Область техники
Настоящее изобретение относится к молекулам антител, которые связываются и способны оказывать агонистическое действие как на CD137, так и на OX40. Указанные молекулы антител содержат связывающий сайт на основе области, определяющей комплементарность (CDR), для CD137 и антигенсвязывающий сайт для OX40, который расположен в константном домене молекулы антитела. Молекулы антител согласно настоящему изобретению можно применять, например, в лечении заболеваний, таких как рак и инфекционные заболевания.
Уровень техники
Иммунная система млекопитающих представляет собой тонко сбалансированную систему, функции которой иногда нарушаются заболеваниями, такими как разные виды рака. Рецепторы контрольных точек играют важнейшую роль в ответе иммунной системы на заболевание, оказывая либо костимулирующие, либо коингибирующие эффекты, баланс которых определяет направление иммунного ответа (Pardoll, 2012). Коингибиторы ингибируют пролиферацию Т-клеток и индуцируют высвобождение противовоспалительных цитокинов. Они подавляют воспаление и предотвращают повреждение органа/ткани в результате чрезмерной иммунной реакции. Костимуляторы, с другой стороны, стимулируют клональное размножение Т-клеток, дифференцировку и выживание эффекторных клеток, чтобы облегчить развитие защитного иммунного ответа.
Один проверенный подход к иммунотерапии рака запускает распознавание и уничтожение опухолевых клеток иммунной системой путем нацеливания на эти рецепторы контрольных точек антител, которые либо блокируют функцию коингибирующих рецепторов, либо индуцируют активность костимулирующих рецепторов (Pardoll, 2012). Антитела, которые блокируют активность коингибирующих рецепторов, показали хорошую клиническую активность и в настоящее время одобрены для лечения рака (Larkin et al., 2015). Антитела, которые индуцируют активность костимулирующих рецепторов, продемонстрировали значительную перспективность в доклинических модельных системах (Moran et al., 2013; Schaer et al., 2014), и несколько агентов в настоящее время проходят клинические исследования (Mayes et al., 2018; Melero et al., 2013). Эти антитела также называют агонистическими антителами, поскольку они предназначены для имитации лигандов таких костимулирующих рецепторов.
Несколько костимулирующих рецепторов Т-клеток являются представителями суперсемейства рецепторов TNF, большого семейства белков, вовлеченных в функции как иммунных, так и неиммунных клеток, которые экспрессируются на поверхности клетки (Bremer, 2013). Структурный анализ комплексов, образованных между рецепторами семейства TNF и их когнатными лигандами, свидетельствует о том, что в большинстве случаев имеет место стехиометрия тример к тримеру, и лиганды семейства TNFR, как правило, экспрессируются на поверхности клетки в виде тримеров (Wajant, 2015). Предложенная модель активации TNFR заключается в том, что взаимодействие с тримерным лигандом индуцирует тримеризацию мономерных рецепторов и инициирует передачу сигнала. Это позволяет заранее предположить, что представители семейства TNFR экспрессируются в виде мономеров и только взаимодействие с лигандом индуцирует образование тримеров рецептора. Эта модель недавно была подвергнута критике (Vanamee & Faustman, 2018), и ассоциация этих мономеров в структуры более высокого порядка в отсутствие взаимодействия с лигандом все еще остается предметом спора. Существование заранее собранных димеров рецептора или даже неактивных тримеров, которые нуждаются в дополнительной кластеризации нескольких рецепторных комплексов, могло бы объяснить более низкую активность некоторых растворимых, только тримерных лигандов TNF, по сравнению с их связанными с мембраной формами, которые могут образовывать суперкластеры лигандов и индуцировать суперкластеры рецепторов TNF, что индуцирует более высокие уровни активации рецептора (Müller et al., 2008). Эта теория также согласуется с наблюдением, что TNFR-специфичные антитела, как правило, не обладают или обладают только низкой агонистической активностью и нуждаются во вторичном перекрестном связывании комплексов антитело-рецептор TNF, чтобы индуцировать достаточную кластеризацию и активацию рецептора, имитируя посредством этого суперкластеры лиганда TNF (Wajant, 2015).
Вторичное перекрестное связывание комплексов антитело-рецептор TNF может быть достигнуто in vitro с помощью перекрестно связывающих агентов, таких как белок A или G или вторичных антител, нацеленных на константные домены агонистических антител, специфичных в отношении рецептора TNF (Vanamee & Faustman, 2018; Wajant, 2015). Однако in vivo для такого вторичного перекрестного связывания требуется взаимодействие с рецепторами Fc-гамма, присутствующими на поверхности иммунных клеток, таких как макрофаги, NK-клетки или B-клетки. Взаимодействие антител с рецепторами Fc-гамма является сложным, поскольку у человека имеется 6 рецепторов Fc-гамма с разными профилями экспрессии и аффинностью в отношении 4 изотипов IgG человека (Bruhns et al., 2009). Было показано, что рецепторы Fc-гамма требуются для оптимальной противоопухолевой активности агонистических антител, нацеленных на мишени из суперсемейства рецепторов TNF in vivo (Bulliard et al., 2013; Bulliard et al., 2014). Однако зависимость агонистических антител к TNFR от опосредуемого рецептором Fc-гамма перекрестного связывания для индукции сильной активации рецепторов, вероятно, ограничит их общую активность in vivo по нескольким причинам: 1) связанные с антителами клетки будут нуждаться во взаимодействии с клетками, экспрессирующими рецептор Fc гамма, в транс-условиях, и частота этого взаимодействия будет ограничивать активацию TNFR-экспрессирующих клеток; 2) аффинность рецепторов Fc-гамма в отношении IgG человека, как правило, значительно ниже по сравнению с аффинностью типичного терапевтического антитела в отношении его мишени (микромолярный диапазон в сравнении с наномолярным диапазоном, соответственно); и 3) рецепторы Fc-гамма опосредуют эффекторные функции антител, такие как АЗКЦ (антителозависимая клеточная цитотоксичность) и АЗКФ (антителозависимый клеточный фагоцитоз), и, следовательно, потенциально способны устранять именно те клетки, для активации которых предназначены агонистические антитела (Mayes et al., 2018).
Двухвалентные биспецифичные антитела, которые используют один из когнатных антигенов для перекрестного связывания агониста рецептора TNF, представляют собой альтернативный вариант для перекрестного связывания, опосредуемого рецептором Fc-гамма. Эффект перекрестного связывания антитела будет возникать в результате связывания с представителем семейства рецепторов TNF и другим рецептором, экспрессируемым на поверхности клетки, либо на одной и той же клетке, в цис-условиях, либо на другой клетке, в транс-условиях. Этот механизм перекрестного связывания антитела затем будет приводить к суперкластеризации рецептора TNF при условии, что вторая мишень экспрессируется на высоких уровнях, имитируя суперкластеры лиганда TNF. Подход к разработке агонистических антител к TNFR на основе биспецифичных антител имеет несколько теоретических преимуществ в сравнении с моноспецифичными агонистическими антителами: 1) агонистическое действие на TNFR может быть направлено к определенным иммунным клеткам в микросреде опухоли и на периферии путем нацеливания на второй антиген, такой как рецептор контрольной точки или ассоциированный с опухолью антиген, в качестве второго вида специфичности биспецифичного антитела; 2) аффинность перекрестно связывающих доменов связывания биспецифичного антитела может быть разработана так, чтобы превышать аффинность антитела в отношении рецепторов Fc-гамма, что делает перекрестное связывание более эффективным; 3) эффекторные функции антител могут быть селективно отключены с использованием мутаций, что гарантирует отсутствие истощения клеток, предназначенных для активации; 4) агонистическое действие на два отдельных рецептора TNF может быть достигнуто в одной молекуле двойного агониста путем комбинирования активации разных иммунных клеток с получением более сильной стимуляции иммунного ответа; 5) нацеливание на совместно экспрессируемые рецепторы может привести к активации отдельной клетки в цис-условиях без потребности во взаимодействии двух клеток.
Несколько из представителей семейства рецепторов TNF имеют перекрывающиеся профили экспрессии в иммунных клетках. В частности, OX40, CD137, GITR и CD27 экспрессируются на активированных Т-клетках, и совместная экспрессия OX40 и CD137 была подтверждена экспериментально (Ma et al., 2005).
OX40 преимущественно экспрессируется на активированных Т-клетках, включая CD4+ Т-клетки, CD8+ Т-клетки, Т-хелперы типа 1 и 2 (Th1 и Th2) и регуляторные Т-клетки (Treg), а также экспрессируется на активированных природных клетках-киллерах (NK-клетках). Взаимодействие OX40 с его лигандом, OX40-лигандом (OX40L), экспрессируемым на антигенпрезентирующих клетках (АПК), повышает клональное размножение, дифференцировку и выживаемость Т-клеток и усиливает образование Т-клеток памяти (Croft et al., 2009). Стимуляция OX40 может оказывать непосредственный эффект на Т-клетки, стимулируя их пролиферацию и выживание, или косвенный эффект за счет усиленной выработки воспалительных цитокинов, таких как IL2 и ИФНγ. Передача сигнала OX40 также может модулировать функцию Treg, несмотря на то, что на этих клетках он устраняет их супрессорную активность (Takeda et al., 2004). Было обнаружено, что при раке OX40 экспрессируется на инфильтрирующих опухоль Т-клетках от пациентов, страдающих раком головы и шеи, меланомой и разными видами колоректального рака, при этом высокие уровни OX40-положительных лимфоцитов коррелируют с лучшей выживаемостью (Petty et al., 2002; Vetto et al., 1997). Доклинические исследования агонистических антител к OX40 на мышах продемонстрировали терапевтическую эффективность в нескольких моделях сингенных опухолей, но эффективность нацеливания на OX40 в качестве монотерапии была различной и, по-видимому, коррелирует с иммуногенностью опухоли (Kjærgaard et al., 2000). Это согласуется с представлением о том, что экспрессия OX40 на опухолеспецифических Т-клетках потребует достаточного прайминга, который, вероятно, не будет обеспечиваться слабоиммуногенными опухолями. В определенных сингенных моделях было установлено, что противоопухолевая активность антитела к OX40, OX86, обусловлена его способностью истощать внутриопухолевые Treg, которые экспрессируют высокие уровни OX40, зависимым от рецептора Fc-гамма образом (Bulliard et al., 2014).
В настоящее время агонистические антитела к OX40 проходят клинические исследования для лечения рака, при этом большинство из них показывает хорошие профили безопасности, но ограниченную клиническую активность (Curti et al., 2013). Изотип, выбранный для этих антител, варьирует, но несколько исследуемых лекарственных средств представляют собой антитела IgG1 человека, задействующие рецептор Fc-гамма, которые предназначены для потенциального истощения Treg в качестве механизма действия. Отсутствие явной клинической активности этих антител побудило провести исследования комбинации агонистических антител к OX40 с несколькими другими видами терапии, включая ингибирование PD1/PD-L1 или CTLA4, терапию против VEGF и ингибитор тирозинкиназы акситиниб.
Было продемонстрировано, что этот механизм действия на основе истощения Treg очень эффективен в доклинических моделях, и несколько рецепторов могут быть мишенями для элиминации Treg, таких как GITR (Bulliard et al., 2014) и CTLA4 (Simpson et al., 2013). Однако в клинических условиях не было показано, что антитела, нацеленные на эквивалентные рецепторы у человека, обладают такими же уровнями противоопухолевой эффективности (Glisson et al., 2016; Tran et al., 2017). Причины этого неясны, но более низкие уровни клеток, экспрессирующих рецептор Fc-гамма, таких как макрофаги, в опухолях человека по сравнению с моделями сингенных опухолей на мышах (Milas et al., 1987) отчасти могут объяснить отсутствие экстраполяции механизма действия этих антител на клинические условия. Другие причины могут заключаться в разных уровнях экспрессии этих маркеров в Treg человека по сравнению с Treg мыши (Aspeslagh et al., 2016).
CD137 также экспрессируется на активированных Т-клетках, включая CD4+, CD8+, Th1, Th2 и Treg, но его профиль экспрессии также включает В-клетки, природные клетки-киллеры (NK-клетки), природные T-клетки-киллеры (NKT-клетки) и дендритные клетки (ДК) (Bartkowiak & Curran, 2015). Как и в случае OX40, взаимодействие CD137 с его лигандом запускает активацию внутриклеточных сигнальных путей, которые приводят к выживанию, пролиферации и индукции цитотоксической активности Т-клеток. Стимуляция CD137 преимущественно стимулирует CD8+ Т-клетки по сравнению с CD4+ Т-клетками и приводит к их пролиферации, выживанию и цитотоксической эффекторной функции за счет выработки воспалительных цитокинов, а также способствует дифференцировке и поддержанию CD8+ Т-клеток памяти. Также было продемонстрировано, что CD137 специфически экспрессируется на подгруппах инфильтрирующих опухоль лимфоцитов (TIL), реактивных в отношении опухоли (Weigelin et al., 2016), что частично обосновывает его агонистическое связывание in vivo и его применение в отборе TIL для адоптивного переноса. Монотерапия CD137 является эффективной в нескольких доклинических моделях иммуногенных опухолей, таких как MC38, CT26 и В-клеточные лимфомы. Однако, в случае еще более эффективного лечения развившихся опухолей, связывание CD137 в комбинации с другими агентами, такими как химиотерапия, цитокины и другие регуляторы контрольных точек, показало более благоприятные эффекты в уменьшении роста опухоли (Bartkowiak & Curran, 2015). В доклинических моделях нацеливание на CD137 с помощью агонистических антител также ассоциировано с воспалением печени и трансаминитом, который возникает в результате повышенного накопления CD8+ Т-клеток, зависимого от выработки IL27 миелоидными клетками (Bartkowiak et al., 2018).
В настоящее время агонистические антитела к CD137 проходят клинические исследования для лечения рака, однако клинический прогресс замедлился из-за ограничивающего дозу выраженного воспаления печени, вероятно, сходного с тем, который наблюдается у мышей (Sanchez-Paulete et al., 2016). Урелумаб (BMS-663513) представлял собой первое агонистическое антитело к CD137, включенное в клинические исследования, и показал признаки клинической активности до того, как исследования были прекращены из-за фатальной гепатотоксичности при дозах выше 1 мг/кг (Segal et al., 2017). Он представляет собой антитело IgG4 человека, которое способно активировать CD137 в отсутствие перекрестного связывания (US 8137667 B2), хотя активность повышается при перекрестном связывании, как и ожидалось в соответствии с теорией полной активации рецептора, опосредуемой суперкластеризацией. Напротив, разные виды ограничивающей дозу токсичности не были обнаружены при применении утомилумаба (PF-05082566) при тестировании до 10 мг/кг (Tolcher et al., 2017). Он представляет собой антитело IgG2 человека и способен активировать CD137 только при перекрестном связывании (US 8337850 B2). Дополнительные клинические исследования обоих антител осуществляются в настоящее время, при этом оба антитела тестируют как в виде монотерапии, так и в комбинации с лучевой терапией и химиотерапией, а также существующими нацеленными и иммуноонкологическими видами терапии. Из-за гепатотоксичности, наблюдаемой при применении урелумаба, это антитело необходимо было дозировать при очень низких уровнях, и ранние признаки клинической активности еще не наблюдались на этих уровнях.
Несколько биспецифичных молекул, нацеленных как на CD137, так и на OX40, находятся на ранней стадии разработки рядом компаний. Нацеливание стимуляции CD137 на опухоль тестируется Macrogenics с использованием HER2- и EphA2-нацеленных молекул-агонистов CD137 DART, Roche с использованием FAPalpha- или CD20-нацеленных гибридных белков лиганда CD137 и Pieris Pharmaceuticals с использованием HER2-нацеленных молекул-агонистов CD137 антикалинов. Двойное нацеливание на OX40 и CTLA4 тестируется Aligator Biosciences для специфичного истощения внутриопухолевых Treg, которые, как ожидается, экспрессируют высокие уровни обеих мишеней.
Было показано, что совместная стимуляция OX40 и CD137 in vivo стимулирует как CD4+, так и CD8+ Т-клетки и индуцирует цитотоксическую функцию как контактировавших с антигеном, так и не контактировавших с антигеном сторонних (bystander) CD4+ Т-клеток (Qui et al., 2011). Представляется интересным, что двойная совместная стимуляция была способна индуцировать трансплантированные CD4+ Т-клетки с уменьшением роста опухоли у мышей с иммунодефицитом, которым инокулировали модель сингенной меланомной опухоли (B16-F10), это подчеркивает способность этой терапии индуцировать противоопухолевую активность CD4+ Т-клеток (Qui et al., 2011). В настоящее время осуществляется клиническое исследование фазы I с повышением дозы с целью исследования эффекта комбинирования агониста OX40 (PF-04518600) с агонистом CD137 (утомилумаб - PF-05082566) (NCT02315066) для оценки безопасности этой комбинации, также в настоящее время осуществляется клиническое исследование фазы Ib/II комбинирования этих же агонистов TNFR с блокадой PD-1 за счет авелумаба (NCT02554812). В этих исследованиях будет рассматриваться комбинация простых моноспецифичных агонистических антител, которые будут нуждаться в перекрестном связывании рецепторов Fc-гамма для их агонистического действия, и, следовательно, может быть недооценена клиническая активность нацеливания на эти рецепторы в комбинации.
Двойная совместная стимуляция OX40 и CD137 также была недавно протестирована на мышах с использованием подхода на основе биспецифичного антитела путем химической конъюгации двух существующих антител против OX40 и CD137 (Ryan et al., 2018). Молекула, названная OrthomAb, была способна индуцировать пролиферацию CD4+ и CD8+ Т-клеток, а также выработку воспалительных цитокинов IL-2 и ИФНγ in vitro. OrthomAb также был способен уменьшать рост опухоли в модели сингенной меланомной опухоли in vivo (B16-F10). Прогнозируется, что двухвалентный биспецифичный характер OrthomAb обеспечит эффективное перекрестное связывание молекул при связывании с обеими мишенями, что приведет к кластеризации рецепторов OX40 и CD137 и, следовательно, к активации Т-клеток. Эти результаты подтверждают подход на основе биспецифичного антитела к нацеливанию на OX40 и CD137 в одной молекуле. Способ изготовления молекулы OrthomAb приводит к получению множества молекул более высокого порядка, а также целевых димеров антитела, которые необходимо дополнительно очищать с помощью нескольких раундов этапов гель-фильтрации. Маловероятно, что такой способ изготовления сделает этот подход конкурентоспособным для чего-либо, за исключением исследовательского инструмента для проверки конкретных комбинаций мишеней. Кроме того, структура этого биспецифичного антитела, в которой две большие макромолекулы удерживаются вместе небольшим химическим линкером, вероятно, будет нестабильной in vivo, и отсутствуют фармакокинетические данные, направленные на решение этой проблемы. К сожалению, противоопухолевый эффект OrthomAb in vivo сравнивали только с активностью агонистических антител либо к OX40, либо к CD137, а не с их комбинацией, в результате неясно, оказывала ли молекула эффект из-за своей биспецифичности или из-за того, что OrthomAb ведет себя как комбинация агонистических антител к OX40 и к CD137 в виде отдельных агентов.
Таким образом, установлена целесообразность комбинирования агонистического действия на представителей семейства рецепторов TNF, OX40 и CD137, в одной двухвалентной, биспецифичной и стабильной молекуле, которая является перспективной для выполнения суперкластеризации OX40 и CD137, независимой от рецептора Fc-гамма, активируя посредством этого как CD4+, так и CD8+ T-клетки для вызова эффективного противоопухолевого иммунного ответа. На основании доклинических данных о комбинации, полученных с использованием моноклональных антител, нацеленных либо на путь OX40, либо на путь CD137, эта молекула также является перспективной в качестве партнера по комбинации для усиления эффекта стандартных видов терапии рака, чтобы обеспечить пользу для пациента.
Изложение сущности изобретения
Авторы настоящего изобретения установили, что молекулы антител, которые связываются как с CD137, так и с OX40 и которые способны индуцировать кластеризацию и передачу сигнала OX40 и/или CD137 при связывании с обеими мишенями, являются высокоэффективными в активации иммунных клеток, например, в микросреде опухоли. Кроме того, авторы настоящего изобретения установили, что ограничение активации CD137 местами, в которых совместно экспрессируются CD137 и OX40, будет высокоэффективным в активации иммунных клеток, не вызывая разные виды токсичности, ассоциированной с известными молекулами агонистов CD137. Ожидается, что это можно будет применять, например, в иммунотерапии для лечения рака и других заболеваний.
Как описано выше в разделе уровня техники, полагают, что первоначальное лигирование лиганда OX40 или лиганда CD137 с OX40 или CD137, соответственно, инициирует цепочку событий, которая приводит к тримеризации рецептора с последующей кластеризацией и активацией рецептора и впоследствии инициацией эффективной противоопухолевой активности Т-клеток. Таким образом, ожидается, что для достижения эффективной активации OX40 или CD137 терапевтическим агентом несколько мономеров рецептора должны быть соединены вместе так, чтобы имитировать связывание тримерным лигандом.
Авторы настоящего изобретения выделили молекулы антител, которые содержат антигенсвязывающий сайт на основе области, определяющей комплементарность (CDR), для CD137 и антигенсвязывающий сайт для OX40, расположенный в константном домене молекулы антитела. Авторы настоящего изобретения показали, что такие молекулы антител способны одновременно связывать обе мишени, когда обе мишени совместно экспрессируются. В данном случае совместная экспрессия включает ситуации, когда CD137 и OX40 экспрессируются на одной и той же клетке, например, иммунной клетке, и ситуации, когда CD137 и OX40 экспрессируются на разных клетках, например, двух разных иммунных клетках, расположенных рядом друг с другом в микросреде опухоли. Таким образом, полагают, что молекулы антител согласно настоящему изобретению способны связываться в цис-условиях с обеими мишенями, экспрессируемыми на одной клетке, а также способны связываться в транс-условиях с двумя мишенями, экспрессируемыми на разных клетках.
Авторы настоящего изобретения также показали, что молекула антитела, которая содержит антигенсвязывающий сайт для CD137 на основе CDR и антигенсвязывающий сайт для OX40, расположенный в константном домене молекулы антитела, способна двухвалентно связываться с обеими мишенями. В частности, авторы настоящего изобретения показали, что, когда такой молекуле антитела обеспечивали условия для связывания с OX40 и CD137, а полученные комплексы были перекрестно связаны и подвергнуты масс-спектрометрическому анализу, было показано, что 19% комплексов содержат две группы OX40 и две группы CD137, это свидетельствует о том, что молекула антитела была двухвалентно связана с обеими мишенями.
Кроме того, авторы настоящего изобретения показали, что, когда эти молекулы антител связаны с обеими мишенями, они способны индуцировать кластеризацию и передачу сигнала OX40 и CD137 in vitro. На основании этого способа действия, такие молекулы антител назвали «двойными агонистами», т. е. молекулы антител способны индуцировать передачу сигнала за счет рецепторов в результате перекрестного связывания путем двойного связывания как с OX40, так и с CD137.
Как продемонстрировано в примерах, OX40 преимущественно экспрессируется на CD4+ Т-клетках, а CD137 преимущественно экспрессируется на CD8+ Т-клетках. Авторы настоящего изобретения продемонстрировали, что молекулы антител способны индуцировать агонистическое действие на OX40 на CD4+ Т-клетках. В этих случаях, как полагают, молекула антитела связывается с CD137 за счет своего антигенсвязывающего домена на основе CDR для перекрестного связывания молекулы антитела, а домен связывания антигена OX40, в то же время, способен связываться, кластеризовать и активировать OX40, экспрессируемый на CD4+ Т-клетках. Аналогичным образом, авторы настоящего изобретения продемонстрировали, что молекулы антител способны индуцировать агонистическое действие на CD137 на CD8+ Т-клетках. В этих случаях, как полагают, молекула антитела связывается с OX40 за счет своего домена связывания антигена OX40 для перекрестного связывания молекулы антитела, а домен связывания антигена OX40, в то же время, способен связываться, кластеризовать и активировать CD137, экспрессируемый на CD8+ Т-клетках.
Кроме того, авторы настоящего изобретения показали, что молекулы антител, содержащие два антигенсвязывающих сайта, подробно описанные выше, и модифицированные, чтобы уменьшить или устранить связывание с рецепторами Fcγ, были способны индуцировать передачу сигнала за счет рецепторов при совместной экспрессии CD137 и OX40, это указывает на то, что агонистическое действие возникало без потребности в перекрестном связывании рецепторами Fcγ. Поскольку для активности молекулы антитела согласно настоящему изобретению не требуется перекрестное связывание, опосредуемое рецептором Fcγ, ожидается, что передача сигнала за счет рецепторов OX40 или CD137 будет локализована в местах, в которых присутствуют обе мишени, например, в микросреде опухоли. Таким образом, молекула антитела способна автономно запускать агонистическое действие на основании экспрессии обеих специфичных мишеней и без потребности в дополнительных перекрестно связывающих агентах.
Кроме того, поскольку для АЗКЦ требуется связывание с рецептором Fcγ, ожидается, что уменьшение связывания с рецепторами Fcγ также приведет к уменьшению АЗКЦ так, что иммунные клетки-мишени не будут истощены молекулами антител согласно настоящему изобретению. Авторы настоящего изобретения сочли это важным, поскольку молекулы антител были разработаны для активации иммунных клеток, экспрессирующих CD137 и/или OX40, чтобы стимулировать иммунный ответ. Таким образом, истощение этих иммунных клеток не является желательным. Авторы настоящего изобретения продемонстрировали, что молекулы антител, обладающие свойствами, определенными в настоящей заявке, были способны активировать и индуцировать пролиферацию иммунных клеток, в частности, Т-клеток, которые экспрессируют CD137 и/или OX40.
Авторы настоящего изобретения также показали, что молекулы антител, содержащие антигенсвязывающие сайты CD137 и OX40, как подробно описано выше, были способны подавлять рост опухоли in vivo у мышей. Кроме того, более эффективное подавление роста опухоли наблюдали при применении молекул биспецифичных антител по сравнению с комбинацией двух молекул моноспецифичных антител, где одна из молекул антител содержала антигенсвязывающий сайт для CD137 на основе CDR, а другая молекула содержала антигенсвязывающий сайт для OX40 на основе CDR, это свидетельствует о том, что одновременное связывание и агонистическое действие на OX40 и CD137 приводит к улучшенной противоопухолевой эффективности. Кроме того, было показано, что молекулы антител способны индуцировать полную регрессию опухоли и формирование защитной иммунологической памяти против повторной стимуляции опухолевыми клетками в модели опухоли CT26 на мышах. Таким образом, ожидается, что молекулы антител согласно настоящему изобретению проявят эффективность в лечении рака у пациентов-людей. Поскольку эти молекулы антител устраняли АЗКЦ-активность, таким образом, ожидается, что они подавляют рост опухоли путем агонистического действия на иммунные клетки-мишени без значительного истощения этих полезных Т-клеток (клеток памяти и эффекторных клеток).
Как наблюдали в исследованиях in vivo на мышах, активация и пролиферация Т-клеток, индуцированная молекулами антител, описанными в настоящей заявке, представляла собой системный, а не локализованный в опухоли эффект. Кроме того, увеличение пролиферации и активации CD4+ и CD8+ Т-клеток центральной памяти в периферической крови и эффекторных CD4+ и CD8+ Т-клеток памяти наблюдали в предварительном исследовании по определению диапазона доз у яванских макак, которым вводили молекулу антитела согласно настоящему изобретению. Таким образом, помимо нацеливания на Т-клетки в микросреде опухоли, ожидается, что мишенями молекулы антитела будут Т-клетки памяти в периферической крови, экспрессирующие OX40 и CD137, чтобы запустить размножение Т-клеток, реактивных в отношении опухоли, которые затем будут обеспечивать их противоопухолевый эффект.
Таким образом, в дополнение к собственно очагу существующей опухоли, анатомическое место, пораженное опухолью, также может рассматриваться, как включающее места где-либо еще в организме, например, лимфатические узлы на периферии, в которых возникают специфичные в отношении опухоли иммунные ответы.
Как объяснялось в разделе уровня техники выше, клиническая разработка молекул агонистов CD137 была приостановлена, по меньшей мере частично, поскольку лечение было ассоциировано либо с ограничивающим дозу выраженным воспалением печени (урелумаб), либо с низкой клинической эффективностью (утомилумаб).
Не ограничиваясь какой-либо теорией, полагают, что Т-клетки, присутствующие в печени, могут быть способны к активации молекулами агонистов CD137, что приводит к воспалению печени. Было показано, что CD8+ Т-клетки стимулируют воспаление и апоптоз печени после сепсиса/вирусной инфекции (Wesche-Soldato et al., 2007). Было показано, что терапия агонистическим антителом к CD137 у мышей приводит к CD137-зависимой инфильтрации Т-клеток в печень (Dubrot J et al., 2010). В совокупности результаты этих исследований указывают на то, что агонистические антитела к CD137 с высокой активностью, такие как урелумаб, могут вызывать инфильтрацию активированных CD8+ Т-клеток в печень, что приводит к воспалению печени. Активность утомилумаба могла быть слишком низкой для обнаружения этого эффекта. В качестве альтернативы, ограничивающая дозу токсичность для печени, наблюдаемая при лечении урелумабом, может быть обусловлена конкретным эпитопом, связанным этим антителом.
Авторы настоящего изобретения провели обширную программу отбора для выделения молекул антител, которые связывают димерный CD137 человека с высокой аффинностью, т. е. ожидается, что они будут связывать CD137 с высокой авидностью. С учетом используемого протокола отбора ожидается, что молекулы антител будут связываться с мономерным CD137 с более низкой аффинностью, чем аффинность, наблюдаемая в отношении димерного CD137.
«Аффинность», упоминаемая в настоящей заявке, может относиться к силе связывающего взаимодействия между молекулой антитела и ее когнатным антигеном, измеренной с помощью KD. Как легко поймет квалифицированный специалист, если молекула антитела способна образовывать несколько связывающих взаимодействий с антигеном (например, если молекула антитела способна двухвалентно связывать антиген и необязательно антиген является димерным), на аффинность, измеренную с помощью KD, также может влиять авидность, при этом авидность относится к общей силе комплекса антитело-антиген.
Экспрессия CD137 иммунными клетками, такими как Т-клетки, положительно регулируется при активации. Не ограничиваясь какой-либо теорией, полагают, что из-за высокой экспрессии CD137 на активированных иммунных клетках CD137 будет находиться в форме димеров, тримеров и мультимеров более высокого порядка на поверхности таких клеток. Напротив, наивные иммунные клетки, такие как наивные Т-клетки, экспрессируют низкие или незначительные уровни CD137 на своей клеточной поверхности, и, следовательно, любой присутствующий CD137, вероятно, будет в мономерной форме. Таким образом, ожидается, что молекулы антител, которые связываются с CD137 с высокой авидностью, будут преимущественно связываться с активированными иммунными клетками, такими как активированные Т-клетки, в отличие от наивных иммунных клеток.
Таким образом, с учетом изложенного выше ожидается, что молекулы антител согласно настоящему изобретению будут в значительной степени неспособны активировать CD137 в отсутствие перекрестного связывания за счет связывания с OX40. Кроме того, как описано выше, авторы настоящего изобретения разработали молекулы антител, в которых перекрестное связывание, опосредуемое рецептором Fcγ, было уменьшено или устранено, при этом ожидается, что это позволит избежать активации CD137 в местах, в которых совместная экспрессия OX40 является незначительной или отсутствует. Было показано, что устранение связывания рецептора Fcγ не влияет на противоопухолевую активность молекулы антитела. Не ограничиваясь какой-либо теорией, полагают, что такие молекулы антител будут проявлять уменьшенную токсичность при введении пациентам. Полагают, что это связано со значительным ограничением активации CD137 местами, в которых OX40 и CD137 совместно экспрессируются на уровнях, достаточных для запуска кластеризации и активации CD137. Авторы настоящего изобретения показали, что в предварительном исследовании по определению диапазона доз на яванских макаках дозы молекулы антитела согласно настоящему изобретению до 30 мг/кг хорошо переносились.
Авторы настоящего изобретения показали, что молекулы антител согласно настоящему изобретению способны индуцировать низкие уровни кластеризации и активации OX40 даже в отсутствие перекрестного связывания. В отличие от агонистических антител к CD137, агонистические антитела к OX40 не проявили никаких видов ограничивающей дозу токсичности (DLT) в клинических условиях, и поэтому не ожидается, что агонистическая активность в отношении OX40 в отсутствие перекрестного связывания будет представлять проблему для клинического лечения. Напротив, в зависимости от состояния, подлежащего лечению, низкий уровень агонистической активности молекул антител в отношении OX40 в отсутствие перекрестного связывания может представлять собой преимущество. Не ограничиваясь какой-либо теорией, полагают, что молекулы антител, содержащие антигенсвязывающий сайт для OX40, с этим свойством можно применять в случае лечения рака путем индукции ограниченной активации и размножения Т-клеток, реактивных в отношении опухоли, в отсутствие перекрестного связывания, что приводит к большему пулу Т-клеток, реактивных в отношении опухоли, которые затем могут быть дополнительно активированы перекрестно связанными молекулами Fcab в микросреде опухоли.
Другое преимущество молекул антител согласно настоящему изобретению, которые были модифицированы для уменьшения или устранения связывания с рецепторами Fcγ, заключается в том, что эти молекулы антител обладают противоопухолевой активностью, которая не опирается на истощение OX40-экспрессирующих регуляторных Т-клеток (Treg). Treg локализованы на периферии, потенциально являются защитными и могут уменьшать влияние аутоиммунитета, который может быть вызван сверхстимуляцией иммунной системы (Vignali DA et al., 2008). Таким образом, было выдвинуто предположение, что истощение Treg может оказывать существенный эффект на уменьшение роста опухоли в моделях на мышах (Bulliard et al., 2014; Simpson et al., 2013). Однако имеются немногочисленные доказательства того, что истощение Treg в опухолях человека может быть достигнуто с помощью АЗКЦ, и, если истощение Treg действительно происходит у человека, по-видимому, это не приводит к такой значительной противоопухолевой активности, как наблюдаемая в моделях на мышах (Powell et al., 2007; Nizar S et al., 2009; Glisson BS et al., 2016; Tran B et al., 2017). Таким образом, если молекула антитела существенно не истощает Treg, но все еще обладает противоопухолевой активностью, это может указывать на то, что молекула антитела обладает противоопухолевой активностью, которая не зависит от истощения Treg, опосредуемого рецептором Fcγ.
Кроме того, было показано, что молекулы антител способны связываться с высокой аффинностью как с CD137 человека и яванской макаки, так и с OX40 человека и яванской макаки. Такая перекрестная реактивность представляет собой преимущество, поскольку она позволяет выполнять дозирование и тестирование безопасности молекул антител у яванских макак во время доклинической разработки.
Еще один признак молекул антител, идентифицированных авторами настоящего изобретения, заключается в том, что антигенсвязывающий сайт для CD137 и антигенсвязывающий сайт для OX40 оба содержатся в самой структуре антитела. В частности, молекулы антител не нуждаются в гибридизации других белков с молекулой антитела за счет линкеров или других средств, чтобы получить молекулу, которая может двухвалентно связываться с обеими своими мишенями. Это обеспечивает ряд преимуществ. В частности, молекулы антител, идентифицированные авторами настоящего изобретения, могут быть получены с использованием способов, аналогичных тем, которые применяют для получения стандартных антител, поскольку они не содержат никаких дополнительных гибридизованных частей. Также ожидается, что структура приведет к улучшенной стабильности антитела, поскольку линкеры могут разрушаться со временем, что приводит к гетерогенной популяции молекул антител. Антитела в популяции, имеющие только один гибридизованный белок, могут быть неспособны действовать как двойной агонист и передавать сигнал за счет рецепторов в результате перекрестного связывания путем связывания как с OX40, так и с CD137. Расщепление или разрушение линкера может происходить до введения или после введения терапевтического средства индивидууму (например, посредством ферментативного расщепления или рН in vivo индивидуума), что приводит к уменьшению его эффективности во время циркуляции у индивидуума. Поскольку в молекулах антител, идентифицированных авторами настоящего изобретения, нет линкеров, ожидается, что молекулы антител сохранят одно и то же количество связывающих сайтов как до, так и после введения. Кроме того, структура молекул антител, идентифицированных авторами настоящего изобретения, также является предпочтительной с точки зрения иммуногенности молекул, поскольку введение гибридных белков или линкеров или того и другого может индуцировать иммуногенность при введении молекул индивидууму, что приводит к уменьшению эффективности терапевтического средства.
Таким образом, согласно настоящему изобретению предложено:
[1] Молекула антитела, которая связывается с CD137 и OX40, содержащая
(a) антигенсвязывающий сайт на основе области, определяющей комплементарность (CDR), для CD137; и
(b) антигенсвязывающий сайт для OX40, расположенный в домене CH3 указанной молекулы антитела;
причем указанный антигенсвязывающий сайт на основе CDR содержит CDR 1-6, представленные в:
(i) SEQ ID NO 1, 2, 3, 4, 5 и 6, соответственно [FS30-10-16];
(ii) SEQ ID NO 1, 2, 16, 4, 5 и 6, соответственно [FS30-10-3];
(iii) SEQ ID NO 1, 2, 21, 4, 5 и 6, соответственно [FS30-10-12];
(iv) SEQ ID NO 25, 26, 27, 4, 5 и 28, соответственно [FS30-35-14]; или
(v) SEQ ID NO 33, 34, 35, 4, 5 и 36, соответственно [FS30-5-37]; и
при этом указанный антигенсвязывающий сайт для OX40 содержит первую последовательность, вторую последовательность и третью последовательность, расположенные в структурных петлях AB, CD и EF домена CH3, соответственно, причем указанная первая, вторая и третья последовательность имеют последовательность, указанные под SEQ ID NO 51, 52 и 53, соответственно [FS20-22-49].
[2] Молекула антитела, которая связывается с CD137 и OX40, содержащая
(a) антигенсвязывающий сайт на основе области, определяющей комплементарность (CDR), для CD137; и
(b) антигенсвязывающий сайт для OX40, расположенный в домене CH3 указанной молекулы антитела;
причем указанный антигенсвязывающий сайт на основе CDR содержит CDR 1-6, представленные в:
(i) SEQ ID NO 7, 8, 9, 10, 11 и 6, соответственно [FS30-10-16];
(ii) SEQ ID NO 7, 8, 17, 10, 11 и 6, соответственно [FS30-10-3];
(iii) SEQ ID NO 7, 8, 22, 10, 11 и 6, соответственно [FS30-10-12];
(iv) SEQ ID NO 29, 30, 31, 10, 11 и 28, соответственно [FS30-35-14]; или
(v) SEQ ID NO 37, 38, 39, 10, 11 и 36, соответственно [FS30-5-37]; и
при этом указанный антигенсвязывающий сайт для OX40 содержит первую последовательность, вторую последовательность и третью последовательность, расположенные в структурных петлях AB, CD и EF указанного домена CH3, соответственно, причем первая, вторая и третья последовательность имеют последовательность, указанные под SEQ ID NO 51, 52 и 53, соответственно [FS20-22-49].
[3] Молекула антитела по п. [1] или [2], характеризующаяся тем, что:
(i) указанная первая последовательность расположена в положениях с 14 по 18 домена CH3 указанной молекулы антитела;
(ii) указанная вторая последовательность расположена в положениях с 45.1 по 77 домена CH3 указанной молекулы антитела; и/или
(iii) указанная третья последовательность расположена в положениях с 93 по 101 домена CH3 указанной молекулы антитела; и
при этом указанная нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[4] Молекула антитела по любому из пп. [1]-[3], характеризующаяся тем, что указанная молекула антитела содержит последовательность домена CH3, указанную под SEQ ID NO: 54 [FS20-22-49].
[5] Молекула антитела по любому из пп. [1]-[4], характеризующаяся тем, что указанная молекула антитела содержит CDR 1-6, представленные в любом из (i)-(iv) пп. [1] или [2].
[6] Молекула антитела по любому из пп. [1]-[5], характеризующаяся тем, что указанная молекула антитела содержит CDR 1-6, представленные в любом из (i)-(iii) пп. [1] или [2].
[7] Молекула антитела по любому из пп. [1]-[6], характеризующаяся тем, что указанная молекула антитела содержит CDR 1-6, представленные в (i) пп. [1] или [2].
[8] Молекула антитела по любому из пп. [1]-[7], характеризующаяся тем, что указанная молекула антитела содержит вариабельный домен тяжелой цепи (VH) и/или вариабельный домен легкой цепи (VL), предпочтительно домен VH и домен VL.
[9] Молекула антитела по любому из пп. [1]-[8], характеризующаяся тем, что указанная молекула антитела содержит тяжелую цепь иммуноглобулина и/или легкую цепь иммуноглобулина, предпочтительно тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина.
[10] Молекула антитела по пп. [8] или [9], характеризующаяся тем, что указанная молекула антитела содержит домен VH и/или домен VL, предпочтительно домен VH и домен VL, представленные в:
(i) SEQ ID NO 12 и 14, соответственно [FS30-10-16];
(ii) SEQ ID NO 18 и 14, соответственно [FS30-10-3];
(iii) SEQ ID NO 23 и 14, соответственно [FS30-10-12];
(iv) SEQ ID NO 170 и 172, соответственно [FS30-35-14]; или
(v) SEQ ID NO 40 и 42, соответственно [FS30-5-37];
[11] Молекула антитела по п. [10], характеризующаяся тем, что указанная молекула антитела содержит указанный домен VH и домен VL, представленные в любом из (i)-(iv) п. [10].
[12] Молекула антитела по пп. [10] или [11], характеризующаяся тем, что указанная молекула антитела содержит указанный домен VH и VL, представленные в любом из (i)-(iii) п. [10].
[13] Молекула антитела по любому из пп. [10]-[12], характеризующаяся тем, что указанная молекула антитела содержит указанный домен VH и домен VL, представленные в (i) п. [10].
[14] Молекула антитела по любому из пп. [1]-[13], характеризующаяся тем, что указанная молекула антитела представляет собой молекулу IgG1 человека.
[15] Молекула антитела по любому из пп. [1]-[14], характеризующаяся тем, что указанная молекула антитела содержит тяжелую цепь и легкую цепь антитела:
(i) FS20-22-49AA/FS30-10-16, представленные в SEQ ID NO 95 и 97, соответственно;
(ii) FS20-22-49AA/FS30-10-3, представленные в SEQ ID NO 99 и 97, соответственно;
(iii) FS20-22-49AA/FS30-10-12, представленные в SEQ ID NO 103 и 97, соответственно;
(iv) FS20-22-49AA/FS30-35-14, представленные в SEQ ID NO 105 и 107, соответственно; или
(v) FS20-22-49AA/FS30-5-37, представленные в SEQ ID NO 109 и 111, соответственно.
[16] Молекула антитела по п. [15], характеризующаяся тем, что указанная молекула антитела содержит указанную легкую цепь и тяжелую цепь, представленные в любом из (i)-(iv) п. [15].
[17] Молекула антитела по п. [15], характеризующаяся тем, что указанная молекула антитела содержит указанную легкую цепь и тяжелую цепь, представленные в любом из (i)-(iii) п. [15].
[18] Молекула антитела по п. [15], характеризующаяся тем, что указанная молекула антитела содержит указанную легкую цепь и тяжелую цепь, представленные в (i) п. [15].
[19] Молекула антитела по любому из пп. [1]-[18], характеризующаяся тем, что указанная молекула антитела связывает CD137 человека и OX40 человека.
[20] Молекула антитела по п. [19], характеризующаяся тем, что указанный CD137 человека состоит из последовательности, представленной в SEQ ID NO: 127, или содержит ее.
[21] Молекула антитела по п. [19] или [20], характеризующаяся тем, что указанный OX40 человека состоит из последовательности, представленной в SEQ ID NO: 130, или содержит ее.
[22] Молекула антитела по любому из пп. [1]-[21], характеризующаяся тем, что указанная молекула антитела связывает CD137 яванской макаки и OX40 яванской макаки.
[23] Молекула антитела по п. [22], характеризующаяся тем, что указанный CD137 яванской макаки состоит из последовательности, представленной в SEQ ID NO: 129, или содержит ее.
[24] Молекула антитела по п. [23] или [24], характеризующаяся тем, что указанный OX40 яванской макаки состоит из последовательности, представленной в SEQ ID NO: 131, или содержит ее.
[25] Молекула антитела по любому из пп. [5]-[7], [11]-[13] и [16]-[18], характеризующаяся тем, что указанная молекула антитела связывает CD137 человека и OX40 человека, и аффинность (KD), с которой указанная молекула антитела связывает CD137 человека, находится в пределах 2-кратной аффинности (KD), с которой указанная молекула антитела связывает OX40 человека.
[26] Молекула антитела по любому из пп. [19]-[25], характеризующаяся тем, что указанная молекула антитела способна одновременно связываться с CD137 человека и OX40 человека.
[27] Молекула антитела по любому из пп. [1]-[26], характеризующаяся тем, что указанная молекула антитела способна активировать OX40 на иммунной клетке в присутствии CD137, экспрессируемого на поверхности клетки.
[28] Молекула антитела по любому из пп. [1]-[27], характеризующаяся тем, что связывание указанной молекулы антитела с OX40 на иммунной клетке и с CD137 вызывает кластеризацию OX40 на указанной иммунной клетке.
[29] Молекула антитела по любому из пп. [1]-[28], характеризующаяся тем, что указанная молекула антитела способна активировать CD137 на иммунной клетке в присутствии OX40, экспрессируемого на поверхности клетки.
[30] Молекула антитела по любому из пп. [1]-[29], характеризующаяся тем, что связывание указанной молекулы антитела с CD137 на иммунной клетке и с OX40 вызывает кластеризацию CD137 на указанной иммунной клетке, и при этом OX40 экспрессируется на той же иммунной клетке или на отдельной клетке.
[31] Молекула антитела по любому из пп. [27]-[30], характеризующаяся тем, что указанная иммунная клетка представляет собой Т-клетку.
[32] Молекула антитела по любому из пп. [1]-[31], характеризующаяся тем, что указанная молекула антитела была модифицирована, чтобы уменьшить или устранить связывание домена CH2 указанной молекулы антитела с одним или более рецепторами Fcγ.
[33] Молекула антитела по любому из пп. [1]-[32], характеризующаяся тем, что указанная молекула антитела не связывается с одним или более рецепторами Fcγ.
[34] Молекула антитела по п. [32] или [33], характеризующаяся тем, что указанный рецептор Fcγ выбран из группы, состоящей из: FcγRI, FcγRIIa, FcγRIIb и FcγRIII.
[35] Молекула антитела по любому из пп. [1]-[34], характеризующаяся тем, что указанная молекула антитела способна индуцировать пролиферацию Т-клеток.
[36] Конъюгат, содержащий молекулу антитела по любому из пп. [1]-[35] и биоактивную молекулу.
[37] Конъюгат, содержащий молекулу антитела по любому из пп. [1]-[36] и обнаруживаемую метку.
[38] Молекула или молекулы нуклеиновой кислоты, кодирующие молекулу антитела по любому из пп. [1]-[35].
[39] Молекула или молекулы нуклеиновой кислоты, кодирующие молекулу антитела по любому из пп. [1]-[4], [8]-[10], [14]-[15] и [19]-[35], которые характеризуются тем, что указанная молекула (молекулы) нуклеиновой кислоты содержит (ат) последовательность нуклеиновой кислоты тяжелой цепи и/или последовательность нуклеиновой кислоты легкой цепи:
(i) FS20-22-49AA/FS30-10-16, представленные в SEQ ID NO 96 и 98, соответственно;
(ii) FS20-22-49AA/FS30-10-3, представленные в SEQ ID NO 100 и 102, соответственно;
(iii) FS20-22-49AA/FS30-10-12, представленные в SEQ ID NO 104 и 102, соответственно;
(iv) FS20-22-49AA/FS30-35-14, представленные в SEQ ID NO 106 и 108, соответственно; или
(v) FS20-22-49AA/FS30-5-37, представленные в SEQ ID NO 110 и 112, соответственно.
[40] Вектор или векторы, содержащие молекулу или молекулы нуклеиновой кислоты по любому из пп. [38]-[39].
[41] Рекомбинантная клетка-хозяин, содержащая молекулу (молекулы) нуклеиновой кислоты по любому из пп. [38]-[39], или вектор (ы) по п. [40].
[42] Способ получения молекулы антитела по любому из пп. [1]-[35], включающий культивирование рекомбинантной клетки-хозяина по п. [41] в условиях для получения указанной молекулы антитела.
[43] Способ по п. [42], дополнительно включающий выделение и/или очистку указанной молекулы антитела.
[44] Фармацевтическая композиция, содержащая молекулу антитела или конъюгат по любому из пп. [1]-[37] и фармацевтически приемлемое вспомогательное вещество.
[45] Молекула антитела или конъюгат по любому из пп. [1]-[37] для применения в способе лечения организма человека или животного с помощью терапии.
[46] Способ лечения заболевания или нарушения у индивидуума, включающий введение указанному индивидууму терапевтически эффективного количества молекулы антитела или конъюгата по любому из пп. [1]-[37].
[47] Молекула антитела или конъюгат для применения по п. [45], характеризующиеся тем, что указанная молекула антитела или конъюгат предназначены для применения в лечении рака или инфекционного заболевания у индивидуума.
[48] Способ по п. [46], который характеризуется тем, что указанное заболевание или нарушение представляет собой рак или инфекционное заболевание у индивидуума.
[49] Применение молекулы антитела или конъюгата по любому из пп. [1]-[37] в получении лекарственного средства для лечения рака или инфекционного заболевания.
[50] Молекула антитела или конъюгат для применения по п. [47], способ по п. [48] или применение молекулы антитела или конъюгата по п. [49], характеризующиеся тем, что указанный рак представляет собой солидный рак, причем необязательно указанный солидный рак выбран из группы, состоящей из меланомы, рака мочевого пузыря, рака головного мозга, рака молочной железы, рака яичников, рака легких, колоректального рака, рака шейки матки, рака печени, рака головы и шеи, рака поджелудочной железы, рака почек и рака желудка.
[51] Молекула антитела или конъюгат для применения по п. [47], способ по п. [48] или применение молекулы антитела или конъюгата по п. [49], характеризующиеся тем, что указанное инфекционное заболевание представляет собой персистирующую вирусную инфекцию, причем необязательно указанная персистирующая вирусная инфекция выбрана из группы, состоящей из вируса иммунодефицита человека (ВИЧ), вируса Эпштейна-Барр, цитомегаловируса, вируса гепатита В, вируса гепатита С, вируса ветряной оспы.
[52] Молекула антитела или конъюгат для применения по п. [47], способ по п. [48] или применение молекулы антитела или конъюгата по п. [49], характеризующиеся тем, что указанное инфекционное заболевание представляет собой персистирующую бактериальную инфекцию, причем необязательно указанная персистирующая бактериальная инфекция представляет собой персистирующую инфекцию Staphylococcus aureus, Hemophilus influenza, Mycobacterium tuberculosis, Mycobacterium leprae, Helicobacter pylori, Treponema pallidum, Enterococcus faecalis или Streptococcus pneumoniae.
[53] Молекула антитела или конъюгат для применения по п. [47], способ по п. [48] или применение молекулы антитела или конъюгата по п. [49], характеризующиеся тем, что указанное инфекционное заболевание представляет собой персистирующую грибковую инфекцию, причем необязательно указанная персистирующая грибковая инъекция представляет собой персистирующую инфекцию Candida, например, Candida albicans, Cryptococcus (gattii и neoformans), Talaromyces (Penicillium) marneffe, Microsporum, например, Microsporum audouinii, и Trichophyton tonsurans.
[54] Молекула антитела или конъюгат для применения по п. [47], способ по п. [48] или применение молекулы антитела или конъюгата по п. [49], характеризующиеся тем, что указанное инфекционное заболевание представляет собой персистирующую паразитарную инфекцию, причем необязательно указанная персистирующая паразитарная инъекция представляет собой персистирующую инфекцию Plasmodium, такую как Plasmodium falciparum, или Leishmania, такую как Leishmania donovani.
[55] Молекула антитела или конъюгат для применения по любому из пп. [45], [47] и [50]-[54], характеризующиеся тем, что указанное лечение включает введение указанной молекулы антитела или конъюгата индивидууму в комбинации со вторым терапевтическим средством.
[56] Способ по пп. [46], [48] и [50]-[54], характеризующийся тем, что указанный способ дополнительно включает введение терапевтически эффективного количества второго терапевтического средства индивидууму.
[57] Молекула антитела или конъюгат для применения в способе лечения рака у индивидуума по пп. [47] или [50], характеризующиеся тем, что указанный способ включает введение указанному индивидууму указанной молекулы антитела или конъюгата в комбинации с антителом, которое связывает PD-1 или PD-L1.
Краткое описание чертежей
Фигура 1 показывает выравнивание последовательностей доменов CH3 Fcab FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-49, FS20-22-85, FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 и FS20-31-115, а также Fcab дикого типа (WT). Указаны положения структурных петель AB, CD и EF, а также любые аминокислотные замены, делеции (обозначены тильдой «~») или вставки, присутствующие в доменах CH3 Fcab по сравнению с последовательностью WT. Показаны номера остатков в соответствии с системами нумерации IMGT, IMGT экзонов (последовательная нумерация), ЕС и Kabat.
Фигура 2 показывает активность МАТ к CD137 и МАТ2 к OX40/CD137 в анализе активации Т-клеток, экспрессирующих CD137 человека, в присутствии и в отсутствие перекрестного связывания. Фигуры 2A и B показывают высвобождение IL2 в присутствии повышающихся концентраций МАТ к CD137 и в присутствии (Фигура 2A) или в отсутствие (Фигура 2B) перекрестно связывающего антитела. G1AA/20H4.9 проявило активность в присутствии и в отсутствие перекрестно связывающего антитела, в то время как активность антител G1AA/MOR7480.1 и G1AA/FS30-10-16 была обнаружена только в присутствии перекрестно связывающего антитела. Фигура 2C и D показывают высвобождение IL-2 в присутствии повышающихся концентраций МАТ к CD137 FS30 в формате МАТ2, содержащем Fcab к OX40 человека (FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14) в присутствии (Фигура 2C) или в отсутствие (Фигура 2D) перекрестно связывающего агента. Были включены следующие контроли: антитело к CD137 G2/MOR7480.1 (положительный контроль); МАТ к OX40 G1/11D4 и МАТ2 FS20-22-49AA/4420 (отрицательные контроли); МАТ к FITC G1/4420 (отрицательный контроль изотипа). Фигура 2C показывает, что имело место зависящее от концентрации увеличение активации клеток DO11.10-hCD137, о чем свидетельствует увеличение высвобождения IL-2 мыши в присутствии перекрестно связанного положительного контроля МАТ (G2/MOR7480.1) и МАТ2 к CD137 FS30 (FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14), но не в присутствии МАТ и МАТ2 для отрицательного контроля (G1/4420, FS20-22-49AA/4420 и G1/11D4). Фигура 2D показывает, что в отсутствие перекрестного связывания положительный контроль G2/MOR7480.1, МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14, а также отрицательные контроли G1/4420, FS20-22-49AA/4420 и G1/11D4 показали отсутствие или слабую активацию Т-клеток, о чем свидетельствуют измеренные низкие начальные уровни IL-2.
Фигура 3 показывает активность МАТ к CD137, Fcab к OX40 и МАТ2 к OX40/CD137 в анализах с использованием стафилококкового энтеротоксина A (SEA). Высвобождение IL-2 измеряли в присутствии указанных МАТ/МАТ2, а также в присутствии и в отсутствие перекрестно связывающих агентов (FITC-декстран для МАТ к FITC и контрольных имитационных МАТ2 к OX40/FITC, а также антитело к CH2 человека для всех других протестированных молекул). Фигура 3A показывает высвобождение IL-2 в присутствии МАТ G1/4420 (против FITC; контроль изотипа), G1AA/MOR7480.1 (против CD137), G1AA/FS30-10-16 (против CD137), G1AA/20H4.9 (против CD137), G1AA/11D4 (против OX40), FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) и FS20-22-49AA/4420 плюс G1AA/FS30-10-16 в комбинации, а также МАТ2 FS20-22-49AA/FS30-10-16, в концентрации 3,7 нМ. Результаты показывают, что только МАТ2 к OX40/CD137 увеличивало активацию Т-клеток в отсутствие искусственных перекрестно связывающих агентов по сравнению с контролем изотипа, в то время как OX40-нацеленные антитела G1AA/11D4 и FS20-22-49AA/4420 и антитело к CD137 G1AA/20H4.9 показали увеличение активации Т-клеток только в присутствии искусственных перекрестно связывающих агентов по сравнению с контролем изотипа, а антитела к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 не проявили статистически достоверной активности даже в присутствии искусственного перекрестно связывающего агента. Фигура 3B показывает высвобождение IL-2 в присутствии МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в повышающихся концентрациях в присутствии и в отсутствие искусственного перекрестно связывающего агента (антитела к CH2 человека). Результаты показывают, что активация Т-клеток, индуцированная МАТ2 к OX40/CD137 в отсутствие антитела к CH2 человека, была сопоставима с активацией, которую тестировали в присутствии указанного искусственного перекрестно связывающего агента. Фигура 3C и D показывают высвобождение IL-2 в присутствии повышающихся концентраций МАТ и МАТ2 в присутствии (Фигура 3D) и в отсутствие (Фигура 3C) искусственного перекрестно связывающего агента (FITC-декстран для МАТ к FITC и контрольных имитационных МАТ2 к OX40/FITC и антитело к CH2 человека для всех других протестированных молекул). Контроли представляли собой: G1/4420 (против FITC), G1/11D4 (против OX40), G2/MOR7480.1 (против CD137), G1/11D4 плюс G2/MOR7480.1 в комбинации и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC). Результаты показывают, что имело место зависящее от концентрации увеличение активации Т-клеток, когда OX40 был связан контролями G1/11D4, как по отдельности, так и при дозировании в комбинации с МАТ к CD137 G2/MOR7480.1 и FS20-22-49A/4420, когда они были перекрестно связаны. МАТ2 к OX40/CD137 имело сравнимую активность в присутствии и в отсутствие искусственного перекрестно связывающего агента, и активность была сходна с активностью перекрестно связанного Fcab к OX40 (FS20-22-49AA/4420 Xlink). Незначительную активность наблюдали при использовании только контрольного антитела к CD137 (G2/MOR7480.1) как с перекрестным связыванием, так и без него.
Фигура 4 показывает активность МАТ к CD137, Fcab к OX40 и МАТ2 к OX40/CD137 в анализах активации общей популяции Т-клеток человека. Высвобождение IL-2 измеряли в присутствии указанных МАТ/МАТ2, а также в присутствии и в отсутствие перекрестно связывающих агентов (FITC-декстран для МАТ к FITC и контрольных имитационных МАТ2 к OX40/FITC, а также антитело к CH2 человека для всех других протестированных молекул). Фигура 4A показывает высвобождение IL-2 в присутствии МАТ и МАТ2 в концентрации 3,7 нМ. Результаты показывают, что только МАТ2 к OX40/CD137 увеличивало активацию Т-клеток в отсутствие искусственных перекрестно связывающих агентов. OX40-нацеленные антитела G1AA/11D4 и FS20-22-49AA/4420 и антитело к CD137 G1AA/20H4.9 показали увеличенную активацию Т-клеток только в присутствии перекрестно связывающих агентов. Активность не была обнаружена для антител к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 даже в присутствии искусственного перекрестно связывающего агента, что подтверждает результаты анализа с SEA, представленные на Фигуре 3A. Фигура 4B показывает высвобождение IL-2, индуцированное повышающимися концентрациями МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в присутствии и в отсутствие искусственного перекрестно связывающего агента (антитела к CH2 человека). МАТ2 к OX40/CD137 имело сравнимую активность в присутствии и в отсутствие искусственного перекрестно связывающего агента. Фигура 4C показывает высвобождение IL-2 в присутствии повышающихся концентраций МАТ2 к OX40/CD137 и контролей в отсутствие искусственных перекрестно связывающих агентов, в то время как Фигура 4D показывает высвобождение IL-2 в присутствии повышающихся концентраций контролей G1/4420, G1/11D4, G2/MOR7480.1 и FS20-22-49AA/4420 в виде отдельных агентов в присутствии искусственного перекрестно связывающего агента (FITC-декстран или антитело к CH2 человека, в зависимости от ситуации). Результаты показывают, что МАТ2 к OX40/CD137 обладало активностью в субнаномолярном или одноразрядном наномолярном диапазоне в отсутствие искусственного перекрестно связывающего агента. Как и ожидалось, контроль G1/4420 не обладал активностью, вне зависимости от присутствия перекрестно связывающего агента. Без присутствия перекрестно связывающего агента контроли G1/11D4, FS20-22-49AA/4420, G2/MOR7480.1 и комбинация G1/11D4 и G2/MOR7480.1 обладали незначительной активностью или не обладали ей. При перекрестном связывании антителом к CH2 человека или FITC-декстраном контроли против OX40 и против CD137 в виде отдельных агентов проявили увеличение активации Т-клеток, зависящее от концентрации, демонстрируя таким образом, что анализ был способен обнаруживать передачу сигнала за счет как рецепторов OX40, так и рецепторов CD137 на Т-клетках.
Фигура 5 показывает активность МАТ2 к OX40/CD137 человека в анализах активации CD4+ и CD8+ Т-клеток. Фигура 5A и B показывают высвобождение IL-2 в анализе активации CD4+ Т-клеток в присутствии повышающихся концентраций МАТ и МАТ2, как указано. МАТ и МАТ2 тестировали в присутствии (Фигура 5B) или в отсутствие (Фигура 5A) искусственных перекрестно связывающих агентов (FITC-декстран для МАТ к FITC и контрольных имитационных МАТ2 к OX40/FITC, а также антитело к CH2 человека для всех других протестированных молекул). Результаты показывают, что МАТ2 к OX40/CD137 было способно активировать CD4+ Т-клетки в отсутствие искусственного перекрестно связывающего агента. CD4+ Т-клетки были активированы перекрестно связанными контролями против OX40 G1AA/11D4 и FS20-22-49AA/4420 (по отдельности и в комбинации с G1AA/FS30-10-16), но не контролями против CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 в виде отдельных агентов. Контроль против OX40 FS20-22-49AA/4420 также показал низкий уровень активности в присутствии CD4+ Т-клеток без перекрестного связывания, который значительно повышался при перекрестном связывании антитела. Таким образом, было показано, что Fcab к OX40, общий для обоих имитационного МАТ2 FS20-22-49AA/4420 и МАТ2 FS20-22-49AA/FS30-10-16, был способен активировать CD4+ Т-клетки за счет агонистического действия на OX40, когда антитела были перекрестно связаны с помощью искусственного перекрестно связывающего агента или связывания Fab с CD137. Фигура 5C и D показывают высвобождение IL-2 в анализе активации CD8+ Т-клеток в присутствии повышающихся концентраций МАТ и МАТ2, как указано. МАТ и МАТ2 тестировали в присутствии (Фигура 5D) или в отсутствие (Фигура 5C) искусственных перекрестно связывающих агентов (для более подробной информации см. подпись к Фигурам 5A и B). Результаты показывают, что МАТ2 к OX40/CD137 было способно активировать CD8+ Т-клетки в отсутствие искусственного перекрестно связывающего агента. Активацию CD8+ Т-клеток наблюдали как для контролей против CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 (по отдельности и в комбинации с FS20-22-49AA/4420), так и для контролей против OX40 FS20-22-49AA/4420 и, в меньшей степени, G1AA/11D4 в присутствии искусственного перекрестно связывающего агента. Таким образом, было показано, что плечи Fab к CD137, общие как для контрольного МАТ G1AA/FS30-10-16, так и для МАТ2 FS20-22-49AA/FS30-10-16, способны оказывать агонистическое действие на CD137, экспрессируемый на CD8+ Т-клетках, когда антитела были перекрестно связаны либо с помощью искусственного перекрестно связывающего агента, либо связывания Fcab с OX40, в то время как Fcab к OX40, общий как для имитационного МАТ2 FS20-22-49AA/4420, так и для МАТ2 FS20-22-49AA/FS30-10-16, был способен активировать CD8+ Т-клетки за счет агонистического действия на OX40, когда антитела были перекрестно связаны с помощью искусственного перекрестно связывающего агента или связывания Fab с CD137. Фигура 5E и F показывают высвобождение IL-2 в анализе активации CD4+ и CD8+ Т-клеток, соответственно, в присутствии МАТ/МАТ2 в концентрации 3,7 нМ и в присутствии или в отсутствие искусственного перекрестно связывающего агента (для более подробной информации см. подпись к Фигурам 5A и B). Фигура 5E показывает, что МАТ2 к OX40/CD137 было способно активировать CD4+ Т-клетки в отсутствие искусственного перекрестно связывающего агента. CD4+ Т-клетки были активированы перекрестно связанными контролями против OX40 G1AA/11D4 и FS20-22-49AA/4420, но не контролями против CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 в виде отдельных агентов. Контроль против OX40 FS20-22-49AA/4420 также показал низкий уровень активности без перекрестного связывания, который значительно повышался при перекрестном связывании антитела. Таким образом, было показано, что Fcab к OX40, общий как для имитационного МАТ2 FS20-22-49AA/4420, так и для МАТ2 FS20-22-49AA/FS30-10-16, был способен активировать CD4+ Т-клетки за счет агонистического действия на OX40, когда антитела были перекрестно связаны с помощью искусственного перекрестно связывающего агента или связывания Fab с CD137. Фигура 5F показывает, что МАТ2 к OX40/CD137 было способно активировать CD8+ Т-клетки в отсутствие искусственного перекрестно связывающего агента. Активацию CD8+ Т-клеток наблюдали для контролей против CD137 G1AA/20H4.9 и G1AA/FS30-10-16 (по отдельности и в комбинации с FS20-22-49AA/4420) в присутствии искусственного перекрестно связывающего агента, но не для контроля против CD137 G1AA/MOR7480.1 или перекрестно связанных контролей против OX40 G1AA/11D4 и FS20-22-49AA/4420. Активацию CD8+ Т-клеток также наблюдали для контроля против CD137 G1AA/20H4.9 в отсутствие искусственного перекрестно связывающего агента. Таким образом, было показано, что плечи Fab к CD137, общие как для контрольного МАТ G1AA/FS30-10-16, так и для МАТ2 FS20-22-49AA/FS30-10-16, были способны оказывать агонистическое действие на CD137, экспрессируемый на CD8+ Т-клетках, когда антитела были перекрестно связаны либо с помощью искусственного перекрестно связывающего агента, либо связывания Fcab с OX40.
Фигура 6 показывает, что CD4+ Т-клетки экспрессируют более низкие уровни CD137 и более высокие уровни OX40, чем CD8+ Т-клетки. На графике показано среднее геометрическое значение интенсивности флуоресценции (GMFI) CD4+ или CD8+ Т-клеток, обработанных G1AA/MOR7480.1 или G1AA/11D4. Связывание G1AA/MOR7480.1 с CD137 является мерой экспрессии CD137, а связывание G1AA/11D4 с OX40 является мерой экспрессии OX40.
Фигура 7 показывает активность МАТ и МАТ2 к CD137 мыши в анализе активации Т-клеток. Фигура 7A и B показывают высвобождение IL-2 в присутствии повышающихся концентраций МАТ2, которое связывает рецепторы OX40 мыши и CD137 мыши (FS20m-232-91AA/Lob12.3), а также контрольных антител, в отсутствие (Фигура 7A) и в присутствии (Фигура 7B) искусственного перекрестно связывающего агента (антитела к CH2 человека или FITC-декстрана, в зависимости от ситуации). Контроли представляли собой антитела G1/4420 (против FITC), G1AA/OX86 (против mOX40), G1AA/Lob12.3 (против mCD137), G1AA/OX86 плюс G1AA/Lob12.3 в комбинации и FS20m-232-91AA/4420 (имитационное МАТ2 к mOX40/FITC). Результаты показывают, что в отсутствие перекрестно связывающего агента контроли G1AA/OX86, FS20m-232-91AA/4420, G1AA/Lob12.3 и комбинация G1AA/OX86 и G1AA/Lob12.3 не обладали активностью. При перекрестном связывании антителом к CH2 человека или FITC-декстраном контроли G1AA/OX86, FS20m-232-91AA/4420 и G1AA/OX86 плюс G1AA/Lob12.3 проявили зависимое от концентрации увеличение активации Т-клеток. Незначительное увеличение активности наблюдали для контроля G1AA/Lob12.3 при перекрестном связывании. МАТ2 к OX40/CD137 проявило хорошую активность независимо от присутствия искусственного перекрестно связывающего агента. Фигура 7C и D показывают активность различных антител к CD137 мыши (G1AA/Lob12.3 и G1AA/3H3) в отсутствие (Фигура 7C) или в присутствии (Фигура 7D) перекрестно связывающего антитела (клон MK1A6) в CD3-стимулированных клетках DO11.10-mCD137. Активность G1AA/3H3 наблюдали в присутствии и в отсутствие перекрестно связывающего антитела, в то время как активность антитела G1AA/Lob12.3 наблюдали только в присутствии перекрестно связывающего антитела. Таким образом, антитело G1AA/3H3 называют «независимым от перекрестного связывания», а антитело G1AA/Lob12.3 называют «зависимым от перекрестного связывания».
Фигура 8 показывает конкурентный анализ для тестирования активности клона МАТ2 к OX40/CD137 человека FS20-22-49AA/FS30-10-16 в присутствии 100-кратного избытка имитационного МАТ2, нацеленного на OX40 человека (FS20-22-49AA/4420), антитела к CD137 человека (G1AA/FS30-10-16) или их комбинации. Данные для двух повторов показаны как среднее значение плюс или минус стандартное отклонение среднего значения (СО). Статистический анализ выполняли с помощью одностороннего дисперсионного анализа и критерия Даннета для множественных сравнений. Звездочки над планками погрешностей представляют достоверное различие по сравнению с образцами, обработанными контролем изотипа (G1/4420) (***p<0,0002). Результаты показывают, что активность МАТ2 к OX40/CD137 была значительно уменьшена при превосходящей конкуренции со стороны как имитационного МАТ2 FS20-22-49AA/4420 за связывание с OX40, так и МАТ G1AA/FS30-10-16 за связывание с CD137, по сравнению с тем, когда МАТ2 к OX40/CD137 могло связываться с обоими рецепторами в отсутствие антитела к OX40 и антитела к CD137. Комбинация OX40-нацеленного имитационного МАТ2 FS20-22-49AA/4420 и МАТ к CD137 G1AA/FS30-10-16 дополнительно уменьшала активность МАТ2 к OX40/CD137. Эти результаты показывают, что для того чтобы МАТ2 к OX40/CD137 индуцировало активацию Т-клеток за счет кластеризации и агонистического действия на OX40 и CD137 требуется двойное связывание МАТ2 с обоими рецепторами.
Фигура 9 показывает конкурентный анализ для тестирования активности МАТ2 к OX40/CD137 мыши FS20m-232-91AA/Lob12.3 в присутствии 100-кратного избытка либо OX40-нацеленного имитационного МАТ2 к FS20m-232-91AA/4420, либо МАТ к CD137 G1/Lob12.3, либо отрицательного контроля МАТ G1AA/4420 (против FITC). Результаты показывают, что активность МАТ2 была значительно уменьшена при превосходящей конкуренции со стороны МАТ G1/Lob12.3 за связывание с CD137, а также уменьшилась до низкого уровня при превосходящей конкуренции со стороны имитационного МАТ2 FS20m-232-91AA/4420 за связывание с OX40, по сравнению с тем, когда МАТ2 могло связываться с обоими рецепторами в отсутствие антитела к OX40 и антитела к CD137. Как и ожидалось, сходный уровень активности наблюдали для МАТ2 в присутствии избытка МАТ для отрицательного контроля, как и в его отсутствие, и антител к OX40 и к CD137. Эти результаты показывают, что для того чтобы МАТ2 индуцировало активацию Т-клеток за счет кластеризации и агонистического действия на OX40 и CD137 требуется двойное связывание МАТ2 с обоими рецепторами.
Фигура 10 показывает противоопухолевую активность МАТ2 к OX40/CD137 мыши в модели сингенной опухоли CT26. На Фигуре 10A показаны средние объемы опухолей CT26 (плюс или минус стандартная ошибка среднего значения) мышей Balb/c, получавших G1/OX86 (положительный контроль против OX40 без мутации LALA), G1/Lob12.3 (положительный контроль против CD137 без мутации LALA), G1/4420 (контроль IgG), комбинацию G1/OX86 и G1/Lob12.3, комбинацию МАТ к OX40 G1AA/OX86 и МАТ к CD137 G1AA/Lob12.3 (оба с мутацией LALA), FS20m-232-91/Lob12.3 (МАТ2 к OX40/CD137 без мутации LALA) и FS20m-232-91AA/Lob12.3 (МАТ2 к OX40/CD137 с мутацией LALA). Результаты показывают, что лечение МАТ2 к OX40/CD137 как с мутацией LALA, так и без нее (FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3, соответственно) приводило к уменьшению роста опухоли по сравнению с лечением антителом к OX40 G1/OX86, антителом к CD137 G1/Lob12.3, комбинацией этих двух антител (G1/OX86 плюс G1/Lob12.3) и комбинацией LALA-содержащих антител к OX40 и к CD137 (G1AA/OX86 плюс G1AA/Lob12.3). Фигура 10B показывает объемы опухолей (с течением времени) отдельных мышей, несущих опухоль CT26, которые получали путем внутрибрюшинной инъекции 3 мг/кг контроля изотипа (клон G1AA/4420), имитационного МАТ2 к mOX40/FITC (клон FS20m-232-91AA/4420), МАТ к mCD137 (клон G1AA/Lob12.3), комбинации имитационного МАТ2 к mOX40/FITC и МАТ к mCD137 или МАТ2 к mOX40/CD137 (клон FS20m-232-91AA/Lob12.3). Горизонтальные пунктирные линии указывают положение 0 мм3 на оси ординат. В качественном анализе МАТ2 к mOX40/CD137 и комбинация имитационного МАТ2 к mOX40/FITC и МАТ к mCD137 ингибировали рост опухоли CT26 в подгруппе животных. Фигура 10C показывает средние объемы опухолей (плюс или минус стандартная ошибка среднего значения) мышей, несущих опухоль CT26, индивидуально представленных на Фигуре 10B. Группа, получавшая МАТ2 к mOX40/CD137, имела отсроченную раннюю фазу роста опухоли (дни 10-22) по сравнению с группой контроля изотипа. МАТ к mCD137 и имитационное МАТ2 к mOX40/FITC не влияли на скорости ранней фазы роста опухоли ни в виде отдельных агентов, ни в комбинации. Фигура 10D показывает график выживаемости Каплана-Мейера тех же мышей, несущих опухоль CT26, представленных на Фигуре 10B и 10C. Анализ выживаемости показывает, что лечение МАТ2 к mOX40/CD137, но не МАТ к mCD137 и имитационным МАТ2 к mOX40/FITC, как в виде отдельных агентов, так и в комбинации, привело к статистически достоверному увеличению выживаемости по сравнению с контролем изотипа. (Попарное сравнение выполняли с использованием лог-рангового критерия (Мантела-Кокса); ****p≤0,0001, ns = статистически недостоверно).
Фигура 11 показывает противоопухолевую активность МАТ2 к OX40/CD137 мыши в модели сингенной опухоли B16-F10. Мыши получали FS20m-232-91AA/Lob12.3 (МАТ2 к OX40/CD137) или G1/4420 (контроль IgG). На график нанесен средний объем опухоли плюс или минус стандартная ошибка среднего значения. Результаты показывают, что МАТ2 к OX40/CD137 было способно значительно уменьшать рост опухоли в сингенной модели B16-F10 по сравнению с мышами, получавшими контрольное антитело G1/4420.
Фигура 12 показывает активность МАТ2 к OX40/CD137 в комбинации с антителом к PD-1 или антителом к PD-L1 в анализе с SEA. Тестируемое МАТ2 представляло собой FS20-22-49AA/FS30-10-16. Контроли представляли собой G1/4420 (против FITC), G1AA/S1 (против PD-L1; Фигура 12A), G1AA/5C4 (против PD-1; Фигура 12B), протестированные либо в присутствии, либо в отсутствие МАТ2 FS20-22-49AA/FS30-10-16. Результаты показывают зависимое от концентрации увеличение активации Т-клеток в присутствии FS20-22-49AA/FS30-10-16, а также что добавление G1AA/S1 или G1AA/5C4 к МАТ2 FS20-22-49AA/FS30-10-16 увеличивало высвобождение IL-2 (максимальный ответ) по сравнению с Т-клетками, обработанными МАТ2 по отдельности. При обработке Т-клеток контрольными антителами по отдельности активность не наблюдалась. Статистический анализ между группами G1/4420 плюс FS20-22-49AA/FS30-10-16 и G1AA/S1 плюс FS20-22-49AA/FS30-10-16 (Фигура 12A) или G1/4420 плюс FS20-22-49AA/FS30-10-16 и G1AA/5C4 плюс FS20-22-49AA/FS30-10-16 (Фигура 12B) выполняли с использованием двустороннего дисперсионного анализа и критерия Тьюки для множественных сравнений, при этом звездочки указывают p-значение (*p<0,032, **p<0,0021, ***p<0,0002, ****p<0,0001).
Фигура 13 показывает противоопухолевую активность МАТ2 к OX40/CD137 мыши и антагониста PD-1 в модели опухоли CT26 на мышах, которые тестировали по отдельности и в комбинации. Показаны объемы опухолей у мышей, несущих опухоль CT26, получавших (Фигура 13A) комбинацию антител для контроля изотипа (G1AA/4420 и mIgG1/4420), (Фигура 13B) антитело к PD-1 мыши, (Фигура 13C) МАТ2 к OX40/CD137 мыши (МАТ2 FS20m-232-91AA/Lob12.3) или (Фигура 13D) комбинацию антитела к PD-1 мыши и МАТ2 к OX40/CD137 мыши FS20m-232-91AA/Lob12.3. Доля мышей с регрессировавшими опухолями (определяется как объем опухоли меньше или равный 62,5 мм3) по окончании исследования, через 60 дней после инокуляции клеток, показана для каждой группы лечения. Результаты показывают, что комбинация антагонистического антитела к PD-1 и FS20m-232-91AA/Lob12.3 привела к самой высокой доле животных, 7 из 15 (47%), с ответом в виде полной регрессии опухоли (Фигура 13D). Мыши, подвергнутые обработке антителом к PD-1 (Фигура 13B) или FS20m-232-91AA/Lob12.3 (Фигура 13C) в виде отдельных агентов, показали 0% и 7% регрессию опухоли в конце исследования, соответственно. Фигура 13E показывает график выживаемости Каплана-Мейера мышей, несущих опухоль CT26, получавших лечение, как описано для Фигур 13A-D. Анализ выживаемости показал, что комбинация FS20m-232-91AA/Lob12.3 и антитела к PD-1 привела к статистически достоверному преимуществу в выживаемости по сравнению с антителами для контроля изотипа (лог-ранговый критерий (Мантела-Кокса), p<0,0001). Не наблюдали значительных различий в выживаемости для обработок отдельным агентом в сравнении с антителами для контроля изотипа.
Фигура 14 показывает дозозависимую противоопухолевую активность МАТ2 к OX40/CD137 мыши в модели сингенной опухоли CT26. Фигура 14A показывает объемы опухолей мышей, несущих опухоль CT26, получавших путем внутрибрюшинной (в/б) инъекции либо 10 мг/кг антитела для контроля изотипа (G1AA/4420), либо 0,1, 0,3, 1, 3 или 10 мг/кг FS20m-232-91AA/Lob12.3. Доля мышей с регрессировавшими опухолями (определяется как объем опухоли меньше или равный 62,5 мм3) по окончании исследования, через 67 дней после инокуляции клеток, показана для каждой группы лечения (см. верхнюю правую часть каждого графика). Результаты показывают, что 0,3, 1, 3 или 10 мг/кг FS20m-232-91AA/Lob12.3 привели к регрессии опухоли у 4% (1/25), 4% (1/25), 8% (2/25) и 4% (1/25) животных в конце исследования, соответственно. Регрессия опухоли не наблюдалось ни у одного из животных в группах контроля изотипа и 0,1 мг/кг FS20m-232-91AA/Lob12.3. Фигура 14B показывает график выживаемости Каплана-Мейера мышей, несущих опухоль CT26, получавших лечение, как описано для Фигуры 14A. Анализ выживаемости показал, что FS20m-232-91AA/Lob12.3 при всех протестированных уровнях дозы привело к статистически достоверному преимуществу в выживаемости по сравнению с контролем изотипа. Сравнение групп 1 и 3 мг/кг и групп 3 и 10 мг/кг не выявило статистического различия в выживаемости. Попарное сравнение выполняли между каждой группой и контролем изотипа 10 мг/кг, если не указано иное, с использованием лог-рангового критерия (Мантела-Кокса); *p≤0,05, ***p≤0,0005, ****p≤0,0001, ns = статистически недостоверно.
Фигура 15 показывает сравнение противоопухолевой эффективности антител МАТ2 к OX40/CD137, содержащих различные клоны Fab к CD137, в модели сингенной опухоли CT26. Фигура 15A показывает средние объемы опухолей CT26 у мышей BALB/c, получавших G1/4420 (контроль IgG), FS20m-232-91AA/Lob12.3 (МАТ2 к OX40/CD137 с клоном агониста CD137 Lob12.3, зависимым от перекрестного связывания) и FS20m-232-91AA/3H3 (МАТ2 к OX40/CD137 с клоном агониста CD137 3H3, независимым от перекрестного связывания). Показаны средние объемы опухолей плюс или минус стандартная ошибка среднего значения. Результаты показывают, что лечение любым из антител МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3 или FS20m-232-91AA/3H3) привело к уменьшению роста опухоли по сравнению с лечением антителом для контроля изотипа (G1/4420) и что отсутствуют различия в уровне уменьшения у мышей, получавших FS20m-232-91AA/Lob12.3 или FS20m-232-91AA/3H3. Фигура 15B показывает график выживаемости Каплана-Мейера мышей, несущих опухоль CT26, получавших лечение, как описано для Фигуры 15A. Анализ выживаемости показал, что лечение любым из МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3 или FS20m-232-91AA/3H3) привело к статистически достоверному преимуществу в выживаемости по сравнению с лечением антителом для контроля изотипа (лог-ранговый критерий (Мантела-Кокса); p<0,05), при этом не наблюдали различий между мышами, получавшими любое из МАТ2 к OX40/CD137.
Подробное описание
Аспекты и варианты реализации настоящего изобретения далее будут описаны со ссылкой на прилагаемые фигуры. Дополнительные аспекты и варианты реализации будут понятны специалистам в данной области техники. Все документы, упомянутые в данном тексте, включены в настоящую заявку посредством ссылки.
Настоящее изобретение относится к молекулам антител, которые связываются как с CD137, так и с OX40. В частности, молекулы антител согласно настоящему изобретению содержат антигенсвязывающий сайт на основе CDR для CD137 и антигенсвязывающий сайт для OX40, расположенный в константном домене молекулы антитела. Термины «CD137» и «OX40» могут относиться к CD137 человека и OX40 человека, CD137 мыши и OX40 мыши и/или CD137 яванской макаки и OX40 яванской макаки, если контекст не требует иного. Предпочтительно термины «CD137» и «OX40» относятся к CD137 человека и OX40 человека, если контекст не требует иного.
Термин «молекула антитела» описывает иммуноглобулин, будь то природный или полученный частично или полностью синтетическим путем. Молекула антитела может быть человеческой или гуманизированной, предпочтительно человеческой. Молекула антитела предпочтительно представляет собой молекулу моноклонального антитела. Примеры антител представляют собой изотипы иммуноглобулинов, такие как иммуноглобулин G, и их изотипические подклассы, такие как IgG1, IgG2, IgG3 и IgG4, а также их фрагменты. Молекула антитела может быть выделенной в том смысле, что она не содержит примеси, такие как антитела, способные связывать другие полипептиды и/или компоненты сыворотки.
В настоящей заявке термин «молекула антитела» включает, таким образом, фрагменты антител, при условии, что указанные фрагменты содержат антигенсвязывающий сайт на основе CDR для CD137 и антигенсвязывающий сайт для OX40, расположенный в константном домене. Таким образом, если контекст не требует иного, в настоящей заявке термин «молекула антитела» эквивалентен «молекула антитела или ее фрагмент».
Для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела, можно применять моноклональные и другие антитела и использовать методики технологии рекомбинантной ДНК. Такие методики могут включать введение CDR или вариабельных областей и/или последовательностей константных доменов, обеспечивающих антигенсвязывающий сайт для OX40, в другой иммуноглобулин. Введение CDR одного иммуноглобулина в другой иммуноглобулин описано, например, в EP-A-184187, GB 2188638A и EP-A-239400. Аналогичные методики можно применять для соответствующих последовательностей константных доменов. В качестве альтернативы, гибридома или другая клетка, продуцирующая молекулу антитела, может быть подвергнута генетической мутации или другим изменениям, которые могут изменять специфичность связывания продуцируемых антител или могут не изменять ее.
Поскольку антитела можно модифицировать различными способами, термин «молекула антитела» следует толковать как охватывающий фрагменты антител, производные, функциональные эквиваленты и гомологи антител, включая любой полипептид, содержащий связывающий домен иммуноглобулина, будь то природный или полностью или частично синтетический. Таким образом, включены химерные молекулы, содержащие связывающий домен иммуноглобулина, или его эквивалент, гибридизованный с другим полипептидом. Клонирование и экспрессия химерных антител описаны в EP-A- 0120694 и EP-A-0125023.
Примером фрагмента антитела, содержащего как последовательности CDR, так и домен CH3, является минитело, которое содержит scFv, присоединенный к домену CH3 (Hu et al., 1996).
Молекула антитела согласно настоящему изобретению связывается с CD137 и OX40. Связывание в данном случае может относиться к специфичному связыванию. Термин «специфичный» может относиться к ситуации, в которой молекула антитела не будет показывать какое-либо значительное связывание с молекулами, отличными от ее партнера (партнеров) по специфичному связыванию, в настоящей заявке CD137 и OX40. Термин «специфичный» также применим, если молекула антитела специфична в отношении определенных эпитопов, таких как эпитопы на CD137 и OX40, которые несет ряд антигенов, и в этом случае молекула антитела будет способна связываться с различными антигенами, несущими эпитоп. Согласно предпочтительному варианту реализации молекула антитела согласно настоящему изобретению не связывается или не показывает какое-либо значительное связывание с TNFRSF1A, TNFRSF1B, GITR, NGFR, CD40 и/или DR6.
Антитела и способы их конструирования и применения хорошо известны в данной области техники и описаны, например, в Holliger and Hudson 2005. Для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела, можно применять моноклональные и другие антитела и использовать методики технологии рекомбинантной ДНК. Такие методики могут включать введение CDR или вариабельных областей одной молекулы антитела в другую молекулу антитела (EP-A-184187, GB 2188638A и EP-A-239400).
Антигенсвязывающий сайт на основе CDR представляет собой антигенсвязывающий сайт в вариабельной области антитела. Антигенсвязывающий сайт на основе CDR может быть образован тремя CDR, такими как три CDR вариабельного домена легкой цепи (VL) или три CDR вариабельного домена тяжелой цепи (VH). Предпочтительно антигенсвязывающий сайт на основе CDR образован шестью CDR, тремя CDR VL и тремя CDR VH. Вклад различных CDR в связывание антигена может варьироваться в разных антигенсвязывающих сайтах.
Три CDR домена VH антигенсвязывающего сайта могут быть расположены внутри домена VH иммуноглобулина, а три CDR домена VL могут быть расположены внутри домена VL иммуноглобулина. Например, антигенсвязывающий сайт на основе CDR может быть расположен в вариабельной области антитела.
Молекула антитела может иметь один или предпочтительно более одного, например, два, антигенсвязывающих сайта на основе CDR для первого антигена. Таким образом, молекула антитела может содержать один домен VH и один домен VL, но предпочтительно содержит два домена VH и два домена VL, то есть две пары доменов VH/VL, как в случае природных молекул IgG, в качестве примера.
Антигенсвязывающий сайт на основе CDR может содержать три CDR VH или три CDR VL, предпочтительно три CDR VH и три CDR VL антитела FS30-10-16, FS30-10-3, FS30-10-12 или FS30-35-14, или FS30-5-37, предпочтительно антитела FS30-10-16.
Последовательности доменов VH и VL этих антител представлены далее:
(i) последовательности доменов VH и VL для SEQ ID NO FS30-10-16 показаны в SEQ ID NO 12 и 14, соответственно;
(ii) последовательности доменов VH и VL для SEQ ID NO FS30-10-3 показаны в SEQ ID NO 18 и 14, соответственно;
(iii) последовательности доменов VH и VL для SEQ ID NO FS30-10-12 показаны в SEQ ID NO 23 и 14, соответственно;
(iv) последовательности доменов VH и VL для SEQ ID NO FS30-35-14 показаны в SEQ ID NO 170 и 172, соответственно; и
(v) последовательности доменов VH и VL для SEQ ID NO FS30-5-37 показаны в SEQ ID NO 40 и 42, соответственно.
Квалифицированный специалист легко определит последовательности CDR из последовательностей доменов VH и VL антител, представленных выше. Последовательности CDR могут быть определены, например, в соответствии с Kabat (Kabat et al., 1991) или международной информационной системой ImMunoGeneTics (IMGT) (Lefranc et al., 2015).
Последовательности CDR1, CDR2 и CDR3 домена VH молекулы антитела в соответствии с нумерацией IMGT могут представлять собой последовательности, расположенные в положениях 27-38, 56-65 и 105-117 домена VH молекулы антитела, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VH молекулы антитела в соответствии с нумерацией Kabat могут представлять собой последовательности, расположенные в положениях 31-35, 50-65 и 95-102 домена VH, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VL молекулы антитела в соответствии с нумерацией IMGT могут представлять собой последовательности, расположенные в положениях 27-38, 56-65 и 105-117 домена VL, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VL молекулы антитела в соответствии с нумерацией Kabat могут представлять собой последовательности, расположенные в положениях 24-34, 50-56 и 89-97 домена VL, соответственно.
Например, молекула антитела может содержать последовательность CDR1, CDR2 и CDR3 домена VH из:
(i) SEQ ID NO 1, 2 и 3, соответственно [FS30-10-16];
(ii) SEQ ID NO 1, 2 и 16, соответственно [FS30-10-3];
(iii) SEQ ID NO 1, 2 и 21, соответственно [FS30-10-12];
(iv) SEQ ID NO 25, 26 и 27, соответственно [FS30-35-14]; или
(v) SEQ ID NO 33, 34 и 35, соответственно [FS30-5-37],
причем указанные последовательности CDR определены в соответствии со схемой нумерации ImMunoGeneTics (IMGT).
Молекула антитела может содержать последовательность CDR1, CDR2 и CDR3 домена VH из:
(i) SEQ ID NO 7, 8 и 9, соответственно [FS30-10-16];
(ii) SEQ ID NO 7, 8 и 17, соответственно [FS30-10-3];
(iii) SEQ ID NO 7, 8 и 22, соответственно [FS30-10-12];
(iv) SEQ ID NO 29, 30 и 31, соответственно [FS30-35-14]; или
(v) SEQ ID NO 37, 38 и 39, соответственно [FS30-5-37],
причем указанные последовательности CDR определены в соответствии со схемой нумерации Kabat.
Например, молекула антитела может содержать последовательность CDR1, CDR2 и CDR3 домена VL из:
(i) SEQ ID NO 4, 5 и 6, соответственно [FS30-10-16];
(ii) SEQ ID NO 4, 5 и 6, соответственно [FS30-10-3];
(iii) SEQ ID NO 4, 5 и 6, соответственно [FS30-10-12];
(iv) SEQ ID NO 4, 5 и 28, соответственно [FS30-35-14]; или
(v) SEQ ID NO 4, 5 и 36, соответственно [FS30-5-37],
причем указанные последовательности CDR определены в соответствии со схемой нумерации ImMunoGeneTics (IMGT).
Например, молекула антитела может содержать последовательность CDR1, CDR2 и CDR3 домена VL из:
(i) SEQ ID NO 10, 11 и 6, соответственно [FS30-10-16];
(ii) SEQ ID NO 10, 11 и 6, соответственно [FS30-10-3];
(iii) SEQ ID NO 10, 11 и 6, соответственно [FS30-10-12];
(iv) SEQ ID NO 10, 11 и 28, соответственно [FS30-35-14]; или
(v) SEQ ID NO 10, 11 и 36, соответственно [FS30-5-37],
причем указанные последовательности CDR определены в соответствии со схемой нумерации Kabat.
Последовательности VH и VL антител FS30-10-16, FS30-10-3 и FS30-10-12 идентичны, за исключением остатка в положении 109 VH в соответствии со схемой нумерации IMGT (остаток 97 VH в соответствии со схемой нумерации Kabat). Таким образом, молекула антитела может содержать последовательности CDR1, CDR2 и CDR3 домена VH и/или последовательности CDR1, CDR2 и CDR3 домена VL, последовательность домена VH и/или последовательность домена VL, антитела FS30-10-16, причем указанная молекула антитела необязательно содержит аминокислотную замену в положении 109 тяжелой цепи в соответствии со схемой нумерации IMGT (остаток 97 тяжелой цепи в соответствии со схемой нумерации Kabat), при этом указанный остаток в указанном положении предпочтительно выбран из группы, состоящей из аспарагина (N), треонина (T) и лейцина (L).
Антигенсвязывающий сайт на основе CDR может содержать домены VH или VL, предпочтительно домены VH и VL, антитела FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14 или FS30-5-37, предпочтительно антитела FS30-10-16, FS30-10-3, FS30-10-12 или FS30-35-14, более предпочтительно антитела FS30-10-16, FS30-10-3 или FS30-10-12, наиболее предпочтительно антитела FS30-10-16.
Домен VH антител FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14 и FS30-5-37 может иметь последовательность, указанные под SEQ ID NO 12, 18, 23, 170 и 40, соответственно. Домен VL антител FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14 и FS30-5-37 может иметь последовательность, указанные под SEQ ID NO 14, 14, 14, 172 и 42, соответственно.
Молекула антитела согласно настоящему изобретению содержит антигенсвязывающий сайт для OX40, расположенный в константном домене молекулы антитела. Константный домен может представлять собой домен CL, CH1, CH2, CH3 или CH4, предпочтительно константный домен представляет собой домен CH1, CH2 или CH3, более предпочтительно домен CH2 или CH3, наиболее предпочтительно домен CH3.
Положения аминокислотных остатков константного домена пронумерованы в настоящей заявке в соответствии со схемой нумерации ImMunoGeneTics (IMGT), если не указано иное. Схема нумерации IMGT описана в Lefranc et al., Dev. Comp. Immunol., 29, 185-203 (2005).
Антигенсвязывающий сайт для OX40 может содержать первую, вторую и третью последовательность, расположенные в первой, второй и третьей структурной петле константного домена, соответственно. Конструирование структурных петель константного домена антитела для создания антигенсвязывающих сайтов для антигенов-мишеней известно в данной области техники и описано, например, в Wozniak-Knopp et al., 2010, и публикациях патентов №№ WO2006/072620 и WO2009/132876. Предпочтительно первая, вторая и третья структурные петли представляют собой структурные петли AB, CD и EF домена CH3 молекулы антитела, соответственно. В домене CH3 структурные петли AB, CD и EF расположены в остатках 11-18, 43-78 и 92-101 домена CH3, соответственно. Модификация последовательностей структурных петель константных доменов антител для создания новых антигенсвязывающих сайтов описана, например, в WO2006/072620 и WO2009/132876.
Согласно предпочтительному варианту реализации антигенсвязывающий сайт для OX40 молекулы антитела содержит первую, вторую и третью последовательность:
(i) FS20-22-49, указанные под SEQ ID NO 51, 52 и 53, соответственно;
(ii) FS20-22-38, указанные под SEQ ID NO 51, 59 и 60, соответственно;
(iii) FS20-22-41, указанные под SEQ ID NO 51, 52 и 60, соответственно;
(iv) FS20-22-47, указанные под SEQ ID NO 51, 52 и 65, соответственно; или
(v) FS20-22-85, указанные под SEQ ID NO 51, 52 и 68, соответственно.
Антигенсвязывающий сайт для OX40 может содержать последовательности структурных петель AB, CD и EF FS20-22-49, FS20-22-38, FS20-22-41, FS20-22-47 или FS20-22-85, причем указанные структурные петли AB, CD и EF представляют собой последовательности, расположенные в остатках 11-18, 43-78 и 92-101 домена CH3, соответственно, и домен CH3 FS20-22-49, FS20-22-38, FS20-22-41, FS20-22-47 или FS20-22-85 представлен в SEQ ID NO: 54, 61, 63, 66 и 69, соответственно.
Согласно более предпочтительному варианту реализации антигенсвязывающий сайт для OX40 молекулы антитела содержит первую, вторую и третью последовательность FS20-22-49, указанные под SEQ ID NO 51, 52 и 53, соответственно. Например, антигенсвязывающий сайт для OX40 может содержать последовательности структурных петель AB, CD и EF FS20-22-49, указанные под SEQ ID NO 56, 57 и 58, соответственно.
Если антигенсвязывающий сайт для OX40 молекулы антитела содержит первую, вторую и третью последовательность FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-49 или FS20-22-85, указанная первая, вторая и третья последовательность предпочтительно расположены в положениях с 14 по 18, с 45.1 по 77 и с 93 по 101 домена CH3 молекулы антитела, соответственно.
Если антигенсвязывающий сайт для OX40 содержит последовательности структурных петель AB, CD и EF FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-49 или FS20-22-85, указанные последовательности структурных петель AB, CD и EF предпочтительно расположены в положениях с 11 по 18, с 43 по 78 и с 92 по 101 домена CH3 молекулы антитела, соответственно.
Молекула антитела может дополнительно содержать лейцин (L) в положении 91 домена CH3 молекулы антитела. В частности, молекула антитела, содержащая антигенсвязывающий сайт для OX40, содержащий первую, вторую и третью последовательность FS20-22-85, может содержать лейцин в положении 91 домена CH3 молекулы антитела.
Согласно альтернативному варианту реализации антигенсвязывающий сайт для OX40 молекулы антитела содержит первую, вторую и третью последовательность:
(i) FS20-31-58, указанные под SEQ ID NO 71, 72 и 73, соответственно;
(ii) FS20-31-66, указанные под SEQ ID NO 71, 72 и 76, соответственно;
(iii) FS20-31-94, указанные под SEQ ID NO 79, 80 и 81, соответственно;
(iv) FS20-31-102, указанные под SEQ ID NO 84, 85 и 76, соответственно;
(v) FS20-31-108, указанные под SEQ ID NO 84, 88 и 89, соответственно; или
(vi) FS20-31-115, указанные под SEQ ID NO 84, 92 и 89, соответственно.
Антигенсвязывающий сайт для OX40 может содержать последовательности структурных петель AB, CD и EF FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 или FS20-31-115, причем указанные структурные петли AB, CD и EF представляют собой последовательности, расположенные в остатках 11-18, 43-78 и 92-101 домена CH3, соответственно, и домен CH3 FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 или FS20-31-115 представлен в SEQ ID NO: 54, 61, 63, 66 и 69, соответственно.
Если антигенсвязывающий сайт для OX40 молекулы антитела содержит первую, вторую и третью последовательность FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 или FS20-31-115, указанная первая, вторая и третья последовательность предпочтительно расположены в положениях с 14 по 18, с 45.1 по 77 и с 92 по 101 домена CH3 молекулы антитела, соответственно.
Если антигенсвязывающий сайт для OX40 содержит последовательности структурных петель AB, CD и EF FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 или FS20-31-115, указанные последовательности структурных петель AB, CD и EF предпочтительно расположены в положениях с 11 по 18, с 43 по 78 и с 92 по 101 домена CH3 молекулы антитела, соответственно.
В качестве альтернативы нумерации IMGT положения аминокислотных остатков в константном домене, включая положение аминокислотных последовательностей, замен, делеций и вставок, описанных в настоящей заявке, могут быть пронумерованы в соответствии с нумерацией IMGT экзонов (также называемой последовательной нумерацией), нумерацией ЕС или нумерацией Kabat. Соответствие между нумерацией IMGT, нумерацией IMGT экзонов, нумерацией ЕС и нумерацией Kabat положений остатков домена CH3 показано на Фигуре 1.
Таким образом, например, в том случае, когда настоящая заявка относится к первой, второй и третьей последовательности, расположенным в положениях с 14 по 18, с 45.1 по 77 и с 93 по 101 домена CH3 молекулы антитела, соответственно, причем указанные положения остатков пронумерованы в соответствии со схемой нумерации IMGT, указанная первая, вторая и третья последовательность расположены в положениях с 18 по 22, с 46 по 50 и с 74 по 82 домена CH3, при этом указанные положения остатков пронумерованы в соответствии со схемой нумерацией IMGT экзонов, как показано на Фигуре 1.
Согласно одному варианту реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-49, FS20-22-85, FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 или FS20-31-115, причем указанная последовательность домена CH3 FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-49, FS20-22-85, FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 и FS20-31-115 представлена в SEQ ID NO 54, 61, 63, 66, 69, 74, 77, 82, 86, 90 и 93, соответственно.
Согласно предпочтительному варианту реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 FS20-22-49, представленной в SEQ ID NO 54.
Домен CH3 молекулы антитела необязательно может содержать дополнительный остаток лизина (К) на самом C-конце последовательности домена CH3.
Кроме того, молекула антитела согласно настоящему изобретению может содержать домен CH2 молекулы иммуноглобулина G, такой как домен CH2 молекулы IgG1, IgG2, IgG3 или IgG4. Предпочтительно молекула антитела согласно настоящему изобретению содержит домен CH2 молекулы IgG1. Домен CH2 может иметь последовательность, указанную под SEQ ID NO: 48.
Домен CH2 молекулы антитела может содержать одну или более мутаций, которые уменьшают или устраняют связывание домена CH2 с одним или более рецепторами Fcγ, такими как FcγRI, FcγRIIa, FcγRIIb, FcγRIII, и/или комплементом. Авторы настоящего изобретения допускают, что уменьшение или устранение связывания с рецепторами Fcγ будет уменьшать или устранять АЗКЦ, опосредуемую молекулой антитела. Аналогичным образом, ожидается, что уменьшение или устранение связывания с комплементом приведет к уменьшению или устранению комплементзависимой цитотоксичности (КЗЦ), опосредуемой молекулой антитела. Мутации для уменьшения или устранения связывания домена CH2 с одним или более рецепторами Fcγ и/или комплементом известны в данной области техники (Wang et al., 2018). Эти мутации включают «мутацию LALA», описанную в Bruhns et al., 2009 и Hezareh et al., 2001, которая включает замену остатков лейцина в положениях 1.3 и 1.2 IMGT домена CH2 аланином (L1.3A и L1.2A). В качестве альтернативы, получение a-гликозильных антител посредством мутации консервативного сайта N-связанного гликозилирования путем мутации аспарагина (N) в положении 84.4 IMGT домена CH2 на аланин, глицин или глутамин (N84.4A, N84.4G или N84.4Q), как известно, также уменьшает эффекторную функцию IgG1 (Wang et al., 2018). В качестве другой альтернативы, активация комплемента (связывание C1q) и АЗКЦ, как известно, уменьшаются посредством мутации пролина в положении 114 IMGT домена CH2 на аланин или глицин (P114A или P114G) (Idusogie et al., 2000; Klein et al., 2016). Эти мутации также можно комбинировать для того чтобы получить молекулы антител с дополнительно уменьшенной активностью АЗКЦ или КЗЦ или без нее.
Таким образом, молекула антитела может содержать домен CH2, причем указанный домен CH2 содержит:
(i) остатки аланина в положениях 1.3 и 1.2; и/или
(ii) аланин или глицин в положении 114; и/или
(iii) аланин, глутамин или глицин в положении 84.4;
при этом указанная нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Согласно предпочтительному варианту реализации молекула антитела содержит домен CH2, причем указанный домен CH2 содержит:
(i) остаток аланина в положении 1.3; и
(ii) остаток аланина в положении 1.2;
при этом указанная нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Например, домен CH2 может иметь последовательность, указанную под SEQ ID NO: 49.
Согласно альтернативному предпочтительному варианту реализации молекула антитела содержит домен CH2, причем указанный домен CH2 содержит:
(i) остаток аланина в положении 1.3;
(ii) остаток аланина в положении 1.2; и
(iii) аланин в положении 114;
при этом указанная нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Например, домен CH2 может иметь последовательность, указанную под SEQ ID NO: 50.
Согласно предпочтительному варианту реализации молекула антитела, которая связывается с CD137 и OX40, содержит
(а) антигенсвязывающий сайт на основе CDR для CD137; и
(b) антигенсвязывающий сайт для OX40, расположенный в домене CH3 молекулы антитела;
причем указанный антигенсвязывающий сайт на основе CDR содержит три CDR VH и три CDR VL (CDR 1-6) антитела FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14, или FS30-5-37, предпочтительно FS30-10-16, FS30-10-3 или FS30-10-12, более предпочтительно FS30-10-16 или FS30-10-3, наиболее предпочтительно FS30-10-16; и
причем указанный антигенсвязывающий сайт для OX40 содержит первую последовательность, вторую последовательность и третью последовательность, расположенные в структурных петлях AB, CD и EF домена CH3, соответственно, при этом указанная первая, вторая и третья последовательности имеют последовательность FS20-22-49, указанные под SEQ ID NO 51, 52 и 53, соответственно.
Согласно другому предпочтительному варианту реализации молекула антитела, которая связывается с CD137 и OX40, содержит
(а) антигенсвязывающий сайт на основе CDR для CD137; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 54 [FS20-22-49];
причем указанный антигенсвязывающий сайт на основе CDR содержит три CDR VH и три CDR VL (CDR 1-6) антитела FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14, или FS30-5-37, предпочтительно FS30-10-16, FS30-10-3 или FS30-10-12, более предпочтительно FS30-10-16 или FS30-10-3, наиболее предпочтительно FS30-10-16.
Согласно другому предпочтительному варианту реализации молекула антитела, которая связывается с CD137 и OX40, содержит
(а) домен VH и домен VL, содержащие антигенсвязывающий сайт на основе CDR для CD137; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 54 [FS20-22-49];
причем указанный домен VH и VL содержит, имеет или состоит из VH и VL антитела FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14 или FS30-5-37, предпочтительно FS30-10-16, FS30-10-3 или FS30-10-12, более предпочтительно FS30-10-16 или FS30-10-3, наиболее предпочтительно FS30-10-16.
Согласно другому предпочтительному варианту реализации молекула антитела, которая связывается с CD137 и OX40, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела:
(i) FS20-22-49AA/FS30-10-16, представленных в SEQ ID NO 95 и 97, соответственно;
(ii) FS20-22-49AA/FS30-10-3, представленных в SEQ ID NO 99 и 97, соответственно;
(iii) FS20-22-49AA/FS30-10-12, представленных в SEQ ID NO 103 и 97, соответственно;
(iv) FS20-22-49AA/FS30-35-14, представленных в SEQ ID NO 105 и 107, соответственно; или
(v) FS20-22-49AA/FS30-5-37, представленных в SEQ ID NO 109 и 111, соответственно;
причем указанная молекула антитела предпочтительно содержит легкую цепь и тяжелую цепь, представленные в (i)-(iv), более предпочтительно содержит легкую цепь и тяжелую цепь, представленные в (i)-(iii), наиболее предпочтительно содержит легкую цепь и тяжелую цепь, представленные в (i).
Молекулы антител согласно настоящему изобретению также могут содержать варианты первой, второй или третьей последовательности, последовательности структурной петли AB, CD или EF, домена CH3, домена CH2, домена CH2 и CH3, CDR, домена VH, домена VL, последовательностей легкой цепи и/или тяжелой цепи, раскрытых в настоящей заявке. Подходящие варианты могут быть получены с помощью способов изменения последовательности, или мутации, и скрининга. Согласно предпочтительному варианту реализации молекула антитела, содержащая один или более вариантов последовательностей, сохраняет одну или более функциональных характеристик исходной молекулы антитела, таких как специфичность связывания и/или аффинность связывания в отношении CD137 и OX40. Например, молекула антитела, содержащая один или более вариантов последовательностей, предпочтительно связывается с CD137 и/или OX40 с такой же аффинностью или с более высокой аффинностью, чем (исходная) молекула антитела. Исходная молекула антитела представляет собой молекулу антитела, которая не содержит аминокислотную (ые) замену (замены), делецию (делеции) и/или вставку (вставки), которые были включены в вариант молекулы антитела.
Например, молекула антитела согласно настоящему изобретению может содержать первую, вторую или третью последовательность, последовательность структурной петли AB, CD или EF, домен CH3, домен CH2, домен CH2 и CH3, CDR, домен VH, домен VL, последовательность легкой цепи и/или тяжелой цепи, которая имеет по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичность последовательности со структурной петлей, доменом CH3, доменом CH2, доменом CH2 и CH3, CDR, доменом VH, доменом VL, последовательностью легкой цепи или тяжелой цепи, раскрытыми в настоящей заявке.
Согласно предпочтительному варианту реализации молекула антитела согласно настоящему изобретению содержит последовательность домена CH3, которая имеет по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичность последовательности с последовательностью домена CH3, представленной в SEQ ID NO: 54 [FS20-22-49].
Согласно другому предпочтительному варианту реализации молекула антитела имеет или содержит последовательность домена CH2, которая имеет по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,1%, по меньшей мере 99,2%, по меньшей мере 99,3%, по меньшей мере 99,4%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% идентичность последовательности с последовательностью домена CH2, представленной в SEQ ID NO: 48 или 49.
Идентичность последовательности обычно определяется со ссылкой на алгоритм GAP (пакет Wisconsin GCG, Accelerys Inc, Сан-Диего, США). GAP использует алгоритм Нидлмана и Вунша для выравнивания двух полных последовательностей, увеличивая до максимального количество совпадений и уменьшая до минимального количество пропусков. Обычно используются параметры по умолчанию со штрафом за создание пропуска равным 12 и штрафом за расширение пропуска равным 4. Использование GAP может быть предпочтительным, но могут использоваться другие алгоритмы, например, BLAST (который использует метод Altschul et al. (1990)), FASTA (который использует метод Pearson and Lipman (1988)), или алгоритм Смита-Уотермана (Smith and Waterman (1981), или программа TBLASTN, Altschul et al. 1990, выше, обычно с применением параметров по умолчанию. В частности, можно использовать алгоритм psi-Blast (Altschul et al., 1997).
Молекула антитела согласно настоящему изобретению также может содержать первую, вторую или третью последовательность, последовательность структурной петли AB, CD или EF, домен CH3, домен CH2, домен CH2 и CH3, CDR, домен VH, домен VL, легкую цепь и/или тяжелую цепь, которая имеет одно или более изменений аминокислотной последовательности (добавление, делецию, замену и/или вставку аминокислотного остатка), предпочтительно 20 изменений или менее, 15 изменений или менее, 10 изменений или менее, 5 изменений или менее, 4 изменения или менее, 3 изменения или менее, 2 изменения или менее или 1 изменение по сравнению с первой, второй или третьей последовательностью, последовательностью структурной петли AB, CD или EF, доменом CH3, доменом CH2, доменом CH2 и CH3, Fcab, CDR, доменом VH, доменом VL, последовательностью легкой цепи или тяжелой цепи, раскрытыми в настоящей заявке. В частности, изменения могут быть сделаны в одном или более каркасных участках молекулы антитела за пределами последовательностей доменов VH и VL и/или в одном или более каркасных участках домена CH3. Например, изменения могут присутствовать в домене CH3 за пределами последовательностей, описанных в настоящей заявке как первая, вторая и третья последовательности или как последовательности структурных петель AB, CD или EF.
Согласно предпочтительному варианту реализации молекула антитела согласно настоящему изобретению может содержать последовательность домена CH3 с одним или более изменениями аминокислотной последовательности (добавлением, делецией, заменой и/или вставкой аминокислотного остатка), предпочтительно 20 изменениями или менее, 15 изменениями или менее, 10 изменениями или менее, 5 изменениями или менее, 4 изменениями или менее, 3 изменениями или менее, 2 изменениями или менее или 1 изменением по сравнению с последовательностью домена CH3, представленной в SEQ ID NO: 54, 61, 63, 66, 69, 74, 77, 82, 86, 90 или 93.
Согласно другому предпочтительному варианту реализации молекула антитела содержит последовательность домена CH2 с одним или более изменениями аминокислотной последовательности (добавлением, делецией, заменой и/или вставкой аминокислотного остатка), предпочтительно 20 изменениями или менее, 15 изменениями или менее, 10 изменениями или менее, 5 изменениями или менее, 4 изменениями или менее, 3 изменениями или менее, 2 изменениями или менее или 1 изменением по сравнению с последовательностью домена CH2, представленной в SEQ ID NO: 48 или 49.
Согласно предпочтительным вариантам реализации, в которых одна или более аминокислот заменены другой аминокислотой, замены могут представлять собой консервативные замены, например, в соответствии со следующей Таблицей. Согласно некоторым вариантам реализации аминокислоты в одной и той же категории в средней колонке заменяют друг другом, т. е., например, неполярную аминокислоту заменяют другой неполярной аминокислотой. Согласно некоторым вариантам реализации аминокислоты в одной строке в крайней правой колонке заменяют друг другом.
Согласно некоторым вариантам реализации замена (ы) может (могут) быть функционально консервативной (ыми). Другими словами, в некоторых вариантах реализации замена может не влиять (или может не влиять в значительной степени) на одно или более функциональных свойств (например, аффинность связывания) молекулы антитела, содержащей замену, по сравнению с эквивалентной незамещенной молекулой антитела.
Молекула антитела предпочтительно связывается с CD137 человека и OX40 человека. Предпочтительно молекула антитела способна одновременно связываться с CD137 человека и OX40 человека, причем CD137 человека и OX40 человека совместно экспрессируются. Совместная экспрессия в данном случае включает ситуации, в которых CD137 и OX40 экспрессируются на одной и той же клетке, например, иммунной клетке, такой как Т-клетка, и ситуации, в которых CD137 и OX40 экспрессируются на разных клетках, например, на двух разных иммунных клетках, расположенных рядом друг с другом в микросреде опухоли. Таким образом, полагают, что молекулы антител согласно настоящему изобретению способны связываться с обеими мишенями на одной клетке в цис-условиях, а также способны связываться с двумя мишенями, экспрессируемыми на разных клетках, в транс-условиях.
Молекула антитела предпочтительно связывается с димерным CD137 человека с аффинностью (KD) 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,4 нМ или 0,3 нМ или с более высокой аффинностью. Предпочтительно молекула антитела связывается с CD137 человека с аффинностью (KD) 0,3 нМ или с более высокой аффинностью. Молекула антитела может связывать димерный CD137 с более высокой аффинностью, чем мономерный CD137. CD137 человека может иметь последовательность, например, указанную под SEQ ID NO: 127.
Молекула антитела предпочтительно связывается с димерным OX40 человека с аффинностью (KD) 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,4 нМ или 0,3 нМ или с более высокой аффинностью. Предпочтительно молекула антитела связывается с OX40 человека с аффинностью (KD) 0,3 нМ или с более высокой аффинностью. Молекула антитела может связывать димерный OX40 с более высокой аффинностью, чем мономерный OX40. OX40 человека может иметь последовательность, например, указанную под SEQ ID NO: 130.
Молекула антитела предпочтительно связывается с CD137 яванской макаки и OX40 яванской макаки. Связывание с CD137 и OX40 яванской макаки, а также с CD137 и OX40 человека является благоприятным, поскольку оно позволяет тестировать эффективность и токсичность молекулы антитела на яванских макаках перед введением человеку. Предпочтительно молекула антитела способна одновременно связываться с CD137 яванской макаки и OX40 яванской макаки, при этом CD137 яванской макаки и OX40 яванской макаки совместно экспрессируются.
Молекула антитела предпочтительно связывается с димерным CD137 яванской макаки с аффинностью (KD) 10 нМ, 9 нМ, 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,4 нМ, или 0,3 нМ, или с более высокой аффинностью. Предпочтительно молекула антитела связывается с димерным CD137 яванской макаки с аффинностью (KD) 0,3 нМ или с более высокой аффинностью. CD137 яванской макаки может иметь последовательность, например, указанную под SEQ ID NO: 129.
Молекула антитела предпочтительно связывается с димерным OX40 яванской макаки с аффинностью (KD) 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2,5 нМ, 2 нМ, 1,5 нМ или 1 нМ или с более высокой аффинностью. Предпочтительно молекула антитела связывается с OX40 яванской макаки с аффинностью (KD) 1 нМ или с более высокой аффинностью. OX40 яванской макаки может иметь последовательность, например, указанную под SEQ ID NO: 131.
Молекула антитела предпочтительно связывается с димерным OX40 яванской макаки с аффинностью (KD), которая находится в пределах 10-кратной, 9-кратной, 8-кратной, 7-кратной, 6-кратной или 5-кратной аффинности (KD), с которой молекула антитела связывается с димерным OX40 человека. Предпочтительно молекула антитела связывается с димерным OX40 яванской макаки с аффинностью (KD), которая находится в пределах 5-кратной аффинности (KD), с которой молекула антитела связывается с димерным OX40 человека.
Молекула антитела предпочтительно связывается с димерным CD137 яванской макаки с аффинностью (KD), которая находится в пределах 30-кратной, 20-кратной, 10-кратной, 5-кратной, 4-кратной, 3-кратной или 2-кратной аффинности (KD), с которой молекула антитела связывается с димерным CD137 человека. Предпочтительно молекула антитела связывается с димерным CD137 яванской макаки с аффинностью (KD), которая находится в пределах 2-кратной аффинности (KD), с которой молекула антитела связывается с димерным CD137 человека.
Как описано в Примерах в настоящей заявке, полагают, что сходство в связывании с антигенами человека и яванской макаки может представлять собой преимущество, поскольку можно надеяться, что поведение МАТ2 в исследованиях на яванских макаках можно будет экстраполировать на человека. Полагают, что это является благоприятным для проведения исследований эффективности и токсичности, осуществляемых с использованием молекулы антитела, на яванских макаках, которые могут иметь прогностическую ценность в отношении эффективности и токсичности молекулы антитела у человека.
Молекула антитела предпочтительно связывается с димерным CD137 человека с аффинностью (KD), которая находится в пределах 10-кратной, 9-кратной, 8-кратной, 7-кратной, 6-кратной или 5-кратной аффинности (KD), с которой молекула антитела связывается с димерным OX40 человека. Предпочтительно молекула антитела связывается с димерным CD137 человека с аффинностью (KD), которая находится в пределах 2-кратной аффинности (KD), с которой молекула антитела связывается с димерным OX40 человека.
Молекула антитела предпочтительно связывается с димерным CD137 яванской макаки с аффинностью (KD), которая находится в пределах 10-кратной, 9-кратной, 8-кратной, 7-кратной, 6-кратной или 5-кратной аффинности (KD), с которой молекула антитела связывается с димерным OX40 яванской макаки.
Как описано в Примерах в настоящей заявке, полагают, что молекула антитела, имеющая сходную аффинность в отношении связывания с обеими мишенями, т. е. CD137 и OX40, может обеспечивать преимущество, поскольку молекула антитела с большей вероятностью будет связываться с клетками, которые экспрессируют обе мишени.
Аффинность связывания молекулы антитела с когнатным антигеном, таким как OX40 человека, CD137 человека, OX40 яванской макаки или CD137 яванской макаки, может быть определена с помощью поверхностного плазмонного резонанса (SPR), такого как Biacore, в качестве примера. Аффинность связывания молекулы антитела с OX40 или CD137, экспрессируемыми на поверхности клетки, может быть определена с помощью проточной цитометрии.
Было показано, что молекулы антител обладают несколькими видами активности в отношении связывания лиганда. Например, молекула антитела может быть способна блокировать, может быть неспособна блокировать или может быть способна частично блокировать связывание CD137L с CD137.
Предпочтительно молекула антитела может быть способна блокировать, может быть неспособна блокировать или может быть способна частично блокировать связывание CD137L с CD137. Более предпочтительно молекула антитела способна частично блокировать связывание CD137L с CD137.
Предпочтительно молекула антитела способна индуцировать передачу сигнала OX40 и/или CD137 в результате перекрестного связывания путем двойного связывания как с OX40, так и с CD137, когда две мишени совместно экспрессируются. На основании такого способа действия такие молекулы антител называют «двойными агонистами», т. е. молекулы антител способны индуцировать передачу сигнала за счет рецепторов в результате перекрестного связывания путем двойного связывания как с OX40, так и с CD137. Таким образом, предпочтительно молекула антитела способна вызывать двойное агонистическое действие, когда оба OX40 и CD137 совместно экспрессируются. Как описано в настоящей заявке, ожидается, что такие двойные агонисты обеспечат преимущество. Например, полагают, что такой двойной агонист может быть способен вызывать более сильную стимуляцию иммунного ответа, поскольку он может сочетать активацию разных иммунных клеток, например, сочетать активность CD8+ и CD4+ Т-клеток с помощью связывания с обеими мишенями на разных клетках в транс-условиях. В качестве другого примера, полагают, что такой двойной агонист может быть способен приводить к активации одной клетки, совместно экспрессирующей обе мишени, без потребности во взаимодействии двух клеток, путем связывания с обеими мишенями в цис-условиях.
Более предпочтительно двойной агонист должен быть способен запускать агонистическое действие автономно путем одновременного связывания со своими специфичными мишенями (OX40 и CD137) и без потребности в дополнительном перекрестном связывании, например, перекрестно связывающих агентов или рецепторов Fcγ. Как описано в настоящей заявке, ожидается, что такая автономная активность будет обеспечивать преимущество, поскольку она будет ограничена местами, в которых совместно экспрессируются обе мишени, и, следовательно, ожидается, что она уменьшит токсичность, потенциально ассоциированную с активацией CD137, в местах, в которых совместная экспрессия OX40 является незначительной или отсутствует.
Способность молекулы антитела активировать Т-клетки может быть измерена с помощью анализа активации Т-клеток. Т-клетки высвобождают IL-2 при активации. Таким образом, анализ активации Т-клеток может измерять высвобождение IL-2 для определения уровня активации Т-клеток, индуцированной молекулой антитела.
Например, способность молекулы антитела активировать Т-клетки определяют путем измерения концентрации молекулы антитела, требуемой для достижения полумаксимального высвобождения IL-2 Т-клетками в анализе активации Т-клеток. Далее она называется ЭК50.
Согласно предпочтительному варианту реализации молекула антитела имеет ЭК50 в анализе активации Т-клеток, которая находится в пределах 50-кратной, 40-кратной, 30-кратной, 20-кратной, 10-кратной или 5-кратной ЭК50 FS20-22-49AA/FS30-10-16 в этом же анализе, причем FS20-22-49AA/FS30-10-16 состоит из тяжелой цепи, имеющей SEQ ID NO: 95, и легкой цепи, имеющей SEQ ID NO: 97.
Например, молекула антитела может иметь ЭК50 в анализе активации Т-клеток 30 нМ или менее, 25 нМ или менее, 20 нМ или менее, 14 нМ или менее, 10 нМ или менее, 5 нМ или менее, 4 нМ или менее, 3 нМ или менее, 2 нМ или менее, 1,5 нМ, 1 нМ или 0,5 нМ или менее, предпочтительно 1,5 нМ или менее, более предпочтительно 1 нМ или менее при перекрестном связывании.
В дополнение или альтернативно, способность молекулы антитела активировать Т-клетки может быть определена путем измерения максимальной концентрации IL-2, высвобождаемого Т-клетками в анализе активации Т-клеток в присутствии молекулы антитела.
Согласно предпочтительному варианту реализации максимальная концентрация IL-2, высвобождаемого Т-клетками в анализе активации Т-клеток в присутствии молекулы антитела, находится в пределах 20% или 10% от максимальной концентрации IL-2, высвобождаемого Т-клетками в присутствии FS20-22-49AA/FS30-10-16 в этом же анализе, причем FS20-22-49AA/FS30-10-16 состоит из тяжелой цепи, имеющей SEQ ID NO: 95, и легкой цепи, имеющей SEQ ID NO: 97.
Анализ активации Т-клеток предпочтительно содержит Т-клетки, совместно экспрессирующие OX40 и CD137. Согласно предпочтительному варианту реализации анализ активации Т-клеток не содержит какие-либо агенты, способные перекрестно связывать молекулы антител, отличные от CD137 и OX40.
Анализ активации Т-клеток может представлять собой анализ Т-клеток, описанный в настоящей заявке, такой как анализ общей популяции Т-клеток, анализ CD4+ Т-клеток или анализ CD8+ Т-клеток, описанные в Примерах в настоящей заявке.
Например, анализ активации Т-клеток может представлять собой анализ высвобождения IL-2 на основе Т-клеток, выделенных из мононуклеарных клеток периферической крови (МКПК). Анализ активации CD4+ T-клеток или анализ активации CD8+ T-клеток может представлять собой анализ высвобождения IL-2 на основе CD4+ T-клеток или CD8+ T-клеток, выделенных из МКПК человека, соответственно. Как объясняется в Примерах в настоящей заявке, молекула антитела, которая способна активировать Т-клетки как в анализе CD4+ Т-клеток, так и в анализе CD8+ Т-клеток, способна активировать как OX40, так и CD137 (также называется «двойной агонист»). Например, анализ активации Т-клеток может включать выделение МКПК человека из конусов для истощения лейкоцитов. Способы выделения МКПК известны в данной области техники и описаны в примерах в настоящей заявке. Затем Т-клетки могут быть выделены из МКПК. Способы выделения Т-клеток (всех Т-клеток, CD4+ Т-клеток или CD8+ Т-клеток) из МКПК также известны в данной области техники и описаны в Примерах в настоящей заявке.
Анализ активации может включать получение требуемого количества Т-клеток, например, в экспериментальных средах, таких как среда для Т-клеток. Требуемое количество Т-клеток может быть получено в концентрации 1,0×106 клеток/мл. Затем Т-клетки можно стимулировать с использованием подходящего реагента для активации Т-клеток, который обеспечивает сигналы, требуемые для активации Т-клеток. Например, реагент для активации Т-клеток может представлять собой реагент, содержащий CD3 и CD28, такой как гранулы, содержащие CD3 и CD28. Выделенные Т-клетки можно инкубировать в течение ночи с реагентом для активации Т-клеток, чтобы активировать Т-клетки. После этого активированные Т-клетки можно промыть, чтобы отделить Т-клетки от реагента для активации Т-клеток, и повторно суспендировать в среде для Т-клеток в подходящей концентрации, такой как 2,0×106 клеток/мл. Затем активированные Т-клетки могут быть добавлены в планшеты, покрытые антителом к CD3 человека.
Подходящее разведение каждой тестируемой молекулы антитела может быть приготовлено и добавлено в лунки. Затем Т-клетки можно инкубировать при 37°C, 5% CO2 в течение 24 часов с тестируемым антителом. Супернатанты могут быть собраны и проанализированы для определения концентрации IL-2 в супернатанте. Способы определения концентрации IL-2 в растворе известны в данной области техники и описаны в примерах в настоящей заявке. Концентрация IL-2 человека может быть нанесена на график в зависимости от логарифмической концентрации молекулы антитела. Полученные кривые могут быть аппроксимированы с использованием уравнения зависимости ответа от log(агониста).
Молекула антитела может быть конъюгирована с биоактивной молекулой или обнаруживаемой меткой. В этом случае молекула антитела может называться конъюгатом. Такие конъюгаты можно применять в лечении заболеваний, описанных в настоящей заявке.
Например, биоактивная молекула может представлять собой модулятор иммунной системы, такой как цитокин, предпочтительно цитокин человека. Например, цитокин может представлять собой цитокин, который стимулирует активацию и/или пролиферацию Т-клеток. Примеры цитокинов для конъюгации с молекулой антитела включают IL-2, IL-10, IL-12, IL-15, IL-21, ГМ-КСФ и ИФН-гамма.
В качестве альтернативы, биоактивная молекула может представлять собой ловушку лиганда, такую как ловушка лиганда цитокина, например TGF-бета или IL-6.
В качестве альтернативы, биоактивная молекула может представлять собой терапевтический радиоизотоп.
Радиоиммунотерапия используется в лечении рака, в качестве примера. Терапевтические радиоизотопы, подходящие для радиоиммунотерапии, известны в данной области техники и включают иттрий-90, йод-131, висмут-213, астат-211, лютеций 177, рений-188, медь-67, актиний-225 и йод-125 и тербий-161.
Подходящие обнаруживаемые метки, которые могут быть конъюгированы с молекулами антител, известны в данной области техники и включают радиоизотопы, такие как йод-125, йод-131, иттрий-90, индий-111 и технеций-99; флуорохромы, такие как флуоресцеин, родамин, фикоэритрин, техасский красный и производные цианинового красителя, например Cy7 и Alexa750; хромогенные красители, такие как диаминобензидин; латексные гранулы; ферментные метки, такие как пероксидаза хрена; люминофорные или лазерные красители со спектрально изолированными характеристиками поглощения или испускания; и химические группы, такие как биотин, которые могут быть обнаружены за счет связывания со специфичной когнатной обнаруживаемой группой, например, меченым авидином.
Молекула антитела может быть конъюгирована с биоактивной молекулой или обнаруживаемой меткой с помощью любой подходящей ковалентной или нековалентной связи, такой как дисульфидная или пептидная связь. Если биоактивная молекула представляет собой цитокин, цитокин может быть присоединен к молекуле антитела с помощью пептидного линкера. Подходящие пептидные линкеры известны в данной области техники и могут представлять собой от 5 до 25, от 5 до 20, от 5 до 15, от 10 до 25, от 10 до 20 или от 10 до 15 аминокислот в длину.
Согласно некоторым вариантам реализации биоактивная молекула может быть конъюгирована с молекулой антитела с помощью расщепляемого линкера. Линкер может обеспечивать высвобождение биоактивной молекулы из молекулы антитела в месте терапии. Линкеры могут включать амидные связи (например, пептидные линкеры), дисульфидные связи или гидразоны. Пептидные линкеры, например, могут быть расщеплены сайт-специфичными протеазами, дисульфидные связи могут быть расщеплены восстанавливающей средой цитозоля, а гидразоны могут быть расщеплены посредством кислотно-опосредуемого гидролиза.
Согласно настоящему изобретению также предложена выделенная молекула или молекулы нуклеиновой кислоты, кодирующие молекулу антитела согласно настоящему изобретению. Квалифицированный специалист легко получит такие молекулы нуклеиновой кислоты с использованием способов, хорошо известных в данной области техники.
Молекула или молекулы нуклеиновой кислоты могут содержать, например, последовательность, указанную под SEQ ID NO: 55 или 113, 62, 64, 67, 70, 75, 78, 83, 87, 91 или 94, которые кодируют домены CH3 FS20-22-49, FS20-22-38, FS20-22-41, FS20-22-47, FS20-22-85, FS20-31-58, FS20-31-66, FS20-31-94, FS20-31-102, FS20-31-108 и FS20-31-115, соответственно. Например, молекула или молекулы нуклеиновой кислоты могут содержать последовательность, указанную под SEQ ID NO: 55 или 113, обе из которых кодируют домен CH3 FS20-22-49. Согласно некоторым вариантам реализации молекула или молекулы нуклеиновой кислоты содержат последовательность, указанную под SEQ ID NO: 113, которая кодирует домен CH3 FS20-22-49. Предпочтительно молекула или молекулы нуклеиновой кислоты содержат последовательность, указанную под SEQ ID NO: 55, которая кодирует домен CH3 FS20-22-49.
Молекула или молекулы нуклеиновой кислоты могут кодировать домен VH и/или домен VL, предпочтительно домен VH и домен VL антитела FS30-10-16, FS30-10-3, FS30-10-12, FS30-35-14 или FS30-5-37, предпочтительно антитела FS30-10-16, FS30-10-3, FS30-10-12 или FS30-35-14, более предпочтительно антитела FS30-10-16, FS30-10-3 или FS30-10-12, наиболее предпочтительно антитела FS30-10-16. В настоящей заявке описаны последовательности доменов VH и VL этих антител.
Например, молекула (ы) нуклеиновой кислоты может (могут) содержать:
(i) последовательность нуклеиновой кислоты домена VH антитела FS30-10-16, указанную под SEQ ID NO: 13, и/или последовательность нуклеиновой кислоты домена VL антитела FS30-10-16, указанную под SEQ ID NO: 15; или
(ii) последовательность нуклеиновой кислоты домена VH антитела FS30-10-3, указанную под SEQ ID NO: 19, и/или последовательность нуклеиновой кислоты домена VL антитела FS30-10-3, указанную под SEQ ID NO: 20;
(iii) последовательность нуклеиновой кислоты домена VH антитела FS30-10-12, указанную под SEQ ID NO: 24, и/или последовательность нуклеиновой кислоты домена VL антитела FS30-10-12, указанную под SEQ ID NO: 20;
(iv) последовательность нуклеиновой кислоты домена VH антитела FS30-35-14, указанную под SEQ ID NO: 171, и/или последовательность нуклеиновой кислоты домена VL антитела FS30-35-14, указанную под SEQ ID NO: 32; или
(v) последовательность нуклеиновой кислоты домена VH антитела FS30-5-37, указанную под SEQ ID NO: 41, и/или последовательность нуклеиновой кислоты домена VL антитела FS30-5-37, указанную под SEQ ID NO: 43.
Молекула или молекулы нуклеиновой кислоты могут кодировать тяжелую цепь и/или легкую цепь, предпочтительно тяжелую цепь и легкую цепь антитела FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-35-14 или FS20-22-49AA/FS30-5-37, предпочтительно антитела FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 или FS20-22-49AA/FS30-35-14, более предпочтительно антитела FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-10-3 или FS20-22-49AA/FS30-10-12, наиболее предпочтительно FS20-22-49AA/FS30-10-16. В настоящей заявке описаны последовательности доменов VH и VL этих антител.
Например, молекула (молекулы) нуклеиновой кислоты может (могут) содержать:
(i) последовательность нуклеиновой кислоты тяжелой цепи антитела FS20-22-49AA/FS30-10-16, указанную под SEQ ID NO: 96, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS20-22-49AA/FS30-10-16, указанную под SEQ ID NO: 98; или
(ii) последовательность нуклеиновой кислоты тяжелой цепи антитела FS20-22-49AA/FS30-10-3, указанную под SEQ ID NO: 100, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS20-22-49AA/FS30-10-3, указанную под SEQ ID NO: 102;
(iii) последовательность нуклеиновой кислоты тяжелой цепи антитела FS20-22-49AA/FS30-10-12, указанную под SEQ ID NO: 104, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS20-22-49AA/FS30-10-12, указанную под SEQ ID NO: 102;
(iv) последовательность нуклеиновой кислоты тяжелой цепи антитела FS20-22-49AA/FS30-35-14, указанную под SEQ ID NO: 106, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS20-22-49AA/FS30-35-14, указанную под SEQ ID NO: 108; или
(v) последовательность нуклеиновой кислоты тяжелой цепи антитела FS20-22-49AA/FS30-5-37, указанную под SEQ ID NO: 110, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS20-22-49AA/FS30-5-37, указанную под SEQ ID NO: 112.
Если нуклеиновая кислота кодирует домен VH и VL, или тяжелую и легкую цепь, молекулы антитела согласно настоящему изобретению, два домена или цепи могут кодироваться на двух отдельных молекулах нуклеиновой кислоты.
Выделенную молекулу нуклеиновой кислоты можно применять для экспрессии молекулы антитела согласно настоящему изобретению. Нуклеиновая кислота обычно будет обеспечена в форме рекомбинантного вектора для экспрессии. Таким образом, согласно другому аспекту настоящего изобретения предложен вектор, содержащий нуклеиновую кислоту, описанную выше. Могут быть выбраны или сконструированы подходящие векторы, содержащие подходящие регуляторные последовательности, включая промоторные последовательности, фрагменты терминаторов, последовательности полиаденилирования, последовательности энхансеров, маркерные гены и другие последовательности, в зависимости от ситуации. Предпочтительно вектор содержит соответствующие регуляторные последовательности для управления экспрессией нуклеиновой кислоты в клетке-хозяине. Векторы могут представлять собой плазмиды, вирусные векторы, например, фаг, или фагмиду, в зависимости от ситуации.
Молекула нуклеиновой кислоты или вектор, описанные в настоящей заявке, могут быть введены в клетку-хозяина. Методики введения нуклеиновой кислоты или векторов в клетки-хозяева хорошо установлены в данной области техники, и можно применять любую подходящую методику. Ряд клеток-хозяев, подходящих для получения рекомбинантных молекул антител, известен в данной области техники и включает клетки-хозяева бактерий, дрожжей, насекомых или млекопитающих. Предпочтительная клетка-хозяин представляет собой клетку млекопитающего, такую как клетка CHO, NS0 или HEK, например, клетку HEK293.
Согласно другому аспекту настоящего изобретения предложен способ получения молекулы антитела согласно настоящему изобретению, включающий экспрессию нуклеиновой кислоты, кодирующей молекулу антитела, в клетке-хозяине и необязательно выделение и/или очистку молекулы антитела, полученной таким способом. Способы культивирования клеток-хозяев хорошо известны в данной области техники. Способ может дополнительно включать выделение и/или очистку молекулы антитела. Методики очистки рекомбинантных молекул антител хорошо известны в данной области техники и включают, например, ВЭЖХ, FPLC или аффинную хроматографию, например, с использованием белка A или белка L. Согласно некоторым вариантам реализации очистка может быть выполнена с использованием аффинной метки на молекуле антитела. Способ также может включать изготовление молекулы антитела в виде фармацевтической композиции, необязательно с фармацевтически приемлемым вспомогательным веществом или другим веществом, описанным ниже. Как объяснялось выше, CD137 и OX40 оба экспрессируются на клетках иммунной системы, включая Т-клетки. Например, OX40 экспрессируется на клетках иммунной системы, включая активированные Т-клетки, в частности, CD4+ Т-клетки, CD8+ Т-клетки, хелперные Т-клетки (Th1) типа 1, хелперные Т-клетки типа 2 (Th2), регуляторные Т-клетки (Treg) и инфильтрирующие опухоль Т-клетки, а также активированные природные клетки-киллеры (NK-клетки). CD137 экспрессируется на клетках иммунной системы, включая Т-клетки, в частности, CD8+ Т-клетки, В-клетки, NK-клетки и инфильтрирующие опухоль лимфоциты (TIL). CD137 экспрессируется на более низком уровне на CD4+ Т-клетках, чем на CD8+ Т-клетках (см. Пример 14 и Фигуру 6), но также было показано, что он вовлечен в индукцию пролиферации и активации некоторых подгрупп CD4+ Т-клеток (Wen et al., 2002).
Было показано, что активация OX40 играет роль в усилении активации Т-клеток, клональном размножении Т-клеток, дифференцировке и выживании Т-клеток, а также в выработке Т-клеток памяти. Было показано, что активация CD137 играет роль в усилении пролиферации, выживаемости и цитотоксической эффекторной функции CD8+ Т-клеток, а также в дифференцировке CD8+ Т-клеток и поддержании CD8+ Т-клеток памяти. Также было продемонстрировано, что активация CD137 усиливает АЗКЦ, опосредуемую NK-клетками, а также пролиферацию, выживаемость B-клеток и выработку ими цитокинов.
С учетом активности OX40 и CD137 в усилении иммунного ответа молекулы агонистов OX40 и CD137 были изучены применительно к лечению рака, также ожидается, что они найдут применение в лечении инфекционных заболеваний.
Таким образом, молекулы антител, описанные в настоящей заявке, можно применять для терапевтических приложений, в частности, в лечении рака и инфекционных заболеваний.
Молекулу антитела, описанную в настоящей заявке, можно применять в способе лечения организма человека или животного. Согласно связанным аспектам настоящего изобретения предложена;
(i) молекула антитела, описанная в настоящей заявке, для применения в качестве лекарственного средства,
(ii) молекула антитела, описанная в настоящей заявке, для применения в способе лечения заболевания или нарушения,
(iii) применение молекулы антитела, описанной в настоящей заявке, в изготовлении лекарственного средства для применения в лечении заболевания или нарушения; и,
(iv) способ лечения заболевания или нарушения у индивидуума, причем указанный способ включает введение индивидууму терапевтически эффективного количества молекулы антитела, описанной в настоящей заявке.
Индивидуум может представлять собой пациента, предпочтительно пациента-человека.
Лечение может представлять собой любое лечение или терапию, при которых достигается некоторый целевой терапевтический эффект, например, ингибирование или отсрочка прогрессирования состояния, и включает уменьшение скорости прогрессирования, замедление скорости прогрессирования, улучшение состояния, излечение или ремиссию (будь то частичную или полную) состояния, предотвращение, улучшение, отсрочку, ослабление или купирование одного или более симптомов и/или признаков состояния или увеличение продолжительности выживаемости индивидуума или пациента сверх ожидаемой в отсутствие лечения.
Также включено лечение в качестве профилактической меры (например, профилактики). Например, индивидуума, восприимчивого или подверженного риску возникновения или повторного возникновения заболевания, такого как рак, можно лечить, как описано в настоящей заявке. Такое лечение может предотвратить или отсрочить возникновение или повторное возникновение заболевания у индивидуума.
Описанный способ лечения может включать введение по меньшей мере одного дополнительного средства лечения индивидууму в дополнение к молекуле антитела. Таким образом, молекула антитела, описанная в настоящей заявке, может быть введена индивидууму по отдельности или в комбинации с одним или более другими средствами лечения. Если молекулу антитела вводят индивидууму в комбинации с другим средством лечения, дополнительное средство лечения может быть введено индивидууму одновременно, последовательно или отдельно от введения молекулы антитела. Если дополнительное средство лечение вводят одновременно с молекулой антитела, молекула антитела и дополнительное средство лечения могут быть введены индивидууму в виде комбинированного препарата. Например, дополнительная терапия может представлять собой известную терапию или терапевтический агент для заболевания, подлежащего лечению.
Несмотря на то, что молекула антитела может быть введена отдельно, молекулы антител обычно будут введены в виде фармацевтической композиции, которая может содержать по меньшей мере один компонент в дополнение к молекуле антитела. Таким образом, согласно другому аспекту настоящего изобретения предложена фармацевтическая композиция, содержащая молекулу антитела, описанную в настоящей заявке. Также предложен способ, включающий изготовление молекулы антитела в виде фармацевтической композиции.
Фармацевтические композиции могут содержать, в дополнение к молекуле антитела, фармацевтически приемлемое вспомогательное вещество, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области техники. В настоящей заявке термин «фармацевтически приемлемый» относится к соединениям, материалам, композициям и/или лекарственным формам, которые, в рамках здравого медицинского заключения, подходят для применения в контакте с тканями субъекта (например, человека) без чрезмерной токсичности, раздражения, аллергической реакции или другой проблемы или осложнения, соизмеримо с разумным соотношением польза/риск. Каждый носитель, вспомогательное вещество и т. д. также должен быть «приемлемым» в плане совместимости с другими ингредиентами состава. Конкретный характер носителя или другого материала будет зависеть от пути введения, который может представлять собой инфузию, инъекцию или любой другой подходящий путь, как обсуждается ниже.
Для парентерального, например, подкожного или внутривенного введения, например, путем инъекции, фармацевтическая композиция, содержащая молекулу антитела, может быть в форме парентерально приемлемого водного раствора, который не содержит пирогены и имеет подходящий pH, изотоничность и стабильность. Специалисты с соответствующими навыками в данной области техники легко смогут приготовить подходящие растворы с использованием, например, изотонических носителей, таких как хлорид натрия для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости можно применять консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки, включая буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, такие как аскорбиновая кислота и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3'-пентанол и м-крезол); низкомолекулярные полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагины, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как твин™, плюроновые агенты™ или полиэтиленгликоль (ПЭГ).
Согласно некоторым вариантам реализации молекулы антител могут быть обеспечены в лиофилизированной форме для восстановления перед введением. Например, лиофилизированные молекулы антител могут быть восстановлены в стерильной воде и смешаны с физиологическим солевым раствором перед введением индивидууму.
Введение может быть осуществлено в «терапевтически эффективном количестве», которого достаточно, чтобы показать пользу для индивидуума. Фактическое вводимое количество, а также скорость и продолжительность введения будут зависеть от характера и степени тяжести того, что лечат, конкретного индивидуума, которого лечат, клинического состояния индивидуума, причины нарушения, места доставки композиции, типа молекулы антитела, способа введения, графика введения и других факторов, известных практикующим врачам. Назначение лечения, например, решения по дозировке и т. д., находится в сфере ответственности врачей общей практики и других врачей и может зависеть от степени тяжести симптомов и/или прогрессирования заболевания, которое лечат. Соответствующие дозы молекул антител хорошо известны в данной области техники (Ledermann et al., 1991; Bagshawe et al., 1991). Могут быть использованы конкретные дозировки, указанные в настоящей заявке или в Physician's Desk Reference (2003), соответствующие вводимой молекуле антитела. Терапевтически эффективное количество или подходящая доза молекулы антитела может быть определена путем сравнения активности in vitro и активности in vivo в модели на животных. Известны способы экстраполяции эффективных дозировок у мышей и других тестируемых животных на человека. Точная доза будет зависеть от ряда факторов, включая размер и расположение области, подлежащей лечению, а также конкретного характера молекулы антитела.
Типичная доза антитела находится в диапазоне от 100 мкг до 1 г для разных видов системного применения и от 1 мкг до 1 мг для разных видов местного применения. Может быть введена более высокая начальная нагрузочная доза с последующей одной или более чем одной более низкими дозами. Это доза для однократного лечения взрослого индивидуума, которую можно пропорционально скорректировать для детей и младенцев, а также скорректировать для других форматов антител пропорционально молекулярной массе.
Лечение можно повторять с ежедневными, дважды в неделю, еженедельными или ежемесячными интервалами по усмотрению врача. График лечения для индивидуума может зависеть от фармакокинетических и фармакодинамических свойств композиции антитела, пути введения и характера состояния, которое лечат.
Лечение может быть периодическим, и период между введениями может составлять приблизительно две недели или более, например, приблизительно три недели или более, приблизительно четыре недели или более, приблизительно один раз в месяц или более, приблизительно пять недель или более или приблизительно шесть недель или более. Например, лечение можно осуществлять каждые две-четыре недели или каждые четыре-восемь недель. Подходящие составы и пути введения описаны выше.
Согласно предпочтительному варианту реализации молекула антитела, описанная в настоящей заявке, может быть предназначена для применения в способе лечения рака.
Рак может характеризоваться аномальной пролиферацией злокачественных раковых клеток. При упоминании конкретного типа рака, такого как рак молочной железы, это относится к аномальной пролиферации злокачественных клеток соответствующей ткани, такой как ткань молочной железы. Вторичный рак, который локализован в молочной железе, но являющийся результатом аномальной пролиферации злокачественных клеток другой ткани, такой как ткань яичника, не является раком молочной железы, упоминаемым в настоящей заявке, а раком яичников.
Рак может представлять собой первичный или вторичный рак. Таким образом, молекула антитела, описанная в настоящей заявке, может быть предназначена для применения в способе лечения рака у индивидуума, причем указанный рак представляет собой первичную опухоль и/или метастаз опухоли.
Раковая опухоль, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, может содержать TIL, которые экспрессируют OX40 и/или CD137, например, на их клеточной поверхности. Согласно одному варианту реализации опухоль могла быть определена как содержащая TIL, которые экспрессируют один или оба из OX40 и CD137. Способы определения экспрессии антигена на поверхности клетки известны в данной области техники и включают, например, проточную цитометрию.
Например, рак, подлежащий лечению с применением молекулы антитела, описанной в настоящей заявке, может быть выбран из группы, состоящей из лейкозов, таких как острый миелолейкоз (AML), хронический миелолейкоз (CML), острый лимфобластный лейкоз (ALL) и хронический лимфоцитарный лейкоз (ХЛЛ); лимфом, таких как лимфома Ходжкина, неходжкинская лимфома и множественная миелома; и разных видов солидного рака, такого как саркомы (например, саркомы мягких тканей), рак кожи (например, карцинома из клеток Меркеля), меланома, рак мочевого пузыря (например, уротелиальная карцинома мочевого пузыря), рак головного мозга (например, мультиформная глиобластома), рак молочной железы, рак матки/эндометрия, рак яичников (например, серозная цистаденома яичников), рак предстательной железы, рак легких (например, немелкоклеточная карцинома легких (NSCLC), такая как плоскоклеточная карцинома легких, и мелкоклеточный рак легких (SCLC)), колоректальный рак (например, колоректальная аденокарцинома), рак шейки матки (например, плоскоклеточный рак шейки матки и аденокарцинома канала шейки матки), рак печени (например, гепатоцеллюлярная карцинома), рак головы и шеи (например, плоскоклеточная карцинома головы и шеи), рак пищевода (например, карцинома пищевода), рак поджелудочной железы, рак почек (например, почечно-клеточный рак), рак надпочечников, рак желудка (например, аденокарцинома желудка), рак яичка (например, герминогенные опухоли яичка), рак желчного пузыря и желчевыводящих путей (например, холангиокарцинома), рак щитовидной железы, рак тимуса, рак костей и церебральный рак.
Согласно предпочтительному варианту реализации рак, подлежащий лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой солидный рак.
Более предпочтительно рак, подлежащий лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой солидный рак, выбранный из группы, состоящей из меланомы, рака мочевого пузыря, рака головного мозга, рака молочной железы, рака яичников, рака легких, колоректального рака, рака шейки матки, рака печени, рака головы и шеи, рака поджелудочной железы, рака почек и рака желудка.
Согласно другому предпочтительному варианту реализации рак, подлежащий лечению с применением молекулы антитела, описанной в настоящей заявке, может представлять собой рак, который отвечает на лечение одним или более ингибиторами контрольных точек, такими как антитело, которое связывает PD-1, PD-L1 или CTLA4. Полагают, что такие опухоли имеют более высокие уровни TIL и/или более высокую мутационную нагрузку опухоли, чем опухоли, которые не отвечают на терапию ингибитором контрольной точки. Такие опухоли также называют «теплыми» (warm) или «горячими» (hot) опухолями.
Примеры таких опухолей включают плоскоклеточную карциному головы и шеи (HNSCC), меланому, рак легких (такой как плоскоклеточный рак легких, аденокарциному легких, немелкоклеточную карциному легких [NSCLC] или мелкоклеточную карциному легких [SCLC]), рак предстательной железы, рак шейки матки, рак мочевого пузыря, рак молочной железы, рак щитовидной железы, рак почек, колоректальный рак (MSI или MSS; например, колоректальную аденокарциному), рак пищевода, неходжкинскую лимфому (NHL), рак желудка, рак эндометрия, рак поджелудочной железы, рак яичников, гепатоцеллюлярную карциному, мезотелиому и уротелиальный рак. Согласно предпочтительному варианту реализации рак представляет собой рак желудка. Рак также может представлять собой рак, который ранее не лечили химиотерапевтическим или радиотерапевтическим агентом, т. е. индивидуум, подлежащий лечению, может представлять собой пациента, страдающего раком, который не получал лечения химиотерапевтическим или радиотерапевтическим агентом для лечения рассматриваемого рака. Согласно предпочтительному варианту реализации молекула антитела, описанная в настоящей заявке, предназначена для применения в способе лечения рака, который отвечает на один или более ингибиторов контрольных точек иммунного ответа у индивидуума, причем указанный способ включает лечение пациента молекулой антитела в комбинации с агентом, который ингибирует взаимодействие между PD-1 и PD-L1.
В качестве альтернативы, рак, подлежащий лечению с применением молекулы антитела, описанной в настоящей заявке, может представлять собой рак, такой как рак поджелудочной железы или рак предстательной железы, который не отвечает на лечение одним или более ингибиторами контрольных точек, такими как антитело, которое связывает PD-1, PD-L1 или CTLA4. Такие опухоли также называют «холодными» (cold) опухолями.
Авторы настоящего изобретения показали, что опухоли, которые не отвечали на лечение только антителом к PD-1 или антителом к PD-L1, отвечали на лечение антителом к PD-1 или антителом к PD-L1 в комбинации с молекулой антитела, описанной в настоящей заявке. Таким образом, молекула антитела согласно настоящему изобретению может быть предназначена для применения в способе лечения рака у индивидуума, причем указанный рак не отвечает или рефрактерен к лечению одним или более ингибиторами контрольных точек по отдельности, и при этом указанный способ включает введение индивидууму молекулы антитела в комбинации с агентом, который ингибирует взаимодействие между PD-1 и PD-L1. Также предусмотрен способ лечения рака у индивидуума, причем указанный рак не отвечает или рефрактерен к лечению одним или более ингибиторами контрольных точек по отдельности, и при этом указанный способ включает введение молекулы антитела индивидууму в комбинации с агентом, который ингибирует взаимодействие между PD-1 и PD-L1.
Не ограничиваясь какой-либо теорией, полагают, что лечение рака, который не отвечает на лечение с применением одного или более ингибиторов контрольных точек по отдельности, химиотерапии, лучевой терапии, иммунотерапевтического агента, такого как иммуностимулирующий агент, или противоопухолевой вакцины приведет к гибели раковых клеток, что, в свою очередь, приведет к увеличению TIL в опухоли и более высокой экспрессии иммуносупрессорных рецепторов, что, в свою очередь, сделает рак восприимчивым к лечению ингибиторами контрольных точек, т. е. превратит «холодную» опухоль в «теплую» опухоль. Таким образом, молекула антитела согласно настоящему изобретению может быть предназначена для применения в способе лечения рака у индивидуума, причем указанный рак не отвечает или рефрактерен к лечению одним или более ингибиторами контрольных точек по отдельности, и при этом указанный способ включает введение индивидууму молекулы антитела в комбинации с химиотерапевтическим, радиотерапевтическим или иммуностимулирующим агентом или противораковой вакциной и необязательно агентом, который ингибирует взаимодействие между PD-1 и PD-L1. Также предусмотрен способ лечения рака у индивидуума, причем указанный рак не отвечает или рефрактерен к лечению одним или более ингибиторами контрольных точек по отдельности, и при этом указанный способ включает введение молекулы антитела индивидууму в комбинации с химиотерапевтическим, радиотерапевтическим или иммуностимулирующим агентом или противораковой вакциной и необязательно агентом, который ингибирует взаимодействие между PD-1 и PD-L1. Согласно предпочтительному варианту реализации агент, который ингибирует взаимодействие между PD-1 и PD-L1, представляет собой антитело, которое связывает PD-1 или PD-L1.
Применительно к раку лечение может включать ингибирование роста рака, включая полную ремиссию рака, и/или ингибирование метастазирования рака, а также ингибирование рецидива рака. Рост рака обычно относится к любому из ряда показателей, которые указывают на изменение внутри рака до более развитой формы. Таким образом, показатели для измерения ингибирования роста рака включают уменьшение выживаемости раковых клеток, уменьшение объема или морфологии опухоли (например, как определено с использованием компьютерной томографии (КТ), сонографии или другого метода визуализации), отсроченный рост опухоли, разрушение сосудистой сети опухоли, улучшение результатов кожного теста на гиперчувствительность замедленного типа, увеличение активности противораковых иммунных клеток или других противораковых иммунных ответов, а также уменьшение уровней опухолеспецифических антигенов. Активация или усиление иммунных ответов на раковые опухоли у индивидуума может улучшить способность индивидуума противостоять росту рака, в частности, росту рака, уже присутствующего у субъекта, и/или уменьшить склонность к росту рака у индивидуума.
Применительно к лечению рака молекула антитела, описанная в настоящей заявке, может быть введена индивидууму в комбинации с другой противораковой терапией или терапевтическим агентом, таким как противораковая терапия или терапевтический агент, который, как было показано, является подходящим, или потенциально подходящим, для лечения рассматриваемого рака. Например, молекула антитела может быть введена индивидууму в комбинации с химиотерапевтическим агентом, лучевой терапией, радионуклидом, иммунотерапевтическим агентом, противоопухолевой вакциной, онколитическим вирусом, терапией с адоптивным переносом клеток (ACT), такой как адоптивная терапия NK-клетками или терапия Т-клетками, экспрессирующими химерный антигенный рецептор (CAR), аутологичными TIL или гамма/дельта Т-клетками, или агентом для гормональной терапии. Молекула антитела, описанная в настоящей заявке, также может быть введена индивидууму в комбинации с адъювантом или неоадъювантом, таким как неоадъювантная гормональная терапия, антиангиогенным агентом, таким как антитело к VEGF или антитело к VEGFR2, цитотоксическим агентом.
Не ограничиваясь какой-либо теорией, полагают, что молекула антитела, описанная в настоящей заявке, может действовать как адъювант в противораковой терапии. В частности, полагают, что введение молекулы антитела индивидууму в комбинации с химиотерапией или лучевой терапией, например, запустит более сильный иммунный ответ против рака, чем тот, который достигается при применении химиотерапии или лучевой терапии по отдельности.
Один или более химиотерапевтических агентов для введения в комбинации с молекулой антитела, описанной в настоящей заявке, могут быть выбраны из группы, состоящей из: таксанов, цитотоксических антибиотиков, ингибиторов тирозинкиназы, ингибиторов PARP, ингибиторов фермента B-Raf, ингибиторов MEK, ингибиторов c-MET, ингибиторов VEGFR, ингибиторов PDGFR, алкилирующих агентов, аналогов платины, аналогов нуклеозидов, антифолатов, производных талидомида, антинеопластических химиотерапевтических агентов и других. Таксаны включают доцетаксел, паклитаксел и наб-паклитаксел; цитотоксические антибиотики включают актиномицин, блеомицин и антрациклины, такие как доксорубицин, митоксантрон и валрубицин; ингибиторы тирозинкиназы включают эрлотиниб, гефитиниб, акситиниб, PLX3397, иматиниб, кобемитиниб и траметиниб; ингибиторы PARP включают пирапариб; ингибиторы фермента B-Raf включают вемурафениб и дабрафениб; алкилирующие агенты включают дакарбазин, циклофосфамид и темозоломид; аналоги платины включают карбоплатин, цисплатин и оксалиплатин; аналоги нуклеозидов включают азацитидин, капецитабин, флударабин, фторурацил и гемцитабин; антифолаты включают метотрексат и пеметрексед. Другие химиотерапевтические агенты, подходящие для применения в настоящем изобретении, включают дефактиниб, энтиностат, эрибулин, иринотекан и винбластин. Химиотерапевтический агент для введения в комбинации с молекулой антитела, описанной в настоящей заявке, может представлять собой фторпиримидин. Например, если рак, подлежащий лечению, представляет собой HER2-отрицательный рак, такой как HER2-отрицательный рак желудка, молекула антитела, описанная в настоящей заявке, может быть введена в комбинации с платиной, аналогом платины и фторпиримидином. Если рак, подлежащий лечению, представляет собой HER2-положительный рак, например, HER2-положительный рак желудка, молекула антитела, описанная в настоящей заявке, может быть введена в комбинации с платиной или аналогом платины, фторпиримидином и трастузумабом.
Предпочтительные терапевтические агенты для введения с молекулой антитела, описанной в настоящей заявке, представляют собой доксорубицин, митоксантрон, циклофосфамид, цисплатин и оксалиплатин.
Лучевая терапия для введения в комбинации с молекулой антитела, описанной в настоящей заявке, может представлять собой дистанционную лучевую терапию или брахитерапию.
Радионуклиды для введения с молекулой антитела, описанной в настоящей заявке, могут быть выбраны из группы, состоящей из: иттрия-90, йода-131, висмута-213, астата-211, лютеция-177, рения-188, меди-67, актиния-225, йода-125 и тербия-161.
Иммунотерапевтический агент для введения в комбинации с молекулой антитела, описанной в настоящей заявке, может представлять собой молекулу терапевтического антитела, нуклеотид, цитокин или терапию на основе цитокина. Например, молекула терапевтического антитела может связываться с иммунной регуляторной молекулой, например, с молекулой ингибитора контрольной точки или иммунной костимулирующей молекулой, рецептором врожденной иммунной системы или опухолевым антигеном, например, опухолевым антигеном поверхности клетки или растворимым опухолевым антигеном. Примеры иммунных регуляторных молекул, с которыми может связываться молекула терапевтического антитела, включают CTLA-4, LAG-3, TIGIT, TIM-3, VISTA, лиганд запрограммированной гибели клеток 1 (PD-L1), белок запрограммированной гибели клеток 1 (PD-1), CD47, CD73, CSF-1R, KIR, CD40, HVEM, IL-10 и CSF-1. Примеры рецепторов врожденной иммунной системы, с которыми может связываться молекула терапевтического антитела, включают TLR1, TLR2, TLR4, TLR5, TLR7, TLR9, RIG-I-подобные рецепторы (например, RIG-I и MDA-5) и STING. Примеры опухолевых антигенов, с которыми может связываться молекула терапевтического антитела, включают HER2, EGFR, CD20 и TGF-бета.
Авторы настоящего изобретения показали, что введение молекулы антитела согласно настоящему изобретению в комбинации с антителом к PD-1 или антителом к PD-L1 привело к усилению активации Т-клеток и регрессии опухоли в модели опухоли на мышах по сравнению с лечением либо молекулой антитела согласно настоящему изобретению, либо антителом к PD-1 или антителом к PD-L1 по отдельности. Не ограничиваясь какой-либо теорией, эти результаты свидетельствуют о том, что введение молекулы антитела согласно настоящему изобретению в комбинации с агентом, способным ингибировать взаимодействие между PD-1 и PD-L1, приводит к усилению противоопухолевых эффектов и что такое комбинированное введение может быть подходящим для лечения опухолей, которые являются рефрактерными или резистентными к монотерапии антителом к PD-1 или антителом к PD-L1 или рецидивировали после нее.
Таким образом, молекула антитела согласно настоящему изобретению может быть предназначена для применения в способе лечения рака у индивидуума, причем указанный способ включает введение молекулы антитела в комбинации с агентом, который способен ингибировать взаимодействие между PD-1 и PD-L1. Также предложен агент, способный ингибировать взаимодействие между PD1 и PD-L1, такой как молекула антитела, которая связывает PD-1 или PD-L1, для применения в способе лечения рака у индивидуума, причем указанный способ включает введение агента, который способен ингибировать взаимодействие между PD-1 и PD-L1, в комбинации с антителом согласно настоящему изобретению. Способ лечения рака у индивидуума, включающий введение индивидууму терапевтически эффективного количества молекулы антитела согласно настоящему изобретению и терапевтически эффективного количества агента, который способен ингибировать взаимодействие между PD-1 и PD-L1.
Согласно предпочтительному варианту реализации агент, который способен ингибировать взаимодействие PD-1 и PD-L1, представляет собой молекулу антитела, которая связывает PD-1 или PD-L1. Антитела, которые связываются с PD-1, известны в данной области техники и включают ниволумаб (5C4) и пембролизумаб. Известные антитела, которые связываются с PD-L1, включают YW243.55.S1, дурвалумаб, атезолизумаб и авелумаб. Молекула антитела согласно настоящему изобретению может быть предназначена для введения с одним из этих известных антител к PD-1 или к PD-L1 или с другим антителом к PD-1 или к PD-L1. Получение альтернативных антител, которые связываются с PD-1 или PD-L1, находится в пределах возможностей квалифицированного специалиста с использованием обычных способов.
Нуклеиновая кислота для введения в комбинации с молекулой антитела, описанной в настоящей заявке, может представлять собой миРНК.
Цитокины или терапия на основе цитокина могут быть выбраны из группы, состоящей из: IL-2, пролекарства конъюгированного IL2, ГМ-КСФ, IL-7, IL-12, IL-9, IL-15, IL-18, IL-21 и интерферона I типа.
Противоопухолевые вакцины для лечения рака были внедрены в клинику и подробно обсуждаются в научной литературе (такой как Rosenberg, 2000). В основном затрагиваются стратегии стимуляции ответа иммунной системы на различные клеточные маркеры, экспрессируемые аутологичными или аллогенными раковыми клетками, путем использования этих клеток в качестве способа вакцинации, как с гранулоцитарно-макрофагальным колониестимулирующим фактором (ГМ-КСФ), так и без него. ГМ-КСФ вызывает сильный ответ при презентации антигена и особенно хорошо работает при применении с указанными стратегиями.
Молекула антитела, описанная в настоящей заявке, также может быть введена индивидууму, страдающему раком, в частности, индивидууму, страдающему раком желудка, в комбинации с рамуцирумабом и/или паклитакселом; иринотеканом и доцетакселом или паклитакселом; или пембролизумабом. Лечение молекулой антитела, описанной в настоящей заявке, в комбинации с пембролизумабом является предпочтительным в лечении MSI-H и/или dMMR рака желудка.
Учитывая активность OX40 и CD137 в усилении иммунного ответа, ожидается, что молекулы двойных агонистов OX40 и CD137 найдут применение в лечении инфекционных заболеваний. Таким образом, согласно другому предпочтительному варианту реализации молекула антитела, описанная в настоящей заявке, может быть предназначена для применения в способе лечения инфекционного заболевания, такого как острое или персистирующее инфекционное заболевание.
Не ограничиваясь какой-либо теорией, полагают, что молекулы агонистов OX40 и CD137 могут быть способны усиливать иммунный ответ против острого инфекционного заболевания, вызванного патогеном, путем индукции быстрой инфильтрации и активации врожденных иммунных клеток, таких как нейтрофилы и моноциты, облегчая посредством этого устранение патогена, вызвавшего острое инфекционное заболевание. Таким образом, согласно другому варианту реализации молекула антитела, описанная в настоящей заявке, может быть предназначена для применения в способе лечения острого инфекционного заболевания, такого как острое бактериальное заболевание. Согласно предпочтительному варианту реализации острое инфекционное заболевание представляет собой острое бактериальное заболевание, вызванное инфекцией грамположительной бактерией, такой как бактерия из рода Listeria, Streptococcus pneumoniae или Staphylococcus aureus.
Инфекционные заболевания обычно устраняются иммунной системой, но некоторые инфекции персистируют в течение длительных периодов времени, например, месяцев или лет, и иммунная система неэффективно борется с ними. Такие инфекции также называют персистирующими или хроническими инфекциями.
Предпочтительно молекулу антитела, описанную в настоящей заявке, применяют для лечения персистирующего инфекционного заболевания, такого как персистирующая вирусная, бактериальная, грибковая или паразитарная инфекция, предпочтительно персистирующая вирусная или бактериальная инфекция.
Согласно предпочтительному варианту реализации персистирующая вирусная инфекция, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой персистирующую инфекцию: вирусом иммунодефицита человека (ВИЧ), вирусом Эпштейна-Барр, цитомегаловирусом, вирусом гепатита B, вирусом гепатита C или вирусом ветряной оспы
Согласно предпочтительному варианту реализации персистирующая бактериальная инфекция, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой персистирующую инфекцию: Staphylococcus aureus, Hemophilus influenza, Mycobacterium tuberculosis, Mycobacterium leprae, Helicobacter pylori, Treponema pallidum, Enterococcus faecalis или Streptococcus pneumoniae.
Было описано, что агонистическое действие на CD137 является благоприятным применительно к лечению инфекций грамположительными бактериями. Таким образом, согласно предпочтительному варианту реализации персистирующая бактериальная инфекция, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой персистирующую инфекцию грамположительной бактерией. Согласно более предпочтительному варианту реализации персистирующая бактериальная инфекция представляет собой персистирующую инфекцию грамположительной бактерией, выбранной из группы, состоящей из Staphylococcus aureus, Mycobacterium leprae, Enterococcus faecalis и Streptococcus pneumoniae.
Согласно предпочтительному варианту реализации персистирующая грибковая инфекция, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой персистирующую инфекцию: Candida, например, Candida albicans, Cryptococcus (gattii и neoformans), Talaromyces (Penicillium) marneffe, Microsporum, например, Microsporum audouinii и Trichophyton tonsurans.
Согласно предпочтительному варианту реализации персистирующая паразитарная инфекция, подлежащая лечению с применением молекулы антитела, описанной в настоящей заявке, представляет собой персистирующую инфекцию: Plasmodium, например, Plasmodium falciparum, или Leishmania, например, Leishmania donovani.
Применительно к лечению персистирующего инфекционного заболевания молекула антитела может быть введена индивидууму в комбинации со второй терапией или терапевтическим агентом, которая (ый), как было показано, является подходящей (им), или, как ожидается, будет подходящей (им), для лечения рассматриваемого патогена. Например, молекула антитела может быть введена индивидууму в комбинации с иммунотерапевтическим агентом. Иммунотерапевтический агент для введения в комбинации с молекулой антитела, описанной в настоящей заявке, может представлять собой молекулу терапевтического антитела. Например, молекула терапевтического антитела может связываться с рецептором врожденной иммунной системы. Примеры рецепторов врожденной иммунной системы, с которыми может связываться молекула терапевтического антитела, включают TLR1, TLR2, TLR4, TLR5, TLR7, TLR9, RIG-I-подобные рецепторы (например, RIG-I и MDA-5) и STING.
Если молекулу антитела применяют для предотвращения инфекционного заболевания, молекула антитела может быть введена в комбинации с вакциной против рассматриваемого патогена. Не ограничиваясь какой-либо теорией, полагают, что молекула антитела, описанная в настоящей заявке, может действовать как адъювант при вакцинации. В частности, полагают, что введение молекулы антитела индивидууму в комбинации с вакциной вызовет более сильный иммунный ответ против патогена, чем тот, который достигается при применении вакцины по отдельности.
Применительно к лечению персистирующего инфекционного заболевания лечение может включать устранение инфекции, уменьшение патогенной нагрузки индивидуума, предотвращение рецидива инфекции. Например, лечение может включать предупреждение, улучшение, отсрочку, ослабление или купирование одного или более симптомов и/или признаков персистирующей инфекции. В качестве альтернативы, лечение может включать предотвращение инфекционного заболевания.
Признаки, раскрытые в предшествующем описании, или в прилагаемой формуле изобретения, или на прилагаемых чертежах, выраженные в их конкретных формах или в контексте средств для выполнения раскрытой функции, или способ или технологический способ получения раскрытых результатов, в зависимости от обстоятельств, можно применять, по отдельности или в любой комбинации таких признаков, для реализации настоящего изобретения в его различных формах.
Несмотря на то, что настоящее изобретение было описано во взаимосвязи с примерными вариантами реализации, описанными выше, многие эквивалентные модификации и варианты будут понятны специалистам в данной области техники при рассмотрении настоящего раскрытия. Соответственно, примерные варианты реализации настоящего изобретения, изложенные выше, считаются иллюстративными и неограничивающими. Различные изменения могут быть внесены в описанные варианты реализации, не отступая от сущности и объема настоящего изобретения.
Во избежание каких-либо сомнений, любые теоретические объяснения, представленные в настоящей заявке, предназначены для улучшения понимания читателя. Авторы настоящего изобретения не хотят быть связанными ни одним из этих теоретических объяснений.
Любые заголовки разделов, используемые в настоящей заявке, предназначены только для организационных целей и не должны рассматриваться как ограничивающие описанный предмет.
По всему тексту описания, включая прилагаемую формулу изобретения, если контекст не требует иного, слова «содержать» и «включать», и варианты, такие как «содержит», «содержащий» и «включающий», следует понимать как подразумевающие включение указанного целого числа или этапа либо группы целых чисел или этапов, но не исключение любого другого целого числа или этапа либо группы целых чисел или этапов.
Следует отметить, что в описании и прилагаемой формуле изобретения формы единственного числа (соответственно, «а», «an» и «the» в исходном тексте на английском языке) включают множественное число, если контекст явно не диктует иное. Диапазоны могут быть выражены в настоящей заявке как от «приблизительно» одного конкретного значения и/или до «приблизительно» другого конкретного значения. При выражении такого диапазона другой вариант реализации включает от одного конкретного значения и/или до другого конкретного значения. Аналогичным образом, когда значения выражены как приближения, с использованием антецедента «приблизительно», следует понимать, что конкретное значение образует другой вариант реализации. Термин «приблизительно» по отношению к числовому значению является необязательным и обозначает, например, +/- 10%.
Примеры
Цель авторов настоящего изобретения заключалась в получении МАТ2, которые были способны оказывать агонистическое действие как на OX40, так и на CD137 в отсутствие искусственных перекрестно связывающих агентов или перекрестного связывания, опосредуемого рецептором Fcγ, и которые были способны вызывать усиленный иммунный ответ против заболеваний, таких как рак. В данном случае МАТ2 представляет собой молекулу антитела, которая содержит антигенсвязывающий сайт на основе CDR, который связывает CD137, и связывающий сайт для OX40, расположенный в домене CH3 молекулы антитела.
Для достижения этой цели авторы настоящего изобретения сначала использовали способы отбора и созревания аффинности для идентификации Fcab, которые были способны связывать OX40 и индуцировать активацию Т-клеток у человека и мыши, соответственно (см. Примеры 2 и 3). Затем авторы настоящего изобретения ввели антигенсвязывающий сайт для OX40 из этих Fcab в формат МАТ2 и показали, что несколько из этих «имитационных» МАТ2 к OX40 человека были способны связывать OX40 человека и яванской макаки с высокой аффинностью и активировать Т-клетки при перекрестном связывании (см. Пример 4). Из них клон FS20-22-49 показал самое большое увеличение агонистической активности при перекрестном связывании, а также имел самую низкую ЭК50 в отношении его агонистической активности в присутствии перекрестного связывания и поэтому был приоритетно взят в качестве антигенсвязывающего сайта для OX40 для разработки рассматриваемого МАТ2.
Для того чтобы разработать антигенсвязывающий сайт на основе CDR, который связывает и способен оказывать агонистическое действие на CD137, авторы настоящего изобретения использовали способы отбора для идентификации моноклональных антител (МАТ), которые могли связывать CD137 человека и были способны активировать Т-клетки только при перекрестном связывании (см. Пример 5). CDR из этих идентифицированных МАТ впоследствии клонировали в МАТ2, которые содержали антигенсвязывающий сайт для OX40 FS20-22-49. CDR этих МАТ2 оптимизировали по последовательности для того чтобы получить следующие МАТ2: FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-35-14 и FS20-22-49AA/FS30-5-37 (см. Пример 6). Было продемонстрировано, что все эти МАТ2 имеют высокий уровень специфичности в отношении CD137 человека и были способны активировать CD137 при перекрестном связывании в анализе активации Т-клеток (см. Пример 7). Ни одно из МАТ2 не показало какой-либо значительной способности активировать CD137 в отсутствие перекрестного связывания.
После того, как было установлено, что антигенсвязывающий сайт для OX40 FS20-22-49AA в выбранном МАТ2 был способен связывать и активировать OX40 при перекрестном связывании и что, отдельно, антигенсвязывающие сайты для CD137 на основе CDR из FS30-10-3, FS30-10-12, FS30-10-16, FS30-35-14 и FS30-5-37 были способны связывать и активировать CD137 при перекрестном связывании, авторы настоящего изобретения попытались продемонстрировать, что МАТ2, содержащие эти антигенсвязывающие домены, были способны активировать как OX40, так и CD137 (также называется «двойное агонистическое действие»). Такой двойной агонист может быть способен i) связываться с OX40 для перекрестного связывания МАТ2 и связываться, кластеризовать и активировать (оказывать агонистическое действие) CD137, и ii) связываться с CD137 для перекрестного связывания МАТ2 и связываться, кластеризовать и активировать (оказывать агонистическое действие) OX40. Важно отметить, что двойной агонист должен быть способен автономно запускать двойное агонистическое действие, основываясь на экспрессии специфичных мишеней (OX40 и CD137) и без потребности в дополнительных перекрестно связывающих агентах.
Авторы настоящего изобретения продемонстрировали, что тестируемые молекулы МАТ2 были способны связывать CD137 человека, OX40 человека, CD137 яванской макаки и OX40 яванской макаки (см. Пример 8) и что тестируемые молекулы МАТ2 были способны одновременно связываться с CD137 человека и OX40 человека (см. Пример 9). Авторы настоящего изобретения показали, что мутация «LALA» в домене CH2 МАТ2 уменьшала их связывание с рецепторами Fcγ и что клон МАТ2 FS20-22-49AA/FS30-10-16 был неспособен индуцировать активацию АЗКЦ в биотесте АЗКЦ (см. Пример 10).
Авторы настоящего изобретения также показали, что тестируемые молекулы МАТ2 к OX40/CD137 связывались с экспрессируемыми клетками OX40 и CD137 человека и яванской макаки без наблюдаемого неспецифичного связывания (см. Пример 11).
Затем авторы настоящего изобретения продемонстрировали, что тестируемые молекулы МАТ2 к OX40/CD137, содержащие мутацию LALA, были способны индуцировать активацию Т-клеток в отсутствие искусственных перекрестно связывающих агентов в анализе активации Т-клеток с использованием стафилококкового энтеротоксина А (SEA; см. Пример 12). Авторы настоящего изобретения также продемонстрировали, что тестируемые молекулы МАТ2 могут индуцировать активацию Т-клеток в отсутствие искусственных перекрестно связывающих агентов в анализе активации общей популяции Т-клеток и что эта активность зависит от одновременного связывания МАТ2 как с OX40, так и с CD137 (см. Примеры 13 и 16). Авторы настоящего изобретения дополнительно подтвердили, что МАТ2 FS20-22-49AA/FS30-10-16 было способно активировать эти рецепторы в CD4+ и CD8+ Т-клетках, соответственно, в отсутствие перекрестного связывания (см. Пример 14).
Поскольку МАТ2 к OX40/CD137 человека не связывалось с белками мыши, чтобы протестировать способность МАТ2 к OX40/CD137 вызывать противоопухолевый ответ, опосредуемый Т-клетками, получали подобное МАТ2, нацеленное на OX40 мыши и CD137 мыши, как с мутацией LALA, так и без нее (обозначено как FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3, соответственно). Авторы настоящего изобретения показали, что МАТ2 FS20m-232-91AA/Lob12.3 может индуцировать активацию Т-клеток без каких-либо дополнительных перекрестно связывающих агентов и что эта активность зависит от одновременного связывания МАТ2 как с OX40, так и с CD137 (см. Примеры 15 и 16).
Авторы настоящего изобретения демонстрируют, что МАТ2 FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3 обладают противоопухолевой эффективностью in vivo в модели сингенной опухоли CT26 (см. Пример 17). Авторы настоящего изобретения дополнительно демонстрируют, что МАТ2 FS20m-232-91AA/Lob12.3 оказывает влияние на циркулирующие Т-клетки, увеличивая частоту активированных и пролиферирующих Т-клеток (см. Примеры 18 и 19). Авторы настоящего изобретения продемонстрировали, что МАТ2 FS20m-232-91AA/Lob12.3 обладает противоопухолевой эффективностью in vivo в модели сингенной опухоли B16-F10 (см. Пример 20).
Авторы настоящего изобретения осуществили аналитическую характеристику и предварительную оценку стабильности МАТ2 (см. Пример 21). Все пять протестированных МАТ2 показали благоприятные аналитические характеристики и благоприятную стабильность.
Авторы настоящего изобретения продемонстрировали, что комбинация МАТ2 FS20-22-49AA/FS30-10-16 с антителом к PD-L1 или антителом к PD-1 в анализе активации Т-клеток с использованием SEA может привести к увеличению максимальной активности Т-клеток in vitro свыше активности, наблюдаемой для МАТ2 к OX40/CD137 по отдельности. Авторы настоящего изобретения также показали, что лечение комбинацией МАТ2 FS20m-232-91AA/Lob12.3 и антитела к PD-1 in vivo в модели опухоли CT26 на мышах было способно приводить к увеличению противоопухолевой активности, обеспечивать преимущество в выживаемости и усиливать фармакодинамическую модуляцию пролиферирующих Т-клеток и NK-клеток по сравнению с лечением любым отдельным агентом (см. Пример 22).
Авторы настоящего изобретения продемонстрировали, что МАТ2 FS20m-232-91AA/Lob12.3 обладает дозозависимой противоопухолевой активностью in vivo в модели сингенной опухоли CT26 до определенного уровня дозы и что эта активность сохранялась при более высоких уровнях дозы. Авторы настоящего изобретения также показали, что МАТ2 FS20m-232-91AA/Lob12.3 может индуцировать формирование защитной иммунологической памяти у мышей с «полным ответом» и защищать от повторной инокуляции клеток CT26 (см. Пример 23). Авторы настоящего изобретения продемонстрировали, что МАТ2 FS20m-232-91AA/Lob12.3 оказывает влияние на циркулирующие Т-клетки, значительно увеличивая частоту пролиферирующих (Ki67+) CD4+ и CD8+ Т-клеток при варьирующихся уровнях дозы (см. Пример 24). Авторы настоящего изобретения также показали, что МАТ2 FS20m-232-91AA/Lob12.3 способно увеличивать частоту активированных (CD69+) и пролиферирующих (Ki67+) CD8 Т-клеток и что истощение CD4 Т-клеток оказывает неблагоприятный эффект на периферический фармакодинамический ответ, опосредуемый МАТ2 FS20m-232-91AA/Lob12.3 (см. Пример 25). Авторы настоящего изобретения показали, что МАТ2 FS20-22-49AA/FS30-10-16 имело сходную функциональную активность в анализе первичных МКПК яванских макак по сравнению с эквивалентным анализом у человека и что МАТ2 хорошо переносилось у яванских макак в дозах до 30 мг/кг, и что оно было способно индуцировать связанное с лекарственным средством увеличение пролиферации и активации CD4+ и CD8+ Т-клеток центральной памяти и эффекторных CD4+ и CD8+ Т-клеток памяти и NK-клеток у яванских макак (см. Пример 26).
Авторы настоящего изобретения также показали, что при исследовании на мышах BALB/c МАТ2 FS20m-232-91AA/Lob12.3 индуцировало умеренное и кратковременное увеличение уровней инфильтрации и пролиферации Т-клеток в печени по сравнению с агонистом CD137, независимым от перекрестного связывания, который индуцировал повышенную и устойчивую инфильтрацию, пролиферацию и активацию Т-клеток в печени (см. Пример 27). Наконец, в модели сингенной опухоли CT26 на мышах авторы настоящего изобретения показали отсутствие различий в росте опухолей или выживаемости между мышами, получавшими либо МАТ2 FS20m-232-91AA/Lob12.3, либо МАТ2 к OX40/CD137, содержащее тот же Fcab к OX40, спаренный с клоном Fab к CD137, независимым от перекрестного связывания, несмотря на способность клона Fab к CD137, независимого от перекрестного связывания, индуцировать повышенные уровни и пролиферацию Т-клеток по сравнению с зависимым от перекрестного связывания клоном антитела к CD137 Lob12.3 из МАТ2 FS20m-232-91AA/Lob12.3 (см. Пример 28).
Эти эксперименты более подробно описаны в следующих Примерах.
Пример 1 - Отбор и характеристика антигена
Способы отбора и скрининга, использованные для идентификации МАТ2, которые способны связывать и оказывать агонистическое действие как на OX40, так и на CD137, требуют использования различных антигенов OX40 и CD137. Получение этих антигенов более подробно описано ниже.
1.1 Антигены OX40
Антигены OX40, использованные для отбора Fcab, специфичных в отношении OX40 человека и мыши, и для тестирования перекрестной реактивности отобранных Fcab с OX40 яванской макаки, либо получали собственными силами, либо получали из коммерческих источников, как описано ниже.
1.1.1 Получение рекомбинантных растворимых антигенов OX40 человека, яванской макаки и мыши
Для получения рекомбинантных растворимых димерных антигенов OX40 внеклеточный домен OX40 гибридизовали с Fc мыши, что улучшило растворимость и стабильность антигена. В частности, внеклеточный домен соответствующего OX40 (человека, яванской макаки или мыши) клонировали в вектор pFUSE-mIgG2aFc2 (Invivogen, номер по каталогу pfuse-mg2afc2) с использованием рестрикционных ферментов EcoRI-HF и BglII с получением антигенов с доменом Fc IgG2a мыши на C-конце. Рекомбинантные антигены OX40 затем получали путем кратковременной экспрессии в клетках HEK293-6E (Национальный исследовательский совет Канады) и очищали с использованием колонок MabSelect SuRe™ с белком A (GE Healthcare, 11003494) с последующей гель-проникающей хроматографией (SEC), чтобы гарантировать, что полученный антиген представлял собой единственную молекулу и не содержал агрегатов.
Чтобы получить биотинилированные варианты рекомбинантных антигенов OX40, антигены биотинилировали с использованием набора EZ-Link™ Sulfo-NHS-SS-Biotin (Thermo Fisher Scientific, номер по каталогу 21331) в соответствии с протоколом производителя. Биотинилированный антиген OX40 использовали для экспериментов по отбору, описанных ниже, но не для измерений аффинности связывания. Очистку биотинилированных антигенов OX40 выполняли в два этапа с использованием обессоливающей колонки PD-10 (GE Healthcare, 17-0851-01), а затем центрифужной колонки Amicon 30k (Millipore, UFC903024) в соответствии с инструкциями производителя. Биофизические свойства рекомбинантных антигенов характеризовали с помощью анализа методом SE-HPLC, чтобы гарантировать отсутствие агрегатов, и с помощью электрофореза в ДСН-ПААГ (PAGE), чтобы подтвердить размер молекул. Определение размера с помощью PAGE показало, что растворимые антигены были димерными, поскольку их оцененная молекулярная масса была вдвое больше, чем прогнозируемая молекулярная масса мономера. Рекомбинантные антигены также анализировали с помощью анализа сдвига электрофоретической подвижности в геле, который показал, что степень биотинилирования была выше 90%. Твердофазный иммуноферментный анализ (ИФА) и поверхностный плазмонный резонанс (SPR) использовали, чтобы подтвердить, что биотинилированные рекомбинантные антигены OX40 человека (hOX40-mFc), мыши (mOX40-mFc) и яванской макаки (cOX40-mFc) могут быть связаны OX40-специфичными антителами (антитело 11D4 [Европейский патент № 2242771] для OX40 человека и яванской макаки; поликлональное овечье антитело к OX40 человека для OX40 яванской макаки [R&D Systems, номер по каталогу AF3388]; антитело ACT35 для OX40 человека [Biolegend, номер по каталогу 35002] и антитело OX86 для OX40 мыши [Biolegend, номер по каталогу 119408]). Эти антигены перечислены в Таблице 2 ниже.
1.1.2 Получение линий клеток, экспрессирующих OX40 человека, яванской макаки и мыши
OX40 человека, яванской макаки и мыши (последовательности см. в Таблице 1) клонировали в вектор pLVX-EF1a-IRES-puro (Clontech, номер по каталогу 631253) с использованием рестрикционных ферментов SpeI-HF и NotI-HF. Затем векторы трансформировали в линию клеток Lenti-X 293T (Clontech, номер по каталогу 632180) вместе со смесью для упаковки Lenti-X HTX (Clontech, номер по каталогу 631249), чтобы получить лентивирус. Затем лентивирус использовали для трансдукции клеток DO11.10 (National Jewish Health). Клетки, сверхэкспрессирующие OX40, отбирали путем инкубации клеток с 5 мкг/мл пуромицина (Life Technologies, номер по каталогу A11113803) в течение приблизительно 2 недель с последующим клонированием линии клеток путем последовательного разведения. Экспрессию OX40 линиями клеток тестировали с помощью проточной цитометрии с использованием флуоресцентномеченых OX40-специфичных антител (OX86; ACT35; и поликлонального овечьего антитела к OX40 человека, как описано в Примере 1.1.1 и Таблице 2). Отбирали линии клеток, экспрессирующие OX40 человека (DO11.10-hOX40), мыши (DO11.10-mOX40) или яванской макаки (DO11.10-cOX40), в которых все клетки показали по меньшей мере в 10 раз более высокие значения флуоресценции, чем нетрансдуцированные клетки в анализе методом проточной цитометрии. Эти линии клеток перечислены в Таблице 2 ниже.
Таблица 1: Последовательности OX40
(№ по каталогу)
1.1.3 Коммерчески доступные антигены OX40
Тестировали несколько коммерчески доступных антигенов OX40.
Рекомбинантный His-меченый внеклеточный домен OX40 человека получали от SinoBiologicals (номер по каталогу 10481-H08H-50). Однако анализ этого антигена методом SE-HPLC показал, что менее 50% антигена было в мономерной, неагрегированной форме. Поэтому этот антиген не использовали в последующем анализе.
Рекомбинантный OX40 человека/Fc человека (hOX40-hFc) и рекомбинантный OX40 мыши/Fc человека (mOX40-hFc), которые содержали домен Fc IgG1 человека на С-конце, получали от R&D Systems (hOX40-hFc: номер по каталогу 3388-OX-050; mOX40-hFc: номер по каталогу 1256-OX-050) и биотинилировали собственными силами. Биофизические свойства этих растворимых антигенов характеризовали с помощью анализа методом SE-HPLC, чтобы гарантировать отсутствие агрегатов, и с помощью PAGE, чтобы проверить размер молекул. Определение размера с помощью PAGE показало, что растворимые антигены были димерными, поскольку их оцененная молекулярная масса в два раза превышала ожидаемую массу для мономерного антигена. Растворимые антигены также анализировали с помощью анализа сдвига электрофоретической подвижности в геле, который показал, что степень биотинилирования была выше 90%. ИФА и SPR использовали, чтобы подтвердить, что биотинилированные рекомбинантные антигены OX40 человека (hOX40-hFc) и мыши (mOX40-hFc) могут быть связаны OX40-специфичными антителами (11D4; ACT35; и OX86, как описано в Примере 1.1.1 и Таблице 2 ниже).
Таблица 2: Антигены OX40
1.2 Антигены CD137
Антигены CD137, использованные для отбора МАТ, специфичных в отношении CD137 человека, и для тестирования перекрестной реактивности отобранных Fcab с OX40 яванской макаки, либо получали собственными силами, либо получали из коммерческих источников, как описано ниже.
1.2.1 Получение рекомбинантных растворимых антигенов CD137 человека и яванской макаки
Поскольку было установлено, что несколько коммерчески доступных рекомбинантных антигенов непригодны для применения, например, из-за неприемлемых уровней агрегатов, присутствующих при тестировании, следующие рекомбинантные димерные и мономерные антигены (Таблица 3) получали собственными силами для применения в отборах, скрининге и дальнейшей характеристике МАТ к CD137.
Таблица 3: Рекомбинантные антигены CD137 человека и яванской макаки
Мономерный антиген получали путем клонирования ДНК, кодирующей внеклеточный домен CD137 человека вместе с последовательностью Avi и шестью C-концевыми остатками гистидина, в модифицированные векторы pFUSE (Invivogen, номер по каталогу pfuse-mg2afc2) с использованием рестрикционных ферментов EcoRI-HF и BamHI-HF. Векторы трансфецировали в клетки HEK293-6E, и экспрессированный CD137 очищали с использованием колонки с никелем HisTrap™ excel (GE Healthcare, 17-3712-06) и гель-проникающей хроматографии (SEC), чтобы гарантировать, что антиген представлял собой единственную молекулу и не содержал агрегатов.
Для получения димерных антигенов конструкции ДНК, кодирующие внеклеточный домен CD137 человека или яванской макаки (cyno), гибридизованный с доменом Fc mIgG2a вместе с последовательностью Avi, клонировали в модифицированные векторы pFUSE и трансфецировали в клетки HEK293-6E. Рекомбинантный CD137 очищали с использованием колонки MabSelect SuRe™ с белком A (GE Healthcare, 11003494) и гель-проникающей хроматографии (SEC), чтобы гарантировать, что антиген представлял собой единственную молекулу и не содержал агрегатов.
Биотинилированные варианты димерных и мономерных антигенов CD137 получали с использованием набора для реакции биотин-протеинлигаза биотина BirA (Avidity LLC, BirA500) с получением мономерного антигена CD137, меченного одной молекулой биотина, и димерных антигенов CD137, меченных двумя молекулами биотина, по одной на каждый из двух мономеров. В частности, 3 мг антигена CD137 смешивали с 7,8 мкл смеси фермента BirA до молярного соотношения фермента и субстрата 1:50. Добавки затем добавляли в соответствии с рекомендациями производителя (142 мкл Biomix A, 142 мкл Biomix B, 142 мкл биотина), и реакционную смесь инкубировали в течение двух часов при комнатной температуре. Для поддержания целостности биотинилированных антигенов буфер в реакционной смеси немедленно заменяли ДФСБ с использованием фильтров Amicon 30 мкм.
Антигены CD137 дополнительно очищали с помощью SEC, чтобы гарантировать удаление фермента BirA и получение конечного монодисперсного белкового препарата высокого качества без высокомолекулярных агрегатов. В частности, антигены из одной и той же производственной партии смешивали и анализировали их стабильность и чистоту с помощью гель-проникающей высокоэффективной жидкостной хроматографии (SE-HPLC), электрофореза в полиакриламидном геле с ДСН (SDS-PAGE) и гель-проникающей хроматографии с многоугловым рассеянием света (SEC-MALS). Полное биотинилирование белков подтверждали с помощью электрофореза в ДСН-ПААГ со сдвигом электрофоретической подвижности при связывании со стрептавидином. Связывание рекомбинантных антигенов CD137 человека с антителом к CD137 человека, 20H4.9, в качестве положительного контроля (патент США № 7288638), in vitro подтверждали с помощью поверхностного плазмонного резонанса (SPR), а связывание с клетками DO11.10, экспрессирующими лиганд CD137 человека, подтверждали с помощью проточной цитометрии. Связывание рекомбинантного антигена CD137 яванской макаки с клетками DO11.10, экспрессирующими лиганд CD137 яванской макаки, подтверждали с помощью проточной цитометрии. Чтобы гарантировать максимально высокую чистоту антигенов CD137, используемых в протоколах отбора, выполняли тщательную характеристику белков антигенов, чтобы гарантировать, что процентное содержание присутствующих агрегатов белка не превышало 2%.
1.2.2 Получение линий клеток, экспрессирующих CD137 человека, яванской макаки и мыши
Клетки DO11.10 (National Jewish Health), экспрессирующие полноразмерный CD137 человека или яванской макаки, обозначенные «DO11.10-hCD137» и «DO11.10-cCD137», соответственно (см. Таблицу 4), получали, чтобы презентировать антиген в его наиболее природной конформации во время отбора и для дальнейшей характеристики отобранных МАТ к CD137.
Также получали клетки DO11.10, экспрессирующие полноразмерный CD137 мыши, обозначенные «DO11.10-mCD137», чтобы определить связывание МАТ2 к OX40/CD137 мыши с CD137 мыши, экспрессируемым клетками (см. Пример 11.2).
Лентивирусную трансдукцию использовали для получения клеток DO11.10, сверхэкспрессирующих рецепторы CD137 человека, яванской макаки или мыши, с использованием системы для упаковки Lenti-X HTX (Clontech, 631249). Вектор экспрессии Lenti-X (pLVX) (Clontech, 631253), содержащий кДНК, кодирующую CD137 человека (SEQ ID NO: 126), CD137 яванской макаки (SEQ ID NO: 128) или CD137 мыши (SEQ ID NO: 164), котрансфецировали со смесью для упаковки Lenti-X HTX в линию клеток Lenti-X 293T (Clontech, 632180), чтобы получить вирус. Затем линию клеток DO11.10 трансдуцировали этими лентивирусными векторами.
Экспрессию CD137 человека, яванской макаки и мыши на этих клетках подтверждали с использованием проточной цитометрии на основании связывания с клетками антител к CD137 (20H4.9, MOR7480.1 (публикация патента № US 2012/0237498 A1) и Lob12.3 (Университет Саутгемптона), соответственно) в качестве положительного контроля.
Таблица 4: Антигены CD137 человека и яванской макаки, экспрессируемые на поверхности клетки
Пример 2 - Отбор и характеристика Fcab к OX40 человека
2.1 Наивный отбор Fcab к OX40 человека
Для отбора Fcab, специфичных в отношении OX40 человека, из наивных фаговых библиотек в качестве антигенов использовали как рекомбинантный биотинилированный растворимый димерный OX40 человека (hOX40-mFc; см. Таблицу 2), так и экспрессируемый клетками OX40 человека (DO11.10-hOX40). Клетки, экспрессирующие OX40 человека, использовали в дополнение к рекомбинантному биотинилированному растворимому димерному OX40 человека в некоторых протоколах отбора, чтобы гарантировать, что отобранные Fcab были способны связываться с OX40 в его природной конформации на поверхности клетки.
Конструировали шесть наивных фаговых библиотек, экспонирующих домен CH3 (нумерация IMGT 1.4-130), содержащий частично рандомизированные петли AB (остатки с 14 по 18 в соответствии со схемой нумерации IMGT) и петли EF (остатки с 92 по 101 в соответствии со схемой нумерации IMGT) в домене CH3. Одна из шести библиотек дополнительно содержала клоны со вставкой либо двух, либо четырех аминокислот (кодируемых двумя или четырьмя кодонами NNK) в положении 101 в петле EF домена CH3 (вставленные остатки пронумерованы с 101.1 по 101.4 в соответствии со схемой нумерации IMGT).
Все шесть библиотек подвергали трем раундам отбора с использованием рекомбинантного биотинилированного растворимого димерного OX40 человека (hOX40-mFc; см. Таблицу 2). Все шесть библиотек также подвергали последующей кампании отбора с использованием hOX40-mFc в первом раунде отбора, а затем OX40 человека, экспрессируемого клетками (DO11.10-hOX40 в двух последующих раундах отбора; см. Таблицу 2).
2133 клона, идентифицированных после третьего раунда отбора, из шести библиотек подвергали скринингу с помощью ИФА для определения связывания с OX40 человека. В результате идентифицировали 32 уникальных положительных связывающих элемента, которые субклонировали и экспрессировали как растворимые Fcab (состоящие из усеченного шарнира [SEQ ID NO: 101], домена CH2 и CH3) в клетках HEK Expi293 (Fcab, клонированные в вектор pTT5 [Национальный исследовательский совет Канады], трансфецированный с использованием набора для трансфекции ExpiFectamine 293 [Life Technologies, A14524] в клетки Expi293F [Life Technologies, A14527]).
Тестировали способность 32 уникальных Fcab связывать OX40 человека, экспрессируемый клетками (DO11.10-hOX40). 15 из 32 Fcab, подвергнутых скринингу, показали связывание с клетками DO11.10-hOX40, и ЭК50 для этих взаимодействий варьировалась от 0,1 до 62 нМ. 15 Fcab, которые показали связывание с DO11.10-hOX40, тестировали с использованием репортерного анализа NF-κB человека, разработанного собственными силами, который тестирует активацию сигнального пути NF-κB. Шесть из 15 Fcab показали увеличение активности при перекрестном связывании с антителом к Fc человека в репортерном анализе NF-κB человека, это свидетельствует о том, что эти Fcab будут способны активировать передачу сигнала OX40. Fcab, обозначенные FS20-22 и FS20-31, показали высокие уровни активности в этом анализе, и их активность увеличивалась при перекрестном связывании Fcab с МАТ к CH2 человека (клон MK1A6 (Jefferis et al., 1985 и Jefferis et al., 1992), полученный собственными силами). Они были отобраны для созревания аффинности.
2.2 Созревание аффинности Fcab к OX40 человека
Библиотеки для созревания аффинности FS20-22 и FS20-31 создавали путем рандомизации пяти остатков в петле AB (остатки с 14 по 18) или пяти остатков в петле CD (остатки с 45.1 по 77) домена CH3 с использованием рандомизированных праймеров от ELLA Biotech, используя эквимолярное распределение аминокислот, исключая цистеины, или путем рандомизации частей петли EF (остатки с 92 по 94 и с 97 по 101 домена CH3 (все остатки пронумерованы в соответствии со схемой нумерации IMGT).
1410 Fcab из выходных продуктов созревания аффинности подвергали скринингу с помощью ИФА для определения связывания с OX40 человека, и 204 уникальных положительных связывающих элемента идентифицировали, субклонировали и экспрессировали как растворимые Fcab в клетках HEK Expi293, как описано в Примере 2.1 выше.
Скорости диссоциации растворимых Fcab при связывании с hOX40-mFc измеряли с использованием Biacore 3000 (GE Healthcare) в отсутствие и в присутствии перекрестного связывания против CH2 с использованием клона МАТ к CH2 человека, MK1A6 (см. Пример 2.1). Fcab с улучшенными скоростями диссоциации по сравнению с соответствующим исходным Fcab подвергали дальнейшему скринингу, чтобы определить связывание с OX40 человека, экспрессируемым клетками, и активность в анализе активации Т-клеток человека, разработанном собственными силами. Все Fcab связывали OX40 человека, экспрессируемый клетками. 10 Fcab из линии FS20-22 и 18 Fcab из линии FS20-31, показавшие высокие уровни активности в анализе активации Т-клеток человека, были отобраны для перестановки петель, как описано ниже.
Для линии FS20-22 получали две библиотеки с переставленными петлями путем перестановки трех петель CD, шести петель EF и либо исходной петли AB, либо петли AB с созревшей аффинностью. Для линии FS20-31 получали одну библиотеку с переставленными петлями, содержащую четыре петли AB, семь петель CD и семь петель EF.
Последовательности с перестановками экспрессировали в виде растворимых Fcab в клетках HEK Expi293, как описано в Примере 2.1 выше, и подвергали скринингу для определения связывания с биотинилированным антигеном hOX40-mFc с использованием стрептавидиновых биосенсоров Dip and Read™ (Pall FortéBio, 18-5050) на системе Octet QKe (Pall FortéBio). Fcab с улучшенной скоростью диссоциации при связывании с hOX40-mFc по сравнению с исходным Fcab секвенировали с получением 35 уникальных Fcab из линии FS20-22 и 62 из линии FS20-31. Идентифицированные уникальные Fcab тестировали для определения связывания с антигеном hOX40-mFc в присутствии и в отсутствие перекрестного связывания CH2 с использованием клона МАТ к CH2 человека MK1A6 с помощью прибора Biacore 3000 (GE Healthcare).
Для линии FS20-22 18 Fcab отобрали для экспрессии в формате имитационного (4420 LALA) МАТ2 и дальнейшей характеристики на основании самой низкой скорости диссоциации с перекрестным связыванием CH2 при связывании с hOX40-mFc, наибольшего различия в скорости диссоциации между скоростью диссоциации без перекрестного связывания и скоростью диссоциации с перекрестным связыванием CH2 при связывании с hOX40-mFc, а также силы связывания с hOX40-mFc, как указано выше. Для линии FS20-31 девять Fcab с самой низкой скоростью диссоциации при связывании с hOX40-mFc с перекрестным связыванием CH2 и девять Fcab с самой низкой скоростью диссоциации при связывании с hOX40-mFc без перекрестного связывания CH2 отобрали для экспрессии и дальнейшей характеристики в формате имитационного (4420 LALA) МАТ2. Поскольку несколько Fcab были общими для обеих групп из девяти Fcab, из линии FS20-31 отобрали дополнительные Fcab, которые показали низкие скорости диссоциации при связывании с hOX40-mFc в отсутствие перекрестного связывания CH2, чтобы общее количество Fcab из этой линии для экспрессии и дальнейшей характеристики в формате имитационного МАТ2 составило 18. Используя данные из анализа активации Т-клеток, идентифицировали еще шесть Fcab из линии FS20-22 и восемь Fcab из линии FS20-31, которые показали высокую активность в этом анализе и которые поэтому также экспрессировали в формате имитационного (4420 LALA) МАТ2 и дополнительно характеризовали (см. Пример 4).
Пример 3 - Отбор и характеристика Fcab к OX40 мыши
3.1. Наивный отбор Fcab к OX40 мыши
Для отборов использовали наивную дрожжевую библиотеку, экспонирующую домены CH1-CH3 IgG1 человека, которая содержала рандомизации в петле AB (остатки 11-18 в соответствии со схемой нумерации IMGT) и петле EF (остатки 92-101 в соответствии со схемой нумерации IMGT) домена CH3 и включала рандомизированную вставку из пяти остатков между остатками 16 и 17 (в соответствии со схемой нумерации IMGT) петли AB. Дрожжи инкубировали с биотинилированным рекомбинантным OX40 мыши, гибридизованным с доменом Fc IgG человека (mOX40-hFc; Таблица 2), и сортировали с помощью MACS с использованием гранул, покрытых стрептавидином. Затем выполняли три раунда отборов с помощью FACS с использованием понижающихся концентраций биотинилированного mOX40-hFc в присутствии пятикратного молярного избытка hFc. Клетки окрашивали стрептавидином-аллофикоцианином (APC) (BD Bioscience, 349024) или антителом к биотину-APC (Miltenyi Biotec, 130-090-856) и сортировали с использованием сортировщика клеток FACSAria (BD Bioscience). 182 индивидуальных Fcab из обогащенных популяций подвергали скринингу для определения связывания антигена, и два уникальных положительных связывающих элемента субклонировали и экспрессировали в виде растворимых Fcab, как описано ранее в Примере 2.1. Связывание Fcab с mOX40-hFc характеризовали с помощью ИФА, а активность характеризовали в репортерном анализе NF-κB мыши, разработанном собственными силами. Только один Fcab, FS20m-232, был активен в репортерном анализе NF-κB и показал связывание с клетками, экспрессирующими OX40 мыши, поэтому этот Fcab отобрали для созревания аффинности.
3.2 Созревание аффинности Fcab к mOX40
Три библиотеки фагового дисплея для созревания аффинности конструировали путем рандомизации семи остатков в петле AB (остатки 15-16.5 в соответствии со схемой нумерации IMGT) (Библиотека 1), шести остатков в петле CD (остатки 45.1-78 в соответствии со схемой нумерацией IMGT) (Библиотека 2) или пяти остатков в петле EF (остатки 92-94 и 97-98 в соответствии со схемой нумерации IMGT) (Библиотека 3) Fcab FS20m-232 с использованием рандомизированных праймеров от ELLA Biotech, используя эквимолярное распределение аминокислот, за исключением цистеина. Три раунда отбора выполняли на библиотеках для созревания аффинности с использованием рекомбинантного биотинилированного mOX40-mFc, попеременно захваченного на покрытых стрептавидином (ThermoFisher Scientific, 11205D) и покрытых нейтравидином (ThermoFisher Scientific, 14203 и A2666) гранулах Dynabeads. Для идентификации связывающих элементов с высокой аффинностью использовали понижение концентраций антигена с 50 нМ (Раунд 1) до 10 нМ (Раунд 2) и до 1 нМ (Раунд 3). 1655 индивидуальных фагов из третьего раунда отбора подвергали скринингу с помощью ИФА фага для определения связывания с mOX40-mFc, и 98 уникальных положительных связывающих элементов идентифицировали, субклонировали и экспрессировали в виде растворимых Fcab в клетках HEK Expi293, как описано в Примере 2.1. Fcab подвергали дальнейшему скринингу для определения связывания с клетками и активности в репортерном анализе NF-κB мыши. Для перестановки петель были отобраны наиболее активные Fcab.
Получали библиотеку с переставленными петлями, содержащую 27 петель CD (все 26 уникальных последовательностей, идентифицированных в результате созревания аффинности, и последовательность WT), переставленных с 37 петлями EF (которые имеют наилучшее связывание с OX40 мыши в ИФА фага и последовательность WT), при этом все клоны с перестановками содержали петлю AB Fcab FS20m-232. 750 последовательностей с перестановками экспрессировали как растворимые Fcab (содержащие усеченный шарнир) в клетках HEK Expi293, как описано выше. Супернатанты НЕК, содержащие Fcab, подвергали скринингу для определения улучшенных скоростей диссоциации путем измерения связывания Fcab с биотинилированным mOX40-mFc (Таблица 2) с использованием стрептавидиновых биосенсоров Dip and Read™ (Pall FortéBio, 18-5050) на системе Octet QKe (Pall FortéBio). 11 уникальных Fcab с рандомизированной петлей AB и 60 уникальных Fcab с рандомизированной петлей EF субклонировали и экспрессировали как растворимые Fcab в клетках HEK Expi293, как описано выше. Эти Fcab дополнительно подвергали скринингу вместе с 43 Fcab с перестановками с самыми низкими скоростями диссоциации для определения связывания клеток и активности в анализе активации Т-клеток мыши. Fcab FS20m-232-91 имел самую низкую скорость диссоциации при связывании с биотинилированным mOX40-mFc и самую высокую активность в анализе активации Т-клеток мыши при перекрестном связывании клоном МАТ к CH2 человека MK1A6 и поэтому был отобран в качестве (суррогатного) Fcab мыши для применения в последующих экспериментах.
Пример 4 - Конструирование, экспрессия и характеристика Fcab к OX40 в формате МАТ2
4.1 Конструирование и экспрессия имитационного МАТ2
«Имитационные» МАТ2, содержащие Fcab к OX40 человека и к OX40 мыши, идентифицированные выше, получали для того чтобы обеспечить возможность характеристики этих Fcab в формате МАТ2. Имитационные МАТ2 получали из Fcab к OX40 и вариабельных областей антитела к FITC 4420 (Bedzyk et al., 1989 и Bedzyk et al., 1990) в остове IgG1 человека (см. SEQ ID NO: 114, SEQ ID NO: 115 и SEQ ID NO: 116 для более подробной информации) или вариабельных областей антитела к лизоциму куриного яичного белка D1.3 (HEL) (Braden et al., 1996) в остове IgG1 человека (см. SEQ ID NO: 117 и SEQ ID NO: 118 для более подробной информации) путем замены доменов CH3 антитела к FITC и антитела к HEL доменами CH3 Fcab к OX40 в сайтах XhoI и BamHI, присутствующих в последовательности немодифицированного домена CH3 IgG1 человека. Имитационное МАТ2 содержало легкую цепь МАТ к FITC 4420 (SEQ ID NO: 116) или МАТ к HEL D1.3 (SEQ ID NO: 118), соответственно, а также содержало мутацию LALA в домене CH2 тяжелой цепи, чтобы уменьшить взаимодействие с рецептором Fc-гамма и потенциальное перекрестное связывание, индуцированное рецептором Fc-гамма. Наличие мутации LALA в имитационных МАТ2 и МАТ2, упомянутых в этих примерах, обозначено суффиксом «AA» на конце части Fcab названий их клонов.
Имитационные МАТ2 получали путем кратковременной экспрессии в клетках HEK293-6E и очищали с использованием колонок MabSelect SuRe™ с белком A.
4.2 Аффинность связывания Fcab к OX40 человека в формате имитационного МАТ2 с экспрессируемым клетками OX40 человека и яванской макаки
Аффинность Fcab к OX40 человека в формате имитационного (4420 LALA) МАТ2 в отношении связывания с экспрессируемым клетками OX40 человека или яванской макаки (клетки DO11.10, экспрессирующие либо OX40 человека [DO11.10-hOX40], либо OX40 яванской макаки [DO11.10-cOX40]; см. Таблицу 2) измеряли с помощью проточной цитометрии. Неспецифичное связывание также оценивали с помощью проточной цитометрии путем тестирования связывания с клетками HEK, не экспрессирующими OX40.
Разведения имитационного (4420 LALA) МАТ2 и контрольного МАТ (2× конечная концентрация) готовили с тремя повторами в 1× ДФСБ (Gibco, 14190-094). Суспензии клеток DO11.10-hOX40 или DO11.10-cOX40, или HEK готовили в ФСБ + 2% БСА (Sigma, A7906) и высевали при 4×106 клеток/мл с 50 мкл/лунку в 96-луночные планшеты с V-образным дном (Costar, 3897). По 50 мкл разведений имитационного (4420 LALA) МАТ2 или контрольного МАТ (МАТ к OX40 человека, 11D4) добавляли в лунки, содержащие клетки (конечный объем 100 мкл), и инкубировали при 4°C в течение 1 часа. Планшеты промывали и затем добавляли 100 мкл/лунку вторичного антитела (антитело к Fc-488 человека, Jackson ImmunoResearch, 109-546-098), разведенного 1:1000 в ФСБ + 2% БСА, и инкубировали в течение 30 минут при 4°C в темноте. Планшеты промывали и содержимое повторно суспендировали в 100 мкл ФСБ, содержащего DAPI (Biotium, номер по каталогу 40043) при 1 мкг/мл. Планшеты считывали с использованием проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали, и измеряли флуоресценцию в канале FITC (488 нм/530/30). Данные аппроксимировали с использованием уравнения зависимости ответа от log(агониста) в программном обеспечении GraphPad Prism.
Fcab (все протестированные в формате имитационного [4420 LALA] МАТ2) и положительный контроль МАТ к OX40 человека, 11D4, в остове IgG1 человека и содержащий мутацию LALA в домене CH2 тяжелой цепи (G1AA/11D4; SEQ ID NO 173 и 175), связывались с OX40 человека с диапазоном значений аффинности. Тестировали способность пяти клонов из линии FS20-22 и шести клонов из линии FS20-31 связывать экспрессируемый клетками OX40 человека и яванской макаки; значения аффинности связывания этих клонов представлены в Таблице 5.
Таблица 5: Аффинность связывания Fcab к OX40 в формате имитационного (4420 LALA) МАТ2 с экспрессируемым клетками OX40 человека или яванской макаки
4.3 Активация OX40 in vitro с помощью Fcab к OX40 в формате имитационного МАТ2
Активированные Т-клетки экспрессируют OX40 на своей клеточной поверхности. Связывание тримерного лиганда OX40 с OX40 приводит к тримеризации рецептора. Поскольку лиганд OX40 экспрессируется в виде кластеров на клеточной поверхности антигенпрезентирующих клеток, взаимодействие между лигандом OX40 и OX40 приводит к кластеризации OX40, которая, как известно, имеет существенное значение для передачи сигнала OX40 и последующей активации Т-клеток. Антитела, которые оказывают агонистическое действие на OX40, должны имитировать эту кластеризующую активность лиганда OX40. В случае моноспецифичных антител к OX40 рецепторы Fc-гамма связываются с доменами Fc антител и перекрестно связывают их, что приводит к кластеризации OX40.
Fcab к OX40 человека и к OX40 мыши в формате имитационного (4420) МАТ2, содержащего мутацию LALA, описанные выше, тестировали в анализах активации Т-клеток для определения их способности активировать OX40, экспрессируемый на Т-клетках, при перекрестном связывании Fcab в присутствии перекрестно связывающего агента. Анализ активации Т-клеток человека для тестирования Fcab к OX40 человека в формате имитационного (4420 LALA) МАТ2 включал выделение Т-клеток из мононуклеарных клеток периферической крови (МКПК) человека и тестирование высвобождения IL-2, который является маркером активации Т-клеток. Анализы проводили способом, аналогичным тому, который описан ниже в Примере 13, и включали использование клона МАТ к CH2 человека, MK1A6, или FITC-декстрана (Sigma) для перекрестного связывания антитела для положительного контроля (11D4) или Fcab в формате имитационного (4420 LALA) МАТ2, соответственно.
Fcab к OX40 человека в формате имитационного (4420 LALA) МАТ2 при перекрестном связывании мишенью Fab (FITC-декстран) показали диапазон значений активности в анализе активации Т-клеток. Все Fcab были способны костимулировать Т-клетки в присутствии антитела к CD3 и индуцировать выработку IL2 человека. Fcab из линий FS20-22 и FS20-31 проявили активность как при перекрестном связывании, так и без него. В частности, Fcab из этих линий обладали активностью в отсутствие перекрестно связывающего агента, которая значительно повышалась при перекрестном связывании. Поскольку эти Fcab обладают высокой перекрестной реактивностью с OX40 яванской макаки (сравнимой со связыванием OX40 человека), возможно проведение токсикологических исследований на этом виде. Из всех клонов в линии FS20-22 клоны FS20-22-41, FS20-22-47, FS20-22-49 и FS20-22-85 имели самые низкие значения ЭК50 в отношении их агонистической активности при перекрестном связывании и поэтому являются предпочтительными клонами из этой линии. Из них клон FS20-22-49 показал самое высокое увеличение агонистической активности при перекрестном связывании, а также имел самую низкую ЭК50 в отношении своей агонистической активности в присутствии перекрестного связывания и поэтому является предпочтительным клоном.
Как описано выше, цель авторов настоящего изобретения заключалась в получении МАТ2, которые способны оказывать агонистическое действие как на OX40, так и на CD137 в отсутствие дополнительных перекрестно связывающих агентов. Упомянутые выше эксперименты демонстрируют, что Fcab FS20-22-49 способен активировать OX40 в присутствии дополнительного перекрестно связывающего агента. Для получения двойного агониста, который не требует дополнительных перекрестно связывающих агентов, авторы настоящего изобретения решили получить антитела к CD137 с намерением использовать CDR из этих антител в конечной молекуле МАТ2, нацеленной на OX40 и CD137.
Пример 5 - Отбор и характеристика антител к CD137 человека
Синтетические наивные фагмидные библиотеки, экспонирующие домен Fab зародышевых линий человека с рандомизацией в CDR1, CDR2 и CDR3 (MSM Technologies), использовали для наивных отборов МАТ к CD137 человека с рекомбинантными и экспрессируемыми на поверхности клеток антигенами CD137, описанными в Примере 1.2.
Библиотеки Fab отбирали в трех раундах с использованием Dynabeads со стрептавидином (Thermo Fisher Scientific, 11205D) и связанного с Dynabeads связывающего белка, нейтравидина (Thermo Fisher Scientific, 31000), чтобы выделить фаг, связанный с биотинилированным CD137-mFc-Avi человека или CD137-Avi-His человека. Чтобы гарантировать связывание Fab с CD137, экспрессируемым на поверхности клеток, выходные продукты первого раунда из отборов с использованием рекомбинантного антигена CD137 также подвергали двум дополнительным раундам отборов с использованием клеток DO11.10-hCD137 и четвертому раунду с использованием клеток DO11.10-cCD137.
Приблизительно 2200 клонов из выходных продуктов раундов 3 и 4 подвергали скринингу с помощью ИФА фага для определения связывания с CD137-mFc-Avi человека и яванской макаки. Биотинилированный mFc включали в качестве отрицательного контроля. Вариабельные области положительных клонов (клоны с сигналом связывания CD137 по меньшей мере в 4 раза превышающим сигнал связывания с mFc) секвенировали, что привело к идентификации 36 уникальных комбинаций последовательностей VH/VL. Идентифицированные последовательности происходили из обоих отборов с использованием рекомбинантного антигена CD137 и антигена CD137, экспрессируемого на поверхности клетки, при этом несколько клонов выделили с использованием обеих стратегий отбора. На основании ИФА фага 22 из 36 клонов были перекрестно реактивными в отношении яванской макаки (cyno), однако поскольку чувствительность ИФА фага могла быть недостаточной для обнаружения слабых связывающих элементов, перекрестно реактивных в отношении яванской макаки, все 36 клонов приоритетно взяли для получения в формате молекул IgG1. Для каждого клона домены VH и VL индивидуально клонировали в вектор экспрессии pTT5 (Национальный исследовательский совет Канады), содержащий либо домены CH1, CH2 (с мутацией LALA в домене CH2) и CH3, либо домены CL, соответственно. Полученный pTT5-FS30 VH с мутацией LALA (AA) и векторы pTT5-FS30 VL кратковременно совместно трансфецировали в клетки HEK293-6E. Двадцать восемь клонов экспрессировались в виде растворимых молекул IgG1. Их очищали с помощью колонок MabSelect SuRe™ с белком A и подвергали дальнейшему тестированию.
Связывание МАТ к CD137 анализировали с помощью ИФА с использованием CD137-mFc-Avi человека и яванской макаки. Из 28 протестированных клонов 10 показали дозозависимое связывание с CD137-mFc-Avi человека и отсутствие связывания с OX40-mFc-Avi человека, mFc или стрептавидином. Четыре клона в этой группе, FS30-5, FS30-10, FS30-15 и FS30-16, были перекрестно реактивными в отношении CD137-mFc-Avi яванской макаки. Из-за небольшого количества полученных клонов, перекрестно реактивных в отношении яванской макаки, дополнительные клоны подвергали скринингу и экспрессировали, как описано выше. В результате был выделен один дополнительный связывающий элемент FS30-35, перекрестно реактивный в отношении яванской макаки.
МАТ к CD137 человека FS30-5, FS30-10, FS30-15 и FS30-16 тестировали для определения связывания с клетками, экспрессирующими CD137 человека или яванской макаки (DO11.10-hCD137 или DO11.10-cCD137), с использованием проточной цитометрии. Неспецифичное связывание также оценивали путем тестирования связывания с клетками DO11.10 и клетками HEK293, не экспрессирующими CD137. Значения аффинности связывания сравнивали со значениями аффинности двух МАТ для положительного контроля, MOR7480.1 (патент США № 2012/0237498) и 20H4.9 (патент США № 7288638), вариабельные домены которых были клонированы и экспрессированы в формате IgG1 человека, содержащем мутацию LALA в домене CH2 (формат G1AA).
Было обнаружено, что клоны FS30-5, FS30-10, FS30-15 и FS30-16 связываются с рецепторами CD137 человека и яванской макаки, экспрессируемыми на поверхности клетки, со значениями ЭК50 в диапазоне 0,15-0,57 нМ, что сравнимо с МАТ для положительного контроля. Связывание с исходными клетками DO11.10 или HEK293 не было обнаружено, что свидетельствует о специфичности связывания. В этих клетках не было обнаружено связывание положительного контроля, антитела к CD137 20H4.9, с CD137 яванской макаки. Опубликованные данные (патент США № 7288638) показывают, что 20H4.9 в формате IgG1 действительно связывается с CD137 яванской макаки на МКПК яванской макаки, индуцированных PMA (форболмиристатацетатом). Авторы настоящего изобретения показали, что 20H4.9 в формате G1AA связывался с рекомбинантным CD137 яванской макаки, но аффинность была намного ниже, чем в отношении CD137 человека (данные не показаны), что может объяснять наблюдаемое отсутствие связывания антитела с клетками DO11.10-cCD137.
Чтобы определить биофизические характеристики МАТ FS30 их подвергали гель-проникающей хроматографии (SEC) и анализировали процентное содержание мономерной фракции. Все четыре протестированных МАТ FS30 показали профиль с одним пиком и содержали >97% мономеров. Высокий уровень мономерного белка позволил продолжить тестирование функциональной активности.
Функциональную активность МАТ к CD137 затем анализировали в анализе активации первичных Т-клеток. МАТ к CD137 индуцируют агонистическое действие in vivo путем вовлечения рецепторов Fcγ, вызывая посредством этого кластеризацию МАТ и рецептора CD137. Чтобы имитировать максимальную способность МАТ кластеризовать поверхностные молекулы рецептора CD137, МАТ FS30 перекрестно связывали с использованием антитела к CH2 человека (клон MK1A6, полученный собственными силами) перед анализом. Активацию Т-клеток сравнивали с МАТ, не связанными перекрестно. Антитело D1.3 к лизоциму куриного яичного белка (HEL) в остове IgG1 человека с мутацией LALA (G1AA/HelD1.3) включали в качестве отрицательного контроля.
При перекрестном связывании МАТ FS30-5, FS30-10, FS30-15 и FS30-16 показали сильную активность в анализе активации Т-клеток со значениями ЭК50 менее 10 нМ и максимальным уровнем IL-2 (Emax), сходным с положительным контролем МАТ к CD137 (МАТ к CD137 MOR7480.1, 5637 hIL-2 пг/мл; и МАТ к CD137 20H4.9, 10232 hIL-2 пг/мл). Emax МАТ FS30-6 (1512 hIL-2 пг/мл) было значительно ниже, чем у положительных контролей и других МАТ FS30, это указывает на более низкий общий уровень активации Т-клеток. В отличие от положительного контроля, МАТ к CD137 20H4.9, который проявил активность в отсутствие перекрестного связывания (выработка hIL-2 3174 пг/мл), МАТ FS30 не проявили активность (без перекрестного связывания, о чем свидетельствуют измеренные фоновые уровни ответа IL-2).
Пример 6 - Конструирование и экспрессия МАТ2, нацеленного на OX40 человека и CD137 человека
Получали МАТ2, содержащие Fcab к OX40 человека, спаренный с Fab к CD137 человека. Fcab FS20-22-49, нацеленный на OX40 человека, отобрали для спаривания с Fab, нацеленными на CD137, вследствие его более высокой активности в анализах Т-клеток (см. Пример 4.3).
6.1 Экспрессия и характеристика МАТ в формате МАТ2
Получали молекулы МАТ2, которые состояли из молекулы IgG1, содержащей CDR клона FS30-5, FS30-10, FS30-15, FS30-16 или FS30-35 и содержащей мутацию LALA в домене CH2, а также связывающий сайт рецептора OX40 человека из FS20-22-49 в домене CH3. Эти молекулы МАТ2 получали путем замены домена VH МАТ2 к OX40 человека, FS20-22-49AA/HelD1.3, соответствующими доменами VH клонов FS30 и совместной трансфекции полученного VH с соответствующей легкой цепью МАТ FS30. Мутация LALA в домене CH2 молекулы IgG1 сохранялась в полученных молекулах МАТ2. Эти молекулы МАТ2 обозначили FS20-22-49AA/FS30-5, FS20-22-49AA/FS30-10, FS20-22-49AA/FS30-15, FS20-22-49AA/FS30-16 и FS20-22-49AA/FS30-35. МАТ2 получали путем кратковременной экспрессии в клетках HEK293-6E и очищали с использованием колонок MabSelect SuRe™ с белком A.
CD137 принадлежит к суперсемейству рецепторов фактора некроза опухоли (TNFRSF) рецепторов цитокинов (Moran et al., 2013). Чтобы проанализировать специфичность связывающего сайта Fab к CD137 пяти молекул МАТ2, связывание МАТ2 с CD137 человека и пятью близкородственными представителями TNFRSF человека (TNFRSF1A, TNFRSF1B, GITR, NGFR и CD40) тестировали с использованием SPR. Цель заключалась в том, чтобы продемонстрировать 1000-кратную специфичность, показав отсутствие связывания МАТ2 с близкородственными антигенами в концентрации 1 мкМ, при этом показав связывание с рецепторами CD137 в концентрации 1 нМ.
В то время как МАТ2 FS20-22-49AA/FS30-5, FS20-22-49AA/FS30-10, FS20-22-49AA/FS30-16 и FS20-22-49AA/FS30-35 показали высокий уровень специфичности (близко к 1000-кратному), МАТ2 FS20-22-49AA/FS30-15 показало неспецифичное связывание со всеми пятью тестируемыми близкородственными представителями TNFRSF. Неспецифичное связывание, проявленное этим клоном, было в среднем приблизительно в 5-10 раз ниже, чем связывание с рецепторами CD137 в той же концентрации, и был сделан вывод, что это обусловлено связывающим сайтом Fab молекулы МАТ2, поскольку МАТ FS30-15 показало такой же профиль связывания при тестировании связывания с теми же пятью представителями TNFRSF, близкородственными CD137. На основании этих данных клон FS30-15 был исключен из дальнейших кампаний отбора.
6.2 Оптимизация последовательности МАТ к CD137
Несмотря на то, что МАТ к CD137, FS30-5, FS30-10, FS30-16 и FS30-35, показали высокую аффинность и специфичность в отношении CD137, а также активность в анализе активации Т-клеток, они содержали один или более потенциальных сайтов посттрансляционных модификаций (PTM) в петлях CDR. Было принято решение дополнительно модифицировать эти клоны, чтобы попытаться идентифицировать аминокислотные остатки, которые могут быть заменены в этих сайтах с сохранением или улучшением связывания и активности. Идентифицированные потенциальные сайты PTM включали остатки метионина в CDR3 VH (положение согласно Kabat M100D и M100H в FS30-5, M97 в FS30-10, M100A в FS30-16 и M100F в FS30-35), потенциальный мотив изомеризации аспартата в CDR2 VH (положение Kabat D54G55 в FS30-16) и потенциальный сайт дезамидирования в CDR3 VL (положение Kabat Q90G91 в FS30-16).
Сайт-направленный мутагенез осуществляли с использованием пяти клонов МАТ2 FS20-22-49AA/FS30 в качестве матриц и праймеров, которые содержали вырожденный кодон NNK в сайтах, кодирующих остатки метионина, аспартата или глицина, чтобы обеспечить все возможные замены аминокислот. Были исключены остатки цистеина и аминокислоты, способные приводить к образованию новых потенциальных мотивов PTM. Клоны экспрессировали и подвергали скринингу для определения связывания с клетками DO11.10-hCD137. Клоны со сходным (в пределах двух раз) или улучшенным связыванием при 10 нМ по сравнению с исходными клонами МАТ2 отбирали для экспрессии в масштабе 30-50 мл, очищали на колонках с белком А и подвергали скринингу в анализе активации Т-клеток с использованием клеток DO11.10-hCD137 и антитела к CH2 человека, MK1A6, в качестве перекрестно связывающего агента.
Клетки DO11.10-hCD137 промывали один раз в ФСБ и повторно суспендировали в среде для клеток DO11.10 (среда RPMI (Life Technologies) с 10% фетальной бычьей сыворотки (ФБС (Life Technologies)) и 5 мкг/мл пуромицина (Life Technologies, A11113803)) в концентрации 1,0×106 клеток/мл. 96-луночные планшеты с плоским дном покрывали антителом к CD3 мыши (Thermo Fisher Scientific, клон 17A2) путем инкубации с 0,1 мкг/мл антитела к CD3 мыши, разведенного в ФСБ, в течение 2 часов при 37°C, 5% CO2 и затем дважды промывали ФСБ. Клетки DO11.10-hCD137 добавляли в планшеты при 1×105 клеток/лунку. Разведение 2 мкМ каждого тестируемого антитела готовили в ДФСБ (Gibco) и дополнительно разводили в соотношении 1:10 в среде для клеток DO11.10 (30 мкл + 270 мкл), чтобы получить разведение 200 нМ. Перекрестно связывающий агент MK1A6 добавляли в лунки в молярном соотношении 1:1 с образцами тестируемого антитела, подлежащими перекрестному связыванию. В 96-луночном планшете готовили последовательные разведения каждого антитела или смеси антитело/перекрестно связывающий агент. По 100 мкл разведенного антитела или смеси антитело/перекрестно связывающий агент добавляли к клеткам DO11.10-hCD137 на планшете. Клетки инкубировали при 37°C, 5% CO2 в течение 72 часов. Супернатанты собирали и анализировали с помощью набора для ИФА IL-2 мыши (eBioscience или R&D Systems) в соответствии с инструкциями производителя. Планшеты считывали при 450 нм, используя считывающее устройство для планшетов с программным обеспечением Gen5, BioTek. Значения поглощения 630 нм вычитали из значений поглощения 450 нм (Коррекция). Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 мыши (mIL-2) наносили на график в зависимости от логарифмической концентрации антитела, и полученные кривые аппроксимировали с использованием уравнения зависимости ответа от log(агониста) в GraphPad Prism.
Для каждого из клонов идентифицировали ограниченное количество аминокислот, которые сохраняли или улучшали связывание с CD137 на поверхности клетки, для замены остатка метионина в CDR3 тяжелой цепи. Клон МАТ2 FS20-22-49AA/FS30-16 содержал три потенциальных сайта PTM и мутация каждого из них приводила к небольшому уменьшению аффинности связывания. При их комбинировании в одной молекуле уменьшение связывания было аддитивным (данные не показаны), и, следовательно, этот клон не использовали в дальнейшем. Было обнаружено несколько мутаций, которые улучшили связывание с CD137 и функциональную активность по сравнению с соответствующим исходным клоном. Было обнаружено, что три мутантных клона МАТ2, все происходящие из клона МАТ2 FS20-22-49AA/FS30-10, обладают улучшенной аффинностью связывания и функциональной активностью. Эти МАТ2 содержали либо остаток аспарагина, либо остаток треонина, либо остаток лейцина, заменяющий остаток метионина в положении 97 в исходном МАТ2 FS20-22-49AA/FS30-10, и были обозначены FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16, соответственно. Несмотря на то, что значения ЭК50 для мутантных клонов, происходящих из исходного клона МАТ2 FS20-22-49AA/FS30-35, не показали улучшения функциональной активности по сравнению с исходным клоном, один мутантный клон, обозначенный FS20-22-49AA/FS30-35-14, который содержал остаток аланина, заменяющий остаток метионина в положении 100F в исходном клоне, однако показал улучшенное связывание. В случае исходного клона МАТ2 FS20-22-49AA/FS30-5 оба остаток метионина в положении 100D и остаток метионина в положении 100H были заменены, соответственно, остатком изолейцина и остатком лейцина в той же молекуле с получением мутантного клона МАТ2, обозначенного FS20-22-49AA/FS30-5-37. Клоны FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-35-14 и FS20-22-49AA/FS30-5-37 отобрали для дальнейшей характеристики.
6.3 Анализы блокирования лиганда CD137 человека
Для активации рецептора CD137 требуется взаимодействие CD137-CD137L. Агонистические антитела к CD137 могут запускать активацию CD137, имитируя взаимодействие лиганда, что потенциально блокирует связывание лиганда, или запуская кластеризацию и активацию рецепторов без нарушения связывания лиганда. Если антитело потенциально имитирует CD137L, оно может блокировать взаимодействие рецептора и лиганда. В данной области техники известно, что MOR7480.1 блокирует взаимодействие лиганд/рецептор (US 2012/0237498), в то время как ранее сообщалось, что антитело 20H4.9 не блокирует взаимодействие между CD137 и его лигандом (патент США № 7288638).
Способность клонов МАТ2 к CD137 человека FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14 блокировать взаимодействие CD137-CD137L тестировали с использованием метода на основе ИФА. МАТ к OX40 11D4 (Европейский патент № 2242771) в формате IgG1 (G1/11D4; SEQ ID NO 174 и 175) использовали в качестве изотипического/отрицательного контроля; МАТ2 FS20-22-49AA/4420, содержащее клон Fcab к OX40 FS20-22-49AA и область Fab антитела к FITC 4420, использовали в качестве МАТ2 для отрицательного контроля для связывания OX40; и МАТ к CD137 G1/MOR7480.1 (SEQ ID NO 119 и 120) и G1/20H4.9 (SEQ ID NO 121 и 122) использовали в качестве положительных контролей для связывания CD137 и активности блокирования лиганда.
В частности, 96-луночные планшеты Maxisorp покрывали рекомбинантным антигеном CD137-mFc-Avi человека в концентрации 1 мкг/мл в ФСБ в течение ночи при 4°C. На следующий день планшеты промывали ФСБТ (ФСБ + 0,05% твин20™) и блокировали ФСБ + 1% БСА (Sigma, A3059-500G) в течение 1 часа при комнатной температуре с перемешиванием. После блокирования планшеты снова промывали ФСБТ. Разведение 100 нМ каждого тестируемого антитела готовили в ФСБ + 1% БСА, добавляли в планшеты, покрытые CD137, и инкубировали в течение 1 часа при комнатной температуре с перемешиванием. После этой инкубации планшеты промывали ФСБТ и затем инкубировали с 20 нг/мл CD137L-His (R&D Systems, 2295-4L-025/CF) в ФСБ в течение 1 часа при комнатной температуре с перемешиванием. Затем планшеты промывали ФСБТ, а затем инкубировали с вторичным антителом к His (R&D Systems, MAB050H) в разведении 1:1000 в ФСБ в течение 1 часа при комнатной температуре с перемешиванием. Планшеты затем промывали ФСБТ и инкубировали с реагентом для обнаружения TMB (Thermo Fisher Scientific, 002023) до появления синего окрашивания в лунках с положительным контролем, после этого реакцию останавливали добавлением 2 н H2SO4. Планшеты считывали при 450 нм, используя считывающее устройство для планшетов с программным обеспечением Gen5, BioTek. Значения поглощения 630 нм вычитали из значений поглощения 450 нм (Коррекция). Значения поглощения после вычитания наносили на график в зависимости от логарифмической концентрации антитела, и полученные кривые аппроксимировали с использованием уравнения зависимости ответа от log(ингибитора) в GraphPad Prism. Значения нормировали, устанавливая контрольные МАТ G1/11D4 и G1/MOR7480.1 как значения блокирования 0% и 100%, соответственно. Данные анализировали с использованием одностороннего дисперсионного анализа и критерия Холма-Сидака для множественных сравнений с использованием GraphPad Prism.
Для пяти протестированных клонов МАТ2 к CD137 человека наблюдали ряд значений активности блокирования. FS20-22-49AA/FS30-5-37 показало, как и антитела для положительного контроля, полное ингибирование взаимодействия рецептор-лиганд. Все клоны МАТ2, содержащие области Fab из МАТ к CD137 линии FS30-10 (т. е. FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16), ингибировали взаимодействие между CD137 и CD137L на 49-54% и поэтому считались частичными блокаторами. Вследствие только частичного блокирования взаимодействия между CD137 и CD137L, возможно, эти МАТ не могут полностью ингибировать природное взаимодействие CD137L с его рецептором так, что некоторая передача сигнала CD137 все еще может происходить за счет этого механизма, даже если одно из этих антител связано. Клон FS20-22-49AA/FS30-35-14, как и молекула МАТ2 FS20-22-49AA/4420 для отрицательного контроля, не был способен значительно ингибировать взаимодействие рецептор-лиганд и поэтому считался неблокирующим.
Подытоживая, результаты этого анализа на основе ИФА показали, что панель протестированных МАТ к CD137 показала несколько видов способности блокирования лиганда, включая полную, частичную и отсутствующую блокирующую активность. Каждый из клонов FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14 показал блокирующую активность, которая отличалась от таковой МАТ к CD137 в качестве положительных контролей. Поскольку был идентифицирован ряд видов активности блокирования лиганда, тестировали функциональную активность каждого из антител.
Способность клонов FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16 блокировать взаимодействие CD137-CD137L дополнительно тестировали с использованием клеточного метода. Наблюдали ряд видов активности блокирования, при этом FS20-22-49AA/FS30-5-37 показало, как и антитело для положительного контроля (G1/MOR7480.1), использованное в этом анализе, полное ингибирование взаимодействия рецептор-лиганд. Все три клона МАТ2, содержащие области Fab из МАТ к CD137 линии FS30-10 (т. е. FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16), ингибировали взаимодействие между CD137 и CD137L на 46-76% и поэтому считались частичными блокаторами. Таким образом, результаты этого анализа были аналогичны результатам анализа блокирования на основе ИФА и показали, что панель протестированных МАТ к CD137 проявила несколько видов способности блокирования лиганда, от полной до частичной блокирующей активности. Каждый из клонов FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16 показал блокирующую активность, которая отличалась от таковой антитела для положительного контроля.
Пример 7 - Специфичность связывания и функциональная активность клонов МАТ и МАТ2 в анализе активации Т-клеток, экспрессирующих CD137 человека
7.1 Специфичность связывания клонов МАТ2
CD137 и OX40 принадлежат к суперсемейству рецепторов фактора некроза опухоли (TNFRSF) рецепторов цитокинов (Moran et al., 2013). Для анализа специфичности Fab к CD137, а также сайта связывания Fcab к OX40 пяти молекул МАТ2 связывание МАТ2 FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16, FS20-22-49AA/FS30-35-14 и FS20-22-49AA/FS30-5-37 с CD137 человека, OX40 человека и шестью близкородственными представителями TNFRSF человека тестировали с использованием поверхностного плазмонного резонанса (SPR). Цель состояла в том, чтобы продемонстрировать 1000-кратную специфичность, показав отсутствие связывания МАТ2 с близкородственными антигенами в концентрации 1 мкМ, при этом показав связывание с рецепторами CD137 и OX40 в концентрации 1 нМ. МАТ к CD137 MOR7480.1 и МАТ к OX40 11D4 использовали в качестве положительных контролей.
В общих чертах, CD137-mFc-Avi человека (Таблица 3), OX40-mFc (Таблица 2), рекомбинантный TNFRSF1A-Fc человека, рекомбинантный TNFRSF1B-Fc человека, рекомбинантный GITR-Fc человека, рекомбинантный NGFR-Fc человека, рекомбинантный CD40-Fc человека или рекомбинантный DR6-Fc человека иммобилизовали на проточных ячейках на чипах CM5 при прибл. 1000 RU. Проточную ячейку 1 оставили для иммобилизации холостого образца. Пять МАТ2 разводили до 1 мкМ и 1 нМ в 1× буфере HBS-EP (GE Healthcare, код продукта BR100188), пропускали над чипом в течение 3 минут, а затем позволяли диссоциировать в течение 4 минут. Для регенерации использовали 30-секундное впрыскивание 10 мМ глицина, pH=1,5. МАТ для положительного контроля впрыскивали при 50-100 нМ, чтобы продемонстрировать покрытие каждого антигена. Уровни связывания определяли в конце фазы ассоциации и сравнивали.
Все из отобранных МАТ2 показали высокий уровень специфичности в отношении рецепторов CD137 и OX40 человека, сходный или более высокий, чем у положительных контролей MOR7480.1 и 11D4, соответственно.
7.2 Функциональная активность агонистических антител к CD137 в анализе активации Т-клеток, экспрессирующих CD137 человека
Чтобы установить активность различных агонистических антител к CD137, использовали анализ активации Т-клеток с использованием клеток DO11.10-hCD137. Тестировали агонистические антитела к CD137 G1AA/MOR7480.1 (SEQ ID NO: 125 и 120), G1AA/20H4.9 (SEQ ID NO: 165 и 122) и G1AA/FS30-10-16 (SEQ ID NO: 154 и 97), а также антитело к FITC 4420 в формате IgG1 (G1/4420; SEQ ID NO: 115 и 116) в качестве отрицательного контроля изотипа. Молекулы антител тестировали как в присутствии, так и в отсутствие перекрестно связывающего антитела к CH2 человека MK1A6 (см. Пример 2.1). Выработку IL-2 мыши использовали в качестве меры активации Т-клеток.
Клетки DO11.10-hCD137 один раз промывали в ФСБ и повторно суспендировали в среде для клеток DO11.10 (среда RPMI (Life Technologies) с 10% ФБС (Life Technologies) и 5 мкг/мл пуромицина (Life Technologies, A11113803)) в концентрации 1,0×106 клеток/мл. 96-луночные планшеты с плоским дном покрывали антителом к CD3 мыши (Thermo Fisher Scientific, клон 17A2) путем инкубации с 0,1 мкг/мл антитела к CD3 мыши, разведенного в ФСБ, в течение 2 часов при 37°C, 5% CO2, а затем дважды промывали ФСБ. Клетки DO11.10-hCD137 добавляли в планшеты при 1×105 клеток/лунку. Разведение 2 мкМ каждого тестируемого антитела готовили в ДФСБ (Gibco) и дополнительно разводили 1:10 в среде для клеток DO11.10 (30 мкл + 270 мкл), чтобы получить разведение 200 нМ. Перекрестно связывающий агент MK1A6 добавляли в лунки в молярном соотношении 1:1 с тестируемыми антителами, где это было необходимо. В 96-луночном планшете готовили последовательные разведения антитела или смеси антитело/перекрестно связывающее антитело. По 100 мкл разведенного антитела или смеси антитело/перекрестно связывающее антитело добавляли к клеткам DO11.10-hCD137 на планшете. Клетки инкубировали при 37°C, 5% CO2 в течение 72 часов. Супернатанты собирали и анализировали с помощью набора для ИФА IL-2 мыши (eBioscience или R&D Systems) в соответствии с инструкциями производителя. Планшеты считывали при 450 нм, используя считывающее устройство для планшетов с программным обеспечением Gen5, BioTek. Значения поглощения 630 нм вычитали из значений поглощения 450 нм (Коррекция). Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 мыши (mIL-2) наносили на график в зависимости от логарифмической концентрации антитела, и полученные кривые аппроксимировали с использованием уравнения зависимости ответа от log(агониста) в GraphPad Prism.
Результаты анализа показаны на Фигуре 2C и D. Антитела к CD137 различались по их потребности в перекрестно связывающем антителе для осуществления своей активности, при этом все три антитела к CD137 показали зависимое от концентрации увеличение выработки IL-2 в присутствии перекрестно связывающего антитела, но только антитело G1AA/20H4.9 проявило активность в отсутствие перекрестно связывающего антитела. Таким образом, G1AA/MOR7480.1 и G1AA/FS30-10-16 требовали добавления перекрестно связывающего антитела, т. е. их активность была «зависимой от перекрестного связывания», в то время как G1AA/20H4.9 проявило активность как в присутствии, так и в отсутствие перекрестно связывающего антитела, т. е. его активность была «независимой от перекрестного связывания».
7.3 Функциональная активность клонов МАТ2 в анализе активации Т-клеток, экспрессирующих CD137 человека
Функциональную активность отобранных клонов МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16 тестировали в анализе активации Т-клеток с использованием клеток DO11.10-hCD137. Антитело к FITC 4420 в формате IgG1 (G1/4420; SEQ ID NO: 115 и 116) использовали в качестве отрицательного контроля изотипа; МАТ к OX40 G1/11D4 (SEQ ID NO 174 и 175) и клон МАТ2 FS20-22-49AA/4420 (SEQ ID NO 123 и 116) использовали в качестве отрицательных контролей; и антитело к CD137 MOR7480.1 как в формате IgG1 (G1/MOR7480.1; SEQ ID NO 119 и 120), так и IgG2 (G2/MOR7480.1; SEQ ID NO 124 и 120), причем формат IgG2 представляет собой формат, в котором антитело было протестировано в клинических исследованиях (Gopal et al., 2017; Tolcher et al., 2017), использовали в качестве положительного контроля. Молекулы МАТ и МАТ2 перекрестно связывали с антителом к CH2 человека, MK1A6 (см. Пример 2.1), и в одном эксперименте изучали активность молекул МАТ и МАТ2, не связанных перекрестно. Выработку IL-2 мыши использовали в качестве меры активации Т-клеток. Эксперимент выполняли, как описано в Примере 7.2.
При перекрестном связывании все пять отобранных клонов МАТ2 показали сильную активность в анализе активации Т-клеток со средними значениями ЭК50 менее 15 нМ и средними значениями Emax в диапазоне приблизительно 16000-20000 пг/мл IL-2 (Таблица 6 и Фигура 2А). В отсутствие перекрестного связывания активность тестируемых клонов МАТ2 не наблюдалась (Фигура 2B). Активность антитела для положительного контроля MOR7480.1 наблюдали только при перекрестном связывании (ЭК50 3,3 нМ и Emax 12575 пг/мл для G1/MOR7480.1 и ЭК50 2,4 нМ и Emax 8547 пг/мл для G2/MOR7480.1). Комбинация отсутствия активности перекрестно связанного МАТ к OX40 (G1/11D4) и низких фоновых сигналов, наблюдаемых для не связанных перекрестно молекул МАТ2, содержащих Fcab к OX40, показывает, что результаты этого анализа представляют собой считывание только активности CD137, наиболее вероятно, из-за высоких уровней экспрессии рецептора CD137 и необнаруживаемых уровней экспрессии рецептора OX40 клетками DO11.10 (данные не показаны).
Таблица 6: Активность МАТ2 в анализе активации Т-клеток, экспрессирующих CD137 человека
(среднее n=2)
N/A: неприменимо, поскольку низкий сигнал не позволил достоверно определить ЭК50/Emax
NM: не измеряли
Таким образом, МАТ2, содержащее CDR моноклональных антител к CD137 человека FS30-5-37, FS30-10-3, FS30-10-12, FS30-10-16 и FS30-35-14, показало сильную активность в отношении способности активировать CD137 в анализе активации Т-клеток DO11.10-hCD137 при перекрестном связывании. В отсутствие перекрестного связывания не наблюдали значимую активность. Эти МАТ2 содержат домен CH3 из Fcab к OX40 человека FS20-22-49, который также показал высокую активность при перекрестном связывании в анализе Т-клеток (см. Пример 4.3). МАТ2, полученные с мутацией LALA, обозначили FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16.
Эти МАТ2 отобрали для дальнейшего анализа, чтобы определить, способны ли они действовать как двойной агонист, который может автономно оказывать агонистическое действие как на OX40, так и на CD137, основываясь на экспрессии специфичных мишеней и без потребности в дополнительных перекрестно связывающих агентах.
Пример 8 - Аффинность связывания МАТ2 в отношении OX40 и CD137 человека и яванской макаки
Для определения аффинности в отношении CD137 чип CM5 Biacore (GE Healthcare) покрывали антителом к Fc человека с использованием набора Human Antibody Capture (GE Healthcare) в соответствии с условиями производителя до поверхностной плотности приблизительно 4000 RU. Образцы тестируемых антител (МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16, положительный контроль к CD137 G1/MOR7480.1 и отрицательный контроль к hOX40 G1/11D4) захватывали приблизительно до 80 RU. CD137 человека или яванской макаки (hCD137-mFc-Avi или cCD137-mFc-Avi) пропускали над чипом в диапазоне концентраций в ряде трехкратных разведений, начиная с 200 нМ, при скорости потока 70 мкл/мин. Время ассоциации составило 2 мин, а время диссоциации составило 8 мин. Рабочий буфер представлял собой HBS-EP (GE Healthcare, BR100188). Проточные ячейки регенерировали путем впрыскивания 3 М хлорида магния при скорости потока 30 мкл/мин в течение 30 секунд.
Для определения аффинности в отношении OX40 чип CM5 Biacore покрывали антителом к Fab человека с использованием набора Human Fab Capture (GE Healthcare, 28958325) в соответствии с условиями производителя до поверхностной плотности приблизительно 8000 RU. Образцы тестируемых антител (МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16, G1/MOR7480.1 (отрицательный контроль) и G1/11D4 (положительный контроль)) захватывали приблизительно до 80 RU, а затем антиген OX40 человека или яванской макаки (hOX40-mFc или cOX40-mFc) пропускали над чипом в диапазоне концентраций в ряде трехкратных разведений, начиная с 200 нМ, при скорости потока 70 мкл/мин. Время ассоциации составило 2 мин, а время диссоциации составило 8 мин. Рабочий буфер представлял собой HBS-EP. Проточные ячейки регенерировали путем впрыскивания глицин-HCl при pH=2,1 при скорости потока 30 мкл/мин в течение 30 секунд.
Данные анализировали путем вычитания двух эталонов против проточной ячейки, которая была намеренно оставлена холостой (без связывания антитела). Кинетики связывания аппроксимировали с использованием модели Ленгмюра 1:1, чтобы получить скорости ассоциации (ka) и диссоциации (kd) связывания. Равновесные константы связывания (KD) рассчитывали путем деления скорости диссоциации на скорость ассоциации для каждого образца. Анализ данных выполняли с помощью программного обеспечения BiaEvaluation, версия 3.2. Результаты представлены в Таблице 7.
Таблица 7: Аффинность связывания МАТ2 с CD137 и OX40 человека и яванской макаки, определенная с помощью SPR
NB - связывание не обнаружено. NM - Не измеряли.
Значения аффинности связывания для МАТ2 к OX40/CD137 показывают, что эти молекулы связываются с высокой аффинностью с обоими рецепторами. Аффинность этих молекул в отношении OX40 человека является сходной, как и следовало ожидать, поскольку все эти молекулы имеют общий Fcab к OX40. Аффинность в отношении OX40 яванской макаки находится в пределах 5-кратной аффинности в отношении OX40 человека. Аффинность в отношении CD137 человека колеблется в пределах 4-0,2 нМ, а перекрестная реактивность с CD137 яванской макаки также варьируется, поскольку Fab к CD137 различаются в каждой молекуле. FS20-22-49AA/FS30-10-16 имеет более высокую аффинность в отношении CD137 человека, а также сходную аффинность в отношении CD137 яванской макаки. Сходство в связывании с антигенами человека и яванской макаки может представлять собой преимущество, поскольку можно надеяться, что поведение МАТ2 в исследованиях на яванских макаках может быть экстраполировано на человека.
FS20-22-49AA/FS30-10-16 также имеет сходную аффинность в отношении OX40 человека и CD137 человека, поэтому ожидается, что МАТ2 должно одинаково хорошо связываться с обеими мишенями при их совместной экспрессии.
Ожидается, что МАТ2, которое связывается с OX40 и CD137 и одновременно запускает кластеризацию и активацию обеих мишеней, будет действовать как двойной агонист. Известно, что оба OX40 и CD137 присутствуют на Т-клетках (Ma, et al., 2005). Не ограничиваясь какой-либо теорией, полагают, что МАТ2, имеющее сходную аффинность в отношении связывания с обеими мишенями, может обеспечивать преимущество в качестве двойного агониста, поскольку МАТ2 с большей вероятностью будет связываться с клетками, которые экспрессируют обе мишени. МАТ2, которое преимущественно связывало одну мишень со значительно более высокой аффинностью, чем другую, может быть неспособно действовать как двойной агонист, поскольку оно может преимущественно связываться с клетками, которые не экспрессируют обе мишени.
Пример 9 - Одновременное связывание МАТ2 с OX40 и CD137
9.1 Одновременное связывание МАТ2 с OX40 человека и CD137 человека
Способность МАТ2 к OX40/CD137 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3 и FS20-22-49AA/FS30-10-16 одновременно связываться с OX40 и CD137 тестировали с помощью SPR на Biacore 3000. G1/MOR7480.1 использовали в качестве контроля. Биотинилированный CD137 человека (hCD137-mFc-Avi-Bio) разводили до 100 нМ в буфере HBS-EP и иммобилизовали на стрептавидиновом (SA) чипе (GE Healthcare, BR100032) до поверхностной плотности приблизительно 1000 RU, в соответствии с инструкциями производителя, и проточную ячейку активировали и деактивировали без какого-либо белка, иммобилизованного для вычитания фона. Антитела, разведенные до 100 нМ в буфере HBS-EP, впрыскивали совместно с 100 нМ OX40 человека (hOX40-mFc) или буфером HBS-EP при скорости потока 30 мкл/мин. Для каждого этапа связывания за диссоциацией наблюдали в течение 3 минут. Сенсорный чип регенерировали после каждого цикла путем впрыскивания 15 мкл глицина 2.5 (GE Healthcare) при скорости потока 30 мкл/мин. Все протестированные МАТ2 были способны одновременно связываться с OX40 и CD137. Контрольное МАТ, G1/MOR7480.1, связывалось только с CD137.
9.2 Одновременное связывание МАТ2, нацеленного на рецептор мыши, с OX40 мыши и CD137 мыши
МАТ2, содержащее Fcab к OX40 мыши с Fab к CD137 мыши, получали для тестирования его способности одновременно связываться с OX40 мыши и CD137 мыши. Fcab, нацеленный на OX40 мыши, FS20m-232-91, отобрали из-за его более высокой активности в анализах Т-клеток, а Fab антитела к CD137 мыши Lob12.3 (Taraban et al., 2002) в формате изотипа IgG1 человека (G1/Lob12.3; Университет Саутгемптона) отобрали для спаривания с Fcab FS20m-232-91, поскольку он показал хорошее клеточное связывание с клетками, экспрессирующими CD137 мыши, и широко используется в литературе как агонистическое антитело к CD137 с активностью in vitro и in vivo. МАТ2, содержащее домен CH3 FS20m-232-91 и Fab антитела к CD137 мыши Lob12.3 и мутацию LALA, обозначили «FS20m-232-91AA/Lob12.3», в то время как МАТ2, содержащее домен CH3 FS20m-232-91 и Fab антитела к CD137 мыши Lob12.3 без мутации LALA, обозначили «FS20m-232-91/Lob12.3».
Способность МАТ2 FS20m-232-91AA/Lob12.3 одновременно связываться с двумя его мишенями тестировали с помощью SPR на приборе BIAcore 3000 (GE Healthcare). G1/Lob12.3 использовали в качестве положительного контроля. Рекомбинантный CD137 мыши (mCD137-hFc; R&D Systems, номер по каталогу 937-4B-050) разводили до 200 нМ в ацетате натрия с pH=5,0 (GE Healthcare) и иммобилизовали на чипе CM5 Biacore до поверхностной плотности приблизительно 1000 RU, в соответствии с инструкциями производителя, и проточную ячейку активировали и деактивировали без какого-либо белка, иммобилизованного для вычитания фона. МАТ2 и положительный контроль, разведенный до 100 нМ в буфере HBS-EP, впрыскивали совместно с 100 нМ OX40 человека (mOX40-mFc) или буфером HBS-EP при скорости потока 30 мкл/мин. Для каждого этапа связывания за диссоциацией наблюдали в течение 3 минут. Сенсорный чип регенерировали после каждого цикла с помощью 30-секундного впрыскивания водного раствора глицин-HCl при pH=1,7 при скорости потока 20 мкл/мин. МАТ2 было способно одновременно связываться с OX40 и CD137. МАТ G1/Lob12.3 связывалось только с CD137.
Пример 10 - Связывание МАТ2 с рецепторами Fcγ
Как известно из литературы, агонистические антитела, нацеленные на представителей семейства TNFR, требуют перекрестного связывания за счет рецепторов Fcγ, чтобы запустить кластеризацию и активацию мишени для активности in vivo (Wajant, 2015). Однако это может быть нежелательно для антитела, которое предусмотрено как двойной агонист. В связи с этим было принято решение уменьшить способность МАТ2 связываться с рецепторами Fcγ путем вставки мутации LALA.
Антитела изотипа IgG1 человека способны связываться с рецепторами Fcγ. В результате они могут индуцировать эффекторную функцию, такую как антителозависимая клеточная цитотоксичность (АЗКЦ), клеток, экспрессирующих мишень, когда они связываются с рецепторами Fcγ, что приводит к лизису клеток. Поскольку предусмотренный механизм МАТ2 к OX40/CD137 заключается в активации клеток, экспрессирующих OX40 и CD137, без их уничтожения, целевым является уменьшение АЗКЦ, индуцированной МАТ2. Поскольку МАТ2 к OX40/CD137 также предназначены для функционирования в качестве двойных агонистов, предусмотренный механизм их действия заключается в передаче сигнала за счет рецепторов в результате перекрестного связывания путем двойного связывания как с OX40, так и с CD137, когда они совместно экспрессируются на одной и той же клетке или экспрессируются на разных клетках, и поэтому для их функционирования не требуется способность перекрестно связываться за счет рецепторов Fcγ.
Кроме того, известно, что CD137-нацеленные антитела были токсичны для печени в клинических условиях (Segal et al., 2017) и, несмотря на то, что механизм токсичности не известен, возможно, он основан на FcγR-опосредуемом перекрестном связывании антител к CD137 и активации CD137-экспрессирующих клеток в печени или на периферии. Предотвращение агонистического действия на CD137 за счет FcγR-опосредуемого перекрестного связывания может уменьшить риск любой токсичности МАТ2 к OX40/CD137 согласно настоящему изобретению, поскольку эти молекулы будут перекрестно связываться только за счет двойного связывания с OX40 и CD137.
Связывание, согласно SPR, использовали, чтобы подтвердить, что наличие мутации LALA в МАТ2 FS20-22-49AA/FS30-10-16 уменьшило аффинность связывания в отношении рецепторов Fcγ, в частности hFcγR1 (R&D Systems, номер по каталогу 1257-FC-050/CF), hFcγR2a (R&D Systems, номер по каталогу 1330-CD-050/CF), hFcγR2b (R&D Systems, номер по каталогу 1460-CD-050/CF) и hFcγR3a (R&D Systems, номер по каталогу 4325-FC-050/CF). МАТ к hOX40 G1AA/11D4 и G1/11D4 (с мутацией LALA и без нее, соответственно) и МАТ к CD137 G1AA/20H4.9 и G1/20H4.9 (с мутацией LALA и без нее, соответственно), все в формате изотипа hIgG1, а также МАТ к hCD137 G4/20H4.9 в формате изотипа hIgG4 использовали в качестве контрольных антител. Связывание тестировали на приборе Biacore 3000 (GE Healthcare). Чип SA (GE Healthcare, номер по каталогу BR100398) покрывали биотинилированными his-мечеными антигенами OX40 человека (BPS Bioscience, номер по каталогу 71310) и CD137 человека (получен собственными силами) в концентрации 2 мкМ. OX40 человека и CD137 человека наносили на отдельные проточные ячейки, в то время как другую проточную ячейку оставили холостой для вычитания фона. Условия регенерации были определены как 12 мкл водного 10 мМ глицин-HCl при pH=2,0 при скорости потока 20 мкл/мин. Антитела (см. Таблицу 8) и FcγR человека (см. Таблицу 8) разводили до 100 нМ (антитела) или 500 нМ (FcγR человека) в HBS-P (0,01 М HEPES, рН=7,4, 0,15 М NaCl, 0,005% об./об. Surfactant P20, GE Healthcare, BR-1003-68) и совместно впрыскивали при скорости потока 20 мкл/мин, а затем наблюдали за диссоциацией в течение 5 мин.
Анализ данных выполняли с использованием программного обеспечения BiaEvaluation, версия 3.2 RC1, путем сравнения с холостой проточной ячейкой и сопоставления кривых после ассоциации антитела. Значения для ответа связывания в конце фазы ассоциации получали путем вычитания абсолютного ответа в конце фазы ассоциации FcγR из абсолютного ответа в конце фазы ассоциации антитела, чтобы нормировать эффект связывания антител с рецепторами OX40 и CD137.
Измерение значений ответа связывания в конце фазы диссоциации FcγRI выполняли, чтобы продемонстрировать эффект мутации LALA на увеличение скорости диссоциации для связывания FcγRI в отсутствие полного устранения связывания с этим FcγR. Эти значения получали путем вычитания абсолютного ответа в конце фазы ассоциации FcγR из абсолютного ответа в конце фазы диссоциации FcγR. Значения для антител к CD137 получали из проточной ячейки, покрытой антигеном CD137-his, значения для антител к OX40 получали из проточной ячейки, покрытой антигеном OX40-his, а значения для МАТ2 к OX40/CD137 получали как из проточной ячейки, покрытой антигеном OX40-his, так и из проточной ячейки, покрытой антигеном CD137-his. Результаты представлены в Таблице 8.
Таблица 8: Ответ связывания антител с рецепторами Fcγ человека согласно SPR
МАТ2 и контрольные антитела без мутации LALA все связывались с каждым из рецепторов Fcγ, как и ожидалось, в обоих форматах IgG1 и IgG4. МАТ2 и контрольные антитела в формате IgG1, содержащие мутацию LALA, показали значительно уменьшенное связывание в конце фазы ассоциации (прямая реакция) с каждым из тестируемых рецепторов Fcγ, за исключением FcγRI, по сравнению с контрольными антителами в формате IgG1 без мутации LALA и контрольным антителом в формате IgG4. Скорость прямой реакции связывания высокоаффинного рецептора FcγR, FcγRI, с LALA-содержащими антителами hIgG1 лишь незначительно уменьшилась по сравнению с антителами IgG1, не содержащими LALA, таким образом, она не изменялась в значительной степени в результате введения мутации. Однако для FcγRI скорость диссоциации была выше для антител, содержащих мутацию LALA, чем для антител без LALA, о чем свидетельствует большее уменьшение ответа связывания в конце фазы диссоциации FcγRI (более 200 RU для каждого из LALA-содержащих антител по сравнению с менее чем 60 RU для антител, не содержащих LALA).
В целом, МАТ2 к OX40/CD137, содержащее мутацию LALA, уменьшало связывание с рецепторами Fcγ по сравнению с IgG1 человека дикого типа, аналогичным образом с другими LALA-содержащими антителами hIgG1, и на более низком уровне, чем контрольное антитело IgG4. Поскольку связывание с рецептором Fcγ необходимо для АЗКЦ-активности, ожидается, что уменьшение связывания с рецепторами Fcγ, вызванное мутацией LALA, также приведет к уменьшению АЗКЦ так, что клетки-мишени не будут истощены связыванием МАТ2. Это считается важным, поскольку МАТ2 к OX40/CD137 являются агонистическими антителами и поэтому истощение клеток-мишеней нежелательно, поскольку это клетки представляют собой клетки, для стимуляции которых предназначено МАТ2.
FcγRIIIa экспрессируется на иммунных эффекторных клетках, таких как природные клетки-киллеры (NK-клетки), и было показано, что он играет важную роль в качестве посредника АЗКЦ (Chan et al., 2015). Чтобы определить, приведет ли уменьшенное связывание МАТ2 FS20-22-49AA/FS30-10-16 с FcγRIIIa, подтвержденное данными SPR, к низкой или незначительной активации пути АЗКЦ, биотест АЗКЦ выполняли с использованием модифицированных клеток Jurkat, экспрессирующих FcγRIIIa, в качестве эффекторных клеток, и клеток Raji, сверхэкспрессирующих либо OX40 человека, либо CD137 человека, в качестве клеток-мишеней. Было обнаружено, что МАТ2 не индуцирует активацию АЗКЦ ни в OX40-экспрессирующих, ни в CD137-экспрессирующих клетках Raji, по сравнению с ответами, наблюдаемыми для антител для отрицательного и положительного контроля, использованных в анализе.
Известно, что для осуществления своей агонистической активности другие агонистические антитела полагаются на перекрестное связывание антител с рецептором Fcγ для образования структур более высокого порядка (Stewart et al., 2014; Wajant, 2015), что приводит к кластеризации и активации рецепторов на поверхности клетки. Поскольку для активности МАТ2 согласно настоящему изобретению не требуется перекрестное связывание, опосредуемое рецептором Fcγ, агонистическое действие на клетки будет локализовано в местах, где присутствуют обе мишени. Поскольку мутация LALA в МАТ2 к OX40/CD137 приводит к уменьшенному связыванию с рецепторами Fcγ, не ожидается, что активация, запускаемая перекрестным связыванием с рецептором Fcγ, будет возможна за счет связывания только CD137. Следовательно, маловероятно, что МАТ2 активируют CD137-экспрессирующие клетки в отсутствие экспрессии OX40. Поскольку существует известный риск токсичности для печени, ассоциированный с нацеливанием на CD137 у человека (например, как наблюдали при лечении урелумабом (BMS-663513) (Segal et al., 2017), можно надеяться, что уменьшение вероятности индуцированного рецептором Fcγ перекрестного связывания МАТ2, содержащего мутацию LALA, уменьшит шансы возникновения такой токсичности для печени после лечения МАТ2, поскольку CD137 будет активирован только в местах, в которых также экспрессируется OX40. Существующая на сегодняшний день теория CD137-индуцированной токсичности для печени свидетельствует о том, что миелоидные клетки, экспрессирующие CD137, представляют собой тип клеток, ответственный за воспаление печени, наблюдаемое у мышей, которых лечили агонистами CD137 (Bartkowiak, et al., 2018).
Известно, что макрофаги экспрессируют FcγRI, который потенциально может опосредовать перекрестное связывание антитела, нацеленного на CD137, однако нет данных, что эти клетки экспрессируют OX40. Таким образом, МАТ2 согласно настоящему изобретению, содержащее мутацию LALA, теоретически не должно быть способно активировать макрофаги печени, которые экспрессируют CD137, но не OX40. Полагают, что это уменьшит риск токсичности МАТ2 к OX40/CD137 согласно настоящему изобретению для печени по сравнению как с агонистом CD137, который требует перекрестного связывания с рецептором Fcγ для осуществления активности, так и агонистом CD137, который не требует перекрестного связывания для осуществления активности. В случае OX40, несмотря на то, что в отсутствие перекрестного связывания наблюдалась некоторая остаточная активность Fcab, которая может привести к некоторой активации OX40 в отсутствие связывания CD137, поскольку на сегодняшний день разные виды ограничивающей дозу токсичности не были зарегистрированы в клинических исследованиях с применением агонистов OX40, это не считается риском.
Пример 11 - Связывание МАТ2 с клетками, экспрессирующими OX40 или CD137
11.1. Связывание МАТ2 с клетками, экспрессирующими OX40 или CD137 человека или яванской макаки
Аффинность связывания МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12 и FS20-22-49AA/FS30-10-16 в отношении экспрессируемых клетками OX40 и CD137 человека или яванской макаки определяли с помощью проточной цитометрии. Разведения (2× конечная концентрация) этих антител МАТ2 и контрольных антител G1/4420 (FITC), G1/11D4 (OX40), G1/MOR7480.1 (CD137) и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) (все в формате изотипа IgG1) готовили в 1×ДФСБ (Gibco, 14190-094). Суспензии клеток DO11.10-hOX40, DO11.10-cOX40, DO11.10-hCD137, DO11.10-cCD137 или HEK готовили в ФСБ + 2% БСА (Sigma, A7906) и высевали при 4×106 клеток/мл с 50 мкл/лунку в 96-луночных планшетах с V-образным дном (Costar, 3897). По 50 мкл разведений антител добавляли в лунки, содержащие клетки (конечный объем 100 мкл), и инкубировали при 4°C в течение 1 часа. Планшеты промывали и затем добавляли 100 мкл/лунку вторичного антитела (антитело к Fc-488 человека, Jackson ImmunoResearch, 109-546-098), разведенного 1:1000 в ФСБ + 2% БСА, и инкубировали в течение 30 минут при 4°C в темноте. Планшеты промывали и содержимое повторно суспендировали в 100 мкл ФСБ, содержащего DAPI (Biotium, номер по каталогу 40043) при 1 мкг/мл. Планшеты анализировали с использованием проточного цитометра Canto II (BD Bioscience), а данные анализировали с использованием FlowJo. Мертвые клетки идентифицировали по их более высокой флуоресценции в канале УФ (405 нм/450/50) и исключали из анализа. Среднее геометрическое значение интенсивности флуоресценции (GMFI) в канале FITC (488 нм/530/30) использовали в качестве меры связывания антитела. Данные GMFI аппроксимировали с использованием уравнения зависимости ответа от log(агониста) (три параметра) в программном обеспечении GraphPad Prism, чтобы получить значения ЭК50.
Таблица 9: Аффинность связывания МАТ2 к OX40/CD137 в отношении клеток DO11.10, экспрессирующих OX40 или CD137 человека или яванской макаки, по данным проточной цитометрии.
(нМ)
(нМ)
(нМ)
(нМ)
(нМ)
NB: связывание не обнаружено.
Результаты подтверждают, что протестированные МАТ2 к OX40/CD137 связываются с OX40 и CD137 человека и яванской макаки, экспрессируемыми на клетках DO11.10. МАТ2 и положительные контроли (МАТ к OX40 человека, G1/11D4, в остове IgG1 человека; и МАТ к CD137 человека G1/MOR7480.1, в остове IgG1 человека) связывались с обоими OX40 и CD137 человека и яванской макаки с диапазоном значений аффинности (см. Таблицу 9). Не наблюдали перекрестную реактивность с другими белками, экспрессируемыми на поверхности линии клеток HEK, поскольку ни для одного из тестируемых антител не было обнаружено связывание с этой линией клеток. Таким образом, МАТ2 к OX40/CD137 специфично связывалось с OX40 человека и CD137 человека, при этом неспецифичное связывание не наблюдали.
11.2 Связывание МАТ2 FS20-22-49AA/FS30-10-16 и его составных частей с клетками, экспрессирующими OX40 или CD137 человека или яванской макаки
Чтобы сравнить аффинность МАТ2 FS20-22-49AA/FS30-10-16 и его составных частей, то есть Fcab к OX40 (в формате имитационного МАТ2 к OX40/FITC; FS20-22-49AA/4420) и Fab к CD137 (в формате IgG1; FS30-010-016), в отношении экспрессируемых клетками OX40 и CD137 человека или яванской макаки использовали тот же способ, что и описанный в Примере 11.1. Однако в этом эксперименте, вместо использования клеток HEK для анализа неспецифичного связывания, использовали нетрансдуцированные клетки DO11.10. Антитело к FITC G1/4420 использовали в качестве контроля. Эксперимент повторяли трижды, чтобы повысить надежность рассчитанных значений ЭК50. Средние значения ЭК50 для протестированных молекул показаны в Таблице 10.
Таблица 10: Аффинность связывания МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 и его составных частей с клетками DO11.10, экспрессирующими OX40 или CD137 человека или яванской макаки, определенная с помощью проточной цитометрии.
Сред. ± СО
(нМ)
Сред. ± СО
(нМ)
Сред. ± СО
(нМ)
Сред. ± СО
(нМ)
Сред. ± СО
(нМ)
Сред.: среднее значение; СО: стандартное отклонение; NB: связывание не обнаружено.
Результаты подтверждают, что МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) связывается с OX40 и CD137 человека и яванской макаки, экспрессируемыми на клетках DO11.10, с субнаномолярной аффинностью, и что компонент Fcab к OX40 из МАТ2 связывается с OX40 человека и яванской макаки с аффинностью, сравнимой с таковой МАТ2 к OX40/CD137, и что компонент Fab к CD137 из МАТ2 связывается с CD137 человека и яванской макаки с аффинностью, сравнимой с таковой МАТ2 к OX40/CD137. Неспецифичное связывание с нетрансдуцированными клетками DO11.10 не наблюдали ни для МАТ2 к OX40/CD137, ни для его составных частей, ни для антитела для контроля изотипа (G1/4420). Результаты свидетельствуют о том, что аффинность МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 и Fcab к OX40 FS20-22-49AA в отношении OX40 яванской макаки, экспрессируемого клетками, выше (о чем свидетельствуют более низкие значения ЭК50), чем наблюдали ранее (Пример 11.1 и Таблица 9), и сходна с результатами определения аффинности с помощью SPR (Пример 8 и Таблица 7). Поскольку средние значения ЭК50, более подробно представленные в Таблице 10, получены в трех независимых экспериментах, они лучше представляют аффинность тестируемых молекул в отношении OX40 и CD137 человека и яванской макаки, экспрессируемых на клетках DO11.10.
11.3 Связывание МАТ2 с клетками, экспрессирующими OX40 или CD137 мыши
Аффинность связывания МАТ2 FS20m-232-91AA/Lob12.3 в отношении экспрессируемых клетками OX40 и CD137 мыши определяли с использованием проточной цитометрии. Разведения (2× конечная концентрация) FS20m-232-91AA/Lob12.3 и контрольных антител G1/4420 (FITC), G1/Lob12.3 (CD137), G1/OX86 (OX40) и FS20m-232-91AA/HEL D1.3 (имитационное МАТ2 к OX40/HEL) готовили в 1×ДФСБ (Gibco, 14190-094). Суспензии клеток DO11.10-mOX40, DO11.10-mCD137 или HEK готовили в ФСБ + 2% БСА (Sigma, A7906) и высевали при 4×106 клеток/мл с 50 мкл/лунку в 96-луночные планшеты с V-образным дном (Costar, 3897). По 50 мкл разведений антител добавляли в лунки, содержащие клетки (конечный объем 100 мкл), и инкубировали при 4°C в течение 1 часа. Планшеты промывали и затем добавляли 100 мкл/лунку вторичного антитела (антитело к Fc-488 человека, Jackson ImmunoResearch, 109-546-098), разведенного 1:1000 в ФСБ + 2% БСА, и инкубировали в течение 30 минут при 4°C в темноте. Планшеты промывали и содержимое повторно суспендировали в 100 мкл ФСБ, содержащего DAPI (Biotium, номер по каталогу 40043) при 1 мкг/мл. Планшеты анализировали с использованием проточного цитометра Canto II (BD Bioscience), а данные анализировали с использованием FlowJo. Мертвые клетки идентифицировали по их более высокой флуоресценции в канале УФ (405 нм/450/50) и исключали из анализа. Среднее геометрическое значение интенсивности флуоресценции (GMFI) в канале FITC (488 нм/530/30) использовали в качестве меры связывания антитела. Данные GMFI аппроксимировали с использованием уравнения зависимости ответа от log(агониста) (три параметра) в программном обеспечении GraphPad Prism, чтобы получить значения ЭК50. Результаты представлены в Таблице 11.
Таблица 11: Аффинность связывания МАТ2 к OX40/CD137 мыши в отношении клеток DO11.10, экспрессирующих OX40 или CD137 мыши, определенная с помощью проточной цитометрии.
(нМ)
(нМ)
NB: связывание не обнаружено.
Результаты подтверждают, что МАТ2 FS20m-232-91AA/Lob12.3 связывается с OX40 и CD137 мыши, экспрессируемыми на клетках DO11.10. МАТ2 и положительные контроли (МАТ к OX40 мыши, OX86, в остове IgG1 человека; и МАТ к CD137 мыши Lob12.3 в остове IgG1 человека) связывались с OX40 и/или CD137 мыши с диапазоном значений аффинности (см. Таблицу 11). Не наблюдали перекрестную реактивность с другими белками, экспрессируемыми на поверхности линии клеток HEK, поскольку ни для одного из тестируемых антител не было обнаружено связывание с этой линией клеток.
Таким образом, МАТ2 к OX40/CD137 мыши специфично связывались с OX40 мыши и CD137 мыши без наблюдаемого неспецифичного связывания.
Пример 12 - Активность МАТ2 к OX40/CD137, нацеленных на совместно экспрессируемые рецепторы, в анализе с использованием стафилококкового энтеротоксина A (SEA)
Экспрессия OX40 на инфильтрирующих опухоль лимфоцитах, вероятно, будет сопровождаться экспрессией CD137, поскольку эти две молекулы часто совместно экспрессируются на активированных Т-клетках (Ma et al., 2005). Агонистическое действие на OX40 и CD137 с помощью МАТ2, нацеленного на эти два совместно экспрессируемых рецептора, может индуцировать пролиферацию и выработку воспалительных цитокинов предварительно активированными Т-клетками.
Для полной активации Т-клеткам требуются два сигнала, первый сигнал, который является антигенспецифичным и обеспечивается посредством рецептора Т-клеток (TCR), который взаимодействует с молекулами главного комплекса гистосовместимости (ГКГС), экспонирующими пептидный антиген на мембране антигенпрезентирующих клеток (АПК), и второй, антиген-неспецифичный сигнал - костимулирующий сигнал, который обеспечивается посредством взаимодействия между костимулирующими молекулами, экспрессируемыми на мембране АПК и Т-клетки.
Чтобы протестировать активность МАТ2 к OX40/CD137, разработали анализ активации Т-клеток с использованием суперантигена стафилококкового энтеротоксина A (SEA) в качестве первого сигнала. SEA перекрестно связывает молекулы ГКГС класса II на поверхности АПК и TCR Т-клеток, обеспечивая посредством этого первый сигнал для активации Т-клеток. Для своей полной активации Т-клетки также должны получить второй, костимулирующий, сигнал от контрольных молекул или МАТ2, перекрестно связанных соответствующим образом. Этот анализ выполняется с использованием выделенных МКПК из крови и должен более точно отражать события, ожидаемые in vivo, по сравнению с анализом, который выполняют с использованием выделенных Т-клеток.
Анализ SEA-стимуляции использовали для установления активности различных агонистических антител к OX40 и к CD137 и антитела МАТ2 к OX40/CD137 в присутствии или в отсутствие искусственных перекрестно связывающих агентов, чтобы сравнить различные клоны МАТ2 к OX40/CD137 и установить типичное значение ЭК50 для клона МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в группе из 10 доноров МКПК.
12.1 Активность агонистических антител к OX40 и к CD137 в отношении SEA-стимулированных МКПК
Чтобы установить чувствительность анализа с SEA к различным агонистическим антителам к OX40 и к CD137, в данном анализе тестировали активность антитела МАТ2 (FS22-20-49AA/FS30-10-16) и контрольных антител, перечисленных в Таблице 12. G1/4420 (против FITC), G1AA/MOR7480.1 (против CD137), G1AA/FS30-10-16 (против CD137), G1AA/20H4.9 (против CD137), G1AA/11D4 (против OX40) и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) использовали в качестве контролей. Выработку IL-2 использовали как меру активации Т-клеток.
Таблица 12: Подробная информация о протестированных антителах и МАТ2
Мононуклеарные клетки периферической крови (МКПК) выделяли из конусов для истощения лейкоцитов (Государственная служба донорства и трансплантации Национальной службы здравоохранения Великобритании (NHS Blood and Transplant service)), побочного продукта донорства тромбоцитов. В общих чертах, содержимое лейкоцитарного конуса смывали ФСБ и наносили на градиент фиколла (GE Lifesciences, номер по каталогу 17144002). МКПК выделяли путем центрифугирования и выделения клеток, которые не пересекали градиент фиколла. МКПК дополнительно промывали ФСБ, а оставшиеся красные клетки крови лизировали путем добавления 10 мл буфера для лизиса красных клеток крови (eBioscience) в соответствии с инструкциями производителя. МКПК подсчитывали и повторно суспендировали до 2,0×106 клеток/мл в среде для Т-клеток (среда RPMI (Life Technologies) с 10% ФБС (Life Technologies), 1× пенициллин/стрептомицин (Life Technologies), пируват натрия (Gibco), 10 мМ Hepes (Gibco), 2 мМ L-глутамина (Gibco) и 50 мкМ 2-меркаптоэтанола (Gibco)). Затем к МКПК добавляли SEA (Sigma, номер по каталогу S9399) при 200 нг/мл, и клетки добавляли в планшеты при 2×105 клеток/лунку (100 мкл/лунку).
Разведения 2 мкМ каждого тестируемого антитела (более подробная информация представлена в Таблице 12) готовили в ДФСБ (Gibco) и дополнительно разводили 1:10 в среде для T-клеток (30 мкл + 270 мкл), чтобы получить разведения 200 нМ. Искусственные перекрестно связывающие агенты (антитело к CH2 человека (клон MK1A6, полученное собственными силами) или FITC-декстран (Sigma) (см. Таблицу 12) добавляли в лунки в молярном соотношении 1:1 с тестируемыми антителами, где это необходимо. В 96-луночном планшете готовили последовательные разведения тестируемых антител, и по 100 мкл смеси разведенных антител добавляли к активированным Т-клеткам на планшете.
Клетки инкубировали при 37°C, 5% CO2 в течение 120 часов. Супернатанты собирали, и высвобождение IL-2 измеряли с использованием набора для ИФА IL-2 человека (eBioscience или R&D Systems) в соответствии с инструкциями производителя. Планшеты считывали при 450 нм, используя считывающее устройство для планшетов с программным обеспечением Gen5, BioTek. Значения поглощения 630 нм вычитали из значений поглощения 450 нм (Коррекция). Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 человека (hIL-2) наносили на график в зависимости от логарифмической концентрации тестируемых антител, и полученные кривые аппроксимировали с использованием уравнения зависимости ответа от log(агониста) в GraphPad Prism. В Таблице 13 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в анализе с SEA в присутствии или в отсутствие перекрестного связывания с искусственными перекрестно связывающими агентами. Фигура 3А показывает уровни высвобождения IL-2, индуцированного тестируемыми антителами в одной концентрации (3,7 нМ) в анализе с SEA. Для этого анализа была выбрана концентрация, при которой эти антитела индуцировали самые высокие уровни выработки IL-2. Статистический анализ выполняли с помощью двустороннего дисперсионного анализа и критерия Тьюки для множественных сравнений. Звездочки над планками погрешностей представляют достоверное различие по сравнению с образцами, обработанными контролем изотипа (G1/4420) (*p<0,032, **p<0,0021, ***p<0,0002, ****p<0,0001). Фигура 3B показывает графики высвобождения IL-2, индуцированного МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) в присутствии или в отсутствие искусственного перекрестно связывающего агента в анализе с SEA.
Таблица 13: Анализ с SEA с использованием агонистических антител и МАТ2 к OX40 и CD137
NAD = активность не обнаружена.
Результаты показывают, что только МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) было способно повышать уровни IL-2 в отсутствие искусственных перекрестно связывающих агентов и что добавление искусственного перекрестно связывающего агента не увеличивало активность МАТ2 к OX40/CD137, применительно как к ЭК50, так и к максимальному ответу. Активность OX40-нацеленных антител G1AA/11D4 и FS20-22-49AA/4420 и антитела к CD137 G1AA/20H4.9 наблюдали только в присутствии искусственных перекрестно связывающих агентов, а для антител к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 статистически достоверная активность не была обнаружена, по сравнению с контролем изотипа, даже в присутствии искусственного перекрестно связывающего агента. Антитело к OX40 G1AA/11D4 индуцировало более высокие уровни IL-2, чем антитела к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16, и уровень IL-2, сравнимый с таковым для антитела к CD137 G1AA/20H4.9, несмотря на наблюдение, что антитело G1AA/11D4 обладает большей эффективностью, чем антитело G1AA/20H4.9, о чем свидетельствует его существенно более низкое значение ЭК50. Эти результаты свидетельствуют о том, что анализ с SEA более чувствителен к агонистическому действию на OX40, чем к агонистическому действию на CD137. Возможно, это связано с тем, что OX40 преимущественно экспрессируется на CD4+ Т-клетках, а CD137 преимущественно экспрессируется на CD8+ Т-клетках (Croft, 2014 и собственные данные, показанные на Фигуре 6), а также поскольку в МКПК человека, как правило, больше CD4+ Т-клеток, чем CD8+ Т-клеток.
12.2 Активность различных клонов МАТ2 к OX40/CD137 в отношении SEA-стимулированных МКПК
Активность пяти различных клонов МАТ2 к OX40/CD137 тестировали в анализе с SEA. Подробная информация о МАТ2 и контрольных антителах, использованных в анализе, представлена в Таблице 14. G1/4420 (против FITC), G1/11D4 (против OX40), G2/MOR7480.1 (против CD137), G1/11D4 плюс G2/MOR7480.1 в комбинации и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) использовали в качестве контролей. Анализ выполняли, как описано в Примере 12.1.
Таблица 14: Подробная информация о протестированных антителах и МАТ2
В Таблице 15 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в анализе с SEA в присутствии или в отсутствие перекрестного связывания с искусственными перекрестно связывающими агентами. Фигура 3C и D показывают графики высвобождения IL-2 для анализа с SEA.
Таблица 15: Анализ с SEA с использованием МАТ2, нацеленных на OX40 и CD137
NAD = активность не обнаружена
На Фигуре 3C и D и в Таблице 15 показано, что выработка IL-2 не наблюдалась при использовании антитела к FITC G1/4420, как не связанного перекрестно, так и перекрестно связанного, или при использовании не связанного перекрестно антитела к OX40 (G1/11D4 по отдельности или в комбинации с G2/MOR7480.1), как и ожидалось. IL-2 вырабатывался Т-клетками, когда OX40 был активирован посредством связывания антитела к OX40, в качестве положительного контроля, но только в присутствии искусственного перекрестно связывающего агента (ЭК50 0,13 нМ для G1/11D4 по отдельности и ЭК50 0,11 нМ в комбинации с G2/MOR7480.1). Fcab, нацеленный на OX40, в формате имитационного МАТ2 (4420 LALA) FS20-22-49AA/4420 проявил некоторую агонистическую активность в отсутствие перекрестного связывания в этом анализе (ЭК50 8,53 нМ), но при перекрестном связывании посредством связывания плеч Fab с FITC-декстраном имел повышенную активность, о чем свидетельствует уменьшение ЭК50 (0,31 нМ) и увеличение максимального количества вырабатываемого IL-2 (максимальный ответ), о чем свидетельствует повышенная выработка IL-2.
Активность не наблюдали при использовании перекрестно связанного CD137-нацеленного антитела G2/MOR7480.1 по отдельности, и активность комбинации OX40-нацеленного антитела G1/11D4 и CD137-нацеленного антитела G2/MOR7480.1 при перекрестном связывании была сходна с активностью перекрестно связанного OX40-нацеленного антитела G1/11D4 по отдельности.
В этом анализе активации Т-клеток с SEA активность пяти клонов МАТ2 к OX40/CD137 (см. Таблицу 15) была сопоставимой, независимо от присутствия искусственного перекрестно связывающего агента. Активность МАТ2 к OX40/CD137 в присутствии искусственного перекрестно связывающего агента также была сопоставима с активностью перекрестно связанного имитационного МАТ2 FS20-22-49AA/4420. Результаты анализа с SEA показывают, что МАТ2 к OX40/CD137 способны передавать сигнал за счет OX40, без потребности в искусственных перекрестно связывающих агентах, в результате перекрестного связывания, обеспеченного путем связывания плеч Fab к CD137 МАТ2.
Несмотря на то, что в этом анализе не была обнаружена активность перекрестно связанного CD137-нацеленного антитела G2/MOR7480.1, ожидается, что CD137 экспрессировался на Т-клетках на уровне, обеспечивающем возникновение перекрестного связывания МАТ2. Предполагается, что экспрессия была на уровне, при котором каждый из пяти клонов МАТ2, при связывании с CD137, также мог связываться с OX40 и запускать его активацию в гораздо большей степени, в сравнении с низким уровнем активности, индуцированным имитационным МАТ2 FS20-22-49AA/4420, не связанным перекрестно.
Активация Т-клеток, наблюдаемая при использовании МАТ2 к OX40/CD137 в отсутствие искусственного перекрестно связывающего агента, также свидетельствует о том, что эти молекулы будут способны активировать Т-клетки, в которых как OX40, так и CD137 экспрессируются in vivo.
12.3 Активность клона МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в отношении SEA-стимулированных МКПК от 10 доноров МКПК
Клон МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 тестировали в анализе с SEA с использованием МКПК от 10 различных доноров, чтобы установить точные значения ЭК20, ЭК30 и ЭК50 для его активности. Анализ выполняли, как описано в Примере 12.1, в отсутствие искусственного перекрестно связывающего агента. Средние значения плюс или минус стандартное отклонение среднего значения (СО) рассчитывали на основании исходных данных для каждого донора. Для расчета значений ЭК50 исходные данные аппроксимировали к логистической функции (4 параметра: максимум (top), минимум (bottom), коэффициент Хилла (Hill slope) и ЭК50 (EC50)):
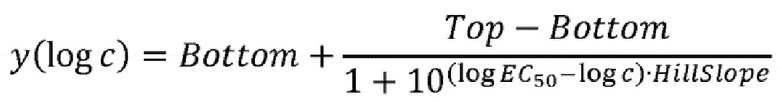
Ось ординат показывает измеренный ответ (уровни IL-2) как функцию log10(c), где c обозначает концентрацию тестируемого изделия.
Оценка каждого параметра из аппроксимации имеет стандартную ошибку, которая указывает на точность этой оценки. Поскольку разные доноры и/или технические повторы для данного эксперимента будут давать разные оценки параметров и разные уровни точности (в зависимости, например, от качества данных в каждом случае), параметры от каждого донора и/или технических повторов включали во взвешенное среднее значение. Веса определяли как величину, обратную квадрату стандартной ошибки параметра, предполагая нормальность распределения параметра.
Кроме того, значения log10(ЭК20) и log10(ЭК30) рассчитывали путем аппроксимации данных к аналогичным уравнениям:
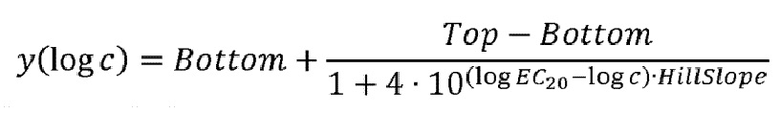
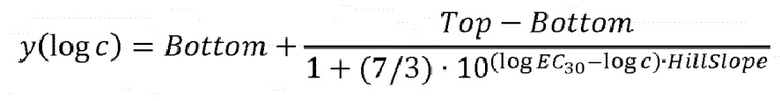
Все логистические аппроксимации выполняли с использованием GraphPad Prism, а взвешенное усреднение выполняли с помощью Microsoft Excel. Формулы, используемые для расчета взвешенного среднего значения и стандартной ошибки взвешенного среднего значения, приведены ниже:
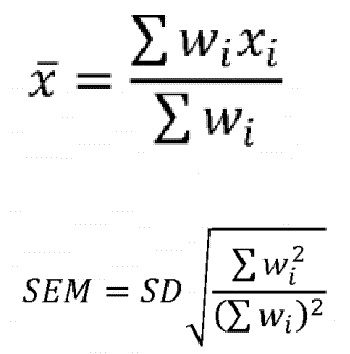
причем взвешенное стандартное отклонение рассчитывали как:
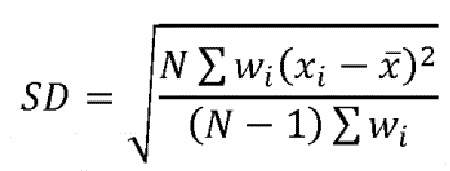
Значения ЭК20, ЭК30 и ЭК50 для высвобождения IL-2, наблюдаемого для МАТ2 к OX40/CD137 в анализе с SEA, показаны в Таблице 16.
Таблица 16: Значения ЭК20, ЭК30 и ЭК50 для МАТ2 к OX40/CD137 в анализе с SEA
Эти результаты показывают, что МАТ2 к OX40/CD137 имеет сравнимую активность с МКПК от различных доноров.
Пример 13 - Активность МАТ2 к OX40/CD137 человека в анализе активации общей популяции Т-клеток
В анализе активации Т-клеток с SEA, описанном в Примере 12, использовали МКПК и суперантиген SEA, чтобы стимулировать Т-клетки. Для оценки эффекта агонистов OX40 и CD137 на выделенные Т-клетки разрабатывали анализ активации Т-клеток. В этом анализе Т-клетки выделяли и стимулировали с использованием антитела к CD3, иммобилизованного на пластиковой поверхности. Иммобилизованное антитело к CD3 способно кластеризовать TCR Т-клеток, обеспечивая первый сигнал, требуемый для активации Т-клеток, в то время как тестируемые молекулы обеспечивали второй сигнал.
Анализ стимуляции Т-клеток использовали, чтобы установить активность различных агонистических антител к OX40 и CD137 и антитела МАТ2 к OX40/CD137 в присутствии или в отсутствие перекрестно связывающих агентов, для сравнения различных клонов МАТ2 к OX40/CD137, и чтобы установить типичное значение ЭК50 для клона МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в группе из девяти доноров МКПК.
13.1 Активность агонистических антител к OX40 и CD137 в анализе активации общей популяции Т-клеток
Чтобы установить чувствительность анализа активации Т-клеток к различным агонистическим антителам к OX40 и CD137, в данном анализе тестировали активность антитела МАТ2 (FS20-22-49AA/FS30-10-16) и контрольных антител, перечисленных в Таблице 17. G1/4420 (против FITC), G1AA/MOR7480.1 (против CD137), G1AA/FS30-10-16 (против CD137), G1AA/20H4.9 (против CD137), G1AA/11D4 (против OX40) и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) использовали в качестве контролей. Выработку IL-2 использовали как меру активации Т-клеток.
Таблица 17: Подробная информация о протестированных антителах и МАТ2
МКПК человека выделяли, как описано в Примере 12.1. Затем Т-клетки выделяли из МКПК с использованием набора для выделения общей популяции Т-клеток II (Miltenyi Biotec Ltd) в соответствии с инструкциями производителя.
Активатор Т-клеток человека CD3/CD28 Dynabeads™ (Life Technologies, 11452D) повторно суспендировали путем встряхивания. Гранулы дважды промывали средой для Т-клеток (среда RPMI (Life Technologies) с 10% ФБС (Life Technologies), 1× пенициллин/стрептомицин (Life Technologies), пируват натрия (Gibco), 10 мМ Hepes (Gibco), 2 мМ L-глутамина (Gibco) и 50 мкМ 2-меркаптоэтанола (Gibco)).
Требуемое количество Т-клеток в концентрации 1,0×106 клеток/мл в среде для Т-клеток стимулировали промытым активатором Т-клеток человека CD3/CD28 Dynabeads™ при соотношении клеток и гранул 2:1 в колбе Т-25 (Sigma) и инкубировали в течение ночи при 37°C, 5% CO2, чтобы активировать Т-клетки. Активированные Т-клетки отмывали от Dynabeads и повторно суспендировали в среде для Т-клеток в концентрации 2,0×106 клеток/мл. 96-луночные планшеты с плоским дном покрывали антителом к CD3 человека путем инкубации с 2,5 мкг/мл антитела к CD3 человека (R&D Systems, клон UHCT1), разведенного в ФСБ, в течение 2 часов при 37°C, 5% CO2, а затем дважды промывали ФСБ. Активированные Т-клетки добавляли в планшеты при 2×105 клеток/лунку.
Готовили разведения 2 мкМ каждого тестируемого антитела (более подробная информация представлена в Таблице 17) и добавляли в лунки в молярном соотношении 1:1 с перекрестно связывающим агентом (антитело к CH2 человека (клон MK1A6, полученное собственными силами) или FITC-декстран (Sigma) (см. Таблицу 17)), где необходимо, как описано выше в Примере 12.1. В 96-луночном планшете готовили последовательные разведения тестируемых антител и по 100 мкл смеси разведенных антител добавляли к активированным Т-клеткам на планшете.
Т-клетки инкубировали при 37°C, 5% CO2 в течение 72 часов. Затем собирали супернатанты, измеряли высвобождение IL-2 и получали данные, как описано в Примере 12.1. В Таблице 18 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в анализе активации Т-клеток в присутствии или в отсутствие перекрестного связывания перекрестно связывающими агентами. Фигура 4А показывает уровни высвобождения IL-2, индуцированного тестируемыми антителами в одной концентрации (3,7 нМ) в анализе активации Т-клеток. Для этого анализа выбрали концентрацию, при которой эти антитела индуцировали самые высокие уровни выработки IL-2. Статистический анализ выполняли с помощью двустороннего дисперсионного анализа и критерия Тьюки для множественных сравнений. Звездочки над планками погрешностей представляют достоверное различие по сравнению с образцами, обработанными контролем изотипа (G1/4420) (*p<0,032, **p<0,0021, ***p<0,0002, ****p<0,0001). Фигура 4B показывает графики высвобождения IL-2, индуцированного МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) в присутствии или в отсутствие перекрестно связывающего агента в анализе активации Т-клеток.
Таблица 18: Анализ активации Т-клеток с использованием агонистических антител и МАТ2 к OX40 и CD137
NAD - активность не обнаружена.
Результаты показывают, что только МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) было способно повышать уровни IL-2 в отсутствие искусственных перекрестно связывающих агентов и что добавление искусственного перекрестно связывающего агента не увеличивало активность МАТ2 к OX40/CD137 в отношении как ЭК50, так и максимального ответа. Активность OX40-нацеленных антител G1AA/11D4 и FS20-22-49AA/4420 и антитела к CD137 G1AA/20H4.9 наблюдали только в присутствии искусственных перекрестно связывающих агентов, а активность антител к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 не была обнаружена даже в присутствии искусственного перекрестно связывающего агента. Агонистическое антитело к OX40 FS20-22-49AA/4420 индуцировало более высокие уровни IL-2, чем все три агонистических антитела к CD137. Антитело к OX40 G1AA/11D4 индуцировало более высокие уровни IL-2, чем антитела к CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16, и уровень IL-2, сравнимый с таковым для антитела к CD137 G1AA/20H4.9, несмотря на наблюдение, что антитело G1AA/11D4 обладает большей эффективностью, чем антитело G1AA/20H4.9, о чем свидетельствует его более низкое значение ЭК50. Эти результаты свидетельствуют о том, что этот анализ активации Т-клеток более чувствителен к агонистическому действию на OX40, чем к агонистическому действию на CD137. Как предполагается в Примере 12.1, возможно, это связано с тем, что OX40 преимущественно экспрессируется на CD4+ Т-клетках, а CD137 преимущественно экспрессируется на CD8+ Т-клетках (Croft, 2014 и собственные данные, показанные на Фигуре 6), а также поскольку в МКПК человека, как правило, больше CD4+ Т-клеток, чем CD8+ Т-клеток.
13.2 Анализ активности агонистических антител к OX40 и к CD137 в отношении множества цитокинов в анализе активации общей популяции Т-клеток
Чтобы лучше понять эффект стимуляции OX40 и CD137 на анализ активации Т-клеток, анализировали уровни множества цитокинов. Использовали антитела и антитело МАТ2 (FS20-22-49AA/FS30-10-16), а также контрольные антитела, перечисленные в Таблице 19. Контрольные антитела G1/4420 (против FITC), G1AA/FS30-10-16 (против CD137) и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) тестировали в присутствии искусственных перекрестно связывающих агентов, а МАТ2 к OX40/CD137 тестировали в отсутствие искусственного перекрестно связывающего агента. Все антитела использовали в одной концентрации (10 нМ). Анализ выполняли, как описано в Примере 13.1.
Таблица 19: Подробная информация о протестированных антителах и МАТ2
Уровни цитокинов IL-2, IL-6, IL12p70, IL-13, TNFα, ИФНγ и IL-10 в супернатантах, собранных после инкубации, затем определяли с использованием набора Pro-inflammatory V-plex (MSD, K15049D-1) в соответствии с инструкциями производителя. Результаты показали, что МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) и перекрестно связанное антитело, нацеленное на OX40 (FS20-22-49AA/4420), увеличивали высвобождение цитокинов IL-2, IL-6, IL-12p70, IL-13 и TNFα и уменьшали высвобождение IL-10 Т-клетками. Активность антитела к CD137 (G1AA/FS30-10-16) не была обнаружена.
13.3 Активность различных клонов МАТ2 к OX40/CD137 в анализе активации общей популяции Т-клеток
Подробная информация о молекулах, протестированных в этом анализе, и их соответствующих перекрестно связывающих агентах, где это применимо, представлена в Таблице 20 ниже. G1/4420 (против FITC), G1/11D4 (против OX40), G2/MOR7480.1 (против CD137), G1/11D4 плюс G2/MOR7480.1 в комбинации и FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) использовали в качестве контролей. Все молекулы тестировали в отсутствие искусственного перекрестно связывающего агента. Контроли G1/4420, G1/11D4, G2/MOR7480.1 и FS20-22-49AA/4420 в виде отдельных агентов дополнительно тестировали в присутствии искусственного перекрестно связывающего агента. Анализ выполняли, как описано в Примере 13.1.
Таблица 20: Подробная информация о протестированных антителах и МАТ2
В Таблице 21 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые для всех молекул, протестированных в анализе активации Т-клеток в отсутствие перекрестного связывания. В Таблице 22 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые для контролей G1/4420, G1/11D4, G2/MOR7480.1 и FS20-22-49AA/4420 в виде отдельных агентов, дополнительно протестированных в присутствии перекрестно связывающих агентов. Фигура 4C и D показывают графики высвобождения IL-2 для анализа активации Т-клеток.
Таблица 21: Анализ активации Т-клеток с использованием МАТ2, нацеленного на совместно экспрессируемые рецепторы, в отсутствие перекрестно связывающего агента
NAD = активность не обнаружена
ND = не определено
Таблица 22: Контроли в виде отдельных агентов в присутствии перекрестно связывающего агента
NAD = активность не обнаружена
ND = не определено
В Таблице 21 и на Фигуре 4C показано, что не связанные перекрестно МАТ2 к OX40/CD137 обладали активностью (значения ЭК50, варьирующиеся от 0,2019 до 1,201 нМ) и, следовательно, были способны связываться с обеими мишенями, что приводило к кластеризации одной или обеих из них с индукцией активации Т-клеток. Выработку IL-2 не наблюдали при использовании не связанного перекрестно или перекрестно связанного антитела к FITC G1/4420, как ожидалось, или при использовании не связанного перекрестно антитела к OX40 (G1/11D4 по отдельности или в комбинации с G2/MOR7480.1). Т-клетки вырабатывали IL-2, когда рецептор OX40 был мишенью антитела к OX40 в качестве положительного контроля в присутствии перекрестно связывающего агента (ЭК50 0,05 нМ для G1/11D4 по отдельности и ЭК50 0,02 нМ в комбинации с G2/MOR7480.1).
Fcab, нацеленный на OX40, в формате имитационного МАТ2 (4420 LALA) FS20-22-49AA/4420 обладал некоторой агонистической активностью в отсутствие перекрестного связывания (ЭК50 5,02 нМ и максимальный ответ 1508 пг/мл hIL-2), как наблюдалось в анализе с SEA, и активность была дополнительно усилена, когда имитационное МАТ2 было перекрестно связано путем связывания его плеч Fab с FITC-декстраном.
Активность не наблюдали при использовании не связанного перекрестно антитела к CD137 G2/MOR7480.1 по отдельности, но при перекрестном связывании оно было способно индуцировать активацию Т-клеток, это свидетельствует о том, что, в отличие от анализа активации Т-клеток с SEA (Пример 12), этот анализ способен измерять передачу сигнала CD137 под действием данного клона к CD137, а также передачу сигнала OX40, подтвержденную выше. Разница в активности, наблюдаемая для этого перекрестно связанного антитела по сравнению с тем же клоном к CD137 в формате IgG1 (G1AA/MOR7480.1) в Примере 13.1, для которого активность не была обнаружена ни в отсутствие, ни в присутствии искусственного перекрестно связывающего агента, может быть объяснена вариабельностью доноров Т-клеток, когда одни доноры могут лучше отвечать на стимуляцию CD137, чем другие.
В анализе активации Т-клеток, экспрессирующих CD137 человека, с использованием клеток DO11.10-hCD137, описанных в Примере 7.1, тестируемые МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14) и контроль G2/MOR7480.1 эффективно индуцировали выработку IL-2. Таким образом, предполагается, что плечи Fab к CD137 из МАТ2 к OX40/CD137 также способны оказывать агонистическое действие на CD137, экспрессируемый Т-клетками, с получением обнаруживаемого сигнала IL-2 в анализе активации первичных Т-клеток в настоящем примере.
13.4 Активность клона МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 в анализе активации общей популяции Т-клеток с использованием Т-клеток от девяти доноров МКПК
Клон МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16 тестировали в анализе активации Т-клеток с использованием МКПК от девяти различных доноров, чтобы установить точные значения ЭК20, ЭК30 и ЭК50 для его активности. Анализ выполняли, как описано в Примере 13.1, в отсутствие искусственного перекрестно связывающего агента.
Средние значения плюс или минус стандартное отклонение среднего значения (СО) рассчитывали на основании исходных данных, как описано в Примере 12.3, для каждого донора. Значения ЭК20, ЭК30 и ЭК50 для высвобождения IL-2, наблюдаемые для МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) в анализе Т-клеток, также рассчитывали, как описано в Примере 12.3, и представили в Таблице 23.
Таблица 23: Значения ЭК20, ЭК30 и ЭК50 МАТ2 к OX40/CD137 в анализе активации Т-клеток
Эти результаты показывают, что МАТ2 к OX40/CD137 имеет сопоставимую активность в отношении Т-клеток от разных доноров.
Пример 14 - Активность МАТ2 к OX40/CD137 человека в анализах активации CD4+ и CD8+ Т-клеток
Т-клетки можно подразделить на CD4+ и CD8+ Т-клетки в соответствии с их функцией в иммунной системе. CD4+ Т-клетки называются хелперными Т-клетками и вырабатывают цитокины, которые модулируют иммунный ответ, а CD8+ Т-клетки называются киллерными Т-клетками и непосредственно устраняют клетки-мишени. Было обнаружено, что на CD4+ T-клетках экспрессия OX40 выше, чем экспрессия CD137, и, наоборот, на CD8+ T-клетках экспрессия CD137 выше, чем экспрессия OX40 (Croft, 2014 и см. Фигуру 6). Несмотря на это различие в уровнях экспрессии, как CD4+, так и CD8+ Т-клетки совместно экспрессируют два рецептора (Ma et al., 2005).
Для дальнейшего изучения активности МАТ2 к OX40/CD137 в этих двух популяциях Т-клеток выделяли CD4+ и CD8+ Т-клетки, чтобы протестировать способность молекул, перечисленных в Таблице 24 ниже, активировать каждую популяцию Т-клеток в отдельных анализах активации CD4+ и CD8+ Т-клеток. В этом анализе совместную экспрессию OX40 и CD137 использовали для определения перекрестного связывания МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16. G1/4420 (против FITC), G1AA/11D4 (против OX40), G1AA/MOR7480.1 (против CD137) G1AA/FS30-10-16 (против CD137), FS20-22-49AA/4420 (имитационное МАТ2 к OX40/FITC) и FS20-22-49AA/4420 плюс G1AA/FS30-10-16 в комбинации использовали в качестве контролей. Выработку IL-2 использовали как меру активации Т-клеток.
Таблица 24: Подробная информация о протестированных антителах и МАТ2
Чтобы выделить CD4+ и CD8+ Т-клетки человека, сначала выделяли МКПК, как описано в Примере 13.1. CD4+ и CD8+ Т-клетки затем по отдельности выделяли из МКПК с использованием, соответственно, набора для выделения CD4+ Т-клеток (человека) (Miltenyi Biotec, 130-096-533) и набора для выделения CD8+ Т-клеток (человека) (Miltenyi Biotec, 130-096-495) в соответствии с инструкциями производителя.
CD4+ или CD8+ Т-клетки активировали в течение ночи в необходимом количестве в концентрации 1,0×106 клеток/мл в среде для Т-клеток с использованием активатора T-клеток человека CD3/CD28 Dynabeads, как описано в Примере 13.1.
Активированные CD4+ или CD8+ Т-клетки отмывали от Dynabeads и повторно суспендировали в среде для T-клеток в концентрации 2,0×106 клеток/мл. 96-луночные планшеты с плоским дном покрывали антителом к CD3 человека путем инкубации с 2,5 мкг/мл (для анализа активации CD4+ Т-клеток) или 10 мкг/мл (для анализа активации CD8+ Т-клеток) антитела к CD3 человека (R&D Systems, клон UHCT1), разведенного в ФСБ, в течение 2 часов при 37°C, 5% CO2, а затем дважды промывали ФСБ. Затем активированные CD4+ или CD8+ Т-клетки добавляли в соответствующие планшеты при 2×105 клеток/лунку.
Готовили разведения 2 мкМ каждого тестируемого антитела (более подробная информация представлена в Таблице 24) и добавляли в лунки в молярном соотношении 1:1 с перекрестно связывающим агентом (антитело к CH2 человека или FITC-декстран (Sigma) (см. Таблицу 24)), где требовалось, как описано в Примере 6. В 96-луночном планшете готовили последовательные разведения тестируемых антител и по 100 мкл смеси разведенных антител добавляли к активированным CD4+ или CD8+ Т-клеткам на соответствующих планшетах.
Т-клетки инкубировали при 37°C, 5% CO2 в течение 72 часов. Собирали супернатанты, измеряли высвобождение IL-2 и получали данные, как описано в Примере 12.1. В Таблице 25 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в отдельных анализах активации Т-клеток в присутствии или в отсутствие перекрестного связывания с использованием перекрестно связывающих агентов. Фигуры 5A-C показывают графики высвобождения IL-2 для анализа активации CD4+ или CD8+ Т-клеток, соответственно.
После сбора супернатантов Т-клетки промывали в ФСБ и окрашивали вторичным антителом к Fc человека, меченным Alexa Fluor 488 (Jackson Immunoresearch, номер по каталогу 109-546-098), разведенным 1:1000 в ФСБ, в течение 1 часа при 4°C. Затем клетки промывали один раз ФСБ и повторно суспендировали в 100 мкл/лунку ФСБ с DAPI (Biotium, номер по каталогу 89139-054) при 1 мкг/мл. После этого клетки анализировали на проточном цитометре BD FACSCanto II (BD Biosciences). Фигура 6 показывает среднее геометрическое значение интенсивности флуоресценции в канале 488 либо CD4+, либо CD8+ Т-клеток, обработанных G1AA/MOR7480.1 или G1AA/11D4.
Таблица 25: Анализ активации CD4+ и CD8+ Т-клеток с использованием МАТ2, нацеленных на совместно экспрессируемые рецепторы
NAD = активность не обнаружена
ND = не определено
В Таблице 25 и на Фигуре 5B показано, что CD4+ Т-клетки могут быть активированы перекрестно связанными контролями против OX40 G1AA/11D4 и FS20-22-49AA/4420 (как по отдельности, так и в комбинации с G1AA/FS30-10-16), но не под действием контролей против CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 в виде отдельных агентов. С другой стороны, Фигура 5C показывает, что CD8+ Т-клетки были активированы обоими контролями против CD137 G1AA/MOR7480.1 и G1AA/FS30-10-16 при перекрестном связывании, а также перекрестно связанными контролями против OX40 G1AA/11D4 и FS20-22-49AA/4420, несмотря на то, что уровень ответа на контроль против CD137 G1AA/FS30-10-16 в виде отдельного агента был выше, чем на оба контроля против OX40 в виде отдельных агентов. Как наблюдали в анализе с SEA (Пример 12.2) и анализе активации общей популяции Т-клеток человека (Пример 13.3), Fcab к OX40 в формате имитационного МАТ2 (FS20-22-49AA/4420) показал некоторую активность в отсутствие перекрестного связывания в присутствии CD4+ Т-клеток, и эта активность увеличивалась, когда антитело было перекрестно связано. МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) проявило активность в присутствии как CD4+, так и CD8+ Т-клеток в отсутствие перекрестного связывания, как и ожидалось на основании предыдущих результатов (см. Примеры 12 и 13).
Фигура 6 показывает, что CD4+ Т-клетки экспрессируют более низкие уровни CD137 и более высокие уровни OX40, чем CD8+ Т-клетки. Связывание G1AA/MOR7480.1 с CD137 является мерой экспрессии CD137, а связывание G1AA/11D4 с OX40 является мерой экспрессии OX40.
Анализ Т-клеток с выделенными CD4+ и CD8+ Т-клетками повторяли в соответствии с тем же протоколом, что и описанный выше, но с Т-клетками, выделенными из МКПК от другого донора, и с добавлением антитела к CD137 G1AA/20H4.9 (см. Таблицу 24). В соответствии с результатами, показанными на Фигурах 5A-D, Фигуры 5E и 5F показывают, что CD8+ Т-клетки отвечают в более значительной степени на агонистическое действие на CD137, а CD4+ Т-клетки отвечают в более значительной степени на агонистическое действие на OX40. Антитела к OX40 (G1AA/11D4 и Fcab к OX40 в формате имитационного МАТ2 FS20-22-49AA/4420) при перекрестном связывании активировали CD4+ Т-клетки, но не CD8+ Т-клетки, а антитела к CD137 (G1AA/20H4.9 и G1AA/FS30-10-16) при перекрестном связывании активировали CD8+ Т-клетки, но не CD4+ Т-клетки. Антитело G1AA/20H4.9 также активировало CD8+ Т-клетки в отсутствие перекрестно связывающего антитела, что согласуется в результатами, полученными в анализе клеток DO11.10-hCD137, описанном в Примере 7.1. В этом повторном эксперименте антитело G1AA/MOR7480.1 не активировало CD8+ Т-клетки при перекрестном связывании. Некоторые доноры МКПК могут быть более восприимчивыми к костимуляции CD137, чем другие, и разные результаты, полученные в этом эксперименте, могут быть обусловлены этой природной изменчивостью.
Эти данные показывают, что CD4+ Т-клетки более чувствительны к активации за счет агонистического действия на OX40, чем CD8+ Т-клетки, и, наоборот, что CD8+ Т-клетки более чувствительны к активации за счет агонистического действия на CD137, чем CD4+ Т-клетки. Это коррелирует с сообщенными различиями в уровнях экспрессии рецепторов OX40 и CD137 на CD4+ T-клетках и CD8+ T-клетках, причем первые экспрессируют более высокие уровни OX40, чем CD137, а вторые экспрессируют более высокие уровни CD137, чем OX40. Активность перекрестно связанного контрольного антитела к CD137 G1AA/FS30-10-16, плечи Fab которого присутствуют в МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16, в присутствии CD8+ Т-клеток демонстрирует, что МАТ2 обладает способностью активировать рецептор CD137 при перекрестном связывании путем связывания его Fcab с OX40. Кроме того, активность перекрестно связанного Fcab к OX40 в формате имитационного МАТ2 (FS20-22-49AA/4420), который также присутствует в МАТ2 к OX40/CD137 FS20-22-49AA/FS30-10-16, в присутствии CD4+ Т-клеток показывает, что МАТ2 обладает способностью активировать рецептор OX40 при перекрестном связывании путем связывания его плеч Fab с CD137. Таким образом, можно сделать вывод, что МАТ2 FS20-22-49AA/FS30-10-16 может действовать как двойной агонист путем активации CD4+ Т-клеток за счет агонистического действия на OX40 и CD8+ T-клеток за счет агонистического действия на CD137 и в меньшей степени на OX40. Активация OX40 под действием МАТ2 происходит за счет его Fcab и увеличивается путем перекрестного связывания МАТ2 при связывании с CD137 за счет его плеч Fab, в то время как активация CD137 происходит за счет связывания его плеч Fab с CD137 и перекрестного связывания МАТ2 при связывании с OX40 за счет его Fcab.
Пример 15 - Активность МАТ2 к OX40/CD137 мыши и антител к CD137 мыши в анализах активации Т-клеток
Поскольку МАТ2 к OX40/CD137 человека не связываются с белками мыши, чтобы протестировать способность МАТ2 к OX40/CD137 вызывать противоопухолевый ответ, опосредуемый Т-клетками, получали подобный реагент, нацеленный на OX40 мыши и CD137 мыши (см. Пример 8.2).
15.1 Активность МАТ2 к OX40/CD137 мыши в анализе активации общей популяции Т-клеток
Чтобы протестировать, может ли МАТ2 к OX40/CD137 мыши (FS20m-232-91AA/Lob12.3), нацеленное на эти два совместно экспрессируемых рецептора, индуцировать выработку воспалительных цитокинов предварительно активированными Т-клетками, разрабатывали анализ активации Т-клеток мыши. Антитела G1/4420 (против FITC), G1AA/OX86 (против mOX40), G1AA/Lob12.30 (против mCD137), G1AA/OX86 и G1AA/Lob12.3 в комбинации, а также FS20m-232-91AA/4420 (имитационное МАТ2 к mOX40/FITC) использовали в качестве контролей (более подробная информация представлена в Таблице 26), и выработку IL-2 использовали как меру стимуляции Т-клеток.
Таблица 26: Подробная информация о протестированных антителах и МАТ2
Для выделения Т-клеток селезенки собирали у самок мышей Balb/C в возрасте 4-8 недель (Charles River). Мышей гуманно умерщвляли, и выделяли селезенки путем диссекции. Спленоциты выделяли путем проталкивания селезенок через сито для клеток 70 мкм (Corning), используя внутреннюю часть пластикового шприца объемом 5 мл. Сито для клеток промывали 10 раз 1 мл фосфатно-солевого буфера Дульбекко (ДФСБ) (Gibco) и элюат собирали в пробирку объемом 50 мл. Красные клетки крови, присутствующие в элюате, лизировали посредством добавления 10 мл буфера для лизиса красных клеток крови (eBioscience) в соответствии с инструкциями производителя. Т-клетки выделяли из спленоцитов, присутствующих в элюате, с использованием набора для выделения общей популяции Т-клеток II (мышь) (Miltenyi Biotec Ltd) в соответствии с инструкциями производителя, а затем активировали и использовали в протоколе, по существу аналогичном анализу активации Т-клеток человека, описанному в Примере 13.1, но с использованием активатора Т-клеток мыши CD3/CD28 Dynabeads (Life Technologies) для активации Т-клеток, антитела к CD3 мыши (Biolegend, клон 145-2C11) для покрытия планшетов и набора для ИФА IL-2 мыши (eBioscience или R&D systems) для измерения высвобождения IL-2.
В Таблице 27 показаны значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в анализе активации Т-клеток в присутствии тестируемых МАТ2 и МАТ. Фигура 7A и B показывают типичные графики высвобождения IL-2 для анализа активации Т-клеток.
Таблица 27: Анализ активации Т-клеток с использованием МАТ2, нацеленных на совместно экспрессируемые рецепторы
NAD = активность не обнаружена
В Таблице 27 и на Фигуре 7B показано, что увеличение активации Т-клеток происходит, когда рецептор OX40 является мишенью, а антитела к OX40 перекрестно связаны. Активацию Т-клеток не наблюдали при использовании перекрестно связанного или не связанного перекрестно антитела к FITC G1/4420, как и ожидалось, или при использовании не связанного перекрестно антитела к OX40 (G1AA/OX86 по отдельности или в комбинации с G1AA/Lob12.3). Т-клетки вырабатывали IL-2, когда рецептор OX40 являлся мишенью антитела к OX40 G1AA/OX86 в присутствии перекрестно связывающего агента (ЭК50 2,41 нМ для G1AA/OX86 по отдельности и ЭК50 1,72 нМ в комбинации с G1AA/Lob12.3).
OX40-нацеленный Fcab в формате имитационного МАТ2 (FS20m-232-91AA/4420) не обладал агонистической активностью в отсутствие перекрестного связывания, но при перекрестном связывании путем связывания плеч Fab с FITC-декстраном показал эффективную активацию Т-клеток. Когда OX40-нацеленный Fcab спаривали с Fab к CD137 (Lob12.3), МАТ2 проявило активность в отношении Т-клеток в отсутствие каких-либо дополнительных перекрестно связывающих агентов. Это указывает на то, что МАТ2 перекрестно связано путем связывания с совместно экспрессируемыми рецепторами на поверхности одной и той же клетки.
Минимальную активность наблюдали при использовании перекрестно связанного CD137-нацеленного антитела G1AA/Lob12.3 по отдельности, а активность комбинации OX40-нацеленного антитела G1AA/OX86 и CD137-нацеленного антитела G1AA/Lob12.3 при перекрестном связывании была сопоставима с таковой перекрестно связанного OX40-нацеленного антитела G1AA/OX86, это свидетельствует о низкой чувствительности анализа для обнаружения агонистического действия на CD137 с помощью Lob12.3. Эти данные противоречат анализу Т-клеток человека, описанному в Примере 13.3, в котором для перекрестно связанного контроля против CD137 G2/MOR7480.1 наблюдали более сильный CD137-специфичный сигнал (максимальный ответ высвобождения IL-2). Это различие в функциональной активности, наблюдаемое для контрольных антител к CD137 мыши и к CD137 человека, может быть связано с их разной аффинностью в отношении их соответствующих мишеней CD137. Это также может отражать источник клеток (МКПК человека в сравнении со спленоцитами мыши) или тонкие различия между биологией мишени в системах мыши и человека.
Эти данные показывают, что МАТ2 к OX40/CD137 FS20m-232-91AA/Lob12.3 может индуцировать активацию Т-клеток без каких-либо дополнительных перекрестно связывающих агентов за счет одновременного связывания обоих рецепторов.
Поскольку молекулы МАТ2 к OX40/CD137 человека не обладают перекрестной реактивностью с белками мыши и МАТ2 к OX40/CD137 мыши функционально сопоставимы с соединениями-кандидатами человека в параллельных экспериментальных системах in vitro, молекулы против белков мыши считаются подходящими суррогатами для того чтобы сделать вывод о способности МАТ2 к OX40/CD137 индуцировать противоопухолевый иммунитет in vivo.
15.2 Активность антител к CD137 мыши в анализе активации Т-клеток, экспрессирующих CD137 мыши
Поскольку в анализе с общей популяции Т-клеток из Примера 15.1 активность клона Fab к CD137 (Lob12.3) МАТ2 к OX40/CD137 мыши была незначительной или отсутствовала, чтобы установить активность различных агонистических антител к CD137 выполняли анализ активации Т-клеток с использованием клеток DO11.10-mCD137. Тестировали агонистические антитела к CD137 G1AA/Lob12.3 (см. Таблицу 26) и G1AA/3H3 (SEQ ID NO: 166 и 167), а также антитело к FITC 4420 в формате IgG1 (G1/4420; SEQ ID NO 115 и 116) в качестве отрицательного контроля изотипа. Молекулы МАТ тестировали как в присутствии, так и в отсутствие перекрестно связывающего антитела к CH2 человека, MK1A6 (см. Пример 2.1). Выработку IL-2 мыши использовали в качестве меры активации Т-клеток.
Анализ выполняли, как описано в Примере 6.2, но с использованием клеток DO11.10-mCD137 вместо клеток DO11.10-hCD137. Планшеты считывали при 450 нм, используя считывающее устройство для планшетов с программным обеспечением Gen5 (BioTek). Значения поглощения 630 нм вычитали из значений поглощения 450 нм (Коррекция). Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 мыши (mIL-2) наносили на график в зависимости от логарифмической концентрации антитела, и полученные кривые аппроксимировали с использованием уравнения зависимости ответа от log(агониста) в GraphPad Prism.
Результаты показаны на Фигуре 7C и D. Антитела к CD137 различались по своей потребности в перекрестно связывающем антителе для индукции активности. В то время как было обнаружено, что G1AA/Lob12.3 требует добавления перекрестно связывающего антитела для активности, т. е. его активность зависела от перекрестного связывания, G1AA/3H3 проявило активность как в присутствии, так и в отсутствие перекрестно связывающего антитела и, таким образом, имело независимую от перекрестного связывания активность.
Пример 16 - Для активности МАТ2 к OX40/CD137 требуется двойное связывание OX40 и CD137.
16.1 МАТ2 к OX40/CD137 человека
МАТ2, нацеленное на OX40/CD137, проявило активность в отсутствие дополнительных перекрестно связывающих агентов в анализах с использованием SEA (Пример 12), общей популяции Т-клеток человека (Пример 13) и CD4+ и CD8+ Т-клеток человека (Пример 14), в которых Т-клетки совместно экспрессировали OX40 и CD137. Чтобы протестировать, требуется ли одновременное связывание с двумя рецепторами для активности МАТ2 к OX40/CD137, выполняли конкурентный анализ с Т-клетками, чтобы оценить способность МАТ2 FS20-22-49AA/FS30-10-16 активировать выделенные T-клетки в присутствии 100-кратного избытка OX40-нацеленного имитационного МАТ2 FS20-22-49AA/4420, МАТ к CD137 G1AA/FS30-10-16, комбинации имитационного МАТ2 FS20-22-49AA/4420 плюс МАТ G1AA/FS30-10-16 или МАТ для контроля изотипа G1/4420. Выработку IL-2 использовали как меру активации Т-клеток.
Т-клетки выделяли, как описано в Примере 13. Затем выделенные Т-клетки активировали, и планшеты покрывали антителом к CD3, как описано в Примере 13. К активированным Т-клеткам добавляли 2 нМ МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) и добавляли в планшеты при 2×105 клеток/лунку в 100 мкл. Таким образом, конечная концентрация МАТ2 к OX40/CD137 составила 1 нМ.
Разведения 2 мкМ каждого тестируемого антитела готовили в ДФСБ (Gibco) и дополнительно разводили 1:10 в среде для Т-клеток (30 мкл + 270 мкл) с получением разведений 200 нМ, и по 100 мкл каждого разведенного антитела добавляли к активированным Т-клеткам на планшете.
Т-клетки инкубировали, супернатанты собирали и высвобождение IL-2 измеряли, как описано в Примере 13. Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Статистический анализ выполняли с использованием одностороннего дисперсионного анализа и критерия Даннета для множественных сравнений с использованием программного пакета GraphPad Prism.
Фигура 8 показывает высвобождение IL-2 для конкурентного анализа. Активность МАТ2 значительно уменьшалась при превосходящей конкуренции со стороны как имитационного МАТ2 FS20-22-49AA/4420 за связывание с OX40, так и МАТ G1AA/FS30-10-16 за связывание с CD137, по сравнению с тем, когда МАТ2 было способно связываться с обоими рецепторами в отсутствие антител к OX40 и к CD137. Комбинация OX40-нацеленного имитационного МАТ2 FS20-22-49AA/4420 и МАТ к CD137 G1AA/FS30-10-16 дополнительно уменьшала активность МАТ2 к OX40/CD137. Эти результаты показывают, что для того чтобы МАТ2 индуцировало активацию Т-клеток за счет кластеризации и агонистического действия на OX40 и CD137, требуется двойное связывание МАТ2 с обоими рецепторами.
16.2 МАТ2 к OX40/CD137 мыши
МАТ2, нацеленное на OX40/CD137 мыши, проявляет активность в отсутствие дополнительных перекрестно связывающих агентов в анализе Т-клеток, когда Т-клетки совместно экспрессируют два рецептора. Чтобы протестировать, требуется ли одновременное связывание с двумя рецепторами для активности МАТ2 к OX40/CD137, выполняли конкурентный анализ для оценки способности МАТ2 FS20m-232-91AA/Lob12.3 активировать выделенные Т-клетки в присутствии 100-кратного избытка либо OX40-нацеленного имитационного МАТ2 FS20m-232-91AA/4420, либо МАТ к CD137 G1/Lob12.3, либо МАТ G1AA/4420 (FITC) в качестве отрицательного контроля. Т-клетки выделяли, как описано в Примере 15.1. Затем выделенные Т-клетки активировали, и планшеты покрывали антителом к CD3, как описано в Примере 13.1 (анализ активации общей популяции Т-клеток человека), однако использовали активатор Т-клеток мыши CD3/CD28 Dynabeads (Life Technologies) для активации Т-клеток и антитело к CD3 мыши (Biolegend, клон 145-2C11) для покрытия планшетов. К активированным Т-клеткам добавляли 2 нМ МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3) и добавляли в планшеты при 2×105 клеток/лунку.
Разведения 2 мкМ каждого тестируемого антитела готовили в ДФСБ (Gibco) и дополнительно разводили 1:10 в среде для Т-клеток (30 мкл + 270 мкл) с получением разведений 200 нМ, и по 100 мкл каждого разведенного антитела добавляли к активированным Т-клеткам на планшете.
Т-клетки инкубировали, супернатанты собирали и высвобождение IL-2 измеряли, как описано в Примере 12.1, однако использовали набор для ИФА IL-2 мыши (eBioscience или R&D systems), чтобы измерить высвобождение IL-2. Стандартная кривая для расчета концентрации цитокина была основана на аппроксимации с использованием четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Статистический анализ выполняли с использованием одностороннего дисперсионного анализа и критерия Даннета для множественных сравнений с использованием программного пакета GraphPad Prism. Фигура 9 показывает типичный график высвобождения IL-2 для конкурентного анализа.
Фигура 9 показывает, что имеет место уменьшение величины выработки IL-2, индуцированной МАТ2 к OX40/CD137, когда антитела, конкурирующие за связывание OX40 или CD137, вводят в избытке. В качестве конкурирующих антител использовали составные части МАТ2 (Fcab в формате имитационного МАТ2 (4420) и Fab без Fcab), чтобы гарантировать нацеливание на один и тот же эпитоп. Добавление этих конкурирующих антител уменьшало величину высвобождения IL-2, индуцированного МАТ2 к OX40/CD137, это свидетельствует о том, что молекула нуждается в двойном связывании для осуществления своей активности. Это показывает, что активность МАТ2 к OX40/CD137 зависит от одновременного связывания как OX40, так и CD137, что приводит к кластеризации и агонистическому действию на оба рецептора.
Пример 17 - Активность МАТ2 к OX40/CD137 в модели сингенной опухоли CT26
17.1 Сравнение противоопухолевой активности МАТ2 к OX40/CD137 с мутацией LALA или без нее
Модель сингенной колоректальной опухоли CT26 на мышах Balb/c использовали для тестирования противоопухолевой активности МАТ2 к OX40/CD137 мыши in vivo. Ранее было показано, что модель опухоли CT26 чувствительна к агонистическим антителам как к OX40, так и к CD137 (Sadun et al., 2008), и ожидается, что инфильтрирующие опухоль лимфоциты (TIL), выделенные из опухолей CT26, экспрессируют как OX40, так и CD137. Протестированные антитела подробно описаны в Таблице 28.
Таблица 28: Подробная информация о протестированных антителах и МАТ2
Способность МАТ2, с мутацией LALA или без нее (FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3, соответственно), ингибировать рост опухоли сравнивали с контролем изотипа МАТ G1/4420 (против FITC), МАТ G1/OX86 (контроль против OX40 без мутации LALA) или G1/Lob12.3 (контроль против CD137 без мутации LALA) в виде отдельных агентов, комбинацией G1/OX86 плюс G1/Lob12.3 или комбинацией G1AA/OX86 (МАТ к OX40 с мутацией LALA) плюс G1AA/Lob12.3 (МАТ к CD137 с мутацией LALA).
Самок мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая акклиматизировали в течение одной недели перед началом исследования. Всем животным вводили микрочип и присваивали уникальный идентификатор. В каждой когорте было по 12 мышей. Линию клеток карциномы толстой кишки CT26 (ATCC, CRL-2638) размножали, создавали банк, а затем подвергали предварительному скринингу в IDEXX Bioresearch для обнаружения патогенов с использованием протокола IMPACT I, и было показано, что линия не содержит патогенов. Клетки CT26 (приблизительно 3-5×106) размораживали из хранилища при -150°C и добавляли к 20 мл DMEM (Gibco, 61965-026) с 10% фетальной сыворотки теленка (ФСТ) (Gibco, 10270-106) в колбе для тканевой культуры T175. Мышей анестезировали с использованием изофлурана (Abbott Laboratories), и каждое животное получило 1×106 клеток, введенных путем подкожной инъекции в левый бок для создания опухолей. На день 10 после инокуляции опухолевых клеток опухоли измеряли и мышей рандомизировали в исследуемые когорты на основании объема опухоли. Любых мышей, которые не имели опухолей в этот момент, исключали из исследования.
В течение 24 часов до инъекции антитела анализировали с помощью профилирования SEC-HPLC и проверяли на наличие примесей. Антитела разводили до конечной концентрации 0,1 мг/мл в ФСБ и вводили путем внутрибрюшинной инъекции (в/б) при 200 мкл/мышь с получением конечной дозы 1 мг/кг для мыши массой 20 г. Инъекции выполняли в дни 13, 15 и 17 (три дозы каждые два дня) после инокуляции опухоли. Состояние здоровья животных проверяли под анестезией три раза в неделю, в течение этого времени выполняли точные измерения опухолей. Измерения объема опухоли выполняли с помощью штангенциркулей, чтобы определить самую длинную ось и самую короткую ось. Для расчета объема опухоли использовали следующую формулу:
L×(S2)/2
(Где L = самая длинная ось; S = самая короткая ось)
Исследование останавливали на день 27, когда объем опухоли достиг гуманной конечной точки в соответствии с Законом Соединенного Королевства о животных (научные процедуры) и Директивой ЕС EU86/609.
Для статистического анализа объемы опухолей анализируются на логарифмической шкале с использованием смешанной модели. Каждую пару обработок, представляющих интерес, аппроксимировали с использованием отдельной модели. Модель представляет собой:
Log10(объем)=A+B×(день-день начала)+ ε
A и B представляют собой точку пересечения и наклон, соответственно; они различны для каждой мыши и включают фиксированный эффект для группы и случайный эффект для животного:
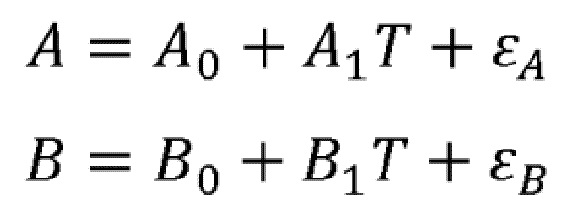
T представляет собой фиктивную переменную, представляющую группу лечения, со значением 0 в одной группе и 1 в другой. Случайные эффекты распределяются в соответствии с нормальным распределением:
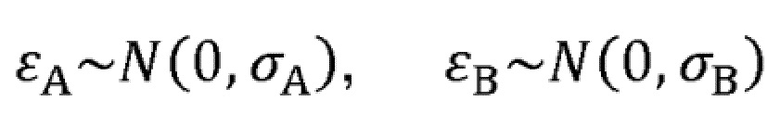
где 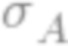 и
и 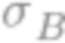 представляют собой стандартные отклонения изменчивости между животными в точке пересечения и наклона, соответственно. Изменчивость между животными также обычно имеет нормальное распределение со стандартным отклонением σ:
представляют собой стандартные отклонения изменчивости между животными в точке пересечения и наклона, соответственно. Изменчивость между животными также обычно имеет нормальное распределение со стандартным отклонением σ:
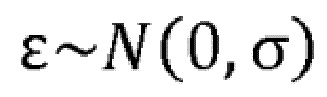
Для каждой пары обработок модель, приведенную выше, аппроксимировали к данным. Для A1 и B1 рассчитывали (двустороннее) p-значение для отличия от нуля; p-значение ниже 0,05 является статистически достоверным доказательством различия между группами лечения.
Результаты показаны на Фигуре 10А. На график нанесены средние объемы опухолей CT26 плюс или минус стандартная ошибка среднего значения. Результаты показывают, что обработка МАТ2 к OX40/CD137 как с мутацией LALA, так и без нее (FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3, соответственно) привела к уменьшению роста опухоли по сравнению с обработкой контролем против OX40 (G1/OX86), контролем против CD137 (G1/Lob12.3), комбинацией этих двух антител (G1/OX86 + G1/Lob12.3) или комбинацией LALA-содержащих антител к OX40 и к CD137 (G1AA/OX86 + G1AA/Lob12.3).
Результаты свидетельствуют о статистически достоверном противоопухолевом эффекте МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3 и FS20-232-91/Lob12.3) по сравнению с контрольным антителом (G1/4420). Активность комбинации нацеленных на OX40 и CD137 антител (G1/OX86 плюс G1/Lob12.3 или G1AA/OX86 плюс G1AA/Lob12.3) не подавляла в значительной степени рост опухоли, как и контроли в виде отдельных агентов (G1/OX86 или G1/Lob12.3).
Ожидается, что введение мутации LALA в область Fc остова IgG1 человека МАТ2 к OX40/CD137 предотвратит АЗКЦ и АЗКФ OX40- или CD137-экспрессирующих клеток, а также опосредуемое рецептором Fcγ перекрестное связывание МАТ2 при связывании либо с OX40, либо с CD137 на клетках, экспрессирующих эти рецепторы. Следовательно, полагают, что активность МАТ2 FS20m-232-91AA/Lob12.3 будет запущена за счет совместного связывания OX40 и CD137, что приводит к передаче сигнала за счет одного или обоих рецепторов, а не за счет Fc-опосредуемой эффекторной функции или перекрестного связывания, опосредуемого рецептором Fcγ. Ожидается, что впоследствии это приведет к активации OX40- и CD137-экспрессирующих Т-клеток, что в конечном итоге приведет к опосредуемой Т-клетками противоопухолевой активности.
Эти результаты демонстрируют, что антитело МАТ2 к OX40/CD137 обладает противоопухолевой эффективностью in vivo против опухоли, которая, как ожидается, содержит TIL, экспрессирующие OX40 и CD137, это свидетельствует о том, что активация OX40 и CD137 in vivo, опосредуемая биспецифичным связыванием OX40 и CD137 с помощью МАТ2 к OX40/CD137, эффективно контролирует рост опухоли.
Как описано в разделе уровня техники выше, при применении агонистического антитела к CD137 в клинических условиях наблюдалась токсичность для печени (Segal et al., 2017). Механизм этого токсического эффекта полностью не определен, но исследования на доклинических моделях подчеркнули роль CD137-экспрессирующих миелоидных клеток, которые вырабатывают IL-27 в ответ на агонистические антитела к CD137 (Bartkowiak et al., 2018). Роль рецепторов Fcγ в механизме токсичности для печени не была исследована, но одним их возможных объяснений наблюдаемой токсичности является то, что совместная экспрессия CD137 и рецепторов Fcγ в миелоидных клетках может привести к перекрестному связыванию агонистических антител к CD137 на этих клетках с запуском выработки воспалительных цитокинов. Таким образом, в случае если опосредуемое рецептором Fcγ перекрестное связывание молекулы может привести к какой-либо активации клеток, экспрессирующих CD137 в отсутствие OX40, в местах, удаленных от микросреды опухоли или периферии, считали желательным включение мутации LALA в молекулу антитела к OX40/CD137, являющегося двойным агонистом, согласно настоящему изобретению. Таким образом, при конструировании молекулы антитела, являющегося двойным агонистом, которая стимулирует Т-клетку, экспрессирующую как OX40, так и CD137, путем одновременного связывания обеих мишеней, но которая не активирует CD137-экспрессирующие клетки за счет опосредуемого рецептором Fcγ перекрестного связывания в отсутствие OX40 из-за наличия мутации LALA в молекуле, полагают, что молекула антитела согласно настоящему изобретению, вероятно, будет иметь пониженную потенциальную токсичность в клинических условиях.
Еще одна причина включения мутации LALA в молекулу антитела согласно настоящему изобретению состоит в том, что она позволяет избежать опосредуемого рецептором Fcγ уничтожения OX40- и CD137-экспрессирующих клеток, для активации которых предназначена молекула, чтобы подавить рост опухоли. Механизм действия агонистических антител к OX40 в некоторых доклинических моделях опухолей был описан и заключается в истощении Treg, опосредуемом рецептором Fcγ, в микросреде опухоли, а введение в эти молекулы мутаций, подавляющих функцию рецептора Fcγ, нарушало их противоопухолевую активность (Bulliard et al., 2014). Несмотря на то, что эффект мутации LALA может заключаться в сохранении полезных иммунных клеток, для активации которых предназначена молекула антитела согласно настоящему изобретению, совместно с отсутствием истощения Treg-клеток, следует отметить, что OX40-нацеленные антитела IgG1 человека, разработанные для вызова того же механизма истощения опухолевых Treg, как и наблюдаемый в доклинических моделях опухолей, не проявили аналогичную способность контролировать рост опухоли (Glisson et al., 2016). Другие молекулы, разработанные для истощения Treg, также не показали высоких уровней клинической активности (Powell et al., 2007; Tran et al., 2017). Отсутствие клинической экстраполяции эффектов истощения Treg, наблюдаемых в моделях сингенных опухолей на мышах, может быть связано с более низкими уровнями клеток, экспрессирующих рецептор Fcγ, в микросреде опухоли (Milas et al., 1987), различиями в биологии Treg у человека и мыши (Liu et al., 2016) или другими неизвестными факторами (Stewart et al., 2014).
Включение мутации LALA в МАТ2 FS20m-232-91AA/Lob12.3 неожиданно не ухудшило его противоопухолевую активность в модели CT26, это свидетельствует о том, что оно имеет независимый от рецептора Fcγ механизм действия, который не опирается на взаимодействие с клетками, экспрессирующими рецептор Fcγ. Отсутствие наблюдаемого истощения опухолевых Treg и индукция сильной пролиферации Т-клеток в крови под действием этого МАТ2, содержащего мутацию LALA, в исследовании «механизма действия», описанном в Примере 19, обеспечивает дополнительную поддержку независимого от рецептора Fcγ механизма действия двойного агониста МАТ2 к OX40/CD137, описанного в настоящей заявке. Учитывая низкую клиническую активность, наблюдаемую при использовании антител, которые опираются на взаимодействие с рецептором Fcγ для осуществления их активности, независимый от рецептора Fcγ механизм действия молекулы антитела согласно настоящему изобретению, как ожидается, приведет к большей эффективности в клинических условиях.
17.2 Сравнение противоопухолевой активности МАТ2 к OX40/CD137 и его составных частей Fcab и Fab
В анализе активации общей популяции Т-клеток мыши (Пример 15) МАТ2 к OX40/CD137 мыши (FS20m-232-91AA/Lob12.3) проявило активность in vitro в отсутствие дополнительных перекрестно связывающих агентов, в отличие от моноспецифичных контрольных антител G1AA/Lob12.3 (МАТ к mCD137) и FS20m-232-91AA/4420 (имитационного МАТ2 к mOX40/FITC), путем одновременного связывания обоих рецепторов CD137 и OX40 (Пример 16.2). После анализа активации общей популяции Т-клеток противоопухолевую активность FS20m-232-91AA/Lob12.3 сравнивали с активностью его составных частей, то есть Fcab FS20m-232-91AA в формате имитационного (против FITC) МАТ2 (FS20m-232-91AA/4420) и моноспецифичного МАТ к CD137 мыши без Fcab (G1AA/Lob12.3) в виде отдельных агентов или в комбинации, или контроля изотипа (G1AA/4420) в модели опухоли CT26.
Придерживаясь того же способа, как и описанный в Примере 17.1, опухоли CT26 формировали подкожно у самок мышей BALB/c. На день 10 после инокуляции клеток CT26 мышей, несущих опухоли, рандомизировали в исследуемые когорты по 25 мышей на группу и вводили антитела.
Антитела разводили до конечной концентрации 0,3 мг/мл в ФСБ и вводили путем внутрибрюшинной инъекции в объеме 200 мкл каждой мыши с получением конечной дозы 3 мг/кг для мыши массой 20 г (фиксированная доза 60 мкг каждого антитела). Инъекции выполняли один раз каждые два дня (Q2D) для получения в общей сложности трех доз, начиная со дня 10 после инокуляции опухоли. Объемы опухолей определяли с помощью измерений штангенциркулем, как описано ранее. Исследование прекращали через 64 дня после инокуляции клеток, животных исключали из исследования, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии.
Данные по объему опухоли в зависимости от времени для отдельных животных показаны на Фигуре 10B, а средние результаты, показанные на Фигуре 10C, свидетельствуют о том, что МАТ2 FS20m-232-91AA/Lob12.3 ингибировало скорость роста опухоли CT26 на более ранним этапе (между днями 10 и 22) по сравнению с антителом для контроля изотипа (G1AA/4420). Не наблюдали явное ингибирование роста опухоли в когортах, обработанных МАТ к CD137 мыши, имитационным МАТ2 к OX40/FITC мыши или их комбинацией.
Придерживаясь того же метода смешанной модели, как и описанный ранее, анализ данных по объему опухоли до дня 22 (после инокуляции клеток, Таблица 29) показал, что FS20m-232-91AA/Lob12.3 привело к статистически достоверному (p=0,003) уменьшению средней скорости роста опухоли по сравнению с контролем изотипа. Для сравнения, лечение МАТ к CD137 мыши, имитационным МАТ2 к OX40/FITC мыши или их комбинацией не привело к значительно различающимся скоростям роста опухоли по сравнению с контролем изотипа. Сравнение скоростей роста опухоли на протяжении всего периода исследования (64 дня) с использованием метода смешанной модели показало статистически достоверные уменьшения скоростей роста опухоли во всех группах лечения по сравнению с контролем изотипа (анализ не показан).
Таблица 29: Попарное сравнение средних скоростей роста опухоли CT26 с использованием анализа модели со смешанными эффектами
[0,279, 0,340]
[0,244, 0,339]
[0,279, 0,340]
[0,235, 0,327]
[0,279, 0,340]
[0,237, 0,316]
[0,279, 0,340]
[0,164, 0,247]
ns = статистически недостоверно; TGR = скорость роста опухоли; ДИ = доверительный интервал
ПРИМЕЧАНИЕ: чтобы сравнить скорости роста опухоли на раннем этапе, в модели со смешанными эффектами использовали данные по объему опухоли за первые 22 дня после инокуляции. Для каждого попарного сравнения по меньшей мере одна из групп, включенных в расчет p-значений, содержит более 50% скоростей роста опухоли, которые достоверно не соответствуют логарифмическому нормальному распределению.
Анализ выживаемости с использованием лог-рангового критерия (Мантела-Кокса) показал, что FS20m-232-91AA/Lob12.3 привело к статистически достоверному улучшению выживаемости по сравнению с контролем изотипа (p≤0,0001) (Фигура 10D). Мыши, несущие опухоль, получавшие либо МАТ к CD137 мыши, либо имитационное МАТ2 к OX40 мыши/FITC, либо их комбинацию, не имели статистически достоверных различий в выживаемости по сравнению с контролем изотипа.
Подытоживая, результаты демонстрируют, что МАТ2 FS20m-232-91AA/Lob12.3 обладало большей и неэквивалентной противоопухолевой активностью по сравнению с комбинацией его составных частей Fcab и Fab или одной из составных частей по отдельности.
Пример 18 - Фармакодинамический ответ МАТ2 к OX40/CD137 в модели сингенной опухоли CT26
18.1 Сравнение фармакодинамического ответа МАТ2 к OX40/CD137 и контрольных МАТ к OX40 и к CD137
Фармакодинамический ответ суррогатного МАТ2 к OX40/CD137 оценивали у мышей, несущих сингенные опухоли CT26. С этой целью у мышей, несущих CT26, которым инокулировали МАТ2 FS20m-232-91AA/Lob12.3, контроль изотипа (G1/4420), контроль против OX40 мыши (G1/OX86) в виде отдельного агента, контроль против CD137 мыши (G1/Lob12.3) в виде отдельного агента или комбинацию этих контролей против OX40 и против CD137 (G1/OX86 плюс G1/Lob12.3), брали образцы крови в течение определенного времени и анализировали с помощью проточной цитометрии для определения активации Т-клеток и маркеров пролиферации.
Придерживаясь того же протокола, как и описанный в Примере 17, самок мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая подготавливали к началу исследования и инокулировали линию клеток карциномы толстой кишки CT26 (ATCC, CRL-2638). На день 10 после инокуляции опухолевых клеток опухоли измеряли и мышей рандомизировали в исследуемые когорты по 10 мышей на группу на основании объема опухоли. Любых мышей, которые не имели опухолей к этому моменту, исключали из исследования.
Антитела анализировали и проверяли на наличие примесей, как описано ранее, разводили до конечной концентрации 0,1 мг/мл в ФСБ и вводили путем инъекции при 200 мкл/мышь с получением конечной дозы 1 мг/кг для мыши массой 20 г. Антитела вводили мышам путем внутрибрюшинной (в/б) инъекции в дни 10, 12 и 14 после инокуляции опухоли.
Кровь собирали из хвостовой вены в пробирки, содержащие ЭДТА, за 1 час до дозирования в день 10, в день 11 (через 24 часа после введения первой дозы), в день 15 (через 24 часа после введения третьей дозы) и путем сердечной пункции в день 17 и день 24. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (eBioscience, номер по каталогу 00-4300-54) в соответствии с инструкциями производителя. Клетки окрашивали для проточной цитометрии с использованием либо красителя 1 (CD4-E450 (клон GK1.5), Ki67-FITC (клон SolA15), Foxp3-PE (клон FJK-16s), CD69-PECy5 (клон H1.2F3), CD3-PECy7 (клон 145-2C11), CD8-APC (клон 53-6.7), фиксируемый краситель для оценки жизнеспособности 780, все поставлены eBioscience; и CD45-V500 (клон 30-F11), поставленный BD Bioscience), либо красителя 2 (CD49b-E450 (клон DX5), F4/80-PE (клон 6F12), CD69-PECy5 (клон H1.2F3), CD19-PECy7 (клон 1D3), CD3-APC (клон 145-2C11) и фиксируемый краситель для оценки жизнеспособности 780, все поставлены eBioscience; CD45-V500 (клон 30-F11), поставленный BD Bioscience; и антитело к hFc-488 (поликлональное), поставленное Jackson ImmunoResearch) в присутствии блока Fc (eBioscience, номер по каталогу 14-0161-86 при 1:100). Затем клетки промывали один раз ФСБ и образцы, окрашенные красителем 2, повторно суспендировали в 200 мкл ФСБ и анализировали на FACS Canto II. Для образцов, окрашенных красителем 1, клетки первоначально окрашивали 100 мкл смеси антител 1 (все, кроме антитела к Ki67 и антитела к Foxp3) в течение 30 минут при 4°C. Затем клетки фиксировали и пермеабилизировали с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. В общих чертах, 200 мкл раствора для фиксации добавляли в каждую лунку и оставляли на ночь в темноте при 4°C. Затем клетки промывали в 200 мкл буфера для пермеабилизации. Клетки затем снова центрифугировали и повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 в присутствии блока Fc (все в разведении 1:100) и инкубировали 30 минут в темноте при 4°C. Затем клетки промывали один раз буфером для пермеабилизации и повторно суспендировали в 200 мкл ФСБ. После этого клетки анализировали в цитометре BD FACS CantoII. Данные анализировали с помощью FlowJoX, Excell и GraphPad Prism. Активацию и пролиферацию Т-клеток, наблюдаемую с течением времени, определяли для всех Т-клеток, а также субпопуляций CD4+ и CD8+.
Этот эксперимент показал, что МАТ2 к OX40/CD137 оказывало влияние на циркулирующие Т-клетки, увеличивая частоту активированных Т-клеток (CD45+ CD3+ CD69+) и CD4+ Т-клеток (CD45+ CD3+ CD4+ CD69+), а также пролиферирующих Т-клеток (CD45+ CD3+ Ki67+), CD4+ Т-клеток (CD45+ CD3+ CD4+ Ki67+) и CD8+ Т-клеток (CD45+ CD3+ CD8+ Ki67+) по сравнению со всеми группами, получавшими контроль, а также увеличивая частоту активированных CD8+ Т-клеток (CD45+ CD3+ CD8+ CD69+) по сравнению с обработкой либо контролем против OX40, либо контролем против CD137 по отдельности, либо контролем изотипа. Сходное увеличение частоты активированных CD8+ Т-клеток (CD45+ CD3+ CD8+ CD69+) наблюдали для контрольной группы, получавшей комбинацию контрольных МАТ к OX40 и к CD137. Эти результаты согласуются с результатами, наблюдаемыми in vitro, в которых МАТ2 к OX40/CD137 также показало увеличение активации Т-клеток, измеренное на основании выработки IL-2, который также известен как цитокин, участвующий в пролиферации Т-клеток.
18.2 Сравнение фармакодинамического ответа МАТ2 к OX40/CD137 и его составных частей Fcab и Fab
Периферический фармакодинамический ответ МАТ2 к OX40/CD137 мыши (FS20m-232-91AA/Lob12.3) сравнивали с таковым его составных частей, в частности, Fcab FS20m-232-91AA в формате имитационного (4420) МАТ2 (FS20m-232-91AA/4420) и моноспецифичного МАТ к CD137 мыши без Fcab (G1AA/Lob12.3) в виде отдельных агентов или в комбинации, или контроля изотипа (G1AA/4420) в модели опухоли CT26.
В аналогичном исследовании, описанном в Примере 17.2, на день 16 после инокуляции клеток CT26 у 10 мышей на группу из хвостовых вен брали образцы крови и собирали в пробирки, содержащие ЭДТА. Придерживаясь тех же методов, как и описанные в Примере 18.1, красные клетки крови лизировали, оставшиеся клетки затем окрашивали красителем для оценки жизнеспособности с последующим окрашиванием поверхности реагентами, перечисленными в Примере 22.2.2 (за исключением использования в этом исследовании клона антитела к CD4 мыши GK1.5 (BD Bioscience, номер по каталогу 563790) вместо клона антитела к CD4 RM4-5), за исключением антитела к Ki67 и антитела к Foxp3, в присутствии блока Fc. Затем клетки фиксировали и пермеабилизировали в течение ночи с помощью набора для окрашивания eBioscience Foxp3 (eBioscience) в соответствии с инструкциями производителя. Затем клетки внутриклеточно окрашивали антителом к Ki67 и антителом к Foxp3. После промывания клетки затем анализировали с использованием проточного цитометра BD Fortessa. Анализ данных выполняли с использованием программного обеспечения FlowJo, Excel и GraphPad Prism 7.
Было обнаружено, что FS20m-232-91AA/Lob12.3 значительно увеличивает доли Ki67+ CD4+ эффекторных (в % от общего количества CD4+ Foxp3- клеток) и Ki67+ CD8+ периферических Т-клеток (в % от общего количества CD8+ клеток) в крови по сравнению с обработкой контролем изотипа. МАТ к CD137 мыши и имитационное МАТ2 FS20m-232-91AA/4420, либо в виде отдельных агентов, либо в комбинации, также были способны индуцировать значительное повышение уровней пролиферирующих Ki67+ CD4+ эффекторных Т-клеток и Ki67+ CD8+ Т-клеток по сравнению с мышами, получавшими контроль изотипа. Однако повышение уровней пролиферирующих Ki67+ CD8+ Т-клеток после дозирования FS20m-232-91AA/Lob12.3 было значительно больше, чем наблюдаемое для моноклонального антитела к CD137 мыши по отдельности, имитационного МАТ2 FS20m-232-91AA/4420 по отдельности или их комбинации.
Подытоживая, эти результаты демонстрируют, что МАТ2 FS20m-232-91AA/Lob12.3 было способно индуцировать усиленный периферический фармакодинамический ответ в отношении увеличения частоты пролиферирующих Ki67+ CD8+ Т-клеток по сравнению с комбинацией его составных частей Fcab и Fab или любой составной частью по отдельности.
Пример 19 - Механизм действия МАТ2 к OX40/CD137 в модели сингенной опухоли CT26
Модель сингенной опухоли CT26 использовали для определения механизма действия (MOA) противоопухолевой активности МАТ2 к OX40/CD137 мыши in vivo. Ранее было показано, что модель сингенной опухоли CT26 чувствительна к агонистическим антителам к OX40 и к CD137 (Sadun et al., 2008), и ожидается, что инфильтрирующие опухоль лимфоциты (TIL), выделенные из опухолей CT26, экспрессируют как OX40, так и CD137. Протестированные антитела подробно описаны в Таблице 30.
Таблица 30. Подробная информация о протестированных антителах и МАТ2
Способность МАТ2, с мутацией LALA или без нее (FS20m-232-91AA/Lob12.3 и FS20m-232-91/Lob12.3, соответственно), активировать и индуцировать пролиферацию Т-клеток в крови и опухоли сравнивали с МАТ для контроля изотипа G1/4420 (против FITC), МАТ G1/OX86 (контроль против OX40 без мутации LALA) или G1/Lob12.3 (контроль против CD137 без мутации LALA) в виде отдельных агентов, комбинацией G1/OX86 плюс G1/Lob12.3 или комбинацией G1AA/OX86 (МАТ к OX40 с мутацией LALA) плюс G1AA/Lob12.3 (МАТ к CD137 с мутацией LALA).
Самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая обеспечивали период покоя в течение одной недели перед началом исследования. Всем животным вводили микрочип и присваивали уникальный идентификатор. В каждой когорте было по 5 мышей. Линию клеток карциномы толстой кишки CT26 (ATCC, CRL-2638) первоначально размножали, сохраняли, а затем подвергали предварительному скринингу в IDEXX Bioresearch для обнаружения патогенов с использованием протокола IMPACT I, и было показано, что линия не содержит патогенов. Клетки CT26 (приблизительно 3-5×106) размораживали из хранилища -150°C и добавляли к 20 мл DMEM (Gibco, 61965-026) с 10% ФСТ (Gibco, 10270-106) в колбе для тканевой культуры T175. Мышей анестезировали с использованием изофлурана (Abbott Laboratories), и каждое животное получило 1×106 клеток, введенных путем подкожной инъекции в левый бок. На день 10 после инокуляции опухолевых клеток состояние здоровья мышей контролировали, опухоли измеряли с помощью штангенциркулей, и мышей рандомизировали в исследуемые когорты на основании объема опухоли. Любых мышей, которые не имели опухолей в этот момент, исключали из исследования.
Антитела, вводимые путем инъекции, анализировали в течение 24 часов до инъекции с помощью профилирования SEC-HPLC и проверяли на наличие примесей. Антитела разводили до конечной концентрации 0,1 мг/мл в ФСБ и вводили путем инъекции при 200 мкл/мышь с получением конечной дозы 1 мг/кг для мыши массой 20 г. Антитела вводили мышам путем внутрибрюшинной (в/б) инъекции в дни 10, 12 и 14 после инокуляции опухоли. Измерения объема опухоли проводили три раза в неделю с помощью штангенциркулей для определения самой длинной оси и самой короткой оси опухоли. Через семь дней после введения третьей дозы (день 21 после инокуляции опухоли) мышей умерщвляли, опухоли выделяли путем диссекции и собирали кровь путем сердечной пункции.
Опухоли диссоциировали с использованием набора для диссоциации опухолей, мышь (Miltenyi 130-096-730), в соответствии с инструкциями производителя. В общих чертах, смесь ферментов готовили путем добавления 2,35 мл RPMI 1640, 100 мкл фермента D, 50 мкл фермента R и 12,5 мкл фермента A на опухоли, и каждую опухоль помещали в пробирку gentleMACS™ C, эту пробирку помещали на диссоциатор gentleMACS™ и запускали программу m_TDK_1, а затем инкубировали в течение 1 часа при 37°C при встряхивании (200 об./мин). Полученную суспензию клеток пропускали через сито, используя сито для клеток 70 мкм (Corning, номер по каталогу 352350), центрифугировали (10 минут при 1500 об./мин), промывали один раз в ФСБ и повторно суспендировали в 5 мл ФСБ.
Кровь собирали с помощью сердечной пункции в пробирки, содержащие ЭДТА. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (eBioscience, номер по каталогу 00-4300-54) в соответствии с инструкциями производителя.
Клетки, выделенные из опухолей и крови, окрашивали для проточной цитометрии с использованием следующей панели антител и реагентов (Краситель 1): CD4-E450 (клон GK1.1), Ki67-FITC (клон SolA15), Foxp3-PE (FJK-16s), CD69-PECy5 (клон H1.2F3), CD3-PECy7 (клон 145-2C11), CD8-APC (клон 53-6.7), фиксируемый краситель для оценки жизнеспособности 780 и блок Fc (клон 93), все поставленные eBioscience; и CD45-V500 (клон 30-F11), поставленный BD Bioscience. Клетки промывали в ФСБ и затем инкубировали с 100 мкл смеси антител 1 (все, кроме антитела к Ki67 и антитела к Foxp3) в течение 30 минут при 4°C. После этого клетки промывали ФСБ, а затем фиксировали и пермеабилизировали с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. В общих чертах, 200 мкл раствора для фиксации добавляли в каждую лунку и оставляли на ночь в темноте при 4°C. Затем клетки промывали в 200 мкл буфера для пермеабилизации. Клетки затем снова центрифугировали и повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 в присутствии блока Fc (все в разведении 1:100) и инкубировали 30 минут в темноте при 4°C. После этого клетки промывали один раз буфером для пермеабилизации и повторно суспендировали в 200 мкл ФСБ. Затем клетки анализировали в цитометре BD FACS CantoII.
Данные анализировали с помощью FlowJoX, Excell и GraphPad Prism. Статистический анализ для сравнения групп выполняли с использованием однофакторного дисперсионного анализа, а затем критерия Тьюки для множественных сравнений каждой пары с использованием программного пакета GraphPad Prism.
Определяли частоту Т-клеток (CD45+CD3+), пролиферирующих Т-клеток (CD45+ CD3+ Ki67+) и регуляторных Т-клеток (CD45+ CD3+ CD4+ Foxp3+) в крови или опухолях мышей, которым были инокулированы клетки CT26, после обработки МАТ2 к OX40/CD137 или контролями. МАТ2 FS20m-232-91AA/Lob12.3 показало статистически достоверное увеличение пролиферирующих Т-клеток, а также увеличение Treg в крови по сравнению с контролем изотипа (G1/4420). МАТ2 FS20m-232-91AA/Lob12.3, как правило, увеличивало частоту Т-клеток в опухоли.
Наблюдали статистически достоверное уменьшение уровней Treg в опухоли у мышей, получавших антитело к CD137 G1/Lob12.3 и комбинацию этого антитела к CD137 с антителом к OX40 G1/OX86, по сравнению с обработкой контролем изотипа. Однако при введении мутации LALA в антитело к OX40 и антитело к CD137 обработка комбинацией этих антител (G1AA/OX86 плюс G1AA/Lob12.3) больше не уменьшала уровни Treg в опухолях. МАТ2 к OX40/CD137, содержащее мутацию LALA (FS20m-232-91AA/Lob12.3), не уменьшало уровни Treg, в то время как вариант IgG1 человека дикого типа МАТ2 к OX40/CD137 без мутации LALA (FS20m-232-91/Lob12.3) действительно показал статистически достоверное уменьшение уровней Treg в опухоли.
Эти данные демонстрируют, что введение мутации LALA в IgG1 человека устраняет способность МАТ2 к OX40/CD137 истощать Treg и, следовательно, что противоопухолевая активность, наблюдаемая при использовании LALA-варианта МАТ2 к OX40/CD137 в формате IgG1 человека (FS20m-232-91AA/Lob12.3), не зависит от истощения Treg. Кроме того, было обнаружено, что МАТ2 FS20m-232-91AA/Lob12.3 индуцирует пролиферацию Т-клеток на периферии в оцененный момент времени, что, как ожидается, приведет к размножению пула Т-клеток, вызывающих противоопухолевый иммунный ответ. Эти данные свидетельствуют о том, что LALA-содержащее МАТ2 к OX40/CD137 в формате IgG1 человека может обладать противоопухолевой активностью при разных видах рака в отсутствие связывания МАТ2 с рецепторами Fcγ, которые могут быть распространенными в опухоли или могут не быть распространенными.
Пример 20 - Активность МАТ2 к OX40/CD137 мыши в модели сингенной опухоли B16-F10
Модель сингенной опухоли B16-F10 использовали для тестирования противоопухолевой активности МАТ2 к OX40/CD137 мыши (FS20m-232-91AA/Lob12.3) in vivo. Антитело G1/4420 использовали в качестве контроля в исследовании. Ранее не было показано, что модель сингенной опухоли B16-F10 чувствительна к агонистическим антителам к OX40 или к CD137 (Hirschhorn-Cymerman et al., 2009; Wilcox et al., 2002). Однако ожидается, что инфильтрирующие опухоль лимфоциты (TIL), выделенные из опухолей B16-F10, экспрессируют как OX40, так и CD137.
Самок мышей C57BL/6 (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая акклиматизировали в течение одной недели перед началом исследования. Всем животным вводили микрочип и присваивали уникальный идентификатор. В каждой когорте было по 10 мышей. Линию клеток карциномы толстой кишки B16-F10 (ATCC, номер по каталогу CRL-6475) первоначально размножали, сохраняли, а затем подвергали предварительному скринингу в IDEXX Bioresearch для обнаружения патогенов с использованием протокола IMPACT I, и было показано, что линия не содержит патогенов.
Клетки B16-F10 размораживали из хранилища при -150°C и добавляли к 20 мл DMEM (Gibco, 61965-026) с 10% ФСТ (Gibco, 10270-106) в колбе для тканевой культуры T175. Каждое животное получило 1×106 клеток путем подкожной инъекции в левый бок. Через 7-8 дней после инокуляции опухолевых клеток мышей, которые не имели опухолей в этот момент, исключали из исследования. Антитела анализировали и проверяли на наличие примесей, как описано ранее, перед инъекцией в конечной концентрации 0,1 мг/мл в ФСБ в объеме 200 мкл/мышь с получением конечной дозы 1 мг/кг для мыши массой 20 г. Каждая мышь получала антитела путем внутрибрюшинной (в/б) инъекции в дни 8, 10 и 12 после инокуляции опухоли. Объемы опухолей определяли путем измерения с использованием штангенциркулей (как описано в Примере 17), и выполняли дозирование любого лекарственного средства, назначенное на рассматриваемый день.
Мышей умерщвляли, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии. Статистический анализ роста опухоли выполняли с использованием статистического анализа смешанной модели, описанного в Примере 17. Результаты исследования показаны на Фигуре 11.
МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3) проявило значительную противоопухолевую активность по сравнению с контрольными животными, которым вводили путем инъекции контрольное антитело (G1/4420). Этот результат был неожиданным, поскольку ранее было показано, что эта модель нечувствительна к стимуляции OX40 или CD137 (Hirschhorn-Cymerman et al., 2009; Wilcox et al., 2002). Важно отметить, что активность, наблюдаемая для МАТ2 к OX40/CD137, была в присутствии мутации LALA и, следовательно, не зависела от истощения опухолевых Treg. Это свидетельствует о том, что MOA МАТ2 к OX40/CD137 приводит к противоопухолевой активности в различных моделях сингенных опухолей, даже в тех, которые имеют более низкие уровни иммунного инфильтрата, таких как B16-F10.
Пример 21 - Аналитическая характеристика и предварительная оценка стабильности МАТ2 к OX40/CD137
21.1 Экспрессия, очистка и аналитическая характеристика МАТ2
МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14 получали в лабораторном масштабе и характеризовали с помощью стандартных аналитических методов с использованием SE-HPLC и SDS-PAGE.
Последовательности ДНК, кодирующие МАТ2, кратковременно экспрессировали в HEK293-6E (Национальный исследовательский совет Канады). Через 5 дней культуральные жидкости собирали и очищали на предварительно заполненных колонках MabSelect с белком A с использованием прибора AKTAxpress (и то, и другое от GE Healthcare). Уравновешивание колонок осуществляли в 50 мМ Трис-HCl, 250 мМ NaCl, pH=7,0, с последующей загрузкой собранной культуральной жидкостью. Затем смолу подвергали промывке с использованием 50 мМ трис-HCl, 250 мМ NaCl при pH=7,0, а затем элюировали МАТ2 с использованием буфера при pH=3,5. Буфер МАТ2 заменяли на предварительный буфер для изготовления состава с использованием обессоливающих колонок PD-10 (GE Healthcare, номер продукта 17085101).
SE-HPLC выполняли на системе для ВЭЖХ Agilent серии 1100 (Agilent), снабженной колонкой TSK-GEL SUPERSW3000 с внутренним диаметром 4,6 мм × 30,0 см (Tosoh Bioscience), с использованием 20 мМ фосфата натрия, 200 мМ хлорида натрия, pH=6,8 в качестве подвижной фазы. Количественное определение процентного содержания мономера выполняли с использованием программного обеспечения Chemstation (Agilent). Результаты анализа SE-HPLC обобщены в Таблице 31.
Таблица 31. Аналитическая характеристика с помощью SE-HPLC
Анализ с помощью электрофореза в ДСН-ПААГ выполняли с использованием 4-12% бис-трис гелей для белков NuPAGE® Novex® и 1× буфера для разделения MOPS (Thermo Fisher Scientific), по существу в соответствии с инструкциями производителя. Для невосстанавливающего электрофореза в ДСН-ПААГ образцы подвергали воздействию алкилирующего реагента N-этилмалеимида (Sigma-Aldrich) перед этапом денатурации, и 2-меркаптоэтанол был исключен из смеси для денатурации. Полосы белка визуализировали с помощью Coomassie InstantBlue (Expedeon).
Все пять МАТ2 показали благоприятные параметры аналитической характеристики после очистки на белке A с чистотой мономера выше 95% при определении с помощью SE-HPLC. Анализ с помощью электрофореза в ДСН-ПААГ выявил профили полос белка, типичные для рекомбинантного IgG1. Таким образом, в невосстанавливающих условиях одна полоса мигрировала в область, соответствующую ожидаемой молекулярной массе, а в восстанавливающих условиях две полосы мигрировали близко к маркерам молекулярной массы 51 кДа и 28 кДа, соответствующим тяжелой цепи и легкой цепи, соответственно. Фрагментация не наблюдалось (данные не показаны).
21.2 Предварительная оценка стабильности МАТ2 к OX40/CD137
Выполняли предварительную оценку стабильности МАТ2 FS20-22-49AA/FS30-5-37, FS20-22-49AA/FS30-10-3, FS20-22-49AA/FS30-10-12, FS20-22-49AA/FS30-10-16 и FS20-22-49AA/FS30-35-14. Перед тем как подвергнуть МАТ2 предварительной оценке стабильности, их дополнительно очищали с помощью гель-проникающей хроматографии (SEC) с использованием колонки Superdex HiLoad 26/600 200 пг (GE Healthcare), уравновешенной предварительным буфером для изготовления состава. Образцы для оценки стабильности хранили при 5°C и анализировали через 2 и 4 недели с помощью стандартных аналитических методов с использованием SE-HPLC и капиллярного электрофореза с додецилсульфатом натрия (CE-SDS).
SE-HPLC выполняли на системе ВЭЖХ Agilent серии 1100 (Agilent), снабженной колонкой TSK-GEL SUPERSW3000 с внутренним диаметром 4,6 мм × 30,0 см (Tosoh Bioscience), с использованием 20 мМ фосфата натрия, 200 мМ хлорида натрия, pH=6,8 в качестве подвижной фазы. Сбор данных и количественное определение содержания мономера выполняли с использованием программного обеспечения Chemstation (Agilent). Результаты обобщены в Таблице 32.
После хранения при 5°C в течение 4 недель содержание мономера, определенное с помощью SE-HPLC для всех протестированных МАТ2, оставалось сопоставимым (в пределах ± 0,9%) с исходным материалом (T = 0). Таким образом, все протестированные МАТ2 проявили благоприятный профиль стабильности.
Таблица 32: Анализ стабильности с помощью SE-HPLC
T=0
T= 2 недели при 5°C
T= 4 недели при 5°C
Анализ CE-SDS выполняли на системе для капиллярного электрофореза Bioanalyzer 2100 (Agilent) в соответствии с рекомендациями производителя. Для создания восстанавливающих условий при CE-SDS добавляли DTT, и образцы денатурировали при 70°C в течение 5 минут. Сбор данных и количественную оценку в процентах материала тяжелой цепи и легкой цепи выполняли с использованием программного обеспечения Expert 2100 (Agilent). Чистоту в процентах рассчитывали как сумму процентного содержания материала тяжелой цепи и процентного содержания материала легкой цепи. Результаты анализа обобщены в Таблице 33.
Чистота всех протестированных МАТ2, определенная как сумма процентного содержания материала тяжелой цепи и материала легкой цепи с помощью CE-SDS в восстанавливающих условиях, также оставалась сопоставимой (в пределах ± 1,0%) с исходным материалом. Таким образом, все протестированные МАТ2 также показали благоприятную стабильность.
Таблица 33. Анализ стабильности с помощью CE-SDS
T=0
T= 2 недели при 5°C
T= 4 недели при 5°C
Пример 22 - Активность МАТ2 к OX40/CD137 в комбинации с антителом к PD-1 или антителом к PD-L1
Известно, что экспрессия PD-L1 на антигенпрезентирующих клетках (например, дендритных клетках, макрофагах, B-клетках), опухолевых клетках и на клетках в микросреде опухоли ингибирует активацию, пролиферацию, а также эффекторные и цитотоксические функции Т-клеток посредством взаимодействия с PD-1. Было показано, что блокирование этого взаимодействия с использованием моноклональных антител к PD-1 или к PD-L1 приводит к увеличению показателей выживаемости у пациентов с несколькими типами рака.
Однако в некоторых опухолях антитела к PD-L1 и к PD-1 оказывают незначительный эффект или не оказывают его. Авторы настоящего изобретения протестировали комбинацию МАТ2 к OX40/CD137 с антителом к PD-L1 или антителом к PD-1 в исследованиях in vitro и in vivo, чтобы установить, может ли использование комбинации привести к улучшенному эффекту по сравнению с использованием МАТ2 к OX40/CD137, антитела к PD-L1 или антитела к PD-1 по отдельности.
22.1 Активность МАТ2 к OX40/CD137 в комбинации с блокадой PD-1 или PD-L1 в анализе с использованием стафилококкового энтеротоксина A (SEA)
Активность МАТ2 к OX40/CD137 тестировали в анализе активации Т-клеток с использованием суперантигена стафилококкового энтеротоксина A (SEA) в качестве первого сигнала, как описано в Примере 12 выше. Чтобы протестировать эффект МАТ2 к OX40/CD137 на активность стимуляции Т-клеток в комбинации с блокированием взаимодействия между PD-1 и PD-L1, блокирующие антитела к PD-1 или к PD-L1 комбинировали с МАТ2 к OX40/CD137 в анализе с SEA.
Антитела и МАТ2, использованные в анализе с SEA, перечислены в Таблице 34 ниже. Тестировали G1/4420 (против FITC) в комбинации с МАТ2 FS20-22-49AA/FS30-10-16, G1AA/S1 (против PD-L1), G1AA/5C4 (против PD-1) по отдельности или в комбинации с МАТ2 FS20-22-49AA/FS30-10-16. Выработку интерлейкина-2 (IL-2) использовали в качестве меры активации Т-клеток.
Таблица 34: Подробная информация о протестированных антителах и МАТ2
FS30-10-16
Последовательности вариабельных доменов антител 5C4 и YW243.55.S1 (S1) также раскрыты в US 8008449 B2 и US 2013/0045202 A1, соответственно.
Выделяли МКПК и выполняли анализ с SEA, по существу как описано в Примере 12.1 выше. G1/4420 использовали в качестве контроля изотипа, и в анализах не применяли перекрестно связывающие агенты.
Активность МАТ2 к OX40/CD137 (FS20-22-49AA/FS30-10-16) в комбинации либо с антителом к PD-L1 (G1AA/S1), либо с антителом к PD-1 (G1AA/5C4) сравнивали с активностью МАТ2 FS20-22-49AA/FS30-10-16 плюс контроль изотипа (G1/4420) или с активностью антитела к PD-L1 (G1AA/S1) или антитела к PD-1 (G1AA/5C4), или контроля изотипа (G1/4420) по отдельности. Значения ЭК50 и максимальный ответ высвобождения IL-2, наблюдаемые в анализе с SEA, показаны в Таблице 35. Фигура 12A и B показывают графики высвобождения IL-2 для анализа с SEA.
Таблица 35: Анализ с SEA с использованием МАТ2, нацеленных на OX40 и CD137, в комбинации с антителами, блокирующими взаимодействие между PD-1 или PD-L1
NAD = активность не обнаружена
Как и ожидалось, активность не была обнаружена при использовании контроля изотипа (G1/4420). Аналогичным образом, блокирование взаимодействия между PD-1 и PD-L1 по отдельности не обладало активностью в этом анализе. Однако комбинирование стимуляции рецепторов OX40 и CD137 (с помощью МАТ2 к OX40/CD137) с блокадой взаимодействия между PD-1 и PD-L1 (с помощью либо антитела к PD-L1, либо антитела к PD-1) привело к увеличению максимальной активности Т-клеток, измеренной на основании максимальной выработки IL-2, свыше той, которая наблюдалась при использовании МАТ2 к OX40/CD137 по отдельности. Увеличение максимальной активности Т-клеток, наблюдаемое при комбинировании МАТ2 к OX40/CD137 с антителом к PD-L1 или антителом к PD-1, было сходным.
22.2 Противоопухолевая активность и фармакодинамический ответ введения МАТ2 к OX40/CD137 мыши и антагониста PD-1 в модели опухоли CT26 на мышах
Модель опухоли CT26 на мышах использовали для установления противоопухолевой активности и фармакодинамического ответа комбинации FS20m-232-91AA/Lob12.3 и антагонистического антитела к PD-1 (клон RMP1-14 IgG1 мыши) по сравнению с любым агентом по отдельности.
22.2.1 Оценка противоопухолевой активности
Придерживаясь того же протокола, как и описанный в Примере 17, самок мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г подготавливали к началу исследования и инокулировали линию клеток карциномы толстой кишки CT26. Через 10 дней после инокуляции опухолевых клеток опухоли измеряли, любых мышей, которые не имели опухолей, исключали из исследования, а оставшихся мышей рандомизировали на 4 группы лечения (Таблица 36) по 15 животных на группу. Животным вводили путем внутрибрюшинной инъекции: (1) комбинацию антител для контроля изотипа 1 мг/кг G1AA/4420 и 10 мг/кг mIgG1/4420 (Absolute Antibodies, клон 4420, номер по каталогу Ab00102-1.1), (2) 10 мг/кг антитела к PD-1 мыши (Absolute Antibodies, клон RMP1-14 IgG1 мыши, номер по каталогу Ab00813-1.1), (3) 1 мг/кг МАТ2 FS20m-232-91AA/Lob12.3, или (4) 10 мг/кг антитела к PD-1 мыши и 1 мг/кг МАТ2 FS20m-232-91AA/Lob12.3 в ФСБ. Животные получали внутрибрюшинные (в/б) инъекции G1AA/4420 или FS20m-232-91AA/Lob12.3 один раз каждые 2 дня, в общей сложности 3 дозы, начиная со дня 10 после инокуляции опухоли. mIgG1/4420 или антитело к PD-1 мыши дозировали в/б один раз каждые 4 дня, в общей сложности 4 дозы, начиная со дня 10 после инокуляции опухоли. Объемы опухолей определяли с помощью измерений штангенциркулем (как описано в Примере 17). Исследование прекращали через 60 дней после инокуляции опухолевых клеток, животных исключали из исследования, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии. Группы лечения, протестированные молекулы, дозы и схема дозирования обобщены в Таблице 36.
Таблица 36. Сводная информация по группам лечения и протестированным молекулам
Q4D
Антитело к PD-1 мыши mIgG1
Q4D
Как показано на Фигуре 13A-D, комбинация антагонистического антитела к PD-1 и 1 мг/кг FS20m-232-91AA/Lob12.3 привела к самой высокой доле животных, 7 из 15 (47%), с ответом в виде полной регрессии опухоли (определяемым как объем опухоли ≤62,5 мм3) по окончании исследования (Фигура 13D). Антитела для контроля изотипа (Фигура 13A), антитело к PD-1 (Фигура 13B) в виде отдельного агента и 1 мг/кг FS20m-232-91AA/Lob12.3 (Фигура 13C) показали 0%, 0% и 7% регрессию опухоли по окончании исследования, соответственно.
Анализ выживаемости показал, что комбинация FS20m-232-91AA/Lob12.3 с антителом к PD-1 привела к статистически достоверному преимуществу в выживаемости по сравнению с антителами для контроля изотипа (лог-ранговый критерий (Мантела-Кокса), p<0,0001) (Фигура 13E). Не наблюдали значительных различий в выживаемости между обработками любым агентом по отдельности в сравнении с антителами для контроля изотипа. Эти результаты демонстрируют, что в этой модели блокада ингибиторного пути PD-1/PD-L1 с помощью антагониста и двойного агонистического действия на OX40 и CD137 с применением МАТ2 к OX40/CD137 была способна увеличивать противоопухолевую активность и обеспечивать преимущество в выживаемости по сравнению с отдельными агентами.
22.2.2 Оценка периферического фармакодинамического ответа
В исследовании, описанном в Примере 22.2.1, также исследовали способность антагониста PD-1 модулировать фармакодинамический ответ на FS20m-232-91AA/Lob12.3 и сравнивали с обработкой отдельным агентом. Через 6 дней после начала дозирования (16 дней после инокуляции опухолевых клеток) кровь из хвостовых вен 6 случайно отобранных мышей, несущих опухоль CT26, из групп лечения 1, 2, 3 и 5 (Таблица 36) собирали в пробирки, содержащие ЭДТА. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (Miltenyi Biotech, №130-094-183) в соответствии с инструкциями производителя. Клетки окрашивали для анализа методом проточной цитометрии с использованием реагентов CD4-BUV395 (клон RM4-5), CD8-BUV737 (клон 53-6.7), CD44-BV510 (клон IM7) и CD3e-BV786 (клон 145-2C11), все поставлены BD Bioscience; CD69-FITC (клон H1.2F3), NKp46-PE (клон 29A1.4), PD-1-APC (клон J43), CD45-Alexa700 (клон 30-F11) и фиксируемого красителя для оценки жизнеспособности 780, все поставлены eBioscience; и CD62L-BV421 (клон MEL-14), поставленный Biolegend, в присутствии блока Fc (eBioscience, номер по каталогу 14-0161-86 при 1:50) в течение 30 минут при 4°C. Затем клетки фиксировали и пермеабилизировали в течение ночи с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. Клетки повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 (Ki67-PE-Cy7 (клон SolA15) и Foxp3-PerCP-Cy5.5 (клон FJK-16s), оба поставлены eBioscience) и инкубировали в течение 30 минут при комнатной температуре в темноте. Затем клетки дважды промывали буфером для пермеабилизации и повторно суспендировали в ФСБ + 0,5% БСА. После этого клетки анализировали в проточном цитометре BD Fortessa. Анализ данных выполняли в программном обеспечении FlowJo, Excel и GraphPad Prism 7.
Частоты пролиферирующих Ki67+ CD4+ Т-клеток (от общего количества CD45+ CD3+ CD4+), Ki67+ CD8+ Т-клеток (от общего количества CD45+ CD3+ CD8+) и Ki67+ NKp46+ NK-клеток (от общего количества CD45+ CD3- NKp46+) определяли с помощью анализа методом проточной цитометрии, как описано выше. По сравнению с контролями изотипа, FS20m-232-91AA/Lob12.3 индуцировало статистически достоверное увеличение пролиферирующих Ki67+ CD4+ и Ki67+ CD8+ Т-клеток, подтверждая предыдущие результаты (Пример 18), и пролиферирующих Ki67+ NK-клеток (непараметрический критерий Манна-Уитни для попарного сравнения; p≤0,005 для всех трех популяций иммунных клеток). Антагонистическое антитело к PD-1 в виде отдельного агента не оказало заметного эффекта на три популяции иммунных клеток по сравнению с контролями изотипа. Комбинация 1 мг/кг FS20m-232-91AA/Lob12.3 и антитела к PD-1 привела к статистически достоверно более высоким уровням пролиферирующих Ki67+ CD4+ Т-клеток, Ki67+ CD8+ Т-клеток и Ki67+ NKp46+ NK-клеток по сравнению как с отдельным агентом, так и с контролями изотипа (p≤0,005 для всех статистически достоверных сравнений, за исключением эффекта комбинации на уровни пролиферирующих Ki67+ CD4+ Т-клеток по сравнению с FS20m-232-91AA/Lob12.3 по отдельности, для которого p≤0,05).
Эффекты антитела к PD-1, МАТ2 FS20m-232-91AA/Lob12.3 в виде отдельных агентов и комбинации антитела к PD-1 и FS20m-232-91AA/Lob12.3 на PD-1-экспрессирующие Т-клетки периферической крови (CD4+ и CD8+ Т-клетки) и NK-клетки также определяли с помощью анализа методом проточной цитометрии, как описано выше. Антитело к PD-1 и FS20m-232-91AA/Lob12.3 в виде отдельных агентов увеличивали долю PD-1-экспрессирующих CD4+ и CD8+ Т-клеток по сравнению с контролем изотипа с более высокой медианной частотой PD-1+ клеток после обработки FS20m-232-91AA/Lob12.3 по сравнению антителом к PD-1 по отдельности. FS20m-232-91AA/Lob12.3 по отдельности увеличивало частоту PD-1+ NK-клеток по сравнению с контролями изотипа. Комбинация привела к статистически достоверно более высоким уровням PD-1-экспрессирующих CD4+ и CD8+ Т-клеток (но не NK-клеток) по сравнению как с отдельным агентом, так и контролями изотипа (непараметрический критерий Манна-Уитни для попарного сравнения; p≤0,005 для всех статистически достоверных сравнений, за исключением эффекта комбинации на частоту PD-1-экспрессирующих CD4+ Т-клеток по сравнению с FS20m-232-91AA/Lob12.3 по отдельности, для которого p≤0,05).
В соответствии с результатами оценки противоопухолевой активности одновременная блокада ингибиторного пути PD-1/PD-L1 с помощью антагониста и двойного агонистического действия на OX40 и CD137 при применении МАТ2 к OX40/CD137 привела к усилению фармакодинамической модуляции пролиферирующих Т-клеток и NK-клеток, что поддерживает использование комбинированного подхода для запуска противоопухолевого иммунитета.
Подытоживая, блокирование оси PD-1/PD-L1 при одновременном агонистическом действии на OX40 и CD137 приводит к увеличению эффекта в сравнении с блокированием PD-1/PD-L1 по отдельности. В частности, комбинация антитела к PD-1 или антитела к PD-L1 с МАТ2 к OX40/CD137 привела к улучшенному эффекту в сравнении с ответом при использовании одного из антител по отдельности. В анализе с SEA, описанном в Примере 22.1, ни одно из протестированных антител к PD-1 или к PD-L1 не проявило какой-либо активности по сравнению с МАТ2 к OX40/CD137, которое имело ЭК50 0,1474 нМ. Однако при тестировании МАТ2 к OX40/CD137 в комбинации с антителом к PD-1 или антителом к PD-L1 значения ЭК50 составили 0,2373 нМ и 0,5961 нМ, соответственно. Кроме того, максимальный ответ IL-2, вызванный любой комбинацией, более чем в два раза превышал ответ МАТ2 к OX40/CD137 по отдельности. Эти данные in vitro демонстрируют, что в системе, в которой не обнаружена активность антитела к PD-L1 или антитела к PD-1, комбинирование любого из этих антител с МАТ2 к OX40/CD137 приводит к значительному улучшению активности.
Эту активность in vitro дополнительно подтверждали с помощью тестирования in vivo МАТ2 к OX40/CD137 мыши, по отдельности или в комбинации с антителом к PD-1, в модели опухоли CT26, как описано в Примере 22.2. Результаты этого исследования показали, что к концу исследования опухоли отсутствовали у большего числа животных из группы, получавшей комбинацию антитела к PD-1 с МАТ2 к OX40/CD137, по сравнению с МАТ2 к OX40/CD137 или антителом к PD-1 по отдельности (когда отсутствие опухоли не было обнаружено ни у одного из животных). Кроме того, также наблюдали статистически достоверные преимущества в выживаемости (Фигура 13E), и фармакодинамическая модуляция пролиферирующих Т-клеток и NK-клеток усиливалась с помощью обработки комбинацией по сравнению как с МАТ2, так и антителом к PD-1 по отдельности.
Поскольку ни в исследованиях in vitro, ни в исследованиях in vivo не наблюдали активность антител к PD-1 или к PD-L1, но значительные улучшения наблюдали при дозировании любого из них с МАТ2 к OX40/CD137, это может свидетельствовать о том, что МАТ2 к OX40/CD137 в комбинации с таким антителом приведет к усилению противоопухолевой эффективности, а также что такая комбинация может быть подходящей для лечения опухолей, которые являются невосприимчивыми, например, рефрактерными или резистентными к монотерапии антителом к PD-1 или антителом к PD-L1 или рецидивировали после нее.
Пример 23 - Дозозависимая противоопухолевая активность МАТ2 к OX40/CD137 мыши в модели сингенной опухоли CT26 и формирование защитной иммунологической памяти против повторной стимуляции опухолевыми клетками CT26
Для оценки взаимосвязи между дозой и противоопухолевой активностью суррогатного МАТ2 к OX40/CD137 в модели сингенной колоректальной опухоли CT26 на мышах оценивали пять различных уровней дозы от 0,1 до 10 мг/кг.
Придерживаясь того же протокола, как и описанный в Примере 17, самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая вводили путем подкожной инъекции клетки карциномы толстой кишки CT26 в левый бок каждого животного. Через 10 дней после инокуляции опухолевых клеток опухоли измеряли и животных без развившейся опухоли исключали из исследования. Оставшихся мышей рандомизировали на шесть экспериментальных групп по 25 животных на группу.
Антитело для контроля изотипа (G1AA/4420) и суррогатное МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3) фильтровали и разводили в ФСБ перед инъекцией. Каждому животному вводили путем внутрибрюшинной инъекции разведенное антитело в объеме 200 мкл на одно введение с получением конечной дозы 10 мг/кг G1AA/4420 или 0,1, 0,3, 1, 3 или 10 мг/кг FS20m-232-91AA/Lob12.3 на введение для мыши массой 20 г. Инъекции выполняли один раз каждые два дня (Q2D), в общей сложности три дозы, начиная со дня 10 после инокуляции опухоли. Объемы опухолей определяли с помощью измерений штангенциркулем, как описано в Примере 17. Исследование прекращали через 67 дней после инокуляции опухолевых клеток, животных исключали из исследования, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии.
Объемы опухолей с течением времени для отдельных животных, получавших либо G1AA/4420, либо FS20m-232-91AA/Lob12.3 при различных уровнях дозы, показаны на Фигуре 14A. Уровни дозы 0,3, 1, 3 или 10 мг/кг FS20m-232-91AA/Lob12.3 привели к полной регрессии опухоли (определяемой как ≤62,5 мм3 на день 60) у 4% (1/25), 4% (1/25), 8% (2/25) и 4% (1/25) животных на группу, соответственно. Ни одно из животных в группах контроля изотипа и 0,1 мг/кг суррогатного МАТ2 не имело полной регрессии опухоли.
Попарные сравнения средних скоростей роста опухоли между группами, обработанными FS20m-232-91AA/Lob12.3 и G1AA/4420, выполняли с использованием статистического анализа смешанной модели, как описано в Примере 17, и для всех протестированных уровней дозы (0,1, 0,3, 1, 3 и 10 мг/кг) наблюдали статистически достоверные различия (p<0,01) по сравнению с контролем изотипа (Таблица 37). FS20m-232-91AA/Lob12.3 уменьшало среднюю скорость роста опухоли (TGR) дозозависимым образом при дозировании от 0,1 до 3 мг/кг (среднее значение log(TGR) от 0,255529 до 0,156767, соответственно). Среднее значение TGR для 3 мг/кг FS20m-232-91AA/Lob12.3 статистически не отличалось от такового для уровня дозы 1 мг/кг (p = 0,18). Однако увеличение уровня дозы до 10 мг/кг привело к более высокой TGR по сравнению с группой, получавшей дозу 3 мг/кг (p<0,001).
Таблица 37: Попарное сравнение средних скоростей роста опухоли CT26 с использованием статистического анализа смешанной модели
Сокращения: ns = статистически недостоверно; TGR = скорость роста опухоли
Примечание: для каждого попарного сравнения по меньшей мере одна из групп, включенных в вычисление p-значений, содержит более 50% скоростей роста опухоли, достоверно не соответствующих логарифмическому нормальному распределению.
Анализ выживаемости с использованием лог-рангового критерия (Мантела-Кокса) показал, что FS20m-232-91AA/Lob12.3 при всех протестированных уровнях дозы приводило к статистически достоверному преимуществу в выживаемости по сравнению с контролем изотипа (Фигура 14B). Сравнение групп 1 мг/кг и 3 мг/кг не показало статистического различия в выживаемости.
Подытоживая, объем опухоли и данные о выживаемости, показанные на Фигурах 14A и B и в Таблице 37, подтверждают результаты Примера 17, что суррогатное МАТ2 к OX40/CD137 может вызывать противоопухолевую активность in vivo в модели опухоли CT26 на мышах. Кроме того, наблюдаемая противоопухолевая активность дозозависимо увеличивалась от 0,1 мг/кг до 1 мг/кг и поддерживалась при протестированных более высоких уровнях дозы (3 мг/кг и 10 мг/кг).
Чтобы протестировать, может ли суррогатное МАТ2 к OX40/CD137 индуцировать защитную иммунологическую память против опухолевых клеток CT26, животным, которые имели полную регрессию опухоли (животные с полным ответом) из исследования с определением диапазона доз настоящего примера, описанного выше, повторно инокулировали подкожно 1×105 клеток CT26 на день 84 после первой инокуляции клеток. Не получавшим лечения мышам BALB/c, не несущим опухоли, также инокулировали клетки CT26 в качестве контрольной группы. Объемы опухолей контролировали, как описано выше. Исследование прекращали на день 137 после первой инокуляции клеток, животных исключали из исследования, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии. В конце исследования в контрольной группе выжило 0% (0/4) мышей, в то время как, напротив, выжило 100% (4/4) животных с полным ответом. Эти результаты показывают, что в подгруппе мышей суррогатное МАТ2 к OX40/CD137 может индуцировать полную регрессию опухоли и формирование защитной иммунологической памяти против повторной стимуляции клетками CT26.
Пример 24 - Дозозависимый фармакодинамический ответ МАТ2 к OX40/CD137 мыши в модели опухоли CT26 на мышах
Взаимосвязь между уровнями дозы, частотой дозирования и периферическим фармакодинамическим ответом суррогатного МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3) оценивали с использованием модели сингенной колоректальной опухоли CT26 на мышах. Сравнивали однократные внутрибрюшинные (в/б) инъекции FS20m-232-91AA/Lob12.3 при различных уровнях дозы 1, 3, 10 или 30 мг/кг или три в/б инъекции FS20m-232-91AA/Lob12.3 при 1 мг/кг, вводимые один раз каждые 2 дня (Q2D). Фармакодинамический ответ FS20m-232-91AA/Lob12.3, в частности, эффект суррогатного МАТ2 на циркулирующие Т-клетки, оценивали с помощью анализа методом проточной цитометрии подгрупп иммунных клеток в крови, как описано в Примере 18.
Придерживаясь того же протокола, как и описанный в Примере 17, самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая вводили путем подкожной инъекции клетки карциномы толстой кишки CT26 в левый бок каждого животного. Через 10 дней после инокуляции опухолевых клеток опухоли измеряли и животных без разившейся опухоли исключали из исследования. Оставшихся мышей рандомизировали на шесть групп лечения по шесть животных на группу.
Антитело для контроля изотипа (G1AA/4420) и FS20m-232-91AA/Lob12.3 фильтровали и разводили в ФСБ перед инъекцией. Каждому животному внутрибрюшинно вводили разведенное антитело в объеме 200 мкл на введение с получением конечной дозы 30 мг/кг G1AA/4420 или 1, 3, 10 или 30 мг/кг FS20m-232-91AA/Lob12.3 на введение для мыши массой 20 г. Животные получили либо однократную в/б инъекцию G1AA/4420 (при 30 мг/кг), либо FS20m-232-91AA/Lob12.3 (при 1, 3, 10 или 30 мг/кг), либо в общей сложности 3 дозы FS20m-232-91AA/Lob12.3 (при 1 мг/кг на дозу), вводимые один раз каждые два дня (Q2D), начиная со дня 10 после инокуляции опухоли. Объемы опухолей определяли с помощью измерений штангенциркулем, как описано в Примере 17. Животных исключали из исследования через шесть дней после начала дозирования (16 дней после инокуляции клеток).
Кровь собирали в пробирки, содержащие ЭДТА, путем сердечной пункции. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (Miltenyi Biotech, №130-094-183) в соответствии с инструкциями производителя. Клетки окрашивали для анализа методом проточной цитометрии с использованием реагентов CD4-BUV395 (клон RM4-5), CD8-BUV737 (клон 53-6.7), CD44-BV510 (клон IM7) и CD3e-BV786 (клон 145-2C11), все поставлены BD Bioscience); CD69-FITC (клон H1.2F3), NKp46-PE (клон 29A1.4), CD45-Alexa700 и фиксируемого красителя для оценки жизнеспособности 780, все поставлены eBioscience; и CD62L-BV421 (клон MEL-14), поставленный Biolegend, в присутствии блока Fc (eBioscience, номер по каталогу 14-0161-86). Затем клетки фиксировали и пермеабилизировали в течение ночи с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. Клетки повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Gzmb, антителом к Ki67 и антителом к Foxp3 (Gzmb-AF647 (клон GB11), поставленный Biolegend, и Ki67-PE-Cy7 (клон SolA15) и Foxp3-PerCP-Cy5.5 (клон FJK-16s), оба поставлены eBioscience) и инкубировали в течение 30 минут в темноте при комнатной температуре. Затем клетки дважды промывали буфером для пермеабилизации и повторно суспендировали в ФСБ плюс 0,5% БСА. После этого клетки анализировали в проточном цитометре BD Fortessa. Анализ данных выполняли с использованием программного обеспечения FlowJo, Excel и GraphPad Prism 7.
Частоты Ki67+ CD8+ (от общего числа CD8+) и Ki67+ CD4+ (от общего числа CD4+) пролиферирующих Т-клеток в периферической крови, через шесть дней после введения первой дозы, определяли с помощью анализа методом проточной цитометрии. Статистически достоверное увеличение частот пролиферирующих Ki67+ CD4+ Т-клеток наблюдали при однократных дозах 1 и 10 мг/кг FS20m-232-91AA/Lob12.3 по сравнению с контролем изотипа. Статистически достоверное увеличение частот пролиферирующих Ki67+ CD8+ Т-клеток наблюдали при однократных дозах 1, 3 и 10 мг/кг FS20m-232-91AA/Lob12.3 по сравнению с контролем изотипа.
Пролиферирующие Ki67+ CD8+ Т-клетки, как правило, достигали максимума при уровне однократной дозы 1 мг/кг, в то время как пролиферирующие Ki67+ CD4+ Т-клетки, как правило, достигали максимума при уровнях дозы 1 и 10 мг/кг. Повышение уровня дозы до 30 мг/кг не привело к значительному эффекту на Ki67+ CD8+ и Ki67+ CD4+ Т-клетки по сравнению с контролем изотипа. Следует отметить, что при любом из уровней дозы не было обнаружено явных клинических наблюдений или потери массы тела.
Сравнение группы многократного дозирования (FS20m-232-91AA/Lob12.3 при 1 мг/кг Q2D, три дозы) и группы однократной дозы 1 мг/кг не выявило статистической достоверности в уровнях Ki67+ CD8+ и Ki67+ CD4+ T-клеток (непарный критерий Манна-Уитни, p = 0,4848 и p = 0,0931, соответственно). Эти данные свидетельствуют о том, что многократное дозирование, по меньшей мере в течение шестидневного периода, оцениваемого в этом исследовании, не обеспечило дополнительный эффект на периферическую фармакодинамическую модуляцию Ki67+.
В соответствии с результатами Примера 18 этот эксперимент показывает, что суррогатное МАТ2 к OX40/CD137 оказывает эффект на циркулирующие Т-клетки, значительно увеличивая частоту пролиферирующих (Ki67+) CD8+ Т-клеток при уровнях дозы от 1 мг/кг до 10 мг/кг и пролиферирующих (Ki67+) CD4+ Т-клеток при уровнях дозы 1 мг/кг и 10 мг/кг.
Пример 25 - Эффект истощения CD4 Т-клеток на фармакодинамический ответ МАТ2 к OX40/CD137 мыши в модели опухоли CT26 на мышах
Ранее было показано, что у мышей комбинация костимулирующих антител, нацеленных на CD137 и OX40, синергически усиливает клональное размножение специфичных CD8+ Т-клеток, по сравнению с любым агентом по отдельности, после введения стафилококкового энтеротоксина А (Lee et al., 2004). В отношении механизма действия Lee et al. продемонстрировали, что CD4 T-клетки играют роль в запуске усиленного ответа специфичных CD8+ T-клеток. Чтобы протестировать, требуются ли CD4 Т-клетки хозяина для активации и пролиферации периферических CD8+ Т-клеток в ответ на лечение суррогатным МАТ2 к OX40/CD137, или вносят ли они вклад в эти процессы, использовали модель опухоли CT26 на мышах и антитело, истощающее CD4 Т-клетки мыши.
Придерживаясь того же протокола, как и описанный в Примере 17, самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г вводили путем подкожной инъекции в левый бок каждого животного клетки карциномы толстой кишки CT26. На седьмой день животных рандомизировали в группы лечения, по пять животных на группу на каждую временную точку.
Антитела анализировали и проверяли на наличие примесей, как описано ранее. Антитело для контроля изотипа (G1/4420) и суррогатное МАТ2 к OX40/CD137 (FS20m-232-91AA/Lob12.3) разводили до конечной концентрации 0,1 мг/мл в ФСБ. Антитело к CD4 мыши (GK1.5; BioXCell, номер по каталогу BE0003-1) разводили до конечной концентрации 1 мг/мл в ФСБ. Каждое животное получило разведенное антитело в объеме 200 мкл на введение с получением конечной дозы либо 1 мг/кг (G1/4420 или FS20m-232-91AA/Lob12.3), либо 10 мг/кг (GK1.5) для мыши массой 20 г. G1/4420 и FS20m-232-91AA/Lob12.3 вводили животным путем внутрибрюшинных (в/б) инъекций в дни 10, 12 и 14 после инокуляции клеток. В/б инъекции GK1.5 вводили в дни 8, 9, 11, 13 и 15.
Животных исключали из исследования на день 16 после инокуляции клеток, и ткани собирали для анализа методом проточной цитометрии. Кровь собирали в пробирки, содержащие ЭДТА, путем сердечной пункции. Придерживаясь того же протокола, как и описанный в Примере 19, красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (Miltenyi Biotech, №130-094-183) в соответствии с инструкциями производителя, и опухоли диссоциировали, используя набор для диссоциации опухолей, мышь (Miltenyi Biotech, 130-096-730), и диссоциатор gentleMACS (Miltenyi Biotech) в соответствии с инструкциями производителя. Полученную суспензию опухолевых клеток пропускали через сито, используя сито для клеток 70 мкм (Corning, номер по каталогу 352350), промывали и повторно суспендировали в ФСБ. Суспензию клеток из селезенок готовили путем проталкивания селезенок через сито для клеток 70 мкм (Corning), лизиса красных клеток крови путем инкубации в буфере для лизиса красных клеток крови (Milteny Biotech), промывки оставшихся спленоцитов и повторного суспендирования их в ФСБ.
Вначале клетки окрашивали реагентами CD4-E450 (клон GK1.5), CD69-PE-Cy5 (клон H1.2F3), CD3-PE-Cy7 (клон 145-2C11), CD8-APC (клон 53-6.7) и фиксируемым красителем для оценки жизнеспособности 780, все поставленные eBioscience; и CD45-V500 (клон 30-F11), поставленный BD Bioscience, в присутствии блока Fc (eBioscience, номер по каталогу 14-0161-86). Затем клетки фиксировали и пермеабилизировали с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. Клетки повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 (Ki67-FITC (клон SolA15) и Foxp3-PE (клон FJK-16s), оба поставлены eBioscience) в присутствии блока Fc (все 1:100) и инкубировали в течение 30 минут в темноте при 4°C. После этого клетки промывали один раз буфером для пермеабилизации и повторно суспендировали в 200 мкл ФСБ. Клетки анализировали на цитометре BD FACSCanto II. Анализ данных выполняли с использованием программного обеспечения FlowJo, Excel и GraphPad Prism. Попарное сравнение между группами лечения выполняли с использованием двустороннего критерия Манна-Уитни в программном обеспечении GraphPad Prism.
Обработка FS20m-232-91AA/Lob12.3 по отдельности индуцировала статистически достоверное увеличение доли активированных CD69+ и пролиферирующих Ki67+ CD8+ Т-клеток в крови и селезенке, а также пролиферирующих Ki67+ CD8+ Т-клеток в опухоли по сравнению с животными, получившими контроль изотипа.
Комбинирование FS20m-232-91AA/Lob12.3 с антителом GK1.5, истощающим CD4+ T-клетки, также привело к статистически достоверному увеличению пролиферирующих Ki67+ CD8+ T-клеток в крови по сравнению с контролем изотипа, но это увеличение было значительно ниже, чем то, которое наблюдали у животных, получавших FS20m-232-91AA/Lob12.3 в виде отдельного агента. Не было обнаружено статистически достоверных различий в уровнях пролиферирующих CD8+ Т-клеток в селезенке и опухолевых тканях после обработки FS20m-232-91AA/Lob12.3 по отдельности по сравнению с обработкой FS20m-232-91AA/Lob12.3 плюс антитело GK1.5, истощающее CD4+ Т-клетки.
Индуцированное FS20m-232-91AA/Lob12.3 увеличение активированных CD69+ CD8+ Т-клеток в крови ингибировалось антителом GK1.5, поскольку не было обнаружено статистически достоверных различий между группой контроля изотипа и группой FS20m-232-91AA/Lob12.3 плюс истощение CD4 (медиана 1,6% и 2,33% от общего количества CD8 Т-клеток, соответственно). Сравнение группы FS20m-232-91AA/Lob12.3 в виде отдельного агента и группы FS20m-232-91AA/Lob12.3 плюс истощение CD4 показало, что частота активированных CD8+ Т-клеток была значительно уменьшена в селезенке (медианная частота 29,3% в сравнении с 6,45% без истощения и с ним, соответственно) и в опухоли (медианная частота 86,4% в сравнении с 66,5%, без истощения и с ним, соответственно).
В соответствии с предыдущими результатами, описанными в Примере 18, суррогатное МАТ2 к OX40/CD137 увеличивало частоту активированных (CD69+) и пролиферирующих (Ki67+) CD8 Т-клеток, и результаты настоящего исследования показывают, что истощение CD4+ Т-клеток оказывало отрицательный эффект на периферический фармакодинамический ответ, опосредуемый МАТ2 к OX40/CD137. Более того, данные свидетельствуют о потенциальном взаимодействии CD4+ и CD8+ T-клеток в качестве посредников активности МАТ2 к OX40/CD137 in vivo, и что CD4+ T-клетки могут требоваться для оптимальной совместной стимуляции CD8+ T-клеточного иммунитета in vivo при применении МАТ2 к OX40/CD137.
Пример 26 - Функциональная активность МАТ2 к OX40/CD137 в анализе на клетках яванской макаки и фармакодинамический ответ на МАТ2 к OX40/CD137 и его переносимость у яванских макак
26.1 Функциональная активность МАТ2 к OX40/CD137 в анализе на клетках яванской макаки
Анализ первичных МКПК, аналогичный анализу первичных Т-клеток, описанному в Примере 13, но с использованием МКПК вместо выделенных активированных Т-клеток, выполняли, чтобы установить относительную эффективность МАТ2 FS20-22-49AA/FS30-10-16 против белков человека в отношении эндогенно экспрессируемых рецепторов человека и яванской макаки. В общих чертах, МКПК яванской макаки или человека выделяли и стимулировали антителом к CD3 в виде покрытия в присутствии повышающихся концентраций МАТ2 FS20-22-49AA/FS30-10-16 или контроля изотипа в течение трех (яванская макака) или четырех (человек) дней, при этом высвобождение IL-2 служило мерой активации Т-клеток.
Было обнаружено, что функциональная активность МАТ2 в отношении МКПК яванской макаки (средняя ЭК50 = 0,28±0,15 нМ) была сходна с активностью, наблюдаемой в эквивалентном анализе для человека (средняя ЭК50 = 0,26±0,1 нМ; IL-2). Таким образом, яванские макаки считаются фармакологически целесообразным видом для исследований токсичности МАТ2.
26.2 Переносимость МАТ2 к OX40/CD137 и фармакодинамический ответ на него у яванских макак
Предварительное исследование по определению диапазона доз проводили для оценки переносимости МАТ2 к OX40/CD137 человека FS20-22-49AA/FS30-10-16 и для оценки потенциальных фармакодинамических изменений долей основных популяций лейкоцитов, а также индукции пролиферации и активации подгрупп специфичных Т-клеток в ответ на FS20-22-49AA/FS30-10-16 у яванских макак.
В общих чертах, МАТ2 FS20-22-49AA/FS30-10-16 вводили яванским макакам путем внутривенной инфузии в виде однократной дозы или в виде повторных доз. Стандартные токсикологические параметры, такие как масса тела, потребление пищи, клинические наблюдения, гематология и биохимический состав крови, оценивали на протяжении всего исследования для оценки переносимости.
МАТ2 FS20-22-49AA/FS30-10-16 хорошо переносилось до 30 мг/кг при еженедельном дозировании, как было определено на основании результатов клинической биохимии и гистопатологии.
В соответствии с результатами исследования по оценке эффекта МАТ2 к OX40/CD137 мыши на циркулирующие Т-клетки в модели сингенной опухоли CT26 на мышах (Пример 18), связанное с лекарственным средством увеличение пролиферации и активации клеток наблюдали в отношении CD4+ и CD8+ Т-клеток центральной памяти и эффекторных CD4+ и CD8+ Т-клеток памяти, а также NK-клеток, как было измерено на основании повышенной экспрессии Ki67 и, в некоторой степени, CD69.
В совокупности эти результаты убедительно указывают, что МАТ2 FS20-22-49AA/FS30-10-16 против белков человека обладает сильной фармакологической активностью in vivo у яванских макак и хорошо переносится до 30 мг/кг. Кроме того, фармакодинамические данные, полученные в этом исследовании, соответствуют данным, наблюдаемым для суррогатного МАТ2 к OX40/CD137 в фармакодинамическом исследовании на мышах, описанном в Примере 18, и обеспечивают дополнительные доказательства ожидаемой противоопухолевой эффективности и переносимости МАТ2, связывающего OX40 и CD137, такого как МАТ2 FS20-22-49AA/FS30-10-16, у пациентов-людей, страдающих раком.
Пример 27 - Фармакология МАТ2 к OX40/CD137 в печени у мышей BALB/c
В доклинических моделях на мышах было показано, что агонистические антитела к CD137 индуцируют повышенную инфильтрацию Т-клеток в печень, а одно агонистическое антитело к CD137 индуцировало токсичность для печени в дозах выше 1 мг/кг в клинических условиях (Dubrot et al., 2010; Segal et al., 2017). Таким образом, эффекты МАТ2 к OX40/CD137 мыши исследовали на мышах BALB/c, чтобы определить, имеется ли повышенная инфильтрация Т-клеток в печень по сравнению с агонистическими антителами к CD137. Кровь и ткани селезенки использовали в качестве контролей и исследовали уровни Т-клеток, а также пролиферацию и активацию Т-клеток. Подробная информация о протестированных антителах представлена в Таблице 38.
Таблица 38: Подробная информация о протестированных антителах и МАТ2
Способность МАТ2 (FS20m-232-91AA/Lob12.3) увеличивать, активировать и индуцировать пролиферацию Т-клеток в крови, селезенке и печени сравнивали с контрольными МАТ в виде отдельных агентов (G1/OX86, G1/Lob12.3, G1/3H3 и G1/4420) и контрольной комбинацией (G1/OX86 и G1/Lob12.3). Самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая обеспечивали период покоя в течение одной недели перед началом исследования. Всем животным вводили микрочип и присваивали уникальный идентификатор. В каждой когорте было по 6 мышей.
В течение 24 часов до инъекции антитела анализировали с помощью профилирования SEC-HPLC и проверяли на наличие примесей. Антитела разводили до конечной концентрации 1 мг/мл в ФСБ и вводили путем внутрибрюшинной (в/б) инъекции при 200 мкл/мышь с получением конечной дозы 10 мг/кг для мышей массой 20 г. Инъекции выполняли в дни 0, 2 и 4 (одна доза каждые два дня) исследования. Через семь и четырнадцать дней после введения третьей дозы по 3 мыши на группу умерщвляли, селезенку и печень выделяли путем диссекции, а кровь собирали путем сердечной пункции.
Печени и селезенки диссоциировали с использованием наборов для диссоциации Miltenyi (печень - Miltenyi, 130-105-807; селезенка - Miltenyi, 130-095-926) в соответствии с инструкциями производителя. Полученную суспензию клеток пропускали через сито, используя сито для клеток 70 мкм (Corning, номер по каталогу 352350), центрифугировали (10 минут при 1500 об./мин), промывали один раз в ФСБ и повторно суспендировали в 5 мл ФСБ.
Кровь собирали в пробирки, содержащие ЭДТА, путем сердечной пункции. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (eBioscience, номер по каталогу 00-4300-54) в соответствии с инструкциями производителя.
Клетки, выделенные из опухолей и крови, окрашивали для проточной цитометрии с использованием панели антител и реагентов, подробно описанных в Примере 19 (Краситель 1). Клетки промывали в ФСБ и затем инкубировали с 100 мкл смеси антител 1 (все, за исключением антитела к Ki67 и антитела к Foxp3) в течение 30 минут при 4°C. Затем клетки промывали ФСБ и после этого фиксировали и пермеабилизировали с помощью набора для окрашивания eBioscience Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. В общих чертах, 200 мкл раствора для фиксации добавляли в каждую лунку и оставляли на ночь в темноте при 4°C. Клетки затем промывали в 200 мкл буфера для пермеабилизации. После этого клетки снова центрифугировали и повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 в присутствии блока Fc (все в разведении 1:100) и инкубировали в течение 30 минут в темноте при 4°С. Затем клетки промывали один раз буфером для пермеабилизации и повторно суспендировали в 200 мкл ФСБ. После этого клетки анализировали в проточном цитометре BD FACSCanto II.
Данные анализировали с помощью программного обеспечения FlowJoX, Excel и GraphPad Prism. Статистический анализ для сравнения групп выполняли с использованием однофакторного дисперсионного анализа, а затем критерия Тьюки для множественных сравнений каждой пары с использованием программного пакета GraphPad Prism. Данные выражали как процент от исходной популяции.
Результаты показали, что независимое от перекрестного связывания агонистическое антитело к CD137 (G1/3H3) индуцировало повышенные уровни Т-клеток в печени, селезенке и крови как на 7, так и на 14 день, и что эти Т-клетки показали повышенные уровни пролиферации и активации, по сравнению с антителом для контроля изотипа (G1/4420). Зависимое от перекрестного связывания агонистическое антитело к CD137 (G1/Lob12.3) не показало значительного повышения уровней, пролиферации или активации Т-клеток в печени, селезенке или крови. Агонистическое антитело к OX40 (G1/OX86) не индуцировало повышенные уровни Т-клеток ни в одной из тканей, но показало повышенные уровни пролиферации Т-клеток в печени, селезенке и крови на 7 день исследования, которые вернулись к уровням для контроля изотипа ко дню 14. Комбинация агонистического антитела к OX40 и зависимого от перекрестного связывания агонистического антитела к CD137 (G1/OX86 и G1/Lob12.3) показала повышение уровней инфильтрации Т-клеток в печени на день 7, увеличение пролиферации Т-клеток в печени на день 7 и в селезенке (недостоверно) и крови в дни 7 и 14, и увеличение активации Т-клеток в печени и крови на день 14 и в селезенке в дни 7 и 14. МАТ2 к OX40/CD137 показало повышение уровней инфильтрации Т-клеток в печени (недостоверно) и уровней Т-клеток в крови на день 7, которые вернулись к уровням для контроля изотипа ко дню 14, и увеличение пролиферации Т-клеток в печени (недостоверно), селезенке и крови на день 7, которые также вернулись к уровням для контроля изотипа ко дню 14. Эти результаты указывают, что только независимый от перекрестного связывания агонист CD137 (G1/3H3) индуцировал повышенную и устойчивую инфильтрацию, пролиферацию и активацию Т-клеток в печени, а также в селезенке и крови, и свидетельствуют о том, что молекулы антител, нацеленных на OX40/CD137, согласно настоящему изобретению могут иметь более низкий риск гепатотоксичности по сравнению с независимыми от перекрестного связывания агонистическими антителами к CD137. Эти результаты указывают на возможность ассоциации между независимым от перекрестного связывания агонистическим действием на CD137, индуцированным клоном 3H3, и повышенным воспалением печени, вызванным Т-клетками, наблюдаемым для этого независимого от перекрестного связывания клона в данном исследовании.
Пример 28 - Сравнение антител МАТ2 к OX40/CD137, содержащих различные клоны Fab к CD137, в модели сингенной опухоли CT26
В Примере 27 наблюдали, что независимое от перекрестного связывания агонистическое антитело к CD137 (G1/3H3) индуцирует повышенные и устойчивые уровни инфильтрации, пролиферации и активации Т-клеток у мышей BALB/c. Чтобы протестировать, обладает ли эта повышенная активность благоприятной противоопухолевой активностью в случае МАТ2 к OX40/CD137, модель сингенной опухоли CT26 использовали для сравнения активности двух различных МАТ2 к OX40/CD137 мыши in vivo, одно из которых - агонист CD137, который представляет собой зависимый от перекрестного связывания клон Lob1.23, а другое - агонист CD137, который представляет собой независимый от перекрестного связывания клон 3H3. Ранее было показано, что модель сингенной опухоли CT26 чувствительна к агонистическим антителам к OX40 и к CD137, а инфильтрирующие опухоль лимфоциты (TIL), выделенные из опухолей CT26, экспрессируют как OX40, так и CD137.
28.1 Противоопухолевая активность антител МАТ2 к OX40/CD137, содержащих различные клоны Fab к CD137, в модели сингенной опухоли CT26
Противоопухолевую активность двух различных МАТ2 к OX40/CD137, FS20m-232-91AA/3H3 (SEQ ID NO: 169 и 167) и FS20m-232-91AA/Lob12.3 (см. Таблицу 38), определяли in vivo в модели сингенной опухоли CT26 на мышах и сравнивали с активностью антитела для контроля изотипа (G1/4420; см. Таблицу 38). Кроме того, уровни пролиферации и активации Т-клеток, индуцированные в крови двумя МАТ2 к OX40/CD137, анализировали и сравнивали с уровнями, индуцированными антителом для контроля изотипа. Самкам мышей BALB/c (Charles River) в возрасте 8-10 недель и массой приблизительно 20 г каждая обеспечивали период покоя в течение одной недели перед началом исследования. Всем животным вводили микрочип и присваивали уникальный идентификатор. В каждой когорте было по 10 мышей. Линию клеток карциномы толстой кишки CT26 (ATCC, CRL-2638) первоначально размножали, сохраняли, а затем подвергали предварительному скринингу в IDEXX Bioresearch для обнаружения патогенов с использованием протокола IMPACT I, и было показано, что линия не содержит патогенов. Клетки CT26 (приблизительно 3-5×106) размораживали из хранилища при -150°C и добавляли к 20 мл DMEM (Gibco, 61965-026) с 10% ФСТ (Gibco, 10270-106) в колбе для тканевой культуры Т175. Мышей анестезировали с использованием изофлурана (Abbott Laboratories), и каждое животное получило 1×106 клеток, введенных путем подкожной инъекции в левый бок. На день 10 после инокуляции опухолевых клеток у мышей контролировали состояние здоровья и рост опухоли, сортировали и рандомизировали в исследуемые когорты. Любых мышей, которые не имели опухолей в этот момент, исключали из исследования.
В течение 24 часов до инъекции антитела анализировали с помощью профилирования SEC-HPLC и проверяли на наличие примесей. Антитела разводили до конечной концентрации 0,1 мг/мл в ФСБ и вводили путем внутрибрюшинной (в/б) инъекции при 200 мкл/мышь с получением конечной дозы 1 мг/кг для мыши массой 20 г. Инъекции выполняли в дни 10, 12 и 14 (одна доза каждые два дня) после инокуляции опухоли. Животных подвергали скринингу для определения состояния здоровья под анестезией три раза в неделю слепым методом, в это время проводили точные измерения опухолей. Объемы опухолей определяли с помощью измерений штангенциркулем (как описано в Примере 17). Исследование прекращали через 35 дней после инокуляции опухолевых клеток, и животных исключали из исследования, когда были достигнуты гуманные конечные точки, основанные на объеме опухоли и состоянии. Группы лечения, протестированные молекулы, дозы и схема дозирования обобщены в Таблице 39. Объемы опухолей на день 21 подвергали статистическому анализу с помощью двустороннего дисперсионного анализа и критерия Тьюки для множественных сравнений с использованием программного обеспечения GraphPad Prism. Статистический анализ выживаемости выполняли с помощью лог-рангового критерия (Мантела-Кокса) с использованием программного обеспечения GraphPad Prism.
Таблица 39: Обобщенные данные по группам лечения и протестированным молекулам
Как показано на Фигуре 15A и 15B, лечение одним из двух антител МАТ2 к OX40/CD137 отсрочило рост опухоли и увеличило выживаемость по сравнению с лечением антителом для контроля изотипа. Не было обнаружено различий в росте опухоли или выживаемости между мышами, получавшими МАТ2 FS20m-232-91AA/3H3 и МАТ2 FS20m-232-91AA/Lob12.3, соответственно. Эти данные свидетельствуют о том, что, несмотря на повышенную активацию и пролиферацию Т-клеток, наблюдаемую для независимого от перекрестного связывания агониста CD137 (G1/3H3), как описано в Примере 27, не наблюдали повышенную противоопухолевую активность МАТ2 к OX40/CD137, в котором клон Fab к CD137 представляет собой независимый от перекрестного связывания клон 3H3 (FS20m-232-91AA/3H3), по сравнению с МАТ2 к OX40/CD137, в котором клон Fab к CD137 представляет собой зависимый от перекрестного связывания клон Lob12.3 (FS20m-232-91AA/Lob12.3).
28.2 Оценка периферического фармакодинамического ответа МАТ2 к OX40/CD137, содержащего различные клоны Fab к CD137, в модели сингенной опухоли CT26
В расширении исследования, описанного выше в Примере 28.1, через пять дней после введения третьей дозы (т. е. на день 19 после инокуляции опухоли) кровь из хвостовой вены пяти мышей собирали в пробирки, содержащие ЭДТА. Красные клетки крови некоагулированной крови дважды лизировали в буфере для лизиса красных клеток крови (eBioscience, номер по каталогу 00-4300-54) в соответствии с инструкциями производителя.
Клетки, выделенные из крови, окрашивали для проточной цитометрии с использованием панели антител и реагентов, подробно описанных в Примере 19 (Краситель 1). Клетки промывали в ФСБ и затем инкубировали с 100 мкл смеси антител 1 (все, кроме антитела к Ki67 и антитела к Foxp3) в течение 30 минут при 4°C. После этого клетки промывали ФСБ, а затем фиксировали и пермеабилизировали с помощью набора для окрашивания Foxp3 (eBioscience, номер по каталогу 00-5523-00) в соответствии с инструкциями производителя. В общих чертах, 200 мкл раствора для фиксации добавляли в каждую лунку и оставляли на ночь в темноте при 4°C. Затем клетки промывали в 200 мкл буфера для пермеабилизации. Клетки затем снова центрифугировали и повторно суспендировали в 100 мкл буфера для пермеабилизации с антителом к Ki67 и антителом к Foxp3 в присутствии блока Fc (все в разведении 1:100) и инкубировали в течение 30 минут в темноте при 4°C. После этого клетки промывали один раз буфером для пермеабилизации и повторно суспендировали в 200 мкл ФСБ. Затем клетки анализировали в цитометре BD FACS CantoII.
Данные анализировали с использованием программного обеспечения FlowJoX, Excel и GraphPad Prism. Статистический анализ для сравнения групп выполняли с использованием однофакторного дисперсионного анализа, а затем критерия Тьюки для множественных сравнений каждой пары с использованием программного пакета GraphPad Prism.
FS20m-232-91AA/3H3 индуцировало статистически достоверное повышение уровней Т-клеток в крови по сравнению как с антителом для контроля изотипа (G1/4420), так и с FS20m-232-91AA/Lob12.3. Повышенные уровни Т-клеток, индуцированные FS20m-232-91AA/3H3, сопровождались статистически достоверным уменьшением относительного процента CD4+ Т-клеток и статистически достоверным увеличением относительного процента CD8+ Т-клеток по сравнению с относительными процентами этих типов клеток, наблюдаемыми для контроля изотипа G1/4420 и МАТ2 FS20m-232-91AA/Lob12.3. Оба антитела МАТ2 к OX40/CD137 также индуцировали пролиферацию CD4+ и CD8+ Т-клеток, но уровни, индуцированные FS20m-232-91AA/3H3, были значительно выше, чем уровни, индуцированные МАТ2 FS20m-232-91AA/Lob12.3. МАТ2 FS20m-232-91AA/3H3 индуцировало повышенные уровни активированных CD4+ Т-клеток по сравнению с контролем изотипа. Изменения уровней активированных Т-клеток и активированных CD8+ Т-клеток у мышей, получавших FS20m-232-91AA/Lob12.3 или FS20m-232-91AA/3H3, по сравнению с когортой, получавшей контроль изотипа, были незначительными и статистически недостоверными, как и изменения уровней активированных CD4+ Т-клеток у мышей, получавших FS20m-232-91AA/Lob12.3. Эти результаты свидетельствуют о том, что независимый от перекрестного связывания клон агониста CD137 3H3 активен применительно к МАТ2 к OX40/CD137 и способен индуцировать повышенные уровни и пролиферацию Т-клеток по сравнению с зависимым от перекрестного связывания клоном агониста CD137 Lob12.3 применительно к МАТ2 к OX40/CD137, и, следовательно, согласуются с повышенными уровнями и пролиферацией Т-клеток, индуцированными клоном 3H3 в виде моноклонального антитела (МАТ), как наблюдали в исследовании на мышах BALB/c, описанном в Примере 27.
Вместе с данными о противоопухолевой активности эти результаты свидетельствуют об отсутствии дополнительного преимущества с точки зрения противоопухолевого ответа от повышенных уровней и пролиферации Т-клеток, индуцированных независимым от перекрестного связывания агонистом CD137 применительно к МАТ2 к OX40/CD137. Эти результаты, вместе с результатами Примера 27, в котором повышенное воспаление печени, вызванное Т-клетками, наблюдали для независимого от перекрестного связывания агонистического действия на CD137, индуцированного клоном 3H3, свидетельствуют о том, что использование МАТ2 к OX40/CD137, агонистическое действие на CD137 которого зависит от связывания с OX40, может обеспечить безопасный и эффективный путь стимуляции иммунной системы для борьбы с раком.
Перечень последовательностей
Аминокислотные последовательности CDR МАТ FS30-10-16 (IMGT)
CDR1 VH - GFTFSSYD (SEQ ID NO: 1)
CDR2 VH- IDPTGSKT (SEQ ID NO: 2)
CDR3 VH - ARDLLVYGFDY (SEQ ID NO: 3)
CDR1 VL - QSVSSSY (SEQ ID NO: 4)
CDR2 VL - GAS (SEQ ID NO: 5)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотные последовательности CDR МАТ FS30-10-16 (Kabat)
CDR1 VH- SYDMS (SEQ ID NO: 7)
CDR2 VH - DIDPTGSKTDYADSVKG (SEQ ID NO: 8)
CDR3 VH - DLLVYGFDY (SEQ ID NO: 9)
CDR1 VL - RASQSVSSSYLA (SEQ ID NO: 10)
CDR2 VL - GASSRAT (SEQ ID NO: 11)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотная последовательность вариабельного домена тяжелой цепи МАТ FS30-10-16 (SEQ ID NO: 12)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EVQLLESGGGLVQPGGSLRLSCAAS GFTFSSYD MSWVRQAPGKGLEWVSD IDPTGSKT DYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC ARDLLVYGFDY WGQGTLVTVSS
Последовательность нуклеиновой кислоты вариабельного домена тяжелой цепи МАТ FS30-10-16 (SEQ ID NO: 13)
GAAGTTCAGCTGCTGGAATCTGGCGGCGGATTGGTTCAACCTGGCGGCTCTCTGAGACTGTCTTGTGCCGCTTCCGGCTTCACCTTCTCCAGCTACGACATGTCCTGGGTCCGACAGGCTCCTGGCAAAGGACTGGAATGGGTGTCCGACATCGACCCCACCGGCTCTAAGACCGACTACGCCGATTCTGTGAAGGGCAGATTCACCATCAGCCGGGACAACTCCAAGAACACCCTGTACCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCAGAGATCTGCTGGTGTACGGCTTCGACTATTGGGGCCAGGGCACACTGGTCACCGTGTCCTCT
Аминокислотная последовательность вариабельного домена легкой цепи МАТ FS30-10-16 (SEQ ID NO: 14)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EIVLTQSPGTLSLSPGERATLSCRAS QSVSSSY LAWYQQKPGQAPRLLIY GAS SRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC QQSYSYPVT FGQGTKVEIK
Последовательность нуклеиновой кислоты вариабельного домена легкой цепи МАТ FS30-10-16 (SEQ ID NO: 15)
GAGATCGTGCTGACCCAGTCTCCTGGCACACTGTCACTGTCTCCAGGCGAGAGAGCTACCCTGTCCTGTAGAGCCTCTCAGTCCGTGTCCTCCTCTTACCTGGCCTGGTATCAGCAGAAGCCTGGACAGGCTCCCCGGCTGTTGATCTACGGCGCTTCTTCTAGAGCCACAGGCATCCCTGACCGGTTCTCCGGATCTGGCTCTGGCACCGATTTCACCCTGACCATCTCTCGGCTGGAACCCGAGGATTTCGCCGTGTACTACTGCCAGCAGTCCTACAGCTACCCCGTGACCTTTGGCCAGGGCACCAAGGTGGAAATCAAG
Аминокислотные последовательности CDR МАТ FS30-10-3 (IMGT)
CDR1 VH - GFTFSSYD (SEQ ID NO: 1)
CDR2 VH - IDPTGSKT (SEQ ID NO: 2)
CDR3 VH - ARDLNVYGFDY (SEQ ID NO: 16)
CDR1 VL - QSVSSSY (SEQ ID NO: 4)
CDR2 VL - GAS (SEQ ID NO: 5)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотные последовательности CDR МАТ FS30-10-3 (Kabat)
CDR1 VH - SYDMS (SEQ ID NO: 7)
CDR2 VH - DIDPTGSKTDYADSVKG (SEQ ID NO: 8)
CDR3 VH - DLNVYGFDY (SEQ ID NO: 17)
CDR1 VL - RASQSVSSSYLA (SEQ ID NO: 10)
CDR2 VL - GASSRAT (SEQ ID NO: 11)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотная последовательность вариабельного домена тяжелой цепи МАТ FS30-10-3 (SEQ ID NO: 18)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EVQLLESGGGLVQPGGSLRLSCAAS GFTFSSYD MSWVRQAPGKGLEWVSD IDPTGSKT DYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC ARDLNVYGFDY WGQGTLVTVSS
Последовательность нуклеиновой кислоты вариабельного домена тяжелой цепи МАТ FS30-10-3 (SEQ ID NO: 19)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCAATGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGT
Аминокислотная последовательность вариабельного домена легкой цепи МАТ FS30-10-3 (SEQ ID NO: 14)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EIVLTQSPGTLSLSPGERATLSCRAS QSVSSSY LAWYQQKPGQAPRLLIY GAS SRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC QQSYSYPVT FGQGTKVEIK
Последовательность нуклеиновой кислоты вариабельного домена легкой цепи МАТ FS30-10-3 (SEQ ID NO: 20)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATCTTATTCTTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотные последовательности CDR МАТ FS30-10-12 (IMGT)
CDR1 VH - GFTFSSYD (SEQ ID NO: 1)
CDR2 VH - IDPTGSKT (SEQ ID NO: 2)
CDR3 VH - ARDLTVYGFDY (SEQ ID NO: 21)
CDR1 VL - QSVSSSY (SEQ ID NO: 4)
CDR2 VL - GAS (SEQ ID NO: 5)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотные последовательности CDR МАТ FS30-10-12 (Kabat)
CDR1 VH - SYDMS (SEQ ID NO: 7)
CDR2 VH - DIDPTGSKTDYADSVKG (SEQ ID NO: 8)
CDR3 VH - DLTVYGFDY (SEQ ID NO: 22)
CDR1 VL - RASQSVSSSYLA (SEQ ID NO: 10)
CDR2 VL - GASSRAT (SEQ ID NO: 11)
CDR3 VL - QQSYSYPVT (SEQ ID NO: 6)
Аминокислотная последовательность вариабельного домена тяжелой цепи МАТ FS30-10-12 (SEQ ID NO: 23)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EVQLLESGGGLVQPGGSLRLSCAAS GFTFSSYD MSWVRQAPGKGLEWVSD IDPTGSKT DYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC ARDLTVYGFDY WGQGTLVTVSS
Последовательность нуклеиновой кислоты вариабельного домена тяжелой цепи МАТ FS30-10-12 (SEQ ID NO: 24)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCACGGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGT
Аминокислотная последовательность вариабельного домена легкой цепи МАТ FS30-10-12 (SEQ ID NO: 14)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EIVLTQSPGTLSLSPGERATLSCRAS QSVSSSY LAWYQQKPGQAPRLLIY GAS SRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC QQSYSYPVT FGQGTKVEIK
Последовательность нуклеиновой кислоты вариабельного домена легкой цепи МАТ FS30-10-12 (SEQ ID NO: 20)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATCTTATTCTTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотные последовательности CDR МАТ FS30-35-14 (IMGT)
CDR1 VH - GFTFSAYN (SEQ ID NO: 25)
CDR2 VH - ISPYGGAT (SEQ ID NO: 26)
CDR3 VH - ARNLYELSAYSYGADY (SEQ ID NO: 27)
CDR1 VL - QSVSSSY (SEQ ID NO: 4)
CDR2 VL - GAS (SEQ ID NO: 5)
CDR3 VL - QQYYYSSPIT (SEQ ID NO: 28)
Аминокислотные последовательности CDR МАТ FS30-35-14 (Kabat)
CDR1 VH - AYNIH (SEQ ID NO: 29)
CDR2 VH - DISPYGGATNYADSVKG (SEQ ID NO: 30)
CDR3 VH - NLYELSAYSYGADY (SEQ ID NO: 31)
CDR1 VL - RASQSVSSSYLA (SEQ ID NO: 10)
CDR2 VL - GASSRAT (SEQ ID NO: 11)
CDR3 VL - QQYYYSSPIT (SEQ ID NO: 28)
Аминокислотная последовательность вариабельного домена тяжелой цепи МАТ FS30-35-14 (SEQ ID NO: 170)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EVQLLESGGGLVQPGGSLRLSCAAS GFTFSAYN IHWVRQAPGKGLEWVSD ISPYGGAT NYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC ARNLYELSAYSYGADY WGQGTLVTVSS
Последовательность нуклеиновой кислоты вариабельного домена тяжелой цепи МАТ FS30-35-14 (SEQ ID NO: 171)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTGCCTATAATATCCATTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTTCTCCGTATGGTGGCGCGACCAACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAAACCTCTACGAGTTGAGCGCTTACTCTTACGGGGCGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGTCG
Аминокислотная последовательность вариабельного домена легкой цепи МАТ FS30-35-14 (SEQ ID NO: 172)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EIVLTQSPGTLSLSPGERATLSCRAS QSVSSSY LAWYQQKPGQAPRLLIY GAS SRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC QQYYYSSPIT FGQGTKVEIK
Последовательность нуклеиновой кислоты вариабельного домена легкой цепи МАТ FS30-35-14 (SEQ ID NO: 32)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATATTATTATTCTTCTCCTATCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотные последовательности CDR МАТ FS30-5-37 (IMGT)
CDR1 VH - GFTFSSYA (SEQ ID NO: 33)
CDR2 VH - ISGSGGST (SEQ ID NO: 34)
CDR3 VH - ARSYDKYWGSSIYSGLDY (SEQ ID NO: 35)
CDR1 VL - QSVSSSY (SEQ ID NO: 4)
CDR2 VL - GAS (SEQ ID NO: 5)
CDR3 VL - QQYYSYYPVT (SEQ ID NO: 36)
Аминокислотные последовательности CDR МАТ FS30-5-37 (Kabat)
CDR1 VH - SYAMS (SEQ ID NO: 37)
CDR2 VH - AISGSGGSTYYADSVKG (SEQ ID NO: 38)
CDR3 VH - SYDKYWGSSIYSGLDY (SEQ ID NO: 39)
CDR1 VL - RASQSVSSSYLA (SEQ ID NO: 10)
CDR2 VL - GASSRAT (SEQ ID NO: 11)
CDR3 VL - QQYYSYYPVT (SEQ ID NO: 36)
Аминокислотная последовательность вариабельного домена тяжелой цепи МАТ FS30-5-37 (SEQ ID NO: 40)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EVQLLESGGGLVQPGGSLRLNCAAS GFTFSSYA MSWVRQAPGKGLEWVSA ISGSGGST YYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYC ARSYDKYWGSSIYSGLDY WGQGTLVTVSS
Последовательность нуклеиновой кислоты вариабельного домена тяжелой цепи МАТ FS30-5-37 (SEQ ID NO: 41)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAATTGCGCGGCCAGTGGCTTTACCTTCAGTAGCTATGCCATGAGCTGGGTGCGTCAGGCGCCGGGCAAAGGTCTGGAATGGGTTAGCGCGATTAGCGGTAGTGGCGGTAGCACGTACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGATCTTACGACAAATACTGGGGTTCTTCTATTTACTCTGGCTTGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGT
Аминокислотная последовательность вариабельного домена легкой цепи МАТ FS30-5-37 (SEQ ID NO: 42)
CDR согласно нумерации IMGT (жирный курсив), CDR согласно нумерации Kabat (подчеркнутый курсив)
EIVLTQSPGTLSLSPGERATLSCRAS QSVSSSY LAWYQQKPGQAPRLLIY GAS SRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYC QQYYSYYPVT FGQGTKVEIK
Последовательность нуклеиновой кислоты вариабельного домена легкой цепи МАТ FS30-5-37 (SEQ ID NO: 43)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATATTATTCTTATTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAA
Аминокислотные последовательности структурных петель домена CH3 WT
Петля WT AB - RDELTKNQ (SEQ ID NO: 44)
Петля WT CD - SNGQPENNY (SEQ ID NO: 45)
Петля WT EF - DKSRWQQGNV (SEQ ID NO: 46)
Аминокислотная последовательность домена CH3 WT (SEQ ID NO: 47)
Петли AB, CD и EF подчеркнуты
GQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTP
PVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность домена CH2 (SEQ ID NO: 48)
APELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAK
Аминокислотная последовательность домена CH2 с мутацией LALA (SEQ ID NO: 49)
Мутация LALA подчеркнута
APEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAK
Аминокислотная последовательность домена CH2 с мутацией LALA и мутацией P114A (SEQ ID NO: 50)
Мутация LALA подчеркнута; P114A мутация выделена жирным шрифтом и подчеркнута
APEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALAAPIEKTISKAK
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-22-49
Первая последовательность FS20-22-49 - YWDQE (SEQ ID NO: 51)
Вторая последовательность FS20-22-49 - DEQFA (SEQ ID NO: 52)
Третья последовательность FS20-22-49 - QYRWNPADY (SEQ ID NO: 53)
Аминокислотная последовательность домена CH3 Fcab FS20-22-49 (SEQ ID NO: 54)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTT
PPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-49 (SEQ ID NO: 55)
GGCCAGCCTAGGGAACCCCAGGTTTACACCTTGCCTCCAAGCCGGGACGAGTACTGGGATCAAGAGGTGTCCCTGACCTGCCTCGTGAAGGGCTTCTACCCTTCCGATATCGCCGTGGAATGGGAGAGCAATGGCGACGAGCAGTTCGCCTACAAGACAACCCCTCCTGTGCTGGACTCCGACGGCTCATTCTTTCTGTACTCCAAGCTGACAGTGGACCAGTACAGATGGAACCCCGCCGACTACTTCTCTTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACACAGAAGTCCCTGTCTCTGTCCCCTGGC
Аминокислотные последовательности последовательностей петель AB, CD и EF домена CH3 Fcab FS20-22-49
Петля AB FS20-22-49 - RDEYWDQE (SEQ ID NO: 56)
Петля CD FS20-22-49 - SNGDEQFAY (SEQ ID NO: 57)
Петля EF FS20-22-49 - DQYRWNPADY (SEQ ID NO: 58)
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-22-38
Первая последовательность FS20-22-38 - YWDQE (SEQ ID NO: 51)
Вторая последовательность FS20-22-38 - AEKYQ (SEQ ID NO: 59)
Третья последовательность FS20-22-38 - QYRWNPGDY (SEQ ID NO: 60)
Аминокислотная последовательность домена CH3 Fcab FS20-22-38 (SEQ ID NO: 61)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGAEKYQYKTT
PPVLDSDGSFFLYSKLTVDQYRWNPGDYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-38 (SEQ ID NO: 62)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGCAGAAAAATACCAGTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAACCCAGGCGACTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-22-41
Первая последовательность FS20-22-41 - YWDQE (SEQ ID NO: 51)
Вторая последовательность FS20-22-41 - DEQFA (SEQ ID NO: 52)
Третья последовательность FS20-22-41 - QYRWNPGDY (SEQ ID NO: 60)
Аминокислотная последовательность домена CH3 Fcab FS20-22-41 (SEQ ID NO: 63)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTT
PPVLDSDGSFFLYSKLTVDQYRWNPGDYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-41 (SEQ ID NO: 64)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAACCCAGGCGACTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGA
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-22-47
Первая последовательность FS20-22-47 - YWDQE (SEQ ID NO: 51)
Вторая последовательность FS20-22-47 - DEQFA (SEQ ID NO: 52)
Третья последовательность FS20-22-47 - QYRWSPGDY (SEQ ID NO: 65)
Аминокислотная последовательность домена CH3 Fcab FS20-22-47 (SEQ ID NO: 66)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTT
PPVLDSDGSFFLYSKLTVDQYRWSPGDYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-47 (SEQ ID NO: 67)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAGTCCGGGTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-22-85
Первая последовательность FS20-22-85 - YWDQE (SEQ ID NO: 51)
Вторая последовательность FS20-22-85 - DEQFA (SEQ ID NO: 52)
Третья последовательность FS20-22-85 - QYRWNPFDD (SEQ ID NO: 68)
Аминокислотная последовательность домена CH3 Fcab FS20-22-85 (SEQ ID NO: 69)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTT
PPVLDSDGSFFLYSKLTLDQYRWNPFDDFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-85 (SEQ ID NO: 70)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCTTGGATCAGTATAGGTGGAATCCGTTTGATGATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-58
Первая последовательность FS20-31-58 - YYSGE (SEQ ID NO: 71)
Вторая последовательность FS20-31-58 - QPEND (SEQ ID NO: 72)
Третья последовательность FS20-31-58 - PYWRWGSPRT (SEQ ID NO: 73)
Аминокислотная последовательность домена CH3 Fcab FS20-31-58 (SEQ ID NO: 74)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYYSGEVSLTCLVKGFYPSDIAVEWESNGQPENDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGSPRTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-58 (SEQ ID NO: 75)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTACTCTGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACGACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCTTATTGGAGGTGGGGTAGTCCGCGTACTTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-66
Первая последовательность FS20-31-66 - YYSGE (SEQ ID NO: 71)
Вторая последовательность FS20-31-66 - QPEND (SEQ ID NO: 72)
Третья последовательность FS20-31-66 - PYWRWGVPRT (SEQ ID NO: 76)
Аминокислотная последовательность домена CH3 Fcab FS20-31-66 (SEQ ID NO: 77)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEYYSGEVSLTCLVKGFYPSDIAVEWESNGQPENDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGVPRTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-66 (SEQ ID NO: 78)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTACTCTGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACGACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCGTATTGGAGGTGGGGTGTTCCGCGTACTTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-94 Fcab
Первая последовательность FS20-31-94 - WEHGE (SEQ ID NO: 79)
Вторая последовательность FS20-31-94 - IREHD (SEQ ID NO: 80)
Третья последовательность FS20-31-94 - PYWRWGGPGT (SEQ ID NO: 81)
Аминокислотная последовательность домена CH3 Fcab FS20-31-94 Fcab (SEQ ID NO: 82)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEWEHGEVSLTCLVKGFYPSDIAVEWESNGIREHDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGGPGTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-94 Fcab (SEQ ID NO: 83)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTGGGAACATGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGATCAGAGAACATGATTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCATATTGGAGGTGGGGCGGCCCAGGCACCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-102
Первая последовательность FS20-31-102 - WASGE (SEQ ID NO: 84)
Вторая последовательность FS20-31-102 - QPEVD (SEQ ID NO: 85)
Третья последовательность FS20-31-102 - PYWRWGVPRT (SEQ ID NO: 76)
Аминокислотная последовательность домена CH3 Fcab FS20-31-102 (SEQ ID NO: 86)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEWASGEVSLTCLVKGFYPSDIAVEWESNGQPEVDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGVPRTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-102 (SEQ ID NO: 87)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTGGGCATCTGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCAGAAGTTGATTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCGTATTGGAGGTGGGGTGTTCCGCGTACTTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-108
Первая последовательность FS20-31-108 - WASGE (SEQ ID NO: 84)
Вторая последовательность FS20-31-108 - EKEID (SEQ ID NO: 88)
Третья последовательность FS20-31-108 - PYWRWGAKRT (SEQ ID NO: 89)
Аминокислотная последовательность домена CH3 Fcab FS20-31-108 (SEQ ID NO: 90)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEWASGEVSLTCLVKGFYPSDIAVEWESNGEKEIDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGAKRTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-108 (SEQ ID NO: 91)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTGGGCATCTGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGAAAAAGAAATCGATTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCGTATTGGAGGTGGGGTGCTAAGCGTACTTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Аминокислотные последовательности последовательностей структурных петель домена CH3 Fcab FS20-31-115
Первая последовательность FS20-31-115 - WASGE (SEQ ID NO: 84)
Вторая последовательность FS20-31-115 - EQEFD (SEQ ID NO: 92)
Третья последовательность FS20-31-115 - PYWRWGAKRT (SEQ ID NO: 89)
Аминокислотная последовательность домена CH3 Fcab FS20-31-115 (SEQ ID NO: 93)
Первая, вторая и третья последовательности подчеркнуты
GQPREPQVYTLPPSRDEWASGEVSLTCLVKGFYPSDIAVEWESNGEQEFDYKTTP
PVLDSDGSFFLYSKLTVPYWRWGAKRTFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS20-31-115 (SEQ ID NO: 94)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTGGGCATCTGGTGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGAACAGGAATTCGATTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGCCGTATTGGAGGTGGGGTGCTAAGCGTACTTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/FS30-10-16 с мутацией LALA (SEQ ID NO: 95)
Вариабельный домен (курсив), мутация LALA (подчеркнутый жирный шрифт)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLLVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи FS20-22-49AA/FS30-10-16 с мутацией LALA (SEQ ID NO: 96)
GAAGTTCAGCTGCTGGAATCTGGCGGCGGATTGGTTCAACCTGGCGGCTCTCTGAGACTGTCTTGTGCCGCTTCCGGCTTCACCTTCTCCAGCTACGACATGTCCTGGGTCCGACAGGCTCCTGGCAAAGGACTGGAATGGGTGTCCGACATCGACCCCACCGGCTCTAAGACCGACTACGCCGATTCTGTGAAGGGCAGATTCACCATCAGCCGGGACAACTCCAAGAACACCCTGTACCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCAGAGATCTGCTGGTGTACGGCTTCGACTATTGGGGCCAGGGCACACTGGTCACCGTGTCCTCTGCTTCTACCAAGGGACCCAGCGTGTTCCCTCTGGCTCCTTCCAGCAAGTCTACCTCTGGCGGAACAGCTGCTCTGGGCTGCCTGGTCAAGGACTACTTTCCTGAGCCTGTGACCGTGTCTTGGAACTCTGGCGCTCTGACATCTGGCGTGCACACCTTTCCAGCAGTGCTGCAGTCCTCCGGCCTGTACTCTCTGTCCTCTGTCGTGACCGTGCCTTCCAGCTCTCTGGGAACCCAGACCTACATCTGCAATGTGAACCACAAGCCTTCCAACACCAAGGTGGACAAGAAGGTGGAACCCAAGTCCTGCGACAAGACCCACACCTGTCCTCCATGTCCTGCTCCAGAAGCTGCTGGCGGCCCTTCCGTGTTTCTGTTCCCTCCAAAGCCTAAGGACACCCTGATGATCTCTCGGACCCCTGAAGTGACCTGCGTGGTGGTGGATGTGTCTCACGAGGACCCAGAAGTGAAGTTCAATTGGTACGTGGACGGCGTGGAAGTGCACAACGCCAAGACCAAGCCTAGAGAGGAACAGTACAACTCCACCTACAGAGTGGTGTCCGTGCTGACCGTGCTGCACCAGGATTGGCTGAACGGCAAAGAGTACAAGTGCAAGGTGTCCAACAAGGCCCTGCCTGCTCCTATCGAAAAGACCATCTCCAAGGCCAAGGGCCAGCCTAGGGAACCCCAGGTTTACACCTTGCCTCCAAGCCGGGACGAGTACTGGGATCAAGAGGTGTCCCTGACCTGCCTCGTGAAGGGCTTCTACCCTTCCGATATCGCCGTGGAATGGGAGAGCAATGGCGACGAGCAGTTCGCCTACAAGACAACCCCTCCTGTGCTGGACTCCGACGGCTCATTCTTTCTGTACTCCAAGCTGACAGTGGACCAGTACAGATGGAACCCCGCCGACTACTTCTCTTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACACAGAAGTCCCTGTCTCTGTCCCCTGGC
Аминокислотная последовательность легкой цепи FS30-10-16 (SEQ ID NO: 97)
Вариабельный домен (курсив)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQSYSYPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи FS30-10-16 (SEQ ID NO: 98)
GAGATCGTGCTGACCCAGTCTCCTGGCACACTGTCACTGTCTCCAGGCGAGAGAGCTACCCTGTCCTGTAGAGCCTCTCAGTCCGTGTCCTCCTCTTACCTGGCCTGGTATCAGCAGAAGCCTGGACAGGCTCCCCGGCTGTTGATCTACGGCGCTTCTTCTAGAGCCACAGGCATCCCTGACCGGTTCTCCGGATCTGGCTCTGGCACCGATTTCACCCTGACCATCTCTCGGCTGGAACCCGAGGATTTCGCCGTGTACTACTGCCAGCAGTCCTACAGCTACCCCGTGACCTTTGGCCAGGGCACCAAGGTGGAAATCAAGCGTACGGTGGCCGCTCCCAGCGTGTTCATCTTCCCCCCAAGCGACGAGCAGCTGAAGAGCGGCACCGCCAGCGTGGTGTGTCTGCTGAACAACTTCTACCCCAGGGAGGCCAAGGTGCAGTGGAAGGTGGACAACGCCCTGCAGAGCGGCAACAGCCAGGAGAGCGTCACCGAGCAGGACAGCAAGGACTCCACCTACAGCCTGAGCAGCACCCTGACCCTGAGCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGTGAGGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGAGCTTCAACAGGGGCGAGTGC
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/FS30-10-3 с мутацией LALA (SEQ ID NO: 99)
Вариабельный домен (курсив), мутация LALA (подчеркнутый жирный шрифт)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLNVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи FS20-22-49AA/FS30-10-3 с мутацией LALA (SEQ ID NO: 100)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCAATGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность легкой цепи FS30-10-3 (SEQ ID NO: 97)
Вариабельный домен (курсив)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQSYSYPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи FS30-10-3 (SEQ ID NO: 102)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATCTTATTCTTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACTGTGGCCGCTCCTAGCGTGTTCATTTTTCCGCCATCCGACGAGCAGCTCAAGTCCGGCACCGCCTCCGTGGTCTGCCTGCTCAACAACTTCTACCCTCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAATGCCCTGCAGTCCGGAAACTCGCAGGAAAGCGTGACTGAACAGGACTCCAAGGACTCCACCTATTCACTGTCCTCGACTCTGACCCTGAGCAAGGCGGATTACGAAAAGCACAAAGTGTACGCATGCGAAGTGACCCACCAGGGTCTTTCGTCCCCCGTGACCAAGAGCTTCAACAGAGGAGAGTGT
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/FS30-10-12 с мутацией LALA (SEQ ID NO: 103)
Вариабельный домен (курсив), мутация LALA (подчеркнутый жирный шрифт)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLTVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи FS20-22-49AA/FS30-10-12 с мутацией LALA (SEQ ID NO: 104)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCACGGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность легкой цепи FS30-10-12 (SEQ ID NO: 97)
Вариабельный домен (курсив)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQSYSYPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи FS30-10-12 (SEQ ID NO: 102)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATCTTATTCTTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACTGTGGCCGCTCCTAGCGTGTTCATTTTTCCGCCATCCGACGAGCAGCTCAAGTCCGGCACCGCCTCCGTGGTCTGCCTGCTCAACAACTTCTACCCTCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAATGCCCTGCAGTCCGGAAACTCGCAGGAAAGCGTGACTGAACAGGACTCCAAGGACTCCACCTATTCACTGTCCTCGACTCTGACCCTGAGCAAGGCGGATTACGAAAAGCACAAAGTGTACGCATGCGAAGTGACCCACCAGGGTCTTTCGTCCCCCGTGACCAAGAGCTTCAACAGAGGAGAGTGT
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/FS30-35-14 с мутацией LALA (SEQ ID NO: 105)
Вариабельный домен (курсив), мутация LALA (подчеркнутый жирный шрифт)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSAYNIHWVRQAPGKGLEWVSDISPYGGATNYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARNLYELSAYSYGADYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи FS20-22-49AA/FS30-35-14 с мутацией LALA (SEQ ID NO: 106)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTGCCTATAATATCCATTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTTCTCCGTATGGTGGCGCGACCAACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAAACCTCTACGAGTTGAGCGCTTACTCTTACGGGGCGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGTCGGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность легкой цепи FS30-35-14 (SEQ ID NO: 107)
Вариабельный домен (курсив)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYYYSSPITFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи FS30-35-14 (SEQ ID NO: 108)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATATTATTATTCTTCTCCTATCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACTGTGGCCGCTCCTAGCGTGTTCATTTTTCCGCCATCCGACGAGCAGCTCAAGTCCGGCACCGCCTCCGTGGTCTGCCTGCTCAACAACTTCTACCCTCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAATGCCCTGCAGTCCGGAAACTCGCAGGAAAGCGTGACTGAACAGGACTCCAAGGACTCCACCTATTCACTGTCCTCGACTCTGACCCTGAGCAAGGCGGATTACGAAAAGCACAAAGTGTACGCATGCGAAGTGACCCACCAGGGTCTTTCGTCCCCCGTGACCAAGAGCTTCAACAGAGGAGAGTGT
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/FS30-5-37 с мутацией LALA (SEQ ID NO: 109)
Вариабельный домен (курсив), мутация LALA (подчеркнутый жирный шрифт)
EVQLLESGGGLVQPGGSLRLNCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARSYDKYWGSSIYSGLDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи FS20-22-49AA/FS30-5-37 с мутацией LALA (SEQ ID NO: 110)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAATTGCGCGGCCAGTGGCTTTACCTTCAGTAGCTATGCCATGAGCTGGGTGCGTCAGGCGCCGGGCAAAGGTCTGGAATGGGTTAGCGCGATTAGCGGTAGTGGCGGTAGCACGTACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGATCTTACGACAAATACTGGGGTTCTTCTATTTACTCTGGCTTGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность легкой цепи FS30-5-37 (SEQ ID NO: 111)
Вариабельный домен (курсив)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQYYSYYPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи FS30-5-37 (SEQ ID NO: 112)
GAAATTGTGCTGACCCAGTCTCCGGGCACGTTATCTCTGAGCCCTGGTGAGCGCGCCACTCTGTCATGCCGGGCTTCTCAAAGTGTTAGCAGTAGCTACCTGGCGTGGTATCAGCAAAAACCGGGCCAGGCCCCGCGTCTGCTGATTTACGGTGCATCCAGCCGTGCCACCGGCATTCCAGATCGTTTTTCCGGTAGTGGTTCTGGGACGGACTTCACTCTGACAATCTCACGCCTGGAACCGGAGGATTTTGCGGTGTATTACTGCCAGCAATATTATTCTTATTATCCTGTCACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACTGTGGCCGCTCCTAGCGTGTTCATTTTTCCGCCATCCGACGAGCAGCTCAAGTCCGGCACCGCCTCCGTGGTCTGCCTGCTCAACAACTTCTACCCTCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAATGCCCTGCAGTCCGGAAACTCGCAGGAAAGCGTGACTGAACAGGACTCCAAGGACTCCACCTATTCACTGTCCTCGACTCTGACCCTGAGCAAGGCGGATTACGAAAAGCACAAAGTGTACGCATGCGAAGTGACCCACCAGGGTCTTTCGTCCCCCGTGACCAAGAGCTTCAACAGAGGAGAGTGT
Альтернативная последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-49 (SEQ ID NO: 113)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи МАТ к FITC G1AA/4420, содержащего мутацию LALA (SEQ ID NO: 114)
Положения CDR подчеркнуты. Положение мутации LALA выделено жирным шрифтом.
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи МАТ к FITC G1/4420 без мутации LALA (SEQ ID NO: 115)
Положения CDR подчеркнуты.
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ 4420 (SEQ ID NO: 116)
Домен VL (курсив)
DVVMTQTPLSLPVSLGDQASISCRSSQSLVHSNGNTYLRWYLQKPGQSPKVLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYFCSQSTHVPWTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи антитела G1AA/HelD1.3 с мутацией LALA (SEQ ID NO: 117)
QVQLQESGPGLVRPSQTLSLTCTVSGSTFSGYGVNWVRQPPGRGLEWIGMIWGDGNTDYNSALKSRVTMLVDTSKNQFSLRLSSVTAADTAVYYCARERDYRLDYWGQGSLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность легкой цепи МАТ HelD1.3 (SEQ ID NO: 118)
Домен VL (курсив)
DIQMTQSPASLSASVGETVTITCRASGNIHNYLAWYQQKQGKSPQLLVYNAKTLADGVPSRFSGSGSGTQYSLKINSLQPEDFGSYYCQHFWSTPRTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи G1/MOR7480.1 (SEQ ID NO: 119)
Домен VH (курсив)
EVQLVQSGAEVKKPGESLRISCKGSGYSFSTYWISWVRQMPGKGLEWMGKIYPGDSYTNYSPSFQGQVTISADKSISTAYLQWSSLKASDTAMYYCARGYGIFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ G1/MOR7480.1, G1AA/MOR7480.1 и G2/MOR7480.1 (SEQ ID NO: 120)
Домен VL (курсив)
SYELTQPPSVSVSPGQTASITCSGDNIGDQYAHWYQQKPGQSPVLVIYQDKNRPSGIPERFSGSNSGNTATLTISGTQAMDEADYYCATYTGFGSLAVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Аминокислотная последовательность тяжелой цепи G1/20H4.9 (SEQ ID NO: 121)
Домен VH (курсив)
QVQLQQWGAGLLKPSETLSLTCAVYGGSFSGYYWSWIRQSPEKGLEWIGEINHGGYVTYNPSLESRVTISVDTSKNQFSLKLSSVTAADTAVYYCARDYGPGNYDWYFDLWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ G1/20H4.9 и G1AA/20H4.9 (SEQ ID NO: 122)
Домен VL (курсив)
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQRSNWPPALTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи FS20-22-49AA/4420 (с мутацией LALA) (SEQ ID NO: 123)
Домен VH (курсив); мутация LALA (жирный шрифт и подчеркнуто)
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи G2/MOR7480.1 (SEQ ID NO: 124)
Домен VH (курсив)
EVQLVQSGAEVKKPGESLRISCKGSGYSFSTYWISWVRQMPGKGLEWMGKIYPGDSYTNYSPSFQGQVTISADKSISTAYLQWSSLKASDTAMYYCARGYGIFDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSNFGTQTYTCNVDHKPSNTKVDKTVERKCCVECPPCPAPPVAGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTFRVVSVLTVVHQDWLNGKEYKCKVSNKGLPAPIEKTISKTKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPMLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи G1AA/MOR7480.1 (SEQ ID NO: 125)
Домен VH (курсив) LALA (жирный шрифт и подчеркнуто)
EVQLVQSGAEVKKPGESLRISCKGSGYSFSTYWISWVRQMPGKGLEWMGKIYPGDSYTNYSPSFQGQVTISADKSISTAYLQWSSLKASDTAMYYCARGYGIFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность CD137 человека (SEQ ID NO: 126)
Внеклеточный домен (курсив); трансмембранный и внутриклеточный домены (жирный шрифт)
LQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQ IISFFLALTSTALLFLLFFLTLRFSVVKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
Аминокислотная последовательность внеклеточного домена CD137 человека (SEQ ID NO: 127)
LQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQ
Аминокислотная последовательность CD137 яванских макак (SEQ ID NO: 128)
Внеклеточный домен (курсив); трансмембранный и внутриклеточный домены (жирный шрифт)
LQDLCSNCPAGTFCDNNRSQICSPCPPNSFSSAGGQRTCDICRQCKGVFKTRKECSSTSNAECDCISGYHCLGAECSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSATPPAPAREPGHSPQ IIFFLALTSTVVLFLLFFLVLRFSVVKRSRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
Аминокислотная последовательность внеклеточного домена CD137 яванской макаки (SEQ ID NO: 129)
LQDLCSNCPAGTFCDNNRSQICSPCPPNSFSSAGGQRTCDICRQCKGVFKTRKECSSTSNAECDCISGYHCLGAECSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSATPPAPAREPGHSPQ
Аминокислотная последовательность внеклеточного домена OX40 человека (SEQ ID NO: 130)
LHCVGDTYPSNDRCCHECRPGNGMVSRCSRSQNTVCRPCGPGFYNDVVSSKPCKPCTWCNLRSGSERKQLCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPATQPQETQGPPARPITVQPTEAWPRTSQGPSTRPVEVPGGRA
Аминокислотная последовательность внеклеточного домена OX40 яванской макаки (SEQ ID NO: 131)
LHCVGDTYPSNDRCCQECRPGNGMVSRCNRSQNTVCRPCGPGFYNDVVSAKPCKACTWCNLRSGSERKQPCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPPTQPQETQGPPARPTTVQPTEAWPRTSQRPSTRPVEVPRGPA
Аминокислотная последовательность DO11.10-hOX40 и рецептора OX40 человека (SEQ ID NO: 132)
LHCVGDTYPSNDRCCHECRPGNGMVSRCSRSQNTVCRPCGPGFYNDVVSSKPCKPCTWCNLRSGSERKQLCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPATQPQETQGPPARPITVQPTEAWPRTSQGPSTRPVEVPGGRAVAAILGLGLVLGLLGPLAILLALYLLRRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKI
Аминокислотная последовательность DO11.10-mOX40 и рецептора OX40 мыши (SEQ ID NO: 133)
VTARRLNCVKHTYPSGHKCCRECQPGHGMVSRCDHTRDTLCHPCETGFYNEAVNYDTCKQCTQCNHRSGSELKQNCTPTQDTVCRCRPGTQPRQDSGYKLGVDCVPCPPGHFSPGNNQACKPWTNCTLSGKQTRHPASDSLDAVCEDRSLLATLLWETQRPTFRPTTVQSTTVWPRTSELPSPPTLVTPEGPAFAVLLGLGLGLLAPLTVLLALYLLRKAWRLPNTPKPCWGNSFRTPIQEEHTDAHFTLAKI
Аминокислотная последовательность DO11.10-cOX40 и рецептора OX40 яванской макаки (SEQ ID NO: 134)
KLHCVGDTYPSNDRCCQECRPGNGMVSRCNRSQNTVCRPCGPGFYNDVVSAKPCKACTWCNLRSGSERKQPCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPPTQPQETQGPPARPTTVQPTEAWPRTSQRPSTRPVEVPRGPAVAAILGLGLALGLLGPLAMLLALLLLRRDQRLPPDAPKAPGGGSFRTPIQEEQADAHSALAKI
Аминокислотная последовательность OX40 человека-mFc (SEQ ID NO: 135)
Лидерная последовательность IL-2 (подчеркнуто), внеклеточный домен OX40 (курсив), домен Fc IgG2a мыши (жирный шрифт)
MYRMQLLSCIALSLALVTNS LHCVGDTYPSNDRCCHECRPGNGMVSRCSRSQNTVCRPCGPGFYNDVVSSKPCKPCTWCNLRSGSERKQLCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPATQPQETQGPPARPITVQPTEAWPRTSQGPSTRPVEVPGGRAVA GSPRGPTIKPCPPCKCPAPNLLGGPSVFIFPPKIKDVLMISLSPIVTCVVVDVSEDDPDVQISWFVNNVEVHTAQTQTHREDYNSTLRVVSALPIQHQDWMSGKEFKCKVNNKDLPAPIERTISKPKGSVRAPQVYVLPPPEEEMTKKQVTLTCMVTDFMPEDIYVEWTNNGKTELNYKNTEPVLDSDGSYFMYSKLRVEKKNWVERNSYSCSVVHEGLHNHHTTKSFSRTPGK
Аминокислотная последовательность OX40 мыши-mFc (SEQ ID NO: 136)
Лидерная последовательность IL-2 (подчеркнуто), внеклеточный домен OX40 (курсив), домен Fc IgG2a мыши (жирный шрифт)
MYRMQLLSCIALSLALVTNS VTARRLNCVKHTYPSGHKCCRECQPGHGMVSRCDHTRDTLCHPCETGFYNEAVNYDTCKQCTQCNHRSGSELKQNCTPTQDTVCRCRPGTQPRQDSGYKLGVDCVPCPPGHFSPGNNQACKPWTNCTLSGKQTRHPASDSLDAVCEDRSLLATLLWETQRPTFRPTTVQSTTVWPRTSELPSPPTLVTPEGPA GSPRGPTIKPCPPCKCPAPNLLGGPSVFIFPPKIKDVLMISLSPIVTCVVVDVSEDDPDVQISWFVNNVEVHTAQTQTHREDYNSTLRVVSALPIQHQDWMSGKEFKCKVNNKDLPAPIERTISKPKGSVRAPQVYVLPPPEEEMTKKQVTLTCMVTDFMPEDIYVEWTNNGKTELNYKNTEPVLDSDGSYFMYSKLRVEKKNWVERNSYSCSVVHEGLHNHHTTKSFSRTPGK
Аминокислотная последовательность OX40 яванской макаки-mFc (SEQ ID NO: 137)
Лидерная последовательность IL-2 (подчеркнуто), внеклеточный домен OX40 (курсив), домен Fc IgG2a мыши (жирный шрифт)
MYRMQLLSCIALSLALVTNS LHCVGDTYPSNDRCCQECRPGNGMVSRCNRSQNTVCRPCGPGFYNDVVSAKPCKACTWCNLRSGSERKQPCTATQDTVCRCRAGTQPLDSYKPGVDCAPCPPGHFSPGDNQACKPWTNCTLAGKHTLQPASNSSDAICEDRDPPPTQPQETQGPPARPTTVQPTEAWPRTSQRPSTRPVEVPRGPAVAAI GSPRGPTIKPCPPCKCPAPNLLGGPSVFIFPPKIKDVLMISLSPIVTCVVVDVSEDDPDVQISWFVNNVEVHTAQTQTHREDYNSTLRVVSALPIQHQDWMSGKEFKCKVNNKDLPAPIERTISKPKGSVRAPQVYVLPPPEEEMTKKQVTLTCMVTDFMPEDIYVEWTNNGKTELNYKNTEPVLDSDGSYFMYSKLRVEKKNWVERNSYSCSVVHEGLHNHHTTKSFSRTPGK
Аминокислотная последовательность последовательности CD137 человека для применения с рекомбинантным антигеном CD137-mFc-Avi (SEQ ID NO: 138)
SLQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQ
Аминокислотная последовательность последовательности CD137 яванской макаки для применения с рекомбинантными антигенами CD137-mFc-Avi и CD137-Avi-His (SEQ ID NO: 139)
SLQDLCSNCPAGTFCDNNRSQICSPCPPNSFSSAGGQRTCDICRQCKGVFKTRKECSSTSNAECDCISGYHCLGAECSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSATPPAPAREPGHSPQ
Аминокислотная последовательность последовательности CD137 мыши для применения с рекомбинантным антигеном CD137-mFc-Avi (SEQ ID NO: 140)
AVQNSCDNCQPGTFCRKYNPVCKSCPPSTFSSIGGQPNCNICRVCAGYFRFKKFCSSTHNAECECIEGFHCLGPQCTRCEKDCRPGQELTKQGCKTCSLGTFNDQNGTGVCRPWTNCSLDGRSVLKTGTTEKDVVCGPPVVSFSPSTTISVTPEGGPGGHSLQVL
Аминокислотная последовательность mFc-Avi для применения с рекомбинантными антигенами CD137-mFc-Avis (SEQ ID NO: 141)
Домен Fc мыши (курсив) метка Avi (жирный шрифт)
PRGPTIKPCPPCKCPAPNLEGGPSVFIFPPKIKDVLMISLSPIVTCVVVDVSEDDPDVQISWFVNNVEVHTAQTQTHREDYNSTLRVVSALPIQHQDWMSGKAFACAVNNKDLPAPIERTISKPKGSVRAPQVYVLPPPEEEMTKKQVTLTCMVTDFMPEDIYVEWTNNGKTELNYKNTEPVLDSDGSYFMYSKLRVEKKNWVERNSYSCSVVHEGLHNHHTTKSFSRTPGK GGGLNDIFEAQKIEWHE
Аминокислотная последовательность усеченной шарнирной области Fcab (SEQ ID NO: 101)
TCPPCP
Альтернативная последовательность нуклеиновой кислоты домена CH3 Fcab FS20-22-49 (SEQ ID NO: 143)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49/FS30-5-37 (без LALA) (SEQ ID NO: 144)
EVQLLESGGGLVQPGGSLRLNCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARSYDKYWGSSIYSGLDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность ДНК тяжелой цепи FS20-22-49/FS30-5-37 (без LALA) (SEQ ID NO: 145)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAATTGCGCGGCCAGTGGCTTTACCTTCAGTAGCTATGCCATGAGCTGGGTGCGTCAGGCGCCGGGCAAAGGTCTGGAATGGGTTAGCGCGATTAGCGGTAGTGGCGGTAGCACGTACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGATCTTACGACAAATACTGGGGTTCTTCTATTTACTCTGGCTTGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAACTGCTGGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49/FS30-10-3 (без LALA) (SEQ ID NO: 146)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLNVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность ДНК тяжелой цепи FS20-22-49/FS30-10-3 (без LALA) (SEQ ID NO: 147)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCAATGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAACTGCTGGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49/FS30-10-12 (без LALA) (SEQ ID NO: 148)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLTVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность ДНК тяжелой цепи FS20-22-49/FS30-10-12 (без LALA) (SEQ ID NO: 149)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCACGGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAACTGCTGGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49/FS30-10-16 (без LALA) (SEQ ID NO: 150)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLLVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность ДНК тяжелой цепи FS20-22-49/FS30-10-16 (без LALA) (SEQ ID NO: 151)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTAGTTACGATATGAGCTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTGATCCGACTGGTAGCAAGACCGACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAGACCTCTTGGTGTACGGGTTCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGAGTGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAACTGCTGGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи FS20-22-49/FS30-35-14 (без LALA) (SEQ ID NO: 152)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSAYNIHWVRQAPGKGLEWVSDISPYGGATNYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARNLYELSAYSYGADYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEYWDQEVSLTCLVKGFYPSDIAVEWESNGDEQFAYKTTPPVLDSDGSFFLYSKLTVDQYRWNPADYFSCSVMHEALHNHYTQKSLSLSPG
Последовательность ДНК тяжелой цепи FS20-22-49/FS30-35-14 (без LALA) (SEQ ID NO: 153)
GAAGTGCAACTGCTGGAGTCCGGTGGTGGTCTGGTACAGCCGGGTGGTTCTCTGCGTCTGAGTTGCGCGGCCAGTGGCTTTACCTTCAGTGCCTATAATATCCATTGGGTGCGTCAGGCTCCGGGCAAAGGTCTGGAATGGGTTAGCGATATTTCTCCGTATGGTGGCGCGACCAACTATGCGGATAGCGTGAAAGGCCGTTTTACCATTTCTCGCGACAACAGCAAGAACACGCTGTACCTGCAGATGAACTCACTGCGTGCCGAAGATACGGCCGTGTATTACTGTGCGAGAAACCTCTACGAGTTGAGCGCTTACTCTTACGGGGCGGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCGTCGGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAACTGCTGGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGTACTGGGACCAGGAAGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGGATGAACAGTTCGCATACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCAGTATAGGTGGAATCCTGCTGATTATTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Аминокислотная последовательность тяжелой цепи МАТ G1AA/FS30-10-16 (с LALA) (SEQ ID NO: 154)
EVQLLESGGGLVQPGGSLRLSCAASGFTFSSYDMSWVRQAPGKGLEWVSDIDPTGSKTDYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCARDLLVYGFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ G1AA/FS30-10-16 (SEQ ID NO: 97)
EIVLTQSPGTLSLSPGERATLSCRASQSVSSSYLAWYQQKPGQAPRLLIYGASSRATGIPDRFSGSGSGTDFTLTISRLEPEDFAVYYCQQSYSYPVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи МАТ G1AA/OX86 (с LALA) (SEQ ID NO: 155)
QVQLKESGPGLVQPSQTLSLTCTVSGFSLTGYNLHWVRQPPGKGLEWMGRMRYDGDTYYNSVLKSRLSISRDTSKNQVFLKMNSLQTDDTAIYYCTRDGRGDSFDYWGQGVMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ G1/OX86 и G1AA/OX86 (SEQ ID NO: 156)
DIVMTQGALPNPVPSGESASITCRSSQSLVYKDGQTYLNWFLQRPGQSPQLLTYWMSTRASGVSDRFSGSGSGTYFTLKISRVRAEDAGVYYCQQVREYPFTFGSGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи FS20m-232-91AA/4420 (с LALA) (SEQ ID NO: 157)
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELFDPMYYYNQVSLTCLVKGFYPSDIAVEWESNGEPLWDYKTTPPVLDSDGSFFLYSKLTVWRDRWEDGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность легкой цепи FS20m-232-91AA/4420 (SEQ ID NO: 116)
DVVMTQTPLSLPVSLGDQASISCRSSQSLVHSNGNTYLRWYLQKPGQSPKVLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYFCSQSTHVPWTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность CD137 человека-Avi-His (SEQ ID NO: 158)
Внеклеточный домен CD137 (жирный шрифт); метка Avi (курсив); метка His (подчеркнуто)
SLQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQGS GGGLNDIFEAQKIEWHE HHHHHH
Аминокислотная последовательность тяжелой цепи МАТ G1/OX86 (без LALA) (SEQ ID NO: 159)
QVQLKESGPGLVQPSQTLSLTCTVSGFSLTGYNLHWVRQPPGKGLEWMGRMRYDGDIYYNSVLKSRLSISRDTSKNQVFLKMNSLQTDDTAIYYCTRDGRGDSFDYWGQGVMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи МАТ к PD-1 G1AA/5C4 (SEQ ID NO: 160)
Вариабельный домен (жирный шрифт)
QVQLVESGGGVVQPGRSLRLDCKASGITFSNSGMHWVRQAPGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность легкой цепи МАТ к PD-1 G1AA/5C4 (SEQ ID NO: 161)
Вариабельный домен (жирный шрифт)
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQSSNWPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи МАТ к PD-L1 G1AA/S1 (SEQ ID NO: 162)
Вариабельный домен (жирный шрифт)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность легкой цепи МАТ к PD-L1 G1AA/S1 (SEQ ID NO: 163)
Вариабельный домен (жирный шрифт)
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLFTPPTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность CD137 мыши (SEQ ID NO: 164)
Внеклеточный домен (курсив); трансмембранный и внутриклеточный домены (жирный шрифт)
VQNSCDNCQPGTFCRKYNPVCKSCPPSTFSSIGGQPNCNICRVCAGYFRFKKFCSSTHNAECECIEGFHCLGPQCTRCEKDCRPGQELTKQGCKTCSLGTFNDQNGTGVCRPWTNCSLDGRSVLKTGTTEKDVVCGPPVVSFSPSTTISVTPEGGPGGHSLQVL TLFLALTSALLLALIFITLLFSVLKWIRKKFPHIFKQPFKKTTGAAQEEDACSCRCPQEEEGGGGGYEL
Аминокислотная последовательность тяжелой цепи МАТ G1AA/20H4.9 (SEQ ID NO: 165)
Домен VH (курсив)
QVQLQQWGAGLLKPSETLSLTCAVYGGSFSGYYWSWIRQSPEKGLEWIGEINHGGYVTYNPSLESRVTISVDTSKNQFSLKLSSVTAADTAVYYCARDYGPGNYDWYFDLWGRGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи МАТ G1AA/3H3 (SEQ ID NO: 166)
Домен VH (курсив)
EMQLVESGGGLVQPGRSMKLSCAGSGFTLSDYGVAWVRQAPKKGLEWVAYISYAGGTTYYRESVKGRFTISRDNAKSTLYLQMDSLRSEDTATYYCTIDGYGGYSGSHWYFDFWGPGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность легкой цепи МАТ G1AA/3H3 и G1/3H3 и МАТ2 FS20m-232-91AA/3H3 (SEQ ID NO: 167)
Домен VL (курсив)
DIQMTQSPSLLSASVGDRVTLNCRTSQNVYKNLAWYQQQLGEAPKLLIYNANSLQAGIPSRFSGSGSGTDFTLTISSLQPEDVATYFCQQYYSGNTFGAGTNLELKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Аминокислотная последовательность тяжелой цепи МАТ G1/3H3 (SEQ ID NO: 168)
Домен VH (курсив)
EMQLVESGGGLVQPGRSMKLSCAGSGFTLSDYGVAWVRQAPKKGLEWVAYISYAGGTTYYRESVKGRFTISRDNAKSTLYLQMDSLRSEDTATYYCTIDGYGGYSGSHWYFDFWGPGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность тяжелой цепи FS20m-232-91AA/3H3 (с LALA) (SEQ ID NO: 169)
Домен VH (курсив)
EMQLVESGGGLVQPGRSMKLSCAGSGFTLSDYGVAWVRQAPKKGLEWVAYISYAGGTTYYRESVKGRFTISRDNAKSTLYLQMDSLRSEDTATYYCTIDGYGGYSGSHWYFDFWGPGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELFDPMYYYNQVSLTCLVKGFYPSDIAVEWESNGEPLWDYKTTPPVLDSDGSFFLYSKLTVWRDRWEDGNVFSCSVMHEALHNHYTQKSLSLSPGK
Аминокислотная последовательность тяжелой цепи МАТ к OX40 G1AA/11D4 (SEQ ID NO: 173)
Домен VH (курсив)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYSMNWVRQAPGKGLEWVSYISSSSSTIDYADSVKGRFTISRDNAKNSLYLQMNSLRDEDTAVYYCARESGWYLFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELRFYQVSLTCLVKGFYPSDIAVEWESNGQPDIFPNGLNYKTTPPVLDSDGSFFLYSKLTVPYPSWLMGTRFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность тяжелой цепи МАТ к OX40 G1/11D4 (SEQ ID NO: 174)
Домен VH (курсив)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYSMNWVRQAPGKGLEWVSYISSSSSTIDYADSVKGRFTISRDNAKNSLYLQMNSLRDEDTAVYYCARESGWYLFDYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELRFYQVSLTCLVKGFYPSDIAVEWESNGQPDIFPNGLNYKTTPPVLDSDGSFFLYSKLTVPYPSWLMGTRFSCSVMHEALHNHYTQKSLSLSPG
Аминокислотная последовательность легкой цепи МАТ к OX40 G1AA/11D4 и G1/11D4 (SEQ ID NO: 175)
Домен VL (курсив)
DIQMTQSPSSLSASVGDRVTITCRASQGISSWLAWYQQKPEKAPKSLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYNSYPPTFGGGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Литературные источники
Все документы, упомянутые в настоящем описании, полностью включены в настоящую заявку посредством ссылки.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J. Mol. Biol. 215(3), 403-10 (1990).
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25(17), 3389-402 (1997).
Aspeslagh S, Postel-Vinay S, Rusakiewicz S, Soria JC, Zitvogel L, Marabelle A. Rationale for anti-OX40 cancer immunotherapy. Eur. J. Cancer 52, 50-66 (2016).
Bagshawe KD, Sharma SK, Springer CJ, Antoniw P, Rogers GT, Burke PJ, Melton R. Antibody-enzyme conjugates can generate cytotoxic drugs from inactive precursors at tumor sites. Antibody, Immunoconjugates and Radiopharmaceuticals 4, 915-22 (1991).
Bartkowiak T, Curran MA. 4-1BB agonists: multi-potent potentiators of tumor immunity. Front. Oncol. 5, 117 (2015).
Bartkowiak T, Jaiswal AR, Ager CR, Chin R, Chen C-H, Budhani P, Ai M, Reilley MJ, Sebastian MM, Hong DS, Curran MA. Activation of 4-1BB on liver myeloid cells triggers hepatitis via an interleukin-27 dependent pathway. Clin. Cancer Res. 24(5), 1138-51 (2018).
Bedzyk WD, Johnson LS, Riordan GS, Voss EW Jr. Comparison of variable region primary structures within an anti-fluorescein idiotype family. J. Biol. Chem. 264(3), 1565-69 (1989).
Bedzyk WD, Weidner KM, Denzin LK, Johnson LS, Hardman KD, Pantoliano MW, Asel ED, Voss EW Jr. Immunological and structural characterization of a high affinity anti-fluorescein single-chain antibody. J. Biol. Chem. 265(30), 18615-20 (1990).
Braden BC, Fields BA, Ysern X, Goldbaum FA, Dall'Acqua W, Schwarz FP, Poljak RJ, Mariuzza RA. Crystal structure of the complex of the variable domain of antibody D1.3 and turkey egg white lysozyme: a novel conformational change in antibody CD3-L3 selects for antigen. J. Mol. Biol. 257(5), 889-94 (1996).
Bremer E. Targeting of the Tumor Necrosis Factor Receptor Superfamily for Cancer Immunotherapy. ISRN Oncol. 2013, 1-25 (2013).
Bruhns P, Iannascoli B, England P, Mancardi DA, Fernandez N, Jorieux S, Daëron M. Specificity and affinity of human Fcγ receptors and their polymorphic variants for human IgG subclasses. Blood 113(16), 3716-25 (2009).
Bulliard Y, Jolicoeur R, Windman M, Rue SM, Ettenberg S, Knee DA, Wilson DS, Dranoff G, Brogdon JL. Activating Fc γ receptors contribute to the antitumor activities of immunoregulatory receptor-targeting antibodies. J. Exp. Med. 210(9), 1685-93 (2013)
Bulliard Y, Jolicoeur R, Zhang J, Dranoff G, Wilson NS, Brogdon JL. OX40 engagement depletes intratumoral Tregs via activating FcγRs, leading to antitumor efficacy. Immunol. Cell Biol. 92(6), 475-80 (2014).
Chan RC, Ong EZ, Mok DZ, Ooi EE. Fc receptors and their influence on efficacy of therapeutic antibodies for treatment of viral diseses. Expert Rev. Anti-infect. Ther. 13(11), 1351-60 (2015).
Croft M, So T, Duan W, Soroosh P. The significance of OX40 and OX40L to T-cell biology and immune disease. Immunol. Rev. 229(1), 173-91 (2009).
Croft M. The TNF family in T cell differentiation and function - unanswered questions and future directions. Semin. Immunol. 26(3), 183-90 (2014).
Curti BD, Kovacsovics-Bankowski M, Morris N, Walker E, Chisholm L, Floyd K, Walker J, Gonzalez I, Meeuwsen T, Fox BA, Moudgil T, Miller W, Haley D, Coffey T, Fisher B, Delanty-Miller L, Rymarchyk N, Kelly T, Crocenzi T, Bernstein E, Sanborn R, Urba WJ, Weinberg AD. OX40 is a potent immune-stimulating target in late-stage cancer patients. Cancer Res. 73(24), 7189-98 (2013).
Dubrot J, Milheiro F, Alfaro C, Palazón A, Martinez-Forero I, Perez-Gracia JL, Morales-Kastresana A, Romero-Trevejo JL, Ochoa MC, Hervás-Stubbs S, Prieto J, Jure-Kunkel M, Chen L, Melero I. Treatment with anti-CD137 mAbs cause intense accumulations of liver T cells without selective antitumor immunotherapeutic effects in this organ. Cancer Immunol. Immunother. 59(8), 1223-33 (2010).
Glisson BS, Leidner R, Ferris RL, Powderly J, Rizvi N, Norton JD, Burton J, Lanasa MC, Patel SP. Phase 1 study of MEDI0562, a humanized OX40 agonist monoclonal antibody (mAb), in adult patients (pts) with advanced solid tumours. Ann. Oncol. 27(suppl_6), 1052PD (2016).
Gopal A, Levy R, Houot, R, Patel S, Hatake K, Popplewell L, Chen Y, Davis C, Huang, B, Cesari R, Thall A, Woolfson A, Bartlett N. A phase I study of utomilumab (PF-05082566), a 4-1BB/CD137 agonist, in combination with ritixumab in patients with CD20+ non-Hodgkin’s lymphoma. Hematol. Oncol. 35, 260 (2017).
Jefferis R, Reimer CB, Skvaril F, de Lange G, Ling NR, Lowe J, Walker MR, Phillips DJ, Aloisio CH, Wells TW. Evaluation of monoclonal antibodies having specificity for human IgG sub-classes: results of an IUIS/WHO collaborative study. Immunol. Lett.1, 223-52 (1985).
Jefferis R, Reimer CB, Skvaril F, de Lange GG, Goodall DM, Bentley TL, Phillips DJ, Vlug A, Harada S, Radl J. Evaluation of monoclonal antibodies having specificity for human IgG subclasses: results of the 2nd IUIS/WHO collaborative study. Immunol. Lett. 31(2),143-68 (1992).
Hezareh M, Hessell AJ, Jensen RC, van de Winkel JG, Parren PW. Effector function activities of a panel of mutants of a broadly neutralizing antibody against human immunodeficiency virus type 1. J. Virol. 75(24), 12161-8 (2001).
Hirschhorn-Cymerman D, Rizzuto GA, Merghoub T, Cohen AD, Avogadri F, Lesokhin AM, Weinberg AD, Wolchok JD, Houghton AN. OX40 engagement and chemotherapy combination provides potent antitumor immunity with concomitant regulatory T cell apoptosis. J. Exp. Med. 206(5), 1103-16 (2009).
Holliger P, Hudson PJ. Engineered antibody fragments and the rise of single domains. Nat. Biotechnol. 23(9), 1126-36 (2005).
Hu S, Shively L, Raubitschek A, Sherman M, Williams LE, Wong JY, Shively JE, Wu AM. Minibody: A novel engineered anti-carcinoembryonic antigen antibody fragment (single-chain Fv-CH3) which exhibits rapid, high-level targeting of xenografts. Cancer Res. 56(13), 3055-61 (1996).
Idusogie EE, Presta LG, Gazzano-Santoro H, Totpal K, Wong PY, Ultsch M, Meng YG, Mulkerrin MG. Mapping of the C1q binding site on rituxan, a chimeric antibody with a human IgG1 Fc. J. Immunol. 164(8), 4178-84 (2000).
Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequences of Proteins of Immunological Interest, 5th ed. NIH Publication No. 91-3242. Washington, D.C.: U.S. Department of Health and Human Services (1991).
Kjaergaard J, Tanaka J, Kim JA, Rothchild K, Weinberg A, Shu S. Therapeutic efficacy of OX-40 receptor antibody depends on tumor immunogenicity and anatomic site of tumor growth. Cancer Res. 60(19), 5514-21 (2000).
Klein C, Schaefer W, Regula JT. The use of CrossMAb technology for the generation of bi- and multispecific antibodies. MAbs 8(6), 1010-20 (2016).
Ledermann JA, Begent RH, Massof C, Kelly AM, Adam T, Bagshawe KD. A phase-I study of repeated therapy with radiolabelled antibody to carcinoembryonic antigen using intermittent or continuous administration of cyclosporin A to supress the immune response. Int. J. Cancer 47(5), 659-64 (1991).
Lee SJ, Myers L, Muralimohan G, Dari J, Qiao Y, Li Z, Mittler RS, Vella AT. 4-1BB and OX40 dual costimulation synergistically stimulate primary specific CD8 T cells for robust effector function. J. Immunol. 173(5), 3002-12 (2004).
Lefranc MP, Pommié C, Kaas Q, Duprat E, Bosc N, Guiraudou D, Jean C, Ruiz M, Da Piédade I, Rouard M, Foulquier E, Thouvenin V, Lefranc G. IMGT unique numbering for immunoglobulin and T cell receptor constant domains and Ig superfamily C-like domains. Dev. Comp. Immunol. 29(3), 185-203 (2005).
Lefranc MP, Giudicelli V, Duroux P, Jabado-Michaloud J, Folch G, Aouinti S, Carillon E, Duvergey H, Houles A, Paysan-Lafosse T, Hadi-Saljoqi S, Sasorith S, Lefranc G, Kossida S. IMGT®, the international ImMunoGeneTics information system® 25 years on. Nucleic Acids Res. 43(Database issue), D413-22 (2015).
Liu C, Workman CJ, Vignali DA. Targeting regulatory T cells in tumors. FEBS J. 283(14), 2731-48 (2016).
Ma BY, Mikolajczak SA, Danesh A, Hosiawa KA, Cameron CM, Takaori-Kondo A, Uchiyama T, Kelvin DJ, Ochi A. The expression and the regulatory role of OX40 and 4-1BB heterodimer in activated human T cells. Blood 106(6), 2002-10 (2005).
Mayes PA, Hance KW, Hoos A. The promise and challenges of immune agonist antibody development in cancer. Nat. Rev. Drug Discov. 17, 509-27 (2018).
Melero I, Hirschhorn-Cymerman D, Morales-Kastresana A, Sanmamed MF, Wolchok JD. Agonist antibodies to TNFR molecules that costimulate T and NK cells. Clin. Cancer Res. 19(5), 1044-53 (2013).
Milas L, Wike J, Hunter N, Volpe J, Basic I. Macrophage content of murine sarcomas and carcinomas: associations with tumor growth parameters and tumor radiocurability. Cancer Res. 47(4), 1069-75 (1987).
Miller L, Rymarchyk N, Kelly T, Crocenzi T, Bernstein E, Sanborn R, Urba WJ, Weinberg AD. OX40 is a potent immune-stimulating target in late-stage cancer patients. Cancer Res. 73(24), 7189-98 (2013).
Moran AE, Kovacsovics-Bankowski M, Weinberg AD. The TNFRs OX40, 4-1BB, and CD40 as targets for cancer immunotherapy. Curr. Opin. Immunol. 25(2), 230-7 (2013).
Müller N, Wyzgol A, Münkel S, Pfizenmaier K, Wajant H. Activity of soluble OX40 ligand is enhanced by oligomerization and cell surface immobilization. FEBS J. 275(9), 2296-304 (2008).
Nizar S, Copier J, Meyer B, Bodman-Smith M, Galustian C, Kumar D, Dalgleish A. T-regulatory cell modulation: the future of cancer immunotherapy? Br. J. Cancer 100(11), 1697-703 (2009).
Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat. Rev. Cancer 12(4), 252-264 (2012).
Pearson WR, Lipman DJ. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. U.S.A. 85(8), 2444-8 (1988).
Petty JK, He K, Corless CL, Vetto JT, Weinberg AD. Survival in human colorectal cancer correlates with expression of the T-cell costimulatory molecule OX-40 (CD134). Am. J. Surg. 183(5), 512-8 (2002).
Powell DJ Jr, de Vries CR, Allen T, Ahmadzadeh M, Rosenberg SA. Inability to mediate prolonged reduction of regulatory T cells after transfer of autologous CD25-depleted PBMC and interleukin-2 after lymphodepleting chemotherapy. J. Immunother. 30(4), 438-47 (2007).
Qui HZ, Hagymasi AT, Bandyopadhyay S, St Rose M-C, Ramanarasimhaiah R, Menoret A, Mittler RS, Gordon SM, Reiner SL, Vella AT, Adler AJ. CD134 plus CD137 dual costimulation induces Eomesodermin in CD4 T cells to program cytotoxic Th1 differentiation. J. Immunol. 187(7), 3555-64 (2011).
Rosenberg S. Development of Cancer Vaccines. ASCO Educational Book Spring: 60-62 (2000).
Ryan JM, Mittal P, Menoret A, Svedova J, Wasser JS, Adler AJ, Vella AT. A novel biologic platform elicits profound T cell costimulatory activity and antitumor immunity in mice. Cancer Immunol. Immunother. 67(4), 605-13 (2018).
Sadun RE, Hsu WE, Zhang N, Nien YC, Bergfeld SA, Sabzevari H, Lutsiak ME, Khawli L, Hu P, Epstein AL. Fc-mOX40L fusion protein produces complete remission and enhanced survival in 2 murine tumour models. J. Immunother. 31(3), 235-45 (2008).
Sanchez-Paulete AR, Labiano S, Rodriguez-Ruiz ME, Azpilikueta A, Etxeberria I, Bolaños E, Lang V, Rodriguez M, Aznar MA, Jure-Kunkel M, Melero I. Deciphering CD137 (4-1BB) signaling in T-cell costimulation for translation into successful cancer immunotherapy. Eur. J. Immunol. 46(3), 513-22 (2016).
Schaer DA, Hirschhorn-Cymerman D, Wolchok JD. Targeting tumor-necrosis factor receptor pathways for tumor immunotherapy. J. Immunother. Cancer 2, 7 (2014).
Segal NH, Logan TF, Hodi FS, McDermott D, Melero I, Hamid O, Schmidt H, Robert C, Chiarion-Sileni V, Ascierto PA, Maio M, Urba WJ, Gangadhar TC, Suryawanshi S, Neely J, Jure-Kunkel M, Krishnan S, Kohrt H, Sznol M, Levy R. Results from an integrated safety analysis of urelumab, an agonist anti-CD137 monoclonal antibody. Clin. Cancer Res. 23(8), 1929-36 (2017).
Simpson TR., Li F, Montalvo-Ortiz W, Sepulveda MA, Bergerhoff K, Arce F, Roddie C, Henry JY, Yagita H, Wolchok JD, Peggs KS, Ravetch JV, Allison JP, Quezada SA. Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma. J. Exp. Med. 210(9), 1695-710 (2013).
Smith TF, Waterman MS. Identification of common molecular subsequences. J. Mol. Biol. 147(1), 195-7 (1981).
Stewart R, Hammond SA, Oberst M, Wilkinson RW. The role of Fc gamma receptors in the activity of immunomodulatory antibodies for cancer. J. Immunother. Cancer 2(29) (2014).
Takeda I, Ine S, Killeen N, Ndhlovu LC, Murata K, Satomi S, Sugamura K, Ishii N. Distinct roles for the OX40-OX40 ligand interaction in regulatory and nonregulatory T cells. J. Immunol. 172(6), 3580-9 (2004).
Taraban VY, Rowley TF, O’Brien L, Chan HT, Haswell LE, Green MH, Tutt AL, Glennie MJ, Al-Shamkhani A. Expression and costimulatory effects of the TNF receptor superfamily members CD134 OX40) and CD137 (4-1BB), and their role in the generation of anti-tumor immune responses. Eur. J. Immunol. 32(12), 3617-27 (2002).
Tolcher AW, Sznol M, Hu-Lieskovan S, Papadopoulos K P, Patnaik A, Rasco DW, Di Gravio D, Huang B, Gambhire D, Chen Y, Thall AD, Pathan N, Schmidt EV, Chow LQM. Phase Ib study of utomilumab (PF-05082566), a 4-1BB/CD137 agonist, in combination with pembrolizumab (MK-3475) in patients with advanced solid tumors. Clin. Cancer Res. 23(18), 5349-57 (2017).
Tran B, Carvajal RD, Marabelle A, Patel SP, LoRusso P, Ramussen E, Juan G, Upreti VV, Ngarmchamnanrith G, Schöffski P. Dose escalation results from a first-in-human, phase 1 study of the glucocorticoid-induced TNF receptor-related protein (GITR) agonist AMG 228 in patients (Pts) with advanced solid tumours. J. Clin. Oncol. 35(15_suppl), 2521 (2017).
Vanamee ÉS, Faustman DL. Structural principles of tumor necrosis factor superfamily signaling. Sci. Signal. 11(511), 1-12 (2018).
Vetto JT, Lum S, Morris A, Sicotte M, Davis J, Lemon M, Weinberg A. Presence of the T-cell activation marker OX-40 on tumor infiltrating lymphocytes and draining lymph node cells from patients with melanoma and head and neck cancers. Am. J. Surg. 174(3), 258-65 (1997).
Vignali DA, Collison LW, Workman CJ. How regulatory T cells work. Nat. Rev. Immunol. 8(7), 523-32 (2008).
Wajant H. Principles of antibody-mediated TNF receptor activation. Cell Death Differ. 22(11), 1727-41 (2015).
Wang X, Mathieu M, Brezski RJ. IgG Fc engineering to modulate antibody effector functions. Protein Cell 9(1), 63-73 (2018).
Weigelin B, Bolaños E, Rodriguez-Ruiz ME, Martinez-Forero I, Friedl P, Melero I. Anti-CD137 monoclonal antibodies and adoptive T cell therapy: a perfect marriage? Cancer Immunol. Immunother. 65(5), 493-7 (2016).
Wen T, Bukczynski J, Watts TH. 4-1BB ligand-mediated costimulation of human T cells induces CD4 and CD8 T cell expansion, cytokine production, and the development of cytolytic effector function. J. Immunol. 168(10), 4897-906 (2002).
Wesche-Soldato DE, Chung CS, Gregory SH, Salazar-Mather TP, Ayala CA, Ayala A. CD8+ T cells promote inflammation and apoptosis in the liver after sepsis: role of Fas-FasL. Am. J. Pathol. 171(1), 87-96 (2007).
Wilcox RA, Flies DB, Zhu G, Johnson AJ, Tamada K, Chapoval AI, Strome SE, Pease LR, Chen L. Provision of antigen and CD137 signaling breaks immunological ignorance, promoting regression of poorly immunogenic tumors. J. Clin. Invest. 109(5), 651-9 (2002).
Wozniak-Knopp G, Bartl S, Bauer A, Mostageer M, Woisetschläger M, Antes B, Ettl K, Kainer M, Weberhofer G, Wiederkum S, Himmler G, Mudde GC, Rüker F. Introducing antigen-binding sites in structural loops of immunoglobulin constant domains: Fc fragments with engineered HER2/neu-binding sites and antibody properties. Protein Eng. Des. Sel. 23(4), 289-97 (2010).
--->
Перечень последовательностей
<110> F-STAR BETA LIMITED
<120> Молекулы антител, которые связываются с CD137 и OX40
<130> 007508237
<140> PCT/EP2019/068796
<141> 2019-07-12
<150> GB 1811407.4
<151> 2018-07-12
<150> GB 1818281.6
<151> 2018-11-09
<150> GB 1902598.0
<151> 2019-02-26
<160> 175
<170> PatentIn версии 3.5
<210> 1
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH из МАТ FS30-10-16 (IMGT)
<400> 1
Gly Phe Thr Phe Ser Ser Tyr Asp
1 5
<210> 2
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-10-16 (IMGT)
<400> 2
Ile Asp Pro Thr Gly Ser Lys Thr
1 5
<210> 3
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-16 (IMGT)
<400> 3
Ala Arg Asp Leu Leu Val Tyr Gly Phe Asp Tyr
1 5 10
<210> 4
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL МАТ FS30-10-16 (IMGT)
<400> 4
Gln Ser Val Ser Ser Ser Tyr
1 5
<210> 5
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL МАТ FS30-10-16 (IMGT)
<400> 5
Gly Ala Ser
1
<210> 6
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ FS30-10-16 (IMGT)
<400> 6
Gln Gln Ser Tyr Ser Tyr Pro Val Thr
1 5
<210> 7
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ FS30-10-16 (Kabat)
<400> 7
Ser Tyr Asp Met Ser
1 5
<210> 8
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-10-16 (Kabat)
<400> 8
Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 9
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-16 (Kabat)
<400> 9
Asp Leu Leu Val Tyr Gly Phe Asp Tyr
1 5
<210> 10
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL МАТ FS30-10-16 (Kabat)
<400> 10
Gly Ala Ser Ser Arg Ala Thr
1 5
<210> 11
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL МАТ FS30-10-16 (Kabat)
<400> 11
Gln Gln Ser Tyr Ser Tyr Pro Val Thr
1 5
<210> 12
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-16
<400> 12
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Leu Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 13
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-16
<400> 13
gaagttcagc tgctggaatc tggcggcgga ttggttcaac ctggcggctc tctgagactg
60
tcttgtgccg cttccggctt caccttctcc agctacgaca tgtcctgggt ccgacaggct
120
cctggcaaag gactggaatg ggtgtccgac atcgacccca ccggctctaa gaccgactac
180
gccgattctg tgaagggcag attcaccatc agccgggaca actccaagaa caccctgtac
240
ctgcagatga actccctgag agccgaggac accgccgtgt actactgtgc cagagatctg
300
ctggtgtacg gcttcgacta ttggggccag ggcacactgg tcaccgtgtc ctct
354
<210> 14
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-10-16
<400> 14
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Tyr Ser Tyr Pro
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 15
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-10-16
<400> 15
gagatcgtgc tgacccagtc tcctggcaca ctgtcactgt ctccaggcga gagagctacc
60
ctgtcctgta gagcctctca gtccgtgtcc tcctcttacc tggcctggta tcagcagaag
120
cctggacagg ctccccggct gttgatctac ggcgcttctt ctagagccac aggcatccct
180
gaccggttct ccggatctgg ctctggcacc gatttcaccc tgaccatctc tcggctggaa
240
cccgaggatt tcgccgtgta ctactgccag cagtcctaca gctaccccgt gacctttggc
300
cagggcacca aggtggaaat caag
324
<210> 16
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-3 (IMGT)
<400> 16
Ala Arg Asp Leu Asn Val Tyr Gly Phe Asp Tyr
1 5 10
<210> 17
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-3 (Kabat)
<400> 17
Asp Leu Asn Val Tyr Gly Phe Asp Tyr
1 5
<210> 18
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-3
<400> 18
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Asn Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 19
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-3
<400> 19
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
aatgtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagt
354
<210> 20
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-10-3
<400> 20
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatcttatt cttatcctgt cacgttcggc
300
caagggacca aggtggaaat caaa
324
<210> 21
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-12 (IMGT)
<400> 21
Ala Arg Asp Leu Thr Val Tyr Gly Phe Asp Tyr
1 5 10
<210> 22
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-10-12 (Kabat)
<400> 22
Asp Leu Thr Val Tyr Gly Phe Asp Tyr
1 5
<210> 23
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-12
<400> 23
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Thr Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 24
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-10-12
<400> 24
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
acggtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagt
354
<210> 25
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ FS30-35-14 (IMGT)
<400> 25
Gly Phe Thr Phe Ser Ala Tyr Asn
1 5
<210> 26
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-35-14 (IMGT)
<400> 26
Ile Ser Pro Tyr Gly Gly Ala Thr
1 5
<210> 27
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-35-14 (IMGT)
<400> 27
Ala Arg Asn Leu Tyr Glu Leu Ser Ala Tyr Ser Tyr Gly Ala Asp Tyr
1 5 10 15
<210> 28
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ FS30-35-14 (IMGT)
<400> 28
Gln Gln Tyr Tyr Tyr Ser Ser Pro Ile Thr
1 5 10
<210> 29
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ FS30-35-14 (Kabat)
<400> 29
Ala Tyr Asn Ile His
1 5
<210> 30
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-35-14 (Kabat)
<400> 30
Asp Ile Ser Pro Tyr Gly Gly Ala Thr Asn Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 31
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-35-14 (Kabat)
<400> 31
Asn Leu Tyr Glu Leu Ser Ala Tyr Ser Tyr Gly Ala Asp Tyr
1 5 10
<210> 32
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-35-14
<400> 32
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatattatt attcttctcc tatcacgttc
300
ggccaaggga ccaaggtgga aatcaaa
327
<210> 33
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ FS30-5-37 (IMGT)
<400> 33
Gly Phe Thr Phe Ser Ser Tyr Ala
1 5
<210> 34
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-5-37 (IMGT)
<400> 34
Ile Ser Gly Ser Gly Gly Ser Thr
1 5
<210> 35
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-5-37 (IMGT)
<400> 35
Ala Arg Ser Tyr Asp Lys Tyr Trp Gly Ser Ser Ile Tyr Ser Gly Leu
1 5 10 15
Asp Tyr
<210> 36
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ FS30-5-37 (IMGT)
<400> 36
Gln Gln Tyr Tyr Ser Tyr Tyr Pro Val Thr
1 5 10
<210> 37
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ FS30-5-37 (Kabat)
<400> 37
Ser Tyr Ala Met Ser
1 5
<210> 38
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ FS30-5-37 (Kabat)
<400> 38
Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 39
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ FS30-5-37 (Kabat)
<400> 39
Ser Tyr Asp Lys Tyr Trp Gly Ser Ser Ile Tyr Ser Gly Leu Asp Tyr
1 5 10 15
<210> 40
<211> 125
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-5-37
<400> 40
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Asn Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Tyr Asp Lys Tyr Trp Gly Ser Ser Ile Tyr Ser Gly Leu
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 41
<211> 375
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-5-37
<400> 41
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
aattgcgcgg ccagtggctt taccttcagt agctatgcca tgagctgggt gcgtcaggcg
120
ccgggcaaag gtctggaatg ggttagcgcg attagcggta gtggcggtag cacgtactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagatcttac
300
gacaaatact ggggttcttc tatttactct ggcttggact actggggcca gggaaccctg
360
gtcaccgtct cgagt
375
<210> 42
<211> 109
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-5-37
<400> 42
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Ser Tyr Tyr
85 90 95
Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 43
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-5-37
<400> 43
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatattatt cttattatcc tgtcacgttc
300
ggccaaggga ccaaggtgga aatcaaa
327
<210> 44
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля AB домена CH3 WT
<400> 44
Arg Asp Glu Leu Thr Lys Asn Gln
1 5
<210> 45
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD домена CH3 WT
<400> 45
Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1 5
<210> 46
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля EF домена CH3 WT
<400> 46
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
1 5 10
<210> 47
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 WT
<400> 47
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 48
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2
<400> 48
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 49
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2 с мутацией LALA
<400> 49
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 50
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2 с мутацией LALA и мутацией P114A
<400> 50
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 51
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-49
<400> 51
Tyr Trp Asp Gln Glu
1 5
<210> 52
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-49
<400> 52
Asp Glu Gln Phe Ala
1 5
<210> 53
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-49
<400> 53
Gln Tyr Arg Trp Asn Pro Ala Asp Tyr
1 5
<210> 54
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-49
<400> 54
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln
35 40 45
Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr Arg Trp Asn Pro Ala
65 70 75 80
Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 55
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-49
<400> 55
ggccagccta gggaacccca ggtttacacc ttgcctccaa gccgggacga gtactgggat
60
caagaggtgt ccctgacctg cctcgtgaag ggcttctacc cttccgatat cgccgtggaa
120
tgggagagca atggcgacga gcagttcgcc tacaagacaa cccctcctgt gctggactcc
180
gacggctcat tctttctgta ctccaagctg acagtggacc agtacagatg gaaccccgcc
240
gactacttct cttgctccgt gatgcacgag gccctgcaca accactacac acagaagtcc
300
ctgtctctgt cccctggc
318
<210> 56
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля AB домена CH3 Fcab FS20-22-49
<400> 56
Arg Asp Glu Tyr Trp Asp Gln Glu
1 5
<210> 57
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля CD домена CH3 Fcab FS20-22-49
<400> 57
Ser Asn Gly Asp Glu Gln Phe Ala Tyr
1 5
<210> 58
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Петля EF домена CH3 Fcab FS20-22-49
<400> 58
Asp Gln Tyr Arg Trp Asn Pro Ala Asp Tyr
1 5 10
<210> 59
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-38
<400> 59
Ala Glu Lys Tyr Gln
1 5
<210> 60
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-38
<400> 60
Gln Tyr Arg Trp Asn Pro Gly Asp Tyr
1 5
<210> 61
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-38
<400> 61
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Ala Glu Lys
35 40 45
Tyr Gln Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr Arg Trp Asn Pro Gly
65 70 75 80
Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 62
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-38
<400> 62
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggcaga aaaataccag tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtggatc agtataggtg gaacccaggc
240
gactatttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccgggt
318
<210> 63
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-41
<400> 63
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln
35 40 45
Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr Arg Trp Asn Pro Gly
65 70 75 80
Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 64
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-41
<400> 64
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggatga acagttcgca tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtggatc agtataggtg gaacccaggc
240
gactatttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccggga
318
<210> 65
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-47
<400> 65
Gln Tyr Arg Trp Ser Pro Gly Asp Tyr
1 5
<210> 66
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-47
<400> 66
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln
35 40 45
Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr Arg Trp Ser Pro Gly
65 70 75 80
Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 67
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-47
<400> 67
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggatga acagttcgca tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtggatc agtataggtg gagtccgggt
240
gattatttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 68
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-22-85
<400> 68
Gln Tyr Arg Trp Asn Pro Phe Asp Asp
1 5
<210> 69
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-85
<400> 69
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln
35 40 45
Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Leu Asp Gln Tyr Arg Trp Asn Pro Phe
65 70 75 80
Asp Asp Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 70
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-85
<400> 70
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggatga acagttcgca tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accttggatc agtataggtg gaatccgttt
240
gatgatttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 71
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-58
<400> 71
Tyr Tyr Ser Gly Glu
1 5
<210> 72
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-58
<400> 72
Gln Pro Glu Asn Asp
1 5
<210> 73
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-58
<400> 73
Pro Tyr Trp Arg Trp Gly Ser Pro Arg Thr
1 5 10
<210> 74
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS20-31-58
<400> 74
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Tyr Ser Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Ser Pro
65 70 75 80
Arg Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 75
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS20-31-58
<400> 75
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactactct
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atgggcagcc ggagaacgac tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgcctt attggaggtg gggtagtccg
240
cgtactttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccgggt
318
<210> 76
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-66
<400> 76
Pro Tyr Trp Arg Trp Gly Val Pro Arg Thr
1 5 10
<210> 77
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS20-31-66
<400> 77
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Tyr Tyr Ser Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Val Pro
65 70 75 80
Arg Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 78
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS20-31-66
<400> 78
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactactct
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atgggcagcc ggagaacgac tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgccgt attggaggtg gggtgttccg
240
cgtactttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccgggt
318
<210> 79
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-94 Fcab
<400> 79
Trp Glu His Gly Glu
1 5
<210> 80
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-94 Fcab
<400> 80
Ile Arg Glu His Asp
1 5
<210> 81
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-94 Fcab
<400> 81
Pro Tyr Trp Arg Trp Gly Gly Pro Gly Thr
1 5 10
<210> 82
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-94 Fcab
<400> 82
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Trp Glu His Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Ile Arg Glu
35 40 45
His Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Gly Pro
65 70 75 80
Gly Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 83
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-94 Fcab
<400> 83
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtgggaacat
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atgggatcag agaacatgat tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgccat attggaggtg gggcggccca
240
ggcaccttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 84
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-102
<400> 84
Trp Ala Ser Gly Glu
1 5
<210> 85
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-102
<400> 85
Gln Pro Glu Val Asp
1 5
<210> 86
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-102
<400> 86
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Trp Ala Ser Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Val Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Val Pro
65 70 75 80
Arg Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 87
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-102
<400> 87
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtgggcatct
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atgggcagcc agaagttgat tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgccgt attggaggtg gggtgttccg
240
cgtactttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccgggt
318
<210> 88
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-108
<400> 88
Glu Lys Glu Ile Asp
1 5
<210> 89
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-108
<400> 89
Pro Tyr Trp Arg Trp Gly Ala Lys Arg Thr
1 5 10
<210> 90
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-108
<400> 90
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Trp Ala Ser Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Glu Lys Glu
35 40 45
Ile Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Ala Lys
65 70 75 80
Arg Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 91
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-108
<400> 91
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtgggcatct
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggaaaa agaaatcgat tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgccgt attggaggtg gggtgctaag
240
cgtactttct catgctccgt gatgcatgag gctctgcaca accactacac acagaagagc
300
ctctccctgt ctccgggt
318
<210> 92
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab
FS20-31-115
<400> 92
Glu Gln Glu Phe Asp
1 5
<210> 93
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-115
<400> 93
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Trp Ala Ser Gly Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Glu Gln Glu
35 40 45
Phe Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Pro Tyr Trp Arg Trp Gly Ala Lys
65 70 75 80
Arg Thr Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 94
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-31-115
<400> 94
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtgggcatct
60
ggtgaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggaaca ggaattcgat tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtgccgt attggaggtg gggtgctaag
240
cgtactttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 95
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-16 с мутацией LALA
<400> 95
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Leu Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 96
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-16 с мутацией LALA
<400> 96
gaagttcagc tgctggaatc tggcggcgga ttggttcaac ctggcggctc tctgagactg
60
tcttgtgccg cttccggctt caccttctcc agctacgaca tgtcctgggt ccgacaggct
120
cctggcaaag gactggaatg ggtgtccgac atcgacccca ccggctctaa gaccgactac
180
gccgattctg tgaagggcag attcaccatc agccgggaca actccaagaa caccctgtac
240
ctgcagatga actccctgag agccgaggac accgccgtgt actactgtgc cagagatctg
300
ctggtgtacg gcttcgacta ttggggccag ggcacactgg tcaccgtgtc ctctgcttct
360
accaagggac ccagcgtgtt ccctctggct ccttccagca agtctacctc tggcggaaca
420
gctgctctgg gctgcctggt caaggactac tttcctgagc ctgtgaccgt gtcttggaac
480
tctggcgctc tgacatctgg cgtgcacacc tttccagcag tgctgcagtc ctccggcctg
540
tactctctgt cctctgtcgt gaccgtgcct tccagctctc tgggaaccca gacctacatc
600
tgcaatgtga accacaagcc ttccaacacc aaggtggaca agaaggtgga acccaagtcc
660
tgcgacaaga cccacacctg tcctccatgt cctgctccag aagctgctgg cggcccttcc
720
gtgtttctgt tccctccaaa gcctaaggac accctgatga tctctcggac ccctgaagtg
780
acctgcgtgg tggtggatgt gtctcacgag gacccagaag tgaagttcaa ttggtacgtg
840
gacggcgtgg aagtgcacaa cgccaagacc aagcctagag aggaacagta caactccacc
900
tacagagtgg tgtccgtgct gaccgtgctg caccaggatt ggctgaacgg caaagagtac
960
aagtgcaagg tgtccaacaa ggccctgcct gctcctatcg aaaagaccat ctccaaggcc
1020
aagggccagc ctagggaacc ccaggtttac accttgcctc caagccggga cgagtactgg
1080
gatcaagagg tgtccctgac ctgcctcgtg aagggcttct acccttccga tatcgccgtg
1140
gaatgggaga gcaatggcga cgagcagttc gcctacaaga caacccctcc tgtgctggac
1200
tccgacggct cattctttct gtactccaag ctgacagtgg accagtacag atggaacccc
1260
gccgactact tctcttgctc cgtgatgcac gaggccctgc acaaccacta cacacagaag
1320
tccctgtctc tgtcccctgg c
1341
<210> 97
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-10-16 (
<400> 97
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Tyr Ser Tyr Pro
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 98
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-10-16 (
<400> 98
gagatcgtgc tgacccagtc tcctggcaca ctgtcactgt ctccaggcga gagagctacc
60
ctgtcctgta gagcctctca gtccgtgtcc tcctcttacc tggcctggta tcagcagaag
120
cctggacagg ctccccggct gttgatctac ggcgcttctt ctagagccac aggcatccct
180
gaccggttct ccggatctgg ctctggcacc gatttcaccc tgaccatctc tcggctggaa
240
cccgaggatt tcgccgtgta ctactgccag cagtcctaca gctaccccgt gacctttggc
300
cagggcacca aggtggaaat caagcgtacg gtggccgctc ccagcgtgtt catcttcccc
360
ccaagcgacg agcagctgaa gagcggcacc gccagcgtgg tgtgtctgct gaacaacttc
420
taccccaggg aggccaaggt gcagtggaag gtggacaacg ccctgcagag cggcaacagc
480
caggagagcg tcaccgagca ggacagcaag gactccacct acagcctgag cagcaccctg
540
accctgagca aggccgacta cgagaagcac aaggtgtacg cctgtgaggt gacccaccag
600
ggcctgtcca gccccgtgac caagagcttc aacaggggcg agtgc
645
<210> 99
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-3 с мутацией LALA
<400> 99
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Asn Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 100
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-3 с мутацией LALA
<400> 100
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
aatgtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagtgctagc
360
actaagggcc cgtcggtgtt cccgctggcc ccatcgtcca agagcacatc agggggtacc
420
gccgccctgg gctgccttgt gaaggattac tttcccgagc ccgtcacagt gtcctggaac
480
agcggagccc tgacctccgg agtgcatact ttcccggctg tgcttcagtc ctctggcctg
540
tactcattgt cctccgtggt caccgtccct tcgtcctccc tgggcaccca gacctatatc
600
tgtaatgtca accataagcc ctcgaacacc aaggtcgaca agaaggtcga gccgaagtcg
660
tgcgacaaga ctcacacttg cccgccttgc ccagccccgg aagctgccgg tggtccttcg
720
gtgttcctct tcccgcccaa gccgaaggat accctgatga tctcacggac ccccgaagtg
780
acctgtgtgg tggtggacgt gtcccacgag gacccggaag tgaaattcaa ttggtacgtg
840
gatggagtgg aagtgcacaa cgccaagacc aagccacggg aagaacagta caactctacc
900
taccgcgtgg tgtccgtgct cactgtgctg caccaagact ggctgaacgg gaaggagtac
960
aagtgcaaag tgtccaacaa ggcgctgcct gccccaattg agaaaactat ctcgaaagcc
1020
aagggacagc ctcgagaacc acaggtgtac accctgcccc catcccggga tgagtactgg
1080
gaccaggaag tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg
1140
gagtgggaga gcaatgggga tgaacagttc gcatacaaga ccacgcctcc cgtgctggac
1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg atcagtatag gtggaatcct
1260
gctgattatt tctcatgctc cgtgatgcat gaggctctgc acaaccacta cactcagaag
1320
agcttgtccc tgtcgcccgg a
1341
<210> 101
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Усеченная шарнирная область Fcab
<400> 101
Thr Cys Pro Pro Cys Pro
1 5
<210> 102
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-10-3
<400> 102
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatcttatt cttatcctgt cacgttcggc
300
caagggacca aggtggaaat caaacgtact gtggccgctc ctagcgtgtt catttttccg
360
ccatccgacg agcagctcaa gtccggcacc gcctccgtgg tctgcctgct caacaacttc
420
taccctcgcg aagctaaggt ccagtggaag gtcgacaatg ccctgcagtc cggaaactcg
480
caggaaagcg tgactgaaca ggactccaag gactccacct attcactgtc ctcgactctg
540
accctgagca aggcggatta cgaaaagcac aaagtgtacg catgcgaagt gacccaccag
600
ggtctttcgt cccccgtgac caagagcttc aacagaggag agtgt
645
<210> 103
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-12 с мутацией LALA
<400> 103
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Thr Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 104
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-10-12 с мутацией LALA
<400> 104
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
acggtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagtgctagc
360
actaagggcc cgtcggtgtt cccgctggcc ccatcgtcca agagcacatc agggggtacc
420
gccgccctgg gctgccttgt gaaggattac tttcccgagc ccgtcacagt gtcctggaac
480
agcggagccc tgacctccgg agtgcatact ttcccggctg tgcttcagtc ctctggcctg
540
tactcattgt cctccgtggt caccgtccct tcgtcctccc tgggcaccca gacctatatc
600
tgtaatgtca accataagcc ctcgaacacc aaggtcgaca agaaggtcga gccgaagtcg
660
tgcgacaaga ctcacacttg cccgccttgc ccagccccgg aagctgccgg tggtccttcg
720
gtgttcctct tcccgcccaa gccgaaggat accctgatga tctcacggac ccccgaagtg
780
acctgtgtgg tggtggacgt gtcccacgag gacccggaag tgaaattcaa ttggtacgtg
840
gatggagtgg aagtgcacaa cgccaagacc aagccacggg aagaacagta caactctacc
900
taccgcgtgg tgtccgtgct cactgtgctg caccaagact ggctgaacgg gaaggagtac
960
aagtgcaaag tgtccaacaa ggcgctgcct gccccaattg agaaaactat ctcgaaagcc
1020
aagggacagc ctcgagaacc acaggtgtac accctgcccc catcccggga tgagtactgg
1080
gaccaggaag tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg
1140
gagtgggaga gcaatgggga tgaacagttc gcatacaaga ccacgcctcc cgtgctggac
1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg atcagtatag gtggaatcct
1260
gctgattatt tctcatgctc cgtgatgcat gaggctctgc acaaccacta cactcagaag
1320
agcttgtccc tgtcgcccgg a
1341
<210> 105
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-35-14 с мутацией LALA
<400> 105
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr
20 25 30
Asn Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Ser Pro Tyr Gly Gly Ala Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Tyr Glu Leu Ser Ala Tyr Ser Tyr Gly Ala Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Gln Tyr Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 106
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-35-14 с мутацией LALA
<400> 106
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt gcctataata tccattgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat atttctccgt atggtggcgc gaccaactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagaaacctc
300
tacgagttga gcgcttactc ttacggggcg gactactggg gccagggaac cctggtcacc
360
gtctcgtcgg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc
420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc
480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt
540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc
600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag
660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct
720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca
780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa
840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa
900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg
960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa
1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc
1080
cgggatgagt actgggacca ggaagtcagc ctgacctgcc tggtcaaagg cttctatccc
1140
agcgacatcg ccgtggagtg ggagagcaat ggggatgaac agttcgcata caagaccacg
1200
cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggatcag
1260
tataggtgga atcctgctga ttatttctca tgctccgtga tgcatgaggc tctgcacaac
1320
cactacactc agaagagctt gtccctgtcg cccgga
1356
<210> 107
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-35-14
<400> 107
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Tyr Ser Ser
85 90 95
Pro Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 108
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-35-14
<400> 108
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatattatt attcttctcc tatcacgttc
300
ggccaaggga ccaaggtgga aatcaaacgt actgtggccg ctcctagcgt gttcattttt
360
ccgccatccg acgagcagct caagtccggc accgcctccg tggtctgcct gctcaacaac
420
ttctaccctc gcgaagctaa ggtccagtgg aaggtcgaca atgccctgca gtccggaaac
480
tcgcaggaaa gcgtgactga acaggactcc aaggactcca cctattcact gtcctcgact
540
ctgaccctga gcaaggcgga ttacgaaaag cacaaagtgt acgcatgcga agtgacccac
600
cagggtcttt cgtcccccgt gaccaagagc ttcaacagag gagagtgt
648
<210> 109
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-5-37 с мутацией LALA
<400> 109
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Asn Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Tyr Asp Lys Tyr Trp Gly Ser Ser Ile Tyr Ser Gly Leu
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Tyr Trp Asp
355 360 365
Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln Phe Ala Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Gln Tyr Arg Trp Asn Pro Ala Asp Tyr Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 110
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/FS30-5-37 с мутацией LALA
<400> 110
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
aattgcgcgg ccagtggctt taccttcagt agctatgcca tgagctgggt gcgtcaggcg
120
ccgggcaaag gtctggaatg ggttagcgcg attagcggta gtggcggtag cacgtactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagatcttac
300
gacaaatact ggggttcttc tatttactct ggcttggact actggggcca gggaaccctg
360
gtcaccgtct cgagtgctag cactaagggc ccgtcggtgt tcccgctggc cccatcgtcc
420
aagagcacat cagggggtac cgccgccctg ggctgccttg tgaaggatta ctttcccgag
480
cccgtcacag tgtcctggaa cagcggagcc ctgacctccg gagtgcatac tttcccggct
540
gtgcttcagt cctctggcct gtactcattg tcctccgtgg tcaccgtccc ttcgtcctcc
600
ctgggcaccc agacctatat ctgtaatgtc aaccataagc cctcgaacac caaggtcgac
660
aagaaggtcg agccgaagtc gtgcgacaag actcacactt gcccgccttg cccagccccg
720
gaagctgccg gtggtccttc ggtgttcctc ttcccgccca agccgaagga taccctgatg
780
atctcacgga cccccgaagt gacctgtgtg gtggtggacg tgtcccacga ggacccggaa
840
gtgaaattca attggtacgt ggatggagtg gaagtgcaca acgccaagac caagccacgg
900
gaagaacagt acaactctac ctaccgcgtg gtgtccgtgc tcactgtgct gcaccaagac
960
tggctgaacg ggaaggagta caagtgcaaa gtgtccaaca aggcgctgcc tgccccaatt
1020
gagaaaacta tctcgaaagc caagggacag cctcgagaac cacaggtgta caccctgccc
1080
ccatcccggg atgagtactg ggaccaggaa gtcagcctga cctgcctggt caaaggcttc
1140
tatcccagcg acatcgccgt ggagtgggag agcaatgggg atgaacagtt cgcatacaag
1200
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg
1260
gatcagtata ggtggaatcc tgctgattat ttctcatgct ccgtgatgca tgaggctctg
1320
cacaaccact acactcagaa gagcttgtcc ctgtcgcccg ga
1362
<210> 111
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-5-37
<400> 111
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Ser Tyr Tyr
85 90 95
Pro Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 112
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS30-5-37
<400> 112
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatattatt cttattatcc tgtcacgttc
300
ggccaaggga ccaaggtgga aatcaaacgt actgtggccg ctcctagcgt gttcattttt
360
ccgccatccg acgagcagct caagtccggc accgcctccg tggtctgcct gctcaacaac
420
ttctaccctc gcgaagctaa ggtccagtgg aaggtcgaca atgccctgca gtccggaaac
480
tcgcaggaaa gcgtgactga acaggactcc aaggactcca cctattcact gtcctcgact
540
ctgaccctga gcaaggcgga ttacgaaaag cacaaagtgt acgcatgcga agtgacccac
600
cagggtcttt cgtcccccgt gaccaagagc ttcaacagag gagagtgt
648
<210> 113
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Альтернативный домен CH3 Fcab FS20-22-49
<400> 113
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggatga acagttcgca tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtggatc agtataggtg gaatcctgct
240
gattatttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 114
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ к FITC G1AA/4420, содержащего мутацию LALA
<400> 114
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 115
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ к FITC G1/4420 без мутации LALA
<400> 115
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 116
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ 4420
<400> 116
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Arg Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 117
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь антитела G1AA/HelD1.3 с мутацией LALA
<400> 117
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr Phe Ser Gly Tyr
20 25 30
Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp Gly Gln Gly Ser Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 118
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ HelD1.3
<400> 118
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Gly Asn Ile His Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val
35 40 45
Tyr Asn Ala Lys Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His Phe Trp Ser Thr Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 119
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/MOR7480.1
<400> 119
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 120
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ G1/MOR7480.1, G1AA/MOR7480.1 и G2/MOR7480.1
<400> 120
Ser Tyr Glu Leu Thr Gln Pro Pro Ser Val Ser Val Ser Pro Gly Gln
1 5 10 15
Thr Ala Ser Ile Thr Cys Ser Gly Asp Asn Ile Gly Asp Gln Tyr Ala
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Val Leu Val Ile Tyr
35 40 45
Gln Asp Lys Asn Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Gly Thr Gln Ala Met
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Ala Thr Tyr Thr Gly Phe Gly Ser Leu
85 90 95
Ala Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys
100 105 110
Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln
115 120 125
Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly
130 135 140
Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly
145 150 155 160
Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala
165 170 175
Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser
180 185 190
Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val
195 200 205
Ala Pro Thr Glu Cys Ser
210
<210> 121
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/20H4.9
<400> 121
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 122
<211> 216
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ G1/20H4.9 и G1AA/20H4.9
<400> 122
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Arg Ser Asn Trp Pro Pro
85 90 95
Ala Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val
100 105 110
Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys
115 120 125
Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg
130 135 140
Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn
145 150 155 160
Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser
165 170 175
Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys
180 185 190
Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr
195 200 205
Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 123
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20-22-49AA/4420 (с мутацией LALA)
<400> 123
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 124
<211> 441
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G2/MOR7480.1
<400> 124
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe
180 185 190
Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro
210 215 220
Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 125
<211> 445
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1AA/MOR7480.1
<400> 125
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Arg Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Ser Thr Tyr
20 25 30
Trp Ile Ser Trp Val Arg Gln Met Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Lys Ile Tyr Pro Gly Asp Ser Tyr Thr Asn Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Ile Ser Ala Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Leu Gln Trp Ser Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Gly Ile Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 126
<211> 232
<212> БЕЛОК
<213> CD137 Homo sapiens
<400> 126
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Ser Phe Phe Leu Ala Leu Thr Ser Thr Ala Leu
165 170 175
Leu Phe Leu Leu Phe Phe Leu Thr Leu Arg Phe Ser Val Val Lys Arg
180 185 190
Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro
195 200 205
Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu
210 215 220
Glu Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 127
<211> 163
<212> БЕЛОК
<213> Внеклеточный домен CD137 Homo sapiens
<400> 127
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 128
<211> 231
<212> БЕЛОК
<213> CD137 яванской макаки
<400> 128
Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Phe Phe Leu Ala Leu Thr Ser Thr Val Val Leu
165 170 175
Phe Leu Leu Phe Phe Leu Val Leu Arg Phe Ser Val Val Lys Arg Ser
180 185 190
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
195 200 205
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
210 215 220
Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 129
<211> 163
<212> БЕЛОК
<213> Внеклеточный домен CD137 яванской макаки
<400> 129
Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 130
<211> 186
<212> БЕЛОК
<213> Внеклеточный домен OX40 Homo sapiens
<400> 130
Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His
1 5 10 15
Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln
20 25 30
Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val
35 40 45
Ser Ser Lys Pro Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly
50 55 60
Ser Glu Arg Lys Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg
65 70 75 80
Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp
85 90 95
Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala
100 105 110
Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln
115 120 125
Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro
130 135 140
Ala Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr
145 150 155 160
Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr
165 170 175
Arg Pro Val Glu Val Pro Gly Gly Arg Ala
180 185
<210> 131
<211> 186
<212> БЕЛОК
<213> Внеклеточный домен OX40 яванской макаки
<400> 131
Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys Gln
1 5 10 15
Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys Asn Arg Ser Gln
20 25 30
Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val
35 40 45
Ser Ala Lys Pro Cys Lys Ala Cys Thr Trp Cys Asn Leu Arg Ser Gly
50 55 60
Ser Glu Arg Lys Gln Pro Cys Thr Ala Thr Gln Asp Thr Val Cys Arg
65 70 75 80
Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp
85 90 95
Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala
100 105 110
Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln
115 120 125
Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro
130 135 140
Pro Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Thr Thr
145 150 155 160
Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln Arg Pro Ser Thr
165 170 175
Arg Pro Val Glu Val Pro Arg Gly Pro Ala
180 185
<210> 132
<211> 249
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> DO11.10-hOX40 и рецептор OX40 человека
<400> 132
Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys His
1 5 10 15
Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys Ser Arg Ser Gln
20 25 30
Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val Val
35 40 45
Ser Ser Lys Pro Cys Lys Pro Cys Thr Trp Cys Asn Leu Arg Ser Gly
50 55 60
Ser Glu Arg Lys Gln Leu Cys Thr Ala Thr Gln Asp Thr Val Cys Arg
65 70 75 80
Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val Asp
85 90 95
Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln Ala
100 105 110
Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu Gln
115 120 125
Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro Pro
130 135 140
Ala Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Ile Thr
145 150 155 160
Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln Gly Pro Ser Thr
165 170 175
Arg Pro Val Glu Val Pro Gly Gly Arg Ala Val Ala Ala Ile Leu Gly
180 185 190
Leu Gly Leu Val Leu Gly Leu Leu Gly Pro Leu Ala Ile Leu Leu Ala
195 200 205
Leu Tyr Leu Leu Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala His Lys
210 215 220
Pro Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln Ala
225 230 235 240
Asp Ala His Ser Thr Leu Ala Lys Ile
245
<210> 133
<211> 253
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> DO11.10-mOX40 и рецептор OX40 мыши
<400> 133
Val Thr Ala Arg Arg Leu Asn Cys Val Lys His Thr Tyr Pro Ser Gly
1 5 10 15
His Lys Cys Cys Arg Glu Cys Gln Pro Gly His Gly Met Val Ser Arg
20 25 30
Cys Asp His Thr Arg Asp Thr Leu Cys His Pro Cys Glu Thr Gly Phe
35 40 45
Tyr Asn Glu Ala Val Asn Tyr Asp Thr Cys Lys Gln Cys Thr Gln Cys
50 55 60
Asn His Arg Ser Gly Ser Glu Leu Lys Gln Asn Cys Thr Pro Thr Gln
65 70 75 80
Asp Thr Val Cys Arg Cys Arg Pro Gly Thr Gln Pro Arg Gln Asp Ser
85 90 95
Gly Tyr Lys Leu Gly Val Asp Cys Val Pro Cys Pro Pro Gly His Phe
100 105 110
Ser Pro Gly Asn Asn Gln Ala Cys Lys Pro Trp Thr Asn Cys Thr Leu
115 120 125
Ser Gly Lys Gln Thr Arg His Pro Ala Ser Asp Ser Leu Asp Ala Val
130 135 140
Cys Glu Asp Arg Ser Leu Leu Ala Thr Leu Leu Trp Glu Thr Gln Arg
145 150 155 160
Pro Thr Phe Arg Pro Thr Thr Val Gln Ser Thr Thr Val Trp Pro Arg
165 170 175
Thr Ser Glu Leu Pro Ser Pro Pro Thr Leu Val Thr Pro Glu Gly Pro
180 185 190
Ala Phe Ala Val Leu Leu Gly Leu Gly Leu Gly Leu Leu Ala Pro Leu
195 200 205
Thr Val Leu Leu Ala Leu Tyr Leu Leu Arg Lys Ala Trp Arg Leu Pro
210 215 220
Asn Thr Pro Lys Pro Cys Trp Gly Asn Ser Phe Arg Thr Pro Ile Gln
225 230 235 240
Glu Glu His Thr Asp Ala His Phe Thr Leu Ala Lys Ile
245 250
<210> 134
<211> 250
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> DO11.10-cOX40 и рецептор OX40 яванской макаки
<400> 134
Lys Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp Arg Cys Cys
1 5 10 15
Gln Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys Asn Arg Ser
20 25 30
Gln Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr Asn Asp Val
35 40 45
Val Ser Ala Lys Pro Cys Lys Ala Cys Thr Trp Cys Asn Leu Arg Ser
50 55 60
Gly Ser Glu Arg Lys Gln Pro Cys Thr Ala Thr Gln Asp Thr Val Cys
65 70 75 80
Arg Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys Pro Gly Val
85 90 95
Asp Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly Asp Asn Gln
100 105 110
Ala Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys His Thr Leu
115 120 125
Gln Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp Arg Asp Pro
130 135 140
Pro Pro Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala Arg Pro Thr
145 150 155 160
Thr Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln Arg Pro Ser
165 170 175
Thr Arg Pro Val Glu Val Pro Arg Gly Pro Ala Val Ala Ala Ile Leu
180 185 190
Gly Leu Gly Leu Ala Leu Gly Leu Leu Gly Pro Leu Ala Met Leu Leu
195 200 205
Ala Leu Leu Leu Leu Arg Arg Asp Gln Arg Leu Pro Pro Asp Ala Pro
210 215 220
Lys Ala Pro Gly Gly Gly Ser Phe Arg Thr Pro Ile Gln Glu Glu Gln
225 230 235 240
Ala Asp Ala His Ser Ala Leu Ala Lys Ile
245 250
<210> 135
<211> 442
<212> БЕЛОК
<213> OX40-mFc Homo sapiens
<400> 135
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp
20 25 30
Arg Cys Cys His Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys
35 40 45
Ser Arg Ser Gln Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr
50 55 60
Asn Asp Val Val Ser Ser Lys Pro Cys Lys Pro Cys Thr Trp Cys Asn
65 70 75 80
Leu Arg Ser Gly Ser Glu Arg Lys Gln Leu Cys Thr Ala Thr Gln Asp
85 90 95
Thr Val Cys Arg Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys
100 105 110
Pro Gly Val Asp Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly
115 120 125
Asp Asn Gln Ala Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys
130 135 140
His Thr Leu Gln Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp
145 150 155 160
Arg Asp Pro Pro Ala Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala
165 170 175
Arg Pro Ile Thr Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln
180 185 190
Gly Pro Ser Thr Arg Pro Val Glu Val Pro Gly Gly Arg Ala Val Ala
195 200 205
Gly Ser Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys
210 215 220
Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
225 230 235 240
Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys
245 250 255
Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp
260 265 270
Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg
275 280 285
Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln
290 295 300
His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val Asn Asn
305 310 315 320
Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly
325 330 335
Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu
340 345 350
Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met
355 360 365
Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu
370 375 380
Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe
385 390 395 400
Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn
405 410 415
Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr
420 425 430
Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
435 440
<210> 136
<211> 447
<212> БЕЛОК
<213> OX40 мышиных-mFc
<400> 136
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Val Thr Ala Arg Arg Leu Asn Cys Val Lys His Thr
20 25 30
Tyr Pro Ser Gly His Lys Cys Cys Arg Glu Cys Gln Pro Gly His Gly
35 40 45
Met Val Ser Arg Cys Asp His Thr Arg Asp Thr Leu Cys His Pro Cys
50 55 60
Glu Thr Gly Phe Tyr Asn Glu Ala Val Asn Tyr Asp Thr Cys Lys Gln
65 70 75 80
Cys Thr Gln Cys Asn His Arg Ser Gly Ser Glu Leu Lys Gln Asn Cys
85 90 95
Thr Pro Thr Gln Asp Thr Val Cys Arg Cys Arg Pro Gly Thr Gln Pro
100 105 110
Arg Gln Asp Ser Gly Tyr Lys Leu Gly Val Asp Cys Val Pro Cys Pro
115 120 125
Pro Gly His Phe Ser Pro Gly Asn Asn Gln Ala Cys Lys Pro Trp Thr
130 135 140
Asn Cys Thr Leu Ser Gly Lys Gln Thr Arg His Pro Ala Ser Asp Ser
145 150 155 160
Leu Asp Ala Val Cys Glu Asp Arg Ser Leu Leu Ala Thr Leu Leu Trp
165 170 175
Glu Thr Gln Arg Pro Thr Phe Arg Pro Thr Thr Val Gln Ser Thr Thr
180 185 190
Val Trp Pro Arg Thr Ser Glu Leu Pro Ser Pro Pro Thr Leu Val Thr
195 200 205
Pro Glu Gly Pro Ala Gly Ser Pro Arg Gly Pro Thr Ile Lys Pro Cys
210 215 220
Pro Pro Cys Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Ile Phe Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser
245 250 255
Pro Ile Val Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp
260 265 270
Val Gln Ile Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln
275 280 285
Thr Gln Thr His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser
290 295 300
Ala Leu Pro Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys
305 310 315 320
Cys Lys Val Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile
325 330 335
Ser Lys Pro Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro
340 345 350
Pro Pro Glu Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met
355 360 365
Val Thr Asp Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn
370 375 380
Gly Lys Thr Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn
405 410 415
Trp Val Glu Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu
420 425 430
His Asn His His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
435 440 445
<210> 137
<211> 444
<212> БЕЛОК
<213> OX40 яванской макаки-mFc
<400> 137
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Leu His Cys Val Gly Asp Thr Tyr Pro Ser Asn Asp
20 25 30
Arg Cys Cys Gln Glu Cys Arg Pro Gly Asn Gly Met Val Ser Arg Cys
35 40 45
Asn Arg Ser Gln Asn Thr Val Cys Arg Pro Cys Gly Pro Gly Phe Tyr
50 55 60
Asn Asp Val Val Ser Ala Lys Pro Cys Lys Ala Cys Thr Trp Cys Asn
65 70 75 80
Leu Arg Ser Gly Ser Glu Arg Lys Gln Pro Cys Thr Ala Thr Gln Asp
85 90 95
Thr Val Cys Arg Cys Arg Ala Gly Thr Gln Pro Leu Asp Ser Tyr Lys
100 105 110
Pro Gly Val Asp Cys Ala Pro Cys Pro Pro Gly His Phe Ser Pro Gly
115 120 125
Asp Asn Gln Ala Cys Lys Pro Trp Thr Asn Cys Thr Leu Ala Gly Lys
130 135 140
His Thr Leu Gln Pro Ala Ser Asn Ser Ser Asp Ala Ile Cys Glu Asp
145 150 155 160
Arg Asp Pro Pro Pro Thr Gln Pro Gln Glu Thr Gln Gly Pro Pro Ala
165 170 175
Arg Pro Thr Thr Val Gln Pro Thr Glu Ala Trp Pro Arg Thr Ser Gln
180 185 190
Arg Pro Ser Thr Arg Pro Val Glu Val Pro Arg Gly Pro Ala Val Ala
195 200 205
Ala Ile Gly Ser Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys
210 215 220
Lys Cys Pro Ala Pro Asn Leu Leu Gly Gly Pro Ser Val Phe Ile Phe
225 230 235 240
Pro Pro Lys Ile Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val
245 250 255
Thr Cys Val Val Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile
260 265 270
Ser Trp Phe Val Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr
275 280 285
His Arg Glu Asp Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro
290 295 300
Ile Gln His Gln Asp Trp Met Ser Gly Lys Glu Phe Lys Cys Lys Val
305 310 315 320
Asn Asn Lys Asp Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro
325 330 335
Lys Gly Ser Val Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu
340 345 350
Glu Glu Met Thr Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp
355 360 365
Phe Met Pro Glu Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr
370 375 380
Glu Leu Asn Tyr Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Tyr Phe Met Tyr Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu
405 410 415
Arg Asn Ser Tyr Ser Cys Ser Val Val His Glu Gly Leu His Asn His
420 425 430
His Thr Thr Lys Ser Phe Ser Arg Thr Pro Gly Lys
435 440
<210> 138
<211> 164
<212> БЕЛОК
<213> Последовательность CD137 Homo sapiens для применения с
рекомбинантным антигеном CD137-mFc-Avi
<400> 138
Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp
1 5 10 15
Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser
20 25 30
Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly
35 40 45
Val Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
65 70 75 80
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
85 90 95
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
115 120 125
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
130 135 140
Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
145 150 155 160
His Ser Pro Gln
<210> 139
<211> 164
<212> БЕЛОК
<213> Последовательность CD137 яванской макаки для применения с
рекомбинантными антигенами CD137-mFc-Avi и CD137-Avi-His
<400> 139
Ser Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp
1 5 10 15
Asn Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser
20 25 30
Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly
35 40 45
Val Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
Asp Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys
65 70 75 80
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
85 90 95
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
115 120 125
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
130 135 140
Pro Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
145 150 155 160
His Ser Pro Gln
<210> 140
<211> 165
<212> БЕЛОК
<213> Последовательность CD137 мышиных для применения с
рекомбинантным антигеном CD137-mFc-Avi
<400> 140
Ala Val Gln Asn Ser Cys Asp Asn Cys Gln Pro Gly Thr Phe Cys Arg
1 5 10 15
Lys Tyr Asn Pro Val Cys Lys Ser Cys Pro Pro Ser Thr Phe Ser Ser
20 25 30
Ile Gly Gly Gln Pro Asn Cys Asn Ile Cys Arg Val Cys Ala Gly Tyr
35 40 45
Phe Arg Phe Lys Lys Phe Cys Ser Ser Thr His Asn Ala Glu Cys Glu
50 55 60
Cys Ile Glu Gly Phe His Cys Leu Gly Pro Gln Cys Thr Arg Cys Glu
65 70 75 80
Lys Asp Cys Arg Pro Gly Gln Glu Leu Thr Lys Gln Gly Cys Lys Thr
85 90 95
Cys Ser Leu Gly Thr Phe Asn Asp Gln Asn Gly Thr Gly Val Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Arg Ser Val Leu Lys Thr Gly
115 120 125
Thr Thr Glu Lys Asp Val Val Cys Gly Pro Pro Val Val Ser Phe Ser
130 135 140
Pro Ser Thr Thr Ile Ser Val Thr Pro Glu Gly Gly Pro Gly Gly His
145 150 155 160
Ser Leu Gln Val Leu
165
<210> 141
<211> 249
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> mFc-Avi для применения с рекомбинантными антигенами
CD137-mFc-Avi
<400> 141
Pro Arg Gly Pro Thr Ile Lys Pro Cys Pro Pro Cys Lys Cys Pro Ala
1 5 10 15
Pro Asn Leu Glu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro Lys Ile
20 25 30
Lys Asp Val Leu Met Ile Ser Leu Ser Pro Ile Val Thr Cys Val Val
35 40 45
Val Asp Val Ser Glu Asp Asp Pro Asp Val Gln Ile Ser Trp Phe Val
50 55 60
Asn Asn Val Glu Val His Thr Ala Gln Thr Gln Thr His Arg Glu Asp
65 70 75 80
Tyr Asn Ser Thr Leu Arg Val Val Ser Ala Leu Pro Ile Gln His Gln
85 90 95
Asp Trp Met Ser Gly Lys Ala Phe Ala Cys Ala Val Asn Asn Lys Asp
100 105 110
Leu Pro Ala Pro Ile Glu Arg Thr Ile Ser Lys Pro Lys Gly Ser Val
115 120 125
Arg Ala Pro Gln Val Tyr Val Leu Pro Pro Pro Glu Glu Glu Met Thr
130 135 140
Lys Lys Gln Val Thr Leu Thr Cys Met Val Thr Asp Phe Met Pro Glu
145 150 155 160
Asp Ile Tyr Val Glu Trp Thr Asn Asn Gly Lys Thr Glu Leu Asn Tyr
165 170 175
Lys Asn Thr Glu Pro Val Leu Asp Ser Asp Gly Ser Tyr Phe Met Tyr
180 185 190
Ser Lys Leu Arg Val Glu Lys Lys Asn Trp Val Glu Arg Asn Ser Tyr
195 200 205
Ser Cys Ser Val Val His Glu Gly Leu His Asn His His Thr Thr Lys
210 215 220
Ser Phe Ser Arg Thr Pro Gly Lys Gly Gly Gly Leu Asn Asp Ile Phe
225 230 235 240
Glu Ala Gln Lys Ile Glu Trp His Glu
245
<210> 142
<400> 142
000
<210> 143
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS20-22-49
<400> 143
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gtactgggac
60
caggaagtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
120
tgggagagca atggggatga acagttcgca tacaagacca cgcctcccgt gctggactcc
180
gacggctcct tcttcctcta cagcaagctc accgtggatc agtataggtg gaatcctgct
240
gattatttct catgctccgt gatgcatgag gctctgcaca accactacac tcagaagagc
300
ttgtccctgt cgcccgga
318
<210> 144
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-5-37 (без LALA)
<400> 144
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Asn Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Ser Gly Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Tyr Asp Lys Tyr Trp Gly Ser Ser Ile Tyr Ser Gly Leu
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
325 330 335
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Tyr Trp Asp
355 360 365
Gln Glu Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Asp Glu Gln Phe Ala Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Lys Leu Thr Val Asp Gln Tyr Arg Trp Asn Pro Ala Asp Tyr Phe Ser
420 425 430
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Pro Gly
450
<210> 145
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-5-37 (без LALA)
<400> 145
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
aattgcgcgg ccagtggctt taccttcagt agctatgcca tgagctgggt gcgtcaggcg
120
ccgggcaaag gtctggaatg ggttagcgcg attagcggta gtggcggtag cacgtactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagatcttac
300
gacaaatact ggggttcttc tatttactct ggcttggact actggggcca gggaaccctg
360
gtcaccgtct cgagtgctag cactaagggc ccgtcggtgt tcccgctggc cccatcgtcc
420
aagagcacat cagggggtac cgccgccctg ggctgccttg tgaaggatta ctttcccgag
480
cccgtcacag tgtcctggaa cagcggagcc ctgacctccg gagtgcatac tttcccggct
540
gtgcttcagt cctctggcct gtactcattg tcctccgtgg tcaccgtccc ttcgtcctcc
600
ctgggcaccc agacctatat ctgtaatgtc aaccataagc cctcgaacac caaggtcgac
660
aagaaggtcg agccgaagtc gtgcgacaag actcacactt gcccgccttg cccagccccg
720
gaactgctgg gtggtccttc ggtgttcctc ttcccgccca agccgaagga taccctgatg
780
atctcacgga cccccgaagt gacctgtgtg gtggtggacg tgtcccacga ggacccggaa
840
gtgaaattca attggtacgt ggatggagtg gaagtgcaca acgccaagac caagccacgg
900
gaagaacagt acaactctac ctaccgcgtg gtgtccgtgc tcactgtgct gcaccaagac
960
tggctgaacg ggaaggagta caagtgcaaa gtgtccaaca aggcgctgcc tgccccaatt
1020
gagaaaacta tctcgaaagc caagggacag cctcgagaac cacaggtgta caccctgccc
1080
ccatcccggg atgagtactg ggaccaggaa gtcagcctga cctgcctggt caaaggcttc
1140
tatcccagcg acatcgccgt ggagtgggag agcaatgggg atgaacagtt cgcatacaag
1200
accacgcctc ccgtgctgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg
1260
gatcagtata ggtggaatcc tgctgattat ttctcatgct ccgtgatgca tgaggctctg
1320
cacaaccact acactcagaa gagcttgtcc ctgtcgcccg ga
1362
<210> 146
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-3 (без LALA)
<400> 146
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Asn Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 147
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-3 (без LALA)
<400> 147
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
aatgtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagtgctagc
360
actaagggcc cgtcggtgtt cccgctggcc ccatcgtcca agagcacatc agggggtacc
420
gccgccctgg gctgccttgt gaaggattac tttcccgagc ccgtcacagt gtcctggaac
480
agcggagccc tgacctccgg agtgcatact ttcccggctg tgcttcagtc ctctggcctg
540
tactcattgt cctccgtggt caccgtccct tcgtcctccc tgggcaccca gacctatatc
600
tgtaatgtca accataagcc ctcgaacacc aaggtcgaca agaaggtcga gccgaagtcg
660
tgcgacaaga ctcacacttg cccgccttgc ccagccccgg aactgctggg tggtccttcg
720
gtgttcctct tcccgcccaa gccgaaggat accctgatga tctcacggac ccccgaagtg
780
acctgtgtgg tggtggacgt gtcccacgag gacccggaag tgaaattcaa ttggtacgtg
840
gatggagtgg aagtgcacaa cgccaagacc aagccacggg aagaacagta caactctacc
900
taccgcgtgg tgtccgtgct cactgtgctg caccaagact ggctgaacgg gaaggagtac
960
aagtgcaaag tgtccaacaa ggcgctgcct gccccaattg agaaaactat ctcgaaagcc
1020
aagggacagc ctcgagaacc acaggtgtac accctgcccc catcccggga tgagtactgg
1080
gaccaggaag tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg
1140
gagtgggaga gcaatgggga tgaacagttc gcatacaaga ccacgcctcc cgtgctggac
1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg atcagtatag gtggaatcct
1260
gctgattatt tctcatgctc cgtgatgcat gaggctctgc acaaccacta cactcagaag
1320
agcttgtccc tgtcgcccgg a
1341
<210> 148
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-12 (без LALA)
<400> 148
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Thr Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 149
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-12 (без LALA)
<400> 149
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
acggtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagtgctagc
360
actaagggcc cgtcggtgtt cccgctggcc ccatcgtcca agagcacatc agggggtacc
420
gccgccctgg gctgccttgt gaaggattac tttcccgagc ccgtcacagt gtcctggaac
480
agcggagccc tgacctccgg agtgcatact ttcccggctg tgcttcagtc ctctggcctg
540
tactcattgt cctccgtggt caccgtccct tcgtcctccc tgggcaccca gacctatatc
600
tgtaatgtca accataagcc ctcgaacacc aaggtcgaca agaaggtcga gccgaagtcg
660
tgcgacaaga ctcacacttg cccgccttgc ccagccccgg aactgctggg tggtccttcg
720
gtgttcctct tcccgcccaa gccgaaggat accctgatga tctcacggac ccccgaagtg
780
acctgtgtgg tggtggacgt gtcccacgag gacccggaag tgaaattcaa ttggtacgtg
840
gatggagtgg aagtgcacaa cgccaagacc aagccacggg aagaacagta caactctacc
900
taccgcgtgg tgtccgtgct cactgtgctg caccaagact ggctgaacgg gaaggagtac
960
aagtgcaaag tgtccaacaa ggcgctgcct gccccaattg agaaaactat ctcgaaagcc
1020
aagggacagc ctcgagaacc acaggtgtac accctgcccc catcccggga tgagtactgg
1080
gaccaggaag tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg
1140
gagtgggaga gcaatgggga tgaacagttc gcatacaaga ccacgcctcc cgtgctggac
1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg atcagtatag gtggaatcct
1260
gctgattatt tctcatgctc cgtgatgcat gaggctctgc acaaccacta cactcagaag
1320
agcttgtccc tgtcgcccgg a
1341
<210> 150
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-16 (без LALA)
<400> 150
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Leu Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Gln Tyr
405 410 415
Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 151
<211> 1341
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-10-16 (без LALA)
<400> 151
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt agttacgata tgagctgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat attgatccga ctggtagcaa gaccgactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagagacctc
300
ttggtgtacg ggttcgacta ctggggccag ggaaccctgg tcaccgtctc gagtgctagc
360
actaagggcc cgtcggtgtt cccgctggcc ccatcgtcca agagcacatc agggggtacc
420
gccgccctgg gctgccttgt gaaggattac tttcccgagc ccgtcacagt gtcctggaac
480
agcggagccc tgacctccgg agtgcatact ttcccggctg tgcttcagtc ctctggcctg
540
tactcattgt cctccgtggt caccgtccct tcgtcctccc tgggcaccca gacctatatc
600
tgtaatgtca accataagcc ctcgaacacc aaggtcgaca agaaggtcga gccgaagtcg
660
tgcgacaaga ctcacacttg cccgccttgc ccagccccgg aactgctggg tggtccttcg
720
gtgttcctct tcccgcccaa gccgaaggat accctgatga tctcacggac ccccgaagtg
780
acctgtgtgg tggtggacgt gtcccacgag gacccggaag tgaaattcaa ttggtacgtg
840
gatggagtgg aagtgcacaa cgccaagacc aagccacggg aagaacagta caactctacc
900
taccgcgtgg tgtccgtgct cactgtgctg caccaagact ggctgaacgg gaaggagtac
960
aagtgcaaag tgtccaacaa ggcgctgcct gccccaattg agaaaactat ctcgaaagcc
1020
aagggacagc ctcgagaacc acaggtgtac accctgcccc catcccggga tgagtactgg
1080
gaccaggaag tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg
1140
gagtgggaga gcaatgggga tgaacagttc gcatacaaga ccacgcctcc cgtgctggac
1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg atcagtatag gtggaatcct
1260
gctgattatt tctcatgctc cgtgatgcat gaggctctgc acaaccacta cactcagaag
1320
agcttgtccc tgtcgcccgg a
1341
<210> 152
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-35-14 (без LALA)
<400> 152
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr
20 25 30
Asn Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Ser Pro Tyr Gly Gly Ala Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Tyr Glu Leu Ser Ala Tyr Ser Tyr Gly Ala Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Tyr Trp Asp Gln Glu
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Asp Glu Gln Phe Ala Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Gln Tyr Arg Trp Asn Pro Ala Asp Tyr Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 153
<211> 1356
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность тяжелой цепи
FS20-22-49/FS30-35-14 (без LALA)
<400> 153
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt gcctataata tccattgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat atttctccgt atggtggcgc gaccaactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagaaacctc
300
tacgagttga gcgcttactc ttacggggcg gactactggg gccagggaac cctggtcacc
360
gtctcgtcgg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc
420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc
480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt
540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc
600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag
660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaactg
720
ctgggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca
780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa
840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa
900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg
960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa
1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc
1080
cgggatgagt actgggacca ggaagtcagc ctgacctgcc tggtcaaagg cttctatccc
1140
agcgacatcg ccgtggagtg ggagagcaat ggggatgaac agttcgcata caagaccacg
1200
cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggatcag
1260
tataggtgga atcctgctga ttatttctca tgctccgtga tgcatgaggc tctgcacaac
1320
cactacactc agaagagctt gtccctgtcg cccgga
1356
<210> 154
<211> 447
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1AA/FS30-10-16 (с LALA)
<400> 154
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Asp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Asp Pro Thr Gly Ser Lys Thr Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Leu Val Tyr Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 155
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1AA/OX86 (с LALA)
<400> 155
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Gly Tyr
20 25 30
Asn Leu His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Met Arg Tyr Asp Gly Asp Thr Tyr Tyr Asn Ser Val Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Asn Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Thr
85 90 95
Arg Asp Gly Arg Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Val Met
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 156
<211> 219
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ G1/OX86 и G1AA/OX86
<400> 156
Asp Ile Val Met Thr Gln Gly Ala Leu Pro Asn Pro Val Pro Ser Gly
1 5 10 15
Glu Ser Ala Ser Ile Thr Cys Arg Ser Ser Gln Ser Leu Val Tyr Lys
20 25 30
Asp Gly Gln Thr Tyr Leu Asn Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Thr Tyr Trp Met Ser Thr Arg Ala Ser Gly Val Ser
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Tyr Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Arg Ala Glu Asp Ala Gly Val Tyr Tyr Cys Gln Gln Val
85 90 95
Arg Glu Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 157
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20m-232-91AA/4420 (с LALA)
<400> 157
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Phe Asp Pro Met Tyr Tyr Tyr Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Glu Pro Leu Trp Asp Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Trp Arg Asp Arg Trp Glu Asp Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 158
<211> 189
<212> БЕЛОК
<213> Homo sapiens CD137-Avi-His
<400> 158
Ser Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp
1 5 10 15
Asn Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser
20 25 30
Ser Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly
35 40 45
Val Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys
50 55 60
Asp Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys
65 70 75 80
Glu Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys
85 90 95
Asp Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg
100 105 110
Pro Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly
115 120 125
Thr Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser
130 135 140
Pro Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly
145 150 155 160
His Ser Pro Gln Gly Ser Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala
165 170 175
Gln Lys Ile Glu Trp His Glu His His His His His His
180 185
<210> 159
<211> 446
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1/OX86 (без LALA)
<400> 159
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Gly Tyr
20 25 30
Asn Leu His Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Met
35 40 45
Gly Arg Met Arg Tyr Asp Gly Asp Ile Tyr Tyr Asn Ser Val Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Arg Asp Thr Ser Lys Asn Gln Val Phe Leu
65 70 75 80
Lys Met Asn Ser Leu Gln Thr Asp Asp Thr Ala Ile Tyr Tyr Cys Thr
85 90 95
Arg Asp Gly Arg Gly Asp Ser Phe Asp Tyr Trp Gly Gln Gly Val Met
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 160
<211> 443
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ к PD-1 G1AA/5C4
<400> 160
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro
210 215 220
Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro
225 230 235 240
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
245 250 255
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
260 265 270
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
275 280 285
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
290 295 300
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
305 310 315 320
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
325 330 335
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
340 345 350
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
355 360 365
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
370 375 380
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
385 390 395 400
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
405 410 415
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
420 425 430
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440
<210> 161
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ к PD-1 G1AA/5C4
<400> 161
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 162
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ к PD-L1 G1AA/S1
<400> 162
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 163
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ к PD-L1 G1AA/S1
<400> 163
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Phe Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 164
<211> 233
<212> БЕЛОК
<213> CD137 мышиных
<400> 164
Val Gln Asn Ser Cys Asp Asn Cys Gln Pro Gly Thr Phe Cys Arg Lys
1 5 10 15
Tyr Asn Pro Val Cys Lys Ser Cys Pro Pro Ser Thr Phe Ser Ser Ile
20 25 30
Gly Gly Gln Pro Asn Cys Asn Ile Cys Arg Val Cys Ala Gly Tyr Phe
35 40 45
Arg Phe Lys Lys Phe Cys Ser Ser Thr His Asn Ala Glu Cys Glu Cys
50 55 60
Ile Glu Gly Phe His Cys Leu Gly Pro Gln Cys Thr Arg Cys Glu Lys
65 70 75 80
Asp Cys Arg Pro Gly Gln Glu Leu Thr Lys Gln Gly Cys Lys Thr Cys
85 90 95
Ser Leu Gly Thr Phe Asn Asp Gln Asn Gly Thr Gly Val Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Arg Ser Val Leu Lys Thr Gly Thr
115 120 125
Thr Glu Lys Asp Val Val Cys Gly Pro Pro Val Val Ser Phe Ser Pro
130 135 140
Ser Thr Thr Ile Ser Val Thr Pro Glu Gly Gly Pro Gly Gly His Ser
145 150 155 160
Leu Gln Val Leu Thr Leu Phe Leu Ala Leu Thr Ser Ala Leu Leu Leu
165 170 175
Ala Leu Ile Phe Ile Thr Leu Leu Phe Ser Val Leu Lys Trp Ile Arg
180 185 190
Lys Lys Phe Pro His Ile Phe Lys Gln Pro Phe Lys Lys Thr Thr Gly
195 200 205
Ala Ala Gln Glu Glu Asp Ala Cys Ser Cys Arg Cys Pro Gln Glu Glu
210 215 220
Glu Gly Gly Gly Gly Gly Tyr Glu Leu
225 230
<210> 165
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1AA/20H4.9
<400> 165
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Gly Gly Tyr Val Thr Tyr Asn Pro Ser Leu Glu
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Asp Tyr Gly Pro Gly Asn Tyr Asp Trp Tyr Phe Asp Leu Trp Gly
100 105 110
Arg Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 166
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1AA/3H3
<400> 166
Glu Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Met Lys Leu Ser Cys Ala Gly Ser Gly Phe Thr Leu Ser Asp Tyr
20 25 30
Gly Val Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Tyr Ala Gly Gly Thr Thr Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Ile Asp Gly Tyr Gly Gly Tyr Ser Gly Ser His Trp Tyr Phe Asp
100 105 110
Phe Trp Gly Pro Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 167
<211> 213
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ G1AA/3H3 и G1/3H3 и МАТ2 FS20m-232-91AA/3H3
<400> 167
Asp Ile Gln Met Thr Gln Ser Pro Ser Leu Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Leu Asn Cys Arg Thr Ser Gln Asn Val Tyr Lys Asn
20 25 30
Leu Ala Trp Tyr Gln Gln Gln Leu Gly Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asn Ala Asn Ser Leu Gln Ala Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Phe Cys Gln Gln Tyr Tyr Ser Gly Asn Thr
85 90 95
Phe Gly Ala Gly Thr Asn Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 168
<211> 454
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1/3H3
<400> 168
Glu Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Met Lys Leu Ser Cys Ala Gly Ser Gly Phe Thr Leu Ser Asp Tyr
20 25 30
Gly Val Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Tyr Ala Gly Gly Thr Thr Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Ile Asp Gly Tyr Gly Gly Tyr Ser Gly Ser His Trp Tyr Phe Asp
100 105 110
Phe Trp Gly Pro Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
355 360 365
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
370 375 380
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
385 390 395 400
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
405 410 415
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
420 425 430
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
435 440 445
Ser Leu Ser Pro Gly Lys
450
<210> 169
<211> 459
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS20m-232-91AA/3H3 (с LALA)
<400> 169
Glu Met Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Ser Met Lys Leu Ser Cys Ala Gly Ser Gly Phe Thr Leu Ser Asp Tyr
20 25 30
Gly Val Ala Trp Val Arg Gln Ala Pro Lys Lys Gly Leu Glu Trp Val
35 40 45
Ala Tyr Ile Ser Tyr Ala Gly Gly Thr Thr Tyr Tyr Arg Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Thr Leu Tyr
65 70 75 80
Leu Gln Met Asp Ser Leu Arg Ser Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Ile Asp Gly Tyr Gly Gly Tyr Ser Gly Ser His Trp Tyr Phe Asp
100 105 110
Phe Trp Gly Pro Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys
115 120 125
Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly
130 135 140
Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro
145 150 155 160
Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr
165 170 175
Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val
180 185 190
Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn
195 200 205
Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro
210 215 220
Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
225 230 235 240
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
245 250 255
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
260 265 270
Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
275 280 285
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn
290 295 300
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
305 310 315 320
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
325 330 335
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
340 345 350
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Phe Asp Pro
355 360 365
Met Tyr Tyr Tyr Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
370 375 380
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Glu Pro Leu
385 390 395 400
Trp Asp Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
405 410 415
Phe Leu Tyr Ser Lys Leu Thr Val Trp Arg Asp Arg Trp Glu Asp Gly
420 425 430
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
435 440 445
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 170
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-35-14
<400> 170
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ala Tyr
20 25 30
Asn Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Asp Ile Ser Pro Tyr Gly Gly Ala Thr Asn Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asn Leu Tyr Glu Leu Ser Ala Tyr Ser Tyr Gly Ala Asp Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 171
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен тяжелой цепи МАТ FS30-35-14
<400> 171
gaagtgcaac tgctggagtc cggtggtggt ctggtacagc cgggtggttc tctgcgtctg
60
agttgcgcgg ccagtggctt taccttcagt gcctataata tccattgggt gcgtcaggct
120
ccgggcaaag gtctggaatg ggttagcgat atttctccgt atggtggcgc gaccaactat
180
gcggatagcg tgaaaggccg ttttaccatt tctcgcgaca acagcaagaa cacgctgtac
240
ctgcagatga actcactgcg tgccgaagat acggccgtgt attactgtgc gagaaacctc
300
tacgagttga gcgcttactc ttacggggcg gactactggg gccagggaac cctggtcacc
360
gtctcgtcg
369
<210> 172
<211> 109
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-35-14
<400> 172
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Gly Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Tyr Ser Ser
85 90 95
Pro Ile Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 173
<211> 327
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Вариабельный домен легкой цепи МАТ FS30-35-14
<400> 173
gaaattgtgc tgacccagtc tccgggcacg ttatctctga gccctggtga gcgcgccact
60
ctgtcatgcc gggcttctca aagtgttagc agtagctacc tggcgtggta tcagcaaaaa
120
ccgggccagg ccccgcgtct gctgatttac ggtgcatcca gccgtgccac cggcattcca
180
gatcgttttt ccggtagtgg ttctgggacg gacttcactc tgacaatctc acgcctggaa
240
ccggaggatt ttgcggtgta ttactgccag caatattatt attcttctcc tatcacgttc
300
ggccaaggga ccaaggtgga aatcaaa
327
<210> 174
<211> 452
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ к OX40 G1/11D4
<400> 174
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ser Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Tyr Ile Ser Ser Ser Ser Ser Thr Ile Asp Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Ser Gly Trp Tyr Leu Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Arg Phe Tyr Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Asp Ile Phe Pro Asn Gly Leu Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Pro Tyr Pro Ser Trp Leu Met Gly Thr Arg Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 175
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ к OX40 G1AA/11D4 и G1/11D4
<400> 175
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Glu Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТ PD-L1 и CD137 | 2019 |
|
RU2826084C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ PD-L1 и LAG-3 | 2017 |
|
RU2784388C2 |
| МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2793167C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ МЕЗОТЕЛИН И CD137 | 2019 |
|
RU2815066C2 |
| ИММУНОАКТИВИРУЮЩАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2722788C2 |
| АНТИ-ОХ40 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2753493C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| ПОЛИВАЛЕНТНЫЕ И ПОЛИСПЕЦИФИЧЕСКИЕ ОХ40-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2773052C2 |
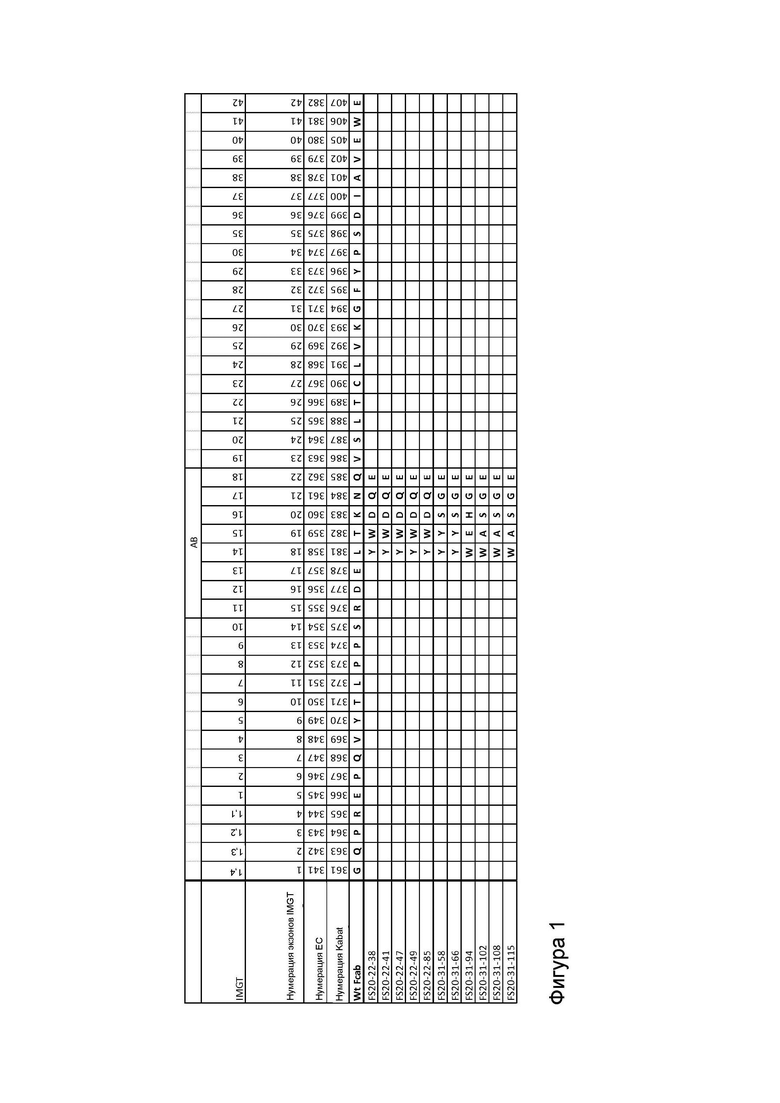

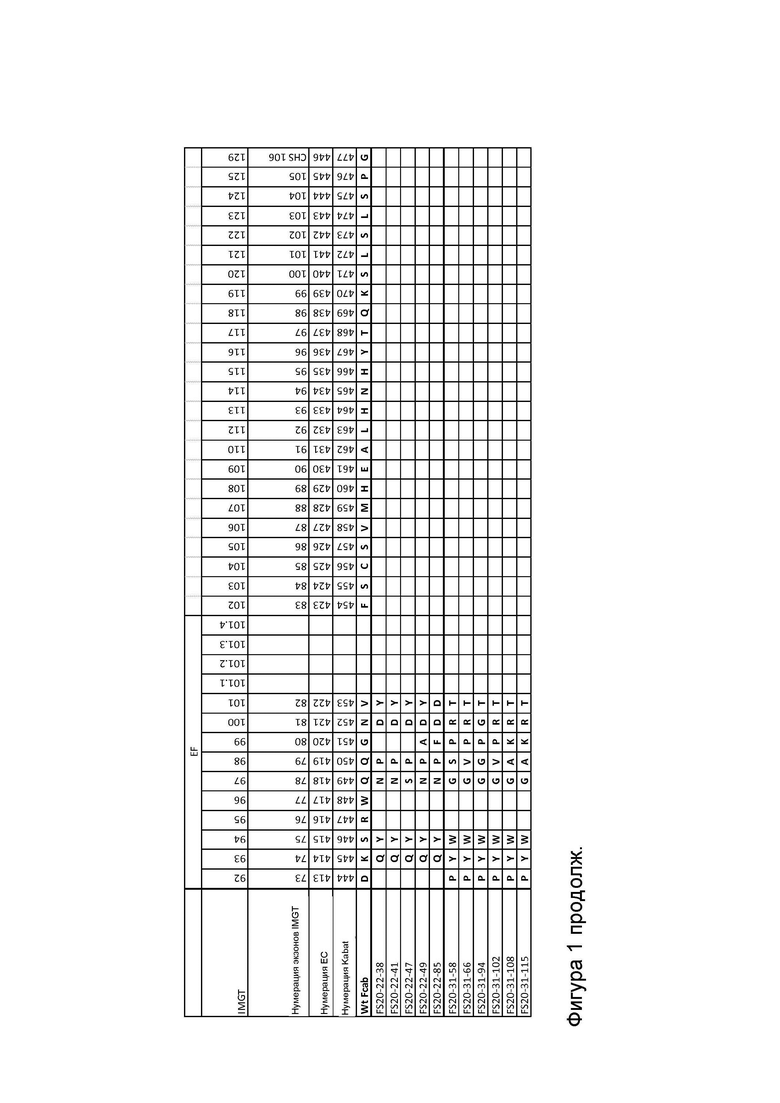
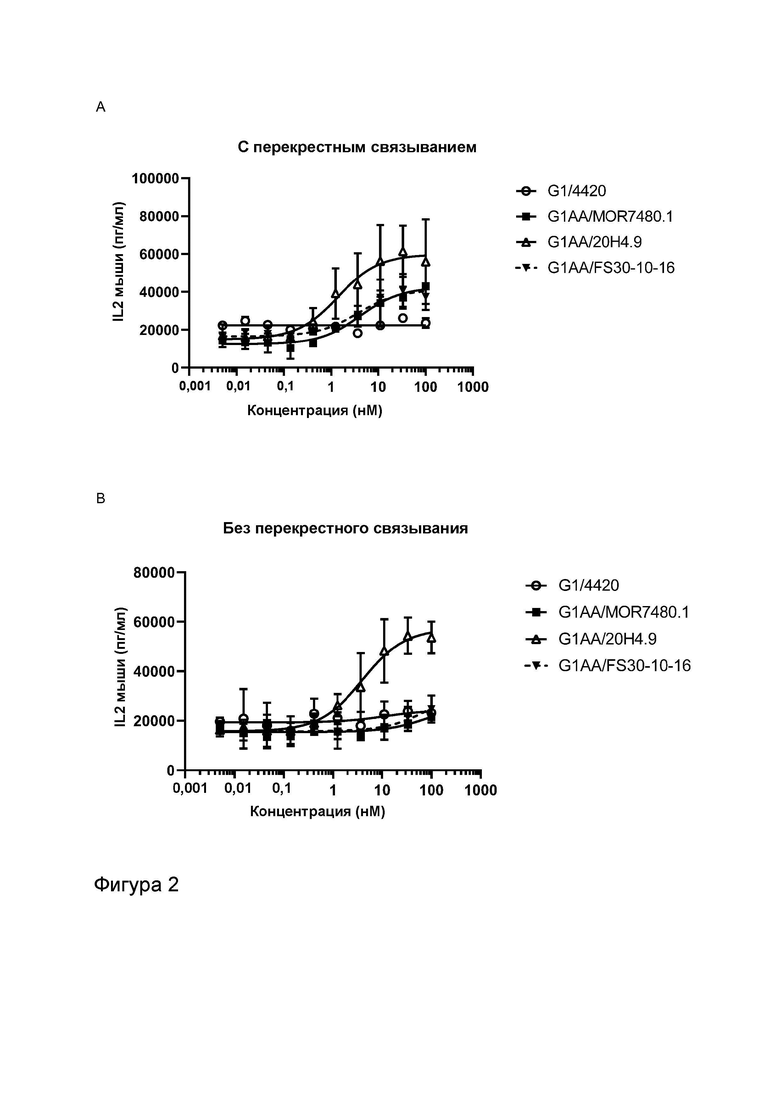
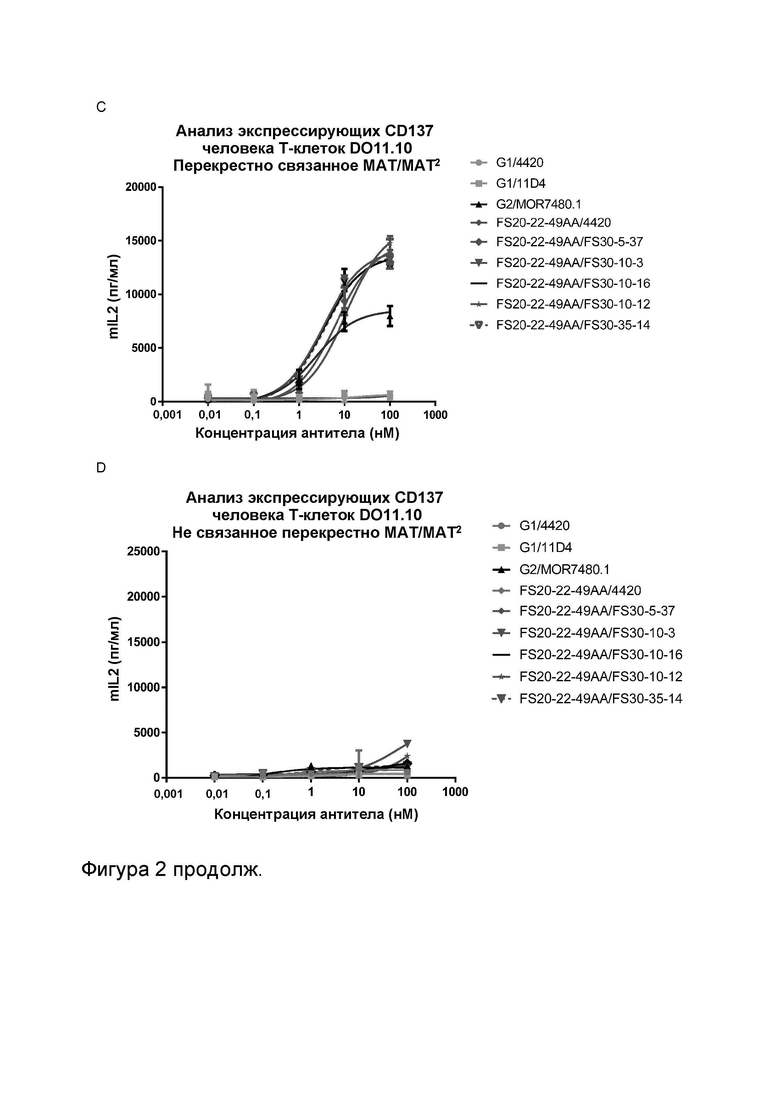
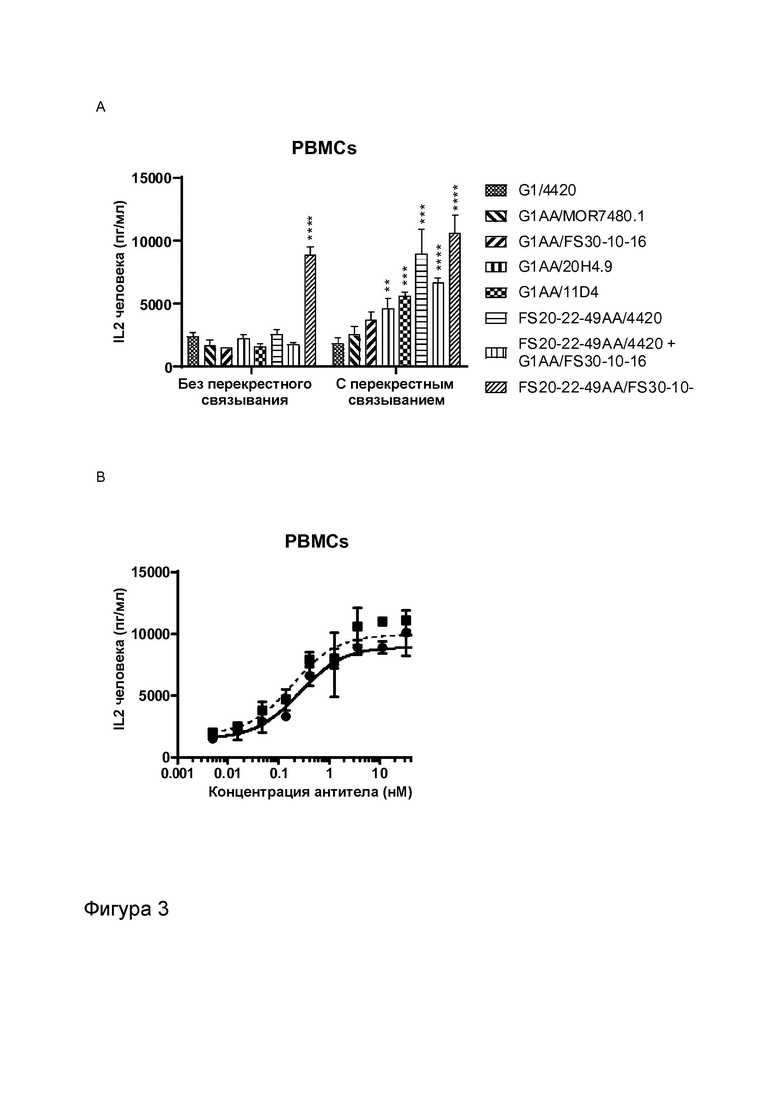
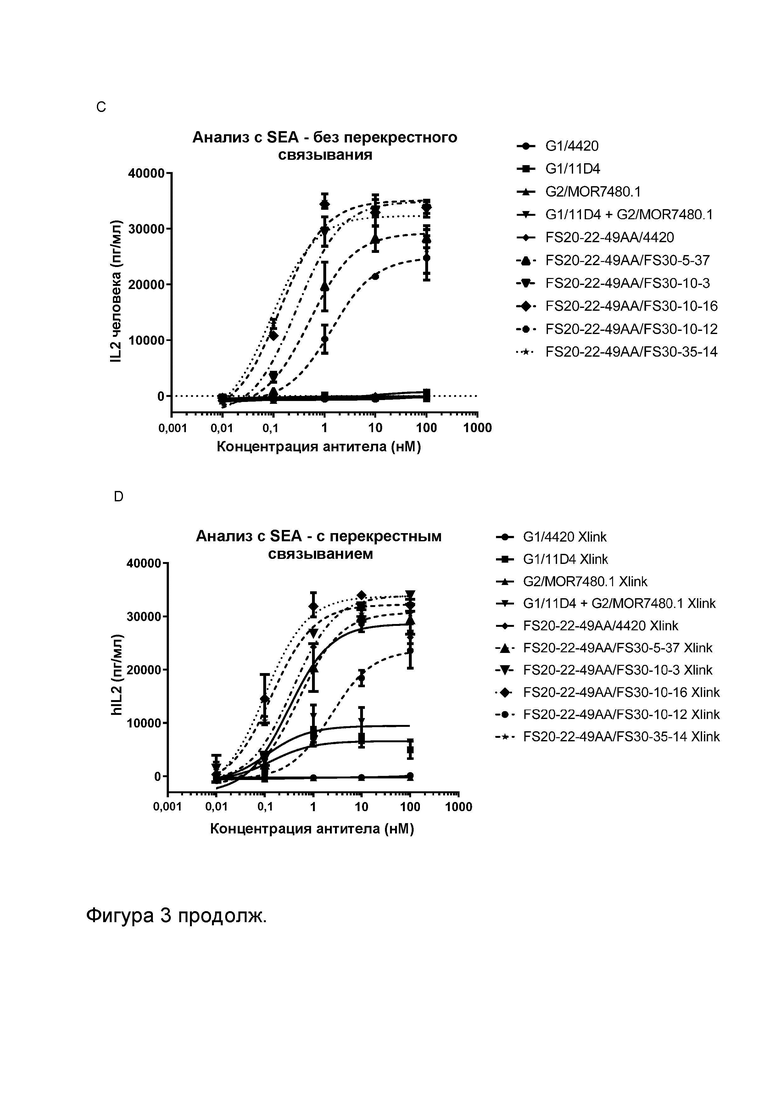
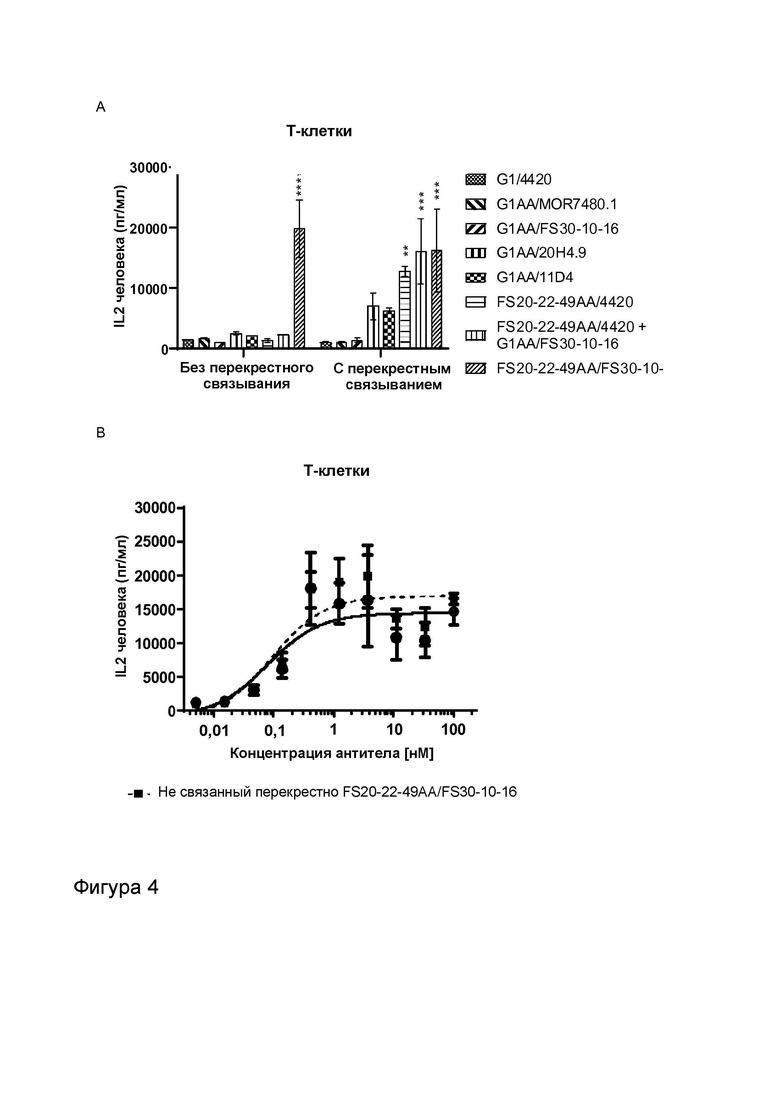
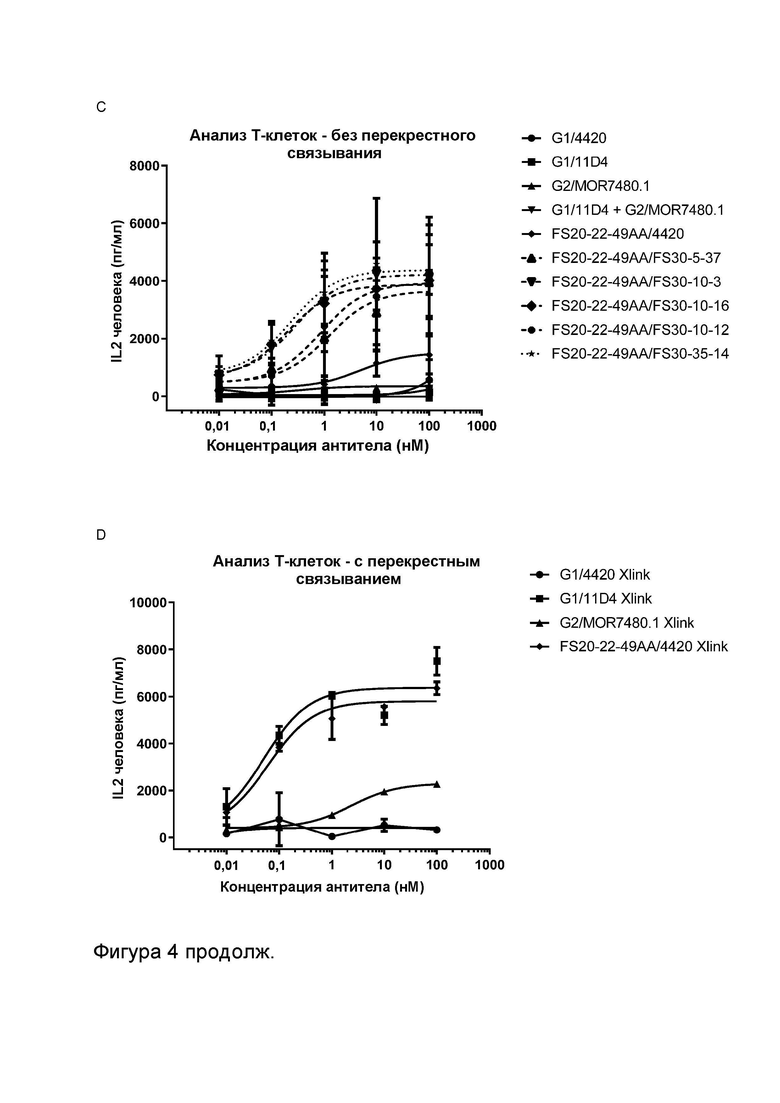
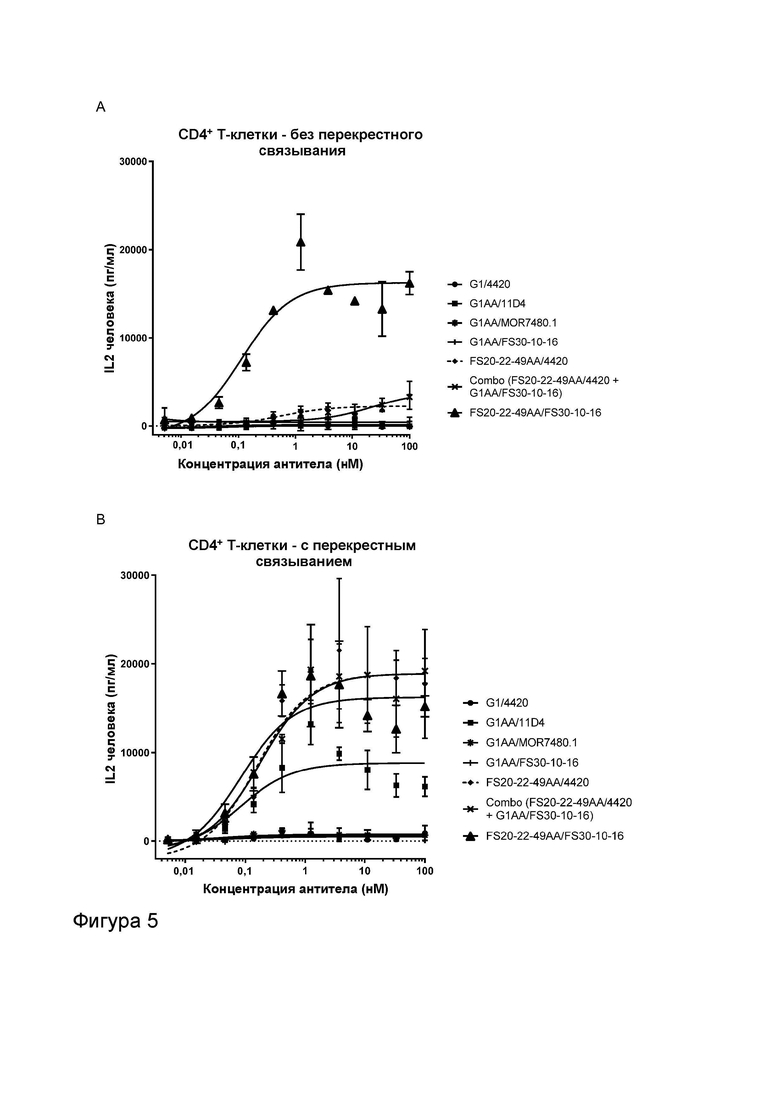
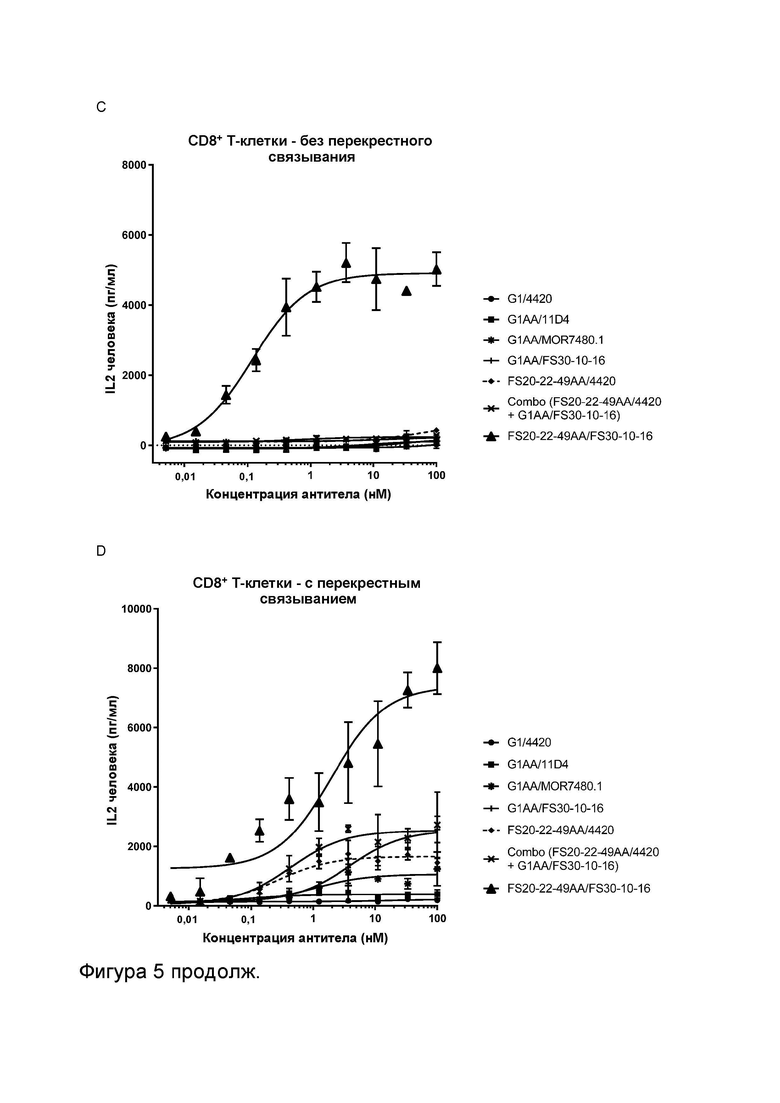
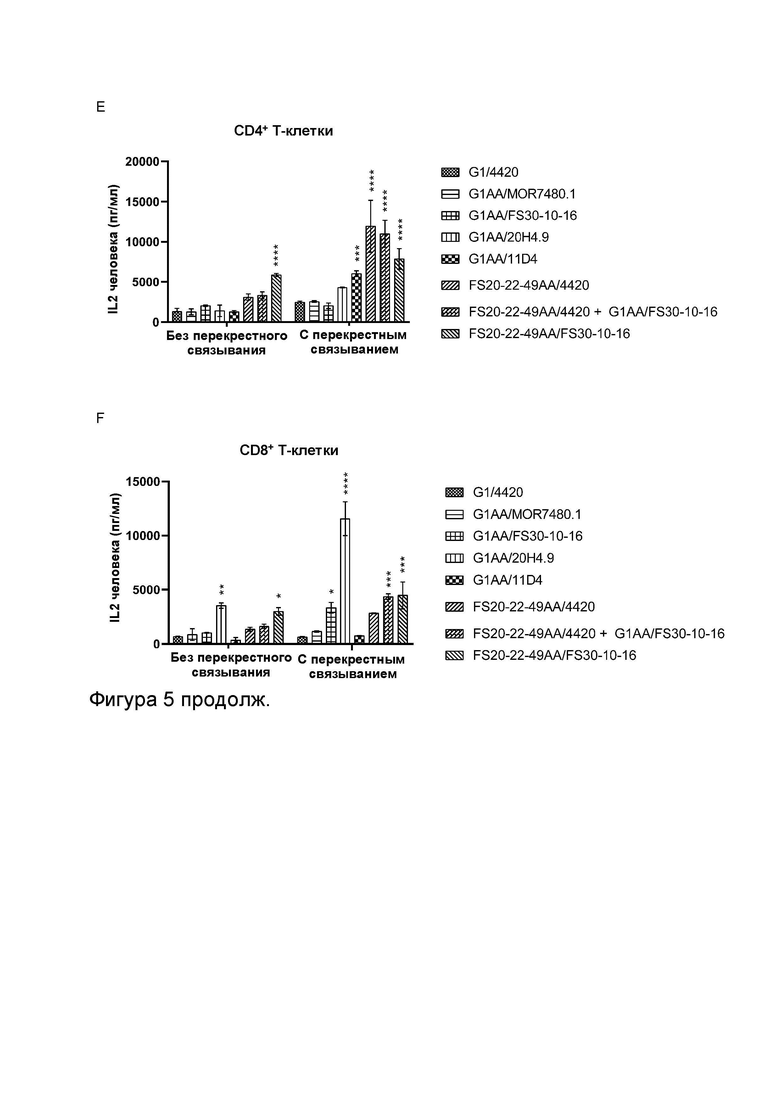
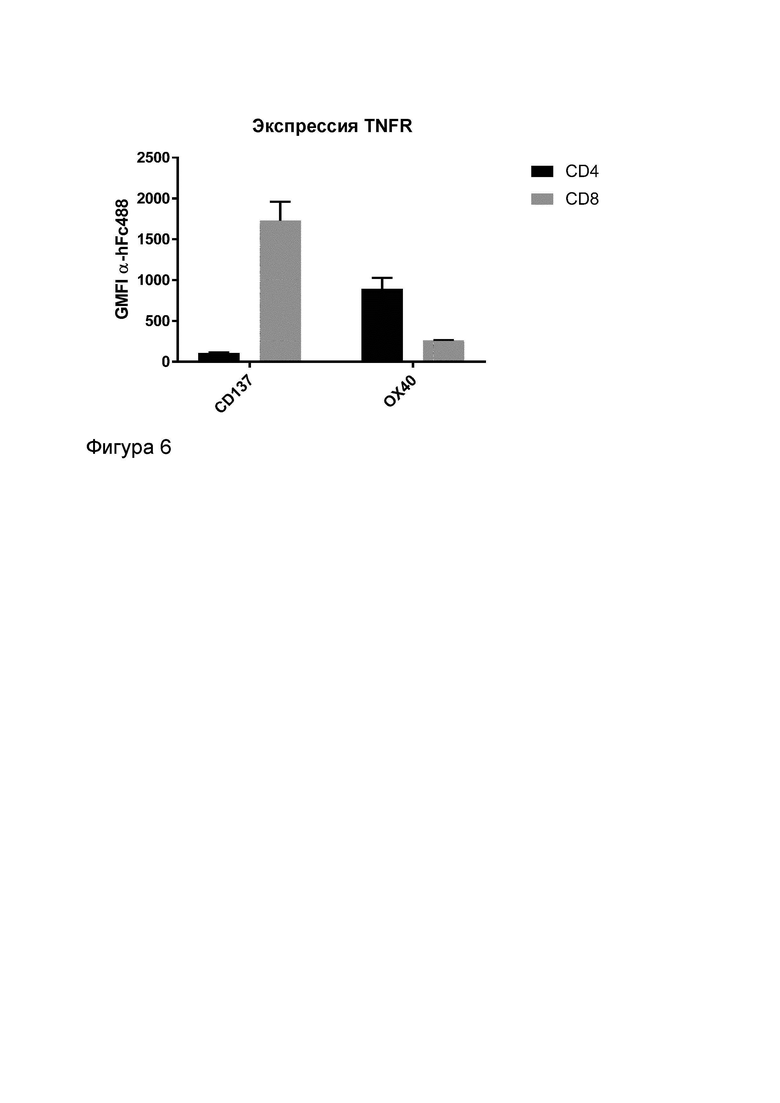
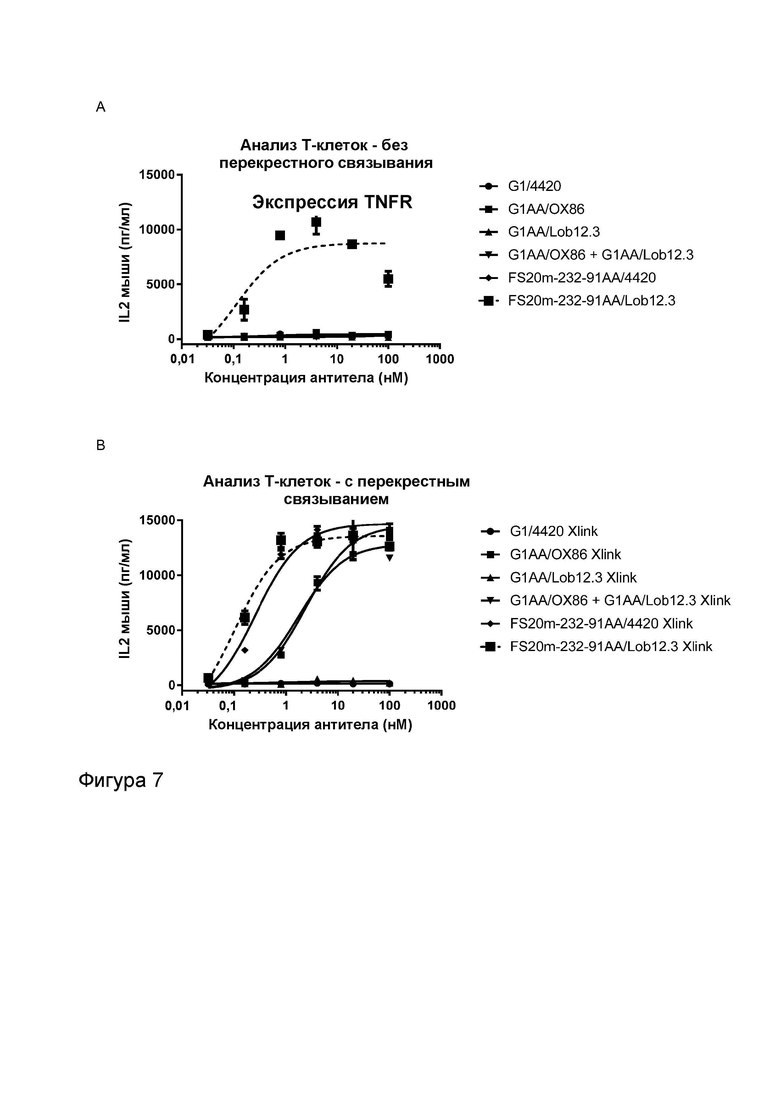
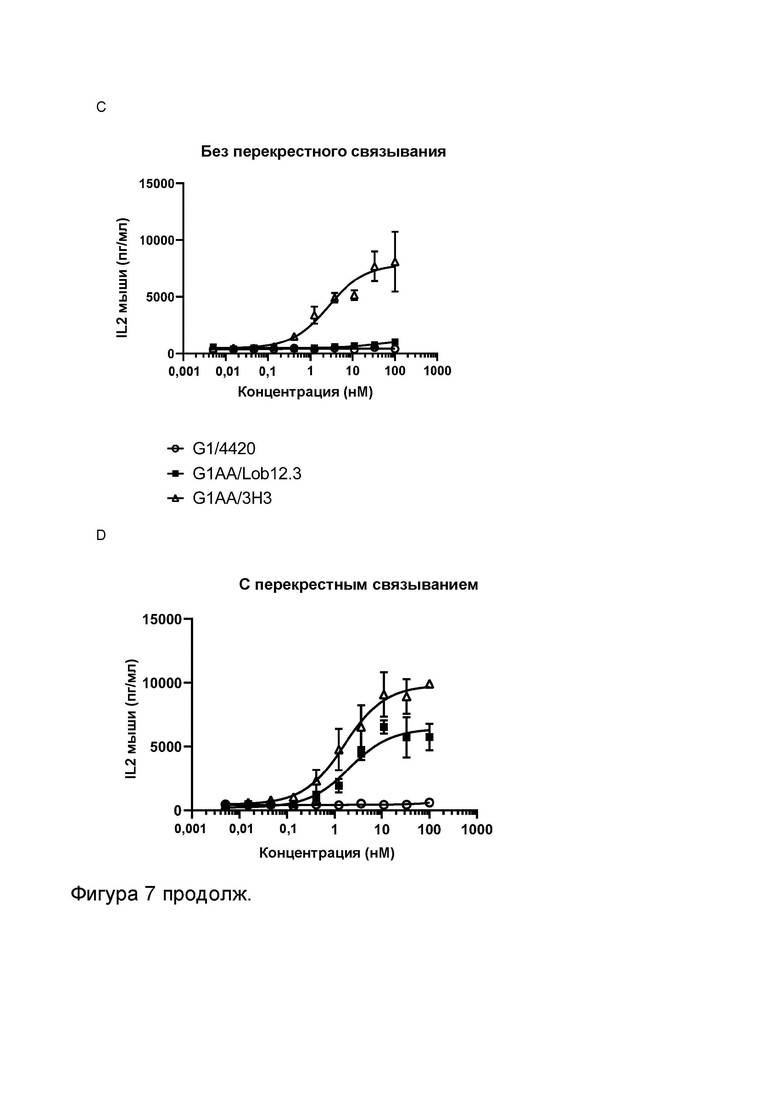
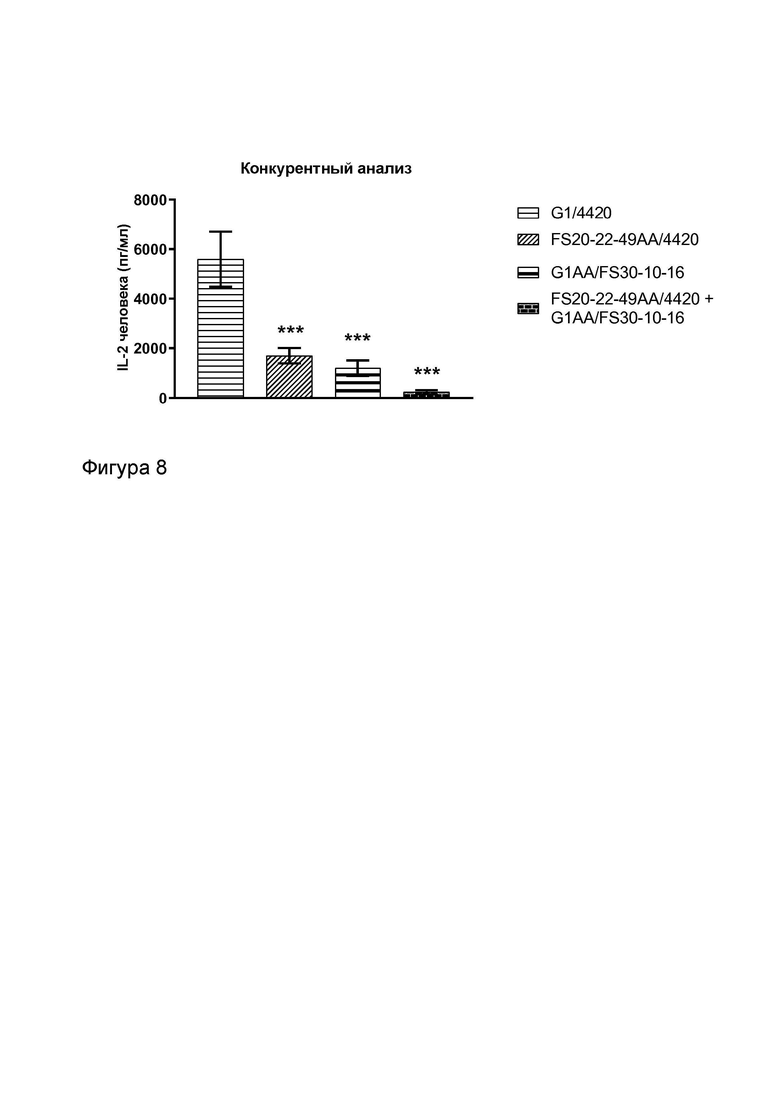
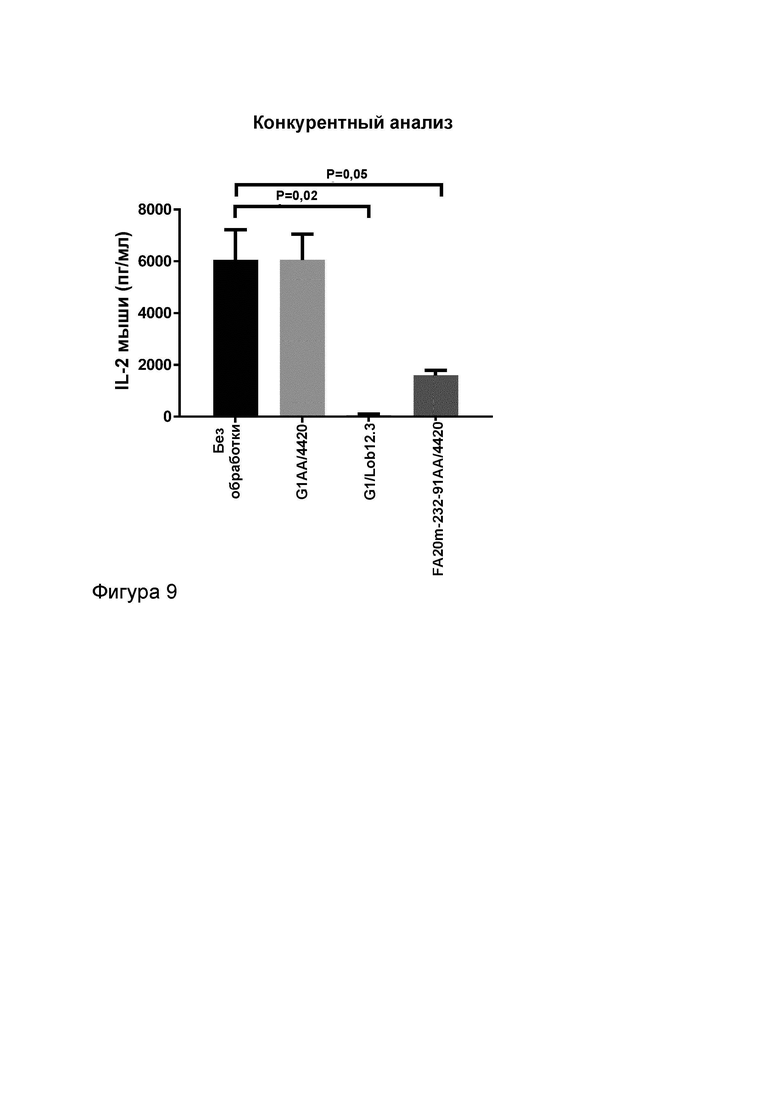
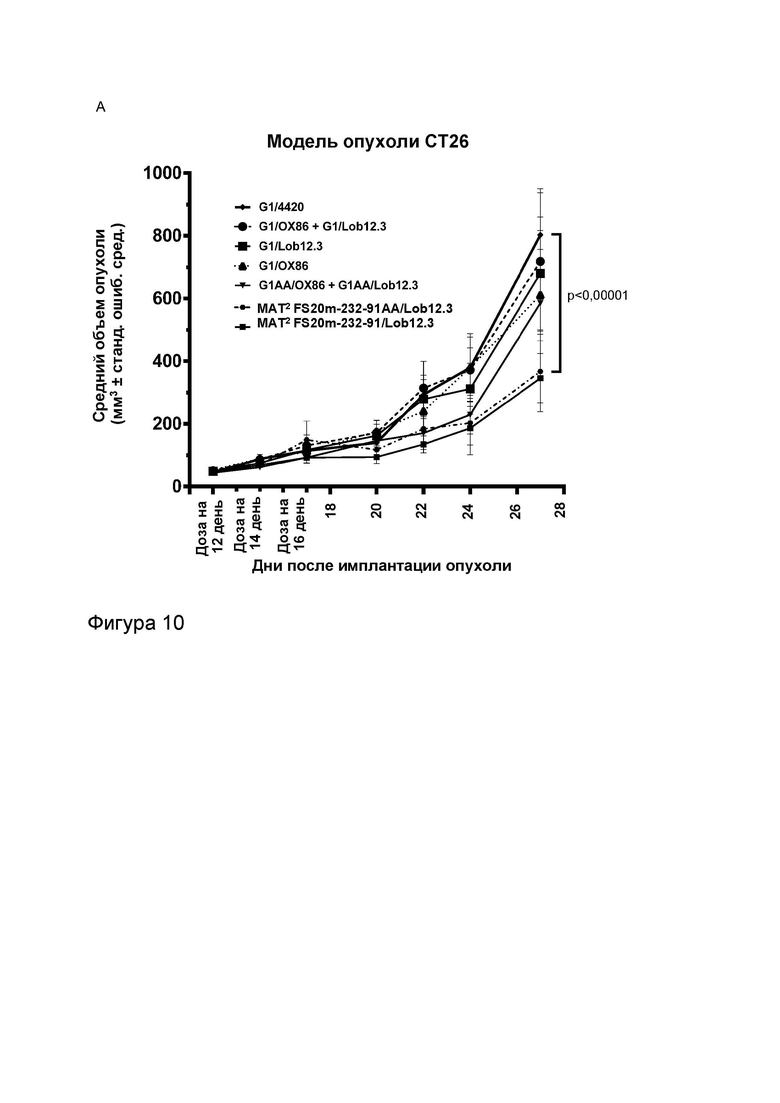
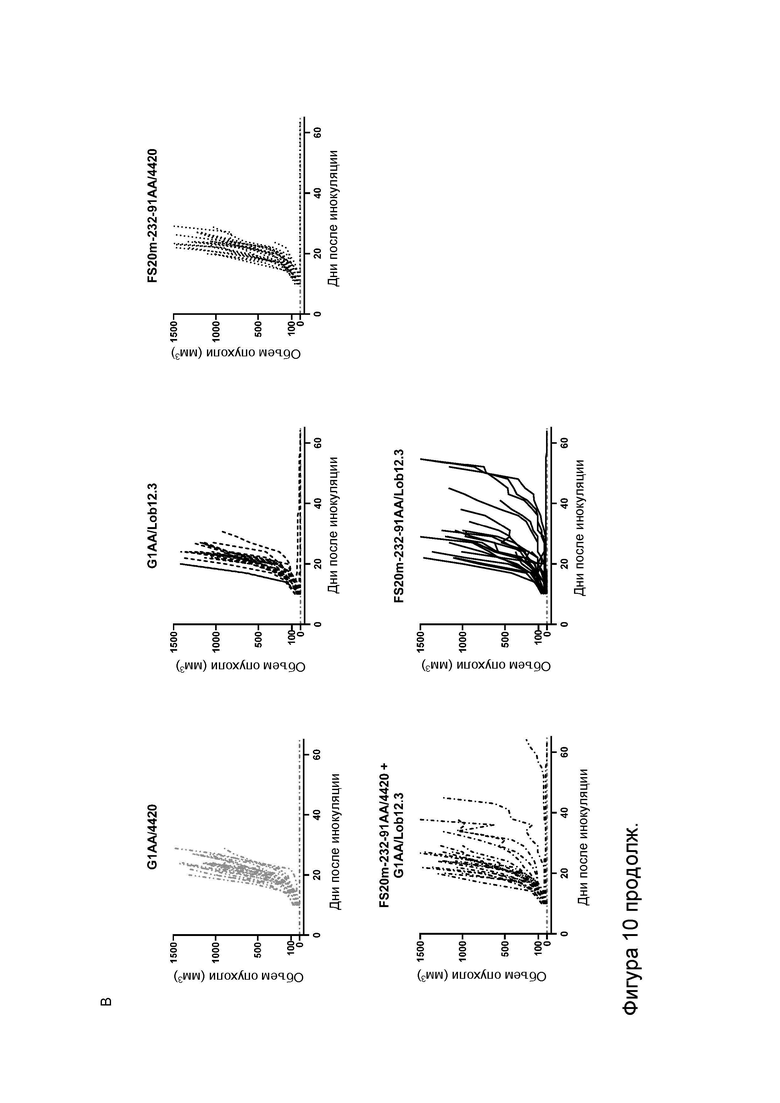
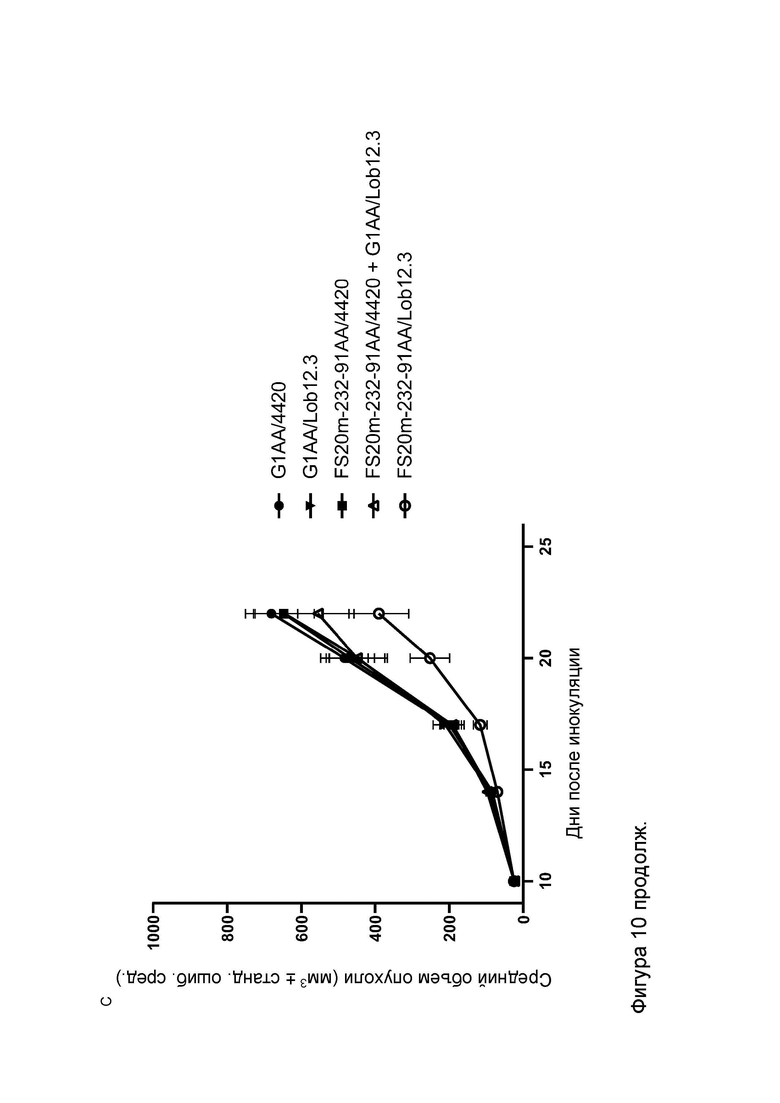
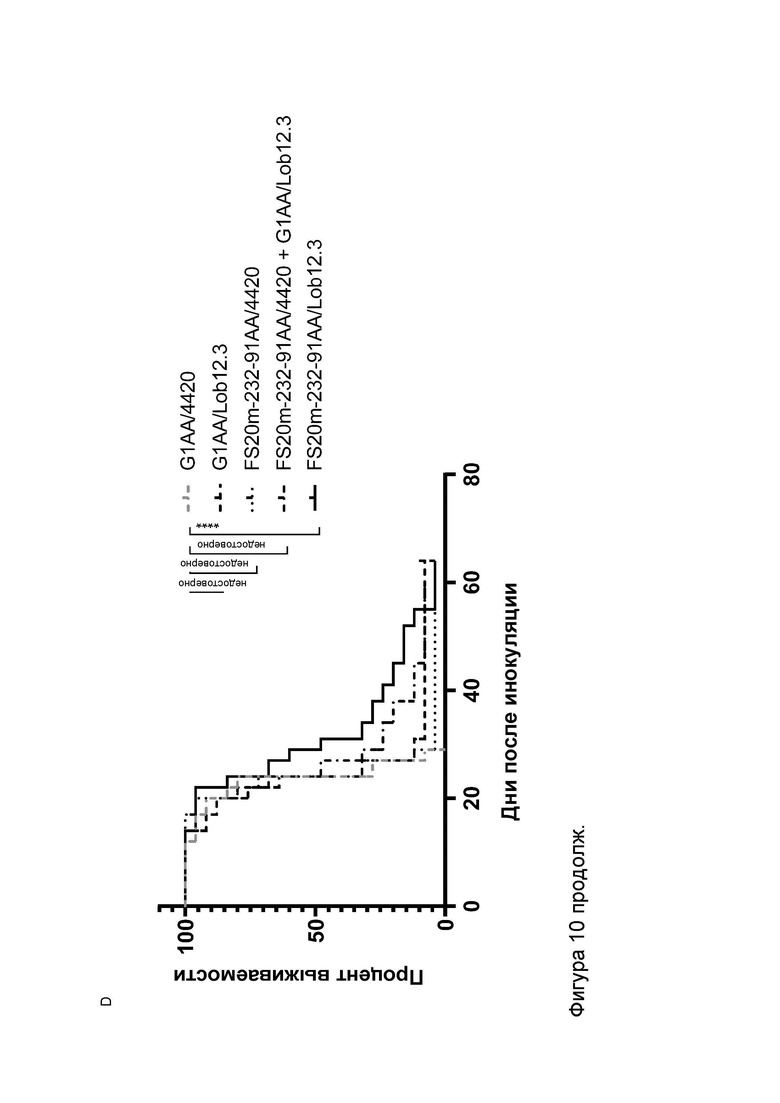
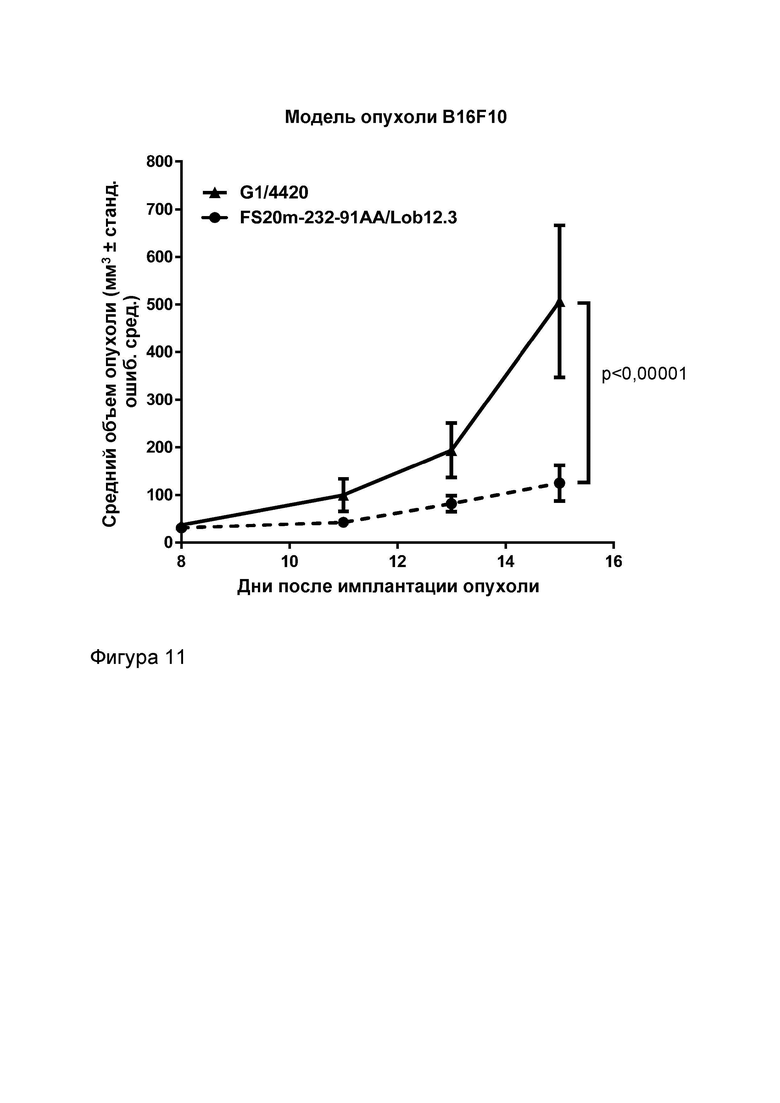
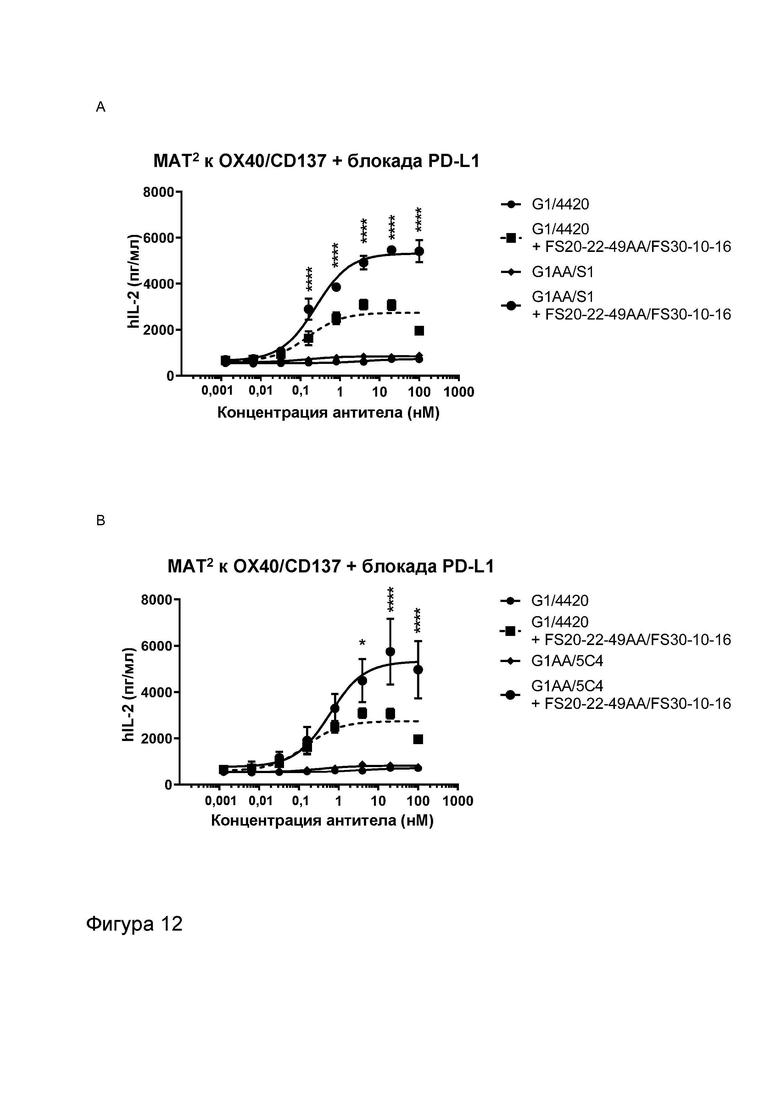
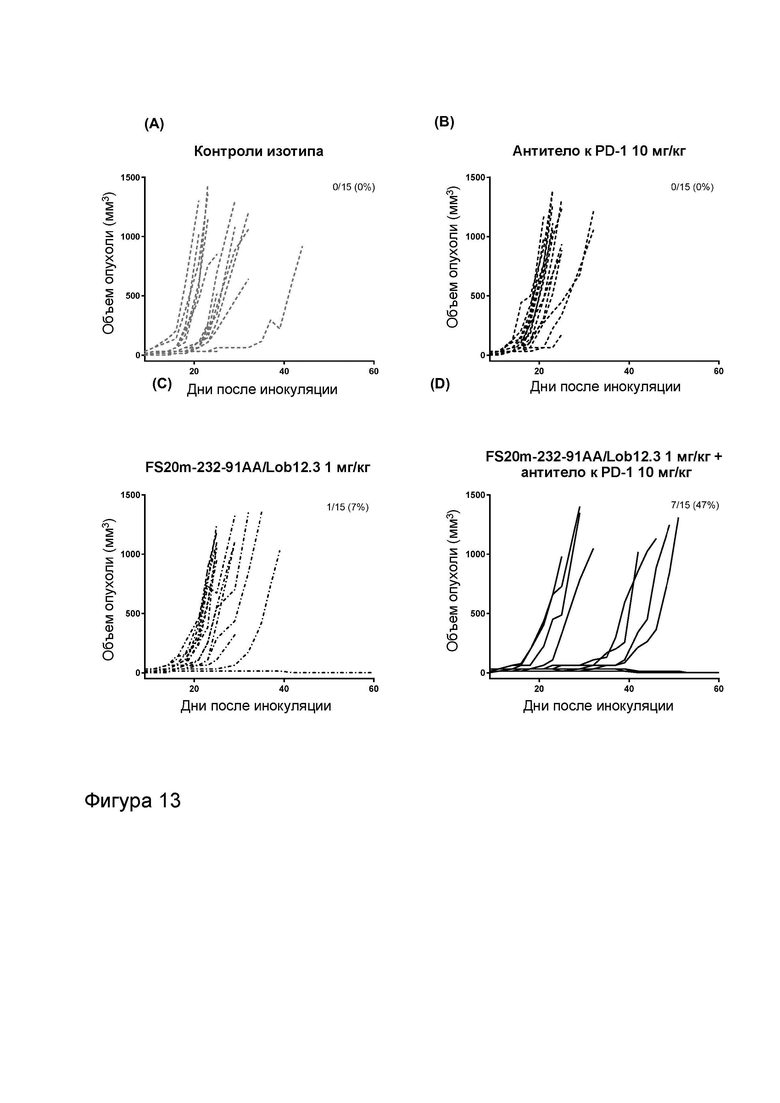
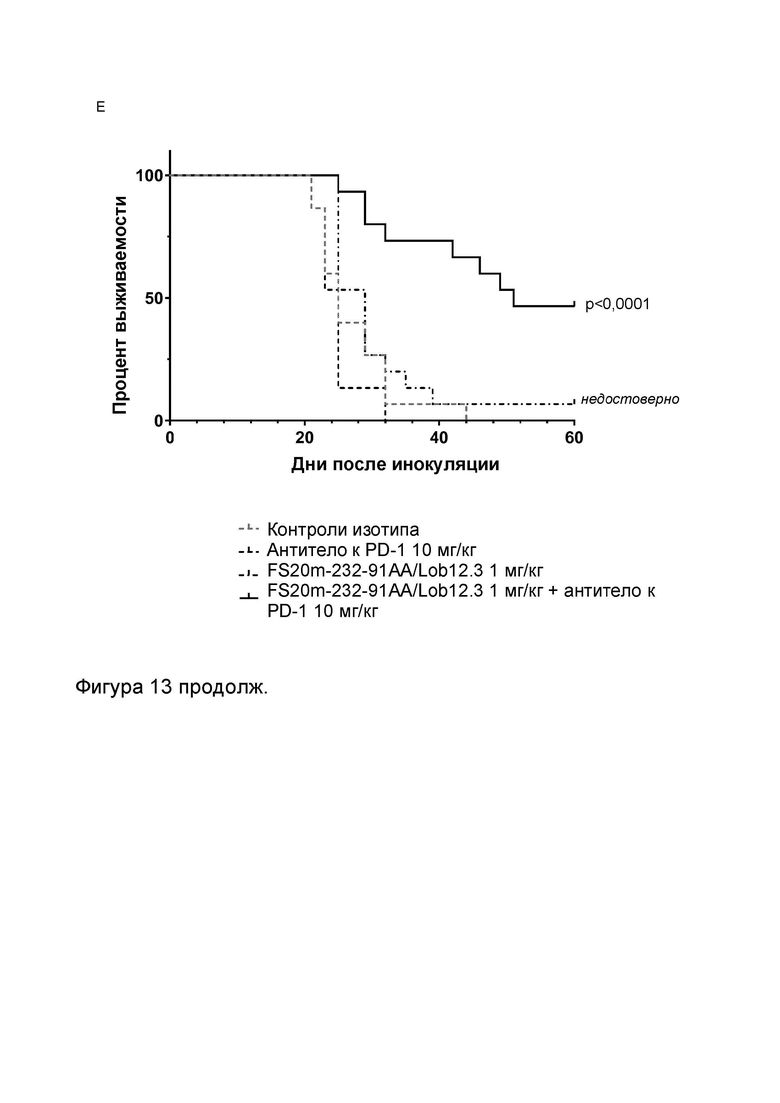
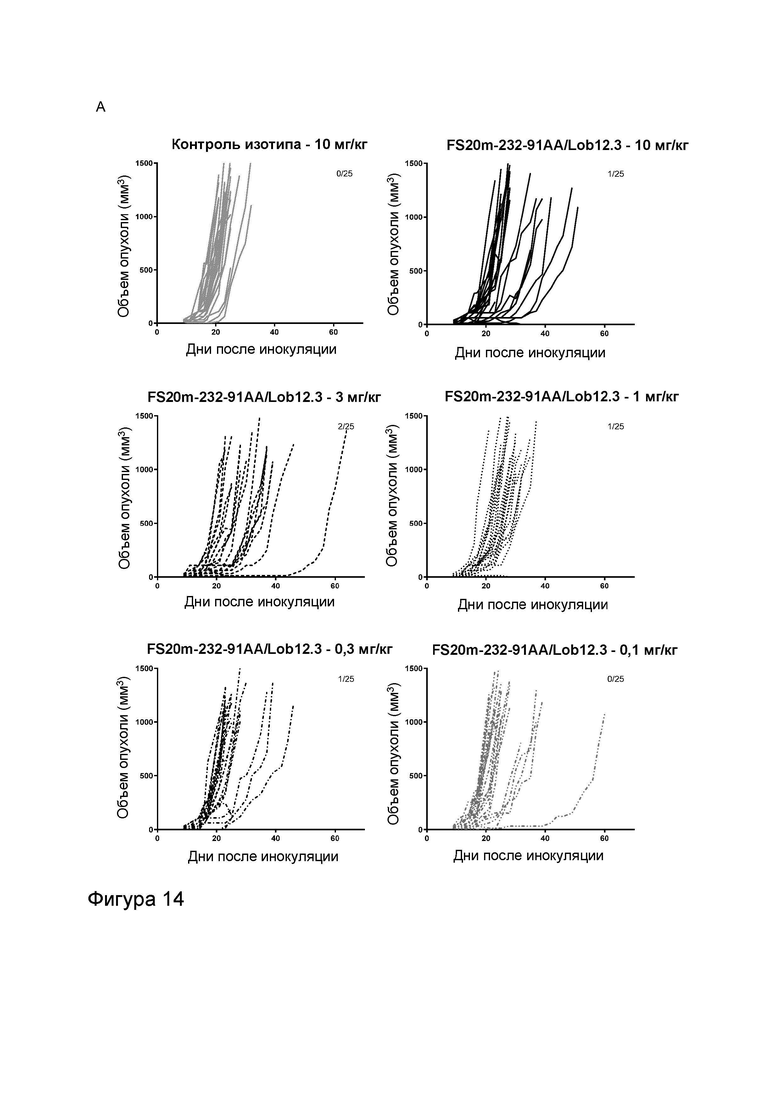
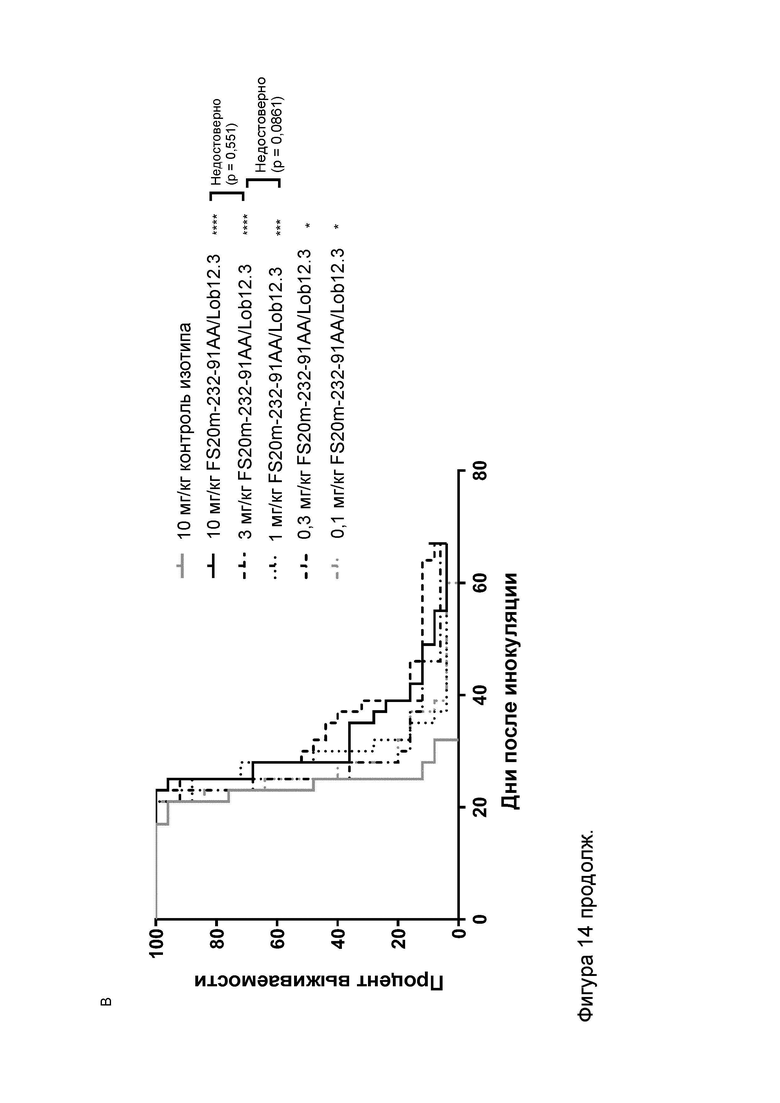
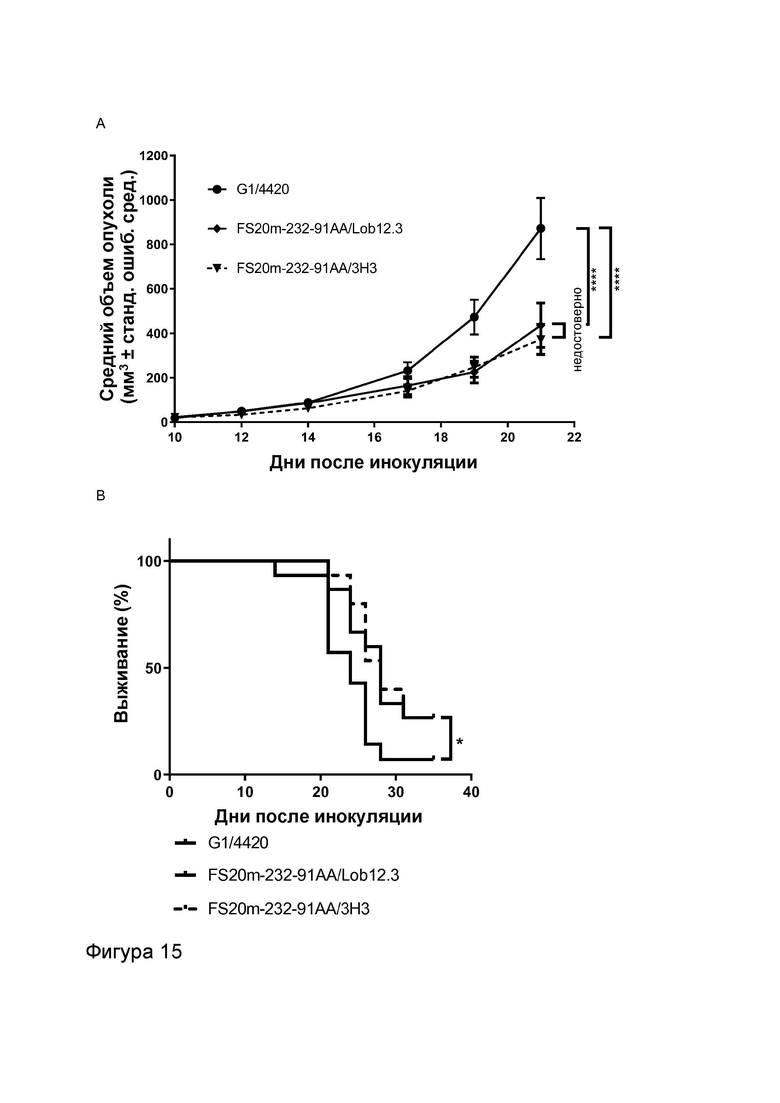
Изобретение относится к области биотехнологии. Предложены молекулы антител, которые связываются с CD137 и OX40 и способны оказывать агонистическое действие как на CD137, так и на OX40. Указанные молекулы антител содержат связывающий сайт на основе CDR для CD137 и антигенсвязывающий сайт для OX40, который расположен в константном домене молекулы антитела. Также предложена фармацевтическая композиция, включающая указанные антитела. Также изобретение относится к нуклеиновым кислотам, кодирующим антитела, векторам экспрессии и клеткам-хозяевам, содержащим указанные нуклеиновые кислоты, способу получения антитела. Изобретение обеспечивает высокоэффективную активацию иммунных клеток с пониженным риском токсичности и может применяться в лечении рака. 11 н. и 14 з.п. ф-лы, 15 ил., 39 табл., 28 пр.
1. Молекула антитела, которая связывается с CD137 и OX40, содержащая:
(a) антигенсвязывающий сайт на основе области, определяющей комплементарность (CDR), для CD137; и
(b) антигенсвязывающий сайт для OX40, расположенный в домене CH3 указанной молекулы антитела;
причем указанный антигенсвязывающий сайт на основе CDR содержит VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации ImMunoGeneTics (IMGT), представленные в:
(i) SEQ ID NO: 1, 2, 3, 4, 5 и 6 соответственно;
(ii) SEQ ID NO: 1, 2, 16, 4, 5 и 6 соответственно;
(iii) SEQ ID NO: 1, 2, 21, 4, 5 и 6 соответственно;
(iv) SEQ ID NO: 25, 26, 27, 4, 5 и 28 соответственно; или
(v) SEQ ID NO: 33, 34, 35, 4, 5 и 36 соответственно; или
причем указанный антигенсвязывающий сайт на основе CDR содержит VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации по Kabat, представленные в:
(vi) SEQ ID NO: 7, 8, 9, 10, 11 и 6 соответственно;
(vii) SEQ ID NO: 7, 8, 17, 10, 11 и 6 соответственно;
(viii) SEQ ID NO: 7, 8, 22, 10, 11 и 6 соответственно;
(ix) SEQ ID NO: 29, 30, 31, 10, 11 и 28 соответственно; или
(x) SEQ ID NO: 37, 38, 39, 10, 11 и 36 соответственно; и
при этом указанный антигенсвязывающий сайт для OX40 содержит первую последовательность, вторую последовательность и третью последовательность, расположенные в структурных петлях AB, CD и EF домена CH3 соответственно, причем указанная первая, вторая и третья последовательности имеют последовательности, указанные под SEQ ID NO: 51, 52 и 53 соответственно.
2. Молекула антитела по п. 1, отличающаяся тем, что:
(i) указанная первая последовательность расположена в положениях с 14 по 18 домена CH3 указанной молекулы антитела;
(ii) указанная вторая последовательность расположена в положениях с 45.1 по 77 домена CH3 указанной молекулы антитела; и/или
(iii) указанная третья последовательность расположена в положениях с 93 по 101 домена CH3 указанной молекулы антитела; и
при этом указанная нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
3. Молекула антитела по любому из пп. 1 или 2, отличающаяся тем, что указанная молекула антитела содержит последовательность домена CH3, указанную под SEQ ID NO: 54.
4. Молекула антитела по п. 3, отличающаяся тем, что указанный домен CH3 дополнительно содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
5. Молекула антитела по любому из пп. 1-4, отличающаяся тем, что указанная молекула антитела содержит домен VH и домен VL, представленные в:
(i) SEQ ID NO: 12 и 14 соответственно;
(ii) SEQ ID NO: 18 и 14 соответственно;
(iii) SEQ ID NO: 23 и 14 соответственно;
(iv) SEQ ID NO: 170 и 172 соответственно; или
(v) SEQ ID NO: 40 и 42 соответственно.
6. Молекула антитела по любому из пп. 1-5, отличающаяся тем, что указанная молекула антитела содержит последовательности тяжелой цепи и легкой цепи антитела:
(i) согласно SEQ ID NO: 95 и 97 соответственно;
(ii) согласно SEQ ID NO: 99 и 97 соответственно;
(iii) согласно SEQ ID NO: 103 и 97 соответственно;
(iv) согласно SEQ ID NO: 105 и 107 соответственно; или
(v) согласно SEQ ID NO: 109 и 111 соответственно.
7. Молекула антитела по п. 6, отличающаяся тем, что указанный домен CH3 в тяжелой цепи содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
8. Молекула антитела по любому из пп. 1-7, отличающаяся тем, что указанная молекула антитела содержит:
(i) VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации IMGT, указанные под SEQ ID NO: 1, 2, 3, 4, 5 и 6 соответственно;
(ii) VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации по Kabat, указанные под SEQ ID NO: 7, 8, 9, 10, 11 и 6 соответственно;
(iii) домен VH и домен VL, указанные под SEQ ID NO: 12 и 14 соответственно; и/или
(iv) тяжелую цепь и легкую цепь, указанные под SEQ ID NO: 95 и 97 соответственно.
9. Молекула антитела по п. 8, отличающаяся тем, что указанная молекула антитела содержит тяжелую цепь и легкую цепь, указанные под SEQ ID NO: 95 и 97 соответственно.
10. Молекула антитела по п. 9, отличающаяся тем, что домен CH3 тяжелой цепи содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
11. Молекула антитела по любому из пп. 1-10, отличающаяся тем, что указанная молекула антитела связывает CD137 человека и OX40 человека, при этом необязательно указанная молекула антитела способна одновременно связываться с CD137 человека и OX40 человека.
12. Молекула антитела по любому из пп. 1-11, отличающаяся тем, что:
(i) указанная молекула антитела способна активировать OX40 на иммунной клетке в присутствии CD137, экспрессируемого на поверхности клетки, и/или указанная молекула антитела способна активировать CD137 на иммунной клетке в присутствии OX40, экспрессируемого на поверхности клетки;
(ii) связывание указанной молекулы антитела с OX40 на иммунной клетке и с CD137 вызывает кластеризацию OX40 на указанной иммунной клетке и/или связывание указанной молекулы антитела с CD137 на иммунной клетке и с OX40 вызывает кластеризацию CD137 на указанной иммунной клетке; и/или
(iii) указанная молекула антитела была модифицирована, чтобы уменьшить или устранить связывание домена CH2 указанной молекулы антитела с одним или более рецепторами Fcγ.
13. Молекула нуклеиновой кислоты, кодирующая молекулу антитела по любому из пп. 1-12.
14. Комбинация молекул нуклеиновой кислоты, кодирующая молекулу антитела по любому из пп. 1-12.
15. Вектор для экспрессии молекулы или молекул нуклеиновой кислоты в клетке-хозяине, содержащий молекулу нуклеиновой кислоты по п. 13 или комбинацию молекул нуклеиновой кислоты по п. 14.
16. Комбинация векторов для экспрессии молекулы или молекул нуклеиновой кислоты в клетке-хозяине, содержащих молекулу нуклеиновой кислоты по п. 13 или комбинацию молекул нуклеиновой кислоты по п. 14.
17. Рекомбинантная клетка-хозяин для получения молекулы антитела по любому из пп. 1-12, содержащая молекулу нуклеиновой кислоты по п. 13, или комбинацию молекул нуклеиновой кислоты по п. 14, или вектор по п. 15, или комбинацию векторов по п. 16.
18. Способ получения молекулы антитела по любому из пп. 1-12, включающий культивирование рекомбинантной клетки-хозяина по п. 17 в условиях для получения указанной молекулы антитела, дополнительно включающий выделение указанной молекулы антитела.
19. Способ по п. 18, дополнительно включающий очистку указанной молекулы антитела.
20. Фармацевтическая композиция для лечения заболевания или нарушения, ассоциированного с экспрессией CD137 и OX40, содержащая молекулу антитела по любому из пп. 1-12 в терапевтически эффективном количестве и фармацевтически приемлемое вспомогательное вещество.
21. Применение молекулы антитела по любому из пп. 1-12 для лечения рака у индивидуума.
22. Применение по п. 21, отличающееся тем, что указанное лечение включает введение указанной молекулы антитела указанному индивидууму в комбинации со вторым терапевтическим средством.
23. Применение по п. 22, отличающееся тем, что указанное второе терапевтическое средство представляет собой антитело, которое связывает PD-1 или PD-L1.
24. Молекула антитела, которая связывается с CD137 и OX40, причем указанная молекула антитела содержит последовательность легкой цепи, указанную под SEQ ID NO: 97, и последовательность тяжелой цепи, указанную под SEQ ID NO: 95.
25. Молекула антитела, которая связывается с CD137 и OX40, причем указанная молекула антитела содержит последовательность легкой цепи, указанную под SEQ ID NO: 97, и последовательность тяжелой цепи, указанную под SEQ ID NO: 95, и причем домен CH3 тяжелой цепи содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
| WO 2016185016 A1, 24.11.2016 | |||
| WO 2017077085 A2, 11.05.2017 | |||
| LEE S.-J | |||
| et al., 4-1BB and OX40 Dual Costimulation Synergistically Stimulate Primary Specific CD8 T Cells for Robust Effector Function, THE JOURNAL OF IMMUNOLOGY, 2004, vol | |||
| Джино-прядильная машина | 1922 |
|
SU173A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| КАДАГИДЗЕ З.Г | |||
| и др., Рецепторы лимфоцитов, регулирующие иммунный ответ - ключ к | |||
