Изобретение относится к области биотехнологии, в частности к генной инженерии, и касается нового штамма бактерий, который используется для получения рекомбинантного интерферона гамма (далее ИНФ-гамма), применяемого в медицинской промышленности.
Известно, что интерферон-гамма (далее - ИНФ-гамма) является полифункциональным белком, вырабатываемым клетками иммунной системы, обладает иммуностимулирующей, противоопухолевой и антивирусной активностями. Создание микроорганизмов-продуцентов человеческого ИНФ-гамма позволило получать биологически активный рекомбинантный белок в количествах, необходимых как для проведения широких структурно-функциональных исследований, так и для использования в медицинской практике. В настоящее время созданы лекарственные препараты на основе рекомбинантного ИНФ-гамма для лечения иммунных, онкологических и тяжелых вирусных заболеваний. В связи с этим разработка новых высокопродуктивных штаммов и эффективных способов получения рекомбинантного ИНФ-гамма является актуальной.
В природе, интерферон-гамма - это цитокин, вырабатываемый Т-лимфоцитами и естественными клетками-киллерами, который существует в виде гомодимера из двух нековалентно связанных полипептидных субъединиц. Зрелый гамма-интерферон человека представляет собой полипептид длиной 143 аминокислотных остатка и не содержит остатков цистеина Предшественник включает 166 аминокислотных остатков /Rinderkneht Е., O'Connor В.Н., Rodriguez Н. Natural human interferon-gamma: Complele aminoasids sequence and determination of sites of glycosilation. J. Biol. Chem. 1984. V.259. P.6790-6797; "Structure of the human immune interferon gene." Gray P.W., Goeddel D.V. Nature 298:859-863(1982). EP 0077670, A2, A61K 35/14, 1981; EP 0089676, A2, C07K 14/57,1983;).
Биологически активной формой ИФН-гамма является нековалентно связанный димер, тепловая денатурация которого происходит при 52°С и сопровождается инактивацией и необратимым агрегированием. С-концевые остатки не участвуют в образовании третичной структуры белка и их делеция вплоть до 20 аминокислот не приводит к потере биологической активности (Slodowski О., Bohm J., Schone В., Otto В. Carboxy-terminal truncated rhuIFN-g with a substitution of Gln 133 or Ser 132 to leucine leads to higher biological activity than in the wild type. Eur. J. Biochem. 1991. V.202. P.1133-1140).
Известно о различных естественных или мутированных формах полипептидов субъединиц ИНФ-гамма, включая форму, содержащую N-концевую последовательность из аминокислот Cys-Tyr-Cys, форму, содержащую N-концевой метионин и различные укороченные с С-конца формы, содержащие 127-134 аминокислотных остатка. Показано, что удаление 1-15 аминокислотных остатков с С-конца не вызывает потери активности молекулы белка. К тому же известно о гетерогенности С-конца человеческого интерферона гамма (RU, 2296130, C1, C07K 14/57, 2002).
Наиболее близким аналогом к заявляемому изобретению является патент (RU, 2214832, А 61 K 38/21, 2002), где описана рекомбинантная плазмидная ДНК, кодирующая синтез белка интерферона гамма, состоящего из 144 аминокислотных остатков. Рекомбинантная плазмидная ДНК pGIF315 размером 3,15 тысяч пар нуклеотидов, кодирующая в клетках Escherichia coli рекомбинантный человеческий гамма-интерферон размером 144 аминокислотных остатка с молекулярной массой 16,9 кДа, характеризуется следующими свойствами: содержит между двумя сайтами рестриктазы EcoRI промотор А2 и между сайтами рестриктаз EcoRI и KpnI промотор A3 фага Т7, между сайтами рестриктаз KpnI и XbaI рибосомосвязывающий сайт, между сайтами рестриктаз XbaI и BgIII ген, кодирующий синтез рекомбинантного человеческого гамма-интерферона, между двумя сайтами рестриктазы XbaI терминатор транскрипции, имеет ген устойчивости к ампициллину. Способ получения гамма-интерферона, представленный в данном аналоге, включает: трансформацию подходящего штамма E. coli плазмидой pGIF315, культивирование клеток трансформированного штамма в обычной среде, лизис клеток и отмывку агрегированного белка, растворение ИФН-гамма в 7М растворе мочевины, хроматографию на катионообменнике с одновременной ренатурацией.
Недостатками ближайшего аналога является то, что для получения биомассы, проводят трансформацию клеток Escherichia coli штамма SG 200-50 рекомбинантной плазмидой перед каждой ферментацией, штамм - продуцент на основе данной плазмиды не создан, а недостатком известного способа является то, что проведение ренатурации одновременно с хроматографией не позволяет получать приемлемый выход правильно свернутой формы рекомбинантного белка.
Также из уровня техники известны и другие способы получения интерферона гамма.
Так, известно много способов получения рекомбинантного ИНФ-гамма человека, но все они трудоемки, дорогостоящи, описаны для выделения аналитических количеств белка и трудно масштабируемы. (US, 4604284, A61K 37/02, 05.08.86; US, 4751078, A61K 45/02,14.06.88; US, 4617378, C07K 15/26, 14.10.86)
Известен способ, позволяющий получить интактный нативный ИНФ-гамма, который включает несколько стадий: химический лизис клеток штамма-продуцента E. coli и одновременную солюбилизацию ИНФ-гамма из телец включения раствором 7М гуанидинхлорида; удаление нерастворившихся компонентов центрифугированием; 10-кратное разбавление целевой фракции буфером рН 9,5 и отделение образующегося осадка белков центрифугированием; фракционирование белков на силикагеле (NuGel-952 АС); и, наконец, аффинная хроматография на сорбенте с моноклональными антителами. (US, 4681930, C07K 15/26, 1987) Полученный продукт находится в растворе 20 мМ натрий-фосфатного буфера, содержащего 1М NaCl и 1М гуанидинхлорид (или 50% этиленгликоль), и представляет собой белок с молекулярной массой 18000, гомогенный по электорофорезу. Выход из 25 г клеток составляет 8 мг (25-32% от содержания в клеточном экстракте). Биологическая активность препарата по ингибированию цитопатического эффекта составляет 107 ед/мг белка.
Недостатком этого способа является то, что одновременное разрушение клеток и солюбилизация не дают возможности использовать одно из преимуществ образования телец включения, заключающееся в легком отделении балластных водорастворимых белков бактериальной цитоплазмы от нерастворимого целевого продукта простым центрифугированием. Использование иммунного сорбента для очистки белка очень дорого, нетехнологично, трудно масштабируемо. Недостатком иммуносорбции в качестве способа очистки гамма-интерферона является использование дорогостоящих, высокоспецифичных моноклональных антител, приготовление которых требует особой технологии. Кроме того, существует опасность загрязнения конечного продукта трансформирующими факторами и вирусами, происходящими от гибридомных линий. В результате могут возникать конформационные изменения гамма-интерферона (денатурация), что приводит к снижению его биологической активности. Поэтому требуется дополнительный сложный анализ конечного продукта на отсутствие онкогенного материала родительской миеломной клетки, используемой при получении моноклональных антител.
Для очистки рекомбинантного ИНФ-гамма так же используют известный способ очистки, включающий осаждение полиэтиленимином, хроматографию на колонке с четвертичным аминоэтилом (QAE), хроматографию на колонке с фенилсефарозой, осаждение сульфатом аммония, хроматографию на колонке с Сефадексом G-100 и диализ.(US,4751078, А, C07K 14/57, 1988).
Известен и другой способ очистки, основанный на серии хроматографических процедур, использующих три свойства ИНФ-гамма - высокую изоэлектрическую точку, отсутствие остатков цистеина и склонность к самоассоциации. (Arakawa, Т., Alton, K.N. and Hsu, R.Y. Preparation and characterization of recombinant DNA-derived human interferon-g. J. Biol. Chem. 260, 14 435-14 439,1985) При этом, не упоминается название какой-либо смолы, кроме Sephadex G-50, в качестве заключительной стадии очистки, которую необходимо проводить в 1 М мочевины, чтобы избежать адсорбции белка в колонке.
Так же известен способ, где используют центрифугирование в градиенте плотности для выделения тел включения и только одну колонну с сорбентом для гель-фильтрациии. (Vanderbrock, K, Martens Е, Andrea S.D. and Billiau, Refolding and single step purification of Porcine interferon-gamma from Escherichia coli inclusion bodies: conditions for reconstitution of dimeric IFN-g. Eur. J. Biochem. 215,481-486, 1993) Эти способы страдают от проблем, связанных с процессом расширения масштабов.
Недостатком иммуносорбции как способа очистки ИНФ-гамма является использование дорогостоящих, высокоспецифичных моноклональных антител, получение которых требует отдельной технологии и существует опасность загрязнения конечного продукта трансформирующими факторами и вирусами, происходящими из гибридомных линий. (US,5278286A, C07K 1/113, 1994). Кроме того, рекомбинантный ИНФ-гамма, как и многие другие рекомбинантные белки, может быть легко агрегирован в растворах, которые не содержат денатурирующих или хаотропных агентов.
Так же известен способ получения гамма-интерферона, предусматривающий ренатурацию за счет десятикратного разведения водой ИНФ-гамма, растворенного в 7М растворе мочевины, а затем фракционирование и осаждение ИФН-гамма сульфатом аммония. (RU, 2097428, C1,C12N 15/23, 1996). Однако, это не технологично, так как операции с большими объемами растворов требуют большего расхода реагентов (того же сульфата аммония) и использования крупномасштабного оборудования. Кроме того, хроматографию осуществляют на анионообменнике, а ИФН-гамма имеет положительный заряд при рН от 0 до 10, поэтому интерферон гамма не может сорбироваться на положительно заряженные анионообменники. Кроме того, осаждение сульфатом аммония вызывает частичную необратимую денатурацию белка.
Известен способ, позволяющий получать большой выход правильно свернутого белка, но поскольку получение молекулярных шаперонов отнимает много времени, а для облегчения рефолдинга белка необходимы большие количества шаперонов при масштабировании производства этот способ не приемлем. (Yong-Gui Gao, Yi-Xin Guan, Shan-Jing Yao, Biotechnol. Prog. 2003, 19(3):915-20).
Наиболее близким аналогом в отношении заявляемого объекта штамм - является штамм С 600 Е. coli, который является штаммом-продуцентом ИФН-гамма человека Е. coli T3g., куда введена рекомбинантная плазмида pTTgKm2 (RU, 2097428, C1, C12N 15/23, 1996). Описанный штамм Е. coli T3g продуцирует белок, состоящий из 144 аминокислотных остатков. Рекомбинантная плазмида pTTgKm2 сконструирована на основе плазмиды pUC 19, имеющей ген устойчивости к канамицину, триптофановый промотор, вслед за ним ген человеческого гамма-интерферона и терминаторы транскрипции rrnBT1T2. Недостатком аналога является использование в качестве штамма-продуцента рекомбинантного человеческого интерферона гамма штамма С 600 Е. coli, который не позволяет получить сстабильный синтезируемый рекомбинантный белкок с высоким выходим. Недостатком является также то, что экспрессия гена человеческого интерферона-гамма с триптофанового промотора запускается в средах с дефицитом триптофана. В обычных питательных средах работа триптофанового промотора репрессирована. При этом в компонентах обычных питательных сред (таких как дрожжевой экстракт, гидролизаты казеина: триптон, пептон) содержание триптофана не ограничивается, поэтому поиск или приготовление сред с пониженным содержанием триптофана усложняет технологию и удорожает себестоимость этапа культивирования.
Таким образом, задачей настоящего изобретения является создание штамма -продуцента белка интерферона гамма на основе сконструированной плазмиды, направляющей синтез интерферона гамма с целью повышения эффективности и снижения трудоемкости способа получения, увеличения выхода и чистоты белка ИНФ-гамма, имеющего аминокислотную последовательность из 144 аминокислотных остатков.
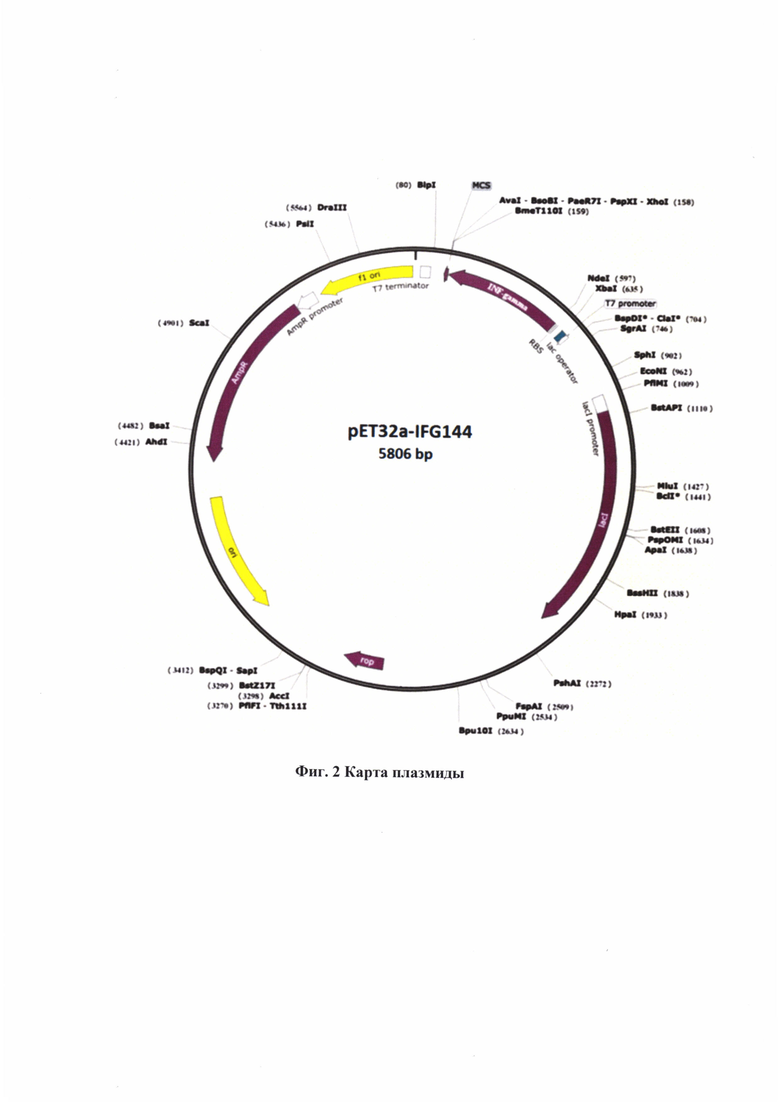
Поставленная задача достигается за счет создания рекомбинантной плазмиды pET32a-IFG144 размером 5806 пар оснований (п.о.), обеспечивающей синтез белка, имеющего аминокислотную последовательность SEQ ID NO: 1 размером 16,9 кДа и содержащей структурные элементы: промотор Т7 и оператор 1асО, синтетическую нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок ИНФ-гамма, ген устойчивости к антибиотику ампициллину (AmpR) и бактериальный промотор гена устойчивости к ампициллину (AmpR промотор) для проведения отбора рекомбинантных клеток, ориджин репликации бактериофага fl (fl ori), ген лактозного репрессора LacI и бактериальный промотор гена лактозного репрессора (LacIpromoter).
Еще одним заявляемым объектом является штамм Escherichia coli JM109/pET32a-IFG144- продуцент белка ИНФ-гамма, полученный трансформацией рекомбинантной плазмидой рЕТ32а-IFG144.
Предложенный штамм имеет ряд существенных преимуществ по сравнению с известными из уровня техники, в т.ч. ранее указанными источниками.
Т.к. белок интерферон гамма используется при производстве лекарственных средств, надо учитывать, что для обеспечения постоянства свойств лекарственного препарата, содержащего допустимые примеси в определенном диапазоне, должен соблюдаться целый ряд требований при ферментации, экспрессии белка и очистке.
Получение для ферментации трансформационной смеси, как в ближайшем аналоге (RU, 2214832, A61K 38/21, 2002),, а не штамма, не позволяет проводить процесс в строго определенных условиях, описанных в стандартных операционных процедурах, что сказывается на качестве получаемого белка, соответственно это сказывается и на лекарственном препарате. Наращивание биомассы происходит до оптической плотности OD600=4 опт.ед. Наращивание биомассы с использованием заявляемого штамма JM109/pET32a-IFG144 происходит до OD600=(30-35) опт.ед Что касается штамма-продуцента ИФН-гамма человека Е. coli T3g., то недостатком является экспрессия гена человеческого интерферона-гамма с триптофанового промотора, который запускается в средах с дефицитом триптофана, что влечет за собой поиск или приготовление сред с пониженным содержанием триптофана усложняет технологию и удорожает себестоимость этапа культивирования. При этом количество белка составляет 7 г/л, в заявляемом изобретении - не менее 40 г/л.
Таким образом, создание штамма-продуцента Escherichia coli JM109/pET32a-IFG144 позволяет получить эталонную культуру, заложить на хранение рабочую культуру клеток штамма и проводить процесс ферментации при стандартизованных условиях. В заявляемом изобретении выход биомассы с 1 л культуральной жидкости составляет от 40 до 60 г. А в вышеуказанных патентах получают биомассу 5-7 г/л. Соответственно, эффективность заявляемой ферментации увеличилась в 6-10 раз.
Так же объектом настоящего изобретения является способ получения белка ИНФ-гамма, имеющего аминокислотную последовательность SEQ ID NO: 1, включающий культивирование штамма-продуцента Escherichia coli JM109/pET32a-IFG144, при этом после 3 -5 часов культивирования проводят индукцию биосинтеза рекомбинантного белка в клетках Escherichia coli JM109/pET32a-IFG144 изопропил-β-D-1-тиогалактопиранозидом и выдерживают 4-6 часов, с получением биомассы клеток штамма-продуцента, продуцирующего белок ИНФ-гамма, выделение из полученной биомассы клеток штамма-продуцента белка ИНФ-гамма, последующую очистку выделенного белка ИНФ-гамма, путем катионнообменной хроматографии в денатурирующих условиях, ренатурацию белка ИНФ-гамма путем последовательного дробного добавления денатурированного белка в буфер для ренатурации, содержащий L-аргинин и очистку ренатурированного белка ИНФ-гамма катионообменной хроматографией, с получением белка с чистотой не менее 97%.
При этом, культивирование клеток Escherichia coli JM109/рЕТ32а-IFG144 можно проводить в ростовой среде на протяжении по меньшей мере 8 часов, но не более 10 часов
А ренатурацию денатурированного белка можно проводить в присутствии L-аргинина с концентрацией 0,3М-0,5М, при этом ренатурацию предпочтительно осуществлять дробным добавлением денатурированного белка через каждые 1-2 часа инкубации.
Настоящее изобретение относится к белку ИНФ-гамма, имеющему аминокислотную последовательность SEQ ID NO: 1.
Изобретение иллюстрируется следующими графическими фигурами.
На Фиг. 1 представлены SEQ ID NO: 1 и SEQ ID NO: 2 (аминокислотная последовательность ИНФ-гамма и нуклеотидная последовательность синтетического фрагмента, кодирующая ИНФ-гамма)
На Фиг. 2 изображена Карта плазмиды, содержащая:
fl ori - ориджин репликации бактериофага fl,
AmpR - ген резистентности к ампициллину (ген β-лактамазы),
Ori - ориджин репликации colE1,
Т7 promoter - промотор бактериофага Т7,
INF-gamma - ген белка IFG144,
Т7 terminator - терминатор бактериофага Т7,
LacI - ген лактозного репрессора LacI.
В заявляемом техническом решении поставленная задача решается посредством конструирования рекомбинантной плазмиды pET32a-IFG144 для экспрессии указанного белка, создания высокопродуктивного бактериального штамма-продуцента Escherichia coli JM109/pET32a-IFG144, продуцирующего белок ИНФ-гамма, а также способа получения белка, позволяющего получать белок ИНФ-гамма с высоким выходом и высокой степенью чистоты.
Так же в заявляемом техническом решении предложен упрощенный способ получения интерферона гамма, состоящий из культивирования клеток штамма-продуцента в обычной среде, лизис клеток, одностадийную отмывку агрегированного белка, растворение ИФН-гамма в 7 М растворе мочевины, хроматографию на катионообменнике в денатурирующих условиях, ренатурацию, очистку на катионообменнике.
Преимущество способа, согласно изобретению, состоит в том, что за счет высокоэффективной ренатурации увеличивается чистота получаемого продукта, а также увеличивается выход получаемого продукта, Неспецифическое агрегирование сильно влияет на конечный выход правильно свернутого интерферона гамма, обладающего большой гидрофобностью и не имеющего дисульфидных связей. Реакции агрегации ИНФ-гамма, которые происходят во время ренатурации белков, могут быть подавлены веществами, которые дестабилизируют неправильно свернутые или неправильно связанные молекулы. Среди этих веществ денатурирующие вещества, используемые в неденатурирующих концентрациях, например, мочевина и L-аргинин, который является мощным подавителем агрегации. Мы наблюдали минимум 2-кратное увеличение выхода, а также удельной активности при проведении ренатурации с аргинином по сравнению с ренатурацией в его отсутствии. Присутствие 0,3-0,5 М L-аргинина в буфере для ренатурации позволяет по меньшей мере трижды последовательно добавлять (каждые 1-2 ч) денатурированный белок в буфер, тем самым увеличивая выход функционального белка. Частично свернутые белковые молекулы, по-видимому, защищены от неспецифической агрегации, поэтому денатурированный ИНФ-гамма добавляют поэтапно, позволяя каждой добавленной аликвоте достичь своего защищенного состояния перед добавлением следующей. Этот метод 'импульсной ренатурации' позволяет эффективно использовать ренатурационный буфер за счет значительного уменьшения требуемых объемов реакции и сокращения времени процесса.
Также преимуществом способа является использование ионообменной хроматографии на катионообменнике, что снижает содержание эндотоксинов в конечной пробе до клинически приемлемых пределов (менее 0,4 ЕС на мг). Известно, что большинство эндотоксинов имеют сильно отрицательный заряд и, таким образом, в наших хроматографических условиях не будут связываться с СМ-сефарозной колонкой, в результате чего получается гамма-интерферон, не содержащий эндотоксинов.
Таким образом, получен рекомбинантный человеческий интерферон гамма с выходом при ренатурации более 95%, чистотой не менее 97%. Также белок соответствует следующим параметрам:
- Мономер интерферона гамма должен имеет молекулярную массу (16,9±0,2) кДа. Положение основной полосы мономера интерферона гамма на электрофореграмме испытуемой субстанции соответствует положению основной полосы мономера стандартного образца интерферона гамма CRS
- Положение основной полосы иммунокомплекса интерферона гамма с специфическими моноклональными антителами к интерферону гамма должно соответствовать положению полосы иммунокомплекса стандартного образца интерферона гамма (CRS) с специфическими моноклональными антителами к интерферону гамма (в имуноблотинге).
- Не более 3 мкг/мл остаточных белков клетки-хозяина
- Остаточная ДНК штамма-продуцента составляет не более 100 пкг/106 ME
- Бактериальные эндотоксины составляют не более 0,5 ЕЭ/106 ME
- Удельная биологическая активность не менее 1,8 × 106 ЕДна 1 мг белка.
Заявленный способ не предусматривает применения моноклональных антител для очистки рекомбинантного протеина, в связи с тем, чтобы избежать загрязнение получаемого продукта, кроме того используются недорогие технические средства для хроматографии, которые широко применяются на практике, и имеется возможность автоматизированного широкомасштабного производства с использованием ионообменника, который может быть регенерирован и стерилизован при многократном промышленном использовании.
Техническим результатом заявленного изобретения является повышение эффективности получения белка интерферона гамма, а именно, увеличение выхода белка из биомассы штамма с чистотой конечного продукта не менее 97% за счет создания рекомбинантного штамма Escherichia coli - продуцента ИНФ-гамма, полученного на основе сконструированной рекомбинантной плазмиды, что обеспечивает повышенный выход ИНФ - гамма за счет увеличения количества получаемой в ходе культивирования биомассы в 6-10 раз.
Указанный технический результат достигается созданием штамма на основе сконструированной рекомбинантной плазмиды рЕТ32а-IFG144 и способом получения белка с использованием такого штамма.
Рекомбинантная плазмида pET32a-IFG144 для экспрессии белка ИНФ-гамма, имеет аминокислотную последовательность SEQ ID NO: 1, длину 5806 пар оснований и состоит из следующих ключевых генетических элементов:
- промотора Т7 и оператора 1асО;
- синтетической нуклеотидной последовательности, представленной в SEQ ID NO: 2, кодирующей белок IFG144;
- гена устойчивости к антибиотику ампициллину (AmpR) и бактериального промотора гена устойчивости к ампициллину (AmpRpromoter);
- гена лактозного репрессора LacI и бактериального промотора гена лактозного репрессора (LacIpromoter);
- ориджина репликации бактериофага fl (fl ori).
В качестве вектора экспрессии выбрана плазмида рЕТ32а. Для получения плазмиды по изобретению последовательность SEQ ID NO: 2, полученную полным нуклеотидным синтезом, встраивают в плазмидный вектор рЕТ-32а по сайтам рестрикции XhoI и NdeI.
Структура указанной плазмидной ДНК (плазмиды), состоящей из указанных ключевых генетических элементов, представлена на Фиг. 2.
В вышеуказанной плазмиде ген AmpR предназначен для селекции стабильных клеток Escherichia coli. Бактериальный промотор гена устойчивости к ампициллину (AmpRpromoter) предназначен для его экспрессии.
Ориджин репликации бактериофага f1 /Analysis of Genes and Genomes, John Wiley&Sons, 2004, S. 140/ широко используется для создания экспрессионных векторов.
Продуктом трансляции синтетической последовательности SEQ ID NO: 2 является полипептид последовательности SEQ ID NO: 1, представляющий интерферон гамма.
После клонирования синтезированной последовательности в составе вектора, полученной рекомбинантной плазмидой была проведена трансформация бактериальных клеток Escherichia coli JM109.
Исходным материалом для создания штамма-продуцента по изобретению является известный из уровня техники штамм JM109. Генотип: endA1, recA1, gyrA96, thi, hsdR17 (rk-, mk+), relA1, supE44, Δ(lac-proAB), [F' traD36, proAB, lacIqZΔM15]. (Yanisch-Perron, J. Vieira, and J. Messing. Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene, 33(1): 103-19, 1985).
Экспрессионной плазмидой длиной 5806 п. о., состоящей из ключевых генетических элементов, расположенных друг относительно друга так, как представлено на Фиг. 2, трансформируют клетки штамма JM109.
Для введения плазмиды в клетки E.coli JM109 могут использоваться методы трансформации, известные из уровня техники, например, электропорация, метод с использованием полиэтиленгликоля, кальций-хлоридный метод. Также для целей настоящего изобретения для введения в клетки Е. coli JM109 плазмиды, представленной на Фиг. 2, могут использоваться и другие методы трансформации, не упомянутые в настоящем описании в явном виде, которые известны в уровне техники в настоящее время или будут созданы впоследствии. При осуществлении конкретных воплощений настоящего изобретения специалист, основываясь на существующем уровне знаний, может выбрать наиболее оптимальный метод трансформации клеток.
Для лучшего понимания сущности предлагаемого изобретения ниже приведены примеры его осуществления. Все стандартные генно-инженерные и микробиологические манипуляции, а также амплификацию и секвенирование ДНК проводили по известным методикам [Маниатис Т., Фрич Э, Сэмбрук Дж. Молекулярное клонирование, М.: Мир, 1984; Клонирование ДНК. Методы. Под ред. Д. Гловера, Пер. с англ., Москва, Мир, 1988; Saiki R.K., Gelfand D.H., Stoffel S., Sharf S.J., Higuehi R., Horn G.T., Mullis K.B., Eriich H.A. Science, 1988. V. 239, №4839, p. 487-491; Sanger F., Nicklen S., Coulson A.R. Proc. Nat. Acad. Sci. USA, 1977, V. 74, p. 5463-5467.].
Пример 1. Конструирование экспрессионной плазмиды
Химический синтез олигонуклеотидов выполняют твердофазным фосфоамидитным методом на ДНК-синтезаторе ASM-102U (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5'-диметокситритил-N-ацил-2'-дезоксинуклеозид-3'-O-(β-цианэтил-диизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500  ), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Используют синтетический цикл стандартного фосфоамидитного метода.
), к которому через 3'-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Используют синтетический цикл стандартного фосфоамидитного метода.
Для приготовления вектора ДНК плазмиды рЕТ32а (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера 20 мМ Трис-ацетата, 10 мМ ацетата магния, 50 мМ ацетатат калия, 100 мкг/мл БСА рестриктазой XhoI (10 ед. акт.), а затем - в 40 мкл буфера 100 мМ NaCl, 50 мМ Трис-HCl, 10 мМ MgCl2, 100 мкг/мл БСА рестриктазой NdeI (10 ед.акт.) в течение 1 ч при 37°С. Векторный фрагмент после электрофореза в 15% агарозном геле вырезают из геля и переносят в 200 мкл буфера NT, растворяют при 50°С в течении 5-10 мин и наносят на колонку NucleoSpinExtractII. Промывают буфером NT 3 и элюируют 50 мкл буфера NE.
Олигонуклеотидную последовательность SEQ ID NO: 2, кодирующую белок SEQ ID NO: 1, получают полным нуклеотидным синтезом. Полученную синтетическую последовательность SEQ ID NO: 2 обрабатывают в 40 мкл буфера 20 мМ Трис-ацетата, 10 мМ ацетата магния, 50 мМ ацетатат калия, 100 мкг/мл БСА рестриктазой XhoI (10 ед. акт.), а затем - в 40 мкл буфера 100 мМ NaCl, 50 мМ Трис-HCl, 10 мМ MgCl2, 100 мкг/мл БСА рестриктазой NdeI (10 ед.акт.) в течение 1 ч при 37°С. Синтетический фрагмент после электрофореза в 15% агарозном геле вырезают из геля и переносят в 200 мкл буфера NT, растворяют при 50°С в течении 5-10 мин и наносят на колонку NucleoSpinExtractII. Промывают буфером NT 3 и элюируют 50 мкл буфера NE.
Описанный выше полученный синтетический фрагмент в количестве 2 пмоль прибавляют к раствору 1 мкг, полученного из ДНК плазмиды рЕТ32а, описанного выше векторного фрагмента в 10 мкл буфера (20 мМ Трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреитол) и лигируют с помощью 10 ед.акт. Т4-ДНК-лигазы в течение 12 ч при 10°С.
Помещают на лед пробирки с компетентными клетками (XL1-Blue, Евроген) до полного размораживания содержимого из расчета одна пробирка на трансформацию. Аккуратно перемешивают суспензию клеток легким встряхиванием. Добавляют в каждую пробирку полученную после обработки Т4-ДНК-лигазой алкивоту реакционной смеси, аккуратно перемешивают содержимое легким встряхиванием. Инкубируют пробирки во льду в течение 20-30 мин. Переносят пробирки в водяную баню (плюс 42°С) на 30-45 сек. Быстро переносят пробирки из водяной бани в лед и инкубируют в течение 3-5 мин. Добавляют не менее 3-х объемов предварительно подогретой до 37°С среды SOB и инкубируют при 37°С в течение 40-60 мин в орбитальном шейкере-инкубаторе (Multitron, Infors) при скорости 225-250 об/мин. Высеивают содержимое пробирок на чашки Петри диамтером 60 мм (Перинт).
Аликвоту полученной плазмидной ДНК используют для трансформации компетентных клеток E. coli JM109.
Трансформанты высевают на чашки с агаризованной средой LB, в которую добавляют ампициллин до конечной концентрации ампициллина 50 мкг/мл. Из клонов выделяют ДНК плазмиды pET32a-IFG144. Скрининг рекомбинантов проводят с помощью секвенирования.
Пример 2. Получение штамма-продуцента E.coli JM109/pET32a-IFG144 и определение уровня его продуктивности
Плазмидой pET32a-IFG144 трансформируют компетентные клетки штамма E. coli JM109 и высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Предпочтительно (без ограничения), для введения указанной плазмиды в клетки штамма Е. coli JM109 используют метод электропорации, известный из уровня техники /Tamara Kleber-Janke, Wolf-Meinhard Becker, Use of Modified JM109 Escherichia coli Cells for High-Level Expression of Recombinant Peanut Allergens Affected by Poor Codon Usage, Protein Expression and Purification, Volume 19, Issue 3,2000, 419-424/ и включенный в настоящее описание посредством ссылки. Отдельно локализованную колонию трижды пересевают на чашки с LB-агаром, содержащим 100 мкг/мл ампициллина. Полученной моноклоновой культурой инокулируют 5 мл жидкой среды LB с ампициллином и инкубируют в течение ночи при интенсивном встряхивании при 37°С. Полученный штамм-продуцент E.coli JM109/pET32a-IFG144 хранят в 20% глицерине при минус 40°С.
Для определения уровня индуцируемой экспрессии белка ночную культуру засевают в разведении 1:50 в 5 мл жидкой среды LB, содержащей 100 мкг/мл ампициллина, и растят до мутности 0,8 при 37°С на качалке при 200 об/мин. К культуре добавляют изопропил-β-D-1-тиогалактоприанозид до концентрации 1,0 мМ и продолжают инкубацию в тех же условиях в течение 3 час. Клетки собирают центрифугированием, осадок суспендируют в буфере, содержащем 62,5 мМ Трис-HCl, рН 6,8, 3% додецилсульфата натрия, 5% 2-меркаптоэтанола, 10% глицерина и 0,01% бромфенолового синего и прогревают 3 мин на кипящей водяной бане. Полученный лизат клеток анализируют электрофорезом в 15% полиакриламидном геле с додецилсульфатом натрия. Гель окрашивают Coomassie R-250, сканируют и проводят его денситометрию. По данным денситометрии содержание белка составляет не менее 30% от общего белка клетки.
Новый штамм Escherichia coli JM109/pET32a-IFG144 является продуцентом белка, содержащего аминокислотную последовательность интерферона гамма и характеризуется следующими признаками:
Морфологические признаки: клетки мелкие, палочковидной формы, грамотрицательные, неспороносные, размером 1×3,5 мкм, подвижные, с хорошо различимыми тельцами включения после индукции синтеза белка.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые. Край ровный, диаметр колоний 1-3 мм, консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением.
Физиолого-биохимические признаки: клетки растут при температуре от 4 до 42°С, оптимум рН 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт. В качестве источника углерода при росте на минимальной среде используют глицерин, углеводы, аминокислоты.
Устойчивость к антибиотикам: клетки штамма-продуцента проявляют устойчивость к ампициллину (до 500 мг/мл), обусловленную наличием в плазмиде гена β-лактамазы (bla).
Стабильность плазмиды в штамме. При поддержании клеток в течение нескольких месяцев на агаризованной среде LB, содержащей ампициллин, не наблюдаются потери или перестройки плазмиды, влияющие на экспрессию белка.
В новом штамме ИНФ-гамма после индуцированной экспрессии накапливается в виде телец включения, и его содержание составляет не менее 30% от общего белка клетки.
Пример 3. Культивирование штамма-продуцента
Приготовление ночной культуры штамма Escherichia coli JM109/ pET32a-IFG144 Для приготовления ночной культуры штамма-продуцента интерферона гамма было разморожено 3 криопробирки из рабочего банка, содержащих культуру штамма объемом 1,5 мл, размороженная культура была полностью стерильно перенесена в качалочные колбы объемом 750 мл с 250 мл питательной среды, содержащие комплексную пептонную среду LB32 г/л, калия гидрофосфат тригидрат 6,3 г/л, калия дигидрофосфат 2,4 г/л, магния сульфат 0,5 г/л, глюкозы моногидрат 8,8 г/л и ампициллин (100 мкг/мл), и инкубировалась при 37°С и 200 об/мин в течение 20 ч. Параметры ночной культуры после инкубации: OD600 - 3.38; рН 5.28.
Приготовление инокулята для засева культуры штамма-продуцента в ферментер.
Для приготовления инокулята для засева культуры в ферментер ночная культура объемом по 125 мл была засеяна в 6 качалочных колб объемом 2000 мл с 1000 мл питательной среды, содержащей комплексную пептонную среду LB32 г/л, калия гидрофосфаттригидрат 6,3 г/л, калия дигидрофосфат 2,4 г/л, магния сульфат 0,5 г/л, глюкозы моногидрат 2,2 г/л и ампициллин (100 мкг/мл), и инкубировалась при 37°С и 200 об/мин в течение 4 ч. После инкубации содержимое всех 6 колб было перенесено в две стерильные емкости объемом 3 л для засева в ферментер. Параметры инокулята после инкубации: OD600 - 2.62; рН 6,36
Ферментация ИНФ-гамма в кулътуралъной жидкости объемом 200 л.
Для ферментацииукультурой штамма-продуцента в ферментере с номинальным объемом 300 л в объеме среды 200 л в ферментер засевают полученный инокулят с общим объемом 6 л. Питательная среда для культивирования, содержащая комплексную пептонную среду LB 45 г/л, калия гидрофосфаттригидрат 8,8 г/л, калия дигидрофосфат 3,4 г/л, магния сульфат 1,1 г/л, глюкозы моногидрат 0,5 г/л и ампициллин (100 мкг/мл). Для предотвращения образования пены в питательную среду был добавлен пеногаситель «Лапрол ПД-1» в концентрации 0,3 г/л.
Перед внесением инокулята в питательную среду был добавлен раствор ампициллина до конечной концентрации 100 мкг/мл.
Наращивание биомассы продуцента в ферментере проводилят при 37°С в течение 3 часов до оптической плотности OD600=(4,9-5,1).
Далее для синтеза белка в культуральную жидкость вносили индуктор 1ас-оперона изопропил-β-D-1-тиогалактопиранозид (ИПТГ) и дополнительную порцию антибиотика и проводили культивирование до фазы замедления роста в течение 5 ч. Конечная оптическая плотность культуральной жидкости составляла OD600=31,2 оптических единиц. В течение индукции 1ас-промотора в культуральную жидкость вносили раствор 50% глюкозы. Для сбора биомассы штамма-продуцента IFN-гамма охлаждали культуральную жидкость до 15°С и подавали на сепаратор для отделения клеток от жидкости. Итого было собрано 8,1 кг биомассы штамма-продуцента, которые ресуспендировали в 40 л воды деминерализованной с помощью погружной мешалки. Затем довели объем суспензии до 80 л. Параметры суспензии: рН - 6,6, t- 6,7°С. Полученную суспензию центрифугировали на проточной центрифуге Сера Z81G при 16000 об/мин в течение 1 ч. Супернатант слили в приемник для утилизации. Масса полученного осадка составила 7,8 кг.Ресуспендированный в воде осадок в объеме 80 л гомогенизировали на гомогенизаторе GAULIN при давлении 600 бар с охлаждением суспензии. Далее дезинтеграт был отцентрифугирован на центрифуге СЕРА Z81G при 16000 об/мин. Масса полученного осадка тел включений ИНФ-гамма составляет 1920 г. Осадок хранят при минус 70°С.
Пример 4. Культивирование штамма-продуцента
Приготовление ночной культуры штамма Escherichia coli JM109/pET32a-IFG144 Для приготовления ночной культуры штамма-продуцента интерферона гамма было разморожено 3 криопробирки из рабочего банка, содержащих культуру штамма объемом 1,5 мл, размороженная культура была полностью стерильно перенесена в качалочные колбы объемом 750 мл с 250 мл питательной среды, содержащие комплексную пептонную среду LB32 г/л, калия гидрофосфат тригидрат 6,3 г/л, калия дигидрофосфат 2,4 г/л, магния сульфат 0,5 г/л, глюкозы моногидрат 8,8 г/л и ампициллин (100 мкг/мл), и инкубировалась при 37°С и 200 об/мин в течение 20 ч. Параметры ночной культуры после инкубации: OD600 - 3.38; рН 5.28.
Приготовление инокулята для засева культуры штамма-продуцента в ферментер.
Для приготовления инокулята для засева культуры в ферментер ночная культура объемом по 125 мл была засеяна в 6 качалочных колб объемом 2000 мл с 1000 мл питательной среды, содержащей комплексную пептонную среду LB 32 г/л, калия гидрофосфаттригидрат 6,3 г/л, калия дигидрофосфат 2,4 г/л, магния сульфат 0,5 г/л, глюкозы моногидрат 2,2 г/л и ампициллин (100 мкг/мл), и инкубировалась при 37°С и 200 об/мин в течение 4 ч. После инкубации содержимое всех 6 колб было перенесено в две стерильные емкости объемом 3 л для засева в ферментер. Параметры инокулята после инкубации: OD600 - 2.62; рН 6,36.
Ферментация ИНФ-гамма в культуральной жидкости объемом 200 л.
Для ферментацииукультурой штамма-продуцента в ферментере с номинальным объемом 300 л в объеме среды 200 л в ферментер засевают полученный инокулят с общим объемом 6 л. Питательная среда для культивирования, содержащая комплексную пептонную среду LB 45 г/л, калия гидрофосфаттригидрат 8,8 г/л, калия дигидрофосфат 3,4 г/л, магния сульфат 1,1 г/л, глюкозы моногидрат 0,5 г/л и ампициллин (100 мкг/мл). Для предотвращения образования пены в питательную среду был добавлен пеногаситель «Лапрол ПД-1» в концентрации 0,3 г/л.
Перед внесением инокулята в питательную среду был добавлен раствор ампициллина до конечной концентрации 100 мкг/мл.
Наращивание биомассы продуцента в ферментере проводят при 37°С в течение 4 часов до оптической плотности OD600=(4,9-5,1).
Далее для синтеза белка в культуральную жидкость вносили индуктор 1ас-оперона изопропил-β-D-1-тиогалактопиранозид (ИПТГ) и дополнительную порцию антибиотика и проводили культивирование до фазы замедления роста в течение 6 ч. Конечная оптическая плотность культуральной жидкости составляла OD600=35 оптических единиц. В течение индукции 1ас-промотора в культуральную жидкость вносили раствор 50% глюкозы. Для сбора биомассы штамма-продуцента IFN-гамма охлаждали культуральную жидкость до 15°С и подавали на сепаратор для отделения клеток от жидкости. Итого было собрано 9 кг биомассы штамма-продуцента, которые ресуспендировали в 40 л воды деминерализованной с помощью погружной мешалки. Затем довели объем суспензии до 80 л. Параметры суспензии: рН - 6,6, t- 6,7°С. Полученную суспензию центрифугировали на проточной центрифуге Сера Z81G при 16000 об/мин в течение 1 ч. Супернатант слили в приемник для утилизации. Масса полученного осадка составила 8,5 кг. Ресуспендированный в воде осадок в объеме 80 л гомогенизировали на гомогенизаторе GAULIN при давлении 600 бар с охлаждением суспензии. Далее дезинтеграт был отцентрифугирован на центрифуге СЕРА Z81G при 16000 об/мин. Масса полученного осадка тел включений ИНФ-гамма составляет 2000 г. Осадок хранят при минус 70°С.
Пример 5. Способ получения белка ИНФ-гамма Растворение телец включений
100 г осадка телец включений по Примеру 3 в центрифужных стаканах ресуспендируют в 50 мМ натрий ацетатном буфере рН 6,0 с 7М мочевиной и переносят в блендер. Доливают буфер до (0,80±0,01) л и перемешивают в течение 10-15 мин. Переносят раствор в мерную емкость, доводят объем до (3,00±0,01) л тем же буфером. Раствор постоянно перемешивают на магнитной мешалке со скоростью 200 об⋅мин-1, Инкубируют при температуре не выше плюс 25°С, в течение от 16 до 20 часов. Раствор белков, входящих в состав телец включений, разливают по центрифужным стаканам вместимостью 250 мл.. Центрифугируют материал в течение 45 минут при температуре плюс 4°С и скорости вращения ротора 14000 об⋅мин-1. Полученный после центрифугирования раствор белков, входящих в состав телец включений, фильтруют на фильтровальной установке с фильтром 0,45 мкм.
Проведение хроматографической очистки белка на сорбенте SepraPrepS
Раствор белка в денатурирующих условиях очищают на колонке объемом 2 л, упакованной сорбентом SepraPrep S и уравновешенной буфером 50 мМ ацетата аммония, 7 М мочевина, рН 7,5. Для очистки используют хроматографическую систему QuantaSep 1000 или любой другой хроматограф низкого давления. После нанесения белка колонку промывают тем же буфером с 0,1М NaCl и пропускают 4 л буфера 50 мМ ацетата аммония, 7 М мочевина, рН 7,5, 0,2М NaCl и 5 л буфера 50 мМ ацетата аммония, 7 М мочевина, рН 7,5, 0,3 М NaCl.
На дисплее компьютера отображается хроматограмма процесса с показаниями электропроводности и оптической плотности при длине волны 280 нм. Раствор белков с оптической плотностью до 0,3 о.е., после выхода из хроматографа, отбрасывают. Раствор десорбированых белков с оптической плотностью от 0,3 о.е. и выше, собирают фракциями по (0,40±0,05) л в емкости для сбора фракций. Фракции собирают до снижения оптической плотности при длине волны 280 нм до (0,20±0,05) оптических единиц.
Из каждой фракции белка отбирают пробу и проводят электрофорез в 17,5% ПААГ - ДНС. После получения результатов электрофореза фракции, содержащие белок с молекулярной массой (16,9±0,2) кДа, объединяют. Промеряют объем полученного раствора интерферона гамма в мочевине, отбирают пробу объемом 3,0 мл и проводят измерения на спектрофотометре при длинах волн 310 и 280 нм. Определяют концентрацию белка в растворе по формуле:
(OD280-OD310)/ε, где
ε - коэффициент экстинкции, равный 0,64 (мл⋅мг-1⋅см-1)
Итоговые показатели раствора интерферона гамма в мочевине должны удовлетворять следующим требованиям:
Количество белка со стадии - не менее 2,2 г;
концентрация белка - не менее 0,8 мг/мл;
рН - 7,2 - 7,5;
объем - от 2,5 до 3,5 л;
Ренатурации белка ИНФ-гамма
Для проведения ренатурации используют стеклянный реактор фирмы Simax с перемешивающим устройством и системой охлаждения. В очищенный реактор загружают 27 л 0,05М аммоний ацетатный буфер с 0,1М хлористым натрием, содержащий 0,5М L-аргинин, рН (7,3±0,1) и охлаждают до плюс (12±1)°С.
Через фильтр с порами 0,22 мкм внутрь реактора с охлажденным буфером подают со скоростью (50±5) мл/мин раствор интерферона гамма в мочевине. При этом, после того, как пройдет (1,0±0,2) л насос останавливают и продолжают процесс через 2 часа. Затем продолжают подачу мочевинного раствора. Проводят остановку подачи после прохождения каждого литра раствора интерферона гамма.
После того, как весь раствор интерферона гамма в мочевине внесен в реактор, ренатурат инкубируют при работающей мешалке в течение 1 часа. С помощью системы охлаждения температуру раствора в реакторе поддерживают от 12 до 14°С. Общее время ренатурации составляет 6 часов.
Раствор ренатурированного интерферона гамма должен быть прозрачным или слегка опалесцирующим, бесцветным. рН раствора от 7,0 до 7,5.
Очистка сорбента CM-SepharoseFF
Со скоростью 200-250 мл/мин из реактора на колонку с сорбентом CM-Sepharose FF подают раствор ренатурированного интерферона гамма. После окончания процесса нанесения белка подают на колонку (5,00±0,02) л 0,1М фосфатного буфера рН 7,4. После того, как показания оптической плотности при 280 нм снизятся до (0,20±0,05) оптических единиц начинают элюцию целевого белка. Элюируют белок градиентом 0,1M.NaCl→0,1M.NaCl в 0,1М фосфатном буфере рН 7,4.
При оптической плотности при 280 нм от 0,3 о.е. и выше собирают фракции по (0,5±0,1) л. Фракции собирают до снижения оптической плотности при 280 нм до (0,20±0,05) оптических единиц. Из каждой фракции белка отбирают пробу и проводят электрофорез в 17,5% ПААГ - ДНС.
После получения результатов электрофореза материал, содержащий белок с молекулярной массой (16,9±0,2) кДа, объединяют.
Показатели раствора интерферона гамма очищенного должны удовлетворять следующим требованиям:
количество белка - не менее 2,1 г;
концентрация белка - не менее 0,7 мг/мл;
рН - от 6,5 до 7,5;
Определение активности
Определение проводят биологическим методом в соответствии с ГФ РФ, ОФС.1.7.2.0002.15 «Биологические методы испытания препаратов интерферона с использованием культур клеток».
Специфическую активность определяют в реакции подавления интерфероном цитопатического действия вируса на культуре перевиваемых клеток, чувствительных к интерферону гамма.
Эталонный штамм «Индиана» вируса везикулярного стоматита получен из Подразделения Института вирусологии им. Д.И. Ивановского ФГБУ «НИЦЭМ им. Н.Ф. Гамалеи» Минздрава России (ГКБ29/1) с инфекционным титром от 10 до 10 ТЦД50/мл.
Противовирусную активность испытуемого образца определяют в сравнении с противовирусной активностью международного стандартного образца (МСО) интерферона гамма рекомбинантного человеческого (NIBSC, Cod 87/586). Степень выраженности противовирусной защиты оценивается в ряду параллельных разведений стандартного и анализируемого образов для одинаковой дозы вируса. Полученная активность интерферона гамма выражается в международных единицах.
Подготовка клеточной культуры. Одну криопробирку с клетками тест-культуры достают из сосуда Дьюара с жидким азотом и помещают в водяную баню до полного оттаивания суспензии (не более чем на 2 минуты) при температуре (37±2)°С. В ламинарном боксе в асептических условиях содержимое пробирки переносят в одноразовую стерильную центрифужную пробирку объемом 15 мл, содержащую 6 мл поддерживающей среды. После центрифугирования при 2000 об/мин 10 мин, надосадочную жидкость сливают, к осадку добавляют 10 мл ростовой среды (см. примечание 3), осадок ресуспендируют и переносят суспензию во флакон площадью 25 см2 для культивирования. Флакон помещают в СО2-инкубатор для культивирования в стандартных условиях (при температуре (37±2)°С, и (5,0±0,5) % углекислого газа) на 48-72 часа до образования монослоя.
После формирования монослоя на поверхности дна культурального флакона, проводят процедуру пассирования клеток, т.е. переноса части клеток монослоя во второй культуральный флакон со средой роста. Перенос клеток необходимо производить, не допуская образования плотного слоя для того, чтобы клетки находились в логарифмической фазе роста.
Из флакона со сформированным клеточным монослоем удаляют среду роста и трижды промывают его 5-6 мл 0,25% раствора Трипсин-ЭДТА, прогретым до температуры (37±2)°С. Последнюю внесенную во флакон порцию раствора удаляют не полностью, а оставляют около 1 мл и равномерно распределяют по всей поверхности монослоя. Флакон с клетками помещают в СО2-инкубатор до тех пор, пока клетки полностью не отслоятся от дна культурального флакона. Вносят 2 мл ростовой среды, суспендируют клетки пипетированием, переносят в чашку Петри и отбирают пробу для подсчета количества клеток в камере Горяева. Переносят 1-1,5 мл клеточной суспензии в новый флакон и добавляют ростовую среду 5-6 мл до посевной концентрации клеток 100 тыс.клеток/мл. После переноса отслоившихся клеток в новый флакон со средой роста, их оба помещают в стандартные условия культивирования на 24-48 часов. При образовании на дне флаконов монослоя, один флакон используют для повторного переноса клеток и определения специфической активности, а с другим повторяют процедуру снятия клеток для закладки в рабочий банк клеток. Перед каждой процедурой работы с клетками при помощи инвертированного микроскопа оценивают морфологию клеток и чистоту культуры в культуральном флаконе.
При отсутствии посторонней микрофлоры и нормальной морфологии клеток определяют культуру, как пригодную к использованию.
Для анализа используют клетки тест-культуры, прошедшие не менее 3-х и не более 30 пассажей после размораживания криопробирки рабочего банка. Для анализа берут клетки в логарифмической фазе роста.
Для определения противовирусной активности испытуемого раствора субстанции готовят 4 рабочих 96-луночных и 1 контрольный планшет с культурой клеток для определения титра вируса. Для этого во все планшеты вносят суспензию клеток и инкубируют 24-48 часов в стандартных условиях культивирования до образования полного, но не плотного монослоя.
После чего из лунок рабочих планшетов удаляют культуральную среду. Во все лунки рабочих планшетов вносят по 100 мкл поддерживающей среды кроме лунок 1 ряда.
Из ампулы МСО интерферона гамма рекомбинантного человеческого ((NIBSC, Cod 87/586) активностью 250 МЕ/ампула готовят раствор с активностью 50 МЕ/мл.
Пробирка МСО-50 будет содержать рабочий раствор стандартного образца с активностью 50 МЕ/мл.
Приготовление испытуемых образцов ИО-50
Пробирку с аликвотой интерферона гамма размораживают при комнатной температуре в течение 30 мин. Переносят 200 мкл в стерильную пробирку типа Эппендорф вместимостью 1,5 мл с 1000 мкл поддерживающей среды, тщательно перемешивают (разведение 1:6). Далее готовят последовательные десятикратные разведения испытуемого образца в поддерживающей среде до 1:60000.
В лунки A1, B1, С1 и D1 рабочих планшетов вносят по 200 мкл предварительно разведенного до 50 МЕ/мл стандарта (МСО-50), а в лунки E1, F1, G1, H1 вносят по 200 мкл предварительно разведенного исследуемого образца (ИО-50) в разведении 1:60000. На каждом рабочем планшете оставляют 10 ряд без интерферона для контроля целостности клеточного монослоя (клеточный контроль, К-).
В лунки с контрольными клетками вносят по 0,1 мл поддерживающей среды.
Рабочие и контрольный планшет инкубируют в течение 22-26 часов в СО2-инкубаторе при стандартных условиях культивирования.
Визуально определяют степень разведения вируса, при которой монослой оказывается пораженным в 50% лунок. Концентрация вируса в этих лунках будет составлять 1 ТЦД50.
Рассчитывают титр вируса (Тв)по формуле:
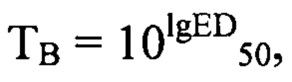
значение степени вычисляют по формуле Спирмена-Кербера:
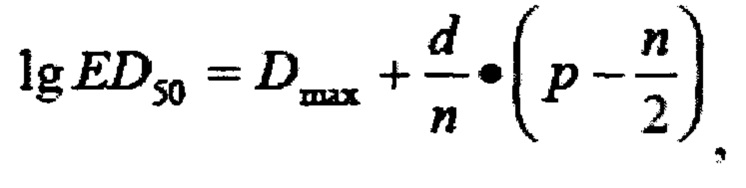
где:
lg ED50 - десятичный логарифм титра вируса;
Dmax - десятичный логарифм разведения, ниже которого произошла 100% гибель клеток (+);
d - десятичный логарифм шага разведения (=1,0);
n - число лунок, приходящееся на каждое разведение вируса (=6);
р - число лунок, давших гибель (+) в разведении, ниже которого произошла 100% гибель клеток.
Титр вируса должен быть не менее 106 ТЦЦ50/мл или 105 ТЦД50/0,1 мл.
Учитывая полученный титр вируса, рассчитывают его рабочую дозу, которая должна составлять 100 ТЦД50/на лунку при постановке теста на определение противовирусной активности. Т.е., для получения вируссодержащего материала с концентрацией 100 ТЦД,50/0,1 мл, исходную суспензию вируса разводят в (значение титра/100) раз.
После определения титра вируса и приготовления его рабочей дозы (РДВ), содержащей 100 ТЦД50 в 0,1 мл (К+), готовят десятикратные разведения РДВ, содержащие 10-1, 10-2 и 10-3 РДВ в 0,1 мл.
Из лунок рабочих планшет удаляют среду культивирования и вносят во все лунки (кроме контрольных лунок К-) по 0,1 мл РДВ. В лунки А11-Н11 и А12-Н12 рабочих планшет вносят по 0,1 мл разведения РДВ согласно рис. 1. В лунки А10-Н10 вносят по 0,1 мл поддерживающей среды для контроля клеточного монослоя (К-).
Планшеты инкубируют в течение 22-26 часов в СО2-инкубаторе при стандартных условиях культивирования до проявления цитопатического действия в 50% лунок в разведении РДВ, соответствующем 1 ТЦД50 в 0,1 мл.
Учет результатов визуальным методом За титр интерферона принимают величину, обратную разведению препарата, при котором клеточная культура в 50% лунок оказалась полностью защищенной от цитопатического действия вируса.
Для каждого рабочего планшета рассчитывается титр испытуемого образца (ТИОi) и стандартного образца (ТМСОi) по формулам:
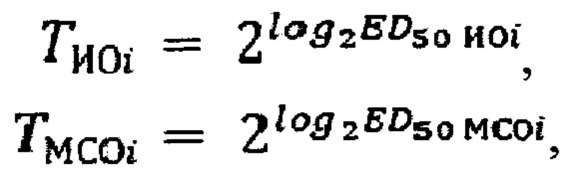
где i - номер планшета.
Значение степени вычисляют по формуле:
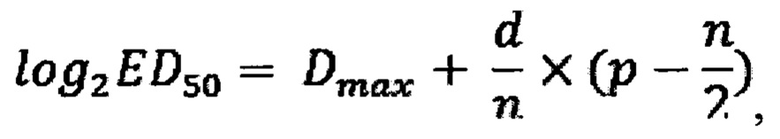
где:
Dmax - двоичный логарифм разведения, ниже которого произошла 100% защита;
d - двоичный логарифм шага разведений (=1,0);
n - количество лунок на каждую дозу;
р - количество лунок, дающих защиту (-) в DmaxH последующих разведениях Специфическую противовирусную активность образца в МЕ/мл для каждого планшета вычисляют по формуле:
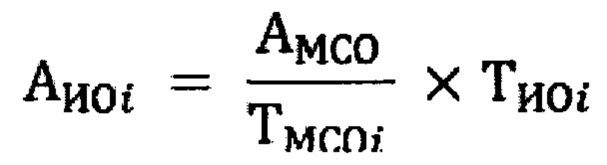
где:
AИОi - противовирусная активность интерферона в исследуемом образце в МЕ/мл,
АМСО - противовирусная активность восстановленного из лиофилизата международного стандартного образца в МЕ/мл,
TИОi - титр испытуемого образца,
TМСОi - титр международного стандартного образца.
Специфическую активность испытуемого образца вычисляют как среднее значение активности по 4 рабочим планшетам по формуле:
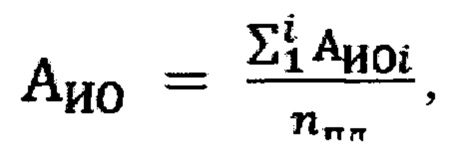
где:
АИО - средняя специфическая противовирусная активность образца в МЕ/мл в планшетах;
nпл - количество планшетов.
Удельная активность
Не менее 18000000 ME активности препарата на 1 мг белка.
Расчет удельной активности (X) проводят по формуле:
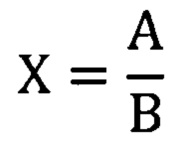
А- специфическая активность образца субстанции, МЕ/мл;
где
В- концентрация белка в образце субстанции, мг/мл.
Получен интерферон гамма с чистотой 98% и удельной биологической активностью 1,8 × 106 ЕД на 1 мг белка
Пример 6. Способ получения белка ИНФ-гамма
Растворение телец включений
100 г осадка телец включений по Примеру 3 в центрифужных стаканах ресуспендируют в 50 мМ натрий ацетатном буфере рН 6,0 с 7М мочевиной и переносят в блендер. Доливают буфер до (0,80±0,01) л и перемешивают в течение 10-15 мин. Переносят раствор в мерную емкость, доводят объем до (3,00±0,01) л тем же буфером. Раствор постоянно перемешивают на магнитной мешалке со скоростью 200 об⋅мин-1. Инкубируют при температуре не выше плюс 25°С, в течение от 16 до 20 часов. Раствор белков, входящих в состав телец включений, разливают по центрифужным стаканам вместимостью 250 мл.. Центрифугируют материал в течение 45 минут при температуре плюс 4°С и скорости вращения ротора 14000 об⋅мин-1. Полученный после центрифугирования раствор белков, входящих в состав телец включений, фильтруют на фильтровальной установке с фильтром 0,45 мкм.
Проведение хроматографической очистки белка на сорбенте SepraPrepS
Раствор белка в денатурирующих условиях очищают на колонке объемом 2 л, упакованной сорбентом SepraPrep S и уравновешенной буфером 50 мМ ацетата аммония, 7 М мочевина, рН 7,5. Для очистки используют хроматографическую систему QuantaSep 1000 или любой другой хроматограф низкого давления. После нанесения белка колонку промывают тем же буфером с 0,1М NaCl, и пропускают 4 л буфера 50 мМ ацетата аммония, 7 М мочевина, рН 7,5, 0,2М NaCl и 5 л буфера 50 мМ ацетата аммония, 7 М мочевина, рН 7,5, 0,3 М NaCl.
На дисплее компьютера отображается хроматограмма процесса с показаниями электропроводности и оптической плотности при длине волны 280 нм. Раствор белков с оптической плотностью до 0,3 о.е., после выхода из хроматографа, отбрасывают. Раствор десорбированых белков с оптической плотностью от 0,3 о.е. и выше, собирают фракциями по (0,40±0,05) л в емкости для сбора фракций. Фракции собирают до снижения оптической плотности при длине волны 280 нм до (0,20±0,05) оптических единиц.
Из каждой фракции белка отбирают пробу и проводят электрофорез в 17,5% ПААГ - ДНС. После получения результатов электрофореза фракции, содержащие белок с молекулярной массой (16,9±0,2) кДа, объединяют. Промеряют объем полученного раствора интерферона гамма в мочевине, отбирают пробу объемом 3,0 мл и проводят измерения на спектрофотометре при длинах волн 310 и 280 нм. Определяют концентрацию белка в растворе по формуле:
(OD280 - OD310)/ε, где
ε - коэффициент экстинкции, равный 0,64 (мл⋅мг-1⋅см-1)
Итоговые показатели раствора интерферона гамма в мочевине должны удовлетворять следующим требованиям:
Количество белка со стадии - не менее 2,2 г;
концентрация белка - не менее 0,8 мг/мл;
рН - 7,2 - 7,5;
объем - от 2,5 до 3,5 л;
Ренатурация белка ИНФ-гамма
Для проведения ренатурации используют стеклянный реактор фирмы Simax с перемешивающим устройством и системой охлаждения. В очищенный реактор загружают 27 л 0,05М аммоний ацетатный буфер с 0,1М хлористым натрием, содержащий 0,3M L-аргинин, рН (7,3±0,1) и охлаждают до плюс (12±1) С.
Через фильтр с порами 0,22 мкм внутрь реактора с охлажденным буфером подают со скоростью (50±5) мл/мин раствор интерферона гамма в мочевине. При этом, после того, как пройдет (1,0±0,2) л насос останавливают и продолжают процесс через 1 час. Затем продолжают подачу мочевинного раствора. Проводят остановку подачи после прохождения каждого литра раствора интерферона гамма.
После того, как весь раствор интерферона гамма в мочевине внесен в реактор, ренатурат инкубируют при работающей мешалке в течение 1 часа. С помощью системы охлаждения температуру раствора в реакторе поддерживают от 12 до 14°С. Общее время ренатурации составляет 3 часа.
Раствор ренатурированного интерферона гамма должен быть прозрачным или слегка опалесцирующим, бесцветным. рН раствора от 7,0 до 7,5.
Очистка сорбента CM-SepharoseFF
Со скоростью 200-250 мл/мин из реактора на колонку с сорбентом CM-Sepharose FF подают раствор ренатурированного интерферона гамма. После окончания процесса нанесения белка подают на колонку (5,00±0,02) л 0,1 М фосфатного буфера рН 7,4. После того, как показания оптической плотности при 280 нм снизятся до (0,20±0,05) оптических единиц начинают элюцию целевого белка. Элюируют белок градиентом 0,1M.NaCl→0,1M.NaCl в 0,1 М фосфатном буфере рН 7,4.
При оптической плотности при 280 нм от 0,3 о.е. и выше собирают фракции по (0,5±0,1) л. Фракции собирают до снижения оптической плотности при 280 нм до (0,20±0,05) оптических единиц. Из каждой фракции белка отбирают пробу и проводят электрофорез в 17,5% ПААГ - ДНС.
После получения результатов электрофореза материал, содержащий белок с молекулярной массой (16,9±0,2) кДа, объединяют.
Показатели раствора интерферона гамма очищенного должны удовлетворять следующим требованиям:
количество белка - не менее 2,1 г;
концентрация белка - не менее 0,7 мг/мл;
рН - от 6,5 до 7,5;
Определяли активность, как описано в примере 4.
Получен интерферон гамма с чистотой 97% и удельной биологической активностью 1,76 × 106 ЕД на 1 мг белка.

Изобретение относится к области биотехнологии, в частности к генной инженерии, и касается нового штамма бактерий, который используется для получения рекомбинантного интерферона гамма (далее ИНФ-гамма), применяемого в медицинской промышленности. Рекомбинантная плазмида рЕТ32а-IFG144 - размером 5806 пар оснований (п.о.) обеспечивает синтез белка, имеющего аминокислотную последовательность SEQ ID NO: 1, размером 16,9 кДа и содержит структурные элементы: промотор Т7 и оператор lacO, синтетическую нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок ИНФ-гамма, ген устойчивости к антибиотику ампициллину (AmpR) и бактериальный промотор гена устойчивости к ампициллину (AmpR промотор) для проведения отбора рекомбинантных клеток, ориджин репликации бактериофага fl (fl ori), ген лактозного репрессора LacI и бактериальный промотор гена лактозного репрессора (LacIpromoter). Изобретение позволяет повысить эффективность получения белка интерферона гамма, а именно увеличить выход белка из биомассы штамма с чистотой конечного продукта не менее 97% за счет создания рекомбинантного штамма Escherichia coli - продуцента ИНФ-гамма, полученного на основе сконструированной рекомбинантной плазмиды, что обеспечивает повышенный выход ИНФ-гамма за счет увеличения количества получаемой в ходе культивирования биомассы в 6-10 раз. 3 н. и 3 з.п. ф-лы, 2 ил.
1. Рекомбинантная плазмида рЕТ32а-IFG144 - размером 5806 пар оснований (п.о.), обеспечивающая синтез белка, имеющего аминокислотную последовательность SEQ ID NO: 1, размером 16,9 кДа и содержащая структурные элементы: промотор Т7 и оператор lacO, синтетическую нуклеотидную последовательность SEQ ID NO: 2, кодирующую белок ИНФ-гамма, ген устойчивости к антибиотику ампициллину (AmpR) и бактериальный промотор гена устойчивости к ампициллину (AmpR промотор) для проведения отбора рекомбинантных клеток - ориджин репликации бактериофага fl (fl ori), ген лактозного репрессора LacI и бактериальный промотор гена лактозного репрессора (LacIpromoter).
2. Штамм Escherichia coli JM109/pET32a-IFG144 - продуцент белка интерферона гамма, полученный трансформацией родительского штамма Е. coli JM109 рекомбинантной плазмидой рЕТ32а-IFG144 по п. 1.
3. Способ получения ИНФ-гамма, имеющего аминокислотную последовательность SEQ ID NO: 1, включающий культивирование штамма-продуцента Escherichia coli JM109/рЕТ32а-IFG144 по п. 2, при этом после 3-5 часов культивирования проводят индукцию биосинтеза рекомбинантного белка в клетках Escherichia coli JM109/рЕТ32а-IFG144 изопропил-β-D-1-тиогалактопиранозидом и выдерживают 4-6 часов с получением биомассы клеток штамма-продуцента, продуцирующего белок ИНФ-гамма, выделение из полученной биомассы клеток штамма-продуцента белка ИНФ-гамма, последующую очистку выделенного белка ИНФ-гамма путем катионообменной хроматографии в денатурирующих условиях, ренатурацию белка ИНФ-гамма путем последовательного дробного добавления денатурированного белка в буфер для ренатурации, содержащий L-аргинин, и очистку ренатурированного белка ИНФ-гамма катионообменной хроматографией с получением белка с чистотой не менее 97%.
4. Способ по п. 3, отличающийся тем, что культивирование клеток Escherichia coli JM109/рЕТ32а-IFG144 проводят в ростовой среде на протяжении по меньшей мере 8 часов, но не более 10 часов.
5. Способ по п. 3, отличающийся тем, что ренатурацию денатурированного белка проводят в присутствии L-аргинина с концентрацией 0,3-0,5М.
6. Способ по п. 3, отличающийся тем, что ренатурацию проводят дробным добавлением денатурированного белка через каждые 1-2 часа инкубации.

