Область техники
Настоящее изобретение может быть включено в область новых терапевтических способов лечения атаксии Фридрейха. В частности, настоящая заявка относится к новым продуктам для генотерапии. Эти продукты могут быть использованы для лечения атаксии Фридрейха.
Предпосылки к созданию изобретения
Атаксия Фридрейха (FRDA; OMIM # 229300) представляет собой редкое наследственное нейродегенеративное заболевание, которое вызывает прогрессирующее повреждение нервной системы, проявляющееся различными симптомами - от нарушений походки и речи до болезней сердца или кардиомиопатии. Это заболевание было названо в честь врача Николауса Фридрейха, который первым описал его в 1860-х годах. Атаксия, ассоциировання с нарушением координации движений, таким как неуклюжие движения и нестабильность, известна как атаксия Фридрейха, вызываемая дегенерацией нервной ткани в спинном мозге и нервах, которые контролируют мышечные движения рук и ног. Спинной мозг сжимается вместе с нервными клетками, что приводит к потере части их миелиновой оболочки. Атаксия Фридрейха, несмотря на то, что она встречается редко, является наиболее распространенной наследственной атаксией, обнаруживающейся у 1 на 30000-50000 новорожденных с распространенностью 2-4 на 100000. Оба пола страдают этой болезнью одинаково. Симптомы обычно начинаются в возрасте от 5 до 15 лет, но в редких случаях, они могут появляться уже в возрасте 18 месяцев или даже в возрасте 50 лет. Первым обнаруживаемым симптомом обычно является затруднение ходьбы или атаксия походки. Атаксия постепенно прогрессирует и медленно распространяется на руки, а затем и на все туловище. В качестве ранних признаков могут наблюдаться деформации стопы, как «полая стопа», непроизвольное складывание пальцев ног или деформация типа молоткообразного пальца стопы. Со временем, мышцы начинают ослабевать и истощаться, особенно на ступнях, голени и руках, и развиваются деформации. Другие симптомы включают потерю сухожильных рефлексов, особенно в коленях и лодыжках. Часто происходит постепенная потеря чувствительности в конечностях, которая может распространиться на другие части тела. При этом развивается дизартрия, в результате чего пациент быстро устает. Особенно распространены быстрые, ритмичные и непроизвольные движения глаз, называемые нистагмом. У большинства людей с атаксией Фридрейха развивается сколиоз, который, в тяжелой форме, может влиять на дыхание.
Другими симптомами, которые могут наблюдаться, являются боль в груди, одышка и сердцебиение. Эти симптомы являются результатом различных форм болезней сердца, которые часто сопровождают атаксию Фридрейха, таких как гипертрофическая кардиомиопатия, фиброз миокарда или сердечная недостаточность. Также часто наблюдаются нарушения сердечного ритма, такие как тахикардия и сердечная блокада. Приблизительно у 20% людей с атаксией Фридрейха развивается непереносимость углеводов, а у 30% - сахарный диабет. Некоторые люди, страдающие таким заболеванием, теряют слух или зрение. Скорость прогрессирования заболевания у разных людей варьируется. Обычно, через 10-20 лет после появления первых симптомов, человек вынужден передвигаться в инвалидной коляске, а на более поздних стадиях заболевания, люди полностью становятся инвалидами. Это, предположительно, может влиять на продолжительность жизни. Многие люди с атаксией Фридрейха умирают в зрелом возрасте из-за связанных с ней болезней сердца, самой распространенной причиной летального исхода. Однако некоторые люди с менее выраженными симптомами атаксии Фридрейха иногда доживают до 60 или 70 лет.
Нейровизуализация, такая как МРТ, показывает нормальное состояние организма на ранних стадиях заболевания, хотя при прогрессировании этого заболевания наблюдается вариабельная атрофия шейного отдела спинного мозга и мозжечка. Это часто указывает на атрофию верхней ножки. Электрофизиологические исследования показывают скорость проводимости, превышающую 40 м/с, в отсутствии или при снижении потенциалов сенсорного действия и в отсутствии рефлекса H. На невропатологическом уровне отмечаются атрофия спинного мозга, задних корешков, а иногда и мозжечка, и гипертрофия сердца. Нейродегенерация идентифицируется в периферических сенсорных нервах с прогрессирующей потерей сенсорных нейронов больших спинномозговых узлов, дегенерацией дорсального столба, трансинаптической дегенерацией нейронов в позвоночнике Кларка и спиноцеребеллярных волокнах и пирамидных путях и атрофией ядер тонкого пучка и клинообразного пучка. Вторичные поражения могут включать атрофию зубчатого ядра в мозжечке, поражающую крупные глутаматергические нейроны, и атрофию клеток Бетца и кортикоспинальных путей.
В 1996 году причина атаксии Фридрейха была идентифицирована как молекулярный дефект в гене FXN, расположенном на хромосоме 9 (полоса 9q21). Мутация состоит из аномального гомозиготного увеличнеия уровней триплета GAA, расположенного внутри последовательности ALU в первом интроне гена FXN. Приблизительно 98% пациентов с FRDA имеют 2 хромосомы с числом повторов GAA от 201 до 1700 (чаще всего от 600 до 900) в каждой из них. Однако у 2-5% пациентов с FRDA наблюдается увеличение уровней GAA и точечная мутация в гене FXN при комбинированной гетерозиготности (Таблица 1). В настоящее время описано более 17 различных мутаций в гене FXN, способных вызывать FRDA. Увеличение уровней GAA при FRDA является очень нестабильным во время мейоза и препятствует транскрипции гена FXN, что приводит к аномально низким уровням мРНК фратаксина. Увеличение уровней GAA может приводить к сайленсингу транскрипции гена FXN и, таким образом, отменить экспрессию фратаксина посредством образования тройных структур ДНК или гибридов ДНК-РНК, или тех и других. Совсем недавно было высказано предположение, что экспансия GAA будет блокировать переход от инициации к повышению уровня транскрипции из-за образования гетерохроматин-подобных структур рядом с областью гиперэкспансии GAA. Также было высказано предположение, что экспансия GAA будет приводить к эпигенетическому метилированию сайтов CpG, расположенных в 5’-области выше гена FXN, и к их сайленсингу. Таким образом, вследствие мутации GAA в гене FXN, при FRDA наблюдается дефицит белка фратаксина.
Ген FXN кодирует небольшой консервативный митохондриальный белок, который содержит 210 аминокислот (а.к.) в форме предшественника, с молекулярной массой 23135 Да, названной фратаксином (Q16595). Эта форма белка-предшественника содержит N-концевую последовательность переноса, которая направляет ее в митохондриальный матрикс, где митохондриальная пептидаза превращает ее в различные более мелкие изоформы (FXN42-210, FXN56-210, FXN81-210, FXN78-210) зрелого белка фратаксина, представляющего собой FXN81-210 размером 130 а.к. и 14,2 кДа, который является наиболее распространенным. У человека, фратаксин детектируется в митохондриальном матриксе в комбинации с его внутренней мембраной в тканях широкого ряда, а его наибольшие уровни обнаруживаются в ткани сердца, спинном мозге и в делящихся лимфобластах, и, что примечательно, самые низкие уровни наблюдаются в мозжечке. До настоящего времени фратаксин не был обнаружен в коре головного мозга. Поскольку был идентифицирован ген, ответственный за это заболевание и было получено несколько животных-моделей, то было высказано предположение, что фратаксин обладает различными функциями, такими как митохондриальный гомеостаз железа, запасание железа, реакция на окислительный стресс, биогенез кластеров Fe-S, модуляция митохондриальной аконитазной активности и регуляция окислительного фосфорилирования. При FRDA, дефицит фратаксина, вызываемый гомозиготной экспансией GAA в гене FXN, приводит к недостаточному биосинтезу кластеров железа-серы, необходимых для транспорта электронов в митохондрии и аконитазной сборки, и тем самым, к нарушению регуляции митохондриальной функции и аккумуляции митохондриального железа за счет изменений его гомеостаза. Фратаксин также модулирует ДНК-связывающую способность белка аконитазы 1 (ACO1), которая помимо ее участия в цикле лимонной кислоты в митохондриальном матриксе, регулирует поглощение и утилизацию клеточного железа.
Несколько животных и клеток с моделями атаксии Фридрейха были созданы посредством генетических манипуляций. Создание AF-модели у мыши, которая как можно точно имитировала бы человеческое заболевание, было трудной задачей, и в настоящее время существуют 8 различных мышиных моделей FRDA. К сожалению, ни одна из них не обладает комбинацией всех фенотипических симптомов FA у одного и того же животного, таких как поражения спинномозговых узлов (DRG) и зубчатых ядер мозжечка, хотя они могут оказаться полезными для изучения отдельных аспектов молекулярного нейродегенеративного процесса при FRDA и для оценки различных методов лечения при конкретных симптомах. В доклиническом лечении наиболее часто используются мыши Prp-CreERT и YG8R. У мышей Prp-CreERT, в частности, развивается мозжечковая и прогрессирующая сенсорная атаксия, наиболее важные неврологические функции атаксии Фридрейха. Гистологические исследования этих животных показали аномалии спинного мозга и спинномозговых узлов с отсутствием двигательной невропатии, которая является отличительным признаком заболевания у человека, а также дефекты арборизации в клетках Пуркинье мозжечка. Напротив, у YG8R-трансгенных мышей в отсутствие эндогенного мышиного белка фратаксина наблюдается медленно прогрессирующая патология FRDA. В отличие от человека, эти мыши не имеют сердечной недостаточности.
Среди вирусных и невирусных векторов, используемых в генотерапии для лечения патологий у человека, векторы, происходящие от аденоассоциированного вируса (AAV), продемонстрировали важные клинические преимущества и пролонгированную экспрессию у животных с моделями болезни Гоше, болезни Фабри, болезни Помпе, метахроматической лейкодистрофии, болезни Нимана-Пика А и мукополисахаридоза I, II, III A, III B, IV и VII и т.п. Внутривенное введение AAV-векторов животным с моделями болезней лизосомного запасания приводит к увеличению активности ферментов в 16 раз по сравнению с нормальными их значениями в крови, печени, селезенке, почках и в мышцах, что делает их очень полезными для лечения патологий этого типа. За последние 10 лет, AAV-векторы были наиболее предпочтительными для большинства клинических испытаний, проводимых для лечения патологий центральной и периферической нервной системы (http://www.abedia.com/wiley/index.html). При этом важно отметить, что до настоящего времени, никаких побочных эффектов при введении этих векторов не наблюдалось, а поэтому такие результаты являются весьма перспективными. Таким образом, некоторые факторы делают AAV идеальными носителями для доставки генов в центральную нервную систему (ЦНС). Аденоассоциированные вирусные векторы, содержащие последовательность фратаксина, были использованы для лечения FRDA у мышей (Gérard et al., 2014. Mol Ther Methods Clin Dev. 1: 14044; Chapdelaine et al., 2016. Gene Ther. 23 (7): 606-614; Tremblay et al., 2015. Mol Ther. 23 (Suppl. 1): pS153; WO 2016/150964). Однако, AAV-векторы, раскрытые в предшествующих работах, либо чрезмерно, либо недостаточно экспрессируют фратаксин и не достигают всех клеток-мишеней и нейронов, пораженных FRDA, что делает их непригодными для эффективного лечения FRDA, а, в частности, ее неврологических симптомов. Таким образом, необходимо создать AAV-вектор, который экспрессировал бы белок фратаксин в терапевтически эффективном количестве.
В настоящее время не существует какой-либо эффективной терапии для лечения FRDA, а поэтому необходимо разработать новые терапевтические средства для лечения FRDA. Целью настоящего изобретения является разработка подходящей терапии для лечения FRDA.
Чертежи
Фигура 1. Оценка влияния человеческих промоторов SYN (синапсина) и FXN (фратаксина; 1255 п.о.) на экспрессию белка фратаксина (FXN). Для тестирования уровней экспрессии человеческого белка фратаксина (FXN) под контролем фрагмента эндогенного промотора (phFXN) и нейронного промотора синапсина (phSYN), по сравнению с экспрессией под контролем в высокой степени экспрессирующегося конститутивного промотора CMV, человеческие клетки нейробластомы SH-SY5Y трансфецировали «пустым» вектором (дорожка 1) или конструкциями, кодирующими человеческий синапсин (phSYN) (дорожка 3), или 1255 п.о.-фрагментом промотора фратаксина (phFXN) (дорожка 4). Конструкции 1.2 и 1.3, полученные с использованием phSYN или phFXN (1255 п.о.) для экспрессии кодирующей последовательности человеческого FXN (hFXN CDS), не приводили к экспрессии рекомбинантного белка фратаксина (rFXN). НА=гемагглютининовая метка.
Фигура 2. Оценка влияния человеческих промоторов SYN (синапсина), NSE (нейрон-специфической энолазы) и FXN (фратаксина; 1255 п.о.) вместе с последовательностью WPRE на экспрессию белка фратаксина (FXN). Было оценено влияние фрагмента длиной 320 п.о. 5’-конца последовательностей FXN и WPRE на экспрессию FXN из промотора phSYN, ранее показанного на фиг. 1 (дорожки 3, 4 и 8), и phNSE (дорожка 6). Различные конструкции были трансфецированы в клетки НЕК, и лизаты были проанализированы через 48 часов после трансфекции. Эффективность последовательностей WPRE в стабилизации РНК наблюдалась при использовании конструкции, экспрессирующей FXN из промотора CMV (дорожка 2). Однако, добавление и комбинация вышеописанных регуляторных элементов не приводит к экспрессии FXN из phSYN, hpFXN или phNSE. HA - гемагглютининовая метка; iFXN - промежуточная форма FXN; mFXN - зрелая форма FXN; WPRE - посттранскрипционный регуляторный элемент вируса гепатита сурка.
Фигура 3. Оценка влияния человеческих промоторов SYN (синапсина), NSE (нейрон-специфической энолазы) и FXN (фратаксина; 1255 п.о.) вместе с последовательностями энхансера CMV и WPRE на экспрессию белка фратаксина (FXN). Клетки HEK и нейробластомы мыши N2a трансфецировали конструкциями, содержащими энхансер CMV вместе с промоторами SYN, NSE или FXN (1255 п.о.). Комбинация регуляторных элементов, показанная выше, не приводила к экспрессии FXN из промоторов phSYN, phFXN или phNSE. HA - гемагглютининовая метка; iFXN - промежуточная форма FXN; mFXN - зрелая форма FXN; WPRE - посттранскрипционный регуляторный элемент вируса гепатита сурка.
Фигура 4. Оценка влияния человеческих промоторов SYN (синапсина), NSE (нейрон-специфической энолазы) и FXN (фратаксина; 1255 п.о.) вместе с последовательностью Козака (5’) и WPRE (3’) на экспрессию белка фратаксина (FXN). Различные конструкции трансфецировали в клетки НЕК, и лизаты анализировали через 48 часов после трансфекции. Эффективность последовательностей WPRE в стабилизации РНК наблюдалась при использовании конструкции, экспрессирующей FXN из промотора CMV (дорожка 1 и 2). Последующее увеличение уровней экспрессии FXN путем добавления последовательности Козака и последовательностей 105 нуклеотидов между промотором и кодирующей последовательностью FXN показано на дорожках 2 и 3. Однако, добавление и комбинация показанных выше регуляторных элементов не приводит к экспрессии FXN из phSYN, phFXN или phNSE. HA - гемагглютининовая метка; iFXN - промежуточная форма FXN; mFXN - зрелая форма FXN; pFXN - форма предшественника; WPRE - посттранскрипционный регуляторный элемент вируса гепатита сурка.
Фигура 5. Оценка влияния комбинации определенного линкера, расположенного между человеческими промоторами PGK1 (фосфоглицераткиназы 1), NSE (нейрон-специфической энолазы), SYN (синапсина) или FXN (фратаксина; 1255 п.о.) и кодирующей области (CDS) FXN вместе с последовательностями Козака (5’) и WPRE (3’) на экспрессию белка фратаксина (FXN). Различные представленные здесь конструкции были трансфецированы либо в клетки N2a (A, B и E), либо в клетки HEK (C и D), и лизаты были проанализированы через 48 часов после трансфекции. Экспрессия FXN была последовательно детектирована из phPGK, но не из промотора phNSE или phFXN (1255 п.о.) при использовании линкера в 105 п.о. между промотором и кодирующей областью (A, C и D: дорожка 4; B: дорожка 5 и E: полоса 8). HA - гемагглютининовая метка; iFXN - промежуточная форма FXN; mFXN - зрелая форма FXN; pFXN - форма предшественника; WPRE - посттранскрипционный регуляторный элемент вируса гепатита сурка.
Фигура 6. Экспрессия люциферазы (LUC) под контролем человеческого промотора PGK1 (фосфоглицераткиназы 1) после интратекальной инъекции вектора rAAV9 (4,6×1012 ВГ/кг) у мышей YG8R Tg/- и у мышей дикого типа. Экспрессию люциферазы детектировали после внутрибрюшинной инъекции 150 мкл D-люциферина мыши с массой 150 мг/кг через 3,5 месяца после интратекальной инъекции вектора (A). Органы иссекали и субстрат люциферазы добавляли в свежем виде перед фиксацией изображения (относительные масштабы показаны на левой и правой панелях (B)).
Фигура 7. Экспрессия мРНК фратаксина in vivo у 7-месячных мышей, которым интратекально вводили rAAV9-PGK1-FXN. Уровни мРНК фратаксина количественно определяли с помощью кол.ОТ-ПЦР в тканях печени, сердца, спинномозговых узлах поясничного отдела (DRG-L), поясничного отдела спинного мозга (SC-L), грудного отдела спинного мозга (SC-T), шейного отдела спинного мозга (SC-C), мозжечка (Cb) и головного мозга (фронтальной области C1) 7-месячных гемизиготных трансгенных (Tg/-) или гомозиготных (Tg/Tg) мышей YG8R. Гемизиготным трансгенным мышам YG8R (Tg/-) вводили либо rAAV-Null (показано черным), либо rAAV9-PGK1-FXN (показано светло-серым), а гомозиготным трансгенным мышам с FRDA (Tg/Tg) (показано темно-серым) инъекции не вводили. В отличие от гемизиготных мышей YG8R (Tg/-), несущих две тандемные копии человеческого гена FXN с повторами тринуклеотидной последовательности ~82 и ~190 GAA в одной из хромосом, гомозиготные мыши YG8R содержали две тандемные копии в каждой из хромосом. У мышей YG8R, которые являются гомозиготными или гемизиготными по человеческому FXN, не экспрессирался эндогенный Fxn, поскольку они имеют генетический фон с нокаутом мышиного Fxn. Гомозиготные мыши YG8R имели бóльшую степень регуляции экспрессии мРНК человеческого FXN, поскольку они содержит больше копий человеческого трансгена, и кроме того, имеют нормальные функции. Мышам в возрасте 2,5 месяцев вводили интратекальные инъекции, и праймеры, специфичные к человеческому FXN, использовали для количественной оценки мРНК FXN в различных тканях. Уровни общей мРНК FXN были нормализованы по уровням, детектируемым у гемизиготных мышей YG8R (Tg/-), обработанных rAAV9-Null для каждой из проанализированных тканей. Через пять месяцев после интратекальной инъекции вектора rAAV9-PGK1-FXN, уровни мРНК FXN были выше (приблизительно в 1,3 и 3,2 раза) у гемизиготных мышей YG8R, которым инъецировали rAAV9-PGK1-FXN (Tg/-) в тестируемой дозе, но не выше, чем уровни у гомозиготных мышей YG8R (Tg/Tg) во всех тканях, по сравнению с уровнями у мышей Tg/-, обработанных AAV-Null. А) Относительные уровни мРНК человеческого FXN у самок и самцов мышей n=6 (3 самки и 3 самца). Кратные различия между мышами YG8R с моделью атаксии Фридрейха, которым инъецировали контрольный AAV-Null, по сравнению с мышами YG8R, которым инъецировали rAAV9-PGK1-FXN, были статистически значимыми в печени, в спинномозговых узлах и в спинном мозге (p=0,019, p=0,004 и p=0,024; обозначено звездочкой *) с тенденцией к увеличению во всех других указанных тканях. Большая вариабельность наблюдалось у самцов в используемой дозе, но не у самок. B) Относительные уровни мРНК hFXN у самок мышей YG8R. Статистически значимые кратные различия уровней фратаксина были обнаружены во всех исследуемых тканях, обозначенных звездочками и перечисленных ниже в Таблице. Кратные увеличения уровней фратаксина у мышей, которым инъецировали rAAV9-PGK1-FXN, никогда не превышали уровней, обнаруженных у мышей Tg/Tg YG8R. Статистическая значимость, р <0,05*, р <0,005**.
Фигура 8. Экспрессия in vivo рекомбинантного белка фратаксина из вектора rAAV9-hPGK1-FXN после интратекальной инъекции. Лизаты культивируемых клеток N2a трансфецировали пустым вектором или вектором, экспрессирующим фратаксин, который использовали в качестве позитивного контроля (фиг. 8А и 8В: дорожки 1 и 2). Эндогенный белок FXN в клетках N2a и у мышей дикого типа детектировали с использованием антитела против FXN (A и B). Человеческий белок FXN, происходящий от трансгена у мышей YG8R, а также человеческий рекомбинантный белок FXN у инъецированных мышей обнаруживаись как в печени, так и в головном мозге мышей (А и В). Инкубирование блотов с анти-FXN антителом в течение одного часа позволило выявить человеческий белок FXN, а инкубирование в течение ночи (o/n) выявило мышиный FXN у мышей дикого типа и человеческий FXN у гемизиготных (Tg/-) и гомозиготных мышей (Tg/Tg). Уровни экспрессии рекомбинантного человеческого белка FXN, экспрессируемого после инъекции rAAV9-FXN, были аналогичны уровням эндогенного мышиного белка FXN у мышей дикого типа. HA - гемагглютининовая метка; iFXN - промежуточная форма FXN; mFXN - зрелая форма FXN; WPRE - посттранскрипционный регуляторный элемент вируса гепатита сурка.
Фигура 9. Восстановление рефлекса поджимания лап у мышей YG8R в течение определенного периода времени после инъекции вектора rAAV9-FXN. Поджимание задних конечностей оценивали по шкале от 0 до 3 (см. Методы), по степени тяжести поражения у гемизиготных (Tg/-) мышей и у мышей дикого типа, которым инъецировали векторы rAAV9-FXN или rAAV9-Null. Представлены общие результаты для самок и самцов (n=10, 5 самцов и 5 самок в каждой группе). Изменения рефлекса поджимания обнаруживались уже в возрасте 4 месяцев у гемизиготных мышей YG8R. После интратекальной инъекции вектора rAAV9-FXN, рефлекс поджимания, по-видимому, нормализуется, на что указывает восстановление этого неврологического патологического фенотипа с течением времени. Звездочками (*) показаны значимые различия между значениями для мышей дикого типа и мышей Tg/-, которым инъецировали rAAV9-Null, а знак фунта (#) обозначают значимые различия между значениями, полученными для мышей Tg/-, которым инъецировали rAAV9-Null, и мышей Tg/-, которым инъецировали rAAV9-FXN. *, #: P <0,05.
Фигура 10. Экспрессия in vivo рекомбинантного белка фратаксина у мышей, которым инъецировали rAAV9-hPGK1-FXN, приводит к восстанавлению электрофизиологических свойств на разных расстояниях от раздражителя (D1-D4) каудального нерва. Звездочки (*) обозначают значимые различия между мышами дикого типа и мышами Tg/- YG8R с FRDA, обработанными rAAV9-Null, а знак фунта (#) обозначает значимые различия между мышами, обработанными rAAV9-Null, и мышами, обработанными rAAV9-FXN YG8R. Интратекальная инъекция 2,5-месячным мышам вектора rAAV9-FXN предотвращает или снижает дефекты нервной проводимости у мышей с моделью FRDA (YG8R). Мыши дикого типа показаны зеленым (n=10); мыши Tg/-YG8R с FRDA, обработанные вектором rAAV9-Null, показаны серым (n=10); мыши Tg/-YG8R с FRDA, обработанные вектором rAAV9-FXN, показаны синим (n=10). Измерения были сделаны на расстоянии 1 см, 2 см, 3 см и 4 см от кончика хвоста, и эти величины были обозначены как значения для участков D1, D2, D3 и D4, соответственно *, #: P <0,05.
Фигура 11. Сохранение спинномозговых узлов у мышей YG8R с моделью FRDA, обработанных rAAV9-FXN. Диаметр спинномозговых узлов поясничного отдела, по-видимому, сохранялся, а также сохранялась морфология митохондрий у мышей с моделью FRDA, обработанных rAAV9-FXN, по сравнению с теми же мышами, обработанными вирусом rAAV9-Null.
Сущность изобретения
Настоящее изобретение относится к вектору на основе аденоассоциированного вируса (AAV), содержащему нуклеиновую кислоту, где нуклеиновая кислота включает: (i) последовательность нуклеиновой кислоты, кодирующую фратаксин; (ii) промотор фосфоглицерат-киназы (PGK); и (iii) посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE). Настоящее изобретение также относится к нуклеиновой кислоте, содержащей: (i) последовательность нуклеиновой кислоты, кодирующую фратаксин; (ii) промотор PGK; и (iii) WPRE; к клонирующему вектору, содержащему эту нуклеиновую кислоту и к вектору для переноса, содержащему эту нуклеиновую кислоту. Кроме того, настоящее изобретение охватывает применение нуклеиновой кислоты, клонирующего вектора или вектора для переноса в целях получения вектора AAV согласно изобретению. Настоящее изобретение также относится к фармацевтической композиции и к применению AAV-вектора, нуклеиновой кислоты или фармацевтической композиции в качестве лекарственного средства, а в частности, в качестве лекарственного средства для лечения FRDA.
Подробное описание изобретения
Определения
Термины «лечение» и «терапия», используемые в настоящей заявке, относятся к набору гигиенических, фармакологических, хирургических и/или физических средств, используемых для лечения и/или облегчения заболевания и/или его симптомов в целях устранения проблем со здоровьем. Термины «лечение» и «терапия» включают профилактические и лечебные методы, поскольку они направлены на поддержание и/или восстановление здоровья человека или животного. Независимо от происхождения симптомов, заболевания и инвалидности, введение подходящего лекарственного средства для облегчения заболевания и/или устранения проблем со здоровьем должно интерпретироваться как форма лечения или терапии в контексте настоящей заявки.
Термин «терапевтически эффективное количество» означает количество вещества, которое обладает терапевтическим действием и которое способно излечивать FRDA.
Термины «индивидуум», «пациент» или «субъект» используются в настоящей заявке как синонимы и никоим образом не должны рассматриваться как ограничение изобретения. «Индивидуум», «пациент» или «субъект» могут иметь любой возраст, пол и физическое состояние.
Используемый здесь термин «фармацевтически приемлемый носитель» или «фармацевтически приемлемый разбавитель» означает любые и все растворители, дисперсионные среды, агенты для нанесения покрытия, антибактериальные и противогрибковые агенты, изотонические агенты и агенты, замедляющие всасывание, которые являются совместимыми с фармацевтическим введением. Использование таких сред и агентов для фармацевтически активных веществ хорошо известно специалистам. Подходящие носители, наполнители или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают следующие соединения, не ограничивающие объема настоящего изобретения, а именно: дополнительные забуферивающие агенты; консерванты; сорастворители; антиоксиданты, включая аскорбиновую кислоту и метионин; хелатообразующие агенты, такие как EDTA; металлокомплексы (например, комплексы Zn-белок); биоразлагаемые полимеры, такие как сложные полиэфиры; солеобразующие противоионы, такие как натрий, многоатомные спирты ряда сахаров; аминокислоты, такие как аланин, глицин, глутамин, аспарагин, гистидин, аргинин, лизин, орнитин, лейцин, 2-фенилаланин, глутаминовая кислота и треонин; органические сахара или спирты ряда сахаров, такие как лактит, стахиоза, манноза, сорбоза, ксилоза, рибоза, рибит, миоинизитоза, миоинизит, галактоза, галактит, глицерин, циклитолы (например, инозит), полиэтиленгликоль; серусодержащие восстановители, такие как мочевина, глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, α-монотиоглицерин и тиосульфат натрия; низкомолекулярные белки, такие как альбумин человеческой сыворотки, альбумин бычьей сыворотки, желатин или другие иммуноглобулины; и гидрофильные полимеры, такие как поливинилпирролидон.
Термин «фратаксин» означает белок, который кодируется геном FXN, и который обычно присутствует в митохондриях. Подробную информацию о белке можно найти в базе данных UniProtKB под регистрационным номером Q16595.
Термин «промотор» означает последовательность ДНК, с которой может связываться РНК-полимераза с инициацией транскрипции. Последовательность может также содержать сайты связывания с различными белками, которые регулируют транскрипцию, такими как факторы транскрипции. Промоторная последовательность может состоять из различных промоторных фрагментов (различных или одинаковых фрагментов), которые локализованы рядом друг с другом в последовательности ДНК и могут быть разделены линкерами или спейсерами. Такие промоторы называются химерными промоторами. В предпочтительном варианте осуществления изобретения, термин «промотор» означает промотор фосфоглицераткиназы (PGK).
Термин «посттранскрипционный регуляторный элемент» означает последовательность ДНК, которая, при ее транскрипции, создает третичную структуру, усиливающую или ингибирующую экспрессию белка.
Термин «функционально присоединенный» относится к двум или более последовательностям нуклеиновой кислоты, которые связаны таким образом, что одна последовательность нуклеиновой кислоты влияет на другую. Так, например, промотор PGK и WPRE функционально присоединены к последовательности нуклеиновой кислоты, кодирующей фратаксин, в результате чего уровни экспрессии фратаксина регулируются промотором PGK и WPRE.
Термин «функциональный вариант» означает нуклеиновые кислоты или аминокислотные последовательности, которые отличаются в одном или более положениях от родительской последовательности нуклеиновой кислоты или аминокислотной последовательности, где эти различия могут представлять собой добавления, делеции и/или замены нуклеиновых кислот или аминокислотных остатков; и которые все еще являются функциональными, а поэтому могут быть использованы для лечения FRDA. Специалист в данной области может определить функциональные варианты путем поиска гомологии с помощью программы поиска BLAST или путем исследования вариабельности белка или гена в популяции.
Термин «аденоассоциированный вирус» означает небольшой вирус, который инфицирует человека и приматов некоторых других видов. «Вектор» представляет собой любой носитель, который может быть использован для искусственного переноса чужеродного генетического материала в клетку. Таким образом, «AAV-вектор» означает рекомбинантный AAV, который переносит нуклеиновую кислоту в клетку.
AAV-вектор
В своем первом аспекте настоящее изобретение относится к вектору на основе аденоассоциированного вируса (AAV), содержащему нуклеиновую кислоту, где нуклеиновая кислота включает: (i) последовательность нуклеиновой кислоты, кодирующую фратаксин; (ii) промотор фосфоглицерат-киназы (PGK); и (iii) посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE), где (ii) и (iii) функционально присоединены к последовательности (i) и регулируют ее экспрессию.
SEQ ID NO: 1 означает следующую последовательность:
gaattccggggttggggttgcgccttttccaaggcagccctgggtctgcgcagggacgcggctgctctgggcgtggttccgggaaacgcagcggcgccgaccctgggtctcgcacattcttcacgtccgttcgcagcgtcacccggatcttcgccgctacccttgtgggccccccggcgacgcttcctgctccgcccctaagtcgggaaggttccttgcggttcgcggcgtgccggacgtgacaaacggaagccgcacgtctcactagtaccctcgcagacggacagcgccagggagcaatggcagcgcgccgaccgcgatgggctgtggccaatagcggctgctcagcagggcgcgccgagagcagcggccgggaaggggcggtgcgggaggcggggtgtggggcggtagtgtgggccctgttcctgcccgcgcggtgttccgcattctgcaagcctccggagcgcacgtcggcagtcggctccctcgttgaccgaatcaccgacctctctccccag.
В предпочтительном варианте осуществления изобретения промотор PGK содержит SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75% идентична последовательности SEQ ID NO: 1. Предпочтительно, промотор PGK содержит SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 1. Более предпочтительно, промотор PGK представляет собой SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 1.
Идентичность двух последовательностей может быть определена стандартными методами. Так, например, она может быть определена с использованием стандартных алгоритмов выравнивания, известных в данной области, таких как BLAST (Altschul et al., 1990. J. Mol. Biol. 215 (3): 403-10). В предпочтительном варианте осуществления изобретения, идентичность двух последовательностей определяют с использованием BLAST.
SEQ ID NO: 2 означает следующую последовательность:
TCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG.
В предпочтительном варианте осуществления изобретения, WPRE содержит SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 2. Предпочтительно, WPRE содержит SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 2. Более предпочтительно, WPRE представляет собой SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 2.
SEQ ID NO: 3 означает следующую последовательность:
ATGTGGACTCTCGGGCGCCGCGCAGTAGCCGGCCTCCTGGCGTCACCCAGCCCGGCCCAGGCCCAGACCCTCACCCGGGTCCCGCGGCCGGCAGAGTTGGCCCCACTCTGCGGCCGCCGTGGCCTGCGCACCGACATCGATGCGACCTGCACGCCCCGCCGCGCAAGTTCGAACCAGAGAGGTCTCAACCAGATTTGGAATGTCAAAAAGCAGAGTGTCTATTTGATGAATTTGAGGAAATCTGGAACTTTGGGCCACCCAGGCTCTCTAGATGAGACCACCTATGAAAGACTAGCAGAGGAAACGCTGGACTCTTTAGCAGAGTTTTTTGAAGACCTTGCAGACAAGCCATACACGTTTGAGGACTATGATGTCTCCTTTGGGAGTGGTGTCTTAACTGTCAAACTGGGTGGAGATCTAGGAACCTATGTGATCAACAAGCAGACGCCAAACAAGCAAATCTGGCTATCTTCTCCATCCAGTGGACCTAAGCGTTATGACTGGACTGGGAAAAACTGGGTGTACTCCCACGACGGCGTGTCCCTCCATGAGCTGCTGGCCGCAGAGCTCACTAAAGCCTTAAAAACCAAACTGGACTTGTCTTCCTTGGCCTATTCCGGAAAAGATGCTT.
В предпочтительном варианте осуществления изобретения последовательность нуклеиновой кислоты, кодирующая фратаксин, содержит SEQ ID NO: 3 или последовательность, которая, по меньшей мере на 75% идентична SEQ ID NO: 3 и представляет собой функциональный вариант фратаксина. Предпочтительно, последовательность нуклеиновой кислоты, кодирующая фратаксин, содержит SEQ ID NO: 3 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или на 99% идентична SEQ ID NO: 3. Более предпочтительно, последовательность нуклеиновой кислоты, кодирующая фратаксин, представляет собой SEQ ID NO: 3 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 3.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит линкер между промотором и последовательностью нуклеиновой кислоты, кодирующей фратаксин, где линкер состоит из SEQ ID NO: 6 или включает SEQ ID NO: 6, или последовательность, которая по меньшей мере на 75% идентична последовательности SEQ ID NO 6. Предпочтительно, линкер состоит из SEQ ID NO: 6, или включает SEQ ID NO: 6, или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 6.
SEQ ID NO: 6 означает следующую последовательность:
TGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGCCGCCACC.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит последовательность Козака, и эта последовательность Козака также функционально присоединена к последовательности нуклеиновой кислоты, кодирующей фратаксин, и регулирует ее экспрессию. Последовательность Козака представляет собой последовательность, которая встречается в эукариотической мРНК и имеет консенсусную последовательность gccRccAUGG, где R представляет собой пурин; строчными буквами обозначены основания, которые наиболее часто встречаются в этих положениях, где, тем не менее, они могут варьироваться; а прописными буквами показаны высококонсервативные основания.
SEQ ID NO: 4 означает следующую последовательность:
GAATTCCGGGGTTGGGGTTGCGCCTTTTCCAAGGCAGCCCTGGGTCTGCGCAGGGACGCGGCTGCTCTGGGCGTGGTTCCGGGAAACGCAGCGGCGCCGACCCTGGGTCTCGCACATTCTTCACGTCCGTTCGCAGCGTCACCCGGATCTTCGCCGCTACCCTTGTGGGCCCCCCGGCGACGCTTCCTGCTCCGCCCCTAAGTCGGGAAGGTTCCTTGCGGTTCGCGGCGTGCCGGACGTGACAAACGGAAGCCGCACGTCTCACTAGTACCCTCGCAGACGGACAGCGCCAGGGAGCAATGGCAGCGCGCCGACCGCGATGGGCTGTGGCCAATAGCGGCTGCTCAGCAGGGCGCGCCGAGAGCAGCGGCCGGGAAGGGGCGGTGCGGGAGGCGGGGTGTGGGGCGGTAGTGTGGGCCCTGTTCCTGCCCGCGCGGTGTTCCGCATTCTGCAAGCCTCCGGAGCGCACGTCGGCAGTCGGCTCCCTCGTTGACCGAATCACCGACCTCTCTCCCCAGTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGCCGCCACCATGTGGACTCTCGGGCGCCGCGCAGTAGCCGGCCTCCTGGCGTCACCCAGCCCGGCCCAGGCCCAGACCCTCACCCGGGTCCCGCGGCCGGCAGAGTTGGCCCCACTCTGCGGCCGCCGTGGCCTGCGCACCGACATCGATGCGACCTGCACGCCCCGCCGCGCAAGTTCGAACCAGAGAGGTCTCAACCAGATTTGGAATGTCAAAAAGCAGAGTGTCTATTTGATGAATTTGAGGAAATCTGGAACTTTGGGCCACCCAGGCTCTCTAGATGAGACCACCTATGAAAGACTAGCAGAGGAAACGCTGGACTCTTTAGCAGAGTTTTTTGAAGACCTTGCAGACAAGCCATACACGTTTGAGGACTATGATGTCTCCTTTGGGAGTGGTGTCTTAACTGTCAAACTGGGTGGAGATCTAGGAACCTATGTGATCAACAAGCAGACGCCAAACAAGCAAATCTGGCTATCTTCTCCATCCAGTGGACCTAAGCGTTATGACTGGACTGGGAAAAACTGGGTGTACTCCCACGACGGCGTGTCCCTCCATGAGCTGCTGGCCGCAGAGCTCACTAAAGCCTTAAAAACCAAACTGGACTTGTCTTCCTTGGCCTATTCCGGAAAAGATGCTTTGCCCACCTAGGGATCGGATCCCCGGGTACCGAGCTCGAATTCTGCAGATATCCAGCACACTTTGCCTTTCTCTCCACAGGTGTCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG.
В предпочтительном варианте осуществления изобретения, последовательность нуклеиновой кислоты, которая содержит (i), (ii) и (iii), представляет собой SEQ ID NO: 4 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 4. Предпочтительно, нуклеиновая кислота, которая содержит (i), (ii) и (iii), представляет собой SEQ ID NO: 4 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 4.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит сигнал PolyA. Предпочтительно, сигнал PolyA имеет длину по меньшей мере 50, 100, 150 или 228 п.о. Более предпочтительно, сигнал PolyA имеет длину по меньшей мере 228 п.о. Наиболее предпочтительно, сигнал PolyA имеет длину 228 п.о.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит одну или более последовательностей инвертированных концевых повторов (ITR). Предпочтительно, нуклеиновая кислота содержит две последовательности ITR. Более предпочтительно, последовательности ITR фланкируют остальные компоненты нуклеиновой кислоты.
SEQ ID NO: 5 означает следующую последовательность:
CTGGCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGGTCGCCCGGCCTCAGTGAGCGAGCGAGCGCGCAGAGAGGGAGTGGCCAACTCCATCACTAGGGGTTCCTGAATTCCGGGGTTGGGGTTGCGCCTTTTCCAAGGCAGCCCTGGGTCTGCGCAGGGACGCGGCTGCTCTGGGCGTGGTTCCGGGAAACGCAGCGGCGCCGACCCTGGGTCTCGCACATTCTTCACGTCCGTTCGCAGCGTCACCCGGATCTTCGCCGCTACCCTTGTGGGCCCCCCGGCGACGCTTCCTGCTCCGCCCCTAAGTCGGGAAGGTTCCTTGCGGTTCGCGGCGTGCCGGACGTGACAAACGGAAGCCGCACGTCTCACTAGTACCCTCGCAGACGGACAGCGCCAGGGAGCAATGGCAGCGCGCCGACCGCGATGGGCTGTGGCCAATAGCGGCTGCTCAGCAGGGCGCGCCGAGAGCAGCGGCCGGGAAGGGGCGGTGCGGGAGGCGGGGTGTGGGGCGGTAGTGTGGGCCCTGTTCCTGCCCGCGCGGTGTTCCGCATTCTGCAAGCCTCCGGAGCGCACGTCGGCAGTCGGCTCCCTCGTTGACCGAATCACCGACCTCTCTCCCCAGTGGCTAACTAGAGAACCCACTGCTTACTGGCTTATCGAAATTAATACGACTCACTATAGGGAGACCCAAGCTGGCTAGCGTTTAAACTTAAGCTTGGCCGCCACCATGTGGACTCTCGGGCGCCGCGCAGTAGCCGGCCTCCTGGCGTCACCCAGCCCGGCCCAGGCCCAGACCCTCACCCGGGTCCCGCGGCCGGCAGAGTTGGCCCCACTCTGCGGCCGCCGTGGCCTGCGCACCGACATCGATGCGACCTGCACGCCCCGCCGCGCAAGTTCGAACCAGAGAGGTCTCAACCAGATTTGGAATGTCAAAAAGCAGAGTGTCTATTTGATGAATTTGAGGAAATCTGGAACTTTGGGCCACCCAGGCTCTCTAGATGAGACCACCTATGAAAGACTAGCAGAGGAAACGCTGGACTCTTTAGCAGAGTTTTTTGAAGACCTTGCAGACAAGCCATACACGTTTGAGGACTATGATGTCTCCTTTGGGAGTGGTGTCTTAACTGTCAAACTGGGTGGAGATCTAGGAACCTATGTGATCAACAAGCAGACGCCAAACAAGCAAATCTGGCTATCTTCTCCATCCAGTGGACCTAAGCGTTATGACTGGACTGGGAAAAACTGGGTGTACTCCCACGACGGCGTGTCCCTCCATGAGCTGCTGGCCGCAGAGCTCACTAAAGCCTTAAAAACCAAACTGGACTTGTCTTCCTTGGCCTATTCCGGAAAAGATGCTTTGCCCACCTAGGGATCGGATCCCCGGGTACCGAGCTCGAATTCTGCAGATATCCAGCACACTTTGCCTTTCTCTCCACAGGTGTCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTGGAATTCGAGCTCGGTACGATCAGCTGATCAGCCTCGACTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGCTGGCGCGCTCGCTCGCTCACTGAGGCCGCCCGGGCAAAGCCCGGGCGTCGGGCGACCTTTGGTCGCCCGGCCTCAGTGAGCGAGCGAGCGCGCAGAGAGGGAGTGGCCAACTCCATCACTAGGGGTTCCT.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота представляет собой SEQ ID NO: 5 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 5. Предпочтительно, нуклеиновая кислота представляет собой SEQ ID NO: 5 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 5.
В предпочтительном варианте осуществления изобретения вектор позволяет экспрессировать терапевтически эффективное количество фратаксина у пациента, страдающего атаксией Фридрейха. Таким образом, вектор доставляет нуклеиновую кислоту в клетки пациента, где эта нуклеиновая кислота затем экспрессирует фратаксин в терапевтически эффективном количестве.
В предпочтительном варианте осуществления изобретения AAV-вектор представляет собой AAV-вектор серотипа 9, то есть, вектор AAV-9.
Нуклеиновая кислота
Во втором своем аспекте настоящее изобретение относится к нуклеиновой кислоте, содержащей: (i) последовательность нуклеиновой кислоты, кодирующую фратаксин; (ii) промотор PGK; и (iii) WPRE; где (ii) и (iii) функционально присоединены к последовательности (i) и регулируют ее экспрессию.
В предпочтительном варианте осуществления изобретения промотор PGK содержит SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75% идентична последовательности SEQ ID NO: 1. Предпочтительно, промотор PGK содержит SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 1. Более предпочтительно, промотор PGK представляет собой SEQ ID NO: 1 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 905, 915, 92%, 95%, 97%, 98% или 99% идентична последовательности SEQ ID NO: 1.
В предпочтительном варианте осуществления изобретения WPRE содержит SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 2. Предпочтительно, WPRE содержит SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 2. Более предпочтительно, WPRE представляет собой SEQ ID NO: 2 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 2.
В предпочтительном варианте осуществления изобретения последовательность нуклеиновой кислоты, кодирующая фратаксин, содержит SEQ ID NO: 3 или последовательность, которая, по меньшей мере на 75% идентична SEQ ID NO: 3 и является функциональным вариантом фратаксина. Предпочтительно, последовательность нуклеиновой кислоты, кодирующая фратаксин, содержит SEQ ID NO: 3 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или на 99% идентична SEQ ID NO: 3. Более предпочтительно, последовательность нуклеиновой кислоты, кодирующая фратаксин, представляет собой SEQ ID NO: 3 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 3.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит линкер между промотором и последовательностью нуклеиновой кислоты, кодирующей фратаксин, где линкер состоит из SEQ ID NO:6 или содержит SEQ ID NO: 6, или последовательность, которая по меньшей мере на 75% идентична последовательности SEQ ID NO 6. Предпочтительно, линкер состоит из SEQ ID NO:6 или содержит SEQ ID NO: 6 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 6.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит последовательность Козака, и эта последовательность Козака также функционально присоединена к последовательности нуклеиновой кислоты, кодирующей фратаксин, и регулирует ее экспрессию.
В предпочтительном варианте осуществления изобретения последовательность нуклеиновой кислоты, которая содержит (i), (ii) и (iii), представляет собой SEQ ID NO: 4 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 4. Предпочтительно, нуклеиновая кислота, которая содержит (i), (ii) и (iii), представляет собой SEQ ID NO: 4 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 4.
В предпочтительном варианте осуществления изобретения, нуклеиновая кислота дополнительно содержит сигнал PolyA. Предпочтительно, сигнал PolyA имеет длину по меньшей мере 50, 100, 150 или 228 п.о. Более предпочтительно, сигнал PolyA имеет длину по меньшей мере 228 п.о. Наиболее предпочтительно, сигнал PolyA имеет длину 228 п.о.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота дополнительно содержит одну или более последовательностей инвертированных концевых повторов (ITR). Предпочтительно, нуклеиновая кислота содержит две последовательности ITR. Более предпочтительно, последовательности ITR фланкируют остальные компоненты нуклеиновой кислоты.
В предпочтительном варианте осуществления изобретения нуклеиновая кислота представляет собой SEQ ID NO: 5 или последовательность, которая по меньшей мере на 75% идентична SEQ ID NO: 5. Предпочтительно, нуклеиновая кислота представляет собой SEQ ID NO: 5 или последовательность, которая по меньшей мере на 75%, 80%, 85%, 89%, 90%, 91%, 92%, 95%, 97%, 98% или 99% идентична SEQ ID NO: 5.
Клонирующий вектор
В своем третьем аспекте настоящее изобретение относится к клонирующему вектору, который содержит нуклеиновую кислоту в соответствии с любым из ранее описанных вариантов осуществления изобретения, и дополнительные элементы нуклеиновой кислоты для стимуляции репликации клонирующего вектора в бактериальной клетке.
Термин «клонирующий вектор» означает любой вектор, который является подходящм для клонирования, и который обычно включает рестрикционные сайты, ориджин репликации для размножения бактерий и селективный маркер.
Клонирующий вектор согласно изобретению содержит нуклеиновую кислоту согласно изобретению и может быть предпочтительно использован для получения вектора для переноса или AAV-вектора согласно изобретению.
Вектор для переноса
В своем четвертом аспекте настоящее изобретение относится к вектору для переноса, который содержит нуклеиновую кислоту в соответствии с любым из ранее описанных вариантов осуществления изобретения, и дополнительные элементы нуклеиновой кислоты для стимуляции интеграции или транспозиции вектора для переноса в AAV-вектор, предпочтительно, вектор AAV-9.
Термин «вектор для переноса» означает вектор, который является подходящим для интеграции или транспозиции в AAV-векторе. Таким образом, вектор для переноса, по существу, позволяет встраивать генетическую информацию в AAV-вектор.
Вектор для переноса согласно изобретению включает нуклеиновую кислоту согласно изобретению и может быть использован для получения AAV-вектора согласно изобретению.
Применение нуклеиновой кислоты, клонирующего вектора и вектора для переноса
В своем пятом аспекте настоящее изобретение относится к применению нуклеиновой кислоты согласно изобретению, клонирующего вектора согласно изобретению или вектора для переноса согласно изобретению в целях получения AAV-вектора согласно любому из ранее описанных вариантов изобретения.
Фармацевтическая композиция
В своем шестом аспекте настоящее изобретение относится к фармацевтической композиции, содержащей AAV-вектор согласно изобретению или нуклеиновую кислоту согласно изобретению и фармацевтически приемлемый носитель или разбавитель.
Описанная здесь фармацевтическая композиция может также содержать и другие вещества. Такими веществами являются, но не ограничиваются ими, криопротекторы, лиопротекторы, поверхностно-активные вещества, наполнители, антиоксиданты и стабилизаторы. В некоторых вариантах осуществления изобретения, фармацевтическая композиция может быть лиофилизована.
Используемый здесь термин «криопротектор» включает агенты, которые обеспечивают резистентность AAV-вектора к стрессам, вызванным замораживанием, преимущественно, благодаря удалению AAV-вектора с поверхности. Криопротекторы могут также обеспечивать защиту при первичной и вторичной сушке и длительном хранении продукта. Неограничивающими примерами криопротекторов являются сахара, такие как сахароза, глюкоза, трегалоза, маннит, манноза и лактоза; полимеры, такие как декстран, гидроксиэтилированный крахмал и полиэтиленгликоль; поверхностно-активные вещества, такие как полисорбаты (например, PS-20 или PS-80); и аминокислоты, такие как глицин, аргинин, лейцин и серин. Обычно используется криопротектор, обладающий низкой токсичностью в биологических системах.
В одном варианте осуществления изобретения лиопротектор добавляют в описанную здесь фармацевтическую композицию. Используемый здесь термин «лиопротектор» включает агенты, которые обеспечивают стабильность AAV-вектора во время сушки замораживанием или во время дегидратации (при первичных и вторичных циклах сушки замораживанием) посредством создания аморфной стеклообразной матрицы и связывания с поверхностью AAV-вектора посредством водородных связей с последующей заменой молекул воды, которые удаляются в процессе сушки. Это позволяет минимизировать разложение продукта во время цикла лиофилизации и сообщать продукту длительную стабильность. Неограничивающими примерами лиопротекторов являются сахара, такие как сахароза или трегалоза; аминокислота, такая как моноглутамат натрия, некристаллический глицин или гистидин; метиламин, такой как бетаин; лиотропная соль, такая как сульфат магния; полиол, такой как трехатомные спирты или многоатомные спирты ряда сахаров, например, глицерин, эритрит, глицерол, арабит, ксилит, сорбит и маннит; пропиленгликоль; полиэтиленгликоль; плюроники; и их комбинации. Количество лиопротектора, добавляемого в фармацевтическую композицию, обычно представляет собой количество, которое не приводит к недопустимому уровню разложения штамма, если фармацевтическая композиция является лиофилизованной.
В некоторых вариантах осуществления изобретения фармацевтическая композиция включает наполнитель. Используемый здесь термин «наполнитель» включает агенты, которые обеспечивают нужную структуру лиофилизованного продукта без непосредственного взаимодействия с фармацевтическим продуктом. Наполнители, помимо их сообщения продукту фармацевтически элегантного вида, могут также придавать продукту полезные свойства в отношении изменения температуры сжатия, обеспечения защиты при замораживании-оттаивании и повышения стабильности штамма в течение длительного периода хранения. Неограничивающими примерами наполнителей являются маннит, глицин, лактоза и сахароза. Наполнители могут быть кристаллическими (например, глицин, маннит или хлорид натрия) или аморфными (например, декстран, гидроксиэтилированный крахмал) и обычно используются в композициях в количестве от 0,5% до 10%.
В описанную здесь фармацевтическую композицию могут быть также включены и другие фармацевтически приемлемые носители, наполнители или стабилизаторы, которые описаны в руководстве Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980), при условии, что они не будут оказывать негативного воздействия на нужные свойства фармацевтической композиции. Используемый здесь термин «фармацевтически приемлемый носитель» означает любые и все растворители, дисперсионные среды, агенты для покрытия, антибактериальные и противогрибковые агенты, изотонические агенты и агенты, замедляющие всасывание, которые являются совместимыми с фармацевтическим введением. Использование таких сред и агентов для фармацевтически активных веществ хорошо известно специалистам. Подходящие носители, наполнители или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают следующие соединения, не ограничивающие объема настоящего изобретения, а именно: дополнительные забуферивающие агенты; консерванты; сорастворители; антиоксиданты, включая аскорбиновую кислоту и метионин; хелатообразующие агенты, такие как EDTA; металлокомплексы (например, комплексы Zn-белок); биоразлагаемые полимеры, такие как сложные полиэфиры; солеобразующие противоионы, такие как натрий, многоатомные спирты ряда сахаров; аминокислоты, такие как аланин, глицин, глутамин, аспарагин, гистидин, аргинин, лизин, орнитин, лейцин, 2-фенилаланин, глутаминовая кислота и треонин; органические сахара или спирты ряда сахаров, такие как лактит, стахиоза, манноза, сорбоза, ксилоза, рибоза, рибит, миоинизитоза, миоинизит, галактоза, галактит, глицерин, циклитолы (например, инозит), полиэтиленгликоль; серусодержащие восстановители, такие как мочевина, глутатион, тиоктовая кислота, тиогликолят натрия, тиоглицерин, α-монотиоглицерин и тиосульфат натрия; низкомолекулярные белки, такие как альбумин человеческой сыворотки, альбумин бычьей сыворотки, желатин или другие иммуноглобулины; и гидрофильные полимеры, такие как поливинилпирролидон.
Фармацевтическая композиция может быть приготовлена для перорального, подъязычного, трансбуккального, внутривенного, внутримышечного, подкожного, внутрибрюшинного, конъюнктивального, ректального, трансдермального, интратекального и местного введения и/или введения путем ингаляции. В предпочтительном варианте осуществления изобретения, фармацевтическая композиция может представлять собой раствор, который является подходящим для внутривенного, внутримышечного, конъюнктивального, трансдермального, внутрибрюшинного и/или подкожного введения. В другом варианте осуществления изобретения, фармацевтическая композиция может представлять собой раствор, который является подходящим для подъязычного и трансбуккального введения и/или введения путем ингаляции. В альтернативном варианте осуществления изобретения, фармацевтическая композиция может представлять собой гель или раствор, который является подходящим для интратекального введения. В альтернативном варианте осуществления изобретения, фармацевтическая композиция может представлять собой аэрозоль, подходящий для введения путем ингаляции. В предпочтительном варианте осуществления изобретения, фармацевтическая композиция может быть приготовлена для интратекального введения.
Фармацевтическая композиция может дополнительно содержать обычные эксципиенты и носители, которые известны специалистам в данной области. Для твердых фармацевтических композиций могут быть использованы стандартные нетоксичные твердые носители, которые включают, например, фармацевтические сорта маннита, лактозы, крахмала, стеарата магния, натрий-содержащего сахарина, талька, целлюлозы, глюкозы, сахарозы, карбоната магния и т.п. В целях приготовления раствора для инъекций, фармацевтическая композиция может дополнительно содержать криопротекторы, лиопротекторы, поверхностно-активные вещества, наполнители, антиоксиданты, стабилизаторы и фармацевтически приемлемые носители. Для аэрозольного введения, фармацевтические композиции обычно поставляются в тонкодисперсной форме вместе с поверхностно-активным веществом и пропеллентом. Само собой разумеется, что поверхностно-активное вещество должно быть нетоксичным и, по существу, растворимым в пропелленте. Такими репрезентативными агентами являются сложные эфиры или неполные сложные эфиры жирных кислот, содержащих от 6 до 22 атомов углерода, таких как капроевая, октановая, лауриновая, пальмитиновая, стеариновая, линолевая, линоленовая, олестериновая и олеиновая кислоты с алифатическим многоатомным спиртом или его циклическим ангидридом. Могут быть использованы смешанные сложные эфиры, такие как смешанные или природные глицериды. При желании, может быть также включен носитель, такой как, например, лецитин для интраназальной доставки. В случае суппозиториев, могут быть включены традиционные связующие вещества и носители, например, полиалкиленгликоли или триглицериды.
Применение в медицине
В своем седьмом аспекте настоящее изобретение относится к AAV-вектору согласно изобретению, к нуклеиновой кислоте согласно изобретению или к фармацевтической композиции согласно изобретению для их применения в качестве лекарственного средства. В своем восьмом аспекте, настоящее изобретение относится к AAV-вектору согласно изобретению, к нуклеиновой кислоте согласно изобретению или к фармацевтической композиции согласно изобретению для их применения в целях лечении атаксии Фридрейха.
В предпочтительном варианте осуществления изобретения AAV-вектор согласно изобретению, нуклеиновую кислоту согласно изобретению или фармацевтическую композицию согласно изобретению вводят интратекально, внутримышечно, интрацеребрально или интрацеребровентрикулярно, предпочтительно, интратекально или внутримышечно.
В предпочтительном варианте осуществления изобретения AAV-вектор согласно изобретению, нуклеиновую кислоту согласно изобретению или фармацевтическую композицию согласно изобретению вводят в дозе по меньшей мере 1×109 векторных геномов/кг массы тела, предпочтительно 1×1010, 1×1011 или 1×1012 векторных геномов/кг массы тела. Более предпочтительно вводить по меньшей мере или приблизительно 4,6×1012 векторных геномов/кг массы тела.
Примеры
Пример 1: Конструирование плазмиды
Кодирующую последовательность изоформы 1 человеческого фратаксина (hFXN) присоединяли к гемагглютининовой метке (HA) и клонировали в экспрессионный вектор pcDNA3.1 с использованием набора для клонирования In-Fusion® HD (Clontech). Эта гибридная система была использована во всех стадиях клонирования. Несколько конструкций было получено путем присоединения промоторов CMV или человеческих (h) промоторов (p) синапсина (phSYN), нейрон-специфической энолазы (phNSE), 1255 п.о. промотора FXN (phFXN1255) или изоформы 1 человеческой фосфоглицераткиназы (phPGK1) к кодирующей области гена FXN. Дополнительные регуляторные элементы, такие как энхансер CMV и последовательности Козака, были присоединены у 5’-конца, а последовательность элемента, отвечающего на вирус гепатита сурка (WPRE), был присоединены у 3’-конца. Все эти конструкции, полученные с использованием экспрессионных векторов, перечислены в Таблице 2.
Плазмиды, содержащие ту же самую комбинацию регуляторных элементов, также получали для инициации экспрессии люциферазы путем замены кодирующей последовательности FXN на последовательность, кодирующую люциферазу светляка (LUC), которая была амплифицирована из вектора pGL3-LUC (Promega), также с использованием набора для клонирования In-Fusion® HD (Clontech). Эти плазмиды перечислены в Таблице 3.
Пример 2. Конструирование и получение рекомбинантного аденоассоциированного вирусного вектора.
Экспрессионные кластеры плазмиды pcDNA3.1-phPGK-kFXN-HA-WPRE и плазмиды pcDNA3.1-phPGK-kLUC-HA-WPRE были клонированы в сайты SnaBI-MfeI рекомбинантного вектора AAV-9 (вектора rAAV9-phPGK1-FXN-HA-WPRE и вектора rAAV9-phPGK-LUC-WPRE). Кроме того, был создан контрольный нулевой вектор без кодирующей последовательности FXN (AAV2/9-null). Все три конструкции и вирусные частицы были получены в лаборатории по продуцированию векторов в Центре Биотехнологии и Генотерапии животных (Universitat Autònoma de Barcelona). Полученные конечные титры составляли 1,4×1013 ВГ/мл для AAV9-phPGK-FXN-HA-WPRE, 9,8×1012 ВГ/мл для AAV9-phPGK-LUC-WPRE и 6×1012 ВГ/мл для AAV2/9-null.
Пример 3: Оптимизация экспрессии фратаксина
1. Клеточная культура, экспрессия in vitro и анализ на ген-репортер люциферазы.
Мышиные клетки нейробластомы (N2a), человеческие клетки нейробластомы (SH-SY5Y) и клетки почек человеческого эмбриона (HEK 293) культивировали в модифицированной по способу Дульбекко среде Игла (DMEM), содержащей 10% фетальную бычью сыворотку (Sigma), 2 мМ глютамина, 50 мкг/мл пенициллина/стрептомицина (Life technologies) при 70% конфлюэнтности в 10 см-чашках для культивирования. Трансфекцию осуществляли с использованием липофектамина 2000 (Life technologies) для клеток N2a и SH-SY5Y и фосфата кальция для HEK 293, с использованием только 4 мкг плазмидных ДНК помимо 0,25 мкг EGFP. После трансфекции, среду заменяли свежей культуральной средой DMEM для клеток HEK и средой, содержащей Neurobasal, добавку B27, 10 мкМ ретиноевой кислоты, 2 мМ глутамина, 50 мкг/мл пенициллина/стрептомицина для клеток N2a и SH-SY5Y. Через 48 часов после замены среды для культивирования клеток, клетки, трансфецированные экспрессионными плазмидами, перечисленными в Таблице 2, собирали и хранили при -80°C до дальнейшего использования. В целях проведения анализа на ген-репортер люциферазы, плазмиды, перечисленные в Таблице 3, трансфецировали, как указано выше, и клетки повторно высевали в 384-луночные планшеты, и люциферазную активность определяли с помощью набора для анализа на два репортера люциферазы в соответствии с инструкциями производителя (Promega).
2. Электрофорез в ДСН-ПААГ и иммуноблоттинг
Белки экстрагировали из культивируемых клеток путем гомогенизации в буфере для лизиса RIPA: 10 мМ Трис-HCl, рН 7,4, 140 мМ NaCl, 0,1% дезоксихолат натрия, 1% Тритона Х-100, 1% ДСН, 2 мМ этилендиаминтетрауксусной кислоты (EDTA), 25 мМ фторида натрия (NaF), 2,5 мМ NaVO3 и коктейля ингибиторов белка (Roche). Концентрацию белка определяли с помощью анализа на белок DC-BioRad (BioRad). Общий экстракт белка смешивали с 4 × буфером для загрузки образцов белка (Li-Cor Biosciences), содержащим 1 мМ DTT, и разделяли с помощью электрофореза в 15% акриламидном геле при постоянной силе тока 20 мА до переноса на PVDF-мембраны. В качестве «первых» антител использовали анти-FXN антитело PAC 2518 (любезно предоставленное Dr. Grazia Isaya, Mayo Clinic, Rochester MN), анти-FXN антитело (1G2, Merck Millipore), антитело против метки HA (клон 16B12, MMS-101P, Covance) и антитело против бета-актина (AC15, Sigma). «Вторые» антитела, конъюгированные с инфракрасным красителем, представляли собой антимышиные IRDye-800CW и антикроличьи IRDye 700CW (Li-Cor Biosciences), и иммунореактивность определяли с использованием программы-анализатора Odyssey v2.1 (Li-Cor Biosciences).
3. Результаты
Как можно видеть на фигурах 1-5 и в Таблицах 2-3, получение конструкции, содержащей промотор hPGK1, линкерные последовательности (определенной длины и определенного состава) и WPRE, позволило экспрессировать фратаксин в клетках. Человеческой промотор PGK1 представляет собой метаболически регулируемый промотор, экспрессируемый в физиологически релевантных тканях, что делает его хорошим кандидатом в качестве промотора для фратаксина. Однако, было не ясно, могут ли регуляторные элементы или линкеры с определенной последовательностью/длиной экспрессировать последовательности, кодирующие фратаксин. Результаты, полученные авторами с использованием различных комбинаций элементов и последовательностей, показали, что экспрессия белка из последовательностей, кодирующих человеческий фратаксин, требует включения функционально присоединенного линкера между промотором hPGK1 и кодирующими последовательностями FXN, поскольку конструкции, имеющие длину приблизительно 105 нуклеотидов, позволяют экспрессировать FXN, а линкеры длиной приблизительно 35 нуклеотидов не выполняют такую функцию даже при добавлении последовательности-стабилизатора РНК WPRE. Интересно отметить, что такое требование преъявлялось к линкеру только при использовании промотора hPGK1, но не других протестированных промоторов (то есть, hSYN). Кроме того, уровни белка фратаксина, экспрессируемые из этого вектора, были гораздо ниже, чем уровни, полученные от векторов, индуцируемых промотором CMV или CAG, даже в отсутствие последовательностей WPRE, и, таким образом, эти уровни были более сопоставимы с эндогенными уровнями. Один из недостатков предыдущих подходов заключается в том, что для достижения экспрессии фратаксина использовались в высокой степени индуцибельные промоторы, такие как промоторы CAG или CMV, которые давали очень высокие уровни экспрессии белка, способные вызывать клеточный стресс и перекрывать некоторые защитные эффекты. Представленный здесь вектор может индуцировать экспрессию белка фратаксина в соответствующей нервной системе и в периферических тканях в количествах в пределах физиологически функционального диапазона.
Пример 4: Данные in viv o
1. Животные
Мыши YG8R с моделью атаксии Фридрейха, полученные Пуком и коллегами (Pook et al. 2001. Neurogenetics. 3 (4):185-93; Virmouni et al., 2014. PLoS ONE. 9 (9): e107416), и используемые в этом исследовании, были взяты из фондов лаборатории Джексона (регистрационный номер 012253). Эти трансгенные мыши с моделью человеческого FXN были нокаутированы по эндогенному гену мышиного фратаксина (Fxn -/-) и содержали человеческий трансген YAC от исходного YG8 (несущего две тандемные копии человеческого гена FXN приблизительно с 82 и со 190 тринуклеотидными повторяющимися последовательностями GAA). Мыши содержались в SPF-подобных условиях в клетках для содержания мышей IVC NexGen (Аллентаун, штат Нью-Джерси) с 12-часовым циклом день-ночь и с регулируемыми отрицательным давлением, температурой и влажностью, а также со свободным доступолм к воде и облученному корму для грызунов Teklad с общим содержанием белка 18% (Envigo). Группа мышей YG8R была получена в Научно-исследовательском институте здравоохранения Germans Trias i Pujol (IGTP). Во всех экспериментах использовали гемизиготных самок и самцов YG8R (Tg/-) и мышей дикого типа C57Bl/6.
Мыши, гемизиготные по мутантному гену человеческого FXN (YG8R), были генотипированы, как сообщалось ранее (29,36). Вкратце, геномную ДНК экстрагировали из хвоста мыши YG8R с помощью набора для очистки ДНК, взятой из хвоста мыши Maxwell® 16 (Promega). Число копий трансгенов определяли для каждой мыши с помощью количественной ПЦР в реальном времени (кол.ПЦР) на устройстве LightCycler® 480 (Roche) с использованием фермента SYBR Premix Ex Taq II (РНКазы Tli H Plus) (Takara) и нижеследующих праймеров для трансгена человеческого фратаксина: hFXN_Tg_FW: 5’-GAACTTCAAATTAGTTCCCCTTTCTTC-3’(SEQ ID NO: 7), hFXN_Tg_RV: 5’-CACAGCCATTCTTTGGGTTTC-3’ (SEQ ID NO: 8); и внутреннего контроля, аполипопротеина B-100 изоформы X1: IC_FW: 5’-CACGTGGGCTCCAGCATT-3’ (SEQ ID NO: 9), IC_RV: TCACCAGTCATTTCTGCCTTTG (SEQ ID NO: 10). Анализ осуществляли путем проведения термоциклов в следующих условиях: 95°C в течение 5 минут и 40 циклов при 95°C в течение 20 секунд и 60°С в течение 30 секунд, 72°C в течение 30 секунд и, наконец, 1 цикл при 72°C в течение 2 минут. Образцы анализировали с тремя повторностями для каждого представляющего интерес гена, а уровни трансгена определяли методом Ct (ΔΔCt). Число копий трансгеннов оценивали по сравнению с данными Ct для контрольного образца с известным числом копий. Длину повторов GAA определяли с помощью ПЦР-амплификации GAA с использованием LA taq (Invitrogen) и нижеследующих праймеров: GAA-F: 5’-GGGATTGGTT-GCCAGTGCTT-AAAAGTTAG-3’ (SEQ ID NO: 11) и GAA-R: 5’-GATCTAAGGACCATCATGG-CCACACTTGCC-3’ (SEQ ID NO: 12). Продукты ПЦР разделяли в 1,5% агарозных гелях с помощью электрофореза при 100 В в течение 3 часов и анализировали размеры полос. Число повторов GAA затем определяли путем вычитания 451 п.о. (фланкирующих неповторяющиеся ДНК) из размера продукта ПЦР и деления полученного размера повторов пар оснований на 3.
Все процедуры на животных проводились в соответствии с нормами, утывержденными ЕС и одобренными местными Регуляторными Органами и соответствующими местными Комитетами по этике.
2. Введение AAV
Гемизиготных мышей YG8R и 10-недельных мышей дикого типа анестезировали путем внутрибрюшинной инъекции кетамина (10 мг/кг массы тела; Imalgene 500; Rhône-Merieux, Lyon, France) и ксилазина (1 мг/кг массы тела; Rompun; Bayer). Интратекальное введение AAV9-phPGK-FXN-HA-WPRE, AAV9-phPGK-LUC-WPRE или AAV2/9-null проводили в поясничную область. После открытия бокового отдела позвоночника путем иссечения паравертебральных мышц, вирусные векторы медленно вводили в CSF с помощью иглы калибра 33 и шприца Гамильтона между поясничными позвонками L3 и L4. Соответствующий доступ к интратекальному пространству был подтвержден движением хвоста животного. После этого, на мышцы и кожу накладывали швы. Количество различных вирусных векторов, вводимое каждой мыши, составляло 4,6×10-12 ВГ/кг массы мыши.
3. Биораспределение вектора с помощью люциферазной визуализации.
Для оценки in vivo биораспределения вектора AAV9-phPGK-LUC-WPRE, мышам в возрасте 10 недель (2,5 месяца) интратекально вводили вектор, а еще через 3,5 месяца мышам вводили одну внутрибрюшинную инъекцию раствора субстрата D-люциферина в количестве 150 мг/кг массы мыши. Мышей анестезировали через 15 минут после введения субстрата путем ингаляционной анестезии (изофлуран 4% для индуцирования и 2% для поддержания). Анестезированных мышей содержали в темной камере Perkin Elmer Ivis Lumina II (Caliper Life Sciences, Germany) для регистрации эмиссии фотонов. Изображения анализировали с помощью программы визуализации Living Imaging (Xenogen Corporation, CA, EUA), где время интеграции составляло 1 мин, а поле зрения=12,5 см. Для оценки ex vivo, ткани и органы мышей извлекали через 15 минут после введения субстрата. Ткани и органы помещали в прозрачные чашки, и изображения фиксировали с помощью программы визуализации Living Imaging (Xenogen Corporation, CA, EUA), как указано выше. Результаты можно видеть на фигуре 6.
4. кол.ОТ-ПЦР
РНК экстрагировали из 30 мг свежезамороженной мышиной ткани с использованием набора RNeasy Mini (Qiagen). Анализ RIN на РНК и количественную оценку РНК проводили на рабочей установке Agilent 2200 TapeStation (Agilent). РНК подвергали обратной транскрипции в кДНК (25 нг/мкл) с использованием набора реагентов PrimeScriptTM RT (Takara) в тепловых условиях: 37°C в течение 15 минут, 85°C в течение 5 секунд. кол.ОТ-ПЦР проводили с использованием кДНК, взятой из мышиных тканей, для оценки уровней экспрессии фратаксина в многолуночном планшета на устройстве LightCycler480 (Roche Diagnostics). Реакционные смеси содержали общий объем 10 мкл, состоящий из 0,1 мМ каждого праймера, 10 нг кДНК и 5 мкл универсальной смеси TaqMan Universal Master Mix II, без UNG (Thermofisher Scientific). Праймер-зонд для детектирования человеческого фратаксина был предварительно сконструирован Bio-rad (dHsaCPE5031641, Bio-rad), а праймеры для гена «домашнего хозяйства» бета-2-микроглобулина (B2m) для образцов мышей были предварительно сконструированы IDT (Mm.PT.39a.22214835; IDT). Анализ осуществляли путем проведения термоциклов в следующих условиях: 50°С в течение 2 минут, 95°C в течение 10 минут и 40 циклов при 95°C в течение 15 секунд и 60°С в течение 1 минуты. Образцы анализировали с тремя повторностями для каждого представляющего интерес гена, а уровни трансгена определяли методом Ct (ΔΔCt). Результаты можно видеть на фигуре 7.
5. Электрофорез в ДСН-ПААГ и иммуноблоттинг
Белки экстрагировали из мышиной ткани путем гомогенизации в буфере для лизиса RIPA: 10 мМ Трис-HCl, рН 7,4, 140 мМ NaCl, 0,1% дезоксихолат натрия, 1% Тритона Х-100, 1% ДСН, 2 мМ этилендиаминтетрауксусной кислоты (EDTA), 25 мМ фторида натрия (NaF), 2,5 мМ NaVO3 и коктейля ингибиторов белка (Roche). Концентрацию белка определяли с помощью анализа на белок DC-BioRad (BioRad). Общий экстракт белка смешивали с 4× буфером для загрузки образцов белка (Li-Cor Biosciences), содержащим 1 мМ DTT, и разделяли с помощью электрофореза в 15% акриламидном геле при постоянной силе тока 20 мА до переноса на PVDF-мембраны. В качестве «первых» антител использовали анти-FXN антитело PAC 2518 (любезно предоставленное Dr. Grazia Isaya, Mayo Clinic, Rochester MN), анти-FXN антитело (1G2, Merck Millipore), антитело против метки HA (клон 16B12, MMS-101P, Covance) и антитело против бета-актина (AC15, Sigma). «Вторые» антитела, конъюгированные с инфракрасным красителем, представляли собой антимышиные IRDye-800CW и антикроличьи IRDye 700CW (Li-Cor Biosciences), и иммунореактивность определяли с использованием программы-анализатора Odyssey v2.1 (Li-Cor Biosciences). Результаты можно видеть на фигуре 8.
6. Поджимание лап
Было показано, что поджимание задних конечностей является показателем прогрессирования заболевания у ряда мышей с моделями нейродегенерации. Каждую мышь поднимали за хвост над любыми окружающими предметами. Положение задних конечностей наблюдалось в течение 10 секунд и оценивалось следующим образом: если задние конечности были последовательно вытянуты наружу от брюшины, то присваивался балл 0. Если одна задняя конечность была притянута к животу с более, чем 50% запаздыванием по времени, то присваивался балл 1. Если обе задние конечности были частично притянуты к животу с более, чем 50% запаздыванием по времени, то присваивался балл 2. Если задние конечности были полностью прижаты и касались живота с более, чем 50% запаздыванием по времени, то присваивался балл 3. Рефлекс поджимания конечностей оценивали каждые 2 месяца у мышей в возрасте от 4 месяцев. Результаты можно видеть на фигуре 9.
7. Электрофизиология
Амплитуду (мкВ) и скорость нервной проводимости (м/с) измеряли в каудальном нерве хвоста мыши. У животных под ингаляционной анестезией (2% изофлураном) иглы с электродом-раздражителем располагались подкожно в 4 различных точках стимуляции (1 см, 2 см, 3 см, 4 см от кончика хвоста), в то время как острие иглы регистрации было зафиксировано на расстоянии 6 см от кончика хвоста. Значения регистрировали с помощью 2-канального EMG/PE N-EP на аппарате Medelec Synergy (Viasys, EUA). Во время электрофизиологических испытаний температуру кожи животных поддерживали выше 32°C. Мышей оценивали каждые 2 месяца с 4-месячного возраста. Результаты можно видеть на фигуре 10.
8. Результаты
Авторы предполагают, что лечение AAV-вектором согласно изобретению будет эффективным для FRDA, поскольку AAV может экспрессироваться в физиологически релевантных тканях, и на уровнях, демонстрирующих процесс, сходный с процессом для эндогенного белка, и может значительно улучшить электрофизиологические свойства пораженных нейронов и неврологические симптомы у мышей с моделью FRDA (YG8R). На фигуре 6 показано, что ген люциферазы в векторе AAV9 под контрлем тех же самых регуляторных элементов, как и FXN в векторе AAV9-hPGK1-FXN, при интратекальном введении, экспрессируется по всему спинному мозгу, головному мозгу, а также в периферических тканях. Кроме того, экспрессия люциферазы под контролем того же самого вектора и регуляторных элементов остается постоянной в течение по меньшей мере приблизительно 7 месяцев после инъекции (данные не приводятся). Поскольку пониженные уровни фратаксина лежат в основе большинства, если не всех, симптомов FRDA, и отдельные индивидуумы-носители с мутацией, экспрессирующие уровни фратаксина ≥25%, не имеют симптомов FRDA, то небольшое увеличение экспрессии FXN должно предотвращать или в значительной степени ослаблять симптомы у пациентов с FRDA. Однако, очень высокие уровни экспрессии FXN или любого другого белка могут индуцировать клеточные эффекты и, в конечном итоге, ослаблять защитные эффекты или даже обострять симптомы. Следовательно, особый интерес представляет разработка FXN-экспрессирующего вектора, который дает уровни, почти такие же, как и уровни эндогенного белка FXN. Следовательно, авторами был разработан вектор (AAV9-hPGK1-FXN), который позволяет экспрессировать FXN под контролем метаболически регулируемого промотора на физиологически значимых уровнях, и доставлять его интратекально 10-недельным гемизиготным мышам YG8R (Tg/-). Более высокие уровни мРНК FXN были детектированы у гемизиготных мышей YG8R (Tg/-) через 3,5 месяца после интратекальной инъекции вектора AAV9-PGK1-FXN по сравнению с неинъецированными мышами, как в печени, так и в грудном отделе спинного мозга (в 0,5 и 2 раза соответственно; Фигура 7). Эти результаты были такими же или ниже, чем у гомозиготных мышей YG8R, у которых уровни в спинном мозге были выше в 0,5 и 3 раза, соответственно, по сравнению с гемизиготными мышами YG8R. Это указывает на то, что FXN-кодирующий вектор, инъецированный в интратекальную область L3-L4, поглощался и экспрессировался в нейронах грудного отдела спинного мозга, либо посредством обратного транспорта, либо посредством диффузии через спинномозговую жидкость. Авторы также отметили, что интратекально вводимый вектор AAV9 также способен преодолевать гематоэнцефалический барьер и достигать периферических тканей, таких как печень, сердце и т.п. (Фигура 6), и способен экспрессировать мРНК hFXN после инъекции этого вектора (Фигура 7).
Также были анализированы уровни экспрессии белка FXN из полученного рекомбинантного вектора. Хотя используемое антитело обладает более высокой аффинностью к человеческому белку FXN, на что указывает белковая структура, наблюдаемая после инкубирования блота с «первым» анти-FXN антителом в течение 1 часа по сравнению с инкубированием в течение ночи, однако, на фигуре 8 показано, что уровни белка FXN, экспрессируемого из вектора rAAV9-PGK1-FXN, по всей видимости, похожи на уровни мышиного FXN у мышей дикого типа (фигура 8А, дорожки 3 и 4 и фигура 8В, вторая панель, дорожка 3 по сравнению с дорожкой 4 и дорожка 5 по сравнению с дорожками 6 и 7). Примечательно то, что авторы смогли обнаружить рекомбинантный белок FXN в области двигательных нейронв коры головного мозга (C1) у мышей (фиг. 8B, дорожка 7). Это указывает на то, что доставка полученного авторами вектора путем интратекальной инъекции в поясничный отдел спинного мозга приводит к распределению вектора по областям ЦНС, относящимся к двигательной системе. Кроме того, рекомбинантный белок, по-видимому, процессируется по механизму, аналогичному механизму процессинга белка FXN дикого типа, причем, его промежуточная и зрелая формы являются наиболее распространенными процессированными формами.
Авторы также проанализировали фенотип гемизиготных мышей YG8R (Tg/-), обработанных вектором rAAV9-PGK1-FXN, по сравнению с фенотипом мышей, обработанных вектором rAAV9-null, с точки зрения неврологического рефлекса поджимания лап и электрофизиологических свойств каудального нерва. Было показано, что рефлекс поджимания наблюдается при некоторых нейродегенеративных расстройствах. Этот рефлекс, по-видимому, включает сенсорные, а также спинномозговые моторные пути, регулирующие движения передней и задней конечностей. В частности, было показано участие кортико-мозжечково-ретикулярных путей. Поэтому, чтобы определить влияние обработки вектора rAAV9-hPGK1-FXN на эти пути у YG8R-мышей, авторами была проведена количественная оценка рефлекса поджимания в различные периоды времени после интратекальной инъекции вектора (см. Методы). На фигуре 9 показано, что у мышей YG8R, обработанных векторами AAV9-null и AAV9-FXN, наблюдаются дефекты в поджимании лап уже в возрасте четырех месяцев (через 1,5 месяца после интратекальной инъекции). Однако, со временем, этот рефлекс поджимания лап нормализуется у AAV-FXN-обработанных мышей, и уже больше напоминает эффект у мышей дикого типа, даже в более зрелом возрасте свыше 10 месяцев (через 6,5 месяцев после инъекции). Это говорит о том, что аномалии в сенсорной и кортико-мозжечково-ретикулярной системах у мышей с моделью FRDA наблюдаются на ранних стадиях и в значительной степени снижаются или устраняются при обработке вектором AAV-FXN.
Для более точного определения влияния обработки на сенсорные нейроны, авторами была проведена оценка электрофизиологических свойств (амплитуды и скорости) каудального нерва у мышей YG8R, обработанных вектором AAV-FXN, по сравнению со свойствами у мышей, обработанных контрольным вестором AAV9-null. На фигуре 10 показано, что на всех расстояниях от кончика хвоста (1-4 см) не было отмечено каких либо значимых различий между мышами дикого типа, обработанным нулевым вектором, или гемизиготными мышами YG8R, обработанными нулевым вектором и вектором AAV-FXN в возрасте четырех месяцев (через 1,5 месяца после обработки). Однако, начиная с 6-месячного возраста, у мышей YG8R (Tg/-), обработанных вектором AAV9-null, наблюдалось значимое снижение амплитуды, а мыши, обработанные AAV9-FXN, были более похожи на мышей дикого типа даже в более позднем возрасте, а именно, 13 месяцев (через 9,5 месяцев после обработки). Никаких изменений в скорости не обнаруживалось (данные не приводятся). Эти данные указывают на то, что обработка вектором AAV-hPGK1-FXN позволяла сохранить функцию каудального нерва у мышей YG8R, возможно, благодаря предотвращению потери аксонов из пораженных нейронов.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> FUNDACIÓ INST D'INVESTIGACIÓ EN CIÈNCIES DE LA SALUT GERMANS TRIAS I
PUJOL
<120> Векторы для лечения атаксии Фридрейха
<130> 903 551
<160> 12
<170> BiSSAP 1.3.6
<210> 1
<211> 518
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Промотор PGK
<400> 1
gaattccggg gttggggttg cgccttttcc aaggcagccc tgggtctgcg cagggacgcg 60
gctgctctgg gcgtggttcc gggaaacgca gcggcgccga ccctgggtct cgcacattct 120
tcacgtccgt tcgcagcgtc acccggatct tcgccgctac ccttgtgggc cccccggcga 180
cgcttcctgc tccgccccta agtcgggaag gttccttgcg gttcgcggcg tgccggacgt 240
gacaaacgga agccgcacgt ctcactagta ccctcgcaga cggacagcgc cagggagcaa 300
tggcagcgcg ccgaccgcga tgggctgtgg ccaatagcgg ctgctcagca gggcgcgccg 360
agagcagcgg ccgggaaggg gcggtgcggg aggcggggtg tggggcggta gtgtgggccc 420
tgttcctgcc cgcgcggtgt tccgcattct gcaagcctcc ggagcgcacg tcggcagtcg 480
gctccctcgt tgaccgaatc accgacctct ctccccag 518
<210> 2
<211> 597
<212> ДНК
<213> Искусственная последовательность
<220>
<223> WPRE
<400> 2
tcgacaatca acctctggat tacaaaattt gtgaaagatt gactggtatt cttaactatg 60
ttgctccttt tacgctatgt ggatacgctg ctttaatgcc tttgtatcat gctattgctt 120
cccgtatggc tttcattttc tcctccttgt ataaatcctg gttgctgtct ctttatgagg 180
agttgtggcc cgttgtcagg caacgtggcg tggtgtgcac tgtgtttgct gacgcaaccc 240
ccactggttg gggcattgcc accacctgtc agctcctttc cgggactttc gctttccccc 300
tccctattgc cacggcggaa ctcatcgccg cctgccttgc ccgctgctgg acaggggctc 360
ggctgttggg cactgacaat tccgtggtgt tgtcggggaa gctgacgtcc tttccatggc 420
tgctcgcctg tgttgccacc tggattctgc gcgggacgtc cttctgctac gtcccttcgg 480
ccctcaatcc agcggacctt ccttcccgcg gcctgctgcc ggctctgcgg cctcttccgc 540
gtcttcgcct tcgccctcag acgagtcgga tctccctttg ggccgcctcc ccgcctg 597
<210> 3
<211> 631
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Фратаксин
<400> 3
atgtggactc tcgggcgccg cgcagtagcc ggcctcctgg cgtcacccag cccggcccag 60
gcccagaccc tcacccgggt cccgcggccg gcagagttgg ccccactctg cggccgccgt 120
ggcctgcgca ccgacatcga tgcgacctgc acgccccgcc gcgcaagttc gaaccagaga 180
ggtctcaacc agatttggaa tgtcaaaaag cagagtgtct atttgatgaa tttgaggaaa 240
tctggaactt tgggccaccc aggctctcta gatgagacca cctatgaaag actagcagag 300
gaaacgctgg actctttagc agagtttttt gaagaccttg cagacaagcc atacacgttt 360
gaggactatg atgtctcctt tgggagtggt gtcttaactg tcaaactggg tggagatcta 420
ggaacctatg tgatcaacaa gcagacgcca aacaagcaaa tctggctatc ttctccatcc 480
agtggaccta agcgttatga ctggactggg aaaaactggg tgtactccca cgacggcgtg 540
tccctccatg agctgctggc cgcagagctc actaaagcct taaaaaccaa actggacttg 600
tcttccttgg cctattccgg aaaagatgct t 631
<210> 4
<211> 1934
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Фратаксин, PGK и кластер WPRE
<400> 4
gaattccggg gttggggttg cgccttttcc aaggcagccc tgggtctgcg cagggacgcg 60
gctgctctgg gcgtggttcc gggaaacgca gcggcgccga ccctgggtct cgcacattct 120
tcacgtccgt tcgcagcgtc acccggatct tcgccgctac ccttgtgggc cccccggcga 180
cgcttcctgc tccgccccta agtcgggaag gttccttgcg gttcgcggcg tgccggacgt 240
gacaaacgga agccgcacgt ctcactagta ccctcgcaga cggacagcgc cagggagcaa 300
tggcagcgcg ccgaccgcga tgggctgtgg ccaatagcgg ctgctcagca gggcgcgccg 360
agagcagcgg ccgggaaggg gcggtgcggg aggcggggtg tggggcggta gtgtgggccc 420
tgttcctgcc cgcgcggtgt tccgcattct gcaagcctcc ggagcgcacg tcggcagtcg 480
gctccctcgt tgaccgaatc accgacctct ctccccagtg gctaactaga gaacccactg 540
cttactggct tatcgaaatt aatacgactc actataggga gacccaagct ggctagcgtt 600
taaacttaag cttggccgcc accatgtgga ctctcgggcg ccgcgcagta gccggcctcc 660
tggcgtcacc cagcccggcc caggcccaga ccctcacccg ggtcccgcgg ccggcagagt 720
tggccccact ctgcggccgc cgtggcctgc gcaccgacat cgatgcgacc tgcacgcccc 780
gccgcgcaag ttcgaaccag agaggtctca accagatttg gaatgtcaaa aagcagagtg 840
tctatttgat gaatttgagg aaatctggaa ctttgggcca cccaggctct ctagatgaga 900
ccacctatga aagactagca gaggaaacgc tggactcttt agcagagttt tttgaagacc 960
ttgcagacaa gccatacacg tttgaggact atgatgtctc ctttgggagt ggtgtcttaa 1020
ctgtcaaact gggtggagat ctaggaacct atgtgatcaa caagcagacg ccaaacaagc 1080
aaatctggct atcttctcca tccagtggac ctaagcgtta tgactggact gggaaaaact 1140
gggtgtactc ccacgacggc gtgtccctcc atgagctgct ggccgcagag ctcactaaag 1200
ccttaaaaac caaactggac ttgtcttcct tggcctattc cggaaaagat gctttgccca 1260
cctagggatc ggatccccgg gtaccgagct cgaattctgc agatatccag cacactttgc 1320
ctttctctcc acaggtgtcg acaatcaacc tctggattac aaaatttgtg aaagattgac 1380
tggtattctt aactatgttg ctccttttac gctatgtgga tacgctgctt taatgccttt 1440
gtatcatgct attgcttccc gtatggcttt cattttctcc tccttgtata aatcctggtt 1500
gctgtctctt tatgaggagt tgtggcccgt tgtcaggcaa cgtggcgtgg tgtgcactgt 1560
gtttgctgac gcaaccccca ctggttgggg cattgccacc acctgtcagc tcctttccgg 1620
gactttcgct ttccccctcc ctattgccac ggcggaactc atcgccgcct gccttgcccg 1680
ctgctggaca ggggctcggc tgttgggcac tgacaattcc gtggtgttgt cggggaagct 1740
gacgtccttt ccatggctgc tcgcctgtgt tgccacctgg attctgcgcg ggacgtcctt 1800
ctgctacgtc ccttcggccc tcaatccagc ggaccttcct tcccgcggcc tgctgccggc 1860
tctgcggcct cttccgcgtc ttcgccttcg ccctcagacg agtcggatct ccctttgggc 1920
cgcctccccg cctg 1934
<210> 5
<211> 2458
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ITR фратаксина, PGK, полиA-ITR WPRE
<400> 5
ctggcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc gggcgacctt 60
tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca actccatcac 120
taggggttcc tgaattccgg ggttggggtt gcgccttttc caaggcagcc ctgggtctgc 180
gcagggacgc ggctgctctg ggcgtggttc cgggaaacgc agcggcgccg accctgggtc 240
tcgcacattc ttcacgtccg ttcgcagcgt cacccggatc ttcgccgcta cccttgtggg 300
ccccccggcg acgcttcctg ctccgcccct aagtcgggaa ggttccttgc ggttcgcggc 360
gtgccggacg tgacaaacgg aagccgcacg tctcactagt accctcgcag acggacagcg 420
ccagggagca atggcagcgc gccgaccgcg atgggctgtg gccaatagcg gctgctcagc 480
agggcgcgcc gagagcagcg gccgggaagg ggcggtgcgg gaggcggggt gtggggcggt 540
agtgtgggcc ctgttcctgc ccgcgcggtg ttccgcattc tgcaagcctc cggagcgcac 600
gtcggcagtc ggctccctcg ttgaccgaat caccgacctc tctccccagt ggctaactag 660
agaacccact gcttactggc ttatcgaaat taatacgact cactataggg agacccaagc 720
tggctagcgt ttaaacttaa gcttggccgc caccatgtgg actctcgggc gccgcgcagt 780
agccggcctc ctggcgtcac ccagcccggc ccaggcccag accctcaccc gggtcccgcg 840
gccggcagag ttggccccac tctgcggccg ccgtggcctg cgcaccgaca tcgatgcgac 900
ctgcacgccc cgccgcgcaa gttcgaacca gagaggtctc aaccagattt ggaatgtcaa 960
aaagcagagt gtctatttga tgaatttgag gaaatctgga actttgggcc acccaggctc 1020
tctagatgag accacctatg aaagactagc agaggaaacg ctggactctt tagcagagtt 1080
ttttgaagac cttgcagaca agccatacac gtttgaggac tatgatgtct cctttgggag 1140
tggtgtctta actgtcaaac tgggtggaga tctaggaacc tatgtgatca acaagcagac 1200
gccaaacaag caaatctggc tatcttctcc atccagtgga cctaagcgtt atgactggac 1260
tgggaaaaac tgggtgtact cccacgacgg cgtgtccctc catgagctgc tggccgcaga 1320
gctcactaaa gccttaaaaa ccaaactgga cttgtcttcc ttggcctatt ccggaaaaga 1380
tgctttgccc acctagggat cggatccccg ggtaccgagc tcgaattctg cagatatcca 1440
gcacactttg cctttctctc cacaggtgtc gacaatcaac ctctggatta caaaatttgt 1500
gaaagattga ctggtattct taactatgtt gctcctttta cgctatgtgg atacgctgct 1560
ttaatgcctt tgtatcatgc tattgcttcc cgtatggctt tcattttctc ctccttgtat 1620
aaatcctggt tgctgtctct ttatgaggag ttgtggcccg ttgtcaggca acgtggcgtg 1680
gtgtgcactg tgtttgctga cgcaaccccc actggttggg gcattgccac cacctgtcag 1740
ctcctttccg ggactttcgc tttccccctc cctattgcca cggcggaact catcgccgcc 1800
tgccttgccc gctgctggac aggggctcgg ctgttgggca ctgacaattc cgtggtgttg 1860
tcggggaagc tgacgtcctt tccatggctg ctcgcctgtg ttgccacctg gattctgcgc 1920
gggacgtcct tctgctacgt cccttcggcc ctcaatccag cggaccttcc ttcccgcggc 1980
ctgctgccgg ctctgcggcc tcttccgcgt cttcgccttc gccctcagac gagtcggatc 2040
tccctttggg ccgcctcccc gcctggaatt cgagctcggt acgatcagct gatcagcctc 2100
gactgtgcct tctagttgcc agccatctgt tgtttgcccc tcccccgtgc cttccttgac 2160
cctggaaggt gccactccca ctgtcctttc ctaataaaat gaggaaattg catcgcattg 2220
tctgagtagg tgtcattcta ttctgggggg tggggtgggg caggacagca agggggagga 2280
ttgggaagac aatagcaggc atgctgggga tgcggtgggc tctatggctg gcgcgctcgc 2340
tcgctcactg aggccgcccg ggcaaagccc gggcgtcggg cgacctttgg tcgcccggcc 2400
tcagtgagcg agcgagcgcg cagagaggga gtggccaact ccatcactag gggttcct 2458
<210> 6
<211> 105
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 6
tggctaacta gagaacccac tgcttactgg cttatcgaaa ttaatacgac tcactatagg 60
gagacccaag ctggctagcg tttaaactta agcttggccg ccacc 105
<210> 7
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hFXN_Tg_FW
<400> 7
gaacttcaaa ttagttcccc tttcttc 27
<210> 8
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> hFXN_Tg_RV
<400> 8
cacagccatt ctttgggttt c 21
<210> 9
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IC_FW
<400> 9
cacgtgggct ccagcatt 18
<210> 10
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> IC_RV
<400> 10
tcaccagtca tttctgcctt tg 22
<210> 11
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> GAA-F
<400> 11
gggattggtt gccagtgctt aaaagttag 29
<210> 12
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> GAA-R
<400> 12
gatctaagga ccatcatggc cacacttgcc 30
<---



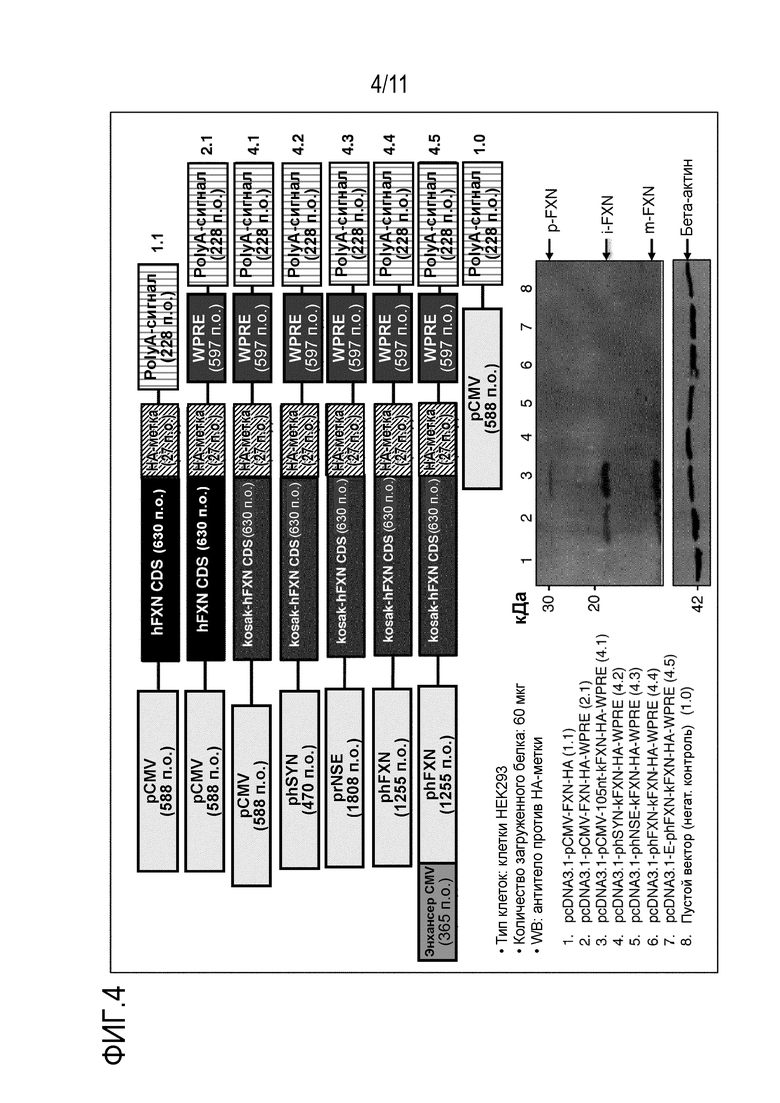

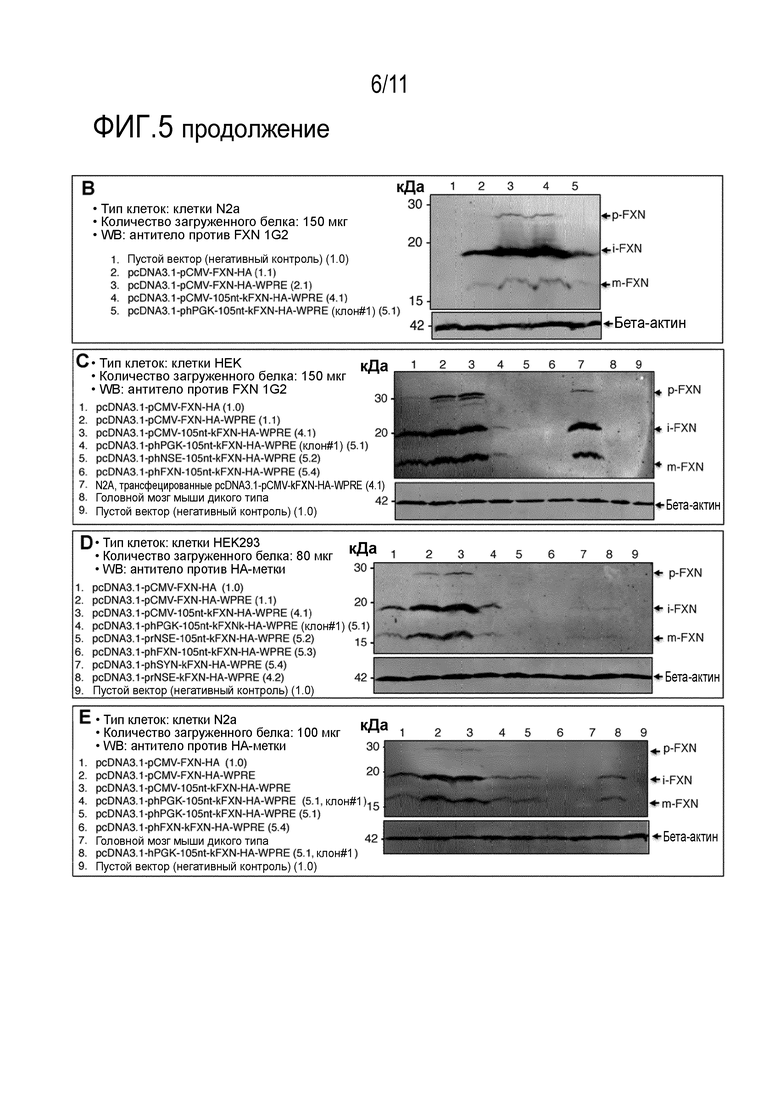

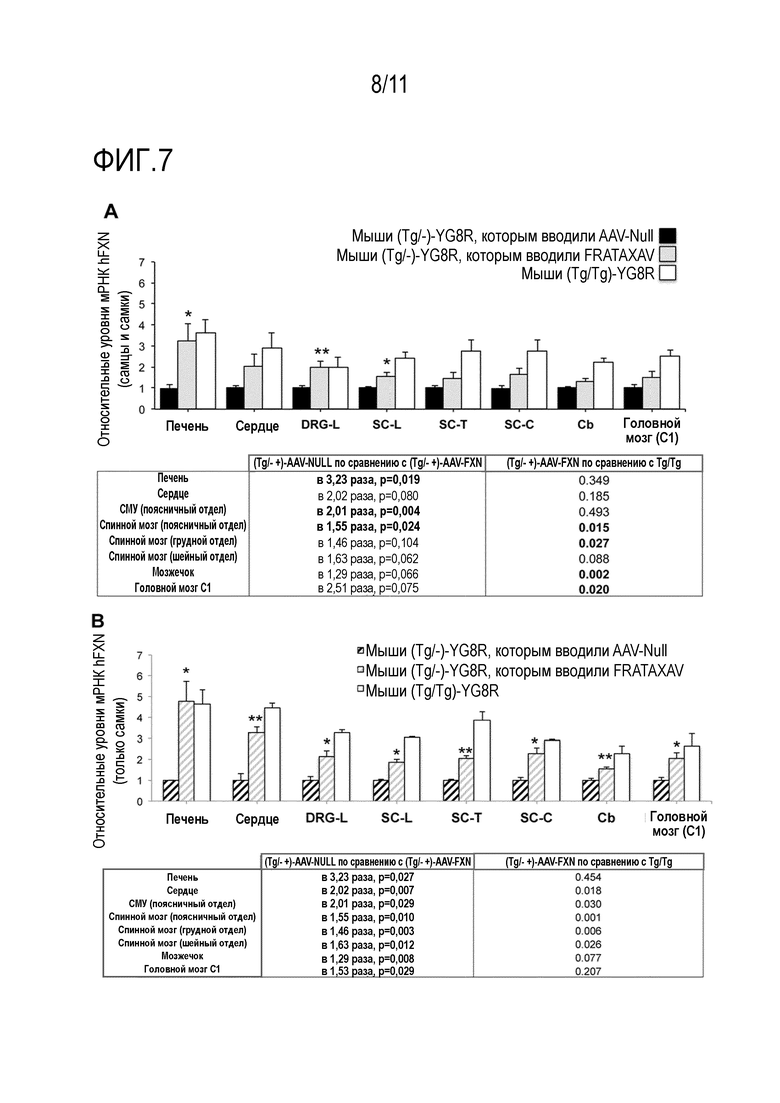



Настоящее изобретение относится к биотехнологии, а именно к аденоассоциированному вирусному вектору (AAV), используемому в генотерапии для лечения атаксии Фридрейха. Аденоассоциированный вирусный вектор (AAV) экспрессии содержит нуклеиновую кислоту, которая включает (i) последовательность нуклеиновой кислоты, кодирующую фратаксин; (ii) промотор фосфоглицерат-киназы (PGK), состоящий из SEQ ID NО:1; и (iii) посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE). (ii) и (iii) функционально присоединены к последовательности (i) и регулируют ее экспрессию. Между (i) и (ii) присутствует функционально присоединенный линкер. Указанный линкер состоит из SEQ ID NO: 6, а AAV-вектор представляет собой AAV-вектор серотипа 9. Представлен аденоассоциированный вирусный вектор (AAV) клонирования. Представлена фармацевтическая композиция для лечения атаксии Фридрейха, содержащая эффективное количество AAV-вектора, применение AAV-вектора в качестве лекарственного средства и в лечении атаксии Фридрейха. Конструкция AAV9 вектора способна обеспечивать промежуточные уровни фратаксина in vitro и in vivo, которые более сопоставимы с эндогенными уровнями. 5 н. и 7 з.п. ф-лы, 3 табл., 11 ил., 4 пр.
1. Аденоассоциированный вирусный вектор (AAV) экспрессии, пригодный для лечения атаксии Фридрейха, содержащий нуклеиновую кислоту, где нуклеиновая кислота включает:
(i) последовательность нуклеиновой кислоты, кодирующую фратаксин;
(ii) промотор фосфоглицерат-киназы (PGK), состоящий из SEQ ID NО:1; и
(iii) посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE),
где (ii) и (iii) функционально присоединены к последовательности (i) и регулируют ее экспрессию, где между (i) и (ii) присутствует функционально присоединенный линкер, и где указанный линкер состоит из SEQ ID NO: 6, и где AAV-вектор представляет собой AAV-вектор серотипа 9.
2. Вектор по п.1, где последовательность нуклеиновой кислоты, кодирующая фратаксин, содержит SEQ ID NO: 3 или последовательность, которая по меньшей мере на 90% идентична последовательности SEQ ID NO: 3 и представляет собой функциональный вариант фратаксина.
3. Вектор по п.1, где последовательность нуклеиновой кислоты, кодирующая фратаксин, состоит из SEQ ID NO: 3 или последовательности, которая по меньшей мере на 98% идентична последовательности SEQ ID NO: 3 и представляет собой функциональный вариант фратаксина.
4. Вектор по любому из пп. 1-3, где WPRE состоит из SEQ ID NO: 2 или последовательности, которая по меньшей мере на 90% идентична последовательности SEQ ID NO: 2.
5. Вектор по любому из пп. 1-3, где WPRE состоит из SEQ ID NO: 2 или последовательности, которая по меньшей мере на 99% идентична последовательности SEQ ID NO: 2.
6. Вектор по п.1, где последовательность нуклеиновой кислоты, кодирующая фратаксин, состоит из SEQ ID NО: 3, а WPRE состоит из SEQ ID NО: 2 или последовательности, которая по меньшей мере на 90% идентична SEQ ID NO: 2.
7. Вектор по п.1, где последовательность нуклеиновой кислоты, кодирующая фратаксин, состоит из SEQ ID NО: 3, а WPRE состоит из SEQ ID NО: 2 или последовательности, которая по меньшей мере на 99% идентична SEQ ID NO: 2.
8. Вектор по п.1, где последовательность нуклеиновой кислоты, которая включает (i), (ii), (iii) и линкер, состоит из SEQ ID NO: 4 и где AAV-вектор представляет собой AAV-вектор серотипа 9.
9. Вектор клонирования для переноса, который включает нуклеиновую кислоту, содержащую:
(i) последовательность нуклеиновой кислоты, кодирующую фратаксин;
(ii) промотор PGK, состоящий из SEQ ID NО:1, и
(iii) WPRE,
где (ii) и (iii) функционально присоединены к последовательности (i) и регулируют ее экспрессию, где между (i) и (ii) присутствует функционально присоединенный линкер, и где указанный линкер состоит из SEQ ID NO: 6, и где вектор для переноса также содержит дополнительные элементы нуклеиновой кислоты для стимуляции интеграции или транспозиции вектора для переноса в AAV-вектор серотипа 9.
10. Фармацевтическая композиция для лечения атаксии Фридрейха, содержащая эффективное количество AAV-вектора по любому из пп. 1-8 и фармацевтически приемлемый носитель или разбавитель.
11. Применение AAV-вектора по любому из пп. 1-8 в качестве лекарственного средства.
12. Применение AAV-вектора по любому из пп. 1-8 в лечении атаксии Фридрейха.
| Токарный резец | 1924 |
|
SU2016A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| MATTAR C N et al, Systemic gene delivery following intravenous administration of AAV9 to fetal and neonatal mice and late-gestation nonhuman primates, The Faseb journal, vol.29, no 9, 01.09.2015, p 3876-3888, DOI: 10.1096/fj.14-269092, abstract | |||
| Устройство для анализа звукозаписей взрывов при сейсмической разведке | 1938 |
|
SU60581A1 |
| ВЕКТОРЫ И ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2011 |
|
RU2588667C2 |

