Область техники
Настоящее изобретение относится к пропуску экзонов, индуцируемому производными пептидно-нуклеиновых кислот с хорошим проникновением внутрь клетки и сильным сродством к нуклеиновой кислоте, и испрашивает преимущество приоритета по предварительной заявке США с № 62/440929, поданной 30 декабря 2016 г., которая включена сюда посредством ссылки в полном объеме.
Предпосылки создания настоящего изобретения
Олигонуклеотиды использовались для разнообразных биологических целей, включая антисмысловое ингибирование экспрессии генов, ПЦР (полимеразную цепную реакцию), диагностический анализ с использованием генного чипа и т.д. Поскольку олигонуклеотиды взаимодействуют с нуклеиновой кислотой, в том числе ДНК и РНК, специфическим для последовательности образом, они применимы для предсказуемой модуляции биологических процессов с участием ДНК или РНК внутри клетки. Олигонуклеотиды с хорошим проникновением внутрь клетки способны модулировать такие биологические процессы в клетке предсказуемым с учетом последовательности образом.
Белки в качестве мишеней лекарственных средств: Белки опосредуют разнообразные клеточные функции. Неудивительно, что большинство продаваемых в настоящее время лекарственных средств проявляют терапевтическую активность благодаря модулированию функций белка(ов). Например, нестероидный противовоспалительный препарат аспирин ингибирует ферменты, называемые циклооксигеназами, благодаря своей противовоспалительной активности. Лозартан связывается с трансмембранным рецептором, называемым рецептором ангиотензина II, благодаря своей антигипертензивной активности. Росиглитазон селективно активирует внутриклеточный рецептор, называемый активируемым пероксисомными пролифераторами рецептором γ (PPAR γ), с вызовом своей противодиабетической активности. Этанерцепт представляет собой слитый белок, который связывается с цитокином, называемым фактором-α некроза опухолей (TNF-α), и нейтрализует биологическую активность TNF-α благодаря своей противоревматической активности. Герцептин представляет собой моноклональное антитело для лечения рака молочной железы путем селективного связывания с erbB2, сверхэкспрессируемым в некоторых типах клеток рака молочной железы.
Пре-мРНК: Генетическая информация переносится на ДНК (содержащей в качестве сахара 2-дезоксирибозу нуклеиновой кислота), которая транскрибируется с продуцированием пре-мРНК (предшественника информационной рибонуклеиновой кислоты) в ядре. Пре-мРНК млекопитающих обычно состоит из экзонов и интронов, и экзон и интрон взаимосвязаны друг с другом. Экзоны и интроны нумеруются, как показано на фиг. 1А.
Сплайсинг пре-мРНК в мРНК. В ядре пре-мРНК превращается в мРНК после делеции интронов и лигирования экзонов с помощью ряда сложных реакций, которые все вместе называются «сплайсингом», как схематически показано на фиг. 1B [Ann. Rev. Biochem. 72(1), 291-336 (2003); Nature Rev. Mol. Cell Biol. 6(5), 386-398 (2005); Nature Rev. Mol. Cell Biol. 15(2), 108-121 (2014)].
Сплайсинг инициируется при образовании «сплайсомного Е-комплекса» (т.е. раннего сплайсомного комплекса) между пре-мРНК и адаптерными факторами сплайсинга. В «сплайсомном Е-комплексе» U1 связывается с местом соединения экзона N и интрона N, а U2AF35 связывается с местом соединения интрона N и экзона (N+1). Таким образом, место соединения экзона/интрона или интрона/экзона имеет решающее значение для образования раннего сплайсомного комплекса. «Сплайсомный Е-комплекс» превращается в «сплайсомный А-комплекс» при дополнительном комплексообразовании с U2. «Сплайсомный А-комплекс» затем подвергается ряду сложных реакций, чтобы удалить или сплайсировать интрон для соединения соседних экзонов.
Альтернативный сплайсинг и вариант сплайсинга (сплайс-вариант): Все экзоны пре-мРНК не всегда сохраняются для образования «полноразмерной» мРНК во время сплайсинга. Некоторые экзоны делетируются, или сплайсируются, с образованием вариантных мРНК, т.е. «вариантов сплайсинга». Таким образом, пре-мРНК может быть «альтернативно сплайсирована» для получения множества вариантов сплайсинга.
Об альтернативном сплайсинге в клетках млекопитающих впервые было сообщено в 1981 году в случае гена, кодирующего кальцитонин [Nature vol 290(5801), 63-65 (1981); Proc. Natl. Acad. Sci. USA vol 79(6), 1717-1721 (1982)]. Ген состоит из 6 экзонов, и мРНК для кальцитонина образуется при пропуске экзона 5 и экзона 6. Между тем, пропуск экзона 4 приводит к образованию варианта мРНК, кодирующего связанный с геном кальцитонина пептид (CGRP).
Альтернативный сплайсинг, по-видимому, полностью зависит от клеток и условий, которым клетки подвергаются. Благодаря альтернативному сплайсингу множество белков продуцируются с одного гена. Альтернативный сплайсинг позволяет животным генерировать большие разнообразия белков для их размера генома. У людей 95% генов с множеством экзонов подвергаются, по оценкам, альтернативному сплайсингу [Nature Genetics vol 40(12), 1413-1415 (2008)].
Варианты сплайсинга и биологические функции. Варианты сплайсинга обнаруживаются как спонтанно возникающие зависящим от типа клетки или ткани образом и кодируют белки, обладающие биологическими профилями, часто отличающимися от профилей полноразмерного белка.
Андрогеновый рецептор (AR) был бы хорошим примером генов, дающих множество вариантов сплайсинга [Int. J. Biol. Sci. vol 7(6), 815-822 (2011)]. Пре-мРНК для AR состоит из 8 экзонов плюс 4 криптических экзона (криптические экзоны представлены в виде затенения на диаграмме ниже). Существует по крайней мере семь вариантов сплайсинга мРНК для AR.
Вариант 1 мРНК для AR состоит из экзонов с 1 по 8, соединенных последовательно, и кодирует полноразмерный белок AR, как показано на фиг. 2А. В случае варианта 3 мРНК для AR экзоны с 4 по 8 сплайсируются (т.е. делетируются). Следовательно, вариант 3 мРНК для AR кодирует усеченный белок AR (AR3), в котором отсутствует лиганд-связывающий домен (LBD), присутствующий в полноразмерном белке.
Полноразмерный белок AR становится функционально активным после образования комплекса с андрогеном, таким как тестостерон или дигидротестостерон (DHT). Между тем, усеченный белок AR3 является функционально активным даже в отсутствие андрогена. В опухолях предстательной железы, устойчивых к терапии в виде андрогенной абляции, часто обнаруживалась увеличение экспрессии белка AR3. Таким образом, эндогенное образование варианта белка AR3 можно рассматривать как естественный процесс отбора клеток рака предстательной железы, чтобы избежать терапии в виде андрогенной абляции.
Индуцируемый гипоксией фактор 1α (HIF-1α) представляет собой субъединицу фактора транскрипции, называемого индуцируемым гипоксией фактором 1 (HIF-1), и кодируется геном HIF1A. HIF-1α активируется в ответ на гипоксию (т.е. низкий уровень кислорода) и, следовательно, может рассматриваться как датчик клеточного кислорода [Proc. Natl. Acad. Sci. USA, vol 92, 5510-5514 (1995)]. HIF-1α индуцирует транскрипцию более 60 генов, в том числе VEGF и EPO. HIF-1α способствует образованию новых кровеносных сосудов через VEGF [Exp. Mol. Med. vol 36, 1-12 (2004)]. Солидные опухоли испытывают гипоксию из-за ограниченного кровоснабжения и повышают экспрессию HIF-1α для выживания в условиях гипоксии.
Белок HIF-1α состоит из различных доменов для его функциональной активности в качестве активатора транскрипции. Он содержит основной элемент спираль-петля-спираль (bHLH) и два PAS-домена [для PAS-домена, ср. Curr. Biol. vol 7(11), R674-677 (1997); Eur. J. Biochem. vol 271(6), 1198-1208 (2004)]. HIF-1α обладает осуществляющим кислород-зависимую деградацию доменом (ODD), который служит в качестве датчика кислорода и, как известно, имеет решающее значение для стабильности белка HIF-1α.
Существует по крайней мере шесть вариантов белка HIF-1α, кодируемых шестью вариантами сплайсинга мРНК HIF-1α, как показано на фиг. 2B [Exp. Mol. Med. vol 36, 1-12 (2004)]. мРНК полноразмерного HIF-1α (HIF-1αFL) схожа с мРНК HIF-1α дикого типа (HIF-1αWT), за исключением дополнительных трех оснований (UAG) между экзоном 1 и экзон 2 вследствие альтернативного сплайсинга. Экзон 14 делетирован или пропущен в HIF-1α736. В HIF-1α736 отсутствует С-концевой активирующий домен (CAD). Известно, что как HIF-1αFL, так и HIF-1α736 активируют промотор VEGF при гипоксии. Между тем, HIF-1α557 (HIF-1αZ) и HIF-1α516 функционируют как доминантно-негативная изоформа HIF-1α. При раке молочной железы вариант сплайсинга мРНК HIF-1αFL отражает стадию прогрессирования рака и связан с плохим прогнозом [BMC Medicine vol 8(44), 1-12 (2010)].
Как показано на примере андрогенового рецептора и белка HIF-1α, варианты сплайсинга играют важную роль в формировании физиологического разнообразия конкретного гена млекопитающего. Природа самопроизвольно создает варианты сплайсинга для поддержания гомеостаза, а также для реагирования на физиологическую динамику.
Синтез белка на рибосомах: Интроны пре-мРНК ферментативно сплайсируются с образованием мРНК (информационной рибонуклеиновой кислоты), которая затем транслоцируется в цитозольный компартмент. В цитозоле комплекс трансляционного аппарата, называемый рибосомой, связывается с мРНК и осуществляет синтез белка, по мере того как он сканирует генетическую информацию, закодированную вдоль мРНК [Biochemistry vol 41, 4503-4510 (2002); Cancer Res. vol 48, 2659-2668 (1988)].
Кодон: Во время синтеза белка на рибосомах каждая аминокислота кодируется триадой в последовательности мРНК. Например, «AUG», «UUA», «CCC» и «AGA» кодируют «метионин», «лейцин», «пролин» и «аргинин», соответственно. Такие триады называются «кодонами». Учитывая, что в мРНК существуют 4 мономера: A, G, U и C, существуют 64 (4×4 x 4=64) возможных кодона. Некоторые кодоны соответствуют «стоп»-сигналу для синтеза белка на рибосомах. «UGA», «UAA» и «UAG» являются кодонами для «стоп»-сигнала. Синтез белка на рибосомах терминируется, когда рибосомный аппарат распознает «стоп-кодон» при сканировании вдоль мРНК.
Антисмысловой олигонуклеотид (ASO). Олигонуклеотид, связывающийся с мРНК или пре-мРНК специфическим для последовательности образом (т.е. комплементарно), называется «антисмысловым олигонуклеотидом» (ASO). ASO, прочно связывающийся с мРНК, может блокировать синтез белков на рибосомах. Также, ASO, прочно связывающийся с пре-мРНК, может вмешиваться в процесс сплайсинга и давать варианты сплайсинга мРНК.
Антисмысловое ингибирование сплайсинга. Сплайсинг пре-мРНК начинается после образования «сплайсомного Е-комплекса» (т.е. E-комплекса). Как схематически представлено на фиг. 3, SR-белки (т.е. богатые серином и аргинином белки) связываются с областями «экзонного энхансера сплайсинга» (ESE) и помогают рекрутировать U1 и U2AF35 для связывания с «5'-сайтом сплайсинга» и «3'-сайтом сплайсинга, соответственно [Biochem. Cell Biol. vol 77(4), 277-291 (1999); Curr. Opin. Cell Biol. vol 13(3), 302-309 (2001)].
В принципе, ASO может стерически ингибировать образование «сплайсомного Е-комплекса» путем связывания с определенной областью пре-мРНК, которая является критической для образования E-комплекса. Образование E-комплекса ингибируется или блокируется, если ASO прочно связывается с «5'-сайтом сплайсинга», «3'-сайтом сплайсинга» или областью ESE.
Поскольку мРНК кодирует белок в соответствии с его последовательностью, вариант сплайсинга мРНК кодирует белок, отличный от белка, кодируемого «исходной» или «полноразмерной» мРНК. Таким образом, антисмысловое ингибирование сплайсинга является эффективным терапевтическим вариантом выбора за счет кодирования варианта белка(ов), проявляющего биологические свойства, отличные от свойств белка, кодируемого «исходной» или «полноразмерной» мРНК.
Сдвиг рамки считывания, индуцируемый антисмысловым ингибированием сплайсинга: Часть «кодирующей последовательности ДНК» (CDS) для мРНК HIF-1α человека [NCBI код для мРНК: NM_001530] представлена на фиг. 4А в качестве примера для иллюстрации «сдвига рамки считывания» (т.е. вне рамки считывания), индуцируемого антисмысловым ингибированием сплайсинга. На CDS (т.е. желтой полосе) отображены кодон и экзон (т.е. зеленая стрелка). Следует отметить, что T (т.е. тимин) в CDS должен быть заменен на U (т.е. урацил) в мРНК или пре-мРНК.
Если экзон 3 делетируется в результате антисмыслового ингибирования сплайсинга, 3'-конец экзона 2 непосредственно связывается с 5'-концом экзона 4. Тогда место соединения экзона 2 и экзона 4 будет прочитываться «...-GAT-GCT-(G-TTT)-GAA-CTA-...», как показано на фиг. 4B (ср. левую диаграмму). Между двумя соседними кодонами полноразмерной мРНК существуют четыре нуклеотида. Делеция экзона 3 выводит кодоны, начиная с экзона 4, из рамки считывания. Таким образом, делеция экзона 3 вызывает «сдвиг рамки считывания» кодонов.
Если экзон 3 и экзон 4 делетируются одновременно в результате антисмыслового ингибирования сплайсинга, 3'-конец экзона 2 соединяется с 5'-концом экзона 5. Тогда место соединения экзона 2 и экзона 5 будет прочитываться «...-GAT-GCT-(G-GC)-CTT-GTC-...», как показано на фиг. 4B (ср. правую диаграмму). Между двумя соседними кодонами полноразмерной мРНК существуют три нуклеотида. Двойная делеция экзона 3 и экзона 4 помещает кодоны, начиная с экзона 5, в рамку считывания, т.е. без сдвига рамки считывания.
Сдвиг рамки считывания дает кодоны, отличные от «исходных» кодонов, и часто порождает кодон преждевременной терминации (PTC), как показано на фиг. 4C для случая делеции экзона 3 в мРНК для HIF-1α. Пропуск экзона, вызывающий сдвиг рамки считывания, предназначен для получения усеченного с С-конца фрагмента белка из-за преждевременной терминации синтеза белка на рибосомах. Такой фрагмент белка может демонстрировать физиологические свойства, отличные от «исходного» или «полноразмерного» белка. Таким образом, антисмысловое ингибирование сплайсинга может быть эффективным терапевтическим вариантом выбора для гена-мишени заболевания.
Обнаружение пропуска экзонов с помощью вложенной ОТ-ПЦР: мРНК варианта сплайсинга, индуцированного ASO, часто детектируется с помощью ПЦР (полимеразная цепная реакция). Если ASO индуцирует пропуск экзона 4 длиной 150 п.н., как показано на фиг. 5А, существуют две возможные мРНК, получаемые из пре-мРНК-мишени ASO, т.е. полноразмерная мРНК и вариант сплайсинга мРНК, в котором отсутствует экзон 4. В случае индукции с помощью ASO полного пропуска экзона 4 (т.е. 100%) клетки, обработанные ASO, дают только ПЦР-продукт, меньший на 150 п.н. ПЦР-продукта в виде полноразмерной мРНК. Относящаяся к ПЦР-продукту в случае пропуска экзона полоса отбирается и подвергается секвенированию с целью подтверждения того, что полоса для ПЦР-продукта действительно произошла от варианта сплайсинга мРНК.
Оценка выхода пропуска экзонов методом ПЦР. В литературе выход или эффективность пропуска экзонов обычно оценивали путем сравнения интенсивности в геле полосы для ПЦР-продукта в виде мРНК варианта сплайсинга с интенсивностью полосы для полноразмерной мРНК. Такая оценка теоретически верна в целом, если только полноразмерная мРНК и мРНК варианта сплайсинга обладают сопоставимой стабильностью в клетках, а также во время процедур анализа, принятых для детектирования ПЦР. Однако, принимая во внимание, что стабильность мРНК является совокупным результатом эволюции на протяжении миллиарда лет, маловероятно, что варианты сплайсинга мРНК будут демонстрировать такую же стабильность, как и полноразмерная мРНК.
Также можно предположить, что относительное обогащение мРНК варианта сплайсинга и полноразмерной мРНК может сильно различаться в зависимости от ПЦР-праймеров условий ПЦР и метода детектирования ПЦР. Недавно цифровая кПЦР была применена для оценки выхода пропуска экзонов в мРНК для дистрофина у мышей mdx, получавших дистрофин-специфический ASO или морфолино-, или 2'-OMe-PTO (фосфоротиоатный). Выход пропуска экзонов, оцененный с помощью цифровой кПЦР, значительно отличался от выходов, оцененных с помощью традиционных методов, таких как вложенная кПЦР [Lab. Investigation, vol 90, 1396-1402 (2010)]. Исследование с помощью цифровой кПЦР пропуска экзонов в миобластах и фибробластах у являющихся людьми пациентов с DMD предполагает, что цифровая кПЦР является выбором для надежного обнаружения продуктов пропуска экзонов с высокой чувствительностью [PLoS One 0162467, September 09 (2016)].
Принимая во внимание, что кажущийся выход пропуска экзонов, как правило, меняется в зависимости от способа и условия ПЦР-анализа, помимо ПЦР-анализа может потребоваться дополнительная проверка выхода пропуска экзонов с помощью анализа экспрессии белка или функциональных анализов для гена-мишени.
Усиление по принципу обратной связи транскрипции с помощью EIciRNA: Интронный лариат образуется как побочный продукт при сплайсинге пре-мРНК. Пропуск экзона дает не только мРНК варианта сплайсинга, но также и состоящую из экзона(ов) и интрона(ов) кольцевую РНК (EIciRNA), как показано на фиг. 5B, в которой экзон 3 и экзон 4 сращены с образованием лариата (структуры типа «лассо»), состоящего из интронов, экзона 3 и экзона 4. Первоначально образованный лариат, т.е. EIciRNA ①, может подвергаться дополнительному сплайсингу с образованием вторичного лариата, определяемого как EIciRNA ②.
Эти лариаты EIciRNA сохраняют последовательность 5'-сайта сплайсинга «экзона 4» и способны рекрутировать «малый ядерный рибонуклеарный белок U1 (snRNP U1)». Затем snRNP U1 рекрутирует РНК-полимеразу II, которая может активировать транскрипцию пре-мРНК. Транскрипция пре-мРНК может увеличиваться, если EIciRNA накапливаются сверх порогового уровня в ядре. Таким образом, EIciRNA часто могут функционировать в качестве регулятора в цепи обратной связи транскрипции, когда пропуск экзонов происходит чрезмерно [Nature Struct. Mol. Biol. vol 22(3), 256-264 (2015)].
Неприродные олигонуклеотиды: ДНК или РНК-олигонуклеотид склонен к деградации эндогенными нуклеазами, что ограничивает их терапевтическое применение. До настоящего времени был разработан и интенсивно исследован ряд неприродных (т.е. не встречающихся в природе) олигонуклеотидов [Clin. Exp. Pharmacol. Physiol. vol 33, 533-540 (2006)]. Многие из них демонстрируют расширенную метаболическую стабильность по сравнению с ДНК и РНК. На фиг. 6А представлены химические структуры некоторых репрезентативных неприродных олигонуклеотидов. Эти олигонуклеотиды предсказуемо связываются с комплементарной нуклеиновой кислотой, как и ДНК или РНК.
Фосфоротиоатный олигонуклеотид (PTO): PTO является аналогом ДНК, в котором один из атомов кислорода в фосфате остова заменен атомом серы в каждом мономере. Такое небольшое структурное изменение сделало PTO сравнительно устойчивым к деградации нуклеазами [Ann. Rev. Biochem. vol 54, 367-402 (1985)].
Отражая структурное сходство между остовами PTO и ДНК, они оба плохо проходят сквозь клеточную мембрану в большинстве типов клеток млекопитающих. Однако в случае некоторых типов клеток, экспрессирующих переносчик(и) для ДНК, ДНК и PTO демонстрируют сравнительно хорошую клеточную проницаемость. Известно, что системно вводимые PTO легко распределяются в печени и почках благодаря избыточной экспрессии переносчиков ДНК [Nucleic Acids Res. vol 25, 3290-3296 (1997)].
Для улучшения клеточной проницаемости PTO in vitro широко применялась липофекция. Однако липофекция физически изменяет клеточную мембрану, вызывает цитотоксичность и, следовательно, не будет идеальной для длительного терапевтического применения.
За последние 30 лет PTO и варианты PTO были клинически оценены для лечения раков, иммунологических нарушений, нарушений обмена веществ и так далее [Biochemistry vol 41, 4503-4510 (2002); Clin. Exp. Pharmacol. Physiol. vol 33, 533-540 (2006)]. Многие из таких кандидатов на антисмысловые лекарственные препараты не были успешно разработаны частично из-за плохой клеточной проницаемости PTO. Для преодоления плохой клеточной проницаемости, PTO должен вводиться в высокой дозе для терапевтической активности. Однако известно, что PTO вызывают ограничивающую дозу токсичность, включая увеличенное время коагуляции, активацию комплемента, тубулярную нефропатию, активацию клеток Купфера и иммунную стимуляцию, в том числе спленомегалию, лимфоидную гиперплазию и инфильтрацию мононуклеарных клеток [Clin. Exp. Pharmacol. Physiol. vol 33, 533-540 (2006)].
Было обнаружено, что многие антисмысловые PTO проявляют клиническую активность при заболеваниях со значительным вкладом со стороны печени или почек. Мипомерсен является аналогом PTO, который ингибирует синтез апоВ-100, белка, участвующего в транспорте холестерина ЛПНП. Мипомерсен проявлял клиническую активность в определенной популяции пациентов с атеросклерозом благодаря его преимущественному распределению в печени. [Circulation vol 118(7), 743-753 (2008)]. ISIS-113715 является аналогом PTO, ингибирующим синтез белка тирозинфосфатазы 1B (PTP1B) и, как было обнаружено, проявляет терапевтическую активность у пациентов с диабетом II типа [Curr. Opin. Mol. Ther. vol 6, 331-336 (2004)].
2'-О-алкил-РНК: 2'-О-алкил-РНК представляет собой аналог РНК, в котором 2'-гидроксигруппа в рибозном кольце замещена алкилоксигруппой. 2'-О-алкил-РНК проявляет сродство к РНК сильнее, чем PTO или ДНК. Кроме того, 2'-О-алкил-РНК демонстрирует увеличенную метаболическую стабильность в терапевтических целях. Однако 2'-О-алкил-РНК демонстрирует плохую мембранную проницаемость, что ограничивает терапевтические возможности.
Замкнутая нуклеиновая кислота (LNA): В LNA рибозное кольцо в остове РНК структурно ограничено для увеличения сродства связывания с РНК или ДНК. Таким образом, LNA может рассматриваться как ДНК или РНК с высоким сродством [Biochemistry vol 45, 7347-7355 (2006)]. Тем не менее, LNA также демонстрирует плохую клеточную проницаемость, как и ДНК или РНК.
Гибридный олигонуклеотид остова ДНК или РНК: РТО и 2-О-алкил-РНК часто сливают в один олигонуклеотид. Благодаря 2'-О-алкил-РНК части такой гибридный олигонуклеотид обладает более высоким сродством к РНК, чем РТО-олигонуклеотид с той же последовательностью. Аналогично, LNA и PTO часто сливают в один олигонуклеотид, и гибридный олигонуклеотид обладает более высоким сродством к РНК, чем PTO-олигонуклеотид с той же последовательностью. Однако такие гибридные олигонуклеотиды также демонстрирует плохую клеточную проницаемость.
Морфолино-фосфородиамидатный олигонуклеотид (PMO): В PMO модули остова ДНК в виде фосфата и 2-дезоксирибозы заменены на фосфородиамидат и морфолин, соответственно [Appl. Microbiol. Biotechnol. vol 71, 575-586 (2006)]. Хотя остов ДНК заряжен отрицательно, основ PMO не заряжен. Таким образом, связь между PMO и мРНК свободна от электростатического отталкивания между остовами и, как правило, является более сильной, чем связь между ДНК и мРНК. Поскольку PMO заметно отличается от ДНК по структуре остова, PMO не будет распознаваться печеночным переносчиком(ами), распознающим ДНК или РНК. Однако PMO не проникает легко через клеточную мембрану.
Пептидо-нуклеиновая кислота (ПНК): ПНК представляет собой полипептид с остовом из N-(2-аминоэтил)глицина в качестве единицы и была открыта Dr. Nielsen и его коллегами [Science vol 254, 1497-1500 (1991)]. На фиг. 6B показана химическая структура и номенклатура для прототипной (т.е. немодифицированной) ПНК.
Подобно ДНК и РНК, ПНК также селективно связывается с комплементарной нуклеиновой кислотой [Nature (London) vol 365, 566-568 (1992)]. При связывании с комплементарной нуклеиновой кислотой N-конец ПНК эквивалентен 5'-концу ДНК или РНК, а С-конец ПНК эквивалентен 3'-концу ДНК или РНК.
Как и PMO, остов ПНК не заряжен. Таким образом, связь между ПНК и РНК, как правило, является сильнее, чем связь между ДНК и РНК. Поскольку ПНК заметно отличается от ДНК по химической структуре, ПНК не будет распознаваться печеночным переносчиком(ами), распознающим ДНК, и будет демонстрировать профиль распределения в ткани, отличный от профиля распределения ДНК или PTO. Однако ПНК также плохо проникает сквозь клеточную мембрану млекопитающего [Adv. Drug Delivery Rev. vol 55, 267-280 (2003)].
Мышечная дистрофия Дюшенна (DMD): DMD представляет собой сопровождающееся истощением мышечной ткани заболевание, которое поражает одного из приблизительно 3500 новорожденных детей мужского пола [Lancet Neurol. vol 9, 77-93 (2010)]. Пациенты с DMD постепенно теряют мышечную функцию и умирают от сердечной или дыхательной недостаточности до достижения 30-летнего возраста. У многих пациентов с DMD ген дистрофина мутирован с образованием мРНК дистрофина с кодоном преждевременной терминации (PTC) и экспрессирует усеченный нефункциональный дистрофин, в котором отсутствует С-концевая часть [HumanMol. Genetics vol. 12 (8), 907-914 (2003); и ссылки там].
Популярный подход к лечению DMD состоит в том, чтобы пропустить экзон, имеющий РТС, в мРНК дистрофина с использованием ASO, и кодировать белок варианта сплайсинга с С-концом, который часто называют полноразмерным дистрофином.
Пропуск экзона-23 в мРНК дистрофина у мышей MDX. Мышь Mdx является мутантом с PTC в экзоне 23 пре-мРНК дистрофина и широко применялась в качестве модели DMD человека на животном [FEBS J. vol 280(17), 4177-4186 (2013)]. ASO, комплементарно выбирающие своей целью (нацеливающиеся на) пре-мРНК для дистрофина мыши, были оценены в отношении их способности к индукции пропуска экзона 23 [Artificial DNA: PNA & XNA vol 2(1), 6-15 (2011)]. В этом отношении мышь mdx послужила хорошей модельной системой для оценки класса олигонуклеотидов в отношении их способности к индукции пропуска экзонов.
20-мерный 2'-OMe-PTO (2'-О-метилфосфоротиоатный) ASO, полностью комплементарный месту соединения экзона 23 и интрона 23 (т.е. 5'-сайту сплайсинга экзона 23), был местно инъецирован в мышцу мыши mdx в дозе приблизительно 10 мкг/кг в композиции с амфифильным агентом для трансфекции F127 и увеличивал экспрессию полноразмерного дистрофина в мышечной ткани при инъекции согласно иммуногистохимическому анализу (IHC) и Вестерн-блоттингом для полноразмерного дистрофина. Эти результаты IHC и Вестерн-блоттинга указывают на то, что экзон 23 был пропущен в результате местной инъекции ASO. ASO обладает способностью к 18-мерному комплементарному перекрыванию с 5'-концом интрона 23 и 2-мерному комплементарному перекрыванию с 3'-концом экзона 23 [Nature Med. vol 9(8), 1009-1014 (2003)].
Другой 20-мерный 2'-OMe-PTO ASO, полностью комплементарный месту соединения экзона 23 и интрона 23 (т.е. 5'-сайту сплайсинга экзона 23), был оценен в отношении его способности к индукции пропуска экзона 23. 20-мерный ASO комплементарно нацеливается на место соединения экзона 23 и интрона 23 и обладает способностью к 18-мерному комплементарному перекрыванию с 5'-концом интрона 23 и 2-мерному комплементарному перекрыванию с 3'-концом экзона 23. Инкубация в течение 96 часов мышиных миобластов с 2 или 4 мкМ ASO индуцировала пропуск экзона 23, что подтверждается вложенной ОТ-ПЦР. Пропуск экзона 23 также был идентифицирован с помощью ОТ-ПЦР у мышей mdx, которые получили две внутримышечные инъекции 2,9 нмоль ASO. Пропуск экзона 23 был обнаружен в мышечной ткани мышей mdx, которым подкожно вводили 2'-OMe-PTO ASO в дозе 50 мг/кг. 20-мерный 2'-FPS (2'-фторфосфоротиоатный) ASO, имеющий ту же последовательность, что и вышеупомянутый 2'-OMe-PTO ASO, также индуцировал пропуск экзона 23 в мышечных миобластах, подобно 2'-OMe-PTO ASO. Однако 2'-FPS ASO не индуцировал пропуск экзона 23 у мышей mdx после внутримышечных или подкожных инъекций [Mol. Ther. Nucl. Acids vol 4, e265 (2015)].
20-мерную пептидо-нуклеиновую кислоту (ПНК), комплементарно нацеливающуюся на место соединения экзона 23 и интрона 23, был оценен в отношении ее способности к индукции пропуска экзона 23 у мышей MDX. 20-мерная ПНК ASO обладает способностью к 18-мерному комплементарному перекрыванию с 5'-концом интрона 23 и 2-мерному комплементарному перекрыванию с 3'-концом экзона 23. 20-мерная ПНК в концентрации 250 нМ индуцировала делецию экзона 23 в клетках H2D мышей MDX, в соответствии с анализом с помощью вложенной ОТ-ПЦР. После внутримышечной инъекции от 5 до 20 мкг (в дозе приблизительно от 0,25 до 2 мг/кг) мышам mdx 20-мерная ПНК индуцировала пропуск экзона 23 в мышечной ткани в месте инъекции. Был сделан вывод, что эффективность пропуска экзонов в случае 20-мерной ПНК выше, чем в случае вышеупомянутых 2'-OMe-PTO ASO у мышей mdx. 20-мерную ПНК ковалентно конъюгировали с различными проникающими в клетки пептидами (СРР) для увеличения клеточной проницаемости. Эти конъюгаты ПНК-CPP и немодифицированная ПНК сравнительно индуцировали пропуск экзона 23 в клетках, а также в мышечной ткани в месте инъекции [Mol. Ther. vol 16(1), 38-45 (2008)].
25-мерный PMO ASO, полностью комплементарный месту соединения экзона 23 и интрона 23 (т. е. 5'-сайту сплайсинга экзона 23), был оценен в отношении его способности к индукции пропуска экзона 23 у мышей MDX. 25-мерный ASO обладает способностью к 18-мерному комплементарному перекрыванию с интроном 23 и 7-мерному комплементарному перекрыванию с экзоном 23. 25-мерный PMO индуцировал пропуск экзона-23 у мышей MDX после многократных внутривенных инъекций по 2 мг на животное (в дозе приблизительно 100 мг/кг) [Nat. Med. vol 12(2), 175-177 (2006)]. 25-мерный PMO был ковалентно конъюгирован с различными проникающими в клетки пептидами (CPP) для увеличения клеточной проницаемости. Эти конъюгаты PMO-CPP индуцировали пропуск экзона 23 в мышцах при однократном внутривенном введении в дозе 3 мг/кг [Human Mol. Genet. vol 18(22), 4405-4414 (2009)].
Пропуск экзона 46 в мРНК дистрофина в миобластах у являющегося человеком пациента с DMD: 2'-OMe-PTO ASO были сконструированы для комплементарного нацеливания на область экзонного энхансера сплайсинга (ESE) внутри экзона 46 в пре-мРНК дистрофина человека и оценены на эффективность пропуска экзона 46 в клетках миобластов, полученных от являющегося человеком пациента с DMD, у которого отсутствует экзон 45 в мРНК дистрофина. Клетки трансфицировали ASO в концентрации 1 мкМ с помощью липофекции и инкубировали в течение 24 часов до экстракции РНК для вложенной ОТ-ПЦР для обнаружения пропуска экзона 46. Некоторые из протестированных ASO индуцировали пропуск экзона 46 [Human Mol. Genet. vol 10(15), 1547-1554 (2001)].
Пропуск экзона 51 в мРНК дистрофина у пациентов с DMD: Drisapersen (PRO051 или GSK24022968) представляет собой 20-мерный 2'-OMe-PTO, сконструированный для комплементарного нацеливания на область ESE внутри экзона 51 в пре-мРНК дистрофина человека, и был оценен на терапевтическую активность у являющихся людьми пациентов с DMD. На основании оценки биопсии мышечной ткани с помощью вложенной ПЦР, Drisapersen индуцировал пропуск экзона 51 у пациентов с DMD, подкожно получавших от 2 до 6 мг/кг в неделю, хотя эффективность пропуска экзона не была высокой [N. Engl. J. Med. vol 364, 1513-1522 (2011)]
Eteplirsen (AVI-4658) представляет собой 30-мерный PMO, сконструированный для комплементарного нацеливания на ESE внутри экзона 51 в пре-мРНК дистрофина человека, и был оценен на его терапевтическую активность у пациентов с DMD. На основании оценки биопсии мышечных тканей с помощью IHC (иммуногистохимического анализа) на полноразмерный дистрофин, Eteplirsen индуцировал пропуск экзона 51 у пациентов с DMD, получающих от 2 до 20 мг/кг в неделю путем внутривенной инфузии [Lancet vol 378(9791), 595-605 (2011)].
Пропуск экзона 27 в мРНК для APOB в клетках HepG2: Аполипопротеин B (APOB) составляет неотъемлемую часть липопротеиновых частиц. мРНК для APOB состоит из 29 экзонов. 2'-OMe-РНК APOB-специфические ASO были сконструированы для нацеливания на 3'-сайт сплайсинга экзона 27, 5'-сайт сплайсинга экзона 27 или как на 3'-сайт сплайсинга, так и на 5'-сайт сплайсинга. Нацеливающийся на 3'-сайт сплайсинга ASO (3'-SS ASO) обладает способностью к 15-мерному перекрыванию с интроном 26 и 5-мерному перекрыванию с экзоном 27 [BMC Mol. Biol. 2007, 8:3., 8:3. опубликовано 17 января 2007 г.]. Нацеливающийся 5'-сайт сплайсинга ASO (5'-SS ASO) обладает способностью к 5-мерному перекрыванию с экзоном 27 и 15-перекрыванию с интроном 27. 40-мерный 2'-OMe-РНК ASO был сконструирован путем ковалентного слияния 3'-SS ASO с 5'-SS ASO. Таким образом, 40-мерный ASO способен взаимодействовать одновременно с 3'-сайтом сплайсинга, а также с 5'-сайтом сплайсинга.
ASO были оценены в отношении их способности к индукции пропуска экзона 27 в клетках HepG2 с помощью липофекции. Интересно отметить, что как 3'-SS ASO, так и 5'-SS ASO не смогли индуцировать пропуск экзона 27 в клетках HepG2 в дозе 25-250 нМ. Между тем, 40-мерный ASO продемонстрировал заметный уровень пропуска экзона 27 в зависимости от дозы от 25 до 250 нМ. Вполне вероятно, что 15-мерное комплементарное перекрывание 2'-OMe-РНК только с интронной частью сайта сплайсинга было недостаточным для эффективного ингибирования образования раннего сплайсомного комплекса. Было бы желательно более прочное связывание с сайтом сплайсинга, охватывающим экзон 27 пре-мРНК для APOB, чтобы индуцировать пропуск экзона в результате эффективного ингибирования образования раннего сплайсомного комплекса в клетках HepG2.
Альтернативный сплайсинг пре-мРНК Bel-x: BCL2L1 (Bcl-x) представляет собой ген человека, кодирующий Bcl-xL или Bcl-xS посредством альтернативного сплайсинга. 18-мерный 2'-OMe-PTO ASO был сконструирован для нацеливания на 5'-сайт сплайсинга экзона 2 и обладает способностью к 16-мерному комплементарному перекрыванию с экзоном 2 и 2-мерному перекрыванию с интроном 2. При липофекции в концентрации 80-400 нМ, ASO способствовал клеточной продукции Bcl-xS посредством альтернативного сплайсинга в группе раковых клеток, включая MCF7, PC3, Dul45, HeLa и MDA MB231 [J. Biol. Chem. vol 277(51), 49374-49382 (2002)].
Бесклеточная коррекция сплайсинга in vitro в пре-мРНК для β-глобина. Талассемия представляет собой наследственное заболевание крови, вызванное аномальным образованием гемоглобина. Редкая мутация IVS2705, обнаруженная у пациентов с большой талассемией, несет точечную мутацию [T→G] в положении нуклеотида 705 в интроне 2 гена β-глобина человека. Мутация IVS2705 создает дополнительный 5'-сайт сплайсинга и активирует криптический 3'-сайт сплайсинга в положении 579 интрона. Мутация IVS2705 индуцирует альтернативный сплайсинг со вставкой 127 нуклеотидов, т.е. нуклеотидов 579-705 интрона, между экзоном 2 и экзоном 3 [J. Biol. Chem. vol 260, 16332-16337 (1985)]
17-мерный 2'-OMe-РНК ASO, полностью комплементарный криптическому 5'-сайту сплайсинга мутанта IVS270, был оценен в отношении его способности к коррекции аберрантного сплайсинга в бесклеточной системе сплайсинга in vitro. ASO обладает способностью к 8-мерному перекрыванию с интроном и 9-мерному перекрыванию с криптическим экзоном. ASO эффективно корректировал аберрантный сплайсинг в диапазоне от 0,12 до 2 мкМ с выходом мРНК без криптического экзона, происходящего от интрона 2 [Proc. Natl. Acad. Sci. USA vol 90, 8673-8677 (1993)]. Система сплайсинга in vitro является бесклеточной и, следовательно, не требует какого-либо средства доставки для индукции пропуска экзона. ASO индуцировал пропуск экзона в концентрации 120 нМ в бесклеточной системе сплайсинга. Если бы ASO обладал более сильным сродством к 5'-сайту сплайсинга, экзон-пропускающая активность была бы большей. С целью увеличения экзон-пропускающей активности желательно использовать ASO, обладающий сильным сродством к 5'-сайту сплайсинга.
Коррекция сплайсинга пре-мРНК для люциферазы в клетках HeLa pLuc/705 с помощью 2'-OMe-РНК: pLuc/705 представляет собой ген люциферазы, модифицированный так, интрон 2 мутанта IVS2705 β-глобина человека был встроен между нуклеотидами 1368 и 1369. Клетки HeLa pLuc/705 стабильно экспрессируют модифицированный ген люциферазы pLuc/705. Модифицированные клетки HeLa экспрессируют мРНК для люциферазы с криптическим экзоном между нуклеотидами 1368 и 1369 и, следовательно, кодируют нефункциональный вариант белка люциферазы.
17-мерный 2'-ОМе-РНК-олигонуклеотид, комплементарно нацеливающийся на криптический 5'-сайт сплайсинга мутанта IVS270 (обладающий способностью к 8-мерному перекрыванию с интроном и 9-мерному перекрыванию с криптическим экзоном), был оценен в отношении его способности к коррекции аберрантного сплайсинга модифицированной пре-мРНК люциферазы в клетках HeLa pLuc/705. После липофекции в концентрации 20-500 нМ 17-мерный ASO восстанавливал активность люциферазы в клетках дозозависимым образом. Было обнаружено методом ОТ-ПЦР, что криптический экзон сплайсируется при обработке ASO. Экзон-пропускающая активность наблюдалась при концентрации 20 нМ или выше [Biochemistry vol 37, 6235-6239 (1998)].
Коррекция сплайсинга пре-мРНК для люциферазы в клетках HeLa pLuc/705 с помощью ПНК: 17-мерные производные ПНК, комплементарно нацеливающиеся на криптический 5'-сайт сплайсинга мутанта IVS2705 (обладающие способностью к 8-мерному перекрыванию с интроном и 9-мерному перекрыванию с криптическим экзоном) были оценены в отношении их способности корректировать аберрантный сплайсинг модифицированной пре-мРНК для люциферазы в клетках HeLa pLuc/705. Были сконструированы производные, которые содержали различное количество фосфонатных групп, ковалентно конъюгированных с N-концом последовательности ПНК [Nucl. Acids Res. vol 30 (13), 4424-4432 (2008)]. Ковалентное конъюгирование фосфонатных фрагментов с ПНК было введено для облегчения трансфекции в клетку путем липофекции.
При липофекции в концентрации 2,5-60 нМ ПНК ASO восстанавливали активность люциферазы в клетке дозозависимым образом. Было обнаружено методом ОТ-ПЦР, что криптический экзон сплайсируется при обработке ASO. ПНК ASO с большим количеством фосфонатных групп, прикрепленных к ним, проявили более высокую активность и эффективность в сплайсинге криптического экзона. ПНК ASO с 12 фосфонатными группами проявил эффективность пропуска экзонов, составляющую 81% при 2,5 нМ.
Наблюдаемая субнаномолярная экзон-пропускающая активность ПНК ASO намного больше активности 17-мерного 2'-Ome-РНК ASO. [Biochemistry vol 37, 6235-6239 (1998)]. ПНК была бы очень полезна для эффективной индукции пропуска экзонов в случае модификации должным образом для доставки в клетку.
Пропуск экзона в пре-мРНК для FOLH1 с помощью 2'-Ome-PTO: Специфический для предстательной железы мембранный антиген (PSMA) является продуктом гена фолатгидролазы (FOLH1) и экспрессируется в высокой степени в злокачественных тканях предстательной железы. 2'-OMe-PTO ASO, нацеливающиеся на пре-мРНК для FOLH1, оценивали в отношении их способности к индукции пропуска экзонов в клетках рака предстательной железы LNCap после трансфекции с помощью липофекции [Oligonucleotides, vol. 16, 186-175 (2006)].
SSO1 представляет собой 18-мерный ASO, нацеливающийся на 5'-сайт сплайсинга экзона 1, и обладает способностью к 16-мерному комплементарному перекрыванию с экзоном 1 и 2-мерному перекрыванию с интроном 1. SS06 и SSO18 представляют собой 18-мерные ASO, комплементарно нацеливающиеся на экзон 6 и экзон 18, соответственно.
SSO1 индуцировал альтернативный сплайсинг с IC50, составляющей приблизительно 400 нМ. SS06 индуцировал пропуск экзона 6 с IC50, составляющей приблизительно 4 нМ. SSOl8 индуцировал пропуск экзона 18 с IC50, составляющей приблизительно 4 нМ.
Интересно отметить, что SS06 и SS018, нацеливающиеся на внутриэкзонную область (т.е. экзонный сайт энхансера сплайсинга), индуцировали пропуск экзона гораздо сильнее, чем SSO1, нацеливающийся на 5'-сайт сплайсинга. Было установлено, что нацеливание на область ESE с помощью 2'-OMe-PTO ASO более эффективно, чем нацеливание на сайт сплайсинга в этом конкретном примере.
Альтернативный сплайсинг пре-мРНК IL-5Rα с помощью 2'-О-MOE-РНК: 2'-О-MOE-РНК (2'-O-метоксиэтил-РНК) ASO, комплементарно нацеливающиеся на пре-мРНК для IL-5Rα мыши, оценивали в отношении их способности индуцировать альтернативный сплайсинг (т.е. пропуск экзонов) в клетках BCL1 после трансфекции путем электропорации. [Mol. Pharmacol. vol 58, 380-387 (2000)].
ASO были разработаны путем сканирования на комплементарность сканирования различных областей экзона 9 и сайтов сплайсинга, фланкирующих экзон 9. 20-мерный ASO, полностью комплементарный 3'-сайту сплайсинга (3' SS) экзона 9 с 4-мерным перекрыванием с интроном 8, заметно индуцировал альтернативный сплайсинг в концентрации 10 мкМ. ASO, нацеливающиеся на внутриэкзонные области экзона 9, индуцировали альтернативный сплайсинг в концентрации 10 мкМ с эффективностью, сравнимой с таковой 3' SS ASO. Все протестированные ASO индуцировали альтернативный сплайсинг, что указывает на то, что экзон 9 и его сайты сплайсинга очень чувствительны к пропуску экзона. 3'-сайт сплайсинга был более чувствительным, чем 5'-сайт сплайсинга.
20-мерные ASO также были сконструированы для комплементарного нацеливания на сайты сплайсинга, фланкирующие экзон 8 с 4-мерным перекрыванием с интроном. ASO индуцировали пропуск экзона 8 в концентрации 10 мкМ, хотя 3' SS ASO был более эффективным, чем 5' SS ASO.
Считается, что микромолярная экзон-пропускающая активность 2'-О-МОЕ-РНК ASO при электропорации является очень низкой по сравнению с наномолярной экзон-пропускающей активностью 2'-OMe-PTO ASO, нацеливающихся на пре-мРНК для FOLH1, при липофекции. [Oligonucleotides, vol. 16, 186-175 (2006)] Липофекция будет более эффективной, чем электропорация, для трансфекции олигонуклеотидов с отрицательно заряженным остовом в клетку.
Пропуск экзона 10 в пре-мРНК тау с помощью 2'-О-MOE PTO: 5'-сайт сплайсинга экзона 10 в пре-мРНК тау имеет 18-мерную последовательность, склонную к образованию петли на стебле, и не подходит для образования сплайсомного Е-комплекса. Поэтому экзон 10 пре-мРНК тау очень склонен к пропуску.
2'-О-MOE PTO ASO, нацеливающиеся либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 10 тау, были оценены в отношении их способности усиливать пропуск экзона 10 [J. Biol. Chem. vol 276(46), 42986-42993 (2001)]. E10α представляет собой 18-мерный ASO, комплементарно нацеливающийся на 3'-сайт сплайсинга. E10α обладает способностью к 10-мерному перекрыванию с интроном 9 и 8-мерному перекрыванию с экзоном 10. E10β представляет собой 21-мерный ASO, комплементарно нацеливающийся на 5'-сайт сплайсинга. E10β обладает способностью к 8-мерному перекрыванию с экзоном 10 и 13-мерному перекрыванию с интроном 10.
После трансфекции в клетки COS-1 с помощью липофекции, E10α и E10β индуцировали пропуск экзона 10 с IC50=2-5 нМ. В клетках РС12, трансфицированных с помощью электропорации, ASO индуцировали пропуск экзона 10 с микромолярной IC50.
Пропуск экзона 2 в пре-мРНК для MyD88 с помощью 2'-О-MOE-РНК ASO: MyD88 является адаптерным белком, участвующим в IL-1R- и TLR-индуцированной активации NF-κB. 2-мерные 2'-О-метоксиэтил (2'-О-MOE)-РНК ASO были сконструированы для комплементарного нацеливания либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 2 в пре-мРНК для MyD88 человека. Были сконструированы 20-мерные ASO, которые обладали способностью к 0-, 5-, 10-, 15- или 20-мерному перекрыванию с либо 5'-концом интрона 1 (т.е. 3'-сайтом сплайсинга экзона 2), либо 3'-концом интрона 2 (т.е. 5'-сайтом сплайсинга экзона 2). ASO были оценены в отношении их способности к индукции пропуска экзона 2 в клетках A549 после трансфекции с помощью липофекции [J. Immunol, vol 176, 3652-3661 (2006)].
Из всех ASO ASO, обладающий способностью к 20-мерному перекрыванию с интроном 1 в 3'-сайте сплайсинга экзона 2, индуцировал пропуск экзона 2 наиболее сильно и эффективно. Наблюдаемая IC50 для пропуска экзона 2 составляла от 50 до 100 нМ. ASO, нацеливающиеся на 5'-сайт сплайсинга, были не так эффективны, как ASO, нацеливающиеся на 3'-сайт сплайсинга. Среди ASO, нацеливающихся на 5'-сайт сплайсинга, самым сильным ASO был ASO, обладающий способностью к 20-мерному перекрыванию с 3'-концом экзона 2.
Среди 2'-О-МОЕ-РНК ASO, сконструированных также для комплементарного нацеливания либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 2 в пре-мРНК для MyD88 мыши, ASO, обладающий способностью к 20-мерному перекрыванию с 5'-концом экзона 2, наиболее сильно индуцировал пропуск экзона 2 в клетках RAW 264.7, трансфицированных с помощью липофекции.
Самый сильный ASO в мышиных клетках вводили два раза в неделю в течение 2 недель в дозе 50 мг/кг. Имело место значительное снижение мРНК для MyD88 на 60-85% в кишечнике, жировой ткани и печени. 50 мг/кг является большой дозой, которая могла бы вызвать типичные неблагоприятные эффекты терапии олигонуклеотидами с фосфатно-рибозным остовом. Существует острая необходимость в значительном увеличении экзон-пропускающей активности, если 2'-О-МОЕ-РНК ASO должны проявлять терапевтическую активность, не вызывая типичные неблагоприятные эффекты.
Восстановление экзона 7 в SMN2 с помощью Nusinersen: Спинальная мышечная атрофия (SMA) представляет собой опасное для жизни редкое заболевание, вызываемое делецией или потерей функции гена SMN1 (выживания двигательного нейрона 1). У людей есть паралогичный ген SMN2, который имеет идентичную кодирующую последовательность, за исключением 11 нуклеотидов. SNP (однонуклеотидный полиморфизм) C→T в экзоне 7 SMN2 индуцирует пропуск экзона 7, и получающаяся в результате сплайс-вариантная мРНК кодирует вариант белка SMN2, быстро метаболизируемый. Поэтому мутант SMN2 не способен компенсировать функциональную нехватку белка SMN1, что приводит к вспышке SMA. [Neurology vol 86, 890-897 (2016)]
Nusinersen (Spiranza™) представляет собой 18-мерный 2'-О-МОЕ-РНК ASO, комлементарно нацеливающийся на область сайленсера сплайсинга в интроне 7 SMN2. Поскольку Nusinersen стерически блокирует связывание белка-сайленсера сплайсинга, экзон 7 сохраняется или восстанавливается с получением полноразмерного белка SMN2. Nusinersen восстанавливает обычный процесс сплайсинга путем связывания с областью сайленсера сплайсинга, расположенной в интроне 7 SMN2.
Nusinersen был одобрен Управлением по контролю качества продуктов питания и лекарственных средств (FDA) США в 2016 году для лечения SMA. Nusinersen вводится интратекально в дозе 12 мг один раз в три месяца или шесть месяцев. Nusinersen находится в спинном мозге с полупериодом существования от 135 до 177 дней в спинномозговой жидкости (CSF) [Nusinersen US Label, FDA, December 2016].
Терапевтическая активность индуцирующих пропуск экзонов олигонуклеотидных терапий: Как и в приведенных ранее в этом документе в качестве примера случаях, олигонуклеотиды с фосфатным остовом индуцируют пропуск экзонов с наномолярной активностью в клетках, трансфицированных с помощью липофекции, но с микромолярной активностью в клетках, обработанных «голым» олигонуклеотидом.
Микромолярная экзон-пропускающая активность в пре-мРНК MyD88 была переведена в терапевтическую дозу, составляющую 10 мг/кг или выше при системном введении в виде «голого» олигонуклеотида у мышей [J. Immunol, vol 176, 3652-3661 (2006)]. В такой высокой терапевтической дозе олигонуклеотиды с фосфатным остовом чувствительны к иммунологическим неблагоприятным событиям. Таким образом, было бы очень желательно разработать способ или препарат для заметного улучшения терапевтической дозы.
Drisapersen, 20-мерный 2'-ОМе-PTO, сконструированный для индукции пропуска экзона 51 в пре-мРНК для дистрофина человека, индуцировал пропуск экзона 51 у пациентов с DMD, подкожно получавших ASO в дозе от 2 до 6 мг/кг в неделю в виде голого олигонуклеотид, хотя эффективность пропуска экзонов не была высокой [N. Engl. J. Med. vol 364, 1513-1522 (2011)]. Была обеспокоенность по поводу увеличения терапевтической дозы Drisapersen из-за ограничивающей дозу токсичности.
ПНК и PMO обладают нейтральным остовом, не распознаются иммунными системами (особенно Toll-подобными рецепторами) и будут свободны от иммунологических реакций, обычно наблюдаемых в случае олигонуклеотидов с фосфатным остовом.
Eteplirsen (AVI-4658), 30-мерный PMO, разработанный для индукции пропуска экзона 51 в пре-мРНК для дистрофина человека, хорошо переносился пациентами с DMD, получавшими ASO путем внутривенной инфузии от 2 до 20 мг/кг в неделю [Lancet vol 378(9791), 595-605 (2011)]. Недавно Eteplirsen получил ускоренное одобрение FDA США для применения для пациентов с DMD.
Несмотря на то, что Nusinersen является ASO со способностью к восстановлению экзонов вместо пропуска экзонов, разрешенная терапевтическая доза, составляющая 12 мг раз в три месяца, является весьма привлекательной. Считается, что эффективное поглощение нейронами после интратекальной инъекции в значительной степени ответственно за активность Nusinersen.
Клиническая разработка терапии олигонуклеотидом с фосфатным остовом была критически затруднена из-за ограничивающей дозу токсичности, включая иммунологическую токсичность через активацию Toll-подобных рецепторов или активацию комплемента, тканеспецифическую токсичность в печени или почках. Повышая терапевтическую активность in vivo, можно преодолеть такие ограничивающие дозу токсичности.
Производство олигонуклеотидов очень дорого. Текущий уровень терапевтической дозы для человека от 100 мг до 2 г в неделю переводится в стоимость API (активного фармацевтического ингредиента) от 100 до 2000 долларов США в неделю, если стоимость производства API щедро предполагается равной 1000 долларов США за грамм. На самом деле, стоимость производства API для терапии олигонуклеотидами намного превышает 1000 долларов США за грамм. Таким образом, у заинтересованных сторон в сфере здравоохранения будет существенная потребность в значительном увеличении терапевтической активности с целью обеспечения терапии олигонуклеотидами по умеренной ежегодной стоимости лечения для постоянного применения.
Хорошая клеточная проницаемость олигонуклеотида. Клеточная мембрана представляет собой липидный бислойный барьер, образовавшийся в процессе эволюции на протяжении миллиарда лет. Клеточная мембрана действительно функционирует как большой барьер для одноцепочечных антисмысловых олигонуклеотидов размером от 4 до 10 кДа. Доставка в клетку таких ASO путем прямого проникновения через клеточную мембрану практически невозможна. Существуют и другие пути клеточного поглощения одноцепочечных олигонуклеотидов. Чтобы привести некоторые пути, опосредованный переносчиками эндоцитоз в гепатоцитах, наблюдаемый в случае Miromersen, нацеливающегося на ApoB 100, поглощение нейронами (вероятно, эндоцитоз), наблюдаемое в случае Nusinersen, GalNac (N-ацетилгалактозамин)-опосредованное поглощение клетками и т.д. Однако такие пути поглощения клетками сильно зависят от тканей и вряд ли применимы в целом к большинству типов тканей [Nature Biotechnol. vol 35(3), 222-229 (2017)].
Олигонуклеотид с фосфатным остовом можно было бы правильно приготовить в виде композиции, чтобы он обладал хорошей клеточной проницаемостью, и для такого приготовленного в виде композиции олигонуклеотида можно бы предсказать, что он будет проявлять лучшую терапевтическую активность in vivo, чем «голый» (т.е. без композиции) олигонуклеотид. Учитывая проявление олигонуклеотидами с фосфатным остовом наномолярной экзон-пропускающей активности чаще всего в клетках, если они были трансфицированы с помощью липофекции, терапевтическая активность in vivo олигонуклеотидов, приготовленных в виде композиции так, чтобы они обладали хорошей клеточной проницаемостью, была бы заметно улучшена, поскольку будет предписываться наномолярная экзон-пропускающая активность in vitro. Таким образом, хорошая клеточная проницаемость будет иметь решающее значение для in vivo терапевтической активности олигонуклеотидов, индуцирующих пропуск экзонов. Тем не менее, разработка композиции, обеспечивающей хорошую доставку в ткани, остается огромной технической проблемой в области терапии олигонуклеотидами.
Модифицированные нуклеотидные основания ПНК для хорошей клеточной проницаемости и высокого сродства. Как упоминалось ранее, были сконструированы производные ПНК, которые содержат различное количество фосфонатных групп, ковалентно конъюгированных для облегчения доставки в клетку с помощью липофекции. Было обнаружено, что такие ПНК ASO проявляют субаномолярную экзон-пропускающую активность в клетках HeLa при липофекции. [Nucl. Acids Res. vol 30(13), 4424-4432 (2008)]. Эта субнаномолярная активность является значительно большей экзон-пропускающей активности, наблюдаемой у ASO с фосфатным остовом. Таким образом, ПНК будет полезна для эффективной индукции пропуска экзонов в случае правильной доставки в клетку.
ПНК была сделана в высокой степени проходимой через клеточную мембрану млекопитающего путем введения модифицированных нуклеотидных оснований вместе с катионным липидом или его эквивалентом, ковалентно присоединенным к ним. Химическая структура таких модифицированных нуклеотидных оснований показана на фиг. 6С. Было обнаружено, что такие модифицированные нуклеотидные основания цитозина, аденина и гуанина предсказуемо гибридизуются с гуанином, тимином и цитозином, соответственно. [PCT-заявка с № PCT/KR2009/001256; EP2268607; US8680253]
Включение таких модифицированных нуклеотидных оснований в ПНК имитирует ситуации с липофекцией. При липофекции молекулы олигонуклеотидов оборачиваются или образуют комплексы с катионными молекулами липидов, такими как липофектамин, и такие комплексы липофектамин/олигонуклеотид, как правило, довольно легко проникают через клеточную мембрану по сравнению с голыми молекулами олигонуклеотидов.
В дополнение к хорошей мембранной проницаемости было обнаружено, что эти производные ПНК обладают сверхсильным сродством к комплементарной нуклеиновой кислоте. Например, включение от 4 до 5 модифицированных нуклеотидных оснований в 11-13-мерные производные ПНК легко дает увеличение Tm на 20οC или больше при образовании дуплекса с комплементарной ДНК.
Было обнаружено, что такие производные ПНК очень чувствительны к ошибочному спариванию пары оснований (несоответствию по одному основанию). Ошибочное спаривание пары оснований приводило к уменьшению Tm на 11-22οC в зависимости от типа модифицированного основания, а также последовательности ПНК.
Учитывая хорошую мембранную проницаемость и сверхвысокое сродство к нуклеиновой кислоте, производные ПНК с такими модифицированными нуклеотидными основаниями были бы применимы для эффективной индукции пропуска экзонов.
Краткое описание фигур
Фиг. 1А. Иллюстрация нумерации интронов и экзонов в пре-мРНК.
Фиг. 1B. Краткая иллюстрация в виде схемы процесса сплайсинга.
Фиг. 2А. Варианты сплайсинга мРНК для AR, кодирующие варианты белков AR.
Фиг. 2B. Варианты сплайсинга мРНК для HIF-1α, кодирующие варианты белков HIF-α.
Фиг. 3. Схематическое изображение биологических процессов, участвующих в образовании сплайсомного раннего комплекса.
Фиг. 4А. Часть CDS, считываемой с мРНК для HIF-1α человека.
Фиг. 4В. Последовательности мест экзон-экзонных соединений в вариантах сплайсинга HIF-1α, в которых отсутствуют экзон 3 (слева) и экзоны 3-4 (справа), иллюстрирующие сдвиг рамки считывания (вне рамки считывания) и нахождение в рамке считывания, соответственно.
Фиг. 4C. Приводимый в качестве примера сдвиг рамки считывания, дающий PTC.
Фиг. 5А. Схематическое изображение вложенной ОТ-ПЦР для обнаружения пропуска экзонов.
Фиг. 5B. Схематическое изображение образования EIciRNA во время пропуска экзонов.
Фиг. 6А. Химическая структура для репрезентативных неприродных олигонуклеотидов.
Фиг. 6B. Химические структуры и сокращенное обозначение прототипной ПНК.
Фиг. 6C. Модифицированные нуклеотидные основания, разработанные для увеличения мембранной проницаемости ПНК.
Фиг. 7. Примеры природных или неприродных (модифицированных) нуклеотидных оснований, выбираемых для производного пептидо-нуклеиновой кислоты формулы I.
Фиг. 8А. Примеры замещенных или незамещенных алкильных радикалов, выбираемых для соединения формулы I.
Фиг. 8В. Примеры замещенных или незамещенных алкилацильных и замещенных или незамещенных арилацильных радикалов, выбираемых для соединения формулы I.
Фиг. 8С. Примеры выбираемых для соединения формулы I радикалов: замещенного алкиламино, замещенного ариламино, замещенного или незамещенного арила, замещенного или незамещенного алкилсульфонила, замещенного или незамещенного арилсульфонила, замещенного или незамещенного алкилфосфонила и замещенного или незамещенного арилфосфонила.
Фиг. 8D. Примеры выбираемых для соединения формулы I радикалов: замещенного или незамещенного алкилоксикарбонила, замещенного или незамещенного арилоксикарбонила, замещенного или незамещенного алкиламинокарбонила и замещенного или незамещенного ариламинокарбонила.
Фиг. 8E. Примеры выбираемых для соединения формулы I радикалов: замещенного или незамещенного алкилокситиокарбонила, замещенного или незамещенного алкиламинотиокарбонила, замещенного или незамещенного ариламинотиокарбонила, замещенного или незамещенного алкилокситиокарбонила и замещенного или незамещенного арилокситиокарбонила.
Фиг. 9. Химических структуры мономеров ПНК с природным или модифицированным нуклеотидным основанием.
Фиг. 10. Химические структуры аббревиатур N-концевых или C-концевых заместителей.
Фиг. 11. Химическая структура 14-мерного ПНК-производного (N→C)Fethoc-GA(5)A-C(102)TT-A(5)TC-CTA(5)-C(102)T-NH2
Фиг. 12. Химическая структура 15-мерного ПНК-производного (N→C)Fmoc-Val-CTC(102)-A(5)TC-CTA(6)-C(103)TT-AA(202)C-NH2.
Фиг. 13. Химические структуры мономеров Fmoc-ПНК, используемых для синтеза производных ПНК этого изобретения.
Фиг. 14. Типичный цикл удлинения мономера, принятый при твердофазном синтезе пептидов.
Фиг. 15А. Хроматограмма, полученная при HPLC c C18-обращенной фазой, для «HIF-ASO 1» перед очисткой.
Фиг. 15B. Хроматограмма, полученная при HPLC c C18-обращенной фазой, для «HIF-ASO 1» после очистки с помощью HPLC.
Фиг. 16. ESI-TOF масс-спектр «HIF-ASO 1», очищенного с помощью препаративной HPLC c C18-обращенной фазой.
Фиг. 17А. Положения-мишени экзон-специфических праймеров, используемых в HIF-1α-специфической вложенной ПЦР для обнаружения пропуска экзонов, индуцируемого «HIF-ASO 2» в клетках HeLa.
Фиг. 17B. Данные электрофореза продуктов HIF-1α-специфической вложенной ПЦР в клетках HeLa, обработанных «HIF-ASO 2» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ.
Фиг. 17C. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзона 2 HIF-1α.
Фиг. 18А. Данные Вестерн-блоттинга HIF-1α в клетках HeLa, обработанных «HIF-ASO 2» в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ, 1 аМ или 10 аМ в течение 24 часов.
Фиг. 18B. Относительные уровни экспрессии белка HIF-1α, приведенные к β-актину, в клетках HeLa, обработанных «HIF-ASO 2», в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ, 1 аМ или 10 аМ в течение 24 часов («усы» согласно среднеквадратической ошибке).
Фиг. 18C. HIF-1α-специфическая вложенная кПЦР с использованием SYBR Green в клетках HeLa, обработанных «HIF-ASO 2» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 18D. HIF-1α-специфическая вложенная кПЦР с использованием зонда TaqMan в клетках HeLa, обработанных «HIF-ASO 2» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 19А. Данные электрофореза продуктов HIF-1α-специфической вложенной ПЦР в клетках HeLa, обработанных «HIF-ASO 6» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ.
Фиг. 19B. Данные Вестерн-блоттинга HIF-1α в клетках HeLa, обработанных «HIF-ASO 6» в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ или 1 аМ в течение 24 часов.
Фиг. 19C. Уровни экспрессии HIF-1α, приведенные к β-актину, в клетках HeLa, обработанных «HIF-ASO 6» в концентрации 0 цM (отрицательный контроль), 10 цM, 100 цМ или 1 аМ в течение 24 часов («усы» согласно среднеквадратической ошибке).
Фиг. 20А. Данные вложенной кПЦР с использованием SYBR Green в клетках HeLa, обработанных «HIF-ASO 6» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 20B. Данные вложенной кПЦР с использованием зонда TaqMan в клетках HeLa, обработанных «HIF-ASO 6» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 21A. Данные электрофореза продуктов HIF-1α-специфической вложенной кПЦР в клетках HeLa, обработанных «HIF-ASO 1» в концентрации 0 (отрицательный контроль), 1, 3, 10, 30 или 100 аМ (слева)); и данные секвенирования по Сэнгеру ПЦР-продукта, приписываемого пропуску экзонов 2-3 (справа).
Фиг. 21B. Данные Вестерн-блоттинга HIF-1α в клетках HeLa, обработанных «HIF-ASO 1» в течение 72 часов в концентрации 0 цМ (отрицательный контроль), 100 цМ, 300 цМ, 1 аМ, 3 аМ, 10 аМ, 30 аМ, 100 аМ или 300 аМ.
Фиг. 21C. Уровни экспрессии HIF-1α, приведенные к β-актину, в клетках HeLa, обработанных «HIF-ASO 1» в течение 72 часов в концентрации 0 цМ (отрицательный контроль), 100 цМ, 300 цМ, 1 аМ, 3 аМ, 10 аМ, 30 аМ, 100 аМ или 300 аМ.
Фиг. 22А. Данные HIF-1α-специфической вложенной кПЦР, полученные в клетках HeLa, обработанных «HIF-ASO 12» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ (слева), вместе с данными секвенирования по Сэнгеру полосы для пропуска экзонов (справа).
Фиг. 22B. Данные Вестерн-блоттинга HIF-1α в клетках HeLa, обработанных «HIF-ASO 12» в концентрации 0 (отрицательный контроль), 0,01, 0,1, 1 или 10 аМ.
Фиг. 22C. Данные HIF-1α-специфической вложенной кПЦР, полученные в клетках HeLa, обработанных «HIF-ASO 12» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 23А. Электрофоретический анализ продуктов AR-специфической вложенной ПЦР в клетках MCF7, обработанных «AR-ASO 1» в течение 3 часов в концентрации 0 (отрицательный контроль), 3, 30, 300 или 3000 аМ.
Фиг. 23B. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов 4-5.
Фиг. 23C. Данные Вестерн-блоттинга AR в клетках MCF7, обработанных «AR-ASO 1» в течение 48 часов в концентрации 0 цМ (отрицательный контроль, т.е. N/C), 10 цМ, 30 цМ, 100 цМ, 300 цМ, 1 аМ, 3 аМ, 10 аМ или 30 аМ.
Фиг. 24А. Данные кПЦР с использованием SYBR Green для уровней экзонов 4-6 AR в клетках MCF7, обработанных «AR-ASO 1» в течение 5 часов в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 24B. Данные кПЦР с использованием SYBR Green для уровней экзонов 4-6 AR в клетках MCF7, обработанных «AR-ASO 5» в течение 5 часов в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 24C. Данные кПЦР с использованием анализа TaqMan для мРНК AR в клетках MCF7, обработанных «AR-ASO 5» в течение 24 часов в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 25А. Необработанные данные Вестерн-блоттинга, полученные в случае кожи в месте инъекции. NC, 1p, 10p, 100p и 1000p относятся к группе отрицательного контроля, группе лечения 1, 10, 100 и 1000 пмоль/кг ASO, соответственно.
Фиг. 25B. Необработанные данные Вестерн-блоттинга, полученные в случае кожи в месте без инъекции. NC, 1p, 10p, 100p и 1000p относятся к группе отрицательного контроля, группе лечения 1, 10, 100 и 1000 пмоль/кг ASO, соответственно.
Фиг. 26А. Уровень экспрессии белка AR по группам, а также по субъектам в месте инъекции (слева) и в месте без инъекции (справа) (** для р<0,01 и * для р<0,05)
Фиг. 26B. Средний уровень экспрессии белка AR по группам в месте инъекции (слева) и в месте без инъекции (справа). (** для р<0,01 и * для р<0,05)
Фиг. 27А. Электрофоретический анализ продуктов AR- специфической вложенной ПЦР в клетках MCF7, обработанных «AR-ASO 1» в течение 3 часов в концентрации 0 (отрицательный контроль), 30, 100 или 1000 аМ.
Фиг. 27B. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзона 5.
Фиг. 28А. Электрофоретический анализ продуктов SCN9A-специфической вложенной ПЦР в клетках PC3, обработанных «SCN-ASO 7» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ.
Фиг. 28B. Данные секвенирования по Сэнгеру продуктов вложенной ПЦР, приписываемых пропуску экзона 4 (вверху) и экзонов 4-5 (внизу), соответственно.
Фиг. 29А. Данные SCN9A-специфической вложенной кПЦР в клетках РС3, обработанных «SCN-ASO 7» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 29B. Данные SCN9A-специфической вложенной кПЦР в клетках РС3, обработанных «SCN-ASO 3» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 29C. Данные SCN9A-специфической вложенной кПЦР в клетках РС3, обработанных «SCN-ASO 8» в течение 24 часов в концентрации 0 (отрицательный контроль), 10 или 100 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 30А. Результаты анализа с использованием CoroNa в клетках PC3, обработанных «SCN-ASO 7» в течение 30 часов в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
Фиг. 30B. Результаты анализа с использованием CoroNa в клетках PC3, обработанных «SCN-ASO 3» в течение 30 часов в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
Фиг. 30C. Результаты анализа с использованием CoroNa в клетках PC3, обработанных «SCN-ASO 8» в течение 30 часов в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
Фиг. 31А. Данные электрофореза продуктов SCN9A-специфической вложенной ОТ-ПЦР в клетках PC3, обработанных «ASO 27» в течение 24 часов в концентрации 0 (отрицательный контроль), 10 или 100 цМ.
Фиг. 31B. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов 4-5.
Фиг. 31C. Данные электрофореза продуктов SCN9A-специфической ОТ-ПЦР в клетках РС3, обработанных «ASO 27» в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 аМ.
Фиг. 32А. Данные SCN9A-специфической кПЦР путем одностадийного синтеза кДНК в клетках РС3, обработанных «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 0,1, 1 или 10 аМ в течение 24 часов («усы» согласно среднеквадратической ошибке).
Фиг. 32B. Данные SCN9A-специфической кПЦР путем синтеза кДНК с использованием случайного гексамера в клетках РС3, обработанных «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 0,1, 1 или 10 аМ в течение 24 часов («усы» согласно среднеквадратической ошибке).
Фиг. 33A. Записи интенсивности флуоресценции в среднем клеток DRG L5 крысы (стимулированных перевязкой L5/L6), обработанных «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
Фиг. 33B. Записи интенсивности флуоресценции в среднем клеток DRG L5 крысы (без перевязки L5/L6), обработанных «SCN-ASO 27» при 0 (отрицательный контроль), 100 или 1000 цМ.
Фиг. 34A. Данные Вестерн-блоттинга для экспрессии белка Nav1.7 в нервных клетках DRG (стимулированных перевязкой L5/L6), обработанных «SCN-ASO 30» в течение 24 часов в концентрации 0 (т.е. отрицательный контроль), 10, 100 или 1000 цМ.
Фиг. 34B. Натриевый ток согласно анализу с использованием патч-клампа вручную в нервных клетках DRG (стимулированных перевязкой L5/L6), обработанных «SCN-ASO 30» в течение 4 часов в концентрации 0 (отрицательный контроль) и 100 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 35А. Данные SCN9A-специфической кПЦР путем одностадийного синтеза кДНК в нервных клетках DR5 L5 крысы, обработанных «SCN-ASO 30» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 30, 100 или 300 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 35В. Данные SCN9A-специфической кПЦР путем синтеза кДНК с использованием случайного гексамера в нервных клетках DRG L5, обработанных «SCN-ASO 30» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 30, 100 или 300 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 36. Обратное развитие аллодинии, вызванной DPNP, у крыс, которым подкожно вводили носитель (PBS, отрицательный контроль), «SCN-ASO 7» 100 пмоль/кг, «SCN-ASO 8» 100 пмоль/кг, «SCN-ASO 21» «100 пмоль/кг», «SCN-ASO 35» 100 пмоль/кг, «SCN-ASO 36» 100 пмоль/кг или «SCN-ASO 37» 100 пмоль/кг («усы» согласно среднеквадратической ошибке).
Фиг. 37А. Данные электрофореза продуктов вложенной ПЦР (способ А), полученных с мышечными тканями от мышей MDX, которым подкожно вводили только с носителем (отрицательный контроль), 1000 пмоль/кг «DMD-ASO 1» или 1000 пмоль/кг «DMD-ASO 4», 2 раза в день (BID) в течение 3 дней.
Фиг. 37B. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзона 23.
Фиг. 38А. Данные электрофореза продуктов вложенной ПЦР (способ В), полученных с мышечными тканями от мышей MDX, которым подкожно вводили только носитель (отрицательный контроль), 1000 пмоль/кг «DMD-ASO 1» или 1000 пмоль/кг «DMD-ASO 4», 2 раза в день (BID) в течение 3 дней.
Фиг. 38B. Данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов 21-23.
Фиг. 38C. Данные электрофореза продуктов вложенной ПЦР (способ А), полученных с образцами трицепса у мышей mdx, которым подкожно вводили «DMD-ASO 1» в дозе 0 (отрицательный контроль) или 10 пмоль/кг, BID в течение 5 дней.
Фиг. 39А. Оценки в тесте с вращающимся стержнем у мышей mdx, получавших носитель (отрицательный контроль), 100 пмоль/кг «DMD-ASO 1» или 1000 пмоль/кг «DMD-ASO 1» (усы» согласно среднеквадратической ошибке и * для p <0,05)
Фиг. 39B. Оценки силы захвата у мышей mdx, которым постоянно вводили «DMD-ASO 1» в дозе 0 (отрицательный контроль), 10, 50 или 200 пмоль/кг («усы» согласно среднеквадратической ошибке и * для p<0,05)
Фиг. 40. IHC-изображения полноразмерного дистрофина, объединенные с окрашиванием DAPI, в мышечной ткани мышей mdx, которым вводили «DMD-ASO 1» в дозе 0 (отрицательный контроль) или 200 пмоль/кг, 2 раза в неделю в течение 30 недель.
Фиг. 41. Относительные уровни экспрессии полноразмерного белка дистрофина в скелетных мышцах мышей mdx, которым постоянно вводили «DMD-ASO 1» в дозе 0 (отрицательный контроль), 10, 50 или 200 пмоль/кг. Уровень экспрессии приведен к уровню экспрессии у мышей WT («усы» согласно среднеквадратической ошибке, * для p<0,05 и ** p<0,01)
Фиг. 42. Данные электрофореза для продуктов вложенной ПЦР, полученных с образцами скелетных мышц, отобранных у мышей mdx на неделе 7.
Фиг. 43. Гистопатологические изменения согласно окрашиванию H&E трицепса мышей C57BL/6 (отрицательный контроль WT) и мышей mdx, которым постоянно вводили «DMD-ASO 1» в дозе 0 (отрицательный контроль mdx), 10, 50 или 200 пмоль/кг.
Фиг. 44А. Пройденные расстояния на беговой дорожке у мышей C57BL/6 (отрицательный контроль WT) и мышей mdx, постоянно подвергавшихся воздействию «DMD ASO2» в дозе 0 (отрицательный контроль mdx), 10 пмоль/кг или 30 пмоль/кг («усы» согласно среднеквадратической ошибке, * для p<0,05, ** для p<0,01 и *** для p<0,001)
Фиг. 44B. Уровни креатинкиназы в сыворотке у мышей C57BL/6 (отрицательный контроль WT) и мышей mdx, постоянно подвергавшихся воздействию «DMD ASO2» в дозе 0 (отрицательный контроль mdx), 10 пмоль/кг или 30 пмоль/кг («усы» согласно среднеквадратической ошибке, ** для p<0,01 и **** для p<0,0001)
Фиг. 44C. Уровни миоглобина в сыворотке у мышей C57BL/6 (отрицательный контроль WT) и мышей mdx, постоянно подвергавшихся воздействию «DMD ASO2» в дозе 0 (отрицательный контроль mdx), 10 пмоль/кг или 30 пмоль/кг («усы» согласно среднеквадратической ошибке, *** для p <0,001 и **** для p <0,0001)
Фиг. 45A. Данные Вестерн-блоттинга с исследованием на наличие полноразмерных дистрофинов в образцах скелетных мышц от мышей дикого типа (отрицательный контроль WT) или мышей mdx, постоянно подвергавшихся воздействию «DMD-ASO 2» в дозе 0 (отрицательный контроль mdx), 10 или 30 пмоль/кг.
Фиг. 45B. Уровни креатинкиназы в сыворотке у мышей WT (отрицательный контроль WT), мышей mdx без воздействия ASO и мышей mdx, которым подкожно вводили 50 ммоль/кг «DMD-ASO 1», 10 ммоль/кг «DMD-ASO 2» или 10 мкмоль/кг «DMD-ASO 6», 2 раза в неделю в течение 66 недель («усы» согласно среднеквадратической ошибке и ** для р<0,01)
Фиг. 45C. Уровни миоглобина в сыворотке у мышей WT (отрицательный контроль WT), мышей mdx без воздействия ASO и мышей mdx, которым подкожно вводили 50 ммоль/кг «DMD-ASO 1», 10 ммоль/кг «DMD-ASO 2» или 10 мкмоль/кг «DMD-ASO 6», 2 раза в неделю в течение 66 недель («усы» согласно среднеквадратической ошибке, * для p<0,05 и *** для p<0,001)
Фиг. 46A. Данные электрофореза для продуктов IDO-1-специфической вложенной ПЦР в клетках SKOV3, обработанных «IDO-ASO 1» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ (левая диаграмма), и данные секвенирования по Сэнгеру полосы для ПЦР-продукта в случае пропуска экзонов (правая диаграмма).
Фиг. 46B. Результаты анализа секреции кинуренина в клетках SKOV3, обработанных «IDO-ASO 1» в концентрации 0 цМ (отрицательный контроль) или от 10 цМ до 1 фМ («усы» согласно среднеквадратической ошибке, и * для p<0,05)
Фиг. 47A. Данные электрофореза продуктов IDO-1- специфической вложенной ПЦР в клетках SKOV3, обработанных «IDO-ASO 5» в концентрации 0 (отрицательный контроль), 1, 3, 10, 30 или 100 аМ.
Фиг. 47B. Данные секвенирования по Сэнгеру полос для ПЦР-продуктов, приписываемых пропуску экзонов 2-4 и экзонов 2-6.
Фиг. 47C. Данные электрофореза продуктов IDO-1-специфической вложенной ПЦР в клетках SKOV3, обработанных «IDO-ASO 6» в концентрации 0 (отрицательный контроль), 1, 3, 10, 30 или 100 аМ (диаграмма слева), а также данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов 2-5 (правая диаграмма).
Фиг. 48А. Электрофоретический анализ продуктов SNAP25-специфической вложенной ПЦР в клетках PC12, обработанных 0 (отрицательный контроль), 10, 100 или 1000 цМ «SNAP-ASO 3» (левая диаграмма), и данные секвенирования полосы, относящейся к ПЦР-продукту в случае пропуска экзонов 5-7.
Фиг. 48B. Изменения уровня полноразмерной мРНК для SNAP25 крысы в клетках РС12, обработанных «SNAP-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 48C. Изменения уровня полноразмерной мРНК для SNAP25 крысы в клетках РС12, обработанных «SNAP-ASO 1» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ («усы» согласно среднеквадратической ошибке).
Фиг. 49A. Данные Вестерн-блоттинга SNAP25 (верхняя диаграмма) и относительные уровни экспрессии SNAP25, приведенные к β-актину, (нижняя диаграмма) в клетках РС12, обработанных «SNAP-ASO 3» в течение 48 часов в концентрации 0 цМ (отрицательный контроль), 1 цМ, 10 цМ, 30 цМ, 100 цМ, 300 цМ, 1 аМ, 3 аМ или 10 аМ.
Фиг. 49B. Данные Вестерн-блоттинга SNAP25 в клетках РС12, обработанных «SNAP-ASO 1» в концентрации 0 (отрицательный контроль), 0,1 или 1 аМ либо в течение 48 часов, либо в течение 72 часов.
Фиг. 50А. Данные Вестерн-блоттинга SNAP25 (верхняя диаграмма) и относительные уровни экспрессии SNAP25, приведенные к β-актину, (нижняя диаграмма) в клетках SiMa, обработанных «SNAP-ASO 3» в течение 48 часов в концентрации 0 цМ (отрицательный контроль), 1 цМ, 10 цМ, 100 цМ, 1 аМ, 10 аМ или 100 аМ.
Фиг. 50B. Изменения уровня полноразмерной мРНК для SNAP25 человека в клетках SiMa, обработанных «SNAP-ASO 3» в концентрации 0 цМ (отрицательный контроль), 1 цМ, 10 цМ, 100 цМ, 1 аМ, 10 аМ или 100 аМ («усы» согласно среднеквадратической ошибке)
Фиг. 51. IHC-изображения SNAP25 для образцов кожи мышей, которым местно вводили «SNAP-ASO 1» в концентрации 0 (отрицательный контроль), 1, 10 или 100 фМ, BID в течение 4 дней.
Фиг. 52А. Электрофоретический анализ продуктов вложенной ПЦР в клетках меланомы мыши B16F10, обработанных «TYR-ASO 4» в концентрации 0 (отрицательный контроль), 1, 10 или 1000 аМ.
Фиг. 52B. Секвенирование по Сэнгеру ПЦР-продукта, приписываемого пропуску экзонов 2-3.
Фиг. 52C. Изменения уровня полноразмерной мРНК для TYR согласно кПЦР в клетках меланомы мыши B16F10, обработанных «TYR-ASO 4» в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 аМ («усы» согласно среднеквадратической ошибке)
Фиг. 53A. Данные Вестерн-блоттинга TYR в клетках B16F10, обработанных «TYR-ASO 4» в течение 24 часов в концентрации 0 (отрицательный контроль), 0,01, 0,1, 1 или 10 аМ.
Фиг. 53B. Изменения содержания меланина в клетках меланомы мыши B16F10, обработанных либо «TYR-ASO 4» в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 аМ, либо 10 мкг/мл или 100 мкг/мл арбутина («усы» согласно среднеквадратической ошибке, * для p<0,05, ** для p<0,01 и *** для p<0,001)
Фиг. 53C. Изменения уровня полноразмерной мРНК для TYR согласно кПЦР в первичных меланоцитах эпидермиса человека, обработанных «TYR-ASO 1» в концентрации 0 цМ (отрицательный контроль), 1 цМ, 100 цМ или 10 аМ («усы» согласно среднеквадратической ошибке)
Фиг. 54А. Электрофоретический анализ продуктов вложенной ПЦР в клетках Jurkat, обработанных «PD-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ.
Фиг. 54B. Секвенирование по Сэнгеру ПЦР-продуктов, приписываемых пропуску экзона 2 (слева) и экзона 3 (справа), соответственно.
Фиг. 55А. Изменения уровня мРНК для PD-1 человека согласно вложенной КПЦР в клетках Jurkat, обработанных «PD-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ («усы» согласно среднеквадратической ошибке, ** для p<0,01 и * для p<0,05)
Фиг. 55B. Изменения уровня мРНК для IL-2 человека согласно кПЦР в клетках Jurkat, обработанных «PD-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ («усы» согласно среднеквадратической ошибке, ** для p<0,01 и * для p<0,05)
Фиг. 56A. Изменения уровня мРНК для PD-1 человека согласно вложенной КПЦР в клетках Jurkat, обработанных «PD-ASO 1» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ. («усы» согласно среднеквадратической ошибке и * для p<0,05)
Фиг. 56B. Ингибирование роста меланомы В16F10 у мышей C57BL/6, которым подкожно вводили «PD-ASO 2» в дозе 2, 10 или 50 пмоль/кг, 2 раза в неделю («усы» согласно среднеквадратической ошибке и * для р<0,05)
Краткое изложение сущности настоящего изобретения
Настоящим изобретением обеспечивается производное пептидо-нуклеиновой кислоты, представленное формулой I, или его фармацевтически приемлемая соль:
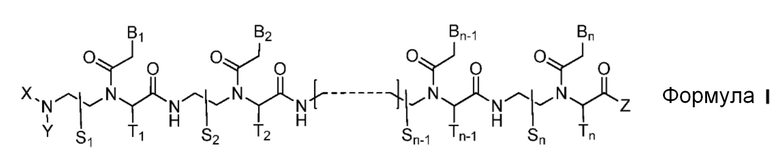
где
n является целым числом между 10 и 25;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn независимо представляют собой дейтеридо [D], гидридо [H], замещенный или незамещенный алкил или замещенный или незамещенный арил;
радикалы X и Y независимо представляют собой гидридо, формил [H-C(=O)-], аминокарбонил [NH2-C(=O)-], аминотиокарбонил [NH2-C(=S)-], замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный арилоксикарбонил, замещенный или незамещенный алкиламинокарбонил, замещенный или незамещенный ариламинокарбонил, замещенный или незамещенный алкиламинотиокарбонил, замещенный или незамещенный ариламинотиокарбонил, замещенный или незамещенный алкилокситиокарбонил, замещенный или незамещенный арилокситиокарбонил, замещенный или незамещенный алкилсульфонил, замещенный или незамещенный арилсульфонил, замещенный или незамещенный алкилфосфонил или замещенный или незамещенный арилфосфонил;
радикал Z представляет собой гидридо, гидрокси, замещенный или незамещенный алкилокси, замещенный или незамещенный арилокси, незамещенный амино [-NH2], замещенный или незамещенный алкиламино, замещенный или незамещенный ариламино, замещенный или незамещенный алкил или замещенный или незамещенный арил;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований; и
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований с замещенным или незамещенным аминорадикалом, ковалентно связанным с фрагментом нуклеотидного основания.
В некоторых вариантах осуществления соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, которая состоит из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не представляет собой [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
В некоторых вариантах осуществления соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с сайтом сплайсинга-мишенью в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не включает [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
Соединение формулы I эффективно индуцирует пропуск экзона-мишени пре-мРНК-мишени, дает вариант(ы) сплайсинга мРНК, в котором отсутствует экзон-мишень, и, следовательно, применимо для модулирования функциональной активности гена, с которого идет транскрибирование пре-мРНК-мишени.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящим изобретением обеспечивается производное пептидо-нуклеиновой кислоты, представленное формулой I, или его фармацевтически приемлемая соль:

где
n является целым числом между 10 и 25;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn независимо представляют собой дейтеридо [D], гидридо [H], замещенный или незамещенный алкил или замещенный или незамещенный арил;
радикалы X и Y независимо представляют собой гидридо, формил [H-C(=O)-], аминокарбонил [NH2-C(=O)-], аминотиокарбонил [NH2-C(=S)-], замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный арилоксикарбонил, замещенный или незамещенный алкиламинокарбонил, замещенный или незамещенный ариламинокарбонил, замещенный или незамещенный алкиламинотиокарбонил, замещенный или незамещенный ариламинотиокарбонил, замещенный или незамещенный алкилокситиокарбонил, замещенный или незамещенный арилокситиокарбонил, замещенный или незамещенный алкилсульфонил, замещенный или незамещенный арилсульфонил, замещенный или незамещенный алкилфосфонил или замещенный или незамещенный арилфосфонил;
радикал Z представляет собой гидридо, гидрокси, замещенный или незамещенный алкилокси, замещенный или незамещенный арилокси, незамещенный амино [-NH2], замещенный или незамещенный алкиламино, замещенный или незамещенный ариламино, замещенный или незамещенный алкил или замещенный или незамещенный арил;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований; и
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований с замещенным или незамещенным аминорадикалом, ковалентно связанным с фрагментом нуклеотидного основания.
В некоторых вариантах осуществления соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, которая состоит из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не представляет собой [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
В некоторых вариантах осуществления соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с сайтом сплайсинга-мишенью в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не включает [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
Соединение формулы I эффективно индуцирует пропуск экзона-мишени пре-мРНК-мишени, дает вариант(ы) сплайсинга мРНК, в котором отсутствует экзон-мишень, и, следовательно, применимо для модулирования функциональной активности гена, с которого идет транскрибирование пре-мРНК-мишени.
Принятое для описания соединения формулы I условие, что «n» является целым числом между 10 и 25», буквально означает, что «n» представляет собой целое число, выбираемое из 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23 и 24».
Было подсчитано, что во всем геноме человека будет 26564 гена [In Silico Biol. vol 4, 387-393 (2004)]. Учитывая, что в среднем в каждом гене имеется приблизительно 8,8 экзонов и 7,8 интронов, в геноме человека возможно 368122 сайтов сплайсинга [{(8,2×2) - 2} x 25564=368122]. Поскольку существует 1048576 (т.е. 410) возможных последовательностей для 10-мерной пре-мРНК, даже 10-мерные производные ПНК будут демонстрировать достаточный уровень специфичности для сайта сплайсинга-мишени. Однако 5'-конец и 3'-конец каждого интрона являются высококонсервативными, чтобы иметь последовательность, начинающуюся с [(5'→3')│GU-], и последовательность, заканчивающуюся [(5'→3')-AG│], соответственно, где «│» обозначает место соединения интрон-экзон или экзон-интрон. Поэтому прогнозируется, что соединение формулы I с длиной олигомера, равной 11 мер или более, (т.е. n представляет собой целое число больше 10) будет демонстрировать достаточный уровень специфичности для сайта сплайсинга-мишени.
Соединение формулы I прочно связывается с комплементарной нуклеиновой кислотой, как показано в известном уровне техники [PCT/KR2009/001256]. Например, включение от 4 до 5 модифицированных (т.е. неприродных или неприродных) нуклеотидных оснований в 11-13-мерные производные ПНК формулы I легко дает увеличение Tm при образовании дуплекса с комплементарной ДНК на 20οC или больше. Соединение настоящего изобретения обладает сильным сродством к комплементарной РНК, как и к комплементарной ДНК. Таким образом, предпочтительно, чтобы соединение этого изобретения было как можно более коротким во избежание эффектов вне мишени, возникающих в результате связывания указанного соединения с другими последовательностями пре-мРНК с небольшим количеством несоответствий (ошибочных спариваний оснований). Таким образом, длина олигомера указанного соединения ограничена короче 25-мера.
Соединение формулы I очень чувствительно к несоответствию по одному основанию, как показано в предшествующем уровне техники [PCT/KR2009/001256]. Например, несоответствие по одному основанию приводило к уменьшению Tm на 11-22οC в зависимости от типа модифицированного основания, а также последовательности ПНК. Однако из-за сильного сродства к РНК соединение этого изобретения все еще прочно связывается с последовательностью сайта сплайсинга-мишени, имеющей одно или два несоответствия, и эффективно индуцирует пропуск экзона-мишени.
Соединение формулы I прочно связывается либо с 3'-сайтом сплайсинга, либо с 5'-сайтом сплайсинга в пре-мРНК-мишени, в зависимости от его последовательности.
В случае, когда соединение связывается с 3'-сайтом сплайсинга, указанное соединение обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью в 3'-сайте сплайсинга-мишени, состоящей из 7-мера из интрона-мишени и 7-мера из экзона-мишени. Таким образом, 3'-сайт сплайсинга однозначно определяется как место соединения 3'-конца интрона-мишени и 5'-конца экзона-мишени.
В случае, когда соединение связывается с 5'-сайтом сплайсинга, указанное соединение обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью в 5'-сайте сплайсинга-мишени, состоящей из 7-мера из экзона-мишени и 7-мера из интрона-мишени. Таким образом, 5'-сайт сплайсинга однозначно определяется как место соединения 3'-конца экзона-мишени и 5'-конца интрона-мишени.
14-мерная последовательность, описывающая соединение формулы I, нацеливающееся на 3'-сайт сплайсинга, иллюстрируется 3'-сайтом сплайсинга, охватывающим место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α (индуцируемого гипоксией фактора 1α) человека, считываемой с гена HIF1A человека [ссылочная последовательность NCBI: NG_029606.1]. 40-мерная последовательность 3'-сайта сплайсинга, состоящая из 20-мера из интрона 1 и 20-мера из экзона 2, прочитывается [(5'→3') uucuuguuguuguuaaguag│GAUAAGUUCUGAACGUCGAA], в которой последовательности интрона и экзона обозначены, соответственно, строчными и заглавными буквами, а место соединения интрона 1 и экзона 2 обозначено «│». Таким образом, 14-мерная последовательность 3'-сайта сплайсинга, состоящая из 7-мера из интрона 1 HIF-1α и 7-мера из экзона 2 HIF-1α, прочитывается [(5'→3')uaaguag│GAUAAGU]. В этом 3'-сайте сплайсинга интрон- и экзон-мишени представляют собой интрон 1 и экзон 2 HIF-1α, соответственно.
Вышеуказанная 40-мерная последовательность пре-мРНК была предоставлена для однозначной идентификации 3'-сайта сплайсинга экзона 2 в пре-мРНК HIF-1α человека, поскольку количества экзонов часто варьируют в зависимости от транскриптов мРНК. На протяжении этого изобретения сайт сплайсинга-мишень указанного соединения ПНК однозначно идентифицируется везде, где это применимо, путем одновременного указания номера экзона-мишени и последовательности пре-мРНК, содержащей сайт сплайсинга-мишень.
14-мерная последовательность, описывающая соединение формулы I, нацеливающееся на 5'-сайт сплайсинга, иллюстрируется 5'-сайтом сплайсинга, охватывающим место соединения экзона 2 и интрона 2 в пре-мРНК HIF-1α человека. 40-мерная последовательность 5'-сайта сплайсинга, состоящая из 20-мера из экзона 2 и 20-мера из интрона 2, прочитывается [(5'→3') GAGGAAACUUCUGGAUGCUG│gugaguuauuuuuacacagggu], в которой последовательности экзона и интрона обозначены заглавными и строчными буквами, соответственно, а место соединения экзона 2 и интрона 2 обозначено «│». Таким образом, 14-мерная последовательность 5'-сайта сплайсинга, состоящая из 7-мера из экзона 2 HIF-1α и 7-мера из интрона 2 HIF-1α, прочитывается [(5'→3')GAUGCUG│gugaguu]. В этом 5'-сайте сплайсинга экзон- и интрон-мишени представляют собой экзон 2 и интрон 2 HIF-1α, соответственно.
Соединение формулы I прочно связывается с сайтом сплайсинга-мишенью в пре-мРНК-мишени и препятствует образованию «сплайсомного раннего комплекса», включающего сайт сплайсинга-мишень соединения. Указанное соединение прочно связывается либо с 3'-сайтом сплайсинга, либо с 5'-сайтом сплайсинга в пре-мРНК-мишени в зависимости от нуклеотидной последовательности указанного соединения. Поскольку соединение этого изобретения стерически ингибирует образование «сплайсомного раннего комплекса», экзон-мишень сплайсируется с получением варианта(ов) сплайсинга мРНК, в котором отсутствует экзон-мишень. Следовательно, соединение настоящего изобретения эффективно индуцирует пропуск экзона-мишени.
Химические структуры природных (т.е. встречающихся в природе) или неприродных (т.е. не встречающихся в природе) нуклеотидных оснований, принятых для описания производного ПНК формулы I, приведены в качестве примера на фиг.7. Природные или неприродные нуклеотидные основания этого изобретения включают, но без ограничения ими, нуклеотидные основания, представленные на фиг. 7. Предоставление таких природных или неприродных нуклеотидных оснований является иллюстрацией разнообразия допустимых нуклеотидных оснований и, следовательно, не должно интерпретироваться как ограничение объема настоящего изобретения нуклеотидными основаниями, представленными на фиг. 7. Специалист в данной области техники может легко определить природное нуклеотидное основание, комплементарное каждому из неприродных нуклеотидных оснований, приведенных в качестве примеров на фиг. 7. Поэтому специалист в данной области техники может однозначно определить комплементарность между соединением формулы I и последовательностью мРНК-мишенью.
Принятые для описания производного ПНК формулы I заместители представлены в качестве примеров на фиг. 8А-8Е. На фиг. 8А представлены примеры замещенных или незамещенных алкильных радикалов. Замещенные или незамещенные алкилацильные и замещенные или незамещенные арилацильные радикалы представлены в качестве примеров на фиг. 8В. Фиг. 8С иллюстрирует примеры следующих радикалов: замещенного алкиламино, замещенного ариламино, замещенного или незамещенного арила, замещенного или незамещенного алкилсульфонила, замещенного или незамещенного арилсульфонила, замещенного или незамещенного алкилфосфонила и замещенного или незамещенного арилфосфонила. На фиг. 8D представлены примеры следующих радикалов: замещенного или незамещенного алкилоксикарбонила, замещенного или незамещенного арилоксикарбонила, замещенного или незамещенного алкиламинокарбонила и замещенного или незамещенного ариламинокарбонила. На фиг. 8E представлены примеры следующих радикалов: замещенного или незамещенного алкилокситиокарбонила, замещенного или незамещенного алкиламинотиокарбонила, замещенного или незамещенного ариламинотиокарбонила, замещенного или незамещенного алкилокситиокарбонила и замещенного или незамещенного арилокситиокарбонила. Предоставление таких примерных заместителей предназначено для иллюстрации разнообразия допустимых заместителей и, следовательно, не должно интерпретироваться как ограничение объема настоящего изобретения заместителями, приведенными в качестве примеров на фиг. 8А-8Е. Поскольку специалист в данной области может без труда понять, что олигонуклеотидная последовательность является определяющим фактором для специфического для последовательности связывания олигонуклеотида с последовательностью пре-мРНК-мишенью по сравнению с заместителями на N-конце или С-конце, существует больше разнообразных заместителей, допустимых для указанного соединения этого изобретения, чем те заместители, которые представлены в качестве примеров на фиг. 8А-8Е.
Соединение формулы I обладает хорошей клеточной проницаемостью и может быть легко доставлено в клетку в виде «голого» (т.е. без добавления адъюванта(ов) для увеличения доставки в клетку) олигонуклеотида, как показано в предшествующем уровне техники [PCT/KR2009/001256]. Таким образом, соединение этого изобретения эффективно индуцирует пропуск экзона-мишени в пре-мРНК-мишени с получением варианта(ов) сплайсинга мРНК, в котором отсутствует экзон-мишень, в клетках, обработанных соединением формулы I в виде «голого» олигонуклеотида.
Соединение формулы I не требует каких-либо средств или композиций для доставки в клетку, чтобы эффективно индуцировать пропуск экзона-мишени в клетках. В этом отношении соединение настоящего изобретения сильно отличается от других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Учитывая сильное сродство к РНК и хорошую клеточную проницаемость, соединение формулы I легко индуцирует пропуск экзона-мишени в клетках с помощью субфемтомолярной антисмысловой активности. До настоящего времени субфемтомолярная активность антисмыслового пропуска экзонов никогда не описывалась и не реализовывалась с другими классами олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д. Даже субнаномолярная активность антисмыслового пропуска экзонов редко документировалась с другими классами олигонуклеотидов. Субнаномолярная активность антисмыслового пропуска экзонов описана в случае ПНК ASO, сконструированных так, что они имеют различное количество фосфонатных групп, ковалентно конъюгированных с N-концом последовательности ПНК для облегчения липофекции для трансфекции в клетку [Nucl. Acids Res. vol 30(13), 4424-4432 (2008)]. Как указывалось ранее в этом документе, активность антисмыслового пропуска экзонов in vitro является наномолярной или микромолярной даже в условиях принудительной доставки в клетку, такой как липофекция, электропорация и т.д. В этом отношении соединение формулы I явно отличается от других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Для того чтобы молекула олигонуклеотида связывалась с комплементарной ей последовательностью в пре-мРНК, эта молекула должна быть растянута или развернута для комплементарного связывания с последовательностью пре-мРНК-мишенью. Молекулы олигонуклеотидов, как правило, агрегируют или остаются свернутыми (например, как шпилька) из-за их высокой склонности к образованию межмолекулярных или внутримолекулярных водородных связей между нуклеотидными основаниями. Таким образом, будет существовать дополнительный энергетический барьер развертывания против антисмыслового пропуска экзона с помощью широко исследованных олигонуклеотидов, включая ДНК, РНК, PTO, 2'-ОМе-PTO, 2'-ОМе-РНК, 2'-ОМОЕ-РНК, LNA, PMO, ПНК и т.д. Олигонуклеотиды обычно количественно определяли путем УФ-абсорбции после инкубации при >90οС в водном буфере, чтобы максимально развернуть молекулы олигонуклеотидов.
Производное ПНК формулы I обладает групповыми положительными зарядами, распределенными по всей олигонуклеотидной цепи, при физиологическом рН благодаря нескольким основным аминогруппам, ковалентно связанным с модифицированными нуклеотидными основаниями в нем. Групповые положительные заряды позволяют соединению формулы I оставаться развернутым или вытянутым из-за электростатического отталкивания между соседними положительными зарядами на одной и той же олигонуклеотидной цепи. Производное формулы I имеет слабую склонность к агрегации с другой молекулой(ами) формулы I. Таким образом, соединение формулы I, как правило, остается структурно готовым (т.е. вытянутым) для комплементарного связывания с последовательностью-мишенью в пре-мРНК-мишени. Структурная готовность также важна для быстрого совмещения олигонуклеотида формулы I с последовательностью пре-мРНК-мишенью по мере транскрибирования пре-мРНК-мишени с ДНК. Таким образом, считается, что структурная готовность в сочетании с сильным сродством сводятся к сильному сродству с получением субфемтомолярной активности антисмыслового пропуска экзона соединения формулы I. В этом отношении соединение формулы I сильно отличается от других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Благодаря хорошей клеточной проницаемости, производное ПНК формулы I может системно вводиться в виде «голого» олигонуклеотида, чтобы эффективно индуцировать пропуск экзона в ткани(ях)-мишени. Соединение формулы I не требует композиции или адъюванта для увеличения доставки в ткань-мишень для вызова желаемой терапевтической активности. Соединение формулы I растворяют просто в PBS (фосфатно-солевом буфере) или физиологическом растворе и системно вводят, чтобы без усилий вызвать терапевтическую активность в ткани(ях)-мишени.
Учитывая субфемтомолярную экзон-пропускающую активность в клетках, обработанных в виде «голого» олигонуклеотида, производное ПНК настоящего изобретения проявляет терапевтическую активность in vivo часто в системной дозе, составляющей 1 мкг/кг или менее. Такая сильная терапевтическая активность никогда не была получена с другими классами олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д. Поскольку стоимость производства олигонуклеотида является, как правило, очень высокой, сверхвысокая активность является большим преимуществом для реализации умеренной стоимости лечения, особенно для пациентов с хроническим заболеванием. В этом отношении соединение формулы I сильно отличается от других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Благодаря хорошей клеточной проницаемости производное ПНК настоящего изобретения легко доставляется местно или трансдермально для вызова терапевтической активности в месте введения. Соединение этого изобретения не нуждается в сильной или инвазивной композиции для вызова намеченной местной терапевтической активности. Производное ПНК формулы I легко доставляется трансдермально в виде «голого» олигонуклеотида. Вследствие сверхсильной экзон-пропускающей активности указанное соединение проявляет терапевтическую активность при местном или трансдермальном введении субпикомолярного раствора олигонуклеотидов. Местная или трансдермальная доставка в виде «голого» олигонуклеотида была чрезвычайно сложной задачей для других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д. В этом отношении соединение формулы I явно отличается от других классов олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Соединение формулы I может использоваться в сочетании с фармацевтически приемлемой кислотой или основанием, включая, но без ограничения этим, гидроксид натрия, гидроксид калия, соляную кислоту, метансульфоновую кислоту, лимонную кислоту, трифторуксусную кислоту и т.д.
Производное ПНК формулы I или его фармацевтически приемлемая соль может вводиться субъекту в комбинации с фармацевтически приемлемым адъювантом, включая, но без ограничения этим, лимонную кислоту, соляную кислоту, винную кислоту, стеариновую кислоту, полиэтиленгликоль, полипропиленгликоль, этанол, изопропанол, бикарбонат натрия, дистиллированную воду, консервант(ы) и т.д.
Соединение настоящего изобретения можно системно вводить субъекту в терапевтически эффективной дозе в диапазоне от 1 фмоль/кг до более чем 1 нмоль/кг, которая может варьировать в зависимости от схемы дозирования, условий или ситуаций субъекта и т.д. Соединение настоящего изобретения можно местно вводить субъекту в терапевтически эффективной концентрации в диапазоне от 1 аМ до более чем 1 нМ, которая может варьировать в зависимости от схемы дозирования, условий или ситуаций субъекта и т.д.
Предпочтительным является производное ПНК формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 10 и 25;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn независимо представляют собой дейтеридо, гидридо, замещенный или незамещенный алкил или замещенный или незамещенный арил;
радикалы X и Y независимо представляют собой гидридо, формил, аминокарбонил, аминотиокарбонил, замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный арилоксикарбонил, замещенный или незамещенный алкиламинокарбонил, замещенный или незамещенный ариламинокарбонил, замещенный или незамещенный алкиламинотиокарбонил, замещенный или незамещенный ариламинотиокарбонил, замещенный или незамещенный алкилокситиокарбонил, замещенный или незамещенный арилокситиокарбонил, замещенный или незамещенный алкилсульфонил, замещенный или незамещенный арилсульфонил, замещенный или незамещенный алкилфосфонил или замещенный или незамещенный арилфосфонил;
радикал Z представляет собой гидридо, гидрокси, замещенный или незамещенный алкилокси, замещенный или незамещенный арилокси, незамещенный амино, замещенный или незамещенный алкиламино, замещенный или незамещенный ариламино, замещенный или незамещенный алкил или замещенный или незамещенный арил;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV:
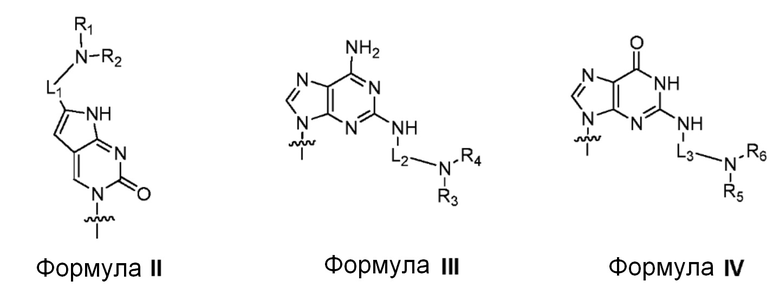
где
радикалы R1, R2, R3, R4, R5 и R6 независимо выбирают из гидридо и замещенного или незамещенного алкила;
L1, L2 и L3 представляют собой ковалентный линкер, представленный формулой V, ковалентно связывающий основную аминогруппу с фрагментом нуклеотидного основания:
где
Q1 и Qm являются замещенным или незамещенным метиленовым (-CH2-) радикалом, и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из замещенного или незамещенного метиленового радикала, радикала кислорода (-O-), радикала серы (-S-) и замещенного или незамещенного аминорадикала [-N(H)- или -N(заместитель)-]; и
m является целым числом между 1 и 15.
Неприродные нуклеотидные основания формулы II, формулы III и формулы IV эквивалентны цитозину, аденину и гуанину, соответственно, для комплементарного спаривания оснований с пре-мРНК, как показано в предшествующем уровне техники [PCT/KR2009/001256].
Принятое для описания соединения формулы I условие, что «m» является целым числом между 1 и 15», буквально означает, что «m» является целым числом, выбираемым из 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13 и 14».
Представляет интерес ПНК-олигомер формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 11 и 23;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn независимо представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо, аминокарбонил, замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный алкиламинокарбонил или замещенный или незамещенный арилсульфонил;
радикал Z представляет собой незамещенный амино или замещенный или незамещенный алкиламино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 независимо выбирают из гидридо и замещенного или незамещенного алкила;
Q1 и Qm являются замещенным или незамещенным метиленовым радикалом, и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из замещенного или незамещенного метиленового радикала, радикала кислорода и аминорадикала; и
m является целым числом между 1 и 11.
Представляет особый интерес производное ПНК формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 11 и 21;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn независимо представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо, замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный алкиламинокарбонил или замещенный или незамещенный арилсульфонил;
радикал Z представляет собой незамещенный амино или замещенный или незамещенный алкиламино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 независимо выбирают из гидридо и замещенного или незамещенного алкила;
Q1 и Qm являются метиленовым радикалом, и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала, радикала кислорода и аминорадикала; и
m является целым числом между 1 и 11.
Представляет большой интерес ПНК-олигомер формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 11 и 19;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо, замещенный или незамещенный алкил, замещенный или незамещенный арил, замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил, замещенный или незамещенный алкилоксикарбонил, замещенный или незамещенный алкиламинокарбонил или замещенный или незамещенный арилсульфонил;
радикал Z представляет собой незамещенный амино или замещенный или незамещенный алкиламино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R3 и R5 представляют собой гидридо, и радикалы R2, R4 и R6 независимо выбирают из гидридо и замещенного или незамещенного алкила;
Q1 и Qm являются метиленовым радикалом, и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 9.
Представляет большой интерес производное ПНК формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 12 и 19;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино или замещенный или незамещенный алкиламино;
B1, B2, …, Bn-1 и Bn независимо выбирают из аденина, тимина, гуанина, цитозина и неприродных нуклеотидных оснований;
по крайней мере пять из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4,R5 и R6 представляют собой гидридо;
Q1 и Qm являются метиленовым радикалом, и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 9.
Представляет наибольший интерес производное ПНК формулы I или его фармацевтически приемлемая соль:
где
n является целым числом между 12 и 18;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикал X представляет собой гидридо;
радикал Y представляет собой замещенный или незамещенный алкилацил, замещенный или незамещенный арилацил или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино или замещенный или незамещенный алкиламино;
B1, B2, …, Bn-1 и Bn независимо выбирают из аденина, тимина, гуанина, цитозина и неприродных нуклеотидных оснований;
по крайней мере пять из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4,R5 и R6 представляют собой гидридо;
L1 представляет собой -(CH2)2-O-(CH2)2-, -CH2-O-(CH2)2-, -CH2-O-(CH2)3-, -CH2-O-(CH2)4-, -CH2-O-(CH2)5-, -CH2-O-(CH2)6- или -CH2-O-(CH2)7-, при этом правый конец непосредственно связан с основной аминогруппой; и
L2 и L3 независимо выбирают из -(CH2)2-, -(CH2)3-, -(CH2)4-, -(CH2)5-, -(CH2)6-, -(CH2)7-, -(CH2)8-, -(CH2)2-O-(CH2)2-, -(CH2)3-O-(CH2)2- и -(CH2)2-O-(CH2)3-, при этом правый конец непосредственно связан с основной аминогруппой.
Соединения формулы I могут обозначаться сокращенно, как описано в предшествующем уровне техники [PCT/KR2009/001256; EP2268607; US8680253]. Ниже приведены примеры таких аббревиатур, использованных для описания производных ПНК формулы I, нацеливающихся на 3'-сайт сплайсинга, охватывающий место соединение интрона 1 и экзона 2 в пре-мРНК HIF-1α человека, считываемой с гена HIF1A (ссылочная последовательность NCBI: NG_029606.1):
(N→C)Fethoc-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-NH2;
(N→C)Fmoc-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-NH2;
(N→C)H-CA(5)G-AA(5)C-TTA(5)-TCC(1O3)-TA(5)-NH2;
(N→C)Ac-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-NH2;
(N→C)Piv-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-NH2;
(N→C)Бензоил-CA(5)G(2O3)-AA(5)C-TTA(4)-TCC(1O2)-TA(5)-NH2;
(N→C)н-пропил-CA(5)G-AA(5)C-TTA(5)-TCC(2O2)-TA(5)-NH2;
(N→C)Бензил-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-NH2;
(N→C)п-толуолсульфонил-CA(5)G-AA(5)C-TTA(2O2)-TCC(1O2)-TA(5)-NH2;
(N→C)[N-(2-фенилэтил)амино]карбонил-CA(5)G(3)-AA(5)C-TTA(3)-TCC(1O2)-TA(5)-NH2;
(N→C)Fethoc-Lys-Leu-CA(5)G(2O2)-AA(5)C-TTA(8)-TCC(1O2)-TA-Lys-NH2;
(N→C)N-фенил-N-Me-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-Lys-NH2;
(N→C)Piv-HEX-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-Lys-NH2;
(N→C)FAM-HEX-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)-Lys-NH2;
(N→C)Fethoc-GA(5)A-C(1O2)TT-A(5)TC-CTA(5)-C(1O2)T-NH2;
(N→C)Fethoc-Arg-GA(5)A-C(1O2)TT-A(5)TC-CTA(5)-C(1O2)T-Gly-NH2;
(N→C)Fethoc-Val-GA(5)A-CTT-A(6)TC-CTA(5)-C(2O2)T-Gly-Lys-NH2;
(N→C)Fethoc-C(1O5)TT-A(6)TC-CTA(6)-C(1O2)TT-AA(6)C-NH2;
(N→C)Fmoc-Val-CTC(1O2)-A(5)TC-CTA(6)-C(1O3)TT-AA(2O2)C-NH2; и
(N→C)Fethoc-TTC(1O5)-AG(5)A-A(4)CT-TA(5)T-CC(2O2)T-A(6)CT-TA(6)-NH2:
где,
A, G, T и C представляют собой мономеры ПНК с природным нуклеотидным основанием: аденином, гуанином, тимином и цитозином, соответственно;
C(pOq), A(p), A(pOq), G(p) и G(pOq) представляют собой мономеры ПНК с неприродным нуклеотидным основанием, представленным формулой VI, формулой VII, формулой VIII, формулой IX и формулой X, соответственно:

где
p и q представляют собой целые числа; и
аббревиатуры для заместителей на N- и C-концах конкретно определены следующим образом: «Fmoc-» - это аббревиатура для «[(9-флуоренил)метилокси]карбонила-»; «Fethoc-» для «[2-(9-флуоренил)этил-1-окси]карбонила»; «Ac-» для «ацетила-»; «Бензоил-» для «бензолкарбонила-»; «Piv-» для «пивалила-»; «н-пропил-» для «1-(н-пропила)-»; «H-» для «гидридо-»группы; «п-толуолсульфонил» для «(4-метилбензол)-1-сульфонила-»; «-Lys-» для аминокислотного остатка «лизин»; «-Val-» для аминокислотного остатка «валин»; «-Leu-» для аминокислотного остатка «лейцин»; «-Arg-» для аминокислотного остатка «аргинин»; «-Gly-» для аминокислотного остатка «глицин»; «[N-(2-фенилэтил)амино] карбонил-» для «[N-1-(2-фенилэтил)амино]карбонила-»; «Бензил-» для «1-(фенил)метила-»; «Фенил-» для «фенила-»; «Me-» для «метила-»; «-HEX-» для «6-амино-1-гексаноила-»; «FAM-» для «5 или 6-флуоресцеин-карбонил-(изомерной смеси)»; и «-NH2» для незамещенной «-амино»группы.
На фиг. 9 в совокупности представлены химические структуры мономеров ПНК, сокращенно обозначенные как A, G, T, C, C(pOq), A(p), A(pOq), G(p) и G(pOq). Как обсуждалось в предшествующем уровне техники [PCT/KR2009/001256], C(pOq) рассматривается как модифицированный мономер ПНК, эквивалентный «цитозину» из-за его предпочтительной гибридизации с «гуанином». A(p) и A (pOq) принимаются за модифицированные мономеры ПНК, действующие как «аденин» из-за их сильного сродства к «тимину». Также G(p) и G(pOq) считаются модифицированными мономерами ПНК, эквивалентными «гуанину» вследствие их продуктивного спаривания оснований с «цитозином».
На фиг. 10 однозначно представлены химические структуры для ряда сокращений заместителей, используемых для разнообразия N-конца или С-конца производного ПНК формулы I в этом изобретении. Предоставление N-концевых или С-концевых групп на фиг. 10 в качестве примеров предназначено для иллюстрации разнообразия допустимых заместителей для N-конца или С-конца производного ПНК формулы I и, следовательно, не должно интерпретироваться как ограничение объема N-концевых или С-концевых групп для соединения этого изобретения. Специалист в данной области техники может легко понять, что олигонуклеотидная последовательность является основным вкладчиком в специфическое для последовательности взаимодействие с пре-мРНК-последовательностью-мишенью.
С целью иллюстрации аббревиатур, принятых для таких производных ПНК, химическая структура 14-мерного производного ПНК, сокращенно обозначенного как «(N→C)Fethoc-GA(5)A-C(1O2)TT-A(5)TC-CTA(5)-C(1O2)T-NH2», представлена на фиг. 11.
В качестве еще одной иллюстрации, химическая структура 15-мерного производного ПНК, сокращенно обозначенного как ((N→C)Fmoc-Val-CTC(1O2)-A(5)TC-CTA(6)-C(1O3)TT-AA(2O2)C-NH2», представлена на фиг. 12.
Соединение формулы I должно удовлетворять требованию обладать способностью к «по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона в пре-мРНК»-мишени. Если соединение формулы I нацеливается, например, на 3'-сайт сплайсинга, охватывающий место соединения интрона 1 и экзона 2, в пре-мРНК для HIF-1α человека, 3'-сайт сплайсинга однозначно определяется 30-мерной последовательностью пре-мРНК для HIF-1α человека [(5'→3')guuguuguuaaguag│GAUAAGUUCUGAACG]. Тогда 14-мерная последовательность являющегося мишенью 3'-сайта сплайсинга HIF-1α прочитывается [(5'→3')uaaguag│GAUAAGU].
15-мерный HIF-1α-специфический ASO с последовательностью «(N→C)Fethoc-CA(5)G-AA(5)C-TTA(5)-TCC(1O2)-TA(5)C-NH2» является эквивалентным последовательности ДНК «(5'→3')CAG-AAC-TTA-TCC-TAC» для комплементарного связывания с пре-мРНК HIF-1α человека. 15-мерный ASO обладает способностью к 15-мерному комплементарному перекрыванию (т.е. является полностью комплементарным) с 3'-сайтом сплайсинга, охватывающим место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК [(5'→3')guuguuguuaaguag│GAUAAGUUCUGAACG]. ПНК ASO обладает способностью к 11-мерному комплементарному перекрыванию (т.е. 4-меров из интрона 1 и 7-меров из экзона 2) с 14-мерной последовательностью пре-мРНК для HIF-1α [(5'→3')uaaguag│GAUAAGU]. Таким образом, 15-мерный специфический для HIF-1α ПНК ASO условиям комплементарного перекрывания, требуемым для соединения формулы I.
Еще один 15-мерный HIF-1α-специфический ASO с последовательностью «(N→C)Fethoc-CTC(1O2)-A(6)TC-CTA(6)-C(1O2)TT-AA(6)C-NH2» является эквивалентным последовательности ДНК «(5'→3')CTC-ATC-CTA-CTT-AAC» для комплементарного связывания с пре-мРНК для HIF-1α. 15-мерный ПНК ASO обладает единственным несоответствием с 3'-сайтом сплайсинга, охватывающим место соединения интрона 1 и экзона 2, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для HIF-1α [(5'→3')guuguuguuaaguag│GAU"A"AGUUCUGAACG], в которой одно несоответствие отмечено знаком кавычки (""). 15-мерная ПНК обладает способностью к 12-мерному комплементарному перекрыванию с 14-мерной последовательностью 3'-сайта сплайсинга, принятой для описания соединения формулы I, как отмечено «жирным шрифтом» и «подчеркиванием» в [(5'→3')uaaguag│GAU"A"AGU], в которой одно несоответствие отмечено знаком кавычки (""). Несмотря на одно несоответствие, 15-мерный HIF-1α-ASO удовлетворяет условиям комплементарного перекрывания, требуемым для соединения формулы I.
В случае, когда соединение формулы I нацеливается, например, на 5'-сайт сплайсинга, охватывающий место соединения экзона 4 и интрона 4 в пре-мРНК для SCN9A человека, 5'-сайт сплайсинга однозначно определяется 30-мерной последовательностью пре-мРНК для SCN9A [(5'→3')CGUCAUUGUUUUUGC│guaaguacuuucagc], считываемой с гена SCN9A человека (доступ к которой получен из ссылочной последовательности NCBI: NC_000002.12). Тогда 14-мерная последовательность 5'-сайта сплайсинга SCN9A-мишени прочитывается[(5'→3')UUUUUGC│guaagua].
16-мерный SCN9A-специфический ASO с последовательностью «(N→C)Fethoc-AC(1O2)T-TA(5)CG(6)CA-A(5)AA(5)-AC(1O2)AA(5)-NH2» является эквивалентным последовательности ДНК «(5'→3')ACT-TAC-GCA-AAA-ACA-A» для комплементарного связывания с пре-мРНК для SCN9A человека. 16-мерная ПНК обладает способностью к 16-мерному комплементарному перекрыванию (т.е. является полностью комплементарной) с 5'-сайтом сплайсинга, охватывающим место соединение экзона 4 и интрона 4 в пре-мРНК для SCN9A человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для SCN9A [(5'→3')CGUCAUUGUUUUUGC│guaaguacuuucagc]. 16-мерный SCN9A-ASO обладает способностью к 13-мерному комплементарному перекрыванию с 14-мерной последовательностью 5'-сайта сплайсинга, как отмечено «жирным шрифтом» и «подчеркиванием» в [(5'→3')UUUUUGC│guaagua]. 16-мерный SCN9A-ASO условиям комплементарного перекрывания, требуемым для соединения формулы I.
ПОДРОБНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Общие процедуры получения ПНК-олигомеров
ПНК-олигомеры синтезировали с помощью твердофазного синтеза пептидов (SPPS) на основе Fmoc-химии в соответствии со способом, описанным в предшествующем уровне техники [патент США 6133444; WO 96/40685] или с незначительными модификациями. Используемым в этом изобретении твердым носителем был H-Rink Amide-ChemMatrix, приобретенный у PCAS BioMatrix Inc. (Quebec, Канада). Мономеры Fmoc-ПНК с модифицированным нуклеотидным основанием синтезировали, как описано в предшествующем уровне техники [PCT/KR 2009/001256] или с незначительными модификациями.
Химические структуры мономеров Fmoc-ПНК с модифицированным нуклеотидным основанием, используемых в этом изобретении, представлены на фиг. 13. Мономеры Fmoc-ПНК, представленные на фиг. 13, должны быть приняты в качестве примеров и, следовательно, не должны приниматься для ограничения объема настоящего изобретения. Специалист в данной области техники может легко понять, что, например, для таких мономеров Fmoc-ПНК, используемых для синтеза производного ПНК формулы I, возможно большое количество вариантов защитных групп.
ПНК-олигомеры очищали с помощью HPLC (ВЭЖХ) с C18-обращенной фазой (вода/ацетонитрил или вода/метанол с 0,1% TFA) и характеризовали с помощью масс-спектрометрии (MS), включая MALDI-TOF/MS и ESI-TOF/MS.
На фиг. 14 представлен типичный цикл удлинения мономера, принятый в SPPS этого изобретения, и ниже приведены подробности синтеза. Однако для специалиста в данной области техники должно представляться явно возможным множество небольших вариаций для проведения таких реакций SPPS в автоматическом синтезаторе пептидов или в ручном синтезаторе пептидов. Соответствующие стадии реакции SPPS представлены ниже в качестве примерных процедур реакции.
[Активация смолы H-Rink-ChemMatrix] 0,01 ммоль (приблизительно 20 мг смолы) смолы ChemMatrix в 1,5 мл 20% пиперидина/диметилформамида (DMF) встряхивали в пробирке Libta Tube® в течение 20 мин, и реакционный раствор отфильтровывали. Смолу промывали в течение 30 сек, последовательно, каждый раз с использованием 1,5 мл метиленхлорида (МС), 1,5 мл DMF, 1,5 мл МС, 1,5 мл DMF и 1,5 мл МС. Полученные в результате свободные аминогруппы на твердом носители подвергались реакции сочетания либо с мономером Fmoc-ПНК, либо с Fmoc-защищенным производным аминокислоты.
[DeFmoc] Смолу встряхивали в 1,5 мл 20% пиперидина/DMF в течение 7 мин, и раствор DeFmoc отфильтровывали. Смолу промывали в течение 30 сек, последовательно, каждый раз с использованием 1,5 мл MC, 1,5 мл DMF, 1,5 мл MC, 1,5 мл DMF и 1,5 мл MC. Полученные в результате свободные аминогруппы на твердом носителе немедленно подвергали реакции сочетания с мономером Fmoc-ПНК.
[Сочетание с мономером Fmoc-ПНК] Свободные аминогруппы на твердом носителе подвергали реакции сочетания с мономером Fmoc-ПНК следующим образом. 0,04 ммоль мономера Fmoc-ПНК, 0,05 ммоль HBTU [2-(1H-бензотриазол-1-ил)-1,1,3,3-тетраметилурония гексафторфосфат] и 10 ммоль DIEA (N,N-диизопропилэтиламин) инкубировали в течение 2 мин в 1 мл безводного DMF и добавляли к смоле со свободными аминогруппами. Раствор смолы встряхивали в течение 1 часа, и реакционную среду отфильтровывали. Затем смолу промывали в течение 30 сек, последовательно, каждый раз с использованием 1,5 мл MC, 1,5 мл DMF и 1,5 мл MC.
[Кэппирование] После реакции сочетания непрореагировавшие свободные аминогруппы подвергали кэппированию встряхиванием в течение 5 мин в 1,5 мл раствора для кэппирования (5% уксусного ангидрида и 6% 2,6-лейтидина в DMF). Затем раствор для кэппирования отфильтровывали, и смолу промывали в течение 30 сек, последовательно, каждый раз с использованием 1,5 мл MC, 1,5 мл DMF и 1,5 мл MC.
[Введение «Fethoc-радикала» в N-конец] «Fethoc-радикал» вводили в N-конец путем взаимодействия свободных аминогрупп в смоле с «Fethoc-OSu» в обычных основных условиях сочетания. Химическая структура «Fethoc-OSu» [CAS № 179337-69-0, C20H17NO5, М.м. 351,36] представляется следующим образом.

[Отщепление от смолы] ПНК-олигомеры, связанные со смолой, отщепляли от смолы путем встряхивания смолы в течение 3 часов в 1,5 мл раствора для расщепления (2,5% триизопропилсилана и 2,5% воды в трифторуксусной кислоте). Смолу отфильтровывали, и фильтрат концентрировали при пониженном давлении или продуванием газообразного азота над раствором. Полученный в результате остаток растирали с диэтиловым эфиром, и полученный осадок собирали фильтрованием для очистки с помощью HPLC c обращенной фазовй.
[Анализ и очистка с помощью HPLC] После отщепления от смолы неочищенный продукт ПНК очищали с помощью HPLC с С18-обращенной фазой, элюируя смесью вода/ацетонитрил или вода/метанол (градиентный способ), содержащей 0,1% TFA. Фиг. 15А и фиг. 15В представляют собой типичные хроматограммы, полученные при HPLC для «HIF-ASO 1» до и после очистки с помощью HPLC, соответственно. Олигомерная последовательность «HIF-ASO 1» представлена в таблице 1.
Примеры синтеза производных ПНК формулы I
Производные ПНК получали в соответствии с описанными выше процедурами синтеза с небольшими модификациями или без них. Производные ПНК этого изобретения сконструированы для нацеливания на сайт сплайсинга в пре-мРНК, включая, но без ограничения этим, пре-мРНК для HIF-1α человека или мыши, пре-мРНК для андрогенового рецептора человека или мыши (AR), пре-мРНК для SCN9A человека или крысы, пре-мРНК для дистрофина мыши, пре-мРНК для тирозиназы человека или мыши, пре-мРНК для SNAP25 человека или мыши, пре-мРНК для IDO1 человека, пре-мРНК для PD-1 человека или мыши и т.д. Предоставление таких производных ПНК, нацеливающихся на сайт сплайсинга в ряде пре-мРНК, является примером производных ПНК формулы I, и его не следует интерпретировать как ограничение объема настоящего изобретения теми сайтами сплайсинга-мишенями, которые приведены в качестве примеров.
В таблице 1 представлены производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление HIF-1α-специфических ASO в таблице 1 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения производными ПНК, указанными в таблице 1.
Таблица 1. Производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
Фиг. 15А представляет собой хроматограмму, полученную при HPLC с неочищенным продуктом «HIF-ASO 1». Неочищенный продукт очищали с помощью препаративной HPLC с С18-обращенной фазой. Фиг. 15В представляет собой хроматограмму, полученную при HPLC с очищенным продуктом «HIF-ASO 1». Степень чистоты HIF-ASO 1 заметно увеличивалась после очистки с помощью препаративной HPLC. На фиг. 16 представлен ESI-TOF/MS спектр, полученный с очищенным продуктом «HIF-ASO 1». Предоставление данных анализа для «HIF-ASO 1» предназначено для иллюстрации того, как производные ПНК формулы I были очищены и идентифицированы в настоящем изобретении, и не должно интерпретироваться как ограничение объема этого изобретения.
В таблице 2 представлены производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 3 и экзона 4 в пре-мРНК для HIF-1α человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление HIF-1α-специфического ASO в таблице 2 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения HIF-1α-ASO, указанными в таблице 2.
Таблица 2. Производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 3 и экзона 4 в пре-мРНК для HIF-1α человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 3 представлены производные ПНК, комплементарно нацеливающиеся на 5'-сайт сплайсинга, охватывающий место соединения экзона 5 и интрона 5 в пре-мРНК для андрогенового рецептора человека (AR), считываемой с гена AR человека (доступ к которой получен из ссылочной последовательности NCBI: NC_000023.11), наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление AR-ASO в Таблице 3 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения AR-ASO, указанными в Таблице 3.
Таблица 3. Производные ПНК, комплементарно нацеливающиеся на 5'-сайт сплайсинга, охватывающий место соединения экзона 5 и интрона 5 в пре-мРНК для АР человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 4 представлены производные ПНК, комплементарно нацеливающиеся на 5'-сайт сплайсинга, охватывающий место соединения экзона 4 и интрона 4 в пре-мРНК для SCN9A (натриевого канала подтипа 9A) человека, считываемой с гена SCN9A человека (доступ к которой получен из ссылочной последовательности NCBI: NC_000002.12), наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление SCN9A-специфических ASO в таблице 4 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения ASO, указанными в таблице 4.
Таблица 4. Производные ПНК, комплементарно нацеливающиеся на 5'-сайт сплайсинга, охватывающий место соединения экзона 4 и интрона 4 в пре-мРНК SCN9A человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 5 представлены производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 3 и экзона 4 в пре-мРНК для SCN9A человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление SCN9A-специфических ASO в таблице 5 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения SCN9A-ASO, указанными в таблице 5.
Таблица 5. Производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения экзона интрона 3 и экзона 4 в пре-мРНК для SCN9A человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 6 представлены производные ПНК, комплементарно нацеливающиеся на конкретный сайт сплайсинга в пре-мРНК для SCN9A человека или крысы, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление SCN9A-специфических ASO в таблице 6 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения SCN9A-ASO, указанными в таблице 6.
Таблица 6. Производные ПНК, комплементарно нацеливающиеся на конкретный сайт сплайсинга (SS) в пре-мРНК для SCN9A человека или крысы, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
& Крыса
a)SS означает сайт сплайсинга; b)теоретическая точная масса; c)наблюдаемая точная масса
«SCN-ASO 34» представляет собой 16-мерный ASO, полностью комплементарный 5'-сайту сплайсинга, охватывающему место соединения экзона 2 и интрона 2 в пре-мРНК для SCN9A человека. «SCN-ASO 34» обладает способностью к 11-мерному комплементарному перекрыванию с экзоном 2 и 5-мерному комплементарному перекрыванию с интроном 2, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК для SCN9A человека[(5'→3')GAUUUUAGUACACUC│auauccuuuu].
«SCN-ASO 35» представляет собой 16-мерный ASO, комплементарно нацеливающийся на 5'-сайт сплайсинга, охватывающий место соединения экзона 2 и интрона 2 в пре-мРНК для SCN9A «крысы». «SCN-ASO 35» обладает способностью к 11-мерному комплементарному перекрыванию с экзоном 2 и 5-мерному комплементарному перекрыванию с интроном 2, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК для SCN9A крысы [(5'→3')GAUCUUAGUGCACUC│auauccuuuc], считываемой с геномной ДНК крысы [доступ к которой получен из ссылочной последовательности NCBI: NC_005102.3]. «SCN-ASO 35» имеет одно несоответствие с пре-мРНК для SCN9A человека, отмеченное знаком кавычки ("") в 25-мерной последовательности пре-мРНК [(5'→3')GAUUUUAGU"A"CACUC│auauccuuuu].
«SCN-ASO 36» представляет собой 15-мерный ASO, комплементарно нацеливающийся на 5'-сайт сплайсинга, охватывающий место соединения экзона 7 и интрона 7 в пре-мРНК для SCN9A человека. «SCN-ASO 36» обладает способностью к 11-мерному комплементарному перекрыванию с экзоном 7 и 4-мерному комплементарному перекрыванию с интроном 7, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК для SCN9A человека [(5 →3')CAGCACAGAUUCAGG│guauguaaua]. Последовательность-мишень «SCN-ASO 43» консервативна в пре-мРНК для SCN9A крысы.
«SCN-ASO 37» представляет собой 14-мерный ASO, комплементарно нацеливающийся на 3'-сайт сплайсинга, охватывающий место соединения интрона 14 и экзона 15 в пре-мРНК для SCN9A человека. «SCN-ASO 37» обладает способностью к 3-мерному комплементарному перекрыванию с интроном 14 и 11-мерному комплементарному перекрыванию с экзоном 15, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК для SCN9A человека [(5→3')uugcuuuuag│CUCCGAGUCUUCAAG]. Последовательность-мишень ASO консервативна в пре-мРНК крысы SCN9A и отмечена жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК крысы [(5'→3')uuauuucuag│CUCCGAGUCUUCAAG].
В таблице 7 представлены производные ПНК, комплементарно нацеливающиеся на либо 3'-, либо 5'-сайт сплайсинга экзона 23 в пре-мРНК дистрофина мыши, считываемой с геномной ДНК мыши [доступ к которой получен из ссылочной последовательности NCBI: NC_000086.7], наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление специфических для дистрофина ASO в таблице 7 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения специфическими для дистрофина ASO, указанными в Таблице 7.
Таблица 7. Производные ПНК, комплементарно нацеливающиеся на либо 3'-, либо 5'-сайт сплайсинга экзона 23 в пре-мРНК дистрофина мыши, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)SS означает сайт сплайсинга; b)теоретическая точная масса; c)наблюдаемая точная масса
В таблице 8 представлены производные ПНК, комплементарно нацеливающиеся на сайт сплайсинга в пре-мРНК для индоламин-2,3-диоксигеназы (IDO1) человека или мыши, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Последовательности пре-мРНК IDO-1 человека и мыши были считаны с геномной ДНК человека [доступ к которой получен из ссылочной последовательности NCBI: NC_000008.11] и с геномной ДНК мыши [доступ к которой получен из ссылочной последовательности NCBI: NC_000074], соответственно. Предоставление специфических для IDO1 ASO в таблице 8 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения IDO1-ASO, указанными в таблице 8.
Таблица 8. Производные ПНК, комплементарно нацеливающиеся на конкретный сайт сплайсинга (SS) в пре-мРНК для IDO1 человека или мыши, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)SS означает сайт сплайсинга; b)теоретическая точная масса; c)наблюдаемая точная масса
В таблице 9 представлены производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга «экзона 7» в пре-мРНК для SNAP25 человека, считываемой с гена SNAP25 человека [ссылочная последовательность NCBI: NG_029626.1], наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии. Предоставление специфических для SNAP25 ASO в Таблице 9 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения SNAP25-ASO, указанными в Таблице 9.
Таблица 9. Производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 6 и экзона 7 в пре-мРНК для SNAP25 человека, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 10 представлены производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 1 и экзона 2 в пре-мРНК для тирозиназы (TYR) человека, считываемой с гена TYR человека [ссылочная последовательность NCBI: NG_0008748], или пре-мРНК для TYR мыши, считываемой с геномной ДНК мыши [доступ к которой получен из ссылочной последовательности NCBI: NC_000073]. Предоставление TYR-специфических ASO в таблице 10 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения TYR-ASO, указанными в таблице 10.
Таблица 10. Производные ПНК, комплементарно нацеливающиеся на 3'-сайт сплайсинга, охватывающий место соединения интрона 1 и экзона 2 в пре-мРНК для TYR человека или мыши, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
ASO 4
a)теоретическая точная масса; b)наблюдаемая точная масса
В таблице 11 представлены производные ПНК, комплементарно нацеливающиеся либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 2 в пре-мРНК для PD-1 человека, считываемой с гена PDCD1 человека [ссылочная последовательность NCBI: NG_012110], или пре-мРНК для PD-1 мыши, считываемой с геномной ДНК мыши [доступ к которой получен из ссылочной последовательности NCBI: NC_000067]. Предоставление ASO, специфических PD-1, в таблице 11 является примером производных ПНК формулы I и не должно интерпретироваться как ограничение объема настоящего изобретения PD-1-ASO, указанными в таблице 11.
Таблица 11. Производные ПНК, комплементарно нацеливающиеся либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 2 в пре-мРНК для PD-1 человека или мыши, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
a)SS означает сайт сплайсинга; b)теоретическая точная масса; c)наблюдаемая точная масса
Сродство модельных производных ПНК к комплементарной РНК или ДНК
10-мерные производные ПНК, содержащие модифицированные нуклеотидные основания, были получены в качестве модельных соединений для демонстрации сильного сродства соединений ПНК формулы I к РНК, а также к ДНК. Эти модельные соединения ПНК были получены в соответствии с процедурами синтеза, предусмотренными в настоящем изобретении, или с незначительными модификациями. 10-мерные производные ПНК представлены в таблице 12, наряду с данными о структурных характеристиках, полученными с помощью масс-спектрометрии.
Таблица 12. 10-мерные производные ПНК в качестве модельных соединений для демонстрации сильного сродства соединений ПНК формулы I к РНК или ДНК.
a)теоретическая точная масса; b)наблюдаемая точная масса
10-мерные ПНК-олигомеры в таблице 12 были оценены в отношении их сродства связывания с комплементарной 10-мерной РНК или ДНК путем измерения значений Tm, как описано ниже.
Смешанный раствор 4 мкМ 10-мерного ПНК-олигомера и 4 мкМ комплементарной 10-мерной ДНК или РНК в 4 мл водного буфера (pH 7,16, 10 мМ фосфата натрия, 100 мМ NaCl) в 15 мл полипропиленовой пробирке Falcon инкубировали при 90οС в течение минуты и медленно охлаждали до температуры окружающей среды. Затем раствор переносили в кварцевую УФ-кювету объемом 4 мл и подвергали измерению Tm при 260 нм в спектрофотометре УФ/видимой области спектра, как описано в предшествующем уровне техники [PCT/KR2009/001256], или с небольшими модификациями. ДНК и РНК для измерения Tm были приобретены у Bioneer (www.bioneer.com, Dajeon, Республика Корея) и использовались без дальнейшей очистки.
В таблице 13 представлены значения Tm (не скорректированные), измеренные между модельными ПНК-олигомерами и комплементарной ДНК или РНК. «ПНК 10-1», эталонный ПНК-олигомер без модифицированного нуклеотидного основания, давал значения Tm 51 и 55οС на фоне комплементарной ДНК и РНК, соответственно. Модельные ПНК-олигомеры, содержащие модифицированное нуклеотидное основание(я), как правило, демонстрировали более высокое значение Tm вместе с большим включением модифицированных нуклеотидных оснований. «ПНК 10-7» продемонстрировал Tm=69οС также на фоне комплементарной ДНК и РНК, что позволяет предположить, что модельные ПНК-олигомеры связываются с комплементарной им ДНК и РНК с сопоставимым сродством.
Таблица 13. Значения Tm между 10-мерной ПНК и комплементарной ДНК или РНК.
a) Значение Tm - значение Tm «ПНК 10-1»
Сродство производных ПНК к 10-мерной комплементарной ДНК
Производные ПНК формулы I оценивали в отношении их сродства к 10-мерным ДНК, комплементарно нацеливающимся на N-конец или С-конец. Сродство оценивали по значению Tm для дуплекса между ПНК и 10-мерной комплементарной ДНК. Дуплекс между производными ПНК и ДНК с полной комплементарностью обычно демонстрирует значения Tm слишком высокие, чтобы их можно было надежно определить в водном буферном растворе. Водный буферный раствор, как правило, выкипает во время измерения Tm.
Наблюдаемые значения Tm (не скорректированные) для производных ПНК формулы I были очень высокими в случае комплементарного связывания с 10-мерной ДНК и представлены в таблице 14. Например, «AR-ASO 1» продемонстрировал значение Tm=86,1οС для дуплекса с 10-мерной комплементарной ДНК, нацеливающейся на 10-мер на N-конце ПНК, как отмечено «жирным шрифтом» и «подчеркиванием» в [(N→C)Fethoc-C(1O2)TT-A(5)CC-A(5)GG-C(1O2)AA(5)-G-NH2]. Между тем, «AR-ASO 1» продемонстрировал значение Tm=81,3οС для дуплекса с 10-мерной комплементарной ДНК, нацеливающейся на 10-мер на С-конце ПНК, как отмечено «жирным шрифтом» и «подчеркиванием» в [(N→C)Fethoc-C(1O2)TT-A(5)CC-A(5)GG-C(1O2)AA(5)-G-NH2].
Таблица 14. Значения Tm между ПНК и 10-мерной комплементарной ДНК, мишенью которой является либо N-конец, либо С-конец ПНК.
Примеры in vitro активности HIF-1α-специфических ASO
Производные ПНК формулы I, комплементарно нацеливающиеся на 3'-сайт сплайсинга либо экзона 2, либо экзона 4 в пре-мРНК для HIF-1α (индуцируемого гипоксией фактор 1α) человека, оценивали в отношении их HIF-1α-антисмысловой экзон-пропускающей активности в клетках HeLa. Биологические примеры для этих HIF-1α-ASO приведены в качестве примеров с целью иллюстрации того, что пропуск экзона эффективно индуцируется соединением формулы I, нацеливающимся на сайт сплайсинга в пре-мРНК-мишени, и, следовательно, не должны интерпретироваться как ограничение объема настоящего изобретение HIF-1α-специфическими ASO.
HIF-1α-пример 1. Пропуск экзонов, индуцируемый «HIF-ASO 2».
«HIF-ASO 2», указанный в таблице 1, представляет собой 14-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга экзона 2 в пре-мРНК для HIF-1α человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')uguuaaguag│GAUAAGUUCU], где символ «│» обозначает место соединения интрона и экзона. «HIF-ASO 2» обладает способностью к 5-мерному комплементарному перекрыванию с интроном 1 и 9-мерному комплементарному перекрыванию с экзоном 2.
«HIF-ASO 2» оценивали с помощью HIF-1α-специфической вложенной ПЦР в отношении его способности к индукции пропуска экзона 2 мРНК для HIF-1α человека в клетках HeLa. Используемые процедуры приведены ниже.
[Клеточная культура и обработка ASO] Клетки HeLa (кат. № CCL-2, ATCC) выращивали в 60 мм чашке для культивирования, содержащей 5 мл среды EMEM с добавлением 10% FBS (фетальной бычьей сыворотки), 1% стрептомицина/пенициллина, 1% L-глютамина и 1% пирувата натрия в атмосфере с 5% CO2 при 37οC. Клетки обрабатывали «HIF-ASO 2» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ. ASO последовательно разводили до соответствующей концентрации в DDW и аликвотировали в культуральную чашку.
[Экстракция РНК] Через 5 часов тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [HIF-экзон 1_прямой: (5'→3')CTTGCCTTTCCTTCTCTTCT; HIF-экзон 8_обратный: (5'→3')AACCCAGACATATCCACC] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 55οC и 1 мин при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК подвергали реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [HIF-экзон 1n_прямой: (5→3')TGAAGACATCGCGGGGAC; HIF-экзон 5n_обратный: (5→3')TTTTTCACAAGG-CCATTTCT] в соответствии со следующими режимами циклирования: 95οC в течение 5 мин с последующими 39 циклами, каждый состоящий из 30 сек при 95οC, 40 сек при 50οC и 50 сек при 72οC.
Наборы экзон-специфических праймеров для одностадийной RT-PCR и амплификации с помощью вложенной ПЦР схематично суммированы на фиг. 17А.
[Идентификация продукта «пропуска экзона 2»] ПЦР-продукты подвергали электрофоретическому разделению в 2% агарозном геле вместе со смесью маркеров размера. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру. Наблюдаемые полосы для ПЦР-продуктов соответствовали полноразмерной мРНК (т.е. без пропуска экзона) и варианту сплайсинга, в котором отсутствует экзон 2, как показано на фиг. 17B. Клетки, обработанные ASO, давали сильную полосу для ПЦР-продукта размером, приписываемым пропуску экзона 2. Клетки без обработки ASO (т.е. отрицательного контроля) также давали ПЦР-продукт, соответствующий пропуску экзона 2, что наводит на мысль, что экзон 2 самопроизвольно делетируется в определенной степени. Однако интенсивность полосы, соответствующей пропуску экзона, была намного выше в клетках, обработанных ASO, чем в клетках без обработки ASO. Таким образом, «HIF-ASO 2» способствовал пропуску экзона 2 в клетках HeLa. Данные секвенирования для полосы, соответствующей пропуску экзона, представлены на фиг. 17C, где показана последовательность мРНК для места соединения экзона 1 и экзона 3.
[Количество клеток, на которые воздействует одна молекула ASO] «HIF-ASO 2» индуцировал пропуск экзона 2 даже в концентрации 10 цМ. Приблизительно 30 молекул ASO в концентрации 10 цМ (т.е. 10×10-21 М) существует в 5 мл культуральной среды в 60-мм чашке для культивирования. Учитывая, что приблизительно 30 молекул ASO индуцировали пропуск в приблизительно 100000 клеток HeLa в 60-мм чашке для культивирования, каждая молекула ASO, по оценкам, воздействовала или контролировала пропуск экзона в среднем приблизительно в 3000 клеток HeLa. Поэтому полагают, что каждая молекула ASO быстро перемещалась вокруг большого количества клеток для выполнения предназначенной для нее роли по пропуску экзона.
HIF-1α-пример 2. Ингибирование экспрессии белка HIF-1α в клетках HeLa с помощью «HIF-ASO 2».
«HIF-ASO 2» оценивали в отношении его способности ингибировать экспрессию белка HIF-1α в клетках HeLa, как описано ниже.
[Клеточная культура и обработка ASO] Клетки HeLa, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «HIF-ASO 2» в концентрации 0 цМ (отрицательный контроль), 10 цM, 100 цМ, 1 аМ или 10 аМ.
[Обработка CoCl2 и лизис клеток] Через 24 часа после обработки ASO чашки для культивирования, за исключением чашки без обработки ASO, обрабатывали 200 мкМ CoCl2 в течение еще 3 часов для повышения уровня белка HIF-1α путем подавления активности пролилгидроксилаз (PHD). Затем клетки дважды промывали 1 мл холодного PBS и подвергали лизису на льду с использованием 200 мкл 1X буфера RIPA (кат. № 9806, Cell Signaling Tech) с добавлением 1% SDS и 1X смеси ингибиторов протеиназ (Complete Mini, Roche). Каждый лизат собирали в 1,5-мл пробирку Эппендорфа, смешивали с 100 мкл 5X буфера для образцов и кипятили в течение 5 мин при 100οC. Лизаты подвергали электрофоретическому разделению в 8% SDS-ПААГ для электрофореза и переносили на мембрану из PVDF 0,45 мкм. Мембрану исследовали с использованием антитела против HIF-1α (кат. № 610958, BD Biosciences) и антитела против β-актина (кат. № sc4778, Santa Cruz).
[Ингибирование экспрессии белка HIF-1α]. На фиг. 18А представлены данные Вестерн-блоттинга HIF-1α, полученные в клетках HeLa, обработанных «HIF-ASO 2». В то время как не было обнаружено полосы для HIF-1α с лизатом клеток без обработки CoCl2, лизаты клеток, обработанных CoCl2, давали сильную полосу для HIF-1α. На фиг. 18В представлены интенсивности отдельных полос для HIF-1α, приведенные е интенсивности отдельной полосы для β-актина с помощью денситометрии. Экспрессия HIF-1α постепенно снижалась по мере повышения концентрации «HIF-ASO 2». Наблюдаемое снижение составило приблизительно 75% при 10 аМ «HIF-ASO 2».
HIF-1α-пример 3. кПЦР с использованием SYBR Green для мРНК для HIF-1α в клетках HeLa, обработанных «HIF-ASO 2».
«HIF-ASO 2» оценивали с помощью вложенной кПЦР в отношении его способности ингибировать экспрессию полноразмерной мРНК для HIF-1α в клетках HeLa следующим образом.
[Клеточная культура и обработка ASO] Клетки HeLa, выращенные в 60 мм чашке для культивирования, содержащей 5 мл среды, обрабатывали «ASO 2» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ (2 чашки для культивирования на каждую концентрацию ASO).
[Экстракция РНК] Через 3 часа после обработки ASO тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК MiniBEST» (кат. № 9767, Takara) в соответствии с инструкциями производителя.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [HIF-экзон 1_прямой: (5'→3')CTTGCCTTTCCTTCTCTTCT; HIF-экзон 8_обратный: (5'→3')AACCCAGACATATCCACC] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 55οC и 1 мин при 72οC.
[Вложенная кПЦР] 1 мкл кДНК, разведенной в 100 раз, подвергали реакция ПЦР в режиме реального времени в объеме 20 мкл на фоне следующих наборов экзон-специфических праймеров: [HIF-экзон 2n_прямой (5'→3')CTTGCTCATCAGTTGCCACTTC; HIF-экзон 2n_обратный (5'→3')AAGTTTCCT-CACACGCAAATAG; HIF-экзон 3n_прямой (5'→3')GAAAGCACAGATGAATTGC; HIF-экзон 3n_обратный (5'→3')TCATGTCACCATCATCTGT; HIF-экзон 4n_прямой (5'→3')CTAACTGGACACAGTGTGTTTG; HIF-экзон 4n_обратный (5'→3')TCTGTGTGTAAGCATTTCTCTC; HIF-экзон 5n_прямой (5'→3')GCCTTGTGAAAAAGGGTAAAG; HIF-экзон 5n_обратный (5'→3')CCATGTTGCAGACTTTATGT]. Реакции ПЦР исследовали с использованием SYBR Green (Takara, Япония) в соответствии со следующими режимами циклирования: 95οC в течение 3 мин с последующими 40 циклами, каждый в течение 5 сек при 95οC и 30 сек при 60οC.
[Изменения уровней экзона в мРНК для HIF-1α]. Индивидуальные уровни экзона в образцах, обработанных ASO, были приведены к каждому индивидуальному уровню экзона без обработки ASO. Наблюдаемые относительные индивидуальные уровни экзона представлены на фиг. 18C. Все индивидуальные уровни экзона значимо снижались на 60-80% и 50-70% в клетках, обработанных «HIF-ASO 2» в концентрации 10 цМ и 100 цМ, соответственно. Однако индивидуальные уровни экзона, полученные с клетками, обработанными «HIF-ASO 2» в концентрации 1000 цМ (т.е. 1 аМ), не отличались от уровней экзона в клетках без обработки ASO. Хотя еще предстоит выяснить, почему увеличенные уровни экзона возвращались к уровням отрицательного контроля, когда концентрация ASO увеличивалась до 1000 цМ. Тем не менее, обратный характер зависимости ответа от дозы на фиг. 18C сравним с обратным характером зависимости от дозы пропуска экзона в «HIF-1α-примере 1». (ср. фиг. 17A).
HIF-1α-пример 4. кПЦР с использованием зонда TaqMan для мРНК HIF-1α в клетках HeLa, обработанных «HIF-ASO 2».
«HIF-ASO 2» оценивали с помощью вложенной кПЦР в отношении его способности ингибировать экспрессию полноразмерной мРНК для HIF-1α в клетках HeLa, как описано в «HIF-1α-примере 3», если не указано иное.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [HIF-экзон l_прямой(2): (5'→3')CGCGAACGACAAGAAAAA; HIF-экзон 8_обратный(2): (5'→3')CTGTGGTGACTTGTCCTTT] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 20 циклов, каждый состоящий из 30 сек при 94οC, 40 сек при 51οC и 50 сек при 72οC.
[Вложенная кПЦР] 1 мкл кДНК, разведенной в 100 раз, подвергали реакция ПЦР в режиме реального времени в объеме 20 мкл, используя зонд TaqMan (Hs00936371_ml, Thermo Fisher), сконструированный для обнаружения места соединения экзона 1 и экзона 2 HIF-1α человека в соответствии со следующими режимами циклирования: 95οC в течение 3 мин с последующими 40 циклами, каждый состоящий из 10 сек при 95οC и 30 сек при 60οC.
[Изменения уровня мРНК для HIF-1α] Уровень полноразмерной мРНК в образцах, обработанных ASO, приводили к уровню мРНК без обработки ASO. Наблюдаемые приведенные уровни мРНК представлены на фиг. 18D. Уровень полноразмерной мРНК для HIF-1α значимо (по t-критерию Стьюдента) снижался на 65% и 55% в клетках, обработанных «HIF-ASO 2» в концентрации 100 цМ и 1000 цМ, соответственно. Уровень полноразмерной мРНК оставался неизменным в клетках, обработанных «ASO 2» в концентрации 10 цМ.
HIF-1α-пример 5. Пропуск экзонов, индуцируемый «HIF-ASO 6».
«HIF-ASO 6», указанный в таблице 1, представляет собой 17-мерный ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α-1 человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')uguuaaguag|GAUAAGUUCU]. «HIF-ASO 6» обладает способностью к 7-мерному комплементарному перекрыванию с интроном 1 и 10-мерному комплементарному перекрыванию с экзоном 2.
«HIF-ASO 6» оценивали с помощью HIF-1α-специфической вложенной ПЦР в отношении его способности к индукции пропуска экзона 2 мРНК для HIF-1α человека в клетках HeLa в соответствии с процедурами, описанными в «HIF-1α-примере 1», если не указано иное.
Продукты ПЦР подвергали электрофоретическому разделению в 2% агарозном геле, и результаты электрофореза представлены на фиг. 19А. Пропуск экзона 2 был устойчивым при всех концентрациях обработки «HIF-ASO 6». «HIF-ASO 6» индуцировал пропуск экзона 2 более эффективно, чем «HIF-ASO 2». Полоса для ПЦР-продукта в виде полноразмерной мРНК HIF-1α исчезала почти полностью при всех концентрациях «HIF-ASO 6». Между тем, в клетках, обработанных «HIF-ASO 2» в диапазоне концентраций от 10 до 1000 цМ, оставался значительный уровень полноразмерной мРНК для HIF-1α [ср. фиг. 17А]
«HIF-ASO 6» обладает способностью к более комплементарному перекрыванию с 3'-сайтом сплайсинга экзона 2, чем «HIF-ASO 2», что ответственно за более высокую эффективность пропуска экзона, наблюдаемую с «HIF-ASO 6». Более прочное связывание ASO с сайтом сплайсинга-мишенью, по-видимому, более эффективно индуцирует пропуск экзона-мишени.
HIF-1α-пример 6. Ингибирование экспрессии белка HIF-1α в клетках HeLa с помощью «HIF-ASO 6».
«HIF-ASO 6» оценивали в отношении его способности снижать экспрессию HIF-1α в клетках HeLa в соответствии с процедурами, описанными в «HIF-1α-примере 2», если не указано иное. Фиг. 19В - данные Вестерн-блоттинга, полученные для клеток HeLa, обработанных «HIF-ASO 6» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ в течение 24 часов. На фиг. 19C представлены интенсивности отдельных полос для HIF-1α, приведенные к интенсивности отдельной полосы для β-актина с помощью денситометрии. Экспрессия белка HIF-1α снижалась приблизительно на 45 ~ 55% в клетках, обработанных «HIF-ASO 6».
HIF-1α-пример 7. кПЦР с использованием SYBR Green для мРНК HIF-1α в клетках HeLa, обработанных «HIF-ASO 6».
«HIF-ASO 6» оценивали в отношении его способности вызывать изменение мРНК для HIF-1α в клетках HeLa с помощью вложенной кПЦР в соответствии с процедурами, описанными в «HIF-1α-примере 4», если не указано иное.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [HIF-экзон 1_прямой(2): (5'→3')CGCGAACGACAAGAAAAA; HIF-экзон 8(2)_обратный: (5'→3')CTGTGGTGACTTGTCCTTT] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οC, 40 сек при 51οC и 50 сек при 72οC.
[Изменения уровней экзона в мРНК для HIF-1α] Индивидуальные уровни экзона, приведенные к индивидуальным уровням экзона без обработки ASO, представлены на фиг. 20(A). Уровни экзона значимо (t-критерий Стьюдента) снижались на 35%, приблизительно 30% и приблизительно 45% в клетках, обработанных «HIF-ASO 6» в концентрации 10, 100 и 1000 цМ, соответственно.
HIF-1α-пример 8. кПЦР с использованием зонда TaqMan для мРНК HIF-1α в клетках HeLa, обработанных «ASO 6».
«HIF-ASO 6» оценивали с помощью вложенной кПЦР в отношении его способности ингибировать экспрессию полноразмерной мРНК для HIF-1α в клетках HeLa, как описано в «HIF-1α-примере 4», если не указано иное.
[Изменения уровня полноразмерной мРНК для HIF-1α] Уровень полноразмерной мРНК в образцах, обработанных ASO, приводили к уровню мРНК без обработки ASO. Наблюдаемые относительные уровни мРНК представлены на фиг. 20(B). Уровень полноразмерной мРНК HIF-1α значимо (t-критерий Стьюдента) снижался приблизительно на 60% и 80% в клетках, обработанных «HIF-ASO 6» в концентрации 100 цМ и 1000 цМ (1 аМ), соответственно. Однако уровень полноразмерной мРНК оставался неизменным в клетках, обработанных «HIF-ASO 6» в концентрации 10 цМ.
HIF-1α-пример 9. Пропуск экзонов, индуцируемый «HIF-ASO 1».
«HIF-ASO 1» представляет собой 14-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 1 и экзона 2 в пре-мРНК для HIF-1α человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 23-мерной последовательности пре-мРНК [(5'→3')uguuaaguag|GAUAAGUUCUGAA]. «HIF-ASO 1» обладает способностью к 3-мерному перекрыванию с интроном 1 и 11-мерному перекрыванию с экзоном 2.
«HIF-ASO 1» оценивали в отношении его способности к индукции пропуска экзонов в мРНК для HIF-1α, как описано в «HIF-1α-примере 1», если не указано иное. Клетки HeLa обрабатывали «HIF-ASO 1» в концентрации 0 (отрицательный контроль), 1, 3, 10, 30 или 100 аМ. Через 24 часа тотальную РНК экстрагировали и подвергали HIF-1α-специфической вложенной ПЦР для детектирования пропуска экзонов.
[Данные пропуска экзонов] На фиг. 21А представлены данные электрофореза, полученные с продуктами вложенной ПЦР, а также данные секвенирования по Сэнгеру ПЦР-продукта, приписываемого пропуску экзонов 2-3. Уровень полноразмерной мРНК имел тенденцию к снижению по мере увеличения концентрации ASO с 1 до 100 аМ. Пропуск экзона 2 был доминирующим в случае «HIF-ASO 1» в концентрации 3 аМ и 10 аМ. Однако пропуск экзонов 2-3 становился чрезмерным, когда концентрация ASO была увеличена до 100 аМ. ПЦР-продукт в случае пропуска экзонов 2-3 был однозначно подтвержден секвенированием по Сэнгеру [ср. фиг. 21 справа]
HIF-1α-пример 10. Ингибирование экспрессии белка HIF-1α в клетках HeLa с помощью «HIF-ASO 1».
«HIF-ASO 1» оценивали в отношении его способности ингибировать экспрессию белка HIF-1α в клетках HeLa в соответствии с процедурами, описанными в «HIF-1α-примере 2», если не указано иное. В этом примере клетки HeLa обрабатывали «HIF-ASO 1» в концентрации 0 цМ (отрицательный контроль), 100 цМ, 300 цМ, 1 аМ, 3 аМ, 10 аМ, 30 аМ, 100 аМ или 300 аМ в течение 72 часов до подавления активности PHD путем инкубации с 200 мкМ CoCl2 в течение 3 часов. Использовали 4 чашки для культивирования для отрицательного контроля, т.е. 0 цМ «HIF-ASO 1».
На фиг. 21B представлены данные Вестерн-блоттинга HIF-1α, полученные с лизатами клеток HeLa. Уровень белка HIF-1α был значительно выше в лизатах отрицательного контроля, чем во всех лизатах клеток, обработанных «HIF-ASO 1». На фиг. 21C представлены интенсивности отдельных полос для HIF-1α, приведенные к интенсивности полосы для β-актина с помощью денситометрии. Экспрессия HIF-1α в клетках HeLa снижалась на 40-80% в результате 72-часовой инкубации с «HIF-ASO 1» в концентрации 0,1-300 аМ.
HIF-1α-пример 11. Пропуск экзонов, индуцируемый «HIF-ASO 12».
«HIF-ASO 12», указанный в таблице 2, представляет собой 15-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 3 и экзона 4 в пре-мРНК для HIF-1α человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')uguuuacag|UUUGAACTAAC]. «HIF-ASO 12» обладает способностью к 6-мерному перекрыванию с интроном 3 и 9-мерному перекрыванию с экзоном 4.
«HIF-ASO 12» оценивали с помощью HIF-1α-специфической вложенной ПЦР в отношении его способности к индукции пропуска экзона 4 в мРНК для HIF-1α человека в HeLa. Клетки HeLa инкубировали с «HIF-ASO 12» в течение 6 часов, а затем подвергали экстракции тотальной РНК в соответствии с протоколом, описанным в «HIF-1α-примере 1», если не указано иное.
На фиг. 22А представлены данные HIF-1α-специфической вложенной ПЦР в клетках HeLa, обработанных «HIF-ASO 12». Полоса, приписываемая пропуску экзонов 2-4, была обнаружена во всех продуктах ПЦР клеток, обработанных ASO, хотя и не в продуктах ПЦР необработанных клеток (ср. левую диаграмму). Интенсивность полосы, приписываемой пропуску экзонов, была наибольшей при концентрации 100 цМ «HIF-ASO 12». Интенсивность полосы для полноразмерной мРНК снизилась больше всего при 100 цМ «HIF-ASO 12». Пропуск экзонов 2-4 был подтвержден секвенированием по Сэнгеру, как показано на правой диаграмме.
HIF-1α-пример 12. Ингибирование экспрессии белка HIF-1α в клетках HeLa с помощью "HIF-ASO 12".
«HIF-ASO 12» оценивали в отношении его способности ингибировать экспрессию HIF-1α в клетках HeLa, как описано в «HIF-1α-примере 2», если не указано иное.
[Обработка ASO] Клетки HeLa обрабатывали «HIF-ASO 12» в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ или 1 аМ. Использовали 3 чашки для культивирования для отрицательного контроля. Через 21 час клетки обрабатывали 200 мкМ CoCl2, за исключением одной чашки отрицательного контроля. Через 3 часа все клетки подвергали лизису на льду следующим образом. Клетки дважды промывали 1 мл холодного PBS и затем подвергали лизису с использованием 200 мкл 2X буфера для образцов Lammeli (24 мМ Трис-HCl, 20% глицерина, 0,8% SDS, 0,04% бромфенолового синего, 2% β-меркаптоэтанола) для минимизации деградация HIF-1α. Каждый лизат собирали в 1,5 мл пробирке Эппендорфа и кипятили в течение 5 мин. Затем лизаты подвергали Вестерн-блоттингу в 8% SDS-ППАГ для электрофореза.
На фиг. 22В представлены данные Вестерн-блоттинга, показывающие, что интенсивность полосы для HIF-1α явно снижалась в клетках, обработанных «HIF-ASO 12» в концентрации 1 и 10 аМ.
HIF-1α-пример 13: кПЦР с использованием SYBR Green для мРНК для HIF-1α в клетках HeLa, обработанных «HIF-ASO 12».
«HIF-ASO 12» оценивали с помощью HIF-1α-специфической вложенной кПЦР в отношении его способности вызывать изменение мРНК для HIF-1α в клетках HeLa, как описано в «HIF-1α-примере 12», если не указано иное. Клетки HeLa обрабатывали «HIF-ASO 12» в течение 6 часов, а затем подвергали тотальной экстракции РНК.
На фиг. 22C представлены данные кПЦР, в которых уровни экзонов 2 и 3 в мРНК значимо (по t-критерию Стьюдента) снижались на 70~80% в клетках, обработанных ASO. Результаты кПЦР соответствуют пропуску экзонов 2-4, индуцированному «HIF-ASO 12» (ср. «Пример 11 HIF-1α).
Примеры in vitro и in vivo активности AR-ASO.
Производные ПНК формулы I, комплементарно нацеливающиеся на 5'-сайт сплайсинга, охватывающий место соединения экзона 5 и интрона 5 в пре-мРНК андрогенового рецептора (AR) человека, оценивали в отношении их AR-антисмысловой экзон-пропускающей активности в клетках, а также у мышей. Биологические примеры для этих AR-ASO приведены в качестве примеров с целью иллюстрации того, что пропуск экзона эффективно индуцируется соединением формулы I, нацеливающимся на сайт сплайсинга в пре-мРНК-мишени, и, следовательно, не должны интерпретироваться как ограничение объема настоящего изобретение AR-ASO.
AR-пример 1. Пропуск экзонов, индуцируемый «AR-ASO 1».
«AR-ASO 1», указанный в таблице 3, представляет собой 13-мерный ASO, полностью комплементарный области в 5'-сайте сплайсинга, охватывающей место соединения экзона 5 и интрона 5 в пре-мРНК для андрогенового рецептора (AR) человека. «AR-ASO 1» комплементарно связывается с 13-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')GCCUUGCCUG|guaaggaaaa]. «AR-ASO 1» обладает способностью к 8-мерному перекрыванию с экзоном 5 и 5-мерному перекрыванию с интроном 5.
«AR-ASO 1» оценивали с помощью AR-специфической вложенной ПЦР в отношении его способности к индукции пропуска экзона 5 в мРНК для AR человека в клетках MCF7 следующим образом.
[Клеточная культура и обработка ASO] Клетки MCF7 (кат. № HTB-22, ATCC) поддерживали в среде EMEM с добавлением 10% FBS, 1% стрептомицина/пенициллина и 0,01 мг/мл бычьего инсулина в атмосфере с 5% CO2 при 37οС. Клетки, выращенные в 60-мм чашке для культивирования, обрабатывали «AR-ASO 1» в течение 3 часов в концентрации 0 (отрицательный контроль), 3, 30, 300 или 3000 аМ (т.е. 3 фМ).
[Экстракция РНК] Тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 100 нг РНК-матрицы использовали в реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) и набором экзон-специфических праймеров [AR-экзон 3_прямой: (5'→3')TGGGTGTCACTATGGAGC; и AR-экзон 9_обратный: (5'→3')GGGT GTGGAAATAGATGGG] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 39 циклов, каждый состоящий из 30 сек при 94οС, 30 сек при 55οС и 1 мин при 72οС.
[Амплификация с помощью вложенной ПЦР] В процессе амплификации использовался уникальный метод амплификации (уточняемый как повышение температуры отжига в каждом цикле), который работал эффективно и специфически в температурном диапазоне, а не при одной конкретной температуре отжига (т.е. в случае обычного метода ПЦР). 1 мкл кДНК дополнительно амплифицировали в 20-мкл реакции вложенной ПЦР с использованием набора экзон-специфических праймеров [AR-экзон 3_прямой: (5'→3')TGGGTGTCACTATGGAGC; и AR-экзон 7n_обратный: (5'→3')GGGGTGATTTGGAGCCAT] в соответствии со следующими режимами циклирования: начальные 10 циклов [каждый состоящий из 94οС C в течение 30 сек, 47οС в течение 40 сек (+ 0,5οС в каждом цикле), 72οС в течение 40 сек] с последующими 20 циклами [каждый состоящий из 94οС в течение 30 сек, 50οС в течение 30 сек и 72οС в течение 40 сек].
[Идентификация продуктов пропуска экзонов] ПЦР-продукты подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру. На фиг. 23А показаны три полосы связанных с обработкой ПЦР-продуктов, приписываемых вариантам сплайсинга мРНК для AR, в которых отсутствует экзон 5. Было обнаружено, что «AR-ASO 1» индуцирует пропуск экзона 5, экзонов 4-5 и экзонов 4-6, хотя соотношение продуктов пропуска, по-видимому, зависит от концентрации ASO. На фиг. 23B представлены фактические данные секвенирования для полосы, приписываемой пропуску экзонов 4-5, на фиг. 23 A.
AR-пример 2. Ингибирование экспрессии белка AR в клетках MCF7 с помощью «AR-ASO 1».
Клетки MCF7 в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «AR-ASO 1» в концентрации 0 цМ (отрицательный контроль) или от 10 цМ до 30 аМ. Было 4 чашки для культивирования для отрицательного контроля. Через 48 часов клетки дважды промывали холодным PBS и затем подвергали лизису с использованием 200 мкл 1Х буфера для лизиса клеток (кат. № 9803, Cell Signaling Tech) с добавлением 1Х смеси ингибиторов протеаз (кат. № P8340, Sigma). Лизаты собирали в 1,5-мл пробирку Эппендорфа. 200 каждого лизата смешивали с 100 мкл 3X буфера для образцов и кипятили в течение 5 мин. 20 мкл каждого лизата (4 отрицательных контроля и 8 образцов обработки ASO) подвергали электрофоретическому разделению на 8% SDS-ПААГ для электрофореза и переносили на мембрану из PVDF. Мембрану исследовали с использованием антитела против АР (кат. № 5153, Cell Signaling Tech) и антитела против β-актина (кат. № sc4778, Santa Cruz). На фиг. 23C представлены данные Вестерн-блоттинга AR, полученные в клетках MCF7, обработанных «AR-ASO 1» в концентрации от 0 цМ (4 отрицательных контроля) до 30 аМ. Интенсивность полосы AR (размером 120K) в случае лизатов, обработанных ASO, была слабее интенсивности соседних с ними лизатов отрицательного контроля.
AR-пример 3. кПЦР с использованием SYBR Green для мРНК AR в клетках MCF7, обработанных «AR-ASO 1».
[Обработка ASO и экстракция РНК] Клетки MCF7 в 5 мл культуральной среды обрабатывали «AR-ASO 1» в концентрации 0 цМ (отрицательный контроль) или от 1 цМ до 1 аМ. (2 чашки для культивирования на концентрацию). Через 5 часов тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК MiniBEST» в соответствии с инструкциями производителя (кат. № 9767, Takara).
[Синтез кДНК с использованием олигоdT] 500 нг РНК-матрицы подвергали синтезу кДНК на фоне «олиго-dT» в соответствии с инструкциями производителя (кат. № 6110A, Takara).
[Первая ПЦР] Затем кДНК подвергали 1-ой ПЦР на фоне набора экзон-специфических праймеров [AR-экзон 3_прямой: (5'→3')TGGGTGTCACTATGGAGC; и AR-экзон 9_обратный: (5'→3')GGGTGTGGAAATAGATGGG] в соответствии со следующими режимами циклирования: 94οС в течение 2 мин с последующими 15 циклами, каждый состоящий из 15 сек при 94οС, 30 сек при 55οС и 2 мин при 72οС.
[Вложенная ПЦР] Продукты 1-ой ПЦР разводили в 2000 раз, и 1 мкл каждого разведенного ПЦР-продукта подвергали реакция ПЦР в режиме реального времени в объеме 20 мкл на фоне наборов экзон-специфических праймеров [AR-экзон 4_прямой(q): (5'→3')GACCATGTTTTGCCCATTG; AR-экзон 4_обратный(q): (5'→3')GGCTCTTTTGAAGAAGACC; AR-экзон 5_прямой(q): (5'→3')GAAACAGAAGTACCTGTGC; AR-экзон 5_обратный(q): (5'→3')GTCATCCCTGCTTCATAAC; AR-экзон 6_прямой(q): (5'→3')CGGAAGCTGAAGAAACTTG; AR-экзон 6_обратный(q): (5'→3')CACTTGACCACGTGTACAAG]. Реакции ПЦР исследовали с использованием SYBR Green (Takara, Япония). Режимы циклирования: 95οС в течение 3 мин с последующими 40 циклами, каждый состоящий из 5 сек при 95οС и 30 сек при 60οС. Уровни экзона постепенно, но значимо снижались по мере увеличения дозы с 1 до 100 цМ. Снижения составляли 40~50% в клетках, обработанных «AR-ASO 1» в концентрации 100 цМ [ср. фиг. 24А]. Однако уровни экзона возвращались близко к уровням отрицательного контроля в клетках, обработанных «AR-ASO 1» в концентрации 1 ам.
AR-пример 4. кПЦР с использованием SYBR Green для мРНК AR в клетках MCF7, обработанных «AR-ASO 5».
«AR-ASO 5», указанный в таблице 3, представляет собой 12-мерный ASO, полностью комплементарный области в 5'-сайте сплайсинга, охватывающей место соединения экзона 5 и интрона 5 в пре-мРНК для AR человека. «AR-ASO 5» комплементарно связывается с 12-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')GCCUUGCCUG|guaaggaaaal. «AR-ASO 5» обладает способностью к 7-мерному перекрыванию с экзоном 5 и 5-мерному перекрыванию с интроном 5.
«AR-ASO 5» оценивали в отношении его способности вызывать изменения уровней экзона в мРНК для AR с помощью кПЦР в соответствии с способами, описанными в «AR-примере 3». Как показано на фиг. 24B, уровни экзона в AR значимо (t-критерий Стьюдента) снижались приблизительно на 60~80% в клетках, обработанных «AR-ASO 5» в концентрации 1-1000 цМ.
В отличие от случая «AR-ASO 1» (ср. «AR-пример 3»), не было отскока в уровнях информационного экзона в концентрации 1000 цМ «AR-ASO 5».
AR-пример 5. кПЦР с использованием зонда TaqMan для мРНК AR в клетках MCF7, обработанных «AR-ASO 5».
«AR-ASO 5» оценивали в отношении его способности снижать экспрессию мРНК AR человека с помощью кПЦР с введением в использование зонда TaqMan.
Клетки MCF7 обрабатывали «AR-ASO 5» в концентрации от 0 цМ (отрицательный контроль) до 1 аМ. (2 чашки на концентрацию). Через 24 часа тотальную РНК экстрагировали с помощью «Универсального набора для экстракции РНК MiniBEST» в соответствии с инструкциями производителя (кат. № 9767, Takara).
400 нг РНК-матрицы подвергали синтезу кДНК с помощью набора для одностадийной ОТ-ПЦР (Invitrogen) на фоне набора экзон-специфических праймеров [AR-экзон 3_прямой: (5'→3')TGGGTGTCACTATGGAGC; и AR-экзон 9_обратный: (5'→3')GGGTGTGGAAATAGATGGG] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οС, 30 сек при 50οС и 1 мин при 72οС.
1 мкл каждого раствора кДНК, разведенного в 50 раз, подвергали ПЦР-реакции в режиме реального времени в объеме 20 мкл на фоне набора экзон-специфических праймеров [AR-экзон 4_прямой(q2): (5'→3')TTGTCCATCTTGTCGTCTT; и AR-экзон 5_обратный(q2): (5'→3') CCTCTCCTTCCTCCTGTA] в соответствии со следующими режимами циклирования: 95οС в течение 3 мин с последующими 40 циклами, каждый состоящий из 15 сек при 95οС и 30 сек при 60οС. Реакцию кПЦР контролировали с использованием зонда TaqMan [(5'→3')TTTCTTCAG-ZEN-CTTCCGGGCTC-3IABkFQ]. Зонд TaqMan был сконструирован для исследования места соединения экзона 4 и экзона 5 в полноразмерной мРНК для AR.
На фиг. 24C представлены данные кПЦР с использованием зонда TaqMan. Относительная экспрессия полноразмерной мРНК для AR значимо (t-критерий Стьюдента) снижалась приблизительно на 50-70% в клетках, обработанных «AR-ASO 5» в концентрации от 1 цМ до 1 аМ.
AR-пример 6. Ингибирование экспрессии белка AR в коже мышей, которым подкожно вводили «AR-ASO-5».
Мишенью «AR-ASO 5» является последовательность пре-мРНК AR, консервативная у людей и мышей. «AR-ASO 5» оценивали в отношении его способности ингибировать экспрессию белка AR в коже мышей после однократного подкожного введения следующим образом.
[Удаление волос и группирование]. В день 0 самцов мышей C57BL/6 в возрасте 7 недель анестезировали с помощью золетила/ромпуна, и волосы на спине стригли машинкой для стрижки волос и удаляли с помощью карбоваска. В день 5 мышей с безупречной (т.е. чистой) эпиляцией отбирали и случайным образом распределяли по пяти группам: 0 пмоль/кг (только носитель, отрицательный контроль), 1 пмоль/кг, 10 пмоль/кг, 100 пмоль/кг, и 1000 пмоль/кг «AR-ASO 5» (6 животных в каждой группе).
[Раствор для инъекций ASO и введение] Водный исходный раствор «AR-ASO 5» последовательно разводили в PBS с добавлением 0,1% Tween 80 для приготовления растворов «AR-ASO 5» с концентрацией 0 нМ (только носитель, отрицательный контроль), 0,5 нМ, 5 нМ, 50 нМ или 500 нМ. В день 5 отдельным мышам в группе каждой дозы делали подкожно в затылок (т.е. около шеи) однократную инъекцию испытуемого препарата в дозе 2 мл/кг.
[Приготовление образцов кожи]. В день 10 животных умерщвляли для получения образцов кожи из места инъекции и бедра в качестве места без инъекции. Образцы кожи замораживали в жидком азоте сразу после отбора. Каждый образец кожи был микронизирован при сохранении образца замороженным жидким азотом. Микронизированные образцы подвергали лизису с использованием буфера RIPA с добавлением 1% SDS. Лизаты смешивали с 5X буфером для образцов и кипятили в течение 5 мин.
[Вестерн-блоттинг AR]. Лизаты подвергали Вестерн-блоттингу AR на мембране из PVDF. Всего в каждый 10% ПААГ для электрофореза загружали 10 лизатов, используя два отдельных лизата из каждой группы. Белок AR (120 кДа) исследовали с использованием поликлонального антитела против AR (N-20, sc-816, Santa Cruz).
[Количественный анализ экспрессии белка AR] Каждую полосу для AR на одной мембране из PVDF приводили к отдельной полосе для β-актина. Среднюю интенсивность полосы для AR (приведенную к β-актину) двух образцов группы отрицательного контроля (т.е. без обработки ASO) использовали для приведения интенсивностей полос для AR других 8 образцов на той же мембране из PVDF. Такое двойное приведение применялось к двум другим мембранам из PVDF для количественного анализа экспрессии белка AR в отдельных образцах с помощью денситометрии. Все уровни экспрессии AR после двойного приведения отдельных образцов были объединены для статистического анализа с помощью t-критерия Стьюдента по отношению к уровню экспрессии без обработки ASO.
[Ингибирование экспрессии белка AR]. На фиг. 25A и 25B представлены данные Вестерн-блоттинга AR, полученные с образцами кожи из места инъекции и места без инъекции, соответственно.
На фиг. 26А представлен уровень экспрессии белка AR по группам, а также по субъектам. Наблюдалась большая степень вариабельности между субъектами в экспрессии белка AR как в месте инъекции, так и в месте без инъекции. Однако экспрессия AR имела тенденцию к снижению по мере увеличения дозы ASO.
На фиг. 26B представлен средний уровень экспрессии AR по группам, приведенный к группе отрицательного контроля. В месте инъекции экспрессия белка AR значимо снижалась приблизительно на 35% в группе введения 1000 пмоль/кг «AR-ASO 5». В месте без инъекции экспрессия белка AR значимо снижалась приблизительно на 40% в группах введения 100 и 1000 пмоль/кг.
Ингибирование экспрессии белка AR, наблюдаемое в коже, удаленной от места инъекции, показывает, что ASO может легко распространяться в ткани, удаленные от места введения, через системный кровоток после подкожной инъекции. Результаты ex vivo были предоставлены для иллюстрации системного поражения мишени после подкожной инъекции производного ПНК формулы I, и поэтому не должны интерпретироваться как ограничивающие объем настоящего изобретения.
AR-пример 7. Пропуск экзонов, индуцируемый «AR-ASO 1» (2).
Морфология клеток MCF7 менялась в зависимости от пассажа, плотности клеток и условий культивирования. Клетки MCF на ранних пассажах, как правило, росли относительно быстро и демонстрировали колонии в форме бугорка. Клетки MCF7 на более поздних пассажах, вероятно, будут расти медленно и образовывать плоские эпителиальные колонии. Однако поддержание клеток MCF7 для демонстрации морфологии бугорковой формы было сложной задачей.
«AR-ASO 1» оценивали в отношении его способности к пропуску экзона в клетках MCF7, в целом демонстрирующих морфологию бугорковой формы, как описано в «AR-примере 1», если не указано иное.
[Обработка ASO] Клетки MCF7 обрабатывали «AR-ASO 1» в течение 3 часов в концентрации 0 (отрицательный контроль), 30, 100 или 1000 аМ (т.е. 1 фМ). (2 чашки для культивирования на каждую концентрацию ASO).
[Экстракция РНК]. Тотальную РНК экстрагировали с использованием набора для мини-приготовления Rneasy (кат. № 74104, Qiagen) в соответствии с инструкциями производителя. 500 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл.
[Результаты пропуска экзонов]. На фиг. 27А представлены данные электрофореза, полученные с продуктами вложенной ОТ-ПЦР. Пропуск экзона 5 (подтвержденный секвенированием по Сэнгеру: ср. фиг. 27B) был отчетливо доминирующим в клетках, обработанных ASO, что будет контрастировать со случаем «AR-примера 1» (ср. фиг. 23A).
Примеры in vitro и in vivo активности SCN9A-ASO.
Производные ПНК формулы I, комплементарно нацеливающиеся на множество сайтов сплайсинга в пре-мРНК для SCN9A (натриевого канала подтипа 9A) человека, оценивали в отношении их SCN9A-антисмысловой и экзон-пропускающей активности в клетках, а также у животных. Биологические примеры для этих SCN9A-ASO приведены в качестве примеров с целью иллюстрации того, что пропуск экзона эффективно индуцируется соединением формулы I, нацеливающимся на сайт сплайсинга в пре-мРНК-мишени, и, следовательно, не должны интерпретироваться как ограничение объема настоящего изобретение SCN9A-ASO.
SCN9A-пример 1. Пропуск экзонов, индуцируемый «SCN-ASO 7».
«SCN-ASO 7», указанный в таблице 4, представляет собой 16-мерный ASO, полностью комплементарный области в 5'-сайте сплайсинга, охватывающей место соединения экзона 4 и интрона 4 в пре-мРНК SCN9A человека, считываемой с гена SCN9A человека (доступ к которой получен из ссылочной последовательности NCBI: NC_000002.12). «SCN-ASO 7» комплементарно связывается с 16-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')UUGUUUUUGC|guaaguacuu]. «SCN-ASO 7» обладает способностью к 10-мерному перекрыванию с экзоном 4 и 6-мерному перекрыванию с интроном 4.
Учитывая, что клетки PC3, как известно, в избытке экспрессируют пре-мРНК для SCN9A человека [Br. J. Pharmacol, vol. 156, 420-431 (2009)], «SCN-ASO 7» оценивали с помощью SCN9A-специфической вложенной ПЦР в отношении его способности к индукции пропуска экзона 4 в пре-мРНК для SCN9A человека в клетках РС3 следующим образом.
[Клеточная культура и обработка ASO] Клетки PC3 (кат. № CRL-1435, ATCC) поддерживали в среде Хэма F-12K с добавлением 10% FBS, 1% стрептомицина/пенициллина, 1% L-глютамина и 1% натрия пирувата в атмосфере с 5% CO2 при 37οС.
Клетки РС3, выращенные в 60-мм чашке для культивирования, содержащей 5 мл среды, обрабатывали «SCN-ASO 7» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ.
[Экстракция РНК] После 18-часовой инкубации клетки РС3 обрабатывали 100 мкг/мл циклогексимида в течение еще 6 часов, чтобы заморозить трансляцию на рибосомах. Затем тотальную РНК экстрагировали из клеток с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы использовали для реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) и набора экзон-специфических праймеров [SCN-экзон 2_прямой: (5'→3')CTTTCTCCTTTCAGTCCTCT и SCN-экзон 9_обратный: (5'→3')CGTCTGTTGGTAAAGGTTTT] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 40 циклов, каждый состоящий из 30 сек при 94οС, 30 сек при 55οС и 2 мин при 72οС.
[Амплификация с помощью вложенной ПЦР] 1 мкл раствора кДНК (разведенного в 100 раз) подвергали амплификации с помощью вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [SCN-экзон 3n_прямой: (5'→3')GGACCAAAAATGTCGAGTATTT; и SCN-экзон 8_обратный: (5'→3')GCTAAGAAGGCCCAGCTGAA] в соответствии со следующими режимами циклирования: 95οС в течение 5 мин с последующими 35 циклами, каждый состоящий из 30 сек при 95οС, 30 сек при 50οС и 1 мин при 72οС.
Следует отметить, что праймер «SCN-экзон 3n_прямой» нацелен на место соединения экзона 3 и экзона 5 для эффективного исследования делеции экзона 4, хотя 22-мерный праймер все еще обладает способностью к 18-мерному комплементарному перекрыванию с местом соединения экзона 3 и экзона 4. Таким образом, «SCN-экзон 3n_прямой» распознает «место соединения экзона 3 и экзона 5» более селективно, чем «место соединения экзона 3 и экзона 4», встречаемое в полноразмерной мРНК для SCN9A. Последовательность праймера была сконструирована для обнаружения вариантов сплайсинга SCN9A, в которых отсутствует экзон 4, с большей чувствительностью, чем полноразмерную мРНК для SCN9A. Такой праймер для пропуска экзонов будет полезен для обнаружения вариантов сплайсинга мРНК, обладающих низкой метаболической стабильностью, поскольку полноразмерная мРНК, как правило, демонстрирует хорошую метаболическую стабильность, достигнутую в ходе эволюции на протяжении миллиардов лет.
[Идентификация продуктов пропуска экзона] Продукты вложенной ПЦР подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру. Пропуск экзона 4 был заметно сильным в клетках РС3, обработанных 1 аМ «SCN ASO 7», хотя пропуск экзона 4 был также заметен при концентрации 10 и 100 цМ, как показано на фиг. 28А. Полоса для пропуска экзона была однозначно подтверждена секвенированием по Сэнгеру, как показано на фиг. 28B.
SCN9A-пример 2. кПЦР с использованием SYBR Green для мРНК SCN9A в клетках РС3, обработанных «SCN-ASO 7».
«SCN-ASO 7» оценивали в отношении их способности ингибировать экспрессию мРНК для SCN9A человека в клетках РС3 с помощью кПЦР на фоне набора экзон-специфических праймеров следующим образом.
[Клеточная культура и обработка ASO] Клетки PC3, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, инкубировали с «SCN-ASO 7» в течение 24 часов в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ (2 чашки для культивирования для каждой концентрации)
[Экстракция РНК] Тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК MiniBEST» (кат. № 767, Takara) в соответствии с инструкциями производителя.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 20 мкл с использованием набора для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) и на фоне набора экзон-специфических праймеров [SCN-экзон 2_прямой: (5'→3')CTTTCTCCTTTCAGTCCTCT; и SCN-экзон 9_обратный: (5'→3')TTGCCTGGTTCTGTTCTT]. Режимы циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 15 сек при 94οС, 30 сек при 55οС и 2 мин при 72οС.
[Амплификация с помощью вложенной кПЦР] 1 мкл каждого раствора кДНК, разведенного в 50 раз, подвергали ПЦР-реакции в режиме реального времени в объеме 20 мкл на фоне наборов экзон-специфических праймеров [SCN-экзон 4_прямой: (5'→3')GTACACTTTTACTGGAATATATATAC; SCN-экзон 4_обратный: (5'→3')AATGACGACAAAATCCAGC; SCN-экзон 5_прямой: (5'→3')GTATTTAACAGAATTTGTAAACCT; SCN-экзон 5_обратный: (5'→3')CTGGGATTACAGAAATAGTTTTCA; SCN-экзон 6_прямой: (5'→3')GAAGACAATTGTAGGGGC; SCN-экзон 6_обратный: (5'→3')GTCTTCTTCACTCTCTAGGG]. Реакции ПЦР исследовали с использованием SYBR Green (Takara, Japan). Режимы циклирования: 95οС в течение 30 сек с последующими 40 циклами, каждый состоящий из 5 сек при 95οС и 30 сек при 60οС.
[Результаты кПЦР] Индивидуальный уровень экзонов в клетках, обработанных ASO, приводили к уровню экзонов в клетках отрицательного контроля (т.е. без обработки ASO). На фиг. 29(A) суммируются результаты кПЦР. Уровни экспрессии экзонов 4-6 значимо снижались приблизительно на 70%, 40% и 20~30% при концентрации 10, 100 и 1000 цМ, соответственно.
SCN9A-пример 3. кПЦР с использованием SYBR Green для мРНК SCN9A в клетках РС3, обработанных «SCN-ASO 3».
«SCN-ASO 3», указанный в таблице 4, представляет собой 14-мерный ASO, нацеливающийся на 5'-сайт сплайсинга, охватывающий место соединения экзона 4 и интрона 4 в пре-мРНК для SCN9A человека. «SCN-ASO 3» комплементарно связывается с 12-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→ 3')UUGUUUUUGC|gua"ag"uacuu], в которой два несоответствия отмечены знаком кавычки (""). «SCN-ASO 3» обладает способностью к 7-мерному комплементарному перекрыванию с экзоном 4 и 5-мерному комплементарному перекрыванию с интроном 4.
«SCN-ASO 3» оценивали в отношении его способности ингибировать экспрессию полноразмерной мРНК для SCN9A в клетках PC3 в соответствии с протоколом, описанным в «SCN9A-примере 2», если не указано иное.
Результаты кПЦР представлены, как показано на фиг. 29B. Уровни экспрессии экзонов 4-6 знамо снижались на >90%, приблизительно 70% и приблизительно 80% при концентрации 10, 100 и 1000 цМ, соответственно. Как и в случае с «SCN-ASO 7», при 10 цМ демонстрировалось наибольшее ингибирование полноразмерной мРНК для SCN9A.
SCN9A-пример 4. кПЦР с использованием SYBR Green для мРНК SCN9A в клетках PC3, обработанных «SCN-ASO 8».
«SCN-ASO 8», указанный в таблице 4, представляет собой 17-мерный ASO, нацеливающийся на область в 5'-сайте сплайсинга, охватывающую место соединения экзона 4 и интрона 4 в пре-мРНК для SCN9A человека. «SCN-ASO 8» комплементарно связывается с 15-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК [(5'→3')UUGUUUUUGC|gua"ag"uacuu], в которой несоответствие помечено знаком кавычки (""). «SCN-ASO 8» обладает способностью к 10-мерному комплементарному перекрыванию с экзоном 4 и 5-мерному комплементарному перекрыванию с интроном 4 в пре-мРНК для SCN9A человека.
«SCN-ASO 8» оценивали в отношении его способности ингибировать экспрессию полноразмерной мРНК для SCN9A в клетках PC3 в соответствии с протоколом, описанным в «SCN9A-примере 2», если не указано иное.
Результаты кПЦР представлены, как показано на фиг. 29C. Уровни экспрессии экзонов 4-6 значимо снижались на 50~70% и 70~80% при концентрации 10 и 100 цМ, соответственно.
SCN9A-пример 5. Ингибирование натриевого тока согласно анализу с использованием CoroNa в клетках РС3, обработанных «SCN-ASO 7».
Натриевый ток в клетках измеряется патч-клампом (фиксацией потенциалов). Когда ионы натрия поступают в клетку, внутриклеточный уровень ионов натрия увеличивается. Внутриклеточный уровень натрия может быть измерен с использованием красителя, чувствительного к ионам натрия. «CoroNa Green» представляет собой краситель с хелатором ионов натрия типа краун-эфира. При хелатировании с ионом натрия «CoroNa Green» испускает зеленую флуоресценцию. «CoroNa Green» использовался для опосредованного измерения внутриклеточного уровня натрия. Было обнаружено, что уровень натрия, измеренный с использованием «CoroNa Green», хорошо коррелирует с током ионов натрия, измеренным фиксацией потенциалов, генерированных ионами натрия [Proc. Natl. Acad. Sci. USA vol 106(38), 16145-16150 (2009)]
Известно, что клетки РС3 экспрессируют мРНК для SCN9A человека в избытке, хотя есть и другие сопутствующим образом экспрессируемые подтипы SCN [Br. J. Pharmacol. vol 156, 420-431 (2009)]. Таким образом, ингибирование экспрессии мРНК для SCN9A может привести к значительному уменьшению натриевого тока в клетках РС3, если ток ионов натрия по подтипу натриевых каналов Nav1.7 занимает заметную часть от общего тока ионов натрия в клетках РС3. Следует отметить, что мРНК SCN9A кодирует подтип натриевых каналов Nav1.7.
«SCN-ASO 7» оценивали в отношении его способности ингибировать ток ионов натрия в клетках РС3 с использованием «CoroNa Green» следующим образом.
[Обработка ASO] Клетки PC3, выращенные в 35-мм чашке для культивирования, содержащей 2 мл среды F-12K, обрабатывали «SCN-ASO 7» в концентрации 0 цМ (отрицательный контроль), 100 цМ или 1 аМ.
[Анализ с использованием CoroNa] Спустя 30 часов клетки промывали 2 мл HBSS (сбалансированным солевым раствором Хенкса, кат. № 14025-092, Life Technologies), а затем вносили 2 мл свежего HBSS. Затем клетки обрабатывали 5 мкМ «CoroNa Green» (кат. № C36676, Life Technologies) при 37οС. Через 30 мин клетки дважды промывали 2 мл HBSS, и вносили 2 мл свежего HBSS. Чашку для культивирования устанавливали на флуоресцентный микроскоп Olympus, снабженный цифровой видеокамерой, для непрерывной фиксации зеленых флуоресцентных изображений клеток. Клетки резко подвергали обработке 100 мМ NaCl, и затем изменения флуоресцентных изображениях клеток регистрировали в цифровой форме в течение 3 мин. В среднем было приблизительно 4 клеток в каждом кадре. Интенсивности флуоресценции каждой отдельной клетки отслеживались с точностью до секунды. Записи интенсивностей внутриклеточной флуоресценции отдельных клеток накладывали и усредняли в каждый момент времени. Записи в среднем от отдельных клеток при каждой концентрации ASO наносили на графики, как показано на фиг. 30А с использованием программы ImageJ (версии 1.50i, NIH). Среднюю запись интенсивности флуоресценции брали в качестве индивидуальной записи внутриклеточной концентрации натрия для клеток, обработанных «SCN-ASO 7» в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
[Результаты анализа с использованием CoroNa] Наблюдаемые записи интенсивности внутриклеточной флуоресценции суммированы на фиг. 29B. Записи (кривые) интенсивности флуоресценции для клеток, обработанных 1000 цМ «SCN-ASO 7», располагались ниже записей для клеток без обработки ASO. Средняя интенсивность флуоресценции клеток без обработки ASO составляла 81,86 (произвольных единиц) в момент времени 100 сек. Между тем, средняя интенсивность флуоресценции клеток, обработанных 1000 цМ «SCN-ASO 7», составляла 51,47 (произвольных единиц) в момент времени 100 сек. Таким образом, 30-часовая инкубация с 1000 цМ «SCN-ASO 7» индуцировала значимое снижение активности натриевого канала на 37% (р<0,05 по t-критерию Стьюдента) в клетках РС3. Принимая во внимание, что клетки РС3 экспрессируют различные подтипы потенциалозависимого натриевого канала (VGSC), снижение на 37% считается заметным для ингибирования экспрессии Nav1.7 с помощью «SCN-ASO 7». Не было заметного уменьшения натриевого тока в клетках, обработанных 100 цМ «SCN-ASO 7».
SCN9A-пример 6. Ингибирование натриевого тока согласно анализу с использованием CoroNa в клетках РС3, обработанных «SCN-ASO 3».
«SCN-ASO 3» оценивали в отношении его способности ингибировать натриевый ток в клетках РС3 с использованием «CoroNa Green» в соответствии с протоколом, описанным в «SCN9A-пример 5», если не указано иное.
Наблюдаемые записи интенсивности флуоресценции клеток представлены на фиг. 30B. Средняя запись (кривая) интенсивности флуоресценции располагалась ниже в клетках, обработанных «SCN-ASO 3», чем в клетках без обработки ASO. Средняя интенсивность флуоресценции клеток без обработки ASO составляла 89,32 (произвольных единиц) в момент времени 100 сек. Между тем, средняя интенсивность флуоресценции клеток, обработанных 1000 цМ «SCN-ASO 3», составляла 61,36 (произвольных единиц) в момент времени 100 сек. Таким образом, 1000 цМ «SCN-ASO 3» значимо (р<0,01) уменьшал натриевый ток на 31% в клетках РС3. Уменьшение, вызванное 100 цМ «SCN-ASO 3», составило 18%, хотя и не имело значимости.
SCN9A-пример 7. Ингибирование натриевого тока в клетках РС3, обработанных «SCN-ASO 8».
«SCN-ASO 8» оценивали в отношении его способности ингибировать натриевый ток в клетках РС3 с использованием «CoroNa Green» в соответствии с протоколом, описанным в «SCN9A-примере 3», если не указано иное.
Наблюдаемые записи интенсивности флуоресценции клеток представлены на фиг. 30C. Средняя запись (кривая) интенсивности флуоресценции располагалась ниже в клетках, обработанных «SCN-ASO 8», чем в клетках без обработки ASO. Средняя интенсивность клеточной флуоресценции клеток без обработки ASO составляла 130,32 (произвольных единиц) в момент времени 100 сек. Между тем, средняя интенсивность флуоресценции клеток, обработанных 1000 цМ «SCN-ASO 8», составляла 89,7 (произвольная единица) в момент времени 100 сек. Таким образом, 1000 цМ «SCN-ASO 8» значимо (р<0,001) уменьшал натриевый ток на 31% в клетках РС3. Уменьшение, вызванное 100 мкМ «SCN-ASO 8», составило 30% (р<0,001).
SCN9A-пример 8. Пропуск экзонов, индуцируемый «SCN-ASO 27» в клетках РС3 (A).
«SCN-ASO 27», указанный в таблице 5, представляет собой 14-мерный ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения «интрона 3» и «экзона 4» в пре-мРНК для SCN9A человека. 14-мерная последовательность-мишень в 3'-сайте сплайсинга отмечена «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК для SCN9A [(5'→3') uuguguuuag|GUACACUUUU]. «SCN-ASO 27» обладает способностью к 6-мерному перекрыванию с «интроном 3» и 8-мерному перекрыванию с «экзоном 4».
«SCN-ASO 27» оценивали в отношении его способности к индукции пропуска «экзона 4» в клетках РС3, как описано в «SCN9A-примере 1», если не указано иное.
[Клеточная культура и обработка ASO] Клетки PC3, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 1, 10 или 100 цМ.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [SCN-экзон 2n_прямой: (5'→3')CCACCGGACTGGACCAAAAA; и SCN-экзон 9n_обратный: (5'→3')GCTAAGAAGGCCCAGCTGAA] в соответствии со следующими режимами циклирования: 95οС в течение 2 мин с последующими 34 циклами, каждый состоящий из 30 сек при 95οС, 30 сек при 55οС и 1 мин при 72οС.
[Идентификация продуктов пропуска экзонов] На фиг. 31А представлены данные электрофореза продуктов вложенной ПЦР, в которых клетки, обработанные «SCN-ASO 27», давали сильную полосу для ПЦР-продукта, приписываемого пропуску экзонов 4-5. Однако интенсивность полосы для ПЦР-продукта в виде полноразмерной мРНК для SCN9A была сильнее в образцах обработки 10 цМ и 100 цМ ASO, чем в образцах отрицательного контроля. Странный характер зависимости ответа от дозы во вложенной ПЦР мог быть обусловлен активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) кольцевой РНК (EIciRNA)», накапливаемой во время пропуска экзонов с помощью «SCN-ASO 27» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)]. С помощью секвенирования по Сэнгеру было подтверждено, что ПЦР-продукт в случае пропуска экзонов соответствует пропускам экзонов 4-5 (ср. фиг. 31B)
SCN9A-пример 9. Пропуск экзонов, индуцируемый «SCN-ASO 27» в клетках РС3 (B).
«SCN-ASO 27» оценивали в отношении его способности к индукции пропуска «экзона 4» в клетках РС3, как описано в «SCN9A-примере 8», если не указано иное. В этом эксперименте клетки РС3 обрабатывали «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 1, 10, 100 и 1000 аМ в течение 24 часов.
[Амплификация с помощью вложенной ПЦР]. Вложенную реакцию ПЦР проводили на фоне набора праймеров [SCN-экзон 3/6_прямой: (5'→3')GGACCAAAAATGTCGAGCCT; и SCN-экзон 9n_обратный: (5'→3') GCTAAGAAGGCCCAGCTGAA], сконструированных для селективной амплификации продукта, имеющего последовательность места соединения экзона 3 и экзона 6.
Следует отметить, что последовательность праймера «SCN-экзон 3/6_прямой» нацелена на место соединения экзона 3 и экзона 6 для исследования пропуска экзонов 4-5, хотя 20-мерный праймер все еще сохраняет способность к 17-мерному комплементарному перекрыванию с местом соединения экзона 3 и экзона 4. Таким образом, последовательность праймера «SCN-экзон 3/6_прямой» распознает «место соединения экзона 3 и экзона 6» более селективно, чем «место соединения экзона 3 и экзона 4», встречаемое в полноразмерной мРНК для SCN9A. Последовательность праймеров была сконструирована для обнаружения варианта сплайсинга SCN9A, в котором отсутствуют экзоны 4-5, с большей чувствительностью, чем полноразмерную мРНК для SCN9A. Такой праймер для пропуска экзонов был бы полезен для обнаружения вариантов сплайсинга мРНК с низкой метаболической стабильностью, поскольку полноразмерная мРНК, как правило, демонстрирует хорошую метаболическую стабильность, достигнутую в ходе эволюции на протяжении миллиардов лет.
На фиг. 31C представлены данные электрофореза продуктов вложенной ПЦР, в которых клетки, обработанные «SCN-ASO 27», давали сильную полосу для ПЦР-продукта, приписываемого пропуску экзонов 4-5, что было подтверждено секвенированием по Сэнгеру.
SCN9A-пример 10. кПЦР с помощью одностадийного синтеза кДНК для мРНК SCN9A в клетках РС3, обработанных «SCN-ASO 27».
«SCN-ASO 27» оценивали с помощью SCN9A-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК для SCN9A человека в клетках РС3, как описано в «SCN9A-примере 2», если не указано иное.
[Обработка ASO] Клетки PC3 обрабатывали «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 0,1, 1 или 10 аМ в течение 24 часов (2 чашки для культивирования на каждую концентрацию ASO).
[Синтез кДНК с помощью одностадийной ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР (Invitrogen, США) на фоне набора экзон-специфических праймеров [SCN-экзон 2_прямой: (5'→3')CTTTCTCCTTTCAGTCCTCT; и SCN-экзон 8/9_обратный: (5'→3')CGTCTGTTGGTAAAGGTTTT] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οС, 30 сек при 55οС и 2 мин при 72οС.
[Амплификация с помощью вложенной кПЦР] 1 мкл каждого раствора кДНК, разведенного в 100 раз, подвергали реакции ПЦР в режиме реального времени в объеме 20 мкл на фоне набора экзон-специфических праймеров [SCN-экзон 3_прямой: (5'→3')TGACCATGAATAACCCAC; и SCN-экзон 4_обратный(2): (5'→3')GCAAGGATTTTTACAAGT] в соответствии со следующими режимами циклирования: 95οС в течение 30 сек с последующими 40 циклами, каждый состоящий из 5 сек при 95οС и 30 сек при 60οС. Реакцию кПЦР контролировали с использованием TaqMan-зонда [(5'→3')5,6-FAM-GGACCAAAA-Zen-ATGTCGAGTACAC-3IABkFQ], нацеленного на место соединения экзона 3 и экзона 4 в полноразмерной мРНК для SCN9A.
Уровень полноразмерной мРНК для SCN9A значимо (по t-критерию Стьюдента) снижался в клетках, обработанных «SCN-ASO 27», приблизительно на 35-45%, как показано на фиг. 32A.
SCN9A-пример 11. кПЦР с помощью синтеза кДНК с использованием случайных гексамеров для мРНК SCN9A в клетках РС3, обработанных «SCN-ASO 27».
«SCN-ASO 27» оценивали с помощью SCN9A-специфической кПЦР в отношении его способности к вызову изменений уровня мРНК для SCN9A человека в клетках PC3, как описано в «SCN9A-примере 10», если не указано иное. кДНК синтезировали с использованием случайных гексамеров и подвергали реакции SCN9A-специфической кПЦР с использованием зонда TaqMan.
Уровень полноразмерной мРНК для SCN9A значимо (по t-критерию Стьюдента) снижался в клетках, обработанных «SCN-ASO 27», приблизительно на 50-60%, как показано на фиг. 32B.
SCN9A-пример 12. Ингибирование натриевого тока согласно анализу с использованием CoroNa в SNL-активированных клетках L5 DRG крысы с помощью «SCN-ASO 27».
«SCN-ASO 27» представляет собой 14-мерный SCN9A-ASO, полностью комплементарный пре-мРНК для SCN9A человека, но имеет одно несоответствие с пре-мРНК для SCN9A крысы, считываемой с геномной ДНК крысы [ссылочная последовательность NCBI: NC_000002.12]. «SCN-ASO 27» обладает способностью к 13-мерному комплементарному перекрыванию и имеет одно несоответствие с пре-мРНК для SCN9A крысы, как отмечено «жирным шрифтом» и «подчеркиванием» в 20мерной последовательности пре-мРНК крысы [(5'→3')uuuc"c"uuuag|GUACACUUUU], в которой одно несоответствие отмечено знаком кавычки ("").
«SCN-ASO 27» оценивали в отношении его способности ингибировать ток ионов натрия в клетках DRG (ганглиях задних корешков) крыс с использованием «CoroNa Green» следующим образом.
[Перевязка спинномозговых нервов] Перевязка спинномозговых нервов (SNL) вызывает невропатию в ганглиях задних корешков (DRG) и спинном мозге и широко использовалась в качестве модели невропатических болей [Pain vol 50 (3), 355-363 (1992)]. Однако в зависимости от того, как перевязывается спинномозговой нерв(ы), может быть несколько вариаций SNL. Степень и продолжительность невропатии в DRG, по-видимому, варьируются в зависимости от того, как перевязывается спинномозговой нерв(ы) [Pain vol 43 (2), 205-218 (1990)]. Двойная перевязка спинномозгового нерва L5 и L6 (т.е. «перевязка L5/L6») вызывает более тяжелую и дольше сохраняющуюся невропатию, чем только перевязка спинномозгового нерва L5 (т. е. «перевязка L5»).
[Хирургическое вмешательство SNL с помощью перевязки L5/L6]. В день 0 самцов крыс SD в возрасте 6 недель анестезировали с помощью золетила/ромпуна. Затем спинномозговой нерв L5 и L6 (левая сторона) обнажали и туго перевязывали. Мышцы и кожу закрывали и обрезали с использованием соответствующих асептических процедур. У крыс время от времени проверяли чувствительность по бальной оценке фон Фрея в течение 4 недель.
[Приготовление нервных клеток DRG]. В день 31 крысу с низкой оценкой фон Фрея умерщвляли для извлечения как левого (перевязанная сторона), так и правого (не перевязанная сторона) DRG. DRG погружали в 0,5 мл PBS сразу после извлечения. Клетки DRG готовили следующим образом в соответствии с процедурами, раскрытыми в литературе [Methods Mol Biol. vol 846, 179-187 (2012); PLOS One vol 8(4); e60558 (2013)].
 DRG погружали в пробирку Эппендорфа объемом 1,5 мл, содержащую 0,2 мл 0,125% коллагеназы (коллагеназы типа IV, кат. № C5138-100MG, Sigma) в HBSS (сбалансированном солевом растворе Хенкса, кат. № 14025-092, Life Technologies), нарезали ножницами на мелкие кусочки и инкубировали в течение 20 мин в термостате при 37οC при 5% CO2 и относительной влажности 95%;
DRG погружали в пробирку Эппендорфа объемом 1,5 мл, содержащую 0,2 мл 0,125% коллагеназы (коллагеназы типа IV, кат. № C5138-100MG, Sigma) в HBSS (сбалансированном солевом растворе Хенкса, кат. № 14025-092, Life Technologies), нарезали ножницами на мелкие кусочки и инкубировали в течение 20 мин в термостате при 37οC при 5% CO2 и относительной влажности 95%;  добавляли 50 мкл 0,25% трипсина/ЭДТА в пробирку Эппендорфа, которую хранили в термостате еще 10 мин,
добавляли 50 мкл 0,25% трипсина/ЭДТА в пробирку Эппендорфа, которую хранили в термостате еще 10 мин,  в пробирку Эппендорфа вносили 1 мл полной среды DMEM, и ее подвергали осаждению центрифугированием при 600хg в течение 5 мин,
в пробирку Эппендорфа вносили 1 мл полной среды DMEM, и ее подвергали осаждению центрифугированием при 600хg в течение 5 мин,  полученный осадок суспендировали в 4 мл среды Neurobasal-А (среды Neurobasal®, кат № 21103-049, Gibco), дополненной 2X В-27 (В-27®, не содержащей сыворотки добавкой, кат № 17504-044, Gibco), 1X пенициллином/стрептомицином, 1X L-глютамином, и 1 мл клеточной суспензии осторожно высевали на покрытое ламинином покровное стекло (кат. № GG-25-1,5-ламинин, Neuvitro), помещаемое в 35 мм-чашку для культивирования;
полученный осадок суспендировали в 4 мл среды Neurobasal-А (среды Neurobasal®, кат № 21103-049, Gibco), дополненной 2X В-27 (В-27®, не содержащей сыворотки добавкой, кат № 17504-044, Gibco), 1X пенициллином/стрептомицином, 1X L-глютамином, и 1 мл клеточной суспензии осторожно высевали на покрытое ламинином покровное стекло (кат. № GG-25-1,5-ламинин, Neuvitro), помещаемое в 35 мм-чашку для культивирования;  через один день после посева в чашку осторожно вносили еще 1 мл свежей среды Neurobasal-A;
через один день после посева в чашку осторожно вносили еще 1 мл свежей среды Neurobasal-A;  через два дня после посева среду заменяли на 2 мл свежей среды Neurobasal-A с добавлением 1 мкМ Ara-C (кат. № C1768-100MG, Sigma) для селективного подавления роста клеток, отличных от нервных клеток DRG;
через два дня после посева среду заменяли на 2 мл свежей среды Neurobasal-A с добавлением 1 мкМ Ara-C (кат. № C1768-100MG, Sigma) для селективного подавления роста клеток, отличных от нервных клеток DRG;  через четыре дня после посева среду снова заменяли 2 мл свежей среды Neurobasal-A с добавлением 1 мкМ Ara-C; и
через четыре дня после посева среду снова заменяли 2 мл свежей среды Neurobasal-A с добавлением 1 мкМ Ara-C; и  через пять или шесть дней после посева нервные клетки DRG обрабатывали «SCN-ASO 27».
через пять или шесть дней после посева нервные клетки DRG обрабатывали «SCN-ASO 27».
[Обработка ASO и анализ с использованием CoroNa] Нервные клетки L5 DRG либо с перевязкой L5/L6, либо без перевязки L5/L6 обрабатывали «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 100 или 1000 цМ. Через 30 часов клетки промывали 2 мл HBSS, а затем вносили 2 мл свежего HBSS. Затем клетки обрабатывали 5 мкМ «CoroNa Green» при 37οС. Через 30 мин клетки дважды промывали 2 мл HBSS, и вносили 2 мл свежего HBSS. Чашку для культивирования устанавливали на флуоресцентный микроскоп, снабженный камерой с зарядовой связью, для непрерывной фиксации зеленых флуоресцентных изображений клеток. Клетки резко подвергали обработке 10 мМ NaCl, и затем изменения интенсивности флуоресценции клеток регистрировали в цифровой форме в течение 300 сек. В случае каждой фиксации изображения было от 4 до 5 клеток на кадр. Интенсивности флуоресценции каждой отдельной клетки отслеживались с точностью до секунды. Записи интенсивностей внутриклеточной флуоресценции отдельных клеток усреднялись с использованием программы ImageJ (версии 1.50i, NIH), и записи в среднем представлены на фиг. 33А и 33В для клеток с «перевязкой L5/L6» и без «перевязки L5/L6», соответственно. Среднюю интенсивность флуоресценции брали в качестве индивидуальной записи внутриклеточной концентрации натрия для клеток, обработанных «SCN-ASO 27» в концентрации 0 (отрицательный контроль), 100 или 1000 цМ.
[Результаты анализа с использованием CoroNa] В клетках, стимулированных перевязкой L5/L6 (ср. фиг. 33А), 30-часовое воздействие «SCN-ASO 27» в концентрации 100 цМ или 1 аМ давало значимое (по t-критерию Стьюдента) снижение средней интенсивности клеточной флуоресценции на 80-85% в момент времени 150 сек.
В клетках без перевязки L5/L6 (ср. фиг. 33B) 30-часовое воздействие «SCN-ASO 27» в концентрации 1 аМ давало снижение интенсивности флуоресценции приблизительно на 50%. В случае нестимулированных клеток, подвергнутых воздействию «SCN-ASO 27» в концентрации 100 цМ, не было снижения интенсивности флуоресценции. Интенсивность флуоресценции клеток без перевязки L5/L6 была значительно меньше, чем у клеток, стимулированных перевязкой L5/L6, что позволяет предположить, что L5/L6 индуцируют заметную активацию функции натриевого канала Nav1.7.
Известно, что нервные клетки DRG без невропатической стимуляции экспрессируют различные подтипы VGSC, в том числе Nav1.7, Nav1.8, Nav1.2 и т.д. Подтип Nav1.7 демонстрирует ограниченный вклад в общий натриевый ток в нервных клетках DRG без стимуляции [Nature Comm. vol 3, Article Number 791: DOI:10.1038/ncomms1795 (2012)]. Нервные клетки DRG без перевязки L5/L6 могут демонстрировать ограниченный вклад в натриевый ток из подтипа Nav1.7.
Между тем известно, что нервные клетки усиливают экспрессию Nav1.7 в ответ на сохраняющуюся невропатию [J Biol Chem. vol 279(28), 29341-29350 (2004); J Neurosci. vol 28(26), 6652-6658 (2008)]. «SCN-ASO 27» в концентрации и 100 цМ, и 1 аМ ингибировал натриевый ток на 80-85% в нервных клетках, стимулированных «перевязкой L5/L6». Более высокая степень ингибирования натриевого тока с помощью «SCN-ASO 27» в клетках DRG с «перевязкой L5/L6» согласуется с активацией Nav1.7 в нервных клетках из-за хронической невропатии.
SCN9A-пример 13. Ингибирование экспрессии белка Nav1.7 в нервных клетках DRG L5 с помощью «SCN-ASO 30».
«SCN-ASO 30» представляет собой 14-мерный ASO, полностью комплементарный пре-мРНК для SCN9A крысы, тогда как «SCN-ASO 27» представляет собой 14-мерный ASO, полностью комплементарный пре-мРНК для SCN9A человека. «SCN-ASO 30» имеет одно несоответствие с «SCN-ASO 27» на С-конце. «SCN-ASO 30» против пре-мРНК для SCN9A крысы может служить хорошим модельным ASO для «SCN-ASO 27» против пре-мРНК для SCN9A человека.
«SCN-ASO 30» оценивали в отношении его способности ингибировать экспрессию белка Nav1.7 в нервных клетках DRG крысы, как описано ниже.
[Приготовление нервных клеток DRG] Самцов крыс SD (возрастом 7 недель) подвергали тугой «перевязке L5/L6». Через 7 дней 4 крыс анестезировали с помощью золетила/ромпуна для взятия образца DRG L5 с перевязанной стороны. DRG объединяли и обрабатывали для получения нервных клеток DRG, как описано в «SCN9A-примере 12».
[Обработка ASO] Нервные клетки DRG обрабатывали «SCN-ASO 30» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ в течение 24 часов, а затем подвергали лизису для Вестерн-блоттинга на фоне антитела против С-конца белка Nav1.7 (кат. № ab85015, Abeam). β-актин исследовали для сравнения.
[Ингибирование экспрессии Nav1.7]. На фиг. 34A представлены данные Вестерн-блоттинга, полученные в случае нервных клетках DRG, обработанных «SCN-ASO 30» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ. Все лизаты давали сильную полосу в положении, соответствующем 170 К, которую можно было бы приписать фрагменту или метаболиту полноразмерного белка Nav1.7. Полоса для полноразмерного белка Nav1.7 была обнаружена в положении, соответствующем 220~240 К, только с лизатами отрицательного контроля и 10 цМ «SCN-ASO 30». Таким образом, экспрессия Nav1.7 была заметно ингибирована в нервных клетках DRG крысы после 24-часовой инкубации с «SCN-ASO 30» в концентрации 100 и 1000 цМ.
SCN9A-пример 14. Ингибирование натриевого тока в нервных клетках DRG L5 крысы с помощью «SCN-ASO 30».
«SCN-ASO 30» оценивали в отношении его способности ингибировать натриевый ток в нервных клетках DRG L5 крысы, стимулированных перевязкой L5/L6, как указано ниже.
[Приготовление нервных клеток DRG] Самцов крыс SD (возрастом 6 недель) подвергали тугой «перевязке L5/L6». Через 7 дней крыс анестезировали с помощью золетила/ромпуна для извлечения DRG L5 с перевязанной стороны. Нервные клетки DRG L5 готовили следующим образом: DRG погружали в пробирку Эппендорфа объемом 1,5 мл, содержащую 0,2 мл 0,125% коллагеназы в HBSS, нарезали ножницами на мелкие кусочки и инкубировали в течение 20 мин в термостате при 37ο C при 5% CO2 и относительной влажности 95%; добавляли 50 мкл 0,25% трипсина/ЭДТА в пробирку Эппендорфа, и пробирку Эппендорфа хранили в термостате еще 10 мин, в пробирку Эппендорфа вносили 1 мл полной среды DMEM, и ее подвергали осаждению центрифугированием при 600хg в течение 5 мин, затем полученный осадок суспендировали в 4 мл среды Neurobasal-А (среды Neurobasal®, кат № 21103-049, Gibco), дополненной 2X В-27 (В-27®, не содержащей сыворотки добавкой, кат № 17504-044, Gibco), 1X пенициллином/стрептомицином, 1X L-глютамином; суспензию клеток DRG переносили на приблизительно один часа в закрытом виде в 15-мл пробирку фирмы Falcon, содержащую приблизительно 15 мл среды Neurobasal-A; 0,5 мл суспензии клеток осторожно высевали на покрытое ламинином покровное стекло, помещенное в лунку 24-луночного планшета для культивирования; клетки, высеянные в планшет для культивирования, инкубировали в термостате с CO2 при 37οC в течение 2 часов для прикрепления клеток к покровному стеклу, а затем обрабатывали «SCN-ASO 30» в концентрации 0 (отрицательный контроль) или 100 цМ в течение 4 часов в термостате; и нервные клетки DRG подвергали измерению натриевого тока с помощью анализа с использованием патч-клампа вручную на устройстве для фиксации потенциалов, генерированных ионами натрия (Axopatch 200B Amplifier, Axon Instruments).
[Результаты анализа с помощью патч-клампа] На фиг. 34В представлены данные о натриевом токе, приведенные к размеру клеток. После инкубации с 100 цМ «SCN-ASO 30» в течение 4 часов натриевый ток значимо (p<0,01 по t-критерию Стьюдента) уменьшался приблизительно на 90% в нервных клетках DRG, экспрессирующих чувствительные к тетродотоксину натриевые каналы, т.е. в нервных клетках малых размер (N=4 клетки на группу).
SCN9A-пример 15. кПЦР с помощью одностадийного синтеза кДНК для мРНК SCN9A в клетках DRG крысы, обработанных «SCN-ASO 30».
«SCN-ASO 30» оценивали с помощью SCN9A-специфической вложенной кПЦР в отношении его способности ингибировать экспрессию мРНК для SCN9A в клетках DRG крысы следующим образом.
[Приготовление клеток DRG L5] Самца крысы SD в возрасте 4 недель анестезировали с помощью золетила/ромпуна для извлечения DRG L5. Образцы DRG были объединены и обработаны для приготовления клеток DRG L5, как описано в «SCN9A-пример 12».
[Обработка ASO] Клетки DRG крысы обрабатывали «SCN-ASO 30» в концентрации 0 (отрицательный контроль), 10, 30, 100, 300 или 1000 цМ (1 чашка для культивирования на каждую концентрацию ASO)
[Экстракция РНК и синтез кДНК с помощью одностадийной ПЦР] Через 24 часа тотальную РНК экстрагировали из клеток с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя. 200 нг РНК-матрицы использовали для реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР (Invitrogen, США) на фоне набора экзон-специфических праймеров [SCN-экзон 2(3)_прямой: (5'→3')CAATCTTCCGTTTCAACGCC; и SCN-экзон 10_обратный: (5'→3') ACCACAGCCAGGATCAAGTT] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οС, 30 сек при 55οС и 2 мин при 72οС.
[Амплификация с помощью вложенной кПЦР] 1 мкл каждого раствора кДНК (дубликат на каждую концентрацию), разведенного в 100 раз, подвергали реакции ПЦР в режиме реального времени в объеме 20 мкл с использованием зонда TaqMan (кат. № Rn01514993_mH, ThermoFisher), нацеленным на место соединения экзона 3 и экзон 4 SCN9A, в соответствии со следующими режимами циклирования: 95οС в течение 30 сек с последующими 40 циклами, каждый состоящий из 5 сек при 95οС и 30 сек при 60οС.
На фиг. 35A представлены данные кПЦР. Уровень экспрессии полноразмерной мРНК для SCN9A значимо (по t-критерию Стьюдента) снижался в клетках, обработанных «SCN-ASO 30», приблизительно на 45~60%, хотя на каждую концентрацию ASO была одна чашка для культивирования.
SCN9A-пример 16. кПЦР с помощью синтеза кДНК с использованием случайных гексамеров для мРНК SCN9A в клетках DRG крысы, обработанных «SCN-ASO 30».
«SCN-ASO 30» оценивали с помощью SCN9A-специфической кПЦР в отношении его способности ингибировать экспрессию мРНК для SCN9A в клетках DRG L5 крысы. Тотальную РНК получали, как описано в «SCN9A-примере 15», и подвергали синтезу кДНК с использованием случайных гексамеров. Растворы кДНК (дубликат для каждой концентрации ASO) разводили в 100 раз, и 1 мкл каждого разведенного ПЦР-продукта подвергали реакции ПЦР в режиме реального времени в объеме 20 мкл с использованием зонда TaqMan, нацеленного на место соединения экзона 3 и экзона 4 SCN9A, в соответствии со следующими режимами циклирования: 95οС в течение 30 сек последующими 40 циклами, каждый состоящий из 5 сек при 95οС и 30 сек при 60οС.
Растворы кДНК также подвергали амплификации с помощью кПЦР для мРНК GAPDH. Значения Ct мРНК для SCN9A приводили к значениям Ct мРНК для GAPDH.
На фиг. 35B представлены данные SCN9A-специфической кПЦР, приведенные к GAPDH. Уровень экспрессии полноразмерной мРНК для SCN9A значимо (t-критерий Стьюдента) снижался в клетках, обработанных «SCN-ASO 30», приблизительно на 45~75%.
SCN9A-пример 17. Обратное развитие аллодинии с помощью SCN9A-ASO у крыс с периферической невропатической болью, вызванной диабетом.
Ген SCN9A кодирует α-субъединицу подтипа Nav1.7 VGSC. Существует очень небольшое количество индивидуумов, которые не испытывают сильных болей, но являются нормальными по другим сенсорным функциям. У таких индивидуумов было обнаружено, что ген SCN9A мутирован с кодированием нефункционального подтипа Nav1.7 [Nature vol 444, 894-898 (2006)]. Это было названо «каналопатией SCN9A». Поведенческие фенотипы каналопатии SCN9A у человека довольно часто воспроизводились у мышей с нокаутом SCN9A [PLOS One 9(9): e105895 (2014)]. Таким образом, SCN9A-ASO формулы I могут проявлять болеутоляющую активность в моделях боли на животном, сопровождающей увеличении экспрессии Nav1.7.
«SCN-ASO 7», «SCN-ASO 8», «SCN-ASO 21», «SCN-ASO 35», «SCN-ASO 36» и «SCN-ASO 37» были оценены в отношении их способности обращать вспять развитие аллодинии у крыс с вызванной диабетом периферической невропатической болью (DPNP). В этом примере шесть SCN9A-ASO, нацеливающихся всего на пять сайтов сплайсинга, были оценены в отношении их способности к обратному развитию аллодинии, вызванной диабетической невропатией у крыс.
[Индукция DPNP и группирование]. Диабет был вызван у крыс путем внутрибрюшинной инъекции стерптозотоцина в дозе 60 мг/кг в день 0. В день 10 крыс с DPNP случайным образом распределяли по 6 группам: отрицательного контроля (только носитель), «SCN-ASO 7» 100 пмоль/кг, «SCN-ASO 8» 100 пмоль/кг, «SCN-ASO 21» 100 пмоль/кг, «SCN-ASO 35» 100 пмоль/кг, «SCN-ASO 36» 100 пмоль/кг, и «SCN-ASO 37» 100 пмоль/кг. Животных группировали на основе бальных оценок по фон Фрею отдельных животных в день 10. (N=8~9 на группу). Аллодинию оценивали с использованием набора микрофиламентов (Touch Test®) в соответствии с методом «вверх и вниз» [J Neurosci. Methods vol 53 (1), 55-63 (1994)]
[Лечение ASO и оценка по фон Фрею] Растворы ASO для инъекций готовили путем последовательного разведения водных исходных растворов SCN9A-ASO до 100 нМ в PBS (фосфатно-солевом буфере). Животным вводили подкожно ASO в дозе 1 мл/кг в дни 11, 13, 15, 17 и 19. Оценку по фон Фрею проводили через 2 часа после введения дозы в дни 11, 13, 15, 17 и 19. Дополнительно оценку по фон Фрею проводили в дни 21 и 23 с целью оценки продолжительности терапевтической активности после введения конечной дозы. Ежедневные оценки по фон Фрею оценивали на статистическую значимость с помощью t-критерия Стьюдента относительно группы отрицательного контроля (только носитель, т.е. PBS).
[Терапевтическая активность] Полученные оценки по фон Фрею суммированы на фиг. 36. Развитие аллодинии было значительно обращено вспять с помощью всех ASO, за исключением «SCN-ASO 36» и «SCN-ASO 37», хотя «SCN-ASO 37» продемонстрировал четкую тенденцию терапевтической активности (р-значение=0,057 в день 19). Терапевтическая активность, как правило, постепенно увеличивалась при повторном введении дозы. Максимальная терапевтическая эффективность, основанная на оценках по фон Фрею в день 19, составила приблизительно 76% (значимая), 61% (значимая), 93% (значимая), 52% (значимая), 0% и 22% (незначимая, p-значение=0,05X) для «SCN-ASO 7», «SCN-ASO 8», «SCN-ASO 21», «SCN-ASO 35», «SCN-ASO 36» и «SCN-ASO 37», соответственно.
«SCN-ASO 7», «SCN-ASO 8», «SCN-ASO 21» и «SCN-ASO 35» обладают способностью к 5-мерному комплементарному перекрыванием с интроном-мишенью. В то же время, «SCN-ASO 36» и «SCN-ASO 37» обладают способностью к 4-мерному и 3-мерному комплементарному перекрыванию с интроном-мишенью, соответственно. Хотя существует ряд факторов, влияющих на терапевтическую эффективность, величина комплементарного перекрывания с интроном-мишенью, по-видимому, влияет на терапевтическую эффективность.
В этом примере антисмысловая активность in vivo наблюдалась в случае SCN9A-ASO, нацеливающихся на 4 из 5 сайтов сплайсинга. Процент совпадений, составляющий 80%, считается очень высоким, учитывая, что терапевтическая активность in vivo зависит от различных факторов, включая клеточную антисмысловую активность, увеличение молекул лекарственного средства в ткани-мишени, фармакокинетический период полувыведения и т.д. Таким образом, соединение формулы I предсказуемо модулирует экспрессию являющегося его мишенью гена.
Учитывая молекулярную массу «SCN-ASO 7» (ср. таблицу 4), 100 пмоль/кг переводятся в терапевтическую дозу, составляющую приблизительно 0,53 мкг/кг. Субаттомолярная экзон-пропускающая активность in vitro «SCN-ASO 7», как полагают, в значительной степени ответственна за сверхсильную терапевтическую активность in vivo, составляющую приблизительно 0,53 мкг/кг у крыс с диабетической невропатией. Еще более удивительно, что ASO вводили в виде «голого» олигонуклеотида. Такая сильная терапевтическая активность in vivo никогда не была реализована с другими классами олигонуклеотидов, включая ДНК, РНК, PTO, 2'-OMe-PTO, 2'-OMe-РНК, 2'-OMOE-РНК, LNA, PMO, ПНК и т.д.
Примеры активности in vivo и ex vivo DMD-ASO.
Мышечная дистрофия Дюшенна (DMD) представляет собой опасное для жизни моногенное редкое заболевание с мышечной дегенерацией. Пациенты с DMD не кодируют полноразмерный белок дистрофина из-за PTC (кодона преждевременной терминации), возникающего в результате точечной мутации или делеции экзона(ов).
Мышь Mdx (C57BL/10ScSn-Dmdmdx/J. Jackson Lab) является мутантной мышью с точечной мутацией в экзоне 23 гена дистрофина, которая приводит к PTC. Мыши Mdx кодируют усеченную форму дистрофина без С-концевой части. Поскольку С-концевая часть связывается с экстраклеточным матриксом (ЕСМ), усеченная форма теряет предназначенную ей роль, заключающуюся в прочном связывании мышечных волокон с ЕСМ. Следовательно, мыши MDX постепенно теряют мышечную целостность и силу с возрастом.
Мыши Mdx широко использовались в качестве модели DMD у человека на животных. Дистрофин-специфические ASO, мишенью которых является экзон 23 дистрофина мыши, были исследованы с целью исключения PTC путем пропуска экзона 23.
Производные ПНК формулы I, комплементарно нацеливающийся на 3'- или 5'-сайт сплайсинга экзона 23 в пре-мРНК дистрофина мыши, оценивали в отношении их способности к индукции пропуска экзона 23 дистрофина у мышей MDX. Биологические примеры, представленные здесь, служат для иллюстрации способности к пропуску экзонов у специфических для дистрофина ASO в качестве примеров соединения формулы I, и поэтому их не следует интерпретировать как ограничение объема настоящего изобретения ASO, специфическими для дистрофина.
DMD-пример 1. Пропуск экзонов, индуцируемый «DMD-ASO 1» и «DMD-ASO 4» у мышей Mdx (способ A вложенной ПЦР).
«DMD-ASO 1», указанный в таблице 7, представляет собой 13-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 22 и экзона 23 в пре-мРНК дистрофина мыши. «DMD-ASO 1» комплементарно связывается с 13-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиваем» в 25-мерной последовательности [(5'→3')uaauuuugag|GCUCUGCAAAGTTCT]. «DMD-ASO 1» обладает способностью к 8-мерному перекрыванию с интроном 22 и 5-мерному перекрыванию с экзоном 23.
«DMD-ASO 4», указанный в таблице 7, представляет собой 17-мерный ASO, полностью комплементарный области в 3'-сайте сплайсига, охватывающей место соединения интрона 22 и экзона 23 в пре-мРНК для дистрофина мыши. «DMD-ASO 4» комплементарно связывается с 17-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности [(5'→3') uaauuuugag|GCUCUGCAAAGTTCT]. «DMD-ASO 4» обладает способностью к 5-мерному перекрыванию с интроном 22 и 12-мерному перекрыванию с экзоном 23.
«DMD-ASO 1» и «DMD-ASO 4» были оценены в отношении их способности к индукции пропуска экзона в мышцах мышей MDX при подкожном введении следующим образом.
[Лечение ASO и отбор образцов мышечных тканей] Инъекционные растворы готовили путем разведения водного маточного раствора «DMD-ASO 1» или «DMD-ASO 4» в PBS до 500 нМ. Самцам мышей mdx подкожно вводили только носитель (отрицательный контроль), «DMD-ASO 1» или «DMD-ASO 4» в дозе 2 мл/кг, 2 раза в день (BID) в течение 3 дней. Через один день после последней дозы животных анестезировали с помощью золетила/ромпуна и умерщвляли для отбора образцов мышечных тканей, включая сердце, диафрагму, икроножную мышцу, четырехглавую мышцу и трицепс.
[Экстракция РНК]. Образцы мышц гомогенизировали путем измельчения в пробирке, содержащейся на льду, и подвергали экстракции тотальной РНК с использованием 1 мл реагента тризола (Invitrogen) на приблизительно 100 мг мышечной ткани.
[Синтез кДНК с помощью одностадийной ОТ-ПЦР] 500 нг РНК-матрицы использовали в реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) и набор экзон-специфических праймеров [DMD-экзон 21_прямой: (5'→3') CAAAGAGAAAGAGCTACAGACA; и DMD-экзон 25_обратный: (5'→3') CTGGGCTGAATTGTTTGAAT] в соответствии со следующими режимами циклирования: 50οС в течение 30 мин и 94οС в течение 2 мин, после чего следовали 40 циклов, каждый состоящий из 30 сек при 94οС, 1 мин при 58οС и 2 мин при 72οС.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в 20-мкл реакции вложенной ПЦР (кат. № K2612, Bioneer) на фоне набора праймеров [DMD-экзон 22n_прямой: (5'→3')ATCCAGCAGTCAGAAAGCAAA; и DMD-экзон 25n_обратный: (5'→3')ACTAAAAGTCTGCATTGT] в соответствии со следующими режимами циклирования: 95οС в течение 5 мин с последующими 39 циклами, каждый состоящий из 30 сек при 95οС, 40 сек при 50οС и 50 сек при 72οС.
[Идентификация продукта пропуска экзона] ПЦР-продукты подвергали электрофоретическому разделению на 2% агарозном геле, как показано на фиг. 37А. Пропуск экзона 23 был обнаружен только у животных, получавших «DMD-ASO 1» и «DMD- ASO 4». Хотя пропуск экзона 23 был обнаружен только в четырехглавой мышце и икроножной мышце. «DMD-ASO 1», по-видимому, более эффективен, чем «DMD-ASO 4».
Субъект в качестве отрицательного контроля (т.е. без лечения ASO) давал полосу для ПЦР-продукта, приписываемого пропуску экзонов 22-23 в четырехглавой мышце. Пропуск экзонов 22-23 приводит к сдвигу рамки считывания, и вариант сплайсинга мРНК дистрофина, в котором отсутствуют экзоны 22-23, обречен на кодирование усеченного дистрофина без С-концевой части.
Полосы целевого размера были собраны и проанализированы с помощью секвенирования по Сэнгеру, и пропуск экзона 23, индуцированный «DMD-ASO 1» и «DMD-ASO 4», был подтвержден (ср. фиг. 37B). Полосы для полноразмерных продуктов ПЦР (т.е. без пропуска) и для пропуска экзонов 22-23 были подтверждены секвенированием, хотя данные секвенирования для пропуска экзонов 22-23 не предоставлены.
DMD-пример 2. Пропуск экзонов, индуцируемый «DMD-ASO 1» и «DMD-ASO 4» у мышей Mdx (способ В вложенной ПЦР).
Образцы РНК, полученные в «DMD-примере 1», подвергали одностадийному синтезу кДНК с использованием набора экзон-специфических праймеров [DMD-экзон 20_прямой: (5'→3')CAGAATTCTGCCAATTGCTGAG; и DMD-экзон 26_обратный:(5'→3')TTCTTCAGCTTGTGTCATCC]. Образцы кДНК затем анализировали с помощью вложенной ПЦР на фоне другого набора экзон-специфических праймеров [DMD-экзон 20n_прямой: (5'→3')CCCAGTCTACCACCCTATCAGAGC; и DMD-экзон 26n_обратный: (5'→3')CCTGCCTTTAAGGCTTCCTT].
Результаты вложенной ОТ-ПЦР представлены, как показано на фиг. 38A и фиг. 38B. У животных, которым вводили ASO, (N=2 на группу) были обнаружены пропуски экзонов 21-23 и экзонов 22-25. В то время как пропуск экзонов 21-23 происходит в рамке считывания (т.е. без сдвига рамки считывания), пропуск экзонов 22-25 происходит вне рамки считывания. Полоса для ПЦР-продукта, приписываемого пропуску экзонов 21-23, была однозначно подтверждена секвенированием по Сэнгеру (ср. фиг. 38В)
Профили пропуска экзонов менялись в зависимости от способа ПЦР, указанного выше. Таким образом, профили пропуска экзонов следует интерпретировать с осторожностью.
DMD-пример 3. Пропуск экзонов, индуцируемый «DMD-ASO 1» у мышей Mdx (способ A вложенной ПЦР).
Мыши Mdx подкожно получали «DMD-ASO 1» в дозе 0 (отрицательный контроль) или 10 пмоль/кг, 2 раза в день в течение 5 дней (2 животных в группе). Через один день после конечной дозы животных умерщвляли для отбора образцов ткани. Образцы трицепса оценивали на пропуск экзона 23 в соответствии с способом вложенной ОТ-ПЦР, описанным в «DMD-примере 1».
Результаты вложенной ОТ-ПЦР суммированы на фиг. 38C. Хотя одно животное в группе отрицательного контроля давало полосу для ПЦР-продукта в случае пропуска экзонов 22-23, вариант сплайсинга мРНК, в котором отсутствуют экзоны 22-23, находится вне рамки считывания. Полоса для ПЦР-продукта в случае пропуска экзона 23 была обнаружена только у одного животного в группе введения ASO. Таким образом, DMD-ASO 1 индуцировал пропуск экзона 23, как и было запланировано.
DMD-пример 4. Улучшение мышечной функции согласно тесту с вращающимся стержнем у мышей Mdx, которым подкожно вводили «DMD-ASO 1».
Пропуск экзона 23 у мышей mdx удаляет PTC в экзоне 23, и вариант сплайсинга мРНК, в котором отсутствует экзон 23, находится в рамке считывания и, следовательно, кодирует вариант белка с С-концевой частью, связывающейся с ЕСМ. Поэтому полноразмерный вариант белка дистрофина частично восстанавливает физиологические функции исходного полноразмерного дистрофина или дистрофина дикого типа.
Учитывая возможность пропуска экзонов, «DMD-ASO 1» оценивали на мышах MDX в отношении его способности улучшать мышечную функцию согласно тесту с вращающимся стержнем, как описано ниже.
[Группирования] Самцов мышей MDX в возрасте 6 недель готовили к тесту с вращающимся стержнем в течение 2 недель, а затем случайным образом распределяли по 3 группам: 0 (отрицательный контроль), 100 и 1000 пмоль/кг «DMD-ASO 1» - на основе оценок индивидуумов в тесте с вращающимся стержнем в день 0, т.е. в день группирования. (N=10 на группу)
[Лечение ASO] Растворы для инъекций готовили путем последовательного разведения ASO до 20 нМ и 200 нМ в PBS. Животным вводили подкожно носитель (PBS, отрицательный контроль), 20 нМ «DMD-ASO 1» (100 пмоль/кг) или 200 нМ «DMD-ASO 1» (1000 пмоль/кг) в дозе 5 мл/кг, 3X в неделю в течение периода с дня 0 по день 21.
[Тест с вращающимся стержнем и статистический анализ] Мышей подвергали тесту с вращающимся стержнем на аппарате с вращающимся стержнем (модель № 47650, Ugo Basile) со схемой ускорения от 4 об/мин до 45 об/мин в течение 60 сек. Время до падения (т.е. период, в течение которого животное оставалось на вращающемся стержне), оценивалось для каждого отдельного животного. Статистическая значимость оценивалась с помощью t-критерия Стьюдента относительно группы отрицательного контроля.
[Улучшение мышечной функции]. На фиг. 39A суммированы оценки в тесте с вращающимся стержнем по группам. Оценки в тесте вращающимся стержнем (т.е. время до падения) оставались на прежнем уровне в среднем приблизительно от 70 до 120 секунд в группе отрицательного контроля. Между тем, мышечная функция в группе введения 1000 пмоль/кг постепенно и заметно улучшалась до дня 12, а затем оставалась неизменной при оценке в тесте с вращающимся стержнем в среднем от 210 до 230 секунд. Мышечная функция была значимо улучшена в дни 12, 14 и 19 при введении 1000 пмоль/кг ASO. Мышечная функция в группе введения 100 пмоль/кг показала сильную склонность к улучшению, но не была значимой.
DMD-пример 5. Улучшение мышечной функции согласно тесту на захват лапами и мышечной целостности у мышей Mdx, которым постоянно вводили «DMD-ASO 1».
«DMD-ASO 1» оценивали при постоянном введении мышам mdx в отношении его способности к улучшению мышечной функции согласно тесту на захват лапами, к индукции пропуска экзонов, к повышению экспрессии полноразмерного дистрофина (т.е. кодируемого белка дистрофина с С-концом) согласно IHC и к улучшению целостности мышц согласно гистопатологии с окрашиванием H&E, как описано ниже.
[Группирование и лечение ASO] Самцов мышей MDX (возрастом 7 недель) случайным образом распределяли по 4 группам: 0 (отрицательный контроль MDX), 10, 50 и 200 пмоль/кг «DMD-ASO 1» на основании индивидуальных оценок силы захвата с помощью теста на захват лапами («прогулки по приподнятой перекладине») (N=12 на группу). Вспомогательная группа из 12 самцов мышей C57BL/6 (возрастом 7 недель) была включена в это исследование в качестве группы отрицательного контроля дикого типа для уровня экспрессии полноразмерного дистрофина.
Группа введения 50 и 200 пмоль/кг получала подкожно «DMD-ASO 1» в растворенном виде в PBS, 2 раза в неделю до конечного умерщвления на неделе 43. Между тем, группа введения 10 пмоль/кг получала подкожно «DMD-ASO 1» сначала 2 раза в неделю в течение недели с 0 по 8 и 3 раза в неделю после этого, чтобы увеличить воздействие ASO.
[Мышечная функция согласно тесту на захват лапами] Мышечную функцию оценивали еженедельно по силе захвата на измерителе силы захвата (кат. № 47200, Ugo Basile) в соответствии с литературой [J. Appl. Physiol, vol 106 (4), 1311-1324 (2009)] Оценки силы захвата по группам оценивали на статистическую значимость с помощью t-критерия Стьюдента относительно группы отрицательного контроля MDX.
На фиг. 39В суммированы оценки наблюдаемой силы захвата по группам в течение неделей с 0 по 30. В течение первых 11 недель после введения первой дозы не было заметных изменений силы захвата между группами введения ASO и группой отрицательного контроля. Сила захвата группы отрицательного контроля MDX достигла максимума=приблизительно 107 г на неделе 3, постепенно снижалась в течение нескольких недель, а затем оставалась относительно неизменной - приблизительно от 75 до 95 г. Сила захвата в группах введения ASO начинала постепенно улучшаться с недели 10 или около того. В группах лечения сила захвата, как правило, возрастала на 30~50% по сравнению с группой отрицательного контроля MDX.
Следует отметить, что 2-3 животных в каждой группе случайным образом отбирали и умерщвляли на неделях 7, 13, 21 и 30 для оценки с помощью IHC (иммуногистохимического анализа) или вложенной ПЦР (см. ниже)
[Оценка с помощью IHC скелетных мышц по сравнению с полноразмерным дистрофином] На неделях 7, 13 и 21 двух животных в каждой группе выбирали случайным образом и умерщвляли для извлечения мышечной ткани. На неделе 30 умерщвляли 3 животных в каждой группе.
Мышечные ткани, отобранные на неделе 7, подвергались IHC в отношении полноразмерного дистрофина путем изготовления срезов из замороженных тканей. Мышечные ткани, отобранные на неделях 13, 21 и 30, подвергались иммуноокрашиванию с использованием парафинового блока. IHC с использованием парафинового блока давал изображения лучшего качества, чем IHC с изготовлением срезов из замороженных тканей.
Образцы мышц подвергали иммуноокрашиванию последовательно с использованием первого антитела против С-конца дистрофина мыши (кат. № sc-816, Santa Cruz) в разведении 1:100, второго антитела против IgG (кат. № BA-1100, Vector) в разведении 1:200, а затем Dylight 594-стептавидина (кат. № SA-5594, Vector, CA, США) в разведении 1:200 для мечения красной флуоресценцией. IHC-изображения фиксировали на слайд-сканере Zeiss (на неделях 13, 21 и 30) или на флуоресцентном микроскопе Olympus (на неделе 7). Дополнительно проводилось окрашивание DAPI.
На фиг. 40 представлен репрезентативный набор IHC-изображений полноразмерного дистрофина по группам для образцов мышц, полученных на неделе 30. Группа отрицательного контроля дикого типа (WT) давала отличительные и угловые характеры экспрессии дистрофина, отражающие естественную структуру пучков мышечных волокон. В группе отрицательного контроля мышей mdx не было большой степени окрашивания полноразмерного дистрофина. Между тем, сильные и угловые характеры окрашивания дистрофина наблюдались в скелетных мышцах в группе введения 200 пмоль/кг. Хотя структуры пучков мышечных волокон были размытыми у мышей MDX по сравнению с мышами WT, экспрессия полноразмерного дистрофина заметно увеличилась у животных, получавших ASO.
IHC-изображения дистрофина подвергали количественному анализу в отношении экспрессии полноразмерного дистрофина путем цифровой оценки интенсивности красной флуоресценции в каждом отдельном IHC-изображении, используя программу ImageJ (NIH). Индивидуальные оценки флуоресценции объединяли по группам и типу мышц для статистической оценки относительно группы отрицательного контроля дикого типа.
На фиг. 41 суммированы изменения уровня относительной экспрессии полноразмерного белка дистрофина у мышей MDX. Экспрессия полноразмерного дистрофина, как правило, больше увеличивалась при большей дозе ASO и большей продолжительности лечения. На неделе 30 экспрессия полноразмерного дистрофина в группе введения 200 пмоль/кг достигала >80% от таковой в группе отрицательного контроля WT, тогда как экспрессия в группе отрицательного контроля mdx составляла менее 20% от таковой в группе отрицательного контроля дикого типа.
[Вложенная ПЦР для обнаружения пропуска экзонов (способ B)] Образцы мышц гомогенизировали путем измельчения в пробирке, содержащейся на льду, и подвергали экстракции тотальной РНК с использованием 1 мл реагента тризола (Invitrogen) на приблизительно 100 мг мышечной ткани. Тотальную РНК оценивали на пропуск экзонов с помощью вложенной ПЦР, как описано в «DMD-примере 2».
На фиг. 42 представлены данные электрофореза продуктов вложенной ПЦР, полученных с использованием скелетных мышц, отобранных на неделе 7. ПЦР-продукты Δэкзонов 21-23 и Δэкзонов 21-24 в рамке считывания были обнаружены в образцах мышц групп лечения ASO, но полностью отсутствовали в группе отрицательного контроля MDX.
[Гистопатология с использованием окрашивания H&E] Воспаление и дегенерация мышц являются признаками симптомов МДД. Образцы скелетных мышц подвергали гистопатологической оценке путем окрашивания H&E. На фиг. 43 представлен репрезентативный набор изображений окрашивания H&E для трицепса по группам и моментам времени отбора образцов.
В группе отрицательного контроля WT мышечная структура была плотной во все моменты времени отбора образцов. Не было никаких предположений о мышечном воспалении в мышцах мышей дикого типа во все моменты времени.
В группе отрицательного контроля mdx мышечная структура показала четкую картину постепенной дегенерации с возрастом. Особенно на неделе 30 мышечные пучки не были связаны между собой и имели тенденцию к дегенерации в округлую форму. Также наблюдалось массивная инфильтрация воспалительных клеток, о чем свидетельствуют пятна в виде синих точек, особенно на неделях 13 и 21.
В группе введения 200 пмоль/кг рыхлая мышечная структура на неделе 7 постепенно восстанавливалась до плотной структуры. Заметная инфильтрация воспалительных клеток на неделе 7 постепенно исчезала с возрастом. Таким образом, дегенерация и воспаление мышц у мышей MDX были обращены вспять при постоянном введении ASO в дозе 200 пмоль/кг, что соответствовало увеличению экспрессии полноразмерного дистрофина в группе введения 200 пмоль/кг.
Согласно гистопатологическим данным степень тяжести дегенерации в группах введения 10 и 50 пмоль/кг была меньше таковой в отрицательном контроле mdx, но больше, чем в группе введения 200 пмоль/кг. Таким образом, увеличению экспрессии полноразмерного дистрофина, вызванного воздействием ASO, в значительной степени согласуется с гистопатологическими данными.
[Прочие данные] Было три случая незапланированного умерщвления или смерти (недели 26, 39 и 43) в группе отрицательного контроля MDX из-за большой массы лимфомы мышечной ткани, наиболее вероятно развившейся вследствие хронического мышечного воспаления. Не было случаев лимфомы во всех группах лечения ASO.
[Сравнение с другим дистрофин-специфическим ASO] Eteplirsen (exondys 51) представляет собой антисмысловой олигонуклеотид PMO, сконструированный для индукции пропуска экзона 51 в пре-мРНК для дистрофина человека. Недавно Управление по контролю качества продуктов питания и лекарственных средств (FDA) США выдало ускоренное одобрение применения Eteplirsen для популяции пациентов с DMD, которым требуется пропуск экзона 51. Рекомендуемая доза Eteplirsen - внутривенная инъекция 30 мг/кг в неделю.
Подкожная доза 200 пмоль/кг «DMD-ASO 1» соответствует приблизительно 1 мкг/кг. Дистрофин-специфический ASO формулы I является приблизительно в 30000 раз более эффективным, чем PMO ASO, хотя существуют различия в видах и экзоне между двумя типами ASO. Беспрецедентно сверхсильная экзон-пропускающая активность производного ПНК формулы I снова была переведена в сверхсильную in vivo терапевтическую активность для этого трудно поддающегося лечению редкого заболевания.
DMD-пример 6. Улучшение мышечной функции по пройденному расстоянию у мышей MDX, которым постоянно вводили «DMD-ASO 2».
«DMD ASO 2», указанный в таблице 7, представляет собой 17-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 22 и экзона 23 в пре-мРНК дистрофина мыши. «DMD-ASO 2» комплементарно связывается с 17-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием, как в 20-мерной последовательности пре-мРНК дистрофина мыши [(5'→3')uaauuuugag|GCUCUGCAAA] «DMD-ASO 2» обладает способностью к 8-мерному перекрыванию с интроном 22 и 9-мерному комплементарному перекрыванию с экзоном 23.
«DMD-ASO 2» постоянно вводили мышам mdx для оценки его способности улучшать мышечную функцию по пройденному расстоянию на беговой дорожке и подавлять деградацию мышц по уровню креатинкиназы (CK) и миоглобина в сыворотке.
[Животные и группирование] Самцов мышей MDX (в возрасте 6 недель) случайным образом распределяли по трем группам: отрицательного контроля MDX (без лечения ASO), «DMD-ASO 2» 10 пмоль/кг и «DMD-ASO 2» 30 мг/кг в расчете на вес тела. (N=16 на группу). 12 самцов мышей C57BL/6 (в возрасте 6 недель) приняли в качестве группы отрицательного контроля дикого типа (WT).
[Растворы для инъекций и лечение ASO] Водный исходный раствор «DMD-ASO 2» разводили либо в PBS, либо в PBS с добавлением 0,1% Tween 80 для приготовления инъекционных растворов 20 и 60 нМ «DMD-ASO 2» для 10 пмоль/кг и 30 пмоль/кг «DMD-ASO 2», соответственно. Считалось, что добавление 0,1% Tween 80 к раствору для инъекций PBS необходимо для предотвращения прилипания молекул ASO к пластиковым инъекционным пробиркам, наконечникам пипеток и шприцам.
[Пройденное расстояние на беговой дорожке] Во время первых 30 недель после группирования животным вводили инъекционные растворы без Tween 80 2 раза в неделю в дозе 2 мл/кг. Начиная с недели 7, животных еженедельно подвергали ходьбе на беговой дорожке (Model # LE8710, PanLab). Однако во время первых 30 недель группы лечения ASO не продемонстрировали каких-либо значимых увеличений пройденного расстояния по сравнению с группой отрицательного контроля MDX.
С целью эффективного увеличения дозы ASO животным вводили инъекционные растворы, дополненные Tween 80, 2 раза в неделю с недели 36. Между периодами лечения был период вымывания (т.е. без введения дозы ASO), составляющий 5 недель.
На фиг. 44A показаны пройденные расстояния на беговой дорожке по группам на неделях с 43 по 48. Среднее пройденное расстояние группы отрицательного контроля mdx постепенно, но быстро уменьшалось с приблизительно 250 метров на неделе 43 до 130 метров на неделе 48. Среднее пройденное расстояние групп лечения ASO было заметно больше пройденного расстояния для группы отрицательного контроля MDX.
На неделях с 46 по 48 группа введения 10 пмоль/кг продемонстрировала среднее пройденное расстояние 240-280 метров, что было значительно больше, чем пройденное расстояние для группы отрицательного контроля MDX. Между тем, группа введения 30 пмоль/кг продемонстрировала пройденное расстояние, составляющее приблизительно 180-230 метров, на неделях с 46 по 48. В случае группы введения 10 пмоль/кг, по сравнению с группой введения 30 пмоль/кг, отмечалась тенденция к увеличению пройденного расстояния. Обратная зависимость от дозы ответа в виде пройденного расстояния могла бы предполагать естественный выбор отличного экзона(ов) при более высокой дозе ASO, как это наблюдалось в «HIF-1α-примере 9». Интересно, что группа отрицательного контроля WT и группа введения 30 ммоль/кг продемонстрировали сопоставимые пройденные расстояния на неделях с 44 по 47.
[Конечное умерщвление] На неделе 48 животных подвергали конечному умерщвлению. Образцы крови анализировали на уровень в сыворотке СК (с помощью набора для анализа активности креатинкиназы, кат. № abl55901, Abeam) и миоглобина (с помощью набора для ELISA миоглобина, кат. № ab210965, Abeam) для оценки степени деградации мышц в соответствии с инструкциями производителя. Мышечные ткани были проанализированы с помощью Вестерн-блоттинга на полноразмерный дистрофин.
[Уровни в сыворотке СК и миоглобина] На фиг. 44B представлены наблюдаемые уровни CK в сыворотке по группам. Отражая мышечную слабость мышей mdx, все группы мышей mdx давали намного более высокие активности СК в сыворотке, чем группа отрицательного контроля WT. Например, уровень CK в сыворотке группы отрицательного контроля MDX был приблизительно в 54 раза выше, чем уровень в группе отрицательного контроля WT. Уровни CK в сыворотке в группе введения 10 и 30 пмоль/кг были ниже, чем уровень в группе отрицательного контроля MDX на 58% и 38%, соответственно. Разница в уровнях CK в сыворотке между группой введения 10 пмоль/кг и группой отрицательного контроля MDX была значимой.
На фиг. 44C представлены наблюдаемые уровни миоглобина в сыворотке по группам. Отражая мышечную слабость мышей mdx, все группы мышей mdx давали намного более высокие уровни миоглобина в сыворотке, чем уровень в группе отрицательного контроля WT. Например, уровень миоглобина в сыворотке в группе отрицательного контроля MDX был приблизительно в 22 раза выше, чем этот уровень в группе отрицательного контроля WT. Уровни миоглобина в сыворотке в группе введения 10 и 30 пмоль/кг были значительно ниже, чем уровень в группе отрицательного контроля MDX на 67% и 58%, соответственно.
Наблюдаемые данные по сывороточным биомаркерам мышечной деградации в целом соответствуют зависимости от дозы пройденного расстояние, представленной на фиг. 44А.
[Экспрессия полноразмерного дистрофина в трицепсе, оцениваемая с помощью Вестерн-блоттинга] После гомогенизации при температуре жидкого азота образцы мышц (трицепса) подвергали лизису в буфере RIPA с добавлением 1% SDS. Концентрацию белка в каждом лизате определяли с помощью анализа BCA относительно стандарта BSA. 50 мг белка каждого лизата подвергали электрофоретическому разделению в 8% ПААГ для электрофореза. Затем мембрану из PVDF исследовали с использованием антитела против С-конца дистрофина (кат. № ab 154168, Abeam).
На фиг. 45А представлены наблюдаемые данные Вестерн-блоттинга. Полноразмерный дистрофин размером 427К был обнаружен не во всех образцах. Вместо этого образцы мышц WT давали белки дистрофина меньшего размера: 170К, 130К и 117К, среди которых наиболее обогащенной была полоса размером 130К.
В случае групп MDX были обнаружены три полосы для дистрофина. Группы лечения 10 и 30 пмоль/кг давали заметно более сильную полосу 117К, чем группа отрицательного контроля MDX, а также контрольная группа WT. Интенсивность полосы 130К была значительно выше в группе лечения 10 пмоль/кг, чем в группе отрицательного контроля MDX.
DMD-пример 7. Длительная оценка мышей MDX, которым вводили «DMD-ASO 1», «DMD-ASO 2» или «DMD-ASO 6».
«DMD ASO 6», указанный в таблице 7, представляет собой 18-мерный ASO, полностью комплементарный области в 5'-сайте сплайсинга, охватывающей место соединения экзона 23 и интрона 23 в пре-мРНК для дистрофина мыши. «DMD-ASO 6» комплементарно перекрывается с 18-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК для дистрофина мыши [(5'→3')AAAAUUUCAG|guaagccgagguuug]. «DMD-ASO 6» обладает способностью к 8-мерному перекрыванию с экзоном 23 и 10-мерному перекрыванию с интроном 23.
«DMD-ASO 1», «DMD-ASO 2» и «DMD-ASO 6» оценивали на предмет их физиологических эффектов у самцов мышей MDX при длительном подкожном введении. В ходе этой оценке животные не подвергались физическим тестам, требующим мышечного стресса, чтобы чрезмерная мышечная стимуляция не нарушала транскрипцию гена дистрофина.
[Животные и группирование] Самцов мышей mdx (в возрасте 6 недель) случайным образом распределяли по 4 группам: отрицательного контроля mdx (без воздействия ASO), «DMD-ASO 1», 50 пмоль/кг, «DMD-ASO 2» 10 мг/кг и «DMD-ASO 6» 10 мг/кг в расчете на вес тела (N=12~13 на группу). 12 мышей-самцов C57BL/6 (6 недель) приняли в качестве группы отрицательного контроля дикого типа (WT).
[Инъекционные растворы и воздействие ASO] Водные исходные растворы ASO последовательно разводили в PBS для приготовления инъекционных растворов 25 нМ «DMD-ASO 1» для 50 пмоль/кг «DMD-ASO 1», 5 нМ «DMD-ASO 2» для «DMD-ASO 2» 10 пмоль/кг и 5 нМ «DMD-ASO 6» для «DMD-ASO 6» 10 пмоль/кг. Животным вводили подкожно инъекционные растворы в дозе 2 мл/кг, 2 раза в неделю.
[Незапланированная смерть или умерщвление] Мыши MDX, получавшие ASO, как правило, демонстрировали более длительное время жизни, чем группа отрицательного контроля MDX, что указывает на терапевтическую активность ASO.
В группе отрицательного контроля MDX было три случая незапланированной смерти или умерщвления: один случай каждый раз на неделях 42, 58 и 61. В группе лечения «DMD-ASO 1» произошли две преждевременных смерти, одна на неделе 38, а другая - на неделе 61. В случае группы лечения «DMD-ASO 2» одна смерть произошла на неделе 56, а другая на неделе 62. В группе лечения «DMD-ASO 6» произошли три преждевременных смерти: одна каждый раз на неделях 55, 65 и 66.
[Конечное умерщвление] Всех выживших животных умерщвляли для забора крови на неделе 66 после группирования. Образцы крови подвергали анализам ELISA на креатинкиназу (CK) в сыворотке и миоглобин в сыворотке, как описано в «DMD-примере 6».
[Уровни в сыворотке СК и миоглобина] На фиг. 45B представлены наблюдаемые уровни CK в сыворотке по группам. Отражая мышечную слабость мышей mdx, все группы мышей mdx давали намного более высокие активности СК в сыворотке, чем группа отрицательного контроля WT. Например, уровень CK в сыворотке группы отрицательного контроля MDX был приблизительно в 17 раз выше, чем уровень в группе отрицательного контроля WT. Уровни CK в сыворотке в группах лечения ASO, как правило, были ниже, чем уровень в группе отрицательного контроля MDX, что указывает на терапевтическую активность, проявляемую ASO. Однако различия не были значимыми.
На фиг. 45C представлены наблюдаемые уровни миоглобина в сыворотке по группам. Отражая мышечную слабость мышей MDX, все группы мышей MDX давали намного более высокий уровень миоглобина в сыворотке, чем группа отрицательного контроля WT. Например, уровень миоглобина в сыворотке в группе отрицательного контроля MDX был приблизительно в 26 раз выше, чем уровень в группе отрицательного контроля WT. Уровни миоглобина в сыворотке в группах лечения «DMD-ASO 1», «DMD-ASO 2» и «DMD-ASO 6» были ниже, чем уровень в группе отрицательного контроля MDX на 43%, 49% и 68%, соответственно. Разница между отрицательным контролем MDX и группой лечения «DMD-ASO 6» была значимой. Сывороточные биомаркеры мышечной целостности косвенно подтверждают, что «DMD-ASO 6» индуцирует пропуск экзона 23 дистрофина и дает функционально активный полноразмерный дистрофин(ы) у мышей MDX.
Примеры активности in vitro IDO1-ASO
Производные ПНК формулы I в таблице 8 были сконструированы для комплементарного нацеливания на различные сайты сплайсинга в пре-мРНК IDO1 человека. IDO1-ASO оценивали на экзон-пропускающую активность в клетках SKOV3. Учитывая, что IDO1 катализирует деградацию L-триптофана до N-формилкинуренина. IDO1-ASO оценивали в отношении их функциональной способности ингибировать продукцию кинуренина. Биологические примеры, представленные здесь, служат для иллюстрации экзон-пропускающей активности IDO1-ASO в качестве примеров соединения формулы I, и поэтому их не следует интерпретировать как ограничение объема настоящего изобретения IDO1-ASO.
IDO1-пример 1. Пропуск экзонов, индуцируемый «IDO-ASO 1».
«IDO-ASO 1», указанный в таблице 8, представляет собой 13-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 6 и экзона 7 в пре-мРНК IDO1 человека. «IDO-ASO 1» комплементарно нацеливается на 13-мерную последовательность, как отмечено «жирный шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК IDOl человека [(5'→3')uuuguuuuag|GUAAUUCCUA1. «IDO-ASO 1» обладает способностью к 5-мерному перекрыванию с интроном 6 и 8-мерному перекрыванию с экзоном 7.
«IDO-ASO 1» оценивали в отношении его способности к индукции пропуска экзона в клетках SKOV3 (кат. № HTB-77, ATCC) с помощью IDO1-специфической вложенной ПЦР следующим образом.
[Клеточная культура и обработка ASO] Клетки SKOV3 (аденокарциномы яичника человека) субкультивировали в 60-мм чашке для культивирования, содержащей 5 мл модифицированной среды 5C McCoy, дополненной 10% FBS, 1% стрептомицина/пенициллина, 1% L-глютамина и 1% пирувата натрия, в атмосфере с 5% CO2 при 37οC и обрабатывали «IDO-ASO 1» в течение 48 часов в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ или 1 аМ.
[Экстракция РНК и синтез кДНК с помощью одностадийной ОТ-ПЦР] Тотальную РНК экстрагировали из клеток с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями изготовителя. 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [IDO-экзон 2_прямой: (5'→3')TTCATTGCTAAACATCTGCC; и IDO-экзон 10_обратный: (5'→ 3')TGAAAGGACAAACTCACGGA] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 40 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 50οC и 1 мин при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в 20-мкл реакции вложенной ПЦР (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [IDO-экзон 4_прямой: (5'→3')CCTTACTGCCAACTCTCC; и IDO-экзон 9_обратный: (5'→3')CTGCTTTGGCCTGCACTG] в соответствии со следующими режимами циклирования: 95οC в течение 5 мин с последующими 30 циклами, каждый состоящий из 30 сек при 95οC, 30 сек при 50οC и 1 мин при 72οC.
[Идентификация продукта пропуска экзона] ПЦР-продукты подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру.
На фиг. 46А представлены данные электрофореза продуктов вложенной ПЦР (левая диаграмма) и данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов 6-7 (справа). Пропуск экзонов 6-7 был обнаружен в экстракте РНК клеток, обработанных 100 цМ «IDO-ASO 1». Полоса для пропуска экзонов не была обнаружена в экстрактах РНК клеток, обработанных ASO в концентрации 10 цМ или 1 аМ, скорее всего из-за слабой стабильности варианта сплайсинга мРНК IDO1, в котором отсутствуют экзоны 6-7. Хотя интенсивность полноразмерной мРНК для IDO-1 снижалась в клетках, обработанных ASO в концентрации 10 или 100 цМ, интенсивность полноразмерной мРНК увеличивалась в клетках, обработанных ASO в концентрации 1 аМ. Наблюдаемое увеличение уровня полноразмерной мРНК при 1 аМ еще предстоит выяснить. Это может быть артефактом во время реакций ПЦР или может быть связано с (транзиторной) активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) кольцевой РНК (EIciRNA)», накапливаемой во время пропуска экзона с помощью «IDO-ASO 1» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)]. Данные секвенирования по Сэнгеру (правая диаграмма) однозначно продемонстрировали пропуск экзонов 6-7, индуцированный «IDO-ASO 1» в клетках SKOV3.
IDO1-пример 2. Антисмысловая функциональная активность «IDO-ASO 1».
Функциональную активность «IDO-ASO 1» оценивали в отношении его способности ингибировать секрецию кинуренина в клетках SKOV3 следующим образом.
[Анализ секреции кинуренина] Клетки SKOV3, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «IDO-ASO 1» в концентрации 0 цМ (отрицательный контроль) или от 10 цМ до 1 фМ (3 чашки на каждую концентрацию). Клетки обрабатывали ASO вместе с 10 нг/мл γ-интерферона для увеличения секреции кинуренина. Спустя 24 часа 200 мкл культуральной среды отбирали из каждой чашки для культивирования и смешивали с 100 мкл 30% трихлоруксусной кислоты. Смесь встряхивали и подвергали центрифугированию при 8000хg в течение 5 мин. 75 мкл полученного супернатанта смешивали с 75 мкл реагента Эрлиха (0,8% п-диметиламинобензальдегида в уксусной кислоте), и смесь подвергали количественному анализу на кинуренин при 490 нм на считывающем устройстве для ELISA. [PLOS One 5 (8): e63301 (2013)]
На фиг. 46B представлены результаты анализа кинуренина. За исключением клеток, обработанных 1 аМ «IDO-ASO 1», секреция кинуренина значимо (t-критерий Стьюдента) снижалась в клетках, обработанных ASO в концентрации 10 цМ - 1 фМ. Секреция кинуренина снижалась приблизительно на 40% в клетках, обработанных 1 фМ «IDO-ASO 1».
IDO1-пример 3. Пропуск экзонов, индуцируемый «IDO-ASO 5».
«IDO-ASO 5», указанный в таблице 8, представляет собой 13-мерный ASO, полностью комплементарный области в 5'-сайте сплайсинга, охватывающей место соединения экзона 3 и интрона 3 в пре-мРНК для IDO1 человека. «IDO-ASO 5» комплементарно перекрывается с 13-мерной последовательностью, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК для IDO1 человека [(5'→3')UGUCCGUAAG|guuuggagau]. «IDO-ASO 5» обладает способностью к 8-мерному комплементарному перекрыванию с экзоном 3 и 5-мерному комплементарному перекрыванию с интроном 3.
«IDO-ASO 5» оценивали в отношении его способности к индукции пропуска экзонов в клетках SKOV3 с помощью IDO1-специфической вложенной ОТ-ПЦР, как описано в «IDO1-примере 1», если не указано иное.
[Обработка ASO] Клетки SKOV3, выращенные в 60-мм чашке для культивирования, обрабатывали «IDO-ASO 5» в концентрации 0 (отрицательный контроль), 1, 3, 10, 30 или 100 аМ в течение 48 часов.
[Синтез кДНК с помощью одностадийной ПЦР] 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [IDO-экзон 1_прямой: (5'→3')AAAACTCCTGGACAATCAGT; и IDO-экзон 8_обратный: (5'→3')ACTTGAAGGGCTTTCTCC] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 40 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 52οC и 40 сек при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) с использованием набора экзон-специфических праймеров [IDO-экзон 1n_прямой: (5'→3')TATTGATGAAGAAGTGGG; и IDO-экзон 8n_обратный: (5'→3')GTTCACATGATCGTGGATTTG] в соответствии со следующими режимами циклирования: 95οC в течение 5 мин с последующими 30 циклами, каждый состоящий из 30 сек при 95οC, 40 сек при 52οC и 40 сек при 72οC.
[Данные о продуктах вложенной ПЦР] На фиг. 47А представлены данные электрофореза продуктов вложенной ПЦР. Клетки, обработанные «IDO-ASO 5» в концентрации 30 и 100 аМ, явно давали варианты сплайсинга мРНК, в которых отсутствовали экзоны 2-4 и экзоны 2-6. Интенсивность полосы для полноразмерной мРНК снижалась в клетках, обработанных ASO, хотя интенсивность слегка увеличивалась в клетках, обработанных ASO в концентрации 10 аМ. На фиг. 47B представлены данные секвенирования по Сэнгеру вариантов сплайсинга мРНК, в которых отсутствуют экзоны 2-4 и экзоны 2-6.
IDO1-пример 4. Пропуск экзонов, индуцируемый «IDO-ASO 6».
«IDO-ASO 6», указанный в таблице 8, представляет собой 13-мерный ASO, полностью комплементарный области в 3'-сайте сплайсинга, охватывающей место соединения интрона 3 и экзона 4 в пре-мРНК для IDO1 человека. «IDO-ASO 6» комплементарно нацеливается на 13-мерную последовательность, как отмечено «жирным шрифтом» и «подчеркиванием» в 20-мерной последовательности пре-мРНК для IDO1 человека [(5'→3')uuuuaaucag|GUCUUGCCAA]. «IDO-ASO 6» обладает способностью к 5-мерному комплементарному перекрыванию с интроном 3 и 8-мерному комплементарному перекрыванию с экзоном 4.
«IDO-ASO 6» оценивали в отношении его способности к индукции пропуска экзонов в клетках SKOV3 с помощью IDO1-специфической вложенной ПЦР, как описано в «IDO1-примере 3», если не указано иное.
[Данные о продуктах вложенной ПЦР] На фиг. 47C представлены данные электрофореза продуктов вложенной ПЦР (левая диаграмма), а также данные секвенирования по Сэнгеру полосы для ПЦР-продукта, приписываемого пропуску экзонов (правая диаграмма). Клетки, обработанные «IDO-ASO 6» в диапазоне от 1 до 30 аМ, явно давали один вариант сплайсинга мРНК, в котором отсутствовали экзоны 2-5, хотя полоса для пропуска экзонов не была обнаружена в клетках, обработанных ASO в концентрации 100 аМ. Интенсивность полосы для полноразмерной мРНК была больше в клетках, обработанных ASO, чем в клетках без обработки ASO, что могло быть связано с активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) кольцевой РНК (EIciRNA)", накапливаемой во время пропуска экзона с помощью «IDO-ASO 1» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)].
Примеры активности in vitro и ex vivo SNAP25-ASO.
SNAP25 (связанный с синаптосомой белок с М.м. 25 кДа) представляет собой белок SNARE, участвующий в экзоцитозе нейромедиаторов в клетках двигательных нейронов. Ботулинический токсин А (Botox™) расщепляет SNAP25 для проявления его знаменитой активности против морщин. Производные ПНК формулы I в таблице 9 были сконструированы для комплементарного нацеливания на 3'-сайт сплайсинга экзона 7 в пре-мРНК SNAP25 человека. SNAP25-ASO оценивали в отношении SNAP25-антисмысловой активности в клетках SiMa (нейробластомы человека) и клетках РС12 крысиного происхождения, а также в отношении их способности ингибировать экспрессию SNAP25 в коже мышей при местном введении. Биологические примеры, представленные здесь, служат для иллюстрации экзон-пропускающей активности SNAP25-АSO в качестве примеров для соединения формулы I, и поэтому их не следует интерпретировать как ограничение объема настоящего изобретения SNAP25-ASO.
SNAP25-пример 1. Пропуск экзонов в клетках РС12, обработанных «SNAP-ASO 3».
«SNAP-ASO 3», указанный в таблице 9, представляет собой 14-мерный ASO, полностью комплементарный 14-мерной последовательности в 3'-сайте сплайсинга, охватывающем место соединения «интрона 6» и «экзона 7» в пре-мРНК для SNAP25 человек. «SNAP-ASO 3» комплементарно перекрывается с 14-мерной последовательностью пре-мРНК, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК [(5'→3') cucuuuggaucccag|GGUAACAAAUGAUGC1. «SNAP-ASO 3» обладает способностью к 7-мерному перекрыванию с «интроном 6» и еще одному 7-мерному перекрыванию с «экзоном 7».
«SNAP-ASO 3» оценивали в отношении его способности к индукции пропуска экзонов в клетках РС12 (кат. № CRL-1721, ATCC), хотя «SNAP-ASO 3» имеет одно несоответствие с 3'-сайтом сплайсинга «экзона 7» в пре-мРНК для SNAP25 крысы, считываемой с геномной ДНК крысы [доступ к которой получен из ссылочной последовательности NCBI: NC_005012]. 14-мерный ASO обладает способностью к 13-мерному комплементарному перекрыванию с пре-мРНК для SNAP25 крысы, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК [(5'→3')ugg"c"ucccag|GGUAACAAACGAUGC], в которой единственное несоответствие отмечено знаком кавычки ("").
[Клеточная культура и обработка ASO] Клетки РС12 поддерживали в среде RPMI 1640 с добавлением 5% FBS, 10% лошадиной сыворотки, 1% стрептомицина/пенициллина, 1% L-глютамина и 1% пирувата натрия в атмосфере с 5% CO2 при 37οC. Клетки, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «SNAP-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ.
[Экстракция РНК и синтез кДНК с помощью одностадийной ПЦР] После инкубации с «SNAP-ASO 3» в течение 42 часов клетки обрабатывали 100 мкг/мл циклогексимида в течение еще 6 часов, чтобы заморозить трансляцию на рибосомах. Затем тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя. 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [SNAP-экзон 1_прямой: (5'→3') ATGGCCGAGGACGCAGACA; и SNAP-экзон 14_обратный: (5'→3') AGCATCTTTGTTGCACGTTG] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 40 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 50οC и 1 мин при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК подвергали реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [SNAP-экзон 1_прямой: (5'→3')ATGGCCGAGGACGCAGACA; SNAP-экзон 14n_обратный: (5'→3')TTGTTGGAGTCAGCGCCT] в соответствии со следующими режимами циклирования: 95οC в течение 2 мин с последующими 34 циклами, каждый состоящий из 30 сек при 95οC, 30 сек при 55οC и 1 мин при 72οC.
[Идентификация продуктов пропуска экзонов] ПЦР-продукты подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру.
На фиг. 48A представлены данные электрофореза ПЦР-продуктов, в которых образец обработки 10 цМ ASO давал слабую, полосу для ПЦР-продукта, приписываемого пропуску экзонов 5-7 (ср. левую диаграмму). Даже если клетки обрабатывали циклогексимидом для дестабилизации полноразмерной мРНК путем замораживания трансляции на рибосомах, была обнаружена только слабая полоса для пропуска экзонов. Таким образом, вариант сплайсинга мРНК SNAP25, приписываемый пропуску экзонов 5-7, вероятно, демонстрирует плохую метаболическую стабильность в клетках по сравнению с полноразмерной мРНК. ПЦР-продукт пропуска экзонов, на основании данных секвенирования, соответствовал пропуску экзонов 5-7, как показано на фиг. 48А. (см. правую диаграмму) Учитывая, что ПЦР-продукт, приписываемый пропуску экзона 6, наблюдался независимо от концентрации ASO, считается, что пропуск экзона 6 происходит спонтанно.
Интенсивность полосы для полноразмерной мРНК SNAP25 больше всего снижалась в клетках, обработанных 10 цМ «SNAP-ASO 3». Интенсивность полосы для полноразмерной мРНК постепенно увеличивалась до уровня отрицательного контроля (т.е. без обработки ASO) по мере увеличения концентрации ASO с 10 до 1000 цМ. Обратный характер зависимости ответа от дозы в данных вложенной ПЦР может быть обусловлен активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) круговой РНК (EIciRNA)», накапливаемой во время пропуска экзона с помощью «SNAP-ASO 3» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)].
SNAP25-пример 2. кПЦР для мРНК SNAP25 в клетках РС12, обработанных «SNAP-ASO 3».
«SNAP-ASO 3» оценивали с помощью SNAP25-специфической вложенной кПЦР в отношении его способности к вызову изменений уровня мРНК для SNAP25 крысы в клетках РС12 следующим образом.
[Клеточная культура и обработка ASO] Клетки РС12, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «SNAP-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 цМ (2 чашки для культивирования на каждую концентрацию ASO)
[Экстракция РНК и синтез кДНК с помощью одностадийной ОТ-ПЦР] После инкубации с «SNAP-ASO 3» в течение 42 часов клетки обрабатывали 100 мкг/мл циклогексимида в течение еще 6 часов до заморозки трансляции на рибосомах. Затем тотальную РНК экстрагировали из клеток с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя. 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР (Invitrogen, США) на фоне набора экзон-специфических праймеров [SNAP-экзон 1_прямой: (5'→3')ATGGCCGAGGACGCAGACAA; и SNAP-экзон 14_обратный: (5'→3')AGCATCTTTGTTGCACGTTG] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 20 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 55οC и 1 мин при 72οC.
[Амплификация с помощью вложенная кПЦР] 1 мкл каждого раствора кДНК, разведенного в 100 раз, подвергали реакции ПЦР в режиме реального времени в объеме 20 мкл на фоне набора экзон-специфических праймеров [SNAP-экзон 7q_прямой: (5'→3')ATGGATGAAAACCTAGAGC; и SNAP-экзон 8q_обратный: (5'→3')CTTCCCAGCATCTTTGTT] в соответствии со следующими режимами циклирования: 95οC в течение 3 мин с последующими 40 циклами, каждый состоящий из 10 сек при 95οC и 30 сек при 60οC. Реакцию кПЦР отслеживали с помощью зонда Taqman [[5'→3')5,6-FAM-CAGCCTTCT-ZEN-CCATGATCCT-3IABkFQ], нацеленного на место соединения экзона 7 и экзона 8, для количественного анализа полноразмерной мРНК для SNAP25.
На фиг. 48B представлены данные кПЦР, в которых уровень полноразмерной мРНК значимо (t-критерий Стьюдента) снижался в клетках, обработанных «SNAP-ASO 3» в концентрации 10 цМ и 100 цМ, приблизительно на 50% и 20%, соответственно. Однако уровень полноразмерной мРНК в клетках, обработанных 1000 цМ «SNAP-ASO 3», был немного выше, чем уровень в клетках без обработки ASO (т.е. отрицательном контроле).
Обратный характер зависимости ответа от дозы в данных кПЦР в значительной степени согласуется с характером зависимости от дозы ответа уровня полноразмерной мРНК во время пропуска экзона, описанной в «SNAP25-примере 1», что предполагает повышение уровня транскрипции при увеличении дозы ASO с 10 до 1000 цМ. Таким образом, 13-мерное комплементарное перекрывание с пре-мРНК для SNAP25 крысы было недостаточным для подавления активации транскрипции, индуцированной EIciRNA, накапливаемой во время пропуска экзонов.
SNAP25-пример 3. кПЦР для мРНК SNAP25 в клетках РС12, обработанных «SNAP-ASO 1».
«SNAP-ASO 1», указанный в таблице 8, представляет собой 16-мерный ASO, полностью комплементарный 16-мерной последовательности 3'-сайта сплайсинга, охватывающего место соединения интрона 6 и экзона 7 в пре-мРНК для SNAP25 человека. «SNAP-ASO 1» комплементарно перекрывается с 16-мерной последовательностью-мишенью, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК человека [(5'→3')cucuuuggaucccag|GGUAACAAUGAUGC]. «SNAP-ASO 1» обладает способностью к 6-мерному перекрыванию с интроном 6 и 10-мерному перекрыванию с экзоном 7. Однако с мРНК для SNAP25 крысы ASO имеет одно несоответствие, как отмечено «жирным шрифтом» и «подчеркиванием» в 25-мерной последовательности пре-мРНК [(5'→3')uggcucccag|GGUAACAAA"C"GAUGC1. в которой одно несоответствие отмечено знаком кавычки ("").
«SNAP-ASO 1» оценивали с помощью SNAP25-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК для SNAP25 крысы в клетках РС12, как описано в «SNAP25-примере 2», если не указано иное.
На фиг. 48C представлены данные кПЦР, в которых уровень полноразмерной мРНК значимо (t-критерий Стьюдента) снижался в клетках, обработанных «SNAP-ASO 1» в концентрации 10 цМ, 100 цМ и 1000 цМ приблизительно на 50%, 40% и 70%, соответственно.
Как и в случае с «SNAP-ASO 3», обратный характер зависимости ответа от дозы был частично воспроизведен с «SNAP-ASO 1» при увеличении дозы с 10 до 100 цМ. Однако, принимая во внимание, что уровень полноразмерной мРНК еще больше снижался при увеличении концентрации ASO до 1000 цМ, эффективность пропуска экзонов в случае «SNAP-ASO 1», по-видимому, выше, чем в случае «SNAP-ASO 3». 15-мерное комплементарное перекрывание «SNAP-ASO 1» с пре-мРНК крысы будет отвечать за более высокую эффективность пропуска экзонов.
SNAP25-пример 4. Ингибирование экспрессии белка SNAP25 в клетках РС12 с помощью «SNAP-ASO 3».
«SNAP-ASO 3» оценивали в отношении его способности ингибировать экспрессию белка SNAP25 в клетках РС12 следующим образом.
Клетки РС12 выращивали в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, и обрабатывали «SNAP-ASO 3» в концентрации 0 цМ (отрицательный контроль), 1 цМ, 10 цМ, 30 цМ, 100 цМ, 300 цМ, 1 аМ, 3 аМ или 10 аМ в течение 48 часов. Использовали 4 чашки для культивирования для отрицательного контроля, чтобы компенсировать возможные технические артефакты во время анализа с помощью Вестерн-блоттинга.
[Лизис клеток] Затем клетки подвергали лизису на льду с использованием 200 мкл 1X буфера RIPA (кат. № 9806, Cell Signaling Tech) с добавлением 1% SDS и 1X смеси ингибиторов протеиназ (Complete Mini, Roche). Лизаты собирали в 1,5 мл пробирку Эппендорфа, смешивали со 100 мкл 5X буфера для образцов и кипятили в течение 5 мин.
[Вестерн-блоттинг] Лизаты подвергали электрофоретическому разделению в 4-15% градиентном геле TGX-ПААГ для электрофореза (кат. № 456-1086, Bio-Rad), а затем переносили на мембрану из PVDF 0,45 мкм. Мембрану исследовали с использованием антитела против SNAP25 (кат. № S9684, Sigma) и антитела против β-актина (кат. № A3845, Sigma).
На фиг. 49А представлены данные Вестерн-блоттинга SNAP25, полученные с лизатами клеток РС12 (верхняя диаграмма), а также относительные уровни экспрессии SNAP25, приведенные к β-актина с помощью денситометрии (нижняя диаграмма). Уровень белка SNAP25 снижался на 10-60% в клетках, обработанных «SNAP-ASO 3». Уровень экспрессии отрицательного контроля (т.е. 0 цМ «SNAP-ASO 3») представляет собой средний уровень экспрессии 4 образцов.
SNAP25-пример 5. Ингибирование экспрессии белка SNAP25 в клетках РС12 с помощью «SNAP-ASO 1».
«SNAP-ASO 1» оценивали в отношении его способности ингибировать экспрессию белка SNAP25 в клетках РС12, как описано в «SNAP25-примере 4», если не указано иное. Клетки РС12 обрабатывали «SNAP-ASO 1» в концентрации 0 (отрицательный контроль), 100 или 1000 цМ либо в течение 48 часов, либо в течение 72 часов. (Одна чашка для культивирования для каждой концентрации ASO).
На фиг. 49B представлены данные Вестерн-блоттинга для обработки ASO в течение 48 часов (слева) и 72 часа (справа). «SNAP-ASO 1» значительно ингибировал экспрессию белка SNAP25 в клетках РС12 в оба момента времени.
SNAP25-пример 6. Ингибирование экспрессии белка SNAP25 в клетках SiMa с помощью «SNAP-ASO 3».
«SNAP-ASO 3» оценивали в отношении его способности ингибировать экспрессию белка SNAP25 в клетках нейробластомы человека SiMa следующим образом.
[Клеточная культура и обработка ASO] Клетки SiMa (кат. № ACC164, DSMZ) поддерживали в среде RPMI 1640 с добавлением 10% FBS, 1% стрептомицина/пенициллина, 1% L-глютамина и 1% пирувата натрия в атмосфере с 5% CO2 при 37οC. Клетки SiMa выращивали в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, и обрабатывали в течение 48 часов «SNAP-ASO 3» в концентрации 0 цМ (отрицательный контроль), от 1 цМ до 100 аМ. Для отрицательного контроля использовали 3 культуральных чашки, чтобы компенсировать возможные технические артефакты во время анализа с помощью Вестерн-блоттинга.
[Лизис клеток] Клетки подвергали лизису на льду с использованием 200 мкл 1X буфера RIPA (кат. № 9806, Cell Signaling Tech) с добавлением 1% SDS и 1X смеси ингибиторов протеиназ (Complete Mini, Roche). Затем лизаты собирали в 1,5 мл пробирку Эппендорфа, смешивали со 100 мкл 5X буфера для образцов и кипятили в течение 5 мин.
[Вестерн-блоттинг] Лизаты подвергали электрофоретическому разделению в 12% SDS-ПААГ для электрофореза и переносили на мембрану из поливинилидендифторида (PVDF) 0,2 мкм. Мембрану исследовали с использованием антитела против SNAP25 (кат. № ab41455, Sigma) и антитела против β-актина (кат. № A3845, Sigma).
На фиг. 50А представлены данные Вестерн-блоттинга SNAP25, полученные с лизатами клеток SiMa (верхняя диаграмма), а также относительные уровни экспрессии SNAP25, приведенные к β-актину с помощью денситометрии (нижняя диаграмма). Уровень белка SNAP25 снижался на 40-50% в клетках, обработанных «SNAP-ASO 3».
SNAP25-пример 7. кПЦР для мРНК SNAP25 в клетках SiMa, обработанных «SNAP-ASO 3».
«SNAP-ASO 3» оценивали с помощью SNAP25-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК SNAP25 человека в клетках SiMa следующим образом.
[Клеточная культура и обработка ASO] Клетки SiMa выращивали в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, и обрабатывали «SNAP-ASO 3» в концентрации 0 цМ (отрицательный контроль), 1 цМ, 10 цМ, 100 цМ, 1 аМ, 10 аМ или 100 аМ (2 чашки для культивирования для каждой концентрации ASO)
[Экстракция РНК и синтез кДНК] Тотальную РНК экстрагировали из клеток с использованием набора для мини-приготовления RNeasy (кат. № 74106, Qiagen) в соответствии с инструкциями производителя. 200 нг РНК-матрицы подвергали 25-мкл реакции обратной транскрипции с использованием набора для синтеза первой цепи кДНК PrimeScript (кат. № 6110B, Takara) на фоне случайных гексамеров.
[Амплификация с помощью кПЦР]. ПЦР-реакции контролировали с использованием зонда Taqman [(5'→3')56-FAM-CGGCTTCAT-ZEN-CCGCAGGGTAACAA-3IABkFQ], нацеленного на место соединения экзона 6 и экзона 7, на фоне набора экзон-специфических праймеров. [SNAP-экзон 6_прямой: (5'→3')GACGAACGGGAGCAGATG; и SNAP-экзон 8_обратный (2): (5'→3')ATCTCATTGCCC-ATATCCAGG]. Режимы циклирования: 95οC в течение 3 мин с последующими 40 циклами: 15 сек при 95οC и 30 сек при 60οC.
На фиг. 50B представлены данные кПЦР, в которых уровень полноразмерной мРНК для SNAP25 человека значимо (t-критерий Стьюдента) снижался в клетках, обработанных «SNAP-ASO 3» в концентрации 1 цМ, 100 цМ, 1 аМ и 100 аМ, на 20-40%. Клетки, обработанные ASO в концентрации 100 аМ, продемонстрировали самое сильное ингибирование, составляющее 40%.
SNAP25-пример 8. Ингибирование экспрессии белка SNAP25 в коже мыши, которой местно вводили «SNAP-ASO 1».
«SNAP-ASO 1» представляет собой 16-мерный ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения интрона 6 и экзона 7 в пре-мРНК для SNAP25 мыши, считываемой с геномной ДНК мыши [доступ к которым осуществляется из ссылочной последовательности NCBI: NC_000068]. «SNAP-ASO 1» оценивали в отношении его способности ингибировать экспрессию белка SNAP25 в коже при местном введении следующим образом.
[Стрижка и группировка] В день 0 8 самок мышей C57BL/6 (возрастом 5 недель) анестезировали с помощью золетила/ромпуна, и стрижка была выполнена на спине (приблизительно 3 см × 4 см) машинкой для стрижки. Мышей случайным образом распределяли по 4 группам, т.е. группе без лечения ASO (отрицательный контроль) и 3 группам лечения 1 фМ, 10 фМ и 100 фМ «SNAP-ASO 1». (2 животных на группу)
[Местное введение] Растворы для местного применения готовили путем последовательного разведения водного исходного раствора «SNAP-ASO 1» в 30% (в объемном отношении) водно-этанольном растворе с добавлением 3% (в объемном отношении) глицерина до 0, 1, 10 и 100 фМ «SNAP-ASO 1». Каждому животному местно вводили приблизительно 1000 мкл раствора для местного применения в кожу спины с удалением волос с использованием ватного тампона два раза в день в дни 0-4.
[Отбор образцов кожи] Днем в день 4 животных анестезировали с помощью золетила/ромпуна для отбора образцов части кожи, подвергнутой местному лечению ASO. Образцы кожи затем подвергали IHC против белка SNAP25, как описано ниже.
[SNAP25-IHC] Образцы кожи были подвергнуты изготовлению срезов из замороженных тканей и иммуноокрашиванию последовательно с использованием первого антитела против SNAP25 (кат. № ab41455, Abeam) в разведении 1:200, второго антитела против IgG (кат. № BA-1100, Vector) в разведении 1:200, а затем Dylight 594-стептавидина (кат. № SA-5594, Vector, CA, США) в разведении 1:200 для мечения красной флуоресценцией. Антитело против SNAP25 исследует С-конец белка SNAP25. IHC-изображения получали на слайд-сканере Zeiss для оценки экспрессии белка SANP25. Окрашивание DAPI проводили для визуализации микроструктуры кожи.
На фиг. 51 представлен репрезентативный набор IHC-изображений SNAP25 по группам. В группе отрицательного контроля экспрессия белка SNAP25 была высокой в мышечном слое прямо под дермой. Полагают, что экспрессия белка SNAP25 в мышечном слое происходит из-за экспрессии белка SNAP25 в аксонах двигательных нейронов, внедренных в мышечный слой. Экспрессия белка SNAP25 в мышечном слое постепенно снижалась с увеличением дозы. Наиболее заметное снижение наблюдалось в группе лечения 100 фМ.
Ингибирование экспрессии полноразмерного белка SNAP25 в коже согласно IHC, по-видимому, сильнее этого ингибирования согласно Вестерн-блоттингу, наблюдаемого в клетках РС12 (ср. «SNAP25-пример 5»). Активация транскрипции с помощью EIciRNA в первичных клетках, если таковые имеются, по-видимому, не столь выражена, как положительная регуляция, наблюдаемая в раковых клетках, включая клетки PC12 и SiMa.
Примеры активности in vitro TYR-ASO.
Тирозиназа (TYR) представляет собой фермент, участвующий в меланогенезе или пигментации кожи. Производные ПНК формулы I в таблице 10 были сконструированы для комплементарного нацеливания на 3'-сайт сплайсинга экзона 2 в пре-мРНК для TYR человека или мыши. TYR-ASO оценивали в отношении TYR-антисмысловой экзон-пропускающей активности в меланоцитах человека, а также в клетках B16F10 (меланомы мыши). Биологические примеры, представленные здесь, служат для иллюстрации экзон-пропускающей активности TYR-ASO в качестве примеров соединения формулы I, и поэтому их не следует интерпретировать как ограничение объема настоящего изобретения TYR ASO.
TYR-пример 1. Пропуск экзонов, индуцируемый «TYR-ASO 4» в клетках B 16F10.
«TYR-ASO 4», указанный в таблице 10, представляет собой 13-мерный TYR-ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения интрона 1 и экзона 2 в TYR мыши, как отмечено «жирным шрифтом» и «подчеркиваем» в 30-мерной. последовательности пре-мРНК для TYR мыши [[5'→3')aauuguuuuucacag|AUCAUUUGUAGCAGA]. Между тем, «TYR-ASO 4» имеет 4 несоответствия с 3'-сайтом сплайсинга, охватывающим место соединения интрона 1 и экзона 2 в пре-мРНК TYR человека, отмеченных знаком кавычки ("") в 30-мерной последовательности пре-мРНК [(5'→3')ggguguuuug"u"acag|AU "UG"U"C"UGUAGCCGA].
«TYR-ASO 4» оценивали в отношении его способности к индукции пропуска экзона 2 TYR мыши в клетках меланомы B16F10 следующим образом. «TYR-ASO 4» может служить хорошим заместителем «TYR-ASO 1», который является полностью комплементарным пре-мРНК для TYR человека.
[Клеточная культура и обработка ASO] Клетки меланомы мыши B16F10 (кат. № CRL-6475, ATCC) поддерживали в DMEM (модифицированной по Дульбекко минимальной питательной среде Игла) с добавлением 10% FBS, 1% стрептомицина/пенициллина и 0,01 мг/мл бычьего инсулина. Клетки B16F10, выращенные в 60-мм чашке для культивирования, содержащей 5 мл DMEM, инкубировали в течение 5 часов с «TYR-ASO 4» в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 аМ.
[Экстракция РНК и синтез кДНК с помощью одностадийной ПЦР] Тотальную РНК экстрагировали с использованием «Универсального набора для экстракции РНК» (кат. № 9767, Takara) в соответствии с инструкциями производителя. 200 нг РНК-матрицы использовали для реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen) на фоне набора экзон-специфических праймеров [TYR-экзон 1_прямой: (5'→3')GTAAGTTTGGATTTGGGG; и TYR-экзон 4_обратный: (5'→3')AGAGCGGTATGAAAGGAA] в соответствии со следующими режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 52οC и 40 сек при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [TYR-экзон 1n_прямой: (5'→3')GAGAACTAACTGGGGATGA; и TYR-экзон 4n_обратный: (5'→3')CGATAGGTGCATTGGCTT] в соответствии со следующими режимами циклирования: 95οC в течение 5 мин с последующими 30 циклами, каждый состоящий из 30 сек при 95οC, 30 сек при 52οC и 40 сек при 72οC.
[Идентификация продуктов пропуска экзонов] ПЦР-продукты подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру.
На фиг. 52A представлены данные электрофореза ПЦР-продуктов. Клетки без обработки ASO давали две полосы для ПЦР-продуктов, одну для полноразмерной мРНК TYR, а другую для варианта сплайсинга мРНК TYR, в котором отсутствуют экзоны 2 и 3, что предполагает спонтанный пропуск экзонов 2-3. Однако клетки, обработанные «TYR-ASO 4» в концентрации 1-1000 аМ, по существу давали только вариант сплайсинга мРНК TYR, в котором отсутствуют экзоны 2 и 3. Таким образом, «TYR-ASO 4» увеличивает склонность к пропуску экзонов 2-3 в клетках меланомы B16F10.
ПЦР-продукт в случае пропуска экзонов, согласно секвенированию, представляет собой вариант сплайсинга мРНК, в котором отсутствуют экзоны 2-3, как показано на фиг. 52В.
TYR-пример 2. кПЦР для мРНК TYR в клетках B16F10, обработанных «TYR-ASO 4».
«TYR-ASO 4» оценивали с помощью TYR-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК TYR мыши в клетках B16F10 следующим образом.
[Клеточная культура и обработка ASO] Клетки B16F10, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «TYR-ASO 4» в концентрации 0 (отрицательный контроль), 1, 10, 100 или 1000 аМ. (2 чашки для культивирования для каждой дозы)
[Экстракция РНК и синтез кДНК с помощью одностадийной ПЦР]. Тотальную РНК экстрагировали и подвергали синтезу кДНК, как описано в «TYR-примере 1».
[Амплификация с помощью вложенной кПЦР] 1 мкл каждого раствора кДНК, разведенного в 100 раз, подвергали реакции ПЦР в режиме реального времени в объеме 20 мкл на фоне набора зондов Taqman, нацеленных на место соединения экзона 2 и экзона 3 (кат. № Mm00495818_ml, Thermo Fisher Scientific), в соответствии со следующими режимами циклирования: 95οC в течение 3 мин с последующими 30 циклами, каждый состоящий из 10 сек при 95οC и 30 сек при 60οC.
[Статистический анализ] Эксперимент с вложенной кПЦР повторяли независимо четыре раза, и индивидуальные уровни мРНК в каждом эксперименте приводили к уровню мРНК без обработки ASO. Уровни мРНК, полученные из всех 4 отдельных экспериментов, объединяли для статистического анализа с помощью t-критерия Стьюдента. Таким образом, количество образцов РНК составляет 8 на каждую концентрацию ASO.
На фиг. 52C представлены объединенные данные кПЦР, в которых уровень полноразмерной мРНК значимо (t-критерий Стьюдента) снижался приблизительно на 40% в клетках, обработанных «TYR-ASO 4» в концентрации 1-1000 аМ.
TYR-пример 3. Ингибирование экспрессии белка TYR с помощью «TYR-ASO 4» в клетках B16F10.
«TYR-ASO 4» оценивали в отношении его способности ингибировать экспрессию белка TYR в клетках B16F10, как описано ниже.
Клетки B16F10, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «TYR-ASO 4» в течение 24 часов в концентрации 0 цМ (отрицательный контроль), 10 цМ, 100 цМ, 1 аМ или 10 аМ и подвергали лизису с использованием 200 мкл 1X буфера для лизиса клеток (кат. № 9803, Cell Signaling Tech), дополненного 1Х смесью ингибиторов (кат. № P8340, Sigma). 200 мкл каждого лизата смешивали с 100 мкл 5X буфера для образцов и кипятили при 100οC в течение 5 мин. 20 мкл каждого лизата подвергали электрофоретическому разделению в 4-15% градиентом TGX-геле (кат. № 456-1086, Bio-Rad) и переносу белка на мембрану из PVDF 0,45 мкм. Мембрану исследовали с использованием антитела против TYR (кат. № 9319, Cell Signaling Tech) и антитела против β-актина (кат. № a3845, Sigma).
На фиг. 53А представлены данные Вестерн-блоттинга TYR, полученные с лизатами клеток B16F10. Уровень белка TYR был значительно выше в лизатах отрицательного контроля, чем в лизатах клеток, обработанных «TYR-ASO 4».
TYR-пример 4. Ингибирование меланогенеза с помощью «TYR-ASO 4» в клетках B16F10.
«TYR-ASO 4» оценивали в отношении его способности ингибировать меланогенез в клетках B16F10, как описано ниже.
Клетки B16F10, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали либо «TYR-ASO 4» в концентрации 0 (отрицательный контроль) или 1-1000 аМ, либо 10 или 100 мкг/мл арбутина в качестве положительного контроля (2 чашки для культивирования для каждой дозы). Через 24 часа клетки подвергали лизису с использованием 200 мкл 1 н NaOH. Каждый лизат собирали в 1,5 мл пробирку Эппендорфа и хранили в течение ночи при комнатной температуре. Содержание меланина в каждом лизате определяли по оптической плотности при 475 нм на считывающем устройстве для ELISA. Эксперимент повторяли четыре раза, используя клетки при разных пассажах. Четыре набора данных о содержании меланина объединяли для статистического анализа с помощью t-критерия Стьюдента относительно содержания меланина без обработки (отрицательный контроль).
На фиг. 53B суммированы изменения содержания меланина в клетках B16F10 после 24-часовой инкубации либо с «TYR ASO 4», либо с арбутином. Содержание меланина значимо уменьшалось приблизительно на 15% и 25% в клетках, обработанных 10 мкг/мл и 100 мкг/мл арбутина, соответственно. В случае клеток, обработанных «TYR-ASO 4», содержание меланина значимо уменьшалось приблизительно на 15% без значительной зависимости от дозы. Ингибиторная активность «TYR-ASO 4» была сравнима с таковой 10 мкг/мл арбутина.
TYR-пример 5. кПЦР для мРНК TYR в меланоцитах человека, обработанных «TYR-ASO 1».
«TYR-ASO 1», указанный в таблице 10, представляет собой 13-мерный TYR-ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения интрона 1 и экзона 2 в TYR человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для TYR человека [(5'→3')ggguguuuuguacag|AUUGUCUGUAGCCGA].
«TYR-ASO 1» оценивали с помощью TYR-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК TYR в первичных меланоцитах эпидермиса человека следующим образом.
[Клеточная культура и обработка ASO] Первичные меланоциты эпидермиса (кат. № PCS-200-013, ATCC) поддерживали в базальной среде для клеток кожи (кат. № PCS-200-030, ATCC), дополненной компонентом из набора для роста меланоцитов у взрослых (кат № PCS-200-042, ATCC). Меланоциты, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали «TYR-ASO 1» в концентрации 0 цМ (отрицательный контроль), 1 цМ, 100 цМ или 10 аМ (3 чашки для культивирования для каждой концентрации).
[Экстракция РНК и синтез кДНК с помощью одностадийной ПЦР] После инкубации с «TYR-ASO 1» в течение 5 часов, тотальную РНК экстрагировали с использованием набора для мини-приготовления RNeasy (кат. № 74106, Qiagen) в соответствии с инструкциями производителя. 200 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл, используя набор для одностадийной ОТ-ПЦР Super Script® с Taq-полимеразой Platinum® (кат. № 10928-042, Invitrogen), на фоне набора экзон-специфических праймеров [TYR-экзон 1_прямой (2): (5'→3')CTCTTTGTCTGGATGCATT; и TYR-экзон 5_обратный: (5'→3')CTGTGGTAATCCTCTTTCT] в соответствии со следующими указанными режимами циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 15 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 50οC и 1 мин при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК дополнительно амплифицировали в реакции вложенной ПЦР в объеме 20 мкл (кат. № K2612, Bioneer) на фоне набора экзон-специфических праймеров [TYR-экзон 2n_прямой: (5'→3')GATAAAGCTGCCAATTTC; и TYR-экзон 3n_обратный: (5'→3')TTGTGCATGCTGCTTTGA] на фоне зонда Taqman [(5'→3')5,6-FAM-CACTGG-ZEN-AAGGATTTGCTAGTCCAC-3IABkFQ]. Режимы циклирования: 95οC в течение 3 мин с последующими 40 циклами, каждый состоящий из 10 сек при 95οC и 30 сек при 60οC.
На фиг. 53C представлены данные кПЦР, в которых уровень полноразмерной мРНК для TYR снижался приблизительно на 30% в меланоцитах человека, обработанных «TYR-ASO 1» в концентрации от 1 цМ до 10 аМ. Наблюдаемые снижения были значимыми (t-критерий Стьюдента) в клетках, обработанных «TYR-ASO 1» в концентрации 1 цМ и 10 аМ.
Примеры биологической активности PD-1-ASO
PD-1, также известный как белок 1 запрограммированной клеточной гибели или CD279, представляет собой рецептор на клеточной поверхности, экспрессируемый в иммунных клетках. PD-1 является иммунным белком контрольной точки, участвующим в подавлении иммунного ответа. Моноклональные антитела против PD-1, такие как ниволумаб и пембролизумаб, использовались для лечения солидных опухолей путем усиления иммунного ответа.
PD-1-ASO формулы I в таблице 11 были сконструированы для комплементарного нацеливания либо на 3'-сайт сплайсинга, либо на 5'-сайт сплайсинга экзона 2 в пре-мРНК для PD-1 человека или мыши. PDO-1-ASO оценивали в отношении антисмысловой экзон-пропускающей активности в клетках Jurkat, а также в отношении противоопухолевой активности у мышей дикого типа с имплантированной сингенной опухолью. Биологические примеры, представленные здесь, служат для иллюстрации экзон-пропускающей активности PD-1-ASO в качестве примеров соединения формулы I, и поэтому их не следует интерпретировать как ограничение объема настоящего изобретения PD-1-ASO.
PD-1-пример 1. Пропуск экзона, индуцируемый «PD-ASO 32 в клетках Jurkat.
«PD-ASO 3», указанный в таблице 11, представляет собой 14-мерный PD-1-ASO, полностью комплементарный 5'-сайту сплайсинга, охватывающему место соединения экзона 2 и интрона 2 в пре-мРНК PD-1 человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для PD-1 человека [(5'→3')AGCUCAGGGUGACAG|gugcggccucggagg]. «PD-ASO 3» обладает способностью к 9-мерному перекрыванию с экзоном 2 и 5-мерному перекрыванию с интроном 2.
«PD-ASO 3» оценивали в отношении его способности к индукции пропуска экзона 2 PD-1 человека в клетках Jurkat следующим образом.
[Клеточная культура и обработка ASO] Клетки Jurkat (кат. № TIB-152, ATCC) поддерживали в RPMI-1640 с добавлением 10% FBS и 1% стрептомицина/пенициллина. Клетки Jurkat, выращенные в 60-мм чашке для культивирования, содержащей 5 мл культуральной среды, обрабатывали в течение 5 часов «PD-ASO 5» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ.
[Экстракция РНК и синтез кДНК с помощью одностадийной ОТ-ПЦР] Тотальную РНК экстрагировали с использованием набора для мини-приготовления Rneasy (Qiagen, США) в соответствии с протоколом производителя. 500 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР (Invitrogen, США) на фоне набора экзон-специфических праймеров [PD-экзон 1_прямой: (5'→3')GTCGTCTGGGCGGTGCTACAAC; и PD-экзон 5_обратный: (5'→3')GGGTGTGGAA-ATAGATGGG]. Режимы циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 30 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 53οC и 1 мин при 72οC.
[Амплификация с помощью вложенной ПЦР] 1 мкл кДНК, разведенной в 100 раз, дополнительно амплифицировали в реакции вложенной ПЦР в объеме 20 мкл (Invitrogen, США), используя следующие режимы циклирования: 20 сек при 95οC, 30 сек при 56οC и 40 сек при 72οC в течение 40 циклов на фоне набора экзон-специфических праймеров [PD-экзон 1n_прямой: (5'→3')GGCTGGCGGCCAGGATGGTTC; и PD-экзон 5n_обратный: (5'→ 3')GAAAGAC AATGGTGGC ATACTCC].
[Идентификация продуктов пропуска экзонов] Полученные продукты вложенной ПЦР подвергали электрофоретическому разделению в 2% агарозном геле. Полосы целевого размера собирали и анализировали с помощью секвенирования по Сэнгеру.
На фиг. 54A представлены данные электрофореза ПЦР-продуктов. Клетки без обработки ASO давали только полосу для продукта в виде полноразмерной мРНК. Между тем, наблюдались три полосы ПЦР-продуктов, вновь образованных в клетках, обработанных PD-1-ASO. Из трех полос для ПЦР-продуктов полоса размером приблизительно 470 п.н. (обозначенная как «неспецифическая» на фиг. 54А) не была вариантом сплайсинга мРНК PD-1 с пропуском экзона в соответствии с анализом секвенированием по Сэнгеру.
Клетки, обработанные «PD-ASO 3» в концентрации 10 и 100 аМ, давали полосу для ПЦР-продукта, соответствующего пропуску экзона 2, согласно секвенированию по Сэнгеру. Однако в клетках, обработанных 1000 аМ «PD-ASO 3», ПЦР-продукт в случае пропуска экзона 2 исчез, и уровень полноразмерной мРНК был выше уровня полноразмерной мРНК в клетках, обработанных более низкими концентрациями ASO. Два ПЦР-продукта, приписываемых пропуску экзона 2 и экзона 3, были подтверждены секвенированием по Сэнгеру (ср. фиг. 54B). Обратный характер зависимости ответа от дозы в данных вложенной ПЦР мог быть обусловлен активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) круговой РНК» (EIciRNA)», накапливаемой во время пропуска экзона с помощью «PD-ASO 3» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)].
PD-1-пример 2. кПЦР для мРНК PD-1 в клетках Jurkat, обработанных «PD-ASO 3».
«PD-ASO 3» оценивали с помощью PD-1-специфической вложенной кПЦР в отношении его способности к вызову изменений уровня мРНК для PD-1 человека в клетках Jurkat следующим образом.
[Клеточная культура и обработка ASO] Клетки Jurkat, выращенные в 60-мм чашке для культивирования, активировали с помощью антитела против CD3 (кат. № 16-0037, eBioscience) в дозе 1 мкг/мл и антитела против CD28 (кат. № 16-0289, eBioscience) в дозе 0,5 мкг/мл в течение 48 часов. Затем культуральную среду заменяли свежей средой и обрабатывали «PD-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ в течение 24 часов (4 чашки для культивирования для каждой концентрации ASO)
[Экстракция РНК и синтез кДНК с помощью одностадийной ОТ-ПЦР]. Тотальную РНК экстрагировали с использованием набора для мини-приготовления RNAeasy (Qiagen, США) в соответствии с протоколом производителя. 500 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для одностадийной ОТ-ПЦР (Invitrogen, США) на фоне набора экзон-специфических праймеров [PD-экзон 1_прямой: (5'→3')GTCGTCTGGGCGGTGCTACAAC; и PD-экзон 5_обратный: (5'→3')GGGTGTGGAA-ATAGATGGG]. Режимы циклирования: 50οC в течение 30 мин и 94οC в течение 2 мин, после чего следовали 30 циклов, каждый состоящий из 30 сек при 94οC, 30 сек при 53οC и 1 мин при 72οC.
[Вложенная кПЦР с использованием SYBR] 1 мкл каждого раствора кДНК, разведенного в 100 раз, подвергали амплификациях с помощью вложенной кПЦР в объеме 20 мкл на фоне набора экзон-специфических праймеров [PD-экзон 2_прямой: (5'→3')ACAACGCCACCTTCACCTGC; и PD-экзон 2_обратный: (5'→3')GCCAGCTTGTCCGTCTGGTTG] в соответствии со следующими режимами циклирования: 20 сек при 95οC, 30 сек при 56οC и 40 сек при 72οC в течение 40 циклов. Реакцию кПЦР исследовали с использованием SYBR (Bio-Rad, США).
На фиг. 55A представлены данные кПЦР, в которых уровень мРНК PD-1 значимо (t-критерий Стьюдента) снижался на >80% в клетках, обработанных «PD-ASO 3» в концентрации 10 и 100 аМ. В случае клеток, обработанных ASO в концентрации 1000 аМ, уровень мРНК PD-1 возвращался к приблизительно 55% от уровня отрицательного контроля. Возвращение уровня мРНК при 1000 аМ согласуется с обратным характером зависимости ответа от дозы, наблюдаемым в «PD-1-примере 1» (ср. фиг. 54A).
PD-1-пример 3. кПЦР для мРНК IL-2 в клетках Jurkat, обработанных «PD-ASO 3».
Известно, что подавление активности PD-1 усиливает экспрессию интерлейкина 2 (IL-2) [Am. J. Clin. Oncol. Vol 39(1), 98-106 (2016)]. «PD-ASO 3» оценивали с помощью IL-2-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК для IL-2 человека в клетках Jurkat следующим образом.
[Клеточная культура и обработка ASO] Клетки Jurkat, выращенные в 60-мм чашке для культивирования, активировали с помощью антитела против CD3 (кат. № 16-0037, eBioscience) в дозе 1 мкг/мл и антитела против CD28 (кат. № 16-0289, eBioscience) в дозе 0,5 мкг/мл в течение 48 часов. Затем культуральную среду заменяли свежей средой и обрабатывали «PD-ASO 3» в концентрации 0 (отрицательный контроль), 10, 100 или 1000 аМ в течение 24 часов (4 чашки для культивирования на каждую концентрацию ASO).
[Экстракция РНК и синтез кДНК] Тотальную РНК экстрагировали с использованием набора для мини-приготовления RNAeasy (Qiagen, США) в соответствии с протоколом производителя. 500 нг РНК-матрицы подвергали реакции обратной транскрипции в объеме 25 мкл с использованием набора для синтеза первой цепи кДНК PrimeScript™ (Takara, Япония) в соответствии с протоколом производителя.
[кПЦР с использованием SYBR] 1 мкл каждого раствора кДНК подвергали амплификации с помощью кПЦР в объеме 20 мкл на фоне набора праймеров, нацеленных на мРНК для IL-2 [IL-2_прямой: (5'→3')GTCACAAACAGTGCACCTAC; и IL-2_обратный: (5'→3')GGTGAGTTTGGGATTCTTGTA], в соответствии со следующими режимами циклирования: 20 сек при 95οC, 30 сек при 56οC и 40 сек при 72οC в течение 40 циклов. Реакцию кПЦР исследовали с использованием SYBR (Bio-Rad, США).
На фиг. 55B представлены данные IL-2-специфической кПЦР, в которых уровень мРНК для IL-2 значимо (t-критерий Стьюдента) увеличивался приблизительно на 140%, 120% и 40% в клетках, обработанных «PD-ASO 3» в концентрации 10, 100 и 1000 аМ, соответственно. Обратный характер зависимости ответа от дозы согласуется с обратным характером зависимости ответа от дозы, наблюдаемым в «PD-1-примере 1» и «PD-1-примере 2» (ср. фиг. 54A и фиг. 55A)
PD-1-пример 4. кПЦР для мРНК PD-1 в клетках Jurkat, обработанных «PD-ASO 1».
«PD-ASO 1», указанный в таблице 11, представляет собой 14-мерный PD-1-ASO, полностью комплементарный 3'-сайту сплайсинга, охватывающему место соединения интрона 1 и экзона 2 в пре-мРНК для PD-1 человека, как отмечено «жирным шрифтом» и «подчеркиваем» в 30-мерной последовательности пре-мРНК для PD-1 человека [(5'→3')cucuccaucucucag|ACUCCCCAGACAGGC1. «PD-ASO 1» обладает способностью к 5-мерному перекрыванию с интроном 1 и 9-мерному перекрыванию с экзоном 2.
«PD-ASO 1» оценивали с помощью PD-1-специфической вложенной кПЦР в отношении его способности вызывать изменения уровня мРНК для PD-1 человека в клетках Jurkat, как описано в «PD-1-примере 2», если не указано иное.
На фиг. 56А представлены данные кПЦР, в которых уровень мРНК для PD-1 значимо (t-критерий Стьюдента) снижался приблизительно на 60% в клетках, обработанных «PD-ASO 1» в концентрации 100 и 1000 аМ. В отличие от случая с «PD-ASO 3», «PD-ASO 1» не заставил явно предположить обратный характер зависимости ответа от дозы.
PD-1-пример 5. Противоопухолевая активность «PD-ASO 2» против меланомы B16F10 у мышей C57BL/6.
«PD-ASO 2», указанный в таблице 11, представляет собой 16-мерный PD-1-ASO, полностью комплементарный 5'-сайту сплайсинга, охватывающему место соединения экзона 2 и интрона 2 в пре-мРНК для PD-1 мыши, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для PD-1 мыши [(5'→3')AGCUCGUGGUAACAG|gugaggcuaguagaa]. «PD-ASO 2» обладает способностью к 10-мерному перекрыванию с экзоном 2 и 6-мерному перекрыванию с интроном 2.
Между тем, «PD-ASO 2» имеет четыре несоответствия с пре-мРНК для PD-1 человека, как отмечено «жирным шрифтом» и «подчеркиванием» в 30-мерной последовательности пре-мРНК для PD-1 человека [(5'→3')AGCUC"AG"GGU"G"ACAG|gug"c"ggccucggagg], в которой четыре несоответствия отмечены знаком кавычки (""). «PD-ASO 2» может рассматриваться как ASO-заместитель «PD-ASO 3» для пре-мРНК PD-1 человека.
«PD-ASO 2» оценивали в отношении его противоопухолевой активности у самцов мышей C57BL/6 (возрастом 4 недели), которым инъецировали клетки меланомы B16F10, как указано ниже.
[Инокуляция клеток меланомы B16F10] Клетки меланомы мыши B16F10 (кат. № CRL-6475, ATCC) поддерживали в 150-мм чашке для культивирования, содержащей DMEM, дополненную 10% FBS, 1% стрептомицина/пенициллина и 0,01 мг/мл бычьего инсулина. В день 0 приблизительно 1×105 клеток B16F10, суспендированных в 50 мкл PBS, инъецировали каждому животному в правый бок сзади.
[Группирование и обработка ASO] В день 3 животных случайным образом распределяли по весу по 4 группам: 0 (отрицательный контроль), 2, 10 и 50 пмоль/кг «PD-ASO 2». (N=15 на группу)
Инъекционные растворы готовили путем последовательного разведения водного маточного раствора «PD-ASO 2» в PBS до 0 нМ (только PBS), 0,4 нМ, 2 нМ и 12,5 нМ «PD-ASO 2» для отрицательного контроля, группы лечения 2, 10 и 50 пмоль/кг «PD-ASO 2», соответственно.
Животным вводили подкожно инъекционный раствор в дозе 5 мл/кг, 2 раза в неделю в дни 3-17.
[Противоопухолевая активность] Противоопухолевую активность оценивали по изменениям объема опухоли между каждой группой лечения ASO и группой отрицательного контроля.
На фиг. 56B представлены наблюдаемые объемы опухолей по группам в дни 0-19. Рост опухоли в группе лечения 2 пмоль/кг значимо ингибировался в дни 10-19. Наблюдаемое ингибирование в 19 день составляло приблизительно 55% (приблизительно 375 мм3 и 850 мм3 в случае группы лечения 2 пмоль/кг и группы отрицательного контроля, соответственно). Противоопухолевая активность в группе лечения 50 пмоль/кг была сравнима с таковой в группе лечения 2 пмоль/кг в дни 10-17. Однако противоопухолевая активность в группе лечения 50 пмоль/кг исчезала в день 19. Противоопухолевая активность в группе лечения 10 пмоль/кг была минимальной без значимости на протяжении всего периода.
Странный характер зависимости ответа от дозы может быть обусловлен активацией транскрипции, вызванной «состоящей из экзона(ов) и интрона(ов) круговой РНК (EIciRNA)», накапливаемой во время пропуска экзона с помощью «PD-ASO 2» [Nature Struc. Mol. Biol, vol. 22 (3), 256-264 (2015)]. Однако учитывая, что ниволумаб, препарат моноклональных антител против PD-1, одобренный FDA США, продемонстрировал характер зависимости ответа от дозы в формы колокола у пациентов с опухолями [J. Clin. Oncol. Vol 33 (18), 2013-2020 (2015)], странный характер зависимости ответа от дозы «PD-ASO 2» может быть обусловлен внутренней фармакологией ингибирования PD-1.
В день 19 животных умерщвляли для измерения веса опухоли по группам. Средний вес опухоли составлял приблизительно 0,35 г и 1,20 г для группы лечения 2 пмоль/кг и группы отрицательного контроля, соответственно. Таким образом, рост опухоли согласно весу был значимо ингибирован в группе лечения 2 пмоль/кг приблизительно на 70% (р<0,01 по t-критерию Стьюдента).
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ К HIF-1-АЛЬФА | 2017 |
|
RU2753517C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ ДЛЯ SNAP25 | 2017 |
|
RU2766701C2 |
| АНТИСМЫСЛОВОЕ ДЛЯ SCN9A ОБЕЗБОЛИВАЮЩЕЕ СРЕДСТВО | 2018 |
|
RU2762293C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ ТИРОЗИНАЗЫ | 2018 |
|
RU2778786C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ ПРОТИВ SCN9A | 2017 |
|
RU2748834C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ АНДРОГЕНОВОГО РЕЦЕПТОРА | 2017 |
|
RU2753966C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ АЦЕТИЛ-СОА-КАРБОКСИЛАЗЫ-2 | 2019 |
|
RU2807629C2 |
| АНТИСМЫСЛОВЫЕ ОЛИГОНУКЛЕОТИДЫ МАТРИКСНОЙ МЕТАЛЛОПРОТЕИНАЗЫ-1 | 2019 |
|
RU2802418C2 |
| Система направленного изменения сплайсинга в гене MARK2 | 2023 |
|
RU2810907C1 |
| НАПРАВЛЯЕМОЕ АНТИСМЫСЛОВЫМ ОЛИГОНУКЛЕОТИДОМ УДАЛЕНИЕ САЙТОВ ПРОТЕОЛИТИЧЕСКОГО РАСЩЕПЛЕНИЯ, МУТАЦИИ HCHWA-D И УВЕЛИЧЕННОГО ЧИСЛА ТРИНУКЛЕОТИДНЫХ ПОВТОРОВ | 2014 |
|
RU2692634C2 |


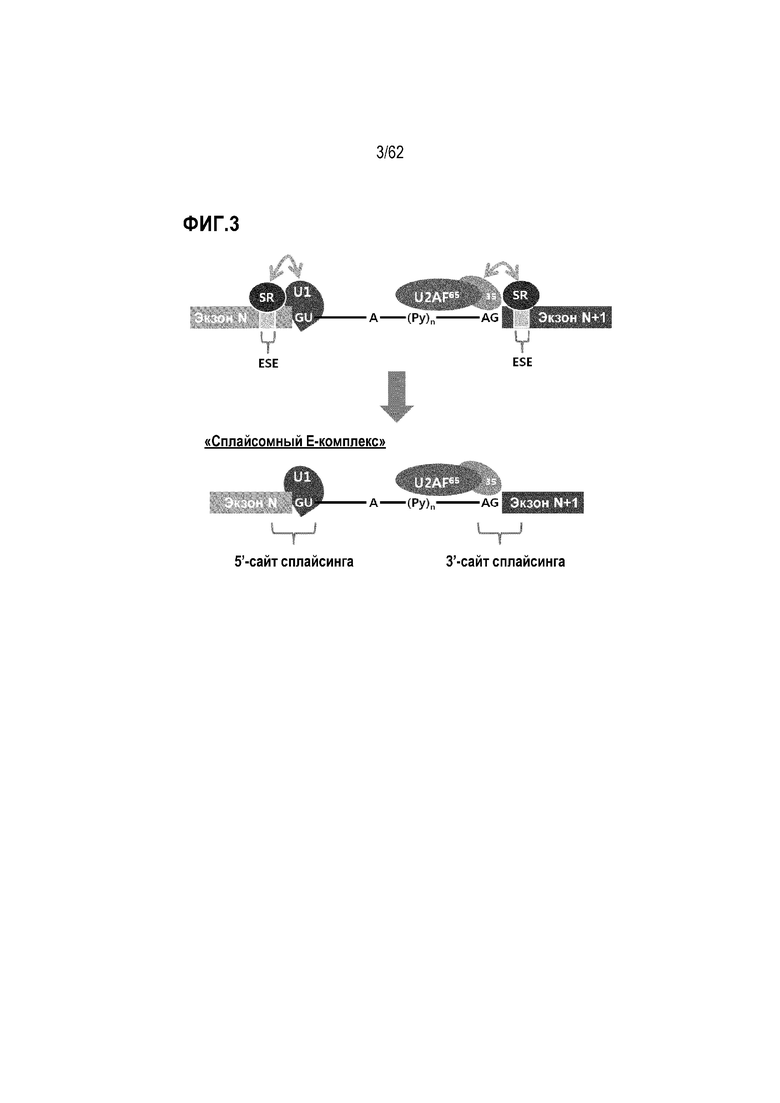
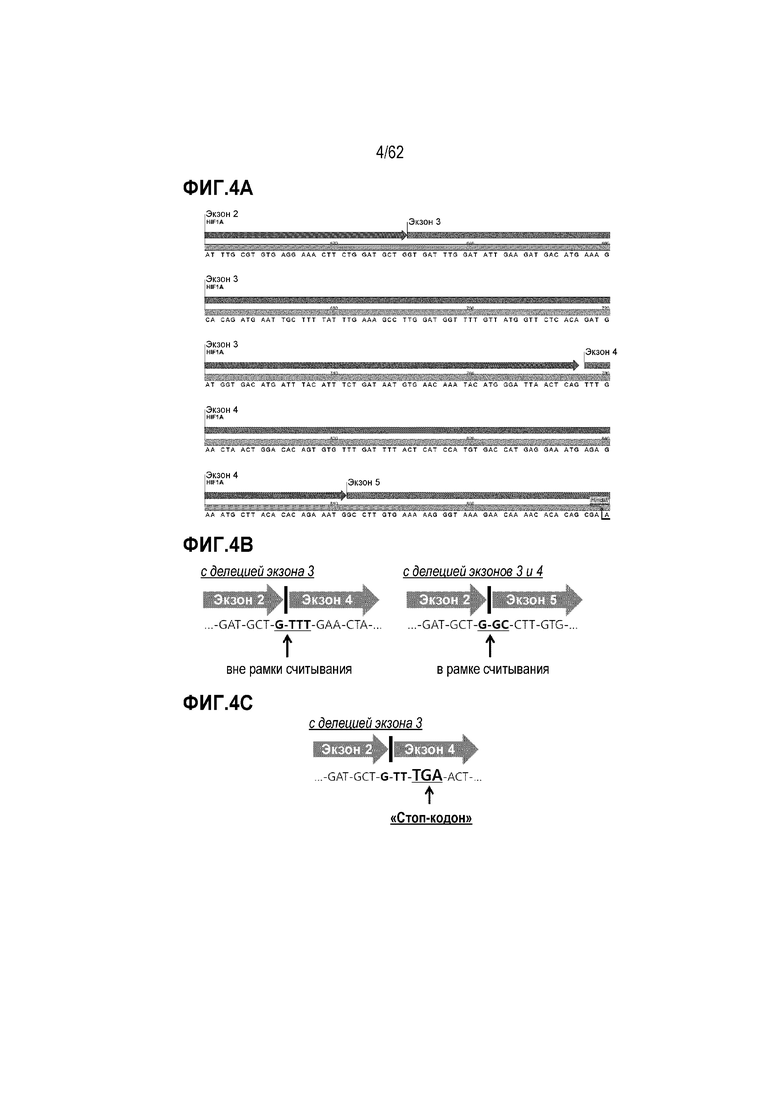



















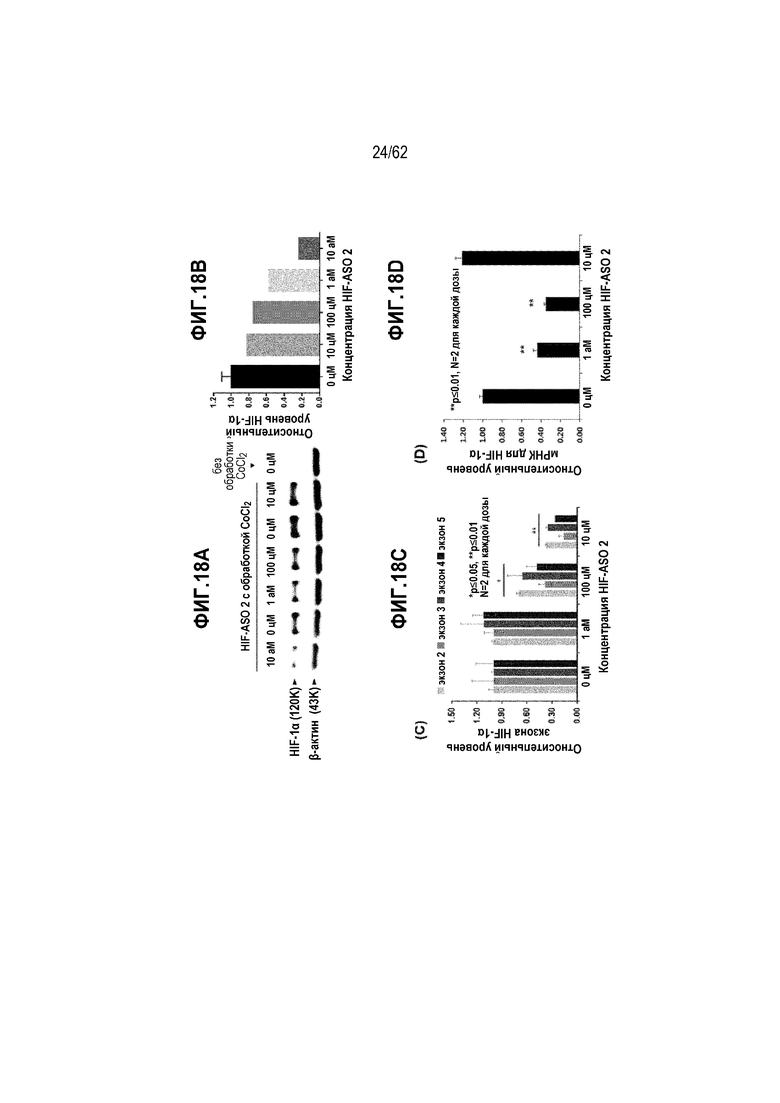



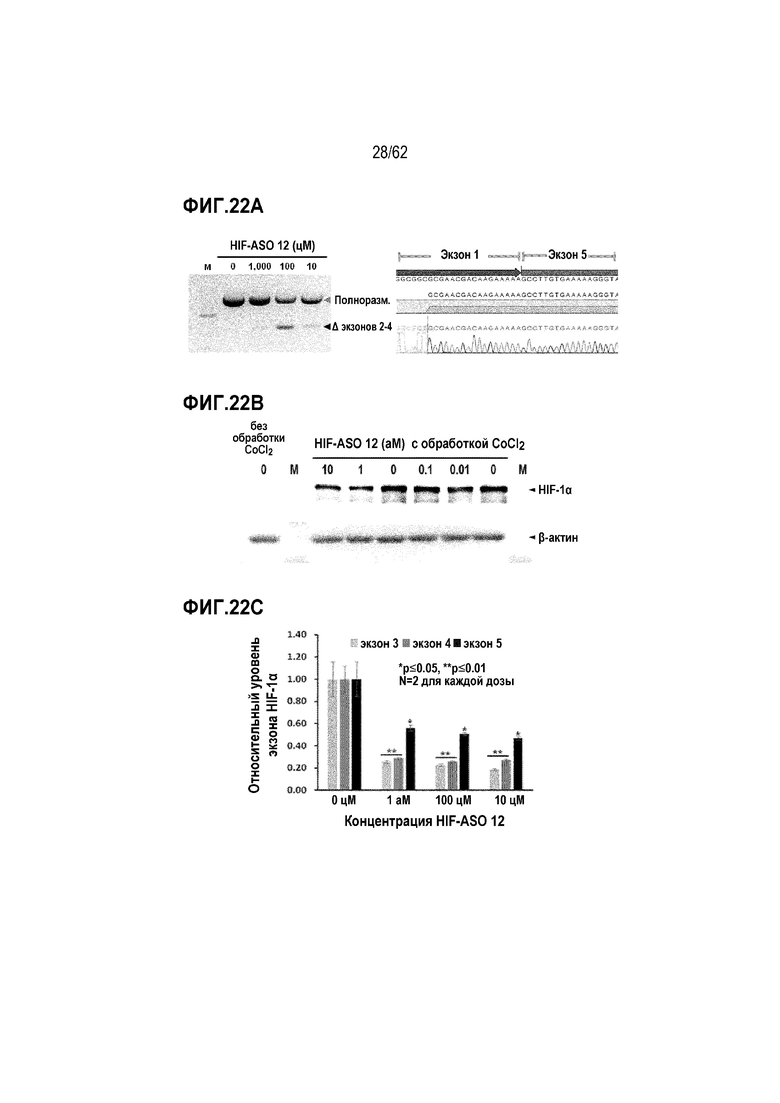
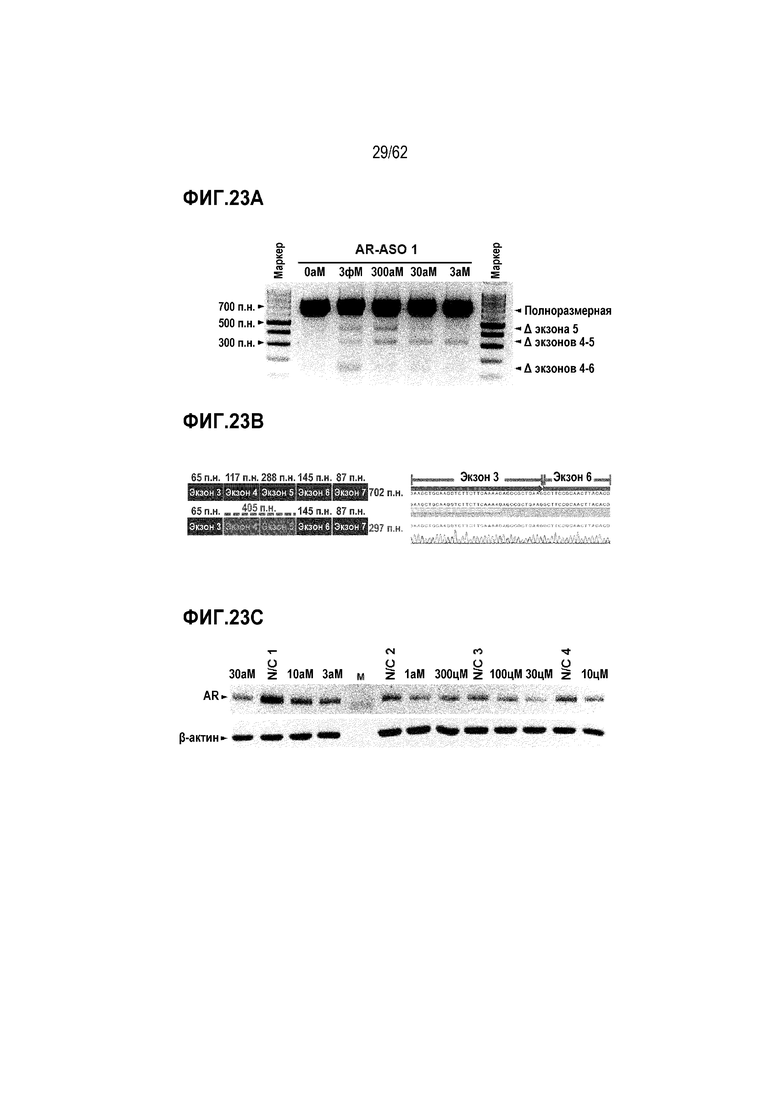
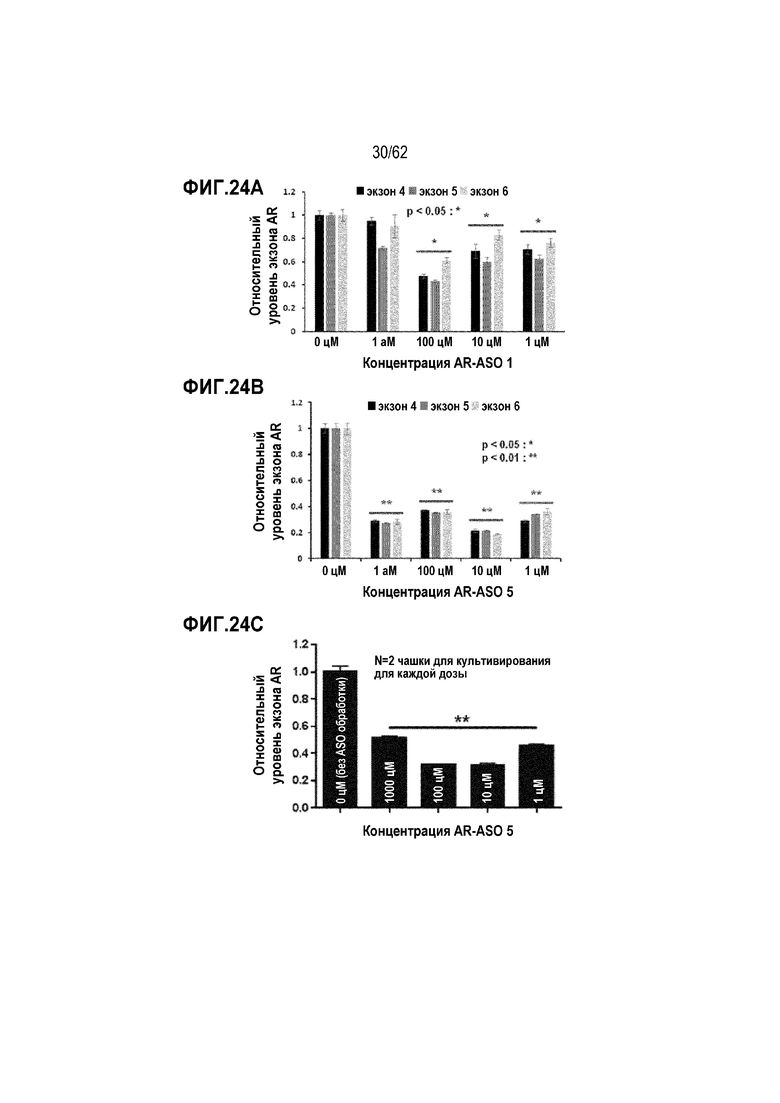





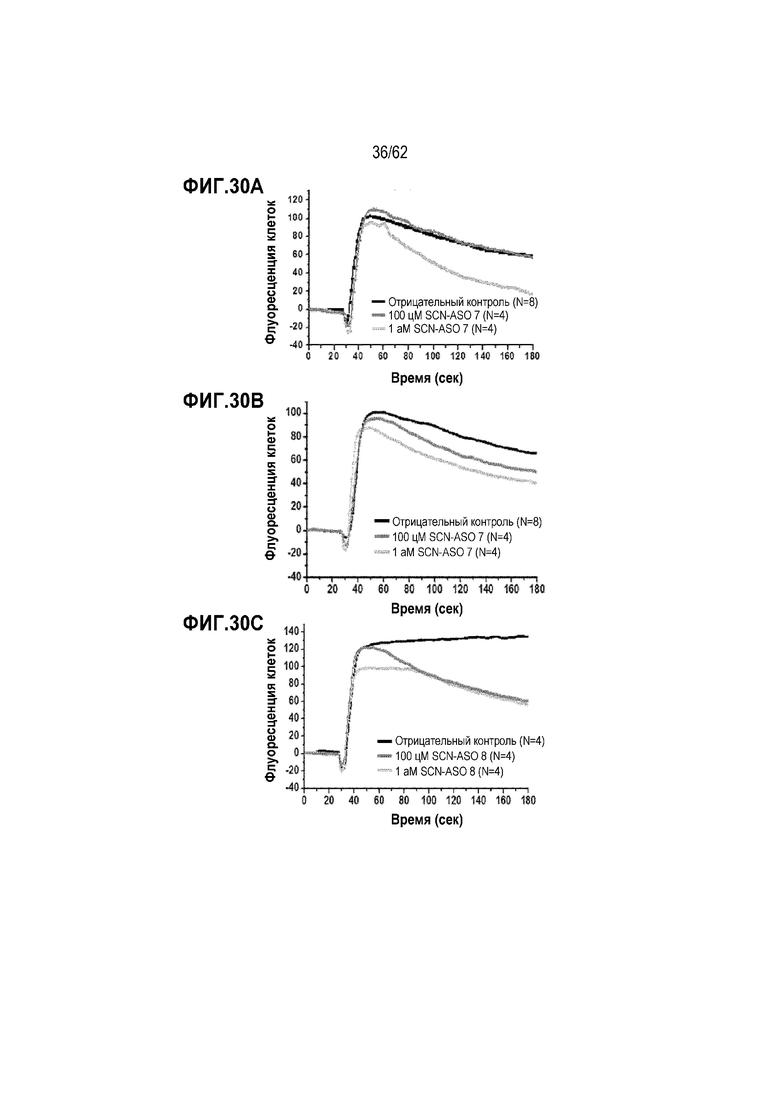

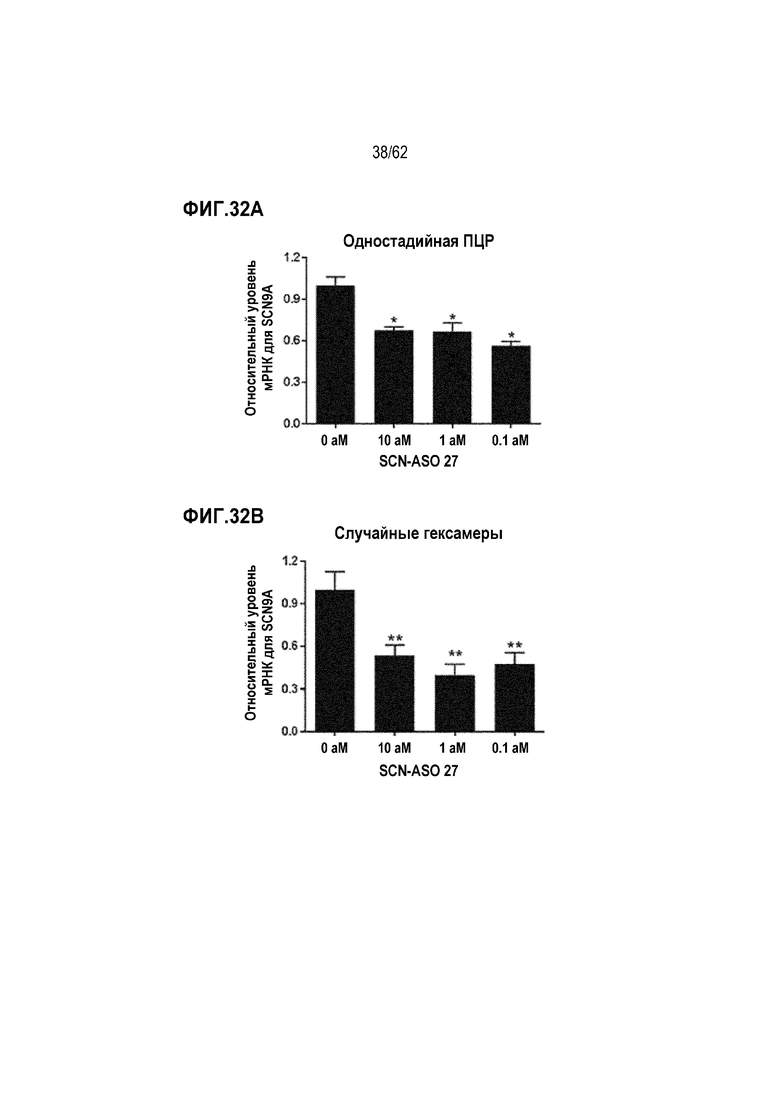


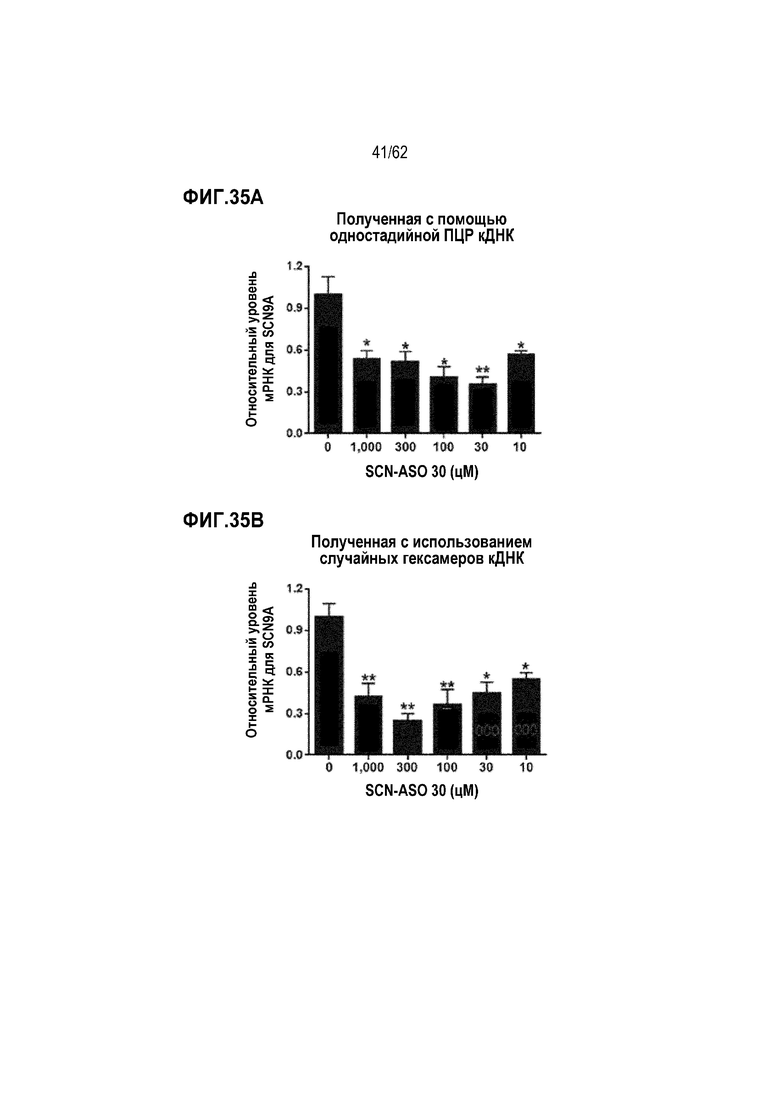
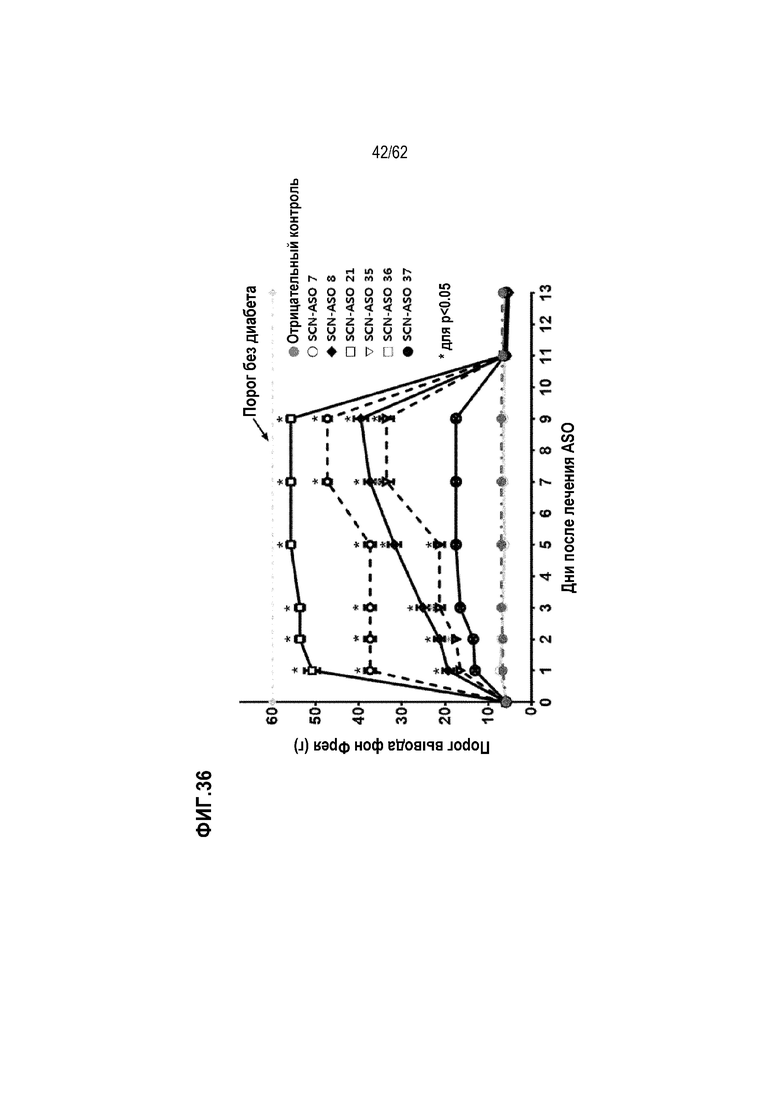

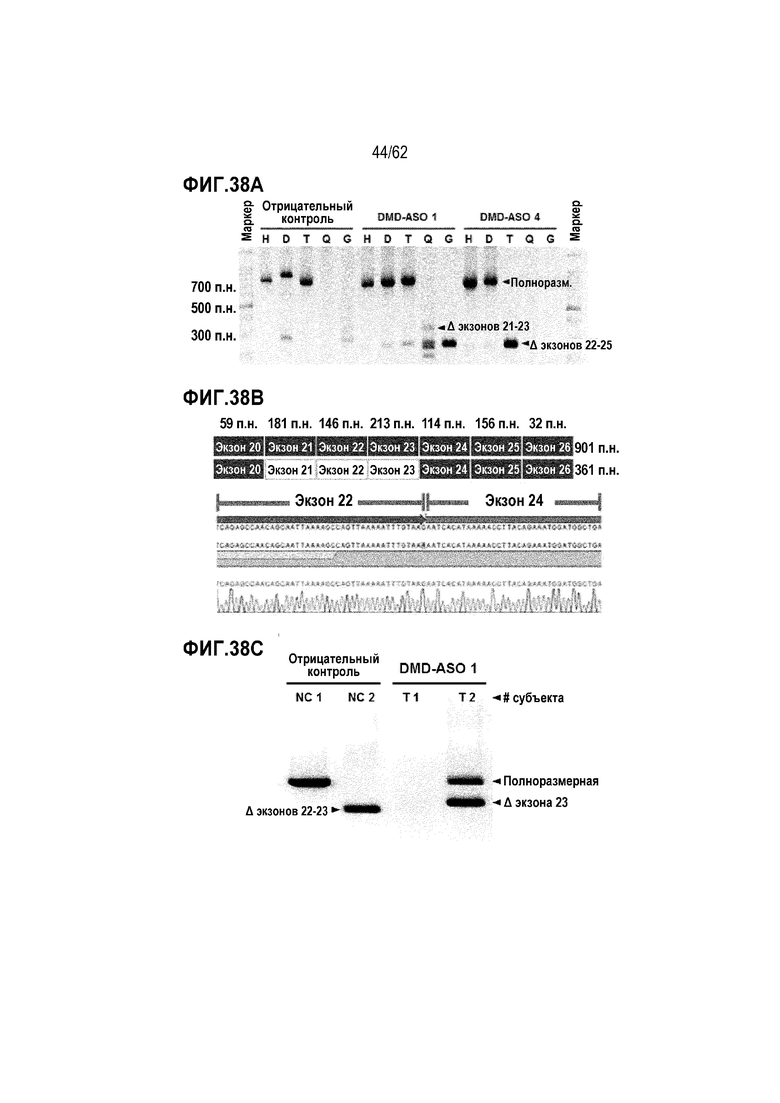
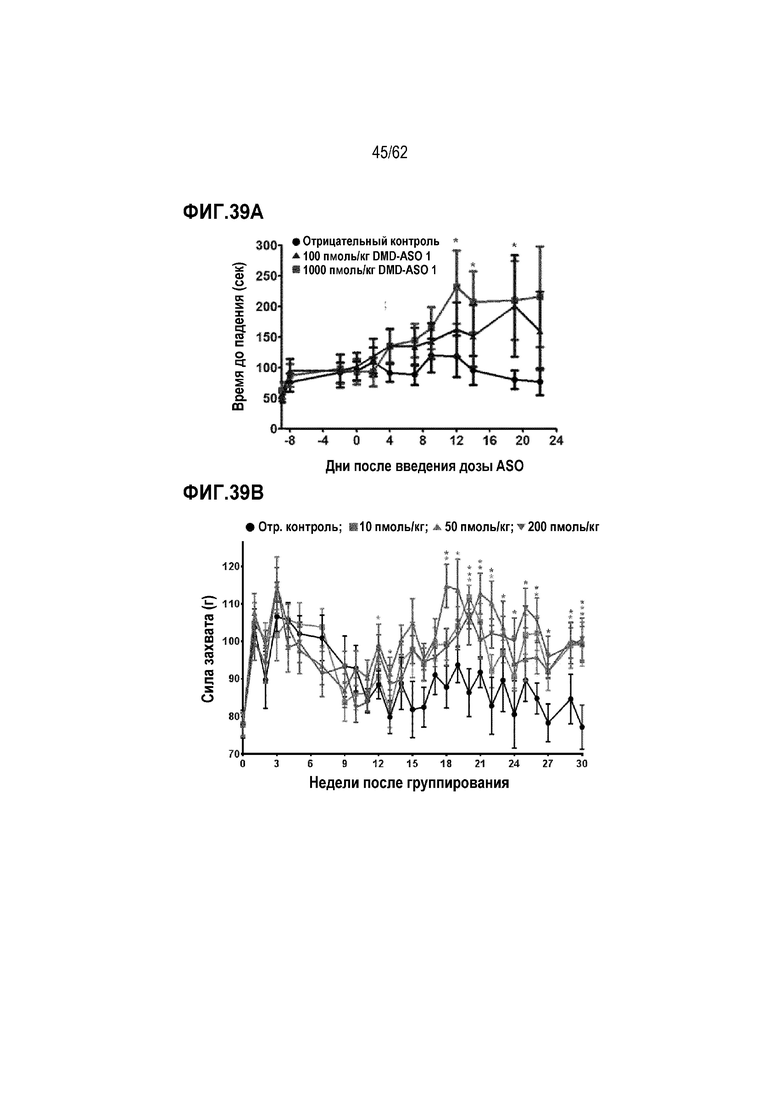


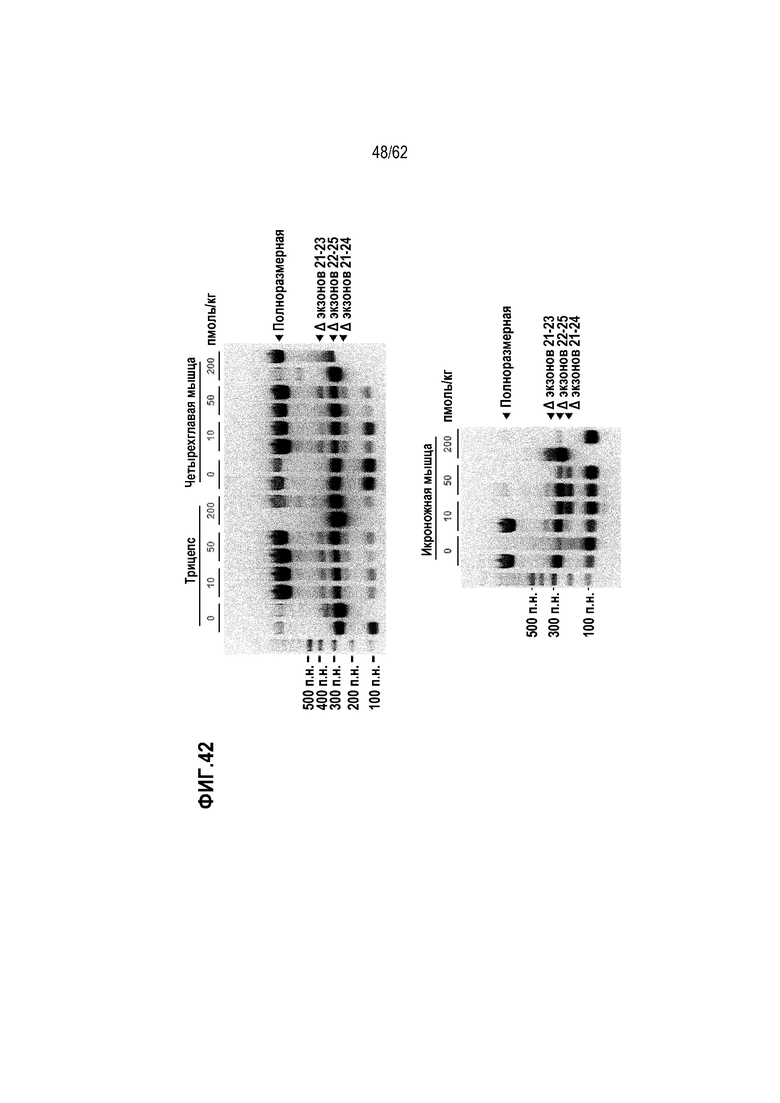

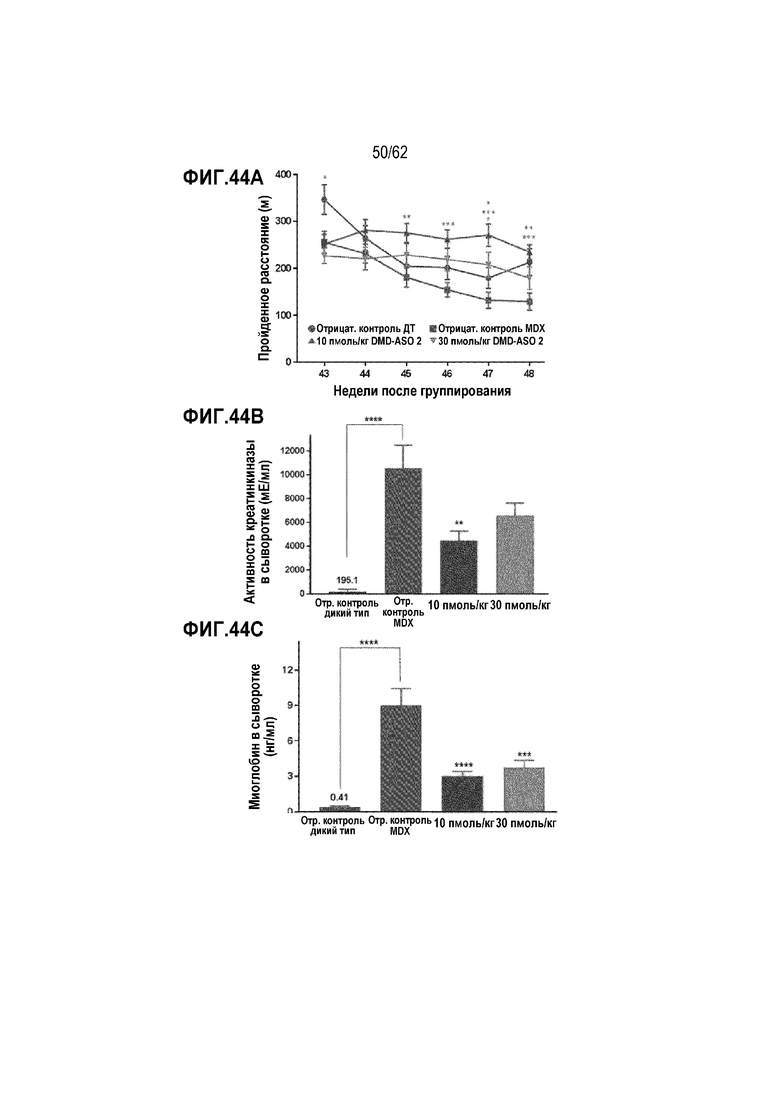





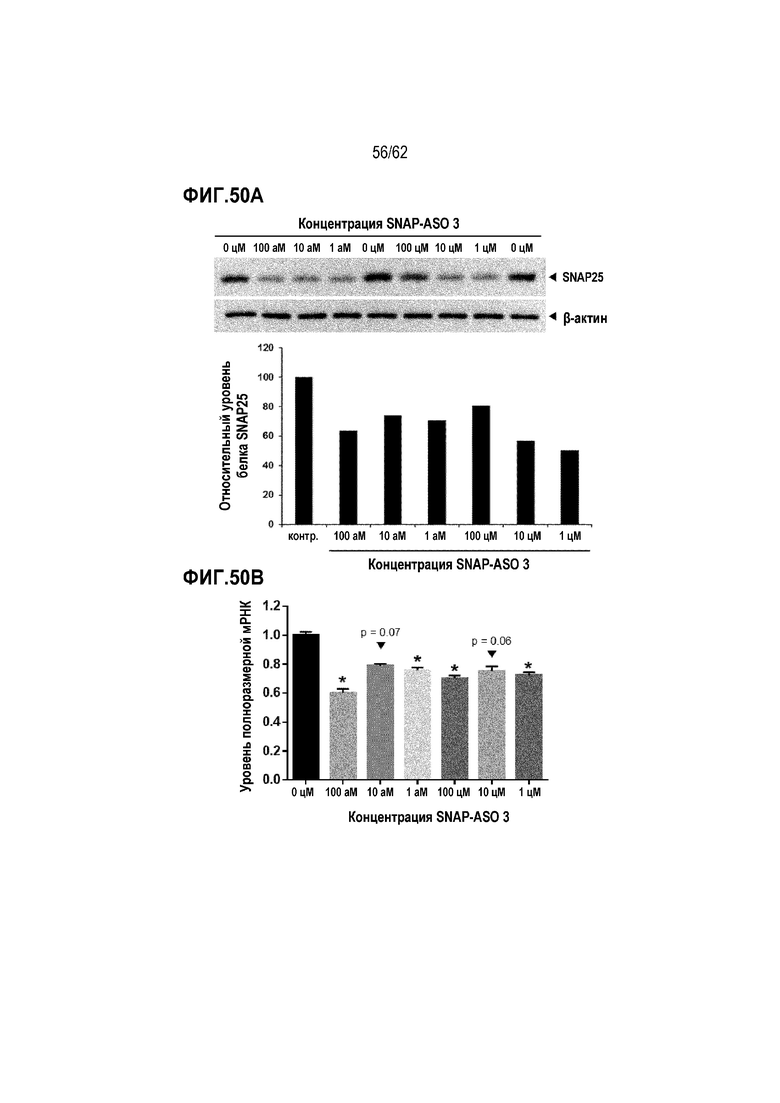






Изобретение относится к области биотехнологии, конкретно к новым производным пептидо-нуклеиновых кислот, и может быть применимо в медицине. Изобретение раскрывает молекулу пептидо-нуклеиновой кислоты, способную к специфическому прочному связыванию с сайтом сплайсинга в пре-мРНК-мишени. В силу улучшенной способности проникать через клеточную мембрану, а также сильного сродства к РНК-мишени указанная пептидо-нуклеиновая кислота способна индуцировать пропуск экзонов в клетках. Изобретение может быть применимо в качестве альтернативы антисмысловым олигонуклеотидам при вакцинации и при лечения различных заболеваний. 9 н. и 10 з.п. ф-лы, 66 пр., 14 табл., 56 ил.
1. Производное пептидо-нуклеиновой кислоты, представленное формулой I, или его фармацевтически приемлемая соль, для индукции пропуска экзона в пре-мРНК-мишени:
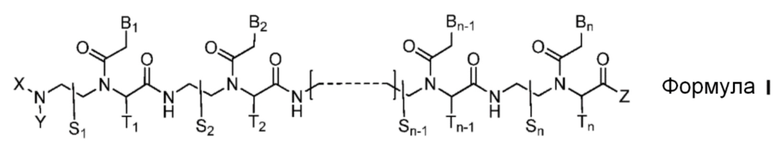
где n является целым числом между 10 и 25;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо [H];
радикалы X и Y независимо представляют собой гидридо или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино [-NH2];
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований; и
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV:

где радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
L1, L2 и L3 представляют собой ковалентный линкер, представленный формулой V, ковалентно связывающий основную аминогруппу с фрагментом нуклеотидного основания:

где Q1 и Qm являются замещенным или незамещенным метиленовым (-CH2-) радикалом и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из замещенного или незамещенного метиленового радикала и радикала кислорода (-O-); и
m является целым числом между 1 и 15.
2. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль,
где n является целым числом между 11 и 23;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
Q1 и Qm являются замещенным или незамещенным метиленовым радикалом и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из замещенного или незамещенного метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 11.
3. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль,
где n является целым числом между 11 и 21;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени или частично комплементарным последовательности пре-мРНК-мишени с одним или двумя несоответствиями;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
Q1 и Qm являются метиленовым радикалом и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 11.
4. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль,
где n является целым числом между 11 и 19;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино;
B1, B2, …, Bn-1 и Bn независимо выбирают из природных нуклеотидных оснований, включая аденин, тимин, гуанин, цитозин и урацил, и неприродных нуклеотидных оснований;
по крайней мере четыре из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
Q1 и Qm являются метиленовым радикалом и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 9.
5. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль,
где n является целым числом между 12 и 19;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикалы X и Y независимо представляют собой гидридо или замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино;
B1, B2, …, Bn-1 и Bn независимо выбирают из аденина, тимина, гуанина, цитозина и неприродных нуклеотидных оснований;
по крайней мере пять из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
Q1 и Qm являются метиленовым радикалом и Qm непосредственно связан с основной аминогруппой;
Q2, Q3,… и Qm-1 независимо выбирают из метиленового радикала и радикала кислорода; и
m является целым числом между 1 и 9.
6. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль,
где n является целым числом между 12 и 18;
соединение формулы I обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, состоящей из 7-мера из интрона и 7-мера из экзона, в указанной пре-мРНК-мишени;
соединение формулы I является полностью комплементарным последовательности пре-мРНК-мишени;
радикалы S1, S2, …, Sn-1, Sn, T1, T2, …, Tn-1 и Tn представляют собой гидридо;
радикал X представляет собой гидридо;
радикал Y представляет собой замещенный или незамещенный алкилоксикарбонил;
радикал Z представляет собой незамещенный амино;
B1, B2, …, Bn-1 и Bn независимо выбирают из аденина, тимина, гуанина, цитозина и неприродных нуклеотидных оснований;
по крайней мере пять из B1, B2, …, Bn-1 и Bn независимо выбирают из неприродных нуклеотидных оснований, представленных формулой II, формулой III или формулой IV;
радикалы R1, R2, R3, R4, R5 и R6 представляют собой гидридо;
L1 представляет собой -(CH2)2-O-(CH2)2-, -CH2-O-(CH2)2-, -CH2-O-(CH2)3-, -CH2-O-(CH2)4-, -CH2-O-(CH2)5-, -CH2-O-(CH2)6- или -CH2-O-(CH2)7-, при этом правый конец непосредственно связан с основной аминогруппой; и
L2 и L3 независимо выбирают из -(CH2)2-, -(CH2)3-, -(CH2)4-, -(CH2)5-, -(CH2)6-, -(CH2)7-, -(CH2)8-, -(CH2)2-O-(CH2)2-, -(CH2)3-O-(CH2)2- и -(CH2)2-O-(CH2)3-, при этом правый конец непосредственно связан с основной аминогруппой.
7. Производное пептидо-нуклеиновой кислоты по п. 1 или его фармацевтически приемлемая соль, где радикал X представляет собой гидридо; и радикал Y представляет собой замещенный или незамещенный алкилоксикарбонил.
8. Способ индукции в клетках пропуска экзона-мишени в пре-мРНК-мишени, включающий приведение клеток в контакт с производным пептидно-нуклеиновой кислоты по любому из пп. 1-7.
9. Способ по п. 8, где указанные клетки приводят в контакт с указанным производным пептидно-нуклеиновой кислоты в концентрации 1 пМ или менее в водном растворе.
10. Способ индукции у субъекта пропуска экзона-мишени в пре-мРНК-мишени, включающий введение субъекту производного пептидо-нуклеиновой кислоты по любому из пп. 1-7.
11. Способ ингибирования в клетках функциональной активности гена, с которого транскрибируется пре-мРНК-мишень, включающий приведение клеток в контакт с производным пептидо-нуклеиновой кислоты по любому из пп. 1-7.
12. Способ по п. 11, где указанные клетки приводят в контакт с указанным производным пептидно-нуклеиновой кислоты в концентрации 1 пМ или менее в водном растворе.
13. Способ ингибирования у субъекта функциональной активности гена, с которого транскрибируется пре-мРНК-мишень, включающий введение производного пептидо-нуклеиновой кислоты по любому из пп. 1-7.
14. Производное пептидо-нуклеиновой кислоты по любому из пп. 1-7, которое обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с 14-мерной последовательностью сайта сплайсинга-мишени, которая состоит из 7-мера из интрона и 7-мера из экзона, в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не представляет собой [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
15. Производное пептидо-нуклеиновой кислоты по любому из пп. 1-7, которое обладает способностью к по крайней мере 10-мерному комплементарному перекрыванию с сайтом сплайсинга-мишенью в пре-мРНК-мишени, причем последовательность сайта сплайсинга-мишени не включает [(5'→3')UUGCCUGGUAAGGA] в пре-мРНК для андрогенового рецептора человека, [(5'→3')UUUUUGCGUAAGUA] в пре-мРНК для SCN9A человека, [(5'→3')UAAGUAGGAUAAGU] в пре-мРНК для HIF-1α человека, [(5'→3')AUCCCAGGGUAACA] в пре-мРНК для SNAP25 человека, [(5'→3')UGUUUAGGUACACU] в пре-мРНК для SCN9A человека или [(5'→3')UGUACAGAUUGUCU] в пре-мРНК для тирозиназы человека.
16. Применение производного пептидо-нуклеиновой кислоты по любому из пп. 1-7 в производстве лекарственного средства для индукции у субъекта пропуска экзона-мишени в пре-мРНК-мишени.
17. Применение производного пептидо-нуклеиновой кислоты по любому из пп. 1-7 в производстве лекарственного средства для ингибирования у субъекта функциональной активности гена, с которого транскрибируется пре-мРНК-мишень.
18. Применение производного пептидо-нуклеиновой кислоты по любому из пп. 1-7 для индукции у субъекта пропуска экзона-мишени в пре-мРНК-мишени.
19. Применение производного пептидо-нуклеиновой кислоты по любому из пп. 1-7 для ингибирования у субъекта функциональной активности гена, с которого транскрибируется пре-мРНК-мишень.
| WO 2009113828 A2, 17.09.2009 | |||
| US 6617422 B1, 09.09.2003 | |||
| ПРОИЗВОДНЫЕ ПЕПТИДО-НУКЛЕИНОВЫХ КИСЛОТ С ХОРОШЕЙ КЛЕТОЧНОЙ ПЕНЕТРАЦИЕЙ И СИЛЬНОЙ АФФИННОСТЬЮ К НУКЛЕИНОВОЙ КИСЛОТЕ | 2009 |
|
RU2564032C2 |
| SIWKOWSKI A.M | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Nucl.Ac | |||
| Res | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| PEACEY E | |||
| et al.: Targeting a pre-mRNA structure with bipartite antisense molecules | |||

