Область техники
[0001] Настоящее изобретение относится к получению анти-Orai1 антитела, обладающего более высокой активностью, для лечения расстройств и заболеваний.
Уровень техники
[0002] Активируемые высвобождением кальция кальциевые каналы (CRAC) являются разновидностью депо-управляемых каналов (SOC), которые открываются в ответ на истощение запасов кальция, находящихся во внутриклеточном эндоплазматическом ретикулуме, и которые отвечают за проникновение внеклеточного кальция в определенные не чувствительные к раздражению клетки, в частности клетки иммунной системы, включая T-лимфоциты и тучные клетки, что приводит к активации этих клеток. Ингибиторы активности CRAC-каналов известны из уровня техники, и их идентификация и терапевтический потенциал описаны в Feske et al. (Непатентная литература 1 и 2).
[0003] Orai1 (CRACM1: модулятор 1 активируемых высвобождением кальция кальциевых каналов, трансмембранный белок 142A: TMEM142A), проходящий четыре раза через мембрану трансмембранный белок, состоящий из 301 аминокислотного остатка, был идентифицирован в качестве компонента порообразующей субъединицы CRAC-каналов, который образует гомотетрамер (Патентная литература 1 и Непатентная литература 3-5). Семейство генов Orai включает в себя ген Orai1 человека вместе с генами Orai2 и Orai3 человека, каждый из которых имеет 90% или выше гомологии с геном Orai1 человека (Непатентная литература 6). Также сообщалось, что в некоторых случаях существует возможность образования гетеротетрамера или гетерогексамера, включающего в себя белок Orai2 и/или Orai3 и белок Orai1 (Непатентная литература 7 и 8). Orai1 состоит из N-концевой и C-концевой цитоплазматических областей, которые связаны с белком STIM-1 (стромальная взаимодействующая молекула 1) или STIM-2, который представляет собой белок, воспринимающий истощение запасов кальция во внутриклеточном эндоплазматическом ретикулуме, и 4 трансмембранных доменов, домен первой внеклеточной петли состоит из приблизительно 20 аминокислотных остатков, а домен второй внеклеточной петли состоит из приблизительно 40 аминокислотных остатков (Непатентная литература 9). Последовательность ДНК и аминокислотная последовательность Orai1 доступны в публичной базе данных, и могут иметь регистрационные номера, например, Nos. NM_032790 и NP_116179 (NCBI).
[0004] Было обнаружено, что врожденное нарушение функции гена Orai1 человека устраняет активность CRAC-каналов и блокирует ответ организма на иммуногены, что приводит к развитию тяжелых иммунодефицитных состояний. Таким образом, было доказано, что молекулярная функция Orai1 является существенной для активации CRAC-каналов (Непатентная литература 10 и 11). Таким образом, блокирующие функцию антитела, специфичные по отношению к молекуле Orai1, могут служить в качестве ингибиторов активности CRAC-каналов.
[0005] В свете информации, указывающей на то, что ингибиторы активности CRAC-каналов могут применяться для лечения пациентов с иммунологическими заболеваниями, аллергическими заболеваниями, воспалительными заболеваниями, при отторжении трансплантата клеток или органов, тромбозе, раке и т.д. (Непатентная литература 12), были предприняты попытки получить анти-Orai1 антитела с целью ингибировать молекулярную функцию Orai1, и проводилось изучение их действия (Патентная литература 2 и 3 и НеПатентная литература 13 и 14). Хотя в этой литературе указывается на то, что каждое антитело в отдельности ингибирует активацию T-лимфоцитов, их ингибирующая активность не является пока достаточно сильной. Клиническое применение этих антител в качестве биопрепаратов, направленных на Orai1, не может удовлетворить медицинские потребности из-за необходимости введения высоких доз или частого введения, ограничений по способу введения или тому подобного.
Список литературы
Патентная литература
[0006]
Патентная литература 1: Публикация Международной заявки No. WO07/081804
Патентная литература 2: Публикация Международной заявки No. WO11/063277
Патентная литература 3: Публикация Международной заявки No. WO13/091903
Непатентная литература
[0007]
Непатентная литература 1: Feske S, Nature Rev. Immunol. 7, p. 690-702 (2007)
Непатентная литература 2: Derler I, et al., Expert Opin. Drug Discovery 3 (7), p. 787-800 (2008)
Непатентная литература 3: Prakriya M, et al., Nature 443, p. 230-233 (2006)
Непатентная литература 4: Vig M, et al., Science 312, p. 1220-1223 (2006)
Непатентная литература 5: Park CY, et al., Cell 136, p. 876-890 (2008)
Непатентная литература 6: Mercer JC, et al., J. Biol. Chem. 281, p. 24979-24990 (2006)
Непатентная литература 7: Gwack Y, et al., J. Biol. Chem. 282, p. 16232-16243 (2007)
Непатентная литература 8: Hou X, et al., Science 338, p. 1308-1313 (2012)
Непатентная литература 9: Vig M, et al., Curr. Biol. 16, p. 2073-2079 (2006)
Непатентная литература 10: Feske S, Nature 441, p. 179-185 (2006)
Непатентная литература 11: McCarl CA, et al., J. Allergy Clin. Immunol. 124, p. 1311-1318 (2009)
Непатентная литература 12: McCarl CA, et al., J. Immunol. 185, p. 5845-5858 (2010)
Непатентная литература 13: Lin F, et al., J. Pharmacol. Exp. Ther. 345, p. 225-238 (2013)
Непатентная литература 14: Cox JH, et al., PLOS ONE 8 (12), e82944 (2013)
Сущность изобретения
Техническая задача
[0008] Целью настоящего изобретения является получение терапевтического и/или профилактического агента для применения при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях, тромбозе, раке и т.д.
Решение задачи
[0009] Авторами настоящего изобретения были получены крысиные анти-Orai1 антитела с целью поиска веществ, обладающих терапевтическим и/или профилактическим эффектом при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях, тромбозе или раке. Полученные крысиные анти-Orai1 антитела были гуманизированы, и CDRs, каркасные области и вариабельные области этих гуманизированных антител были сконструированы. Таким образом, настоящее изобретение было выполнено.
[0010] В частности, настоящее изобретение включает в себя следующие аспекты:
(1) Антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 104, 106, 107 и 108, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 109 или 110; и
последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 93, 94 и 95, CDRL2 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 96, 97 и 98, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(2) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(3) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 94, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 97, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(4) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 95, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 98, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(5) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 107, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(6) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 108, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(7) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1), где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 104, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 109; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
(8) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(2), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(9) Антитело или антигенсвязывающий фрагмент этого антитела по п.(8), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(10) Антитело или антигенсвязывающий фрагмент этого антитела по п.(8), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 70, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(11) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(3), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 58.
(12) Антитело или антигенсвязывающий фрагмент этого антитела по п.(11), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 58.
(13) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(4), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 60.
(14) Антитело или антигенсвязывающий фрагмент этого антитела по п.(13), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 60.
(15) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(5), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 64, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(16) Антитело или антигенсвязывающий фрагмент этого антитела по п.(15), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 64, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(17) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(6), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 66, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(18) Антитело или антигенсвязывающий фрагмент этого антитела по п.(17), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 66, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(19) Антитело или антигенсвязывающий фрагмент этого антитела по п.(1) или п.(7), где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 43, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(20) Антитело или антигенсвязывающий фрагмент этого антитела по п.(19), где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 43, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 235 в аминокислотной последовательности, представленной SEQ ID NO: 56.
(21) Антигенсвязывающий фрагмент антитела по любому из п.п. (1)-(20), где этот антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
(22) Антитело по любому из п.п. (1)-(8), (11), (13), (15), (17) и (19), где это антитело представляет собой scFv.
(23) Фармацевтическая композиция, содержащая, по меньшей мере, любое одно антитело или антигенсвязывающий фрагмент этого антитела по п.п. (1)-(22).
(24) Фармацевтическая композиция по п.(23), где эта фармацевтическая композиция представляет собой терапевтический и/или профилактический агент, применяемый при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях или раке, антитромбоцитарный или антитромботический активатор, или ингибитор активации клеток, экспрессирующих Orai1.
(25) Фармацевтическая композиция по п.(24), где отторжения трансплантата представляют собой реакции отторжения и реакции по типу «хозяин против трансплантата» при трансплантации органа или ткани, таких как сердце, почка, печень, костный мозг или кожа, и реакцию по типу «трансплантат против хозяина», вызванную трансплантацией гемопоэтических клеток (костного мозга, периферической крови, пуповинной крови и т.д.).
(26) Фармацевтическая композиция по п.(24), где иммунные заболевания представляют собой заболевания соединительной ткани или опорно-двигательного аппарата (ревматоидный артрит, анкилозирующий спондилит, системная красная волчанка, склеродермия, полимиозит, дерматомиозит и т.д.), гематологические заболевания (апластическая анемия, идиопатическая тромбоцитопеническая пурпура и т.д.), заболевания желудочно-кишечного тракта (болезнь Крона, неспецифический язвенный колит и т.д.), неврологические заболевания (рассеянный склероз, миастения гравис и т.д.), офтальмологические заболевания (увеит и т.д.), сосудистые заболевания (болезнь Бехчета, гранулематоз Вегенера и т.д.), кожные заболевания (псориаз, пузырчатка, лейкодермия и т.д.), эндокринные заболевания (сахарный диабет первого типа, аутоиммунный тиреоидит, болезнь Грейвcа, тиреоидит Хашимото и т.д.) и тому подобное, аллергические заболевания представляют собой атопический дерматит, астму, анафилаксию, анафилактоидную реакцию, алиментарную аллергию, ринит, отит среднего уха, лекарственную аллергию, аллергическую реакцию на укусы насекомых, аллергическую реакцию на растения, аллергию на латекс, конъюнктивит, крапивницу и тому подобное, и воспалительные заболевания представляют собой воспалительные заболевания почек (гломерулонефрит, нефроз и т.д.), воспалительные заболевания легких (хроническое обструктивное заболевание легких, муковисцидоз, интерстициальная пневмония и т.д.), воспалительные заболевания кишечника (неспецифический язвенный колит, илеит и т.д.), воспалительные заболевания печени (аутоиммунный гепатит, вирусный гепатит и т.д.), воспалительные заболевания сердца (миокардит, ишемическая болезнь сердца, атеросклероз и т.д.), воспалительные заболевания кожи (контактный дерматит, экзема и т.д.), воспалительные заболевания глаз (трахома, эндофтальмит и т.д.), воспалительные заболевания центральной нервной системы (менингит, энцефаломиелит, аутоиммунный энцефалит и т.д.), воспалительные заболевания суставов (артрит, остеоартрит и т.д.), системные воспаления (сепсис, кровотечения, гиперчувствительность, симптомы шока, связанные с химиотерапией рака или тому подобным, и т.д.), и тому подобное.
(27) Фармацевтическая композиция по п.(24), где рак представляет собой рак молочной железы, рак легкого, рак кожи, лейкоз и тому подобное, и случаи, при которых антитромбоцитарная или антитромботическая активность используется для лечения и/или предотвращения заболевания, представляют собой инфаркт миокарда, инсульт, ишемическую болезнь сердца, тромбоз и тому подобное, и случаи, при которых ингибирование активации клеток, экспрессирующих Orai1, используется для лечения и/или предотвращения заболевания, представляют собой тучноклеточный лейкоз, мастоцитоз, базофильный лейкоз, эндометриоз, миопатию с тубулярными агрегатами, синдром Шторморкена, ревматоидный артрит, анкилозирующий спондилоартрит, атопический дерматит и тому подобное.
(28) Полинуклеотид, кодирующий антитело или антигенсвязывающий фрагмент этого антитела по любому из п.п. (1)-(22).
(29) Вектор, содержащий полинуклеотид по п.(28).
(30) Трансформированная клетка-хозяин, содержащая полинуклеотид по п.(28).
(31) Трансформированная клетка-хозяин, содержащая вектор по п.(29).
(32) Способ получения антитела или антигенсвязывающего фрагмента этого антитела по любому из п.п. (1)-(22), включающий в себя этап культивирования клетки-хозяина по п.(30) или п.(31) и очистку антитела от продукта культивирования.
(33) Антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 50%, составляет 80 нг/мл или меньше.
(34) Антитело или антигенсвязывающий фрагмент этого антитела по п.(33), где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 50%, составляет 10 нг/мл или меньше.
(35) Антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество IL-2, высвобождаемое из мононуклеарных клеток периферической крови (МКПК) человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 100 нг/мл или меньше.
(36) Антитело или антигенсвязывающий фрагмент этого антитела по п.(35), где концентрация, при которой количество IL-2, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 20 нг/мл или меньше.
(37) Антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 800 нг/мл или меньше.
(38) Антитело или антигенсвязывающий фрагмент этого антитела по п.(37), где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 40 нг/мл или меньше.
Полезный эффект изобретения
[0011] В соответствии с настоящим изобретением может быть получен терапевтический и/или профилактический агент для применения при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях, тромбозе или раке, механизм действия которого основан на ингибировании активности CRAC-каналов.
Краткое описание чертежей
[0012]
[Фигура 1] На Фигуре 1 представлена диаграмма, показывающая, что R118 и R198 каждое связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T зависимым от концентрации образом.
[Фигура 2] На Фигуре 2 представлена диаграмма, показывающая, что R118 и R198 каждое ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187.
[Фигура 3] На Фигуре 3 представлена диаграмма, показывающая, что cR118 и cR198 каждое связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T зависимым от концентрации образом.
[Фигура 4] На Фигуре 4 представлена диаграмма, показывающая, что cR118 и cR198 каждое ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187.
[Фигура 5] На Фигуре 5 представлена диаграмма, показывающая, что гуманизированные анти-человеческий Orai1 антитела hR198_H1/L1, hR198_H2/L2, hR198_H3/L3 и hR198_H4/L4 каждое связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T зависимым от концентрации образом, как и исходное антитело cR118 или cR198.
[Фигура 6] На Фигуре 6 представлена диаграмма, показывающая, что гуманизированные анти-человеческий Orai1 антитела каждое ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187.
[Фигура 7] На Фигуре 7 представлена диаграмма, показывающая, что клоны с мутантным Fab LCDR60, LCDR67, LCDR83, CE151, PE057, HCDR046, HCDR047, HEP087, HEP124 и HEP237, полученные с помощью рибосомного дисплея, каждый сильно связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T, по сравнению с антителом hR198_H4/L4-Fab, имеющим исходный Fab.
[Фигура 8] На Фигуре 8 представлена диаграмма, показывающая, что при созревании аффинности антител hR198_H3/LG1, hR198_HG1/LG1, hR198_HG1/LG2, hR198_HG1/LG3, hR198_HG2/LG1 и hR198_HG3/LG1, каждое из них связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T на уровне эквивалентном или более сильном, чем исходное антитело hR198_H3/L3 или hR198_H4/L4.
[Фигура 9] На Фигуре 9 представлена диаграмма, показывающая, что при созревании аффинности каждое из антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187.
[Фигура 10] На Фигуре 10 представлена диаграмма, показывающая, что каждое из антител 10F8, 14F74 и 17F6 связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T.
[Фигура 11] На Фигуре 11 представлена диаграмма, показывающая, что каждое из антител hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1 и 5H3.1 связывается с трансфицированными pcDNA3.1-hOrai1 клетками HEK293T.
[Фигура 12] На Фигуре 12 представлена диаграмма, показывающая, что каждое из анти-Orai1 моноклональных антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187.
[Фигура 13] На Фигуре 13 представлена диаграмма, показывающая, полумаксимальную ингибирующую концентрацию (ИК50) и 80% от ингибирующей концентрации (ИК80) для высвобождения IL-2 из клеток Юрката для антител hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6.
[Фигура 14] На Фигуре 14 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую вариабельную область легкой цепи антитела R118, и аминокислотную последовательность этой вариабельной области.
[Фигура 15] На Фигуре 15 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи антитела R118, и аминокислотную последовательность этой вариабельной области.
[Фигура 16] На Фигуре 16 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую вариабельную область легкой цепи антитела R198, и аминокислотную последовательность этой вариабельной области.
[Фигура 17] На Фигуре 17 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую вариабельную область тяжелой цепи антитела R198, и аминокислотную последовательность этой вариабельной области.
[Фигура 18] На Фигуре 18 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь человеческого гибридного антитела cR118, и аминокислотную последовательность этой легкой цепи.
[Фигура 19] На Фигуре 19 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь человеческого гибридного антитела cR198, и аминокислотную последовательность этой легкой цепи.
[Фигура 20] На Фигуре 20 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь человеческого гибридного антитела cR118, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 21] На Фигуре 21 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь человеческого гибридного антитела cR198, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 22] На Фигуре 22 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_L1, и аминокислотную последовательность этой легкой цепи.
[Фигура 23] На Фигуре 23 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_L2, и аминокислотную последовательность этой легкой цепи.
[Фигура 24] На Фигуре 24 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_L3, и аминокислотную последовательность этой легкой цепи.
[Фигура 25] На Фигуре 25 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_L4, и аминокислотную последовательность этой легкой цепи.
[Фигура 26] На Фигуре 26 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H1, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 27] На Фигуре 27 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H2, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 28] На Фигуре 28 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H3, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 29] На Фигуре 29 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H4, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 30] На Фигуре 30 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_LG1, и аминокислотную последовательность этой легкой цепи.
[Фигура 31] На Фигуре 31 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_LG2, и аминокислотную последовательность этой легкой цепи.
[Фигура 32] На Фигуре 32 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь типа hR198_LG3, и аминокислотную последовательность этой легкой цепи.
[Фигура 33] На Фигуре 33 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_HG1, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 34] На Фигуре 34 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_HG2, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 35] На Фигуре 35 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_HG3, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 36] На Фигуре 36 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H4-LALA, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 37] На Фигуре 37 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_HG1-LALA, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 38] На Фигуре 38 представлена диаграмма, показывающая, SEQ ID NOs, соответствующие последовательностям CDR, содержащимся в каждой из легких цепей антител cR118, cR198, hR198_L1 - hR198_L4 и hR198_LG1 - hR198_LG3 и в каждой из тяжелых цепей антител cR118, cR198, hR198_H1 - hR198_H4, hR198_HG1 - hR198_HG3, hR198_H4-LALA и hR198_HG1-LALA.
[Фигура 39] На Фигуре 39 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь антитела 2C1.1, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 40] На Фигуре 40 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь антитела 2C1.1, и аминокислотную последовательность этой легкой цепи.
[Фигура 41] На Фигуре 41 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь антитела 5H3.1, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 42] На Фигуре 42 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь антитела 5H3.1, и аминокислотную последовательность этой легкой цепи.
[Фигура 43] На Фигуре 43 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь антитела 10F8, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 44] На Фигуре 44 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь антитела 10F8, и аминокислотную последовательность этой легкой цепи.
[Фигура 45] На Фигуре 45 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь антитела 14F74, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 46] На Фигуре 46 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь антитела 14F74, и аминокислотную последовательность этой легкой цепи.
[Фигура 47] На Фигуре 47 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь антитела 17F6, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 48] На Фигуре 48 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую легкую цепь антитела 17F6, и аминокислотную последовательность этой легкой цепи.
[Фигура 49] На Фигуре 49 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H0, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 50] На Фигуре 50 представлена диаграмма, показывающая, нуклеотидную последовательность, кодирующую тяжелую цепь типа hR198_H5, и аминокислотную последовательность этой тяжелой цепи.
[Фигура 51] На Фигуре 51 представлена диаграмма, показывающая, что каждое из анти-Orai1 моноклональных антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из МКПК человека, обработанных ФМА и A23187.
[Фигура 52] На Фигуре 52 представлена диаграмма, показывающая, полумаксимальную ингибирующую концентрацию (ИК50) и 80% от ингибирующей концентрации (ИК80) для высвобождения IL-2 из МКПК человека для антител hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6.
[Фигура 53] На Фигуре 53 представлена диаграмма, показывающая, что каждое из анти-Orai1 моноклональных антител ингибирует, зависимым от концентрации образом, высвобождение ИФН-γ из МКПК человека, обработанных ФМА и A23187.
[Фигура 54] На Фигуре 54 представлена диаграмма, показывающая, полумаксимальную ингибирующую концентрацию (ИК50) и 80% от ингибирующей концентрации (ИК80) для высвобождения ИФН-γ из МКПК человека для антител hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6.
[Фигура 55] На Фигуре 55 представлена диаграмма, показывающая, что антитело hR198_HG1/LG1 ингибирует снижение массы тела, связанное с реакцией по типу «трансплантат против хозяина», которая развивается при трансплантации МКПК человека мышам с тяжелым комбинированным иммунодефицитом.
[Фигура 56] На Фигуре 56 представлена диаграмма, показывающая, что антитело hR198_HG1/LG1 подавляет пассивную кожную анафилактическую реакцию, индуцированную у мышей с активным Orai1 человека.
[Фигура 57] На Фигуре 57 представлена диаграмма, показывающая, что антитело hR198_HG1/LG1 подавляет гиперчувствительную реакцию замедленного типа, индуцированную у мышей с активным Orai1 человека, в каждый момент времени: через 6 часов (A), 24 часа (B) и 48 часов (C) после введения антигена.
[Фигура 58] На Фигуре 58 представлена диаграмма, показывающая, вязкость, измеренную при концентрации 90 мг/мл, 120 мг/мл и 150 мг/мл антител hR198_HG1/LG1, hR198_H3/LG1, hR198_HG1-LALA/LG1 и 2C1.1.
Описание вариантов осуществления изобретения
[0013] В настоящем описании термин «ген» включает в себя не только ДНК, но и мРНК, кДНК и кРНК.
[0014] В настоящем описании термин «полинуклеотид» используется в том же значении, что и нуклеиновая кислота, и также включает в себя ДНК, РНК, зонды, олигонуклеотиды и праймеры.
[0015] В настоящем описании термин «полипептид» и термин «белок» используются как взаимозаменяемые.
[0016] В настоящем описании термин «РНК фракция» относится к фракции, содержащей РНК.
[0017] В настоящем описании термин «клетка» включает в себя клетки внутри отдельного животного и культивируемые клетки.
[0018] В настоящем описании термин «Orai1» используется в том же значении, что и белок Orai1.
[0019] В настоящем описании термин «антигенсвязывающий фрагмент антитела» означает частичный фрагмент антитела, обладающий активностью связывания по отношению к антигену, и включает в себя Fab, F(ab')2, Fv, scFv, диатела, линейные антитела и мультиспецифичные антитела, образованные из фрагментов антител, и т.д. Антигенсвязывающий фрагмент антитела также включает в себя Fab', который представляет собой моновалентный фрагмент вариабельных областей антитела, полученный путем обработки F(ab')2 в восстанавливающих условиях. Однако антигенсвязывающий фрагмент антитела не ограничивается этими молекулами, при условии, что этот антигенсвязывающий фрагмент обладает способностью связываться с антигеном. Такой антигенсвязывающий фрагмент включает в себя не только фрагмент, полученный путем обработки полноразмерной молекулы белка антитела соответствующим ферментом, но также белок, продуцируемый соответствующими клетками-хозяевами с использованием генно-инженерного гена антитела.
[0020] Известно, что каждая из тяжелых и легких цепей молекулы антитела имеет три определяющих комплементарность области (CDRs). Эти определяющие комплементарность области также называются гипервариабельными доменами. Эти области расположены в вариабельных областях тяжелой и легкой цепей антитела. Эти участки имеют особенно высоко вариабельную первичную структуру и разделены на три положения в соответствующих первичных структурах тяжелой и легкой полипептидных цепей. В настоящем описании определяющие комплементарность области антитела обозначаются как CDRH1, CDRH2 и CDRH3 с амино-конца аминокислотной последовательности тяжелой цепи для определяющих комплементарность областей тяжелой цепи, и как CDRL1, CDRL2 и CDRL3 с амино-конца аминокислотной последовательности легкой цепи для определяющих комплементарность областей легкой цепи. Эти участки расположены близко друг к другу в трехмерной структуре и определяют специфичность к антигену, с которым они связываются.
[0021] В настоящем изобретении термин «гибридизация в жестких условиях» означает гибридизацию в условиях, включающих в себя гибридизацию при температуре 68°C в коммерчески доступном растворе для гибридизации ExpressHyb Hybridization Solution (Clontech Laboratories, Inc.), или гибридизацию при температуре 68°C в присутствии от 0,7 до 1,0 М NaCl с использованием иммобилизующего ДНК фильтра с последующей отмывкой при температуре 68°C с использованием раствора SSC, имеющего от 0,1- до 2-кратную концентрацию (имеющий 1-кратную концентрацию раствор SSC состоит из 150 мМ NaCl и 15 мМ цитрата натрия), что позволяет проводить идентификацию или гибридизацию в эквивалентных им условиях.
[0022] В настоящем изобретении термин «реакция по типу «хозяин против трансплантата»» относится к гипериммунному состоянию реципиента, наблюдаемому после трансплантации органа, а также к происходящему в результате этого повреждению трансплантированного органа.
[0023] В настоящем изобретении термин «реакция по типу «трансплантат против хозяина»» относится к симптомам, возникающим в результате иммунологической атаки реципиента трансплантированными клетками после трансплантации гемопоэтических клеток.
[0024]
1. Orai1
Используемый в настоящем изобретении белок Orai1 может быть непосредственно очищен из T-лимфоцитов или тучных клеток человека, не являющегося человеком млекопитающего (например, морской свинки, крысы, мыши, кролика, свиньи, овцы, коровы или обезьяны) или курицы, или он может быть использован в составе фракции клеточных мембран, полученной из клеток. В альтернативном варианте, Orai1 может быть синтезирован in vitro, или получен путем продуцирования клетками-хозяевами в результате генных манипуляций. В частности, при таких генных манипуляциях кДНК Orai1 встраивают в вектор, который обеспечивает экспрессию этой кДНК, и затем Orai1 может быть синтезирован в растворе, содержащем фермент, субстрат и энергетическое вещество, необходимое для транскрипции и трансляции, или он может быть экспрессирован путем трансформации различных прокариотических или эукариотических клеток-хозяев для получения белка.
[0025] Нуклеотидная последовательность кДНК Orai1 человека зарегистрирована под регистрационным номером No. NM_032790 в регистрационной базе GenBank. Нуклеотидная последовательность кДНК мышиного Orai1 зарегистрирована под регистрационным номером No. NM_175423 в регистрационной базе GenBank. Нуклеотидная последовательность, кодирующая Orai1 человека, приведена в SEQ ID NO: 1 в Списке последовательностей, и аминокислотная последовательность Orai1 человека, приведена в SEQ ID NO: 2 в Списке последовательностей. Orai1 также именуется модулятор 1 активируемых высвобождением кальция кальциевых каналов (CRACM1) или трансмембранный белок 142A (TMEM142A), все эти наименования относятся к одной и той же молекуле.
[0026] кДНК Orai1 может быть получена при помощи так называемого метода ПЦР, который включает в себя проведение полимеразной цепной реакции (далее именуемой «ПЦР») (Saiki, R.K., et al., Science, (1988) 239, 487-49), например, используя библиотеку кДНК из органов, экспрессирующих мРНК Orai1, в качестве матрицы с праймерами, специфично амплифицирующими кДНК Orai1.
[0027] Термин «кДНК Orai1» также включает в себя полинуклеотид, который в жестких условиях гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности, кодирующей человеческий или мышиный Orai1, и который кодирует белок, обладающий биологической активностью, эквивалентной Orai1. Термин «кДНК Orai1» также включает в себя кДНК варианта сплайсинга, который транскрибируется с локуса гена человеческого или мышиного Orai1, или полинуклеотид, который в жестких условиях гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной этому варианту сплайсинга, и который кодирует белок, обладающий биологической активностью, эквивалентной Orai1.
[0028] Термин «Orai1» также включает в себя белок, который состоит из аминокислотной последовательности, полученной из аминокислотной последовательности человеческого или мышиного Orai1, или его аминокислотной последовательности, лишенной сигнальной последовательности путем замены, делеции или добавления 1, 2 или 3, или 4 или 5 аминокислот, и который обладает биологической активностью, эквивалентной Orai1. Термин «Orai1» также включает в себя белок, который состоит из аминокислотной последовательности, кодируемой вариантом сплайсинга, транскрибируемым с локуса гена человеческого или мышиного Orai1, или аминокислотной последовательности, полученной из этой аминокислотной последовательности путем замены, делеции или добавления 1, 2 или 3, или 4 или 5 аминокислот, и который обладает биологической активностью, эквивалентной Orai1.
[0029]
2. Получение анти-Orai1 антитела
Антитело против Orai1 в соответствии с настоящим изобретением может быть получено при помощи общеизвестных способов путем иммунизации животного Orai1 или произвольным пептидом, выбранным из аминокислотной последовательности Orai1, и сбора и очистки антитела, продуцируемого in vivo. Биологические виды для Orai1, используемого в качестве антигена, не ограничиваются человеком, и животное может быть иммунизировано Orai1, полученным от не являющегося человеком животного, такого как мышь или крыса. В этом случае полученное антитело, связывающееся с гетерологичным Orai1, может быть проверено на перекрестное реагирование с Orai1 человека, чтобы выбрать антитело, применимое для лечения заболеваний человека. Антиген Orai1 может быть получен с использованием клетки-хозяина, продуцирующей Orai1 в результате генных манипуляций. В частности, получают вектор, который обеспечивает экспрессию гена Orai1, и этот вектор переносят в клетку-хозяин для экспрессии этого гена. Экспрессируемый Orai1 может быть очищен.
[0030] Антитело против Orai1 в соответствии с настоящим изобретением также может быть получено при помощи метода ДНК-иммунизации. Метод ДНК-иммунизации представляет собой подход, который включает в себя трансфекцию животного (например, мыши или крысы) экспрессирующей антиген плазмидой, и экспрессию этого антигена в животном, чтобы индуцировать иммунитет против этого антигена. Трансфекция включает в себя способ непосредственной инъекции плазмиды в мышцу, способ инъекции комплекса плазмиды и липосомы, полиэтиленимина или тому подобного в вену, подход с использованием вирусного вектора, инъекцию при помощи генной пушки частиц золота, прикрепленных к плазмиде, гидродинамический способ быстрой инъекции раствора плазмиды в большом количестве в вену, и тому подобное. Метод, носящий название электропорация in vivo, который включает в себя применение электропорации к области, предназначенной для внутримышечной инъекции плазмиды, известен в качестве подхода, повышающего уровень эффективности способа трансфекции путем инъекции экспрессионной плазмиды в мышцу (Aihara H, Miyazaki J., Nat Biotechnol. 1998 Sep; 16 (9): 867-70 or Mir LM, Bureau MF, Gehl J, Rangara R, Rouy D, Caillaud JM, Delaere P, Branellec D, Schwartz B, Scherman D., Proc Natl Acad Sci USA. 1999 Apr 13; 96 (8): 4262-7). Этот подход также повышает уровень экспрессии путем обработки мышцы гиалуронидазой перед внутримышечной инъекцией плазмиды (McMahon JM1, Signori E, Wells KE, Fazio VM, Wells DJ., Gene Ther. 2001 Aug; 8 (16): 1264-70).
[0031] Моноклональное антитело также может быть получено способами, известными из уровня техники (например, Kohler and Milstein, Nature (1975) 256, p. 495-497; and Kennet, R. ed., Monoclonal Antibodies, p. 365-367, Plenum Press, N.Y. (1980)), путем слияния продуцирующих антитело клеток, которые продуцируют антитело против Orai1, с клетками миеломы, для получения гибридом. Конкретные примеры такого способа описаны в публикациях Международных патентных заявок Nos. WO09/48072 (опубликована 16 апреля 2009) и WO10/117011 (опубликована 14 октября 2010).
[0032] Фактические примеры полученного таким образом крысиного анти-человеческий Orai1 антитела могут включать в себя антитела R118 и R198. Аминокислотная последовательность вариабельной области легкой цепи антитела R118 приведена в SEQ ID NO: 11 в Списке последовательностей, и последовательность вариабельной области тяжелой цепи антитела R118 приведена в SEQ ID NO: 13 в Списке последовательностей. Аминокислотная последовательность вариабельной области легкой цепи антитела R198 приведена в SEQ ID NO: 15 в Списке последовательностей, и последовательность вариабельной области тяжелой цепи антитела R198 приведена в SEQ ID NO: 17 в Списке последовательностей.
[0033] Антитело настоящего изобретения включает в себя описанное выше моноклональное антитело против Orai1, а также рекомбинантное антитело, искусственно сконструированное с целью, например, уменьшения гетерогенной антигенности по отношению к человеку, например, гибридного антитела и гуманизированного антитела, человеческого антитела и тому подобного. Эти антитела могут быть получены при помощи известных способов.
[0034] Примеры гибридного антитела могут включать в себя гибридное антитело, содержащее вариабельные области и константные области антител, полученных из разных видов, например, вариабельные области антитела, полученного от мыши или крысы, соединенные с константными областями, полученными от человека (см. Proc. Natl. Acad. Sci. U.S.A., 81, 6851-6855, (1984)).
[0035] Примеры гибридного антитела, полученного из крысиного анти-человеческий Orai1 антитела R118 могут включать в себя антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положения с 21 по 126 последовательности SEQ ID NO: 23, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положения с 20 по 136 последовательности SEQ ID NO: 27. Один пример такого полученного из R118 гибридного антитела может включать в себя антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 23, и тяжелой цепи, состоящей из аминокислотных остатков в положения с 20 по 466 последовательности SEQ ID NO: 27. В настоящем описании это антитело именуется «cR118» или «антитело cR118».
[0036] Примеры гибридного антитела, полученного из крысиного анти-человеческий Orai1 антитела R198 могут включать в себя антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положения с 21 по 126 последовательности SEQ ID NO: 25, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положения с 20 по 136 последовательности SEQ ID NO: 29. Один пример такого полученного из R198 гибридного антитела может включать в себя антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 25, и тяжелой цепи, состоящей из аминокислотных остатков в положения с 20 по 466 последовательности SEQ ID NO: 29. В настоящем описании это антитело именуется «cR198» или «антитело cR198».
[0037] Вышеописанная последовательность гибридного антитела против Orai1 может быть искусственно сконструирована для получения гуманизированного антитела в виде рекомбинантного гена антитела с целью, например, уменьшения гетерогенной антигенности по отношению к человеку. Антитело настоящего изобретения включает в себя антитело, чьи CDRs представляют собой сконструированные CDRs гуманизированного антитела. Эти антитела могут быть получены с использованием известных способов.
[0038] Примеры гуманизировнного антитела могут включать в себя антитело, содержащее определяющие комплементарность области (CDRs) по-отдельности привитые на антитело человеческого происхождения (см. Nature (1986) 321, p. 522-525), и антитело, содержащее последовательности CDR, а также аминокислотные остатки части каркасной области, привитые на человеческое антитело (публикация Международной патентной заявки No. WO90/07861).
[0039] Гуманизировнное антитело, полученное из антитела cR118 или cR198, сохраняет все шесть последовательностей CDR, полученных из cR118 или cR198, и обладает активностью по ингибированию активации T-лимфоцитов. Вариабельная область легкой цепи гуманизированного антитела сохраняет любую одну из CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 92 (RASQSIGNSLS), и CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 116 (RASQSISNSLS), CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96 (STSTLES), и любую одну из CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 99 (LQFATFPDT), и CDRL3, представленной в SEQ ID NO: 100 (LQFATYPDT). Также вариабельная область тяжелой цепи гуманизированного антитела сохраняет любую одну из CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102 (AYYIS), и CDRH1, представленной в SEQ ID NO: 103 (SYYIS), любую одну из CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 104 (YIDMGNGRTNYNARFKG), CDRH2, представленной в SEQ ID NO: 105 (YVDMGNGRTNYNEKFKG), и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 109 (DSNWGVDY). Эти аминокислотные последовательности CDRs также показаны на Фигуре 38.
[0040] Примеры антитела, имеющего предпочтительную комбинацию CDRs, могут включать в себя антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 92, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 100, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 103, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 105, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 109, и антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 92, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 99, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 104, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 109.
[0041] Предпочтительные примеры гуманизированного антитела могут включать в себя антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 31, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 39, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 33, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 41, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 35, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 43, и антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 37, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 45.
[0042] Более предпочтительные примеры такого антитела могут включать в себя антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 31, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 39, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 33, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 41, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 35, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 43, и антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 37, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 45.
[0043] Антитело настоящего изобретения может представлять собой антитело с повышенной способностью связываться с Orai1, полученное при помощи дополнительного изменения описанного выше гуманизированного антитела. Такой подход получил название созревание аффинности. Конкретные примеры этого способа могут включать в себя метод рибосомного дисплея. Метод рибосомного дисплея представляет собой способ, который включает в себя использование трехчастного комплекса белка, связанного с мРНК, несущей генетическую информацию о нем, через рибосому, и выделение последовательности гена, кодирующего белок, который связывается с целевой молекулой (Stafford RL. et al., Protein Eng. Des. Sel. 2014 (4): 97-109).
[0044] Вариабельная область легкой цепи антитела, полученного при помощи генной инженерии описанным выше способом, сохраняет любую одну из CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 93 (RASQSIGGSLS), CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 94 (HASQNIGGSLS), и CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 95 (HASRNIGGSLS), любую одну из CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96 (STSTLES), CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 97 (LTSTLDW), и CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 98 (LTSSLDW), и CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101 (LQFAIFPDS). Также вариабельная область тяжелой цепи антитела, полученного при помощи генной инженерии, сохраняет CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102 (AYYIS), любую одну из CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 104 (YIDMGNGRTNYNARFKG), CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 106 (YIDMGNGRTDYNARFKG), CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 107 (YIDMGNGRTDYNGRFKG), и CDRH2, представленной в SEQ ID NO: 108 (YIDMGNGRTDYNMRFKG), и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 109 (DSNWGVDY), или CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110 (DSNWGADY). Эти аминокислотные последовательности CDRs также показаны на Фигуре 38.
[0045] Примеры антитела, имеющего предпочтительную комбинацию CDRs, могут включать в себя антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110, антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 94, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 97, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110, антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 95, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 98, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110, антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 107, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110, антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 108, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 110, и антитело, содержащее CDRL1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 96, CDRL3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 101, CDRH1, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 104, и CDRH3, состоящую из аминокислотной последовательности, представленной SEQ ID NO: 109.
[0046] Предпочтительные примеры содержащего генно-инженерные CDR антитела могут включать в себя антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 56, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 58, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 60, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 56, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 64, антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 56, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 66, и антитело, состоящее из легкой цепи, содержащей вариабельную область легкой цепи, состоящую из аминокислотных остатков в положениях с 21 по 126 последовательности SEQ ID NO: 56, и тяжелой цепи, содержащей вариабельную область тяжелой цепи, состоящую из аминокислотных остатков в положениях с 20 по 136 последовательности SEQ ID NO: 43.
[0047] Предпочтительные примеры антитела, содержащего описанные выше вариабельную область легкой цепи и вариабельную область тяжелой цепи, могут включать в себя антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 56, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 58, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 60, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 62, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 56, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 64, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 56, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 66, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 56, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 43.
[0048] Чтобы избежать цитотоксичности в отношении нормальных клеток, экспрессирующих Orai1 человека, желательно, чтобы антитело обладало низкой эффекторной активностью. Известно, что эффекторная активность различается у антител различных субклассов. Отмечены следующие характеристики, например, IgG4 обладает низкой АЗКЦ и КЗЦ активностью, а IgG2 обладает КЗЦ активностью, но низкой АЗКЦ активностью. На основе этих свойств можно получить антитело с пониженной АЗКЦ и КЗЦ активностью путем замены константных областей IgG1 константными областями IgG2 или IgG4. Также возможно получить антитело IgG1 с пониженной АЗКЦ и КЗЦ активностью путем частичной замены последовательностей константных областей IgG1 последовательностями, характерными для IgG2 или IgG4. В качестве одного примера, в работе Marjan Hezareh et al., Journal of Virology, 75 (24): 12161-12168 (2001) показано, что АЗКЦ и КЗЦ активность IgG1 уменьшается путем замены каждого из остатков лейцина в положениях 234 и 235 (положения обозначены EU-индексом, как описано у Kabat et al.) IgG1 остатком аланина.
[0049] Примеры тяжелой цепи анти-Orai1 антитела, полученного вышеописанным способом, могут включать в себя последовательность тяжелой цепи, приведенную в SEQ ID NO: 68 или 70. Тяжелая цепь, приведенная в SEQ ID NO: 68 или 70, может быть объединена с каждой последовательностью легкой цепи, описанной в настоящем изобретении, и использована в качестве терапевтического антитела. Конкретные примеры легкой цепи, подходящей для объединения, могут включать в себя легкую цепь, описанную в последовательностях SEQ ID NO: 31, 33, 35, 37, 56, 58 или 60. Примеры антитела, имеющего предпочтительную комбинацию легкой цепи и тяжелой цепи, могут включать в себя антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 56, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 70, антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 58, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 70, и антитело, состоящее из легкой цепи, состоящей из аминокислотных остатков в положениях с 21 по 234 последовательности SEQ ID NO: 60, и тяжелой цепи, состоящей из аминокислотных остатков в положениях с 20 по 466 последовательности SEQ ID NO: 70.
[0050] Известно, что карбокситерминальные остатки лизина удаляются в тяжелых цепях антител, продуцируемых в культивируемых клетках млекопитающих (Journal of Chromatography A, 705: 129-134 (1995)). Также известно, что два карбокситерминальных аминокислотных остатка (глицин и лизин) удаляются в тяжелых цепях, и остаток пролина, вновь расположенный на карбоксильном конце, подвергается амидированию (Analytical Biochemistry, 360: 75-83 (2007)). Однако, такая делеция и модификация последовательностей тяжелой цепи не оказывает влияния на способность антитела связываться с антигеном и эффекторные функции (активация комплемента, эффект антителозависимой клеточной цитотоксичности и т.д.) антитела. Таким образом, антитело настоящего изобретения также включает в себя антитело, модифицированное таким образом. Примеры такого антитела включают в себя делеционный мутант, полученный путем делеции 1 или 2 аминокислот на карбоксильном конце тяжелой цепи, и амидированную форму этого делеционного мутанта (например, тяжелая цепь, амидированная по остатку пролина на карбоксильном конце). Тем не менее, мутант с делецией на карбоксильном конце тяжелой цепи антитела настоящего изобретения не ограничивается вышеописанными типами, при условии, что делеционный мутант сохраняет способность связываться с антигеном и эффекторные функции. Две тяжелые цепи, образующие антитело настоящего изобретения, могут быть любым типом тяжелой цепи, выбранным из группы, состоящей из полноразмерных тяжелых цепей и тяжелых цепей делеционных мутантов, описанных выше, или могут представлять собой комбинацию любых двух типов, выбранных из них. Количественное соотношение каждого делеционного мутанта может определяться типом и условиями культивирования клеток млекопитающих, продуцирующих антитело настоящего изобретения. Примеры основного компонента антитела настоящего изобретения могут включать в себя две тяжелые цепи, обе из которых не имеют одного карбокситерминального аминокислотного остатка. В частности, тяжелая цепь, состоящая из аминокислотных остатков в положениях с 20 по 465 в последовательности тяжелой цепи, представленной в каждой из последовательностей SEQ ID NOs: 27, 29, 39, 41, 43, 45, 62, 64, 66, 68 и 70 в Списке последовательностей, или тяжелая цепь, состоящая из аминокислотных остатков в положениях с 20 по 464 в этих последовательностях, также могут быть использованы в антителе настоящего изобретения.
[0051] Антитела, полученные этими способами, могут быть оценены по их активности связывания с антигеном, с целью выбора подходящего антитела. Один пример другого показателя для сравнения свойств антител может включать в себя стабильность антитела. Дифференциальная сканирующая калориметрия (ДСК) представляет собой способ, способный быстро и точно измерить серединную точку термической денатурации (Tm), которая является хорошим показателем относительной структурной стабильности белков. Значения Tm могут быть измерены при помощи ДСК и сопоставлены для определения отличительной термостабильности. Известно, что стабильность антитела при хранении до некоторой степени коррелирует с термостабильностью этого антитела (Lori Burton, et al., Pharmaceutical Development and Technology (2007) 12, p. 265-273). Подходящее антитело может быть выбрано по показателю термостабильности. Примеры других показателей для выбора антитела могут включать в себя высокий выход в соответствующих клетках-хозяевах и низкую агрегацию в водном растворе. Например, так как не всегда верно, что антитело, имеющее самый высокий выход, обладает самой высокой термостабильностью, то антитело, наиболее подходящее для введения человеку, должно выбираться при помощи комплексной оценки на основе вышеупомянутых показателей.
[0052] Способ получения одноцепочечного иммуноглобулина путем соединения последовательностей полноразмерных тяжелой и легкой цепей антитела через соответствующий линкер также известен (Lee, H-S, et al., Molecular Immunology (1999) 36, p. 61-71; and Schirrmann, T. et al., mAbs (2010), 2 (1), p. 73-76). Такой одноцепочечный иммуноглобулин может быть димеризован, чтобы таким образом поддерживать структуру и активность, аналогичные структуре и активности антитела, которое изначально представляет собой тетрамер. Также антитело настоящего изобретения может представлять собой антитело, которое имеет одну вариабельную область тяжелой цепи и не имеет последовательности легкой цепи. Такое антитело, получившее название однодоменное антитело (sdAb) или наноантитело, в природе наблюдается у верблюдов и лам, и имеются сообщения, что они сохраняют способность связываться с антигеном (Muyldemans S. et al., Protein Eng. (1994) 7 (9), 1129-35; and Hamers-Casterman C. et al., Nature (1993) 363 (6428), 446-8). Эти антитела могут рассматриваться как одна из разновидностей антигенсвязывающего фрагмента антитела настоящего изобретения.
[0053] Антителозависимая клеточная цитотоксичность антитела настоящего изобретения может быть повышена путем корректировки модификации углеводной цепи, связанной с антителом. Например, способы, описанные в Международных патентных заявках WO99/54342, WO00/61739 и WO02/31140 известны в качестве примеров такой методики корректировки модификации углеводной цепи антитела, хотя эта методика не ограничивается описанием в этих источниках.
[0054] В случае получения антитела путем временного выделения гена антитела и последующего переноса этого гена в соответствующего хозяина, соответствующий хозяин может использоваться в сочетании с экспрессионным вектором. Конкретные примеры гена антитела могут включать в себя комбинацию гена, кодирующего последовательность тяжелой цепи, и гена, кодирующего последовательность легкой цепи, антитела, описанного в настоящем изобретении. Для трансформации клеток-хозяев ген последовательности тяжелой цепи и ген последовательности легкой цепи могут быть встроены в один и тот же экспрессионный вектор, или могут быть встроены в разные экспрессионные векторы. В случае использования эукариотических клеток-хозяев, клетки животных, клетки растений или клетки эукариотических микроорганизмов могут быть использованы. Примеры клеток животных могут включать в себя клетки млекопитающих, например, клетки обезьяны COS (Gluzman, Y., Cell (1981) 23, p. 175-182, ATCC CRL-1650), мышиные фибробласты NIH3T3 (ATCC No. CRL-1658) и дигидрофолатредуктаза дефицитные линии (Urlaub, G. and Chasin, L.A., Proc. Natl. Acad. Sci. U.S.A. (1980) 77, p. 4126-4220) клеток яичника китайского хомячка (клетки CHO, ATCC CCL-61). В случае использования прокариотических клеток, их примеры могут включать в себя E. coli и Bacillus subtilis. Целевой ген антитела переносят в эти клетки путем трансформации, и трансформированные клетки культивируют in vitro для получения антитела. Такой способ культивирования может отличаться выходом в зависимости от последовательности антитела. Антитело, которое легко получать в качестве лекарственного средства, может быть выбрано из числа антител, имеющих эквивалентную активность связывания, используя в качестве показателя его выход.
[0055] Примеры изотипа антитела настоящего изобретения могут включать в себя без ограничений IgG (IgG1, IgG2, IgG3 и IgG4), IgM, IgA (IgA1 и IgA2), IgD и IgE. Изотип может представлять собой предпочтительно IgG или IgM, более предпочтительно IgG1 или IgG2.
[0056] Антитело настоящего изобретения может представлять собой антигенсвязывающий фрагмент антитела, имеющий антигенсвязывающий сайт этого антитела, или его модифицированную форму. Фрагмент антитела может быть получен путем обработки антитела протеолитическим ферментом, таким как папаин, или путем экспрессии генно-инженерного гена антитела в соответствующих культивируемых клетках. Среди таких фрагментов антитела, фрагмент, который полностью или частично сохраняет функции, присущие полноразмерной молекуле антитела, может именоваться антигенсвязывающий фрагмент антитела.
[0057] Примеры функций антитела, как правило, могут включать в себя антигенсвязывающую активность, активность, нейтрализующую активность антигена, активность, повышающую активность антигена, активность антителозависимой клеточной цитотоксичности, активность комплементзависимой цитотоксичности и активность комплементзависимой клеточной цитотоксичности. Функция, которой обладает антигенсвязывающий фрагмент антитела настоящего изобретения, представляет собой Orai1-связывающую активность, и, предпочтительно, активность ингибирования активации T-лимфоцитов, более предпочтительно, активность ингибирования продуцирования IL-2 и/или интерферона γ T-лимфоцитами.
[0058] Примеры фрагмента антитела могут включать в себя Fab, F(ab')2, Fv, одноцепочечный Fv (scFv), содержащие тяжелую и легкую цепь Fvs, соединенные через соответствующий линкер, диатела, линейные антитела и мультиспецифичные антитела, образованные из фрагментов антител. Фрагмент антитела также включает в себя Fab', который представляет собой моновалентный фрагмент вариабельных областей антитела, полученный путем обработки F(ab')2 в восстанавливающих условиях.
[0059] Антитело настоящего изобретения может представлять собой мультиспецифичное антитело, обладающее специфичностью в отношении, по меньшей мере, двух различных типов антигенов. Такая молекула обычно связывается с двумя типами антигенов (т.е. биспецифичное антитело). Термин «мультиспецифичное антитело» в соответствии с настоящим изобретением включает в себя антитело, обладающее специфичностью в отношении большего количества типов (например, 3 типов) антигенов.
[0060] Мультиспецифичное антитело настоящего изобретения может представлять собой антитело, состоящее из полноразмерного антитела, или может представлять собой фрагмент такого антитела (например, F(ab')2 биспецифичное антитело). Биспецифичное антитело может быть получено путем соединения тяжелой и легкой цепей (HL пары) двух типов антител, или может быть получено путем слияния гибридом, продуцирующих различные моноклональные антитела, для получения слитых клеток, продуцирующих биспецифичное антитело (Millstein et al., Nature (1983) 305, p. 537-539).
[0061] Антитело настоящего изобретения может представлять собой одноцепочечное антитело (одноцепочечный Fv, также именуемый «scFV»). scFV получают путем связывания вариабельных областей тяжелой и легкой цепей антитела через полипептидный линкер (Pluckthun, The Pharmacology of Monoclonal Antibodies, 113 (Rosenberg and Moore ed., Springer Verlag, New York, p. 269-315 (1994); and Nature Biotechnology (2005), 23, p. 1126-1136). Кроме того, biscFv фрагмент, полученный путем связывания двух scFVs через полипептидный линкер, также может быть использован в качестве биспецифичного антитела.
[0062] Способ получения scFV хорошо известен из уровня техники (см., например, Патенты США Nos. 4946778, 5260203, 5091513 и 5455030). В этом scFV вариабельная область тяжелой цепи и вариабельная область легкой цепи соединены через линкер, предпочтительно, полипептидный линкер, который не позволяет им образовывать конъюгат (Huston, J.S. et al., Proc. Natl. Acad. Sci. U.S.A. (1988), 85, p. 5879-5883). Вариабельная область тяжелой цепи и вариабельная область легкой цепи в scFV могут быть получены из различных антител. Например, произвольный одноцепочечный пептид, состоящий из 12-19 остатков используется в качестве полипептидного линкера, который соединяет эти вариабельные области.
[0063] Для того чтобы получить ДНК, кодирующую scFV, каждая часть ДНК, кодирующая целую или желаемую аминокислотную последовательность в последовательностях ДНК, кодирующих тяжелую цепь или вариабельную область тяжелой цепи антитела, и ДНК, кодирующих легкую цепь или вариабельную область легкой цепи антитела, используется в качестве матрицы и амплифицируется при помощи ПЦР с использованием праймеров, фланкирующих оба конца этой матрицы. Затем ДНК, кодирующую полипептидный линкер, дополнительно амплифицируют в комбинации с парой праймеров, фланкирующих оба конца этой ДНК, таким образом, чтобы полученнный фрагмент мог быть соединен своими концами с ДНК тяжелой и легкой цепей, соответсвенно.
[0064] После получения ДНК, кодирующей scFv, экспрессионный вектор, содержащий ДНК, и хозяин, трансформированный этим экспрессионным вектором, могут быть получены стандартными способами. Кроме того, хозяин может быть использован для получения scFv в соответствии со стандартными способами. Эти фрагменты антитела могут быть продуцированы хозяином таким же образом, как описано выше путем получения и экспрессии гена.
[0065] Антитело настоящего изобретения может быть мультимеризировано, чтобы таким образом повысить его аффинность к антигену. Антитела одного и того же типа могут быть мультимеризированы, или несколько антител, распознающих несколько эпитопов, соответственно, одного и того же антигена могут быть мультимеризированы. Примеры способа мультимеризации этих антител могут включать в себя связывание двух scFvs с CH3 доменом IgG, связывание их со стрептавидином и введение мотива «спираль-поворот-спираль».
[0066] Антитело настоящего изобретения может представлять собой поликлональное антитело, которое представляет собой смесь нескольких типов анти-Orai антител, отличающихся по аминокислотной последовательности. Один пример поликлонального антитела может включать в себя смесь нескольких типов антител, отличающихся по CDRs. Антитело, полученное путем культивирования смеси клеток, продуцирующих различные антитела, с последующей очисткой от культуры, может быть использовано в качестве такого поликлонального антитела (см. WO2004/061104).
[0067] Антитело, конъюгированное с любой из различных молекул, таких полиэтиленгликоль (ПЭГ), также может быть использовано в качестве модифицированной формы антитела.
[0068] Антитело настоящего изобретения также может представлять собой любой из конъюгатов, образованный этими антителами с другими лекарственными препаратами (иммуноконъюгаты). Примеры такого антитела могут включать в себя антитело, конъюгированное с радиоактивным материалом или химическим соединением, обладающим фармакологическим действием (Nature Biotechnology (2005) 23, p. 1137-1146).
[0069] Полученное антитело может быть очищено до однородного состояния. Обычные способы разделения и очистки белков могут быть использованы для разделения и очистки антител. Антитело может быть отделено и очищено при помощи соответствующим образом выбранного или комбинированного подхода(ов), например, колоночной хроматографии, фильтрации через фильтр, ультрафильтрации, высаливания, диализа, препаративного электрофореза в полиакриламидном геле и/или изоэлектрического фокусирования (Strategies for Protein Purification and Characterization: A Laboratory Course Manual, Daniel R. Marshak et al. eds., Cold Spring Harbor Laboratory Press (1996); and Antibodies: A Laboratory Manual. Ed Harlow and David Lane, Cold Spring Harbor Laboratory (1988)), хотя способ разделения и очистки не ограничивается вышеперечисленными способами.
[0070] Примеры такой хроматографии могут включать в себя аффинную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, гельфильтрационную хроматографию, хроматографию с обращенной фазой и адсорбционную хроматографию. Эти хроматографические подходы могут быть осуществлены с использованием жидкостной хроматографии, такой как ВЭЖХ или ЖХБР. Примеры колонки для использования в аффинной хроматографии могут включать в себя колонки с белком A и колонки с белком G. Примеры колонок с белком A могут включать в себя Hyper D, POROS, и Sepharose F.F. (Pharmacia Corp.). Также антитело может быть очищено с использованием его активности связывания с антигеном, используя антиген, иммобилизованный на носителе.
[0071]
3. Анти-Orai1 антитело, содержащее лекарственный препарат
Антитело, нейтрализующее биологическую активность Orai1, может быть получено из числа анти-Orai1 антител, полученных способами, описанными в предыдущем параграфе «2. Получение анти-Orai1 антитела». Такое антитело, нейтрализующее биологическую активность Orai1, может ингибировать биологическую активность Orai1 in vivo, т.е. активацию активируемых высвобождением кальция кальциевых каналов (CRAC каналов) клеток, сверхэкспрессирующих Orai1, типичными представителями которых являются T-лимфоциты и тучные клетки, и, таким образом, оно может фармацевтически применяться в качестве терапевтического и/или профилактического агента для заболеваний, вызываемых активацией CRAC каналов этих клеток.
[0072] Заболевания и состояния, вызываемые активацией CRAC каналов, включают в себя отторжение трансплантата, иммунологические заболевания, аллергические заболевания, воспалительные заболевания, тромбоз и рак и т.д.
[0073] Примеры лечения и/или предотвращения отторжения трансплантата включают в себя лечение и/или предотвращение реакции отторжения и реакции по типу «хозяин против трансплантата» при трансплантации органа или ткани, таких как сердце, почка, печень, костный мозг или кожа, и реакцию по типу «трансплантат против хозяина», вызванную трансплантацией гемопоэтических клеток (костного мозга, периферической крови, пуповинной крови и т.д.).
[0074] Примеры иммунных заболеваний включают в себя заболевания соединительной ткани или опорно-двигательного аппарата (ревматоидный артрит, анкилозирующий спондилит, системная красная волчанка, склеродермия, полимиозит, дерматомиозит и т.д.), гематологические заболевания (апластическая анемия, идиопатическая тромбоцитопеническая пурпура и т.д.), заболевания желудочно-кишечного тракта (болезнь Крона, неспецифический язвенный колит и т.д.), неврологические заболевания (рассеянный склероз, миастения гравис и т.д.), офтальмологические заболевания (увеит и т.д.), сосудистые заболевания (болезнь Бехчета, гранулематоз Вегенера и т.д.), кожные заболевания (псориаз, пузырчатка, лейкодермия и т.д.), эндокринные заболевания (сахарный диабет первого типа, аутоиммунный тиреоидит, болезнь Грейвcа, тиреоидит Хашимото и т.д.).
[0075] Примеры аллергических заболеваний включают в себя атопический дерматит, астму, анафилаксию, анафилактоидную реакцию, алиментарную аллергию, ринит, отит среднего уха, лекарственную аллергию, аллергическую реакцию на укусы насекомых, аллергическую реакцию на растения, аллергию на латекс, конъюнктивит и крапивницу.
[0076] Примеры воспалительных заболеваний включают в себя воспалительные заболевания почек (гломерулонефрит, нефроз и т.д.), воспалительные заболевания легких (хроническое обструктивное заболевание легких, муковисцидоз, интерстициальная пневмония и т.д.), воспалительные заболевания кишечника (неспецифический язвенный колит, илеит и т.д.), воспалительные заболевания печени (аутоиммунный гепатит, вирусный гепатит и т.д.), воспалительные заболевания сердца (миокардит, ишемическая болезнь сердца, атеросклероз и т.д.), воспалительные заболевания кожи (контактный дерматит, экзема и т.д.), воспалительные заболевания глаз (трахома, эндофтальмит и т.д.), воспалительные заболевания центральной нервной системы (менингит, энцефаломиелит, аутоиммунный энцефалит и т.д.), воспалительные заболевания суставов (артрит, остеоартрит и т.д.) и системные воспаления (сепсис, кровотечения, гиперчувствительность, симптомы шока, связанные с химиотерапией рака или тому подобным, и т.д.).
[0077] Примеры случаев, при которых антитромбоцитарная или антитромботическая активность используется для лечения и/или предотвращения заболевания, включают в себя инфаркт миокарда, инсульт, ишемическую болезнь сердца и тромбоз.
[0078] Примеры лечения рака включают в себя рак молочной железы, рак легкого, рак кожи и лейкоз.
[0079] Другие примеры заболеваний, которые можно лечить при помощи антитела настоящего изобретения включают в себя: патологические состояния, связанные с тучными клетками и базофилами, такие как тучноклеточный лейкоз, мастоцитоз, базофильный лейкоз и эндометриоз; и миопатию с тубулярными агрегатами, синдром Шторморкена, ревматоидный артрит, анкилозирующий спондилоартрит и атопический дерматит, которые развиваются, или риск развития которых увеличивается при генетической гиперфункции CRAC каналов.
[0080] Примеры анти-Orai1 антитела в качестве лекарственного средства могут включать в себя гуманизированные антитела, полученные из антитела R118 и/или антитела R198, и их формы со сконструированными CDR.
[0081] Нейтрализующая активность in vitro анти-Orai1 антитела в отношении биологической активности Orai1 может быть измерена на основе, например, ингибирующей активности в отношении активации T-лимфоцитов, экспрессирующих Orai1. Например, анти-Orai1 антитело добавляют в различных концентрациях к полученной из T-лимфоцитов человека клеточной линии клеток Юрката, и ингибирующая активность в отношении высвобождения IL-2 из клеток Юрката, стимулированных ФМА и A23187, может быть измерена. Также анти-Orai1 антитело добавляют в различных концентрациях к мононуклеарным клеткам периферической крови человека (МКПК), и ингибирующая активность в отношении высвобождения IL-2 и интерферона γ из МКПК, стимулированных ФМА и A23187, может быть измерена.
[0082] Экспериментально доказанным и широко признанным фактом считается, что клетками, ответственными за развитие реакции по типу «трансплантат против хозяина», являются T-лимфоциты. Когда трансплантированные T-лимфоциты распознают реципиента в качестве чужеродного вещества, самовоспроизводство, основанное на интерлейкине 2 (IL-2), продуцируемом ими самими, и высвобождение воспалительных цитокинов, таких как интерферон гамма (ИФН-γ), путем активации вызывает системную иммунологическую воспалительную реакцию, приводящую к развитию реакции по типу «трансплантат против хозяина». Таким образом, ингибирование продуцирования этих цитокинов приводит к предотвращению или лечению реакции по типу «трансплантат против хозяина», при этом пригодность анти-Orai1 антитела в качестве терапевтического лекарственного средства может быть определена с использованием его ингибирующей активности в качестве показателя.
[0083] Предпочтительные примеры антитела настоящего изобретения могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 50%, составляет 80 нг/мл или меньше. Более предпочтительные примеры этого антитела могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 50%, составляет 10 нг/мл или меньше. Антитело настоящего изобретения также включает в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 80%, составляет 60000 нг/мл или меньше, и антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из клеток Юрката, обработанных ФМА и A23187, ингибируется на 80%, составляет 200 нг/мл или меньше.
[0084] Альтернативные предпочтительные примеры антитела настоящего изобретения могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество IL-2, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 100 нг/мл или меньше. Более предпочтительные примеры этого антитела могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 20 нг/мл или меньше. Антитело настоящего изобретения также включает в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 80%, составляет 17000 нг/мл или меньше, и антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество IL-2, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 80%, составляет 400 нг/мл или меньше.
[0085] Другие альтернативные предпочтительные примеры антитела настоящего изобретения могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 800 нг/мл или меньше. Более предпочтительные примеры этого антитела могут включать в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 50%, составляет 40 нг/мл или меньше. Антитело настоящего изобретения также включает в себя антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 80%, составляет 300000 нг/мл или меньше, и антитело или антигенсвязывающий фрагмент этого антитела, где концентрация, при которой количество ИФН-γ, высвобождаемое из МКПК человека, обработанных ФМА и A23187, ингибируется на 80%, составляет 2000 нг/мл или меньше.
[0086] Терапевтическое или профилактическое действие анти-Orai1 антитела на реакцию по типу «трансплантат против хозяина» может быть подтверждено in vivo у лабораторного животного, например, путем измерения ингибирования снижения массы тела у мышей с трансплантированными МКПК человека в модели реакции по типу «трансплантат против хозяина». В частности мышей NOG или NSG, которые являются мышами с тяжелым комбинированным иммунодефицитом, облучают рентгеновскими лучами в дозе 2,0 Гр. На следующий день 200 мкл раствора ФСБ, содержащего от 15000000 до 50000000 клеток МКПК человека на мл, трансплантировали каждой мыши через хвостовую вену. Группу облученных рентгеновскими лучами мышей, не получавших МКПК человека, использовали в качестве группы, в которой не было реакции по типу «трансплантат против хозяина», а группу мышей с трансплантированными МКПК человека, получавших только раствор инертного вещества для антитела, использовали в качестве группы, в которой имела место реакция по типу «трансплантат против хозяина». Перед или после трансплантации МКПК человека, анти-Orai1 антитело вводили или внутривенно в хвостовую вену или внутрибрюшинно для измерения ингибирующего действия на снижение массы в группе, в которой имела место реакция по типу «трансплантат против хозяина».
[0087] Кроме того, терапевтическое или профилактическое влияние анти-Orai1 антитела на различные заболевания также может быть подтверждено следующими системами оценки in vivo с использованием мышей, в чей геном встроен ген Orai1 человека.
[0087] Для влияния на дерматит, раствор белкового антигена внутрибрбшинно или внутривенно вводили мышам, а через две недели этот антиген вводили им в качестве бустера. Затем тот же раствор белкового антигена, как описано выше, повторно наносили на ухо или спину от 3 до 6 раз с интервалами от 3 дней до 2 недель, чтобы вызвать дерматит. Влияние на дерматит оценивали путем сравнения толщины ушной раковины, видимых признаков дерматита на спине, концентраций антитела, цитокинов или биомаркеров сыворотки в крови или тканях, и активности роста, способности продуцировать цитокины или поверхностных антигенов клеток, полученных из кожи, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга, между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0088] Для влияния на зуд, мазь, содержащую клещевой антиген, повторно наносили на ушную раковину или спину от 3 до 6 раз с интервалами от 3 дней до 2 недель, или гаптен наносили на ушную раковину, область живота или спину с частотой от одного раза в сутки до одного раза в неделю, или вызывающее зуд вещество вводили внутрикожно в ушную раковину, подкожно в спину или интратекальным способом. Зудящее действие, индуцируемое таким образом, количественно определяли путем измерения количества событий почесывания, используя для этого магниты, прикрепленные к тыльной поверхности обеих лап мышей, и аппарат для измерения зудящего действия, или путем изучения паталогических признаков на коже, в периферической крови или тканях спинного мозга. Влияние на зуд оценивали путем сравнения полученных результатов между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0090] Для влияния на псориаз, имиквимод наносили на обе стороны или одну сторону ушной раковины и на бритую спину, или суспензию зимозана вводили внутрибрюшинно, или цитокин, такой как IL-23, вводили внутрикожно в одну сторону ушной раковины мыши. Псориатический дерматит, индуцированный таким способом, количественно определяли путем изучения массы и толщины участка воспаления, активности миелопероксидазы нейтрофилов, инфильтрированных в этот участок, анализа с использованием проточной цитометрии инфильтрированных клеток, генетического анализа, измерения концентрации цитокинов и т.д. Влияние на псориаз оценивали путем сравнения полученных результатов между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0091] Для влияния на рассеянный склероз, раствор миелинового олигодендроцитарного гликопротеина или его частичного пептидного антигена эмульгировали путем смешивания с водным раствором полного адъюванта Фрейнда в равных количествах. Эту эмульсию внутримышечно вводили в бок или в область живота мыши. Затем в хвостовую вену вводили водный раствор коклюшного токсина. Через несколько дней раствор коклюшного токсина снова вводили в хвостовую вену, чтобы индуцировать экспериментальный энцефаломиелит. Симптомы паралича, которые развивались через 1-2 недели эксперимента и распространялись от хвоста к задним конечностям и передним конечностям, оценивали путем наружного наблюдения. Влияние на рассеянный склероз оценивали путем сравнения полученных результатов между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0092] Для влияния на артрит, эмульсию, полученную путем смешивания бычьего коллагена типа II с полным адъювантом Фрейнда, внутрикожно вводили в основание хвоста мыши. Через 2-3 недели осуществляли такое же введение, как указано выше. Затем влияние на артрит оценивали путем сравнения степени опухания суставов, концентрации антитела, цитокинов или биомаркеров сыворотки в крови или тканях, активности роста, способности продуцировать цитокины или поверхностных антигенов клеток, полученных из кожи, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга, и т.д. между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0093] Для влияния на колит, тринитробензолсульфоновую кислоту вводили в кишечник мышей, которых не кормили в течение 24 часов, или мышей, которые имели свободный доступ к водному раствору, содержащему от 1 до 10% декстрана сульфата натрия, из поилки в течение периода времени от 4 суток до 2 недель, или CD4+CD25-CD45RBhi T-лимфоциты, собранные и очищенные из лимфатических узлов и селезенки мышей, в чей геном встроен ген Orai1 человека, внутрибрюшинно трансплантировали Rag2-/- мышам. Влияние на колит, индуцированный таким способом, оценивали путем сравнения массы тела в течение периода наблюдения, степени утолщения кишечного тракта, числа и размера полипов и патологических признаков, что определяли при помощи аутопсии после завершения эксперимента, концентрации антитела, цитокинов или биомаркеров сыворотки в крови или тканях, активности роста, способности продуцировать цитокины или поверхностных антигенов клеток, полученных из кишечного тракта, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга, и т.д. между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0094] Для влияния на системную красную волчанку, яичный альбумин или другой нестимулирующий антиген внутрибрюшинно или подкожно вводили максимум 15 раз с интервалами от 5 до 10 дней, чтобы вызвать симптомы, сходные с симптомами системной красной волчанки. Влияние на системную красную волчанку оценивали путем сравнения концентрации антитела, цитокинов или биомаркеров сыворотки в крови или тканях, отбираемых в течение периода наблюдения, титра антитела или концентрации биомаркера в моче, симптомов, индуцированных у не получавших лечение мышей путем трансплантации клеток, полученных из таких органов, как селезенка или лимфатические узлы, собранных после завершения эксперимента, и т.д. между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0095] Для влияния на гепатит, D-галактозамин внутрибрюшинно вводили мышам или в чистом виде, или в комбинации с липополисахаридом, или конканавалин А внутривенно вводили в чистом виде в хвостовую вену. Влияние на гепатит, индуцированный таким способом, оценивали путем сравнения концентрации АсТ и АлТ в крови, отбираемой в интервале времени от 1 часа до 1 недели после введения вызывающего гепатит вещества, и гистопатологических состояний поражений печени между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0096] Для влияния на выживаемость трансплантата при трансплантации клеток костного мозга, или профилактического действия на реакцию по типу «трансплантат против хозяина», клетки костного мозга, полученные из бедренной кости или большой берцовой кости мышей, в чей геном встроен ген Orai1 человека, трансплантировали облученным рентгеновскими лучами мышам-реципиентам или в чистом виде или в комбинации с клетками селезенки, полученными из селезенок мышей, в чей геном встроен ген Orai1 человека. Влияние на выживаемость трансплантата или профилактическое действие оценивали путем сравнения изменения массы тела или выживаемости в течение 4-16 недель, концентрации антитела, цитокинов, биомаркеров сыворотки в крови ли тканях в конце эксперимента, и поверхностных антигенов, активности роста или способности продуцировать цитокины у клеток, полученных из кишечного тракта, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга, между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0097] Для влияния на выживаемость трансплантата при трансплантации органа, кожу, полученную с основания хвоста мыши-хозяина, прикрепляли к кожно-туловищной мышце, разрезая кожу на спине мыши, в чей геном встроен ген Orai1 человека. Размер трансплантата и его статус приживления оценивали в течение 4-16 недель после трансплантации. Влияние на выживаемость трансплантата оценивали путем сравнения результатов между группой, получавшей анти-Orai1 антитело, и группой, не получавшей анти-Orai1 антитело.
[0098] Полученное таким образом антитело, которое нейтрализует биологическую активность Orai1, применимо в качестве лекарственного средства, в частности, антитела для предотвращения или лечения отторжения трансплантата, иммунологических заболеваний, аллергических заболеваний, воспалительных заболеваний, тромбоза или рака и т.д.
[0099] В качестве одного примера, анти-Orai1 антитело может быть введено или само по себе, или в комбинации с, по меньшей мере, одним дополнительным терапевтическим агентом, для лечения или предотвращения отторжения трансплантата, иммунологических заболеваний, аллергических заболеваний, воспалительных заболеваний, тромбоза или рака. Примеры дополнительного терапевтического агента, который может вводиться в комбинации с анти-Orai1 антителом, могут включать в себя без ограничений антифолаты, ингибиторы кальциневрина, стероиды коры надпочечников, антитимоцитарные глобулины, антиметаболиты нуклеиновых кислот, ингибиторы синтеза нуклеиновых кислот, биопрепараты, направленно воздействующие на антигены клеточной поверхности, и биопрепараты, направленно воздействующие на цитокины или рецепторы цитокинов.
[0100] Конкретные примеры этих терапевтических агентов могут включать в себя метотрексат в качестве антифолата; циклоспорин и такролимус в качестве ингибиторов кальциневрина; метилпреднизолон в качестве стероида коры надпочечников; циклофосфамид в качестве ингибитора синтеза нуклеиновых кислот; цетбулин, лимфоглобулин и тимоглобулин в качестве антитимоцитарных глобулинов; мофетила микофенолат в качестве антиметаболита нуклеиновых кислот; алемтузумаб и ритуксимаб в качестве биопрепарата, направленно воздействующего на антигены клеточной поверхности; и инфликсимаб, этанерцепт и ритуксимаб в качестве биопрепарата, направленно воздействующего на цитокины или рецепторы цитокинов.
[0101] В зависимости от условий отторжения трансплантата, иммунологических заболеваний, аллергических заболеваний, воспалительных заболеваний, тромбоза или рака, или от цели их лечения и/или предотвращения, может вводиться 2, 3 или более типов дополнительных терапевтических агентов, и эти дополнительные терапевтические агенты могут находиться в составе одного и того же препарата и вводиться в одно и то же время. Дополнительный терапевтический агент и анти-Orai1 антитело могут находиться в составе одного и того же препарата и вводиться в одно и то же время. В качестве альтернативы, анти-Orai1 антитело и дополнительный терапевтический агент могут находиться в составе разных препаратов и вводиться в одно и то же время. Кроме того, дополнительный терапевтический агент и анти-Orai1 антитело могут вводиться раздельно в разное время. В частности, терапевтический агент, содержащий анти-Orai1 антитело или антигенсвязывающий фрагмент этого антитела, в качестве активного ингредиента, может вводиться после введения дополнительного терапевтического агента, или дополнительный терапевтический агент может вводиться после введения терапевтического агента, содержащего анти-Orai1 антитело или антигенсвязывающий фрагмент этого антитела, в качестве активного ингредиента. В случае введения для генной терапии, ген белкового терапевтического агента и ген анти-Orai1 антитела могут быть встроены после отдельных промоторных областей или после одной промоторной области и введены в отдельные векторы или в один и тот же вектор.
[0102] Анти-Orai1 антитело или его фрагмент может быть конъюгировано с терапевтическим агентом для получения специализированного конъюгата лекарственного средства, описанного в M.C. Garnet, «Targeted drug conjugates: principles and progress», Advanced Drug Delivery Reviews, (2001) 53, 171-216. Для этой цели применима молекула антитела, а также любой фрагмент антитела, при условии, что этот фрагмент антитела не полностью утрачивает способность распознавать T-лимфоциты. Примеры этого фрагмента могут включать в себя фрагменты Fab, F(ab')2 и Fv. В этом качестве антитело и фрагмент могут быть использованы в настоящем изобретении. Способ конъюгации между анти-Orai1 антителом или фрагментом этого антитела и терапевтическим агентом может представлять собой любой из различных способов, описанных в M.C. Garnet, «Targeted drug conjugates: principles and progress», Advanced Drug Delivery Reviews, (2001) 53, 171-216, G.T. Hermanson, «Bioconjugate Techniques», Academic Press, California (1996), Putnam and J. Kopecek, «Polymer Conjugates with Anticancer Activity», Advances in Polymer Science (1995) 122, 55-123, и т.д. Конкретные примеры таких способов могут включать в себя способ, в котором анти-Orai1 антитело и терапевтический агент являются конъюгированы непосредственно химическим способом, или через спейсер, такой как олигопептид, и способ, в котором анти-Orai1 антитело и терапевтический агент являются конъюгированы через соответствующий носитель лекарственного средства. Примеры носителя лекарственного средства включают в себя липосомы и водорастворимые полимеры. Примеры такого опосредованного этими носителями лекарственного средства способа более конкретно могут включать в себя способ, в котором антитело и терапевтический агент заключены в липосому путем конъюгации липосомы и антитела, и способ, в котором терапевтический агент конъюгирован с водорастворимым полимером (химическим соединением, имеющим молекулярную массу порядка от 1000 до 100000) непосредственно химическим способом, или через спейсер, такой как олигопептид, в то время как антитело конъюгировано с водорастворимым полимером. Конъюгация антитела (или фрагмента) с терапевтическим агентом или носителем лекарственного средства (например, липосомой или водорастворимым полимером) может быть проведена способом, хорошо известным специалистам в данной области техники, например, способом, описанным в G.T. Hermanson, «Bioconjugate Techniques», Academic Press, California (1996), и Putnam and J. Kopecek, «Polymer Conjugates with Anticancer Activity», Advances in Polymer Science (1995) 122, 55-123. Включение терапевтического агента в липосому может быть проведено способом, хорошо известным специалистам в данной области техники, например, способом, описанным в D.D. Lasic, «Liposomes: From Physics to Applications», Elsevier Science Publishers B.V., Amsterdam (1993), и т.д. Конъюгация терапевтического агента с водорастворимым полимером может быть проведена способом, хорошо известным специалистам в данной области техники, например, способом, описанным в D. Putnam and J Kopecek, «Polymer Conjugates with Anticancer Activity», Advances in Polymer Science (1995) 122, 55-123. Конъюгат антитела (или фрагмента) и белкового терапевтического агента (или фрагмента) может быть получен любым из описанных выше способов, а также способами генной инженерии, хорошо известными специалистам в данной области техники.
[0103] Настоящее изобретение также относится к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество анти-Orai1 антитела и фармацевтически приемлемые разбавитель, вспомогательное вещество, солюбилизатор, эмульгатор, консервант и/или добавку.
[0104] Настоящее изобретение также относится к фармацевтической композиции, содержащей терапевтически и/или профилактически эффективное количество анти-Orai1 антитела, терапевтически и/или профилактически эффективное количество, по меньшей мере, одного терапевтического агента и фармацевтически приемлемые разбавитель, вспомогательное вещество, солюбилизатор, эмульгатор, консервант и/или добавку. Примеры терапевтического агента могут включать в себя без ограничений вышеперечисленные антифолаты, ингибиторы кальциневрина, стероиды коры надпочечников, антитимоцитарные глобулины, антиметаболиты нуклеиновых кислот, ингибиторы синтеза нуклеиновых кислот, биопрепараты, направленно воздействующие на антигены клеточной поверхности, и биопрепараты, направленно воздействующие на цитокины или рецепторы цитокинов.
[0105] Предпочтительно, чтобы фармацевтически приемлемые вещества, используемые в фармацевтической композиции настоящего изобретения, были нетоксичными для реципиента это фармацевтической композиции, предпочтительно, это относится к дозе или вводимой концентрации.
[0106] Фармацевтическая композиция настоящего изобретения может содержать фармацевтические материалы для изменения или поддержания значения pH, осмотического давления, вязкости, прозрачности, цвета, изотоничности, стерильности, стабильности, растворимости, длительности высвобождения, всасываемости или впитываемости. Примеры этих фармацевтических материалов могут включать в себя без ограничений следующие вещества: аминокислоты, такие как глицин, аланин, глутамин, аспарагин, аргинин и лизин; противомикробные агенты; антиокисданты, такие как аскорбиновая кислота, сульфат натрия и бисульфит натрия; буферы, такие как фосфатный, цитратный или боратный буферы, растворы бикарбоната натрия и Трис-HCl; наполнители, такие как маннит и глицин; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (ЭДТА); комплексообразующие агенты, такие как кофеин, поливинилпирролидин, β-циклодекстрин и гидроксипропил-β-циклодекстрин; объемообразующие агенты, такие как глюкоза, манноза и декстрин; другие углеводы, такие как моносахариды и дисахариды; красители; корригирующие вещества; разбавители; эмульгаторы; гидрофильные полимеры, такие как поливинилпирролидин; низкомолекулярные полипептиды; солеобразующие противоионы; антисептики, такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота и перекись водорода; растворители, такие как глицерин, пропиленгликоль и полиэтиленгликоль; сахароспирты, такие как маннит и сорбит; суспендирующие агенты; поверхностно-активные вещества, такие как сложный эфир сорбита, полисорбаты, такие как полисорбат 20 и полисорбат 80, тритон, трометамин, лецитин и холестерин; повышающие стабильность вещества, такие как сахароза и сорбит; повышающие эластичность вещества, такие как хлорид натрия, хлорид калия, маннит и сорбит; транспортные агенты; вспомогательные вещества; и/или фармацевтические добавки. Количество этих добавленных фармацевтических материалов предпочтительно составляет от 0,01 до 100 раз, в частности, от 0,1 до 10 раз от массы анти-Orai1 антитела. Подходящая рецептура для фармацевтической композиции в препарате может быть соответствующим образом определена специалистами в данной области техники, в зависимости от подлежащего лечению заболевания, применяемого способа введения и т.д.
[0107] Вспомогательное вещество или носитель в фармацевтической композиции может быть жидким или твердым. Подходящие вспомогательные вещества или носители могут представлять собой другие материалы, обычно используемые в воде для инъекций, физиологическом растворе, искусственной спинномозговой жидкости и парентеральном введении. Нейтральный физиологический раствор или физиологический раствор, содержащий сывороточный альбумин, может быть использован в качестве носителя. Фармацевтическая композиция может содержать Трис-буфер, имеющий значение pH от 7,0 до 8,5, ацетатный буфер, имеющий значение pH от 4,0 до 5,5, или цитратный буфер, имеющий значение pH от 3,0 до 6,2. Эти буферы также могут содержать сорбит или другие химические соединения. Примеры фармацевтической композиции настоящего изобретения могут включать в себя фармацевтическую композицию, содержащую анти-Orai1 антитело, и фармацевтическую композицию, содержащую анти-Orai1 антитело и, по меньшей мере, один терапевтический агент. Фармацевтическую композицию настоящего изобретения получают в виде сублимированного продукта или жидкости, в форме лекарственного препарата, имеющего выбранную рецептуру и требуемую степень чистоты. Фармацевтическая композиция, содержащая анти-Orai1 антитело, или фармацевтическая композиция, содержащая анти-Orai1 антитело и, по меньшей мере, один терапевтический агент, также может быть получена в виде сублимированного продукта с использованием подходящего вспомогательного вещества, такого как сахароза.
[0108] Фармацевтическая композиция настоящего изобретения также может быть получена для парентерального введения, или может быть получена для всасывания в пищеварительном тракте при пероральном способе введения. Рецептура и концентрация препарата может быть определена в соответствии со способом введения. Чем выше аффинность анти-Orai1 антитела, содержащегося в фармацевтической композиции настоящего изобретения, по отношению к Orai1, т.е. чем ниже константа диссоциации (значение Kd) для Orai1, тем ниже доза анти-Orai1 антитела, которая может имметь эффективность у человека. Таким образом, доза фармацевтической композиции настоящего изобретения для человека также может быть определена на основе этого результата. Доза для введения человеческого анти-Orai1 антитела человеку составляет приблизительно от 0,1 до 100 мг/кг, которая может быть введена один раз в течение периода времени от 1 до 180 дней.
[0109] Примеры формы фармацевтической композиции настоящего изобретения могут включать в себя инъекции, включая внутривенные капельницы, суппозитории, трансназальные препараты, сублингвальные препараты и препараты, всасывающиеся через кожу.
[0110] Хотя большинство одобренных для медицинского применения препаратов антител вводится внутривенно, подкожное введение часто является более предпочтительным в медицинской практике. В этом случае объем ограничивается 1,0-1,5 мл. Таким образом, в зависимости от дозы, требуется раствор антитела, имеющий высокую концентрацию. Однако так как более высокая концентрация повышает вязкость раствора лекарственного средства, может возникнуть ситуация, при которой раствор лекарственного средства не сможет быть введен при помощи иглы для инъекции, имеющей стандартно используемую толщину, по причине своей высокой вязкости. Иными словами, в случае выбора инъекции в качестве формы фармацевтической композиции, низкая вязкость является важным свойством, которому следует уделять наибольшее внимание. Таким образом, при выборе подходящего антитела в качестве показателя может использоваться вязкость.
Примеры
[0111] Далее настоящее изобретение будет подробно описано со ссылкой на Примеры. Однако не предполагается, что настоящее изобретение ограничивается ими. Каждая процедура, связанная с манипуляциями с генами, в приведенных ниже Примерах проводилась в соответствии со способами, описанными в «Molecular Cloning» (Sambrook, J., Fritsch, E.F. and Maniatis, T., Cold Spring Harbor Laboratory Press, 1989), или с использованием коммерчески доступных реагентов или наборов реагентов в соответствии с инструкциями производителя этих коммерческих продуктов, если не указано иное.
[0112]
Пример 1. Получение крысиного анти-человеческий Orai1 антитела
1)-1 Иммунизация
1)-1-1 Конструирование вектора, экспрессирующего Orai1 человека, (pcDNA3.1-hOrai1)
Клон с полной ОРС (Clone No. IOH40869, Life Technologies Corp.), клонированной в pDONR221 (Life Technologies Corp.) приобретали и использовали в качестве последовательности гена Orai1 человека. Эта последовательность гена показана в SEQ ID NO: 1 в Списке последовательностей, и аминокислотная последовательность показана в SEQ ID NO: 2 в Списке последовательностей. Также pcDNA3.1(+) модифицировали в принимающий вектор с использованием системы преобразования вектора Gateway (Life Technologies Corp.) для получения pcDNA3.1-DEST. Последовательность гена рекомбинировали с pcDNA3.1-DEST с последовательностью att с использованием смеси Gateway LR Clonase Enzyme (Life Technologies Corp.) для получения вектора, экспрессирующего Orai1 человека, pcDNA3.1-hOrai1. Набор реагентов EndoFree Plasmid Giga (Qiagen N.V.) использовали для крупномасштабного получения вектора, экспрессирующего Orai1 человека.
[0113]
1)-1-2 Иммунизация крыс
Самок крыс линии WKY/Izm (Japan SLC, Inc.) использовали для иммунизации. Сначала, обе задние конечности каждой крысы предварительно обрабатывали гиалуронидазой (Sigma-Aldrich Corp.), и затем pcDNA3.1-hOrai1 внутримышечно вводили в эти области. Затем проводили электропорацию in vivo в этих областях, используя ECM830 (BTX) и двухигольный электрод. После повторения аналогичной электропорации in vivo один раз каждые две недели, от крыс получали лимфатические узлы, и использовали их в получении гибридом.
[0114]
1)-2 Получение гибридом
Клетки лимфатического узла при действии электрического тока сливали с клетками мышиной миеломы SP2/0-ag14, используя систему для получения гибридом Hybrimune (Cyto Pulse Sciences Inc.). Слитые клетки разбавляли селективной средой ClonaCell-HY Selection Medium D (StemCell Technologies Inc.) и культивировали. Обнаруженные колонии гибридом выделяли для получения моноклональных гибридом. Каждую выделенную колонию гибридомы культивировали, и полученный супернатант культуры гибридом использовали для выявления гибридомы, продуцирующей анти-человеческий Orai1 антитело.
[0115]
1)-3 Первичный скрининг при помощи ИФА на клетках
1)-3-1 Получение клеток, экспрессирующих ген антигена, для ИФА на клетках
Клетки HEK293 получали в концентрации 7,5×105 клеток/мл в среде DMEM, содержащей 10% ФБС. Клетки трансфицировали pcDNA3.1-hOrai1 или pcDNA3.1-DEST в качестве контроля в соответствии с процедурами трансфекции, используя липофектамин 2000, распределяли по 50 мкл на лунку в 96-луночный планшет (Corning Inc.) и культивировали при температуре 37°C в течение двух ночей в атмосфере с 5% содержанием CO2 в среде DMEM, содержащей 10% ФБС. Полученные трансфицированные клетки использовали в прикрепленном состоянии в ИФА на клетках.
[0116]
1)-3-2 ИФА на клетках
После удаления культурального супернатанта из клеток HEK293, трансфицированных экспрессионным вектором, полученным в Примере 1)-1-1, каждый супернатант культуры гибридом добавляли к трансфицированным pcDNA3.1-hOrai1 или pcDNA3.1-DEST клеткам HEK293, и планшет инкубировали при температуре 4°C в течение 1 часа. Клетки в лунках один раз промывали ФСБ, содержащим 5% ФБС. Затем к ним добавляли полученное от кролика анти-крысиный IgG антитело, конъюгированное с пероксидазой, (Sigma-Aldrich Corp.), разбавленное в 500 раз ФСБ, содержащим 5% ФБС, и планшет инкубировали при температуре 4°C в течение 1 часа. Клетки в лунках 6 раз промывали ФСБ, содержащим 5% ФБС. Затем к ним добавляли раствор хромогена ОФД (раствор ОФД (о-фенилендиамина дигидрохлорид (Wako Pure Chemicals Industries, Ltd.) и H2O2, растворенные в концентрациях 0,4 мг/мл и 0,6% (по объему), соответственно, в 0,05 M тринатрийцитрате и 0,1 M динатрия гидрофосфат додекагидрате, pH 4,5)) в количестве 25 мкл на лунку. Цветную реакцию проводили при периодическом помешивании и останавливали путем добавления 1 M HCl в количестве 25 мкл на лунку. Затем измеряли поглощение при длине волны 490 нм, используя планшет-ридер (ENVISION; PerkinElmer, Inc.). Для выбора гибридомы, продуцирующей антитело, специфично связывающееся с Orai1 человека, экспрессированным на поверхности клеточной мембраны, гибридомы, которые давали культуральный супернатант, имеющий более высокое значение поглощения для клеток HEK293, трансфицированных вектором, экспрессирующим pcDNA3.1-hOrai1, по сравнению с контрольными клетками HEK293, трансфицированными pcDNA3.1-DEST, отбирали в качестве гибридом, положительных по анти-человеческий Orai1 антителу.
[0117]
1)-4 Вторичный скрининг при помощи проточной цитометрии
1)-4-1 Получение клеток, экспрессирующих ген антигена, для анализа при помощи проточной цитометрии
Клетки HEK293T инокулировали в количестве 5×104 клеток/см2 в колбу на площадь 225 см2 и культивировали в течение ночи при температуре 37°C в течение двух ночей в атмосфере с 5% содержанием CO2 в среде DMEM, содержащей 10% ФБС. На следующий день клетки HEK293T трансфицировали pcDNA3.1-hOrai1 или pcDNA3.1-DEST в качестве контроля, используя липофектамин 2000, и затем культивировали при температуре 37°C в течение двух ночей в атмосфере с 5% содержанием CO2. На следующий день клетки HEK293T, трансфицированные экспрессионным вектором, обрабатывали TrypLE Express (Life Technologies Corp.), промывали DMEM, содержащей 10% ФБС, и затем суспендировали в ФСБ, содержащем 5% ФБС. Полученную суспензию клеток использовали в анализе при помощи проточной цитометрии.
[0118]
1)-4-2 Анализ при помощи проточной цитометрии
Специфичность связывания с Orai1 человека для антитела, продуцируемого каждой гибридомой, подтвержденную как положительная при помощи ИФА на клетках в Примере 1)-3, дополнительно подтверждали при помощи проточной цитометрии. Каждую суспензию клеток HEK293T, полученную в Примере 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T, трансфицированные pcDNA3.1-DEST, или клетки HEK293T, трансфицированные pcDNA3.1-hOrai1, суспендировали путем добавления супернатанта культуры гибридом и инкубировали при температуре 4°C в течение 1 часа. Клетки один раз промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-крысиного IgG, конъюгированного с ФИТЦ, (Sigma-Aldrich Corp.), разбавленного в 500 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 1 часа. Клетки три раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 2 мкл/мл 7-аминоактиномицина D (Molecular Probes, Inc.), с последующей детекцией при помощи проточного цитометра (FC500; Beckman Coulter Inc.). Данные анализировали с использованием программы Flowjo (Tree Star Inc.). После исключения 7-аминоактиномицин D положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ наносили на график в виде гистограммы. Гибридомы, которые дали образец, имеющий сдвиг к более сильной интенсивности флуоресценции на гистограмме клеток HEK293, трансфицированных pcDNA3.1-hOrai1, по сравнению с гистограммой интенсивности флуоресценции контрольных клеток HEK293T, трансфицированных pcDNA3.1-DEST, выделяли в виде гибридом, продуцирующих анти-человеческий Orai1 антитело. В результате было получено 225 гибридом, имеющих значительный сдвиг.
[0119]
1)-5 Изотипирование антитела
Антитела R118 и R198, которые предположительно активно связываются с Orai1 человека, были выбраны из крысиных анти-человеческий Orai1 антител, продуцируемых гибридомами, полученными в пункте 1)-4, и изотипированы. Их изотипы определяли с использованием тестового набора реагентов для изотипирования крысиных моноклональных антител (AbD Serotec). В результате было установлено, что оба крысиных анти-человеческий Orai1 антитела R118 и R198 представляют собой IgG2a и κ цепи.
[0120]
1)-6 Получение крысиного анти-человеческий Orai1 антитела
Каждое крысиное анти-человеческий Orai1 моноклональное антитело очищали из супернатанта культуры гибридом.
[0121] Сначала гибридому, продуцирующую R118 и R198, выращивали в достаточном количестве с селективной средой ClonaCell-HY Selection Medium E. Затем среду заменяли средой Hybridoma SFM (Life Technologies Corp.), дополненной 20% ФБС со сверхнизким содержанием IgG (Life Technologies Corp.), после чего культивировали в течение 5 суток. Этот культуральный супернатант собирали и стерилизовали через фильтр с размером пор 0,45 мкм.
[0122] Антитело очищали из супернатанта культуры гибридом при помощи аффинной хроматографии на белке G (при температуре от 4 до 6°C) в один этап. Этап замены буфера после очистки при помощи аффинной хроматографии на белке G проводили при температуре от 4 до 6°C. Сначала супернатант культуры гибридом наносили на колонку, заполненную белком G (GE Healthcare Bio-Sciences Corp.), уравновешенную ФСБ. После введения в колонку всего объема культурального супернатанта, колонку промывали ФСБ в объеме, в 2 или более раз превышающем объем колонки. Затем, фракции, содержащие антитело, собирали элюированием 0,1 М водным раствором гидрохлорида глицина (pH 2,7). Собранные фракции получали при значении pH от 7,0 до 7,5 путем добавления 1 М Трис-HCl (pH 9,0), и затем буфер заменяли ФСБ с использованием ультрафильтрационного центробежного концентратора VIVASPIN 20 (отсечение по молекулярной массе: UF30K, Sartorius Japan K.K., при температуре от 4 до 6°C), повышая при этом концентрацию антитела до 0,2 мг/мл или выше. На последнем этапе, раствор антитела фильтровали через фильтр Minisart-Plus (Sartorius Japan K.K.) и использовали в качестве очищенного образца.
[0123]
Пример 2. Оценка крысиного анти-человеческий Orai1 антитела in vitro
2)-1 Оценка способности крысиного анти-человеческий Orai1 антитела к связыванию при помощи проточной цитометрии
Для того чтобы оценить специфичность связывания с Orai1, суспензию клеток HEK293T, трансфицированных pcDNA3.1-DEST, или суспензию клеток HEK293T, трансфицированных pcDNA3.1-hOrai1, полученную способом, описанным в пункте 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T суспендировали путем добавления крысиного анти-человеческий Orai1 моноклонального антитела R118 или R198, полученного в пункте 1)-6, или контрольного крысиного антитела IgG (Beckman Coulter Inc.), и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-крысиного IgG, конъюгированного с ФИТЦ, разбавленного в 320 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида (Life Technologies Corp.), с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). R118 и R198 не связывались с клетками HEK293T, трансфицированными pcDNA3.1-DEST, и, как показано на Фигуре 1, каждое из них связывалось только с клетками HEK293T, трансфицированными pcDNA3.1-hOrai1, зависимым от концентрации образом, свидетельствуя о том, что каждое из этих антител специфично связывается с Orai1 человека. С другой стороны, такое связывание не наблюдалось в случае контрольного крысиного антитела IgG.
[0124]
2)-2 Ингибирующее действие крысиного анти-человеческий Orai1 антитела на активацию T-лимфоцитов
T-лимфоцитов человека клеточную линию клеток Юрката получали в концентрации 1,5×106 клеток/мл в среде RPMI1640 (Life Technologies Corp.), содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина (Life Technologies Corp.), инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали крысиным анти-человеческий Orai1 антителом R118 или R198 или контрольным крысиным антителом IgG, которые добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут в атмосфере с 5% содержанием CO2. Затем добавляли 100 нг/мл ФМА (Sigma-Aldrich Corp.) и 1 мкг/мл A23187 (Sigma-Aldrich Corp.) в количестве 10 мкл на лунку (конечная концентрация: 10 нг/мл ФМА и 100 нг/мл A23187) и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию интерлейкина 2 (IL-2), содержащегося в супернатанте, измеряли при помощи ИФА (R&D systems, Inc.). На Фигуре 2 показано, что каждое из полученных анти-человеческий Orai1 моноклональных антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187. R118 и R198 каждое ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката. С другой стороны, такое ингибирование не наблюдалось в случае контрольного крысиного антитела IgG.
[0125]
Пример 3. Определение нуклеотидной последовательности кДНК, кодирующих вариабельные области крысиного анти-человеческий Orai1 антитела
3)-1 Синтез кДНК
Клеточный лизат (50 мМ Трис-HCl (pH 7,5), 250 мМ LiCl, 5 мМ ЭДТА (pH 8), 0,5% додецилсульфата лития (LiDS), 2,5 мМ дитиотреитол (ДТТ)) гибридом, продуцирующих каждое антитело R118 или R198, смешивали с магнитными частицами (Dynabeads mRNA DIRECT Kit, Invitrogen Corp.), связанными с олигонуклеотидом dT25, так чтобы мРНК связывалась с этими магнитными частицами. Затем магнитные частицы один раз промывали каждым из следующих растворов: мРНК промывающий раствор A (10 мМ Трис-HCl (pH 7,5), 0,15 М LiCl, 1 мМ ЭДТА, 0,1% LiDS, 0,1% Тритон X-100) и раствор для синтеза кДНК (50 мМ Трис-HCl (pH 8,3), 75 мМ KCl, 3 мМ MgCl2, 5 мМ ДТТ, 0,5 мМ dNTP, 0,2% Тритон X-100, 1,2 единицы ингибитора РНКазы (Life Technologies Corp.)), с последующим синтезом кДНК с использованием раствора для синтеза кДНК, дополненного 12 единицами обратной транскриптазы SuperScript III (Life Technologies Corp.). Затем магнитные частицы промывали реакционным раствором для реакции 3'-конца (50 мМ фосфата калия, 4 мМ MgCl2, 0,5 мМ dGTP, 0,2% Тритон X-100, 1,22 единицы ингибитора РНКазы (Life Technologies Corp.)) с последующим осуществлением реакции 3'-конца с использованием реакционного раствора, дополненного 48 единицами рекомбинантной терминальной трансферазы (F. Hoffmann-La Roche, Ltd.).
[0126]
3)-2 Амплификация и секвенирование фрагментов генов вариабельной области тяжелой и легкой цепи иммуноглобулина
Магнитные частицы промывали раствором TE (10 мМ Трис-HCl (pH 7,5), 1 мМ ЭДТА, 0,1% Тритон X-100), и затем гены тяжелой и легкой цепи иммуноглобулина крысы амплифицировали при помощи 5'-RACE ПЦР. В частности, магнитные частицы переносили реакционный раствор для ПЦР (0,2 мкМ праймер, 0,2 мМ dNTP, 0,25 единиц HS ДНК-полимеразы PrimeSTAR (Takara Bio Inc.)) и подвергали 35 циклам реакции, каждый из которых включал в себя 94°C в течение 30 секунд - 68°C в течение 90 секунд. Используемые наборы праймеров были следующими:
[0127]
Набор праймеров для тяжелой цепи
5'-GCTAGCGCTACCGGACTCAGATCCCCCCCCCCCCCDN-3' (Nhe-polyC-S) (SEQ ID NO: 3)
5'-TCACTGAGCTGGTGAGAGTGTAGAGCCC-3' (rIgγ-AS1) (SEQ ID NO: 4)
5'-TCACCGAGCTGCTGAGGGTGTAGAGCCC-3' (rIgγ-AS2) (SEQ ID NO: 5)
Набор праймеров для легкой цепи
5'-GCTAGCGCTACCGGACTCAGATCCCCCCCCCCCCCDN-3' (Nhe-polyC-S) (SEQ ID NO: 6) (такой же, как для тяжелой цепи)
5'-TCAGTAACACTGTCCAGGACACCATCTC-3' (rIgκ-AS) (SEQ ID NO: 7).
Фрагменты, амплифицированные при помощи ПЦР-реакции, секвенировали для анализа их нуклеотидных последовательностей. Олигонуклеотид, имеющий нуклеотидную последовательность 5'-CTGGCTCAGGGAAATAGCC-3' (rIgγ-seq) (SEQ ID NO: 8), использовали в качестве праймера для секвенирования тяжелой цепи, и олигонуклеотид, имеющий нуклеотидную последовательность 5'-TCCAGTTGCTAACTGTTCC-3' (rIgκ-seq) (SEQ ID NO: 9), использовали в качестве праймера для секвенирования легкой цепи.
Анализ секвенирования проводили на аппарате для анализа последовательностей генов («ABI PRISM 3700 DNA Analyzer; Applied Biosystems, Inc.» или «Applied Biosystems 3730xl Analyzer; Applied Biosystems, Inc.»). Систему для циклического секвенирования Dye Terminator с ДНК-полимеразой AmpliTaq (Life Technologies Corp.) и GeneAmp 9700 (Applied Biosystems, Inc.) использовали в реакции секвенирования.
[0128] Определенные нуклеотидные последовательности, кодирующие вариабельные области тяжелой и легкой цепи R118 и R198 приведены в SEQ ID NOs: 10 (R118 легкая цепь) и 12 (R118 тяжелая цепь) и SEQ ID NOs: 14 (R198 легкая цепь) и 16 (R198 тяжелая), соответственно, в Списке последовательностей. Аминокислотные последовательности этих вариабельных областей приведены в SEQ ID NOs: 11 (R118 легкая цепь) и 13 (R118 тяжелая цепь) и SEQ ID NOs: 15 (R198 легкая цепь) и 17 (R198 тяжелая цепь), соответственно, в Списке последовательностей. SEQ ID NOs: 10 и 11 показаны на Фигуре 14. SEQ ID NOs: 12 и 13 показаны на Фигуре 15. SEQ ID NOs: 14 и 15 показаны на Фигуре 16. SEQ ID NOs: 16 и 17 показаны на Фигуре 17.
[0129]
Пример 4. Получение человеческого гибридного анти-человеческий Orai1 антитела
4)-1 Конструирование вектора pCMA-LK, экспрессирующего гибридную и гуманизированную легкую цепь
Фрагмент размером приблизительно 5,4 т.п.н., полученный расщеплением плазмиды pcDNA3.3-TOPO/LacZ (Life Technologies Corp.) ферментами рестрикции XbaI и PmeI, лигировали с фрагментом ДНК (приведенным в SEQ ID NO: 18 в Списке последовательностей), содержащим последовательность, кодирующую сигнал секреции κ цепи человека и константную область κ цепи человека, с использованием набора реагентов In-Fusion Advantage PCR cloning kit (Clontech Laboratories, Inc.) для получения pcDNA3.3/LK.
[0130] pcDNA3.3/LK использовали в качестве матрицы для ПЦР с набором праймеров, приведенным ниже. Полученный фрагмент размером приблизительно 3,8 т.п.н. фосфорилировали и затем подвергали самолигированию для получения вектора pCMA-LK, экспрессирующего гибридную и гуманизированную легкую цепь, который содержал сигнальную последовательность, сайт клонирования и последовательность, кодирующую константную область κ цепи человека, после промотора ЦМВ.
Набор праймеров
5'-TATACCGTCGACCTCTAGCTAGAGCTTGGC-3' (SEQ ID NO: 19; праймер 3.3-F1)
5'-GCTATGGCAGGGCCTGCCGCCCCGACGTTG-3' (SEQ ID NO: 20; праймер 3.3-R1).
4)-2 Конструирование вектора pCMA-G1, экспрессирующего гибридную и гуманизированную тяжелую цепь IgG1
Фрагмент ДНК, полученный путем расщепления pCMA-LK рестриктазами XbaI и PmeI для удаления последовательности, кодирующей сигнал секреции κ цепи и константную область κ цепи человека, лигировали с фрагментом ДНК (приведенным в SEQ ID NO: 21 в Списке последовательностей), содержащим последовательность, кодирующую сигнальную последовательность тяжелой цепи человека и аминокислоты константной области IgG1 человека, с использованием набора реагентов In-Fusion Advantage PCR cloning kit для получения вектора pCMA-G1, экспрессирующего гибридную и гуманизированную тяжелую цепь IgG1, который содержал сигнальную последовательность, сайт клонирования и последовательность, кодирующую константную область тяжелой цепи IgG1 человека, после промотора ЦМВ.
[0131]
4)-3 Конструирование вектора, экспрессирующего легкую цепь человеческого гибридного анти-человеческий Orai1 антитела
4)-3-1 Конструирование вектора cR118_L, экспрессирующего легкую цепь человеческого гибридного антитела R118
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область легкой цепи R118, полученную в Примере 3)-2, (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, размером приблизительно 0,7 т.п.н., полученный путем расщепления фрагмента ДНК, содержащего последовательность, кодирующую вариабельную область легкой цепи R118, ферментами рестрикции XbaI и PmeI, встраивали в фрагмент ДНК размером приблизительно 3,4 т.п.н., полученный путем расщепления универсального вектора pCMA-LK для экспрессии гибридной и гуманизированной легкой цепи антитела теми же ферментами рестрикции, которые указаны выше, с использованием реагента Ligation High ver. 2 (Toyobo Co., Ltd.), для конструирования вектора, экспрессирующего легкую цепь человеческого гибридного антитела R118 (cR118_L). Полученный экспрессионный вектор был обозначен как «pCMA-LK/cR118_L». Нуклеотидная последовательность, кодирующая легкую цепь человеческого гибридного антитела cR118, и аминокислотная последовательность этой легкой цепи приведены в SEQ ID NOs: 22 и 23 (Фигура 18), соответственно, в Списке последовательностей.
[0132]
4)-3-1 Конструирование вектора cR198_L, экспрессирующего легкую цепь человеческого гибридного антитела R198
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область легкой цепи R198, полученную в Примере 3)-2, (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, размером приблизительно 0,7 т.п.н., полученный путем расщепления фрагмента ДНК, содержащего последовательность, кодирующую вариабельную область легкой цепи R198, ферментами рестрикции XbaI и PmeI, встраивали в фрагмент ДНК размером приблизительно 3,4 т.п.н., полученный путем расщепления универсального вектора pCMA-LK для экспрессии гибридной и гуманизированной легкой цепи антитела теми же ферментами рестрикции, которые указаны выше, с использованием реагента Ligation High ver. 2 (Toyobo Co., Ltd.), для конструирования вектора, экспрессирующего легкую цепь человеческого гибридного антитела R198 (cR198_L). Полученный экспрессионный вектор был обозначен как «pCMA-LK/cR198_L». Нуклеотидная последовательность, кодирующая легкую цепь человеческого гибридного антитела cR198, и аминокислотная последовательность этой легкой цепи приведены в SEQ ID NOs: 24 и 25 (Фигура 19), соответственно, в Списке последовательностей.
[0134]
4)-4-2 Конструирование вектора cR198_H, экспрессирующего тяжелую цепь человеческого гибридного антитела R198
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область тяжелой цепи R198, полученную в Примере 3)-2, (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, размером приблизительно 0,3 т.п.н., полученный путем расщепления фрагмента ДНК, содержащего последовательность, кодирующую вариабельную область тяжелой цепи R198, ферментом рестрикции BlpI, встраивали в фрагмент ДНК размером приблизительно 4,5 т.п.н., полученный путем расщепления вектора pCMA-G1 для экспрессии гибридной и гуманизированной тяжелой цепи IgG1 тем же ферментом рестрикции, который указан выше, с использованием реагента Ligation High ver. 2, для конструирования вектора, экспрессирующего тяжелую цепь человеческого гибридного антитела R198 (cR198_H). Полученный экспрессионный вектор был обозначен как «pCMA-G1/cR198_H». Нуклеотидная последовательность, кодирующая тяжелую цепь человеческого гибридного антитела cR198, и аминокислотная последовательность этой тяжелой цепи приведены в SEQ ID NOs: 28 и 29 (Фигура 21), соответственно, в Списке последовательностей.
[0135]
4)-5 Получение человеческого гибридного анти-человеческий Orai1 антитела
4)-5-1 Продуцирование человеческого гибридного анти-человеческий Orai1 антитела
Клетки FreeStyle 293F (Life Technologies Corp.) субкультивтровали и культивировали в соответствии с инструкцией производителя.
[0136] 107 клеток FreeStyle 293F (Life Technologies Corp.), находящихся в логарифмической фазе роста, инокулировали в 30 мл флакон (Thermo Fisher Scientific Inc.) в объеме 9 мл, полученном путем разбавления экспрессионной средой FreeStyle 293 (Life Technologies Corp.), и затем культивировали при встряхивании при 135 об/мин при температуре 37°C в течение 1 часа в инкубаторе с атмосферой, содержащей 8% CO2. 30 мкг полиэтиленимина (Polysciences #24765) растворяли в 500 мкл среды Opti-Pro SFM (Life Technologies Corp.). Затем каждый вектор, экспрессирующий тяжелую цепь человеческого гибридного анти-человеческий Orai1 антитела, (4 мкг) и каждый вектор, экспрессирующий легкую цепь человеческого гибридного анти-человеческий Orai1 антитела, (6 мкг), полученные с использованием набора реагентов QIAGEN Plasmid Maxi (Qiagen N.V.), суспендировали в 500 мкл среды Opti-Pro SFM. 500 мкл смешанного раствора экспрессионный вектор/среда Opti-Pro SFM добавляли к 500 мкл смешанного раствора полиэтиленимин/среда Opti-Pro SFM и эту смесь осторожно перемешивали, затем оставляли на 5 минут и затем добавляли к клеткам FreeStyle 293F. Затем клетки культивировали при встряхивании при 95 об/мин при температуре 37°C в течение 5-7 суток в инкубаторе с атмосферой, содержащей 8% CO2, и полученный культуральный супернатант фильтровали через фильтр Millex (Millipore Corp.) с размером пор 0,22 мкм.
[0137] Человеческое гибридное антитело R118, полученное путем комбинации pCMA-G1/cR118_H и pCMA-LK/cR118_L, было обозначено как «cR118». Аналогично, человеческое гибридное антитело R198, полученное путем комбинации pCMA-G1/cR198_H и pCMA-LK/cR198_L, было обозначено как «cR198».
[0138] 4)-5-2 Очистка человеческого гибридного анти-человеческий Orai1 антитела
Каждый культуральный супернатант, полученный в пункте 4)-5-1, при помощи аффинной хроматографии с белком A в один этап. Сначала, 10 мл культурального супернатанта наносили на колонку MabSelectSuRe (GE Healthcare Bio-Sciences Corp.), уравновешенную ФСБ. После ввода в колонку всего культурального раствора, колонку промывали 7 мл ФСБ. Затем проводили элюирование 5 мл 2 М раствора аргинин-HCl (pH 4,0), и элюат подвергали замене буфера на 4 мл гистидинового буфера (25 мМ гистидина, 5% сорбита, pH 6,0) с использованием колонки обессоливания PD-10 (GE Healthcare Bio-Sciences Corp.). На последнем этапе раствор концентрировали приблизительно в 100 мкл, используя фильтр Amicon Ultracel 30K (отсечение по молекулярной массе: 30 K, Millipore Corp.), и использовали в качестве очищенного образца.
[0139]
Пример 5. In vitro активность человеческого гибридного анти-человеческий Orai1 антитела
5)-1 Антигенсвязывающая активность человеческого гибридного анти-человеческий Orai1 антитела, определенная при помощи проточной цитометрии
Для оценки специфичности связывания с Orai1 человека, суспензию клеток HEK293T, трансфицированных pcDNA3.1-DEST, или суспензию клеток HEK293T, трансфицированных pcDNA3.1-hOrai1, полученные способом, описанным в пункте 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T суспендировали путем добавления человеческого гибридного анти-человеческий Orai1 антитела cR118 или cR198, полученного в пункте 4)-5, или контрольного человеческого антитела IgG (Jackson ImmunoResearch Laboratories, Inc.), и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-человеческого IgG, конъюгированного с ФИТЦ (Jackson ImmunoResearch Laboratories, Inc.), разбавленного в 100 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида (Invitrogen Corp.), с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). cR118 и cR198 не связывались с клетками HEK293T, трансфицированными pcDNA3.1-DEST, и, как показано на Фигуре 3, каждое из них связывалось только с клетками HEK293T, трансфицированными pcDNA3.1-hOrai1, зависимым от концентрации образом, свидетельствуя о том, что каждое из этих антител специфично связывается с Orai1 человека. С другой стороны, такое связывание не наблюдалось в случае контрольного человеческого антитела IgG.
[0140]
5)-2 Ингибирующее действие человеческого гибридного анти-человеческий Orai1 антитела на активацию T-лимфоцитов
T-лимфоцитов человека клеточную линию клеток Юрката получали в концентрации 1,5×106 клеток/мл в среде RPMI1640, содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина, инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали человеческим гибридным анти-человеческий Orai1 антителом cR118 или cR198, исходным антителом R198 или контрольным человеческим антителом IgG, которые добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут в атмосфере с 5% содержанием CO2. Затем добавляли 100 нг/мл ФМА и 1 мкг/мл A23187 в количестве 10 мкл на лунку и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию IL-2, содержащегося в супернатанте, измеряли при помощи ИФА. На Фигуре 4 показано, что каждое из полученных человеческих гибридных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187. cR118 и cR198 каждое ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, и их ингибирующее действие эквивалентно действию исходного антитела R198. С другой стороны, такое ингибирование не наблюдалось в случае контрольного человеческого антитела IgG.
[0141]
Пример 6. Дизайн гуманизированного варианта hR198 человеческого гибридного анти-человеческий Orai1 антитела cR198
6)-1 Молекулярное моделирование вариабельных областей антитела R198
Молекулярное моделирование вариабельных областей антитела cR198 осуществляли способом, широко известным как гомологичное моделирование (Methods in Enzymology, 203, 121-153, (1991)). Вариабельные области R198, определенные выше, сравнивали с первичными последовательностями (доступными трехмерными структурами, полученными при помощи рентгеноструктурного анализа), вариабельных областей иммуноглобулина человека, зарегистрированными в Protein Data Bank (Nuc. Acid Res. 28, 235-242 (2000)). В результате, антитело 1AJ7 было выбрано в качестве антитела, имеющего самую высокую гомологию по последовательности с вариабельной областью легкой цепи cR198. Также антитело 1XGY было выбрано в качестве антитела, имеющего самую высокую гомологию по последовательности с вариабельной областью тяжелой цепи cR198. Трехмерные структуры каркасных областей получали в виде «каркасной модели» путем комбинирования координат 1AJ7 и 1XGY, соответствующих легкой цепи и тяжелой цепи cR198. CDRs антитела cR198 обозначили как кластеры 11A, 7A, 9A, 10A и 10A для CDRL1, CDRL2, CDRL3, CDRH1 и CDRH2, соответственно, согласно классификации Thornton et al. (J. Mol. Biol., 263, 800-815, (1996)). Его CDRH3 был классифицирован как k(6)- в соответствии с правилом H3 (FEBS letter, 399, 1-8 (1996)). Затем типичную конформацию каждой CDR включали в каркасную модель.
[0142] На последнем этапе проводили расчет энергии для исключения нежелательного межатомного контакта, чтобы получить возможные молекулярные модели вариабельных областей cR198 с точки зрения энергии. Эти процедуры проводили с использованием коммерчески доступной программы предсказания трехмерной структуры белков Prime и программы поиска конформаций MacroModel (Schrodinger, LLC).
[0143]
6)-2 Дизайн аминокислотной последовательности гуманизированного антитела R198
Гуманизированное антитело R198 конструировали способом, широко известным как прививка CDR (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)). Антитело-акцептор выбирали на основе гомологии аминокислотных последовательностей в каркасных областях.
[0144] Последовательности каркасных областей cR198 сравнивали с последовательностями всех каркасных областей человека, зарегистрированными в базе данных Kabat (Nuc. Acid Res., 29, 205-206 (2001)) аминокислотных последовательностей антител. В результате антитело 1C10'CL было выбрано в качестве акцептора, благодаря его 71% гомологии по последовательности в части каркасных областей. Аминокислотные остатки каркасных областей 1C10'CL были выравнены с аминокислотными остатками каркасных областей cR198 с целью идентификации положений аминокислот, которые не совпадают между собой. Положения этих остатков анализировали, используя трехмерную модель cR198, сконструированную выше. Затем донорные остатки, которые должны быть привиты на акцептор, выбирали в соответствии с критериями, представленными Queen et al. (Proc. Natl. Acad. Sci. USA 86, 10029-10033 (1989)).
[0145] Некоторые выбранные таким образом донорные остатки переносили в акцепторное антитело с целью конструирования гуманизированной последовательности R198, как описано в Примерах ниже.
[0146] Кроме того, от 1 до 5 аминокислотных остатков в каждой CDR или FR антитела cR198 заменяли аминокислотными остатками антитела cR118, чтобы получить CDR сконструированную гуманизированную последовательность R198, как описано в Примерах ниже.
[0147]
6)-3 Дизайн легкой цепи hR198_L гуманизированного антитела R198
6)-3-1 Легкая цепь типа hR198_L1:
Гуманизированную легкую цепь антитела R198, сконструированную путем замены остатка треонина в аминокислотном положении 30 остатком серина, остатка пролина в аминокислотном положении 32 остатком серина, остатка лейцина в аминокислотном положении 35 остатком валина, остатка глутаминовой кислоты в аминокислотном положении 37 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 42 остатком треонина, остатка аспарагиновой кислоты в аминокислотном положении 61 остатком глицина, остатка глицина в аминокислотном положении 62 остатком лизина, остатка серина в аминокислотном положении 63 остатком аланина, остатка валина в аминокислотном положении 64 остатком пролина, остатка серина в аминокислотном положении 92 остатком треонина, остатка серина в аминокислотном положении 94 остатком треонина, остатка треонина в аминокислотном положении 96 остатком серина, остатка глутаминовой кислоты в аминокислотном положении 99 остатком глутамина, остатка серина в аминокислотном положении 100 остатком пролина, остатка треонина в аминокислотном положении 120 остатком глутамина, остатка лейцина в аминокислотном положении 124 остатком валина, остатка лейцина в аминокислотном положении 126 остатком изолейцина, остатка аргинина в аминокислотном положении 127 остатком лизина и остатка аланина в аминокислотном положении 129 остатком треонина в последовательности легкой цепи cR198, приведенной в SEQ ID NO: 25 в Списке последовательностей, обозначили как «легкая цепь типа hR198_L1».
[0148] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L1, приведена в SEQ ID NO: 30 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_L1 приведена в SEQ ID NO: 31 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 30 и 31 также показаны на Фигуре 22. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0149]
6)-3-2 Легкая цепь типа hR198_L2:
Гуманизированную легкую цепь антитела R198, сконструированную путем замены остатка треонина в аминокислотном положении 30 остатком серина, остатка пролина в аминокислотном положении 32 остатком серина, остатка лейцина в аминокислотном положении 35 остатком валина, остатка глутаминовой кислоты в аминокислотном положении 37 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 42 остатком треонина, остатка аспарагиновой кислоты в аминокислотном положении 61 остатком глицина, остатка глицина в аминокислотном положении 62 остатком лизина, остатка серина в аминокислотном положении 63 остатком аланина, остатка серина в аминокислотном положении 92 остатком треонина, остатка серина в аминокислотном положении 94 остатком треонина, остатка треонина в аминокислотном положении 96 остатком серина, остатка глутаминовой кислоты в аминокислотном положении 99 остатком глутамина, остатка серина в аминокислотном положении 100 остатком пролина, остатка треонина в аминокислотном положении 120 остатком глутамина, остатка лейцина в аминокислотном положении 124 остатком валина, остатка лейцина в аминокислотном положении 126 остатком изолейцина, остатка аргинина в аминокислотном положении 127 остатком лизина и остатка аланина в аминокислотном положении 129 остатком треонина в последовательности легкой цепи cR198, приведенной в SEQ ID NO: 25 в Списке последовательностей, обозначили как «легкая цепь типа hR198_L2».
[0150] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L2, приведена в SEQ ID NO: 32 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_L2 приведена в SEQ ID NO: 33 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 32 и 33 также показаны на Фигуре 23. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0151]
6)-3-3 Легкая цепь типа hR198_L3:
Гуманизированную легкую цепь антитела R198, сконструированную путем замены остатка треонина в аминокислотном положении 30 остатком серина, остатка пролина в аминокислотном положении 32 остатком серина, остатка лейцина в аминокислотном положении 35 остатком валина, остатка глутаминовой кислоты в аминокислотном положении 37 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 42 остатком треонина, остатка аспарагиновой кислоты в аминокислотном положении 61 остатком глицина, остатка глицина в аминокислотном положении 62 остатком лизина, остатка серина в аминокислотном положении 63 остатком аланина, остатка валина в аминокислотном положении 64 остатком пролина, остатка серина в аминокислотном положении 92 остатком треонина, остатка серина в аминокислотном положении 94 остатком треонина, остатка треонина в аминокислотном положении 96 остатком серина, остатка глутаминовой кислоты в аминокислотном положении 99 остатком глутамина, остатка серина в аминокислотном положении 100 остатком пролина, остатка тирозина в аминокислотном положении 114 остатком фенилаланина, остатка треонина в аминокислотном положении 120 остатком глутамина, остатка лейцина в аминокислотном положении 124 остатком валина, остатка лейцина в аминокислотном положении 126 остатком изолейцина, остатка аргинина в аминокислотном положении 127 остатком лизина и остатка аланина в аминокислотном положении 129 остатком треонина в последовательности легкой цепи cR198, приведенной в SEQ ID NO: 25 в Списке последовательностей, обозначили как «легкая цепь типа hR198_L3».
[0152] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L3, приведена в SEQ ID NO: 34 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_L3 приведена в SEQ ID NO: 35 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 34 и 35 также показаны на Фигуре 24. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0153]
6)-3-4 Легкая цепь типа hR198_L4:
Гуманизированную легкую цепь антитела R198, сконструированную путем замены остатка треонина в аминокислотном положении 30 остатком серина, остатка пролина в аминокислотном положении 32 остатком серина, остатка лейцина в аминокислотном положении 35 остатком валина, остатка глутаминовой кислоты в аминокислотном положении 37 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 42 остатком треонина, остатка аспарагиновой кислоты в аминокислотном положении 61 остатком глицина, остатка глицина в аминокислотном положении 62 остатком лизина, остатка серина в аминокислотном положении 63 остатком аланина, остатка серина в аминокислотном положении 92 остатком треонина, остатка серина в аминокислотном положении 94 остатком треонина, остатка треонина в аминокислотном положении 96 остатком серина, остатка глутаминовой кислоты в аминокислотном положении 99 остатком глутамина, остатка серина в аминокислотном положении 100 остатком пролина, остатка тирозина в аминокислотном положении 114 остатком фенилаланина, остатка треонина в аминокислотном положении 120 остатком глутамина, остатка лейцина в аминокислотном положении 124 остатком валина, остатка лейцина в аминокислотном положении 126 остатком изолейцина, остатка аргинина в аминокислотном положении 127 остатком лизина и остатка аланина в аминокислотном положении 129 остатком треонина в последовательности легкой цепи cR198, приведенной в SEQ ID NO: 25 в Списке последовательностей, обозначили как «легкая цепь типа hR198_L4».
[0154] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L4, приведена в SEQ ID NO: 36 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_L4 приведена в SEQ ID NO: 37 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 36 и 37 также показаны на Фигуре 25. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0155]
6)-4 Дизайн тяжелой цепи hR198_H гуманизированного антитела R198
6)-4-1 Тяжелая цепь типа hR198_H1:
Гуманизированную тяжелую цепь антитела R198, сконструированную путем замены остатка глутамина в аминокислотном положении 24 остатком валина, остатка лейцина в аминокислотном положении 30 остатком валина, остатка аланина в аминокислотном положении 31 остатком лизина, остатка серина в аминокислотном положении 35 остатком аланина, остатка метионина в аминокислотном положении 37 остатком валина, остатка изолейцина в аминокислотном положении 39 остатком валина, остатка изолейцина в аминокислотном положении 56 остатком валина, остатка лизина в аминокислотном положении 57 остатком аргинина, остатка треонина в аминокислотном положении 59 остатком аланина, остатка треонина в аминокислотном положении 60 остатком пролина, остатка лизина в аминокислотном положении 86 остатком аргинина, остатка серина в аминокислотном положении 95 остатком треонина, остатка фенилаланина в аминокислотном положении 99 остатком тирозина, остатка глутамина в аминокислотном положении 101 остатком глутаминовой кислоты, остатка треонина в аминокислотном положении 106 остатком аргинина, остатка пролина в аминокислотном положении 107 остатком серина, остатка аспарагиновой кислоты в аминокислотном положении 108 остатком глутаминовой кислоты, остатка серина в аминокислотном положении 110 остатком треонина, остатка валина в аминокислотном положении 130 остатком треонина и остатка метионина в аминокислотном положении 131 остатком лейцина в последовательности тяжелой цепи cR198, приведенной в SEQ ID NO: 29 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_H1».
[0156] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H1, приведена в SEQ ID NO: 38 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_H1 приведена в SEQ ID NO: 39 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 136 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 38 и 39 также показаны на Фигуре 26. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0157]
6)-4-2 Тяжелая цепь типа hR198_H2:
Гуманизированную тяжелую цепь антитела R198, сконструированную путем замены остатка глутамина в аминокислотном положении 24 остатком валина, остатка лейцина в аминокислотном положении 30 остатком валина, остатка аланина в аминокислотном положении 31 остатком лизина, остатка серина в аминокислотном положении 35 остатком аланина, остатка метионина в аминокислотном положении 37 остатком валина, остатка изолейцина в аминокислотном положении 39 остатком валина, остатка лизина в аминокислотном положении 57 остатком аргинина, остатка треонина в аминокислотном положении 59 остатком аланина, остатка треонина в аминокислотном положении 60 остатком пролина, остатка лизина в аминокислотном положении 86 остатком аргинина, остатка серина в аминокислотном положении 95 остатком треонина, остатка фенилаланина в аминокислотном положении 99 остатком тирозина, остатка глутамина в аминокислотном положении 101 остатком глутаминовой кислоты, остатка треонина в аминокислотном положении 106 остатком аргинина, остатка пролина в аминокислотном положении 107 остатком серина, остатка аспарагиновой кислоты в аминокислотном положении 108 остатком глутаминовой кислоты, остатка серина в аминокислотном положении 110 остатком треонина, остатка валина в аминокислотном положении 130 остатком треонина и остатка метионина в аминокислотном положении 131 остатком лейцина в последовательности тяжелой цепи cR198, приведенной в SEQ ID NO: 29 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_H2».
[0158] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H2, приведена в SEQ ID NO: 40 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_H2 приведена в SEQ ID NO: 41 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 136 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 40 и 41 также показаны на Фигуре 27. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0159]
6)-4-3 Тяжелая цепь типа hR198_H3:
Гуманизированную тяжелую цепь антитела R198, сконструированную путем замены остатка глутамина в аминокислотном положении 24 остатком валина, остатка лейцина в аминокислотном положении 30 остатком валина, остатка аланина в аминокислотном положении 31 остатком лизина, остатка серина в аминокислотном положении 35 остатком аланина, остатка метионина в аминокислотном положении 37 остатком валина, остатка изолейцина в аминокислотном положении 39 остатком валина, остатка серина в аминокислотном положении 50 остатком аланина, остатка изолейцина в аминокислотном положении 56 остатком валина, остатка лизина в аминокислотном положении 57 остатком аргинина, остатка треонина в аминокислотном положении 59 остатком аланина, остатка треонина в аминокислотном положении 60 остатком пролина, остатка изолейцина в аминокислотном положении 67 остатком валина, остатка валина в аминокислотном положении 70 остатком изолейцина, остатка глутаминовой кислоты в аминокислотном положении 81 остатком аланина, остатка лизина в аминокислотном положении 82 остатком аргинина, остатка лизина в аминокислотном положении 86 остатком аргинина, остатка серина в аминокислотном положении 95 остатком треонина, остатка фенилаланина в аминокислотном положении 99 остатком тирозина, остатка глутамина в аминокислотном положении 101 остатком глутаминовой кислоты, остатка треонина в аминокислотном положении 106 остатком аргинина, остатка пролина в аминокислотном положении 107 остатком серина, остатка аспарагиновой кислоты в аминокислотном положении 108 остатком глутаминовой кислоты, остатка серина в аминокислотном положении 110 остатком треонина, остатка валина в аминокислотном положении 130 остатком треонина и остатка метионина в аминокислотном положении 131 остатком лейцина в последовательности тяжелой цепи cR198, приведенной в SEQ ID NO: 29 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_H3».
[0160] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H3, приведена в SEQ ID NO: 42 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_H3 приведена в SEQ ID NO: 43 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 136 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 42 и 43 также показаны на Фигуре 28. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0161] 6)-4-4 Тяжелая цепь типа hR198_H4:
Гуманизированную тяжелую цепь антитела R198, сконструированную путем замены остатка глутамина в аминокислотном положении 24 остатком валина, остатка лейцина в аминокислотном положении 30 остатком валина, остатка аланина в аминокислотном положении 31 остатком лизина, остатка серина в аминокислотном положении 35 остатком аланина, остатка метионина в аминокислотном положении 37 остатком валина, остатка изолейцина в аминокислотном положении 39 остатком валина, остатка серина в аминокислотном положении 50 остатком аланина, остатка лизина в аминокислотном положении 57 остатком аргинина, остатка треонина в аминокислотном положении 59 остатком аланина, остатка треонина в аминокислотном положении 60 остатком пролина, остатка изолейцина в аминокислотном положении 67 остатком валина, остатка валина в аминокислотном положении 70 остатком изолейцина, остатка глутаминовой кислоты в аминокислотном положении 81 остатком аланина, остатка лизина в аминокислотном положении 82 остатком аргинина, остатка лизина в аминокислотном положении 86 остатком аргинина, остатка серина в аминокислотном положении 95 остатком треонина, остатка фенилаланина в аминокислотном положении 99 остатком тирозина, остатка глутамина в аминокислотном положении 101 остатком глутаминовой кислоты, остатка треонина в аминокислотном положении 106 остатком аргинина, остатка пролина в аминокислотном положении 107 остатком серина, остатка аспарагиновой кислоты в аминокислотном положении 108 остатком глутаминовой кислоты, остатка серина в аминокислотном положении 110 остатком треонина, остатка валина в аминокислотном положении 130 остатком треонина и остатка метионина в аминокислотном положении 131 остатком лейцина в последовательности тяжелой цепи cR198, приведенной в SEQ ID NO: 29 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_H4».
[0162] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H4, приведена в SEQ ID NO: 44 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_H4 приведена в SEQ ID NO: 45 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 136 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 44 и 45 также показаны на Фигуре 29. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0163]
Пример 7. Получение гуманизированного анти-человеческий Orai1 антитела
7)-1 Конструирование вектора, экспрессирующего легкую цепь hR198_L гуманизированного анти-человеческий Orai1 антитела
7)-1-1 Конструирование вектора, экспрессирующего hR198_L1
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_L1, соответствующую нуклеотидам в положениях с 38 по 402 нуклеотидной последовательности hR198_L1, представленной SEQ ID NO: 30 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Синтезированный фрагмент ДНК расщепляли ферментами рестрикции AvaI и EcoRV и встраивали в соответствующий сайт вектора pCMA-LK, экспрессирующего легкую цепь гибридного и гуманизированного антитела, расщепленного теми же ферментами рестрикции, которые указаны выше, для конструирования экспрессионного вектора hR198_L1. Полученный экспрессионный вектор обозначили как «pCMA-LK/hR198_L1».
[0164]
7)-1-2 Конструирование вектора, экспрессирующего hR198_L2
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_L2, соответствующую нуклеотидам в положениях с 38 по 402 нуклеотидной последовательности hR198_L2, представленной SEQ ID NO: 32 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_L2, амплифицировали с синтезированным фрагментом ДНК в качестве матрицы с использованием ДНК-полимеразы KOD -Plus- (Toyobo Co., Ltd.), расщепляли ферментом рестрикции BsiWI и встраивали в соответствующий сайт вектора pCMA-LK, экспрессирующего легкую цепь гибридного и гуманизированного антитела, расщепленного ферментом рестрикции BsiWI, для конструирования экспрессионного вектора hR198_L2. Полученный экспрессионный вектор обозначили как «pCMA-LK/hR198_L2».
[0165]
7)-1-3 Конструирование векторов, экспрессирующих hR198_L3 и hR198_L4
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_L4, соответствующую нуклеотидам в положениях с 38 по 402 нуклеотидной последовательности hR198_L4, представленной SEQ ID NO: 36 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий hR198_L4, конструировали тем же способом, что и в Примере 7)-1-2. Полученный экспрессионный вектор обозначили как «pCMA-LK/hR198_L4».
[0166] Мутацию для замены остатка валина в аминокислотном положении 64 остатком пролина вводили с использованием pCMA- LK/hR198_L4 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза QuikChange (Stratagene Corp.). Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 34 в Списке последовательностей, обозначили как «pCMA- LK/hR198_L3».
[0167]
7)-2 Конструирование вектора, экспрессирующего тяжелую цепь hR198_H гуманизированного анти-человеческий Orai1 антитела
7)-2-1 Конструирование векторов, экспрессирующих hR198_H1 и hR198_H2
Фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_H0, соответствующую нуклеотидам в положениях с 36 по 425 нуклеотидной последовательности hR198_H0, представленной SEQ ID NO: 111 в Списке последовательностей, синтезировали в качестве кандидата для гуманизации тяжелой цепи анти-Orai1 антитела hR198 (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_H0, амплифицировали с синтезированным фрагментом ДНК в качестве матрицы с использованием ДНК-полимеразы KOD -Plus- (Toyobo Co., Ltd.) и встраивали в соответствующий сайт вектора pCMA-G1, экспрессирущего тяжелую цепь гибридного и гуманизированного антитела типа IgG1, расщепленного ферментом рестрикции BlpI, с использованием набора реагентов для клонирования ПЦР продуктов In-Fusion HD (Clontech Laboratories, Inc.) для конструирования экспрессионного вектора hR198_H0. Полученный экспрессионный вектор обозначили как «pCMA-G1/hR198_H0». Аминокислотная последовательность hR198_H0 приведена в SEQ ID NO: 112.
[0168] Затем мутацию для замены остатка триптофана в аминокислотном положении 66 остатком тирозина вводили с использованием pCMA-G1/hR198_H0 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus- (Toyobo Co., Ltd.). Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 40 в Списке последовательностей, обозначили как «pCMA-G1/hR198_H2».
[0169] Затем мутацию для замены остатка изолейцина в аминокислотном положении 56 остатком валина вводили с использованием pCMA-G1/hR198_H2 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза QuikChange (Stratagene Corp.) и набора праймеров, указанного ниже. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 38 в Списке последовательностей, обозначили как «pCMA-G1/hR198_H1».
[0170]
7)-2-2 Конструирование векторов, экспрессирующих hR198_H3, hR198_H4 и hR198_H5
Фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_H5, соответствующую нуклеотидам в положениях с 36 по 425 нуклеотидной последовательности hR198_H5, представленной SEQ ID NO: 113 в Списке последовательностей, синтезировали в качестве кандидата для гуманизации тяжелой цепи анти-Orai1 антитела hR198 (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий hR198_H5, конструировали тем же способом, что и в Примере 7)-2-1. Полученный экспрессионный вектор обозначили как «pCMA-G1/hR198_H5». Аминокислотная последовательность hR198_H5 приведена в SEQ ID NO: 114.
[0171] Затем мутации для замены остатка триптофана в аминокислотном положении 66 и остатка изолейцина в аминокислотном положении 67 остатком тирозина и остатком валина, соответственно, вводили с использованием pCMA-G1/H5 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 44 в Списке последовательностей, обозначили как «pCMA-G1/hR198_H4».
[0172] Затем мутацию для замены остатка изолейцина в аминокислотном положении 56 остатком валина вводили тем же способом, что и в Примере 7)-2-1 с использованием pCMA-G1/hR198_H4 в качестве матрицы. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 44 в Списке последовательностей, обозначили как «pCMA-G1/hR198_H3».
[0173] Получение гуманизированного анти-человеческий Orai1 антитела hR198
Клетки FreeStyle 293F трансфицировали каждым вектором, экспрессирующим тяжелую цепь гуманизированного анти-человеческий Orai1 антитела, и каждым вектором, экспрессирующим легкую, цепь гуманизированного анти-человеческий Orai1 антитела, тем же способом, что и в пункте 4)-5-1, для получения культурального супернатанта, содержащего антитело.
[0174] Гуманизированные анти-человеческий Orai1 антитела, полученные путем комбинации pCMA-G1/hR198_H1 и pCMA-LK/hR198_L1, комбинации pCMA-G1/hR198_H2 и pCMA-LK/hR198_L2, комбинации pCMA-G1/hR198_H3 и pCMA-LK/hR198_L3, и комбинации pCMA-G1/hR198_H4 и pCMA-LK/hR198_L4 обозначили как «hR198_H1/L1», «hR198_H2/L2», «hR198_H3/L3» и «hR198_H4/L4», соответственно.
[0175] Каждый полученный культуральный супернатант очищали при помощи аффинной хроматографии с белком A тем же способом, что и в пункте 4)-5-2, для получения образца очищенного антитела.
[0176]
Пример 8. In vitro активность гуманизированного анти-человеческий Orai1 антитела
8)-1 Антигенсвязывающая активность гуманизированного анти-человеческий Orai1 антитела, определенная при помощи проточной цитометрии
Для оценки специфичности связывания с Orai1 человека, суспензию клеток HEK293T, трансфицированных pcDNA3.1-DEST, или суспензию клеток HEK293T, трансфицированных pcDNA3.1-hOrai1, полученные способом, описанным в пункте 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T суспендировали путем добавления гуманизированного анти-человеческий Orai1 антитела hR198_H1/L1, hR198_H2/L2, hR198_H3/L3 или hR198_H4/L4, полученного в пункте 7)-5, или исходного антитела cR118 или cR198, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-человеческого IgG, конъюгированного с ФИТЦ, разбавленного в 100 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида, с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). Гуманизированные анти-человеческий Orai1 антитела hR198_H1/L1, hR198_H2/L2, hR198_H3/L3 и hR198_H4/L4 не связывались с клетками HEK293T, трансфицированными pcDNA3.1-DEST, и, как показано на Фигуре 5, каждое из них связывалось только с клетками HEK293T, трансфицированными pcDNA3.1-hOrai1, зависимым от концентрации образом, как в случае исходного антитела cR118 или cR198, свидетельствуя о том, что каждое из этих антител специфично связывается с Orai1 человека.
[0177]
8)-2 Ингибирующее действие гуманизированного анти-человеческий Orai1 антитела на активацию T-лимфоцитов
T-лимфоцитов человека клеточную линию клеток Юрката получали в концентрации 1,5×106 клеток/мл в среде RPMI1640, содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина, инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали гуманизированным анти-человеческий Orai1 антителом hR198H1/L1, hR198H2/L2, hR198H3/L3 или hR198H4/L4, или человеческим гибридным анти-человеческий Orai1 антителом cR118 или cR198, которые добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут в атмосфере с 5% содержанием CO2. Затем добавляли 100 нг/мл ФМА и 1 мкг/мл A23187 в количестве 10 мкл на лунку и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию IL-2, содержащегося в супернатанте, измеряли при помощи ИФА. На Фигуре 6 показано, что каждое из гуманизированных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187. Каждое из гуманизированных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, и ингибирующее действие hR198H1/L1 и hR198H2/L2 эквивалентно действию человеческих гибридных анти-человеческий Orai1 антител cR118 или cR198. С другой стороны, hR198H3/L3 и hR198H4/L4 обладают ингибирующей активностью в более низкой концентрации, чем концентрация антител cR118 или cR198.
[0178]
Пример 9. Идентификация повышающей активность мутации при помощи рибосомного дисплея
9)-1 Получение библиотек H цепи и L цепи
Библиотеку, содержащую мутированные H цепи и L цепи, конструировали с гуманизированным анти-человеческий Orai1 антителом hR198_H4/L4 в качестве матрицы и подвергали идентификации повышающей активность мутации при помощи рибосомного дисплея.
9)-1-1 Получение библиотеки H цепи
Чтобы случайным образом изменить область гена, проводили 80 циклов ПЦР с геном hR198_H4 в качестве матрицы, используя приведенный ниже набор праймеров и ДНК-полимеразу (Toyobo Co., Ltd.).
Набор праймеров
5'-ATGCAAGTCCAACTGGTTCAATC-3' (SEQ ID NO: 46; праймер Orai1 HF)
5'-TGACGGAGCCAGCGGGAAGAC-3' (SEQ ID NO: 47; праймер Orai1 CH FR).
Затем проводили перекрывающуюся ПЦР с каждым случайным образом мутированным геном H цепи, 5'-UTR сайтом, содержащим T7 промотор, и фрагментом гена TolA, содержащим 5' добавленную c-Myc последовательность и 3' добавленную SecM последовательность, в качестве матрицы, используя приведенный ниже набор праймеров, для получения библиотеки H цепи.
Набор праймеров
5'-CAGGAAACAGCTATGACCATG-3' (SEQ ID NO: 48; праймер M13 rev long)
5'-CTCGAGTTATTCATTAGGTGAGGCGTTGAGG-3' (SEQ ID NO: 49; праймер SecM Stop R).
9)-1-2 Получение библиотеки L цепи
Чтобы случайным образом изменить область гена, проводили 80 циклов ПЦР с геном hR198_L4 в качестве матрицы, используя приведенный ниже набор праймеров и ДНК-полимеразу.
Набор праймеров
5'-ATGGACATTCAACTGACCCAAAGC-3' (SEQ ID NO: 50; праймер Orai1 Lc F)
5'-GATAAAAACACTCGGGGCCGCCAC-3' (SEQ ID NO: 51; праймер Orai1 CL-FR).
Так же как, как описано в пункте 9)-1-1 проводили перекрывающуюся ПЦР с каждым случайным образом мутированным геном L цепи и двумя вышеописанными фрагментами генов в качестве матрицы для получения библиотеки L цепи.
[0179]
9)-1-3 Получение фрагмента гена H цепи
Область гена H цепи амплифицировали при помощи ПЦР с геном hR198_H4 в качестве матрицы, используя набор праймеров, приведенный ниже, ДНК-полимеразу KOD -Plus-.
Набор праймеров
5'-ATGCAAGTCCAACTGGTTCAATC-3' (SEQ ID NO: 46; праймер Orai1 HF)
5'-TCATTATTTGTCATCGTCATCTTTATAGTCGAATTCTTCGCCACGATTAAAGGATTTGGTGAC-3' (SEQ ID NO: 52; праймер Orai1 HR-FLAG R).
Затем проводили перекрывающуюся ПЦР с фрагментом гена, 5'-UTR сайтом, содержащим T7 промотор, в качестве матрицы, используя приведенный ниже набор праймеров, для получения фрагмента гена H цепи.
Набор праймеров
5'-CAGGAAACAGCTATGACCATG-3' (SEQ ID NO: 48; праймер M13 rev long)
5'-TCATTATTTGTCATCGTCATCTTTATAGTCGAATTCTTCGCCACGATTAAAGGATTTGGTGAC-3' (SEQ ID NO: 52; праймер Orai1 HR-FLAG R).
9)-1-4 Получение фрагмента гена L цепи
Область гена L цепи амплифицировали при помощи ПЦР с геном hR198_L4 в качестве матрицы, используя набор праймеров, приведенный ниже, ДНК-полимеразу KOD -Plus-.
5'-ATGGACATTCAACTGACCCAAAGC-3' (SEQ ID NO: 50; праймер Orai1 Lc F)
5'-TCATTATTTGTCATCGTCATCTTTATAGTCGAATTCTTCGCCACGATTAAAGGATTTGGTGAC-3' (SEQ ID NO: 53; праймер Orai1 CL-FLAG R).
Затем, таким же образом, как описано в пункте 9)-1-3, проводили перекрывающуюся ПЦР для получения фрагмента гена L цепи.
Набор праймеров
5'-CAGGAAACAGCTATGACCATG-3' (SEQ ID NO: 48; праймер M13 rev long)
5'-TCATTATTTGTCATCGTCATCTTTATAGTCGAATTCTTCGCCACGATTAAAGGATTTGGTGAC-3' (SEQ ID NO: 53; праймер Orai1 CL-FLAG R).
9)-1-5 Получение мРНК
Каждую мРНК синтезировали с библиотек и фрагментов генов, полученных в Примерах 9)-1-1 по 9)-1-4, в качестве матриц, используя набор реагентов T7 RiboMax Express Large Scale RNA Production System (Promega Corp.).
[0180]
9)-2 Скрининг при помощи рибосомного дисплея
Рибосомный дисплей Fabs H цепи получали путем комбинации библиотеки H цепи и фрагмента гена L цепи, и рибосомный дисплей Fabs L цепи получали путем комбинации библиотеки L цепи и фрагмента гена H цепи. 20 пмоль библиотеки мРНК H цепи (или L цепи) и 100 пмоль рибосом добавляли к реакционному раствору PUREfrex (GeneFrontier Corp.), и эту смесь инкубировали при температуре 30°C в течение 45 минут. Аналогичным образом, 40 пмоль библиотеки мРНК L цепи (или H цепи) и 200 пмоль рибосом добавляли к реакционному раствору PUREfrex, и эту смесь инкубировали при температуре 30°C в течение 45 минут. Затем посттрансляционные реакционные растворы библиотеки H цепи (или L цепи) и L цепи (или H цепи) смешивали и допонительно инкубировали при температуре 30°C в течение 90 минут для получения рибосомного дисплея Fabs H цепи (или L цепи). Затем реакцию останавливали путем охлаждения до 4°C. Затем к реакционному раствору добавляли антиген, и смесь осторожно перемешивали при температуре 4°C в течение 1 часа, так, чтобы каждый Fab связался с антигеном. Используемый антиген представлял собой фиксированный в формалине образец клеток CHO, постоянно экспрессирующих Orai1 человека, полученных с использованием pcDNA3.1-hOrai1, полученного в пункте 1)-1-1, или биотин-ПЭГилированного пептида петлевой области Orai1 человека (Sigma-Aldrich Corp.), показанной ниже.
[0181]
Биотин-ПЭГилированный пептид петлевой области Orai1 человека
Биотин-ПЭГ-SGSGFLPLKKQPGQPRPTSKPPASGAAANVSTSGITPGQAAAIASTTI (SEQ ID NO: 115)
Рибосомный дисплей Fabs, связанных с антигеном, выделяли с использованием магнитных частиц Nonolink Streptavidin (SoluLink, Inc.). Затем антиген промывали 50 мМ Трис-HCl (pH 7,4), 150 мМ NaCl, 15 мМ Mg(OAc)2, 0,05% Твин 20 и 1 мг/мл РНК дрожжей, и 50 мМ Трис-HCl (pH 7,4), 150 мМ NaCl, 15 мМ Mg(OAc)2 и 0,05% Твин 20. Затем 50 мМ Трис-HCl (pH 7,4), 150 мМ NaCl, 15 мМ Mg(OAc)2 и 50 мМ ЭДТА добавляли к антигену, и эту смесь оставляли при комнатной температуре на 10 минут, после чего при помощи центрифугирования получали супернатант, содержащий мРНК. кДНК синтезировали из выделенной мРНК с использованием набора реагентов для синтеза кДНК Transcriptor High Fidelity cDNA Synthesis Kit (F. Hoffmann-La Roche, Ltd.), и затем амплифицировали в виде ДНК при помощи ПЦР с использованием приведенного ниже набора праймеров и ДНК-полимеразы KOD -Plus-. мРНК синтезировали с кДНК в качестве матрицы и затем подвергали скринингу таким же способом, как указано выше. Цикл скринига проводили несколько раз для выбора генов антитела, связывающегося с антигеном с высокой степенью эффективности.
Набор праймеров
5'-ATGGACATTCAACTGACCCAAAGC-3' (SEQ ID NO: 50; праймер Orai1-LcF)
5'-CAGATCCTCCTCAGAGATCAGCTTCTGCTC-3' (SEQ ID NO: 54; праймер Myc-R).
9)-3 Получение белка Fab
Отобранные гены субклонировали в векторы для экспрессии в E.coli и скринировали при помощи ИФА на клетках для поиска клонов с улучшенной активностью связывания с клетками CHO, постоянно экспрессирующими Orai1 человека. Сначала каждую отобранную ДНК расщепляли ферментами рестрикции EcoRV и XhoI и встраивали в вектор для экспрессии Fab (GeneFrontier Corp.), расщепленный теми же ферментами, которые указаны выше, этот вектор затем переносили в E.coli BL21 (DE3). Полученные таким образом трансформанты культивировали при температуре 37°C в течение 4-5 часов в 150 мкл карбенициллина/0,1% глюкозы/2×YT на лунку в 96-луночном круглодонном планшете. Затем планшет охлаждали до 4°C, и затем добавляли в него IPTG с конечной концентрацией 0,5 мМ с последующим культивированием при встряхивании в течение ночи при температуре 30°C. Затем бактериальные клетки выделяли при помощи центрифугирования и затем к ним добавляли лизирующий буфер (2,5 мг/мл лизоцима, 100 Ед. ДНКазы I) с последующим встряхиванием при комнатной температуре в течение 60 минут. Затем при помощи центрифугирования отделяли супернатант для получения образцов Fab.
[0182]
9)-4 Скрининг при помощи ИФА на клетках
Клетки, экспрессирующие Orai1 человека, культивировали в 384-луночном планшете до образования ими сплошного слоя. Затем планшет промывали промывочным буфером (ФСБ(-), 20 мМ MgSO4, 2,5% ФБС). Затем в этот планшет добавляли образцы Fab и встряхивали планшет при температуре 4°C в течение 1 часа. После четырехкратного промывания промывочным буфером добавляли меченое пероксидазой анти-человеческий F(ab')2 козье антитело (Jackson ImmunoResearch Laboratories, Inc.) и встряхивали планшет при температуре 4°C в течение 30 минут. Планшет три раза промывали промывочным буфером и затем три раза промывали ФСБ(-) и 20 мМ MgSO4. Затем добавляли хромогенный реагент (0,4 мг/мл тетраметилбензидина, 200 мМ ацетата натрия (pH 3,4), 0,01% водный раствор перекиси водорода), и встряхивали планшет при комнатной температуре в течение 15 минут. Затем добавляли 2Н HCl, после чего измеряли OD450. Отбирали образец Fab, который имел более высокое значение OD450 при связывании с клетками CHO, постоянно экспрессирующими Orai1 человека, по сравнению с контрольными клетками CHO.
[0183]
9)-5 Определение антигенсвязывающей активности Fab анти-человеческий Orai1 антитела при помощи проточной цитометрии
Для оценки специфичности связывания с Orai1 человека, суспензию клеток HEK293T, трансфицированных pcDNA3.1-DEST, или суспензию клеток HEK293T, трансфицированных pcDNA3.1-hOrai1, полученные способом, описанным в пункте 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T суспендировали путем добавления мутированных клонов легкой цепи LCDR60, LCDR67, LCDR83, CE151 или PE057, или мутированных клонов тяжелой цепи HCDR046, HCDR047, HEP087, HEP124 или HEP237, отобранных в пункте 9)-4, или исходного Fab антитела hR198_H4/L4-Fab, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-человеческого IgG, конъюгированного с ФИТЦ, разбавленного в 100 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида, с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). Мутированные клоны легкой цепи LCDR60, LCDR67, LCDR83, CE151 и PE057, и мутированные клоны тяжелой цепи HCDR046, HCDR047, HEP087, HEP124 и HEP237 не связывались с клетками HEK293T, трансфицированными pcDNA3.1-DEST, как и исходный Fab антитела hR198_H4/L4-Fab, и, как показано на Фигуре 7, каждый из них имел тенденцию связываться с Orai1 человека в клетках HEK293T, трансфицированных pcDNA3.1-hOrai1, на уровне эквивалентном или превышающем уровень исходного Fab антитела.
[0184]
Пример 10 Получение созревания аффинности гуманизированного анти-человеческий Orai1 антитела
100 или более типов сконструированных hR198_H3/L3 антител получали путем переноса некоторых повышающих активность мутаций, обнаруженных в мутированных клонах легкой цепи LCDR60, LCDR67, LCDR83, CE151 и PE057, и мутированных клонах тяжелой цепи HCDR046, HCDR047, HEP087, HEP124 и HEP237, отобранных в Примере 9), в hR198_H3/L3, и оценивали их на предмет аффинности связывания, активности in vitro, продуктивности и гетерогенной антигенности по отношению к человеку. В результате, были отобраны антитела, приведенные ниже.
[0185]
10)-1 Дизайн созревания аффинности гуманизированного анти-человеческий Orai1 антитела
10)-1-1 Легкая цепь типа hR198_LG1:
Гуманизированную легкую цепь R198, сконструированную путем замены остатка аспарагина в аминокислотном положении 51 остатком глицина, остатка треонина в аминокислотном положении 113 остатком изолейцина и остатка треонина в аминокислотном положении 117 остатком серина в последовательности легкой цепи hR198_L3, приведенной в SEQ ID NO: 35 в Списке последовательностей, обозначили как «легкая цепь типа hR198_LG1».
[0186] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG1, приведена в SEQ ID NO: 55 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_LG1 приведена в SEQ ID NO: 56 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 55 и 56 также показаны на Фигуре 30. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0187]
10)-1-2 Легкая цепь типа hR198_LG2:
Гуманизированную легкую цепь R198, сконструированную путем замены остатка аргинина в аминокислотном положении 44 остатком гистидина, остатка серина в аминокислотном положении 48 остатком аспарагина, остатка аспарагина в аминокислотном положении 51 остатком глицина, остатка серина в аминокислотном положении 70 остатком лейцина, остатка глутаминовой кислоты в аминокислотном положении 75 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 76 остатком триптофана, остатка треонина в аминокислотном положении 113 остатком изолейцина и остатка треонина в аминокислотном положении 117 остатком серина в последовательности легкой цепи hR198_L3, приведенной в SEQ ID NO: 35 в Списке последовательностей, обозначили как «легкая цепь типа hR198_LG2».
[0188] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG2, приведена в SEQ ID NO: 57 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_LG2 приведена в SEQ ID NO: 58 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 57 и 58 также показаны на Фигуре 31. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0189]
10)-1-3 Легкая цепь типа hR198_LG3:
Гуманизированную легкую цепь R198, сконструированную путем замены остатка аргинина в аминокислотном положении 44 остатком гистидина, остатка глутамина в аминокислотном положении 47 остатком аргинина, остатка серина в аминокислотном положении 48 остатком аспарагина, остатка аспарагина в аминокислотном положении 51 остатком глицина, остатка серина в аминокислотном положении 70 остатком лейцина, остатка треонина в аминокислотном положении 73 остатком серина, остатка глутаминовой кислоты в аминокислотном положении 75 остатком аспарагиновой кислоты, остатка серина в аминокислотном положении 76 остатком триптофана, остатка треонина в аминокислотном положении 113 остатком изолейцина и остатка треонина в аминокислотном положении 117 остатком серина в последовательности легкой цепи hR198_L3, приведенной в SEQ ID NO: 35 в Списке последовательностей, обозначили как «легкая цепь типа hR198_LG3».
[0190] Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG3, приведена в SEQ ID NO: 59 в Списке последовательностей. Нуклеотиды в положениях с 61 по 702 кодируют зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 61 по 378 кодируют вариабельную область. Аминокислотная последовательность легкой цепи типа hR198_LG3 приведена в SEQ ID NO: 60 в Списке последовательностей. Аминокислоты в положениях с 21 по 234 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 21 по 126 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 59 и 60 также показаны на Фигуре 32. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0191]
10)-1-4 Тяжелая цепь типа hR198_HG1:
Гуманизированную тяжелую цепь R198, сконструированную путем замены остатка аспарагина в аминокислотном положении 78 остатком аспарагиновой кислоты и остатка валина в аминокислотном положении 123 остатком аланина в последовательности тяжелой цепи hR198_H3, приведенной в SEQ ID NO: 43 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_HG1».
[0192] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG1, приведена в SEQ ID NO: 61 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_HG1 приведена в SEQ ID NO: 62 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 135 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 61 и 62 также показаны на Фигуре 33. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0193]
10)-1-5 Тяжелая цепь типа hR198_HG2:
Гуманизированную тяжелую цепь R198, сконструированную путем замены остатка валина в аминокислотном положении 48 остатком изолейцина, остатка аспарагина в аминокислотном положении 78 остатком аспарагиновой кислоты, остатка аланина в аминокислотном положении 81 остатком глицина и остатка валина в аминокислотном положении 123 остатком аланина в последовательности тяжелой цепи hR198_H3, приведенной в SEQ ID NO: 43 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_HG2».
[0194] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG2, приведена в SEQ ID NO: 63 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_HG2 приведена в SEQ ID NO: 64 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 135 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 63 и 64 также показаны на Фигуре 34. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0195]
10)-1-6 Тяжелая цепь типа hR198_HG3:
Гуманизированную тяжелую цепь R198, сконструированную путем замены остатка валина в аминокислотном положении 48 остатком изолейцина, остатка аспарагина в аминокислотном положении 78 остатком аспарагиновой кислоты, остатка аланина в аминокислотном положении 81 остатком метионина и остатка валина в аминокислотном положении 123 остатком аланина в последовательности тяжелой цепи hR198_H3, приведенной в SEQ ID NO: 43 в Списке последовательностей, обозначили как «тяжелая цепь типа hR198_HG3».
[0196] Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG3, приведена в SEQ ID NO: 65 в Списке последовательностей. Нуклеотиды в положениях с 58 по 1398 кодируют зрелую тяжелую цепь, образованную путем отщепления сигнальной последовательности. Нуклеотиды в положениях с 58 по 408 кодируют вариабельную область. Аминокислотная последовательность тяжелой цепи типа hR198_HG3 приведена в SEQ ID NO: 66 в Списке последовательностей. Аминокислоты в положениях с 20 по 466 представляют собой зрелую легкую цепь, образованную путем отщепления сигнальной последовательности. Аминокислоты в положениях с 20 по 135 представляют собой вариабельную область. Обе последовательности SEQ ID NOs: 65 и 66 также показаны на Фигуре 35. Каждая последовательность CDR и ее соответствующий SEQ ID NO показаны на Фигуре 38.
[0197]
10)-2 Получение экспрессионного вектора созревания аффинности гуманизированного анти-человеческий Orai1 антитела
10)-2-1 Конструирование вектора, экспрессирующего легкую цепь типа hR198_LG1
Мутации для замены остатка аспарагина в аминокислотном положении 51 остатком глицина, остатка треонина в аминокислотном положении 113 остатком изолейцина и остатка треонина в аминокислотном положении 117 остатком серина вводили с использованием pCMA-LK/hR198_L3 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 55 в Списке последовательностей, обозначили как «pCMA-LK/hR198_LG1».
[0198]
10)-2-2 Конструирование векторов, экспрессирующих легкую цепь типа hR198_LG2 и LG3
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область hR198_LG3, соответствующую нуклеотидам в положениях с 38 по 402 нуклеотидной последовательности hR198_LG3, представленной SEQ ID NO: 59 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий hR198_H5, конструировали тем же способом, что и в Примере 7)-2-1. Полученный экспрессионный вектор обозначили как «pCMA-G1/hR198_H5». Синтезированный фрагмент ДНК расщепляли ферментами рестрикции EcoRV и AvaI и встраивали в pCMA-LK/hR198_LG1, расщепленный теми же ферментами, которые указаны выше, для получения экспрессионного вектора hR198_LG3. Полученный экспрессионный вектор обозначили как «pCMA-LK/hR198_LG3».
[0199] Мутации для замены остатка аргинина в аминокислотном положении 47 остатком глутамина и остатка серина в аминокислотном положении 73 остатком треонина вводили с использованием pCMA-LK/hR198_LG3 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 57 в Списке последовательностей, обозначили как «pCMA-LK/hR198_LG2».
[0200]
10)-2-3 Конструирование вектора, экспрессирующего тяжелую цепь типа hR198_HG1
Мутации для замены остатка аспарагина в аминокислотном положении 78 остатком аспарагиновой кислоты и остатка валина в аминокислотном положении 123 остатком аланина вводили с использованием pCMA-G1/hR198_H3 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 61 в Списке последовательностей, обозначили как «pCMA-G1/hR198_HG1».
[0201]
10)-2-4 Конструирование вектора, экспрессирующего тяжелую цепь типа hR198_HG2
Мутации для замены остатка валина в аминокислотном положении 48 остатком изолейцина и остатка аланина в аминокислотном положении 81 остатком глицина вводили с использованием pCMA-G1/hR198_HG1 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 63 в Списке последовательностей, обозначили как «pCMA-G1/hR198_HG2».
[0202]
10)-2-5 Конструирование вектора, экспрессирующего тяжелую цепь типа hR198_HG3
Мутацию для замены остатка глицина в аминокислотном положении 81 остатком метионина вводили с использованием pCMA-G1/hR198_HG2 в качестве матрицы при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-. Полученный экспрессионный вектор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 65 в Списке последовательностей, обозначили как «pCMA-G1/hR198_HG3».
[0203]
10)-3 Получение созревания аффинности гуманизированного анти-человеческий Orai1 антитела
Клетки FreeStyle 293F трансфицировали каждым вектором, экспрессирующим тяжелую цепь гуманизированного анти-человеческий Orai1 антитела, и каждым вектором, экспрессирующим легкую, цепь гуманизированного анти-человеческий Orai1 антитела, получеными в пункте 10)-2, тем же способом, что и в пункте 4)-5-1, для получения культурального супернатанта, содержащего антитело.
[0204] Гуманизированные анти-человеческий Orai1 антитела, полученные путем комбинации взятых в качестве матрицы pCMA-G1/hR198_H3 или pCMA-G1/hR198_HG1, pCMA-G1/hR198_HG2 или pCMA-G1/hR198_HG3, содержащих мутацию(и) с pCMA-LK/hR198_LG1, pCMA-LK/hR198_LG2 или pCMA-LK/hR198_LG3 обозначили как «hR198_H3/LG1», «hR198_HG1/LG1», «hR198_HG1/LG2», «hR198_HG1/LG3», «hR198_HG2/LG1» и «hR198_HG3/LG1», соответственно.
[0205] Каждый полученный культуральный супернатант очищали при помощи аффинной хроматографии с белком A тем же способом, что и в пункте 4)-5-2, для получения образца очищенного антитела.
[0206]
Пример 11. In vitro активность антитела при созревании аффинности
11)-1 Оценка способности к связыванию антитела при созревании аффинности, выполненная при помощи проточной цитометрии
Для оценки специфичности связывания с Orai1 человека, суспензию клеток HEK293T, трансфицированных pcDNA3.1-DEST, или суспензию клеток HEK293T, трансфицированных pcDNA3.1-hOrai1, полученные способом, описанным в пункте 1)-4-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T суспендировали путем добавления антитела с созреванием аффинности hR198_H3/LG1, hR198_HG1/LG1, hR198_HG1/LG2, hR198_HG1/LG3, hR198_HG2/LG1 или hR198_HG3/LG1, полученного в пункте 10)-3, или исходного антитела hR198_H3/L3 или hR198_H4/L4, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-человеческого IgG, конъюгированного с ФИТЦ, разбавленного в 100 раз ФСБ, содержащим 5% ФБС, и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида, с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). Антитела с созреванием аффинности hR198_H3/LG1, hR198_HG1/LG1, hR198_HG1/LG2, hR198_HG1/LG3, hR198_HG2/LG1 и hR198_HG3/LG1 не связывались с клетками HEK293T, трансфицированными pcDNA3.1-DEST, как в случае исходного антитела hR198_H3/L3 или hR198_H4/L4, и, как показано на Фигуре 8, каждое из них имело тенденцию связываться с Orai1 человека в клетках HEK293T, трансфицированных pcDNA3.1-hOrai1, на уровне эквивалентном или превышающем уровень исходного антитела.
[0207]
11)-2 Ингибирующее действие антитела при созревании аффинности на активацию T-лимфоцитов
T-лимфоцитов человека клеточную линию клеток Юрката получали в концентрации 1,5×106 клеток/мл в среде RPMI1640, содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина, инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали антителом при созревании аффинности hR198_H3/LG1, hR198_HG1/LG1, hR198_HG1/LG2, hR198_HG1/LG3, hR198_HG2/LG1 или hR198_HG3/LG1, или гуманизированным анти-человеческий Orai1 антителом hR198_H3/L3 или hR198_H4/L4, которые добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут в атмосфере с 5% содержанием CO2. Затем добавляли 100 нг/мл ФМА и 1 мкг/мл A23187 в количестве 10 мкл на лунку и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию IL-2, содержащегося в супернатанте, измеряли при помощи ИФА. На Фигуре 9 показано, что каждое из антител при созревании аффинности ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187. Каждое из антител при созревании аффинности ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, и все они имеют более высокую ингибирующую активность по сравнению с гуманизированными анти-человеческий Orai1 антителами H3/L3 и H4/L4, которые являются исходными антителами.
[0208]
Пример 12. Получение сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью
Сконструированную последовательность константной области hR198_HG1-LALA, полученную из hR198_HG1 путем замены двух аминокислотных остатков в константной области, конструировали как описано в Примерах ниже.
12)-1 Дизайн вектора, экспрессирующего тяжелую цепь типа LALA
Чтобы избежать эффекта цитотоксичности в отношении нормальных клеток, экспрессирующих Orai1 человека, желательно, чтобы антитело обладало низкой эффекторной активностью. Известно, что эффекторная активность различается у антител различных субклассов. Отмечены следующие характеристики, например, IgG4 обладает низкой АЗКЦ и КЗЦ активностью, а IgG2 обладает КЗЦ активностью, но низкой АЗКЦ активностью. На основе этих свойств можно получить антитело IgG1 с пониженной АЗКЦ и КЗЦ активностью путем частичной замены последовательностей константных областей IgG1 последовательностями, характерными для IgG2 или IgG4. В качестве одного примера, в работе Marjan Hezareh et al., Journal of Virology, 75 (24): 12161-12168 (2001) показано, что АЗКЦ и КЗЦ активность IgG1 уменьшается путем замены каждого из остатков лейцина в положениях 234 и 235 (положения обозначены EU-индексом, как описано у Kabat et al.) IgG1 остатком аланина. Таким образом, тяжелую цепь гуманизированного анти-человеческий Orai1 антитела, полученного путем замены остатка лейцина в аминокислотном положении 253 остатком аланина и остатка лейцина в аминокислотном положении 254 остатком аланина в последовательности тяжелой цепи hR198_HG1, полученной в пункте 10)-1, обозначили как «тяжелая цепь типа hR198_HG1-LALA».
[0209]
12)-2-1 Конструирование вектора, экспрессирующего тяжелую цепь типа LALA
12)-2-1 Конструирование вектора, экспрессирующего тяжелую цепь типа hR198_HG1-LALA
Мутации вводили с использованием в качестве матрицы полученного в пункте 7)-2 вектора pCMA-G1/hR198_H4, содержащего тяжелую цепь типа hR198_H4, при помощи набора реагентов для сайт-направленного мутагенеза KOD -Plus-, для конструирования вектора, экспрессирующего тяжелую цепь типа hR198_H4-LALA. Полученный экспрессионный вектор обозначили как «pCMA-G1-LALA/hR198_H4». Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H4-LALA, и аминокислотная последовательность этой тяжелой цепи представлены в SEQ ID NOs: 67 и 68 (Фигура 36), соответсвенно, в Списке последовательностей.
[0210] Фрагмент ДНК, размером приблизительно 0,6 т.п.н., содержащий последовательность, кодирующую вариабельную область антитела, получали путем расщепления pCMA-G1/hR198_HG1, полученного в пункте 10)-2, ферментами рестрикции PstI и XbaI и встраивали в фрагмент ДНК размером приблизительно 4,2 т.п.н., полученный путем расщепления pCMA-G1-LALA/hR198_H4 теми же ферментами рестрикции, которые указаны выше с использованием реагента Ligation High ver. 2, для конструирования вектора, экспрессирующего тяжелую цепь типа hR198_HG1-LALA. Полученный экспрессионный вектор был обозначен как «pCMA-G1-LALA/hR198_HG1». Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG1-LALA, и аминокислотная последовательность этой тяжелой цепи приведены в SEQ ID NOs: 69 и 70 (Фигура 37), соответственно, в Списке последовательностей.
[0211]
12)-3 Получение сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью
12)-3-1 Продуцирование сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью
Клетки FreeStyle 293F (Life Technologies Corp.) субкультивтровали и культивировали в соответствии с инструкцией производителя. 1,2×109 клеток FreeStyle 293F (Life Technologies Corp.), находящихся в логарифмической фазе роста, инокулировали в 3 л колбу Фернбаха Эрленмейера (Corning Inc.), разбавляли экспрессионной средой FreeStyle 293 (Life Technologies Corp.) до концентрации 1,0×106 клеток/мл, и затем культивировали при встряхивании при 90 об/мин при температуре 37°C в течение 1 часа в инкубаторе с атмосферой, содержащей 8% CO2. 3,6 мг полиэтиленимина (Polysciences #24765) растворяли в 20 мл среды Opti-Pro SFM (Life Technologies Corp.). Затем каждый вектор, экспрессирующий легкую цепь, (0,8 мг) и каждый вектор, экспрессирующий тяжелую цепь, (0,4 мг), полученные с использованием набора реагентов PureLink HiPure Plasmid (Life Technologies Corp.) добавляли к 20 мл среды Opti-Pro SFM (Life Technologies Corp.). 20 мкл смешанного раствора экспрессионный вектор/среда Opti-Pro SFM добавляли к 20 мл смешанного раствора полиэтиленимин/среда Opti-Pro SFM и эту смесь осторожно перемешивали, затем оставляли на 5 минут и затем добавляли к клеткам FreeStyle 293F. Клетки культивировали при встряхивании при 90 об/мин при температуре 37°C в течение 7 суток в инкубаторе с атмосферой, содержащей 8% CO2, и полученный культуральный супернатант фильтровали через одноразовый мембранный фильтр Millex (ADVANTEC #CCS-045-E1H). hR198_HG1/LG1 получали путем комбинации pCMA-G1/hR198_HG1 и pCMA-LK/hR198_LG1, и hR198_HG1-LALA/LG1 получали путем комбинации pCMA-G1-LALA/hR198_HG1 и pCMA-LK/hR198_LG1.
12)-3-2 Двухстадийная очистка сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью
Каждое антитело очищали из культурального супернатанта, полученного в Примере 12)-3-1, в два этапа при помощи аффинной хроматографии на белке A (при температуре от 4 до 6°C) и керамического гидроксиапатита (при комнатной температуре). Этап замены буфера после очистки при помощи аффинной хроматографии на белке A проводили и после очистки при помощи керамического гидроксиапатита проводили при температуре от 4 до 6°C. Культуральный супернатант наносили на колонку MabSelectSuRe (GE Healthcare Bio-Sciences Corp., HiTrap column), уравновешенную ФСБ. После введения в колонку всего объема культурального супернатанта, колонку промывали ФСБ в объеме, в 2 или более раз превышающем объем колонки. Затем, фракции, содержащие антитело, собирали элюированием 2 М раствором гидрохлорида аргинина (pH 4,0). Буферы фракций заменяли ФСБ при помощи диализа (Thermo Fisher Scientific Inc., Slide-A-Lyzer Dialysis Cassette), и затем разбавляли в 5 раз буфером, содержащим 5 мМ фосфата натрия/50 мМ MES (pH 7,0). Полученный раствор антитела наносили на колонку с керамическим гидроксиапатитом (Bio-Rad Laboratories, Inc., Bio-Scale CHT Type-1 Hydroxyapatite Column), уравновешенную буфером 5 мМ NaPi/50 мМ MES/30 мМ NaCl (pH 7,0). Фракции, содержащие антитело, собирали при помощи элюирования с линейным градиентом концентрации хлорида натрия. Буферы фракций заменяли HBSor (25 мМ гистидин/5% сорбит, pH 6,0) путем диализа (Thermo Fisher Scientific Inc., Slide-A-Lyzer Dialysis Cassette). Раствор концентрировали до концентрации IgG 5 мг/мл или выше с использованием ультрафильтрационного центробежного концентратора VIVASPIN 20 (отсечение по молекулярной массе: UF30K, Sartorius Japan K.K., при температуре 4°C) и использовали в качестве очищенного образца.
[0213]
Пример 13. Получение векторов, экспрессирующих человеческое анти-человеческий Orai1 антитело 2C1.1 и 5H3.1
Антитела 2C1.1 и 5H3.1 получали на основе аминокислотных последовательностей легкой и тяжелой цепей, описанных в Международной патентной заявке WO2011063277A1.
13)-1 Конструирование вектора pCMA-G2, экспрессирующего тяжелую цепь гибридного и гуманизированного антитела типа IgG2
Фрагмент ДНК, полученный путем расщепления pCMA-LK ферментами рестрикции XbaI и PmeI для удаления последовательности, кодирующей сигнал секреции κ цепи человека и константную область κ цепи человека, лигировали с фрагментом ДНК (приведенным в SEQ ID NO: 71 в Списке последовательностей), содержащим последовательность, кодирующую сигнал секреции тяжелой цепи человека и аминокислоты константной области IgG2 человека, с использованием набора реагентов In-Fusion Advantage PCR cloning kit (Clontech Laboratories, Inc.) для конструирования вектора pCMA-G2, экспрессирующего тяжелую цепь гибридного и гуманизированного антитела типа IgG2, содержащего сигнальную последовательность, сайт клонирования и последовательность, кодирующую константную область тяжелой цепи IgG2 человека, после промотора ЦМВ.
[0214]
13)-2 Конструирование вектора, экспрессирующего тяжелую цепь антитела 2C1.1
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область тяжелой цепи антитела 2C1.1, соответствующую нуклеотидам в положениях с 36 по 434 в нуклеотидной последовательности, кодирующей тяжелую цепь антитела 2C1.1, которая представлена в SEQ ID NO: 72 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область тяжелой цепи антитела 2C1.1, амплифицировали с синтезированным фрагментом ДНК в качестве матрицы с использованием ДНК-полимеразы KOD -Plus- и встраивали в соответствующий сайт вектора pCMA-G2, экспрессирующего тяжелую цепь гибридного и гуманизированного антитела типа IgG2, расщепленного ферментом рестрикции BlpI, с использованием набора реагентов In-Fusion HD PCR cloning kit для конструирования вектора, экспрессирующего тяжелую цепь антитела 2C1.1. Полученный экспрессионный вектор обозначили как «pCMA-G2/2C1.1».
[0215] Аминокислотная последовательность тяжелой цепи антитела 2C1.1 приведена в SEQ ID NO: 73 в Списке последовательностей.
[0216]
13)-3 Конструирование вектора, экспрессирующего легкую цепь антитела 2C1.1
Синтезировали фрагмент ДНК, содержащий последовательности, кодирующие вариабельную область и константную область (λ цепь) легкой цепи антитела 2C1.1, соответствующие нуклеотидам в положениях с 38 по 739 в нуклеотидной последовательности, кодирующей легкую цепь антитела 2C1.1, которая представлена в SEQ ID NO: 74 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, содержащий последовательности, кодирующие вариабельную область и константную область легкой цепи антитела 2C1.1, амплифицировали с синтезированным фрагментом ДНК в качестве матрицы с использованием ДНК-полимеразы KOD -Plus- и встраивали в соответствующий сайт вектора pCMA-LK, экспрессирующего легкую цепь гибридного и гуманизированного антитела, расщепленного ферментами рестрикции BsiWI и PmeI, с использованием набора реагентов In-Fusion HD PCR cloning kit для конструирования вектора, экспрессирующего легкую цепь антитела 2C1.1. Полученный экспрессионный вектор обозначили как «pCMA-L/2C1.1»
[0217] Аминокислотная последовательность легкой цепи антитела 2C1.1 приведена в SEQ ID NO: 75 в Списке последовательностей.
[0218]
13)-4 Конструирование вектора, экспрессирующего тяжелую цепь антитела 5H3.1
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область тяжелой цепи антитела 5H3.1, соответствующую нуклеотидам в положениях с 36 по 434 в нуклеотидной последовательности, кодирующей тяжелую цепь антитела 5H3.1, которая представлена в SEQ ID NO: 76 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий тяжелую цепь антитела 5H3.1, конструировали тем же способом, что и в Примере 13)-2. Полученный экспрессионный вектор обозначили как «pCMA-G2/5H3.1».
[0219] Аминокислотная последовательность тяжелой цепи антитела 5H3.1 приведена в SEQ ID NO: 77 в Списке последовательностей.
[0220]
13)-5 Конструирование вектора, экспрессирующего легкую цепь антитела 5H3.1
Синтезировали фрагмент ДНК, содержащий последовательность, кодирующую вариабельную область легкой цепи антитела 5H3.1, соответствующую нуклеотидам в положениях с 38 по 742 в нуклеотидной последовательности, кодирующей легкую цепь антитела 5H3.1, которая представлена в SEQ ID NO: 76 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий легкую цепь антитела 5H3.1, конструировали тем же способом, что и в Примере 13)-3. Полученный экспрессионный вектор обозначили как «pCMA-L/5H3.1».
[0221] Аминокислотная последовательность легкой цепи антитела 5H3.1 приведена в SEQ ID NO: 79 в Списке последовательностей.
[0222]
13)-6 Получение антител 2C1.1 и 5H3.1
13)-6-1 Продуцирование антител 2C1.1 и 5H3.1
Каждое антитело продуцировали тем же способом, что и в Примере 12)-3-1. Антитело 2C1.1 продуцировали путем комбинации pCMA-G2/2C1.1 и pCMA-L/2C1.1, и антитело 5H3.1 продуцировали путем комбинации pCMA-G2/5H3.1 и pCMA-L/5H3.1.
[0223]
13)-6-2 Двухстадийная очистка антител 2C1.1 и 5H3.1
Каждое антитело очищали от культурального супернатанта, полученного в Примере 13)-6-1, тем же способом, что и в Примере 12)-3-2.
[0224]
Пример 14. Получение мышиных анти-человеческий Orai1 антител 10F8, 14F74 и 17F6
Антитела 10F8, 14F74 и 17F6 получали на основе аминокислотных последовательностей легкой и тяжелой цепей, описанных в Международной патентной заявке WO2013091903A1.
14)-1 Конструирование вектора, экспрессирующего тяжелую цепь антитела 10F8
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую тяжелую цепь антитела 10F8, которая представлена в SEQ ID NO: 80 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Фрагмент ДНК, содержащий последовательность, кодирующую тяжелую цепь антитела 10F8, амплифицировали с синтезированным фрагментом ДНК в качестве матрицы с использованием ДНК-полимеразы KOD -Plus- и встраивали в сайт, из которого последовательность, кодирующую сигнал секреции κ цепи и константную область κ цепи человека, удаляли путем расщепления вектора pCMA-LK, экспрессирующего легкую цепь гибридного и гуманизированного антитела, расщепленного ферментами рестрикции XbaI и PmeI, с использованием набора реагентов In-Fusion HD PCR cloning kit для конструирования вектора, экспрессирующего тяжелую цепь антитела 10F8. Полученный экспрессионный вектор обозначили как «pCMA/10F8H».
[0225] Аминокислотная последовательность тяжелой цепи антитела 10F8 приведена в SEQ ID NO: 81 в Списке последовательностей.
[0226]
14)-2 Конструирование вектора, экспрессирующего легкую цепь антитела 10F8
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую легкую цепь антитела 10F8, которая представлена в SEQ ID NO: 82 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Синтезированный фрагмент ДНК встраивали в сайт, из которого последовательность, кодирующую сигнал секреции κ цепи и константную область κ цепи человека, удаляли путем расщепления вектора pCMA-LK, экспрессирующего легкую цепь гибридного и гуманизированного антитела, расщепленного ферментами рестрикции XbaI и PmeI, с использованием набора реагентов In-Fusion HD PCR cloning kit для конструирования вектора, экспрессирующего легкую цепь антитела 10F8. Полученный экспрессионный вектор обозначили как «pCMA/10F8L».
[0227] Аминокислотная последовательность легкой цепи антитела 10F8 приведена в SEQ ID NO: 83 в Списке последовательностей.
[0228]
14)-3 Конструирование вектора, экспрессирующего тяжелую цепь антитела 14F74
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую тяжелую цепь антитела 14F74, которая представлена в SEQ ID NO: 84 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий тяжелую цепь антитела 14F74, конструировали тем же способом, что и в Примере 14)-1. Полученный экспрессионный вектор обозначили как «pCMA/14F74H».
[0229] Аминокислотная последовательность тяжелой цепи антитела 14F74 приведена в SEQ ID NO: 85 в Списке последовательностей.
[0230]
14)-4 Конструирование вектора, экспрессирующего легкую цепь антитела 14F74
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую легую цепь антитела 14F74, которая представлена в SEQ ID NO: 86 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий легкую цепь антитела 14F74, конструировали тем же способом, что и в Примере 14)-2. Полученный экспрессионный вектор обозначили как «pCMA/14F74L».
[0231] Аминокислотная последовательность легкой цепи антитела 14F74 приведена в SEQ ID NO: 87 в Списке последовательностей.
[0232]
14)-5 Конструирование вектора, экспрессирующего тяжелую цепь антитела 17F6
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую тяжелую цепь антитела 17F6, которая представлена в SEQ ID NO: 88 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий тяжелую цепь антитела 17F6, конструировали тем же способом, что и в Примере 14)-1. Полученный экспрессионный вектор обозначили как «pCMA/17F6H».
[0233] Аминокислотная последовательность тяжелой цепи антитела 17F6 приведена в SEQ ID NO: 89 в Списке последовательностей.
[0234]
14)-6 Конструирование вектора, экспрессирующего легкую цепь антитела 17F6
Синтезировали фрагмент ДНК, содержащий нуклеотидную последовательность, кодирующую легую цепь антитела 17F6, которая представлена в SEQ ID NO: 90 в Списке последовательностей (сервис GeneArt Artificial Gene Synthesis). Вектор, экспрессирующий легкую цепь антитела 17F6, конструировали тем же способом, что и в Примере 14)-2. Полученный экспрессионный вектор обозначили как «pCMA/17F6L».
[0235] Аминокислотная последовательность легкой цепи антитела 17F6 приведена в SEQ ID NO: 91 в Списке последовательностей.
[0236]
14)-7 Получение антител 10F8, 14F74 и 17F6
14)-7-1 Продуцирование антител 10F8, 14F74 и 17F6
Каждое антитело продуцировали тем же способом, что и в Примере 12)-3-1. Антитело 10F8 продуцировали путем комбинации pCMA/10F8H и pCMA/10F8L. Антитело 14F74 продуцировали путем комбинации pCMA/14F74H и pCMA/14F74L. Антитело 17F6 продуцировали путем комбинации pCMA/17F6H и pCMA/17F6L.
[0237]
14)-7-2 Двухстадийная очистка антител 10F8, 14F74 и 17F6
Каждое антитело очищали от культурального супернатанта, полученного в Примере 14)-7-1, тем же способом, что и в Примере 12)-3-2.
[0238]
Пример 15. Сравнение активности in vitro сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью с другими анти-человеческий Orai1 антителами
15)-1 Определение антигенсвязывающей активности анти-человеческий Orai1 антитела при помощи проточной цитометрии
Суспензию клеток HEK293T, трансфицированных способом, описанным в пункте 1)-4-2, каждым вектором, экспрессирующим Orai1 человека, сконструированным в пункте 1)-1-1, центрифугировали для удаления супернатанта. Затем клетки HEK293T, трансфицированные pcDNA3.1-hOrai1, или клетки HEK293T, трансфицированные pcDNA3.1-DEST, суспендировали путем добавления hR198_HG1/LG1 или hR198_HG1/LG1-LALA, полученных в пункте 12)-3, 2C1.1 или 5H3.1, полученных в пункте 13)-6, 10F8, 14F74 или 17F6, полученных в пункте 14)-7, или контрольного антитела IgG человека или контрольного антитела IgG мыши в качестве контроля и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, затем суспендировали путем добавления анти-человеческого IgG, конъюгированного с ФИТЦ, разбавленного в 100 раз ФСБ, содержащим 5% ФБС, для человеческих антител, или анти-мышиного IgG, конъюгированного с ФИТЦ, (Cappel Laboratories, Inc.) для мышиных антител и инкубировали при температуре 4°C в течение 30 минут. Клетки два раза промывали ФСБ, содержащим 5% ФБС, и затем ресуспендировали в ФСБ, содержащим 5% ФБС и 1 мкг/мл пропидиум иодида, с последующей детекцией при помощи проточного цитометра (FC500). Данные анализировали с использованием программы Flowjo. После исключения пропидиум иодид положительных мертвых клеток путем гейтирования, интенсивность флуоресценции ФИТЦ живых клеток наносили на график в виде гистограммы для вычисления средней интенсивности флуоресценции (СИФ). hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6 не связывались с клетками HEK293T, и, как показано на Фигуре 10 (человеческие антитела) и Фигуре 11 (мышиные антитела), каждое из них связывалось с клетками HEK293T, трансфицированными pcDNA3.1-hOrai1, указывая на то, что все эти антитела специфично связываются с Orai1 человека. С другой стороны, такое связывание не наблюдалось в случае контрольного антитела IgG мыши.
[0239]
15)-2 Ингибирующее действие анти-человеческий Orai1 антитела на активацию линии T-лимфоцитов
T-лимфоцитов человека клеточную линию клеток Юрката получали в концентрации 1,5×106 клеток/мл в среде RPMI1640, содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина, инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 или 17F6, которые добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут в атмосфере с 5% содержанием CO2. Затем добавляли 100 нг/мл ФМА и 1 мкг/мл A23187 в количестве 10 мкл на лунку (конечная концентрация: 10 нг/мл ФМА и 100 нг/мл A23187) и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию интерлейкина 2 (IL-2), содержащегося в супернатанте, измеряли при помощи ИФА. На Фигуре 12 показано, что каждое из полученных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из клеток Юрката, обработанных ФМА и A23187. На Фигуре 13 показана полумаксимальная ингибирующая концентрация (ИК50) и 80% от ингибирующей концентрации (ИК80) hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6 с концентрацией IL-2 в отсутствии антитела, определенной как 100%. ИК50 антител предшествующего уровня техники составила 80 нг/мл или выше, в то время как ИК50 типичных антител настоящего изобретения составила 10 нг/мл или ниже. ИК80 антител предшествующего уровня техники составила 60000 нг/мл или выше, в то время как ИК80 типичных антител настоящего изобретения составила 200 нг/мл или ниже.
[0240]
15)-3 Ингибирующее действие на мононуклеарные клетки периферической крови сконструированной формы антитела с созреванием аффинности с пониженной эффекторной активностью и других анти-человеческий Orai1 антител
Мононуклеарные клетки периферической крови (МКПК) человека приобретали в виде замороженного продукта в компании Cellular Technology Ltd. и использовали после оттаивания в соответствии с инструкцией производителя. МКПК получали в концентрации 2,0×106 клеток/мл в среде RPMI1640, содержащей 10% ФБС, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина, инокулировали в количестве 80 мкл на лунку в 96-луночный планшет для культивирования клеток и предварительно обрабатывали каждым анти-человеческий Orai1 антителом которое добавляли в количестве 10 мкл на лунку при температуре 37°C в течение 60 минут. Затем добавляли 100 нг/мл ФМА и 1 мкг/мл A23187 в количестве 10 мкл на лунку и хорошо перемешивали, после чего культивировали при температуре 37°C в течение приблизительно 16 часов в атмосфере с 5% содержанием CO2. Планшет хорошо перемешивали и затем центрифугировали при 600 g в течение 3 минут. Концентрацию интерлейкина 2 (IL-2) и интерферона γ (ИФНγ), содержащихся в супернатанте, измеряли при помощи ИФА (Mabtech AB). На Фигуре 51 показано, что каждое из полученных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из МКПК человека, обработанных ФМА и A23187. На Фигуре 52 показана полумаксимальная ингибирующая концентрация (ИК50) и 80% от ингибирующей концентрации (ИК80) hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6 с концентрацией IL-2 в отсутствии антитела, определенной как 100%. ИК50 антител предшествующего уровня техники составила 100 нг/мл или выше, в то время как ИК50 типичных антител настоящего изобретения составила 20 нг/мл или ниже. ИК80 антител предшествующего уровня техники составила 17000 нг/мл или выше, в то время как ИК80 типичных антител настоящего изобретения составила 400 нг/мл или ниже. На Фигуре 53 показано, что каждое из полученных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение ИФНγ из МКПК человека, обработанных ФМА и A23187. На Фигуре 51 показано, что каждое из полученных анти-человеческий Orai1 антител ингибирует, зависимым от концентрации образом, высвобождение IL-2 из МКПК человека, обработанных ФМА и A23187. На Фигуре 54 показана полумаксимальная ингибирующая концентрация (ИК50) и 80% от ингибирующей концентрации (ИК80) hR198_HG1/LG1, hR198_HG1-LALA/LG1, 2C1.1, 5H3.1, 10F8, 14F74 и 17F6 с концентрацией ИФНγ в отсутствии антитела, определенной как 100%. ИК50 антител предшествующего уровня техники составила 800 нг/мл или выше, в то время как ИК50 типичных антител настоящего изобретения составила 40 нг/мл или ниже. ИК80 антител предшествующего уровня техники составила 300000 нг/мл или выше, в то время как ИК80 типичных антител настоящего изобретения составила 2000 нг/мл или ниже.
[0241]
Пример 16. In vivo активность hR198_HG1/LG1
16)-1 Влияние введения hR198_HG1/LG1 в мышиной модели реакции по типу «трансплантат против хозяина», вызванной трансплантацией МКПК человека
Известно, что реакция, подобная реакции по типу «трансплантат против хозяина» у человека, может быть индуцирована путем трансплантации МКПК человека мышам NSG (NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJ), которые являются мышами с тяжелым комбинированным иммунодефицитом (Clinical and Experimental Immunology, 157: 104-118 (2009)). 17 шестинедельных самцов мышей NSG, приобретенных у Charles River Laboratories Japan, Inc., облучали рентгеновскими лучами в дозе 2,0 Гр (аппарат для рентгеновского облучения Hitachi MBR-1520R-4). Затем мышей разделяли на группы. 1 группа включала в себя двух мышей, и 3 группы включали в себя по 5 мышей каждая. hR198_HG1/LG1 в концентрации 3 мг/мл и 6 мг/л в HBSor (25 мМ гистидин/5% сорбит, pH 6,0) внутривенно вводили в хвостовую вену в количестве 10 мл/кг, т.е. 30 мг/кг и 60 мг/кг мышам двух групп (n=5). Только HBSor вводили еще одной группе (n=5), служившей контрольной группой, получавшей только носитель. На следующий день замороженные МКПК человека (Cellular Technology Ltd.) оттаивали с использованием антиагрегатора CTL (Cellular Technology Ltd.) по протоколу фирмы-производителя, и 3000000 клеток МКПК человека суспендировали в 200 мкл ФСБ и трансплантировали мышам трех групп (n=5), при этом МКПК человека не трансплантировали мышам первой группы (n=2), которую наблюдали во времени как облученную рентгеновскими лучами контрольную группу. Через 0, 7, 14 и 21 суток после облучения рентгеновскими лучами hR198_HG1/LG1 или HBSor вводили в той же дозе, которая указана выше, группам, получавшим их ранее. Массу тела каждой мыши измеряли в дни 0, 1, 4, 7, 9, 10, 11 и 13 и затем каждый день. Изменение массы тела вычисляли в процентах от массы тела в день 0, принятой за 100%. Изменение средней массы тела в каждой группе показано на Фигуре 55. Снижение средней массы тела в контрольной группе, получавшей носитель, начинало наблюдаться примерно с дня 16. Эксперимент прекращали в день 21, когда средняя масса тела в контрольной группе, получавшей носитель, опустилась ниже 80%. В этот момент времени облученная рентгеновскими лучами контрольная группа, которая не получала МКПК человека, не обнаруживала снижение массы тела. Мыши, получавшие hR198_HG1/LG1 в количестве 30 мг/кг и 60 мг/кг, не снижали своей массы тела, и их средняя масса тела была эквивалентна средней массе тела мышей, которым не трансплантировали МКПК человека. Ожидается, что hR198_HG1/LG1 (анти-Orai1 антитело), котрое заметно подавляло сиптомы реакции по типу «трансплантат против хозяина», вызванной активацией МКПК человека в этой системе, будет оказывать терапевтическое и/или профилактическое действие на реакцию по типу «трансплантат против хозяина» у человека.
[0242]
16)-2 Оценка in vivo активности hR198_HG1/LG1 с использованием мышей, в чей геном встроен ген Orai1 человека.
16)-2-1 Получение мышей с геном Orai1 человека
Мышей с геном Orai1 человека, у которых аминокислотная последовательность домена внеклеточной петли мышиного Orai1 была заменена последовательностью Orai1 человека, получали в Институте иммунологии Co., Ltd. стандартным способом получения генно-инженерной мыши. В кратком изложении, направленный вектор для введения гена Orai1 человека конструировали путем замены последовательности ДНК в BAC-клоне, кодирующей область, содержащую локус мышиного гена Orai1, локусом гена Orai1 человека при введении в него гена устойчивости к неомицину, фланкированного последовательностями loxP. Мышиные клетки ES трансфицировали этим направленным вектором для получения устойчивой линии G418. Линию клеток ES, содержащую специфично рекомбинироанный целевой локус гена, отбирали при помощи гибридизации по Саузерну. Селекционный маркер удаляли при помощи трансфекции экспрессирующего Cre вектора, и полученную линию клеток ES использовали для получения гибридных мышей F1. Генотипирование проводили при помощи гибридизации по Саузерну для отбора гетерозиготных мутантных особей из числа мышей F1. Отобранные гетерозиготные мутантные особи F1 спаривались для получения мышей с геном Orai1 человека в виде гомозиготных мутантов F2.
[0243]
16)-2-2 Подавляющее действие hR198_HG1/LG1 на пассивную кожную анафилактическую (ПКА) реакцию
ПКА реакцию у мышей вызывали стандартным способом. Восьминедельных мышей с геном Orai1 человека получали в Институте иммунологии Co., Ltd., или потомство мышей дикого типа содержали в удерживающих приспособлениях. Затем hR198_HG1/LG1 в концентрации 6 мг/л в HBSor внутривенно вводили мышам в хвостовую вену в количестве 10 мл/кг, т.е. 60 мг/кг. Только HBSor вводили группе, служившей контрольной группой, получавшей только носитель. На следующий день 10 мкл моноклонального анти-OVA IgE (Chondrex, Inc.), доведенного до концентрации 10 мкг/мл физиологическим раствором внутрикожно вводили в ушную раковину каждой мыши под ингаляционной анестезией изофлураном (Pfizer Japan Inc.). Через 24 часа физиологический раствор, содержащий 2 мг/мл овальбумина (OVA) (альбумин белка куриного яйца, Sigma-Aldrich Corp.) и 20 мг/мл красителя Эванс синий (Merck KGaA) внутривенно вводили в количестве 5 мг/кг OVA и 100 мг/кг красителя Эванс синий в хвостовую вену. Через 30 минут каждую мышь умерщвляли путем кровопускания под глубокой анестезией изофлураном, ее ушную раковину отрезали и погружали в 0,5 мл ДМСО с последующей экстракцией Эванса синего (37°C, 72 часа). Раствор ДМСО, содержащий экстрагированный краситель Эванс синий, переносили в количестве 200 мкл на лунку в 96-луночный микропланшет и измеряли OD650, используя планшет-ридер (Molecular Devices Corp., SpectraMax M5e). Поглощение экстравазированного красителя Эванс синий определяли способом расчета, приведенным ниже, и указывали со средним значением для группы, получавшей только носитель, (контроль) взятым за 100%. Поглощение раствора ДМСО определяли как холостую пробу. %=(OD650 образца - OD650 холостой пробы)/(OD650 контроля - OD650 холостой пробы). На Фигуре 56 показано, что hR198_HG1/LG1 подавляет ПКА реакцию, индуцированную у мышей с геном Orai1 человека. Мышиная ПКА реакция является базовой модельной системой аллергии немедленного типа, вызывающей анафилаксию, которая представляет собой аллергическую реакцию I типа у человека, и известно, что эта система пригодна для оценки антиаллергических лекарственных средств (Archives internationales de pharmacodynamie et de therapie, 165: 92-102 (1967); и International archives of allergy and applied immunology, 78: 113-117 (1985)). Ожидается, что hR198_HG1/LG1 (анти-Orai1 антитело), обладающее подтвержденной в этой системе ингибирующей активностью против IgE зависимой дегрануляции тучных клеток, будет оказывать терапевтическое и/или профилактическое действие при бронхиальной астме, аллергическом рините и атопическом дерматите, обнаруженное у существующих ингибиторов дегрануляции тучных клеток, и аналогичных аллергических заболеваниях I типа у людей, например, при аллергической астме.
[0244]
16)-2-3 Подавляющее действие hR198_HG1/LG1 на гиперчувствительную реакцию замедленного типа (ГЗТ)
ГЗТ реакцию у мышей вызывали стандартным способом. Восьминедельных мышей с геном Orai1 человека получали в Институте иммунологии Co., Ltd., или потомство мышей дикого типа содержали в удерживающих приспособлениях. Затем hR198_HG1/LG1 в концентрации 6 мг/л в HBSor внутривенно вводили мышам в хвостовую вену в количестве 10 мл/кг, т.е. 60 мг/кг. Только HBSor вводили группе, служившей контрольной группой, получавшей только носитель. На следующий день 50 мкл эмульсии, полученной путем смешивания в равных количествах мБСА (бычий сывороточный альбумин Sigma-Aldrich Corp.), разбавленного до концентрации 5 мг/мл физиологическим раствором, и полного адъюванта Фрейнда (Difco Laboratories), подкожно вводили в каждую подмышку каждой мыши для иммунизации. Через 6 дней hR198_HG1/LG1 снова внутривенно вводили в хвостовую вену в количестве 60 мг/кг. На следующий день мБСА, разбавленный до концентрации 0,5 мг/мл физиологическим раствором, и физиологический раствор каждый внутрикожно вводили в заднюю лапу под ингаляционной анестезией изофлураном. Через 6, 24 и 48 часов после введения опухание лапы измеряли при помощи измерительного прибора с круговой шкалой. Зависимое от времени изменение толщины припухлости лапы вычисляли, используя опухание в 0 часов, принятым за 0 (10-2 мм), и определенное среднее значение в контрольной группе, получавшей носитель, принятое за 100%. На Фигуре 57 показано, что hR198_HG1/LG1 подавляет ГЗТ реакцию, индуцированную у мышей с геном Orai1 человека, в каждый момент времени через 6 часов (A), 24 часа (B) и 48 часов (C) после введения. Мышиная ПКА реакция является системой для наблюдения за формированием и ответом при Т-клеточно зависимой иммунизации, и хорошо известно, что лекарственное средство, проявляющее активность в этой системе, было создано в качестве мощного иммуносупрессора (Clinical and Experimental Immunology, 52: 599-606 (1983)). Ожидается, что hR198_HG1/LG1 (анти-Orai1 антитело), обладающее подтвержденной в этой системе супрессивной активностью против Т-клеточной иммунизации, будет оказывать терапевтическое и/или профилактическое действие на реакции организма или заболевания, вызываемые активностью у человека, например, отторжение трансплантата, иммунологические заболевания и воспалительные заболевания.
[0245]
16)-2-4 Изучение подавляющей активности анти-Orai1 антитела против дерматита
Дерматит индуцировали любым из следующих способов: 0,5 мл раствора антигена альбумина внутрибрюшинно вводили мышам, а через 2 недели указанное выше количество антигена вводили им в качестве бустера. Затем раствор антигена альбумина повторно наносили на ухо (20 мкл) или спину (100 мкл) от 3 до 6 раз с интервалами от 3 дней до 2 недель. В альтернативном варианте, мазь, содержащую клещевой антиген, повторно наносили на ухо (20 мкл) или спину (100 мкл) от 3 до 6 раз с интервалами от 3 дней до 2 недель. Гаптен пикрилхлорид или динитрофторбензол получали в соответствии с протоколом и наносили на ухо (20 мкл) или спину (100 мкл) однин или два раза в неделю в течение 8 недель максимум. В альтернативном варианте, вызывающее кожную реакцию вещество гистамин, химическое соединение 40/80, 5- (и 6-) сукцинимидиловый сложный эфир карбоксифлуоресцеин диацетата, флуоресцеинизотиоцианат или бомбезинподобный пептид вводили в ухо (20 мкл), спину (100 мкл) или спинной мозг (5 мкл). Анти-Orai1 антитело вводили внутривенно или подкожно за одни сутки до индуцирования дерматита, или за 1-4 часа до индуцирования дерматита. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. После индуцирования дерматита измерение толщины ушной раковины с использованием толщиномера с циферблатным индикатором или шкалы оценки макроскопических признаков дерматита производили с течением времени. После завершения тестового периода, проводили количественное определение концентрации антитела, цитокинов или биомаркеров сыворотки в крови или тканях, исследование активности роста, способности продуцировать цитокины или поверхностных антигенов клеток, полученных из кожи, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга, гистопатологический анализ и т.д. для определения подавляющей активности анти-Orai1 антитела против дерматита.
[0246]
16)-2-5 Изучение подавляющей активности анти-Orai1 антитела против псориаза
В случае использования имиквимода, псориатический дерматит индуцировали путем нанесения его на обе стороны или одну сторону ушной раковины (5-30 мг) и на бритую спину (50-100 мг). В альтернативном варианте дерматит индуцировали путем внутрибрюшинного введения 200 мкл суспензию зимозана с концентрацией 10 мг/мл в фосфатно-буферном растворе. В случае использования в качестве проглогистического вещества цитокина (IL-23 и т.д.), псориатический дерматит индуцировали путем внутрикожного введения 20-50 мкл раствора, содержащего 0,1-2 мкг цитокина, в одну сторону ушной раковины мыши под глубокой анестезией 1-5% изофлурана. Анти-Orai1 антитело вводили внутривенно или подкожно за одни сутки до индуцирования дерматита, или за 1-4 часа до индуцирования дерматита. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. После индуцирования дерматита измерение толщины ушной раковины с использованием толщиномера с циферблатным индикатором или шкалы оценки макроскопических признаков дерматита производили с течением времени. После завершения тестового периода, проводили исследование массы участка воспаления, активности миелопероксидазы нейтрофилов, инфильтрированных в этот участок, анализ инфильтрированных клеток при помощи проточной цитометрии, генетический анализ, измерение концентрации цитокинов и т.д. для определения подавляющей активности анти-Orai1 антитела против псориаза.
[0247]
16)-2-6 Изучение подавляющей активности анти-Orai1 антитела против рассеянного склероза
Раствор миелинового олигодендроцитарного гликопротеина или его частичного пептидного антигена доводили до концентрации 4 мг/мл физиологическим раствором и эмульгировали путем смешивания в равных количествах с полным адъювантом Фрейнда, доведенным до концентрации 8 мг/мл физиологическим раствором. 200 мкл этого смешанного раствора внутрикожно вводили в бок или в область живота мыши. Непосредственно после этого 100 мкл водного раствора коклюшного токсина с концентрацией 2 мкг/мл вводили эотй мыши в хвостовую вену. Через 2 дня вышеупомянутый раствор коклюшного токсина снова вводили в хвостовую вену, чтобы индуцировать экспериментальный энцефаломиелит. Через 1-2 недели эксперимента развивался энцефаломиелит, и паралич от хвоста к задним конечностям и передним конечностям. Степень этого паралича оценивали путем наружного наблюдения за движением конечностей и хвоста. Анти-Orai1 антитело вводили внутривенно или подкожно за одни сутки до индуцирования энцефаломиелита, или за 1-4 часа до индуцирования энцефаломиелита. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. Определяли влияние введения анти-Orai1 антитела на рассеянный склероз.
[0248]
16)-2-7 Изучение подавляющей активности анти-Orai1 антитела против артрита,
100 мкл эмульсии, полученную путем смешивания бычьего коллагена типа II (концентрация 2 мг/мл) с полным адъювантом Фрейнда в объемном соотношении 1:1,3, внутрикожно вводили в основание хвоста мыши при помощи 1мл шприца и туберкулиновой иглы. Через 2-3 недели осуществляли такое же введение, как указано выше. Затем опухание суставов конечностей оценивали макроскопически или при помощи толщиномера с циферблатным индикатором. После завершения тестового периода измеряли концентрацию антитела, цитокинов или биомаркеров сыворотки в крови или тканях, активность роста, способность продуцировать цитокины или поверхностные антигены клеток, полученных из кожи, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга и т.д. Анти-Orai1 антитело вводили внутривенно или подкожно за одни сутки до индуцирования энцефаломиелита, или за 1-4 часа до индуцирования энцефаломиелита. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. Определяли влияние введения анти-Orai1 антитела на артрит.
[0249]
16)-2-8 Изучение подавляющей активности анти-Orai1 антитела против колита
Лимфоциты, собранные и очищенные из лимфатических узлов и селезенки мышей, в чей геном встроен ген Orai1 человека, выделяли при помощи клеточного сортера с использованием анти-CD4 антитела GK1.5, анти-CD25 антитела PC61.5 и анти-CD45R антитела C363.16A (все от eBioscience). При помощи проточного цитометра подтверждали, что клетки, полученные этим способом, представляют собой CD4+CD25-CD45RBhi T-лимфоциты, имеющие чистоту 95% или выше. Затем 500000 клеток внутрибрюшинно трансплантировали 12-16-недельным Rag2-/- мышам. Затем измерение массы тела и наблюдение таких симптомов, как диарея, осуществляли в течение 12 недель. После завершения периода наблюдения степень утолщения кишечного тракта, число и размер полипов и наличие или отсутствие патологических признаков определяли при помощи аутопсии. Также проводили количественое определение концентрации антитела, цитокинов или биомаркеров сыворотки в крови или тканях, и анализировали активность роста, способность продуцировать цитокины, поверхностные антигены или тому подобное клеток, полученных из кишечного тракта, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга. Анти-Orai1 антитело вводили внутривенно или подкожно за одни сутки до трансплантации клеток, или за 1-4 часа до трансплантации клеток. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. Определяли влияние введения анти-Orai1 антитела на колит.
[0250]
16)-2-9 Изучение влияния анти-Orai1 антитела на систему трансплантации клеток костного мозга
Бедренную кость или большую берцовую кость извлекали из мыши, в чей геном встроен ген Orai1 человека, и клетки костного мозга извлекали из эпифиза путем инъекции от 1 до 5 мл среды RPMI1640, содержащей 10% ФБС, с использованием шприца и иглы. После пропускания клеток через фильтр, клетки костного мозга выделяли при помощи центрифугирования, и их концентрацию регулировали таким образом, чтобы 10000000 клеток костного мозга содержалось в 200 мкл инъекционного буфера, содержащего 10 мМ HEPES, 0,5 мМ ЭДТА и 0,5% пенициллина/стрептомицина. Одновременно с этим клетки селезенки получали из селезенки мыши, в чей геном встроен ген Orai1 человека, стандартным способом. 10-13-недельных мышей линии BALB/c в качестве реципиентов, облучали рентгеновскими лучами в дозе от 2,0 до 8,0 Гр. Затем 4000000 клеток селезенки донора добавляли к 200 мкл клеток костного мозга донора, и этот раствор клеточной смеси вводили в хвостовую вену реципиента. Затем изменение массы тела и выживаемость оценивали в течение 4-16 недель. После окончания эксперимента измеряли концентрацию антитела, цитокинов или биомаркеров сыворотки в крови ли тканях мышей-реципиентов, и поверхностные антигены, активность роста или способность продуцировать цитокины у клеток, полученных из кишечного тракта, периферической крови, тимуса, селезенки, лимфатических узлов или костного мозга. Анти-Orai1 антитело вводили внутривенно или подкожно мышам-реципиентам за одни сутки до трансплантации костного мозга, или за 1-4 часа до трансплантации костного мозга. Затем частоту введения устанавливали в интервале от 7 до 28 дней для продолжения введения антитела. Определяли влияние анти-Orai1 антитела на приживаемость трансплантированных клеток и число случаев реакции по типу «трансплантат против хозяина».
[0251]
Пример 17. Сравнение физико-химических свойств между антителом с созреванием аффинности и другими анти-Orai1 антителами
Измерение вязкости с использованием вискозиметра m-VROC
hR198_HG1/LG1 или hR198_H3/LG1, полученные в пункте 10)-3, hR198_HG1-LALA/LG1, полученное в пункте 12)-3, или 2C1.1, полученное в пункте 13)-6, концентрировали до 150 мг/мл или выше при помощи концентратора VIVAPORE 5 (Sartorius Japan K.K.), и затем получали в концентрациях 90, 120 и 150 мг/мл в HBSor. Вязкость при температуре 25°C измеряли три раза для каждой концентрации с использованием вискозиметра m-VROC (RheoSense, Inc.) для расчета средней вязкости (мПа⋅с). Результаты показаны на Фигуре 58 и в Таблице 1. Вязкость hR198_HG1/LG1, hR198_H3/LG1 и hR198_HG1-LALA/LG1 при всех концентрациях была ниже, чем вязкость 2C1.1. В частности, при 150 мг/мл, вязкость hR198_HG1/LG1, hR198_H3/LG1 и hR198_HG1-LALA/LG1 была 11,4, 10,0 и 10,8 мПа⋅с, соответственно, в то время как вязкость 2C1.1 составила 21,0 мПа⋅с, демонстрируя, что антитела с созреванием аффинности имеют низкую вязкость даже при высокой концентрации по сравнению с 2C1.1.
[0252]
Таблица 1
Промышленная применимость
[0253]
Гуманизированное анти-Orai1 антитело настоящего изобретения обладает более сильным действием, ингибирующим активность T-лимфоцитов, по сравнению с действием антител, известных из уровня техники, и оно может служить в качестве терапевтического или профилактического агента при отторжении трансплантата и т.д.
Свободный текст списка последовательностей
[0254]
SEQ ID NO: 1: Последовательность гена Orai1 человека
SEQ ID NO: 2: Аминокислотная последовательность Orai1 человека
SEQ ID NO: 3: ПЦР праймер Nhe-polyC-S
SEQ ID NO: 4: ПЦР праймер rIgγ-AS1
SEQ ID NO: 5: ПЦР праймер rIgγ-AS2
SEQ ID NO: 6: ПЦР праймер Nhe-polyC-S
SEQ ID NO: 7: ПЦР праймер rIgκ-AS
SEQ ID NO: 8: Секвенирующий праймер rIgγ-seq
SEQ ID NO: 9: Секвенирующий праймер rIgκ-seq
SEQ ID NO: 10: Нуклеотидная последовательность, кодирующая легкую цепь R118
SEQ ID NO: 11: Аминокислотная последовательность легкую цепь R118
SEQ ID NO: 12: Нуклеотидная последовательность, кодирующая тяжелую цепь R118
SEQ ID NO: 13: Аминокислотная последовательность тяжелой цепи R118
SEQ ID NO: 14: Нуклеотидная последовательность, кодирующая легкую цепь R198
SEQ ID NO: 15: Аминокислотная последовательность легкой цепи R198
SEQ ID NO: 16: Нуклеотидная последовательность, кодирующая тяжелую цепь R198
SEQ ID NO: 17: Аминокислотная последовательность тяжелой цепи R198
SEQ ID NO: 18: Фрагмент ДНК, содержащий последовательность, кодирующую сигнал секреции κ цепи человека и константную область κ цепи человека
SEQ ID NO: 19: ПЦР праймер 3.3-F1
SEQ ID NO: 20: ПЦР праймер 3.3-R1
SEQ ID NO: 21: Фрагмент ДНК, содержащий последовательность, кодирующую сигнальную последовательность тяжелой цепи человека и аминокислоты константной области IgG1 человека
SEQ ID NO: 22: Нуклеотидная последовательность, кодирующая легкую цепь человеческого гибридного cR118
SEQ ID NO: 23: Аминокислотная последовательность легкой цепи человеческого гибридного cR118
SEQ ID NO: 24: Нуклеотидная последовательность, кодирующая легкую цепь человеческого гибридного cR198
SEQ ID NO: 25: Аминокислотная последовательность легкой цепи человеческого гибридного cR198
SEQ ID NO: 26: Нуклеотидная последовательность, кодирующая тяжелую цепь человеческого гибридного cR118
SEQ ID NO: 27: Аминокислотная последовательность тяжелой цепи человеческого гибридного cR118
SEQ ID NO: 28: Нуклеотидная последовательность, кодирующая тяжелую цепь человеческого гибридного cR198
SEQ ID NO: 29: Аминокислотная последовательность тяжелой цепи человеческого гибридного cR198
SEQ ID NO: 30: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L1
SEQ ID NO: 31: Аминокислотная последовательность легкой цепи типа hR198_L1
SEQ ID NO: 32: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L2
SEQ ID NO: 33: Аминокислотная последовательность легкой цепи типа hR198_L2
SEQ ID NO: 34: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L3
SEQ ID NO: 35: Аминокислотная последовательность легкой цепи типа hR198_L3
SEQ ID NO: 36: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_L4
SEQ ID NO: 37: Аминокислотная последовательность легкой цепи типа hR198_L4
SEQ ID NO: 38: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H1
SEQ ID NO: 39: Аминокислотная последовательность тяжелой цепи типа hR198_H1
SEQ ID NO: 40: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H2
SEQ ID NO: 41: Аминокислотная последовательность тяжелой цепи типа hR198_H2
SEQ ID NO: 42: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H3
SEQ ID NO: 43: Аминокислотная последовательность тяжелой цепи типа hR198_H3
SEQ ID NO: 44: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H4
SEQ ID NO: 45: Аминокислотная последовательность тяжелой цепи типа hR198_H4
SEQ ID NO: 46: ПЦР праймер Orai1 HF
SEQ ID NO: 47: ПЦР праймер Orai1 CH FR
SEQ ID NO: 48: ПЦР праймер M13 rev long
SEQ ID NO: 49: ПЦР праймер SecM Stop R
SEQ ID NO: 50: ПЦР праймер Orai1 Lc F
SEQ ID NO: 51: ПЦР праймер Orai1 CL-FR
SEQ ID NO: 52: ПЦР праймер Orai1 HR-FLAG R
SEQ ID NO: 53: ПЦР праймер Orai1 CL-FLAG R
SEQ ID NO: 54: ПЦР праймер Myc-R
SEQ ID NO: 55: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG1
SEQ ID NO: 56: Аминокислотная последовательность легкой цепи типа hR198_LG1
SEQ ID NO: 57: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG2
SEQ ID NO: 58: Аминокислотная последовательность легкой цепи типа hR198_LG2
SEQ ID NO: 59: Нуклеотидная последовательность, кодирующая легкую цепь типа hR198_LG3
SEQ ID NO: 60: Аминокислотная последовательность легкой цепи типа hR198_LG3
SEQ ID NO: 61: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG1
SEQ ID NO: 62: Аминокислотная последовательность тяжелой цепи типа hR198_HG1
SEQ ID NO: 63: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG2
SEQ ID NO: 64: Аминокислотная последовательность тяжелой цепи типа hR198_HG2
SEQ ID NO: 65: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG3
SEQ ID NO: 66: Аминокислотная последовательность тяжелой цепи типа hR198_HG3
SEQ ID NO: 67: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H4-LALA
SEQ ID NO: 68: Аминокислотная последовательность тяжелой цепи типа hR198_H4-LALA
SEQ ID NO: 69: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_HG1-LALA
SEQ ID NO: 70: Аминокислотная последовательность тяжелой цепи типа hR198_HG1-LALA
SEQ ID NO: 71: Фрагмент ДНК, содержащий последовательность, кодирующую сигнальную последовательность тяжелой цепи человека и аминокислоты константной области IgG2 человека
SEQ ID NO: 72: Нуклеотидная последовательность, кодирующая тяжелую цепь антитела 2C1.1
SEQ ID NO: 73: Аминокислотная последовательность тяжелой цепи антитела 2C1.1
SEQ ID NO: 74: Нуклеотидная последовательность, кодирующая легкую цепь антитела 2C1.1
SEQ ID NO: 75: Аминокислотная последовательность легкой цепи антитела 2C1.1
SEQ ID NO: 76: Нуклеотидная последовательность, кодирующая тяжелую цепь антитела e 5H3.1
SEQ ID NO: 77: Аминокислотная последовательность тяжелой цепи антитела 5H3.1
SEQ ID NO: 78: Нуклеотидная последовательность, кодирующая легкую цепь антитела 5H3.1
SEQ ID NO: 79: Аминокислотная последовательность легкой цепи антитела 5H3.1
SEQ ID NO: 80: Нуклеотидная последовательность, кодирующая тяжелую цепь антитела 10F8
SEQ ID NO: 81: Аминокислотная последовательность тяжелой цепи антитела 10F8
SEQ ID NO: 82: Нуклеотидная последовательность, кодирующая легкую цепь антитела 10F8
SEQ ID NO: 83: Аминокислотная последовательность легкой цепи антитела 10F8
SEQ ID NO: 84: Нуклеотидная последовательность, кодирующая тяжелую цепь антитела 14F74
SEQ ID NO: 85: Аминокислотная последовательность тяжелой цепи антитела 14F74
SEQ ID NO: 86: Нуклеотидная последовательность, кодирующая легкую цепь антитела 14F74
SEQ ID NO: 87: Аминокислотная последовательность легкой цепи антитела 14F74
SEQ ID NO: 88: Нуклеотидная последовательность, кодирующая тяжелую цепь антитела 17F6
SEQ ID NO: 89: Аминокислотная последовательность тяжелой цепи антитела 17F6
SEQ ID NO: 90: Нуклеотидная последовательность, кодирующая легкую цепь антитела 17F6
SEQ ID NO: 91: Аминокислотная последовательность легкой цепи антитела 17F6
SEQ ID NO: 92: Крысиная CDRL1 (которая была получена из R198 и использовалась для легких цепей типа hR198_L1 to hR198_L4)
SEQ ID NO: 93: Сконструированная CDRL1 (которая использовалась для легкой цепи типа hR198_LG1)
SEQ ID NO: 94: Сконструированная CDRL1 (которая использовалась для легкой цепи типа hR198_LG2)
SEQ ID NO: 95: Сконструированная CDRL1 (которая использовалась для легкой цепи типа hR198_LG3)
SEQ ID NO: 96: Крысиная CDRL2 (которая была общей для R118 и R198 и использовалась для легких цепей типа hR198_L1 - hR198_L4 и hR198_LG1)
SEQ ID NO: 97: Сконструированная CDRL2 (которая использовалась для легкой цепи типа hR198_LG2)
SEQ ID NO: 98: Сконструированная CDRL2 (которая использовалась для легкой цепи типа hR198_LG3)
SEQ ID NO: 99: Крысиная CDRL3 (которая была получена из R118 и использовалась для легких цепей типа hR198_L3 и hR198_L4)
SEQ ID NO: 100: Крысиная CDRL3 (которая была получена из R198 и использовалась для легких цепей типа hR198_L1 и hR198_L2)
SEQ ID NO: 101: Сконструированная CDRL3 (которая использовалась для легких цепей типа hR198_LG1 to hR198_LG3)
SEQ ID NO: 102: Крысиная CDRH1 (которая была получена из R118 и использовалась для тяжелых цепей типа hR198_H3, hR198_H4, и hR198_HG1 - hR198_ HG3)
SEQ ID NO: 103: Крысиная CDRH1 (которая была получена из R198 и использовалась для тяжелых цепей типа hR198_H1 и hR198_H2)
SEQ ID NO: 104: Крысиная CDRH2 (которая была получена из R118 и использовалась для тяжелых цепей типа hR198_H3 и hR198_H4)
SEQ ID NO: 105: Крысиная CDRH2 (которая была получена из R198 и использовалась для тяжелых цепей типа hR198_H1 и hR198_H2)
SEQ ID NO: 106: Сконструированная CDRH2 (которая использовалась для тяжелой цепи типа hR198_HG1)
SEQ ID NO: 107: Сконструированная CDRH2 (которая использовалась для тяжелой цепи типа hR198_HG2)
SEQ ID NO: 108: Сконструированная CDRH2 (которая использовалась для тяжелой цепи типа hR198_HG3)
SEQ ID NO: 109: Крысиная CDRH3 (которая была общей для R118 и R198 и использовалась для тяжелой цепи типа hR198_H1 - hR198_H4)
SEQ ID NO: 110: Сконструированная CDRH3 (которая использовалась для тяжелых цепей типа hR198_HG1 to hR198_HG3)
SEQ ID NO: 111: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H0
SEQ ID NO: 112: Аминокислотная последовательность тяжелой цепи типа hR198_H0
SEQ ID NO: 113: Нуклеотидная последовательность, кодирующая тяжелую цепь типа hR198_H5
SEQ ID NO: 114: Аминокислотная последовательность тяжелой цепи типа hR198_H5
SEQ ID NO: 115: Последовательность биотин-ПЭГилированного пептида петлевой области Orai1 человека
SEQ ID NO: 116: Крысиная CDRL1 (которая была получена из R118, но не использовалась для гуманизированных антител)
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Daiichi Sankyo Co., Ltd.
<120> Анти Orai1 антитела
<130> FP1522
<160> 116
<170> Патентная версия 3.5
<210> 1
<211> 906
<212> ДНК
<213> Homo sapiens
<400> 1
atgcatccgg agcccgcccc gcccccgagc cgtagcagtc ccgagcttcc cccaagcggc 60
ggcagcacca ccagcggcag ccgccggagc cgccgccgca gcggggacgg ggagcccccg 120
ggggccccgc caccgccgcc gtccgccgtc acctacccgg actggatcgg ccagagttac 180
tccgaggtga tgagcctcaa cgagcactcc atgcaggcgc tgtcctggcg caagctctac 240
ttgagccgcg ccaagcttaa agcctccagc cggacctcgg ctctgctctc cggcttcgcc 300
atggtggcaa tggtggaggt gcagctggac gctgaccacg actacccacc ggggctgctc 360
atcgccttca gtgcctgcac cacagtgctg gtggctgtgc acctgtttgc gctcatgatc 420
agcacctgca tcctgcccaa catcgaggcg gtgagcaacg tgcacaatct caactcggtc 480
aaggagtccc cccatgagcg catgcaccgc cacatcgagc tggcctgggc cttctccacc 540
gtcattggca cgctgctctt cctagctgag gtggtgctgc tctgctgggt caagttcttg 600
cccctcaaga agcagccagg ccagccaagg cccaccagca agccccccgc cagtggcgca 660
gcagccaacg tcagcaccag cggcatcacc ccgggccagg cagctgccat cgcctcgacc 720
accatcatgg tgcccttcgg cctgatcttt atcgtcttcg ccgtccactt ctaccgctca 780
ctggttagcc ataagaccga ccgacagttc caggagctca acgagctggc ggagtttgcc 840
cgcttacagg accagctgga ccacagaggg gaccaccccc tgacgcccgg cagccactat 900
gcctag 906
<210> 2
<211> 301
<212> Белок
<213> Homo sapiens
<400> 2
Met His Pro Glu Pro Ala Pro Pro Pro Ser Arg Ser Ser Pro Glu Leu
1 5 10 15
Pro Pro Ser Gly Gly Ser Thr Thr Ser Gly Ser Arg Arg Ser Arg Arg
20 25 30
Arg Ser Gly Asp Gly Glu Pro Pro Gly Ala Pro Pro Pro Pro Pro Ser
35 40 45
Ala Val Thr Tyr Pro Asp Trp Ile Gly Gln Ser Tyr Ser Glu Val Met
50 55 60
Ser Leu Asn Glu His Ser Met Gln Ala Leu Ser Trp Arg Lys Leu Tyr
65 70 75 80
Leu Ser Arg Ala Lys Leu Lys Ala Ser Ser Arg Thr Ser Ala Leu Leu
85 90 95
Ser Gly Phe Ala Met Val Ala Met Val Glu Val Gln Leu Asp Ala Asp
100 105 110
His Asp Tyr Pro Pro Gly Leu Leu Ile Ala Phe Ser Ala Cys Thr Thr
115 120 125
Val Leu Val Ala Val His Leu Phe Ala Leu Met Ile Ser Thr Cys Ile
130 135 140
Leu Pro Asn Ile Glu Ala Val Ser Asn Val His Asn Leu Asn Ser Val
145 150 155 160
Lys Glu Ser Pro His Glu Arg Met His Arg His Ile Glu Leu Ala Trp
165 170 175
Ala Phe Ser Thr Val Ile Gly Thr Leu Leu Phe Leu Ala Glu Val Val
180 185 190
Leu Leu Cys Trp Val Lys Phe Leu Pro Leu Lys Lys Gln Pro Gly Gln
195 200 205
Pro Arg Pro Thr Ser Lys Pro Pro Ala Ser Gly Ala Ala Ala Asn Val
210 215 220
Ser Thr Ser Gly Ile Thr Pro Gly Gln Ala Ala Ala Ile Ala Ser Thr
225 230 235 240
Thr Ile Met Val Pro Phe Gly Leu Ile Phe Ile Val Phe Ala Val His
245 250 255
Phe Tyr Arg Ser Leu Val Ser His Lys Thr Asp Arg Gln Phe Gln Glu
260 265 270
Leu Asn Glu Leu Ala Glu Phe Ala Arg Leu Gln Asp Gln Leu Asp His
275 280 285
Arg Gly Asp His Pro Leu Thr Pro Gly Ser His Tyr Ala
290 295 300
<210> 3
<211> 37
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Nhe-polyC-S
<220>
<221> прочий признак
<222> (37)..(37)
<223> n представляет собой a, c, g, или t
<400> 3
gctagcgcta ccggactcag atcccccccc cccccdn 37
<210> 4
<211> 28
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер rIg gamma-AS1
<400> 4
tcactgagct ggtgagagtg tagagccc 28
<210> 5
<211> 28
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер rIG gamma-AS2
<400> 5
tcaccgagct gctgagggtg tagagccc 28
<210> 6
<211> 37
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Nhe-polyC-S
<220>
<221> прочий признак
<222> (37)..(37)
<223> n представляет собой a, c, g, или t
<400> 6
gctagcgcta ccggactcag atcccccccc cccccdn 37
<210> 7
<211> 28
<212> ДНК
<213> искусственный
<220>
<223> rIg kappa-AS
<400> 7
tcagtaacac tgtccaggac accatctc 28
<210> 8
<211> 19
<212> ДНК
<213> искусственный
<220>
<223> rIG gamma-seq
<400> 8
ctggctcagg gaaatagcc 19
<210> 9
<211> 19
<212> ДНК
<213> искусственный
<220>
<223> rIG kappa-seq
<400> 9
tccagttgct aactgttcc 19
<210> 10
<211> 405
<212> ДНК
<213> Rattus rattus
<220>
<221> прочий признак
<222> (382)..(382)
<223> n представляет собой a, c, g, или t
<220>
<221> прочий признак
<222> (403)..(403)
<223> n представляет собой a, c, g, или t
<400> 10
atgaaaatga cgacacctgc tcagttcctt gggcttctgt tgctctggtt tccaggtgcc 60
aggtgtgaca tccagttgac ccagtctcca tccacattgc ctgcatccct gggagagaga 120
gtcaccatca gttgcagagc aagtcagagt attagcaata gtttaagctg gtttcaacag 180
aaaccagatg gaactgttaa acgcctgatc tattctacat ccactttaga atctggtgtc 240
ccatcaaggt tcagtggcag tgggtctggg acagattatt ctctctccat caccagtctt 300
gagtctgaag attttgcaat gtattactgt ctacagtttg ctacttttcc ggacacgttt 360
ggaactggga ccaaactgga antgagacgg gctgatgctg canca 405
<210> 11
<211> 135
<212> Белок
<213> Rattus rattus
<220>
<221> прочий признак
<222> (128)..(128)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> прочий признак
<222> (135)..(135)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 11
Met Lys Met Thr Thr Pro Ala Gln Phe Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Phe Pro Gly Ala Arg Cys Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Pro Ala Ser Leu Gly Glu Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Ser Ile Ser Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Asp Gly
50 55 60
Thr Val Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Ser
85 90 95
Ile Thr Ser Leu Glu Ser Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln
100 105 110
Phe Ala Thr Phe Pro Asp Thr Phe Gly Thr Gly Thr Lys Leu Glu Xaa
115 120 125
Arg Arg Ala Asp Ala Ala Xaa
130 135
<210> 12
<211> 468
<212> ДНК
<213> Rattus rattus
<220>
<221> прочий признак
<222> (467)..(467)
<223> n представляет собой a, c, g, или t
<400> 12
atggaatgga actgggtctt tctcctcctc ctgtcagtaa ctgcagaagt ccagtcccag 60
gtccagctgc agcagtctgg agcggagctg gcaaagcctg gctcctcagt gaagatttcc 120
tgcaaggctt ccggctacac cgtcaccgcc tattatataa gttggataag gcagacgatt 180
ggacagggcc ttgagtatgt tggatatatt gacatgggaa atggaaggac taactacaat 240
gcgaggttca agggcaaggc cacattgact gtggacaaat cctccagcac agccttcatg 300
caactcagca gcctgacacc tgacgactct gcggtctatt actgtgcaag ggactccaac 360
tggggggttg attactgggg ccaaggagtc atggtcacag tctcctcagc tgaaacaaca 420
gccccatctg tctatccact ggctcctgga actgcttctc aaaagtna 468
<210> 13
<211> 156
<212> Белок
<213> Rattus rattus
<220>
<221> прочий признак
<222> (156)..(156)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 13
Met Glu Trp Asn Trp Val Phe Leu Leu Leu Leu Ser Val Thr Ala Glu
1 5 10 15
Val Gln Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Ile Arg Gln Thr Ile Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Phe Met Gln Leu Ser Ser Leu Thr Pro Asp Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Glu Thr Thr Ala Pro Ser Val
130 135 140
Tyr Pro Leu Ala Pro Gly Thr Ala Ser Gln Lys Xaa
145 150 155
<210> 14
<211> 408
<212> ДНК
<213> Rattus rattus
<220>
<221> прочий признак
<222> (403)..(403)
<223> n представляет собой a, c, g, или t
<400> 14
atgaaaatga cgacacctgc tcagttcctt gggcttctgt tgctctggtt tccaggtgcc 60
aggtgtgaca tccagttgac ccagtctcca tccacattgc ctgcatctct gggagagaga 120
gtcaccatca gttgcagagc aagtcagagt attggcaata gtttaagctg gtttcagcag 180
aaaccagatg gatctgttaa acgcctgatc tactctacat ccactttaga atctggtgtc 240
ccatcaaggt tcagtggcag tgggtctggg acagattatt ctctctccat caccagtctt 300
gagtctgaag attttgcaat gtattactgt ctacagtttg ctacttatcc ggacacgttt 360
ggaactggga ccaaactgga actgagacgg gctgatgctg cancaact 408
<210> 15
<211> 136
<212> Белок
<213> Rattus rattus
<220>
<221> прочий признак
<222> (135)..(135)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 15
Met Lys Met Thr Thr Pro Ala Gln Phe Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Phe Pro Gly Ala Arg Cys Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr
20 25 30
Leu Pro Ala Ser Leu Gly Glu Arg Val Thr Ile Ser Cys Arg Ala Ser
35 40 45
Gln Ser Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Asp Gly
50 55 60
Ser Val Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Ser
85 90 95
Ile Thr Ser Leu Glu Ser Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln
100 105 110
Phe Ala Thr Tyr Pro Asp Thr Phe Gly Thr Gly Thr Lys Leu Glu Leu
115 120 125
Arg Arg Ala Asp Ala Ala Xaa Thr
130 135
<210> 16
<211> 471
<212> ДНК
<213> Rattus rattus
<220>
<221> прочий признак
<222> (469)..(469)
<223> n представляет собой a, c, g, или t
<400> 16
atggaatgga actgggtctt tctcttcctc ctgtcagtaa ctgcagaagt ccagtcccag 60
gtccagctgc agcagtctgg agcggagctg gcaaagcctg gctcctcaat gaagatttcc 120
tgcaaggctt ccggctaccc cgtcaccagc tattatataa gttggataaa gcagacgact 180
ggacagggcc ttgagtatat tggatatgtt gacatgggaa atggacggac taactacaat 240
gagaagttca agggcaaggc cacattgact gtagacaaat cctccagcac agccttcatg 300
caactcagca gcctgacacc tgacgactct gcggtctatt actgtgcaag ggactccaac 360
tggggggttg attactgggg ccaaggagtc atggtcacag tctcctcagc tgaaacaaca 420
gccccatctg tctatccact ggctcctgga actgctctca aaagtaacnc c 471
<210> 17
<211> 157
<212> Белок
<213> Rattus rattus
<220>
<221> прочий признак
<222> (157)..(157)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 17
Met Glu Trp Asn Trp Val Phe Leu Phe Leu Leu Ser Val Thr Ala Glu
1 5 10 15
Val Gln Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys
20 25 30
Pro Gly Ser Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ser Tyr Tyr Ile Ser Trp Ile Lys Gln Thr Thr Gly Gln Gly Leu
50 55 60
Glu Tyr Ile Gly Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Phe Met Gln Leu Ser Ser Leu Thr Pro Asp Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Glu Thr Thr Ala Pro Ser Val
130 135 140
Tyr Pro Leu Ala Pro Gly Thr Ala Leu Lys Ser Asn Xaa
145 150 155
<210> 18
<211> 449
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая сигнал секреции легкой цепи человека и константную область каппа цепи человека
<400> 18
gcctccggac tctagagcca ccatggtgct gcagacccag gtgttcatct ccctgctgct 60
gtggatctcc ggcgcgtacg gcgatatcgt gatgattaaa cgtacggtgg ccgccccctc 120
cgtgttcatc ttccccccct ccgacgagca gctgaagtcc ggcaccgcct ccgtggtgtg 180
cctgctgaat aacttctacc ccagagaggc caaggtgcag tggaaggtgg acaacgccct 240
gcagtccggg aactcccagg agagcgtgac cgagcaggac agcaaggaca gcacctacag 300
cctgagcagc accctgaccc tgagcaaagc cgactacgag aagcacaagg tgtacgcctg 360
cgaggtgacc caccagggcc tgagctcccc cgtcaccaag agcttcaaca ggggggagtg 420
ttaggggccc gtttaaacgg gggaggcta 449
<210> 19
<211> 30
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер 3.3-F1
<400> 19
tataccgtcg acctctagct agagcttggc 30
<210> 20
<211> 30
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер 3.3-R1
<400> 20
gctatggcag ggcctgccgc cccgacgttg 30
<210> 21
<211> 1132
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая сигнальную последовательность тяжелой цепи человека и аминокислоты константной области IgG1 человека
<400> 21
gcctccggac tctagagcca ccatgaaaca cctgtggttc ttcctcctgc tggtggcagc 60
tcccagatgg gtgctgagcc aggtgcaatt gtgcaggcgg ttagctcagc ctccaccaag 120
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 180
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 240
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 300
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 360
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 420
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 480
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 540
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 600
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 660
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 720
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 780
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 840
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 900
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 960
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1020
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1080
tccctgtctc ccggcaaatg agatatcggg cccgtttaaa cgggggaggc ta 1132
<210> 22
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая cR118_L
<400> 22
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccagt tgacccagtc tccatccaca ttgcctgcat ccctgggaga gagagtcacc 120
atcagttgca gagcaagtca gagtattagc aatagtttaa gctggtttca acagaaacca 180
gatggaactg ttaaacgcct gatctattct acatccactt tagaatctgg tgtcccatca 240
aggttcagtg gcagtgggtc tgggacagat tattctctct ccatcaccag tcttgagtct 300
gaagattttg caatgtatta ctgtctacag tttgctactt ttccggacac gtttggaact 360
gggaccaaac tggaactgag acgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 23
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность cR118_L
<400> 23
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Pro
20 25 30
Ala Ser Leu Gly Glu Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Ser
35 40 45
Ile Ser Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Asp Gly Thr Val
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Ser Ile Thr
85 90 95
Ser Leu Glu Ser Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Phe Pro Asp Thr Phe Gly Thr Gly Thr Lys Leu Glu Leu Arg Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 24
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая cR198_L
<400> 24
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gacatccagt tgacccagtc tccatccaca ttgcctgcat ctctgggaga gagagtcacc 120
atcagttgca gagcaagtca gagtattggc aatagtttaa gctggtttca gcagaaacca 180
gatggatctg ttaaacgcct gatctactct acatccactt tagaatctgg tgtcccatca 240
aggttcagtg gcagtgggtc tgggacagat tattctctct ccatcaccag tcttgagtct 300
gaagattttg caatgtatta ctgtctacag tttgctactt atccggacac gtttggaact 360
gggaccaaac tggaactgag acgggctgtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 25
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность cR198_L
<400> 25
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Pro
20 25 30
Ala Ser Leu Gly Glu Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Asp Gly Ser Val
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Ser Leu Ser Ile Thr
85 90 95
Ser Leu Glu Ser Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Tyr Pro Asp Thr Phe Gly Thr Gly Thr Lys Leu Glu Leu Arg Arg
115 120 125
Ala Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 26
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая cR118_H
<400> 26
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtccagctgc agcagtctgg agcggagctg gcaaagcctg gctcctcagt gaagatttcc 120
tgcaaggctt ccggctacac cgtcaccgcc tattatataa gttggataag gcagacgatt 180
ggacagggcc ttgagtatgt tggatatatt gacatgggaa atggaaggac taactacaat 240
gcgaggttca agggcaaggc cacattgact gtggacaaat cctccagcac agccttcatg 300
caactcagca gcctgacacc tgacgactct gcggtctatt actgtgcaag ggactccaac 360
tggggggttg attactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 27
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность cR118_H
<400> 27
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Ile Arg Gln Thr Ile Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Phe Met Gln Leu Ser Ser Leu Thr Pro Asp Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 28
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая cR198_H
<400> 28
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtccagctgc agcagtctgg agcggagctg gcaaagcctg gctcctcaat gaagatttcc 120
tgcaaggctt ccggctaccc cgtcaccagc tattatataa gttggataaa gcagacgact 180
ggacagggcc ttgagtatat tggatatgtt gacatgggaa atggacggac taactacaat 240
gagaagttca agggcaaggc cacattgact gtagacaaat cctccagcac agccttcatg 300
caactcagca gcctgacacc tgacgactct gcggtctatt actgtgcaag ggactccaac 360
tggggggttg attactgggg ccaaggagtc atggtcacag tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 29
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность cR198_H
<400> 29
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Lys
20 25 30
Pro Gly Ser Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ser Tyr Tyr Ile Ser Trp Ile Lys Gln Thr Thr Gly Gln Gly Leu
50 55 60
Glu Tyr Ile Gly Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Phe Met Gln Leu Ser Ser Leu Thr Pro Asp Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Val Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 30
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_L1
<400> 30
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgta gagccagcca gagcatcggc aacagcctga gctggttcca gcagaaaccc 180
ggcaaggccc ccaagcggct gatctacagc accagcaccc tggaaagcgg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccacct accccgacac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 31
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_L1
<400> 31
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Tyr Pro Asp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 32
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_L2
<400> 32
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgta gagccagcca gagcatcggc aacagcctga gctggttcca gcagaaaccc 180
ggcaaggccg tgaagcggct gatctacagc accagcaccc tggaaagcgg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccacct accccgacac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 33
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_L2
<400> 33
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Val
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Tyr Pro Asp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 34
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_L3
<400> 34
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgta gagccagcca gagcatcggc aacagcctga gctggttcca gcagaaaccc 180
ggcaaggccc ccaagcggct gatctacagc accagcaccc tggaaagcgg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccacct tccccgacac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 35
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_L3
<400> 35
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Phe Pro Asp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 36
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_L4
<400> 36
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgta gagccagcca gagcatcggc aacagcctga gctggttcca gcagaaaccc 180
ggcaaggccg tgaagcggct gatctacagc accagcaccc tggaaagcgg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccacct tccccgacac ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 37
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_L4
<400> 37
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Asn Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Val
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Thr Phe Pro Asp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 38
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H1
<400> 38
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccagc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacat cggctatgtg gacatgggca acggccggac caactacaac 240
gagaagttca agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 39
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H1
<400> 39
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ser Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Ile Gly Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 40
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H2
<400> 40
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccagc tactacatca gctggatcag acaggcccca 180
ggccagggcc tggaatacat cggctatgtg gacatgggca acggccggac caactacaac 240
gagaagttca agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 41
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H2
<400> 41
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ser Tyr Tyr Ile Ser Trp Ile Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Ile Gly Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 42
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H3
<400> 42
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac caactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 43
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H3
<400> 43
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 44
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H4
<400> 44
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctggatcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac caactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 45
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H4
<400> 45
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Ile Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 46
<211> 23
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 HF
<400> 46
atgcaagtcc aactggttca atc 23
<210> 47
<211> 21
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 CH FR
<400> 47
tgacggagcc agcgggaaga c 21
<210> 48
<211> 21
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер M13 rev long
<400> 48
caggaaacag ctatgaccat g 21
<210> 49
<211> 31
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер SecM Stop R
<400> 49
ctcgagttat tcattaggtg aggcgttgag g 31
<210> 50
<211> 24
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 Lc F
<400> 50
atggacattc aactgaccca aagc 24
<210> 51
<211> 24
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 CL-FR
<400> 51
gataaaaaca ctcggggccg ccac 24
<210> 52
<211> 63
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 HR-FLAG R
<400> 52
tcattatttg tcatcgtcat ctttatagtc gaattcttcg ccacgattaa aggatttggt 60
gac 63
<210> 53
<211> 63
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Orai1 CL -FLAG R
<400> 53
tcattatttg tcatcgtcat ctttatagtc gaattcttcg ccacgattaa aggatttggt 60
gac 63
<210> 54
<211> 30
<212> ДНК
<213> искусственный
<220>
<223> ПЦР праймер Myc-R
<400> 54
cagatcctcc tcagagatca gcttctgctc 30
<210> 55
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_LG1
<400> 55
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgta gagccagcca gagcatcggc ggcagcctga gctggttcca gcagaaaccc 180
ggcaaggccc ccaagcggct gatctacagc accagcaccc tggaaagcgg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccatct tccccgacag ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 56
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_LG1
<400> 56
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser
35 40 45
Ile Gly Gly Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Arg Leu Ile Tyr Ser Thr Ser Thr Leu Glu Ser Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Ile Phe Pro Asp Ser Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 57
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_LG2
<400> 57
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgtc acgccagcca gaacatcggc ggcagcctga gctggttcca gcagaaaccc 180
ggcaaggccc ccaagcggct gatctacctg accagcaccc tggactgggg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccatct tccccgacag ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 58
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_LG2
<400> 58
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys His Ala Ser Gln Asn
35 40 45
Ile Gly Gly Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Arg Leu Ile Tyr Leu Thr Ser Thr Leu Asp Trp Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Ile Phe Pro Asp Ser Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 59
<211> 705
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_LG3
<400> 59
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
gatatccagc tgacccagag ccctagcagc ctgtctgcca gcgtgggcga cagagtgacc 120
atcacctgtc acgccagccg gaacatcggc ggcagcctga gctggttcca gcagaaaccc 180
ggcaaggccc ccaagcggct gatctacctg accagcagcc tggactgggg cgtgcccagc 240
agattttctg gcagcggctc cggcaccgac tacaccctga caatcagcag cctgcagccc 300
gaggacttcg ccatgtacta ctgcctgcag ttcgccatct tccccgacag ctttggccag 360
ggcaccaagg tggaaatcaa gcgtacggtg gccgccccct ccgtgttcat cttccccccc 420
tccgacgagc agctgaagtc cggcaccgcc tccgtggtgt gcctgctgaa taacttctac 480
cccagagagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg gaactcccag 540
gagagcgtga ccgagcagga cagcaaggac agcacctaca gcctgagcag caccctgacc 600
ctgagcaaag ccgactacga gaagcacaag gtgtacgcct gcgaggtgac ccaccagggc 660
ctgagctccc ccgtcaccaa gagcttcaac aggggggagt gttag 705
<210> 60
<211> 234
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_LG3
<400> 60
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser
20 25 30
Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys His Ala Ser Arg Asn
35 40 45
Ile Gly Gly Ser Leu Ser Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
50 55 60
Lys Arg Leu Ile Tyr Leu Thr Ser Ser Leu Asp Trp Gly Val Pro Ser
65 70 75 80
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser
85 90 95
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Tyr Cys Leu Gln Phe Ala
100 105 110
Ile Phe Pro Asp Ser Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
115 120 125
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
130 135 140
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
145 150 155 160
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
165 170 175
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
180 185 190
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
195 200 205
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
210 215 220
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 61
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_HG1
<400> 61
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac cgactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgccg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 62
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_HG1
<400> 62
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Ala Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 63
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_HG2
<400> 63
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc catcaccgcc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac cgactacaac 240
ggccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgccg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 64
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_HG2
<400> 64
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Ile
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn
65 70 75 80
Gly Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Ala Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 65
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_HG3
<400> 65
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc catcaccgcc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac cgactacaac 240
atgcggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgccg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 66
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_HG3
<400> 66
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Ile
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn
65 70 75 80
Met Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Ala Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 67
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H4-LALA
<400> 67
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctggatcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac caactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaagccg cggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 68
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H4-LALA
<400> 68
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Ile Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 69
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_HG1-LALA
<400> 69
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctgggtcag acaggcccca 180
ggccagggcc tggaatacgt gggctacatc gacatgggca acggccggac cgactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgccg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaagccg cggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 70
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_HG1-LALA
<400> 70
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Tyr Val Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Ala Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 71
<211> 1118
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая сигнальную последовательность тяжелой цепи человека и аминокислоты константной области IgG2 человека
<400> 71
gcctccggac tctagagcca ccatgaaaca cctgtggttc ttcctcctgc tggtggcagc 60
tcccagatgg gtgctgagcc aggtgcaatt gtgcaggcgg ttagctcagc cagcaccaag 120
ggcccttccg tgttccctct ggccccttgt agccgttcca ccagcgagtc caccgccgcc 180
cttggctgtc tggtgaagga ctacttccct gagcctgtga ccgtgagctg gaactccgga 240
gcccttacca gcggcgtgca caccttccct gccgtgctgc agtccagcgg cctttactcc 300
ctgagctccg tggtgaccgt gcctagctcc aacttcggca cccaaaccta cacctgtaac 360
gtggaccaca agcctagcaa caccaaggtg gacaagaccg tggagcgtaa gtgttgtgtg 420
gagtgtcctc cttgtcctgc ccctcctgtg gccggacctt ccgtgttcct tttccctcct 480
aagcctaagg acaccctgat gatcagccgt acccctgagg tgacctgtgt ggtggtggac 540
gtgtcccacg aggaccctga ggtgcagttc aactggtacg tggacggcgt ggaggtgcac 600
aacgccaaga ccaagcctcg tgaggagcaa ttcaacagca ccttccgtgt ggtgtccgtg 660
cttaccgtgg tgcaccaaga ctggctgaac ggcaaggagt acaagtgtaa ggtgagcaac 720
aagggacttc ctgcccctat cgagaagacc atctccaaga ccaagggcca acctcgtgag 780
cctcaagtgt acacccttcc tcctagccgt gaggagatga ccaagaacca agtgtccctt 840
acctgtctgg tgaagggctt ctaccctagc gacatcgccg tggagtggga gtccaacgga 900
caacctgaga acaactacaa gaccacccct cctatgcttg acagcgacgg ctccttcttc 960
ctgtacagca agctgaccgt ggacaagtcc cgttggcaac aaggcaacgt gttcagctgt 1020
tccgtgatgc acgaggccct gcacaaccac tacacccaaa agagcctttc cctgagccct 1080
ggaaagtgat atcgggcccg tttaaacggg ggaggcta 1118
<210> 72
<211> 1395
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь 2C1.1
<400> 72
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgc agcagtgggg agccggcctg ctgaagccca gcgagacact gagcctgacc 120
tgcgctgtgt acggaggcag cttcagcggc tactactggt cctggatccg gcagccccct 180
ggcaagggcc tggaatggat cggcgagatc gaccacagcg gcagcaccaa ctacaacccc 240
gccctgaagt cccggctgac catcagcgtg gacaccagca agaaccagtt cagcctgaag 300
ctgagcagcg tgacagccgc cgacaccgcc gtgtactact gtgccagagc cggcagcggc 360
ggctacgagg attggttcga tccttggggc cagggcaccc tggtgaccgt cagctcagcc 420
agcaccaagg gcccttccgt gttccctctg gccccttgta gccgttccac cagcgagtcc 480
accgccgccc ttggctgtct ggtgaaggac tacttccctg agcctgtgac cgtgagctgg 540
aactccggag cccttaccag cggcgtgcac accttccctg ccgtgctgca gtccagcggc 600
ctttactccc tgagctccgt ggtgaccgtg cctagctcca acttcggcac ccaaacctac 660
acctgtaacg tggaccacaa gcctagcaac accaaggtgg acaagaccgt ggagcgtaag 720
tgttgtgtgg agtgtcctcc ttgtcctgcc cctcctgtgg ccggaccttc cgtgttcctt 780
ttccctccta agcctaagga caccctgatg atcagccgta cccctgaggt gacctgtgtg 840
gtggtggacg tgtcccacga ggaccctgag gtgcagttca actggtacgt ggacggcgtg 900
gaggtgcaca acgccaagac caagcctcgt gaggagcaat tcaacagcac cttccgtgtg 960
gtgtccgtgc ttaccgtggt gcaccaagac tggctgaacg gcaaggagta caagtgtaag 1020
gtgagcaaca agggacttcc tgcccctatc gagaagacca tctccaagac caagggccaa 1080
cctcgtgagc ctcaagtgta cacccttcct cctagccgtg aggagatgac caagaaccaa 1140
gtgtccctta cctgtctggt gaagggcttc taccctagcg acatcgccgt ggagtgggag 1200
tccaacggac aacctgagaa caactacaag accacccctc ctatgcttga cagcgacggc 1260
tccttcttcc tgtacagcaa gctgaccgtg gacaagtccc gttggcaaca aggcaacgtg 1320
ttcagctgtt ccgtgatgca cgaggccctg cacaaccact acacccaaaa gagcctttcc 1380
ctgagccctg gaaag 1395
<210> 73
<211> 465
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность тяжелой цепи 2C1.1
<400> 73
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys
20 25 30
Pro Ser Glu Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe
35 40 45
Ser Gly Tyr Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu
50 55 60
Glu Trp Ile Gly Glu Ile Asp His Ser Gly Ser Thr Asn Tyr Asn Pro
65 70 75 80
Ala Leu Lys Ser Arg Leu Thr Ile Ser Val Asp Thr Ser Lys Asn Gln
85 90 95
Phe Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr
100 105 110
Tyr Cys Ala Arg Ala Gly Ser Gly Gly Tyr Glu Asp Trp Phe Asp Pro
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val
210 215 220
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys
225 230 235 240
Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys
465
<210> 74
<211> 739
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая легкую цепь 2C1.1
<400> 74
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
cagtccgtgc tgacccagcc tccttccgtg tctggcgccc ctggccagag agtgaccatc 120
agctgtaccg gcagcagcag caacatcgga gccggctaca acgtgcactg gtatcagcag 180
ttcccccgga ccgaccccaa gctgctgatc tacgtgtaca acatccggcc cagcggcgtg 240
cccgaccggt tttctggcag cagaagcggc acaagcgcca gcctggccat caccggcctg 300
cagaccgagg acgaggccga ctactactgc cagagctacg acagcagcct gagcggcgtg 360
gtgttcggcg gaggcaccaa gctgacagtg ctgggccagc ccaaggccaa ccccaccgtg 420
accctgttcc ccccaagcag cgaggaactg caggccaaca aggccaccct ggtgtgcctg 480
atcagcgact tctaccctgg cgccgtgaca gtggcctgga aggccgatgg atctcccgtg 540
aaggccggcg tggaaaccac caagcccagc aagcagagca acaacaaata cgccgccagc 600
agctacctga gcctgacccc cgagcagtgg aagtcccacc ggtcctacag ctgccaggtg 660
acacacgagg gcagcaccgt ggaaaagacc gtggccccca ccgagtgcag ctaggggccc 720
gtttaaacgg gggaggcta 739
<210> 75
<211> 237
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность легкой цепи 2C1.1
<400> 75
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly
20 25 30
Ala Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn
35 40 45
Ile Gly Ala Gly Tyr Asn Val His Trp Tyr Gln Gln Phe Pro Arg Thr
50 55 60
Asp Pro Lys Leu Leu Ile Tyr Val Tyr Asn Ile Arg Pro Ser Gly Val
65 70 75 80
Pro Asp Arg Phe Ser Gly Ser Arg Ser Gly Thr Ser Ala Ser Leu Ala
85 90 95
Ile Thr Gly Leu Gln Thr Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser
100 105 110
Tyr Asp Ser Ser Leu Ser Gly Val Val Phe Gly Gly Gly Thr Lys Leu
115 120 125
Thr Val Leu Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro
130 135 140
Pro Ser Ser Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu
145 150 155 160
Ile Ser Asp Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp
165 170 175
Gly Ser Pro Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln
180 185 190
Ser Asn Asn Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu
195 200 205
Gln Trp Lys Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly
210 215 220
Ser Thr Val Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
225 230 235
<210> 76
<211> 1395
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь 5H3.1
<400> 76
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaacctg gcgctagcgt gaaggtgtcc 120
tgcaaggcca gcggctacac cttcaccgac tactacatga actgggtgcg ccaggctcca 180
ggacagggcc tggaatggat gggctggatc aaccccaaca gcggcggcac caaatacgcc 240
cagaaattcc agggcagagt gaccatgacc cgggacacca gcatccggac cgcctacatg 300
gaactgagcc ggctgagaag cgacgacacc gccgtgtact actgcgccag agagtacggc 360
ggcaacagcg attggttcga cccctggggc cagggcaccc tggtgaccgt cagctcagcc 420
agcaccaagg gcccttccgt gttccctctg gccccttgta gccgttccac cagcgagtcc 480
accgccgccc ttggctgtct ggtgaaggac tacttccctg agcctgtgac cgtgagctgg 540
aactccggag cccttaccag cggcgtgcac accttccctg ccgtgctgca gtccagcggc 600
ctttactccc tgagctccgt ggtgaccgtg cctagctcca acttcggcac ccaaacctac 660
acctgtaacg tggaccacaa gcctagcaac accaaggtgg acaagaccgt ggagcgtaag 720
tgttgtgtgg agtgtcctcc ttgtcctgcc cctcctgtgg ccggaccttc cgtgttcctt 780
ttccctccta agcctaagga caccctgatg atcagccgta cccctgaggt gacctgtgtg 840
gtggtggacg tgtcccacga ggaccctgag gtgcagttca actggtacgt ggacggcgtg 900
gaggtgcaca acgccaagac caagcctcgt gaggagcaat tcaacagcac cttccgtgtg 960
gtgtccgtgc ttaccgtggt gcaccaagac tggctgaacg gcaaggagta caagtgtaag 1020
gtgagcaaca agggacttcc tgcccctatc gagaagacca tctccaagac caagggccaa 1080
cctcgtgagc ctcaagtgta cacccttcct cctagccgtg aggagatgac caagaaccaa 1140
gtgtccctta cctgtctggt gaagggcttc taccctagcg acatcgccgt ggagtgggag 1200
tccaacggac aacctgagaa caactacaag accacccctc ctatgcttga cagcgacggc 1260
tccttcttcc tgtacagcaa gctgaccgtg gacaagtccc gttggcaaca aggcaacgtg 1320
ttcagctgtt ccgtgatgca cgaggccctg cacaaccact acacccaaaa gagcctttcc 1380
ctgagccctg gaaag 1395
<210> 77
<211> 465
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность тяжелой цепи 5H3.1
<400> 77
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asp Tyr Tyr Met Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Trp Ile Asn Pro Asn Ser Gly Gly Thr Lys Tyr Ala
65 70 75 80
Gln Lys Phe Gln Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Arg
85 90 95
Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Glu Tyr Gly Gly Asn Ser Asp Trp Phe Asp Pro
115 120 125
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
130 135 140
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
145 150 155 160
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
165 170 175
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
180 185 190
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
195 200 205
Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val
210 215 220
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys
225 230 235 240
Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro
245 250 255
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
260 265 270
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
275 280 285
Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
290 295 300
Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val
305 310 315 320
Val Ser Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu
325 330 335
Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys
340 345 350
Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
355 360 365
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
370 375 380
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
385 390 395 400
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu
405 410 415
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
420 425 430
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
435 440 445
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455 460
Lys
465
<210> 78
<211> 742
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая легкую цепь 5H3.1
<400> 78
atggtgctgc agacccaggt gttcatctcc ctgctgctgt ggatctccgg cgcgtacggc 60
cagtccgtgc tgacccagcc tccttccgtg tctggcgccc ctggccagag agtgaccatc 120
agctgtaccg gcagcagcag caacatcgga gctggatacg acgtgcactg gtatcagcag 180
ctgcccggca ccgcccccaa gctgctgatc tacggcaaca gcaaccggcc cagcggcgtg 240
cccgatagat tcagcggcag caagagcggc accagcgcca gcctggccat taccggactg 300
caggccgagg acgaggccga ctactactgc cagagctacg acaaccggct gagcgacagc 360
gtggtgatcg gcggaggcac caagctggcc gtgcagggac agcccaaggc caaccccacc 420
gtgaccctgt tccccccaag cagcgaggaa ctgcaggcca acaaggccac cctggtgtgc 480
ctgatcagcg acttctaccc tggcgccgtg acagtggcct ggaaggccga tggatctccc 540
gtgaaggccg gcgtggaaac caccaagccc agcaagcaga gcaacaacaa atacgccgcc 600
agcagctacc tgagcctgac ccccgagcag tggaagtccc accggtccta cagctgccag 660
gtgacacacg agggcagcac cgtggaaaag accgtggccc ccaccgagtg cagctagggg 720
cccgtttaaa cgggggaggc ta 742
<210> 79
<211> 238
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность легкой цепи 5H3.1
<400> 79
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly
20 25 30
Ala Pro Gly Gln Arg Val Thr Ile Ser Cys Thr Gly Ser Ser Ser Asn
35 40 45
Ile Gly Ala Gly Tyr Asp Val His Trp Tyr Gln Gln Leu Pro Gly Thr
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Gly Asn Ser Asn Arg Pro Ser Gly Val
65 70 75 80
Pro Asp Arg Phe Ser Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala
85 90 95
Ile Thr Gly Leu Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser
100 105 110
Tyr Asp Asn Arg Leu Ser Asp Ser Val Val Ile Gly Gly Gly Thr Lys
115 120 125
Leu Ala Val Gln Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe
130 135 140
Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys
145 150 155 160
Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala
165 170 175
Asp Gly Ser Pro Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys
180 185 190
Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro
195 200 205
Glu Gln Trp Lys Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu
210 215 220
Gly Ser Thr Val Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
225 230 235
<210> 80
<211> 1433
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь 10F8
<400> 80
ccagcctccg gactctagag ccaccatgaa gcacctgtgg ttctttctgc tgctggtggc 60
cgctcccaga tgggtgctgt ctcaggtgca gctgcagcag tctggcgccg aactcgtgcg 120
gcctggaagc agcgtgaaga tcagctgcaa ggccagcggc tacgccttcc ggtcctactg 180
gatgaactgg gtcaagcaga ggccaggcca gggcctggaa tggatcggcc acatctatcc 240
cggcgacggc gacaccaact acaacggcaa gttcaagggc aaggccaccc tgaccgccga 300
caagagcagc agcacagcct acatgcagct gtccagcctg accagcgagg acagcgccgt 360
gtacctgtgt ggcagaggcg gcacaaccgt ggtggtggat tattggggcc agggcaccac 420
actgaccgtg tccagcgcca agaccacccc cccatctgtg tatcctctgg cccctggatc 480
tgccgcccag accaacagca tggtcaccct gggctgcctc gtgaagggct acttccctga 540
gcctgtgacc gtgacctgga acagcggctc tctgtctagc ggcgtgcaca cctttccagc 600
cgtgctgcag agcgacctgt acaccctgag cagctccgtg accgtgccta gcagcacctg 660
gcctagcgag acagtgacct gcaacgtggc ccaccctgcc agctctacca aggtggacaa 720
gaaaatcgtg ccccgggact gcggctgcaa gccctgtatc tgtaccgtgc ccgaggtgtc 780
ctccgtgttc atcttcccac ccaagcccaa ggacgtgctg accatcaccc tgacacccaa 840
agtgacatgt gtggtggtgg acatcagcaa ggacgacccc gaggtgcagt tcagttggtt 900
cgtggacgac gtggaagtgc acacagccca gacccagccc agagaggaac agttcaacag 960
caccttcaga agcgtgtccg agctgcccat catgcaccag gactggctga acggcaaaga 1020
attcaagtgc agagtgaaca gcgccgcctt ccctgccccc atcgagaaaa ccatctccaa 1080
gaccaagggc agacccaagg ccccccaggt gtacacaatc cccccaccca aagaacagat 1140
ggccaaggac aaggtgtccc tgacctgcat gatcaccgat ttcttcccag aggacatcac 1200
cgtggaatgg cagtggaacg gccagcccgc cgagaactac aagaacaccc agcctatcat 1260
ggacaccgac ggcagctact tcgtgtacag caagctgaac gtgcagaagt ccaactggga 1320
ggccggcaac accttcacct gtagcgtgct gcacgagggc ctgcacaatc accacaccga 1380
gaagtccctg tcccacagcc ccggcaaatg agtttaaacg ggggaggcta act 1433
<210> 81
<211> 461
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность тяжелой цепи 10F8
<400> 81
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe
35 40 45
Arg Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly His Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn
65 70 75 80
Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Leu Cys Gly Arg Gly Gly Thr Thr Val Val Val Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Ala Lys Thr Thr Pro Pro Ser
130 135 140
Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met Val
145 150 155 160
Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val Pro
195 200 205
Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His Pro
210 215 220
Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys Gly
225 230 235 240
Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe Ile
245 250 255
Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro Lys
260 265 270
Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val Gln
275 280 285
Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr Gln
290 295 300
Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu Leu
305 310 315 320
Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys Arg
325 330 335
Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
340 345 350
Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro Pro
355 360 365
Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile Thr
370 375 380
Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly Gln
385 390 395 400
Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp Gly
405 410 415
Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp Glu
420 425 430
Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His Asn
435 440 445
His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
450 455 460
<210> 82
<211> 770
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая легкую цепь 10F8
<400> 82
ccagcctccg gactctagag ccaccatggt gctgcagacc caggtgttca tcagcctgct 60
gctgtggatc agcggcgcct acggcgacat cgtgatgagc cagagcccta gcagcctggc 120
cgtgtctgcc ggcgagaaag tgaccatgag ctgcaagagc agccagtccc tgctgaacag 180
ccggacccgg aagaactacc tggcctggta tcagcagaag cccggccagt cccccaagct 240
gctgatctac tgggccagca ccagagaaag cggcgtgccc gatagattca ccggcagcgg 300
ctctggcacc gacttcaccc tgacaatcag cagcgtgcag gccgaggacc tggctgtgta 360
ctactgcaag cagagctaca acctgccctg gaccttcggc ggaggcacca agctggaaat 420
caagagagcc gacgccgctc ccaccgtgtc catctttcca cctagcagcg agcagctgac 480
cagcggcgga gctagcgtcg tgtgcttcct gaacaacttc taccccaagg acatcaacgt 540
gaagtggaag atcgacggca gcgagcggca gaacggcgtg ctgaatagct ggaccgacca 600
ggacagcaag gactccacct acagcatgtc cagcaccctg accctgacca aggacgagta 660
cgagcggcac aacagctaca catgcgaggc cacccacaag accagcacct cccccatcgt 720
gaagtccttc aaccggaacg agtgctgagt ttaaacgggg gaggctaact 770
<210> 83
<211> 240
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность легкой цепи 10F8
<400> 83
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala
20 25 30
Val Ser Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg
65 70 75 80
Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr
100 105 110
Tyr Cys Lys Gln Ser Tyr Asn Leu Pro Trp Thr Phe Gly Gly Gly Thr
115 120 125
Lys Leu Glu Ile Lys Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe
130 135 140
Pro Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys
145 150 155 160
Phe Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile
165 170 175
Asp Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln
180 185 190
Asp Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr
195 200 205
Lys Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His
210 215 220
Lys Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
225 230 235 240
<210> 84
<211> 1436
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь 14F74
<400> 84
ccagcctccg gactctagag ccaccatgaa gcacctgtgg ttctttctgc tgctggtggc 60
cgctcccaga tgggtgctgt ctcaggtgca gctgcagcag tctggcgccg aactcgtgcg 120
gcctggaagc agcgtgaaga tcagctgcaa ggccagcggc tacgccttca gcagctactg 180
gatgaactgg gtcaagcagc ggccaggcca gggcctggaa tggatcggcc atatctatcc 240
cggcgacggc gacaccaact acaacggcaa gttcaagggc aaggccaccc tgaccgccga 300
caagagcagc agcacagcct acatgcagct gagcggcctg accagcgagg acagcgccgt 360
gtacttctgc gccagaagcg gcagactgag attcgccatg gactactggg gccagggcac 420
cagcgtgaca gtgtctagcg ccaagaccac cccccccagc gtgtaccctc tggctcctgg 480
atctgccgcc cagaccaaca gcatggtcac cctgggctgc ctcgtgaagg gctacttccc 540
tgagcctgtg accgtgacct ggaacagcgg ctctctgtct agcggcgtgc acacctttcc 600
agccgtgctg cagagcgacc tgtacaccct gagcagctcc gtgaccgtgc ctagcagcac 660
ctggcctagc gagacagtga cctgcaacgt ggcccaccct gccagctcta ccaaggtgga 720
caagaaaatc gtgccccggg actgcggctg caagccctgt atctgtaccg tgcccgaggt 780
gtccagcgtg ttcatcttcc cacccaagcc caaggacgtg ctgaccatca ccctgacacc 840
caaagtgacc tgtgtggtgg tggacatcag caaggacgac cccgaggtgc agttcagttg 900
gttcgtggac gacgtggaag tgcacacagc ccagacccag cccagagagg aacagttcaa 960
cagcaccttc agaagcgtgt ccgagctgcc catcatgcac caggactggc tgaacggcaa 1020
agaattcaag tgcagagtga acagcgccgc cttccctgcc cccatcgaga aaaccatctc 1080
caagaccaag ggcagaccca aggcccctca ggtgtacaca atccccccac ccaaagaaca 1140
gatggccaag gacaaggtgt ccctgacctg catgatcacc gatttcttcc cagaggacat 1200
caccgtggaa tggcagtgga acggccagcc cgccgagaac tacaagaaca cccagcctat 1260
catggacacc gacggcagct acttcgtgta cagcaagctg aacgtgcaga agtccaactg 1320
ggaggccggc aacaccttca cctgtagcgt gctgcacgag ggcctgcaca atcaccacac 1380
cgagaagtcc ctgtcccaca gccccggcaa atgagtttaa acgggggagg ctaact 1436
<210> 85
<211> 462
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность тяжелой цепи 14F74
<400> 85
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe
35 40 45
Ser Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly His Ile Tyr Pro Gly Asp Gly Asp Thr Asn Tyr Asn
65 70 75 80
Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Gly Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ala Arg Ser Gly Arg Leu Arg Phe Ala Met Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
130 135 140
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
145 150 155 160
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
195 200 205
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
210 215 220
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
225 230 235 240
Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe
245 250 255
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
260 265 270
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
275 280 285
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
290 295 300
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
305 310 315 320
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
325 330 335
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
355 360 365
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
370 375 380
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
385 390 395 400
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
405 410 415
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
420 425 430
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
435 440 445
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
450 455 460
<210> 86
<211> 767
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая легкую цепь 14F74
<400> 86
ccagcctccg gactctagag ccaccatggt gctgcagacc caggtgttca tcagcctgct 60
gctgtggatc agcggcgcct acggcgacat cgtgatgagc cagagcccta gcagcctggc 120
cgtgtctgcc ggcgagaaag tgaccatgag ctgcaagagc agccagtccc tgctgaacag 180
ccggacccgg aagaactacc tggcctggta tcagcagaag cccggccagt cccccaagct 240
gctgatctac tgggccagca ccagagaaag cggcgtgccc gatagattca ccggcagcgg 300
ctctggcacc gacttcaccc tgacaatcag cagcgtgcag gccgaggacc tggctgtgta 360
ctactgcaag cagagctaca acctgcggac cttcggcgga ggcaccaagc tggaaatcca 420
gagagccgac gccgctccca ccgtgtccat ctttccacct agcagcgagc agctgaccag 480
cggcggagct agcgtcgtgt gcttcctgaa caacttctac cccaaggaca tcaacgtgaa 540
gtggaagatc gacggcagcg agcggcagaa cggcgtgctg aatagctgga ccgaccagga 600
cagcaaggac tccacctaca gcatgtccag caccctgacc ctgaccaagg acgagtacga 660
gcggcacaac agctacacat gcgaggccac ccacaagacc agcacctccc ccatcgtgaa 720
gtccttcaac cggaacgagt gctgagttta aacgggggag gctaact 767
<210> 87
<211> 239
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность легкой цепи 14F74
<400> 87
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala
20 25 30
Val Ser Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg
65 70 75 80
Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr
100 105 110
Tyr Cys Lys Gln Ser Tyr Asn Leu Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Gln Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro
130 135 140
Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe
145 150 155 160
Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp
165 170 175
Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp
180 185 190
Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys
195 200 205
Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys
210 215 220
Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
225 230 235
<210> 88
<211> 1436
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая тяжелую цепь 17F6
<400> 88
ccagcctccg gactctagag ccaccatgaa gcacctgtgg ttctttctgc tgctggtggc 60
cgctcccaga tgggtgctgt ctcaggtgca gctgcagcag tctggcgccg aactcgtgcg 120
gcctggaagc agcgtgaaga tcagctgcaa ggccagcggc tacgccttca gcagctactg 180
gatgaactgg gtcaagcagc ggccaggcca gggcctggaa tggatcggcc atatctatcc 240
cggcgacgcc gacaccaact acaacggcaa gttcaagggc aaggccaccc tgaccgccga 300
caagagcagc agcacagcct acatgcacct gtccagcctg accagcgagg acagcgccgt 360
gtacttctgc agccggcagc tgggcttcag atacgccatg gactattggg gccagggcac 420
cagcgtgacc gtgtctagcg ccaagaccac cccccctagc gtgtaccctc tggcccctgg 480
atctgccgcc cagaccaaca gcatggtcac cctgggctgc ctcgtgaagg gctacttccc 540
tgagcctgtg accgtgacct ggaacagcgg ctctctgtct agcggcgtgc acacctttcc 600
agccgtgctg cagagcgacc tgtacaccct gagcagctcc gtgacagtgc ccagctctac 660
ctggcccagc gagacagtga cctgcaacgt ggcccaccct gccagcagca ccaaggtgga 720
caagaaaatc gtgccccggg actgcggctg caagccctgt atctgtaccg tgcccgaggt 780
gtccagcgtg ttcatcttcc cacccaagcc caaggacgtg ctgaccatca ccctgacacc 840
caaagtgacc tgtgtggtgg tggacatcag caaggacgac cccgaggtgc agttcagttg 900
gttcgtggac gacgtggaag tgcacacagc ccagacccag cccagagagg aacagttcaa 960
cagcaccttc agaagcgtgt ccgagctgcc catcatgcac caggactggc tgaacggcaa 1020
agaattcaag tgcagagtga acagcgccgc cttccctgcc cccatcgaga aaaccatctc 1080
caagaccaag ggcagaccca aggcccccca ggtgtacaca atccccccac ccaaagaaca 1140
gatggccaag gacaaggtgt ccctgacctg catgatcacc gatttcttcc cagaggacat 1200
caccgtggaa tggcagtgga acggccagcc cgccgagaac tacaagaaca cccagcctat 1260
catggacacc gacggcagct acttcgtgta cagcaagctg aacgtgcaga agtccaactg 1320
ggaggccggc aacaccttca cctgtagcgt gctgcacgag ggcctgcaca atcaccacac 1380
cgagaagtcc ctgtcccaca gccccggcaa atgagtttaa acgggggagg ctaact 1436
<210> 89
<211> 462
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность тяжелой цепи 17F6
<400> 89
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala Phe
35 40 45
Ser Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly His Ile Tyr Pro Gly Asp Ala Asp Thr Asn Tyr Asn
65 70 75 80
Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met His Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Phe Cys Ser Arg Gln Leu Gly Phe Arg Tyr Ala Met Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Ser Val Thr Val Ser Ser Ala Lys Thr Thr Pro Pro
130 135 140
Ser Val Tyr Pro Leu Ala Pro Gly Ser Ala Ala Gln Thr Asn Ser Met
145 150 155 160
Val Thr Leu Gly Cys Leu Val Lys Gly Tyr Phe Pro Glu Pro Val Thr
165 170 175
Val Thr Trp Asn Ser Gly Ser Leu Ser Ser Gly Val His Thr Phe Pro
180 185 190
Ala Val Leu Gln Ser Asp Leu Tyr Thr Leu Ser Ser Ser Val Thr Val
195 200 205
Pro Ser Ser Thr Trp Pro Ser Glu Thr Val Thr Cys Asn Val Ala His
210 215 220
Pro Ala Ser Ser Thr Lys Val Asp Lys Lys Ile Val Pro Arg Asp Cys
225 230 235 240
Gly Cys Lys Pro Cys Ile Cys Thr Val Pro Glu Val Ser Ser Val Phe
245 250 255
Ile Phe Pro Pro Lys Pro Lys Asp Val Leu Thr Ile Thr Leu Thr Pro
260 265 270
Lys Val Thr Cys Val Val Val Asp Ile Ser Lys Asp Asp Pro Glu Val
275 280 285
Gln Phe Ser Trp Phe Val Asp Asp Val Glu Val His Thr Ala Gln Thr
290 295 300
Gln Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Ser Val Ser Glu
305 310 315 320
Leu Pro Ile Met His Gln Asp Trp Leu Asn Gly Lys Glu Phe Lys Cys
325 330 335
Arg Val Asn Ser Ala Ala Phe Pro Ala Pro Ile Glu Lys Thr Ile Ser
340 345 350
Lys Thr Lys Gly Arg Pro Lys Ala Pro Gln Val Tyr Thr Ile Pro Pro
355 360 365
Pro Lys Glu Gln Met Ala Lys Asp Lys Val Ser Leu Thr Cys Met Ile
370 375 380
Thr Asp Phe Phe Pro Glu Asp Ile Thr Val Glu Trp Gln Trp Asn Gly
385 390 395 400
Gln Pro Ala Glu Asn Tyr Lys Asn Thr Gln Pro Ile Met Asp Thr Asp
405 410 415
Gly Ser Tyr Phe Val Tyr Ser Lys Leu Asn Val Gln Lys Ser Asn Trp
420 425 430
Glu Ala Gly Asn Thr Phe Thr Cys Ser Val Leu His Glu Gly Leu His
435 440 445
Asn His His Thr Glu Lys Ser Leu Ser His Ser Pro Gly Lys
450 455 460
<210> 90
<211> 767
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая легкую цепь 17F6
<400> 90
ccagcctccg gactctagag ccaccatggt gctgcagacc caggtgttca tcagcctgct 60
gctgtggatc agcggcgcct acggcgacat cgtgatgagc cagagcccta gcagcctggc 120
cgtgtctgcc ggcgagaaag tgaccatgag ctgcaagagc agccagtccc tgctgaacag 180
ccggacccgg aagaactacc tggcctggta tcagcagaag cccggccagt cccccaagct 240
gctgatctac tgggccagca ccagagaaag cggcgtgccc gatagattca ccggcagcgg 300
ctctggcacc gacttcaccc tgacaatcag cagcgtgcag gccgaggacc tggctgtgta 360
ctactgcaag cagagctaca acctgcggac cttcggcgga ggcaccaagc tggaaatcaa 420
gagagccgac gccgctccca ccgtgtccat ctttccacct agcagcgagc agctgaccag 480
cggcggagct agcgtcgtgt gcttcctgaa caacttctac cccaaggaca tcaacgtgaa 540
gtggaagatc gacggcagcg agcggcagaa cggcgtgctg aatagctgga ccgaccagga 600
cagcaaggac tccacctaca gcatgtccag caccctgacc ctgaccaagg acgagtacga 660
gcggcacaac agctacacat gcgaggccac ccacaagacc agcacctccc ccatcgtgaa 720
gtccttcaac cggaacgagt gctgagttta aacgggggag gctaact 767
<210> 91
<211> 239
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность легкой цепи 17F6
<400> 91
Met Val Leu Gln Thr Gln Val Phe Ile Ser Leu Leu Leu Trp Ile Ser
1 5 10 15
Gly Ala Tyr Gly Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala
20 25 30
Val Ser Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Leu Leu Asn Ser Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg
65 70 75 80
Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr
100 105 110
Tyr Cys Lys Gln Ser Tyr Asn Leu Arg Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys Arg Ala Asp Ala Ala Pro Thr Val Ser Ile Phe Pro
130 135 140
Pro Ser Ser Glu Gln Leu Thr Ser Gly Gly Ala Ser Val Val Cys Phe
145 150 155 160
Leu Asn Asn Phe Tyr Pro Lys Asp Ile Asn Val Lys Trp Lys Ile Asp
165 170 175
Gly Ser Glu Arg Gln Asn Gly Val Leu Asn Ser Trp Thr Asp Gln Asp
180 185 190
Ser Lys Asp Ser Thr Tyr Ser Met Ser Ser Thr Leu Thr Leu Thr Lys
195 200 205
Asp Glu Tyr Glu Arg His Asn Ser Tyr Thr Cys Glu Ala Thr His Lys
210 215 220
Thr Ser Thr Ser Pro Ile Val Lys Ser Phe Asn Arg Asn Glu Cys
225 230 235
<210> 92
<211> 11
<212> Белок
<213> Rattus rattus
<400> 92
Arg Ala Ser Gln Ser Ile Gly Asn Ser Leu Ser
1 5 10
<210> 93
<211> 11
<212> Белок
<213> искусственный
<220>
<223> CDRL1 of hR198_LG1
<400> 93
Arg Ala Ser Gln Ser Ile Gly Gly Ser Leu Ser
1 5 10
<210> 94
<211> 11
<212> Белок
<213> искусственный
<220>
<223> CDRL1 of hR198_LG2
<400> 94
His Ala Ser Gln Asn Ile Gly Gly Ser Leu Ser
1 5 10
<210> 95
<211> 11
<212> Белок
<213> искусственный
<220>
<223> CDRL1 of hR198_LG3
<400> 95
His Ala Ser Arg Asn Ile Gly Gly Ser Leu Ser
1 5 10
<210> 96
<211> 7
<212> Белок
<213> Rattus rattus
<400> 96
Ser Thr Ser Thr Leu Glu Ser
1 5
<210> 97
<211> 7
<212> Белок
<213> искусственный
<220>
<223> CDRL2 of hR198_LG2
<400> 97
Leu Thr Ser Thr Leu Asp Trp
1 5
<210> 98
<211> 7
<212> Белок
<213> искусственный
<220>
<223> CDRL2 of hR198_LG3
<400> 98
Leu Thr Ser Ser Leu Asp Trp
1 5
<210> 99
<211> 9
<212> Белок
<213> Rattus rattus
<400> 99
Leu Gln Phe Ala Thr Phe Pro Asp Thr
1 5
<210> 100
<211> 9
<212> Белок
<213> Rattus rattus
<400> 100
Leu Gln Phe Ala Thr Tyr Pro Asp Thr
1 5
<210> 101
<211> 9
<212> Белок
<213> искусственный
<220>
<223> CDRL3 of hR198_LG1 to LG3
<400> 101
Leu Gln Phe Ala Ile Phe Pro Asp Ser
1 5
<210> 102
<211> 5
<212> Белок
<213> Rattus rattus
<400> 102
Ala Tyr Tyr Ile Ser
1 5
<210> 103
<211> 5
<212> Белок
<213> Rattus rattus
<400> 103
Ser Tyr Tyr Ile Ser
1 5
<210> 104
<211> 17
<212> Белок
<213> Rattus rattus
<400> 104
Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn Ala Arg Phe Lys
1 5 10 15
Gly
<210> 105
<211> 17
<212> Белок
<213> Rattus rattus
<400> 105
Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 106
<211> 17
<212> Белок
<213> искусственный
<220>
<223> CDRH2 of hR198_HG1
<400> 106
Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn Ala Arg Phe Lys
1 5 10 15
Gly
<210> 107
<211> 17
<212> Белок
<213> искусственный
<220>
<223> CDRH2 of hR198_HG2
<400> 107
Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn Gly Arg Phe Lys
1 5 10 15
Gly
<210> 108
<211> 17
<212> Белок
<213> искусственный
<220>
<223> CDRH2 of hR198_HG3
<400> 108
Tyr Ile Asp Met Gly Asn Gly Arg Thr Asp Tyr Asn Met Arg Phe Lys
1 5 10 15
Gly
<210> 109
<211> 8
<212> Белок
<213> Rattus rattus
<400> 109
Asp Ser Asn Trp Gly Val Asp Tyr
1 5
<210> 110
<211> 8
<212> Белок
<213> искусственный
<220>
<223> CDRH3 of hR198_HG1 to HG3
<400> 110
Asp Ser Asn Trp Gly Ala Asp Tyr
1 5
<210> 111
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H0
<400> 111
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccagc tactacatca gctggatcag acaggcccca 180
ggccagggcc tggaatggat cggctatgtg gacatgggca acggccggac caactacaac 240
gagaagttca agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 112
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H0
<400> 112
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ser Tyr Tyr Ile Ser Trp Ile Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Val Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Glu Lys Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 113
<211> 1401
<212> ДНК
<213> искусственный
<220>
<223> Нуклеотидная последовательность, кодирующая hR198_H5
<400> 113
atgaaacacc tgtggttctt cctcctgctg gtggcagctc ccagatgggt gctgagccag 60
gtgcagctgg tgcagtctgg cgccgaagtg aagaaaccag gcgccagcgt gaaggtgtcc 120
tgcaaggcct ctggctaccc cgtgaccgcc tactacatca gctggatcag acaggcccca 180
ggccagggcc tggaatggat cggctacatc gacatgggca acggccggac caactacaac 240
gcccggttta agggcagagc caccctgacc gtggacaaga gcaccagcac cgcctacatg 300
gaactgagca gcctgcggag cgaggacacc gccgtgtact actgcgccag agacagcaac 360
tggggcgtgg actattgggg ccagggcaca ctcgtgaccg tcagctcagc ctccaccaag 420
ggcccaagcg tcttccccct ggcaccctcc tccaagagca cctctggcgg cacagccgcc 480
ctgggctgcc tggtcaagga ctacttcccc gaacccgtga ccgtgagctg gaactcaggc 540
gccctgacca gcggcgtgca caccttcccc gctgtcctgc agtcctcagg actctactcc 600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac 660
gtgaatcaca agcccagcaa caccaaggtg gacaagagag ttgagcccaa atcttgtgac 720
aaaactcaca catgcccacc ctgcccagca cctgaactcc tggggggacc ctcagtcttc 780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 900
gtggaggtgc ataatgccaa gacaaagccc cgggaggagc agtacaacag cacgtaccgg 960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggc 1080
cagccccggg aaccacaggt gtacaccctg cccccatccc gggaggagat gaccaagaac 1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 1200
gagagcaatg gccagcccga gaacaactac aagaccaccc ctcccgtgct ggactccgac 1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcagggcaac 1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacaccca gaagagcctc 1380
tccctgtctc ccggcaaatg a 1401
<210> 114
<211> 466
<212> Белок
<213> искусственный
<220>
<223> Аминокислотная последовательность hR198_H5
<400> 114
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1 5 10 15
Val Leu Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Val
35 40 45
Thr Ala Tyr Tyr Ile Ser Trp Ile Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asp Met Gly Asn Gly Arg Thr Asn Tyr Asn
65 70 75 80
Ala Arg Phe Lys Gly Arg Ala Thr Leu Thr Val Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Ser Asn Trp Gly Val Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 115
<211> 48
<212> Белок
<213> Homo sapiens
<400> 115
Ser Gly Ser Gly Phe Leu Pro Leu Lys Lys Gln Pro Gly Gln Pro Arg
1 5 10 15
Pro Thr Ser Lys Pro Pro Ala Ser Gly Ala Ala Ala Asn Val Ser Thr
20 25 30
Ser Gly Ile Thr Pro Gly Gln Ala Ala Ala Ile Ala Ser Thr Thr Ile
35 40 45
<210> 116
<211> 11
<212> Белок
<213> Rattus rattus
<400> 116
Arg Ala Ser Gln Ser Ile Ser Asn Ser Leu Ser
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
| АНТИТЕЛО ПРОТИВ SIRPα | 2019 |
|
RU2791002C2 |
| АНТИТЕЛО К TIGIT И ЕГО ИСПОЛЬЗОВАНИЕ | 2019 |
|
RU2786434C2 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| АНТИТЕЛО К CLDN-18.2 И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2829997C1 |
| Композиции и способы активации интегринов | 2020 |
|
RU2836262C1 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛА ПРОТИВ СИГНАЛ-РЕГУЛЯТОРНОГО БЕЛКА АЛЬФА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2771964C2 |
| СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800923C2 |
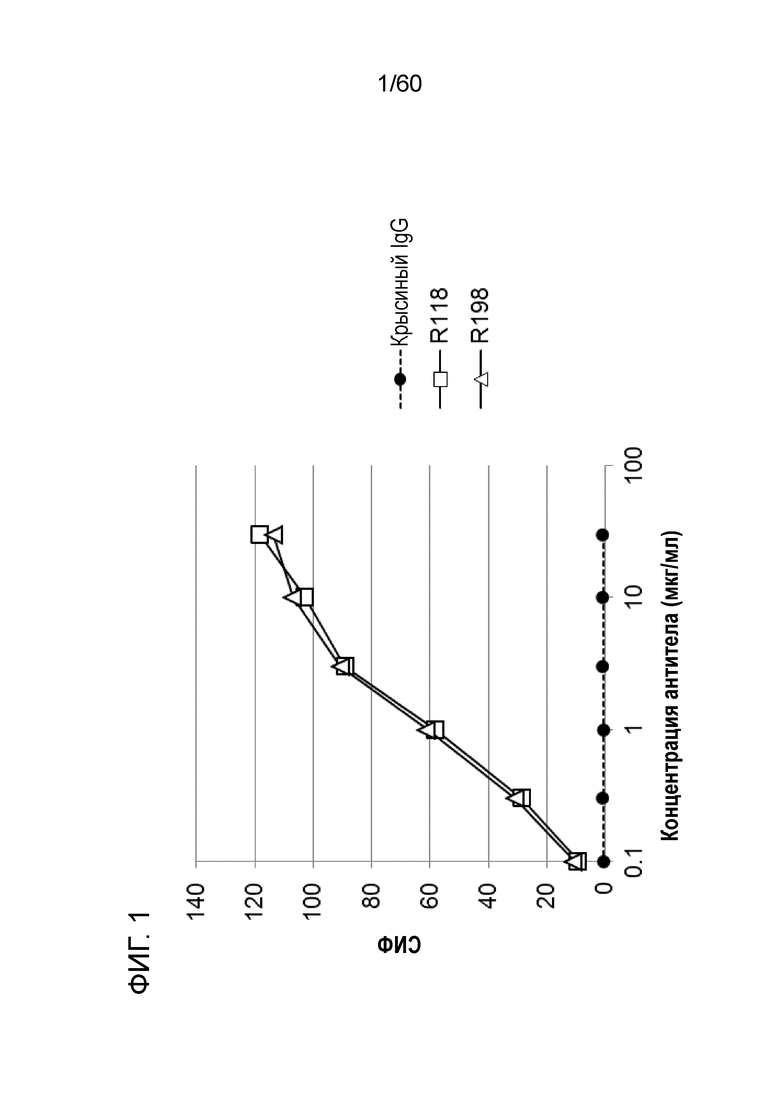
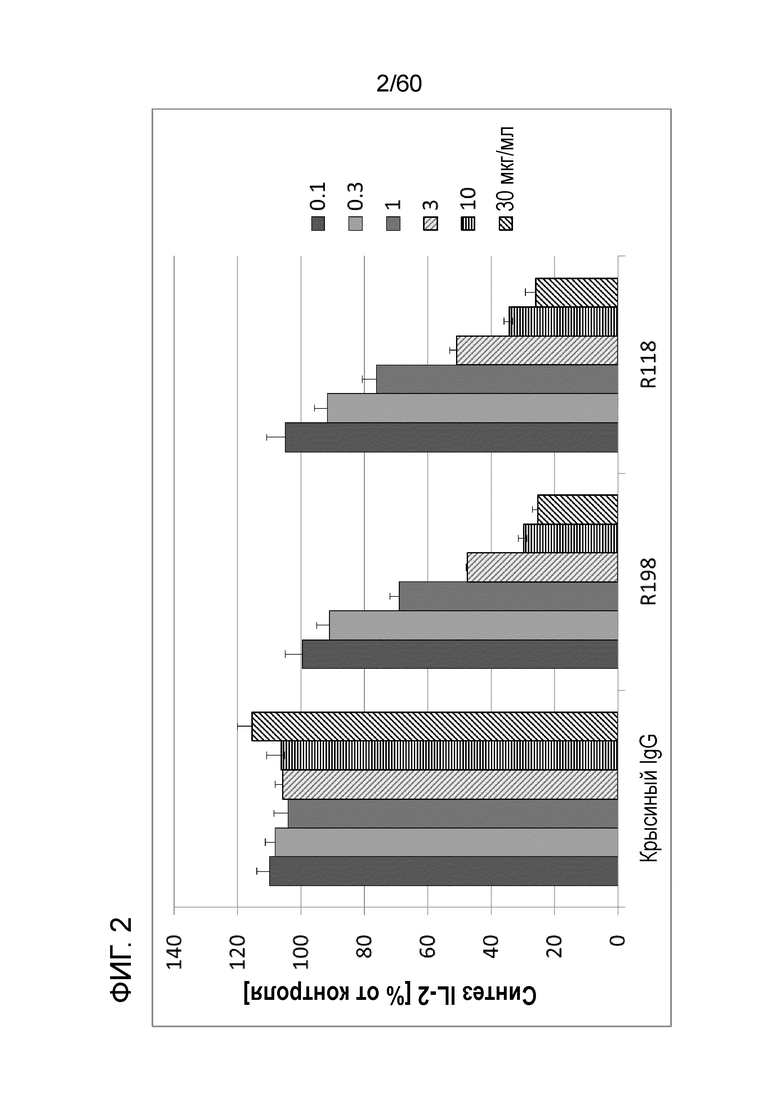
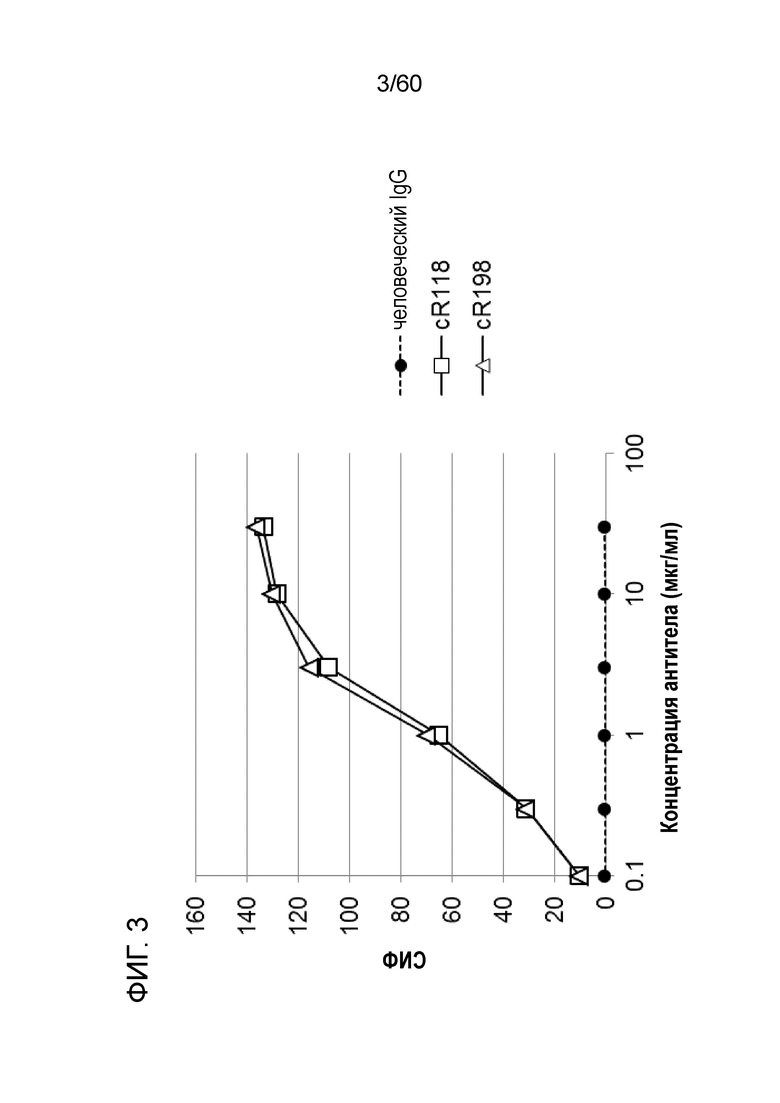
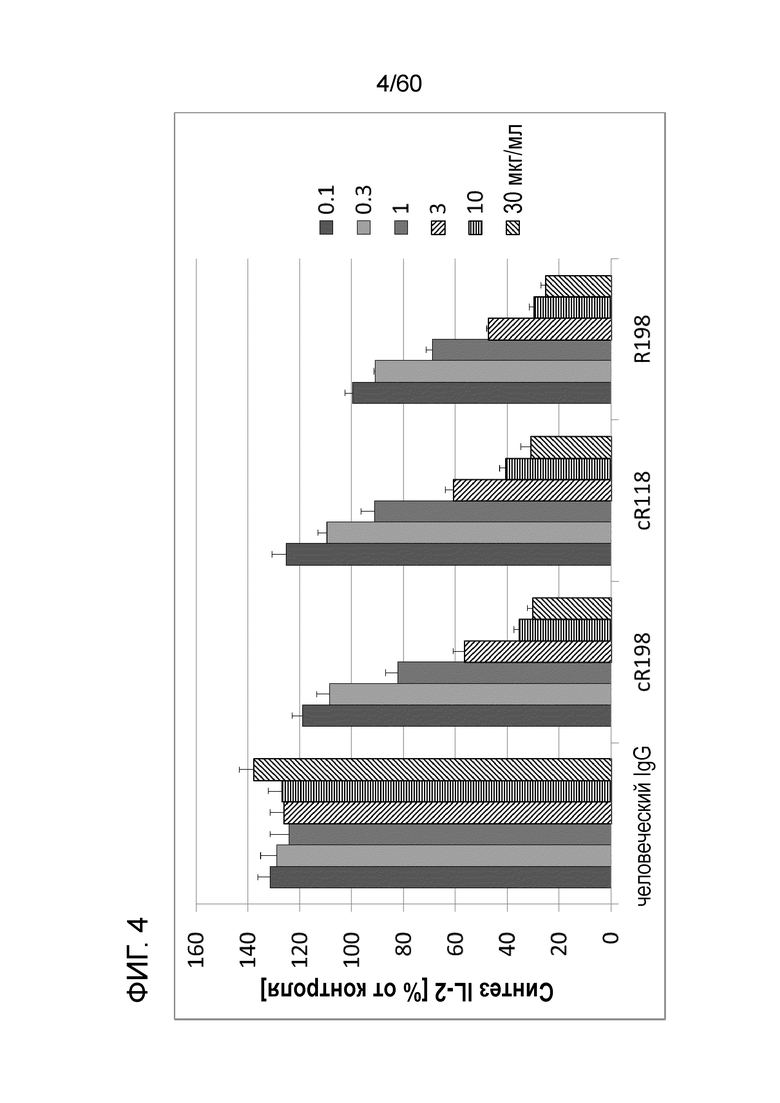
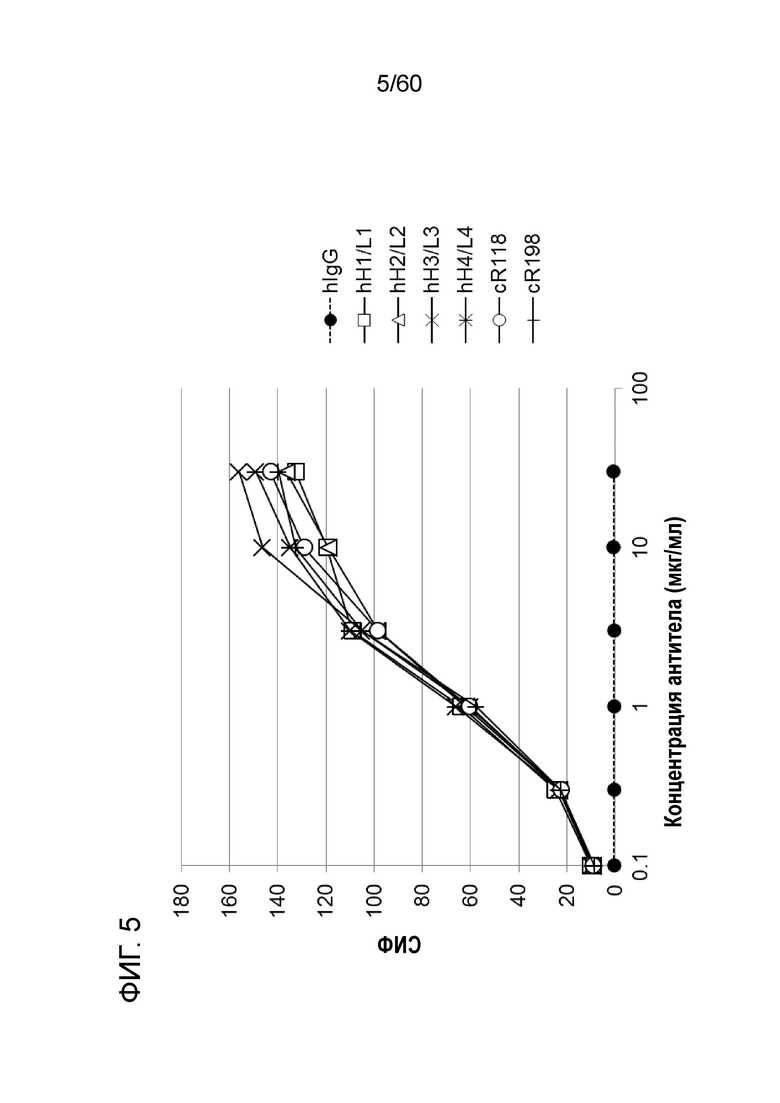
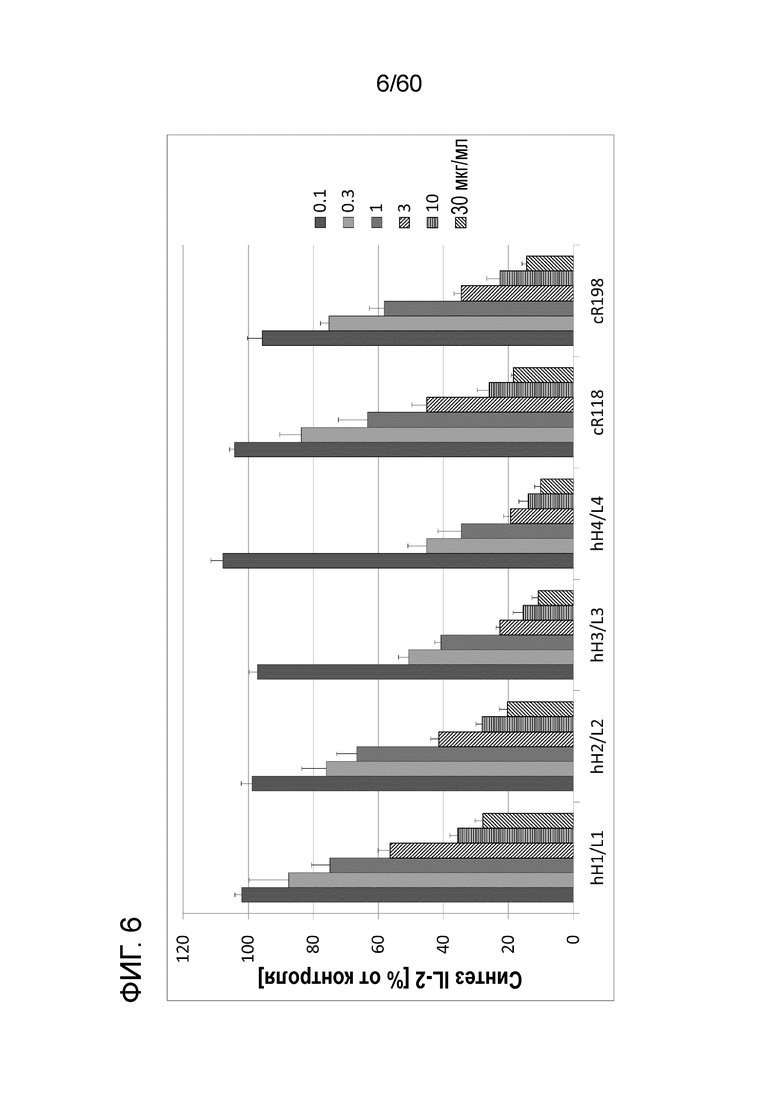
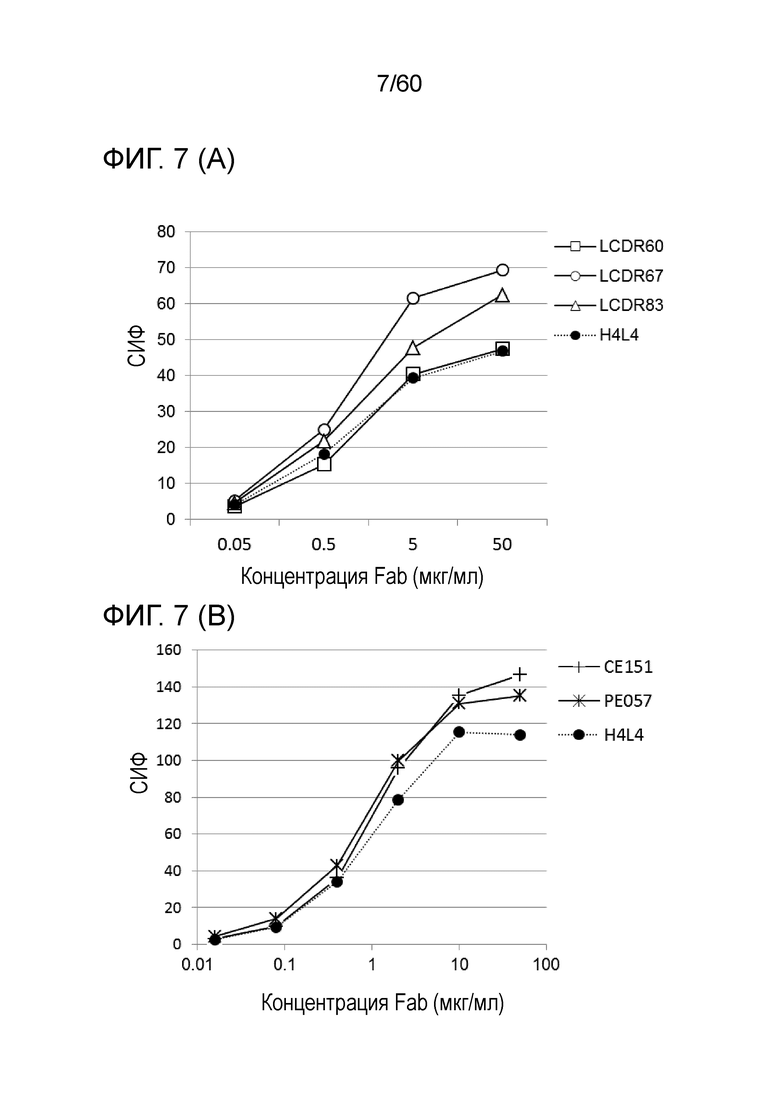
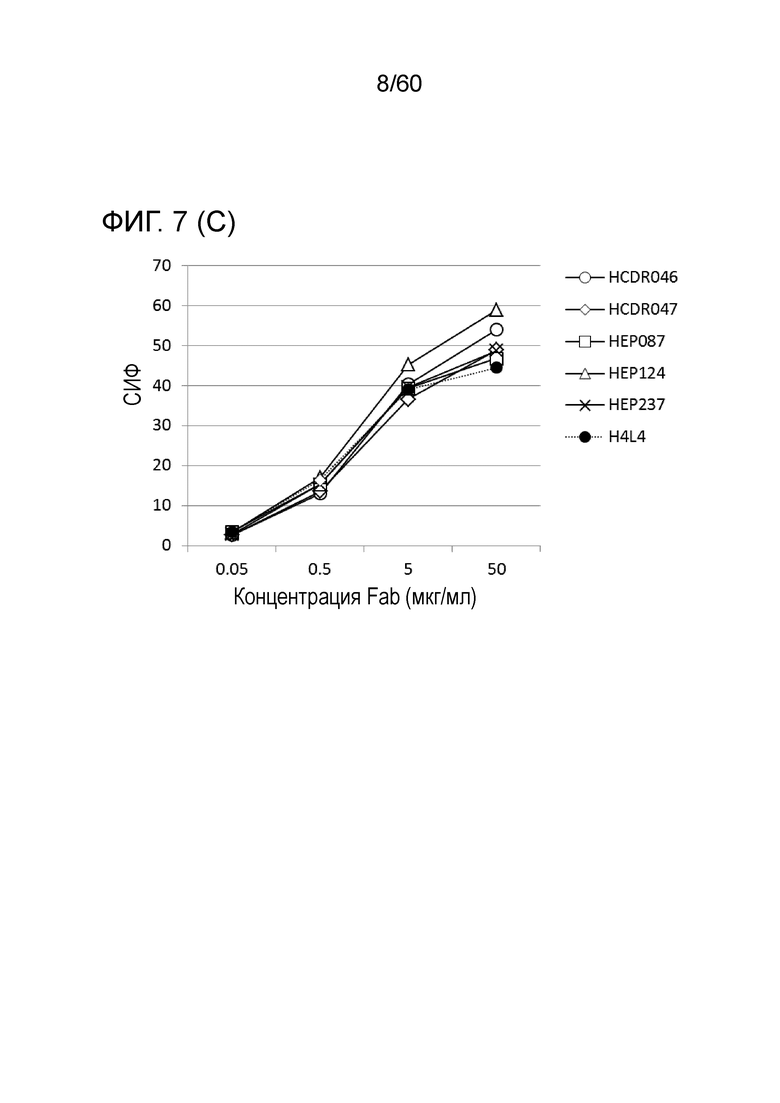
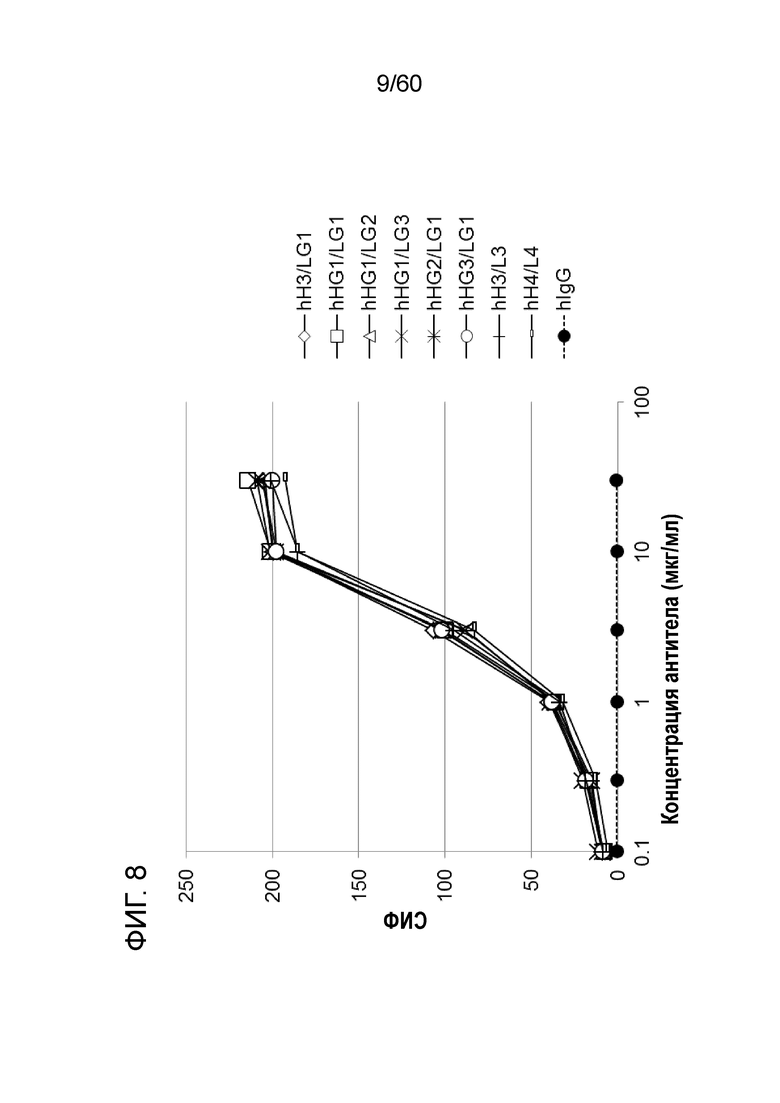
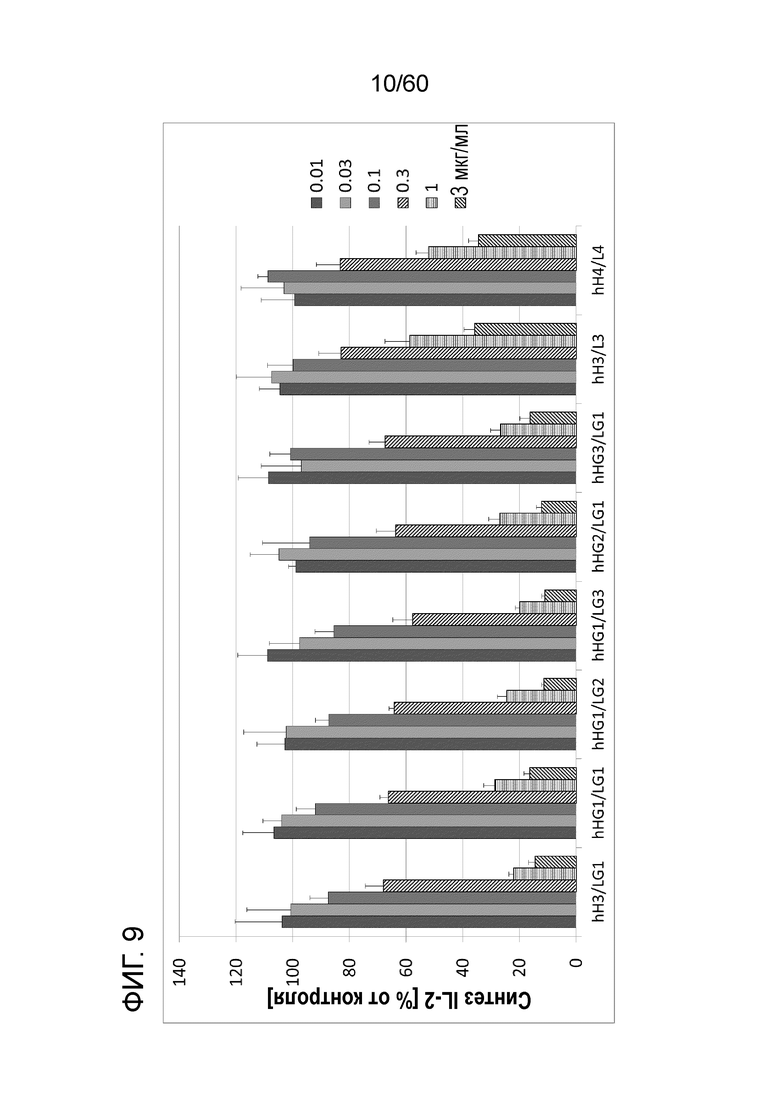
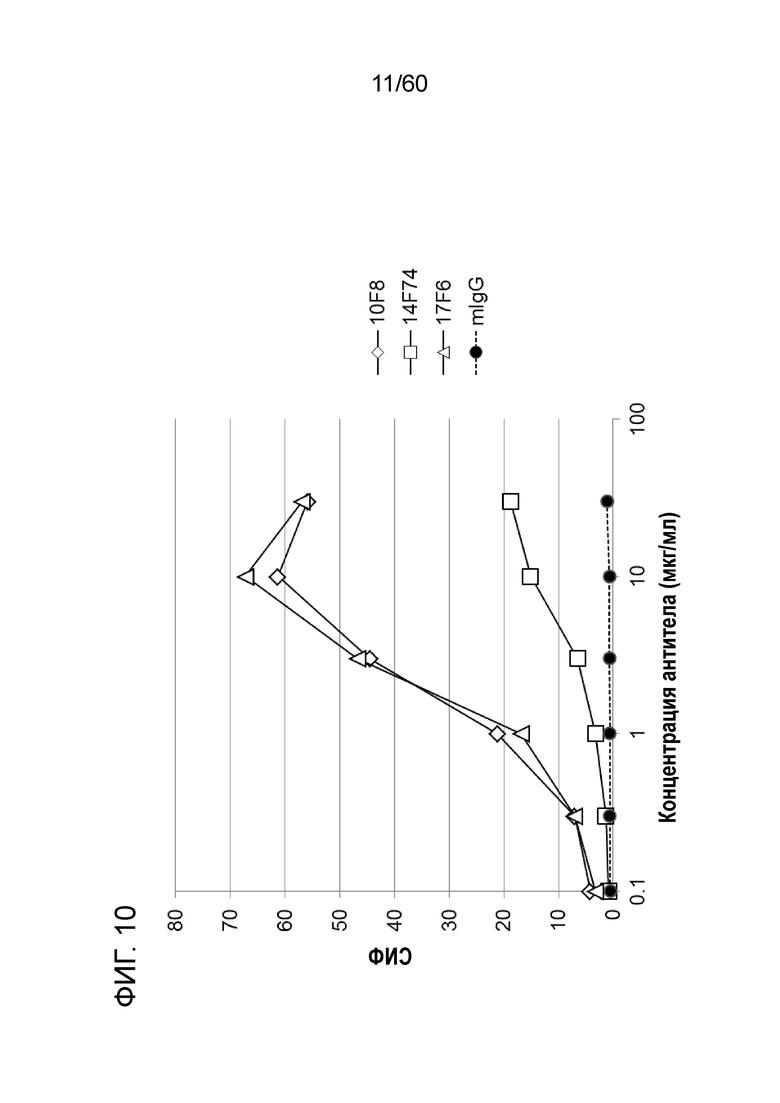
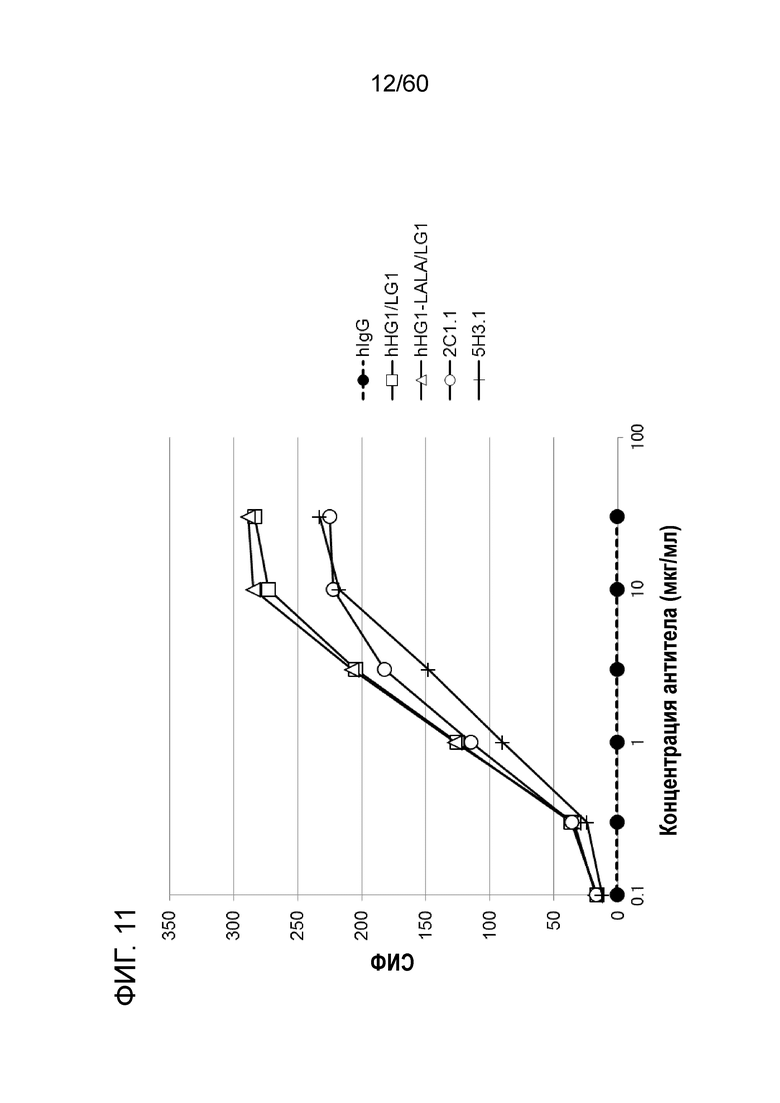
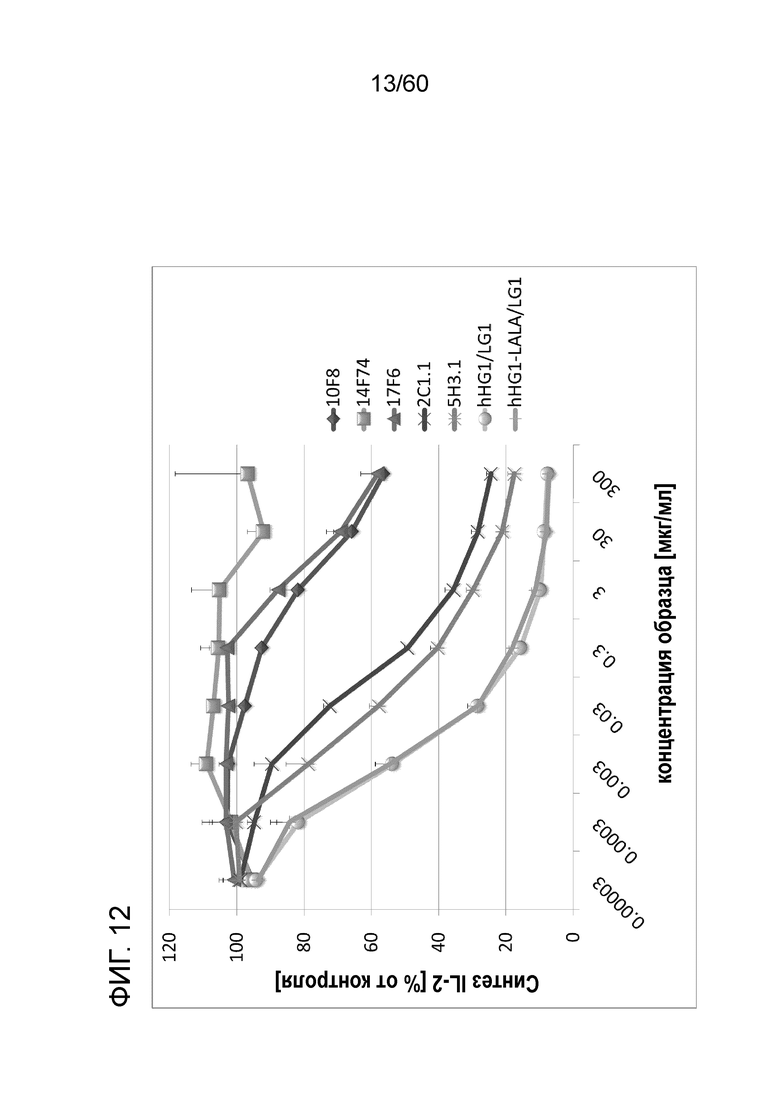
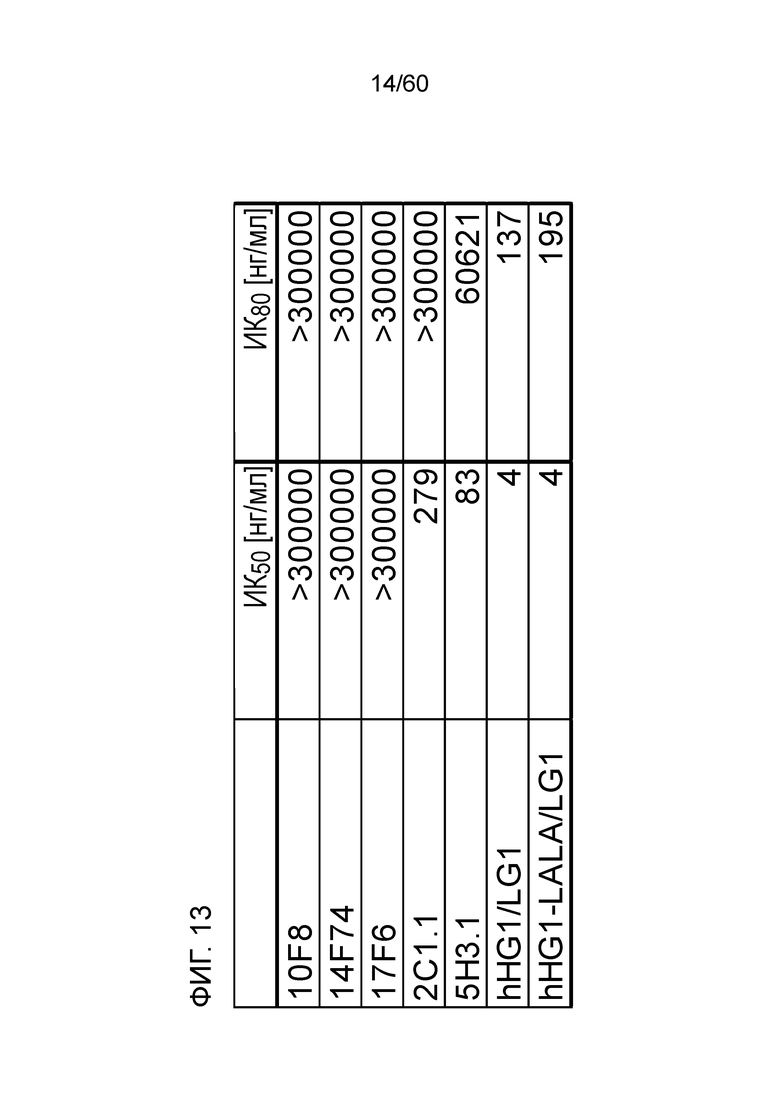
























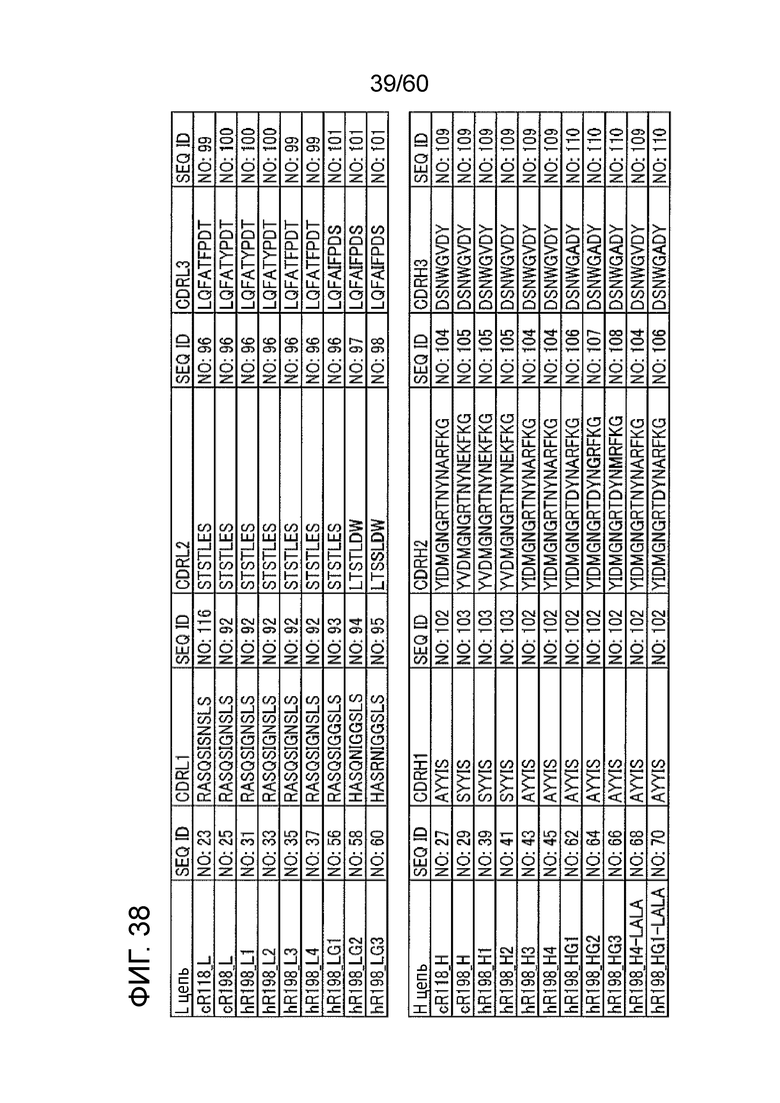












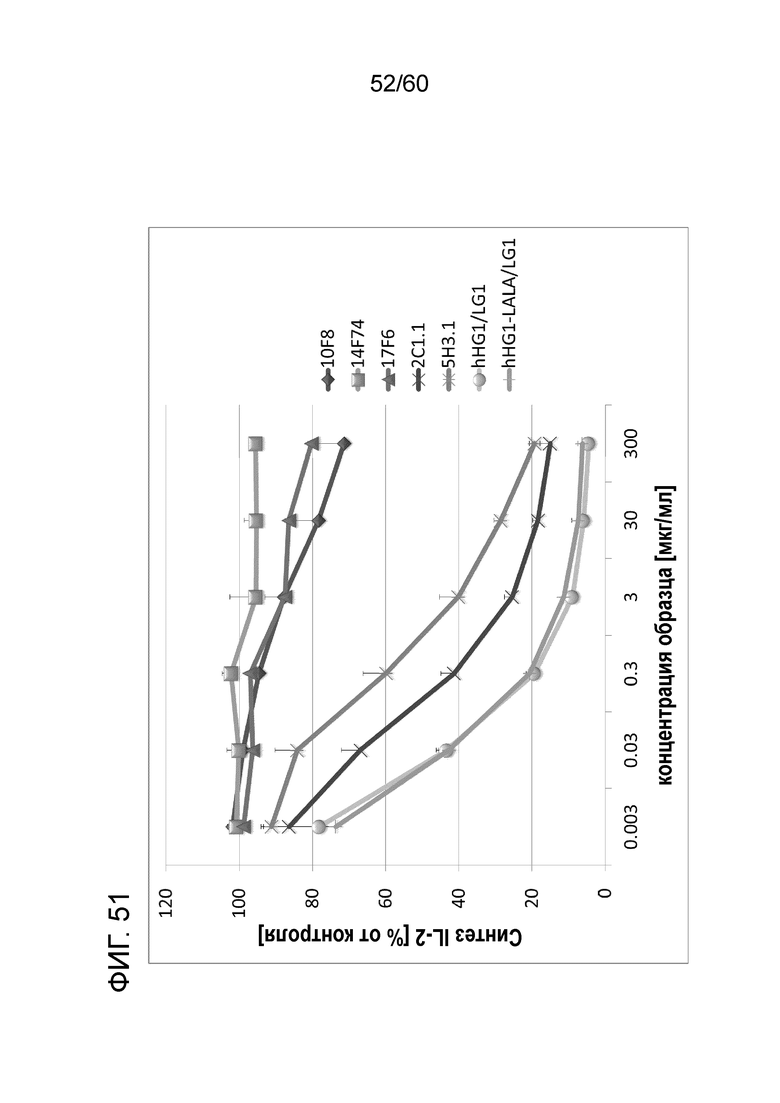
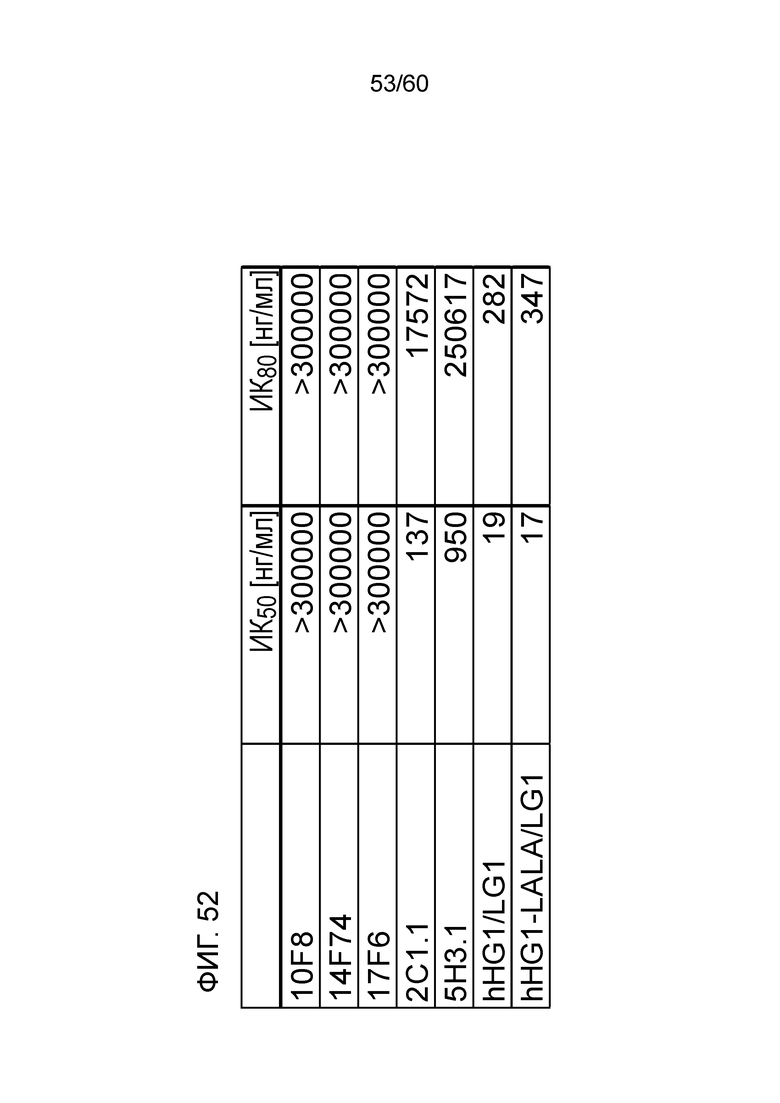
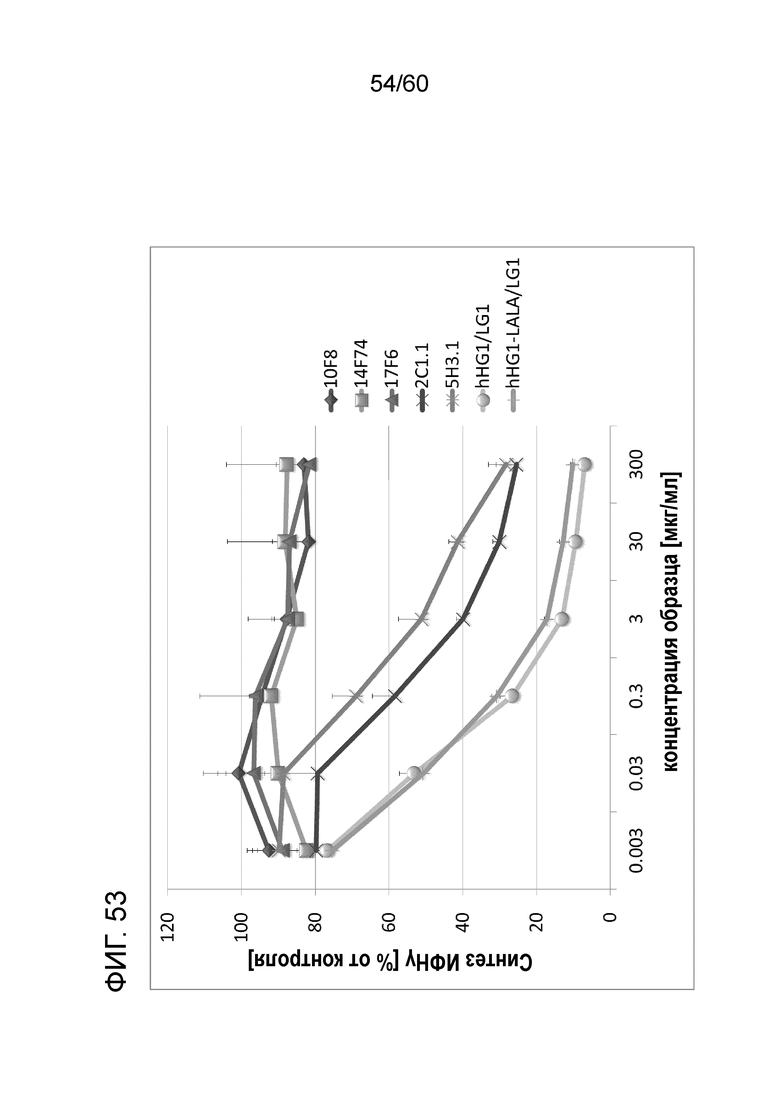
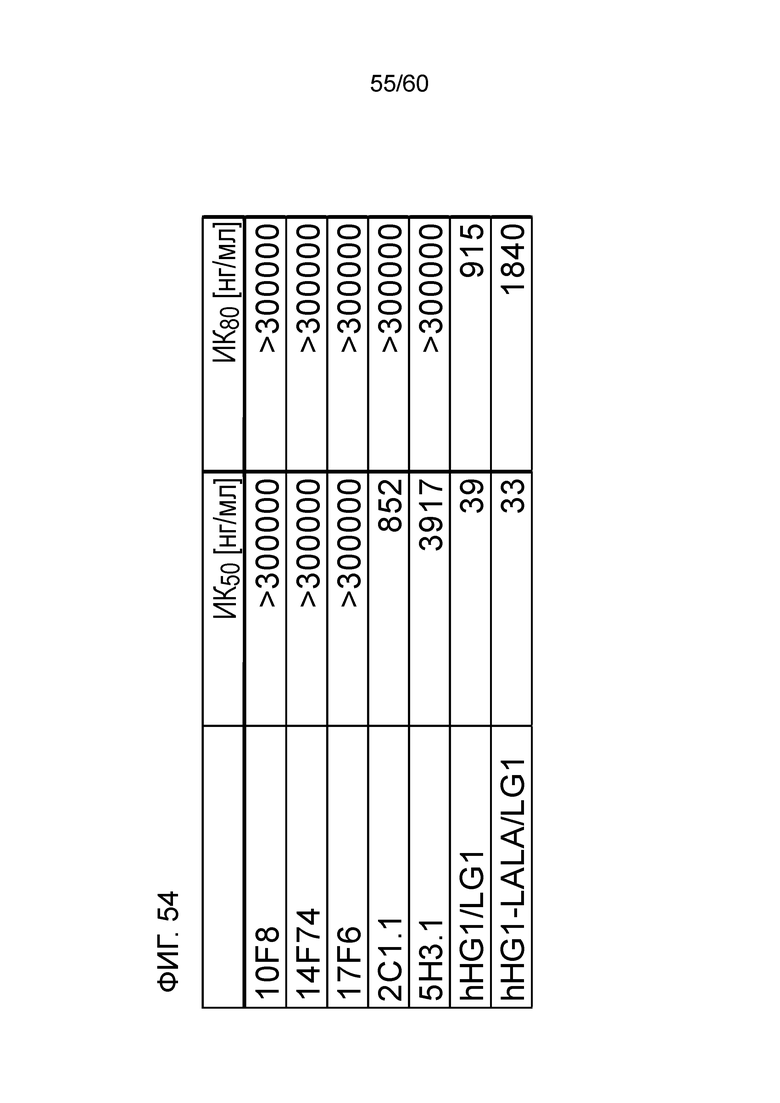
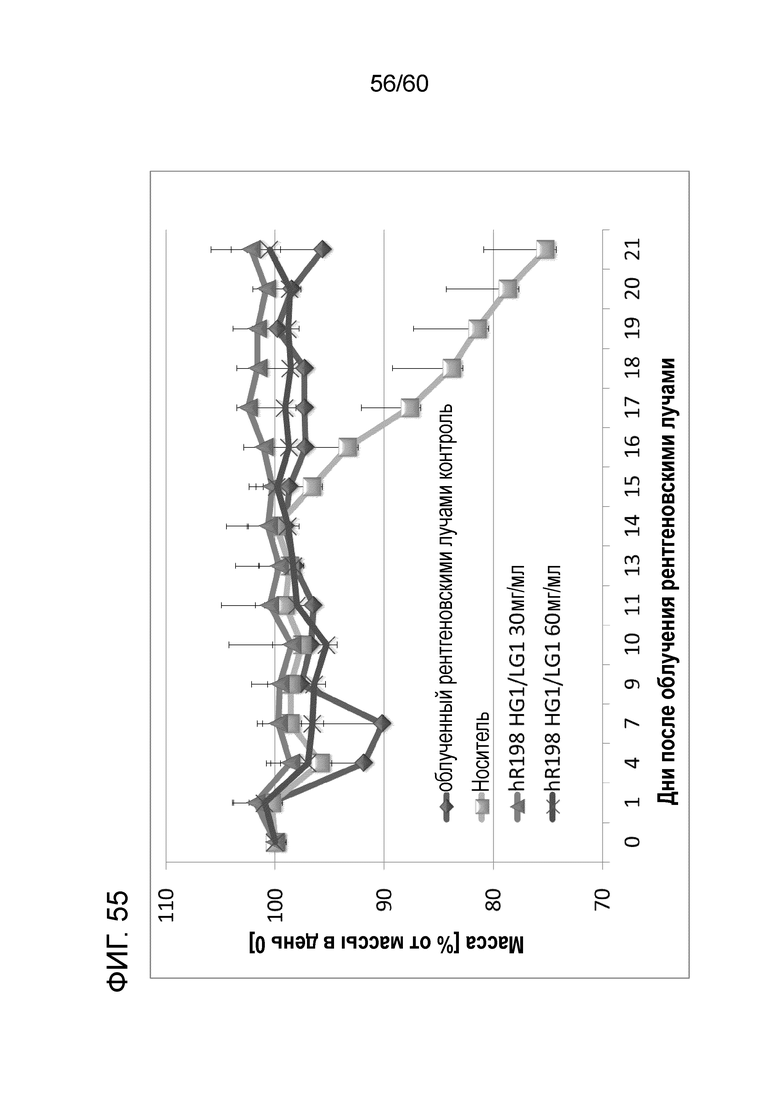
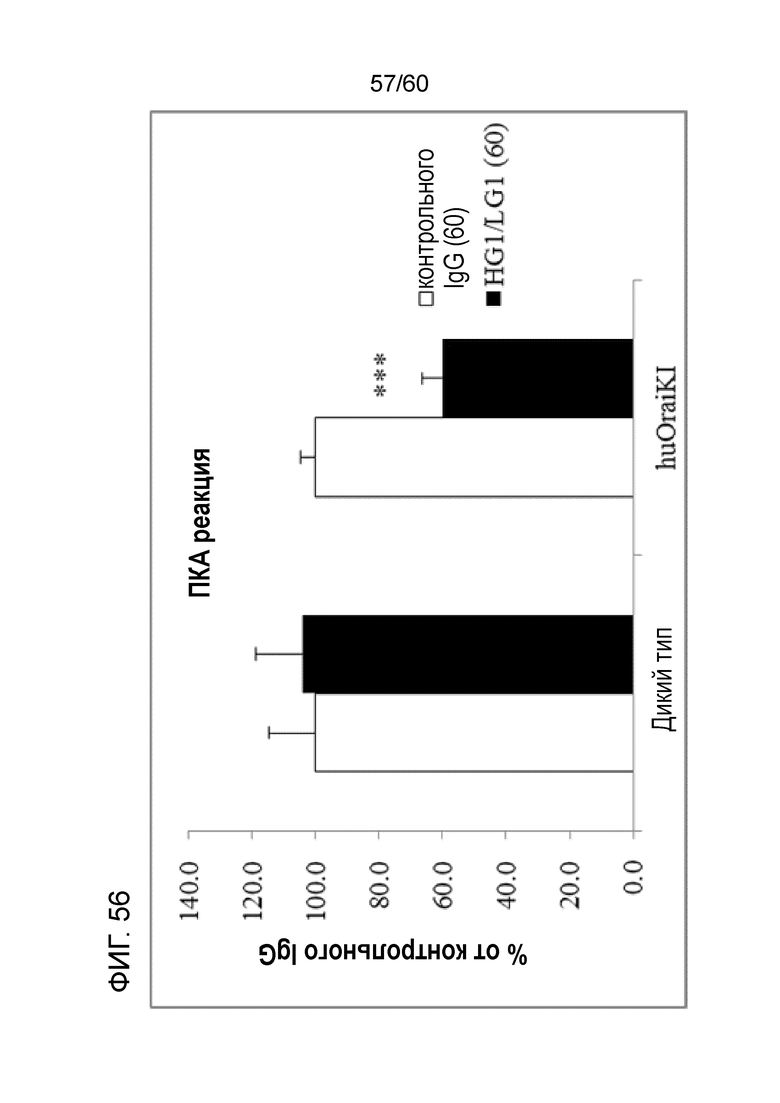
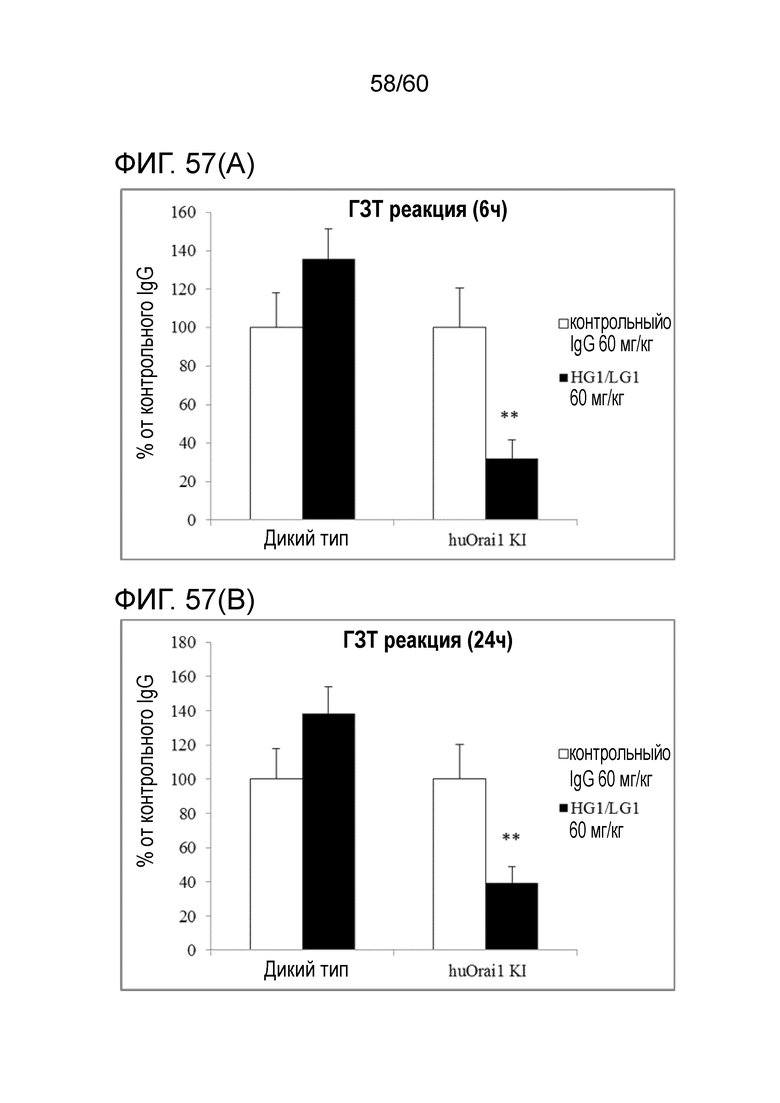
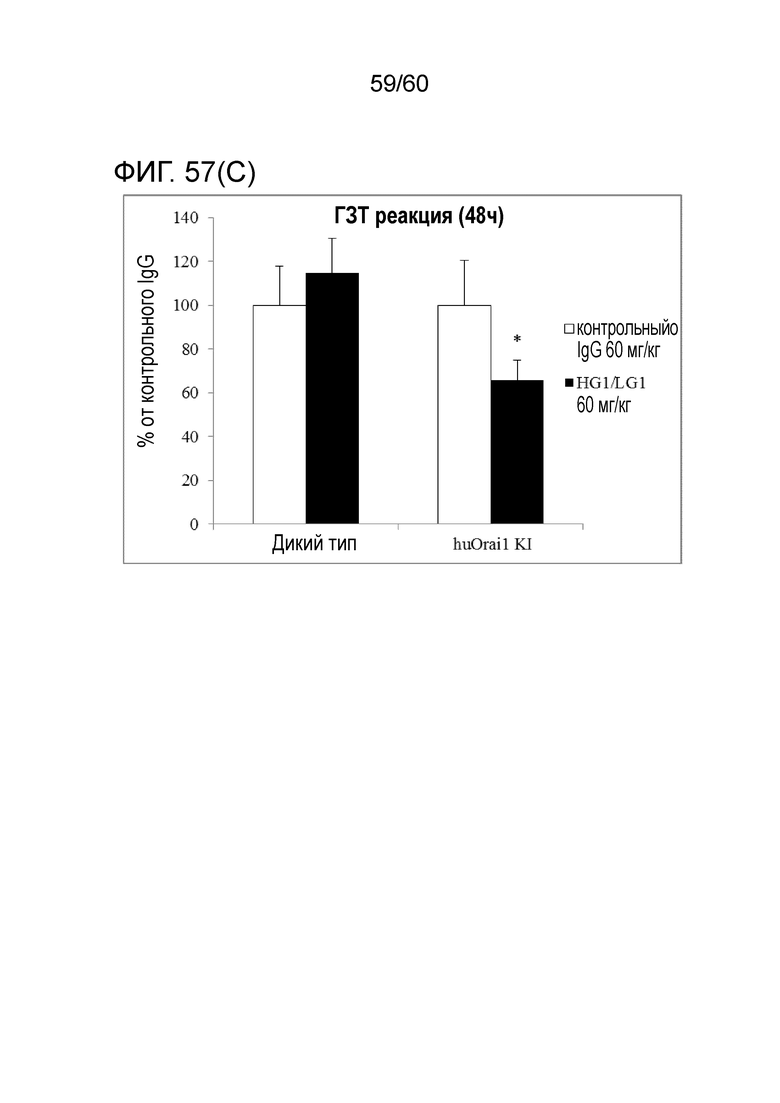
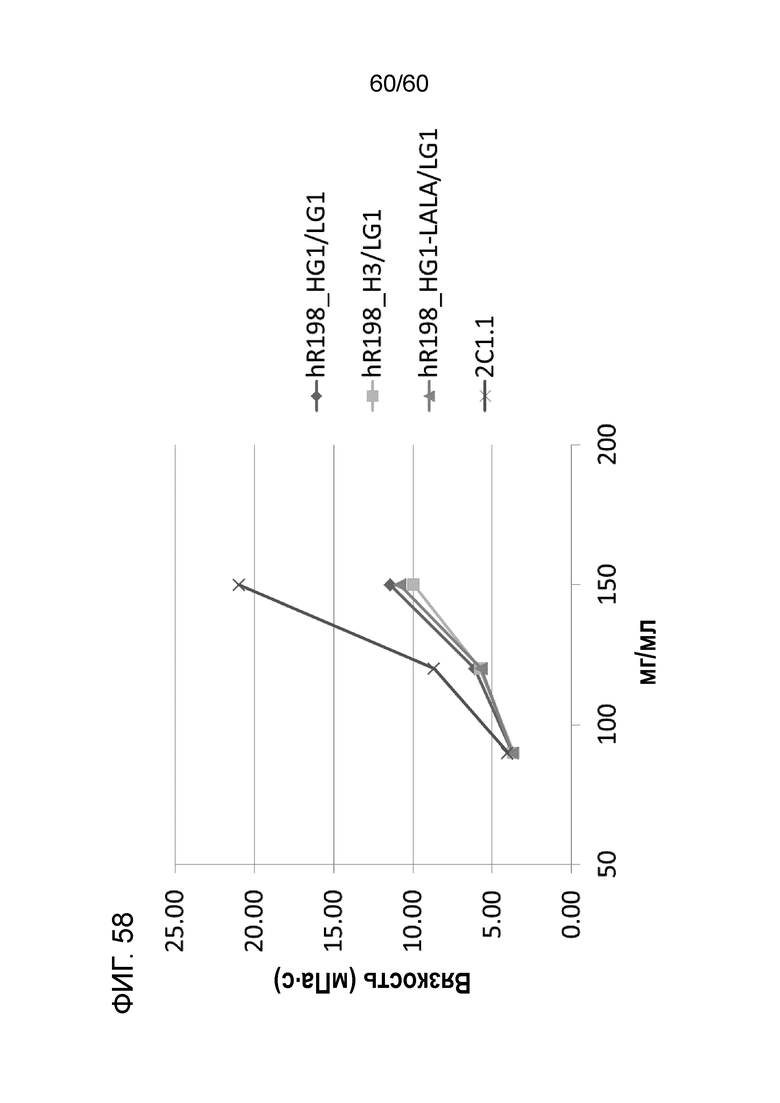
Настоящее изобретение относится к получению терапевтического и/или профилактического агента для применения при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях, тромбозе, раке и т.д., направлено воздействующего на Orai1 человека. Настоящее изобретение относится к фармацевтической композиции, содержащей антитело, которое специфично распознает Orai1 человека и обладает активностью, ингибирующей активацию T-лимфоцитов. 7 н. и 24 з.п. ф-лы, 58 ил., 1 табл., 17 пр.
1. Антитело или антигенсвязывающий фрагмент этого антитела, которое специфично связывается с аминокислотной последовательностью, представленной SEQ ID NO: 2, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 104, 106, 107 и 108, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 109 или 110; и
последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 93, 94 и 95, CDRL2 состоит из аминокислотной последовательности, представленной любой из SEQ ID NOs: 96, 97 и 98, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
2. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
3. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 94, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 97, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
4. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 106, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 95, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 98, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
5. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 107, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
6. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 108, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 110; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
7. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где последовательность тяжелой цепи содержит вариабельную область, имеющую CDRH1, CDRH2 и CDRH3, где CDRH1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 102, CDRH2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 104, и CDRH3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 109; и последовательность легкой цепи содержит вариабельную область, имеющую CDRL1, CDRL2 и CDRL3, где CDRL1 состоит из аминокислотной последовательности, представленной SEQ ID NO: 93, CDRL2 состоит из аминокислотной последовательности, представленной SEQ ID NO: 96, и CDRL3 состоит из аминокислотной последовательности, представленной SEQ ID NO: 101.
8. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
9. Антитело или антигенсвязывающий фрагмент этого антитела по п. 8, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
10. Антитело или антигенсвязывающий фрагмент этого антитела по п. 8, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 70, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
11. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 58.
12. Антитело или антигенсвязывающий фрагмент этого антитела по п. 11, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 58.
13. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 60.
14. Антитело или антигенсвязывающий фрагмент этого антитела по п. 13, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 62, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 60.
15. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 64, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
16. Антитело или антигенсвязывающий фрагмент этого антитела по п. 15, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 64, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
17. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 66, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
18. Антитело или антигенсвязывающий фрагмент этого антитела по п. 17, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 66, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 234 в аминокислотной последовательности, представленной SEQ ID NO: 56.
19. Антитело или антигенсвязывающий фрагмент этого антитела по п. 1, где это антитело содержит последовательность вариабельной области тяжелой цепи, состоящую из аминокислотных остатков от положения 20 до положения 136 в аминокислотной последовательности, представленной SEQ ID NO: 43, и последовательность вариабельной области легкой цепи, состоящую из аминокислотных остатков от положения 21 до положения 126 в аминокислотной последовательности, представленной SEQ ID NO: 56.
20. Антитело или антигенсвязывающий фрагмент этого антитела по п. 19, где это антитело состоит из последовательности тяжелой цепи, состоящей из аминокислотных остатков от положения 20 до положения 465 или от положения 20 до положения 466 в аминокислотной последовательности, представленной SEQ ID NO: 43, и последовательности легкой цепи, состоящей из аминокислотных остатков от положения 21 до положения 235 в аминокислотной последовательности, представленной SEQ ID NO: 56.
21. Антигенсвязывающий фрагмент антитела по любому из пп. 1-20, где этот антигенсвязывающий фрагмент выбран из группы, состоящей из Fab, F(ab')2, Fab' и Fv.
22. Антитело по любому из пп. 1-8, 11, 13, 15, 17 и 19, где это антитело представляет собой scFv.
23. Фармацевтическая композиция для применения в терапии, содержащая эффективное количество по меньшей мере любого одного антитела или антигенсвязывающего фрагмента этого антитела по пп. 1-22, где эта фармацевтическая композиция представляет собой терапевтический и/или профилактический агент, применяемый при отторжениях трансплантата, иммунологических заболеваниях, аллергических заболеваниях, воспалительных заболеваниях или раке, антитромбоцитарный или антитромботический активатор или ингибитор активации клеток, экспрессирующих Orai1.
24. Фармацевтическая композиция по п. 23, где отторжения трансплантата представляют собой реакции отторжения и реакции по типу «хозяин против трансплантата» при трансплантации органа или ткани, таких как сердце, почка, печень, костный мозг или кожа, и реакцию по типу «трансплантат против хозяина», вызванную трансплантацией гемопоэтических клеток (костного мозга, периферической крови, пуповинной крови и т.д.).
25. Фармацевтическая композиция по п. 23, где иммунные заболевания представляют собой заболевания соединительной ткани или опорно-двигательного аппарата (ревматоидный артрит, анкилозирующий спондилит, системная красная волчанка, склеродермия, полимиозит, дерматомиозит и т.д.), гематологические заболевания (апластическая анемия, идиопатическая тромбоцитопеническая пурпура и т.д.), заболевания желудочно-кишечного тракта (болезнь Крона, неспецифический язвенный колит и т.д.), неврологические заболевания (рассеянный склероз, миастения гравис и т.д.), офтальмологические заболевания (увеит и т.д.), сосудистые заболевания (болезнь Бехчета, гранулематоз Вегенера и т.д.), кожные заболевания (псориаз, пузырчатка, лейкодермия и т.д.), эндокринные заболевания (сахарный диабет первого типа, аутоиммунный тиреоидит, болезнь Грейвcа, тиреоидит Хашимото и т.д.) и тому подобное, аллергические заболевания представляют собой атопический дерматит, астму, анафилаксию, анафилактоидную реакцию, алиментарную аллергию, ринит, отит среднего уха, лекарственную аллергию, аллергическую реакцию на укусы насекомых, аллергическую реакцию на растения, аллергию на латекс, конъюнктивит, крапивницу и тому подобное, и воспалительные заболевания представляют собой воспалительные заболевания почек (гломерулонефрит, нефроз и т.д.), воспалительные заболевания легких (хроническое обструктивное заболевание легких, муковисцидоз, интерстициальная пневмония и т.д.), воспалительные заболевания кишечника (неспецифический язвенный колит, илеит и т.д.), воспалительные заболевания печени (аутоиммунный гепатит, вирусный гепатит и т.д.), воспалительные заболевания сердца (миокардит, ишемическая болезнь сердца, атеросклероз и т.д.), воспалительные заболевания кожи (контактный дерматит, экзема и т.д.), воспалительные заболевания глаз (трахома, эндофтальмит и т.д.), воспалительные заболевания центральной нервной системы (менингит, энцефаломиелит, аутоиммунный энцефалит и т.д.), воспалительные заболевания суставов (артрит, остеоартрит и т.д.), системные воспаления (сепсис, кровотечения, гиперчувствительность, симптомы шока, связанные с химиотерапией рака или тому подобным, и т.д.), и тому подобное.
26. Фармацевтическая композиция по п. 23, где рак представляет собой рак молочной железы, рак легкого, рак кожи, лейкоз и тому подобное, и случаи, при которых антитромбоцитарная или антитромботическая активность используется для лечения и/или предотвращения заболевания, представляют собой инфаркт миокарда, инсульт, ишемическую болезнь сердца, тромбоз и тому подобное, и случаи, при которых ингибирование активации клеток, экспрессирующих Orai1, используется для лечения и/или предотвращения заболевания, представляют собой тучноклеточный лейкоз, мастоцитоз, базофильный лейкоз, эндометриоз, миопатию с тубулярными агрегатами, синдром Шторморкена, ревматоидный артрит, анкилозирующий спондилоартрит, атопический дерматит и тому подобное.
27. Полинуклеотид, кодирующий антитело или антигенсвязывающий фрагмент этого антитела по любому из пп. 1-22.
28. Вектор экспрессии, содержащий полинуклеотид по п. 27.
29. Трансформированная клетка-хозяин, содержащая полинуклеотид по п. 27, для продуцирования антитела или антигенсвязывающего фрагмента антитела по любому из пп. 1-22.
30. Трансформированная клетка-хозяин, содержащая вектор по п. 28, для продуцирования антитела или антигенсвязывающего фрагмента антитела по любому из пп. 1-22.
31. Способ получения антитела или антигенсвязывающего фрагмента этого антитела по любому из пп. 1-22, включающий в себя этап культивирования клетки-хозяина по п. 29 или 30 и очистку антитела от продукта культивирования.
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| LIN FF et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО-АНТАГОНИСТ ПРОТИВ CD40 ЧЕЛОВЕКА | 2007 |
|
RU2491295C2 |

