Область техники
Настоящее изобретение относится к фармацевтической композиции для профилактики или лечения аритмии сердца. Конкретно, настоящее изобретение относится к фармацевтической композиции, включающий в качестве активного ингредиента белок CCN5 или кодирующую его нуклеотидную последовательность.
Предпосылки изобретения
Аритмия сердца представляет собой заболевание сердца, вызванное нарушениями ритма сердца и эффективных сокращений сердца, возникающими из-за проблем с электрическими импульсами в сердце. По определению сердечная аритмия классифицируется на предсердную аритмию и желудочковую аритмию в зависимости от места ее возникновения. В частности, предсердная аритмия классифицируется на фибрилляцию предсердий, предсердную тахикардию и дисфункцию синусового узла; а желудочковая аритмия классифицируется на желудочковую тахикардию и фибрилляцию желудочков (Swarminathan PD, et al. Circ Res 2012; 110:1661-1677).
Аритмия сердца вызывает значительную заболеваемость и смертность, особенно в развитых странах, а остановка сердца является одной из основных причин смерти в развитых странах (Mozaffarian D, et al. Circulation 2015; 131: e29-e322). В частности, желудочковая аритмия является основной причиной внезапной сердечной смерти, а другие факторы риска сердечных заболеваний ускоряют и усиливают желудочковую аритмию (Roberts-Thomson KC, et al. Nat Rev Cardiol 2011; 8: 311-321). Кроме того, в классификации аритмии сердца фибрилляция предсердий является наиболее распространенной аритмией с возрастающей заболеваемостью и значительно увеличивает частоту таких заболеваний, как сердечная аритмия и инсульт (Andrada D, et al. Circ Res 2014; 114; 1453-1458).
Виды терапии для лечения аритмии сердца включают антиаритмические препараты, катетерную аблацию, имплантируемый кардиовертер-дефибриллятор для лечения желудочковой аритмии. Однако эти виды терапии обладают ограниченным терапевтическим действием. Блокада ионных каналов, которая является основным механизмом методов лечения аритмии, имеет ограничения при длительном лечении и профилактике аритмии. В клинических исследованиях, связанных с подавлением сердечной аритмии, в процессе лечения экстрасистолии было показано, что антиаритмические препараты увеличивают смертность от сердечно-сосудистых заболеваний у пациентов с инфарктом миокарда. Общим побочным эффектом используемых в настоящее время антиаритмических средств является риск развития аритмии (Camm J, Int Cardiol 2012; 155: 363-371).
Между тем, исследования показали, что CaMKII (Ca2+/кальмодулин-зависимая протеинкиназа II) играет центральную роль в электрических аспектах сердечной аритмии. Повышенная активность CaMKII приводит к гиперактивации ионных каналов, дефектам внутриклеточного гомеостаза Ca2+ и повреждению тканей, способствуя тем самым аритмии. Таким образом, ингибирование активности CaMKII может быть использовано как эффективный способ лечения аритмии.
Для создания эффективного средства для лечения сердечной аритмии существует необходимость разработки такого средства для лечения посредством понимания его патологического механизма действия и обнаружения новых терапевтических мишеней.
Соответственно, настоящее изобретение предназначено для решения задачи по разработке средства для лечения, нацеленного одновременно на CaMKII, которая является ключевой мишенью электрической дисфункции, и на фиброз, который является основной причиной структурной дисфункции, в качестве средства для лечения при начале и протекании аритмии и при продолжительном ухудшении симптомов аритмии.
Описание изобретения
Техническая задача
В связи с этим авторы настоящего изобретения провели исследование для разработки эффективного средства для лечения аритмии сердца, и в результате установили, что фармацевтическая композиция, которая содержит ген, кодирующий белок CCN5 или его фрагмент, ингибирует патологическую активность CaMKII и ингибирует активность миофибробластов, тем самым разработав настоящее изобретение. Кроме того, авторы настоящего изобретения установили, что фармацевтическая композиция, которая содержит ген, кодирующий белок CCN5 и белок SERCA2a, проявляет синергетический терапевтический эффект в отношении электрической дисфункции в экспериментах с использованием модели сердечной аритмии на животных, тем самым разработав настоящее изобретение.
Решение задачи
В одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В другом аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В еще одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения субъекту фармацевтической композиции по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая белок CCN5 в качестве активного ингредиента.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения белка CCN5 субъекту.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения субъекту генетической конструкции, содержащей нуклеотидную последовательность, кодирующую белок CCN5.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения субъекту вектора экспрессии, несущего нуклеотидную последовательность, кодирующую белок CCN5.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения субъекту рекомбинантного вируса, содержащего нуклеотидную последовательность, кодирующую белок CCN5.
Положительные эффекты изобретения
Фармацевтическая композиция для профилактики или лечения аритмии сердца по настоящему изобретению подавляет патологическую активность CaMKII, которая вызывает электрическую дисфункцию сердца, основную причину предсердной аритмии и желудочковой аритмии, так что электрические функции сердца восстанавливаются, и ингибирует активность миофибробластов, которые вызывают структурную дисфункцию. Следовательно, фармацевтическая композиция по настоящему изобретению может быть эффективно использована для профилактики или лечения аритмии сердца.
Краткое описание фигур
На фиг. 1 показано схематическое изображение структуры вектора pTR-CMV-CCN5.
На фиг. 2 показано схематическое изображение структуры вектора pTR-CMV-SERCA2a-P2A-CCN5.
На фиг. 3a показано графическое отображение концептуальной модели экспериментов на животных с использованием мышей дикого типа или мышей CCN5 TG для определения подавляющего действия белка CCN5 на фиброз сердца.
На фиг. 3b показаны фотографии, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и окрашивания тканей предсердий трихромом по Массону.
На фиг. 3c показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий, окрашивания тканей предсердий трихромом по Массону и количественной оценки степени фиброза окрашенных тканей предсердия (*: p <0,05; **: p <0,01).
На фиг. 3d показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК α-SMA в тканях предсердий (**: p <0,01).
На фиг. 3e показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК Collagen I в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 3f показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК TGF-β1 в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 3g показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК IL-1β в тканях предсердий (*: p <0,05).
На фиг. 3h показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК RANTES в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 3i показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК F4/80 в тканях предсердий (**: p <0,01).
На фиг. 3j показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них тканей предсердий и определения посредством qRT-PCR уровня экспрессии мРНК MCP-1 в тканях предсердий (**: p <0,01).
На фиг. 4a показано графическое отображение концептуальной модели экспериментов на животных с использованием мышей дикого типа или мышей CCN5 TG для определения подавляющего действия белка CCN5 на фибрилляцию предсердий.
На фиг. 4b показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них сердца, применения к нему электрической стимуляции для индукции фибрилляции предсердий и получения электрокардиограммы.
На фиг. 4c показаны результаты, полученные путем введения ангиотензина II мышам дикого типа или мышам CCN5 TG в течение 14 дней, извлечения из них сердца и определения частоты фибрилляции предсердий, индуцируемой при воздействии на сердце электрической стимуляции для индукции фибрилляции предсердий.
На фиг. 5a показано графическое отображение концептуальной модели экспериментов с использованием клеток HL-1 для определения подавляющего действия белка CCN5 на фибрилляцию предсердий.
На фиг. 5b показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем определения посредством вестерн-блоттинга экспрессии белков, p-CaMKII (Thr286), CaMKII, pRyR2 (Ser2808), pRyR2 (Ser2814), RyR2, калсеквестрина 2, Na+/Ca+ обменника 2 (NCX2) и GAPDH в клетках HL-1.
На фиг. 5c показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов, а затем вычисление значений p-CaMKII (Thr286)/CaMKII в клетках HL-1 (**: p <0,01).
На фиг. 5d показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений pRyR2 (Ser2808)/RyR2 в клетках HL-1 (**: p <0,01).
На фиг. 5e показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений pRyR2 (Ser2814)/RyR2 в клетках HL-1 (*: p <0,05; **: p <0,01).
На фиг. 5f показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений калсеквестрин2/GAPDH в клетках HL-1 (*: p <0,05; **: p <0,01).
На фиг. 5g показаны результаты, полученные путем обработки клеток HL-1 ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений NCX2/GAPDH в клетках HL-1 (**: p <0,01).
На фиг. 6a показано графическое отображение концептуальной модели экспериментов с использованием фибробластов предсердий крысы для определения подавляющего действия белка CCN5 на фибрилляцию предсердий.
На фиг. 6b показаны фотографии, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем окрашивания культивированных фибробластов предсердий крысы флуоресцентной иммунохимией.
На фиг. 6c показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем определения посредством вестерн-блоттинга экспрессии белков, α-SMA, Collagen I, TGF-β1 и α-тубулина в фибробластах предсердий.
На фиг. 6d показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений α-SMA/α-тубулин в фибробластах предсердий (*: p <0,05; **: p <0,01).
На фиг. 6e показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений Collagen I/α-тубулин в фибробластах предсердий (*: p <0,05; **: p <0,01).
На фиг. 6f показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем вычисления значений TGF-β1/α-тубулин в фибробластах предсердий (*: p <0,05; **: p <0,01).
На фиг. 6g показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем определения посредством qRT-PCR уровня экспрессии мРНК α-SMA в тканях предсердий (*: p <0,05).
На фиг. 6h показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем определения посредством qRT-PCR уровня экспрессии мРНК Collagen I в тканях предсердий (**: p <0,01).
На фиг. 6i показаны результаты, полученные путем обработки фибробластов предсердий крысы ангиотензином II и одновременно CM-Con или CM-CCN5, проведения культивирования в течение 48 часов и затем определения посредством qRT-PCR уровня экспрессии мРНК TGF-β1 в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 7a показано графическое отображение концептуальной модели экспериментов на животных с использованием мышей с индуцированным фиброзом предсердий для определения терапевтического эффекта белка AAV-CCN5 на фиброз предсердий.
На фиг. 7b показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции, и определения экспрессии белка CCN5 посредством вестерн-блоттинга.
На фиг. 7c показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения уровней экспрессии белка и мРНК CCN5.
На фиг. 7d показаны фотографии, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции, и окрашивания тканей предсердий трихромом по Массону.
На фиг. 7e показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции, окрашивания тканей предсердий трихромом по Массону и количественная оценка фиброза в окрашенных тканях предсердия (*: p <0,05).
На фиг. 7f показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений α-SMA/18s рРНК в тканях предсердий (*: p <0,05).
На фиг. 7g показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений Collagen I/18s рРНК в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 7h показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений TGF-β1/18s рРНК в тканях предсердий (**: p <0,01).
На фиг. 7i показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений IL-1β/18s рРНК в тканях предсердий (*: p <0,05).
На фиг. 7j показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений RANTES/18s рРНК в тканях предсердий (**: p <0,01).
На фиг. 7k показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений F4/80/18s рРНК в тканях предсердий (*: p <0,05; **: p <0,01).
На фиг. 7l показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5, извлечения из них тканей предсердий через 4 недели после инъекции и определения посредством qRT-PCR значений MCP-1/18s рРНК в тканях предсердий (**: p <0,01).
На фиг. 8a показано графическое отображение концептуальной модели экспериментов на животных c использованием мышей с индуцированным фиброзом предсердий для определения подавляющего фиброз предсердий действия белка AAV-CCN5.
На фиг. 8b показаны данные электрокардиограммы, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и получения электрокардиограммы через 4 недели после инъекции.
На фиг. 8c показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и наблюдения частоты возникновения аритмии с помощью электрической стимуляции через 4 недели после инъекции.
На фиг. 8d показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и наблюдения интенсивности электрической стимуляции, необходимой для индукции аритмии через 4 недели после инъекции.
На фиг. 8e показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и определения потенциала действия Ca2+ через 4 недели после инъекции.
На фиг. 8f показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и определения продолжительности потенциала действия 50 (APD50) и продолжительности потенциала действия 75 (APD75) через 4 недели после инъекции.
На фиг. 8g показаны результаты, полученные путем введения ангиотензина II мышам дикого типа в течение 14 дней, инъецирования им AAV-Control или AAV-CCN5 и определения скорости деполяризации через 4 недели после инъекции (*: p <0,05).
На фиг. 9a показаны фракция укорочения и изменения в весе тела в течение 6 недель у мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 9b показано оптическое картирование, полученное применением электрической стимуляции в 10 Гц к правому желудочку (RV) мышей дикого типа и мышей с индуцированной желудочковой аритмией и фотографирования.
На фиг. 9c показано оптическое картирование, полученное применением электрической стимуляции 20 Гц к правому желудочку (RV) мышей дикого типа и мышей с индуцированной желудочковой аритмией и фотографирования.
На фиг. 9d показаны результаты, полученные путем определения потенциала действия Ca2+ у мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 9e показаны результаты, полученные путем определения продолжительности потенциала действия 50 (APD50) и продолжительности потенциала действия 75 (APD75) у мышей дикого типа и мышей с индуцированной желудочковой аритмией (*: p <0,05).
На фиг. 9f показаны результаты, полученные путем определения дисперсии продолжительности потенциала действия у мышей дикого типа и мышей с индуцированной желудочковой аритмией (*: p <0,05).
На фиг. 9g показаны скорость проводимости и скорость деполяризации у мышей дикого типа и мышей с индуцированной желудочковой аритмией (***: p <0,001).
На фиг. 10a показаны изменения в потенциале действия Ca2+ в зависимости от обработки ISO у мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 10b показаны изменения в картине реполяризации Ca2+ в зависимости от обработки ISO в правом желудочке мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 10c показаны изменения в продолжительности потенциала действия 75 (APD75) в зависимости от обработки ISO у мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 10d показано Ca2+ оптическое картирование, полученное после обработки с помощью ISO мышей дикого типа и мышей с индуцированной желудочковой аритмией.
На фиг. 10e показаны результаты, полученные после обработки с помощью ISO мышей дикого типа и мышей с индуцированной желудочковой аритмией и затем определения продолжительности потенциала действия 75 (APD75) (*: p <0,05).
На фиг. 10f показаны изменения в скорости деполяризации в зависимости от обработки ISO у мышей дикого типа и мышей с индуцированной желудочковой аритмией (***: p <0,001).
На фиг. 11a показана фракция укорочения в течение 6 недель у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 11b показаны изменения толщины стенки левого желудочка в течение 6 недель у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 11c показано Ca2+ оптическое картирование для мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных ISO и AAV9-CCN5.
На фиг. 11d показано Ca2+ оптическое картирование для мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных ISO и AAV9-SERCA2a-P2A-CCN5.
На фиг. 11e показаны изменения в потенциале действия Ca2+ в зависимости от обработки ISO у мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 11f показана картина реполяризации Ca2+ в левом желудочке мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 11g показаны изменения в продолжительности потенциала действия 75 (APD75) в зависимости от обработки ISO у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5 (*: p <0,05; **: p <0,01; ***: p <0,001).
На фиг. 11h показана скорость проводимости в зависимости от обработки ISO у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5 (*: p <0,05; ***: p <0,001).
На фиг. 11i показана скорость деполяризации в зависимости от обработки ISO у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5 (**: p <0,01; ***: p <0,001).
На фиг. 12a показаны изменения в электрокардиограмме, наблюдаемые в случае, когда электрическая стимуляция применялась к мышам дикого типа, мышам с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышам с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышам с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 12b показана интенсивность электрической стимуляции, требуемая для индукции аритмии у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 12c показана частота аритмии с электрической стимуляцией у мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
На фиг. 13a показаны фотографии, полученные путем извлечения сердца из мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5, и окрашивания сердца трихромом по Массону.
На фиг. 13b показаны результаты, полученные путем определения посредством вестерн-блоттинга экспрессии белков, Nav 1.5, FAP, коннексин 43, MsrA, Kir2.1, тубулина, SERCA2a, Smad7 и CCN5 в тканях сердца мышей дикого типа, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-Control, мышей с индуцированной желудочковой аритмией, инъецированных AAV9-CCN5, и мышей с индуцированной желудочковой аритмией, инъецированных AAV9-SERCA2a-P2A-CCN5.
Лучший способ осуществления изобретения
Далее настоящее изобретение описано подробно.
В одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
Как используется в настоящем документе, термин «белок CCN5» относится к матрицеллюлярному белку, принадлежащему к семейству CCN, который играет различные роли в регуляции клеточных функций, таких как индукция сосудистых заболеваний, ангиогенез, онкогенез, индукция фиброзной болезни, дифференцировка клеток и выживание. Белок CCN5, в отличие от других белков семейства CCN, не имеет С-концевого домена и также его называют WISP-2, HICP, Cop1, CTGF-L или тому подобное. Кроме того, белок CCN5 состоит из одной полипептидной цепи с 250-аминокислотной последовательностью. Благодаря секреторной лидерной последовательности из 22 аминокислот на N-конце белок CCN5 секретируется из клетки и функционирует как сигнальный белок. Таким образом, когда нуклеотидная последовательность экспрессируется в клетке, белок CCN5 может секретироваться из клетки. Согласно настоящему документу нуклеотидная последовательность может быть в виде мРНК.
В частности, белок CCN5 может иметь аминокислотную последовательность, представленную SEQ ID №: 1. Кроме того, нуклеотидная последовательность, кодирующая белок CCN5, может представлять собой последовательность, представленную SEQ ID №: 2 или SEQ ID №: 41.
Кроме того, фрагмент белка CCN5 может представлять собой фрагмент, полученный путем усечения части N-конца и/или C-конца CCN5 дикого типа, в той степени, пока этот фрагмент поддерживает активность белка CCN5. Конкретно, фрагмент белка CCN5 может быть фрагментом, полученным укорочением от 1 до 30, от 1 до 20, от 1 до 10 или от 1 до 5 аминокислот с N-конца или С-конца.
Кроме того, генетическая конструкция может содержать промоторную последовательность, функционально с ней связанную.
Как используется в настоящем документе, термин «функционально связанный» относится к функциональной связи между нуклеотидной регуляторной последовательностью экспрессии (такой как промотор, сигнальная последовательность или набор сайтов связывания транскрипционных факторов) и другими нуклеотидными последовательностями. Регуляторная последовательность регулирует транскрипцию и/или трансляцию других нуклеотидных последовательностей.
В частности, промотор, связанный с нуклеотидной последовательностью, кодирующей белок CCN5 или его фрагмент, может действовать, предпочтительно, в клетках животных и, более предпочтительно, в клетках млекопитающих, для регуляции транскрипции гена CCN5. Промотор включает промоторы, полученные из вирусов млекопитающих, и промоторы, полученные из геномов клеток млекопитающих. Промотор может действовать специфически в клетках сердца.
Промотор может быть любым промотором, выбранным из группы, состоящей из промотора цитомегаловируса (CMV), позднего промотора аденовируса, промотора вируса коровьей оспы 7.5K, промотора SV40, промотора HSV tk, промотора RSV, промотора EF1alpha, металлотионеинового промотора, β-актинового промотора, промотора гена IL-2 человека, промотора гена IFN человека, промотора гена IL-4 человека, промотора гена лимфотоксина человека и промотора гена GM-CSF человека. Однако перечень промоторов этим не ограничивается. В частности, промотор может представлять собой промотор CMV.
Генетическая конструкция может дополнительно содержать нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент. В настоящем документе нуклеотидная последовательность может быть в виде мРНК.
В настоящем документе в генетической конструкции нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент, может находиться в направлении от 5' до 3' в порядке следования «нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент-нуклеотидная последовательность, кодирующая белок CCN5 или его фрагмент». В настоящем документе нуклеотидная последовательность, кодирующая белок CCN5 или его фрагмент, может содержать стоп-кодон.
Кроме того, в генетической конструкции нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент, может находиться в направлении от 5' до 3' в порядке следования «нуклеотидная последовательность, кодирующая белок CCN5 или его фрагмент-нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент». В настоящем документе нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент, может содержать стоп-кодон.
Кроме того, генетическая конструкция может дополнительно содержать саморасщепляющуюся последовательность между нуклеотидной последовательностью, кодирующая белок SERCA2a или его фрагмент, и нуклеотидной последовательностью, кодирующей белок CCN5 или его фрагмент.
Как используется в настоящем документе, термин «белок SERCA2a» относится к белку, который функционирует, чтобы вызвать обратный захват кальция в саркоплазматический ретикулум, используя энергию ATP. Сообщалось, что у пациентов, страдающих сердечной недостаточностью со сниженной фракцией выброса (HFrEF), наблюдается заметно сниженный уровень экспрессии белка SERCA2a. Снижение обратного захвата кальция в саркоплазматический ретикулум, что является следствием снижения экспрессии белка SERCA2a, анормально увеличивает концентрацию кальция в цитоплазме, ослабляет функцию сокращения-расслабления кардиомиоцитов и действует как прямая причина гибели кардиомиоцитов, вызывая образование вредного кислорода, нарушение функции энергетического обмена и тому подобное из-за притока кальция в митохондрии.
В частности, белок SERCA2a может иметь аминокислотную последовательность, представленную SEQ ID №: 3. Кроме того, нуклеотидная последовательность, кодирующая белок SERCA2a может представлять собой последовательность, представленную SEQ ID №: 4 или SEQ ID №: 42.
Кроме того, фрагмент белка SERCA2a может представлять собой фрагмент, полученным путем усечения части N-конца и/или С-конца SERCA2a дикого типа, в той степени, пока этот фрагмент поддерживает активность белка SERCA2a. В частности, фрагмент белка SERCA2a может представлять собой фрагмент, полученный усечением от 1 до 100, от 1 до 50, от 1 до 20 или от 1 до 10 аминокислот с N-конца или С-конца.
Саморасщепляющаяся последовательность может представлять собой нуклеотидную последовательность, кодирующую пептид 2A, полученный из тешовируса 1 свиней, вируса Thosea asigna, вируса ринита лошадей A или вируса ящура. В частности, саморасщепляющаяся последовательность может представлять собой нуклеотидную последовательность, кодирующую пептид 2A, полученный из тешовируса 1 свиней. Кроме того, саморасщепляющаяся последовательность может представлять собой нуклеотидную последовательность, представленную SEQ ID №: 6.
Нуклеотидная последовательность, кодирующая пептид 2A, полученный из тешовируса 1 свиней, может представлять собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID №: 5. Кроме того, нуклеотидная последовательность, кодирующая аминокислотную последовательность, представленную SEQ ID №: 5, может представлять собой нуклеотидную последовательность, представленную SEQ ID №: 6.
Нуклеотидная последовательность, кодирующая пептид 2A, полученный из вируса Thosea asigna, может представлять собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID №: 7. Кроме того, нуклеотидная последовательность, кодирующая аминокислотную последовательность, представленную SEQ ID №: 7, может представлять собой нуклеотидную последовательность, представленную SEQ ID №: 8.
Нуклеотидная последовательность, кодирующая пептид 2A, полученный из вируса ринита лошадей A может представлять собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID №: 9. Кроме того, нуклеотидная последовательность, кодирующая аминокислотную последовательность, представленную SEQ ID №: 9, может представлять собой нуклеотидную последовательность, представленную SEQ ID №: 10.
Нуклеотидная последовательность, кодирующая пептид 2A, полученный из вируса ящура, может представлять собой нуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную SEQ ID №: 11. Кроме того, нуклеотидная последовательность, кодирующая аминокислотную последовательность, представленную SEQ ID №: 11, может представлять собой нуклеотидную последовательность, представленную SEQ ID №: 12.
Когда SERCA2a-P2A-CCN5, вариант генетической конструкции, экспрессируется в клетке, белок SERCA2a может быть встроен в мембрану саркоплазматического ретикулума, и белок CCN5 может секретироваться из клетки. Кроме того, когда CCN5-P2a-SERCA2a, вариант генетической конструкции по настоящему изобретению, экспрессируется в клетке, белок SERCA2a может быть встроен в мембрану саркоплазматического ретикулума, и белок CCN5 может секретироваться из клетки.
Как используется в настоящем документе, термин «аритмия сердца» относится к заболеванию, при котором в сердце не продолжается регулярное сокращение из-за плохой генерации электрической стимуляции в сердце или плохой передачи стимуляции, и сердцебиение становится анормально быстрым, медленным или нерегулярным.
Аритмия сердца классифицируется на предсердную аритмию и желудочковую аритмию в зависимости от места ее возникновения. Предсердная аритмия может включать фибрилляцию предсердий, предсердную тахикардию или дисфункцию синусового узла. Желудочковая аритмия может включать желудочковую тахикардию или фибрилляцию желудочков.
Кроме того, генетическая конструкция по настоящему изобретению может быть доставлена в клетку с использованием липосом. Липосомы образуются автоматически из фосфолипидов, диспергированных в водной фазе, и липосомы, содержащие нуклеотидную последовательность, кодирующую белок CCN5, и/или нуклеотидную последовательность, кодирующую белок SERCA2a, дают возможность нуклеотидной последовательности, кодирующей белок CCN5, и/или нуклеотидной последовательности, кодирующей белок SERCA2a, быть доставленными в клетку посредством такого механизма, как эндоцитоз, адсорбция на поверхности клетки или слияние с плазматической мембраной клетки.
В другом аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
Белок CCN5 является таким, как описанный выше при описании фармацевтической композиции для профилактики или лечения аритмии сердца, которая содержит генетическую конструкцию в качестве активного ингредиента.
Как используется в настоящем документе, термин «вектор экспрессии» относится к рекомбинантному вектору, способному экспрессировать белок-мишень в мишеневой клетке-хозяине, причем рекомбинантный вектор представляет собой генетическую конструкцию, которая содержит важные регуляторные элементы, функционально связанные с вставкой гена, так что вставка гена экспрессируется.
Кроме того, вектор экспрессии может содержать сигнальную последовательность для секреции слитого полипептида, так что облегчается выделение белка из клеточной культуры. Специфические сигналы инициации также могут потребоваться для эффективной трансляции вставленной последовательности нуклеиновых кислот. Эти сигналы содержат стартовый кодон ATG и смежные последовательности. В некоторых случаях должны предоставляться экзогенные трансляционные регуляторные сигналы, которые могут содержать стартовый кодон ATG. Эти экзогенные трансляционные регуляторные сигналы и стартовые кодоны могут иметь различные природные и синтетические источники. Эффективность экспрессии может быть повышена путем введения соответствующего элемента, усиливающего транскрипцию или трансляцию.
Кроме того, вектор экспрессии может дополнительно содержать нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент. В настоящем документе в векторе экспрессии нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент, может находиться в направлении от 5' до 3', в порядке следования «нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент-нуклеотидная последовательность, кодирующая белок CCN5 или его фрагмент».
Вектор экспрессии может дополнительно содержать саморасщепляющуюся последовательность между нуклеотидом, кодирующим белок SERCA2a или его фрагмент, и нуклеотидом, кодирующим белок CCN5 или его фрагмент. Саморасщепляющаяся последовательность является такой, как описанная выше при описании фармацевтической композиции для профилактики или лечения аритмии сердца, которая содержит генетическую конструкцию в качестве активного ингредиента.
Вектор экспрессии может содержать нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент, и/или нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент, по настоящему изобретению. В настоящем документе используемый вектор конкретно не ограничен при условии, что он может продуцировать белок CCN5 и/или белок SERCA2a по настоящему изобретению. Вектор экспрессии может быть любым, выбранным из группы, состоящей из плазмидных векторов и космидных векторов.
Плазмидный вектор может включать, но не ограничивается ими, коммерчески доступные плазмиды, такие как pUC18, pBAD и pIDTSAMRT-AMP.
Аритмия сердца является такой, как описано выше при описании фармацевтической композиции для предотвращения или лечения сердечной аритмии, содержащей генетическую конструкцию в качестве активного ингредиента.
В еще одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая в качестве активного ингредиента рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
Белок CCN5 является таким, как описано выше при описании фармацевтической композиции для профилактики или лечения аритмии сердца, которая содержит генетическую конструкцию в качестве активного ингредиента.
Вирус может представлять собой любой вектор, выбранный из группы, состоящей из аденовируса, аденоассоциированного вируса (AAV), ретровируса, лентивируса, вируса простого герпеса, вируса коровьей оспы и тому подобное. В частности, вирус может представлять собой, но не ограничивается этим, аденоассоциированный вирусом.
Аденовирус широко используется в качестве вектора для переноса генов из-за его среднего генома, простоты манипулирования, высокого титра, широкого диапазона клеток-мишеней и превосходной инфекционности. Его геном окружен от 100 до 200 п.н. инвертированного концевого повтора (ITR), который является важным цис-элементом для репликации и упаковки ДНК. Области E1 (E1A и E1B) генома кодируют белки, которые участвуют в репликации вирусной ДНК.
Из аденовирусных векторов широко используются неспособные к репликации аденовирусы, в которых отсутствуют области E1. С другой стороны, область E3 удаляется из обычных векторов аденовируса, чтобы предоставить участок для вставки чужеродного гена.
Таким образом, ген CCN5 по настоящему изобретению может быть встроен в удаленные области E1 (область E1A и/или область E1B, предпочтительно, область E1B) или область E3. В частности, ген белка CCN5 может быть встроен в область E3.
Между тем, нуклеотидная последовательность-мишень, подлежащая доставке в клетку, может быть встроена в удаленные области E1 (область E1A и/или область E1B, предпочтительно, область E1B) или область E3, предпочтительно, область E3. Кроме того, мишеневая нуклеотидная последовательность также может быть экспрессирована бицистронной системой экспрессии, связанной внутренним сайтом входа в рибосому (IRES), такой как ген белка «промотор-мишеневая нуклеотидная последовательность-поли A последовательность-IRES-CCN5».
Кроме того, поскольку до приблизительно 105% генома дикого типа может быть упаковано в аденовирус, около 2 т.п.н. может быть дополнительно упаковано в аденовирус. Таким образом, чужеродная последовательность, которая должна быть встроена в аденовирус, может быть дополнительно связана с аденовирусным геномом.
Аденовирус имеет 42 различных серотипа и подгруппы от А до F. Из них аденовирус типа 5, принадлежащий к подгруппе С, подходит для получения аденовирусного вектора по настоящему изобретению. Биохимическая и генетическая информация об аденовирусе типа 5 является хорошо известной.
Чужеродные гены, доставляемые аденовирусом, реплицируются так же, как и эписомы, и таким образом, имеют очень низкую генотоксичность для клеток-хозяев.
Ретровирус широко используется в качестве вектора переноса генов, поскольку ретровирус способен встраивать свой ген в геном хозяина и доставлять большое количество чужеродного генетического материала, и имеет широкий спектр клеток, которые он может инфицировать.
Для создания ретровирусного вектора ген CCN5 и мишеневая нуклеотидная последовательность, подлежащая доставке, встраиваются в ретровирусный геном вместо ретровирусной последовательности для получения вируса, не способного к репликации. Чтобы получить вирионы, конструируют упаковочную клеточную линию, которая содержит гены gag, pol и env и не имеет длинного концевого повтора (LTR) и последовательности ψ. Когда рекомбинантная плазмида, которая содержит ген CCN5, мишеневую нуклеотидную последовательность, подлежащую доставке, LTR и последовательность ψ, вводится в клеточную линию, последовательность ψ обеспечивает продуцирование РНК-транскрипта рекомбинантной плазмиды. Этот транскрипт упакован в вирус, и вирус секретируется в среду. Среду, содержащую рекомбинантные ретровирусы, собирают, обогащают и используют в качестве системы доставки генов.
Аденоассоциированный вирус (AAV) подходит в качестве системы доставки генов по настоящему изобретению, поскольку он способен инфицировать неделящиеся клетки и обладает способностью инфицировать различные типы клеток. Детали конструкции и использования векторов AAV описаны в патентах США №№5139941 и 4797368.
Обычно вирус AAV получают путем совместной трансформации плазмиды, содержащей последовательность гена-мишени (ген CCN5 и мишеневую нуклеотидную последовательность, которая должна быть доставлена), которая фланкирована двумя концевыми повторами AAV и плазмидой экспрессии, содержащей кодирующую последовательность AVV дикого типа, в которой отсутствуют концевые повторы.
Для доставки в клетку гена CCN5 и мишеневой нуклеотидной последовательности, подлежащей доставке, также можно использовать векторы, полученные из вируса коровьей оспы, лентивируса или вируса простого герпеса.
Кроме того, вирус может дополнительно содержать промоторную последовательность, функционально связанную с нуклеотидной последовательностью. Функционально связанная промоторная последовательность является такой, как описано выше при описании фармацевтической композиции для профилактики или лечения аритмии сердца, которая содержит генетическую конструкцию в качестве активного ингредиента.
Кроме того, рекомбинантный вирус может дополнительно содержать нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент. В настоящем документе в рекомбинантном вирусе нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент, может находиться в направлении от 5' до 3' в порядке следования «нуклеотидная последовательность, кодирующая белок SERCA2a или его фрагмент-нуклеотидная последовательность, кодирующая белок CCN5 или его фрагмент».
Кроме того, рекомбинантный вирус может содержать саморасщепляющуюся последовательность между нуклеотидной последовательностью, кодирующей белок SERCA2a или его фрагмент, и нуклеотидной последовательностью, кодирующей белок CCN5 или его фрагмент. Саморасщепляющаяся последовательность является такой, как описано выше при описании фармацевтической композиции для профилактики или лечения аритмии сердца, которая содержит генетическую конструкцию в качестве активного ингредиента.
Аритмия сердца является такой, как описано выше.
В настоящем изобретении способ введения фармацевтической композиции, которая содержит в качестве активного ингредиента вирус, который содержит генетическую конструкцию, может осуществляться в соответствии со способами вирусного инфицирования, известными в данной области. Кроме того, по настоящему изобретению, когда генетическая конструкция в качестве активного ингредиента содержится в рекомбинантной голой молекуле ДНК или плазмиде, для введения гена в клетки может быть использован метод микроинъекции, метод липосом-опосредованной трансфекции, метод обработки DEAE-декстраном и метод генетической бомбардировки.
Фармацевтически приемлемый носитель, который должен содержаться в фармацевтической композиции по настоящему изобретению, является одним из традиционно используемых для ее составления, и его примеры включают, но не ограничиваются ими, лактозу, декстрозу, сахарозу, сорбит, маннит, крахмал, аравийскую камедь, фосфат кальция, альгинат, желатин, силикат кальция, микрокристаллическую целлюлозу, поливинилпирролидон, целлюлозу, воду, сироп, метилцеллюлозу, метилгидроксибензоат, пропилгидроксибензоат, тальк, стеарат магния и минеральное масло.
Фармацевтическая композиция может включать помимо перечисленных выше ингредиентов смазывающее вещество, смачивающий агент, подсластитель, ароматизатор, эмульгатор, суспендирующий агент, консервант и тому подобное. Подходящие фармацевтически приемлемые носители и составы подробно описаны в обзоре Remington's Pharmaceutical Sciences (19th ed., 1995).
Лекарственная форма фармацевтической композиции может варьироваться в зависимости от способа применения и может быть изготовлена в виде инъекций.
Дозу фармацевтической композиции по настоящему изобретению желательно определять с учетом возраста, пола, состояния пациента, степени поглощения активных веществ организмом, степени инактивации и препаратов, используемых в комбинации; и фармацевтическая композиция может вводиться в количестве от 0,0001 мг/кг (веса тела) до 100 мг/кг (веса тела) в расчете на белок CCN5.
Дозу фармацевтической композиции по настоящему изобретению желательно определять с учетом возраста, пола, состояния пациента, степени поглощения активных веществ организмом, степени инактивации и препаратов, используемых в комбинации; и когда фармацевтическая композиция представляет собой вирус, фармацевтическая композиция может вводиться в количестве от 1,0×103 до 1,0×1020 вирусных геномов в день в расчете на взрослого. В частности, фармацевтическая композиция по настоящему изобретению может быть введена в количестве от 1,0×103 до 1,0×1020, от 1,0×108 до 1,0×1016, от 1,0×1012 до 1,0×1015 или от 1,0×1013 до 1,0×014 вирусных геномов в день в расчете на взрослого.
Кроме того, когда фармацевтическая композиция представляет собой плазмидный вектор, фармацевтическая композиция может вводиться в концентрации от 0,1 мкг/1 мкл до 1 мг/1 мкл в день в расчете на взрослого. Кроме того, когда фармацевтическая композиция представляет собой плазмидный вектор, доза может включать 0,1 мл, 1 мл, 2 мл, 3 мл, 4 мл, 5 мл, 6 мл, 7 мл, 8 мл, 9 мл, 10 мл или выше, и включает все значения и диапазоны между ними.
В еще одном аспекте настоящего изобретения предложена фармацевтическая композиция для профилактики или лечения аритмии сердца, включающая белок CCN5 в качестве активного ингредиента. Фармацевтическая композиция может дополнительно включать белок SERCA2a. Белок CCN5 и белок SERCA2a являются такими, как описано выше.
Фармацевтическая композиция по настоящему изобретению вводится парентерально, и парентеральное введение включает внутривенную инъекцию, подкожную инъекцию, внутримышечную инъекцию, внутрибрюшинную инъекцию, трансдермальное введение, способ прямой инъекции в ткань и тому подобное.
Как используется в настоящем документе, термин «приемлемый носитель» относится к некоторым или ко всем следующим веществам и включает вещества, подходящие для конкретной дозы: растворители, разбавители, жидкие несущие среды, диспергаторы, суспензионные адъюванты, поверхностно-активные вещества, изотонические агенты, загустители, эмульгаторы, консерванты твердые связующие, смазочные материалы или тому подобное. В работе Alfanso R. Gennaro, Remingtonʼs Pharmaceutical Sciences, 19th edition, 1995, Macna Publishing Co. Easton, PA, представлены различные носители для использования в фармацевтических композициях с известными методиками и композициями. Примеры фармацевтически приемлемых носителей включают, но не ограничиваются ими, следующие: глюкоза, сахароза, крахмал, такой как кукурузный крахмал и картофельный крахмал, целлюлоза и ее производные, такие как натрий-карбоксиметилцеллюлоза, этилцеллюлоза и ацетат целлюлозы; трагакант в виде порошка; солод; желатин; тальк; по усмотрению производителя состава могут содержаться инертные вспомогательные вещества, такие как масло какао, воск для суппозиториев, арахисовое масло, хлопковое масло, сафлоровое масло, кунжутное масло, оливковое масло и соевое масло; гликоли, такие как пропиленгликоль; сложные эфиры, такие как этилолеат и этиллаурат; агар; буферы, такие как гидроксид магния и гидроксид алюминия; альгиновая кислота; апирогенная дистиллированная вода; изотонический солевой раствор; раствор Рингера; этиловый спирт и забуференная фосфатом вода, лаурилсульфат натрия и стеарат магния, красители, антиадгезивы, покрывающие агенты, подсластители, ароматизаторы и отдушки, антиоксиданты и тому подобное.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий стадию введения субъекту фармацевтической композиции по настоящему изобретению.
Фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента генетическую конструкцию, содержащую нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент. Кроме того, фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент. Кроме того, фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В данном случае субъектом может быть млекопитающее, предпочтительно, человек. Конкретно, субъектом может быть человек или другое млекопитающее, которое страдает или может подвергаться риску аритмии сердца.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий введение субъекту белка CCN5. Кроме того, способ может дополнительно включать введение субъекту белка SERCA2a.
В данном случае субъектом может быть млекопитающее, предпочтительно, человек. Конкретно, субъектом может быть человек или другое млекопитающее, которое страдает или может подвергаться риску аритмии сердца.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий введение субъекту генетической конструкции, которая содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент, и генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент.
В данном случае субъектом может быть млекопитающее, предпочтительно, человек. Конкретно, субъектом может быть человек или другое млекопитающее, которое страдает или может подвергаться риску аритмии сердца.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий введение субъекту вектора экспрессии, несущего нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент, и вектора экспрессии, несущего нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В данном случае субъектом может быть млекопитающее, предпочтительно, человек. Конкретно, субъектом может быть человек или другое млекопитающее, которое страдает или может подвергаться риску аритмии сердца.
В еще одном аспекте настоящего изобретения предложен способ профилактики или лечения аритмии сердца, включающий введение субъекту рекомбинантного вируса, который содержит нуклеотидную последовательность, кодирующую белок SERCA2a или его фрагмент, и рекомбинантного вируса, который содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
В данном случае субъектом может быть млекопитающее, предпочтительно, человек. Конкретно, субъектом может быть человек или другое млекопитающее, которое страдает или может подвергаться риску аритмии сердца.
В еще одном аспекте настоящего изобретения предложено применение фармацевтической композиции по настоящему изобретению для профилактики или лечения аритмии сердца.
В еще одном аспекте настоящего изобретения предложено применение фармацевтической композиции по настоящему изобретению для изготовления лекарственного препарата для профилактики или лечения аритмии сердца.
Фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент. Кроме того, фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент. Кроме того, фармацевтическая композиция может представлять собой фармацевтическую композицию для профилактики или лечения аритмии сердца, которая содержит в качестве активного ингредиента рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5 или его фрагмент.
Метод осуществления изобретения
Далее настоящее изобретение описано подробно с помощью примеров. Однако следующие экспериментальные примеры и примеры предназначены только для иллюстрации настоящего изобретения, и настоящее изобретение не ограничивается следующими препаративными примерами и примерами.
Препаративный пример 1. Конструирование AAV9-CCN5 и AAV9-SERCA2a-P2A-CCN5
Генетическую конструкцию pTR-CMV-CCN5 сконструировали для экспрессии белка CCN5. Кроме того, генетическую конструкцию pTR-CMV-SERC2a-P2A-CCN5 сконструировали одновременной экспрессии белка CCN5 и белка SERCA2a (фиг. 1 и 2).
В генетической конструкции фрагмент SERCA2a состоит из последовательности кДНК белка SERCA2a человека. Следующий связанный фрагмент P2A представляет собой сайт саморасщепления, полученный из тешовируса 1 свинец, и состоит из нуклеотидной последовательности, кодирующей 22 аминокислоты. Наконец, фрагмент CCN5 состоит из последовательности кДНК человеческого белка CCN5.
Рекомбинантную плазмиду pTR-CMV-SERCA2a-P2A-CCN5 конструировали, удаляя часть люциферазы из вектора pTR-CMV-люциферазы и вставляя вместо нее генетическую конструкцию SERCA2a-P2A-CCN5. Белок, продуцируемый рекомбинантной плазмидой, подразделяли на фрагмент SERCA2a и фрагмент CCN5 путем саморасщепления между 21-й аминокислотой глицином и 22-й аминокислотой пролином в сайте P2A. Часть SERCA2a может оставаться в мембране эндоплазматического ретикулума и выполнять свою встраиваемую функцию. Кроме того, фрагмент CCN5 может мигрировать в эндоплазматический ретикулум и затем секретироваться из клетки в форме, в которой расщепляется сигнальный пептид, тем самым выполняя свою встраиваемую функцию.
Ген CCN5 человека клонировали в вектор pds-AAV2-EGFP для конструирования аденоассоциированного вируса (AAV, серотип 9). Чтобы улучшить упаковку вируса и эффективность доставки вируса, в процессе конструирования вектора удаляли AAV последовательность eGFP. Рекомбинантный AAV конструировали с использованием клеток 293T. Частицы AAV в клеточной культуре собирали и осаждали сульфатом аммония. Полученный продукт очищали ультрацентрифугированием с использованием градиента йодиксанола. Частицы AAV обогащали путем осуществления нескольких разбавлений и обогащений таким образом, чтобы йодиксанол заменялся на раствор Рингер-лактата, используя центрифугирования. Концентрацию AAV определяли количественно с использованием количественной ОТ-PCR и SDS-PAGE.
Экспериментальный метод 1. Создание экспериментальной модели и введение гена
Экспериментальный метод 1.1. Создание мышиной модели фибрилляции предсердий путем инфузии ангиотензина II в мышиную модель избыточной экспрессией CCN5
Для экспериментов использовали самцов мышей C57BL6 WT (дикий тип, черная окраска шерсти) и трансгенных (TG) мышей, у которых была специфическая для сердца сверхэкспрессия CCN5.
Мышей CCN5-TG получали путем субклонирования мышиного гена CCN5 в вектор pNC (Clontech, США), содержащий промотор α-MHC, который индуцирует специфическую для сердце экспрессию гена, и введения полученного продукта в оплодотворенные яйца C57BL/6 с использованием техники микроинъекции. Дополнительно, для приобретения и поддержания значительной линии мышей секвенирование было поручено компании Macrogen Inc. (Южная Корея). Для идентификации присутствия трансгена CCN5 в геноме мыши использовали саузерн-блоттинг.
Все используемые мыши были мышами в возрасте от 8 до 10 недель весом от 20 до 25 г. Мышей анестезировали путем внутрибрюшинной инъекции кетамина (95 мг/кг) и ксилазина (5 мг/кг), а мерцательную аритмию индуцировали подкожной инфузией ангиотензина II. В данном случае ангиотензин II вводили подкожно в течение 14 дней в концентрации 3,0 мг/кг в день с использованием небольшого осмотического насоса (Alzet 1002, Alzet).
Экспериментальный метод 1.2. Создание мышиной модели фибрилляции предсердий посредством инфузии ангиотензина II и инъекции вируса
8-10-недельных мышей B6C3F1 (серый окрас шерсти) анестезировали путем внутрибрюшинной инъекции кетамина (95 мг/кг) и ксилазина (5 мг/кг), и фибрилляцию предсердий индуцировали путем подкожной инфузии ангиотензина II. В данном методе ангиотензин II инфузировали подкожно в течение 2 недель в концентрации 3 мг/кг в день с использованием небольшого осмотического насоса. Через 2 недели после индукции фибрилляции предсердий инфузией ангиотензина II каждой мыши через хвостовую вену инъецировали 1×1011 вирусных геномов (vgs) от AAV9-Control или AAV9-CCN5.
Экспериментальный метод 1.3. Создание мышиной модели желудочковой аритмии посредством инфузии ангиотензина II и введения гена вируса
8-10-недельных мышей B6C3F1 анестезировали путем внутрибрюшинной инъекции кетамина (95 мг/кг) и ксилазина (5 мг/кг), и желудочковую аритмию индуцировали путем подкожной инфузии ангиотензина II. Ангиотензин II инфузировали подкожно в течение 2 недель в концентрации 3 мг/кг в день с использованием небольшого осмотического насоса. Через 2 недели после индукции желудочковой аритмии инфузией ангиотензина II каждой мыши через хвостовую вену инъецировали 1×1011 vgs от AAV9-Control, AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5.
Экспериментальный метод 2. Окрашивание тканей
У животных брали ткани сердца и затем фиксировали 10% (масс/объем) формалином при комнатной температуре в течение 5 дней. Затем промывали PBS. Каждый образец погружали в парафин, и блок образца ткани разрезали на секции толщиной 7 мкм.
Для измерения степени фиброза осуществляли окрашивание трихромом по Массону. Ткань в месте, где прогрессировал фиброз, окрашивается в синий цвет, а нормальная ткань окрашивается в красный цвет. Степень фиброза выражали путем расчета части, в которой фиброз происходил во всей ткани. Это наблюдали под оптическим микроскопом и анализировали с использованием программы Aperio Imagescope (Leica Biosystems).
Экспериментальный метод 3. Определение уровня экспрессии мРНК с помощью PCR в реальном времени
У животных брали ткани сердца и затем из них извлекали мРНК для проведения qRT-PCR. PCR в реальном времени проводили с использованием набора для PCR QuantiTect SYBR Green в реальном времени (Qiagen Ltd). Посредством этого был проанализирован их уровень транскрипции. РНК выделяли из ткани сердца с использованием тризола (Gibco BRL) и из нее синтезировали кДНК. Количественные условия PCR в реальном времени были следующими: 37 циклов при 94°С в течение 10 секунд, при 57°С в течение 15 секунд, при 72°С в течение 5 секунд. Данные о праймерах, использованных в эксперименте, приведена в таблице 1.
[Таблица 1]
Экспериментальный метод 4. Электрофизиологический экспериментальный метод
Экспериментальный метод 4.1. Измерение электрокардиограммы в модели мерцательной аритмии с использованием мышей CCN5 TG
Для исследований электрофизиологии предсердий эксперименты проводились ex vivo путем подключения удаленного мышиного сердца к системе Лангендорфа. Удаленное сердце перфузировали буфером Кребса-Хенселейта (118 мМ NaCl, 4,7 мМ KCl, 1,2 мМ MgSO4, 1,25 мМ CaCl2, 1,2 мМ KH2PO4, 25 мМ NaHCO3, 11 мМ глюкозы) и 95% O2/5% газообразного CO2, поддерживали температуру 37°C и давление 60 мм рт.ст. Перед применением электрической стимуляции удаленное сердце стабилизировали в течение 10 минут путем подключения к системе Лангендорфа. Затем серебряный биполярный электрод с тефлоновым покрытием помещали в правое предсердие, левое предсердие и левый желудочек. Чтобы вызвать фибрилляцию предсердий, 2-секундную импульсную стимуляцию выполняли три раза с использованием автоматического стимулятора. В первой 2-секундной импульсной стимуляции применялось стимулирование с длительностью цикла 40 мс с длительностью импульса 5 мс. После применения стимула стабилизацию проводили в течение 3 минут. Второй 2-секундный пакет был применен с длительностью цикла 20 мс с длительностью импульса 5 мс, и стабилизацию проводили снова в течение 3 минут. Последний 2-секундный пакет был применен с длительностью цикла 20 мс с длительностью импульса 10 мс. Было установлено, что предсердие, которое показало нерегулярный интервал R-R в течение не менее 1 секунды и показало нерегулярный, быстрый ритм, имело фибрилляцию предсердий.
Экспериментальный метод 4.2. Измерение электрокардиограммы у мышей с фибрилляцией предсердий, которым вводили AAV9-CCN5
Эксперименты выполнялись ex vivo путем подключения извлеченного мышиного сердца к системе Лангендорфа, как описано в экспериментальном методе 4.1. В частности, сначала мышам вводили гепарин для предотвращения свертывания крови, и мышей анестезировали, используя 100% изофлуран (Forane, USP, Baxter Healthcare Corporation). После этого сердце мыши извлекали и подключали к системе Лангендорфа. Через аорту в сердце устанавливали канюлю, перфузировали раствором Тироде (NaCl 130 мМ, NaHCO3 24 мМ, KCl 4 мМ, MgCl2 1 мМ, CaCl2 1,8 мМ, KH2PO4 1,2 мМ, C6H12O6 5,6 мМ, 1% альбумина) при потоке от 1,5 до 2,0 мин-1 и 95% O2/5% газообразного CO2, поддерживая температуру 38±1°C, pH от 7,3 до pH 7,5 и давление от 60 мм до 70 мм рт.ст. (датчик давления BP-1, World Precision Instruments). Для минимизации механического сокращения сердца использовали электромеханическую развязку (5 мМ блеббистатина, Sigma Aldrich, США).
Изготовленный на заказ кардиостимулятор Ag-AgCl помещали в правое предсердие, а другой электрод прикрепляли как к эпикардиальной поверхности, так и к сердечной перегородке. Передняя поверхность сердца использовалась для непрерывного отображения электрической активности в левом предсердии полувертикальным способом (Mightex BioLED Light, модуль управления источником, серия BLS).
Для определения аритмии проводили объемную электрокардиографию. Усиление и фильтрацию нижних частот выполняли при 150 Гц с использованием электронного усилителя для непрерывной записи объемной электрокардиограммы сердца. Кроме того, проводилась цифровая выборка с частотой 1 кГц (BioPac Systems MP150). Чтобы идентифицировать возникновение персистирующей аритмии с помощью электрических и оптических сигналов, стимуляцию начинали с основной частоты 7 Гц (PCL=140 мс) в правом предсердии с постепенным увеличением частоты при применении длительности стимуляции 2 мс с частотой от 2 Гц до 3 Гц.
Экспериментальный метод 4.3. Экспериментальный метод оптического картирования на мышиных моделях фибрилляции предсердий и желудочковой аритмии
Когда через 20-30 минут после начала обратной перфузии сердца сердцебиение достигало устойчивого состояния от 4 Гц до 5 Гц, сердце окрашивали, вводя в непосредственной близости от аорты 0,3 мл 15 мкМ чувствительного к напряжению красителя (Di-4-ANEPPS, Invitrogen, Thermofisher Scientific). Для возбуждения флуорофоров на длине волны 530 нм использовали монохроматический свет (Mightex, BioLED). Излучаемые длины волн отфильтровывали через фильтр длинных полос (>590 нм) и проецировали с пространственным разрешением 87,5 мкм, частотой кадров 1 кГц и ПЗС-матрицей 80×80 пикселей (SciMeasure, SciMeasure Analytical Systems, США) на 3× общее увеличение.
Экспериментальный метод 5. Вестерн-блоттинг
Используемые клетки и сердце, полученные в настоящих экспериментах, были приготовлены с использованием гомогенизированного буфера RIPA (0,1% (мас./объем) SDS, 50 мМ Трис-HCl (pH 7,4), 150 мМ NaCl, 1% (мас./объем) NP-40, 0,5% (мас./объем) дезоксихолата натрия) с добавлением коктейля ингибиторов протеазы широкого спектра действия (Calbiochem). Белки разделяли по размеру, используя гели SDS-PAGE, и переносили на мембрану PVDF (Millipore). После блокирования 5% (мас./объем) обезжиренным молоком в течение 1 часа и промывания TBST подготовленной мембране давали возможность реагировать с p-CaMKII, CaMKII, Na+/Ca+ обменником 2 (NCX2), RyR2 (Santa Cruz), pRyR2 (Ser2808), pRyR2 (Ser2814) (Badrilla), GAPDH (лабораторный), α-тубулином, TGF-β1, α-SMA (Sigma), Collagen I (Rockland), NaV 1.5, коннексином 43, Kir2.1 (Alomone labs), белком активации фибробластов (FAP), митохондриальным пептидом, метионинсульфоксидредуктазой (MsrA), тубулином (Abcam), SERCA2a (21st Century Biochemicals), Smad7 (Invitrogen) и CCN5 (Genescipt) антителами. Затем мембрану инкубировали с вторичным антителом, конъюгированным с пероксидазой хрена (Jackson ImmunoResearch, West Grove, PA, USA) и разрабатывали с использованием хемилюминесцентного субстрата (Dogen). Блоты сканировали и количественно определяли с использованием программного обеспечения LAS.
Экспериментальный метод 6. Выделение и культура фибробластов предсердий
Для выделения фибробластов из предсердий белых крыс использовали сердце белых крыс Sprague Dawley (SD). Левое предсердие измельчали на отдельные клетки путем расщепления ткани с использованием раствора коллагеназы. Клетки, собранные из предсердия, сначала центрифугировали при 50×g в течение 3 минут. Полученный таким образом супернатант собирали и снова центрифугировали при 500×g в течение 10 минут. Полученный таким образом клеточный слой культивировали с использованием культуры DMEM, содержащей 10% эмбриональной бычьей сыворотки (FBS) и 1% антибиотиков. Через 2-3 дня культуру подвергали обработке 100 нМ ангиотензина II и одновременно контрольной кондиционированной средой (CM-Con) или CCN5-содержащей кондиционированной средой (CM-CCN5). Через 48 часов эксперимент прекращали. Для ясных экспериментальных результатов в качестве фибробластов левого предсердия использовали только фибробласты левого предсердия.
Экспериментальный метод 7. Приготовление CM-CCN5
Для получения CM-CCN5 использовали плазмиду pcDNA3.1-CCN5HA. Клетки HEK293 распределяли по 5×105 клеток в 60-мм культуральной чашке и стабилизировали в течение одного дня. Затем проводили трансфекцию pcDNA3.1-CCN5HA с использованием липофектамина (Invitrogen). Через 4 часа проводили обмен среды для удаления липофектамина. Затем культивирование проводили в течение 24 часов, и полученная культуральная среда называлась CM-CCN5.
Экспериментальный метод 8. Флуоресцентная иммунохимия
15000 клеток распределяли на 16-мм покровном стекле и инкубировали в течение ночи для стабилизации. Затем клетки подвергали обработке 100 нМ ангиотензином II и CM-Con или CM-CCN5 и проводили инкубацию. Полученные клетки фиксировали 4% (мас./объем) раствором параформальдегида, придавая клеточной мембране проницаемость, используя 0,5% раствор Triton X-100, а затем блокировали 5% (мас./объем) раствором BSA. Затем осуществляли взаимодействие с антителом против α-SMA (Sigma), и в качестве вторичного антитела использовали конъюгированное с Alexa Fluor 488 антитело (Invitrogen). Ядра окрашивали с использованием красителя Hoechst. Для клеток, которые были подвергнуты иммунохимии, использовали конфокальный микроскоп Fluoview FV 1000.
Экспериментальный метод 9. Измерение функции миокарда с помощью эхокардиографии
Мышей анестезировали путем внутрибрюшинной инъекции кетамина (95 мг/кг) и ксилазина (5 мг/кг) и проводили эхокардиографию. Запись выполнялась с помощью 2-мерной визуализации и функции слежения в М-режиме, были определены фракция укорочения и соотношение размеров желудочков (GE Vivid Vision).
Экспериментальный пример 1. Выявление терапевтического эффекта белка CCN5 на мышиной модели фибрилляции предсердий
Экспериментальный пример 1.1. Выявление эффекта подавления фибрилляции предсердий белком CCN5
Мышам WT и мышам CCN5 TG, полученным способом, описанным в экспериментальном методе 1.1, инфузировали подкожно ангиотензин II в течение 14 дней в концентрации 3,0 мг/кг в день с использованием небольшого осмотического насоса (Alzet 1002, Alzet). Спустя 2 недели у мышей извлекали сердце. Окрашивание тканей осуществляли способом, описанным в экспериментальном методе 2, для определения степени фиброза (фиг. 3a).
В результате в контрольной группе мышей, которым вводили ангиотензин II, было обнаружено, что коллаген накапливался примерно в 8% тканей предсердия; с другой стороны, в группе мышей CCN5 TG, которым вводили ангиотензин II, накопление коллагена наблюдалось примерно в 4% тканей предсердия, что указывает на значительное уменьшение (фиг. 3b и 3c).
Кроме того, чтобы идентифицировать изменения, происходящие в экспрессии мРНК, из левого предсердия мыши выделяли мРНК и проводили qRT-PCR способом, описанным в экспериментальном методе 3.
В результате, у маркерных генов фиброза сердца, α-SMA, Collagen I и TGF-β1 и маркерных генов воспалительного ответа, IL-1β, RANTES (регулируется при активации, нормальные Т-клетки экспрессируются и секретируются), F4/80, белка 1 хемоаттрактанта моноцитов (МСР-1) экспрессия их мРНК была повышена в контрольной группе мышей, которым вводили ангиотензин II; с другой стороны, экспрессия этих маркерных генов была значительно снижена в группе мышей CCN5 TG, которым вводили ангиотензин II (фиг. 3d-3j). На основании этих результатов было установлено, что CCN5 эффективно подавляет фиброз предсердий, вызванный ангиотензином II.
Экспериментальный пример 1.2. Выявление эффекта подавления фибрилляции предсердий CCN5
Для исследований электрофизиологии предсердий эксперименты выполнялись ex vivo путем подключения удаленного сердца мыши к системе Лангендорфа таким же образом, как в эксперименте 4.1. В результате у мышей WT, которым вводили ангиотензин II, фибрилляция предсердий была индуцирована у 4 из 6 животных. Однако у мышей CCN5 TG, которым вводили ангиотензин II, у всех 4 животных были показаны нормальные результаты электрокардиограммы даже после электростимуляции (фиг. 4b и 4c). На основании этих результатов было установлено, что CCN5 ингибирует фибрилляцию предсердий, вызванную ангиотензином II.
Экспериментальный пример 1.3. Выявление регуляции внутриклеточной концентрации Ca2+ с помощью CCN5
Предположение, что CCN5 регулирует Ca2+ в кардиомиоцитах, было исследовано с использованием клеток HL-1 (Sigma), которые являются клеточной линией предсердных кардиомиоцитов мыши. Кроме того, для выявления его регулирующего влияния на концентрацию Ca2+в культивируемых клетках HL-1 использовали вестерн-блоттинг.
В экспериментах на клеточном уровне для выявления эффекта CCN5 использовали CM-CCN5, полученный способом, описанным в экспериментальном примере 8. Клетки HL-1 подвергали обработке ангиотензином II и одновременно CM-Con (контроль) или CM-CCN5 (экспериментальная группа). Через 48 часов с помощью вестерн-блоттинга контролировали изменения уровня белка (фиг. 5a).
В результате было обнаружено, что фосфорилирование CaMKII (Thr287) возрастало, когда клетки HL-1 подвергались обработке 400 нМ ангиотензином II, тогда как фосфорилирование CaMKII снижалось в клетках, которые подвергались одновременной обработке CM-CCN5. В клетках HL-1, которые были подвергнуты обработке ангиотензином II, было индуцировано гиперфосфорилирование рианодинового рецептора 2 саркоплазматического ретикулума (RyR2) на Ser2808 и Ser2814; однако такое гиперфосфорилирование ингибировалось CCN5. Повышенная экспрессия Na+/Ca+ обменника 2 (NCX2), вызванная ангиотензином II, также снижалась с помощью CCN5. Уровень экспрессии калсеквестрина 2, связывающего кальций белка в саркоплазматической сети, снижался ангиотензином II, тогда как его экспрессия повышалась в клетках, которые были подвергнуты одновременной обработке CM-CCN5 (фиг. 5b-5g). На основании этих результатов было установлено, что CCN5 непосредственно регулирует кардиомиоциты и, таким образом, участвует в фибрилляции предсердий.
Экспериментальный пример 1.4. Выявление эффекта подавления фиброза предсердий с помощью CCN5 in vitro
Для выделения фибробластов из предсердия использовали сердце белых крыс Sprague Dawley (SD). Фибробласты предсердий выделяли способом, описанным в экспериментальном методе 6. Через 2-3 дня выделенные фибробласты предсердий обрабатывали 100 нМ ангиотензина II одновременно с контрольной кондиционированной средой (CM-Con) или CCN5-содержащей кондиционированной средой (CM-CCN5), приготовленный в экспериментальном примере 8. Через 48 часов эксперимент прекращали (фиг. 6а). Фибробласты, которые обрабатывали ангиотензином II и CM-Con или CM-CCN5, подвергались флуоресцентной иммуноцитохимии таким же образом, как и в экспериментальном методе 8.
В результате фибробласты, которые были подвергнуты обработке 100 нМ ангиотензином II, дифференцировались в миофибробласты и экспрессировали специфичный для миофибробластов маркерный белок α-SMA, тогда как фибробласты, которые были подвергнуты одновременной обработке CM-CCN5, вообще не экспрессировали α-SMA (фиг. 6b).
Кроме того, для выявления эффекта ингибирования фиброза предсердий культивируемые фибробласты предсердий подвергали вестерн-блоттингу в соответствии со способом, описанным в экспериментальном методе 5. В результате фибробласты, которые обрабатывали ангиотензином II, проявляли повышенную экспрессию белков α-SMA, Collagen I и TGF-β1, тогда как фибробласты, которые подвергались одновременной обработке CM-CCN5 демонстрировали заметно сниженную экспрессию этих белков (фиг. 6c-6f).
Кроме того, для выявления уровня экспрессии мРНК культивируемых фибробластов предсердий, эффект подавления фиброза предсердий контролировали с помощью qRT-PCR. В результате было установлено, что в фибробластах, которые обрабатывали ангиотензином II и одновременно с CM-CCN5, уровни экспрессии мРНК α-SMA, Collagen I и TGF-β1 были снижены до контрольного уровня (фиг. 6g-6i). На основании этих результатов было установлено, что CCN5 не только регулирует фиброз и, таким образом, ингибирует фибрилляцию предсердий, но также непосредственно регулирует кардиомиоциты и, таким образом, участвует в фибрилляции предсердий.
Экспериментальный пример 1.5. Выявление эффекта подавления фиброза предсердий AAV-CCN5 на мышиной модели фибрилляции предсердий
Таким же образом, как и в экспериментальном методе 1.2, ангиотензин II инфузировали 8-10-недельным мышам в концентрации 3 мг/кг/день. В момент времени 2 недели в хвостовую вену инъецировали AAV9-Control (контроль) или AAV9-CCN5 (сравнительная группа) в количестве 5×1011 вирусных геномов (vgs). Через 4 недели ткань предсердия подвергали окрашиванию трихромом по Массону и молекулярному анализу (фиг. 7a).
Сначала, для определения того, хорошо ли вирус AAV9 был экспрессирован специфическим для сердца способом, для выявления экспрессии CCN5 в ткани предсердия использовали вестерн-блоттинг. В результате было установлено, что белок CCN5 сверхэкспрессировал в группе мышей, которым инъецировали AAV9-CCN5. Кроме того, из результатов, полученных путем идентификации уровня экспрессии мРНК CCN5 в ткани предсердия, было обнаружено, что мРНК CCN5 аналогичным образом увеличивалась путем инъекции AAV9-CCN5 (фиг. 7b и 7c).
Из результатов, полученных путем определения степени накопления коллагена в предсердии посредством окрашивания трихромом, было установлено, что у мышей, инъецированных ангиотензином II и затем AAV9-Control, коллаген накапливался примерно в 6% ткани предсердия, тогда как в группе мышей, инъецированных AAV9-CCN5, степень накопления коллагена снижалась примерно до 3% (фиг. 7d и 7e).
Кроме того, для определения уровней экспрессии мРНК маркерных генов, связанных с фиброзом, α-SMA, Collagen I и TGF-β1, и маркерных генов, связанных с воспалением, IL-1β, RANTES, F4/80 и MCP-1, проводили qRT-PCR с использованием ткани предсердия таким же образом, как в экспериментальном методе 3. В результате было обнаружено, что AAV9-CCN5 значительно уменьшал эти маркерные гены, уровень экспрессии которых был увеличен ангиотензином II (фиг. 7f-7l). На основании этих результатов было установлено, что даже на мышиной модели со специфической для сердца сверхэкспрессией CCN5 с использованием AAV9-CCN5, индуцированный ангиотензином II фиброз предсердий эффективно подавлялся.
Экспериментальный пример 1.6. Выявление эффекта подавления фибрилляции предсердий, проявляемого AAV-CCN5
Мышиная модель была получена таким же образом, как в экспериментальном методе 4.2, и кардиофизиологические эксперименты были выполнены в конце 6 недели (фиг. 8a). Сначала была проверена электрокардиограмма, и в результате было установлено, что в группе мышей, которым инъецировали AAV9-Control, функция предсердия была ослаблена ангиотензином II и, таким образом, фибрилляция предсердий постоянно индуцировалась после электростимуляции. С другой стороны, было замечено, что в группе мышей, которым инъецировали AAV9-CCN5, сердцебиение возвращалось к нормальному ритму после электростимуляции. Даже в экспериментах, в которых измеряли потенциал действия Ca2+ (оптический сигнал), группа, инъецированная AAV9-Control, постоянно демонстрировала нарушения проводимости после стимуляции, тогда как группа, инъецированная AAV9-CCN5, демонстрировала нормальную предсердную активацию после стимуляции (фиг. 8b и 8c).
В результате анализа электрокардиограммы в группе мышей, которым инъецировали AAV9-Control, фибрилляция предсердий была индуцирована у 4 из 5 мышей после электростимуляции, тогда как в группе мышей, которым инъецировали AAV9-CCN5, у 2 из 5 мышей были предсердные аномалии, а остальные показали нормальную предсердную активацию (фиг. 8d).
При проведении экспериментов с электрокардиограммой, когда для стимуляции фибрилляции предсердий применялась электрическая стимуляция, контрольной (плацебо) группе мышей требовалась стимуляция примерно 28 Гц для индукции фибрилляции предсердий; однако в контрольной группе мышей, которым инъецировали ангиотензин II и AAV9-Control, предсердная фибрилляция в достаточной степени индуцировалась даже при такой низкой стимуляции, как примерно 20 Гц. С другой стороны, было обнаружено, что в группе мышей, которым индуцировалась только тогда, когда стимуляция была повышена до примерно 25 Гц, что было аналогично уровню в группе плацебо (фиг. 8e).
Как указывалось в экспериментальном методе 4.2, электрическая активность в левом предсердии была картирована для графического представления потенциала действия Ca2+. График анализировался в каждый момент продолжительности потенциала действия 50 (APD50) и продолжительности потенциала действия 75 (APD75). В результате было установлено, что в группе, которой инъецировали AAV9-CCN5, продолжительности потенциала действия проявлялись с такой же скоростью, что и в группе плацебо, по сравнению с группой, которой вводили AAV9-Control (фиг. 8f).
Это указывает на скорость, требуемую для деполяризации потенциала действия, и было обнаружено, что в группе мышей, которым инъецировали AAV9-CCN5, скорость, необходимая для деполяризации, которая была замедлена ангиотензином II, была восстановлена до уровня, подобного плацебо (фиг. 8g). На основании этих результатов было установлено, что даже в группе мышей, у которых CCN5 сверхэкспрессировал посредством инъекции AAV9-CCN5, фибрилляция предсердий, индуцированная ангиотензином II, надежно подавлялась.
Экспериментальный пример 2. Определение терапевтического эффекта AAV-CCN5 и AAV9-SERCA2a-P2A-CCN5 на мышиной модели желудочковой аритмии
Экспериментальный пример 2.1. Создание мышиной модели желудочковой аритмии
Мышиная модель желудочковой аритмии была создана способом,, описанным в экспериментальном методе 1.3. Используемые самцы мышей B6C3F1 WT (дикий тип, шкурка серого цвета) представляли собой 8-10-недельных мышей весом от 20 до 25 г. Ангиотензин II инфузировали подкожно в течение 2 недель в концентрации 3 мг/кг. Спустя 2 недели с помощью эхокардиографии выявляли изменения функции сердца. Идентифицированные мыши были отобраны случайным образом и инъецированы только вектором CCN5 (AAV9-CCN5) или комбинацией векторов CCN5 и SERCA2a (AAV9-SERCA2a-P2A-CCN5) в количестве 5×1011 вирусных геномов (vgs). Затем этих мышей содержали еще 4 недели для окончательной функциональной оценки.
Сначала, чтобы установить, что модель желудочковой аритмии у мышей была хорошо продуцирована ангиотензином II, функцию сердца в контрольной группе мышей (плацебо) и в группе сравнительных мышей (AngII) проверяли с помощью эхокардиографии. В результате у группы мышей, которым подкожно инъецировали ангиотензин II, наблюдалось снижение фракция укорочения по сравнению с контролем (плацебо). Тем не менее две группы не показали большой разницы с точки зрения веса мыши (фиг. 9a).
Для определения желудочковой тахикардии, оптическое картирование выполняли, применяя к правому желудочку (RV) электрическую стимуляцию в 10 Гц (фиг. 9b) или электрическую стимуляцию в 20 Гц (фиг. 9c), таким же образом, что и в экспериментальном методе 4.3. Чем больше электрическая стимуляция, тем более выраженная разница наблюдалась между экспериментальной группой и контролем. В группе мышей, которым вводили ангиотензин II, когда применяли электрическую стимуляцию в 20 Гц, канал ионов кальция или тому подобное не был локально восстановлен в своем первоначальном состоянии, и, таким образом, наблюдался разрыв в передаче стимуляции. Потенциал действия кальция был представлен графически и проиллюстрирован на фиг. 9d. График анализировался в каждый момент времени продолжительности потенциала действия 50 (APD50) и продолжительности потенциала действия 75 (APD75). В результате было обнаружено, что более быстрая продолжительность потенциала действия была достигнута инъекцией ангиотензина II (фиг. 9e). Кроме того, в группе, принимавшей ангиотензин II, наблюдалось значительное снижение скорости продолжительность потенциала действия (фиг. 9f). В результате анализа скорости проводимости и скорости, необходимой для деполяризации потенциала действия, было обнаружено, что такие скорости были значительно снижены ангиотензином II (фиг. 9g). На основании этих результатов было установлено, что мышиная модель желудочковой аритмии достоверно продуцировалась ангиотензином II.
Экспериментальный пример 2.2. Влияние на антагонистов β-адренергического рецептора на мышиной модели желудочковой аритмии
Изопротеренол (ISO) представляет собой лекарственное средство, которое клинически воздействует на β-адренергические рецепторы и, таким образом, увеличивает сократимость миокарда, тем самым увеличивая сердечный выброс. На мышиной модели с индуцированной ангиотензином II желудочковой аритмией был установлен потенциал действия Ca2+ в желудочке, когда изопротеренол использовался для стимуляции β-рецепторов (фиг. 10a). Электрическая стимуляция в 20 Гц была применена к правому желудочку, и результирующая картина реполяризации Ca2+ показана на диаграмме.
В результате было установлено, что в мышиной модели, инъецированной ангиотензином II, картина реполяризации кальция была медленнее даже при обработке ISO (фиг. 10b). В результате анализа продолжительности потенциала действия 75 (APD75), было обнаружено, что в мышиной модели, инъецированной ангиотензином II, обработка ISO не давала различий в степени продолжительности потенциала действия (фиг. 10c).
Чтобы определить желудочковую тахикардию, оптическое картирование выполняли, применяя к правому желудочку (RV) электрическую стимуляцию в 20 Гц, так же, как в экспериментальном методе 4.3. В результате в группе мышей с ангиотензином II, обработанных ISO, был отмечен разрыв в проводящем блоке со стимуляцией по сравнению с контрольной группой мышей, обработанной ISO (фиг. 10d). В результате анализа продолжительности потенциала действия 75 (APD75) по сравнению с группой плацебо, мыши, обработанные ангиотензином II, продемонстрировали значительно увеличенную продолжительность потенциала действия, но показали незначительные изменения при обработке ISO (фиг. 10e). В результате анализа скорости проводимости было установлено, что группа мышей, которым вводили ангиотензин II, показала гораздо более медленную скорость по сравнению с плацебо, и обработка ISO не привела к восстановлению скорости проводимости (фиг. 10f). Из этих результатов было обнаружено, что инъекция ангиотензина II делала β-адренергические рецепторы нечувствительными к стимулу.
Экспериментальный пример 2.3. Определение терапевтического эффекта AAV9-CCN5 и AAV9-SERCA2a-P2A-CCN5 на мышиной модели желудочковой аритмии
Модель желудочковой аритмии была надежно получена экспериментальным методом, как описано в экспериментальном примере 2.3. Ангиотензин II вводили мышам в течение 2 недель, а в конце недели 2 вектор AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5 вводили в хвостовую вену мыши. Через 6 недель эксперимент прекращали и проводили эксперименты, связанные с желудочковой аритмией.
Эхокардиографию проводили для определения сердечной функции, пораженной AAV9-CCN5 и AAV9-SERCA2a-P2A-CCN5 на модели мышей с индуцированной ангиотензином II желудочковой аритмией. Инъекция AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5 вызывала фракцию укорочения, которая было уменьшена ангиотензином II, чтобы показать фракцию укорочения, которая является уровнем плацебо. Кроме того, в результате анализа толщины стенки левого желудочка (LVSd) было установлено, что толщина стенки левого желудочка, которая была уменьшена ангиотензином II, восстанавливалась почти до нормального уровня AAV9-SERCA2a-P2A-CCN5 (фиг. 11a и 11b).
Потенциал действия Ca2+ был показан с помощью оптического картирования. Было обнаружено, что при воздействии на желудочек электрической стимуляцией в 20 Гц проводимость регулярно распространялась в группе мышей, которым вводили AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5, независимо от обработки ISO (фиг. 11c и 11d). Потенциал действия кальция был представлен графически и проиллюстрирован на фиг. 11e.
Для соответствующих групп плацебо, AngII, AngII+AAV9-CCN5, AngII+AAV9-SERCA2a-P2A-CCN5 проводили обработку ISO и анализ выполняли путем построения карт реполяризации. В результате было установлено, что инъекция AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5 вызывала реполяризацию, которая была замедлена инъекцией ангиотензина II, чтобы проявлять реполяризацию, имеющую сходный характер с плацебо (фиг. 11f).
В результате анализа продолжительности потенциала действия 75 (APD75) Ca2+, было обнаружено, что продолжительность потенциала действия, которая увеличивалась ангиотензином II, уменьшалась путем инъекции AAV9-CCN5 (фиг. 11g) и надежно снижалась до уровня плацебо с помощью AAV9-SERCA2a-P2A-CCN5. Кроме того, было установлено, что скорость проводимости и скорости деполяризации, которые замедлялись ангиотензином II, нормализовались путем инъекции AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5 (фиг. 11h и 11i). На основании этих результатов было установлено, что AAV9-CCN5 и AAV9-SERCA2a-P2A-CCN5 были эффективными при лечении желудочковой аритмии, индуцированной ангиотензином II.
Кроме того, желудочковая аритмия, вызванная запрограммированной электрической стимуляцией (PES), была идентифицирована в модели с инъекцией ангиотензина II (фиг. 12a). В результате анализа частоты стимуляции, чтобы вызвать желудочковую аритмию, желудочковая аритмия была вызвана даже низкой частотой стимуляции в группе мышей AngII, тогда как желудочковая аритмия была вызвана частотой стимуляции, которая была равна или больше, чем уровень группы плацебо, в группе мышей, которым инъецировали AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5. В результате анализа частоты возникновения желудочковой аритмии желудочковая аритмия возникла с частотой эпизодов 5/5 в группе мышей AngII со скоростью эпизодов 1/5 в группе мышей AngII+AAV9-CCN5 и с частотой эпизодов 0/5 в группе мышей AngII+AAV9-SERCA2a-P2A-CCN5 (фиг. 12b и 12c).
Поскольку аритмия сердца тесно связана с фиброзом сердца, фиброз сердца определяли окрашиванием трихромом по Массону. В результате было обнаружено, что фиброз сердца, индуцированной ангиотензином II, был значительно уменьшен в сердце, в которое вводили AAV9-CCN5 или AAV9-SERCA2a-P2A-CCN5 (фиг. 13a).
Кроме того, это явление также вплотную ассоциируется с белками, связанными с каналом, который регулируют электрические импульсы сердца. Таким образом, экспрессию белков NaV1.5 и коннексин 43 в ткани сердца определяли с помощью вестерн-блоттинга. В результате было установлено, что снижение экспрессии связанных с каналом белков NaV1.5 и коннексин 43, которое было вызвано ангиотензином II, восстанавливалось с помощью AAV9-SERCA2a-P2A-CCN5 (фиг. 13b). На основании этих результатов было установлено, что генная терапия с использованием только вектора CCN5 или совместной экспрессии SERCA2A и CCN5 также может подавлять возникновение желудочковой тахикардии, индуцированной ангиотензином II.
--->
Перечень последовательностей
<110> BethphaGen Inc.
<120> ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ
АРИТМИИ СЕРДЦА
<130> PCB807072BPG
<150> KR 2017/127550
<151> 2017-09-29
<160> 42
<170> KoPatentIn 3.0
<210> 1
<211> 250
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Arg Gly Thr Pro Lys Thr His Leu Leu Ala Phe Ser Leu Leu Cys
1 5 10 15
Leu Leu Ser Lys Val Arg Thr Gln Leu Cys Pro Thr Pro Cys Thr Cys
20 25 30
Pro Trp Pro Pro Pro Arg Cys Pro Leu Gly Val Pro Leu Val Leu Asp
35 40 45
Gly Cys Gly Cys Cys Arg Val Cys Ala Arg Arg Leu Gly Glu Pro Cys
50 55 60
Asp Gln Leu His Val Cys Asp Ala Ser Gln Gly Leu Val Cys Gln Pro
65 70 75 80
Gly Ala Gly Pro Gly Gly Arg Gly Ala Leu Cys Leu Leu Ala Glu Asp
85 90 95
Asp Ser Ser Cys Glu Val Asn Gly Arg Leu Tyr Arg Glu Gly Glu Thr
100 105 110
Phe Gln Pro His Cys Ser Ile Arg Cys Arg Cys Glu Asp Gly Gly Phe
115 120 125
Thr Cys Val Pro Leu Cys Ser Glu Asp Val Arg Leu Pro Ser Trp Asp
130 135 140
Cys Pro His Pro Arg Arg Val Glu Val Leu Gly Lys Cys Cys Pro Glu
145 150 155 160
Trp Val Cys Gly Gln Gly Gly Gly Leu Gly Thr Gln Pro Leu Pro Ala
165 170 175
Gln Gly Pro Gln Phe Ser Gly Leu Val Ser Ser Leu Pro Pro Gly Val
180 185 190
Pro Cys Pro Glu Trp Ser Thr Ala Trp Gly Pro Cys Ser Thr Thr Cys
195 200 205
Gly Leu Gly Met Ala Thr Arg Val Ser Asn Gln Asn Arg Phe Cys Arg
210 215 220
Leu Glu Thr Gln Arg Arg Leu Cys Leu Ser Arg Pro Cys Pro Pro Ser
225 230 235 240
Arg Gly Arg Ser Pro Gln Asn Ser Ala Phe
245 250
<210> 2
<211> 750
<212> ДНК
<213> Homo sapiens
<400> 2
atgagaggca caccgaagac ccacctcctg gccttctccc tcctctgcct cctctcaaag 60
gtgcgtaccc agctgtgccc gacaccatgt acctgcccct ggccacctcc ccgatgcccg 120
ctgggagtac ccctggtgct ggatggctgt ggctgctgcc gggtatgtgc acggcggctg 180
ggggagccct gcgaccaact ccacgtctgc gacgccagcc agggcctggt ctgccagccc 240
ggggcaggac ccggtggccg gggggccctg tgcctcttgg cagaggacga cagcagctgt 300
gaggtgaacg gccgcctgta tcgggaaggg gagaccttcc agccccactg cagcatccgc 360
tgccgctgcg aggacggcgg cttcacctgc gtgccgctgt gcagcgagga tgtgcggctg 420
cccagctggg actgccccca ccccaggagg gtcgaggtcc tgggcaagtg ctgccctgag 480
tgggtgtgcg gccaaggagg gggactgggg acccagcccc ttccagccca aggaccccag 540
ttttctggcc ttgtctcttc cctgccccct ggtgtcccct gcccagaatg gagcacggcc 600
tggggaccct gctcgaccac ctgtgggctg ggcatggcca cccgggtgtc caaccagaac 660
cgcttctgcc gactggagac ccagcgccgc ctgtgcctgt ccaggccctg cccaccctcc 720
aggggtcgca gtccacaaaa cagtgccttc 750
<210> 3
<211> 997
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Met Glu Asn Ala His Thr Lys Thr Val Glu Glu Val Leu Gly His Phe
1 5 10 15
Gly Val Asn Glu Ser Thr Gly Leu Ser Leu Glu Gln Val Lys Lys Leu
20 25 30
Lys Glu Arg Trp Gly Ser Asn Glu Leu Pro Ala Glu Glu Gly Lys Thr
35 40 45
Leu Leu Glu Leu Val Ile Glu Gln Phe Glu Asp Leu Leu Val Arg Ile
50 55 60
Leu Leu Leu Ala Ala Cys Ile Ser Phe Val Leu Ala Trp Phe Glu Glu
65 70 75 80
Gly Glu Glu Thr Ile Thr Ala Phe Val Glu Pro Phe Val Ile Leu Leu
85 90 95
Ile Leu Val Ala Asn Ala Ile Val Gly Val Trp Gln Glu Arg Asn Ala
100 105 110
Glu Asn Ala Ile Glu Ala Leu Lys Glu Tyr Glu Pro Glu Met Gly Lys
115 120 125
Val Tyr Arg Gln Asp Arg Lys Ser Val Gln Arg Ile Lys Ala Lys Asp
130 135 140
Ile Val Pro Gly Asp Ile Val Glu Ile Ala Val Gly Asp Lys Val Pro
145 150 155 160
Ala Asp Ile Arg Leu Thr Ser Ile Lys Ser Thr Thr Leu Arg Val Asp
165 170 175
Gln Ser Ile Leu Thr Gly Glu Ser Val Ser Val Ile Lys His Thr Asp
180 185 190
Pro Val Pro Asp Pro Arg Ala Val Asn Gln Asp Lys Lys Asn Met Leu
195 200 205
Phe Ser Gly Thr Asn Ile Ala Ala Gly Lys Ala Met Gly Val Val Val
210 215 220
Ala Thr Gly Val Asn Thr Glu Ile Gly Lys Ile Arg Asp Glu Met Val
225 230 235 240
Ala Thr Glu Gln Glu Arg Thr Pro Leu Gln Gln Lys Leu Asp Glu Phe
245 250 255
Gly Glu Gln Leu Ser Lys Val Ile Ser Leu Ile Cys Ile Ala Val Trp
260 265 270
Ile Ile Asn Ile Gly His Phe Asn Asp Pro Val His Gly Gly Ser Trp
275 280 285
Ile Arg Gly Ala Ile Tyr Tyr Phe Lys Ile Ala Val Ala Leu Ala Val
290 295 300
Ala Ala Ile Pro Glu Gly Leu Pro Ala Val Ile Thr Thr Cys Leu Ala
305 310 315 320
Leu Gly Thr Arg Arg Met Ala Lys Lys Asn Ala Ile Val Arg Ser Leu
325 330 335
Pro Ser Val Glu Thr Leu Gly Cys Thr Ser Val Ile Cys Ser Asp Lys
340 345 350
Thr Gly Thr Leu Thr Thr Asn Gln Met Ser Val Cys Arg Met Phe Ile
355 360 365
Leu Asp Arg Val Glu Gly Asp Thr Cys Ser Leu Asn Glu Phe Thr Ile
370 375 380
Thr Gly Ser Thr Tyr Ala Pro Ile Gly Glu Val His Lys Asp Asp Lys
385 390 395 400
Pro Val Asn Cys His Gln Tyr Asp Gly Leu Val Glu Leu Ala Thr Ile
405 410 415
Cys Ala Leu Cys Asn Asp Ser Ala Leu Asp Tyr Asn Glu Ala Lys Gly
420 425 430
Val Tyr Glu Lys Val Gly Glu Ala Thr Glu Thr Ala Leu Thr Cys Leu
435 440 445
Val Glu Lys Met Asn Val Phe Asp Thr Glu Leu Lys Gly Leu Ser Lys
450 455 460
Ile Glu Arg Ala Asn Ala Cys Asn Ser Val Ile Lys Gln Leu Met Lys
465 470 475 480
Lys Glu Phe Thr Leu Glu Phe Ser Arg Asp Arg Lys Ser Met Ser Val
485 490 495
Tyr Cys Thr Pro Asn Lys Pro Ser Arg Thr Ser Met Ser Lys Met Phe
500 505 510
Val Lys Gly Ala Pro Glu Gly Val Ile Asp Arg Cys Thr His Ile Arg
515 520 525
Val Gly Ser Thr Lys Val Pro Met Thr Ser Gly Val Lys Gln Lys Ile
530 535 540
Met Ser Val Ile Arg Glu Trp Gly Ser Gly Ser Asp Thr Leu Arg Cys
545 550 555 560
Leu Ala Leu Ala Thr His Asp Asn Pro Leu Arg Arg Glu Glu Met His
565 570 575
Leu Glu Asp Ser Ala Asn Phe Ile Lys Tyr Glu Thr Asn Leu Thr Phe
580 585 590
Val Gly Cys Val Gly Met Leu Asp Pro Pro Arg Ile Glu Val Ala Ser
595 600 605
Ser Val Lys Leu Cys Arg Gln Ala Gly Ile Arg Val Ile Met Ile Thr
610 615 620
Gly Asp Asn Lys Gly Thr Ala Val Ala Ile Cys Arg Arg Ile Gly Ile
625 630 635 640
Phe Gly Gln Asp Glu Asp Val Thr Ser Lys Ala Phe Thr Gly Arg Glu
645 650 655
Phe Asp Glu Leu Asn Pro Ser Ala Gln Arg Asp Ala Cys Leu Asn Ala
660 665 670
Arg Cys Phe Ala Arg Val Glu Pro Ser His Lys Ser Lys Ile Val Glu
675 680 685
Phe Leu Gln Ser Phe Asp Glu Ile Thr Ala Met Thr Gly Asp Gly Val
690 695 700
Asn Asp Ala Pro Ala Leu Lys Lys Ala Glu Ile Gly Ile Ala Met Gly
705 710 715 720
Ser Gly Thr Ala Val Ala Lys Thr Ala Ser Glu Met Val Leu Ala Asp
725 730 735
Asp Asn Phe Ser Thr Ile Val Ala Ala Val Glu Glu Gly Arg Ala Ile
740 745 750
Tyr Asn Asn Met Lys Gln Phe Ile Arg Tyr Leu Ile Ser Ser Asn Val
755 760 765
Gly Glu Val Val Cys Ile Phe Leu Thr Ala Ala Leu Gly Phe Pro Glu
770 775 780
Ala Leu Ile Pro Val Gln Leu Leu Trp Val Asn Leu Val Thr Asp Gly
785 790 795 800
Leu Pro Ala Thr Ala Leu Gly Phe Asn Pro Pro Asp Leu Asp Ile Met
805 810 815
Asn Lys Pro Pro Arg Asn Pro Lys Glu Pro Leu Ile Ser Gly Trp Leu
820 825 830
Phe Phe Arg Tyr Leu Ala Ile Gly Cys Tyr Val Gly Ala Ala Thr Val
835 840 845
Gly Ala Ala Ala Trp Trp Phe Ile Ala Ala Asp Gly Gly Pro Arg Val
850 855 860
Ser Phe Tyr Gln Leu Ser His Phe Leu Gln Cys Lys Glu Asp Asn Pro
865 870 875 880
Asp Phe Glu Gly Val Asp Cys Ala Ile Phe Glu Ser Pro Tyr Pro Met
885 890 895
Thr Met Ala Leu Ser Val Leu Val Thr Ile Glu Met Cys Asn Ala Leu
900 905 910
Asn Ser Leu Ser Glu Asn Gln Ser Leu Leu Arg Met Pro Pro Trp Glu
915 920 925
Asn Ile Trp Leu Val Gly Ser Ile Cys Leu Ser Met Ser Leu His Phe
930 935 940
Leu Ile Leu Tyr Val Glu Pro Leu Pro Leu Ile Phe Gln Ile Thr Pro
945 950 955 960
Leu Asn Val Thr Gln Trp Leu Met Val Leu Lys Ile Ser Leu Pro Val
965 970 975
Ile Leu Met Asp Glu Thr Leu Lys Phe Val Ala Arg Asn Tyr Leu Glu
980 985 990
Pro Ala Ile Leu Glu
995
<210> 4
<211> 2991
<212> ДНК
<213> Homo sapiens
<400> 4
atggagaacg cgcacaccaa gacggtggag gaggtgctgg gccacttcgg cgtcaacgag 60
agtacggggc tgagcctgga acaggtcaag aagcttaagg agagatgggg ctccaacgag 120
ttaccggctg aagaaggaaa aaccttgctg gaacttgtga ttgagcagtt tgaagacttg 180
ctagttagga ttttattact ggcagcatgt atatcttttg ttttggcttg gtttgaagaa 240
ggtgaagaaa caattacagc ctttgtagaa ccttttgtaa ttttactcat attagtagcc 300
aatgcaattg tgggtgtatg gcaggaaaga aatgctgaaa atgccatcga agcccttaag 360
gaatatgagc ctgaaatggg caaagtgtat cgacaggaca gaaagagtgt gcagcggatt 420
aaagctaaag acatagttcc tggtgatatt gtagaaattg ctgttggtga caaagttcct 480
gctgatataa ggttaacttc catcaaatct accacactaa gagttgacca gtcaattctc 540
acaggtgaat ctgtctctgt catcaagcac actgatcccg tccctgaccc acgagctgtc 600
aaccaagata aaaagaacat gctgttttct ggtacaaaca ttgctgctgg gaaagctatg 660
ggagtggtgg tagcaactgg agttaacacc gaaattggca agatccggga tgaaatggtg 720
gcaacagaac aggagagaac accccttcag caaaaactag atgaatttgg ggaacagctt 780
tccaaagtca tctcccttat ttgcattgca gtctggatca taaatattgg gcacttcaat 840
gacccggttc atggagggtc ctggatcaga ggtgctattt actactttaa aattgcagtg 900
gccctggctg tagcagccat tcctgaaggt ctgcctgcag tcatcaccac ctgcctggct 960
cttggaactc gcagaatggc aaagaaaaat gccattgttc gaagcctccc gtctgtggaa 1020
acccttggtt gtacttctgt tatctgctca gacaagactg gtacacttac aacaaaccag 1080
atgtcagtct gcaggatgtt cattctggac agagtggaag gtgatacttg ttcccttaat 1140
gagtttacca taactggatc aacttatgca cctattggag aagtgcataa agatgataaa 1200
ccagtgaatt gtcaccagta tgatggtctg gtagaattag caacaatttg tgctctttgt 1260
aatgactctg ctttggatta caatgaggca aagggtgtgt atgaaaaagt tggagaagct 1320
acagagactg ctctcacttg cctagtagag aagatgaatg tatttgatac cgaattgaag 1380
ggtctttcta aaatagaacg tgcaaatgcc tgcaactcag tcattaaaca gctgatgaaa 1440
aaggaattca ctctagagtt ttcacgtgac agaaagtcaa tgtcggttta ctgtacacca 1500
aataaaccaa gcaggacatc aatgagcaag atgtttgtga agggtgctcc tgaaggtgtc 1560
attgacaggt gcacccacat tcgagttgga agtactaagg ttcctatgac ctctggagtc 1620
aaacagaaga tcatgtctgt cattcgagag tggggtagtg gcagcgacac actgcgatgc 1680
ctggccctgg ccactcatga caacccactg agaagagaag aaatgcacct tgaggactct 1740
gccaacttta ttaaatatga gaccaatctg accttcgttg gctgcgtggg catgctggat 1800
cctccgagaa tcgaggtggc ctcctccgtg aagctgtgcc ggcaagcagg catccgggtc 1860
atcatgatca ctggggacaa caagggcact gctgtggcca tctgtcgccg catcggcatc 1920
ttcgggcagg atgaggacgt gacgtcaaaa gctttcacag gccgggagtt tgatgaactc 1980
aacccctccg cccagcgaga cgcctgcctg aacgcccgct gttttgctcg agttgaaccc 2040
tcccacaagt ctaaaatcgt agaatttctt cagtcttttg atgagattac agctatgact 2100
ggcgatggcg tgaacgatgc tcctgctctg aagaaagccg agattggcat tgctatgggc 2160
tctggcactg cggtggctaa aaccgcctct gagatggtcc tggcggatga caacttctcc 2220
accattgtgg ctgccgttga ggaggggcgg gcaatctaca acaacatgaa acagttcatc 2280
cgctacctca tctcgtccaa cgtcggggaa gttgtctgta ttttcctgac agcagccctt 2340
ggatttcccg aggctttgat tcctgttcag ctgctctggg tcaatctggt gacagatggc 2400
ctgcctgcca ctgcactggg gttcaaccct cctgatctgg acatcatgaa taaacctccc 2460
cggaacccaa aggaaccatt gatcagcggg tggctctttt tccgttactt ggctattggc 2520
tgttacgtcg gcgctgctac cgtgggtgct gctgcatggt ggttcattgc tgctgacggt 2580
ggtccaagag tgtccttcta ccagctgagt catttcctac agtgtaaaga ggacaacccg 2640
gactttgaag gcgtggattg tgcaatcttt gaatccccat acccgatgac aatggcgctc 2700
tctgttctag taactataga aatgtgtaac gccctcaaca gcttgtccga aaaccagtcc 2760
ttgctgagga tgcccccctg ggagaacatc tggctcgtgg gctccatctg cctgtccatg 2820
tcactccact tcctgatcct ctatgtcgaa cccttgccac tcatcttcca gatcacaccg 2880
ctgaacgtga cccagtggct gatggtgctg aaaatctcct tgcccgtgat tctcatggat 2940
gagacgctca agtttgtggc ccgcaactac ctggaacctg caatactgga g 2991
<210> 5
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из
тешовируса 1 свиней
<400> 5
Gly Ser Gly Ala Thr Asn Phe Ser Leu Leu Lys Gln Ala Gly Asp Val
1 5 10 15
Glu Glu Asn Pro Gly Pro
20
<210> 6
<211> 66
<212> ДНК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из
тешовируса 1 свиней
<400> 6
ggaagcggag ctactaactt cagcctgctg aagcaggctg gagacgtgga ggagaaccct 60
ggacct 66
<210> 7
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из вируса Thosea asigna
<400> 7
Gly Ser Gly Glu Gly Arg Gly Ser Leu Leu Thr Cys Gly Asp Val Glu
1 5 10 15
Glu Asn Pro Gly Pro
20
<210> 8
<211> 63
<212> ДНК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из
вируса Thosea asigna
<400> 8
ggaagcggag agggcagagg aagtctgcta acatgcggtg acgtcgagga gaatcctgga 60
cct 63
<210> 9
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из
вируса ринита лошадей A (ERAV)
<400> 9
Gly Ser Gly Gln Cys Thr Asn Tyr Ala Leu Leu Lys Leu Ala Gly Asp
1 5 10 15
Val Glu Ser Asn Pro Gly Pro
20
<210> 10
<211> 69
<212> ДНК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из
вируса ринита A лошадей (ERAV)
<400> 10
ggaagcggac agtgtactaa ttatgctctc ttgaaattgg ctggagatgt tgagagcaac 60
cctggacct 69
<210> 11
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из FMDV
2A
<400> 11
Gly Ser Gly Val Lys Gln Thr Leu Asn Phe Asp Leu Leu Lys Leu Ala
1 5 10 15
Gly Asp Val Glu Ser Asn Pro Gly Pro
20 25
<210> 12
<211> 75
<212> ДНК
<213> Искусственная последовательность
<220>
<223> саморасщепляющаяся последовательность, полученная из FMDV
2A
<400> 12
ggaagcggag tgaaacagac tttgaatttt gaccttctca agttggcggg agacgtggag 60
tccaaccctg gacct 75
<210> 13
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для альфа-SMA мыши
<400> 13
cccacccaga gtggagaa 18
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для альфа-SMA мыши
<400> 14
acatagctgg agcagcgtct 20
<210> 15
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для Collagen I мыши
<400> 15
catgttcagc tttgtggacc t 21
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для Collagen I мыши
<400> 16
gacgctgact tcagggatgt 20
<210> 17
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для TGF-бета 1 мыши
<400> 17
tggagcaaca tgtggaactc 20
<210> 18
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для TGF-бета 1 мыши
<400> 18
cagcagccgg ttaccaag 18
<210> 19
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для IL-1 бета мыши
<400> 19
tccaggatga ggacatgatg agca 24
<210> 20
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для IL-1 бета мыши
<400> 20
gaacgtcaca cacaccagca ggtta 25
<210> 21
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для RANTES мыши
<400> 21
tgcagaggac tctgagacag c 21
<210> 22
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для RANTES мыши
<400> 22
gagtggtgtc cgagccata 19
<210> 23
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для F4/80 мыши
<400> 23
cctggacgaa tcctgtgaag 20
<210> 24
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для F4/80 мыши
<400> 24
ggtgggacca cagagagttg 20
<210> 25
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для MCP-1 мыши
<400> 25
catccacgtg ttggctca 18
<210> 26
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для MCP-1 мыши
<400> 26
gatcatcttg ctggtgaatg agt 23
<210> 27
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для CCN5 мыши
<400> 27
atacaggtgc caggaaggtg 20
<210> 28
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для CCN5 мыши
<400> 28
gttggatact cgggtggcta 20
<210> 29
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для GAPDH мыши
<400> 29
ctcatgacca cagtccatgc 20
<210> 30
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для GAPDH мыши
<400> 30
ttcagctctg ggatgacctt 20
<210> 31
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для 18s рРНК мыши
<400> 31
gtaacccgtt gaaccccatt 20
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для 18s рРРК мыши
<400> 32
ccatccaatc ggtagtagcg 20
<210> 33
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для альфа-SMA крысы
<400> 33
tctgtctcta gcacacaact gtgaatg 27
<210> 34
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для альфа-SMA крысы
<400> 34
ttgacaggcc agggctagaa ggg 23
<210> 35
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для Collagen I крысы
<400> 35
aatgcacttt tggtttttgg tcacgt 26
<210> 36
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для Collagen I крысы
<400> 36
cagcccactt tgccccaacc c 21
<210> 37
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для TGF-бета 1 крысы
<400> 37
tgttcgcgct ctcggcagtg 20
<210> 38
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для TGF-бета 1 крысы
<400> 38
cggatggcct cgatgcgctt 20
<210> 39
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Прямой праймер для GAPDH крысы
<400> 39
acccagccca gcaaggatac tg 22
<210> 40
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Обратный праймер для GAPDH крысы
<400> 40
attcgagaga agggagggct ccc 23
<210> 41
<211> 753
<212> ДНК
<213> Homo sapiens
<400> 41
atgagaggca caccgaagac ccacctcctg gccttctccc tcctctgcct cctctcaaag 60
gtgcgtaccc agctgtgccc gacaccatgt acctgcccct ggccacctcc ccgatgcccg 120
ctgggagtac ccctggtgct ggatggctgt ggctgctgcc gggtatgtgc acggcggctg 180
ggggagccct gcgaccaact ccacgtctgc gacgccagcc agggcctggt ctgccagccc 240
ggggcaggac ccggtggccg gggggccctg tgcctcttgg cagaggacga cagcagctgt 300
gaggtgaacg gccgcctgta tcgggaaggg gagaccttcc agccccactg cagcatccgc 360
tgccgctgcg aggacggcgg cttcacctgc gtgccgctgt gcagcgagga tgtgcggctg 420
cccagctggg actgccccca ccccaggagg gtcgaggtcc tgggcaagtg ctgccctgag 480
tgggtgtgcg gccaaggagg gggactgggg acccagcccc ttccagccca aggaccccag 540
ttttctggcc ttgtctcttc cctgccccct ggtgtcccct gcccagaatg gagcacggcc 600
tggggaccct gctcgaccac ctgtgggctg ggcatggcca cccgggtgtc caaccagaac 660
cgcttctgcc gactggagac ccagcgccgc ctgtgcctgt ccaggccctg cccaccctcc 720
aggggtcgca gtccacaaaa cagtgccttc tag 753
<210> 42
<211> 2994
<212> ДНК
<213> Homo sapiens
<400> 42
atggagaacg cgcacaccaa gacggtggag gaggtgctgg gccacttcgg cgtcaacgag 60
agtacggggc tgagcctgga acaggtcaag aagcttaagg agagatgggg ctccaacgag 120
ttaccggctg aagaaggaaa aaccttgctg gaacttgtga ttgagcagtt tgaagacttg 180
ctagttagga ttttattact ggcagcatgt atatcttttg ttttggcttg gtttgaagaa 240
ggtgaagaaa caattacagc ctttgtagaa ccttttgtaa ttttactcat attagtagcc 300
aatgcaattg tgggtgtatg gcaggaaaga aatgctgaaa atgccatcga agcccttaag 360
gaatatgagc ctgaaatggg caaagtgtat cgacaggaca gaaagagtgt gcagcggatt 420
aaagctaaag acatagttcc tggtgatatt gtagaaattg ctgttggtga caaagttcct 480
gctgatataa ggttaacttc catcaaatct accacactaa gagttgacca gtcaattctc 540
acaggtgaat ctgtctctgt catcaagcac actgatcccg tccctgaccc acgagctgtc 600
aaccaagata aaaagaacat gctgttttct ggtacaaaca ttgctgctgg gaaagctatg 660
ggagtggtgg tagcaactgg agttaacacc gaaattggca agatccggga tgaaatggtg 720
gcaacagaac aggagagaac accccttcag caaaaactag atgaatttgg ggaacagctt 780
tccaaagtca tctcccttat ttgcattgca gtctggatca taaatattgg gcacttcaat 840
gacccggttc atggagggtc ctggatcaga ggtgctattt actactttaa aattgcagtg 900
gccctggctg tagcagccat tcctgaaggt ctgcctgcag tcatcaccac ctgcctggct 960
cttggaactc gcagaatggc aaagaaaaat gccattgttc gaagcctccc gtctgtggaa 1020
acccttggtt gtacttctgt tatctgctca gacaagactg gtacacttac aacaaaccag 1080
atgtcagtct gcaggatgtt cattctggac agagtggaag gtgatacttg ttcccttaat 1140
gagtttacca taactggatc aacttatgca cctattggag aagtgcataa agatgataaa 1200
ccagtgaatt gtcaccagta tgatggtctg gtagaattag caacaatttg tgctctttgt 1260
aatgactctg ctttggatta caatgaggca aagggtgtgt atgaaaaagt tggagaagct 1320
acagagactg ctctcacttg cctagtagag aagatgaatg tatttgatac cgaattgaag 1380
ggtctttcta aaatagaacg tgcaaatgcc tgcaactcag tcattaaaca gctgatgaaa 1440
aaggaattca ctctagagtt ttcacgtgac agaaagtcaa tgtcggttta ctgtacacca 1500
aataaaccaa gcaggacatc aatgagcaag atgtttgtga agggtgctcc tgaaggtgtc 1560
attgacaggt gcacccacat tcgagttgga agtactaagg ttcctatgac ctctggagtc 1620
aaacagaaga tcatgtctgt cattcgagag tggggtagtg gcagcgacac actgcgatgc 1680
ctggccctgg ccactcatga caacccactg agaagagaag aaatgcacct tgaggactct 1740
gccaacttta ttaaatatga gaccaatctg accttcgttg gctgcgtggg catgctggat 1800
cctccgagaa tcgaggtggc ctcctccgtg aagctgtgcc ggcaagcagg catccgggtc 1860
atcatgatca ctggggacaa caagggcact gctgtggcca tctgtcgccg catcggcatc 1920
ttcgggcagg atgaggacgt gacgtcaaaa gctttcacag gccgggagtt tgatgaactc 1980
aacccctccg cccagcgaga cgcctgcctg aacgcccgct gttttgctcg agttgaaccc 2040
tcccacaagt ctaaaatcgt agaatttctt cagtcttttg atgagattac agctatgact 2100
ggcgatggcg tgaacgatgc tcctgctctg aagaaagccg agattggcat tgctatgggc 2160
tctggcactg cggtggctaa aaccgcctct gagatggtcc tggcggatga caacttctcc 2220
accattgtgg ctgccgttga ggaggggcgg gcaatctaca acaacatgaa acagttcatc 2280
cgctacctca tctcgtccaa cgtcggggaa gttgtctgta ttttcctgac agcagccctt 2340
ggatttcccg aggctttgat tcctgttcag ctgctctggg tcaatctggt gacagatggc 2400
ctgcctgcca ctgcactggg gttcaaccct cctgatctgg acatcatgaa taaacctccc 2460
cggaacccaa aggaaccatt gatcagcggg tggctctttt tccgttactt ggctattggc 2520
tgttacgtcg gcgctgctac cgtgggtgct gctgcatggt ggttcattgc tgctgacggt 2580
ggtccaagag tgtccttcta ccagctgagt catttcctac agtgtaaaga ggacaacccg 2640
gactttgaag gcgtggattg tgcaatcttt gaatccccat acccgatgac aatggcgctc 2700
tctgttctag taactataga aatgtgtaac gccctcaaca gcttgtccga aaaccagtcc 2760
ttgctgagga tgcccccctg ggagaacatc tggctcgtgg gctccatctg cctgtccatg 2820
tcactccact tcctgatcct ctatgtcgaa cccttgccac tcatcttcca gatcacaccg 2880
ctgaacgtga cccagtggct gatggtgctg aaaatctcct tgcccgtgat tctcatggat 2940
gagacgctca agtttgtggc ccgcaactac ctggaacctg caatactgga gtag 2994
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ СЕРДЕЧНОЙ НЕДОСТАТОЧНОСТИ | 2018 |
|
RU2788124C2 |
| СПОСОБЫ ЛЕЧЕНИЯ БОЛЕЗНИ ДАНОНА И ДРУГИХ НАРУШЕНИЙ АУТОФАГИИ | 2017 |
|
RU2777571C2 |
| ОПТИМИЗИРОВАННЫЕ ГЕНЫ И ЭКСПРЕССИОННЫЕ КАССЕТЫ МИНИ-ДИСТРОФИНА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2753194C2 |
| Синергетическое действие SMN1 и miR-23a при лечении спинальной мышечной атрофии | 2021 |
|
RU2839898C2 |
| СКОНСТРУИРОВАННЫЕ АДЕНОАССОЦИИРОВАННЫЕ (AAV) ВЕКТОРЫ ДЛЯ ЭКСПРЕССИИ ТРАНСГЕНА | 2020 |
|
RU2839774C2 |
| ВАРИАНТЫ КИСЛОЙ АЛЬФА-ГЛЮКОЗИДАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2017 |
|
RU2780410C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2809246C2 |
| ГЕНОТЕРАПЕВТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА | 2020 |
|
RU2807158C2 |
| Способ получения модифицированного капсида аденоассоциированного вируса | 2021 |
|
RU2831732C1 |
| Выделенный модифицированный белок VPI капсида аденоассоциированного вируса 9 серотипа (AAV9), капсид и вектор на его основе | 2021 |
|
RU2825667C2 |



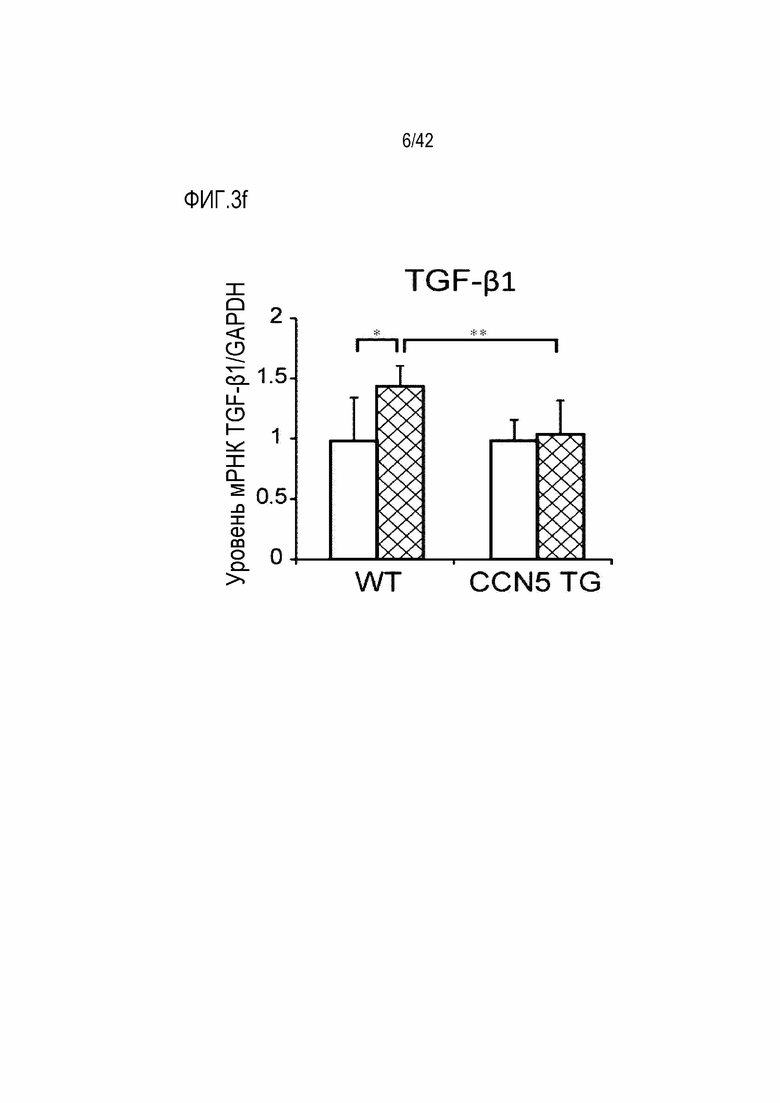

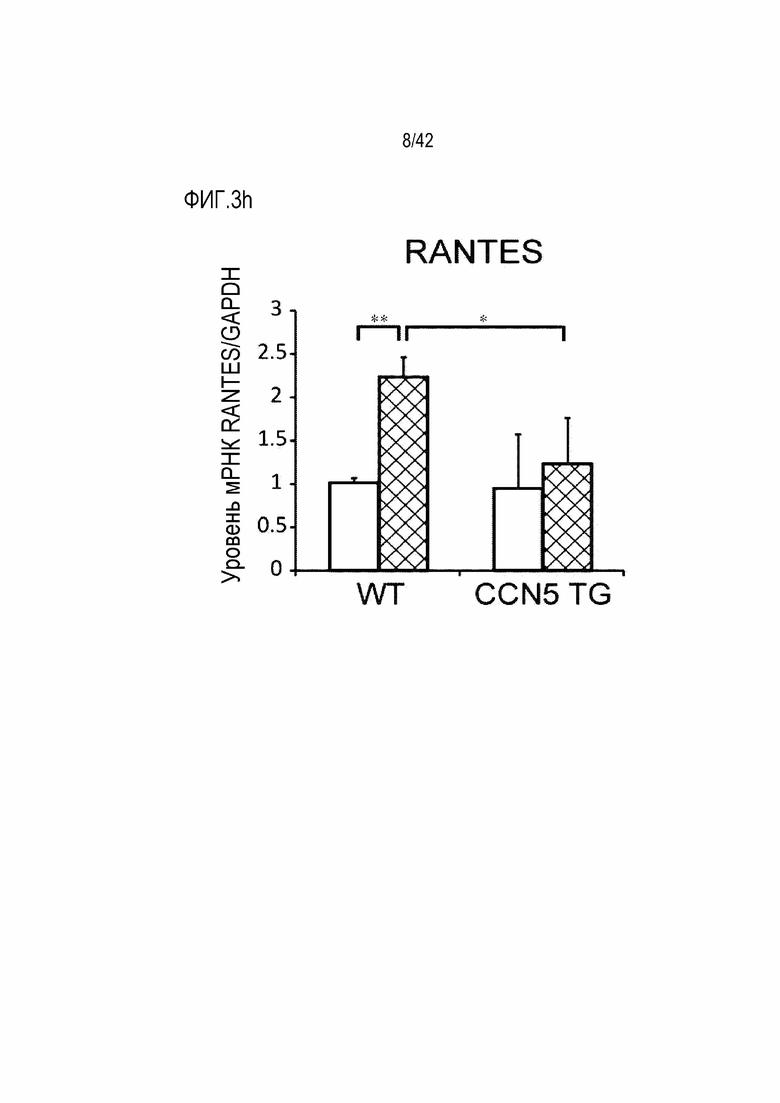
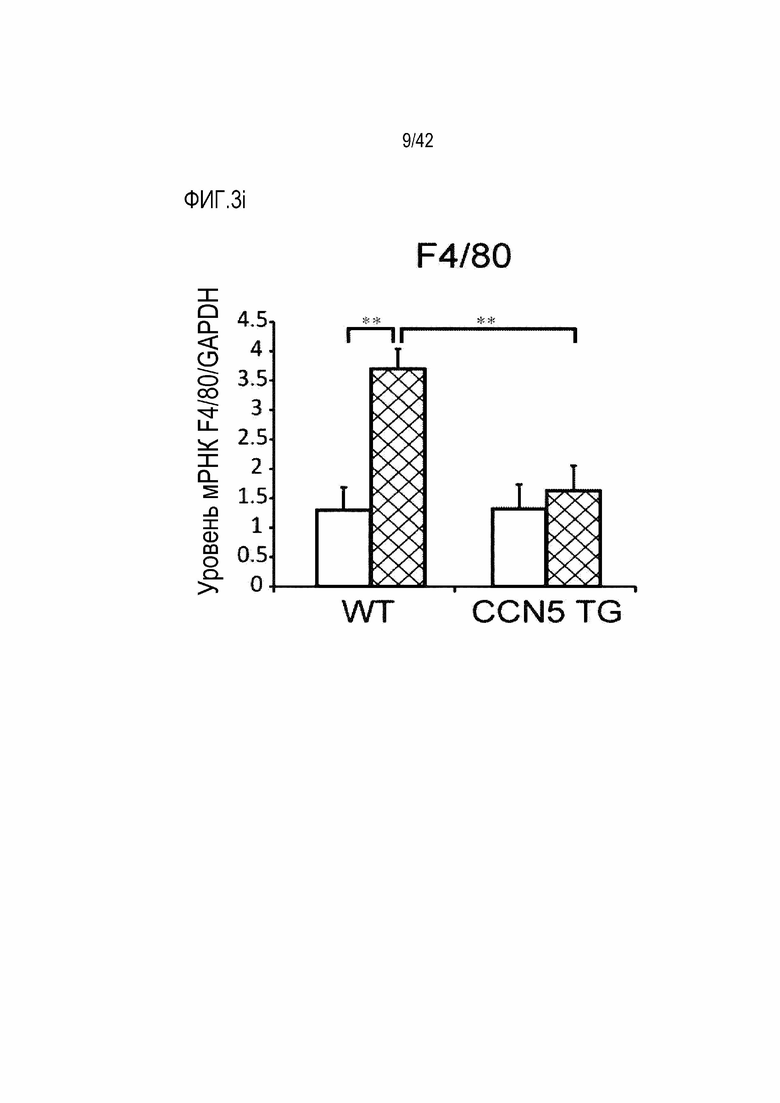
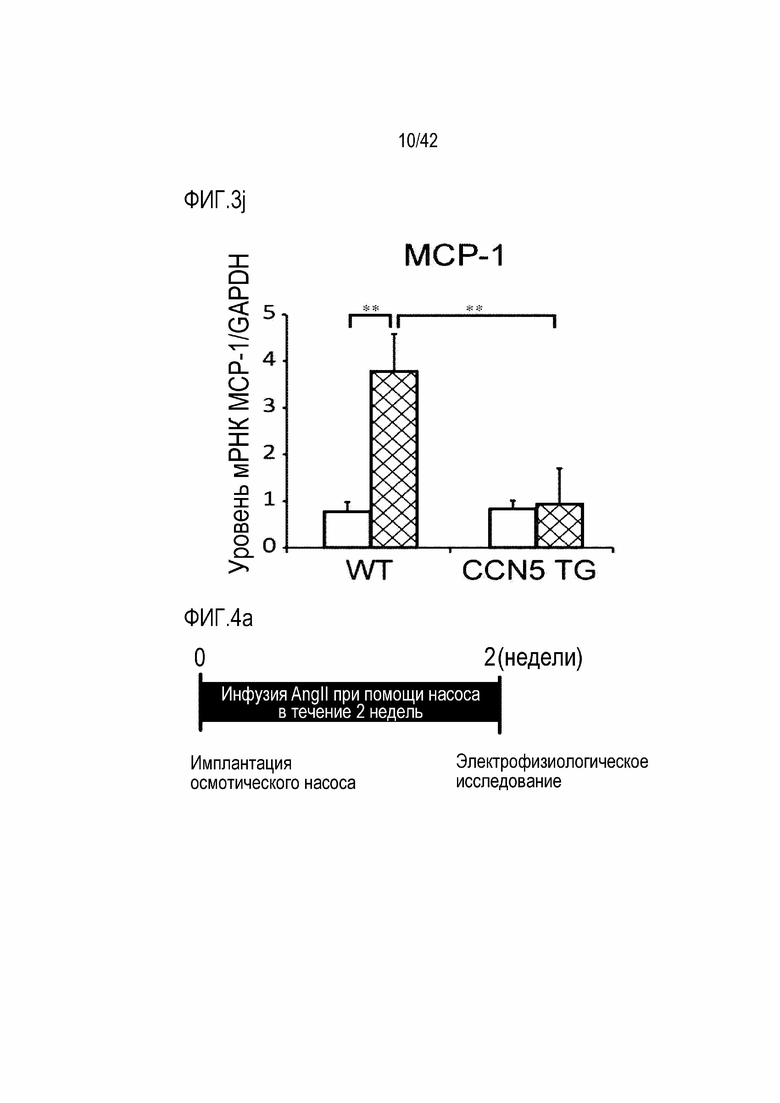
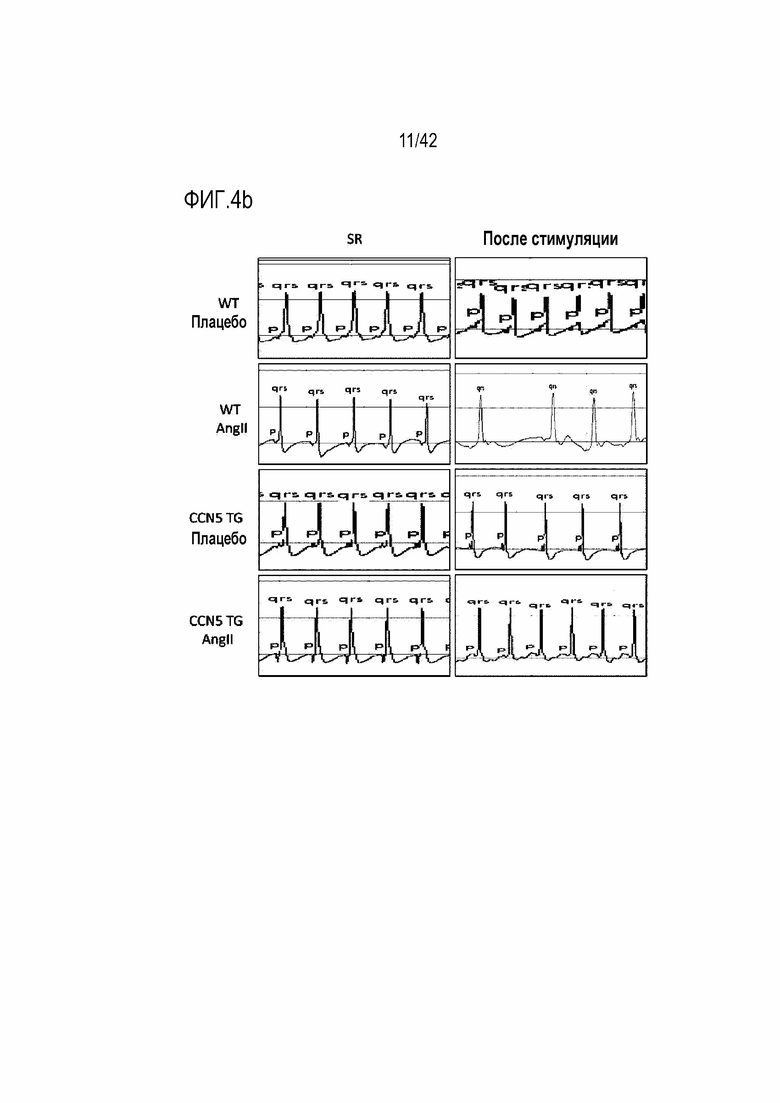
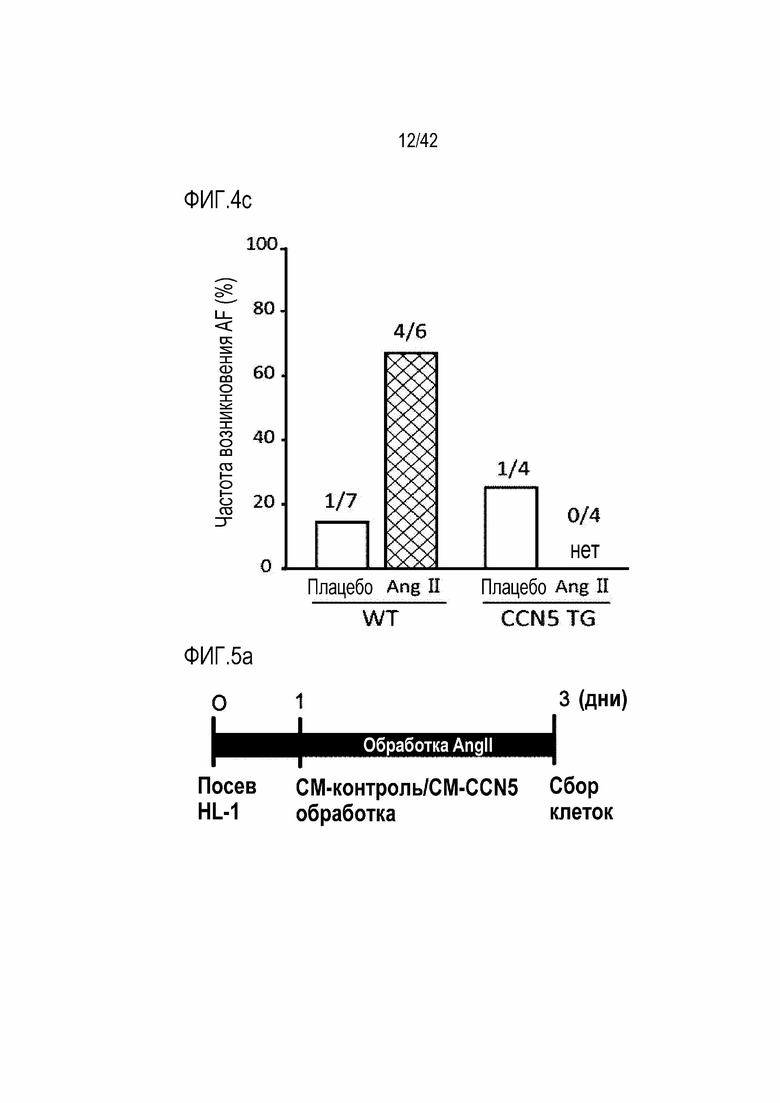
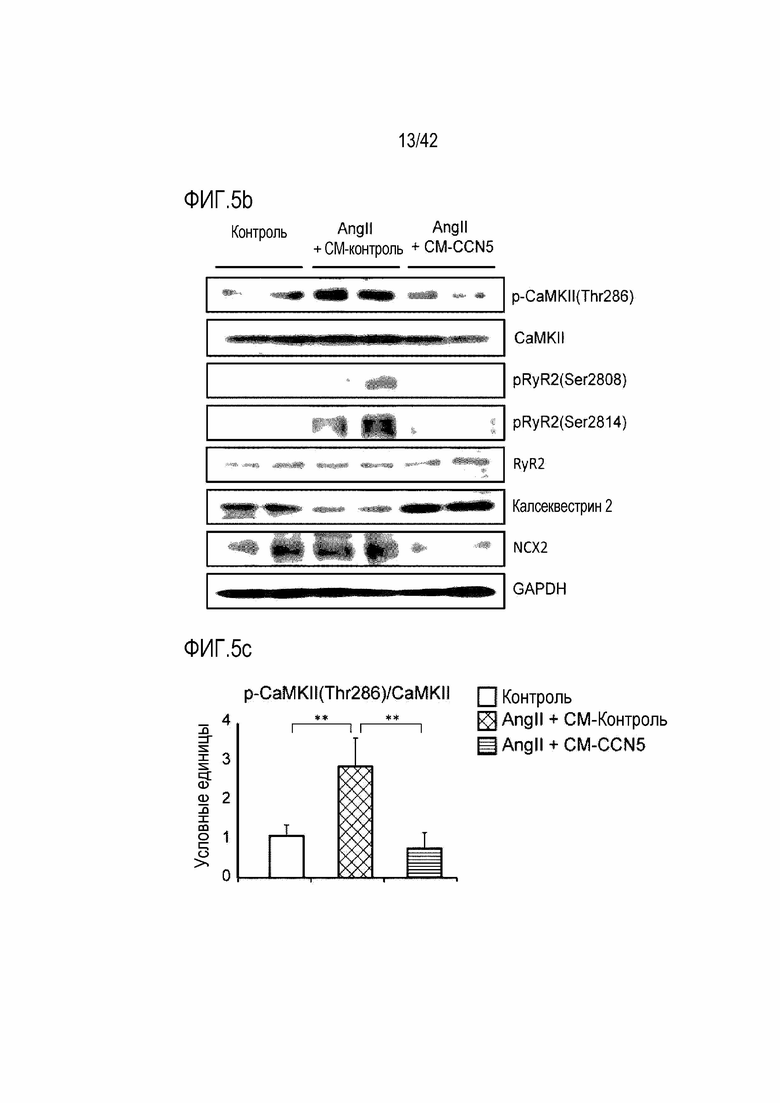
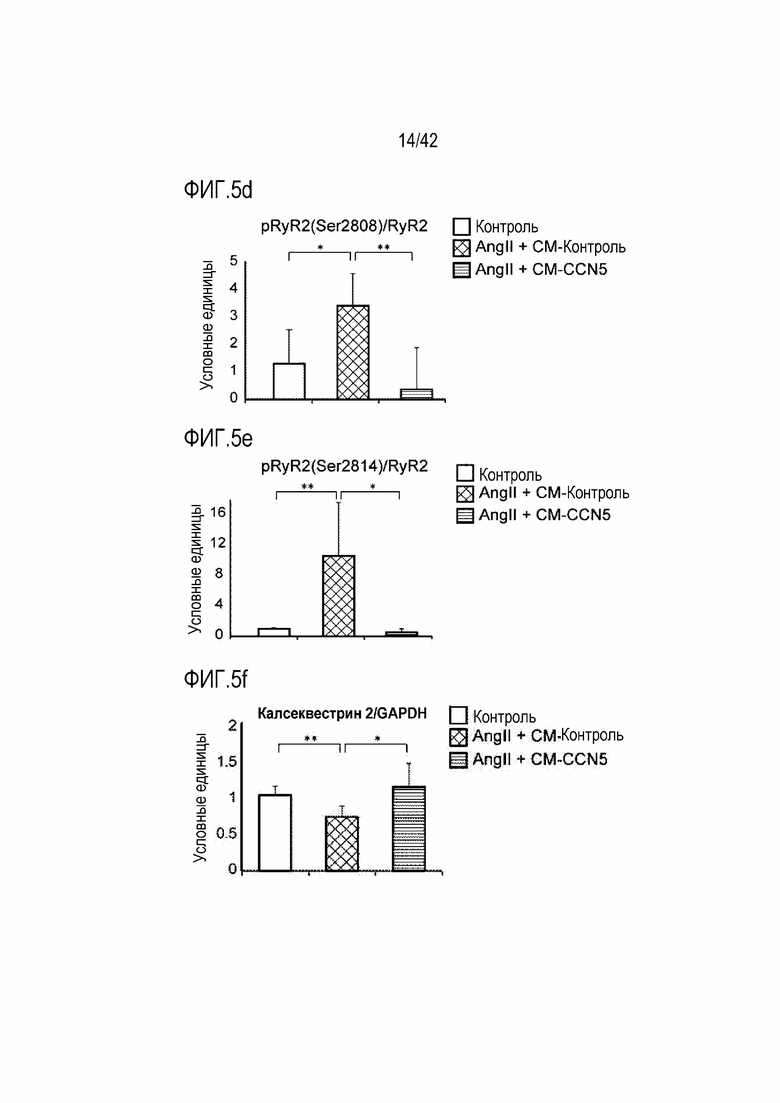
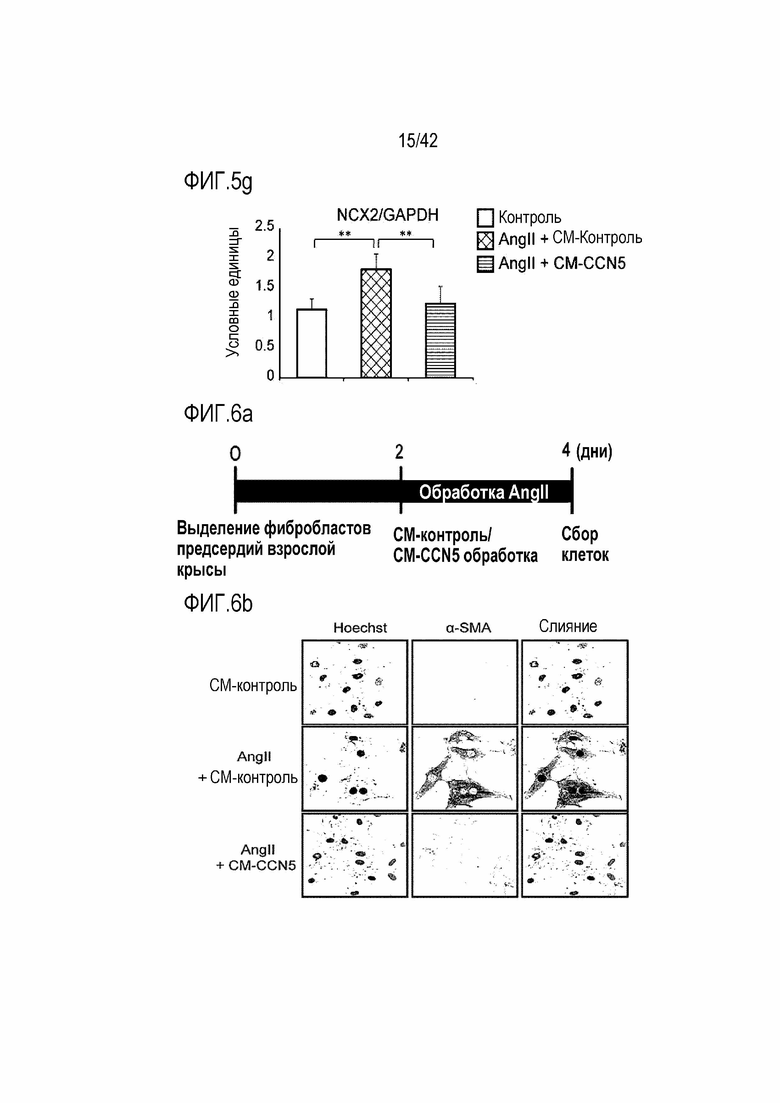

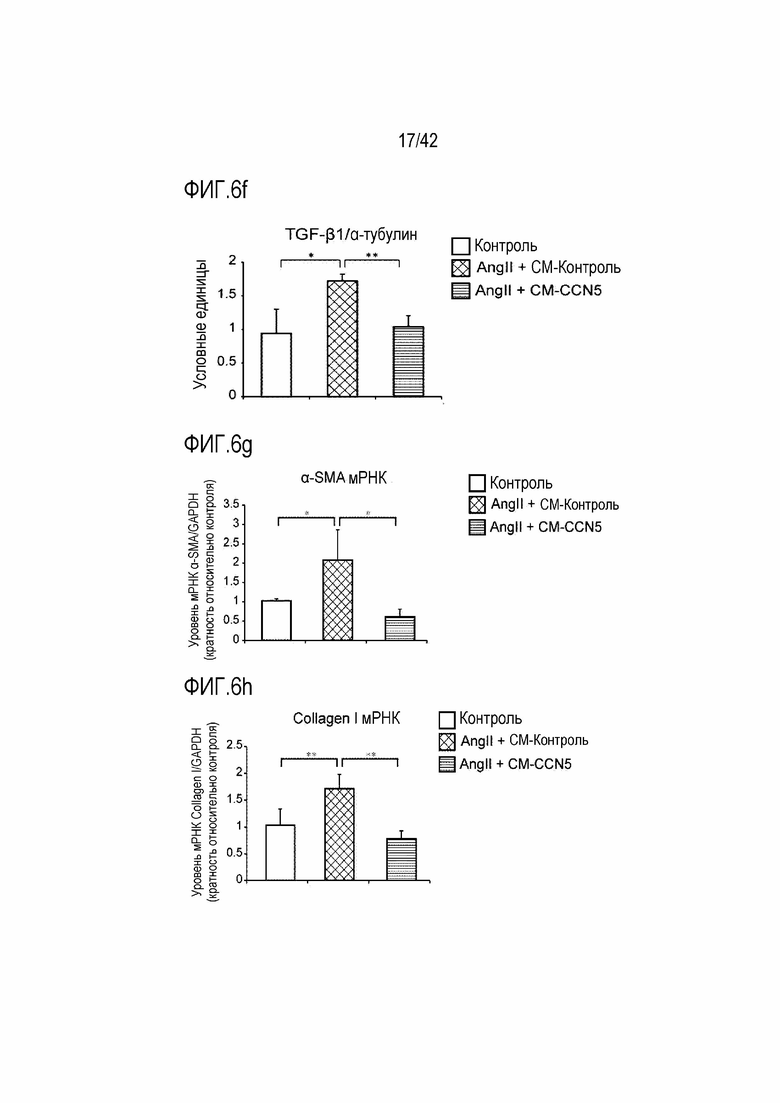
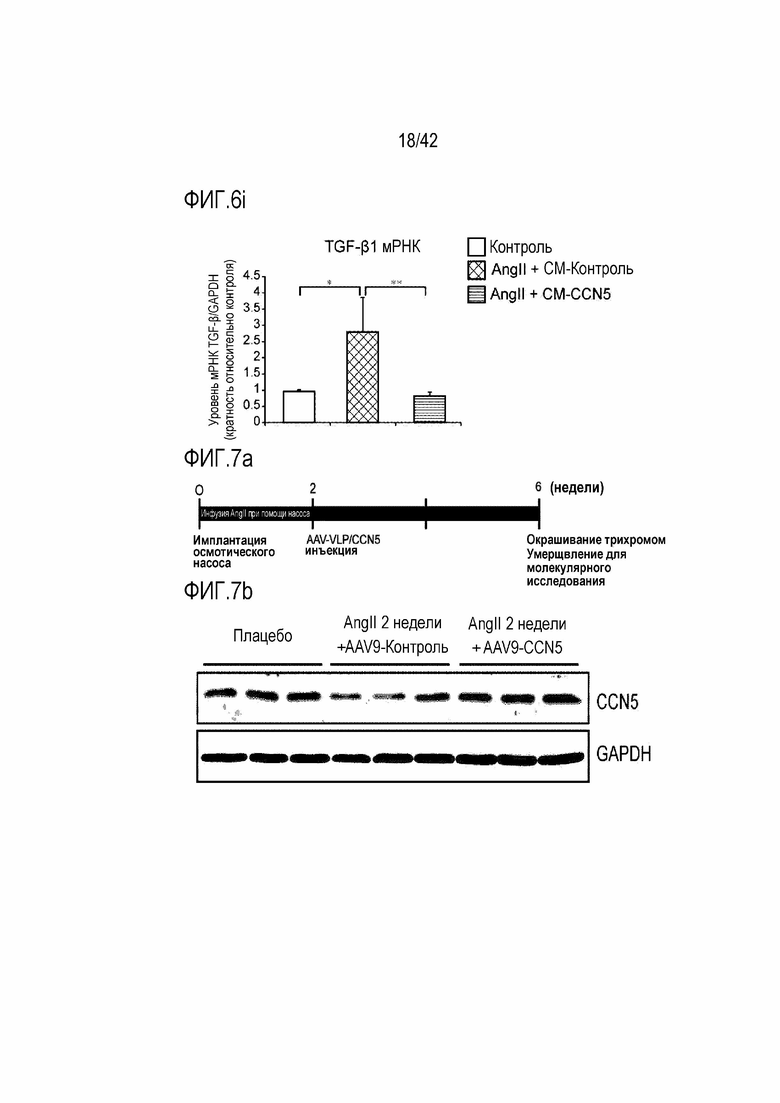
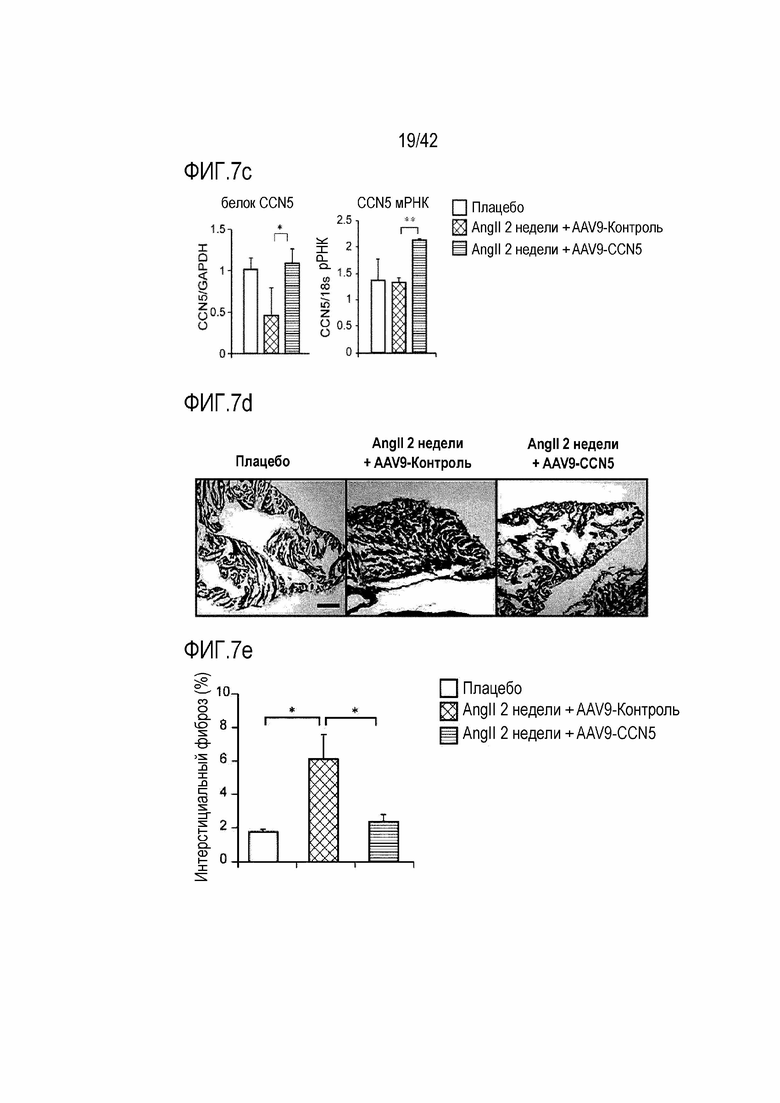
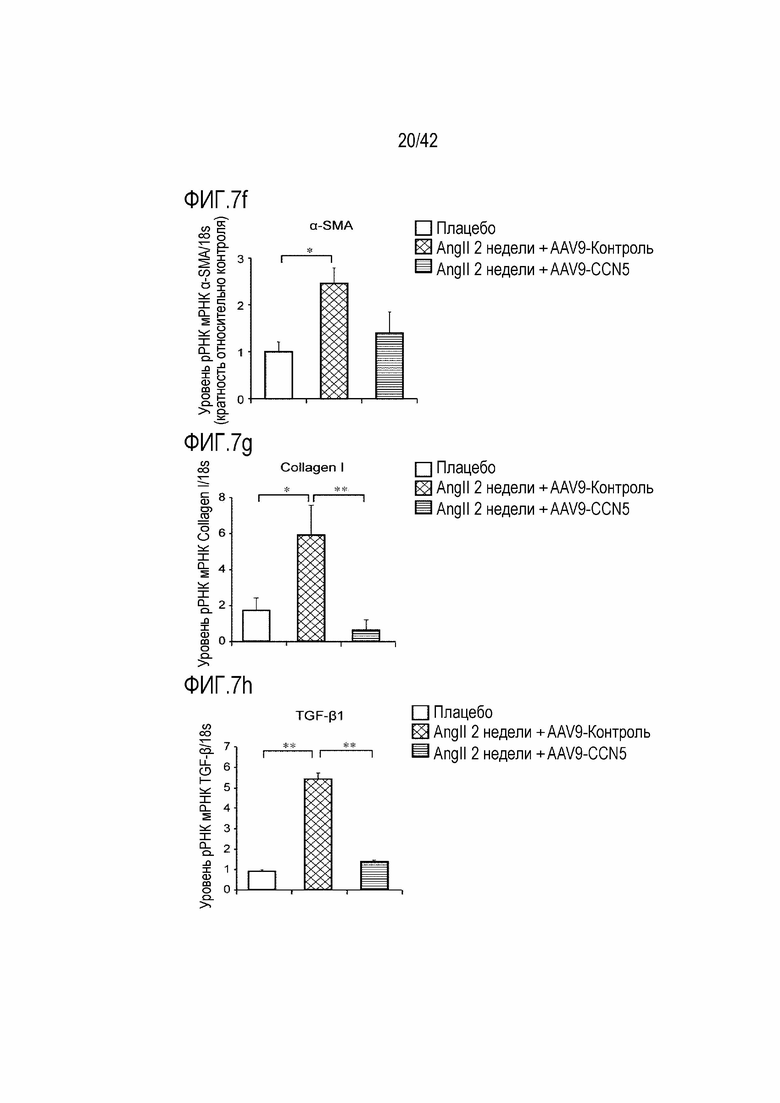
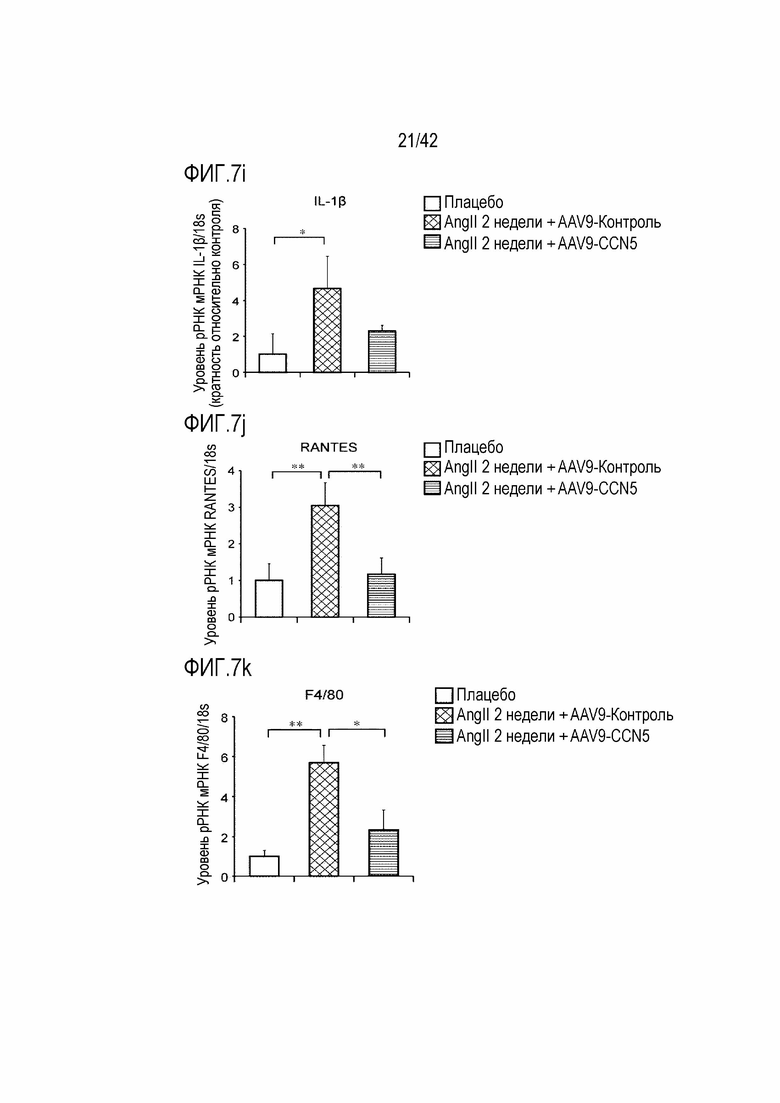
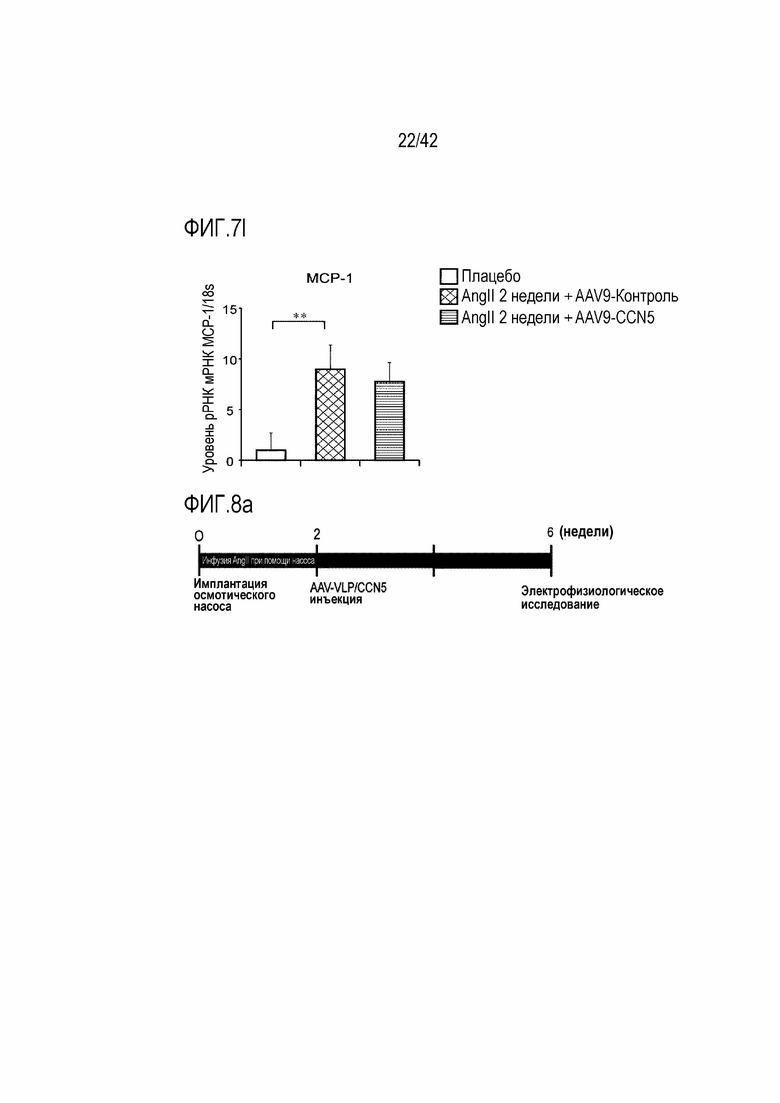
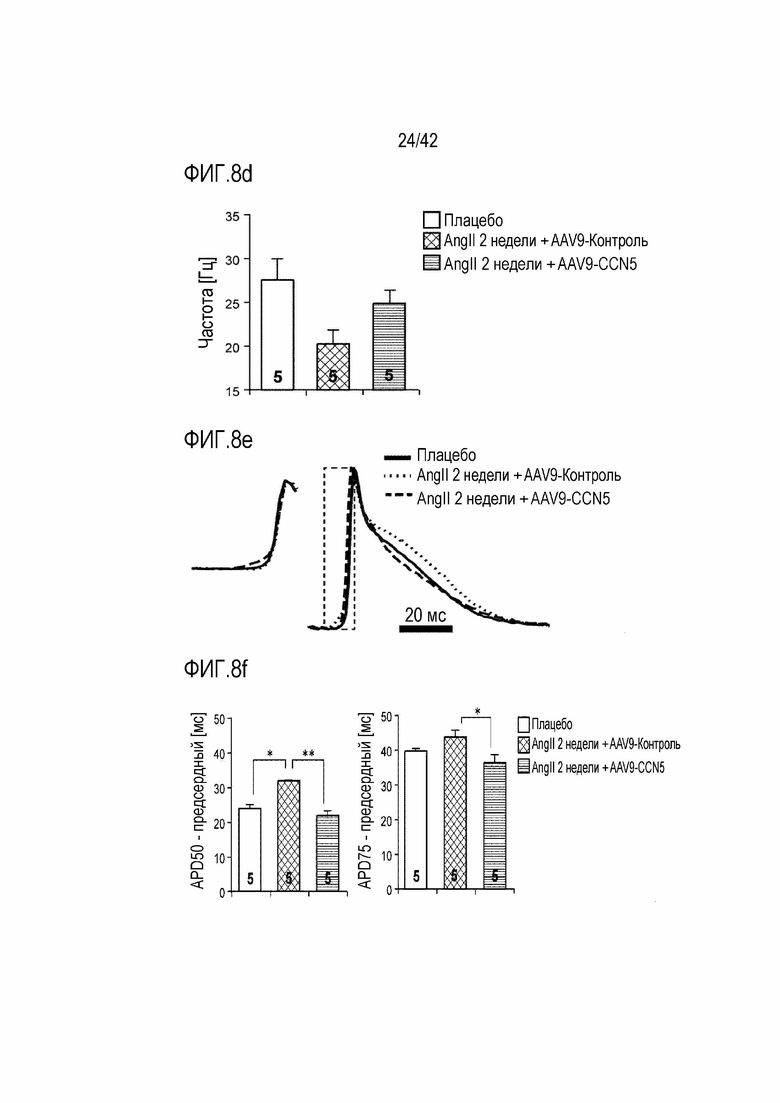
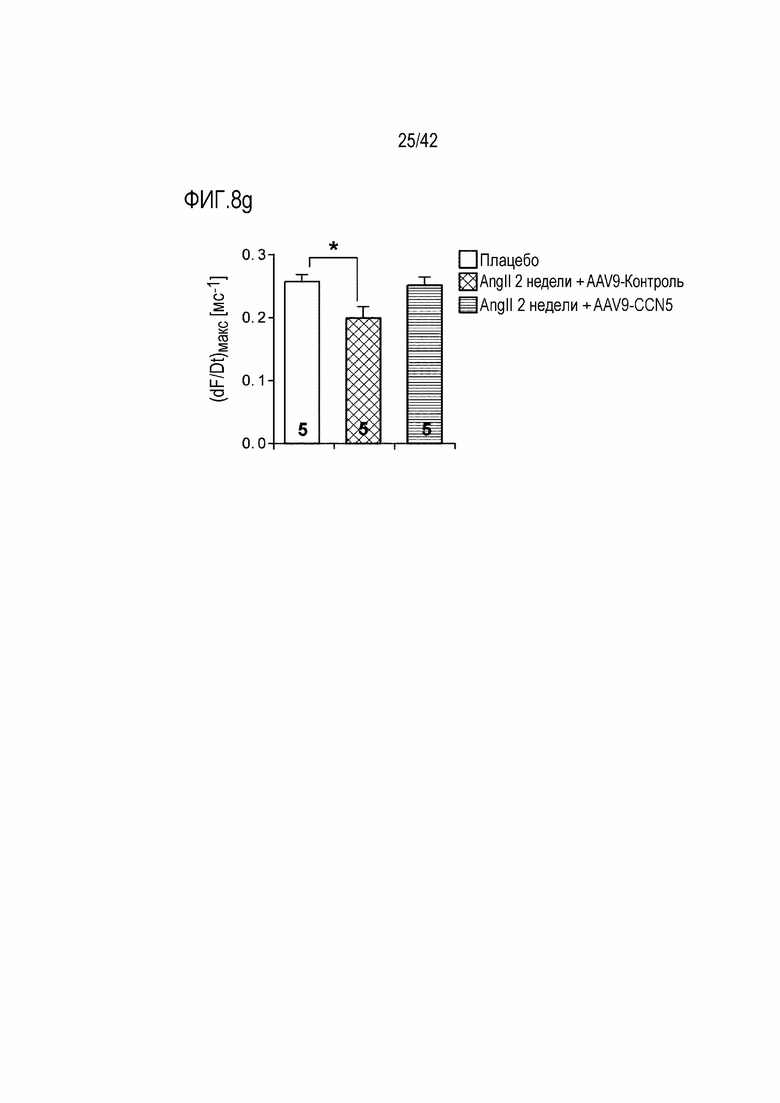
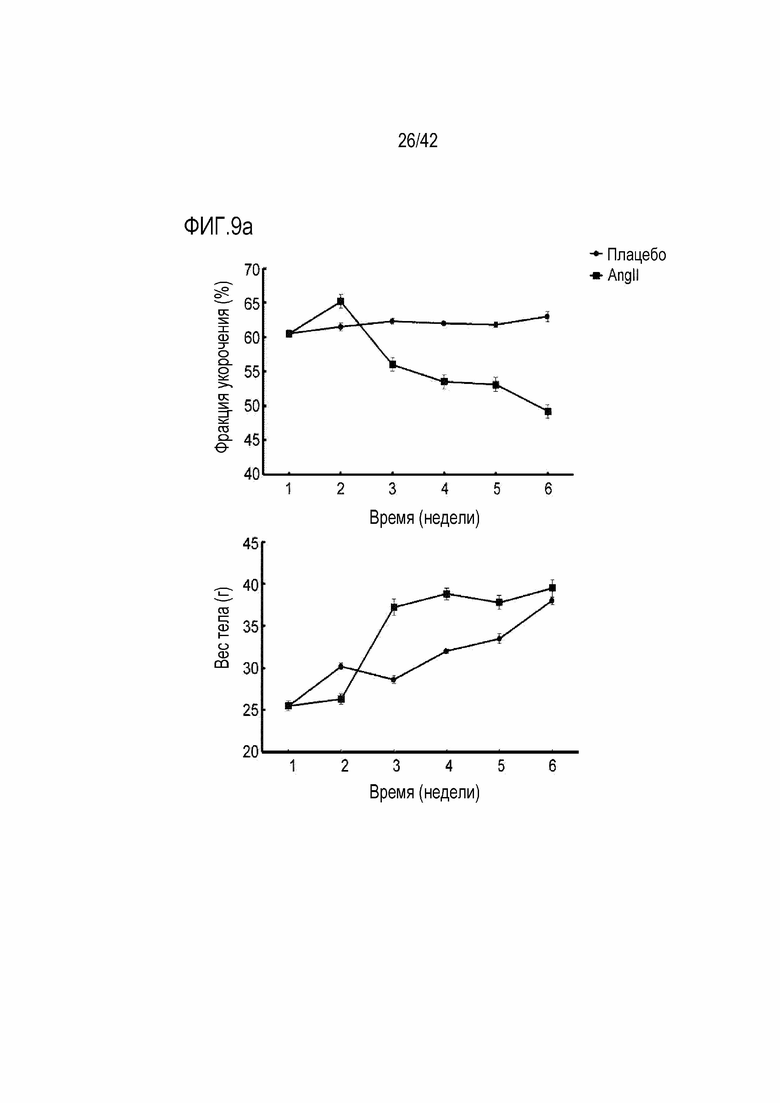
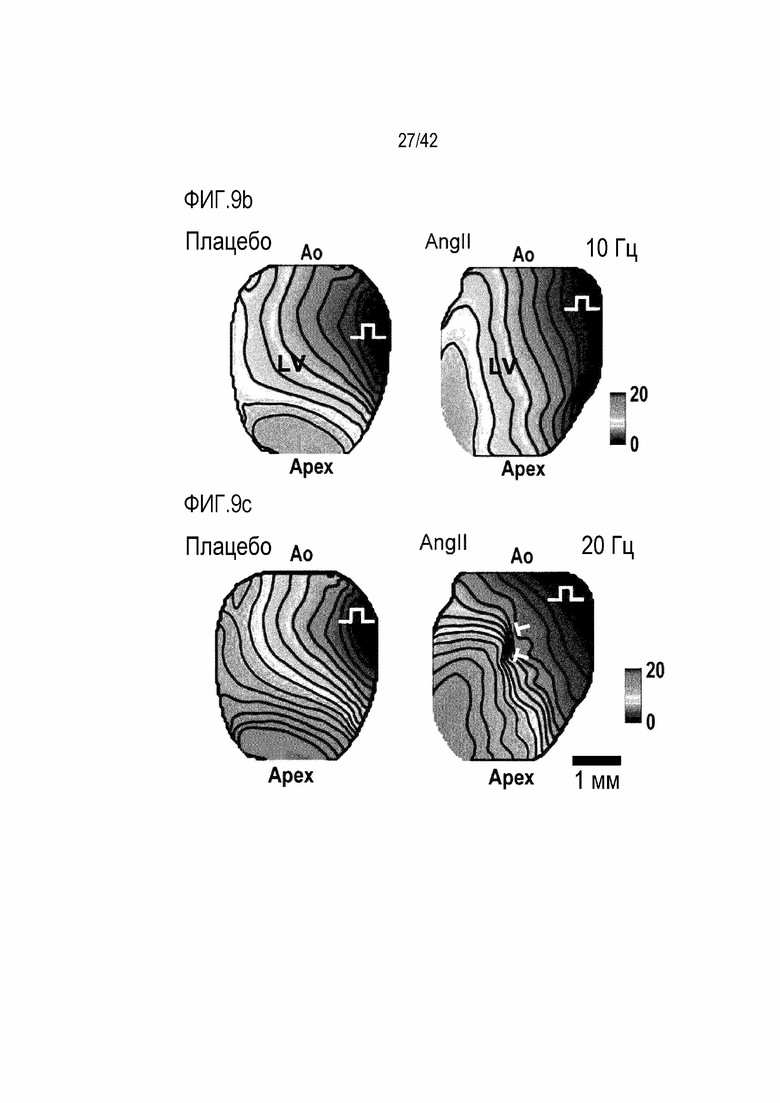
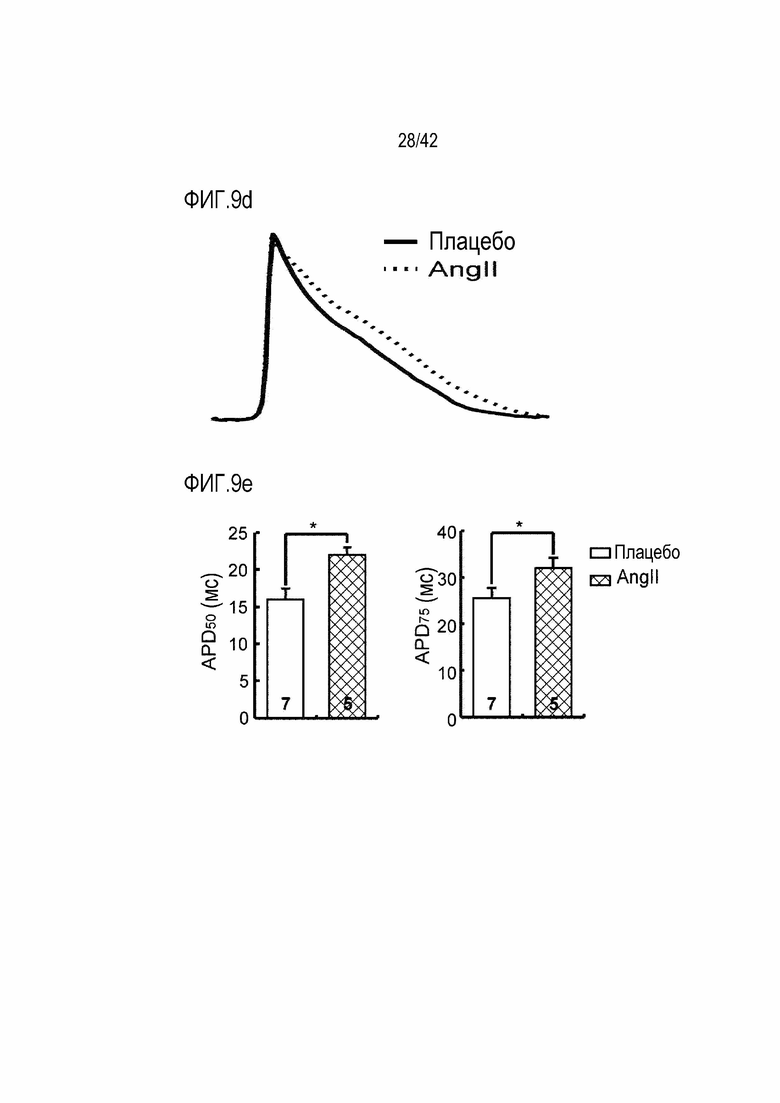
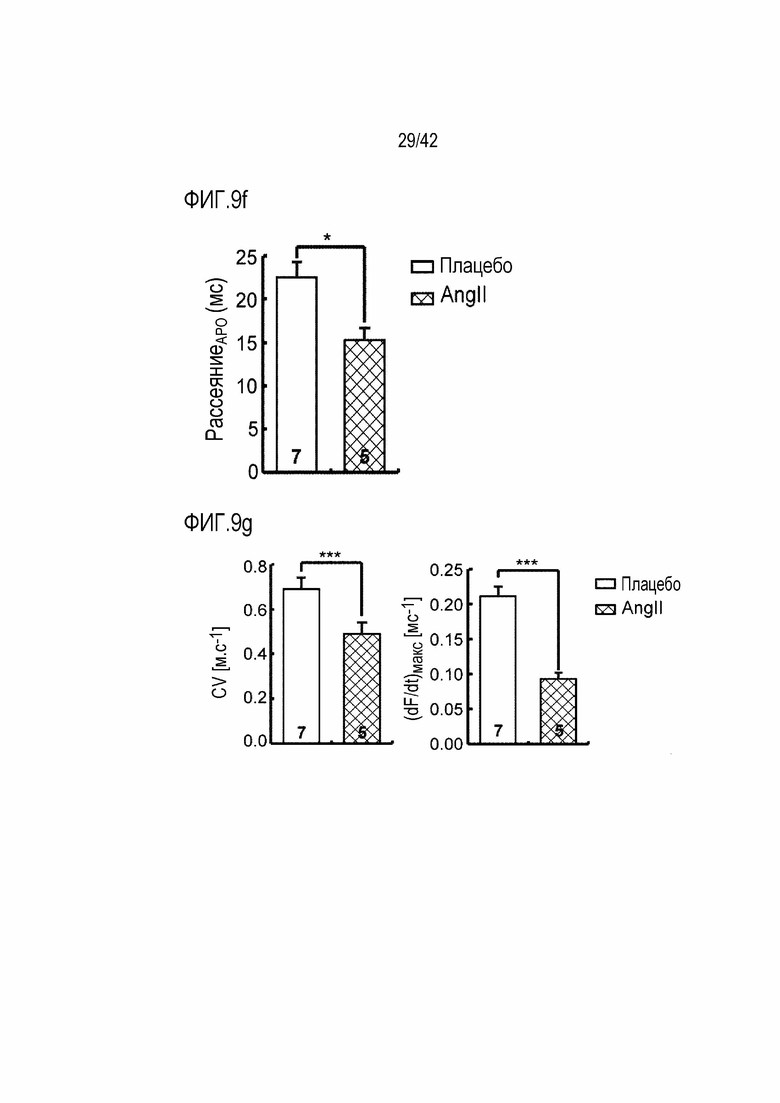
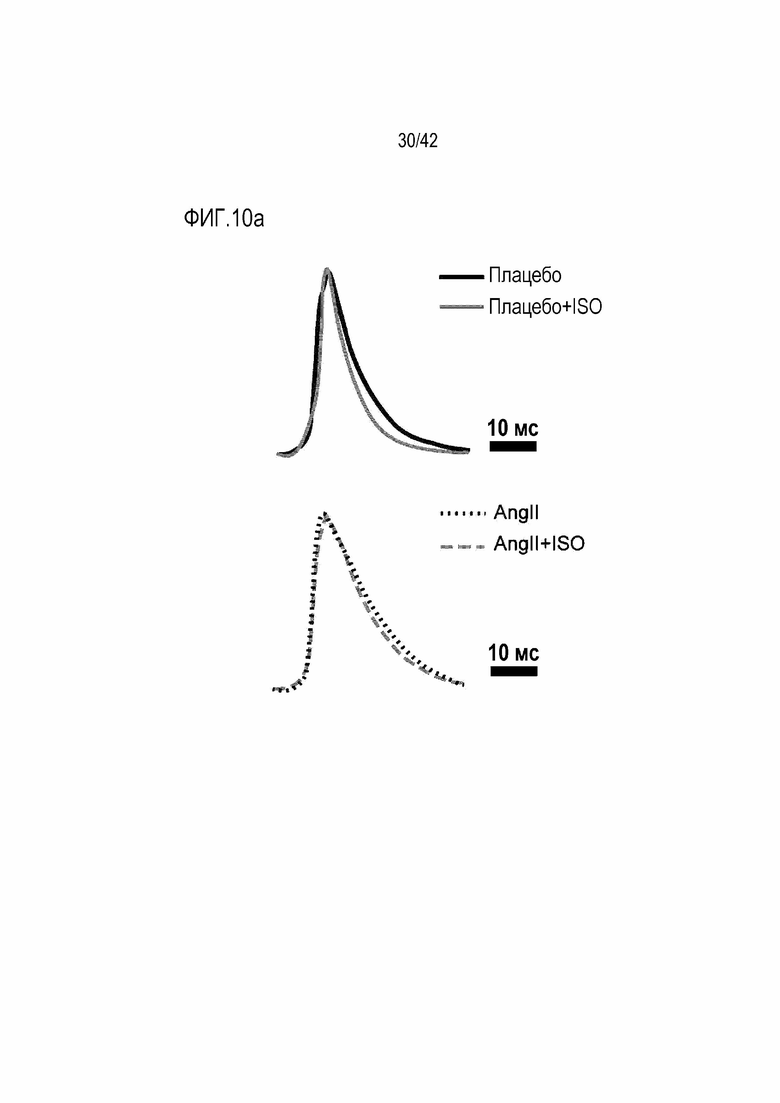

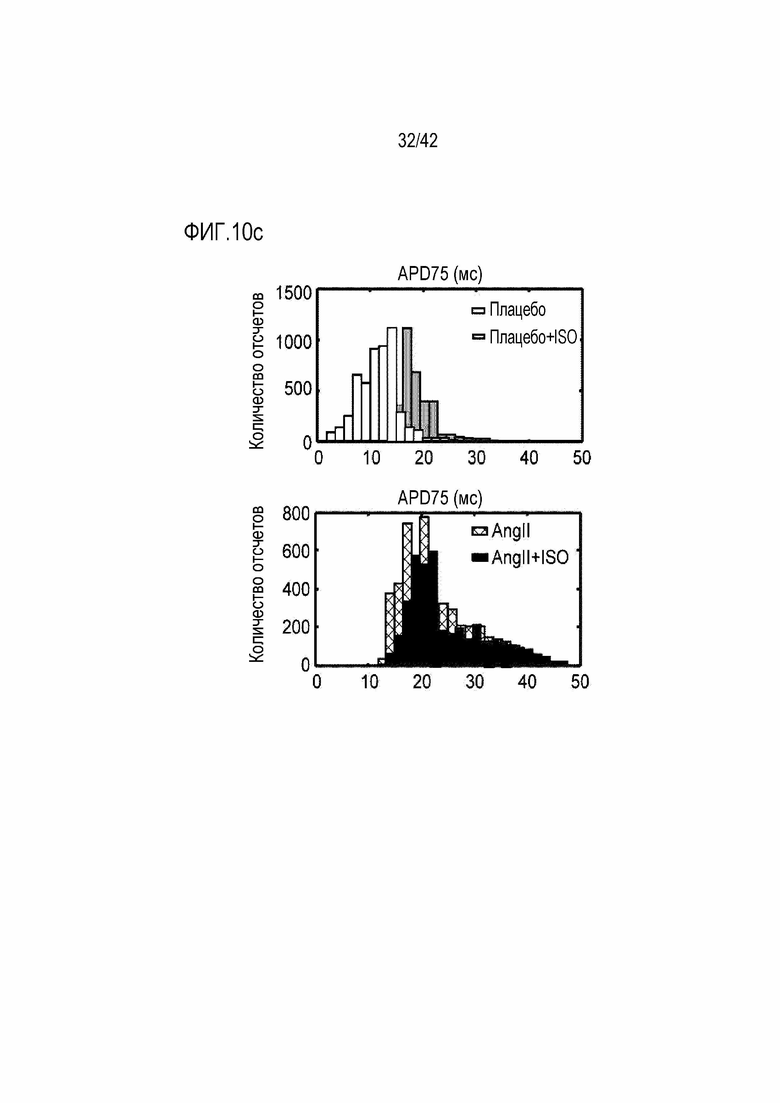
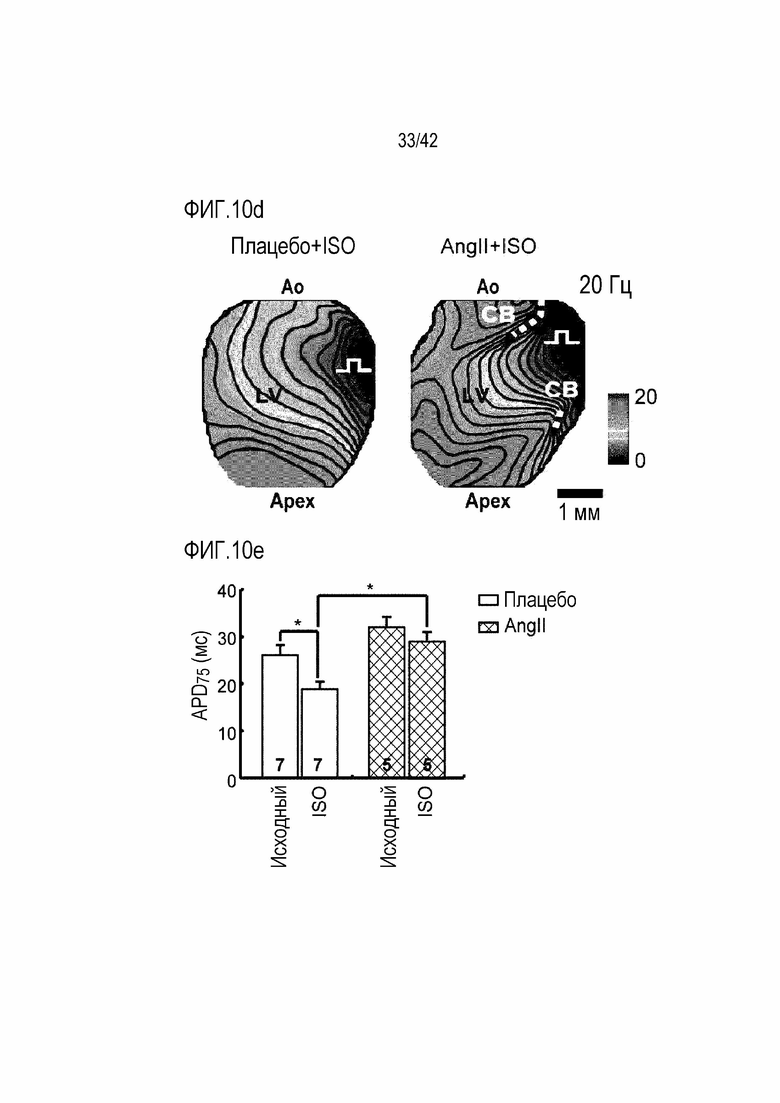
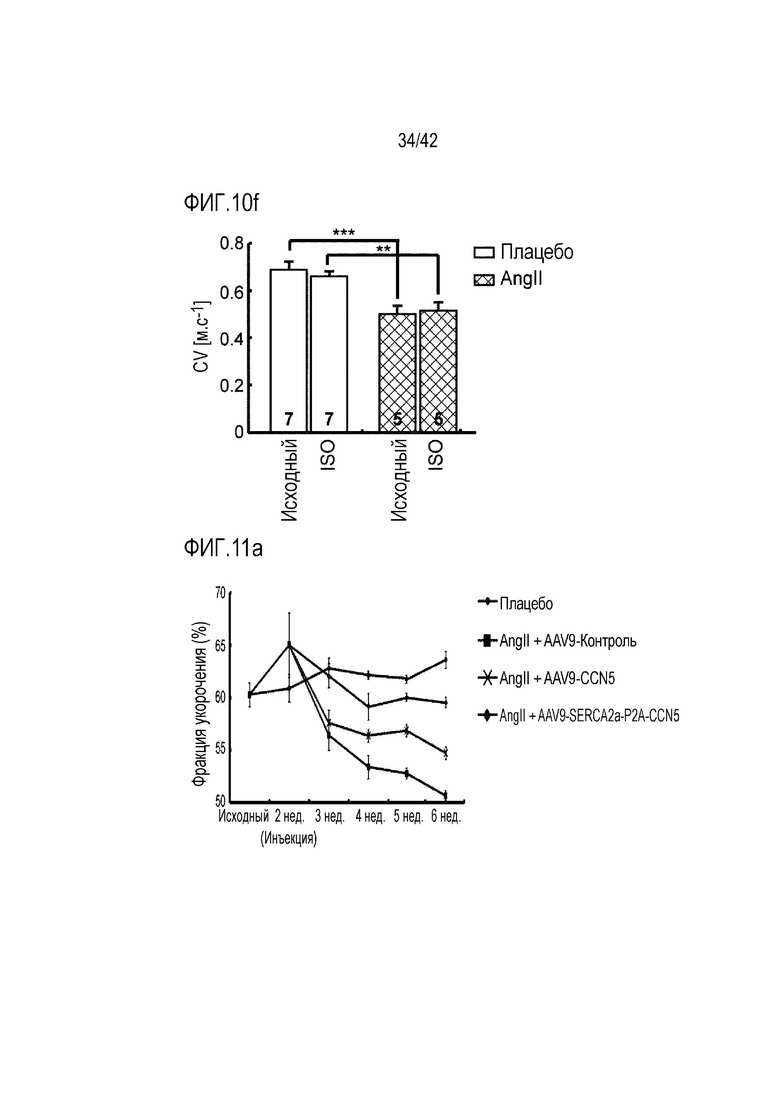
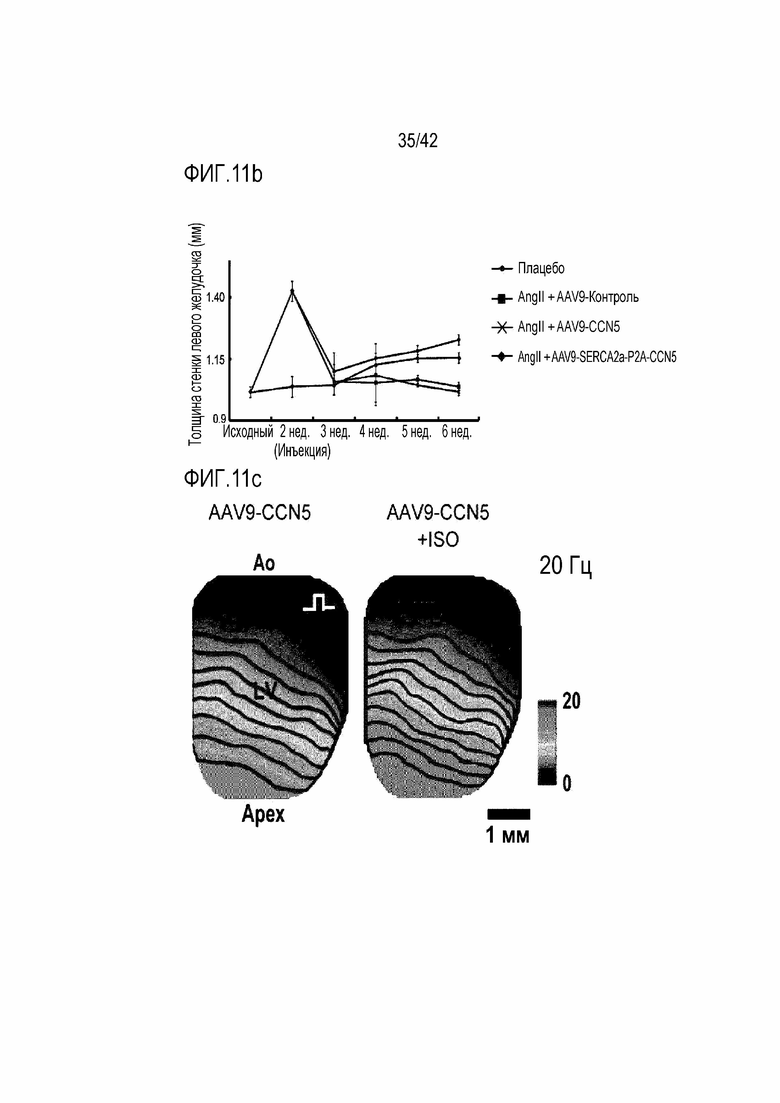
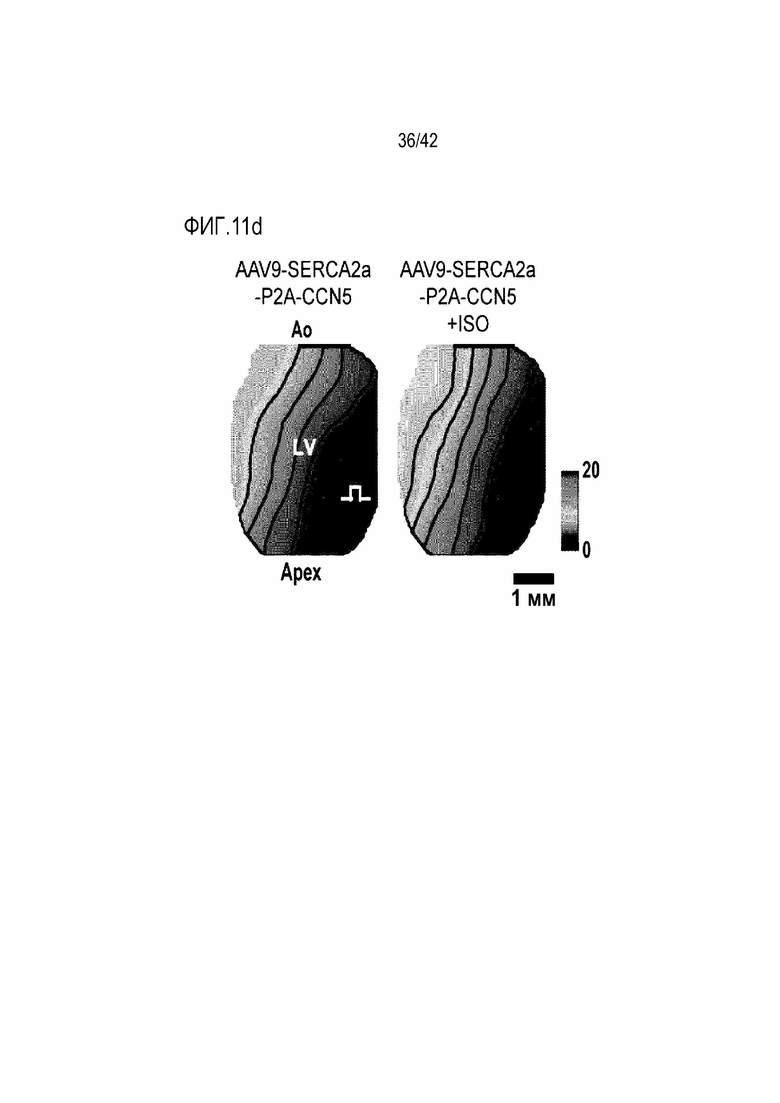
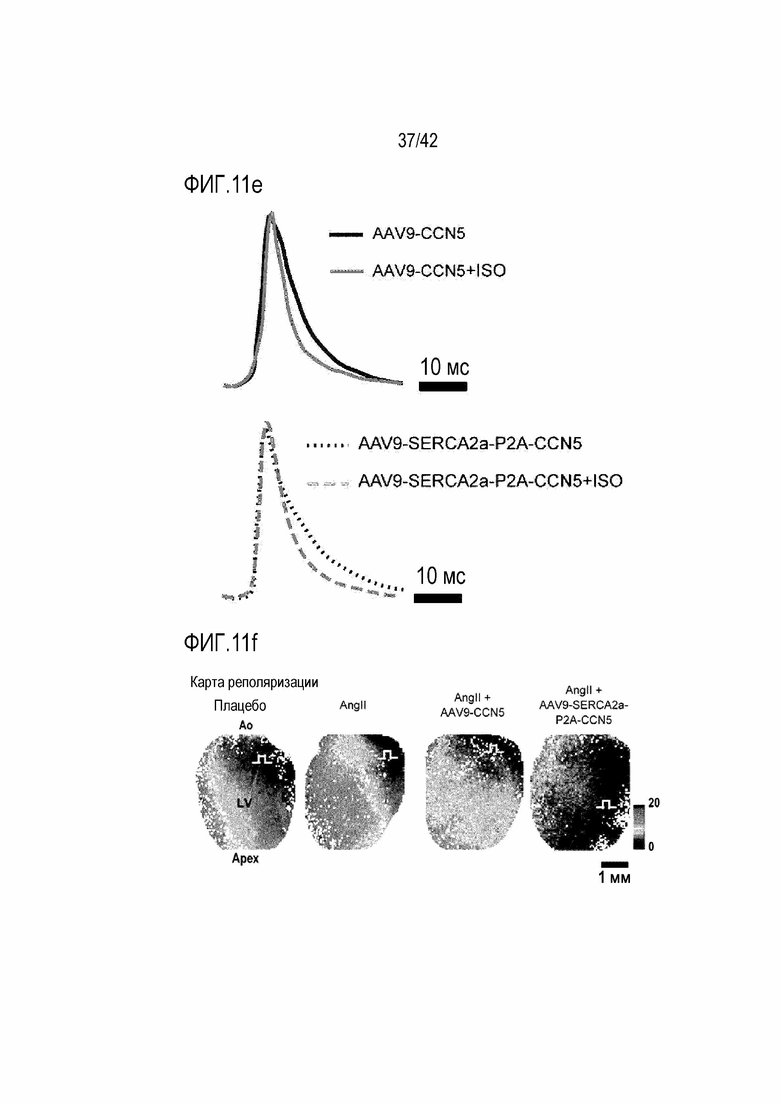
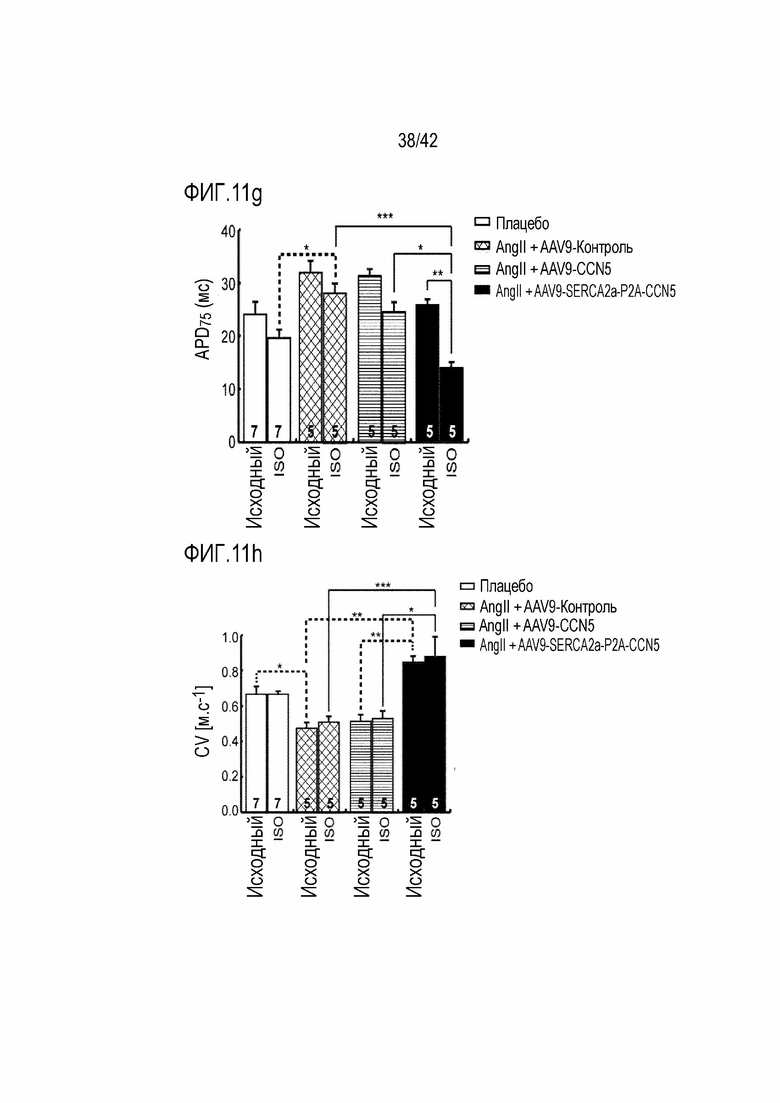


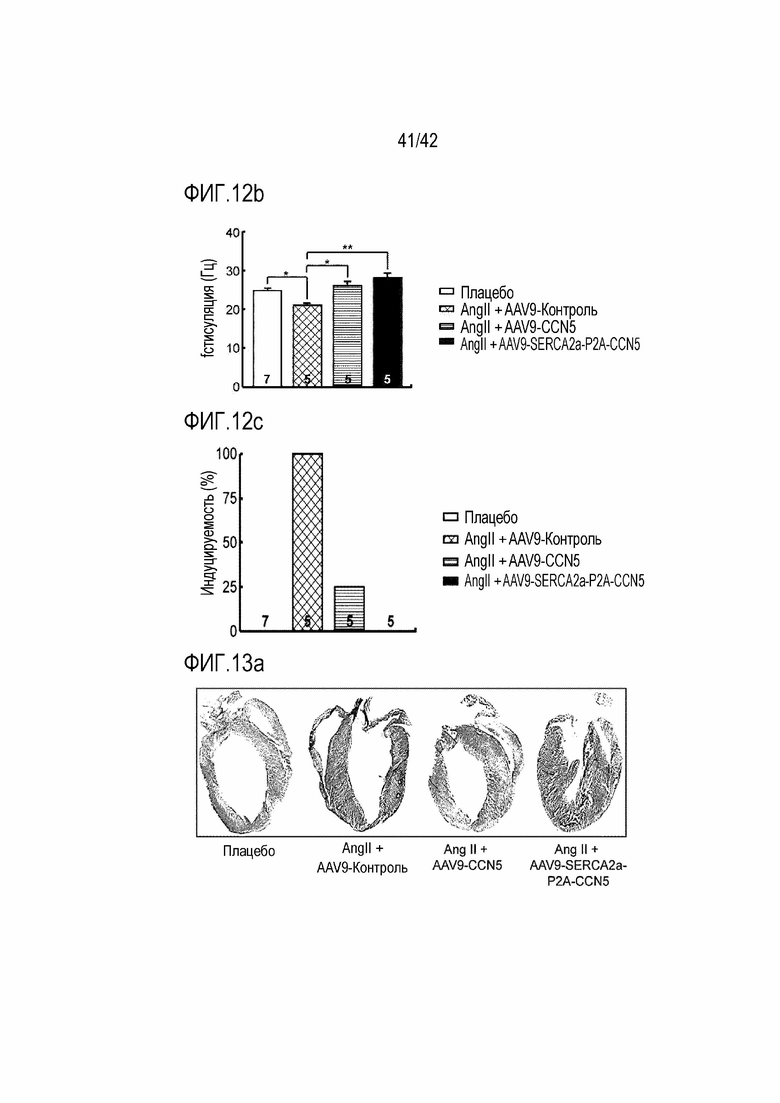

Настоящее изобретение относится к биотехнологии. Предложено применение фармацевтической композиции, включающей в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца. Применение фармацевтической композиции, включающей в качестве активного ингредиента вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца. Применение фармацевтической композиции, включающей в качестве активного ингредиента рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца. Применение фармацевтической композиции, включающей белок CCN5 и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца. Применение фармацевтической композиции, содержащей в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, или вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, или рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для изготовления лекарственного препарата для профилактики или лечения аритмии сердца. Предложен способ профилактики или лечения аритмии сердца, включающий введение субъекту белка CCN5, способ профилактики или лечения аритмии сердца, включающий введение фармацевтической композиции, содержащей в качестве активного ингредиента генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, или вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, или рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель. Изобретение может быть эффективно использовано для профилактики или лечения аритмии сердца. 7 н. и 19 з.п. ф-лы, 1 табл., 2 пр., 77 ил.
1. Применение фармацевтической композиции, включающей в качестве активного ингредиента:
генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца.
2. Применение по п.1, где белок CCN5 имеет аминокислотную последовательность, представленную SEQ ID №: 1.
3. Применение по п.1, где нуклеотидная последовательность, кодирующая белок CCN5, представляет собой последовательность, представленную SEQ ID №: 2.
4. Применение по п.1, где генетическая конструкция содержит промоторную последовательность, функционально с ней связанную.
5. Применение по п.4, где промотор представляет собой любой промотор, выбранный из группы, состоящей из промотора CMV, позднего промотора аденовируса, промотора вируса коровьей оспы 7.5K, промотора SV40, промотора HSV tk, промотора RSV, промотора EF1alpha, металлотионеинового промотора, бета-актинового промотора, промотора гена IL-2 человека, промотора гена IFN человека, промотора гена IL-4 человека, промотора гена лимфотоксина человека и промотора гена GM-CSF человека.
6. Применение по п.1, где генетическая конструкция дополнительно содержит нуклеотидную последовательность, кодирующую белок SERCA2a.
7. Применение по п.6, где в генетической конструкции нуклеотидная последовательность, кодирующая белок SERCA2a, содержится в направлении от 5' до 3' в порядке следования «нуклеотидная последовательность, кодирующая белок SERCA2a -нуклеотидная последовательность, кодирующая белок CCN5».
8. Применение по п.7, где генетическая конструкция содержит саморасщепляющуюся последовательность, расположенную между нуклеотидной последовательностью, кодирующей белок SERCA2a, и нуклеотидной последовательностью, кодирующей белок CCN5.
9. Применение по п.8, где саморасщепляющаяся последовательность представляет собой нуклеотидную последовательность, кодирующую пептид 2A, полученный из тешовируса 1 свиней, вируса Thosea asigna, вируса ринита лошадей A или вируса ящура.
10. Применение по п.8, где саморасщепляющаяся последовательность представляет собой нуклеотидную последовательность, кодирующую пептид 2A, полученный из тешовируса 1 свиней.
11. Применение по п.8, где саморасщепляющаяся последовательность представляет собой нуклеотидную последовательность, представленную SEQ ID №: 6.
12. Применение по п.1, где аритмия сердца представляет собой предсердную аритмию или желудочковую аритмию.
13. Применение фармацевтической композиции, включающей в качестве активного ингредиента:
вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца.
14. Применение по п.13, где вектор экспрессии дополнительно содержит нуклеотидную последовательность, кодирующую белок SERCA2a.
15. Применение по п.14, где в векторе экспрессии нуклеотидная последовательность, кодирующая белок SERCA2a, содержится в направлении от 5' до 3', в порядке следования «нуклеотидная последовательность, кодирующая белок SERCA2a-нуклеотидная последовательность, кодирующая белок CCN5».
16. Применение по п.15, где вектор экспрессии дополнительно содержит саморасщепляющуюся последовательность, расположенную между нуклеотидной последовательностью, кодирующей белок SERCA2a, и нуклеотидной последовательностью, кодирующей белок CCN5.
17. Применение по п.13, где вектор экспрессии представляет собой любой вектор экспрессии, выбранный из группы, состоящей из плазмидного вектора и космидного вектора.
18. Применение фармацевтической композиции, включающей в качестве активного ингредиента:
рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца.
19. Применение по п.18, где вирус представляет собой любой вирус, выбранный из группы, состоящей из аденовируса, аденоассоциированных вирусов (AAV), ретровируса, лентивируса, вируса простого герпеса и вируса коровьей оспы.
20. Применение по п.18, где рекомбинантный вирус дополнительно содержит нуклеотидную последовательность, кодирующую белок SERCA2a.
21. Способ профилактики или лечения аритмии сердца, включающий:
введение фармацевтической композиции, содержащей в качестве активного ингредиента:
генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, или вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, или рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и
фармацевтически приемлемый носитель.
22. Применение фармацевтической композиции, включающей белок CCN5 и фармацевтически приемлемый носитель, для профилактики или лечения аритмии сердца.
23. Применение по п.22, где фармацевтическая композиция дополнительно содержит белок SERCA2a.
24. Способ профилактики или лечения аритмии сердца, включающий введение субъекту белка CCN5.
25. Способ по п.24, дополнительно включающий введение субъекту белка SERCA2a.
26. Применение фармацевтической композиции, содержащей в качестве активного ингредиента:
генетическую конструкцию, которая содержит нуклеотидную последовательность, кодирующую белок CCN5, или вектор экспрессии, несущий нуклеотидную последовательность, кодирующую белок CCN5, или рекомбинантный вирус, который содержит нуклеотидную последовательность, кодирующую белок CCN5, и
фармацевтически приемлемый носитель, для изготовления лекарственного препарата для профилактики или лечения аритмии сердца.
| KR 1020110105957 A, 28.09.2011 | |||
| AI X, Ca2+/Ca1 modul in - Dependent Protein kinase vodulates cardiac ryanodine receptor phosphorylation and Sarcoplasmic reticulum Ca2+ leak in heart failure, Circulation research, 2005, 97, p.p | |||
| Прибор для распиливания дерева при помощи проволоки, накаливаемой электрическим током | 1925 |
|
SU1314A1 |
| KR 1020170056460 A, 23.05.2017 | |||
| WO 2006007444 A2, 19.01.2006 | |||
| АПАРЦИН Е.К | |||
| и др | |||
| Методы доставки | |||

