Притязания на приоритет
В настоящей заявке испрашивается преимущество предварительной заявки на патент США с регистрационным номером 62/825703, поданной 28 марта 2019 г. Все вышеизложенное содержание включено в настоящее описание посредством ссылки.
Исследования или разработки, спонсируемые Федеральными органами
Настоящее изобретение было разработано при поддержке правительства в соответствии с грантами № AG047336 и DC017117, выданными Национальным Институтом Здравоохранения. Правительство имеет определенные права на данное изобретение.
Область техники
В настоящем изобретении описаны сконструированные векторы AAV для экспрессии трансгена, например, в ЦНС, ПНС, во внутреннем ухе, в сердце или в сетчатке глаза, и способы их применения. Также представлены способы обнаружения новых сконструированных векторов AAV, которые опосредуют экспрессию трансгена в клетках нужного типа.
Предпосылки создания изобретения
Хотя было продемонстрировано, что векторы AAV9 обладают значительной способностью к доставке в ЦНС после системного введения, что послужило успешному лечению детей со спинальной мышечной атрофией1, однако, системная инъекция высоких доз векторов AAV может привести к индуцированию Т-клеточного ответа, который может удалять трансдуцированные клетки2. Сообщалось, что у обезьян, которым были введены высокие системные дозы AAV9-подобного вектора, развивалась токсичность, которая приводила к гибели животных, ассоциированной с системным воспалением3.. Недавно проведенное клиническое испытание фазы I с использованием высоких доз AAV9 для лечения мышечной дистрофии было приостановлено FDA из-за иммунной реакции после инфузии вектора у одного пациента. Причина, требующая введения высоких доз, связана с относительно низкой эффективностью AAV, созданного на основе копий генома вектора, для обеспечения адекватной экспрессии трансгена в значительном количестве клеток-мишеней. Таким образом, разработка новых капсидов AAV, которые обеспечивали бы более эффективную трансдукцию при более низких дозах, должна обеспечить лучшую терапевтическую эффективность при одновременном решении проблем безопасности, таких как иммунотоксичность.
Сущность изобретения
В настоящем изобретении описаны способы, в которых используется векторный геном аденоассоциированного вируса (AAV) с двухкомпонентным экспрессионным кластером для идентификации новых клонов вируса. Первый компонент представляет собой кластер Cre-рекомбиназы, находящийся под контролем представляющеого интерес промотора. Второй компонент представляет собой промотор AAV для инициации экспрессии сконструированного капсидного гена, клонированного «в цис-ориентации» по отношению к первому компоненту вирусного генома. Вирусные векторы отбирают для экспрессии трансгена (высокочувствительной экспрессии Cre) с использованием клеток, которые экспрессируют репортерный ген (например, белок, флуоресцирующий в зеленом диапазона спектра) с вышерасположенным loxP/стоп-сайтом, что тем самым позволяет предотвратить экспрессию репортера до тех пор, пока Cre, доставляемый вектором AAV, не будет удалять стоп-сайт. Могут быть выделены клетки, позитивные по репортерному гену, и полученные последовательности капсида AAV будут иметь более высокую вероятность опосредования эффективной экспрессии трансгена. В настоящей заявке также описаны сконструированные вирусные последовательности, которые обеспечивают эффективную экспрессию в центральной нервной системе (ЦНС) и в периферической нервной системе (ПНС), в сердце, в печени и во внутреннем ухе.
Таким образом, в настоящей заявке описаны капсидные белки AAV, содержащие аминокислотную последовательность, которая включает по меньшей мере четыре смежных аминокислоты последовательности STTLYSP (SEQ ID NO: 1) или FVVGQSY (SEQ ID NO: 2). В некоторых вариантах осуществления изобретения, капсидные белки AAV содержат аминокислотную последовательность, которая включает по меньшей мере пять смежных аминокислот последовательности STTLYSP (SEQ ID NO: 1) или FVVGQSY (SEQ ID NO: 2). В некоторых вариантах осуществления изобретения, капсидные белки AAV содержат аминокислотную последовательность, которая включает по меньшей мере шесть смежных аминокислот последовательности STTLYSP (SEQ ID NO: 1) или FVVGQSY (SEQ ID NO: 2). Альтернативно, капсидные белки AAV содержат аминокислотную последовательность, которая включает по меньшей мере четыре, пять или шесть смежных аминокислот последовательностей, представленных на фиг. 2A или 7C (SEQ ID NO: 17-150).
В некоторых вариантах осуществления изобретения, AAV представляет собой AAV9.
В некоторых вариантах осуществления изобретения, капсидные белки AAV содержат VP1 AAV9.
В некоторых вариантах осуществления изобретения, последовательность встраивают в капсид в положении, соответствующем аминокислотам 588 и 589 SEQ ID NO: 6, в пограничной области VP1/VP2 (аминокислота 138) или в любом сайте между 583-590.
Настоящее изобретение также относится к нуклеиновым кислотам, кодирующим капсидный белок AAV, как описано в настоящей заявке.
Кроме того, настоящее изобретение относится к AAV, содержащим описанные здесь капсидные белки и предпочтительно не содержащим капсидный белок VP1, VP2 или VP3 дикого типа. В некоторых вариантах осуществления изобретения, AAV дополнительно содержат трансген, а предпочтительно, терапевтический трансген.
Кроме того, настоящее изобретение относится к способам доставки трансгена в клетку, например, в клетку in vivo или ex vivo/in vitro. Эти способы включают контактирование клетки с описанным здесь AAV. В некоторых вариантах осуществления изобретения, клетка представляет собой нейрон (необязательно нейрон спинномозговых узлов или нейрон спирального узла), астроцит, кардиомиоцит или миоцит, астроцит, глиальную клетку, внутреннюю волосковую клетку, внешнюю волосковую клетку, поддерживающую клетку, фиброцит внутреннего уха, фоторецепторы, интернейроны, узлы сетчатки или пигментный эпителий сетчатки.
В некоторых вариантах осуществления изобретения, клетка находится в живом организме, например, у млекопитающего, а предпочтительно, у человека. В некоторых вариантах осуществления изобретения, клетка находится в ткани, выбранной из ткани головного мозга, спинного мозга, спинномозновых узлов, сердца, внутреннего уха, глаза или мышцы, а также их комбинации. В некоторых вариантах осуществления изобретения, индивидуум страдает болезнью Альцгеймера; болезнью Паркинсона; Х-сцепленной адренолейкодистрофией; болезнью Канавана; болезнью Нимана-Пика; спинальной мышечной атрофией; болезнью Гентингтона; болезнью, ассоциированной с коннексином-26; болезнью Ашера типа 3A; болезнью Ашера типа 2D; потерей слуха, ассоциированной с волосковыми клетками; потерей слуха, ассоциированной с волосковыми клетками (DFNB7/11); потерей слуха, ассоциированной с внутренними волосковыми клетками (DFNB9); болезнью Ашера типа 1F; болезнью Ашера типа 1B; пигментным ретинитом (RP; несиндромным); врожденным амаврозом Лебера; наследственной невропатией зрительного нерва Лебера; синдромом Ашера (RP; синдромом, ассоциированным с глухотой); мышечной дистрофией Дюшенна; васкулопатией после аллотрансплантации; или гемофилией А и В. Описанные здесь способы и композиции могут быть использованы для лечения этих состояний путем введения терапевтически эффективного количества AAV, несущего терапевтический трансген и достаточного для ослабления, снижения риска или задержки начала развития одного или более симптомов указанного состояния.
В некоторых вариантах осуществления изобретения, клетка находится в головном мозге индивидуума, и AAV вводят парентерально; интрацеребрально или интратекально.
В некоторых вариантах осуществления изобретения, интратекальную доставку осуществляют посредством поясничной инъекции, интрацистернальной инъекции или интрапаренхимальной инъекции.
В некоторых вариантах осуществления изобретения, AAV доставляют парентерально, предпочтительно путем внутривенной, внутриартериальной, подкожной, внутрибрюшинной или внутримышечной инъекции.
В некоторых вариантах осуществления изобретения, клетка находится в глазу индивидуума, и AAV вводят путем субретинальной инъекции или инъекции в стекловидное тело.
В некоторых вариантах осуществления изобретения, клетка находится во внутреннем ухе индивидуума, и AAV вводят в улитку путем нанесения поверх мембраны или через мембрану круглого окна, посредством хирургической кохлеостомии путем просверливания отверстия, смежного с круглым окном, в отверстие, находящееся в костном овальном окне или в полукруглый канал.
Настоящее изобретение также относится к библиотеке конструкций AAV, включающей:
(i) последовательность, кодирующую рекомбиназу Cre, под контролем промотора;
(ii) последовательность, кодирующую капсидный белок AAV9 с пептидом, описанным в настоящей заявке, например, с гептамерным пептидом, встроенным между последовательностями, кодирующими аминокислоты (а.к.) 588-589 капсида, под контролем промотора, расположенного ниже кластера Cre. В некоторых вариантах осуществления изобретения, пептид содержит рандомизированную пептидную последовательность или предварительно отобранную пептидную последовательность.
Кроме того, настоящее изобретение относится к библиотекиам, содержащим множество библиотек конструкций, описанных в настоящей заявке. В некоторых вариантах осуществления изобретения, где пептидные последовательности являются рандомизированными, библиотека включает конструкции, имеющие последовательности, кодирующие все возможные варианты гептамера.
Настоящее изобретение также относится к способам идентификации сконструированного капсида, который опосредует экспрессию трансгена в клетке предварительно выбранного типа. Эти способы включают: (а) введение библиотеки согласно пунктам 23 или 24 животному-модели, не являющемуся человеком, а предпочтительно млекопитающему, где клетки животного-модели экспрессируют loxP-фланкированный кластер STOP, расположенный выше репортерной последовательности; (b) выделение клеток предварительно выбранного типа; (c) отбор клеток, в которых экспрессируется репортерная последовательность; (d) выделение по меньшей мере части конструкции библиотек, а предпочтительно, части, содержащей гептамер, из выбранных клеток, в которых репортерная последовательность экспрессируется на стадии (c); и (e) определение идентичности гептамеров в конструкциях библиотек, выделенных на стадии (d), где выделенные гептамеры могут опосредовать экспрессию трансгена в клетках предварительно выбранного типа.
В некоторых вариантах осуществления изобретения, репортерная последовательность кодирует флуоресцентный репортерный белок.
В некоторых вариантах осуществления изобретения, животное-модель является трансгенным по loxP-фланкированному кластеру STOP, расположенному выше репортерной последовательности, или loxP-фланкированный кластер STOP, расположенный выше репортерной последовательности, может экспрессироваться из второй конструкции.
В некоторых вариантах осуществления изобретения, определение идентичности гептамеров в конструкциях библиотеки включает проведение анализа посредством секвенирования ДНК.
В некоторых вариантах осуществления изобретения, эти способы также включают, до и/или после стадии (е): проведение ПЦР для амплификации последовательностей, содержащих последовательности гептамера, и необязательно включающих последовательности полноразмерного капсида, из конструкций библиотек, выделенных на стадии (d); клонирование амплифицированных последовательностей обратно во второй набор векторов библиотек; повторную упаковку второго набора векторов библиотек; и проведение стадий (a) - (d) или (a) - (e) во втором наборе векторов библиотек.
Если это не оговорено особо, то все используемые здесь технические и научные термины имеют свое общепринятое значение, известное специалистам в области, к которой относится изобретение. Описанные здесь способы и материалы предназначены для их использования в настоящем изобретении, однако, могут быть также применены и другие подходящие способы и материалы, известные специалистам в данной области. Материалы, способы и примеры приводятся лишь в иллюстративных целях и не должны рассматриваться как ограничение объема изобретения. Все упомянутые здесь публикации, патентные заявки, патенты, последовательности, записи в базах данных и другие документы в полном объеме включены в настоящее описание посредством ссылки. В случае возникновения противоречий, следует отдать предпочтение определениям, представленным в настоящей заявке.
Другие особенности и преимущества настоящего изобретения будут очевидны из нижеследующего подробного описания и чертежей, а также из формулы изобретения.
Описание чертежей
Фиг. 1А-В. Библиотека iTransduce для отбора новых капсидов AAV, способных эффективно экспрессировать трансген в ткани-мишени. а. Двухкомпонентная система конструкции библиотеки. 1. Cre-рекомбиназа находится под контролем минимального промотора куриного бета-актина (CBA). 2. Капсид AAV9 находится под контролем промотора p41 с рандомизированным гептамерным пептидом, встроенным между аминокислотами 588-589 и клонирован ниже кластера Cre. b. Стратегия отбора. i. Библиотеку iTransduce, состоящую из различных пептидных вставок, экспрессируемых на капсиде (показанных разными цветами), вводят внутривенно трансгенной мыши Ai9 с использованием loxP-фланкированного кластера STOP, расположенного выше репортерного гена tdTomato, встроенного в локус Gt(ROSA)26Sor. Капсиды AAV, способные проникать в представляющую интерес клетку, но не функционально трансдуцировать клетку (без экспрессии Cre), не инициируют экспрессию tdTomato. Капсиды, которые могут опосредовать функциональную трансдукцию (экспрессировать Cre), будут запускать экспрессию tdTomato. ii. Клетки выделяют из представляющего интерес органа (например, головного мозга), и трансдуцированные клетки отбирают по экспрессии tdTomato и, необязательно, по клеточным маркерам. iii. Капсидную ДНК подвергают ПЦР-амплификации из отобранных клеток, клонируют обратно в вектор библиотеки и снова упаковывают для проведения другого раунда отбора. Анализ методом секвенирования ДНК проводят после каждого раунда для мониторинга процесса отбора.
Фиг. 2А-В. Идентификация AAV-S и AAV-F после двух раундов отбора in vivo для трансдукции в головной мозг после системной инъекции. Кольцевые диаграммы показывают частоту определенных пептидных вставок, оцененную посредством секвенирования следующего поколения. а. Таблица векторных последовательностей раунда 2 после получения, но до инъекции (SEQ ID NO: 17-86). b. Кольцевая диаграмма частоты встречаемости пептидов при выделении iTransduce после инъекции в раунде 2 (SEQ ID NO 1-3). «Другие» означает варианты последовательностей, составляющие менее 1% от общего пула (в (а) варианты, выделенные после скрининга в раунде 2, также отмечены более ярким цветом, и составляют менее 1%). * означает стоп-кодон.
Фиг. 3A-F. AAV-F эффективно проникает в головной мозг мышей после системной инъекции. а. Одноцепочечный экспрессионный кластер AAV-GFP, используемый для сравнения трансдукционного потенциала капсидов. ITR, инвертированные концевые повторы; CBA, гибрид энхансера CMV/промотора куриного бета-актина; WPRE, посттранскрипционный регуляторный элемент вируса гепатита сурка; pA, сигналы polyA (полученные как из SV40, так и из бычьего гормона роста). b. Репрезентативные изображения с низким увеличением сагиттальных срезов всего головного мозга мышей C57BL/6 (самцов), трансдуцированных 1×1011 ВГ (низкая доза) AAV9, AAV9-PHP.B, AAV-S или AAV-F. c. Репрезентативные изображения сагиттального среза головного мозга после инъекции 8×1011 ВГ (высокая доза) каждого вектора у самцов C57BL/6. d. Примеры срезов спинного мозга, трансдуцированных каждым из четырех векторов, вводимых внутривенно в более высокой дозе (8×1011 ВГ/мышь). е. Количественное определение нативной экспрессии GFP из каждого вектора по проценту участков, охватываемых флуоресценцией при низких (левая панель) и высоких (правая панель) дозах. f. Мультирегиональное сравнение трансдукции в головной мозг при более высокой дозе. ***, р <0,001; ****, p < 0,0001 после однофакторного дисперсионного анализа ANOVA с критерием множественного сравнения Тьюки (n=3 в каждой группе).
Фиг. 4A-E. Сравнение трансдукции нейронов и астроцитов AAV-векторами и биораспределения этих векторов. Изображения с большим увеличением клеток, трансдуцированных AAV9, AAV-PHP.B, AAV-S и AAV-F (GFP-позитивных) после совместного иммунологического окрашивания маркерами для a. нейронов (NeuN) и b. астроцитов (глутамин-синтетазы, GS), также показаны объединенные клетки с окрашиванием ядра красителем DAPI. c. Стереологическая оценка процента трансдуцированных кортикальных астроцитов и нейронов после внутривенной доставки 1×1011 ВГ каждого вектора. P <0,0001 - однофакторный ANOVA. (*) и (**) означают значимые различия между каждой группой векторов после анализа с использованием критерия множественного сравнения Тьюки (n=3 мыши/группу). d. Биораспределение векторов в головном мозге и в печени, оцененное с помощью кол.ПЦР векторных геномов, и нормализованное по уровням геномной ДНК в присутствии GAPDH (исходной ДНК). е. Трансдукция AAV9, AAV9-PHP.B, AAV-S и AAV-F в периферические органы после внутривенного введения в дозе 8×1011 ВГ самцам C57BL/6. Изображения сетчатки: RPE, пигментный эпителий сетчатки; ONL; внешний ядерный слой; OPL, внешний плексиформный слой; INL, внутренний ядерный слой; IPL, внутренний плексиформный слой; GCL, слой ганглиозных клеток. ****, p <0,0001 (n=3 мыши/группу, однофакторный ANOVA с критерием множественного сравнения Тьюки).
Фиг. 5А-С. AAV-F опосредует высокую эффективность трансдукции у самцов и самок мышей C57BL/6, а также у мышей BALB/c. а. Репрезентативные изображения сигнала GFP в сагиттальных срезах головного мозга самцов и самок мышей (n=3), трансдуцированных AAV-F в дозе 1×1011 ВГ/мышь. b. Сагиттальные срезы головного мозга самцов мышей BALB/c, которым вводили AAV-F (слева) или AAV9-PHP.В (справа) в дозе 1×1011 ВГ/мышь. DAPI представлен как контрастный краситель для окрашивания на GFP в целях визуализации срезов головного мозга, обработанных PHP.B. c. Количественное определение экспрессии эндогенного GFP из каждого вектора по проценту срезов, охватываемых флуоресценцией. **, р <0,01. Непарный t-критерий (n=3 мыши/группу).
Фиг. 6А-В. AAV-F опосредует более высокую эффективность трансдукции, чем AAV9, в нейронах коры головного мозга человека. а. Экспрессия GFP в первичных нейронах плода человека, трансдуцированных AAV-F. Нейроны были помечены антителом против β-тубулина для количественной оценки трансдукции. b. Количественная оценка эффективности трансдукции нейронов человека векторами AAV9, AAV-S и AAV-F. *, р <0,05.
Фиг. 7A-C. Библиотека iTransduce функционально инициирует Cre-рекомбинацию, и ПЦР-амплификация 7-мерных вставок, кодирующих пептид, в гене cap, может быть осуществлена для их выделения из ткани. а. Примеры окрашивания DAB tdTomato в тканях после трансдукции неотобранной библиотекой iTransduce у мышей, трансгенных по Ai9 floxed/STOP tdTomato (правые панели). PBS вводили в качестве контроля (левые панели). Красные стрелки означают примеры трансдуцированных клеток. b. Примеры ПЦР, сохраняющих область гена Cap, содержащую вставку, в различных тканях, включая головной мозг, по сравнению с тканями мышей дикого типа и трансгенных нетрансдуцированных мышей. c. Таблица, иллюстрирующая спектр вариантов, наблюдаемых после первого раунда отбора, с выделенными наиболее частыми вариантами (SEQ ID NO: 87-150). «Другие» означают сгруппированные варианты последовательностей, каждый из которых составляет менее 1% от общего пула. * означает стоп-кодон.
Фиг. 8A-В. Отбор на основе Cre в раунде 2 выявляет компетентные по трансдукции AAV. а. Анализ tdTomato-позитивных клеток методом проточной цитометрии. После разрушения головного мозга мышей, суспензия клеток была проанализирована и отсортирована на наличие tdTomato-позитивных клеток, при этом, стробирование было проведено на основе прямого и бокового рассеяния (FSC, SSC) для исключения нежизнеспособных клеток (общие события) и захвата только отдельных клеток (синглетов), и, наконец, для экспрессии tdTomato (tdTomato+/--клеток). Для отбора tdTomato-позитивных клеток из головного мозга, трансдуцированного библиотекой AAV, стробирование было осуществлено на основе негативного контроля (Ai9 с инъекцией PBS) и позитивного контроля (мышей Ai9, трансдуцированных AAV9-PHP.B, несущей hSyn-Cre-нейрон-специфический кластер). b. Cre-зависимые события рекомбинации были обнаружены после иммунологического окрашивания DAB на tdTomato в печени и в головном мозге после инъекции библиотеки в раунде 2 (по сравнению с инъекциями PBS или AAV9-PHP.B hSyn-Cre). Стрелка указывает на позитивные клетки (астроциты) в головном мозге мышей, которым инъецировали библиотеку AAV.
Фиг. 9А-В. Трансдукция головного мозга с помощью AAV-F и AAV-S после внутривенной доставки низких (1×1011 ВГ) или высоких доз вектора (8×1011 ВГ). Профиль трансдукции в головном мозге после трансдукции AAV9, AAV9-PHP.B, AAV-S и AAV-F у n=3 мышей, демонстрирующий эндогенную (неокрашенную) флуоресценцию GFP в сагиттальных срезах. Мышам вводили либо 1×1011 ВГ (a), либо 8×1011 ВГ (b). Каждый срез для каждой группы был взят от отдельной мыши.
Фиг. 10А-В. (а) AAV-F трансдуцирует несколько подтипов нейронов в головном мозге мыши. Экспрессия GFP, инициируемая AAV-F, была детектирована в широком диапазоне нейронных подтипов в различных областях головного мозга и ЦНС. CamKII, возбуждающие нейроны. GAD67, ингибирующие нейроны. Тирозингидроксолаза (TH), нейроны Пуркинье. Холинацетилтрансфераза (ChAT), двигательные нейроны (белые стрелки представляют примеры трансдуцированных нейронов для каждого подтипа). (b) AAV-F обеспечивает эффективную трансдукцию, в то время как AAV9 не обеспечивает ее при концентрации 1×1011 ВГ/мышь. Репрезентативные 10-кратные изображения мышей на фиг. 3b демонстрируют экспрессию GFP в полосатом теле, в гиппокампе и в мозжечке у мышей, которым инъецировали AAV-F и AAV9.
Фиг. 11. Биораспределение AAV-F после системного введения. Биораспределение AAV-F по сравнению с AAV9 показано в скелетных мышцах, в сердце и в ткани спинного мозга. Биораспределение оценивали с помощью кол.ПЦР векторных геномов, и образцы нормализовали по геномной ДНК GAPDH для равного ввода. n.s., не значимые. * p <0,05 (n=3 мыши/группу, каждый образец, взятый у мышей, оценивали с тремя повторностями с помощью двустороннего t-критерия).
Фиг. 12A-E. Количественное определение пустых капсидов с помощью просвечивающей электронной микроскопии (ТЕМ). а-d. Репрезентативные сегменты изображений на электронных микрофотографиях препаратов AAV9 и AAV-F. Два препарата AAV9 (a, b) и AAV-F (c, d) были количественно определены путем подсчета полных и пустых капсидов на пяти изображениях для каждого препарата (примеры пустых капсидов указаны стрелками). е. Количественная оценка процентного содержания пустого капсида для препаратов AAV9 и AAV-F. n.s.: незначимое (p=0,54, непарный t-критерий).
Фиг. 13. Пролонгированная трансдукция нервных клеток после прямой внутричерепной инъекции AAV-F и AAV-S. Репрезентативные изображения сигнала флуоресценции GFP (и DAPI) на сагиттальных срезах головного мозга мышей после прямых инъекций AAV-F (верхние панели) или AAV-S (нижние панели) в кору головного мозга и в гиппокамп (1,65×1010 и 5,6×1010 ГК/участок инъекции для AAV-F и AAV-S, соответственно). Масштаб: 1000 мкм для изображений всего головного мозга с небольшим увеличением и 200 мкм для изображений коры головного мозга и гиппокампа с еще бόльшим увеличением.
Фиг. 14A-F. Широкое распределение трансдукции в спинном и головном мозге после поясничной интратекальной инъекции вектора AAV-F. Десять микролитров AAV9 (1,25×1011 ВГ) или AAV-F (8,8×1010 ВГ), упаковывающих одноцепочечный экспрессионный кластер AAV-CBA-GFP, вводили интратекально в поясничную область взрослых мышей (n=2/вектор). Через три недели, мышей умерщвляли и спинной мозг и головной мозг анализировали на экспрессию GFP после иммунологического окрашивания на антитела против GFP. (а) Верхние изображения: Полное сканирование коронарного среза головного мозга, иллюстрирующее надежную трансдукцию головного мозга с помощью AAV-F, но не AAV9. Нижние изображения: Полное сканирование срезов спинного мозга с AAV-F и AAV9. Мыши, которым вводили AAV-F, показали очень высокий сигнал GFP. AAV9 показал очень низкую экспрессию у обеих мышей. Все изображения были сделаны с экспозицией 33 мс; для AAV-F было сделано дополнительное изображение с интервалом 4 мс, для лучшего разрешения деталей. Белые контуры границ срезов были включены там, где срезы являются тусклыми. Если не указано конкретно , то масштаб составлял 250 мкм. (b-d) Изображения спинного мозга с большим увеличением, полученных от мышей, обработанных AAV9 (b) и AAV-F (c, d). GFAP указывает на астроцит-специфическое окрашивание, а NeuN - на нейроны. Область спинного мозга показана в правом верхнем углу каждого изображения. Изображения на нижних панелях представляют собой изображения в рамке на верхнем изображении с большим увеличением. (e, f) Изображения головного мозга, трансдуцированного AAV-F, с большим увеличением после интратекальной инъекции. (e) показаны нейроны астроцитов (f), трансдуцированные AAV-F. AAV9 не опосредовал детектируемую трансдукцию головного мозга после интратекальной инъекции. SC, спинной мозг; CC, мозолистое тело, CP, скорлупа хвостатого тела.
Фиг. 15A-D. Флуоресценция GFP после введения AAV-S-CBA-GFP во внутреннее ухо. (a, b): Типичные изображения сенсорного эпителия улитки, преобразованного с помощью AAV-S (увеличение 63×). (c): Трансдукция в спиральном лимбе. (d): Трансдукция в области спирального узла. Z и стрелка указывают на различные слои Z-стэка. OHC: внешние волосковые клетки. IHC: внутренние волосковые клетки.
Фиг. 16А-В. Использование библиотеки iTransduce в нетрансгенных NHP для отбора капсидов AAV, которые эффективно трансдуцируют фиброциты внутреннего уха и спинной мозг. (А) i. Собакоподобным обезьянам (или другим приматам, кроме человека) вводили библиотеку капсидов AAV вместе с AAV9-PHP.B, кодирующей кластер floxed-Stop-tdTomato, под контролем GJB2. ii. AAV9-PHP.B будет селективно экспрессировать tdTomato в фиброцитах (показано затенением), если капсид библиотеки AAV экспрессирует Cre. iii. внутреннее ухо является пораженным. iv. tdТомато-позитивные фиброциты были отсортированы с помощью проточной цитометрии. v. Потенциально функциональные капсиды были ПЦР-амплифицированы из восстановленной ДНК отсортированных клеток, а затем библиотеку повторно упаковывали и проводили еще один раунд отбора. Анализ посредством секвенирования ДНК следующего поколения проводили после каждого раунда для мониторинга процесса отбора. Сокращения: ТМ - покровная мембрана; OC, кортиев орган; SL, спиральная связка. (В). i. Собакоподобным обезьянам (или другим приматам, кроме человека) вводили библиотеку капсидов AAV вместе с AAV9-кодирующим кластером floxed-Stop-mPlum под контролем CBA. ii, iii. AAV9 будет экспрессировать mPlum в спинном мозге (показано затенением), если капсид библиотеки AAV экспрессирует Cre. Спинной мозг является пораженным, а mPlum-позитивные клетки были отсортированы с помощью проточной цитометрии. iv. Потенциально функциональные капсиды были ПЦР-амплифицированы из восстановленной ДНК отсортированных клеток, а затем библиотеку повторно упаковывали и проводили еще один раунд отбора. Анализ посредством секвенирования ДНК следующего поколения проводили после каждого раунда для мониторинга процесса отбора.
Подробное описание изобретения
Перспективным способом эффективной доставки трансгенов в клетки-мишени является способ отбора пула или библиотеки вариантов капсидов вектора AAV in vivo на «выживание наиболее приспособленных»4-8. Способы создания библиотек AAV, в которых используются рандомизированные олигомерные нуклеотиды для встраивания коротких (6-9 аминокислот) рандомизированных пептидов в доступную область на поверхности капсида, продемонстрировали успешную идентификацию новых вариантов капсида AAV с уникальными свойствами, такими как усиленная трансдукция тканей-мишеней9,10. Одним из основных ограничений библиотек AAV является то, что конечное считывание в процессе отбора не всегда позволяет отличить капсиды, которые опосредуют функциональную экспрессию трансгена, от капсидов, которые не обладают такой функцией. Трансдукция AAV представляет собой способ, включающий множество стадий, от связывания с клеточными рецепторами и транспорта в ядро, и до синтеза второй цепи и, наконец, экспрессии генов и белков11. Недавнее усовершенствование стандартного метода получения библиотеки AAV, названного CREATE, позволило сконструировать Cre-чувствительный геном AAV, обеспечивающий селективное выделение капсидов, которые с успехом транспортируется в ядро у трансгенных животных, экспрессирующих Cre12. В настоящей заявке описана система отбора капсидов, один из примеров которой представляет собой iTransduce, где используется активность системы Cre/loxP. Вместо использования Cre-трансгенных мышей был сконструирован AAV для кодирования обоих капсидов с пептидными вставками вместе с кластером для экспрессии Cre. Затем был проведен отбор мышей с Cre-чувствительным флуоресцентным репортером так, чтобы можно было провести отбор капсидов, которые опосредуют весь процесс трансдукции, включая экспрессию трансгена. Отбор библиотеки in vivo позволил идентифицировать капсид AAV, который обеспечивает значительную эффективность трансдукции в ЦНС, и другой капсид, который опосредует трансдукцию во внутреннее ухо.
С использованием системы iTransduce, авторами были выделены два новых капсида AAV, называемых здесь AAV-F и AAV-S, которые опосредуют высокоэффективную экспрессию трансгена в ЦНС (два протестированных штамма) и во внутреннем ухе мыши, соответственно. Капсид AAV-F также опосредует надежную трансдукцию первичных нейронов человека.
Интересно отметить, что путем проведения отбора на основе экспрессии были идентифицированы 3 пептидных клона (STTLYSP (SEQ ID NO: 1), FVVGQSY (SEQ ID NO: 2) и FQPCP* (SEQ ID NO: 3), которые представляют 97% от всех ридов NGS. Поскольку FQPCP* имеет стоп-кодон, то было высказано предположение, что этот геном был перекрестно упакован в другой капсид во время продуцирования, и такой феномен был оттмечен для библиотек AAV15. Это также может иметь место для STTLYSP (SEQ ID NO: 1, «AAV-S») (фиг. 2). AAV-S не является дефектным вектором, поскольку он опосредует надежную трансдукцию в периферические органы (фиг. 4e) и во внутреннее ухо (фиг. 15). Поскольку в данном случае наблюдалась высокая эффективность продуцирования (Таблица 4), то возможно, что AAV-S будет иметь предрасположенность к перекрестной упаковке. С другой стороны, AAV-F (FVVGQSY (SEQ ID NO: 2)) был чрезвычайно эффективным при трансдукции и был одним из двух потенциальных кандидатов на NGS (фиг. 2). Оба этих кандидата обнаруживались на очень низких уровнях в пуле библиотеки в раунде 2 (фиг. 2), а поэтому авторы смогли подтвердить, что их обогащение не было ассоциировано с уже существовавшим смещением.
Поскольку авторами настоящего изобретения была продемонстрирована система iTransduce с применением агностического подхода к типу клеток (всего головного мозга), то неудивительно, что AAV-F обладал высокой степенью тропизма в отношении астроцитов и нейронов, то есть, клеток, которые были трансдуцированы AAV9. В последующих исследованиях, авторы будут комбинировать клетко-специфические промоторы для инициации экспрессии Cre из вектора библиотеки AAV, а также магнитный сортинг клеток для выделения капсидов, которые могут трансдуктировать клетки, невосприимчивые к традиционной трансдукции вектором AAV.
Помимо возможностей системы iTransduce относительно выбора клинических кандидатов на векторы AAV, она может быть использована для идентификации векторов в целях их применения в качестве инструментов для исследования. Недавно идентифицированный капсид AAV-PHP.B послужил эффективным вектором для генетической модификации головного мозга мыши12. Однако, он не трансдуцирует линии дифференцировки мышей BALB/c или мышей, родственных мышам BALB/c13,14,16. Интересно отметить, что после внутривенной инъекции AAV-F наблюдалась надежная трансдукция в головной мозг у мышей BALB/c и C57BL/6. Это указывает на то, что механизм усиленной трансдукции посредством AAV9 отличается между AAV-PHP.B и AAV-F. Это также позволяет использовать AAV-F в качестве эффективного инструмента для исследования ЦНС у стандартных мышей вида BALB/c (labome.com/method/Laboratory-Mice-and-Rats.html). Кроме того, как показано в настоящей заявке, AAV-F может также опосредовать устойчивую экспрессию трансгена в ЦНС как после прямой, так и после интратекальной инъекции ударной дозы, а AAV-S может опосредовать экспрессию трансгена во внутреннем ухе.
Будущие исследования на более крупных животных могут быть проведены для дальнейшего тестирования AAV-F, например, в доклинических исследованиях. Для проверки дозозависимой токсичности AAV-F могут быть проведены исследования с увеличением дозы, как это наблюдалось в случае PHP.B в NHP14. Повторные раунды отбора могут быть проведены у животных различных видов (например, у мышей, а затем у крыс) для еще лучшего обеспечения эффективности трансдукции между видами. Так, например, это может быть осуществлено на мышах, а затем на крысах, трансгенных по floxed-Stop-tdTomato17. Альтернативно, прямой отбор библиотеки iTransduce может быть осуществлен у трансгенных игрунок18,19 или даже у других нетрансгенных приматов, не являющихся человеком (фиг. 16A и B).
Способы идентификации оптимизированных капсидных последовательностей
«Библиотеки вирусных векторов» представляют собой объединенные варианты вирусов, которые под давлением отбора (in vivo или in vitro), могут обеспечивать выделение клонов вирусов, специфичных для представляющих интерес клетки/ткани/органа-мишени. Одним из ограничений современных технологий создания библиотек является то, что многие из клонов вирусов-кандидатов не опосредуют экспрессию трансгена (необходимую конечную функцию вектора). Основная причина такого ограничения заключается в том, что пока еще не было разработано какой-либо стратегии, позволяющей проводить отбор вектора на основе экспрессии трансгена, опосредуемой вектором. В настоящее заявке описаны способы, в которых используется геном вектора аденоассоциированного вируса (AAV) с экспрессионным кластером из двух частей. Первая часть представляет собой кластер Cre-рекомбиназы под контролем представляющего интерес промотора. Вторая часть представляет собой промотор AAV для инициации экспрессии сконструированного капсидного гена, клонированного «в цис-ориентации» по отношению к первой части вирусного генома. Вирусные векторы могут быть отобраны для экспрессии трансгена (высокочувствительной экспрессии Cre) с использованием клеток, которые экспрессируют репортерный ген (например, белок, флуоресцирующий в зеленом диапазона спектра) с вышерасположенным loxP/стоп-сайтом, что тем самым позволяет предотвратить экспрессию репортера до тех пор, пока Cre, доставляемый вектором AAV, не будет удалять стоп-сайт. Могут быть выделены клетки, позитивные по репортерному гену, и восстановленные последовательности капсида AAV будут иметь более высокую вероятность опосредования эффективной экспрессии трансгена.
Таким образом, в настоящей заявке описаны конструкции библиотеки AAV, содержащие: (i) рекомбиназу Cre, находящуюся под контролем промотора, например, минимального промотора куриного бета-актина (CBA); (ii) последовательность капсида AAV9 под контролем промотора (например, промотора p41), вместе с последовательностью, кодирующей пептид, описанный в настоящей заявке, например, рандомизированный гептамерный пептид или выбранный гептамерный пептид, встроенный в капсидный белок ниже кластера Cre. Предпочтительно, пептид встраивают между последовательностями, кодирующими аминокислоты (а.к.) 588-589 капсида, но он также может быть встроен в любой другой участок, при условии, что он не будет негативно влиять на функцию вируса и будет сохранять свою активность в стимуляции инфицирования выбранных клеток, например, в пограничной области VP1/VP2 (аминокислота 138) или в любом сайте между аминокислотами 583-590. Промотор CBA является сильным активным промотором, который инициирует активность Cre в клетках большинства типов. Промотор P41 представляет собой природный AAV-специфичный промотор, который инициирует экспрессию гена Cap. Другими промоторами, которые могут быть использованы, являются, но не ограничиваются ими, промотор синапсина, промотор GFAP, промотор CD68, промотор F4/80, промотор CX3CR1, промотор CD3 или CD4, промотор CMV, промотор, специфичный для печени; и другие промоторы, перечисленные ниже. Конструкции также могут включать стоп-кодон на конце кДНК Cre и на конце ДНК сар. Сигналы poly-A находятся за кластером Cre и кластером сар. Рекомбиназы Cre известны специалистам в данной области, см., например, Van Duyne, Microbiol Spectr. 2015 Feb; 3(1): MDNA3-0014-2014. На Фиг. 1A представлена репрезентативная конструкция библиотеки.
В настоящей заявке также описаны библиотеки (то есть, композиции, содержащие множество конструкций библиотек). Если используются рандомизированные гептамерные последовательности, то предпочтительно, чтобы библиотека включала конструкции с последовательностями, кодирующими все или почти все возможные варианты гептамера.
Способы, проиллюстрированные на фиг. 1B(i), могут включать введение библиотеки, состоящей из различных пептидных вставок, экспрессируемых на капсиде (представленных различными оттенками серого), животному-модели, например, млекопитающему, такому как мышь (например, трансгенная мышь Ai9), кролик, крыса или обезьяна. Животное-модель содержит loxP-фланкированный кластер STOP, расположенный перед репортерной последовательностью, например, флуоресцентной последовательностью репортерного белка, например, репортерным геном tdTomato, необязательно встроенным в локус Gt(ROSA)26Sor. Животное-модель может быть трансгенным, или loxP-фланкированный кластер STOP, расположенный перед репортерной последовательностью, может экспрессироваться из второй конструкции, например, второго AAV, вводимого животному-модели (например, вводимого до, во время или после введения конструкций библиотеки). Любые капсиды AAV, которые проникают в представляющую интерес клетку, но функционально не трансдуцируют клетку (не экспрессируют Cre), не инициируют экспрессию репортера. Капсиды, которые могут опосредовать функциональную трансдукцию (экспрессировать Cre), будут инициировать экспрессию tdTomato. Как показано на фиг. 1B (ii), клетки выделяют из представляющего интерес органа (например, головного мозга, глаза, уха, сетчатки, сердца и т.п.), а затем трансдуцированные клетки могут быть отобраны на экспрессию репортерного гена и, необязательно, клеточных маркеров. Как показано на фиг. 1B(iii), капсидную ДНК получают и анализируют, например, но необязательно, с помощью ПЦР-амплификации последовательностей из отсортированных клеток, с последующим их повторным клонированием обратно в вектор библиотеки и повторной упаковкой для проведения другого раунда отбора. После каждого раунда может быть проведен анализ путем секвенирования ДНК для мониторинга процесса отбора.
Промоторы
Описанные здесь конструкции библиотек включают два промотора: один промотор управляет рекомбиназой Cre, а второй - последовательностью капсида AAV.
Специалистам в данной области известен ряд промоторных последовательностей, включая так называемые «часто встречающиеся» промоторы, которые инициируют экспрессию в клетках большинства типов, например промотор цитомегаловируса (CMV) (необязательно с энхансером CMV), промотор куриного бета-актина (CBA), промотор LTR вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор SV40, промотор дигидрофолатредуктазы, промотор фосфоглицеринкиназы, промотор фосфоглицеринкиназы (PGK), промотор EF1-альфа, промотор убихитина C (UBC), промотор B-глюкуронидазы и энхансер предраннего/раннего гена CMV/промотор CBA.
Экспрессия Cre-рекомбиназы также может быть инициирована тканеспецифическим промотором, например, тканеспецифическим промотором для ЦНС, печени, сердца, улитки, сетчатки или Т-клеток, inter alia. В некоторых вариантах осуществления изобретения, тканеспецифический промотор для ЦНС включает промотор нейронов, макрофагов/микроглиальных клеток и промоторы астроцитов. Специалистам в данной области известен ряд тканеспецифических промоторов, включая промотор синапсина (нейроны), нейрон-специфической энолазы (NSE) (нейроны), MeCP2 (метил-CPG-связывающего белка 2) (нейроны), глиального фибриллярного кислотного белка (GFAP) (астроциты), фактора транскрипции олигодендроцитов 1 (Olig1) (олигодендроциты), CNP (2',3'-циклической нуклеотид-3'-фосфодиэстеразы) (клетки широкого спектра) или CBh (гибридный промотор CBA или интрона MVM и CBA) (клетки широкого спектра). См., например, патент США № 20190032078. Промоторы макрофагов/микроглиальных клеток включают, но не ограничиваются ими, промотор хемокинового рецептора 1 (CX3CR1) с мотивом C-X3-C, промотор CD68, промотор молекулы адаптора, связывающейся с ионами кальция 1 (IBA1), промотор трансмембранного белка 119 (TMEM119), промотор spalt-подобного фактора транскрипции 1 (SALL1), промотор адгезионного рецептора, связанного с G-белком E1 (F4/80), энхансер вируса миелопролиферативной саркомы, промотор с замененным сайтом связывания с праймером d1587rev и с делетированной областью негативного контроля (MND); промотор субъединицы интегрина альфа-M (ITGAM; промотор CD11b-миелоидных клеток (нейтрофилов, моноцитов и макрофагов)). Для экспрессии во внутреннем ухе, промотором может быть, например, PKG, CAG, престин, Atoh1, POU4F3, Lhx3, Myo6, α9AchR, α10AchR, онкомод или промотор myo7A; см. Ryan et al., Adv Otorhinolaryngol. 2009; 66: 99-115.
Репортерные белки
Специалистам в данной области известен ряд репортерных белков, которые включают белок, флуоресцирующий в зеленом диапазоне спектра (GFP), вариант белка, флуоресцирующего в зеленом диапазоне спектра (GFP10), активированный GFP (eGFP), TurboGFP, GFPS65T, TagGFP2, mUKGEmerald GFP, Superfolder GFP, GFPuv, дестабилизированный EGFP (dEGFP), Azami Green, mWasabi, Clover, mClover3, mNeonGreen, NowGFP, Sapphire, T-Sapphire, mAmetrine, фотоактивируемый GFP (PA-GFP), Kaede, Kikume, mKikGR, tdEos, Dendra2, mEosFP2, Dronpa, белок, флуоресцирующий в синем диапазоне спектра (BFP), eBFP2, азурит BFP, mTagBFP, mKalamal, mTagBFP2, shBFP, белок, флуоресцирующий в голубом диапазоне спектра (CFP), eCFP, Cerulian CFP, SCFP3A, дестабилизированный ECFP (dECFP), CyPet, mTurquoise, mTurquoise2, mTFP1, фотопереключаемый CFP2 (PS-CFP2), TagCFP, mTFP1, mMidoriishi-Cyan, аквамариновый, mKeima, mBeRFP, LSS-mKate2, LSS-mKatel, LSS-mOrange, CyOFP1, сандерцианин, белок, флуоресцирующий в красном диапазоне спектра (RFP), eRFP, mRaspberry, mRuby, mApple, mCardinal, mStable, mMaroonl, mGarnet2, tdTomato, mTangerine, mStrawberry, TagRFP, TagRFP657, TagRFP675, mKate2, HcRed, t-HcRed, HcRed-Tandem, mPlum, mNeptune, NirFP, Kindling, белок, флуоресцирующий в дальнем красном диапазоне спектра, белок, флуоресцирующий в желтом диапазоне спектра (YFP), eYFP, дестабилизированный EYFP (dEYFP), TagYFP, Topaz, Venus, SYFP2, mCherry, PA-mCherry, Citrine, mCitrine, Ypet, IANRFP-AS83, mPapayal, mCyRFP1, mHoneydew, mBanana, mOrange, Kusabira Orange, Kusabira Orange 2, mKusabira Orange, mOrange 2, mKOK, mKO2, mGrapel, mGrape2, zsYellow, eqFP611, Sirius, сандерцианин, shBFP-N158S/L173I, белок, флуоресцирующий в ближнем инфракрасном диапазоне спектра, iFP1.4, iRFP713, iRFP670, iRFP682, iRFP702, iRFP720, iFP2.0, mIFP, TDsmURFP, miRFP670, бриллиантовый фиолетовый (BV) 421, BV 605, BV 510, BV 711, BV786, PerCP, PerCP/Cy5.5, DsRed, DsRed2, mRFPl, поциллопорин, Renilla GFP, Monster GFP, paGFP, или фикобилипротеин, или биологически активный вариант или фрагмент любого из них.
Наборы
Настоящее изобретение также относится к описанным здесь наборам, содержащим одну или более конструкций AAV-библиотек с рандомизированными гептамерными последовательностями или без них. Наборы могут также включать конструкцию, содержащую loxP-фланкированный кластер STOP, расположенный перед репортерной последовательностью.
Сконструированные капсидные белки AAV
Способы согласно изобретению позволяют идентифицировать две пептидные последовательности, которые изменяют способность AAV опосредовать экспрессию трансгена в определенных клетках при их встраивании в капсид AAV, например, AAV1, AAV2, AAV8 или AAV9. В некоторых вариантах осуществления изобретения, пептиды содержат последовательности по меньшей мере из 7 аминокислот. В некоторых вариантах осуществления изобретения, аминокислотная последовательность содержит по меньшей мере 4, например, 5, 6 или 7 смежных аминокислот последовательностей (STTLYSP (SEQ ID NO: 1) или FVVGQSY (SEQ ID NO: 2).
Также могут быть использованы пептиды, включающие обратные последовательности, например, PSYLTTS (SEQ ID NO: 4) и YSQGVVF (SEQ ID NO: 5). Альтернативно, пептиды могут содержать по меньшей мере четыре, пять или шесть смежных аминокислот последовательностей, представленных на фиг. 2A или 7C (SEQ ID NO: 17-150).
AAV
Вирусные векторы для использования в способах, наборах и композициях согласно изобретению включают рекомбинантные ретровирусы, аденовирус, аденоассоциированный вирус, альфавирус и лентивирус, предпочтительно содержащие капсидный пептид, как описано в настоящей заявке, и необязательно трансген для экспрессии в ткани-мишени.
Предпочтительной вирусной векторной системой, пригодной для доставки нуклеиновых кислот в способах согласно изобретению, является аденоассоциированный вирус (AAV). AAV представляет собой небольшой вирус без оболочки, имеющий капсид размером 25 нм. Пока не известно или не сообщалось, что какое-либо заболевание связано с этим вирусом дикого типа. AAV имеет геном одноцепочечной ДНК (оцДНК). Было показано, что AAV обнаруживает длительную эписомальную экспрессию трансгена и превосходную экспрессию трансгена в головном мозге, особенно в нейронах. Пространство для экзогенной ДНК ограничено приблизительно 4,7 т.п.о. Вектор AAV, такой как вектор, описанный в Tratschin et al., Mol. Cell. Biol. 5: 3251-3260 (1985), может быть использован для введения ДНК в клетки. Различные нуклеиновые кислоты были введены в клетки различных типов с использованием векторов AAV (см., например, Hermonat et al., Proc. Natl. Acad. Sci. USA 81: 6466-6470 (1984); Tratschin et al., Mol. Cell. Biol. 4: 2072-2081 (1985); Wondisford et al., Mol. Endocrinol. 2: 32-39 (1988); Tratschin et al., J. Virol. 51: 611-619 (1984); и Flotte et al., J. Biol. Chem. 268: 3781-3790 (1993). Существует множество альтернативных вариантов AAV (было клонировано более 100), и варианты AAV были идентифицированы исходя из желаемых свойств. В некоторых вариантах осуществления изобретения, AAV представляет собой AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AV6.2, AAV7, AAV8, rh.8, AAV9, rh.10, rh.39, rh.43 или CSp3; а для использования в ЦНС, в некоторых вариантах осуществления изобретения, AAV представляет собой AAV1, AAV2, AAV4, AAV5, AAV6, AAV8 или AAV9. В качестве одного примера было показано, что AAV9 в некоторой степени эффективно преодолевает гематоэнцефалический барьер. С применением способов согласно изобретению, капсид AAV может быть генетически модифицирован для повышения способности проникать через ГЭБ, или в определенную ткань, посредством встраивания описанной здесь пептидной последовательности в капсидный белок, например, в капсидный белок VP1 AAV9 между аминокислотами 588 и 589.
Репрезентативныая последовательность VP1 капсидного белка AAV9 дикого типа (Q6JC40-1) представляет собой:

Таким образом, настоящее изобретение относится к AAV, который включают одну или более описанных здесь пептидных последовательностей, например, AAV, включающий капсидный белок, содержащий описанную здесь последовательность, например, капсидный белок, содержащий SEQ ID NO: 1 или SEQ ID NO: 2, где в последовательность была встроена пептидная последовательность, например, между аминокислотами 588 и 589.
Репрезентативные последовательности AAV представлены ниже. Встроенные пептидные последовательности выделены жирным шрифтом и подчеркнуты двойным подчеркиванием в белковых последовательностях, и жирным шрифтом и заглавными буквами в последовательностях ДНК.
Последовательность капсидного белка AAV-F
MAADGYLPDWLEDNLSEGIREWWALKPGAPQPKANQQHQDNARGLVLPGYKYLGPGNGLDKGEPVNAADAAALEHDKAYDQQLKAGDNPYLKYNHADAEFQERLKEDTSFGGNLGRAVFQAKKRLLEPLGLVEEAAKTAPGKKRPVEQSPQEPDSSAGIGKSGAQPAKKRLNFGQTGDTESVPDPQPIGEPPAAPSGVGSLTMASGGGAPVADNNEGADGVGSSSGNWHCDSQWLGDRVITTSTRTWALPTYNNHLYKQISNSTSGGSSNDNAYFGYSTPWGYFDFNRFHCHFSPRDWQRLINNNWGFRPKRLNFKLFNIQVKEVTDNNGVKTIANNLTSTVQVFTDSDYQLPYVLGSAHEGCLPPFPADVFMIPQYGYLTLNDGSQAVGRSSFYCLEYFPSQMLRTGNNFQFSYEFENVPFHSSYAHSQSLDRLMNPLIDQYLYYLSKTINGSGQNQQTLKFSVAGPSNMAVQGRNYIPGPSYRQQRVSTTVTQNNNSEFAWPGASSWALNGRNSLMNPGPAMASHKEGEDRFFPLSGSLIFGKQGTGRDNVDADKVMITNEEEIKTTNPVATESYGQVATNHQSAQFVVGQSYAQAQTGWVQNQGILPGMVWQDRDVYLQGPIWAKIPHTDGNFHPSPLMGGFGMKHPPPQILIKNTPVPADPPTAFNKDKLNSFITQYSTGQVSVEIEWELQKENSKRWNPEIQYTSNYYKSNNVEFAVNTEGVYSEPRPIGTRYLTRNL(SEQ ID NO:7)
Последовательность ДНК капсида AAV-F
atggctgccgatggttatcttccagattggctcgaggacaaccttagtgaaggaattcgcgagtggtgggctttgaaacctggagcccctcaacccaaggcaaatcaacaacatcaagacaacgctcgaggtcttgtgcttccgggttacaaataccttggacccggcaacggactcgacaagggggagccggtcaacgcagcagacgcggcggccctcgagcacgacaaggcctacgaccagcagctcaaggccggagacaacccgtacctcaagtacaaccacgccgacgccgagttccaggagcggctcaaagaagatacgtcttttgggggcaacctcgggcgagcagtcttccaggccaaaaagaggcttcttgaacctcttggtctggttgaggaagcggctaagacggctcctggaaagaagaggcctgtagagcagtctcctcaggaaccggactcctccgcgggtattggcaaatcgggtgcacagcccgctaaaaagagactcaatttcggtcagactggcgacacagagtcagtcccagaccctcaaccaatcggagaacctcccgcagccccctcaggtgtgggatctcttacaatggcttcaggtggtggcgcaccagtggcagacaataacgaaggtgccgatggagtgggtagttcctcgggaaattggcattgcgattcccaatggctgggggacagagtcatcaccaccagcacccgaacctgggccctgcccacctacaacaatcacctctacaagcaaatctccaacagcacatctggaggatcttcaaatgacaacgcctacttcggctacagcaccccctgggggtattttgacttcaacagattccactgccacttctcaccacgtgactggcagcgactcatcaacaacaactggggattccggcctaagcgactcaacttcaagctcttcaacattcaggtcaaagaggttacggacaacaatggagtcaagaccatcgccaataaccttaccagcacggtccaggtcttcacggactcagactatcagctcccgtacgtgctcgggtcggctcacgagggctgcctcccgccgttcccagcggacgttttcatgattcctcagtacgggtatctgacgcttaatgatggaagccaggccgtgggtcgttcgtccttttactgcctggaatatttcccgtcgcaaatgctaagaacgggtaacaacttccagttcagctacgagtttgagaacgtacctttccatagcagctacgctcacagccaaagcctggaccgactaatgaatccactcatcgaccaatacttgtactatctctcaaagactattaacggttctggacagaatcaacaaacgctaaaattcagtgtggccggacccagcaacatggctgtccagggaagaaactacatacctggacccagctaccgacaacaacgtgtctcaaccactgtgactcaaaacaacaacagcgaatttgcttggcctggagcttcttcttgggctctcaatggacgtaatagcttgatgaatcctggacctgctatggccagccacaaagaaggagaggaccgtttctttcctttgtctggatctttaatttttggcaaacaaggaactggaagagacaacgtggatgcggacaaagtcatgataaccaacgaagaagaaattaaaactactaacccggtagcaacggagtcctatggacaagtggccacaaaccaccagagtgcccaaTTTGTTGTTGGTCAGAGTTATgcacaggcgcagaccggctgggttcaaaaccaaggaatacttccgggtatggtttggcaggacagagatgtgtacctgcaaggacccatttgggccaaaattcctcacacggacggcaactttcacccttctccgctgatgggagggtttggaatgaagcacccgcctcctcagatcctcatcaaaaacacacctgtacctgcggatcctccaacggccttcaacaaggacaagctgaactctttcatcacccagtattctactggccaagtcagcgtggagatcgagtgggagctgcagaaggaaaacagcaagcgctggaacccggagatccagtacacttccaactattacaagtctaataatgttgaatttgctgttaatactgaaggtgtatatagtgaaccccgccccattggcaccagatacctgactcgtaatctg (SEQ ID NO:8)
Последовательность капсидного белка AAV-S
MAADGYLPDWLEDNLSEGIREWWALKPGAPQPKANQQHQDNARGLVLPGYKYLGPGNGLDKGEPVNAADAAALEHDKAYDQQLKAGDNPYLKYNHADAEFQERLKEDTSFGGNLGRAVFQAKKRLLEPLGLVEEAAKTAPGKKRPVEQSPQEPDSSAGIGKSGAQPAKKRLNFGQTGDTESVPDPQPIGEPPAAPSGVGSLTMASGGGAPVADNNEGADGVGSSSGNWHCDSQWLGDRVITTSTRTWALPTYNNHLYKQISNSTSGGSSNDNAYFGYSTPWGYFDFNRFHCHFSPRDWQRLINNNWGFRPKRLNFKLFNIQVKEVTDNNGVKTIANNLTSTVQVFTDSDYQLPYVLGSAHEGCLPPFPADVFMIPQYGYLTLNDGSQAVGRSSFYCLEYFPSQMLRTGNNFQFSYEFENVPFHSSYAHSQSLDRLMNPLIDQYLYYLSKTINGSGQNQQTLKFSVAGPSNMAVQGRNYIPGPSYRQQRVSTTVTQNNNSEFAWPGASSWALNGRNSLMNPGPAMASHKEGEDRFFPLSGSLIFGKQGTGRDNVDADKVMITNEEEIKTTNPVATESYGQVATNHQSAQSTTLYSPAQAQTGWVQNQGILPGMVWQDRDVYLQGPIWAKIPHTDGNFHPSPLMGGFGMKHPPPQILIKNTPVPADPPTAFNKDKLNSFITQYSTGQVSVEIEWELQKENSKRWNPEIQYTSNYYKSNNVEFAVNTEGVYSEPRPIGTRYLTRNL (SEQ ID NO:9)
Последовательность ДНК капсида AAV-S
atggctgccgatggttatcttccagattggctcgaggacaaccttagtgaaggaattcgcgagtggtgggctttgaaacctggagcccctcaacccaaggcaaatcaacaacatcaagacaacgctcgaggtcttgtgcttccgggttacaaataccttggacccggcaacggactcgacaagggggagccggtcaacgcagcagacgcggcggccctcgagcacgacaaggcctacgaccagcagctcaaggccggagacaacccgtacctcaagtacaaccacgccgacgccgagttccaggagcggctcaaagaagatacgtcttttgggggcaacctcgggcgagcagtcttccaggccaaaaagaggcttcttgaacctcttggtctggttgaggaagcggctaagacggctcctggaaagaagaggcctgtagagcagtctcctcaggaaccggactcctccgcgggtattggcaaatcgggtgcacagcccgctaaaaagagactcaatttcggtcagactggcgacacagagtcagtcccagaccctcaaccaatcggagaacctcccgcagccccctcaggtgtgggatctcttacaatggcttcaggtggtggcgcaccagtggcagacaataacgaaggtgccgatggagtgggtagttcctcgggaaattggcattgcgattcccaatggctgggggacagagtcatcaccaccagcacccgaacctgggccctgcccacctacaacaatcacctctacaagcaaatctccaacagcacatctggaggatcttcaaatgacaacgcctacttcggctacagcaccccctgggggtattttgacttcaacagattccactgccacttctcaccacgtgactggcagcgactcatcaacaacaactggggattccggcctaagcgactcaacttcaagctcttcaacattcaggtcaaagaggttacggacaacaatggagtcaagaccatcgccaataaccttaccagcacggtccaggtcttcacggactcagactatcagctcccgtacgtgctcgggtcggctcacgagggctgcctcccgccgttcccagcggacgttttcatgattcctcagtacgggtatctgacgcttaatgatggaagccaggccgtgggtcgttcgtccttttactgcctggaatatttcccgtcgcaaatgctaagaacgggtaacaacttccagttcagctacgagtttgagaacgtacctttccatagcagctacgctcacagccaaagcctggaccgactaatgaatccactcatcgaccaatacttgtactatctctcaaagactattaacggttctggacagaatcaacaaacgctaaaattcagtgtggccggacccagcaacatggctgtccagggaagaaactacatacctggacccagctaccgacaacaacgtgtctcaaccactgtgactcaaaacaacaacagcgaatttgcttggcctggagcttcttcttgggctctcaatggacgtaatagcttgatgaatcctggacctgctatggccagccacaaagaaggagaggaccgtttctttcctttgtctggatctttaatttttggcaaacaaggaactggaagagacaacgtggatgcggacaaagtcatgataaccaacgaagaagaaattaaaactactaacccggtagcaacggagtcctatggacaagtggccacaaaccaccagagtgcccaaTCTACTACGCTTTATAGTCCTgcacaggcgcagaccggctgggttcaaaaccaaggaatacttccgggtatggtttggcaggacagagatgtgtacctgcaaggacccatttgggccaaaattcctcacacggacggcaactttcacccttctccgctgatgggagggtttggaatgaagcacccgcctcctcagatcctcatcaaaaacacacctgtacctgcggatcctccaacggccttcaacaaggacaagctgaactctttcatcacccagtattctactggccaagtcagcgtggagatcgagtgggagctgcagaaggaaaacagcaagcgctggaacccggagatccagtacacttccaactattacaagtctaataatgttgaatttgctgttaatactgaaggtgtatatagtgaaccccgccccattggcaccagatacctgactcgtaatctg (SEQ ID NO:10).
Последовательности AAV могут быть, например, по меньшей мере на 80, 85, 90, 95, 97 или 99% идентичны эталонной последовательности AAV, представленной в настоящей заявке, и например, могут включать предпочтительно варианты, которые не снижают способность AAV опосредовать экспрессию трансгена в клетке. Для определения процента идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновых кислот, эти последовательности выравнивают для их оптимального сравнения (например, могут быть введены пробелы в одной или обеих первой и второй аминокислотных последовательностях или в последовательностях нуклеиновой кислоты для оптимального выравнивания, при этом, при сравнении, негомологичные последовательности можно не принимать во внимание). В предпочтительном варианте осуществления изобретения, длина эталонной последовательности, выравниваемой для сравнения, составляет по меньшей мере 80% длины эталонной последовательности, а в некоторых вариантах осуществления изобретения, по меньшей мере 90% или 100%. Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих положениях аминокислот или нуклеотидов. Если положение в первой последовательности занято таким же аминокислотным остатком или нуклеотидом, как и соответствующее положение во второй последовательности, то такие молекулы являются идентичными в этом положении (используемый здесь термин «идентичность» аминокислот или нуклеиновых кислот эквивалентен термину «гомология» аминокислот или нуклеиновых кислот). Процент идентичности между двумя последовательностями зависит от числа идентичных положений в обеих последовательностях, а также от количества пробелов и длины каждого пробела, которые необходимо ввести для оптимального выравнивания двух последовательностей.
Сравнение последовательностей и определение процента идентичности между двумя последовательностями может быть осуществлено с использованием математического алгоритма. Так, например, процент идентичности между двумя аминокислотными последовательностями можно определить с помощью алгоритма Нидлмана и Вюнша (Needleman and Wunsch ((1970) J. Mol. Biol. 48: 444-453), который был включен в программу GAP в пакете программного обеспечения GCG (доступном в Интернете на сайте gcg.com), с использованием параметров по умолчанию, например, оценочной матрицы Blossum 62 со штрафом за пробел 12, штрафом за продление пробела 4 и штрафом за пробел со сдвигом рамки считывания 5.
Трансгены
В некоторых вариантах осуществления изобретения, AAV также включает последовательность трансгена (то есть, гетерологичную последовательность), например, трансген, кодирующий терапевтический агент, например, описанный в настоящей заявке или известный специалистам в данной области, или репортерный белок, например, флуоресцентный белок, фермент, который катализирует реакцию с образованием детектируемого продукта, или антиген клеточной поверхности. Трансген предпочтительно связан с последовательностями, которые стимулируют/инициируют экспрессию трансгена в ткани-мишени.
Примеры трансгенов для использования в качестве терапевтических средств включают белок, ингибирующий апоптоз нейронов (NAIP); фактор роста нервных клеток (NGF); фактор роста глиального происхождения (GDNF); фактор роста, происходящий от головного мозга (BDNF); цилиарный нейротропный фактор (CNTF); тирозингидрокслазу (TH); GTP-циклогидролазу (GTPCH); декарбоксилазу аминокислот (AADC); аспартоацилазу (ASPA); факторы крови, такие как β-глобин, гемоглобин, тканевый активатор плазминогена и факторы свертывания крови; колониестимулирующие факторы (CSF); интерлейкины, такие как IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9 и т.п; факторы роста, такие как фактор роста кератиноцитов (KGF), фактор стволовых клеток (SCF), фактор роста фибробластов (FGF, такой как основный FGF и кислотный FGF), фактор роста гепатоцитов (HGF), инсулиноподобные факторы роста (IGF), белок морфогенеза кости (BMP), эпидермальный фактор роста (EGF), фактор дифференцировки и роста-9 (GDF-9), фактор роста, происходящий от гепатомы (HDGF), миостатин (GDF-8), фактор роста нервных клеток (NGF), нейротропины, тромбоцитарный фактор роста (PDGF), тромбопоэтин (TPO), трансформирующий фактор роста альфа (TGF-α), трансформирующий фактор роста бета (TGF-β) и т.п.; растворимые рецепторы, такие как растворимые рецепторы TNF-α, растворимые рецепторы VEGF, растворимые рецепторы интерлейкина (например, растворимые рецепторы IL-1 и растворимые рецепторы IL-1 типа II), растворимые Т-клеточные рецепторы гамма/дельта, лиганд-связывающие фрагменты растворимого рецептора и т.п.; ферменты, такие как α-глюкозидаза, имиглюкараза и β-глюкоцереброзидаза; активаторы ферментов, такие как тканевый активатор плазминогена; хемокины, такие как IP-10, монокин, индуцированный гамма-интерфероном (Mig), Groa/IL-8, RANTES, MIP-1α, MIP-1β, MCP-1, PF-4 и т.п.; ангиогенные агенты, такие как васкулярные эндотелиальные факторы роста (VEGF, например, VEGF121, VEGF165, VEGF-C, VEGF-2), трансформирующий фактор роста бета, основный фактор роста фибробластов, фактор роста, происходящий от глиомы, ангиогенин, ангиогенин-2, и т.п.; антиангиогенные агенты, такие как растворимый рецептор VEGF; белковую вакцину; нейроактивные пептиды, такие как фактор роста нервных клеток (NGF), брадикинин, холецистокинин, гастин, секретин, окситоцин, гонадотропин-высвобождающий гормон, бета-эндорфин, энкефалин, субстанцию P, соматостатин, пролактин, галанин, рилизинг-фактор гормона роста, бомбезин, динорфин, варфарин, нейротензин, мотилин, тиреотропин, нейропептид Y, лютеинизирующий гормон, кальцитонин, инсулин, глюкагоны, вазопрессин, ангиотензин II, тиреотропин-высвобождающий гормон, вазоактивный кишечный пептид, пептид сайленсинга и т.п.; тромболитические агенты; предсердный натрийуретический пептид; релаксин; глиальный фибриллярный кислотный белок; фолликулостимулирующий гормон (FSH); человеческий альфа-1-антитрипсин; фактор ингибирования лейкоза (LIF); трансформирующие факторы роста (TGF); тканевые факторы, лютеинизирующий гормон; факторы активации макрофагов; фактор некроза опухоли (TNF); хемотаксический фактор нейтрофилов (NCF); фактор роста нервных клеток; тканевые ингибиторы металлопротеиназ; вазоактивный кишечный пептид; ангиогенин; ангиотропин; фибрин; гирудин; антагонисты рецептора IL-1; и т.п. Некоторые другие примеры представляющего интерес белка включают цилиарный нейротропный фактор (CNTF); нейротропины 3 и 4/5 (NT-3 и 4/5); нейротропный фактор глиальных клеток (GDNF); ароматическую декарбоксилазу аминокислот (AADC); белки свертывания крови, ассоциированные с гемофилией, такие как фактор VIII, фактор IX, фактор X; дистрофин или нинидистрофин; лизосомную кислую липазу; фенилаланингидроксилазу (PAH); ферменты, ассоциированные с болезнью накопления гликогена, такие как глюкозо-6-фосфатаза, кислая мальтаза, гликоген-деветвящий фермент, мышечная гликогенфосфорилаза, гликогенфосфорилаза печени, мышечная фосфофруктокиназа, фосфорилазокиназа (например, PHKA2), переносчик глюкозы (например, GLUT2), альдолаза А, β-энолаза и гликогенсинтаза; лизосомные ферменты (например, бета-N-ацетилгексозаминидазу A); и любые их варианты.
Трансген может также кодировать антитело, например антитело, ингибирующее иммунную контрольную точку, например, антитело против PD-L1, PD-1, CTLA-4 (белка-4, ассоциированного с цитотоксическим T-лимфоцитом; CD152); LAG-3 (гена активации лимфоцитов 3; CD223); TIM-3 (домена Т-клеточного иммуноглобулина и домена 3 муцина; HAVCR2); TIGIT (Т-клеточного иммунорецептора с доменами Ig и ITIM); B7-H3 (CD276); VSIR (иммунорегуляторного рецептора серии V, также известного как VISTA, B7H5, C10orf54); BTLA 30 (аттенюатора В- и Т-лимфоцитов, CD272); GARP (доменов гликопротеина А с большим количеством повторов); PVRIG (содержащего домен иммуноглобулина, ассоциированный с PVR); или VTCN1 (домена серии V, содержащего ингибитор 1 активации Т-клеток, также известный как B7-H4).
Другие трансгены могут включать небольшие или ингибирующие нуклеиновые кислоты, которые изменяют/снижают экспрессию гена-мишени, например, киРНК, кшРНК, миРНК, антисмысловые олигонуклеотиды или длинные некодирующие РНК, которые изменяют экспрессию гена (см., например, WO2012087983 и US20140142160), или CRISPR Cas9/cas12a и руководящие РНК.
Вирус также может включать одну или более последовательностей, которые стимулируют экспрессию трансгена, например, одну или более промоторных последовательностей; энхансерные последовательности, например, 5'-нетранслируемую область (UTR) или 3'-UTR; сайт полиаденилирования; и/или изоляторные последовательности. В некоторых вариантах осуществления изобретения, промотор представляет собой промотор, специфичный для ткани головного мозга, например, нейрон-специфический промотор или промотор, специфичный к глиальным клеткам. В некоторых вариантах осуществления изобретения, промотор представляет собой промотор гена, выбранный из промотора нейронных ядер (NeuN), глиального фибриллярного кислотного белка (GFAP), MeCP2, аденоматозного кишечного полипоза (APC), молекулы адаптера, связывающегося с ионами кальция 1 (Iba-1), синапсина I (SYN), кальций/кальмодулин-зависимой протеинкиназы II, тубулина альфа I, нейрон-специфической энолазы и бета-цепи тромбоцитарного фактора роста. В некоторых вариантах осуществления изобретения, промотор представляет собой промотор клеток различных типов, например, промотор цитомегаловируса (CMV), бета-глюкуронидазы (GUSB), убихитина C (UBC) или вируса саркомы Рауса (RSV). Также может быть использован элемент посттранскрипционного ответа вируса гепатита сурка (WPRE).
В некоторых вариантах осуществления изобретения, AAV также имеет одну или более дополнительных мутаций, которые улучшают доставку в ткань-мишень, например, в ЦНС, или снижают нацеливание на ткань, не являющуюся мишенью, например, мутации, которые снижают доставку в печень, в том случае, если необходима доставка в ЦНС, сердце или мышцы (например, как описано Pulicherla et al. (2011) Mol Ther 19: 1070-1078); или добавление других пептидов, например, как описано у Chen et al. (2008) Nat Med 15: 1215-1218 или Xu et al., (2005) Virology 341: 203-214 или в патентах США 9102949; 9585971 и 20170166926. См. также публикации Gray and Samulski (2011) «Vector design and considerations for CNS applications» in Gene Vector Design and Application to Treat Nervous System Disorders ed. Glorioso J., editor. (Washington, DC: Society for Neuroscience) 1-9, доступные на сайте sfn.org/~/media/SfN/Documents/Short%20Courses/ 2011%20Short%20Course%20I/2011_SC1_Gray.ashx.
Способы применения
Описанные здесь способы и композиции могут быть применены для доставки любой композиции, например, представляющей интерес последовательности, в ткань, например, в центральную нервную систему (головной мозг), сердце, мышцы, периферическую нервную систему (например, в спинномозговые узлы или в спинной мозг) или во внутреннее ухо или в сетчатку. В некоторых вариантах осуществления изобретения, способы включают доставку в определенные области головного мозга, например, в кору головного мозга, мозжечок, гиппокамп, черное вещество, миндалины. В некоторых вариантах осуществления изобретения, способы включают доставку в поясничный отдел, например, в субарахноидальное пространство или в эпидуральное пространство. В некоторых вариантах осуществления изобретения, способы включают доставку в нейроны, астроциты или глиальные клетки. В некоторых вариантах осуществления изобретения, способы включают доставку во внутренние и/или внешние волосковые клетки, нейроны спиральных узлов, поддерживающие клетки или фиброциты внутреннего уха. В некоторых вариантах осуществления изобретения, способы включают доставку в фоторецепторы, интернейроны, в клетки узлов сетчатки (например, с использованием AAV-F) или в пигментный эпителий сетчатки (RPE) (например, с использованием AAV-S).
В некоторых вариантах осуществления изобретения, способы и композиции, например, AAV, применяют для доставки последовательности нуклеиновой кислоты индивидууму, который страдает заболеванием, например, заболеванием ЦНС; см., например, патенты США 9102949; 9585971; и в заявке на патент США 20170166926. В некоторых вариантах осуществления изобретения, индивидуум страдает заболеваниями, перечисленными в Таблицах 1-3; а в некоторых вариантах осуществления изобретения, векторы используют для доставки терапевтического средства, указанного в Таблицах 1-3, для лечения соответствующих заболеваний, перечисленных в Таблицах 1-3. Терапевтическое средство может быть доставлено в виде нуклеиновой кислоты, например, посредством вирусного вектора, где нуклеиновая кислота кодирует терапевтический белок или другую нуклеиновую кислоту, такую как антисмысловой олигонуклеотид, киРНК, кшРНК и т.п.; или в виде слитого белка/комплекса с пептидом, как описано в настоящей заявке.
Описанные здесь способы и композиции могут быть применены для лечения этих состояний у индивидуума, нуждающегося в этом, путем введения терапевтически эффективного количества AAV, несущего терапевтический трансген, а именно, количества, достаточного для ослабления, снижения риска или задержки начала развития одного или более симптомов указанного состояния.
Таблица 1. Мишени для ЦНС (AAV-F, AAV-S)
Нейроны
Таблица 2. Мишени для внутреннего уха (AAV-S)
Фиброциты/поддерживающие клетки
Волосковые клетки (внутренние и внешние)
Волосковые клетки (внутренние и внешние)
Поддерживающие клетки (для регенерации HC)
Волосковые клетки (внутренние и внешние)
Внутренние волосковые клетки
Волосковые клетки (внутренние и внешние)
Волосковые клетки (внутренние и внешние)
* Синдром Ашера приводит как к глухоте, как указано выше, так и к слепоте из-за пигментного ретинита.
Таблица 3. Мишени для периферической системы (AAV-F, AAV-S)
Фармацевтические композиции и способы введения
Описанные здесь способы включают использование фармацевтических композиций, содержащих AAV, в качестве активного ингредиента.
Фармацевтические композиции обычно включают фармацевтически приемлемый носитель. Используемый здесь термин «фармацевтически приемлемый носитель» включает физиологический раствор, растворители, дисперсионные среды, агенты для нанесения покрытия, антибактериальные и противогрибковые средства, изотонические агенты и агенты, замедляющие всасывание, и т.п., совместимые с введением фармацевтических средств.
Фармацевтические композиции обычно получают так, чтобы они были совместимы с предполагаемым способом введения. Примеры способов введения включают парентеральное, например, внутривенное, внутриартериальное, подкожное, внутрибрюшинное, интратекальное, внутримышечное введение или введение путем инъекции или инфузии. Таким образом, доставка может быть системной или местной. Так, например, для доставки во внутреннее ухо может быть осуществлено введение в улитку поверх или через мембрану круглого окна, посредством хирургической кохлеостомии путем просверливания отверстия рядом с круглым окном, через отверстие в овальном окне кости или через полукруглый канал. (см., например, Kim et al., Mol Ther Methods Clin Dev. 2019 Jan 11: 13: 197-204; Ren et al., Front Cell Neurosci. 2019; 13: 323); а для доставки в сетчатку могут быть введены субретинальные инъекции или инъекции в стекловидное тело (см., например, Ochakovski et al., Front Neurosci. 2017; 11: 174; Xue et al., Eye (Lond). 2017 Sep; 31 (9): 1308-1316).
Методы получения подходящих фармацевтических композиций известны специалистам в данной области, см., например, руководство Remington: The Science and Practice of Pharmacy, 21st ed., 2005; и справочники из серии «Drugs and the Pharmaceuticals Sciences: a Series of Textbooks and Monographs» (Dekker, NY). Так, например, растворы или суспензии, используемые для парентерального введения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, жирные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатообразующие агенты, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетаты, цитраты или фосфаты, и агенты для регуляции тоничности, такие как хлорид натрия или декстроза. pH может быть скорректирован с помощью кислот или оснований, таких как соляная кислота или гидроксид натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или флаконы для многократных доз, изготовленные из стекла или пластика.
Фармацевтические композиции, подходящие для инъекций, могут включать стерильные водные растворы (если они являются водорастворимыми) или дисперсии и стерильные порошки, приготовленные в виде стерильных растворов или дисперсий для немедленного приема в виде инъекций. Для внутривенного введения, подходящими носителями являются физиологический раствор, бактериостатическая вода, Cremophor EL ™ (BASF, Parsippany, NJ) или забуференный фосфатом физиологический раствор (PBS). Во всех случаях, композиция должна быть стерильной и жидкой до такой степени, чтобы ее можно было легко забрать с помощью шприца. Она должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего действия микроорганизмов, таких как бактерии и грибы. Носитель может представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Подходящую текучесть можно поддерживать, например, за счет использования покрытия, такого как лецитин, путем поддержания необходимого размера частиц в случае дисперсии и за счет использования поверхностно-активных веществ. Предотвращение действия микроорганизмов может быть достигнуто с помощью различных антибактериальных и противогрибковых средств, например, парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимерозала и т.п. Во многих случаях, в композицию предпочтительно включать изотонические агенты, например сахара, многоатомные спирты, такие как маннит и сорбит; и хлорид натрия. Длительное всасывание композиций для инъекций может быть достигнуто путем включения в композицию агента, замедляющего всасывание, например, моностеарата алюминия и желатина.
Стерильные растворы для инъекций могут быть приготовлены путем включения активного соединения в нужном количестве в соответствующий растворитель с одним из ингредиентов или с комбинацией вышеперечисленных ингредиентов, если это необходимо, с последующей стерилизацией фильтрованием. Обычно дисперсии приготавливают путем включения активного соединения в стерильный носитель, который содержит оснόвную дисперсионную среду и другие необходимые ингредиенты из числа ингредиентов, перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций, предпочтительными методами приготовления являются вакуумная сушка и сушка вымораживанием, в результате чего, из раствора путем стерильной фильтрации получают порошок, состоящий из активного ингредиента и любого дополнительного желаемого ингредиента.
В одном варианте осуществления изобретения, терапевтические соединения получают вместе с носителями, которые будут защищать терапевтические соединения от их быстрого выведения из организма, например, в виде состава с контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Могут быть использованы биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Такие составы могут быть приготовлены стандартными методами, либо они могут быть закуплены, например, у Alza Corporation и Nova Pharmaceuticals, Inc. Липосомные суспензии (включая липосомы, нацеленные на выбранные клетки под действием моноклональных антител против клеточных антигенов) могут быть также использованы в качестве фармацевтически приемлемых носителей. Они могут быть приготовлены методами, известными специалистам в данной области, например, как описано в патенте США № 4522811.
Фармацевтические композиции могут быть включены в набор, контейнер, упаковку или дозатор вместе с инструкциями по их применению. Так, например, набор может включать композиции, включающие AAV, содержащий описанный здесь пептид.
ПРИМЕРЫ
Настоящее изобретение далее описано в нижеследующих примерах, которые не должны рассматриваться как ограничение объема изобретения, изложенного в формуле изобретения.
Материалы и методы
Если это не оговорено особо, то в приведенных ниже примерах были использованы нижеследующие материалы и методы.
Конструирование библиотеки AAV.
Плазмида iTransduce: pAAV-CBA-Cremut-p41-Cap9del
Авторами была сконструирована плазмида остова библиотеки iTransduce, названная pAAV-CBA-Cremut-p41-Cap9del и содержащая два экспрессионных кластера в цис-ориентации: 1) CBA-Cremut, в который авторами была введена мутантная кДНК Cre (CCG → CCT, кодирующую аминокислоту Pro15 для удаления изначально присутствующего сайта AgeI) под контролем часто встречающегося промотора CBA, 2) p41-Cap9del, состоящий из гена капсида AAV9 под контролем промотора p41 AAV5 (остатки 1680-1974 GenBank AF085716.1) и последовательности сплайсинга гена rep AAV2 (аналогичного гену, описанному со ссылкой на 12).
Мутантная кДНК Cre (Cremut), фланкированная рестрикционными сайтами KpnI и SalI, и фрагмент p41-CAP9(del)-polyA были синтезированы с помощью GenScript и клонированы в остов puC57. Сначала авторами была сконструирована плазмида pAAV-CBA-Cremut-polyA путем субклонирования мутантной кДНК Cre (с фрагментом KpnI-blunt, SalI) вместо eGFP-WPRE в остове pAAV-CBA-WPRE согласно изобретению (линеаризованном с помощью NcoI-blunt/SalI, который удаляет изначально присутствующий фрагмент eGFP-WPRE). Затем авторами был введен фрагмент p41-Cap9del, несущий мутацию K449R в последовательности Cap9, позволяющую создать уникальный сайт XbaI, где была удалена последовательность Cap размером 447 п.о. между рестрикционными сайтами XbaI и AgeI.
Плазмида для генерирования рандомизированных 7-мерных пептидных сар-фрагментов и субклонирования фрагментов CAP9, полученных с помощью ПЦР: pUC57-Cap9-XbaI/KpnI/AgeI.
Сначала авторами была получена 447 п.о.-область капсидной последовательности AAV9 между сайтами XbaI/AgeI (последовательность, которая отсутствует в pAAV-CBA-Cremut-p41-Cap9del), синтезированная Genscript (Piscataway, NJ) и субклонированная в pUC57-Kan. Эта последовательность капсида имеет такую же делецию сайта EarI в этой 447 п.о.-области сар AAV9 дикого типа, что позволяет удалить рестрикционный гидролизат из AAV9 дикого типа, как описано Deverman et al. 2015. Так же был создан уникальный сайт KpnI. Фрагмент сар pUC57-Cap9-XbaI/KpnI/AgeI использовали для создания исходной библиотеки рандомизированных 21-мерных нуклеотидных последовательностей (кодирующих 7-мерные пептиды), встроенных между нуклеотидами, кодирующими аминокислоты 588 и 589 VP1 AAV9. Авторами была использована стратегия, аналогичная стратегии, описанной Deverman et al. (2015). Вкратце, pUC57-Cap9-XbaI/KpnI/AgeI служила в качестве матрицы для амплификации ДНК сар и встраивания рандомизированных 21-мерных последовательностей с использованием прямого и обратного праймера. Информация о праймерах: XF-удлиненный праймер (5'-GTACTATCTCTCTAGAACtattaacggttc-3'; SEQ ID NO: 11) и обратный праймер 588iRev 5'-(GTATTCCTTGGTTTTGAACCCAACCGGTCTGCGCCTGTGCXMNNMNNMNNMNNMNNMN
NMNNTTGGGCACTCTGGTGGTTTGTG-3'; SEQ ID NO: 12), где повтор МНН означает рандомизированные 21-мерные нуклеотиды (закупленные у IDT). XF-удлиненный праймер и 588iRev использовали в ПЦР с использованием полимеразы Phusion (NEB) и pUC57-Cap9-XbaI/KpnI/AgeI в качестве матрицы. 447 п.о.-ПЦР-продукт гидролизовали ферментами XbaI и AgeI в течение ночи при 37°С, а затем продукт очищали на геле (Qiagen). Аналогичным образом, pAAV-CBA-Cremut-p41-Cap9del расщепляли XbaI и AgeI и очищали на геле. Затем проводили реакцию лигирования (1 час при комнатной температуре) с ДНК-лигазой Т4 (NEB) с использованием молярного отношения вставки сар к вектору, составляющего 3:1. Затем лигированная плазмида была названа pAAV-CBA-Cremut-p41-Cap9-7mer, и эта плазмида содержала пул из плазмид с рандомизированными 7-мерными пептидами, встроенными в ген сар между нуклеотидами, кодирующих 588 и 589 VP1 AAV9.
Эту плазмиду (pUC57-Cap9-XbaI/KpnI/AgeI) также использовали в качестве плазмиды-реципиента согласно изобретению для субклонирования фрагментов CAP9, амплифицированных с помощью ПЦР из ткани головного мозга. Авторами был удален расположенный выше сайт KpnI в плазмиде pUC57 путем гидролиза ферментами SacI и NsiI и лигирования. Это позволило создать уникальный сайт KpnI во фрагменте капсида. См. ниже.
Плазмида для экспрессии rep.
Авторами была сконструирована плазмида для экспрессии rep, названная pAR9-Cap9-stop/AAP/Rep, с применением стратегии, аналогичной стратегии, описанной Deverman et al.12. Полноразмерную кДНК синтезировали с помощью Genscript и клонировали в плазмиду pUC57-Kan. Стоп-кодоны встраивали вместо старт-кодонов для VP1, VP2, VP3, так, чтобы не продуцировались родительские капсиды AAV9, но сохранялась экспрессия AAP и rep.
Создание и очистка библиотеки AAV.
Для каждого продуцирования, авторы засевали 15-сантиметровые чашки для культивирования тканей 1,5×107 клеток 293T/чашку. На следующий день, клетки трансфецировали аденовирусной хелперной плазмидой (pAdΔF6, 26 мкг на чашку), плазмидой rep (pAR9-Cap9-stop/ААР/Rep, 12 мкг на чашку) и ITR-фланкированной библиотекой AAV (pAAV-CBA-Cre-mut/p41-Cap9-7mer, 1 мкг на чашку) методом с использованием фосфата кальция, для индуцирования продуцирования AAV. На следующий день после трансфекции, среду заменяли на DMEM, содержащую 2% FBS. AAV очищали от клеточного лизата путем ультрацентрифугирования в градиенте плотности йодиксанола. Замену буфера на PBS проводили с использованием центрифужных колонок ZEBA (7K MWCO; Thermo Fisher Scientific), а последующее концентрирование проводили на центрифужных устройствах для ультрафильтрации Amicon Ultra с отсечкой молекулярной массы 100 кДа (Millipore). Векторы хранили при -80°C до их использования. Затем авторами была проведена количественная оценка геномных копий AAV (ВГ) в препаратах AAV с помощью кол.ПЦР TaqMan с использованием праймеров и зондов, специфичных к последовательности ITR20,21 .
Секвенирование библиотеки следующего поколения.
Секвенирование следующего поколения было осуществлено на пуле библиотеки плазмид AAV9, с последующей упаковкой капсидов. Секвенирование также осуществляли после ПЦР-восстановления фрагмента сар (либо из ткани головного мозга, либо из выделенных tdTomato-позитивных клеток, отобранных с помощью проточной цитометрии). Для каждого раунда отбора, вирусную ДНК, соответствующую области, содержащей вставку, амплифицировали с помощью ПЦР с использованием набора для ПЦР Phusion High-Fidelity PCR от New England Biolabs (прямой праймер: 5'-AATCCTGGACCTGCTATGGC-3' (SEQ ID NO: 13), обратный праймер: 5'-TGCCAAACCATACCCGGAAG-3' (SEQ ID NO: 14)). ПЦР-амплификацию проводили с использованием полимеразы Q5 (New England Biolabs). Уникальные адаптеры штрих-кода были гибридизованы с каждым образцом, и образцы секвенировали на Illumina Miseq (150 п.о.- ридов). Было проанализировано приблизительно 50-100000 ридов на образец. Файлы с полученными последовательностями качественно подтверждали с помощью FastQC (bioinformatics.babraham.ac.uk/projects/fastqc/) и анализировали с помощью программы, написанной специально на Python. Вкратце, последовательности были количественно проанализированы на присутствие или отсутствие вставки, а затем последовательности, содержащие вставку, сравнивали с базовой эталонной последовательностью, и безошибочные риды заносили в таблицу на основе встречаемости каждой обнаруженной уникальной вставки. Вставки были транслированы и нормализованы.
Животные.
Все эксперименты на животных были одобрены Подкомитетом Массачусетской клиники общего профиля по уходу за животными в соответствии с руководствами, принятыми Национальным Институтом Здравоохранения по уходу и использованию лабораторных животных. Авторы использовали взрослых мышей (8-10-недельных) Ai9 (типа № 007909), C57BL/6 (типа № 000664) и BALB/c (типа № 000651), взятых из лаборатории Джексона, Бэр-Харбор, штат Мэн. Всех животных подвергали эвтаназии через три недели после инъекции, проводили транскардиальную перфузию, а затем ткани собирали и либо фиксировали в 4% параформальдегиде в PBS и быстро замораживали в жидком азоте, либо диссоциировали для проточной цитометрии.
In vivo отбор капсидов, обладающих тропизмом по отношению к головному мозгу.
Мышам Ai9 внутривенно (в хвостовую вену) вводили дозу ВГ, указанную в разделе Результаты, и через 3 недели после инъекции, мышей умерщвляли и собирали ткани.
Мышей подвергали глубокой анестезии изофлуораном и подвергали декапитации. В первом раунде, головной мозг быстро препарировали и брали два коронарных среза (толщиной 2 мм). Один срез использовали для экстракции полноразмерной ДНК головного мозга (DNeasy Blood and Tissue Kits, Qiagen, Hilden, Germany). Другой коронарный срез фиксировали в 4% PFA и заливали парафином для проведения иммуногистологического анализа (tdtomato-позитивные клетки детектировали после каждого раунда отбора путем окрашивания DAB с использованием кроличьих антител против RFP от Rockland Immunochemicals). Для раунда 2, ткань мозга затем разрезали лезвием бритвы на кусочки размером 1 мм3, и нервные клетки выделяли путем диссоциации папаином (Papain Dissociation System, Worthington) в соответствии с инструкциями производителя. После диссоциации, миелин удаляли (с использованием сфер для удаления миелина II у человека, мышей и крыс от Miltenyi) и tdtomato-позитивные клетки отсортировывали с помощью клеточного сортера S3eTM Cell Sorter (Bio-Rad). Клетки были отсортированы путем первой серии стробирования для исключения клеточного дебриса и отбора только синглетов. Суспензии клеток, взятых у мышей Ai9 с инъекцией AAV9-PHP.B-Cre (позитивный контроль), и мышей Ai9 с инъекцией PBS (негативный контроль) использовали для проведения серии стробирования в целях отбора tdTomato-позитивных и негативных клеток. После отбора, tdTomato-позитивные клетки сразу осаждали центрифугированием, и ДНК экстрагировали с помощью набора для экстракции ДНК ARCTURUS PicoPure (ThermoFisher).
После экстракции ДНК, вставки Cap9 (содержащие 21-мерную последовательность, кодирующую 7-мерные пептиды) амплифицировали с использованием следующих праймеров: Cap9_Kpn/Age_For: 5'-AGCTACCGACAACAACGTGT-3' (SEQ ID NO: 15) и Cap9_Kpn/Age_Rev: 5'-AGAAGGGTGAAAGTTGCCGT-3' (SEQ ID NO: 16) (с помощью набора для ПЦР с высокой достоверностью Phusion, New England Biolabs). Затем ампликоны очищали (с помощью набора Monarch для ПЦР и очистки ДНК, New England Biolabs), гидролизовали ферментами KpnI, AgeI и BanII, а фрагменты Cap9 KpnI-AgeI (144 п.о.) очищали на агарозном геле (с помощью набора для гель-экстракции ДНК Monarch, New England Biolabs) с последующим лигированием в плазмиду pUC57-Cap9-XbaI/AgeI/KpnI (линеаризованную с использованием KpnI и AgeI и дефосфорилированную инозит-фосфатазой теленка, New England Biolabs). Продукты лигирования трансформировали в электрокомпетентные бактерии DH5alpha (New England Biolabs), и все трансформированные бактерии культивировали в течение ночи в среде LB с ампициллином. Плазмиду pUC57-Cap9-XbaI/AgeI/KpnI очищали с помощью набора Maxi-Prep (Qiagen). Плазмиду гидролизовали ферментами XbaI/AgeI для высвобождения сар-фрагмента размером 447 п.о., который затем очищали на геле и лигировали с аналогичным образом разрезанной плазмидой pAAV-CBA-Cre-mut/p41-Cap9del для проведения последующего раунда получения библиотеки AAV.
Плазмиды rep/cap AAV, содержащие вставки пептидов AAV-F и AAV-S для получения вектора.
Для получения плазмид rep/cap, кодирующих капсиды AAV9, несущие представляющую интерес пептидную вставку для продуцирования векторов, кодирующих представляющий интерес трансген (например, GFP), авторами был проведен гидролиз плазмиды rep/cap AAV9 ферментами BsiWI и BaeI, которые удаляют фрагмент, фланкирующий аминокислоту 588 VP3 в сайте для встраивания пептидной последовательности. Затем авторами был заказан у компании Integrated DNA Technologies (IDT, Coralville, IA) фрагмент дцДНК размером 997 п.о., который содержит перекрывающиеся гомологичные ветви Гибсона с BsiWI/BaeI-разрезанным AAV9, а также 21-мерную нуклеотидную последовательность, кодирующую представляющий интерес пептид в одной рамке считывания после аминокислоты 588 VP3. И наконец, авторами была проведена сборка Гибсона с использованием смеси Gibson Assembly® Master Mix (NEB, Ipswich, МA) для лигирования вставки, содержащей пептид, в плазмиду rep/cap AAV9.
Векторы AAV для анализа на трансдукцию.
Для каждого продуцирования, авторы засевали 15-сантиметровые чашки для культивирования тканей 1,5×107 клеток 293T/чашку. На следующий день, клетки трансфецировали аденовирусной хелперной плазмидой (pAdΔF6, 26 мкг на чашку), плазмидой rep/сар (AAV9, ААВ-F, ААВ-S;, 12 мкг на чашку) и ITR-фланкированной плазмидой трансгенного кластера (одноцепочечной AAV-CBA-GFP-WPRE22, 10 мкг/чашку) для индуцирования продуцирования AAV. На следующий день после трансфекции, среду заменяли на DMEM, содержащую 2% FBS. AAV очищали от клеточного лизата путем ультрацентрифугирования в градиенте плотности йодиксанола. Замену буфера на PBS проводили с использованием центрифужных колонок ZEBA (7K MWCO; Thermo Fisher Scientific), а последующее концентрирование проводили на центрифужных устройствах для ультрафильтрации Amicon Ultra с отсечкой молекулярной массы 100 кДа (Millipore). Векторы хранили при -80°C до их использования. Затем авторами была проведена количественная оценка геномных копий AAV в препаратах AAV с помощью кол.ПЦР TaqMan с использованием праймеров и зондов, специфичных к последовательности BGH-polyА23.
Эвтаназия животных и сбор тканей
Мышам (линии, указанной на каждой фигуре) медленно вводили через боковую хвостовую вену 200 мкл тестируемого вектора AAV, разведенного в стерильном PBS (низкая доза: 4×1012 ВГ/кг и высокая доза: 3,2×1012 ВГ/кг), осторожно зажимая место инъекции пальцем до остановки кровотечения. Через три недели после инъекции, мышей подвергали эвтаназии и транскардиальной перфузии стерильным холодным забуференным фосфатом физиологическиим раствором (PBS). Затем головной мозг был продольно рассечен на два полушария. Одно полушарие подвергали последующей фиксации в 15% глицерине/4% параформальдегиде, разведенном в PBS в течение 48 часов, а затем в 30% глицерине для криоконсервации в течение еще 48-72 часов. Для группы с высокими дозами, небольшой кусочек сердца, мышцы (икроножной мышцы) и сетчатки также обрабатывали для проведения иммуногистологического анализа. Авторами было получено 3 независимых препарата AAV-S, AAV-F и AAV9 (Таблица I). Результаты трансдукции у мышей были получены от одного препарата каждого вектора, однако, авторами были воспроизведены эти результаты еще в двух независимых экспериментах.
Иммуногистология и визуализация с большим увеличением популяций нервных и нейронных клеток, трансдуцированных AAV-CBA-GFP
Коронарные плавающие срезы (40 мкм) вырезали с помощью микротома-криостата. После смывания глицерина в трис-забуференном физиологическом растворе (TBS), криосрезы делали проницаемыми с использованием 0,5% тритона X-100 (AmericanBio) в TBS в течение 30 минут при комнатной температуре, и блокировали 5% нормальной козьей сывороткой (или нормальной ослиной сывороткой) и 0,05% тритона в TBS в течение 1 часа при комнатной температуре. Первые антитела инкубировали в течение ночи при 4°C в 2,5% NGS и 0,05% тритоне в TBS, а вторые антитела, конъюгированные с Alexa Fluor 488 или Cy3 (Jackson ImmunoResearch laboratories, Baltimore, USA), инкубировали в течение 1 часа на следующий день. Первые антитела, используемые в этом исследовании, представляли собой: куриные анти-GFP антитела (Aves Labs, Tigard, USA), мышиные анти-NeuN антитела (EMD Millipore, Burlington, USA); кроличьи антитела против глутамин- синтетазы (Abcam, Cambridge, USA); кроличьи анти-Olig2 антитела (EMD Millipore, Burlington, USA); кроличьи анти-Iba1 антитела (Wako, Japan); кроличьи анти-CamKII антитела (Abcam, Cambridge, USA); мышиные анти-GAD67 антитела (EMD Millipore, Burlington, USA); кроличьи анти-ChAT антитела (EMD Millipore, Burlington, USA); мышиные антитела против кальбиндина (Abcam, Cambridge, USA) и кроличьи анти-TH антитела (Novus Biologicals, Littleton, USA). Срезы заполняли заливочной средой Vectashield с DAPI (Vector Laboratories, Burlingame, USA).
Для идентификации нервных клеток, трансдуцированных каждым вектором, и для исследования различных подтипов нейронов, на которые нацелен AAV-F, использовали эпифлуоресцентный микроскоп Zeiss Axio Imager Z, оснащенный программным обеспечением AxioVision и объективом 60× для получения изображений с высоким разрешением, иллюстрирующих совместную локализацию между GFP и каждым клеточным маркером.
Визуализация и количественная оценка общего охвата сигнала GFP
Для количественной оценки общего нативного сигнала флуоресценции GFP в срезе головного мозга и печени был использован роботизированный сканер предметных стекол Virtual Slide Microcope VS120 (Olympus) для получения изображения всей партии предметных стекол за один раз с помощью объектива Olympus UPLSAPO 10×. Для снижения вариабельности было получено изображение всей партии предметных стекол за один раунд. Начальное время экспозиции для GFP было настроено так, чтобы флуоресцентный сигнал не был чрезмерно насыщенным для всей экспериментальной группы и оставался неизменным на протяжении всего сканирования партии. Порядок предметных стекол был рандомизирован и оставался неизвестным до окончания статистического анализа. Программное обеспечение Olympus cellSens Standard затем использовали для анализа процентного охвата GFP в каждом отделе головного мозга. Представляющую интерес область (ROI) первоначально определяли с использованием инструмента «ROI-polygon», и количественно оценивали GFP-позитивную область в этой исходной ROI, а затем проводили аналогичное детектирование порога обнаружения на канале GFP для всех проанализированных предметных стекол (один порог был установлен на аналогичном уровне для анализа всех срезов головного мозга мышей, а другой порог применялся для анализа всех срезов печени мышей и всех срезов головного мозга крыс). Затем вычисляли процент GFP-позитивной области по отношению к общей поверхности ROI. Сигнал аутофлуоресценции учитывался в проводимом авторами анализе, поскольку ими был устанавлен порог интенсивности флуоресценции eGFP выше уровня аутофлуоресценции (при этом, следует убедиться, что учитывался только сигнал от клеток, трансдуцированных AAV-GFP). Кроме того, авторами была изображена каждая область ROI для каждого среза головного мозга, но при этом, они избегали самых крайних участков срезов, так как эти участки также могут иметь высокий уровень аутофлуоресценции. Желудочковое пространство также было исключено из анализа. И наконец, было оценено три технических повтора (срезы головного мозга) на каждую мышь, и все измерения были сделаны вслепую вплоть до оконачания анализа.
Количественный анализ процентного содержания трансдуцированных астроцитов и нейронов на основе стереологии
Исследования на основе стереологии были осуществлены как описано ранее24,25, после совместного окрашивания срезов головного мозга на GFP и NeuN (маркер нейронов) или GFP и GS (глутаминсинтетазу, маркер всех астроцитов). Авторы не включили в этот анализ микроглиальные клетки и олигодендроциты, так как клетки этих типов не трансдуцировались в заметном количестве. Стереологическая оценка процента нейронов и астроцитов, трансдуцированных AAV, была проведена вслепую после идентификации первоначально введенного вектора на подвижном предметном столике эпифлуоресцентного микроскопа Olympus BX51, оснащенного цифровой камерой CCD DP70, флуоресцентной лампой X-Cite и соответствующего программного обеспечения для проведения стереологический исследований CAST версии 2.3.1.5 (Olympus, Tokyo, Japan). Кора головного мозга была первоначально очерчена под 4-кратным объективом. Случайная выборка отобранной области была определена с помощью оптического диссекторного зонда с помощью программного обеспечения CAST. Чтобы оценить процентное содержание астроцитов или нейронов, трансдуцированных AAV9, AAV9-PHP.B, AAV-S и AAV-F, был проведен подсчет клеток на основе стереологического анализа под объективом 20×, с меандром выборки 10% для поверхности коры головного мозга в случае AAV с «высокой трансдукцией», и 20% для AAV с «низкой трансдукцией» (с учетом редко встречающихся GFP-позитивных клеток в этих случаях). Для каждой выборки оценивали общее количество астроцитов (GS-позитивных клеток) или нейронов (NeuN-позитивных клеток) и для каждой из этих популяций определяли процент GFP-позитивных клеток. Учитывали только глиальные клетки и нейроны с DAPI-позитивным ядром в пределах данной выборки.
Количественная оценка векторного генома в головном мозге и печени
Одно полушарие головного мозга и небольшой кусочек печени были заморожены для выделения генома AAV в целях анализа биораспределения векторного генома. Для свежезамороженных образцов головного мозга и печени авторы выделили геномную ДНК и ДНК вектора AAV из 10 мг ткани с помощью набора DNeasy для выделения ДНК из крови и ткани (Qiagen) в соответствии с инструкциями производителя. Количественный анализ ДНК проводили с использованием спектрофотометра NanoDrop ND-1000 (Thermo Scientific). Затем, с использованием 50 нг геномной ДНК в качестве матрицы, авторами была проведена кол.ПЦР Taqman с использованием зонда и праймеров, специфичных к области polyА кластера для экспрессии трансгена (такой же анализ был проведен для титрования очищенных векторов AAV). Дла гарантии одинакового ввода геномной ДНК для каждого образца была проведена отдельная кол.ПЦР для каждого образца с использованием набора зондов и праймеров Taqman, которые обнаруживают геномную ДНК GAPDH (Thermo Fisher Scientific, Assay ID Mm01180221_g1; gene symbol Gm12070). Для каждого органа/ткани, число копий векторных геномов AAV было скорректировано для каждого образца, с учетом любых различий в значениях Ct GAPDH по следующей формуле: (число копий векторных геномов AAV)/(2ΔCt). Значение Δ Ct рассчитывали по следующей формуле: значение Ct GAPDH для представляющего интерес образца - среднее значение Ct GAPDH для образца, который имел наименьшее количество GAPDH (наивысшее значение Ct). Данные выражали в виде геномов вектора AAV на 50 нг геномной ДНК.
Трансдукция человеческих нейронов :
Первичные стволовые нервные клетки плода человека (NSC) были получены из Лаборатории по исследованию врожденных дефектов (Вашингтонский Университет, Сиэтл, Вашингтон) с соблюдением всех требований в соответствии с этическими принципами NIH. Процедура выделения была подробно описана ранее26 с небольшими изменениями. Вкратце, ткань головного мозга инкубировали в буфере с 0,25% трипсином, ДНКазой (90 единиц/мл), разведенном в сбалансированном солевом растворе Хэнкса (HBSS) в течение 45 минут. Ткань титровали, переносили в термоинактивированную фетальную бычью сыворотку при 4°C и центрифугировали при 500× g в течение 20 минут. Осадок ресуспендировали в полной среде NSC, состоящей из реагента x-Vivo 15 (без фенолового красного и гентамицина; Lonza) с добавлением 10 мкг основного фактора роста фибробластов (Life Technologies), 100 мкг эпидермального фактора роста (Life Technologies), 5 мкг фактора ингибирования лейкоза (EMD Millipore), 60 нг/мл N-ацетилцистеина (Sigma-Aldrich), 4 мл добавки, а именно, фактора, ответственного за выживание нервных клеток-1 (Lonza), 5 мл добавки 100× N-2 (Life Technologies), 100 единиц пенициллина, 100 мкг/мл стрептомицина (Life Technologies) и 2,5 мкг/мл фунгизона (Life Technologies). Затем супернатанты фильтровали через сетчатый фильтр для клеток 40 мкм (Corning Life Science). Нейросферы диаметром более 40 мкм подвергали диссоциации с использованием аккутазы (10 минут). Среда для дифференцировки нейронов состояла из 1× нейробазальной среды, 2% добавки B-27, не содержащей сыворотки, и 2 мМ добавки GlutaMAX-I (все от Invitrogen) с добавлением человеческого рекомбинантного нейротропного фактора головного мозга (BDNF) (10 нг/мл; Peprotech).
Дифференцирующиеся NSC культивировали на предметных стеклах в камере со средой для дифференцировки в течение 2 недель, а затем обрабатывали указанным вектором AAV, кодирующим GFP (было добавлено 7×109 ВГ/лунку, 150 ВГ/клетку). Через одну неделю после трансдукции, клетки фиксировали 4% параформальдегидом и делали их проницаемыми с использованием 0,05% Triton X-100 (Sigma-Aldrich) в 1×забуференном фосфатом физиологическом растворе (PBS; Invitrogen). Клетки окрашивали первым моноклональным антителом (TU-20) против нейрон-специфического β-тубулина класса III (1:50; Abcam). Вторые антитела, конъюгированные с Alexa Fluor 594 (разведение 1:200; Invitrogen), добавляли в течение 1 часа, а затем добавляли DAPI в течение 30 минут. Затем на предметные стекла наносили обесцвечивающий реагент ProLong (Invitrogen). Максимальные проекции изображения были получены из захваченных Z-стэков на конфокальном микроскопе Nikon A1R.
Z-стэки были загружены в Imaris, а затем для преобразования изображений в объеме 3D использовали поверхностный модуль. После этого подсчитывали GFP+-нейроны (в зеленом канале 1) и нейроны, позитивные по β-тубулину класса III (в красном канале 2). С использованием модуля солокализации Imaris, была определена популяция нейронов, дважды позитивных по вышеуказанным компонентам.
Статистический анализ
Статистический анализ данных проводили с использованием программного обеспечения GraphPad Prism (версия 8.00). Для различных групп AAV9, AAV9-PHP.B, AAV-S и AAV-F был проведен анализ с использованием однофакторного критерия ANOVA, а затем с использованием критерия множественного сравнения Тьюки. Статистически значимым считалось значение p <0,05. Результаты представлены как среднеее±ср. кв. ош. Были проведены подобные анализы на биораспределение геномов AAV в головном мозге и в печени, а также на трансдукцию человеческих нейронов.
Прямая стереотаксическая инъекция векторов
Взрослых мышей C57BL/6 анестезировали внутрибрюшинной инъекцией кетамина/ксилазина (100 мг/кг и 50 мг/кг массы тела, соответственно) и помещали на стереотаксическую рамку (Kopf Instruments, Tujunga, USA). Инъекции векторов делали в кору головного мозга (соматосенсорную кору) и в гиппокамп. В общей сложности вводили 3 мкл вирусной суспензии (1,65×1010 и 5,6×1010 ГК на участок инъекции для AAV-F и AAV-S, соответственно, со скоростью 0,15 мкл/минуту) острой иглой 33-го калибра 10 мкл-шприца Гамильтона (Sigma-Aldrich, St. Louis, USA). Стереотаксические координаты участков инъекции рассчитывали по брегме (координаты коры головного мозга: переднезадний участок -1 мм, медиолатеральный участок ±1 мм и дорсовентральный участок -0,8 мм; координаты гиппокампа: переднезадний участок -2 мм, медиолатеральный участок ±1,7 мм и дорсовентральный участок -2,5 мм).
Интратекальное введение ударной дозы (ударной IТ-дозы)
Взрослых мышей C57BL/6 подвергали анестезии изофлураном. После того, как кожа в поясничной области была выбрита и очищена, через кожу был сделан срединный сагиттальный разрез 3-4 см, обнажающий мышцы и позвоночник. Катетер вводили между областью позвоночника L4-L5 и подсоединяли к газонепроницаемому шприцу Гамильтона с помощью стальной иглы 33-го калибра. Десять микролитров векторов AAV9-CBA-GFP (1,25×1011 ВГ) или AAV-F-CBA-GFP (8,8×1010 ВГ) медленно вводили со скоростью 2 мкл/мин. Мышей умерщвляли через три недели после инъекции.
Для получения изображений с малым увеличением всех срезов спинного и головного мозга было проведено окрашивание в течение ночи анти-GFP антителом (Invitrogen, номер по каталогу G10362, разведение 1:250) с последующим окрашиванием вторым антителом и визуализацией, как показано на фигуре 3.
Для иммунологического окрашивания клеток головного и спинного мозга различных типов, фиксированные срезы тканей промывали PBS, блокировали козьей сывороткой и повышали проницаемость с помощью 0,3% тритона в PBS. Целевые антитела разводили в блокирующем буфере и инкубировали при 4°С в течение ночи.
Первые антитела:
анти-GFP антитело: номер по каталогу ab1218 (abcam): разведение 1:1000
анти- GFAP антитело: номер по каталогу catz0334 (Dako); разведение 1:500
анти-NeuN антитело: номер по каталогу ab177487 (abcam); разведение 1:300
После инкубирования в течение ночи, предметные стекла тщательно промывали PBS с 0,1% твином. Затем добавляли вторые антитела, конъюгированные с флуорохромом (разведение 1:500), и инкубировали в течение 1 часа при комнатной температуре. После промывки, предметные стекла промывали PBS и закрепляли с использованием заливочного раствора DAPI (ThermoFisher, номер по каталогу P36931). Предметные стекла визуализировали на конфокальном лазерном сканирующем микроскопе Zeiss LSM 800.
Просвечивающая электронная микроскопия
Сетки с углеродным покрытием (Electron Microscopy Sciences, EMS) были сделаны гидрофильными путем воздействия на них тлеющего разряда 25 мА в течение 20 сек. Предварительно, 5 мкл каждого вектора AAV адсорбировали на сетку в течение 1 минуты и окрашивали 1% уранилацетатом (EMS # 22400) в течение 20 сек. Сетки оценивали в TecnaiG2 Spirit BioTWIN и получали изображения с помощью камеры AMT 2k с зарядовой связью (CCD). Исследования проводили в лаборатории электронной микроскопии Гарвардской Медицинской Школы. Подсчет проводили следующим образом: были сделаны 5 репрезентативных изображений каждого препарата вектора; а затем все полные и пустые капсиды были подсчитаны на счетчике в Photoshop (CS6). Процент пустых капсидов вычисляли для каждого изображения и по полученным данным строили график.
Пример 1. Конструирование библиотеки AAV iTransduce на основе экспрессии
Сначала, авторами была сконструирована плазмида библиотеки AAV, которая состояла из ITR-фланкированного экспрессионного кластера AAV2, состоящего из рекомбиназы Cre, происходящей от куриного бета-актина (CBA), и капсида AAV9, находящегося под контролем промотора p41 (схематически показано на фиг. 1a). Псевдорандомизированные нуклеотиды из 21 основания встраивали между нуклеотидами VP1 AAV9, кодирующими аминокислоты 588/589, с помощью ПЦР. Перед упаковкой вируса, эту плазмидную библиотеку секвенировали путем низкоуровневого секвенирования следующего поколения (NGS), и было подтверждено наличие 21-мерных вставок в подавляющем большинстве плазмид и отсутствие смещения вариантов (данные не приводятся). Затем капсидную библиотеку упаковывали и проводили NGS, чтобы убедиться, что процесс создания вектора сохранял разнообразие, достаточное для проведения отбора. Библиотека iTransduce основана на каждом уникальном капсиде, несущем как свой собственный ген сар, так и конструкцию, экспрессирующую Cre (фиг. 1b). Трансгенным мышам (Ai9), несущим кластер floxed-STOP tdTomato, внутривенно вводили библиотеку AAV (фиг. 1b-i). Капсиды, которые успешно трансдуцируют клетки, обеспечивают экспрессию tdTomato в любом органе-мишени или в клетках любого типа (независимо от доступности специфических линий трансгенных мышей Cre); и эти tdTomato-позитивные клетки могут быть затем подвергнуты сортингу с помощью проточной цитометрии из представляющей интерес ткани (необязательно, вместе с клетко-специфическими маркерами, фиг. 1b-ii). Вирусная ДНК, выделенная из этих клеток, должна соответствовать вариантам капсида, которые могут эффективно преодолевать все внеклеточные и внутриклеточные биологические барьеры для экспрессии трансгена (фиг. 1b-iii).
Пример 2. Отбор новых вариантов капсида AAV9 на основе экспрессии трансгена для идентификации функциональных капсидов.
Для тестирования стратегии библиотеки iTransduce, авторы проводили отбор для подтверждения концепции в целях выделения вариантов капсида AAV, способных трансдуцировать клетки головного мозга после системной инъекции. 1,27×1011 векторных геномов (5×1012 ВГ/кг) библиотеки инъецировали внутривенно в хвостовую вену одному взрослому самцу и одной самке мыши Ai9, и через три недели после инъекции, мышей умерщвляли; а затем брали срезы печени, головного мозга, селезенки и почек и проводили иммунологическое окрашивание tdTomato. Авторы легко обнаружили tdTomato-позитивные клетки (вероятно, гепатоциты) в печени с разбросом tdTomato-позитивных клеток (как нейронов, так и астроцитов) в головном мозге, а также в других органах (фиг. 7a). Авторами было также проверено, можно ли сохранить ДНК сар, содержащую вставку из 21 основания, с помощью ПЦР, и были обнаружены специфические полосы в 10 органах/тканях у мышей, которым инъецировали библиотеку, но не у контрольных, неинъецированных мышей (фиг. 7b). Затем оставшуюся ткань головного мозга самок и самцов мышей объединили и экстрагировали ДНК, сначала из всей ткани головного мозга для первого раунда отбора. После этого амплифицировали ДНК сар, содержащую 21-мерную вставку, и эту ДНК анализировали на идентичность и число ридов с помощью NGS. После этого было обнаружено значительное обогащение популяции библиотеки специфическими пептидами, собранной из ткани головного мозга после одного раунда отбора, по сравнению с неотобранной библиотекой и с вариантами, выделенными из печени (фиг. 7c, данные не приводятся). Затем была выделена область вирусной ДНК, содержащая вставку, и эта область была повторно клонирована обратно в остов плазмиды AAV с последующей повторной упаковкой капсидов («библиотека, обогащенная капсидами головного мозга») для второго раунда отбора.
Для второго раунда отбора, двум мышам Ai9 (одному самцу и одной самке) инъецировали библиотеку, полученную в первом раунде (доза 1,91×1010 ВГ, 7,64×1011 ВГ/кг), и мышей умерщвляли через три недели. Перед инъекцией, последовательность, содержащая модифицированную область, была амплифицирована и секвенирована с помощью NGS для гарантии того, что ранее имеющееся смещение не было внесено в пул векторов (фиг. 2). Ткань головного мозга диссоциировали для получения клеточной суспензии в целях проведения сортинга tdTomato-позитивных клеток с помощью проточной цитометрии. Авторами был проведен клеточный сортинг 3834 tdTomato-позитивных клеток (0,043% от исходной клеточной суспензии) с помощью проточной цитометрии, и была подтверждена успешная трансдукция (фиг. 8a-b). Вирусную ДНК из отсортированных клеток tdTomato амплифицировали и секвенировали, как это было проведено ранее (фиг. 2). Вирусная ДНК, выделенная из tdTomato-позитивных клеток, обнаруживала 97% ридов, представленных всего тремя пептидами, STTLYSP, FVVGQSY и FQPCP* (где * означает стоп-кодон) (фиг. 2b). Авторами были выбраны два из них, STTLYSP (названный AAV-S) и FVVGQSY (названный AAV-F) для функциональной оценки in vivo (авторы не проводили оценку FQPCP*, поскольку он, вероятно, был продуктом перекрестной упаковки). Как видно на фиг. 2, обе эти последовательности обнаруживались в библиотеке второго раунда на низких уровнях (~ 0,4% ридов для каждого варианта), но были сильно обогащены в головном мозге после отбора.
Пример 3. Капсид AAV-F опосредует эффективную экспрессию трансгена в ЦНС мыши.
Чтобы проверить, могут ли эти пептиды, экспрессируемые на капсиде AAV, опосредовать эффективную экспрессию трансгена с помощью вектора AAV, авторами были созданы капсиды (AAV-S и AAV-F), упаковывающие одноцепочечный кластер для экспрессии GFP под контролем промотора CBA (фиг. 3а). Для сравнения был включен родительский вектор AAV9 и AAV9-PHP.B, то есть, наиболее широко изученный вариант AAV9 с 7-мерной пептидной вставкой (TLAVPFK), генерируемой благодаря направленной эволюции12. Все векторы продуцировались на достаточно высоком уровне и давали несколько более низкую эффективность продуцирования, чем AAV9 (Таблица 4).
Таблица 4. Эффективность продуцирования капсидов AAV *
* Все капсиды упакованы в одноцепочечный кластер с трансгеном AAV-CBA-GFP-WPRE, фланкированным ITR AAV2.
Взрослым самцам мышей линии C57BL/6J в боковую хвостовую вену инъецировали низкую дозу или высокую дозу (1×1011 ВГ и 8×1011 ВГ вектора, соответственно, приблизительно 4×1012 и 3,2×1013 ВГ/кг) одного из следующих векторов: AAV9, AAV9-PHP.B, AAV-S и AAV-F (n=3 каждый). Через три недели после инъекции, мышей умерщвляли и органы собирали для эндогенного (неокрашенного) флуоресцентного анализа на GFP. Авторами была проведена количественная оценка процента охвата сигнала GFP в серийных сагиттальных срезах головного мозга (на каждое животное было проанализировано 3 среза). Примечательно то, что AAV-F продемонстрировал 119-кратное (p <0,0001) и 68-кратное (p=0,0004) увеличение охвата флуоресценции GFP по сравнению с родительским вектором AAV9 при 1×1011 ВГ и 8×1011 ВГ, соответственно (фиг. 3b, c, e, f; фиг.9A-B). AAV9 и AAV-S отображали аналогичные уровни охвата GFP (фиг. 3b, c, e, f). AAV9-PHP.B давал немного более высокий охват GFP при низкой дозе по сравнению с AAV-F, в то время как аналогичные уровни охвата GFP наблюдались при дозе 8×1011 ВГ (фиг. 3b, c, e, f). Подобно AAV9-PHP.B, AAV-F проникал в спинной мозг с превосходной эффективностью (фиг. 3d). Большинство областей головного мозга были эффективными мишенями для AAV-F, и устойчивый сигнал GFP наблюдался в коре головного мозга, в гиппокампе, в полосатом теле, в мозжечке и в обонятельной луковице (фиг. 3f).
Чтобы получить подробную информацию о типах клеток, на которые нацелены AAV-S и AAV-F, авторы затем проводили серию иммунологических окрашиваний на GFP и маркеры нейронов (NeuN), астроцитов (глутаминсинтетаза, GS), микроглиальных клеток (Iba-1) и олигодендроцитов (Olig2). AAV-F и AAV-S, аналогичные двум другим эталонным векторам, в основном, трансдуцировали нейроны и астроциты (оказалось, что ни один из вариантов эффективно не трансдуцировал микроглиальные или олигодендроглиальные клетки, фиг. 4a, b). Стереологическое количественное определение нейронов и астроцитов в коре головного мозга при дозе 1×1011 ВГ подтвердило потенциал эффективной трансдукции AAV-F по сравнению со стандартным AAV9 в 65 раз в астроцитах и в 171 раз в нейронах, в то время как различие между AAVS и AAV9 было незначимым (процент GFP-позитивных астроцитов составлял 0,63%±0,24% для AAV9 и 0,36±0,15% для AAV-S, соответственно, а процент GFP-позитивных нейронов составлял 0,039%±0,0,02% для AAV9 и 0,029±0,002%) для AAV-S; все±перед числами означают стандартную ошибку среднего, SEM). Следует отметить, что AAV-F нацелен в значительной степени на астроциты (40,78±0,73%) в отличие от AAV9-PHP.B (28,21±0,25%), и обратное справедливо для нейронов (6,67±0,5% для AAV-F и 10,59±0,16% для AAV9-PHP.B, фиг.4c), что наводит на мысль о немного отличающемся тропизме между этими двумя векторами у мышей. Кроме того, AAV-F трансдуцировал нейроны различных подтипов, включая возбуждающие (CamKII-позитивные) и ингибирующие (GAD67-позитивные) нейроны коры головного мозга, допаминергические нейроны в полосатом теле (экспрессирующие тирозингидроксилазу, TH), нейроны Пуркинье в мозжечке (позитивные по кальбиндину) и двигательные нейроны в спинном мозге (экспрессирующие маркер холинацетил-трансферазы, ChAT, фиг. 10a). В соответствии со стереологическими показателями коры головного мозга (фиг. 4c) и изображениями для высокой дозы AAV-F по сравнению с AAV9 (фиг. 3F), авторами была обнаружена эффективная трансдукция нейронов и астроцитов ААV-F, а не AAV9 в дозе 1×1011 ВГ/мышь в полосатом теле, в гиппокампе и в мозжечке (фиг. 10b).
Для того, чтобы лучше понять, соответствуют ли высокие уровни экспрессии трансгена GFP в головном мозге под действием AAV-F более высоким уровням геномов AAV в головном мозге, авторами были выделены вектор и мышиная геномная ДНК из печени и головного мозга и была проведена кол.ПЦР для дозы 8×1011 ВГ/мышь. AAV-F показал 20-кратное увеличение (p <0,0001) геномов AAV в головном мозге по сравнению с AAV9 (фиг. 4d). Как сообщалось, AAV9-PHP.B имел гораздо большее (25-кратное) количество геномов AAV в головном мозге по сравнению с AAV9, в то время как AAV-S имел низкий уровень в соответствии с данными флуоресценции GFP (фиг. 3). PHP.B показал уровни экспрессии в печени, которые были немного ниже, чем уровни AAV9 и AAV-F (но не AAV-S; фиг. 4d). Уровни AAV-F в печени были такими же, как и уровни AAV9, а AAV-S показал более низкую, но незначительную тенденцию к снижению. Авторами было также оценено биораспределение AAV-F по сравнению с AAV9 в нескольких других органах, а именно в скелетных мышцах, в сердце и в спинном мозге (фиг. 11). Какого-либо существенного различия между двумя векторами в мышцах и в сердце не наблюдалось (хотя наблюдалась тенденция к повышению уровня экспрессии AAV-F в сердце). Как и следовало ожидать исходя из экспрессии в головном мозге, AAV-F показал значительно более высокие уровни экспрессии в спинном мозге по сравнению с AAV9 (p <0,05, t-критерий).
Для исследования экспрессии трансгена в периферических органах после системной инъекции, авторами была проанализирована флуоресценция GFP в печени, в сердце, в скелетных мышцах и в сетчатке (фиг. 4e). Неудивительно, что все векторы эффективно проникали в печень. В сердце и скелетных мышцах, AAV-S давал большее количество ярких сигналов GFP/срез, чем AAV9 и AAV-F. Все капсиды опосредовали низкую трансдукцию в сетчатке, как и ожидалось при внутривенной доставке. Интересно отметить, что хотя AAV-9 и AAV-S не проникали в нейронную сетчатку, однако, соответствующая экспрессия наблюдалась в пигментном эпителии сетчатки (RPE). AAV9-PHP.B, и AAV-F проходили в несколько слоев сетчатки, в первую очередь, в слой ганглиозных клеток (GCL). Экспрессия GFP также наблюдалась во внутреннем ядерном слое (INL), при этом значительная экспрессия проявлялась во внешнем плексиформном слое (OPL), что может указывать на биполярную или ингибирующую горизонтальную трансдукцию клеток.
Поскольку трансдукция векторами AAV у мышей может существенно различаться в зависимости от пола и линии мышей, то авторами было проведено исследование для того, чтобы подтвердить, может ли наиболее эффективный капсид, AAV-F, эффективно проникать в головной мозг после системной инъекции самкам C57BL/6, а также самцам BALB/c. Мышам вводили 1×1011 ВГ (4×1012 ВГ/кг), в результате чего наблюдалась устойчивая и эффективная трансдукция, не зависящая от линии или пола (фиг. 5a-c). Эти результаты контрастировали с отсутствием эффективности AAV9-PHP.B в отношении трансдукции центральной нервной системы после системной доставки мышам BALB/c, как сообщалось ранее (фиг.5b, c)13,14.
Хотя очистка в градиенте плотности йодиксанола удаляет большую часть пустых капсидов, однако, чтобы проверить, может ли избыток пустых капсидов в AAV-F объяснить его повышенное биораспределение в головном мозге по сравнению с AAV9, авторами была проведена просвечивающая электронная микроскопия (ТЕМ) на двух отдельных препаратах AAV-F, с последующим их сравнением их с двумя независимыми препаратами AAV9. Как видно на фиг. 12A-E, не наблюдалось какого-либо значительного различия в уровнях пустого капсида между AAV-F и AAV9 (среднее 4,63±1,99% против 5,2±2,18% пустых капсидов для AAV9 и AAV-F, соответственно, среднее±стандартное отклонение, p=0,54, непарный t-критерий).
Затем авторами был проведен анализ чтобы определить, может ли AAV-F использоваться в качестве вектора для трансдукции в ЦНС другими способами введения. Сначала были протестированы AAV-S и AAV-F на опосредование экспрессии трансгена в головном мозге после прямой инъекции в гиппокамп взрослым мышам C57BL/6. Авторами было обнаружено, что в обоих капсидах достигалась широкая экспрессия GFP после прямой инъекции, главным образом, в нейронах (фиг. 13). Интратекальная инъекция векторов AAV для трансдукции в спинной мозг показала перспективность лечения этого органа. Одним из недостатков является ограниченное распространение вектора в мозг после инъекции вектора в поясничную область. Авторами было проведено сравнение AAV9 и AAV-F после интратекальной инъекции ударной дозы вектора в поясничную область спинного мозга у взрослых мышей C57BL/6. Через три недели после инъекции, мышей умерщвляли и анализировали спинной мозг и головной мозг. Примечательно, что AAV-F приводил к гораздо более интенсивной экспрессии GFP по всему спинному мозгу по сравнению с AAV9, и при этом, трансдукция наблюдалась как в белом, так и в сером веществе. Между тем, трансдукция AAV9 ограничивалась, в основном, белым веществом. Поразительно, но была также обнаружена трансдукции астроцитов и нейронов в головном мозге мышей, которым инъецировали AAV-F, но не AAV9 (фиг. 14).
Пример 4. AAV-F опосредует усиленную трансдукцию человеческих нейронов.
Поскольку отбор проводили на мышах, то был проведен анализ для того, чтобы определить, транслируются ли устойчивые свойства трансдукции AAV-F в клетки человека. Первичные нейроны, полученные из стволовых клеток человека, трансдуцировали равными дозами AAV9, AAV-S и AAV-F, все из которых кодировали GFP. Через неделю их фиксировали, окрашивали β-тубулином класса III и анализировали на процентное содержание GFP-позитивных нейронов. Примечательно, что AAV-F трансдуцировал 62% нейронов, что в 3 раза выше, чем для AAV9 (p <0,05) (фиг. 6a, b). AAV-S давал небольшое, но статистически значимое (p <0,05) повышение эффективности трансдукции по сравнению с AAV9 (фиг. 6a).
Пример 5. AAV-S проникает во внутреннее ухо с высокой эффективностью
В последние годы были разработаны новые векторы AAV, предназначенные для высокоэффективного нацеливания на органы и клетки определенных типов. Во многих из них используются 7-мерные пептидные вставки, которые изменяют свойства трансдукции конкретного серотипа AAV. Однако, эти векторы часто можно использовать для трансдукции других тканей, включая органы, трудно поддающиеся лечению, такие как внутреннее ухо. Трансдукция определенных типов клеток внутреннего уха, таких как волосковые клетки, все еще остается проблемой, поскольку многие традиционные векторы AAV не могут соответствующим образом нацеливаться на волосковые клетки одного класса, а именно, внешние волосковые клетки (OHC).
Как отмечалось выше, один вектор (AAV-F) показал большие перспективы и высокую экспрессию в ЦНС после системной инъекции, тогда как другой (AAV-S) не обнаруживал таких свойств. Хотя системно вводимый AAV-S не может эффективно преодолевать гематоэнцефалический барьер, однако, он проникает в различные ткани, включая сердце, печень и мышцы. После прямой инъекции в головной мозг, AAV-S показал высокую эффективность локальной трансдукции в нейронах даже при относительно низких дозах. Чтобы проверить его трансдукционные свойства во внутреннем ухе, авторами был получен AAV-S, кодирующий одноцепочечный экспрессионный кластер, инициирующий продуцирование EGFP под контролем промотора CBA, и этот вектор был введен новорожденным (P1) мышам через круглое окно. Авторами было обнаружено, что AAV-S очень хорошо проникает в волосковые клетки улитки. Как внутренние, так и внешние волосковые клетки трансдуцировались с эффективностью до 100% и 99% (фиг. 15a, b) при тестируемой дозе (2×1010 ВГ). Авторы также наблюдали значительную трансдукцию спирального лимба и спиральных узлов (фиг. 15c, d). В целом, как показано в настоящей заявке, AAV-S может быть использован для генотерапии внутреннего уха.
Пример 6. Использование системы iTransduce для отбора нетрансгенных взрослых приматов в целях выделения капсидов, которые эффективно проникают в фиброциты.
2-3 раунда отбора на капсиды AAV, нацеленные на фиброциты, проводили у приматов, не являющихся человеком, например, у собакоподобных обезьян. Для идентификации селективности к фиброцитам, отбор капсида AAV, компетентного по трансдукции, осуществляли путем совместной инъекции библиотеки iTransduce AAV вместе с AAV9-PHP.B, который кодирует кластер GJB2-floxed-STOP-tdTomato (размер соответствует размеру капсида AAV). Во внутреннем ухе NHP, AAV9-PHP.B-CBA-GFP проникает во множество клеток улитки, включая фиброциты, HC и область нейронов спиральных узлов. В этой стратегии отбора (см. Фиг. 16A), экспрессия tdTomato ограничивается фиброцитами под контролем промотора GJB2, по существу, создавая внутреннее ухо трансгенного NHP. Капсиды AAV, которые проникают в фиброциты и включают ген tdTomato, отбирают с помощью проточной цитометрии из диссоциированной улитки, и капсидную ДНК, сохраняемую для NGS, клонируют для идентификации пептидных последовательностей, которые обеспечивают AAV-опосредованную экспрессию в фиброцитах. После обогащения отдельными пептидами проводят глубокое секвенирование, как описано выше, чтобы получить информацию о том, когда следует останавливать дополнительные раунды отбора (вероятно, когда конкретный пептид представляет > 25% ридов).
Альтернативно или дополнительно, 2-3 раунда отбора на капсиды AAV, нацеленные на клетки спинного мозга, проводят у приматов, не являющихся человеком, например, собакоподобных обезьян. Для идентификации селективности к спинному мозгу, отбор капсидов AAV, компетентных по трансдукции, осуществляли путем совместной инъекции библиотеки iTransduce AAV вместе с AAV9, который кодирует кластер CBA-floxed-STOP-mPlum (размер соответствует размеру капсида AAV). В спинном мозге NHP, AAV9 проникает в клетки, включая нейроны и астроциты. В этой стратегии отбора (см. Фиг. 16В), капсиды AAV, которые проникают в клетки спинного мозга и запускают флуоресценцию mPlum, отбирают с помощью проточной цитометрии из диссоциированного спинного мозга, и капсидную ДНК, сохраняемую для NGS, клонируют для идентификации пептидных последовательностей, которые обеспечивают AAV-опосредованную экспрессию в клетках спинного мозга. После обогащения отдельными пептидами проводят глубокое секвенирование, как описано выше, чтобы получить информацию о том, когда следует останавливать дополнительные раунды отбора (вероятно, когда конкретный пептид представляет > 25% ридов).
После идентификации капсидных клонов-кандидатов, капсиды были превращены, как описано ранее, в векторы, кодирующие кластер GFP. Затем капсиды тестировали на трансдукцию клеток-мишеней после прямой инъекции через мембрану круглого окна (RMW) (например, как показано на фиг. 16A) или интратекальной инъекции (например, как показано на фиг. 16B).
Ссылки на литературу
1. Mendell, J.R., S. Al-Zaidy, R. Shell, W.D. Arnold, L.R. Rodino-Klapac, T.W. Prior, L. Lowes, L. Alfano, K. Berry, K. Church, J.T. Kissel, S. Nagendran, J. L'Italien, D.M. Sproule, C. Wells, J.A. Cardenas, M.D. Heitzer, A. Kaspar, S. Corcoran, L. Braun, S. Likhite, C. Miranda, K. Meyer, K.D. Foust, A.H.M. Burghes, and B.K. Kaspar, Single-Dose Gene-Replacement Therapy for Spinal Muscular Atrophy. N Engl J Med, 2017. 377(18): p. 1713-1722.
2. Manno, C.S., G.F. Pierce, V.R. Arruda, B. Glader, M. Ragni, J.J. Rasko, M.C. Ozelo, K. Hoots, P. Blatt, B. Konkle, M. Dake, R. Kaye, M. Razavi, A. Zajko, J. Zehnder, P.K. Rustagi, H. Nakai, A. Chew, D. Leonard, J.F. Wright, R.R. Lessard, J.M. Sommer, M. Tigges, D. Sabatino, A. Luk, H. Jiang, F. Mingozzi, L. Couto, H.C. Ertl, K.A. High, and M.A. Kay, Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host immune response. Nat Med, 2006. 12(3): p. 342-7.
3. Hinderer, C., N. Katz, E.L. Buza, C. Dyer, T. Goode, P. Bell, L.K. Richman, and J.M. Wilson, Severe Toxicity in Nonhuman Primates and Piglets Following High-Dose Intravenous Administration of an Adeno-Associated Virus Vector Expressing Human SMN. Hum Gene Ther, 2018. 29(3): p. 285-298.
4. Paulk, N.K., K. Pekrun, G.W. Charville, K. Maguire-Nguyen, M.N. Wosczyna, J. Xu, Y. Zhang, L. Lisowski, B. Yoo, J.G. Vilches-Moure, G.K. Lee, J.B. Shrager, T.A. Rando, and M.A. Kay, Bioengineered Viral Platform for Intramuscular Passive Vaccine Delivery to Human Skeletal Muscle. Mol Ther Methods Clin Dev, 2018. 10: p. 144-155.
5. Paulk, N.K., K. Pekrun, E. Zhu, S. Nygaard, B. Li, J. Xu, K. Chu, C. Leborgne, A.P. Dane, A. Haft, Y. Zhang, F. Zhang, C. Morton, M.B. Valentine, A.M. Davidoff, A.C. Nathwani, F. Mingozzi, M. Grompe, I.E. Alexander, L. Lisowski, and M.A. Kay, Bioengineered AAV Capsids with Combined High Human Liver Transduction In Vivo and Unique Humoral Seroreactivity. Mol Ther, 2018. 26(1): p. 289-303.
6. Gray, S.J., B.L. Blake, H.E. Criswell, S.C. Nicolson, R.J. Samulski, T.J. McCown, and W. Li, Directed evolution of a novel adeno-associated virus (AAV) vector that crosses the seizure-compromised blood-brain barrier (BBB). Mol Ther, 2010. 18(3): p. 570-8.
7. Tse, L.V., K.A. Klinc, V.J. Madigan, R.M. Castellanos Rivera, L.F. Wells, L.P. Havlik, J.K. Smith, M. Agbandje-McKenna, and A. Asokan, Structure-guided evolution of antigenically distinct adeno-associated virus variants for immune evasion. Proc Natl Acad Sci U S A, 2017. 114(24): p. E4812-E4821.
8. Koerber, J.T., N. Maheshri, B.K. Kaspar, and D.V. Schaffer, Construction of diverse adeno-associated viral libraries for directed evolution of enhanced gene delivery vehicles. Nat Protoc, 2006. 1(2): p. 701-6.
9. Korbelin, J., T. Sieber, S. Michelfelder, L. Lunding, E. Spies, A. Hunger, M. Alawi, K. Rapti, D. Indenbirken, O.J. Muller, R. Pasqualini, W. Arap, J.A. Kleinschmidt, and M. Trepel, Pulmonary Targeting of Adeno-associated Viral Vectors by Next-generation Sequencing-guided Screening of Random Capsid Displayed Peptide Libraries. Mol Ther, 2016. 24(6): p. 1050-1061.
10. Sallach, J., G. Di Pasquale, F. Larcher, N. Niehoff, M. Rubsam, A. Huber, J. Chiorini, D. Almarza, S.A. Eming, H. Ulus, S. Nishimura, U.T. Hacker, M. Hallek, C.M. Niessen, and H. Buning, Tropism-modified AAV vectors overcome barriers to successful cutaneous therapy. Mol Ther, 2014. 22(5): p. 929-39.
11. Berry, G.E. and A. Asokan, Cellular transduction mechanisms of adeno-associated viral vectors. Curr Opin Virol, 2016. 21: p. 54-60.
12. Deverman, B.E., P.L. Pravdo, B.P. Simpson, S.R. Kumar, K.Y. Chan, A. Banerjee, W.L. Wu, B. Yang, N. Huber, S.P. Pasca, and V. Gradinaru, Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat Biotechnol, 2016. 34(2): p. 204-9.
13. Matsuzaki, Y., M. Tanaka, S. Hakoda, T. Masuda, R. Miyata, A. Konno, and H. Hirai, Neurotropic Properties of AAV-PHP.B Are Shared among Diverse Inbred Strains of Mice. Mol Ther, 2019. 27(4): p. 700-704.
14. Hordeaux, J., Q. Wang, N. Katz, E.L. Buza, P. Bell, and J.M. Wilson, The Neurotropic Properties of AAV-PHP.B Are Limited to C57BL/6J Mice. Mol Ther, 2018. 26(3): p. 664-668.
15. Nonnenmacher, M., H. van Bakel, R.J. Hajjar, and T. Weber, High capsid-genome correlation facilitates creation of AAV libraries for directed evolution. Mol Ther, 2015. 23(4): p. 675-82.
16. Hordeaux, J., Y. Yuan, P.M. Clark, Q. Wang, R.A. Martino, J.J. Sims, P. Bell, A. Raymond, W.L. Stanford, and J.M. Wilson, The GPI-Linked Protein LY6A Drives AAV-PHP.B Transport across the Blood-Brain Barrier. Mol Ther, 2019. 27(5): p. 912-921.
17. Igarashi, H., K. Koizumi, R. Kaneko, K. Ikeda, R. Egawa, Y. Yanagawa, S. Muramatsu, H. Onimaru, T. Ishizuka, and H. Yawo, A Novel Reporter Rat Strain That Conditionally Expresses the Bright Red Fluorescent Protein tdTomato. PLoS One, 2016. 11(5): p. e0155687.
18. Park, J.E., X.F. Zhang, S.H. Choi, J. Okahara, E. Sasaki, and A.C. Silva, Generation of transgenic marmosets expressing genetically encoded calcium indicators. Sci Rep, 2016. 6: p. 34931.
19. Sasaki, E., H. Suemizu, A. Shimada, K. Hanazawa, R. Oiwa, M. Kamioka, I. Tomioka, Y. Sotomaru, R. Hirakawa, T. Eto, S. Shiozawa, T. Maeda, M. Ito, R. Ito, C. Kito, C. Yagihashi, K. Kawai, H. Miyoshi, Y. Tanioka, N. Tamaoki, S. Habu, H. Okano, and T. Nomura, Generation of transgenic non-human primates with germline transmission. Nature, 2009. 459(7246): p. 523-7.
20. D'Costa, S., V. Blouin, F. Broucque, M. Penaud-Budloo, A. Francois, I.C. Perez, C. Le Bec, P. Moullier, R.O. Snyder, and E. Ayuso, Practical utilization of recombinant AAV vector reference standards: focus on vector genomes titration by free ITR qPCR. Mol Ther Methods Clin Dev, 2016. 5: p. 16019.
21. Aurnhammer, C., M. Haase, N. Muether, M. Hausl, C. Rauschhuber, I. Huber, H. Nitschko, U. Busch, A. Sing, A. Ehrhardt, and A. Baiker, Universal real-time PCR for the detection and quantification of adeno-associated virus serotype 2-derived inverted terminal repeat sequences. Hum Gene Ther Methods, 2012. 23(1): p. 18-28.
22. Gyorgy, B., E.J. Meijer, M.V. Ivanchenko, K. Tenneson, F. Emond, K.S. Hanlon, A.A. Indzhykulian, A. Volak, K.D. Karavitaki, P.I. Tamvakologos, M. Vezina, V.K. Berezovskii, R.T. Born, M. O'Brien, J.F. Lafond, Y. Arsenijevic, M.A. Kenna, C.A. Maguire, and D.P. Corey, Gene Transfer with AAV9-PHP.B Rescues Hearing in a Mouse Model of Usher Syndrome 3A and Transduces Hair Cells in a Non-human Primate. Mol Ther Methods Clin Dev, 2019. 13: p. 1-13.
23. Maguire, C.A., L. Balaj, S. Sivaraman, M.H. Crommentuijn, M. Ericsson, L. Mincheva-Nilsson, V. Baranov, D. Gianni, B.A. Tannous, M. Sena-Esteves, X.O. Breakefield, and J. Skog, Microvesicle-associated AAV vector as a novel gene delivery system. Mol Ther, 2012. 20(5): p. 960-71.
24. Hudry, E., C. Martin, S. Gandhi, B. Gyorgy, D.I. Scheffer, D. Mu, S.F. Merkel, F. Mingozzi, Z. Fitzpatrick, H. Dimant, M. Masek, T. Ragan, S. Tan, A.R. Brisson, S.H. Ramirez, B.T. Hyman, and C.A. Maguire, Exosome-associated AAV vector as a robust and convenient neuroscience tool. Gene Ther, 2016. 23(4): p. 380-92.
25. Dashkoff, J., E.P. Lerner, N. Truong, J.A. Klickstein, Z. Fan, D. Mu, C.A. Maguire, B.T. Hyman, and E. Hudry, Tailored transgene expression to specific cell types in the central nervous system after peripheral injection with AAV9. Mol Ther Methods Clin Dev, 2016. 3: p. 16081.
26. Watters, A.K., S. Rom, J.D. Hill, M.K. Dematatis, Y. Zhou, S.F. Merkel, A.M. Andrews, J. Cena, R. Potula, A. Skuba, Y.J. Son, Y. Persidsky, and S.H. Ramirez, Identification and dynamic regulation of tight junction protein expression in human neural stem cells. Stem Cells Dev, 2015. 24(12): p. 1377-89.
27. Akil, O., Dyka, F., Calvet, C., Emptoz, A., Lahlou, G., Nouaille, S., De Monvel, J. B., Hardelin, J. P., Hauswirth, W. W., Avan, P., Petit, C., Safieddine, S., and Lustig, L. R. (2019). Dual AAV-mediated gene therapy restores hearing in a DFNB9 mouse model. Proc. Natl. Acad. Sci. U. S. A. doi:10.1073/pnas.1817537116.
28. Al-Zaidy, S. A., Kolb, S. J., Lowes, L., Alfano, L. N., Shell, R., Church, K. R., Nagendran, S., Sproule, D. M., Feltner, D. E., Wells, C., Ogrinc, F., Menier, M., L’Italien, J., Arnold, W. D., Kissel, J. T., Kaspar, B. K., and Mendell, J. R. (2019). AVXS-101 (Onasemnogene Abeparvovec) for SMA1: Comparative Study with a Prospective Natural History Cohort. J. Neuromuscul. Dis. doi:10.3233/JND-190403.
29. Caron, N. S., Southwell, A. L., Brouwers, C. C., Cengio, L. D., Xie, Y., Black, H. F., Anderson, L. M., Ko, S., Zhu, X., van Deventer, S. J., Evers, M. M., Konstantinova, P., and Hayden, M. R. (2020). Potent and sustained huntingtin lowering via AAV5 encoding miRNA preserves striatal volume and cognitive function in a humanized mouse model of Huntington disease. Nucleic Acids Res. doi:10.1093/nar/gkz976.
30. Cehajic-Kapetanovic, J., Xue, K., Martinez-Fernandez de la Camara, C., Nanda, A., Davies, A., Wood, L. J., Salvetti, A. P., Fischer, M. D., Aylward, J. W., Barnard, A. R., Jolly, J. K., Luo, E., Lujan, B. J., Ong, T., Girach, A., Black, G. C. M., Gregori, N. Z., Davis, J. L., Rosa, P. R., Lotery, A. J., Lam, B. L., Stanga, P. E., and MacLaren, R. E. (2020). Initial results from a first-in-human gene therapy trial on X-linked retinitis pigmentosa caused by mutations in RPGR. Nat. Med. doi:10.1038/s41591-020-0763-1.
31. Christine, C. W., Bankiewicz, K. S., Van Laar, A. D., Richardson, R. M., Ravina, B., Kells, A. P., Boot, B., Martin, A. J., Nutt, J., Thompson, M. E., and Larson, P. S. (2019). Magnetic resonance imaging-guided phase 1 trial of putaminal AADC gene therapy for Parkinson’s disease. Ann. Neurol. doi:10.1002/ana.25450.
32. Duan, D. (2018). Systemic AAV Micro-dystrophin Gene Therapy for Duchenne Muscular Dystrophy. Mol. Ther. doi:10.1016/j.ymthe.2018.07.011.
33 Eichler, F., Duncan, C., Musolino, P. L., Orchard, P. J., De Oliveira, S., Thrasher, A. J., Armant, M., Dansereau, C., Lund, T. C., Miller, W. P., Raymond, G. V., Sankar, R., Shah, A. J., Sevin, C., Gaspar, H. B., Gissen, P., Amartino, H., Bratkovic, D., Smith, N. J. C., Paker, A. M., Shamir, E., O’Meara, T., Davidson, D., Aubourg, P., and Williams, D. A. (2017). Hematopoietic stem-cell gene therapy for cerebral adrenoleukodystrophy. N. Engl. J. Med. doi:10.1056/NEJMoa1700554.
34. Gong, Y., Mu, D., Prabhakar, S., Moser, A., Musolino, P., Ren, J. Q., Breakefield, X. O., Maguire, C. A., and Eichler, F. S. (2015). Adenoassociated virus serotype 9-mediated gene therapy for X-linked adrenoleukodystrophy. Mol. Ther. doi:10.1038/mt.2015.6.
35. Griciuc, A., Serrano-Pozo, A., Parrado, A. R., Lesinski, A. N., Asselin, C. N., Mullin, K., Hooli, B., Choi, S. H., Hyman, B. T., and Tanzi, R. E. (2013). Alzheimer’s disease risk gene cd33 inhibits microglial uptake of amyloid beta. Neuron. doi:10.1016/j.neuron.2013.04.014.
36. György, B., Meijer, E. J., Ivanchenko, M. V., Tenneson, K., Emond, F., Hanlon, K. S., Indzhykulian, A. A., Volak, A., Karavitaki, K. D., Tamvakologos, P. I., Vezina, M., Berezovskii, V. K., Born, R. T., O’Brien, M., Lafond, J. F., Arsenijevic, Y., Kenna, M. A., Maguire, C. A., and Corey, D. P. (2019). Gene Transfer with AAV9-PHP.B Rescues Hearing in a Mouse Model of Usher Syndrome 3A and Transduces Hair Cells in a Non-human Primate. Mol. Ther. - Methods Clin. Dev. doi:10.1016/j.omtm.2018.11.003.
37. Hughes, M. P., Smith, D. A., Morris, L., Fletcher, C., Colaco, A., Huebecker, M., Tordo, J., Palomar, N., Massaro, G., Henckaerts, E., Waddington, S. N., Platt, F. M., and Rahim, A. A. (2018). AAV9 intracerebroventricular gene therapy improves lifespan, locomotor function and pathology in a mouse model of Niemann-Pick type C1 disease. Hum. Mol. Genet. doi:10.1093/hmg/ddy212.
38. Isgrig, K., Shteamer, J. W., Belyantseva, I. A., Drummond, M. C., Fitzgerald, T. S., Vijayakumar, S., Jones, S. M., Griffith, A. J., Friedman, T. B., Cunningham, L. L., and Chien, W. W. (2017). Gene Therapy Restores Balance and Auditory Functions in a Mouse Model of Usher Syndrome. Mol. Ther. doi:10.1016/j.ymthe.2017.01.007.
39. Keskin, S., Brouwers, C. C., Sogorb-Gonzalez, M., Martier, R., Depla, J. A., Vallès, A., van Deventer, S. J., Konstantinova, P., and Evers, M. M. (2019). AAV5-miHTT Lowers Huntingtin mRNA and Protein without Off-Target Effects in Patient-Derived Neuronal Cultures and Astrocytes. Mol. Ther. - Methods Clin. Dev. doi:10.1016/j.omtm.2019.09.010.
40. Leone, P., Shera, D., McPhee, S. W. J., Francis, J. S., Kolodny, E. H., Bilaniuk, L. T., Wang, D. J., Assadi, M., Goldfarb, O., Goldman, H. W., Freese, A., Young, D., During, M. J., Samulski, R. J., and Janson, C. G. (2012). Long-term follow-up after gene therapy for canavan disease. Sci. Transl. Med. doi:10.1126/scitranslmed.3003454.
41. Lopes, V. S., and Williams, D. S. (2015). Gene therapy for the retinal degeneration of usher syndrome caused by mutations in MYO7A. Cold Spring Harb. Perspect. Med. doi:10.1101/cshperspect.a017319.
42. Maguire, A. M., Russell, S., Wellman, J. A., Chung, D. C., Yu, Z. F., Tillman, A., Wittes, J., Pappas, J., Elci, O., Marshall, K. A., McCague, S., Reichert, H., Davis, M., Simonelli, F., Leroy, B. P., Wright, J. F., High, K. A., and Bennett, J. (2019). Efficacy, Safety, and Durability of Voretigene Neparvovec-rzyl in RPE65 Mutation-Associated Inherited Retinal Dystrophy: Results of Phase 1 and 3 Trials. Ophthalmology. doi:10.1016/j.ophtha.2019.06.017.
43. Millington-Ward, S., Chadderton, N., O’Reilly, M., Palfi, A., Goldmann, T., Kilty, C., Humphries, M., Wolfrum, U., Bennett, J., Humphries, P., Kenna, P. F., and Farrar, G. J. (2011). Suppression and Replacement Gene Therapy for Autosomal Dominant Disease in a Murine Model of Dominant Retinitis Pigmentosa. Mol. Ther. 19, 642-649. doi:10.1038/mt.2010.293.
44. Mookherjee, S., Hiriyanna, S., Kaneshiro, K., Li, Y., Li, W., Qian, H., Li, T., Colosi, P., Swaroop, A., Wu, Z., Li, L., and Khanna, H. (2015). Long-term rescue of cone photoreceptor degeneration in retinitis pigmentosa 2 (RP2)-knockout mice by gene replacement therapy. Hum. Mol. Genet. doi:10.1093/hmg/ddv354.
45. Nathwani, A. C. (2019). Gene therapy for hemophilia. Hematol. (United States). doi:10.1182/hematology.2019000007.
46. Nist-Lund, C. A., Pan, B., Patterson, A., Asai, Y., Chen, T., Zhou, W., Zhu, H., Romero, S., Resnik, J., Polley, D. B., Géléoc, G. S., and Holt, J. R. (2019). Improved TMC1 gene therapy restores hearing and balance in mice with genetic inner ear disorders. Nat. Commun. doi:10.1038/s41467-018-08264-w.
47. Remes, A., Franz, M., Zaradzki, M., Borowski, C., Frey, N., Karck, M., Kallenbach, K., Müller, O. J., Wagner, A. H., and Arif, R. (2020). AAV-mediated TIMP-1 overexpression in aortic tissue reduces the severity of allograft vasculopathy in mice. J. Hear. Lung Transplant. doi:10.1016/j.healun.2020.01.1338.
48. Tan, F., Chu, C., Qi, J., Li, W., You, D., Li, K., Chen, X., Zhao, W., Cheng, C., Liu, X., Qiao, Y., Su, B., He, S., Zhong, C., Li, H., Chai, R., and Zhong, G. (2019). AAV-ie enables safe and efficient gene transfer to inner ear cells. Nat. Commun. doi:10.1038/s41467-019-11687-8.
49. Wan, X., Pei, H., Zhao, M., Yang, S., Hu, W., He, H., Ma, S., Zhang, G., Dong, X., Chen, C., Wang, D., and Li, B. (2016). Efficacy and Safety of rAAV2-ND4 Treatment for Leber’s Hereditary Optic Neuropathy. Sci. Rep. 6, 1-10. doi:10.1038/srep21587.
50. Yang, S., Ma, S.-Q., Wan, X., He, H., Pei, H., Zhao, M.-J., Chen, C., Wang, D.-W., Dong, X.-Y., Yuan, J.-J., and Li, B. (2016). Long-term outcomes of gene therapy for the treatment of Leber’s hereditary optic neuropathy. EBioMedicine 10, 258-268. doi:10.1016/j.ebiom.2016.07.002.
51. Yu, W., Mookherjee, S., Chaitankar, V., Hiriyanna, S., Kim, J. W., Brooks, M., Ataeijannati, Y., Sun, X., Dong, L., Li, T., Swaroop, A., and Wu, Z. (2017). Nrl knockdown by AAV-delivered CRISPR/Cas9 prevents retinal degeneration in mice. Nat. Commun. doi:10.1038/ncomms14716.
52. Zhang, W., Kim, S. M., Wang, W., Cai, C., Feng, Y., Kong, W., and Lin, X. (2018). Cochlear Gene Therapy for Sensorineural Hearing Loss: Current Status and Major Remaining Hurdles for Translational Success. Front. Mol. Neurosci. doi:10.3389/fnmol.2018.00221.
Другие варианты осуществления изобретения
Следует отметить, что хотя настоящее изобретение было подробно описано выше, однако, такое описание приводится лишь в целях иллюстрации и не рассматривается как ограничение объема изобретения, который определяется прилагаемой формулой изобретения. Другие аспекты, преимущества и модификации входят в объем нижеследующей формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> THE GENERAL HOSPITAL CORPORATION
PRESIDENT AND FELLOWS OF HARVARD COLLEGE
<120> СКОНСТРУИРОВАННЫЕ АДЕНОАССОЦИИРОВАННЫЕ (AAV) ВЕКТОРЫ
ДЛЯ ЭКСПРЕСИИ ТРАНСГЕНА
<130> 29539-0390WO1
<140> PCT/US2020/025720
<141> 2020-03-30
<150> 62/825,703
<151> 2019-03-28
<160> 149
<170> PatentIn version 3.5
<210> 1
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 1
Ser Thr Thr Leu Tyr Ser Pro
1 5
<210> 2
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 2
Phe Val Val Gly Gln Ser Tyr
1 5
<210> 3
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 3
Phe Gln Pro Cys Pro
1 5
<210> 4
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 4
Pro Ser Tyr Leu Thr Thr Ser
1 5
<210> 5
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 5
Tyr Ser Gln Gly Val Val Phe
1 5
<210> 6
<211> 736
<212> PRT
<213> Аденоассоциированный вирус 9
<400> 6
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ala Gln Ala Gln
580 585 590
Thr Gly Trp Val Gln Asn Gln Gly Ile Leu Pro Gly Met Val Trp Gln
595 600 605
Asp Arg Asp Val Tyr Leu Gln Gly Pro Ile Trp Ala Lys Ile Pro His
610 615 620
Thr Asp Gly Asn Phe His Pro Ser Pro Leu Met Gly Gly Phe Gly Met
625 630 635 640
Lys His Pro Pro Pro Gln Ile Leu Ile Lys Asn Thr Pro Val Pro Ala
645 650 655
Asp Pro Pro Thr Ala Phe Asn Lys Asp Lys Leu Asn Ser Phe Ile Thr
660 665 670
Gln Tyr Ser Thr Gly Gln Val Ser Val Glu Ile Glu Trp Glu Leu Gln
675 680 685
Lys Glu Asn Ser Lys Arg Trp Asn Pro Glu Ile Gln Tyr Thr Ser Asn
690 695 700
Tyr Tyr Lys Ser Asn Asn Val Glu Phe Ala Val Asn Thr Glu Gly Val
705 710 715 720
Tyr Ser Glu Pro Arg Pro Ile Gly Thr Arg Tyr Leu Thr Arg Asn Leu
725 730 735
<210> 7
<211> 743
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 7
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Phe Val Val Gly
580 585 590
Gln Ser Tyr Ala Gln Ala Gln Thr Gly Trp Val Gln Asn Gln Gly Ile
595 600 605
Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro
610 615 620
Ile Trp Ala Lys Ile Pro His Thr Asp Gly Asn Phe His Pro Ser Pro
625 630 635 640
Leu Met Gly Gly Phe Gly Met Lys His Pro Pro Pro Gln Ile Leu Ile
645 650 655
Lys Asn Thr Pro Val Pro Ala Asp Pro Pro Thr Ala Phe Asn Lys Asp
660 665 670
Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val
675 680 685
Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro
690 695 700
Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys Ser Asn Asn Val Glu Phe
705 710 715 720
Ala Val Asn Thr Glu Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr
725 730 735
Arg Tyr Leu Thr Arg Asn Leu
740
<210> 8
<211> 2229
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 8
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgcttggc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaatttgtt gttggtcaga gttatgcaca ggcgcagacc 1800
ggctgggttc aaaaccaagg aatacttccg ggtatggttt ggcaggacag agatgtgtac 1860
ctgcaaggac ccatttgggc caaaattcct cacacggacg gcaactttca cccttctccg 1920
ctgatgggag ggtttggaat gaagcacccg cctcctcaga tcctcatcaa aaacacacct 1980
gtacctgcgg atcctccaac ggccttcaac aaggacaagc tgaactcttt catcacccag 2040
tattctactg gccaagtcag cgtggagatc gagtgggagc tgcagaagga aaacagcaag 2100
cgctggaacc cggagatcca gtacacttcc aactattaca agtctaataa tgttgaattt 2160
gctgttaata ctgaaggtgt atatagtgaa ccccgcccca ttggcaccag atacctgact 2220
cgtaatctg 2229
<210> 9
<211> 743
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полипептид
<400> 9
Met Ala Ala Asp Gly Tyr Leu Pro Asp Trp Leu Glu Asp Asn Leu Ser
1 5 10 15
Glu Gly Ile Arg Glu Trp Trp Ala Leu Lys Pro Gly Ala Pro Gln Pro
20 25 30
Lys Ala Asn Gln Gln His Gln Asp Asn Ala Arg Gly Leu Val Leu Pro
35 40 45
Gly Tyr Lys Tyr Leu Gly Pro Gly Asn Gly Leu Asp Lys Gly Glu Pro
50 55 60
Val Asn Ala Ala Asp Ala Ala Ala Leu Glu His Asp Lys Ala Tyr Asp
65 70 75 80
Gln Gln Leu Lys Ala Gly Asp Asn Pro Tyr Leu Lys Tyr Asn His Ala
85 90 95
Asp Ala Glu Phe Gln Glu Arg Leu Lys Glu Asp Thr Ser Phe Gly Gly
100 105 110
Asn Leu Gly Arg Ala Val Phe Gln Ala Lys Lys Arg Leu Leu Glu Pro
115 120 125
Leu Gly Leu Val Glu Glu Ala Ala Lys Thr Ala Pro Gly Lys Lys Arg
130 135 140
Pro Val Glu Gln Ser Pro Gln Glu Pro Asp Ser Ser Ala Gly Ile Gly
145 150 155 160
Lys Ser Gly Ala Gln Pro Ala Lys Lys Arg Leu Asn Phe Gly Gln Thr
165 170 175
Gly Asp Thr Glu Ser Val Pro Asp Pro Gln Pro Ile Gly Glu Pro Pro
180 185 190
Ala Ala Pro Ser Gly Val Gly Ser Leu Thr Met Ala Ser Gly Gly Gly
195 200 205
Ala Pro Val Ala Asp Asn Asn Glu Gly Ala Asp Gly Val Gly Ser Ser
210 215 220
Ser Gly Asn Trp His Cys Asp Ser Gln Trp Leu Gly Asp Arg Val Ile
225 230 235 240
Thr Thr Ser Thr Arg Thr Trp Ala Leu Pro Thr Tyr Asn Asn His Leu
245 250 255
Tyr Lys Gln Ile Ser Asn Ser Thr Ser Gly Gly Ser Ser Asn Asp Asn
260 265 270
Ala Tyr Phe Gly Tyr Ser Thr Pro Trp Gly Tyr Phe Asp Phe Asn Arg
275 280 285
Phe His Cys His Phe Ser Pro Arg Asp Trp Gln Arg Leu Ile Asn Asn
290 295 300
Asn Trp Gly Phe Arg Pro Lys Arg Leu Asn Phe Lys Leu Phe Asn Ile
305 310 315 320
Gln Val Lys Glu Val Thr Asp Asn Asn Gly Val Lys Thr Ile Ala Asn
325 330 335
Asn Leu Thr Ser Thr Val Gln Val Phe Thr Asp Ser Asp Tyr Gln Leu
340 345 350
Pro Tyr Val Leu Gly Ser Ala His Glu Gly Cys Leu Pro Pro Phe Pro
355 360 365
Ala Asp Val Phe Met Ile Pro Gln Tyr Gly Tyr Leu Thr Leu Asn Asp
370 375 380
Gly Ser Gln Ala Val Gly Arg Ser Ser Phe Tyr Cys Leu Glu Tyr Phe
385 390 395 400
Pro Ser Gln Met Leu Arg Thr Gly Asn Asn Phe Gln Phe Ser Tyr Glu
405 410 415
Phe Glu Asn Val Pro Phe His Ser Ser Tyr Ala His Ser Gln Ser Leu
420 425 430
Asp Arg Leu Met Asn Pro Leu Ile Asp Gln Tyr Leu Tyr Tyr Leu Ser
435 440 445
Lys Thr Ile Asn Gly Ser Gly Gln Asn Gln Gln Thr Leu Lys Phe Ser
450 455 460
Val Ala Gly Pro Ser Asn Met Ala Val Gln Gly Arg Asn Tyr Ile Pro
465 470 475 480
Gly Pro Ser Tyr Arg Gln Gln Arg Val Ser Thr Thr Val Thr Gln Asn
485 490 495
Asn Asn Ser Glu Phe Ala Trp Pro Gly Ala Ser Ser Trp Ala Leu Asn
500 505 510
Gly Arg Asn Ser Leu Met Asn Pro Gly Pro Ala Met Ala Ser His Lys
515 520 525
Glu Gly Glu Asp Arg Phe Phe Pro Leu Ser Gly Ser Leu Ile Phe Gly
530 535 540
Lys Gln Gly Thr Gly Arg Asp Asn Val Asp Ala Asp Lys Val Met Ile
545 550 555 560
Thr Asn Glu Glu Glu Ile Lys Thr Thr Asn Pro Val Ala Thr Glu Ser
565 570 575
Tyr Gly Gln Val Ala Thr Asn His Gln Ser Ala Gln Ser Thr Thr Leu
580 585 590
Tyr Ser Pro Ala Gln Ala Gln Thr Gly Trp Val Gln Asn Gln Gly Ile
595 600 605
Leu Pro Gly Met Val Trp Gln Asp Arg Asp Val Tyr Leu Gln Gly Pro
610 615 620
Ile Trp Ala Lys Ile Pro His Thr Asp Gly Asn Phe His Pro Ser Pro
625 630 635 640
Leu Met Gly Gly Phe Gly Met Lys His Pro Pro Pro Gln Ile Leu Ile
645 650 655
Lys Asn Thr Pro Val Pro Ala Asp Pro Pro Thr Ala Phe Asn Lys Asp
660 665 670
Lys Leu Asn Ser Phe Ile Thr Gln Tyr Ser Thr Gly Gln Val Ser Val
675 680 685
Glu Ile Glu Trp Glu Leu Gln Lys Glu Asn Ser Lys Arg Trp Asn Pro
690 695 700
Glu Ile Gln Tyr Thr Ser Asn Tyr Tyr Lys Ser Asn Asn Val Glu Phe
705 710 715 720
Ala Val Asn Thr Glu Gly Val Tyr Ser Glu Pro Arg Pro Ile Gly Thr
725 730 735
Arg Tyr Leu Thr Arg Asn Leu
740
<210> 10
<211> 2229
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
полинуклеотид
<400> 10
atggctgccg atggttatct tccagattgg ctcgaggaca accttagtga aggaattcgc 60
gagtggtggg ctttgaaacc tggagcccct caacccaagg caaatcaaca acatcaagac 120
aacgctcgag gtcttgtgct tccgggttac aaataccttg gacccggcaa cggactcgac 180
aagggggagc cggtcaacgc agcagacgcg gcggccctcg agcacgacaa ggcctacgac 240
cagcagctca aggccggaga caacccgtac ctcaagtaca accacgccga cgccgagttc 300
caggagcggc tcaaagaaga tacgtctttt gggggcaacc tcgggcgagc agtcttccag 360
gccaaaaaga ggcttcttga acctcttggt ctggttgagg aagcggctaa gacggctcct 420
ggaaagaaga ggcctgtaga gcagtctcct caggaaccgg actcctccgc gggtattggc 480
aaatcgggtg cacagcccgc taaaaagaga ctcaatttcg gtcagactgg cgacacagag 540
tcagtcccag accctcaacc aatcggagaa cctcccgcag ccccctcagg tgtgggatct 600
cttacaatgg cttcaggtgg tggcgcacca gtggcagaca ataacgaagg tgccgatgga 660
gtgggtagtt cctcgggaaa ttggcattgc gattcccaat ggctggggga cagagtcatc 720
accaccagca cccgaacctg ggccctgccc acctacaaca atcacctcta caagcaaatc 780
tccaacagca catctggagg atcttcaaat gacaacgcct acttcggcta cagcaccccc 840
tgggggtatt ttgacttcaa cagattccac tgccacttct caccacgtga ctggcagcga 900
ctcatcaaca acaactgggg attccggcct aagcgactca acttcaagct cttcaacatt 960
caggtcaaag aggttacgga caacaatgga gtcaagacca tcgccaataa ccttaccagc 1020
acggtccagg tcttcacgga ctcagactat cagctcccgt acgtgctcgg gtcggctcac 1080
gagggctgcc tcccgccgtt cccagcggac gttttcatga ttcctcagta cgggtatctg 1140
acgcttaatg atggaagcca ggccgtgggt cgttcgtcct tttactgcct ggaatatttc 1200
ccgtcgcaaa tgctaagaac gggtaacaac ttccagttca gctacgagtt tgagaacgta 1260
cctttccata gcagctacgc tcacagccaa agcctggacc gactaatgaa tccactcatc 1320
gaccaatact tgtactatct ctcaaagact attaacggtt ctggacagaa tcaacaaacg 1380
ctaaaattca gtgtggccgg acccagcaac atggctgtcc agggaagaaa ctacatacct 1440
ggacccagct accgacaaca acgtgtctca accactgtga ctcaaaacaa caacagcgaa 1500
tttgcttggc ctggagcttc ttcttgggct ctcaatggac gtaatagctt gatgaatcct 1560
ggacctgcta tggccagcca caaagaagga gaggaccgtt tctttccttt gtctggatct 1620
ttaatttttg gcaaacaagg aactggaaga gacaacgtgg atgcggacaa agtcatgata 1680
accaacgaag aagaaattaa aactactaac ccggtagcaa cggagtccta tggacaagtg 1740
gccacaaacc accagagtgc ccaatctact acgctttata gtcctgcaca ggcgcagacc 1800
ggctgggttc aaaaccaagg aatacttccg ggtatggttt ggcaggacag agatgtgtac 1860
ctgcaaggac ccatttgggc caaaattcct cacacggacg gcaactttca cccttctccg 1920
ctgatgggag ggtttggaat gaagcacccg cctcctcaga tcctcatcaa aaacacacct 1980
gtacctgcgg atcctccaac ggccttcaac aaggacaagc tgaactcttt catcacccag 2040
tattctactg gccaagtcag cgtggagatc gagtgggagc tgcagaagga aaacagcaag 2100
cgctggaacc cggagatcca gtacacttcc aactattaca agtctaataa tgttgaattt 2160
gctgttaata ctgaaggtgt atatagtgaa ccccgcccca ttggcaccag atacctgact 2220
cgtaatctg 2229
<210> 11
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 11
gtactatctc tctagaacta ttaacggttc 30
<210> 12
<211> 84
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<220>
<221> модифицированное_основание
<222> (41)..(41)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (43)..(44)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (46)..(47)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (49)..(50)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (52)..(53)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (55)..(56)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (58)..(59)
<223> a, c, t, g, неизвестные или другие
<220>
<221> модифицированное_основание
<222> (61)..(62)
<223> a, c, t, g, неизвестные или другие
<400> 12
gtattccttg gttttgaacc caaccggtct gcgcctgtgc nmnnmnnmnn mnnmnnmnnm 60
nnttgggcac tctggtggtt tgtg 84
<210> 13
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 13
aatcctggac ctgctatggc 20
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 14
tgccaaacca tacccggaag 20
<210> 15
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 15
agctaccgac aacaacgtgt 20
<210> 16
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
праймер
<400> 16
agaagggtga aagttgccgt 20
<210> 17
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 17
cagcctcgtc ttactgagct t 21
<210> 18
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 18
Gln Pro Arg Leu Thr Glu Leu
1 5
<210> 19
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 19
agtcatttta gtaattctat g 21
<210> 20
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 20
Ser His Phe Ser Asn Ser Met
1 5
<210> 21
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 21
ccgccttcta cggggaatcc t 21
<210> 22
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 22
Pro Pro Ser Thr Gly Asn Pro
1 5
<210> 23
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 23
aatagtcttg gggtgatgga t 21
<210> 24
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 24
Asn Ser Leu Gly Val Met Asp
1 5
<210> 25
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 25
cctcctatgg ctacgcctca g 21
<210> 26
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 26
Pro Pro Met Ala Thr Pro Gln
1 5
<210> 27
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 27
ccgcatactc gggctgagcc t 21
<210> 28
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 28
Pro His Thr Arg Ala Glu Pro
1 5
<210> 29
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 29
gtgaatcaga ctaattattc t 21
<210> 30
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 30
Val Asn Gln Thr Asn Tyr Ser
1 5
<210> 31
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 31
catgcgccga ggctggttca g 21
<210> 32
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 32
His Ala Pro Arg Leu Val Gln
1 5
<210> 33
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 33
tggagtaagg agtcgaatca t 21
<210> 34
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 34
Trp Ser Lys Glu Ser Asn His
1 5
<210> 35
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 35
actactgctg gtccgactga g 21
<210> 36
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 36
Thr Thr Ala Gly Pro Thr Glu
1 5
<210> 37
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 37
tcgtggccgc agtcgccttc g 21
<210> 38
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 38
Ser Trp Pro Gln Ser Pro Ser
1 5
<210> 39
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 39
acttgtgctg ttccggatcg g 21
<210> 40
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 40
Thr Cys Ala Val Pro Asp Arg
1 5
<210> 41
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 41
cctcgtgcgg agcctcggac g 21
<210> 42
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 42
Pro Arg Ala Glu Pro Arg Thr
1 5
<210> 43
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 43
cctactgcgg gtgtgtatct g 21
<210> 44
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 44
Pro Thr Ala Gly Val Tyr Leu
1 5
<210> 45
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 45
cagttgagta atacgacgtt g 21
<210> 46
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 46
Gln Leu Ser Asn Thr Thr Leu
1 5
<210> 47
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 47
cggatggata tggctgtgag g 21
<210> 48
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 48
Arg Met Asp Met Ala Val Arg
1 5
<210> 49
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 49
cggccgccgc ctgcgactcc t 21
<210> 50
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 50
Arg Pro Pro Pro Ala Thr Pro
1 5
<210> 51
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 51
cagtatctgt ttcatatttt t 21
<210> 52
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 52
Gln Tyr Leu Phe His Ile Phe
1 5
<210> 53
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 53
ggtgctggga ctcttacttc t 21
<210> 54
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 54
Gly Ala Gly Thr Leu Thr Ser
1 5
<210> 55
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 55
cctctttcgc gtgttgcttt g 21
<210> 56
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 56
Pro Leu Ser Arg Val Ala Leu
1 5
<210> 57
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 57
actactgcta agacgactgg g 21
<210> 58
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 58
Thr Thr Ala Lys Thr Thr Gly
1 5
<210> 59
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 59
tcgctgccgg ctactcatgt g 21
<210> 60
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 60
Ser Leu Pro Ala Thr His Val
1 5
<210> 61
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 61
agtgagaagg tgattcggag t 21
<210> 62
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 62
Ser Glu Lys Val Ile Arg Ser
1 5
<210> 63
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 63
aagtcgacta tgcggaagcc g 21
<210> 64
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 64
Lys Ser Thr Met Arg Lys Pro
1 5
<210> 65
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 65
gtgtcgaagc agaatacgac t 21
<210> 66
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 66
Val Ser Lys Gln Asn Thr Thr
1 5
<210> 67
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 67
gctgatatga cggggcctac t 21
<210> 68
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 68
Ala Asp Met Thr Gly Pro Thr
1 5
<210> 69
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 69
cctactcgtg cgactcctat t 21
<210> 70
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 70
Pro Thr Arg Ala Thr Pro Ile
1 5
<210> 71
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 71
tcttggcctg ggctgttgct g 21
<210> 72
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 72
Ser Trp Pro Gly Leu Leu Leu
1 5
<210> 73
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 73
tggaggctgg agcagttgaa g 21
<210> 74
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 74
Trp Arg Leu Glu Gln Leu Lys
1 5
<210> 75
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 75
cttccgagtt ctccggtttt t 21
<210> 76
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 76
Leu Pro Ser Ser Pro Val Phe
1 5
<210> 77
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 77
cctcagcggc atcagaatga t 21
<210> 78
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 78
Pro Gln Arg His Gln Asn Asp
1 5
<210> 79
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 79
tcgattccgt ttcggtatcc g 21
<210> 80
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 80
Ser Ile Pro Phe Arg Tyr Pro
1 5
<210> 81
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 81
cctgatggtc cgcaggctct g 21
<210> 82
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 82
Pro Asp Gly Pro Gln Ala Leu
1 5
<210> 83
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 83
tcgactacgt ggaagtctga t 21
<210> 84
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 84
Ser Thr Thr Trp Lys Ser Asp
1 5
<210> 85
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 85
ttgcagaaga tggctgttcc g 21
<210> 86
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 86
Leu Gln Lys Met Ala Val Pro
1 5
<210> 87
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 87
acggggtgta agtgttcgca g 21
<210> 88
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 88
Thr Gly Cys Lys Cys Ser Gln
1 5
<210> 89
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 89
catctgcctg ttatgaatga g 21
<210> 90
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 90
His Leu Pro Val Met Asn Glu
1 5
<210> 91
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 91
gtttcgttga attatactgc t 21
<210> 92
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 92
Val Ser Leu Asn Tyr Thr Ala
1 5
<210> 93
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 93
cctcctcctc ctccgcctgt g 21
<210> 94
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 94
Pro Pro Pro Pro Pro Pro Val
1 5
<210> 95
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 95
agttctactg ctagtcgtcg t 21
<210> 96
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 96
Ser Ser Thr Ala Ser Arg Arg
1 5
<210> 97
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 97
ctgcttgtga cgtgtccgga t 21
<210> 98
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 98
Leu Leu Val Thr Cys Pro Asp
1 5
<210> 99
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 99
tcgttgctgc gttatgctcc g 21
<210> 100
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 100
Ser Leu Leu Arg Tyr Ala Pro
1 5
<210> 101
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 101
cagccgctgc gtccgaatct g 21
<210> 102
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 102
Gln Pro Leu Arg Pro Asn Leu
1 5
<210> 103
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 103
gctttggata ctcatcattg g 21
<210> 104
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 104
Ala Leu Asp Thr His His Trp
1 5
<210> 105
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 105
cggcctactc gtctttctcc g 21
<210> 106
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 106
Arg Pro Thr Arg Leu Ser Pro
1 5
<210> 107
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 107
gatgcttggt cggcttctat t 21
<210> 108
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 108
Asp Ala Trp Ser Ala Ser Ile
1 5
<210> 109
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 109
tataagtgtt ggccgaggag t 21
<210> 110
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 110
Tyr Lys Cys Trp Pro Arg Ser
1 5
<210> 111
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 111
aagtatctga ctaatctttc t 21
<210> 112
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 112
Lys Tyr Leu Thr Asn Leu Ser
1 5
<210> 113
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 113
tcgcggcctc cgccgtctct g 21
<210> 114
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 114
Ser Arg Pro Pro Pro Ser Leu
1 5
<210> 115
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 115
taggcgccga ttctttagtt g 21
<210> 116
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 116
atgcctcctt ctcctccggg t 21
<210> 117
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 117
Met Pro Pro Ser Pro Pro Gly
1 5
<210> 118
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 118
ccgtttcctc agatgctttt t 21
<210> 119
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 119
Pro Phe Pro Gln Met Leu Phe
1 5
<210> 120
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 120
cctcaggcgt ctcatgttaa t 21
<210> 121
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 121
Pro Gln Ala Ser His Val Asn
1 5
<210> 122
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 122
gagtattgtt cttagattcc t 21
<210> 123
<211> 4
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 123
Glu Tyr Cys Ser
1
<210> 124
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 124
tcgtagaatt tgcgggtgtt g 21
<210> 125
<211> 1
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 125
Ser
1
<210> 126
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 126
tattatcatt cgctgtcgtc g 21
<210> 127
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 127
Tyr Tyr His Ser Leu Ser Ser
1 5
<210> 128
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 128
aagcattctc agcggcggaa t 21
<210> 129
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 129
Lys His Ser Gln Arg Arg Asn
1 5
<210> 130
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 130
tagactaatg tgtggattaa t 21
<210> 131
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 131
actaagacgc ctgagcatta g 21
<210> 132
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 132
Thr Lys Thr Pro Glu His
1 5
<210> 133
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 133
gggccggagg ggccgcgtct g 21
<210> 134
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 134
Gly Pro Glu Gly Pro Arg Leu
1 5
<210> 135
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 135
tgtacttcta atctgcatgg g 21
<210> 136
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 136
Cys Thr Ser Asn Leu His Gly
1 5
<210> 137
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 137
gttattactc atctgactga t 21
<210> 138
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 138
Val Ile Thr His Leu Thr Asp
1 5
<210> 139
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 139
gattctaatc attttggtcc g 21
<210> 140
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 140
Asp Ser Asn His Phe Gly Pro
1 5
<210> 141
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 141
ccgagtcagc agacttagat g 21
<210> 142
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 142
Pro Ser Gln Gln Thr
1 5
<210> 143
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 143
tttgttgctg attgggagcc t 21
<210> 144
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 144
Phe Val Ala Asp Trp Glu Pro
1 5
<210> 145
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 145
ctgatgcaga ctaatattac g 21
<210> 146
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 146
Leu Met Gln Thr Asn Ile Thr
1 5
<210> 147
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
олигонуклеотид
<400> 147
ctggtggttc ctaatcgtga t 21
<210> 148
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 148
Leu Val Val Pro Asn Arg Asp
1 5
<210> 149
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Описание искусственной последовательности: Синтетический
пептид
<400> 149
Thr Leu Ala Val Pro Phe Lys
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения модифицированного капсида аденоассоциированного вируса | 2021 |
|
RU2831732C1 |
| Выделенный модифицированный белок VPI капсида аденоассоциированного вируса 9 серотипа (AAV9), капсид и вектор на его основе | 2021 |
|
RU2825667C2 |
| ГЕНОТЕРАПЕВТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ВИЛЬСОНА | 2020 |
|
RU2807158C2 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ЧАСТИЦЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2811426C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ С МОДИФИЦИРОВАННЫМ ТРОПИЗМОМ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НАЦЕЛЕННОГО ВВЕДЕНИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА В КЛЕТКИ ЧЕЛОВЕКА | 2018 |
|
RU2809246C2 |
| Синергетическое действие SMN1 и miR-23a при лечении спинальной мышечной атрофии | 2021 |
|
RU2839898C2 |
| ВЕКТОРЫ AAV, НАЦЕЛЕННЫЕ НА ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ | 2015 |
|
RU2727015C2 |
| СПОСОБЫ ВЫЯВЛЕНИЯ AAV | 2017 |
|
RU2771622C2 |
| МОДУЛЯЦИЯ АКТИВНОСТИ REP БЕЛКА ПРИ ПОЛУЧЕНИИ ДНК С ЗАМКНУТЫМИ КОНЦАМИ (ЗКДНК) | 2020 |
|
RU2812850C2 |
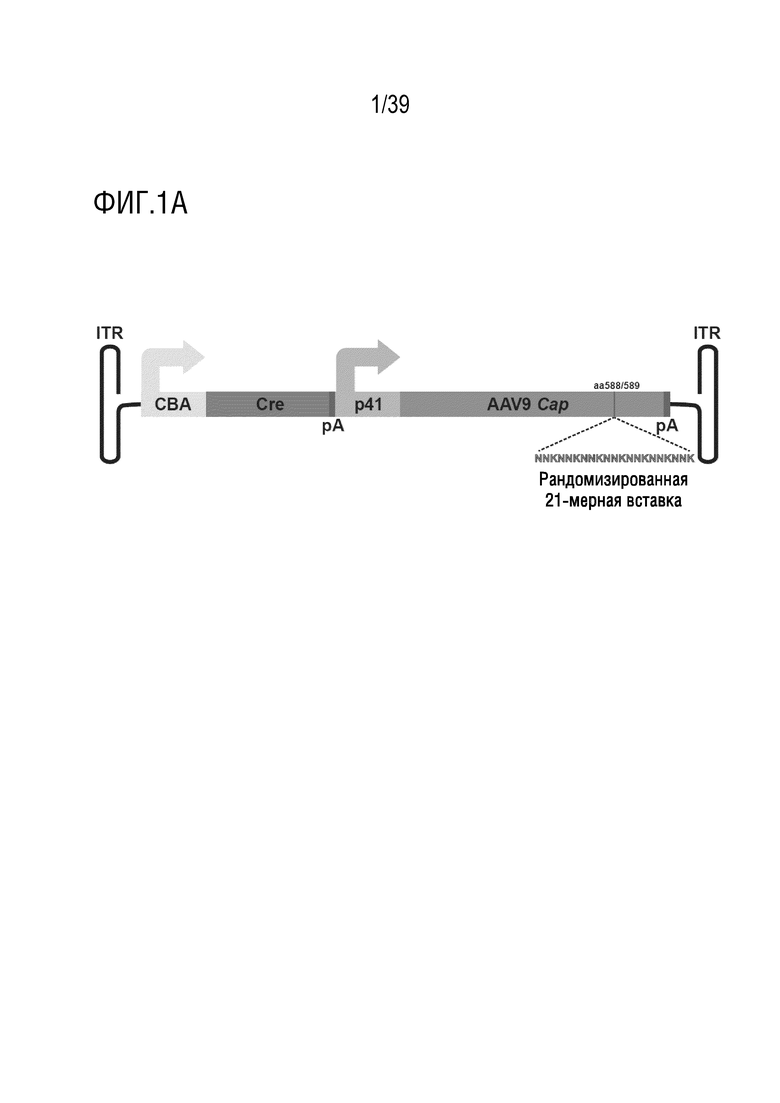
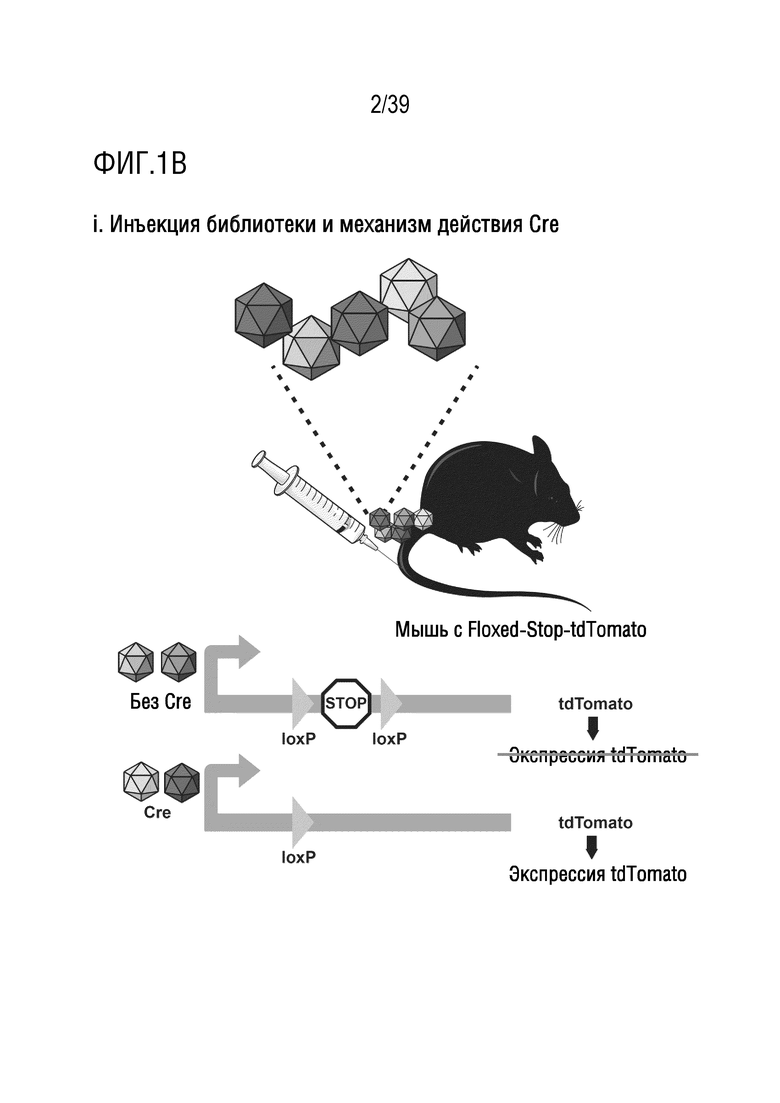
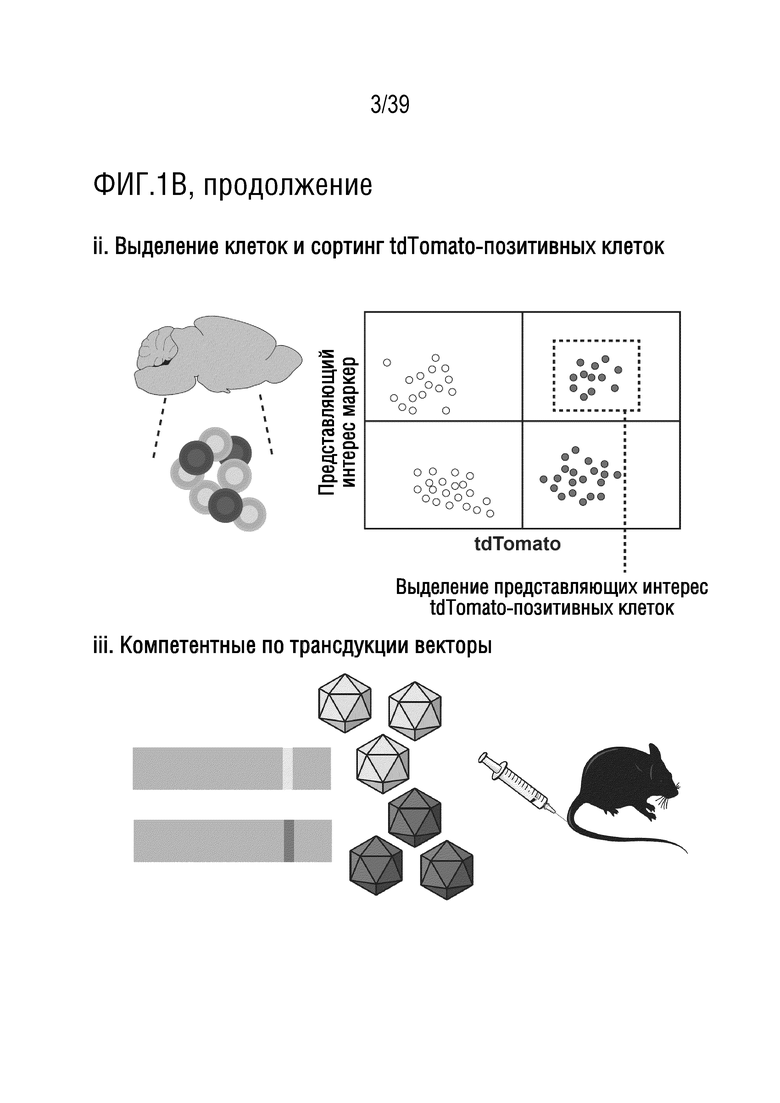

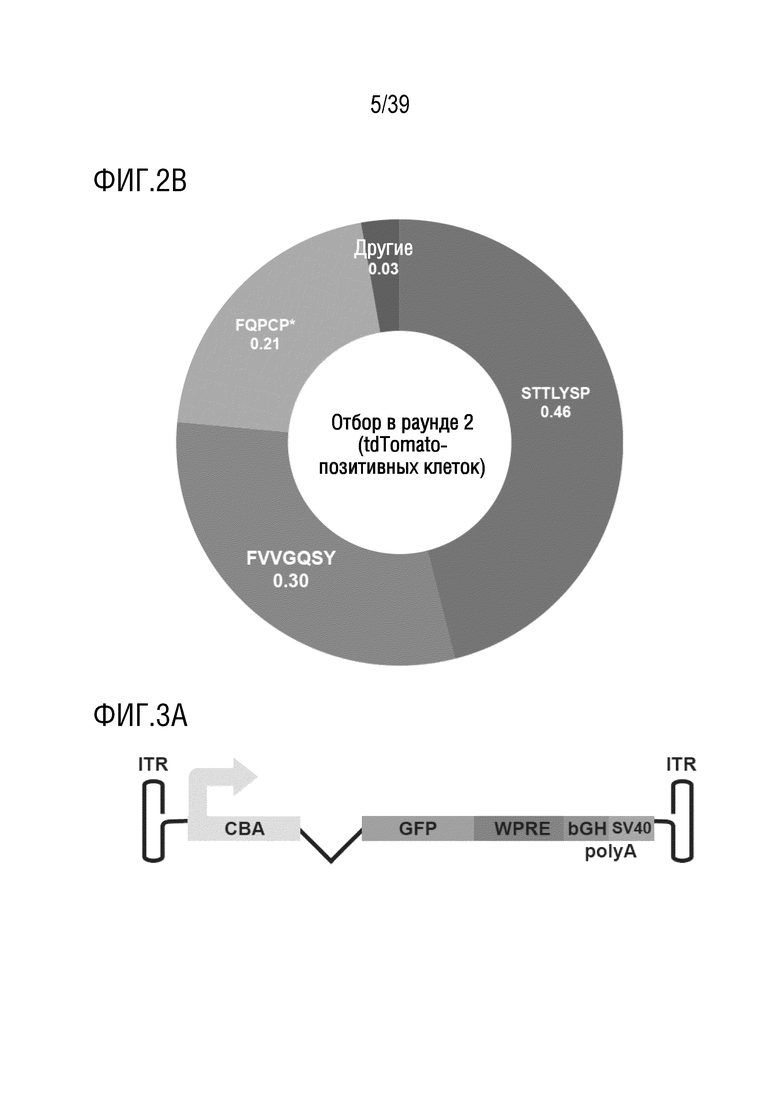
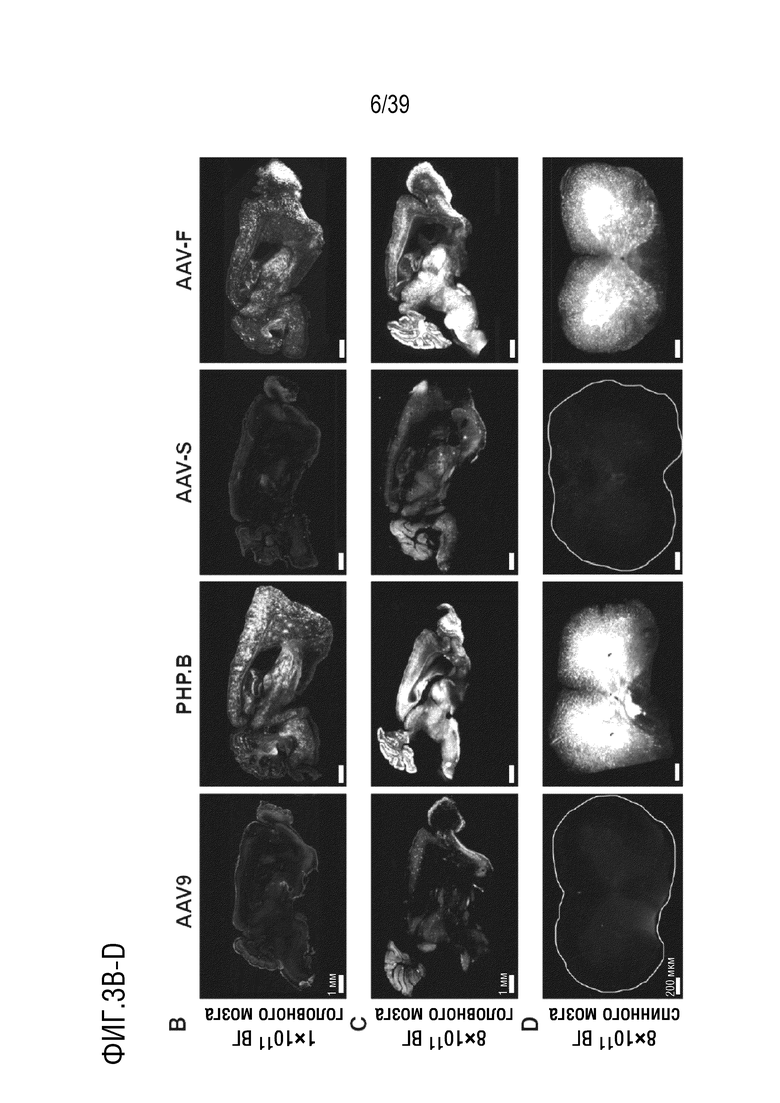
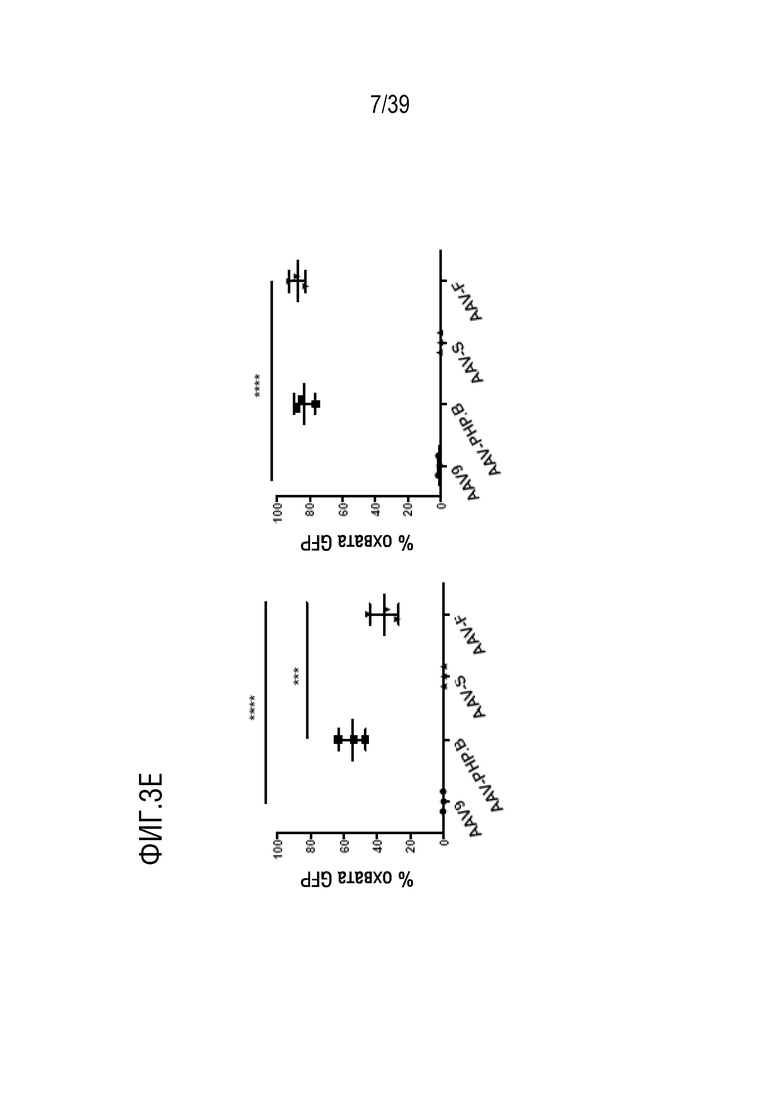
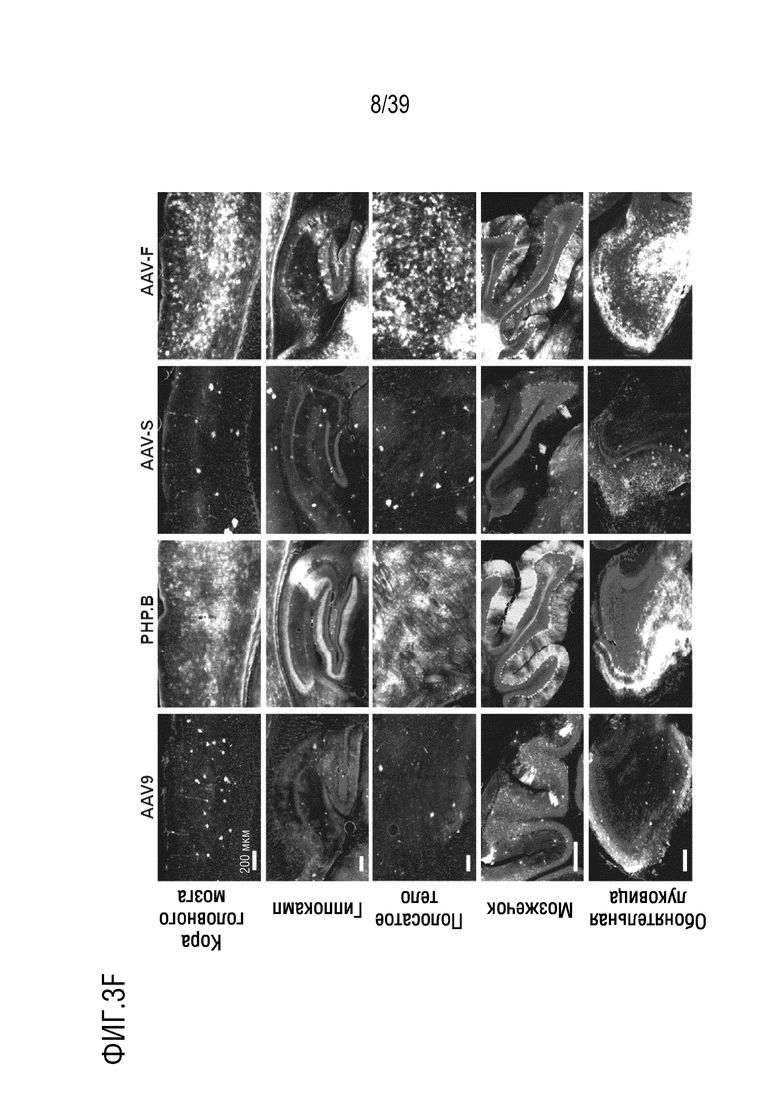

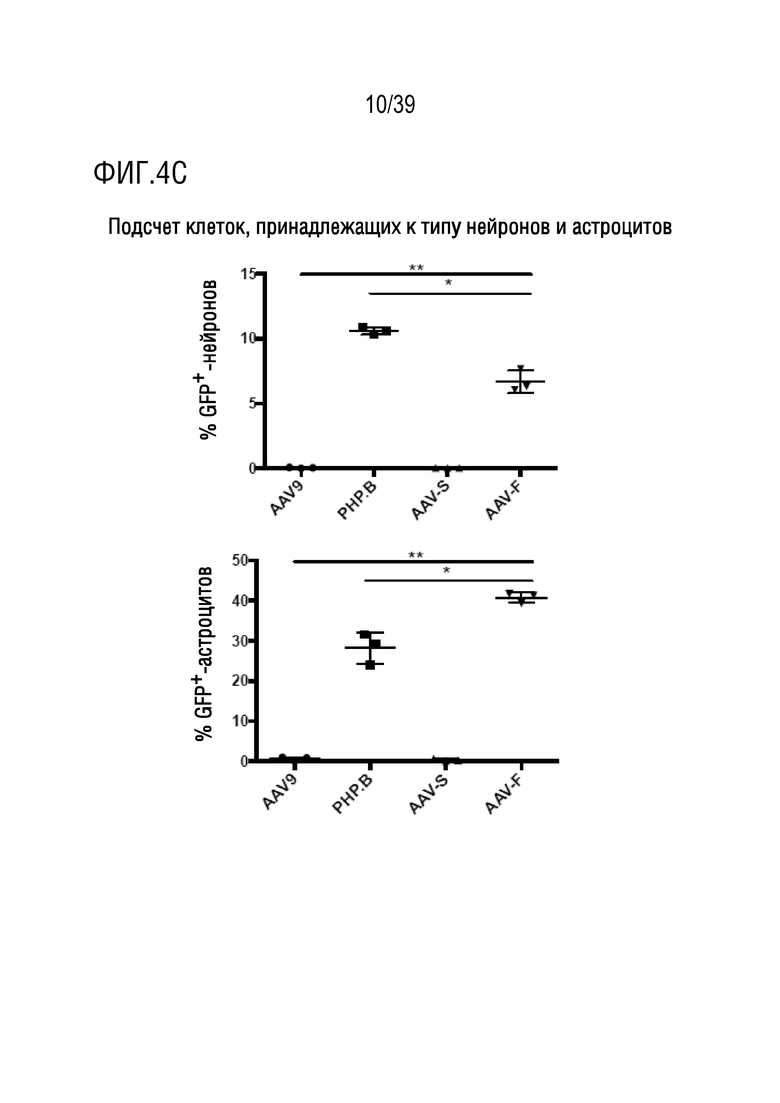
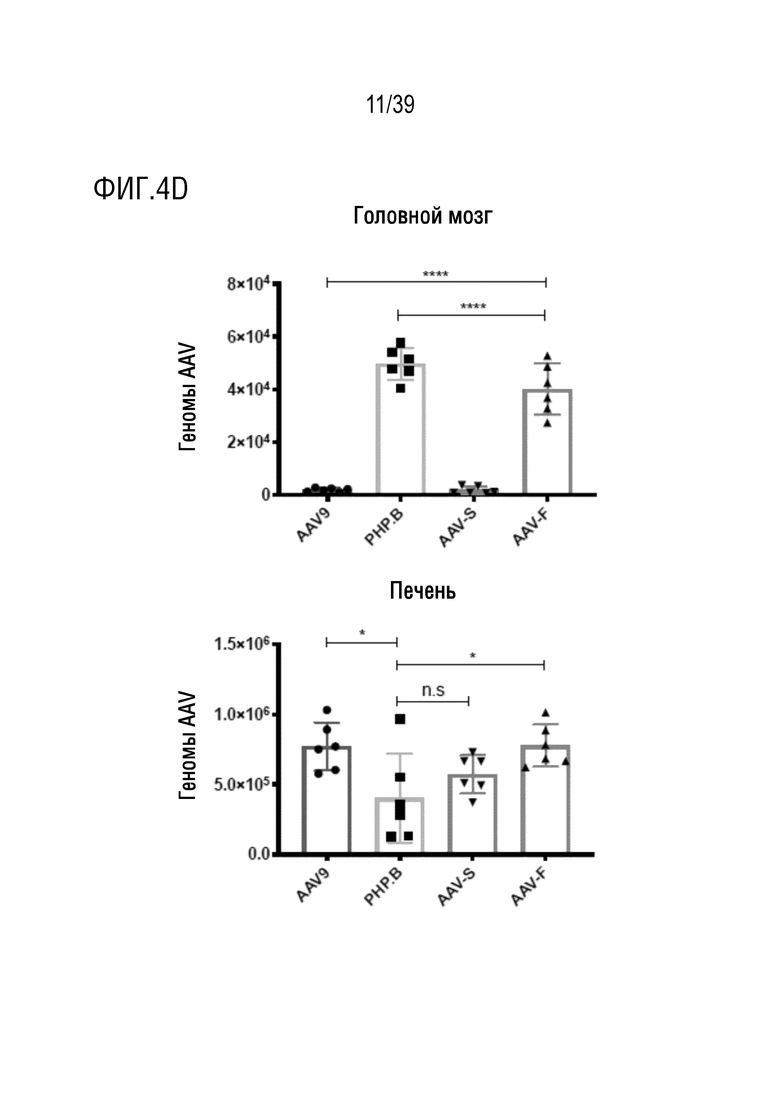

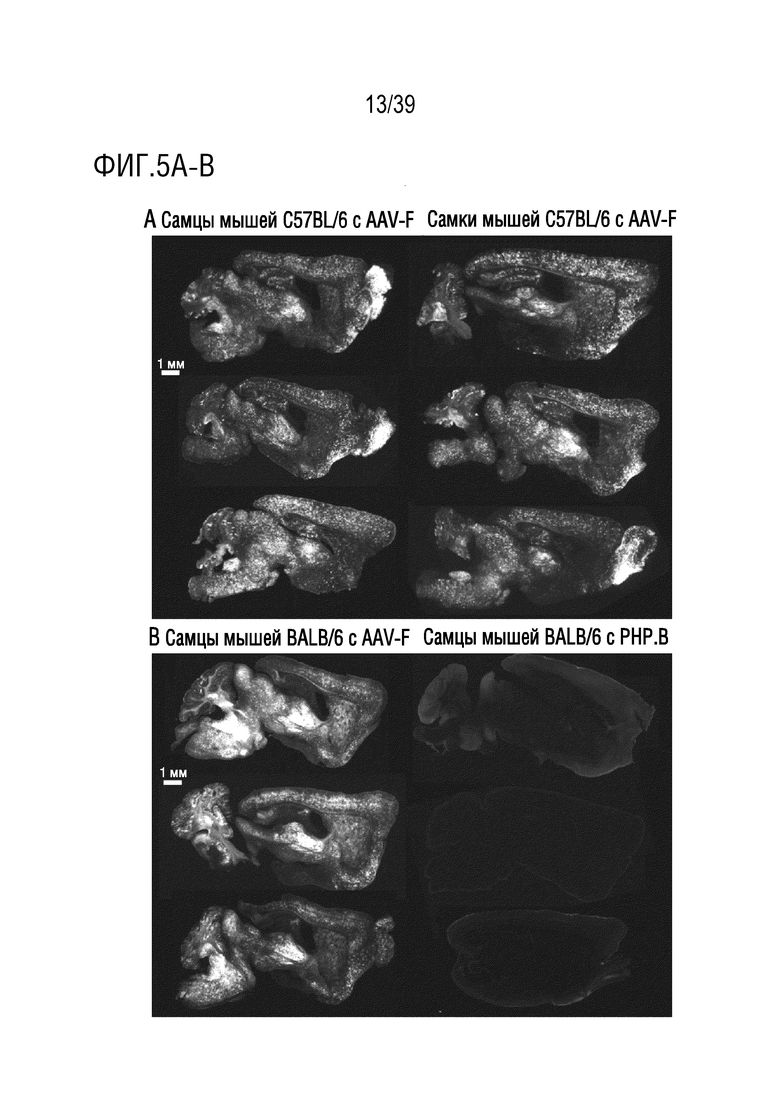
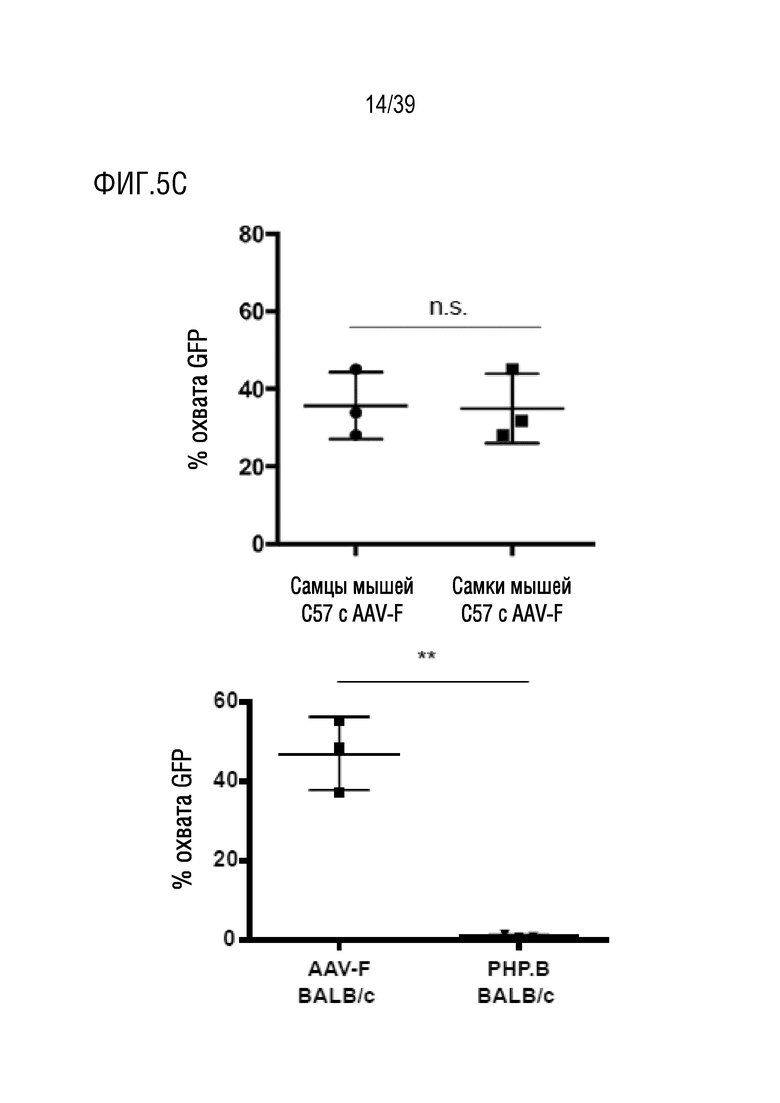
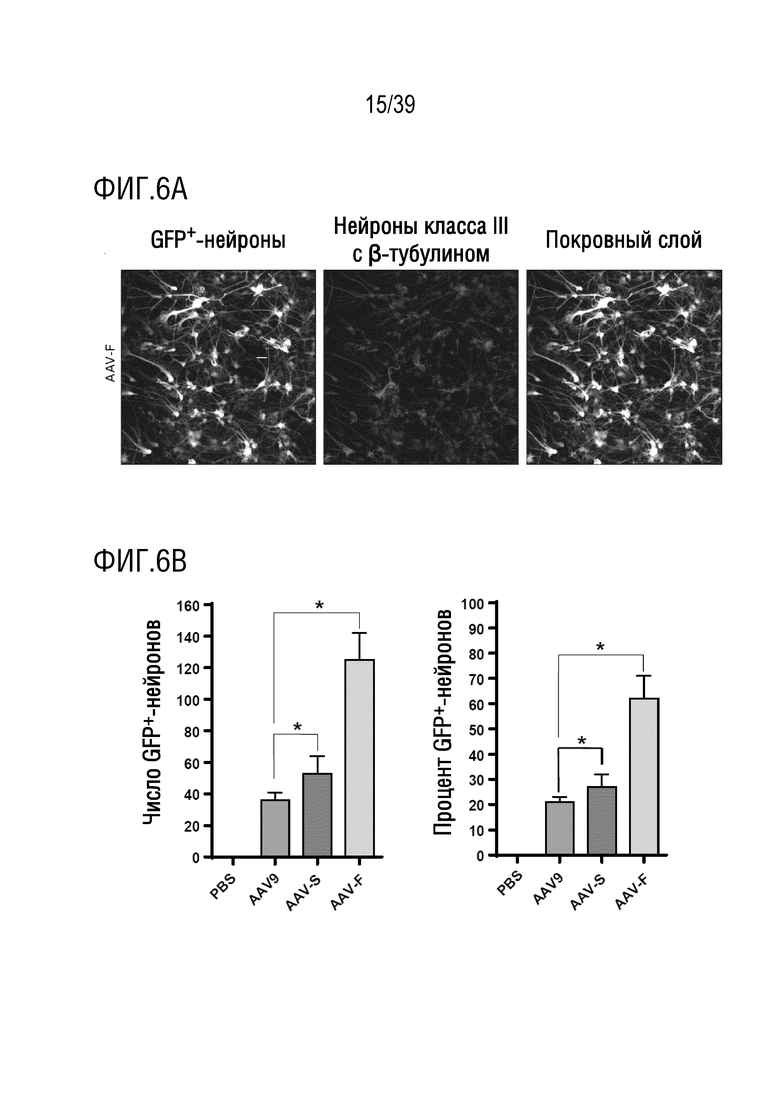


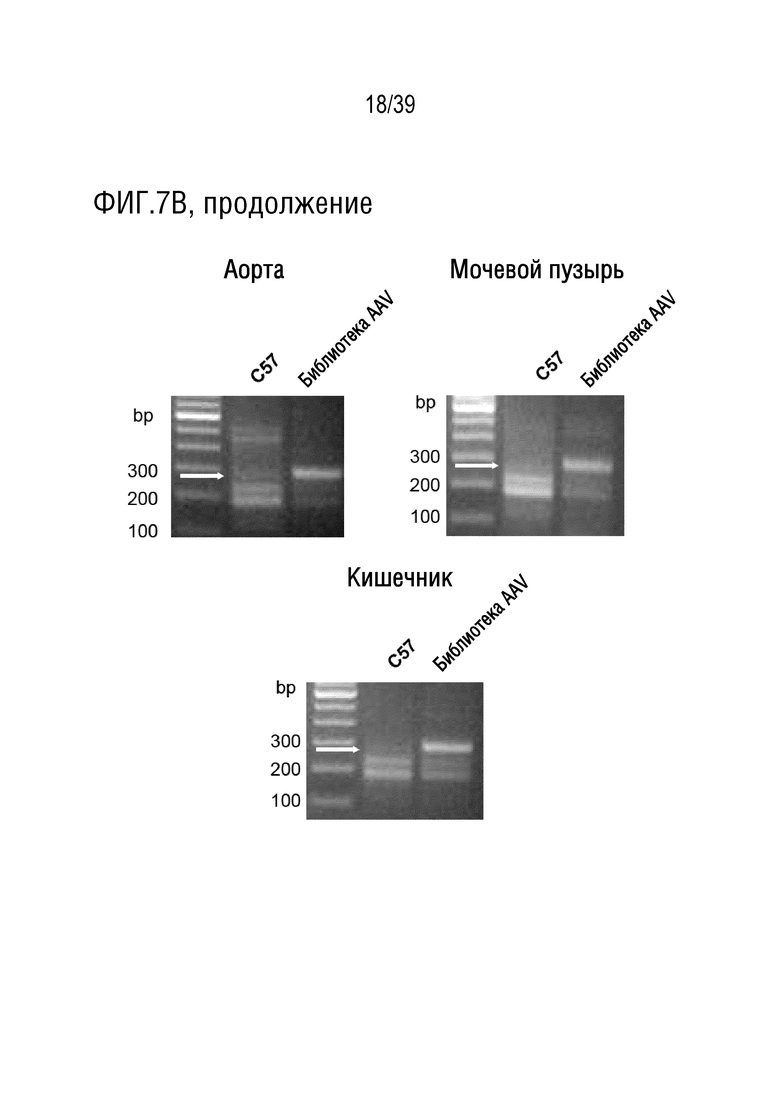


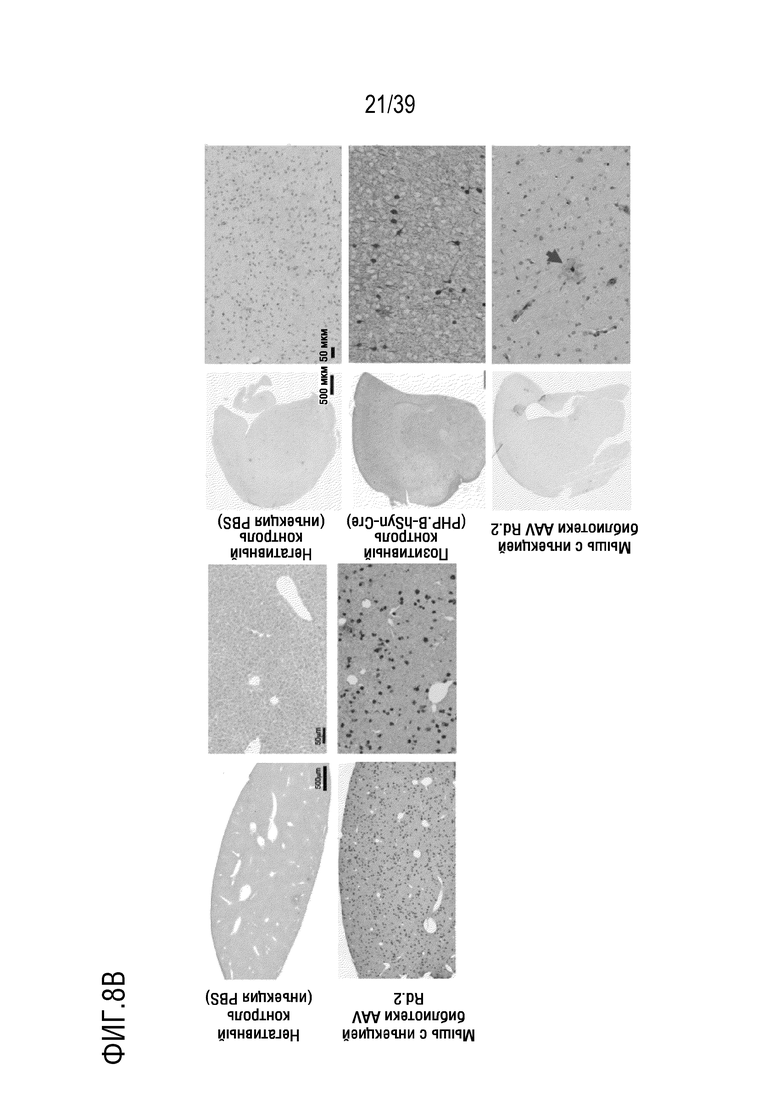

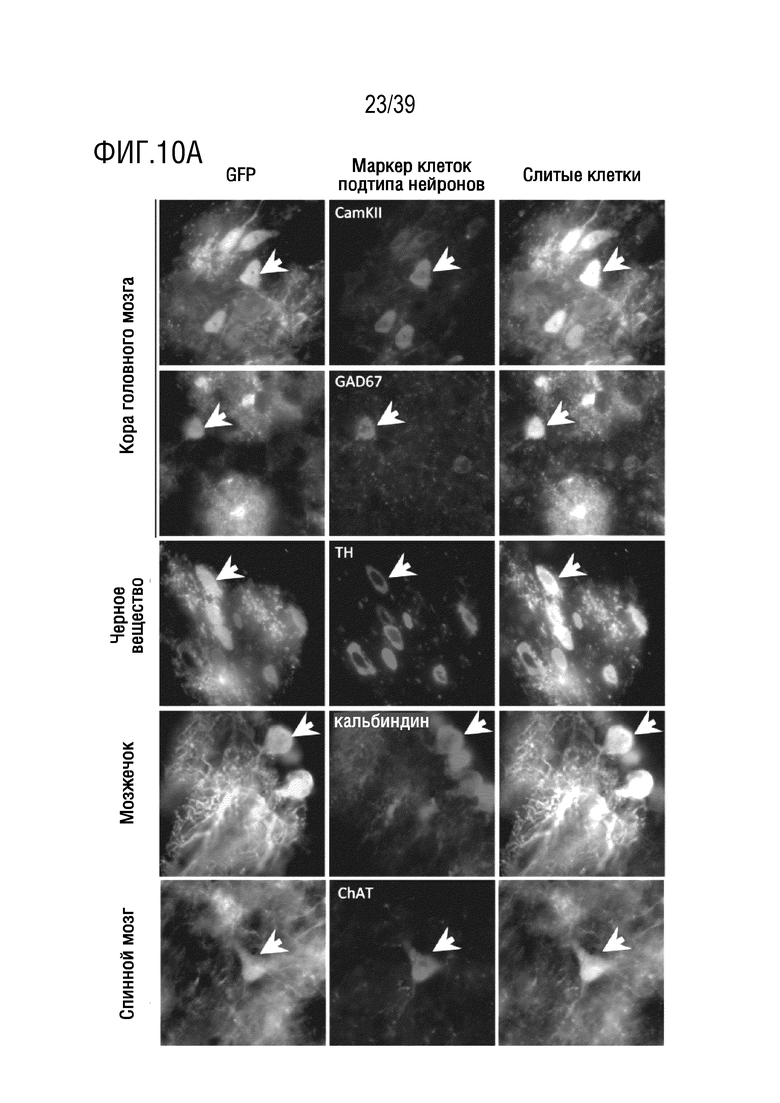
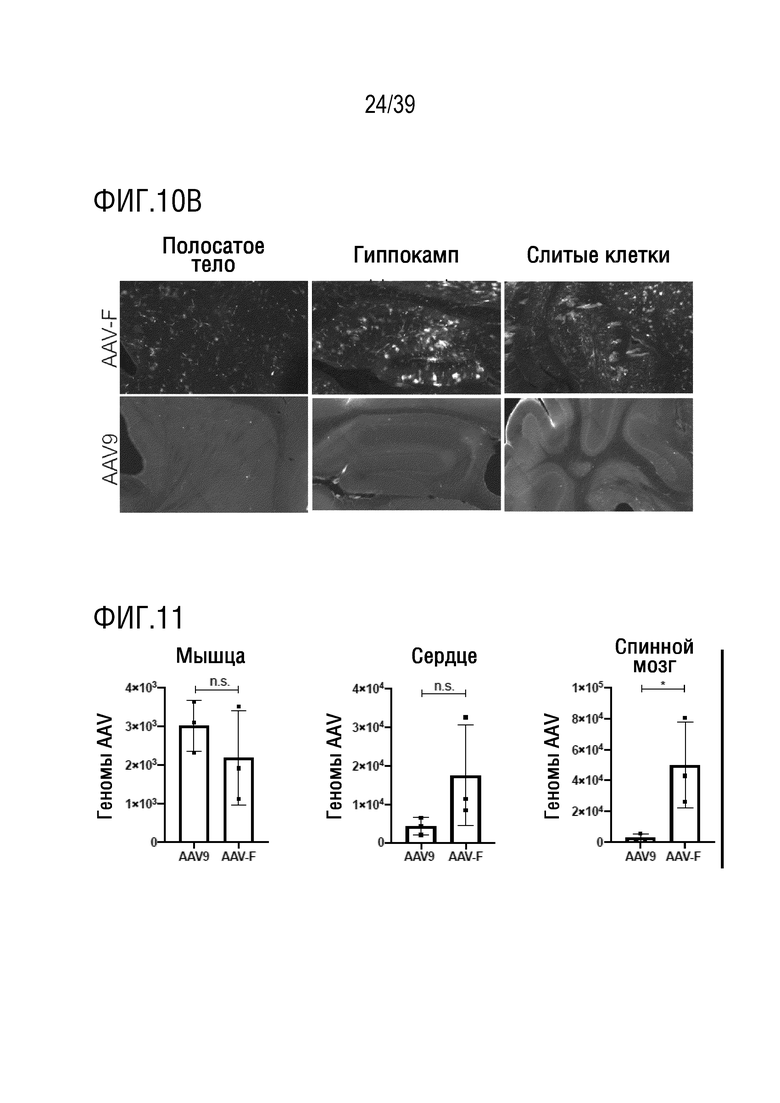

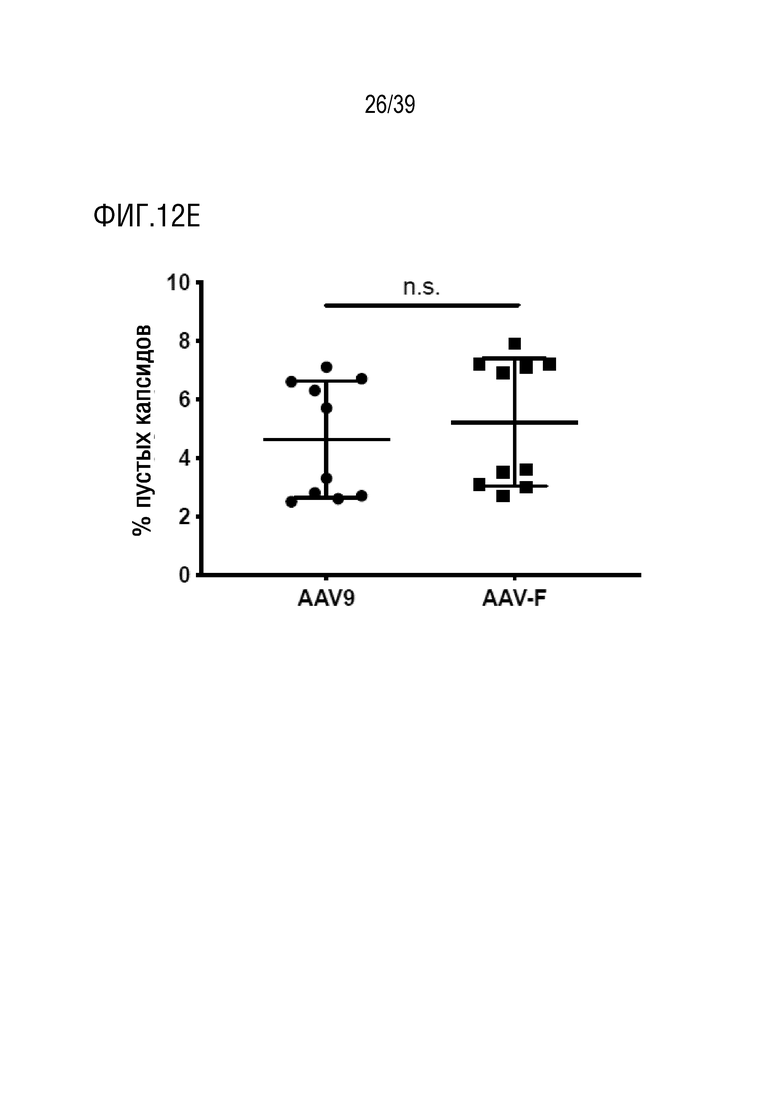



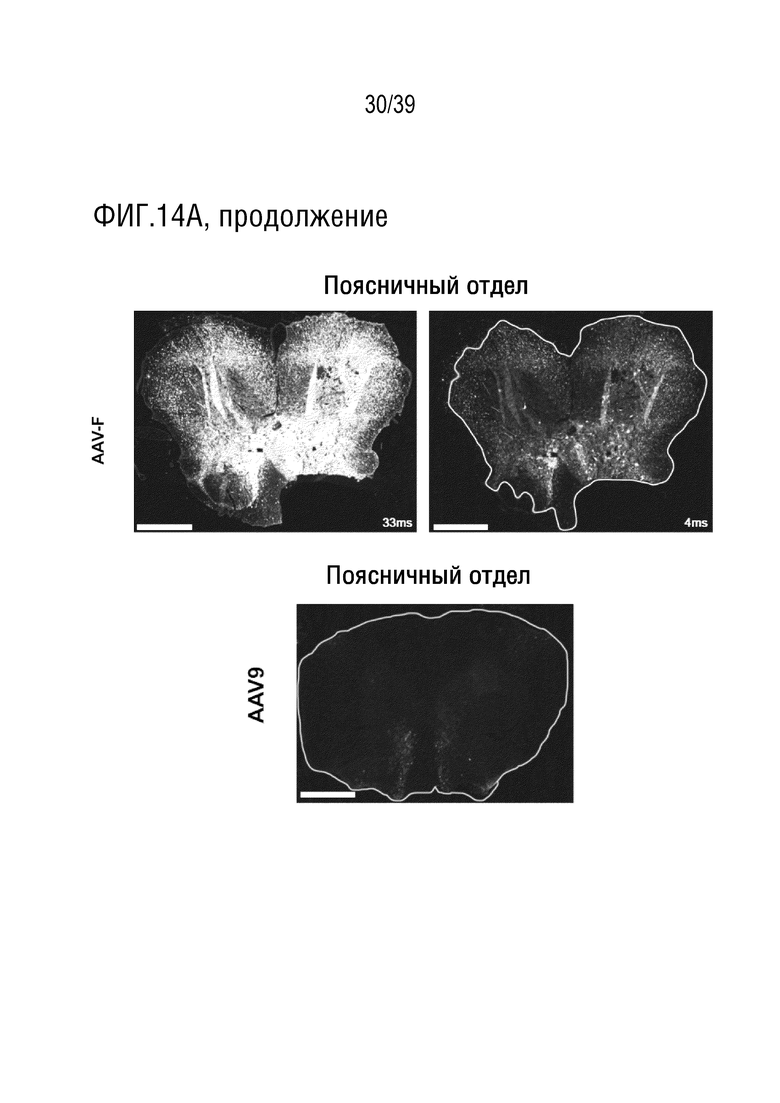
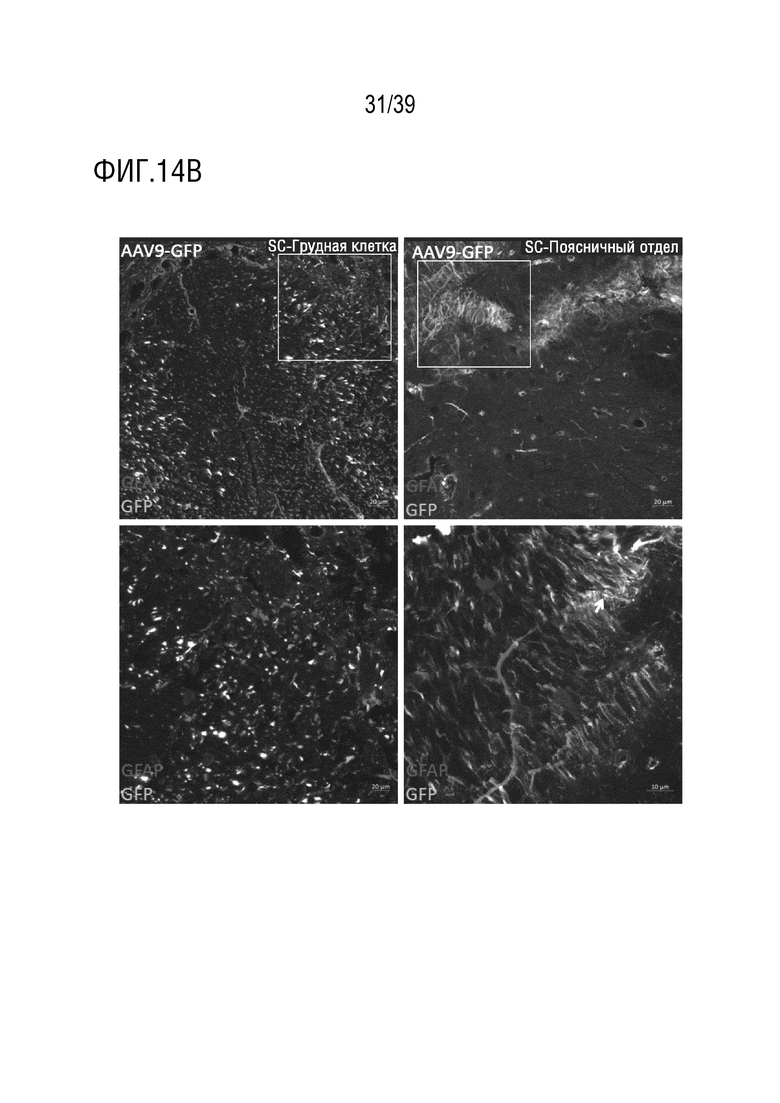




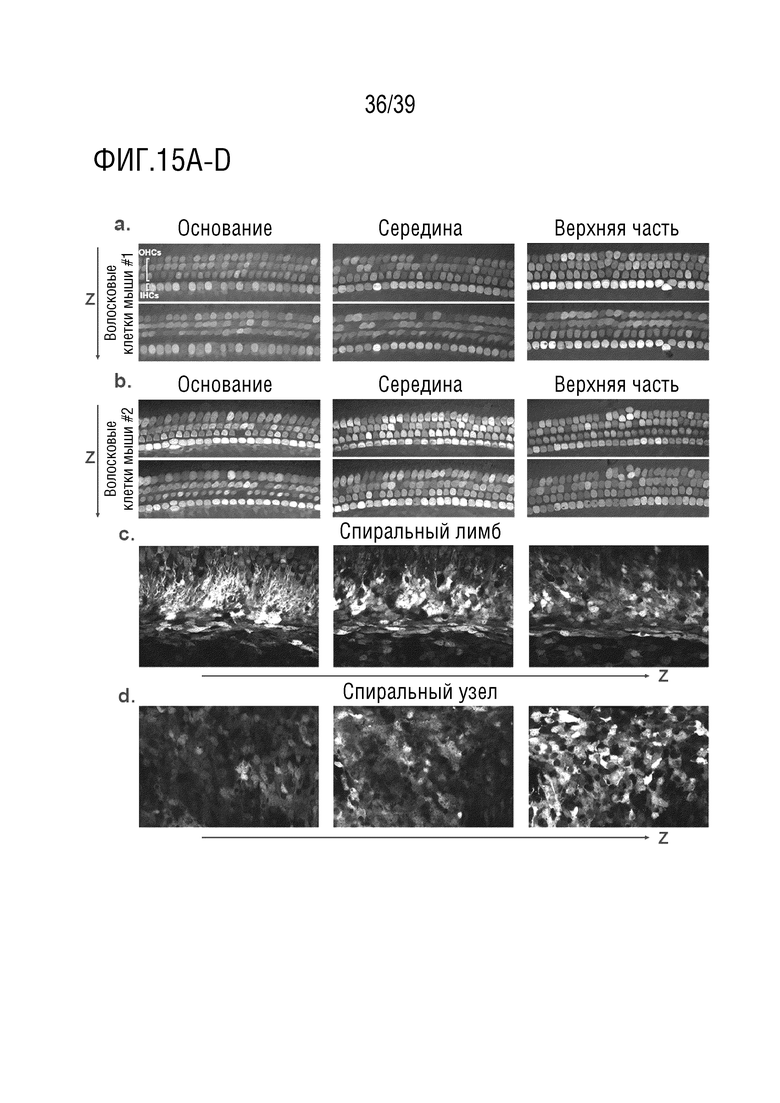

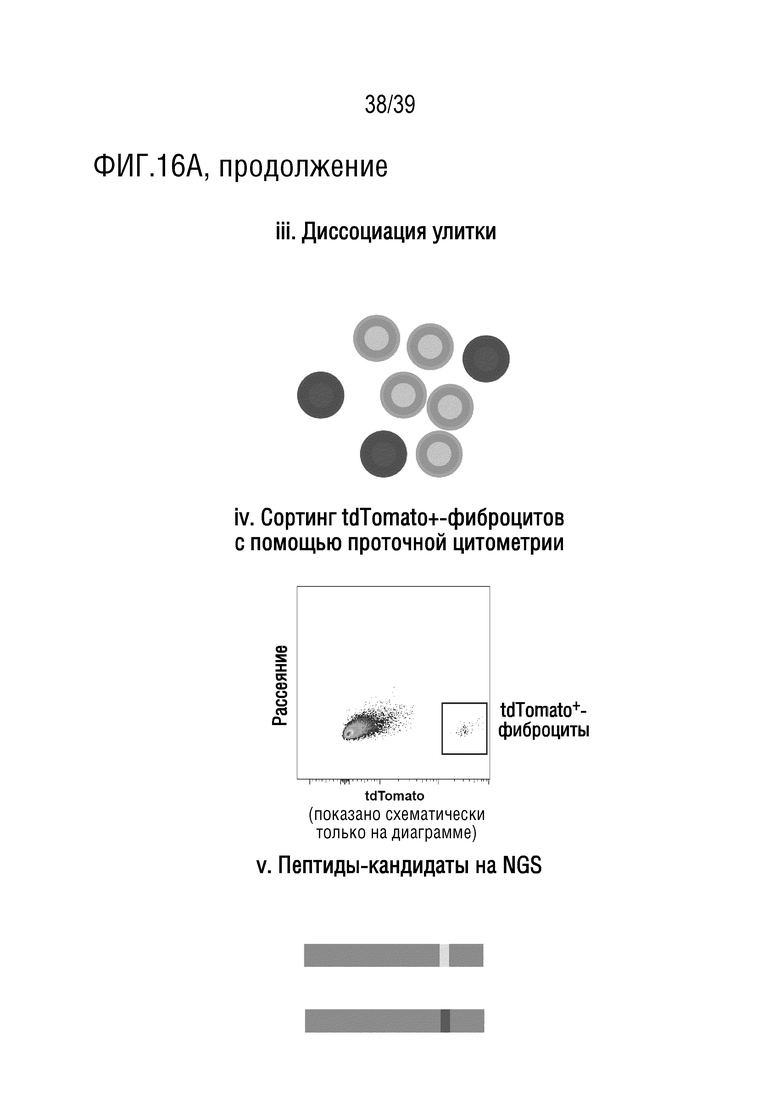
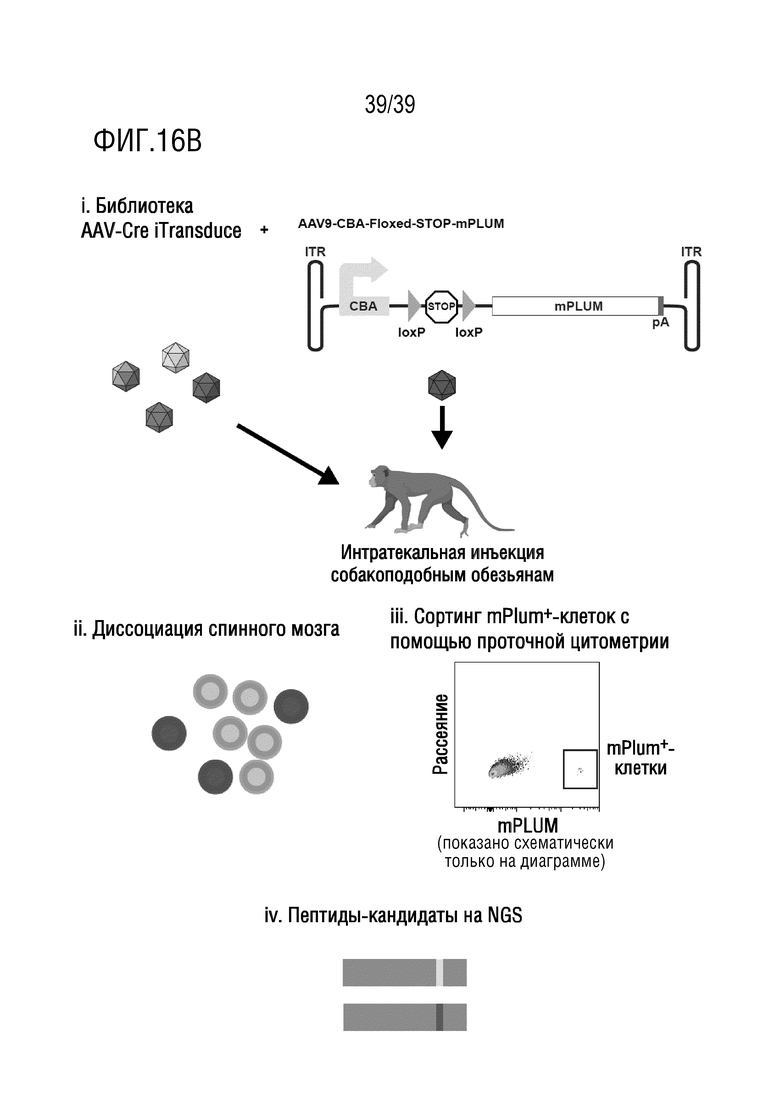
Изобретение относится к биотехнологии. Описан капсидный белок AAV для использования в векторе AAV, где капсидный белок AAV содержит аминокислотную последовательность, которая содержит по меньшей мере семь смежных аминокислот последовательности STTLYSP (SEQ ID NO:1). Представлена нуклеиновая кислота, кодирующая описанный капсидный белок AAV. Раскрыт вектор AAV для использования при доставке нуклеиновой кислоты в клетку, где вектор AAV содержит описанный капсидный белок AAV. Представлен способ доставки трансгена в клетку, где способ включает приведение клетки в контакт с раскрытым вектором AAV. Изобретение позволяет экспрессировать трансген, например, в ЦНС, ПНС, во внутреннем ухе, в сердце или в сетчатке. Также изобретение позволяет экспрессировать трансген в клетках нужных типов. 4 н. и 18 з.п. ф-лы, 9 ил., 4 табл., 6 пр.
1. Капсидный белок AAV для использования в векторе AAV, где капсидный белок AAV содержит аминокислотную последовательность, которая содержит по меньшей мере семь смежных аминокислот последовательности STTLYSP (SEQ ID NO:1).
2. Капсидный белок AAV по п.1, где AAV представляет собой AAV9.
3. Капсидный белок AAV по п.1 или 2, содержащий VP1 AAV9.
4. Капсидный белок AAV по п.3, где последовательность, которая содержит по меньшей мере семь последовательных аминокислот последовательности STTLYSP (SEQ ID NO:1), встроена в положение, соответствующее аминокислотам 588 и 589 последовательности SEQ ID NO:6.
5. Нуклеиновая кислота, кодирующая капсидный белок AAV по любому из пп.1-4.
6. Вектор AAV для использования при доставке нуклеиновой кислоты в клетку, где вектор AAV содержит капсидный белок AAV по любому из пп.1-4.
7. Вектор AAV по п.6, где AAV не содержит капсидный белок VP1, VP2 или VP3 дикого типа.
8. Вектор AAV по п.6 или 7, дополнительно содержащий трансген.
9. Вектор AAV по п.8, где трансген представляет собой терапевтический трансген.
10. Способ доставки трансгена в клетку, где способ включает приведение клетки в контакт с вектором AAV по п.8 или 9.
11. Способ по п.10, где клетка представляет собой нейрон, кардиомиоцит, миоцит, астроцит, глиальную клетку, внутреннюю волосковую клетку, внешнюю волосковую клетку, поддерживающую клетку, фиброцит внутреннего уха, фоторецепторную клетку, интернейрон, ганглиональную клетку сетчатки или клетку пигментного эпителия сетчатки.
12. Способ по п.11, где нейрон является нейроном спинномозговых ганглиев или нейроном спиральных ганглиев.
13. Способ по любому из пп.10-12, где клетка присутствует у живого индивидуума.
14. Способ по п.13, где индивидуум является млекопитающим.
15. Способ по п.13 или 14, где клетка присутствует в ткани, выбранной из ткани головного мозга, спинного мозга, спинномозговых ганглиев, сердца, внутреннего уха, глаза, мышцы, и их комбинаций.
16. Способ по любому из пп.13-15, где индивидуум страдает болезнью Альцгеймера; болезнью Паркинсона; Х-сцепленной адренолейкодистрофией; болезнью Канавана; болезнью Нимана-Пика; спинальной мышечной атрофией; болезнью Гентингтона; болезнью, ассоциированной с коннексином-26; болезнью Ашера типа 3A; болезнью Ашера типа 2D; потерей слуха, ассоциированной с волосковыми клетками; потерей слуха, ассоциированной с волосковыми клетками (DFNB7/11); потерей слуха, ассоциированной с внутренними волосковыми клетками (DFNB9); болезнью Ашера типа 1F; болезнью Ашера типа 1B; пигментным ретинитом (RP; несиндромным); врожденным амаврозом Лебера; наследственной невропатией зрительного нерва Лебера; синдромом Ашера (RP; синдромом, ассоциированным с глухотой); мышечной дистрофией Дюшенна; васкулопатией после аллотрансплантации; гемофилией А; или гемофилией В.
17. Способ по любому из пп.13-16, где клетка находится в головном мозге индивидуума, и AAV вводят посредством парентеральной доставки, интрацеребральной доставки или интратекальной доставки.
18. Способ по п.17, где интратекальную доставку осуществляют путем поясничной инъекции, интрацистернальной инъекции или интрапаренхимальной инъекции.
19. Способ по любому из пп.10-17, где AAV вводят парентерально.
20. Способ по п.19, где AAV добавляют внутривенно, внутриартериально, подкожно, внутрибрюшинно или внутримышечно.
21. Способ по любому из пп.13-16, где клетка находится в глазу индивидуума, и AAV вводят путем субретинальной инъекции или инъекции в стекловидное тело.
22. Способ по любому из пп.13-16, где клетка находится во внутреннем ухе индивидуума, и AAV вводят в улитку путем нанесения на мембрану круглого окна или через нее, через хирургически просверленную кохлеостому, расположенную рядом с круглым окном, через отверстие в костном овальном окне или через полукруглый канал.
| WO 2018022608 A2, 01.02.2018 | |||
| ВИРИОНЫ АДЕНОАССОЦИИРОВАННОГО ВИРУСА С ВАРИАНТНЫМ КАПСИДОМ И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2611202C2 |
| RU 2018128780 A, 05.12.2018. | |||

