УРОВЕНЬ ТЕХНИКИ
Область техники
Настоящее изобретение относится к области биологии и, в частности, относится к способу оценки токсинообразующей способности афлатоксигенного штамма.
Описание предшествующего уровня техники
Афлатоксин представляет собой наиболее токсичный микотоксин, обнаруженный к настоящему времени. Если в качестве примера взять афлатоксин В1, то его токсичность в 10 раз превышает таковую для цианида калия и в 68 раз превышает таковую для мышьяка. Афлатоксин классифицируется Международной онкологической организацией как канцероген класса I. Афлатоксин может легко загрязнить зерновые продукты и маслопродукты, такие как арахис, кукурузу, рис и т.д., а также многие растительные продукты, такие как грецкие орехи, фисташки и сырье для китайской медицины. Наблюдалось много случаев отравления людей и скота афлатоксином в Китае и других странах. Согласно последнему отчету Международного агентства по изучению рака (IARC), только в развивающихся странах приблизительно 500 миллионов человек подвергаются риску воздействия афлатоксина. Китай является регионом с высоким уровнем загрязнения афлатоксином. Результаты учета, проводимого Министерством сельского хозяйства в течение многих лет, показывают, что заражение афлатоксином основных сельскохозяйственных продуктов в Китае постепенно увеличивается, а содержание токсинов на сильно загрязненных территориях превышает предельное содержание в сотни раз. Хотя высокотоксичные микотоксигенные штаммы составляют менее 20%, они стали серьезной скрытой опасностью, угрожающей качеству и безопасности продукции растениеводства.
Афлатоксин в основном вырабатывается такими грибами, как Aspergillus flavus и Aspergillus parasiticus. Исследования показали, что способность различных штаммов Aspergillus flavus вырабатывать афлатоксин, то есть их токсинообразующая способность, может различаться в сотни раз. Штаммы, обладающие токсинообразующей способностью, являются основным источником высокого загрязнения. Однако до сих пор отсутствует эффективный способ успешного выявления штаммов, обладающих токсинообразующей способностью, с применением специфичности. В настоящее время существуют, главным образом, два способа оценки токсинообразующей способности штаммов Aspergillus flavus: один заключается в оценке токсинообразующей способности штаммов путем измерения выхода афлатоксина, вырабатываемого штаммами после некоторого периода времени культивирования штамма. Во-первых, способ этого типа занимает много времени, поскольку штаммы необходимо выделить, а затем культивировать, и затем проводить оценку путем измерения содержания афлатоксина. А во-вторых, на биосинтез афлатоксина влияет множество сложных факторов. Кроме того, выход афлатоксина сильно варьируется от одной партии к другой партии одних и тех же штаммов, и результат токсинообразующей способности не подходит для использования при описании характеристик выхода, присущих штамму, что влияет на точность и надежность результатов этого способа в отношении выявления и оценки. Способ другого типа представляет собой оценку токсинообразующей способности штаммов путем измерения транскрипционной активности генов, связанных с токсинообразующей способностью. Существует литература, посвященная выявлению гена Nor-1 для оценки токсинообразующей способности. Однако недостатком способа этого типа является то, что в естественном состоянии имеют место дефекты в генах токсигенности, отличных от гена Nor-1, что приводит к тому, что даже в случае выявления экспрессии гена Nor-1 афлатоксин не образуется в силу его свойств, а это ведет к ложным результатам таких способов.
Вкратце, объективная и точная оценка токсинообразующей способности афлатоксигенного штамма представляют собой нерешенную проблему для практикующих специалистов во всем мире.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Ввиду недостатков предшествующего уровня техники настоящее изобретение направлено на предоставление способа оценки токсинообразующей способности афлатоксигенного штамма.
Для достижения цели настоящего изобретения техническое решение, принятое согласно настоящему изобретению, представляет собой следующее.
Способ оценки токсинообразующей способности афлатоксигенного штамма включает следующие стадии, предусматривающие измерение выхода афлатоксина и транскрипционной активности гена Nor-1 штаммов Aspergillus flavus для получения отношения выхода афлатоксина к транскрипционной активности гена Nor-1, оценку токсинообразующей способности афлатоксигенного штамма в зависимости от отношения выхода афлатоксина к транскрипционной активности гена Nor-1.
Согласно изложенному выше решению способ измерения выхода афлатоксина включает следующие стадии: культивирование штамма Aspergillus flavus, сбор спор Aspergillus flavus для культивирования со встряхиванием, после культивирования фильтрацию с получением фильтрата и измерение концентрации афлатоксина в фильтрате.
Согласно изложенному выше решению среда, используемая для культивирования штамма Aspergillus flavus, представляет собой среду CDA, а условия культивирования являются следующими: культивирование при 28°C и влажности 90% в течение 10 дней.
Среда, используемая для встряхиваемой культуры спор Aspergillus flavus, представляет собой жидкую среду с картофельной декстрозой, а условия культивирования являются следующими: культивирование со встряхиванием при 28°C и 200 об/мин в течение 96 часов.
Согласно изложенному выше решению применяют способ иммуноаффинной очистки в сочетании с высокоэффективной жидкостной хроматографией для измерения концентрации афлатоксина в фильтрате после культивирования со встряхиванием спор Aspergillus flavus.
Согласно изложенному выше решению способ измерения транскрипционной активности гена Nor-1 включает следующие стадии: культивирование штамма Aspergillus flavus, сбор спор Aspergillus flavus для культивирования со встряхиванием, после завершения культивирования фильтрацию гифов Aspergillus flavus и получение высушенных микроорганизмов в результате сушки, а затем используют способ измерения транскрипционной активности гена Nor-1 для измерения транскрипционной активности гена Nor-1 в высушенных микроорганизмах.
Согласно изложенному выше решению показатели выхода афлатоксина и транскрипционной активности гена Nor-1 также можно получить посредством способа одновременного выявления на основе RT-PCR. Конкретные стадии включают в себя следующее.
(1) Построение S-образной кривой для количественного определения афлатоксина: на стадии иммунной реакции, если количество нанесенного в качестве покрытия моноклонального антитела к афлатоксину является постоянным, используют препараты стандарта афлатоксина для конкуренции с фагом с представленным на поверхности антиидиотипическим наноантителом к афлатоксину за связывание с моноклональным антителом к афлатоксину. После завершения иммунной реакции фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, связанным с моноклональным антителом к афлатоксину, элюируют. Препараты стандарта афлатоксина соответствуют разному количеству элюированного фага. Из содержащегося в элюате фага высвобождаются молекулы ДНК во время нагревания в ходе ПЦР, и при этом высвободившиеся молекулы ДНК используют в качестве мишеней для амплификации в реакции RT-PCR. Каждый элюат подвергают реакции амплификации в RT-PCR с использованием набора, и после завершения реакции амплификации получают разные значения Ct. Логарифм концентрации афлатоксина используют в качестве абсциссы, а значение Ct используют в качестве ординаты, и для получения S-образной кривой для количественного определения афлатоксина выполняют регрессионный анализ.
(2) Построение кривой RT-PCR для транскрипционной активности гена Nor-1: серийно разводят образцы с известным числом копий фрагмента ДНК гена Nor-1 Tq-nor1 с получением разного числа копий, а затем используют набор для проведения реакции RT-PCR-амплификации для каждого числа копий Tq-nor1 отдельно. После реакции амплификации получают разные значения Ct. Логарифм числа копий Tq-nor1 используют в качестве абсциссы, а значение Ct используют в качестве ординаты для выполнения регрессионного анализа с получением при этом кривой для количественного определения транскрипции гена Nor-1.
(3) Осуществление культивирования штамма Aspergillus flavus с получением спор Aspergillus flavus, сбор спор Aspergillus flavus для культивирования со встряхиванием. После завершения культивирования фильтруют раствор для культивирования штамма от мицелия грибов Aspergillus flavus. После разбавления раствора для культивирования штамма в некотором соотношении используют его для замены препаратов стандарта афлатоксина в иммунной реакции, проводимой на стадии (1), для осуществления реакции, основанной на иммунной конкуренции. После конкурентной реакции фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, связанным с моноклональным антителом к афлатоксину, элюируют, и молекулы ДНК, высвобождаемые содержащимися в элюате фагами, используют в качестве матрицы амплификации для количественного определения афлатоксина в реакции одновременной RT-PCR-амплификации. Кроме того, после высушивания мицелия грибов Aspergillus flavus общую РНК выделяют и подвергают обратной транскрипции с получением cDNA, и cDNA разбавляют в некотором соотношении и используют в качестве матрицы для амплификации гена Nor-1 в реакции одновременной RT-PCR-амплификации.
(4) Бактериофаг V2-5 и cDNA используют в качестве матриц для проведения реакции одновременной RT-PCR-амплификации. После реакции амплификации получают два значения Ct, и эти два значения Ct соответственно сопоставляют с S-образной кривой для количественного определения афлатоксина и кривой для количественного определения транскрипции гена Nor-1, при этом выполняют преобразование для получения показателей концентрации афлатоксина и транскрипционной активности гена Nor-1 с определением отношения выхода афлатоксина к транскрипционной активности гена Nor-1.
Согласно изложенному выше решению фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину представляет собой фаг VHH 2-5, а моноклональное антитело к афлатоксину представляет собой моноклональное антитело к афлатоксину 1С11.
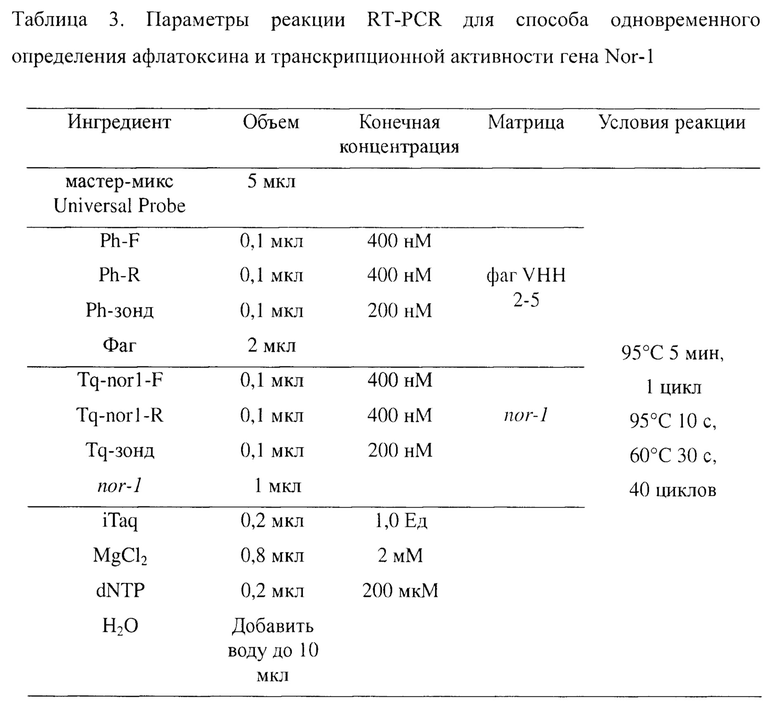
Согласно изложенному выше решению в реакционной системе, предусматривающей реакцию одновременной RT-PCR-амплификации, конечная концентрация прямых и обратных праймеров Ph-F, Ph-R, Tq-nor1-F и Tq-nor1-R составляет от 300 нМ до 400 нМ; флуоресцентный зонд: конечная концентрация Ph-зонда и Tq-зонда составляет от 200 нМ до 400 нМ; конечная доза ДНК-полимеразы: от 0,5 Ед до 1,0 Ед; конечная концентрация MgCl2: от 1 мМ до 2 мМ; конечная концентрация dNTP: от 200 мкМ до 400 мкМ.
Согласно изложенному выше решению нуклеотидная последовательность прямого праймера Ph-F представлена под SEQ ID N0:1; нуклеотидная последовательность обратного праймера Ph-R представлена под SEQ ID NO: 2; нуклеотидная последовательность Ph-зонда, представляющего собой флуоресцентный зонд, представлена под SEQ ID NO: 3; нуклеотидная последовательность прямого праймера Tq-nor1-F представлена под SEQ ID NO: 4; нуклеотидная последовательность обратного праймера Tq-nor1-R представлена под SEQ ID NO: 5; нуклеотидная последовательность Tq-зонда, представляющего собой флуоресцентный зонд, представлена под SEQ ID NO: 6.
Согласно изложенному выше решению реакционная система, предусматривающая реакцию одновременной RT-PCR-амплификации, включает 5 мкл мастер-микса Universal Probe для qPCR, 0,1 мкл Ph-F, 0,1 мкл Ph-R, 0,1 мкл Ph-зонда, 2 мкл фаговой матрицы, 0,1 мкл Tq-nor1-F, 0,1 мкл Tq-nor1-R, 0,1 мкл Tq-зонда, 1 мкл матрицы гена Nor-1, 0,2 мкл ДНК-полимеразы, 0,8 мкл MgCl2, 0,2 мкл dNTP, при этом добавляют Н2О для доведения до 10 мкл.
Согласно изложенному выше решению условия реакции одновременной RT-PCR-амплификации являются следующими: 95°C, 5 мин; 95°C, 10 с, 60°C, 30 с, 40 циклов.
Согласно изложенному выше решению диапазон концентраций афлатоксина на S-образной кривой для количественного определения афлатоксина составляет от 33,33 нг/мл до 1,69 пг/мл, и нижний предел выявления (LOD) афлатоксина составляет 0,018 нг/мл. На кривой для количественного определения транскрипции гена Nor-1 число копий гена Nor-1 находится в диапазоне от 102 до 108.
Согласно изложенному выше решению токсинообразующую способность афлатоксигенного штамма обозначают как Y, и отношение выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1) обозначают как X. Формула для оценки токсинообразующей способности Aspergillus flavus представляет собой Y=10,14Х - 16,20. Для высокотоксигенного штамма токсинообразующая способность составляет >150 нг/мл; умеренная токсинообразующая способность: 50< токсинообразующая способность <150 нг/мл; низкая токсинообразующая способность соответствует токсинообразующей способности <50 нг/мл; отсутствие выработки токсина: 0. Указанные выше значения подставляют в формулу для расчета диапазона оценки токсинообразующей способности, причем AFT/Nor-1 > 16,4 указывает на высокотоксигенный штамм; 6,5 < AFT/Nor-1 < 16,4 указывает на умеренно токсигенный штамм; 0<AFT/Nor-1 < 6,5 указывает на низкотоксигенный штамм; AFT/Nor-1=0 указывает на нетоксигенный штамм.
Полезный эффект настоящего изобретения заключается в следующем.
(1) В исследовании согласно настоящему изобретению было доказано, что отношение выхода афлатоксина к транскрипционной активности гена Nor-1 характеризуется высокой относительной стабильностью. Путем построения модели оценки токсинообразующей способности штаммов Aspergillus flavus можно получить уравнение регрессии для отношения токсинообразующей способности Aspergillus flavus и выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1). Путем определения отношения AFT/Nor-1 можно быстро и точно осуществлять оценку токсинообразующей способности штаммов Aspergillus flavus, что имеет большое значение для раннего предупреждения, предотвращения и контроля загрязнения афлатоксином.
(2) В настоящем изобретении предусматривается способ одновременного обнаружения на основе RT-PCR для одновременного выявления токсинообразующей способности штаммов Aspergillus flavus и транскрипционной активности гена Nor-1. Существует четкая линейная взаимосвязь между токсинообразующей способностью штаммов Aspergillus flavus и транскрипционной активностью гена Nor-1, полученными при помощи способа одновременного выявления на основе RT-PCR согласно настоящему изобретению, при этом количественное определение афлатоксина проводили с помощью ВЭЖХ, а также проводили количественное определение транскрипционной активности гена Nor-1 при помощи Nanodrop. Следовательно, результат, полученный для отношения AFT/Nor-1 (токсинообразующая способность штамма Aspergillus flavus/транскрипционная активность гена Nor-1), полученного с использованием способа одновременного выявления на основе RT-PCR, является надежным и точным и может использоваться в качестве показателя идентификации для оценки токсинообразующей способности штамма Aspergillus flavus.
(3) В способе одновременного выявления на основе RT-PCR токсинообразующей способности штамма Aspergillus flavus и транскрипционной активности гена Nor-1, описанном в данном документе, необходимое количество реагентов меньше и стоимость ниже, и, следовательно, можно достичь высокой производительности выявления. Способ одновременного выявления на основе RT-PCR обеспечивает упрощение режима анализа, оптимизацию процедуры и структуры эксперимента, а также обеспечивает платформу для выявления и теоретическую основу для одновременного анализа афлатоксина и других низкомолекулярных соединений, связанных с путем его синтеза.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
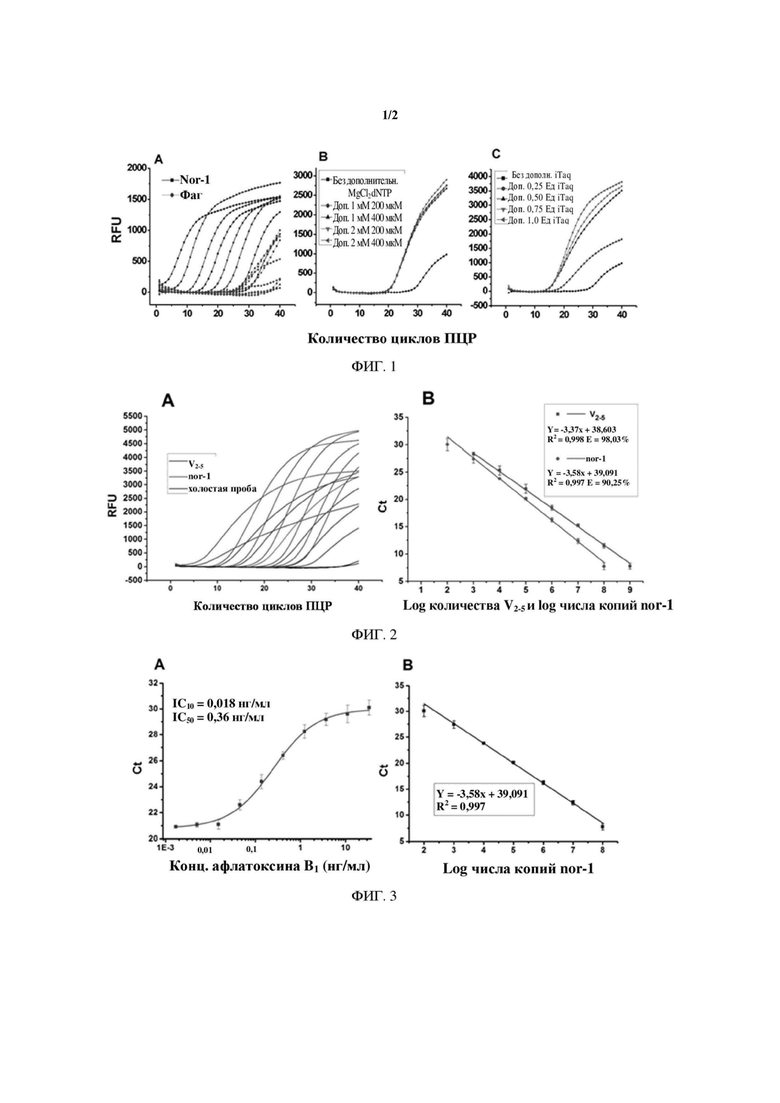
На ФИГ. 1 показана оптимизация концентрации ДНК-полимеразы, dNTP и MgCl2 в одновременной реакции RT-PCR.
На ФИГ. 2 показана оценка эффективности одновременной RT-PCR-амплификации.
ФИГ. 3 представляет собой стандартную кривую количественного анализа для количественного определения афлатоксина В1 и транскрипции гена Nor-1 посредством способа на основе одновременной RT-PCR.
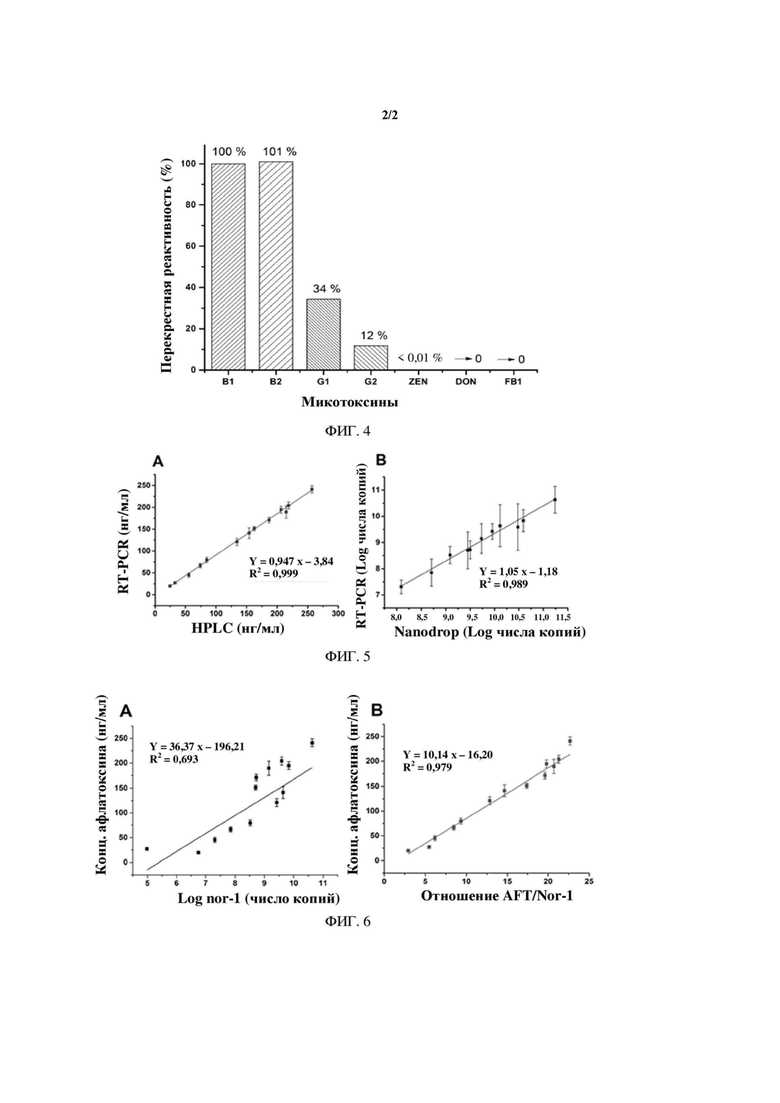
На ФИГ. 4 показана перекрестная реактивность при количественном определении в одновременной RT-PCR для афлатоксина B1, В2, G1, G2, ZEN, DON, FBI.
На ФИГ. 5 показано сравнение результатов количественного определения посредством одновременной RT-PCR, ВЭЖХ и Nanodrop.
На ФИГ. 6 показана корреляционная взаимосвязь (А) между выходом афлатоксина и величиной экспрессии гена Nor-1, а также корреляционная взаимосвязь (В) между выходом афлатоксина и отношением AFT/Nor-1.
ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Для того чтобы сделать настоящее изобретение более доступным для понимания, содержание настоящего изобретения будет дополнительно разъяснено ниже в связи с вариантами осуществления, но при этом содержание настоящего изобретения не ограничено следующими вариантами осуществления.
Вариант осуществления 1
1. В настоящем исследовании доказано, что отношение выхода афлатоксина к транскрипционной активности гена Nor-1 характеризуется высокой относительной стабильностью.
Взвешивают 1 г NaNO3, 1 г K2HPO4, 0,5 г MgSO4⋅7H2O, 0,5 г KCl, 0,01 г FeSO4, 30 г глюкозы и 20 г порошкообразного агара. Разбавляют их деионизированной водой до общего объема 1000 мл и стерилизуют их высокотемпературным паром при 121°C в течение 30 мин для получения среды CDA. Штамм Aspergillus flavus культивируют на твердой среде CDA при 28°C и влажности 90% в течение 10 дней, а затем культуральный планшет промывают 20% Tween-20 для получения раствора со спорами Aspergillus flavus. Используют способ подсчета при помощи гемоцитометра, раствор спор Aspergillus flavus равномерно встряхивают с помощью вихревого встряхивателя и споры Aspergillus flavus в растворе подсчитывают под микроскопом.
В колбу Эрленмейера объемом 50 мл помещают 15 мл жидкой среды с картофельной декстрозой, автоклавируют жидкую среду с картофельной декстрозой при 121°C в течение 30 минут. После этого на основании результатов подсчета добавляют раствор со спорами Aspergillus flavus до тех пор, пока конечная концентрация не составит 1×105 спор на мл, и культивируют со встряхиванием при 28°C и 200 об/мин в течение 96 ч. Культуральный бульон фильтруют с помощью двухслойной фильтровальной бумаги с получением фильтрата (сохраняют для последующего использования) и мицелия грибов Aspergillus flavus. Для получения мицелия грибов Aspergillus flavus отжимают лишнюю воду фильтровальной бумагой и высушивают в сушильной камере при 65°C в течение 12 часов для получения высушенных микроорганизмов. После охлаждения до комнатной температуры хранят при -70°C для последующего использования. Фильтрат, полученный при фильтровании, хранят при 4°C для последующего использования.
Для измерения концентрации афлатоксина в фильтрате (хранящемся для последующего использования, как описано выше) после культивирования со встряхиванием спор Aspergillus flavus применяют стандартный способ иммуноаффинной очистки в сочетании с высокоэффективной жидкостной хроматографией, а затем для измерения транскрипционной активности гена Nor-1 в высушенных микроорганизмах, хранящихся для последующего использования, применяют обычный способ измерения транскрипционной активности гена Nor-1.
С применением той же процедуры, что описана выше, семь штаммов Aspergillus flavus культивируют в разные периоды времени до и после тестирования двух партий. Результаты измерения показаны в таблице 1.
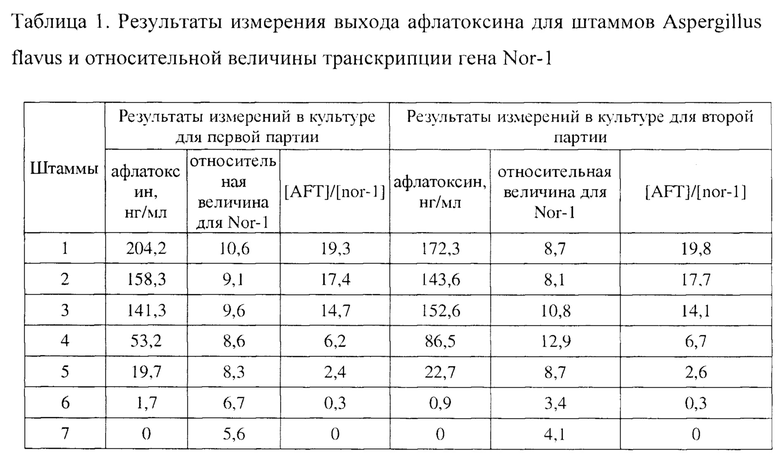
Согласно результатам измерений в таблице 1 наблюдается большая разница между концентрацией афлатоксина в двух партиях, и поэтому выход афлатоксина сам по себе не может служить для оценки токсинообразующей способности штаммов Aspergillus flavus. В некоторых случаях, когда относительная величина транскрипции гена Nor-1 для штаммов Aspergillus flavus является высокой, токсинообразующая способность оказывается неожиданно низкой. Кроме того, нетоксигенные штаммы также могут транскрибировать ген Nor-1, и, следовательно, одна только величина транскрипции гена Nor-1 не может служить для оценки токсинообразующей способности штаммов Aspergillus flavus. В таблице 1 в связи с отношением, полученным путем деления концентрации афлатоксина на относительную величину транскрипции гена Nor-1, то есть значением [AFT]/[nor-1] в таблице 1, обнаруживается строгая закономерность. Являются сопоставимыми не только порядки величин токсинообразующей способности 7 штаммов, полученных из двух партий культуры, но также значение [AFT]/[nor-1] для каждого штамма является относительно стабильным и, таким образом, позволяет оценить токсинообразующую способность штаммов Aspergillus flavus.
Из данных в таблице 1 следует, что если для оценки токсинообразующей способности штаммов Aspergillus flavus применяют отношение концентрации афлатоксина к относительной величине транскрипции гена Nor-1, точность и надежность такого подхода значительно лучше, чем у подхода с использованием только концентрации афлатоксина или подхода с использованием только относительной величины транскрипции гена Nor-1.
Вариант осуществления 2. Разработка способа одновременного выявления на основе RT-PCR для получения показателей выхода афлатоксина и транскрипционной активности гена Nor-1
Используют известное антиидиотипическое наноантитело к афлатоксину, представленное на поверхности фага VHH 2-5, и фрагмент ДНК гена Nor-1, Tq-nor1, для разработки способа одновременного выявления на основе RT-PCR выхода афлатоксина и транскрипционной активности гена Nor-1. Приведенный выше результат выявления используется в качестве научной основы для определения и оценки токсинообразующей способности штаммов Aspergillus flavus, обеспечивая аргументацию способа быстрого определения и оценки токсинообразующей способности штаммов Aspergillus flavus. Конкретные стадии являются следующими.
Представленное на поверхности фага VHH 2-5 антиидиотипическое наноантитело к афлатоксину разработано Центром контроля качества Института исследований масличных культур Китайской академии сельскохозяйственных наук и опубликовано в журнале «Yanru Wang; Peiwu Li; Zuzana Majkova; Candace RS Bever; Нее Too Kim; Qi Zhang; Julie E. Dechant; Shirley J. Gee; Bruce D. Hammock; Isolation of Alpaca Anti-Idiotypic Heavy-Chain Single-Domain Antibody for the Aflatoxin Immunoassay; Analytical Chemistry, 2013, 8298-8303». Фаг, используемый в этом примере, предварительно хранят в Е. coli ER2738 в лаборатории и получают путем амплификации. Способ амплификации является следующим.
Отбирают случайным образом одну колонию из планшета с отдельными колониями ER2738, содержащими фаг VHH 2-5 с наноантителами, вносят ее в жидкую среду, содержащую 1 мл SB-ампициллина, и инкубируют в течение ночи при 225 об/мин во встряхивателе с постоянной температурой при 37°C. Добавляют вышеуказанный образованный в течение ночи культуральный бульон к 100 мл жидкой среды с SB-ампициллином при 225 об/мин и 37°C и культивируют его до OD600=0,5-0,6. Добавляют 1,5 мл фага-помощника М13К07 (титр 1×1011 - 1×1012 БОЕ/мл) в раствор культивируемых бактерий и оставляют при 37°C в течение 30 мин. Добавляют канамицин с конечной концентрацией 70 мкг/мл в бульон при 37°C и 225 об/мин и встряхивают культуру в течение ночи. Полученный в течение ночи культуральный бульон центрифугируют при 4°C и 10000 об/мин в течение 15 минут, отбирают супернатант и переносят его в чистую пробирку для центрифугирования. Добавляют 1/4 объема PEG/NaCl и оставляют выдерживаться на льду в течение 2 часов. Центрифугируют при 10000 об/мин в течение 30 минут при 4°C, повторно суспендируют в 2 мл 0,5% BSA/PBS, а затем центрифугируют при 12000 об/мин в течение 5 минут. Супернатант отбирают, отфильтровывают его с помощью фильтра с размером пор 0,22 мкм и смешивают его с равным объемом стерильного глицерина, отбирают аликвоту и измеряют титр. Способ измерения титра является следующим.
Случайным образом отбирают одну колонию из планшета с отдельными колониями ER2738, инокулируют ее в жидкую среду LB, содержащую 0,04 мг/мл тетрациклина, культивируют в течение ночи во встряхивателе с постоянной температурой при 37°C и 225 об/мин. Отбирают 20 мкл вышеупомянутого бульона, который культивировали в течение ночи, и добавляют его к 2 мл среды SB. Его инкубируют при 225 об/мин и 37°C до OD600≈1. Для постепенного разбавления фага, титр которого необходимо проверить, используют жидкую среду LB. Отбирают 10 мкл каждого разведения и добавляют его к 100 мкл бульона ER2738 с OD600≈1 и оставляют его без изменения при 37°C в течение 20 минут. Осуществляют инфицирование фагом Е. coli; распределяют бульон с инфицированной Е. coli на чашке с LB-ампициллином, помещают ее вертикально в инкубатор с постоянной температурой 37°C на 30 минут, а затем переворачивают культуру на ночь. Отбирают приблизительно 100 отдельных колоний для подсчета на чашке и рассчитывают титр фага по следующей формуле:

Способ получения фрагмента ДНК гена Nor-1, Tq-nor1. В колбу Эрленмейера объемом 50 мл помещают 15 мл жидкой среды с картофельной декстрозой, автоклавируют ее при 121°C в течение 30 минут и добавляют раствор со спорами штамма N73 Aspergillus flavus до конечной концентрации 1×105 мл-1. После культивирования со встряхиванием при 28°C, 200 об/мин в течение 96 часов используют двухслойную фильтровальную бумагу для выжимания раствора для культивирования из мицелия грибов, высушивают его при 65°C в течение 12 часов, быстро замораживают жидким азотом и измельчают в порошок. Точно отвешивают 200 мг порошка гифов, используют набор для выделения ДНК (DNeasy Plant Mini Kit) для выделения геномной ДНК штамма N73 в соответствии с прилагаемыми к набору инструкциями. Используют следующие праймеры (последовательность Nor1-F представлена под SEQIDNO:7; последовательность Nor1-R представлена под SEQIDNO:8) для амплификации продукта фрагмента гена Nor-1 размером 400 п.о. и используют набор для выделения из геля E.Z.N.A.T.M (OMEGA) для очистки фрагмента ДНК размером 400 п.о. в соответствии с инструкциями для получения Tq-nor1.
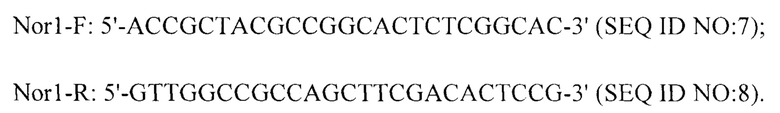
1. Разработка способа одновременного выявления на основе RT-PCR для выхода афлатоксина и транскрипционной активности гена Nor-1
1. Разработка последовательностей праймеров и зондов для одновременного выявления на основе RT-PCR выхода афлатоксина и транскрипционной активности гена Nor-1
Фрагмент ДНК Tq-nor1 фрагмента ДНК и ген Nor-1, высвобождаемый фагом VHH 2-5 с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, выбирают в качестве матрицы для одновременной амплификации на основе RT-PCR. Определяют отношение выхода афлатоксина к транскрипционной активности гена Nor-1 с помощью результатов одновременной амплификации на основе RT-PCR.
Исходя из последовательности, кодирующей наноантитело, и последовательности гена Nor-1 фага VHH 2-5 с представленным на поверхности антиидиотипическим наноантителом, получают последовательность прямого и обратного праймеров (Ph-F, Ph-R) для афлатоксина и зонда (Ph-зонда), а также последовательность прямого и обратного праймеров (Tq-nor1-F, Tq-nor1-R) для Tq-nor-1 и зонда (Tq-зонда). Затем для анализа праймеров используют программное обеспечение Oligo7.0 для выполнения проверки в соответствии с принципами и руководствами для разработки праймеров и зондов для дуплексной реакции RT-PCR.
Принцип проверки праймеров и зонда. Значение ТМ для всех праймеров должно быть установлено на один и тот же или аналогичный уровень, а значение ТМ для всех зондов должно быть как можно более схожим и превышать значение ТМ для праймера приблизительно на 5-10°C. Поскольку реакция является одновременной реакцией в одной и той же системе, необходимо убедиться, что все праймеры и зонды не будут образовывать димеры. Выполняют поиск с помощью BLAST с целью удостовериться, что праймеры и зонды специфичны для предполагаемой мишени.
Было установлено, что следующие последовательности праймеров и зондов соответствуют указанным выше принципам и требованиям, как подытожено в таблице 2.

2. Параметры реакции способа одновременного выявления на основе RT-PCR для определения выхода афлатоксина и транскрипционной активности гена Nor-1
Параметры реакции одновременной RT-PCR отличаются от параметров реакции RT-PCR, в которой проводят амплификацию одного гена, поскольку необходимо тщательно учитывать взаимодействие между реакционными системами. Во-первых, с помощью RT-PCR амплифицируют два одноцепочечных специфичных фрагмента ДНК: фрагмент фага с антиидиотипическим наноантителом к афлатоксину и фрагмент ДНК гена Nor-1. Компоненты для реакции смешивают непосредственно, без добавления других компонентов, а затем проводят двойную реакцию RT-PCR, результат которой показан как изображение А на ФИГ. 1. Из фигуры следует, что амплификация молекул ДНК фага VHH 2-5 в значительной степени подавляется. В дуплексной реакции RT-PCR эффективность амплификации и целевая последовательность различных амплифицированных продуктов могут быть разными. Амплификация образцов с низкой эффективностью амплификации или с низким содержанием последовательностей-мишеней может подавляться высокоамплифицируемым образцом или целевой последовательностью с более высоким содержанием.
По этой причине в настоящем изобретении оптимизируют условия реакции RT-PCR для способа одновременного выявления выхода афлатоксина и транскрипционной активности гена Nor-1. В настоящем изобретении оптимизированы значения концентрации ДНК-пол им ер азы, MgCl2 и dNTP. Результат, показанный как изображение В на ФИГ. 1, демонстрирует, что если концентрация фага VHH 2-5 составляет 106 БОЕ/мл, пороговое значение циклов Ct уменьшается после добавления dNTP и MgCl2, и кривая амплификации появляется при более низком значении количества циклов. Таким образом, показано, что амплификация молекул ДНК фага VHH 2-5 улучшается. На изображении С на ФИГ. 1 показано, что когда количество ДНК-полимеразы повышают с 0,25 Ед до 1,0 Ед, эффективность амплификации фага также в значительной степени улучшается. Следовательно, исходя из принципа, что чем раньше достигается пороговое значение циклов Ct, тем ближе кривая амплификации к S-образной форме в экспоненциальной фазе, при этом предпочтительный диапазон доз ДНК-полимеразы, MgCl2 и dNTP согласно настоящему изобретению составляет:
ДНК-полимераза: от 0,5 Ед до 1,0 Ед; MgCl2: от 1 мМ до 2 мМ; dNTP: от 200 мкМ до 400 мкМ.
Согласно приведенным выше результатам исследования, настоящее изобретение обеспечивает окончательные оптимизированные параметры реакции амплификации, как показано в таблице 3.
3. Эффективность амплификации согласно способу одновременного выявления на основе RT-PCR для выхода афлатоксина и транскрипционной активности гена Nor-1
Результаты одновременной амплификации на основе RT-PCR последовательно разведенного фага и последовательно разведенного Nor-1 показаны на кривой одновременной амплификации на основе RT-PCR (RFU, относительная единица флуоресценции), как показано на изображении А на ФИГ. 2. Изображение В на ФИГ. 2 представляет собой стандартную кривую эффективности амплификации, полученную из кривой амплификации. Угол наклона стандартной кривой эффективности амплификации фага VHH 2-5 составляет -3,37. Расчеты производили с использованием формулы для расчета эффективности амплификации 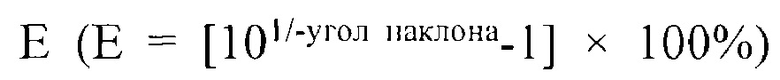 , причем если эффективность амплификации Е составляла 98,03%, обнаруживаемый диапазон концентрации фага VHH 2-5 составлял от 109 БОЕ/мл до 103 БОЕ/мл. Если число копий фрагмента ДНК Tq-nor1 гена Nor-1 составляло от 102 копий до 108 копий, эффективность амплификации гена Nor-1 по аналогии составляла 90,25%. Эффективность амплификации Е соответствовала требуемому диапазону от 90% до 105% и коэффициент корреляции R2 стандартной кривой эффективности амплификации составлял >0,99. Таким образом, оптимизированная система одновременной RT-PCR может использоваться для одновременной и эффективной амплификации фага VHH 2-5 и гена Nor-1.
, причем если эффективность амплификации Е составляла 98,03%, обнаруживаемый диапазон концентрации фага VHH 2-5 составлял от 109 БОЕ/мл до 103 БОЕ/мл. Если число копий фрагмента ДНК Tq-nor1 гена Nor-1 составляло от 102 копий до 108 копий, эффективность амплификации гена Nor-1 по аналогии составляла 90,25%. Эффективность амплификации Е соответствовала требуемому диапазону от 90% до 105% и коэффициент корреляции R2 стандартной кривой эффективности амплификации составлял >0,99. Таким образом, оптимизированная система одновременной RT-PCR может использоваться для одновременной и эффективной амплификации фага VHH 2-5 и гена Nor-1.
2. Построение стандартной кривой для количественного анализа и оценка способа на основе одновременной RT-PCR
1. S-образная стандартная кривая для количественного определения афлатоксина
1.1. Иммунная реакция
(1) Нанесение в качестве покрытия. Использовали PBS для разбавления коммерчески доступного моноклонального антитела к афлатоксину 1С11 (секретируется штаммом 1С11 клеток гибридомы с номером депонирования ССТСС №: С201013, номер заявки на патент CN201010245095.5, для которого доступны соответствующие отчеты) до 1,0 мкг/мл. Использовали микропипетку для добавления его в 96-луночный микропланшет по 100 мкл на лунку, инкубировали его в течение ночи при 4°C (-12 ч), промывали планшет 3 раза с помощью PBST; блокировали его: блокировали каждую лунку блокирующим раствором, 300 мкл на лунку, инкубировали при 37°C в течение 45 минут, промывали планшет 3 раза с помощью PBST.
(2) Блокировка. Блокировали блокирующим раствором, 300 мкл на лунку, инкубировали при 37°C в течение 45 минут, 3 раза промывали планшет с помощью PBST.
(3) Конкуренция. Разбавляли препарат стандарта афлатоксина В1 100% чистым метанолом до концентрации 200 нг/мл, затем разбавляли стандартный раствор афлатоксина В1 3-кратным градиентом 10% (об./об.) смеси метанол/PBS так, чтобы диапазон концентраций составлял от 33,33 нг/мл до 1,69 пг/мл. Затем смешивали фаг VHH 2-5 с известной концентрацией (1,0×1010 КОЕ/мл) с 50 мкл афлатоксина В1 с серийной концентрацией, добавляли 100 мкл смеси в 96-луночный микротитровальный планшет. После инкубации при 37°C в течение 1 часа промывали планшет с помощью PBST 10 раз.
(4) Элюирование. Вводили 90 мкл элюата фага в каждую лунку, оставляли его на теплой бане при 37°C на 15 минут, аккуратно захватывали микропипеткой и переносили элюат, содержащий фагмиду.
(5) Нейтрализация. Смешивали 90 мкл извлеченного раствора с 10 мкл нейтрализующего раствора для нейтрализации смеси, причем объем добавляемого нейтрализующего раствора регулировали в соответствии с фактическим значением рН элюента и нейтрализующего раствора с целью обеспечения нейтрализации смеси.
1.2. Построение стандартной кривой. На стадии иммунной реакции при условии, что количество нанесенного в качестве покрытия моноклонального антитела к афлатоксину 1С11 является фиксированным, афлатоксин будет конкурировать с фагом VHH 2-5 за связывание с 1С11. Чем выше концентрация афлатоксина, тем меньше вероятность связывания фага с 1С11 и меньше величина связывания. После завершения иммунной реакции фаг, связанный с 1С11, элюировали. Количество фагов в элюате зависело от концентрации афлатоксина. Фагмида в элюате высвобождала молекулы ДНК во время процесса нагревания в реакции ПЦР. Высвободившиеся молекулы ДНК использовали в качестве мишени для амплификации в реакции RT-PCR. После реакции амплификации программное обеспечение системы количественного анализа флуоресценции предоставило значения Ct для амплификации фагов с разным количеством в разных элюатах, соответствующие разным количествам афлатоксина. Значения Ct, полученные при серийном разбавлении афлатоксина в концентрациях от 33,33 нг/мл до 1,69 пг/мл до логарифма концентрации афлатоксина, подвергали четырехпараметрической логистической регрессии с использованием программного обеспечения Origin Pro 8.0, и результат использовали для построения S-образной стандартной кривой для количественного определения афлатоксина, как показано на изображении А на ФИГ. 3.
2. Построение стандартной кривой для количественного определения транскрипции гена Nor-1
Использовали штамм Aspergillus flavus. В этом примере использовали депонированный в исследовательском центре штамм Aspergillus flavus N73, который является высокотоксигенным штаммом и использовался для получения Tq-nor1 в данном исследовании. Для других штаммов Aspergillus flavus можно использовать описанный выше «способ получения фрагмента ДНК Tq-nor1 гена Nor-1» для амплификации фрагмента ДНК Tq-nor1 размером 400 п.о. Оба они могут быть использованы в качестве представляющих интерес штаммов для построения стандартной кривой для количественного определения транскрипции гена Nor-1. После использования спектрофотометра (NanoDrop 2000, Thermo Scientific, США) для определения концентрации Tq-nor1 рассчитывали число копий Tq-nor1. Tq-nor1 серийно разводили (от 102 до 108 копий) в виде образца с известным количеством, а затем осуществляли RT-PCR-амплификацию. Для выполнения регрессионного анализа использовали программное обеспечение Origin Pro 8.0, в котором значение логарифма числа копий серийного разведения стандартного образца Tq-nor1 является абсциссой, а значение Ct является ординатой. На изображении В на ФИГ. 3 показана стандартная кривая для количественного определения транскрипции гена Nor-1.
Стандартная кривая для количественного определения афлатоксина показана на изображении А на ФИГ. 3. На фигуре представлено, что предел выявления LOD (выраженный при помощи IC10) для количественного определения афлатоксина В1 посредством одновременной RT-PCR составляет 0,018 нг/мл. Следовательно, разработанное количественное определение афлатоксина В1 посредством RT-PCR характеризуется высокой чувствительностью. Кроме того, на изображении В на ФИГ. 3 показано, что с помощью одновременной RT-PCR можно количественно определять число копий гена Nor-1 в диапазоне от 102 до 108, что полностью доказывает, что разработанная одновременная RT-PCR имеет заметные преимущества при низкоуровневом абсолютном количественном определении гена Nor-1.
3. Измерение перекрестной реактивности афлатоксина при одновременном выявлении на основе RT-PCR
Препараты стандарта афлатоксина В2, G1, G2, зеараленона (ZEN), дезоксиниваленола (DON), фумонизина (FBI) разводили в серии концентраций. После их конкуренции с фагом VHH 2-5 в конкурентной иммунной реакции фаг, связанный с моноклональным антителом 1С11 на дне микропланшета, элюировали для проведения одновременной амплификации на основе RT-PCR с Tq-nor1. Значение Ct, полученное при амплификации, соответствует логарифмическому значению различных концентраций токсина, которое использовали для построения стандартной кривой. Вычисляли значение IC50 и рассчитывали перекрестную реактивность в соответствии с формулой расчета перекрестной реактивности % CR=(IC50 AFB1/IC50 апалита)×100. Как показано на ФИГ. 4, показатели перекрестной реактивности для афлатоксина В1, В2, G1 и G2 составляли 100%, 101%, 34% и 12% соответственно. Из этого следует, что при помощи данного способа можно измерять общее количество афлатоксина. В частности, показатели перекрестной реактивности афлатоксина G1 и G2 составляли 34% и 12% соответственно, но это не влияет на применение способа при определении токсинообразующей способности Aspergillus flavus, поскольку штаммы Aspergillus flavus вырабатывают только афлатоксин группы В. Перекрестная реактивность афлатоксина В2 составляет 101%. Разработанный способ на основе RT-PCR для количественного определения афлатоксина В2 имеет более высокую чувствительность, что в некоторых условиях повышает надежность разработанного способа на основе RT-PCR для определения токсинообразующей способности Aspergillus flavus по той причине, что штаммы Aspergillus flavus могут вырабатывать как афлатоксин В1, так и афлатоксин В2. Следовательно, при использовании разработанного способа на основе RT-PCR для определения токсинообразующей способности штаммов Aspergillus flavus получают оценку общего выхода В1 и В2.
4. Проверка выявления добавки с помощью одновременной RT-PCR
Препарат стандарта афлатоксина В1 и фрагмент ДНК Tq-nor-1 гена Nor-1 одновременно добавляли к 2 мл чистой среды PDB. Встряхивали их и хорошо перемешивали, а после равномерного перемешивания помещали смешанный раствор при 4°C без доступа света в течение 4 дней. Через 4 дня смесь разбавляли в 20 раз, и содержание афлатоксина В1 и Tq-nor-1 в смеси одновременно определяли с помощью RT-PCR. Получали 3 повторности для внутригруппового эксперимента группу в один день и 3 повторности для межгруппового эксперимента в разные дни. Результаты выявления добавки показаны в таблице 4.
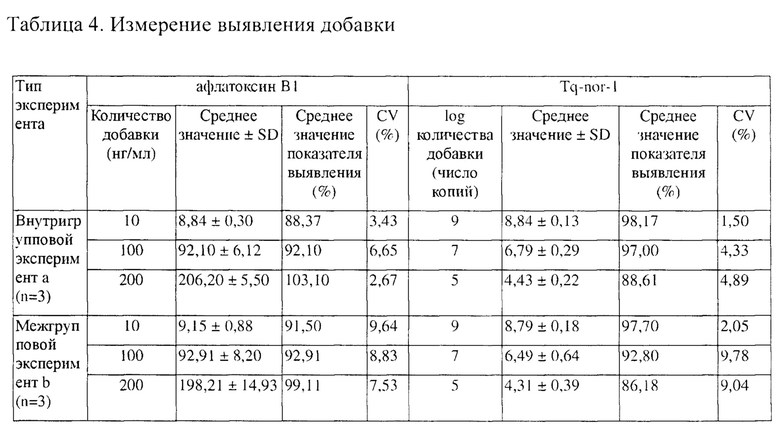
Степень выявления добавки афлатоксина В1 составляла от 88,37% до 103,10%, и степень выявления добавки фрагмента ДНК Tq-nor-1 гена Nor-1 составляла от 86,18% до 98,17%. Этот результат демонстрирует, что разработанная одновременная RT-PCR имеет надежную повторяемость и воспроизводимость результатов при выявлении и анализе реальных образцов.
5. Применение способа на основе одновременной RT-PCR для количественной оценки уровня экспрессии гена Nor-1 и токсинообразующей способности штаммов Aspergillus flavus
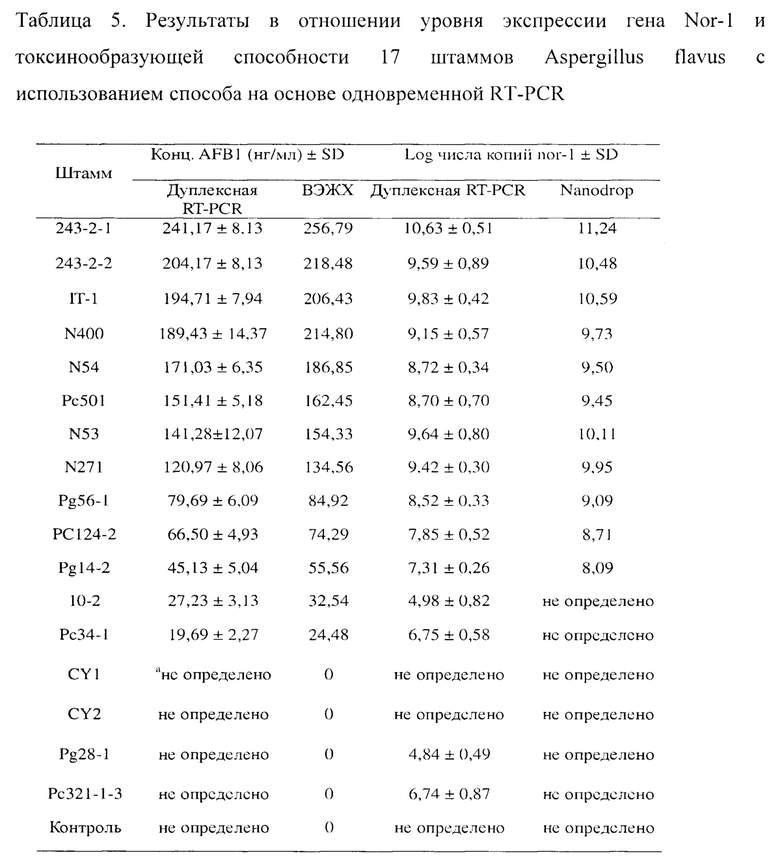
В этом исследовании отбирали 17 штаммов Aspergillus flavus с разной токсинообразующей способностью. Помещали 15 мл жидкой среды с картофельной декстрозой в колбу Эрленмейера объемом 50 мл, автоклавировали ее при 121°C в течение 30 минут и добавляли раствор со спорами Aspergillus flavus до конечной концентрации 1×105 мл-1. После культивирования со встряхиванием культуры при 28°C и 200 об/мин в течение 96 часов фильтровали культуральный бульон с помощью двухслойной фильтровальной бумаги для получения культурального бульона штамма и мицелия грибов Aspergillus flavus. С помощью фильтровальной бумаги отжимали лишнюю воду из мицелия грибов Aspergillus flavus и высушивали его в течение 12 часов при 65°C в сушильной камере, охлаждали до комнатной температуры, измельчали в порошок с жидким азотом и точно отвешивали по 0,20 мг измельченного гриба на образец. При проведении выделения общей РНК следовали инструкциям к набору (RNeasy Plant Mini Kit) для выделения РНК. Затем использовали набор для обратной транскрипции QuantiTect для синтеза cDNA. Раствор cDNA разводили в 100-1000 раз и использовали вместо Tq-nor1 в системе одновременной амплификации на основе RT-PCR и использовали в качестве одной из матриц амплификации при выполнении одновременной амплификации на основе RT-PCR для измерения транскрипционной активности гена Nor-1. После того, как культуральную среду штамма разбавляли в 10 раз с помощью 10% (вес/об.) BSA/PBS, ее использовали вместо препарата стандарта афлатоксина, используемого в иммунной реакции, для участия в реакции иммунной конкуренции. После конкурентной реакции фагмиду в элюате использовали в качестве другой матрицы в системе одновременной амплификации на основе RT-PCR и выполняли одновременную амплификацию на основе RT-PCR для измерения токсинообразующей способности штаммов. Для количественной оценки выработки токсина и уровня экспрессии гена Nor-1 для 17 штаммов Aspergillus flavus использовали способ на основе одновременной RT-PCR, и результаты показаны в таблице 5.
Для проверки точности результатов со ссылкой на способ национального стандарта GB5009.22-2016 применяли способ ВЭЖХ для определения количества токсина, вырабатываемого штаммами Aspergillus flavus. В то же время экспрессию гена Nor-1 конкретного штамма количественно определяли с помощью спектрофотометра (Nanodrop). Количественные результаты представлены в таблице 5. Полученный результат сравнивали с результатом, полученным в способе на основе одновременной RT-PCR, и результат сравнения показан на изображении А на ФИГ. 5 и изображении В на ФИГ. 5. На изображении А на ФИГ. 5 показан результат сравнения RT-PCR и ВЭЖХ в отношении количественного определения афлатоксина. Полученное уравнение линейной регрессии имеет вид Y=0,947X - 3,84, и анализ линейной регрессии дает хорошую корреляцию (R2=0,999). На изображении В на ФИГ. 5 показан результат сравнения RT-PCR и Nanodrop в отношении количественного определения транскрипции гена Nor-1. Уравнение линейной регрессии, полученное этим способом, имеет вид Y=1,05Х-1,18, а коэффициент корреляции R2=0,989. Результаты сравнения различных способов выявления демонстрируют, что количественные результаты разработанной одновременной RT-PCR надежны и могут использоваться для одновременного анализа токсинообразующей способности штаммов Aspergillus flavus и транскрипционной активности гена Nor-1.
Вариант осуществления 3
1. Применение транскрипционной активности Nor-1 и AFT/Nor-1 (отношение токсинообразующей способности к транскрипционной активности Nor-1) для оценки токсинообразующей способности Aspergillus flavus соответственно
Путем сравнения и анализа результатов было обнаружено, что если токсинообразующую способность Aspergillus flavus оценивают исключительно по транскрипционной активности Nor-1, результаты определения являются ненадежными. Например, логарифмические значения числа копий экспрессии гена Nor-1 в штаммах Aspergillus flavus Pel24-2 и Рс34-1 составляют 7,85±0,52 и 6,75±0,58 соответственно. Уровни экспрессии гена Nor-1 этих двух штаммов эквивалентны. Однако выработка афлатоксина штаммом Pel24-2 составляет 66,5±4,93 нг/мл, а выработка афлатоксина штаммом Рс34-1 составляет всего 19,69±2,27 нг/мл. Кроме того, штаммы CY1, CY2, Pg28-1 и Рс321-1-3 не вырабатывают афлатоксин. При этом уровни экспрессии гена Nor-1 штаммами Pg28-1 и Рс321-1-3 даже выше, чем у некоторых штаммов, вырабатывающих афлатоксин.
Однако более надежно оценивать токсинообразующую способность Aspergillus flavus с использованием отношения выработки афлатоксина к уровню экспрессии гена Nor-1. Для того чтобы дополнительно изучить корреляционную взаимосвязь между токсинообразующей способностью штаммов Aspergillus flavus, уровнем экспрессии гена nor-1, а также AFT/Nor-1, токсинообразующую способность штаммов принимают за ординату, а логарифм величины экспрессии гена Nor-1 и отношение AFT/Nor-1 используют в качестве абсциссы для построения графика. Корреляционная взаимосвязь между токсинообразующей способностью штаммов Aspergillus flavus и величиной экспрессии гена Nor-1 показана на изображении А на ФИГ. 6. Уравнение линейной регрессии имеет вид: у=36,37 х-196,21, а коэффициент корреляции, полученный с помощью анализа линейной регрессии, составляет R2=0,693. При этом корреляционная взаимосвязь между токсинообразующей способностью Aspergillus flavus и отношением AFT/Nor-1 показана на изображении В на ФИГ. 6. Уравнение линейной регрессии имеет вид: у=10,14 х - 16,20, а коэффициент корреляции, полученный с помощью анализа линейной регрессии, составляет R2=0,979. Имеет место очень хорошая корреляция между токсинообразующей способностью штаммов Aspergillus flavus и отношением AFT/Nor-1.
В свете вышеизложенного в настоящем изобретении предусмотрено, что уравнение для определения токсинообразующей способности штаммов Aspergillus flavus имеет вид: у=10,14 х - 16,20, где X представляет AFT/Nor-1, а Y представляет токсинообразующую способность. Для высокотоксигенных штаммов токсинообразующая способность составляет >150 нг/мл; умеренная токсинообразующая способность: 50 <токсинообразующая способность <150 нг/мл; низкая токсинообразующая способность: токсинообразующая способность <50 нг/мл; отсутствие выработки токсина: 0; затем рассчитывают диапазон определения токсинообразующей способности в соответствии с уравнением регрессии.
Высокотоксигенные штаммы: AFT/Nor-1 > 16,4;
умеренно токсигенные штаммы: 6,5<AFT/Nor-1<16,4;
низкотоксигенные штаммы: 0<AFT/Nor-1<6,5;
нетоксигенные штаммы: AFT/Nor-1=0.
Проверка стабильности AFT/Nor-1 (отношение токсинообразующей способности к транскрипционной активности Nor-1) в связи с результатом определения токсинообразующей способности
2. Проверка стабильности AFT/Nor-1 (отношение токсинообразующей способности к транскрипционной активности Nor-1) в связи с результатом определения токсинообразующей способности
Для дополнительной проверки стабильности отношения AFT/Nor-1 при определении результатов токсинообразующей способности, проводят внутригрупповые и межгрупповые экспериментальные анализы соответственно для пяти штаммов Aspergillus flavus с точки зрения токсинообразующей способности в отношении афлатоксина и транскрипционной активности гена Nor-1. Во внутригрупповом эксперименте получают по 5 повторностей одновременно в один и тот же день, и среднее значение для 5 повторностей используют в качестве токсинообразующей способности и транскрипционной активности гена Nor-1 для конкретной партии штаммов. Для межгруппового эксперимента определяют три разные даты, каждую дату определяют как партию и всего получают 3 партии. Результаты показаны в таблице 6.
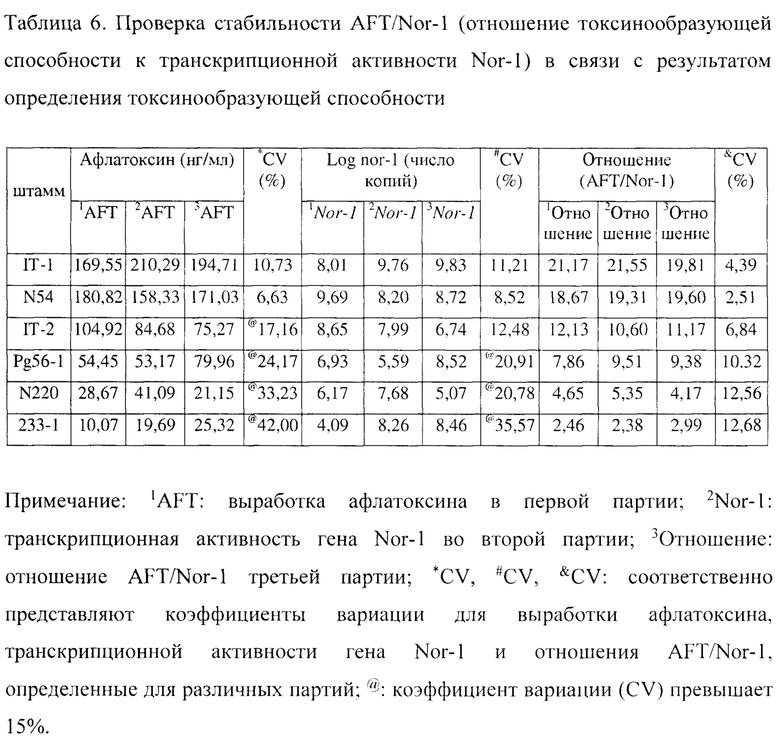
Величина афлатоксинообразующей способности и транскрипционной активности гена Nor-1 в таблице представляют собой среднее значение для внутригрупповых экспериментов с 5 повторностями.
Исходя из таблицы можно легко обнаружить, что существует большая разница между выходом афлатоксина, вырабатываемым разными партиями культур, и транскрипционной активностью гена Nor-1. При рассмотрении штамма 233-1 в качестве примера, показатели выхода афлатоксина, определенные для трех партий, составляют 10,07, 19,69 и 25,32 нг/мл соответственно, и коэффициент вариации между различными партиями составляет 42,00%. Кроме того, для показателей выхода афлатоксина, вырабатываемого штаммами IT-2, Pg56-1-2, N200 и 233-1, все внутригрупповые коэффициенты вариации между различными партиями превышают 17,00%. Из этого следует, что оценка токсинообразующей способности штаммов только на основании полученного показателя выхода афлатоксина ненадежна. По аналогии, исходя из межгрупповых показателей транскрипционной активности гена Nor-1 для разных партий, можно обнаружить, что имеют место большие различия в транскрипционной активности гена Nor-1, что соответственно доказывает, что определение и оценка токсинообразующей способности штаммов исключительно по транскрипционной активности гена Nor-1 являются ненадежными. Однако межгрупповые коэффициенты вариации для отношения AFT/Nor-1 составляют менее 13%, что обеспечивает лучшую стабильность.
Таким образом, способ определения и оценки токсинообразующей способности токсигенных штаммов, продуцирующих афлатоксин, предусмотренный в настоящем изобретении, то есть определение отношения AFT/Nor-1, является более точным и надежным способом определения и оценки токсинообразующей способности штаммов Aspergillus flavus.
Очевидно, что вышеупомянутые варианты осуществления являются просто примерами, изложенными для более ясного описания, и не предназначены для ограничения реализации настоящего изобретения. Для специалистов в данной области техники другие изменения или изменения в различных формах могут быть внесены на основе приведенного выше описания. Перечисление в данном документе всех способов реализации нецелесообразно и не представляется возможным. Следовательно, очевидные изменения или модификации настоящего изобретения все еще находятся в пределах объема правовой охраны настоящего изобретения.
Настоящее изобретение относится к области биологии и, в частности, относится к оценке токсинообразующей способности афлатоксигенного штамма. Способ включает измерение выхода афлатоксина и транскрипционной активности гена Nor-1 штаммов Aspergillus flavus для получения отношения выхода афлатоксина к транскрипционной активности гена Nor-1 и оценку токсинообразующей способности афлатоксигенного штамма. Токсинообразующую способность афлатоксигенного штамма обозначают как Y, отношение выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1) обозначают как X с расчетом по формуле Y = 10,14X - 16,20. При этом для высокотоксигенного штамма токсинообразующая способность составляет > 150 нг/мл; для умеренно токсигенного штамма: 50 < токсинообразующая способность < 150 нг/мл; для низкотогсигенного штамма: токсинообразующая способность составляет < 50 нг/мл. Подставляют указанные выше значения в формулу для получения диапазона оценки токсинообразующей способности с оценкой токсинообразующей способности афлатоксигенного штамма в соответствии с отношением выхода афлатоксина к транскрипционной активности гена Nor-1(AFT/Nor-1). Причем AFT/Nor-1>16,4 указывает на высокотоксигенный штамм; 6,5<AFT/Nor-1<16,4 указывает на умеренно токсигенный штамм; 0<AFT/Nor-1<6,5 указывает на низкотоксигенный штамм. Отношение выхода афлатоксина к транскрипционной активности гена Nor-1(AFT/Nor-1) получают посредством одного из двух предложенных способов. Изобретение обеспечивает быструю и точную оценку токсинообразующей способности штамма Aspergillus flavus, так что данный способ имеет важное значение для раннего предупреждения, предотвращения и контроля загрязнения афлатоксином. Кроме того, разработанный способ одновременного выявления на основе RT-PCR для показателей выхода афлатоксина и транскрипционной активности гена Nor-1 с результатом AFT/Nor-1 является надежным и точным и может использоваться в качестве индекса идентификации для определения токсинообразующей способности штамма Aspergillus flavus. 5 з.п. ф-лы, 6 ил., 6 табл.
1. Способ оценки токсинообразующей способности афлатоксигенного штамма, отличающийся тем, что включает измерение выхода афлатоксина и транскрипционной активности гена Nor-1 штаммов Aspergillus flavus для получения отношения выхода афлатоксина к транскрипционной активности гена Nor-1 и оценку токсинообразующей способности афлатоксигенного штамма в зависимости от отношения выхода афлатоксина к транскрипционной активности гена Nor-1,
при этом токсинообразующую способность афлатоксигенного штамма обозначают как Y, и отношение выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1) обозначают как X, при этом формула для оценки токсинообразующей способности Aspergillus flavus представляет собой Y = 10,14Х - 16,20, при этом для высокотоксигенного штамма токсинообразующая способность составляет > 150 нг/мл; для умеренно токсигенного штамма: 50 < токсинообразующая способность < 150 нг/мл; для низкотогсигенного штамма: токсинообразующая способность составляет < 50 нг/мл; при этом подставляют указанные выше значения в формулу для получения диапазона оценки токсинообразующей способности с оценкой токсинообразующей способности афлатоксигенного штамма в соответствии с отношением выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1), причем AFT/Nor-1>16,4 указывает на высокотоксигенный штамм; 6,5<AFT/Nor-1<16,4 указывает на умеренно токсигенный штамм; 0<AFT/Nor-1<6,5 указывает на низкотоксигенный штамм;
при этом отношение выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1) получают посредством одного из следующих способов:
способ 1: культивирование штаммов Aspergillus flavus, сбор спор Aspergillus flavus для культивирования со встряхиванием, после завершения культивирования фильтрация с получением фильтрата и измерение концентрации афлатоксина в фильтрате; фильтрация мицелия грибов Aspergillus flavus для высушивания с получением высушенных микроорганизмов, измерение транскрипционной активности гена Nor-1 в высушенных микроорганизмах, таким образом получая отношение выхода афлатоксина к транскрипционной активности гена Nor-1 (AFT/Nor-1);
способ 2: показатели выхода афлатоксина и транскрипционной активности гена Nor-1 также можно получить посредством способа одновременного выявления на основе RT-PCR, при этом указанный способ включает следующие стадии:
(1) построение S-образной кривой для количественного определения афлатоксина: на стадии реакции иммунной конкуренции, если количество нанесенного в качестве покрытия моноклонального антитела к афлатоксину является постоянным, используют препараты стандарта афлатоксина для конкуренции с представленным на поверхности фага антиидиотипическим наноантителом к афлатоксину за связывание с моноклональным антителом к афлатоксину, после завершения реакции иммунной конкуренции фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, связанным с моноклональным антителом к афлатоксину, элюируют, при этом препараты стандарта афлатоксина соответствуют разному количеству элюированного фага с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, при этом во время нагревания в ходе ПЦР из содержащегося в элюате фага высвобождаются молекулы ДНК, и высвободившиеся молекулы ДНК используют в качестве мишеней для амплификации в реакции RT-PCR, при этом каждый элюат подвергают реакции одновременной RT-PCR-амплификации с использованием набора и после завершения реакции амплификации получают разные значения Ct, при этом логарифм концентрации афлатоксина используют в качестве абсциссы, а значение Ct используют в качестве ординаты, и для получения S-образной кривой для количественного определения афлатоксина проводят регрессионный анализ;
(2) построение кривой RT-PCR для транскрипционной активности гена Nor-1, предусматривающее серийное разведение образцов с известным числом копий фрагмента ДНК Tq-nor1 гена Nor1 с получением разного числа копий, а затем с использованием набора проведение реакции одновременной RT-PCR-амплификации для каждого числа копий Tq-nor1 отдельно, после завершения реакции амплификации получение разных значений Ct, использование логарифма числа копий Tq-nor1 в качестве абсциссы, а значения Ct в качестве ординаты для выполнения регрессионного анализа с получением при этом кривой для количественного определения транскрипции гена Nor-1;
(3) осуществление культивирования штаммов Aspergillus flavus с получением спор Aspergillus flavus, при этом используют споры Aspergillus flavus для культивирования со встряхиванием, после завершения культивирования фильтруют раствор для культивирования штамма от мицелия грибов Aspergillus flavus; после разбавления раствора для культивирования штамма в некотором соотношении его используют для замены препаратов стандарта афлатоксина в иммунной реакции, проводимой на стадии (1), для осуществления реакции, основанной на иммунной конкуренции, после реакции иммунной конкуренции фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину, связанным с моноклональным антителом к афлатоксину, элюируют, и молекулы ДНК, высвобождаемые содержащимися в элюате фагами, используют в качестве матрицы для количественной амплификации афлатоксина в реакции одновременной RT-PCR-амплификации; дополнительно после высушивания мицелия грибов Aspergillus flavus общую РНК выделяют и подвергают обратной транскрипции с получением cDNA, и cDNA разбавляют в некотором соотношении и используют в качестве матрицы для количественной амплификации гена Nor-1 в реакции одновременной RT-PCR-амплификации;
(4) использование молекул ДНК, высвобождаемых фагом, и cDNA в качестве матриц для одновременной реакции RT-PCR-амплификации, при этом после реакции амплификации получают два значения Ct, и эти два значения Ct соответственно сопоставляют с S-образной кривой для количественного определения афлатоксина и кривой для количественного определения транскрипции гена Nor-1, при этом выполняют преобразование для получения концентрации афлатоксина и транскрипционной активности гена Nor-1 с определением отношения выхода афлатоксина к транскрипционной активности гена Nor-1.
2. Способ оценки токсинообразующей способности афлатоксигенного штамма по п. 1, отличающийся тем, что культуральная среда, используемая для культивирования штаммов Aspergillus flavus, представляет собой среду CDA, а условия культивирования являются следующими: культивирование при 28°С и влажности 90% в течение 10 дней; среда, используемая для культивирования со встряхиванием спор Aspergillus flavus, представляет собой жидкую среду с картофельной декстрозой, а условия культивирования являются следующими: культивирование со встряхиванием в течение 96 ч при 28°С и 200 об/мин.
3. Способ оценки токсинообразующей способности афлатоксигенного штамма по п. 1, отличающийся тем, что применяют способ иммуноаффинной очистки в сочетании с высокоэффективной жидкостной хроматографией для измерения концентрации афлатоксина в фильтрате после культивирования со встряхиванием спор Aspergillus flavus.
4. Способ оценки токсинообразующей способности афлатоксигенного штамма по п. 1, отличающийся тем, что фаг с представленным на поверхности антиидиотипическим наноантителом к афлатоксину представляет собой фаг VHH 2-5, а моноклональное антитело к афлатоксину представляет собой моноклональное антитело к афлатоксину 1С11.
5. Способ оценки токсинообразующей способности афлатоксигенного штамма по п. 1, отличающийся тем, что в реакционной системе, предусматривающей реакцию одновременной RT-PCR-амплификации, конечная концентрация прямых и обратных праймеров Ph-F, Ph-R, Tq-nor1-F и Tq-nor1-R составляет от 300 до 400 нМ; флуоресцентный зонд: конечная концентрация Ph-зонда и Tq-зонда составляет от 200 до 400 нМ; конечная доза ДНК-полимеразы: от 0,5 до 1,0 Ед; конечная концентрация MgCl2: от 1 до 2 мМ; конечная концентрация dNTP: от 200 до 400 мкМ.
6. Способ оценки токсинообразующей способности афлатоксигенного штамма по п. 1, отличающийся тем, что реакционная система, предусматривающая реакцию одновременной RT-PCR-амплификации, содержит 5 мкл мастер-микса Universal Probe для qPCR, 0,1 мкл Ph-F, 0,1 мкл Ph-R, 0,1 мкл Ph-зонда, 2 мкл фаговой матрицы, 0,1 мкл Tq-nor1-F, 0,1 мкл Tq-nor1-R, 0,1 мкл Tq-зонда, 1 мкл матрицы гена Nor-1, 0,2 мкл ДНК-полимеразы, 0,8 мкл MgCl2, 0,2 мкл dNTP, при этом добавляют H2O для доведения до 10 мкл.
| СПОСОБ КОЛИЧЕСТВЕННОГО ОПРЕДЕЛЕНИЯ АФЛАТОКСИНА В1 МЕТОДОМ ДИФФЕРЕНЦИАЛЬНОЙ ВОЛЬТАМПЕРОМЕТРИИ | 2013 |
|
RU2534732C1 |
| CN 1556391 A, 22.12.2004 | |||
| CN 108593832 A, 28.09.2018 | |||
| Abdel-Hadi, A., Carter, D., and Magan, N | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| J | |||
| Appl | |||
| Microbiol | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Mohammad | |||

