)-(d3); и
(d5) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по любому из пунктов (d1)-(d4).
[0085]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#110 антитела, включают антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137; антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149; или антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151.
[0086]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#110 антитела, включают антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:135, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:137; антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:149; или антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:151.
[0087]
Примеры гуманизированного антитела, произведенного из rat_CD147_#131 антитела, включают гуманизированное антитело против CD147 или антигенсвязывающий фрагмент антитела, имеющие следующие области (g) и (h):
(g) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих областей (g1)-(g3):
(g1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139;
(g2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (g1); и
(g3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (g1) или (g2), и
(h) вариабельную область легкой цепи, выбранную из группы, состоящей из следующих областей (h1)-(h3):
(h1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141;
(h2) аминокислотной последовательности, имеющей идентичность, по меньшей мере, 95% более с последовательностью каркасной области вне каждой CDR последовательности в последовательности по пункту (h1); и
(h3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой CDR последовательности в последовательности по пункту (h1) или (h2).
[0088]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#131 антитела, включают антитело, содержащее вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139, и вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141.
[0089]
Предпочтительные примеры гуманизированного антитела, произведенного из rat_CD147_#131 антитела, включают антитело, содержащее тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:139, и легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:141.
[0090]
Предпочтительно, чтобы приведенные выше гуманизированное антитело, произведенное из rat_CD147_#84 антитела, гуманизированное антитело, произведенное из rat_CD147_#101 антитела, гуманизированное антитело, произведенное из rat_CD147_#110 антитела или гуманизированное антитело, произведенное из rat_CD147_#131 антитела активировали p38MAPK сигнальную трансдукцию и/или SMAD4 сигнальную трансдукцию посредством CD147.
[0091]
Примеры антитела по настоящему изобретению могут дополнительно включать человеческие антитела. Термин "человеческое антитело против CD147" относится к человеческому антителу, имеющему только нуклеотидную последовательность гена человеческого антитела, произведенного из хромосомы. Такое человеческое антитело против CD147 может быть получено, например, методом с использованием мышей, продуцирующих человеческое антитело, имеющих фрагмент хромосомы человека, включающий гены тяжелых цепей и легких цепей человеческого антитела (смотрите публикации Tomizuka, K. et al., Nature Genetics (1997)16, p.133-143; Kuroiwa, Y. et al., Nucl. Acids Res. (1998) 26, p.3447-3448; Yoshida, H. et al., Animal Cell Technology: Basic and Applied Aspects vol. 10, p.69-73 (Kitagawa, Y., Matsuda, T. and Iijima, S. eds.), Kluwer Academic Publishers, 1999; Tomizuka, K. et al., Proc. Natl. Acad. Sci. USA (2000) 97, p.722-727 etc.).
[0092]
В частности, такие продуцирующие человеческое антитело мыши могут быть получены продуцированием животных с рекомбинантным геном путем разрушения генных локусов тяжелой цепи и легкой цепи эндогенного иммуноглобулина, и затем введения генных локусов тяжелой цепи и легкой цепи иммуноглобулина человека с помощью вектора, такого как вектор искусственной хромосомы человека (HAC) или вектор искусственной хромосомы мыши (MAC), вместо разрушенных локусов; и затем создают рекомбинантных животных путем скрещивания этих животных.
[0093]
Эукариотические клетки трансформируют методами рекомбинации генов с помощью кДНК, кодирующей каждую из тяжелой цепи и легкой цепи человеческого антитела, преимущественно, с помощью вектора, включающего кДНК, и затем трансформированные клетки, продуцирующие человеческое моноклональное антитело с рекомбинантным геном, культивируют, в результате чего из культуральной надосадочной жидкости может быть получено антитело. В данном случае, в качестве клеток-хозяина могут быть использованы эукариотические клетки, предпочтительно, клетки CHO, клетки млекопитающих, такие как лимфоциты и миеломные клетки.
[0094]
Кроме того, также известно получение человеческого антитела методами фагового дисплея, выбранного из библиотеки человеческих антител (смотрите публикации Wormstone, I. M. et al., Investigative Ophthalmology & Visual Science. (2002)43(7), p.2301-2308; Carmen, S. et al., Briefings in Functional Genomics and Proteomics (2002), 1(2), p.189-203; Siriwardena, D. et al., Ophthalmology (2002)109(3), p.427-431, etc.).
[0095]
Например, может быть использован метод фагового дисплея, в соответствии с которым вариабельную область человеческого антитела экспрессируют на поверхности фага в виде одноцепочечного антитела (scFv), и затем выбирают фаг, связывающийся с антигеном (Nature Biotechnology (2005), 23, (9), p.1105-1116). Анализируют ген фага, выбранного на основе его связывания с антигеном, для того чтобы можно было определить последовательность ДНК, кодирующую вариабельную область человеческого антитела, связывающуюся с антигеном. После того как выявлена последовательность ДНК одноцепочечного вариабельного фрагмента scFv, связывающегося с антигеном, приготавливают экспрессирующий вектор, имеющий эту последовательность, и затем его вводят в соответствующую клетку-хозяина для экспрессии, и в результате может быть получено человеческое антитело (патентные документы WO92/01047, WO92/20791, WO93/06213, WO93/11236, WO93/19172, WO95/01438, WO95/15388, публикация Annu. Rev. Immunol. (1994)12, p.433-455, Nature Biotechnology (2005) 23(9), p.1105-1116).
[0096]
Примеры антитела по настоящему изобретению также включают антитело, имеющее такой же эпитоп, как эпитоп антитела, предлагаемого в настоящем изобретении. Его примеры включают антитело, имеющее такой же эпитоп, как эпитоп, по меньшей мере, одного из LN22R8, 2P10F2, rat_CD147_#84, rat_CD147_#101, rat_CD147_#110 и rat_CD147_#131.
[0097]
LN22R8 и 2P10F2 по настоящему изобретению распознавали эпитоп, обозначаемый как mu3 на фигуре 3 (человеческий CD147v1: DALPGQKTEFKVDSDDQ (SEQ ID NO:143), обезьяний CD147: DTLPGQKTDFEVDSDDL (SEQ ID NO:144)). Примеры антитела по настоящему изобретению также включают антитело, которое распознает или связывается с последовательностью, включающей аминокислотную последовательность, в которой 1 или несколько, предпочтительно, от 1 до 3, более предпочтительно, 1 или 2 аминокислоты удалены, заменены или добавлены в последовательности SEQ ID NO:143 или SEQ ID NO:144, или SEQ ID NO:143 или SEQ ID NO:144. Предпочтительно, чтобы антитело активировало сигнальную трансдукцию посредством CD147.
[0098]
Примеры антитела по настоящему изобретению включают антитело, которое распознает такой же эпитоп, как антитело rat_CD147_#110, предпочтительно, гуманизированное #110H1L4. Предпочтительно, чтобы антитело активировало сигнальную трансдукцию посредством CD147. Результаты анализа эпитопа гуманизированного антитела #110H1L4 описаны в примере 17.
[0099]
Подходящее антитело может быть выбрано в результате оценки активности связывания с антигеном, проведение которой известно специалистам в данной области. Константа диссоциации антитела и антигена (CD147) может быть измерена с помощью системы Biacore T200 (GE Healthcare Bioscience) с применением в качестве метода детекции поверхностного плазмонного резонанса (SPR). Например, антитело при соответствующей концентрации и аналит взаимодействуют с иммобилизированным антигеном в качестве лиганда, измеряют его связывание и диссоциацию, и в результате чего могут быть определены константа скорости ассоциации ka1, константа скорости диссоциации kd1 и константа диссоциации (KD; KD=kd1/ka1).
[0100]
Прибор, используемый для оценки активности связывания с CD147, не ограничивается системой Biacore T200. Активность связывания с CD147 может быть также оценена с помощью прибора с использованием в качестве метода детекции поверхностного плазмонного резонанса (SPR), например, прибора KinExA (Sapidyne Instruments) с использованием в качестве метода детекции анализа кинетического исключения, системы BLItz (Pall) с использованием в качестве метода детекции биослойную интерферометрию или ELISA (твердофазный иммуноферментный анализ).
[0101]
Примером другого критерия для сравнения свойств антител может служить стабильность антител. Дифференциальная сканирующая калориметрия (DSC) является методом, позволяющим быстро и точно измерить среднюю температуру термического разворачивания (Tm), служащую в качестве хорошего критерия относительной структурной стабильности белков. Измеряют величины Tm методом DSC, сравнивают величины Tm, и затем может быть проведены сравнение различий термической стабильности. Известно, что стабильность свойств антитела при хранении коррелирует в определенной степени с термической стабильностью антитела (Lori Burton, et al., Pharmaceutical Development and Technology (2007)12, p.265-273). И поэтому, подходящее антитело может быть выбрано с использованием в качестве критерия его термическую стабильность. Примеры других критериев для выбора антитела могут включать высокий выход в соответствующих клетках-хозяина и низкую склонность к агрегации в водном растворе. Например, так как антитело, характеризующееся высоким выходом, не всегда характеризуется высокой термической стабильностью, то оптимальное антитело для введения людям следует выбирать путем всестороннего исследования на основе упомянутых выше критериев.
[0102]
Также известен метод получения одноцепочечного иммуноглобулина, который включает связывание полноразмерных последовательностей тяжелой цепи и легкой цепи антитела с соответствующим линкером (Lee, H-S, et al., Molecular Immunology (1999)36, p.61-71; Shirrmann, T. et al., mAbs (2010), 2, (1) p.1-4). Такой одноцепочечный иммуноглобулин может сохранять структуру и активность, аналогичную структуре и активности антитела, которое исходно представляет собой тетрамер в результате димеризации. Антитело по настоящему изобретению может также представлять собой антитело, имеющее одиночную вариабельную область тяжелой цепи, но не имеющее последовательность легкой цепи. Такое антитело называют однодоменным антителом (sdAb) или нанотелом, и оно в действительности обнаруживается у верблюдов или у лам, и у него сохраняется способность к связыванию антигена (Muyldemans S. et al., Protein Eng. (1994)7(9), 1129-35, Hamers-Casterman C. et al., Nature (1993) 363 (6428) 446-8). Упомянутое выше антитело может также рассматриваться в качестве типа антигенсвязывающего фрагмента антитела по настоящему изобретению.
[0103]
Антителозависимая цитотоксичность может быть повышена путем регулирования гликозилирования сахарной цепи, связывающейся с антителом по настоящему изобретению. Примеры известных методов регулирования гликозилирования антител включают, но этим не ограничивая, методы, описанные в патентных документах WO99/54342, WO2000/61739 и WO2002/31140.
[0104]
Когда антитело получают путем выделения гена антитела и затем введения выделенного гена в соответствующую клетку-хозяина, такая соответствующая клетка-хозяин и экспрессирующий вектор могут быть использованы в комбинации.
[0105]
Конкретные примеры гена антитела могут включать комбинации генов, кодирующих последовательности тяжелых цепей антитела, и генов, кодирующих последовательности легких цепей, описанных в изобретении. После трансформации клеток-хозяина, последовательность тяжелой цепи гена и последовательность легкой цепи гена могут быть введены в один и тот же экспрессирующий вектор, или они могут быть введены в различные экспрессирующие векторы. Когда в качестве клеток-хозяина используют эукариотические клетки, могут быть использованы клетки животных, клетки растений и эукариотические микроорганизмы. Примеры клеток животных могут включать клетки млекопитающих, например, клетки обезьян, то есть, клетки COS (Gluzman, Y. Cell (1981) 23, p.175-182, ATCC CRL-1650), линии дигидрофолатредуктаза-дефицитных клеток фибробластов мыши NIH3T3 (ATCC, CRL-1658) или клетки яичника китайского хомячка (клетки CHO, ATCC CCL-61) (Urlaub, G. and Chasin, L. A. Proc. Natl. Acad. Sci. USA. (1980) 77, p.4126-4220). Когда используют прокариотические клетки, их примеры могут включать Escherichia coli и Bacillus subtilis. Соответствующий ген антитела вводят в эти клетки путем трансформации, и затем трансформированные таким образом клетки культивируют in vitro, получая в результате антитело. Упомянутые выше методы культивирования могут характеризоваться различным выходом, зависящим от последовательности антитела. Поэтому, антитело, которое легко получить в качестве лекарственного средства, может быть выбрано путем использования в качестве критерия выхода, на основе которого проводит выбор между антителами, имеющими эквивалентную активность связывания.
[0106]
На изотип антитела по настоящему изобретению не накладывают конкретных ограничений, и его примеры могут включать IgG (IgG1, IgG2, IgG3, IgG4), IgM, IgA (IgA1, IgA2), IgD или IgE. Предпочтительно, чтобы такой изотип представлял собой IgG или IgM, и предпочтительно, чтобы другим его примером являлся IgG.
[0107]
Человеческий IgG1 проявляет очень сильные эффекторные функции, включающие комплементзависимую цитотоксичность (CDC) в результате опосредования фиксации комплемента и антителозависимую цитотоксичность из существующих 4 типов подклассов человеческого IgG (Bruggemann et al., J. Exp. Med., 1351-1361, 1987). Когда молекула, которая сверхэкспрессируется при раке, таргетирована терапевтическим антителом, человеческий IgG1 используют фактически в качестве изотипа IgG, который проявляет терапевтические эффекты путем промотирования индукции гибели раковых клеток вследствие цитотоксичности, опосредованной эффекторными функциями (например, трастузумаб и ритуксимаб). Однако, сообщалось, что человеческое IgG1 антитело, таргетирующее HLA-DR с его естественной комплементзависимой цитотоксичностью (CDC), вызывало гибель яванских макаков после его введения. Использование такого антитело в качестве лекарственного средства, которое таргетирует молекулу, которая также экспрессируется в нормальных органах, вызывает опасение по поводу того, что эффекторные функции могут индуцировать тяжелую побочную реакцию (Tawara, T., J. Immunology, 2008, 2294-2298). Когда IgG1 используют в качестве изотипа антитела по настоящему изобретению, IgG1 антитело может иметь мутацию, и, в частности, эффекторные функции могут быть скорректированы путем замены некоторых аминокислотных остатков константных областей (смотрите патентные документы WO88/007089, WO94/28027, WO94/29351). Примеры варианта IgG1 с ослабленными эффекторными функциями включают IgG1 LALA (IgG1-L234A, L235A) и 1gG1 LAGA (IgG1-L235A, G237A).
[0108]
Человеческий IgG2 имеет очень слабые эффекторные функции, включающие комплементзависимую цитотоксичность (CDC) в результате опосредования фиксация комплемента и антителозависимую цитотоксичность из существующих 4 типов подклассов человеческого IgG (Bruggemann et al., J. Exp. Med., 1351-1361, 1987). Когда молекула, которая экспрессируется в нормальных органах, таргетирована терапевтическим антителом, IgG2 используют в качестве одного из форматов IgG для предотвращения токсичности, обусловленной цитотоксичностью, опосредованной эффекторными функциями (Denosumab, Evolocumab, Brodalumab).
[0109]
Человеческий IgG4 имеет очень слабые эффекторные функции, включающие комплементзависимую цитотоксичность (CDC) в результате опосредования фиксация комплемента и антителозависимую цитотоксичность из существующих 4 типов подклассов человеческого IgG (Bruggemann et al., J. Exp. Med., 1351-1361, 1987). Когда молекула, которая экспрессируется в нормальных органах, таргетирована терапевтическим антителом, IgG4 используют в качестве одного из форматов IgG для предотвращения токсичности, обусловленной цитотоксичностью, опосредованной эффекторными функциями (ниволумаб). Когда IgG4 используют в качестве изотипа антитела по настоящему изобретению, подавляется расщепление, отличительная черта IgG4, и увеличивается период полувыведения путем замены некоторых аминокислотных остатков константных областей (смотрите публикацию Molecular Immunology, 30, 1 105-108 (1993)).
[0110]
Когда IgG4 используют в качестве изотипа антитела по настоящему изобретению, IgG4 антитело может иметь мутации. Примеры мутации варианта IgG4 включают (Proc Natl Acad Sci USA. 1969, 63(1), 78-85; Kabat et al., Sequences of proteins of immunological interest, 1991 Fifth edition) замену (F234A) фенилаланина в положении 234 в соответствии с системой нумерации аминокислот в белке на аланин и замену (L235A) лейцина в положении 235 в соответствии с системой нумерации аминокислот в белке на аланин (Parekh et al., mAbs, 310-318, 2012). Такую мутацию антитела называют мутацией FALA. IgG4P FALA дополнительно снижает эффекторные функции путем замены двух аминокислотных остатков, требуемых для взаимодействия с FcγRs (например, FcγRI, FcγRII или FcγRIII), существующих в домене CH2, на аланин. Так как образование связи S-S между тяжелыми цепями антитела является нестабильной в IgG4, мутацию для ускорения образования связи S-S между тяжелыми цепями антитела вводят в IgG4 для повышения стабильности. Примеры такой мутации включают замену (S228P) серина в положении 228 в соответствии с системой нумерации аминокислот в белке на пролин (ANGAL et al., Molecular Immunology, 105-108, 1993). Такую мутацию антитела называют мутацией Pro. В константной области антитела по настоящему изобретению, обе описанные выше мутация FALA и мутация Pro могут быть введены одновременно (Vafa et al., Methods, 65, 114-126, 2014). Тяжелую цепь IgG4, имеющую мутацию FALA, называют тяжелой цепью типа "IgG4FALA", тяжелую цепь IgG4, имеющую мутацию Pro, называют тяжелой цепью типа "IgG4P", и тяжелую цепь IgG4, имеющую обе мутации FALA и Pro называют тяжелой цепью типа "IgG4PFALA".
[0111]
Константная область тяжелой цепи антитела состоит из областей CH1, шарнира, CH2 и CH3, где CH1 определяется как находящаяся в диапазоне положений от 118 до 215 в соответствии с системой нумерации аминокислот в белке, шарнир определяется как находящийся в диапазоне положений от 216 до 230 в соответствии с системой нумерации аминокислот в белке, CH2 определяется как находящаяся в диапазоне положений от 231 до 340 в соответствии с системой нумерации аминокислот в белке, и CH3 определяется как находящаяся в диапазоне положений от 341 до 446, в соответствии с системой нумерации аминокислот в белке. В соответствии с системой нумерации аминокислот в белке, пролин, полученный в результате замены серина в положении 228, аланин, полученный в результате замены фенилаланина в положении 234, и аланин, полученный в результате замены лейцина в положении 235, соответствует пролину в положении 248, аланину в положении 254 и аланину в положении 255, соответственно, в последовательности SEQ ID NO:100, представляющей аминокислотную последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#84; пролин в положении 245, аланин в положении 251 и аланин в положении 252, соответственно, в последовательности SEQ ID NO:108, представляющей аминокислотную последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#101; и пролин в положении 244, аланин в положении 250 и аланин в положении 251, соответственно, в последовательности SEQ ID NO:108, представляющей аминокислотную последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#110.
[0112]
Предпочтительные примеры изотипа антитело по настоящему изобретению включают IgG1, IgG2, IgG4, IgG4P или IgG4PFALA, в частности, его предпочтительные примеры включают IgG2, IgG4P или IgG4PFALA, и его еще более предпочтительные примеры включают IgG2 или IgG4P.
[0113]
Антитело по настоящему изобретению может также представлять собой антигенсвязывающий фрагмент антитела, который имеет антигенсвязывающий сайт антитела или его модифицированного продукта. Антитело обрабатывают протеазой, такой как папаин или пепсин, или ген антитела модифицируют методами генной инженерии и затем полученный продукт экспрессируют в подходящих культивируемых клетках, в результате чего может быть получен фрагмент антитела. Из этих фрагментов антитела, фрагменты, сохраняющие все или часть функций молекулы полноразмерного антитела, могут называться антигенсвязывающим фрагментом антитела. Примером функций антитела является активация связанной с антигеном сигнальной трансдукции.
[0114]
CD147 также экспрессируется в клеточных элементах крови, в том числе в эритроцитах, и в здоровых органах, существенно важных для сохранения жизни (Spring, et al., Eur. J. Immunol., 1997, 891-897), и, поэтому, противоопухолевый эффект, оказываемый с использованием антитела с сопровождающими его эффекторными функциями, характеризуется высокой степенью риска возникновения нежелательного побочного эффекта. Действительно, сообщалось, что эритроциты чувствительны к эффекторным функциям (ADCC, CDC, ADCP), возникающим вследствие их связывания с антителом (Flegel, W., Transfusion, 2015, S47-S58), и известно, что повышенное связывание in vivo антитела с эритроцитами вызывает аутоиммунную гемолитическую анемию (Gibson, J., Aust. N. Z. J. Med., 1988. 625-637). Антитело по настоящему изобретению в качестве терапевтического антитела, направленного против CD147, который также экспрессируется в нормальных клетках, характеризуется тем, что любой один или все виды токсичности ADCC, ADCP и CDC, которые вызывают серьезные побочные эффекты, обнаруживаются при низких уровнях или вообще не обнаруживаются.
[0115]
Авторами настоящего изобретения впервые было обнаружено, что антитело против человеческого CD147 проявляет противоопухолевую активность, не зависящую от эффекторных функций антитела, путем активации клеточной сигнальной трансдукции посредством CD147. В настоящем изобретении, антитело сохраняет функции активности связывания с CD147 и/или активации CD147. Предпочтительно, чтобы антитело по настоящему изобретению активировало связанные с нисходящим сигнальным путем молекулы, опосредующие CD147, например, FAK, MEK, Erk, JAK/STAT, AKT или MAP киназу (MAPK), или активировало связанные с нисходящим сигнальным путем молекулы, расположенные ниже его. Более предпочтительно, чтобы антитело по настоящему изобретению активировало молекулы, расположенные ниже MAPK или MAPK. Предпочтительным примером MAPK является p38MAPK. Примеры сигнальных молекул, расположенных ниже MAPK, включают HSP27, cxcl8 и SMAD (например, SMAD2, SMAD3 или SMAD4, предпочтительно, SMAD4). Примеры "активации CD147" включают повышение уровня экспрессии p38MAPK мРНК, повышение уровня экспрессии белка p38MAPK, фосфорилирование p38MAPK, фосфорилирование HSP27 (например, фосфорилирование Ser82 в HSP27 или фосфорилирование Ser15 в HSP27), повышение уровня экспрессии cxcl8 мРНК, повышение уровня экспрессии белка cxcl8, повышение уровня экспрессии rhoB мРНК или повышение уровня экспрессии белки rhoB через активацию сигнала SMAD. Примеры "активации CD147" включают, преимущественно, повышение уровня экспрессии белка p38MAPK, фосфорилирование p38MAPK, фосфорилирование HSP27 (например, фосфорилирование Ser82 в HSP27 или фосфорилирование Ser15 в HSP27), повышение уровня экспрессии cxcl8 мРНК и повышение уровня экспрессии rhoB мРНК через активацию сигнала SMAD.
Известно, что SMAD2 или SMAD3 фосфорилируются рецептором TGFβ, когда TGFβ связывается с рецептором TGFβ (TGFBR1/2), в результате чего образуется гетеротример с SMAD4. Известно, что после его импорта в ядро, гетеротример связывается с доменом регуляции транскрипции, имеющем SMAD ДНК связывающую последовательность на хромосоме (Smad связывающий элемент: SBE), и затем положительно или отрицательно контролирует экспрессию мРНК гена, регулирующего последующие звенья сигнальных каскадов (Miyazono, The Official Journal of the Japan Geriatrics Society, 1999, 162-166). Поэтому, считается, что активация SMAD4 требует присутствия SMAD2 или SMAD3. SMAD2, SMAD3 и SMAD4 отрицательно контролирует уровень экспрессии KLF5 зависящем от TGFβ образом (David et al., Cell, 2016, 164(5), 1015-1030). В клетках SMAD4-дефицитного рака поджелудочной железы, гасятся сигналы для подавления экспрессии гена KLF5 через SMAD2, SMAD3 и SMAD4, и в результате экспрессируется белок KLF5. Известно, что когда теряется SMAD4 и экспрессируется KLF5, подавляется TGFβ-зависимая система сигнализации гибели клеток (SOX4-зависимая) (упомянутая выше публикация David et al, Cell).
[0116]
Авторами настоящего изобретения было обнаружено, что антитело против человеческого CD147 по настоящему изобретению фосфорилирует p38MAPK (фигура 21), фосфорилирует HSP27 (фигура 20) и повышает экспрессию cxcl8 (фигура 22(b) и фигура 23(b)). Поэтому, активация CD147 антителом по настоящему изобретению может быть подтверждена путем подтверждения изменения экспрессии гена, экспрессии белка или фосфорилирования, по меньшей мере, одной из этих молекул до и после введения антитела.
[0117]
Авторами настоящего изобретения было обнаружено, что антитело против человеческого CD147 по настоящему изобретению проявляет противоопухолевую активность в модели рака поджелудочной железы, экспрессирующего белок SMAD4 (фигура 25); антитело против человеческого CD147 по настоящему изобретению проявляет частичную (приблизительно до 30%) противоопухолевую активность в модели рака поджелудочной железы, таком как BxPC-3, характеризующемся мутацией гена SMAD4 и отсутствием мутации SMAD4 (фигура 24); и, после введения антитела против человеческого CD147 по настоящему изобретению, в опухолях индуцируются расположенные ниже молекулы SMAD2, SMAD3 и SMAD4 (rhoB, фигура 22(c) и фигура 23(c)). Поэтому, до и после введения антитела по настоящему изобретению, подтверждают присутствие или отсутствие изменения экспрессии гена rhoB или экспрессии белка rhoB, следовательно, если антитело по настоящему изобретению опосредует активацию CD147, то это может быть подтверждено. Геномная последовательность, экспрессия гена или экспрессия белка SMAD4 в образцах пациента измеряют методом, известным специалистам в данной области; пациентов, у которых экспрессируется SMAD4, отбирают в качестве тех пациентов, которым будут вводить антитело по настоящему изобретению, и, поэтому, им может быть введено антитело по настоящему изобретению.
[0118]
Известно, что уровень экспрессии KLF5 будет высоким в SMAD4-отрицательной модели, такой как BxPC-3 (David et al., Cell, 2016, 164(5), 1015-1030). Чувствительность к антителу против человеческого CD147 по настоящему изобретению снижалось с 91% до 20% в KLF5-экспрессирующей MIA PaCa-2 модели (пример 26). Соответственно, авторы настоящего изобретения считают, что экспрессия KLF5 подавляет SMAD2-, SMAD3- и SMAD4-зависимый сигнальный путь гибели клеток, индуцируемый антителом против CD147. В связи с тем, что уровни экспрессии KLF5 являются низкими при раке печени, остром лимфобластном лейкозе (ALL), лимфоме, гастроинтестинальной стромальной опухоли (GIST), раке кожи, саркоме, остром миелолейкозе (AML) или раке почки, то ожидают, что антитело против человеческого CD147 по настоящему изобретению будет эффективным для многих пациентов, страдающих от этих видов рака. Экспрессия гена или экспрессия белка KLF5 в образцах пациента измеряют методом, известным специалистам в данной области, пациентов, характеризующихся пониженными уровнями экспрессии KLF5 или отсутствием экспрессии KLF5, отбирают в качестве тех пациентов, которым будут вводить антитело по настоящему изобретению, и, поэтому, им может быть введено антитело по настоящему изобретению. Степень снижения экспрессии KLF5 в этом случае может быть определена методом, известным специалистам в данной области, и путем проведения соответствующих клинических испытаний. Например, пациентов, для которых могут быть достигнуты эффекты, и пациентов, для которых эффекты не могут быть достигнуты, сравнивают по уровню экспрессии KLF5, в результате чего определяют соответствующий пороговый уровень.
[0119]
Примеры фрагментов антитела, могут включать Fab, F(ab')2, Fv, или одноцепочечный Fv (scFv), в которых тяжелая цепь Fv и легкая цепь Fv соединены с помощью соответствующего линкера, диатело (диатела), линейное антитело и полиспецифическое антитело, образованное из фрагментов антитела. Примеры фрагментов антитела включают Fab', которое представляет собой моновалентный фрагмент вариабельной области антитела, и который получают путем обработки F(ab')2 в восстановительных условиях.
[0120]
Кроме того, антитело по настоящему изобретению может также представлять собой полиспецифическое антитело, имеющее специфичность, по меньшей мере, к двум различным типам антигенов. Такая молекула обычно связывается с двумя типами антигенов (то есть, биспецифическое антитело), и примеры "полиспецифического антитела" в настоящем изобретении включают антитело, имеющее специфичность к трем или более (например, 3 типам) антигенам.
[0121]
Полиспецифическое антитело по настоящему изобретению может представлять собой полноразмерное антитело или фрагменты такого антитела (например, F(ab')2 биспецифическое антитело). Биспецифическое антитело может быть получено путем связывания тяжелой цепи и легкой цепи (HL пара) двух типов антител или путем гибридизации гибридом, продуцирующих различные моноклональные антитела, друг с другом, с получением слившихся клеток, продуцирующих биспецифическое антитело (Millstein et al., Nature (1983) 305, P. 537-539).
[0122]
Антитело по настоящему изобретению может представлять собой одноцепочечное антитело (описываемое так же как scFv). Одноцепочечное антитело получают путем соединения вариабельной области тяжелой цепи антитела и вариабельной области легкой цепи с помощью полипептидного линкера (Pluckthun, The Pharmacology of Monoclonal Antibodies, 113 (Ed., Rosenberg and Moore, Springer Verlag, New York, p.269-315 (1994), Nature Biotechnology (2005), 23, p.1126-1136). BiscFv фрагмент, полученный путем соединения двух scFvs с помощью полипептидного линкера, может также быть использован в качестве биспецифического антитела.
[0123]
Методы получения одноцепочечного антитела хорошо известны (например, смотрите патентные документы U.S. Patent No. 4946778, U.S. Patent No. 5260203, U.S. Patent No. 5091513 и U.S. Patent No. 5455030.). В scFv, вариабельная область тяжелой цепи и вариабельная область легкой цепи соединены с помощью линкера, который не образует конъюгата, предпочтительно, с помощью полипептидного линкера (Huston, J. S. et al., Proc. Natl. Acad. Sci. USA. (1988), 85, p.5879-5883). Вариабельная область тяжелой цепи и вариабельная область легкой цепи в scFv могут быть произведены из одного и того же антитела или из разных антител. В качестве полипептидного линкера для соединения вариабельных областей, используют, например, произвольный одноцепочечный пептид, состоящий из от 12 до 19 остатков.
[0124]
ДНК, кодирующая scFv, может быть получена путем амплификации методом полимеразной цепной реакции (PCR), используя в качестве матрицы, матрицу, выбранную из ДНК, кодирующей упомянутую выше тяжелую цепь или вариабельную область тяжелой цепи антитела, и ДНК, кодирующей легкую цепь или вариабельную область легкой цепи, ДНК, кодирующей полноразмерную последовательность каждой области, или части ДНК, кодирующей ее требуемую аминокислотную последовательность, и пары праймеров, определяющих каждый ее конец, и затем амплифицирующих, используя ДНК, кодирующую часть полипептидного линкера, и пары праймеров, определяющих конец, в результате чего эти концы присоединяются к тяжелой цепи и легкой цепи, соответственно, в комбинации.
[0125]
После того, как получена ДНК, кодирующая scFv, может быть получен экспрессирующий вектор, содержащий ДНК, и клетки-хозяина, трансформированные с помощью экспрессирующего вектора стандартным методом, и может быть получена scFv стандартным методом, используя клетки-хозяина. Эти фрагменты антитела могут быть продуцированы клетками-хозяина после получения его гена и экспрессии этого гена таким же способом, как описано выше.
[0126]
Антитело по настоящему изобретению может быть полимеризовано с целью повышения его аффинности к антигену. Антитело, подвергаемое полимеризации, может представлять собой единственный тип антитела или множество антител, распознающих множество эпитопов одного и того же антигена. Примеры метода полимеризации антитела могут включать связывание IgG CH3 домена с двумя scFvs, связывание с стрептавидином и введение мотива спираль-петля-спираль.
[0127]
Антитело по настоящему изобретению может также представлять собой поликлональное антитело, которое является смесью множества типов антител против CD147, отличающихся аминокислотной последовательностью. Примером поликлонального антитела может быть смесь множества типов антител, отличающихся CDR. Что касается такого поликлонального антитела, то культивируют смесь клеток, продуцирующих различные антитела, и антитело, очищенное от культуры клеток, может быть использовано (смотрите патентный документ WO2004/061104).
[0128]
Антитело по настоящему изобретению может представлять собой антитело, имеющее от 80% до 99% идентичности (или гомологичности) с упомянутой выше тяжелой цепью и/или легкой цепью антитела. Путем использования комбинации последовательностей, характеризующихся высокой гомологичностью с упомянутой выше аминокислотной последовательностью тяжелой цепи и аминокислотной последовательностью легкой цепи, может быть выбрано антитело, имеющее антигенсвязывающую способность, характеризующееся активацией CD147, предпочтительно, активацией MAPK, и активацией расположенных ниже молекул сигнального пути MAPK, эквивалентными характеристикам каждого упомянутого выше антитела. Такая гомологичность составляет обычно 80% или более гомологичности, предпочтительно, 90% или более гомологичности, более предпочтительно, 95% или более гомологичности, и наиболее предпочтительно, 99% или более гомологичности. Путем использования комбинации аминокислотной последовательности тяжелой цепи и/или легкой цепи с аминокислотной последовательностью, в которой заменены, удалены и/или добавлены от 1 до нескольких аминокислотных остатков, может быть выбрано антитело, обладающее различными эффектами, эквивалентными эффектам каждого упомянутого выше антитела. Число заменяемых, удаляемых и/или добавляемых аминокислотных остатков составляет обычно 10 или менее аминокислотных остатков, предпочтительно, от 5 до 6 или менее аминокислотных остатков, более предпочтительно, от 2 до 3 или менее аминокислотных остатков, и, наиболее предпочтительно, 1 аминокислотный остаток. Кроме того, известно, что удаляют карбоксильный концевой остаток лизина тяжелой цепи антитела, продуцированного культивируемыми клетками млекопитающего (Tsubaki et al., Int. J. Biol. Macromol, 139-147, 2013). Однако, удаление и модификация этих последовательностей тяжелой цепи не влияют на способность антитела связываться с антигеном и эффекторные функции (например, на активацию комплемента и антителозависимую цитотоксичность). Поэтому, настоящее изобретение включает антитело, подвергнутое модификации, и его примеры могут включать антитело, в котором удалены 1 или 2 аминокислоты на карбоксильном конце тяжелой цепи, и антитело, подвергнутое амидированию (например, тяжелая цепь, в которой амидируют остаток пролина на карбоксильном конце). Следует отметить, что дефицитная по карбоксильному концу тяжелая цепь антитела по настоящему изобретению не ограничивается упомянутыми выше типами, при условии, что в случае других типов поддерживается способность связывания с антигеном и функция активации расположенных ниже CD147 связанных с передачей сигналов молекул. Две тяжелые цепи, которые образовывают антитело по настоящему изобретению, могут представлять собой любой один тип тяжелой цепи, выбранный из группы, состоящей из полноразмерных тяжелых цепей и упомянутых выше дефицитных тяжелых цепей, или комбинации их двух типов. На содержание дефицитных по карбоксильному концу тяжелых цепей может влиять тип и условия культивирования клеток млекопитающего, продуцирующих антитело по настоящему изобретению, и примером такого антитела может служить случай, при котором каждая из двух тяжелых цепей, являющихся основными компонентами антитело по настоящему изобретению, имеет удаление одного аминокислотного остатка на карбоксильном конце.
[0129]
Гомологичность между двумя типами аминокислотной последовательности может быть определена, используя алгоритм BLAST версии 2.2.2 с параметрами по умолчанию (Altschul, Stephen F., Thomas L. Madden, Alejandro A. Schaffer, Jinghui Zhang, Zheng Zhang, Webb Miller, and David J. Lipman (1997), "Gapped BLAST and PSI-BLAST: a new generation of protein database search programs", Nucleic acids Res. 25: 3389-3402). Алгоритмом BLAST можно также воспользоваться через доступ к сайту в интернете www.ncbi.nlm.nih.gov/blast. Следует отметить, что с помощью алгоритма Blast рассчитывают два типа процентов идентичности (или идентичностей) и положительный результат (положительные результаты). Упомянутый первым процент представляет собой величину, полученную из совпадающих аминокислотных остатков при сравнении двух типов аминокислотной последовательности, в которой предполагается наличие гомологичности, и упомянутый вторым процент представляет собой численную величину, полученную с учетом обнаружения в химической структуре аналогичных друг другу аминокислотных остатков. Величина идентичности при совпадении аминокислотных остатков обозначают в изобретении величиной гомологичности.
[0130]
В качестве модифицированных продуктов антитела могут быть использованы антитела, связанные с различными молекулами, такими как полиэтиленгликоль (PEG).
[0131]
Антитело по настоящему изобретению может также представлять собой иммуноконъюгат, образованный таким антителом и другим лекарственным средством. Примеры такого антитела могут включать антитело, связанное с радиоактивным веществом или с соединением, обладающим фармакологическими эффектами (Nature Biotechnology (2005) 23, p.1137-1146).
[0132]
Полученное таким образом антитело может быть очищено до однородного состояния. Для выделения и очистки антитела, могут быть использованы методы выделения и очистки, применяемые для обычных белков. Например, антитело может быть выделено и очищено путем соответствующего выбора или комбинирования методов колоночной хроматографии, фильтрации через фильтр, ультрафильтрации, высаливания, диализа, препаративного электрофореза в полиакриламидном геле, изоэлектрического фокусирования (Strategies for Protein Purification and Characterization: A Laboratory Course Manual, Daniel R. Marshak et al. eds., Cold Spring Harbor Laboratory Press (1996); Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), но методы выделения и очистки не ограничиваются этими примерами.
[0133]
Примеры хроматографии могут включать аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гель-проникающую хроматографию, хроматографию с обращенной фазой и адсорбционную хроматографию.
[0134]
Эти виды хроматографии могут быть осуществлены с использованием жидкостной хроматографии, такой как высокоэффективная жидкостная хроматография (HPLC) и жидкостная хроматография быстрого разрешения (FPLC).
[0135]
Примеры колонки, используемой для аффинной хроматографии, могут включать колонку Protein A и колонку Protein G.
[0136]
Примеры колонки Protein A могут включать Hyper D, POROS, Sepharose F. F. (GE Healthcare).
[0137]
Антитело может быть также очищено путем использования носителя, на котором иммобилизирован антиген и который характеризуется активностью связывания с антигеном.
[0138]
(Лекарственное средство, включающее антитело против CD147)
Антитело против CD147 по настоящему изобретению может быть получено из числа антител против CD147, полученных методом, описанным в приведенном выше разделе "Получение антитело против CD147". Полученное таким методом антитело может быть использовано в качестве средства для лечения и/или предотвращения опухоли и/или рака. Антитело против CD147 по настоящему изобретению имеет очень высокую противоопухолевую активность и может применяться в качестве средства для лечения опухоли или рака. Антитело против CD147 по настоящему изобретению проявляет очень высокую противоопухолевую активность в отношении гемцитабин-резистентных раковых клеток и раковых клеток, имеющих низкую восприимчивость к сорафенибу. Антитело против CD147 по настоящему изобретению проявляет значительно более высокую активность в отношении клеток хронического миелолейкоза, чем иматиниб.
[0139]
Примеры опухолей, которые могут быть подвергнуты лечению с использованием антитела против CD147 по настоящему изобретению или лекарственного средства, включающего антитело, конкретно не ограничиваются, при условии, что эти опухоли являются опухолями, экспрессирующими CD147, и они включают, предпочтительно, рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль(GIST), лейкоз (например, острый миелолейкоз (AML), хронический миелолейкоз (CML), или хронический лимфоцитарный лейкоз (CLL) или острый лимфоцитарный лейкоз (ALL)), лимфому или злокачественную лимфому (например, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL)), и, более предпочтительно, рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
[0140]
Примеры опухолей, которые могут быть подвергнуты лечению с использованием антитела по настоящему изобретению или лекарственного средства, включающего антитело, включают SMAD-положительные CD147-экспрессирующие опухоли. Примеры SMAD-положительных CD147-экспрессирующих опухолей включают SMAD-положительный CD147-экспрессирующий рак печени и рак поджелудочной железы. Антитело против CD147 по настоящему изобретению или лекарственное средство, включающее антитело, предпочтительно вводить пациентам, у которых была подтверждена экспрессия CD147 и/или SMAD. Примеры SMAD включают, предпочтительно, SMAD2, SMAD3 и/или SMAD4, и более предпочтительно, SMAD4. Предпочтительно, совместно с подтверждением экспрессии SMAD4, также подтверждать экспрессию, по меньшей мере, одного из SMAD2 и SMAD3.
[0141]
В качестве варианта, примеры опухоли, которая может быть подвергнута лечению с помощью антитела по настоящему изобретению или лекарственного средства, включающего антитело, включают опухоль, характеризующуюся пониженной экспрессией KLF5 или отсутствием экспрессии KLF5. Примеры опухоли, характеризующейся пониженной экспрессией KLF5 или отсутствием экспрессии KLF5, включают рак печени, острый лимфоцитарный лейкоз (ALL), лимфому, гастроинтестинальную стромальную опухоль (GIST), рак кожи, саркому, острый миелолейкоз (AML) и рак почки. Антитело против CD147 по настоящему изобретению или лекарственное средство, включающее антитело, предпочтительно вводить пациенту, у которого было подтверждено наличие пониженной экспрессии KLF5 или отсутствие экспрессии KLF5.
[0142]
Антитело против CD147 по настоящему изобретению может также быть введено вместе с 2, 3 или более другими терапевтическими средствами, в зависимости от цели лечения. Эти другие терапевтические средства могут быть инкапсулированы в одном и том же препарате и введены одновременно. Другие терапевтические средства и антитело против CD147 могут быть также инкапсулированы в одном и том же препарате и, следовательно, могут быть введены одновременно. Антитело против CD147 и другие терапевтические средства могут быть раздельно инкапсулированы в препаратах и затем могут быть введены одновременно. Кроме того, другие лекарственные средства и антитело против CD147 могут также быть введены раздельно, в частности, лекарственные средства могут быть введены до или после введения антитела против CD147. В частности, после введения других терапевтических средств, вводят терапевтическое средство, включающее антитело против CD147 или антигенсвязывающий фрагмент антитела в качестве активного компонента, или после введения терапевтического средства, включающего антитело против CD147 или антигенсвязывающий фрагмент антитела в качестве активного компонента, могут быть введены другие терапевтические средства.
[0143]
В настоящем изобретении также предлагается фармацевтическая композиция, включающая терапевтически и/или профилактически эффективное количество антитела против CD147 и фармацевтически приемлемый разбавитель, носитель, солюбилизатор, эмульгатор, консервант и/или вспомогательное вещество.
[0144]
В настоящем изобретении также предлагается фармацевтическая композиция, включающая терапевтически и/или профилактически эффективное количество антитела против CD147, терапевтически и/или профилактически эффективное количество, по меньшей мере, одного противоопухолевого терапевтического средства, и фармацевтически приемлемый разбавитель, носитель, солюбилизатор, эмульгатор, консервант и/или вспомогательное вещество.
[0145]
Вещества, используемые для приготовления препарата, которые являются приемлемыми в фармацевтической композиции по настоящему изобретению, являются, преимущественно, нетоксичными для субъекта, которому вводят фармацевтическую композицию, при предпочтительной дозе и концентрации для введения.
[0146]
Фармацевтическая композиция по настоящему изобретению может содержать вещества для ее приготовления, которые позволяют изменять или поддерживать pH, осмотическое давление, вязкость, прозрачность, цвет, изотоничность, стерильность, стабильность, растворимость, скорость пролонгированного высвобождения, всасываемость, проницаемость и другие подобные свойства. Примеры веществ, используемых для приготовления препарата, могут включать, но этим не ограничивая, следующие вещества: аминокислоты, такие как глицин, аланин, глутамин, аспарагин, аргинин или лизин, антимикробные средства, антиоксиданты, такие как аскорбиновая кислота, сульфат натрия или гидросульфит натрия, буферные вещества, такие как фосфатный, цитратный или боратный буфер, гидрокарбонат натрия и раствор Трис-хлористоводородная кислота (Tris-HCl), наполнители, такие как маннит и глицин, хелатообразующие вещества, такие как этилендиаминтетрауксусная кислота (EDTA), комплексообразующие вещества, такие как кофеин, поливинилпирролизин, β-циклодекстрин и гидроксипропил-β- циклодекстрин, увеличивающие объем средства, такие как глюкоза, манноза или декстрин, углеводы, такие как моносахарид и дисахарид, окрашивающие вещества, вещества, корригирующие вкус и запах лекарственного средства, разбавители, эмульгаторы, гидрофильные полимеры, такие как поливинилпирролизин, низкомолекулярные полипептиды, солеобразующие противоионы, бензалкония хлорид, бензойную кислоту, салициловую кислоту, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, антисептики, такие как сорбиновая кислота или пероксид водорода, глицерин, растворители, такие как пропиленгликоль или полиэтиленгликоль, сахарные спирты, такие как маннит или сорбитан, суспензии, эфиры сорбитана, полисорбаты, такие как полисорбат 20 и полисорбат 80, тритон, трометамин, поверхностно-активные вещества, такие как лецитин или холестерин, стабилизаторы, такие как сахароза и сорбит, хлорид натрия, хлорид калия, усилители эластичности, такие как маннит и сорбит, транспортирующие средства, вспомогательные вещества и/или фармацевтические адъюванты. Предпочтительно, чтобы такое вещество, используемое для приготовления препарата, добавляли в количестве большем, чем масса антитела против CD147, в 0,01-100 раз, и, в частности, в 0,1-10 раз. Состав подходящей фармацевтической композиции в препарате может быть соответствующим образом определен специалистами в данной области в соответствии с подвергаемым лечению заболеванием, применяемым способом введения и другими подобными параметрами.
[0147]
Вспомогательное вещество и носитель в фармацевтической композиции может представлять собой жидкость или твердое вещество. Соответствующее вспомогательное вещество и соответствующий носитель могут представлять собой воду или физиологический раствор для инъекции, искусственную спинномозговую жидкость или другие вещества, которые обычно используют для парентерального введения. В качестве носителя может быть также использован нейтральный физиологический раствор или физиологический раствор, содержащий сывороточный альбумин. Фармацевтическая композиция может содержать Трис-буфер с pH 7,0-8,5, ацетатный буфер с pH 4,0-5,5 или цитратный буфер с pH 3,0-6,2. Эти буферы могут содержать сорбит и другие соединения. Примеры фармацевтической композиции по настоящему изобретению могут включать фармацевтическую композицию, содержащую антитело против CD147, и фармацевтическую композицию, содержащую антитело против CD147 и, по меньшей мере, одно противоопухолевое терапевтическое средство. Фармацевтическую композицию по настоящему изобретению приготавливают в форме лекарственного средства, имеющего выбранный состав и требуемую чистоту, лиофилизированного продукта или жидкости. Фармацевтическая композиция, включающая антитело против CD147, и фармацевтическая композиция, включающая антитело против CD147 и, по меньшей мере, одно противораковое терапевтическое средство, может быть также приготовлено в форме лиофилизированного продукта, в котором используют соответствующее вспомогательное вещество, такое как сахароза.
[0148]
Фармацевтическая композиция по настоящему изобретению может быть также приготовлена для парентерального введения или для перорального введения для всасывания в желудочно-кишечном тракте. Состав и концентрация препарата могут быть определены с учетом способа введения. При введении антитела по настоящему изобретению человеку, может быть введено приблизительно от 0,1 до 100 мг/кг антитела один раз или несколько раз в течение от 1 до 180 дней. Однако, дозу и частоту введения следует обычно определять с учетом пола, массы тела и возраста пациента, симптомов, тяжести заболевания, побочного действия и других подобных параметров, и поэтому, приведенные выше примеры не являются ограничениями для дозы и методов.
[0149]
Примеры лекарственной формы фармацевтической композиции по настоящему изобретению могут включать инъекционные препараты, в том числе препараты для инфузии, суппозитории, назальные препараты, сублингвальные препараты и препараты для трансдермального всасывания. Примеры способа введения включают пероральное введение или парентеральное введение. Примеры парентерального введения включают внутривенное, интраартериальное, внутримышечное, ректальное, трансмукозальное или интрамукозальное, и интрадермальное введения.
[0150]
Антитело по настоящему изобретению или антигенсвязывающий фрагмент антитела, комплексное лекарственное средство, которое их содержит, биспецифическое антитело, которое их содержит, или фармацевтическая композиция, которая их содержит, могут предлагаться в комбинации с биомаркером для отбора пациентов, которым они могут быть введены. Такое антитело или фармацевтическая композиция могут также предлагаться в виде набора в комбинации со средствами для детекции биомаркера, или такое антитело или фармацевтическая композиция могут предлагаться по отдельности с биомаркером. Путем использования биомаркера, антитело или фармацевтическая композиция по настоящему изобретению могут быть введены группе пациентов, в отношении которых предполагается, что введение антитела по настоящему изобретению позволит достигать высокой эффективности.
[0151]
Настоящее изобретение относится к способу предсказания восприимчивости к противораковому лечению, включающему использование биологических образцов, взятых у больных раком пациентов, измерение экспрессии SMAD4 или экспрессии KLF5, содержащихся в биологических образцах, и определение пациентов, у которых выявляют SMAD4, или пациентов, у которых выявляют пониженную экспрессию KLF5 или отсутствие экспрессии KLF5, в качестве критерия наличия восприимчивости к противораковому лечению с помощью антитела по настоящему изобретению или функционального фрагмента антитела, или фармацевтической композиции по настоящему изобретению; к способу отбора пациентов для противоракового лечения, включающему использование биологических образцов, взятых у больных раком пациентов, обнаружение экспрессии SMAD4 или экспрессии KLF5 в биологических образцах, и отбор пациентов, у которых выявляют SMAD4, или пациентов, у которых выявляют пониженную экспрессию KLF5 или отсутствие экспрессии KLF5, в качестве субъектов для противоракового лечения с помощью антитела по настоящему изобретению или функционального фрагмента антитела, или фармацевтической композиции по настоящему изобретению; к способу лечения рака, включающему использование биологических образцов, взятых у больных раком пациентов, обнаружение экспрессии SMAD4 или экспрессии KLF5 в биологических образцах, и введение антитела по настоящему изобретению или функционального фрагмента антитела, или фармацевтической композиции по настоящему изобретению пациентам, у которых выявляют SMAD4, или пациентов, у которых выявляют пониженную экспрессию KLF5 или отсутствие экспрессии KLF5; или к набору для определения восприимчивости к противораковому лечению с помощью антитела по настоящему изобретению или функционального фрагмента антитела, или фармацевтической композиции по настоящему изобретению, включающему, по меньшей мере, средства для выявления экспрессии SMAD4 или экспрессии KLF5 в биологических образцах, взятых у больных раком пациентов.
[0152]
Используемый в изобретении термин "биологический образец (биологические образцы)" относится к ткани, жидкости и клеткам, взятым у индивидуума, и к их смеси, и его примеры могут включать, но этим не ограничивая, биоптат опухоли, спинномозговые жидкости, плевральные жидкости, интраперитонеальные жидкости, лимфу, срезы кожи, кровь, мочу, кал, мокроту, органы дыхания, кишечник, урогенитальные органы, слюну, грудное молоко, органы пищеварения и собранные из них клетки. Примеры "биологического образца (биологических образцов)" включают предпочтительно образец, содержащий раковые клетки, более предпочтительно, ткань или клетки, полученные путем иссечения или биопсии, или клетки, полученные из плевральной жидкости или интраперитонеальной жидкости. Более предпочтительными биологическими образцами являются образцы, содержащие раковые клетки или раковую ткань.
[0153]
"Экспрессия SMAD4" может быть выявлена или измерена, используя геномную последовательность, экспрессию гена или экспрессию белка SMAD4, методом, известным специалистам в данной области. Примеры такого метода включают секвенирование РНК, микроматричный анализ, секвенирование генома и иммуноанализ.
[0154]
"Экспрессия KLF5" может быть выявлена или измерена, используя геномную последовательность, экспрессию гена или экспрессию белка KLF5, методом, известным специалистам в данной области. Примеры такого метода включают иммуногистохимию (IHC), секвенирование РНК, микроматричный анализ, секвенирование генома и иммуноанализ. Термин "пониженная экспрессия KLF5" означает, что при сравнении с контрольной величиной (например, с уровнем экспрессии у здорового субъекта или уровнем экспрессии в доброкачественной ткани этого же пациента), обнаруживается, что уровень экспрессии является более низким, чем уровень экспрессии в случае контроля. В качестве варианта, степень снижения экспрессии KLF5, при которой может быть подтверждена восприимчивость к противораковому лечению, являющаяся результатом использования антитела или фармацевтической композиции по настоящему изобретению, может быть определена с помощью метода, известного специалистам в данной области и путем проведения соответствующих клинических испытаний. Например, пациентов, в отношении которых был достигнут положительный эффект лечения, и пациентов, в отношении которых не был достигнут положительный эффект лечения, сравнивают по уровню экспрессии KLF5, на основании чего устанавливают соответствующую пороговую величину. Поэтому, например, термин "пониженная экспрессия KLF5" относится к уровню экспрессии, который ниже определенной таким образом пороговой величины.
Примеры
[0155]
Далее, настоящее изобретение конкретно описывается с помощью примеров, но настоящее изобретение не ограничивается приводимыми далее примерами. Кроме того, эти примеры никоим образом не следует истолковывать в качестве ограничений. В следующих далее примерах, методики, относящиеся к генной инженерии, проводили в соответствии с методами, описанными в монографии "Molecular Cloning" (Authors: Sambrook, J., Fritsch, E. F., and Maniatis, T., published by Cold SpringHarbor Laboratory Press in 1989), с другими методами, описанными в руководствах по экспериментам и используемыми специалистами в данной области, или с инструкциями к производимых промышленностью продуктам в случае использования производимых промышленностью реагентов или наборов, если не указано иное. Кроме того, реагенты, растворители и исходные материалы, которые конкретно не упомянуты в этом описании изобретения, могут быть без проблем приобретены у фирм-поставщиков. В этих примерах, в качестве человеческих клеток поджелудочной железы линии MIA PaCa-2 использовали клетки из Американской коллекции типовых культур (ATCC), номер по каталогу CRL-1420, и в качестве человеческих клеток рака поджелудочной железы линии PANC-1 использовали клетки из ATCC, номер по каталогу CRL-1469.
[0156]
(Пример 1) Продукция мышиных и крысиных антител путем клеточной иммунизации
1)-1 Продукция CD147 экспрессирующего вектора
Клон IOH3378 (Invitrogen, Thermo Fisher Scientific K.K.) коммерчески доступного гена человеческого CD147 (BSG вариант2/CD147v2) клонировали в экспрессирующий вектор для клеток млекопитающего pcDNA-DEST40 (Invitrogen, Thermo Fisher Scientific K.K.), используя смесь ферментов Gateway LR Clonase, для продукции человеческого CD147v2 экспрессирующего вектора (pcDNA-DEST40-CD147v2).
[0157]
Приобретали экспрессирующий вектор для клеток млекопитающего pCMV6-XL5-hBSGv1 (OriGene Technologies, Inc., Cat. SC303059) коммерчески доступного гена человеческого CD147 (BSG variant1/CD147v1) и использовали его в качестве человеческого CD147v1 экспрессирующего вектора.
[0158]
В качестве яванского макака CD147 экспрессирующего вектора, приобретали pCMV3-cynoBSG (Sino Biological Inc., Cat. CG90636-UT).
[0159]
В качестве мышиного CD147v2 экспрессирующего вектора, приобретали pCMV3-mBSGv2 (Sino Biological Inc., Cat. MG50332-UT).
[0160]
1)-2 Продукция мышиных гибридом
Использовали мышей линии BALB/cAnNCrlCrlj (CHARLES RIVER LABORATORIES JAPAN, INC.) в возрасте от 4 до 6 недель. 5 × 106 клеток LNCaP (ATCC, CRL-1740), выделенных с использованием реагента Versene (поставляемого фирмой Thermo Fisher SCIENTIFIC K.K.) на день 0, 7, 15 и 24, суспендировали в PBS, и суспензию вводили подкожно в области спины. 5 × 106 этих же клеток вводили внутривенно на день 31, и в этот же день собирали селезенку и использовали ее для продукции гибридом. Для того чтобы продуцировать гибридомы, клетки селезенки подвергали слиянию с мышиными миеломными клетками P3X63Ag8U.1 (ATCC, CRL-1597), используя PEG4000 (Immuno-Biological Laboratories Co., Ltd.). Для выделения и культивирования гибридом использовали среды ClonaCell-HY MediumD (STEMCELL Technologies Inc.) и ClonaCell-HY MediumE (STEMCELL Technologies Inc.).
[0161]
1)-3 Продукция крысиных гибридом
Использовали крыс линии WKY/Izm (Japan SLC, Inc.) в возрасте 7 недель. Через 13 дней после иммунизации с помощью 1 × 107 клеток линии PANC-1 рака поджелудочной железы человека в ягодицы крыс, собирали клетки подвздошных лимфатических узлов и использовали их для генерации гибридом. Клетки селезенки крыс подвергали слиянию с миеломными клетками линии SP2/0-Ag14 мышей (ATCC, CRL-1581), используя прибор для стимуляции слияния клеток LF301 (BEX CO., LTD.), для того чтобы продуцировать гибридомы. Для выделения и культивирования гибридом использовали среды ClonaCell-HY MediumD (STEMCELL Technologies Inc.) и ClonaCell-HY MediumE (STEMCELL Technologies Inc.).
[0162]
1)-4 Идентификация антигена методом твердофазного иммуноферментного анализа (ELISA)
Использовали гибридный белок человеческий CD147-Fc (Sino Biological Inc., Cat. 10186-H02H) и гибридный белок мышиный CD147-Fc (Sino Biological Inc., Cat.50332-M03H). Гибридный белок человеческий CD147-Fc (Sino Biological Inc., Cat. 10186-H02H) и гибридный белок мышиный CD147-Fc получали путем добавления буферного раствора PBS и растворения на льду до 1 мкг/мл. 100 мкл раствора белка добавляли в 96-луночный планшет (Nunc, Thermo Fisher Scientific K.K., Cat. 442404), затем хранили в течение ночи при 4°C, и в лунки наносили слой гибридного белка CD147-Fc. Удаляли раствор белка, и лунки блокировали с помощью буферного раствора PBS, содержащего 1% BSA (Research Organics, Inc., Cat. 1334A), при 4°C в течение двух часов. Лунки промывали три раза буферным раствором PBS, содержащим 0,05% Tween20 (ATTO CORPORATION, Cat. WSE-7235). Затем, надосадочные жидкости из культуры гибридомы, приготовленной в примерах 1)-2 и 1)-3, разбавляли в 20 раз буферным раствором PBS, и добавляли каждую в лунки, затем инкубировали при комнатной температуре в течение одного часа. Лунки промывали три раза буферным раствором PBS, содержащим 0,05% Tween20 (ATTO CORPORATION, Cat.WSE-7235). После этого, в лунки добавляли 100 мкл anti-rat-Fab2-igG-HRP (Jackson ImmunoResearch Inc., Cat. 112-036-072), разбавленный в 50000 раз буферным раствором PBS, содержащем 1% BSA, затем встряхивали при комнатной температуре в течение 30 минут. Лунки промывали 5 раз буферным раствором PBS, содержащим 0,05% Tween20 (ATTO CORPORATION, Cat. WSE-7235). Вслед за этим, в лунки добавляли 100 мкл окрашивающего реагента из фермента пероксидазы хрена (HRP) (eBioscience, Thermo Fisher SCIENTIFIC K.K., Super AquaBlue ELISA substrate, Cat. 00-4203), затем нагревали при комнатной температуре в течение от 10 до 20 минут, и измеряли поглощение при 405 нм с помощью планшет-ридера (Envision, PerkinElmer, Inc). Рассчитывали среднее значение измеренных величин поглощения для двух или для трех лунок, и антитела, характеризующиеся поглощениями, которые были в два раза или более выше измеренной величины для контрольной лунки, не содержащей первичных антител, определяли как обладающие активностью связывания (+), а антитела, характеризующиеся поглощениями, которые были выше менее, чем в два раза, определяли как не обладающие активностью связывания (-). Результаты представлены в таблице 1. Развитие окраски, специфическое для лунок, в которые наносили слой гибридного белка человеческого CD147-Fc, наблюдали в надосадочных жидкостях из культуры клеток LN22R8, 2P1A6, 2P3A9, 2P8C12, 2P10F2, 2P2D7, 2P2D10 и 2P1B7. Развитие окраски, специфическое для лунок, в которые наносили слой гибридных белков человеческого и мышиного CD147-Fc наблюдали для LN24R7, 2P5F5, 2P6A2 и 2P3G8.
[0163]
[Таблица 1]
[0164]
1)-5 Получение моноклональных антител и определение изотипов антител
Для гибридом, в которых в примере 1)-4 наблюдали продукцию антител против человеческого CD147 и которые могли быть стабильно культивированы, определяли изотипы антител, содержавшихся в надосадочных жидкостях из культуры клеток, используя выпускаемый промышленностью набор реагентов для изотипирования, и результаты приведены в таблице 2. Используя колбу CL-1000 (Becton, Dickinson and Company), эти гибридомы культивировали для получения надосадочных жидкостей из культуры клеток гибридом, содержащих моноклональные антитела.
[0165]
[Таблица 2]
Антитела против CD147, полученные клеточной иммунизацией
[0166]
1)-6 Очистка моноклональных антител
Каждое антитело, полученное в примере 1)-5, очищали от надосадочной жидкости из культуры клеток. Мышиное моноклональное тело против человеческого CD147 очищали с помощью одностадийного процесса аффинной хроматографии на колонке с наполнителем rProtein A (при температуре от 4 до 6°C). После очистки методом аффинной хроматографии на колонке с наполнителем rProtein A, проводили стадию замены буфера при температуре от 4 до 6°C. Предварительно, надосадочную жидкость из культуры клеток наносили на колонку, приведенную в равновесие с PBS и заполненную смолой для очистки антител MabSelectSuRe (поставляемой фирмой GE Healthcare Bioscience). Затем полностью вводили в колонку культуральный раствор, колонку промывали с помощью PBS в количестве, составляющем два или более объема колонки. Далее, проводили элюирование раствором 2M гидрохлорида аргинина (pH 4,0), собирая фракцию, содержащую антитело. Фракцию подвергали замене жидкости на HBSor (25 мM гистидин/5% сорбит/pH 6,0) методом диализа (Thermo Scientific, Slide-A-Lyzer Dialysis Cassette). После концентрирования с использованием устройства для центробежной ультрафильтрации VIVASPIN20 (фракция с молекулярной массой UF10K, Sartorius AG, при 4°C), концентрацию IgG доводили до 4,9 мг/мл. И наконец, проводили фильтрацию с использованием фильтра Minisart-Plus (Sartorius AG) с получением очищенного образца.
[0167]
Крысиное моноклональное антитело против человеческого CD147 очищали с помощью одностадийного процесса аффинной хроматографии на колонке с наполнителем Protein G (при температуре от 4 до 6°C). После очистки методом аффинной хроматографии на колонке с наполнителем Protein G, проводили стадию замены буфера при температуре от 4 до 6°C. Предварительно, надосадочную жидкость из культуры гибридомы наносили на колонку, приведенную в равновесие с PBS и заполненную смолой Protein G (GE Healthcare Bioscience). Затем полностью вводили в колонку культуральный раствор, колонку промывали с помощью PBS в количестве, составляющем два или более объема колонки. Далее, проводили элюирование смесью 0,1M глицин/водный раствор хлористоводородной кислоты (pH 2,7), собирая фракцию, содержащую антитело. Затем к собранной фракции добавляли 1M Tris-HCl (pH 9,0) для доведения величины pH до 7,0-7,5, проводили замену буфера на HBSor (25 мM гистидин/5% сорбит/pH 6,0) и концентрировали с использованием устройства для центробежной ультрафильтрации VIVASPIN20 (фракция с молекулярной массой UF30K, Sartorius AG, при температуре от 4 до 6°C), затем доводили концентрацию антитела до 1 мг/мл или более. И наконец, проводили фильтрацию с использованием фильтра Minisart-Plus (Sartorius AG) с получением очищенного образца.
[0168]
1)-7 Скрининг антител путем измерения их in-vivo противоопухолевой активности
1 × 107 клеток линии PANC-1 поджелудочной железы человека суспендировали в PBS, и суспензию инокулировали подкожно в подмышечную впадину иммунодефицитных мышей линии NOD-scid (CHARLES RIVER LABORATORIES JAPAN, INC., NOD. CB17-Prkdc<scid>/J). Проводили распределение по группам на основе объема опухоли, и несущим злокачественную опухоль мышам вводили интраперитонеально мышиное антитело против CD147 (LN22R8) или крысиное антитело против CD147 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12 или 2P10F2) при 10 мг/кг на день 27, 34 и 41 после инокуляции (n=6). Крысиное антитело против CD147 (2P2D6) вводили интраперитонеально несущим злокачественную опухоль мышам при 10 мг/кг на день 27 и 34 после инокуляции (n=6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0169]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигурах 1(a)-(c). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. На фигуре 1(a) представлены результаты для антитела 2P2D6, антитела 2P3G8 и антитела 2P2D10. Степень ингибирования роста опухоли на день 48 после инокуляции, который был датой проведения последнего измерения, составляла 10%, 45% и 40%, соответственно, для группы, в которой вводили 10 мг/кг. На фигуре 1(b) представлены результаты для антитела LN22R8, антитела 2P1A6 и антитела 2P1B7. Степень ингибирования роста опухоли на день 48 после инокуляции, который был датой проведения последнего измерения, составляла 50%, 26% и 24%, соответственно, для группы, в которой вводили 10 мг/кг. На фигуре 1(c) представлены результаты для антитела 2P8C12 и антитела 2P10F2. Степень ингибирования роста опухоли на день 48 после инокуляции, который был датой проведения последнего измерения, составляла -2% и 62%, соответственно, для группы, в которой вводили 10 мг/кг.
[0170]
1)-8 Анализ межвидовой перекрестной реактивности антитела против CD147
Используя реагент для трансфектирования Lipofectamine 2000 (Thermo Fisher SCIENTIFIC K.K., Cat. 11668-019), pcDNA-DEST40-CD147v2 или pCMV3-cynoBSG, продуцированные в примере 1)-1, трансфектировали в клетки CHO-K1 (ATCC, CCL-61), и через день клетки подвергали обработке мышиным антителом против человеческого CD147 (LN22R8) или крысиным антителом против человеческого CD147 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2 или 2P2D6) при 10 мкг/мл, что давало возможность флуоресцентной детекции связывания каждого антитела с CD147, который экспрессируется клетками CHO-K1, используя мышиное антитело класса IgG-FITC (MP Biomedicals, Inc., Cat. 554936) или крысиное антитело класса IgG-PE (BD Biosciences, Cat. 550767). Флуоресцентная детекция экспрессии CD147 человека и яванского макака в клетках CHO-K1 была возможна в результате связывания выпускаемого промышленностью антитела против CD147 (MEM-M6/1, AbD Serotec, Cat. MCA28822). Измерение упомянутых выше клеток проводили на проточном цитометре (CantoII, BD Biosciences), и результаты представлены на фигурах от 2-1 до 2-3. На графических зависимостях, по вертикальной оси представлено число клеток, а по горизонтальной оси представлена интенсивность флуоресцентных сигналов.
[0171]
Выпускаемое промышленностью антитело против CD147 (MEM-M6/1), мышиное антитело против человеческого CD147 (LN22R8) и крысиные антитела против CD147 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2 и 2P2D6) все проявляли активность в отношении связывания с человеческим CD147, который экспрессируют клетки CHO-K1 (фигуры от 2-1 до 2-3).
[0172]
Выпускаемое промышленностью антитело против CD147 (MEM-M6/1) проявляло активность в отношении связывания с CD147 яванского макака, который экспрессируют клетки CHO-K1, но ни одно из мышиного антитела против человеческого CD147 (LN22R8) и крысиных антител против CD147 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2 и 2P2D6) не проявляло активности в отношении связывания с CD147 яванского макака, который экспрессируют клетки CHO-K1 (фигуры от 2-1 до 2-3).
[0173]
Ни одно из антител против CD147 не проявляло активности в отношении связывания с мышиным CD147, который экспрессируют клетки CHO-K1 (данные не показаны).
[0174]
1)-9 Анализ эпитопа
Продукция экспрессирующих векторов мутировавшего CD147 человека для анализа эпитопа
В результате проведения поиска с использованием алгоритма BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) было рассчитано, что аминокислотные последовательности CD147 яванского макака и CD147 человека являются на 81% идентичными. Поэтому, проводилась оценка противоопухолевых эпитопов с использованием мутантов, в которых различные аминокислотные последовательности были частично перевиты между видами, поскольку было сделано предположение о том, что ограниченное число различий в аминокислотных последовательностях может непосредственно влиять на распознавание связывающих эпитопов антитела против CD147. Аминокислотные последовательности, содержащиеся в hCD147v1 и v2, вместе подвергали процедуре сравнения последовательностей между CD147 яванского макака и CD147 человека и классифицировали на 9 различных аминокислотных областей между видами с формированием областей от mu1 до mu9 (фигура 3). Для продукции CD147 мутантных экспрессирующих векторов и для подтверждения экспрессии на поверхности клеток, была искусственно синтезирована (фирмой GenScript Biotech Corporation) кДНК последовательность человеческого CD147 variant2 с последовательностью FLAG, введенной в N-конец, для продуцирования плазмиды, которую затем вводили в вектор pcDNA3.1, Signal-N-Flag-hCD147v2_pcDNA3.1. Кроме того, было продуцировано (фирмой GenScript Biotech Corporation) 9 типов человека-яванского макака химерных CD147 экспрессирующих векторов, hCD147-mu1_pcDNA3.1, hCD147-mu2_pcDNA3.1, hCD147-mu3_pcDNA3.1, hCD147-mu4_pcDNA3.1, hCD147-mu5_pcDNA3.1, hCD147-mu6_pcDNA3.1, hCD147-mu7_pcDNA3.1, hCD147-mu8_pcDNA3.1 и hCD147-mu9_pcDNA3.1, которые образовывали путем введения ДНК, кодирующей CD147 аминокислотные последовательности яванского макака от mu1 до mu9 в качестве мутаций замены аминокислоты путем замены ДНК в гене CD147 человека той же самой плазмиды.
[0175]
1)-10 Идентификация противоопухолевых эпитопных областей с использованием мутантов
9 типов экспрессирующих векторов CD147 человека, CD147 яванского макака или химерных CD147 человека-яванского макака экспрессирующих векторов вводили в клетки CHO-K1 (ATCC, CCL-61), используя реагент для трансфекции Lipofectamine 2000 (Thermo Scientific, Cat. 11668-019), затем через день подвергали обработке с помощью мышиного антитела против человеческого CD147 (LN22R8) или крысиного антитела против человеческого CD147 (2P1A6, 2P1B7, 2P3G8, 2P2D10, 2P8C12, 2P10F2 или 2P2D6) при 10 мкг/мл, и исследовали связывание антитела против CD147 с экспрессирующими CD147 клетками CHO-K1, используя мышиный IgG-PE (DAKO, Cat. R480) или крысиный IgG-PE (Becton, Dickinson and Company, #550767). Экспрессию белка CD147 подтверждали, используя выпускаемое промышленностью антитело против FLAG (anti-Flag M2, SIGMA, Cat. F4049-.2MG). Измерения проводили на проточном цитометре (CantoII, BD Biosciences), и результаты представлены в таблице 3. Образцы с флуоресцентными сигналами выше в 10 или более раз по сравнению с флуоресцентными сигналами в контрольных клетках, которые не подвергали обработке с помощью первичных антител, характеризовали как положительное связывание (+). Образцы с флуоресцентными сигналами выше менее чем в 10 раз характеризовали как слабоположительное связывание (±). Образцы с флуоресцентными сигналами, не отличавшиеся по интенсивности от флуоресцентных сигналов в контрольных клетках, которые не подвергали обработке с помощью первичных антител, характеризовали как отрицательное связывание (-).
[0176]
Все антитела 2P3G8, 2P10F2, 2P2D10 и LN22R8, обладающие противоопухолевой активностью 40% или более, наблюдаемой в примере 1)-7, теряли активность связывания с CD147 с мутацией mu3. Это позволило предположить, что важным эпитопом для противоопухолевой активности является область m3.
[0177]
[Таблица 3]
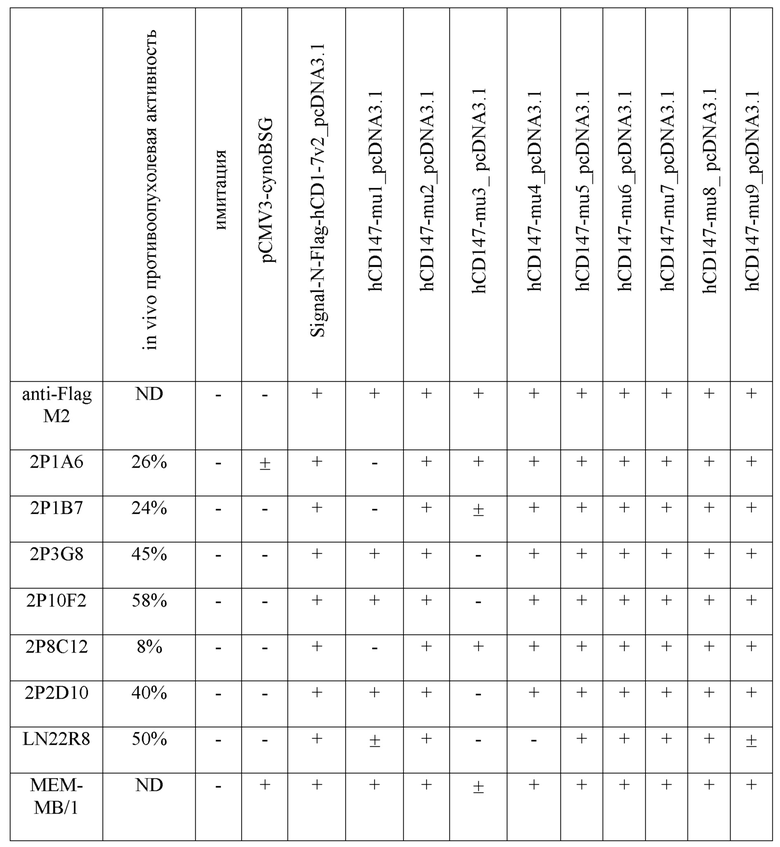
[0178]
1)-11 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антител LN22R8 и 2P10F2
1)-11-1 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела LN22R8
1)-11-1-1 Получение суммарной РНК гибридомы, продуцирующей антитело LN22R8
Для амплификации кДНК, кодирующей вариабельную область антитела LN22R8, получали суммарную РНК, используя реагент TRIzol (Ambion), из гибридом, продуцирующих антитело LN22R8.
[0179]
1)-11-1-2 Амплификация кДНК, кодирующей вариабельную область легкой цепи антитела LN22R8, с помощью 5'-RACE PCR и определение ее последовательности
кДНК, кодирующую вариабельную область легкой цепи, амплифицировали, используя приблизительно 1 мкг суммарной РНК, полученной в примере 1)-11-1-1, и набор реагентов SMARTer RACE 5'/3' Kit (Clontech Laboratories, Inc). В качестве праймеров для амплификации кДНК, кодирующей вариабельную область легкой цепи гена антитела LN22R8, методом PCR использовали смесь универсальных праймеров UPM (Universal Primer A Mix: прилагаемая в наборе реагентов SMARTer RACE 5'/3' Kit) и праймер, сконструированный из последовательности константной области известной мышиной легкой цепи.
[0180]
кДНК, кодирующую вариабельную область легкой цепи, амплифицированную с помощью 5'-RACE PCR, клонировали в плазмиду, и затем подвергали анализу нуклеотидную последовательность кДНК, кодирующей вариабельную область легкой цепи.
[0181]
SEQ ID NO:7 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область легкой цепи антитела LN22R8, определенную таким методом, и SEQ ID NO:8 представляет ее аминокислотную последовательность. SEQ ID NOs: 11, 12 и 13, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела LN22R8.
[0182]
1)-11-1-3 Амплификация кДНК, кодирующей вариабельную область тяжелой цепи антитела LN22R8, с помощью 5'-RACE PCR и определение ее последовательности
кДНК, кодирующую вариабельную область тяжелой цепи, амплифицировали, используя приблизительно 1 мкг суммарной РНК, полученной в примере 1)-11-1-1, и набор реагентов SMARTer RACE 5'/3' Kit (Clontech Laboratories, Inc). В качестве праймеров для амплификации кДНК, кодирующей вариабельную область тяжелой цепи гена антитела LN22R8, методом PCR использовали смесь универсальных праймеров UPM (Universal Primer A Mix: прилагаемая в наборе реагентов SMARTer RACE 5'/3' Kit) и праймер, сконструированный из последовательности константной области известной мышиной тяжелой цепи.
[0183]
кДНК, кодирующую вариабельную область тяжелой цепи, амплифицированную с помощью 5'-RACE PCR, клонировали в плазмиду, и затем подвергали анализу нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи.
[0184]
SEQ ID NO:9 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи антитела LN22R8, определенную таким методом, и SEQ ID NO:10 представляет ее аминокислотную последовательность. SEQ ID NOs: 14, 15 и 16, соответственно, представляют CDRH1, CDRH2 и CDRH3 вариабельной области тяжелой цепи антитела LN22R8.
[0185]
1)-11-2 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела 2P10F2
Использовали те же самые методы, что и в примере 1)-11-1. Однако, в качестве праймеров для амплификации кДНК, кодирующей вариабельную область легкой цепи гена, методом PCR использовали смесь универсальных праймеров UPM (Universal Primer A Mix: прилагаемая в наборе реагентов SMARTer RACE 5'/3' Kit) и праймер, сконструированный из последовательности константной области известной крысиной легкой цепи, и в качестве праймеров для амплификации кДНК, кодирующей вариабельную область тяжелой цепи гена, методом PCR использовали смесь универсальных праймеров UPM (Universal Primer A Mix: прилагаемая в наборе реагентов SMARTer RACE 5'/3' Kit) и праймер, сконструированный из последовательности константной области известной крысиной тяжелой цепи.
[0186]
SEQ ID NO:17 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область легкой цепи антитела 2P10F2, определенной таким образом, и SEQ ID NO:18 представляет ее аминокислотную последовательность. SEQ ID NOs: 21, 22 и 23, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела 2P10F2. SEQ ID NO:19 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи, и SEQ ID NO:20 представляет ее аминокислотную последовательность. SEQ ID NOs: 24, 25 и 26, соответственно, представляют CDRH1, CDRH2 и CDRH3 вариабельной области тяжелой цепи антитела 2P10F2.
[0187]
1)-12 Продукция экспрессирующих векторов человеческого химерное антитело LN22R8
1)-12-1 Конструирование экспрессирующего вектора pCMA-LK человеческой химерной и гуманизированной легкой цепи
Приблизительно 5,4 тысячи пар нуклеотидов фрагмента, полученного путем дигерирования плазмиды pcDNA3.3-TOPO/LacZ (Invitrogen Corp.) с помощью рестрикционных ферментов XbaI и PmeI, связывали с фрагментом ДНК, содержащим сигнальную последовательность человеческой легкой цепи, представленной последовательностью SEQ ID NO:27, и последовательностью ДНК, кодирующей константную область κ цепи человека, используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), с продукцией pcDNA3.3/LK.
[0188]
Из pcDNA 3.3/LK удаляли экспрессирующие неомицин элементы, создавая в результате pCMA-LK.
[0189]
1)-12-2 Конструирование экспрессирующего вектора pCMA-G1 человеческой химерной и гуманизированной тяжелой цепи типа IgG1
Фрагмент ДНК, из которого удаляли сигнальную последовательность легкой цепи и константную область κ цепи человека путем дигерирования pCMA-LK с помощью XbaI и PmeI, связывали с фрагментом ДНК, содержащим сигнальную последовательность тяжелой цепи человека, представленную последовательностью SEQ ID NO:28, и последовательностью ДНК, кодирующей аминокислоты в константной области человеческого IgG1, используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), с получением pCMA-G1.
[0190]
1)-12-3 Конструирование экспрессирующего вектора pCMA-G2 человеческой химерной и гуманизированной тяжелой цепи типа IgG2
Используя фрагмент ДНК, содержащий сигнальную последовательность тяжелой цепи человека, представленную последовательностью SEQ ID NO:29, и последовательность ДНК, кодирующей аминокислоты в константной области человеческого IgG2, конструировали pCMA-G2 таким же методом, как в примере 1)-12-2.
[0191]
1)-12-4 Конструирование экспрессирующего вектора легкой цепи человеческого химерного LN22R8
Используя кДНК, кодирующую вариабельную область легкой цепи LN22R8, полученную в примере 1)-11-1-2, в качестве матрицы, проводили PCR, используя праймер, сконструированный для прямого клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область легкой цепи. Амплифицированный фрагмент ДНК вставляли в том месте, где дигерировали pCMA-LK с помощью рестрикционного фермента BsiWI, используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), в результате чего конструировали экспрессирующий вектор легкой цепи человеческого химерного LN22R8. SEQ ID NO:30 и SEQ ID NO:31, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного LN22R8 и аминокислотную последовательность легкой цепи.
[0192]
1)-12-5 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG1
Используя кДНК, кодирующую вариабельную область тяжелой цепи LN22R8, полученную в примере 1)-11-1-3, в качестве матрицы, проводили PCR, используя праймер, сконструированный для прямого клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), амплифицированный фрагмент ДНК вставляли в том месте, где дигерировали pCMA-G1 с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор тяжелой цепи человеческого химерного LN22R8 типа IgG1. SEQ ID NO:32 и SEQ ID NO:33, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG1 и аминокислотную последовательность тяжелой цепи.
[0193]
1)-12-6 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи LN22R8, полученную в примере 1)-11-1-3, в качестве матрицы, проводили PCR, используя праймер, сконструированный для прямого клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), амплифицированный фрагмент ДНК вставляли в том месте, где дигерировали pCMA-G2 с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор тяжелой цепи человеческого химерного LN22R8 типа IgG2. SEQ ID NO:34 и SEQ ID NO:35, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG2 и аминокислотную последовательность тяжелой цепи.
[0194]
1)-12-7 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG4P
Был синтезирован (Geneart AG) фрагмент ДНК, содержащий последовательность ДНК, кодирующей аминокислотную последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG4P, представленную последовательностью SEQ ID NO:36. Используя синтезированный фрагмент ДНК, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного LN22R8 типа IgG4P таким же методом, как в примере 1)-12-2. SEQ ID NO:37 представляет аминокислотную последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG4P.
[0195]
1)-13 Продукция экспрессирующих векторов человеческого химерного антитела 2P10F2
1)-13-1 Конструирование экспрессирующего вектора pCMA-G1LALA тяжелой цепи человеческого химерного и гуманизированного антитела типа IgG1LALA
Используя сигнальную последовательность человеческой тяжелой цепи, представленной последовательностью SEQ ID NO:38, и фрагмент ДНК, содержащий последовательность ДНК, кодирующей аминокислоты константной области человеческого IgG1LALA, конструировали pCMA-G1LALA таким же методом, как в примере 1)-12-2.
[0196]
1)-13-2 Конструирование экспрессирующего вектора pCMA-G4P тяжелой цепи человеческого химерного гуманизированного антитела типа IgG4P
Используя сигнальную последовательность человеческой тяжелой цепи, представленной последовательностью SEQ ID NO:39, и фрагмент ДНК, содержащий последовательность ДНК, кодирующей аминокислоты константной области человеческого IgG4P, конструировали pCMA-G4P таким же методом, как в примере 1)-12-2.
[0197]
1)-13-3 Конструирование экспрессирующего вектора легкой цепи человеческого химерного 2P10F2
Используя кДНК, кодирующую 2P10F2 вариабельную область легкой цепи, полученную в примере 1)-11-2, в качестве матрицы, конструировали экспрессирующий вектор легкой цепи человеческого химерного 2P10F2 таким же методом, как в примере 1)-12-4. SEQ ID NO:40 и SEQ ID NO:41, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного 2P10F2 и аминокислотную последовательность легкой цепи.
[0198]
1)-13-4 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного 2P10F2 типа IgG1LALA
Используя кДНК, кодирующую вариабельную область тяжелой цепи 2P10F2, полученную в примере 1)-11-2, в качестве матрицы, проводили PCR, используя праймер сконструированный для клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), амплифицированный фрагмент ДНК вставляли в том месте, где дигерировали pCMA-G1LALA с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор тяжелой цепи человеческого химерного 2P10F2 типа IgG1LALA. SEQ ID NO:42 и SEQ ID NO:43, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG1LALA и аминокислотную последовательность тяжелой цепи.
[0199]
1)-13-5 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного 2P10F2 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи 2P10F2, полученную в примере 1)-11-2, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного 2P10F2 типа IgG2 таким же методом, как в примере 1)-12-6. SEQ ID NO:44 и SEQ ID NO:45, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG2 и аминокислотную последовательность тяжелой цепи.
[0200]
1)-13-6 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного 2P10F2 типа IgG4P
Используя кДНК, кодирующую вариабельную область тяжелой цепи 2P10F2, полученную в примере 1)-11-2, в качестве матрицы, проводили PCR, используя праймер, сконструированный для клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), амплифицированный фрагмент ДНК вставляли в том месте, где дигерировали pCMA-G4P с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор тяжелой цепи человеческого химерного 2P10F2 типа IgG4P. SEQ ID NO:46 и SEQ ID NO:47, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG4P и аминокислотную последовательность тяжелой цепи.
[0201]
1)-14 Продукция и получение человеческих химерных антител LN22R8 и 2P10F2
1)-14-1 Продукция человеческих химерных антител LN22R8 и 2P10F2
Клетки FreeStyle 293F (Invitrogen Corp.) пассировали и культивировали в соответствии с инструкцией фирмы. 1,2 × 109 клеток FreeStyle 293F (Invitrogen Corp.) в фазе логарифмического роста высевали в колбе Эрленмейера (дизайн Фернбаха) объемом 3 литра (Corning Incorporated) и разбавляли экспрессионной средой FreeStyle293 (Invitrogen Corp.) до 2,0 × 106 клеток/мл. К 40 мл среды Opti-Pro SFM (Invitrogen Corp.) добавляли 0,24 мг экспрессирующего вектора тяжелой цепи, 0,36 мг экспрессирующего вектора легкой цепи и 1,8 мг полиэтиленимина (Polyscience #24765), затем осторожно перемешивали и дополнительно выдерживали в течение 5 минут. После этого, смесь добавляли к клеткам FreeStyle 293F. После встряхивания культуры при 90 об/мин в инкубаторе при 37°C и 8% CO2 в течение 4 часов, к ней добавляли 600 мл среды EX-CELL VPRO (SAFC Biosciences, Sigma-Aldrich Corporation), 18 мл GlutaMAX I (Gibco) и 30 мл Yeastolate Ultrafiltrate (Gibco), затем встряхивали культуру при 90 об/мин в инкубаторе при 37°C и 8% CO2 в течение 7 дней, и полученную надосадочную жидкость культуры клеток фильтровали через одноразовый капсульный фильтр (Advantec #CCS-045-E1H).
[0202]
Человеческое химерное антитело LN22R8, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG1 и экспрессирующего вектора легкой цепи человеческого химерного LN22R8 обозначали как "LN22R8chIgG1". Человеческое химерное антитело LN22R8, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного LN22R8 обозначали как "LN22R8chIgG2". Человеческое химерное антитело LN22R8, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного LN22R8 обозначали как "LN22R8chIgG4P". Человеческое химерное антитело L2P10F2, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного LN22R8 типа IgG1LALA и экспрессирующего вектора легкой цепи человеческого химерного 2P10F2 обозначали как "2P10F2chIgG1LALA". Человеческое химерное антитело L2P10F2, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного 2P10F2 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного 2P10F2 обозначали как "2P10F2chIgG2". Человеческое химерное антитело L2P10F2, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного 2P10F2 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного 2P10F2 обозначали как "2P10F2chIgG4P".
[0203]
1)-14-2 Очистка человеческих химерных антител LN22R8 и 2P10F2
Каждое антитело очищали от полученной надосадочной жидкости культуры клеток в примере 1)-14-1 с помощью одностадийного процесса аффинной хроматографии на колонке с наполнителем rProtein A. После внесения надосадочной жидкости культуры клеток в колонку, приведенную в равновесие с PBS и заполненную средой MabSelectSuRe (поставляемой фирмой GE Healthcare Bioscience), колонку промывали с помощью PBS, в количестве, которое было в два или более раз больше объема колонки. Далее, проводили элюирование раствором 2M гидрохлорида аргинина (pH 4,0), собирая фракцию, содержащую антитело. Фракцию подвергали замене жидкости на буфер HBSor (25 мM гистидин/5% сорбит/pH 6,0) методом диализа (Thermo Scientific, Slide-A-Lyzer Dialysis Cassette). Антитело концентрировали с использованием устройства для центробежной ультрафильтрации VIVASPIN20 (фракция с молекулярной массой UF10K, Sartorius AG) с доведением концентрации IgG до 1 мг/мл или выше. И наконец, проводили фильтрацию с использованием фильтра Minisart-Plus (Sartorius AG) с получением очищенного образца.
[0204]
1)-15 Опосредованная антителами клеточная цитотоксичность (ADCC) человеческих химерных антител
Опосредованную антителами клеточную цитотоксичность (ADCC) каждого человеческого химерного антитела оценивали с использованием человеческих мононуклеарных клеток периферической крови (PBMC) в качестве эффекторных клеток и линию человеческих клеток поджелудочной железы MIA PaCa-2 в качестве клеток мишеней для ADCC. После обработки клеток MIA PaCa-2, меченых радиоизотопом 51Cr мышиным антителом (LN22R8), крысиным антителом (2P10F2) или человеческими химерными антителами (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA или 2P10F2chIgG4P) при концентрации 0,5 или 5 мкг/мл при 4°C в течение 30 минут, к ним добавляли PBMC, выделенные из периферической крови человека, в соотношении в 20 раз большем, чем клетки MIA PaCa-2, затем культивировали в течение 4 часов при 37°C в присутствии 5% CO2. Используя сцинтилляционный счетчик TopCount NXT v2,53, измеряли 51Cr, высвобождаемый в надосадочную жидкость, с получением a величины суммарного высвобождения. Используя измеренную величину высвобождаемого 51Cr при обработке клеток MIA PaCa-2, меченых 51Cr, с помощью неионного поверхностно-активного вещества Triton-100, в качестве величины максимального высвобождения, и измеренную величину высвобождаемого 51Cr при обработке клеток, обработанных антителом, без добавления PBMC, в качестве величины спонтанного высвобождения, рассчитывали % специфическое высвобождение по приведенной ниже формуле. Результаты представлены на фигуре 6. В качестве образца отрицательного контроля, измеряли таким же методом образец, полученный путем обработки с помощью человеческого IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Inc., Cat. 009-000-003), и эти результаты представлены вместе. Измерения проводили три раза, и рассчитывали и представляли средние значения и стандартные отклонения.
[0205]
% Специфическое высвобождение = (суммарное высвобождение-спонтанное высвобождение)/максимальное высвобождение
В то время как человеческий IgG (hIgG) и мышиное антитело LN22R8 не проявляли опосредованной антителами клеточной цитотоксичности (ADCC), LN22R8chIgG1 проявляло опосредованную антителами клеточную цитотоксичность (ADCC), равную 17,4% при 0,5 мкг/мл и 18,1% при 5 мкг/мл. Опосредованная антителами клеточная цитотоксичность (ADCC) в случае LN22R8chIgG2 и LN22R8chIgG4P была ниже, чем в случае LN22R8chIgG1, и эти значения составляли, соответственно, 3,0% и 2,2% даже при 5 мкг/мл.
[0206]
Крысиное антитело 2P10F2 проявляло опосредованную антителами клеточную цитотоксичность (ADCC), равную 4,8% при 0,5 мкг/мл и 8,4% при 5 мкг/мл. 2P10F2chIgG1LALA проявляло ADCC, равную 4,7% при 0,5 мкг/мл и 2,9% при 5 мкг/мл. 2P10F2chIgG4P проявляло ADCC, равную 3,4% при 0,5 мкг/мл и 1,1% при 5 мкг/мл, которые были ниже, чем показатели для крысиного антитела 2P10F2 и 2P10F2chIgG1LALA. Как известно из литературных источников (Bruggemann et al., J.Exp. Med., 1351-1361, 1987), человеческие химерные антитела с использованием IgG1 субтипа проявляют самую высокую опосредованную антителами клеточную цитотоксичность (ADCC).
[0207]
1)-16 Комплементзависимая цитотоксичность (CDC) человеческих химерных антител
Комплементзависимую цитотоксичность (CDC) каждого антитела против человеческого CD147 оценивали с использованием человеческих клеток поджелудочной железы линии MIA PaCa-2 в качестве клеток-мишеней. В качестве комплемента использовали выпускаемый промышленностью кроличий комплемент (Low Tox-M Rabbit Complement, CEDARLANE LABORATORIES LIMITED, Cat. CL3051). В качестве антитела против человеческого CD147 использовали мышиное антитело (LN22R8), крысиное антитело (2P10F2) или человеческие химерные антитела (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA или 2P10F2chIgG4P). В качестве антитела для отрицательного контроля комплементзависимой цитотоксичности (CDC) использовали человеческий IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Inc., Cat. 009-000-003). После обработки антитела при 4°C и при концентрации 0, 0,1, 1 или 10 мкг/мл в течение одного часа, к нему добавляли кроличий комплемент до конечной концентрации 7,5%, затем нагревали при 37°C в присутствии 5% CO2 в течение трех часов. После этого, измеряли межклеточный АТФ, содержащийся в живых клетках, с помощью анализа жизнеспособности клеток CellTiter-Glo Lumimescent Cell Viability Assay (Promega Corp., Cat. G7572). Измеряли сигналы люминесценции, полученные при проведении анализа CellTiter-Glo Lumimescent Cell Viability Assay, используя многоканальный ридер EnVision 2104 Multilabel Reader (PerkinElmer Co., Ltd). Измерение проводили три раза, и рассчитывали средние значения и стандартные отклонения. Сигналы люминесценции, полученные от неподвергнутых обработке клеток, принимали за 100%, и любое антителозависимое и комплементзависимое уменьшение сигналов люминесценции принимали за комплементзависимую цитотоксичность (CDC). Результаты представлены на фигуре 7.
[0208]
Только в случае мышиного антитела (LN22R8) и крысиного антитела (2P10F2) обнаруживалась зависимая от концентрации комплементзависимая цитотоксичность (CDC) при сравнении с hIgG в качестве отрицательного контроля. В случае LN22R8, количество живых клеток при 10 мкг/мл максимально снижалось до 41,1%. В случае 2P10F2, количество живых клеток при 10 мкг/мл максимально снижалось до 53,5%.
[0209]
В случае человеческих химерных антител (LN22R8chIgG1, LN22R8chIgG2, LN22R8chIgG4P, 2P10F2chIgG1LALA и 2P10F2chIgG4P), не обнаруживалось очевидной комплементзависимой цитотоксичности (CDC) при сравнении с hIgG в качестве отрицательного контроля.
[0210]
1)-17 Антителозависимый клеточный фагоцитоз (ADCP), вызываемый человеческими химерными антителами
Сообщалось, что человеческие IgG антитела вызывают гибель раковых клеток в результате индуцирования фагоцитоза (ADCP) антителозависимыми моноцитами или макрофагами через взаимодействие с мышиными Fcγ рецепторами (Overdijk et al., Journal of Immunology, 1-9, 2012). Антителозависимый клеточный фагоцитоз (ADCP) каждого человеческого химерного антитела оценивали путем использования RAW264.7 (ATCC, TIB-71) в качестве эффекторных клеток и клеток поджелудочной железы человека линии PANC-1 или линии MIA PaCa-2 в качестве клеток-мишеней для ADCP. После обработки клеток-мишеней для ADCP, меченых с помощью мини-набора реагентов PKH67 Green Fluorescent Cell Linker Mini Kit for General Cell Membrane Labeling (SIGMA, Cat. MINI67-1KIT), и человеческих химерных антител (LN22R8chIgG1, LN22R8chIgG2 или LN22R8chIgG4P) при концентрации 20 мкг/мл при 4°C в течение одного часа, к ним добавляли клетки RAW264.7, меченые с помощью набора реагентов PKH26 Red Fluorescent Cell Linker Kit for General Cell Membrane Labeling (SIGMA, Cat. PKH26GL-1KT), в количестве в 5 раз превышающее количество клеток-мишеней для ADCP, затем нагревали при 37°C в присутствии 5% CO2 в течение трех часов. Используя проточный цитометр (Becton, Dickinson and Company, CantoII), измеряли долю PKH26-положительных клеток, которые переносились в PKH67 сигнал-положительные клетки в результате фагоцитоза. В качестве образца отрицательного контроля, измеряли таким же способом образец, полученный обработкой человеческого IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Inc., Cat. 009-000-003). Измерение проводили три раза, и рассчитывали средние значения и стандартные отклонения. На фигуре 8(a) представлены результаты для PANC-1, и на фигуре 8(b) представлены результаты для MIA PaCa-2, соответственно.
[0211]
В случае использования клеток PANC-1 в качестве клеток-мишеней для ADCP, LN22R8chIgG1 проявляло высокий антителозависимый клеточный фагоцитоз (ADCP) порядка 9,2%, а LN22R8chIgG4P проявляло высокий антителозависимый клеточный фагоцитоз (ADCP) порядка 9,0%, по сравнению с человеческим IgG (5,5%). LN22R8chIgG2 не проявляло антителозависимого клеточного фагоцитоза (ADCP) при 5,9%.
[0212]
Аналогичная тенденция была обнаружена также в случае использования клеток MIA PaCa-2 в качестве клеток-мишеней для ADCP, в которых LN22R8chIgG1 проявляло высокий антителозависимый клеточный фагоцитоз (ADCP) порядка 6,6%, и LN22R8chIgG4P проявляло высокий антителозависимый клеточный фагоцитоз (ADCP) порядка 6,1%, по сравнению с человеческим IgG (3,6%). LN22R8chIgG2 не проявляло антителозависимого клеточного фагоцитоза (ADCP) при 3,6%.
[0213]
1)-18 Измерение in-vivo противоопухолевой активности человеческих химерных антител
5 × 106 клеток линии MIA PaCa-2 поджелудочной железы человека суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию инокулировали подкожно в подмышечную впадину самок иммунодефицитных мышей в возрасте от 4 до 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 5-7 дней после инокуляции, и несущим злокачественную опухоль мышам вводили интраперитонеально мышиное антитело (LN22R8) и три типа человеческих химерных антител (LN22R8chIgG1, LN22R8chIgG2 и LN22R8chIgG4P) антитела LN22R8 против человеческого CD147 при 1 мг/кг, 3 мг/кг или 10 мг/кг (n=5). Крысиное антитело (2P10F2) и два типа человеческих химерных антител (2P10F2chIgG2 и 2P10F2chIgG4P) антитела 2P10F2 против человеческого CD147 вводили интраперитонеально несущим злокачественную опухоль мышам при 10 мг/кг (n=5-6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0214]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигурах 9-1(a)-(d) и фигурах 9-2(e)-(g). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли.
[0215]
Как в мышином антителе, так и в трех типах человеческих химерных антител, LN22R8 проявляло дозозависимую противоопухолевую активность. В группе, в которой вводили человеческое химерное антитело LN22R8chIgG4P в дозе 10 мг/кг, наблюдали полный регресс опухоли через 18 дней после инокуляции у 5 мышей из 5 мышей, и возобновление роста опухоли не обнаруживалось даже через 41 день после инокуляции в момент завершения эксперимента. В других группах введения антитела LN22R8, возобновление роста опухоли обнаруживалось у некоторых или у всех мышей.
[0216]
В крысином антителе и в двух типах человеческих химерных антител, обнаруживалась противоопухолевая активность 2P10F2. В группе, в которой вводили 2P10F2chIgG4P в дозе 10 мг/кг, наблюдали полный регресс опухоли через 21 дней после инокуляции у 6 мышей из 6 мышей.
[0217]
Было сделано предположение, что противоопухолевая активность проявляется на основе нового механизма воздействия на CD147, независимо от иммунитета мыши, так как мышиное антитело LN22R8 и крысиное антитело 2P10F2, которые являются антителами против человеческого CD147, сконфигурированные для распознания одного и того же эпитопного сайта, сохраняют противоопухолевую активность 90% или более не только в человеческом химерном антителе chIgG1, имеющем эффекторные функции, зависящие от иммунной системы мыши, такие как опосредованная антителами клеточная цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP) и комплементзависимая цитотоксичность (CDC), или в человеческом химерном антителе chIgG4P, имеющем антителозависимый клеточный фагоцитоз (ADCP), но также и в человеческом химерном антителе chIgG2, которое почти не проявляет никаких эффекторных функций.
[0218]
1)-19 Противоопухолевая активность человеческого химерного антитела против CD147 у мышей линии NOG
Мыши линии NOG (NOD/Shi-scid, IL-2Rnull), получаемые путем гибридизации нокаута γ цепи рецептора IL-2, которая является общим доменом рецепторов цитокинов, с мышами линии NOD-scid, характеризующихся дефицитом T-клеток и B-клеток, имеют недостаточную активность естественных клеток-киллеров (NK) и комплемента помимо дефицита T- и B-клеток и характеризуются пониженными функциями макрофагов и дендритных клеток, вследствие чего находятся в крайне тяжелом состоянии иммунной недостаточности (Ito, Blood, 3175-3182, 2002). На модели подкожной имплантации MIA PaCa-2 проводили исследование возможности влияния такого тяжелого состояния недостаточности иммунной системы мышей на противоопухолевую активность антитела против CD147.
[0219]
5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащим 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензии инокулировали подкожно в подмышечную впадину самок мышей линии NOG (NOD/Shi-scid, IL-2RγKO Jic, поставляемых фирмой In-Vivo Science Inc.) в возрасте 7 недель. Проводили распределение по группам на основе объема опухоли через 6 дней после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам человеческое химерное антитело против CD147 (LN22R8chIgG4P) при 10 мг/кг (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0220]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 10. На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли.
[0221]
Согласно полученным результатам для человеческого химерного антитела LN22R8chIgG4P, представленным на фигуре 10, степень ингибирования роста опухоли через 17 дней после инокуляции составляла 99% в группе, в которой вводили 10 мг/кг, и полный регресс опухоли наблюдали у 3 мышей из 5 мышей.
[0222]
Было выдвинуто предположение о возможности проявления противоопухолевой активности вне зависимости от мышиных иммуноцитов, так как человеческое химерное антитело против CD147 (LN22R8chIgG4P) проявляло высокую противоопухолевую активность также в отношении злокачественных опухолей поджелудочной железы, образованных у мышей линии NOG, которые помимо дефицита T- и B-клеток имеют недостаточную активность естественных клеток-киллеров (NK) и комплемента.
[0223]
(Пример 2) Продукция обезьяньего кросс-реактивного крысиного антитела путем иммунизации с помощью белка CD147
Антитела против человеческого CD147, проявляющие высокую противоопухолевую активность, полученные в примере 1, не проявляют кросс-реактивность с CD147 мыши, крысы и яванского макака. Была сделана попытка получить антитело против CD147, проявляющее кросс-реактивность с CD147 яванского макака, используя каждое антитело, полученное в примере 1.
[0224]
2)-1 Иммунизация
Для иммунизации использовали самок крыс линии WKY/Izm (Japan SLC, Inc.). Смесь рекомбинантного человеческого базиджина (BSG), антигена His-концевого белка (фирмы Creative BioMart) и полного адъюванта Фрейнда (фирмы Wako Pure Chemical Industries, Ltd.) вводили в основание хвоста крысы, собирали лимфатические узлы и селезенку крысы и использовали их для продукции гибридом.
[0225]
2)-2 Продукция гибридом
Клетки лимфатического узла или клетки селезенки подвергали электрослиянию с мышиными миеломными клетками линии SP2/0-ag14 (ATCC, No. CRL-1581), используя прибор LF301 Cell Fusion Unit (BEX CO., LTD.), и слившиеся клетки разбавляли средой ClonaCell-HY Selection Medium D (StemCell Technologies Inc.) и культивировали. Собирали появляющиеся колонии гибридомы, в результате чего получали моноклональные гибридомы. Каждую из собранных колоний гибридомы культивировали, и антитела против CD147, которые продуцируют гибридомы, подвергали скринингу, используя гибридомы, получаемые в надосадочной жидкости культуры клеток.
[0226]
2)-3 Скрининг антител методом проточной цитометрии
Для выбора продуцируемого гибридомами антитела, сконфигурированного для связывания с раковыми клетками человека и проявляющего активность связывания с CD147 человека и CD147 яванского макака, проводили скрининг активности связывания антитела, используя проточный цитометр. В качестве раковых клеток человека использовали CD147 положительные клетки поджелудочной железы человека линии MIA PaCa-2. Клетки CHO-K1 (CHO-K1-hCD147v2 и CHO-K1-cynoCD147), экспрессирующие CD147 человека или яванского макака, использовали таким же методом, как в примере 1)-8, для подтверждения активности связывания с CD147 человека или яванского макака. Равное количество содержащей гибридому надосадочной жидкости культуры клеток добавляли к суспензии MIA PaCa-2, CHO-K1-hCD147v2 или CHO-K1-cynoCD147, затем проводили реакцию при 4°C в течение одного часа или более. После этого, клетки промывали с помощью PBS, содержащим 5% FBS, для того чтобы можно было провести флуоресцентное обнаружение связывание каждого антитела с клетками, используя анти-крысиный IgG-PE (BD Biosciences, Cat. 550767). Измеряли флуоресцентные сигналы клеток, используя проточный цитометр (CantoII, BD Biosciences), и рассчитывали отношение этих флуоресцентных сигналов к флуоресцентным сигналам в образце отрицательного контроля (клетки без добавления культурального раствора с гибридомами). Часть этих результатов представлена в таблице 4.
[0227]
Используя производимый промышленностью набор реагентов для изотипирования крысиного антитела (Bio-Rad Laboratories, RMT1), определяли изотип антитела, содержащегося в надосадочной жидкости культуры клеток. Результаты представлены в таблице 4.
[0228]
2P10F2, полученное в примере 1, проявляло активность связывания с MIA PaCa-2 и CHO-K1-hCD147v2, и не проявляло активности связывания с CHO-K1-cynoCD147. Rat_CD147_#84 (которое может быть обозначено в этом описании как r#84), rat_CD147_#131 (которое может быть обозначено в этом описании как r#131), rat_CD147_#110 (которое может быть обозначено в этом описании как r#110) и rat_CD147_#101 (которое может быть обозначено в этом описании как r#101) проявляли активность связывания с MIA PaCa-2, CHO-K1-hCD147v2 и CHO-K1-cynoCD147. В результате, были получены крысиные антитела против CD147, проявляющие кросс-реактивность с CD147 яванского макака.
[0229]
[Таблица 4]
[0230]
2)-4 Получение крысиного моноклонального антитела с использованием сыворотки с низким содержанием IgG
Моноклональные антитела против человеческого CD147, проявляющие кросс-реактивность с CD147 яванского макака, очищали от содержащей гибридому надосадочной жидкости культуры клеток. Сначала, выращивали достаточное количество гибридом, продуцирующих антитело rat_CD147_#131, в среде ClonaCell-HY Selection Medium E, и после этого среду заменяли на среду Hybridoma SFM (Life Technologies), к которой добавляли Ultra Low IgG FBS (Life Technologies) до 20%, затем культивировали в течение 7 дней. Эту надосадочную жидкость культуры клеток собирали и пропускали через фильтр с размером пор 0,45 мкм с целью стерилизации.
[0231]
2)-5 Получение крысиного моноклонального тела в культуре с высокой плотностью клеток
Культивировали гибридомы rat_CD147_#84, rat_CD147_#101 или rat_CD147_#110, используя колбу CL-1000 (Becton, Dickinson and Company), с целью получения в надосадочной жидкости культуры клеток гибридомы, содержащей каждое моноклональное антитело.
[0232]
2)-6 Очистка моноклонального антитела
Антитело, полученное в примере 2)-4 и примере 2)-5, очищали от надосадочной жидкости культуры клеток таким же методом, как в примере 1)-6.
[0233]
2)-7 Скрининг антитела путем измерения in-vivo противоопухолевой активности
5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащим 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC.). Проводили распределение по группам на основе объема опухоли через 6-8 дней после инокуляции, и вводили интраперитонеально яванский макак-кросс-реактивное крысиное антитело #84, #101 или #110 против CD147 при 10 мг/кг несущим злокачественную опухоль мышам через 8 дней и 15 дней после инокуляции (n=5). Аналогичным методом, вводили интраперитонеально PBS мышам контрольной группы. Яванский макак-кросс-реактивное крысиное антитело #131 против CD147 вводили интраперитонеально несущим злокачественную опухоль мышам при 10 мг/кг через 6 дней после инокуляции (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0234]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигурах 11(a)-(d). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. Степень ингибирования роста опухоли через 28 дней после инокуляции r#84 составляла 95% в группе дозирования 10 мг/кг (фигура 11(a)). Степень ингибирования роста опухоли через 15 дней после инокуляции r#101 составляла 37% в группе дозирования 10 мг/кг, но наблюдалось возобновление роста опухоли через 28 дней после инокуляции (фигура 11(b)). Степень ингибирования роста опухоли через 15 дней после инокуляции r#110 составляла 51% в группе дозирования 10 мг/кг, но наблюдалось возобновление роста опухоли через 28 дней после инокуляции (фигура 11(c)). Степень ингибирования роста опухоли через 16 дней после инокуляции r#131 составляла 50% в группе дозирования 10 мг/кг (фигура 11(d)). В результате получали крысиное антитело r#84, которое сильно ингибирует рост опухоли в клетках MIA PaCa-2, и крысиные антитела r#101, r#110 и r#131, которые частично ингибируют рост опухоли.
[0235]
-[0236]
2)-8 Эпитопный анализ: 2P10F2chIgG4P-конкурентный твердофазный иммуноферментный анализ (ELISA)
Применительно к эпитопному анализу обезьяньих-кросс-реактивных крысиных антител против CD147, проводили исследование возможности каждого антитела ингибировать активность связывания с CD147 рекомбинантного белка 2P10F2chIgG4P методом конкурентного твердофазного иммуноферментного анализа (ELISA). Человеческий CD147-Fc гибридный белок (Sino Biological Inc., 10186-H02H) растворяли в PBS до концентрации 2 мкг/мл, и 50 мкл этого раствора добавляли в 96-луночный планшет (Thermo Fisher SCIENTIFIC K.K., Cat. 43454), затем выдерживали при 4°C. После чего удаляли раствор белка, добавляли 300 мкл PBS, содержащего 1% BSA, затем нагревали при комнатной температуре в течение одного часа. В качестве конкурирующих антител добавляли в 96-луночный планшет 25 мкл CD147 крысиных антител r#84, r#101, r#110, r#131 или 2P10F2, разбавленных с помощью PBS, содержащего 1% BSA, до 20 или 60 мкг/мл, или с помощью PBS, содержащего 1% BSA, затем нагревали при комнатной температуре в течение двух часов. Добавляли в 96-луночный планшет 25 мкл 2P10F2chIgG4P, разбавленного с помощью PBS, содержащего 1% BSA, до 20 нг/мл, затем нагревали при комнатной температуре в течение двух часов. 96-луночный планшет промывали два раза с помощью PBS, содержащего 0,05% Tween 20 (BIO RAD, Cat. 170-6531). В 96-луночный планшет добавляли 50 мкл мышиного моноклонального HP6025 античеловеческого IgG4 Fc (HRP) (Abcam plc., Cat. ab99823), разбавленного в 2000 раз с помощью PBS, содержащего 1% BSA, затем нагревали при комнатной температуре в течение одного часа. 96-луночный планшет промывали три раза с помощью PBS, содержащего 0,05% Tween 20 (BIO RAD, Cat. 170-6531). В 96-луночный планшет добавляли 50 мкл субстрата Super AquaBlue ELISA Substrate (eBioscience, Thermo Fisher SCIENTIFIC K.K., 00-4203-58), затем нагревали при комнатной температуре в течение 20 минут. Измеряли поглощение при 405 нм 96-луночного планшета, используя многоканальный ридер EnVision 2104 Multilabel Reader (PerkinElmer Co., Ltd). Измеренную величину для лунки, не содержащей конкурирующих антител, использовали в качестве контроля, и рассчитывали снижение поглощения, вызванное каждым конкурирующим антителом, в процентах (%). Результаты представлены на фигуре 12. Измерения проводили для трех лунок, и представлены средние значения.
[0237]
Аналогично крысиному антителу 2P10F2, антитела r#84, r#101 и r#131 ингибировали активность связывания 2P10F2chIgG4P на 90% или более, и в результате чего было сделано предположение, что их сайты распознавания антитела были близки к сайту распознавания 2P10F2chIgG4P. При этом, r#110 не ингибировало связывание крысиного антитела 2P10F2chIgG4P. Можно допустить, что связывание крысиного антитела 2P10F2chIgG4P не могло быть ингибировано вследствие того, что его сайт распознавания антитела отстоял далеко от сайта распознаванияr#110, или активность связывания r#110 была низкой.
[0238]
(Пример 3) Клонирование обезьяньего-кросс-реактивного крысиного антитела и продукция человеческого химерного антитела
3)-1 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область крысиных антител против CD147, для клонирования
3)-1-1 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела rat_CD147_#84
Определение проводили таким же методом, как в примере 1)-11-2. SEQ ID NO:48 представляет нуклеотидную последовательность кДНК, кодирующей определенную вариабельную область легкой цепи антитела rat_CD147_#84, и SEQ ID NO:49 представляет ее аминокислотную последовательность. SEQ ID NO:50 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область ее тяжелой цепи, и SEQ ID NO:51 представляет ее аминокислотную последовательность. SEQ ID NOs: 52, 53 и 54, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела rat_CD147_#84. SEQ ID NOs: 55, 56 и 57, соответственно, представляют CDRH1, CDRH2 и CDRH3 вариабельной области тяжелой цепи антитела rat_CD147_#84.
[0239]
3)-1-2 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела rat_CD147_#101
Определение проводили таким же методом, как в примере 1)-11-2. SEQ ID NO:58 представляет нуклеотидную последовательность кДНК, кодирующей определенную вариабельную область легкой цепи антитела rat_CD147_#101, и SEQ ID NO:59 представляет ее аминокислотную последовательность. SEQ ID NO:60 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи, и SEQ ID NO:61 представляет ее аминокислотную последовательность. SEQ ID NOs: 62, 63 и 64, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела rat_CD147_#101. SEQ ID NOs: 65, 66 и 67, соответственно, представляют CDRH1, CDRH2, и CDRH3 вариабельной области тяжелой цепи антитела rat_CD147_#101.
[0240]
3)-1-3 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела rat_CD147_#110
Определение проводили таким же методом, как в примере 1)-11-2. SEQ ID NO:68 представляет нуклеотидную последовательность кДНК, кодирующей определенную вариабельную область легкой цепи антитела rat_CD147_#110, и SEQ ID NO:69 представляет ее аминокислотную последовательность. SEQ ID NO:70 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи, и SEQ ID NO:71 представляет ее аминокислотную последовательность. SEQ ID NOs: 72, 73 и 74, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела rat_CD147_#110. SEQ ID NOs: 75, 76 и 77, соответственно, представляют CDRH1, CDRH2 и CDRH3 вариабельной области тяжелой цепи антитела rat_CD147_#110.
[0241]
3)-1-4 Определение нуклеотидной последовательности кДНК, кодирующей вариабельную область антитела rat_CD147_#131
Определение проводили таким же методом, как в примере 1)-11-2. SEQ ID NO:78 представляет нуклеотидную последовательность кДНК, кодирующей определенную вариабельную область легкой цепи антитела rat_CD147_#131, и SEQ ID NO:79 представляет ее аминокислотную последовательность. SEQ ID NO:80 представляет нуклеотидную последовательность кДНК, кодирующей вариабельную область тяжелой цепи, и SEQ ID NO:81 представляет ее аминокислотную последовательность. SEQ ID NOs: 82, 83 и 84, соответственно, представляют CDRL1, CDRL2 и CDRL3 вариабельной области легкой цепи антитела rat_CD147_#131. SEQ ID NOs: 85, 86 и 87, соответственно, представляют CDRH1, CDRH2 и CDRH3 вариабельной области тяжелой цепи антитела rat_CD147_#131.
[0242]
3)-2 Продукция экспрессирующих векторов человеческого химерного антитела
3)-2-1 Продукция экспрессирующего вектора человеческого химерного антитела rat_CD147_#84
3)-2-1-1 Конструирование экспрессирующего вектора pCMA-G4PFALA тяжелой цепи человеческого химерное и гуманизированного антитела типа IgG4PFALA
Используя сигнальную последовательность человеческой тяжелой цепи, представленной последовательностью SEQ ID NO:88, и фрагмент ДНК, содержащий последовательность ДНК, кодирующую аминокислоты в константной области человеческого IgG4PFALA, конструировали pCMA-G4PFALA таким же методом, как в примере 1)-12-2.
[0243]
3)-2-1-2 Конструирование экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84
Используя кДНК, кодирующую вариабельную область легкой цепи rat_CD147_#84, полученную в примере 3)-1-1, в качестве матрицы, конструировали экспрессирующий вектор легкой цепи человеческого химерного rat_CD147_#84 таким же методом, как в примере 1)-12-4. SEQ ID NO:89 и SEQ ID NO:90, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного rat_CD147_#84 и аминокислотную последовательность легкой цепи.
[0244]
3)-2-1-3 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#84 полученную в примере 3)-1-1, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1 таким же методом, как в примере 1)-12-5. SEQ ID NO:91 и SEQ ID NO:92, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1 и аминокислотную последовательность тяжелой цепи.
[0245]
3)-2-1-4 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#84, полученную в примере 3)-1-1, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2 таким же методом, как в примере 1)-12-6. SEQ ID NO:93 и SEQ ID NO:94, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2 и аминокислотную последовательность тяжелой цепи.
[0246]
3)-2-1-5 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#84, полученную в примере 3)-1-1, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P таким же методом, как в примере 1)-13-6. SEQ ID NO:95 и SEQ ID NO:96, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P и аминокислотную последовательность тяжелой цепи.
[0247]
3)-2-1-6 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#84, полученную в примере 3)-1-1, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA таким же методом, как в примере 1)-13-4. SEQ ID NO:97 и SEQ ID NO:98, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA и аминокислотную последовательность тяжелой цепи.
[0248]
3)-2-1-7 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4PFALA
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#84, полученную в примере 3)-1-1, в качестве матрицы, проводили PCR, используя праймер, сконструированный для клонирования In-fusion, в результате чего амплифицировали фрагмент ДНК, содержащий кДНК, кодирующую вариабельную область тяжелой цепи. Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), амплифицированный фрагмент ДНК вставляли в том месте, где pCMA-G4PFALA дигерировали с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4PFALA. SEQ ID NO:99 и SEQ ID NO:100, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4PFALA и аминокислотную последовательность тяжелой цепи.
[0249]
3)-2-2 Продукция экспрессирующего вектора человеческого химерного антитела rat_CD147_#101
3)-2-2-1 Конструирование экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#101
Используя кДНК, кодирующую вариабельную область легкой цепи rat_CD147_#101, полученную в примере 3)-1-2, в качестве матрицы, конструировали экспрессирующий вектор легкой цепи человеческого химерного rat_CD147_#101 таким же методом, как в примере 1)-12-4. SEQ ID NO:101 и SEQ ID NO:102, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного rat_CD147_#101 и аминокислотную последовательность легкой цепи.
[0250]
3)-2-2-2 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#101, полученную в примере 3)-1-2, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG2 таким же методом, как в примере 1)-12-6. SEQ ID NO:103 и SEQ ID NO:104, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG2 и аминокислотную последовательность тяжелой цепи.
[0251]
3)-2-2-3 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4P
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#101, полученную в примере 3)-1-2, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4P таким же методом, как в примере 1)-13-6. SEQ ID NO:105 и SEQ ID NO:106, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4P и аминокислотную последовательность тяжелой цепи.
[0252]
3)-2-2-4 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4PFALA
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#101, полученную в примере 3)-1-2, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4PFALA таким же методом, как в примере 3)-2-1-7. SEQ ID NO:107 и SEQ ID NO:108, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4PFALA и аминокислотную последовательность тяжелой цепи.
[0253]
3)-2-3 Продукция экспрессирующего вектора человеческого химерного антитела rat_CD147_#110
3)-2-3-1 Конструирование экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#110
Используя кДНК, кодирующую вариабельную область легкой цепи rat_CD147_#110, полученную в примере 3)-1-3, в качестве матрицы, конструировали экспрессирующий вектор легкой цепи человеческого химерного rat_CD147_#110 таким же методом, как в примере 1)-12-4. SEQ ID NO:109 и SEQ ID NO:110, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного rat_CD147_#110 и аминокислотную последовательность легкой цепи.
[0254]
3)-2-3-2 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#110, полученную в примере 3)-1-3, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG2 таким же методом, как в примере 1)-12-6. SEQ ID NO:111 и SEQ ID NO:112, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG2 и аминокислотную последовательность тяжелой цепи.
[0255]
3)-2-3-3 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4P
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#110, полученную в примере 3)-1-3, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4P таким же методом, как в примере 1)-13-6. SEQ ID NO:113 и SEQ ID NO:114, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4P и аминокислотную последовательность тяжелой цепи.
[0256]
3)-2-3-4 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4PFALA
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#110, полученную в примере 3)-1-3, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4PFALA таким же методом, как в примере 3)-2-1-7. SEQ ID NO:115 и SEQ ID NO:116, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4PFALA и аминокислотную последовательность тяжелой цепи.
[0257]
3)-2-4 Продукция экспрессирующего вектора человеческого химерного антитела rat_CD147_#131
3)-2-4-1 Конструирование экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#131
Используя кДНК, кодирующую вариабельную область легкой цепи rat_CD147_#131, полученную в примере 3)-1-4, в качестве матрицы, конструировали экспрессирующий вектор легкой цепи человеческого химерного rat_CD147_#131 таким же методом, как в примере 1)-12-4. SEQ ID NO:117 и SEQ ID NO:118, соответственно, представляют нуклеотидную последовательность легкой цепи человеческого химерного rat_CD147_#131 и аминокислотную последовательность легкой цепи.
[0258]
3)-2-4-2 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG2
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#131, полученную в примере 3)-1-4, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG2 таким же методом, как в примере 1)-12-6. SEQ ID NO:119 и SEQ ID NO:120, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG2 т аминокислотную последовательность тяжелой цепи.
[0259]
3)-2-4-3 Конструирование экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG4P
Используя кДНК, кодирующую вариабельную область тяжелой цепи rat_CD147_#131, полученную в примере 3)-1-4, в качестве матрицы, конструировали экспрессирующий вектор тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG4P таким же методом, как в примере 1)-13-6. SEQ ID NO:121 и SEQ ID NO:122, соответственно, представляют нуклеотидную последовательность тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG4P и аминокислотную последовательность тяжелой цепи.
[0260]
3)-3 Получение человеческого химерного антитела
3)-3-1 Продукция человеческого химерного антитела из обезьяньего-кросс-реактивного крысиного антитела
Антитело продуцировали таким же методом, как в примере 1)-14-1.
[0261]
Человеческое химерное антитело rat_CD147_#84, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1 и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84, обозначали как "#84chIgG1". Человеческое химерное антитело rat_CD147_#84, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84, обозначали как "#84chIgG2". Человеческое химерное антитело rat_CD147_#84, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84, обозначали как "#84chIgG4P". Человеческое химерное антитело rat_CD147_#84, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84, обозначали как "#84chIgG1LALA". Человеческое химерное антитело rat_CD147_#84, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4PFALA и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#84, обозначали как "#84chIgG4PFALA".
[0262]
Человеческое химерное антитело rat_CD147_#101, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#101, обозначали как "#101chIgG2". Человеческое химерное антитело rat_CD147_#101, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#101, обозначали как "#101chIgG4P". Человеческое химерное антитело rat_CD147_#101, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#101 типа IgG4PFALA и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#101, обозначали как "#101chIgG4PFALA".
[0263]
Человеческое химерное антитело rat_CD147_#110, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#110, обозначали как "#101chIgG2". Человеческое химерное антитело rat_CD147_#110, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#110, обозначали как "#110chIgG4P". Человеческое химерное антитело rat_CD147_#110, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#110 типа IgG4PFALA и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#110, обозначали как "#110chIgG4PFALA".
[0264]
Человеческое химерное антитело rat_CD147_#131, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG2 и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#131, обозначали как "#131chIgG2". Человеческое химерное антитело rat_CD147_#131, полученное путем комбинации экспрессирующего вектора тяжелой цепи человеческого химерного rat_CD147_#131 типа IgG4P и экспрессирующего вектора легкой цепи человеческого химерного rat_CD147_#131, обозначали как "#131chIgG4P".
[0265]
3)-3-2 Очистка человеческого химерного антитела обезьяньих-кросс-реактивных крысиных антител
Антитела очищали от надосадочной жидкости культуры клеток, полученные в примере 3)-3-1, таким же методом, как в примере 1)-14-2.
[0266]
(Пример 4) In-vivo противоопухолевая активность человеческих химерных антител
5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5-6 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 5-6 дней после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам яванский макак-кросс-реактивное человеческое химерное антитело против CD147 (#84chIgG1, #84chIgG1LALA, #84chIgG2, #84chIgG4P или #84chIgG4PFALA) в дозе 1, 3 или 10 мг/кг через один день после распределения мышей по группам (n=5). Яванский макак-кросс-реактивное человеческое химерное антитело против CD147 (#101chIgG2, #101chIgG4P, #101chIgG4PFALA, #110chIgG2, #110chIgG4P, #110chIgG4PFALA, #131chIgG2 или #131chIgG4P) вводили интраперитонеально несущим злокачественную опухоль мышам в дозе 3 или 10 мг/кг через один день после распределения мышей по группам (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0267]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигурах 13-1(a)-(d), фигурах 13-2(e)-(h), фигурах 13-3 (i)-(l) и фигурах 13-4 (m)-(n). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли.
[0268]
Человеческое химерное антитело #84 проявляло дозозависимую противоопухолевую активность во всех субтипах IgG. В группе введения 10 мг/кг #84chIgG4P, в которой наблюдалась самая высокая противоопухолевая активность, через 20 дней после инокуляции наблюдали полный регресс опухоли у 5 мышей из 5 мышей.
[0269]
Человеческое химерное антитело #101 проявляло дозозависимую противоопухолевую активность во всех субтипах IgG. В группе введения 10 мг/кг 101chIgG4P, в которой наблюдалась самая высокая противоопухолевая активность, через 20 дней после инокуляции наблюдали полный регресс опухоли у 4 мышей из 5 мышей.
[0270]
Человеческое химерное антитело #110 проявляло дозозависимую противоопухолевую активность во всех субтипах IgG. В группе введения 10 мг/кг 110chIgG4P, в которой наблюдалась самая высокая противоопухолевая активность, через 22 дня после инокуляции наблюдали полный регресс опухоли у 3 мышей из 5 мышей.
[0271]
Человеческое химерное антитело #131 проявляло дозозависимую противоопухолевую активность во всех субтипах IgG4P. Отсутствовало различие в противоопухолевой активности, наблюдаемой в группах введения #131chIgG2 в дозах 3 мг/кг и 10 мг/кг.
[0272]
Все человеческие химерные антитела проявляли эффект ингибирования роста опухоли от 68 до 100% при дозе 10 мг/кг, независимо от субтипа человеческого IgG каждого антитела. Было сделано предположение, что яванский макак-кросс-реактивные человеческие химерные антитела #084, #101, #110 и #131 против CD147 человека также не зависели от иммунной системы мыши, аналогично антителу LN22R8 или 2P10F2 против человеческого CD147, полученному путем клеточной иммунизации, и проявляли противоопухолевую активность на основе нового механизма воздействия на CD147.
[0273]
(Пример 5) Оценка активности связывания человеческого химерного антитела с CD147
Измеряли константу диссоциации для связывания #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P или #110chIgG4P, продуцированных в примере 3)-3-1, с CD147 человека с использованием системы Biacore T200 (поставляемой фирмой GE Healthcare Bioscience). Антитело против IgG (Fc) человека иммобилизовали на сенсорном чипе, используя набор для иммобилизации антител Human Antibody Capture Kit (поставляемый фирмой GE Healthcare Bioscience), затем захватывали человеческое химерное антитело в качестве лиганда и ассоциировали антиген в качестве аналита. Использовали HBS-EP+ (поставляемый фирмой GE Healthcare Bioscience) в качестве подвижного буфера и CM5 (поставляемый фирмой GE Healthcare Bioscience) в качестве сенсорного чипа. Затем 1 мкг/мл или 2 мкг/мл человеческого химерного антитела добавляли на чип при 10 мкл/минута в течение 60 секунд, ассоциировали последовательные разведения белка CD147 в качестве антигена (от 0,25 до 4 мкг/мл в отношении #131chIgG4P или от 0,5 до 8 мкг/мл в отношении #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P или #110chIgG4P) при скорости 30 мкл/минута в течение 120 секунд, затем проводили последующий мониторинг фазы диссоциации 120 секунд, 300 секунд или 600 секунд. В этом примере, используемый белок CD147 экспрессировали в Escherichia coli, очищали в две стадии аффинной хроматографией на никель-агарозе и гель-проникающей хроматографией (SEC), и затем расщепляли метку. В качестве регенерирующего раствора добавляли раствор 3M хлорида магния (поставляемый фирмой GE Healthcare Bioscience) при скорости 20 мкл/минута в течение 30 секунд. Используя модель связывания 1:1, рассчитывали константу скорости связывания ka, константу скорости диссоциации kd и константу диссоциации (KD; KD=kd/ka). Результаты приведены в таблице 5.
[0274]
[Таблица 5]
Константа диссоциации для связывания человеческого химерного антитела с CD147 человека
[0275]
(Пример 6) Продукция гуманизированного антитела
6)-1 Конструирование гуманизированного антитела
6)-1-1 Молекулярное моделирование вариабельной области
Использовали метод, известный как гомологичное моделирование (Methods in Enzymology, 203, 121-153, (1991)). Используя коммерчески доступную программу для анализа трехмерной структуры белка Discovery Studio (поставляемую фирмой Dassault Systemes), идентифицировали гомологические последовательности вариабельных областей из зарегистрированных структур в Базе данных белковых структур (Nuc. Acids Res. 35, D301-D303 (2007)). Создавали трехмерные модели, используя идентифицированные тяжелые цепи, легкие цепи и структуры поверхности раздела тяжелых и легких цепей в качестве матриц.
[0276]
6)-1-2 Метод конструирования гуманизированного антитела
Антитело гуманизировали путем прививки CDR (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). Выбирали имеющих высокую гомологичность консенсусную последовательность человеческой каппа цепи подгруппы 1 и консенсусную последовательность человеческой каппа цепи подгруппы 3, определенных в публикации KABAT et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, available from Panasonic Corporation Institutes of Health, Bethesda, MD. (1991), в качестве акцепторов для каркасных областей легких и тяжелых цепей антитела rat_CD147_#84, соответственно. Выбирали имеющих высокую гомологичность консенсусную последовательность человеческой каппа цепи подгруппы 1, определенную в публикации KABAT et al., и IGHV3-30*05 и IGHJ3*01 человеческой гамма цепи, определенные в системе IMGT (THE INTERNATIONAL IMMUNOGENETICS INFORMATION SYSTEM (R)), в качестве акцепторов для каркасных областей легких и тяжелых цепей антитела rat_CD147_#101, соответственно. Выбирали имеющих высокую гомологичность IGKV1-39*01 и IGKJ4*01 человеческой каппа цепи, определенной в IMGT, и IGHV1-2*02 и IGHJ6*01 человеческой гамма цепи, определенной в IMGT, в качестве акцепторов для легких и тяжелых цепей антитела rat_CD147_#110, соответственно. Выбирали имеющих высокую гомологичность IGKV1-39*01 и IGKJ2*01 человеческой каппа цепи, определенной в IMGT, и IGHV3-30*05 и IGHJ6*01 человеческой гамма цепи, определенной в IMGT, в качестве акцепторов для легких и тяжелых цепей антитела rat_CD147_#131, соответственно. Донорные остатки, импортируемые в акцепторы, выбирали на основе анализа трехмерных моделей с использованием критерия, предложенного в публикации Queen et al., Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989), или других критериев.
[0277]
6)-1-3 Гуманизация тяжелой цепи rat_CD147_#84
Аминокислотные остатки акцептора, представленные на фигуре 14(a), импортировали в вариабельную область тяжелой цепи rat_CD147_#84, в результате конструируя вариабельную область тяжелой цепи #84H1h гуманизированного антитела.
[0278]
Тяжелые цепи гуманизированного антитела конструировали путем соединения сконструированной выше вариабельной области с константными областями гамма цепи человеческого IgG2 и IgG4P, и обозначали как #84H1hIgG2 и #84H1hIgG4P, соответственно. SEQ ID NO:123 представляет полноразмерную аминокислотную последовательность #84H1hIgG2. SEQ ID NO:124 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:123. SEQ ID NO:125 представляет полноразмерную аминокислотную последовательность #84H1hIgG4P. SEQ ID NO:126 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:125.
[0279]
6)-1-4 Гуманизация легкой цепи rat_CD147_#84
Аминокислотные остатки акцептора, представленные на фигуре 14(b), импортировали в вариабельную область легкой цепи rat_CD147_#84, в результате конструируя вариабельную область тяжелой цепи #84L2h гуманизированного антитела.
[0280]
Гуманизированное антитело легкой цепи конструировали путем соединения сконструированной выше вариабельной области с константной область каппа цепи, и обозначали как #84L2h. SEQ ID NO:127 представляет полноразмерную аминокислотную последовательность легкой цепи #84L2h. SEQ ID NO:128 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:127.
[0281]
6)-1-5 Гуманизированное антитело #84
Гуманизированное антитело #84H1L2hIgG2 конструировали путем комбинации сконструированных выше тяжелой цепи #84H1hIgG2 и легкой цепи #84L2h. Кроме того, гуманизированное антитело #84H1L2hIgG4P конструировали путем комбинации тяжелой цепи #84H1hIgG4P и легкой цепи #84L2h.
[0282]
6)-1-6 Гуманизация тяжелой цепи rat_CD147_#101
Аминокислотные остатки акцептора, представленные на фигуре 15(a), импортировали в вариабельную область тяжелой цепи rat_CD147_#101, в результате конструируя вариабельную область тяжелой цепи #101H1h гуманизированного антитела. Тяжелые цепи гуманизированного антитела конструировали путем соединения сконструированной выше вариабельной области с константными областями гамма цепи человеческого IgG2 и IgG4P, и обозначали как #101H1hIgG2 и #101H1hIgG4P, соответственно. SEQ ID NO:129 представляет полноразмерную аминокислотную последовательность #101H1hIgG2. SEQ ID NO:130 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:129. SEQ ID NO:131 представляет полноразмерную аминокислотную последовательность #101H1hIgG4P. SEQ ID NO:132 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:131.
[0283]
6)-1-7 Гуманизация легкой цепи rat_CD147_#101
Аминокислотные остатки акцептора, представленные на фигуре 15(b), импортировали в вариабельную область легкой цепи rat_CD147_#101, в результате конструируя вариабельную область тяжелой цепи #101L2h гуманизированного антитела. В сконструированной выше вариабельной области, конструировали легкую цепь гуманизированного антитела, соединенную с константной областью каппа цепи человека, и обозначали как #101L2h. SEQ ID NO:133 представляет полноразмерную аминокислотную последовательность легкой цепи #101L2h. SEQ ID NO:134 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:133.
[0284]
6)-1-8 Гуманизированное антитело #101
Гуманизированное антитело #101H1L2hIgG2 конструировали путем комбинации сконструированных выше тяжелой цепи #101H1hIgG2 и легкой цепи #101L2h. Кроме того, гуманизированное антитело #101H1L2hIgG4P конструировали путем комбинации тяжелой цепи #101H1hIgG4P и легкой цепи #101L2h.
[0285]
6)-1-9 Гуманизация тяжелой цепи rat_CD147_#110
Аминокислотные остатки акцепторов, представленные на фигурах 16(a) и (b), каждый импортировали в вариабельную область тяжелой цепи rat_CD147_#110, в результате конструируя вариабельные области тяжелых цепей #110H1h и #110H13h гуманизированного антитела.
[0286]
Тяжелые цепи гуманизированного антитела конструировали путем соединения сконструированной выше вариабельной области с константной областью гамма цепь человеческого IgG4P, и обозначали как #110H1hIgG4P и #110H13hIgG4P, соответственно. SEQ ID NO:135 представляет полноразмерную аминокислотную последовательность #110H1hIgG4P, и SEQ ID NO:147 представляет полноразмерную аминокислотную последовательность #110H13hIgG4P. SEQ ID NO:136 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:135, и SEQ ID NO:148 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:147.
[0287]
6)-1-10 Гуманизация легкой цепи rat_CD147_#110
Аминокислотные остатки акцепторов, представленные на фигурах 16(c), (d) и (e), каждый импортировали в вариабельную область легкой цепи rat_CD147_#110, в результате конструируя вариабельные области легких цепей #110L4h, #110L2h и #110L12h гуманизированного антитела.
[0288]
Легкие цепи гуманизированного антитела конструировали путем соединения сконструированной выше вариабельной области с константной областью каппа цепи человека, и обозначали как #110L4h, #110L2h и #110L12h, соответственно. SEQ ID NO:137 представляет полноразмерную аминокислотную последовательность легкой цепи #110L4h, SEQ ID NO:149 представляет полноразмерную аминокислотную последовательность легкой цепи #110L2h, и SEQ ID NO:151 представляет полноразмерную аминокислотную последовательность легкой цепи #110L12h. SEQ ID NO:138 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:137, SEQ ID NO:150 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:149, и SEQ ID NO:152 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:151.
[0289]
6)-1-11 Гуманизированное антитело #110
Гуманизированное антитело #110H1L4hIgG4P конструировали путем комбинации сконструированных выше тяжелой цепи #110H1hIgG4P и легкой цепи #110L4h. Кроме того, гуманизированное антитело #110H13L2hIgG4P конструировали путем комбинации тяжелой цепи #110H13hIgG4P и легкой цепи #110L2h, и гуманизированное антитело #110H13L12hIgG4P конструировали путем комбинации тяжелой цепи #110H13hIgG4P и легкой цепи #110L12h.
[0290]
6)-1-12 Гуманизация тяжелой цепи rat_CD147_#131
Аминокислотные остатки акцептора, представленные на фигуре 17(a), импортировали в вариабельную область тяжелой цепи rat_CD147_#131, в результате конструируя вариабельную область тяжелая цепь #131H2h гуманизированного антитела.
[0291]
Тяжелую цепь гуманизированного антитела конструировали путем соединения сконструированной выше вариабельной области с константной областью гамма цепи человеческого IgG2, и обозначали как #131H2hIgG2. SEQ ID NO:139 представляет полноразмерную аминокислотную последовательность #131H2hIgG2. SEQ ID NO:140 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:139.
[0292]
6)-1-13 Гуманизация легкой цепи rat_CD147_#131
Аминокислотные остатки акцептора, представленные на фигуре 17(b), импортировали в вариабельную область легкой цепи rat_CD147_#131, в результате конструируя вариабельную область тяжелой цепи #131L2h гуманизированного антитела.
[0293]
Гуманизированное антитело легкой цепи конструировали путем соединения сконструированной выше вариабельной области с константной область каппа цепи, и обозначали как #131L2h. SEQ ID NO:141 представляет полноразмерную аминокислотную последовательность легкой цепи #131L2h. SEQ ID NO:142 представляет нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:141.
[0294]
6)-1-14 Гуманизированное антитело #131
Гуманизированное антитело #131H2L2hIgG2 конструировали путем комбинации сконструированных выше тяжелой цепи #131H2hIgG2 и легкой цепи #131L2h.
[0295]
6)-2 Конструирование экспрессирующих векторов легкой цепи гуманизированного антитела
6)-2-1 Конструирование экспрессирующего вектора #84L2h
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 37-402 нуклеотидной последовательности #84L2h представленной последовательностью SEQ ID NO:128 (Geneart AG). Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), синтезированный фрагмент ДНК вставляли в то же самое место, где дигерировали pCMA-LK с помощью рестрикционного фермента BsiWI, в результате чего конструировали экспрессирующий вектор #84L2h.
[0296]
6)-2-2 Конструирование экспрессирующего вектора #101L2h
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 37-402 нуклеотидной последовательности #101L2h, представленной последовательностью SEQ ID NO:134 (Geneart AG). Экспрессирующий вектор #101L2h конструировали таким же методом, как в примере 6)-2-1.
[0297]
6)-2-3 Конструирование экспрессирующих векторов #110L4h, #110L2h и #110L12h
Синтезировали фрагменты ДНК, представленные нуклеотидами в положениях 37-402 нуклеотидных последовательностей #110L4h, #110L2h и #110L12h, представленных последовательностями SEQ ID NO:138, SEQ ID NO:150 и SEQ ID NO:152 (GeneArt AG). Конструировали экспрессирующие векторы #110L4h, #110L2h и #110L12h таким же методом, как в примере 6)-2-1.
[0298]
6)-2-4 Конструирование экспрессирующего вектора #131L2h
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 37-402 нуклеотидной последовательности #131L2h, представленной последовательностью SEQ ID NO:142 (GeneArt AG). Экспрессирующий вектор #131L2h конструировали таким же методом, как в примере 6)-2-1.
[0299]
6)-3 Конструирование экспрессирующих векторов тяжелой цепи гуманизированного антитела
6)-3-1 Конструирование экспрессирующего вектора #84H1hIgG2
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 36-437 нуклеотидной последовательности #84H1hIgG2, представленной последовательностью SEQ ID NO:124 (GeneArt AG). Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), синтезированный фрагмент ДНК вставляли в тоже самое место, где дигерировали pCMA-G2 с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор #84H1hIgG2.
[0300]
6)-3-2 Конструирование экспрессирующего вектора #84H1hIgG4P
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 36-437 нуклеотидной последовательности #84H1hIgG4P, представленной последовательностью SEQ ID NO:126 (GeneArt AG). Используя набор реагентов для клонирования In-Fusion HD PCR (Clontech Laboratories, Inc.), синтезированный фрагмент ДНК вставляли в тоже самое место, где дигерировали pCMA-G4P с помощью рестрикционного фермента BlpI, в результате чего конструировали экспрессирующий вектор #84H1hIgG4P.
[0301]
6)-3-3 Конструирование экспрессирующего вектора #101H1hIgG2
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 36-428 нуклеотидной последовательности #101H1hIgG2, представленной последовательностью SEQ ID NO:130 (GeneArt AG). Экспрессирующий вектор #101H1hIgG2 конструировали таким же методом, как в примере 6)-3-1.
[0302]
6)-3-4 Конструирование экспрессирующего вектора #101H1hIgG4P
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 36-428 нуклеотидной последовательности #101H1hIgG4P, представленной последовательностью SEQ ID NO:132 (GeneArt AG). Экспрессирующий вектор #101H1hIgG4P конструировали таким же методом, как в примере 6)-3-2.
[0303]
6)-3-5 Конструирование экспрессирующих векторов #110H1hIgG4P и #110H13hIgG4P
Синтезировали фрагменты ДНК, представленные нуклеотидами в положениях 36-425 нуклеотидных последовательностей #110H1hIgG4P и #110H13hIgG4P, представленных последовательностью SEQ ID NO:136 и SEQ ID NO:148 (GeneArt AG). Экспрессирующие векторы #110H1hIgG4P и #110H13hIgG4P конструировали таким же методом, как в примере 6)-3-2.
[0304]
6)-3-6 Конструирование экспрессирующего вектора #131H2hIgG2
Синтезировали фрагмент ДНК, представленный нуклеотидами в положениях 36-431 нуклеотидной последовательности #131H2hIgG2, представленной последовательностью SEQ ID NO:140 (GeneArt AG). Экспрессирующий вектор #131H2hIgG2 конструировали таким же методом, как в примере 6)-3-1.
[0305]
6)-4 Получение гуманизированного антитела
6)-4-1 Продукция гуманизированного антитела
Антитело продуцировали таким же методом, как в примере 1)-14-1. Получали целый ряд гуманизированных антител путем комбинации экспрессирующего вектора H цепи и экспрессирующего вектора L цепи, соответствующей комбинациям H цепи и L цепи, приведенным в примере 6)-1-5, примере 6)-1-8, примере 6)-1-11 и примере 6)-1-14.
[0306]
6)-4-2 Одностадийная очистка гуманизированного антитела
Антитело очищали от надосадочной жидкости культуры клеток, полученной в примере 6)-4-1, одностадийным процессом аффинной хроматографии на колонке с наполнителем rProtein A. После внесения надосадочной жидкости культуры клеток в колонку, приведенную в равновесие с PBS и заполненную средой MabSelectSuRe (поставляемой фирмой GE Healthcare Bioscience), колонку промывали с помощью PBS, в количестве, которое было в два или более раз больше объема колонки. Далее, проводили элюирование раствором 2M гидрохлорида аргинина (pH 4,0), собирая фракцию, содержащую антитело. Фракцию подвергали замене буфера на PBS методом диализа (Thermo Scientific, Slide-A-Lyzer Dialysis Cassette). Антитело концентрировали с использованием устройства для центробежной ультрафильтрации VIVASPIN20 (фракция с молекулярной массой UF10K, Sartorius AG) с доведением концентрации IgG до 1 мг/мл или выше. И наконец, проводили фильтрацию с использованием фильтра Minisart-Plus (Sartorius AG) с получением очищенного образца.
[0307]
6)-4-3 Двухстадийная очистка гуманизированного антитела
Надосадочную жидкость культуры клеток, полученную в примере 6)-4-1, очищали двухстадийным процессом аффинной хроматографии на колонке с наполнителем rProtein A и на колонке с наполнителем из керамического гидроксиапатита. После внесения надосадочной жидкости культуры клеток в колонку, приведенную в равновесие с PBS и заполненную средой MabSelectSuRe (поставляемой фирмой GE Healthcare Bioscience), колонку промывали с помощью PBS, в количестве, которое было в два или более раз больше объема колонки. Далее, проводили элюирование раствором 2M гидрохлорида аргинина (pH 4,0). Фракцию, содержащую антитело, подвергали замене буфера на PBS методом диализа (Thermo Scientific, Slide-A-Lyzer Dialysis Cassette), затем разбавляли в 5 раз с помощью буфера 5 мM фосфат натрия/50 мM MES/pH 7.0, и вносили в колонку, наполненную керамическим гидроксиапатитом (Japan Bio-Rad Laboratories, Inc., Bio-Scale CHT Type-1 Hydroxyapatite Column), приведенную в равновесие с буфером 5 мM NaPi/50 мM MES/30 мM NaCl/pH 7.0. Проводили элюирование с линейным градиентом хлорида натрия с получением фракции, содержащей антитело. Фракцию подвергали замене буфера на буфер HBSor (25 мM гистидин/5% сорбит/pH 6.0) методом диализа (Thermo Scientific, Slide-A-Lyzer Dialysis Cassette). Антитело концентрировали с использованием устройства для центробежной ультрафильтрации VIVASPIN20 (фракция с молекулярной массой UF10K, Sartorius AG) с доведением концентрации IgG до 25 мг/мл. И наконец, проводили фильтрацию с использованием фильтра Minisart-Plus (Sartorius AG) с получением очищенного образца.
[0308]
(Пример 7) Измерение активностей гуманизированного антитела
7)-1 Оценка активности связывания гуманизированного антитела с CD147
Измеряли константу диссоциации для связывания гуманизированных антител #84H1L2hIgG2, #84H1L2hIgG4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIgG4P или #131H2L2hIgG2 против человеческого CD147, продуцированных в примере 6)-4-3, с CD147 человека, используя систему Biacore T200 (поставляемую фирмой GE Healthcare Bioscience). Антитело против IgG (Fc) человека иммобилизовали на сенсорном чипе, используя набор для иммобилизации антител Human Antibody Capture Kit (поставляемый фирмой GE Healthcare Bioscience), затем захватывали гуманизированное антитело в качестве лиганда и ассоциировали антиген в качестве аналита. Использовали HBS-EP+ (поставляемый фирмой GE Healthcare Bioscience) в качестве подвижного буфера и CM5 (поставляемый фирмой GE Healthcare Bioscience) в качестве сенсорного чипа. Затем 1 мкг/мл гуманизированного антитела добавляли на чип при 10 мкл/минута в течение 60 секунд, ассоциировали последовательные разведения антигена, используемого в примере 5) при скорости 30 мкл/минута в течение 120 секунд (от 0,0625 до 1 мкг/мл в отношении #101H1L2hIgG2 и #101H1L2hIgG4P, и от 0,25 до 4 мкг/мл в отношении #84H1L2hIgG2, #84H1L2hIgG4P, #110H1L4hIgG4P, #110H13L2hIgG4P, #110H13L12hIgG4P и #131H2L2hIgG2), затем проводили последующий мониторинг фазы диссоциации 300 секунд. Добавляли раствор 3M хлорида магния (поставляемого фирмой GE Healthcare Bioscience) в качестве регенерирующего раствора добавляли при скорости 20 мкл/минута в течение 30 секунд. Используя модель связывания 1:1, рассчитывали константу скорости связывания ka, константу скорости диссоциации kd и константу диссоциации (KD; KD=kd/ka).
[0309]
Результаты представлены в таблице.
[0310]
[Таблица 6]
Константа диссоциации для связывания гуманизированного антитела с CD147 человека
[0311]
7)-2 Противоопухолевая активность гуманизированных антител против CD147 в ксенотрансплантатной модели рака поджелудочной железы
5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самок мышей в возрасте 4 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли, вводили интраперитонеально гуманизированное антитело против CD147 (#84H1L2hIgG2, #84H1L2IgG4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIgG4P или #131H2L2hIgG2), продуцированное в примере 6)-4-2, несущим злокачественную опухоль мышам в дозе 3 мг/кг или 10 мг/кг через 7 дней после инокуляции (n=5). В качестве контрольного лекарственного средства, вводили интраперитонеально гемцитабин (поставляемый фирмой Eli Lilly Japan K.K.), который является стандартным терапевтическим средством при раке поджелудочной железы, несущим злокачественную опухоль мышам в дозе 400 мг/кг через 7 и 14 дней после инокуляции (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0312]
Результаты представлены на фигурах 18(a)-(d). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли.
[0313]
В то время как степень ингибирования роста опухоли при применении контрольного лекарственного средства составляла 71% и не обнаруживалось исчезновение опухоли, все гуманизированные антитела против CD147 проявляли более мощную противоопухолевую активность в группах введения 3 или 10 мг/кг, чем гемцитабин. В группах введения 10 мг/кг #84H1L2hIgG4P, #101H1L2hIgG2, #101H1L2hIgG4P или #110H1L4hIgG4P наблюдалась мощная противоопухолевая активность с исчезновением опухоли.
[0314]
(Пример 8) In-vitro активация сигнала p38MAPK антителом против CD147
8)-1 Индукция фосфорилирования p38MAPK человеческим химерным антителом против CD147 человека в клетках рака поджелудочной железы линии PANC-1
Сообщалось, что активация CD147 промотирует фосфорилирование p38MAPK (Lim et al., FEBS Letters, 88-92, 1998) (Li et., al. J. Hepatology, 1378-1389, 2015). Для того чтобы исследовать влияние антитела против CD147, проявляющего противоопухолевую активность, на сигнал p38MAPK, оценивали фосфорилирование p38MAPK в клетках линии PANC-1, подвергнутых обработке с помощью человеческого химерного антитела LN22R8chIgG4P против CD147 при 10 мкг/мл в течение 15 минут, используя методы анализа Simple Western assays (ProteinSimple Japan K.K., Wes). В качестве контрольного образца, анализировали подобным образом клетки линии PANC-1, подвергнутые обработке человеческим IgG (Jackson ImmunoResearch Inc., Cat. 009-000-003) при 10 мкг/мл. Для детекции p38MAPK, использовали p38 MAPK кроличьего mAb (Cell Signaling Technology, Inc., Cat. #9212). Для детекции фосфорилированного p38MAPK, использовали P-p38 MAPK (T180/Y182) (D3F9) XP кроличьего mAb (Cell Signaling Technology, Inc., Cat. #4511S). На фигуре 19 представлено отношение сигнала фосфорилированной p38MAPK к детектируемому сигналу p38MAPK. Образцы, обработанные антителом, измеряли два раза, и на графиках приведены средние значения.
[0315]
В группе, подвергнутой обработке антителом LN22R8chIgG4P, наблюдалось увеличение сигнала фосфорилированной MAPK.
[0316]
8)-2 Опосредованное антителом против CD147 фосфорилирование HSP27, который является молекулой, расположенной ниже по потоку от сигнала p38
Сообщалось, что HSP27 фосфорилируется в результате активации p38MAPK (Landry et al., Biochem. Cell Biol., 703-707, 1995). Для подтверждения того, что фосфорилирование p38, индуцированное человеческим химерным антителом LN22R8 против CD147, в действительности промотирует активацию сигнала p38, оценивали фосфорилирование в обработанных антителом образцах 8)-1, используя методы анализа Simple Western assays (ProteinSimple Japan K.K., Wes). Для детекции HSP27, использовали антитело против HSP27 (R&D systems, Cat. AF15801). Для детекции фосфорилированного HSP27, использовали Phospho-HSP27 (Ser82) антитело (Cell Signaling Technology, Inc., Cat. 2401S) или антитело против HSP27 (phospho Ser15) (Abcam plc., Cat. ab76313).
[0317]
На фигуре 20(a) или фигуре 20(b) приведено отношение сигнала Ser82 или Ser15 фосфорилированного HSP27 к детектируемому сигналу HSP27, соответственно. Обработанные антителом образцы оценивали в двух параллельных анализах, и на графиках приведены средние значения.
[0318]
В группе, подвергнутой обработке антителом LN22R8chIgG4P, наблюдалось увеличение сигнала фосфорилированного HSP27. Так как фосфорилирование и активация HSP27 расположены ниже по потоку и индуцируются в результате фосфорилирования p38, которое в свою очередь индуцируется человеческим химерным антителом против CD147, было обнаружено, что человеческое химерное антитело LN22R8IgG4P против CD147 человека индуцирует активацию сигнала p38MAPK. Аналогично этому, активацию сигнала p38MAPK также наблюдали в клетках рака поджелудочной железы линии MIA PaCa-2.
[0319]
(Пример 9) Активация сигнала p38 антителом против CD147 в ткани ксенотрансплантатной опухоли
Проводили исследование возможности активации сигнала p38MAPK антителом против CD147 in vitro в опухолях, сформированных подкожно у мышей, в клетках подкожных опухолей линии MIA PaCa-2. 5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 5 дней после инокуляции, и вводили интраперитонеально человеческое химерное антитело LN22R8chIgG4P против CD147 человека несущим злокачественную опухоль мышам при 10 мг/кг на следующий день после распределения на группы. Отбирали образцы опухоли через 6, 24, 48, 72 часа после введения антитела, замораживали с использованием сухого льда и хранили в замороженном виде. Для приготовления образцов из замороженных тканей опухоли (n=3), использовали прибор для автоматической гомогенизации тканей с нагревателями gentleMACS Octo Dissociator with Heaters (Miltenyi Biotec K.K.). Полученные лизаты опухолей подвергали анализу методом Simple Western assays (ProteinSimple Japan K.K., Wes).
[0320]
На фигуре 21(a) представлены средние значения детектируемого сигнала p38MAPK со стандартными ошибками. На фигуре 21(b) представлены средние значения детектируемого сигнала фосфорилированного p38MAPK со стандартными ошибками. На фигуре 21(c) представлены средние значения отношения сигнала фосфорилированного p38MAPK к сигналу p38MAPK вместе со стандартными ошибками. В случае LN22R8chIgG4P, фосфорилированный P38MAPK увеличивался в период от 6 часов до 72 часов после введения. Отношение сигнала фосфорилированного p38MAPK достигало максимального значения через 6 часов после введения и затем снижалось по мере роста p38MAPK в период от 24 часов до 72 часов.
[0321]
Так как увеличение фосфорилирования p38 и увеличение фосфорилирования HSP27, обусловленное введением химерного антитела против CD147, наблюдалось также в подкожных опухолях мышей, было обнаружено, что происходит активация сигнала p38.
[0322]
(Пример 10) Индукция молекул, таких как CXCL8, расположенных ниже по потоку от p38MAPK
Сообщалось, что активация p38MAPK вызывает индукцию CXCL8 через стабилизацию мРНК (Hoffmann et al., J. Leukoc. Biol., 847-855, 2002) и активацию сигнала SMAD3/4 (Leovonen et al., PLOS ONE, e57474, 2013). Проводили исследование РНК, извлеченной из подкожных опухолей мышей после введения антитела, методом количественной PCR с целью определения, была ли индуцирована экспрессия гена CXCL8 в опухолях MIA PaCa-2 после введения антитела. Аналогично этому, ген RHOB (Vasilaki et al., FASEB Journal, 891-905, 2010), который, как сообщалось, индуцируется путем активации сигнального пути SMAD, также исследовали на наличие изменений вследствие введения антитела против CD147. В качестве эндогенных контрольных генов, измеряли экспрессию гена импортина (ipo8) и гена ТАТА-связывающего белка (tbp). После введения человеческого химерного антитела против CD147 человека, после отбора через 72 часа образцов опухолей MIA PaCa-2 и затем обработки с помощью РНК (Qiagen N.V., Cat. 76104), извлекали РНК, используя набор RNeasy Mini Kit (250) (Qiagen N.V., Cat. 74106). В качестве человеческого химерного антитела против CD147 человека, использовали LN22R8chIgG1, LN22R8chIgG2 или LN22R8chIgG4P, продуцированные в примере 1)-14. Для проведения количественной RT-PCR, использовали универсальный набор EXPRESS One-Step SuperScript qRT-PCR kit Universal (Thermofisher scienticic, Cat. 11781-01K), и импортин (ipo8) (Thermo Fisher Scientific, Cat. Hs00183533_m1), ТАТА-связывающий белок (tbp) (Thermo Fisher Scientific, Cat. Hs00427621_m1), представитель семейства Ras homolog B (rhoB) (Thermo Fisher Scientific, Cat. hs00269660_s1) и интерлейкин-8 (Thermo Fisher Scientific, Cat. Hs00174103_m1) использовали в качестве зондов для количественного определения генов. Ген-специфические увеличения флуоресцентных сигналов после проведения реакции PCR измеряли с использованием амплификатора ABIPrism 7500 (Applied Biosystems). На фигуре 22(a) представлены средние значения (n=3) отношения экспрессии генов ipo8/tbp вместе со стандартными отклонениями. На фигуре 22(b) представлены средние значения (n=3) отношения экспрессии генов cxcl8/tbp вместе со стандартными отклонениями. На фигуре 22(c) представлены средние значения (n=3) отношения экспрессии генов rhoB/tbp вместе со стандартными отклонениями.
[0323]
После введения LN22R8chIgG1, LN22R8chIgG2 или LN22R8chIgG4P, экспрессия гена ipo8 не изменялась, но наблюдалась индукция экспрессии cxcl8 и rhoB в группе введения человеческого химерного антитела против CD147 человека. Индукция экспрессии cxcl8 и rhoB находилась в соответствии с величиной противоопухолевой активности человеческого химерного антитела, приведенного в примере 1)-18, что позволяет предположить, что индукция экспрессии обоих генов, возможно, может коррелировать с противоопухолевой активностью.
[0324]
(Пример 11) Индукция генов cxcl8 и rhoB путем усиления противоопухолевой активности в результате конверсии крысиного антитела против CD147 человека в человеческое химерное антитело
Противоопухолевую активность крысиного антитела rat_CD147_#110 против CD147 человека усиливали путем химеризации, приведенной в примере 2 и примере 7. Для индукции cxcl8 и rhoB, наблюдаемой в опухолях после введения человеческого химерного антитела LN22R8chIgG1, LN22R8chIgG2 или LN22R8chIgG4P, rat_CD147_#110 сравнивали с химерным антителом #110chIgG4P таким же способом, как в примере 10.
[0325]
На фигуре 23(a) представлены средние значения (n=3) отношения экспрессии генов ipo8/tbp вместе со стандартными отклонениями.
[0326]
На фигуре 23(b) представлены средние значения (n=3) отношения экспрессии генов cxcl8/tbp вместе со стандартными отклонениями.
[0327]
На фигуре 23(c) представлены средние значения (n=3) отношения экспрессии генов rhoB/tbp вместе со стандартными отклонениями.
[0328]
Экспрессия гена ipo8 не изменялась вследствие введения антитела. Не смотря на то, что не наблюдалось изменений экспрессии генов cxcl8 и rhoB вследствие введения rat_CD147_#110, так же как в группе, не подвергнутой обработке антителом, тем не менее, наблюдалась индукция cxcl8 и rhoB, вызываемая введением #110chIgG4P. Было сделано предположение, что индукция обоих генов являлась показателем, который коррелировал с противоопухолевой активностью антитела против CD147.
[0329]
(Пример 12) Проведение оценки с использованием экспериментальной модели SMAD4-отрицательного рака поджелудочной железы на мышах
Фактор транскрипции SMAD4 известен в качестве одной из молекул, важной для активации сигнального пути SMAD (Zang, et al., Current Biology, 270-276, 1997). Известно, что сигнальный путь SMAD частично поврежден вследствие генетических дефектов SMAD4 при некоторых типов рака поджелудочной железы (Hahn, et al., Science, 350-353, 1996). Проводили исследование наличия противоопухолевой активности у антитела против CD147 в клетках рака поджелудочной железы линии BxPC-3 с дефицитом SMAD4 и с частично ингибированным сигнальным путем SMAD (Yasutome et al., Clin. Exp. Metastasis, 461-473, 2005). 2,5 × 106 клеток рака поджелудочной железы линии BxPC-3 (ATCC, Cat. CRL-1687) суспендировали в PBS, содержащем 100% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самок мышей в возрасте 6 недель линии BALB/c-nu (CAnN. Cg-Foxn1nu/CrlCrlj, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 8 дней после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам мышиное антитело LN22R8 против CD147 человека или крысиное антитело 2P10F2 против CD147 человека в дозе 10 мг/кг через 8, 15 и 22 дня после инокуляции (n=5). В качестве контрольного лекарственного средства, вводили несущим злокачественную опухоль мышам через хвостовую вену гемцитабин (Eli Lilly Japan K.K., Gemzar (R)), который является терапевтическим средством при раке поджелудочной железы, в дозе 200 мг/кг через 8, 15, 22 дня после инокуляции (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0330]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 24. На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. Опухоль BxPC-3 была резистентна к антителу LN22R8, антителу 2P10F2 и гемцитабину.
[0331]
(Пример 13) Приобретение восприимчивости к воздействию антитела против CD147 путем восстановления функции SMAD4
Сообщалось, что сигнал SMAD может быть восстановлен в результате восстановления функции SMAD4 в клетках некоторых линий рака поджелудочной железы. Поэтому, проводили исследование способности сигнала SMAD, который восстанавливали путем восстановления функции SMAD4, повышать восприимчивость к воздействию антитела против CD147.
[0332]
13)-1 Генерация клеток, стабильно экспрессирующих SMAD4
Используя набор Retro-X™ Q vector kit, генерировали линию клеток BxPC-3, стабильно экспрессирующую SMAD4. Ген SMAD4 человека, полученный искусственным синтезом, вводили в сайт клонирования плазмиды pQCXIP, содержащейся в наборе в качестве ретровирусного вектора (Takara Bio Inc., Retro-X™ Q Vector Set, Cat. 631516), с формированием ретровирусного вектора, экспрессирующего SMAD4. Используя систему ретровирусной сборки Retro-X Universal Packaging System (Takara Bio Inc., Cat. 631530), ретровирусный вектор, экспрессирующий SMAD4, вводили в BxPC-3, и клетки BxPC-3, в которых ретровирус был введен в хромосому путем вирусного инфицирования и которые в результате этого стали резистентными к пуромицину и SMAD4-положительными, отбирали, используя пуромицин (Takara Bio Inc., Cat. 631306), для использования в качестве SMAD4-положительных клеток BxPC-3, BxPC-3-SMAD4. Вводили таким же способом ретровирусный вектор плазмиды pQCXIP, и клетки BxPC-3, которые становились резистентными к пуромицину, использовали в качестве BxPC-3-имитации. Эксперимент по ретровирусному инфицированию проводили два раза с продуцированием партии 1 и партии 2 BxPC-3-имитации и BxPC-3-SMAD4.
[0333]
13)-2 Подтверждение экспрессии CD147 и SMAD4
Проводили исследование клеток BxPC-3 (ATCC, Cat. CRL-1687), и клеток BxPC-3-имитации и BxPC-3-SMAD4, продуцированных в примере 13)-1, с использованием методов анализа Simple Western assays (ProteinSimple Japan K.K., Wes). В качестве образца SMAD4-положительного контроля, использовали клетки MIA PaCa-2. Для детекции SMAD4, использовали антитело против SMAD4 (R&D systems, Cat. AF2097). Для детекции GAPDH, использовали антитело против GAPDH (Abfrontier, Cat. LF-MA0026). Для детекции CD147, использовали антитело против CD147 (Abcam, Cat. Ab108317).
[0334]
Результаты представлены на фигуре 25(a). Клетки MIA PaCa-2 были SMAD4-положительными и CD147-положительными. Клетки BxPC-3 были SMAD4-отрицательными и CD147-положительными. Клетки BxPC-3-имитации были SMAD4-отрицательными и CD147-положительными с отсутствием наблюдаемого влияния ретровирусного инфицирования. Как партия 1, так и партия 2 клеток BxPC-3-SMAD4 с введенным ретровирусным вектором, экспрессирующим SMAD4, были SMAD4-положительными. Экспрессия SMAD4 партии 2 была высокой, и, поэтому, партию 2 использовали в качестве BxPC-3-SMAD4 в последующих экспериментах.
[0335]
13)-3 Восприимчивость клеток опухоли BxPC-3, стабильно экспрессирующих SMAD4, к воздействию человеческого химерного антитела против CD147
2,5 × 106 клеток BxPC-3-имитация или BxPC-3-SMAD4 суспендировали в PBS, содержащем 100% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии BALB/c-nu (CAnN. Cg-Foxn1nu/CrlCrlj, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 6 дней после инокуляции BxPC-3-имитация, и через 3 дня после инокуляции BxPC-3-SMAD4, соответственно, вводили интраперитонеально несущим злокачественную опухоль мышам в дозе 10 мг/кг человеческое химерное антитело LN22R8chIgG или LN22R8chIgG4 против человеческого CD147 в день распределения по группам и через 7, 14, 21, 28 дней после распределения по группам (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0336]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 25(b) и фигуре 25(c). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. Опухоль BxPC-3-имитация была резистентной или проявляла низкую восприимчивость к воздействию антитела LN22R8chIgG2 или LN22R8chIgG4P. Опухоль BxPC-3-SMAD4 проявляла частичную восприимчивость к антителу LN22R8chIgG2 или LN22R8chIgG4P.
[0337]
13)-4 Внутриопухолевые изменения сигнала p38 вследствие экспрессии SMAD4
Сообщалось, что сигнал p38 усиливается в результате экспрессии SMAD4 в клетках SMAD4-отрицательного рака поджелудочной железы (Chen et al., B. M. C., 1471-2407, 2014). Проводили исследование изменения p38MAPK и фосфорилированного p38MAPK в опухоли BxPC-3-SMAD4 вследствие введения человеческого химерного антитела против CD147 человека (через 72 часа после введения антитела), используя методы анализа Simple Western assays (ProteinSimple Japan K.K., Wes), таким же образом, как в примере 13)-1. Вводили LN22R8chIgG4P в качестве человеческого химерного антитела против CD147 человека несущим подкожную опухоль MIA PaCa-2 мышам в дозе 10 мг/кг таким же методом, как в примере 1)-18. Для приготовления образцов (n=3) из тканей опухолей, использовали прибор для автоматической гомогенизации тканей с нагревателями gentleMACS Octo Dissociator with Heaters (Miltenyi Biotec K.K.).
[0338]
На фигуре 26(a) представлены в графической форме средние значения измеренных величин p38 MAPK для трех образцов опухолей вместе со стандартными ошибками. На фигуре 26(b) представлены в графической форме средние значения измеренных величин фосфорилированного p38 MAPK для трех образцов опухолей вместе со стандартными ошибками.
[0339]
В опухолях BxPC-3-SMAD4, количество p38 и фосфорилированного p38 увеличивалось в два раза. Через 72 часа после введения человеческого химерного антитела против CD147, наблюдалось частичное снижение p38 и частичное увеличение фосфорилированного p38.
[0340]
Было обнаружено, что вследствие экспрессии SMAD4 в опухолевых клетках рака поджелудочной железы линии BxPC-3 увеличивался сигнал p38. Весьма вероятно, что увеличение SMAD4-зависимого сигнала p38 способствовало увеличению восприимчивости к воздействию человеческого химерного антитела LN22R8chIgG4P против CD147.
[0341]
(Пример 14) Противоопухолевая активность химерного антитела против CD147 при гемцитабин-резистентном раке поджелудочной железы
Исследовали противоопухолевую активность антитела против CD147 в модели гемцитабин-резистентного рака поджелудочной железы. 5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самок мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, CHARLES RIVER LABORATORIES JAPAN, INC). Гемцитабин (Eli Lilly Japan K.K.), который является стандартным терапевтическим средством при раке поджелудочной железы, вводили интраперитонеально в дозе 400 мг/кг через 6 дней после инокуляции, и через неделю проводили распределение мышей, у которых подтверждался рост гемцитабин-резистентных опухолей, по группам на основе размера опухоли. Гемцитабин вводили интраперитонеально в дозе 400 мг/кг контрольной группе (n=5) через 13 дней после распределения на группы и через 20 дней после инокуляции. В группе комбинированного введения антитела против CD147 и гемцитабина (n=5), человеческое химерное антитело LN22R8chIgG4P против CD147 вводили интраперитонеально в дозе 10 мг/кг через 13 дней после распределения на группы и через 20 дней после инокуляции в сочетании с гемцитабином, вводимым интраперитонеально в дозе 400 мг/кг через 13 дней после распределения на группы и через 20 дней после инокуляции. Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
[0342]
На фигуре 27 в графической форме представлены средние значения и стандартные ошибки измеренных изменений объема опухоли. В графической форме также продемонстрирован рост опухоли в группе, в которой не вводили гемцитабин и антитело (n=5).
[0343]
В то время как средний размер опухоли в группе введения гемцитабина в качестве контрольного лекарственного средства составлял 1269 мм3 через 28 дней после инокуляции и не наблюдалось никакого регресса опухолей, средний размер опухоли в группе, в которой в комбинации вводили человеческое химерное антитело LN22R8chIgG4P против CD147, составлял 15 мм3, и наблюдался регресс опухоли у 3 мышей из 5 мышей. Эти результаты показали, что существует возможность проявления восприимчивости к воздействию антитела против CD147 у растущих злокачественных опухолей поджелудочной железы, резистентных к гемцитабину.
[0344]
(Пример 15) Противоопухолевая активность гуманизированного антитела против CD147 в модели рака печени
15)-1 Экспрессия CD147 и SMAD4 в клетках линии Hep G2
Исследовали экспрессию CD147 и SMAD4 в клетках рака печени линии HepG2 (ATCC, Cat. HB-8065) таким же методом, как в примере 13)-2. В качестве контрольных образцов, которые били CD147-положительными, использовали MIA PaCa-2 и BxPC-3 (ATCC, Cat. CRL-1687). MIA PaCa-2 использовали в качестве контрольного образца, который был SMAD4-положительным, и BxPC-3 использовали в качестве контрольного образца, который был SMAD4-отрицательным.
[0345]
Определяли отношение детектируемого сигнала к GAPDH, и результаты представлены ниже. Было обнаружено, что клетки Hep G2 являются CD147-положительными и SMAD4-положительными.
[0346]
Отношение сигналов CD147/GAPDH
MIA PaCa-2 0,481
BxPC-3 0,481
Hep G2 2,944
[0347]
Отношение сигналов SMAD4/GAPDH
MIA PaCa-2 0,329
BxPC-3 0,003
Hep G2 0,723
[0348]
15)-2 Подтверждение экспрессии CD147 в клетках Hep G2 с использованием проточного цитометра
CD147, экспрессировавшийся на поверхности клеток HepG2 (ATCC, Cat. HB-8065), анализировали на проточном цитометре. Для подтверждения экспрессии CD147 человека, использовали мышиное IgG1 антитело MEM-M6/1-APC против CD147 человека, меченое с помощью APC (Thermofisher, Cat. MA1-10104), в качестве выпускаемого промышленностью антитела против человеческого CD147. В качестве контрольного мышиного антитела изотипа IgG1, использовали mIgG1-APC (Miltenyi Biotec K.K., Cat. 130-092-214). MEM-M6/1-APC добавляли к суспензии клеток HepG2 в количестве 1/10, затем обрабатывали при 4°C в течение 30 минут. Клетки промывали буферным раствором PBS, содержащим 5% FBS, затем проводили измерения, используя проточный цитометр (CantoII, BD Biosciences). Результаты представлены на фигуре 28(a).
[0349]
Было обнаружено, что клетки Hep G2, покрашенные с помощью MEM-M6/1-APC, проявляли сильный флуоресцентный сигнал, из чего следовало, что они экспрессировали CD147.
[0350]
15)-3 Активация p38 гуманизированным антителом против CD147 в клетках рака печени
Для исследования влияния антитела против CD147 на p38MAPK в клетках рака печени HepG2, оценивали фосфорилирование P38 в клетках HepG2 (ATCC, Cat. HB-8065), обработанных с помощью человеческого химерного антитела (LN22R8chIgG4P) против CD147 человека или с помощью гуманизированного антитела против CD147 человека (#84H1L2hIgG2, #84H1L2hIgG4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIgG4P или #131H2L2hIgG2), продуцированного в примере 6)-4-2, при дозе 10 мкг/мл в течение 15 минут, используя методы анализа Simple Western assays (ProteinSimple Japan K.K., Wes). В качестве контрольного образца, проводили анализ аналогичным методом клеток HepG2, обработанных человеческим IgG (hIgG, Jackson ImmunoResearch Inc., 009-000-003) при дозе 10 мкг/мл. Для детекции p38MAPK, использовали p38 MAPK кроличьего mAb (Cell Signaling Technology, Inc., Cat. #9212). Для детекции фосфорилированного p38MAPK, использовали P-p38 MAPK (T180/Y182) (D3F9) XP кроличьего mAb (Cell Signaling Technology, Inc., #4511S). На фигуре 28(b) представлено отношение сигнала фосфорилированного p38MAPK к детектируемого сигналу p38MAPK.
[0351]
Наблюдалось увеличение сигнала фосфорилированного p38 MAPK в два или более раз по сравнению с группой, обработанной человеческим IgG, обусловленное обработкой с помощью LN22R8chIgG4P, #84H1L2hIgG2, #84H1L2hIgG4P, #101H1L2hIgG2, #101H1L2hIgG4P, #110H1L4hIgG4P или #131H2L2hIgG2, и было обнаружено, что фосфорилирование p38MAPK в клетках рака печени, а также в клетках рак поджелудочной железы, индуцируется антителом против человеческого CD147.
[0352]
15)-4 Сравнение противоопухолевой активности гуманизированного антитела против CD147 и сорафениба при раке печени
Исследовали противоопухолевую активность человеческого химерного антитела против CD147 человека и гуманизированного антитела в клетках печени человека линии HepG2 (ATCC, Cat. HB-8065), которые были CD147-положительными и SMAD4-положительными и в которых наблюдалось фосфорилирование p38, обусловленное воздействием антитела против человеческого CD147.
[0353]
5 × 106 клеток печени человека линии HepG2 суспендировали в PBS, содержащем 50% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 4 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CHARLES RIVER LABORATORIES JAPAN, INC). Проводили распределение по группам на основе объема опухоли через 9 дней после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам человеческое химерное антитело против CD147 (LN22R8chIgG4P) или гуманизированное антитело против CD147 (#84H1L2hIgG2, #84H1L2hIgG4P или #110H1L4hIgG4P), продуцированное в примере 6)-4-2, при 1 мг/кг или 10 мг/кг (n=5). В качестве контрольного лекарственного средства, сорафениб (таблетки нексавар® 200 мг, Bayer AG), который является терапевтическим средством при раке печени, растворяли в этаноле с эмульгатором PEG-35 касторовое масло (Cremophor EL, NACALAI TESQUE, INC., Cat. 09727-14) в соответствии с инструкцией (Chang et.al., Cancer Chem.Thera.Pharm., 2007), прилагаемой к таблеткам нексавар®, и раствор перорально вводили несущим злокачественную опухоль мышам при 30 мг/кг или 90 мг/кг через 9, 10, 11, 12, 13, 16, 17 и 18 дней после инокуляции (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0354]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены в таблице 7 и на фигурах 29(a)-(d). На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. В результате введения сорафениба, наблюдалось частичное ингибирование роста опухоли, но опухоль не исчезала. Все антитела LN22R8chIgG4P, #84H1L2hIgG2, #84H1L2hIgG4P и #110H1L4hIgG4P проявляли высокую противоопухолевую активность с исчезновением опухоли.
[0355]
[Таблица 7]
(мм3)
qD1×9-один раз в сутки на протяжении 9 дней
po-перорально
ip-интраперитонеально
[0356]
(Пример 16) Воздействие на T-клетки и мононуклеарные клетки периферической крови (PBMC)
Сообщалось, что экспрессия CD147 увеличивается в CD4-положительных и CD8-положительных T-клетках с активацией T-клеток (Hu et al., J. Cell. Mol. Med., 2132-2143, 2010), и некоторые антитела против CD147 обладают эффектом индуцирования активации T-клеток и ингибирования роста (Koch et al., Int. Immunology, 777-786, 1999; Chiampanichayakul et al., Immunology 167-178, 2006). Исследовали влияние антитела против CD147 человека, проявляющего мощную противоопухолевую активность, которая не зависит от эффекторной функции, на лимфоциты периферической крови (PBL), содержащие T-клетки.
[0357]
16)-1 Увеличение экспрессии CD147 путем стимуляции CD3/CD28 мононуклеарных клеток периферической крови (PBMC)
Исследовали возможность увеличение экспрессии CD147 с активацией T-клеток, используя PBMC человека. PBMC человека культивировали в среде RPMI1640, содержащей 10% FBS, при 37°C в присутствии 5% CO2. Рост индуцировали путем добавления активатора Dynabeads Human T-Activator CD3/CD28 (CD3/CD28 сферы, Thermo Fisher SCIENTIFIC K.K., Cat. 1131D) в процессе культивирования, и через 4 дня проводили анализ, используя проточный цитометр, с целью исследования изменений экспрессии CD147. Для подтверждения экспрессии CD147 человека, использовали мышиное IgG1 антитело против CD147 человека MEM-M6/1-APC, меченое с помощью APC (CD147-APC, Thermofisher, Cat. MA1-10104) в качестве выпускаемого промышленностью антитела против CD147 человека. В качестве контрольного мышиного антитела изотипа IgG1, использовали mIgG1-APC (Miltenyi Biotec K.K., Cat. 130-092-214). Для детекции CD3, CD4 и CD8 T-клеток, содержавшихся в PBMC человека, использовали антитело против CD3 человека APC/Fire™ 750 (поставляемое фирмой BioLegend, Inc., Cat. 344840), антитело против CD4 человека PerCP/Cy5.5 (поставляемое фирмой BioLegend, Inc., Cat. 344608), и антитело против CD8 человека Brilliant Violet 510 (поставляемое фирмой BioLegend, Inc., Cat. 344732).
[0358]
На фигуре 30 представлены результаты связывания CD147-APC в CD3- и CD4-положительных клетках и CD3- и CD8-положительных клетках. В случае, когда CD3- и CD4-положительные клетки и CD3- и CD8-положительные клетки стимулировали с помощью CD3/CD28 сфер, подтверждалось увеличение экспрессии CD147, и подтверждалось увеличение экспрессии CD147 на поверхности мембраны T-клеток с активацией Т-клеток.
[0359]
16)-2 Оценка воздействия антитела против человеческого CD147 на рост мононуклеарных клеток периферической крови человека
Исследовали воздействие антитела против человеческого CD147 на рост мононуклеарных клеток периферической крови человека (PBMC). В качестве антитела против человеческого CD147, использовали 2P10F2chIgG4P. После нанесения флуоресцентной метки на PBMC, используя набор CellVue Claret Far Red Fluorescent Cell Linker Kit (sigma, Cat. MIDCLARET-1KT), клетки культивировали в среде RPMI1640, содержащей 10% FBS, при 37°C в присутствии 5% CO2. Для исследования влияния на рост, в процессе культивирования добавляли 2P10F2chIgG4P (10 мкг/мл), когда индуцировали рост путем добавления IL-2 и Dynabeads Human T-Activator CD3/CD28 (CD3/CD28 сферы, Thermo Fisher SCIENTIFIC K.K., Cat. 1131D). Флуоресцентные сигналы PBMC, которые уменьшались вследствие деления клеток, измеряли с использованием проточного цитометра (CantoII, BD Biosciences) через 3 дня и 5 дней после культивирования. Результаты представлены на фигуре 31.
[0360]
Регистрировали флуоресцентные сигналы PBMC, которые уменьшались вследствие деления клеток 3 дня и 5 дней после культивирования в результате добавления сфер CD3/CD28. В случае добавления IL-2, 2P10F2chIgG4P или IL-2 и 2P10F2chIgG4P в процессе культивирования, не обнаруживалось уменьшения наблюдаемых флуоресцентных сигналов. Было сделано предположение, что человеческое химерное антитело 2P10F2chIgG4P против CD147 человека, которое проявляет высокую противоопухолевую активность в экспериментальной модели на мышах рака поджелудочной железы человека, приведенной в примере 1)-18, не влияет на рост PBMC.
[0361]
16)-3 Оценка влияния антитела против человеческого CD147 на продукцию цитокинов лимфоцитами периферической крови человека
В качестве антитела против человеческого CD147, использовали человеческое химерное антитело #84chIgG1, #84chIgG2, #84chIgG4P, #84chIgG1LALA, #84chIgG4PFALA, #101chIgG4P или #110chIgG4P. Лимфоциты периферической крови (PBL) получали из периферической крови человека с использованием Ficoll-Paque PLUS (GE Healthcare Japan). На 96-луночный планшет наносили 10 мкг/мл человеческого химерного антитела. Использовали IgG человека (hIgG, Jackson ImmunoResearch Inc., Cat. 009-000-003) в качестве антитела для отрицательного контроля и активатор Dynabeads Human T-Activator CD3/CD28 (CD3/CD28-сферы, Thermo Fisher SCIENTIFIC K.K., Cat. 1131D) в качестве антитела для положительного контроля, которое индуцирует активацию T-клеток и индукцию цитокинов. 1×106 клеток PBL добавляли в лунки с нанесенным антителом, и через 24 часа измеряли цитокины (IL2, TNFα и INFγ) в среде. В лунки с добавленными PBL непосредственно добавляли активатор Dynabeads Human T-Activator CD3/CD28, и через 24 часа измеряли цитокины (IL2, TNFα и INFγ) в среде таким же способом. Для измерения IL2 использовали набор реагентов для иммунологического анализа Quantikine ELISA Human IL-2 (R&D systems, Cat. D2050). Для измерения TNFα использовали набор для иммунологического анализа Amersham TNF-α Human, Biotrak Easy ELISA (GE Healthcare Japan, Cat. RPN5967). Для измерения INFγA использовали набор реагентов для иммунологического анализа Human IFN-γ ELISA development kit (Mabtech, Cat. 3420-1H-6). Каждое измерение проводили три раза, и рассчитывали средние значения и стандартные отклонения. Результаты представлены на фигуре 32.
[0362]
Было подтверждено, что количество всех измеренных и продуцированных из PBL цитокинов (IL2, TNFα и INFγ) увеличивалось только в культуре в лунках с добавленными CD3/CD28-сферами, служившими в качестве положительного контроля, и количество цитокинов не увеличивалось в культуре в лунках с нанесенным любым одним из человеческих химерных антител против CD147 человека, относительно hIgG, который служил в качестве отрицательного контроля.
[0363]
(Пример 17) Рентгеноструктурный анализ комплекса гуманизированный #110Fab'-CD147
17)-1 Кристаллизация комплекса
Гуманизированное #110H1L4hIgG4P расщепляли пепсином, и полученный фрагмент Fab'2 восстанавливали с помощью дитиотреитола, затем алкилировали с помощью иодоацетамида с получением фрагмента Fab'. Смесь фрагмента Fab' и hCD147v2 (22-205), используемого в примере 5), подвергали гель-проникающей хроматографии, используя колонку Superdex 10/300GL Increase (GE Healthcare), с получением фракции комплекса. Комплекс заменяли буферным раствором (10 мM Tris HCl pH 7,5, 50 мM NaCl), используя AmiconUltra15 MWCO 10K (фирмы Millipore Corporation) с концентрированием до 13 г/л. Раствор комплекса подвергали кристаллизации методом диффузии из паровой фазы. Раствор, образующийся в результате добавления равного количества преципитирующего раствора (0,1 M малонат натрия, pH 7,0, 12% (масса/объем) полиэтиленгликоль 3350) к 0,5 мкл раствора белка, помещали в закрытый контейнер, содержащий 0,05 мл преципитирующего раствора, так чтобы эти два раствора не контактировали друг с другом, затем выдерживали при 25°C. Полученный приблизительно через одну неделю кристалл размером 0,15 мм × 0,15 мм × 0,3 мм погружали в преципитирующий раствор с добавленным полиэтиленгликолем 400 с получением 30% (масса/объем), и затем замораживали с помощью жидкого азота. Данные рентгеноструктурного анализа получали, используя канал синхротронного излучения PF-BL17A синхротрона Photon Factory, High Energy Accelerator Research Organization (KEK) (Ibaraki, Japan). Интенсивность дифракции количественно определяли из дифракционной картины, полученной с использованием программного обеспечения Mosflm (CCP4: Collaborative Computational Project No. 4) для определения структурного фактора кристалла. Пространственная группа кристалла представляла собой P21, и элементарная ячейка кристалла имела следующие параметры: a=64,96 Å, b=93,37 Å, c=98,31 Å, альфа=гамма=90, и бета=90,89).
[0364]
17)-2 Структурный анализ комплекса
Для определения фазы, проводили молекулярное замещение, используя координаты пространственной структуры полученного структурного фактора и модель гомологичности фрагмента Fab', и известную структуру CD147 человека (PDBID: 3b5h). Для расчета использовали программное обеспечение Phaser (CCP4: Collaborative Computational Project No.4). Кристалл содержал два комплекса в асимметричной ячейке. Структуру уточняли, используя программное обеспечение Refmac5 (CCP4: Collaborative Computational Project No.4), и модель корректировали, используя программное обеспечение Coot. Эту операцию повторяли с получением конечной величины расхождения R 23% и свободной величины расхождения R 28% с разрешение 2,3 Å. Конечная модель включала два фрагмента Fab' гуманизированного #110H1L4 и hCD147v2, связывающихся с каждым из них. Для одного hCD147v2, наблюдалась электронная плотность, соответствующая аминокислотным остаткам 23-203, но для другого, электронная плотность, соответствующая домену 1, не была отчетливой, и наблюдалась только электронная плотность, соответствующая аминокислотным остаткам 103-202.
[0365]
17)-3 Идентификация эпитопа
Аминокислотные остатки hCD147v2, которые присутствовали на расстоянии менее 4 Å от фрагмента Fab', связывающего поверхность гуманизированного #110H1L4 в двух комплексах, содержавшихся вместе в асимметричной ячейке, были следующими: Arg106, Lys108, Ala109, Val110, Lys127, Ser128, Glu129, Ser130, Val131, Pro132, Pro133, Val134, Gln164 и Gly165. На фигуре 41 представлена ленточная модель и вся поверхность комплексов, и на фигуре 42 показано взаимодействие между hCD147v2 и гуманизированным #110H1L4.
[0366]
(Пример 18) Противоопухолевое действие гуманизированного антитела против CD147 в модели рака желудка
Оценивали противоопухолевую активность человеческого химерного антитела и гуманизированного антитела против CD147 человека в клетках рака желудка линии KATO III (ATCC, Cat. HTB-103), по поводу которых было подтверждено, используя проточный цитометр, что они являются CD147-положительными.
[0367]
5 × 106 клеток рака желудка человека линии KATO III суспендировали в 100% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CLEA Japan, Inc). Проводили распределение по группам на основе объема опухоли через 3 дня после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам человеческое химерное антитело против CD147 (LN22R8chIgG4P) или гуманизированное антитело против CD147 (#110H1L4hIgG4P), продуцированные в примере 6)-4-2, при 10 мг/кг в течение каждого из 7 дней после распределения мышей по группам (n=6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0368]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 43. На графиках также показаны средние значения и стандартные ошибки изменений объема опухоли. В то время как средний объем опухоли мышей через 24 дня после инокуляции составлял в группе, не подвергавшейся обработке, 290 мм3, в группе введения LN22R8chIgG4p средний объем опухоли составлял 199 мм3, а в группе введения #110H1L4hIgG4P средний объем опухоли составлял 134 мм3. Рассчитанная противоопухолевая активность составляла 31% в группе введения LN22R8chIgG4p и 54% в группе введения #110H1L4hIgG4P.
[0369]
(Пример 19) Противоопухолевое действие гуманизированного антитела против CD147 в модели хронического миелолейкоза
Оценивали противоопухолевую активность гуманизированного антитела против CD147 человека на клетках хронического миелолейкоза человека линии KU812 (ATCC, Cat. CRL-2099) по поводу которых было подтверждено, используя проточный цитометр, что они являются CD147-положительными.
[0370]
5 × 106 клеток хронического миелолейкоза линии KU812 суспендировали в PBS, содержащим 50% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CLEA Japan, Inc). Проводили распределение по группам на основе объема опухоли через 3 дня после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам гуманизированное антитело против CD147 (#110H1L4hIgG4P), продуцированное в примере 6)-4-2, при 10 мг/кг каждый день в течение 7 дней после распределения по группам (n=5). В качестве контрольного лекарственного средства, перорально вводили несущим злокачественную опухоль мышам в форме раствора в дистиллированной воде с концентрацией 9 мг/мл иматиниб (AstaTech, Inc., Cat. #63168), который является стандартным терапевтическим средством при хроническом миелолейкозе, в дозе 90 мг/кг (ежедневное введение, за исключением суббот и воскресений; введение через 4, 7, 8, 9, 10, 11, 14, 15, 16, 17, 18, 21, 22, 23, 24 дня после инокуляции). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0371]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 44. На графиках показаны средние значения и стандартные ошибки изменений объема опухоли. В то время как средний объем опухоли мышей через 25 дней после инокуляции составлял в группе, не подвергавшейся обработке, 627 мм3, в группе введения иматиниба средний объем опухоли составлял 328 мм3, а в группе введения #110H1L4hIgG4P средний объем опухоли составлял 6 мм3. Рассчитанная противоопухолевая активность составляла 48% в группе введения иматиниба и 97% в группе введения #110H1L4hIgG4P. Только лишь в группе введения #110H1L4hIgG4P, наблюдался полный регресс опухоли в 4 примерах из 5 примеров.
[0372]
(Пример 20) Противоопухолевое действие гуманизированного антитела против CD147 в модели рака толстой
Оценивали противоопухолевую активность каждого гуманизированного антитела на клетках рака толстой кишки человека линии SW620 (ATCC, Cat. CCL-227), по поводу которых было подтверждено, используя проточный цитометр, что они являются CD147-положительными.
[0373]
5 × 106 клеток рака толстой кишки человека линии SW620 суспендировали в 100% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CLEA Japan, Inc). Проводили распределение по группам на основе объема опухоли через 3 дня после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам человеческое химерное антитело против CD147 (LN22R8chIgG4P) или гуманизированное антитело против CD147 (#084H1L2hIgG4P или #110H1L4hIgG4P), продуцированные в примере 6)-4-2, при 10 мг/кг каждый день в течение 7 дней после распределения по группам (n=5). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0374]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 45. На графиках показаны средние значения и стандартные ошибки изменений объема опухоли. В то время как средний объем опухоли мышей через 21 день после инокуляции составлял в группе, не подвергавшейся обработке, 1302 мм3, в группе введения #084H1L2hIgG4P средний объем опухоли составлял 709 мм3, а в группе введения #110H1L4hIgG4P средний объем опухоли составлял 403 мм3. Рассчитанная противоопухолевая активность составляла 46% в группе введения #084H1L2hIgG4P и 69% в группе введения #110H1L4hIgG4P.
[0375]
(Пример 21) Противоопухолевая активность гуманизированного антитела против CD147 в модели рака почки 786-O
Оценивали противоопухолевую активность каждого гуманизированного антитела на клетках рака почки человека линии 786-O, по поводу которых было подтверждено, используя проточный цитометр, что они являются CD147-положительными. 5 × 106 клеток рака почки человека линии 786-O суспендировали в 50% Matrigel (Corning Inc., Cat. 354234), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CLEA Japan, Inc). Проводили распределение по группам на основе объема опухоли через 3 дня после инокуляции, и вводили интраперитонеально несущим злокачественную опухоль мышам человеческое химерное антитело против CD147 (LN22R8chIgG4P) или гуманизированное антитело против CD147 (#084H1L2hIgG4P или #110H1L4hIgG4P), продуцированные в примере 6)-4-2, при 10 мг/кг каждый день в течение 7 дней после распределения по группам в общей сложности 4 раза (n=6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0376]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 46. На графиках показаны средние значения и стандартные ошибки изменений объема опухоли. В то время как средний объем опухоли мышей через 31 день после инокуляции составлял в группе, не подвергавшейся обработке, 918 мм3, в группе введения #084H1L2hIgG4P средний объем опухоли составлял 224 мм3, а в группе введения #110H1L4hIgG4P средний объем опухоли составлял 379 мм3. Рассчитанная противоопухолевая активность составляла 76% в группе введения #084H1L2hIgG4P и 59% в группе введения #110H1L4hIgG4P.
[0377]
(Пример 22) Противоопухолевое действие гуманизированного антитела против CD147 в модели острого миелолейкоза (AML)
Оценивали противоопухолевую активность гуманизированного антитела на клетках AML человека линии OCI-AML3 (DSMZ, Cat. ACC 582), по поводу которых было подтверждено, используя проточный цитометр, что они являются CD147-положительными.
[0378]
5 × 106 клеток AML человека линии OCI-AML3 суспендировали в 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 5 недель линии NOD-scid (NOD. CB17-Prkdc<scid>/J, поставляемых фирмой CLEA Japan, Inc). Проводили распределение по группам на основе объема опухоли через 3 дня после инокуляции, и вводили путем инъекции в хвостовую вену несущим злокачественную опухоль мышам гуманизированное антитело против CD147 (#110H1L4hIgG4P), продуцированное в примере 6)-4-2, при 10 мг/кг каждый день в течение 7 дней после распределения по группам (n=6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
[0379]
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
Результаты представлены на фигуре 47. На графиках показаны средние значения и стандартные ошибки изменений объема опухоли. В то время как средний объем опухоли мышей через 21 день после инокуляции составлял в группе, не подвергавшейся обработке, 1533 мм3, в группе введения #110H1L4hIgG4P средний объем опухоли составлял 394 мм3. Рассчитанная противоопухолевая активность в группе введения #110H1L4hIgG4P составляла 74%.
[0380]
(Пример 23) Измерение действия гуманизированных антител против CD147, имеющих различные активности связывания
5 × 106 клеток поджелудочной железы человека линии MIA PaCa-2 суспендировали в PBS, содержащем 50% GFR-Matrigel (Corning Inc., Cat. 354230), и суспензию подкожно инокулировали в подмышечную впадину самкам мышей в возрасте 4 недель линии Nude (BALB/c Slc-nu/nu, поставляемых фирмой Japan SLC, Inc). Проводили распределение по группам на основе объема опухоли, и три типа гуманизированных антител против CD147 (#110H1L4hIgG4P, #110H13L02hIgG4P и #110H13L12hIgG4P; в таблице 6 приведены их активности связывания), имеющих различные активности связывания с белком CD147, продуцированных вы примере 6)-4-2, каждое вводили в хвостовую вену несущим злокачественную опухоль мышам в дозе 10 мг/кг через 7 дней после инокуляции (n=6). В качестве контрольного лекарственного средства, вводили в хвостовую вену несущим злокачественную опухоль мышам гемцитабин (поставляемый фирмой Eli Lilly Japan K.K.), который является стандартным терапевтическим средством при раке поджелудочной железы, в дозе 400 мг/кг через 3 и 10 дней после инокуляции (n=6). Измеряли два раза в неделю большую ось и малую ось инокулированной опухоли с помощью штангенциркуля с электронным цифровым отсчетом (фирмы MITUTOYO CORPORATION), и рассчитывали объем опухоли по следующей формуле.
Объем опухоли (мм3) = 1/2 × малая ось (мм) × малая ось (мм) × большая ось (мм)
[0381]
Результаты представлены на фигуре 48. На графиках показаны средние значения и стандартные ошибки изменений объема опухоли.
[0382]
В то время как степень ингибирования роста опухоли в случае введения гемцитабина в качестве контрольного лекарственного средства составляла 66%, все гуманизированные антитела против CD147 в группе введения в дозе 10 мг/кг проявляли более высокую противоопухолевую активность, чем гемцитабин.
[0383]
(Пример 24) Сравнение антител против CD147
24)-1 Оценка активности связывания существующих антител против CD147 с антигеном
Очищенные антитела получали на основе последовательностей 4A5, 5F6, описанных в патентном документе WO2010/036460, и PPAT-082-03, описанной в патентном документе WO2017/061602. Измеряли константу диссоциации для связывания 4A5, 5F6 или PPAT-082-03 с белком CD147, используя систему Biacore T200 (поставляемую фирмой GE Healthcare Bioscience). Антитело против IgG (Fc) человека иммобилизовали на сенсорном чипе, используя набор для иммобилизации антител Human Antibody Capture Kit (поставляемый фирмой GE Healthcare Bioscience), затем захватывали человеческое химерное антитело в качестве лиганда и ассоциировали антиген в качестве аналита. Использовали HBS-EP+ (поставляемый фирмой GE Healthcare Bioscience) в качестве подвижного буфера и CM5 (поставляемый фирмой GE Healthcare Bioscience) в качестве сенсорного чипа. Затем 1 мкг/мл конкурирующего антитела добавляли на чип при 10 мкл/минута в течение 60 секунд, добавляли последовательное разведение антигена (0,5-8 мкг/мл), используемого в примере 5, при скорости 30 мкл/минута в течение 120 секунд, затем проводили последующий мониторинг фазы диссоциации 300 секунд. В качестве регенерирующего раствора добавляли раствор 3M хлорида магния (поставляемый фирмой GE Healthcare Bioscience) при скорости 20 мкл/минута в течение 30 секунд. Используя для анализа данных модель связывания 1:1, рассчитывали константу скорости связывания ka, константу скорости диссоциации kd и константу диссоциации (KD; KD=kd/ka). Константу диссоциации для связывания гуманизированного антитела против CD147 (#084H1L2hIgG4P или #110H1L4hIgG4P) с белком CD147 рассчитывали методом, описанным в примере 7)-1.
[0384]
В таблице 8 приведена информация по рассчитанной константе диссоциации, эффекторная функция и эпитопная область.
[0385]
[Таблица 8]
CDC(-)
ADCP(±)
CDC(-)
ADCP(+)
CDC(+)
ADCP(+)
CDC(-)
ADCP(+)
CDC(-)
ADCP(+)
[0386]
24)-2 Конкурирующий твердофазный иммуноферментный анализ (ELISA) с существующим антителом против CD147
Используя гуманизированное антитело против CD147 (#084H1L2hIgG4P или #110H1L4hIgG4P), продуцированные в примере 6)-4-2, оценивали активность связывания с рекомбинантным человеческим CD147/Fc (Sino Biological Inc., Cat. 10186-H02H) методом конкурирующего твердофазного иммуноферментного анализа (ELISA). В качестве конкурирующих антител, использовали антитела 4A5 и 5F6, полученные в примере 24)-1, и антитело PPAT-082-03. В качестве конкурирующего антитела для отрицательного контроля, использовали человеческий IgG (Jackson ImmunoResearch Inc., Cat. 130093). В качестве конкурирующего антитела для положительного контроля, использовали антитело #84H1L2hIgG2 применительно к антителу #084H1L2hIgG4P, и антитело #110chIgG2 применительно к антителу #110H1L4hIgG4P.
[0387]
Рекомбинантный человеческий CD147/Fc, разбавленный с помощью PBS до 2 мкг/мл, добавляли в 96-луночный планшет (Thermo Scientific, Cat. 43454) при 50 мкл/лунка, затем выдерживали в течение ночи при 4°C. После удаления раствора белка, добавляли 300 мкл PBS, содержащего 1% BSA, затем нагревали при комнатной температуре в течение одного часа. После удаления раствора, добавляли 25 мкл раствора существующего антитела (0, 0,2, 2 или 20 мкг/мл), затем нагревали при комнатной температуре в течение двух часов. Добавляли в каждую лунку 25 мкл гуманизированного антитела против CD147 (#084H1L2hIgG4P или #110H1L4hIgG4P) при концентрации 20 нг/мл, затем нагревали при комнатной температуре в течение двух часов. После двухкратного промывания с помощью PBS, содержащего 0,05% Tween20 (Bio-Rad Laboratories, Inc., Cat. 170-6531), добавляли 50 мкл анти-человеческого IgG4-HRP (Abcam plc., Cat.ab99823), разбавленного в 2000 раз с помощью PBS, содержащего 1% BSA, затем нагревали при комнатной температуре в течение одного часа. После троекратного промывания с помощью PBS, содержащего 0,05% Tween20 (Bio-Rad Laboratories, Inc., Cat. 170-6531), в достаточной степени удаляли промывающий раствор и добавляли 50 мкл раствора субстрата HRP (eBioscience, Thermo Fisher SCIENTIFIC K.K., Cat.00-4203-58), затем нагревали при комнатной температуре в течение времени от 15 до 20 минут. После этого, измеряли поглощение при 405 нм на планшет-ридере (название модели: EnVision2104, фирмы PerkinElmer, Inc).
[0388]
Результаты представлены в таблице 8 и на фигурах 49(a)-(b).
[0389]
Связывание #084H1L2hIgG4P с рекомбинантным человеческим CD147/Fc ингибировалось в результате присутствия существующего антитела против CD147 при 1-10 мкг/мл в присутствии hIgG. Связывание #110H1L4hIgG4P с рекомбинантным человеческим CD147/Fc не ингибировалось в результате присутствия существующего антитела против CD147 при 1-10 мкг/мл в присутствии hIgG, но ингибировалось в результате присутствия антитела #110chIgG2. Было обнаружено, что эпитоп #084H1L2hIgG4P конкурировал со связыванием существующего антитела против CD147, но на эпитоп #110H1L4hIgG4P не оказывало влияние связывание существующего антитела против CD147. В таблице 8 приведена информация по эпитопу антитела H110H1L4h, представленного в примере 17)-3. Эпитопную область #084H1L2h оценивали по результатам испытаний конкурирующего связывания (Пример 2)-8) с антителом 2P10F2, эпитом которого оценивали путем проведения анализа с использованием мутантов CD147 из примера 1)-9. Результаты приведены в таблице 8.
[0390]
24)-3 Оценка опосредованной антителами клеточной цитотоксичности (ADCC) антител против CD147
Опосредованную антителами клеточную цитотоксичность (ADCC) антитела против CD147 оценивали в соответствии с методом, использованным в примере 1)-15. Условия, отличные от условий в методе, используемом в примере 1)-15, включали проведение оценки с использованием клеток HepG2 (ATCC, Cat. HB-8065) в качестве клеток-мишеней для ADCC, и использование # 110H1L4hIgG4P, # 084H1L2hIgG4P, 4A5, 5F6 и PPAT-082-03 в качестве антител против CD147 при концентрации 1 мкг/мл. Измерение проводили в трех параллельных опытах, и рассчитывали средние значения и стандартные отклонения. В случае обнаружения 51Cr в 5% или более клеток, опосредованную антителами клеточную цитотоксичность (ADCC) определяли как положительную (+). В случае обнаружения 51Cr в менее чем 5% клеток, опосредованную антителами клеточную цитотоксичность (ADCC) определяли как отрицательную (-). Результаты приведены в таблице 8.
[0391]
Опосредованная антителами клеточная цитотоксичность (ADCC) не обнаруживалась в случае #110H1L4hIgG4P и #084H1L2hIgG4P, и, поэтому ADCC определяли как отрицательную (-). В случае 4A5, 5F6 и PPAT-082-03, 51Cr обнаруживали в 5% или более клеток, и, поэтому ADCC определяли как положительную (+). #110H1L4hIgG4P и #084H1L2hIgG4P характеризовались отрицательной ADCC, и, поэтому, можно ожидать, что они будут индуцировать более низкий уровень гибели нормальных клеток, таких как клетки крови, экспрессирующие CD147, в организме человека, чем 4A5, 5F6 и PPAT-082-03, которые характеризовались положительной опосредованной антителами клеточной цитотоксичностью (ADCC).
[0392]
24)-4 Оценка комплементзависимой цитотоксичности (CDC) антител против CD147
Комплементзависимую цитотоксичность (CDC) антител против CD147 оценивали в соответствии с методом, использованным в примере 1)-16. Условия, отличные от условий в методе, используемом в примере 1)-16, включали проведение измерений с использованием клеток печени человека линии HepG2 (ATCC, Cat. HB-8065) в качестве клеток-мишеней и #110H1L4hIgG4P, #084H1L2hIgG4P, 4A5, 5F6 и PPAT-082-03 в качестве антител против CD147 человека, и добавление кроличьего комплемента до конечной концентрации 10%. Измерение проводили в трех параллельных опытах, и рассчитывали средние значения и стандартные отклонения. Антитела, для которых наблюдали 30% или более антителозависимой комплементзависимой цитотоксичности (CDC), определяли как имеющие положительную комплементзависимую цитотоксичность (CDC) и обозначали ее в таблице как CDC (+). Результаты представлены в таблице 8. Только антитело 4A5 из антител против человеческого CD147 проявляла положительную комплементзависимую цитотоксичность (CDC). #110H1L4hIgG4P, #084H1L2hIgG4P, 5F6 и PPAT-082-03 характеризовались отрицательной CDC, и можно ожидать, что они будут индуцировать более низкий уровень гибели нормальных клеток, таких как клетки крови, экспрессирующие CD147, в организме человека, чем 4A5, которое имело положительную комплементзависимую цитотоксичность (CDC).
24)-5 Оценка антителозависимого клеточного фагоцитоза (ADCP) антител против CD147
Антитело против CD147 измеряли в соответствии с методом, используемым в пример 1)-17. Условия, отличные от условий в методе, используемом в примере 1)-17, включали проведение измерения антителозависимого клеточного фагоцитоза (ADCP) с использованием клеток печени человека линии HepG2 (ATCC, Cat. HB-8065) в качестве клеток-мишеней и добавление #110H1L4hIgG4P, #084H1L2hIgG4P, 4A5, 5F6 и PPAT-082-03 в качестве антител против CD147 при концентрации 1 мкг/мл и равного количества меченых клеток RAW264.7 к клеткам-мишеням для ADCP. Измерение проводили в трех параллельных опытах, и рассчитывали средние значения и стандартные отклонения. Увеличение антителозависимого клеточного фагоцитоза (ADCP) менее чем у 10% в группе, подвергнутой обработке с помощью человеческого IgG, определяли как слегка положительный (±), и увеличение активности у 10% или более определяли как положительный (+). Результаты представлены в таблице 8.
[0393]
#110H1L4hIgG4P имело антителозависимый клеточный фагоцитоз (ADCP) менее чем 10%, и антителозависимый клеточный фагоцитоз (ADCP) определяли как ±. Антитела #084H1L2hIgG4P, 4A5, 5F6 и PPAT-082-03 имели антителозависимый клеточный фагоцитоз (ADCP) 10% или более, и антителозависимый клеточный фагоцитоз (ADCP) определяли как +. #110H1L4hIgG4P, которое распознает CD147-D2, имело более низкий антителозависимый клеточный фагоцитоз (ADCP), чем другие антитела против CD147, которые распознают CD147-D1. После связывания антитело против CD147 с CD147, Fс-фрагмент антитела должен быть распознан рецептором Fcγ, экспрессируемым на макрофагах или мононуклеарных клетках для ADCP. Однако существует вероятность того, что антителозависимый клеточный фагоцитоз (ADCP) является низким вследствие того, что эпитоп антитела #110 находится близко к поверхности клеток, и рецептору Fcγ более сложно распознать Fс-фрагмент антитела, связанного к CD147, служащий в качестве антигена, чем другие антитела, которые распознают CD147-D1. Можно ожидать, что #110H1L4hIgG4P, имеющее слегка положительный антителозависимый клеточный фагоцитоз (ADCP), будет с меньшей вероятностью индуцировать гибель нормальных клеток, таких как клетки крови, экспрессирующие CD147, в организме человека, чем #084H1L2hIg4P, 4A5, 5F6 и PPAT-082-03, которые имеют положительный антителозависимый клеточный фагоцитоз (ADCP).
[0394]
(Пример 25)
25)-1 Агрегация клеток системы крови под воздействием антитела против CD147
Сообщалось, что некоторые антитела против CD147 индуцируют агрегацию клеток крови (Kasinrerk, et al., Immunology 1999, 96 (2) p184-192). Агрегация клеток крови может потенциально быть причиной серьезной токсичности для крови (Doll, C., et al., 1994, Curr. Opin. Oncol., 345-350), и агрегация клеток крови представляет собой свойство, которое является нежелательным для терапевтического антитела. Исследовали различия в активности в отношении агрегации между антителами против CD147. В качестве антител против CD147, оценивали #110H1L4hIgG4P, #084H1L2hIgG4P, 4A5, 5F6 и PPAT-082-03. В качестве антитела для отрицательного контроля, использовали человеческий IgG (hIgG, ChromPure Human IgG, Jackson ImmunoResearch Laboratories, Inc., Cat. 009-000-003). В 96-луночный планшет с U-образным дном (Sumitomo Bakelite Co., Ltd., Cat. MS-9096U) добавляли клетки HEL92.1.7 (поставляемые из ATCC, Cat. #TIB-180) со средой RPMI1640 (Thermo Fisher SCIENTIFIC K.K., Cat. 11875-093), содержащей 10% FBS (HyClone, GE Healthcare, Cat. SH30084.03), при 1600 клеток/80 мкл на лунку, затем культивировали в течение 4 часов в присутствии 5% CO2 при влажности 95% и 37°C. Добавляли в каждую лунку 20 мкл раствора антитела против CD147 (150 мкг/мл, 50 мкг/мл) до конечной концентрации 30 или 10 мкг/мл. После культивирования в течение 2 часов в присутствии 5% CO2 при влажности 95% и 37°C, проводили наблюдение под микроскопом.
[0395]
В то время как при добавлении человеческого IgG или #110H1L4hIgG4P агрегация клеток не наблюдалась, клетки агрегировали в присутствии антитела #084H1L2hIgG4P, 4A5, 5F6 или PPAT-082-03, и наблюдалось перекрывание клеточных масс в центре планшета. Было обнаружено, что #110H1L4hIgG4P, которое распознает CD147-D2, не имеет активности в отношении агрегации клеток крови и, поэтому, оно отличалось от других антител CD147, которые распознают домен CD147-D1. Так как антитела против CD147, включающие #084, которые имеют активность в отношении агрегации клеток, могут потенциально вызывать токсичность, такую как образование тромба в результате агрегации клеток крови, при введении людям, то желательно предотвращать или снижать такие побочные эффекты путем комбинированного использования подкожной инъекции гепарина в дозе, используемой для лечения тромбозов, или низкомолекулярного гепарина или антиагрегантных лекарственных средств.
[0396]
25)-2 Оценка риска проявления синдрома высвобождения цитокинов
В случае некоторых антител, таких как OKT3 и TGN1412, введение терапевтических антител вызывает увеличение цитокинов в крови в результате активации иммуноцитов, в силу этого вызывая серьезный синдром высвобождения цитокинов (Gaston, R., Kidney International, 1991, 141-148; Suntharalingam, G., et al., N. Engl. J. Med. 2006, 1018-1028). Сообщалось, что некоторые антитела против CD147 воздействуют на иммуноциты, обладающие способностью увеличивать продукцию гамма-интерферона или интерлейкин-4 (Hu, J., et al., J. Cell. Mol. Med., 2010, 2132-2143). Токсичность антител, обусловленная синдромом высвобождения цитокинов, может быть предсказана путем проведения анализа на высвобождение цитокинов с использованием периферической крови (Vessillier, S. et al., J. Immunolol. Methods, 2015, 43-52). В связи с этим, проводили оценку риска проявления синдрома высвобождения цитокинов с помощью анализа на высвобождение цитокинов с использованием периферической крови. В качестве антител против CD147, использовали #110H1L4hIgG4P, #110chIgG4PFALA, #084H1L2hIgG4P и #084H1L2hIgG2, и в качестве антител для сравнения, использовали бевацизумаб (Genentech, Inc.), трастузумаб (Roche Pharma AG), алемтузумаб (Sanofi K.K.) и антитело против CD3 человека (BioLegend Cat. No317326). Для всех подвергнутых оценке антител против CD147 не было обнаружено ускорения роста клеток в мононуклеарных клетках периферической крови человека (проводили оценку каждого антитела на 6 донорах), и их влияние на высвобождение цитокинов (TNFα, INF-γ, IL-2, IL-6, IL-8, IL-10 и MIP-1α) было меньше, чем в случае бевацизумаба, который характеризуется низким риском проявления синдрома высвобождения цитокинов. Для антитела против CD3 (OKT3), были обнаружены ускорение роста клеток и ускорение высвобождения цитокинов (TNFα, INF-γ, IL-2, IL-6, IL-8, IL-10 и MIP-1α). Было показано, что #110H1L4hIgG4P, #110chIgG4PFALA, #084H1L2hIgG4P и #084H1L2hIgG2 не индуцируют высвобождения цитокинов, которое может потенциально вызывать синдром высвобождения цитокинов.
[0397]
25)-3 Оценка безопасности антитела против CD147 у обезьян
Сообщалось, что некоторые мышиные антитела против CD147 при введении мышам ингибируют функции CD147, индуцируют агрегацию эритроцитов в селезенке и снижают количество эритроцитов в периферической крови, вызывая в результате анемию (Coste, I. et al., Blood, 2001, 3984-3988). Так как полученные в настоящем изобретении антитела против CD147, такие как #110H1L4hIgG4P, не проявляют связывание с мышиным CD147, то нельзя было проводить оценку безопасности антител на мышах. Поэтому, оценку безопасности проводили путем введения яванским макакам антитела #110H1L4hIgG4P, которое является антителом против CD147, и по поводу которого было подтверждено, что оно имеет активность связывания с CD147 человека и обезьяны в экспериментах с использованием проточного цитометра. #110H1L4hIgG4P вводили внутривенно яванским макакам (одному самцу и одной самке) в разовой дозе 99,2 мг/кг, которая является максимальной дозой. В результате, не было обнаружено проявления серьезной токсичности (изменений массы тела и потребления пищи или патологогистологических изменений) ни в период наблюдения в течение 15 дней после введения, ни в результате патологогистологического исследования по завершению периода наблюдения. #110H1L4hIgG4P не проявляло токсичности в отношении яванских макаков, тем самым указывая на то, что оно может быть использовано для лечения рака у человека.
[0398]
(Пример 26) Влияние KLF5 на восприимчивость к антителу против CD147
Сигнал смерти EMT (эпителиально-мезенхимального перехода) известен как сигнал SMAD2/SMAD3/SMAD4-зависимой гибели клеток в раковых клетках, и сообщалось, что в SMAD4-отрицательных раковых клетках обнаруживается повышенная экспрессия белка фактора транскрипции KLF5, которая в нормальных условиях снижается под воздействием сигнального пути SMAD, и это уменьшает сигнал смерти EMT в SMAD4-отрицательных раковых клетках (David, C., Cell, 2016, 1015-1030). Так как антитело против CD147 по настоящему изобретению активирует сигнальный путь SMAD и проявляет противоопухолевую активность на SMAD4-положительных клетках, то считалось, что оно может индуцировать зависимую от сигнала SMAD гибель клеток. Поэтому, проводили исследование возможности влияния KLF5 на восприимчивость к противоопухолевой активности антител против CD147.
[0399]
26)-1 Продукция линии клеток, экспрессирующих KLF5
В соответствии с методом, используемым в примере 13, продуцировали клетки линии MIA PaCa-2, стабильно экспрессирующие KLF5. Последовательности SEQ ID NOs: 145 и 146, соответственно, представляют аминокислотную последовательность и нуклеотидную последовательность KLF5 человека. Продуцировали ретровирусный вектор pQCXIP с последовательностью (Ref seq. ID: NM_001730.4) гена KLF5, содержащейся в нем (GenScript Biotech Corporation, Cat. OHu21278C), и использовали ретровирусный вектор для продукции ретровируса. Ретровирус вводили в хромосому путем инфицирования вирусом, и отбирали клетки MIA PaCa-2, которые становились резистентными к пуромицину и положительными по KLF5, для образования KLF5-положительных клеток MIA PaCa-2, клеток MIA PaCa-2-KLF5. Ретровирусный вектор pQCXIP инфицировали аналогичным методом с образованием пуромицин-резистентных клеток MIA PaCa-2, MIA PaCa-2-имитация.
[0400]
26)-2 Подтверждение экспрессии KLF5
Экспрессии KLF5 в клетках MIA PaCa-2-mock и MIA PaCa-2-KLF5 подтверждали, используя проточный цитометр. В соответствии с методом примера 13-2, было подтверждено, что уровень экспрессии KLF5 в MIA PaCa-2-KLF5 был выше, чем в MIA PaCa-2-имитация. Для детекции KLF5, использовали антитело против KLF5 (Cell Signaling Technology, Inc., Cat. #51586).
26)-3 Восприимчивость опухоли MIA PaCa-2, экспрессирующей KLF5, к гуманизированному антителу против CD147
В соответствии с методом примера 7)-2, сравнивали восприимчивости опухолей MIA PaCa-2-KLF5 и MIA PaCa-2-имитация к гуманизированному антителу #110H1L4hIgG4P против CD147. Гуманизированное антитело (#110H1L4hIgG4P) против CD147, продуцированное в примере 6)-4-2, вводили в хвостовую вену мышам, несущим злокачественную опухоль, в дозе 1 мг/кг через 3 дня после инокуляции клеток (n=6). Антитело вводили через 7 дней аналогичным способом. Несущим злокачественную опухоль мышам контрольной группы вводили через хвостовую вену аналогичным способом буферный раствор PBS (n=6). Результаты представлены на фигуре 50.
[0401]
Средний объем опухоли MIA PaCa-2-имитация снижался до 9% от среднего объема опухоли в контрольной группе через 14 дней после введения гуманизированного антитела против CD147, проявляя в результате восприимчивость к антителу против CD147. Средний объем опухоли MIA PaCa-2-KLF5 составлял 80% от среднего объема опухоли в контрольной группе, проявляя в результате низкую восприимчивость к антителу против CD147. Было обнаружено, что SMAD сигнал-зависимая противоопухолевая активность антитела против CD147 подавляется экспрессией KLF5.
Промышленная применимость
[0402]
В настоящем изобретении предлагается специфичное антитело против CD147, которое активирует CD147 и проявляет высокую противоопухолевая активность. В настоящем изобретении предлагается антитело, проявляющее высокую противоопухолевую активность, независимую от эффекторных функций. Антитело по настоящему изобретению проявляет активность в клетках рака печени, которая несомненно выше активности сорафениба, который применяют в качестве одного из стандартных лекарственных средств для лечения рака печени. Антитело по настоящему изобретению проявляет активность в клетках рака поджелудочной железы, которая значительно выше активности гемцитабина, который применяют в качестве одного из стандартных лекарственных средств для лечения рака поджелудочной железы. Антитело по настоящему изобретению проявляет активность в клетках хронического миелолейкоза, которая значительно выше активности иматиниба, который применяют в качестве одного из стандартных лекарственных средств для лечения хронического миелолейкоза. Антитело по настоящему изобретению имеет противоопухолевую активность, которая проявляется независимо от эффекторных функций и вызывает меньше проблем, связанных с его безопасностью, и, поэтому, в изобретении предлагается антитело против CD147, гарантирующее высокую безопасность при его применении. CD147 экспрессируется не только в опухолевых клетках, но также и в клетках крови. Однако, антитело по настоящему изобретению не воздействует на T- клетки и PBMC и не зависит от эффекторных функций, в результате чего антитело имеет преимущество, связанное с меньшим количеством проблем, связанных с его безопасностью, при его разработке в качестве противоопухолевого средства. В настоящем изобретении предлагается фармацевтическая композиция, включающая упомянутое выше антитело, а также способ лечения опухолей с применением антитела и/или фармацевтической композиции.
Свободный текст, содержащий перечень последовательностей
[0403]
SEQ ID NO:1: Аминокислотная последовательность варианта 1 человеческого CD147
SEQ ID NO:2: Нуклеотидная последовательность варианта 1 человеческого CD147
SEQ ID NO:3: Аминокислотная последовательность варианта 2 человеческого CD147
SEQ ID NO:4: Нуклеотидная последовательность варианта 2 человеческого CD147
SEQ ID NO:5: Аминокислотная последовательность SMAD4 человека
SEQ ID NO:6: Нуклеотидная последовательность SMAD4 человека
SEQ ID NO:7: Нуклеотидная последовательность вариабельной области легкой цепи LN22R8
SEQ ID NO:8: Аминокислотная последовательность вариабельной области легкой цепи LN22R8
SEQ ID NO:9: Нуклеотидная последовательность вариабельной области тяжелой цепи LN22R8
SEQ ID NO:10: Аминокислотная последовательность вариабельной области тяжелой цепи LN22R8
SEQ ID NO:11: Аминокислотная последовательность CDRL1 LN22R8
SEQ ID NO:12: Аминокислотная последовательность CDRL2 LN22R8
SEQ ID NO:13: Аминокислотная последовательность CDRL3 LN22R8
SEQ ID NO:14: Аминокислотная последовательность CDRH1 LN22R8
SEQ ID NO:15: Аминокислотная последовательность CDRH2 LN22R8
SEQ ID NO:16: Аминокислотная последовательность CDRH3 LN22R8
SEQ ID NO:17: Нуклеотидная последовательность вариабельной области легкой цепи 2P10F2
SEQ ID NO:18: Аминокислотная последовательность вариабельной области легкой цепи 2P10F2
SEQ ID NO:19: Нуклеотидная последовательность вариабельной области тяжелой цепи 2P10F2
SEQ ID NO:20: Аминокислотная последовательность вариабельной области тяжелой цепи 2P10F2
SEQ ID NO:21: Аминокислотная последовательность CDRL1 2P10F2
SEQ ID NO:22: Аминокислотная последовательность CDRL2 2P10F2
SEQ ID NO:23: Аминокислотная последовательность CDRL3 2P10F2
SEQ ID NO:24: Аминокислотная последовательность CDRH1 2P10F2
SEQ ID NO:25: Аминокислотная последовательность CDRH2 2P10F2
SEQ ID NO:26: Аминокислотная последовательность CDRH3 2P10F2
SEQ ID NO:27: ДНК, включающая последовательность ДНК, кодирующую сигнальную последовательность легкой цепи человека и константную область каппа цепи человека
SEQ ID NO:28: ДНК, включающая последовательность ДНК, кодирующую аминокислоту сигнальной последовательности тяжелой цепи человека и константной области человеческого IgG1
SEQ ID NO:29: ДНК, включающая последовательность ДНК, кодирующую аминокислоту сигнальной последовательности тяжелой цепи человека и константной области человеческого IgG2
SEQ ID NO:30: Нуклеотидная последовательность легкой цепи человеческого химерного LN22R8
SEQ ID NO:31: Аминокислотная последовательность легкой цепи человеческого химерного LN22R8
SEQ ID NO:32: Нуклеотидная последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG1
SEQ ID NO:33: Аминокислотная последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG1
SEQ ID NO:34: Нуклеотидная последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG2
SEQ ID NO:35: Аминокислотная последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG2
SEQ ID NO:36: ДНК, включающая последовательность ДНК, кодирующую аминокислотную последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG4P
SEQ ID NO:37: Аминокислотная последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG4P
SEQ ID NO:38: ДНК, включающая последовательность ДНК, кодирующую аминокислоту сигнальной последовательности тяжелой цепи человека и константой области IgG1LALA
SEQ ID NO:39: ДНК, включающая последовательность ДНК, кодирующую аминокислоту сигнальной последовательности тяжелой цепи человека и константной области IgG4P
SEQ ID NO:40: Нуклеотидная последовательность легкой цепи человеческого химерного 2P10F2
SEQ ID NO:41: Аминокислотная последовательность легкой цепи человеческого химерного 2P10F2
SEQ ID NO:42: Нуклеотидная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG1LALA
SEQ ID NO:43: Аминокислотная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG1LALA
SEQ ID NO:44: Нуклеотидная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG2
SEQ ID NO:45: Аминокислотная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG2
SEQ ID NO:46: Нуклеотидная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG4P
SEQ ID NO:47: Аминокислотная последовательность тяжелой цепи человеческого химерного 2P10F2 типа IgG4P
SEQ ID NO:48: Нуклеотидная последовательность вариабельной области легкой цепи rat_CD147_#84(r#84)
SEQ ID NO:49: Аминокислотная последовательность вариабельной области легкой цепи rat_CD147_#84(r#84)
SEQ ID NO:50: Нуклеотидная последовательность вариабельной области тяжелой цепи rat_CD147_#84(r#84)
SEQ ID NO:51: Аминокислотная последовательность вариабельной области тяжелой цепи rat_CD147_#84(r#84)
SEQ ID NO:52: Аминокислотная последовательность CDRL1 rat_CD147_#84(r#84)
SEQ ID NO:53: Аминокислотная последовательность CDRL2 rat_CD147_#84(r#84)
SEQ ID NO:54: Аминокислотная последовательность CDRL3 rat_CD147_#84(r#84)
SEQ ID NO:55: Аминокислотная последовательность CDRH1 rat_CD147_#84(r#84)
SEQ ID NO:56: Аминокислотная последовательность CDRH2 rat_CD147_#84(r#84)
SEQ ID NO:57: Аминокислотная последовательность CDRH3 rat_CD147_#84(r#84)
SEQ ID NO:58: Нуклеотидная последовательность вариабельной области легкой цепи rat_CD147_#101(r#101)
SEQ ID NO:59: Аминокислотная последовательность вариабельной области легкой цепи rat_CD147_#101(r#101)
SEQ ID NO:60: Нуклеотидная последовательность вариабельной области тяжелой цепи rat_CD147_#101(r#101)
SEQ ID NO:61: Аминокислотная последовательность вариабельной области тяжелой цепи rat_CD147_#101(r#101)
SEQ ID NO:62: Аминокислотная последовательность CDRL1 rat_CD147_#101(r#101)
SEQ ID NO:63: Аминокислотная последовательность CDRL2 rat_CD147_#101(r#101)
SEQ ID NO:64: Аминокислотная последовательность CDRL3 rat_CD147_#101(r#101)
SEQ ID NO:65: Аминокислотная последовательность CDRH1 rat_CD147_#101(r#101)
SEQ ID NO:66: Аминокислотная последовательность CDRH2 rat_CD147_#101(r#101)
SEQ ID NO:67: Аминокислотная последовательность CDRH3 rat_CD147_#101(r#101)
SEQ ID NO:68: Нуклеотидная последовательность вариабельной области легкой цепи rat_CD147_#110(r#110)
SEQ ID NO:69: Аминокислотная последовательность вариабельной области легкой цепи rat_CD147_#110(r#110)
SEQ ID NO:70: Нуклеотидная последовательность вариабельной области тяжелой цепи rat_CD147_#110(r#110)
SEQ ID NO:71: Аминокислотная последовательность вариабельной области тяжелой цепи rat_CD147_#110(r#110)
SEQ ID NO:72: Аминокислотная последовательность CDRL1 rat_CD147_#110(r#110)
SEQ ID NO:73: Аминокислотная последовательность CDRL2 rat_CD147_#110(r#110)
SEQ ID NO:74: Аминокислотная последовательность CDRL3 rat_CD147_#110(r#110)
SEQ ID NO:75: Аминокислотная последовательность CDRH1 rat_CD147_#110(r#110)
SEQ ID NO:76: Аминокислотная последовательность CDRH2 rat_CD147_#110(r#110)
SEQ ID NO:77: Аминокислотная последовательность CDRH3 rat_CD147_#110(r#110)
SEQ ID NO:78: Нуклеотидная последовательность вариабельной области легкой цепи rat_CD147_#131(r#131)
SEQ ID NO:79: Аминокислотная последовательность вариабельной области легкой цепи rat_CD147_#131(r#131)
SEQ ID NO:80: Нуклеотидная последовательность вариабельной области тяжелой цепи rat_CD147_#131(r#131)
SEQ ID NO:81: Аминокислотная последовательность вариабельной области тяжелой цепи rat_CD147_#131(r#131)
SEQ ID NO:82: Аминокислотная последовательность CDRL1 rat_CD147_#131(r#131)
SEQ ID NO:83: Аминокислотная последовательность CDRL2 rat_CD147_#131(r#131)
SEQ ID NO:84: Аминокислотная последовательность CDRL3 rat_CD147_#131(r#131)
SEQ ID NO:85: Аминокислотная последовательность CDRH1 rat_CD147_#131(r#131)
SEQ ID NO:86: Аминокислотная последовательность CDRH2 rat_CD147_#131(r#131)
SEQ ID NO:87: Аминокислотная последовательность CDRH3 rat_CD147_#131(r#131)
SEQ ID NO:88: ДНК, включающая последовательность ДНК, кодирующую аминокислоту сигнальной последовательности тяжелой цепи человека и константной области человеческого IgG4PFALA
SEQ ID NO:89: Нуклеотидная последовательность легкой цепи человеческого химерного rat_CD147_#84
SEQ ID NO:90: Аминокислотная последовательность легкой цепи человеческого химерного rat_CD147_#84
SEQ ID NO:91: Нуклеотидная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1
SEQ ID NO:92: Аминокислотная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1
SEQ ID NO:93: Нуклеотидная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2
SEQ ID NO:94: Аминокислотная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG2
SEQ ID NO:95: Нуклеотидная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P
SEQ ID NO:96: Аминокислотная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4P
SEQ ID NO:97: Нуклеотидная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA
SEQ ID NO:98: Аминокислотная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG1LALA
SEQ ID NO:99: Нуклеотидная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа IgG4PFALA
SEQ ID NO:100: Аминокислотная последовательность тяжелой цепи человеческого химерного rat_CD147_#84 типа
SEQ ID NO:101: Нуклеотидная последовательность легкой цепи человеческого химерного rat_CD147_#101
SEQ ID NO:102: Аминокислотная последовательность легкой цепи человеческого химерного rat_CD147_#101
SEQ ID NO:103: Нуклеотидная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#101
SEQ ID NO:104: Аминокислотная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#101
SEQ ID NO:105: Нуклеотидная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#101
SEQ ID NO:106: Аминокислотная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#101
SEQ ID NO:107: Нуклеотидная последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#101
SEQ ID NO:108: Аминокислотная последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#101
SEQ ID NO:109: Нуклеотидная последовательность легкой цепи человеческого химерного rat_CD147_#110
SEQ ID NO:110: Аминокислотная последовательность легкой цепи человеческого химерного rat_CD147_#110
SEQ ID NO:111: Нуклеотидная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#110
SEQ ID NO:112: Аминокислотная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#110
SEQ ID NO:113: Нуклеотидная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#110
SEQ ID NO:114: Аминокислотная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#110
SEQ ID NO:115: Нуклеотидная последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#110
SEQ ID NO:116: Аминокислотная последовательность тяжелой цепи IgG4PFALA человеческого химерного rat_CD147_#110
SEQ ID NO:117: Нуклеотидная последовательность легкой цепи человеческого химерного rat_CD147_#131
SEQ ID NO:118: Аминокислотная последовательность легкой цепи человеческого химерного rat_CD147_#131
SEQ ID NO:119: Нуклеотидная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#131
SEQ ID NO:120: Аминокислотная последовательность тяжелой цепи IgG2 человеческого химерного rat_CD147_#131
SEQ ID NO:121: Нуклеотидная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#131
SEQ ID NO:122: Аминокислотная последовательность тяжелой цепи IgG4P человеческого химерного rat_CD147_#131
SEQ ID NO:123: Аминокислотная последовательность #84H1hIgG2
SEQ ID NO:124: Нуклеотидная последовательность #84H1hIgG2
SEQ ID NO:125: Аминокислотная последовательность #84H1hIgG4P
SEQ ID NO:126: Нуклеотидная последовательность #84H1hIgG4P
SEQ ID NO:127: Аминокислотная последовательность #84L2h
SEQ ID NO:128: Нуклеотидная последовательность #84L2h
SEQ ID NO:129: Аминокислотная последовательность #101H1hIgG2
SEQ ID NO:130: Нуклеотидная последовательность #101H1hIgG2
SEQ ID NO:131: Аминокислотная последовательность #101H1hIgG4P
SEQ ID NO:132: Нуклеотидная последовательность #101H1hIgG4P
SEQ ID NO:133: Аминокислотная последовательность #101L2h
SEQ ID NO:134: Нуклеотидная последовательность #101L2h
SEQ ID NO:135: Аминокислотная последовательность #110H1hIgG4P
SEQ ID NO:136: Нуклеотидная последовательность #110H1hIgG4P
SEQ ID NO:137: Аминокислотная последовательность #110L4h
SEQ ID NO:138: Нуклеотидная последовательность #110L4h
SEQ ID NO:139: Аминокислотная последовательность #131H2hIgG2
SEQ ID NO:140: Нуклеотидная последовательность #131H2hIgG2
SEQ ID NO:141: Аминокислотная последовательность #131L2h
SEQ ID NO:142: Нуклеотидная последовательность #131L2h
SEQ ID NO:143: область mu3 CD147v1 человека
SEQ ID NO:144: область mu3 CD147 яванского макака
SEQ ID NO:145: Аминокислотная последовательность KLF5 человека
SEQ ID NO:146: Нуклеотидная последовательность KLF5 человека
SEQ ID NO:147: Аминокислотная последовательность #110H13hIgG4P
SEQ ID NO:148: Нуклеотидная последовательность #110H13hIgG4P
SEQ ID NO:149: Аминокислотная последовательность #110L2h
SEQ ID NO:150: Нуклеотидная последовательность #110L2h
SEQ ID NO:151: Аминокислотная последовательность #110L12h
SEQ ID NO:152: Нуклеотидная последовательность #110L12h
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ДАЙИТИ САНКИО КОМПАНИ, ЛИМИТЕД
<120> Антитело против CD147
<130> DSPCT-FP1821
<150> JP2017-145701
<151> 2017-07-27
<160> 152
<170> PatentIn version 3.5
<210> 1
<211> 385
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Ala Ala Ala Leu Phe Val Leu Leu Gly Phe Ala Leu Leu Gly Thr
1 5 10 15
His Gly Ala Ser Gly Ala Ala Gly Phe Val Gln Ala Pro Leu Ser Gln
20 25 30
Gln Arg Trp Val Gly Gly Ser Val Glu Leu His Cys Glu Ala Val Gly
35 40 45
Ser Pro Val Pro Glu Ile Gln Trp Trp Phe Glu Gly Gln Gly Pro Asn
50 55 60
Asp Thr Cys Ser Gln Leu Trp Asp Gly Ala Arg Leu Asp Arg Val His
65 70 75 80
Ile His Ala Thr Tyr His Gln His Ala Ala Ser Thr Ile Ser Ile Asp
85 90 95
Thr Leu Val Glu Glu Asp Thr Gly Thr Tyr Glu Cys Arg Ala Ser Asn
100 105 110
Asp Pro Asp Arg Asn His Leu Thr Arg Ala Pro Arg Val Lys Trp Val
115 120 125
Arg Ala Gln Ala Val Val Leu Val Leu Glu Pro Gly Thr Val Phe Thr
130 135 140
Thr Val Glu Asp Leu Gly Ser Lys Ile Leu Leu Thr Cys Ser Leu Asn
145 150 155 160
Asp Ser Ala Thr Glu Val Thr Gly His Arg Trp Leu Lys Gly Gly Val
165 170 175
Val Leu Lys Glu Asp Ala Leu Pro Gly Gln Lys Thr Glu Phe Lys Val
180 185 190
Asp Ser Asp Asp Gln Trp Gly Glu Tyr Ser Cys Val Phe Leu Pro Glu
195 200 205
Pro Met Gly Thr Ala Asn Ile Gln Leu His Gly Pro Pro Arg Val Lys
210 215 220
Ala Val Lys Ser Ser Glu His Ile Asn Glu Gly Glu Thr Ala Met Leu
225 230 235 240
Val Cys Lys Ser Glu Ser Val Pro Pro Val Thr Asp Trp Ala Trp Tyr
245 250 255
Lys Ile Thr Asp Ser Glu Asp Lys Ala Leu Met Asn Gly Ser Glu Ser
260 265 270
Arg Phe Phe Val Ser Ser Ser Gln Gly Arg Ser Glu Leu His Ile Glu
275 280 285
Asn Leu Asn Met Glu Ala Asp Pro Gly Gln Tyr Arg Cys Asn Gly Thr
290 295 300
Ser Ser Lys Gly Ser Asp Gln Ala Ile Ile Thr Leu Arg Val Arg Ser
305 310 315 320
His Leu Ala Ala Leu Trp Pro Phe Leu Gly Ile Val Ala Glu Val Leu
325 330 335
Val Leu Val Thr Ile Ile Phe Ile Tyr Glu Lys Arg Arg Lys Pro Glu
340 345 350
Asp Val Leu Asp Asp Asp Asp Ala Gly Ser Ala Pro Leu Lys Ser Ser
355 360 365
Gly Gln His Gln Asn Asp Lys Gly Lys Asn Val Arg Gln Arg Asn Ser
370 375 380
Ser
385
<210> 2
<211> 1158
<212> ДНК
<213> Homo sapiens
<400> 2
atggcggctg cgctgttcgt gctgctggga ttcgcgctgc tgggcaccca cggagcctcc 60
ggggctgccg gcttcgtcca ggcgccgctg tcccagcaga ggtgggtggg gggcagtgtg 120
gagctgcact gcgaggccgt gggcagcccg gtgcccgaga tccagtggtg gtttgaaggg 180
cagggtccca acgacacctg ctcccagctc tgggacggcg cccggctgga ccgcgtccac 240
atccacgcca cctaccacca gcacgcggcc agcaccatct ccatcgacac gctcgtggag 300
gaggacacgg gcacttacga gtgccgggcc agcaacgacc cggatcgcaa ccacctgacc 360
cgggcgccca gggtcaagtg ggtccgcgcc caggcagtcg tgctagtcct ggaacccggc 420
acagtcttca ctaccgtaga agaccttggc tccaagatac tcctcacctg ctccttgaat 480
gacagcgcca cagaggtcac agggcaccgc tggctgaagg ggggcgtggt gctgaaggag 540
gacgcgctgc ccggccagaa aacggagttc aaggtggact ccgacgacca gtggggagag 600
tactcctgcg tcttcctccc cgagcccatg ggcacggcca acatccagct ccacgggcct 660
cccagagtga aggctgtgaa gtcgtcagaa cacatcaacg agggggagac ggccatgctg 720
gtctgcaagt cagagtccgt gccacctgtc actgactggg cctggtacaa gatcactgac 780
tctgaggaca aggccctcat gaacggctcc gagagcaggt tcttcgtgag ttcctcgcag 840
ggccggtcag agctacacat tgagaacctg aacatggagg ccgaccccgg ccagtaccgg 900
tgcaacggca ccagctccaa gggctccgac caggccatca tcacgctccg cgtgcgcagc 960
cacctggccg ccctctggcc cttcctgggc atcgtggctg aggtgctggt gctggtcacc 1020
atcatcttca tctacgagaa gcgccggaag cccgaggacg tcctggatga tgacgacgcc 1080
ggctctgcac ccctgaagag cagcgggcag caccagaatg acaaaggcaa gaacgtccgc 1140
cagaggaact cttcctga 1158
<210> 3
<211> 269
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Met Ala Ala Ala Leu Phe Val Leu Leu Gly Phe Ala Leu Leu Gly Thr
1 5 10 15
His Gly Ala Ser Gly Ala Ala Gly Thr Val Phe Thr Thr Val Glu Asp
20 25 30
Leu Gly Ser Lys Ile Leu Leu Thr Cys Ser Leu Asn Asp Ser Ala Thr
35 40 45
Glu Val Thr Gly His Arg Trp Leu Lys Gly Gly Val Val Leu Lys Glu
50 55 60
Asp Ala Leu Pro Gly Gln Lys Thr Glu Phe Lys Val Asp Ser Asp Asp
65 70 75 80
Gln Trp Gly Glu Tyr Ser Cys Val Phe Leu Pro Glu Pro Met Gly Thr
85 90 95
Ala Asn Ile Gln Leu His Gly Pro Pro Arg Val Lys Ala Val Lys Ser
100 105 110
Ser Glu His Ile Asn Glu Gly Glu Thr Ala Met Leu Val Cys Lys Ser
115 120 125
Glu Ser Val Pro Pro Val Thr Asp Trp Ala Trp Tyr Lys Ile Thr Asp
130 135 140
Ser Glu Asp Lys Ala Leu Met Asn Gly Ser Glu Ser Arg Phe Phe Val
145 150 155 160
Ser Ser Ser Gln Gly Arg Ser Glu Leu His Ile Glu Asn Leu Asn Met
165 170 175
Glu Ala Asp Pro Gly Gln Tyr Arg Cys Asn Gly Thr Ser Ser Lys Gly
180 185 190
Ser Asp Gln Ala Ile Ile Thr Leu Arg Val Arg Ser His Leu Ala Ala
195 200 205
Leu Trp Pro Phe Leu Gly Ile Val Ala Glu Val Leu Val Leu Val Thr
210 215 220
Ile Ile Phe Ile Tyr Glu Lys Arg Arg Lys Pro Glu Asp Val Leu Asp
225 230 235 240
Asp Asp Asp Ala Gly Ser Ala Pro Leu Lys Ser Ser Gly Gln His Gln
245 250 255
Asn Asp Lys Gly Lys Asn Val Arg Gln Arg Asn Ser Ser
260 265
<210> 4
<211> 810
<212> ДНК
<213> Homo sapiens
<400> 4
atggcggctg cgctgttcgt gctgctggga ttcgcgctgc tgggcaccca cggagcctcc 60
ggggctgccg gcacagtctt cactaccgta gaagaccttg gctccaagat actcctcacc 120
tgctccttga atgacagcgc cacagaggtc acagggcacc gctggctgaa ggggggcgtg 180
gtgctgaagg aggacgcgct gcccggccag aaaacggagt tcaaggtgga ctccgacgac 240
cagtggggag agtactcctg cgtcttcctc cccgagccca tgggcacggc caacatccag 300
ctccacgggc ctcccagagt gaaggctgtg aagtcgtcag aacacatcaa cgagggggag 360
acggccatgc tggtctgcaa gtcagagtcc gtgccacctg tcactgactg ggcctggtac 420
aagatcactg actctgagga caaggccctc atgaacggct ccgagagcag gttcttcgtg 480
agttcctcgc agggccggtc agagctacac attgagaacc tgaacatgga ggccgacccc 540
ggccagtacc ggtgcaacgg caccagctcc aagggctccg accaggccat catcacgctc 600
cgcgtgcgca gccacctggc cgccctctgg cccttcctgg gcatcgtggc tgaggtgctg 660
gtgctggtca ccatcatctt catctacgag aagcgccgga agcccgagga cgtcctggat 720
gatgacgacg ccggctctgc acccctgaag agcagcgggc agcaccagaa tgacaaaggc 780
aagaacgtcc gccagaggaa ctcttcctga 810
<210> 5
<211> 552
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Met Asp Asn Met Ser Ile Thr Asn Thr Pro Thr Ser Asn Asp Ala Cys
1 5 10 15
Leu Ser Ile Val His Ser Leu Met Cys His Arg Gln Gly Gly Glu Ser
20 25 30
Glu Thr Phe Ala Lys Arg Ala Ile Glu Ser Leu Val Lys Lys Leu Lys
35 40 45
Glu Lys Lys Asp Glu Leu Asp Ser Leu Ile Thr Ala Ile Thr Thr Asn
50 55 60
Gly Ala His Pro Ser Lys Cys Val Thr Ile Gln Arg Thr Leu Asp Gly
65 70 75 80
Arg Leu Gln Val Ala Gly Arg Lys Gly Phe Pro His Val Ile Tyr Ala
85 90 95
Arg Leu Trp Arg Trp Pro Asp Leu His Lys Asn Glu Leu Lys His Val
100 105 110
Lys Tyr Cys Gln Tyr Ala Phe Asp Leu Lys Cys Asp Ser Val Cys Val
115 120 125
Asn Pro Tyr His Tyr Glu Arg Val Val Ser Pro Gly Ile Asp Leu Ser
130 135 140
Gly Leu Thr Leu Gln Ser Asn Ala Pro Ser Ser Met Met Val Lys Asp
145 150 155 160
Glu Tyr Val His Asp Phe Glu Gly Gln Pro Ser Leu Ser Thr Glu Gly
165 170 175
His Ser Ile Gln Thr Ile Gln His Pro Pro Ser Asn Arg Ala Ser Thr
180 185 190
Glu Thr Tyr Ser Thr Pro Ala Leu Leu Ala Pro Ser Glu Ser Asn Ala
195 200 205
Thr Ser Thr Ala Asn Phe Pro Asn Ile Pro Val Ala Ser Thr Ser Gln
210 215 220
Pro Ala Ser Ile Leu Gly Gly Ser His Ser Glu Gly Leu Leu Gln Ile
225 230 235 240
Ala Ser Gly Pro Gln Pro Gly Gln Gln Gln Asn Gly Phe Thr Gly Gln
245 250 255
Pro Ala Thr Tyr His His Asn Ser Thr Thr Thr Trp Thr Gly Ser Arg
260 265 270
Thr Ala Pro Tyr Thr Pro Asn Leu Pro His His Gln Asn Gly His Leu
275 280 285
Gln His His Pro Pro Met Pro Pro His Pro Gly His Tyr Trp Pro Val
290 295 300
His Asn Glu Leu Ala Phe Gln Pro Pro Ile Ser Asn His Pro Ala Pro
305 310 315 320
Glu Tyr Trp Cys Ser Ile Ala Tyr Phe Glu Met Asp Val Gln Val Gly
325 330 335
Glu Thr Phe Lys Val Pro Ser Ser Cys Pro Ile Val Thr Val Asp Gly
340 345 350
Tyr Val Asp Pro Ser Gly Gly Asp Arg Phe Cys Leu Gly Gln Leu Ser
355 360 365
Asn Val His Arg Thr Glu Ala Ile Glu Arg Ala Arg Leu His Ile Gly
370 375 380
Lys Gly Val Gln Leu Glu Cys Lys Gly Glu Gly Asp Val Trp Val Arg
385 390 395 400
Cys Leu Ser Asp His Ala Val Phe Val Gln Ser Tyr Tyr Leu Asp Arg
405 410 415
Glu Ala Gly Arg Ala Pro Gly Asp Ala Val His Lys Ile Tyr Pro Ser
420 425 430
Ala Tyr Ile Lys Val Phe Asp Leu Arg Gln Cys His Arg Gln Met Gln
435 440 445
Gln Gln Ala Ala Thr Ala Gln Ala Ala Ala Ala Ala Gln Ala Ala Ala
450 455 460
Val Ala Gly Asn Ile Pro Gly Pro Gly Ser Val Gly Gly Ile Ala Pro
465 470 475 480
Ala Ile Ser Leu Ser Ala Ala Ala Gly Ile Gly Val Asp Asp Leu Arg
485 490 495
Arg Leu Cys Ile Leu Arg Met Ser Phe Val Lys Gly Trp Gly Pro Asp
500 505 510
Tyr Pro Arg Gln Ser Ile Lys Glu Thr Pro Cys Trp Ile Glu Ile His
515 520 525
Leu His Arg Ala Leu Gln Leu Leu Asp Glu Val Leu His Thr Met Pro
530 535 540
Ile Ala Asp Pro Gln Pro Leu Asp
545 550
<210> 6
<211> 1659
<212> ДНК
<213> Homo sapiens
<400> 6
atggacaata tgtctattac gaatacacca acaagtaatg atgcctgtct gagcattgtg 60
catagtttga tgtgccatag acaaggtgga gagagtgaaa catttgcaaa aagagcaatt 120
gaaagtttgg taaagaagct gaaggagaaa aaagatgaat tggattcttt aataacagct 180
ataactacaa atggagctca tcctagtaaa tgtgttacca tacagagaac attggatggg 240
aggcttcagg tggctggtcg gaaaggattt cctcatgtga tctatgcccg tctctggagg 300
tggcctgatc ttcacaaaaa tgaactaaaa catgttaaat attgtcagta tgcgtttgac 360
ttaaaatgtg atagtgtctg tgtgaatcca tatcactacg aacgagttgt atcacctgga 420
attgatctct caggattaac actgcagagt aatgctccat caagtatgat ggtgaaggat 480
gaatatgtgc atgactttga gggacagcca tcgttgtcca ctgaaggaca ttcaattcaa 540
accatccagc atccaccaag taatcgtgca tcgacagaga catacagcac cccagctctg 600
ttagccccat ctgagtctaa tgctaccagc actgccaact ttcccaacat tcctgtggct 660
tccacaagtc agcctgccag tatactgggg ggcagccata gtgaaggact gttgcagata 720
gcatcagggc ctcagccagg acagcagcag aatggattta ctggtcagcc agctacttac 780
catcataaca gcactaccac ctggactgga agtaggactg caccatacac acctaatttg 840
cctcaccacc aaaacggcca tcttcagcac cacccgccta tgccgcccca tcccggacat 900
tactggcctg ttcacaatga gcttgcattc cagcctccca tttccaatca tcctgctcct 960
gagtattggt gttccattgc ttactttgaa atggatgttc aggtaggaga gacatttaag 1020
gttccttcaa gctgccctat tgttactgtt gatggatacg tggacccttc tggaggagat 1080
cgcttttgtt tgggtcaact ctccaatgtc cacaggacag aagccattga gagagcaagg 1140
ttgcacatag gcaaaggtgt gcagttggaa tgtaaaggtg aaggtgatgt ttgggtcagg 1200
tgccttagtg accacgcggt ctttgtacag agttactact tagacagaga agctgggcgt 1260
gcacctggag atgctgttca taagatctac ccaagtgcat atataaaggt ctttgatttg 1320
cgtcagtgtc atcgacagat gcagcagcag gcggctactg cacaagctgc agcagctgcc 1380
caggcagcag ccgtggcagg aaacatccct ggcccaggat cagtaggtgg aatagctcca 1440
gctatcagtc tgtcagctgc tgctggaatt ggtgttgatg accttcgtcg cttatgcata 1500
ctcaggatga gttttgtgaa aggctgggga ccggattacc caagacagag catcaaagaa 1560
acaccttgct ggattgaaat tcacttacac cgggccctcc agctcctaga cgaagtactt 1620
cataccatgc cgattgcaga cccacaacct ttagactga 1659
<210> 7
<211> 324
<212> ДНК
<213> Домовая мышь
<400> 7
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 60
ctcacttgtc gggcaagtct ggaaattagt ggtttcttaa gttggcttca gcagaaacca 120
gatggaacta ttaaacgcct gatctacgcc gcatccattt tagattctgg tgtcccaaaa 180
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 240
gaagattttg cagactatta ctgtctacaa tatgctagtt atccgtggac gttcggtgga 300
ggcaccaagc tggaaatcaa acgg 324
<210> 8
<211> 108
<212> БЕЛОК
<213> Домовая мышь
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Leu Glu Ile Ser Gly Phe
20 25 30
Leu Ser Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile Lys Arg Leu Ile
35 40 45
Tyr Ala Ala Ser Ile Leu Asp Ser Gly Val Pro Lys Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Ser
65 70 75 80
Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Ala Ser Tyr Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 9
<211> 366
<212> ДНК
<213> Домовая мышь
<400> 9
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc 60
tcctgcaagg cttctgggta taccttcaca aactatggaa tgaactgggt gaagcaggct 120
ccaggaaagg gtttaaagtg gatgggctgg ataaacacct acactggaga gccaacatat 180
gctgatgact tcaagggacg gtttgccttc tctttggaaa cctctgccag cactgcctat 240
ttgcagatca acaacctcaa aaatgaggac acggctacat atttctgtgc aagagggggg 300
tatggtaact acggggcggg ggctatggac tactggggtc aaggtacctc agtcaccgtc 360
tcctca 366
<210> 10
<211> 122
<212> БЕЛОК
<213> Домовая мышь
<400> 10
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Asp Asp Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Gly Gly Tyr Gly Asn Tyr Gly Ala Gly Ala Met Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
<210> 11
<211> 11
<212> БЕЛОК
<213> Домовая мышь
<400> 11
Arg Ala Ser Leu Glu Ile Ser Gly Phe Leu Ser
1 5 10
<210> 12
<211> 7
<212> БЕЛОК
<213> Домовая мышь
<400> 12
Ala Ala Ser Ile Leu Asp Ser
1 5
<210> 13
<211> 9
<212> БЕЛОК
<213> Домовая мышь
<400> 13
Leu Gln Tyr Ala Ser Tyr Pro Trp Thr
1 5
<210> 14
<211> 10
<212> БЕЛОК
<213> Домовая мышь
<400> 14
Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
1 5 10
<210> 15
<211> 10
<212> БЕЛОК
<213> Домовая мышь
<400> 15
Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr
1 5 10
<210> 16
<211> 13
<212> БЕЛОК
<213> Домовая мышь
<400> 16
Gly Gly Tyr Gly Asn Tyr Gly Ala Gly Ala Met Asp Tyr
1 5 10
<210> 17
<211> 324
<212> ДНК
<213> Rattus norvegicus
<400> 17
gacatccaga tgacacagtc tccagcttcc ctgtctgcat ctctgggaga aactgtctcc 60
atcgaatgtc ttgcaagtga gggcatttcc aatagtttag cgtggtatca gcagaagcca 120
gggaaatctc ctcagctcct gatctatggt gcaagtagct tgcaagacgg ggtcccatca 180
cggttcagtg gcagtggatc tggcacacag tattctctca agatcagcgg catgcaacct 240
gaagatgaag gggtttatta ctgtcaacag ggttacaagt atccattcac gttcggctca 300
gggacgaagt tggaaataaa acgg 324
<210> 18
<211> 108
<212> БЕЛОК
<213> Rattus norvegicus
<400> 18
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Glu Thr Val Ser Ile Glu Cys Leu Ala Ser Glu Gly Ile Ser Asn Ser
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro Gln Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Ser Gly Met Gln Pro
65 70 75 80
Glu Asp Glu Gly Val Tyr Tyr Cys Gln Gln Gly Tyr Lys Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 19
<211> 351
<212> ДНК
<213> Rattus norvegicus
<400> 19
gaggtgcagc ttcaggagtc aggacctggc cttgtgaaac cctcacagtc actctccctc 60
acctgttctg tcactggtta ctccatcact agtaattact ggggctggat ccggaagttc 120
ccaggaaata aaatggagtg gatgggatgc ataacctaca gtggtggcac tagctacaac 180
ccatctctca aaagtcgaat ctccattact agagacacat caaagaatca gttcttcctg 240
cagttgaact ctgtaactac tgaggacaca gccacatatt actgtgcaag ttcctatacc 300
agtggtgacg tcgattactg gggccaagga gtcatggtca cagtctcctc a 351
<210> 20
<211> 117
<212> БЕЛОК
<213> Rattus norvegicus
<400> 20
Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Ser Val Thr Gly Tyr Ser Ile Thr Ser Asn
20 25 30
Tyr Trp Gly Trp Ile Arg Lys Phe Pro Gly Asn Lys Met Glu Trp Met
35 40 45
Gly Cys Ile Thr Tyr Ser Gly Gly Thr Ser Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe Leu
65 70 75 80
Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys Ala
85 90 95
Ser Ser Tyr Thr Ser Gly Asp Val Asp Tyr Trp Gly Gln Gly Val Met
100 105 110
Val Thr Val Ser Ser
115
<210> 21
<211> 11
<212> БЕЛОК
<213> Rattus norvegicus
<400> 21
Leu Ala Ser Glu Gly Ile Ser Asn Ser Leu Ala
1 5 10
<210> 22
<211> 7
<212> БЕЛОК
<213> Rattus norvegicus
<400> 22
Gly Ala Ser Ser Leu Gln Asp
1 5
<210> 23
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 23
Gln Gln Gly Tyr Lys Tyr Pro Phe Thr
1 5
<210> 24
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 24
Gly Tyr Ser Ile Thr Ser Asn Tyr Trp Gly
1 5 10
<210> 25
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 25
Cys Ile Thr Tyr Ser Gly Gly Thr Ser
1 5
<210> 26
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 26
Ser Tyr Thr Ser Gly Asp Val Asp Tyr
1 5
<210> 27
<211> 449
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую сигнальную
последовательность легкой цепи человека и константную область каппа цепи
человека
<400> 27
gcctccggac tctagagcca ccatggtgct gcagacccag gtgttcatct ccctgctgct 60
gtggatctcc ggcgcgtacg gcgatatcgt gatgattaaa cgtacggtgg ccgccccctc 120
cgtgttcatc ttccccccct ccgacgagca gctgaagtcc ggcaccgcct ccgtggtgtg 180
cctgctgaat aacttctacc ccagagaggc caaggtgcag tggaaggtgg acaacgccct 240
gcagtccggg aactcccagg agagcgtgac cgagcaggac agcaaggaca gcacctacag 300
cctgagcagc accctgaccc tgagcaaagc cgactacgag aagcacaagg tgtacgcctg 360
cgaggtgacc caccagggcc tgagctcccc cgtcaccaag agcttcaaca ggggggagtg 420
ttaggggccc gtttaaacgg gggaggcta 449
<210> 28
<211> 1132
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислоту
сигнальной последовательности тяжелой цепи человека и константной
области человеческого IgG1
<400> 28
gcctccggac tctagagcca ccatgaaaca cctgtggttc ttcctcctgc tggtggcagc 60
tcccagatgg gtgctgagcc aggtgcaatt gtgcaggcgg ttagctcagc ctccaccaag 120
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 180
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 240
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 300
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 360
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 420
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 480
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 540
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 600
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 660
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 720
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 780
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 840
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 900
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 960
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1020
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1080
tccctgtctc ccggcaaatg agatatcggg cccgtttaaa cgggggaggc ta 1132
<210> 29
<211> 1114
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислоту
сигнальной последовательности тяжелой цепи человека и константной
области человеческого IgG2
<400> 29
ccagcctccg gactctagag ccaccatgaa acacctgtgg ttcttcctcc tgctggtggc 60
agctcccaga tgggtgctga gccaggtgca attgtgcagg cggttagctc agcctccacc 120
aagggccctt ccgtgttccc tctggcccct tgtagccgtt ccaccagcga gtccaccgcc 180
gcccttggct gtctggtgaa ggactacttc cctgagcctg tgaccgtgag ctggaactcc 240
ggagccctta ccagcggcgt gcacaccttc cctgccgtgc tgcagtccag cggcctttac 300
tccctgagct ccgtggtgac cgtgcctagc tccaacttcg gcacccaaac ctacacctgt 360
aacgtggacc acaagcctag caacaccaag gtggacaaga ccgtggagcg taagtgttgt 420
gtggagtgtc ctccttgtcc tgcccctcct gtggccggac cttccgtgtt ccttttccct 480
cctaagccta aggacaccct gatgatcagc cgtacccctg aggtgacctg tgtggtggtg 540
gacgtgtccc acgaggaccc tgaggtgcag ttcaactggt acgtggacgg cgtggaggtg 600
cacaacgcca agaccaagcc tcgtgaggag caattcaaca gcaccttccg tgtggtgtcc 660
gtgcttaccg tggtgcacca agactggctg aacggcaagg agtacaagtg taaggtgagc 720
aacaagggac ttcctgcccc tatcgagaag accatctcca agaccaaggg ccaacctcgt 780
gagcctcaag tgtacaccct tcctcctagc cgtgaggaga tgaccaagaa ccaagtgtcc 840
cttacctgtc tggtgaaggg cttctaccct agcgacatcg ccgtggagtg ggagtccaac 900
ggacaacctg agaacaacta caagaccacc cctcctatgc ttgacagcga cggctccttc 960
ttcctgtaca gcaagctgac cgtggacaag tcccgttggc aacaaggcaa cgtgttcagc 1020
tgttccgtga tgcacgaggc cctgcacaac cactacaccc aaaagagcct ttccctgagc 1080
cctggaaagt gagtttaaac gggggaggct aact 1114
<210> 30
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь человеческого химерного LN22R8
<400> 30
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccaga tgacccagtc tccatcctcc ttatctgcct ctctgggaga aagagtcagt 120
ctcacttgtc gggcaagtct ggaaattagt ggtttcttaa gttggcttca gcagaaacca 180
gatggaacta ttaaacgcct gatctacgcc gcatccattt tagattctgg tgtcccaaaa 240
aggttcagtg gcagtaggtc tgggtcagat tattctctca ccatcagcag ccttgagtct 300
gaagattttg cagactatta ctgtctacaa tatgctagtt atccgtggac gttcggtgga 360
ggcaccaagc tggaaatcaa acgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 31
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь человеческого химерного LN22R8
<400> 31
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Leu Gly Glu Arg Val Ser Leu Thr Cys Arg Ala Ser Leu Glu
35 40 45
Ile Ser Gly Phe Leu Ser Trp Leu Gln Gln Lys Pro Asp Gly Thr Ile
50 55 60
Lys Arg Leu Ile Tyr Ala Ala Ser Ile Leu Asp Ser Gly Val Pro Lys
65 70 75 80
Arg Phe Ser Gly Ser Arg Ser Gly Ser Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Ser Leu Glu Ser Glu Asp Phe Ala Asp Tyr Tyr Cys Leu Gln Tyr Ala
100 105 110
Ser Tyr Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 32
<211> 1413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелаяй цепь человеческого химерного LN22R8 типа IgG1
<400> 32
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
atccagttgg tgcagtctgg acctgagctg aagaagcctg gagagacagt caagatctcc 120
tgcaaggctt ctgggtatac cttcacaaac tatggaatga actgggtgaa gcaggctcca 180
ggaaagggtt taaagtggat gggctggata aacacctaca ctggagagcc aacatatgct 240
gatgacttca agggacggtt tgccttctct ttggaaacct ctgccagcac tgcctatttg 300
cagatcaaca acctcaaaaa tgaggacacg gctacatatt tctgtgcaag aggggggtat 360
ggtaactacg gggcgggggc tatggactac tggggtcaag gtacctcagt caccgtcagc 420
tcagcctcca ccaagggccc aagcgtcttc cccctggcac cctcctccaa gagcacctct 480
ggcggcacag ccgccctggg ctgcctggtc aaggactact tccccgaacc cgtgaccgtg 540
agctggaact caggcgccct gaccagcggc gtgcacacct tccccgctgt cctgcagtcc 600
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 660
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gagagttgag 720
cccaaatctt gtgacaaaac tcacacatgc ccaccctgcc cagcacctga actcctgggg 780
ggaccctcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 840
cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac 900
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccccggga ggagcagtac 960
aacagcacgt accgggtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 1020
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 1080
tccaaagcca aaggccagcc ccgggaacca caggtgtaca ccctgccccc atcccgggag 1140
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 1200
atcgccgtgg agtgggagag caatggccag cccgagaaca actacaagac cacccctccc 1260
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1320
tggcagcagg gcaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1380
acccagaaga gcctctccct gtctcccggc aaa 1413
<210> 33
<211> 471
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелаяй цепь человеческого химерного LN22R8 типа IgG1
<400> 33
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Gly Gly Tyr Gly Asn Tyr Gly Ala Gly Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr
130 135 140
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
145 150 155 160
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
165 170 175
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
180 185 190
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
195 200 205
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
210 215 220
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
225 230 235 240
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
245 250 255
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
260 265 270
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
275 280 285
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
290 295 300
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
305 310 315 320
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
325 330 335
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
340 345 350
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
355 360 365
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys
370 375 380
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
385 390 395 400
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
405 410 415
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
420 425 430
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
435 440 445
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
450 455 460
Leu Ser Leu Ser Pro Gly Lys
465 470
<210> 34
<211> 1401
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного LN22R8 типа IgG2
<400> 34
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
atccagttgg tgcagtctgg acctgagctg aagaagcctg gagagacagt caagatctcc 120
tgcaaggctt ctgggtatac cttcacaaac tatggaatga actgggtgaa gcaggctcca 180
ggaaagggtt taaagtggat gggctggata aacacctaca ctggagagcc aacatatgct 240
gatgacttca agggacggtt tgccttctct ttggaaacct ctgccagcac tgcctatttg 300
cagatcaaca acctcaaaaa tgaggacacg gctacatatt tctgtgcaag aggggggtat 360
ggtaactacg gggcgggggc tatggactac tggggtcaag gtacctcagt caccgtcagc 420
tcagcctcca ccaagggccc ttccgtgttc cctctggccc cttgtagccg ttccaccagc 480
gagtccaccg ccgcccttgg ctgtctggtg aaggactact tccctgagcc tgtgaccgtg 540
agctggaact ccggagccct taccagcggc gtgcacacct tccctgccgt gctgcagtcc 600
agcggccttt actccctgag ctccgtggtg accgtgccta gctccaactt cggcacccaa 660
acctacacct gtaacgtgga ccacaagcct agcaacacca aggtggacaa gaccgtggag 720
cgtaagtgtt gtgtggagtg tcctccttgt cctgcccctc ctgtggccgg accttccgtg 780
ttccttttcc ctcctaagcc taaggacacc ctgatgatca gccgtacccc tgaggtgacc 840
tgtgtggtgg tggacgtgtc ccacgaggac cctgaggtgc agttcaactg gtacgtggac 900
ggcgtggagg tgcacaacgc caagaccaag cctcgtgagg agcaattcaa cagcaccttc 960
cgtgtggtgt ccgtgcttac cgtggtgcac caagactggc tgaacggcaa ggagtacaag 1020
tgtaaggtga gcaacaaggg acttcctgcc cctatcgaga agaccatctc caagaccaag 1080
ggccaacctc gtgagcctca agtgtacacc cttcctccta gccgtgagga gatgaccaag 1140
aaccaagtgt cccttacctg tctggtgaag ggcttctacc ctagcgacat cgccgtggag 1200
tgggagtcca acggacaacc tgagaacaac tacaagacca cccctcctat gcttgacagc 1260
gacggctcct tcttcctgta cagcaagctg accgtggaca agtcccgttg gcaacaaggc 1320
aacgtgttca gctgttccgt gatgcacgag gccctgcaca accactacac ccaaaagagc 1380
ctttccctga gccctggaaa g 1401
<210> 35
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного LN22R8 типа IgG2
<400> 35
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Gly Gly Tyr Gly Asn Tyr Gly Ala Gly Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr
130 135 140
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser
145 150 155 160
Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
165 170 175
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
180 185 190
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
195 200 205
Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys
210 215 220
Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu
225 230 235 240
Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Lys
465
<210> 36
<211> 1454
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислотную
последовательность тяжелой цепи человеческого химерного LN22R8 типа IgG4P
<400> 36
ccagcctccg gactctagag ccaccatgaa gcacctgtgg ttctttctgc tgctggtggc 60
cgctcccaga tgggtgctgt ctcagatcca gctggtgcag agcggccctg agctgaagaa 120
acccggcgag acagtgaaga tcagctgcaa ggccagcggc tacaccttca ccaactacgg 180
catgaactgg gtcaagcagg cccctggcaa gggcctgaag tggatgggct ggatcaacac 240
ctacaccggc gagcccacct acgccgacga cttcaagggc agattcgcct tcagcctgga 300
aaccagcgcc agcaccgcct acctgcagat caacaacctg aagaacgagg acaccgccac 360
ctacttttgc gccagaggcg gctacggcaa ttacggcgct ggcgccatgg attactgggg 420
ccagggaaca agcgtgaccg tgtccagcgc ctctaccaag ggccctagcg tgttccctct 480
ggccccttgc agcagaagca ccagcgaatc tacagccgcc ctgggctgcc tcgtgaagga 540
ctactttccc gagcccgtga cagtgtcctg gaactctggc gccctgacaa gcggcgtgca 600
cacctttcca gccgtgctgc agagcagcgg cctgtactct ctgagcagcg tcgtgactgt 660
gcccagcagc tctctgggca ccaagaccta cacctgtaac gtggaccaca agcccagcaa 720
caccaaggtg gacaagcggg tggaatctaa gtacggccct ccctgccctc cttgcccagc 780
ccctgaattt ctgggcggac cctccgtgtt cctgttcccc ccaaagccca aggacaccct 840
gatgatcagc cggacccccg aagtgacctg cgtggtggtg gatgtgtccc aggaagatcc 900
cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg cacaacgcca agaccaagcc 960
tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc gtgctgacag tgctgcacca 1020
ggactggctg aacggcaaag agtacaagtg caaggtgtcc aacaagggac tgcccagctc 1080
catcgagaaa accatcagca aggccaaggg ccagccccgc gaaccccagg tgtacacact 1140
gcctccaagc caggaagaga tgaccaagaa ccaggtgtcc ctgacctgtc tcgtgaaagg 1200
cttctacccc tccgatatcg ccgtggaatg ggagagcaac ggccagcccg agaacaacta 1260
caagaccacc ccccctgtgc tggacagcga cggctcattc ttcctgtaca gcagactgac 1320
cgtggacaag agccggtggc aggaaggcaa cgtgttcagc tgcagcgtga tgcacgaggc 1380
cctgcacaac cactacaccc agaagtccct gtctctgagc ctgggcaaat gagtttaaac 1440
gggggaggct aact 1454
<210> 37
<211> 468
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного LN22R8 типа IgG4P
<400> 37
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Asp Asp Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Gly Gly Tyr Gly Asn Tyr Gly Ala Gly Ala Met
115 120 125
Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr
130 135 140
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser
145 150 155 160
Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
165 170 175
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
180 185 190
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
195 200 205
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys
210 215 220
Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu
225 230 235 240
Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Leu Gly Lys
465
<210> 38
<211> 1137
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислоту
сигнальной последовательности тяжелой цепи человека и константой области
IgG1LALA
<400> 38
ccagcctccg gactctagag ccaccatgaa acacctgtgg ttcttcctcc tgctggtggc 60
agctcccaga tgggtgctga gccaggtgca attgtgcagg cggttagctc agcctccacc 120
aagggcccaa gcgtcttccc cctggcaccc tcctccaaga gcacctctgg cggcacagcc 180
gccctgggct gcctggtcaa ggactacttc cccgaacccg tgaccgtgag ctggaactca 240
ggcgccctga ccagcggcgt gcacaccttc cccgctgtcc tgcagtcctc aggactctac 300
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc 360
aacgtgaatc acaagcccag caacaccaag gtggacaaga gagttgagcc caaatcttgt 420
gacaaaactc acacatgccc accctgccca gcacctgaag ccgcgggggg accctcagtc 480
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 540
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac 600
ggcgtggagg tgcataatgc caagacaaag ccccgggagg agcagtacaa cagcacgtac 660
cgggtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag 720
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 780
ggccagcccc gggaaccaca ggtgtacacc ctgcccccat cccgggagga gatgaccaag 840
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 900
tgggagagca atggccagcc cgagaacaac tacaagacca cccctcccgt gctggactcc 960
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggc 1020
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac ccagaagagc 1080
ctctccctgt ctcccggcaa atgagatatc gggcccgttt aaacggggga ggctaac 1137
<210> 39
<211> 1117
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислоту
сигнальной последовательности тяжелой цепи человека и константной области
IgG4P
<400> 39
ccagcctccg gactctagag ccaccatgaa acacctgtgg ttcttcctcc tgctggtggc 60
agctcccaga tgggtgctga gccaggtgca attgtgcagg cggttagctc agcctccacc 120
aagggcccta gcgtgttccc tctggcccct tgtagcagaa gcaccagcga gtctacagcc 180
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 240
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tgcagagcag cggcctgtac 300
tctctgtcca gcgtcgtgac tgtgcccagc agctctctgg gcaccaagac ctacacctgt 360
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 420
cctccctgcc ctccttgccc agcccctgaa tttctgggcg gaccctccgt gttcctgttc 480
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggtg 540
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 600
gtgcacaacg ccaagaccaa gcctagagag gaacagttca acagcaccta ccgggtggtg 660
tccgtgctga cagtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 720
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 780
cgcgaacccc aggtgtacac actgcctcca agccaggaag agatgaccaa gaatcaggtg 840
tccctgacct gtctcgtgaa aggcttctac ccctccgata tcgccgtgga atgggagagc 900
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggactc cgatggctca 960
ttcttcctgt acagcagact gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1020
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1080
agcctgggca aatgagttta aacgggggag gctaact 1117
<210> 40
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь человеческого химерного 2P10F2
<400> 40
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccaga tgacacagtc tccagcttcc ctgtctgcat ctctgggaga aactgtctcc 120
atcgaatgtc ttgcaagtga gggcatttcc aatagtttag cgtggtatca gcagaagcca 180
gggaaatctc ctcagctcct gatctatggt gcaagtagct tgcaagacgg ggtcccatca 240
cggttcagtg gcagtggatc tggcacacag tattctctca agatcagcgg catgcaacct 300
gaagatgaag gggtttatta ctgtcaacag ggttacaagt atccattcac gttcggctca 360
gggacgaagt tggaaataaa acgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 41
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь человеческого химерного 2P10F2
<400> 41
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser
20 25 30
Ala Ser Leu Gly Glu Thr Val Ser Ile Glu Cys Leu Ala Ser Glu Gly
35 40 45
Ile Ser Asn Ser Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ser Pro
50 55 60
Gln Leu Leu Ile Tyr Gly Ala Ser Ser Leu Gln Asp Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Ser
85 90 95
Gly Met Gln Pro Glu Asp Glu Gly Val Tyr Tyr Cys Gln Gln Gly Tyr
100 105 110
Lys Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 42
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG1LALA
<400> 42
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagcttc aggagtcagg acctggcctt gtgaaaccct cacagtcact ctccctcacc 120
tgttctgtca ctggttactc catcactagt aattactggg gctggatccg gaagttccca 180
ggaaataaaa tggagtggat gggatgcata acctacagtg gtggcactag ctacaaccca 240
tctctcaaaa gtcgaatctc cattactaga gacacatcaa agaatcagtt cttcctgcag 300
ttgaactctg taactactga ggacacagcc acatattact gtgcaagttc ctataccagt 360
ggtgacgtcg attactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaagccg cggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaa 1398
<210> 43
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG1LALA
<400> 43
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
20 25 30
Pro Ser Gln Ser Leu Ser Leu Thr Cys Ser Val Thr Gly Tyr Ser Ile
35 40 45
Thr Ser Asn Tyr Trp Gly Trp Ile Arg Lys Phe Pro Gly Asn Lys Met
50 55 60
Glu Trp Met Gly Cys Ile Thr Tyr Ser Gly Gly Thr Ser Tyr Asn Pro
65 70 75 80
Ser Leu Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln
85 90 95
Phe Phe Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr
100 105 110
Tyr Cys Ala Ser Ser Tyr Thr Ser Gly Asp Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 44
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG2
<400> 44
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagcttc aggagtcagg acctggcctt gtgaaaccct cacagtcact ctccctcacc 120
tgttctgtca ctggttactc catcactagt aattactggg gctggatccg gaagttccca 180
ggaaataaaa tggagtggat gggatgcata acctacagtg gtggcactag ctacaaccca 240
tctctcaaaa gtcgaatctc cattactaga gacacatcaa agaatcagtt cttcctgcag 300
ttgaactctg taactactga ggacacagcc acatattact gtgcaagttc ctataccagt 360
ggtgacgtcg attactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggcccttccg tgttccctct ggccccttgt agccgttcca ccagcgagtc caccgccgcc 480
cttggctgtc tggtgaagga ctacttccct gagcctgtga ccgtgagctg gaactccgga 540
gcccttacca gcggcgtgca caccttccct gccgtgctgc agtccagcgg cctttactcc 600
ctgagctccg tggtgaccgt gcctagctcc aacttcggca cccaaaccta cacctgtaac 660
gtggaccaca agcctagcaa caccaaggtg gacaagaccg tggagcgtaa gtgttgtgtg 720
gagtgtcctc cttgtcctgc ccctcctgtg gccggacctt ccgtgttcct tttccctcct 780
aagcctaagg acaccctgat gatcagccgt acccctgagg tgacctgtgt ggtggtggac 840
gtgtcccacg aggaccctga ggtgcagttc aactggtacg tggacggcgt ggaggtgcac 900
aacgccaaga ccaagcctcg tgaggagcaa ttcaacagca ccttccgtgt ggtgtccgtg 960
cttaccgtgg tgcaccaaga ctggctgaac ggcaaggagt acaagtgtaa ggtgagcaac 1020
aagggacttc ctgcccctat cgagaagacc atctccaaga ccaagggcca acctcgtgag 1080
cctcaagtgt acacccttcc tcctagccgt gaggagatga ccaagaacca agtgtccctt 1140
acctgtctgg tgaagggctt ctaccctagc gacatcgccg tggagtggga gtccaacgga 1200
caacctgaga acaactacaa gaccacccct cctatgcttg acagcgacgg ctccttcttc 1260
ctgtacagca agctgaccgt ggacaagtcc cgttggcaac aaggcaacgt gttcagctgt 1320
tccgtgatgc acgaggccct gcacaaccac tacacccaaa agagcctttc cctgagccct 1380
ggaaag 1386
<210> 45
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG2
<400> 45
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
20 25 30
Pro Ser Gln Ser Leu Ser Leu Thr Cys Ser Val Thr Gly Tyr Ser Ile
35 40 45
Thr Ser Asn Tyr Trp Gly Trp Ile Arg Lys Phe Pro Gly Asn Lys Met
50 55 60
Glu Trp Met Gly Cys Ile Thr Tyr Ser Gly Gly Thr Ser Tyr Asn Pro
65 70 75 80
Ser Leu Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln
85 90 95
Phe Phe Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr
100 105 110
Tyr Cys Ala Ser Ser Tyr Thr Ser Gly Asp Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val
225 230 235 240
Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 46
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG4P
<400> 46
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagcttc aggagtcagg acctggcctt gtgaaaccct cacagtcact ctccctcacc 120
tgttctgtca ctggttactc catcactagt aattactggg gctggatccg gaagttccca 180
ggaaataaaa tggagtggat gggatgcata acctacagtg gtggcactag ctacaaccca 240
tctctcaaaa gtcgaatctc cattactaga gacacatcaa agaatcagtt cttcctgcag 300
ttgaactctg taactactga ggacacagcc acatattact gtgcaagttc ctataccagt 360
ggtgacgtcg attactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggccctagcg tgttccctct ggccccttgt agcagaagca ccagcgagtc tacagccgcc 480
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 540
gctctgacaa gcggcgtgca cacctttcca gccgtgctgc agagcagcgg cctgtactct 600
ctgtccagcg tcgtgactgt gcccagcagc tctctgggca ccaagaccta cacctgtaac 660
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 720
ccctgccctc cttgcccagc ccctgaattt ctgggcggac cctccgtgtt cctgttcccc 780
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 840
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 900
cacaacgcca agaccaagcc tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 960
gtgctgacag tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 1020
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1080
gaaccccagg tgtacacact gcctccaagc caggaagaga tgaccaagaa tcaggtgtcc 1140
ctgacctgtc tcgtgaaagg cttctacccc tccgatatcg ccgtggaatg ggagagcaac 1200
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga tggctcattc 1260
ttcctgtaca gcagactgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1320
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgagc 1380
ctgggcaaa 1389
<210> 47
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь человеческого химерного 2P10F2 типа IgG4P
<400> 47
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys
20 25 30
Pro Ser Gln Ser Leu Ser Leu Thr Cys Ser Val Thr Gly Tyr Ser Ile
35 40 45
Thr Ser Asn Tyr Trp Gly Trp Ile Arg Lys Phe Pro Gly Asn Lys Met
50 55 60
Glu Trp Met Gly Cys Ile Thr Tyr Ser Gly Gly Thr Ser Tyr Asn Pro
65 70 75 80
Ser Leu Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln
85 90 95
Phe Phe Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr
100 105 110
Tyr Cys Ala Ser Ser Tyr Thr Ser Gly Asp Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 48
<211> 324
<212> ДНК
<213> Rattus norvegicus
<400> 48
gatgtccaga tgacccagtc tccatcttat cttgctgcgt ctcctggaga aagtgtttcc 60
atcagttgca aggcaagtaa gagcattagc aataatttag cctggtatca ggagaaacct 120
gggaaagcaa ataagcttct tattcactct gggtcaactt tgcaatctgg aactccatcg 180
aggttcagtg gcagtggatc tggtacagat ttcacgctca ccatcagaag cctggagttt 240
gaagattttg cagtctatta ctgtcaacag tataatgaat acccactcac gttcggttct 300
gggaccaagc tggagatcaa acgg 324
<210> 49
<211> 108
<212> БЕЛОК
<213> Rattus norvegicus
<400> 49
Asp Val Gln Met Thr Gln Ser Pro Ser Tyr Leu Ala Ala Ser Pro Gly
1 5 10 15
Glu Ser Val Ser Ile Ser Cys Lys Ala Ser Lys Ser Ile Ser Asn Asn
20 25 30
Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Ala Asn Lys Leu Leu Ile
35 40 45
His Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Ser Leu Glu Phe
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn Glu Tyr Pro Leu
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 50
<211> 363
<212> ДНК
<213> Rattus norvegicus
<400> 50
gaggtgcagc tggtggagtc tgggggaggc ctagtgcagc ctggaaggtc cctgaaactc 60
tcctgtgcag cctcaggatt cactttcagt aactattaca tggcctgggt ccgccaggct 120
ccaaagaagg gtctggagtg ggtcgcaacc attactacca gtggtagcag accttactat 180
ccagactccg tgaaaggccg attcactatc tccagagata atgcaaaaag cagcctatac 240
ctgcaaatga acagtctgaa gtctgaggac acggccactt attactgtgc aagacatatt 300
tattactatg atggttaccc ctttgcttac tggggccaag gcactctggt cactgtctct 360
tca 363
<210> 51
<211> 121
<212> БЕЛОК
<213> Rattus norvegicus
<400> 51
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 52
<211> 11
<212> БЕЛОК
<213> Rattus norvegicus
<400> 52
Lys Ala Ser Lys Ser Ile Ser Asn Asn Leu Ala
1 5 10
<210> 53
<211> 7
<212> БЕЛОК
<213> Rattus norvegicus
<400> 53
Ser Gly Ser Thr Leu Gln Ser
1 5
<210> 54
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 54
Gln Gln Tyr Asn Glu Tyr Pro Leu Thr
1 5
<210> 55
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 55
Gly Phe Thr Phe Ser Asn Tyr Tyr Met Ala
1 5 10
<210> 56
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 56
Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr
1 5 10
<210> 57
<211> 12
<212> БЕЛОК
<213> Rattus norvegicus
<400> 57
His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala Tyr
1 5 10
<210> 58
<211> 324
<212> ДНК
<213> Rattus norvegicus
<400> 58
gacatccaga tgacccagtc tccttcactc ctgtctgcat ctgtgggagg cagaatcact 60
ctcaactgca aagcaagtca gaatctctat aagaacttag cctggtatca gcagaagctt 120
ggagaagctc ccaaactcct gattgataat gcaaacagtt tgcaaacggg catcccatca 180
aggttcagtg gcagtggatc tggtacagat ttcacactca ccatcagcag cctgcagcct 240
gaagatgttg ccacatattt ctgccagcag tattatagcg ggtcgtacac gtttggagct 300
gggaccaagc tggaactgaa acgg 324
<210> 59
<211> 108
<212> БЕЛОК
<213> Rattus norvegicus
<400> 59
Asp Ile Gln Met Thr Gln Ser Pro Ser Leu Leu Ser Ala Ser Val Gly
1 5 10 15
Gly Arg Ile Thr Leu Asn Cys Lys Ala Ser Gln Asn Leu Tyr Lys Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Leu Gly Glu Ala Pro Lys Leu Leu Ile
35 40 45
Asp Asn Ala Asn Ser Leu Gln Thr Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Phe Cys Gln Gln Tyr Tyr Ser Gly Ser Tyr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
100 105
<210> 60
<211> 354
<212> ДНК
<213> Rattus norvegicus
<400> 60
gaggtgcagc tggtggagtc tgatggaggc ttagtgcagc ctggaaggtc cctaaaactc 60
tcctgtgcag cctcaggatt cactttcagt gactattaca tggcctgggt ccgccaggct 120
ccaacgaagg ggctggagtg ggtcgcaacc attagttatg atggtagtaa cacttactat 180
cgagactccg tgaagggccg attcactatc tccagagata atgcaaaaag caccctatac 240
ctgcaaatgg acagtctgag gtctggggac acggccactt attactgtgc aagattttac 300
aacaactact actttgatta ctggggccaa ggagtcatgg tcacagtctc ctca 354
<210> 61
<211> 118
<212> БЕЛОК
<213> Rattus norvegicus
<400> 61
Glu Val Gln Leu Val Glu Ser Asp Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Gly Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly Gln Gly Val
100 105 110
Met Val Thr Val Ser Ser
115
<210> 62
<211> 11
<212> БЕЛОК
<213> Rattus norvegicus
<400> 62
Lys Ala Ser Gln Asn Leu Tyr Lys Asn Leu Ala
1 5 10
<210> 63
<211> 7
<212> БЕЛОК
<213> Rattus norvegicus
<400> 63
Asn Ala Asn Ser Leu Gln Thr
1 5
<210> 64
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 64
Gln Gln Tyr Tyr Ser Gly Ser Tyr Thr
1 5
<210> 65
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 65
Gly Phe Thr Phe Ser Asp Tyr Tyr Met Ala
1 5 10
<210> 66
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 66
Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr
1 5 10
<210> 67
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 67
Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr
1 5
<210> 68
<211> 324
<212> ДНК
<213> Rattus norvegicus
<400> 68
gacatccaga tgacccagac tccatcctcc atgcctgcat ctctgggaga gagagtcacc 60
atcagttgca gagcaagtca gggtattagc aattatctaa actggtatca gcagaaacca 120
gatggaacga ttaaacccct gatctactac acatccaatt tacaatctgg tgtcccatca 180
aggttcagtg gcagtgggtc tgggacagat tattctctca ccatcagcag cctggagcct 240
gaagattttg caatgtattt ctgccaacag tatgatagtt ctcctcggac gttcggtgga 300
ggcaccaagc tggaattgaa acgg 324
<210> 69
<211> 108
<212> БЕЛОК
<213> Rattus norvegicus
<400> 69
Asp Ile Gln Met Thr Gln Thr Pro Ser Ser Met Pro Ala Ser Leu Gly
1 5 10 15
Glu Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Ile Lys Pro Leu Ile
35 40 45
Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Tyr Asp Ser Ser Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys Arg
100 105
<210> 70
<211> 351
<212> ДНК
<213> Rattus norvegicus
<400> 70
caggtacagc tgcagcaatc tggggctgaa ctggtgaagc ctgggtcctc agtgaaaatt 60
tcctgcaagg cttctggcta caccttcacc agtgacttta tgcactggat aaaacagcag 120
cctggaaatg gccttgagtg gattgggtgg atttatcctg gagatggtga tacagagtac 180
aatcaaaagt tcaatgggaa ggcaacactc actgcagaca aatcctccag cacagcctat 240
atgcagctca gcagcctgac atctgaggac tctgcagtct atttctgtgc aaggggacgg 300
gggtatgtta tggatgcctg gggtcaagga gcttcagtca ctgtctcctc a 351
<210> 71
<211> 117
<212> БЕЛОК
<213> Rattus norvegicus
<400> 71
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Asp
20 25 30
Phe Met His Trp Ile Lys Gln Gln Pro Gly Asn Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Asn Gln Lys Phe
50 55 60
Asn Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln Gly Ala Ser
100 105 110
Val Thr Val Ser Ser
115
<210> 72
<211> 11
<212> БЕЛОК
<213> Rattus norvegicus
<400> 72
Arg Ala Ser Gln Gly Ile Ser Asn Tyr Leu Asn
1 5 10
<210> 73
<211> 7
<212> БЕЛОК
<213> Rattus norvegicus
<400> 73
Tyr Thr Ser Asn Leu Gln Ser
1 5
<210> 74
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 74
Gln Gln Tyr Asp Ser Ser Pro Arg Thr
1 5
<210> 75
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 75
Gly Tyr Thr Phe Thr Ser Asp Phe Met His
1 5 10
<210> 76
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 76
Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu
1 5 10
<210> 77
<211> 8
<212> БЕЛОК
<213> Rattus norvegicus
<400> 77
Gly Arg Gly Tyr Val Met Asp Ala
1 5
<210> 78
<211> 324
<212> ДНК
<213> Rattus norvegicus
<400> 78
gatgtccaga tgacccagtc tccatcttat cttgctgcgt ctcctggaga aagtgtttcc 60
atcagttgca aggcaagtaa aagcattaac acatacttag cctggtatca ggagaaacct 120
gggaaaacga ataagcttct tatctactct gggtcaactt tgcaatctgg aactccatcg 180
agattcagtg gcagtggatc tggtacagat ttcacgctca ccatcagaag cctggagcct 240
gaagattttg cagtctacta ctgtcaacag cataatgaat accccttcac gttcggctca 300
gggacgaagt tggaaataaa acgg 324
<210> 79
<211> 108
<212> БЕЛОК
<213> Rattus norvegicus
<400> 79
Asp Val Gln Met Thr Gln Ser Pro Ser Tyr Leu Ala Ala Ser Pro Gly
1 5 10 15
Glu Ser Val Ser Ile Ser Cys Lys Ala Ser Lys Ser Ile Asn Thr Tyr
20 25 30
Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Thr Asn Lys Leu Leu Ile
35 40 45
Tyr Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Asn Glu Tyr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
100 105
<210> 80
<211> 357
<212> ДНК
<213> Rattus norvegicus
<400> 80
gaggtgcagc tggtggagtc tgggggaggc ctagtgcagc ctggaaggtc tctgaaacta 60
tcctgtggag cctctggatt cacattcaat aactactgga tgacctgggt ccgccaggct 120
ccagggaagg ggctggagtg ggttgcatcc attactaaag ctggtggtag cacttactat 180
cgagactctg tgaagggccg attcactatc tccagagata atgcaaaaag caccctatat 240
ctgcaaatga acagtctgag gtctgaggac acggccactt attactgtac aagagaactg 300
ggagagttct atgttatgga tgcctggggt caaggagctt cagtcactgt ctcctca 357
<210> 81
<211> 119
<212> БЕЛОК
<213> Rattus norvegicus
<400> 81
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Lys Leu Ser Cys Gly Ala Ser Gly Phe Thr Phe Asn Asn Tyr
20 25 30
Trp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ser Ile Thr Lys Ala Gly Gly Ser Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Arg Glu Leu Gly Glu Phe Tyr Val Met Asp Ala Trp Gly Gln Gly
100 105 110
Ala Ser Val Thr Val Ser Ser
115
<210> 82
<211> 11
<212> БЕЛОК
<213> Rattus norvegicus
<400> 82
Lys Ala Ser Lys Ser Ile Asn Thr Tyr Leu Ala
1 5 10
<210> 83
<211> 7
<212> БЕЛОК
<213> Rattus norvegicus
<400> 83
Ser Gly Ser Thr Leu Gln Ser
1 5
<210> 84
<211> 9
<212> БЕЛОК
<213> Rattus norvegicus
<400> 84
Gln Gln His Asn Glu Tyr Pro Phe Thr
1 5
<210> 85
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 85
Gly Phe Thr Phe Asn Asn Tyr Trp Met Thr
1 5 10
<210> 86
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 86
Ser Ile Thr Lys Ala Gly Gly Ser Thr Tyr
1 5 10
<210> 87
<211> 10
<212> БЕЛОК
<213> Rattus norvegicus
<400> 87
Glu Leu Gly Glu Phe Tyr Val Met Asp Ala
1 5 10
<210> 88
<211> 1117
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ДНК, включающая последовательность ДНК, кодирующую аминокислоту
сигнальной последовательности тяжелой цепи человека и константной
области человеческого IgG4PFALA
<400> 88
ccagcctccg gactctagag ccaccatgaa acacctgtgg ttcttcctcc tgctggtggc 60
agctcccaga tgggtgctga gccaggtgca attgtgcagg cggttagctc agcctccacc 120
aagggcccta gcgtgttccc tctggcccct tgtagcagaa gcaccagcga gtctacagcc 180
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 240
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tgcagagcag cggcctgtac 300
tctctgtcca gcgtcgtgac tgtgcccagc agctctctgg gcaccaagac ctacacctgt 360
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 420
cctccctgcc ctccttgccc agcccctgaa gccgcgggcg gaccctccgt gttcctgttc 480
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggtg 540
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 600
gtgcacaacg ccaagaccaa gcctagagag gaacagttca acagcaccta ccgggtggtg 660
tccgtgctga cagtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 720
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 780
cgcgaacccc aggtgtacac actgcctcca agccaggaag agatgaccaa gaatcaggtg 840
tccctgacct gtctcgtgaa aggcttctac ccctccgata tcgccgtgga atgggagagc 900
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggactc cgatggctca 960
ttcttcctgt acagcagact gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1020
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1080
agcctgggca aatgagttta aacgggggag gctaact 1117
<210> 89
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#84
<400> 89
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatgtccaga tgacccagtc tccatcttat cttgctgcgt ctcctggaga aagtgtttcc 120
atcagttgca aggcaagtaa gagcattagc aataatttag cctggtatca ggagaaacct 180
gggaaagcaa ataagcttct tattcactct gggtcaactt tgcaatctgg aactccatcg 240
aggttcagtg gcagtggatc tggtacagat ttcacgctca ccatcagaag cctggagttt 300
gaagattttg cagtctatta ctgtcaacag tataatgaat acccactcac gttcggttct 360
gggaccaagc tggagatcaa acggacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 90
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#84
<400> 90
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Val Gln Met Thr Gln Ser Pro Ser Tyr Leu Ala
20 25 30
Ala Ser Pro Gly Glu Ser Val Ser Ile Ser Cys Lys Ala Ser Lys Ser
35 40 45
Ile Ser Asn Asn Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Ala Asn
50 55 60
Lys Leu Leu Ile His Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg
85 90 95
Ser Leu Glu Phe Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Asn
100 105 110
Glu Tyr Pro Leu Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 91
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG1
<400> 91
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtccct gaaactctcc 120
tgtgcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
aagaagggtc tggagtgggt cgcaaccatt actaccagtg gtagcagacc ttactatcca 240
gactccgtga aaggccgatt cactatctcc agagataatg caaaaagcag cctatacctg 300
caaatgaaca gtctgaagtc tgaggacacg gccacttatt actgtgcaag acatatttat 360
tactatgatg gttacccctt tgcttactgg ggccaaggca ctctggtcac tgtcagctca 420
gcctccacca agggcccaag cgtcttcccc ctggcaccct cctccaagag cacctctggc 480
ggcacagccg ccctgggctg cctggtcaag gactacttcc ccgaacccgt gaccgtgagc 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc ccgctgtcct gcagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccctgcccag cacctgaact cctgggggga 780
ccctcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cccgggagga gcagtacaac 960
agcacgtacc gggtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag gccagccccg ggaaccacag gtgtacaccc tgcccccatc ccgggaggag 1140
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tggccagccc gagaacaact acaagaccac ccctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggca acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacc 1380
cagaagagcc tctccctgtc tcccggcaaa 1410
<210> 92
<211> 470
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG1
<400> 92
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 93
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG2
<400> 93
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtccct gaaactctcc 120
tgtgcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
aagaagggtc tggagtgggt cgcaaccatt actaccagtg gtagcagacc ttactatcca 240
gactccgtga aaggccgatt cactatctcc agagataatg caaaaagcag cctatacctg 300
caaatgaaca gtctgaagtc tgaggacacg gccacttatt actgtgcaag acatatttat 360
tactatgatg gttacccctt tgcttactgg ggccaaggca ctctggtcac tgtcagctca 420
gcctccacca agggcccttc cgtgttccct ctggcccctt gtagccgttc caccagcgag 480
tccaccgccg cccttggctg tctggtgaag gactacttcc ctgagcctgt gaccgtgagc 540
tggaactccg gagcccttac cagcggcgtg cacaccttcc ctgccgtgct gcagtccagc 600
ggcctttact ccctgagctc cgtggtgacc gtgcctagct ccaacttcgg cacccaaacc 660
tacacctgta acgtggacca caagcctagc aacaccaagg tggacaagac cgtggagcgt 720
aagtgttgtg tggagtgtcc tccttgtcct gcccctcctg tggccggacc ttccgtgttc 780
cttttccctc ctaagcctaa ggacaccctg atgatcagcc gtacccctga ggtgacctgt 840
gtggtggtgg acgtgtccca cgaggaccct gaggtgcagt tcaactggta cgtggacggc 900
gtggaggtgc acaacgccaa gaccaagcct cgtgaggagc aattcaacag caccttccgt 960
gtggtgtccg tgcttaccgt ggtgcaccaa gactggctga acggcaagga gtacaagtgt 1020
aaggtgagca acaagggact tcctgcccct atcgagaaga ccatctccaa gaccaagggc 1080
caacctcgtg agcctcaagt gtacaccctt cctcctagcc gtgaggagat gaccaagaac 1140
caagtgtccc ttacctgtct ggtgaagggc ttctacccta gcgacatcgc cgtggagtgg 1200
gagtccaacg gacaacctga gaacaactac aagaccaccc ctcctatgct tgacagcgac 1260
ggctccttct tcctgtacag caagctgacc gtggacaagt cccgttggca acaaggcaac 1320
gtgttcagct gttccgtgat gcacgaggcc ctgcacaacc actacaccca aaagagcctt 1380
tccctgagcc ctggaaag 1398
<210> 94
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG2
<400> 94
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
145 150 155 160
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn
210 215 220
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg
225 230 235 240
Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 95
<211> 1401
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG4P
<400> 95
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtccct gaaactctcc 120
tgtgcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
aagaagggtc tggagtgggt cgcaaccatt actaccagtg gtagcagacc ttactatcca 240
gactccgtga aaggccgatt cactatctcc agagataatg caaaaagcag cctatacctg 300
caaatgaaca gtctgaagtc tgaggacacg gccacttatt actgtgcaag acatatttat 360
tactatgatg gttacccctt tgcttactgg ggccaaggca ctctggtcac tgtcagctca 420
gcctccacca agggccctag cgtgttccct ctggcccctt gtagcagaag caccagcgag 480
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 540
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct gcagagcagc 600
ggcctgtact ctctgtccag cgtcgtgact gtgcccagca gctctctggg caccaagacc 660
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 720
aagtacggcc ctccctgccc tccttgccca gcccctgaat ttctgggcgg accctccgtg 780
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 840
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 900
ggcgtggaag tgcacaacgc caagaccaag cctagagagg aacagttcaa cagcacctac 960
cgggtggtgt ccgtgctgac agtgctgcac caggactggc tgaacggcaa agagtacaag 1020
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1080
ggccagcccc gcgaacccca ggtgtacaca ctgcctccaa gccaggaaga gatgaccaag 1140
aatcaggtgt ccctgacctg tctcgtgaaa ggcttctacc cctccgatat cgccgtggaa 1200
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggactcc 1260
gatggctcat tcttcctgta cagcagactg accgtggaca agagccggtg gcaggaaggc 1320
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1380
ctgtctctga gcctgggcaa a 1401
<210> 96
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG4P
<400> 96
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
145 150 155 160
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
210 215 220
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
225 230 235 240
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
275 280 285
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Leu Gly Lys
465
<210> 97
<211> 1410
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG1LALA
<400> 97
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtccct gaaactctcc 120
tgtgcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
aagaagggtc tggagtgggt cgcaaccatt actaccagtg gtagcagacc ttactatcca 240
gactccgtga aaggccgatt cactatctcc agagataatg caaaaagcag cctatacctg 300
caaatgaaca gtctgaagtc tgaggacacg gccacttatt actgtgcaag acatatttat 360
tactatgatg gttacccctt tgcttactgg ggccaaggca ctctggtcac tgtcagctca 420
gcctccacca agggcccaag cgtcttcccc ctggcaccct cctccaagag cacctctggc 480
ggcacagccg ccctgggctg cctggtcaag gactacttcc ccgaacccgt gaccgtgagc 540
tggaactcag gcgccctgac cagcggcgtg cacaccttcc ccgctgtcct gcagtcctca 600
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 660
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagag agttgagccc 720
aaatcttgtg acaaaactca cacatgccca ccctgcccag cacctgaagc cgcgggggga 780
ccctcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 840
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 900
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cccgggagga gcagtacaac 960
agcacgtacc gggtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 1020
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1080
aaagccaaag gccagccccg ggaaccacag gtgtacaccc tgcccccatc ccgggaggag 1140
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1200
gccgtggagt gggagagcaa tggccagccc gagaacaact acaagaccac ccctcccgtg 1260
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg 1320
cagcagggca acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacc 1380
cagaagagcc tctccctgtc tcccggcaaa 1410
<210> 98
<211> 470
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG1LALA
<400> 98
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
145 150 155 160
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
210 215 220
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro
225 230 235 240
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
245 250 255
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
260 265 270
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
275 280 285
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
290 295 300
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
305 310 315 320
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
325 330 335
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
340 345 350
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
355 360 365
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
370 375 380
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
385 390 395 400
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
405 410 415
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
420 425 430
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
435 440 445
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
450 455 460
Ser Leu Ser Pro Gly Lys
465 470
<210> 99
<211> 1401
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG4PFALA
<400> 99
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtccct gaaactctcc 120
tgtgcagcct caggattcac tttcagtaac tattacatgg cctgggtccg ccaggctcca 180
aagaagggtc tggagtgggt cgcaaccatt actaccagtg gtagcagacc ttactatcca 240
gactccgtga aaggccgatt cactatctcc agagataatg caaaaagcag cctatacctg 300
caaatgaaca gtctgaagtc tgaggacacg gccacttatt actgtgcaag acatatttat 360
tactatgatg gttacccctt tgcttactgg ggccaaggca ctctggtcac tgtcagctca 420
gcctccacca agggccctag cgtgttccct ctggcccctt gtagcagaag caccagcgag 480
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 540
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct gcagagcagc 600
ggcctgtact ctctgtccag cgtcgtgact gtgcccagca gctctctggg caccaagacc 660
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 720
aagtacggcc ctccctgccc tccttgccca gcccctgaag ccgcgggcgg accctccgtg 780
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 840
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 900
ggcgtggaag tgcacaacgc caagaccaag cctagagagg aacagttcaa cagcacctac 960
cgggtggtgt ccgtgctgac agtgctgcac caggactggc tgaacggcaa agagtacaag 1020
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1080
ggccagcccc gcgaacccca ggtgtacaca ctgcctccaa gccaggaaga gatgaccaag 1140
aatcaggtgt ccctgacctg tctcgtgaaa ggcttctacc cctccgatat cgccgtggaa 1200
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggactcc 1260
gatggctcat tcttcctgta cagcagactg accgtggaca agagccggtg gcaggaaggc 1320
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1380
ctgtctctga gcctgggcaa a 1401
<210> 100
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#84 типа IgG4PFALA
<400> 100
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Pro
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
145 150 155 160
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
210 215 220
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
225 230 235 240
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
275 280 285
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Leu Gly Lys
465
<210> 101
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#101
<400> 101
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccaga tgacccagtc tccttcactc ctgtctgcat ctgtgggagg cagaatcact 120
ctcaactgca aagcaagtca gaatctctat aagaacttag cctggtatca gcagaagctt 180
ggagaagctc ccaaactcct gattgataat gcaaacagtt tgcaaacggg catcccatca 240
aggttcagtg gcagtggatc tggtacagat ttcacactca ccatcagcag cctgcagcct 300
gaagatgttg ccacatattt ctgccagcag tattatagcg ggtcgtacac gtttggagct 360
gggaccaagc tggaactgaa acggacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 102
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#101
<400> 102
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Leu Leu Ser
20 25 30
Ala Ser Val Gly Gly Arg Ile Thr Leu Asn Cys Lys Ala Ser Gln Asn
35 40 45
Leu Tyr Lys Asn Leu Ala Trp Tyr Gln Gln Lys Leu Gly Glu Ala Pro
50 55 60
Lys Leu Leu Ile Asp Asn Ala Asn Ser Leu Gln Thr Gly Ile Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Val Ala Thr Tyr Phe Cys Gln Gln Tyr Tyr
100 105 110
Ser Gly Ser Tyr Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 103
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG2
<400> 103
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctga tggaggctta gtgcagcctg gaaggtccct aaaactctcc 120
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg ccaggctcca 180
acgaaggggc tggagtgggt cgcaaccatt agttatgatg gtagtaacac ttactatcga 240
gactccgtga agggccgatt cactatctcc agagataatg caaaaagcac cctatacctg 300
caaatggaca gtctgaggtc tggggacacg gccacttatt actgtgcaag attttacaac 360
aactactact ttgattactg gggccaagga gtcatggtca cagtcagctc agcctccacc 420
aagggccctt ccgtgttccc tctggcccct tgtagccgtt ccaccagcga gtccaccgcc 480
gcccttggct gtctggtgaa ggactacttc cctgagcctg tgaccgtgag ctggaactcc 540
ggagccctta ccagcggcgt gcacaccttc cctgccgtgc tgcagtccag cggcctttac 600
tccctgagct ccgtggtgac cgtgcctagc tccaacttcg gcacccaaac ctacacctgt 660
aacgtggacc acaagcctag caacaccaag gtggacaaga ccgtggagcg taagtgttgt 720
gtggagtgtc ctccttgtcc tgcccctcct gtggccggac cttccgtgtt ccttttccct 780
cctaagccta aggacaccct gatgatcagc cgtacccctg aggtgacctg tgtggtggtg 840
gacgtgtccc acgaggaccc tgaggtgcag ttcaactggt acgtggacgg cgtggaggtg 900
cacaacgcca agaccaagcc tcgtgaggag caattcaaca gcaccttccg tgtggtgtcc 960
gtgcttaccg tggtgcacca agactggctg aacggcaagg agtacaagtg taaggtgagc 1020
aacaagggac ttcctgcccc tatcgagaag accatctcca agaccaaggg ccaacctcgt 1080
gagcctcaag tgtacaccct tcctcctagc cgtgaggaga tgaccaagaa ccaagtgtcc 1140
cttacctgtc tggtgaaggg cttctaccct agcgacatcg ccgtggagtg ggagtccaac 1200
ggacaacctg agaacaacta caagaccacc cctcctatgc ttgacagcga cggctccttc 1260
ttcctgtaca gcaagctgac cgtggacaag tcccgttggc aacaaggcaa cgtgttcagc 1320
tgttccgtga tgcacgaggc cctgcacaac cactacaccc aaaagagcct ttccctgagc 1380
cctggaaag 1389
<210> 104
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG2
<400> 104
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Asp Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Gly Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys
225 230 235 240
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 105
<211> 1392
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG4P
<400> 105
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctga tggaggctta gtgcagcctg gaaggtccct aaaactctcc 120
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg ccaggctcca 180
acgaaggggc tggagtgggt cgcaaccatt agttatgatg gtagtaacac ttactatcga 240
gactccgtga agggccgatt cactatctcc agagataatg caaaaagcac cctatacctg 300
caaatggaca gtctgaggtc tggggacacg gccacttatt actgtgcaag attttacaac 360
aactactact ttgattactg gggccaagga gtcatggtca cagtcagctc agcctccacc 420
aagggcccta gcgtgttccc tctggcccct tgtagcagaa gcaccagcga gtctacagcc 480
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 540
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tgcagagcag cggcctgtac 600
tctctgtcca gcgtcgtgac tgtgcccagc agctctctgg gcaccaagac ctacacctgt 660
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 720
cctccctgcc ctccttgccc agcccctgaa tttctgggcg gaccctccgt gttcctgttc 780
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggtg 840
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 900
gtgcacaacg ccaagaccaa gcctagagag gaacagttca acagcaccta ccgggtggtg 960
tccgtgctga cagtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 1020
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 1080
cgcgaacccc aggtgtacac actgcctcca agccaggaag agatgaccaa gaatcaggtg 1140
tccctgacct gtctcgtgaa aggcttctac ccctccgata tcgccgtgga atgggagagc 1200
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggactc cgatggctca 1260
ttcttcctgt acagcagact gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1320
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1380
agcctgggca aa 1392
<210> 106
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG4P
<400> 106
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Asp Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Gly Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
225 230 235 240
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 107
<211> 1392
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG4PFALA
<400> 107
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctga tggaggctta gtgcagcctg gaaggtccct aaaactctcc 120
tgtgcagcct caggattcac tttcagtgac tattacatgg cctgggtccg ccaggctcca 180
acgaaggggc tggagtgggt cgcaaccatt agttatgatg gtagtaacac ttactatcga 240
gactccgtga agggccgatt cactatctcc agagataatg caaaaagcac cctatacctg 300
caaatggaca gtctgaggtc tggggacacg gccacttatt actgtgcaag attttacaac 360
aactactact ttgattactg gggccaagga gtcatggtca cagtcagctc agcctccacc 420
aagggcccta gcgtgttccc tctggcccct tgtagcagaa gcaccagcga gtctacagcc 480
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 540
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tgcagagcag cggcctgtac 600
tctctgtcca gcgtcgtgac tgtgcccagc agctctctgg gcaccaagac ctacacctgt 660
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 720
cctccctgcc ctccttgccc agcccctgaa gccgcgggcg gaccctccgt gttcctgttc 780
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggt 840
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 900
gtgcacaacg ccaagaccaa gcctagagag gaacagttca acagcaccta ccgggtggtg 960
tccgtgctga cagtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 1020
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 1080
cgcgaacccc aggtgtacac actgcctcca agccaggaag agatgaccaa gaatcaggtg 1140
tccctgacct gtctcgtgaa aggcttctac ccctccgata tcgccgtgga atgggagagc 1200
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggactc cgatggctca 1260
ttcttcctgt acagcagact gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1320
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1380
agcctgggca aa 1392
<210> 108
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#101 типа IgG4PFALA
<400> 108
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Asp Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Thr Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asp Ser Leu Arg Ser Gly Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
225 230 235 240
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 109
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#110
<400> 109
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccaga tgacccagac tccatcctcc atgcctgcat ctctgggaga gagagtcacc 120
atcagttgca gagcaagtca gggtattagc aattatctaa actggtatca gcagaaacca 180
gatggaacga ttaaacccct gatctactac acatccaatt tacaatctgg tgtcccatca 240
aggttcagtg gcagtgggtc tgggacagat tattctctca ccatcagcag cctggagcct 300
gaagattttg caatgtattt ctgccaacag tatgatagtt ctcctcggac gttcggtgga 360
ggcaccaagc tggaattgaa acggacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 110
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#110
<400> 110
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Thr Pro Ser Ser Met Pro
20 25 30
Ala Ser Leu Gly Glu Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Ile
50 55 60
Lys Pro Leu Ile Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser
85 90 95
Ser Leu Glu Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Tyr Asp
100 105 110
Ser Ser Pro Arg Thr Phe Gly Gly Gly Thr Lys Leu Glu Leu Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 111
<211> 1386
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG2
<400> 111
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtacagctgc agcaatctgg ggctgaactg gtgaagcctg ggtcctcagt gaaaatttcc 120
tgcaaggctt ctggctacac cttcaccagt gactttatgc actggataaa acagcagcct 180
ggaaatggcc ttgagtggat tgggtggatt tatcctggag atggtgatac agagtacaat 240
caaaagttca atgggaaggc aacactcact gcagacaaat cctccagcac agcctatatg 300
cagctcagca gcctgacatc tgaggactct gcagtctatt tctgtgcaag gggacggggg 360
tatgttatgg atgcctgggg tcaaggagct tcagtcactg tcagctcagc ctccaccaag 420
ggcccttccg tgttccctct ggccccttgt agccgttcca ccagcgagtc caccgccgcc 480
cttggctgtc tggtgaagga ctacttccct gagcctgtga ccgtgagctg gaactccgga 540
gcccttacca gcggcgtgca caccttccct gccgtgctgc agtccagcgg cctttactcc 600
ctgagctccg tggtgaccgt gcctagctcc aacttcggca cccaaaccta cacctgtaac 660
gtggaccaca agcctagcaa caccaaggtg gacaagaccg tggagcgtaa gtgttgtgtg 720
gagtgtcctc cttgtcctgc ccctcctgtg gccggacctt ccgtgttcct tttccctcct 780
aagcctaagg acaccctgat gatcagccgt acccctgagg tgacctgtgt ggtggtggac 840
gtgtcccacg aggaccctga ggtgcagttc aactggtacg tggacggcgt ggaggtgcac 900
aacgccaaga ccaagcctcg tgaggagcaa ttcaacagca ccttccgtgt ggtgtccgtg 960
cttaccgtgg tgcaccaaga ctggctgaac ggcaaggagt acaagtgtaa ggtgagcaac 1020
aagggacttc ctgcccctat cgagaagacc atctccaaga ccaagggcca acctcgtgag 1080
cctcaagtgt acacccttcc tcctagccgt gaggagatga ccaagaacca agtgtccctt 1140
acctgtctgg tgaagggctt ctaccctagc gacatcgccg tggagtggga gtccaacgga 1200
caacctgaga acaactacaa gaccacccct cctatgcttg acagcgacgg ctccttcttc 1260
ctgtacagca agctgaccgt ggacaagtcc cgttggcaac aaggcaacgt gttcagctgt 1320
tccgtgatgc acgaggccct gcacaaccac tacacccaaa agagcctttc cctgagccct 1380
ggaaag 1386
<210> 112
<211> 462
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG2
<400> 112
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Asp Phe Met His Trp Ile Lys Gln Gln Pro Gly Asn Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Asn
65 70 75 80
Gln Lys Phe Asn Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln
115 120 125
Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val
225 230 235 240
Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe
245 250 255
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
260 265 270
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
275 280 285
Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
290 295 300
Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val
305 310 315 320
Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
325 330 335
Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
355 360 365
Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
370 375 380
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
385 390 395 400
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp
405 410 415
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
420 425 430
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
435 440 445
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 113
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG4P
<400> 113
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtacagctgc agcaatctgg ggctgaactg gtgaagcctg ggtcctcagt gaaaatttcc 120
tgcaaggctt ctggctacac cttcaccagt gactttatgc actggataaa acagcagcct 180
ggaaatggcc ttgagtggat tgggtggatt tatcctggag atggtgatac agagtacaat 240
caaaagttca atgggaaggc aacactcact gcagacaaat cctccagcac agcctatatg 300
cagctcagca gcctgacatc tgaggactct gcagtctatt tctgtgcaag gggacggggg 360
tatgttatgg atgcctgggg tcaaggagct tcagtcactg tcagctcagc ctccaccaag 420
ggccctagcg tgttccctct ggccccttgt agcagaagca ccagcgagtc tacagccgcc 480
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 540
gctctgacaa gcggcgtgca cacctttcca gccgtgctgc agagcagcgg cctgtactct 600
ctgtccagcg tcgtgactgt gcccagcagc tctctgggca ccaagaccta cacctgtaac 660
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 720
ccctgccctc cttgcccagc ccctgaattt ctgggcggac cctccgtgtt cctgttcccc 780
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 840
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 900
cacaacgcca agaccaagcc tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 960
gtgctgacag tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 1020
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1080
gaaccccagg tgtacacact gcctccaagc caggaagaga tgaccaagaa tcaggtgtcc 1140
ctgacctgtc tcgtgaaagg cttctacccc tccgatatcg ccgtggaatg ggagagcaac 1200
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga tggctcattc 1260
ttcctgtaca gcagactgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1320
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgagc 1380
ctgggcaaa 1389
<210> 114
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG4P
<400> 114
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Asp Phe Met His Trp Ile Lys Gln Gln Pro Gly Asn Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Asn
65 70 75 80
Gln Lys Phe Asn Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln
115 120 125
Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 115
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG4PFALA
<400> 115
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtacagctgc agcaatctgg ggctgaactg gtgaagcctg ggtcctcagt gaaaatttcc 120
tgcaaggctt ctggctacac cttcaccagt gactttatgc actggataaa acagcagcct 180
ggaaatggcc ttgagtggat tgggtggatt tatcctggag atggtgatac agagtacaat 240
caaaagttca atgggaaggc aacactcact gcagacaaat cctccagcac agcctatatg 300
cagctcagca gcctgacatc tgaggactct gcagtctatt tctgtgcaag gggacggggg 360
tatgttatgg atgcctgggg tcaaggagct tcagtcactg tcagctcagc ctccaccaag 420
ggccctagcg tgttccctct ggccccttgt agcagaagca ccagcgagtc tacagccgcc 480
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 540
gctctgacaa gcggcgtgca cacctttcca gccgtgctgc agagcagcgg cctgtactct 600
ctgtccagcg tcgtgactgt gcccagcagc tctctgggca ccaagaccta cacctgtaac 660
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 720
ccctgccctc cttgcccagc ccctgaagcc gcgggcggac cctccgtgtt cctgttcccc 780
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 840
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 900
cacaacgcca agaccaagcc tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 960
gtgctgacag tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 1020
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1080
gaaccccagg tgtacacact gcctccaagc caggaagaga tgaccaagaa tcaggtgtcc 1140
ctgacctgtc tcgtgaaagg cttctacccc tccgatatcg ccgtggaatg ggagagcaac 1200
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga tggctcattc 1260
ttcctgtaca gcagactgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1320
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgagc 1380
ctgggcaaa 1389
<210> 116
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#110 типа IgG4PFALA
<400> 116
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Asp Phe Met His Trp Ile Lys Gln Gln Pro Gly Asn Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Asn
65 70 75 80
Gln Lys Phe Asn Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln
115 120 125
Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 117
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#131
<400> 117
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatgtccaga tgacccagtc tccatcttat cttgctgcgt ctcctggaga aagtgtttcc 120
atcagttgca aggcaagtaa aagcattaac acatacttag cctggtatca ggagaaacct 180
gggaaaacga ataagcttct tatctactct gggtcaactt tgcaatctgg aactccatcg 240
agattcagtg gcagtggatc tggtacagat ttcacgctca ccatcagaag cctggagcct 300
gaagattttg cagtctacta ctgtcaacag cataatgaat accccttcac gttcggctca 360
gggacgaagt tggaaataaa acggacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 118
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь химерного человек/крыса_CD147_#131
<400> 118
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Val Gln Met Thr Gln Ser Pro Ser Tyr Leu Ala
20 25 30
Ala Ser Pro Gly Glu Ser Val Ser Ile Ser Cys Lys Ala Ser Lys Ser
35 40 45
Ile Asn Thr Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Thr Asn
50 55 60
Lys Leu Leu Ile Tyr Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg
85 90 95
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln His Asn
100 105 110
Glu Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 119
<211> 1392
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#131 типа IgG2
<400> 119
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtctct gaaactatcc 120
tgtggagcct ctggattcac attcaataac tactggatga cctgggtccg ccaggctcca 180
gggaaggggc tggagtgggt tgcatccatt actaaagctg gtggtagcac ttactatcga 240
gactctgtga agggccgatt cactatctcc agagataatg caaaaagcac cctatatctg 300
caaatgaaca gtctgaggtc tgaggacacg gccacttatt actgtacaag agaactggga 360
gagttctatg ttatggatgc ctggggtcaa ggagcttcag tcactgtcag ctcagcctcc 420
accaagggcc cttccgtgtt ccctctggcc ccttgtagcc gttccaccag cgagtccacc 480
gccgcccttg gctgtctggt gaaggactac ttccctgagc ctgtgaccgt gagctggaac 540
tccggagccc ttaccagcgg cgtgcacacc ttccctgccg tgctgcagtc cagcggcctt 600
tactccctga gctccgtggt gaccgtgcct agctccaact tcggcaccca aacctacacc 660
tgtaacgtgg accacaagcc tagcaacacc aaggtggaca agaccgtgga gcgtaagtgt 720
tgtgtggagt gtcctccttg tcctgcccct cctgtggccg gaccttccgt gttccttttc 780
cctcctaagc ctaaggacac cctgatgatc agccgtaccc ctgaggtgac ctgtgtggtg 840
gtggacgtgt cccacgagga ccctgaggtg cagttcaact ggtacgtgga cggcgtggag 900
gtgcacaacg ccaagaccaa gcctcgtgag gagcaattca acagcacctt ccgtgtggtg 960
tccgtgctta ccgtggtgca ccaagactgg ctgaacggca aggagtacaa gtgtaaggtg 1020
agcaacaagg gacttcctgc ccctatcgag aagaccatct ccaagaccaa gggccaacct 1080
cgtgagcctc aagtgtacac ccttcctcct agccgtgagg agatgaccaa gaaccaagtg 1140
tcccttacct gtctggtgaa gggcttctac cctagcgaca tcgccgtgga gtgggagtcc 1200
aacggacaac ctgagaacaa ctacaagacc acccctccta tgcttgacag cgacggctcc 1260
ttcttcctgt acagcaagct gaccgtggac aagtcccgtt ggcaacaagg caacgtgttc 1320
agctgttccg tgatgcacga ggccctgcac aaccactaca cccaaaagag cctttccctg 1380
agccctggaa ag 1392
<210> 120
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#131 типа IgG2
<400> 120
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Gly Ala Ser Gly Phe Thr Phe
35 40 45
Asn Asn Tyr Trp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Lys Ala Gly Gly Ser Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Thr Arg Glu Leu Gly Glu Phe Tyr Val Met Asp Ala Trp
115 120 125
Gly Gln Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys
225 230 235 240
Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr
340 345 350
Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 121
<211> 1395
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#131 типа IgG4P
<400> 121
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgag 60
gtgcagctgg tggagtctgg gggaggccta gtgcagcctg gaaggtctct gaaactatcc 120
tgtggagcct ctggattcac attcaataac tactggatga cctgggtccg ccaggctcca 180
gggaaggggc tggagtgggt tgcatccatt actaaagctg gtggtagcac ttactatcga 240
gactctgtga agggccgatt cactatctcc agagataatg caaaaagcac cctatatctg 300
caaatgaaca gtctgaggtc tgaggacacg gccacttatt actgtacaag agaactggga 360
gagttctatg ttatggatgc ctggggtcaa ggagcttcag tcactgtcag ctcagcctcc 420
accaagggcc ctagcgtgtt ccctctggcc ccttgtagca gaagcaccag cgagtctaca 480
gccgccctgg gctgcctcgt gaaggactac tttcccgagc ccgtgaccgt gtcctggaac 540
tctggcgctc tgacaagcgg cgtgcacacc tttccagccg tgctgcagag cagcggcctg 600
tactctctgt ccagcgtcgt gactgtgccc agcagctctc tgggcaccaa gacctacacc 660
tgtaacgtgg accacaagcc cagcaacacc aaggtggaca agcgggtgga atctaagtac 720
ggccctccct gccctccttg cccagcccct gaatttctgg gcggaccctc cgtgttcctg 780
ttccccccaa agcccaagga caccctgatg atcagccgga cccccgaagt gacctgcgtg 840
gtggtggatg tgtcccagga agatcccgag gtgcagttca attggtacgt ggacggcgtg 900
gaagtgcaca acgccaagac caagcctaga gaggaacagt tcaacagcac ctaccgggtg 960
gtgtccgtgc tgacagtgct gcaccaggac tggctgaacg gcaaagagta caagtgcaag 1020
gtgtccaaca agggcctgcc cagctccatc gagaaaacca tcagcaaggc caagggccag 1080
ccccgcgaac cccaggtgta cacactgcct ccaagccagg aagagatgac caagaatcag 1140
gtgtccctga cctgtctcgt gaaaggcttc tacccctccg atatcgccgt ggaatgggag 1200
agcaacggcc agcccgagaa caactacaag accacccccc ctgtgctgga ctccgatggc 1260
tcattcttcc tgtacagcag actgaccgtg gacaagagcc ggtggcagga aggcaacgtg 1320
ttcagctgct ccgtgatgca cgaggccctg cacaaccact acacccagaa gtccctgtct 1380
ctgagcctgg gcaaa 1395
<210> 122
<211> 465
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь химерного человек/крыса_CD147_#131 типа IgG4P
<400> 122
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Lys Leu Ser Cys Gly Ala Ser Gly Phe Thr Phe
35 40 45
Asn Asn Tyr Trp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Lys Ala Gly Gly Ser Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ser Glu Asp Thr Ala Thr
100 105 110
Tyr Tyr Cys Thr Arg Glu Leu Gly Glu Phe Tyr Val Met Asp Ala Trp
115 120 125
Gly Gln Gly Ala Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr
225 230 235 240
Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys
340 345 350
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
450 455 460
Lys
465
<210> 123
<211> 466
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #84H1hIgG2
<400> 123
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
145 150 155 160
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn
210 215 220
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg
225 230 235 240
Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 124
<211> 1398
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #84H1hIgG2
<400> 124
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgaa 60
gtgcagctgg ttgagtctgg cggaggactg gttcaacctg gcggaagcct gagactgtct 120
tgtgccgcca gcggcttcac cttcagcaac tactatatgg cctgggtccg acaggcccct 180
ggcaaaggac ttgaatgggt gtccaccatc accaccagcg gcagcagacc ttactacgcc 240
gatagcgtga agggcagatt caccatcagc cgggacaaca gcaagaacac cctgtacctg 300
cagatgaaca gcctgagagc cgaggacacc gccgtgtact attgcgccag acacatctac 360
tactacgacg gctacccctt cgcctattgg ggccagggaa cactggtcac agttagctca 420
gcctccacca agggcccttc cgtgttccct ctggcccctt gtagccgttc caccagcgag 480
tccaccgccg cccttggctg tctggtgaag gactacttcc ctgagcctgt gaccgtgagc 540
tggaactccg gagcccttac cagcggcgtg cacaccttcc ctgccgtgct gcagtccagc 600
ggcctttact ccctgagctc cgtggtgacc gtgcctagct ccaacttcgg cacccaaacc 660
tacacctgta acgtggacca caagcctagc aacaccaagg tggacaagac cgtggagcgt 720
aagtgttgtg tggagtgtcc tccttgtcct gcccctcctg tggccggacc ttccgtgttc 780
cttttccctc ctaagcctaa ggacaccctg atgatcagcc gtacccctga ggtgacctgt 840
gtggtggtgg acgtgtccca cgaggaccct gaggtgcagt tcaactggta cgtggacggc 900
gtggaggtgc acaacgccaa gaccaagcct cgtgaggagc aattcaacag caccttccgt 960
gtggtgtccg tgcttaccgt ggtgcaccaa gactggctga acggcaagga gtacaagtgt 1020
aaggtgagca acaagggact tcctgcccct atcgagaaga ccatctccaa gaccaagggc 1080
caacctcgtg agcctcaagt gtacaccctt cctcctagcc gtgaggagat gaccaagaac 1140
caagtgtccc ttacctgtct ggtgaagggc ttctacccta gcgacatcgc cgtggagtgg 1200
gagtccaacg gacaacctga gaacaactac aagaccaccc ctcctatgct tgacagcgac 1260
ggctccttct tcctgtacag caagctgacc gtggacaagt cccgttggca acaaggcaac 1320
gtgttcagct gttccgtgat gcacgaggcc ctgcacaacc actacaccca aaagagcctt 1380
tccctgagcc ctggaaag 1398
<210> 125
<211> 467
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #84H1hIgG4P
<400> 125
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asn Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ser Thr Ile Thr Thr Ser Gly Ser Arg Pro Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg His Ile Tyr Tyr Tyr Asp Gly Tyr Pro Phe Ala
115 120 125
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys
130 135 140
Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu
145 150 155 160
Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
165 170 175
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
180 185 190
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
195 200 205
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn
210 215 220
Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser
225 230 235 240
Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln
275 280 285
Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Leu Gly Lys
465
<210> 126
<211> 1401
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #84H1hIgG4P
<400> 126
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgaa 60
gtgcagctgg ttgagtctgg cggaggactg gttcaacctg gcggaagcct gagactgtct 120
tgtgccgcca gcggcttcac cttcagcaac tactatatgg cctgggtccg acaggcccct 180
ggcaaaggac ttgaatgggt gtccaccatc accaccagcg gcagcagacc ttactacgcc 240
gatagcgtga agggcagatt caccatcagc cgggacaaca gcaagaacac cctgtacctg 300
cagatgaaca gcctgagagc cgaggacacc gccgtgtact attgcgccag acacatctac 360
tactacgacg gctacccctt cgcctattgg ggccagggaa cactggtcac agttagctca 420
gcctccacca agggccctag cgtgttccct ctggcccctt gtagcagaag caccagcgag 480
tctacagccg ccctgggctg cctcgtgaag gactactttc ccgagcccgt gaccgtgtcc 540
tggaactctg gcgctctgac aagcggcgtg cacacctttc cagccgtgct gcagagcagc 600
ggcctgtact ctctgtccag cgtcgtgact gtgcccagca gctctctggg caccaagacc 660
tacacctgta acgtggacca caagcccagc aacaccaagg tggacaagcg ggtggaatct 720
aagtacggcc ctccctgccc tccttgccca gcccctgaat ttctgggcgg accctccgtg 780
ttcctgttcc ccccaaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 840
tgcgtggtgg tggatgtgtc ccaggaagat cccgaggtgc agttcaattg gtacgtggac 900
ggcgtggaag tgcacaacgc caagaccaag cctagagagg aacagttcaa cagcacctac 960
cgggtggtgt ccgtgctgac agtgctgcac caggactggc tgaacggcaa agagtacaag 1020
tgcaaggtgt ccaacaaggg cctgcccagc tccatcgaga aaaccatcag caaggccaag 1080
ggccagcccc gcgaacccca ggtgtacaca ctgcctccaa gccaggaaga gatgaccaag 1140
aatcaggtgt ccctgacctg tctcgtgaaa ggcttctacc cctccgatat cgccgtggaa 1200
tgggagagca acggccagcc cgagaacaac tacaagacca ccccccctgt gctggactcc 1260
gatggctcat tcttcctgta cagcagactg accgtggaca agagccggtg gcaggaaggc 1320
aacgtgttca gctgctccgt gatgcacgag gccctgcaca accactacac ccagaagtcc 1380
ctgtctctga gcctgggcaa a 1401
<210> 127
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #84L2h
<400> 127
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Val Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Lys Ser
35 40 45
Ile Ser Asn Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Asn
50 55 60
Lys Leu Leu Ile His Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn
100 105 110
Glu Tyr Pro Leu Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 128
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #84L2h
<400> 128
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacgttcaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 120
atcacatgca aggccagcaa gagcatcagc aacaacctgg cctggtatca gcagaagccc 180
ggaaaggcca acaagctgct gatccacagc ggcagcacac tgcagtctgg cacccctagc 240
agattttccg gctctggcag cggcaccgat ttcaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctacta ctgccagcag tacaacgagt accctctgac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 129
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #101H1hIgG2
<400> 129
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Gly Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys
225 230 235 240
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
340 345 350
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 130
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #101H1hIgG2
<400> 130
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgaa 60
gtgcagctgg ttgagtctgg cggaggactg gttcagcctg gcagaagcct gagactgtct 120
tgtgccgcca gcggcttcac cttcagcgac tactatatgg cctgggtccg acaggcccct 180
ggcaaaggac ttgaatgggt cgccaccatc agctacgacg gcagcaacac ctactaccgg 240
gacagcgtga agggcagatt caccatctcc agagacaaca gcaagagcac cctgtacctg 300
cagatgaaca gcctgagagc cggcgatacc gccgtgtact actgcgccag attctacaac 360
aactactact tcgactactg gggccagggc accctggtca cagttagctc agcctccacc 420
aagggccctt ccgtgttccc tctggcccct tgtagccgtt ccaccagcga gtccaccgcc 480
gcccttggct gtctggtgaa ggactacttc cctgagcctg tgaccgtgag ctggaactcc 540
ggagccctta ccagcggcgt gcacaccttc cctgccgtgc tgcagtccag cggcctttac 600
tccctgagct ccgtggtgac cgtgcctagc tccaacttcg gcacccaaac ctacacctgt 660
aacgtggacc acaagcctag caacaccaag gtggacaaga ccgtggagcg taagtgttgt 720
gtggagtgtc ctccttgtcc tgcccctcct gtggccggac cttccgtgtt ccttttccct 780
cctaagccta aggacaccct gatgatcagc cgtacccctg aggtgacctg tgtggtggtg 840
gacgtgtccc acgaggaccc tgaggtgcag ttcaactggt acgtggacgg cgtggaggtg 900
cacaacgcca agaccaagcc tcgtgaggag caattcaaca gcaccttccg tgtggtgtcc 960
gtgcttaccg tggtgcacca agactggctg aacggcaagg agtacaagtg taaggtgagc 1020
aacaagggac ttcctgcccc tatcgagaag accatctcca agaccaaggg ccaacctcgt 1080
gagcctcaag tgtacaccct tcctcctagc cgtgaggaga tgaccaagaa ccaagtgtcc 1140
cttacctgtc tggtgaaggg cttctaccct agcgacatcg ccgtggagtg ggagtccaac 1200
ggacaacctg agaacaacta caagaccacc cctcctatgc ttgacagcga cggctccttc 1260
ttcctgtaca gcaagctgac cgtggacaag tcccgttggc aacaaggcaa cgtgttcagc 1320
tgttccgtga tgcacgaggc cctgcacaac cactacaccc aaaagagcct ttccctgagc 1380
cctggaaag 1389
<210> 131
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #101H1hIgG4P
<400> 131
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Tyr Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Thr Ile Ser Tyr Asp Gly Ser Asn Thr Tyr Tyr Arg
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Gly Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Phe Tyr Asn Asn Tyr Tyr Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
225 230 235 240
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
340 345 350
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 132
<211> 1392
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #101H1hIgG4P
<400> 132
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagcgaa 60
gtgcagctgg ttgagtctgg cggaggactg gttcagcctg gcagaagcct gagactgtct 120
tgtgccgcca gcggcttcac cttcagcgac tactatatgg cctgggtccg acaggcccct 180
ggcaaaggac ttgaatgggt cgccaccatc agctacgacg gcagcaacac ctactaccgg 240
gacagcgtga agggcagatt caccatctcc agagacaaca gcaagagcac cctgtacctg 300
cagatgaaca gcctgagagc cggcgatacc gccgtgtact actgcgccag attctacaac 360
aactactact tcgactactg gggccagggc accctggtca cagttagctc agcctccacc 420
aagggcccta gcgtgttccc tctggcccct tgtagcagaa gcaccagcga gtctacagcc 480
gccctgggct gcctcgtgaa ggactacttt cccgagcccg tgaccgtgtc ctggaactct 540
ggcgctctga caagcggcgt gcacaccttt ccagccgtgc tgcagagcag cggcctgtac 600
tctctgtcca gcgtcgtgac tgtgcccagc agctctctgg gcaccaagac ctacacctgt 660
aacgtggacc acaagcccag caacaccaag gtggacaagc gggtggaatc taagtacggc 720
cctccctgcc ctccttgccc agcccctgaa tttctgggcg gaccctccgt gttcctgttc 780
cccccaaagc ccaaggacac cctgatgatc agccggaccc ccgaagtgac ctgcgtggtg 840
gtggatgtgt cccaggaaga tcccgaggtg cagttcaatt ggtacgtgga cggcgtggaa 900
gtgcacaacg ccaagaccaa gcctagagag gaacagttca acagcaccta ccgggtggtg 960
tccgtgctga cagtgctgca ccaggactgg ctgaacggca aagagtacaa gtgcaaggtg 1020
tccaacaagg gcctgcccag ctccatcgag aaaaccatca gcaaggccaa gggccagccc 1080
cgcgaacccc aggtgtacac actgcctcca agccaggaag agatgaccaa gaatcaggtg 1140
tccctgacct gtctcgtgaa aggcttctac ccctccgata tcgccgtgga atgggagagc 1200
aacggccagc ccgagaacaa ctacaagacc accccccctg tgctggactc cgatggctca 1260
ttcttcctgt acagcagact gaccgtggac aagagccggt ggcaggaagg caacgtgttc 1320
agctgctccg tgatgcacga ggccctgcac aaccactaca cccagaagtc cctgtctctg 1380
agcctgggca aa 1392
<210> 133
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #101L2h
<400> 133
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Asn Cys Lys Ala Ser Gln Asn
35 40 45
Leu Tyr Lys Asn Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Leu Leu Ile Asp Asn Ala Asn Ser Leu Gln Thr Gly Ile Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Tyr Tyr
100 105 110
Ser Gly Ser Tyr Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 134
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #101L2h
<400> 134
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga tagagtgacc 120
atcaactgca aggccagcca gaacctgtac aagaacctgg cctggtatca gcagaagccc 180
ggcaaggctc ctaagctgct gatcgacaac gccaacagcc tgcagaccgg cattcccagc 240
agattttctg gcagcggctc cggcaccgat ttcaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctactt ttgccagcag tactacagcg gcagctacac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 135
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #110H1hIgG4P
<400> 135
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Asp Phe Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Asn
65 70 75 80
Gln Lys Phe Gln Gly Arg Val Thr Leu Thr Arg Asp Thr Ser Ile Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln
115 120 125
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 136
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #110H1hIgG4P
<400> 136
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gttcagctgg ttcagtctgg cgccgaagtg aagaaacctg gcgcctctgt gaaggtgtcc 120
tgcaaggcca gcggctacac ctttaccagc gacttcatgc actgggtccg acaggctcca 180
ggacagggac ttgaatggat gggctggatc tatcccggcg acggcgacac cgagtacaac 240
cagaaattcc agggcagagt gaccctgacc agagacacca gcatcagcac cgcctacatg 300
gaactgagcc ggctgagatc cgatgacacc gccgtgtact actgcgccag aggcagaggc 360
tatgtgatgg atgcttgggg ccagggcacc accgttacag ttagctcagc ctccaccaag 420
ggccctagcg tgttccctct ggccccttgt agcagaagca ccagcgagtc tacagccgcc 480
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 540
gctctgacaa gcggcgtgca cacctttcca gccgtgctgc agagcagcgg cctgtactct 600
ctgtccagcg tcgtgactgt gcccagcagc tctctgggca ccaagaccta cacctgtaac 660
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 720
ccctgccctc cttgcccagc ccctgaattt ctgggcggac cctccgtgtt cctgttcccc 780
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 840
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 900
cacaacgcca agaccaagcc tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 960
gtgctgacag tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 1020
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1080
gaaccccagg tgtacacact gcctccaagc caggaagaga tgaccaagaa tcaggtgtcc 1140
ctgacctgtc tcgtgaaagg cttctacccc tccgatatcg ccgtggaatg ggagagcaac 1200
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga tggctcattc 1260
ttcctgtaca gcagactgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1320
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgagc 1380
ctgggcaaa 1389
<210> 137
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #110L4h
<400> 137
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Ile
50 55 60
Lys Pro Leu Ile Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Tyr Asp
100 105 110
Ser Ser Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 138
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #110L4h
<400> 138
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 120
atcacctgta gagccagcca gggcatcagc aactacctga actggtatca gcagaagccc 180
ggcaaggcca tcaagcccct gatctactac accagcaacc tgcagagcgg cgtgcccagc 240
agattttctg gctctggcag cggcaccgac tacaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctactt ttgccagcag tacgacagca gccccagaac ctttggcggc 360
ggaacaaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 139
<211> 464
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #131H2hIgG2
<400> 139
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln
20 25 30
Pro Gly Arg Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Asn Asn Tyr Trp Met Thr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Ser Ile Thr Lys Ala Gly Gly Ser Thr Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Ser
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Gly Thr Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Glu Leu Gly Glu Phe Tyr Val Met Asp Ala Trp
115 120 125
Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
130 135 140
Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr
145 150 155 160
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
195 200 205
Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp
210 215 220
His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Cys
225 230 235 240
Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser
245 250 255
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
260 265 270
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
275 280 285
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
290 295 300
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val
305 310 315 320
Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
325 330 335
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr
340 345 350
Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
355 360 365
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
370 375 380
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
385 390 395 400
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp
405 410 415
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
420 425 430
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
435 440 445
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 140
<211> 1392
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #131H2hIgG2
<400> 140
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gttcagctgg ttgaatctgg tggcggagtg gtgcagcctg gcagatctct gagactgtct 120
tgtgccgcca gcggcttcac cttcaacaac tactggatga cctgggtccg acaggcccct 180
ggcaaaggac ttgaatgggt cgccagcatc accaaggctg gcggctctac ctactacgcc 240
gatagcgtga agggcagatt caccatcagc cgggacaaca gcaagagcac cctgtacctg 300
cagatgaaca gcctgagagc cgagggcaca gccgtgtact actgtacaag agagctgggc 360
gagttctacg tgatggatgc ctggggccag ggcaccacag ttacagttag ctcagcctcc 420
accaagggcc cttccgtgtt ccctctggcc ccttgtagcc gttccaccag cgagtccacc 480
gccgcccttg gctgtctggt gaaggactac ttccctgagc ctgtgaccgt gagctggaac 540
tccggagccc ttaccagcgg cgtgcacacc ttccctgccg tgctgcagtc cagcggcctt 600
tactccctga gctccgtggt gaccgtgcct agctccaact tcggcaccca aacctacacc 660
tgtaacgtgg accacaagcc tagcaacacc aaggtggaca agaccgtgga gcgtaagtgt 720
tgtgtggagt gtcctccttg tcctgcccct cctgtggccg gaccttccgt gttccttttc 780
cctcctaagc ctaaggacac cctgatgatc agccgtaccc ctgaggtgac ctgtgtggtg 840
gtggacgtgt cccacgagga ccctgaggtg cagttcaact ggtacgtgga cggcgtggag 900
gtgcacaacg ccaagaccaa gcctcgtgag gagcaattca acagcacctt ccgtgtggtg 960
tccgtgctta ccgtggtgca ccaagactgg ctgaacggca aggagtacaa gtgtaaggtg 1020
agcaacaagg gacttcctgc ccctatcgag aagaccatct ccaagaccaa gggccaacct 1080
cgtgagcctc aagtgtacac ccttcctcct agccgtgagg agatgaccaa gaaccaagtg 1140
tcccttacct gtctggtgaa gggcttctac cctagcgaca tcgccgtgga gtgggagtcc 1200
aacggacaac ctgagaacaa ctacaagacc acccctccta tgcttgacag cgacggctcc 1260
ttcttcctgt acagcaagct gaccgtggac aagtcccgtt ggcaacaagg caacgtgttc 1320
agctgttccg tgatgcacga ggccctgcac aaccactaca cccaaaagag cctttccctg 1380
agccctggaa ag 1392
<210> 141
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #131L2h
<400> 141
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Val Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Lys Ser
35 40 45
Ile Asn Thr Tyr Leu Ala Trp Tyr Gln Glu Lys Pro Gly Lys Thr Asn
50 55 60
Lys Leu Leu Ile Tyr Ser Gly Ser Thr Leu Gln Ser Gly Thr Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Asn
100 105 110
Glu Tyr Pro Phe Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 142
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #131L2h
<400> 142
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacgttcaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 120
atcacatgca aggccagcaa gagcatcaac acctacctgg cctggtatca agagaagccc 180
ggcaagacca acaagctgct gatctacagc ggcagcacac tgcagagcgg caccccttct 240
agattttccg gctctggcag cggcaccgac ttcaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctacta ctgccagcag cacaacgagt accccttcac ctttggccag 360
ggcaccaagc tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 143
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 143
Asp Ala Leu Pro Gly Gln Lys Thr Glu Phe Lys Val Asp Ser Asp Asp
1 5 10 15
Gln
<210> 144
<211> 17
<212> БЕЛОК
<213> Макак-крабоед
<400> 144
Asp Thr Leu Pro Gly Gln Lys Thr Asp Phe Glu Val Asp Ser Asp Asp
1 5 10 15
Leu
<210> 145
<211> 457
<212> БЕЛОК
<213> Homo sapiens
<400> 145
Met Ala Thr Arg Val Leu Ser Met Ser Ala Arg Leu Gly Pro Val Pro
1 5 10 15
Gln Pro Pro Ala Pro Gln Asp Glu Pro Val Phe Ala Gln Leu Lys Pro
20 25 30
Val Leu Gly Ala Ala Asn Pro Ala Arg Asp Ala Ala Leu Phe Pro Gly
35 40 45
Glu Glu Leu Lys His Ala His His Arg Pro Gln Ala Gln Pro Ala Pro
50 55 60
Ala Gln Ala Pro Gln Pro Ala Gln Pro Pro Ala Thr Gly Pro Arg Leu
65 70 75 80
Pro Pro Glu Asp Leu Val Gln Thr Arg Cys Glu Met Glu Lys Tyr Leu
85 90 95
Thr Pro Gln Leu Pro Pro Val Pro Ile Ile Pro Glu His Lys Lys Tyr
100 105 110
Arg Arg Asp Ser Ala Ser Val Val Asp Gln Phe Phe Thr Asp Thr Glu
115 120 125
Gly Leu Pro Tyr Ser Ile Asn Met Asn Val Phe Leu Pro Asp Ile Thr
130 135 140
His Leu Arg Thr Gly Leu Tyr Lys Ser Gln Arg Pro Cys Val Thr His
145 150 155 160
Ile Lys Thr Glu Pro Val Ala Ile Phe Ser His Gln Ser Glu Thr Thr
165 170 175
Ala Pro Pro Pro Ala Pro Thr Gln Ala Leu Pro Glu Phe Thr Ser Ile
180 185 190
Phe Ser Ser His Gln Thr Ala Ala Pro Glu Val Asn Asn Ile Phe Ile
195 200 205
Lys Gln Glu Leu Pro Thr Pro Asp Leu His Leu Ser Val Pro Thr Gln
210 215 220
Gln Gly His Leu Tyr Gln Leu Leu Asn Thr Pro Asp Leu Asp Met Pro
225 230 235 240
Ser Ser Thr Asn Gln Thr Ala Ala Met Asp Thr Leu Asn Val Ser Met
245 250 255
Ser Ala Ala Met Ala Gly Leu Asn Thr His Thr Ser Ala Val Pro Gln
260 265 270
Thr Ala Val Lys Gln Phe Gln Gly Met Pro Pro Cys Thr Tyr Thr Met
275 280 285
Pro Ser Gln Phe Leu Pro Gln Gln Ala Thr Tyr Phe Pro Pro Ser Pro
290 295 300
Pro Ser Ser Glu Pro Gly Ser Pro Asp Arg Gln Ala Glu Met Leu Gln
305 310 315 320
Asn Leu Thr Pro Pro Pro Ser Tyr Ala Ala Thr Ile Ala Ser Lys Leu
325 330 335
Ala Ile His Asn Pro Asn Leu Pro Thr Thr Leu Pro Val Asn Ser Gln
340 345 350
Asn Ile Gln Pro Val Arg Tyr Asn Arg Arg Ser Asn Pro Asp Leu Glu
355 360 365
Lys Arg Arg Ile His Tyr Cys Asp Tyr Pro Gly Cys Thr Lys Val Tyr
370 375 380
Thr Lys Ser Ser His Leu Lys Ala His Leu Arg Thr His Thr Gly Glu
385 390 395 400
Lys Pro Tyr Lys Cys Thr Trp Glu Gly Cys Asp Trp Arg Phe Ala Arg
405 410 415
Ser Asp Glu Leu Thr Arg His Tyr Arg Lys His Thr Gly Ala Lys Pro
420 425 430
Phe Gln Cys Gly Val Cys Asn Arg Ser Phe Ser Arg Ser Asp His Leu
435 440 445
Ala Leu His Met Lys Arg His Gln Asn
450 455
<210> 146
<211> 1374
<212> ДНК
<213> Homo sapiens
<400> 146
atggctacaa gggtgctgag catgagcgcc cgcctgggac ccgtgcccca gccgccggcg 60
ccgcaggacg agccggtgtt cgcgcagctc aagccggtgc tgggcgccgc gaatccggcc 120
cgcgacgcgg cgctcttccc cggcgaggag ctgaagcacg cgcaccaccg cccgcaggcg 180
cagcccgcgc ccgcgcaggc cccgcagccg gcccagccgc ccgccaccgg cccgcggctg 240
cctccagagg acctggtcca gacaagatgt gaaatggaga agtatctgac acctcagctt 300
cctccagttc ctataattcc agagcataaa aagtatagac gagacagtgc ctcagtcgta 360
gaccagttct tcactgacac tgaagggtta ccttacagta tcaacatgaa cgtcttcctc 420
cctgacatca ctcacctgag aactggcctc tacaaatccc agagaccgtg cgtaacacac 480
atcaagacag aacctgttgc cattttcagc caccagagtg aaacgactgc ccctcctccg 540
gccccgaccc aggccctccc tgagttcacc agtatattca gctcacacca gaccgcagct 600
ccagaggtga acaatatttt catcaaacaa gaacttccta caccagatct tcatctttct 660
gtccctaccc agcagggcca cctgtaccag ctactgaata caccggatct agatatgccc 720
agttctacaa atcagacagc agcaatggac actcttaatg tttctatgtc agctgccatg 780
gcaggcctta acacacacac ctctgctgtt ccgcagactg cagtgaaaca attccagggc 840
atgccccctt gcacatacac aatgccaagt cagtttcttc cacaacaggc cacttacttt 900
cccccgtcac caccaagctc agagcctgga agtccagata gacaagcaga gatgctccag 960
aatttaaccc cacctccatc ctatgctgct acaattgctt ctaaactggc aattcacaat 1020
ccaaatttac ccaccaccct gccagttaac tcacaaaaca tccaacctgt cagatacaat 1080
agaaggagta accccgattt ggagaaacga cgcatccact actgcgatta ccctggttgc 1140
acaaaagttt ataccaagtc ttctcattta aaagctcacc tgaggactca cactggtgaa 1200
aagccataca agtgtacctg ggaaggctgc gactggaggt tcgcgcgatc ggatgagctg 1260
acccgccact accggaagca cacaggcgcc aagcccttcc agtgcggggt gtgcaaccgc 1320
agcttctcgc gctctgacca cctggccctg catatgaaga ggcaccagaa ctga 1374
<210> 147
<211> 463
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #110H13hIgG4P
<400> 147
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Asp Phe Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Trp Ile Tyr Pro Gly Asp Gly Asp Thr Glu Tyr Ala
65 70 75 80
Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Gly Arg Gly Tyr Val Met Asp Ala Trp Gly Gln
115 120 125
Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly Pro
225 230 235 240
Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser Val
245 250 255
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
260 265 270
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
275 280 285
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
290 295 300
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
305 310 315 320
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
325 330 335
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
340 345 350
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
355 360 365
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
370 375 380
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
385 390 395 400
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
405 410 415
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
420 425 430
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
435 440 445
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
450 455 460
<210> 148
<211> 1389
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #110H13hIgG4P
<400> 148
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gttcagctgg ttcagtctgg cgccgaagtg aagaaacctg gcgcctctgt gaaggtgtcc 120
tgcaaggcca gcggctacac ctttaccagc gacttcatgc actgggtccg acaggctcca 180
ggacagggac ttgaatggat gggctggatc tatcccggcg acggcgatac agagtacgcc 240
cagaaattcc agggcagagt gaccatgacc agagacacca gcatcagcac cgcctacatg 300
gaactgagcc ggctgagatc cgatgacacc gccgtgtact actgcgccag aggcagaggc 360
tatgtgatgg atgcttgggg ccagggcacc accgttacag ttagctcagc ctccaccaag 420
ggccctagcg tgttccctct ggccccttgt agcagaagca ccagcgagtc tacagccgcc 480
ctgggctgcc tcgtgaagga ctactttccc gagcccgtga ccgtgtcctg gaactctggc 540
gctctgacaa gcggcgtgca cacctttcca gccgtgctgc agagcagcgg cctgtactct 600
ctgtccagcg tcgtgactgt gcccagcagc tctctgggca ccaagaccta cacctgtaac 660
gtggaccaca agcccagcaa caccaaggtg gacaagcggg tggaatctaa gtacggccct 720
ccctgccctc cttgcccagc ccctgaattt ctgggcggac cctccgtgtt cctgttcccc 780
ccaaagccca aggacaccct gatgatcagc cggacccccg aagtgacctg cgtggtggtg 840
gatgtgtccc aggaagatcc cgaggtgcag ttcaattggt acgtggacgg cgtggaagtg 900
cacaacgcca agaccaagcc tagagaggaa cagttcaaca gcacctaccg ggtggtgtcc 960
gtgctgacag tgctgcacca ggactggctg aacggcaaag agtacaagtg caaggtgtcc 1020
aacaagggcc tgcccagctc catcgagaaa accatcagca aggccaaggg ccagccccgc 1080
gaaccccagg tgtacacact gcctccaagc caggaagaga tgaccaagaa tcaggtgtcc 1140
ctgacctgtc tcgtgaaagg cttctacccc tccgatatcg ccgtggaatg ggagagcaac 1200
ggccagcccg agaacaacta caagaccacc ccccctgtgc tggactccga tggctcattc 1260
ttcctgtaca gcagactgac cgtggacaag agccggtggc aggaaggcaa cgtgttcagc 1320
tgctccgtga tgcacgaggc cctgcacaac cactacaccc agaagtccct gtctctgagc 1380
ctgggcaaa 1389
<210> 149
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #110L2h
<400> 149
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Pro Leu Ile Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp
100 105 110
Ser Ser Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 150
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #110L2h
<400> 150
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 120
atcacctgta gagccagcca gggcatcagc aactacctga actggtatca gcagaagccc 180
ggcaaggccc ctaagcctct gatctactac accagcaacc tgcagagcgg cgtgcccagc 240
agattttctg gctctggcag cggcaccgac tacaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctacta ctgccagcag tacgacagca gccccagaac atttggcgga 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<210> 151
<211> 234
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> #110L12h
<400> 151
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly
35 40 45
Ile Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Pro Leu Ile Tyr Tyr Thr Ser Asn Leu Gln Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asp
100 105 110
Ser Ser Pro Arg Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 152
<211> 702
<212> ДНК
<213> Искусственная последовательность
<220>
<223> #110L12h
<400> 152
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccaga tgacacagag ccctagcagc ctgtctgcca gcgtgggaga cagagtgacc 120
atcacctgta gagccagcca gggcatcagc aactacctga actggtatca gcagaagccc 180
ggcaaggccc ctaagcctct gatctactac accagcaacc tgcagagcgg cgtgcccagc 240
agattttctg gctctggcag cggcaccgac ttcaccctga ccatatctag cctgcagcct 300
gaggacttcg ccacctacta ctgccagcag tacgacagca gccccagaac atttggcgga 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gt 702
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| АНТИТЕЛА К ВИРУСУ ДЕНГЕ, ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ВАРИАНТЫ FC-ОБЛАСТЕЙ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2758596C2 |
| КАНИНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ЧЕЛОВЕЧЕСКОГО CTLA-4 | 2020 |
|
RU2822460C2 |
| АНТИТЕЛО ПРОТИВ ЧЕЛОВЕЧЕСКОГО TLR7 | 2019 |
|
RU2808123C2 |
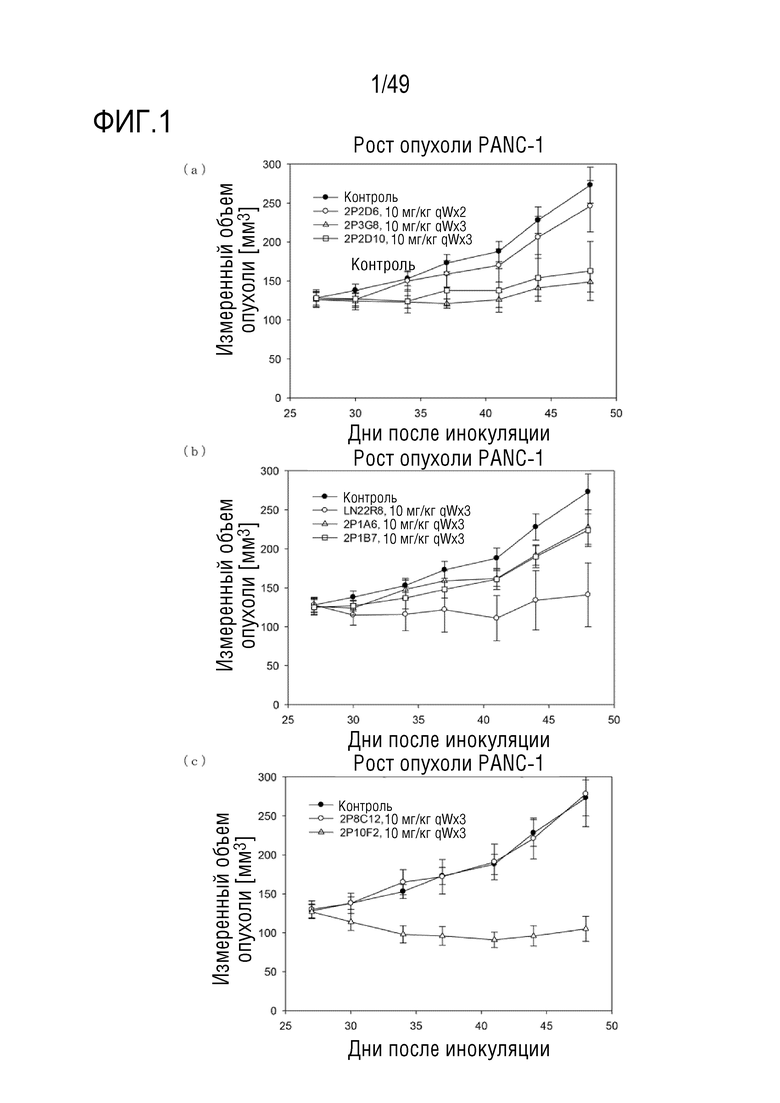







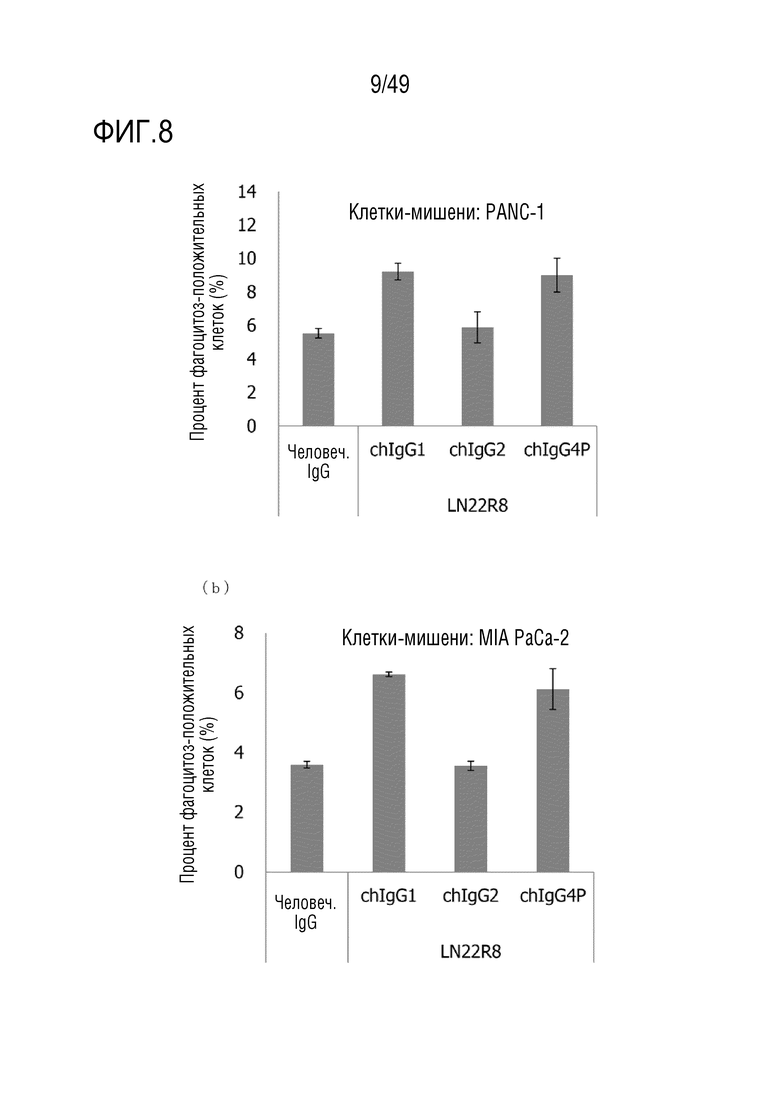














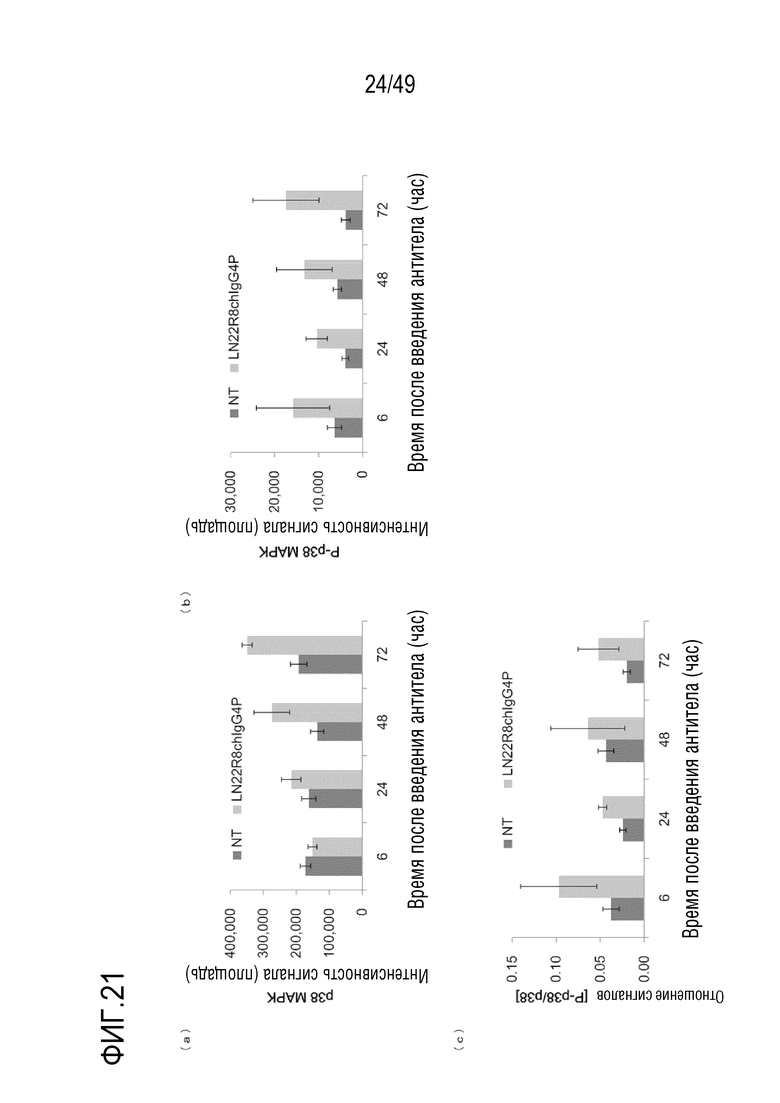


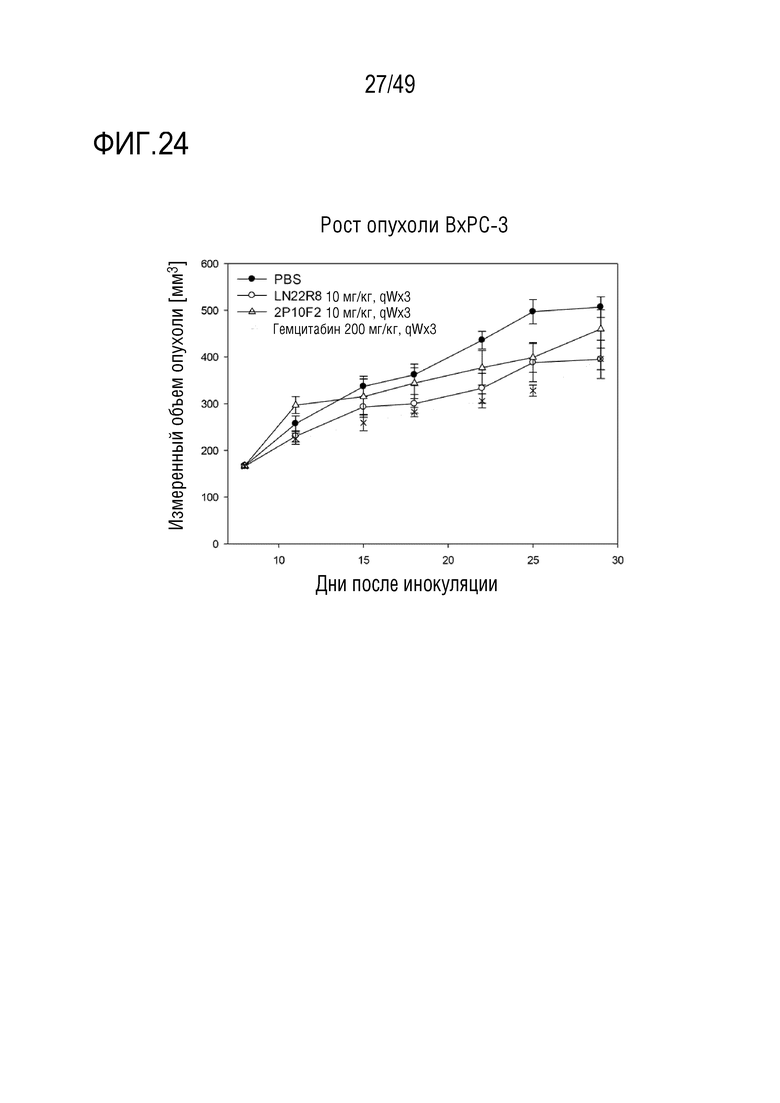





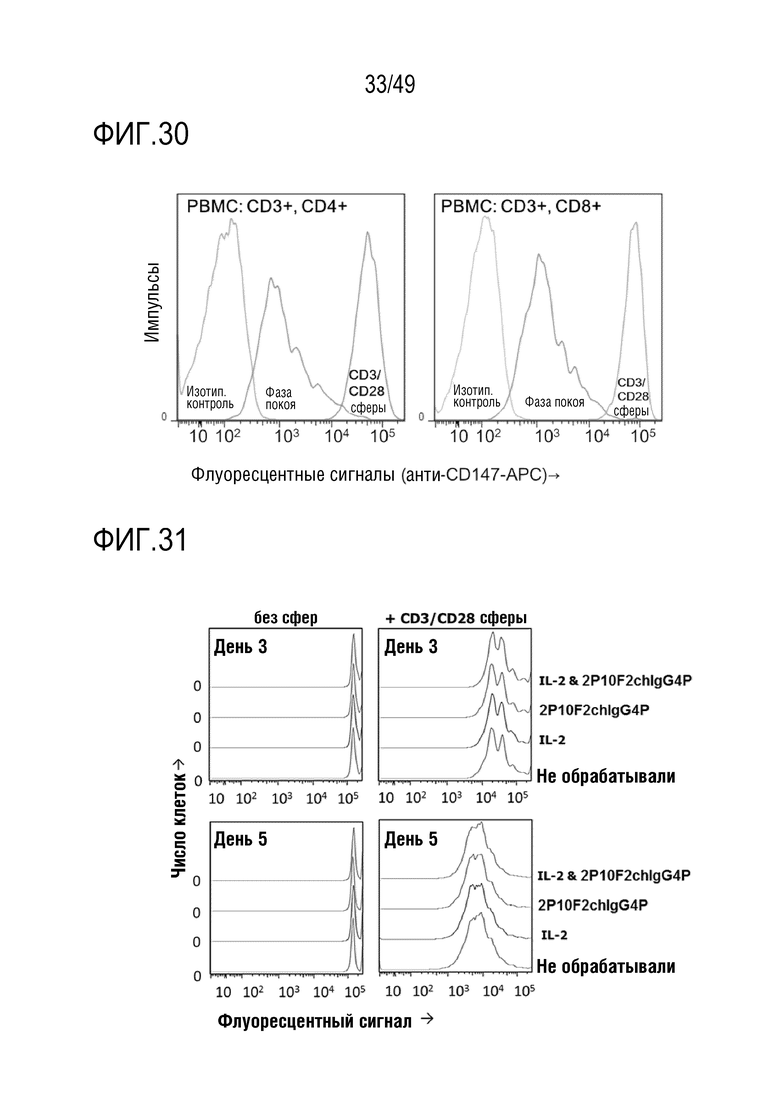









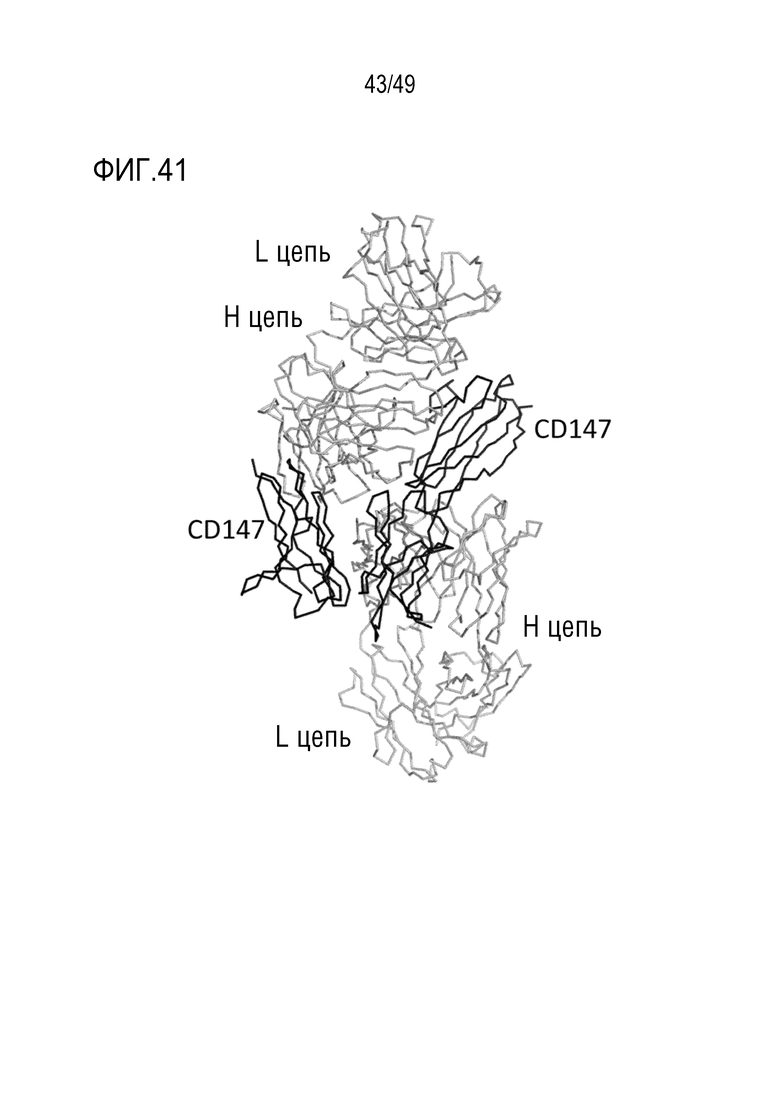


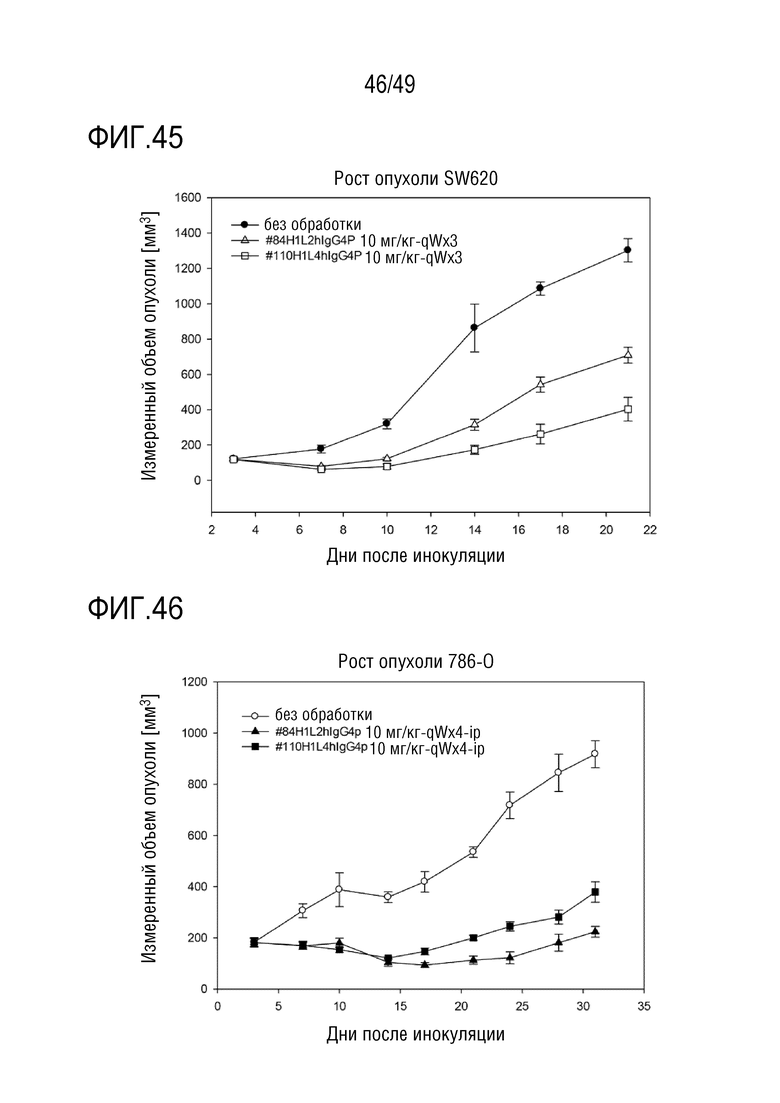

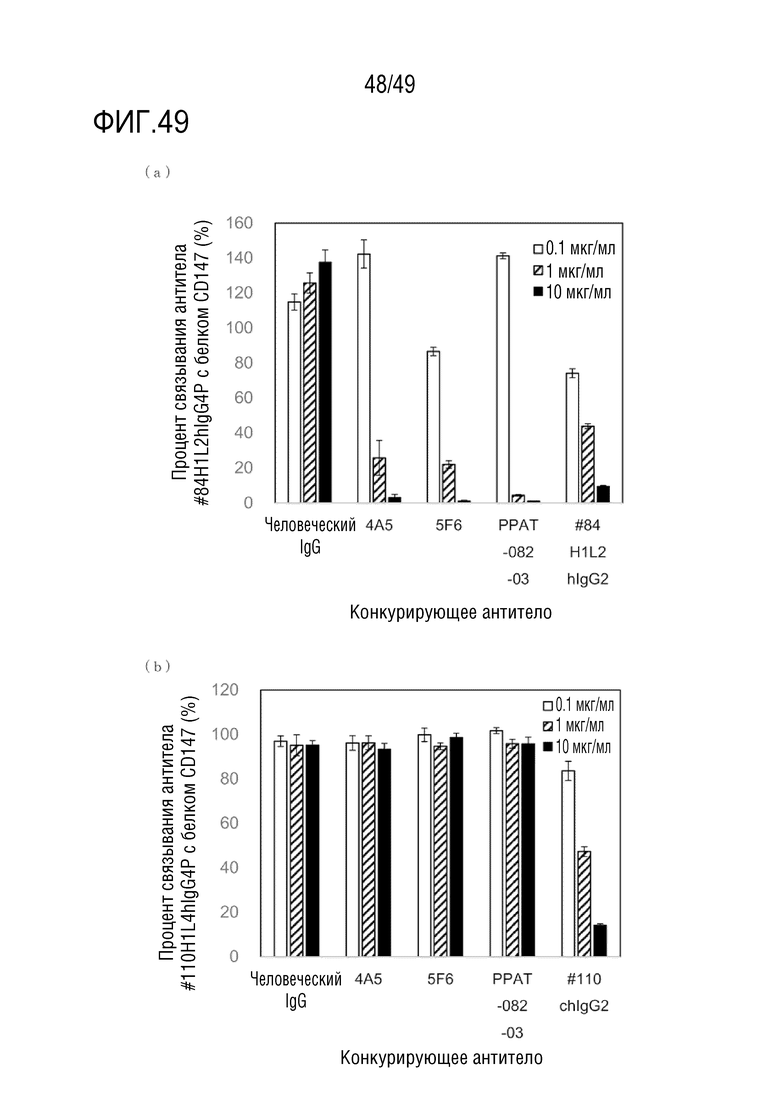

Изобретение относится к области биотехнологии. Описана группа изобретений, включающая антитело против CD147 человека или его антигенсвязывающий фрагмент, которое активирует сигнальную трансдукцию посредством CD147 (варианты), противоопухолевую композицию, содержащую вышеуказанное антитело, где опухоль представляет собой опухоль, экспрессирующую CD147, способ лечения опухоли, экспрессирующей CD147, и применение антитела, или его антигенсвязывающего фрагмента, или композиции для лечения опухоли, где опухоль является опухолью, экспрессирующей CD147. Изобретение расширяет арсенал средств против CD147. 7 н. и 27 з.п. ф-лы, 50 ил., 8 табл., 26 пр.
1. Антитело против CD147 человека или его антигенсвязывающий фрагмент, которое содержит последовательности, указанные в нижеследующих подпунктах (c) и (d), и которое активирует сигнальную трансдукцию посредством CD147:
(c) вариабельную область тяжелой цепи, выбранную из группы, состоящей из нижеследующих подпунктов (c1)-(c4):
(c1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135;
(c2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147;
(c3) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпунктах (c1) или (c2); и
(c4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в любом из подпунктов (c1)-(c3); и
(d) вариабельную область легкой цепи, выбранной из группы, состоящей из нижеследующих последовательностей, указанных в подпунктах (d1)-(d5):
(d1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137;
(d2) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149;
(d3) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151;
(d4) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в любом из подпунктов (d1) и (d3); и
(d5) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в любом из подпунктов (d1)-(d4).
2. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:135; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:137.
3. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:135; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:137.
4. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:149.
5. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:149.
6. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-136 в аминокислотной последовательности SEQ ID NO:147; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:151.
7. Антитело или его антигенсвязывающий фрагмент по п.1, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:147; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:151.
8. Антитело против CD147 человека или его антигенсвязывающий фрагмент, которое содержит последовательности, указанные в нижеследующих подпунктах (а) и (b), и которое активирует сигнальную трансдукцию посредством CD147:
(a) вариабельную область тяжелой цепи, выбранную из группы, состоящей из последовательностей, указанных в нижеследующих подпунктах (a1)-(a4):
(a1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123;
(a2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125;
(a3) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (a1) или (a2); и
(a4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в любом из подпунктов (a1)-(a3); и
(b) вариабельную область легкой цепи, выбранную из группы, состоящей из последовательностей, указанных в нижеследующих подпунктах (b1)-(b3):
(b1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127;
(b2) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности по подпункту (b1); и
(b3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (b1) или (b2).
9. Антитело или его антигенсвязывающий фрагмент по п.8, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:123, или
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-140 в аминокислотной последовательности SEQ ID NO:125; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:127.
10. Антитело или его антигенсвязывающий фрагмент по п.8, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-466 в аминокислотной последовательности SEQ ID NO:123, или
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-467 в аминокислотной последовательности SEQ ID NO:125; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:127.
11. Антитело против CD147 человека или его антигенсвязывающий фрагмент, которое содержит последовательности, указанные в нижеследующих подпунктах (e) и (f), и которое активирует сигнальную трансдукцию посредством CD147:
(e) вариабельную область тяжелой цепи, выбранную из группы, состоящей из следующих последовательностей, указанных в нижеследующих подпунктах (e1)-(e4):
(e1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129;
(e2) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131;
(e3) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (e1) или (e2); и
(e4) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в любом из подпунктов (e1)-(e3); и
(f) вариабельную область легкой цепи, выбранную из группы, состоящей из последовательностей, указанных в нижеследующих подпунктах (f1)-(f3):
(f1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133;
(f2) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (f1); и
(f3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (f1) или (f2).
12. Антитело или его антигенсвязывающий фрагмент по п.11, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:129, или
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-137 в аминокислотной последовательности SEQ ID NO:131; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:133.
13. Антитело или его антигенсвязывающий фрагмент по п.11, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-463 в аминокислотной последовательности SEQ ID NO:129, или
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:131; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:133.
14. Антитело против CD147 человека или его антигенсвязывающий фрагмент, которое содержит последовательности, указанные в нижеследующих подпунктах (g) и (h), и которое активирует сигнальную трансдукцию посредством CD147:
(g) вариабельную область тяжелой цепи, выбранную из группы, состоящей из последовательностей, указанных в нижеследующих подпунктах (g1)-(g3):
(g1) вариабельной области тяжелой цепи, состоящей из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139;
(g2) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (g1); и
(g3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (g1) или (g2); и
(h) вариабельную область легкой цепи, выбранную из группы, состоящей из последовательностей, указанных в нижеследующих подпунктах (h1)-(h3):
(h1) вариабельной области легкой цепи, состоящей из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141;
(h2) аминокислотной последовательности, которая по меньшей мере на 95% или более идентична последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (h1); и
(h3) аминокислотной последовательности, в которой одна или несколько аминокислот удалены, заменены или добавлены в последовательности каркасной области вне каждой последовательности CDR в последовательности, указанной в подпункте (h1) или (h2).
15. Антитело или его антигенсвязывающий фрагмент по п.14, которое содержит:
вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях 20-138 в аминокислотной последовательности SEQ ID NO:139; и
вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях 21-128 в аминокислотной последовательности SEQ ID NO:141.
16. Антитело или его антигенсвязывающий фрагмент по п.14, которое содержит:
тяжелую цепь, состоящую из аминокислотных остатков в положениях 20-464 в аминокислотной последовательности SEQ ID NO:139; и
легкую цепь, состоящую из аминокислотных остатков в положениях 21-234 в аминокислотной последовательности SEQ ID NO:141.
17. Антитело против CD147 человека или его антигенсвязывающий фрагмент по любому из пп.1-16, которое характеризуется пониженной опосредованной антителами клеточной цитотоксичностью (ADCC) или у которого ADCC отсутствует.
18. Антитело против CD147 человека или его антигенсвязывающий фрагмент по любому из пп.1-16, которое характеризуется пониженной комплементзависимой цитотоксичностью (CDC) или у которого CDC отсутствует.
19. Антитело против CD147 человека или его антигенсвязывающий фрагмент по любому из пп.1-16, которое характеризуется пониженным антителозависимым клеточным фагоцитозом (ADCP) или у которого ADCP отсутствует.
20. Противоопухолевая композиция, где композиция содержит по меньшей мере одно антитело или его антигенсвязывающий фрагмент по любому из пп.1-19 и опухоль представляет собой опухоль, экспрессирующую CD147.
21. Композиция по п.20, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, лимфому, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль (GIST), лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
22. Композиция по п.20, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
23. Композиция по п.20, где опухоль представляет собой SMAD4-положительную опухоль или опухоль c пониженным или отсутствующим уровнем экспрессии KLF5.
24. Композиция по любому из пп.20-23, дополнительно содержащая еще одно противоопухолевое средство.
25. Способ лечения опухоли, экспрессирующей CD147, включающий введение пациенту антитела или его антигенсвязывающего фрагмента по любому из пп.1-19 или композиции по любому из пп.20-24.
26. Способ по п.25, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, лимфому, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль (GIST), лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
27. Способ по п.25, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
28. Способ по п.25, где опухоль представляет собой SMAD4-положительную опухоль или опухоль с пониженным уровнем или отсутствием экспрессии KLF5.
29. Способ по любому из пп.25-28, где антитело, или его антигенсвязывающий фрагмент, или композицию вводят в комбинации с еще одним противоопухолевым средством.
30. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-19 или композиции по любому из пп.20-24 для лечения опухоли, где опухоль является опухолью, экспрессирующей CD147.
31. Применение по п.30, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, рак молочной железы, рак матки, рак яичников, рак легкого, лимфому, рак щитовидной железы, рак кожи, рак головы и шеи, саркому, рак предстательной железы, рак мочевого пузыря, рак головного мозга, гастроинтестинальную стромальную опухоль (GIST), лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
32. Применение по п.30, где опухоль представляет собой рак поджелудочной железы, рак печени, рак желудка, рак толстой кишки, рак почки, лейкоз, острый миелолейкоз (AML), хронический миелолейкоз (CML), хронический лимфоцитарный лейкоз (CLL), острый лимфоцитарный лейкоз (ALL), злокачественную лимфому, B-клеточную лимфому, неходжкинскую лимфому или диффузную крупноклеточную В-клеточную лимфому (DLBCL).
33. Применение по п.30, где опухоль представляет собой SMAD4-положительную опухоль или опухоль, имеющую пониженный уровень экспрессии или с отсутствием экспрессии KLF5.
34. Применение по п.30, где антитело, или его антигенсвязывающий фрагмент, или композицию вводят в комбинации с еще одним противоопухолевым средством.
| WO 2010036460 A2, 01.04.2010 | |||
| WO 1999045031 A2, 10.09.1999 | |||
| Qin H | |||
| et al | |||
| Раздвижной паровозный золотник со скользящими по его скалке поршнями и упорными для них шайбами | 1922 |
|
SU147A1 |
| Experimental Cell Research, 2017, 358(2), 279-289 | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| WO 2017061602 A1, 13.04.2017 | |||
| СПОСОБ УПРАВЛЕНИЯ МНОГОЗОННЫМ ВЫПРЯМИТЕЛЕМ ОДНОФАЗНОГО ПЕРЕМЕННОГО ТОКА | 2006 |
|
RU2322749C1 |
| RU 2014131097 A, 20.02.2016. | |||

