Изобретение относится к медицинской вирусологии и может быть использовано в практическом здравоохранении для профилактики заболеваемости гриппом и пневмонии, вызванной Streptococcus pneumoniae.
Грипп - острое респираторное заболевание, вызванное вирусами гриппа, характеризующееся высокой заболеваемостью и смертностью среди населения, что определяет его высокую социально-экономическую значимость. Ежегодно вирусы гриппа вызывают эпидемии, а раз в 10-40 лет - пандемии гриппа, что связано с высокой антигенной изменчивостью вируса [https://www.who.intbulletin/volumes/90/4/12-020412/ru/].
Для человека основную опасность представляют вирусы гриппа A/H1N1, A/H3N2 и В, однако в последние годы штаммы вирусов гриппа подтипов A/H7N9 и A/H5N1 стали представлять потенциальную угрозу возникновения пандемии, поскольку некоторые представители данных сероподтипов преодолели видовой барьер и вызывали вспышки заболеваемости в различных уголках мира [Jonathan S. Nguyen-Van-Tam, Chloe Sellwood. Preparing for a potential A(H7N9) pandemic: lessons from the deployment of A(H1N1) pandemic vaccines. Expert Rev. Vaccines 12(8), 825-828 (2013)].
Зачастую грипп сопровождается вторичной бактериальной пневмонией, чаще всего вызываемой Staphylococcus aureus и Streptococcus pneumoniae. В настоящее время известно, что вирусные инфекции, проявляющиеся одновременно с бактериальной пневмонией, встречаются с частотой 30-50% как среди взрослых, так и среди детей. Все больше данных свидетельствуют о том, что бактериальные инфекции, сопутствующие вирусу гриппа, связаны с более высокой заболеваемостью и более высокой смертностью [Influenza virus-related critical illness: Pathophysiology and epidemiology December 2019 Critical Care 23(1) DOI: 10.1186/s13054-019-2539-х]. Существуют данные, указывающие на то, что во время коинфекции гриппа и пневмококка у человека бактериальное носительство и передача пневмококка увеличиваются [F Ansaldi, D De Florentiis, V Parodi, E Rappazzo, M Coppelli, M Martini, С Alicino, P Durando, G Icardi. Bacterial carriage and respiratory tract infections in subjects > or=60 years during an influenza season: implications for the epidemiology of community acquired pneumonia and influenza vaccine effectiveness. J. Prev. Med. Hyg. 2012 Jun; 53(2): 94-7.]
Вторичная бактериальная пневмония приводит к критическому состоянию, при котором лечение антибиотиками не всегда дает положительный результат. Так, например, во время пандемии вируса гриппа 2009 года смертность среди тяжелобольных с вторичной пневмонией (14-46%) оставалась высокой (95-99%), несмотря на использование рекомендованных β-лактамных антибиотиков [Gustavo Palacios, Mady Hornig., Danie Cisterna, Nazir Savji, Ana Valeria Bussetti, Vishal Kapoor, Jeffrey Hui, Rafal Tokarz, Thomas Briese, Elsa Baumeister, W. Ian Lipkin. Streptococcus pneumoniae Coinfection Is Correlated with the Severity of H1N1 Pandemic Influenza. PLoS One. 2009 Dec 31; 4(12):e8540. doi: 10.1371/journal.pone.0008540].
Разрушение стенки грамположительной бактерии S.pneumoniae под действием таких антибиотиков приводит к высвобождению провоспалительных бактериальных компонентов, усугубляя течение инфекции [
 Sarah М. Heston, Kelli L. Boyd, Elaine I. Tuomanen, Jonathan A. McCullers. Toll-Like Receptor 2 Mediates Fatal Immunopathology in Mice During Treatment of Secondary Pneumococcal Pneumonia Following Influenza. The Journal of Infectious Diseases. 2011 Nov 204(9); 1358-1366. https://doi.org/10.1093/infdis/jir5221.
Sarah М. Heston, Kelli L. Boyd, Elaine I. Tuomanen, Jonathan A. McCullers. Toll-Like Receptor 2 Mediates Fatal Immunopathology in Mice During Treatment of Secondary Pneumococcal Pneumonia Following Influenza. The Journal of Infectious Diseases. 2011 Nov 204(9); 1358-1366. https://doi.org/10.1093/infdis/jir5221.
В России для профилактики пневмококковой инфекции рекомендованы полисахаридные вакцины «Превенар 13», «Синфлорикс», «Пневмо-23». Вакцинация современными полисахаридными вакцинами значительно снижает заболеваемость пневмококком, однако в последние годы появилась тревожная тенденция к увеличению заболеваемости пневмококковыми инфекциями, вызванными штаммами, не входящими в состав вакцины [Sam Mehr, Nicholas Wood. Streptococcus pneumoniae - a review of carriage, infection, serotype replacement and vaccination. J. Paediatric Respiratory Reviews. Dec. 2012 13 (4); 258-264]. Также появились исследования пневмококковой конъюгированной вакцины PCV7, демонстрирующие, что она не снижает заболеваемость во время распространения вируса гриппа и не облегчает тяжесть протекания пневмонии. В то же время вакцинация живыми аттенуированными гриппозными вакцинами может способствовать значительному снижению количества бактериальных заболеваний во время эпидемий гриппа [Michael J. Mina, Keith P. Klugman, Jonathan A. McCullers. Live attenuated influenza vaccine, but not pneumococcal conjugate vaccine, protects against increased density and duration of pneumococcal carriage after influenza infection in pneumococcal colonized mice. J. of infectious diseases, 208(8), October 2013, 1281-1285].
Все вышеизложенное свидетельствует о необходимости разработки комбинированной вакцины против гриппа и пневмонии, вызванной Streptococcus pneumoniae, которая позволит совместить сезонную вакцинацию против гриппа с вакцинацией против сопутствующей пневмококковой инфекции. Такая комбинированная вакцина будет иметь ряд преимуществ. Во-первых, в последнее время среди населения нарастет недоверие к вакцинам и внедрение еще одной дополнительной может привести к добровольным отказам от иммунизации. Вакцинация же хорошо изученной живой гриппозной вакциной с новым защитным свойством может, наоборот, положительно сказаться на отношении населения к вакцинации. Во-вторых, иммунизация такой комбинированной вакциной поможет не только снизить вызванную вирусом гриппа заболеваемость, но и защитить от его возможных осложнений. В-третьих, во время сезонной эпидемии в человеческой популяции не исключено появление вирусов гриппа, антигенно отличающихся от входящих в состав живой гриппозной вакцины вакцинных штаммов, что может снизить эффективность профилактики, но позволит уменьшить тяжесть осложнений и смертность, вызванные вторичной бактериальной инфекцией.
Для разработки комбинированной вакцины необходимо использовать в качестве антигена пневмококка иммуногенный стабильный консервативный белок, что позволит индуцировать протективные уровни антител для борьбы с пневмококковой инфекцией. Наиболее привлекательной мишенью для представления иммунной системе является поверхностный белок S.pneumoniae PspA. Данный белок пневмококка имеет консервативные области [М.J. Jedrzejas. Unveiling molecular mechanisms of bacterial surface proteins: Streptococcus pneumoniae as a model organism for structural studies. J. Cell Mol Life Sci, 64 (21), Nov 2007, 2799-2822] и выполняет, по крайней мере, две функции в вирулентности патогена. PspA является лактоферрин-связывающим белком. Он конкурентно связывается с лактоферрином (hLf) -железо-связывающим гликопротеином, преобладающим в секрете слизистых оболочек человека. После связывания PspA-hLf высвобождается железо, достаточный уровень которого на поверхности слизистых оболочек необходим для роста бактерий [S Hammerschmidt 1, G Bethe, Р Н Remane, G S Chhatwal. Identification of pneumococcal surface protein A as a lactoferrin-binding protein of Streptococcus pneumoniae. J. Infect Immun. Apr 1999, 67(4), 1683-7.]. Кроме того, N-домен белка PspA отрицательно заряжен и выступает на поверхности бактериальной клетки, препятствуя взаимодействию системы комплемента с пневмококком.
Задачей, на решение которой направлено заявляемое изобретение, является получение реассортантного вакцинного штамма для производства комбинированной вакцины против вируса гриппа А и бактериальной пневмонии, вызываемой Streptococcus pneumoniae.
Цель заявляемого изобретения - получить вакцинный штамм вируса гриппа, содержащий модифицированный ген, кодирующий поверхностный белок гемагглютинин, несущий в себе фрагмент белка Streptococcus pneumoniae PspA. Остальные гены соответствуют классическому реассортантному штамму аттенуированной живой гриппозной вакцины, подготовленной на базе отечественного донора аттенуации А/Ленинград/134/17/57.
Получение комбинированного вакцинного штамма. Комбинированный вакцинный штамм ИнфлюБакт-HV/PspA был получен методом обратной генетики, формула генома которого (6:2) соответствует требованиям, предъявляемым к штаммам живой гриппозной вакцины: гены, кодирующие поверхностные белки гемагглютинин (НА) и нейраминидазу (NA), принадлежат потенциально-пандемическому вирусу А/Ануи/1/2013 (H7N9), а гены, кодирующие внутренние белки (РВ2, РВ1, PA, NP, М, NS), принадлежат донору аттенуации А/Ленинград/134/17/57 (H2N2) (табл 1). При этом ген, кодирующий поверхностный белок НА, модифицирован: после сигнального пептида в ген включен фрагмент бактериального гена PspA (160-262 a.o. gb|EHD89266.1| pneumococcal surface protein A [Streptococcus pneumoniae]).

Бактериальная вставка была встроена между НА1 субъединицей НА и сигнальным пептидом SP через гибкие линкеры (AAAPGAA). Так, после трансляции и сборки вирусной частицы, бактериальный пептид PspA экспонирован снаружи вируса, представляя себя клеткам иммунной системы. Гибкие линкеры не нарушают пространственную конфигурацию бактериальной вставки и гемагглютинина.
На фиг. 1 (фрагмент А) представлена пространственная конфигурация молекулы гемагглютинина с бактериальной вставкой PspA. Показан один мономер. Линкер - AAAPGAA. На фрагменте Б показана структурная схема модифицированного белка гемагглютинина НА с бактериальной вставкой PspA. SP - сигнальный пептид, НА1/2 - первая и вторая субъединицы гемагглютинина.
Получение химерного гена гемагглютинина. Этап 1: Подготовка бактериальной составляющей. Фрагмент гена белка пневмококка PspA был любезно предоставлен проф. А.Н. Суворовым (отдел молекулярной микробиологии, ФГБНУ «ИЭМ») в составе экспрессионного вектора рЕТ-24а. С помощью перекрывающейся ПЦР на первом этапе с N и С концов от бактериального фрагмента были внесены гибкие линкеры AAAPGAA с помощью специально подобранных праймеров (табл. 2). Затем слева и справа от линкеров были достроены BsmBI сайты рестрикции (также с помощью перекрывающейся ПЦР).

Этап 2: Подготовка вирусной составляющей. Плазмидная ДНК на основе вектора pCIPolISapIT, кодирующая ген НА, была разработана в отделе вирусологии ФГБНУ «ИЭМ» и ранее была депонирована в коллекцию отдела [Е.А. Stepanova, Т.S. Kotomina, V.A. Matyushenko, Т.A. Smolonogina, V.S. Shapovalova, L.G. Rudenko, and I.N. Isakova-Sivak. Amino Acid Substitutions N123D and N149D in Hemagglutinin Molecule Enhance Immunigenicity of Live Attenuated Influenza H7N9 Vaccine Strain in Experiment. Bulletin of Experimental Biology and Medicine, 2019 March, 166(5), doi:10.1007/s10517-019-04407-1].
В ходе двух реакций ПЦР со специальными праймерами (табл. 3) с С-конца сигнального пептида и N-конца субъединицы НА1 были внесены сайты рестрикции BsmBI. Далее эти два фрагмента были объединены в общей ПЦР с прямым и обратным праймерами для получения полноразмерного гена НА.
Цикл для полимеразной цепной реакции состоял из следующих этапов: 1) Тотальная денатурация ДНК 98°С (30 с) → вход в цикл 2) денатурация ДНК 98°С (10 с) → 3) отжиг праймеров 55°С (30 с) → 4) элонгация одноцепочечной ДНК 72°С (1:30 с) → конец цикла, обратно к шагу 2 (35 циклов) → 5) инактивация полимеразы 72°С (10 мин) → 4°С (хранение). ПЦР проводили с помощью набора Phusion Green High-Fidelity DNA Polymerase (Thermo Scientific) согласно инструкциям производителя.
После каждой реакции ПЦР амплифицированные фрагменты подвергали электрофоретическому разделению в 1% агарозном геле с бромистым этидием, в трис-ацетатном буфер (ТАЕ) в течение 40-50 минут. Все специфические продукты, соответствующие по размеру заданным фрагментам, вырезали из геля и очищали с помощью набора для выделения и очистки ДНК из агарозного геля и реакционных смесей (Cleanup Mini, Евроген, Россия) и секвенировали по методу Сенгера.
С помощью реакции рестрикции и лигирования по BsmBI сайту (CGTCTC(1/5)) фрагмент гена PspA был внесен в ген НА. Для рестрикции использовали эндонуклеазу Esp3I (Fast Digest, Thermo, США). Рестрикцию проводили в течение двух часов при рабочей температуре 37°С, после чего инактивировали фермент в течение 10 минут при 65°С. Рестрикционную смесь очищали от ферментов и буфера с помощью набора Cleanup Mini (Евроген). Лигирование проводили с помощью Т4 ДНК лигазы (Escherichia virus Т4) в течение 1 часа при 22°С.
Далее химерный ген гемагглютинина с бактериальной вставкой H7-PspA (аминокислотная последовательность SEQ ID No:1) встраивали в вектор для обратной генетики вируса гриппа (pCIPolISapIT) для последующей сборки нового комбинированного штамма вируса гриппа. Для этого к 3° и 5° концам химерного гена H7-PspAc помощью перекрывающейся ПЦР вносили сайты рестрикции SapI (табл. 4). Цикл для полимеразной реакции использовали аналогичный описанному выше. Электрофорез для очистки и концентрации амплифицированного гена с внесенными сайтами рестрикциями проводили в стандартных условиях, приведенных выше. Затем проводили рестрикцию вектора pCIPolISapIT и химерного гена H7-PspA по SapI сайтам рестрикции (GCTCTTC(1/4)), в течение 20 минут при 37°С, после чего фермент инактивировали при 65°С в течение 10 мин. Очистку рестрикционной смеси и лигирование проводили аналогично описанному выше способу. Таким образом, была получена плазмида на основе вектора pCIPolISapIT с включенным в нее химерным геном гемагглютинина H7-PspA.
Накопление плазмидной ДНК. Плазмидная ДНК несущая химерный ген гемагглютинина H7-PspA после последнего этапа лигирования была введена в компетентные клетки E.coli путем трансформации тепловым шоком. После трансформации, бактерии были высеяны на чашку Петри с 1% агарозой, питательной средой LB-Medium и 0.1% ампициллином и инкубированы при 37°С в течение 16 ч. Далее с помощью скрининга бактериальных колоний методом ПЦР были отобраны клоны с химерным геном H7-PspA. Отобранные варианты были помещены в жидкую среду и накоплены в течение 16 часов в LB-medim среде при 37°С и 200 rpm/мин. Затем плазмидная ДНК была выделена из культуры E.coli с помощью набора Plasmid Midiprep 2.0 согласно протоколу производителя.
Итоговая концентрация ДНК составила 437 мкг/мл. Вставка в плазмиду была полностью секвенирована, чтобы убедиться в том, что полученный клон содержит целевую последовательность H7-PspA, и что эта последовательность не содержит мутаций, которые могли бы возникнуть при амплификации и клонировании.
Сборку комбинированного вакцинного штамма ИнфлюБакт-H7/PspA осуществляли методом трансфекции клеток Vero необходимым набором плазмидных ДНК. Для этого были подготовлены 8 плазмидных ДНК на основе вектора pCIPolISapIT, содержащих гены донора аттенуации А/Ленинград/134/17/57(H2N2): РВ2, РВ1, PA, NP, M,NS, ген NА от вирулентного штамма А/Ануи/1/2013 (H7N9) (плазмиды разработаны в отделе вирусологии ФГБНУ «ИЭМ») и модифицированный ген гемагглютинина PspA-H7. С помощью электропорации клеток Vero с использованием системы Neon Transfection System (Invitrogen) в режиме двух импульсов по 1150 вольт в течение 20 мс, плазмиды были доставлены в клетки. Клетки после трансфекции инкубировали в течение 6 часов в среде OptiPRO (Gibco) с добавлением 1× Glutamax (Gibco), 5% эмбриональной бычьей сыворотки, при температуре 37°С в атмосфере 5% СО2. Спустя 6 часов среду меняли на бессывороточную среду Opti PRO (Gibco), содержащую 1× Glutamax, 1× антибиотик-антимикотик (Gibco) и 4,0 мкг/мл трипсина, при 33°С и 5% СО2. Спустя 3 дня инкубации клетки были сняты с подложки, ресуспендированы в ростовой среде. Данной трансфекционной смесью были заражены развивающиеся куриные эмбрионы, которые затем были инкубированы в течение 72 часов при 33°С. Наличие вируса в аллантоисной жидкости определяли по реакции гемагглютинации с куриными эритроцитами. Все гены собранного вируса были секвенированы на наличие мутаций и соответствие комбинированному вакцинному штаммуИнфлюБакт-Н7/PspA.
Накопление стока вируса. Сток был накоплен в развивающихся куриных эмбрионах (РКЭ). Для этого РКЭ были заражены 10-кратными разведениями комбинированного вакцинного вируса ИнфлюБакт-Н7/PspA, собранного после трансфекции. Спустя 72 ч инкубации при 33°С, каждый эмбрион был вскрыт, протитрован 2-кратными разведениями с куриными эритроцитами. Вируссодержащая аллантоисная жидкость с наибольшими титрами по реакции гемагглютинации собиралась для подготовки рабочего стока. Объединенный сток был осветлен центрифугированием при 3×103 rpm, 15 минут, 4°С. Вирус был разлит в аликвоты по 1 мл и хранился при -70°С.
Антигенная характеристика вакцинного штамма. Изучение антигенных свойств вакцинного штамма ИнфлюБакт-Н7/PspA показало, что:
- ответственный за антигенную специфичность поверхностный белок вакцинного штамма - гемагглютинин (НА) - в РТГА с гомологичной сывороткой антигенно идентичен потенциально-пандемическому вирусу A/Ануи/1/2013(H7N9), поскольку показатели взаимодействия реассортанта ИнфлюБакт-Н7/PspA и родительского штамма А/17/Ануи/2013/61 (H7N9), подготовленному на основе А/Ануи/1/2013 (H7N9), с крысиной сывороткой к вирусу А/Ануи/1/2013 (H7N9), полностью совпадали (табл. 5). Антигенная идентичность НА вирулентному штамму H7N9 также подтверждалась полногеномным секвенированием.
- второй ответственный за антигенную специфичность поверхностный белок вакцинного штамма - нейраминидаза (NA) - проверен методом полногеномного секвенирования и идентичен вирусу А/Ануи/1/2013 (H7N9).

Генетическую стабильность бактериальной составляющей комбинированного вакцинного штамма ИнфлюБакт-H7/PspA изучали путем определения сохранности химерной генетической конструкции H7-PspA в течение 10 пассажей на развивающихся куриных эмбрионах с использованием аналитического ПЦР-анализа после каждого пассажа, а также секвенирования последнего пассажа. Показано, что данная химерная генетическая конструкция H7-PspA является генетически стабильной, что подтверждено результатами секвенированием 10-го пассажа гемагглютинина комбинированного вакцинного штамма ИнфлюБакт-Н7/PspA.
Генетическую стабильность кодирующих мутаций вакцинного штамма ИнфлюБакт-Н7/PspA изучали сравнением сохранности кодирующих мутаций до и после пятикратного пассирования вакцинного штамма в куриных эмбрионах с использованием метода частичного секвенирования. Показано, что нуклеотидная последовательность вакцинного штамма ИнфлюБакт-Н7/PspA до и после его десятикратного пассирования, осталась неизменной, доказательством чего служат результаты секвенирования, приведенные в таблице 6.
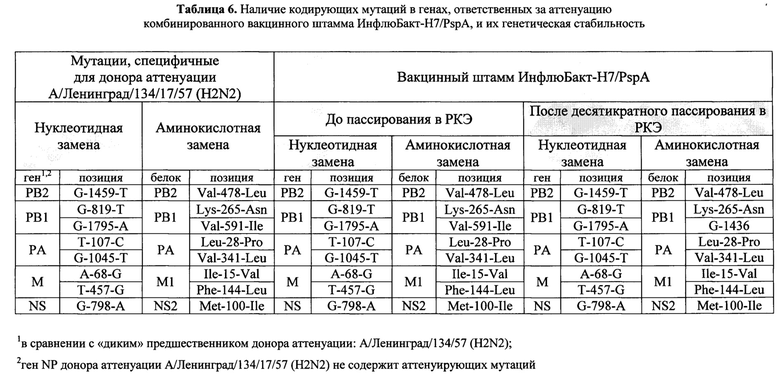
Оценку фенотипических свойств комбинированного вакцинного штамма ИнфлюБакт-Н7/PspA проводили путем его параллельного титрования в РКЭ при разных температурах. Комбинированный вакцинный штамм ИнфлюБакт-Н7/PspA в развивающихся куриных эмбрионах обладает выраженным температурочувствительным и холодоадаптированным фенотипом, идентичным фенотипу донора аттенуации А/Ленинград/134/17/57 (H2N2).
Установлено, что вакцинный вирус является температурочувствительным (ts фенотип) - его инфекционная активность при температуре 38°С составила 2.0±0.6 lg ЭИД50/мл, и холодоадаптированным (са фенотип) - инфекционная активность при пониженной до 26°С температуре инкубации достигала 4,9±0,5 lg ЭИД50/мл, что свидетельствует о его безвредности для человека, поскольку по этим показателям он идентичен донору аттенуации А/Ленинград/134/17/57 (H2N2).
Было показано, что комбинированный вакцинный штамм, несущий бактериальную вставку, имеет незначительно сниженный титр при всех температурах инкубации по сравнению с классическим штаммом с формулой генома 6:2 без вставки А/17/Ануи/2013/61 (H7N9). Результаты фенотипического анализа представлены в таблице 7.
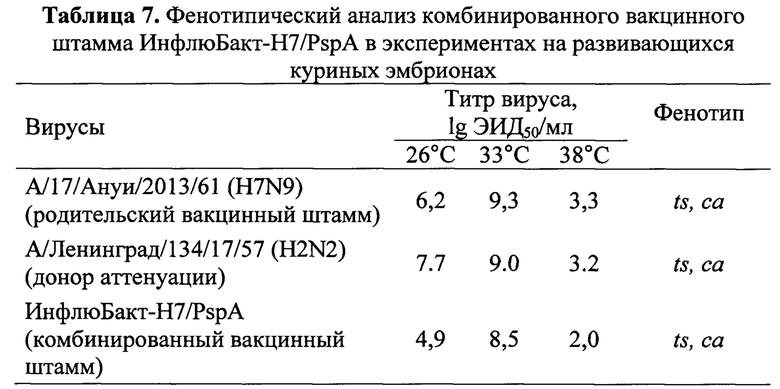
Безвредность для мышей. Доклинические исследования острой токсичности вакцинного штамма ИнфлюБакт-Н7/PspA проводили на белых беспородных мышах в соответствии с действующими правилами [Методические рекомендации «Доклинические испытания эффективности и безопасности новых иммунобиологических лекарственных препаратов». М.: 2010. 39 с; Правила надлежащей лабораторной практики. Приказ Минздрава РФ №199н от 01.04.2016. https://rg.ru/2016/09/02/minzdrav-prikaz199-site-dok.html].
Мышам вводили однократно одну человеческую дозу, соответствующую 0.5 мл, вакцинного вируса внутрибрюшинно в дозе 7.0 lg ЭИД50/мл. Животным контрольной группы вводили внутрибрюшинно физиологический раствор. Ежедневно в течение всего исследования (10 дней) проводился контроль общего состояния каждого животного.
Данные физиологического исследования (подвижность, поведенческие реакции, изменение массы тела) показали, что внутрибрюшинное введение вакцинного штамма не вызывало гибели экспериментальных животных (табл. 8) и не приводило к изменению их массы тела (фиг. 2), внешнего вида, поведения, не отражалось на потреблении ими пищи и воды, что свидетельствует о безвредности вакцинного препарата.

Формирование специфических IgG антител после иммунизации мышей комбинированной вакциной ИнфлюБакт-H7/PspA. Мыши линии BALB/c были иммунизированы двумя дозами комбинированной вакциной ИнфлюБакт-Н7/PspAc интервалом в 21 день. Спустя 3 недели после второй дозы, у мышей ретроорбитально были собраны сыворотки для определения титра и специфичности IgG антител методом иммуноферментного анализа. Было показано, что новый комбинированный вакцинный штамм индуцирует вирус-специфические сывороточные антитела к антигенам вируса гриппа, сопоставимо с классическим вакцинным штаммом А/17/Ануи/2013/61 (H7N9).
Кроме того, в сыворотках мышей, иммунизированных комбинированной вакциной ИнфлюБакт-Н7/PspA, обнаружены специфические IgG антитела к гибридному белку пневмококка PspA-Spr1895-PsaA (табл. 9) [Суворов А.Н., Духовлинов И.В., Орлов А.И., Байгузин Е.Я. Вакцина против пневмонии, вызываемой Streptococcus Pneumoniae, на основе гибридного белка. Патент РФ №2510281, опубл. 27.03.2014].

Кроме того, специфические к вирусу гриппа антитела были обнаружены в сыворотках крови иммунизированных мышей по реакции торможения гемагглютинации (РТГА). Данный метод позволяет определить титр антигемагглютинирующих антител и основан на явлении отсутствия реакции гемагглютинации при добавлении к вирусу специфической сыворотки. По результатам РТГА титр антигемагглютинирующих антител к вирусу гриппа А подтипа H7N9 в группе мышей, иммунизированных комбинированной вакциной ИнфлюБакт-Н7/PspA, был сопоставим с животными, получившими классическую вакцину А/17/Ануи/2013/61 (H7N9) (табл. 10).

Защитная эффективность комбинированной вакцины ИнфлюБакт-H7/PspA в отношении вируса гриппа и пневмококка. Мыши линии BALB/c после двукратной иммунизации были подвержены вирусно-бактериальной инфекции. В качестве патогенных вирусов для проверки защитной эффективности использовали реассортантный штамм А/Шанхай/2/2013 (H7N9) CDC-RG, содержащий поверхностные антигены H7N9, а остальные гены - от адаптированного к мышам вируса A/PR8/34, либо реассортантный штамм A/Philippines/2/1982 (Х-79) (H3N2), содержащий поверхностные антигены H3N2, а остальные гены - от адаптированного к мышам вируса A/PR8/34, в дозе 1×105 ЭИД50. На третий день после заражения вирусом гриппа мыши подвергались инфицированию S.pneumoniae 3 серотипа (штамм 73) в дозе 6,0×104 КОЕ/мышь. Защитную эффективность оценивали по изменению массы тела иммунизированных мышей и выживаемости. Двукратная иммунизация комбинированной вакциной ИнфлюБакт-Н7/PspA частично защищает мышей от вирусно-бактериальной суперинфекции: как при инфекции гомологичном вирусом подтипа H7N9, так и при использовании гетерологичного вируса подтипа H3N2. Выживаемость в группе, иммунизированной комбинированной вакциной ИнфлюБакт-H7/PspA, составила 40%, тогда как выживаемость животных из контрольных групп (PBS и группа классической вакцины А/17/Ануи/2013/61 (H7N9)) составила 0%. На фиг. 3 представлены результаты выживаемости и изменения массы тела мышей при заражении (челлендже) вирусом гриппа подтипа H7N9 либо H3N2 и пневмококком S.pneumoniae 3 серотип (штамм 73). Группа H7N9 6:2 - получившая классическую ЖГВ с формулой генома 6:2А/17/Ануи/2013/61 (H7N9); группа ИнфлюБакт-Н7/PspA-комбинированную вакцину; группа контроля - получившая PBS.
При заражении только вирусом подтипа H7N9 - гомологичным вакцинному вирусу - выживаемость мышей, получивших комбинированную вакцину ИнфлюБакт-H7/PspA, составила 100%, как и в группе, иммунизированной классической ЖГВ А/17/Ануи/2013/61 (H7N9). На фиг. 4 - результаты выживаемости при заражении (челлендже) вирусом гриппа А/Шанхай/2/2013 (H7N9) CDC-RGV1/Е2 мышей, иммунизированных комбинированной вакциной ИнфлюБакт-Н7/PspA, классической ЖГВ А/17/Ануи/2013/61 (H7N9)h неиммунизированных мышей из группы контроля.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА RN9/13-HUMAN A(H6N9) ДЛЯ ОПРЕДЕЛЕНИЯ АНТИТЕЛ К НЕЙРАМИНИДАЗЕ ПРИ ГРИППОЗНОЙ ИНФЕКЦИИ И ВАКЦИНАЦИИ | 2014 |
|
RU2587629C1 |
| Рекомбинантный вакцинный штамм для живой интраназальной вакцины, обеспечивающей сочетанную профилактику гриппозной и коронавирусной инфекций | 2022 |
|
RU2782531C1 |
| ШТАММ ВИРУСА ГРИППА А/17/Ануи/2013/61 (H7N9) ДЛЯ ПРОИЗВОДСТВА ЖИВОЙ ИНТРАНАЗАЛЬНОЙ ГРИППОЗНОЙ ВАКЦИНЫ | 2013 |
|
RU2563351C2 |
| Живая вакцина на основе штамма пробиотиков ENTEROCOCCUS FAECIUM L3 для профилактики инфекции, вызванной STREPTOCOCCUS PNEUMONIE | 2018 |
|
RU2701733C1 |
| ВАКЦИНА ПРОТИВ ПНЕВМОНИИ, ВЫЗЫВАЕМОЙ Streptococcus pneumoniae, НА ОСНОВЕ ГИБРИДНОГО БЕЛКА | 2012 |
|
RU2510281C2 |
| Рекомбинантная плазмидная ДНК pSp-raPLY, кодирующая синтез рекомбинантного атоксичного белка пневмолизина Streptococcus pneumoniae, штамм Escherichia coli M15/pSp-raPLY - продуцент рекомбинантной атоксичной формы пневмолизина и получение указанного белка для разработки вакцинного препарата для профилактики пневмококковых инфекций | 2021 |
|
RU2782598C1 |
| Штамм вируса гриппа A/Shanghai/HK/6:2/2013 (H7N9) для получения инактивированных и живых гриппозных вакцин | 2017 |
|
RU2664460C1 |
| Вакцинный штамм вируса гриппа A/17/Гонконг/2017/75108 (H7N9) для производства потенциально пандемической живой интраназальной гриппозной вакцины | 2018 |
|
RU2685121C1 |
| РЕАССОРТАНТНЫЙ ШТАММ ВИРУСА ГРИППА RN2/57-HUMAN A(H7N2) ДЛЯ ОПРЕДЕЛЕНИЯ АНТИТЕЛ К НЕЙРАМИНИДАЗЕ ПРИ ГРИППОЗНОЙ ИНФЕКЦИИ И ВАКЦИНАЦИИ | 2011 |
|
RU2464312C1 |
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |


Изобретение относится к вирусологии. Описан комбинированный вакцинный штамм ИнфлюБакт-H7/PspA - реассортант, формула генома 6:2 которого соответствует требованиям, предъявляемым к штаммам живой гриппозной вакцины: гены, кодирующие поверхностные белки гемагглютинин (НА) и нейраминидазу (NA), принадлежат потенциально-пандемическому вирусу А/Ануи/1/2013 (H7N9), а гены, кодирующие внутренние белки (РВ2, РВ1, PA, NP, М, NS), принадлежат донору аттенуации А/Ленинград/134/17/57 (H2N2), полученному методом обратной генетики. При этом ген, кодирующий поверхностный белок НА, модифицирован: после сигнального пептида в ген встроен фрагмент бактериального гена PspA (160-262 а.о. gb|EHD89266.1| pneumococcal surface protein A [Streptococcus pneumoniae]). Изобретение может быть использовано в практическом здравоохранении для профилактики заболеваемости гриппом и пневмонии, вызванной Streptococcus pneumoniae. Комбинированный вакцинный штамм ИнфлюБакт-Н7/PspA активно размножается в развивающихся куриных эмбрионах при оптимальной температуре 32°С, характеризуется температурочувствительностью, холодоадаптированностью и безвредностью для лабораторных животных. Обладает высокой генетической стабильностью и иммуногенен как против вируса гриппа, так и против пневмококка. 4 ил., 10 табл.
Штамм вируса гриппа ИнфлюБакт-Н7/PspA для производства комбинированной вакцины против вируса гриппа А и бактериальной пневмонии, вызываемой Streptococcus pneumoniae, полученный на основе штамма А/Ленинград/134/17/57, содержащий модифицированный ген, в который после сигнального пептида включен фрагмент бактериального гена PspA.
| Смолоногина Т | |||
| А., Исакова-Сивак И | |||
| Н | |||
| и др | |||
| Конструирование векторной вакцины на основе холодоадаптированного вируса гриппа для защиты от бактериальной инфекции, вызываемой стрептококками группы В | |||
| Молекулярная генетика, микробиология и вирусология, 2019, том 37, номер 1, стр.25-34 | |||
| CN 110691611 A, 14.01.2020 | |||
| CN 104428417 A, 18.03.2015 | |||
| EA |

