Родственные заявки
На настоящей заявке испрашивается приоритет заявки на патент Великобритании № 1707561.5, поданной 11 мая 2017 года, полное содержание которой включено в настоящее описание посредством ссылки.
Список последовательностей
Настоящая заявка содержит список последовательностей, который был представлен в электронном виде в формате ASCII и в полном объеме включен в настоящее описание посредством ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые связываются с комплексом GARP и TGF-β1, в частности, с комплексом человеческого GARP и человеческого TGF-β1. Эти антитела и антигенсвязывающие фрагменты проявляют комбинацию преимущественных свойств, включая связывание антигена с высокой аффинностью и способность ингибировать высвобождение активного TGF-β из регуляторных Т-клеток. Антитела и антигенсвязывающие фрагменты по настоящему изобретению усовершенствованы по сравнению с антителами предшествующего уровня техники, которые связываются с комплексом GARP и TGF-β1. В частности, антитела и антигенсвязывающие фрагменты по настоящему изобретению относительно устойчивы к дезамидированию, изомеризации и окислению, так что они проявляют повышенную стабильность по сравнению с анти-GARP-TGF-β1-антителами, описанными на предшествующем уровне техники.
Уровень техники
Регуляторные T-клетки (также известные как «Treg» или Foxp3+ регуляторные T-клетки) являются важным компонентом иммунной системы. В частности, Treg играют критическую роль в иммунном гомеостазе, подавляя различные аспекты иммунного ответа. Как следствие их роль в координации иммунного ответа, Treg с нарушенной активностью может привести к развитию различных заболеваний и патологических состояний. В частности, недостаточная функция Treg может быть причиной аутоиммунной патологии, тогда как избыточная активность Treg связана с ингибированием противоопухолевых ответов у пациентов со злокачественным новообразованием.
Белок GARP (преобладающие повторы гликопротеина A) был идентифицирован как высокоэкспрессируемый маркер на поверхности Treg, особенно активированных Treg. GARP представляет трансмембранный белок c молекулярной массой 80 кДа с внеклеточной областью, включающей 20 лейцин-богатых повторов. Они также известны как LRRC32. GARP служит рецептором для TGF-β, особенно латентной формы TGF-β, и необходим для экспрессии латентного TGF-β на клетках Treg (EM Shevach. Expert Opin. Ther. Targets (2016) 21 (2), 191-200).
TGF-β является цитокином, который, как известно, играет роль во многих процессах, включая пролиферацию и дифференцировку клеток, морфогенез тканей, воспаление и апоптоз. Он также был идентифицирован в качестве важного фактора роста, участвующего в развитии злокачественного новообразования, и довольно необычно, был идентифицирован в качестве цитокина с опухоль-стимулирующей и опухоль-подавляющей активностью.
Синтез и активация TGF-β является многоступенчатым процессом, который регулируется на разных уровнях. TGF-β синтезируется в виде димерного предшественника про-TGF-β, где каждая полипептидная цепь состоит из латентно-ассоциированного пептида (LAP) и зрелой области TGF-β. Про-TGF-β подвергается расщеплению ферментом фурином с образованием «латентного TGF-β», неактивной формы, в которой LAP остается нековалентно связанным со зрелой областью TGF-β каждой полипептидной цепи (см. фиг.1). Локализованный на мембране GARP служит для транспорта и якорного закрепления латентного TGF-β на клеточной поверхности Treg, и именно из этого мембраносвязанного комплекса GARP-латентный TGF-β высвобождается активная форма TGF-β. Для объяснения того, как активный TGF-β высвобождается из комплекса GARP-латентный TGF-β на поверхности Treg, были предложены различные механизмы. Однако в настоящее время полагается, что интегрины, в частности, αvβ6 и αvβ8, играют важную роль в регуляции усилий сдвига, необходимых для высвобождения зрелого димера TGF-β.
После высвобождения активный димер TGF-β может функционировать в качестве аутокринного или паракринного медиатора даунстрим сигнальных путей. Считается, что в контексте иммунной системы высвобождение TGF-β из клеток Treg влияет на активность различных эффекторных T-клеток, а также самих Treg (см. фиг. 1). Поскольку Treg играют важную роль в иммуносупрессии, то полагается, что TGF-β, высвобождаемый из Treg и действующий аутокринно, может участвовать в обеспечении супрессии Treg. В частности, полагают, что TGF-β1, происходящий из Treg, играет важную роль в Treg-опосредованной супрессии опухолевого иммунитета.
Учитывая роль TGF-β, происходящего из Treg, в подавлении иммунного ответа в микроокружении опухоли, имеется интерес к нацеливанию на этот путь в качестве альтернативного подхода для иммунотерапии злокачественного новообразования. Например, терапевтические агенты, способные ослабить этот путь, могут служить пригодными инструментами для повышения эффективности противораковых вакцин или других стратегий иммунотерапии злокачественного новообразования, в которых используются возможности иммунной системы организма для лечения злокачественного новообразования.
Cuende et al. (Sci. Transl. Med., 2015 Apr. 22; 7 (284): 284ra56) описывают получение и характеристику двух моноклональных антител (MHG-8 и LHG10), которые связываются с комплексом GARP-TGF-β на Treg и ингибируют продукцию TGF-β. Эти два антитела также описаны и охарактеризованы в международных заявках на патент WO2015/015003 и WO2016/125017. Было показано, что эти антитела способны ингибировать иммуносупрессивную активность человеческих Treg на мышиной модели синдрома «ксеногенный трансплантат против хозяина». Результаты этой работы служат для подтверждения того, что комплекс GARP-TGF-β является интересной терапевтической мишенью для целей модуляции функции Treg и, следовательно, для лечения таких заболеваний, как злокачественное новообразование и аутоиммунные заболевания, где уровень активности Treg играет важную роль. Однако остается необходимость в усовершенствованных антителах к GARP-TGF-β, способных ингибировать высвобождение TGF-β и тем самым модифицировать активность Treg. Настоящее изобретение решает эту проблему, как в настоящем описании описано.
Сущность изобретения
Настоящее изобретение улучшает современное состояние обеспечением новых антител и их антигенсвязывающих фрагментов, которые связываются с комплексом человеческих GARP-TGF-β1. Антитела и антигенсвязывающие фрагменты по настоящему изобретению получены из антитела к GARP-TGF-β1 «LHG-10», описанного в международных заявках на патент WO2015/015003 и WO2016/125017. Последовательности вариабельной области тяжелой цепи и легкой цепи LHG-10 показаны в SEQ ID NO:1 и 2, соответственно, и вариабельная область варианта с перестановленной легкой цепью, LHG-10.6 (также описанного в WO2015/ 015003 и WO2016/125017), показана в SEQ ID NO: 3. Антитела по настоящему изобретению отличаются, в частности, в отношении определенных последовательностей CDR по сравнению с LHG-10 и LHG-10.6, в частности, в отношении последовательностей CDR2 и CDR3 вариабельной области тяжелой цепи. Анти-GARP-TGF-β1-антитела, LHG-10 и LHG-10.6, обладают последовательностью CDR2 тяжелой цепи: RIDPEDGGTKYAQKFQG (SEQ ID NO: 5); и последовательностью CDR3 тяжелой цепи: NEWETVVVGDLMYEYEY (SEQ ID NO: 6), тогда как антитела по настоящему изобретению содержат последовательность CDR2 тяжелой цепи: RIDPEDAGTKYAQKFQG (SEQ ID NO: 12); и последовательность CDR3 тяжелой цепи: YEWETVVVGDLMYEYEY (SEQ ID NO: 13).
Различия в последовательностях CDR2 и CDR3 тяжелой цепи, описанные в настоящем описании, приводят к антителам, которые усовершенствованы по сравнению с антителами предшествующего уровня техники благодаря их повышенной стабильности. Более конкретно, антитела по настоящему изобретению относительно устойчивы к дезамидированию, изомеризации и окислению, так что они проявляют повышенную стабильность. Удивительно, что эти специфические замены в областях CDR2 и CDR3 тяжелой цепи, которые приводят к повышенной стабильности, существенно не снижают аффинность связывания антител с комплексом GARP-TGF-β1. Повышенная стабильность в сочетании с высокоаффинным связыванием с мишенью делает антитела по настоящему изобретению особенно подходящими для клинической разработки в качестве терапевтических агентов, например, в качестве терапевтических агентов для лечения злокачественного новообразования.
В первом аспекте настоящее изобретение обеспечивает антитело или его антигенсвязывающий фрагмент, которые связываются с комплексом человеческих GARP-TGF-β1, где антитело или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи (VH), где:
CDR3 VH содержит аминокислотную последовательность YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит аминокислотную последовательность RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержит аминокислотную последовательность SYYID (SEQ ID NO: 4).
В некоторых вариантах осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которые связываются с комплексом человеческих GARP-TGF-β1, где антитело или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи (VH), где:
CDR3 VH состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4).
Антитело или антигенсвязывающий фрагмент могут дополнительно содержать вариабельную область легкой цепи (VL), где:
CDR3 VL содержит аминокислотную последовательность QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит аминокислотную последовательность GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит аминокислотную последовательность QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления антитело или антигенсвязывающий фрагмент могут дополнительно содержать вариабельную область легкой цепи (VL), где:
CDR3 VL состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL состоит из аминокислотной последовательности GASRLKT (SEQ ID NO:10), и
CDR1 VL состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент содержит:
вариабельную область тяжелой цепи (VH), где:
CDR3 VH содержит аминокислотную последовательность YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит аминокислотную последовательность RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержит аминокислотную последовательность SYYID (SEQ ID NO: 4); и
вариабельную область легкой цепи (VL), где:
CDR3 VL содержит аминокислотную последовательность QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит аминокислотную последовательность GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит аминокислотную последовательность QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент содержит:
вариабельную область тяжелой цепи (VH), где:
CDR3 VH состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4); и
вариабельную область легкой цепи (VL), где:
CDR3 VL состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты включают, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и/или, по меньшей мере, одну вариабельную область легкой цепи (VL), которые представляют собой гуманизированный вариант, вариант зародышевого типа или аффинный вариант верблюжьей области VH или VL.
В некоторых вариантах осуществления обеспечиваются антитела или их антигенсвязывающие фрагменты, которые связываются с комплексом человеческого GARP и человеческого TGF-β1, где антитела или антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, выбранную из следующего:
(i) VH, содержащей или состоящей из аминокислотной последовательности SEQ ID NO: 14; или
(ii) VH, содержащей или состоящей из аминокислотной последовательности, имеющей, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность с SEQ ID NO: 14.
Альтернативно или дополнительно, антитела или антигенсвязывающие фрагменты могут содержать вариабельную область легкой цепи (VL), выбранную из следующего:
(i) VL, содержащей или состоящей из аминокислотной последовательности SEQ ID NO: 15; или
(ii) VL, содержащей или состоящей из аминокислотной последовательности, имеющей, по меньшей мере, 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, 99% идентичность с SEQ ID NO: 15.
Для вариантов осуществления, в которых области антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью последовательности с референсной последовательностью, области VH и/или VL могут сохранять последовательности CDR, идентичные тем, которые присутствуют в референсной последовательности так, что вариабельность присутствует только в областях каркасной области.
В конкретном варианте осуществления обеспечены антитела или их антигенсвязывающие фрагменты, где вариабельная область тяжелой цепи (VH) содержит или состоит из аминокислотной последовательности SEQ ID NO: 14, и вариабельная область легкой цепи (VL) содержит или состоит из аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах осуществления антитела по изобретению включают домен CH1, шарнирную область, домен CH2 и домен CH3 антитела человека, в частности, IgG1, IgG2 человека, IgG3 или IgG4 человека. В некоторых вариантах осуществления антитело включает домен СН3 человеческого IgG4 и включает замену S228P в домене СН3.
Антитела, которые связываются с комплексом GARP-TGF-β1, могут включать, по меньшей мере, одну полноразмерную тяжелую цепь иммуноглобулина и/или, по меньшей мере, одну полноразмерную легкую цепь лямбда или каппа. В некоторых вариантах осуществления антитела содержат тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 17. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 16. В некоторых вариантах осуществления обеспечиваются моноклональные антитела содержащие легкую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 17. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 16, и легкую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью с аминокислотной последовательностью, показанной в SEQ ID NO: 17.
Для вариантов осуществления, в которых тяжелая и/или легкая цепи антител определяются конкретной процентной идентичностью последовательности с референсной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять последовательности CDR, идентичные тем, которые присутствуют в референсной последовательности, так что вариабельность присутствует только за пределами доменов CDR.
Если в настоящей заявке не указано иное, то % идентичность последовательности между двумя аминокислотными последовательностями можно определить сравнением этих двух последовательностей, выровненных оптимальным образом, и в котором сравниваемая аминокислотная последовательность может содержать добавления или делеции относительно референсной последовательности для оптимального выравнивания этих двух последовательностей. Процент идентичности рассчитывается определением числа идентичных положений, для которых аминокислотный остаток является идентичным между двумя последовательностями, делением этого числа идентичных положений на общее число положений в окне сравнения и умножением полученного результата на 100, с получением процента идентичности между этими двумя последовательностями. Например, можно использовать программу BLAST «Последовательности BLAST 2» (Tatusova et al, «Blast 2 sequences - a new tool for comparing protein and nucleotide sequences», FEMS Microbiol Lett. 174:247-250), используются параметры по умолчанию (в частности, такие параметры как «штраф за открытый гэп»: 5 и «штраф за удлинение гэпа»: 2; выбранной матрицей является, например, матрица «BLOSUM 62», предложенная программой), процент идентичности между двумя сравниваемыми последовательностями рассчитывается непосредственно программой.
Каждое из анти-GARP-TGF-β1-антител или их антигенсвязывающих фрагментов, обеспеченных в настоящем описании, может проявлять одно или более из следующих свойств/признаков:
- антитело или антигенсвязывающий фрагмент могут перекрестно реагировать с комплексом GARP-TGF-β1 яванского макака;
- антитело или антигенсвязывающий фрагмент могут связываться с GARP-TGF-β1 человека с высокой аффинностью;
- антитело или антигенсвязывающий фрагмент могут включать область VH и область VL, которые при тестировании в виде Fab-фрагмента демонстрируют скорость диссоциации (Koff) комплекса человеческих GARP и TGF-β1 ниже 5×10-4 с-1;
- антитело или антигенсвязывающий фрагмент могут включать область VH и область VL, которые при тестировании в виде Fab-фрагмента демонстрируют скорость диссоциации (Koff) комплекса человеческих GARP и TGF-β1 в диапазоне от 1×10-6 с-1 до 5×10-4 с-1;
- антитело или антигенсвязывающий фрагмент могут включать область VH и область VL, которые при тестировании в виде mAb демонстрируют KD ниже 1,7×10-9 М;
- антитело или антигенсвязывающий фрагмент могут блокировать или ингибировать высвобождение активного TGF-β1 из регуляторных Т-клеток.
В дополнительных аспектах изобретение также относится к полинуклеотидным молекулам, которые кодируют вышеуказанные антитела и антигенсвязывающие фрагменты, в дополнение к экспрессионным векторам, содержащим полинуклеотиды, клеткам-хозяевам, содержащим векторы, и способам рекомбинантной экспрессии/продуцирования антител, описанных в настоящем описании.
В еще одном аспекте изобретение относится к фармацевтической композиции, содержащей любое из антител к GARP-TGF-β1 или их антигенсвязывающих фрагментов, описанных в настоящем описании, и фармацевтически приемлемый носитель или эксципиент.
Еще один аспект изобретения относится к способам терапевтического лечения с использованием вышеуказанных антител к GARP-TGF-β1 или их антигенсвязывающих фрагментов, в частности, для профилактики и/или лечения расстройств, связанных с TGF-β. В некоторых вариантах осуществления изобретение относится к способам лечения с использованием антител к GARP-TGF-β1 или их антигенсвязывающих фрагментов, где заболевание или патологическое состояние, подлежащее лечению, выбрано из группы, состоящей из воспалительных заболеваний, хронической инфекции, злокачественного новообразования, фиброза, сердечно-сосудистого заболевания, цереброваскулярной болезни и нейродегенеративного заболевания. В некоторых вариантах осуществления антитела к GARP-TGF-β1 или их антигенсвязывающие фрагменты вводят в комбинации с другим лечением в виде части комбинированной терапии. Например, антитела к GARP-TGF-β1 или их антигенсвязывающие фрагменты можно вводить в комбинации с иммунотерапевтическим агентом, необязательно иммуностимулирующим антителом или противоопухолевой вакциной.
Эти и другие варианты осуществления изобретения будут лучше оценены и поняты при рассмотрении в сочетании со следующим описанием и прилагаемыми фигурами. Следует понимать, однако, что нижеследующее описание, в котором описаны различные варианты осуществления изобретения и многочисленные его конкретные детали, представлено в качестве иллюстрации, а не ограничения. Многие замены, модификации, дополнения и/или перестановки могут быть сделаны в пределах объема изобретения, не отступая от его сущности, и изобретение включает все такие замены, модификации, дополнения и/или перестановки.
Краткое описание фигур
На фиг.1 представлена схема, показывающая связывание латентного TGF-β с GARP на поверхности регуляторных Т-клеток. TGF-β продуцируется в виде предшественника «про-TGF-β» и подвергается расщеплению с образованием «латентного-TGF-β», формы, в которой зрелый димер TGF-β остается нековалентно связанным с областью латентно-ассоциированного пептида (LAP) каждого полипептида. Именно эта латентная форма связывается с GARP на поверхности Treg-клеток. Полагается, что интегрины αvβ6 и αvβ8 ответственны за высвобождение зрелого или «активного TGF-β» с поверхности клетки. Эта активная форма может функционировать в качестве паракринного медиатора, вызывая эффекты в различных клетках-мишенях, или может функционировать в качестве аутокринного медиатора, связываясь с рецептором TGF-β на Treg-клетках.
На фиг.2 показана активность связывания с мишенью, измеренная с помощью анализа Biacore™ или поверхностного плазмонного резонанса (SPR) для антител 39B6 IgG1N297Q и 39B6 IgG4S228P в течение 56-дневного периода для образцов, хранившихся при -20°C, 5°C и 37°C в PBS или PBS-Твине (PBSTw). Контрольный образец (который хранили при -20°С) принимали за 100% активность связывания на каждую временную точку.
На фиг. 3 показаны результаты тестирования вариантов антитела 39B6-A в анализе, предназначенном для мониторинга фосфорилирования SMAD2 после активации рецептора TGF-β. Фосфорилирование SMAD2 служит маркером активации сигнального пути TGF-β после связывания TGF-β с его рецептором. Если фосфорилирование SMAD2 снижено, то активность TGF-β ингибирована. На фиг. 3А: вестерн-блоты, показывающие снижение фосфорилирования SMAD2 в присутствии различных концентраций анти-GARP-TGF-β-антител 39B6-A, 39B6-AVE, 39B6-AEE, 39B6-AYE, 39B6-ANR и 39B6-ANK. На фиг. 3В: графическое представление данных, представленных в (А), показывающих процент ингибирования фосфорилирования SMAD2 при различных концентрациях антител.
На фиг.4 показаны результаты тестирования вариантов антитела 39B6-A в анализе, предназначенном для измерения активности TGF-β с помощью репортерного гена люциферазы, конъюгированного с промотором SMAD. Графики показывают процентное ингибирование сигнала люминесценции в присутствии различных концентраций анти-GARP-TGF-β-антител, LHG-10, 39B6-A, 39B6-AVE, 39B6-AEE, 39B6-AYE, 39B6-ANR и 39B6-ANK.
На фиг.5 показано процентное образование агрегатов в течение 56-дневного периода для антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR, хранившихся при 5°C и 37°C. Образование агрегатов контролировали с помощью эксклюзионной хроматографии (SE-HPLC).
На фиг.6 показано процентное образование фрагментов в течение 56-дневного периода для антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR, хранившихся при 37°C. Образование фрагментов контролировали с помощью эксклюзионной хроматографии (SE-HPLC).
На фиг.7 показана процентная площадь мономеров в течение 56-дневного периода для антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR, хранившихся при 5°C и 37°C. Площадь мономеров контролировали с помощью эксклюзионной хроматографии (SE-HPLC).
На фиг. 8 показаны результаты анализа SDS-PAGE образцов антител, хранившихся в течение 56 суток при референсной температуре (-20°C), при 5°C и при 37°C. На фиг. 8А: 39B6-AVE. На фиг. 8B: 39B6-AYE. На фиг. 8C: 39B6-ANK. На фиг. 8D: 39B6-ANR. Маркеры находятся в центре каждого геля. Слева от маркеров 3 образца: (i) контрольный образец; (ii) хранение при 5°C; и (iii) хранение при 37°C, тестированные в невосстанавливающих условиях, и справа от маркеров, 3 образца: (i) контрольный образец; (ii) хранение при 5°C; и (iii) хранение при 37°С, тестированные в восстанавливающих условиях.
На фиг.9 показана активность связывания с мишенью, измеренная анализом Biacore® или SPR для антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR в течение 56-дневного периода для образцов, хранившихся при -20°C, 5°C и 37°C. Контрольный образец (хранение при -20°С) принимали за 100% активность связывания на каждую временную точку.
На фиг.10 показана концентрация белка (мг/мл) для образцов антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR в течение 56-дневного периода для образцов, хранившихся при -20°С (контрольный образец), 5°С и 37°С.
На фиг. 11 показаны результаты анализа SDS-PAGE образцов антител после 10 циклов замораживания-оттаивания. Маркеры находятся в центре геля. Слева от маркеров, 4 образца представляют образцы, анализированные в невосстанавливающих условиях: (i) контрольный образец для 39B6-AVE; (ii) образец с замораживанием-оттаиванием 39B6-AVE; (iii) контрольный образец для 39B6-AYE; и (iv) образец с замораживанием-оттаиванием 39B6-AYE. Справа от маркеров 4 образца представляют образцы, анализированные в восстанавливающих условиях: (i) контрольный образец для 39B6-AVE; (ii) образец с замораживанием-оттаиванием 39B6-AVE; (iii) контрольный образец для 39B6-AYE; и (iv) образец с замораживанием-оттаиванием 39B6-AYE.
На фиг. 12 показана активность связывания с мишенью, измеренная анализом Biacore® или SPR после 10 циклов замораживания-оттаивания для антител 39B6-AVE и 39B6-AYE. Контрольный образец (хранение при -20°С) принимали за 100% активность связывания на каждую временную точку.
На фиг. 13 показана концентрация белка (мг/мл) для образцов антител 39B6-AVE и 39B6-AYE после 10 циклов замораживания-оттаивания.
На фиг. 14 показана активность связывания с мишенью, измеренная анализом Biacore ® или SPR после тестирования термостабильности при температуре от 54,6°C до 71,4°C для антител 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR. Контрольный образец принимали за 100% активность связывания.
На фиг. 15 показаны результаты анализа SDS-PAGE образцов антител после 96 ч вращения. Маркеры находятся в центре геля. Слева от маркеров, 4 образца представляют образцы, анализированные в невосстанавливающих условиях: (i) контрольный образец для 39B6-AVE; (ii) подвергшийся ротационному вращению образец 39B6-AVE; (iii) контрольный образец для 39B6-AYE; и (iv) подвергшийся ротационному вращению образец 39B6-AYE. Справа от маркеров 4 образца представляют образцы, анализированные в восстанавливающих условиях: (i) контрольный образец для 39B6-AVE; (ii) подвергшийся ротационному вращению образец 39B6-AVE; (iii) контрольный образец для 39B6-AYE; и (iv) подвергшийся ротационному вращению образец 39B6-AYE.
На фиг. 16 показана активность связывания с мишенью, измеренная анализом Biacore® или SPR после тестирования ротационной стабильности для mAb 39B6-AVE и 39B6-AYE. Контрольный образец принимали за 100% активность связывания.
На фиг. 17 показана концентрация белка (мг/мл) для образцов mAb 39B6-AVE и 39B6-AYE после тестирования ротационной стабильности.
На фиг. 18 показан относительный уровень дезамидирования и изомеризации в положении N95 в антителах 39B6-ANE, 39B6-ANR и 39B6-ANK в течение 56-дневного периода. Антитела 39B6-AVE и 39B6-AYE не включали в испытание, поскольку у этих антител был удален остаток «N95» из CDR3. Также показана относительная связывающая активность для mAb 39B6-ANE, 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR в течение 56 дней, для образцов, хранившихся при 37°C.
На фиг. 19 показана необходимость в зрелом TGF-β в связывании 39B6-AYE (ARGX-115) с комплексом GARP-TGF-β. Планшеты для ELISA покрывали GARP или анти-GARP Ab, ARGX-115. Для планшетов для ELISA, покрытых GARP, обеспечивали возможность образования комплекса с полноразмерным латентным TGF-β (включая как области LAP, так и зрелого TGF-β) или комплекса с рекомбинантным LAP добавлением соответствующего рекомбинантного белка. Для планшетов для ELISA, покрытых ARGX-115, добавляли GARP и затем добавляли полноразмерный латентный TGF-β или LAP. ARGX-115 был способен связываться с GARP только в присутствии полноразмерного TGF-β. Связывание ARGX-115 с комплексом GARP-LAP отсутствовало. Напротив, анти-LAP-антитело было способным связываться с комплексом GARP-LAP. Это демонстрирует необходимость в зрелом TGF-β для связывания ARGX-115 с комплексом GARP-TGF-β.
На фиг. 20 показана способность антител нейтрализовать активацию TGF-β комплексом GARP-TGF-β с различными мутантными формами TGF-β. Нейтрализующая активность ARGX-115 отменялась в результате мутации R58 в LAP и K338 в зрелом TGF-β.
Подробное описание изобретения
А. Определения
«GARP». GARP (преобладающие повторы гликопротеина A) является членом семейства белков, содержащих лейцин-богатые повторы. Его также называют белком 32, содержащим лейцин-богатые повторы (LRRC32). GARP представляет собой трансмембранный белок с молекулярной массой 80 кДа с внеклеточной областью, состоящей в основном из 20 лейцин-богатых повторов. Полная аминокислотная последовательность варианта 2 транскрипта белка GARP человека (идентификационный номер GenBank NP_001122394.1) представляет:
MRPQILLLLALLTLGLAAQHQDKVPCKMVDKKVSCQVLGLLQVPSVLPPDTETLDLSGNQLRSILASPLGFYTALRHLDLSTNEISFLQPGAFQALTHLEHLSLAHNRLAMATALSAGGLGPLPRVTSLDLSGNSLYSGLLERLLGEAPSLHTLSLAENSLTRLTRHTFRDMPALEQLDLHSNVLMDIEDGAFEGLPRLTHLNLSRNSLTCISDFSLQQLRVLDLSCNSIEAFQTASQPQAEFQLTWLDLRENKLLHFPDLAALPRLIYLNLSNNLIRLPTGPPQDSKGIHAPSEGWSALPLSAPSGNASGRPLSQLLNLDLSYNEIELIPDSFLEHLTSLCFLNLSRNCLRTFEARRLGSLPCLMLLDLSHNALETLELGARALGSLRTLLLQGNALRDLPPYTFANLASLQRLNLQGNRVSPCGGPDEPGPSGCVAFSGITSLRSLSLVDNEIELLRAGAFLHTPLTELDLSSNPGLEVATGALGGLEASLEVLALQGNGLMVLQVDLPCFICLKRLNLAENRLSHLPAWTQAVSLEVLDLRNNSFSLLPGSAMGGLETSLRRLYLQGNPLSCCGNGWLAAQLHQGRVDVDATQDLICRFSSQEEVSLSHVRPEDCEKGGLKNINLIIILTFILVSAILLTTLAACCCVRRQKFNQQYKA (SEQ ID NO: 33).
«TGF-β». TGF-β представляет цитокин, относящийся к суперсемейству факторов роста. Существует три различных изоформы TGF-β (TGF-β1, TGF-β2 и TGF-β3), кодируемых тремя различными генами, но общие структуры изоформ TGF-β очень сходны, с гомологией на уровне 70-80%. Как в настоящем описании используется, термин TGF-β обычно используется для охвата всех трех различных изоформ TGF-β цитокина, если в контексте не указано иное.
Все три изоформы TGF-β кодируются в виде крупных белков-предшественников; TGF-β1 (идентификационный номер GenBank: NM_000660) содержит 390 аминокислот, и TGF-β2 (идентификационный номер GenBank: NM_001135599 и NM_003238) и TGF-β3 (идентификационный номер GenBank: XM_005268028), где каждый содержит 412 аминокислот. Каждый из них имеет N-концевой сигнальный пептид из 20-30 аминокислот, который необходим для секреции из клетки, прообласть (называемую латентно- ассоциированным пептидом или LAP) и С-концевую область из 112-114 аминокислот, которая становится зрелой молекулой TGF-β после ее высвобождения из прообласти в результате протеолитического расщепления. После протеолитического расщепления LAP и зрелый TGF-β остаются нековалентно связанными и образуют молекулу «латентного TGF-β». В этой латентной форме зрелый TGF-β не связывается с рецептором TGF-β посредством LAP. Для того, чтобы проявлять сигнал, зрелый TGF-β должен освободиться от LAP. Зрелый TGF-β, который не связан с LAP, называется активным TGF-β, поскольку он может связываться с рецептором TGF-β и передавать сигнал.
Полноразмерный TGF-β1 имеет следующую аминокислотную последовательность:
MPPSGLRLLPLLLPLLWLLVLTPGRPAAGLSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHCSCDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRRALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS (SEQ ID NO: 34).
LAP имеет следующую аминокислотную последовательность:
LSTCKTIDMELVKRKRIEAIRGQILSKLRLASPPSQGEVPPGPLPEAVLALYNSTRDRVAGESAEPEPEPEADYYAKEVTRVLMVETHNEIYDKFKQSTHSIYMFFNTSELREAVPEPVLLSRAELRLLRLKLKVEQHVELYQKYSNNSWRYLSNRLLAPSDSPEWLSFDVTGVVRQWLSRGGEIEGFRLSAHCSCDSRDNTLQVDINGFTTGRRGDLATIHGMNRPFLLLMATPLERAQHLQSSRHRR (SEQ ID NO: 35).
Зрелый TGF-β1 имеет следующую аминокислотную последовательность:
ALDTNYCFSSTEKNCCVRQLYIDFRKDLGWKWIHEPKGYHANFCLGPCPYIWSLDTQYSKVLALYNQHNPGASAAPCCVPQALEPLPIVYYVGRKPKVEQLSNMIVRSCKCS (SEQ ID NO: 36).
«Комплекс GARP-TGF-β». Как в настоящем описании используется, термин «комплекс GARP-TGF-β» означает нативный комплекс, который образуется, когда латентный TGF-β связывается с GARP, в частности, с GARP, расположенным на поверхности Treg-клеток. Несмотря на то, что в настоящем описании не указано, термин «комплекс GARP-TGF-β» или просто «GARP-TGF-β» предназначен для обозначения комплекса между GARP и латентным TGF-β. Связывание GARP с TGF-β, более конкретно с латентным TGF-β, было охарактеризовано на молекулярном уровне, например, как сообщалось в публикации Wang et al. Mol. Biol. Cell., 2012 Mar; 2 3(6):1129-39. GARP образует дисульфидную связь с Cys4 латентного TGF-β, и также связывается с латентным TGF-β через нековалентные взаимодействия. Во внеклеточном домене GARP имеется 15 остатков Cys, и GARP использует Cys-192 и Cys-331 для образования дисульфидных связей с двумя остатками Cys4 латентного TGF-β. Следовательно, один белок GARP связывается с одним латентным TGF-β-димером.
«Антитело» или «иммуноглобулин». Как в настоящем описании используется, термин «иммуноглобулин» включает полипептид, имеющий комбинацию двух тяжелых и двух легких цепей, независимо от того, обладает он какой-либо соответствующей специфической иммунореактивностью или нет. «Антитела» относятся к таким молекулам, которые обладают значительной известной специфической иммунореактивностью в отношении антигена, представляющего интерес (например, комплекса GARP и TGF-β). Как в настоящем описании используется, термин «анти-GARP-TGF-β-антитела» относится к антителам, которые проявляют иммунологическую специфичность для комплекса GARP-TGF-β1, в частности, комплекса человеческих GARP-TGF-β1 и в некоторых случаях их видовых гомологов. Антитела и иммуноглобулины включают легкие и тяжелые цепи, с или без межцепочечной ковалентной связью между ними. Основные структуры иммуноглобулина у позвоночных относительно хорошо изучены.
Общий термин «иммуноглобулин» включает пять различных классов антител (IgG, IgM, IgA, IgD или IgE), которые можно различить биохимически. Все пять классов антител входят в объем настоящего изобретения. Последующее обсуждение, в общем, будет относиться к классу иммуноглобулиновых молекул IgG. В отношении IgG, то иммуноглобулины обычно содержат два идентичных полипептида легкой цепи с молекулярной массой примерно 23000 дальтон и два идентичных полипептида тяжелой цепи с молекулярной массой 53000-70000 дальтон. Четыре цепи соединены дисульфидными связями в конфигурации «Y», где легкие цепи скрепляют тяжелые цепи, начиная с развилки «Y» и продолжаясь через вариабельную область.
Легкие цепи антитела классифицируются как каппа (κ) или лямбда (λ). Каждый класс тяжелой цепи может быть связан с легкой цепью каппа или лямбда. В общем, легкая и тяжелая цепи ковалентно связаны друг с другом, и «хвостовые» части двух тяжелых цепей связаны друг с другом ковалентными дисульфидными связями или нековалентными связями, когда иммуноглобулины продуцируются гибридомами, B клетками или генно-инженерными клетками-хозяевами. В тяжелой цепи аминокислотные последовательности проходят от N-конца на раздвоенных концах Y-конфигурации к С-концу в нижней части каждой цепи. Специалистам в данной области будет понятно, что тяжелые цепи классифицируются как гамма, мю, альфа, дельта или эпсилон (γ, μ, α, δ, ε) с наличием некоторых подклассов среди них (например, γ1-γ4). Именно природа этой цепи определяет «класс» антитела как IgG, IgM, IgA, IgD или IgE соответственно. Подклассы иммуноглобулинов (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и т.д., хорошо охарактеризованы и, как известно, придают функциональную специализацию. Специалисты в данной области могут легко определить модифицированные варианты каждого из этих классов и изотипов с учетом настоящего раскрытия и, следовательно, они находятся в объеме настоящего изобретения.
Как указано выше, вариабельная область антитела обеспечивает возможность антителу селективно распознавать и специфически связываться с эпитопами на антигенах. Т. е. домен VL и домен VH антитела объединяются, образуя вариабельную область, которая определяет трехмерный антигенсвязывающий сайт. Эта четвертичная структура антитела образует антигенсвязывающий сайт, находящийся на конце каждого плеча Y. Более конкретно, антигенсвязывающий сайт определяется тремя определяющими комплементарность участками (CDR) в каждой из цепей VH и VL.
«Сайт связывания». Как в настоящем описании используется, термин «сайт связывания» включает область полипептида, которая ответственна за избирательное связывание с антигеном-мишенью, представляющим интерес. Связывающие домены содержат, по меньшей мере, один сайт связывания. Типичные связывающие домены включают вариабельную область антитела. Молекулы антител по изобретению могут содержать один сайт связывания или несколько сайтов связывания (например, два, три или четыре).
«Вариабельная область» или «вариабельный домен». Термины «вариабельная область» и «вариабельный домен» используются в настоящем описании взаимозаменяемо и имеют одинаковое значение. Термин «вариабельный» относится к тому факту, что определенные части вариабельных областей VH и VL существенно различаются по последовательности среди антител и обеспечивают связывание и специфичность каждого конкретного антитела для его антигена-мишени. Однако вариабельность неравномерно распределена по вариабельным областям антител. Она сконцентрирована в трех сегментах, называемых «гипервариабельными петлями» в каждой области VL и области VH, которые образуют часть антигенсвязывающего сайта. Первая, вторая и третья гипервариабельные петли домена легкой цепи Vлямбда относятся в настоящем описании к L1 (λ), L2 (λ) и L3 (λ) и могут быть определены как содержащие остатки 24-33 (L1(λ), состоящая из 9, 10 или 11 аминокислотных остатков), 49-53 (L2(λ), состоящая из 3 остатков) и 90-96 (L3(λ), состоящая из 5 остатков) в области VL (Morea et al., Methods, 20: 267-279 (2000). Первая, вторая и третья гипервариабельные петли домена легкой цепи Vкаппа относятся в настоящем описании к L1(κ), L2(κ) и L3(κ) и могут быть определены как содержащие остатки 25-33 (L1(κ), состоящая из 6, 7, 8, 11, 12 или 13 остатков), 49-53 (L2(κ), состоящая из 3 остатков) и 90-97 (L3(κ), состоящая из 6 остатков) в области VL (Morea et al., Methods 20: 267-279 (2000)). Первая, вторая и третья гипервариабельные петли области VH относятся в настоящем описании к H1, H2 и H3 и могут быть определены как содержащие остатки 25-33 (H1, состоящая из 7, 8 или 9 остатков), 52-56 (H2, состоящая из 3 или 4 остатков) и 91-105 (Н3, имеющая разную длину) в области VH (Morea et al., Methods, 20: 267-279 (2000)).
Если не указано иное, то термины L1, L2 и L3, соответственно, относятся к первой, второй и третьей гипервариабельным петлям области VL и охватывают гипервариабельные петли, полученные из изотипов Vкаппа и Vлямбда. Термины H1, H2 и H3 соответственно относятся к первой, второй и третьей гипервариабельным петлям области VH и охватывают гипервариабельные петли, полученные из любого из известных изотипов тяжелой цепи, включая γ, μ, α, δ, ε.
Гипервариабельные петли L1, L2, L3, H1, H2 и H3, каждая может составлять часть «определяющего комплементарность участка» или «CDR», как определено ниже. Термины «гипервариабельная петля» и «определяющий комплементарность участок» не являются полными синонимами, поскольку гипервариабельные петли (HV) определяются на основе структуры, тогда как определяющие комплементарность участки (CDR), определяются на основе вариабельности последовательности (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD., 1983) и границы HV и CDR могут различаться в некоторых областях VH и VL.
CDR в VL и VH обычно можно определить как включающие следующие аминокислоты: остатки 24-34 (LCDR1), 50-56 (LCDR2) и 89-97 (LCDR3) в вариабельной области легкой цепи и остатки 31-35 или 31-35b (HCDR1), 50-65 (HCDR2) и 95-102 (HCDR3) в вариабельной области тяжелой цепи; (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Таким образом, HV могут находиться внутри соответствующих CDR, и определение в настоящем описании «гипервариабельные петли» областей VH и VL следует интерпретировать как также включающее соответствующие CDR, и наоборот, если не указано иное.
Более высококонсервативные участки вариабельных областей называются каркасной областью (FR), как определено ниже. Каждая вариабельная область нативной тяжелой и легкой цепей содержит четыре FR (FR1, FR2, FR3 и FR4 соответственно), в основном принимающих β-складчатую конфигурацию, соединенную тремя гипервариабельными петлями. Гипервариабельные петли в каждой цепи удерживаются вместе в непосредственной близости с помощью FR и, вместе с гипервариабельными петлями из другой цепи, участвуют в образовании антигенсвязывающего сайта антител. Структурный анализ антител выявил взаимосвязь между последовательностью и формой сайта связывания, образованного определяющими комплементарность участками (Chothia et al., J. Mol. Biol., 227: 799-817 (1992)); Tramontano et al., J. Mol. Biol., 215: 175-182 (1990)). Несмотря на их высокую вариабельность последовательности, пять из шести петель принимают только небольшой репертуар конформаций главной цепи, называемых «каноническими структурами». Эти конформации, во-первых, определяются длиной петель и, во-вторых, наличием ключевых остатков в определенных положениях петель и в каркасных областях, которые определяют конформацию посредством их упаковки, водородных связей или способности принимать необычные конформации главной цепи.
«CDR». Как в настоящем описании используется, термин «CDR» или «определяющий комплементарность участок» означает несмежные антигенсвязывающие сайты, находящиеся в вариабельной области полипептидов как тяжелой, так и легкой цепи. Эти указанные области были описаны Kabat et al., J. Biol. Chem., 252, 6609-6616 (1977) и Kabat et al., Sequences of protein of immunological interest. (1991), и Chothia et al., J. Mol. Biol., 196:901-917 (1987) и MacCallum et al., J. Mol. Biol., 262:732-745 (1996), где определения включают перекрывающиеся или субпопуляции аминокислотных остатков при сравнении друг с другом. Аминокислотные остатки, которые охватывают CDR, определенные в каждой из приведенных выше ссылок, приводятся для сравнения. Предпочтительно, термин «CDR» представляет собой CDR, определяемый Kabat на основании сравнений последовательностей.
2Нумерация остатков согласно номенклатуре Chothia et al., приведенной в публикации выше
3Нумерация остатков согласно номенклатуре MacCallum et al., приведенной в публикации выше
«Каркасная область». Как в настоящем описании используется, термин «каркасная область» или «FR-область» включает аминокислотные остатки, которые являются частью вариабельной области, но не являются частью CDR (например, с использованием определения CDR по Kabat). Следовательно, каркасная область вариабельной области имеет длину примерно от 100 до 120 аминокислот, но включает только те аминокислоты, которые находятся за пределами CDR. В конкретном примере вариабельной области тяжелой цепи и CDR, как определено Kabat et al., каркасная область 1 соответствует домену вариабельной области, охватывающей аминокислоты 1-30; каркасная область 2 соответствует домену вариабельной области, охватывающей аминокислоты 36-49; каркасная область 3 соответствует домену вариабельной области, охватывающей аминокислоты 66-94, и каркасная область 4 соответствует домену вариабельной области от аминокислоты 103 до конца вариабельной области. Каркасные области легкой цепи аналогичным образом отделены друг от друга CDR вариабельной области легкой цепи. Аналогичным образом, используя определение CDR по Chothia et al. или McCallum et al. границы каркасной области отделены концами соответствующих CDR, как описано выше. В предпочтительных вариантах осуществления CDR являются такими, как определено Kabat.
Во встречающихся в природе антителах шесть CDR, присутствующих на каждом мономерном антителе, представляют короткие несмежные последовательности аминокислот, которые специфически расположены с образованием антигенсвязывающего сайта, когда антитело принимает свою трехмерную конфигурацию в водной среде. Остальные вариабельные области тяжелой и легкой цепей проявляют меньшую межмолекулярную вариабельность аминокислотных последовательностей и называются каркасными областями. Каркасные области в основном имеют β-складчатую конформацию, и CDR образуют петли, которые соединяют, и в некоторых случаях являются частью β-складчатой структуры. Таким образом, эти каркасные области функционируют с образованием каркаса, который обеспечивает правильную ориентацию шести CDR посредством межцепочечных нековалентных взаимодействий. Антигенсвязывающий сайт, образованный расположенными CDR, определяет поверхность, комплементарную эпитопу на иммунореактивном антигене. Эта комплементарная поверхность способствует нековалентному связыванию антитела с эпитопом иммунореактивного антигена. Специалист в данной области может легко идентифицировать положение CDR.
«Константная область». Как в настоящем описании используется, термин «константная область» относится к части молекулы антитела вне вариабельных доменов или вариабельных областей. Легкие цепи иммуноглобулина имеют единственный домен «константная область», обычно называемый «доменом CL или CL1». Данный домен находится от C-конца до домена VL. Тяжелые цепи иммуноглобулина различаются по константной области в зависимости от класса иммуноглобулина (γ, μ, α, δ, ε). Тяжелые цепи γ, α и δ имеют константную область, состоящую из трех доменов иммуноглобулина (называемых СН1, СН2 и СН3) с гибкой шарнирной областью, разделяющей домены СН1 и СН2. Тяжелые цепи μ и ε имеют константную область, состоящую из четырех доменов (CH1-CH4). Константные домены тяжелой цепи располагаются на С-конце относительно домена VH.
Нумерация аминокислот в тяжелых и легких цепях иммуноглобулина проходит от N-конца на разветвленных концах Y-конфигурации к С-концу в нижней части каждой цепи. Различные системы нумерации используются для определения константных областей тяжелой и легкой цепей иммуноглобулина. В соответствии с системой нумерации ЕС константные области тяжелой цепи молекулы IgG идентифицируются следующим образом: CH1 - аминокислотные остатки 118-215; СН2 - аминокислотные остатки 231-340; СН3 - аминокислотные остатки 341-446. В соответствии с системой нумерации по Kabat константные области тяжелой цепи молекулы IgG идентифицируются следующим образом: CH1 - аминокислотные остатки 114-223; СН2 - аминокислотные остатки 244-360; СН3 - аминокислотные остатки 361-477. «Шарнирная область» включает часть молекулы тяжелой цепи, которая соединяет домен CH1 с доменом CH2. Эта шарнирная область содержит примерно 25 остатков и является гибкой, что позволяет двум N-концевым антигенсвязывающим областям двигаться независимо. Шарнирные области можно подразделить на три отдельных домена: верхний, средний и нижний шарнирные домены (Roux K.H. et al. J. Immunol., 161: 4083-90 1998). Антитела по изобретению, содержащие «полностью человеческую» шарнирную область, могут содержать одну из последовательностей шарнирной области, показанных в таблице 2 ниже.
Последовательности человеческих шарнирных областей
(SEQ ID NO:37)
(SEQ ID NO:38)
(SEQ ID NO:39)
(SEQ ID NO:40)
(SEQ ID NO:41)
(SEQ ID NO:42)
(SEQ ID NO:43)
(SEQ ID NO:44)
(SEQ ID NO:45)
(SEQ ID NO:46)
(SEQ ID NO:47)
(SEQ ID NO:48)
«Фрагмент». Термин «фрагмент», используемый в контексте антител по изобретению, относится к части или участку антитела или цепи антитела, содержащим меньшее число аминокислотных остатков, чем целое или полное антитело или цепь антитела. Термин «антигенсвязывающий фрагмент» относится к полипептидному фрагменту иммуноглобулина или антитела, который связывается с антигеном или конкурирует с интактным антителом (т. е. с интактным антителом, из которого он был получен) за связывание антигена (т.е. специфическое связывание с комплексом GARP-TGF-β). Как в настоящем описании используется, термин «фрагмент» молекулы антитела включает антигенсвязывающие фрагменты антител, например, вариабельную область легкой цепи антитела (VL), вариабельную область тяжелой цепи антитела (VH), одноцепочечное антитело (scFv), F(ab')2-фрагмент, Fab-фрагмент, Fd-фрагмент, Fv-фрагмент, антитело с одним плечом (одновалентное), диатела, триатела, тетратела или любую антигенсвязывающую молекулу, образованную комбинацией, сборкой или конъюгацией таких антигенсвязывающих фрагментов. Как в настоящем описании используется, термин «антигенсвязывающий фрагмент», кроме того, предназначен для охвата фрагментов антител, выбранных из группы, состоящей из монотел, доменных антител и нанотел. Фрагменты могут быть получены, например, посредством химической или ферментативной обработки интактного или полного антитела или цепи антитела или рекомбинантными способами.
«Консервативная аминокислотная замена». «Консервативная аминокислотная замена» представляет собой замену, в которой аминокислотный остаток заменяется аминокислотным остатком, имеющим сходную боковую цепь. В данной области классифицированы семейства аминокислотных остатков, имеющих сходные боковые цепи, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глютамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, остаток заменимой аминокислоты в полипептиде иммуноглобулина может быть заменен другим аминокислотным остатком из того же семейства боковых цепей. В еще одном варианте осуществления цепочка аминокислот может быть заменена структурно подобной цепочкой, которая отличается порядком и/или составом членов семейства боковых цепей.
«Химерный». «Химерный» белок содержит первую аминокислотную последовательность, связанную со второй аминокислотной последовательностью, с которой она не связана естественным образом в природе. Аминокислотные последовательности обычно могут существовать в отдельных белках, которые объединяются в слитый полипептид, или они обычно могут существовать в одном и том же белке, будучи расположенными в новом порядке в слитом полипептиде. Химерный белок может быть создан, например, химическим синтезом или созданием и трансляцией полинуклеотида, в котором пептидные области кодируются в желаемой взаимосвязи. Типичные химерные антитела по изобретению включают слитые белки, содержащие области VH и VL, полученные от верблюда, или их гуманизированные варианты, слитые с константными доменами человеческого антитела, например, человеческого IgG1, IgG2, IgG3 или IgG4.
«Валентность». Как в настоящем описании используется, термин «валентность» относится к числу потенциальных сайтов связывания мишени в полипептиде. Каждый сайт связывания мишени специфически связывается с одной молекулой-мишенью или специфическим сайтом на молекуле-мишени. Когда полипептид содержит более одного сайта связывания мишени, то каждый сайт связывания мишени может специфически связываться с одними и теми же или разными молекулами (например, может связываться с разными лигандами или разными антигенами или разными эпитопами на одном и том же антигене).
«Специфичность». Термин «специфичность» относится к способности связывать (например, иммунореагировать) с конкретной мишенью, например, комплексом GARP-TGF-β1. Полипептид может быть моноспецифическим и содержать один или более сайтов связывания, которые специфически связываются с мишенью, или полипептид может быть мультиспецифическим и содержать два или более сайтов связывания, которые специфически связываются с одной и той же или разными мишенями.
«Синтетический». Как в настоящем описании используется, термин «синтетический» по отношению к полипептидам, включает полипептиды, которые содержат аминокислотную последовательность, которая не встречается в природе. Например, не встречающиеся в природе полипептиды представляют модифицированные формы встречающихся в природе полипептидов (например, содержащих мутацию, такую как добавление, замену или делецию) или которые содержат первую аминокислотную последовательность (которая может встречаться или не встречаться в природе), которая связана в линейной последовательности аминокислот со второй аминокислотной последовательностью (которая может встречаться или не встречаться в природе), с которой она не связана естественным образом в природе.
«Сконструированный». Как в настоящем описании используется, термин «сконструированный» включает манипулирование молекулами нуклеиновых кислот или полипептидов с использованием синтетических методов (например, рекомбинантных методов, пептидного синтеза in vitro, ферментативного или химического сочетания пептидов или некоторых комбинаций этих методов). Предпочтительно антитела по изобретению являются сконструированными, включая, например, гуманизированные и/или химерные антитела, и антитела, которые были сконструированы для улучшения одного или более свойств, таких как связывание антигена, стабильность/период полураспада или эффекторная функция.
«Гуманизирующие замены». Как в настоящем описании используется, термин «гуманизирующие замены» относится к аминокислотным заменам, в соответствии с которыми аминокислотный остаток, находящийся в определенном положении в области VH или VL антитела (например, антитела к GARP-TGF-β, полученного верблюда) заменяется аминокислотным остатком, который находится в эквивалентном положении в референсной области VH или VL человека. Референсная область VH или VL человека может представлять собой область VH или VL, кодируемая генами зародышевой линией человека. Гуманизирующие замены могут быть сделаны в каркасных областях и/или CDR антител, определенных в настоящем описании.
«Гуманизированные варианты». Как в настоящем описании используется, термин «гуманизированный вариант» относится к варианту антитела, которое содержит одну или более «гуманизирующих замен» по сравнению с референсным антителом, где часть референсного антитела (например, область VH и/или область VL или ее части, содержащие, по меньшей мере, один CDR), содержит аминокислоту, происходящую от вида, отличного от человека, и «гуманизирующие замены» имеют место в аминокислотной последовательности, полученной от вида, отличного от человека.
«Варианты зародышевого типа». Как в настоящем описании используется, термин «вариант зародышевого типа» относится к «гуманизированным вариантам», в которых «гуманизирующие замены» приводят к замене одного или более аминокислотных остатков, присутствующих в определенном положении(ях) в области VH или VL антитела (например, антитела к GARP-TGF-β1, полученного от верблюда) аминокислотным остатком, который находится в эквивалентном положении в референсной области VH или VL человека, кодированной генами зародышевой линией человека. Типично, что для любого данного «варианта зародышевого типа» замещающие аминокислотные остатки, замещенные зародышевым вариантом, берутся исключительно или преимущественно из одной области VH или VL, кодированной генами зародышевой линией человека. Термины «гуманизированный вариант» и «вариант зародышевого типа» часто используются в настоящем описании взаимозаменяемо. Введение одной или более «гуманизирующих замен» в области VH или VL, полученной от верблюда (например, от ламы), приводит к получению «гуманизированного варианта» области VH или VL, полученной от верблюда (ламы). Если замещенные аминокислотные остатки получены преимущественно или исключительно из одной последовательности области VH или VL, кодированной генами зародышевой линией человека, то результатом может быть «вариант зародышевой линии человека» области VH или VL, полученной от верблюда (ламы).
«Аффинные варианты». Как в настоящем описании используется, термин «аффинный вариант» относится к варианту антитела, который имеет одно или более изменений в аминокислотной последовательности по сравнению с референсным антителом, где аффинный вариант проявляет измененную аффинность к антигену-мишени по сравнению с референсным антителом. Например, аффинные варианты будут проявлять измененную аффинность к GARP-TGF-β по сравнению с референсным антителом к GARP-TGF-β. Предпочтительно аффинный вариант будет проявлять повышенную аффинность к антигену-мишени по сравнению с референсным антителом. Аффинные варианты обычно имеют одно или более изменений в аминокислотной последовательности в CDR по сравнению с референсным антителом. Такие замены могут привести к замене исходной аминокислоты, присутствующей в данном положении в CDR, другим аминокислотным остатком, который может быть встречающимся в природе аминокислотным остатком или не встречающимся в природе аминокислотным остатком. Аминокислотные замены могут быть консервативными или неконсервативными.
В. Анти-GARP-TGF-β-антитела
Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, которые специфически связываются с комплексом GARP и TGF-β, в частности с комплексом человеческого GARP и человеческого TGF-β. Антитела и антигенсвязывающие фрагменты по настоящему изобретению могут быть определены в отношении структурных и функциональных характеристик, как в настоящем описании описано.
Важно, что антитела к GARP-TGF-β по изобретению усовершенствованы по сравнению с антителами к GARP-TGF-β, описанными ранее, в том отношении, что они демонстрируют повышенную стабильность. В частности, стабильность анти-GARP-TGF-β1-антител, описанных в настоящем описании, повышена по сравнению с антителами, имеющими последовательности CDR тяжелой цепи и легкой цепи референсных антител к GARP-TGF-β, LHG-10 и LHG-10.6, описанных в WO 2015/015003 и WO2016/125017. Такое повышение стабильности достигается без существенного снижения аффинности связывания антител с комплексом GARP-TGF-β по сравнению с референсными антителами LHG10 и LHG10.6.
Антитела по настоящему изобретению отличаются от референсных антител к GARP-TGF-β LHG-10 и LHG-10.6, описанных ранее, особенно в отношении последовательностей CDR2 и CDR3 тяжелой цепи. Более конкретно, антитела к GARP-TGF-β1, LHG-10 и LHG-10.6, содержат последовательность CDR2 тяжелой цепи: RIDPEDGGTKYAQKFQG (SEQ ID NO:5) и последовательность CDR3 тяжелой цепи: NEWETVVVGDLMYEYEY (SEQ ID NO:6), в то время как антитела по настоящему изобретению содержат последовательность CDR2 тяжелой цепи: RIDPEDAGTKYAQKFQG (SEQ ID NO:12) и последовательность CDR3 тяжелой цепи: YEWETVVVGDLMYEYEY (SEQ ID NO:13). Как описано и приведено в настоящем описании в качестве примера, было установлено, что аминокислотные замены G55A и N95Y в последовательностях CDR2 и CDR3 тяжелой цепи, соответственно, повышают стабильность антител посредством снижения дезамидирования, изомеризации и окисления, в то же время, достигая аффинности связывания с комплексом GARP-TGF-β1, примерно эквивалентного для референсных антител.
В первом аспекте настоящее изобретение относится к антителам или их антигенсвязывающим фрагментам, которые связываются с комплексом GARP-TGF-β1 и содержат вариабельную область тяжелой цепи (VH), где:
CDR3 VH содержит или состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит или состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержит или состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4).
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты дополнительно содержат вариабельную область легкой цепи (VL), где:
CDR3 VL содержит или состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит или состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит или состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления обеспечиваются антитела или их антигенсвязывающие фрагменты, которые специфически связываются с комплексом GARP-TGF-β1, где антитела или их антигенсвязывающие фрагменты содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где:
CDR3 VH содержит или состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит или состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержит или состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4),
CDR3 VL содержит или состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит или состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит или состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления обеспечиваются антитела или их антигенсвязывающие фрагменты, которые специфически связываются с комплексом GARP-TGF-β1, где антитела или их антигенсвязывающие фрагменты содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкий цепи (VL), где:
CDR3 VH состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4),
CDR3 VL состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты являются рекомбинантными. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты являются моноклональными.
Термин «антитело» в настоящем описании используется в самом широком смысле и охватывает, но не ограничивается этим, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и полиспецифические антитела (например, биспецифические антитела), при условии, что они проявляют соответствующую иммунологическую специфичность для комплекса GARP-TGF-β1. Как в настоящем описании используется, термин «моноклональное антитело» относится к антителу, полученному из популяции по существу гомогенных антител, т. е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных встречающихся в природе мутаций, которые могут присутствовать в небольших количествах. Моноклональные антитела являются высокоспецифичными, будучи направленными против одного антигенного сайта. Кроме того, в отличие от препаратов обычных (поликлональных) антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов) на антигене, каждое моноклональное антитело направлено против одной детерминанты или эпитопа на антигене.
Настоящее изобретение также охватывает «антигенсвязывающие фрагменты» антител, и такие фрагменты определены в другом месте в данном документе. Фрагменты антител обычно включают часть полноразмерного антитела, обычно его антигенсвязывающий или вариабельный домен. Примеры фрагментов антител включают Fab, Fab', F(ab')2, биспецифические Fab-фрагменты и Fv-фрагменты, линейные антитела, молекулы одноцепочечных антител, вариабельный фрагмент с одной цепью (scFv) и мультиспецифические антитела, образованные из фрагментов антител (см. Holliger and Hudson, Nature Biotechnol., 23: 1126-36 (2005)).
Антитела и антигенсвязывающие фрагменты по настоящему изобретению могут проявлять высокую гомологию с человеком. Уровень гомологии с человеческой последовательностью можно оценить по длине вариабельной области тяжелой цепи (VH) и/или по длине вариабельной области легкой цепи (VL). В контексте настоящего изобретения антитело, содержащее вариабельную область тяжелой цепи (VH) и вариабельную область легкой цепи (VL), можно рассматривать в качестве обладающего высокой гомологией с человеком, если области VH и домены VL, взятые вместе, проявляют, по меньшей мере, 90%, по меньшей мере, 92%, по меньшей мере, 94% или, по меньшей мере, 96% идентичность аминокислотных последовательностей с наиболее подходящими последовательностями VH и VL зародышевой линии человека. В одном варианте осуществления область VH антитела с высокой гомологией с человеком может иметь идентичность аминокислотной последовательности или гомологию последовательности, по меньшей мере, на 90%, по меньшей мере, на 92%, по меньшей мере, на 94% или, по меньшей мере, на 96% с одной или более человеческими областями VH по каркасным областям FR1, FR2, FR3 и FR4. В одном варианте осуществления область VH антитела с высокой гомологией с человеком может содержать одну или более (например, от 1 до 10) аминокислотных последовательностей, несовпадающих по каркасным областям FR1, FR2, FR3 и FR4, по сравнению с наиболее близкой человеческой последовательностью VH.
В еще одном варианте осуществления область VL антитела с высокой с высокой гомологией с человеком может иметь идентичность аминокислотной последовательности или гомологию последовательности, по меньшей мере, на 90%, по меньшей мере, на 92%, по меньшей мере, на 94% или, по меньшей мере, на 96% с одной или более человеческими областями VL по каркасным областям FR1, FR2, FR3 и FR4. В одном варианте осуществления область VL антитела с высокой гомологией с человеком может содержать одну или более (например, от 1 до 10) аминокислотных последовательностей, несовпадающих по каркасным областям FR1, FR2, FR3 и FR4, по сравнению с наиболее близкой человеческой последовательностью VL.
Антитела и антигенсвязывающие фрагменты по настоящему изобретению, имеющие высокую гомологию с человеком, могут включать антитела, содержащие области VH и VL нативных антител, отличных от человеческих, которые имеют достаточно высокую % идентичность последовательности с последовательностями зародышевой линии человека. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты по изобретению представляют гуманизированные варианты или варианты зародышевого типа, антител, отличных от человеческих, например антитела, содержащие области VH и VL обычных верблюжьих антител, сконструированных таким образом, чтобы они представляли гуманизированные варианты или варианты зародышевого типа исходных антител.
Антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи (VH), содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 14, и, необязательно, вариабельную область легкой цепи (VL), содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 14. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 15.
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи (VH), состоящую из аминокислотной последовательности SEQ ID NO: 14. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область легкой цепи (VL), состоящую из аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательности SEQ ID NO: 14, и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 15.
В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область тяжелой цепи (VH), состоящую из аминокислотной последовательности SEQ ID NO: 14, и вариабельную область легкой цепи (VL), состоящую из аминокислотной последовательности SEQ ID NO: 15.
В некоторых вариантах осуществления обеспечиваются моноклональные антитела или их антигенсвязывающие фрагменты, содержащие вариабельную область тяжелой цепи и вариабельную область легкой цепи, где вариабельная область тяжелой цепи содержит последовательность VH, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 14, и/или вариабельную область легкой цепи, содержащую VL, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 15.
Для вариантов осуществления, в которых домены антител или антигенсвязывающих фрагментов определяются конкретной процентной идентичностью последовательности с референсной последовательностью, области VH и/или VL могут сохранять последовательности CDR, идентичные тем, которые присутствуют в референсной последовательности, так что вариабельность присутствует только в каркасных областях. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, содержащие вариабельные области тяжелой цепи и/или вариабельные области легкой цепи, определенные как имеющие конкретный процент идентичности с SEQ ID NO: 14 и 15 соответственно, будут иметь следующие последовательности CDR:
CDR3 VH содержащий или состоящий из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержащий или состоящий из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержащий или состоящий из аминокислотной последовательности SYYID (SEQ ID NO:4),
CDR3 VL содержащий или состоящий из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержащий или состоящий из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержащий или состоящий из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В неограничивающих вариантах осуществления антитела по настоящему изобретению могут содержать домены CH1 и/или домены CL (соответственно из тяжелой цепи и легкой цепи), аминокислотная последовательность которых является полностью или по существу человеческой. В тех случаях, когда антитело или антигенсвязывающий фрагмент по изобретению представляет антитело, предназначенное для терапевтического применения у людей, то обычно для всей константной области антитела или, по меньшей мере, его части, имеется полная или по существу человеческая аминокислотная последовательность. Следовательно, одно или более, или любая комбинация домена CH1, шарнирной области, домена CH2, домена CH3 и домена CL (и домена CH4, если он присутствует) может быть полностью или по существу человеческими в отношении их аминокислотной последовательности.
Преимущественно, домен CH1, шарнирная область, домен CH2, домен CH3 и домен CL (и домен CH4, если он присутствует) все могут иметь полностью или по существу человеческую аминокислотную последовательность. В контексте константной области гуманизированного или химерного антитела или фрагмента антитела термин «по существу человеческая» относится к идентичности аминокислотной последовательности, по меньшей мере, на 90%, или, по меньшей мере, на 92%, или, по меньшей мере, на 95%, или, по меньшей мере, 97% или, по меньшей мере, 99% с человеческой константной областью. Термин «человеческая аминокислотная последовательность» в данном контексте относится к аминокислотной последовательности, которая кодируется геном иммуноглобулина человека, который включает зародышевые, реаранжированные и соматически мутированные гены. Изобретение также предусматривает полипептиды, содержащие константные области «человеческой» последовательности, которые были изменены добавлениями, делециями или заменами одной или более аминокислот относительно человеческой последовательности, за исключением тех вариантов осуществления, где прямо требуется, чтобы присутствовала «полностью человеческая» шарнирная область.
Наличие «полностью человеческой» шарнирной области в анти-GARP-TGF-β1-антителах по изобретению может быть преимущественным как для минимизации иммуногенности, так и для оптимизации стабильности антитела.
Как обсуждается в настоящем описании в другом месте, предполагается, что одна или более аминокислотных замен, вставок или делеций могут быть сделаны в константной области тяжелой и/или легкой цепи, в частности, в Fc-области. Аминокислотные замены могут привести к замене замещенной аминокислоты другой природной аминокислотой или неприродной или модифицированной аминокислотой. Также допускаются другие структурные модификации, например, такие как изменения в паттерне гликозилирования (например, добавлением или делецией сайтов N- или O-связанного гликозилирования).
Анти-GARP-TGF-β1-антитела можно модифицировать в Fc-области для повышения аффинности связывания с неонатальным рецептором FcRn. Повышенную аффинность связывания можно измерить при кислом значении рН (например, примерно от 5,5 до примерно 6,0). Повышенную аффинность связывания также можно измерить при нейтральном рН (например, примерно от 6,9 до примерно 7,4). Под «повышенной аффинностью связывания» подразумевается повышенная аффинность связывания с FcRn относительно немодифицированной Fc-области. Как правило, немодифицированная Fc-область будет обладать аминокислотной последовательностью дикого типа человеческого IgG1, IgG2, IgG3 или IgG4. В таких вариантах осуществления повышенная аффинность связывания с FcRn молекулы антитела, имеющей модифицированную Fc-область, будет измеряться относительно аффинности связывания IgG1, IgG2, IgG3 или IgG4 дикого типа для FcRn.
В некоторых вариантах осуществления один или более аминокислотных остатков в Fc-области можно заменить другой аминокислотой, для повышения связывания с FcRn. Сообщалось о некоторых заменах в Fc, которые увеличивают связывание с FcRn и тем самым улучшают фармакокинетику антител. Такие замены описаны, например, в публикациях Zalevsky et al. (2010) Nat. Biotechnol., 28 (2): 157-9; Zalevsky et al. (2010) Nat. Biotechnol., 28(2):157-9; Hinton et al. (2006) J. Immunol., 176:346-356; Yeung et al. (2009) J. Immunol., 182:7663-7671; Presta L.G. (2008) Curr. Op. Immunol., 20:460-470; и Vaccaro et al. (2005) Nat. Biotechnol., 23(10):1283-88, содержание которых в полном объеме включено в настоящем описании посредством ссылки.
В некоторых вариантах осуществления анти-GARP-TGF-β1-антитела содержат модифицированную Fc-область человеческого IgG, содержащую или состоящую из аминокислотных замен H433K и N434F, где нумерация Fc-областей соответствует системе нумерации ЕС. В дополнительном варианте осуществления антитела к GARP-TGF-β1, описанные в настоящем описании, содержат модифицированную Fc-область человеческого IgG, содержащую или состоящую из аминокислотных замен M252Y, S254T, T256E, H433K и N434F, где нумерация Fc-области соответствует системе нумерации EU.
В некоторых вариантах осуществления анти-GARP-TGF-β1-антитела содержат модифицированную Fc-область человеческого IgG, имеющую до 2, до 3, до 4, до 5, до 6, до 7, до 8, до 9, до 10, до 12, до 15, до 20 замен относительно соответствующей последовательности IgG дикого типа.
В зависимости от предполагаемого применения антитела может быть желательным модифицировать антитело по изобретению в отношении его связывающих свойств с Fc-рецепторами, например, для модуляции эффекторной функции. Например, остаток(и) цистеина могут быть введены в Fc-область, что обеспечивает образование межцепочечных дисульфидных связей в этой области. Полученное таким образом гомодимерное антитело может обладать улучшенной способностью к интернализации и/или повышенной комплемент-опосредованной клеточной цитотоксичностью и антителозависимой клеточной цитотоксичностью (ADCC). См. Caron et al., J. Exp. Med., 176: 1191-1195 (1992) и Shopes B.J. Immunol., 148: 2918-2922 (1992). Изобретение также предусматривает иммуноконъюгаты, содержащие антитело, как в настоящем описании описано, конъюгированное с цитотоксическим агентом, таким как химиотерапевтический агент, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или его фрагменты) или радиоактивный изотоп (т. е. радиоконъюгат). Fc-области также можно сконструировать для увеличения периода полураспада, как описано в публикации Chan and Carter, Nature Reviews: Immunology, Vol.10, p. 301-316, 2010, включенной в настоящем описании посредством ссылки.
В еще одном варианте осуществления Fc-область модифицируется для повышения способности антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или для увеличения аффинности антитела к рецептору Fcγ посредством модификации одной или нескольких аминокислот.
В конкретных вариантах осуществления Fc-область может быть сконструирована таким образом, что отсутствует эффекторная функция. Антитело к GARP-TGF-β1, не обладающее эффекторной функцией Fc, может быть особенно пригодным в качестве агента, блокирующего рецепторы. В определенных вариантах осуществления антитела по изобретению могут иметь Fc-область, полученную из встречающихся в природе изотипов IgG, имеющих сниженную эффекторную функцию, например, IgG4. Fc-области, полученные из IgG4, можно дополнительно модифицировать для повышения терапевтической применимости, например, введением модификаций, которые минимизируют обмен плечами между молекулами IgG4 in vivo. Fc-области, полученные из IgG4, можно модифицировать с включением замены S228P.
В еще одном варианте модифицируется гликозилирование антитела. Например, может быть получено агликозилированное антитело (т. е. антитело не имеет гликозилирования). Гликозилирование можно изменить, например, для повышения аффинности антитела к антигену-мишени. Такие модификации углеводов можно выполнить, например, изменением одного или более сайтов гликозилирования в последовательности антитела. Например, можно сделать одну или более аминокислотных замен, которые приводят к элиминации одного или более сайтов гликозилирования каркасной области вариабельной области, чтобы тем самым элиминировать гликозилирование в этом сайте. Такое агликозилирование может повысить аффинность антитела к антигену.
Также предусматриваются варианты анти-GARP-TGF-β1-антител, имеющих измененный тип гликозилирования, такие как гипофукозилированное антитело, содержащее уменьшенные количества фукозильных остатков, или полностью или частично дефукозилированное антитело (как описано Natsume et al., Drug Design Development и Therapy, Vol.3, p. 7-16, 2009) или антитело, имеющее повышенные бисектные структуры GlcNac. Было показано, что такие измененные паттерны гликозилирования увеличивают ADCC активность антител, обычно приводя к 10-кратному усилению ADCC по сравнению с эквивалентным антителом, содержащим «нативную» Fc-область человека. Такие модификации углеводов можно выполнить, например, экспрессией антитела в клетке-хозяине с измененным ферментативным механизмом гликозилирования (как описано Yamane-Ohnuki and Satoh, mAbs 1: 3, 230-236, 2009). Примерами нефукозилированных антител с повышенной ADCC функцией являются антитела, которые получены с использованием технологии Potelligent® от BioWa Inc.
Антитела, предназначенные для терапевтического применения у людей, обычно относятся к типу IgG, IgM, IgA, IgD или IgE, часто типа IgG, и в этом случае они могут принадлежать к любому из четырех подклассов IgG1, IgG2a и IgG2b, IgG3 или IgG4. Внутри каждого из этих подклассов допускается делать одну или более аминокислотных замен, вставок или делеций в пределах Fc-области или делать другие структурные модификации, например, для усиления или уменьшения Fc-зависимых функций.
В некоторых вариантах осуществления антитела, которые специфически связываются с GARP-TGF-β1, содержат, по меньшей мере, одну иммуноглобулиновую полноразмерную тяжелую цепь и/или, по меньшей мере, одну полноразмерную легкую цепь лямбда или каппа, где тяжелая цепь содержит или состоит из аминокислотной последовательности SEQ ID NO:16, и легкая цепь содержит или состоит из аминокислотной последовательности SEQ ID NO: 17.
В некоторых вариантах осуществления антитела, которые специфически связываются с GARP-TGF-β1, содержат, по меньшей мере, одну иммуноглобулиновую полноразмерную тяжелую цепь и/или, по меньшей мере, одну полноразмерную легкую цепь лямбда или каппа, где тяжелая цепь содержит аминокислотную последовательность SEQ ID NO:16, и легкая цепь содержит аминокислотную последовательность SEQ ID NO: 17.
В некоторых вариантах осуществления антитела, которые специфически связываются с GARP-TGF-β1, содержат, по меньшей мере, одну иммуноглобулиновую полноразмерную тяжелую цепь и/или по меньшей мере одну полноразмерную легкую цепь лямбда или каппа, где тяжелая цепь состоит из аминокислотной последовательности SEQ ID NO:16, и легкая цепь состоит из аминокислотной последовательности SEQ ID NO: 17.
В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 16, и/или легкую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 17.
В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO:16. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие легкую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 17. В некоторых вариантах осуществления обеспечиваются моноклональные антитела, содержащие тяжелую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 16 и легкую цепь, по меньшей мере, с 90%, по меньшей мере, 95%, по меньшей мере, 97%, по меньшей мере, 98% или, по меньшей мере, с 99% идентичностью последовательности с аминокислотной последовательностью, показанной в SEQ ID NO: 17.
Для вариантов осуществления, в которых цепи антител определяются конкретной процентной идентичностью последовательности с референсной последовательностью, тяжелая цепь и/или легкая цепь могут сохранять идентичные последовательности CDR с последовательностями, находящимися в референсной последовательности, так что вариабельность присутствует только вне областей CDR. В частности, антитела или антигенсвязывающие фрагменты, содержащие тяжелые цепи и/или легкие цепи, определенные как имеющие конкретную процентную идентичность с SEQ ID NO: 16 и 17 соответственно, могут иметь следующие последовательности CDR:
CDR3 VH содержащий или состоящий из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержащий или состоящий из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
CDR1 VH содержащий или состоящий из аминокислотной последовательности SYYID (SEQ ID NO: 4),
CDR3 VL содержащий или состоящий из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержащий или состоящий из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержащий или состоящий из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
Связывание с GARP-TGF-β1
Антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются с комплексом GARP и TGF-β1, в частности, с комплексом человеческого GARP и человеческого TGF-β1. Как пояснено в другом месте в данном документе, GARP представляет собой трансмембранный белок, экспрессируемый на поверхности регуляторных Т-клеток, и он функционирует в качестве рецептора для латентной формы TGF-β1. На фиг. 1 приведено схематическое представление комплекса, который образуется между GARP и латентным TGF-β1 на клеточной поверхности регуляторных Т-клеток.
Комплекс GARP-TGF-β1, с которым связываются антитела и антигенсвязывающие фрагменты по настоящему изобретению, представляет собой нативный комплекс GARP-TGF-β1, который образуется на клеточной поверхности между GARP и TGF-β1.
Антитела и антигенсвязывающие фрагменты по настоящему изобретению характеризуются тем, что они связываются с комплексом GARP-TGF-β1, но не связываются с GARP в отсутствии TGF-β1 или латентного TGF-β. Антитела и антигенсвязывающие фрагменты связываются с GARP только в присутствии TGF-β1. В частности, антитела и антигенсвязывающие фрагменты связываются с GARP только в присутствии латентного TGF-β1. Антитела и антигенсвязывающие фрагменты могут блокировать или ингибировать высвобождение активного TGF-β из Treg-клеток.
Поскольку антиген-мишень для антител и их антигенсвязывающих фрагментов по настоящему изобретению представляет собой комплекс, включающий два отдельных белка, то эпитоп, с которым связываются антитела и антигенсвязывающие фрагменты, является конформационным, в отличие от линейного эпитопа. Конформационный эпитоп содержит, по меньшей мере, один остаток из GARP и, по меньшей мере, один остаток из латентного TGF-β1. В предпочтительных вариантах осуществления конформационный эпитоп содержит, по меньшей мере, один остаток из GARP, по меньшей мере, один остаток из латентно-ассоциированного пептида (LAP) латентного TGF-β1 и, по меньшей мере, один остаток из зрелого TGF-β1.
Антитела и антигенсвязывающие фрагменты по настоящему изобретению могут связываться с эпитопом комплекса, образованного человеческим GARP и человеческим TGF-β1, где эпитоп содержит, по меньшей мере, один остаток из GARP, выбранный из Y137, S138, G139, T162 и R163 (со ссылкой на SEQ ID NO:33) и, по меньшей мере, один остаток из TGF-β1. В предпочтительных вариантах осуществления эпитоп содержит, по меньшей мере, остатки GARP Y137, S138, G139, T162 и R163 (со ссылкой на SEQ ID NO: 33).
Эпитоп может содержать, по меньшей мере, один остаток из полипептида TGF-β1 (SEQ ID NO: 34), где необязательно, по меньшей мере, один остаток представляет K338. Эпитоп может содержать, по меньшей мере, один остаток из латентно-ассоциированного пептида (LAP) TGF-β1 и, по меньшей мере, один остаток из зрелого TGF-β1. Эпитоп может содержать R58 из LAP и K338 из зрелого TGF-1 (со ссылкой на SEQ ID NO: 34).
Антитела и антигенсвязывающие фрагменты по настоящему изобретению связываются с комплексом GARP и TGF-β1. Антитела и антигенсвязывающие фрагменты по настоящему изобретению могут дополнительно связываться с комплексом человеческого GARP и человеческого TGF-β2 и/или комплексом человеческого GARP и человеческого TGF-β3.
В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты по изобретению связываются с комплексом GARP и TGF-β1 с высокой аффинностью. Как в настоящем описании используется, термин «аффинность» или «аффинность связывания» следует понимать, исходя из обычного значения, принятого в данной области техники в контексте связывания антител, и отражает силу и/или стабильность связывания между антигеном и сайтом связывания на антителе или его антигенсвязывающем фрагменте.
Аффинность связывания антитела или его антигенсвязывающего фрагмента с его соответствующим антигеном можно определить экспериментально с использованием методик, известных в данной области. Например, методы SPR, такие как BiacoreTM, измеряют аффинность на основе иммобилизации белка-мишени или антигена на биосенсорном чипе, когда антитело или фрагмент антитела пропускают над иммобилизованной мишенью в определенных условиях потока. Эти эксперименты дают значения kon и koff, которые можно перевести в значения KD, где KD является константой равновесия диссоциации антигена с антителом или его фрагментом. Чем ниже значение KD, тем сильнее связывающее взаимодействие между антителом и его антигеном-мишенью.
Как отмечено выше, аффинность антитела можно определить с помощью BiacoreTM или SPR, например, с использованием протокола, описанного в настоящем описании в другом месте. Аффинность антитела или антигенсвязывающего фрагмента к комплексу GARP-TGF-β1, измеренная с помощью BiacoreTM или SPR, можно определить с использованием рекомбинантно экспрессированного комплекса GARP-TGF-β1, как описано, например, в примере 2.
Антитела к GARP-TGF-β1 или их антигенсвязывающие фрагменты по изобретению могут иметь скорость диссоциации (koff) комплекса GARP-TGF-β1 ниже 7×10-4 с-1, ниже 5×10-4 с-1, ниже 3×10-4 с-1, ниже 1,5×10-4 при тестировании в виде Fab. Антитела к GARP-TGF-β1 или их антигенсвязывающие фрагменты по изобретению могут иметь скорость диссоциации (koff) комплекса GARP-TGF-β1 в диапазоне от 1×10-6 с-1 до 5×10-4 с-1, предпочтительно в диапазоне от 1×10-6 с-1 до 3×10-4 с-1, более предпочтительно в диапазоне от 1×10-6 с-1 до 1,5×10-4 с-1.
Антитела к GARP-TGF-β1 по изобретению могут иметь значение KD ниже 5×10-9 М, ниже 2×10-9 М. В предпочтительных вариантах осуществления антитела к GARP-TGF-β1 по изобретению имеют значение KD ниже 1,7×10-9 М.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты, описанные в настоящем описании, которые связываются с комплексом GARP и TGF-β1, могут перекрестно реагировать с одним или несколькими видовыми гомологами GARP и TGF-β1, например, гомологами GARP и TGF-β1 приматов, отличных от человека.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению не реагируют перекрестно с комплексом GARP и TGF-β1 мышиного происхождения. Альтернативно или в дополнение, антитела или антигенсвязывающие фрагменты могут связываться с комплексом GARP-TGF-β1 примата, отличного от человека, в частности, с комплексом GARP-TGF-β1 яванского макака. Перекрестная реактивность с гомологами других видов может быть особенно преимущественной при разработке и испытании терапевтических антител. Например, доклиническое токсикологическое исследование терапевтических антител часто проводится на видах приматов, включая, не ограничиваясь этим, яванский макак. Следовательно, перекрестная реактивность с этими видовыми гомологами может быть особенно преимущественной для разработки антител в качестве кандидатов для применения в клинике.
Повышенная стабильность
Анти-GARP-TGF-β1-антитела по изобретению усовершенствованы по сравнению с анти-GARP-TGF-β1-антителами, описанными ранее, в том отношении, что они демонстрируют повышенную стабильность. В частности, стабильность антител к GARP-TGF-β1 повышена по сравнению с антителами, имеющими последовательности CDR тяжелой цепи и легкой цепи референсного анти-GARP-TGF-β1-антитела, LHG10.6, описанного в WO2015/015003 и WO2016/125017.
Анти-GARP-TGF-β1-антитело LHG10.6 обладает следующей комбинацией последовательностей CDR вариабельной области тяжелой цепи и вариабельной области легкой цепи:
CDR3 тяжелой цепи, состоящего из NEWETVVVGDLMYEYEY (SEQ ID NO: 6),
CDR2 тяжелой цепи, состоящего из RIDPEDGGTKYAQKFQG (SEQ ID NO: 5),
CDR1 тяжелой цепи, состоящего из SYYID (SEQ ID NO: 4),
CDR3 легкой цепи, состоящего из QQYASVPVT (SEQ ID NO: 11),
CDR2 легкой цепи, состоящего из GASRLKT (SEQ ID NO: 10), and
CDR1 легкой цепи, состоящего из QASQSISSYLA (SEQ ID NO: 9).
Как сообщалось в другом месте в данном документе, было установлено, что антитела к GARP-TGF-β1, имеющие последовательности CDR тяжелой цепи и легкой цепи LHG10.6, не обладают стабильностью. В частности, было обнаружено, что варианты моноклонального антитела LHG10.6 зародышевого типа (в другом месте в настоящем документе обозначаемые как mAb 39B6 IgG4 и 39B6 IgG1) демонстрируют тенденцию к снижению активности связывания с мишенью при хранении при 37°C как в PBS, так и в PBS/Твине (см. например, фиг. 2). Такая нестабильность относится за счет, по меньшей мере, частично, изомеризации и дезамидирования в положении N95 HCDR3, т. е. первом остатке CDR3 тяжелой цепи.
Антитела к GARP-TGF-β1 и антигенсвязывающие фрагменты по настоящему изобретению отличаются по своим последовательностям CDR, в частности, по последовательностям CDR тяжелых цепей, так что их стабильность повышается. В частности, последовательность CDR2 тяжелой цепи (HCDR2 или CDR2 VH) модифицирована с включением замены G55A, так что последовательность HCDR2 антител по изобретению содержит HCDR2, представленный RIDPEDAGTKYAQKFQG (SEQ ID NO:12). Кроме того, последовательность CDR3 тяжелой цепи (HCDR3 или CDR3 VH) модифицирована с включением замены N95Y, так что последовательность HCDR3 антител по изобретению содержит HCDR3, представленный YEWETVVVGDLMYEYEY (SEQ ID NO:13). Антитела по настоящему изобретению не подвергаются дезамидированию или изомеризации. Кроме того, неожиданно было установлено, что анти-GARP-TGF-β1-антитела по настоящему изобретению относительно устойчивы к окислению. Эта устойчивость к дезамидированию, изомеризации и окислению коррелирует с повышенной стабильностью антител по изобретению, в частности, с повышенной стабильностью при хранении при температуре 37°С.
Кроме того, антитела и антигенсвязывающие фрагменты по изобретению являются неожиданно преимущественными, поскольку модификации последовательностей CDR2 и CDR3 тяжелой цепи, по сравнению с референсным антителом LHG10.6, существенно не снижают активность связывания с мишенью. Как в настоящем описании показано, антитела против GARP-TGF-β1 по настоящему изобретению относительно стабильны при 37°С и не демонстрируют значительного снижения аффинности связывания с комплексом GARP-TGF-β1 по сравнению с референсным антителом LHG10.6 или его вариантом зародышевого типа (39B6). Как описано в другом месте, в предпочтительных вариантах осуществления антитела к GARP-TGF-β1 по изобретению имеют значение KD ниже 1,7×10-9 М.
Как в настоящем описании отмечалось, не все замены в положении N95 HCDR3, в результате которых удаляется остаток Asn (N), способны повышать стабильность антитела, сохраняя при этом аффинность связывания с комплексом GARP-TGF-β1. Было установлено, что вариант моноклонального антитела зародышевого типа, имеющий замену N95V (называемый в настоящем описании 39B6-AVE), устойчив к дезамидированию и изомеризации, но подвергался значительному окислению при хранении в течение 28 суток при 37°С. Также было обнаружено, что активность связывания этого варианта антитела N95V значительно снижается в течение 56-дневного периода хранения при 37°С. Заявители также обнаружили, что объемные замены в смежном положении 96 в тяжелой цепи (т. е. во втором остатке HCDR3) были неспособны повышать стабильность и сохранить активность высокоаффинного связывания с антигеном. Как в настоящем описании показано в качестве примера, заявители изобретения тестировали два варианта моноклонального антитела зародышевого типа, включающего замены E96K и E96R в домене HCDR3. Вариант E96K (называемый в настоящем описании 39B6-ANK) не подвергался окислению, но подвергался дезамидированию и изомеризации в течение 28-дневного периода хранения при 37°C и подвергался значительному снижению активности связывания в течение 56-дневного периода хранения. Вариант E96R (называемый в настоящем описании 39B6-ANR) подвергался значительному окислению и дезамидированию в течение 28-дневного периода хранения при 37°C и подвергался снижению активности связывания в течение 56-дневного периода хранения.
В предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где:
CDR3 VH содержит или состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит или состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12),
CDR1 VH содержит или состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4),
CDR3 VL содержит или состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит или состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит или состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
В некоторых предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где:
CDR3 VH содержит аминокислотную последовательность YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH содержит аминокислотную последовательность RIDPEDAGTKYAQKFQG (SEQ ID NO: 12),
CDR1 VH содержит аминокислотную последовательность SYYID (SEQ ID NO: 4),
CDR3 VL содержит аминокислотную последовательность QQYASVPVT (SEQ ID NO: 11),
CDR2 VL содержит аминокислотную последовательность GASRLKT (SEQ ID NO: 10), и
CDR1 VL содержит аминокислотную последовательность QASQSISSYLA (SEQ ID NO: 9).
В некоторых предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат, по меньшей мере, одну вариабельную область тяжелой цепи (VH) и, по меньшей мере, одну вариабельную область легкой цепи (VL), где:
CDR3 VH состоит из аминокислотной последовательности YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
CDR2 VH состоит из аминокислотной последовательности RIDPEDAGTKYAQKFQG (SEQ ID NO: 12),
CDR1 VH состоит из аминокислотной последовательности SYYID (SEQ ID NO: 4),
CDR3 VL состоит из аминокислотной последовательности QQYASVPVT (SEQ ID NO: 11),
CDR2 VL состоит из аминокислотной последовательности GASRLKT (SEQ ID NO: 10), и
CDR1 VL состоит из аминокислотной последовательности QASQSISSYLA (SEQ ID NO: 9).
Эти антитела предпочтительно имеют значение KD ниже 1,7×10-9 М.
В особенно предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат вариабельную область тяжелой цепи (VH), содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 14 и необязательно вариабельную область легкой цепи (VL), содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 15.
В особенно предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность SEQ ID NO: 14 и вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность SEQ ID NO: 15.
В особенно предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат вариабельную область тяжелой цепи (VH), состоящую из аминокислотной последовательности SEQ ID NO: 14 и вариабельную область легкой цепи (VL), состоящую из аминокислотной последовательности SEQ ID NO: 15.
В дополнительных предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат тяжелую цепь, содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 16, и необязательно легкую цепь, содержащую или состоящую из аминокислотной последовательности SEQ ID NO: 17.
В еще одних предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:16, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO:17.
В еще одних предпочтительных вариантах осуществления изобретения антитела, которые специфически связываются с комплексом GARP-TGF-β1 и проявляют повышенную стабильность, содержат тяжелую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 16, и легкую цепь, состоящую из аминокислотной последовательности SEQ ID NO: 17.
Полинуклеотиды, кодирующие антитела к GARP-TGF-β1
Изобретение также относится к полинуклеотидным молекулам, содержащим одну или более нуклеотидных последовательностей, кодирующих антитела к GARP-TGF-β1 или антигенсвязывающие фрагменты по изобретению, экспрессионным векторам, содержащим указанные нуклеотидные последовательности по изобретению, функционально связанные с регуляторными последовательностями, которые обеспечивают экспрессию антител или их фрагментов в клетке-хозяине или бесклеточной системе экспрессии, и клеткам-хозяевам или бесклеточным системах экспрессии, содержащим указанные экспрессионные векторы.
В некоторых вариантах осуществления вариабельная область тяжелой цепи и/или вариабельная область легкой цепи анти-GARP-TGF-β1-антител или антигенсвязывающих фрагментов по настоящему изобретению кодируются первой и второй полинуклеотидными последовательностями, где первая и вторая полинуклеотидные последовательности соответствуют аминокислотным последовательностям SEQ ID NO: 18 и 19 соответственно. В некоторых вариантах осуществления полинуклеотиды, кодирующие антитела к GARP-TGF-β1 по изобретению, могут содержать вариантные последовательности, которые кодируют функциональные области VH или VL антитела к GARP-TGF-β1. Вариантные последовательности, кодирующие области VH, могут иметь, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичность последовательности при оптимальном выравнивании с SEQ ID NO:18, и вариантные последовательности, кодирующие области VL, могут иметь, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичность последовательности при оптимальном выравнивании с SEQ ID NO: 19.
В некоторых вариантах осуществления тяжелая цепь и/или легкая цепь анти-GARP-TGF-β1-антител или антигенсвязывающих фрагментов по настоящему изобретению кодируются первой и второй полинуклеотидными последовательностями, где первая и вторая полинуклеотидные последовательности соответствуют аминокислотным последовательностям SEQ ID NO: 20 и 21 соответственно. В некоторых вариантах осуществления полинуклеотиды, кодирующие антитела к GARP-TGF-β1 по изобретению, могут содержать вариантные последовательности, которые кодируют тяжелые цепи или легкие цепи антитела к GARP-TGF-β1. Вариантные последовательности, кодирующие тяжелые цепи, могут иметь, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичность последовательности при оптимальном выравнивании с SEQ ID NO: 20, и вариантные последовательности, кодирующие легкие цепи, могут иметь, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичность последовательности при оптимальном выравнивании с SEQ ID NO: 21.
В этом контексте % идентичность последовательностей между двумя полинуклеотидными последовательностями можно определить сравнением этих двух последовательностей, выровненных оптимальным образом, и в которых сравниваемая полинуклеотидная последовательность может содержать добавления или делеции относительно референсной последовательности для оптимального выравнивания этих двух последовательностей. Процент идентичности рассчитывается определением числа идентичных положений, для которых нуклеотидный остаток является идентичным между двумя последовательностями, делением этого числа идентичных положений на общее число положений в окне сравнения и умножением полученного результата на 100 с получением процента идентичности этих двух последовательностей. Например, можно использовать программу BLAST «Последовательности BLAST 2» (Tatusova et al, «Blast 2 sequences - a new tool for comparing protein and nucleotide sequences», FEMS Microbiol Lett. 174:247-250), доступную на сайте ncbi.nlm.nih.gov/gorf/bl2, в которой используются параметры по умолчанию (в частности, такие параметры как «штраф за открытый гэп»: 5 и «штраф за удлинение гэпа»: 2; например, выбранная матрица представляет матрицу «BLOSUM 62», предложенную программой), при этом процент идентичности двух сравниваемых последовательностей рассчитывается непосредственно программой.
Полинуклеотидные молекулы, кодирующие антитела по изобретению, включают, например, рекомбинантные молекулы ДНК. Термины «нуклеиновая кислота», «полинуклеотид» или «полинуклеотидная молекула», используемые в настоящем описании, взаимозаменяемы и относятся к любой молекуле ДНК или РНК, одно- или двухцепочечной, и, если она одноцепочечная, то к молекуле с комплементарной ей последовательностью. При обсуждении молекул нуклеиновой кислоты последовательность или структура конкретной молекулы нуклеиновой кислоты может быть описана в настоящем описании в соответствии с обычным условным обозначением последовательности в направлении от 5' к 3'. В некоторых вариантах осуществления изобретения нуклеиновые кислоты или полинуклеотиды являются «выделенными». Данный термин, когда используется применительно к молекуле нуклеиновой кислоты, относится к молекуле нуклеиновой кислоты, которая отделена от последовательностей, с которыми она непосредственно ассоциирована в природном геноме организма, в котором она возникла. Например, «выделенная нуклеиновая кислота» может включать молекулу ДНК, вставленную в вектор, такой как плазмидный или вирусный вектор, или интегрированную в геномную ДНК прокариотической или эукариотической клетки или организма-хозяина, не являющегося человеком. Применительно к РНК термин «выделенный полинуклеотид» в основном относится к молекуле РНК, кодированной выделенной молекулой ДНК, как определено выше. Альтернативно, данный термин может относиться к молекуле РНК, которая была очищена/отделена от других нуклеиновых кислот, с которыми она была связана в своем природном состоянии (т. е. в клетках или тканях). Выделенный полинуклеотид (ДНК или РНК) может, кроме того, представлять собой молекулу, полученную непосредственно биологическим или синтетическим способом и отделенную от других компонентов, присутствующих в процессе ее получения.
Для рекомбинантной продукции антитела по изобретению рекомбинантный полинуклеотид, кодирующий его, можно получить (с использованием стандартных методов молекулярной биологии) и вставить в реплицируемый вектор для экспрессии в выбранной клетке-хозяине или бесклеточной системе экспрессии. Подходящими клетками-хозяевами могут быть клетки прокариот, дрожжей или высших эукариот, в частности, клетки млекопитающих. Примерами пригодных линий клеток-хозяев млекопитающих являются следующие: линия CV1 почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); линия почки эмбриона человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol., 36:59 (1977)); клетки почки детеныша хомячка (BHK, ATCC CCL 10); клетки яичника китайского хомячка/-DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA, 77: 4216 (1980)); мышиные клетки Сертоли (TM4, Mather, Biol. Reprod., 23: 243-251 (1980)); клетки мышиной миеломы SP2/0-AG14 (ATCC CRL 1581; ATCC CRL 8287) или NS0 (коллекции культур HPA № 85110503); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крыс buffalo (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383: 44-68 (1982)); клетки MRC 5; клетки FS4; и линия гепатомы человека (Hep G2), а также клеточная линия PERC-6 DSM. Экспрессионные векторы, подходящие для использования в каждой из этих клеток-хозяев, также в целом известны в данной области.
Следует отметить, что термин «клетка-хозяин» обычно относится к культивируемой клеточной линии. Целый организм человека, в который был введен экспрессионный вектор, кодирующий антитело или антигенсвязывающий фрагмент по изобретению, явно исключается из определения «клетки-хозяина».
Получение антител
В дополнительном аспекте изобретение также относится к способу получения антител по изобретению, который включает культивирование клетки-хозяина (или бесклеточной системы экспрессии), содержащей один или более полинуклеотидов (например, экспрессионный вектор), кодирующих антитело, в условиях, которые обеспечивают экспрессию антитела и выделение экспрессированного антитела. Данный процесс рекомбинантной экспрессии можно использовать для крупномасштабного производства антител, в том числе анти-GARP-TGF-β1-антител по изобретению, включая моноклональные антитела, предназначенные для терапевтического применения у людей. Подходящие векторы, клеточные линии и производственные процессы для крупномасштабного производства рекомбинантных антител, подходящих для терапевтического применения in vivo, обычно доступны в данной области и хорошо известны специалисту в данной области.
Фармацевтические композиции
В объем изобретения входят фармацевтические композиции, содержащие одно или комбинацию антител к GARP-TGF-β1 по изобретению или их антигенсвязывающих фрагментов, формулированных с одним или более фармацевтически приемлемыми носителями или эксципиентами. Такие композиции могут включать одно или комбинацию (например, два или более разных) антител к GARP-TGF-β1. Способы формуляции моноклональных антител для терапевтического применения у людей хорошо известны в данной области и их обзор имеется, например, в публикации Wang et al., Journal of Pharmaceutical Sciences, Vol.96, pp1-26, 2007, содержание которой в полном объеме включено в настоящем описании посредством ссылки.
В некоторых вариантах осуществления фармацевтические композиции формулированы для введения индивиду любым подходящим путем введения, включая, не ограничиваясь этим, внутривенное, внутримышечное, внутрикожное, внутрибрюшинное, подкожное, эпидуральное, интраназальное, оральное, ректальное, местное, ингаляционное, буккальное (например, сублингвальное) и трансдермальное введение.
Фармацевтически приемлемые эксципиенты, которые можно использовать в этих композициях, включают, не ограничиваясь этим, ионообменники, оксид алюминия, стеарат алюминия, лецитин, сывороточные белки, такие как человеческий сывороточный альбумин, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, смеси частичных глицеридов насыщенных растительных жирных кислот, воду, соли или электролиты, такие как протамина сульфат, динатрия гидрофосфат, гидрофосфат калия, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, соединения на основе целлюлозы (например натрий карбоксиметилцеллюлоза), полиэтиленгликоль, полиакрилаты, воска, блок-сополимеры полиэтилена-полиоксипропилена, полиэтиленгликоль и ланолин.
Терапевтическая применимость антител к GARP-TGF-β1
Антитела и антигенсвязывающие фрагменты по настоящему изобретению можно использовать в способах лечения, где индивиду, вводят терапевтически эффективное количество антитела к GARP-TGF-β1 или его антигенсвязывающего фрагмента. В некоторых вариантах осуществления антитела и антигенсвязывающие фрагменты по настоящему изобретению можно использовать в способах лечения, где человеку, вводят терапевтически эффективное количество антитела к GARP-TGF-β1 или его антигенсвязывающего фрагмента. Обеспечиваются антитела или их антигенсвязывающие фрагменты, которые связываются с комплексом человеческих GARP и TGF-β1 для применения в качестве лекарственных средств.
Антитела и антигенсвязывающие фрагменты по изобретению связываются с комплексом GARP-TGF-β1 на регуляторных Т-клетках и могут блокировать или ингибировать продукцию или высвобождение активного TGF-β. Следовательно, в дополнительном аспекте изобретения обеспечиваются способы лечения индивидов, имеющих или имеющих подозрение на наличие расстройств, связанных с TGF-β. Такие способы включают введение индивиду, терапевтически эффективного количества антитела к GARP-TGF-β1 по изобретению.
Типичные расстройства, связанные с TGF-β, включают, не ограничиваясь этим, воспалительные заболевания, хроническую инфекцию, злокачественное новообразование, фиброз, сердечно-сосудистое заболевание, цереброваскулярную болезнь (например, ишемический инсульт) и нейродегенеративные заболевания. В некоторых вариантах осуществления расстройство, связанное с TGF-β, представляет собой хроническую инфекцию. В некоторых вариантах осуществления расстройство, связанное с TGF-β, представляет злокачественное новообразование.
Для применения при введении индивиду антитела и антигенсвязывающие фрагменты по изобретению можно формулировать в виде фармацевтических композиций. Композиции можно вводить перорально, парентерально, с помощью ингаляционного спрея, местно, ректально, интраназально, буккально, интравагинально или через имплантированный резервуар. Как в настоящем описании используется, термин «введение» включает, без ограничения, методы подкожной, внутривенной, внутрибрюшинной, внутримышечной, внутрисуставной, интрасиновиальной, интрастернальной, интратекальной, внутрипеченочной, в очаг поражения, интратуморальной и интракраниальной инъекции или инфузии.
Стерильные инъецируемые формы композиций могут представлять водную или масляную суспензию. Такие суспензии можно формулировать в соответствии с методиками, известными в данной области техники, с использованием подходящих диспергирующих или смачивающих агентов и суспендирующих агентов. Стерильный инъекционный препарат также может представлять стерильный инъекционный раствор или суспензию, в нетоксичном приемлемом для парентерального введения разбавителе или растворителе. Среди приемлемых носителей и растворителей, которые можно использовать, находятся вода, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, стерильные нелетучие масла обычно используются в качестве растворителя или суспендирующей среды. Для этой цели можно использовать любое мягкое нелетучее масло, включая синтетические моно- или диглицериды жирных кислот. Жирные кислоты, такие как олеиновая кислота и ее глицеридные производные, пригодны для приготовления инъекционных препаратов, как и природные фармацевтически приемлемые масла, такие как оливковое масло или касторовое масло, особенно в их полиоксиэтилированных вариантах. Эти масляные растворы или суспензии также могут включать длиноцепочечные спиртовые растворители или дисперсанты, такие как карбоксиметилцеллюлоза или аналогичные диспергирующие агенты, которые обычно используются при формуляции фармацевтически приемлемых лекарственных форм, включая эмульсии и суспензии. Другие обычно используемые поверхностно-активные вещества, такие как твины, спаны и другие эмульгирующие агенты или усилители биодоступности, которые обычно используются в изготовлении фармацевтически приемлемых твердых, жидких или других лекарственных форм, также можно использовать для целей формуляции.
Режимы и дозировки для введения антитела или его антигенсвязывающего фрагмента в фармацевтических композициях можно определить в соответствии с известными способами для этих типов продуктов, например, с использованием инструкций производителя. Например, антитело, находящееся в фармацевтической композиции по данному изобретению, можно вводить в концентрации 10 мг/мл в одноразовых флаконах по 100 мг (10 мл) или 500 мг (50 мл). Продукт формулирован для внутривенного (в/в) введения в 9,0 мг/мл хлорида натрия, 7,35 мг/мл дигидрата цитрата натрия, 0,7 г/мл полисорбата 80 и стерильной воде для инъекций. Значение рН доводят до 6,5.
Для клинического применения антитело или антигенсвязывающий фрагмент можно вводить индивиду в одной или нескольких дозах. Для парентеральных путей введения разовая доза антитела или антигенсвязывающего фрагмента может составлять, например, примерно от 0,01 до примерно 100 мг/кг массы тела. В одном варианте осуществления разовая доза антитела или антигенсвязывающего фрагмента может составлять, например, примерно от 0,1 до примерно 50 мг/кг массы тела. В одном варианте осуществления разовая доза антитела или антигенсвязывающего фрагмента может составлять, например, примерно от 1 до примерно 20 мг/кг массы тела. Для многократного дозирования индивидуальные дозы можно вводить одним и тем же или разными путями введения. Также для многократного дозирования индивидуальные дозы могут быть одинаковыми или разными. Например, первая или нагрузочная доза может быть выше, чем последующая доза. Также для многократного дозирования индивидуальные дозы можно вводить по фиксированной схеме или по корректируемой или изменяемой схеме с учетом, например, клинического состояния индивида или клинического ответа. Также для многократного дозирования индивидуальные дозы обычно можно вводить один раз в день, через день, один раз в 3, 4, 5, 6 или 7 дней, один раз в неделю, один раз в две недели, один раз в три или четыре недели или один раз в месяц. Другие схемы также предусмотрены изобретением.
Понятно, что эти дозы и схемы являются примерными и что оптимальную схему и режим можно определить, принимая во внимание такие факторы, как конкретное антитело или антигенсвязывающий фрагмент, которые нужно вводить, заболевание или расстройство, которое нужно лечить, размер, возраст и состояние индивида, подлежащего лечению, путь введения, другие виды лечения, применяемые для индивида, а также аффинность и переносимость конкретного антитела. Такие факторы и соображения дозирования могут быть определены в одном или нескольких клинических испытаниях.
Настоящее изобретение также относится к способам стимуляции иммунной системы у индивида, включающим введение индивиду терапевтически эффективного количества антитела или антигенсвязывающего фрагмента по изобретению. Настоящее изобретение также относится к способам ингибирования иммуносупрессивной функции человеческих Treg у индивида, включающим введение индивиду терапевтически эффективного количества антитела или антигенсвязывающего фрагмента по изобретению.
Настоящее изобретение также обеспечивает способы лечения злокачественного новообразования с использованием антител к GARP-TGF-β1 и антигенсвязывающих фрагментов по изобретению. Такие способы могут включать снижение иммуносупрессии в окружении опухоли у индивида, нуждающегося в этом.
Для вариантов осуществления, в которых антитела к GARP-TGF-β1 или их антигенсвязывающие фрагменты предназначены для применения в способах лечения злокачественного новообразования, антитела или их антигенсвязывающие фрагменты можно вводить в комбинации с одним или более дополнительными способами лечения злокачественного новообразования, например одним или более иммунотерапевтическими агентами.
Для вариантов осуществления, в которых антитела к GARP-TGF-β1 вводят в комбинации с иммунотерапевтическим агентом, указанный иммунотерапевтический агент может представлять противоопухолевую вакцину. Альтернативно, иммунотерапевтический агент может представлять иммуностимулирующее антитело. Не желая связываться с какой-либо теорией, антитела по изобретению, вероятно, повышают эффективность иммунотерапевтического агента посредством предупреждения или ослабления проявления любой иммуносупрессии. В определенных вариантах осуществления комбинация с иммунотерапевтическим агентом может проявлять синергический эффект.
Различные виды злокачественного новообразования можно лечить в соответствии со способами, описанными в настоящем описании, включая, не ограничиваясь этим, адренокортикальную карциному, злокачественное новообразование анального канала, рак мочевого пузыря, опухоль головного мозга, глиому, рак молочной железы, карциноидную опухоль, рак шейки матки, рак толстой кишки, рак эндометрия, рак пищевода, рак внепеченочных желчных протоков, опухоль Юинга, экстракраниальную опухоль из зародышевых клеток, рак глаза, рак желчного пузыря, рак желудка, опухоль из зародышевых клеток, гестационную трофобластическую опухоль, рак головы и шеи, гипофарингеальный рак, карциному островков Лангерганса, рак почки, рак гортани, лейкоз, рак губ и ротовой полости, рак печени, рак легкого, лимфому, меланому, мезотелиому, карциному Меркеля, метастатический плоскоклеточный рак головы и шеи, миелому, новообразование, назофарингеальный рак, нейробластому, рак ротовой полости, рак ротоглотки, остеосаркому, рак яичника, рак поджелудочной железы, рак околоносовых пазух и полости носа, рак паращитовидной железы, рак полового члена, феохромоцитому, рак гипофиза, новообразование из плазматических клеток, рак предстательной железы, рабдомиосаркому, рак прямой кишки, почечноклеточную карциному, рак слюнной железы, рак кожи, саркому Капоши, Т-клеточную лимфому, саркому мягких тканей, рак желудка, рак яичка, тимому, рак щитовидной железы, рак уретры, рак матки, рак влагалища, рак вульвы или опухоль Вильмса.
Подходящие опухолевые антигены для применения в качестве противоопухолевых вакцин, известные в данной области, включают, например: (а) антигены рака яичка, такие как NY-ESO-1, SSX2, SCP1, а также семейства полипептидов RAGE, BAGE, GAGE и MAGE, например, GAGE-1, GAGE-2, MAGE-1, MAGE-2, MAGE-3, MAGE-4, MAGE-5, MAGE-6 и MAGE-12 (которые можно использовать, например, для лечения меланомы, рака легкого, головы и шеи, NSCLC, рака молочной железы, желудочно-кишечного тракта и мочевого пузыря), (b) мутированные антигены, например, p53 (ассоциированный с различными солидными опухолями, например, колоректальным раком, раком легкого, раком головы и шеи), p21/Ras (ассоциированный, например, с меланомой, раком поджелудочной железы и колоректальным раком), CD 4 (ассоциированный, например, с меланомой), MUM 1 (ассоциированный, например, с меланомой), каспазу-8 (ассоциированную, например, с раком головы и шеи), CIA 0205 (ассоциированный, например, с раком мочевого пузыря), HLA-A2-R1701, бета-катенин (ассоциированный, например, с меланомой), TCR (ассоциированный, например, с T-клеточной неходжкинской лимфомой), BCR-abl (ассоциированный, например, с хроническим миелоидным лейкозом), триозофосфатизомеразу, IA 0205, CDC-27 и LDLR-FUT, (c) сверхэкспрессируемые антигены, например, галектин 4 (ассоциированный, например, с колоректальным раком), галектин 9 (ассоциированный, например, с болезнью Ходжкина), протеиназу 3 (ассоциированную, например, с хроническим миелоидным лейкозом), WT 1 (ассоциированный, например, с различными лейкозами), карбоангидразу (ассоциированную, например, с раком почки), альдолазу A (ассоциированную, например, с раком легкого), PRAME (ассоциированный, например, с меланомой), HER-2/neu (ассоциированный, например, с раком молочной железы, толстой кишки, легкого и яичника), альфа-фетопротеин (ассоциированный, например, с гепатомой), SA (ассоциированный, например, с колоректальным раком), гастрин (ассоциированный, например, с раком поджелудочной железы и желудка), каталитический белок теломеразы, MUC-1 (ассоциированный, например, с раком молочной железы и яичника), G-250 (ассоциированный, например, с почечно-клеточным раком), и карциноэмбриональный антиген (ассоциированный, например, с раком молочной железы, раком легкого и раком желудочно-кишечного тракта, таким как колоректальный рак), (d) общие антигены, например, дифференцировочные антигены меланомы-меланоцитов, такие как MART-1/Melan A, gp100, MC1R, рецептор меланоцитостимулирующего гормона, тирозиназа, родственный тирозиназе белок-1/TRP1 и родственный тирозиназе белок-2/TRP2 (ассоциированный, например, с меланомой), (e) антигены, ассоциированные с предстательной железой, такие как PAP, PSA, PSMA, PSH-P1, PSM-P1, PSM-P2, связанные, например, с раком предстательной железы, (f) идиотипы иммуноглобулинов (например, ассоциированные с миеломой и В-клеточными лимфомами) и (g) другие опухолевые антигены, такие как полипептид- и сахаридсодержащие антигены, включая (i) гликопротеины, такие как сиалил-Tn и сиалил-Lex (ассоциированные, например, с раком молочной железы и колоректальным раком), а также различные муцины; гликопротеины могут быть связаны с белком-носителем (например, MUC-1 может быть связан с LH); (ii) липополипептиды (например, MUC-1, связанный с липидной молекулой); (iii) полисахариды (например, Globo H-синтетический гексасахарид), которые могут быть соединены с белками-носителями (например, с KLH), (iv) ганглиозиды, такие как GM2, GM12, GD2, GD3 (ассоциированные, например, с раком мозга, легкого, меланомой), которые также могут быть связана с белками-носителями (например, KLH). Другие опухолевые антигены включают pi 5, Hom/Mel-40, H-Ras, E2A-PRL, H4-RET, IGH-IGK, MYL-RAR, антигены вируса Эпштейна-Барра, EBNA, антигены вируса папилломы человека (HPV), включая E6 и Е7, антигены вируса гепатита В и С, антигены Т-лимфотропного вируса человека, TSP-180, pl85erbB2, pl80erbB-3, c-met, mn-23H1, TAG-72-4, CA 19-9, CA 72- 4, CAM 17.1, NuMa, K-ras, p 16, TAGE, PSCA, CT7, 43-9F, 5T4, 791 Tgp72, бета-ХГЧ, BCA225, BTAA, CA 125, CA 15-3 (CA 27.29/BCAA), CA 195, CA 242, CA-50, CAM43, CD68/KP1, CO-029, FGF-5, Ga733 (EpCAM), HTgp-175, M344, MA-50, MG7-Ag, MOV 18, NB/70K , NY-CO-1, RCAS1, SDCCAG16, TA-90 (белок, связывающий Mac-2/белок, связанный с циклофилином C), TAAL6, TAG72, TLP, TPS и тому подобное. Подходящие иммуностимулирующие антитела включают, не ограничиваясь этим: анти-CTLA-4, анти-PD1, анти-PDL1 и анти-KIR антитела.
В некоторых вариантах осуществления способы лечения злокачественного новообразования, описанные в настоящем описании, включают введение индивиду антитела или антигенсвязывающего фрагмента по изобретению до, одновременно и/или после введения другого противоопухолевого агента или лечения злокачественного новообразования, такого как химиотерапия.
Настоящее изобретение также включает способы профилактики инфекционных заболеваний введением антител или антигенсвязывающих фрагментов по изобретению, для повышения эффективности стратегий вакцинации. Например, способы по изобретению могут включать профилактику инфекционных заболеваний, таких как ВИЧ, малярия или лихорадка Эбола, комбинированным введением антител или антигенсвязывающих фрагментов, как в настоящем описании описано, вместе с вакцинами, специфичными для этих заболеваний.
Наборы
В дополнительном аспекте настоящее изобретение относится к набору, содержащему, по меньшей мере, одно антитело к GARP-TGF-β1 или антигенсвязывающий фрагмент по изобретению.
Термин «набор» предназначен для обозначения любого предмета производства (например, упаковки или контейнера), содержащего, по меньшей мере, один реагент, например антитело или антигенсвязывающий фрагмент по изобретению, для специфического связывания комплекса GARP-TGF-β1. Набор может рекламироваться, распространяться или продаваться в виде комплекта для осуществления способов по настоящему изобретению. Кроме того, любой или все реагенты набора могут быть представлены в контейнерах, которые защищают их от внешней среды, например в герметичных контейнерах. Наборы могут также содержать вкладыш в упаковку, описывающий набор и способы его применения.
Примеры
Изобретение будет дополнительно пояснено со ссылкой на следующие неограничивающие примеры.
Пример 1. Получение антитела зародышевого типа LHG-10.6.
Получение и характеристика антитела «LHG-10» описаны в международных заявках на патент WO2015/015003 и WO2016/125017, содержание которых в полном объеме включено в настоящем описании посредством ссылки. Антитело LHG-10 получали иммунизацией лам клетками HEK293E, сверхэкспрессирующими комплекс человеческих GARP-TGF-β1, и было идентифицировано селекцией и скринингом Fab GARP-TGF-β1. Подход перестановки легких цепей (как описано, например, в международной заявке на патент WO2014/033252) использовали для повышения аффинности моноклонального антитела LHG-10, и вариант «LHG-10.6» (также относящийся в настоящем описании к 17H5) был определен как имеющий улучшенные характеристики связывания. Последовательности VH и VL LHG-10 и варианта с переставленными Vκ LHG-10.6 показаны в таблице 3.
Было установлено, что антитело 17H5 обладает 88,5% идентичностью с наиболее близкой последовательностью зародышевой линии человека (X92343|IGHV1-46*01) по области VH и 86,2% идентичностью с наиболее близкой последовательностью зародышевой линии человека (X59315|IGKV1-39*01) по области VL. Это антитело было подвергнуто превращению в антитело зародышевого типа с использованием 3-стадийного метода:
1 - Создание библиотеки генов с использованием перекрывающихся олигонуклеотидов для синтетического получения генов, кодирующих вариабельную область легкой (VL) и вариабельную область тяжелой (VH) цепи, посредством ПЦР.
2 - Клонирование этой библиотеки генов в вектор (pCB13), содержащий гены, кодирующие человеческий константный домен тяжелой цепи (CH1) и константный домен легкой цепи каппа (Cκ) (конструирование библиотеки).
3 - Селекция функциональных Fab с использованием фагового дисплея и селекция аффинности.
1. Конструирование библиотеки
VH 17H5 сравнивали с самой близкой последовательностью зародышевой линии человека и идентифицировали остатки каркаса, отклоняющиеся от зародышевой линии человека. Этим каркасным остаткам давали возможность мутировать в остаток, имеющийся в V-области человека, также одновременно сохраняя в библиотеке исходный остаток. 12 остатков VH, которым давали возможность мутировать в человеческие аналоги, приведены в таблице ниже. В дополнение к этим мутациям для получения зародышевого типа, сайт изомеризации в CDR2 (D54; D55) и сайт окисления в CDR3 (M100f) подвергали мутации. Эти сайты также показаны в таблице ниже.
Остатки-мишени в области VH 17H5
Для Vk 17H5 сравнение между самой близкой последовательностью человеческой зародышевой линии и каркасными последовательностями привело к идентификации 11 остатков-мишеней. Эти сайты показаны в таблице ниже.
Остатки-мишени в области VL 17H5
Теоретические размеры библиотеки 17H5 зародышевого типа показаны в таблице 6.
2. Конструирование библиотеки генов
Библиотеки генов зародышевой линии были созданы сборкой генов. Синтетические гены VH и VL (два набора), основанные на этом дизайне, получали сборкой на основе ПЦР (Stemmer et al., Gene (1995) 164: 49-530). Перекрывающиеся олигонуклеотиды со специфическими мутациями в определенных положениях собирали с помощью ПЦР. Нуклеотиды были вырожденными для кодирования аминокислоты человека и ламы. Это было сделано для предупреждения полной потери связывания в случае, если остатки ламы были критическими для обеспечения стабильности, фолдинга или высокоаффинного связывания.
Синтетические гены библиотек VH и VH 17H5 сначала клонировали в pCB13-CK1, фагемидный вектор с человеческими доменами Ck и CH1 соответственно. После конструирования этих подбиблиотек VkCk и VHCH1 конечные библиотеки получали лигированием вставки тяжелой цепи в библиотеку легкой цепи. ПЦР колоний проводили для определения процента вставки, и клоны отправляли для секвенирования, для оценки гарантии того, что все желаемые мутации присутствовали в библиотеке. Характеристики различных библиотек приведены в таблице ниже.
Характеристики библиотек 17H5 зародышевого типа
Было установлено, что библиотеки зародышевого типа хорошего качества, и их можно использовать для селекции.
3. Селекция Fab с высокой идентичностью с человеком
Фаговый дисплей использовали для селекции Fab-фрагментов с высокой аффинностью к комплексу человеческих GARP-TGF-β1, как описано ранее в WO 2015/015003, содержание которой в полном объеме включено в настоящем описании посредством ссылки. Вкратце, микротитрационный планшет покрывали O/N при 4°C комплексом GARP-TGF-β1, и на следующий день фаги, экспрессирующие Fab, инкубировали в течение 2 ч в лунках, покрытых различными концентрациями комплекса. Планшеты промывали и специфический фаг элюировали трипсином и использовали для инфицирования клеток TG1. Детали условий селекции приведены в таблице 8.
Детали раундов селекции для библиотеки 17H5 зародышевого типа
После первого раунда селекции обогащение наблюдали только с покрытием нерелевантным белком в концентрации 10 мкг/мл. Во втором раунде обогащение было в 10 раз выше, тогда как количество введенного фага было в 10 раз ниже. Начиная с третьего раунда, жесткость селекции увеличивали посредством повышения продолжительности отмывки и добавления избытка мишени в растворе. Добавляли большой избыток раствора растворимой мишени для предупреждения повторного связывания диссоциированного фага. Даже после 6 раунда, с отмывкой в течение одной недели и 100-кратным избытком растворимой мишени, обогащение все еще наблюдали.
4. Скрининг Fab с высокой идентичностью с человеком
Мастер-планшеты отбирали из раундов селекции 2, 3, 4, 5 и 6 (48 клонов на каждый раунд). Готовили периплазматические экстракты, и скорость диссоциации определяли с помощью SPR (с использованием Biacore). Все клоны имели секвенированные ДНК, и из них были выведены аминокислотные последовательности. Клоны с идентичностью V-области Ig зародышевой линии человека выше 92% приведены в таблице 9. Скорость диссоциации этих клонов измеряли в виде периплазматических экстрактов в анализе SPR (с использованием Biacore) и сравнивали со скоростью диссоциации Fab 17H5.
Аффинность связывания клонов с более чем 92% общей идентичностью с человеком
Эти 5 клонов, имеющих общую идентичность с человеком на уровне > 93% и скорость диссоциации, сравнимую с таковой у родительского Fab 17H5, реклонировали в виде полноразмерных человеческих IgG1 антител. Последовательности CDR, VH и VL этих антител показаны в таблицах 10, 11 и 12.
Последовательность CDR VH для Ab против GARP-TGF-β1
Последовательность VL CDR для Ab против GARP-TGF-β1
Последовательности вариабельных областей для Ab против GARP-TGF-β1
5. Характеристика mAb in vitro с высокой идентичностью остатков каркасной области с человеком
Пять клонов зародышевой линии, переформатированных в человеческий IgG1, получали транзиентной трансфекцией клеток HEK 293E. Все антитела тестировали на связывание с комплексом человеческих GARP-TGF-β1 с использованием SPR, и они показали аффинность связывания, сходную с родительским клоном незародышевой линии (KD в 2-5 раз ниже, чем 17H5). Замена остатка M100f в CDR3 на треонин снижала KD ≈ в 5 раз. Связывающие свойства и характеристики Ab 17H5 зародышевого типа приведены в таблице 13.
Связывающие свойства и характеристики Ab 17H5 зародышевого типа
6. Стабильность mAb 39B6 зародышевого типа
Клон 39B6 отбирали в качестве основного клона зародышевой линии на основании его характеристик связывания. Это антитело получали в двух форматах с дефицитной эффекторной функцией, hIgG1N297Q и hIgG4S228P, и проводили тесты на стабильность. Перед тестированием стабильности аффинность дефицитных по эффекторной функции антител к GARP-TGF-β1 39B6 анализировали BiacoreT200 с использованием биосенсорного чипа CM5, покрытого комплексом GARP-TGF-β1 при 750RU (таблица 14) или покрытого антителами при 1000RU (таблица 15).
Термостабильность антител 39B6 тестировали с использованием следующей схемы.
Подготовка образца mAb: 1 мл mAb (39B6-IgG1 и 39B6-IgG4), свежеприготовленный исходный раствор (5 мг/мл), помещали в стеклянные флаконы со стеклянными завинчивающимися крышками емкостью 2 мл и хранили при 5°C и 37°C. Флаконы сразу же проверяли на отсутствие частиц.
Отрицательный контроль PBS: аликвотные порции 1 мл профильтрованного PBS Дульбекко готовили в стеклянных флаконах емкостью 2 мл в качестве отрицательного контроля PBST. Флаконы сразу же проверяли на отсутствие частиц.
Отрицательный контроль PBSTw: аликвотные 1 мл профильтрованного PBS Дульбекко, содержащего 0,02% Твина 80 (Sigma), готовили в стеклянных флаконах емкостью 2 мл в качестве отрицательного контроля PBSTw. Флаконы сразу же проверяли на отсутствие частиц.
Контроль с высокой агрегацией: аликвотные порции 1 мл собственного антитела готовили с обильной видимой агрегацией в стеклянных флаконах емкостью 2 мл.
Контрольный образец: для каждого антитела аликвотную порцию mAb 60 мкл готовили в стерильных пробирках для ПЦР емкостью 500 мкл, маркировали и хранили при -20°C.
Стабильность антител, хранившихся при разных температурах и в разных условиях (PBS по сравнению с PBSTw), контролировали в течение 56 суток. Влияние хранения на активность связывания с мишенью, измеренное анализом SPR (Biacore™), показано на фиг.2. Как видно из представленных результатов, антитело 39B6 (в обоих форматах с дефицитом эффекторной функции) проявляло тенденцию к снижению активности связывания с мишенью в PBS и PBS/Твине при хранении при 37°С. В противоположность, образцы, хранившиеся при 5°С, не проявляли значительной потери в активности связывания с мишенью во времени.
Пример 2. Разработка антител против GARP-TGF-β1 с повышенной стабильностью
2.1 Получение вариантов 39В6
Было обнаружено, что снижение активности связывания с мишенью, отмеченное выше для антитела 39B6, связано с дезамидированием, имевшим место в положениях 95-96 CDR3 VH. Таким образом, была проведена дальнейшая работа по повышению стабильности 39B6.
Сначала этот клон подвергали модификации VH цепи в области CDR2 для введения замены G55A (39B6-A). Области VH и VL 39B6-A реклонировали в скелет человеческого IgG4S228P, т. е. в формате человеческого антитела с дефицитной эффекторной функцией.
В попытке повысить стабильность антитела 39B6-A IgG4S228P было получено пять вариантов, имеющих мутации в положениях 95 или 96 области CDR3 VH. Эти варианты показаны ниже. Все варианты были получены в дефицитном по эффекторной функции формате IgG4S228P.
Варианты 9B6-A
Антитела фильтровали и концентрировали до 5 мг/мл и хранили в PBS Дульбекко с 0,02% Твина 80 (Sigma). Твин 80 был необходим для стабилизации составов антител.
Аффинность всех антител к GARP-TGF-β1 (включая 39B6-A) тестировали анализом BiacoreT200 с использованием биосенсорного чипа CM5, покрытого комплексом GARP-TGF-β1 при 150RU (таблица 17) или покрытого антителами при 200RU (таблица 18).
Способность блокировать высвобождение активного TGF-β клетками Treg человека анализировали со всеми вариантами по определению уровня фосфорилирования SMAD2, стимулированных CD3/CD28 человеческих Treg в присутствии mAb. Как показано на фиг.3, антитела 39B6-AYE, 39B6-ANK и 39B6-ANR обладали сходной активностью в анализе фосфорилирования SMAD2 по сравнению с исходным антителом 39B6-A. Варианты 39B6-AEE и 39B6-AVE показали явную потерю активности.
Аналогичные результаты были получены с использованием анализа GAGA-luc, где клетки 293T-hGARP транзиентно трансфицировали репортерной плазмидой, в которой люцифераза светлячков находится под контролем SMAD-реактивного промотора (GAGA-luc). Антитела 39B6-AYE, 39B6-ANK и 39B6-ANR обладали сходной активностью в анализе с люциферазой по сравнению с исходным антителом 39B6-A. Варианты 39B6-AEE и 39B6-AVE показали явную потерю активности (см. фиг. 4).
2.2 Исследование стабильности
За исключением 39B6-AEE, остальные варианты тестировали на стабильность с использованием различных подходов, как описано ниже.
Для каждого из различных исследований стабильности антитела тестировали, используя один или более из следующих методов:
- визуальный контроль
- эксклюзионная ВЭЖХ
- SDS-PAGE (восстанавливающие и невосстанавливающие условия)
- аффинность связывания с мишенью с использованием анализа SPR
- концентрация белка.
Протоколы этих методов описаны ниже.
Протокол визуального контроля
Образцы исследовали «вслепую» и оценивали на наличие видимых частиц визуальным контролем флаконов тремя аналитиками. Образцам давали возможность достичь комнатной температуры в течение 30 мин перед анализом. Следующую оценочную систему использовали для оценки:
A: образец прозрачный, видимые частицы отсутствуют
В: очень мало частиц
C: умеренное наличие частиц
D: наличие обильных частиц
F: волокна
Протокол эксклюзионной хроматографии (SE-HPLC)
Система, колонка и пробоподготовка
Колонка, используемая на протяжении всего исследования, представляла собой Xbridge Protein BEH SEC 200A (3,5 мкм, 7,8×30 мм, Waters), которую обычно хранили в 20% EtOH. Предколонку Xbridge Protein BEH 200A соединяли с аналитической колонкой (3,5 мкм, 7,8×3 мм; Waters). Перед каждым использованием колонку уравновешивали PBS Дульбекко в объеме, по меньшей мере, 5CV, и перед удалением из системы ее промывали 5CV воды категории LC. Все растворители фильтровали и дегазировали перед использованием.
Используемая хроматографическая система представляла собой Agilent 1260 Infinity, оснащенная насосом для четырехкомпонентных смесей, автоматическим инжектором, он-лайн дегазатором и DAD-детектором. Колонку не хранили в термостатированном отделении, и образцы анализировали при комнатной температуре. Детектор был настроен на длины волн 280 и 214 нм одновременно (референсная длина волны при 360 нм с отсечением 100 нм). Агрегацию контролировали на канале 214 нм. Сбор данных проводили с помощью программного обеспечения Chemstation (Agilent).
Анализ образцов проводили переносом 25 мкл всех исходных образцов непосредственно из раствора с концентрацией 5 мг/мл в асептических условиях. Образцы центрифугировали при 2000 об/мин в течение 1 мин и 20 мкл осторожно переносили во флаконы из коричневого стекла с завинчивающейся крышкой. Объем инжектирования для каждого условия составлял 5 мкл (25 мкг) со скоростью потока 0,7 мл PBS/мин в течение 25 мин. Каждое инжектирование сопровождали промыванием иглы водой категории LC, и промывку септы проводили в конце анализа каждого образца.
Каждую последовательность временных точек начинали с 2× инжектирований PBS (пустых; инжектируемый объем 5 мкл) с последующим введением 2 мкл стандартной белковой смеси BEH 200 (Waters) и образца mAb с известной агрегацией. Анализировали каждые двенадцать образцов, и вводили пустую пробу. Каждую аналитическую последовательность завершали как начинали, но в обратном порядке (контрольное mAb с известной агрегацией, стандартная белковая смесь BEH и 2× пустая проба).
Метод, использованный для определения % агрегации и % площади мономеров
Использовали следующий протокол:
- Интегрируют все хроматограммы с помощью метода ARGX-115 (основные технические параметры: стандартный режим касательной, чувствительность наклона 2,0, отклонение по высоте 1,7, отклонение по площади 1,0, ширина пика: 0,02)
- Следует убедиться, что все пики интегрированы таким образом, чтобы это совпадало с результатами предыдущих анализов
- Экспортируют результаты интеграции в файл PDF и Excel
- Рассчитывают % общую агрегацию как сумму площадей всех пиков, элюированных до пика мономера, деленную на общую площадь инжектирования и умноженную на 100
- Также рассчитывают % площадь мономера делением площади мономера на общую площадь и умножением на 100. Эта % площадь мономера отражает наличие нерастворимых агрегатов в образце
- Хранят подробные отчеты для мониторинга производительности, эффективности и разрешения аналитической колонки (т.е. симметрия пика, площадь мономера, общая площадь, воспроизводимость времени удерживания и т. д.)
Протокол электрофореза SDS-PAGE
Подготовка образцов
Анализ образцов проводили переносом 25 мкл всех исходных образцов с концентрацией 5 мг/мл в 100 мкл смеси PBS/0,02% Твин 80 (сокращенно обозначенного как PBSTw на протяжении всего этого исследования) в асептических условиях, с получением промежуточной концентрации 1 мг/мл, для проведения электрофореза SDS-PAGE (в восстанавливающих и невосстанавливающих условиях), оценки аффинности связывания анализом SPR и определения концентрации белка. Для анализа образцов SDS-PAGE (Biorad) использовали 4-20% Трис-глициновый буфер, не окрашенные, готовые гели Mini-Protean. Готовили партию 4× концентрированного красителя с восстановителем DTT или без него, и готовили аликвоты для хранения при -20°C. Обычно 5 мкл отбирали из промежуточного разведения с концентрацией 1 мг/мл, и затем добавляли 5 мкл 4× концентрированного красителя (+/- DDT) вместе с 10 мкл mQ. Конечное количество для каждого условия составляло 5 мкг. Образцы кипятили в течение 10 мин при 95°С и затем наносили (20 мкл) на готовые гели. Электрофорез проводили в течение 35 мин в Трис-глициновой буферной системе при постоянном напряжении 200 В. Последующее окрашивание синим (Gentaur) проводили в течение 1 ч. Все гели промывали водой mQ в течение, по меньшей мере, 1 ч.
Определение % полноразмерного Ab
Интенсивность различных полос определяли в системе серии Odyssey v3.0 Li-Cor сканированием белковых гелей с одноканальным детектированием (700 нм) при настройках: смещение фокуса 0,5 мм и «высокое» качество. Яркость и контрастность для каждого анализа устанавливали на 50% с линейным параметром ручного управления 5.
Метод был следующим:
- Сканируют гель и подтверждают, что все полосы на геле окружены узкими прямоугольниками
- Выбирают «экспорт» в настройках для получения необработанных данных по интенсивности всех полос в формате Excel
- Рассчитывают % необработанную интенсивность каждой полосы, присутствующей на дорожке, как необработанную интенсивность полосы, деленную на общую необработанную интенсивность полосы, и умножают на 100
- Для невосстанавливающих условий % полноразмерного Ab определяется как % необработанная интенсивность полосы, которая соответствует полосе ~ 100 кДа
- Для восстанавливающих условий % полноразмерного Ab определяется как сумма % необработанной интенсивности полос, которые соответствуют полосам ~ 25-35 кДа и ~ 55 кДа.
Протокол измерения активности связывания с мишенью анализом Biacore 3000
Подготовка образцов
Из всех образцов с промежуточной концентрацией 1 мг/мл, описанных выше, для анализа SPR готовили дополнительное разведение 1/250, с получением конечной концентрации 4 мкг/мл. Это разведение 1/250 готовили для всех условий двумя стадиями: а) 5 мкл (1 мг/мл) + 120 мкл HBS-EP (буфер для SPR) и b) дополнительное 10-кратное разведение (15 мкл+135 мкл HBS-ЕР). Для каждого антитела и каждой временной точки строили отдельную калибровочную кривую, начиная с образца, замороженного при -80°С. Его анализировали вместе с образцами, использованными для испытания стабильности, для определения наклона каждой кривой после общего подбора стандартов. Эти наклоны использовали для расчета процентной активности образцов в анализе стабильности, установив контрольный образец на 100%.
Определение % активности в анализе Biacore
Для определения активности связывания с мишенью в Biacore сенсорный чип CM5 покрывали комплексом человеческих GARP-TGF-β1 при ~ 4000 RU. Поток устанавливали на 30 мкл/мин с режимом введения «kinject» и двумя введениями для регенерации (1 мМ NaCl, 2,5 мМ глицин, pH 1,5) с 5-минутным интервалом.
Метод проводили следующим образом:
- Открывают кривые в программе BIAEVAL и выбирают значение «2-1» (1: пустой, 2: hGARP-TGF-β1)
- Выбирают одну кривую на время для наложения графика
- Удаляют часть кривой регенерации
- Выбирают базовую линию непосредственно перед введением и трансформируют ось Y на «ноль при медиане выбора».
- Выбирают «Общая подгонка», начиная с 120 с после введения и заканчивают через 155 с
- Строят стандартную кривую в Excel и определяют наклон кривой для каждого антитела
- Эти значения преобразуют в процентную активность установкой контрольного значения (образец, который хранится при -20°C) на 100%.
Протокол определения концентрации белка
Для определения содержания белка для каждого условия хранения использовали систему NanoDrop, измеряя оптическую плотность каждого образца (2 мкл) при 280 нм. Контролем в системе служил PBS, и определение белка проводили измерением оптической плотности при 280 нм каждого образца в трех повторах (промежуточное разбавление с 1 мг/мл). Все значения, полученные при 280 нм, делили на коэффициент 1,51. Измерение оптической плотности пустого образца с PBS проводили после анализа образца для каждого отдельного условия. Все данные были представлены в виде среднего значения для трех измерений для каждого условия хранения.
2.2.1 Исследование термостабильности
Для контроля стабильности антител в различных температурных условиях хранения, следовали следующей схеме:
Подготовка образцов mAb: 1 мл mAb (39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR), свежеприготовленный исходный раствор (с концентрацией 5 мг/мл), помещали в стеклянные флаконы с завинчивающейся стеклянной крышкой емкостью 2 мл и хранили при 5°С и 37°С. Флаконы сразу же проверяли на отсутствие частиц.
Отрицательный контроль PBSTw: аликвотные порции 1 мл профильтрованного PBS Дульбекко, содержащего 0,02% Твина 80 (Sigma), готовили в стеклянных флаконах емкостью 2 мл и использовали в качестве отрицательного контроля, обозначенного PBSTw. Флаконы сразу же проверяли на отсутствие частиц.
Контроль с высокой агрегацией: аликвотные порции 1 мл собственного антитела готовили с обильной видимой агрегацией в стеклянных флаконах емкостью 2 мл.
Контрольный образец: для каждого антитела аликвотные порции 60 мкл mAb готовили в стерильных пробирках для ПЦР емкостью 500 мкл, маркировали и хранили при -20°C.
На разные временные точки пробы mAb отбирали из образцов, соответствующих условиям хранения и тестировали следующим образом:
- Визуальный контроль
- SE-HPLC
- SDS-PAGE (в восстанавливающих и невосстанавливающих условиях)
- аффинность связывания с мишенью в анализе SPR
- концентрация белка
Визуальный контроль
Результаты визуального контроля приведены в таблице 19 ниже. Все mAb находились в PBSTw, что должно оказывать стабилизирующий эффект.
Результаты визуального контроля образцов mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR в течение 56-дневного периода в условиях хранения при 5°C и 37°C
Для условий хранения при 5°C средние оценки для mAb через 56 суток были следующими:
39B6-AYE: «образец прозрачный, видимые частицы отсутствуют»
39B6-AVE и 39B6-ANK: «очень мало частиц»
39B6-ANR: «умеренное наличие частиц».
Буфер PBSTw был оценен как «образец прозрачный, видимые частицы отсутствуют» при 5°C, указывая на то, что наблюдаемые частицы связаны с белками.
Для условий хранения при 37°С средние оценки для mAb через 56 суток были следующими:
39B6-AYE, 39B6-ANK и 39B6-ANR: «очень мало частиц»
39B6-AVE: «образец прозрачный, видимые частицы отсутствуют».
Буфер PBSTw был оценен как «образец прозрачный, видимые частицы отсутствуют» при 37°C, указывая на то, что наблюдаемые частицы связаны с белками.
Анализ с использованием SE-HPLC
Агрегацию и фрагментацию белка измеряли с помощью SE-HPLC.
Результаты агрегации белка для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR, измеренные с помощью SE-HPLC, обобщены в таблице 20 ниже и показаны на фиг.5.
% образование агрегатов, контролируемое SE-HPLC, для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
При хранении при 5°C не наблюдали изменений % уровней агрегатов для mAb 39B6-AYE, 39B6-AVE и 39B6-ANK в течение 56 суток. Наблюдаемые % уровни агрегатов были низкими в пределах от 0,9% до 1,1%. Незначительное увеличение уровней агрегатов наблюдали для mAb 39B6-ANR с 1,1% до 1,4%.
При хранении при 37°С не наблюдали изменения % уровней агрегатов для mAb 39B6-AYE и 39B6-ANK. Для 39B6-ANR наблюдали незначительное увеличение с 1,1% на сутки 0 до 1,5% на сутки 56. Для 39B6-AVE увеличение % агрегатов наблюдали с 1,0% на сутки 0 до 4,4% на сутки 56.
Присутствие пиков фрагментов для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR, как измерено с помощью SE-HPLC, показано в таблице 21 и на фиг.6. Поскольку пики фрагментации наблюдали только при хранении при 37°C через 56 суток, то представлены только эти результаты.
% образование фрагментов, контролируемое анализом SE-HPLC для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR на 56 сутки
Для mAb 39B6-AVE, 39B6-ANK и 39B6-ANR процент пиков фрагментов составляет от 0,2% до 0,3% через 56 суток, тогда как для mAb 39B6-AYE процент пиков фрагментов составляет 5,7%.
Результаты % пика мономеров для всех mAb обобщены в таблице 22.
Площадь мономеров (%), контролируемая эксклюзионной хроматографией для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
SDS-PAGE
Результаты анализа SDS-PAGE для всех антител показаны в таблице 23 и на фиг.8.
Общий процент полноразмерных Ab/тяжелой цепи, измеренный анализом SDS-PAGE и сканированием Odyssey для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
Какой-либо тенденции к деградации не наблюдали ни для одного mAb для температурного режима хранения при 5°C в анализе, проведенном как в восстанавливающих условиях, так и в невосстанавливающих условиях.
При хранении при 37°С все mAb показали одинаковую скорость деградации в анализе, проведенном в невосстанавливающих условиях, за исключением mAb 39B6-AYE. Данное mAb показывает сходную скорость деградации до 28 суток, но демонстрирует более высокую скорость деградации в период между 28 и 56 сутками по сравнению с другими mAb. В анализе в восстанавливающих условиях все mAb показывают одинаковую скорость деградации.
Аффинность связывания с мишенью в анализе Biacore
Образцы тестировали на их активность связывания с мишенью в анализе Biacore измерением наклона на каждую временную точку. Контрольный образец устанавливали на 100% активности. Результаты для всех mAb приведены в таблице 24 и на фиг. 9.
Процентная активность в анализе Biacore для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
Для образцов mAb, хранившихся при 37°С, только антитело 39B6-AYE сохраняло активность связывания с мишенью в течение всего 56-дневного периода. Все другие антитела демонстрировали значительное снижение активности связывания с мишенью в течение 56-дневного периода при хранении при 37°С.
Концентрация белка
Концентрацию белка во всех образцах измеряли для каждого условия на приборе NanoDrop.
В таблице 25 и на фиг.10 показана измеренная концентрация белка для всех mAb.
Концентрация белка (мг/мл) для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
Резюме и заключение по результатам исследования термостабильности
Исследование термостабильности выявило некоторые существенные различия между четырьмя тестированными mAb: агрегацию на уровне 4,4% наблюдали в анализе SE-HPLC для mAb 39B6-AVE через 56 суток при хранении при 37°C и фрагментацию на уровне 5,7% для mAb 39B6-AYE. Однако эта фрагментация для mAb 39B6-AYE не влияла на активность связывания с мишенью при хранении при 37°C в анализе Biacore; через 56 суток этот показатель все еще был таким же благоприятным, как у контрольного образца. Между тем, более низкая активность связывания с мишенью для mAb 39B6-AVE, 39B6-ANK и 39B6-ANR была четко видна при хранении при 37°C через 56 суток.
2.2.2 Исследование стабильности при замерзании-оттаивании
Для контроля стабильности антител в условиях замораживания-оттаивания дизайн опыта был следующим. Аликвотную порцию mAb 1 мл (с концентрацией 5 мг/мл) замораживали, по меньшей мере, на 6 ч при -20°С и оттаивали в течение 1 ч при комнатной температуре. Этот цикл повторяли 9 раз (всего 10 циклов замораживания-оттаивания). Образцы анализировали визуальным контролем, SE-HPLC, SDS-PAGE, оценивали активность связывания с мишенью в анализе Biacore и определяли концентрацию белка. Контрольные образцы, хранившиеся при -20°C, использовали для всех анализов параллельно. Поскольку mAb 39B6-ANR и 39B6-ANK имеют возможный сайт дезамидирования, то они подвергались анализу только визуальным контролем.
Визуальный контроль
Результаты визуального контроля в исследовании стабильности при замораживании-оттаивании приведены в таблице 26. Все mAb находились в PBSTw, что должно было оказывать стабилизирующий эффект.
Визуальный контроль в оценке стабильности при замораживании-оттаивании для mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-AN
Средние оценки для mAb были следующими:
39B6-AVE, 39B6-AYE и 39B-ANK: «очень мало частиц»; и
39B6-ANR: «образец прозрачный, видимые частицы отсутствуют».
Буфер PBSTw был оценен двумя экспертами как «очень мало частиц», и, следовательно, частицы, видимые в образцах 39B6-AVE, 39B6-AYE и 39B-ANK, могут быть не связаны с белками. Можно сделать вывод, что все mAb оставались без изменений после 10 циклов замораживания-оттаивания.
Анализ с помощью SE-HPLC
Агрегацию и фрагментацию белка измеряли с помощью SE-HPLC.
Результаты по оценке агрегации белка в исследовании стабильности при замораживании-оттаивании для mAb 39B6-AVE и 39B6-AYE обобщены в таблице 27.
% образование агрегатов, контролируемое эксклюзионной хроматографией в оценке стабильности при замораживании-оттаивании для mAb 39B6-AVE и 39B6-AYE
Какого-либо изменения процентных уровней агрегатов не наблюдали для mAb после 10 циклов замораживания-оттаивания по сравнению с контрольными образцами.
Площади пика мономеров для всех различных инжектирований также были исследованы. Результаты % площади мономеров для mAb 39B6-AVE и 39B6-AYE обобщены в таблице 28.
% площадь мономеров, контролируемая эксклюзионной хроматографией в оценке стабильности при замораживании-оттаивании для mAb 39B6-AVE и 39B6-AYE
Каких-либо изменений в % площади мономеров не наблюдали для обоих mAb после 10 циклов замораживания-оттаивания по сравнению с контрольными образцами.
Анализ электрофорезом SDS-PAGE
Образцы для оценки стабильности при замораживании-оттаивании анализировали на целостность с использованием электрофореза SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Результаты показаны в таблице 29 и на фиг. 11 для mAb 39B6-AVE и 39B6-AYE.
Общий процент полноразмерных Ab/тяжелой цепи, оцененный анализом SDS-PAGE и сканированием Odyssey в исследовании стабильности при замораживании-оттаивании для mAb 39B6-AVE и 39B6-AYE
Каких-либо изменений не наблюдали для обоих mAb после 10 циклов замораживания-оттаивания.
Анализ связывания с мишенью с использованием Biacore
Образцы тестировали на их активность связывания с мишенью в Biacore измерением наклона после 10 циклов замораживания-оттаивания. Контрольный образец принимали за 100% активность. Результаты для mAb 39B6-AVE и 39B6-AYE показаны на фиг. 12.
Результаты показывают, что после 10 циклов замораживания-оттаивания изменения активности связывания с мишенью не наблюдали.
Анализ концентрации белка
Концентрацию белка в образцах, предназначенных для анализа стабильности при замораживании-оттаивании, измеряли на приборе NanoDrop.
В таблице 30 показана измеренная концентрация белка для mAb 39B6-AVE и 39B6-AYE после 10 циклов замораживания-оттаивания. Кроме того, на фиг. 13 показаны эти результаты в графической форме для обоих mAb.
Концентрация белка (мг/мл) при оценке стабильности к замораживанию-оттаиванию для mAb 39B6-AVE и 39B6-AYE
Заключение
Исследование стабильности при замораживании-оттаивании не выявило каких-либо существенных различий для тестированных mAb 39B6-AVE и 39B6-AYE.
2.2.3 Исследование термостабильности
Для анализа кривых плавления mAb нагревали по схеме, приведенной ниже. После завершения термического цикла в приборе для ПЦР образцы тестировали на аффинность с использованием анализа Biacore.
Используемый протокол был следующим:
1) Аликвотные порции по 1 мл каждого mAb хранили в стеклянных флаконах при -20°C в начале исследования
2) После одной недели хранения их размораживали один раз
3) mAb разводили до 1 мг/мл как обычно, и затем дополнительно разводили до 100 мкг/мл (10-кратное разведение: 200 мкл+1800 мкл PBSTw)
4) Разведенные mAb вносили в планшет для ПЦР (из расчета 50 мкл/лунку)
5) Выдерживали достаточное количество образца и также образца при 5°C в качестве контроля для параллельного анализа
6) 1 ч в приборе для ПЦР при температуре, указанной ниже
7) 2 ч в приборе для ПЦР при 25°C
8) При 4°C в приборе ПЦР
9) Готовили образцы и контроли для анализа при 4 мкг/мл (25× разведение: 168 мкл буфера Biacore+7 мкл образца).
Запускают следующую программу в приборе для градиентной ПЦР:
Протокол анализа Biacore:
- Использовали тот же биосенсорный чип CM5, покрытый комплексом человеческих GARP-TGF-β при ~ 4000 RU
- Открывали кривые в программе оценки BIAE и выбирали значение «2-1» (1: пустой, 2: hGARP-TGF-β)
- Выбирали одну кривую на каждое время для наложения графика
- Удаляли часть кривой регенерации
- Выбирали базовую линию непосредственно перед введением и изменяли ось Y на «ноль при медиане выбора»
- Выбирали «Общая подгонка», начиная с 120 с после введения и заканчивая через 155 с
- Для расчета IC50: наклон для контролей 5°C выравнивали на 100%. Можно было рассчитать процент (ось Y) других температур (ось X).
Оценивали термостабильность четырех mAb, и результаты обобщены на фиг. 14. Температуру плавления рассчитывали как температуру, при которой 50% антитела все еще функционирует. Контрольные образцы, хранившиеся при 5°С, принимали за 100% активность. Температуры плавления приведены в таблице 31.
Температура плавления в исследовании термостабильности
MAb 39B6-ANK и 39B6-ANR показали самые высокие температуры плавления: 69,13°C и 68,55°C соответственно. MAb 39B6-AVE и 39B6-AYE также дали благоприятные температуры плавления, 66,99°C и 66,53°C соответственно. Все температуры плавления для всех mAb можно считать высокими.
Заключение по исследованию термостабильности
Четыре MAb демонстрировали хорошую термостабильность. Температуры плавления были сопоставимы с исходным mAb 39B6-A в предыдущем исследовании стабильности, 66,8°C.
2.2.4 Исследование ротационной стабильности
Для исследования ротационной стабильности протокол был следующим. Аликвотные порции по 1 мл каждого mAb хранили в стеклянных флаконах при -20°C в начале исследования. После одной недели хранения аликвоты размораживали один раз и вращали сверху со скоростью 15 об/мин при комнатной температуре.
Образцы оценивали на наличие частиц на указанные временные точки:
- Часы: 0, 3, 6, 24, 30, 48, 54, 72 и 96
Образцы также анализировали через 96 ч с использованием SE-HPLC, SDS-PAGE, анализа активности связывания с мишенью в Biacore и определения концентрации белка. Контрольные образцы, хранившиеся при -20°C, использовали для всех анализов параллельно. Поскольку mAb 39B6-ANR и 39B6-ANK все еще имеют возможный сайт дезамидирования, то их оценивали только визуальным контролем.
Визуальный контроль
Результаты визуального контроля в исследовании ротационной стабильности приведены в таблице 32. Все mAb находились в PBSTw, что должно было оказывать стабилизирующий эффект.
Результаты визуального контроля в исследовании ротационной стабильности mAb 39B6-AVE, 39B6-AYE, 39B6-ANK и 39B6-ANR
Было установлено, что все mAb имеют среднюю оценку «очень мало частиц», поэтому они остаются относительно интактными после 96 ч вращения. Буфер PBSTw не содержал никаких частиц ни при одном из экспериментальных условий. Это указывает на то, что наблюдаемая оценка «очень мало частиц» связана с белками в образцах антител.
Анализ SE-HPLC
Агрегацию и фрагментацию белка измеряли с помощью SE-HPLC. Результаты оценки агрегации белка в исследовании ротационной стабильности mAb 39B6-AVE и 39B6-AYE приведены в таблице 33.
% образование агрегатов, контролируемое эксклюзионной хроматографией в исследовании ротационной стабильности mAb 39B6-AVE и 39B6-AYE
Каких-либо изменений в % уровнях агрегатов не наблюдали для mAb после ротационного напряжения по сравнению с контрольными образцами.
Площади пика мономеров для всех различных инжектирований также были исследованы. Результаты по % площади мономеров для mAb 39B6-AVE и 39B6-AYE показаны в таблице 34.
% площадь мономеров, контролируемая эксклюзионной хроматографией в исследовании ротационной стабильности mAb 39B6-AVE и 39B6-AYE
Какого-либо изменения в % площади мономеров не наблюдали для mAb после 96 ч вращения по сравнению с контрольными образцами.
Для обоих антител не наблюдали изменения профиля SE-HPLC после 96 ч вращения по сравнению с контрольными образцами.
Анализ электрофорезом SDS-PAGE
Подвергшиеся вращению образцы анализировали на их целостность электрофорезом SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Результаты обобщены в таблице 35 для mAb 39B6-AVE и 39B6-AYE.
Общий процент полноразмерных Ab/тяжелой цепи, оцененный анализом SDS-PAGE и сканированием Odyssey в исследовании ротационной стабильности для mAb 39B6-AVE и 39B6-AYE
Гели SDS-PAGE для обоих mAb после 96 ч вращения представлены на фиг. 15.
Каких-либо изменений не наблюдали ни для одного антитела после 96 ч вращения.
Анализ связывания с мишенью в Biacore
Образцы тестировали на их активность связывания с мишенью в анализе Biacore измерением наклона после 96 ч вращения. Контрольный образец устанавливали на 100% активности. Результаты для mAb 39B6-AVE и 39B6-AYE показаны на фиг. 16.
Анализ концентрации белка
Концентрацию белка в образцах, подвергшихся вращению, измеряли на приборе NanoDrop.
В таблице 36 показана измеренная концентрация белка для mAb 39B6-AVE и 39B6-AYE после 96 ч вращения. Кроме того, на фиг. 17 показано графическое представление этих результатов для обоих mAb.
Концентрация белка (мг/мл) в исследовании ротационной стабильности mAb 39B6-AVE и 39B6-AYE
Потери белка не наблюдали для обоих mAb после 96 ч вращения.
Заключение по результатам исследования ротационной стабильности
Исследование ротационной стабильности не выявило различий между тестированными mAb 39B6-AVE и 39B6-AYE.
2.2.5 Анализ первичной последовательности с помощью пептидного картирования
Образцы из исследования термостабильности, стабильности при замораживании-оттаивании и ротационной стабильности анализировали с использованием методологии RP-HPLC-UV-MS для триптического пептидного картирования в целях выявления модификаций (дезамидирования, изомеризации и окисления) на уровне белка (пептида).
Дезамидирование в CDR3 тяжелой цепи в положениях 95-96 (аминокислоты NE) было сконструировано в mAb 39B6-AVE и 39B6-AYE мутацией положения N95. Данный сайт дезамидирования по-прежнему присутствует в mAb 39B6-ANK и 39B6-ANR, и значительное дезамидирование/изомеризацию обнаруживали через 28 суток при хранении при 5°C и 37°C в исследовании термостабильности, а также в исследовании стабильности при замораживании-оттаивании и ротационной стабильности. Был сделан вывод, что дезамидирование невозможно предупреждать введением объемных положительно заряженных остатков даунстрим от N95.
В положении 100f CDR3 тяжелой цепи находится метионин, который необходим для обеспечения высокой аффинности связывания. Исследование термостабильности, стабильности при замораживании-оттаивании и ротационной стабильности показали, что окисление M100f не происходит в mAb 39B6-AYE и 39B6-ANK, в то время как окисление все еще наблюдается для mAb 39B6-AVE и 39B6-ANR. Это свидетельствует о том, что аминокислоты в положениях 95 и 96 влияют на чувствительность к окислению даунстрим метионина. В таблице 37 представлен обзор уровней окисления и дезамидирования пептидов, покрывающих CDR3 тяжелой цепи, которые были получены триптическим расщеплением.
Степень окисления и дезамидирования пептидов в CDR3 тяжелой цепи
сутки 28
сутки 28
сутки 28
сутки28
Относительный уровень дезамидирования и изомеризации N95, а также относительная активность связывания вариантов, хранившихся при 37°C, представлены на фиг. 18. Также на фиг. 18 показано исходное антитело 39B6-A (обозначенное 39B6-ANE). Поскольку N95 был мутирован в mAb 39B6-AVE и 39B6-AYE, то уровни дезамидирования и изомеризации равны нулю и поэтому не были включены в анализ. Корреляция между дезамидированием N95 и более низкой активностью связывания с мишенью, наблюдаемая для mAb 39B6-ANK и 39B6-ANR, позволяет предположить, что дезамидирование N95 отрицательно влияет на связывание mAb с его мишенью.
Заключение
Вариант анти-GARP-TGF-β1-антитела 39B6-AYE является особенно благоприятным антителом к GARP-TGF-β1 для клинической разработки, потому что оно:
- сохраняет высокую аффинность связывания с его мишенью, комплексом GARP-TGF-β1;
- показывает хорошую активность в анализе фосфорилирования SMAD2;
- не подвергается дезамидированию или изомеризации в CDR3
- не подвергается окислению в CDR3
- показывает высокую гомологию с человека (95%)
- показывает повышенную стабильность по сравнению с 39B6-A, оцененную в разных анализах стабильности.
Последовательности CDR и вариабельной области для 39B6-AYE приведены в таблицах 38 и 39 ниже. Последовательности полноразмерной тяжелой цепи и легкой цепи показаны в таблице 40. Полинуклеотидные последовательности, кодирующие области VH и VL, а также полноразмерные тяжелую и легкую цепи показаны в таблице 41.
Последовательности CDR VH и VL в 39B6-AYE
Последовательности VH и VL в 39B6-AYE
Последовательности тяжелой и легкой цепи в 39B6-AYE
Полинуклеотидные последовательности, кодирующие 39B6-AYE
Пример 3. Испытание партий 39B6-AYE (ARGX-115)
Опытную партию лекарственной субстанции ARGX-115 испытывали на стабильность в течение трехмесячного периода. Испытуемые образцы опытной партии лекарственной субстанции хранили в предназначенных условиях хранения при -70°C, в условиях «ускоренного старения» при +5°C и в стрессовых условиях хранения при +25°C. Опытная партия лекарственной субстанции была формулирована в следующем составе: 10 мМ гистидин/гистидина гидрохлорид, 200 мМ сахарозы, 40 мМ аргинина, 0,03% (мас./об.) полисорбата 80 при рН 6,0 и концентрации белка 20,0±2,0 мг/мл.
Было подтверждено, что опытная партия лекарственной субстанции ARGX-115 является стабильной в течение трех месяцев при хранении в предназначенных условиях хранения при -70°C. Активность связывания в анализе SPR также определяли в качестве показателя стабильности. Активность связывания выражали в процентах от активности связывания контрольного образца, который хранили при -70°С.
Результаты показаны в таблице 42 ниже.
Эти результаты подтверждают, что ARGX-115 стабильно в течение длительного периода хранения.
Пример 4. Характеристика связывания ARGX-115 с комплексом GARP-TGF-β
4.1. Зрелый TGF-β необходим для связывания с ARGX-115
В природе комплекс GARP-TGF-β (GARP в комплексе с латентным TGF-β) образуется в эндоплазматическом ретикулуме посредством ковалентных цистеиновых взаимодействий (дисульфидных мостиков) между GARP и латентным TGF-β. Этот комплекс затем располагается на клеточной поверхности. In vitro комплекс GARP-TGF-β может быть образован из рекомбинантного человеческого GARP и рекомбинантного латентного TGF-β (C33S)-3xstrep-tag человека. Мутантная форма C33S латентного TGF-β не образует ковалентных взаимодействий с GARP (или любым из латентный TGF-β-связывающих белков (LTBP)), подобных присутствующим в нативном комплексе.
Для демонстрации связывания ARGX-115 с образованным in vitro комплексом рекомбинантного GARP и рекомбинантного латентного TGF-β (C33S), планшет для ELISA покрывали рекомбинантным GARP (1 мкг/мл человеческого GARP O/N при 4°C), блокировали с использованием блокирующего агента (казеин-PBS) и латентный TGF-β (5 мкг/мл в течение 1 ч при комнатной температуре) захватывали покрытым GARP. Для ARGX-115 и антитела контрольного изотипа (1 мкг/мл в течение 1 ч при комнатной температуре) обеспечивали возможность связываться с комплексом и определяли с помощью HRP-конъюгированного античеловеческого IgG. Как показано на фиг. 19, ARGX-115 связывался с комплексом GARP-латентный TGF-β (C33S), тогда как для контрольного изотипа такое связывание отсутствовало.
Также было установлено, что анализ «работает» в обратном порядке. Планшет для ELISA покрывали ARGX-115 или антителом контрольного изотипа (1 мкг/мл при 4°C) и блокировали с использованием блокирующего агента (казеин-PBS). Рекомбинантный GARP (5 мкг/мл) захватывался покрытым антителом ARGX-115, планшет отмывали, и рекомбинантный латентный TGF-β (C33S)-3xstrep-tag (5 мкг/мл) захватывался и детектировался с помощью стрептавидина-HRP. Активность HRP детектировали только в присутствии ARGX-115, но не в лунках планшета, покрытого контрольным изотипом.
Для тестирования связывания ARGX-115 с комплексом GARP и латентно-ассоциированного пептида (LAP) TGF-β, был образован комплекс между рекомбинантным GARP и рекомбинантным LAP. Планшет для ELISA покрывали рекомбинантным GARP (1 мкг/мл O/N 4°C), блокировали с использованием блокирующего агента (казеин-PBS), и LAP (5 мкг/мл) захватывали на покрытом рекомбинантном GARP. Связывание LAP детектировали с использованием анти-LAP-HRP. Связывание анти-LAP-HRP показывало, что комплекс GARP-LAP действительно образуется in vitro. ARGX-115, однако, не проявило какого-либо связывания с комплексом GARP-LAP. Кроме того, когда планшет для ELISA покрывали ARGX-115 (1 мкг/мл ON при 4°С), добавляли рекомбинантный GARP (5 мкг/мл) с последующим добавлением LAP (5 мкг/мл), то связывание анти-LAP-HRP не детектировали. Эти результаты подтверждают, что зрелый TGF-β необходим для связывания ARGX-115 с комплексом GARP-TGF-β.
4.2 Влияние мутаций в hTGF-β в комплексе с GARP на нейтрализующую активность ARGX-115
Клетки 293T, стабильно экспрессирующие интегрин αvβ6 (293Tcl.ITGB6), транзиентно трансфицировали смесью из 3 плазмид (смесь плазмид): (i) репортерная плазмида CAGA-luc; (ii) человеческий GARP (pEF-BOS-puro-hGARP); и (iii) человеческий TGF-β дикого типа или мутантный TGF-β (pDisplay). Интегрин αvβ6 является одним из двух интегринов, активирующих TGF-β. Клетки 293Tcl.ITGB6 отделяли и собирали из полуконфлюентных слоев в колбе емкостью 75 см2, подсчитывали и разбавляли до 1E+06 клеток/мл и распределяли по 1 мл на пробирку Эппендорфа. 250 мкл плазмидной смеси добавляли в каждую пробирку для трансфекции. Непосредственно после трансфекции трансфицированные клетки распределяли в 96-луночные оптические планшеты, по 50 мкл (4E+04 клеток) на лунку, содержащие различные тестируемые mAb (ARGX-115, LHG10.6, 1D11 и MHG-8 при 100 мкл/мл). 1D11 представляет собой mAb против активного TGF-β изоформы-1, -2 и -3. MHG-8 и LHG10.6 описаны в WO2015/015003 и WO2016/125017. После инкубации в течение 24 ч при 37°С измеряли активность люциферазы. Значение, полученное при трансфекции мутантным TGF-β, выражали в процентах от значения, полученного для TGF-β дикого типа. Результаты показаны на фиг. 20. Как можно видеть из фиг. 20, нейтрализующая активность ARGX-115, измеренная в отношении мутанта TGF-β, включающего замену R58A, и мутанта TGF-β, включающего замену K338E, была достоверно снижена. Это указывает на то, что эти два остатка в комплексе GARP-TGF-β особенно важны для проявления нейтрализующей активности ARGX-115.
--->
Список последовательностей
<110> АРДЖЕНКС БВБА
<120> АНТИ-GARP-TGF-БЕТА-АНТИТЕЛА
<130> N415743WO
<140> PCT/EP2018/062251
<141> 2018-05-11
<150> GB1707561.5
<151> 2017-05-11
<160> 48
<170> PatentIn version 3.5
<210> 1
<211> 126
<212> БЕЛОК
<213> Llama glama
<400> 1
Glu Val Gln Leu Val Gln Pro Gly Ala Glu Leu Arg Asn Ser Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Gly Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Gln Val Thr Val Ser Ser
115 120 125
<210> 2
<211> 107
<212> БЕЛОК
<213> Llama glama
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Gln Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Ser Phe Thr Leu Thr Ile Ser Gly Leu Glu Ala
65 70 75 80
Glu Asp Ala Gly Thr Tyr Tyr Cys Gln Gln Tyr Asp Ser Leu Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Leu Lys
100 105
<210> 3
<211> 107
<212> БЕЛОК
<213> Llama glama
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Asn Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Ser Phe Thr Leu Thr Ile Ser Gly Leu Glu Ala
65 70 75 80
Glu Asp Ala Gly Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Leu Lys
100 105
<210> 4
<211> 5
<212> БЕЛОК
<213> Llama glama
<400> 4
Ser Tyr Tyr Ile Asp
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Llama glama
<400> 5
Arg Ile Asp Pro Glu Asp Gly Gly Thr Lys Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 6
<211> 17
<212> БЕЛОК
<213> Llama glama
<400> 6
Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu Tyr Glu
1 5 10 15
Tyr
<210> 7
<211> 17
<212> БЕЛОК
<213> Llama glama
<400> 7
Arg Ile Asp Pro Glu Glu Gly Gly Thr Lys Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 8
<211> 17
<212> БЕЛОК
<213> Llama glama
<400> 8
Arg Ile Asp Pro Glu Asp Ala Gly Thr Lys Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 9
<211> 11
<212> БЕЛОК
<213> Llama glama
<400> 9
Gln Ala Ser Gln Ser Ile Ser Ser Tyr Leu Ala
1 5 10
<210> 10
<211> 7
<212> БЕЛОК
<213> Llama glama
<400> 10
Gly Ala Ser Arg Leu Lys Thr
1 5
<210> 11
<211> 9
<212> БЕЛОК
<213> Llama glama
<400> 11
Gln Gln Tyr Ala Ser Val Pro Val Thr
1 5
<210> 12
<211> 17
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 12
Arg Ile Asp Pro Glu Asp Ala Gly Thr Lys Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 13
<211> 17
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 13
Tyr Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu Tyr Glu
1 5 10 15
Tyr
<210> 14
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 14
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Ala Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 15
<211> 107
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 15
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Ser Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 16
<211> 452
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 16
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Ala Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120 125
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr
130 135 140
Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
145 150 155 160
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
165 170 175
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
180 185 190
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr
195 200 205
Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val
210 215 220
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
325 330 335
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Leu Gly
450
<210> 17
<211> 214
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 17
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Ser Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 18
<211> 378
<212> ДНК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 18
caagtccaac ttgtccaacc gggggcggaa gtgcggaagc cgggggcgag cgtgaaagtc 60
tcgtgcaagg catcgggata ccgattcaca tcatattaca tcgactgggt caggcaagcg 120
ccggggcaag ggctggaatg gatggggcgg atcgacccgg aggatgccgg gacgaaatat 180
gcgcaaaaat tccaagggcg cgtcacgatg acggccgaca catcgacgag cacggtatac 240
gtggagctga gctcgctgag gagcgaggac accgcggtat actactgcgc gcgatacgaa 300
tgggagaccg tcgtcgtcgg ggacctgatg tacgaatacg aatactgggg gcaagggacg 360
cttgtcacgg tctcgagc 378
<210> 19
<211> 321
<212> ДНК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 19
gacatccaga tgactcagag cccttccagc ctgagcgcct ctgtgggaga tagagtcacc 60
atcacatgcc aggctagtca gtcaatttct agttacctgg catggtatca gcagaagcct 120
ggccaggcac ctaaaatcct gatctacgga gccagtaggc tgaagacagg ggtgccatct 180
cggttctccg gcagcggatc tgggacatcc tttactctga ccatctcatc cctggagcca 240
gaagacgccg ctacatacta ttgtcagcag tatgcttccg tgcccgtcac attcggtcag 300
ggcactaagg tcgagatcaa g 321
<210> 20
<211> 420
<212> ДНК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 20
caagtccaac ttgtccaacc gggggcggaa gtgcggaagc cgggggcgag cgtgaaagtc 60
tcgtgcaagg catcgggata ccgattcaca tcatattaca tcgactgggt caggcaagcg 120
ccggggcaag ggctggaatg gatggggcgg atcgacccgg aggatgccgg gacgaaatat 180
gcgcaaaaat tccaagggcg cgtcacgatg acggccgaca catcgacgag cacggtatac 240
gtggagctga gctcgctgag gagcgaggac accgcggtat actactgcgc gcgatacgaa 300
tgggagaccg tcgtcgtcgg ggacctgatg tacgaatacg aatactgggg gcaagggacg 360
cttgtcacgg tctcgagcgc tagcaccaag ggcccctccg tgttccccct ggccccttgc 420
<210> 21
<211> 936
<212> ДНК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 21
tcccggtcca cctccgagtc taccgccgct ctgggctgcc tggtgaaaga ctacttcccc 60
gagcctgtga ccgtgagctg gaactctggc gccctgacct ccggcgtgca caccttccct 120
gccgtgctgc aatcctccgg cctgtactcc ctgtcctccg tggtgacagt gccctcctcc 180
agcctgggca ccaagaccta cacctgtaac gtggaccaca agccctccaa caccaaggtg 240
gacaagcggg tggaatctaa atacggccct ccctgccccc cctgccctgc ccctgaattt 300
ctgggcggac cttccgtgtt tctgttcccc ccaaagccca aggacaccct gatgatctcc 360
cggacccccg aagtgacctg cgtggtggtg gacgtgtccc aggaagatcc agaggtgcag 420
ttcaactggt atgttgacgg cgtggaagtg cacaacgcca agaccaagcc cagagaggaa 480
cagttcaact ccacctaccg ggtggtgtcc gtgctgaccg tgctgcacca ggactggctg 540
aacggcaaag agtacaagtg caaggtgtcc aacaagggcc tgccctccag catcgaaaag 600
accatctcca aggccaaggg ccagccccgc gagccccagg tgtacaccct gccccctagc 660
caggaagaga tgaccaagaa ccaggtgtcc ctgacctgtc tggtgaaagg cttctacccc 720
tccgacattg ccgtggaatg ggagtccaac ggccagcccg agaacaacta caagaccacc 780
ccccctgtgc tggactccga cggctccttc ttcctgtact ctcggctgac agtggataag 840
tcccggtggc aggaaggcaa cgtgttctcc tgcagcgtga tgcacgaggc cctgcacaac 900
cactataccc agaagtccct gtccctgagc ctgggc 936
<210> 22
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 22
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Val Arg Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Gly Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 23
<211> 107
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 23
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Ser Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 24
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 24
Glu Val Gln Leu Val Gln Pro Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Glu Gly Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Thr Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 25
<211> 107
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 26
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 26
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Leu Arg Asn Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Gly Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 27
<211> 107
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 28
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 28
Gln Val Gln Leu Val Gln Pro Gly Ala Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Asp Ala Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Phe Thr Ala Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Val Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Thr Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 29
<211> 106
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 29
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Glu Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ile Ser Gly Leu Glu Pro Glu
65 70 75 80
Asp Ala Gly Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 30
<211> 126
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 30
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Arg Phe Thr Ser Tyr
20 25 30
Tyr Ile Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Asp Pro Glu Glu Gly Gly Thr Lys Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ala Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Met Tyr Glu
100 105 110
Tyr Glu Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 31
<211> 107
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Asn Ile Leu Ile
35 40 45
Tyr Gly Ala Ser Arg Leu Lys Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Gly Leu Glu Ala
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ala Ser Val Pro Val
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 32
<211> 17
<212> БЕЛОК
<213> Искуственная
<220>
<223> фрагмент антитела
<400> 32
Asn Glu Trp Glu Thr Val Val Val Gly Asp Leu Thr Tyr Glu Tyr Glu
1 5 10 15
Tyr
<210> 33
<211> 662
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Met Arg Pro Gln Ile Leu Leu Leu Leu Ala Leu Leu Thr Leu Gly Leu
1 5 10 15
Ala Ala Gln His Gln Asp Lys Val Pro Cys Lys Met Val Asp Lys Lys
20 25 30
Val Ser Cys Gln Val Leu Gly Leu Leu Gln Val Pro Ser Val Leu Pro
35 40 45
Pro Asp Thr Glu Thr Leu Asp Leu Ser Gly Asn Gln Leu Arg Ser Ile
50 55 60
Leu Ala Ser Pro Leu Gly Phe Tyr Thr Ala Leu Arg His Leu Asp Leu
65 70 75 80
Ser Thr Asn Glu Ile Ser Phe Leu Gln Pro Gly Ala Phe Gln Ala Leu
85 90 95
Thr His Leu Glu His Leu Ser Leu Ala His Asn Arg Leu Ala Met Ala
100 105 110
Thr Ala Leu Ser Ala Gly Gly Leu Gly Pro Leu Pro Arg Val Thr Ser
115 120 125
Leu Asp Leu Ser Gly Asn Ser Leu Tyr Ser Gly Leu Leu Glu Arg Leu
130 135 140
Leu Gly Glu Ala Pro Ser Leu His Thr Leu Ser Leu Ala Glu Asn Ser
145 150 155 160
Leu Thr Arg Leu Thr Arg His Thr Phe Arg Asp Met Pro Ala Leu Glu
165 170 175
Gln Leu Asp Leu His Ser Asn Val Leu Met Asp Ile Glu Asp Gly Ala
180 185 190
Phe Glu Gly Leu Pro Arg Leu Thr His Leu Asn Leu Ser Arg Asn Ser
195 200 205
Leu Thr Cys Ile Ser Asp Phe Ser Leu Gln Gln Leu Arg Val Leu Asp
210 215 220
Leu Ser Cys Asn Ser Ile Glu Ala Phe Gln Thr Ala Ser Gln Pro Gln
225 230 235 240
Ala Glu Phe Gln Leu Thr Trp Leu Asp Leu Arg Glu Asn Lys Leu Leu
245 250 255
His Phe Pro Asp Leu Ala Ala Leu Pro Arg Leu Ile Tyr Leu Asn Leu
260 265 270
Ser Asn Asn Leu Ile Arg Leu Pro Thr Gly Pro Pro Gln Asp Ser Lys
275 280 285
Gly Ile His Ala Pro Ser Glu Gly Trp Ser Ala Leu Pro Leu Ser Ala
290 295 300
Pro Ser Gly Asn Ala Ser Gly Arg Pro Leu Ser Gln Leu Leu Asn Leu
305 310 315 320
Asp Leu Ser Tyr Asn Glu Ile Glu Leu Ile Pro Asp Ser Phe Leu Glu
325 330 335
His Leu Thr Ser Leu Cys Phe Leu Asn Leu Ser Arg Asn Cys Leu Arg
340 345 350
Thr Phe Glu Ala Arg Arg Leu Gly Ser Leu Pro Cys Leu Met Leu Leu
355 360 365
Asp Leu Ser His Asn Ala Leu Glu Thr Leu Glu Leu Gly Ala Arg Ala
370 375 380
Leu Gly Ser Leu Arg Thr Leu Leu Leu Gln Gly Asn Ala Leu Arg Asp
385 390 395 400
Leu Pro Pro Tyr Thr Phe Ala Asn Leu Ala Ser Leu Gln Arg Leu Asn
405 410 415
Leu Gln Gly Asn Arg Val Ser Pro Cys Gly Gly Pro Asp Glu Pro Gly
420 425 430
Pro Ser Gly Cys Val Ala Phe Ser Gly Ile Thr Ser Leu Arg Ser Leu
435 440 445
Ser Leu Val Asp Asn Glu Ile Glu Leu Leu Arg Ala Gly Ala Phe Leu
450 455 460
His Thr Pro Leu Thr Glu Leu Asp Leu Ser Ser Asn Pro Gly Leu Glu
465 470 475 480
Val Ala Thr Gly Ala Leu Gly Gly Leu Glu Ala Ser Leu Glu Val Leu
485 490 495
Ala Leu Gln Gly Asn Gly Leu Met Val Leu Gln Val Asp Leu Pro Cys
500 505 510
Phe Ile Cys Leu Lys Arg Leu Asn Leu Ala Glu Asn Arg Leu Ser His
515 520 525
Leu Pro Ala Trp Thr Gln Ala Val Ser Leu Glu Val Leu Asp Leu Arg
530 535 540
Asn Asn Ser Phe Ser Leu Leu Pro Gly Ser Ala Met Gly Gly Leu Glu
545 550 555 560
Thr Ser Leu Arg Arg Leu Tyr Leu Gln Gly Asn Pro Leu Ser Cys Cys
565 570 575
Gly Asn Gly Trp Leu Ala Ala Gln Leu His Gln Gly Arg Val Asp Val
580 585 590
Asp Ala Thr Gln Asp Leu Ile Cys Arg Phe Ser Ser Gln Glu Glu Val
595 600 605
Ser Leu Ser His Val Arg Pro Glu Asp Cys Glu Lys Gly Gly Leu Lys
610 615 620
Asn Ile Asn Leu Ile Ile Ile Leu Thr Phe Ile Leu Val Ser Ala Ile
625 630 635 640
Leu Leu Thr Thr Leu Ala Ala Cys Cys Cys Val Arg Arg Gln Lys Phe
645 650 655
Asn Gln Gln Tyr Lys Ala
660
<210> 34
<211> 390
<212> БЕЛОК
<213> Homo sapiens
<400> 34
Met Pro Pro Ser Gly Leu Arg Leu Leu Pro Leu Leu Leu Pro Leu Leu
1 5 10 15
Trp Leu Leu Val Leu Thr Pro Gly Arg Pro Ala Ala Gly Leu Ser Thr
20 25 30
Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg Ile Glu Ala
35 40 45
Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser Pro Pro Ser
50 55 60
Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val Leu Ala Leu
65 70 75 80
Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala Glu Pro Glu
85 90 95
Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr Arg Val Leu
100 105 110
Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys Gln Ser Thr
115 120 125
His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg Glu Ala Val
130 135 140
Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu Leu Arg Leu
145 150 155 160
Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys Tyr Ser Asn
165 170 175
Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro Ser Asp Ser
180 185 190
Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg Gln Trp Leu
195 200 205
Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala His Cys Ser
210 215 220
Cys Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn Gly Phe Thr
225 230 235 240
Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met Asn Arg Pro
245 250 255
Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln His Leu Gln
260 265 270
Ser Ser Arg His Arg Arg Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser
275 280 285
Thr Glu Lys Asn Cys Cys Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys
290 295 300
Asp Leu Gly Trp Lys Trp Ile His Glu Pro Lys Gly Tyr His Ala Asn
305 310 315 320
Phe Cys Leu Gly Pro Cys Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr
325 330 335
Ser Lys Val Leu Ala Leu Tyr Asn Gln His Asn Pro Gly Ala Ser Ala
340 345 350
Ala Pro Cys Cys Val Pro Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr
355 360 365
Tyr Val Gly Arg Lys Pro Lys Val Glu Gln Leu Ser Asn Met Ile Val
370 375 380
Arg Ser Cys Lys Cys Ser
385 390
<210> 35
<211> 249
<212> БЕЛОК
<213> Homo sapiens
<400> 35
Leu Ser Thr Cys Lys Thr Ile Asp Met Glu Leu Val Lys Arg Lys Arg
1 5 10 15
Ile Glu Ala Ile Arg Gly Gln Ile Leu Ser Lys Leu Arg Leu Ala Ser
20 25 30
Pro Pro Ser Gln Gly Glu Val Pro Pro Gly Pro Leu Pro Glu Ala Val
35 40 45
Leu Ala Leu Tyr Asn Ser Thr Arg Asp Arg Val Ala Gly Glu Ser Ala
50 55 60
Glu Pro Glu Pro Glu Pro Glu Ala Asp Tyr Tyr Ala Lys Glu Val Thr
65 70 75 80
Arg Val Leu Met Val Glu Thr His Asn Glu Ile Tyr Asp Lys Phe Lys
85 90 95
Gln Ser Thr His Ser Ile Tyr Met Phe Phe Asn Thr Ser Glu Leu Arg
100 105 110
Glu Ala Val Pro Glu Pro Val Leu Leu Ser Arg Ala Glu Leu Arg Leu
115 120 125
Leu Arg Leu Lys Leu Lys Val Glu Gln His Val Glu Leu Tyr Gln Lys
130 135 140
Tyr Ser Asn Asn Ser Trp Arg Tyr Leu Ser Asn Arg Leu Leu Ala Pro
145 150 155 160
Ser Asp Ser Pro Glu Trp Leu Ser Phe Asp Val Thr Gly Val Val Arg
165 170 175
Gln Trp Leu Ser Arg Gly Gly Glu Ile Glu Gly Phe Arg Leu Ser Ala
180 185 190
His Cys Ser Cys Asp Ser Arg Asp Asn Thr Leu Gln Val Asp Ile Asn
195 200 205
Gly Phe Thr Thr Gly Arg Arg Gly Asp Leu Ala Thr Ile His Gly Met
210 215 220
Asn Arg Pro Phe Leu Leu Leu Met Ala Thr Pro Leu Glu Arg Ala Gln
225 230 235 240
His Leu Gln Ser Ser Arg His Arg Arg
245
<210> 36
<211> 112
<212> БЕЛОК
<213> Homo sapiens
<400> 36
Ala Leu Asp Thr Asn Tyr Cys Phe Ser Ser Thr Glu Lys Asn Cys Cys
1 5 10 15
Val Arg Gln Leu Tyr Ile Asp Phe Arg Lys Asp Leu Gly Trp Lys Trp
20 25 30
Ile His Glu Pro Lys Gly Tyr His Ala Asn Phe Cys Leu Gly Pro Cys
35 40 45
Pro Tyr Ile Trp Ser Leu Asp Thr Gln Tyr Ser Lys Val Leu Ala Leu
50 55 60
Tyr Asn Gln His Asn Pro Gly Ala Ser Ala Ala Pro Cys Cys Val Pro
65 70 75 80
Gln Ala Leu Glu Pro Leu Pro Ile Val Tyr Tyr Val Gly Arg Lys Pro
85 90 95
Lys Val Glu Gln Leu Ser Asn Met Ile Val Arg Ser Cys Lys Cys Ser
100 105 110
<210> 37
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 37
Glu Pro Lys Ser Cys Asp Lys Thr His Thr
1 5 10
<210> 38
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 38
Cys Pro Pro Cys Pro
1 5
<210> 39
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 39
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 40
<211> 12
<212> БЕЛОК
<213> Homo sapiens
<400> 40
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr
1 5 10
<210> 41
<211> 50
<212> БЕЛОК
<213> Homo sapiens
<400> 41
Cys Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys
1 5 10 15
Pro Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro
20 25 30
Arg Cys Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg
35 40 45
Cys Pro
50
<210> 42
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 42
Ala Pro Glu Leu Leu Gly Gly Pro
1 5
<210> 43
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 43
Glu Ser Lys Tyr Gly Pro Pro
1 5
<210> 44
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 44
Cys Pro Ser Cys Pro
1 5
<210> 45
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 45
Ala Pro Glu Phe Leu Gly Gly Pro
1 5
<210> 46
<211> 3
<212> БЕЛОК
<213> Homo sapiens
<400> 46
Glu Arg Lys
1
<210> 47
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 47
Cys Cys Val Glu Cys Pro Pro Pro Cys Pro
1 5 10
<210> 48
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 48
Ala Pro Pro Val Ala Gly Pro
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ РАКА, НАЦЕЛИВАЮЩИЕСЯ НА CD38 И TGF-БЕТА | 2019 |
|
RU2808632C2 |
| ИММУНОКОНЪЮГАТЫ IL2 И МУТАНТНОГО TNF | 2017 |
|
RU2758139C2 |
| МЕЖВИДОВЫЕ АНТИТЕЛА К ЛАТЕНТНОМУ TGF-БЕТА 1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2837609C2 |
| АНТИТЕЛА, НАЦЕЛИВАЮЩИЕСЯ НА КОСТНУЮ ТКАНЬ | 2018 |
|
RU2801206C2 |
| СЛИТЫЙ БЕЛОК, СОДЕРЖАЩИЙ РЕЦЕПТОР TGF-БЕТА, И ЕГО ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2776204C1 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СОРТИЛИНОМ И ПОДАВЛЯЮТ СВЯЗЫВАНИЕ ПРОГРАНУЛИНА | 2016 |
|
RU2735639C2 |
| СЛИТЫЙ БЕЛОК НА ОСНОВЕ АНТИТЕЛА К ЕТА И TGF-β TRAP, СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2832190C2 |
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |

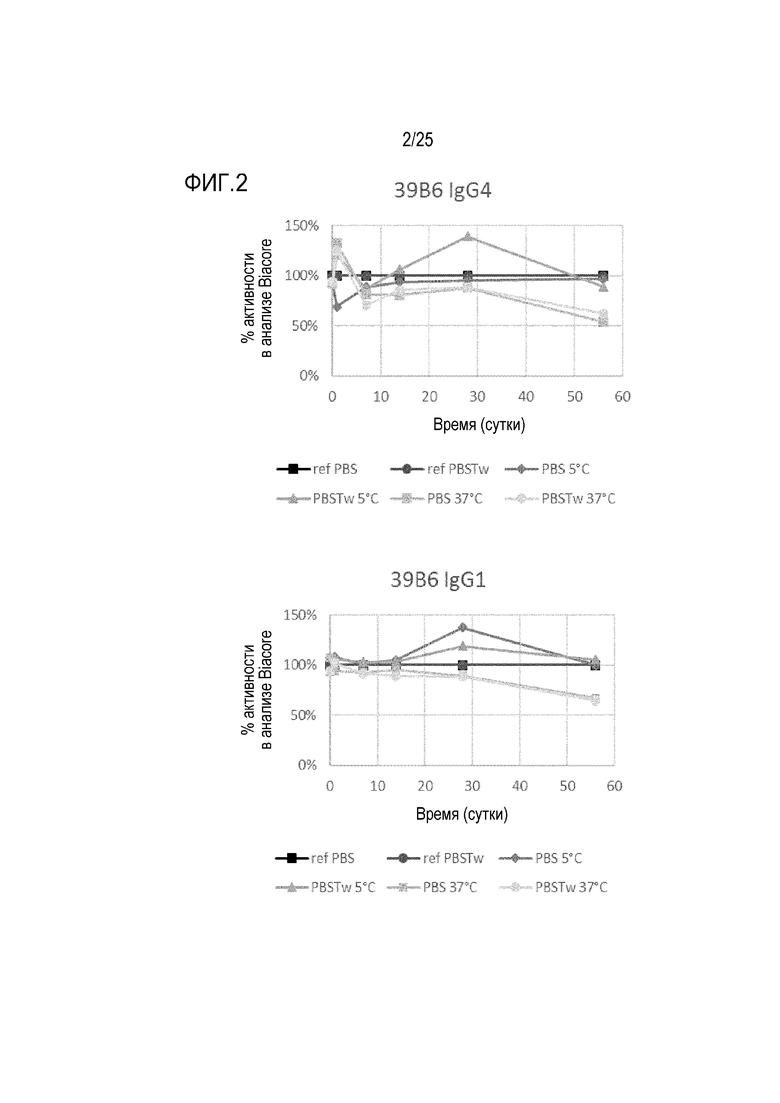






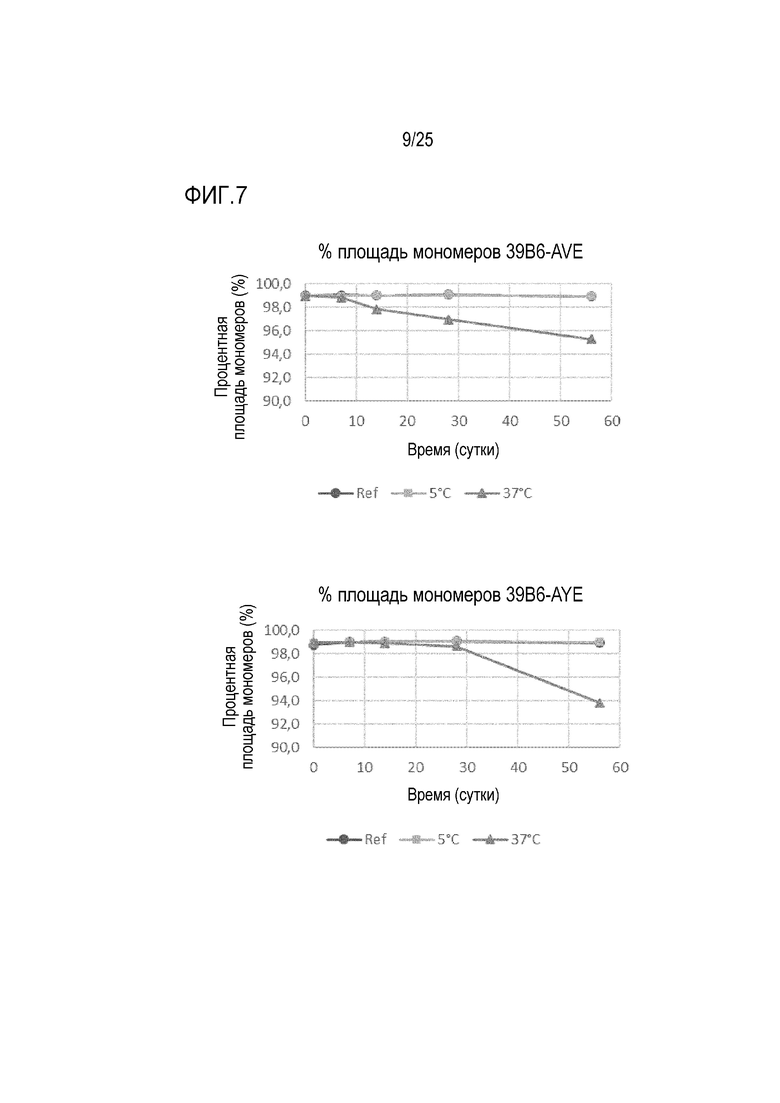
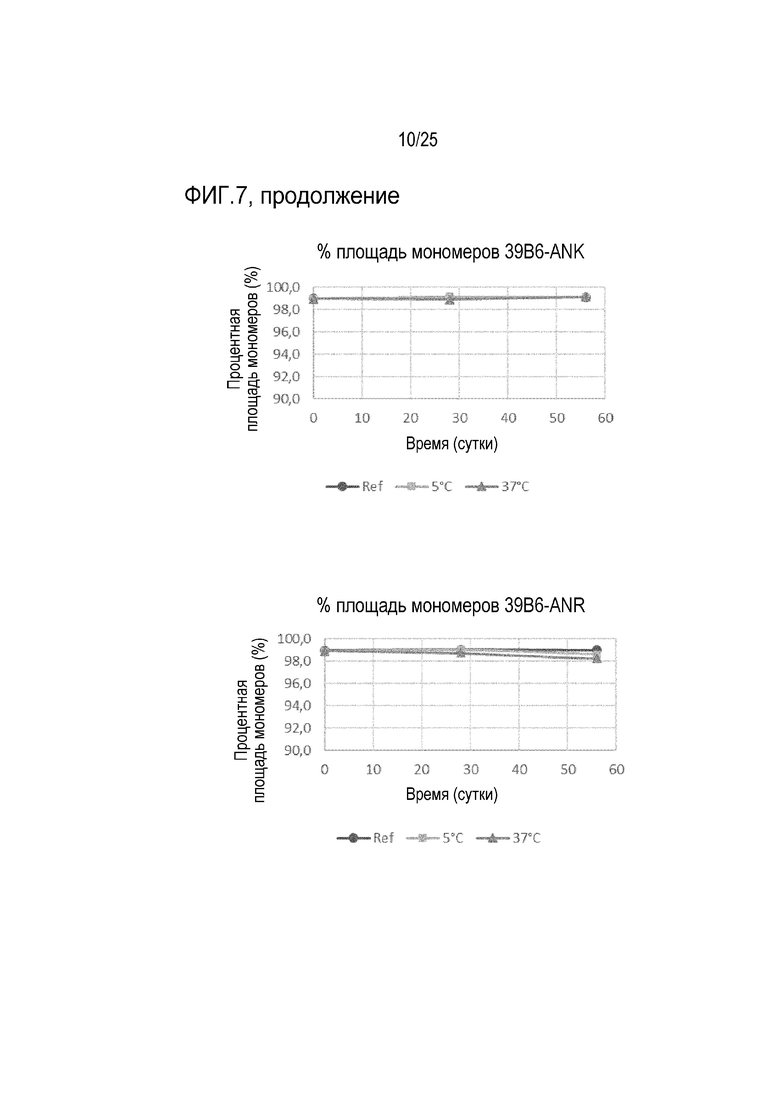


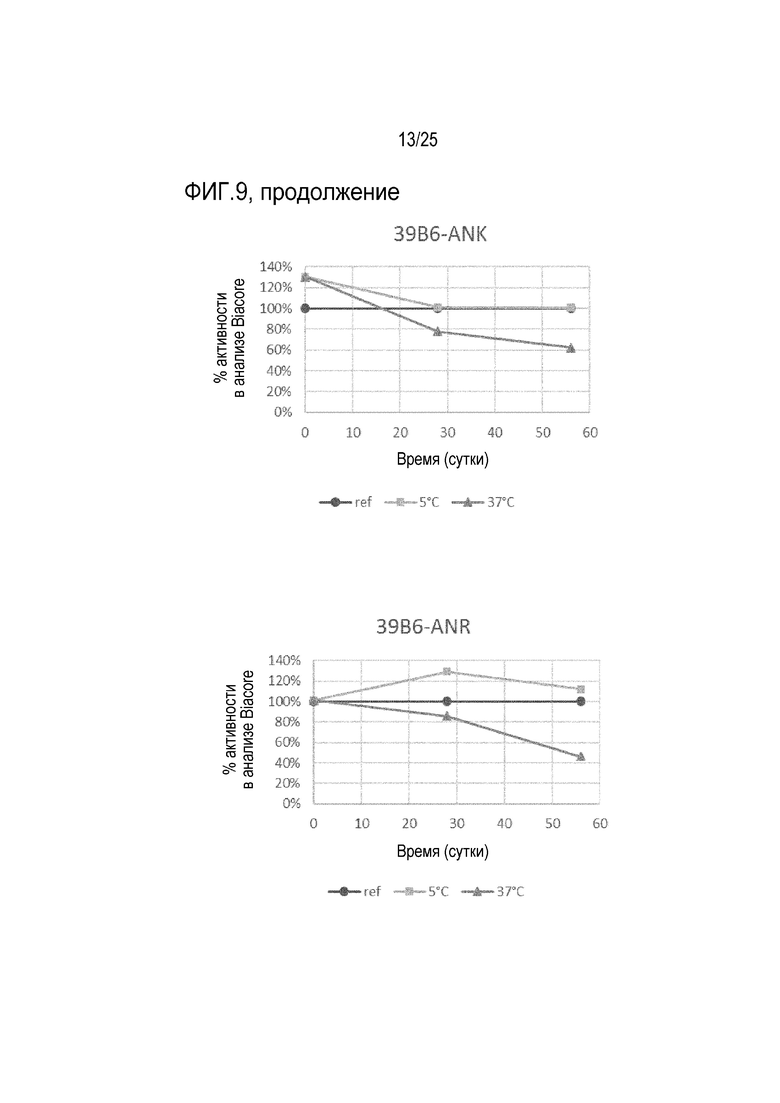






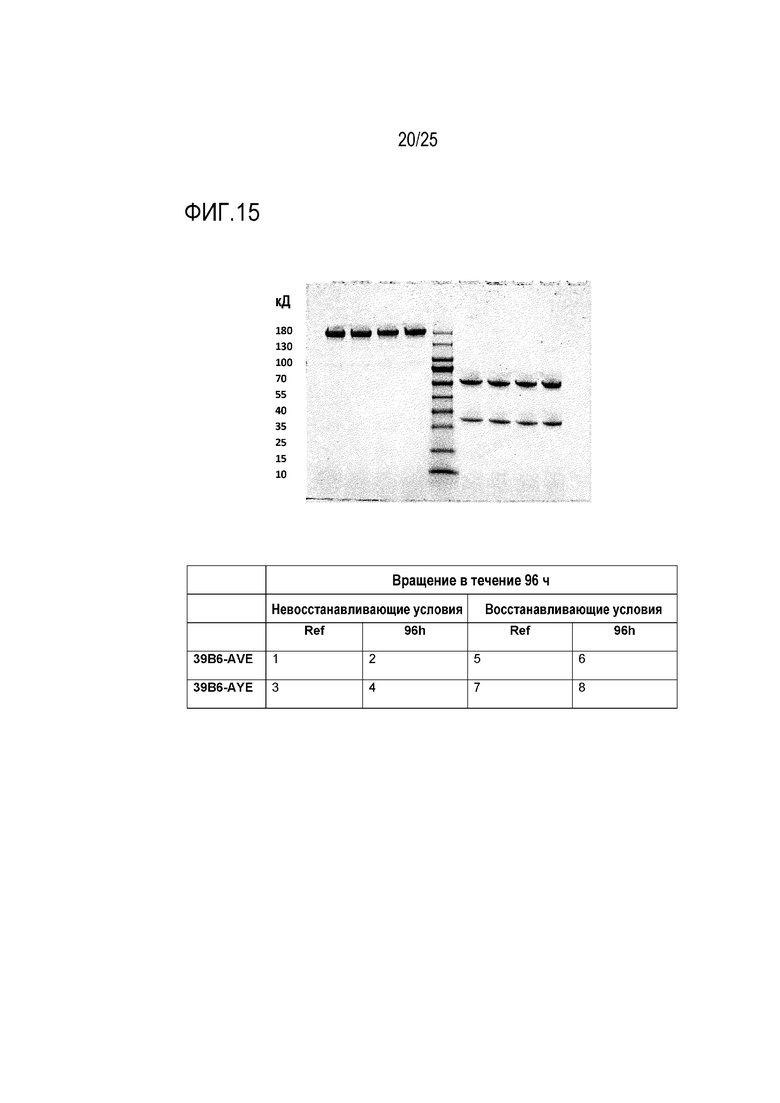





Изобретение относится к области биотехнологии. Предложено антитело, которое связывается с комплексом человеческого GARP и человеческого TGF–β1. Также предложены полинуклеотиды, кодирующие указанное антитело, вектор экспрессии, клетка-хозяин, бесклеточная система экспрессии и способ получения антитела. Изобретение обеспечивает высокую аффинность и способность ингибировать высвобождение активного TGF–β из регуляторных Т–клеток, а также повышенную стабильность антитела вследствие его устойчивости к дезамидированию, изомеризации и окислению. 7 н. и 6 з.п. ф-лы, 20 ил., 43 табл., 4 пр.
1. Антитело, которое связывается с комплексом человеческих GARP и TGF–β1, где антитело содержит вариабельную область тяжелой цепи (VH), где:
VH CDR3 содержит аминокислотную последовательность YEWETVVVGDLMYEYEY (SEQ ID NO: 13),
VH CDR2 содержит аминокислотную последовательность RIDPEDAGTKYAQKFQG (SEQ ID NO: 12), и
VH CDR1 содержит аминокислотную последовательность SYYID (SEQ ID NO: 4); и
вариабельную область легкой цепи (VL), где:
VL CDR3 содержит аминокислотную последовательность QQYASVPVT (SEQ ID NO: 11),
VL CDR2 содержит аминокислотную последовательность GASRLKT (SEQ ID NO: 10), и
VL CDR1 содержит аминокислотную последовательность QASQSISSYLA (SEQ ID NO: 9).
2. Антитело по п. 1, где
вариабельная область тяжелой цепи (VH) содержит аминокислотную последовательность SEQ ID NO: 14, и
вариабельная область легкой цепи (VL) содержит аминокислотную последовательность SEQ ID NO: 15.
3. Антитело по п. 1 или 2, которое содержит домен CH1, шарнирную область, домен CH2 и/или домен CH3 IgG человека.
4. Антитело по п. 3, где IgG человека представляет IgG1.
5. Антитело по п. 3, где IgG человека представляет IgG4.
6. Антитело по п. 5, где IgG4 человека имеет замену S228P в домене СН3.
7. Антитело по любому из пп. 1-3, 5 или 6, где антитело содержит
тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 16, и
легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 17.
8. Выделенный полинуклеотид, который кодирует антитело по любому из пп. 1-7, где полинуклеотид кодирует вариабельную область тяжелой цепи этого антитела и вариабельную область легкой цепи этого антитела.
9. Выделенный полинуклеотид, который кодирует антитело по любому из пп. 1–7, где полинуклеотид, который кодирует тяжелую цепь, содержит последовательность SEQ ID NO: 18, и полинуклеотид, который кодирует легкую цепь, содержит последовательность SEQ ID NO: 19.
10. Вектор экспрессии, содержащий полинуклеотид по п. 8 или 9, функционально связанный с регуляторными последовательностями, которые обеспечивают экспрессию антитела в клетке–хозяине или в бесклеточной системе экспрессии.
11. Клетка–хозяин для получения антитела по любому из пп. 1-7, где клетка содержит вектор экспрессии по п. 10.
12. Бесклеточная система экспрессии для получения антитела по любому из пп. 1-7, где система содержит вектор экспрессии по п. 10.
13. Способ получения рекомбинантного антитела по любому из пп. 1-7, где способ включает:
культивирование клетки–хозяина по п. 11 или бесклеточной системы экспрессии по п. 12 в условиях, которые обеспечивают экспрессию антитела, и
выделение экспрессированного антитела.
| WO 2015015003 A1, 05.02.2015 | |||
| WO 2016125017 A1, 11.08.2016 | |||
| ПРИМЕНЕНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ ДНК, КОТОРАЯ КОДИРУЕТ ПЕПТИД, СПОСОБНЫЙ СВЯЗЫВАТЬСЯ С ТРАНСФОРМИРУЮЩИМ ФАКТОРОМ РОСТА β1 (ВАРИАНТЫ) | 2008 |
|
RU2455358C2 |
| WANG R | |||
| et al., GARP regulates the bioavailability and activation of TGFβ, Molecular biology of the cell, 2012, Vol | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| CUENDE J | |||
| et al., Monoclonal antibodies against GARP/TGF-β1 complexes inhibit the immunosuppressive activity of human | |||

