ОБЛАСТЬ ТЕХНИКИ
Согласно настоящему изобретению предложены молекулы антитела, которые связывают как PD-L1, так и CD137 и способны вызывать агонизм CD137. Молекулы антитела содержат сайт связывания с PD-L1 на основе CDR и сайт связывания антигена CD137, который расположен в константном домене молекулы антитела. Молекулы антитела согласно настоящему изобретению находят применение, например, при лечении заболеваний, таких как рак.
УРОВЕНЬ ТЕХНИКИ
Белок программируемой гибели клеток 1 (PD-1) и его лиганды PD-L1 (CD274, B7-H1) и PD-L2 (B7-DC) доставляют ингибиторные сигналы, которые регулируют равновесие между T-клеточной активацией, толерантностью и иммунопатологией. PD-L1 временно экспрессируется на всех иммунных клетках и некоторых клетках опухоли. PD-L1 является представителем семейства белков B7, и его последовательность аминокислот идентична приблизительно на 20% последовательности B7.1 и B7.2. Последовательность аминокислот PD-L1 человека идентична на 70% и 93% последовательности ортологов PD-L1 мыши и яванского макака, соответственно.
PD-L1 связывается со своим рецептором PD-1 с аффинностью (KD) 770 нМ. PD-1 экспрессируется на активированных T-клетках, B-клетках и миелоидных клетках и модулирует активацию или ингибирование клеточных иммунных ответов. Связывание PD-L1 с PD-1 доставляет ингибиторный сигнал, уменьшающий продукцию цитокинов и пролиферацию T-клеток. Следовательно, экспрессия PD-L1 клетками может опосредовать защиту против уничтожения цитотоксическими T-лимфоцитами (CTL) и является регуляторным механизмом, который сдерживает хронические иммунные ответы во время вирусных инфекций. Рак, как хроническое и провоспалительное заболевание, нарушает этот иммунный защитный путь за счет повышения экспрессии PD-L1, чтобы ускользнуть от иммунного ответа хозяина. В контексте активного иммунного ответа, интерферон-гамма (IFN-γ) также повышает экспрессию PD-L1.
PD-L1 также опосредует подавление иммунного ответа путем взаимодействия с другим белком B7.1 (также известным как CD80), блокируя его способность доставлять один из вторичных сигналов активации T-клеткам посредством CD28. В отношении экспрессии PD-L1 на клетках опухоли и его взаимодействия с B7.1, значение данного конкретного взаимодействия для иммунной устойчивости опухоли все еще неясно.
Было показано, что PD-L1 экспрессируется в большом количестве различных солидных опухолей. Из 654 образцов, изученных в одном исследовании, охватывающем 19 опухолей из различных локализаций, 89 (14%) были положительны по PD-L1 (встречаемость ≥5%). Наибольшую встречаемость положительных по PD-L1 опухолей наблюдали при раке головы и шеи (17/54; 31%), раке шейки матки (10/34; 29%), раке неизвестного первичного происхождения (CUP; 8/29; 28%), мультиформной глиобластоме (GBM; 5/20; 25%), раке мочевого пузыря (8/37; 21%), раке пищевода (16/80; 20%), трижды негативном (TN) раке молочной железы (6/33; 18%) и гепатокарциноме (6/41; 15%) (Grosso и др., 2013). Было показано, что связанная с опухолью экспрессия PD-L1 придает устойчивость к иммунному ответу и потенциально защищает клетки опухоли от опосредованного T-клетками апоптоза.
На сегодняшний день клинические испытания показали клиническую пользу нацеливания на PD-L1 у пациентов и привели к одобрению трех моноклональных антител, нацеленных на PD-L1. Атезолизумаб (MPDL3280A, RG7466, Тецентрик™), гуманизированное антитело IgG1, которое связывает PD-L1, одобрено для терапии первой линии немелкоклеточной карциномы легких (НМРЛ) и первой и второй линии для лечения рака мочевого пузыря после того, как в клинических испытаниях показали частоту объективного ответа (ЧОО) 38% и 43%, соответственно, у пациентов с положительными по PD-L1 опухолями (Iwai и др., 2017). Авелумаб (MSB0010718C, Бавенсио™) представляет собой полностью человеческое антитело IgG1, которое связывается с PD-L1, и оно одобрено для лечения карциномы из клеток Меркеля и как лечение второй линии лечения рака мочевого пузыря, тогда как полностью человеческое антитело IgG1 дурвалумаб (MEDI4736, Имфинзи™) одобрено как лечение второй линии рака мочевого пузыря. Продолжаются дополнительные испытания с данными антителами и другими лекарствами против PD-L1, с целью расширения диапазона типов солидного рака, которые можно лечить, включая колоректальный рак, рак желудка, рак молочной железы, рак головы и шеи, рак поджелудочной железы, рак яичников и почечно-клеточную карциному.
CD137 (4-1BB; TNFRSF9) представляет собой костимулирующую молекулу из суперсемейства рецепторов фактора некроза опухоли (TNFRSF). Широко известно, что экспрессия CD137 повышена на CD8+ T-клетках после активации, и он также может экспрессироваться на активированных CD4+ хелперных T-клетках, B-клетках, регуляторных T-клетках, естественных клетках-киллерах (NK), естественных T-клетках-киллерах (NKT) и дендритных клетках (ДК) (Bartkowiak и Curran, 2015). Главная функциональная роль CD137 в повышении цитотоксичности T-клеток была впервые описана в 1997 г. (Shuford и др., 1997), и вскоре после этого МАТ против CD137 были предложены в качестве противоракового лекарства.
CD137 представляет собой трансмембранный белок с четырьмя внеклеточными богатыми цистеином доменами, называемыми CRD1-4, и цитоплазматической областью, отвечающей за передачу сигналов CD137. Лигандом для CD137 является CD137L. Хотя нет кристаллической структуры для комплекса CD137/CD137L, предсказали, что CD137 образует комплекс тример/тример с CD137L (Won и др., 2010). Связывание CD137L приводит к образованию тримера рецептора и последующей кластеризации множества тримеров рецептора, и приводит к активации каскада передачи сигналов CD137. Данный каскад передачи сигналов передает сигнал выживаемости T-клеткам против вызванной активацией гибели клеток (Hurtado и др., 1997), тем самым играя важную роль в поддержании эффективных T-клеточных иммунных ответов и формировании иммунологической памяти (Bartkowiak & Curran, 2015).
CD137 экспрессируется активированными T-клетками, и его использовали в качестве маркера для идентификации антигенспецифических CD4+ и CD8+ T-клеток (Wolfl и др., 2007; Ye и др., 2014). Обычно экспрессия CD137 выше на CD8+ T-клетках, чем на CD4+ T-клетках (Wen и др., 2002). В случае CD8+ T-клеток, пролиферацию, выживаемость и цитотоксическую эффекторную функцию посредством продукции интерферона гамма и интерлейкина 2 считали следствием перекрестного связывания с CD137. Перекрестное связывание с CD137 также способствует дифференцировке и сохранению CD8+ T-клеток памяти (Chacon и др., 2013). В некоторых субпопуляциях CD4+ T-клеток, перекрестное связывание CD137 аналогично приводит к пролиферации и активации и приводит к высвобождению цитокинов, таких как интерлейкин 2 (Makkouk и др., 2016). Также продемонстрировали, что CD137 экспрессируется на субпопуляциях опухолереактивных инфильтрирующих опухоль лимфоцитов (TIL). Было показано, что монотерапия CD137 эффективна в нескольких иммуногенных доклинических моделях опухоли, таких как MC38, CT26 и B-клеточные лимфомы.
Клиническая разработка МАТ против CD137 шла медленно вследствие ограничивающего дозу воспаления печени высокой степени, связанного с лечением агонистическим антителом против CD137. Урелумаб (BMS-663513), не блокирующее лиганд антитело изотипа IgG4 человека (Chester и др., 2018), было первым антителом против CD137, которое участвовало в клинических испытаниях, но их остановили после того как наблюдали значительную, связанную с воздействием на мишень, зависимую от дозы гепатотоксичность (Chester и др., 2018; Segal и др., 2017; и Segal и др., 2018). Позже возобновили клинические испытания урелумаба для лечения типов солидного рака, в котором лечение урелумабом комбинировали с лучевой терапией (NCT03431948) или с другими терапевтическими антителами, такими как ритуксимаб (NCT01775631), цетуксимаб (NCT02110082), антитело против PD-1 ниволумаб (NCT02253992, NCT02534506, NCT02845323) и комбинация ниволумаба и антитела против LAG-3 BMS986016 (NCT02658981). Тем не менее для уменьшения гепатотоксичности, связанной с лечением урелумабом, дозировку урелумаба в данных испытаниях пришлось ограничить, и результаты по эффективности были разочаровывающими (Chester и др., 2018).
Не наблюдали ограничивающей дозу токсичности для антитела против CD137 утомилумаба (PF-05082566) от Pfizer, антитела изотипа IgG2 человека, в диапазоне доз от 0,03 мг/кг до 10 мг/кг в I фазе клинических испытаний распространенного рака (Chester и др. 2016; Segal и др., 2018). Тем не менее общая частота объективного ответа на данное антитело составляла лишь 3,8% у пациентов с солидными опухолями, потенциально свидетельствуя о том, что утомилумаб обладает меньшей активностью и клинической эффективностью, чем урелумаб, при этом имея более благоприятный профиль безопасности (Chester и др., 2018; Segal и др., 2018). Утомилумаб исследовали в комбинации с лучевой терапией (NCT03217747) или химиотерапией, а также в комбинации с другими методами лечения антителами, включая антитело против PD-L1 - авелумаб (NCT02554812), и антитело против PD-1 -пембролизумаб (NCT02179918), чтобы оценить безопасность, переносимость, ограничивающую дозу токсичность (DLT), максимальную переносимую дозу (MTD) и эффективность различных комбинаций лечения. Эти испытания продолжаются, причем ранние результаты выявили отсутствие DLT для доз вплоть до 5 мг/кг и частоту ответа пациентов, составляющую 26%, для комбинации утомилумаба и пембролизумаба (Tolcher и др., 2016) (Pérez-Ruiz и др., 2017).
Продолжаются клинические испытания, в которых исследуют антитело против CD137 - утомилумаб - в комбинации с антителом против PD-L1 - авелумабом (NCT02554812, NCT3390296). Также исследуют тройные комбинации утомилумаба с авелумабом и другими методами лечения (NCT02554812, NCT03217747, NCT03440567, NCT03414658).
Множество биспецифических молекул, нацеленных на CD137, также находятся на ранней стадии разработки, например, молекулы, нацеленные на CD137, а также FAP-альфа (Link и др., 2018; Reichen и др., 2018), HER2 (Hinner и др., 2015 и WO 2016/177802 A1) или EphA2 (Liu и др., 2017). Также разрабатываются слитые белки CD137L, которые нацелены на опухоли, например, посредством FAP-альфа (Claus и др., 2017). Наиболее перспективное с клинической точки зрения биспецифическое антитело против CD137 представляет собой PRS-343 - биспецифическую молекулу CD137/HER2, которую недавно начали исследовать в I фазе клинических испытаний для лечения ряда солидных опухолей, чтобы оценить его безопасность, переносимость и эффективность (NCT03330561).
Существуют другие подходы к комбинированию активности против CD137 и активности против PD-L1 в биспецифических методах лечения. В одном подходе применяли технологию Biclonics для получения полноразмерного гетеродимерного IgG человека, моновалентно связывающегося с обоими CD137 и PD-L1 (WO2018056821), с получением молекулы, которая может связываться как с CD137, так и с PD-L1 и вызывать агонизм CD137 в присутствии высоких уровней PD-L1. Был описан второй подход с применением слитых sdAb-Fc для нацеливания на оба CD137 и PD-L1 (WO2017123650). При обоих подходах антитела способны связывать CD137 в отсутствие связывания PD-L1 и вызывать низкие уровни агонизма CD137 в отсутствие PD-L1, данный агонизм повышается в присутствии высоких уровней PD-L1. Было описано дополнительное гетеродимерное биспецифическое антитело, моновалентно связывающее как CD137, так и PD-L1 (WO2019/025545 A1), содержащее гуманизированную область связывания с CD137 и область связывания с PD-L1 человека, которая вызывает агонизм CD137 в присутствии некоторых уровней PD-L1.
Современные данные показывают, что в целом монотерапевтическое лечение против PD-L1 приводит к ответу у менее чем 50% страдающих раком пациентов. Таким образом, в данной области остается потребность в дополнительных молекулах, которые могут нацеливаться на PD-L1 и которые найдут применение в терапии рака.
Блокирование PD-1/PD-L1 имеет значительное клиническое обоснование, тем не менее, менее 50% пациентов отвечают на монотерапевтическую схему лечения. Ожидают, что комбинации PD-L1 и дополнительных иммунных модуляторов продемонстрируют улучшенную эффективность. Тем не менее, такие комбинации могут быть связаны с увеличением связанных с лечением нежелательных явлений, и, в результате, эффективность может быть ограничена ограниченным терапевтическим окном. Нацеленные на CD137 агонистические молекулы еще должны продемонстрировать значительные ответы у страдающих раком пациентов, это может быть отчасти следствием относительно низких уровней доз вследствие ограниченного терапевтического коэффициента. Таким образом, в данной области остается потребность в разработке методов лечения, в которых комбинируют блокирование PD-L1 и агонизм CD137 в безопасных и эффективных методах лечения.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В разделе уровня техники выше объясняется, что клиническая разработка молекул-агонистов CD137 задерживалась вследствие того, что лечение было связано либо с ограничивающим дозу воспалением печени высокой степени (урелумаб), либо с низкой клинической эффективностью (утомилумаб).
Авторы настоящего изобретения поняли, что в данной области существует потребность в молекулах-агонистах CD137, которые проявляют высокую активность, причем агонизм можно ограничить в микроокружении опухоли. Такие молекулы можно вводить индивидам в дозах, которые оптимизируют активность и, следовательно, эффективность молекулы, и могут применяться для лечения рака, например, в качестве иммунотерапевтических агентов.
Молекулы антитела согласно настоящему изобретению содержат сайт связывания антигена CD137, который расположен в константном домене молекулы антитела. Авторы настоящего изобретения провели всесторонние процедуры селекции и созревания аффинностии выделили панель молекул, содержащих сайт связывания антигена CD137 (в данной заявке также называемых “Fcab”), которые связываются с димерным CD137 с более высокой аффинностью, чем с мономерным CD137.
«Аффинность» в данной заявке может относиться к силе связывающего взаимодействия между молекулой антитела и распознаваемым ею антигеном, измеряемой KD. Для квалифицированного специалиста будет очевидно, что если молекула антитела способна образовывать множество связывающих взаимодействий с антигеном (например, когда молекула антитела способна связывать антиген бивалентно и необязательно когда антиген димерный), на аффинность, измеренную в виде KD, также может влиять авидность, где авидность относится к общей силе связывания комплекса антитело-антиген.
Экспрессия CD137 T-клетками повышается при активации. Без привязки к теории полагают, что вследствие высокой экспрессии CD137 на активированных T-клетках, CD137 будет находиться в виде димеров, тримеров и мультимеров более высокого порядка на поверхности таких клеток. Наоборот, наивные иммунные клетки, такие как наивные T-клетки, экспрессируют низкие или незначительные уровни CD137 на поверхности клеток, и любой присутствующий CD137, следовательно, вероятно находится в мономерной форме. Следовательно, ожидается, что молекулы антитела, содержащие сайт связывания антигена CD137, которые связываются с димерным или мультимерным CD137 с высокой авидностью, преимущественно будут связываться с активированными иммунными клетками, такими как активированные T-клетки, в противоположность, например, наивным иммунным клеткам.
Полагают, что данные особенности сайта связывания антигена CD137 помогают отличить молекулы антитела согласно настоящему изобретению от известных антител, которые связывают CD137, например, антитела, описанного в WO2018056821. В WO2018056821 описано антитело, содержащее моновалентный связывающий CD137 домен, который связывается с CD137 с высокой аффинностью (низкий нМ диапазон, см. таблицу 6 в WO2018056821). Поскольку данные антитела не различают мономерный и димерный или мультимерный CD137, не ожидается, что у данных антител известного уровня техники будут выявлять такое же предпочтительное связывание с активированными иммунными клетками.
Как описано в разделе «Уровень техники» выше, полагают, изначальное связывание лиганда CD137 с CD137 дает начало цепи событий, которая приводит к тримеризации рецептора, а затем кластеризации рецептора, его активации и последующему запуску сильной противоопухолевой активности T-клеток. Следовательно, ожидается, что для того, чтобы добиться эффективной активации CD137 терапевтическим агентом, несколько мономеров рецептора должны быть соединены мостиком таким образом, который имитирует мостиковое соединение тримерным лигандом.
Утомилумаб представляет собой молекулу IgG2, и его агонистическая активность зависит от перекрестного связывания Fcγ-рецепторами. Урелумаб представляет собой молекулу IgG4 с конститутивной активностью, и для его активности не требуется перекрестное связывание Fcγ-рецепторами, хотя его агонистическая активность усиливается при перекрестном связывании некоторыми Fcγ-рецепторами. Fcγ-рецепторы находятся повсеместно в организме человека. Активность активации иммунных клеток утомилумабом и урелумабом, следовательно, не ограничена конкретными сайтами в организме и, таким образом, может происходить в местах, отличных от микроокружения опухоли, таких как печень.
Авторы настоящего изобретения показали, что для сайта связывания антигена CD137, присутствующего в молекулах антитела согласно настоящему изобретению, требуется перекрестное связывание, чтобы кластеризовать и активировать CD137. Тем не менее следует отметить, что это не является свойственным признаком связывающих CD137 молекул. Точнее, множество связывающих CD137 молекул, выделенных во время программы скрининга, связывались с CD137, но не требовали перекрестного связывания для кластеризации и активации CD137, или вызывали ограниченную кластеризацию и активацию CD137 в отсутствие перекрестного связывания.
Как упоминалось выше, у опосредованного Fcγ-рецептором перекрестного связывания есть недостаток, состоящий в том, что Fcγ-рецепторы находятся повсеместно в организме человека, и, следовательно, активация CD137 не ограничена конкретным местом. Соответственно, авторы настоящего изобретенияввели мутации в домен CH2 Fcab, чтобы уменьшить или устранить связывание с Fcγ-рецептором. Таким образом, в отсутствие перекрестного связывания посредством агента, отличного от Fcγ-рецепторов, молекулы антитела согласно настоящему изобретению не проявляют агонистическую активность в отношении CD137. Кроме того, ожидается, что данные мутации приведут к тому, что молекулы антитела согласно настоящему изобретению будут неспособны вызывать антителозависимую клеточную цитотоксичность, так что молекулы антитела не будут вызывать уничтожение иммунных клеток, которые они активируют.
Авторы настоящего изобретения продемонстрировали, что молекулы антитела, которые содержат сайт связывания антигена CD137, описанный выше, и сайт связывания с PD-L1 на основе CDR, высокоэффективно активируют иммунные клетки в локализациях, где коэкспрессируются CD137 и PD-L1, например, в микроокружении опухоли. Коэкспрессия в данном смысле включает случаи, когда CD137 и PD-L1 экспрессируются на одних и тех же клетках, например, T-клетке, и случаи, когда CD137 и PD-L1 экспрессируются на различных клетках, например, T-клетке и клетке опухоли, соответственно.
Молекулы антитела способны связываться одновременно с CD137 и PD-L1. Таким образом, считают, что в локализациях, где PD-L1 и CD137 коэкспрессируются, связывание молекул антитела с PD-L1 вызывает перекрестное связывание молекул антитела, что, в свою очередь, приводит к кластеризации и активации связанного CD137 на поверхности T-клетки.
Авторы настоящего изобретения продемонстрировали, что при снижении или устранении связывания с Fcγ-рецептором, агонистическая активность молекул антитела зависит от присутствия обоих PD-L1 и CD137. Другими словами, агонистическая активность условная, и, следовательно, ожидается, что молекулы антитела способны активировать иммунные клетки только в локализациях, где присутствует PD-L1, таких как микроокружение опухоли. Ожидается, что данная направленная активация иммунных клеток предпочтительна тем, что позволяет избежать воспаления печени, наблюдаемого при лечении, например, урелумабом.
Действительно, авторы настоящего изобретения демонстрируют, что молекулы антитела, обладающие свойствами, описанными выше, не проявляют сильную гепатотоксичность при введении моделям на мышах в терапевтических дозах. Лишь минимальная патология печени наблюдалась у мышей, которым вводили данные молекулы антитела, которую не считали представляющей сильную гепатотоксичность, о которой ранее сообщали для других агонистических антител против CD137. Предварительные исследования на яванских макаках также показали, что молекулы антитела безопасны и хорошо переносятся вплоть до 30 мг/кг. Без привязки к теории ожидается, что результаты, полученные в данных моделях на животных, можно использовать в клинике для прогнозирования риска гепатотоксичности у пациентов-людей, и, следовательно, что будет низкий риск того, что молекулы антител согласно настоящему изобретению вызовут гепатотоксичность у пациентов-людей, которых будут лечить терапевтическими дозами.
Авторы настоящего изобретения также предоставляют доказательство in vitro того, что уровень агонистической активности в отношении CD137, вызванный молекулой антитела, коррелирует с уровнем экспрессии PD-L1 на поверхности клетки. Авторы настоящего изобретения демонстрируют, что молекула антитела способна агонизировать CD137, даже если присутствует низкий уровень экспрессии PD-L1, и что при повышении уровня PD-L1 в системе также повышается агонистическая активность в отношении CD137. Данный результат дополнительно подтверждает, что агонистическая активность в отношении CD137 зависит от экспрессии PD-L1, и позволяет предположить, что молекулы антитела согласно настоящему изобретению будут проявлять широкий спектр активности по отношению к опухолям, которые экспрессируют различные уровни PD-L1 на поверхности клеток опухоли.
Сайты связывания PD-L1 на основе CDR, описанные выше, способны эффективно блокировать связывание PD-L1 с его рецептором PD-1. PD-1 экспрессируется на активированных T-клетках, B-клетках и миелоидных клетках и модулирует активацию или ингибирование клеточных иммунных ответов. Связывание PD-L1 с PD-1 доставляет ингибиторный сигнал, уменьшая продукцию цитокинов и пролиферацию T-клеток, тем самым тормозя иммунный ответ. При раке взаимодействие PD-L1 на клетке опухоли с PD-1 на T-клетке уменьшает активность T-клетки, чтобы предотвратить атаку иммунной системой клеток опухоли. Следовательно, ожидается, что путем блокирования связывания PD-L1 с PD-1 молекулы антитела согласно настоящему изобретению могут предотвратить такое ускользание клеток опухоли от иммунной системы. Без привязки к теории полагают, что такое эффективное блокирование связывания PD-L1 с PD-1 действует вместе с агонистической активностью в отношении CD137, описанной выше, повышая противоопухолевую эффективность молекулы антитела.
Авторы настоящего изобретения также показали, что такие биспецифические молекулы антитела, содержащие сайт связывания антигена CD137 и сайт связывания с PD-L1 на основе CDR, описанные выше, способны подавлять рост опухоли in vivo. Более того, более эффективное подавление роста опухоли наблюдали для биспецифических молекул антитела по сравнению с комбинацией двух моноспецифических молекул антитела, где одна из молекул антитела содержала сайты связывания антигена PD-L1 на основе CDR и другая молекула содержала сайты связывания антигена CD137 на основе CDR, демонстрируя, что повышенная кластеризация и передача сигналов CD137 и, следовательно, активация T-клеток и соответствующие противоопухолевые эффекты, наблюдается для молекул антитела согласно настоящему изобретению.
Молекулы антитела, содержащие сайт связывания антигена CD137 согласно настоящему изобретению, могут дополнительно быть способны связывать PD-L1 бивалентно, так что молекулы антитела связывают оба CD137 и PD-L1 бивалентно. Ожидают, что это будет преимуществом, так как ожидается, что бивалентное связывание обоих мишеней сделает образование мостика между T-клеткой, экспрессирующей CD137, и клеткой, экспрессирующей PD-L1, более стабильным и, таким образом, продлит время, в течение которого T-клетка локализуется в сайтах, где PD-L1 коэкспрессируется с CD137, например, в микроокружении опухоли, и может оказывать действие на заболевание, например, опухоль. Это служит отличием от большинства обычных форматов биспецифических антител, которые гетеродимерны и связывают каждый целевой антиген моновалентно посредством одного плеча Fab. Ожидают, что такое моновалентное взаимодействие не только менее стабильно, но также менее эффективно вызывает кластеризацию рецепторов TNF, таких как CD137, и/или требует более высокого уровня экспрессии одной или обеих мишеней для индукции такой кластеризации и, следовательно, активации T-клеток. Это подтверждается экспериментами, проведенными авторами настоящего изобретения, в которых показали, что молекулы МАТ2, содержащие бивалентный сайт связывания Fab с PD-L1 и моновалентный сайт связывания с CD137 в одном из доменов CH3 указанной молекулы, вызывали более низкие уровни активации T-клеток, что измеряли по высвобождению IFN-γ, чем МАТ2, связывающее обе мишени бивалентно.
Дополнительным свойством молекул антитела, идентифицированных авторами настоящего изобретения, является то, что как сайт связывания антигена CD137, так и сайт связывания с PD-L1 на основе CDR содержатся в самой структуре антитела. В частности, не требуется, чтобы молекулы антитела были слиты с другими белками посредством линкеров или других средств с образованием молекулы, которая связывается бивалентно с обеими мишенями. Это дает множество преимуществ. В частности, молекулы антитела можно получить, применяя способы, аналогичные таковым, используемым для получения стандартных антител, так как они не содержат никаких дополнительных слитых частей. Также ожидается, что такая структура приведет к улучшенной стабильности антитела, так как линкеры могут разрушаться с течением времени, приводя к гетерогенной популяции молекул антител. Ожидают, что в таких гетерогенных популяциях те антитела в популяции, которые содержат только один слитый с ними белок и, следовательно, связывают одну мишень лишь моновалентно, не вызывают условный агонизм рецепторов TNF, таких как CD137, так же эффективно, как антитела, которые содержат два слитых с ними белка и которые, следовательно, способны связывать обе мишени бивалентно. Расщепление/деградация линкера может происходить перед введением или после введения терапевтического средства пациенту (например, посредством ферментативного расщепления или под действием pH пациента in vivo), таким образом, приводя к снижению его эффективности в кровотоке пациента. Так как в молекулах антитела согласно настоящему изобретению нет линкеров, ожидают, что в указанных молекулах антитела сохраняется одинаковое количество сайтов связывания как до, так и после введения. Более того, структура указанных молекул антитела также предпочтительна с точки зрения иммуногенности указанных молекул, так как встраивание слитых белков или линкеров или обоих может вызывать иммуногенность, когда указанные молекулы вводят пациенту, приводя к снижению эффективности терапевтического средства.
Таким образом, в соответствии с настоящим изобретением предложены:
[1] Молекула антитела, которая связывается с лигандом программируемой гибели клеток (PD-L1) и CD137, содержащая
(a) сайт связывания антигена PD-L1 на основе определяющей комплементарность области (CDR); и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
причем указанный сайт связывания антигена на основе CDR содержит CDR 1 - 6, представленные в:
(i) SEQ ID NO: 1, 2, 3, 4, 5 и 6, соответственно [E12v2];
(ii) SEQ ID NO: 1, 2, 3, 18, 19 и 20, соответственно [E05v2]; или
(iii) SEQ ID NO: 1, 2, 3, 18, 19 и 29, соответственно [G12v2]; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно, причем первая и вторая последовательность имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003] или 79 и 80 [FS22-53-008], соответственно.
[2] Молекула антитела, которая связывается с лигандом программируемой гибели клеток (PD-L1) и CD137, содержащая
(a) сайт связывания антигена PD-L1 на основе определяющей комплементарность области (CDR); и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
причем указанный сайт связывания антигена на основе CDR содержит CDR 1 - 6 согласно IMGT, представленные в:
(i) SEQ ID NO: 7, 8, 9, 10, 11 и 6, соответственно [E12v2];
(ii) SEQ ID NO: 21, 8, 9, 22, 11 и 20, соответственно [E05v2]; или
(iii) SEQ ID NO: 21, 8, 9, 22, 11 и 29, соответственно [G12v2]; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно, причем первая и вторая последовательность имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003] или 79 и 80 [FS22-53-008], соответственно.
[3] Молекула антитела согласно [1] или [2], отличающаяся тем, что молекула антитела содержит CDR 1 - 6, представленные в (i) или (ii) из [1] или [2].
[4] Молекула антитела согласно [1] или [2], отличающаяся тем, что молекула антитела содержит CDR 1 - 6, представленные в (i) из [1] или [2].
[5] Молекула антитела согласно любому из [1] - [4], отличающаяся тем, что молекула антитела содержит вариабельный домен тяжелой цепи (VH) и/или вариабельный домен легкой цепи (VL), предпочтительно домен VH и домен VL.
[6] Молекула антитела согласно любому из [1] - [5], отличающаяся тем, что молекула антитела содержит тяжелую цепь иммуноглобулина и/или легкую цепь иммуноглобулина, предпочтительно тяжелую цепь иммуноглобулина и легкую цепь иммуноглобулина.
[7] Молекула антитела согласно любому из [5] - [6], отличающаяся тем, что молекула антитела содержит домен VH и/или домен VL, предпочтительно домен VH и домен VL, представленные в:
(i) SEQ ID NO: 12 и 14, соответственно [E12v2];
(ii) SEQ ID NO: 23 и 25, соответственно [E05v2]; или
(iii) SEQ ID NO: 23 и 30, соответственно [G12v2].
[8] Молекула антитела согласно [7], отличающаяся тем, что молекула антитела содержит домен VH и домен VL, представленные в (i) или (ii).
[9] Молекула антитела согласно [8], отличающаяся тем, что молекула антитела содержит домен VH и домен VL, представленные в (i).
[10] Молекула антитела согласно любому из [1] - [9], отличающаяся тем, что первая последовательность расположена между положениями 14 и 17 домена CH3 молекулы антитела, где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[11] Молекула антитела согласно [10], отличающаяся тем, что первая последовательность расположена в положениях 15, 16, 16.5, 16.4, 16.3, 16.2, и 16.1 домена CH3 молекулы антитела, где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[12] Молекула антитела согласно любому из [1] - [11], отличающаяся тем, что вторая последовательность расположена в положениях 92 - 98 домена CH3 молекулы антитела, где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[13] Молекула антитела согласно любому из [1] - [12], отличающаяся тем, что молекула антитела дополнительно содержит третью последовательность, расположенную в структурной петле CD домена CH3.
[14] Молекула антитела согласно [13], отличающаяся тем, что третья последовательность расположена в положениях 43 - 78 домена CH3 молекулы антитела, где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[15] Молекула антитела согласно любому из [13] - [14], отличающаяся тем, что третья последовательность имеет последовательность, представленную в SEQ ID NO: 73.
[16] Молекула антитела согласно любому из [1] - [15], отличающаяся тем, что молекула антитела содержит последовательность домена CH3, представленную в:
(i) SEQ ID NO: 115 [FS22-172-003]; или
(ii) SEQ ID NO: 81 [FS22-53-008].
[17] Молекула антитела согласно любому из [1] - [15], отличающаяся тем, что первая и вторая последовательность имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003], соответственно.
[18] Молекула антитела согласно любому из [1] - [15] и [17], отличающаяся тем, что молекула антитела содержит последовательность домена CH3, представленную в SEQ ID NO: 115 [FS22-172-003].
[19] Молекула антитела согласно любому из [1] - [18], отличающаяся тем, что молекула антитела представляет собой молекулу IgG1 человека.
[20] Молекула антитела согласно любому из [1] - [19], отличающаяся тем, что молекула антитела содержит тяжелую цепь и легкую цепь антитела:
(i) FS22-172-003-AA/E12v2, представленные в SEQ ID NO: 134 и 17, соответственно;
(ii) FS22-172-003-AA/E05v2, представленные в SEQ ID NO: 137 и 28, соответственно;
(iii) FS22-172-003-AA/G12v2, представленные в SEQ ID NO: 140 и 33, соответственно;
(iv) FS22-053-008-AA/E12v2, представленные в SEQ ID NO: 143 и 17, соответственно;
(v) FS22-053-008-AA/E05v2, представленные в SEQ ID NO: 146 и 28, соответственно; или
(vi) FS22-053-008-AA/G12v2, представленные в SEQ ID NO: 149 и 33, соответственно.
[21] Молекула антитела согласно [20], отличающаяся тем, что молекула антитела содержит легкую цепь и тяжелую цепь, представленные в любом из (i) - (iv) из [20].
[22] Молекула антитела согласно [20], отличающаяся тем, что молекула антитела содержит легкую цепь и тяжелую цепь, представленные в (i) из [20].
[23] Молекула антитела согласно любому из [20] - [22], отличающаяся тем, что пролин (P) в положении 114 домена CH2 антитела заменен на аланин (A), где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
[24] Молекула антитела согласно любому из [1] - [23], отличающаяся тем, что молекула антитела связывает PD-L1 человека и CD137 человека.
[25] Молекула антитела согласно [24], отличающаяся тем, что PD-L1 содержит или состоит из последовательности, представленной в SEQ ID NO: 180.
[26] Молекула антитела согласно [24] или [25], отличающаяся тем, что CD137 человека содержит или состоит из последовательности, представленной в SEQ ID NO: 186.
[27] Молекула антитела согласно любому из [1] - [26], отличающаяся тем, что молекула антитела способна активировать CD137 на иммунной клетке в присутствии связанного с поверхностью опухолевой клетки PD-L1.
[28] Молекула антитела согласно любому из [1] - [26], отличающаяся тем, что связывание молекулы антитела с CD137 на иммунной клетке и со связанным с поверхностью опухолевой клетки PD-L1 вызывает кластеризацию CD137 на иммунной клетке.
[29] Молекула антитела согласно [27] или [28], отличающаяся тем, что иммунная клетка представляет собой T-клетку, B-клетку, естественную клетку-киллера (NK), естественную T-клетку-киллера (NKT) или дендритную клетку (ДК).
[30] Молекула антитела согласно [29], отличающаяся тем, что иммунная клетка представляет собой T-клетку.
[31] Молекула антитела согласно любому из [1] - [30], отличающаяся тем, что молекула антитела была модифицирована для уменьшения или устранения связывания домена CH2 молекулы антитела с одним или более Fcγ-рецепторами.
[32] Молекула антитела согласно любому из [1] - [31], отличающаяся тем, что молекула антитела не связывается с одним или более Fcγ-рецепторами.
[33] Молекула антитела согласно [31] или [32], отличающаяся тем, что Fcγ-рецептор выбран из группы, состоящей из: FcγRI, FcγRIIa, FcγRIIb и FcγRIII.
[34] Конъюгат, содержащий молекулу антитела согласно любому из [1] - [33] и биоактивную молекулу.
[35] Конъюгат, содержащий молекулу антитела согласно любому из [1] - [33] и детектируемую метку.
[36] Молекула или молекулы нуклеиновых кислот, кодирующие молекулу антитела согласно любому из [1] - [33].
[37] Молекула или молекулы нуклеиновых кислот, кодирующие молекулу антитела согласно любому из [1] - [2], [5] - [7], [10] - [16], [19] - [20] и [23] - [33], отличающиеся тем, что молекулы(-а) нуклеиновых(-ой) кислот(ы) содержат(-ит) последовательность нуклеиновой кислоты тяжелой цепи и/или последовательность нуклеиновой кислоты легкой цепи из:
(i) FS22-172-003-AA/E12v2, представленные в SEQ ID NO: 32 и 39 или 135 и 136, соответственно, предпочтительно в SEQ ID NO: 32 и 39, соответственно;
(ii) FS22-172-003-AA/E05v2, представленные в SEQ ID NO: 138 и 139, соответственно;
(iii) FS22-172-003-AA/G12v2, представленные в SEQ ID NO: 141 и 142, соответственно;
(iv) FS22-053-008-AA/E12v2, представленные в SEQ ID NO: 144 и 145, соответственно;
(v) FS22-053-008-AA/E05v2, представленные в SEQ ID NO: 147 и 148, соответственно; или
(vi) FS22-053-008-AA/G12v2, представленные в SEQ ID NO: 150 и 151, соответственно.
[38] Вектор или векторы, содержащие молекулу или молекулы нуклеиновых кислот согласно любому из [36] - [37].
[39] Рекомбинантная клетка-хозяин, содержащая молекулы(-у) нуклеиновых(-ой) кислот(ы) согласно любому из [36] - [37] или вектор(ы) согласно [38].
[40] Способ получения молекулы антитела согласно любому из [1] - [33], включающий культивирование рекомбинантной клетки-хозяина из [39] в условиях, подходящих для продукции молекулы антитела.
[41] Способ согласно [40], дополнительно включающий выделение и/или очистку молекулы антитела.
[42] Фармацевтическая композиция, содержащая молекулу антитела или конъюгат согласно любому из [1] - [35] и фармацевтически приемлемое вспомогательное вещество.
[43] Молекула антитела или конъюгат согласно любому из [1] - [35] для применения в способе лечения рака у индивида.
[44] Способ лечения рака у индивида, включающий введение индивиду терапевтически эффективного количества молекулы антитела или конъюгата согласно любому из [1] - [35].
[45] Применение молекулы антитела или конъюгата согласно любому из [1] - [35] для получения лекарственного средства для лечения рака.
[46] Молекула антитела или конъюгат для применения, способ или применение согласно любому из [43] - [45], отличающиеся тем, что рак выбран из следующего перечня: меланома, рак мочевого пузыря, рак головного мозга, рак молочной железы, рак яичников, рак легких, колоректальный рак, рак шейки матки, рак печени, рак головы и шеи, рак поджелудочной железы, рак почки, рак желудка и стромальные опухоли желудочно-кишечного тракта (GIST).
[47] Молекула антитела или конъюгат для применения согласно [43], отличающегося тем, что лечение включает введение индивиду молекулы антитела или конъюгата в комбинации со вторым терапевтическим агентом.
[48] Способ согласно [44], отличающийся тем, что способ дополнительно включает введение индивиду терапевтически эффективного количества второго терапевтического агента.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1. На фигурах 1A, B и C показано выравнивание последовательностей доменов CH3 Fcab FS22-053, FS22-053-008, FS22-053-009, FS22-053-010, FS22-053-011, FS22-053-012, FS22-053-013, FS22-053-014, FS22-053-015, FS22-053-016, FS22-053-017, FS22-172, FS22-172-001, FS22-172-002, FS22-172-003, FS22-172-004, FS22-172-005 и FS22-172-006, а также Fcab дикого типа (ДТ). Указаны номера остатков согласно системам нумерации IMGT, IMGT экзона (последовательная нумерация), EU и Кабата.
На фигуре 2 показано, что CD137/PD-L1 МАТ2 FS22-172-003-AA/E12v2 способно связываться с активированными CD4+ (фигура 2A) и CD8+ (фигура 2B) T-клетками. МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 связывало активированные T-клетки со значениями EC50, равными 0,8 нМ в случае CD4+ T-клеток и 0,9 нМ в случае CD8+ T-клеток. Для антитела положительного контроля против PD-L1 человека (G1-AA/E12v2) выявили аффинность, очень сходную с таковой у МАТ2, со значениями EC50, равными 0,8 нМ, для обоих типов клеток. Для антитела положительного контроля против CD137 человека (G1-AA/MOR7480.1) выявили низкий уровень связывания с CD4+ и CD8+ клетками, свидетельствующий о низком уровне экспрессии CD137 на данных типах клеток.
На фигуре 3 показано, что МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 не связывается с клетками CHO, экспрессирующими на повышенном уровне Fcγ-рецептор. МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2, антитело положительного контроля связывания с Fcγ-рецептором G1/4420 и антитело отрицательного контроля отсутствия связывания с Fcγ-рецептором G1-AA/4420 исследовали в анализе связывания с клетками из линий клеток CHO, экспрессирующих на повышенном уровне пять различных Fcγ-рецепторов, экспрессирующих (A) FcgRIIA 131H, (B) FcgRIIA 131E, (C) FcgRIIB, (D) FcgRIIIA 158F, (E) FcgRIIIA 158V, а также (F) линии клеток CHO ДТ, которую использовали в качестве отрицательного контроля. МАТ2 против CD137/PD-L1 человека, которое содержало мутацию LALA в области CH2, не связывалось ни с какими исследованными Fcγ-рецепторами, и оно также не связывалось неспецифично с клетками CHO ДТ (заштрихованные черные треугольники). Антитело положительного контроля связывало Fcγ-рецепторы (заштрихованные черные круги), а антитело отрицательного контроля G1-AA/4420, которое содержало мутацию LALA в области CH2, не связывало Fcγ-рецепторы (незаштрихованные серые квадраты), как и ожидалось.
На фигуре 4 показано высвобождение IL-2 человека (hIL-2) в анализе активации первичных CD8+ T-клеток человека в присутствии МАТ2 против CD137/PD-L1 человека или антитела положительного контроля против CD137 человека G1-AA/20H4.9. Все исследованные МАТ2 запускали кластеризацию CD137 и активацию CD8+ T-клеток, только когда МАТ2 перекрестно связывались клетками HEK с повышенной экспрессией PD-L1 человека, приводя к высвобождению IL-2 человека. Значения EC50 для МАТ2 против CD137/PD-L1 человека были меньше, чем значение EC50 для антитела положительного контроля против CD137 человека - G1-AA/20H4.9, при перекрестном связывании со вторичным антителом против CH2 человека (заштрихованные черные круги, пунктирная линия), свидетельствуя о том, что с МАТ2 добились улучшенной активации T-клеток. Для всех выявили повышение высвобождения hIL-2, и наблюдалось большее высвобождение IL-2 (Emax) с антителом положительного контроля, чем с МАТ2 против CD137/PD-L1 человека.
На фигуре 5 показано высвобождение IL-2 человека (hIL-2) в анализе активации первичных CD8+ T-клеток человека в присутствии МАТ2 против CD137/PD-L1 человека или антитела положительного контроля против CD137 человека - G1-AA/20H4.9. Все исследованные МАТ2 запускали кластеризацию CD137 и активацию CD8+ T-клеток, только когда МАТ2 перекрестно связывались клетками MDA-MD-231, эндогенно экспрессирующими PD-L1 человека, приводя к высвобождению IL-2 человека. Значения EC50 для МАТ2 против CD137/PD-L1 человека были меньше, чем значение EC50 для антитела положительного контроля против CD137 человека - G1-AA/20H4.9, при перекрестном связывании со вторичным антителом против CH2 человека (заштрихованные черные круги, пунктирная линия), свидетельствуя о том, что с МАТ2 добивались лучшей активности. Для всех выявили повышение высвобождения hIL-2, и большее высвобождение IL-2 (Emax) наблюдалось с антителом положительного контроля, чем с МАТ2 против CD137/PD-L1 человека.
На фигуре 6 показано высвобождение IL-2 человека (hIL-2) в анализе активации первичных CD8+ T-клеток человека в присутствии МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 или антитела положительного контроля против CD137 человека - G1-AA/20H4.9. Все исследованные МАТ2 запускали кластеризацию CD137 и активацию CD8+ T-клеток, только когда МАТ2 перекрестно связывались клетками HEK.hPD-L1, которые составляли по меньшей мере 6,5% всей популяции клеток HEK, приводя к высвобождению IL-2 человека. Значения EC50 для МАТ2 против CD137/PD-L1 человека оставались сходными при модулировании процента клеток HEK.hPD-L1, присутствующих в анализе, но максимальная активация (Emax) повышалась в соответствии с возрастанием процента перекрестно связывающих клеток HEK.hPD-L1 в анализе. Это указывало на то, что добивались лучшей максимальной активности с МАТ2, когда в целом 100% клеток HEK экспрессировали PD-L1.
На фигуре 7 показано, что МАТ2 против CD137/PD-L1 человека способны активировать T-клетки. (A) и (B): высвобождение IFN-γ в анализе реакции смешанной культуры первичных лимфоцитов человека (СКЛ) исследовали в присутствии МАТ2 против CD137/PD-L1 человека, антитела положительного контроля против PD-L1 человека G1/S70 или антитела положительного контроля против CD137 человека G1-AA/20H4.9 (перекрестно связанного с антителом против CH2 человека). Для (A) МАТ2 на основе Fcab FS22-053-008 и (B) МАТ2 на основе Fcab FS22-172-003 выявили более высокую активность в данном анализе, чем для любого из антител положительного контроля, свидетельствуя о том, что блокирование PD-L1 и агонизм CD137 в одной и той же молекуле вызывает большую активацию T-клеток, предположительно посредством основанной на PD-L1 кластеризации и активации CD137. (C) Высвобождение IFN-γ в анализе первичных СКЛ человека исследовали в присутствии МАТ2 против CD137/PD-L1 человека, антитела против PD-L1 человека G1-AA/E12v2, антитела положительного контроля против CD137 человека G1-AA/20H4.9 (перекрестно связанного с антителом против CH2 человека) или Fcab против CD137 человека в формате ложного МАТ2 (FS22-172-003-AA/D1.3). В данном анализе выявили более высокую активность МАТ2 FS22-172-003-AA/E12v2 в отношении активации T-клеток, чем у любого из антител положительного контроля или их компонента Fcab в формате ложного МАТ2, свидетельствуя о том, что блокирование PD-L1 и агонизм CD137 в одной и той же молекуле вызывает большую активацию T-клеток, предположительно посредством перекрестного связывания на основе PD-L1, приводящего к кластеризации и активации CD137.
На фигуре 8 показано высвобождение IL-2 в анализе активации T-клеток, где T-клетки были сконструированы для сверхэкспрессии CD137 яванского макака, в присутствии МАТ2 против CD137/PD-L1 человека и антитела положительного контроля против CD137 человека G1-AA/MOR7480.1. Все исследованные МАТ2 запускали кластеризацию и активацию CD137, только когда они были перекрестно связаны с HEK клетками с повышенной экспрессией PD-L1 яванского макака (незаштрихованные и заштрихованные черные и серые символы со сплошными соединяющими линиями), приводя к высвобождению интерлейкина-2 мыши (mIL-2) в анализе активации T-клеток DO11.10. При возрастании концентраций, для антитела положительного контроля против CD137 человека - G1-AA/MOR7480.1, которое, как было показано ранее другими исследователями, перекрестно реагирует с CD137 яванского макака, показали повышение высвобождения mIL-2 (заштрихованные черные круги, пунктирная линия), тем не менее, максимальное высвобождение было значительно меньшим, чем таковое для всех МАТ2 против CD137/PD-L1 человека. Для всех МАТ2 против CD137/PD-L1 человека без клеток HEK с повышенной экспрессией PD-L1, с которыми они могли бы перекрестно связаться, выявили значительно более низкое высвобождение mIL-2 по сравнению с таковым при перекрестном связывании (результаты не представлены).
На фигуре 9 показано высвобождение IL-2 на 2 сутки (фигура 9A) или высвобождение IFN-γ на 6 сутки (фигура 9B) из CD4+ T-клеток яванского макака, культивированных с аллогенными моноцитами яванского макака в присутствии титрованного МАТ2 против CD137/PD-L1 человека FS22-053-008-AA/E12v2 или 172-003-AA/E12v2, или антитела положительного контроля против PD-L1 человека G1-AA/E12v2, или изотипического контрольного антитела G1-AA/HelD1.3.
На фигуре 10 показано высвобождение IL-2 мыши в анализе активации T-клеток с CD137 мыши DO11.10, в котором тестировали перекрестно реагирующие с CD137 мыши и человека Fcab FS22-053-014 и FS22-053-017 в формате ложного МАТ2, при перекрестном связывании с белком L. FS22-053-014 и FS22-053-017, и Fcab против CD137 мыши FS22m-063, все в формате ложного МАТ2 HelD1.3, активировали CD137 при перекрестном связывании с белком L, что приводило к высвобождению mIL-2. Для МАТ положительного контроля против CD137 мыши - Lob12.3 - выявили повышение высвобождения mIL-2, как и ожидалось. Для всех Fcab против CD137 в формате ложного МАТ2 без белка L, с которым они могли бы перекрестно связаться (пунктирные линии), выявили гораздо меньшее высвобождение mIL-2 по сравнению с таковым при перекрестном связывании. Для FS22-053-017 наблюдалась более низкая активность в данном анализе (в 8 раз худшая EC50 по сравнению с FS22-053-014), но все еще выявлялась активность в данном анализе.
На фигуре 11 показано высвобождение IFNγ мыши, которое свидетельствовало об активации CD8+ T-клеток. МАТ2 против CD137/PD-L1 мыши проявило наибольшую эффективность в данном анализе по сравнению с любым из антител положительного контроля против CD137 и PD-L1 человека или комбинацией указанных двух антител. Как ожидалось, для FS22m-063-AA Fcab в формате ложного МАТ2 (FS22m-063-AA/HelD1.3) не выявили никакой активности.
На фигуре 12 показаны измерения объема опухоли в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/4420 (контроль IgG), G1/S70 (положительный контроль против PD-L1), G1-AA/Lob12.3 и G1/Lob12.3 (положительный контроль против CD137 с мутацией LALA и без нее), комбинацией G1/S70 плюс G1-AA/Lob12.3 и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). Средний объем опухоли плюс или минус 95%-ный доверительный интервал наносили на график. FS22m-063-AA/S70 способно значительно уменьшать рост опухоли в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG и МАТ положительного контроля против PD-L1. Статистическая значимость показана попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели. Средний объем опухоли представлен в виде среднего геометрического или арифметического, в соответствующих случаях, на основании проверки нормальности результатов. * P ≤ 0,05; ** P ≤ 0,01; *** P ≤ 0,001; **** P ≤ 0,0001.
На фигуре 13 показаны результаты исследования доза-ответ МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли CT26 мыши. A: показаны измерения объема опухоли в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили 3 дозами G1-AA/4420 (контроля IgG, 10 мг/кг), G1/Lob12.3 (положительного контроля против CD137, 1 мг/кг) и МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70 в 4 различных дозах (0,1 мг/кг, 0,3 мг/кг, 1,0 мг/кг и 10,0 мг/кг). Каждая доза указана вертикальной черной стрелкой на оси x. Средний объем опухоли плюс или минус 95%-ный доверительный интервал наносили на график. FS22m-063-AA/S70 способно значительно уменьшать рост опухоли в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG и МАТ положительного контроля против PD-L1. Статистическая значимость показана попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели. Средний объем опухоли представлен в виде среднего геометрического или арифметического, в соответствующих случаях, на основании проверки нормальности результатов. ** P ≤ 0,01; *** **** P ≤ 0,0001. B: показан график Каплана-Майера для модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили 3 дозами G1-AA/4420 (контроля IgG, ~10 мг/кг) и МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70 в 4 различных дозах: 2 мкг, 6 мкг, 20 мкг и 200 мкг (эквивалентных приблизительно 0,1 мг/кг, 0,3 мг/кг, 1,0 мг/кг и 10,0 мг/кг). FS22m-063-AA/S70 вызывало зависимое от дозы повышение выживаемости в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 14 зависимое от дозы увеличение процентного соотношения CD8+:CD4+ в опухоли и крови мышей, которым вводили раз в два дня (q2d)x3 МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70 при четырех различных уровнях доз, показано на панели A и B и экспрессия Ki67 на CD8+ T-клетках показана на панелях C и D.
На фигуре 15 показаны измерения объема опухоли в модели сингенной опухоли MC38, которую растили подкожно в мышах C57BL/6, которых лечили G1-AA/4420 (изотипический контроль), G1-AA/S70 (положительный контроль против PD-L1), G1-AA/Lob12.3 (положительный контроль против CD137), комбинацией G1-AA/S70 плюс G1-AA/Lob12.3, и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). Средний объем опухоли [в мм3] плюс или минус 95%-ный доверительный интервал наносили на график. FS22m-063-AA/S70 способно значительно уменьшать рост опухоли в модели сингенной опухоли MC38 по сравнению с мышами, которым вводили контроль IgG, мышами, которых лечили МАТ положительного контроля против PD-L1, мышами, которых лечили антителом положительного контроля против CD137. Статистическая значимость показана попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели. **** p-значение ≤ 0,0001.
На фигуре 16 показаны измерения объема опухоли в модели сингенной опухоли B16.F10, которую растили подкожно в мышах C57BL/6, которых лечили G1-AA/4420 (изотипический контроль) и МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70. Обе композиции вводили в дозе 1 мг/кг. Средний объем опухоли [в мм3] плюс или минус 95%-ный доверительный интервал наносили на график. Лечение FS22m-063-AA/S70 значительно уменьшало рост опухоли в модели сингенной опухоли B16.F10 по сравнению с мышами, которым вводили контроль IgG. Статистическая значимость показана попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели. Средний объем опухоли представлен в виде среднего геометрического или арифметического, в соответствующих случаях, на основании проверки нормальности результатов. * P ≤ 0,05; ** P ≤ 0,01; *** P ≤ 0,001; **** P ≤ 0,0001.
На фигуре 17 показаны диаграммы спагетти для отдельных мышей в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили (A) G1-AA/4420 (контроль IgG), (C) G1/S70 (антитело положительного контроля против PD-L1), (B) G1-AA/Lob12.3 и (D) G1/Lob12.3 (антитело положительного контроля против CD137 с мутацией LALA и без нее) или (E) комбинацией G1/S70 плюс G1-AA/Lob12.3 и (F) FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 вызывало значимое ингибирование роста опухоли в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 18 показан график Каплана-Майера для модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/4420 (контроль IgG), комбинацией G1/S70 плюс G1-AA/Lob12.3, и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1), все в дозе 1 мг/кг. FS22m-063-AA/S70 вызывало значимую выживаемость в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 19 показаны измерения объема опухоли в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/HelD1.3 (контроль IgG), G1/S70 (антитело положительного контроля против PD-L1 ), G1-AA/Lob12.3 (антитело положительного контроля против CD137) и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). Средний объем опухоли плюс или минус 95%-ный доверительный интервал наносили на график. FS22m-063-AA/S70 было способно значимо уменьшить рост опухоли в модели сингенной опухоли CT26 по сравнению с мышами, которым вводили контроль IgG, мышами, которых лечили МАТ положительного контроля против PD-L1, и мышами, которых лечили антителом положительного контроля против CD137. Статистическая значимость показана попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели. Средний объем опухоли представлен в виде среднего геометрического или арифметического, в соответствующих случаях, на основании проверки нормальности результатов. * P ≤ 0,05; ** P ≤ 0,01; *** P ≤ 0,001; **** P ≤ 0,0001.
На фигуре 20 показан график Каплана-Майера для модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/HelD1.3 (контроль IgG), G1/S70 (антитело положительного контроля против PD-L1), G1-AA/Lob12.3 (антитело положительного контроля против CD137) и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 вызывало значимую выживаемость в модели сингенной опухоли CT26 по сравнению с мышами, которых лечили контролем IgG, и мышами, которых лечили антителом положительного контроля против CD137.
На фигуре 21 показаны диаграммы спагетти для отдельных мышей в модели сингенной опухоли MC38, которую растили подкожно в мышах C57BL/6, которых лечили 3 дозами: (A) G1-AA/4420 (изотипический контроль), (B) G1-AA/S70 (антитело положительного контроля против PD-L1), (C) G1-AA/Lob12.3 (антитело положительного контроля против CD137), (D) комбинации G1-AA/S70 плюс G1-AA/Lob12.3, и (E) FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 вызывало полное ингибирование роста опухоли, приводя к 100% отсутствию опухолей у мышей, в модели сингенной опухоли MC38 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 22 показан график Каплана-Майера для модели сингенной опухоли MC38, которую растили подкожно в мышах C57BL/6, которых лечили 3 дозами: G1-AA/4420 (изотипический контроль), G1-AA/S70 (антитело положительного контроля против PD-L1), G1-AA/Lob12.3 (антитело положительного контроля против CD137), комбинации G1-AA/S70 плюс G1-AA/Lob12.3 и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 вызывало полную выживаемость в модели сингенной опухоли MC38 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 23 показаны диаграммы спагетти для отдельных мышей в модели сингенной опухоли B16.F10, которую растили подкожно в мышах C57BL/6, которых лечили G1-AA/4420 (изотипический контроль) (фигура 23A) и МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70 (фигура 23B). FS22m-063-AA/S70 вызывало частичное ингибирование роста опухоли в модели сингенной опухоли B16.F10 по сравнению с мышами, которым вводили контроль IgG.
На фигуре 24 показаны результаты связывания МАТ2 против mCD137/PD-L1 с T-клетками ex vivo, определенные с помощью проточной цитометрии. Значения средней интенсивности флуоресценции (MFI) измеряли для вторичного антитела против Fc человека, конъюгированного с Alexa Fluor 488, которое детектировало человеческую Fc-область МАТ2 против mCD137/PDl-L1, связанного с клетками. Положительные клетки, MFI которых была больше, чем таковая для неокрашенного контрольного образца, идентифицировали как положительные по антителу против mCD137/PD-L1, и представленные результаты показывают положительную популяцию в виде процента (A) от всех CD8+ или (B) от всех CD4+ T-клеток из крови или (C) от всех CD8+ или (D) от всех CD4+ T-клеток из опухоли. Данные результаты показывают, что МАТ2 против mCD137/PD-L1 быстро связывает как CD8+, так и CD4+ T-клетки, присутствующие как в опухоли, так и в крови, и что процент популяции положительных T-клеток снижается с течением времени в зависимости от уровня дозы.
На фигуре 25 показаны результаты экспрессии Ki67 T-клетками ex vivo, определенные с помощью проточной цитометрии. Значения средней интенсивности флуоресценции (MFI) измеряли для антитела против Ki67, конъюгированного с PE-Cy7, связанного с клетками. Положительные клетки, MFI которых была больше, чем таковая для неокрашенного контрольного образца, идентифицировали как положительные по экспрессии Ki67, и представленные результаты показывают положительную популяцию в виде процента (A) от всех CD8+ или (B) от всех CD4+ T-клеток из крови или (C) от всех CD8+ или (D) от всех CD4+ T-клеток из опухоли. Данные результаты показывают, что T-клетки в мышах, которым вводят дозу МАТ2 против mCD137/PD-L1, быстро экспрессируют Ki67 как на CD8+, так и на CD4+ T-клетках в крови, и что высокий процент положительных по Ki67 T-клеток уже присутствует в образце из микроокружения опухоли. Наблюдали повышение экспрессии Ki67 с течением времени в зависимости от уровня дозы.
На фигуре 26 показана занятость рецептора PD-L1 по сравнению с соответствующим по времени образцом из мышей, которым вводили дозу контрольного антитела G1-AA/4420, в который затем добавляли 100 нМ МАТ2 против mCD137/PD-L1 ex vivo, чтобы насытить все рецепторы PD-L1, что указывало на 100% уровень занятости рецептора PD-L1 (треугольные символы с пунктирной линией). Свободные рецепторы PD-L1 детектировали, используя значения средней интенсивности флуоресценции (MFI) для конкурирующего антитела против mPD-L1, конъюгированного с Bv605. Положительные клетки, MFI которых была больше, чем таковая для неокрашенного контрольного образца, идентифицировали как положительные по свободному PD-L1. Результаты показывают положительную популяцию в виде процента занятости рецептора PD-L1 по сравнению с насыщенным на 100% образцом (A) CD8+ или (B) CD4+ T-клеток из крови или (C) CD8+ или (D) CD4+ T-клеток из опухоли. Данные результаты демонстрируют, что занятость рецептора PD-L1 быстро достигала 100% в крови перед снижением в соответствии с уровнем дозы с течением времени. Занятость рецептора PD-L1 на T-клетках поддерживалась дольше в микроокружении опухоли, чем в крови.
На фигуре 27 показан фармакокинетический профиль МАТ2 против CD137/PD-L1 в дозе 25, 10, 3, и 1 мг/кг после однократной внутривенной разовой дозы, которую вводили непримированным (наивным) мышам C57BL/6. Показана концентрация МАТ2 против CD137/PD-L1 в мышах с течением времени. На фигуре 27 продемонстрировано, что скорость клиренса МАТ2 против CD137/PD-L1 при каждой дозе сопоставима с клиренсом стандартного IgG человека в мышах.
На фигуре 28 показано, что МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 способно активировать T-клетки в анализе активации T-клеток МКПК с применением (A) МКПК яванского макака или (B) МКПК человека. Высвобождение IFN-γ в анализе МКПК исследовали в присутствии МАТ2 против CD137/PD-L1 человека, антитела положительного контроля против CD137 человека G1-AA/MOR7480.1 (перекрестно связанного с антителом против CH2 человека). Выявили активность МАТ2 в обоих анализах и выявили более высокие уровни активации, чем для антитела положительного контроля, свидетельствуя о том, что блокирование PD-L1 и агонизм CD137 одной и той же молекулой вызывает большую активацию T-клеток, предположительно посредством основанной на PD-L1 кластеризации и активации CD137, как у яванского макака, так и у людей.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Аспекты и варианты реализации настоящего изобретения далее будут обсуждаться со ссылкой на сопроводительные фигуры. Дополнительные аспекты и варианты реализации должны быть очевидны для специалистов в данной области. Все документы, упомянутые в данном тексте, включены в данную заявку посредством ссылки.
Согласно настоящему изобретению предложены молекулы антитела, которые связываются как с PD-L1, так и с CD137. В частности, молекулы антитела согласно настоящему изобретению содержат сайт связывания антигена PD-L1 на основе CDR и сайт связывания антигена CD137, расположенный в константном домене молекулы антитела. Термины “PD-L1” и “CD137” могут относиться к PD-L1 человека и CD137 человека, PD-L1 мыши и CD137 мыши, и/или PD-L1 яванского макака и CD137 яванского макака, если из контекста не следует иное. Предпочтительно термины “PD-L1” и “CD137” относятся к PD-L1 человека и CD137 человека, если из контекста не следует иное.
Термин “молекула антитела” описывает иммуноглобулин, либо природный, либо полученный частично или полностью синтетическим путем. Молекула антитела может быть человеческой или гуманизированной, предпочтительно человеческой. Молекула антитела предпочтительно представляет собой молекулу моноклонального антитела. Примеры антител представляют собой изотипы иммуноглобулинов, такие как иммуноглобулин G, и их изотипические подклассы, такие как IgG1, IgG2, IgG3 и IgG4, а также их фрагменты. Молекула антитела может быть изолированной, в том смысле, что она свободна от примесей, таких как антитела, способные связываться с другими полипептидами и/или компонентами сыворотки.
Термин “молекула антитела” в данной заявке, следовательно, включает фрагменты антитела, при условии, что указанные фрагменты содержат сайт связывания антигена PD-L1 на основе CDR и сайт связывания антигена CD137, расположенный в константном домене. Если из контекста не следует иное, термин “молекула антитела” в данной заявке, следовательно, эквивалентен термину “молекула антитела или ее фрагмент”.
Можно взять моноклональные и другие антитела и использовать методики технологии рекомбинантных ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела. Такие методики могут включать внедрение CDR, или вариабельных областей, и/или последовательностей константного домена, содержащих сайт связывания антигена CD137, в отличный иммуноглобулин. Внедрение CDR одного иммуноглобулина в другой иммуноглобулин описано, например, в EP-A-184187, GB 2188638A или EP-A-239400. Аналогичные методики можно применять для соответствующих последовательностей константного домена. В качестве альтернативы, гибридому или другую клетку, продуцирующую молекулу антитела, можно подвергнуть генетической мутации или другим изменениям, которые могут изменить или могут не изменить специфичность связывания продуцированных антител.
Так как антитела можно модифицировать множеством способов, термин «молекула антитела» следует толковать как включающий фрагменты, производные, функциональные эквиваленты антитела и гомологи антител, включая любой полипептид, содержащий связывающий домен иммуноглобулина, либо природный, либо полностью или частично синтетический. Следовательно, в объем данного термина входят химерные молекулы, содержащие связывающий домен иммуноглобулина или его эквивалент, слитый с другим полипептидом. Клонирование и экспрессия химерных антител описаны в EP-A-0120694 и EP-A-0125023.
Примером фрагмента антитела, содержащего как последовательности CDR, так и домен CH3, является минитело, которое содержит scFv, соединенный с доменом CH3 (Hu и др. (1996), Cancer Res., 56(13):3055-61).
Молекула антитела согласно настоящему изобретению связывается с PD-L1 и CD137. Связывание в данном контексте может относиться к специфическому связыванию. Термин «специфический» может относиться к случаю, когда у молекулы антитела не будут выявлять какое-либо значительное связывание с молекулами, отличными от ее партнера(-ов) по специфическому связыванию, здесь PD-L1 и CD137. Термин “специфический” также применим, когда молекула антитела специфична к конкретным эпитопам, таким как эпитопы на PD-L1 и CD137, которые находятся на множестве антигенов, и в данном случае молекула антитела будет способна связываться с различными антигенами, несущими указанный эпитоп.
Авторы настоящего изобретения продемонстрировали, что у молекул антител, описанных в данной заявке, выявили высокий уровень специфичности к PD-L1 человека и не выявили какого-либо значимого связывания с другими мишенями T-клеток PD-L1, CD80, PD-1 или B7-H3. См. пример 9.3. Таким образом, в предпочтительном варианте реализации молекула антитела не связывается или не проявляет какого-либо значительного связывания с любым, предпочтительно со всеми, из PD-L2, CD80, PD-1 и B7-H3. Авторы настоящего изобретения также продемонстрировали, что для сайта связывания антигена CD137 не выявили какого-либо значительного связывания с рецепторами TNFRSF человека CD40, OX40 и GITR. См. пример 3.6. Таким образом, в более предпочтительном варианте реализации молекула антитела не связывается или не проявляет какого-либо значительного связывания с любым, предпочтительно со всеми, из PD-L2, CD80, PD-1, B7-H3, CD40, OX40 и GITR.
Антитела и способы их конструирования и применения хорошо известны в данной области и описаны, например, в Holliger & Hudson (2005). Можно взять моноклональные и другие антитела и использовать методики технологии рекомбинантных ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела. Такие методики могут включать внедрение CDR или вариабельных областей одной молекулы антитела в отличную молекулу антитела (EP-A-184187, GB 2188638A и EP-A-239400).
Сайт связывания антигена на основе CDR представляет собой сайт связывания антигена в вариабельной области антитела. Сайт связывания антигена на основе CDR может быть образован тремя CDR, такими как три CDR вариабельного домена легкой цепи (VL) или три CDR вариабельного домена тяжелой цепи (VH). Предпочтительно, сайт связывания антигена на основе CDR образован шестью CDR: тремя CDR VL и тремя CDR VH. Вклад различных CDR в связывание антигена может изменяться в различных сайтах связывания антигена.
Три CDR домена VH сайта связывания антигена могут быть расположены внутри домена VH иммуноглобулина и три CDR домена VL могут быть расположены внутри домена VL иммуноглобулина. Например, сайт связывания антигена на основе CDR может быть расположен в вариабельной области антитела.
Молекула антитела может содержать один или предпочтительно более чем один, например, два сайта связывания антигена на основе CDR для связывания с первым антигеном. Молекула антитела, следовательно, может содержать один домен VH и один домен VL, но предпочтительно содержит два домена VH и два домена VL, т.е., две пары доменов VH/VL, как в случае, например, встречающихся в природе молекул IgG.
Сайт связывания антигена на основе CDR может содержать три CDR VH или три CDR VL, предпочтительно три CDR VH и три CDR VL, антитела E12v2, E05v2, G12v2 или lam-G02v3, предпочтительно антитела E12v2, E05v2 или G12v2, более предпочтительно E12v2 или E05v2, наиболее предпочтительно E12v2.
Последовательности доменов VH и VL данных антител описаны далее:
(i) последовательности доменов VH и VL для антитела E12v2 представлены в SEQ ID NO: 12 и 14, соответственно;
(ii) последовательности доменов VH и VL для антитела E05v2 представлены в SEQ ID NO: 23 и 25, соответственно;
(iii) последовательности доменов VH и VL для антитела G12v2 представлены в SEQ ID NO: 23 и 30, соответственно; и
(iv) последовательности доменов VH и VL для антитела lam-G02v3 представлены в SEQ ID NO: 23 и 41, соответственно.
Для специалиста не будет представлять трудностей определение последовательностей CDR в последовательностях доменов VH и VL антител, представленных выше. Последовательности CDR можно, например, определить согласно Кабату (Kabat, E.A и др. (1991). Sequences of Proteins of Immunological Interest, 5ое изд., публикация NIH № 91-3242. Министерство здравоохранения и социального обеспечения США) или международной информационной системе ImMunoGeneTics (IMGT: Lefranc, M.-P. и др. Nucleic Acids Res. 43, D413-22 (2015)).
Последовательности CDR1, CDR2 и CDR3 домена VH молекулы антитела согласно нумерации по Кабату могут представлять собой последовательности, расположенные в положениях 31 - 35, 50 - 65 и 95 - 102 домена VH, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VH молекулы антитела согласно нумерации IMGT могут представлять собой последовательности, расположенные в положениях 27 -38, 56 - 65 и 105 - 117 домена VH молекулы антитела, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VL молекулы антитела согласно нумерации по Кабату могут представлять собой последовательности, расположенные в положениях 24 - 34, 50 - 56 и 89 - 97 домена VL, соответственно.
Последовательности CDR1, CDR2 и CDR3 домена VL молекулы антитела согласно нумерации IMGT могут представлять собой последовательности, расположенные в положениях 27 - 38, 56 - 65 и 105 - 117 домена VL, соответственно.
Молекула антитела может содержать последовательность CDR1 домена VH SYGIS (SEQ ID NO: 1), CDR2 домена VH WISAYSGGTNYAQKLQG (SEQ ID NO: 2) и CDR3 домена VH DLFPTIFGVSYYYY (SEQ ID NO: 3), где последовательности CDR определены согласно схеме нумерации по Кабату.
В предпочтительных вариантах реализации молекула антитела содержит пролин в положении 28 согласно схеме нумерации по Кабату.
Молекула антитела может содержать последовательность CDR1 домена VH GYX1FTSYG (SEQ ID NO: 45), CDR2 домена VH ISAYSGGT (SEQ ID NO: 8) и CDR3 домена VH ARDLFPTIFGVSYYYY (SEQ ID NO: 9);
где X1 представляет собой P или T , предпочтительно где X1 представляет собой P;
и где последовательности CDR определены согласно схеме нумерации IMGT.
Молекула антитела может содержать последовательности CDR1, CDR2 и CDR3 домена VH:
(i) SEQ ID NO: 7, 8 и 9, соответственно [E12v2]; или
(ii) SEQ ID NO: 21, 8 и 9, соответственно [E05v2, G12v2 или lam-G02v3],
где последовательности CDR определены согласно схеме нумерации IMGT.
CDR домена VH могут быть фланкированы каркасными (FW) последовательностями (HFW1, HFW2, HFW3 и HFW4). Домен VH может содержать последовательности HFW1, HFW2, HFW3 и HFW4, представленные в SEQ ID NO: 51, 52, 53 и 54, соответственно, где последовательности FW и CDR определены согласно схеме нумерации по Кабату. В качестве альтернативы, домен VH может содержать последовательности HFW1, HFW2, HFW3 и HFW4, представленные в SEQ ID NO: 55, 56, 57 и 54, соответственно, где последовательности FW и CDR определены согласно схеме нумерации IMGT.
Молекула антитела может содержать последовательность CDR1 домена VL GTSSDVGGYNYVS (SEQ ID NO: 34), CDR2 VL EVTNRPS (SEQ ID NO: 35) и CDR3 VL SSFKRGSTLVV (SEQ ID NO: 36); где последовательности CDR определены согласно схеме нумерации по Кабату. Домен VL может происходить из домена VL лямбда. Например, каждый из CDR домена VL может быть фланкирован каркасными (FW) последовательностями (LFW1, LFW2, LFW3 и LFW4), которые происходят из домена VL лямбда. В частности, домен VL может содержать последовательности LFW1, LFW2, LFW3 и LFW4, представленные в SEQ ID NO: 65, 66, 67 и 68, соответственно, где последовательности FW определены согласно схеме нумерации по Кабату.
Молекула антитела может содержать последовательность CDR1 домена VL RASQSIX2X3RLA (SEQ ID NO: 46), CDR2 VL EASX4X5EX6 (SEQ ID NO: 47) и CDR3 VL QQSYSX7PX8X9T (SEQ ID NO: 48);
где X2 представляет собой S или G, X3 представляет собой N или G; X4 представляет собой T или N; X5 представляет собой S или L; X6 представляет собой T или S; X7 представляет собой T или W; X8 отсутствует или представляет собой R; X9 представляет собой Y, R или V; и
где последовательности CDR и нумерация положений определены согласно схеме нумерации по Кабату.
Предпочтительно, молекула антитела содержит последовательность CDR1 домена VH RASQSIGNRLA (SEQ ID NO: 4), CDR2 VL EASTSET (SEQ ID NO: 5) и CDR3 VL QQSYSTPYT (SEQ ID NO: 6), где последовательности CDR определены согласно схеме нумерации по Кабату. Домен VL может происходить из домена VL каппа. Например, каждый из CDR домена VL может быть фланкирован каркасными (FW) последовательностями (LFW1, LFW2, LFW3 и LFW4), которые происходят из домена VL каппа. В частности, домен VL может содержать последовательности LFW1, LFW2, LFW3 и LFW4, представленные в SEQ ID NO: 58, 59, 60 и 61, соответственно, где последовательности FW определены согласно схеме нумерации по Кабату.
Например, молекула антитела может содержать последовательности CDR1, CDR2 и CDR3 домена VL, представленные в:
(i) SEQ ID NO: 4, 5 и 6, соответственно [E12v2];
(ii) SEQ ID NO: 18, 19 и 20, соответственно [E05v2];
(iii) SEQ ID NO: 18, 19 и 29, соответственно [G12v2]; или
(iv) SEQ ID NO: 34, 35 и 36, соответственно [lam-G02v3],
где последовательности CDR определены согласно схеме нумерации по Кабату.
Молекула антитела может содержать последовательность CDR1 домена VL SSDVGGYNY (SEQ ID NO: 37), CDR2 VL EVT (SEQ ID NO: 38) и CDR3 VL SSFKRGSTLVV (SEQ ID NO: 36), где последовательности CDR определены согласно схеме нумерации IMGT. Домен VL может происходить из домена VL лямбда. Например, каждый из CDR домена VL может быть фланкирован каркасными (FW) последовательностями (LFW1, LFW2, LFW3 и LFW4), которые происходят из домена VL лямбда. В частности, домен VL может содержать последовательности LFW1, LFW2, LFW3 и LFW4, представленные в SEQ ID NO: 69, 70, 71 и 68, соответственно, где последовательности FW определены согласно схеме нумерации IMGT.
В качестве альтернативы, молекула антитела может содержать последовательность CDR1 домена VL QSIX10X11R (SEQ ID NO: 49), CDR2 VL EAS (SEQ ID NO: 11) и CDR3 VL QQSYSX12X13X14X15T (SEQ ID NO: 50);
где X10 представляет собой G или S; X11 представляет собой N или G; X12 отсутствует или представляет собой T; X13 представляет собой T, W или P; X14 представляет собой P или R; X15 представляет собой Y или R; и
где последовательности CDR и нумерация положений определены согласно схеме нумерации IMGT.
Предпочтительно, молекула антитела содержит последовательность CDR1 домена VH QSIGNR (SEQ ID NO: 10), CDR2 VL EAS (SEQ ID NO: 11) и CDR3 VL QQSYSTPYT (SEQ ID NO: 6), где последовательности CDR определены согласно схеме нумерации IMGT. Домен VL может происходить из домена VL каппа. Например, каждый из CDR домена VL может быть фланкирован каркасными (FW) последовательностями (LFW1, LFW2, LFW3 и LFW4), которые происходят из домена VL каппа. В частности, домен VL может содержать последовательности LFW1, LFW2, LFW3 и LFW4, представленные в SEQ ID NO: 62, 63, 64 и 61, соответственно, где последовательности FW определены согласно схеме нумерации IMGT.
Например, молекула антитела может содержать последовательности CDR1, CDR2 и CDR3 домена V, представленные в:
(i) SEQ ID NO: 10, 11 и 6, соответственно [E12v2];
(ii) SEQ ID NO: 22, 11 и 20, соответственно [E05v2];
(iii) SEQ ID NO: 22, 11 и 29, соответственно [G12v2]; или
(iv) SEQ ID NO: 37, 38 и 36, соответственно [lam-G02v3],
где последовательности CDR определены согласно схеме нумерации IMGT.
Сайт связывания антигена на основе CDR может содержать домены VH или VL, предпочтительно домены VH и VL, антитела E12v2, E05v2, G12v2 или lam-G02v3, предпочтительно антитела E12v2, E05v2 или G12v2, более предпочтительно E12v2 или E05v2, наиболее предпочтительно E12v2.
Домен VH антител E12v2, E05v2, G12v2 и lam-G02v3 могут иметь последовательность, представленную в SEQ ID NO: 12, 23, 23 и 23, соответственно. Домен VL антител E12v2, E05v2, G12v2 и lam-G02v3 может иметь последовательность, представленную в SEQ ID NO: 14, 25, 30 и 41, соответственно.
Молекула антитела согласно настоящему изобретению содержит сайт связывания антигена CD137, расположенный в константном домене молекулы антитела. Константный домен может представлять собой домен CL, CH1, CH2, CH3 или CH4, предпочтительно константный домен представляет собой домен CH1, CH2 или CH3, более предпочтительно домен CH2 или CH3, наиболее предпочтительно домен CH3. Сайт связывания антигена CD137 содержит одну или более модифицированных структурных петель в константном домене молекулы антитела. Конструирование структурных петель константного домена антитела для создания сайтов связывания целевых антигенов известно в данной области и описано, например, в Wozniak-Knopp G и др. (2010); WO2006/072620 и WO2009/132876.
Сайт связывания антигена CD137 молекулы антитела может содержать первую и вторую последовательность, предпочтительно первую и вторую последовательность, причем первая и вторая последовательность расположены в структурных петлях AB и EF константного домена, предпочтительно домена CH3, молекулы антитела, соответственно.
В предпочтительном варианте реализации остатки в положениях 95 и 96 домена CH3 молекулы антитела относятся к дикому типу, т.е., предпочтительно представляют собой аргинин (R) и триптофан (W), соответственно. Оба данных остатка расположены в структурной петле EF. Положения аминокислотных остатков пронумерованы здесь согласно схеме нумерации ImMunoGeneTics (IMGT), если не указано иначе. Схема нумерации IMGT описана в Lefranc и др., 2005.
Первая последовательность предпочтительно содержит последовательность PPY (SEQ ID NO: 78).
Последовательность PPY может быть расположена между положениями 10 и 19, предпочтительно между положениями 15 и 17 домена CH3 молекулы антитела. В предпочтительном варианте реализации последовательность PPY расположена в положениях 16, 16.5 и 16.4 домена CH3. В качестве альтернативы последовательность PPY может быть расположена между положениями 16 и 17 домена CH3. В альтернативном предпочтительном варианте реализации последовательность PPY расположена в положениях 16.3, 16.2 и 16.1 домена CH3. В схеме нумерации IMGT вставленные остатки пронумерованы в соответствии с направлением петли, в которой они расположены. Если петля идет “вверх”, то вставленным остаткам присваивают номер остатка, непосредственно предшествующий вставке, при этом номер вставленного остатка в последовательности указывают по возрастанию десятичного знака, например, 16, 16.1, 16.2, 16.3, где присутствуют три мутации после остатка 16. Если петля идет “вниз”, то вставленным остаткам присваивают номер остатка, непосредственно предшествующий вставке, при этом номер вставленного остатка в последовательности указывают по убыванию десятичного знака, например, 16, 16.3, 16.2, 16.1, где снова присутствуют три мутации после остатка 16 (LeFranc и др., 2005 и LeFranc и др. 2015).
В предпочтительном варианте реализации структурная петля AB содержит вставку аминокислоты. Длина вставки может составлять от 1 до 10, от 2 до 9, от 3 до 7, от 4 до 6 или 5 аминокислот. Предпочтительно, длина вставки составляет 5 аминокислот.
Вставка может быть расположена между положениями 10 и 19, предпочтительно между положениями 14 и 17, более предпочтительно между положениями 16 и 17 домена CH3 молекулы антитела. В предпочтительном варианте реализации вставка расположена в положениях с 16.5 по 16.1 домена CH3 молекулы антитела. На фигуре 1 показаны Fcab, содержащие домен CH3, где вставка расположена в положениях с 16.5 по 16.1 домена CH3.
Большинство Fcab, идентифицированных после созревания аффинности, содержали остаток лейцина (L) в положении 97 домена CH3. Множество данных Fcab также содержало остаток аспарагиновой кислоты (D) или остаток глутаминовой кислоты (E) в положениях 98 домена CH3. Оба данных изменения аминокислот расположены в структурной петле EF. Данные результаты позволяют предположить, что один или оба данных остатка могут быть важны для связывания CD137. Таким образом, вторая последовательность предпочтительно содержит последовательность LD или LE, где последовательность LD или LE предпочтительно расположена в положениях 97 и 98 домена CH3 молекулы антитела.
Первая последовательность и вторая последовательность могут представлять собой первую и вторую последовательность домена CH3 из: FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, предпочтительно FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017 или FS22-053-014, более предпочтительно FS22-053-008, FS22-053-009, FS22-053-011 или FS22-053-017, еще более предпочтительно представителя специфического связывания FS22-053-008.
Первая последовательность и вторая последовательность могут представлять собой первую и вторую последовательность домена CH3 из: FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 или FS22-172, предпочтительно FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005 или FS22-172-006, более предпочтительно FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-172-003, FS22-172-002 или FS22-172-004, еще более предпочтительно FS22-053-008 или FS22-172-003, еще более предпочтительно FS22-172-003. В альтернативном предпочтительном варианте реализации первая последовательность и вторая последовательность могут представлять собой первую и вторую последовательность домена CH3 из FS22-053-017.
Последовательность домена CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 и FS22-172 представлена в последовательностях SEQ ID NO: 81, 84, 87, 90, 93, 96, 99, 102, 105, 108, 111, 115, 118, 121, 124, 127, 130 и 132, соответственно.
Первая и вторая последовательность из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 и FS22-172 может представлять собой последовательность между положениями 14 и 17 и положениями 91 и 99 домена CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 и FS22-172, соответственно.
В качестве альтернативы, первая и вторая последовательность из FS22-053-008, FS22-053-010, FS22-053-011, FS22-053-012 и FS22-053-016 может представлять собой последовательность между положениями 14 и 17 и положениями 92 и 99 домена CH3 из FS22-053-008, FS22-053-010, FS22-053-011, FS22-053-012 и FS22-053-016, соответственно.
Первая и вторая последовательность из FS22-053-015 может в качестве альтернативы представлять собой последовательность между положениями 14 и 17 и положениями 92 и 98 домена CH3 из FS22-053-015, соответственно.
Последовательность петли CD молекулы антитела предпочтительно не модифицирована, т.е., относится к дикому типу. Последовательность петли CD, следовательно, предпочтительно имеет последовательность, представленную в SEQ ID NO: 73. Последовательность петли CD предпочтительно расположена в положениях с 43 по 78 домена CH3.
Первая и вторая последовательности могут представлять собой полные последовательности структурных петель AB и EF из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 или FS22-172, соответственно. Определение положения структурных петель AB, CD и EF в последовательности домена CH3, например, в соответствии с системами нумерации IMGT, экзонов IMGT, EU или Кабата, находится в рамках возможностей специалиста и описано в Hasenhindl и др. (2013). В предпочтительном варианте реализации структурные петли AB, CD и EF согласно системе нумерации IMGT расположены между положениями 10 и 19, 42 и 79 и 91 и 102 домена CH3, соответственно. В предпочтительном варианте реализации первая, вторая и третья последовательности, следовательно, представляют собой последовательность между положениями 10 и 19, 42 и 79 и 91 и 102 домена CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 или FS22-172, соответственно.
Таким образом, в одном предпочтительном варианте реализации первая и вторая последовательности сайта связывания антигена CD137 содержат последовательности петель AB и EF, представленные в SEQ ID NO: 171 и 172 [FS22-172-003], соответственно, или последовательности структурных петель AB и EF, представленные в SEQ ID NO: 173 и 174 [FS22-53-008], соответственно. В более предпочтительном варианте реализации первая и вторая последовательности сайта связывания антигена CD137 содержат структурные петли AB и EF, представленные в последовательностях SEQ ID NO: 171 и 172 [FS22-172-003], соответственно.
В альтернативном предпочтительном варианте реализации первая и вторая последовательности сайта связывания антигена CD137 содержат последовательности структурных петель AB и EF, представленные в SEQ ID NO: 173 и 175 [FS22-053-017], соответственно.
В предпочтительном варианте реализации сайт связывания антигена CD137 содержит первую и/или вторую, предпочтительно первую и вторую последовательности, представленные в:
(i) SEQ ID NO: 79 и 80, соответственно [FS22-053-008];
(ii) SEQ ID NO: 79 и 83, соответственно [FS22-053-009];
(iii) SEQ ID NO: 79 и 86, соответственно [FS22-053-011];
(iv) SEQ ID NO: 79 и 89, соответственно [FS22-053-017];
(v) SEQ ID NO: 79 и 92, соответственно [FS22-053-014];
(vi) SEQ ID NO: 79 и 95, соответственно [FS22-053-010];
(vii) SEQ ID NO: 79 и 98, соответственно [FS22-053-012];
(viii) SEQ ID NO: 79 и 101, соответственно [FS22-053-013];
(ix) SEQ ID NO: 79 и 104, соответственно [FS22-053-015];
(x) SEQ ID NO: 79 и 107, соответственно [FS22-053-016];
(xi) SEQ ID NO: 79 и 110, соответственно [FS22-053];
(xii) SEQ ID NO: 113 и 114, соответственно [FS22-172-003];
(xiii) SEQ ID NO: 117 и 114, соответственно [FS22-172-002];
(xiv) SEQ ID NO: 120 и 114, соответственно [FS22-172-004];
(xv) SEQ ID NO: 123 и 114, соответственно [FS22-172-001];
(xvi) SEQ ID NO: 126 и 114, соответственно [FS22-172-005];
(xvii) SEQ ID NO: 129 и 114, соответственно [FS22-172-006]; или
(xviii) SEQ ID NO: 129 и 114, соответственно [FS22-172]; причем первая и вторая последовательности предпочтительно расположены между положениями 14 и 17 и 91 и 99 домена CH3 молекулы антитела, соответственно.
В еще более предпочтительном варианте реализации молекула антитела содержит первую и/или вторую, предпочтительно первую и вторую последовательности, представленные в:
(i) SEQ ID NO: 79 и 80, соответственно [FS22-053-008];
(ii) SEQ ID NO: 79 и 83, соответственно [FS22-053-009];
(iii) SEQ ID NO: 79 и 86, соответственно [FS22-053-011];
(iv) SEQ ID NO: 79 и 89, соответственно [FS22-053-017];
(v) SEQ ID NO: 113 и 114, соответственно [FS22-172-003];
(vi) SEQ ID NO: 117 и 114, соответственно [FS22-172-002]; или
(vii) SEQ ID NO: 120 и 114, соответственно [FS22-172-004].
В еще более предпочтительном варианте реализации сайт связывания антигена CD137 молекулы антитела содержит первую и вторую последовательности, представленные в SEQ ID NO: 113 и 114 [FS22-172-003], соответственно, или первую и вторую последовательности, представленные в SEQ ID NO: 79 и 80 [FS22-53-008], соответственно. В еще более предпочтительном варианте реализации сайт связывания антигена CD137 молекулы антитела содержит первую и вторую последовательности, представленные в SEQ ID NO: 113 и 114 [FS22-172-003], соответственно. Например, сайт связывания антигена CD137 может содержать последовательности структурных петель AB и EF, представленные в SEQ ID NO: 171 и 172 [FS22-172-003], соответственно.
В варианте реализации молекула антитела, в частности, молекула антитела, содержащая первую и/или вторую, предпочтительно первую и вторую, последовательность, представленную в SEQ ID NO: 129 и 114 [FS22-172-006], может содержать лейцин (L) в положении 19 домена CH3 молекулы антитела.
В качестве альтернативы нумерации IMGT, положения аминокислотных остатков, включая положения последовательностей аминокислот, замен, делеций и вставок, описанных в данной заявке, могут быть пронумерованы согласно нумерации экзонов IMGT (также называют последовательной нумерацией), нумерации EU или нумерации по Кабату. Соответствие между нумерацией IMGT, нумерацией экзонов IMGT, нумерацией EU и нумерацией по Кабату положений остатков домена CH3 показано на фигуре 1. Таким образом, например, если в настоящей заявке предложена первая последовательность, расположенная между положениями 14 и 17 домена CH3 клона, соответственно, если положения остатков пронумерованы в соответствии со схемой нумерации IMGT, первая последовательность расположена между положениями 18 и 21 домена CH3, где положения остатков пронумерованы в соответствии со схемой нумерации экзонов IMGT, как показано на фигуре 1. В качестве альтернативы, положение аминокислотных остатков в домене CH3, включая положения последовательностей аминокислот, замен, делеций и вставок в домене CH3, описанные здесь, можно определить, ссылаясь на их положение в последовательности домена CH3 дикого типа, представленной в SEQ ID NO: 75. Соответствие между нумерацией IMGT и последовательностью домена CH3 дикого типа также показано на фигуре 1.
В одном варианте реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 или FS22-172, где последовательность домена CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 и FS22-172 представлена в последовательностях SEQ ID NO: 81, 84, 87, 90, 93, 96, 99, 102, 105, 108, 111, 115, 118, 121, 124, 127, 130 и 132, соответственно.
В предпочтительном варианте реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 из FS22-172-003 или FS22-053-008, представленной в SEQ ID NO: 115 и 81, соответственно. В более предпочтительном варианте реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 из FS22-172-003, представленной в SEQ ID NO: 115.
В альтернативном предпочтительном варианте реализации молекула антитела содержит домен CH3, который содержит, имеет или состоит из последовательности домена CH3 из FS22-053-017, представленной в SEQ ID NO: 90, соответственно.
Домен CH3 молекулы антитела необязательно может содержать дополнительный остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
Кроме того, молекула антитела согласно настоящему изобретению может содержать домен CH2 молекулы иммуноглобулина G, такой как домен CH2 молекулы IgG1, IgG2, IgG3 или IgG4. Предпочтительно молекула антитела согласно настоящему изобретению содержит домен CH2 молекулы IgG1. Домен CH2 может иметь последовательность, представленную в SEQ ID NO: 76.
Известно, что домен CH2 связывается с Fcγ-рецепторами и комплементом. Связывание домена CH2 с Fcγ-рецепторами необходимо для антителозависимой клеточной цитотоксичности (АЗКЦ), хотя связывание с комплементом необходимо для комплементзависимой цитотоксичности (КЗЦ). Домен CH2 молекулы антитела предпочтительно содержит одну или более мутаций, которые уменьшают или устраняют связывание домена CH2 с одним или более Fcγ-рецепторами, такими как FcγRI, FcγRlla, FcγRllb, FcγRIII, и/или с комплементом. Авторы настоящего изобретения предположили, что уменьшение или устранение связывания с Fcγ-рецепторами будет уменьшать или устранять АЗКЦ, опосредованную молекулой антитела. Аналогично, ожидают, что уменьшение или устранение связывания с комплементом уменьшает или устраняет КЗЦ, опосредованную молекулой антитела. Без привязки к теории, ожидают, что это уменьшит или позволит избежать гепатотоксичности, когда молекулу антитела вводят пациенту. Кроме того, ожидают, что уменьшение или устранение связывания с Fcγ-рецепторами полезно, когда молекула антитела содержит второй сайт связывания с антигеном иммунной клетки, когда опосредованное АЗКЦ и/или КЗЦ уничтожение иммунных клеток, связанных молекулой антитела, следует избежать. Мутации для уменьшения или устранения связывания домена CH2 с одним или более Fcγ-рецепторами и/или комплементом известны в данной области (Wang и др., 2018). Данные мутации включают «мутацию LALA», описанную в Bruhns и др., 2009 и Hezareh и др., 2001, которая включает замену остатков лейцина в положениях 1.3 и 1.2 домена CH2 на аланин (L1.3A и L1.2A). В качестве альтернативы, также известно, что получение агликозилированных антител посредством мутации консервативного сайта N-связанного гликозилирования путем мутации аспарагина (N) в положении 84.4 домена CH2 на аланин, глицин или глутамин (N84.4A, N84.4G или N84.4Q) снижает эффекторную функцию IgG1 (Wang и др., 2018). В качестве дополнительной альтернативы, известно, что активация комплемента (связывание C1q) и АЗКЦ уменьшается при мутации пролина в положении 114 домена CH2 на аланин или глицин (P114A или P114G) (Idusogie и др., 2000; Klein и др., 2016). Данные мутации также можно комбинировать для получения молекул антитела с дополнительно сниженной или отсутствующей активностью АЗКЦ или КЗЦ.
Таким образом, молекула антитела может содержать домен CH2, где домен CH2 предпочтительно содержит:
(i) остатки аланина в положениях 1.3 и 1.2; и/или
(ii) аланин или глицин в положении 114; и/или
(iii) аланин, глутамин или глицин в положении 84.4;
где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
В предпочтительном варианте реализации молекула антитела содержит домен CH2, где домен CH2 содержит:
(i) остаток аланина в положении 1.3; и
(ii) остаток аланина в положении 1.2;
где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Например, домен CH2 может иметь последовательность, представленную в SEQ ID NO: 77.
В альтернативном предпочтительном варианте реализации молекула антитела содержит домен CH2, где домен CH2 содержит:
(i) остаток аланина в положении 1.3;
(ii) остаток аланина в положении 1.2; и
(iii) аланин в положении 114;
где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
Например, домен CH2 может иметь последовательность, представленную в SEQ ID NO: 176.
В предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) сайт связывания антигена на основе CDR для PD-L1; и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
причем указанный сайт связывания антигена на основе CDR содержит три CDR VH и три CDR VL (CDR 1 - 6) антитела E12v2, E05v2 или G12v2, предпочтительно E12v2; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно; причем первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003] или 79 и 80 [FS22-53-008], соответственно, предпочтительно где первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003].
Как описано в примерах, молекула антитела, содержащая сайт связывания антигена CD137, содержащий первую последовательность и вторую последовательность, представленную в SEQ ID NO: 79 и 89, соответственно [FS22-053-017], было способно связывать CD137 человека, яванского макака и, неожиданно, мыши. Авторы настоящего изобретения продемонстрировали, что данная молекула антитела проявляла активность в анализах активации T-клеток человека, яванского макака и мыши при перекрестном связывании.
Вальтернативном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) сайт связывания антигена на основе CDR для PD-L1; и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
причем указанный сайт связывания антигена на основе CDR содержит три CDR VH и три CDR VL (CDR 1 - 6) антитела E12v2, E05v2 или G12v2, предпочтительно E12v2; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно, причем первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 79 и 89 [FS22-053-017].
В некоторых вариантах реализации молекула антитела содержит
(a) сайт связывания антигена на основе CDR для PD-L1; и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
причем указанный сайт связывания антигена на основе CDR содержит три CDR VH и три CDR VL (CDR 1 - 6) антитела E12v2, E05v2 или G12v2, предпочтительно E12v2; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно, причем первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003] или 79 и 80 [FS22-53-008], соответственно, предпочтительно где первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 113 и 114 [FS22-172-003], при условии, что молекула антитела не содержит CDR 1 - 6 антитела G12v2, спаренные с сайтом связывания антигена CD137, содержащим первую и вторую последовательности, представленные в SEQ ID NO: 79 и 80 [FS22-053-008].
В дополнительном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) сайт связывания антигена на основе CDR для PD-L1; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 115 [FS22-172-003] или 81 [FS22-53-008], предпочтительно SEQ ID NO: 115 [FS22-172-003];
причем указанный сайт связывания антигена на основе CDR содержит три CDR VH и три CDR VL (CDR 1 - 6) антитела E12v2, E05v2 или G12v2, предпочтительно E12v2.
В дополнительном альтернативном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) сайт связывания антигена на основе CDR для PD-L1; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 90 [FS22-053-017];
причем указанный сайт связывания антигена на основе CDR содержит три CDR VH и три CDR VL (CDR 1 - 6) антитела E12v2, E05v2 или G12v2, предпочтительно E12v2.
В еще одном дополнительном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) домен VH и домен VL, содержащий сайт связывания антигена на основе CDR для PD-L1; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 115 [FS22-172-003] или 81 [FS22-53-008], предпочтительно SEQ ID NO: 115 [FS22-172-003];
где домен VH и VL содержит, имеет или состоит из VH и VL антитела E12v2, E05v2 или G12v2, предпочтительно E12v2.
В еще одном дополнительном альтернативном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) домен VH и домен VL, содержащий сайт связывания антигена на основе CDR для PD-L1; и
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 90 [FS22-053-017];
где домен VH и VL содержит, имеет или состоит из VH и VL антитела E12v2, E05v2 или G12v2, предпочтительно E12v2.
В еще одном дополнительном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит
(a) домен VH и домен VL, содержащий сайт связывания антигена на основе CDR для PD-L1;
(b) домен CH3, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 115 [FS22-172-003] или 81 [FS22-53-008], предпочтительно SEQ ID NO: 115 [FS22-172-003];
(c) домен CH2, который содержит, имеет или состоит из последовательности, представленной в SEQ ID NO: 76, где домен CH2 содержит:
(i) остаток аланина в положении 1.3;
(ii) остаток аланина в положении 1.2;
где нумерация аминокислотных остатков соответствует схеме нумерации IMGT; и
где домен VH и VL содержит, имеет или состоит из VH и VL антитела E12v2, E05v2 или G12v2, предпочтительно E12v2.
В дополнительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит:
(a) домен VH и VL, содержащий сайт связывания антигена на основе CDR для PD-L1, причем указанный сайт связывания антигена на основе CDR содержит CDR 1 - 6; и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела, сайт связывания антигена CD137, содержащий первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3;
где CDR 1 - 6, первая последовательность и вторая последовательность имеют последовательность, представленную в:
(i) SEQ ID NO: 1, 2, 3, 4, 5, 6, 113 и 114, соответственно [FS22-172-003-AA/E12v2];
(ii) SEQ ID NO: 1, 2, 3, 18, 19, 20, 113 и 114, соответственно [FS22-172-003-AA/E05v2];
(iii) SEQ ID NO: 1, 2, 3, 18, 19, 29, 113 и 114, соответственно [FS22-172-003-AA/G12v2];
(iv) SEQ ID NO: 1, 2, 3, 4, 5, 6, 79 и 80, соответственно [FS22-053-008-AA/E12v2]; или
(v) SEQ ID NO: 1, 2, 3, 18, 19, 20, 79 и 80, соответственно [FS22-053-008-AA/E05v2],
или
где домен VH, домен VL, первая последовательность и вторая последовательность имеют последовательность, представленную в:
(i) SEQ ID NO: 12, 14, 113 и 114, соответственно [FS22-172-003-AA/E12v2];
(ii) SEQ ID NO: 23, 25, 113 и 114, соответственно [FS22-172-003-AA/E05v2];
(iii) SEQ ID NO: 23, 30, 113 и 114, соответственно [FS22-172-003-AA/G12v2];
(iv) SEQ ID NO: 12, 14, 79 и 80, соответственно [FS22-053-008-AA/E12v2]; или
(v) SEQ ID NO: 23, 25, 79 и 80, соответственно [FS22-053-008-AA/E05v2]; или
где домен VH, домен VL и домен CH3 имеют последовательность, представленную в:
(i) SEQ ID NO: 12, 14 и 115, соответственно [FS22-172-003-AA/E12v2];
(ii) SEQ ID NO: 23, 25 и 115, соответственно [FS22-172-003-AA/E05v2];
(iii) SEQ ID NO: 23, 30 и 115, соответственно [FS22-172-003-AA/G12v2];
(iv) SEQ ID NO: 12, 14 и 81, соответственно [FS22-053-008-AA/E12v2]; или
(v) SEQ ID NO: 23, 25 и 81, соответственно [FS22-053-008-AA/E05v2].
В еще одном дополнительном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела:
(i) FS22-172-003-AA/E12v2, представленных в последовательностях SEQ ID NO: 134 и 17, соответственно;
(ii) FS22-172-003-AA/E05v2, представленных в последовательностях SEQ ID NO: 137 и 28, соответственно;
(iii) FS22-172-003-AA/G12v2, представленных в последовательностях SEQ ID NO: 140 и 33, соответственно;
(iv) FS22-053-008-AA/E12v2, представленных в последовательностях SEQ ID NO: 143 и 17, соответственно;
(v) FS22-053-008-AA/E05v2, представленных в последовательностях SEQ ID NO: 146 и 28, соответственно; или
(vi) FS22-053-008-AA/G12v2, представленных в последовательностях SEQ ID NO: 149 и 33, соответственно.
В еще более предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела:
(i) FS22-172-003-AA/E12v2, представленных в последовательностях SEQ ID NO: 134 и 17, соответственно;
(ii) FS22-172-003-AA/E05v2, представленных в последовательностях SEQ ID NO: 137 и 28, соответственно;
(iii) FS22-172-003-AA/G12v2, представленных в последовательностях SEQ ID NO: 140 и 33, соответственно;
(iv) FS22-053-008-AA/E12v2, представленных в последовательностях SEQ ID NO: 143 и 17, соответственно; или
(v) FS22-053-008-AA/E05v2, представленных в последовательностях SEQ ID NO: 146 и 28, соответственно.
В еще более предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела:
(i) FS22-172-003-AA/E12v2, представленных в последовательностях SEQ ID NO: 134 и 17, соответственно; или
(iv) FS22-053-008-AA/E12v2, представленных в последовательностях SEQ ID NO: 143 и 17, соответственно.
В еще более предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела FS22-172-003-AA/E12v2, представленных в последовательностях SEQ ID NO: 134 и 17, соответственно.
В еще одном дополнительном альтернативном предпочтительном варианте реализации молекула антитела, которая связывается с PD-L1 и CD137, содержит тяжелую цепь, которая содержит, имеет или состоит из тяжелой цепи и легкой цепи антитела:
(i) FS22-053-017-AA/E12v2, представленных в последовательностях SEQ ID NO: 152 и 17, соответственно;
(ii) FS22-053-017-AA/E05v2, представленных в последовательностях SEQ ID NO: 153 и 28, соответственно; или
(iii) FS22-053-017-AA/G12v2, представленных в последовательностях SEQ ID NO: 154 и 33, соответственно.
Молекулы антитела согласно настоящему изобретению также могут содержать варианты первой, второй или третьей последовательности, последовательности структурной петли AB, CD или EF, последовательности домена CH3, домена CH2, доменов CH2 и CH3, CDR, FW, домена VH, домена VL, легкой цепи и/или тяжелой цепи, описанных в данной заявке. Подходящие варианты можно получить с помощью способов изменения или мутации последовательностей и скрининга. В предпочтительном варианте реализации молекула антитела, содержащая один или более вариантов последовательностей, сохраняет одно или более функциональных свойств исходной молекулы антитела, таких как специфичность связывания и/или аффинность связывания с PD-L1 и CD137. Например, молекула антитела, содержащая один или более вариантов последовательности, предпочтительно связывается с PD-L1 и/или CD137 с такой же аффинностью или большей аффинностью, чем (исходная) молекула антитела. Исходная молекула антитела представляет собой молекулу антитела, которая не содержит замену(-ы), делецию(-и) и/или вставку(-и) аминокислоты, которые были включены в вариант молекулы антитела.
Например, молекула антитела согласно настоящему изобретению может содержать первую, вторую или третью последовательность, последовательность структурной петли AB, CD или EF, последовательности домена CH3, домена CH2, доменов CH2 и CH3, CDR, FW, домена VH, домена VL, легкой цепи и/или тяжелой цепи, которые по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,1%, по меньшей мере на 99,2%, по меньшей мере на 99,3%, по меньшей мере на 99,4%, по меньшей мере на 99,5%, по меньшей мере на 99,6%, по меньшей мере на 99,7%, по меньшей мере на 99,8% или по меньшей мере на 99,9% идентичны последовательностям структурной петли, домена CH3, домена CH2, доменов CH2 и CH3, CDR, FW, домена VH, домена VL, легкой цепи или тяжелой цепи, описанным в данной заявке.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению содержит последовательность домена CH3, которая по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,1%, по меньшей мере на 99,2%, по меньшей мере на 99,3%, по меньшей мере на 99,4%, по меньшей мере на 99,5%, по меньшей мере на 99,6%, по меньшей мере на 99,7%, по меньшей мере на 99,8% или по меньшей мере на 99,9% идентична последовательности домена CH3, представленной в SEQ ID NO: 115 [FS22-172-003] или 81 [FS22-053-008], предпочтительно SEQ ID NO: 115 [FS22-172-003].
В дополнительном предпочтительном варианте реализации молекула антитела имеет или содержит последовательность домена CH2, которая по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,1%, по меньшей мере на 99,2%, по меньшей мере на 99,3%, по меньшей мере на 99,4%, по меньшей мере на 99,5%, по меньшей мере на 99,6%, по меньшей мере на 99,7%, по меньшей мере на 99,8% или по меньшей мере на 99,9% идентична последовательности домена CH2, представленной в SEQ ID NO: 76 или 77.
Идентичность последовательностей обычно определяют на основании алгоритма GAP (пакет программ Wisconsin GCG, Accelerys Inc, Сан-Диего, США). В GAP применяют алгоритм Нидлмана и Вунша для выравнивания двух полноразмерных последовательностей, максимизирования количества совпадений и минимизирования количества гэпов. Как правило, используют параметры по умолчанию, где штраф за открытие гэпа равнен 12 и штраф за продление гэпа равнен 4. Применение GAP может быть предпочтительным, но можно применять другие алгоритмы, например, BLAST (в котором используется способ Altschul и др., 1990), FASTA (в котором используется способ Пирсона и Липмана, 1988), или алгоритм Смита-Уотермана (Smith-Waterman, 1981), или программу TBLASTN, Altschul и др., 1990, выше, как правило, используя параметры по умолчанию. В частности, можно применять алгоритм PSI-BLAST (Altschul и др., 1997).
Молекула антитела согласно настоящему изобретению также может содержать первую, вторую или третью последовательность, последовательность структурной петли AB, CD или EF, домена CH3, домена CH2, доменов CH2 и CH3, CDR, FW, домена VH, домена VL, легкой цепи и/или тяжелой цепи, которая содержит одно или более изменений в последовательности аминокислот (добавление, делецию, замену и/или вставку аминокислотного остатка), предпочтительно 20 изменений или меньше, 15 изменений или меньше, 10 изменений или меньше, 5 изменений или меньше, 4 изменения или меньше, 3 изменения или меньше, 2 изменения или меньше или 1 изменение по сравнению с первой, второй или третьей последовательностью, последовательностью структурной петли AB, CD или EF, последовательностями домена CH3, домена CH2, доменов CH2 и CH3, CDR, FW, домена VH, домена VL, легкой цепи или тяжелой цепи, описанными в данной заявке. В частности, можно внести изменения в одну или более каркасных областей молекулы антитела за пределами последовательностей доменов VH и VL и/или в одну или более каркасных областей домена CH3. Например, изменения могут быть в домене CH3 за пределами последовательностей, описанных в данной заявке как первая, вторая и третья последовательности или как последовательности структурной петли AB, CD или EF.
В предпочтительном варианте реализации молекула антитела согласно настоящему изобретению может содержать последовательность домена CH3 с одним или более изменениями в последовательности аминокислот (добавлением, делецией, заменой и/или вставкой аминокислотного остатка), предпочтительно 20 изменениями или меньше, 15 изменениями или меньше, 10 изменениями или меньше, 5 изменениями или меньше, 4 изменениями или меньше, 3 изменениями или меньше, 2 изменениями или меньше или 1 изменением по сравнению с последовательностью домена CH3, представленной в SEQ ID NO: 81, 84, 87, 90, 93, 96, 99, 102, 105, 108, 111, 115, 118, 121, 124, 127, 130 или 132. В более предпочтительном варианте реализации молекула антитела согласно настоящему изобретению может содержать последовательность домена CH3 с одним или более изменениями в последовательности аминокислот (добавлением, делецией, заменой и/или вставкой аминокислотного остатка), предпочтительно 20 изменениями или меньше, 15 изменениями или меньше, 10 изменениями или меньше, 5 изменениями или меньше, 4 изменениями или меньше, 3 изменениями или меньше, 2 изменениями или меньше или 1 изменением по сравнению с последовательностью домена CH3, представленной в SEQ ID NO: 115 [FS22-172-003] или 81 [FS22-053-008], предпочтительно SEQ ID NO: 115 [FS22-172-003].
В дополнительном предпочтительном варианте реализации молекула антитела содержит последовательность домена CH2 с одним или более изменениями в последовательности аминокислот (добавлением, делецией, заменой и/или вставкой аминокислотного остатка), предпочтительно 20 изменениями или меньше, 15 изменениями или меньше, 10 изменениями или меньше, 5 изменениями или меньше, 4 изменениями или меньше, 3 изменениями или меньше, 2 изменениями или меньше или 1 изменением по сравнению с последовательностью домена CH2, представленной в SEQ ID NO: 76 или 77.
В предпочтительных вариантах реализации, в которых одна или более аминокислот заменены на другую аминокислоту, замены могут представлять собой консервативные замены, например, в соответствии со следующей таблицей. В некоторых вариантах реализации аминокислоты из одной категории в средней колонке заменяют друг на друга, т.е., например, неполярную аминокислоту заменяют на другую неполярную аминокислоту. В некоторых вариантах реализации аминокислоты в одной строке в правой колонке заменяют друг на друга.
В некоторых вариантах реализации замена(-ы) может(-гут) быть функционально консервативной(-ыми). То есть, в некоторых вариантах реализации замена может не влиять (или может по существу не влиять) на одно или более функциональных свойств (например, аффинность связывания) молекулы антитела, содержащей замену, по сравнению с эквивалентной не содержащей заместителей молекулой антитела.
Если молекула антитела содержит вариант первой последовательности, последовательности структурной петли AB, домена CH3 или последовательности тяжелой цепи, описанных в данной заявке, в молекуле антитела предпочтительно сохраняется последовательность PPY между положениями 11 и 19, предпочтительно положениями 15 и 17, домена CH3 молекулы антитела. Кроме того, в молекуле антитела предпочтительно сохраняется вставка, предпочтительно вставка 5 аминокислот между положениями 16 и 17 домена CH3 молекулы антитела. В дополнительном предпочтительном варианте реализации в молекуле антитела предпочтительно сохраняется последовательность в положениях 97 и 98 домена CH3 молекулы антитела.
В качестве дополнения или альтернативы, если молекула антитела содержит вариант последовательности домена CH3, домена CH2 и CH3, легкой цепи или тяжелой цепи, описанных в данной заявке, указанный вариант предпочтительно не содержит каких-либо изменений аминокислот в первой и второй последовательности, расположенной в структурных петлях AB и EF домена CH3 молекулы антитела. Например, вариант может не содержать каких-либо изменений аминокислот в структурных петлях AB и EF домена CH3 молекулы антитела. Кроме того, вариант может не содержать каких-либо изменений аминокислот в структурной петле CD домена CH3 молекулы антитела. То есть, вариант может не содержать каких-либо изменений аминокислот в структурных петлях AB и EF домена CH3 молекулы антитела.
Если молекула антитела содержит вариант последовательности домена VH, домена VL, легкой цепи или тяжелой цепи, описанных в данной заявке, молекула антитела предпочтительно не содержит каких-либо изменений аминокислот в последовательностях CDR. То есть, любые присутствующие изменения аминокислот могут находиться в последовательностях FW, например, HFW1, HFW2, HFW3, HFW4, LFW1, LFW2, LFW3 и/или LFW4. Например, указанный вариант может не содержать каких-либо изменений аминокислот в последовательностях CDR1, CDR2, CDR3, CDR4, CDR5 и/или CDR6.
Молекула антитела предпочтительно связывается с PD-L1 человека и CD137 человека. Предпочтительно, молекула антитела способна одновременно связываться с PD-L1 человека и CD137 человека, причем CD137 человека и PD-L1 человека коэкспрессируются. В данной заявке коэкспрессия означает, что две мишени экспрессируются на поверхности одной клетки или на поверхности двух отдельных клеток. Например, молекула антитела может быть способна связываться с PD-L1 человека и CD137 человека, когда PD-L1 человека и CD137 человека коэкспрессируются на одной клетке, например, иммунной клетке, а также способна связываться с PD-L1 человека и CD137 человека, когда PD-L1 человека и CD137 человека коэкспрессируются на двух отдельных клетках, например, иммунной клетке, экспрессирующей CD137, и на отдельной клетке опухоли, экспрессирующей PD-L1, в микроокружении опухоли.
Молекула антитела предпочтительно связывается с PD-L1 человека с аффинностью (KD), равной 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,4 нМ или 0,3 нМ, или с более высокой аффинностью. Предпочтительно, молекула антитела связывается с PD-L1 человека с аффинностью (KD), равной 0,3 нМ, или с более высокой аффинностью.
PD-L1 человека может, например, иметь последовательность, представленную в SEQ ID NO: 180. PD-L1 человека может, например, представлять собой рекомбинантный PD-L1 человека с меткой Avi (hPD-L1-Avi-His), доступный от Acro Biosystems (номер в каталоге: PD1-H82E5). Рекомбинантный PD-L1 человека может быть биотинилирован.
Молекула антитела предпочтительно связывается с димерным CD137 человека с аффинностью (KD), равной 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 5 нМ, 4 нМ, 3 нМ или 2 нМ, или с более высокой аффинностью. Предпочтительно, молекула антитела связывается с димерным CD137 человека с аффинностью (KD), равной 2 нМ, или с более высокой аффинностью.
В предпочтительном варианте реализации молекула антитела связывается с димерным CD137 с более высокой аффинностью, чем с мономерным CD137. В предпочтительном варианте реализации молекула антитела связывается с димерным CD137 с аффинностью, которая по меньшей мере в 50 раз, в 60 раз, в 70 раз, в 80 раз, в 90 раз, в 100 раз, в 110 раз, в 120 раз, в 130 раз, в 140 раз, в 150 раз, в 160 раз, в 170 раз или в 200 раз выше, чем аффинность молекулы антитела к мономерному CD137.
CD137 человека может, например, иметь последовательность, представленную в SEQ ID NO: 186. Способы получения димерных и мономерных антигенов CD137 описаны в примерах.
Молекула антитела предпочтительно связывается с PD-L1 яванского макака и CD137 яванского макака. Предпочтительно, молекула антитела способна одновременно связываться с PD-L1 яванского макака и CD137 яванского макака, причем PD-L1 яванского макака и CD137 яванского макака экспрессируются на поверхности одной клетки или на поверхности двух отдельных клеток.
В предпочтительном варианте реализации молекула антитела может связываться с PD-L1 яванского макака с аффинностью (KD), равной 8 нМ, 7 нМ, 6 нМ, 5 нМ, 4 нМ, 3 нМ, 2 нМ, 1 нМ, 0,5 нМ, 0,4 нМ или 0,3 нМ, или с более высокой аффинностью. Предпочтительно молекула антитела связывается с PD-L1 яванского макака с аффинностью (KD), равной 1 нМ, или с более высокой аффинностью.
PD-L1 яванского макака может, например, иметь последовательность, представленную в SEQ ID NO: 184.
В предпочтительном варианте реализации молекула антитела может связываться с димерным CD137 яванского макака с аффинностью (KD), равной 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 5 нМ, 4 нМ, 3 нМ или 2 нМ, или с более высокой аффинностью. Предпочтительно молекула антитела связывается с димерным CD137 яванского макака с аффинностью (KD), равной 2 нМ, или с более высокой аффинностью.
Молекула антитела может связываться с PD-L1 человека и PD-L1 яванского макака со сходной аффинностью, и/или связываться с димерным CD137 человека и димерным CD137 яванского макака со сходной аффинностью. Считают, что это предпочтительно для гарантии того, что исследования эффективности и токсичности, проведенные с молекулой антитела на яванских макаках, позволят прогнозировать эффективность и токсичность молекулы антитела у людей.
Таким образом, в предпочтительном варианте реализации молекула антитела связывается с PD-L1 яванского макака с аффинностью, которая не более чем в 10 раз, предпочтительно не более чем в 5 раз ниже или выше, чем аффинность, с которой молекула антитела связывается с PD-L1 человека. В предпочтительном варианте реализации молекула антитела связывается с димерным CD137 яванского макака с аффинностью, которая не более чем в 10 раз, предпочтительно не более чем в 5 раз ниже или выше, чем аффинность, с которой молекула антитела связывает димерный CD137 человека.
Аффинность связывания молекулы антитела с распознаваемым антигеном, таким как PD-L1 человека, CD137 человека, PD-L1 яванского макака или CD137 яванского макака, можно определить с помощью поверхностного плазмонного резонанса (ППР), такого как, например, Biacore. Дополнительные подробности подходящих способов описаны в примерах.
Способность молекулы антитела одновременно связываться с двумя распознаваемыми антигенами, например, PD-L1 человека и CD137 человека или PD-L1 яванского макака и CD137 яванского макака, можно определить с помощью ППР, такого как, например, Biacore. Дополнительные подробности подходящих способов описаны в примерах.
Молекула антитела может быть способна блокировать взаимодействие между PD-L1 и его рецептором, PD-1, предпочтительно PD-L1 человека и PD-1 человека.
Известно, что путь передачи сигналов PD-1/PD-L1 важен для опосредования подавления иммунного ответа. Таким образом, ожидают, что молекулы антитела, которые способны блокировать данный путь, будут предпочтительны в том отношении, что они могут уменьшить подавление иммунного ответа и помочь повысить противоопухолевый ответ. Ингибирование передачи сигналов PD-1/PD-L1 и активация CD137 могут действовать совместно, чтобы повысить противоопухолевую эффективность.
Способность молекулы антитела блокировать связывание PD-L1 с PD-1 можно определить, применяя основанный на клеточной системе биолюминесцентный анализ, например, используя продукт PD-1/PD-L1 Blockade Bioassay, например, от Promega. Способность молекулы антитела блокировать связывание PD-L1 с PD-1 можно определить, применяя твердофазный иммуноферментный анализ (ELISA). Дополнительные подробности данных анализов описаны в примерах.
Способность молекулы антитела блокировать связывание PD-L1 с PD-1, также называемую в данной заявке активностью блокирования PD-1/PD-L1, можно определить относительно молекулы антитела, содержащей или состоящий из тяжелой цепи и легкой цепи антитела 2.14H9OPT, представленного в WO2011/066389 A1, соответственно, или тяжелой цепи и легкой цепи антитела G1/280_02_G02, представленного в последовательности SEQ ID NO: 177 и 178, соответственно.
Например, молекула антитела может проявлять сходный или более высокий уровень активности блокирования PD-1/PD-L1, чем молекула антитела, содержащая или состоящая из тяжелой цепи и легкой цепи антитела 2.14H9OPT, представленного в WO2011/066389 A1, или тяжелой цепи и легкой цепи антитела G1/280_02_G02, представленных в последовательностях SEQ ID NO: 177 и 178, соответственно.
Молекула антитела может проявлять активность блокирования PD-1/PD-L1, которая составляет по меньшей мере 70%, 80% или 90% от активности блокирования PD-1/PD-L1 молекулы антитела, содержащей или состоящей из последовательности тяжелой цепи и последовательности легкой цепи антитела 2.14H9OPT, представленного в WO2011/066389 A1, или тяжелой цепи и легкой цепи антитела G1/280_02_G02, представленных в последовательностях SEQ ID NO: 177 и 178, соответственно.
Молекула антитела может проявлять активность блокирования PD-1/PD-L1, которая составляет от 70% до 130%, от 80% до 120% или от 90% до 110% от активности блокирования PD-1/PD-L1 молекулы антитела, содержащей или состоящей из последовательности тяжелой цепи и последовательности легкой цепи антитела 2.14H9OPT, представленного в WO2011/066389 A1, или тяжелой цепи и легкой цепи антитела G1/280_02_G02, представленных в последовательностях SEQ ID NO: 177 и 178, соответственно.
Выше описано, что молекулы антитела согласно настоящему изобретению способны одновременно связывать PD-L1 и CD137, что приводит к активации (агонизму) CD137. В предпочтительном варианте реализации молекула антитела способна одновременно связываться как с PD-L1 человека, так и с CD137 человека, причем такое связывание вызывает активацию CD137 человека. В дополнительном предпочтительном варианте реализации молекула антитела способна одновременно связываться как с PD-L1 яванского макака, так и с CD137 яванского макака, причем такое связывание вызывает активацию CD137 яванского макака. Типичные способы тестирования одновременного связывания и активации включают анализы активации T-клеток, описанные более подробно ниже.
Способность молекулы антитела активировать T-клетки можно измерить, применяя анализ активации T-клеток. T-клетки высвобождают IL-2 при активации. В анализе активации T-клеток, следовательно, можно измерить высвобождение IL-2, чтобы определить уровень активации T-клеток, вызванный молекулой антитела.
Например, способность молекулы антитела активировать T-клетки определяют путем измерения концентрации молекулы антитела, необходимой для достижения полумаксимального высвобождения IL-2 T-клетками в анализе активации T-клеток. Ее называют EC50 ниже.
В предпочтительном варианте реализации EC50 молекулы антитела в анализе активации T-клеток находится в пределах 50-кратной, 40-кратной, 30-кратной, 20-кратной, 10-кратной, 5-кратной, 4-кратной, 3-кратной или 2-кратной EC50 FS22-172-003-AA/E12v2 в том же анализе, где FS22-172-003-AA/E12v2 состоит из тяжелой цепи с последовательностью SEQ ID NO: 134 и легкой цепи с последовательностью SEQ ID NO: 17.
Например, EC50 молекулы антитела в анализе активации T-клеток может составлять 30 нМ или менее, 25 нМ или менее, 20 нМ или менее, 14 нМ или менее, 10 нМ или менее, 5 нМ или менее, 4 нМ или менее, 3 нМ или менее, 2 нМ или менее, 1,5 нМ или менее, 1 нМ или менее, 0,4 нМ или менее, 0,4 нМ или менее или 0,3 нМ или менее, предпочтительно 1 нМ или менее, более предпочтительно 1,5 нМ или менее, когда молекула антитела перекрестно связана.
В качестве дополнения или альтернативы, способность молекулы антитела активировать T-клетки можно определить путем измерения максимальной концентрации IL-2, высвобожденного T-клетками в анализе активации T-клеток в присутствии молекулы антитела (Emax).
Для молекулы антитела максимальная концентрация IL-2, высвобожденного T-клетками в анализе активации T-клеток в присутствии указанной молекулы антитела, может составлять по меньшей мере 1500 пг/мл, 2000 пг/мл, 2500 пг/мл, 3000 пг/мл или 3250 пг/мл или более, предпочтительно 2500 пг/мл или более.
В предпочтительном варианте реализации максимальная концентрация IL-2, высвобожденного T-клетками в анализе активации T-клеток в присутствии молекулы антитела, находится в пределах 20% или 10% от максимальной концентрации IL-2, высвобожденного T-клетками в присутствии FS22-172-003-AA/E12v2 в том же анализе, причем FS22-172-003-AA/E12v2 состоит из тяжелой цепи с последовательностью SEQ ID NO: 134 и легкой цепи с последовательностью SEQ ID NO: 17.
В анализе активации T-клеток предпочтительно используют T-клетки, экспрессирующие CD137, и клетки, экспрессирующие PD-L1, например, клетки HEK293 с повышенной экспрессией PD-L1 человека (HEK.hPD-L1) можно получить и применять, как описано в примерах. В качестве дополнения или альтернативы, можно применять клетки, экспрессирующие отличный уровень PD-L1, такие как клетки линии клеток аденокарциномы молочной железы человека MDA-MB-231 (ATCC HTB-26) и/или клетки SKBR3. Как описано в примерах, HEK.hPD-L1 экспрессируют высокий уровень PD-L1 человека, клетки MDA-MB-231 экспрессируют средний уровень PD-L1 человека и клетки SKBR3 экспрессируют низкий уровень PD-L1.
В предпочтительном варианте реализации в анализе активации T-клеток не используют какие-либо агенты, способные перекрестно связывать молекулу антитела, отличные от клеток, экспрессирующих CD137 и PD-L1. Примеры агентов, способных перекрестно связывать молекулу антитела, включают антитело против CH2 человека, описанное в примерах.
Анализ активации T-клеток может представлять собой анализ T-клеток, описанный в данной заявке, такой как анализ общей популяции T-клеток, описанный в примерах в настоящей заявке.
Например, анализ активации T-клеток может представлять собой анализ высвобождения IL-2 на основе T-клеток, выделенных из мононуклеарных клеток периферической крови (МКПК) человека. Например, анализ активации T-клеток может включать выделение МКПК человека из конусов (фильтров) для удаления лейкоцитов. Способы выделения МКПК известны в данной области и описаны в примерах в настоящей заявке. T-клетки затем можно выделить из МКПК. Способы выделения T-клеток (всех T-клеток) из МКПК известны в данной области и описаны в примерах в настоящей заявке.
Анализ активации может включать получение необходимого количества T-клеток, например, в экспериментальной среде, такой как среда для T-клеток. Необходимое количество T-клеток можно получить при концентрации 1,0 x 106 клеток/мл. T-клетки затем можно стимулировать, применяя подходящий реагент для активации T-клеток, который дает сигналы, необходимые для активации T-клеток. Например, реагент для активации T-клеток может представлять собой реагент, содержащий CD3 и CD28, такой как гранулы, содержащие CD3 и CD28. Изолированные T-клетки можно инкубировать в течение ночи с реагентом для активации T-клеток, чтобы активировать T-клетки. После этого активированные T-клетки можно промыть, чтобы отделить T-клетки от реагента для активации T-клеток, и ресуспендировать в среде для T-клеток при подходящей концентрации, такой как 2,0 x 106 клеток/мл. Активированные T-клетки затем можно добавить в планшеты, покрытые антителом против CD3 человека.
Клетки (например, клетки HEK.hPD-L1) можно высеять при концентрации, например, 2 x 105 клеток на лунку на покрытые антителом против CD3 культуральные планшеты в среде для культивирования T-клеток. После, например, 4 часов инкубации всю среду для культивирования T-клеток можно удалить и заменить на 100 мкл среды для культивирования T-клеток, содержащей T-клетки, например, при концентрации 5,0 x 105 клеток/мл с итоговой концентрацией 5,0 x 104 клеток/лунку.
Можно получить подходящее разведение каждой исследуемой молекулы антитела и добавить в лунки. T-клетки затем можно инкубировать при 37°C, 5% CO2 в течение 24 часов с исследуемым антителом. Супернатанты можно собрать и проанализировать, чтобы определить концентрацию IL-2 в супернатантах. Способы определения концентрации IL-2 в растворе известны в данной области и описаны в примерах в настоящей заявке. Концентрацию IL-2 человека можно нанести на график в зависимости от log концентрации молекулы антитела. Полученные в результате этого кривые можно аппроксимировать, используя уравнение log (агониста) против ответа.
Молекулу антитела можно конъюгировать с биоактивной молекулой или детектируемой меткой. В данном случае молекулу антитела можно назвать конъюгатом. Такие конъюгаты находят применение для лечения заболеваний, описанных в данной заявке.
Например, биоактивная молекула может представлять собой модулятор иммунной системы, такой как цитокин, предпочтительно цитокин человека. Например, цитокин может представлять собой цитокин, который стимулирует активацию и/или пролиферацию T-клеток. Примеры цитокинов для конъюгирования с молекулой антитела включают IL-2, IL-10, IL-12, IL-15, IL-21, GM-CSF и IFN-γ.
В качестве альтернативы биоактивная молекула может представлять собой ловушку для лиганда, такую как ловушка для лиганда цитокина, например, TGF-бета или IL-6.
В качестве альтернативы биоактивная молекула может представлять собой терапевтический радиоактивный изотоп.
Радиоиммунотерапию применяют, например, для лечения рака. Терапевтические радиоактивные изотопы, подходящие для радиоиммунотерапии, известны в данной области и включают иттрий-90, йод-131, висмут-213, астат-211, лютеций 177, рений-188, медь-67, актиний-225, йод-125 и тербий-161.
Подходящие детектируемые метки, которые можно конъюгировать с молекулам антитела, известны в данной области и включают радиоактивные изотопы, такие как йод-125, йод-131, иттрий-90, индий-111 и технеций-99; флуорохромы, такие как флуоресцеин, родамин, фикоэритрин, техасский красный и производные красителя цианина, например, Cy7 и Alexa750; хромогенные красители, такие как диаминобензидин; гранулы латекса; ферментативные метки, такие как пероксидаза хрена; фосфор или лазерные красители со спектрально изолированными характеристиками поглощения или испускания; и химические молекулы, такие как биотин, которые можно детектировать по связыванию с определенной распознаваемой детектируемой молекулой, например, меченым авидином.
Молекулу антитела можно конъюгировать с биоактивной молекулой или детектируемой меткой посредством любой подходящей ковалентной или нековалентной связи, такой как дисульфидная или пептидная связь. Если биоактивная молекула представляет собой цитокин, то указанный цитокин может быть соединен с молекулой антитела посредством пептидного линкера. Подходящие пептидные линкеры известны в данной области, и их длина может составлять от 5 до 25, от 5 до 20, от 5 до 15, от 10 до 25, от 10 до 20 или от 10 до 15 аминокислот.
В некоторых вариантах реализации биоактивную молекулу можно конъюгировать с молекулой антитела с помощью расщепляемого линкера. Указанный линкер может позволить высвобождение биоактивной молекулы из молекулы антитела в месте терапии. Линкеры могут содержать амидные связи (например, пептидные линкеры), дисульфидные связи или гидразоны. Пептидные линкеры, например, можно расщепить с помощью сайт-специфических протеаз, дисульфидные связи могут расщепиться под действием восстанавливающей среды цитозоля, и гидразоны можно расщепить с помощью опосредованного кислотами гидролиза.
В соответствии с настоящим изобретением также предложена(-ы) изолированная(-ые) молекула или молекулы нуклеиновых кислот, кодирующая(-ие) молекулу антитела согласно настоящему изобретению. Для специалиста не будет представлять трудностей получение таких молекул нуклеиновых кислот с применением способов, хорошо известных в данной области.
Молекула или молекулы нуклеиновых кислот может(-гут), например, содержать последовательность, представленную в SEQ ID NO: 82, 85, 88, 91, 94, 97, 100, 103, 106, 109, 112, 116, 119, 122, 125, 128, 131 или 133, которая кодирует домены CH3 из FS22-053-008, FS22-053-009, FS22-053-011, FS22-053-017, FS22-053-014, FS22-053-010, FS22-053-012, FS22-053-013, FS22-053-015, FS22-053-016, FS22-053, FS22-172-003, FS22-172-002, FS22-172-004, FS22-172-001, FS22-172-005, FS22-172-006 и FS22-172, соответственно. Предпочтительно, молекула или молекулы нуклеиновых кислот содержат последовательность, представленную в SEQ ID NO: 116 или 82, которая кодирует домен CH3 из FS22-172-003 или FS22-053-008, соответственно. Более предпочтительно, молекула или молекулы нуклеиновых кислот содержат последовательность, представленную в SEQ ID NO: 116, которая кодирует домен CH3 из FS22-172-003.
Молекула или молекулы нуклеиновых кислот могут кодировать домен VH и/или домен VL, предпочтительно домен VH и домен VL антитела E12v2, E05v2, G12v2 или lam-G02v3, предпочтительно антитела E12v2, E05v2 или G12v2, более предпочтительно E12v2 или E05v2, наиболее предпочтительно E12v2. Последовательности доменов VH и VL данных антител описаны в данной заявке.
Например, молекулы(-а) нуклеиновых(-ой) кислот(ы) могут(-жет) содержать:
(i)последовательность нуклеиновой кислоты домена VH антитела E12v2, представленную в SEQ ID NO: 13, и/или последовательность нуклеиновой кислоты домена VL антитела E12v2, представленную в SEQ ID NO: 15; или
(ii)последовательность нуклеиновой кислоты домена VH антитела E05v2, представленную в SEQ ID NO: 24, и/или последовательность нуклеиновой кислоты домена VL антитела E05v2, представленную в SEQ ID NO: 26;
(iii)последовательность нуклеиновой кислоты домена VH антитела G12v2, представленную в SEQ ID NO: 24, и/или последовательность нуклеиновой кислоты домена VL антитела G12v2, представленную в SEQ ID NO: 31; или
(v)последовательность нуклеиновой кислоты домена VH антитела lam-G02v3, представленную в SEQ ID NO: 24, и/или последовательность нуклеиновой кислоты домена VL антитела lam-G02v3, представленную в SEQ ID NO: 42.
Молекула или молекулы нуклеиновых кислот могут кодировать тяжелую цепь и/или легкую цепь, предпочтительно тяжелую цепь и легкую цепь антитела FS22-172-003-AA/E12v2, FS22-172-003-AA/E05v2, FS22-172-003-AA/G12v2, FS22-053-008-AA/E12v2, FS22-053-008-AA/E05v2 или FS22-053-008-AA/G12v2, предпочтительно антитела FS22-172-003-AA/E12v2, FS22-172-003-AA/E05v2, FS22-172-003-AA/G12v2, FS22-053-008-AA/E12v2 или FS22-053-008-AA/E05v2, еще более предпочтительно антитела FS22-172-003-AA/E12v2. Последовательности тяжелых цепей и легких цепей данных антител описаны в данной заявке.
Например, молекулы(-а) нуклеиновых(-ой) кислот(ы) могут(-жет) содержать:
(i)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-172-003-AA/E12v2, представленную в SEQ ID NO: 32 или 135, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-172-003-AA/E12v2, представленную в SEQ ID NO: 39 или 136; или
(ii)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-172-003-AA/E05v2, представленную в SEQ ID NO: 138, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-172-003-AA/E05v2, представленную в SEQ ID NO: 139;
(iii)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-172-003-AA/G12v2, представленную в SEQ ID NO: 141, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-172-003-AA/G12v2, представленную в SEQ ID NO: 142;
(iv)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-053-008-AA/E12v2, представленную в SEQ ID NO: 144, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-053-008-AA/E12v2, представленную в SEQ ID NO: 145;
(v)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-053-008-AA/E05v2, представленную в SEQ ID NO: 147, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-053-008-AA/E05v2, представленную в SEQ ID NO: 148; или
(vi)последовательность нуклеиновой кислоты тяжелой цепи антитела FS22-053-008-AA/G12v2, представленную в SEQ ID NO: 150, и/или последовательность нуклеиновой кислоты легкой цепи антитела FS22-053-008-AA/G12v2, представленную в SEQ ID NO: 151.
Если нуклеиновая кислота кодирует домен VH и VL или тяжелую и легкую цепь молекулы антитела согласно настоящему изобретению, указанные два домена или две цепи могут кодироваться на двух отдельных молекулах нуклеиновых кислот.
Изолированную молекулу нуклеиновой кислоты можно применять для экспрессии молекулы антитела согласно настоящему изобретению. Нуклеиновая кислота, как правило, будет предложена в виде рекомбинантного вектора для экспрессии. В другом аспекте настоящего изобретения, следовательно, предложен вектор, содержащий нуклеиновую кислоту, описанную выше. Подходящие векторы можно выбрать или сконструировать, чтобы они содержали подходящие регуляторные последовательности, включая последовательности промотора, фрагменты терминатора, последовательности полиаденилирования, последовательности энхансера, маркерные гены и другие последовательности, при необходимости. Предпочтительно вектор содержит подходящие регуляторные последовательности для запуска экспрессии нуклеиновой кислоты в клетке-хозяине. Векторы могут представлять собой плазмиды, могут быть вирусными, например, фаги или фагмиды, при необходимости.
Молекулу нуклеиновой кислоты или вектор, описанные в данной заявке, можно внедрить в клетку-хозяина. Методики внедрения нуклеиновой кислоты или векторов в клетки-хозяева хорошо отработаны в данной области, и можно применять любую подходящую методику. Множество клеток-хозяев, подходящих для получения рекомбинантных молекул антител, известны в данной области и включают клетки-хозяева из бактерий, дрожжей, насекомых или млекопитающих. Предпочтительная клетка-хозяин представляет собой клетку млекопитающего, такую как клетка CHO, NS0 или HEK, например, клетка HEK293.
В другом аспекте настоящего изобретения предложен способ получения молекулы антитела согласно настоящему изобретению, включающий экспрессию нуклеиновой кислоты, кодирующей молекулу антитела, в клетке-хозяине и необязательно выделение и/или очистку молекулы антитела, полученной таким образом. Способы культивирования клеток-хозяев хорошо известны в данной области. Способ может дополнительно включать выделение и/или очистку молекулы антитела. Методики очистки рекомбинантных молекул антител хорошо известны в данной области и включают, например, ВЭЖХ, жидкостную хроматографию быстрого разрешения (ЖХБР) или аффинную хроматографию, например, с применением белка A или белка L. В некоторых вариантах реализации очистку можно осуществить, применяя аффинную метку на молекуле антитела. Способ также может включать включение молекулы антитела в состав фармацевтической композиции, необязательно с фармацевтически приемлемым вспомогательным веществом или другим веществом, описанным ниже.
Известно, что PD-L1 экспрессируется на множестве раковых клеток и экспрессируется на клетке иммунной системы. CD137 экспрессируется на клетках иммунной системы, включая T-клетки, в частности, CD8+ T-клетки, B-клетки, NK-клетки и инфильтрирующие опухоль лимфоциты (TIL). CD137 экспрессируется на более низком уровне на CD4+ T-клетках, чем на CD8+ T-клетках, но показали, что он также участвует в индукции пролиферации и активации некоторых субпопуляций CD4+ T-клеток.
Было показано, что активация CD137 играет роль в повышении пролиферации, выживаемости и цитотоксической эффекторной функции CD8+ T-клеток, а также дифференцировке CD8+ T-клеток и сохранении CD8+ T-клеток памяти. Также продемонстрировали, что активация CD137 повышает опосредованную NK-клетками АЗКЦ, а также пролиферацию, выживаемость и продукцию цитокинов B-клетками.
В данной заявке подробно описано, что авторы настоящего изобретения продемонстрировали, что молекулы антитела согласно настоящему изобретению способны одновременно связываться с PD-L1 и CD137, чтобы вызывать агонизм CD137. Таким образом, молекула антитела запускает агонизм автономно, на основе экспрессии PD-L1 и CD137, и без потребности в дополнительных перекрестно связывающих агентах. Поскольку PD-L1 экспрессируется на множестве раковых клеток и CD137 экспрессируется на иммунных клетках, где, как известно, он играет роль в повышении пролиферации и выживаемости иммунных клеток, ожидается, что молекулы антитела согласно настоящему изобретению будут способны усиливать иммунный ответ в локализациях, где экспрессируется PD-L1, например, в микроокружении опухоли. Более того, авторы настоящего изобретения показали, что применение молекулы антитела, обладающей данными свойствами, эффективно подавляет рост опухоли в сингенных моделях рака у мышей, и что такие молекулы антитела более эффективны, чем введение двух связывающих молекул, которые связывают PD-L1 и CD137, соответственно.
Молекулы антитела, описанные в данной заявке, следовательно, могут быть полезны для применений в терапии, в частности, для лечения рака.
Молекулу антитела, описанную в данной заявке, можно применять в способе лечения организма человека или животного. В сходных аспектах настоящего изобретения предложены:
(i) молекула антитела, описанная в данной заявке, для применения в качестве лекарственного средства,
(ii) молекула антитела, описанная в данной заявке, для применения в способе лечения заболевания или расстройства,
(iii) применение молекулы антитела, описанной в данной заявке, при изготовлении лекарственного средства для применения для лечения заболевания или расстройства; и,
(iv) способ лечения заболевания или расстройства у индивида, где указанный способ включает введение индивиду терапевтически эффективного количества молекулы антитела, описанной в данной заявке.
Индивид может представлять собой пациента, предпочтительно пациента-человека.
Лечение может представлять собой любое лечение или терапию, при которой достигается некоторый желательный терапевтический эффект, например, ингибирование или задержка прогрессирования состояния, и включает снижение скорости прогрессирования, замедление скорости прогрессирования, снижение выраженности состояния, исцеление или ремиссию (либо частичную или полную) состояния, предотвращение, облегчение, отсрочивание, ослабление или купирование одного или более симптомов и/или признаков состояния или продление выживаемости индивида или пациента сверх ожидаемой в отсутствие лечения.
Лечение как профилактическая мера (т.е., профилактика) также входит в объем данного термина. Например, индивида, предрасположенного или имеющего риск возникновения или повторного возникновения заболевания, такого как рак, можно лечить, как описано в данной заявке. Такое лечение может предотвращать или отсрочивать возникновение или повторное возникновение заболевания у индивида.
Описанный способ лечения может включать введение по меньшей мере одного дополнительного лечения индивиду, вдобавок к молекуле антитела. Молекулу антитела, описанную в данной заявке, следовательно, можно вводить индивиду отдельно или в комбинации с одним или более другими способами лечения. Если молекулу антитела вводят индивиду в комбинации с другим лечением, дополнительное лечение можно вводить индивиду одновременно, последовательно или отдельно от введения молекулы антитела. Если дополнительное лечение вводят одновременно с молекулой антитела, молекулу антитела и дополнительное лечение можно вводить индивиду в виде комбинированного препарата. Например, дополнительная терапия может представлять собой известную терапию или терапевтический агент для заболевания, которое лечат.
Хотя молекулу антитела можно вводить отдельно, молекулы антитела обычно вводят в виде фармацевтической композиции, которая может содержать по меньшей мере один компонент дополнительно к молекуле антитела. В другом аспекте настоящего изобретения, следовательно, предложена фармацевтическая композиция, содержащая молекулу антитела, описанную в данной заявке. Также предложен способ, включающий составление фармацевтической композиции с молекулой антитела.
Фармацевтические композиции могут содержать, дополнительно к молекуле антитела, фармацевтически приемлемое вспомогательное вещество, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области. Термин “фармацевтически приемлемый” в данной заявке относится к соединениям, материалам, композициям и/или лекарственным формам, которые, в рамках здравого медицинского суждения, подходят для применения в контакте с тканями субъекта (например, человека) без избыточной токсичности, раздражения, аллергической реакции или другой проблемы или осложнения, в соответствии с разумным соотношением риск/польза. Каждый носитель, вспомогательное вещество и т.д. также должны быть “приемлемыми” в том смысле, что они совместимы с другими ингредиентами состава. Точная природа носителя или другого материала будет зависеть от пути введения, который может представлять собой инфузию, инъекцию или любой другой подходящий путь, обсуждаемый ниже.
Для парентерального, например, подкожного или внутривенного введения, например, путем инъекции, фармацевтическая композиция, содержащая молекулу антитела, может находиться в виде приемлемого для парентерального введения водного раствора, который апирогенен и имеет подходящий уровень pH, изотоничность и стабильность. Специалисты с соответствующей квалификацией в данной области вполне способны получить подходящие растворы, применяя, например, изотонические среды, такие как хлорид натрия для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости можно использовать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие вспомогательные вещества, включая буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, такие как аскорбиновая кислота и метионин; консерванты (такие как октадецилдиметилбензилхлорид аммония; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3’-пентанол и м-крезол); низкомолекулярные полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагины, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как твин™, плюроники™ или полиэтиленгликоль (ПЭГ).
В некоторых вариантах реализации молекулы антитела могут быть предложены в лиофилизированной форме для восстановления влагосодержания перед введением. Например, можно восстановить влагосодержание лиофилизированных молекул антитела в стерильной воде и смешать с солевым раствором перед введением индивиду.
Введение может быть в «терапевтически эффективном количестве», которого достаточно, чтобы принести пользу индивиду. Фактическое введенное количество и скорость и временной график введения будут зависеть от природы и тяжести состояния, которое лечат, конкретного индивида, которого лечат, клинического состояния индивида, причины расстройства, места доставки композиции, типа молекулы антитела, способа введения, графика введения и других факторов, известных практикующим врачам. Назначение лечения, например, определение дозировки и т.д., находится в рамках компетенции врачей общей практики и других врачей и может зависеть от тяжести симптомов и/или прогрессирования заболевания, которое лечат. Подходящие дозы молекул антитела хорошо известны в данной области (Ledermann и др. (1991) Int. J. Cancer 47: 659-664; и Bagshawe и др. (1991) Antlbody, Immunoconjugates and Radiopharmaceuticals 4: 915-922). Можно применять конкретные дозировки молекулы антитела, которую нужно вводить, указанные в данной заявке или в Physician’s Desk Reference (2003), в соответствующих случаях. Терапевтически эффективное количество или подходящую дозу молекулы антитела можно определить путем сравнения активности in vitro и активности in vivo в модели на животном. Известны способы экстрополяции эффективных дозировок у мышей и других подопытных животных на людей. Точная доза будет зависеть от множества факторов, включая размер и расположение области, которую нужно лечить, и конкретную природу молекулы антитела.
Обычная доза антитела находится в диапазоне от 100 мкг до 1 г для системного применения и от 1 мкг до 1 мг для топического применения. Можно вводить изначально более высокую нагрузочную дозу, а затем одну или более меньших доз. Указанная доза представляет собой дозу разового введения взрослому индивиду, которую можно пропорционально скорректировать для детей и младенцев, а также скорректировать для других форматов антител пропорционально молекулярной массе.
Лечение можно повторять ежедневно, дважды в неделю, еженедельно или ежемесячно по усмотрению врача. Схема лечения индивида может зависеть от фармакокинетических и фармакодинамических свойств композиции антитела, пути введения и природы состояния, которое лечат.
Лечение может быть периодическим, и период между введениями может составлять приблизительно две недели или более, например, приблизительно три недели или более, приблизительно четыре недели или более, приблизительно один месяц или более, приблизительно пять недель или более или приблизительно шесть недель или более. Например, лечение можно вводить раз в две - четыре недели или раз в четыре - восемь недель. Подходящие составы и пути введения описаны выше.
В предпочтительном варианте реализации молекула антитела, описанная в данной заявке, может быть предназначена для применения в способе лечения рака.
Рак можно охарактеризовать аномальной пролиферацией злокачественных раковых клеток. Если упоминается конкретный тип рака, такой как рак молочной железы, то это относится к аномальной пролиферации злокачественных клеток соответствующей ткани, такой как ткань молочной железы. Вторичный рак, который расположен в молочной железе, но который возник в результате аномальной пролиферации злокачественных клеток другой ткани, такой как ткань яичников, не является раком молочной железы, упомянутым в данной заявке, но является раком яичников.
Рак может представлять собой первичный или вторичный рак. Следовательно, молекула антитела, описанная в данной заявке, может быть предназначена для применения в способе лечения рака у индивида, где рак представляет собой первичную опухоль и/или метастаз опухоли.
Опухоль рака, который лечат с применением молекулы антитела, описанной в данной заявке, может содержать TIL, которые экспрессируют CD137, например, на своей поверхности. В одном варианте реализации, возможно, определили, что опухоль содержит TIL, которые экспрессируют CD137. Способы определения экспрессии антигена на поверхности клетки известны в данной области и включают, например, проточную цитометрию.
Например, рак, который лечат с применением молекулы антитела, описанной в данной заявке, можно выбрать из группы, состоящей из лейкозов, таких как острый миелоидный лейкоз (ОМЛ), хронический миелоидный лейкоз (ХМЛ), острый лимфобластный лейкоз (ОЛЛ) и хронический лимфоцитарный лейкоз (ХЛЛ); лимфом, таких как лимфома Ходжкина, неходжкинская лимфома и множественная миелома; и типов солидного рака, таких как саркомы (например, саркомы мягких тканей), рак кожи (например, карцинома из клеток Меркеля), меланома, рак мочевого пузыря (например, уротелиальная карцинома), рак головного мозга (например, мультиформная глиобластома), рак молочной железы, рак матки/эндометрия, рак яичников (например, серозная цистаденома яичников), рак предстательной железы, рак легких (например, немелкоклеточная карцинома легких (НМРЛ) и мелкоклеточный рак легких (МРЛ), колоректальный рак (например, колоректальная аденокарцинома), рак шейки матки (например, плоскоклеточный рак шейки матки и аденокарцинома шейки матки), рак печени (например, печеночно-клеточная карцинома), рак головы и шеи (например, плоскоклеточная карцинома головы и шеи), рак пищевода, рак поджелудочной железы, рак почки (например, почечно-клеточный рак), рак надпочечника, рак желудка (например, аденокарцинома желудка), рак яичка, рак желчного пузыря и желчных протоков (например, холангиокарцинома), рак щитовидной железы, рак тимуса, рак кости и рак головного мозга.
В предпочтительном варианте реализации рак, который лечат с применением молекулы антитела, описанной в данной заявке, представляет собой солидный рак.
Более предпочтительно, рак, который лечат с применением молекулы антитела, описанной в данной заявке, представляет собой солидный рак, выбранный из группы, состоящей из меланомы, рака мочевого пузыря, рака головного мозга, рака молочной железы, рака яичников, рака легких, колоректального рака, рака шейки матки, рака печени, рака головы и шеи, рака поджелудочной железы, рака почки, рака желудка и стромальных опухолей желудочно-кишечного тракта (GIST).
В дополнительном предпочтительном варианте реализации рак, который лечат с применением молекулы антитела, описанной в данной заявке, может представлять собой рак, который отвечает на лечение одним или более ингибиторами контрольных точек иммунного ответа, такими как антитело, которое связывает PD-1, PD-L1 или CTLA4. Считают, что такие опухоли содержат более высокие уровни TIL и/или более высокую мутационную нагрузку опухоли, чем опухоли, которые не чувствительны к терапии ингибитором контрольных точек иммунного ответа. Такие опухоли также называют теплыми или горячими опухолями.
Примеры таких опухолей включают плоскоклеточную карциному головы и шеи (ПКГШ), меланому, рак легких (такой как плоскоклеточный рак легких, аденокарцинома легких, немелкоклеточная карцинома легких (НМРЛ) или мелкоклеточная карцинома легких (МРЛ)), рак предстательной железы, рак шейки матки (такой как плоскоклеточная карцинома шейки матки или эндоцервикальная аденокарцинома), рак мочевого пузыря, рак молочной железы, рак щитовидной железы, рак почки, колоректальный рак (с микросателлитной нестабильностью (MSI) или с микросателлитной стабильностью (MSS); например, колоректальная аденокарцинома), рак пищевода, неходжкинскую лимфому (NHL), рак желудка, рак эндометрия, рак поджелудочной железы, рак яичников, печеночно-клеточную карциному, мезотелиому, уротелиальный рак, карциному из клеток Меркеля и аденокарциному желудка. В предпочтительном варианте реализации рак представляет собой ПКГШ. Рак может дополнительно представлять собой рак, который ранее не лечили химиотерапевтическим или радиотерапевтическим агентом, т.е., индивид, которого лечат, может представлять собой страдающего раком пациента, который не получал лечение обсуждаемого рака химиотерапевтическим или радиотерапевтическим агентом.
В качестве альтернативы, рак, который лечат с применением молекулы антитела, описанной в данной заявке, может представлять собой рак, такой как рак поджелудочной железы или рак предстательной железы, который не отвечает на лечение одним или более ингибиторами контрольных точек иммунного ответа, такими как антитело, которое связывает PD-1, PD-L1 или CTLA4. Следовательно, индивид, которого лечат антителом согласно настоящему изобретению, может представлять собой страдающего раком пациента, такого как пациент, страдающий раком поджелудочной железы или раком предстательной железы, с предшествующим недостаточным ответом на один или более ингибиторов контрольных точек иммунного ответа, таких как антитело против PD-L1 или против PD-1.
Опухоли, которые не отвечают на лечение одним или более ингибиторами контрольных точек иммунного ответа, также называют холодными опухолями. Без привязки к теории, считают, что лечение рака, который не отвечает на лечение одним или более ингибиторами контрольных точек иммунного ответа отдельно, химиотерапией, лучевой терапией, иммунотерапевтическим агентом, таким как иммуностимулирующий агент или противоопухолевая вакцина, приведет к гибели раковой клетки, что, в свою очередь, приведет к повышению количества TIL в опухоли и к более высокому уровню экспрессии иммуносупрессорных рецепторов, что, в свою очередь, сделает рак отвечающим на лечение ингибиторами контрольных точек иммунного ответа, т.е. превратит холодную опухоль в теплую опухоль. Таким образом, молекула антитела согласно настоящему изобретению может быть предназначена для применения в способе лечения рака у индивида, где рак не отвечает или рефрактерный к лечению одним или более ингибиторами контрольных точек иммунного ответа отдельно, и где указанный способ включает введение молекулы антитела индивиду в комбинации с химиотерапевтическим, радиотерапевтическим или иммуностимулирующим агентом или противораковой вакциной. Также предложен способ лечения рака у индивида, где рак не отвечает или рефрактерный к лечению одним или более ингибиторами контрольных точек иммунного ответа отдельно, и где указанный способ включает введение молекулы антитела индивиду в комбинации с химиотерапевтическим, радиотерапевтическим или иммуностимулирующим агентом или противораковой вакциной.
В контексте рака, лечение может включать ингибирование роста рака, включая полную ремиссию рака, и/или ингибирование метастазирования рака, а также ингибирование рецидива рака. Рост рака, как правило, относится к любому из множества показателей, которые свидетельствуют об изменении внутри рака с образованием более развитой его формы. Таким образом, показатели для измерения ингибирования роста рака включают снижение выживаемости клетки рака, уменьшение объема опухоли или морфологии (например, что определяют с применением компьютерной томографии (КТ), эхографии или другого способа визуализации), замедление роста опухоли, разрушение сосудистой сети опухоли, улучшенные показатели при проверке реакции кожной гиперчувствительности замедленного типа, повышение активности противораковых иммунных клеток или других противораковых иммунных ответов и снижение уровней опухоль-специфических антигенов. Активация или усиление иммунных ответов на раковые опухоли у индивида может улучшить способность индивида противостоять росту рака, в частности, росту рака, уже присутствующего у субъекта, и/или снизить склонность к росту рака у индивида.
В контексте лечения рака, молекулу антитела, описанную в данной заявке, можно вводить индивиду в комбинации с другой противораковой терапией или терапевтическим агентом, такими как противораковая терапия или терапевтический агент, которые, как было показано, подходят или потенциально подходят для лечения обсуждаемого рака. Например, молекулу антитела можно вводить индивиду в комбинации с химиотерапевтическим агентом, лучевой терапией, радионуклидом, иммунотерапевтическим агентом, противоопухолевой вакциной, онколитическим вирусом, терапией адоптивным переносом клеток (ACT), такой как терапия адоптивным переносом NK-клеток или терапия T-клетками с химерным антигенным рецептором (CAR), аутологическими TIL или гамма/дельта-T-клетками или агентом для гормональной терапии.
Без привязки к теории, считают, что молекула антитела, описанная в данной заявке, может действовать как адъювант при противораковой терапии. В частности, считают, что введение молекулы антитела индивиду в комбинации с химиотерапией или лучевой терапией, например, будет запускать больший иммунный ответ против рака, чем достижимый при химиотерапии или лучевой терапии отдельно.
Один или более химиотерапевтических агентов для введения в комбинации с молекулой антитела, описанной в данной заявке, можно выбрать из группы, состоящей из: таксанов, цитотоксических антибиотиков, ингибиторов тирозинкиназы, ингибиторов PARP, ингибиторов фермента B-Raf, ингибиторов MEK, ингибиторов c-MET, ингибиторов VEGFR, ингибиторов PDGFR, алкилирующих агентов, аналогов платины, аналогов нуклеозидов, антифолатов, производных талидомида, противоопухолевых химиотерапевтических агентов и других агентов. Таксаны включают доцетаксел, паклитаксел и nab-паклитаксел; цитотоксические антибиотики включают актиномицин, блеомицин и антрациклины, такие как доксорубицин, митоксантрон и валрубицин; ингибиторы тирозинкиназы включают эрлотиниб, гефитиниб, акситиниб, PLX3397, иматиниб, кобиметиниб и траметиниб; ингибиторы PARP включают пирапариб; ингибиторы фермента B-Raf включают вемурафениб и дабрафениб; алкилирующие агенты включают дакарбазин, циклофосфамид и темозоломид; аналоги платины включают карбоплатин, цисплатин и оксалиплатин; аналоги нуклеозидов включают азацитидин, капецитабин, флударабин, фторурацил и гемцитабин и антифолаты включают метотрексат и пеметрексед. Другие химиотерапевтические агенты, подходящие для применения в настоящем изобретении, включают дефактиниб, энтиностат, эрибулин, иринотекан и винбластин.
Предпочтительные терапевтические агенты для введения с молекулой антитела, описанной в данной заявке, представляют собой доксорубицин, митоксантрон, циклофосфамид, цисплатин и оксалиплатин.
Лучевая терапия для введения в комбинации с молекулой антитела, описанной в данной заявке, может представлять собой наружную дистанционную лучевую терапию или брахитерапию.
Радионуклиды для введения с молекулой антитела, описанной в данной заявке, можно выбрать из группы, состоящей из: иттрия-90, йода-131, висмута-213, астата-211, лютеция 177, рения-188, меди-67, актиния-225, йода-125 и тербия-161.
Иммунотерапевтический агент для введения в комбинации с молекулой антитела, описанной в данной заявке, может представлять собой терапевтическую молекулу антитела, нуклеотид, цитокин или терапевтическое средство на основе цитокина. Например, терапевтическая молекула антитела может связываться с регуляторной молекулой иммунного ответа, например, молекулой ингибитора контрольных точек иммунного ответа или костимулирующей молекулой иммунного ответа, рецептором врожденной иммунной системы или антигеном опухоли, например, антигеном поверхности клетки опухоли или растворимым антигеном опухоли. Примеры регуляторных молекул иммунного ответа, с которыми может связываться терапевтическая молекула антитела, включают CTLA-4, LAG-3, TIGIT, TIM-3, VISTA, PD-L1, PD-1, CD47, CD73, CSF-1R, KIR, CD40, HVEM, IL-10 и CSF-1. Примеры рецепторов врожденной иммунной системы, с которыми может связываться терапевтическая молекула антитела, включают TLR4, TLR7, TLR9, RIG-I-подобные рецепторы (например, RIG-I и MDA-5) и STING. Примеры антигенов опухоли, с которыми может связываться терапевтическая молекула антитела, включают HER2, EGFW, CD20 и TGF-бета.
Нуклеотид для введения в комбинации с молекулой антитела, описанной в данной заявке, может представлять собой миРНК.
Цитокины или терапевическое средство на основе цитокинов можно выбрать из группы, состоящей из: IL-2, пролекарства конъюгированного IL-2, GM-CSF, IL-7, IL-12, IL-9, IL-15, IL-18, IL-21 и интерферона I типа.
Обе противоопухолевые вакцины для лечения рака были внедрены в клинику и подробно обсуждаются в научной литературе (такой как Rosenberg, S. 2000, Development of Cancer Vaccines). Это, главным образом, включает стратегии вынуждения иммунной системы отвечать на различные клеточные маркеры, экспрессированные аутологическими или аллогенными раковыми клетками, путем применения этих клеток в качестве способа вакцинации, с колониестимулирующим фактором гранулоцитов-макрофагов (GM-CSF) или без него. GM-CSF вызывает сильный ответ представления антигена и особенно хорошо работает, когда используется с указанными стратегиями.
Признаки, раскрытые в предшествующем описании, или в следующей формуле изобретения, или в сопроводительных фигурах, выраженные в конкретных формах или на примере средств осуществления описанной функции, или способа или процесса получения описанных результатов, в соответствующих случаях, можно, отдельно или в любой комбинации таких признаков, применять для реализации настоящего изобретения в разнообразных его формах.
Хотя настоящее изобретение было описано в сочетании с типичными вариантами реализации, описанными выше, множество эквивалентных модификаций и вариантов будут очевидны специалистам в данной области с учетом настоящего описания. Соответственно, считают, что типичные варианты реализации настоящего изобретения, описанные выше, являются иллюстрирующими и не ограничивающими. Можно внести различные изменения в описанные варианты реализации, не отклоняясь от сущности и объема настоящего изобретения.
Во избежание любых сомнений, любые теоретические объяснения, предложенные в данной заявке, предложены с целью улучшения понимания читателем. Авторы настоящего изобретения не привязываются к каким-либо данным теоретическим объяснениям.
Любые заголовки разделов, используемые в данной заявке, приведены исключительно с организационной целью, и не должны толковаться как ограничивающие описанный объект изобретения.
Во всем тексте настоящего описания, включая следующую формулу изобретения, если контекст не подразумевает иного, формулировка “включают” и “содержат” и ее варианты, такие как “включает”, “включающий” и “содержащий”, следует понимать как поразумевающие включение указанного целого числа или этапа или группы целых чисел или этапов, но не исключение какого-либо другого целого числа или этапа или группы целых чисел или этапов.
Следует отметить, что в настоящем описании и прилагаемой формуле изобретения использование форм единственного числа включает ссылку на множественное число, если в контексте явно не указано иное. Диапазоны в данной заявке могут быть выражены в виде: от “приблизительно” одного конкретного значения и/или до “приблизительно” другого конкретного значения. Когда указан такой диапазон, другой вариант реализации включает от одного конкретного значения и/или до другого конкретного значения. Аналогично, когда значения выражены в виде приблизительных величин, использование предшествующего термина “приблизительно” будет подразумевать, что конкретное значение образует другой вариант реализации. Термин “приблизительно” в отношении числового значения необязателен и означает, например, ±10%.
ПРИМЕРЫ
Пример 1. Характеристика и селекция антигенов.
Для способов селекции и скрининга, применяемых для идентификации МАТ2, которые способны связывать PD-L1 и CD137 и агонизировать CD137, требуется применение различных антигенов PD-L1 и CD137. Получение данных антигенов описано более подробно ниже.
1.1. Рекомбинантные антигены CD137.
Представители суперсемейства рецепторов фактора некроза опухоли (TNFRSF), такие как CD137, известны за склонность к обобразованию мультимеров, которые кластеризуются, когда они связаны со своими лигандами (Croft, M., 2003). Данная склонность к агрегации для функционирования создает трудности при получении растворимых рекомбинантных белков, которые не агрегируют в растворе, для применения для селекции in vitro, такой как фаговый и дрожжевой дисплей, и для идентификации прошедших селекцию белков.
Исследовали некоторые доступные для приобретения рекомбинантные антигены и обнаружили, что большинство не подходят для применения в данных способах селекции вследствие присутствующих уровней агрегатов. Из исследованных антигенов только для биотинилированного, слитого белка секретируемого CD137 человека и hFc (BPS Biosciences, № в каталоге 71171), обозначенного «hCD137-hFc-Avi-BPS» далее в данной заявке, была достаточно низкая агрегация, чтобы он подходил, и его применяли в способах селекции, хотя и с ограниченным успехом (см. пример 2).
Так как большинство доступных для приобретения антигенов считали неподходящими, следующие рекомбинантные димерные и мономерные антигены CD137 (см. таблицу 1), получали своими силами для применения в способах селекции:
Таблица 1. Антигены CD137.
Мономерные антигены получали путем клонирования ДНК, кодирующей внеклеточный домен CD137 человека (SEQ ID NO: 186) или мыши (SEQ ID NO: 188), наряду с последовательностью Avi и шестью C-концевыми остатками гистидина, в модифицированные векторы pFUSE (№ в каталоге Invivogen pfuse-mg2afc2), применяя ферменты рестрикции EcoRI-HF и BamHI-HF. Векторами трансфицировали клетки HEK293-6E (Канадский национальный совет по научным исследованиям) и экспрессированный CD137 очищали, применяя никелевую колонку HisTrap™ excel (GE LifeSciences 29048586) и эксклюзионную хроматографию (SEC), чтобы гарантировать, что антиген из одного вида и не содержал агрегатов.
Для получения димерных антигенов конструкции ДНК, кодирующие внеклеточный домен CD137 человека, мыши или яванского макака, слитый с доменом Fc mIgG2a наряду с последовательностью Avi, клонировали в модифицированные векторы pFUSE и трансфицировали ими клетки HEK293-6E. Рекомбинантный CD137 очищали, применяя колонку с белком A MabSelect SuRe™ (GE Healthcare, 11003494) и эксклюзионную хроматографию (SEC), чтобы гарантировать, что антиген из одного вида и не содержал агрегаты.
Каждый из димерного и мономерного антигенов биотинилировали, применяя набор для реакции биотин-биотин-протеинлигазы BirA (Avidity LLC, BirA500), с получением мономерных антигенов CD137, меченых одной молекулой биотина, и димерных антигенов CD137, меченых двумя молекулами биотина, по одной на каждом из двух мономеров. 3 мг антигена смешивали с 7,8 мкл смеси ферментов BirA до молярного отношения фермента к субстрату 1:50. Затем добавляли вспомогательные вещества в соответствии с рекомендациями производителя (142 мкл Biomix A, 142 мкл Biomix B, 142 мкл биотина) и реакционную смесь инкубировали в течение двух часов при комнатной температуре. Для сохранения целостности биотинилированного белка незамедлительно заменяли буфер реакционной смеси на DPBS (Life Technologies 14190-169), применяя фильтры Amicon с диаметром пор 30 мкм (Merck Millipore UFC503096).
Белки дополнительно очищали с помощью SEC, чтобы гарантировать удаление фермента BirA и получение конечного препарата высокого качества монодиспергированного белка без высокомолекулярных агрегатов. Более подробно, материалы из той же производственной партии смешивали и анализировали стабильность и чистоту с помощью эксклюзионной высокоэффективной жидкостной хроматографии (ЭВЭЖХ), электрофореза в полиакриламидном геле с ДСН (ПААГ/ДСН) и эксклюзионной хроматографии с детекцией многоуглового рассеяния лазерного света (SEC-MALS). Полное биотинилирование белков подтверждали по перемещению стрептавидина в геле ПААГ/ДСН. Подтвердили, что рекомбинантные антигены человека и мыши связываются с антителами положительного контроля против CD137 (20H4.9 (патент США № 7288638)) и Lob12.3 (Саутгемптонский университет), соответственно) in vitro, с помощью поверхностного плазмонного резонанса (ППР) и с клетками DO11.10, экспрессирующими лиганд CD137 человека и мыши, с помощью проточной цитометрии. Клетки инкубировали с антигенами CD137 в течение 1 часа, а затем применяли флуоресцентно меченое антитело против Fc-фрагмента мыши для детектирования связывания клеток. Подтвердили, что рекомбинантный антиген яванского макака связывается с клетками DO11.10 (National Jewish Health), экспрессирующими лиганд CD137 яванского макака, с помощью проточной цитометрии, как описано выше. Для того чтобы гарантировать как можно большую чистоту материалов, применяемых в протоколах селекции, провели полный белковый анализ антигенов, чтобы гарантировать, что присутствие агрегатов белков не превышало 2%.
1.2. Экспрессированные клетками антигены CD137.
Клетки DO11.10 (National Jewish Health), экспрессирующие полноразмерный CD137 мыши (SEQ ID NO: 187) или CD137 человека (SEQ ID NO: 185), обозначенные «DO11.10.mCD137» и «DO11.10.hCD137», соответственно, получали, чтобы представлять антиген в связанной с мембраной конформации, наиболее близкой к его природной форме, для проведения селекции и дополнительного исследования прошедших селекцию Fcab, перечисленных в таблице 2.
Лентивирусную трансдукцию использовали для получения данных клеток DO11.10, экспрессирующих на повышенном уровне рецепторы CD137 человека или мыши, применяя систему упаковки Lenti-X HTX (Clontech, № в каталоге 631249). Вектором экспрессии Lenti-X (pLVX) (Clontech, № в каталоге 631253), содержащим кДНК, кодирующую CD137 человека (SEQ ID NO: 185) или кодирующую CD137 мыши (SEQ ID NO: 187), котрансфицировали со смесью для упаковки Lenti-X HTX линию клеток Lenti-X 293T (Clontech, № в каталоге 632180), чтобы получить вирус. Линию клеток DO11.10 затем трансдуцировали данными лентивирусными векторами.
Экспрессию CD137 человека или CD137 мыши на данных клетках подтверждали с помощью проточной цитометрии по связыванию антител положительного контроля против CD137 20H4.9 и Lob12.3, соответственно, с указанными клетками. Клетки инкубировали с антителами положительного контроля против человека или мыши в течение 1 часа, а затем применяли флуоресцентно меченое детектирующее антитело против Fc человека (Stratech Scientific Ltd, № в каталоге 109-546-098-JIR) для детектирования связывания клеток.
Клетки DO11.10, экспрессирующие CD137 яванского макака (SEQ ID NO: 189), обозначенные «DO11.10.cCD137», также получали, применяя ту же методику лентивирусной трансдукции, и использовали для тестирования перекрестной реактивности Fcab против CD137 человека с CD137 яванского макака. Экспрессию CD137 яванского макака подтверждали с помощью проточной цитометрии по связыванию антитела положительного контроля против CD137 (MOR7480.1, US 2012/0237498 A1) с клетками, как описано ранее.
Таблица 2. Экспрессированный клетками CD137.
1.3. CD4+ и Fc-меченые PD-L1 мыши и человека.
Антигены PD-L1 мыши и человека со слитыми белками получали для применения для селекции и скрининга антител. Антигены экспрессировали либо с C-концевой His-меткой, а также либо с мономерной меткой CD4+, домены 3 и 4 (rCD4) крысы (Brown и Barclay, 1994), димерным доменом Fc IgG1 человека, либо, только для PD-L1 человека, с меткой Avi (с получением hPD-L1-rCD4-His (SEQ ID NO: 195), hPD-L1-Fc-His (SEQ ID NO: 196), mPD-L1-rCD4-His (SEQ ID NO: 197), mPD-L1-Fc-His (SEQ ID NO: 198) и hPD-L1-His-Avi (SEQ ID NO: 199). Получение антигенов в различных форматах позволило удалить связывающие метку молекулы во время последовательных раундов пэннинга фагового дисплея антител. Плазмидами экспрессии, кодирующими антигены, трансфицировали клетки HEK293, как описано у Chapple и др., 2006. Супернатанты собирали через 5 суток после трансфекции, и секретированные антигены очищали с помощью аффинной хроматографии на сефарозе с Ni-NTA (Schofield и др., 2007). Содержащие rCD4 и Fc антигены PD-L1 биотинилировали, применяя реагент EZ-link сульфо-NHS-биотин (Thermo Fisher Scientific, код продукта 21326), следуя рекомендациям производителя. Продукт реакции биотинилирования подвергали гель-фильтрации и собирали мономерную фракцию. Мономерную фракцию использовали для всех жидкофазных селекций фагового дисплея. Среднее количество биотинов на молекулу составляло от 1 до 3 биотинов на мономер PD-L1, что определяли с применением набора для флуоресцентного определения количества биотина (Thermo Fisher Scientific, код продукта 46610).
Пример 2. Селекция и характеристика Fcab против CD137 человека.
2.1. Наивная селекция Fcab против CD137 человека.
Для того чтобы выбрать Fcab, которые связываются с CD137 человека, проводили мероприятия по селекции дрожжевого и фагового дисплея, чтобы максимизировать разнообразие идентифицированных Fcab. Использовали как выставленный на поверхности клетки CD137 человека, так и рекомбинантный димерный CD137 человека, чтобы предоставить разнообразные форматы антигенов, чтобы оказать селекционное давление под действием авидности на димерные или мультимерные белки CD137. Получение Fcab, которые авидно связяваются с комплексами CD137, но не с мономерным CD137 с высокой аффинностью, считают предпочтительным, так как такие Fcab будут преимущественно нацеливаться только на активированные и примированные T-клетки, где повышенная экспрессия CD137 происходит после стимуляции T-клеток. Без привязки к теории, выдвинули предположение, что T-клетки с очень низкими или незначительными уровнями экспрессии CD137 на мембране более вероятно будут содержать CD137 в мономерном состоянии, в отличие от активированных T-клеток с сильно повышенной экспрессией CD137, где большая часть указанных белков будет в димерном, тримерном или более мультимерном состоянии. В результате селекции под действием авидности, Fcab будут преимущественно связываться с активированными T-клетками и не будут хорошо связываться с наивными T-клетками, на которых выставлен только мономерный CD137. В результате селекции авидных к CD137 Fcab, потенциальная нецелевая активация T-клеток будет снижена, а вместе с ней уменьшится токсичность.
Шесть наивных фаговых библиотек, представляющих домен CH3 IgG1 человека, использовали для проведения селекции с помощью фагового дисплея. Все шесть библиотек содержали рандомизированные петли AB (содержащие остатки в положениях 14 - 18 согласно нумерации IMGT) и рандомизированные петли EF (содержащие остатки в положениях 92 - 101 согласно нумерации IMGT). Одна из библиотек содержала клоны со вставкой либо двух, либо четырех аминокислот (кодируемых двумя или четырьмя кодонами NNK) в положении 101 в петле EF (вставленные остатки находятся в положениях 101.4 - 101.1 согласно нумерации IMGT).
3230 клонов фагов подвергали скринингу с помощью ELISA фагов на связывание с димерным рекомбинантным антигеном hCD137, и рекомбинантный Fc использовали в качестве отрицательного контроля. 1140 клонов фагов подвергали скринингу с помощью FACS фагов на специфическое связывание с клетками DO11.10.hCD137. Отдельные выбранные клоны затем секвенировали, и полученным в результате этого 76 уникальным последовательностям присваивали идентификатор клона Fcab и субклонировали их в вектор экспрессии pTT5 (Канадский национальный совет по научным исследованиям), содержащий кассету экспрессии тяжелой цепи HelD1.3 IgG1, с целью экспрессии клонов Fcab в формате МАТ2 (см. пример 3.2).
Четыре наивные дрожжевые библиотеки, представляющие домены с CH1 по CH3 IgG1 человека, использовали для проведения селекции с помощью дрожжевого дисплея. Все четыре библиотеки содержали рандомизированные петли AB (содержащие остатки в положениях 14 - 18 согласно нумерации IMGT) и рандомизированные петли EF (содержащие остатки в положениях 92 - 101 согласно нумерации IMGT) в домене CH3. Две из указанных библиотек дополнительно содержали вставку пяти аминокислотных остатков в положении 16 в петле AB домена CH3 (остатки в положениях 16.5 - 16.1 согласно нумерации IMGT).
2784 дрожжевых отдельных клона, идентифицированных при селекции из библиотеки, подвергали скринингу на связывание антигена, применяя анализ связывания антигена методом проточной цитометрии, который включал инкубацию клеток с биотинилированным рекомбинантным димерным антигеном человека или Fc-фрагментом мыши, чтобы отличить клоны дрожжей, которые связываются с Fc-областью рекомбинантного антигена hCD137. Селекцию повторяли при различных концентрациях антигена и условиях, таких как повышение температуры индукции, снижение строгости селекции или уменьшение количества раундов, чтобы увеличить количество прошедших селекцию клонов. В результате секвенирования прошедших селекцию клонов выявили значительно низкое разнообразие полученных клонов, причем обнаружили только 9 клонов Fcab с уникальными последовательностями: FS22-053, FS22-172, FS22-173, FS22-174, FS22-175, FS22-176, FS22-177, FS22-178 и FS22-179.
2.2. Получение Fcab против CD137 человека в формате “ложного” МАТ2.
Получали “ложные” антитела МАТ2, состоящие из молекул IgG1, содержащих 76 клонов Fcab против CD137 человека, выделенных из прошедших селекцию фагов, и 9 клонов, выделенных из прошедших селекцию дрожжей, чтобы позволить исследовать Fcab в формате МАТ2. Ложные МАТ2 получали путем замены части домена CH3 Fcab, содержащего петли AB, CD и EF, на соответствующую область домена CH3 из антитела против лизоцима куриного яйца HelD1.3. Получение антитела HelD1.3 описано в Tello и др., 1993. Последовательности тяжелой и легкой цепи антитела HelD1.3 представлены в SEQ ID 191 и 159, соответственно. Молекулы ложного МАТ2 получали путем временной экспрессии в клетках HEK293-6E. Чтобы оценить количество полученного белка, проводили количественный анализ содержания белка IgG с помощью интерферометрии биослоев, применяя платформу Octet QKe с биосенсорами для определения количества белка A от PALL (18-5021). Белки очищали с помощью аффинной хроматографии на белке A, применяя колонки mAb SelectSure. Для 53 полученных из фага белков МАТ² против CD137 выявили измерения ниже порога детектирования, и, следовательно, определили, что они не подходят для дальнейшего анализа. 32 МАТ² очищали, применяя колонки MAb SelectSure с белком A (GE Healthcare, 11003494).
Затем исследовали связывание прошедших селекцию данных МАТ2 с рекомбинантным антигеном человека (биотинилированным hCD137-mFc-Avi), применяя интерферометрию биослоев (BLI) с помощью платформы Octet QKe. 12 МАТ² не связывались и 19 связывались с покрытыми CD137 сенсорами (FS22-007, FS22-033, FS22-042, FS22-049, FS22-050, FS22-052, FS22-053, FS22-054, FS22-169, FS22-172, FS22-173, FS22-174, FS22-179, FS22-180, FS22-181, FS22-183, FS22-187, FS22-194, FS22-195).
2.3. Активность прошедших селекцию ложных МАТ2 против CD137 в репортерном анализе NF-κB человека.
Мультимеризация и кластеризация необходимы для передачи сигналов TNFR (Bitra и др., 2017). CD137 кластеризуется и активирует путь передачи сигналов NF-κB, когда он взаимодействует со своим лигандом CD137L. Молекулы агонистов имитируют лиганд при запуске кластеризации и активации CD137, тем самым активируя путь передачи сигналов NF-κB. Известно, что некоторые агонистические антитела могут по своей природе вызывать кластеризацию CD137 при связывании, например, урелумаб, тогда как для других требуется дополнительное перекрестное связывание самого антитела, чтобы вызвать кластеризацию CD137, для такого как утомилумаб (Fisher и др., 2012). Известно, что рецепторы Fc-гамма на эффекторных клетках вызывают такое перекрестное связывание in vivo, хотя оно неэффективно и может происходить вдали от сайта терапевтического интереса. Поскольку ограничивающая дозу токсичность была связана с урелумабом, но не утолимумабом, решили выбрать связывающие CD137 Fcab, у которых нет способности природного агонизма, но выбрать только те, которым необходимо дополнительное перекрестное связывание, чтобы вызвать кластеризацию CD137. Следовательно, разработали анализ, который позволяет детектировать активацию пути передачи сигналов NF-κB в клетке после кластеризации CD137, экспрессированного на поверхности клетки, с помощью перекрестно связанных антител, но для которого показали небольшую активность, когда антитела не были перекрестно связаны. Данный анализ затем применяли для тестирования агонистической функциональной активности 27 клонов Fcab против CD137 в формате ложного МАТ2 и 6 клонов Fcab против CD137 в формате МАТ² против CD20, независимо от того, было или нет обнаружено связывание Fcab с рекомбинантным антигеном в анализе BLI.
Это можно измерить по активации пути передачи сигналов NF-κB в анализе. Белок L применяли в качестве перекрестно связывающего агента, чтобы запустить перекрестное связывание ложного МАТ2 фрагментами Fab в анализе, и измеряли активацию NF-κB.
кДНК, кодирующую CD137 человека (SEQ ID NO: 185), субклонировали в вектор pMSCV-неомицин (Takara Clontech, № в каталоге 634401), применяя ферменты рестрикции EcoRI-HF и XhoI. Линию клеток RetroPack PT67 (Clontech, № в каталоге 631510) использовали для получения ретровирусных частиц, следуя протоколу производителя. Данный ретровирус впоследствии использовали для трансдукции клеток HEK.FRT.luc, которые ранее получали путем трансдукции линии клеток Flp-In T-REx 293 HEK (Life Technologies, R780-07) лентивирусом с репортером Qiagen Cignal Lenti NFkB Reporter (luc) (Qiagen, № в каталоге 336851), содержащим NF-κB-чувствительный промотор, контролирующий экспрессию люциферазы. Данные клетки HEK.FRT.luc.hCD137 использовали для скрининга ложного МАТ², содержащего связывающие CD137 молкулы, идентифицированные в способах селекции.
Получали разведения 2 мкМ каждого ложного МАТ2 в DPBS (Life Technologies, 14190169) и дополнительно разбавляли 1:3 в среде для репортерного анализа клеток (DMEM (Gibco, № в каталоге 61965-026); 10% фетальная телячья сыворотка (ФТС) (Gibco, № в каталоге 10270-106); 1x пенициллин/стрептомицин (Gibco, № в каталоге 15140-122); 15 мкг/мл бластицидина (Melford Laboratories Ltd., № в каталоге B1105); 5 мкг/мл пуромицина (Life technologies, № в каталоге A11113803); 100 мкг/мл зеоцина (InvivoGen, № в каталоге 11006-33-0); 500 мкг/мл генетицина (Life Technologies, № в каталоге 10131-027)). Белок L (Life Technologies, 21189) применяли в качестве искусственного перекрестно связывающего агента и смешивали с молекулами МАТ² в молярном отношении 1:4. После инкубации в течение 24 часов клетки обрабатывали 100 мкл реагента для люциферазного анализа Promega Bio-Glo™ (Promega, № в каталоге G7941), следуя инструкциям производителя, и измеряли люминесценцию со временем накопления сигнала 0,5 секунд на спектрофотометре для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения люминесценции представляют собой меру люциферазы, продуцированной в ответ на активацию пути передачи сигналов NF-κB посредством кластеризации CD137, вызванной перекрестно связанными Fcab. Значения люминесценции наносили на график против log концентрации Fcab, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Подходящие клоны идентифицировали по повышению по меньшей мере в 10 раз сигнала люциферазы при перекрестном связывании с белком L по сравнению с отсутствием перекрестного связывания. Определили, что данные клоны способны вызывать кластеризацию CD137 и дальнейшую активацию последующих путей передачи сигналов. Из всех исследованных клонов два были способны вызывать такое 10-кратное повышение сигнала люциферазы при перекрестном связывании: FS22-053 и FS22-172, хотя EC50 невозможно было определить для любого из них. Оба выбрали для дальнейшего исследования в анализе активации T-клеток DO11.10. Неожиданно, активность не наблюдали для остальных клонов в условиях перекрестного связывания, несмотря на связывание с мишенью CD137, в анализе BLI, возможно, свидетельствуя о том, что они связывались с посторонним эпитопом на CD137 или что аффинность таких клонов была недостаточной для связывания CD137 с достаточной силой, чтобы инициировать каскад передачи сигналов NF-κB.
В целом, хотя исследовали более 30 Fcab, в первичных селекциях идентифицировали только два Fcab (FS22-053 и FS22-172), которые проявляли желательную функцию в репортерном анализе NF-κB при перекрестном связывании и проявляли небольшую активность, когда не были перекрестно связаны.
2.4. Активность прошедшего селекцию ложного МАТ2 против CD137 в анализе активации T-клеток DO11.10.
Кластеризация CD137 посредством молекул-агонистов на активированных T-клетках вызывает активацию T-клеток и последующую передачу сигналов, приводящую к продукции IL-2, но не ограниченую ею. Поскольку обнаружили, что FS22-053 и FS22-172 проявляют активность в репортерном анализе NF-κB, исследовали их способность активировать CD137 в анализе активации T-клеток. Разработали анализ активации T-клеток DO11.10 с применением DO11.10 T-клеток, сконструированных для сверхэкспрессии CD137 человека, и активацию T-клеток оценивали путем измерения высвобождения IL-2.
T-клетки DO11.10 (National Jewish Health) трансдуцировали лентивирусным вектором, разработанным таким образом, чтобы он сверхэкспрессировал CD137 мыши или человека, как описано ранее. Так же, как и FS22-053 и FS22-172, следующие клоны исследовали в данном анализе активации T-клеток DO11.10: FS22-007, FS22-033, FS22-042, FS22-049, FS22-050, FS22-052, FS22-054 (все в формате “ложного” МАТ2 HelD1.3). Получали разведения МАТ2 или МАТ положительного контроля 20H4.9 либо с добавлением, либо без перекрестно связывающего агента рекомбинантного белка L (Life Technologies, 21189) и добавляли к клеткам DO11.10.hCD137 в 96-луночном планшете со скругленным дном лунок, которые покрывали в течение ночи 0,1 мкг/мл антитела против CD3 (клон 17A2, BioLegend, 100208). После инкубации в течение 18 часов, супернатанты собирали и анализировали с помощью набора для ELISA IL-2 мыши (eBioscience, 88-7024-86), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gens, BioTek. Значения поглощения на 570 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gens, BioTek). Концентрацию mlL-2 наносили на график в зависимости от log концентрации МАТ2 или эталонного МАТ, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для клонов FS22-053 и FS22-172 в данном анализе выявили значительно повышенную активность, когда они были перекрестно связаны с белком L. FS22-053 проявил активность, составляющую 126 нМ, когда он не был перекрестно связан, и активность, составляющую 21 нМ при перекрестном связывании (улучшение в 6 раз), тогда как FS22-172 проявил активность, составляющую 950 нМ, когда он не был перекрестно связан, и активность, составляющую 44 нМ при перекрестном связывании (улучшение в 22 раза). В результате оба клона выбрали для созревания аффинности.
В примере 2.4 показано, что FS22-053 и FS22-172 могли запускать кластеризацию и активацию CD137 на поверхности T-клеток DO11.10 при перекрестном связывании белком L в исследованных форматах МАТ².
Пример 3. Созревание аффинности Fcab против CD137 человека и последующий анализ.
Ранее обсуждалось, что два клона выбрали для созревания аффинности на основании их функциональных свойств в репортерном анализе NF-κB, анализе активации T-клеток DO11.10 (FS22-053 и FS22-172).
3.1. Созревание аффинности FS22-053 и FS22-172.
Четыре библиотеки дрожжевого дисплея конструировали из клонов Fcab FS22-053 и FS22-172. Семь остатков (в положениях 15 - 16.1 согласно IMGT) рандомизировали, применяя праймеры ELLA, в петле AB домена CH3 из каждого клона с получением библиотек FS22-053 AB и FS22-172 AB. Пять остатков (в положениях 92 - 94 и 97 - 98 согласно IMGT) рандомизировали, применяя праймеры ELLA, в петле EF домена CH3 с получением библиотек FS22-053 EF и FS22-172 EF.
Для библиотек FS22-053 AB и FS22-053 EF и FS22-172 AB и FS22-172 EF проводили три или четыре раунда селекции на дрожжевых библиотеках, чтобы выбрать клоны с созревшей аффинностью, применяя либо димерный антиген hCD137-mFc-Avi, либо мономерный антиген hCD137-Avi-His. Мономерный антиген применяли, чередуя с димерными антигенами, чтобы гарантировать, что у клонов сохранилась аффинность к антигену, и что они связываются не исключительно за счет авидности. Применение мономерного или димерного антигена, а также используемую концентрацию определяли опытным путем во время каждого раунда с помощью проточной цитометрии, что определяли по тому, наблюдалось ли обогащение против мономерного или димерного антигена в предыдущем раунде. Во всех случаях, когда это возможно, использовали окно сортировки выше исходного, чтобы выделить клоны с созревшей аффинностью по сравнению с исходной молекулой. Селекционное давление повышали до 1 нМ димерного антигена. Во время каждого раунда селекции отдельные клоны наносили пятнами на чашки с агаром, чтобы оценить успех селекции. Каждый клон растили и индуцировали отдельно, а затем его связывание и структурные параметры определяли с помощью проточной цитометрии, применяя биотинилированный димерный антиген, а также структурные маркеры против CH2, как описано ранее. Данному каскаду скрининга следовали, чтобы позволить определение успеха селекции на основании образца клонов из результата селекции и чтобы позволить ранний скрининг отдельных клонов, которые впоследствии можно получить в виде растворимых белков.
Всего 1152 дрожжевых отдельных клона подвергали скринингу на связывание с биотинилированным рекомбинантным антигеном в анализе методом проточной цитометрии связывания антигена, описанном ранее. Селекции библиотеки FS22-053 EF привели к обогащению 138 уникальными последовательностями петли. Аналогичным образом, 30 уникальных последовательностей петли были выделены из библиотеки FS22-172 AB. Библиотеки FS22-053 AB и FS22-172 EF не содержали каких-либо клонов, проявивших какое-либо улучшение связывания относительно исходных клонов. Анализ последовательностей для всех клонов с наилучшим связыванием из библиотек FS22-053 EF и FS22-172 AB выявил консервативный паттерн последовательности PPY в петлях AB.
Присутствие консервативного мотива PPY может вызвать образование более протяженной области связывания путем внедрения более жесткой или выставленной петли, исходя из ограниченной гибкости остатков пролина. В качестве альтернативы, последовательность PPY может представлять собой специфический консервативный мотив, вовлеченный в связывание данных клонов, так как продемонстрировали, что богатые пролином последовательности связываются с ароматическими последовательностями в белках с доменом SH3. Кроме того, поскольку консервативная последовательность PPY прошла селекцию независимо в двух отдельных линиях Fcab, она может быть важна для связывания эпитопа на CD137. Кроме того, консервативный паттерн последовательности LE или LD в петлях EF домена CH3 клонов обеих линий FS22-053 и FS22-172 позволяет предположить, что данный мотив аминокислот в петле EF необходим для улучшенного связывания.
Для того чтобы оценить ход селекции и будет ли необходимость рекомбинировать мутированные петли AB и EF между клонами с созревшей аффинностью, лучшие пять уникальных клонов из библиотеки FS22-053 EF (FS22-053-008, FS22-053-009, FS22-053-010, FS22-053-011, FS22-053-012), ранжированные по проявлению специфического связывания с 10 нМ димерного антигена человека (более чем 30% положительных по APC клеток в анализе связывания методом проточной цитометрии), и лучшие шесть уникальных клонов из библиотеки FS22-172 AB (FS22-172-001, FS22-172-002, FS22-172-003, FS22-172-004, FS22-172- 005, FS22-172-006, для всех выявили более 10% положительных по APC клеток при скрининге с 10 нМ димерного антигена человека в том же анализе) получали в виде ложного МАТ2 (HelD1.3) и модельного МАТ2 (PD-L1), чтобы оценить функциональное и кинетическое улучшение рандомизированных петель.
3.2. Конструирование Fcab против CD137 человека в формате “ложного” и “модельного” МАТ².
16 клонов с созревшей аффинностью, полученных из исходного клона FS22-053 (FS22-053-001 - FS22-053-016), и 6 клонов с созревшей аффинностью, полученных из исходных клонов FS22-172 (FS22-172-001 - FS22-172-006), получали в формате “ложного” и “модельного” МАТ2. Клоны FS22-053-001 - FS22-053-007 далее не исследовали, так как они не экспрессировались в формате МАТ2 на уровне, который позволил бы последующую очистку для дальнейшего тестирования и определения характеристик.
“Ложные” антитела МАТ², содержащие Fcab против CD137 человека в HelD1.3 получали для дальнейшего исследования Fcab с созревшей аффинностью в формате МАТ². Данные МАТ² получали, как описано в примере 2.2.
Также получали “модельное” МАТ2, содержащее Fcab против CD137 человека, а также связывающую PD-L1 область Fab (клон YW243.55.S70 из US 8217149 B2). Их получали способом, аналогичным способу, описанному в примере 2.2, путем замены части CH3 связывающего PD-L1 антитела, содержащего петли AB, CD и EF, на соответствующий участок из Fcab. Данное модельное МАТ² против PD-L1 содержало мутацию LALA в домене CH2 (AA). Известно, что внедрение мутации LALA в домен CH2 IgG1 человека уменьшает связывание с Fc γ-рецептором (Bruhns, P. и др. (2009) и Hezareh M. и др. (2001)).
“Ложное” CD137/HelD1.3 и “модельное” CD137-AA/PD-L1 МАТ² получали путем временной экспрессии в клетках HEK293-6E и очищали, применяя колонки MAb SelectSure с белком A.
3.3. Активность Fcab человека в формате ложного МАТ2 в репортерном клеточном анализе NF-κB человека.
Функциональную активность перечисленных Fcab против CD137 человека с созревшей аффинностью в формате ложного МАТ² (HelD1.3) исследовали в том же люциферазном анализе NF-κB, который описан в примере 2.3. Люминесценцию измеряли со временем накопления сигнала 0,5 секунд на спектрофотометре для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Как ожидалось, ни у одного из Fcab не выявили активность без перекрестного связывания с белком L (-XL). Для всех Fcab против CD137 с созревшей аффинностью выявили значительное улучшение относительно исходного Fcab против CD137, для которого, несмотря на положительность в данном анализе, расчет значения EC50 был не возможен (см. пример 2.3). Для FS22-053-008 и FS22-172-003 выявили лучшую активность из каждого семейства с наименьшей EC50 (26,34 нМ и 32,64 нМ, соответственно) при перекрестном связывании с белком L (+XL).
3.4. Активность Fcab человека с созревшей аффинностью в формате модельного МАТ2 в анализе активации T-клеток DO11.10 человека.
Функциональную активность Fcab человека с созревшей аффинностью в формате модельного МАТ² (PD-L1 LALA) исследовали в анализе активации T-клеток DO11.10, аналогичном анализу, описанному в примере 2.4.
Клетки HEK.mPD-L1 получали путем субклонирования кДНК, кодирующей PD-L1 мыши (SEQ ID NO: 187), в вектор pcDNA5FRT (Life Technologies), используя сайты рестрикции KpnI и NotI, и трансформирования векторами линии клеток Flp-In T-REx 293 (Life Technologies, R780-07), применяя липофектамин 2000 (Life Technologies, 11668-019). Клетки растили в DMEM, содержащей 10% ФБС, 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475) и 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105), в течение 3 - 4 недель до образования колоний стабильно трансформированных клеток. Данные колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma Aldrich, D9891) и исследовали наличие экспрессии PD-L1, применяя конъюгированное с PE антитело против PD-L1 мыши (MIH5) (BD Biosciences, 558091).
Клетки отделяли, применяя буфер для диссоциации клеток, однократно промывали ФБР и 2x105 клеток высевали в лунки 96-луночного планшета, а затем инкубировали с антителом, разбавленным 1:20 в ФБР, в течение 1 часа при 4 °C. Клетки однократно промывали ФБР, а затем измеряли на цитометре Accuri C6 (BD Biosciences), и результаты анализировали, применяя FlowJoX. Экспрессию PD-L1 мыши снова подтверждали.
15 клонов, полученных в примере 3.2, в CD137/PD-L1 МАТ² исследовали в данном анализе активации T-клеток DO11.10. Готовили разведения МАТ2 или МАТ положительного контроля и добавляли либо в клетки DO11.10.hCD137 (7,5x103 клеток на лунку) и HEK.mPD-L1 (2x104 клеток на лунку), либо в клетки DO11.10.hCD137 (7,5x103 клеток на лунку) и HEK, которые не трансдуцировали для экспрессии mPD-L1 (2x104 клеток на лунку), в 96-луночном планшете с плоским дном лунок, который покрывали в течение ночи 0,1 мкг/мл антитела против CD3 (клон 17A2, BioLegend, 100208). После инкубации в течение 18 часов супернатанты собирали и анализировали с помощью набора ELISA IL-2 мыши (eBioscience, 88-7024-86), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gens, BioTek. Значения поглощения на 570 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gens, BioTek). Концентрацию mlL-2 наносили на график в зависимости от log концентрации МАТ2 или эталонного МАТ, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism. Активацию T-клеток детектировали путем измерения высвобождения mIL-2.
Не наблюдали активность T-клеток без перекрестного связывания путем связывания с экспрессирующими PD-L1 клетками. После перекрестного связывания для всех МАТ2 наблюдали сильную активность T-клеток, что видно по высвобождению высоких уровней mIL-2 и субнаномолярным значениям EC50. Для всех клонов, отличных от FS22-053-009 и FS22-172-005, наблюдали EC50 менее 0,3 нМ, то есть они были также хороши, если не лучше чем положительный контроль (МАТ против CD137 человека, G1-AA/20H4.9). Наименьший наблюдаемый Emax, который является мерой максимальной активации T-клеток и может соответствовать большей противоопухолевой активности T-клеток in vivo, составлял 7758 пг/мл, что было выше, чем для положительного контроля (МАТ против CD137 человека, G1-AA/20H4.9).
3.5. Анализ активации первичных CD8+ T-клеток человека.
Способность Fcab активировать CD137 на T-клетках с повышенной экспрессией CD137 была показана в примере 3.3. Для тестирования активности Fcab на клетках, которые не были сконструированы для сверхэкспрессии CD137, был необходим анализ первичных T-клеток человека. Активированные цитотоксические CD8+ T-клетки отвечают за непосредственное уничтожение раковых клеток и экспрессируют CD137 на своей поверхности (Ye и др., 2014). Известно, что кластеризация CD137 необходима для индукции последующей передачи сигналов и дальнейшей активации CD8+ T-клеток. Следовательно анализ активации CD8+ T-клеток применяли, чтобы оценить способность Fcab (в формате МАТ2, подробно описанном ниже) запускать кластеризацию и последующую передачу сигналов CD137. Активацию CD8+ T-клеток определяли по высвобождению hIL-2.
Для того чтобы выделить T-клетки, мононуклеарные клетки периферической крови (МКПК) собирали из конусов для удаления лейкоцитов, побочного продукта процесса донорства тромбоцитов. Вкратце, содержимое конусов для удаления лейкоцитов смывали с помощью ФБР и наносили на градиент фиколла (Sigma-Aldrich, 1440-02). МКПК собирали путем центрифугирования и получали клетки, которые не пересекали градиент фиколла. МКПК дополнительно промывали ФБР и остальные красные кровяные клетки лизировали путем добавления 10 мл 1X лизирующего эритроциты буфера (eBioscience, 00-4300-54), следуя инструкциям производителя. CD8+ T-клетки выделяли из МКПК, присутствующих в элюэнте, применяя набор II для выделения CD8+ T-клеток (Miltenyi Biotec Ltd, 130-096-495), следуя инструкциям производителя.
Инкубацию с антителом против CD3 использовали в качестве первого сигнала, чтобы запустить исходную активацию T-клеток. 96-луночные культуральные планшеты с плоским дном лунок покрывали 8 мкг/мл антитела против CD3 (клон UCHT1, R&D Systems, МАТ100-SP) в ФБР в течение ночи при 4°C. Планшеты затем трижды промывали 200 мкл ФБР.
Для основанного на клеточной системе перекрестного связывания Fcab с созревшей аффинностью против CD137 человека в формате модельного МАТ2 против PD-L1, получали клетки HEK293 с повышенной экспрессией PD-L1 человека (HEK.hPD-L1) по существу как описано в примере 5.3, но путем субклонирования кДНК, кодирующей последовательность PD-L1 человека (SEQ ID NO: 185) вместо PD-L1 мыши. Клетки HEK.hPD-L1 высевали при концентрации 2×105 клеток на лунку на покрытые антителом против CD3 (8 мкг/мл) 96-луночные планшеты с плоским дном лунок в 100 мкл среды для культивирования T-клеток (среда RPMI (Life Technologies, 61870-044) с 10% ФБС (Life Technologies), 1X пенициллином/стрептомицином (Life Technologies, 15140122), 1 мМ пируватом натрия (Gibco, 11360-070), 10 мМ HEPES (Sigma-Aldrich, H0887), 2 мМ L-глутамином (Sigma-Aldrich, G7513) и 50 мкМ 2-меркаптоэтанолом (Gibco, M6250)). После прикрепления клеток HEK.hPD-L1 или клеток HEK, которые не были трансдуцированы для экспрессии hPD-L1, через 4 часа инкубации всю среду для культивирования T-клеток удаляли и заменяли на 100 мкл среды для культивирования T-клеток, содержащей T-клетки при концентрации 5,0×105 клеток/мл, с конечной концентрацией 5,0 x 104 клеток/лунку.
МАТ2 разбавляли в среде для T-клеток при 2X конечной концентрации, начиная с 500 нМ, и осуществляли титрование 1:3. 100 мкл титрованного МАТ2 добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела.
Каждое из антитела положительного контроля против CD137 (G1-AA/20H4.9) и антитела отрицательного контроля изотипа IgG (G1-AA/HelD1.3) разбавляли в среде для T-клеток при 2X конечной концентрации, начиная с 500 нМ, содержащей 500 нМ перекрестно связывающего агента (антитела против CH2 человека, полученного собственными силами (Jefferis и др., 1985 и Jefferis и др., 1992) (mG1/MK1A6)), и осуществляли титрование 1:3. 100 мкл разбавленной смеси антитела положительного контроля/перекрестно связывающего агента или смеси антитела IgG отрицательного контроля/перекрестно связывающего агента добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела.
Анализируемую смесь инкубировали при 37°C, 5% CO2 в течение 72 часов. Супернатанты собирали и анализировали с помощью набора ELISA Ready-SET-Go! для анализа IL-2 человека (eBioscience, № в каталоге 88-7025-88), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения поглощения на 630 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 человека (hIL-2) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для МАТ положительного контроля против CD137 человека 20H4.9 выявили повышение высвобождения hIL-2 с EC50, равной 0,5 нМ, при перекрестном связывании с антителом против hCH2 (mG1/MK1A6). Для всех клонов выявили активность в данном анализе, для большинства выявили хорошую эффективность с субнаномолярными EC50. МАТ2, содержащие Fcab: FS22-053-007, FS22-053-008, FS22-053-010, FS22-053-011, FS22-053-012, FS22-172-003, FS22-172-004, FS22-172-005, - вызывали наибольший ответ T-клеток с наименьшими EC50 в диапазоне 0,19 - 0,49 нМ. Подгруппу МАТ2 (содержащих Fcab: FS22-053-008, FS22-053-011, FS22-053-014, FS22-173-003 и FS22-172-004) также исследовали без перекрестного связывания с PD-L1, экспрессированным на клетках HEK, и выявили отсутствие активности в данном анализе, как и ожидалось. Это подтверждает активность, наблюдаемую в анализе NF-kB и анализе активации T-клеток DO11.10.
3.6. Определение специфичности Fcab против CD137 человека с помощью поверхностного плазмонного резонанса (ППР).
Исследовали специфичность Fcab против CD137 человека по отношению к CD137 человека по сравнению с другими родственными представителями семейства TNFSFR. 8 из указанных Fcab исследовали в формате ложного МАТ2 (HelD1.3) и измеряли с помощью ППР на Biacore T200 (GE Healthcare) путем тестирования связывания с другими рецепторами TNFRSF человека: CD40, OX40 и GITR. Сочетание по аминогруппе (набор для сочетания по аминогруппе, GE Healthcare, BR-1000-50) использовали, чтобы покрыть CD40, GITR и OX40 человека до приблизительно 1000 RU чипы CM5 Biacore (GE Healthcare, № в каталоге 29149603). Готовили разведения Fcab против CD137 человека в формате ложного МАТ2 (FS22-053-008/HelD1.3, FS22-053-009/HelD1.3, FS22-053-010/HelD1.3, FS22-053-011/HelD1.3, FS22-053-012/HelD1.3, FS22-053-014/HelD1.3, FS22-172-003/HelD1.3, FS22-172-004/HelD1.3), начиная с 1 мкМ, в буфере HBS-EP+ (BR100669) и впрыскивали в течение 3 мин при 30 мкл/мин, а затем позволяли диссоциировать в буфер в течение 4 мин. Чип регенерировали путем впрыскивания 10 мМ глицина, pH 2,5, в течение 12 с при 30 мкл/мин. Антитела, специфичные к различным представителям TNFRSF, использовали в качестве положительных контролей, чтобы проверить покрытие чипа Biacore. Из результатов вычитали два опорных сигнала и анализировали, применяя программное обеспечение BIAevaluation 3.2. Fcab не связывались ни с какими из исследованных рецепторов TNFRSF, демонстрируя их специфичность к CD137. В итоге, не ожидают, что Fcab или МАТ2, содержащие сайты связывания антигена из данных Fcab, будут осуществлять нецелевое связывание с чем-либо, отличным от CD137.
3.7. Аффинность связывания Fcab против CD137 человека в формате ложного МАТ2 с CD137 человека, яванского макака и мыши, измеренная с помощью ППР, и получение FS22-053-017.
Аффинность Fcab против CD137 человека (FS22-053-008, FS22-053-011, FS22-053-014, FS22-172-004, FS22-172-004) в формате ложного МАТ2 (см. пример 3.2) к CD137 человека, макака (яванского макака) и мыши измеряли с помощью ППР, чтобы определить, могут ли Fcab быть пригодны для тестирования в исследованиях на животных. Захватывающее антитело против Fab человека иммобилизовали на всех четырех проточных кюветах чипа CM5 серии S (GE Healthcare, № BR-1005-30) до средней поверхностной плотности 6000 RU, следуя рекомендациям производителя (GE Healthcare, набор для захвата Fab человека, № 28958325). Иммобилизация при 25°C и скорость потока 10 мкл/мин позволили добиться среднего конечного ответа 6000 RU. Каждое МАТ² захватывалось до уровня приблизительно 150 RU при впрыскивании 3 мкг/мл раствора МАТ², разбавленного в буфере HBS-EP+ (GE Healthcare, №BR1006-69), в течение 60 секунд при 30 мкл/мин. Затем различные концентрации антигена CD137 человека, яванского макака или мыши (небиотинилированного CD137 человека-mFc-Avi, яванского макака или мыши или CD137-Avi-His человека) в буфере HBS-EP+ пропускали через чип в течение 3 мин при 60 мкл/мин, а затем позволяли диссоциировать в течение 10 минут. После каждой концентрации антигена чип регенерировали путем впрыскивания 10 мМ глицина, pH 2,1, при скорости потока 30 мкл/мин в течение 30 секунд. Буфер HBS-EP+ впрыскивали перед наибольшей концентрацией антигена и после наименьшей концентрации антигена для вычитания опорного сигнала и одну из концентраций произвольно повторно впрыскивали еще раз. Кинетику связывания аппроксимировали моделью Лэнгмюра 1:1, чтобы получить равновесные константы связывания (KD) для каждого образца. Анализ результатов проводили с помощью программного обеспечения BIAevaluation версии 3.2. Результаты представлены в таблице 3.
Анализ результатов выявил улучшенное связывание с CD137 как человека, так и яванского макака всеми клонами с созревшей аффинностью по сравнению с соответствующими исходными молекулами. Аффинность связывания с мономерными антигенами CD137 человека была слабее (по меньшей мере в 100 раз), чем с димерными Fc-слитыми антигенами человека и яванского макака. В примере 2.1 обсуждается, что Fcab подвергали селекции, чтобы они преимущественно связывались с димерными, чем с мономерными формами CD137, и данный результат подтверждает, что стратегия селекции была успешной. Такое кинетическое поведение свидетельствует о том, что они с меньшей вероятностью будут связываться с мономерным CD137, экспрессированным на минимальных уровнях на нестимулированных T-клетках, что приведет к пониженному риску токсичности для печени или системной токсичности, связанной с некоторыми методами лечения моноклональным антителом против CD137.
Полученные результаты также показывают, что Fcab против CD137 человека связывалось с димерным CD137 яванского макака с аффинностью, сопоставимой с таковой к димерному CD137 человека.
Таблица 3
н/п - не применимо, так как низкий сигнал не позволил определить KD.
Также исследовали способность Fcab в формате ложного МАТ2 связываться с димерным CD137 мыши. Ни для одного из клонов не выявили сильного связывания с антигеном мыши (как показано в таблице, где н/п указывает на то, что KD невозможно было рассчитать), за исключением клона FS22-053-014, для которого неожиданно обнаружили KD 24 нМ для антигена мыши. Это было неожиданным, поскольку гомология последовательностей CD137 мыши и CD137 человека составляет менее 57%.
Анализ последовательности FS22-053-014 выявил потенциальный недостаток в последовательности, который мог приводить к посттрансляционной изомеризации аспартата в положении 98 в петле EF его домена CH3. Аспартат (D) в положении 98 мутировали на глутамат (E) с помощью сайт-направленного мутагенеза, применяя набор для мутагенеза QuickChange II (Agilent, № в каталоге 200523), следуя рекомендациям производителя, что позволило получить клон FS22-053-017.
Затем проводили эксперименты, чтобы подтвердить, что данная мутация не влияла отрицательно на аффинность связывания или функциональную активность клона FS22-053-017 по сравнению с клоном FS22-053-014.
Равновесную константу диссоциации (KD) клона Fcab FS22-053-017 сравнивали с таковой для клона FS22-053-014 с помощью ППР на системе Biacore T200, используя антиген CD137 человека-mFc-Avi и PD-L1 мыши-mFc-Avi, соответственно. Полученные результаты показали очень сходные кинетические профили по отношению к антигену человека для обоих клонов Fcab, таким образом, свидетельствуя о том, что мутирование аспартата в положении 98 на глутамат не оказывало отрицательного влияния на связывание с антигеном человека. Результаты также показали, что для FS22-053-017 наблюдалось снижение в 2 раза силы связывания с антигеном CD137 мыши.
Клон Fcab FS22-053-017 субклонировали и экспрессировали в виде “ложного” МАТ² HelD1.3 (см. пример 2.2), а затем сравнивали с клоном FS22-053-014, также в формате МАТ² HelD1.3, в анализе активации T-клеток DO11.10 с CD137 человека, как описано в примере 2.4. G1-AA/20H4.9 использовали в качестве антитела положительного контроля против CD137, а G1-AA/HelD1.3 - в качестве контроля IgG. МАТ² исследовали либо без перекрестного связывания с белком L, либо при перекрестном связывании при отношении 1:4 с белком L.
Результаты показали, что у обоих Fcab в формате ложного МАТ2 FS22-053-17 и Fcab в формате ложного МАТ2 FS22-053-014 была сопоставимая активность при перекрестном связывании с белком L в одном и том же анализе активации T-клеток DO11.10. Следовательно, проведенный мутагенез не оказал отрицательного влияния на функциональную активность. Оба клона в формате ложного МАТ2 не проявляли активность без перекрестного связывания.
3.8. Краткое описание созревания аффинности и исследования Fcab против CD137 человека.
Вкратце, получали Fcab против CD137 с созревшей аффинностью и готовили в формате МАТ2, которые затем исследовали. Для МАТ2, содержащего данные связывающие антиген CD137 домены в домене CH3, выявили высокие уровни активности в репортерном клеточном анализе NF-κB человека, и показали, что эта активность зависит от перекрестного связывания. Для МАТ2, содержащего антигенсвязывающий домен из FS22-053-008 и FS22-172-003 Fcab, показали наилучшую активность в данном анализе.
Кроме того, показали, что когда МАТ2 были получены из данных Fcab, так что они содержали связывающий антиген CD137 домен и связывающую PD-L1 область Fab, а также мутацию LALA в их домене CH2, чтобы уменьшить или устранить связывание с Fcγ-рецепторами, то для указанных МАТ2 выявляли сильную активность T-клеток в анализе активации T-клеток DO11.10 человека, и показали, что эта активность зависит от перекрестного связывания путем связывания с экспрессирующими PD-L1 клетками. Также показали, что МАТ2 обладают зависимой от перекрестного связывания активностью в анализе активации первичных CD8+ T-клеток человека, дополнительно доказывая, что МАТ2, содержащие данные домены с созревшей аффинностью, связывающие антиген CD137, способны эффективно вызывать агонизм CD137, и что данная агонистическая активность наблюдается при условии перекрестного связывания.
Также показали, что МАТ2, содержащее антигенсвязывающий домен антитела против CD137 человека, специфично к CD137 и не связывалось с другими представителями TNFRSF. Показали, что МАТ2 преимущественно связываются с димерным антигеном CD137 человека, чем с мономерным антигеном CD137 человека. Наконец, показали, что данные МАТ2 связываются с димерным CD137 яванского макака с аффинностью, сопоставимой с таковой к димерному CD137 человека.
Неожиданно обнаржили, что один клон Fcab FS22-053-014 также был способен связывать CD137 мыши. Анализ последовательности FS22-053-014 выявил потенциальный недостаток в последовательности, который скорректировали с помощью сайт-направленного мутагенеза с получением клона FS22-053-017. Исследование FS22-053-017 выявило, что у него сохранились аналогичные свойства связывания и аналогичная функциональная активность в анализе активации T-клеток DO11.10 с CD137 человека, как и у FS22-053-014.
После демонстрирования, что для МАТ2, содержащих антигенсвязывающий домен антитела против CD137 человека, требуется перекрестное связывание, чтобы кластеризоваться и активировать CD137, следующей целью было получение МАТ2, которое связывает PD-L1 человека дополнительно к CD137. Выдвинули предположение, что связывание МАТ2 с PD-L1 человека посредством плеч Fab будет вызывать перекрестное связывание указанных молекул антитела, что, в свою очередь, будет приводить к кластеризации и активации CD137, и что такая активация будет зависеть от наличия экспрессии PD-L1 человека.
Для получения МАТ2, которые связывают PD-L1 человека дополнительно к CD137, сначала было необходимо получить МАТ, которые были способны связывать PD-L1. Получение данных МАТ описано ниже.
Пример 4. Выделение наивных МАТ против PD-L1 280_02_G02.
4.1. Селекция
Для селекции клонов против PD-L1 использовали библиотеку фагового дисплея антител человека “IONTAS 1” (IONTAS Ltd.) Гены антитела, используемые для конструирования библиотеки IONTAS 1, получали из лимфоцитов человека (42 донорских лейкоцитных пленок) и одного образца ткани миндалины. Как лейкоцитные пленки, так и ткань миндалины получали при одобрении местным исследовательским этическим комитетом.
Проводили три раунда твердофазной селекции из библиотеки фагового дисплея антител IONTAS 1, применяя антиген, которым непосредственно покрывали полистироловые пробирки Nunc, как описано у Schofield и др., 2007. В первом, втором и третьем раундах селекции использовали PD-L1 человека-Fc-His, PD-L1 мыши-rCD4-His и PD-L1 человека-Fc-His, соответственно (подробности относительно антигена см. в примере 1.3).
Прошедшую селекцию популяцию вариабельных областей тяжелой цепи (VH) антитела против PD-L1 перетасовывали с популяцией наивных вариабельных областей легкой цепи (VL) антитела, как описано у Dyson и др., 2011, и эту перетасованную, восстановленную (спасенную) популяцию фагового дисплея антител использовали в жидкофазной селекции. Вкратце, пэннинг провели с PD-L1 человека-rCD4-His (10 нМ), PD-L1 человека-rCD4-His (200 пМ) и PD-L1 мыши-rCD4-His (10 нМ) в раундах 1, 2 и 3, соответственно, и это привело к получению популяции scFv против PD-L1, названной “Селекция 280”. Данная популяция scFv содержала человеческие и мышиные scFv против PD-L1, что определили с помощью ELISA для поликлонального фага, проведенного, как описано у Dyson и др., 2011, и проявила минимальную перекрестную реактивность с PD-1 человека или с метками rCD4 или Fc.
4.2. Скрининг: ELISA, рекомбинантный анализ блокирования, основанный на клеточной системе анализ блокирования.
4.2.1. ELISA моноклонального scFv.
Популяцию scFv из Селекции 280 из примера 4.1 подвергали скринингу с помощью ELISA, чтобы определить клоны, которые лучше всего связывались с PD-L1 человека. Популяцию scFv субклонировали в вектор растворимого scFv pSANG10, и культуры E.coli, содержащих растворимые scFv, получали, как описано (Martin и др., 2006; Studier, 2005). Растворимые scFv затем использовали в моноклональном ELISA с иммобилизованным PD-L1 человека-rCD4-His, как описано у Schofield и др., 2007. Планшеты Nunc Maxisorp (Thermo Fisher Scientific, 437111) покрывали PD-L1 человека-rCD4-His (5 мкг/мл, ФБР) в течение ночи, блокировали в 2% молоке на ФБР в течение 1 часа, добавляли культуральный супернатант E. coli (разведение 1:2 в 2x 2% молоке на ФБР) и позволяли связываться scFv в течение 1 часа при комнатной температуре. Связанные scFv детектировали с помощью антитела против FLAG M2 (Sigma, F1804), меченого европием. Всего подвергли скринингу 470 клонов, и это привело к идентификации 346 клонов против PD-L1 с сигналом связывания PD-L1 по меньшей мере в 10 раз выше фонового, по сравнению с “пустыми” блокированными лунками, не содержащими антигены. 192 наилучших клона антитела против PD-L1 человека, что оценивали по первичному сигналу ELISA, выбирали для дальнейшего анализа.
4.2.2. Скрининг scFv в анализе блокирования PD-L1/PD-1 на основе ELISA.
Для того чтобы идентифицировать клоны, которые блокировали взаимодействие между PD-L1 и PD-1, провели ELISA для скрининга блокирующих scFv. Вкратце, планшеты Nunc Maxisorp (437111, Thermo Fisher Scientific) покрывали антителом против rCD4 (домены 3 и 4) (MCA1022, OX-68, Bio-Rad) в течение ночи, блокировали в 3% молоке на ФБР и инкубировали с PD-1 человека-rCD4-His (5 мкг/мл в 3% молоке на ФБР) в течение 1 часа при комнатной температуре. PD-L1 человека-Fc-His (50 мкл, 0,2 нМ) предварительно смешивали с культуральным супернатантом E. coli, содержащим scFv. 96-луночные планшеты Nunc трижды промывали ФБР, 0,1% твин™-20 (ФБР-T) и трижды промывали ФБР, затем добавляли смесь PD-L1 человека-Fc-His/scFv и инкубировали в течение 1 часа при комнатной температуре. Планшеты промывали и связанный PD-L1 человека-Fc-His детектировали, применяя антитело козы против Fc-биотин (Jackson ImmunoResearch Laboratories, 109-065-098, 0,1 мкг/мл, 3% молоко на ФБР) и стрептавидин-европий (Perkin Elmer, 1244-360), а затем усиливающий раствор DELFIA (Perkin Elmer, 4001-0010). Из 192 подвергнутых скринингу клонов у 183 выявили по меньшей мере 90% блокирующей активности по сравнению с контролем средой. Указанные 183 идентифицированных клона scFv против PD-L1 подвергали дополнительному скринингу на перекрестную реактивность с PD-L1 мыши в первичном ELISA, как описано в примере 4.2.1 выше, но применяя иммобилизованный PD-L1 мыши-rCD4-His вместо PD-L1 человека. Это позволило идентифицировать 50 перекрестно реагирующих с PD-L1 мыши клонов, которые представляли собой кандидатов для преобразования в формат IgG1.
4.2.3. Преобразование блокирующих клонов scFv против PD-L1 в формат IgG1.
Клоны scFv против PD-L1, которые блокировали взаимодействие между PD-L1 и PD-1, преобразовывали в формат IgG1 путем субклонирования генов VL и VH в плазмиду экспрессии IgG1 pINT3-IgG1 и экспрессировали в HEK293 в масштабе 4 мл, как описано у Chapple и др., 2006. Антитела подвергали аффинной очистке по партиям с помощью гранул сефарозы с белком А (PC-A100) и центрифужных колонок “1-step batch” midi от Proteus (Generon, GEN-1SB08), следуя инструкциям производителя. Диализ очищенных антител проводили с помощью пробирок GeBAflex maxi с отсечкой 8 кДа (Generon, D045). При необходимости антитела концентрировали до 2 мкМ с помощью ультрафильтрации.
4.2.4. Скрининг активности блокирования PD-L1-PD-1 в анализе совместного культивирования с репортерными клетками Jurkat-NFAT.
Функциональную активность очищенных МАТ против PD-L1 затем оценивали в в репортерном скрининговом анализе совместного культивирования. Данный скрининг проводили, применяя линию клеток Jurkat, стабильно экспрессирующую GloResponse NFAT-luc2/PD-1 (Promega, CS187102), и готовые для применения сразу после размораживания клетки, экспрессирующие PD-L1 (Promega, CS178103), в соответствии с инструкциями производителя. PD-L1-клетки высевали в среду HAM’S-F12, содержащую 10% ФБС. На следующие сутки среду удаляли и параллельно репортерные клетки Jurkat-PD-1 (Promega, CS187102) ресуспендировали в среде для анализа (90% RPMI1640, 1% ФБС). В планшет, содержащий прикрепившиеся PD-L1-клетки, добавляли 40 мкл среды для анализа, содержащей различные антитела в 2x концентрации (200 нМ), а затем 40 мкл смеси PD-1-клеток. Планшет инкубировали в течение 6 часов при 37°C, 5% CO2. Реагент BioGlo (Promega, G7940, 80 мкл) добавляли в каждую лунку и выход люциферазы прочитывали, применяя спектрофотометр для прочтения планшетов BMG pherastar. Это позволило идентифицировать антитело G1/280_02_G02 как способное блокировать взаимодействие PD-L1 с PD-1 в анализе совместного культивирования, что определили по повышенной активности люциферазы по сравнению с контролями без антитела. Данную активность подтверждали в анализе совместного культивирования с зависимостью ответа от дозы (диапазон двухкратного разведения концентраций: от 200 до 1,56 нМ) с получением рассчитанной полумаксимальной эффективной концентрации (EC50), равной 4,2 нМ.
4.3. Оптимизация последовательности.
Предварительный анализ последовательности антитела G1/280_02_G02 привел к идентификации потенциального сайта дезамидирования в VH-CDR2, в частности, мотива NG в положениях по Кабату 54 - 55. Так как дезамидирование в данном сайте потенциально могло повлиять на связывание, получали варианты клона, в которых мотив NG был заменен на любой из NA, NS, SG или GG. Данные модификации не приводили к какому-либо значительному снижению аффинности к рекомбинантному PD-L1 или эффективности активности блокирования PD-L1, и вариант клона, содержащий модификацию NS, обозначенный G1/280_02_G02_NS, выбрали для применения для перетасовки легких цепей.
4.4. Краткое описание наивных селекций.
Стратегии селекции фагов позволили идентифицировать более 50 клонов, связывающих PD-L1 человека, с сильной активностью блокирования PD-1/PD-L1 in vitro, а также перекрестной реактивностью по отношению к PD-L1 мыши. В частности, для G1/280_02_G02 выявили эффективную активацию в основанном на клеточной системе репортерном анализе PD-L1, и, следовательно, выбрали его для дополнительной оптимизации.
Пример 5. Получение и исследование содержащих легкую цепь каппа клонов против PD-L1.
Антитело G1/280_02_G02_NS содержит легкую цепь лямбда. Так как большинство моноклональных антител, используемых в клиническом контексте, содержат легкие цепи каппа (Jain и др., 2017), попытались, применяя подход перетасовки цепей, получить клоны, содержащие тяжелую цепь антитела G1/280_02_G02_NS, но спаренные с легкими цепями каппа, которые сохранили аффинность к PD-L1 человека и перекрестную реактивность по отношению к PD-L1 мыши. Библиотеку легких цепей каппа IONTAS™ в плазмиде для фагового дисплея pIONTAS-1 (библиотеку каппа) использовали для получения библиотеки с перетасованными легкими цепями клонов scFv, содержащих тяжелую цепь антитела G1/280_02_G02_NS, соединенную с вариантами легкой цепи.
5.1. Селекции фагов и стратегия скрининга.
Проводили множество селекций фагового дисплея в растворе в трех раундах, применяя биотинилированные антигены PD-L1 человека-rCD4-His и PD-L1 мыши-rCD4-His (подробности относительно антигенов см. в примере 1.3). Селекции проводили путем снижения концентраций антигена в каждом раунде (изменяющихся от 100 до 0,02 нМ) и для каждого раунда селекции использовали контроль “без антигена”.
Шесть результатов селекции выбрали для скрининга: два из раунда 2 (№ 871 и 872) и четыре из раунда 3 (№ 887, 890, 891 и 894), применяя систему экспрессии растворимого scFv, описанную в разделе 1.3.1. Всего 1692 клона растворимого scFv подвергали скринингу на связывание с иммобилизованным антигеном в анализе ELISA (антигеном hu-PD-L1-rCD4-His при концентрации 3 мкг/мл в Dulbecco ФБР, 50 мкл, покрывали планшеты Maxisorb), применяя анализ, описанный в примере 4.2.1 выше, используя усиливающий раствор DELFIA.
Из 1692 подвергнутых скринингу клонов для 1029 клонов получили сигнал более 2000 RFU в анализе DELFIA, что дает показатель эффективности, приблизительно равный 61%. Затем выбрали лучшие 736 клонов и анализировали их, используя вторичный анализ (ранжирование по аффинности), применяя три концентрации антигена hPD-L1-rCD4-His (1,0 нМ, 0,2 нМ и 0,04 нМ). Из 736 подвергнутых скринингу клонов 48 клонов, для которых выявили наибольший сигнал, выбрали для клонирования и экспрессии в формате IgG1. Клоны экспрессировали в клетках Expi293F™ (Fisher Scientific, номер в каталоге 13479756) в масштабе 800 мкл, и культуральные супернатанты собирали на 5е сутки после трансфекции для дальнейшего скрининга в формате IgG1.
5.2. Скрининг с помощью ППР.
Все 48 антител ранжировали по аффинности, применяя ППР (устройство Biacore T200). Для ранжирования, разбавленные супернатанты (1:10 в подвижном буфере, полученном из 1X ФБР и 0,002% твин™-20) иммобилизовали на чипе с белком A (GE healthcare, код продукта: 29127556), и PD-L1 человека-rCD4-His пропускали через полученную поверхность при концентрации 50 нМ. Константы скорости ассоциации (ka) и диссоциации (kd), полученные с применением данного однократного впрыскивания, использовали для определения константы диссоциации (KD). Значения KD для клонов сравнивали с таковыми для клона G1/280_02_G02_NS в формате Ig1 (G1/280_02_G02_NS). Идентифицировали десять клонов с уникальной последовательностью, для которых выявили более высокую аффинность к PD-L1 человека, чем для клона G1/280_02_G02_NS, и, следовательно, их подвергли полному кинетическому анализу вместе с клоном G1/280_02_G02_NS.
Вкратце, эксперименты ППР проводили, применяя устройство BIAcore T200. Антитела из разбавленного культурального супернатанта захватывались чипом с белком A (GE Healthcare, 29127556) в проточной кювете 2 (FC2) при скорости потока 10 мкл/мин с длительностью контакта 60 секунд. Обычно это приводило к захвату 500 - 800 RU антитела. Двухкратные разведения PD-L1-rCD4-His впрыскивали, начиная с 50 нМ, при скорости потока 30 мкл/мин (диапазон концентраций: 50 нМ - 0,05 нМ) в FC1 и FC2. Ассоциацию измеряли в течение 180 секунд и диссоциацию измеряли в течение 300 секунд. Все измерения проводили при 25°C в ФБР, pH 7,4, 0,05 % твин™-20. Кинетические параметры определяли путем вычитания эталонной кюветы и аппроксимирования экспериментальных результатов сенсограммы, предполагая взаимодействие 1:1, применяя программное обеспечение BIAevaluation (GE, BR-1005-97). Полученные результаты аппроксимировали, применяя программное обеспечение BIAevaluation, и рассчитывали соответствующие значения ka, kd и KD. Из десяти исследованных клонов для четырех антител, обозначенных “G1/887_04_E12”, “894_08_A05”, “G1/894_08_E05” и “G1/887_04_G12”, выявили субнаномолярные значения KD, которые были ниже, чем KD для G1/280_02_G02_NS. Результаты определения аффинности, полученные при описанных условиях скрининга, показали, что перетасовка легких цепей каппа, описанная в примере 5.1, позволяла тяжелой цепи антитела G1/280_02_G02_NS образовывать пару не только с легкой цепью лямбда, но также с легкими цепями каппа с получением антител с хорошей, и по существу улучшенной, аффинностью к рекомбинантному PD-L1 человека.
5.3. Характеристика клонов каппа в формате IgG1.
5.3.1. Основанный на клеточной системе анализ блокирования PD-1/PD-L1.
Способность клонов против PD-L1, содержащих легкую цепь каппа, G1/887_04_E12, G1/894_08_E05 и G1/887_04_G12, блокировать взаимодействие между PD-1 и PD-L1 оценивали в основанном на клеточной системе биолюминесцентном анализе, применяя продукт PD-1/PD-L1 Blockade Bioassay (Promega, J1250/J1255), в соответствии с рекомендациями производителя. Активность блокирования сравнивали с таковой для клона G1/280_02_G02_NS.
Вкратце, все антитела очищали, как описано в примере 4.2.3, и исследовали в виде 3-кратных разведений от 100 нМ до 35 пМ (восемь концентраций) в двух повторах. Показали, что все исследованные клоны были эффективными ингибиторами взаимодействия PD-1/PD-L1, причем для трех содержащих легкую цепь каппа клонов G1/887_04_E12, G1/894_08_E05 и G1/884_04_G12 выявили даже лучшие значения IC50, чем для содержащего легкую цепь лямбда клона G1/280_02_G02_NS.
5.3.2. Аффинности.
Связывание МАТ против PD-L1 G1/887_04_E12, G1/887_04_G12 и G1/894_08_E05 с рекомбинантным PD-L1 человека (биотинилированный hPD-L1-Avi-His, Acro Biosystems, PD1-H82E5), яванского макака (cPD-L1-His, Acro Biosystems, PD1-C52H4) и мыши (mPD-L1-His, Acro Biosystems, PD1-M5220) затем измеряли с помощью ППР, применяя обрабатывающее устройство Biacore T200 (GE Healthcare). Аффинности сравнивали с таковыми для клона 280_02_G02_NS в формате IgG1 (G1-AA/280_02_G02_NS; “AA” в названии данного клона означает, что данный клон также содержал мутацию “LALA” в домене CH2).
Вкратце, МАТ против PD-L1, разбавленные в буфере HBS-EP (GE Healthcare, BR100188) до концентрации 2 мкг/мл, впрыскивали отдельно на проточные кюветы 2, 3 и 4 чипа с белком A (GE Healthcare, 29127556) при скорости 30 мкл/мин, чтобы добиться конечного ответа приблизительно 110 RU. Рекомбинантные антигены PD-L1-His человека, яванского макака и мыши, разбавленные в буфере HBS-EP, впрыскивали на проточную кювету 1, 2, 3 или 4, в соответствующих случаях, в диапазоне концентраций от 81 нМ до 0,037 нМ в виде 3-кратных разведений в течение 4 мин при 75 мкл/мин, а затем позволяли диссоциировать в буфер в течение 10 мин. Регенерирования добивались путем впрыскивания 10 мМ глицина-HCL, pH1,5 (GE Healthcare, набор для захвата антитела человека Human Antibody Capture Kit, BR00356) в течение 30 сек при скорости 30 мкл/мин. Результаты после вычитания (проточная кювета 2 - проточная кювета 1, проточная кювета 3 - проточная кювета 1 или проточная кювета 4 - проточная кювета 1) анализировали, применяя программное обеспечение BIAevaluation 3.2 (GE Healthcare), чтобы определить связывание, применяя модель связывания 1:1 с переносом массы, с постоянным показателем преломления (RI), равным 0. Для определения аффинностей для кривых связывания PD-L1 мыши использовали Rmax для соответствующих профилей связывания PD-L1 человека.
Результаты анализа связывания продемонстрировали, что клон G1-AA/280_02_G02_NS и клоны G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 связывались с PD-L1 человека и яванского макака с низкими одноразрядными наномолярными или субнаномолярными аффинностями и полностью перекрестно реагировали с PD-L1 человека/яванского макака. По сравнению с клоном G1-AA/280_02_G02_NS, аффинности связывания клонов G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 с PD-L1 человека были приблизительно в 1,8 - 4,8 раз выше и с PD-L1 яванского макака - в 2,7 - 4,7 раз выше. Аффинности указанных клонов к рекомбинантному PD-L1 мыши были ниже, со значениями KD в диапазоне от 38 до 225 нМ, и наибольшую аффинность наблюдали для клона G1/887_04_E12. Данные результаты показывают, что тяжелую цепь антитела G1-AA/280_02_G02_NS можно спарить с легкими цепями как лямбда, так и каппа с получением антител с хорошими (и в случае спаривания с легкой цепью каппа - субнаномолярными) аффинностями к рекомбинантному PD-L1 человека и яванского макака, а также некоторой, хотя и более низкой, аффинностью к рекомбинантному PD-L1 мыши.
5.3.3. Связывание МАТ против PD-L1 с экспрессированным клеткой PD-L1.
Затем исследовали связывание МАТ против PD-L1 человека - G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 - с клетками HEK293, экспрессирующими PD-L1 человека (клетки HEK.hPD-L1), применяя проточную цитометрию. Неспецифическое связывание также оценивали путем тестирования связывания с исходными клетками HEK293, которые не содержат PD-L1 человека (линия клеток Flp-In T-Rex 293, Life Technologies, R780-07).
cNDA, кодирующий PD-L1 человека (SEQ ID NO: 185), субклонировали в вектор pcDNA™5/FRT (ThermoFisher Scientific, номер в каталоге V601020), используя сайты рестрикции KpnI и NotI, и вектором затем трансформировали линию клеток Flp-In T-Rex 293 (Life Technologies, R780-07), применяя липофектамин 2000 (Life Technologies, 11668-019). Клетки растили в DMEM, содержащей 10% ФБС, 100 мкг/мл гигромицина B (Melford Laboratories Ltd, Z2475) и 15 мкг/мл бластицидина (Melford Laboratories Ltd, B1105) в течение 3 - 4 недель, пока не образовались колонии стабильно трансформированных клеток. Данные колонии амплифицировали в присутствии 1 мкг/мл доксициклина (Sigma Aldrich, D9891) и исследовали экспрессию PD-L1, применяя конъюгированное с PE антитело против PD-L1 человека (MIH1) (BD Biosciences, 557924). Клетки отделяли, применяя буфер для диссоциации клеток, однократно промывали ФБР, высевали при концентрации 2x105 клеток в лунки 96-луночного планшета, а затем инкубировали с антителом, разбавленным 1:20 в ФБР, в течение 1 часа при 4 °C перед повторной промывкой в ФБР и измеряли, применяя цитометр Accuri C6 (BD Biosciences). Результаты анализировали, применяя программное обеспечение FlowJoX. Экспрессию PD-L1 человека обнаружили в указанной линии клеток.
Обнаружили, что клоны G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 связываются с PD-L1 человека на поверхности клетки со значениями EC50 в диапазоне 0,26 - 0,29 нМ. Не наблюдали связывания с исходными клетками HEK293, что свидетельствовало о специфичности связывания. Следовательно, все исследованные клоны МАТ специфично связывались с PD-L1, при этом не наблюдали неспецифического связывания.
5.3.4. Активность МАТ против PD-L1 в анализе реакции смешанной культуры лимфоцитов.
Активность МАТ против PD-L1 исследовали в анализе реакции смешанной культуры лимфоцитов (СКЛ). В анализе СКЛ измеряют клеточный иммунный ответ, который происходит между двумя аллогенными популяциями лимфоцитов (из одного вида, но генетически различными). В данном анализе использовали CD4+ T-клетки из одного донора и дендритные клетки моноцитарного происхождения (иДК) из другого донора. Так как иммунные клетки содержат физиологические уровни регуляторов иммунных контрольных точек, можно использовать анализ СКЛ, чтобы подтвердить, что активация T-клеток усиливается под действием МАТ в системе человека.
Получение размноженных CD4+ T-клеток.
МКПК выделяли из конусов с лейкоцитами путем разделения в градиенте фиколла. CD4+ T-клетки выделяли, применяя набор для выделения CD4+ T-клеток человека (Miltenyi Biotec Ltd, 130-096-533), следуя инструкциям производителя. Гранулы Human T-ActivatorCD3/CD28 Dynabeads (Life Technologies, 11131D) ресуспендировали путем встряхивания на вортексе. Гранулы переносили в стерильную пробирку емкостью 15 мл и добавляли 10 мл RPMI (Life Technologies, 61870044) с 10% ФБС (Life Technologies, 10270106) и 1x пенициллином/стрептомицином (Life Technologies, 15140122), чтобы промыть Dynabeads. Супернатант отбрасывали. Необходимое количество CD4+ T-клеток при концентрации 1,0×106 клеток/мл в RPMI с 10% ФБС, 1x раствором пенициллина/стрептомицина и 50 МЕ/мл рекомбинантного IL-2 человека (Peprotech, 200-02-50μg) при отношении гранул к клеткам 3:1 переносили во флакон T75 (Greiner Bio-one, 690195) и инкубировали при 37°C + 5% CO2. Через 3 суток клетки аккуратно ресуспендировали и подсчитывали. Плотность клеток поддерживали в диапазоне 0,8-1×106 клеток/мл путем добавления свежей среды (RPMI-10% ФБС + 1X раствор пенициллина/стрептомицина + 50 МЕ/мл rhuIL-2) при необходимости. На 7 или 8 сутки гранулы CD3/28 удаляли и CD4+ T-клетки оставляли в покое в течение ночи при концентрации 1×106 клеток/мл свежей среды RPMI-10% ФБС + 1X раствор пенициллина/стрептомицина с уменьшенным количеством 10 МЕ/мл rhuIL-2. Клетки хранили замороженными до момента, когда они потребуются.
Дифференцировка иДК.
Свежие моноциты выделяли из МКПК человека, применяя набор для выделения общей популяции моноцитов человека (Miltenyi Biotec Ltd, 130-096-537), следуя инструкциям производителя. Моноциты дифференцировали с получением иДК, применяя среду для дифференцировки Mo-DC Differentiation Medium, human (Miltenyi Biotec Ltd, 130-094-812), следуя инструкциям производителя.
Размноженные T-клетки размораживали за одни сутки до проведения эксперимента, промывали средой AIM V (Gibco, 12055-091) и инкубировали при 37 °C, 5% CO2 в среде AIM V в течение ночи. МАТ против PD-L1 человека G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 разбавляли до 4x конечной концентрации в трех повторах в 50 мкл среды AIM V в 96-луночных планшетах со скругленным дном лунок (VWR, 734-1797). Антитело против FITC, обозначенное 4420 (Bedzyk и др., 1989 и Bedzyk и др., 1990), содержащее мутацию LALA, включали в анализ в качестве отрицательного контроля. Исследовали серию 3-кратных разведений, начиная от 30 нМ до 0,002 нМ. Как 1×104 клеток иДК, суспендированных в 50 мкл среды AIM V, так и 1×105 размноженных CD4+ T-клеток, суспендированных в 100 мкл среды AIM V, добавляли к разведениям антитела и инкубировали в течение 5 суток при 37°C + 5% CO2. Включали следующие контроли: CD4+ T-клетки отдельно, иДК отдельно, CD4+ T-клетки + иДК и только среда AIM V. Супернатанты собирали, образцы разбавляли (1:25) и концентрации интерферона гамма (IFN-γ) измеряли, применяя набор для анализа ELISA IFN-гамма человека Ready-SET-Go! (Life Technologies, 88-7316-77). Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения поглощения на 630 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IFN-γ человека наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для МАТ против PD-L1 человека G1/894_8_E05, G1/887_4_E12 и G1/887_4_G12 выявили сильную активность в анализе СКЛ со значениями EC50, составляющими менее 0,030 нМ, и максимальным уровнем IFN-γ (Emax) более чем 10000 пг/мл. EC50 указывает на концентрацию МАТ, при которой достигается половина ответа, тогда как Emax представляет собой абсолютное значение, которое указывает на максимальную концентрацию IFN-γ, достигнутую в анализе. Не наблюдалась активность МАТ отрицательного контроля G1-AA/4420, как и ожидалось.
5.4. Активность МАТ против PD-L1 в анализе активации T-клеток DO11.10 мыши.
Так как у МАТ против PD-L1 человека G1/887_04_E12, G1/887_4_G12 и G1/894_08_E05 выявили способность слабо перекрестно реагировать с PD-L1 мыши (см. пример 5.3.2), исследовали их функциональную активность по отношению к PD-L1 мыши в анализе высвобождения интерлейкина-2 (IL-2) на основе линий клеток гибридомы DO11.10 OVA T-лимфоцитов и LK35.2 B-лимфоцитов. Высвобождение IL-2 является маркером активации T-клеток. T-клетки, экспрессирующие эндогенный PD-1 мыши, трансфицировали пустым вектором (pLVX). B-клетки трансфицировали конструкцией PD-L1 мыши.
5.4.1. Получение линий T-клеток с пустым вектором.
Методику лентивирусной трансдукции использовали для получения клеток DO11.10 (National Jewish Health), содержащих пустой лентивирусный вектор pLVX, применяя систему упаковки Lenti-X HTX (Clontech, 631249). Вектором экспрессии Lenti-X (pLVX) (№ в каталоге 631253) котрансфицировали со смесью для упаковки Lenti-X HTX линию клеток Lenti-X 293T (№ в каталоге 632180), чтобы получить вирус. Линию клеток DO11.10 трансдуцировали, применяя лентивирусные частицы, полученные с помощью системы упаковки Lenti-X HTX.
5.4.2. Получение антигенпредставляющих клеток, экспрессирующих на повышенном уровне PD-L1.
Методику лентивирусной трансдукции использовали для получения клеток B-клеточной лимфомы LK35.2 (ATCC, HB-98), экспрессирующих на повышенном уровне PD-L1 мыши, применяя систему упаковки Lenti-X HTX (№ в каталоге 631249). Вектором экспрессии Lenti-X (pLVX) (№ в каталоге 631253), содержащим кДНК, кодирующую PD-L1 мыши (SEQ ID NO: 187), котрансфицировали со смесью для упаковки Lenti-X HTX линию клеток Lenti-X 293T (№ в каталоге 632180), чтобы получить вирус. Линию клеток LK35.2 трансдуцировали, применяя лентивирусные векторы, полученные с помощью системы упаковки Lenti-X HTX.
5.4.3. Анализ активации T-клеток DO11.10 мыши.
Получали разведения МАТ против PD-L1 G1/887_04_E12, G1/887_4_G12 и G1/894_08_E05 или МАТ отрицательного контроля против FITC (G1-AA/4420) в экспериментальных средах (DMEM (Gibco, 61965-026), 10% ФБС (Gibco, 10270-106), 1 мМ пируват натрия (Gibco, 11360-070)). МАТ смешивали 1:1 с 4×105/мл клеток LK35.2 mPD-L1 в экспериментальных средах в присутствии 2,46 мкМ пептида OVA (H-ISQAVHAAHAEINEAGR-OH (SEQ ID NO: 192) (Pepscan)) (100 мкл смеси клеток LK35.2 mPD-L1 (B-клеточная гибридома, трансдуцированная лентивирусным вектором, содержащим mPD-L1 для сверхэкспрессии PD-L1 мыши)/МАТ на лунку в 96-луночном планшете со скругленным дном лунок) и инкубировали при 37°C, 5% CO2 в течение 1 часа. 2×105 клеток DO11.10 pLVX (T-клеточная гибридома DO11.10, трансдуцированная пустым лентивирусным вектором)/мл в объеме 100 мкл экспериментальный среды добавляли к 100 мкл смеси LK35.2 mPD-L1/(МАТ). Клетки затем смешивали перед инкубацией при 37°C, 5% CO2 в течение 24 часов. Супернатанты собирали и анализировали с помощью набора для ELISA IL-2 мыши (eBioscience, 88-7024-88, или R&D Systems, SM2000), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения поглощения на 570 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 мыши наносили на график в зависимости от log концентрации МАТ и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для МАТ против PD-L1 человека выявили значимую активность в анализе активации T-клеток мыши с эффективностью (EC50) в диапазоне 1 - 4,4 нМ. Не наблюдалась активность МАТ отрицательного контроля, как и ожидалось. Из трех исследованных клонов, G1/887_04_E12, у которого выявили наибольшую аффинность к рекомбинантному PD-L1 мыши, также был наиболее эффективным клоном в анализе активации T-клеток. Различия в эффективности были меньше, чем измеренные аффинности, что, вероятно, является следствием высокого уровня сверхэкспрессии PD-L1 мыши на клетках LK35.2 в данном анализе.
5.5. Краткое описание исследования клонов каппа в формате IgG1.
Для МАТ против PD-L1 G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12, содержащих прошедшие селекцию легкие цепи каппа, продемонстрировали перекрестную реактивность по отношению к PD-L1 яванского макака и мыши, выявили специфическое связывание с экспрессированным на поверхности клетки PD-L1 и выявили еще более высокую аффинность к рекомбинантному PD-L1 человека и яванского макака, чем у содержащего легкую цепь лямбда клона G1/280_02_G02. Показали, что МАТ против PD-L1 G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12 являются эффективными активаторами T-клеток человека in vitro и обладают функциональной перекрестной реактивностью по отношению к PD-L1 мыши.
Пример 6. Оптимизация последовательности МАТ против PD-L1.
6.1. Идентификация и удаление потенциальных сайтов дезамидирования белка.
Анализ последовательности клона G1/280_02_G02_NS привел к идентификации последовательности NSNT в петле H-CDR2 (в положениях по Кабату 54 - 57) как потенциального сайта дезамидирования, который, когда он дезамидирован, может влиять на связывание. Тяжелая цепь данного клона сохранялась во всех содержащих легкую цепь каппа клонах, полученных с помощью подхода перетасовки цепей, описанного в примере 5, так что данный потенциальный сайт дезамидирования также присутствовал в клонах G1/887_04_E12, G1/894_08_A05, G1/894_08_E05 и G1/887_4_G12. Используя специфические праймеры, наиболее близкие к последовательности зародышевой линии, последовательность NSNT заменили в четырех содержащих легкую цепь каппа клонах с помощью сайт-направленного мутагенеза на любую из GGST, SGGT или SGNA с получением вариантов идентифицированных клонов. Во время удаления данного потенциального сайта дезамидирования также исследовали роль остатка пролина в положении 28 по Кабату в области VH клона G1/887_04_E12, который был непреднамеренно введен в последовательность данного антитела во время перетасовки легких цепей каппа, путем его обратной замены на остаток треонина, который содержится в клоне G1/280_02_G02_NS. Проводили трансфекцию исходным и поученными вариантами клона (все в формате IgG1) в масштабе 0,8 мл, и культуральные супернатанты, собранные через пять суток после трансфекции, использовали для определения аффинностей клонов к PD-L1 человека и яванского макака-rCD4-His с помощью ППР. PD-L1 яванского макака-rCD4-His получали, как описано в примере 1.3. За исключением вариантов клона, полученных из клона G1/887_04_E12, все варианты клона сохраняли свои субнаномолярные аффинности к PD-L1 человека и яванского макака по сравнению с соответствующим исходным клоном.
Для всех модифицированных клонов, полученных из исходных клонов G1/894_08_A05, G1/894_08_E05 и G1/887_04_G12, выявили субнаномолярные аффинности к PD-L1 человека и яванского макака, сопоставимые с таковыми для соответствующего исходного клона, свидетельствуя о том, что замены GGST, SGGT и SGNA были вполне допустимы. Сильно сниженные аффинности клонов G1/929_01_A01, G1/929_01_A02 и G1/929_01_A03 по сравнению с исходным клоном (G1/887_04_E12) считали вероятным следствием удаления пролина в области VH в положении 28 по Кабату, а не наличия замен GGST, SGGT и SGNA в H-CDR2. Было неожиданно, что данный остаток пролина в клоне G1/887_04_E12 оказался важен для его аффинности к PD-L1. Варианты, полученные из трех исходных клонов G1/887_04_E12, G1/894_08_E05 и G1/887_04_G12, которые содержали замену SGGT в H-CDR2 (положения 54 - 57), а именно клоны G1/929_01_A02, G1/929_01_A08 и G1/929_01_A11, выбрали для дальнейшего исследования на том основании, что замена SGGT была наиболее близка к последовательности зародышевой линии.
Используя сайт-направленный мутагенез, потенциальный сайт дезамидирования (NSNT в положениях по Кабату 54 - 57) в петле H-CDR2 клона G1/280_02_G02_NS также модифицировали на SGGT. Кроме того, дополнительный потенциальный сайт дезамидирования (мотив NS), обнаруженный в положениях по Кабату 31 - 32 в CDR1 легкой цепи лямбда данного клона модифицировали на NY путем мутирования серина 32 (нумерация по Кабату) на тирозин, так как тирозин находится в данном положении в нескольких последовательностях зародышевой линии, таких как IGLV2-8-01, IGLV2-8-02, IGLV2-8-03, IGLV2-11-01, IGLV2-11-02, IGLV2-11-03 и IGLV2-14-01, IGLV2-14-02, IGLV2-14-03, IGLV2-14-04. Комбинация данных модификаций привела к получению содержащего легкую цепь лямбда клона G1/lam-G02v3, который также выбирали для дальнейшего исследования.
Пример 7. Исследование МАТ.
7.1. Клонирование и получение клонов в формате МАТ.
Остаток треонина в положении 28 по Кабату в области VH варианта клона G1/929_01_A02 “SGGT”, идентифицированного в примере 6, мутировали на пролин, который присутствует в том же положении в исходном клоне G1/887_04_E12, с целью улучшения его аффинности к PD-L1 человека и яванского макака. Временную экспрессию в клетках HEK293-6E и очистку с применением колонок MAb SelectSure с белком A использовали для получения данного модифицированного варианта клона и других трех вариантов клонов “SGGT” (G1/929_01_A08, G1/929_01_A11 и G1/lam-G02v3), идентифицированных в примере 6, в формате IgG1 и с мутацией LALA, чтобы позволить тестирование их функциональной активности в отсутствие эффекторной функции. Полученные в результате этого МАТ обозначили G1-AA/E12v2, G1-AA/E05v2, G1-AA/G12v2 и G1-AA/lamG02v3 (называемый либо G1-AA/lamG02v3, либо G1-AA/lambdav3). Последовательности тяжелой и легкой цепи, соответственно, представлены в SEQ ID NO: 16 и SEQ ID NO: 17 для G1-AA/E12v2, SEQ ID NO: 27 и SEQ ID NO: 28 для G1-AA/E05v2, SEQ ID NO: 27 и SEQ ID NO: 33 для G1-AA/G12v2 и SEQ ID NO: 27 и SEQ ID NO: 44 для G1-AA/lam-G02v3.
7.2. Аффинности МАТ к PD-L1 человека и яванского макака.
Для того чтобы определить, влияли ли дополнительные модификации последовательности, присутствующие в G1-AA/lam-G02v3 (а именно, замена NSNT на SGGT в VH-CDR2, NS на NY в VL-CDR1 и мутация LALA) и содержащих легкую цепь каппа МАТ G1-AA/E12v2, G1-AA/E05v2 и G1-AA/G12v2 (а именно, мутация LALA и, только в G1-AA/E12v2, замена треонина на пролин в положении 28 по Кабату в области VH), на кинетику связывания, определяли аффинности данных МАТ против PD-L1 к PD-L1 человека и яванского макака, как описано в примере 5.3.2. Указанные МАТ G1-AA/lam-G02v3, G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2 проявляли аффинности к PD-L1 человека и яванского макака, сходные с таковыми, наблюдаемыми в примере 5.3 для МАТ G1-AA/280_02_G02_NS, G1/894_08_E05, G1/887_04_E12 и G1/887_04_G12, демонстрируя, что на аффинности связывания исследованных указанных МАТ и МАТ2 не оказывали влияния модификация потенциальных сайтов дезамидирования или внедрение мутации LALA. Для МАТ G1-AA/E12v2 выявили наименьшее значение KD из всех четырех исследованных МАТ (0,21 нМ к PD-L1 человека и 0,37 нМ к PD-L1 яванского макака). Результаты представлены в таблице 4.
VH МАТ G1-AA/E12v2 отличается от участка G1/929_01_A02 (пример 6) одним остатком: E12v2 содержит пролин в положении 28 по Кабату, тогда как G1/929_01_A02 содержит треонин в данном положении. G1/929_01_A02 проявляло более чем в 10 раз более низкую аффинность к PD-L1 как человека, так и яванского макака, по сравнению с G1-AA/E12v2, данный результат демонстрирует важность остатка пролина в положении 28 в VH клона E12v2 (номенклатура Кабата) для аффинности к PD-L1 человека и яванского макака.
Таблица 4
KD (нМ)
KD (нМ)
7.3. Активность МАТ к PD-L1 человека в СКЛ.
МАТ против PD-L1 G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G05v2 исследовали в анализе реакции смешанной культуры лимфоцитов (СКЛ), описанном в примере 5.3.4. G1-AA/4420 использовали в качестве отрицательного контроля. Полученные результаты показаны в таблице 5. Для МАТ G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2 выявили сильную активность в анализе СКЛ со значениями EC50, составляющими менее 0,054 нМ, и максимальным уровнем IFN-γ (Emax) более 600 пг/мл (таблица 5). Значения EC50 и особенно Emax значительно отличались от таковых, описанных в примере 5.3.4. Полагают, что данное различие является следствием различий между донорами, так как ответ зависит от аллогенной реакции между T-клетками из одного донора и дендритными клетками моноцитарного происхождения из другого донора. Эффективность МАТ против PD-L1 человека согласовывалась с результатами, описанными в примере 5.3.4, как и ранжирование клонов в порядке эффективности. Как и ожидалось, для МАТ отрицательного контроля G1-AA/4420 не наблюдали активности.
Таблица 5.
7.4. Экспрессия, очистка и аналитическое исследование МАТ против PD-L1.
МАТ G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2 получали в лабораторном масштабе и исследовали с помощью стандартных аналитических способов SEC и дифференциальной сканирующей калориметрии (ДСК).
7.4.1. Экспрессия в лабораторном масштабе и очистка МАТ против PD-L1.
Последовательностями ДНК, кодирующими МАТ G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2, трансфицировали клетки HEK293 6E (Национальный научно-исследовательский совет Канады), применяя PEIpro (Polyplus, Франция). Через 5 суток культуральные супернатанты собирали и очищали на заранее наполненных белком A колонках MabSelect, применяя устройство AKTAxpress (оба от GE Healthcare, Уппсала, Швеция). Уравновешивание колонок проводили с помощью 50 мМ Трис, 250 мМ NaCl при pH 7,0, а затем загружали на них собранный культуральный супернатант. Смолу промывали, используя 50 мМ Трис, 250 мМ NaCl при pH 7,0, и затем следовало элюирование МАТ с применением буфера при pH менее 3,5.
7.4.2. Анализ с помощью эксклюзионной сверхвысокоэффективной жидкостной хроматографии (эксклюзионной СВЭЖХ).
Эксклюзионную СВЭЖХ после очистки проводили в течение 24 часов с момента очистки (материал хранили при 4°C), применяя Acquity H-Class Bio UPLC (Waters Corp., Великобритания), для измерения процента мономеров. Применяли колонку Acquity UPLC BEH200 SEC 1,7 мм (4,6 × 150 мм), подвижная фаза состояла из 250 мМ фосфата натрия, 100 мМ L-аргинина при pH 6,8. Количественный анализ мономеров, низкомолекулярных и высокомолекулярных соединений проводили, применяя программное обеспечение Empower (Waters Corp., Великобритания).
7.4.3. Термостабильность.
Температуру плавления (Tm) G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2 измеряли, применяя дифференциальный сканирующий калориметр (ДСК) Microcal VP-capillary. G1-AA/lam-G02v3 включали в анализ, чтобы оценить различие между содержащими легкую цепь каппа и лямбда МАТ. Образцы измеряли в буфере для образца при концентрации 0,2 мг/мл. Скорость сканирования устанавливали на 60°C/ч и результаты собирали между 35°C и 100°C. Анализ результатов проводили с помощью программного обеспечения Origin 7.0. Так как пики ДСК для Fab и CH3 перекрывались, регистрировали одно значение.
Таблица 6
Краткое описание результатов представлено в таблице 6. Для трех МАТ G1-AA/E05v2, G1-AA/E12v2 и G1-AA/G12v2 выявили благоприятные параметры аналитического исследования: чистота мономера после очистки на белке A составляла более 98%, и обнаружили, что термостабильность конформационного перехода Fab (Tm) была на более высоком уровне, чем для конформационных переходов, обычно регистируемых для IgG1, причем G1-AA/E12v2 оказалось наиболее термически стабильным (Fab/CH3 TM= 81,4 - 84,1°C). Для МАТ с легкой цепью лямбда G1-AA/lamG02v3 выявили боле низкую Tm, чем для трех МАТ, содержащих легкую цепь каппа.
Пример 8. Получение и исследование МАТ2 против CD137/PD-L1 человека.
8.1. Получение МАТ2 против CD137/PD-L1 человека.
Получали антитела МАТ2 CD137/PD-L1, состоящие из молекул IgG1, содержащих 3 клона Fcab против CD137 человека FS22-053-008, FS22-053-017 и FS22-172-003 (см. пример 3), чтобы позволить исследование Fcab в формате МАТ2. МАТ2 CD137/PD-L1 получали путем замены части домена CH3 Fcab, содержащей петли AB, CD и EF, на соответствующий участок домена CH3 из одного из трех связывающих PD-L1 антител (G1-AA/E05v2, G1-AA/E12v2, G1-AA/G12v2; см. пример 7.1). Все поученные в результате этого МАТ2 содержали мутацию LALA. МАТ2 CD137/PD-L1 временно экспрессировали, применяя PEIpro (Polyplus, Франция), в клетках HEK293 6E (NRCC, Канада). Через 5 суток культуральные супернатанты собирали и очищали на заранее наполненных белком A колонках MabSelect, применяя устройство AKTAxpress (оба от GE Healthcare, Уппсала, Швеция). Уравновешивание колонок проводили с помощью 50 мМ Трис, 250 мМ NaCl при pH 7,0, а затем загружали на них собранный культуральный супернатант. Смолу затем подвергали промывке в 50 мМ Трис-HCl, 250 мМ NaCl при pH 7,0, а затем следовало элюирование МАТ2 с применением буфера при pH<3,5.
Чтобы оценить количество полученного белка, проводили количественный анализ содержания белка IgG с помощью интерферометрии биослоев, применяя платформу Octet QKe (ForteBio) с биосенсорами для определения количества белка A от PALL (18-5021). Белки очищали с помощью аффинной хроматографии на белке A, применяя колонки mAb SelectSure. Девять МАТ2 были очищены с применением колонок MAb SelectSure с белком A (GE Healthcare, 11003494). Результаты представлены в таблице 7.
Таблица 7.
Все 9 антител МАТ2 против CD137/PD-L1 получали путем временной экспрессии в HEK 293 6E, и полученные титры и выход после очистки на белке A находились в рамках обычных диапазонов, ожидаемых для лабораторного масштаба. FS22-172-003-AA-Iam/G02v3 также экспрессировали и очищали для дальнейшего тестирования.
8.2. Биофизическое исследование МАТ2 с помощью эксклюзионной хроматографии (SEC) и ЭФ в ПААГ/ДСН.
8.2.1. Анализ с помощью эксклюзионной ВЭЖХ.
После очистки проводили эксклюзионную ВЭЖХ на системе для ВЭЖХ Agilent серии 1100 (Agilent, Великобритания), оборудованной колонкой TSK-GEL SUPERSW3000, внутренний диаметр 4,6 мм x 30,0 см (Tosoh Bioscience), применяя 20 мМ фосфат натрия, 200 мМ хлорид натрия, pH 6,8, в качестве подвижной фазы. Количественный анализ % мономеров провели, применяя программное обеспечение Chemstation (Agilent, Великобритания).
8.2.2. Анализ с помощью капиллярного электрофореза в присутствии додецилсульфата натрия (КЭ-ДСН).
Анализ КЭ-ДСН проводили на системе для капиллярного электрофореза Bioanalyzer 2100 (Agilent, Великобритания), следуя инструкциям производителя. Для восстанавливающих условий добавляли ДТТ и денатурировали образцы при 70° C в течение 5 минут.
Таблица 8.
Сумма % тяжелых цепей + % легких цепей
% мономеров
Краткое описание аналитических результатов представлено в таблице 8. Для всех МАТ2, за исключением FS22-053-008-AA/G12v2, выявили высокую чистоту после очистки с применением белка A, низкий уровень растворимой агрегации и фрагментации, что выявили по % мономеров после белка A > 95%, и высокую чистоту, что выявили с помощью восстанавливающего КЭ-ДСН (>95 %, сумма % тяжелых цепей + % легких цепей). Для FS22-053-008-AA/G12v2 выявили агрегацию после очистки на белке A, и, в результате, его далее не исследовали.
Пример 9. Исследование связывания МАТ2.
9.1. Аффинность связывания МАТ2 против CD137/PD-L1 человека с PD-L1 человека и яванского макака, исследованная с помощью ППР.
Связывание МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/lam-G02v3, FS22-172-003-AA/E05v2, FS22-053-008-AA/E05v2, FS22-172-003-AA/E12v2, FS22-053-008-AA/E12v2 и FS22-172-003-AA/G12v2 с рекомбинантными антигенами PD-L1 человека и яванского макака затем измеряли с помощью ППР, применяя обрабатывающее устройство Biacore T200 (GE Healthcare).
Вкратце, МАТ2 против CD137/PD-L1 человека, разбавленные в буфере HBS-EP (GE Healthcare, BR100188) до концентрации 2 мкг/мл, впрыскивали отдельно на проточные кюветы 2, 3 и 4 чипа с белком A (GE Healthcare, 29127556) при скорости 30 мкл/мин, чтобы добиться конечного ответа приблизительно 110 RU. Использовали два различных рекомбинантных антигена PD-L1 человека, т.е. hPD-L1-His-Avi (подробности относительно антигенов см. в примере 1.3) и биотинилированный hPD-L1-Avi-His (AcroBiosystems, PD-1-H82E5). Антигены человека и яванского макака PD-L1-His (AcroBiosystems, PD-1-C52H4), разбавленные в буфере HBS-EP, впрыскивали на проточную кювету 1, 2, 3 или 4, в соответствующих случаях, в диапазоне концентраций от 81 нМ до 0,037 нМ при 3-кратных разведениях в течение 4 мин при скорости 75 мкл/мин, а затем позволяли диссоциировать в буфер в течение 10 мин. Регенерирования достигали путем впрыскивания 10 мМ глицина-HCL, pH 1,5 (GE Healthcare, набор для захвата антитела человека, BR00356), в течение 30 сек при скорости, составляющей 30 мкл/мин. Результаты после вычитания (проточная кювета 2 - проточная кювета 1, проточная кювета 3 - проточная кювета 1 или проточная кювета 4 - проточная кювета 1) анализировали, применяя программное обеспечение BIAevaluation 3.2 (GE Healthcare), чтобы определить связывание, применяя модель связывания 1:1 с переносом массы, с постоянным показателем преломления (RI), равным 0.
Таблица 9
KD (нМ)
KD (нМ)
Результаты анализа связывания продемонстрировали, что каждое МАТ2 против CD137/PD-L1 человека связывалось с PD-L1 человека и яванского макака с от низкой до субнаномолярной аффинностью, и, таким образом, перекрестно реагировало с антигеном человека/яванского макака (таблица 9). Все клоны МАТ2 связывались с PD-L1 человека и яванского макака одинаково хорошо с эквивалентными значениями KD по отношению к каждому антигену.
9.2. Аффинность связывания МАТ2 против CD137/PD-L1 человека с CD137 человека и яванского макака, определенная с помощью ППР.
Также измеряли связывание МАТ2 против CD137/PD-L1 человека с рекомбинантными димерными антигенами CD137 человека и яванского макака с помощью ППР, применяя обрабатывающее устройство Biacore T200 (GE Healthcare).
Вкратце, 20 мкг/мл антитела против Fab человека (GE Healthcare, набор для захвата Fab человека 28958325) покрывали проточные кюветы 1, 2, 3 и 4 сенсорного чипа CM5 серии S Biacore (GE Healthcare, BR100530), достигая конечного ответа приблизительно 5000 RU. Клоны МАТ2, разбавленные в буфере HBS-EP (GE Healthcare, BR100188) до концентрации 2 - 5 мкг/мл, впрыскивали отдельно на проточные кюветы 2, 3 и 4 при скорости 30 мкл/мин, чтобы добиться ответа, составляющего приблизительно 70 RU (200 RU для FS22-172-003-AA/E12v2). Рекомбинантные димерные антигены CD137 человека-mFc-Avi или CD137 яванского макака-mFc-Avi (подробности относительно антигенов см. в примере 1.1), разбавленные в буфере HBS-EP, впрыскивали на проточную кювету 1, 2, 3 или 4, в соответствующих случаях, в диапазоне концентраций от 81 нМ до 0,037 нМ при 3-кратных разведениях в течение 4 минут при скорости 70 мкл/мин, а затем позволяли диссоциировать в буфер в течение 10 минут. Регенерирования достигали путем впрыскивания 10 мМ глицина pH 2,1 (GE Healthcare, набор для захвата Fab человека 28958325) в течение 60 секунд при скорости 30 мкл/мин. Анализ результатов проводили, как описано в примере 9.1.
Таблица 10
KD (нМ)
KD (нМ)
Результаты анализа связывания продемонстрировали, что клоны МАТ2 против CD137/PD-L1 человека связывались с димерными CD137 человека и яванского макака с низкими наномолярными аффинностями и полностью перекрестно реагировали с антигеном человека/яванского макака (таблица 10). Как ожидалось, данные результаты были сходны с результатами связывания, представленными для данных клонов Fcab в формате “ложного” МАТ2 (таблица 3, пример 3.7). Наибольшую аффинность к димерному CD137 наблюдали для FS22-172-003-AA/E12v2, указанную аффинность впоследствие также исследовали против мономерного CD137-His-Avi, применяя тот же способ, который описан выше. Не обнаружили связывания FS22-172-003-AA/E12v2 с мономерным CD137 вплоть до 81 нМ, иллюстрируя что связывающие области Fcab МАТ2 против CD137/PD-L1 человека обладают низкой аффинностью к мономерному CD137. Данные результаты сходны с таковыми, также наблюдаемыми для Fcab FS22-053-008 против CD137, показывая, что преобразование Fcab в формат МАТ2 приводило к небольшому изменению связывания с CD137.
9.3. Определение специфичности МАТ2 против CD137/PD-L1 человека с помощью ППР.
9.3.1. Специфичность к PD-L1 человека.
Для анализа специфичности МАТ2 против CD137/PD-L1 человека, связывание шести МАТ2 (FS22-172-003-AA/lam-G02v3, FS22-172-003-AA/E05v2, FS22-053-008-AA/E05v2, FS22-172-003-AA/E12v2, FS22-053-008-AA/E12v2 и FS22-172-003-AA/G12v2) с другими T-клеточными мишенями исследовали, применяя ППР. Целью было продемонстрировать специфичность, показав отсутствие связывания МАТ2 с антигенами PD-L2, CD80, PD-1 и B7-H3 при концентрации 1 мкМ.
На проточных кюветах на чипах CM5 иммобилизовали приблизительно 1000 RU любого из человеческих PD-L2-Fc (R&D Biosystems, 1224-PL), CD80-Fc (R&D Biosystems, 140-B1), PD-1-His (R&D Biosystems, 8986-PD), B7-H3-His (F-star, получен своими силами, SEQ ID NO: 193), PD-L1-Fc (R&D Biosystems, 156-B7) и PD-L1-His (AcroBiosystems, PD-1-H83F3). Проточную кювету 1 оставляли без иммобилизации. МАТ2 разбавляли до 1 мкМ и 1 нМ в 1x буфере HBS-EP (GE Healthcare, код продукта BR100188), позволяли протекать через чип в течение 3 мин, а затем позволяли диссоциировать в течение 4 мин. Впрыскивание 10 мМ глицина, pH 1,5, на 30 секунд использовали для регенерирования. Антитела положительного контроля впрыскивали при 50 - 100 нМ, чтобы продемонстрировать покрытие каждым антигеном. Уровни связывания определяли по окончании фазы связывания и сравнивали.
Для всех исследованных МАТ2 выявили высокий уровень специфичности, причем детектировали менее 10 RU МАТ2, связавшегося с указанными четырьмя антигенами, при 1 мкМ по сравнению с диапазоном от 105 до 570 RU ответа связывания, детектированным при 1 нМ, для связывания с любым из человеческих PD-L1-Fc или PD-L1-His. Данные результаты продемонстрировали высокий уровень специфичности МАТ2 против CD137/PD-L1 человека по отношению к PD-L1 при отсутствии связывания с другими перечисленными T-клеточными мишенями.
9.3.2. Специфичность к CD137 человека.
Ожидается, что специфичность связывающей CD137 молекулы Fcab сохраняется в МАТ2 CD137/PD-L1 по сравнению с МАТ2 CD137/HelD1.3 (см. пример 3.6). Специфичность FS22-172-003-AA/E12v2 к представителям семейства TNFR OX40, GITR, CD40 и DR6 (R&D Biosystems) анализировали с помощью ППР, аналогично анализу, описанному в примере 9.3.1. При концентрации 1 мкМ FS22-172-003-AA/E12v2 связывалось менее чем с 7 RU с OX40, GITR, CD40 и DR6 человека по сравнению с 278 RU с CD137 человека-mFc. Данный результат демонстрирует высокий уровень специфичности МАТ2 FS22-172-003-AA/E12v2 к CD137.
9.4. Одновременное связывание МАТ2 против CD137/PD-L1 человека с PD-L1 человека и CD137 человека, определенное с помощью ППР.
Способность МАТ2 FS22-172-003-AA/E12v2 одновременно связываться с CD137 человека и PD-L1 человека-His Avi (подробности относительно антигенов см. в примере 1.3) исследовали с помощью ППР на Biacore T200. G1/S70 использовали в качестве контроля. PD-L1 человека-His, разбавленный до 10 мкг/мл в буферном растворе ацетата натрия, pH 5,0 (GE Healthcare, 18-1069), иммобилизовали на сенсорном чипе CM5 серии S (GE Healthcare, BR100530) до поверхностной плотности приблизительно 1100 RU. Одну проточную кювету активировали и деактивировали без иммобилизованного белка для вычитания фонового сигнала. Антитела, разбавленные до 5 мкг/мл в буфере HBS-EP, захватывались при скорости потока 30 мкл/мин, чтобы достигнуть плотности приблизительно 200 - 300 RU. Связывание CD137 человека-mFc-Avi исследовали при 0 и 100 нМ в течение 5 мин при 30 мкл/мин с последующим этапом диссоциации в течение 2,5 мин. Сенсорный чип регенерировали после каждого цикла с помощью 20 мМ NaOH в течение 60 секунд при скорости потока 30 мкл/мин. FS22-172-003-AA/E12v2 было способно связываться одновременно с обоими PD-L1 человека и CD137 человека, тогда как контрольное G1/S70 связывалось только с PD-L1 человека.
9.5. Аффинность связывания МАТ2 против CD137/PD-L1 человека с экспрессированным на поверхности клетки PD-L1 человека или яванского макака, измеренная с помощью проточной цитометрии.
Чтобы оценить связывание МАТ2 против CD137/PD-L1 человека с PD-L1 человека и яванского макака на поверхности клетки, получали клетки HEK293 с повышенной экспрессией PD-L1 человека. Клетки HEK293 с повышенной экспрессией PD-L1 человека (HEK.hPD-L1) получали, как описано в примере 5.3.3. Клетки HEK293 с повышенной экспрессией PD-L1 яванского макака (HEK.cPD-L1) также получали, как описано в примере 5.3.3, за исключением того, что кДНК, кодирующую PD-L1 яванского макака (SEQ ID NO: 189), субклонировали в вектор pcDNA™5/FRT вместо последовательности PD-L1 человека.
Затем исследовали связывание МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 с клетками HEK293, экспрессирующими PD-L1 человека или яванского макака, применяя проточную цитометрию. Неспецифическое связывание также оценивали путем тестирования связывания с исходными клетками HEK293, которые не содержат PD-L1 человека (линия клеток Flp-In T-Rex 293, Life Technologies, R780-07). Аффинности связывания сравнивали с таковыми для антитела положительного контроля G1-AA/E12v2 (SEQ ID NO: 16 и 17 для тяжелой цепи и легкой цепи, соответственно), вариабельные домены которого были клонированы и экспрессированы в формате IgG1 человека, содержащего мутацию LALA в домене CH2 (формат G1-AA).
Получали суспензии HEK293, HEK.hPD-L1 и HEK.cPD-L1 в буфере для FACS (DPBS (Gibco, 14190-094), содержащая 0,5% БСА (Sigma, A7906)) и высевали при концентрации 1 x 105 клеток/лунку в 100 мкл в 96-луночные планшеты со скругленным дном лунок (VWR, 734-1797). Клетки однократно промывали в буфере для FACS. МАТ2 FS22-172-003-AA/E12v2, ложное МАТ2 FS22-172-003-AA/HelD1.3, МАТ G1-AA/E12v2 и МАТ отрицательного контроля G1-AA/4420 разбавляли (400 нМ - 0,005 нМ, 5-кратные разведения) в 100 мкл буфера для FACS. Промытые клетки ресуспендировали в смеси разбавленного антитела, инкубировали при 4°C в течение 30 минут, а затем однократно промывали в буфере для FACS. Затем добавляли 100 мкл/лунку вторичного антитела (меченого Alexa Fluor 647 антитела козы против IgG человека, Invitrogen, A21445), разбавленного 1:1000 в буфере для FACS, инкубировали в течение 20 мин при 4°C и клетки снова промывали буфером для FACS и ресуспендировали в 100 мкл ФБР, содержащего 7AAD (1:1000, Biotium, 40043), перед проведением анализа с применением проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию в канале APC (638 нм 660/20). Средние геометрические значения интенсивности флуоресценции (GMFI) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Обнаружили, что МАТ2 FS22-172-003-AA/E12v2 и антитело G1-AA/E12v2 связываются с PD-L1 человека на поверхности клетки со значениями EC50 в диапазоне 3,1 - 3,7 нМ и с PD-L1 яванского макака на поверхности клетки со значениями EC50 в диапазоне 0,6 - 0,7 нМ (см. таблицу 11). Не наблюдали связывания с клетками HEK293, в которых отсутствовала сверхэкспрессия PD-L1, что показывает специфичность связывания. Следовательно, исследованное МАТ2 FS22-172-003-AA/E12v2 специфично связывалось с PD-L1, при этом не наблюдали неспецифического связывания. Данные результаты показывают, что специфичность к PD-L1 человека и яванского макака, наблюдаемая для МАТ против PD-L1 (см. пример 5.3.3), сохраняется.
Таблица 11
EC50 (нМ)
EC50 (нМ)
9.6. Аффинность связывания МАТ2 против CD137/PD-L1 человека с экспрессированным на поверхности клетки CD137 человека или яванского макака, измеренная с помощью проточной цитометрии.
Клетки DO11.10 (National Jewish Health), экспрессирующие полноразмерный CD137 человека (SEQ ID NO: 185) или яванского макака (SEQ ID NO: 189), обозначенные «DO11.10.hCD137» и «DO11.10.cCD137», соответственно, получали, чтобы представлять антиген в связанной с мембраной конформации, наиболее близкой к его природной форме, для дальнейшего исследования прошедших селекцию МАТ2 против CD137/PD-L1 человека. Подробности получения клеток DO11.10.hCD137 и DO11.10.cCD137 и подтверждения экспрессии описаны в примере 1.2.
Исследовали связывание МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 с клетками, экспрессирующими CD137 человека или яванского макака (DO11.10.hCD137 или DO11.10.cCD137), применяя проточную цитометрию. Неспецифическое связывание также оценивали путем тестирования связывания с клетками DO11.10, в которых отсутствовала экспрессия CD137. Вариабельные домены MOR7480.1 (патент США № 2012/0237498) клонировали и экспрессировали в формате IgG1 человека, содержащего мутацию LALA в домене CH2 (формат G1-AA), и применяли их в качестве положительного контроля.
Вкратце, готовили суспензии DO11.10, DO11.10.hCD137 или DO11.10.cCD137 в буфере для FACS (DPBS (Gibco, 14190-094), содержащий 0,5% БСА (Sigma, A7906)) и высевали при концентрации 1 x 105 клеток/лунку в 100 мкл в 96-луночные планшеты со скругленным дном лунок (VWR, 734-1797). Клетки однократно промывали в буфере для FACS и МАТ2 FS22-172-003-AA/E12v2, ложное МАТ2 FS22-172-003-AA/HelD1.3, антитело G1-AA/MOR7480.1 и МАТ отрицательного контроля G1-AA/4420 разбавляли (400 нМ - 0,005 нМ, 5-кратные разведения) в 100 мкл буфера для FACS. Промытые клетки ресуспендировали в смеси разбавленного антитела, инкубировали при 4°C в течение 30 минут, а затем однократно промывали в буфере для FACS. Затем добавляли 100 мкл/лунку вторичного антитела (меченого Alexa Fluor 647 антитела козы против IgG человека, Invitrogen, A21445), разбавленного 1:1000 в буфере для FACS, смесь клетки/антитело инкубировали в течение 20 мин при 4°C и клетки затем снова промывали буфером для FACS и ресуспендировали в 100 мкл ФБР, содержащего 7AAD (1:1000, Biotium, 40043), перед проведением анализа с применением проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию в канале APC (638 нм 660/20). Средние геометрические значения интенсивности флуоресценции (GMFI) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Результаты представлены в таблице 12. Обнаружили, что МАТ2 FS22-172-003-AA/E12v2 и ложное МАТ2 FS22-172-003-AA/HelD1.3 связываются с экспрессированными на поверхности клетки рецепторами CD137 человека и яванского макака со значениями EC50 в диапазоне 4,8 - 9,1 нМ, и обнаружили, что МАТ положительного контроля связывается с рецепторами CD137 человека и яванского макака со значениями EC50 1,45 нМ и 4,4 нМ, соответственно. Не наблюдали связывания с клетками DO11.10 или HEK293, которые не сверхэкспрессируют CD137, что подтвердило специфичность связывания МАТ2.
Таблица 12
HelD1.3 (EC50 (нМ))
9.7. Аффинность связывания МАТ2 против CD137/PD-L1 человека с эндогенно экспрессированными CD137 и PD-L1 на активированных первичных T-клетках человека.
Аффинность МАТ2 против CD137/PD-L1 человека к эндогенно экспрессированным CD137 и PD-L1 на активированных первичных T-клетках человека определяли с применением проточной цитометрии. Для того чтобы выделить T-клетки, мононуклеарные клетки периферической крови (МКПК) выделяли из конусов для удаления лейкоцитов, побочного продукта донорства тромбоцитов. Вкратце, содержимое конусов для удаления лейкоцитов смывали с помощью ФБР и наносили на градиент фиколла (Sigma-Aldrich, 1440-02). МКПК выделяли путем центрифугирования и собирали клетки, которые не пересекали градиент фиколла. МКПК дополнительно промывали ФБР и остальные красные кровяные клетки лизировали путем добавления 10 мл 1X лизирующего эритроциты буфера (eBioscience, 00-4300-54), следуя инструкциям производителя. Общую популяцию T-клеток выделяли из МКПК, присутствующих в элюенте, применяя набор II для выделения общей популяции T-клеток (Miltenyi Biotec Ltd, 130-095-130), следуя инструкциям производителя. CD137 быстро экспрессировался до высоких уровней на активированных CD8+ T-клетках, а также на активированных CD4+ T-клетках на более низких уровнях (Xue-Zhong Yu и др., 2013), в связи с этим, общую популяцию T-клеток стимулировали в течение 24 часов, применяя гранулы Human T-ActivatorCD3/CD28 Dynabeads™ (Thermo, 11132D), следуя инструкциям производителя. Гранулы Human T-Activator смывали с T-клеток, используя магнит DynaMagTM-15 (Thermo, 12301D), следуя инструкциям производителя, перед использованием анализа связывания с клетками для тестирования связывания МАТ2 FS22-172-003-AA/E12v2, ложного МАТ2 FS22-172-003-AA/HelD1.3, антитела положительного контроля против PD-L1 человека G1-AA/E12v2, антитела положительного контроля против CD137 человека G1-AA/MOR7480.1, антитело отрицательного контроля G1-AA/4420.
Готовили суспензии стимулированной общей популяции T-клеток человека в буфере для FACS ((DPBS (Gibco, 14190-094), содержащий 0,5% БСА (Sigma, A7906)) и высевали при концентрации 1 x 105 клеток/лунку в 100 мкл в 96-луночные планшеты со скругленным дном лунок (VWR, 734-1797). Клетки однократно промывали в буфере для FACS и МАТ2 FS22-172-003-AA/E12v2, ложное МАТ2 FS22-172-003-AA/HelD1.3, антитело положительного контроля G1-AA/E12v2, антитело положительного контроля G1-AA/20H4.9 и антитело отрицательного контроля G1-AA/4420 разбавляли (80 нМ - 0,005 нМ, 5-кратные разведения) в 100 мкл буфера для FACS. Промытые клетки ресуспендировали в разбавленной смеси антител, содержащей антитело против CD4+ человека, меченое FITC (BD Biosciences, 550628), и антитело против CD8+ человека, меченое eFluor450 (Invitrogen, 48-0087-42), чтобы различить CD4+ и CD8+ T-клетки, инкубировали при 4°C в течение 30 минут, а затем однократно промывали в буфере для FACS. Затем добавляли 100 мкл/лунку вторичного антитела (меченого Alexa Fluor 647 антитела козы против IgG человека, Invitrogen, A21445), разбавленного 1:1000 в буфере для FACS, смесь клетки/антитело инкубировали в течение 20 мин при 4°C, и клетки затем снова промывали буфером для FACS и ресуспендировали в 100 мкл ФБР, содержащего 7AAD (1:1000, Biotium, 40043), перед проведением анализа с применением проточного цитометра Canto II (BD Bioscience). Мертвые клетки исключали и измеряли флуоресценцию в канале APC (638 нм 660/20), чтобы показать связывание исследуемых антител. Средние геометрические значения интенсивности флуоресценции (GMFI) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Обнаружили, что МАТ2 FS22-172-003-AA/E12v2 связывается со стимулированными первичными CD4+ T-клетками и CD8+ T-клетками человека, что видно по значениям MFI на фигуре 2. Антитело положительного контроля против PD-L1 G1-AA/E12v2 связывалось со значениями MFI, сходными с таковыми для МАТ2, как с CD4+, так и с CD8+ T-клетками, подтверждая, что PD-L1 экспрессировался на клетках.
Обнаружили, что G1-AA/MOR7480.1, антитело положительного контроля против CD137, связывается с теми же клетками, которые описаны выше, хотя MFI была гораздо ниже, чем наблюдаемая для любого из FS22-172-003-AA/E12v2 или G1-AA/E12v2. Это, вероятно, является следствием того, что на указанных клетках более низкий уровень экспрессии CD137 по сравнению с экспрессией PD-L1. Кроме того, значение MFI было выше для связывания G1-AA/MOR7480.1 с CD8+ T-клетками, чем с CD4+ T-клетками. Ранее было описано, что ожидали наблюдать более низкий уровень экспрессии CD137 на стимулированных CD4+ T-клетках, чем на CD8+ T-клетках (Xue-Zhong Yu и др., 2013), что объясняет низкий уровень связывания антитела положительного контроля против CD137 MOR7480.1 с CD4+ T-клетками в противоположность CD8+ T-клеткам. Такой низкий уровень экспрессии CD137, наблюдаемый на первичных T-клетках, особенно на CD4+ T-клетках, также, вероятно, объясняет наблюдаемое отсутствие связывания или незначительное связывание Fcab в формате ложного МАТ2 (FS22-172-003-AA/HelD1.3).
9.8. Связывание МАТ2 против CD137/PD-L1 человека с FcRn, определенное с помощью ППР.
Связывание неонатального Fc-рецептора (FcRn) крайне необходимо для рециркуляции антител (Martins и др., 2016). Связывание МАТ2 FS22-172-003-AA/lam-G02v3, FS22-172-003-AA/E12v2 и FS22-053-008-AA/E12v2 с FcRn человека и мыши измеряли с помощью ППР, применяя BIAcore T200 (GE Healthcare). G1-AA/HelD1.3 включали в качестве изотипического контроля IgG без мутаций в домене CH3.
Вкратце, МАТ2 отдельно покрывали проточные кюветы 2, 3 и 4 сенсорного чипа CM5 серии S (GE Healthcare, BR100530), чтобы добиться ответа, приблизительно составляющего 1000 RU. Проточную кювету 1 активировали/деактивировали, но оставляли пустой для использования в качестве контроля. FcRn человека (Acro Biosystems, FCM-H5286) или FcRn мыши (R&D Systems, 9114-Fc) подвергали диализу и разбавляли в буфере MHBS-EP+B, pH 6,0 или pH 7,4 (10 мМ MES, FortéBio,18-5026; 1x HBS-EP, GE Healthcare, BR100669; 0,1 мг/мл БСА, Sigma, A7906), и впрыскивали на проточную кювету 1, 2, 3 и 4 при 2000 - 3000 нМ с 2-кратными разведениями в течение 1 мин при 80 мкл/мин, а затем позволяли диссоциировать в буфер в течение 3 мин. Регенерирование не требовалось. Результаты после вычитания (проточная кювета 2 - проточная кювета 1, проточная кювета 3 - проточная кювета 1 или проточная кювета 4 - проточная кювета 1) анализировали, применяя программное обеспечение Biacore T200 Evaluation 2.0, чтобы определить связывание, используя модель аффинности в равновесном состоянии. Результаты связывания представлены в таблице 13 и демонстрируют, что МАТ2 FS22-172-003-AA/lam-G02v3, FS22-172-003-AA/E12v2 и FS22-053-008-AA/E12v2 связывались с FcRn человека или мыши при pH 6,0 с аффинностями, сходными с таковой для контрольного антитела G1-AA/HelD1.3. Не наблюдали связывания с FcRn человека или мыши при pH 7,4. Данные результаты показали, что МАТ2 сохраняли связывание с FcRn.
Таблица 13
9.9. Связывание МАТ2 против CD137/PD-L1 человека с клетками, экспрессирующими Fcγ-рецепторы, определенное с помощью проточной цитометрии.
Для агонистических антител, нацеленных на представителей TNFRSF, требуется перекрестное связывание посредством Fcγ-рецепторов, чтобы запустить кластеризацию и активацию мишени, чтобы добиться активности in vivo (Li и др., 2013). Тем не менее чтобы обойти потенциальные побочные эффекты, такие как системная активация CD137 вследствие перекрестного связывания при связывании с Fcγ-рецептором и уменьшить вероятность вызвать АЗКЦ иммунных клеток, например, T-клеток, которые экспрессируют обе мишени, было решено уменьшить способность МАТ2 против CD137/PD-L1 человека связываться с Fcγ-рецептором путем введения мутации LALA в домен CH2 (подробности относительно формата LALA см. в примере 3.2). Измерение связывания с клетками с помощью проточной цитометрии с клетками CHO, экспрессирующими на повышенном уровне только FcγRIIA (ECACC, № в каталоге 15042905), FcγRIIA (ECACC, № в каталоге 15042903), FcγRIIIA (ECACC, № в каталоге 15042902), FcγRIIIA (ECACC, № в каталоге 15042901) или FcγRIIB (ECACC, № в каталоге 15042907), использовали для подтверждения того, что наличие мутации LALA в домене CH2 МАТ2 против CD137/PD-L1 человека снизило его аффинность связывания с ранее упомянутыми Fcγ-рецепторами.
Вкратце, готовили суспензии CHO.FcγRIIA,131H, CHO.FcγRIIA,131E, CHO.FcγRIIIA,158F, CHO.FcγRIIIA,158V, CHO.FcγRIIB или CHO.WT в буфере для FACS (DPBS (Gibco, 14190-094), содержащий 0,5% БСА (Sigma, A7906)) и высевали при концентрации 1 x 105 клеток/лунку в 100 мкл в 96-луночные планшеты со скругленным дном лунок (VWR, 734-1797). Клетки однократно промывали в буфере для FACS и МАТ2 FS22-172-003-AA/E12v2, антитело положительного контроля для связывания с Fcγ-рецептором G1/4420 и антитело отрицательного контроля для связывания с Fcγ-рецептором G1-AA/4420 разбавляли (500 нМ - 0,03 нМ, 4-кратные разведения) в 100 мкл буфера для FACS. Промытые клетки ресуспендировали в смеси разбавленного антитела, инкубировали при 4°C в течение 30 минут, а затем однократно промывали в буфере для FACS. Затем добавляли 100 мкл/лунку вторичного антитела (меченого Alexa Fluor 488 антитела козы против IgG человека, Stratech Scientific Ltd, 109-545-098-JIR), разбавленного 1:1000 в буфере для FACS, смесь клетки/антитело инкубировали в течение 20 мин при 4°C и клетки затем снова промывали буфером для FACS и ресуспендировали в 100 мкл ФБР, содержащего 7AAD (1:1000, Biotium, 40043), перед анализом с применением проточного цитометра CytoFLEX (Beckman Coulter). Мертвые клетки исключали и измеряли флуоресценцию в канале FITC (488 нм 525/40). Средние геометрические значения интенсивности флуоресценции (GMFI) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для МАТ2 FS22-172-003-AA/E12v2 выявили отсутствие связывания (или связывание ниже уровня такового для антитела отрицательного контроля G1-AA/4420) со всеми исследованными экспрессирующими Fcγ-рецептор клеткамии (фигура 3 (A) FcgRIIA 131H, (B) FcgRIIA 131E, (C) FcgRIIB, (D) FcgRIIIA 158F, (E) FcgRIIIA 158V). Это свидетельствует о том, что мутация LALA уменьшает связывание с FcγRIIA, FcγRIIIA или FγgRIIB, что также уменьшит перекрестное связывание МАТ2 посредством данных рецепторов in vivo.
Пример 10. Исследование функции МАТ2.
10.1. Активность МАТ2 против CD137/PD-L1 человека в анализе первичных CD8+ T-клеток человека.
Анализ первичных T-клеток человека, описанный в примере 3.5, использовали для тестирования активности МАТ2.
Для основанного на клеточной системе перекрестного связывания МАТ2 против CD137/PD-L1 человека, клетки HEK293 с повышенной экспрессией hPD-L1 (HEK.hPD-L1) получали, по существу как описано в примере 3.5. Клетки HEK.hPD-L1 были сконструированы для экспрессии PD-L1 человека на высоком уровне, и их использовали наряду с клетками MDA-MB-231, которые экспрессируют PD-L1 человека на среднем уровне, а также с клетками SKBR3, которые экспрессируют PD-L1 человека на низком уровне (таблица 14), что количественно анализировали с помощью гранул для определения способности связывать антитела Antibody Binding Capacity (Quantum Simply Cellular, Bans Laboratories, Inc., № в каталоге 816A), следуя инструкциям производителя.
Таблица 14
CD8+ T-клетки выделяли и активировали, применяя антитело против CD3, как описано в примере 3.5. Клетки HEK.hPD-L1 высевали при концентрации 2×105 клеток на лунку на покрытые антителом против CD3 (8 мкг/мл) 96-луночные планшеты с плоским дном лунок в 100 мкл среды для культивирования T-клеток (среда RPMI (Life Technologies, 61870-044) с 10% ФБС (Life Technologies), 1X пенициллин/стрептомицин (Life Technologies, 15140122), 1 мМ пируват натрия (Gibco, 11360-070) и 50 мкМ 2-меркаптоэтанол (Gibco, M6250)). После того как клетки HEK.hPD-L1 или клетки HEK, не трансдуцированные для экспрессии hPD-L1, используемые в качестве контролей, прикрепились через 4 часа инкубации, всю среду для культивирования T-клеток удаляли и заменяли на 100 мкл среды для культивирования T-клеток, содержащей T-клетки при концентрации 5,0×105 клеток/мл, в результате чего получали 5,0×104 клеток/лунку.
Для основанного на клеточной системе перекрестного связывания с более низкими более физиологическими уровнями экспрессии PD-L1, линию клеток аденокарциномы молочной железы человека MDA-MB-231 (ATCC HTB-26) использовали с такой же методикой, как описанная для основанного на клеточной системе HEK.hPD-L1 перекрестного связывания.
МАТ2 разбавляли в среде для T-клеток при 2X конечной концентрации, начиная с 50 нМ, и осуществляли титрование 1:5. 100 мкл титрованного МАТ2 добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела.
Антитело положительного контроля против CD137 (G1-AA/20H4.9) разбавляли в среде для T-клеток при 2X конечной концентрации, начиная с 50 нМ, содержащей 50 нМ перекрестно связывающего агента (антитела против CH2 человека (mG1/MK1A6)), и осуществляли титрование 1:5. 100 мкл разбавленной смеси антитела положительного контроля/перекрестно связывающего агента добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела.
Анализируемую смесь инкубировали при 37°C, 5% CO2 в течение 72 часов. Супернатанты собирали и анализировали с помощью набора ELISA Ready-SET-Go! для анализа IL-2 человека (eBioscience, № в каталоге 88-7025-88), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения поглощения на 630 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IL-2 человека (hIL-2) наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
В таблице 15 показаны значения EC50 и максимальный ответ высвобождением IL-2, наблюдаемый в анализе активации T-клеток в присутствии МАТ2 против CD137/PD-L1 человека, исследованного с основанным на клеточной системе перекрестным связыванием. Для МАТ положительного контроля против CD137 человека 20H4.9 выявили повышение высвобождения hIL-2 с EC50, составляющей 0,21 - 0,31 нМ, при перекрестном связывании с антителом против hCH2 в любом анализе. Для всех клонов выявили активность в данных анализах, каждое проявило эффективность с субнаномолярными EC50. МАТ2, содержащее Fcab FS22-172-003, вызвало наибольший ответ T-клеток с EC50 в диапазоне от 0,19 до 0,31 нМ в анализе на основе HEK.hPD-L1 и 0,01 - 0,02 нМ в анализе на основе MDA-MB-231. Подгруппу МАТ2 (содержащих Fcab FS22-053-008, FS22-053-011, FS22-053-014, FS22-173-003 и FS22-172-004) также исследовали без перекрестного связывания с PD-L1, экспрессированным на клетках HEK, и выявили отсутствие активности в данном анализе, как и ожидалось. На фигуре 4 и фигуре 5 показаны типичные графики высвобождения IL-2 в анализе активации T-клеток для МАТ2, перечисленных в таблице 15.
Линию Fcab FS22-053 и FS22-172 также исследовали в данном анализе, используя экспрессирующую наименьше количество PD-L1 линию клеток аденокарциномы молочной железы человека SK-BR-3 (ATCC HTB-30). Как и ожидалось, более низкий уровень экспрессии PD-L1 приводил к более низкой активации CD8+ T-клеток, в соответствии с двумя предшествующими анализами (результаты не представлены).
Таблица 15
Результаты показали, что МАТ2 могло связываться с клетками, экспрессирующими различные уровни PD-L1. Связывание с экспрессированным клетками PD-L1 приводило к перекрестному связыванию МАТ2, так что они оказывались способны связывать, кластеризовать и активировать CD137 на T-клетках, что измеряли по продукции IL-2. Наблюдаемый уровень активации соответствовал уровню перекрестного связывания посредством связывания PD-L1, причем для МАТ2, связанного с клетками, экспрессирующими более высокие уровни hPD-L1 (HEK.hPD-L1), наблюдали большую активацию, тогда как более низкие уровни экспрессии PD-L1 вызывали меньшую активацию.
10.2. Активность МАТ2 против CD137/PD-L1 человека, перекрестно связанного с различными популяциями клеток HEK.hPD-L1, в анализе активации первичных CD8+ T-клеток человека.
Так как наблюдали, что уровень экспрессии PD-L1 различными линиями клеток влияет на активацию CD137, приняли решение дополнительно исследовать влияние экспрессии PD-L1 на функциональную активность исследуемого МАТ2. Провели анализ, сходный с описанным в примере 10.1, в котором отношение клеток HEK.hPD-L1 к HEK.ДТ изменяли, чтобы модулировать уровень экспрессии PD-L1 человека в подходе на основе популяций.
Клетки HEK.hPD-L1 и клетки HEK, которые не были трансдуцированы для экспрессии hPD-L1 (HEK.ДТ), высевали в различных соотношениях, чтобы получить процент клеток HEK.hPD-L1 от всех присутствующих клеток HEK, перечисленный на фигуре 6 (100%, 50%, 25%, 12,5%, 6,5% или 0%). Клетки HEK высевали при 2×105 всех клеток HEK на лунку на покрытые антителом против CD3 (8 мкг/мл) 96-луночные планшеты с плоским дном лунок в 100 мкл среды для культивирования T-клеток (среда RPMI (Life Technologies, 61870-044) с 10% ФБС (Life Technologies), 1X пенициллин/стрептомицин (Life Technologies, 15140122), 1 мМ пируват натрия (Gibco, 11360-070) и 50 мкМ 2-меркаптоэтанол (Gibco, M6250)). После того как все клетки HEK прикрепились через 4 часа инкубации, всю среду для культивирования T-клеток удаляли и заменяли на 100 мкл среды для культивирования T-клеток, содержащей T-клетки при концентрации 5,0×105 клеток/мл, с получением 5,0×104 клеток/лунку.
Анализ проводили с МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2, но в остальном следовали протоколу, описанному в примере 10.1. Антитело положительного контроля против CD137 G1-AA/20H4.9 включали в анализ с дополнительным перекрестным связыванием посредством антитела против CH2. Антитело G1-AA/4420 использовали в качестве отрицательного контроля.
Результаты на фигуре 6 покывают, что по мере увеличения популяции клеток HEK.hPD-L1, активность МАТ2 также повышалась, что демонстрирует повышение наблюдаемой максимальной концентрации hIL-2. Если 100% присутствующей популяции клеток HEK были HEK.hPD-L1, достигалось максимальное перекрестное связывание и наблюдалась наибольшая максимальная концентрация hIL-2. Для антитела положительного контроля G1-AA/20H4.9 также достигалась сходная максимальная концентрация hIL-2, когда оно было максимально искусственно перекрестно связано антителом против CH2.
Полученные результаты подтверждают наблюдение, что количество PD-L1, присутствующего в системе (показанное тестированием линии клеток, экспрессирующей различные уровни PD-L1 на своей поверхности, или модулированием количества клеток в популяции, на которых выставлен PD-L1), контролирует перекрестное связывание МАТ2 и, таким образом, определяет уровень агонизма CD137, вызванного МАТ2. Тем не менее даже когда присутствовали только низкие уровни PD-L1, МАТ2 было все еще способно агонизировать CD137. Следовательно, ожидается, что МАТ2 обладает активностью, когда в системе экспрессируется PD-L1 и присутствуют активированные T-клетки, которые экспрессируют CD137. Ожидается, что по мере возрастания уровня PD-L1 в системе, оказываемый агонизм также повышается. В противоположность МАТ2, которое было все еще способно вызывать агонизм CD137 в присутствии низких уровней CD137, ожидают, что у молекул CD137/PD-L1, которые связывают CD137 моновалентно, будет низкая эффективность в присутствии низких уровней PD-L1, так как молекулы, связанные с PD-L1, вероятно, будут находиться на слишком большом расстоянии, чтобы запустить кластеризацию CD137, этой проблемы не ожидают, когда молекула может связывать CD137 бивалентно, как в случае МАТ2 согласно настоящему изобретению.
10.3. Активность МАТ2 против CD137/PD-L1 человека в анализе реакции смешанной культуры первичных лимфоцитов человека.
Активность МАТ2 против CD137/PD-L1 человека исследовали в анализе реакции смешанной культуры лимфоцитов (СКЛ) (см. пример 5.3.4). В анализе СКЛ использовали CD4+ T-клетки из одного донора и дендритные клетки моноцитарного происхождения (иДК) из другого донора. Так как иммунные клетки содержат физиологические уровни регуляторов иммунных контрольных точек, можно использовать анализ СКЛ, чтобы подтвердить, что активация T-клеток усиливается посредством МАТ2 в системе человека.
Размноженные CD4+ T-клетки и иДК получали, как описано в примере 5.3.4.
Размноженные T-клетки размораживали за одни сутки до проведения эксперимента, промывали средой AIM V(Gibco, 12055-091) и инкубировали при 37 °C, 5% CO2 в среде AIM V в течение ночи. Антитела против CD137 и против PD-L1 и МАТ2 против CD137/PD-L1 человека, перечисленные в таблице 16, разбавляли до 4x конечной концентрации в трех повторах в 50 мкл среды AIM V в 96-луночных планшетах со скругленным дном лунок (VWR, 734-1797). Антитело против гликопротеина gH цитомегаловируса (CMV), обозначенное MSL109 (описано в WO1994/016730 A1), содержащее мутацию LALA, включали в качестве отрицательного контроля. Исследовали серию 3-кратных разведений, начиная от 30 нМ до 0,002 нМ. Как 1×104 клеток иДК, суспендированных в 50 мкл среды AIM V, так и 1×105 размноженных CD4+ T-клеток, суспендированных в 100 мкл среды AIM V, добавляли в разведения антител и инкубировали в течение 5 суток при 37°C + 5% CO2. В анализ включили следующие отрицательные контроли: CD4+ T-клетки отдельно, иДК отдельно, CD4+ T-клетки + иДК и среда AIM V отдельно. Супернатанты собирали, образцы разбавляли (1:25) и концентрации интерферона гамма (IFN-γ) измеряли, применяя набор для анализа ELISA IFN-гамма человека Ready-SET-Go! (Life Technologies, 88-7316-77). Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gen5, BioTek. Значения поглощения на 630 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gen5, BioTek). Концентрацию IFN-γ человека наносили на график в зависимости от log концентрации антитела, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для антитела против PD-L1 человека G1/S70 выявили сильную активность в анализе СКЛ со значением EC50, составляющим 0,06 нМ, и максимальным уровнем IFN-γ (Emax), составляющим 1135 пг/мл (таблица 16, типичные фигуры 7A и 7B). EC50 указывает на концентрацию МАТ, при которой достигается половина агонистического ответа, тогда как Emax представляет собой абсолютное значение, которое указывает на максимальную концентрацию IFN-γ, достигнутую в анализе. Как и ожидалось, не наблюдали активности для антитела отрицательного контроля G1-AA/MSL109. Антитело положительного контроля против CD137 человека G1-AA/20H4.9 при перекрестном связывании с антителом против hCH2 не вызывало никакой активности, свидетельствуя о том, что экспрессия CD137 может быть низкой, и что блокирование PD-L1 превосходит передачу сигналов CD137 в данном анализе, так как блокирование PD-L1 очень сильное. Неожиданно, для всех МАТ2 против CD137/PD-L1 человека, исследованных в данном анализе, выявили очень эффективные значения EC50, составляющие менее 0,04 нМ, и высокие значения Emax в диапазоне от 2626 до 3327 пг/мл. Это свидетельствует о том, что МАТ2 проявляет более высокую эффективность в данном анализе, чем любое из моноклональных антител против PD-L1 и против CD137.
Таблица 16.
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
Следуя тому же протоколу, который описан выше, снова исследовали активность МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2 в СКЛ, но на этот раз против каждого компонента (Fcab против CD137 человека FS22-172-003 в формате ложного МАТ2 и антитела против PD-L1 G1-AA/E12v2), а также комбинации указанных двух компонентов.
Для антитела компонента против PD-L1 человека G1-AA/E12v2 выявили сильную активность в анализе СКЛ со значением EC50, составляющим 0,08 нМ, и максимальный уровень IFN-γ (Emax), равный 12421 пг/мл (таблица 17; типичная фигура 7C). Не наблюдали активности для антитела отрицательного контроля G1-AA/4420, как и ожидалось, и не наблюдали активности для Fcab против CD137 человека FS22-173-003 в формате ложного МАТ2. Антитело положительного контроля против CD137 человека G1-AA/20H4.9 при перекрестном связывании с антителом против hCH2 не вызывало никакой активности, свидетельствуя о том, что экспрессия CD137 может быть низкой, и что блокирование PD-L1 подавляет передачу сигналов CD137 в данном анализе, так как блокирование PD-L1 очень сильное, что наблюдали выше. Как и ожидалось, для МАТ2 против человеческих CD137/PD-L1 FS22-172-003-AA/E12v2 выявили очень эффективное значение EC50, составляющее 0,07 нМ, и высокое значение Emax, равное 17874 пг/мл. Это свидетельствует о том, что МАТ2 проявляло более высокую эффективность в данном анализе, чем любое из моноклональных антител против PD-L1 и против CD137 отдельно или оба в комбинации. Полученные результаты также показали, что МАТ2 проявляло более высокую эффективность, чем комбинация двух антител, содержащих Fcab и сайты связывания Fab из МАТ2, соответственно.
Таблица 17.
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
10.4. Активность МАТ2 против CD137/PD-L1 человека в анализе активации T-клеток повторной дозой антигена.
Активность МАТ2 против CD137/PD-L1 человека исследовали в анализе активации T-клеток повторной дозой антигена путем измерения иммунного ответа на два различных пула пептидов, содержащих физиологически значимые антигены. Один пул содержал рестриктированные по MHC I класса пептиды из цитомегаловируса, вируса Эпштейна-Барр и вируса гриппа (CEF), который позволял тестировать активацию CD8+ T-клеток, и второй пул содержал рестриктированные по MHC II класса пептиды из цитомегаловируса, вируса Эпштейна-Барр, вируса гриппа и столбнячный токсин (CEFT), который позволял тестировать активацию CD4+ T-клеток. Так как использовали МКПК человека из одного донора и так как иммунные клетки содержали физиологические уровни регуляторов иммунных контрольных точек и стимулировались представлением антигена, а не перекрестным связыванием T-клеточного рецептора антителом против CD3, результаты данного анализа подтверждают, что указанное МАТ2 усиливает запускаемую антигеном активацию T-клеток в системе человека.
Криоконсервированные мононуклеарные клетки периферической крови (МКПК) размораживали и подсчитывали, и 1,9×105 клеток высевали в каждую лунку 96-луночного планшета с плоским дном лунок в 200 мкл среды Roswell Park Memorial Institute (RPMI), содержащей 10% фетальную бычью сыворотку (ФБС) + 1 нг/мл IL-2 + 5 нг/мл IL-7 и либо 1 мкг/мл пула пептидов CEF (Thinkpeptides, № в каталоге PX-CEF-G), либо 1 мкг/мл пула пептидов CEFT (Thinkpeptides, № в каталоге PX-CEFT-G). Клетки инкубировали при 37°C с 5% CO2 в течение 3 суток. Через 3 суток среды восполняли путем добавления 22,5 мкл свежей RPMI, содержащей 10% ФБС, и поддержания 1 нг/мл IL-2 и 1 нг/мл IL-7. Клетки инкубировали при 37°C с 5% CO2 в течение дополнительных 4 суток.
Клетки собирали и объединяли в общий пул, а затем распределяли по 96-луночным планшетам со скругленным дном лунок при концентрации 1×105 клеток на лунку в 150 мкл RPMI, содержащей 10% ФБС + 1 нг/мл IL-2 + 5 нг/мл IL-7 и либо 1 мкг/мл пула пептидов CEF, либо 1 мкг/мл CEFT пула пептидов. МАТ2 разбавляли в той же среде, в которую клетки высевали и которую добавляли к клеткам.
Каждое из антитела положительного контроля против CD137 (G1-AA/20H4.9) и антитела положительного контроля против PD-L1 (G1-AA/E12v2) разбавляли в той же среде, в которую клетки высевали, при 2X конечной концентрации, начиная с 400 нМ, содержащей 400 нМ перекрестно связывающего агента (антитела против CH2 человека (MK1A6)), и осуществляли титрование 1:5. 50 мкл разбавленной смеси любого из антител положительного контроля/перекрестно связывающего агента добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела.
Анализируемую смесь инкубировали при 37°C, 5% CO2 в течение 72 часов. Супернатанты собирали и анализировали с помощью набора MSD U-PLEX (Meso Scale Discovery, № в каталоге K15067L-2), следуя инструкциям производителя. Планшеты прочитывали на электрохемилюминесцентном спектрофотометре MESO QuickPlex SQ 120 для прочтения планшетов, используя разведение образца 1:5. Стандартную кривую для расчета концентрации цитокина рассчитывали, применяя программное обеспечение Workbench 4.0 MSD Discovery, и концентрацию цитокина в супернатанте аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
В таблице 18 и таблице 19 показано, что для антитела против PD-L1 человека G1-AA/E12v2 выявили активность в отношении активации как CD4+, так и CD8+ T-клеток со значением EC50, составляющим 0,06 нМ и 0,04 нМ, соответственно (Emax 65840 пг/мл и 86613 пг/мл, соответственно). EC50 указывает на концентрацию МАТ, при которой достигается половина агонистического ответа, тогда как Emax представляет собой абсолютное значение, которое указывает на максимальную концентрацию IFN-γ, достигнутую в анализе. Для антитела положительного контроля против CD137 человека G1-AA/20H4.9 при перекрестном связывании с антителом против hCH2 выявили активность в отношении активации CD8+ T-клеток со значением EC50, составляющим 0,33 нМ, и значением Emax, равным 116204 пг/мл, но не выявили какой-либо активности в отношении активации CD4+ T-клеток, свидетельствуя о том, что экспрессия CD137 может быть выше на CD8+ T-клетках по сравнению с CD4+ T-клетками. Не наблюдали активности для антитела отрицательного контроля G1-AA/4420, как и ожидалось. Неожиданно, для МАТ2 выявили активацию в обоих анализах активации CD4+ T-клеток и CD8+ T-клеток со значениями EC50, составляющими 0,08 нМ и 0,03 нМ, соответственно. МАТ2 могло связываться с экспрессированным на МКПК PD-L1 и обладало эквивалентной активностью к PD-L1 на обоих типах клеток. Кроме того, связывание с экспрессированным на МКПК PD-L1 приводило к перекрестному связыванию МАТ2, так что они оказывались способны связывать, кластеризовать и активировать CD137 на обоих субпопуляциях T-клеток, что измеряли по высвобождению IFN-γ. Максимальная концентрация IFN-γ, высвобожденного T-клетками, обработанными МАТ2, была выше, чем таковая для любого из антител положительного контроля, со значениями EC50, составляющими 86680 пг/мл и 188242 пг/мл для активации CD4+ T-клеток и активации CD8+ T-клеток, соответственно. Это указывает на то, что МАТ2 обладает более высокой активностью в данном анализе, чем любое из моноклональных антител против PD-L1 и против CD137.
Таблица 18. Значения в анализе повторной дозы антигена для пула пептидов CEFT (активация CD4+ T-клеток).
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
Таблица 19. Значения в анализе повторной дозы антигена для пула пептидов CEF (активация CD8+ T-клеток).
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
10.5. Активность МАТ2 против CD137/PD-L1 человека в отношении блокирования PD-L1 в анализе активации T-клеток DO11.10 мыши.
Активность FS22-172-003-AA/E12v2 исследовали в анализе активации T-клеток DO11.10 мыши, чтобы исследовать активность блокирования контрольной точки иммунного ответа PD-L1. Функциональную активность в отношении PD-L1 человека исследовали в анализе высвобождения IL-2, используя DO11.10 OVA T-клетки и LK35.2 B-клетки гибридомы, как описано в примере 5.4.
Результаты представлены в таблице 20. Для МАТ2 против CD137/PD-L1 человека выявили сильную активность в анализе активации T-клеток DO11.10 мыши со значением EC50, составляющим 1,27 нМ, которая подтверждает, что FS22-172-003-AA/E12v2 может связывать PD-L1 и блокировать взаимодействие PD-L1 с PD-1. Его активность была сходна с таковой у антител положительного контроля против PD-L1 человека G1-AA/E12v2 и G1-AA/S70, свидетельствуя о том, что не происходила утрата активности антитела по отношению к PD-L1 человека, когда G1-AA/E12v2 внедряли в молекулу МАТ2 FS22-172-003-AA/E12v2.
Таблица 20. Значения в анализе активации T-клеток DO11.10 (высвобождение IL-2 мыши).
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
Пример 11. Функциональная перекрестная реактивность МАТ2 против CD137/PD-L1 человека в функциональных анализах для яванского макака.
11.1. Активность МАТ2 против CD137/PD-L1 человека в анализе активации T-клеток DO11.10 яванского макака.
Кластеризация CD137 посредством молекул агонистов на активированных T-клетках вызывает активацию T-клеток и последующую передачу сигналов, приводящую к продукции IL-2, но не ограниченную ею. Способность МАТ2 против CD137/PD-L1 человека активировать клетки DO11.10, экспрессирующие CD137 яванского макака, исследовали в анализе активации T-клеток. Был разработан анализ активации T-клеток DO11.10 с применением T-клеток DO11.10, сконструированных для сверхэкспрессии CD137 яванского макака, и активацию T-клеток оценивали путем измерения высвобождения IL-2.
Получали клетки DO11.10 (National Jewish Health), экспрессирующие полноразмерный CD137 яванского макака (SEQ ID NO: 189), обозначенные «DO11.10.cCD137», и экспрессию CD137 яванского макака подтверждали, как описано в примере 1.2.
Следующие МАТ2 против CD137/PD-L1 человека исследовали в данном анализе активации T-клеток DO11.10: FS22-172-003-AA/lamG02v3, FS22-053-008-AA/E05v2, FS22-053-008-AA/E12v2, FS22-053-008-AA/G12v2, FS22-053-017-AA/E05v2, FS22-053-017-AA/E12v2, FS22-053-017-AA/G12v2, FS22-172-003-AA/E05v2, FS22-172-003-AA/E12v2, FS22-172-003-AA//G12v2. Готовили разведения МАТ2 или МАТ положительного контроля G1-AA/MOR7480.1 либо с перекрестно связывающим антителом против hCH2 (mG1/MK1A6), либо без него, и добавляли к клеткам DO11.10.cCD137 в 96-луночный планшет со скругленным дном лунок, которые покрывали в течение ночи 0,1 мкг/мл антитела против CD3 (клон 17A2, BioLegend, 100208) и засевали 2x105 клеток HEK.cyPD-L1 для применения в качестве основанных на клеточной системе перекрестно связывающих клеток, которые сверхэкспрессируют PD-L1 яванского макака. Клетки, сверхэкспрессирующие PD-L1 яванского макака, получали, как описано в примере 5.3.3, применяя PD-L1 яванского макака, описанный в примере 9.5. После инкубации в течение 18 часов супернатанты собирали и анализировали с помощью набора для ELISA IL-2 мыши (eBioscience, 88-7024-86), следуя инструкциям производителя. Планшеты прочитывали на 450 нм, применяя спектрофотометр для прочтения планшетов, с помощью программного обеспечения Gens, BioTek. Значения поглощения на 570 нм вычитали из таковых на 450 нм (корректировка). Стандартную кривую для расчета концентрации цитокина получали на основе аппроксимирования четырехпараметрической логистической кривой (программное обеспечение Gens, BioTek). Концентрацию mlL-2 наносили на график в зависимости от log концентрации МАТ2 или эталонного МАТ, и полученные в результате этого кривые аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
Для всех МАТ2 против CD137/PD-L1 человека, исследованных в данном анализе, выявили активность в присутствии клеток HEK с повышенной экспрессией PD-L1 яванского макака для перекрестного связывания с МАТ2, свидетельствуя о том, что все МАТ2 способны как связывать PD-L1 яванского макака, так и связывать и кластеризовать CD137 яванского макака, вызывая активацию T-клеток DO11.10, которую считывают по продукции IL-2 (см. таблицу 21 и фигуру 8). Для всех МАТ2 против CD137/PD-L1 человека выявили сходную активность со значениями EC50 в диапазоне от 0,06 до 0,11 нМ и значениями Emax в диапазоне от 8060 до 12300 пг/мл IL-2 только при перекрестном связывании экспрессированным клетками PD-L1 яванского макака. В отсутствие PD-L1 яванского макака, когда анализ проводили с применением клеток HEK, которые не экспрессируют PD-L1 яванского макака, не наблюдалась активность в результате отсутствия перекрестного связывания. Для антитела положительного контроля против CD137 G1-AA/MOR7480.1, которое, как было показано, перекрестно реагирует с CD137 яванского макака (Fisher и др., 2012), показали активность в данном анализе при перекрестном связывании с антителом против CH2, тем не менее, все МАТ2 против CD137/PD-L1 человека были лучше в отношении как более низких значений EC50, так и более высоких значений Emax, свидетельствуя о перекрестной реактивности и лучшей активности в системе, экспрессирующей CD137 яванского макака.
Таблица 21.
(перекрестно связанный)
(перекрестно связанный)
11.2. Активность МАТ2 против CD137/PD-L1 человека в анализе реакции смешанной культуры первичных лейкоцитов яванского макака.
Активность МАТ2 против CD137/PD-L1 человека исследовали в анализе реакции смешанной культуры первичных лейкоцитов (СКЛ) яванского макака. Аналогично анализу СКЛ человека, описанному в примере 10.3, в данном анализе измеряют клеточный иммунный ответ, который происходит между двумя аллогенными популяциями лимфоцитов (из одного вида, но генетически различными). В данном анализе использовали CD4+ T-клетки из одного индивида яванского макака и моноциты CD14+ из другого индивида. Так как иммунные клетки содержат физиологические уровни регуляторов иммунных контрольных точек, можно использовать анализ СКЛ, чтобы подтвердить, что МАТ2 повышает активацию T-клеток в системе яванского макака.
Выделение CD4+ T-клеток.
МКПК выделяли из образцов крови яванского макака путем разделения в градиенте фиколла. CD4+ T-клетки выделяли, применяя набор для выделения CD4+ отличного от человека примата (Miltenyi Biotec Ltd, 130-091-102), следуя инструкциям производителя. Клетки использовали свежими в анализе СКЛ.
Выделение CD14+ моноцитов.
Свежие моноциты выделяли из МКПК яванского макака, применяя набор для выделения CD14 отличного от человека примата (Miltenyi Biotec Ltd, 130-091-097), следуя инструкциям производителя. Моноциты использовали свежими в анализе СКЛ.
Анализ СКЛ.
МАТ2 против CD137/PD-L1 человека FS22-172-003-AA/E12v2, антитело против PD-L1 человека G1-AA/E12v2 и изотипическое контрольное антитело G1-AA/4420 разбавляли до 4x конечной концентрации в трех повторах в 50 мкл среды AIM V в 96-луночных планшетах со скругленным дном лунок (VWR, 734-1797). Антитело против лизоцима куриного яйца (HEL), обозначенное HelD1.3 (описанное в примере 2.2), содержащее мутацию LALA, включали в качестве отрицательного контроля. Исследовали серию 4-кратных разведений, начиная от 300 нМ до 0,02 нМ. Как 7,5×104 клеток моноцитов, суспендированных в 50 мкл среды AIM V, так и 7,5×105 CD4+ T-клеток, суспендированных в 100 мкл среды AIM V, добавляли к разведениям антител и инкубировали в течение 6 суток при 37°C + 5% CO2. В анализ включали следующие отрицательные контроли: CD4+ T-клетки отдельно, CD14+ моноциты отдельно, CD4+ T-клетки + иДК и только среда AIM V. Супернатанты собирали на 2 сутки и измеряли концентрации интерлейкина 2 (IL-2), используя панель магнитных гранул с цитокинами отличного от человека примата MILLIPLEX MAP (Merck Millipore, № в каталоге PRCYTOMAG-40K-01), супернатанты также собирали на 6 сутки и измеряли концентрации интерферона гамма (IFN-γ), используя тот же набор. Образцы анализировали на устройствах Bio Plex 200 от Bio-Rad.
Для обоих МАТ2 против CD137/PD-L1 человека выявили активность в данном анализе, что свидетельствует о том, что указанные МАТ2 способны связывать PD-L1 яванского макака и впоследствии связывать и кластеризовать CD137 яванского макака при уровнях экспрессии, эндогенных для свежевыделенных иммунных клеток яванского макака, приводя к высвобождению цитокинов в результате активации T-клеток (см. фигуру 9).
11.3. Активность МАТ2 против CD137/PD-L1 человека в анализе первичных МКПК яванского макака.
Активность МАТ2 против CD137/PD-L1 человека исследовали в анализе первичных МКПК яванского макака. В данном анализе измеряют клеточный иммунный ответ, который происходит в смешанной популяции МКПК, которую стимулировали МАТ против CD3. В данном анализе использовалвсю популяцию МКПК из одного индивида яванского макака, и, так как иммунные клетки содержат физиологические уровни регуляторов иммунных контрольных точек, анализ МКПК можно использовать для подтверждения того, что МАТ2 повышает активацию T-клеток в системе яванского макака.
Данный эксперимент провели по существу как описано в примере 3.5, со следующими отклонениями: антитело положительного контроля против CD137 (G1-AA/MOR7480.1) разбавляли в той же среде, в которую высевали клетки, при 2X конечной концентрации, начиная с 40 нМ, содержащей 40 нМ перекрестно связывающего агента (антитела против CH2 человека (MK1A6)) и осуществляли титрование 1:4. МАТ2 (FS22-172-003-AA/E12v2) и антитело отрицательного контроля (G1-AA/HelD1.3) разбавляли таким же способом без перекрестно связывающего агента. 100 мкл разбавленной смеси антитело/перекрестно связывающий агент добавляли к клеткам с получением общего объема для анализа, составляющего 200 мкл, и 1X концентрации антитела. Анализируемую смесь инкубировали при 37°C с 5% CO2 в течение 3 суток.
Супернатанты собирали и анализировали с помощью набора MSD V-Plex Proinflammatory Panel 1 (NHP) (Meso Scale Discovery, № в каталоге K15056D-2), следуя инструкциям производителя. Планшеты прочитывали на электрохемилюминесцентном спектрофотометре MESO QuickPlex SQ 120 для прочтения планшетов. Стандартную кривую для расчета концентрации цитокина рассчитывали, применяя программное обеспечение Workbench 4.0 MSD Discovery, и концентрацию цитокина в супернатанте аппроксимировали, используя уравнение log (агониста) против ответа, в GraphPad Prism.
На фигуре 28A показан типичный график высвобождения IFN-γ в результате активации T-клеток в анализе МКПК яванского макака и на фигуре 28B показан типичный график высвобождения IFN-γ в результате активации T-клеток в анализе МКПК человека.
Для антитела против CD137 человека G1-AA/MOR7480.1 при перекрестном связывании с антителом против hCH2 выявили активность в обоих анализах МКПК. Не наблюдали активности для антитела отрицательного контроля G1-AA/HelD1.3, как и ожидалось. Неожиданно, для МАТ2 выявили сходную активацию в обоих анализах МКПК со значениями EC50, равными 0,08 нМ и 0,03 нМ, соответственно. EC50 указывает на концентрацию МАТ, при которой достигается половина агонистического ответа. Следовательно, МАТ2 могли связываться с экспрессированным на МКПК PD-L1, что приводило к перекрестному связыванию МАТ2, так что они оказывались способны связывать, кластеризовать и активировать CD137 на T-клетках из обоих видов, что измеряли по высвобождению IFN-γ. Максимальная концентрация IFN-γ, высвобожденного T-клетками, обработанными МАТ2, была выше, чем таковая для любого из антител положительного контроля, свидетельствуя о том, что МАТ2 обладало более высокой активностью в данном анализе, чем у моноклональных антител против CD137.
Пример 12. Получение Fcab против CD137 мыши.
Для проверки активности МАТ2, содержащих область связывания антигена CD137, в моделях на мышах in vivo, вследствие риска невозможности получения перекрестно реагирующих с антигеном мыши и человека Fcab вследствие низкой гомологии последовательностей CD137 мыши и человека, получали и характеризовали Fcab, которые специфично связывались с CD137 мыши. После получения связывающих CD137 мыши Fcab, неожиданно, затем обнаружили, что FS22-053-14 способно перекрестно реагировать с антигеном мыши и человека.
12.1. Наивная селекция Fcab против CD137 мыши.
Для того чтобы подвергнуть селекции Fcab, которые связываются с CD137 человека, использовали подходы селекции из дрожжевого и фагового дисплея, сходные с таковыми, ранее описанными для селекции Fcab, связывающихся с CD137 человека (см. пример 2.1). Рекомбинантный димерный CD137 мыши или клетки, экспрессирующие полноразмерный CD137 мыши, использовали в качестве антигенов (см. пример 1).
Полученный своими силами антиген mCD137-mFc-Avi и клетки DO11.10, экспрессирующие mCD137 (DO11.10.mCD137), использовали в способах селекции, применяя шесть фаговых библиотек. Все полученные после 3 раунда селекции на рекомбинантных антигенах (576 клонов) и полученные после 3 раунда селекции на клетках (576 клонов) клоны подвергали скринингу с помощью ELISA фагов и скринингу связывания с клетками DO11.10.mCD137. 34 прошедших селекцию клона Fcab субклонировали и получали в виде МАТ² HelD1.3, как описано в примере 2.2.
Четыре наивные дрожжевые библиотеки, представляющие домены CH1 - CH3 IgG1 человека, ранее использованные для селекции Fcab, связывающихся с CD137 человека, использовали для проведения селекции Fcab, связывающихся с CD137 мыши. Всего проводили 53 отдельных раунда селекции, чтобы определить молекулы, связывающие CD137 мыши. Полученный своими силами, рекомбинантный, димерный, биотинилированный антиген CD137 мыши (mCD137-mFc-Avi) использовали для селекции связывающих молекул из наивных дрожжевых библиотек.
12.2. Исследование Fcab против CD137 мыши из наивных селекций.
Специфичность Fcab против CD137 мыши к CD137 мыши исследовали в формате “ложного” МАТ2 HelD1.3 и измеряли с помощью BLI на системе Octet QKe путем тестирования связывания Fcab с другими рецепторами TNFRSF мыши (CD40, OX40, GITR). Стрептавидиновые биосенсоры (PALL ForteBio 18-5021) покрывали 10 нг/мкл рецепторов CD40, GITR, OX40 мыши (все получены от R&D Systems и биотинилированы с применением набора EZ-Link Sulfo-NHS-SS-Biotin от Thermoscientific, № 21328). Fcab против CD137 мыши в формате ложного МАТ2 разбавляли 1:1 в буфере для кинетического анализа (PALL 18-1092) до конечной концентрации по меньшей мере 1 мкМ. Покрытые антигеном сенсоры погружали в растворы МАТ² на 180 секунд, а затем на 180 секунд в 1× буфер для кинетического анализа. Антитела к каждому из рецепторов TNFRSF использовали в качестве положительных контролей. Клоны Fcab FS22m-055, FS22m-063, FS22m-066, FS22m-075, FS22m-135, FS22m-055, FS22m-063, FS22m-066 не связывались ни с одним из исследованных рецепторов TNFRSF, таким образом, демонстрируя их специфичность к CD137 мыши.
Клетки HEK.FRT.luc, экспрессирующие последовательность CD137 мыши (SEQ ID NO: 187), получали, следуя той же методике, которая описана ранее в примере 2.3. МАТ², содержащие ранее прошедшие селекцию Fcab против CD137 мыши, подвергали скринингу, используя данную линию клеток, HEK.FRT.luc.mCD137, согласно способу, описанному в примере 2.3. Исследовали 56 МАТ2, из которых 29 оказались положительны по активности NF-κB. Lob12.3, содержащее Fc IgG1 человека с мутацией LALA (G1-AA/Lob12.3), использовали в качестве МАТ положительного контроля против CD137 мыши, и выявили повышение люминесценции, что подтверждает достоверность анализа. HelD1.3, также содержащее Fc IgG1 человека с мутацией LALA, использовали в качестве отрицательного изотипического контроля IgG человека, чтобы исключить влияние ложного Fab IgG человека в данном анализе. EC50 рассчитывали, когда это было возможно, и МАТ2, которые не достигали плато активности, пренебрегали в пользу МАТ2, для которых выявили классическую сигмоидальную кинетику активности. МАТ2 ранжировали в порядке EC50 и кратности изменения активности при перекрестном связывании с белком L. FS22m-063 выбрали на основании того, что у него была лучшая EC50 при перекрестном связывании (1,44 нМ) и наибольшая кратность изменения активности при перекрестном связывании (в 27 раз).
12.3. Активность FS22m-063, FS22-053-014 и FS22-053-017 Fcab в формате ложного МАТ2 в анализе активации T-клеток DO11.10 с CD137 мыши.
Клон FS22-053-017 (в формате ложного МАТ² HelD1.3) также сравнивали со связывающим CD137 мыши клоном Fcab FS22m-063 (также в формате ложного МАТ² HelD1.3), а также с исходным клоном FS22-053-014 (в формате ложного МАТ² HelD1.3), в анализе активации T-клеток DO11.10 с CD137 мыши, описанном в примере 2.4. Молекулы МАТ² перекрестно связывались с белком L при молярном отношении 4:1 (МАТ2:белок L). Результаты представлены в таблице 22.
Как ожидалось, для всех исследованных указанных молекул выявили активность, что измеряли по высвобождению IL-2 при перекрестном связывании белком L, но не выявили активность в отсуствие перекрестного связывания. FS22m-063, которое прошло селекцию по связыванию с CD137 мыши, обладало наилучшей активностью в данном анализе с EC50, составляющей 0,39 нМ, при перекрестном связывании. Оба FS22-053-14 и FS22-053-017 обладали активностью в анализе, свидетельствуя о том, что данная функция не утрачивалась в результате мутагенеза, хотя для FS22-053-017 обнаружили небольшую утрату активности с EC50, которая была приблизительно в 8 раз хуже, чем FS22-053-14, при перекрестном связывании белком L. На фигуре 10 показано, что перекрестно реагирующие с CD137 человека и мыши Fcab с созревшей аффинностью FS22-053-014 и FS22-053-017 и Fcab против CD137 мыши FS22m-063 в формате ложного МАТ2 HelD1.3 активируют CD137 при перекрестном связывании белком L, приводя к высвобождению mIL-2, в анализе активации T-клеток DO11.10.
Таблица 22.
н/п - не применимо, так как низкий сигнал не позволяет определение EC50.
Пример 13. Исследование in vitro МАТ2 против CD137/PD-L1 мыши.
Для проверки активности МАТ2, содержащего область связывания антигена CD137, в моделах на мышах in vivo, FS22m-063 выбрали как Fcab, для которого выявили наилучшую активность в анализе активации T-клеток, описанную в примере 12.3.
13.1. Конструирование, экспрессия и очистка МАТ2 против CD137/PD-L1 мыши.
Получали МАТ2 против CD137/PD-L1 мыши, содержащие Fcab против CD137 мыши FS22m-063, а также связывающую PD-L1 область Fab (клон YW243.55.S70 из US 8217149 B2). Их получали аналогично способу, описанному в примере 3.2, путем замены части CH3 связывающего PD-L1 антитела, содержащего петли AB, CD и EF, на соответствующий участок из Fcab. Данные модельные МАТ² против PD-L1 содержали мутацию LALA в домене CH2 (AA). Известно, что введение мутации LALA в домен CH2 IgG1 человека уменьшает связывание с Fcγ-рецептором (Bruhns, P. и др. (2009) и Hezareh M. и др. (2001)).
МАТ² FS22m-063-AA/PD-L1 получали путем временной экспрессии в клетках HEK293-6E и очищали, применяя колонки MAb SelectSure с белком A.
13.2. Активность МАТ2 против CD137/PD-L1 мыши в анализе активации первичных OT-1 T-клеток мыши.
Для проверки активности МАТ2 против CD137/PD-L1 мыши на клетках, которые не были сконструированы для сверхэкспрессии CD137, потребовался анализ первичных T-клеток мыши. Активированные цитотоксические CD8+ T-клетки отвечают за непосредственное уничтожение раковых клеток и экспрессируют CD137 на своей поверхности (Ye и др., 2014). Известно, что кластеризация CD137 необходима для индуцирования последующей передачи сигналов и дальнейшей активации CD8+ T-клеток. Следовательно, использовали анализ активации CD8+ T-клеток, чтобы оценить способность МАТ2 запускать кластеризацию и последующую передачу сигналов CD137. Тогда как пытались идентично повторить анализ активации CD8+ T-клеток человека, для CD8+ T-клеток мыши, выделенных из наивных селезенок C57BL/6 или Balb/c, не выявили активирующий потенциал, необходимый для такого анализа. Следовательно, разработали альтернативный анализ активации антигенспецифических CD8+ T-клеток. Активации CD8+ T-клеток добивались путем стимуляции антигеном генетически модифицированных T-клеток OT-1, выделенных из C57BL/6 OT-1 мыши (Jackson Laboratory, № в каталоге 003831), содержащих T-клеточный рецептор, специфичный к пептиду 257 - 264 овальбумина, и определяли ее по высвобождению IL-2.
Для выделения CD8+ T-клеток спленоциты выделяли из свежих селезенок мышей OT-1. Вкратце, каждую селезенку из мыши C57BL/6 OT-1 собирали и хранили в ФБР перед переносом в лунку 6-луночного культурального планшета и механическим разрушением с помощью 2 игл. Разрушенную селезенку пропускали через клеточное сито с диаметром пор 70 мкм и сито промывали ФБР. Суспензию клеток затем осаждали путем центрифугирования, супернатант удаляли и красные кровяные клетки лизировали путем добавления 10 мл 1X лизирующего эритроциты буфера (eBioscience, 00-4300-54), следуя инструкциям производителя. Спленоциты высевали в среде для активации T-клеток (IMDM, 5% ФТС, 50 мкМ 2-меркаптоэтанол, 1X пенициллин/стрептомицин), содержащей 10 нМ пептид SIINFEKL (SEQ ID NO: 194) InvivoGen, № в каталоге vac-sin) в 6-луночные планшеты при концентрации 10×106 клеток на лунку. Планшеты инкубировали в течение 48 часов при 37°C с 5% CO2. Через 48 часов CD8+ T-клетки выделяли, используя набор для выделения CD8+ T-клеток (Miltenyi Biotec, 130-104-075), следуя инструкциям производителя. Изолированные и активированные CD8+ T-клетки высевали в среде (IMDM, 5% ФТС, 50 мкМ 2-меркаптоэтанол, 1X пенициллин/стрептомицин), дополненной 30 Ед./мл IL-2 (Peprotech, AF-200-02), и поддерживали при концентрации менее 1×106 на мл при каждом ежедневном разведении в течение 3 дополнительных суток. Через трое суток размножения клетки затем использовали в следующем анализе.
Инкубацию с клетками B16.F10, которые культивировали в присутствии IFN-γ, чтобы вызвать экспрессию PD-L1, и которые затем сенсибилизировали пептидом SIINFEKL, использовали в качестве первого сигнала для запуска исходной активации T-клеток OT-1, и впоследствие использовали для оценки эффективности МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70.
Вкратце, клетки B16.F10 сначала культивировали в присутствии 20 нг/мл IFN-γ мыши (Peprotech, № в каталоге AF-315-05-100UG), чтобы вызвать экспрессию PD-L1. Данные клетки затем инкубировали с 500 нМ пептида SIINFEKL в течение одного часа при 37°C, перед тем как высеять при концентрации 1,5×104 клеток на лунку в 100 мкл среды в 96-луночный планшет с плоским дном лунок. 2×104 клеток OT-1 на лунку добавляли в клетки B16.F10 в 50 мкл среды. Осуществляли титрование тестируемых антител (FS22m-063-AA/S70, G1-AA/S70, G1-AA/Lob12.3, FS22m-063-AA/HelD1.3) 1:4, начиная со 160 нМ (4X конечная концентрация), и 50 мкл смеси антитела добавляли в каждую лунку, соответственно, с получением конечного анализируемого объема 200 мкл. Анализируемую смесь инкубировали в течение 3 суток при 37°C с 5% CO2. Через 3 суток собирали супернатанты и проводили анализ mIFN-γ с помощью ELISA (eBioscience, № в каталоге 88-7314-88), следуя инструкциям производителя.
На фигуре 11 и в таблице 23 видно, что МАТ2 против CD137/PD-L1 мыши обладает наибольшей эффективностью со значением EC50, составляющим 0,003 нМ. Данный анализ свидетельствует о том, что МАТ2 обладает сильной эффективностью в результате кластеризации CD137, приводящей к повышенному агонизму и активации CD8+ T-клеток.
Таблица 23
н/п означает - не применимо, так как низкий сигнал не позволяет определение EC50.
После того как показали активность in vitro и активность, превосходящую таковую у антител положительного контроля, было желательно тестировать МАТ2 против CD137/PD-L1 мыши в подходящей модели сингенной опухоли мыши in vivo.
Пример 14. Исследование in vivo МАТ2 против CD137/PD-L1 мыши.
14.1. Активность МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли CT26 мыши.
После того как показали, что Fcab FS22m-063 способно запускать кластеризацию и активацию CD137 in vitro, было желательно проверить его способность активировать CD137 in vivo.
Получение Fcab FS22m-063 в формате МАТ2 для тестирования in vivo на мышах.
Получали МАТ2, содержащие Fcab против CD137 мыши FS22m-063 и область Fab, специфичную к PD-L1, применяя методику, аналогичную модельному МАТ2, полученному в примере 3.2, и исследовали противоопухолевую активность in vivo в модели сингенной опухоли CT26 мыши.
Контроли: G1/Lob12.3, G1-AA/Lob12.3, G1/S70, G1-AA/4420.
Контрольные антитела для экспериментов in vivo получали путем соединения вариабельной области тяжелой цепи антитела против PD-L1 S70 (клон YW243.55.S70 из US 8217149 B2) с константной областью IgG1 человека (G1m17), содержащей мутацию LALA, и вариабельную область легкой цепи из антитела S70 соединяли с константной областью (Lm1) человека посредством J-области цепи каппа человека. Антитело против CD137 мыши Lob12.3 (Taraban и др., 2002) в формате IgG1 человека с мутацией LALA в константной области и без нее использовали в качестве антитела положительного контроля против CD137 мыши. МАТ2 получали путем замены домена CH3 конструкции с измененным форматом, описанной выше, на FS22m-063 и обозначили «FS22m-063-AA/S70».
Сингенные модели на мышах общепризнаны в качестве подходящих систем на мышах для тестирования противоопухолевого эффекта ингибирования терапевтических мишеней и всесторонне применяются для проверки разработки лекарств для человека. В данном эксперименте использовали модель сингенной опухоли CT26, так как известно, что опухоли CT26 высоко иммуногены (Lechner и др., 2013) и частично отвечают на монотерапию антителом против CD137 (Kim и др., 2009) и экспрессируют PD-L1 (Kleinovink, 2017).
Самок мышей Balb/c (Charles River) в возрасте 8 - 10 недель и с массой тела 20 - 25 г каждая оставляли в покое на одну неделю перед началом исследования. Всех животных микрочипировали и присваивали им уникальный идентификатор. В каждой группе было по 12 мышей. Линию клеток карциномы толстого кишечника CT26 (S.Rosenberg, NIH) сначала размножали, хранили, а затем подвергали предварительному скринингу с помощью IDEXX Bioresearch на патогены, применяя протокол IMPACT I, и показали, что они свободны от патогенов. Каждое животное получило 0,1×106 клеток, введенных путем подкожной инъекции в левый бок в 100 мкл DMEM. Через 7 суток после прививки клеток опухоли, мышей, у которых не развились опухоли к данному моменту, исключали из исследования.
МАТ2 FS22m-063-AA/S70 и контрольные антитела (G1/Lob12.3, G1-AA/Lob12.3 (положительные контроли против CD137), G1/S70 (положительный контроль против PD-L1), G1-AA/4420 (изотипический контроль)) вводили путем интраперитонеальной инъекции мышам в дозе 20 мкг на мышь (конечная концентрация составляла ~1 мг/кг) в DPBS + 1 мМ аргинин + 0,05 твин 80. Каждая мышь получила молекулу МАТ2 или контрольное антитело или комбинацию двух контрольных антител путем интраперитонеальной (и/п) инъекции 200 мкл на 7, 9 и 11 сутки после прививки опухоли. Проводили точные измерения опухолей, осуществляли введение доз тех лекарственных средств, которые нужно было вводить в указанные сутки, и мышей помещали под пристальное наблюдение на весь оставшийся период исследования. Измерения объема опухоли проводили с помощью кронциркуля, чтобы определить наибольшую ось и наименьшую ось опухоли. Следующую формулу использовали для вычисления объема опухоли:
L X (S2) / 2,
где L = наибольшая ось; S = наименьшая ось.
На фигуре 12 показано, что для МАТ2 FS22m-063-AA/S70 выявили значимое ингибирование роста опухоли по сравнению с мышами, которым вводили некоторые из контрольных антител. Статистическую значимость показали попарно для скоростей роста на протяжении всего времени исследования с применением анализа смешанной модели, сравнивающего все группы. Ни у одной из мышей не выявили признаки выраженной токсичности и все лекарственные средства хорошо переносились, свидетельствуя о том, что биспецифическое нацеливание на CD137 и PD-L1 не вызывает значимой токсичности.
Всех животных, содержащих опухоли размером равным или меньшим чем 62,5 мм3 засчитывали как полностью отвечающих животных (см. таблицу 24). 28% животных, которых лечили G1/Lob12.3 (антителом положительного контроля против CD137 без мутации LALA), считали не имеющими опухолей по окончании исследования, тогда как 7% животных, которых лечили G1-AA/Lob12.3 (антителом положительного контроля против CD137 с мутацией LALA), и 0% мышей, которых лечили FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1), считали не имеющими опухолей. FS22m-063-AA/S70 вызывало значимое ингибирование роста опухоли в модели сингенной опухоли CT26, по сравнению с мышами, которым вводили контроль IgG. Ни одно из животных, которых лечили G1-AA/4420 (контроль IgG) или G1/S70 (положительный контроль против PD-L1), не было свободно от опухоли по окончании исследования. На диаграммах на фигуре 17 показаны измерения роста опухоли в мм3 для каждого животного в динамике.
Таблица 24. Количество и процент не имеющих опухолей мышей по окончании исследования в исследовании сингенной опухоли CT26.
В данном исследовании показали, что у мышей с полностью функциональной иммунной системой, в модели сингенной опухоли CT26 мыши, агонизм CD137, предположительно в результате перекрестного связывания посредством PD-L1, приводит к снижению роста опухоли, предположительно посредством повышенной цитотоксической активности CD8+ T-клеток в опухоли.
Анализ выживаемости (фигура 18 и таблица 25) показал, что FS22m-063-AA/S70 вызывало значимое преимущество в выживаемости по сравнению с изотипическим контролем (G1-AA/4420). В таблице 25 кратко представлена медиана выживаемости в сутках для каждой группы и попарный статистический анализ (логранговый критерий) в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/4420 (контроль IgG), комбинацией G1/S70 плюс G1-AA/Lob12.3 и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 приводило к значимо большей выживаемости по сравнению с мышами, которым вводили контроль IgG. Для FS22m-063-AA/S70 не выявили значимого преимущества в выживаемости по сравнению с комбинацией G1/S70 + G1-AA/Lob12.3.
Таблица 25. Медианы выживаемости животных, которых лечили каждым соединением.
Показав, что у мышей с полностью функциональной иммунной системой, в модели сингенной опухоли CT26 мыши, агонизм CD137, предположительно в результате перекрестного связывания посредством PD-L1, приводит к снижению роста опухоли, предположительно посредством повышенной цитотоксической активности CD8+ T-клеток в опухоли, исследование повторяли со следующими отклонениями.
МАТ2 FS22m-063-AA/S70 и контрольные антитела G1-AA/Lob12.3 (положительный контроль против CD137), G1/S70 (положительный контроль против PD-L1), G1-AA/HelD1.3 (изотипический контроль)) вводили путем интраперитонеальной инъекции мышам в дозе 200 мкг на мышь (конечная концентрация составляла ~10 мг/кг) в DPBS + 1 мМ аргинин + 0,05 твин 80. Каждая мышь получила молекулу МАТ2, или контрольное антитело, или комбинацию двух контрольных антител путем интраперитонеальной (и/п) инъекции 200 мкл на 7, 9 и 11 сутки после прививки опухоли.
Лечение FS22m-063-AA/S70 приводило к значимому ингибированию роста опухоли (представленному в виде среднего объема опухоли на фигуре 19) по сравнению с контрольными антителами.
Анализ выживаемости (фигура 20 и таблица 26) показал, что FS22m-063-AA/S70 вызывало значимое преимущество в выживаемости по сравнению с изотипическим контролем (G1-AA/HelD1.3) и G1/S70. Не наблюдали значимого преимущества в выживаемости относительно G1-AA/Lob12.3
В таблице 26 кратко представлена медиана выживаемости в сутках для каждой группы и попарный статистический анализ (логранговый критерий) в модели сингенной опухоли CT26, которую растили подкожно в мышах Balb/c, которых лечили G1-AA/HelD1.3 (контроль IgG), G1/S70 (антитело положительного контроля против PD-L1), G1-AA/Lob12.3 (антитело положительного контроля против CD137) и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). FS22m-063-AA/S70 вызывало значимую выживаемость в модели сингенной опухоли CT26 по сравнению с мышами, которых лечили контролем IgG, и мышами, которых лечили антителом положительного контроля против CD137.
Таблица 26. Краткое представление медиан выживаемости в сутках для каждой группы и попарный статистический анализ (логранговый критерий) в модели сингенной опухоли CT26.
14.2. Активность доза-ответ МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли CT26 мыши.
Противоопухолевая активность доза-ответ.
Для МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70 выявили противоопухолевую активность в модели сингенной опухоли CT26 в дозе 20 мкг на мышь (~1 мг/кг), которую вводили три раза за каждые двое суток (q2dx3). Для выявления оптимальной дозы МАТ2 FS22m-063-AA/S70 in vivo, исследовали диапазон доз (2, 6, 20 и 200 мкг/мышь, эквивалентных приблизительно 0,1, 0,3, 1 и 10 мг/кг) в модели CT26 при схеме введения доз q2dx3, описанной выше, начиная с 7 суток после прививки клеток опухоли. Антитело отрицательного контроля G1-AA/4420 включали в анализ в дозе 200 мкг на мышь (~10 мг/кг) при той же схеме. Антитело положительного контроля G1/Lob12.3 включали в анализ в субоптимальной дозе 20 мкг на мышь (~1 мг/кг) при той же схеме.
Следуя такому же протоколу, который описан в примере 14.1, самок мышей Balb/c (Charles River) в возрасте 8 - 10 недель и с массой тела 20 - 25 г каждая оставляли в покое на одну неделю перед началом исследования. Всех животных микрочипировали и присваивали им уникальный идентификатор. В каждой группе было по 12 мышей. Линию клеток карциномы толстого кишечника CT26 (ATCC CRL-2638) изначально размножали, хранили, а затем подвергали предварительному скринингу с помощью IDEXX Bioresearch на патогены, применяя протокол IMPACT I, и показали, что они свободны от патогенов. Каждое животное получило 0,1×105 клеток, введенных путем подкожной инъекции в левый бок в 100 мкл DMEM. Через 7 суток после прививки клеток опухоли, мышей, у которых не развились опухоли к данному моменту, исключали из исследования.
На 7 сутки исследования МАТ2 FS22m-063-AA/S70 вводили путем интраперитонеальной инъекции мышам в конечной концентрации согласно диапазону доз, описаному выше. Контрольные антитела (G1/Lob12.3 (антитело положительного контроля против CD137), G1-AA/4420 (изотипический контроль)) вводили путем интраперитонеальной инъекции мышам в конечной концентрации 20 мкг на мышь (~1 мг/кг) и 200 мкг на мышь (~10 мг/кг), соответственно, в DPBS + 1 мМ аргинин + 0,05 твин 80. Каждая мышь получила молекулу МАТ2 или контрольное антитело путем интраперитонеальной (и/п) инъекции 200 мкл на 7, 9 и 11 сутки после прививки опухоли (q2dx3). Проводили точные измерения опухолей, осуществляли введение доз тех лекарственных средств, которые нужно было вводить в указанные сутки, и мышей помещали под пристальное наблюдение на весь оставшийся период исследования. Измерения объема опухоли проводили с помощью кронциркуля, чтобы определить наибольшую ось и наименьшую ось опухоли, как описано в примере 14.1.
На фигуре 13A показано, что для МАТ2 FS22m-063-AA/S70 выявили значимое ингибирование роста опухоли по сравнению с мышами, которым вводили изотипический контроль (G1-AA/4420) при введении в дозе от ~0,3 мг/кг до ~10 мг/кг. Статистическую значимость показали с применением анализа смешанной модели, сравнивающего все группы. Анализ выживаемости, фигура 13B и таблица 27, показал, что МАТ2 FS22m-63-AA/S70 вызывает статистически значимое преимущество в выживаемости по сравнению с изотипическим контролем в сингенной модели CT26 при введении при уровнях доз выше 0,3 мг/кг. Статистическую значимость определяли, применяя логранговый критерий (критерий Кокса-Мантеля). * P ≤ 0,05; ** P ≤ 0,01; *** P ≤ 0,001; **** P ≤ 0,0001. Ни у одной из мышей не выявили признаки выраженной токсичности, и все варианты лечения хорошо переносились.
Таблица 27.
н/з означает не значимо.
В данном исследовании показали, что у мышей с полностью функциональной иммунной системой, в модели сингенной опухоли CT26 мыши, агонизм CD137, предположительно в результате перекрестного связывания посредством PD-L1, приводит к зависимому от дозы снижению роста опухоли и зависимому от дозы повышению выживаемости, предположительно посредством повышенной цитотоксической активности CD8+ T-клеток в опухоли.
14.3. Механизм действия доза-ответ.
Механизм действия МАТ2 против CD137/PD-L1 мыши оценивали в той же модели несущих опухоль CT26 мышей, которая описана в примере 14.2. В крови, селезенке и опухолевой ткани из несущих опухоль CT26 мышей, которых лечили FS22m-063-AA/S70 в четырех различных дозах (2, 6, 20 и 200 мкг/мышь, эквивалентных приблизительно 0,1, 0,3, 1, 10 мг/кг), а также антителом отрицательного контроля G1-AA/4420, исследовали маркеры активации и пролиферации T-клеток, которые, как известно, являются последующими эффектами агонизма CD137 (Fisher и др., 2012).
Следуя тому же протоколу, который описан в данном примере, на 9 сутки (через 48 часов после первой дозы), на 11 сутки (через 48 часов после второй дозы) и на 15 сутки (через 96 часов после третьей и конечной дозы) кровь собирали в содержащие ЭДТА пробирки путем пункции сердца и селезенку и опухолевую ткань собирали путем диссекции. Селезенку и опухолевую ткань измельчали с получением суспензии отдельных клеток с помощью стандартных механических и ферментативных способов. Красные кровяные клетки однократно лизировали в лизирующем эритроциты буфере (eBioscience, № в каталоге 00-4300-54), следуя инструкциям производителя.
Красные кровяные клетки некоагулированной крови лизировали дважды в лизирующем эритроциты буфере (eBioscience, № в каталоге 00-4300-54), следуя инструкциям производителя.
Все образцы затем обрабатывали одинаково. Клетки затем однократно промывали ФБР и образцы окрашивали фиксирующимся красителем для выявления жизнеспособности (Invitrogen, № в каталоге 65-0865-14) при разведении 1:3000 в ФБР, следуя инструкциям производителя. Клетки окрашивали для проведения проточной цитометрии для выявления маркеров поверхности клетки панелью для окрашивания антителами (всеми, кроме антител против Ki67 и FoxP3) (таблица 28 ниже) в присутствии агента для блокирования Fc (eBioscience, № в каталоге 14-0161-86 при 1:50) в течение 30 минут при 4°C. Клетки затем фиксировали и пермеабилизовали с помощью набора для окрашивания Foxp3 eBioscience (eBioscience, № в каталоге 00-5523-00), следуя инструкциям производителя. Клетки ресуспендировали в 100 мкл пермеабилизирующего буфера с антителами Ki67 и Foxp3 в присутствии агента для блокирования Fc (все при разведении 1:100) и инкубировали 30 минут в темноте при комнатной температуре. Клетки затем однократно промывали пермеабилизирующим буфером и ресуспендировали в 200 мкл ФБР + 0,5% БСА. Клетки затем анализировали на проточном цитометре BD Fortessa. Результаты анализировали с помощью FlowJoX, Excel и GraphPad Prism. Нанесенные на график результаты представляют собой численность и пролиферацию T-клеток, наблюдаемые в динамике для субпопуляций CD4+ и CD8+ T-клеток. Результаты представлены в виде процента от исходной популяции, в соответствии с описанием популяции на оси Y.
На фигуре 14B показано, что процентное соотношение CD8+:CD4+ возрастает в пользу CD8+ T-клеток, как в опухоли, так и на периферии, в соответствии с повышением дозы МАТ2 против CD137/PD-L1 мыши, свидетельствуя о том, что лечение вызывает сдвиг равновесия в двух субпопуляциях T-клеток, вероятно в результате увеличения количества CD8+ T-клеток. К 15 суткам маркер пролиферации на CD8+ T-клетках возрастал в соответствии с уровнем дозы лечения, при этом наибольшая пролиферация наблюдалась при наибольшем уровне дозы. Это будет свидетельствовать о том, что CD8+ T-клетки пролиферируют, что является признаком активации и объяснит повышенное количество CD8+ T-клеток, приводящее к увеличению отношения CD8+:CD4+ T-клеток.
В целом, данное исследование показало, что повышение уровня дозы антитела против CD137/PD-L1 мыши приводит к большему количеству CD8+ T-клеток в опухоли. Главная роль CD8+ T-клеток (часто также называемых цитотоксическими лимфоцитами) состоит в уничтожении инфицированных или злокачественных клеток посредством 3 основных механизмов: 1) высвобождения цитокинов, например, TNFα и IFN-γ, 2) продукции и высвобождения содержимого цитотоксических гранул и 3) экспрессии FasL. Присутствие большего количества CD8+ T-клеток в опухоли после лечения антителом против CD137/PD-L1 мыши предположительно приводит к цитотоксической активности посредством данных механизмов против опухоли, приводя к контролю опухоли.
Таблица 28.
н/п означает не применимо.
14.4. Активность МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли MC38 мыши.
Сингенные модели на мышах общепризнаны в качестве подходящих систем на мышах для тестирования противоопухолевого эффекта ингибирования терапевтических мишеней и всесторонне применяются для проверки разработки лекарств для человека. В данном эксперименте использовали модель сингенной опухоли MC38, так как известно, что опухоли MC38 высоко иммуногены и отвечают на монотерапию антителом против CD137 (Kocak и др., 2006) и экспрессируют PD-L1 (Juneja и др., 2017).
Самок мышей C57BL/6 (The Jackson Laboratory) в возрасте 9 - 10 недель и с массой тела по 18 - 24 г каждая оставляли в покое на одну неделю перед началом исследования. Всех животных микрочипировали и присваивали им уникальный идентификатор. В каждой группе было по 12 мышей. Линию клеток карциномы толстого кишечника MC38 (Национальный институт онкологии, США) изначально размножали, хранили, а затем подвергали предварительному скринингу на патогены и показали, что они свободны от патогенов. Каждое животное получило 1×106 клеток, введенных путем подкожной инъекции в правый бок в объеме 100 мкл в бессывороточной культуральной среде (среде Игла, модифицированная по Дульбекко). Через 7 суток после прививки клеток опухоли мышей, у которых не развились опухоли к данному моменту, исключали из исследования.
МАТ2 FS22m-063-AA/S70 и контрольные антитела (G1-AA/Lob12.3 (положительный контроль против CD137), G1-AA/S70 (положительный контроль против PD-L1), G1-AA/4420 (изотипический контроль)) вводили путем интраперитонеальной инъекции мышам в неизменной концентрации, составляющей 20 мкг на дозу, в DPBS + 1 мМ аргинин + 0,05 твин 80. Каждая мышь получила молекулу МАТ2 или контрольное антитело путем интраперитонеальной (и/п) инъекции 200 мкл на 7, 9 и 11 сутки после прививок опухоли. Проводили точные измерения опухолей, осуществляли введение доз тех лекарственных средств, которые нужно было вводить в указанные сутки, и мышей помещали под пристальное наблюдение на весь оставшийся период исследования. Измерения объема опухоли проводили с помощью кронциркуля, чтобы определить наибольшую ось и наименьшую ось опухоли. Следующую формулу использовали для вычисления объема опухоли:
L X (S2) / 2,
где L = наибольшая ось; S = наименьшая ось.
На фигуре 15 показано, что для МАТ2 FS22m-063-AA/S70 выявили значимое ингибирование роста опухоли по сравнению с мышами, которым вводили любое из контрольных антител. Статистическую значимость показали попарно для скоростей роста на протяжении всего времени исследования, применяя анализ смешанной модели, сравнивающий все группы. В таблице 29 показано, что, неожиданно, все мыши, которых лечили МАТ2 FS22m-063-AA/S70, оказались свободны от опухоли по окончании исследования, по сравнению лишь с 4 из 12 мышей, которых лечили комбинацией антител против PD-L1 и против CD137 (G1-AA/S70 + G1-AA/Lob12.3) или антителами против PD-L1 или CD137 отдельно. На диаграммах на фигуре 21 показаны измерения роста опухоли в мм3 для каждого животного в динамике.
Таблица 29.
В данном исследовании показали, что у мышей с полностью функциональной иммунной системой, во второй модели сингенной опухоли MC38, для которой показали субоптимальный ответ на 1 мг/кг МАТ против PD-L1, неожиданно, 1 мг/кг МАТ2 против CD137/PD-L1 вызывал полную регрессию опухоли у 100 % получивших лечение мышей.
В анализе выживаемости (фигура 22 и таблица 30) показали краткое представление медиан выживаемости в сутках для каждой группы лечения, которую лечили 3 дозами G1-AA/4420 (изотипический контроль), G1-AA/S70 (антитело положительного контроля против PD-L1), G1-AA/Lob12.3 (антитело положительного контроля против CD137), комбинации G1-AA/S70 плюс G1-AA/Lob12.3 и FS22m-063-AA/S70 (Fcab против CD137 мыши FS22m-063 в формате модельного МАТ2 против PD-L1). В таблице 30 также показан попарный статистический анализ (логранговый критерий). FS22m-063-AA/S70 вызывало полную выживаемость в модели сингенной опухоли MC38 со 100% выживаемостью животных по окончании исследования (обозначено «не определена»), по сравнению с мышами, которых лечили контролем IgG.
Таблица 30. Медиана выживаемости в модели опухоли MC38.
14.5. Активность МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли B16.F10 мыши.
Модель сингенной опухоли B16.F10 использовали для проверки противоопухолевой активности МАТ2 против CD137/PD-L1 мыши (FS22m-063-AA/S70) in vivo. Данную модель рассматривают как более сложную для лечения терапевтическими антителами (Baird, J. R. и др., 2013). Антитело G1-AA/4420 использовали в качестве изотипического контроля в данном исследовании. Ранее не было показано, что модель сингенной опухоли B16.F10 чувствительна к агонистическим антителам против CD137. Тем не менее инфильтрирующие опухоль лимфоциты (TIL), выделенные из опухолей B16.F10, экспрессируют CD137 (Curran. M. и др., 2013).
Самкам мышей C57BL/6 (Charles River) в возрасте 8 - 10 недель и с массой тела 20 - 25 г каждая позволяли акклиматизироваться в течение одной недели перед началом исследования. Всех животных микрочипировали и присваивали им уникальный идентификатор. В каждой группе было по 10 мышей. Линию клеток меланомы B16.F10 (ATCC, № в каталоге CRL-6475) изначально размножали, хранили, а затем подвергали предварительному скринингу с помощью IDEXX Bioresearch на патогены, применяя протокол IMPACT I, и показали, что они свободны от патогенов. Клетки B16.F10 размораживали после хранения на -150°C и добавляли в 20 мл DMEM (Gibco, 61965-026) с 10% ФТС (Gibco, 10270-106) во флакон для культивирования ткани T175. Каждое животное получило 1×106 клеток, введенных путем подкожной инъекции в левый бок.
20 мкг каждого антитела вводили путем инъекции мышам (~1 мг/кг) в 200 мкл ФБР. Каждая мышь получила антитела путем интраперитонеальной (и/п) инъекции каждые 2 суток по 3 дозы, начиная в сутки 7 после прививки клеток опухоли. Объемы опухоли определяли путем измерения с применением кронциркуля (как описано в примере 14.1) и осуществляли введение доз тех лекарственных средств, которые нужно было вводить в указанные сутки, и мышей помещали под пристальное наблюдение на весь оставшийся период исследования. Объемы опухоли рассчитывали, как описано в примере 14.1.
Исследование останавливали, когда были достигнуты гуманные конечные точки, на основании объема опухоли и состояния. Статистический анализ скоростей роста осуществляли попарно за весь период исследования с применением анализа смешанной модели.
Наблюдалось значимое замедление скорости роста опухоли для МАТ2 против CD137/PD-L1 мыши по сравнению с антителом изотипического контроля G1-AA/4420 (фигура 16). FS22m-063-AA/S70 вызывало частичное ингибирование роста опухоли в модели сингенной опухоли B16.F10 по сравнению с мышами, которым вводили контроль IgG. На диаграммах на фигуре 23 показаны измерения роста опухоли в мм3 для каждого животного в динамике.
В анализе выживаемости (таблица 31) показали краткое представление медиан выживаемости в сутках для каждой группы и попарный статистический анализ (логранговый критерий) в модели сингенной опухоли B16.F10, которые растили подкожно в мышах C57BL/6, которых лечили G1-AA/4420 (изотипический контроль) и МАТ2 против CD137/PD-L1 мыши FS22m-063-AA/S70. FS22m-063-AA/S70 вызывало повышение выживаемости в модели сингенной опухоли B16.F10 по сравнению с мышами, которым вводили контроль IgG.
Таблица 31. Медиана выживаемости в модели сингенной опухоли B16.F10.
логранговый критерий
В данном исследовании показали, что у мышей с полностью функциональной иммунной системой, в общеизвестно трудной для лечения модели сингенной опухоли B16.F10 мыши, агонизм CD137, предположительно в результате перекрестного связывания посредством PD-L1, приводит к снижению роста опухоли, предположительно посредством повышенной цитотоксической активности CD8+ T-клеток в опухоли.
14.6. Фармакология в печени МАТ2 против CD137/PD-L1 мыши в модели несущих сингенную опухоль CT26 мышей.
Лечение МАТ против CD137 урелумабом пациентов с солидной опухолью в исследовательских клинических испытаниях приводило к тяжелым связанным с лечением иммунным явлениям, которые, как было показано, связаны с введенной дозой урелумаба. Эффекты данных иммунных явлений проявляются в печени как сильная гепатотоксичность (Segal, N.H. и др., 2017).
В доклиническом механистическом исследовании, предпринятом на мышах, в котором животным вводили дозу антител с агонистическим действием против CD137, показали сходную гепатотоксичность. В данных исследованиях выявили необходимость T-клеток и CD137 для итоговой гепатотоксичности (Niu, L., и др. 2007 и Dubrot J и др. 2010). Хотя не до конца изучено, также было показано, что взаимодействие между миелоидным и T-клеточным компартментами важно для запуска воспалительного каскада, приводящего к повреждению печени и гепатотоксичности (Bartkowiak, T и др., 2018). Следовательно, данные модели на животных имеют трансляционную значимость для клиники в прогнозировании риска гепатотоксичности у пациентов-людей после введения других агонистов CD137 и, в частности, МАТ2 CD137/PD-L1.
У мышей из исследования сингенной опухоли CT26, описанного в примере 14.1, не выявили явных признаков токсичности после повторных доз МАТ2 FS22m-063-AA/S70, которое обладает способностью к агонизму CD137 при перекрестном связывании PD-L1. Для того чтобы определить, коррелировала ли у данных животных активация иммунной системы, наблюдаемая в результате лечения ~1 мг/кг МАТ2 FS22m-063-AA/S70 (которая коррелировала с противоопухолевой иммунной активностью), с гепатотоксичностью, образцы печени брали при вскрыти для гистологической оценки. Мышей, получивших МАТ2 FS22m-063-AA/S70, и контрольных мышей вскрывали через 4, 7 и 14 суток после последнего введения (по три мыши на группу на момент времени) и образцы печени фиксировали формалином и заключали в парафин. Затем делали срезы печени и подвергали гистопатологической оценке путем окрашивания гематоксилином и эозином и оценки параметров воспаления и повреждения печени сертифицированным патологом. Способы препарирования ткани и гистопатологического окрашивания гематоксилином и эозином хорошо стандартизированы и хорошо известны в данной области.
Использовали систему оценки, чтобы оценить патологию печени на окрашенных гематоксилином и эозином срезах. Оценивали патологию печени, соответствующую гепатоцеллюлярному некрозу, воспалению портального тракта, дегенеративным изменениям в гепатоцитах и повышенному количеству митозов. Встречаемость мышей, у которых выявили минимальные, незначительные, умеренные и выраженные эффекты внутри каждой группы, представлена в таблице 32.
У животных, которых лечили МАТ2 FS22m-063-AA/S70, наблюдалась минимальная патология печени. В частности:
• от минимального до незначительного гепатоцеллюлярного некроза со смешанным инфильтратом лимфоцитов в паренхиме
• от минимального до незначительного количества смешанных воспалительных клеток в перипортальных трактах
• минимальное количество дегенеративных гепатоцитов
• от минимального до выраженного повышенного количества митозов.
Таблица 32. Находки в печени.
Не считали, что данные находки означают сильную гепатотоксичность, которая наблюдалась для других примеров агонистических антител против CD137.
Учитывая важность доклинических исследований на мышах для оценки риска сильной гепатотоксичности у пациентов-людей, которых лечат агентами-агонистами CD137, данные результаты свидетельствуют о том, что МАТ2, которые агонизируют CD137 посредством перекрестного связывания, опосредованного связыванием PD-L1, имеют низкий риск вызывать гепатотоксичность у пациентов-людей, которых лечат терапевтическими дозами.
14.7. Занятость рецептора в опухоли и на периферии и фармакодинамические эффекты МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли CT26 мыши.
В примере 14.3 для МАТ2 против CD137-PD-L1 мыши FS22m-063-AA/S70 выявили способность повышать количество CD8+ T-клеток в опухоли после множества доз. Для того чтобы исследовать уровень связывания мишени на T-клетках, блокирования PD-L1 и влияния FS22m-063-AA/S70 на пролиферацию T-клеток, проводили фармакодинамическое исследование введения однократной дозы на той же модели сингенной опухоли CT26, которая описана в примере 14.1.
Антитело G1-AA/4420 использовали в качестве контроля. МАТ2 против CD137/PD-L1 мыши и контрольные антитела для экспериментов in vivo получали, как описано в примере 14.1.
Следуя такому же протоколу, который описан в примере 14.1, самок мышей Balb/c (Charles River) в возрасте 8 - 10 недель и с массой тела 20 - 25 г каждая оставляли в покое на одну неделю перед началом исследования. Всех животных микрочипировали и присваивали им уникальный идентификатор. Данное исследование включало 3 группы дозирования с введением либо контрольного антитела, либо МАТ2 в одной из двух доз в 8 моментов времени (2 ч, 6 ч, 24 ч, 48 ч, 72 ч, 96 ч, 120 ч, 192 ч). Каждая группа дозирования содержала 64 мыши (по 8 мышей на момент времени). Линию клеток карциномы толстого кишечника CT26 (S.Rosenberg, NIH) сначала размножали, хранили, а затем подвергали предварительному скринингу с помощью IDEXX Bioresearch на патогены, применяя протокол IMPACT I, и показали, что она свободна от патогенов. Каждое животное получило 1×105 клеток, введенных путем подкожной инъекции в левый бок в 100 мкл DMEM. Через 7 суток после прививки клеток опухоли, мышей, у которых не развились опухоли к данному моменту, исключали из исследования.
Каждая мышь получала тестируемый образец посредством внутривенной (в/в) инъекции в объеме 100 мкл на 11 сутки после прививки опухоли. В группах, получивших МАТ2, МАТ2 FS22m-063-AA/S70 вводили путем внутривенной инъекции мышам в дозе либо 20 мкг, либо 200 мкг на мышь (конечная концентрация составляла ~1 мг/кг и ~10 мг/кг, соответственно) и группе, получившей контроль, антитело G1-AA/4420 (изотипический контроль) вводили путем внутривенной инъекции мышам в дозе 200 мкг на мышь (конечная концентрация составляла ~10 мг/кг) в DPBS + 1 мМ аргинин + 0,05 твин 80.
Оценивали захват рецептора в опухоли и периферической крови и фармакодинамический ответ в результате введения дозы МАТ2 против CD137/PD-L1 мыши. Опухолевую ткань и кровь из несущих опухоль CT26 мышей, которых лечили FS22m-063-AA/S70 (МАТ2 против mCD137/PD-L1) в каждой из доз, а также антителом отрицательного контроля G1-AA/4420, исследовали на наличие связанных с МАТ2 против CD137/PD-L1 мыши положительных T-клеток, пролиферацию T-клеток и свободный PD-L1, не связанный МАТ2 против CD137/PD-L1 мыши. Также оценивали общую экспрессию CD137.
Кровь (100 мкл) собирали в покрытые ЭДТА капиллярные трубочки путем кровоизвлечения из хвостовой вены и лизировали дважды в лизирующем эритроциты буфере (eBioscience, № в каталоге 00-4300-54), следуя инструкциям производителя.
Опухолевую ткань собирали путем диссекции и измельчали с получением суспензии отдельных клеток с помощью стандартных механических и ферментативных способов. Все красные кровяные клетки, оставшиеся в измельченной опухолевой ткани, однократно лизировали в лизирующем эритроциты буфере (eBioscience, № в каталоге 00-4300-54), следуя инструкциям производителя.
Клетки однократно промывали ФБР и образцы окрашивали фиксирующимся красителем для выявления жизнеспособности (Invitrogen, № в каталоге 65-0865-14) при разведении 1:3000 в ФБР, следуя инструкциям производителя. Клетки окрашивали на наличие маркеров поверхности клетки для проточной цитометрии панелью для окрашивания антителами (всеми, кроме антител против Ki67 и FoxP3) (таблица 33 ниже) в присутствии агента для блокирования Fc (eBioscience, № в каталоге 14-0161-86 при разведении 1:50) в течение 30 минут при 4°C. Клетки затем фиксировали и пермеабилизовали с помощью набора для окрашивания Foxp3 eBioscience (eBioscience, № в каталоге 00-5523-00), следуя инструкциям производителя. Клетки ресуспендировали в 100 мкл пермеабилизирующего буфера с антителами Ki67 и Foxp3 в присутствии агента для блокирования Fc и инкубировали в течение 30 минут в темноте при комнатной температуре. Клетки затем однократно промывали пермеабилизирующим буфером и ресуспендировали в 200 мкл ФБР + 0,5% БСА. Клетки затем анализировали на проточном цитометре BD Fortessa. Результаты анализировали с помощью FlowJoX, Excel и GraphPad Prism.
На фигуре 24 показан процент CD4+ и CD8+ T-клеток, выделенных либо из крови, либо из опухоли, которые были положительны по антителу против IgG, связанному с МАТ2 mCD137/PD-L1 в образце. На фигуре 24 показано, что T-клетки были положительны по связыванию МАТ2 против mCD137/PD-L1 уже через 2 часа после внутривенного введения. Наблюдалась зависимая от дозы корреляция длительности связывания, причем МАТ2 против mCD137/PD-L1 более не детектировалось через 96 ч на T-клетках, выделенных из крови и опухоли, после введения дозы 1 мг/кг, тогда как МАТ2 против mCD137/PD-L1 все еще детектировалось через 120 ч - 192 ч после введения дозы 10 мг/кг. Как ожидалось, минимальное фоновое окрашивание наблюдали для контрольного антитела. Популяции T-клеток, выделенных из опухоли, которые были положительны по МАТ2 против mCD137/PD-L1, были больше вначале по сравнению с T-клетками, выделенными из крови. Это может свидетельствовать о том, что уровни экспрессии мишени, либо CD137, либо PD-L1, были выше на T-клетках, выделенных из опухоли, так что антитело против mCD137/PD-L1 более эффективно нацеливалось на опухоль по сравнению с T-клетками в крови.
На фигуре 25 показано детектирование Ki67 как маркера пролиферации T-клеток на CD4+ и CD8+ T-клетках, выделенных из крови и опухоли. Как указано на фигуре 25, по сравнению с контрольным антителом, МАТ2 против mCD137/PD-L1 приводило к увеличению встречаемости положительных по Ki67 T-клеток в периферической крови, свидетельствуя о фармакодинамическом (ФД) ответе, т.е., что обе мишени захвачены или были в какой-то момент захвачены, приводя к активации T-клеток, что видно по пролиферативному ответу, при обоих исследованных уровнях дозы. Поскольку инфильтрирующие опухоль T-клетки контактируют с воспалительным окружением, в популяциях T-клеток, выделенных из опухоли, выявляют более высокую встречаемость положительных по Ki67 пролиферирующих T-клеток. Напротив, на исходном уровне встречаемость положительных по Ki67 T-клеток была гораздо ниже в крови. Похоже, как CD4+, так и CD8+ T-клетки отвечали на лечение против mCD137/PD-L1, свидетельствуя о том, что CD137 на обоих типах клеток захватывались МАТ2 и кластеризовались друг с другом путем связывания PD-L1, приводя к передаче сигналов CD137 и активации T-клеток, что приводило к повышенной пролиферации. Данный эффект проявлялся сильнее для CD8+ T-клеток, что соответствует тому, что CD8+ T-клетки экспрессирут более высокие уровни CD137, чем CD4+ T-клетки. Полученные результаты позволяют предположить, что максимального эффекта достигают при 1 мг/кг, хотя продолжительность ответа оказалась большей при 10 мг/кг, что может быть связано с более длительной занятостью рецептора и, следовательно, более длительным захватом мишени при более высоком уровне дозы. Чем продолжительнее занятость рецептора, тем больше ФД ответ. В опухоли T-клетки уже сильно пролиферирующие, и наблюдали коррелирующее с дозой повышение величины пролиферации как на CD4+, так и на CD8+ T-клетках с течением времени.
В образцы из мышей, которых лечили контрольным антителом G1-AA/4420, добавляли 100 нМ МАТ2 против mCD137/PD-L1 и использовали в качестве контроля 100% захвата рецептора PD-L1. Это показали по отсутствию связывания с конкурирующим антителом против mPD-L1 (клон 10F.9G2, см. таблицу 33). Для образцов из опухолей и крови мышей, которых лечили антителом против mCD137/PD-L1, выявили почти полное блокирование PD-L1 при дозе 10 мг/кг на всем протяжении исследования, как показано на фигуре 26, представленное 100% занятостью рецептора PD-L1. В образцах из мышей, которых лечили 1 мг/кг антитела против mCD137/PD-L1, захват рецептора PD-L1 уменьшался приблизительно через 72 ч на T-клетках, присутствующих в крови и опухоли, причем захват рецептора PD-L1 на T-клетках, присутствующих в крови, уменьшался гораздо быстрее, чем на T-клетках, присутствующих в опухоли. Данные результаты свидетельствуют об опухолеспецифическом блокировании PD-L1 с преимуществом как ингибирования системы PD-1/PD-L1, так и сохранения МАТ2 против mCD137/PD-L1 в опухоли в течение более длительного времени, чем в крови, что, как ожидается, позволит полному действию лекарственного средства сохраняться в опухоли, при этом, в то же время, ограничивая побочные действия на периферии.
В целом, в данном исследовании показали, что увеличение дозы антитела против CD137/PD-L1 мыши приводило к активации и пролиферации CD8+ и CD4+ T-клеток в опухоли, что измеряли по количеству клеток, с которыми связывалось МАТ2, захвату МАТ2 рецептора PD-L1 и повышенной экспрессии Ki67 как маркера пролиферации. Активированные и пролиферирующие CD8+ и CD4+ клетки также наблюдали в периферической крови, что было дополнительным маркером активности, вызванной антителом против CD137/PD-L1 мыши. Такая зависимая от дозы активация T-клеток подтверждает ингибирование роста опухоли, ранее наблюдаемое в примере 14 при введении антитела против CD137/PD-L1 мыши.
Главная роль CD8+ T-клеток (часто также называемых цитотоксическими лимфоцитами) состоит в уничтожении инфицированных или злокачественных клеток посредством 3 основных механизмов: 1) высвобождения цитокинов (например, TNFα и IFN-γ), 2) продукции и высвобождения содержимого цитотоксических гранул и 3) экспрессии лиганда Fas. Ожидают, что присутствие большего количества CD8+ T-клеток в опухоли после лечения антителом против CD137/PD-L1 мыши приведет к цитотоксической активности посредством данных механизмов против опухоли, приводя к контролю опухоли. Тогда как CD8+ T-клетки являются главными цитотоксическими T-клетками, которые отвечают за уничтожение клеток опухоли, CD4+ T-клетки играют главную роль в приобретенном иммунитете, узнавая пептиды, представленные молекулами MHC II класса, становлясь активированными и продуцируя IFN-γ и TNFα, оба из которых опосредуют защиту от опухолей (Zanetti, 2015, JI, 2015).
Таблица 33.
н/п означает не применимо.
Пример 15. Фармакокинетика МАТ2 против CD137/PD-L1 человека в мыши.
Для того чтобы определить фармакокинетику МАТ2 против CD137/PD-L1 в мышах, самкам мышей C57BL/6 (см. пример 14.4), но без опухоли MC38, т.е., не имеющим опухоли мышам, вводили 100 мкл МАТ2 против CD137/PD-L1 в 4 дозах (25 мг/кг, 10 мг/кг, 3 мг/кг и 1 мг/кг) и наблюдали за ними в течение 384 часов.
Взятие микропробы приблизительно 20 мкл цельной крови провели через 2, 6, 24, 48, 96 и 384 часа и обработали, чтобы выделить приблизительно 5 мкл сыворотки. Количество МАТ2 против CD137/PD-L1, присутствующее в каждый момент времени, определяли с применением системы Gyrolab xPlore от Gyros Protein Technologies. Проводили сэндвич-анализ, применяя Gyrolab Bioaffy 1000 CD (номер продукта P0004253), с биотинилированным антителом козы против человеческого IgG-(тяжелая и легкая цепь), адсорбированном на иммуносорбенте из обезьяны (Cambridge Bioscience, номер продукта A80-319B), в качестве захватывающего антитела и антитела козы против IgG человека-AlexaFluor® 647 (Cambridge Bioscience, номер продукта 2040-31) в качестве детектирующего антитела. Стандартную кривую, полученную в диапазоне 8000 - 0,07 нг/мл, использовали для определения концентрации образца, причем образцы подвергали разведению в буфере Rexxip AN при необходимости (Gyros, номер продукта P0004994). Другие буферы использовали в соответствии с рекомендациями производителя. Среднюю концентрацию образца из отдельной мыши на момент времени (по три мыши на момент времени) наносили на график (фигура 27).
На фигуре 27 показана фармакокинетика МАТ2 против CD137/PD-L1, демонстрирующая, что МАТ2 обладало конечным клиренсом, сопоставимым с таковым для стандартного IgG человека в мышах (Bergman и др., 1998).
Пример 16. Фармакокинетический/фармакодинамический ответ и переносимость МАТ2 против CD137/PD-L1 человека яванскими макаками.
Проводили предварительное исследование по определению диапазона доз, чтобы оценить фармакокинетический/фармакодинамический (ФК/ФД) ответ и переносимость МАТ2 против CD137/PD-L2 человека яванскими макаками. Вкратце, МАТ2 FS22-172-003-AA/E12v2 вводили яванским макакам путем внутривенной (в/в) инфузии в виде одной дозы или повторных доз. Оценивали стандартные токсикологические параметры, такие как масса тела, потребление пищи, клиническое наблюдение, гематология и биохимический анализ крови, для оценки переносимости на всем протяжении исследования.
Для МАТ2 FS22-172-003-AA/E12v2 выявили конечное время полужизни приблизительно 6 суток и, как правило, оно хорошо переносилось вплоть до дозы 30 мг/кг, вводимой еженедельно, что определили по результатам клинической биохимии и гистопатологии). Повышенные уровни в сыворотке sPD-L1 (признак непосредственного захвата мишени и активации клеток) наблюдали у всех животных на 1 сутки, причем пик достигался через 168 часов после окончания инфузии, после чего уровни снижались в соответствии со снижением системных уровней МАТ2 FS22-172-003-AA/E12v2.
В соответствии с находками в исследовании для оценки ФД ответа для МАТ2 против CD137/PD-L1 мыши в модели сингенной опухоли у мышей (пример 15), также наблюдали связанное с лекарственным средством повышение пролиферации и активации клеток среди CD4+ и CD8+ T-клеток центральной памяти и эффекторной памяти, что измеряли по повышенной экспрессии Ki67. Аналогичную кинетику наблюдали для NK-клеток и умеренного, но временного повышения относительного процента и абсолютных количеств CD4+ регуляторных T-клеток (CD4+ FoxP3+).
В совокупности данные результаты явно свидетельствуют о том, что МАТ2 против антигенов человека FS22-172-003-AA/E12v2 обладает эффективной фармакологической активностью in vivo в яванских макаках и хорошо переносится в дозе до 30 мг/кг. Более того, полученные результаты ФК/ФД соответствуют таковым для суррогатной молекулы (FS22m-063-AA/S70), усиливая обоснование для применения МАТ2 FS22-172-003-AA/E12v2 в условиях клиники.
Перечень последовательностей
Последовательности аминокислот CDR МАТ E12v2 (Кабат)
CDR1 VH - SYGIS (SEQ ID NO: 1)
CDR2 VH - WISAYSGGTNYAQKLQG (SEQ ID NO: 2)
CDR3 VH - DLFPTIFGVSYYYY (SEQ ID NO: 3)
CDR1 VL - RASQSIGNRLA (SEQ ID NO: 4)
CDR2 VL - EASTSET (SEQ ID NO: 5)
CDR3 VL - QQSYSTPYT (SEQ ID NO: 6)
Последовательности аминокислот CDR E12v2 (IMGT)
CDR1 VH - GYPFTSYG (SEQ ID NO: 7)
CDR2 VH - ISAYSGGT (SEQ ID NO: 8)
CDR3 VH - ARDLFPTIFGVSYYYY (SEQ ID NO: 9)
CDR1 VL - QSIGNR (SEQ ID NO: 10)
CDR2 VL - EAS (SEQ ID NO: 11)
CDR3 VL - QQSYSTPYT (SEQ ID NO: 6)
Последовательность аминокислот домена VH МАТ E12v2 (SEQ ID NO: 12)
CDR IMGT (жирный курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты).
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSS
Последовательность нуклеиновой кислоты домена VH МАТ E12v2 (SEQ ID NO: 13)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACCCCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGC
Последовательность аминокислот домена VL МАТ E12v2 (SEQ ID NO: 14)
CDR IMGT (жирный курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты).
DIQMTQSPSTLSASVRDRVIITCRASQSIGNRLAWYQHKPGKAPKLLIYEASTSETGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPYTFGQGTKLEIK
Последовательность нуклеиновой кислоты домена VL МАТ E12v2 (SEQ ID NO: 15)
GACATCCAGATGACGCAGAGCCCGTCTACCCTGTCCGCCTCCGTGAGAGATCGCGTGATCATCACCTGTCGGGCCAGCCAGTCCATCGGAAACCGCTTGGCGTGGTACCAGCACAAGCCTGGGAAGGCTCCGAAGCTGCTCATCTACGAAGCCTCGACTTCGGAGACTGGTGTCCCTAGCCGGTTCAGCGGATCGGGATCAGGGACCGATTTCACTCTGACCATTTCCTCCCTGCAACCCGAGGACTTCGCCACCTACTACTGCCAACAGTCATATTCCACCCCGTACACCTTCGGACAAGGCACCAAGCTCGAAATCAAG
Последовательность аминокислот тяжелой цепи МАТ G1-AA/E12v2 (с LALA) (SEQ ID NO: 16)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность легкой цепи аминокислот МАТ G1-AA/E12v2 (SEQ ID NO: 17)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
DIQMTQSPSTLSASVRDRVIITCRASQSIGNRLAWYQHKPGKAPKLLIYEASTSETGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательности аминокислот CDR МАТ E05v2 (Кабат)
CDR1 VH - SYGIS (SEQ ID NO: 1)
CDR2 VH - WISAYSGGTNYAQKLQG (SEQ ID NO: 2)
CDR3 VH - DLFPTIFGVSYYYY (SEQ ID NO: 3)
CDR1 VL - RASQSISGRLA (SEQ ID NO: 18)
CDR2 VL - EASNLES (SEQ ID NO: 19)
CDR3 VL - QQSYSTPRVT (SEQ ID NO: 20)
Последовательности аминокислот CDR E05v2 (IMGT)
CDR1 VH - GYTFTSYG (SEQ ID NO: 21)
CDR2 VH - ISAYSGGT (SEQ ID NO: 8)
CDR3 VH - ARDLFPTIFGVSYYYY (SEQ ID NO: 9)
CDR1 VL - QSISGR (SEQ ID NO: 22)
CDR2 VL - EAS (SEQ ID NO: 11)
CDR3 VL - QQSYSTPRVT (SEQ ID NO: 20)
Последовательность аминокислот домена VH МАТ E05v2 (SEQ ID NO: 23)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSS
Последовательность нуклеиновой кислоты домена VH МАТ E05v2 (SEQ ID NO: 24)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGC
Последовательность аминокислот домена VL МАТ E05v2 (SEQ ID NO: 25)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
DIQMTQSPSTLSASVGDRVTITCRASQSISGRLAWYQQKPGKAPNLLIYEASNLESGVPSRFSGSGSGTEFTLTISSLQPEDFATYYCQQSYSTPRVTFGQGTKVEIK
Последовательность нуклеиновой кислоты домена VL МАТ E05v2 (SEQ ID NO: 26)
GACATTCAGATGACCCAATCCCCGTCCACGCTGAGCGCCTCCGTCGGTGATCGCGTGACAATCACTTGTCGGGCGTCGCAGTCCATCTCTGGAAGGCTCGCCTGGTACCAGCAGAAGCCTGGAAAGGCTCCCAACCTCCTTATCTACGAAGCCAGCAACCTGGAGTCCGGAGTGCCTAGCCGGTTCAGCGGATCAGGGTCCGGTACCGAGTTCACCCTGACCATTTCCTCGCTCCAACCTGAGGACTTCGCCACCTACTACTGCCAACAGTCCTATTCAACTCCGCGCGTGACCTTCGGCCAGGGCACTAAGGTCGAAATCAAA
Последовательность аминокислот тяжелой цепи МАТ G1-AA/E05v2 (с LALA) (SEQ ID NO: 27)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность аминокислот легкой цепи МАТ G1-AA/E05v2 и МАТ2 FS22-172-003-AA/E05v2 и FS22-053-008-AA/E05v2 (SEQ ID NO: 28)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
DIQMTQSPSTLSASVGDRVTITCRASQSISGRLAWYQQKPGKAPNLLIYEASNLESGVPSRFSGSGSGTEFTLTISSLQPEDFATYYCQQSYSTPRVTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательности аминокислот CDR МАТ G12v2 (Кабат)
CDR1 VH - SYGIS (SEQ ID NO: 1)
CDR2 VH - WISAYSGGTNYAQKLQG (SEQ ID NO: 2)
CDR3 VH - DLFPTIFGVSYYYY (SEQ ID NO: 3)
CDR1 VL - RASQSISGRLA (SEQ ID NO: 18)
CDR2 VL - EASNLES (SEQ ID NO: 19)
CDR3 VL - QQSYSWPRT (SEQ ID NO: 29)
Последовательности аминокислот CDR G12v2 (IMGT)
CDR1 VH - GYTFTSYG (SEQ ID NO: 21)
CDR2 VH - ISAYSGGT (SEQ ID NO: 8)
CDR3 VH - ARDLFPTIFGVSYYYY (SEQ ID NO: 9)
CDR1 VL - QSISGR (SEQ ID NO: 22)
CDR2 VL - EAS (SEQ ID NO: 11)
CDR3 VL - QQSYSWPRT (SEQ ID NO: 29)
Последовательность аминокислот домена VH МАТ G12v2 (SEQ ID NO: 23)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSS
Последовательность нуклеиновой кислоты домена VH МАТ G12v2 (SEQ ID NO: 24)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGC
Последовательность аминокислот домена VL МАТ G12v2 (SEQ ID NO: 30)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
DIQMTQSPSTLSASVGDRVTITCRASQSISGRLAWYQQKPGKAPNLLIYEASNLESGVPSRFSGSGSGTEFTLTISSLQPEDFATYYCQQSYSWPRTFGQGTKVEIK
Последовательность нуклеиновой кислоты домена VL МАТ G12v2 (SEQ ID NO: 31)
GACATTCAGATGACCCAGTCCCCGAGCACGCTGTCCGCAAGCGTGGGGGACAGAGTGACCATCACTTGCCGCGCCTCACAATCCATCAGCGGACGCTTGGCCTGGTACCAGCAGAAGCCCGGAAAGGCCCCAAACCTTCTGATCTACGAAGCCTCGAACCTGGAGTCAGGCGTCCCTTCGCGGTTCTCTGGCTCCGGTTCCGGAACTGAGTTCACCCTCACCATCTCGTCCCTGCAACCGGAAGATTTCGCCACCTACTACTGCCAACAGTCGTACTCCTGGCCCCGGACATTCGGACAGGGAACCAAAGTCGAGATTAAG
Последовательность аминокислот тяжелой цепи МАТ G1-AA/G12v2 (с LALA) (SEQ ID NO: 27)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность аминокислот легкой цепи МАТ G1-AA/G12v2 и МАТ2 FS22-172-003-AA/G12v2 и FS22-053-008-AA/G12v2 (SEQ ID NO: 33)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
DIQMTQSPSTLSASVGDRVTITCRASQSISGRLAWYQQKPGKAPNLLIYEASNLESGVPSRFSGSGSGTEFTLTISSLQPEDFATYYCQQSYSWPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательности аминокислот CDR МАТ lambdav3/lam-G02v3 (Кабат)
CDR1 VH - SYGIS (SEQ ID NO: 1)
CDR2 VH - WISAYSGGTNYAQKLQG (SEQ ID NO: 2)
CDR3 VH - DLFPTIFGVSYYYY (SEQ ID NO: 3)
CDR1 VL - TGTSSDVGGYNYVS (SEQ ID NO: 34)
CDR2 VL - EVTNRPS (SEQ ID NO: 35)
CDR3 VL - SSFKRGSTLVV (SEQ ID NO: 36)
Последовательности аминокислот CDR lambdav3/lam-G02v3 (IMGT)
CDR1 VH - GYTFTSYG (SEQ ID NO: 21)
CDR2 VH - ISAYSGGT (SEQ ID NO: 8)
CDR3 VH - ARDLFPTIFGVSYYYY (SEQ ID NO: 9)
CDR1 VL - SSDVGGYNY (SEQ ID NO: 37)
CDR2 VL - EVT (SEQ ID NO: 38)
CDR3 VL - SSFKRGSTLVV (SEQ ID NO: 36)
Последовательность аминокислот домена VH МАТ lambdav3/lam-G02v3 (SEQ ID NO: 23)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSS
Последовательность нуклеиновой кислоты домена VH МАТ lambdav3/lam-G02v3 (SEQ ID NO: 24)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGC
Последовательность аминокислот домена VL МАТ lambdav3/lam-G02v3 (SEQ ID NO: 41)
CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
QSALTQPASVSGSPGQSITISCTGTSSDVGGYNYVSWYQQFPGKAPKLMIFEVTNRPSGVSDRFSGSKSDNTASLTISGLQAEDEAEYYCSSFKRGSTLVVFGGGTKLTVL
Последовательность нуклеиновой кислоты домена VL МАТ lambdav3/lam-G02v3 (SEQ ID NO: 42)
CAGTCGGCCCTTACTCAACCCGCGTCAGTCTCCGGTAGCCCCGGACAGTCCATCACGATTTCGTGCACCGGAACCAGCAGCGATGTCGGGGGATACAACTACGTGTCCTGGTACCAGCAGTTCCCGGGAAAGGCCCCTAAGCTGATGATCTTCGAAGTCACTAACAGACCTTCCGGAGTGTCGGACCGGTTCTCCGGCTCCAAGTCCGACAACACTGCGAGCCTGACCATCTCGGGCCTGCAAGCCGAGGACGAAGCCGAGTACTACTGTAGCTCATTCAAGCGCGGTTCCACCCTCGTGGTGTTCGGCGGTGGCACTAAGCTCACCGTGCTGGGA
Последовательность аминокислот тяжелой цепи МАТ G1-AA/lambdav3/lam-G02v3 (с LALA) (SEQ ID NO: 27)
Домен VH (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность аминокислот легкой цепи МАТ G1-AA/lambdav3/lam-G02v3 (SEQ ID NO: 44)
VL домен (курсивный шрифт); CDR IMGT (жирный и курсивный шрифт); CDR по Кабату (курсивный шрифт и подчеркнуты)
QSALTQPASVSGSPGQSITISCTGTSSDVGGYNYVSWYQQFPGKAPKLMIFEVTNRPSGVSDRFSGSKSDNTASLTISGLQAEDEAEYYCSSFKRGSTLVVFGGGTKLTVLGQPAAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Альтернативное определение HCDR1 (IMGT) (SEQ ID NO: 45)
GYX1FTSYG
где X1 представляет собой P или T
Альтернативное определение LCDR1 (Кабат) (SEQ ID NO: 46)
RASQSIX2X3RLA
где X2 представляет собой S или G, X3 представляет собой N или G
Альтернативное определение LCDR2 (Кабат) (SEQ ID NO: 47)
EASX4X5EX6
Где X4 представляет собой T или N; X5 представляет собой S или L; X6 представляет собой T или S
Альтернативное определение LCDR3 (Кабат) (SEQ ID NO: 48)
QQSYSX7PX8X9T
Где X7 представляет собой T или W; X8 отсутствует или представляет собой R; X9 представляет собой Y, R или V
Альтернативное определение LCDR1 (IMGT) (SEQ ID NO: 49)
QSIX10X11R
Где X10 представляет собой G или S; X11 представляет собой N или G
Альтернативное определение LCDR3 (IMGT) (SEQ ID NO: 50)
QQSYSX12X13X14X15T
Где X12 отсутствует или представляет собой T; X13 представляет собой T, W или P; X14 представляет собой P или R; X15 представляет собой Y или R
Последовательности аминокислот HFW IgG1 (Кабат)
HFW1 - EVQLVQSGAEVKRPGASVKVSCKASGYX16FT (SEQ ID NO: 51)
Где X16 представляет собой P или T
HFW2 - WVRQAPGQGLEWMG (SEQ ID NO: 52)
HFW3 - RVTMTTDTSTSTAYMELRSLRSDDTAVYYCAR (SEQ ID NO: 53)
HFW4 - WGQGTLVTVSS (SEQ ID NO: 54)
Последовательности аминокислот HFW IgG1 (IMGT)
HFW1 - EVQLVQSGAEVKRPGASVKVSCKAS (SEQ ID NO: 55)
HFW2 - ISWVRQAPGQGLEWMGW (SEQ ID NO: 56)
HFW3 - NYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYC (SEQ ID NO: 57)
HFW4 - WGQGTLVTVSS (SEQ ID NO: 54)
Последовательности аминокислот LFW легкой цепи каппа (Кабат)
LFW1 - DIQMTQSPSTLSASVRDRVIITC (SEQ ID NO: 58)
LFW2 - WYQHKPGKAPKLLIY (SEQ ID NO: 59)
LFW3 - GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO: 60)
LFW4 - FGQGTKLEIK (SEQ ID NO: 61)
Последовательности аминокислот LFW легкой цепи каппа (IMGT)
LFW1 - DIQMTQSPSTLSASVRDRVIITCRAS (SEQ ID NO: 62)
LFW2 - LAWYQHKPGKAPKLLIY (SEQ ID NO: 63)
LFW3 - TSETGVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO: 64)
LFW4 - FGQGTKLEIK (SEQ ID NO: 61)
Последовательности аминокислот LFW легкой цепи лямбда (Кабат)
LFW1 - QSALTQPASVSGSPGQSITISC (SEQ ID NO: 65)
LFW2 - WYQQFPGKAPKLMIF (SEQ ID NO: 66)
LFW3 - GVSDRFSGSKSDNTASLTISGLQAEDEAEYYC (SEQ ID NO: 67)
LFW4 - FGGGTKLTVL (SEQ ID NO: 68)
Последовательности аминокислот LFW легкой цепи лямбда (IMGT)
LFW1 - QSALTQPASVSGSPGQSITISCTGT (SEQ ID NO: 69)
LFW2 - VSWYQQFPGKAPKLMIF (SEQ ID NO: 70)
LFW3 - NRPSGVSDRFSGSKSDNTASLTISGLQAEDEAEYYC (SEQ ID NO: 71)
LFW4 - FGGGTKLTVL (SEQ ID NO: 68)
Последовательности аминокислот структурных петель домена CH3 ДТ
Петля AB ДТ - RDELTKNQ (SEQ ID NO: 72)
Петля CD ДТ - SNGQPENNY (SEQ ID NO: 73)
Петля EF ДТ - DKSRWQQGNV (SEQ ID NO: 74)
Последовательность аминокислот домена CH3 ДТ (SEQ ID NO: 75)
Петли AB, CD и EF подчеркнуты
GQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность аминокислот домена CH2 (SEQ ID NO: 76)
APELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAK
Последовательность аминокислот домена CH2 с мутацией LALA (SEQ ID NO: 77)
Мутация LALA подчеркнута
APEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAK
Последовательность аминокислот домена CH2 с мутацией LALA и мутацией P114A (SEQ ID NO: 176)
Мутация LALA подчеркнута; мутация P114A выделена жирным шрифтом и подчеркнута
APEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALAAPIEKTISKAK
Последовательность аминокислот мотива PPY (SEQ ID NO: 78)
PPY
Последовательности аминокислот структурной петли домена CH3 FS22-053-008
Первая последовательность FS22-053-008 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-008 - DYWRWLE (SEQ ID NO: 80)
Последовательность аминокислот домена CH3 FS22-053-008 (SEQ ID NO: 81)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 FS22-053-008 (SEQ ID NO: 82)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTACTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACTGGAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательности аминокислот структурных петель домена CH3 FS22-053-009
Первая последовательность FS22-053-009 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-009 - EHTRWLD (SEQ ID NO: 83)
Последовательность аминокислот домена CH3 FS22-053-009 (SEQ ID NO: 84)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVEHTRWLDGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 FS22-053-009 (SEQ ID NO: 85)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGAACATACTAGGTGGCTGGATGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-011
Первая последовательность FS22-053-011 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-011 - DYWRWTD (SEQ ID NO: 86)
Последовательность аминокислот домена CH3 Fcab FS22-053-011 (SEQ ID NO: 87)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYWRWTDGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-011 (SEQ ID NO: 88)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACTGGAGGTGGACTGATGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-017
Первая последовательность FS22-053-017 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-017 - YHWRWLE (SEQ ID NO: 89)
Последовательность аминокислот домена CH3 Fcab FS22-053-017 (SEQ ID NO: 90)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYHWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-017 (SEQ ID NO: 91)
GGACAGCCTCGAGAACCACAGGTGTACACTCTGCCCCCTTCACGCGACGAACTCAATCCGCCCTACCTGTTCTCCAACCAAGTCTCCCTGACCTGTCTTGTGAAGGGTTTCTACCCATCCGATATCGCCGTGGAGTGGGAGAGCAACGGACAGCCGGAGAACAACTATAAGACTACCCCGCCTGTGCTGGACTCGGACGGCAGCTTCTTCTTGTACTCCAAACTGACCGTGTACCACTGGCGGTGGCTGGAAGGGAACGTGTTTAGCTGCTCCGTCATGCATGAAGCCCTGCACAACCACTACACCCAGAAGTCCCTCTCGCTCTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-014
Первая последовательность FS22-053-014 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-014 - YHWRWLD (SEQ ID NO: 92)
Последовательность аминокислот домена CH3 Fcab FS22-053-014 (SEQ ID NO: 93)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYHWRWLDGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-014 (SEQ ID NO: 94)
GGACAGCCTCGAGAGCCTCAAGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGTACCATTGGAGGTGGCTGGATGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-010
Первая последовательность FS22-053-010 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-010 - DYMRWLD (SEQ ID NO: 95)
Последовательность аминокислот домена CH3 Fcab FS22-053-010 (SEQ ID NO: 96)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYMRWLDGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-010 (SEQ ID NO: 97)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACATGAGGTGGCTGGATGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-012
Первая последовательность FS22-053-012 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-012 - DHMRWLE (SEQ ID NO: 98)
Последовательность аминокислот домена CH3 Fcab FS22-053-012 (SEQ ID NO: 99)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDHMRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-012 (SEQ ID NO: 100)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCATATGAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-013
Первая последовательность FS22-053-013 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-013 - GYERWLE (SEQ ID NO: 101)
Последовательность аминокислот домена CH3 Fcab FS22-053-013 (SEQ ID NO: 102)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGYERWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-013 (SEQ ID NO: 103)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGTTACGAAAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-015
Первая последовательность FS22-053-015 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-015 - DHWRWLQ (SEQ ID NO: 104)
Последовательность аминокислот домена CH3 Fcab FS22-053-015 (SEQ ID NO: 105)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDHWRWLQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-015 (SEQ ID NO: 106)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGTTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATCATTGGAGGTGGCTGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-053-016
Первая последовательность FS22-053-016 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-016 - DYIRWLN (SEQ ID NO: 107)
Последовательность аминокислот домена CH3 Fcab FS22-053-016 (SEQ ID NO: 108)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYIRWLNGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053-016 (SEQ ID NO: 109)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACATCAGGTGGCTGAACGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 FS22-053
Первая последовательность FS22-053-008 - NPPYLFS (SEQ ID NO: 79)
Вторая последовательность FS22-053-008 - YYNRWQD (SEQ ID NO: 110)
Последовательности аминокислот домена CH3 FS22-053 (SEQ ID NO: 111)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYYNRWQDGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-053 (SEQ ID NO: 112)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGTATTATAACAGGTGGCAGGATGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-003
Первая последовательность FS22-172-003 - PYIIPPY (SEQ ID NO: 113)
Вторая последовательность FS22-172-003 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-003 (SEQ ID NO: 115)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-003 (SEQ ID NO: 116)
GGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCCATACATCATCCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-002
Первая последовательность FS22-172-002 - PFQMPPY (SEQ ID NO: 117)
Вторая последовательность FS22-172-002 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-002 (SEQ ID NO: 118)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELPFQMPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-002 (SEQ ID NO: 119)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCCATTCCAGATGCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-004
Первая последовательность FS22-172-004 - NYIYPPY (SEQ ID NO: 120)
Вторая последовательность FS22-172-004 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-004 (SEQ ID NO: 121)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELNYIYPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-004 (SEQ ID NO: 122)
GGACAGCCTCGAGAGCCTCAAGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACTACATCTACCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGA
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-001
Первая последовательность FS22-172-001 - PFVMPPY (SEQ ID NO: 123)
Вторая последовательность FS22-172-001 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-001 (SEQ ID NO: 124)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELPFVMPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-001 (SEQ ID NO: 125)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCCATTCGTTATGCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-005
Первая последовательность FS22-172-005 - QQVYPPY (SEQ ID NO: 126)
Вторая последовательность FS22-172-005 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-005 (SEQ ID NO: 127)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELQQVYPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-005 (SEQ ID NO: 128)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCAGCAGGTTTACCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172-006
Первая последовательность FS22-172-006 - RKYYPPY (SEQ ID NO: 129)
Вторая последовательность FS22-172-006 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172-006 (SEQ ID NO: 130)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELRKYYPPYNQLSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172-006 (SEQ ID NO: 131)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCGTAAATACTACCCGCCGTACAACCAGCTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательности аминокислот структурных петель домена CH3 Fcab FS22-172
Первая последовательность FS22-172 - RKYYPPY (SEQ ID NO: 129)
Вторая последовательность FS22-172 - GADRWLE (SEQ ID NO: 114)
Последовательность аминокислот домена CH3 Fcab FS22-172 (SEQ ID NO: 132)
Первая и вторая последовательности подчеркнуты
GQPREPQVYTLPPSRDELRKYYPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты домена CH3 Fcab FS22-172 (SEQ ID NO: 133)
GGCCAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCGTAAATACTACCCGCCGTACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACACAGAAGAGCCTCTCCCTGTCTCCGGGT
Последовательность аминокислот тяжелой цепи МАТ2 FS22-172-003-AA/E12v2 с мутацией LALA (SEQ ID NO: 134)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-172-003-AA/E12v2 с мутацией LALA (SEQ ID NO: 135)
GAAGTTCAGCTGGTTCAGTCTGGCGCCGAAGTGAAAAGACCTGGCGCCTCTGTGAAGGTGTCCTGCAAGGCTTCTGGCTACCCCTTTACCTCCTACGGCATCTCCTGGGTCCGACAGGCTCCTGGACAAGGCTTGGAATGGATGGGCTGGATCTCCGCTTATTCCGGCGGCACCAATTACGCCCAGAAACTGCAGGGCAGAGTGACCATGACCACCGACACCTCTACCTCCACCGCCTACATGGAACTGCGGTCCCTGAGATCTGACGACACCGCCGTGTACTACTGCGCCAGAGATCTGTTCCCCACCATCTTCGGCGTGTCCTACTACTACTATTGGGGCCAGGGCACCCTGGTCACCGTGTCCTCTGCTTCTACCAAGGGACCCAGCGTGTTCCCTCTGGCTCCTTCCAGCAAGTCTACCTCTGGCGGAACAGCTGCTCTGGGCTGCCTGGTCAAGGACTACTTTCCTGAGCCTGTGACCGTGTCTTGGAACTCTGGCGCTCTGACATCTGGCGTGCACACCTTTCCAGCTGTGCTGCAGTCCTCCGGCCTGTACTCTCTGTCCTCTGTCGTGACCGTGCCTTCCAGCTCTCTGGGAACCCAGACCTACATCTGCAATGTGAACCACAAGCCTTCCAACACCAAGGTGGACAAGAAGGTGGAACCCAAGTCCTGCGACAAGACCCACACCTGTCCTCCATGTCCTGCTCCAGAAGCTGCTGGCGGCCCTTCCGTGTTTCTGTTCCCTCCAAAGCCTAAGGACACCCTGATGATCTCTCGGACCCCTGAAGTGACCTGCGTGGTGGTGGATGTGTCTCACGAGGACCCAGAAGTGAAGTTCAATTGGTACGTGGACGGCGTGGAAGTGCACAACGCCAAGACCAAGCCTAGAGAGGAACAGTACAACTCCACCTACAGAGTGGTGTCCGTGCTGACCGTGCTGCACCAGGATTGGCTGAACGGCAAAGAGTACAAGTGCAAGGTGTCCAACAAGGCCCTGCCTGCTCCTATCGAAAAGACCATCTCCAAGGCCAAGGGCCAGCCTAGGGAACCCCAGGTTTACACCTTGCCTCCATCTCGGGACGAGCTGCCCTACATCATCCCTCCATACAACCAGGTGTCCCTGACCTGCCTCGTGAAGGGCTTCTACCCTTCCGATATCGCCGTGGAATGGGAGAGCAATGGCCAGCCTGAGAACAACTACAAGACAACCCCTCCTGTGCTGGACTCCGACGGCTCATTCTTTCTGTACTCCAAGCTGACAGTGGGCGCCGACAGATGGCTGGAAGGGAACGTGTTCTCCTGCAGCGTGATGCACGAGGCCCTGCACAATCACTACACACAGAAGTCCCTGTCTCTGTCCCCTGGC
Последовательность аминокислот легкой цепи МАТ2 FS22-172-003-AA/E12v2 и FS22-053-008-AA/E12v2 (SEQ ID NO: 17)
DIQMTQSPSTLSASVRDRVIITCRASQSIGNRLAWYQHKPGKAPKLLIYEASTSETGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQSYSTPYTFGQGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-172-003-AA/E12v2 (SEQ ID NO: 136)
GACATCCAGATGACCCAGTCTCCATCCACACTGTCCGCCTCTGTGCGGGACAGAGTGATCATCACCTGTAGAGCCAGCCAGTCCATCGGCAACAGACTGGCCTGGTATCAGCACAAGCCTGGCAAGGCTCCCAAGCTGCTGATCTACGAGGCCTCCACATCTGAGACAGGCGTGCCCTCTAGATTCTCCGGCTCTGGCTCTGGCACCGACTTTACCCTGACAATCTCCAGCCTGCAGCCTGAGGACTTCGCCACCTACTACTGCCAGCAGTCCTACAGCACCCCTTACACCTTTGGCCAGGGCACCAAGCTGGAAATCAAGCGTACGGTGGCCGCTCCCAGCGTGTTCATCTTCCCCCCAAGCGACGAGCAGCTGAAGAGCGGCACCGCCAGCGTGGTGTGTCTGCTGAACAACTTCTACCCCAGGGAGGCCAAGGTGCAGTGGAAGGTGGACAACGCCCTGCAGAGCGGCAACAGCCAGGAGAGCGTCACCGAGCAGGACAGCAAGGACTCCACCTACAGCCTGAGCAGCACCCTGACCCTGAGCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGTGAGGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGAGCTTCAACAGGGGCGAGTGC
Последовательность аминокислот тяжелой цепи МАТ2 FS22-172-003-AA/E05v2 с мутацией LALA (SEQ ID NO: 137)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-172-003-AA/E05v2 с мутацией LALA (SEQ ID NO: 138)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGCGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCCATACATCATCCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-172-003-AA/E05v2 (SEQ ID NO: 139)
GACATTCAGATGACCCAATCCCCGTCCACGCTGAGCGCCTCCGTCGGTGATCGCGTGACAATCACTTGTCGGGCGTCGCAGTCCATCTCTGGAAGGCTCGCCTGGTACCAGCAGAAGCCTGGAAAGGCTCCCAACCTCCTTATCTACGAAGCCAGCAACCTGGAGTCCGGAGTGCCTAGCCGGTTCAGCGGATCAGGGTCCGGTACCGAGTTCACCCTGACCATTTCCTCGCTCCAACCTGAGGACTTCGCCACCTACTACTGCCAACAGTCCTATTCAACTCCGCGCGTGACCTTCGGCCAGGGCACTAAGGTCGAAATCAAAAGAACCGTGGCAGCCCCATCGGTGTTTATCTTCCCGCCCTCGGACGAACAGCTGAAGTCAGGCACTGCTAGCGTGGTCTGTCTCCTGAACAATTTCTACCCGCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAACGCGCTGCAGTCCGGAAACAGCCAGGAGTCAGTGACCGAGCAGGACTCCAAGGATTCCACTTATTCCCTGTCCTCCACCCTGACTTTGAGCAAGGCCGACTACGAGAAGCACAAAGTGTACGCCTGCGAAGTGACCCATCAAGGGCTTTCGTCGCCCGTGACCAAGAGCTTCAACCGGGGCGAATGC
Последовательность аминокислот (АК) тяжелой цепи МАТ2 FS22-172-003-AA/G12v2 с мутацией LALA (SEQ ID NO: 140)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-172-003-AA/G12v2 с мутацией LALA (SEQ ID NO: 141)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGCGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGCCATACATCATCCCACCATACAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGGCGCAGATAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-172-003-AA/G12v2 (SEQ ID NO: 142)
GACATTCAGATGACCCAGTCCCCGAGCACGCTGTCCGCAAGCGTGGGGGACAGAGTGACCATCACTTGCCGCGCCTCACAATCCATCAGCGGACGCTTGGCCTGGTACCAGCAGAAGCCCGGAAAGGCCCCAAACCTTCTGATCTACGAAGCCTCGAACCTGGAGTCAGGCGTCCCTTCGCGGTTCTCTGGCTCCGGTTCCGGAACTGAGTTCACCCTCACCATCTCGTCCCTGCAACCGGAAGATTTCGCCACCTACTACTGCCAACAGTCGTACTCCTGGCCCCGGACATTCGGACAGGGAACCAAAGTCGAGATTAAGCGGACTGTGGCGGCTCCTAGCGTGTTCATCTTTCCCCCGTCCGACGAACAGCTGAAGTCCGGTACCGCTAGCGTGGTCTGTCTCCTGAACAATTTCTACCCGCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAACGCGCTGCAGTCCGGAAACAGCCAGGAGTCAGTGACCGAGCAGGACTCCAAGGATTCCACTTATTCCCTGTCCTCCACCCTGACTTTGAGCAAGGCCGACTACGAGAAGCACAAAGTGTACGCCTGCGAAGTGACCCATCAAGGGCTTTCGTCGCCCGTGACCAAGAGCTTCAACCGGGGCGAATGC
Последовательность АК тяжелой цепи МАТ2 FS22-053-008-AA/E12v2 с мутацией LALA (SEQ ID NO: 143)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-053-008-AA/E12v2 с мутацией LALA (SEQ ID NO: 144)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACCCCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGCGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTACTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACTGGAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-053-008-AA/E12v2 (SEQ ID NO: 145)
GACATCCAGATGACGCAGAGCCCGTCTACCCTGTCCGCCTCCGTGAGAGATCGCGTGATCATCACCTGTCGGGCCAGCCAGTCCATCGGAAACCGCTTGGCGTGGTACCAGCACAAGCCTGGGAAGGCTCCGAAGCTGCTCATCTACGAAGCCTCGACTTCGGAGACTGGTGTCCCTAGCCGGTTCAGCGGATCGGGATCAGGGACCGATTTCACTCTGACCATTTCCTCCCTGCAACCCGAGGACTTCGCCACCTACTACTGCCAACAGTCATATTCCACCCCGTACACCTTCGGACAAGGCACCAAGCTCGAAATCAAGCGGACTGTCGCCGCACCTTCCGTGTTCATTTTCCCACCCTCCGACGAACAGCTGAAATCGGGTACAGCTAGCGTGGTCTGTCTCCTGAACAATTTCTACCCGCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAACGCGCTGCAGTCCGGAAACAGCCAGGAGTCAGTGACCGAGCAGGACTCCAAGGATTCCACTTATTCCCTGTCCTCCACCCTGACTTTGAGCAAGGCCGACTACGAGAAGCACAAAGTGTACGCCTGCGAAGTGACCCATCAAGGGCTTTCGTCGCCCGTGACCAAGAGCTTCAACCGGGGCGAATGC
Последовательность АК тяжелой цепи FS22-053-008-AA/E05v2 с мутацией LALA (SEQ ID NO: 146)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-053-008-AA/E05v2 с мутацией LALA (SEQ ID NO: 147)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGCGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTACTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACTGGAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-053-008-AA/E05v2 (SEQ ID NO: 148)
GACATTCAGATGACCCAATCCCCGTCCACGCTGAGCGCCTCCGTCGGTGATCGCGTGACAATCACTTGTCGGGCGTCGCAGTCCATCTCTGGAAGGCTCGCCTGGTACCAGCAGAAGCCTGGAAAGGCTCCCAACCTCCTTATCTACGAAGCCAGCAACCTGGAGTCCGGAGTGCCTAGCCGGTTCAGCGGATCAGGGTCCGGTACCGAGTTCACCCTGACCATTTCCTCGCTCCAACCTGAGGACTTCGCCACCTACTACTGCCAACAGTCCTATTCAACTCCGCGCGTGACCTTCGGCCAGGGCACTAAGGTCGAAATCAAAAGAACCGTGGCAGCCCCATCGGTGTTTATCTTCCCGCCCTCGGACGAACAGCTGAAGTCAGGCACTGCTAGCGTGGTCTGTCTCCTGAACAATTTCTACCCGCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAACGCGCTGCAGTCCGGAAACAGCCAGGAGTCAGTGACCGAGCAGGACTCCAAGGATTCCACTTATTCCCTGTCCTCCACCCTGACTTTGAGCAAGGCCGACTACGAGAAGCACAAAGTGTACGCCTGCGAAGTGACCCATCAAGGGCTTTCGTCGCCCGTGACCAAGAGCTTCAACCGGGGCGAATGC
Последовательность АК тяжелой цепи FS22-053-008-AA/G12v2 с мутацией LALA (SEQ ID NO: 149)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDYWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность нуклеиновой кислоты тяжелой цепи МАТ2 FS22-053-008-AA/G12v2 с мутацией LALA (SEQ ID NO: 150)
GAAGTGCAGCTGGTGCAGTCCGGAGCCGAAGTCAAGAGGCCTGGAGCGTCCGTGAAGGTGTCCTGCAAAGCCTCAGGATACACCTTCACTTCGTACGGGATTTCCTGGGTCCGCCAAGCACCGGGTCAAGGCTTGGAGTGGATGGGATGGATCAGCGCGTATTCCGGGGGAACCAACTACGCTCAAAAGCTGCAGGGTCGCGTGACCATGACCACCGATACCTCCACCTCAACGGCCTACATGGAACTGAGATCTCTGCGGAGCGACGACACTGCCGTGTACTACTGTGCCCGGGACCTGTTCCCCACTATCTTCGGAGTGTCGTACTACTACTACTGGGGCCAGGGGACTCTCGTGACCGTGTCGAGCGCTAGCACTAAGGGCCCGTCGGTGTTCCCGCTGGCCCCATCGTCCAAGAGCACATCAGGGGGTACCGCCGCCCTGGGCTGCCTTGTGAAGGATTACTTTCCCGAGCCCGTCACAGTGTCCTGGAACAGCGGAGCCCTGACCTCCGGAGTGCATACTTTCCCGGCTGTGCTTCAGTCCTCTGGCCTGTACTCATTGTCCTCCGTGGTCACCGTCCCTTCGTCCTCCCTGGGCACCCAGACCTATATCTGTAATGTCAACCATAAGCCCTCGAACACCAAGGTCGACAAGAAGGTCGAGCCGAAGTCGTGCGACAAGACTCACACTTGCCCGCCTTGCCCAGCCCCGGAAGCTGCCGGTGGTCCTTCGGTGTTCCTCTTCCCGCCCAAGCCGAAGGATACCCTGATGATCTCACGGACCCCCGAAGTGACCTGTGTGGTGGTGGACGTGTCCCACGAGGACCCGGAAGTGAAATTCAATTGGTACGTGGATGGAGTGGAAGTGCACAACGCCAAGACCAAGCCACGGGAAGAACAGTACAACTCTACCTACCGCGTGGTGTCCGTGCTCACTGTGCTGCACCAAGACTGGCTGAACGGGAAGGAGTACAAGTGCAAAGTGTCCAACAAGGCGCTGCCTGCCCCAATTGAGAAAACTATCTCGAAAGCCAAGGGACAGCCTCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGATGAGCTGAACCCGCCGTACCTGTTCTCTAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTACTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGATTACTGGAGGTGGCTGGAAGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCGCTGCACAACCACTACACTCAGAAGAGCTTGTCCCTGTCGCCCGGT
Последовательность нуклеиновой кислоты легкой цепи МАТ2 FS22-053-008-AA/G12v2 (SEQ ID NO: 151)
GACATTCAGATGACCCAGTCCCCGAGCACGCTGTCCGCAAGCGTGGGGGACAGAGTGACCATCACTTGCCGCGCCTCACAATCCATCAGCGGACGCTTGGCCTGGTACCAGCAGAAGCCCGGAAAGGCCCCAAACCTTCTGATCTACGAAGCCTCGAACCTGGAGTCAGGCGTCCCTTCGCGGTTCTCTGGCTCCGGTTCCGGAACTGAGTTCACCCTCACCATCTCGTCCCTGCAACCGGAAGATTTCGCCACCTACTACTGCCAACAGTCGTACTCCTGGCCCCGGACATTCGGACAGGGAACCAAAGTCGAGATTAAGCGGACTGTGGCGGCTCCTAGCGTGTTCATCTTTCCCCCGTCCGACGAACAGCTGAAGTCCGGTACCGCTAGCGTGGTCTGTCTCCTGAACAATTTCTACCCGCGCGAAGCTAAGGTCCAGTGGAAGGTCGACAACGCGCTGCAGTCCGGAAACAGCCAGGAGTCAGTGACCGAGCAGGACTCCAAGGATTCCACTTATTCCCTGTCCTCCACCCTGACTTTGAGCAAGGCCGACTACGAGAAGCACAAAGTGTACGCCTGCGAAGTGACCCATCAAGGGCTTTCGTCGCCCGTGACCAAGAGCTTCAACCGGGGCGAATGC
Последовательность АК тяжелой цепи FS22-053-017AA/E12v2 с мутацией LALA (SEQ ID NO: 152)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYHWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи FS22-053-017AA/E05v2 с мутацией LALA (SEQ ID NO: 153)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYHWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи FS22-053-017AA/G12v2 с мутацией LALA (SEQ ID NO: 154)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELNPPYLFSNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVYHWRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи FS22-172-003AA/lam-G02v3 с мутацией LALA (SEQ ID NO: 155)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи FS22-172-003AA/S70 с мутацией LALA (SEQ ID NO: 156)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК легкой цепи S70 (SEQ ID NO: 157)
Домен VL (курсивный шрифт);
DIQMTQSPSSLSASVGDRVTITCRASQDVSTAVAWYQQKPGKAPKLLIYSASFLYSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQYLYHPATFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность АК тяжелой цепи FS22-172-003AA/HelD1.3 с мутацией LALA (SEQ ID NO: 158)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
QVQLQESGPGLVRPSQTLSLTCTVSGSTFSGYGVNWVRQPPGRGLEWIGMIWGDGNTDYNSALKSRVTMLVDTSKNQFSLRLSSVTAADTAVYYCARERDYRLDYWGQGSLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELPYIIPPYNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVGADRWLEGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК легкой цепи HelD1.3 (SEQ ID NO: 159)
Домен VL (курсивный шрифт)
DIQMTQSPASLSASVGETVTITCRASGNIHNYLAWYQQKQGKSPQLLVYNAKTLADGVPSRFSGSGSGTQYSLKINSLQPEDFGSYYCQHFWSTPRTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность АК тяжелой цепи FS22m-063AA/S70 с мутацией LALA (SEQ ID NO: 160)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEPYWSYVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVMNYRWELGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи FS22m-063AA/HelD1.3 с мутацией LALA (SEQ ID NO: 161)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
QVQLQESGPGLVRPSQTLSLTCTVSGSTFSGYGVNWVRQPPGRGLEWIGMIWGDGNTDYNSALKSRVTMLVDTSKNQFSLRLSSVTAADTAVYYCARERDYRLDYWGQGSLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDEPYWSYVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVMNYRWELGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи G1-AA/E12v2 с мутацией LALA (SEQ ID NO: 162)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVQSGAEVKRPGASVKVSCKASGYPFTSYGISWVRQAPGQGLEWMGWISAYSGGTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность АК тяжелой цепи G1/S70 (SEQ ID NO: 163)
Домен VH (курсивный шрифт)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность АК тяжелой цепи G1-AA/S70 с мутацией LALA (SEQ ID NO: 164)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDSWIHWVRQAPGKGLEWVAWISPYGGSTYYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCARRHWPGGFDYWGQGTLVTVSAASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность АК тяжелой цепи G1/4420 (SEQ ID NO: 165)
Домен VH (курсивный шрифт)
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность АК легкой цепи 4420 (SEQ ID NO: 166)
Домен VL (курсивный шрифт)
DVVMTQTPLSLPVSLGDQASISCRSSQSLVHSNGNTYLRWYLQKPGQSPKVLIYKVSNRFSGVPDRFSGSGSGTDFTLKISRVEAEDLGVYFCSQSTHVPWTFGGGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательность АК тяжелой цепи G1-AA/4420 с мутацией LALA (SEQ ID NO: 167)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EVKLDETGGGLVQPGRPMKLSCVASGFTFSDYWMNWVRQSPEKGLEWVAQIRNKPYNYETYYSDSVKGRFTISRDDSKSSVYLQMNNLRVEDMGIYYCTGSYYGMDYWGQGTSVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность АК тяжелой цепи G1-AA/HelD1.3 с мутацией LALA (SEQ ID NO: 168)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
QVQLQESGPGLVRPSQTLSLTCTVSGSTFSGYGVNWVRQPPGRGLEWIGMIWGDGNTDYNSALKSRVTMLVDTSKNQFSLRLSSVTAADTAVYYCARERDYRLDYWGQGSLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность АК тяжелой цепи G1-AA/MLS109 с мутацией LALA (SEQ ID NO: 169)
Домен VH (курсивный шрифт); мутация LALA (жирный шрифт и подчеркнута)
EEQVLESGGGLVKPGGSLRLSCAASGFTFSPYSVFWVRQAPGKGLEWVSSINSDSTYKYYADSVKGRFTISRDNAENSIFLQMNSLRAEDTAVYYCARDRSYYAFSSGSLSDYYYGLDVWGQGTTVIVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
Последовательность аминокислот легкой цепи MLS109 (SEQ ID NO: 170)
Домен VL (курсивный шрифт)
DIVMTQSPLSLSVTPGEPASISCRSSQSLLHTNGYNYLDWYVQKPGQSPQLLIYLASNRASGVPDRFSGSGSGTDFTLKISRVETEDVGVYYCMQALQIPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Последовательности аминокислот петель AB и EF домена CH3 FS22-172-003
Петля AB - RDELPYIIPPYNQ (SEQ ID NO: 171)
Петля EF - GADRWLEGNV (SEQ ID NO: 172)
Последовательности аминокислот петель AB и EF домена CH3 FS22-53-008
Петля AB - RDELNPPYLFSNQ (SEQ ID NO: 173)
Петля EF - DYWRWLEGNV (SEQ ID NO: 174)
Последовательности аминокислот петель AB и EF домена CH3 FS22-053-017
Петля AB - RDELNPPYLFSNQ (SEQ ID NO: 173)
Петля EF - YHWRWLEGNV (SEQ ID NO: 175)
Последовательность аминокислот тяжелой цепи G1/280_02_G02 (SEQ ID NO: 177)
Домен VH (курсивный шрифт)
EVQLVQSGAEVKRPGASVKVSCKASGYTFTSYGISWVRQAPGQGLEWMGWISAYNGNTNYAQKLQGRVTMTTDTSTSTAYMELRSLRSDDTAVYYCARDLFPTIFGVSYYYYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Последовательность аминокислот легкой цепи G1/280_02_G02 (SEQ ID NO: 178)
Домен VL (курсивный шрифт)
QSALTQPASVSGSPGQSITISCTGTSSDVGGYNSVSWYQQFPGKAPKLMIFEVTNRPSGVSDRFSGSKSDNTASLTISGLQAEDEAEYYCSSFKRGSTLVVFGGGTKLTVLGQPAAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
Последовательность аминокислот PD-L1 человека (SEQ ID NO: 179)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERTHLVILGAILLCLGVALTFIFRLRKGRMMDVKKCGIQDTNSKKQSDTHLEET
Последовательность аминокислот внеклеточного домена PD-L1 человека (SEQ ID NO: 180)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNER
Последовательность аминокислот PD-L1 мыши (SEQ ID NO: 181)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
FTITAPKDLYVVEYGSNVTMECRFPVERELDLLALVVYWEKEDEQVIQFVAGEEDLKPQHSNFRGRASLPKDQLLKGNAALQITDVKLQDAGVYCCIISYGGADYKRITLKVNAPYRKINQRISVDPATSEHELICQAEGYPEAEVIWTNSDHQPVSGKRSVTTSRTEGMLLNVTSSLRVNATANDVFYCTFWRSQPGQNHTAELIIPELPATHPPQNRTHWVLLGSILLFLIVVSTVLLFLRKQVRMLDVEKCGVEDTSSKNRNDTQFEET
Последовательность аминокислот внеклеточного домена PD-L1 мыши (SEQ ID NO: 182)
FTITAPKDLYVVEYGSNVTMECRFPVERELDLLALVVYWEKEDEQVIQFVAGEEDLKPQHSNFRGRASLPKDQLLKGNAALQITDVKLQDAGVYCCIISYGGADYKRITLKVNAPYRKINQRISVDPATSEHELICQAEGYPEAEVIWTNSDHQPVSGKRSVTTSRTEGMLLNVTSSLRVNATANDVFYCTFWRSQPGQNHTAELIIPELPATHPPQNRTH
Последовательность аминокислот PD-L1 яванского макака (SEQ ID NO: 183)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLTSLIVYWEMEDKNIIQFVHGEEDLKVQHSNYRQRAQLLKDQLSLGNAALRITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLLNVTSTLRINTTANEIFYCIFRRLDPEENHTAELVIPELPLALPPNERTHLVILGAIFLLLGVALTFIFYLRKGRMMDMKKCGIRVTNSKKQRDTQLEETKGPSARFDIPDEIPVIESKPNTLSIVLGTTLTVAMIIVATIFGYRRQKGRLRTLKL
Последовательность аминокислот внеклеточного домена PD-L1 яванского макака (SEQ ID NO: 184)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLTSLIVYWEMEDKNIIQFVHGEEDLKVQHSNYRQRAQLLKDQLSLGNAALRITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLLNVTSTLRINTTANEIFYCIFRRLDPEENHTAELVIPELPLALPPNERTH
Последовательность аминокислот CD137 человека (SEQ ID NO: 185)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
LQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQIISFFLALTSTALLFLLFFLTLRFSVVKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
Последовательность аминокислот внеклеточного домена CD137 человека (SEQ ID NO: 186)
LQDPCSNCPAGTFCDNNRNQICSPCPPNSFSSAGGQRTCDICRQCKGVFRTRKECSSTSNAECDCTPGFHCLGAGCSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSVTPPAPAREPGHSPQ
Последовательность аминокислот CD137 мыши (SEQ ID NO: 187)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
VQNSCDNCQPGTFCRKYNPVCKSCPPSTFSSIGGQPNCNICRVCAGYFRFKKFCSSTHNAECECIEGFHCLGPQCTRCEKDCRPGQELTKQGCKTCSLGTFNDQNGTGVCRPWTNCSLDGRSVLKTGTTEKDVVCGPPVVSFSPSTTISVTPEGGPGGHSLQVLTLFLALTSALLLALIFITLLFSVLKWIRKKFPHIFKQPFKKTTGAAQEEDACSCRCPQEEEGGGGGYEL
Последовательность аминокислот внеклеточного домена CD137 мыши (SEQ ID NO: 188)
VQNSCDNCQPGTFCRKYNPVCKSCPPSTFSSIGGQPNCNICRVCAGYFRFKKFCSSTHNAECECIEGFHCLGPQCTRCEKDCRPGQELTKQGCKTCSLGTFNDQNGTGVCRPWTNCSLDGRSVLKTGTTEKDVVCGPPVVSFSPSTTISVTPEGGPGGHSLQVL
Последовательность аминокислот CD137 яванского макака (SEQ ID NO: 189)
Внеклеточный домен (курсивный шрифт); трансмембранный и внутриклеточный домены (жирный шрифт)
LQDLCSNCPAGTFCDNNRSQICSPCPPNSFSSAGGQRTCDICRQCKGVFKTRKECSSTSNAECDCISGYHCLGAECSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSATPPAPAREPGHSPQIIFFLALTSTVVLFLLFFLVLRFSVVKRSRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
Последовательность аминокислот внеклеточного домена CD137 яванского макака (SEQ ID NO: 190)
LQDLCSNCPAGTFCDNNRSQICSPCPPNSFSSAGGQRTCDICRQCKGVFKTRKECSSTSNAECDCISGYHCLGAECSMCEQDCKQGQELTKKGCKDCCFGTFNDQKRGICRPWTNCSLDGKSVLVNGTKERDVVCGPSPADLSPGASSATPPAPAREPGHSPQ
Последовательность аминокислот тяжелой цепи G1/HelD1.3 (SEQ ID NO: 191)
Домен VH (курсивный шрифт)
QVQLQESGPGLVRPSQTLSLTCTVSGSTFSGYGVNWVRQPPGRGLEWIGMIWGDGNTDYNSALKSRVTMLVDTSKNQFSLRLSSVTAADTAVYYCARERDYRLDYWGQGSLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPG
Пептид OVA (SEQ ID NO: 192)
ISQAVHAAHAEINEAGR
Последовательность аминокислот внеклеточного домена B7-H3 человека (SEQ ID NO: 193)
ILEVQVPEDPVVALVGTDATLCCSFSPEPGFSLAQLNLIWQLTDTKQLVHSFAEGQDQGSAYANRTALFPDLLAQGNASLRLQRVRVADEGSFTCFVSIRDFGSAAVSLQVAAPYSKPSMTLEPNKDLRPGDTVTITCSSYQGYPEAEVFWQDGQGVPLTGNVTTSQMANEQGLFDVHSILRVVLGANGTYSCLVRNPVLQQDAHSSVTITPQRSPTGAVEVQVPEDPVVALVGTDATLRCSFSPEPGFSLAQLNLIWQLTDTKQLVHSFTEGRDQGSAYANRTALFPDLLAQGNASLRLQRVRVADEGSFTCFVSIRDFGSAAVSLQVAAPYSKPSMTLEPNKDLRPGDTVTITCSSYRGYPEAEVFWQDGQGVPLTGNVTTSQMANEQGLFDVHSVLRVVLGANGTYSCLVRNPVLQQDAHGSVTITGQPMT
Пептид SIINFEKL (SEQ ID NO: 194)
SIINFEKL
Последовательность аминокислот PD-L1 человека-rCD4-His (SEQ ID NO: 195)
Сигнальный пептид (подчеркнут); внеклеточный домен PD-L1 (обычный шрифт); C-конец CD4+ крысы (домены 3 и 4) (курсивный шрифт); соединение между антигеном и C-концевым слитым фрагментом, кодирующее сайт рестрикции NotI (жирный шрифт и подчеркнут); C-концевая гексагистидиновая метка (жирный шрифт)
MRIFAVFIFMTYWHLLNAFTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERTAAATSITAYKSEGESAEFSFPLNLGEESLQGELRWKAEKAPSSQSWITFSLKNQKVSVQKSTSNPKFQLSETLPLTLQIPQVSLQFAGSGNLTLTLDRGILYQEVNLVVMKVTQPDSNTLTCEVMGPTSPKMRLILKQENQEARVSRQEKVIQVQAPEAGVWQCLLSEGEEVKMDSKIQVLSKGLNGSHHHHHH
Последовательность аминокислот PD-L1 человека-Fc-His (SEQ ID NO: 196)
Сигнальный пептид (подчеркнут), внеклеточный домен PD-L1 (обычный шрифт), Fc IgG1 человека (курсивный шрифт), соединение между антигеном и C-концевым слитым фрагментом, кодирующее сайт рестрикции NotI (жирный шрифт и подчеркнуто), C-концевая гексагистидиновая метка (жирный шрифт)
MRIFAVFIFMTYWHLLNAFTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERTAAADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGSHHHHHH
Последовательность аминокислот PD-L1 мыши-rCD4-His (SEQ ID NO: 197)
Сигнальный пептид (подчеркнут); внеклеточный домен PD-L1 (обычный шрифт); C-конец CD4+ крысы (домены 3 и 4) (курсивный шрифт); соединение между антигеном и C-концевым слитым фрагментом, кодирующее сайт рестрикции NotI (жирный шрифт и подчеркнуто); C-концевая гексагистидиновая метка (жирный шрифт)
MRIFAGIIFTACCHLLRAFTITAPKDLYVVEYGSNVTMECRFPVERELDLLALVVYWEKEDEQVIQFVAGEEDLKPQHSNFRGRASLPKDQLLKGNAALQITDVKLQDAGVYCCIISYGGADYKRITLKVNAPYRKINQRISVDPATSEHELICQAEGYPEAEVIWTNSDHQPVSGKRSVTTSRTEGMLLNVTSSLRVNATANDVFYCTFWRSQPGQNHTAELIIPELPATHPPQNRTAAATSITAYKSEGESAEFSFPLNLGEESLQGELRWKAEKAPSSQSWITFSLKNQKVSVQKSTSNPKFQLSETLPLTLQIPQVSLQFAGSGNLTLTLDRGILYQEVNLVVMKVTQPDSNTLTCEVMGPTSPKMRLILKQENQEARVSRQEKVIQVQAPEAGVWQCLLSEGEEVKMDSKIQVLSKGLNGSHHHHHH
Последовательность аминокислот PD-L1 мыши-Fc-His (SEQ ID NO: 198)
Сигнальный пептид (подчеркнут), внеклеточный домен PD-L1 (обычный шрифт), Fc IgG1 человека (курсивный шрифт), соединение между антигеном и C-концевым слитым фрагментом, кодирующее сайт рестрикции NotI (жирный шрифт и подчеркнуто), C-концевая гексагистидиновая метка (жирный шрифт)
MRIFAGIIFTACCHLLRAFTITAPKDLYVVEYGSNVTMECRFPVERELDLLALVVYWEKEDEQVIQFVAGEEDLKPQHSNFRGRASLPKDQLLKGNAALQITDVKLQDAGVYCCIISYGGADYKRITLKVNAPYRKINQRISVDPATSEHELICQAEGYPEAEVIWTNSDHQPVSGKRSVTTSRTEGMLLNVTSSLRVNATANDVFYCTFWRSQPGQNHTAELIIPELPATHPPQNRTAAADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGSHHHHHH
Последовательность аминокислот PD-L1 человека-His-Avi (SEQ ID NO: 199)
Внеклеточный домен PD-L1 (обычный шрифт), C-концевая гексагистидиновая метка (курсивный шрифт), метка Avi (жирный шрифт)
FTVTVPKDLYVVEYGSNMTIECKFPVEKQLDLAALIVYWEMEDKNIIQFVHGEEDLKVQHSSYRQRARLLKDQLSLGNAALQITDVKLQDAGVYRCMISYGGADYKRITVKVNAPYNKINQRILVVDPVTSEHELTCQAEGYPKAEVIWTSSDHQVLSGKTTTTNSKREEKLFNVTSTLRINTTTNEIFYCTFRRLDPEENHTAELVIPELPLAHPPNERGSHHHHHHGGGLNDIFEAQKIEWHE
Кодон-оптимизированные последовательности нуклеиновых кислот, кодирующие тяжелую цепь FS22-172-003-AA/E12v2 (SEQ ID NO: 32)
AAGCTTGCCGCCACCATGGAATGGTCCTGGGTGTTCCTGTTCTTCCTGTCCGTGACCACCGGCGTGCACTCTGAAGTTCAGCTGGTTCAGTCTGGCGCCGAAGTGAAAAGACCTGGCGCCTCTGTGAAGGTGTCCTGCAAGGCTTCTGGCTACCCCTTTACCTCCTACGGCATCTCCTGGGTCCGACAGGCTCCTGGACAAGGCTTGGAATGGATGGGCTGGATCTCCGCTTATTCCGGCGGCACCAATTACGCCCAGAAACTGCAGGGCAGAGTGACCATGACCACCGACACCTCTACCTCCACCGCCTACATGGAACTGCGGTCCCTGAGATCTGACGACACCGCCGTGTACTACTGCGCCAGAGATCTGTTCCCCACCATCTTCGGCGTGTCCTACTACTACTATTGGGGCCAGGGCACCCTGGTCACCGTGTCCTCTGCTTCTACCAAGGGACCCAGCGTGTTCCCTCTGGCTCCTTCCAGCAAGTCTACCTCTGGCGGAACAGCTGCTCTGGGCTGCCTGGTCAAGGACTACTTTCCTGAGCCTGTGACCGTGTCTTGGAACTCTGGCGCTCTGACATCTGGCGTGCACACCTTTCCAGCTGTGCTGCAGTCCTCCGGCCTGTACTCTCTGTCCTCTGTCGTGACCGTGCCTTCCAGCTCTCTGGGAACCCAGACCTACATCTGCAATGTGAACCACAAGCCTTCCAACACCAAGGTGGACAAGAAGGTGGAACCCAAGTCCTGCGACAAGACCCACACCTGTCCTCCATGTCCTGCTCCAGAAGCTGCTGGCGGCCCTTCCGTGTTTCTGTTCCCTCCAAAGCCTAAGGACACCCTGATGATCTCTCGGACCCCTGAAGTGACCTGCGTGGTGGTGGATGTGTCTCACGAGGACCCAGAAGTGAAGTTCAATTGGTACGTGGACGGCGTGGAAGTGCACAACGCCAAGACCAAGCCTAGAGAGGAACAGTACAACTCCACCTACAGAGTGGTGTCCGTGCTGACCGTGCTGCACCAGGATTGGCTGAACGGCAAAGAGTACAAGTGCAAGGTGTCCAACAAGGCCCTGCCTGCTCCTATCGAAAAGACCATCTCCAAGGCCAAGGGCCAGCCTAGGGAACCCCAGGTTTACACCTTGCCTCCATCTCGGGACGAGCTGCCCTACATCATCCCTCCATACAACCAGGTGTCCCTGACCTGCCTCGTGAAGGGCTTCTACCCTTCCGATATCGCCGTGGAATGGGAGAGCAATGGCCAGCCTGAGAACAACTACAAGACAACCCCTCCTGTGCTGGACTCCGACGGCTCATTCTTTCTGTACTCCAAGCTGACAGTGGGCGCCGACAGATGGCTGGAAGGGAACGTGTTCTCCTGCAGCGTGATGCACGAGGCCCTGCACAATCACTACACACAGAAGTCCCTGTCTCTGTCCCCTGGCAAG TGATGAATTC
Кодон-оптимизированные последовательности нуклеиновых кислот, кодирующие легкую цепь FS22-172-003-AA/E12v2 (SEQ ID NO: 39)
AAGCTTGCCGCCACCATGTCTGTGCCTACACAGGTTCTGGGACTGCTGCTGCTGTGGCTGACCGACGCCAGATGCGACATCCAGATGACCCAGTCTCCATCCACACTGTCCGCCTCTGTGCGGGACAGAGTGATCATCACCTGTAGAGCCAGCCAGTCCATCGGCAACAGACTGGCCTGGTATCAGCACAAGCCTGGCAAGGCTCCCAAGCTGCTGATCTACGAGGCCTCCACATCTGAGACAGGCGTGCCCTCTAGATTCTCCGGCTCTGGCTCTGGCACCGACTTTACCCTGACAATCTCCAGCCTGCAGCCTGAGGACTTCGCCACCTACTACTGCCAGCAGTCCTACAGCACCCCTTACACCTTTGGCCAGGGCACCAAGCTGGAAATCAAGCGTACGGTGGCCGCTCCCAGCGTGTTCATCTTCCCCCCAAGCGACGAGCAGCTGAAGAGCGGCACCGCCAGCGTGGTGTGTCTGCTGAACAACTTCTACCCCAGGGAGGCCAAGGTGCAGTGGAAGGTGGACAACGCCCTGCAGAGCGGCAACAGCCAGGAGAGCGTCACCGAGCAGGACAGCAAGGACTCCACCTACAGCCTGAGCAGCACCCTGACCCTGAGCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGTGAGGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGAGCTTCAACAGGGGCGAGTGCTGATGAATTC
Ссылочные материалы
Все документы, упомянутые в настящем описании, полностью включены в данную заявку посредством ссылки.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J. Mol. Biol. 215(3), 403-10 (1990).
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25(17), 3389-402 (1997).
Bagshawe KD, Sharma SK, Springer CJ, Antoniw P, Rogers GT, Burke PJ, Melton R. Antibody-enzyme conjugates can generate cytotoxic drugs from inactive precursors at tumor sites. Antibody, Immunoconjugates and Radiopharmaceuticals 4, 915-922 (1991).
Baird JR et al. Immune-mediated regression of established B16F10 melanoma by intratumoral injection of attenuated Toxoplasma gondii protects against rechallenge. J Immunol. 190(1): 469-478 (2013).
Bartkowiak T, Curran MA. 4-1BB Agonists: Multi-Potent Potentiators of Tumor Immunity. Front Oncol. Jun 8;5, 117 (2015).
Bartkowiak T et al. Activation of 4-1BB on Liver Myeloid Cells Triggers Hepatitis via an Interleukin-27-Dependent Pathway. Clin Cancer Res. 24(5), 1138-51 (2018).
Bergman I, Burckart GJ, Pohl CR, Venkataramanan R, Barmada MA, Griffin JA And Cheung NV. Pharmacokinetics of IgG and IgM Anti-Ganglioside Antibodies in Rats and Monkeys After Intrathecal Administration. The Journal of Pharmacology and Experimental Therapeutics 284(1), 111-115 (1998)
Bitra A, Doukov T, Wang J, Picarda G, Benedict CA, Croft M, Zajonc DM. Crystal structure of murine 4-1BB and its interaction with 4-1BBL support a role for galectin-9 in 4-1BB signaling. J Biol Chem. 293(4):1317-1329 (2017).
Brown MH, Barclay AN. Expression of immunoglobulin and scavenger receptor superfamily domains as chimeric proteins with domains 3 and 4 of CD4+ for ligand analysis. Protein Eng. 7(4), 515-21 (1994).
Bruhns P, Iannascoli B, England P, Mancardi DA, Fernandez N, Jorieux S, Daëron M. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood. 113(16):3716-25 (2009).
Chacon JA1, Wu RC, Sukhumalchandra P, Molldrem JJ, Sarnaik A, Pilon-Thomas S, Weber J, Hwu P, Radvanyi L. Co-stimulation through 4-1BB/CD137 improves the expansion and function of CD8(+) melanoma tumor-infiltrating lymphocytes for adoptive T-cell therapy. PLoS One. 8(4):e60031 (2013).
Chapple SD, Crofts AM, Shadbolt SP, McCafferty J, Dyson MR. Multiplexed expression and screening for recombinant protein production in mammalian cells. BMC Biotechnol. 6:49 (2006).
Chester C, Ambulkar S, Kohrt HE. 4-1BB agonism: adding the accelerator to cancer immunotherapy. Cancer Immunol Immunother. 65(10):1243-8 (2016).
Chester C, Sanmamed MF, Wang J, Melero I. Immunotherapy targeting 4-1BB: mechanistic rationale, clinical results, and future strategies. Blood. 131(1):49-57 (2018).
Claus C et al. A novel tumor-targeted 4-1BB agonist and its combination with T-cell bispecific antibodies: an off-the-shelf cancer immunotherapy alternative to CAR T-cells [abstract]. In: Proceedings of the American Association for Cancer Research Annual Meeting 2017. Cancer Res. 77 (2017).
Croft M. Co-stimulatory members of the TNFR family: keys to effective T-cell immunity? Nat Rev Immunol. 3(8):609-20 (2003).
Curran MA, Geiger TL, Montalvo W, Kim M, Reiner SL, Al-Shamkhani A, Sun JC, Allison J. P. Systemic 4-1BB activation induces a novel T cell phenotype driven by high expression of Eomesodermin. J. Exp.Med. 210(4): 743-755 (2013).
Dubrot J et al. Treatment with anti-CD137 mAbs causes intense accumulations of liver T cells without selective antitumor immunotherapeutic effects in this organ. Cancer Immunol Immunother. 59(8):1223-1233 (2010).
Dyson MR, Zheng Y, Zhang C, Colwill K, Pershad K, Kay BK, Pawson T, McCafferty J: Mapping protein interactions by combining antibody affinity maturation and mass spectrometry. Anal. Biochem. 417(1), 25-35 (2011).
Fisher TS et al. Targeting of 4-1BB by monoclonal antibody PF-05082566 enhances T-cell function and promotes anti-tumor activity. Cancer Immunol Immunother. 61(10):1721-33 (2012).
Grosso J, Inzunza D, Wu Q, Simon J, Singh P, Zhang X, Phillips T, Simmons P, Cogswell J. Programmed death-ligand 1 (PD-L1) expression in various tumor types. Journal for Immunotherapy of Cancer. 1(Suppl 1):P53. (2013).
Hasenhindl C, Traxlmayr MW, Wozniak-Knopp G, Jones PC, Stadlmayr G, Rüker F, Obinger C. Stability assessment on a library scale: a rapid method for the evaluation of the commutability and insertion of residues in C-terminal loops of the CH3 domains of IgG1-Fc. Protein Eng. Des. Sel., 26(10), 675-82 (2013).
Hezareh M, Hessell AJ, Jensen RC, van de Winkel JG, Parren PW. Effector function activities of a panel of mutants of a broadly neutralizing antibody against human immunodeficiency virus type 1. J Virol. 75(24):12161-8. (2001).
Hinner MJ, Aiba, RSB., Wiedenmann A, Schlosser C, Allersdorfer A, Matschiner G, Rothe C, Moebius U, Kohrt HE, Olwill SA. Costimulatory T cell engagement via a novel bispecific anti-CD137 /anti-HER2 protein. J. Immunotherapy Cancer 3 (Suppl 2): P187. (2015).
Holliger P, Hudson PJ. Engineered antibody fragments and the rise of single domains. Nat Biotechnol. 23(9):1126-36 (2005).
Hu S, Shively L, Raubitschek A, Sherman M, Williams LE, Wong JY, Shively JE, Wu AM. Minibody: A novel engineered anti-carcinoembryonic antigen antibody fragment (single-chain Fv-CH3) which exhibits rapid, high-level targeting of xenografts. Cancer Res. 56(13):3055-61 (1996).
Hurtado JC, Kim YJ, Kwon BS. Signals through 4-1BB are costimulatory to previously activated splenic T cells and inhibit activation-induced cell death. J Immunol. 15;158(6):2600-9 (1997).
Idusogie EE, Presta LG, Gazzano-Santoro H, Totpal K, Wong PY, Ultsch M, Meng YG, Mulkerrin MG. Mapping of the C1q binding site on rituxan, a chimeric antibody with a human IgG1 Fc. J Immunol. 164(8), 4178-84 (2000).
Jain T et al. Biophysical properties of the clinical-stage antibody landscape. PNAS 114 (5), 944-949 (2017).
Jefferis R, Reimer CB, Skvaril F, de Lange G, Ling NR, Lowe J, Walker MR, Phillips DJ, Aloisio CH, Wells TW. Evaluation of monoclonal antibodies having specificity for human IgG sub-classes: results of an IUIS/WHO collaborative study. Immunol. Lett.1, 223-52 (1985).
Jefferis R, Reimer CB, Skvaril F, de Lange GG, Goodall DM, Bentley TL, Phillips DJ, Vlug A, Harada S, Radl J. Evaluation of monoclonal antibodies having specificity for human IgG subclasses: results of the 2nd IUIS/WHO collaborative study. Immunol. Lett. 31(2),143-68 (1992).
Juneja VR, McGuire KA, Manguso RT, LaFleur MW, Collins N, Haining WN, Freeman GJ, Sharpe AH. PD-L1 on tumor cells is sufficient for immune evasion in immunogenic tumors and inhibits CD8+ T cell cytotoxicity. J Exp Med. 214(4), 895-904 (2017).
Kabat EA, Wu TT, Perry HM, Gottesman KS, Foeller C. Sequences of Proteins of Immunological Interest, 5th ed. NIH Publication No. 91-3242. Washington, D.C.: U.S. Department of Health and Human Services (1991).
Kim YH, Choi BK, Kim KH, Kang SW, Kwon BS. Combination Therapy with Cisplatin and Anti-4-1BB. Cancer Res. 68(18), 7264-7269 (2009).
Klein C, Schaefer W, Regula JT. The use of CrossMAb technology for the generation of bi- and multispecific antibodies. MAbs 8(6), 1010-20 (2016).
Kleinovink JW, Marijt KA, Schoonderwoerd MJA, Hall T, Ossendorp F, Fransen MF. PD-L1 expression on malignant cells is no prerequisite for checkpoint therapy. Oncoimmunology. 6(4), e1294299 (2017).
Kocak E et al. Combination therapy with anti-CTL antigen-4 and anti-4-1BB antibodies enhances cancer immunity and reduces autoimmunity. Cancer Res. 15;66(14), 7276-84 (2006).
Lechner MG et al. Immunogenicity of murine solid tumor models as a defining feature of in vivo behavior and response to immunotherapy. J Immunother. 36(9), 477-89 (2013).
Ledermann JA, Begent RH, Massof C, Kelly AM, Adam T, Bagshawe KD. A phase-I study of repeated therapy with radiolabelled antibody to carcinoembryonic antigen using intermittent or continuous administration of cyclosporin A to supress the immune response. Int. J. Cancer 47(5), 659-64 (1991).
Lefranc MP et al. IMGT®, the international ImMunoGeneTics information system® 25 years on. Nucleic Acids Res. 43(Database issue):D413-22 (2015).
Lefranc MP et al. IMGT unique numbering for immunoglobulin and T cell receptor constant domains and Ig superfamily C-like domains. Dev. Comp. Immunol. 29(3), 185-203 (2005).
Li F. and Ravetech JV. Antitumor activities of agonistic anti-TNFR antibodies require differential FcγRIIB coengagement in vivo. Proc Natl Acad Sci U S A, 110(48), 19501-6 (2013).
Link A et al. Preclinical pharmacology of MP0310: a 4-1BB/FAP bispecific DARPin drug candidate promoting tumor-restricted T cell co-stimulation [abstract]. In: Proceedings of the Annual Meeting of the American Association for Cancer Research; 2018 Apr 14-18; Chicago (IL) Abstract nr 3752 (2018).
Makkouk A, Chester C, Kohrt HE. Rationale for anti-CD137 cancer immunotherapy. Eur J Cancer. 54, 112-119 (2016).
Martins JP, Kennedy PJ, Santos HA, Barrias C, Sarmento B. A comprehensive review of the neonatal Fc receptor and its application in drug delivery. Pharmcol. Ther. 161, 22-39 (2016).
Niu L et al. Cytokine- mediated disruption of lymphocyte trafficking, hemopoiesis, and induction of lymphopenia, anemia, and thrombocytopenia in anti-CD137-treated mice. J Immunol. 178(7), 4194-4213 (2007).
Pérez-Ruiz E, Etxeberria I, Rodriguez-Ruiz, Melero I. Anti-CD137 and PD-1/PD-L1 Antibodies En Route toward Clinical Synergy. Clin. Cancer Res. 23, (18) (2017)
Reichen C et al. FAP-mediated tumor accumulation of a T-cell agonistic FAP/4-1BB DARPin drug candidate analyzed by SPECT/CT and quantitative biodistribution [abstract]. In: Proceedings of the Annual Meeting of the American Association for Cancer Research; 2018 Apr 14-18; Chicago (IL) Abstract nr 3029 (2018).
Schofield DJ et al. Application of phage display to high throughput antibody generation and characterization. Genome Biol. 8(11), R254 (2007).
Segal NH et al. Phase I Study of Single-Agent Utomilumab (PF-05082566), a 4-1BB/CD137 Agonist, in Patients with Advanced Cancer. Clin Cancer Res. 24(8):1816-1823 (2018).
Segal NH et al. Results from an Integrated Safety Analysis of Urelumab, an Agonist Anti-CD137 Monoclonal Antibody. Clin Cancer Res. 23(8):1929-1936 (2017).
Shuford WW et al.l 4-1BB costimulatory signals preferentially induce CD8+ T cell proliferation and lead to the amplification in vivo of cytotoxic T cell responses. J Exp Med. 186(1), 47-55 (1997).
Smith TF, Waterman MS. Identification of common molecular subsequences. J. Mol. Biol. 147(1), 195-7 (1981).
Taraban VY et al. Expression and costimulatory effects of the TNF receptor superfamily members CD134 (OX40) and CD137 (4-1BB), and their role in the generation of anti-tumor immune responses. Eur. J. Immunol. 32, 3617-3627 (2002).
Tello, D, Goldbaum, FA, Mariuzza, RA, Ysern, X, Schwarz, FP, Poljak, RJ. Three-dimensional structure and thermodynamics of antigen binding by anti-lysozyme antibodies, Biochem Soc. Trans., 21(4), 943-6 (1993).
Tolcher AW et al. Phase Ib Study of Utomilumab (PF-05082566), a 4-1BB/CD137 Agonist, in Combination with Pembrolizumab (MK-3475) in Patients with Advanced Solid Tumors. Clin. Cancer Res. 23, 5349-57 (2017)
Wang X, Mathieu M, Brezski RJ. IgG Fc engineering to modulate antibody effector functions. Protein Cell 9(1), 63-73 (2018).
Wen T, Bukczynski J, Watts TH. 4-1BB ligand-mediated costimulation of human T cells induces CD4+ and CD8+ T cell expansion, cytokine production, and the development of cytolytic effector function. J Immunol. 168(10), 4897-906 (2002).
Wolfl M, Kuball J, Ho WY, Nguyen H, Manley TJ, Bleakley M, Greenberg PD. Activation-induced expression of CD137 permits detection, isolation, and expansion of the full repertoire of CD8+ T cells responding to antigen without requiring knowledge of epitope specificities. Blood. 110(1):201-10 (2007).
Won EY, Cha K, Byun JS, Kim DU, Shin S, Ahn B, Kim YH, Rice AJ, Walz T, Kwon BS, Cho HS. The structure of the trimer of human 4-1BB ligand is unique among members of the tumor necrosis factor superfamily. J Biol Chem.285(12), 9202-10 (2010).
Wozniak-Knopp G et al. Introducing antigen-binding sites in structural loops of immunoglobulin constant domains: Fc fragments with engineered HER2/neu-binding sites and antibody properties. Protein Eng Des Sel. 23(4), 289-97 (2010).
Ye Q, Song DG, Poussin M, Yamamoto T, Best A, Li C, Coukos G, Powell DJ Jr. CD137 accurately identifies and enriches for naturally occurring tumor-reactive T cells in tumor. Clin Cancer Res. 20(1):44-55 (2014).
Yu X, Anasetti C. CH 10 - T-cell costimulation in graft-versus-host disease and graft-versus-leukemia effect. Immune Biology of Allogeneic Hematopoietic Stem Cell Transplantation. 195-222 (2013).
Zanetti M. Tapping CD4 T Cells for Cancer Immunotherapy: The Choice of Personalized Genomics. J Immunol. 194:2049-2056 (2015).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> F-STAR BETA LIMITED
<120> МОЛЕКУЛЫ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТ PD-L1 и CD137
<130> 007508260
<140> PCT/EP2019/068793
<141> 2019-07-12
<150> GB 1811405.8
<151> 2018-07-12
<150> GB 1818283.2
<151> 2018-11-09
<150> GB 1902594.9
<151> 2019-02-26
<160> 199
<170> PatentIn версии 3.5
<210> 1
<211> 5
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH МАТ E12v2 (Кабат)
<400> 1
Ser Tyr Gly Ile Ser
1 5
<210> 2
<211> 17
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH МАТ E12v2 (Кабат)
<400> 2
Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu Gln
1 5 10 15
Gly
<210> 3
<211> 14
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH МАТ E12v2 (Кабат)
<400> 3
Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
1 5 10
<210> 4
<211> 11
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL МАТ E12v2(Кабат)
<400> 4
Arg Ala Ser Gln Ser Ile Gly Asn Arg Leu Ala
1 5 10
<210> 5
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL МАТ E12v2 (Кабат)
<400> 5
Glu Ala Ser Thr Ser Glu Thr
1 5
<210> 6
<211> 9
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ E12v2 (Кабат)
<400> 6
Gln Gln Ser Tyr Ser Thr Pro Tyr Thr
1 5
<210> 7
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH E12v2 (IMGT)
<400> 7
Gly Tyr Pro Phe Thr Ser Tyr Gly
1 5
<210> 8
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VH E12v2 (IMGT)
<400> 8
Ile Ser Ala Tyr Ser Gly Gly Thr
1 5
<210> 9
<211> 16
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VH E12v2 (IMGT)
<400> 9
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
1 5 10 15
<210> 10
<211> 6
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL E12v2 (IMGT)
<400> 10
Gln Ser Ile Gly Asn Arg
1 5
<210> 11
<211> 3
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL E12v2 (IMGT)
<400> 11
Glu Ala Ser
1
<210> 12
<211> 123
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> МАТ E12v2
<400> 12
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 13
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТ E12v2
<400> 13
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata ccccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagc 369
<210> 14
<211> 107
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ E12v2
<400> 14
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Arg
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Asn Arg
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Thr Ser Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 15
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ E12v2
<400> 15
gacatccaga tgacgcagag cccgtctacc ctgtccgcct ccgtgagaga tcgcgtgatc 60
atcacctgtc gggccagcca gtccatcgga aaccgcttgg cgtggtacca gcacaagcct 120
gggaaggctc cgaagctgct catctacgaa gcctcgactt cggagactgg tgtccctagc 180
cggttcagcg gatcgggatc agggaccgat ttcactctga ccatttcctc cctgcaaccc 240
gaggacttcg ccacctacta ctgccaacag tcatattcca ccccgtacac cttcggacaa 300
ggcaccaagc tcgaaatcaa g 321
<210> 16
<211> 452
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ G1-AA/E12v2 (с LALA)
<400> 16
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 17
<211> 214
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ G1-AA/E12v2
<400> 17
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Arg
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Asn Arg
20 25 30
Leu Ala Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Thr Ser Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 18
<211> 11
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL МАТ E05v2 (Кабат)
<400> 18
Arg Ala Ser Gln Ser Ile Ser Gly Arg Leu Ala
1 5 10
<210> 19
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL МАТ E05v2 (Кабат)
<400> 19
Glu Ala Ser Asn Leu Glu Ser
1 5
<210> 20
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ E05v2 (Кабат)
<400> 20
Gln Gln Ser Tyr Ser Thr Pro Arg Val Thr
1 5 10
<210> 21
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VH E05v2
<400> 21
Gly Tyr Thr Phe Thr Ser Tyr Gly
1 5
<210> 22
<211> 6
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL E05v2
<400> 22
Gln Ser Ile Ser Gly Arg
1 5
<210> 23
<211> 123
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен VH МАТ E05v2
<400> 23
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 24
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен VH МАТ E05v2
<400> 24
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata caccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagc 369
<210> 25
<211> 108
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ E05v2
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Gly Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Arg
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 26
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ E05v2
<400> 26
gacattcaga tgacccaatc cccgtccacg ctgagcgcct ccgtcggtga tcgcgtgaca 60
atcacttgtc gggcgtcgca gtccatctct ggaaggctcg cctggtacca gcagaagcct 120
ggaaaggctc ccaacctcct tatctacgaa gccagcaacc tggagtccgg agtgcctagc 180
cggttcagcg gatcagggtc cggtaccgag ttcaccctga ccatttcctc gctccaacct 240
gaggacttcg ccacctacta ctgccaacag tcctattcaa ctccgcgcgt gaccttcggc 300
cagggcacta aggtcgaaat caaa 324
<210> 27
<211> 452
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжелой цепи МАТ G1-AA/E05v2
<400> 27
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 28
<211> 215
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот легкой цепи МАТ G1-AA/E05v2 и МАТ2
FS22-172-003-AA/E05v2 и FS22-053-008-AA/E05v2
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Gly Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Arg
85 90 95
Val Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 29
<211> 9
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ G12v2 (Кабат)
<400> 29
Gln Gln Ser Tyr Ser Trp Pro Arg Thr
1 5
<210> 30
<211> 107
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ G12v2
<400> 30
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Gly Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 31
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ G12v2
<400> 31
gacattcaga tgacccagtc cccgagcacg ctgtccgcaa gcgtggggga cagagtgacc 60
atcacttgcc gcgcctcaca atccatcagc ggacgcttgg cctggtacca gcagaagccc 120
ggaaaggccc caaaccttct gatctacgaa gcctcgaacc tggagtcagg cgtcccttcg 180
cggttctctg gctccggttc cggaactgag ttcaccctca ccatctcgtc cctgcaaccg 240
gaagatttcg ccacctacta ctgccaacag tcgtactcct ggccccggac attcggacag 300
ggaaccaaag tcgagattaa g 321
<210> 32
<211> 1456
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированные последовательности нуклеиновых кислот, кодирующие
тяжелую цепь FS22-172-003-AA/E12v2
<400> 32
aagcttgccg ccaccatgga atggtcctgg gtgttcctgt tcttcctgtc cgtgaccacc 60
ggcgtgcact ctgaagttca gctggttcag tctggcgccg aagtgaaaag acctggcgcc 120
tctgtgaagg tgtcctgcaa ggcttctggc taccccttta cctcctacgg catctcctgg 180
gtccgacagg ctcctggaca aggcttggaa tggatgggct ggatctccgc ttattccggc 240
ggcaccaatt acgcccagaa actgcagggc agagtgacca tgaccaccga cacctctacc 300
tccaccgcct acatggaact gcggtccctg agatctgacg acaccgccgt gtactactgc 360
gccagagatc tgttccccac catcttcggc gtgtcctact actactattg gggccagggc 420
accctggtca ccgtgtcctc tgcttctacc aagggaccca gcgtgttccc tctggctcct 480
tccagcaagt ctacctctgg cggaacagct gctctgggct gcctggtcaa ggactacttt 540
cctgagcctg tgaccgtgtc ttggaactct ggcgctctga catctggcgt gcacaccttt 600
ccagctgtgc tgcagtcctc cggcctgtac tctctgtcct ctgtcgtgac cgtgccttcc 660
agctctctgg gaacccagac ctacatctgc aatgtgaacc acaagccttc caacaccaag 720
gtggacaaga aggtggaacc caagtcctgc gacaagaccc acacctgtcc tccatgtcct 780
gctccagaag ctgctggcgg cccttccgtg tttctgttcc ctccaaagcc taaggacacc 840
ctgatgatct ctcggacccc tgaagtgacc tgcgtggtgg tggatgtgtc tcacgaggac 900
ccagaagtga agttcaattg gtacgtggac ggcgtggaag tgcacaacgc caagaccaag 960
cctagagagg aacagtacaa ctccacctac agagtggtgt ccgtgctgac cgtgctgcac 1020
caggattggc tgaacggcaa agagtacaag tgcaaggtgt ccaacaaggc cctgcctgct 1080
cctatcgaaa agaccatctc caaggccaag ggccagccta gggaacccca ggtttacacc 1140
ttgcctccat ctcgggacga gctgccctac atcatccctc catacaacca ggtgtccctg 1200
acctgcctcg tgaagggctt ctacccttcc gatatcgccg tggaatggga gagcaatggc 1260
cagcctgaga acaactacaa gacaacccct cctgtgctgg actccgacgg ctcattcttt 1320
ctgtactcca agctgacagt gggcgccgac agatggctgg aagggaacgt gttctcctgc 1380
agcgtgatgc acgaggccct gcacaatcac tacacacaga agtccctgtc tctgtcccct 1440
ggcaagtgat gaattc 1456
<210> 33
<211> 214
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот легкой цепи МАТ G1-AA/G12v2 и МАТ2
FS22-172-003-AA/G12v2 и FS22-053-008-AA/G12v2
<400> 33
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Gly Arg
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Asn Leu Leu Ile
35 40 45
Tyr Glu Ala Ser Asn Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 34
<211> 14
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL МАТ lambdav3/lam-G02v3 (Кабат)
<400> 34
Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr Asn Tyr Val Ser
1 5 10
<210> 35
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL МАТ lambdav3/lam-G02v3 (Кабат)
<400> 35
Glu Val Thr Asn Arg Pro Ser
1 5
<210> 36
<211> 11
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR3 VL МАТ lambdav3/lam-G02v3 (Кабат)
<400> 36
Ser Ser Phe Lys Arg Gly Ser Thr Leu Val Val
1 5 10
<210> 37
<211> 9
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR1 VL lambdav3/lam-G02v3 (IMGT)
<400> 37
Ser Ser Asp Val Gly Gly Tyr Asn Tyr
1 5
<210> 38
<211> 3
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> CDR2 VL lambdav3/lam-G02v3 (IMGT)
<400> 38
Glu Val Thr
1
<210> 39
<211> 727
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кодон-оптимизированные последовательности нуклеиновых кислот, кодирующие
легкую цепь FS22-172-003-AA/E12v2
<400> 39
aagcttgccg ccaccatgtc tgtgcctaca caggttctgg gactgctgct gctgtggctg 60
accgacgcca gatgcgacat ccagatgacc cagtctccat ccacactgtc cgcctctgtg 120
cgggacagag tgatcatcac ctgtagagcc agccagtcca tcggcaacag actggcctgg 180
tatcagcaca agcctggcaa ggctcccaag ctgctgatct acgaggcctc cacatctgag 240
acaggcgtgc cctctagatt ctccggctct ggctctggca ccgactttac cctgacaatc 300
tccagcctgc agcctgagga cttcgccacc tactactgcc agcagtccta cagcacccct 360
tacacctttg gccagggcac caagctggaa atcaagcgta cggtggccgc tcccagcgtg 420
ttcatcttcc ccccaagcga cgagcagctg aagagcggca ccgccagcgt ggtgtgtctg 480
ctgaacaact tctaccccag ggaggccaag gtgcagtgga aggtggacaa cgccctgcag 540
agcggcaaca gccaggagag cgtcaccgag caggacagca aggactccac ctacagcctg 600
agcagcaccc tgaccctgag caaggccgac tacgagaagc acaaggtgta cgcctgtgag 660
gtgacccacc agggcctgtc cagccccgtg accaagagct tcaacagggg cgagtgctga 720
tgaattc 727
<210> 40
<400> 40
000
<210> 41
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ lambdav3/lam-G02v3
<400> 41
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Glu Val Thr Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Glu Tyr Tyr Cys Ser Ser Phe Lys Arg Gly
85 90 95
Ser Thr Leu Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 42
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен VL МАТ lambdav3/lam-G02v3
<400> 42
cagtcggccc ttactcaacc cgcgtcagtc tccggtagcc ccggacagtc catcacgatt 60
tcgtgcaccg gaaccagcag cgatgtcggg ggatacaact acgtgtcctg gtaccagcag 120
ttcccgggaa aggcccctaa gctgatgatc ttcgaagtca ctaacagacc ttccggagtg 180
tcggaccggt tctccggctc caagtccgac aacactgcga gcctgaccat ctcgggcctg 240
caagccgagg acgaagccga gtactactgt agctcattca agcgcggttc caccctcgtg 300
gtgttcggcg gtggcactaa gctcaccgtg ctggga 336
<210> 43
<400> 43
000
<210> 44
<211> 217
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот легкой цепи МАТ G1-AA/lambdav3/lam-G02v3
<400> 44
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Glu Val Thr Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Glu Tyr Tyr Cys Ser Ser Phe Lys Arg Gly
85 90 95
Ser Thr Leu Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Ala Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 45
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение HCDR1 (IMGT)
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> X = P или T
<400> 45
Gly Tyr Xaa Phe Thr Ser Tyr Gly
1 5
<210> 46
<211> 11
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение LCDR1 (Кабат)
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> X = S или G
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> X = N или G
<400> 46
Arg Ala Ser Gln Ser Ile Xaa Xaa Arg Leu Ala
1 5 10
<210> 47
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение LCDR2 (Кабат)
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> X = T или N
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> X = S или L
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> X = T или S
<400> 47
Glu Ala Ser Xaa Xaa Glu Xaa
1 5
<210> 48
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение LCDR3 (Кабат)
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> X = T или W
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> X = отсутствует или R
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> X = Y, R или V
<400> 48
Gln Gln Ser Tyr Ser Xaa Pro Xaa Xaa Thr
1 5 10
<210> 49
<211> 6
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение LCDR1 (IMGT)
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> X = G или S
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> X = N или G
<400> 49
Gln Ser Ile Xaa Xaa Arg
1 5
<210> 50
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Альтернативное определение LCDR3 (IMGT)
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> X = отсутствует или T
<220>
<221> MISC_FEATURE
<222> (7)..(7)
<223> X = T, W или P
<220>
<221> MISC_FEATURE
<222> (8)..(8)
<223> X = P или R
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> X = Y или R
<400> 50
Gln Gln Ser Tyr Ser Xaa Xaa Xaa Xaa Thr
1 5 10
<210> 51
<211> 30
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW1 IgG1 (Кабат)
<220>
<221> MISC_FEATURE
<222> (28)..(28)
<223> X = P или T
<400> 51
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Xaa Phe Thr
20 25 30
<210> 52
<211> 14
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW2 IgG1 (Кабат)
<400> 52
Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
1 5 10
<210> 53
<211> 32
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW3 IgG1 (Кабат)
<400> 53
Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr Met Glu
1 5 10 15
Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys Ala Arg
20 25 30
<210> 54
<211> 11
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW4 IgG1 (Кабат)
<400> 54
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 55
<211> 25
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW1 IgG1 (IMGT)
<400> 55
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser
20 25
<210> 56
<211> 17
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW2 IgG1 (IMGT)
<400> 56
Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly
1 5 10 15
Trp
<210> 57
<211> 38
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> HFW3 IgG1 (IMGT)
<400> 57
Asn Tyr Ala Gln Lys Leu Gln Gly Arg Val Thr Met Thr Thr Asp Thr
1 5 10 15
Ser Thr Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Arg Ser Asp Asp
20 25 30
Thr Ala Val Tyr Tyr Cys
35
<210> 58
<211> 23
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW1 легкой цепи каппа (Кабат)
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Arg
1 5 10 15
Asp Arg Val Ile Ile Thr Cys
20
<210> 59
<211> 15
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW2 легкой цепи каппа (Кабат)
<400> 59
Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
1 5 10 15
<210> 60
<211> 32
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW3 легкой цепи каппа (Кабат)
<400> 60
Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr
1 5 10 15
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
20 25 30
<210> 61
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW4 легкой цепи каппа (Кабат)
<400> 61
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
1 5 10
<210> 62
<211> 26
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW1 легкой цепи каппа (IMGT)
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Arg
1 5 10 15
Asp Arg Val Ile Ile Thr Cys Arg Ala Ser
20 25
<210> 63
<211> 17
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW2 легкой цепи каппа (IMGT)
<400> 63
Leu Ala Trp Tyr Gln His Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
1 5 10 15
Tyr
<210> 64
<211> 36
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW3 легкой цепи каппа (IMGT)
<400> 64
Thr Ser Glu Thr Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly
1 5 10 15
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala
20 25 30
Thr Tyr Tyr Cys
35
<210> 65
<211> 22
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW1 легкой цепи лямбда (Кабат)
<400> 65
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys
20
<210> 66
<211> 15
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW2 легкой цепи лямбда (Кабат)
<400> 66
Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu Met Ile Phe
1 5 10 15
<210> 67
<211> 32
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW3 легкой цепи лямбда (Кабат)
<400> 67
Gly Val Ser Asp Arg Phe Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser
1 5 10 15
Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu Ala Glu Tyr Tyr Cys
20 25 30
<210> 68
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW4 легкой цепи лямбда (Кабат)
<400> 68
Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
1 5 10
<210> 69
<211> 25
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW1 легкой цепи лямбда (IMGT)
<400> 69
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr
20 25
<210> 70
<211> 17
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW2 легкой цепи лямбда (IMGT)
<400> 70
Val Ser Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu Met Ile
1 5 10 15
Phe
<210> 71
<211> 36
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> LFW3 легкой цепи лямбда (IMGT)
<400> 71
Asn Arg Pro Ser Gly Val Ser Asp Arg Phe Ser Gly Ser Lys Ser Asp
1 5 10 15
Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu Gln Ala Glu Asp Glu Ala
20 25 30
Glu Tyr Tyr Cys
35
<210> 72
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Структурная петля AB домена CH3 ДТ
<400> 72
Arg Asp Glu Leu Thr Lys Asn Gln
1 5
<210> 73
<211> 9
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Структурная петля CD домена CH3 ДТ
<400> 73
Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1 5
<210> 74
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Структурная петля EF домена CH3 ДТ
<400> 74
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
1 5 10
<210> 75
<211> 106
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 ДТ
<400> 75
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 76
<211> 110
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2
<400> 76
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 77
<211> 110
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2 с мутацией LALA
<400> 77
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 78
<211> 110
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2 с мутацией LALA и мутацией P114A
<400> 78
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 79
<211> 3
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PPY
<400> 79
Pro Pro Tyr
1
<210> 80
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 FS22-053-008
<400> 80
Asp Tyr Trp Arg Trp Leu Glu
1 5
<210> 81
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053-008
<400> 81
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Tyr Trp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 82
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053-008
<400> 82
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtactgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggattactgg 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggcgct gcacaaccac 300
tacactcaga agagcttgtc cctgtcgccc ggt 333
<210> 83
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 FS22-053-009
<400> 83
Glu His Thr Arg Trp Leu Asp
1 5
<210> 84
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053-009
<400> 84
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Glu His Thr
65 70 75 80
Arg Trp Leu Asp Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 85
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053-009
<400> 85
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggaacatact 240
aggtggctgg atgggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 86
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-011
<400> 86
Asp Tyr Trp Arg Trp Thr Asp
1 5
<210> 87
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-011
<400> 87
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Tyr Trp
65 70 75 80
Arg Trp Thr Asp Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 88
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-011
<400> 88
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggattactgg 240
aggtggactg atgggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 89
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-017
<400> 89
Tyr His Trp Arg Trp Leu Glu
1 5
<210> 90
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-017
<400> 90
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Tyr His Trp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 91
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-017
<400> 91
ggacagcctc gagaaccaca ggtgtacact ctgccccctt cacgcgacga actcaatccg 60
ccctacctgt tctccaacca agtctccctg acctgtcttg tgaagggttt ctacccatcc 120
gatatcgccg tggagtggga gagcaacgga cagccggaga acaactataa gactaccccg 180
cctgtgctgg actcggacgg cagcttcttc ttgtactcca aactgaccgt gtaccactgg 240
cggtggctgg aagggaacgt gtttagctgc tccgtcatgc atgaagccct gcacaaccac 300
tacacccaga agtccctctc gctctctccg ggt 333
<210> 92
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-014
<400> 92
Tyr His Trp Arg Trp Leu Asp
1 5
<210> 93
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-014
<400> 93
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Tyr His Trp
65 70 75 80
Arg Trp Leu Asp Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 94
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-014
<400> 94
ggacagcctc gagagcctca agtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gtaccattgg 240
aggtggctgg atgggaacgt cttctcatgc tccgtgatgc atgaggcgct gcacaaccac 300
tacactcaga agagcttgtc cctgtcgccc gga 333
<210> 95
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-010
<400> 95
Asp Tyr Met Arg Trp Leu Asp
1 5
<210> 96
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-010
<400> 96
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Tyr Met
65 70 75 80
Arg Trp Leu Asp Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 97
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-010
<400> 97
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggattacatg 240
aggtggctgg atgggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 98
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-012
<400> 98
Asp His Met Arg Trp Leu Glu
1 5
<210> 99
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-012
<400> 99
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp His Met
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 100
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-012
<400> 100
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggatcatatg 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 101
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-013
<400> 101
Gly Tyr Glu Arg Trp Leu Glu
1 5
<210> 102
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-013
<400> 102
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Tyr Glu
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 103
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-013
<400> 103
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggttacgaa 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 104
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-015
<400> 104
Asp His Trp Arg Trp Leu Gln
1 5
<210> 105
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-015
<400> 105
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp His Trp
65 70 75 80
Arg Trp Leu Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 106
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-015
<400> 106
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgttgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggatcattgg 240
aggtggctgc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 107
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-053-016
<400> 107
Asp Tyr Ile Arg Trp Leu Asn
1 5
<210> 108
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-016
<400> 108
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Tyr Ile
65 70 75 80
Arg Trp Leu Asn Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 109
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-053-016
<400> 109
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggattacatc 240
aggtggctga acgggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 110
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 FS22-053
<400> 110
Tyr Tyr Asn Arg Trp Gln Asp
1 5
<210> 111
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053
<400> 111
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Tyr Tyr Asn
65 70 75 80
Arg Trp Gln Asp Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 112
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 FS22-053
<400> 112
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaacccg 60
ccgtacctgt tctctaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gtattataac 240
aggtggcagg atgggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 113
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-003
<400> 113
Pro Tyr Ile Ile Pro Pro Tyr
1 5
<210> 114
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-003
<400> 114
Gly Ala Asp Arg Trp Leu Glu
1 5
<210> 115
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-003
<400> 115
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Pro Tyr Ile Ile Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 116
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-003
<400> 116
ggacagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgccatac 60
atcatcccac catacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggcgct gcacaaccac 300
tacactcaga agagcttgtc cctgtcgccc ggt 333
<210> 117
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-002
<400> 117
Pro Phe Gln Met Pro Pro Tyr
1 5
<210> 118
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-002
<400> 118
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Pro Phe Gln Met Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 119
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-002
<400> 119
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgccattc 60
cagatgccac catacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 120
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-004
<400> 120
Asn Tyr Ile Tyr Pro Pro Tyr
1 5
<210> 121
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-004
<400> 121
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Asn Tyr Ile Tyr Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 122
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-004
<400> 122
ggacagcctc gagagcctca agtgtacacc ctgcccccat cccgggatga gctgaactac 60
atctacccac catacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggcgct gcacaaccac 300
tacactcaga agagcttgtc cctgtcgccc gga 333
<210> 123
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-001
<400> 123
Pro Phe Val Met Pro Pro Tyr
1 5
<210> 124
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-001
<400> 124
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Pro Phe Val Met Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 125
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-001
<400> 125
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgccattc 60
gttatgccac catacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 126
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-005
<400> 126
Gln Gln Val Tyr Pro Pro Tyr
1 5
<210> 127
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-005
<400> 127
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Gln Gln Val Tyr Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 128
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-005
<400> 128
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgcagcag 60
gtttacccac catacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 129
<211> 7
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность структурной петли домена CH3 Fcab FS22-172-006
<400> 129
Arg Lys Tyr Tyr Pro Pro Tyr
1 5
<210> 130
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-006
<400> 130
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Arg Lys Tyr Tyr Pro Pro Tyr Asn Gln Leu Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 131
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172-006
<400> 131
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgcgtaaa 60
tactacccgc cgtacaacca gctcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 132
<211> 111
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172
<400> 132
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Arg Lys Tyr Tyr Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys
20 25 30
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
35 40 45
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
50 55 60
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp
65 70 75 80
Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
85 90 95
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105 110
<210> 133
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Домен CH3 Fcab FS22-172
<400> 133
ggccagcctc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgcgtaaa 60
tactacccgc cgtacaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 120
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 180
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt gggcgcagat 240
aggtggctgg aagggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 300
tacacacaga agagcctctc cctgtctccg ggt 333
<210> 134
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/E12v2 с мутацией LALA
<400> 134
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Pro Tyr Ile Ile
355 360 365
Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 135
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/E12v2 с мутацией LALA
<400> 135
gaagttcagc tggttcagtc tggcgccgaa gtgaaaagac ctggcgcctc tgtgaaggtg 60
tcctgcaagg cttctggcta cccctttacc tcctacggca tctcctgggt ccgacaggct 120
cctggacaag gcttggaatg gatgggctgg atctccgctt attccggcgg caccaattac 180
gcccagaaac tgcagggcag agtgaccatg accaccgaca cctctacctc caccgcctac 240
atggaactgc ggtccctgag atctgacgac accgccgtgt actactgcgc cagagatctg 300
ttccccacca tcttcggcgt gtcctactac tactattggg gccagggcac cctggtcacc 360
gtgtcctctg cttctaccaa gggacccagc gtgttccctc tggctccttc cagcaagtct 420
acctctggcg gaacagctgc tctgggctgc ctggtcaagg actactttcc tgagcctgtg 480
accgtgtctt ggaactctgg cgctctgaca tctggcgtgc acacctttcc agctgtgctg 540
cagtcctccg gcctgtactc tctgtcctct gtcgtgaccg tgccttccag ctctctggga 600
acccagacct acatctgcaa tgtgaaccac aagccttcca acaccaaggt ggacaagaag 660
gtggaaccca agtcctgcga caagacccac acctgtcctc catgtcctgc tccagaagct 720
gctggcggcc cttccgtgtt tctgttccct ccaaagccta aggacaccct gatgatctct 780
cggacccctg aagtgacctg cgtggtggtg gatgtgtctc acgaggaccc agaagtgaag 840
ttcaattggt acgtggacgg cgtggaagtg cacaacgcca agaccaagcc tagagaggaa 900
cagtacaact ccacctacag agtggtgtcc gtgctgaccg tgctgcacca ggattggctg 960
aacggcaaag agtacaagtg caaggtgtcc aacaaggccc tgcctgctcc tatcgaaaag 1020
accatctcca aggccaaggg ccagcctagg gaaccccagg tttacacctt gcctccatct 1080
cgggacgagc tgccctacat catccctcca tacaaccagg tgtccctgac ctgcctcgtg 1140
aagggcttct acccttccga tatcgccgtg gaatgggaga gcaatggcca gcctgagaac 1200
aactacaaga caacccctcc tgtgctggac tccgacggct cattctttct gtactccaag 1260
ctgacagtgg gcgccgacag atggctggaa gggaacgtgt tctcctgcag cgtgatgcac 1320
gaggccctgc acaatcacta cacacagaag tccctgtctc tgtcccctgg c 1371
<210> 136
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ2 FS22-172-003-AA/E12v2
<400> 136
gacatccaga tgacccagtc tccatccaca ctgtccgcct ctgtgcggga cagagtgatc 60
atcacctgta gagccagcca gtccatcggc aacagactgg cctggtatca gcacaagcct 120
ggcaaggctc ccaagctgct gatctacgag gcctccacat ctgagacagg cgtgccctct 180
agattctccg gctctggctc tggcaccgac tttaccctga caatctccag cctgcagcct 240
gaggacttcg ccacctacta ctgccagcag tcctacagca ccccttacac ctttggccag 300
ggcaccaagc tggaaatcaa gcgtacggtg gccgctccca gcgtgttcat cttcccccca 360
agcgacgagc agctgaagag cggcaccgcc agcgtggtgt gtctgctgaa caacttctac 420
cccagggagg ccaaggtgca gtggaaggtg gacaacgccc tgcagagcgg caacagccag 480
gagagcgtca ccgagcagga cagcaaggac tccacctaca gcctgagcag caccctgacc 540
ctgagcaagg ccgactacga gaagcacaag gtgtacgcct gtgaggtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gagcttcaac aggggcgagt gc 642
<210> 137
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/E05v2 с мутацией LALA
<400> 137
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Pro Tyr Ile Ile
355 360 365
Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 138
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/E05v2 с мутацией LALA
<400> 138
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata caccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagcg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc 420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc 480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt 540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc 600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag 660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct 720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca 780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa 840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa 900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg 960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa 1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgccatacat catcccacca tacaaccagg tcagcctgac ctgcctggtc 1140
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1200
aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag 1260
ctcaccgtgg gcgcagatag gtggctggaa gggaacgtct tctcatgctc cgtgatgcat 1320
gaggcgctgc acaaccacta cactcagaag agcttgtccc tgtcgcccgg t 1371
<210> 139
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ2 FS22-172-003-AA/E05v2
<400> 139
gacattcaga tgacccaatc cccgtccacg ctgagcgcct ccgtcggtga tcgcgtgaca 60
atcacttgtc gggcgtcgca gtccatctct ggaaggctcg cctggtacca gcagaagcct 120
ggaaaggctc ccaacctcct tatctacgaa gccagcaacc tggagtccgg agtgcctagc 180
cggttcagcg gatcagggtc cggtaccgag ttcaccctga ccatttcctc gctccaacct 240
gaggacttcg ccacctacta ctgccaacag tcctattcaa ctccgcgcgt gaccttcggc 300
cagggcacta aggtcgaaat caaaagaacc gtggcagccc catcggtgtt tatcttcccg 360
ccctcggacg aacagctgaa gtcaggcact gctagcgtgg tctgtctcct gaacaatttc 420
tacccgcgcg aagctaaggt ccagtggaag gtcgacaacg cgctgcagtc cggaaacagc 480
caggagtcag tgaccgagca ggactccaag gattccactt attccctgtc ctccaccctg 540
actttgagca aggccgacta cgagaagcac aaagtgtacg cctgcgaagt gacccatcaa 600
gggctttcgt cgcccgtgac caagagcttc aaccggggcg aatgc 645
<210> 140
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/G12v2 с мутацией LALA
<400> 140
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Pro Tyr Ile Ile
355 360 365
Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 141
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-172-003-AA/G12v2 с мутацией LALA
<400> 141
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata caccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagcg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc 420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc 480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt 540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc 600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag 660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct 720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca 780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa 840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa 900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg 960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa 1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgccatacat catcccacca tacaaccagg tcagcctgac ctgcctggtc 1140
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1200
aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag 1260
ctcaccgtgg gcgcagatag gtggctggaa gggaacgtct tctcatgctc cgtgatgcat 1320
gaggcgctgc acaaccacta cactcagaag agcttgtccc tgtcgcccgg t 1371
<210> 142
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ2 FS22-172-003-AA/G12v2
<400> 142
gacattcaga tgacccagtc cccgagcacg ctgtccgcaa gcgtggggga cagagtgacc 60
atcacttgcc gcgcctcaca atccatcagc ggacgcttgg cctggtacca gcagaagccc 120
ggaaaggccc caaaccttct gatctacgaa gcctcgaacc tggagtcagg cgtcccttcg 180
cggttctctg gctccggttc cggaactgag ttcaccctca ccatctcgtc cctgcaaccg 240
gaagatttcg ccacctacta ctgccaacag tcgtactcct ggccccggac attcggacag 300
ggaaccaaag tcgagattaa gcggactgtg gcggctccta gcgtgttcat ctttcccccg 360
tccgacgaac agctgaagtc cggtaccgct agcgtggtct gtctcctgaa caatttctac 420
ccgcgcgaag ctaaggtcca gtggaaggtc gacaacgcgc tgcagtccgg aaacagccag 480
gagtcagtga ccgagcagga ctccaaggat tccacttatt ccctgtcctc caccctgact 540
ttgagcaagg ccgactacga gaagcacaaa gtgtacgcct gcgaagtgac ccatcaaggg 600
ctttcgtcgc ccgtgaccaa gagcttcaac cggggcgaat gc 642
<210> 143
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-053-008-AA/E12v2 с мутацией LALA
<400> 143
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Tyr Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 144
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь МАТ2 FS22-053-008-AA/E12v2 с мутацией LALA
<400> 144
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata ccccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagcg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc 420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc 480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt 540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc 600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag 660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct 720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca 780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa 840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa 900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg 960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa 1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgaacccgcc gtacctgttc tctaaccagg tcagcctgac ctgcctggtc 1140
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1200
aactacaaga ccacgcctcc cgtactggac tccgacggct ccttcttcct ctacagcaag 1260
ctcaccgtgg attactggag gtggctggaa gggaacgtct tctcatgctc cgtgatgcat 1320
gaggcgctgc acaaccacta cactcagaag agcttgtccc tgtcgcccgg t 1371
<210> 145
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь МАТ2 FS22-053-008-AA/E12v2 с мутацией LALA
<400> 145
gacatccaga tgacgcagag cccgtctacc ctgtccgcct ccgtgagaga tcgcgtgatc 60
atcacctgtc gggccagcca gtccatcgga aaccgcttgg cgtggtacca gcacaagcct 120
gggaaggctc cgaagctgct catctacgaa gcctcgactt cggagactgg tgtccctagc 180
cggttcagcg gatcgggatc agggaccgat ttcactctga ccatttcctc cctgcaaccc 240
gaggacttcg ccacctacta ctgccaacag tcatattcca ccccgtacac cttcggacaa 300
ggcaccaagc tcgaaatcaa gcggactgtc gccgcacctt ccgtgttcat tttcccaccc 360
tccgacgaac agctgaaatc gggtacagct agcgtggtct gtctcctgaa caatttctac 420
ccgcgcgaag ctaaggtcca gtggaaggtc gacaacgcgc tgcagtccgg aaacagccag 480
gagtcagtga ccgagcagga ctccaaggat tccacttatt ccctgtcctc caccctgact 540
ttgagcaagg ccgactacga gaagcacaaa gtgtacgcct gcgaagtgac ccatcaaggg 600
ctttcgtcgc ccgtgaccaa gagcttcaac cggggcgaat gc 642
<210> 146
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-008-AA/E05v2 с мутацией LALA
<400> 146
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Tyr Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 147
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-008-AA/E05v2 с мутацией LALA
<400> 147
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata caccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagcg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc 420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc 480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt 540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc 600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag 660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct 720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca 780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa 840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa 900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg 960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa 1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgaacccgcc gtacctgttc tctaaccagg tcagcctgac ctgcctggtc 1140
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1200
aactacaaga ccacgcctcc cgtactggac tccgacggct ccttcttcct ctacagcaag 1260
ctcaccgtgg attactggag gtggctggaa gggaacgtct tctcatgctc cgtgatgcat 1320
gaggcgctgc acaaccacta cactcagaag agcttgtccc tgtcgcccgg t 1371
<210> 148
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS22-053-008-AA/E05v2 с мутацией LALA
<400> 148
gacattcaga tgacccaatc cccgtccacg ctgagcgcct ccgtcggtga tcgcgtgaca 60
atcacttgtc gggcgtcgca gtccatctct ggaaggctcg cctggtacca gcagaagcct 120
ggaaaggctc ccaacctcct tatctacgaa gccagcaacc tggagtccgg agtgcctagc 180
cggttcagcg gatcagggtc cggtaccgag ttcaccctga ccatttcctc gctccaacct 240
gaggacttcg ccacctacta ctgccaacag tcctattcaa ctccgcgcgt gaccttcggc 300
cagggcacta aggtcgaaat caaaagaacc gtggcagccc catcggtgtt tatcttcccg 360
ccctcggacg aacagctgaa gtcaggcact gctagcgtgg tctgtctcct gaacaatttc 420
tacccgcgcg aagctaaggt ccagtggaag gtcgacaacg cgctgcagtc cggaaacagc 480
caggagtcag tgaccgagca ggactccaag gattccactt attccctgtc ctccaccctg 540
actttgagca aggccgacta cgagaagcac aaagtgtacg cctgcgaagt gacccatcaa 600
gggctttcgt cgcccgtgac caagagcttc aaccggggcg aatgc 645
<210> 149
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-008-AA/G12v2 с мутацией LALA
<400> 149
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Asp Tyr Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 150
<211> 1371
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-008-AA/G12v2 с мутацией LALA
<400> 150
gaagtgcagc tggtgcagtc cggagccgaa gtcaagaggc ctggagcgtc cgtgaaggtg 60
tcctgcaaag cctcaggata caccttcact tcgtacggga tttcctgggt ccgccaagca 120
ccgggtcaag gcttggagtg gatgggatgg atcagcgcgt attccggggg aaccaactac 180
gctcaaaagc tgcagggtcg cgtgaccatg accaccgata cctccacctc aacggcctac 240
atggaactga gatctctgcg gagcgacgac actgccgtgt actactgtgc ccgggacctg 300
ttccccacta tcttcggagt gtcgtactac tactactggg gccaggggac tctcgtgacc 360
gtgtcgagcg ctagcactaa gggcccgtcg gtgttcccgc tggccccatc gtccaagagc 420
acatcagggg gtaccgccgc cctgggctgc cttgtgaagg attactttcc cgagcccgtc 480
acagtgtcct ggaacagcgg agccctgacc tccggagtgc atactttccc ggctgtgctt 540
cagtcctctg gcctgtactc attgtcctcc gtggtcaccg tcccttcgtc ctccctgggc 600
acccagacct atatctgtaa tgtcaaccat aagccctcga acaccaaggt cgacaagaag 660
gtcgagccga agtcgtgcga caagactcac acttgcccgc cttgcccagc cccggaagct 720
gccggtggtc cttcggtgtt cctcttcccg cccaagccga aggataccct gatgatctca 780
cggacccccg aagtgacctg tgtggtggtg gacgtgtccc acgaggaccc ggaagtgaaa 840
ttcaattggt acgtggatgg agtggaagtg cacaacgcca agaccaagcc acgggaagaa 900
cagtacaact ctacctaccg cgtggtgtcc gtgctcactg tgctgcacca agactggctg 960
aacgggaagg agtacaagtg caaagtgtcc aacaaggcgc tgcctgcccc aattgagaaa 1020
actatctcga aagccaaggg acagcctcga gaaccacagg tgtacaccct gcccccatcc 1080
cgggatgagc tgaacccgcc gtacctgttc tctaaccagg tcagcctgac ctgcctggtc 1140
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 1200
aactacaaga ccacgcctcc cgtactggac tccgacggct ccttcttcct ctacagcaag 1260
ctcaccgtgg attactggag gtggctggaa gggaacgtct tctcatgctc cgtgatgcat 1320
gaggcgctgc acaaccacta cactcagaag agcttgtccc tgtcgcccgg t 1371
<210> 151
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь FS22-053-008-AA/G12v2 с мутацией LALA
<400> 151
gacattcaga tgacccagtc cccgagcacg ctgtccgcaa gcgtggggga cagagtgacc 60
atcacttgcc gcgcctcaca atccatcagc ggacgcttgg cctggtacca gcagaagccc 120
ggaaaggccc caaaccttct gatctacgaa gcctcgaacc tggagtcagg cgtcccttcg 180
cggttctctg gctccggttc cggaactgag ttcaccctca ccatctcgtc cctgcaaccg 240
gaagatttcg ccacctacta ctgccaacag tcgtactcct ggccccggac attcggacag 300
ggaaccaaag tcgagattaa gcggactgtg gcggctccta gcgtgttcat ctttcccccg 360
tccgacgaac agctgaagtc cggtaccgct agcgtggtct gtctcctgaa caatttctac 420
ccgcgcgaag ctaaggtcca gtggaaggtc gacaacgcgc tgcagtccgg aaacagccag 480
gagtcagtga ccgagcagga ctccaaggat tccacttatt ccctgtcctc caccctgact 540
ttgagcaagg ccgactacga gaagcacaaa gtgtacgcct gcgaagtgac ccatcaaggg 600
ctttcgtcgc ccgtgaccaa gagcttcaac cggggcgaat gc 642
<210> 152
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-017AA/E12v2 с мутацией LALA
<400> 152
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Tyr His Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 153
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-017AA/E05v2 с мутацией LALA
<400> 153
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Tyr His Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 154
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-053-017AA/G12v2 с мутацией LALA
<400> 154
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Asn Pro Pro Tyr
355 360 365
Leu Phe Ser Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Tyr His Trp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 155
<211> 457
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-172-003AA/lam-G02v3 с мутацией LALA
<400> 155
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Pro Tyr Ile Ile
355 360 365
Pro Pro Tyr Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
385 390 395 400
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
405 410 415
Leu Tyr Ser Lys Leu Thr Val Gly Ala Asp Arg Trp Leu Glu Gly Asn
420 425 430
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Leu Ser Leu Ser Pro Gly
450 455
<210> 156
<211> 452
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-172-003AA/S70 с мутацией LALA
<400> 156
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Pro Tyr Ile Ile Pro Pro Tyr Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Gly Ala Asp Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 157
<211> 214
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь S70
<400> 157
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 158
<211> 450
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22-172-003AA/HelD1.3 с мутацией LALA
<400> 158
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr Phe Ser Gly Tyr
20 25 30
Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp Gly Gln Gly Ser Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Pro Tyr Ile Ile Pro Pro Tyr Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Gly Ala Asp Arg Trp Leu Glu Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly
450
<210> 159
<211> 214
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь HelD1.3
<400> 159
Asp Ile Gln Met Thr Gln Ser Pro Ala Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Glu Thr Val Thr Ile Thr Cys Arg Ala Ser Gly Asn Ile His Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Gln Gly Lys Ser Pro Gln Leu Leu Val
35 40 45
Tyr Asn Ala Lys Thr Leu Ala Asp Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Gln Tyr Ser Leu Lys Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Gly Ser Tyr Tyr Cys Gln His Phe Trp Ser Thr Pro Arg
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 160
<211> 447
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22m-063AA/S70 с мутацией LALA
<400> 160
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Pro Tyr Trp Ser Tyr Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Met Asn Tyr
405 410 415
Arg Trp Glu Leu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 161
<211> 445
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь FS22m-063AA/HelD1.3 с мутацией LALA
<400> 161
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr Phe Ser Gly Tyr
20 25 30
Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp Gly Gln Gly Ser Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Pro Tyr Trp Ser Tyr Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Met Asn Tyr Arg Trp
405 410 415
Glu Leu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 162
<211> 452
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1-AA/E12v2 с мутацией LALA
<400> 162
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Pro Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Ser Gly Gly Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala
225 230 235 240
Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 163
<211> 448
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/S70
<400> 163
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 164
<211> 448
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1-AA/S70 с мутацией LALA
<400> 164
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 165
<211> 448
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/4420
<400> 165
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 166
<211> 219
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь 4420
<400> 166
Asp Val Val Met Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu Arg Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Val Leu Ile Tyr Lys Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 167
<211> 448
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1-AA/4420 с мутацией LALA
<400> 167
Glu Val Lys Leu Asp Glu Thr Gly Gly Gly Leu Val Gln Pro Gly Arg
1 5 10 15
Pro Met Lys Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met Asn Trp Val Arg Gln Ser Pro Glu Lys Gly Leu Glu Trp Val
35 40 45
Ala Gln Ile Arg Asn Lys Pro Tyr Asn Tyr Glu Thr Tyr Tyr Ser Asp
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Ser Ser
65 70 75 80
Val Tyr Leu Gln Met Asn Asn Leu Arg Val Glu Asp Met Gly Ile Tyr
85 90 95
Tyr Cys Thr Gly Ser Tyr Tyr Gly Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 168
<211> 446
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1-AA/HelD1.3 с мутацией LALA
<400> 168
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr Phe Ser Gly Tyr
20 25 30
Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp Gly Gln Gly Ser Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 169
<211> 460
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1-AA/MLS109 с мутацией LALA
<400> 169
Glu Glu Gln Val Leu Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Ser Val Phe Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asn Ser Asp Ser Thr Tyr Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Glu Asn Ser Ile Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Ser Tyr Tyr Ala Phe Ser Ser Gly Ser Leu Ser Asp
100 105 110
Tyr Tyr Tyr Gly Leu Asp Val Trp Gly Gln Gly Thr Thr Val Ile Val
115 120 125
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
130 135 140
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
145 150 155 160
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
165 170 175
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
180 185 190
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
195 200 205
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
210 215 220
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
225 230 235 240
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
245 250 255
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
260 265 270
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
275 280 285
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
290 295 300
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
305 310 315 320
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
325 330 335
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
340 345 350
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
355 360 365
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
370 375 380
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
385 390 395 400
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
405 410 415
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
420 425 430
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
435 440 445
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455 460
<210> 170
<211> 219
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь MLS109
<400> 170
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Thr
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Val Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Ala Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Thr Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Ile Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 171
<211> 13
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 петли AB FS22-172-003
<400> 171
Arg Asp Glu Leu Pro Tyr Ile Ile Pro Pro Tyr Asn Gln
1 5 10
<210> 172
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 петли EF FS22-172-003
<400> 172
Gly Ala Asp Arg Trp Leu Glu Gly Asn Val
1 5 10
<210> 173
<211> 13
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 петли AB FS22-53-008
<400> 173
Arg Asp Glu Leu Asn Pro Pro Tyr Leu Phe Ser Asn Gln
1 5 10
<210> 174
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 петли EF FS22-53-008
<400> 174
Asp Tyr Trp Arg Trp Leu Glu Gly Asn Val
1 5 10
<210> 175
<211> 10
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH3 петли EF FS22-053-017
<400> 175
Tyr His Trp Arg Trp Leu Glu Gly Asn Val
1 5 10
<210> 176
<211> 110
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Домен CH2 с мутацией LALA и мутацией P114A
<400> 176
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105 110
<210> 177
<211> 452
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/280_02_G02
<400> 177
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Gly Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Ser Ala Tyr Asn Gly Asn Thr Asn Tyr Ala Gln Lys Leu
50 55 60
Gln Gly Arg Val Thr Met Thr Thr Asp Thr Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Leu Phe Pro Thr Ile Phe Gly Val Ser Tyr Tyr Tyr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly
450
<210> 178
<211> 217
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь G1/280_02_G02
<400> 178
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Ser Val Ser Trp Tyr Gln Gln Phe Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Phe Glu Val Thr Asn Arg Pro Ser Gly Val Ser Asp Arg Phe
50 55 60
Ser Gly Ser Lys Ser Asp Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Glu Tyr Tyr Cys Ser Ser Phe Lys Arg Gly
85 90 95
Ser Thr Leu Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly
100 105 110
Gln Pro Ala Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
115 120 125
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
130 135 140
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
145 150 155 160
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
165 170 175
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
180 185 190
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
195 200 205
Lys Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 179
<211> 272
<212> PRT/БЕЛОК
<213> PD-L1 Homo sapiens
<400> 179
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr His Leu Val
210 215 220
Ile Leu Gly Ala Ile Leu Leu Cys Leu Gly Val Ala Leu Thr Phe Ile
225 230 235 240
Phe Arg Leu Arg Lys Gly Arg Met Met Asp Val Lys Lys Cys Gly Ile
245 250 255
Gln Asp Thr Asn Ser Lys Lys Gln Ser Asp Thr His Leu Glu Glu Thr
260 265 270
<210> 180
<211> 220
<212> PRT/БЕЛОК
<213> Внеклеточный домен PD-L1 Homo sapiens
<400> 180
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg
210 215 220
<210> 181
<211> 272
<212> PRT/БЕЛОК
<213> PD-L1 мыши
<400> 181
Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu Asp Leu
20 25 30
Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val Ile Gln
35 40 45
Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn Phe Arg
50 55 60
Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp Pro Ala
115 120 125
Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly Lys Arg
145 150 155 160
Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val Thr Ser
165 170 175
Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys Thr Phe
180 185 190
Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile Ile Pro
195 200 205
Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr His Trp Val Leu
210 215 220
Leu Gly Ser Ile Leu Leu Phe Leu Ile Val Val Ser Thr Val Leu Leu
225 230 235 240
Phe Leu Arg Lys Gln Val Arg Met Leu Asp Val Glu Lys Cys Gly Val
245 250 255
Glu Asp Thr Ser Ser Lys Asn Arg Asn Asp Thr Gln Phe Glu Glu Thr
260 265 270
<210> 182
<211> 221
<212> PRT/БЕЛОК
<213> Внеклеточный домен PD-L1 мыши
<400> 182
Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu Asp Leu
20 25 30
Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val Ile Gln
35 40 45
Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn Phe Arg
50 55 60
Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Cys Cys
85 90 95
Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu Lys Val
100 105 110
Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp Pro Ala
115 120 125
Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro Glu Ala
130 135 140
Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly Lys Arg
145 150 155 160
Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val Thr Ser
165 170 175
Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys Thr Phe
180 185 190
Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile Ile Pro
195 200 205
Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr His
210 215 220
<210> 183
<211> 328
<212> PRT/БЕЛОК
<213> PD-L1 яванского макака
<400> 183
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Thr Ser Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Asn Tyr Arg
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Arg Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Leu Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Ala Asn Glu Ile Phe Tyr Cys Ile
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala Leu Pro Pro Asn Glu Arg Thr His Leu Val
210 215 220
Ile Leu Gly Ala Ile Phe Leu Leu Leu Gly Val Ala Leu Thr Phe Ile
225 230 235 240
Phe Tyr Leu Arg Lys Gly Arg Met Met Asp Met Lys Lys Cys Gly Ile
245 250 255
Arg Val Thr Asn Ser Lys Lys Gln Arg Asp Thr Gln Leu Glu Glu Thr
260 265 270
Lys Gly Pro Ser Ala Arg Phe Asp Ile Pro Asp Glu Ile Pro Val Ile
275 280 285
Glu Ser Lys Pro Asn Thr Leu Ser Ile Val Leu Gly Thr Thr Leu Thr
290 295 300
Val Ala Met Ile Ile Val Ala Thr Ile Phe Gly Tyr Arg Arg Gln Lys
305 310 315 320
Gly Arg Leu Arg Thr Leu Lys Leu
325
<210> 184
<211> 222
<212> PRT/БЕЛОК
<213> Внеклеточный домен PD-L1 яванского макака
<400> 184
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Thr Ser Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Asn Tyr Arg
50 55 60
Gln Arg Ala Gln Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Arg Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Leu Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Ala Asn Glu Ile Phe Tyr Cys Ile
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala Leu Pro Pro Asn Glu Arg Thr His
210 215 220
<210> 185
<211> 232
<212> PRT/БЕЛОК
<213> CD137 Homo sapiens
<400> 185
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Ser Phe Phe Leu Ala Leu Thr Ser Thr Ala Leu
165 170 175
Leu Phe Leu Leu Phe Phe Leu Thr Leu Arg Phe Ser Val Val Lys Arg
180 185 190
Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro
195 200 205
Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu
210 215 220
Glu Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 186
<211> 163
<212> PRT/БЕЛОК
<213> Внеклеточный домен CD137 Homo sapiens
<400> 186
Leu Gln Asp Pro Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Asn Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Arg Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Thr Pro Gly Phe His Cys Leu Gly Ala Gly Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Val Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 187
<211> 233
<212> PRT/БЕЛОК
<213> CD137 мыши
<400> 187
Val Gln Asn Ser Cys Asp Asn Cys Gln Pro Gly Thr Phe Cys Arg Lys
1 5 10 15
Tyr Asn Pro Val Cys Lys Ser Cys Pro Pro Ser Thr Phe Ser Ser Ile
20 25 30
Gly Gly Gln Pro Asn Cys Asn Ile Cys Arg Val Cys Ala Gly Tyr Phe
35 40 45
Arg Phe Lys Lys Phe Cys Ser Ser Thr His Asn Ala Glu Cys Glu Cys
50 55 60
Ile Glu Gly Phe His Cys Leu Gly Pro Gln Cys Thr Arg Cys Glu Lys
65 70 75 80
Asp Cys Arg Pro Gly Gln Glu Leu Thr Lys Gln Gly Cys Lys Thr Cys
85 90 95
Ser Leu Gly Thr Phe Asn Asp Gln Asn Gly Thr Gly Val Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Arg Ser Val Leu Lys Thr Gly Thr
115 120 125
Thr Glu Lys Asp Val Val Cys Gly Pro Pro Val Val Ser Phe Ser Pro
130 135 140
Ser Thr Thr Ile Ser Val Thr Pro Glu Gly Gly Pro Gly Gly His Ser
145 150 155 160
Leu Gln Val Leu Thr Leu Phe Leu Ala Leu Thr Ser Ala Leu Leu Leu
165 170 175
Ala Leu Ile Phe Ile Thr Leu Leu Phe Ser Val Leu Lys Trp Ile Arg
180 185 190
Lys Lys Phe Pro His Ile Phe Lys Gln Pro Phe Lys Lys Thr Thr Gly
195 200 205
Ala Ala Gln Glu Glu Asp Ala Cys Ser Cys Arg Cys Pro Gln Glu Glu
210 215 220
Glu Gly Gly Gly Gly Gly Tyr Glu Leu
225 230
<210> 188
<211> 164
<212> PRT/БЕЛОК
<213> Внеклеточный домен CD137 мыши
<400> 188
Val Gln Asn Ser Cys Asp Asn Cys Gln Pro Gly Thr Phe Cys Arg Lys
1 5 10 15
Tyr Asn Pro Val Cys Lys Ser Cys Pro Pro Ser Thr Phe Ser Ser Ile
20 25 30
Gly Gly Gln Pro Asn Cys Asn Ile Cys Arg Val Cys Ala Gly Tyr Phe
35 40 45
Arg Phe Lys Lys Phe Cys Ser Ser Thr His Asn Ala Glu Cys Glu Cys
50 55 60
Ile Glu Gly Phe His Cys Leu Gly Pro Gln Cys Thr Arg Cys Glu Lys
65 70 75 80
Asp Cys Arg Pro Gly Gln Glu Leu Thr Lys Gln Gly Cys Lys Thr Cys
85 90 95
Ser Leu Gly Thr Phe Asn Asp Gln Asn Gly Thr Gly Val Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Arg Ser Val Leu Lys Thr Gly Thr
115 120 125
Thr Glu Lys Asp Val Val Cys Gly Pro Pro Val Val Ser Phe Ser Pro
130 135 140
Ser Thr Thr Ile Ser Val Thr Pro Glu Gly Gly Pro Gly Gly His Ser
145 150 155 160
Leu Gln Val Leu
<210> 189
<211> 231
<212> PRT/БЕЛОК
<213> CD137 яванского макака
<400> 189
Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln Ile Ile Phe Phe Leu Ala Leu Thr Ser Thr Val Val Leu
165 170 175
Phe Leu Leu Phe Phe Leu Val Leu Arg Phe Ser Val Val Lys Arg Ser
180 185 190
Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met Arg Pro Val
195 200 205
Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu
210 215 220
Glu Glu Gly Gly Cys Glu Leu
225 230
<210> 190
<211> 163
<212> PRT/БЕЛОК
<213> Внеклеточный домен CD137 яванского макака
<400> 190
Leu Gln Asp Leu Cys Ser Asn Cys Pro Ala Gly Thr Phe Cys Asp Asn
1 5 10 15
Asn Arg Ser Gln Ile Cys Ser Pro Cys Pro Pro Asn Ser Phe Ser Ser
20 25 30
Ala Gly Gly Gln Arg Thr Cys Asp Ile Cys Arg Gln Cys Lys Gly Val
35 40 45
Phe Lys Thr Arg Lys Glu Cys Ser Ser Thr Ser Asn Ala Glu Cys Asp
50 55 60
Cys Ile Ser Gly Tyr His Cys Leu Gly Ala Glu Cys Ser Met Cys Glu
65 70 75 80
Gln Asp Cys Lys Gln Gly Gln Glu Leu Thr Lys Lys Gly Cys Lys Asp
85 90 95
Cys Cys Phe Gly Thr Phe Asn Asp Gln Lys Arg Gly Ile Cys Arg Pro
100 105 110
Trp Thr Asn Cys Ser Leu Asp Gly Lys Ser Val Leu Val Asn Gly Thr
115 120 125
Lys Glu Arg Asp Val Val Cys Gly Pro Ser Pro Ala Asp Leu Ser Pro
130 135 140
Gly Ala Ser Ser Ala Thr Pro Pro Ala Pro Ala Arg Glu Pro Gly His
145 150 155 160
Ser Pro Gln
<210> 191
<211> 445
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь G1/HelD1.3
<400> 191
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Arg Pro Ser Gln
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ser Thr Phe Ser Gly Tyr
20 25 30
Gly Val Asn Trp Val Arg Gln Pro Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Met Ile Trp Gly Asp Gly Asn Thr Asp Tyr Asn Ser Ala Leu Lys
50 55 60
Ser Arg Val Thr Met Leu Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Arg Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Glu Arg Asp Tyr Arg Leu Asp Tyr Trp Gly Gln Gly Ser Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
115 120 125
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
130 135 140
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
145 150 155 160
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
165 170 175
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
180 185 190
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
195 200 205
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr
210 215 220
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
225 230 235 240
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
245 250 255
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
260 265 270
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
275 280 285
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
290 295 300
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
305 310 315 320
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
325 330 335
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
340 345 350
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
355 360 365
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
370 375 380
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
385 390 395 400
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
405 410 415
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
420 425 430
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 192
<211> 17
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Пептид OVA
<400> 192
Ile Ser Gln Ala Val His Ala Ala His Ala Glu Ile Asn Glu Ala Gly
1 5 10 15
Arg
<210> 193
<211> 434
<212> PRT/БЕЛОК
<213> Внеклеточный домен B7-H3 Homo sapiens
<400> 193
Ile Leu Glu Val Gln Val Pro Glu Asp Pro Val Val Ala Leu Val Gly
1 5 10 15
Thr Asp Ala Thr Leu Cys Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser
20 25 30
Leu Ala Gln Leu Asn Leu Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu
35 40 45
Val His Ser Phe Ala Glu Gly Gln Asp Gln Gly Ser Ala Tyr Ala Asn
50 55 60
Arg Thr Ala Leu Phe Pro Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu
65 70 75 80
Arg Leu Gln Arg Val Arg Val Ala Asp Glu Gly Ser Phe Thr Cys Phe
85 90 95
Val Ser Ile Arg Asp Phe Gly Ser Ala Ala Val Ser Leu Gln Val Ala
100 105 110
Ala Pro Tyr Ser Lys Pro Ser Met Thr Leu Glu Pro Asn Lys Asp Leu
115 120 125
Arg Pro Gly Asp Thr Val Thr Ile Thr Cys Ser Ser Tyr Gln Gly Tyr
130 135 140
Pro Glu Ala Glu Val Phe Trp Gln Asp Gly Gln Gly Val Pro Leu Thr
145 150 155 160
Gly Asn Val Thr Thr Ser Gln Met Ala Asn Glu Gln Gly Leu Phe Asp
165 170 175
Val His Ser Ile Leu Arg Val Val Leu Gly Ala Asn Gly Thr Tyr Ser
180 185 190
Cys Leu Val Arg Asn Pro Val Leu Gln Gln Asp Ala His Ser Ser Val
195 200 205
Thr Ile Thr Pro Gln Arg Ser Pro Thr Gly Ala Val Glu Val Gln Val
210 215 220
Pro Glu Asp Pro Val Val Ala Leu Val Gly Thr Asp Ala Thr Leu Arg
225 230 235 240
Cys Ser Phe Ser Pro Glu Pro Gly Phe Ser Leu Ala Gln Leu Asn Leu
245 250 255
Ile Trp Gln Leu Thr Asp Thr Lys Gln Leu Val His Ser Phe Thr Glu
260 265 270
Gly Arg Asp Gln Gly Ser Ala Tyr Ala Asn Arg Thr Ala Leu Phe Pro
275 280 285
Asp Leu Leu Ala Gln Gly Asn Ala Ser Leu Arg Leu Gln Arg Val Arg
290 295 300
Val Ala Asp Glu Gly Ser Phe Thr Cys Phe Val Ser Ile Arg Asp Phe
305 310 315 320
Gly Ser Ala Ala Val Ser Leu Gln Val Ala Ala Pro Tyr Ser Lys Pro
325 330 335
Ser Met Thr Leu Glu Pro Asn Lys Asp Leu Arg Pro Gly Asp Thr Val
340 345 350
Thr Ile Thr Cys Ser Ser Tyr Arg Gly Tyr Pro Glu Ala Glu Val Phe
355 360 365
Trp Gln Asp Gly Gln Gly Val Pro Leu Thr Gly Asn Val Thr Thr Ser
370 375 380
Gln Met Ala Asn Glu Gln Gly Leu Phe Asp Val His Ser Val Leu Arg
385 390 395 400
Val Val Leu Gly Ala Asn Gly Thr Tyr Ser Cys Leu Val Arg Asn Pro
405 410 415
Val Leu Gln Gln Asp Ala His Gly Ser Val Thr Ile Thr Gly Gln Pro
420 425 430
Met Thr
<210> 194
<211> 8
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Пептид SIINFEKL
<400> 194
Ser Ile Ile Asn Phe Glu Lys Leu
1 5
<210> 195
<211> 433
<212> PRT/БЕЛОК
<213> Homo sapiens PD-L1-rCD4-His
<400> 195
Met Arg Ile Phe Ala Val Phe Ile Phe Met Thr Tyr Trp His Leu Leu
1 5 10 15
Asn Ala Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asp Leu Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile
50 55 60
Ile Gln Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser
65 70 75 80
Tyr Arg Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Arg Cys Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val
115 120 125
Lys Val Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val
130 135 140
Asp Pro Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr
145 150 155 160
Pro Lys Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser
165 170 175
Gly Lys Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn
180 185 190
Val Thr Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr
195 200 205
Cys Thr Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu
210 215 220
Val Ile Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr Ala
225 230 235 240
Ala Ala Thr Ser Ile Thr Ala Tyr Lys Ser Glu Gly Glu Ser Ala Glu
245 250 255
Phe Ser Phe Pro Leu Asn Leu Gly Glu Glu Ser Leu Gln Gly Glu Leu
260 265 270
Arg Trp Lys Ala Glu Lys Ala Pro Ser Ser Gln Ser Trp Ile Thr Phe
275 280 285
Ser Leu Lys Asn Gln Lys Val Ser Val Gln Lys Ser Thr Ser Asn Pro
290 295 300
Lys Phe Gln Leu Ser Glu Thr Leu Pro Leu Thr Leu Gln Ile Pro Gln
305 310 315 320
Val Ser Leu Gln Phe Ala Gly Ser Gly Asn Leu Thr Leu Thr Leu Asp
325 330 335
Arg Gly Ile Leu Tyr Gln Glu Val Asn Leu Val Val Met Lys Val Thr
340 345 350
Gln Pro Asp Ser Asn Thr Leu Thr Cys Glu Val Met Gly Pro Thr Ser
355 360 365
Pro Lys Met Arg Leu Ile Leu Lys Gln Glu Asn Gln Glu Ala Arg Val
370 375 380
Ser Arg Gln Glu Lys Val Ile Gln Val Gln Ala Pro Glu Ala Gly Val
385 390 395 400
Trp Gln Cys Leu Leu Ser Glu Gly Glu Glu Val Lys Met Asp Ser Lys
405 410 415
Ile Gln Val Leu Ser Lys Gly Leu Asn Gly Ser His His His His His
420 425 430
His
<210> 196
<211> 477
<212> PRT/БЕЛОК
<213> Homo sapiens PD-L1-Fc-His
<400> 196
Met Arg Ile Phe Ala Val Phe Ile Phe Met Thr Tyr Trp His Leu Leu
1 5 10 15
Asn Ala Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu
35 40 45
Asp Leu Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile
50 55 60
Ile Gln Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser
65 70 75 80
Tyr Arg Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Arg Cys Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val
115 120 125
Lys Val Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val
130 135 140
Asp Pro Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr
145 150 155 160
Pro Lys Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser
165 170 175
Gly Lys Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn
180 185 190
Val Thr Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr
195 200 205
Cys Thr Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu
210 215 220
Val Ile Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Thr Ala
225 230 235 240
Ala Ala Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
245 250 255
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
260 265 270
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
275 280 285
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
290 295 300
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
305 310 315 320
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
325 330 335
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
340 345 350
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
355 360 365
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
370 375 380
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
385 390 395 400
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
405 410 415
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
420 425 430
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
435 440 445
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
450 455 460
Leu Ser Pro Gly Lys Gly Ser His His His His His His
465 470 475
<210> 197
<211> 432
<212> PRT/БЕЛОК
<213> PD-L1 мыши-rCD4-His
<400> 197
Met Arg Ile Phe Ala Gly Ile Ile Phe Thr Ala Cys Cys His Leu Leu
1 5 10 15
Arg Ala Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu
35 40 45
Asp Leu Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val
50 55 60
Ile Gln Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn
65 70 75 80
Phe Arg Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Cys Cys Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu
115 120 125
Lys Val Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp
130 135 140
Pro Ala Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro
145 150 155 160
Glu Ala Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly
165 170 175
Lys Arg Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val
180 185 190
Thr Ser Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys
195 200 205
Thr Phe Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile
210 215 220
Ile Pro Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr Ala Ala
225 230 235 240
Ala Thr Ser Ile Thr Ala Tyr Lys Ser Glu Gly Glu Ser Ala Glu Phe
245 250 255
Ser Phe Pro Leu Asn Leu Gly Glu Glu Ser Leu Gln Gly Glu Leu Arg
260 265 270
Trp Lys Ala Glu Lys Ala Pro Ser Ser Gln Ser Trp Ile Thr Phe Ser
275 280 285
Leu Lys Asn Gln Lys Val Ser Val Gln Lys Ser Thr Ser Asn Pro Lys
290 295 300
Phe Gln Leu Ser Glu Thr Leu Pro Leu Thr Leu Gln Ile Pro Gln Val
305 310 315 320
Ser Leu Gln Phe Ala Gly Ser Gly Asn Leu Thr Leu Thr Leu Asp Arg
325 330 335
Gly Ile Leu Tyr Gln Glu Val Asn Leu Val Val Met Lys Val Thr Gln
340 345 350
Pro Asp Ser Asn Thr Leu Thr Cys Glu Val Met Gly Pro Thr Ser Pro
355 360 365
Lys Met Arg Leu Ile Leu Lys Gln Glu Asn Gln Glu Ala Arg Val Ser
370 375 380
Arg Gln Glu Lys Val Ile Gln Val Gln Ala Pro Glu Ala Gly Val Trp
385 390 395 400
Gln Cys Leu Leu Ser Glu Gly Glu Glu Val Lys Met Asp Ser Lys Ile
405 410 415
Gln Val Leu Ser Lys Gly Leu Asn Gly Ser His His His His His His
420 425 430
<210> 198
<211> 476
<212> PRT/БЕЛОК
<213> PD-L1 мыши-Fc-His
<400> 198
Met Arg Ile Phe Ala Gly Ile Ile Phe Thr Ala Cys Cys His Leu Leu
1 5 10 15
Arg Ala Phe Thr Ile Thr Ala Pro Lys Asp Leu Tyr Val Val Glu Tyr
20 25 30
Gly Ser Asn Val Thr Met Glu Cys Arg Phe Pro Val Glu Arg Glu Leu
35 40 45
Asp Leu Leu Ala Leu Val Val Tyr Trp Glu Lys Glu Asp Glu Gln Val
50 55 60
Ile Gln Phe Val Ala Gly Glu Glu Asp Leu Lys Pro Gln His Ser Asn
65 70 75 80
Phe Arg Gly Arg Ala Ser Leu Pro Lys Asp Gln Leu Leu Lys Gly Asn
85 90 95
Ala Ala Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr
100 105 110
Cys Cys Ile Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Leu
115 120 125
Lys Val Asn Ala Pro Tyr Arg Lys Ile Asn Gln Arg Ile Ser Val Asp
130 135 140
Pro Ala Thr Ser Glu His Glu Leu Ile Cys Gln Ala Glu Gly Tyr Pro
145 150 155 160
Glu Ala Glu Val Ile Trp Thr Asn Ser Asp His Gln Pro Val Ser Gly
165 170 175
Lys Arg Ser Val Thr Thr Ser Arg Thr Glu Gly Met Leu Leu Asn Val
180 185 190
Thr Ser Ser Leu Arg Val Asn Ala Thr Ala Asn Asp Val Phe Tyr Cys
195 200 205
Thr Phe Trp Arg Ser Gln Pro Gly Gln Asn His Thr Ala Glu Leu Ile
210 215 220
Ile Pro Glu Leu Pro Ala Thr His Pro Pro Gln Asn Arg Thr Ala Ala
225 230 235 240
Ala Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
245 250 255
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
260 265 270
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
275 280 285
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
290 295 300
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
305 310 315 320
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
325 330 335
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
340 345 350
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
355 360 365
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
370 375 380
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
385 390 395 400
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
405 410 415
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
420 425 430
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
435 440 445
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
450 455 460
Ser Pro Gly Lys Gly Ser His His His His His His
465 470 475
<210> 199
<211> 245
<212> PRT/БЕЛОК
<213> Homo sapiens PD-L1-His-Avi
<400> 199
Phe Thr Val Thr Val Pro Lys Asp Leu Tyr Val Val Glu Tyr Gly Ser
1 5 10 15
Asn Met Thr Ile Glu Cys Lys Phe Pro Val Glu Lys Gln Leu Asp Leu
20 25 30
Ala Ala Leu Ile Val Tyr Trp Glu Met Glu Asp Lys Asn Ile Ile Gln
35 40 45
Phe Val His Gly Glu Glu Asp Leu Lys Val Gln His Ser Ser Tyr Arg
50 55 60
Gln Arg Ala Arg Leu Leu Lys Asp Gln Leu Ser Leu Gly Asn Ala Ala
65 70 75 80
Leu Gln Ile Thr Asp Val Lys Leu Gln Asp Ala Gly Val Tyr Arg Cys
85 90 95
Met Ile Ser Tyr Gly Gly Ala Asp Tyr Lys Arg Ile Thr Val Lys Val
100 105 110
Asn Ala Pro Tyr Asn Lys Ile Asn Gln Arg Ile Leu Val Val Asp Pro
115 120 125
Val Thr Ser Glu His Glu Leu Thr Cys Gln Ala Glu Gly Tyr Pro Lys
130 135 140
Ala Glu Val Ile Trp Thr Ser Ser Asp His Gln Val Leu Ser Gly Lys
145 150 155 160
Thr Thr Thr Thr Asn Ser Lys Arg Glu Glu Lys Leu Phe Asn Val Thr
165 170 175
Ser Thr Leu Arg Ile Asn Thr Thr Thr Asn Glu Ile Phe Tyr Cys Thr
180 185 190
Phe Arg Arg Leu Asp Pro Glu Glu Asn His Thr Ala Glu Leu Val Ile
195 200 205
Pro Glu Leu Pro Leu Ala His Pro Pro Asn Glu Arg Gly Ser His His
210 215 220
His His His His Gly Gly Gly Leu Asn Asp Ile Phe Glu Ala Gln Lys
225 230 235 240
Ile Glu Trp His Glu
245
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ МЕЗОТЕЛИН И CD137 | 2019 |
|
RU2815066C2 |
| МОЛЕКУЛЫ АНТИТЕЛ, КОТОРЫЕ СВЯЗЫВАЮТ CD137 И OX40 | 2019 |
|
RU2817602C2 |
| АНТИТЕЛА К МЕЗОТЕЛИНУ | 2019 |
|
RU2837574C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ PD-L1 и LAG-3 | 2017 |
|
RU2784388C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| АНТИ-CD137 АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2829536C2 |
| МОНОСПЕЦИФИЧЕСКИЕ И БИСПЕЦИФИЧЕСКИЕ БЕЛКИ С РЕГУЛИРОВАНИЕМ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2793167C2 |
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| Иммунотерапия с применением антител, связывающих лиганд 1 белка программируемой смерти клеток (PD-L1) | 2017 |
|
RU2766582C2 |
| PD-L1 специфические антитела | 2017 |
|
RU2756236C2 |



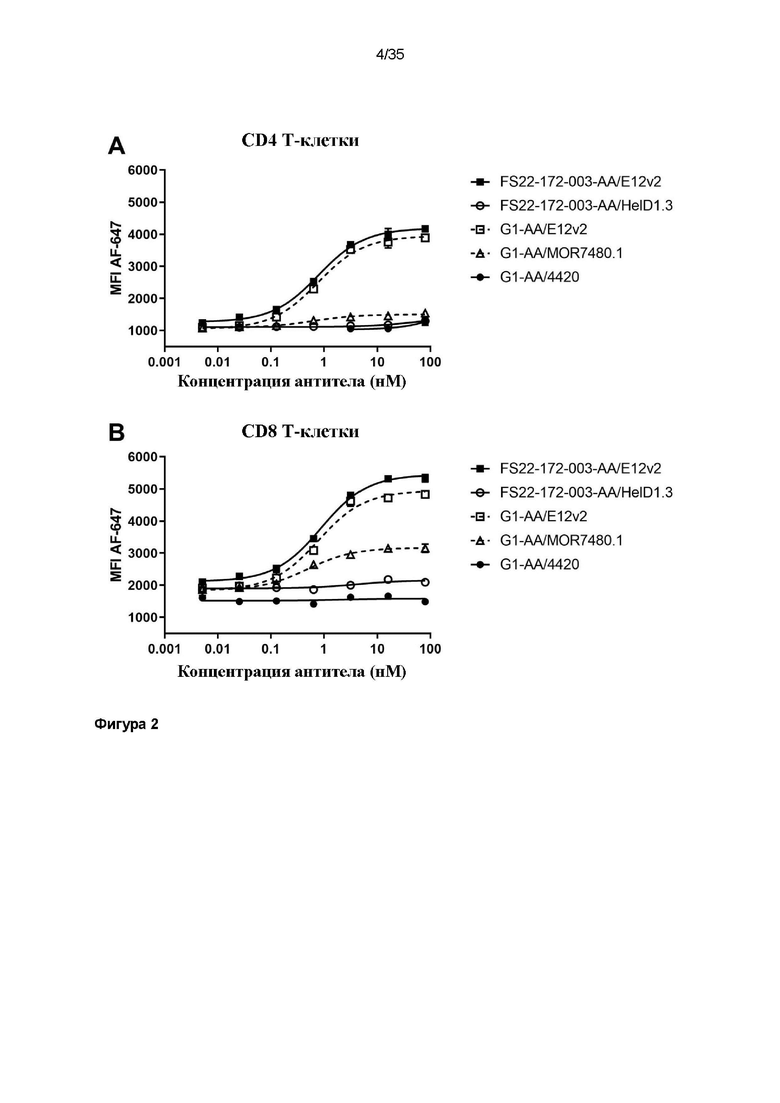
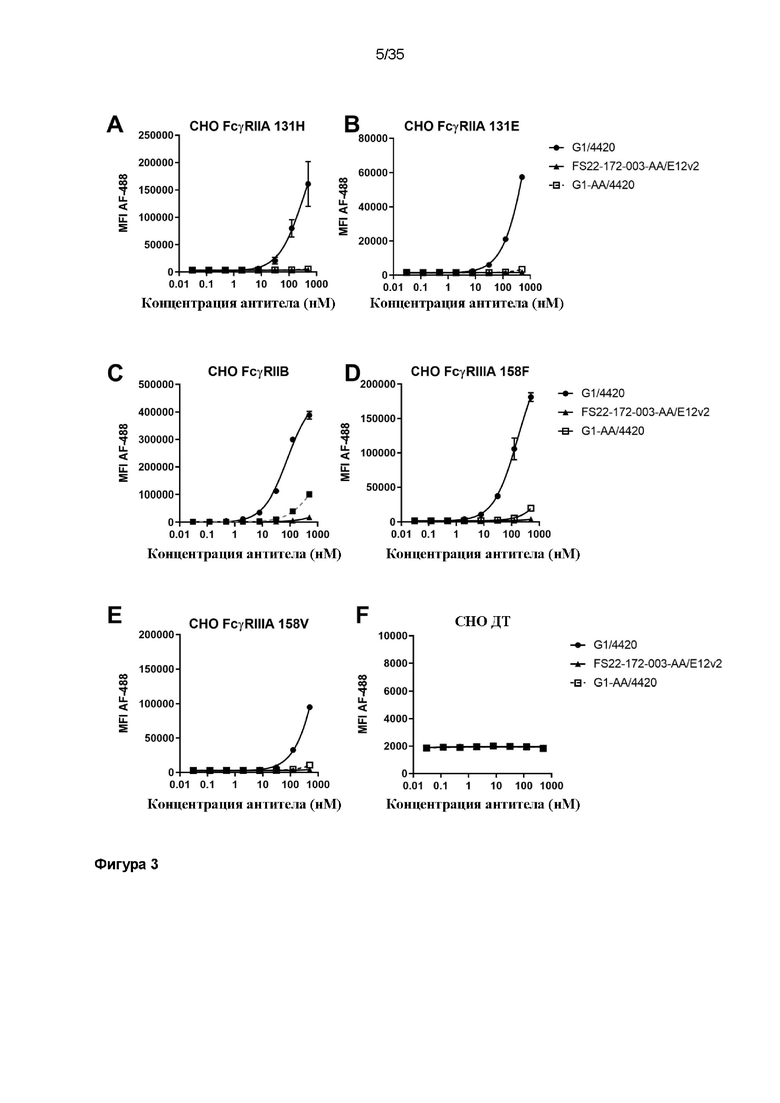
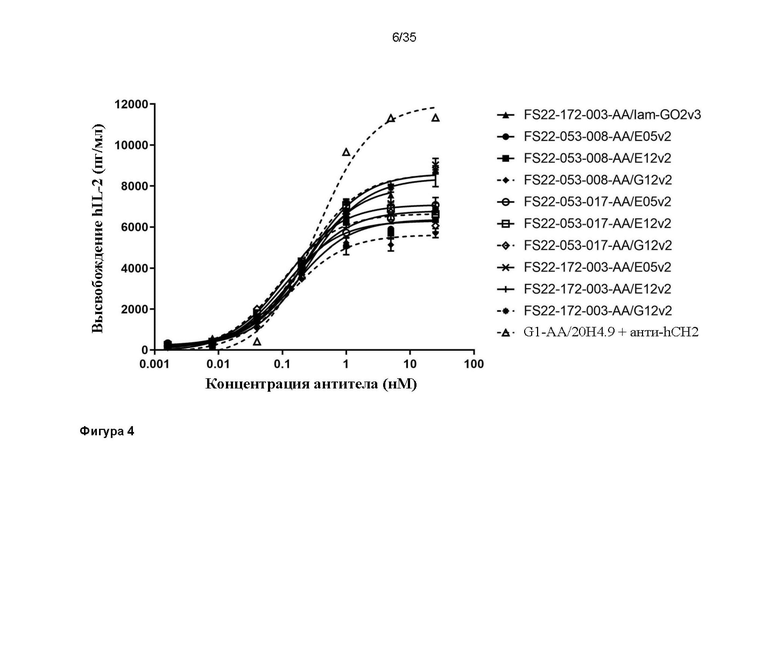
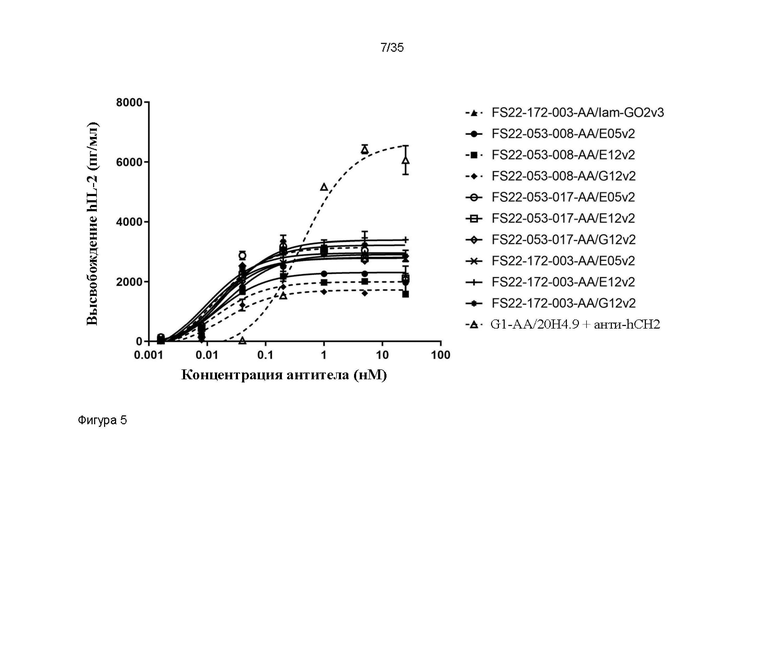
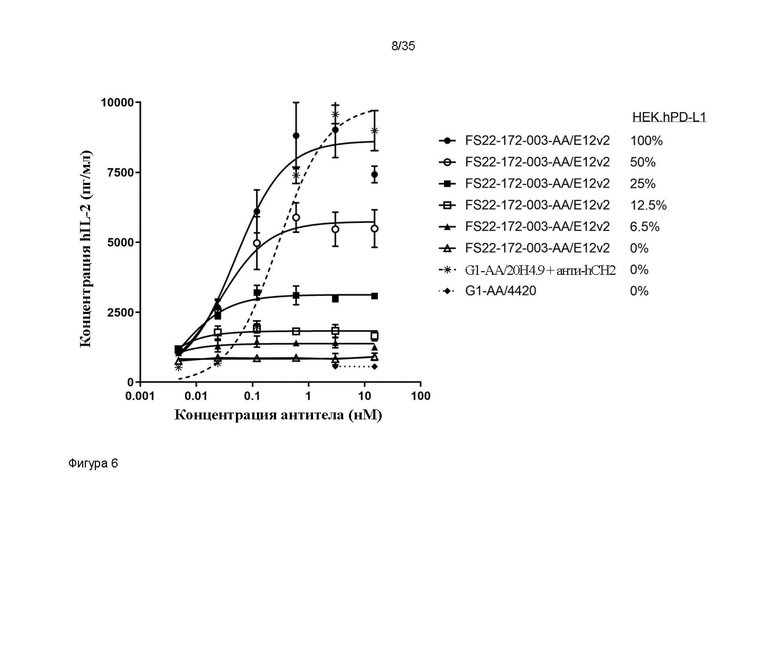
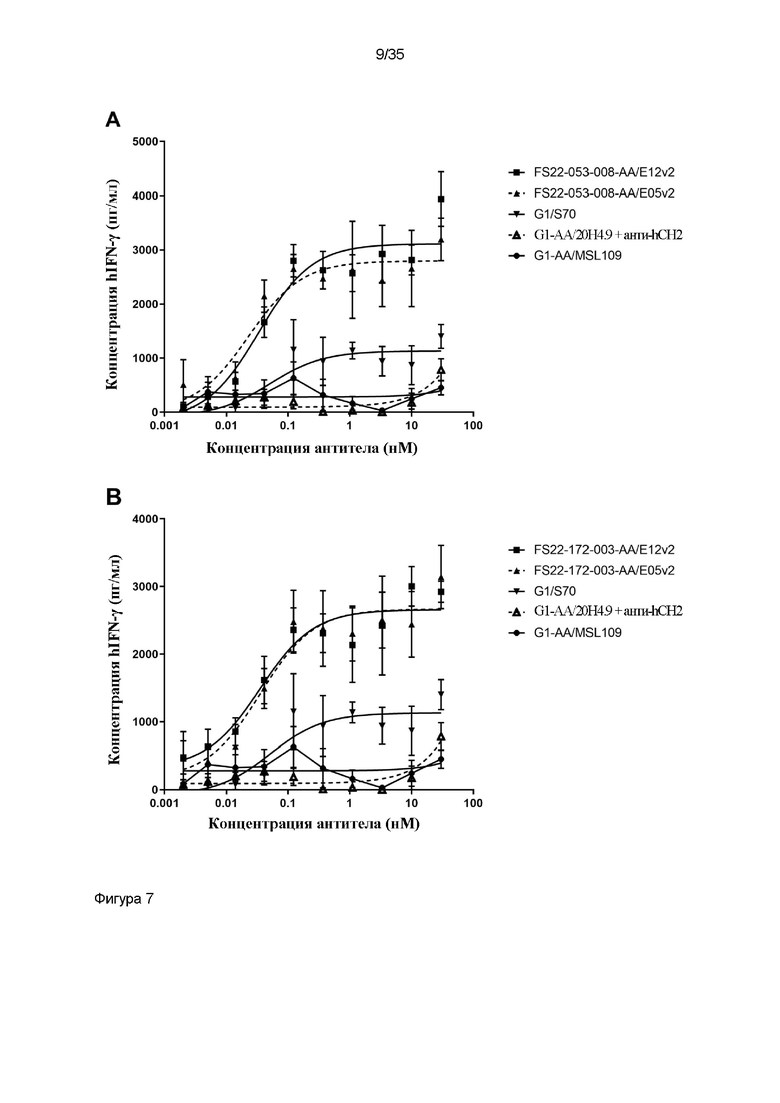
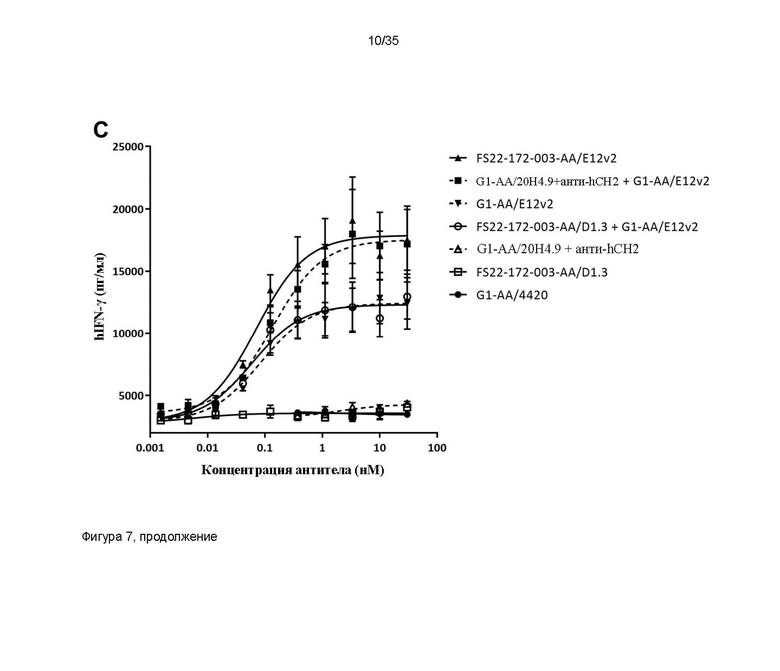
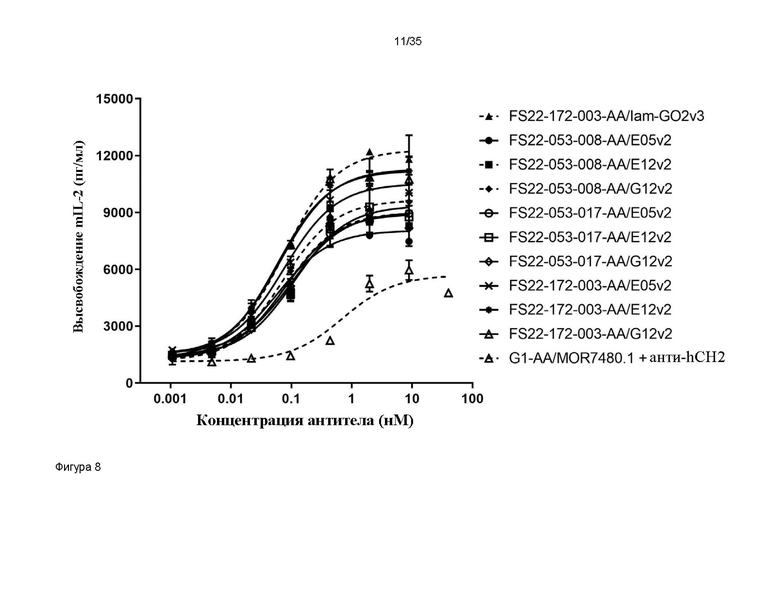
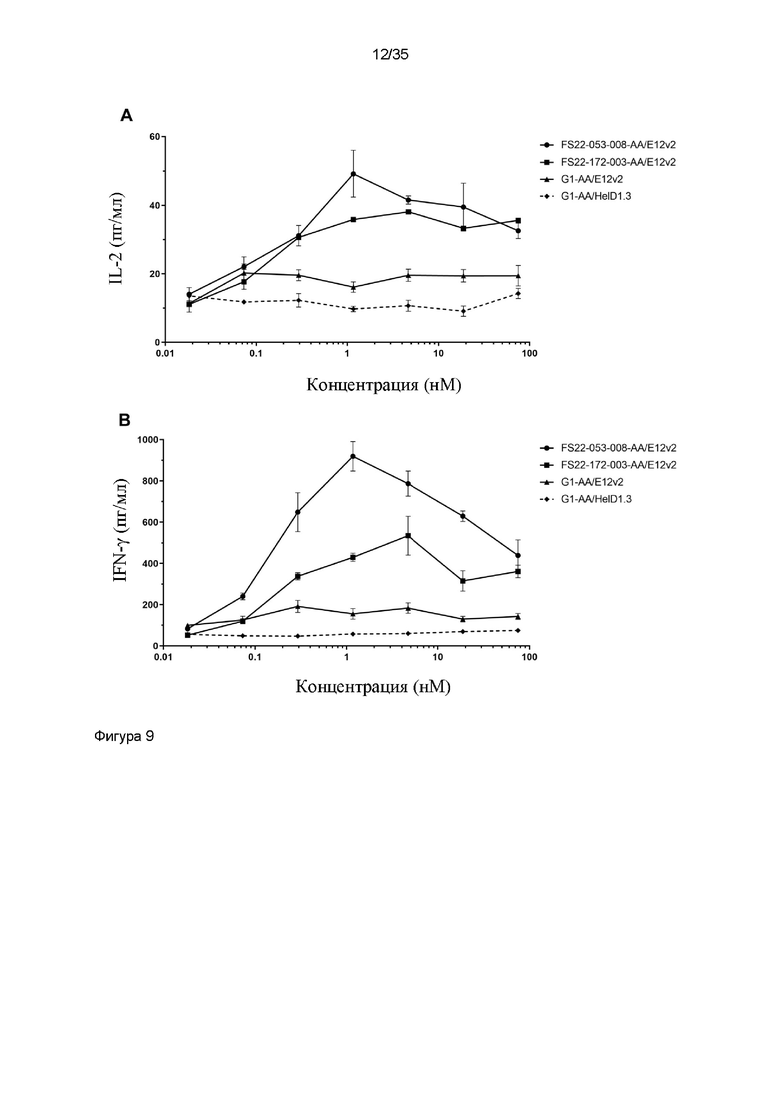
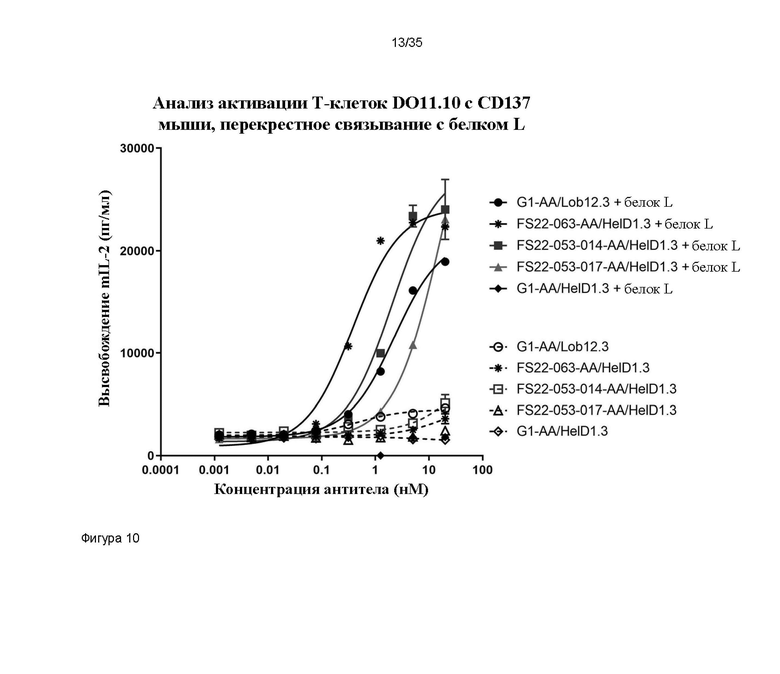
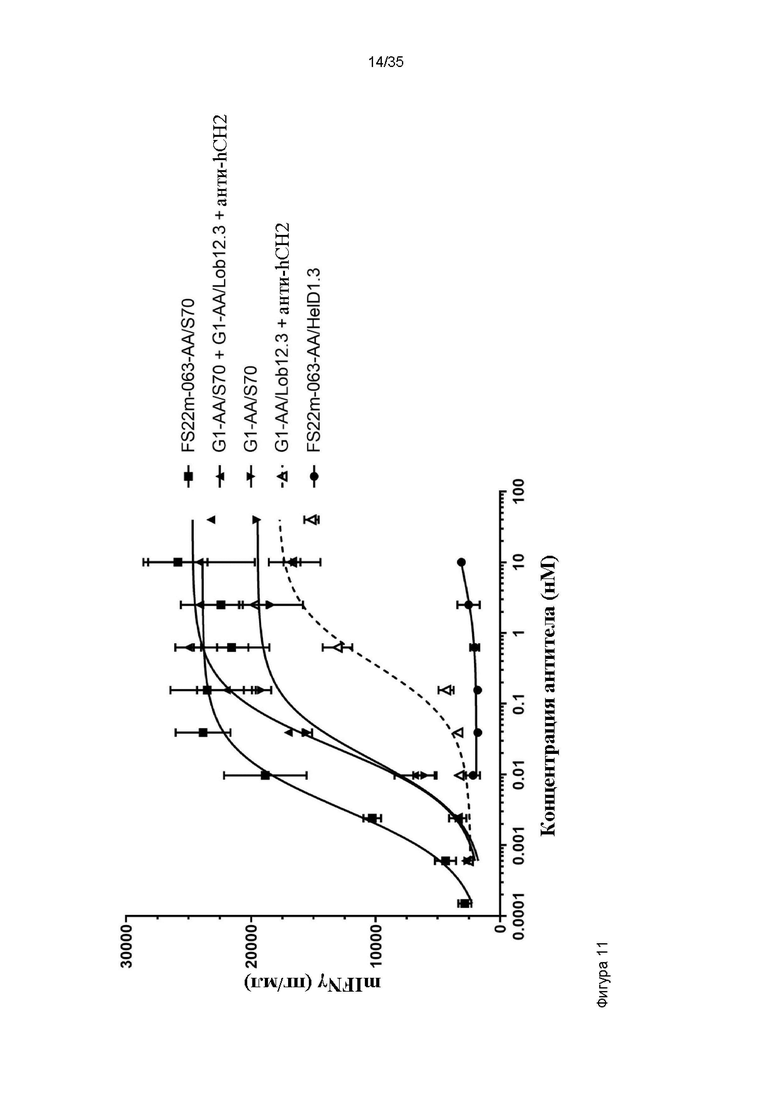
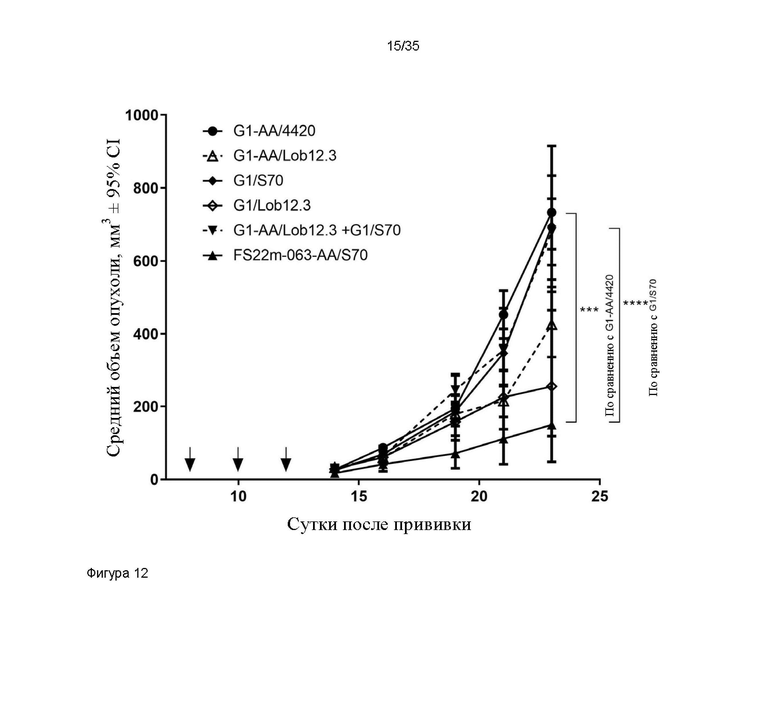
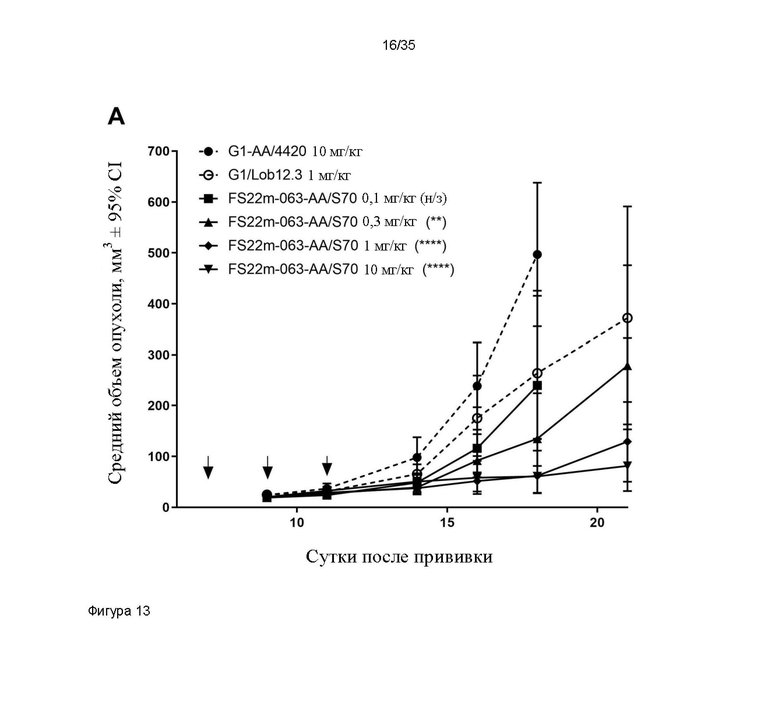
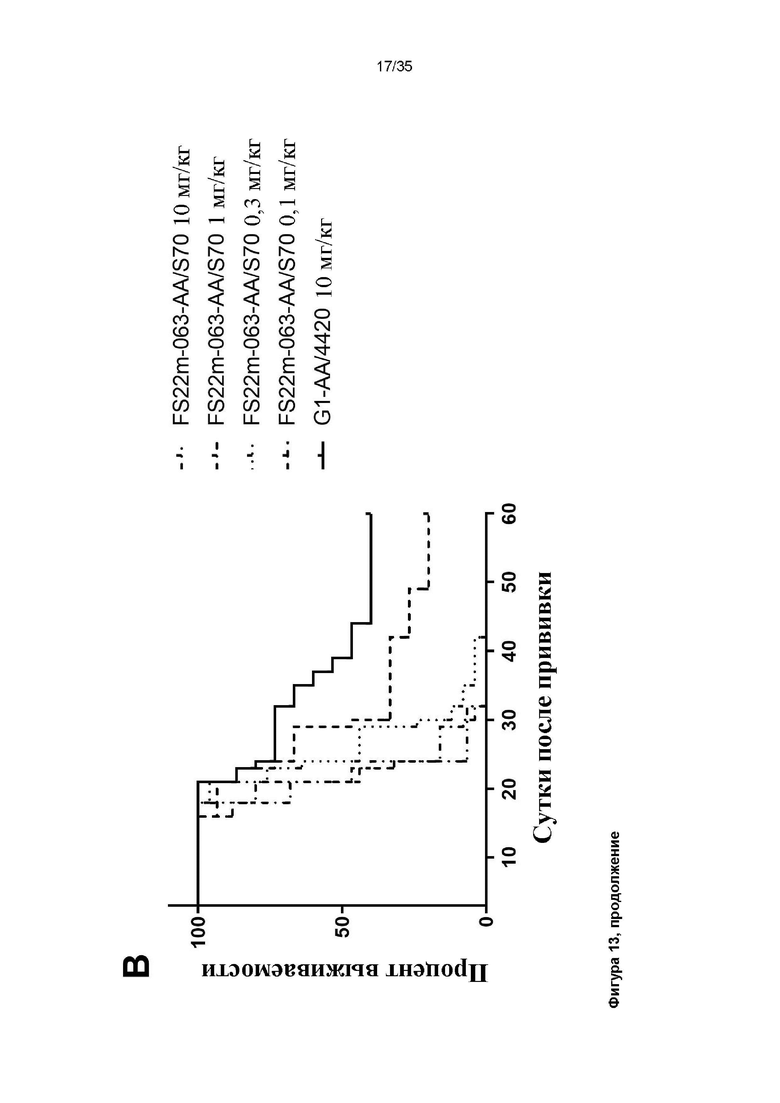
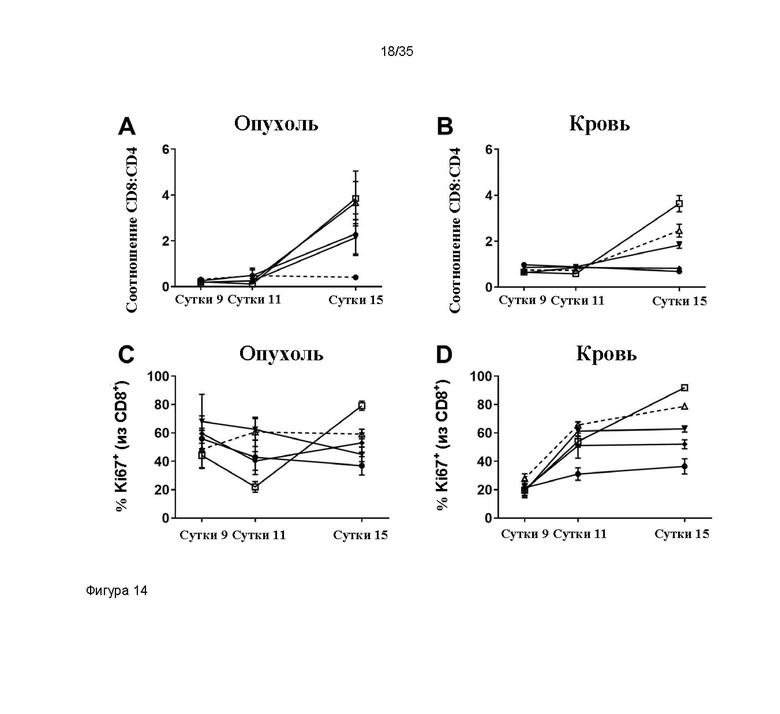
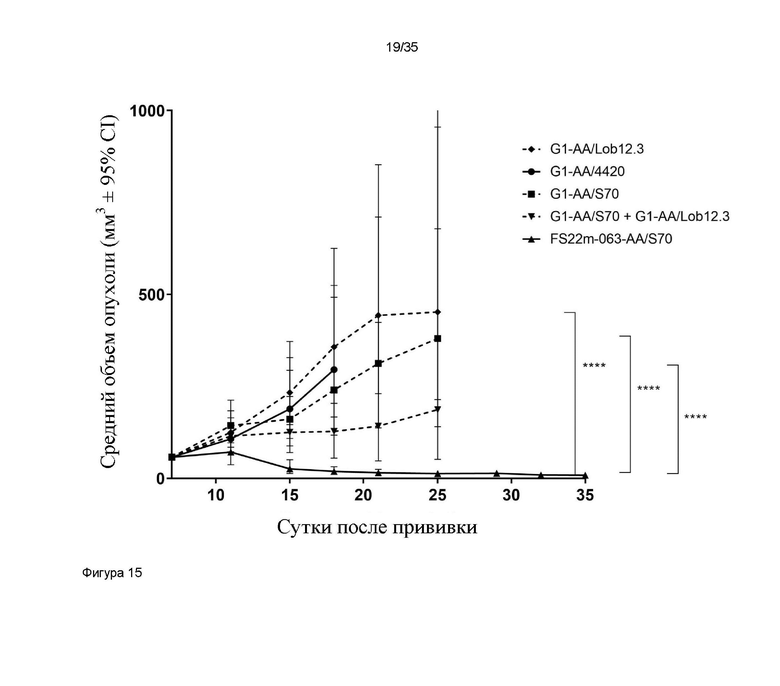
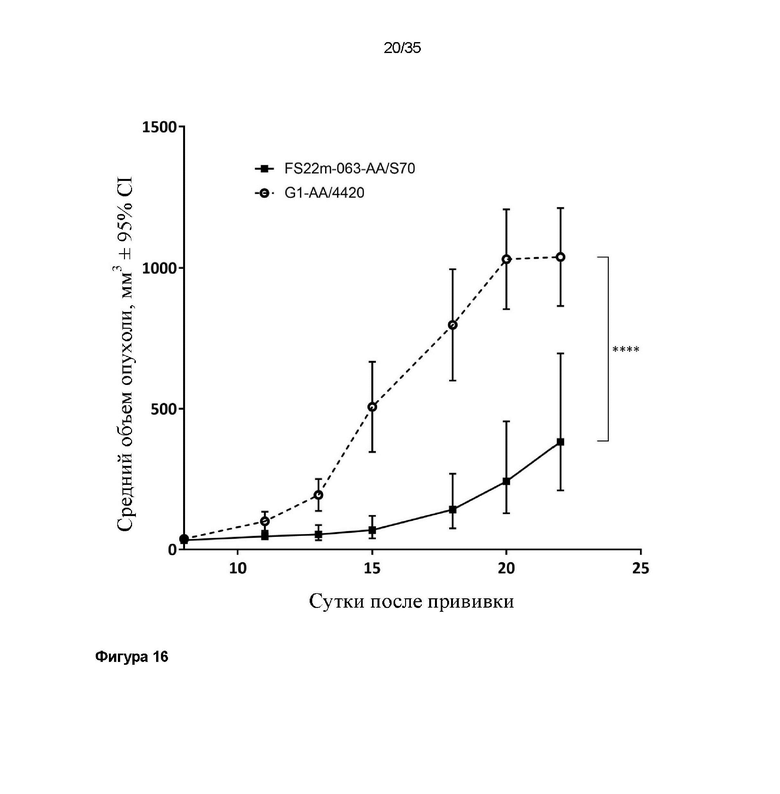
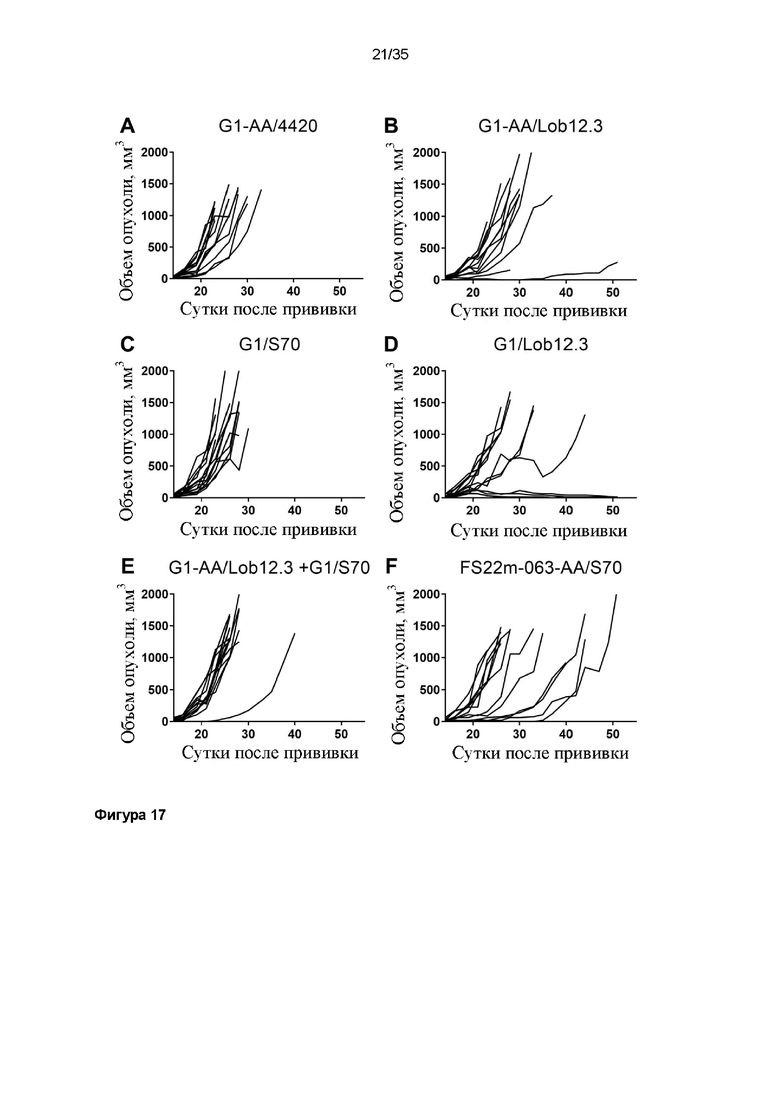
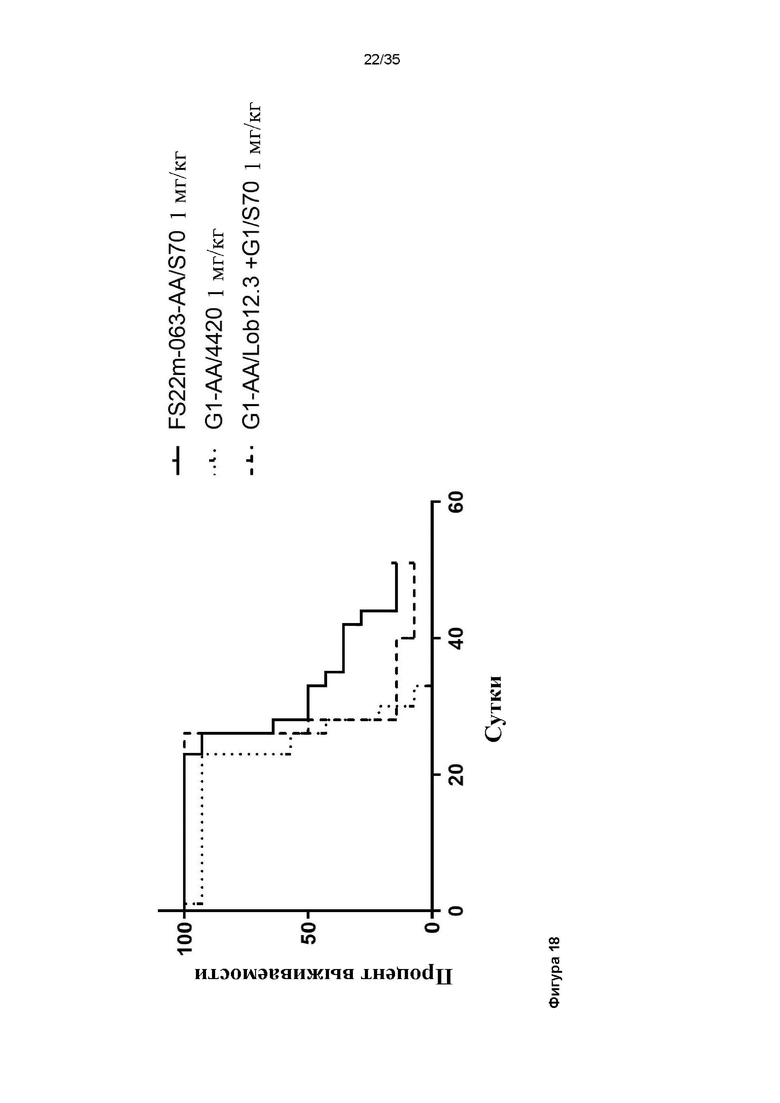
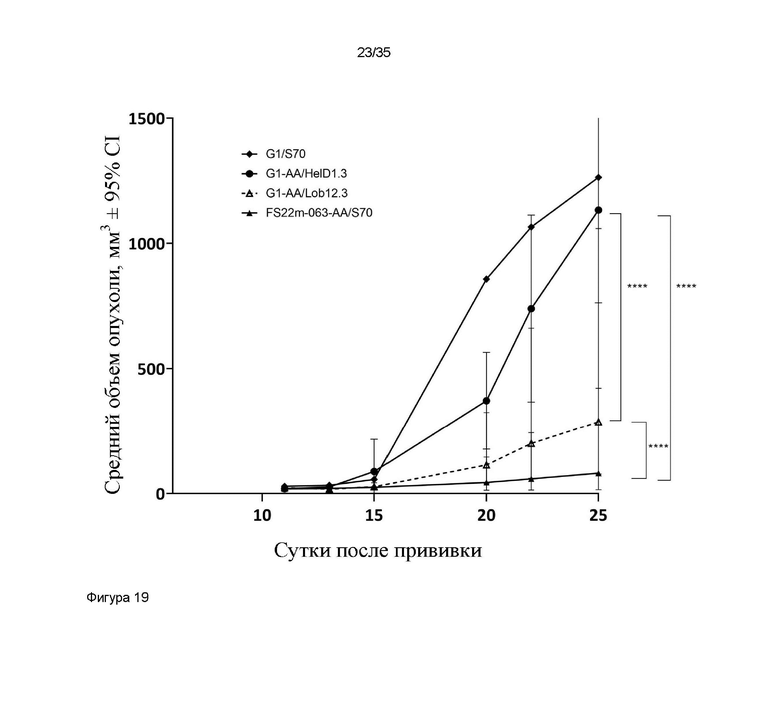
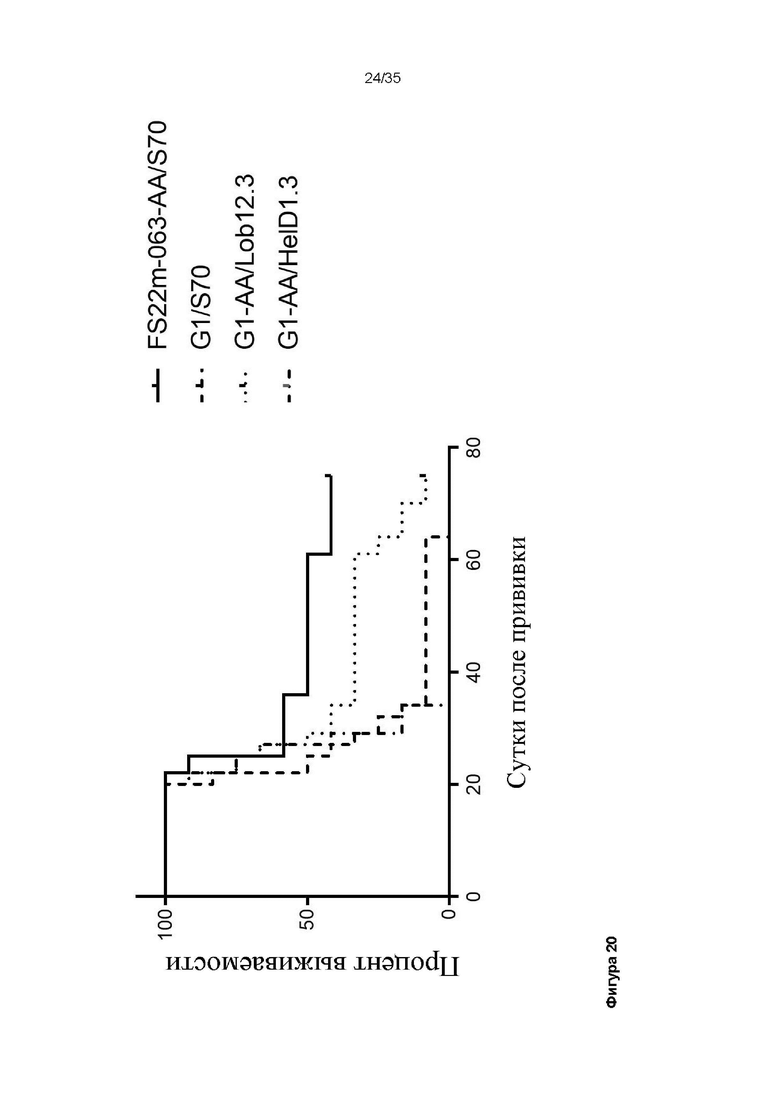
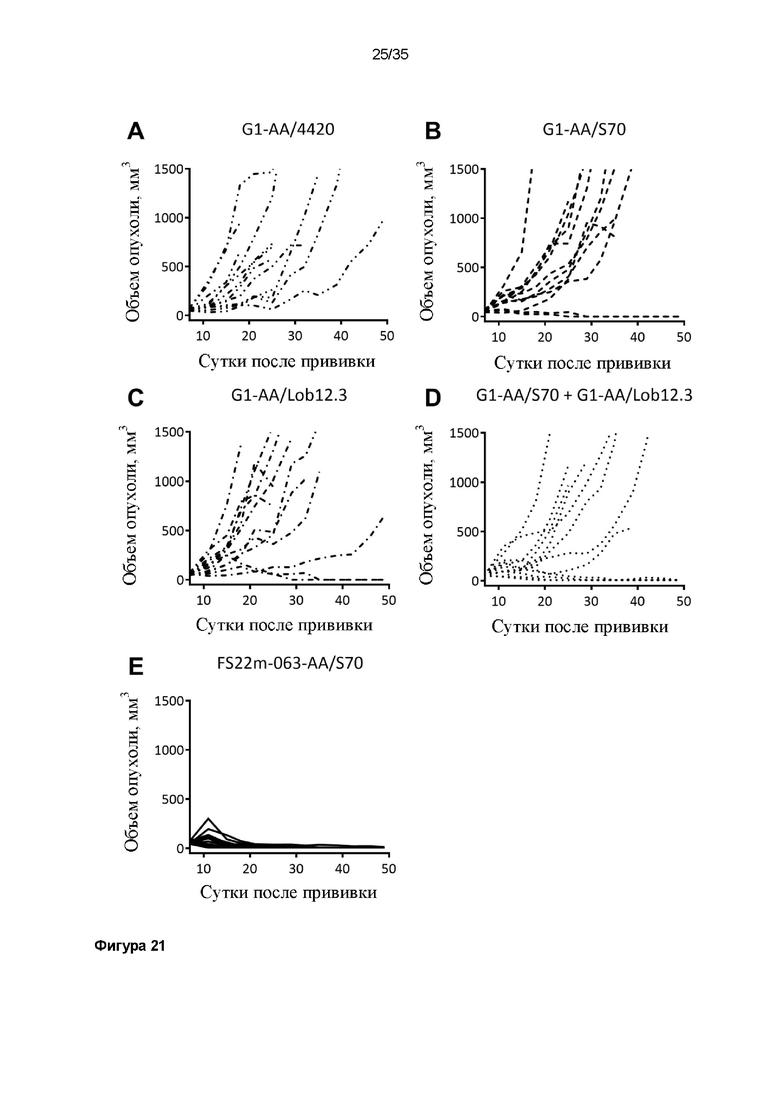
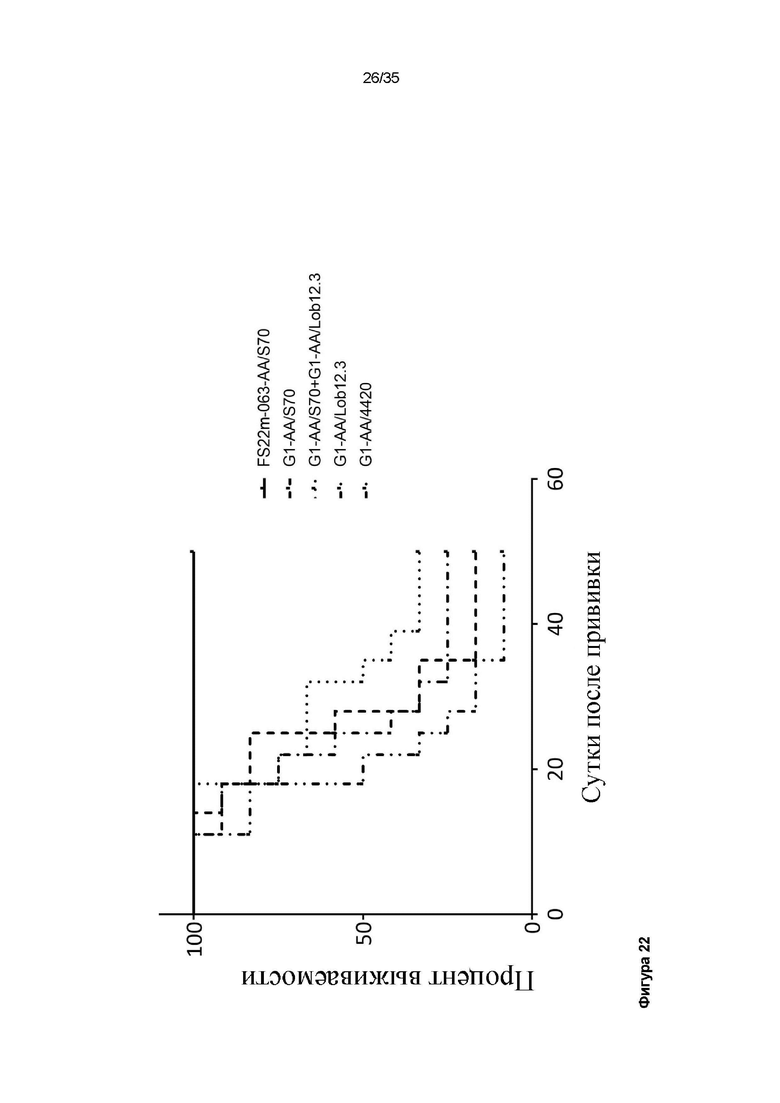
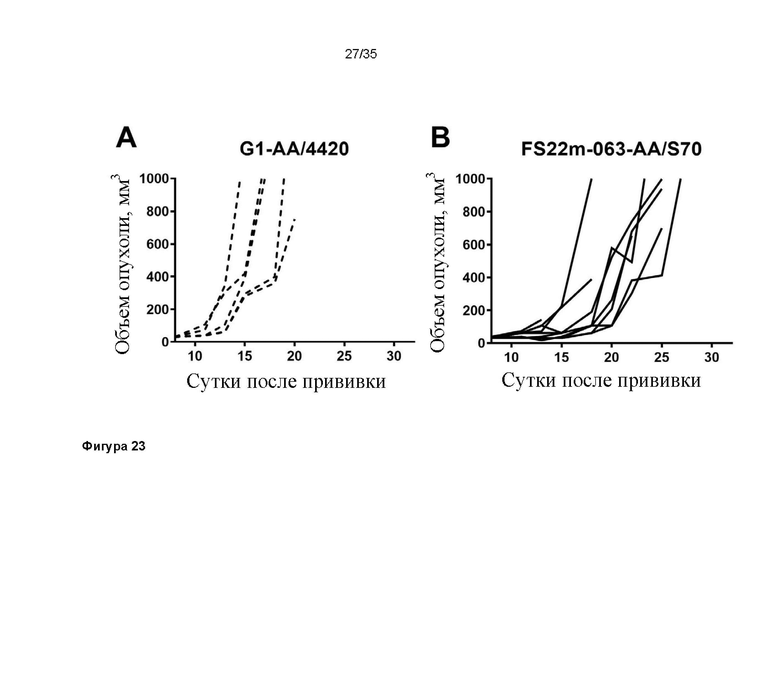
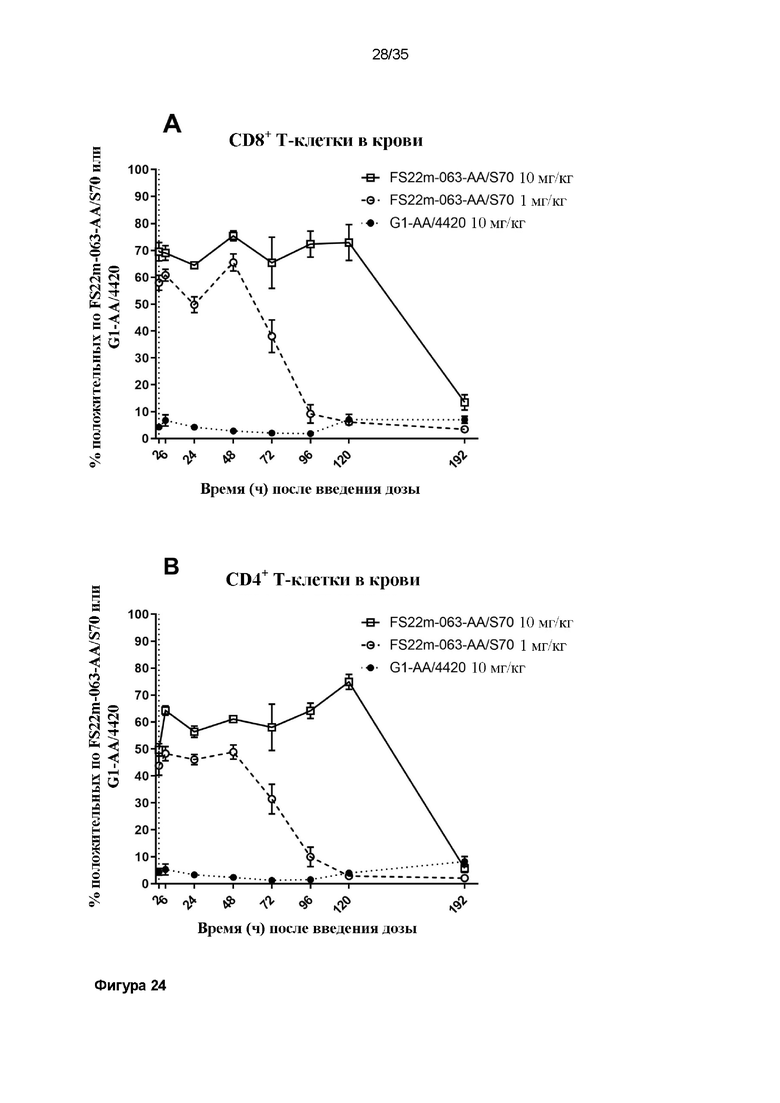
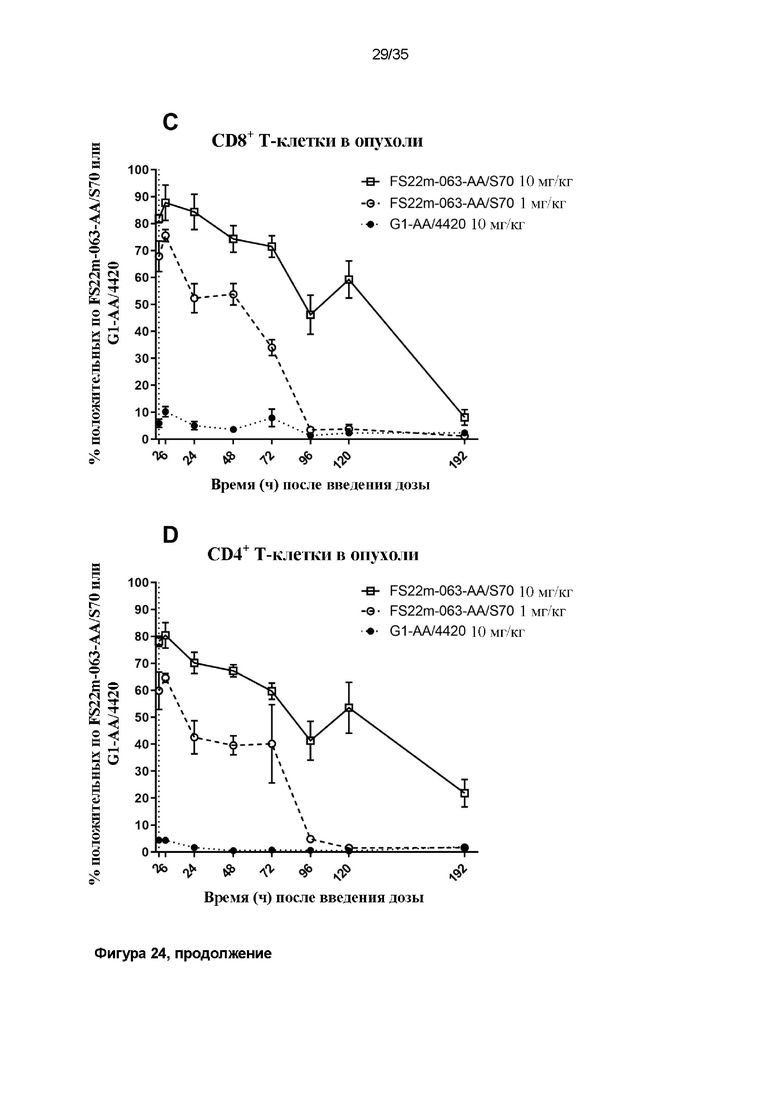
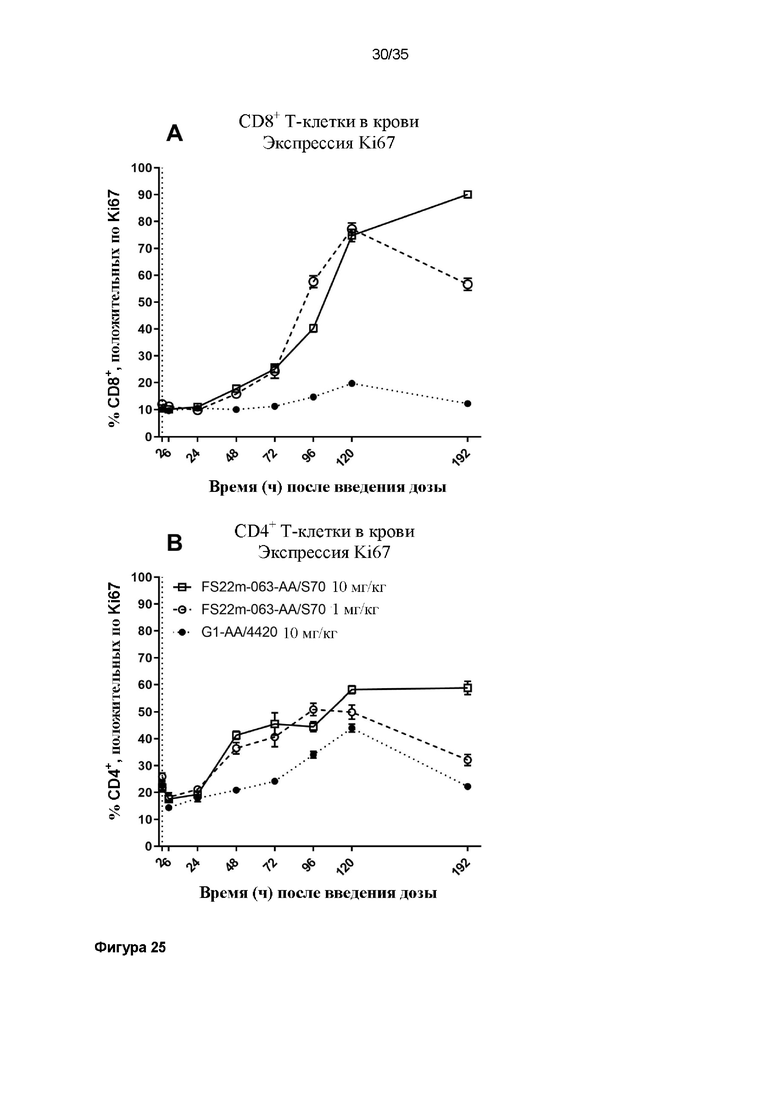
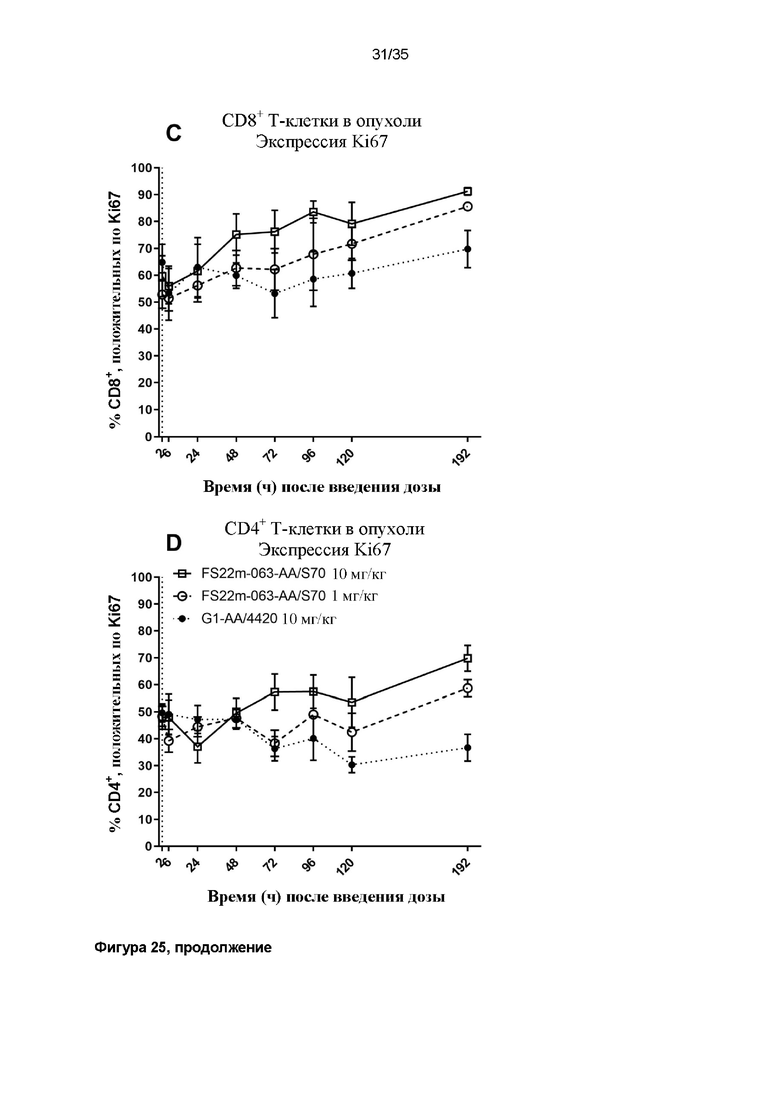
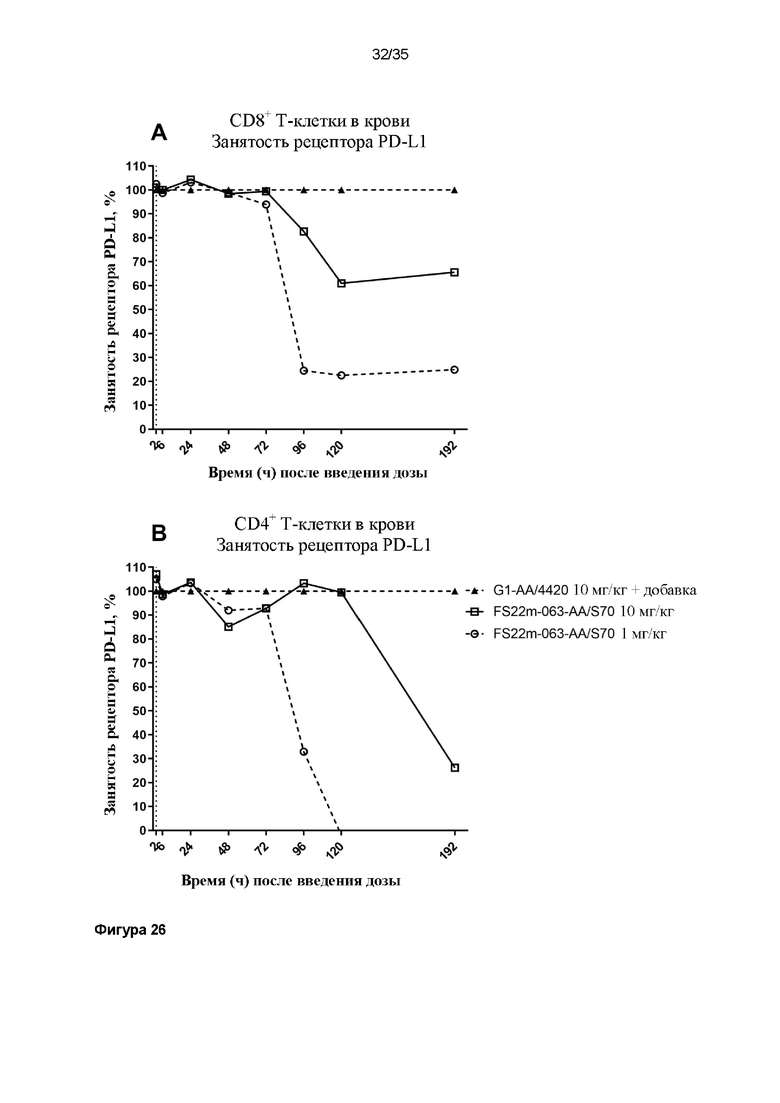
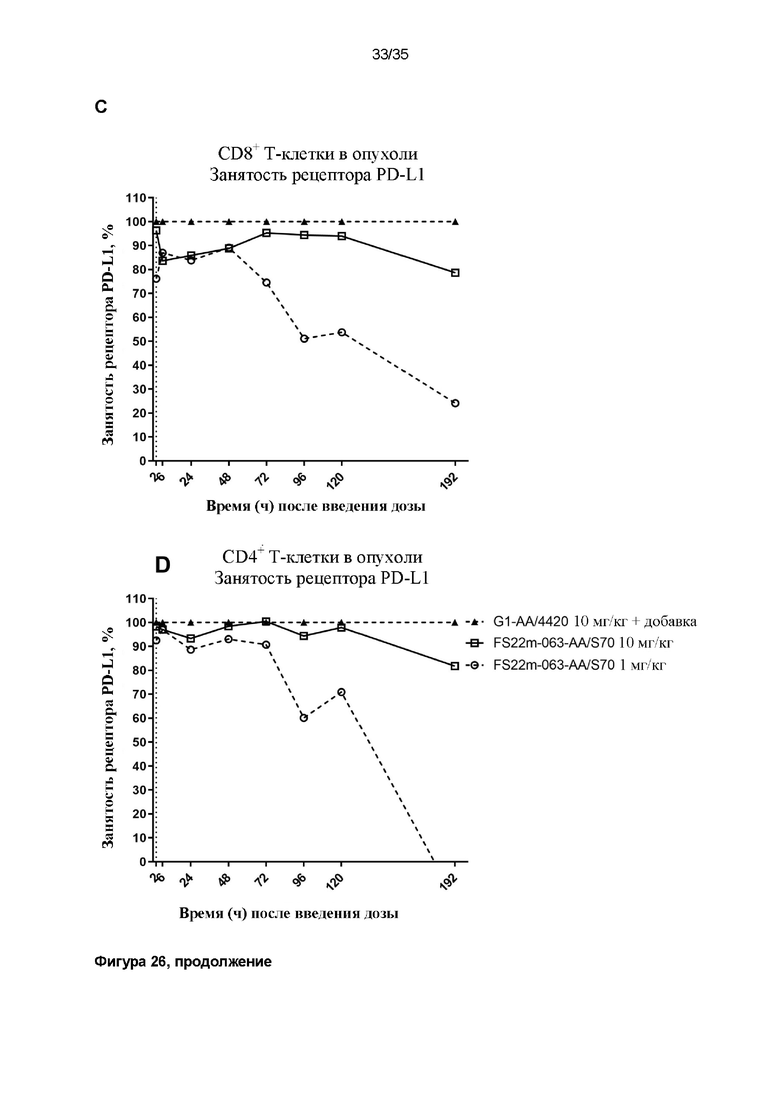
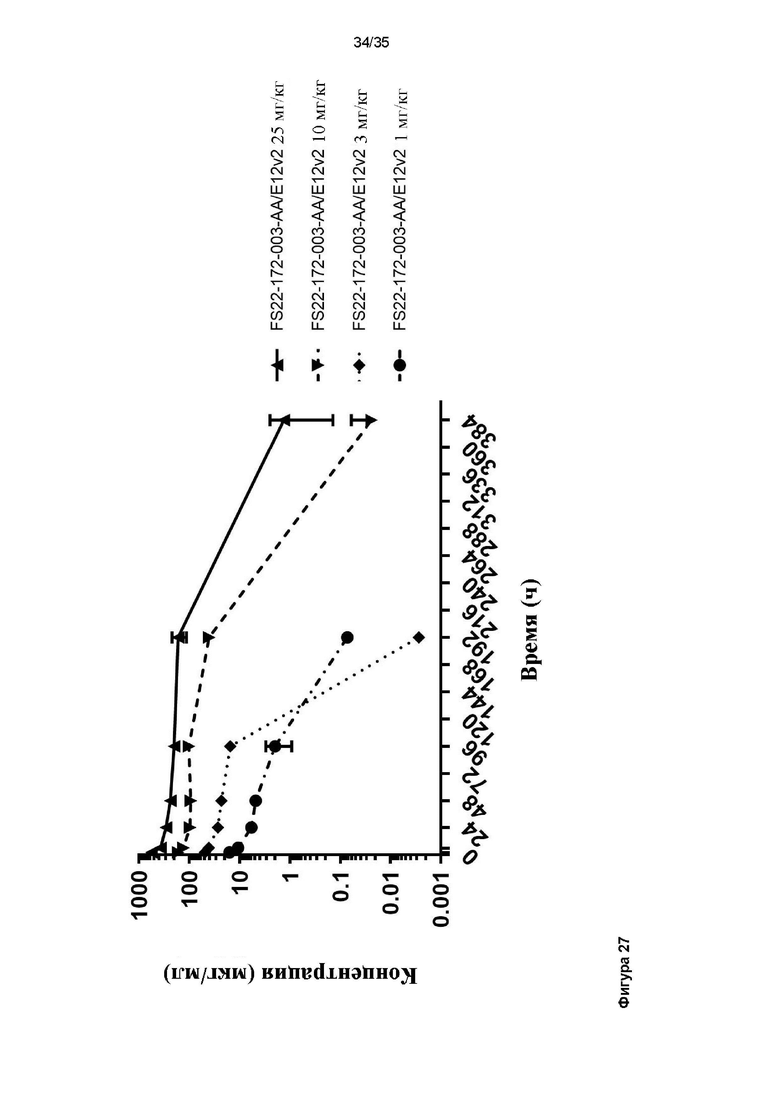
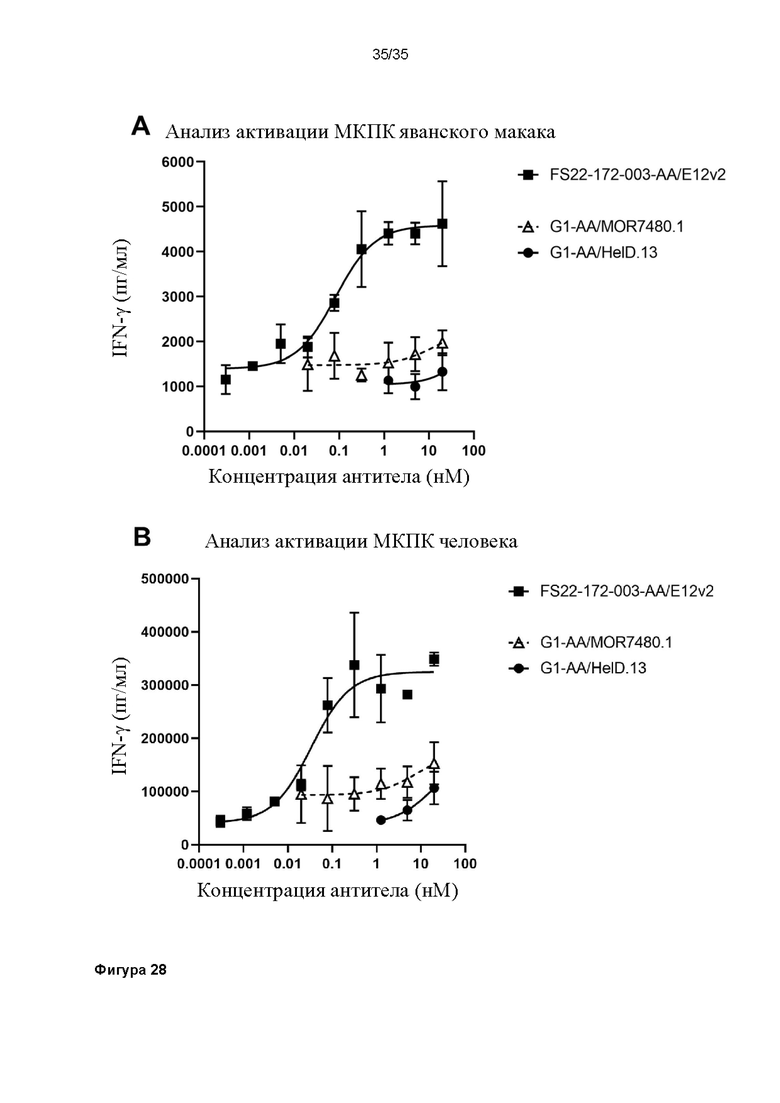
Изобретение относится к области биотехнологии. Предложена молекула антитела, которая связывается с PD-L1 и CD137 и способна вызывать агонизм CD137. Указанная молекула антитела содержит сайт связывания с PD-L1 на основе CDR и сайт связывания CD137, который расположен в домене CH3 молекулы антитела. Также предложены нуклеиновые кислоты, кодирующие молекулы антитела, векторы экспрессии и клетка-хозяин, содержащие указанные нуклеиновые кислоты, способ получения молекулы антитела. Кроме того, изобретение относится к фармацевтической композиции, включающей антитела по изобретению, и к применению указанных антител для лечения рака у индивида. Антитела согласно настоящему изобретению обладают высокой фармакологической активностью и безопасностью и могут найти применение, например, для лечения заболеваний, таких как рак. 9 н. и 10 з.п. ф-лы, 30 ил., 33 табл., 16 пр.
1. Молекула антитела, которая связывается с лигандом программируемой гибели клеток (PD-L1) и CD137, содержащая
(a) сайт связывания антигена PD-L1 на основе определяющих комплементарность областей (CDR); и
(b) сайт связывания антигена CD137, расположенный в домене CH3 молекулы антитела;
где указанный сайт связывания антигена на основе CDR содержит VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации по Кабату, представленные в:
(i) SEQ ID NO: 1, 2, 3, 4, 5 и 6, соответственно;
(ii) SEQ ID NO: 1, 2, 3, 18, 19 и 20, соответственно; или
(iii) SEQ ID NO: 1, 2, 3, 18, 19 и 29, соответственно; или
где указанный сайт связывания антигена на основе CDR содержит VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации ImMunoGeneTics (IMGT), представленные в:
(iv) SEQ ID NO: 7, 8, 9, 10, 11 и 6, соответственно;
(v) SEQ ID NO: 21, 8, 9, 22, 11 и 20, соответственно; или
(vi) SEQ ID NO: 21, 8, 9, 22, 11 и 29, соответственно; и
причем сайт связывания антигена CD137 содержит первую последовательность и вторую последовательность, расположенные в структурных петлях AB и EF домена CH3, соответственно, причем первая и вторая последовательности имеют последовательность, представленную в SEQ ID NO: 113 и 114, SEQ ID NO: 79 и 80 или SEQ ID NO: 79 и 89, соответственно.
2. Молекула антитела по п. 1, отличающаяся тем, что молекула антитела содержит домен VH и домен VL, представленные в:
(i) SEQ ID NO: 12 и 14, соответственно;
(ii) SEQ ID NO: 23 и 25, соответственно; или
(iii) SEQ ID NO: 23 и 30, соответственно.
3. Молекула антитела по п. 1 или 2, отличающаяся тем, что молекула антитела содержит:
(i) VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации по Кабату, представленные в SEQ ID NO: 1, 2, 3, 4, 5 и 6, соответственно;
(ii) VH CDR1, VH CDR2, VH CDR3, VL CDR1, VL CDR2 и VL CDR3, определенные согласно схеме нумерации ImMunoGeneTics (IMGT), представленные в SEQ ID NO: 7, 8, 9, 10, 11 и 6, соответственно; и/или
(iii) домен VH и домен VL, представленные в SEQ ID NO: 12 и 14, соответственно.
4. Молекула антитела по любому из предшествующих пунктов, отличающаяся тем, что:
(i) первая последовательность расположена между положениями 14 и 17 домена CH3 молекулы антитела; и/или
(ii) вторая последовательность расположена между положениями 91 и 99 домена CH3 молекулы антитела; и
где нумерация аминокислотных остатков соответствует схеме нумерации IMGT.
5. Молекула антитела по любому из предшествующих пунктов, отличающаяся тем, что молекула антитела содержит домен CH3, представленный в SEQ ID NO: 115, SEQ ID NO: 81 или SEQ ID NO: 90.
6. Молекула антитела по п. 5, отличающаяся тем, что домен CH3 дополнительно содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
7. Молекула антитела по любому из предшествующих пунктов, отличающаяся тем, что молекула антитела содержит последовательности тяжелой цепи и легкой цепи:
(i) представленные в SEQ ID NO: 134 и 17, соответственно;
(ii) представленные в SEQ ID NO: 137 и 28, соответственно;
(iii) представленные в SEQ ID NO: 140 и 33, соответственно;
(iv) представленные в SEQ ID NO: 143 и 17, соответственно;
(v) представленные в SEQ ID NO: 146 и 28, соответственно;
(vi) представленные в SEQ ID NO: 149 и 33, соответственно,
(vii) представленные в SEQ ID NO: 152 и 17, соответственно;
(viii) представленные в SEQ ID NO: 153 и 28, соответственно; или
(ix) представленные в SEQ ID NO: 154 и 33, соответственно.
8. Молекула антитела по п. 7, отличающаяся тем, что домен CH3 в тяжелой цепи содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
9. Молекула антитела по любому из предшествующих пунктов, отличающаяся тем, что молекула антитела содержит тяжелую цепь и легкую цепь, представленные в SEQ ID NO: 134 и 17, соответственно.
10. Молекула антитела по п. 9, отличающаяся тем, что домен CH3 в тяжелой цепи содержит остаток лизина (K) непосредственно на C-конце последовательности домена CH3.
11. Молекула антитела по любому из предшествующих пунктов, отличающаяся тем, что:
(i) указанная молекула антитела была модифицирована для уменьшения или устранения связывания домена CH2 молекулы антитела с одним или более Fcγ-рецепторами;
(ii) молекула антитела не связывается с одним или более Fcγ-рецепторами; и/или
(iii) связывание молекулы антитела с CD137 на иммунной клетке и со связанным с поверхностью опухолевой клетки PD-L1 вызывает кластеризацию CD137 на иммунной клетке.
12. Молекула нуклеиновой кислоты, кодирующая молекулу антитела по любому из предшествующих пунктов.
13. Комбинация молекул нуклеиновых кислот, кодирующих молекулу антитела по любому из пп. 1-11.
14. Вектор, содержащий молекулу нуклеиновых кислот по п. 12 или комбинацию молекул нуклеиновых кислот по п. 13, для экспрессии указанных молекулы нуклеиновой кислоты или комбинации молекул нуклеиновых кислот в клетке-хозяине.
15. Комбинация векторов, содержащих молекулу нуклеиновой кислоты по п. 12 или комбинацию молекул нуклеиновых кислот по п. 13, для экспрессии указанных молекулы нуклеиновой кислоты или комбинации молекул нуклеиновых кислот в клетке-хозяине.
16. Рекомбинантная клетка-хозяин, содержащая молекулы нуклеиновых кислот по п. 12, или комбинацию молекул нуклеиновой кислоты по п. 13, или вектор по п. 14, или комбинацию векторов по п. 15, для получения рекомбинантных молекул антител.
17. Способ получения молекулы антитела по любому из пп. 1-11, включающий культивирование рекомбинантной клетки-хозяина по п. 16 в условиях, подходящих для продукции молекулы антитела, дополнительно включающий выделение и/или очистку молекулы антитела.
18. Фармацевтическая композиция для лечения заболевания или нарушения, ассоциированного с экспрессией PD-L1 и CD137, содержащая молекулу антитела по любому из пп. 1-11 в эффективном количестве и фармацевтически приемлемое вспомогательное вещество.
19. Применение молекулы антитела по любому из пп. 1-11 для лечения рака у индивида.
| WO 2017123650 A2, 20.07.2017 | |||
| WO 2008003103 A2, 10.01.2008 | |||
| YOSHITARO SHINDO et al., Combination Immunotherapy with 4-1BB Activation and PD-1 Blockade Enhances Antitumor Efficacy in a Mouse Model of Subcutaneous Tumor, Anticancer Research, 2015, Vol | |||
| Скоропечатный станок для печатания со стеклянных пластинок | 1922 |
|
SU35A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| КАДАГИДЗЕ З.Г | |||
| и др., Рецепторы лимфоцитов, регулирующие иммунный ответ - | |||

