ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[1] Данная заявка испрашивает приоритет предварительной заявки на патент США № 62/376298, поданной 17 августа 2016 года, предварительной заявки на патент США № 62/442377, поданной 4 января 2017 года, и предварительной заявки на патент США № 62/502313, поданной 5 мая 2017 года. Каждая из этих предварительных заявок на патент США полностью включена в данный документ посредством ссылки.
ВКЛЮЧЕНИЕ ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[2] Перечень последовательностей, содержащийся в файле с именем "P34494WO00_SEQ.txt", размер которого составляет 293398 байт (измерено в MS-Windows®) и который был создан 17 августа 2017 года, подан в электронном виде и полностью включен в данный документ посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
ОБЛАСТЬ ТЕХНИКИ
[3] Данное описание относится к композициям и способам улучшения признаков, таких как устойчивость к полеганию и повышенный урожай, у однодольных или злаковых растений, включая кукурузу.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[4] Гиббереллины (гиббереллиновые кислоты или GA (GA - gibberellic acid)) представляют собой гормоны растений, которые регулируют ряд основных процессов роста и развития растений. Манипулирование уровнями GA в полукарликовых сортах пшеницы, риса и сорго привело к увеличению урожайности и сокращению полегания посевов этих злаковых культур в течение 20-го века, что в значительной степени привело к Зеленой революции. Тем не менее успешное увеличение урожайности в других злаковых культурах, таких как кукуруза, не было достигнуто с помощью манипулирования путями GA. В самом деле, некоторые мутации в генах пути GA были связаны с различными нетипичными сортами кукурузы, которые несовместимы с урожайностью, что привело исследователей к поиску полукарликовых, высокоурожайных сортов кукурузы посредством манипулирования путем GA.
[5] В данной области техники по-прежнему существует потребность в разработке однодольных или злаковых культур, таких как кукуруза, с повышенной урожайностью и/или устойчивостью к полеганию.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[6] В первом аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом указанная некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA в однодольном или злаковом растении или клетке растения, при этом эндогенный белок оксидазы GA является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 9, 12, 15, 30 или 33, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[7] Во втором аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA20 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 9, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[8] В третьем аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или клетке растения, при этом указанный эндогенный белок оксидазы GA20 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 15, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[9] В четвертом аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA3 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 30 или 33, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[10] В пятом аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA20 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 12, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[11] В шестом аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок в однодольном или злаковом растении или клетке растения, при этом эндогенный белок является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 86, 90, 94, 97, 101, 104, 108, 112, 116, 118, 121, 125, 129, 133 или 136, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении. В дополнительном аспекте данного описания также предложен вектор для трансформации, содержащий конструкцию рекомбинантной ДНК, описанную в данном документе. В дополнительном аспекте данного описания также предложено трансгенное однодольное или злаковое растение, часть растения или клетка растения, содержащая конструкцию рекомбинантной ДНК, описанную в данном документе. В одном аспекте предоставляется трансгенное растение кукурузы, часть растения или клетка растения. В другом аспекте предложен способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата с помощью конструкции рекомбинантной ДНК, описанной в данном документе, и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата. В другом аспекте злаковое растение трансформируется посредством трансформации, опосредованной Agrobacterium или бомбардировкой частицами.
[12] В седьмом аспекте данного описания предложен способ снижения уровня по меньшей мере одной активной молекулы GA в стебле кукурузного или злакового растения, включающий в себя: супрессию одного или более генов оксидазы GA3 или оксидазы GA20 рекомбинантной конструкцией ДНК в одной или более тканях трансгенного злакового или кукурузного растения.
[13] В восьмом аспекте данного описания предложено трансгенное кукурузное или злаковое растение, содержащее конструкцию рекомбинантной ДНК, при этом конструкция рекомбинантной ДНК содержит последовательность транскрибируемой ДНК, кодирующую некодирующую молекулу РНК, которая нацелена по меньшей мере на один эндогенный ген оксидазы GA20 или GA3 для супрессии, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, и при этом трансгенное однодольное или злаковое растение имеет меньшую высоту растения по сравнению с контрольным растением дикого типа.
[14] В девятом аспекте данного описания предложено злаковое растение, содержащее мутацию в эндогенном гене оксидазы GA или около него, введенную методом мутагенеза, при этом уровень экспрессии эндогенного гена оксидазы GA снижается или отсутствует в злаковом растении, и при этом злаковое растение имеет меньшую высоту по сравнению с контрольным растением дикого типа.
[15] В десятом аспекте данного описания предложено кукурузное или злаковое растение, содержащие геномное редактирование, введенное с помощью целевого метода редактирования генома в локус эндогенного гена оксидазы GA или около него, при этом уровень экспрессии эндогенного гена оксидазы GA снижен или отсутствует по сравнению с контрольным растением, и при этом редактированное злаковое растение имеет меньшую высоту по сравнению с контрольным растением.
[16] В одиннадцатом аспекте данного описания предложена композиция, содержащая направляющую РНК, при этом направляющая РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере, 25 последовательным нуклеотидам целевой последовательности ДНК в геномном локусе эндогенного гена оксидазы GA злакового растения или около него. В одном аспекте композиция дополнительно содержит эндонуклеазу, направляемую РНК.
[17] В двенадцатом аспекте данного описания предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу направляющей РНК, при этом молекула направляющей РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или по меньшей мере 25 последовательным нуклеотидам целевой последовательности ДНК в геномном локусе эндогенного гена оксидазы GA кукурузного или злакового растения или около него.
[18] В тринадцатом аспекте данного описания предложена донорная матрица рекомбинантной ДНК содержащая, по меньшей мере, одну гомологичную последовательность, при этом по меньшей мере одна гомологичная последовательность является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК, при этом последовательность целевой ДНК представляет собой геномную последовательность в геномном локусе эндогенного гена оксидазы GA кукурузного или злакового растения или около него.
[19] В четырнадцатом аспекте данного описания предложена донорная матрица рекомбинантной ДНК содержащая два гомологичных плеча, включая первое гомологичное плечо и второе гомологичное плечо, при этом первое гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам первой фланкирующей последовательности ДНК, при этом второе гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам второй фланкирующей последовательности ДНК, и при этом первая фланкирующая последовательность ДНК и вторая фланкирующая последовательность ДНК представляют собой геномные последовательности в геномном локусе эндогенного гена оксидазы GA кукурузного или злакового растения или около него. В одном аспекте дополнительно предоставлена молекула ДНК или вектор, содержащий донорную матрицу рекомбинантной ДНК, описанную в данном документе. В другом аспекте дополнительно предоставлена бактериальная клетка или клетка-хозяин, содержащая донорную матрицу рекомбинантной ДНК, описанную в данном документе. В другом аспекте дополнительно предоставлено кукурузное или злаковое растение, часть растения или клетка растения, содержащая конструкцию рекомбинантной ДНК, описанную в данном документе.
[20] В пятнадцатом аспекте данное описание относится к сконструированной сайт-специфической нуклеазе, которая связывается с целевым сайтом в или рядом с геномным локусом эндогенного гена оксидазы GA кукурузного или злакового растения и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте.
[21] В шестнадцатом аспекте данное описание относится к конструкции рекомбинантной ДНК, содержащей трансген, кодирующий сайт-специфическую нуклеазу, при этом сайт-специфическая нуклеаза связывается с целевым сайтом в геномном локусе эндогенного гена оксидазы GA однодольного или злакового растения, или около него и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте.
[22] В семнадцатом аспекте данное описание относится к способу получения трансгенного кукурузного или злакового растения, включающему в себя: (а) трансформацию по меньшей мере одной клетки эксплантата с помощью донорной матрицы рекомбинантной ДНК, описанной в данном документе, и (b) регенерацию или развитие трансгенного кукурузного или злакового растения из трансформированного эксплантата, при этом трансгенное кукурузное или злаковое растение содержит последовательность вставки донорной матрицы рекомбинантной ДНК.
[23] В восемнадцатом аспекте данное описание относится к способу получения кукурузного или злакового растения, имеющего геномное редактирование в гене эндогенной оксидазы GA или около него, включающему в себя: (а) введение по меньшей мере в одну клетку эксплантата кукурузного или злакового растения сайт-специфической нуклеазы или молекулы рекомбинантной ДНК, содержащей трансген, кодирующий сайт-специфическую нуклеазу, при этом сайт-специфическая нуклеаза связывается с целевым сайтом в геномном локусе эндогенного гена оксидазы GA или около него и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте и (b) регенерацию или развитие редактированного кукурузного или злакового растения по меньшей мере из одной клетки эксплантата, содержащей геномное редактирование в гене эндогенной оксидазы GA или около него редактированного однодольного или злакового растения.
[24] В девятнадцатом аспекте данное описание относится к модифицированному растению кукурузы, имеющему высоту растения менее 2000 мм, менее 1950 мм, менее 1900 мм, менее 1850 мм, менее 1800 мм, менее 1750 мм, менее 1700 мм, менее 1650 мм, менее 1600 мм, менее 1550 мм, менее 1500 мм, менее 1450 мм, менее 1400 мм, менее 1350 мм, менее 1300 мм, менее 1250 мм, менее 1200 мм, менее 1150 мм, менее 1100 мм, менее 1050 мм или менее 1000 мм и один или более из (i) среднего диаметра стебля более 18 мм, более 18,5 мм, более 19 мм, более 19,5 мм, более 20 мм, более 20,5 мм, более 21 мм, более 21,5 мм или более 22 мм, (ii) улучшения устойчивости к полеганию по сравнению с контрольным растением дикого типа, или (iii) повышения устойчивости к засухе по сравнению с контрольным растением дикого типа.
[25] В двадцатом аспекте данное описание относится к модифицированному злаковому растению, имеющему уменьшенную высоту растения по сравнению с контрольным растением дикого типа, и (i) увеличенному диаметру стебля по сравнению с контрольным растением дикого типа, (ii) улучшенной устойчивости к полеганию по сравнению с контрольным растением дикого типа, или (iii) улучшенной устойчивости к засухе по сравнению с контрольным растением дикого типа.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[26] На Фиг. 1 проиллюстрирована уменьшенная высота растений у инбредных растений кукурузы, экспрессирующих конструкцию супрессии оксидазы GA20, через восемь событий трансформации по сравнению с инбредными контрольными растениями;
[27] На Фиг. 2А проиллюстрирована уменьшенная высота растений в среднем по гибридным растениям кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[28] На Фиг. 2В проиллюстрировано изображение гибридного контрольного растения дикого типа (слева) рядом с гибридным растением кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20 (справа), имеющим уменьшенную высоту растения;
[29] На Фиг. 3А проиллюстрирован увеличенный диаметр стебля в среднем по гибридным растениям кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[30] На Фиг. 3В проиллюстрировано изображение поперечного сечения стебля гибридного контрольного растения дикого типа (слева) рядом с поперечным сечением стебля гибридного растения кукурузы, экспрессирующего конструкцию супрессии оксидазы GA20 (справа), имеющего увеличенный диаметр стебля;
[31] На Фиг. 4 проиллюстрирован увеличенный сырой вес початка в среднем по гибридным растениям кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[32] На Фиг. 5 проиллюстрирован увеличенный сырой вес початка в среднем по гибридным растениям кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20, в двух полевых испытаниях по сравнению с гибридными контрольными растениями дикого типа в ответ на случай сильного ветра, который вызвал большее полегание у гибридных контрольных растений;
[33] На Фиг. 6 проиллюстрирован увеличенный уборочный индекс гибридных растений кукурузы, экспрессирующих конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[34] На Фиг. 7 проиллюстрировано увеличение средней оценки урожайности зерна гибридных растений кукурузы, экспрессирующих конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[35] На Фиг. 8 проиллюстрирован увеличенный показатель плодовитости в среднем по гибридным растениям кукурузы, экспрессирующим конструкцию супрессии оксидазы GA20, по сравнению с гибридными контрольными растениями;
[36] На Фиг. 9 проиллюстрировано изменение высоты растений с течением времени на стадиях развития V11 до R1 между трансгенными растениями кукурузы и контролем;
[37] На Фиг. 10 проиллюстрирован график, сравнивающий измерения соотношений стабильных изотопов кислорода (δ18O) как показателя устьичной проводимости и уровня воды в ткани листьев на стадии R5 между трансгенными растениями кукурузы и контролем;
[38] На Фиг. 11 проиллюстрирован график, сравнивающий скорость роста корня во время стадий развития V10 и до R2 между трансгенными и контрольными растениями как в условиях SAP, так и в условиях HD с применением датчиков на разных глубинах почвы, которые обнаруживают изменения в уровнях воды, указывающие на наличие корней на этой глубине;
[39] На Фиг. 12А проиллюстрированы различия в устьичной проводимости утром и днем между трансгенными растениями кукурузы и контролем в нормальных условиях и в условиях засухи в теплице;
[40] На Фиг. 12B проиллюстрированы различия в фотосинтезе утром и днем между трансгенными растениями кукурузы и контролем в нормальных условиях и в условиях засухи в теплице;
[41] На Фиг. 13А проиллюстрированы различия в уровнях экспрессии микроРНК в общей стеблевой ткани или в отдельных сосудистых и несосудистых тканях стебля трансгенных растений кукурузы по сравнению с контролем; а также
[42] На Фиг. 13B проиллюстрированы различия в уровнях экспрессии транскрипта мРНК оксидазы_3 GA20 и оксидазы_5 GA20 в общей стеблевой ткани или в отдельных сосудистых и несосудистых тканях стебля трансгенных растений кукурузы по сравнению с контролем.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
[43] Для облегчения понимания описания, несколько терминов и сокращений, применяемых в данном документе, определены ниже следующим образом:
[44] Термин "и/или" в случае применения в списке из двух или более предметов означает, что любой из перечисленных предметов может применяться сам по себе или в сочетании с любым одним или более перечисленными предметами. Например, выражение "A и/или B" предназначено для обозначения одного или обоих из A и B, то есть одного A, одного B или A и B в комбинации. Выражение "A, B и/или C" предназначено для обозначения одного A, одного B, одного C, A и B в комбинации, A и C в комбинации, B и C в комбинации или A, B и C в комбинации.
[45] Применяемый в данном документе термин "около" предназначен для определения числовых значений, которые он изменяет, и обозначает такое значение как переменную в пределах погрешности. Когда конкретный предел погрешности, такой как стандартное отклонение от среднего значения, не указан, следует понимать, что термин "около" означает тот диапазон, который будет включать указанное значение и диапазон, который будет включен округления в большую или меньшую сторону до этой цифры с учетом значимых цифр.
[46] Термин "злаковое растение" в контексте данного описания относится к однодольному (monocot) культурному растению, которое относится к семейству злаковых Poaceae или Gramineae и которое обычно собирают с целью получения его семян, включая, например, пшеницу, кукурузу, рис, просо, ячмень, сорго, овес и рожь.
[47] Применяемый в данном документе термин "процент идентичности" в отношении двух или более нуклеотидных или белковых последовательностей рассчитывается путем (i) сравнения двух оптимально выровненных последовательностей (нуклеотида или белка) в окне сравнения, (ii) определения количество положений, в которых идентичное основание нуклеиновой кислоты (для нуклеотидных последовательностей) или аминокислотный остаток (для белков) встречается в обеих последовательностях, с целью получения числа совпадающих положений, (iii) деление количества совпадающих положений на общее количество позиций в окне сравнения, а затем (iv) умножение этого коэффициента на 100%, чтобы получить процент идентичности. В целях расчета "процента идентичности" между последовательностями ДНК и РНК урацил (U) последовательности РНК считается идентичным тимину (Т) последовательности ДНК. Если окно сравнения определено как область выравнивания между двумя или более последовательностями (то есть, исключая нуклеотиды на 5' и 3' концах выровненных полинуклеотидных последовательностей или аминокислот на N-конце и С-конце выровненных последовательностей белка, которые не являются идентичными для сравниваемых последовательностей), тогда "процент идентичности" также может упоминаться как "процент идентичности выравнивания". Если "процент идентичности" вычисляется по отношению к эталонной последовательности без указания конкретного окна сравнения, то процент идентичности определяется путем деления числа совпадающих положений в области выравнивания на общую длину эталонной последовательности. Соответственно, для целей данного описания, когда две последовательности (запрашиваемая и предметная) оптимально выровнены (с учетом пробелов в их выравнивании), "процент идентичности" для запрашиваемой последовательности равен количеству идентичных положений между двумя последовательностями поделенному на общее количество положений в запрашиваемой последовательности по всей ее длине (или в окне сравнения), которое затем умножается на 100%.
[48] Признано, что положения остатков белков, которые не являются идентичными, часто отличаются консервативными аминокислотными заменами, где аминокислотные остатки заменены другими аминокислотными остатками с аналогичным размером и химическими свойствами (например, зарядом, гидрофобностью, полярностью и тому подобным), и, следовательно, не могут изменить функциональные свойства молекулы. Когда последовательности отличаются консервативными заменами, процент сходства последовательностей может быть откорректирован в сторону увеличения, чтобы скорректировать консервативную природу неидентичной замены(замен). Считается, что последовательности, которые отличаются такими консервативными заменами, имеют "сходство последовательностей" или "сходство". Таким образом, "процент сходства", как применяется в данном документе в отношении двух или более белковых последовательностей, рассчитывается путем (i) сравнения двух оптимально выровненных белковых последовательностей в окне сравнения, (ii) определения количества положений, в которых один и тот же или подобный аминокислотный остаток встречается в обеих последовательностях, с целью получения числа совпадающих положений, (iii) деления количества совпадающих положений на общее количество положений в окне сравнения (или общую длину эталонного или запрашиваемого белка если окно сравнения не указано), а затем (iv) умножение этого коэффициента на 100%, с целью получения процента сходства. Консервативные аминокислотные замены белков известны в данной области техники.
[49] Для оптимального выравнивания последовательностей с целью вычисления их процента идентичности или сходства в данной области техники известны различные попарные или множественные алгоритмы и программы выравнивания последовательностей, такие как ClustalW или Basic Local Alignment Search Tool® (BLAST®) и тому подобные, которые могут применяться для сравнения идентичности или сходства последовательностей между двумя или более нуклеотидными, или белковыми последовательностями. Не смотря на то, что в данной области техники известны другие способы выравнивания и сравнения, выравнивание между двумя последовательностями (включая диапазоны процента идентичности, описанные выше) может быть таким, как определено алгоритмом ClustalW или BLAST®, см., например, Chenna R. и соавт., ʺMultiple sequence alignment with the Clustal series of programs,ʺNucleic Acids Research 31: 3497-3500 (2003 год); Thompson JD и соавт., ʺClustal W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice,ʺ Nucleic Acids Research 22: 4673-4680 (1994 год); и Larkin MA и соавт., ʺClustal W and Clustal X version 2.0,ʺ Bioinformatics 23: 2947-48 (2007 год); и Altschul, S.F., Gish, W., Miller, W., Myers, E.W. & Lipman, D.J. (1990 год) "Basic local alignment search tool." J. Mol. Biol. 215: 403-410 (1990 год), все содержание и описание которых включено в данный документ посредством ссылки.
[50] В данном документе термин "процент комплементарности", применяется в отношении к двум нуклеотидным последовательностям, является аналогичным концепции процентной идентичности, но относится к проценту нуклеотидов в запрашиваемой последовательности, которая оптимально спаривается основаниями или гибридизуется с нуклеотидами предметной последовательности, когда запрашиваемые и предметные последовательности расположены линейно и оптимально спариваются основаниями без вторичных складывающихся структур, таких как петли, стебли или шпильки. Такой процент комплементарности может быть между двумя цепями ДНК, двумя цепями РНК или цепью ДНК и цепью РНК. "Процент комплементарности" рассчитывается путем (i) оптимального спаривания оснований или гибридизации двух нуклеотидных последовательностей в линейном и полностью вытянутом расположении (то есть, без фолдинга или вторичных структур) через окно сравнения, (ii) определения количества положений этой пары оснований между двумя последовательностями в окне сравнения, с целью получения количества комплементарных положений, (iii) деления количества комплементарных положений на общее количество положений в окне сравнения и (iv) умножения этого коэффициента на 100%, с целью получения процентной комплементарности двух последовательностей. Оптимальное спаривание оснований двух последовательностей может быть определено на основе известных спариваний нуклеотидных оснований, таких как G-C, A-T и A-U, посредством водородных связей. Если "процент комплементарности" вычисляются по отношению к эталонной последовательности без указания конкретного окна сравнения, то процент идентичности определяется путем деления числа дополнительных положений между двумя линейными последовательностями по общей длине эталонной последовательности. Таким образом, для целей данного описания, когда две последовательности (запрашиваемая и предметная) являются оптимально спарены основаниями (с учетом несовпадений или не спаренных основаниями нуклеотидов, но без фолдинга или вторичных структур), "процент комплементарности" для запрашиваемой последовательности равен количеству спаренных основаниями положений между двумя последовательностями, деленному на общее количество положений в запрашиваемой последовательности по всей ее длине (или на количество положений в запрашиваемой последовательности в окне сравнения), которое затем умножается на 100%.
[51] Термин "функционально связанный" относится к функциональной связи между промотором или другим регуляторным элементом и ассоциированной транскрибируемой последовательностью ДНК или кодирующей последовательностью гена (или трансгена), так что промотор, и тому подобное, действует или функционирует, чтобы инициировать, способствовать, воздействовать, вызывать и/или стимулировать транскрипцию и экспрессию ассоциированной транскрибируемой последовательности ДНК или кодирующей последовательности, по меньшей мере, в определенных клетках, тканях, стадии(ях) развития и/или состоянии(ях).
[52] Термин "промотор, экспрессирующийся в растении" относится к промотору, который может инициировать, содействовать, влиять, вызывать и/или стимулировать транскрипцию и экспрессию ассоциированной с ним транскрибируемой последовательности ДНК, кодирующей последовательности или ген в растительной клетке или ткани.
[53] Термин "гетерологичный" в отношении промотора или другой регуляторной последовательности по отношению к ассоциированной полинуклеотидной последовательности (например, транскрибируемой последовательности ДНК или кодирующей последовательности, или гена) представляет собой промотор или регуляторную последовательность, которая функционально не связана с такой ассоциированной полинуклеотидной последовательностью в природе - например, промотор или регуляторная последовательность имеют различное происхождение по сравнению с ассоциированной полинуклеотидной последовательностью, и/или промотор или регуляторная последовательность не встречаются в природе у видов растений, которые должны быть трансформированы промотором или регуляторной последовательностью.
[54] Термин "рекомбинантный" в отношении к полинуклеотидной (ДНК или РНК) молекуле, белку, конструкции, вектору и тому подобному, относится к полинуклеотидной или белковой молекуле или последовательности, искусственной и обычно не встречающейся в природе, и/или присутствующим в контексте, в котором они обычно не встречаются в природе, включая молекулу полинуклеотида (ДНК или РНК), белок, конструкцию и тому подобное, включая комбинацию двух или более последовательностей полинуклеотида или белка, которые не встречаются в природе вместе, таким же образом без вмешательства человека, что и полинуклеотидная молекула, белок, конструкция и тому подобное, включающие в себя по меньшей мере две полинуклеотидные или белковые последовательности, которые функционально связаны, но гетерологичны по отношению друг к другу. Например, термин "рекомбинантный" может относиться к любой комбинации двух или более последовательностей ДНК или белка в одной и той же молекуле (например, плазмиде, конструкции, векторе, хромосоме, белке и тому подобном), где такая комбинация является искусственной и обычно не встречается в природе. Как применяется в данном определении, фраза "обычно не встречается в природе" означает "не встречается в природе без вмешательства человека". Рекомбинантная полинуклеотидная или белковая молекула, конструкция и тому подобное, может содержать полинуклеотидную или белковую последовательность(и), которая (i) отделена от другой полинуклеотидной или белковой последовательности(тей), которые существуют в природе рядом друг с другом, и/или (ii) является смежной (или примыкает к) с другими полинуклеотидными или белковыми последовательностями, которые не находятся в естественной близости друг от друга. Такая рекомбинантная полинуклеотидная молекула, белок, конструкция и тому подобное, также может относиться к полинуклеотидной или белковой молекуле, или последовательности, которая была генетически сконструирована и/или сконструирована вне клетки. Например, рекомбинантная молекула ДНК может содержать любую сконструированную или искусственную плазмиду, вектор и тому подобное, и может включать линейную или кольцевую молекулу ДНК. Такие плазмиды, векторы и тому подобное, могут содержать различные поддерживающие элементы, включая прокариотическую точку начала репликации и селектируемый маркер, а также один или более трансгенов или экспрессионных кассет, возможно, в дополнение к селектируемому маркерному гену растения и тому подобному.
[55] В данном документе термин "выделенный" относится, по меньшей мере, к частичному отделению молекулы от других молекул, обычно связанных с ней в ее естественном состоянии. В одном варианте реализации изобретения термин "выделенный" относится к молекуле ДНК, которая отделена от нуклеиновых кислот, которые обычно фланкируют молекулу ДНК в ее естественном состоянии. Например, молекула ДНК, кодирующая белок, который естественным образом присутствует в бактерии, была бы выделенной молекулой ДНК, если бы она не находилась в ДНК бактерии, в которой естественным образом находится молекула ДНК, кодирующая белок. Таким образом, молекула ДНК, слитая или функционально связанная с одной или более другой молекулой(ами) ДНК, с которыми она не может быть связана в природе, например, в результате рекомбинантной ДНК или методов трансформации растений, считается выделенной в данном документе. Такие молекулы считаются выделенными, даже если они интегрированы в хромосому клетки-хозяина или присутствуют в растворе нуклеиновой кислоты с другими молекулами ДНК.
[56] В данном документе термин "кодирующая область" относится к части полинуклеотида, который кодирует функциональную единицу или молекулу (например, без ограничения, мРНК, белок или некодирующую последовательность или молекулу РНК).
[57] В данном документе термин "модифицированный" в контексте растения, семени растения, части растения, клетки растения и/или генома растения относится к растению, семени растения, части растения, клетки растения и/или геному растения, содержащему сконструированное изменение в уровне экспрессии и/или кодирующей последовательности одного или более гена(ов) оксидазы GA относительно дикого типа или контрольного растения, семени растения, части растения, клетки растения и/или генома растения, например, через (A) трансгенное событие, включающее в себя супрессирующую конструкцию или транскрибируемую последовательность ДНК, кодирующую некодирующую РНК, которая нацелена на один или более генов оксидазы GA3 и/или GA20 для супрессии, или (B) событие редактирования генома или мутацию, влияющую (например, уменьшая или устраняя) на уровень экспрессии или активности одного или более эндогенных генов оксидазы GA3 и/или GA20. Более того, термин "модифицированный" может дополнительно относиться к растению, семени растения, части растения, клетки растения и/или геному растения, имеющему одну или более мутаций, влияющих на экспрессию одного или более эндогенных генов оксидазы GA, таких как один или более эндогенных генов оксидазы GA3 и/или GA20, введенных посредством химического мутагенеза, вставки или исключения транспозона, или любого другого известного метода мутагенеза, или введенных посредством редактирования генома. Поэтому, для ясности, модифицированное растение, семя растения, часть растения, клетка растения и/или геном растения включает мутированное, редактированное и/или трансгенное растение, семя растения, часть растения, клетку растения и/или геном растения, имеющие модифицированный уровень экспрессии, профиль экспрессии и/или кодирующую последовательность одного или более генов оксидазы GA по отношению к дикому типу или контрольному растению, семени растения, части растения, клетке растения и/или геному растения. Модифицированные растения или семена могут содержать различные молекулярные изменения, которые влияют на экспрессию гена(ов) оксидазы GA, таких как ген(ы) оксидазы GA3 и/или GA20, включая генетические и/или эпигенетические модификации. Модифицированные растения, части растений, семена и тому подобное, могут быть подвергнуты мутагенезу, редактированию генома или сайт-направленной интеграции (например, без ограничения, с помощью способов, применяющих сайт-специфические нуклеазы), генетической трансформации (например, без ограничения, с помощью способов трансформации Agrobacterium или бомбардировки микрочастицами) или их комбинации. Такие "модифицированные" растения, семена растений, части растений и клетки растений включают растения, семена растений, части растений и клетки растений, которые являются потомками или получены из "модифицированных" растений, семян растений, частей растений и клеток растений, которые сохраняют молекулярное изменение (например, изменение уровня экспрессии и/или активности) одного или более генов оксидазы GA. Модифицированные семена, представленные в данном документе, могут дать начало модифицированному растению, представленному в данном документе. Модифицированное растение, семя растения, часть растения, клетка растения или геном растения, представленные в данном документе, могут содержать конструкцию рекомбинантной ДНК или вектор, или геном, как указано в данном документе. "Модифицированный продукт растительного происхождения" может представлять собой любой продукт, изготовленный из модифицированного растения, части растения, клетки растения или хромосомы растения, представленных в данном документе, или любой их части или компонента.
[58] Как применяется в данном документе, термин "контрольное растение" (или аналогично "контрольное" семя растения, часть растения, клетка растения и/или геном растения) относится к растению (или семени растения, части растения, клетке растения и/или геному растения) которое применяется для сравнения с модифицированным растением (или модифицированным семенем растения, частью растения, клеткой растения и/или геномом растения) и имеет такой же или сходный генетический фон (например, одинаковые родительские линии, гибридное скрещивание, инбредная линия, тестеры, и тому подобное) что и модифицированное растение (или семя растения, часть растения, клетка растения и/или геном растения), за исключением события(ий) трансгенного и/или геномного редактирования, затрагивающих один или более генов оксидазы GA. Например, контрольное растение может быть инбредной линией, которая является такой же инбредной линией, применяемой для изготовления модифицированного растения, или контрольное растение может быть продуктом того же гибридного скрещивания инбредных родительских линий, что и модифицированное растение, за исключением отсутствия в контрольном растении какого-либо трансгенного события(ий) или события(ий) редактирования генома, затрагивающего один или более генов оксидазы GA. В целях сравнения с модифицированным растением, семенами растения, частью растения, клеткой растения и/или геномом растения, "растение дикого типа" (или аналогичным образом семя растения "дикого типа", часть растения "дикого типа", клетка растения "дикого типа" и/или геном растения "дикого типа") относится к нетрансгенному и геномно-нередактированному контрольному растению, семени растения, части растения, клетке растения и/или геному растения. В данном документе термин "контрольное" растение, семя растения, часть растения, клетка растения и/или геном растения может также представлять собой растение, семя растения, часть растения, клетку растения и/или геном растения, имеющие сходный (но не одинаковый или идентичный) генетический фон с модифицированным растением, семенем растения, частью растения, клеткой растения и/или геномом растения, если он считается достаточно подобным для сравнения характеристик или признаков, подлежащих анализу.
[59] В данном документе термин "целевой сайт" для редактирования генома относится к расположению полинуклеотидной последовательности в геноме растения, которая связывается и расщепляется сайт-специфической нуклеазой, вводящей двухцепочечный разрыв (или одноцепочечный разрыв) в остов нуклеиновой кислоты полинуклеотидной последовательности и/или в ее комплементарную цепь ДНК. Целевой сайт может содержать по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, при по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26, по меньшей мере 27, по меньшей мере 29 или по меньшей мере 30 последовательных нуклеотидов. "Целевой сайт" для РНК-направленной нуклеазы может содержать последовательность либо комплементарной цепи молекулы двухцепочечной нуклеиновой кислоты (ДНК), либо хромосомы в целевом сайте. Сайт-специфическая нуклеаза может связываться с целевым сайтом, например, через некодирующую направляющую РНК (например, без ограничения, CRISPR РНК (crРНК) или одиночную направляющую РНК (sgРНК), как описано далее ниже). Некодирующая направляющая РНК, представленная в данном документе, может быть комплементарной целевому сайту (например, комплементарной либо цепи двухцепочечной молекулы нуклеиновой кислоты, либо хромосоме в целевом сайте). Понятно, что для некодирующей направляющей РНК может не требоваться совершенная идентичность или комплементарность для связывания или гибридизации с целевым сайтом. Например, может допускаться по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7 или по меньшей мере 8 ошибочных спариваний (или более) между целевым сайтом и некодирующей РНК. Термин "целевой сайт" также относится к расположению полинуклеотидной последовательности в геноме растения, которая связана и расщепляется другой сайт-специфической нуклеазой, которая не может направляться некодирующей молекулой РНК, такой как мегануклеаза, нуклеаза с цинковыми пальцами (ZFN) или эффекторная нуклеаза, подобная активаторам транскрипции (TALEN), для введения двухцепочечного разрыва (или одноцепочечного разрыва) в полинуклеотидную последовательность и/или ее комплементарную цепь ДНК. В данном документе термин "целевая область" или "нацеленная область" относится к полинуклеотидной последовательности или области, которая фланкирована двумя или более целевыми сайтами. Без ограничения, в некоторых вариантах реализации изобретения целевая область может быть подвергнута мутации, делеции, вставке или инверсии. В данном документе термин "фланкированный" если применяется для описания целевой области полинуклеотидной последовательности или молекулы, относится к двум или более целевым сайтам полинуклеотидной последовательности или молекулы, окружающей целевую область, с одним целевым сайтом на каждой стороне целевой области. Помимо редактирования генома, термин "целевой сайт" может также применяться в контексте супрессии гена для обозначения части молекулы мРНК (например, "сайта распознавания"), которая комплементарна по меньшей мере части некодирующей молекулы РНК (например, микроРНК, миРНК и тому подобное), кодируемой конструкцией супрессии.
[60] В данном документе термин "донорная молекула", "донорная матрица" или "молекула донорной матрицы" (совместно именуемые "донорная матрица"), которые могут представлять собой рекомбинантную донорную матрицу ДНК, определяется как молекула нуклеиновой кислоты, имеющая матрицу нуклеиновой кислоты или последовательность вставки для сайт-направленной, направленной вставки или рекомбинации в геном растительной клетки посредством репарации одноцепочечного разрыва или двухцепочечного разрыва ДНК в геноме растительной клетки. Например, "донорная матрица" может применяться для сайт-направленной интеграции трансгена или супрессирующей конструкции, или в качестве матрицы для введения мутации, такой как вставка, делеция и тому подобное, в целевой сайт в пределах генома растения. Предложенный в данном документе метод редактирования целевого генома может включать в себя применение одной или более, двух или более, трех или более, четырех или более, или пяти или более донорных молекул или матриц. "Донорская матрица" может представлять собой одноцепочечную или двухцепочечную молекулу ДНК или РНК, или плазмиду. "Инсерционная последовательность" донорной матрицы представляет собой последовательность, предназначенную для целевой инсерции в геном растительной клетки, которая может иметь любую подходящую длину. Например, инсерционная последовательность донорной матрицы может составлять от 2 до 50000, от 2 до 10000, от 2 до 5000, от 2 до 1000, от 2 до 500, от 2 до 250, от 2 до 100, от 2 до 50, от 2 до 30, от 15 до 50, от 15 до 100, от 15 до 500, от 15 до 1000, от 15 до 5000, от 18 до 30, от 18 до 26, от 20 до 26, от 20 до 50, от 20 до 100, от 20 до 250, от 20 до 500, от 20 до 1000, от 20 до 5000, от 20 до 10000, от 50 до 250, от 50 до 500, от 50 до 1000, от 50 до 5000, от 50 до 10000, от 100 до 250, от 100 до 500, от 100 до 1000, от 100 до 5000, от 100 до 10000, от 250 до 500, от 250 до 1000, от 250 до 5000 или от 250 до 10000 нуклеотидов или пар оснований в длину. Донорская матрица также может иметь по меньшей мере одну гомологичную последовательность или гомологичное плечо, такое как два плеча гомологии, для направления интеграции последовательности мутации или вставки в целевой сайт в геноме растения посредством гомологичной рекомбинации, при этом гомологичная последовательность или гомологичное плечо(и) идентичны или комплементарны, или имеют процент идентичности или процент комплементарности последовательности в целевом сайте или около него в геноме растения. В случае, если донорная матрица содержит гомологичное плечо(и) и последовательность вставки, гомологичное плечо(и) будет фланкировать или окружать последовательность вставки донорной матрицы.
[61] Инсерционная последовательность донорной матрицы может содержать один или более генов, или последовательностей, каждая из которых кодирует транскрибированную некодирующую последовательность РНК или мРНК и/или транслированную последовательность белка. Транскрибируемая последовательность или ген донорной матрицы может кодировать белок или некодирующую молекулу РНК. Инсерционная последовательность донорной матрицы может содержать полинуклеотидную последовательность, которая не содержит функциональный ген или полную последовательность гена (например, донорная матрица может просто содержать регуляторные последовательности, такие как последовательность промотора, или только часть гена или кодирующей последовательности) или может не содержать какие-либо идентифицируемые элементы экспрессии гена или любую активно транскрибируемую последовательность гена. Кроме того, донорная матрица может быть линейной или кольцевой и может быть одноцепочечной или двухцепочечной. Донорная матрица может доставляться в клетку в виде "голой" нуклеиновой кислоты (например, посредством бомбардировки микрочастицами), в виде комплекса с одним или более агентами доставки (например, липосомами, белками, полоксамерами, T-цепью, инкапсулированной с белками и тому подобным) или содержаться в бактериальном или вирусном носителе для доставки, таком как, например, Agrobacterium tumefaciens или геминивирус, соответственно. Инсерционная последовательность донорной матрицы, представленная в данном документе, может содержать транскрибируемую последовательность ДНК, которая может быть транскрибирована в молекулу РНК, которая может быть некодирующей и может или не может быть функционально связана с промотором и/или другой регуляторной последовательностью.
[62] В соответствии с некоторыми вариантами реализации изобретения донорная матрица может не содержать инсерционную последовательность, а вместо этого содержать одну или более гомологичных последовательностей, которая(ые) включают в себя одну или более мутаций, таких как вставка, делеция, замена и тому подобное, относительно геномной последовательности на целевом сайте в геноме растения, например, в или около гена оксидазы GA3 или оксидазы GA20 в геноме растения. В альтернативном варианте, донорная матрица может содержать инсерционную последовательность, которая не содержит кодирующую или транскрибируемую последовательность ДНК, при этом инсерционная последовательность применяется для введения одной или более мутаций в целевой сайт в геноме растения, например, в или около гена оксидазы GA3 или оксидазы GA20 в геноме растения.
[63] Донорная матрица, представленная в данном документе, может содержать по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять генов или транскрибируемых последовательностей ДНК. В альтернативном варианте, донорная матрица может не содержать генов. Без ограничения, последовательность гена или транскрибируемая ДНК донорной матрицы может включать в себя, например, ген устойчивости к инсектициду, ген устойчивости к гербицидам, ген эффективности использования азота, ген эффективности использования воды, ген, обеспечивающий пищевую ценность, ДНК-связывающий ген, селектируемый маркерный ген, РНКи или супрессирующую конструкцию, ген фермента сайт-специфической модификации генома, одиночную направляющую РНК системы CRISPR/Cas9, кассету экспрессии на основе геминивируса или векторную систему экспрессии на основе вируса растений. В соответствии с другими вариантами реализации изобретения инсерционная последовательность донорной матрицы может содержать транскрибируемую последовательность ДНК, которая кодирует некодирующую молекулу РНК, которая может нацеливаться на ген оксидазы GA, такой как ген оксидазы GA3 или оксидазы GA20, с целью супрессии. Донорная матрица может содержать промотор, такой как тканеспецифический или тканепредпочтительный промотор, конститутивный промотор или индуцибельный промотор. Донорная матрица может содержать лидерную последовательность, энхансер, промотор, сайт инициации транскрипции, 5'-НТО, один или более экзонов, один или более интронов, сайт терминации транскрипции, область или последовательность, 3'-НТО, и/или сигнал полиаденилирования. Лидерная последовательность, энхансер и/или промотор могут быть функционально связаны с последовательностью гена или транскрибируемой ДНК, кодирующей некодирующую РНК, направляющую РНК, мРНК и/или белок.
[64] В данном документе термин "сосудистый промотор" относится к промотору, экспрессирующемуся в растении, который управляет, вызывает или инициирует экспрессию транскрибируемой последовательности ДНК или трансгена, функционально связанного с таким промотором, в одной или более сосудистой ткани(нях) растения, даже если промотор также экспрессируется в другой несосудистой клетке(ках) или ткани(нях) растения. Такая сосудистая ткань(ни) может содержать одну или более клетку(ки) или ткань(ни) флоэмы, сосудистой паренхимы и/или обкладки сосудисто-волокнистых пучков растения. "сосудистый промотор" отличается от конститутивного промотора тем, что он имеет регулируемый и относительно более ограниченный профиль экспрессии, который включает в себя одну или более сосудистую ткань(ней) растения. сосудистый промотор включает в себя как сосудисто-специфические промоторы, так и сосудисто-предпочтительные промоторы.
[65] В данном документе термин "листовой промотор" относится к промотору, экспрессирующемуся в растении, который управляет, вызывает или инициирует экспрессию транскрибируемой последовательности ДНК или трансгена, функционально связанного с таким промотором, в одной или более листовой ткани(нях) растения, даже если промотор также экспрессируется в другой не листовой клетке(ках) или ткани(нях) растения. Листовой промотор включает в себя как специфичные для листа промоторы, так и предпочтительные для листа промоторы. "Листовой промотор" отличается от сосудистого промотора тем, что он экспрессируется более преимущественно или исключительно в ткани(нях) листьев растения по сравнению с другими тканями растения, тогда как сосудистый промотор экспрессируется предпочтительно в сосудистой ткани(нях), включая сосудистую ткань(ни) вне листа, такую как сосудистая ткань(ни) стебля или стебля и листьев растения.
[66] В данном документе термин "промотор, экспрессирующийся в растении" относится к промотору, который управляет, вызывает или инициирует экспрессию транскрибируемой последовательности ДНК или трансгена, функционально связанных с таким промотором, в одной или более растительных клетках или тканях, таких как одна или более клеток, или ткани кукурузы или злаковых растений.
Описание
[67] Большинство злаковых, производящих зерно, таких как пшеница, рис и сорго, продуцируют как мужские, так и женские структуры в пределах каждого отдельного цветочка метелки (то есть, они имеют единую репродуктивную структуру). Тем не менее, зерновые или кукуруза уникальны среди злаковых культур, поскольку они образуют отдельные мужские (султан) и женские (початок) соцветия. Кукуруза создает полностью сексуально диморфные репродуктивные структуры путем избирательного аборта мужских органов (пыльников) в соцветиях початка и женских органов (семезачаток) в соцветиях султана на ранних стадиях развития. Точно регулируемый синтез и передача сигнала гиббереллина имеет решающее значение для регуляции процесса избирательного аборта, причем женский репродуктивный початок наиболее чувствителен к нарушениям в пути GA. Действительно, фенотип "пыльник початка" является наиболее распространенным репродуктивным фенотипом у мутантов GA кукурузы.
[68] В отличие от кукурузы, мутации в путях синтеза или сигнального пути гиббереллина, которые привели к "зеленой революции" в отношении пшеницы, риса и сорго, оказали незначительное влияние на их репродуктивную структуру, поскольку эти виды сельскохозяйственных культур не подвергаются процессу избирательного аборта зерна, несущего метелку во время развития и, таким образом, не чувствительны к нарушениям в уровнях GA. Те же мутации не применялись на кукурузе, потому что нарушение синтеза и сигнального пути GA неоднократно приводило к драматическим искажениям и маскулинизации початка ("пыльник початка") и стерильности (нарушение развития пыльника и микроспор) в султане, вдобавок к экстремальной карликовости в некоторых случаях. См., например, Chen, Y. и соавт., "The Maize DWARF1 Encodes a Gibberellin 3-Oxidase and Is Dual Localized to the Nucleus and Cytosol," Plant Physiology 166: 2028-2039 (2014 год). Эти мутантные фенотипы GA (нетипичные) в кукурузе привели к значительному сокращению продукции зерна и снижению урожайности. Дополнительно, образование пыльников в початке увеличивает вероятность грибковых инфекций или инфекций, вызванных насекомыми, что снижает качество зерна, производимого на таких мутантных початках. Передовая селекция для разработки полукарликовых линий кукурузы не увенчалась успехом, а репродуктивные отклонения (в том числе экстремальные карликовые) мутантов GA было трудно преодолеть. Таким образом, те же самые мутации в пути GA, которые привели к "зеленой революции" в других злаковых, еще не были успешными в кукурузе.
[69] Несмотря на эти предшествующие трудности в достижении более высоких урожаев зерна в кукурузе посредством манипулирования путем GA, авторы данного изобретения обнаружили способ манипулирования уровнями GA в растениях кукурузы таким образом, чтобы уменьшить общую высоту растения и длину междоузлия стебля и увеличить устойчивость к полеганию, но не вызвать репродуктивных отклонений, ранее связанных с мутациями в пути GA кукурузы. Дополнительные данные указывают на то, что эти невысокого роста или полукарликовые растения кукурузы могут также иметь один или более дополнительных признаков, включая увеличенный диаметр стебля, уменьшенный излом стебля, более глубокие корни, увеличенную площадь листьев, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, повышение эффективности использования воды, снижение содержания антоцианов и площади листьев в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение количества зерна, увеличение массы зерна, увеличение урожайности и/или увеличенный уборочный индекс.
[70] Не ограничиваясь какой-либо теорией, предполагается, что неполная супрессия гена(ов) оксидазы GA20 или GA3 и/или нацеливание на подмножество одного или более генов оксидазы GA может быть эффективным в достижении невысокого роста, полукарликового фенотипа с повышенной устойчивостью к полеганию, но без репродуктивных отклонений в початке. Дополнительно предлагается, не ограничиваясь теорией, что ограничение супрессии гена(ов) оксидазы GA20 и/или GA3 некоторыми тканями, активно продуцирующими GA, такими как сосудистые и/или листовые ткани растения, может быть достаточным для получения растения невысокого роста с повышенной устойчивостью к полеганию, но без значительных дефектов в репродуктивных тканях. Экспрессия элемента супрессии оксидазы GA20 или GA3 тканеспецифическим или тканепредпочтительным способом может быть достаточной и эффективной для получения растений с низкорослым фенотипом, при этом избегая потенциальных дефектов в репродуктивных тканях, которые ранее наблюдались у мутантов GA в кукурузе (например, избегая или ограничивая супрессию гена(ов) оксидазы GA20 в таких репродуктивных тканях). Например, ген(ы) оксидазы GA20 и/или GA3 может быть нацелен на супрессию с применением сосудистого промотора, такого как промотор палочковидного вируса риса тунгро (RTBV - rice tungro bacilliform virus), который управляет экспрессией в сосудистых тканях растений. Как подтверждается в приведенных ниже Примерах, профиль экспрессии промотора RTBV является активным в сосудистых тканях растений кукурузы по сравнению с несосудистыми тканями, что является достаточным для получения полукарликового фенотипа у растений кукурузы, когда он функционально связан с элементом супрессии, нацеленным на ген(ы) оксидазы GA20 и GA3. Снижение уровней активной GA в ткани(ях) кукурузы или злаковых растений, которые производят активную GA, может уменьшить высоту растения и повысить устойчивость к полеганию, а отклонения могут быть предотвращены в этих растениях, если уровни активной GA также не подвергаются значительному воздействию или снижению в репродуктивных тканях, таких как развивающийся женский половой орган или початок растения. Если уровни активной GA могут быть снижены в стебле или междоузлии(ях) кукурузы или злаковых растений без существенного влияния на уровни GA в репродуктивных тканях (например, женских или мужских репродуктивных органах или соцветиях), тогда кукуруза или злаковые растения, имеющие уменьшенную высоту растения и повышенную устойчивость к полеганию, могут быть созданы без дефектов в репродуктивных тканях растения.
[71] Таким образом, в данном документе представлены конструкции рекомбинантной ДНК и трансгенные растения, содержащие элемент или последовательность супрессии оксидазы GA20 или GA3 функционально связанные с промотором, экспрессируемым в растении, который может быть тканеспецифическим или тканепредпочтительным промотором. Такой тканеспецифический или тканепредпочтительный промотор может управлять экспрессией ассоциированного с ним элемента или последовательности супрессии оксидазы GA, в одной или более ткани(нях) растения, активно продуцирующих GA, для супрессии или снижения уровня активных GA, продуцируемых в этой ткани(нях). Такой тканеспецифический или тканепредпочтительный промотор может управлять экспрессией, ассоциированной с ним конструкции супрессии оксидазы GA, или трансгена во время одной или более стадии (стадий) вегетативного развития. Такой тканеспецифический или тканепредпочтительный промотор также может иметь небольшую экспрессию или отсутствие экспрессии в одной или более клетке(ах) или ткани(нях) развивающегося женского полового органа или початка растения с целью избегания возможности возникновения отклонений в этих репродуктивных тканях. Согласно некоторым вариантам реализации изобретения тканеспецифический или тканепредпочтительный промотор представляет собой сосудистый промотор, такой как промотор RTBV. Последовательность промотора RTBV представлена в данном документе как SEQ ID NO: 65, а усеченная версия промотора RTBV дополнительно представлена в данном документе как SEQ ID NO: 66.
[72] Активные или биоактивные гиббереллиновые кислоты (то есть "активные гиббереллины" или "активные GA") известны в данной области техники для данного вида растений, в отличие от неактивных GA. Например, активные GA в кукурузе и высших растениях включают следующее: GA1, GA3, GA4 и GA7. Таким образом, "ткань, продуцирующая активную GA" представляет собой растительную ткань, которая продуцирует одну или более активных GA.
[73] Помимо супрессии генов оксидазы GA20 в тканях растения, продуцирующих активную GA, с помощью промотора сосудистой ткани, было неожиданно обнаружено, что супрессия одних и тех же генов оксидазы GA20 с помощью различных конститутивных промоторов также может вызывать фенотипы низкого, полукарликового роста у кукурузы, но без каких-либо видимых дефектов в початке. Учитывая, что ранее было показано, что мутации в пути GA вызывают нетипичность в репродуктивных тканях, было неожиданно, что конститутивная супрессия оксидазы GA20 не вызывала сходных репродуктивных фенотипов в початке. Таким образом, дополнительно предлагается, что супрессия одного или более генов оксидазы GA20 могла быть осуществлена с применением конститутивного промотора для создания низкорослой, устойчивой к полеганию кукурузы или злакового растения без каких-либо существенных или наблюдаемых репродуктивных отклонений в растении. Другие неожиданные наблюдения были сделаны, когда та же самая конструкция супрессии оксидазы GA20 была экспрессирована в стебле, листьях или репродуктивных тканях. Как описано дополнительно ниже, направленная супрессия тех же генов оксидазы GA20 в тканях стебля или початка растений кукурузы не приводила к фенотипу невысокого, полукарликового роста. Кроме того, нацеленная экспрессия конструкции супрессии оксидазы GA20 непосредственно в репродуктивных тканях развивающегося початка растений кукурузы с помощью промотора женской репродуктивной ткани (початка) не вызывала каких-либо значительных или наблюдаемых отклонений в початке. Тем не менее, экспрессия той же конструкции супрессии оксидазы GA20 в листовых тканях была достаточной для того, чтобы вызвать фенотип умеренного невысокого роста без существенных или наблюдаемых репродуктивных отклонений в растении.
[74] Не ограничиваясь какой-либо теорией, предлагается, что низкорослые, полукарликовые фенотипы в кукурузе и других злаковых растениях могут быть результатом достаточного уровня экспрессии супрессирующей конструкции, нацеленной на определенный ген(ы) оксидазы GA в ткани(нях), продуцирующей активную GA растения. По меньшей мере, для направленной супрессии определенных генов оксидазы GA20 в кукурузе, ограничение профиля экспрессии во избежание репродуктивных тканей початка может быть необязательным для избежания репродуктивных отклонений в развивающемся початке. Тем не менее экспрессия конструкции супрессии оксидазы GA20 на низких уровнях и/или в ограниченном количестве растительных тканей может быть недостаточной для того, чтобы вызвать значительную низкорослость, полукарликовый фенотип. Принимая во внимание, что наблюдаемый полукарликовый фенотип с направленной супрессией оксидазы GA20 является результатом укорочения междоузлий стебля растения, является неожиданным, что супрессия генов оксидазы GA20 по меньшей мере в некоторых тканях стебля была недостаточной, чтобы вызвать укорочение междоузлий и уменьшенную высоту растения. Не ограничиваясь какой-либо теорией, предполагается, что супрессия определенного гена(ов) оксидазы GA в ткани(нях) и/или клетке(ах) растения, где вырабатываются активные GA, и не обязательно в ткани(ах) стебля или междоузлия, может быть достаточной для получения полукарликовых растений, даже несмотря на то, что признак низкорослости обусловлен укорочением междоузлий стебля. Учитывая, что GA могут мигрировать через сосудистую систему растения, предполагается, что манипулирование генами оксидазы GA в растительной ткани(нях), где вырабатываются активные GA, может привести к низкорослому, полукарликовому растению, даже если это может быть в значительной степени достигнуто путем супрессии уровня активных GA, продуцируемых не в тканях стебля (то есть вдали от места действия в стебле, где пониженное удлинение междоузлия приводит к полукарликовому фенотипу). В самом деле, было обнаружено, что супрессия определенных генов оксидазы GA20 в листовых тканях вызывает умеренный полукарликовый фенотип у растений кукурузы. Учитывая, что экспрессия конструкции супрессии оксидазы GA20 с несколькими различными "стеблевыми" промоторами не давала полукарликового фенотипа у кукурузы, следует отметить, что экспрессия той же самой конструкции супрессии оксидазы GA20 с помощью сосудистого промотора была эффективной при последовательном создании полукарликового фенотипа с высокой степенью проявления гена через события и зародышевую плазму. Этот полукарликовый фенотип также наблюдался при экспрессии той же конструкции супрессии оксидазы GA20 с применением других сосудистых промоторов.
[75] В соответствии с вариантами реализации данного изобретения предложены модифицированные злаковые или кукурузные растения, которые имеют, по меньшей мере, один полезный агрономический признак и, по меньшей мере, один женский репродуктивный орган или початок, которые практически или полностью не содержат дефектов. Полезный агрономический признак может включать в себя, например, более короткую высоту растения, более короткую длину междоузлия в одном или более междоузлии(ях), больший (более толстый) диаметр стебля, повышенную устойчивость к полеганию, повышенную устойчивость к засухе, повышенную эффективность использования азота, повышенную эффективность использования воды, более глубокие корни, большую площадь листа, более раннюю сомкнутость полога и/или увеличенную урожайность. Дефекты могут включать мужскую (султан или пыльник) стерильность, уменьшенное количество семян и/или наличие одной или более маскулинизированных или мужских (или подобных мужским) репродуктивных структур в женском органе или початке (например, в пыльнике початка) растения. В данном документе представлен модифицированное злаковое или кукурузное растение, в котором отсутствуют значительные дефекты в репродуктивных тканях растения. Такое модифицированное злаковое или кукурузное растение может иметь женский репродуктивный орган или початок, которые кажутся нормальными по сравнению с контрольным растением или растением дикого типа. Действительно, предлагаются модифицированные злаковые или кукурузные растения, которые содержат, по меньшей мере, один репродуктивный орган или початок, который не имеет или не проявляет, или по существу или полностью лишен отклонений, включая мужскую стерильность, пониженное число зерен, и/или маскулинизированную структуру(ы) в одном или более женских органах или початках. В данном документе женский орган или початок растения, такого как кукуруза, является "по существу свободным" от мужских репродуктивных структур, если мужские репродуктивные структуры отсутствуют или почти отсутствуют в женском органе или початке растения на основании визуального осмотра женского органа или початка на более поздних репродуктивных стадиях. Женский орган или початок растения, такого как кукуруза, является "полностью свободным" от зрелых мужских репродуктивных структур, если мужские репродуктивные структуры отсутствуют или не наблюдаются, или не являются наблюдаемыми в женском органе или початке растения, такого как кукурузное растение, путем визуального осмотра женского органа или початка на более поздних репродуктивных стадиях. Женский орган или початок растения, такого как кукуруза, без существенных отклонений и по существу свободного от мужских репродуктивных структур в початке, может иметь количество зерен на женский орган или початок растения, которое составляет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% от количества зерен на женский орган или початок дикого типа или контрольного растения. Аналогичным образом, женский орган или початок растения, такого как кукуруза, без существенных отклонений и по существу свободного от мужских репродуктивных структур в початке, может иметь среднюю массу зерен на женский орган или початок растения, которая составляет по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99,5%, по меньшей мере 99,6%, по меньшей мере 99,7%, по меньшей мере 99,8% или по меньшей мере 99,9% от средней массы зерен на женский орган или початок дикого типа или контрольного растения. Женский орган или початок растения, такого как кукуруза, который полностью свободен от зрелых мужских репродуктивных структур, может иметь количество зерен на женский орган или початок растения, которое примерно такое же, что и у дикого типа или контрольного растения. Другими словами, репродуктивное развитие женского органа или початка растения может быть нормальным или практически нормальным. Однако количество зерен на женский орган или початок может зависеть от других факторов, которые влияют на использование ресурсов и развитие растения. Действительно, количество зерен на женский орган или початок растения и/или масса зерен на женский орган или початок растения может быть примерно таким же или больше, чем у дикого типа или контрольного растения.
[76] Гормон растений гиббереллин играет важную роль в ряде процессов развития растений, включая прорастание, элонгацию клеток, цветение, эмбриогенез и развитие семян. Определенные биосинтетические ферменты (например, оксидаза GA20 и оксидаза GA3) и катаболические ферменты (например, оксидаза GA2) в пути GA имеют решающее значение для воздействия на уровни активной GA в тканях растений. Таким образом, в дополнение к супрессии некоторых генов оксидазы GA20, дополнительно предлагается, что супрессия гена оксидазы GA3 конститутивным, тканеспецифичным или тканепредпочтительным образом может также продуцировать растения кукурузы, имеющие низкорослый фенотип и повышенную устойчивость к полеганию, с возможной повышенной урожайностью, но без дефектов в початке. Таким образом, в соответствии с некоторыми вариантами реализации изобретения предоставлены конструкции и трансгены, содержащие элемент или последовательность супрессии оксидазы GA3, функционально связанные с конститутивным или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый или листовой промотор. Согласно некоторым вариантам реализации изобретения тканеспецифический или тканепредпочтительный промотор представляет собой сосудистый промотор, такой как промотор RTBV. Тем не менее, другие типы тканеспецифических или тканепредпочтительных промоторов могут потенциально применяться для супрессии оксидазы GA3 в тканях, продуцирующих активные GA кукурузы или злаковых растений для получения полукарликового фенотипа без значительных дефектов.
[77] Любой известный в данной области техники способ супрессии целевого гена можно применять для супрессии гена(ов) оксидазы GA в соответствии с вариантами реализации данного изобретения, включая экспрессию антисмысловых РНК, двухцепочечных РНК (дцРНК) или инвертированных повторов последовательностей РНК, или посредством косупрессии или РНК-интерференции (РНКи) посредством экспрессии малых интерферирующих РНК (миРНК), коротких шпилечных РНК (кшРНК), транс-активных миРНК (та-миРНК) или микро РНК (микроРНК). Кроме того, с целью сайленсинга гена могут применяться смысловые и/или антисмысловые молекулы РНК, которые нацелены на кодирующие и/или некодирующие геномные последовательности или области в пределах или вблизи гена оксидазы GA. Соответственно, любой из этих способов может быть применен для целевой супрессии эндогенного гена(генов) оксидазы (оксидаз) GA20 или оксидазы GA3 тканеспецифическим или тканепредпочтительным способом. См., например, публикации патентных заявок США № 2009/0070898, 2011/0296555 и 2011/0035839, содержание и описание которых включено в данный документ посредством ссылки.
[78] В данном документе термин "супрессия" относится к снижению, уменьшению или устранению уровня экспрессии мРНК и/или белка, кодируемого целевым геном в растении, клетке растения или ткани растения на одной или более стадии(ях) развития растений, по сравнению с уровнем экспрессии такой целевой мРНК и/или белка в контрольном растении, клетке или ткани, или растении, клетке или ткани дикого типа на той же стадии(ях) развития растения. В соответствии с некоторыми вариантами реализации изобретения представлено модифицированное или трансгенное растение, имеющее уровень экспрессии гена оксидазы GA20, который снижается по меньшей мере в одной ткани растения по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25% по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с контрольным растением. В соответствии с некоторыми вариантами реализации изобретения представлено модифицированное или трансгенное растение, имеющее уровень экспрессии гена оксидазы GA3, который снижается по меньшей мере в одной ткани растения по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25% по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с контрольным растением. В соответствии с некоторыми вариантами реализации изобретения представлено модифицированное или трансгенное растение, имеющее уровень экспрессии гена оксидазы GA20, который снижается по меньшей мере в одной ткани растения на 5% - 20%, 5% - 25%, 5% - 30%, 5% - 40%, 5% - 50%, 5% - 60%, 5% - 70%, 5% - 75%, 5% - 80%, 5% - 90%, 5% - 100%, 75% - 100%, 50% - 100%, 50% - 90%, 50% - 75%, 25% - 75%, 30% - 80% или 10% - 75% по сравнению с контрольным растением. В соответствии с некоторыми вариантами реализации изобретения представлено модифицированное или трансгенное растение, имеющее уровень экспрессии гена оксидазы GA3, который снижается по меньшей мере в одной ткани растения на 5% - 20%, 5% - 25%, 5% - 30%, 5% - 40%, 5% - 50%, 5% - 60%, 5% - 70%, 5% - 75%, 5% - 80%, 5% - 90%, 5% - 100%, 75% - 100%, 50% - 100%, 50% - 90%, 50% - 75%, 25% - 75%, 30% - 80% или 10% - 75% по сравнению с контрольным растением. В соответствии с этими вариантами реализации изобретения по меньшей мере одна ткань модифицированного или трансгенного растения, имеющего пониженный уровень экспрессии гена(ов) оксидазы GA20 и/или оксидазы GA3, включает одну или более растительную ткань(и), продуцирующую активную GA, такую как сосудистую и/или листовую ткань(и) растения, в течение одной или более вегетативной стадии(стадий) развития.
[79] В некоторых вариантах реализации изобретения супрессия эндогенного гена оксидазы GA20 или гена оксидазы GA3 является тканеспецифической (например, только в листовой и/или сосудистой ткани). Супрессия гена оксидазы GA20 может быть конститутивной и/или специфичной или предпочтительной для сосудистой или листовой ткани. В других вариантах реализации изобретения супрессия гена оксидазы GA20 или гена оксидазы GA3 является конститутивной, а не тканеспецифической. В соответствии с некоторым вариантам реализации изобретения экспрессия эндогенного гена оксидазы GA20 и/или гена оксидазы GA3 снижается в одном или более типах тканей (например, в листовой и/или сосудистой ткани(нях)) модифицированного или трансгенного растения по сравнению с такой же тканью(тканями) контрольного растения.
[80] В соответствии с вариантами реализации данного изобретения представлена молекула, конструкция или вектор рекомбинантной ДНК, включающие элемент супрессии, нацеленный на ген(ы) оксидазы GA20 или оксидазы GA3, который функционально связан с экспрессируемым в растении конститутивным или тканеспецифическим, или тканепредпочтительным промотором. Элемент супрессии может содержать транскрибируемую последовательность ДНК длиной по меньшей мере 19 нуклеотидов, например, длиной от около 19 нуклеотидов до около 27 нуклеотидов или 19, 20, 21, 22, 23, 24, 25, 26 или 27 нуклеотидов длиной, при этом транскрибируемая последовательность ДНК соответствует, по меньшей мере, части целевого гена оксидазы GA, подлежащего супрессии, и/или комплементарной ей последовательности ДНК. Элемент супрессии может составлять 19-30, 19-50, 19-100, 19-200, 19-300, 19-500, 19-1000, 19-1500, 19-2000, 19-3000, 19-4000 или 19-5000 нуклеотидов в длину. Подавляющий элемент может иметь длину, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22 или по меньшей мере 23 нуклеотида или более (например, по меньшей мере 25, по меньшей мере 30, по меньшей мере 50, по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 1500, по меньшей мере 2000, по меньшей мере 3000, по меньшей мере 4000 или по меньшей мере 5000 нуклеотидов в длину). В зависимости от длины и последовательности элемента супрессии может допускаться одно или более несовпадений последовательностей или некомплементарных оснований без потери супрессии, например, 1, 2, 3, 4, 5, 6, 7, 8 или более несовпадений, если некодирующая молекула РНК, кодируемая элементом супрессии, все еще способна в достаточной степени гибридизоваться и связываться с целевой молекулой мРНК гена(ов) оксидазы GA20 или оксидазы GA3. Действительно, даже более короткие элементы супрессии РНКи в диапазоне от около 19 нуклеотидов до около 27 нуклеотидов в длину могут иметь одно или более несовпадений или некомплементарных оснований, но все же быть эффективными в супрессии целевого гена оксидазы GA. Соответственно, смысловая или антисмысловая последовательность элемента супрессии может быть, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичной соответствующей последовательности по меньшей мере сегмента или части целевого гена оксидазы GA или его комплементарной последовательности, соответственно.
[81] Элемент супрессии или транскрибируемая последовательность ДНК по данному изобретению для целевой супрессии гена(ов) оксидазы GA могут включать одно или более из следующего: (а) последовательность ДНК, которая включает в себя, по меньшей мере, одну антисмысловую последовательность ДНК, которая является антисмысловой или комплементарной, по меньшей мере, одному сегменту или части целевого гена оксидазы GA; (b) последовательность ДНК, которая включает в себя множество копий по меньшей мере одной антисмысловой последовательности ДНК, которая является антисмысловой или комплементарной по меньшей мере одному сегменту или части целевого гена оксидазы GA; (c) последовательность ДНК, которая включает в себя, по меньшей мере, одну смысловую последовательность ДНК, которая включает в себя, по меньшей мере, один сегмент или часть целевого гена оксидазы GA; (d) последовательность ДНК, которая включает в себя множество копий по меньшей мере одной смысловой последовательности ДНК, каждая из которых содержит по меньшей мере один сегмент или часть целевого гена оксидазы GA; (e) последовательность ДНК, которая включает в себя инвертированный повтор сегмента или части целевого гена оксидазы GA и/или транскрибируется в РНК для супрессии целевого гена оксидазы GA путем образования двухцепочечной РНК, при этом транскрибированная РНК включает в себя по меньшей мере одну антисмысловую последовательность ДНК, которая является антисмысловой или комплементарной по меньшей мере к одному сегменту или части целевого гена оксидазы GA и по меньшей мере к одной смысловой последовательности ДНК, которая содержит по меньшей мере один сегмент или часть целевого гена оксидазы GA; (f) последовательность ДНК, которая транскрибируется в РНК с целью супрессии целевого гена оксидазы GA путем образования одной двухцепочечной РНК и включает в себя множество последовательных последовательностей антисмысловой ДНК, каждая из которых является антисмысловой или комплементарной по меньшей мере к одному сегменту или части целевого гена оксидазы GA, и множество последовательных смысловых последовательностей ДНК, каждая из которых содержит по меньшей мере один сегмент или часть целевого гена оксидазы GA; (g) последовательность ДНК, которая транскрибируется в РНК с целью супрессии целевого гена оксидазы GA путем образования множества двойных цепей РНК и включает в себя множество последовательностей антисмысловой ДНК, каждая из которых является антисмысловой или комплементарной по меньшей мере одному сегменту или части целевого гена GA, и множество последовательностей смысловой ДНК, каждая из которых содержит, по меньшей мере, один сегмент или часть целевого гена оксидазы GA, при этом множество антисмысловых сегментов ДНК и множество смысловых сегментов ДНК расположены в виде серии инвертированных повторов; (h) последовательность ДНК, которая включает в себя нуклеотиды, полученные из микроРНК, предпочтительно растительной микроРНК; (i) последовательность ДНК, которая включает в себя предшественник микроРНК, который кодирует искусственную микроРНК, комплементарную по меньшей мере одному сегменту или части целевого гена оксидазы GA; (j) последовательность ДНК, которая включает в себя нуклеотиды миРНК; (k) последовательность ДНК, которая транскрибируется в РНК-аптамер, способный связываться с лигандом; а также (l) последовательность ДНК, которая транскрибируется в РНК-аптамер, способный связываться с лигандом, и ДНК, которая транскрибируется в регуляторную РНК, способную регулировать экспрессию целевого гена оксидазы GA, при этом регуляция целевого гена оксидазы GA зависит от конформации регуляторной РНК, а на конформацию регуляторной РНК аллостерически влияет состояние связывания РНК-аптамера лигандом. Любой из этих элементов супрессии гена, транскрибируемый в одноцепочечную или двухцепочечную РНК, может быть предназначен для супрессии более чем одного целевого гена оксидазы GA, в зависимости от количества и последовательности элемента(ов) супрессии.
[82] Множественные смысловые и/или антисмысловые элементы супрессии для более чем одной цели оксидазы GA могут быть расположены последовательно в тандеме или расположены в виде тандемных сегментов или повторов, таких как тандемно-инвертированные повторы, которые также могут прерываться одной или более спейсерной последовательностью(ами), а последовательность каждого элемента супрессии может быть нацелена на один или более генов оксидазы GA. Кроме того, смысловая или антисмысловая последовательность элемента супрессии может не полностью совпадать или быть не полностью комплементарной целевой последовательности гена оксидазы GA, в зависимости от последовательности и длины элемента супрессии. Даже более короткие элементы супрессии РНКи от около 19 нуклеотидов до около 27 нуклеотидов в длину могут иметь одно или более несовпадений или некомплементарных оснований, но все же быть эффективными в супрессии целевого гена оксидазы GA. Соответственно, смысловая или антисмысловая последовательность элемента супрессии может быть, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичной соответствующей последовательности по меньшей мере сегмента или части целевого гена оксидазы GA или его комплементарной последовательности, соответственно.
[83] Для супрессии антисмысловой последовательностью, транскрибируемая последовательность ДНК или элемент супрессии содержит последовательность, которая является антисмысловой или комплементарной по меньшей мере части или сегменту целевого гена оксидазы GA. Элемент супрессии может содержать множество антисмысловых последовательностей, которые являются комплементарными одной или более частям или сегментам целевого гена(ов) оксидазы GA, или множество копий антисмысловой последовательности, которая комплементарна целевому гену оксидазы GA. Антисмысловая последовательность элемента супрессии может быть, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичной последовательности ДНК, которая является комплементарной к, по меньшей мере, сегменту или части целевого гена оксидазы GA. Другими словами, последовательность антисмыслового элемента супрессии может быть, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной целевому гену оксидазы GA.
[84] Для супрессии гена(ов) оксидазы GA с применением инвертированного повтора или транскрибированной дцРНК, транскрибируемая последовательность ДНК или элемент супрессии могут содержать смысловую последовательность, которая содержит сегмент или часть целевого гена оксидазы GA, и антисмысловую последовательность, которая является комплементарной сегменту или части целевого гена оксидазы GA, при этом смысловые и антисмысловые последовательности ДНК расположены в тандеме. Смысловые и/или антисмысловые последовательности, соответственно, могут быть менее чем на 100% идентичными или комплементарными сегменту, или части целевого гена оксидазы GA, как описано выше. Смысловые и антисмысловые последовательности могут быть разделены спейсерной последовательностью, так что молекула РНК, транскрибируемая с элемента супрессии, образует структуру "стебля", петли или "петли-на-стебле" между смысловой и антисмысловой последовательностями. Вместо этого элемент супрессии может содержать множество смысловых и антисмысловых последовательностей, которые расположены в тандеме, который также может быть разделен одной или более спейсерными последовательностями. Такие элементы супрессии, содержащие множество смысловых и антисмысловых последовательностей, могут быть расположены в виде последовательности смысловых последовательностей, за которыми следует последовательность антисмысловых последовательностей, или в виде последовательности тандемно расположенных смысловых и антисмысловых последовательностей. В альтернативном варианте, одна или более смысловых последовательностей ДНК могут быть экспрессированы отдельно из одной или более антисмысловых последовательностей (то есть, одна или более смысловых последовательностей ДНК могут быть экспрессированы из первой последовательности транскрибируемой ДНК, и одна или более антисмысловых последовательностей ДНК могут быть экспрессированы из второй последовательности транскрибируемой ДНК, при этом первая и вторая последовательности транскрибируемой ДНК экспрессируются в виде отдельных транскриптов).
[85] Для супрессии гена(ов) оксидазы GA с применением микроРНК транскрибируемая последовательность ДНК или элемент супрессии могут содержать последовательность ДНК, полученную из последовательности микроРНК, естественной для вируса или эукариота, такого как животное или растение, или модифицированной или полученной от такой естественной последовательности микроРНК. Такие естественные или полученные естественным образом последовательности микроРНК могут образовывать структуру "самогибридизации" и служить каркасом для предшественника микроРНК (пре-микроРНК), и могут соответствовать стеблевой области нативной последовательности предшественника микроРНК, такой как естественная (или полученная естественным образом) первичная-микроРНК (перв-микроРНК) или последовательность пре-микроРНК. Однако, в дополнение к этим естественным или полученным естественным образом каркасам или предпроцессированными последовательностям микроРНК, сконструированные или синтетические микроРНК по данному варианту реализации изобретения дополнительно содержат последовательность, соответствующую сегменту или части целевого гена(ов) оксидазы GA. Таким образом, в дополнение к предпроцессированным или каркасным последовательностям микроРНК, элемент супрессии может дополнительно содержать смысловую и/или антисмысловую последовательность, которая соответствует сегменту или части целевого гена оксидазы GA, и/или последовательность, которая является комплементарной ей, не смотря на то, что может допускаться одно или более несовпадений последовательности.
[86] Сконструированные микроРНК полезны для супрессии целевого гена с повышенной специфичностью. См., например, Parizotto и соавт., Genes Dev. 18:2237-2242 (2004 год), и публикации патентных заявок США № 2004/0053411, 2004/0268441, 2005/0144669 и 2005/0037988, содержание и описание которых включены в данный документ посредством ссылки. микроРНК представляют собой РНК, не кодирующие белок. Когда молекула предшественника микроРНК расщепляется, образуется зрелая микроРНК, длина которой обычно составляет от около 19 до около 25 нуклеотидов (обычно у растений длина составляет от около 20 до около 24 нуклеотидов), например, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину и имеет последовательность, соответствующую гену, нацеленному на супрессию, и/или его комплементу. Зрелая микроРНК гибридизуется с целевыми транскриптами мРНК и направляет связывание комплекса белков с целевыми транскриптами, которые могут функционировать для ингибирования трансляции и/или приводить к деградации транскрипта, таким образом, негативно регулируя или супрессируя экспрессию целевого гена. Предшественники микроРНК также полезны в растениях для направления поэтапного продуцирования миРНК, транс-активных миРНК (та-миРНК), в процессе, который требует РНК-зависимой РНК-полимеразы с целью супрессии целевого гена. См., например, Allen и соавт., Cell 121:207-221 (2005 год), Vaucheret Science STKE, 2005:pe43 (2005 год) и Yoshikawa и соавт., Genes Dev., 19:2164-2175 (2005 год), содержание и описание которых включены в данный документ посредством ссылки.
[87] микроРНК растений регулируют свои целевые гены путем распознавания и связывания с комплементарной или почти идеально комплементарной последовательностью (сайтом узнавания микроРНК) в целевом транскрипте мРНК с последующим расщеплением транскрипта ферментами РНКазы III, такими как ARGONAUTE1. У растений определенные ошибочные спаривания между данным сайтом распознавания микроРНК и соответствующей зрелой микроРНК, как правило, не допускаются, особенно ошибочные спаривания нуклеотидов в положениях 10 и 11 зрелой микроРНК. Положения в зрелой микроРНК приведены в направлении от 5' до 3'. Совершенная комплементарность между данным сайтом распознавания микроРНК и соответствующей зрелой микроРНК обычно требуется в положениях 10 и 11 зрелой микроРНК. См., например, Franco-Zorrilla и соавт., (2007 год) Nature Genetics, 39:1033-1037; и Axtell и соавт., (2006 год) Cell, 127:565-577.
[88] Многие гены микроРНК (гены MIR) были идентифицированы и обнародованы в базе данных ("miRBase", доступной в сети по адресу microrna.sanger.ac.uk/sequence; также см., Griffiths-Jones и соавт., (2003 год) Nucleic Acids Res., 31:439-441). Сообщалось, что гены MIR встречаются в межгенных областях, как отдельно, так и в кластерах в геноме, но также могут полностью или частично располагаться в интронах других генов (как кодирующих белок, так и не кодирующих белок). Для обзора биогенеза микроРНК, см., Kim (2005 год) Nature Rev. Mol. Cell. Biol., 6:376-385. Транскрипция генов MIR может, по меньшей мере, в некоторых случаях, находиться под промоторным контролем собственного промотора гена MIR. Первичный транскрипт, называемый "перв-микроРНК", может быть довольно большим (несколько тысяч пар оснований) и может быть полицистронным, содержать одну или более пре-микроРНК (структуры "самогибридизации", содержащие расположение "петли-на-стебле", которое процессируется до зрелой микроРНК), а также обычный 5' "кэп" и полиаденилированный хвост мРНК. См., например, Фиг. 1 в Kim (2005 год) Nature Rev. Mol. Cell. Biol., 6:376-385.
[89] Трансгенная экспрессия микроРНК (будь то встречающаяся в природе последовательность или искусственная последовательность) может быть применена для регуляции экспрессии целевого гена или генов микроРНК. Сайты распознавания микроРНК были подтверждены во всех областях мРНК, включая 5'-нетранслируемую область, кодирующую область, интронную область и 3'-нетранслируемую область, что указывает на то, что положение цели микроРНК или участок распознавания относительно кодирующей последовательности могут не обязательно влиять на супрессию (см., например, Jones-Rhoades and Bartel (2004 год)). Mol. Cell, 14:787-799, Rhoades и соавт., (2002 год) Cell, 110:513-520, Allen и соавт., (2004 год) Nat. Genet., 36:1282-1290, Sunkar and Zhu (2004 год) Plant Cell, 16:2001-2019). микроРНК являются важными регуляторными элементами у эукариот, и трансгенная супрессия с помощью микроРНК является полезным инструментом для манипулирования биологическими путями и ответами. Описание нативных микроРНК, их предшественников, участков распознавания и промоторов приведено в публикации заявки на патент США № 2006/0200878, содержание и описание которой включено в данный документ посредством ссылки.
[90] Создание искусственной последовательности микроРНК может быть достигнуто путем замены нуклеотидов в стеблевой области предшественника микроРНК последовательностью, комплементарной предполагаемой цели, как продемонстрировано, например, Zeng и соавт., (2002 год) Mol. Cell, 9:1327-1333. Согласно многим вариантам реализации изобретения, целью может быть последовательность гена оксидазы GA20 или гена оксидазы GA3. Один неограничивающий пример общего способа определения нуклеотидных изменений в нативной последовательности микроРНК для получения сконструированного предшественника микроРНК для представляющей интерес цели включает следующие этапы: (а) Выбор уникальной целевой последовательности из по меньшей мере 18 нуклеотидов, специфичных к целевому гену, например, с применением инструментов выравнивания последовательности, таких как BLAST (см., например, Altschul и соавт., (1990 год) J. Mol. Biol., 215:403-410; Altschul и соавт., (1997 год) Nucleic Acids Res., 25:3389-3402); последовательности кДНК и/или геномной ДНК могут применяться для идентификации ортологов целевого транскрипта и любых потенциальных совпадений со сторонними генами, что позволяет избежать непреднамеренного сайленсинга или супрессии нецелевых последовательностей; (b) Анализ целевого гена на наличие нежелательных последовательностей (например, совпадений с последовательностями из нецелевых видов) и оценка каждой потенциальной целевой последовательности на содержание GC, оценку Рейнольдса (см. Reynolds и соавт., (2004 год) Nature Biotechnol., 22:326-330), и функциональную асимметрию, характеризующуюся отрицательной разницей в свободной энергии ("ΔΔG") (см., Khvorova и соавт., (2003 год) Cell, 115:209-216). Предпочтительно, могут быть выбраны целевые последовательности (например, 19-меры), которые имеют все или большинство из следующих характеристик: (1) оценка Рейнольдса > 4, (2) содержание GC от около 40% до около 60%, (3 ) отрицательную ΔΔG, (4) концевой аденозин, (5) отсутствие идущих подряд 4 или более одних и тех же нуклеотидов; (6) местоположение около 3'-конца целевого гена; (7) минимальные отличия от транскрипта предшественника микроРНК. В одном аспекте некодирующая молекула РНК, применяемая здесь для супрессии целевого гена (например, гена оксидазы GA20 или GA3), предназначена для того, чтобы иметь целевую последовательность, демонстрирующую одну или более, две или более, три или более, четыре или более, или пять или более из вышеуказанных характеристик. Положения на каждом третьем нуклеотиде элемента супрессии могут влиять на эффективность РНКи; например, алгоритм "siExplorer" общедоступен по адресу rna.chem.tu-tokyo.ac.jp/siexplorer.htm (см., Katoh and Suzuki (2007 год) Nucleic Acids Res., 10.1093/nar/gkl1120); (c) Определение обратного комплемента выбранной целевой последовательности (например, 19-мер) для применения при получении модифицированной зрелой миРНК. Относительно последовательности 19-мера, дополнительный нуклеотид в положении 20 может совпадать с выбранной целью или последовательностью распознавания, а нуклеотид в положении 21 может быть либо неспаренным, чтобы предотвратить распространение сайленсинга на целевом транскрипте, либо спаренным с целевой последовательностью, чтобы способствовать распространению сайленсинга на целевом транскрипте; и (d) Трансформация искусственной микроРНК в растение.
[91] В соответствии с вариантами реализации данного изобретения предложена молекула, конструкция или вектор рекомбинантной ДНК, включающие транскрибируемую последовательность ДНК или элемент супрессии, кодирующий молекулу микроРНК или предшественника микроРНК, для целевой супрессии гена(ов) оксидазы GA. Такая транскрибируемая последовательность ДНК и элемент супрессии могут содержать последовательность длиной по меньшей мере 19 нуклеотидов, которая соответствует одному или более гену(ам) оксидазы GA, и/или последовательность, комплементарную одному или более гену(ам) оксидазы GA, хотя могут допускаться один или более несоответствий последовательности или неспаренных нуклеотидов.
[92] Ген(ы) оксидазы GA также может быть супрессирован с применением одной или более малых интерферирующих РНК (миРНК). Путь миРНК включает в себя непоследовательное расщепление более длинного двухцепочечного интермедиата РНК ("РНК-дуплекса") на малые интерферирующие РНК (миРНК). Размер или длина миРНК варьируется от около 19 до около 25 нуклеотидов или пар оснований, но общие классы миРНК включают те, которые содержат 21 или 24 пары оснований. Таким образом, транскрибируемая последовательность ДНК или элемент супрессии могут кодировать молекулу РНК длиной, по меньшей мере, от около 19 до около 25 нуклеотидов (или более), например, по меньшей мере, 19, 20, 21, 22, 23, 24 или 25 нуклеотидов в длину. Таким образом, для супрессии миРНК обеспечивается рекомбинантная молекула, конструкция или вектор ДНК, включающие транскрибируемую последовательность ДНК и элемент супрессии, кодирующий молекулу миРНК для целевой супрессии гена(ов) оксидазы GA. Такая транскрибируемая последовательность ДНК и элемент супрессии могут иметь длину по меньшей мере 19 нуклеотидов и иметь последовательность, соответствующую одному или более гену(ам) оксидазы GA, и/или последовательность, комплементарную одному или более гену(ам) оксидазы GA.
[93] Ген(ы) оксидазы GA также может быть супрессирован с применением одной или более транс-активных малых интерферирующих РНК (та-миРНК). В пути та-миРНК, микроРНК служат для направления поэтапного процессинга первичных транскриптов миРНК в процессе, который требует РНК-зависимой РНК-полимеразы для получения двухцепочечного предшественника РНК. та-миРНК определяются отсутствием вторичной структуры, целевого сайта микроРНК, который инициирует продукцию двухцепочечной РНК, потребностью в DCL4 и РНК-зависимой РНК-полимеразе (RDR6), а также продуцированием множества идеально последовательных малых РНК, длиной ~21 нуклеотид, с идеально подобранными дуплексами с 2-нуклеотидными 3'-"липкими" концами (см., Allen и соавт., (2005 год) Cell, 121:207-221). Размер или длина та-миРНК варьируется от около 20 до около 22 нуклеотидов или пар оснований, но обычно это 21 пара оснований. Таким образом, транскрибируемая последовательность ДНК или элемент супрессии по данному изобретению могут кодировать молекулу РНК длиной, по меньшей мере, от около 20 до около 22 нуклеотидов, например, длиной 20, 21 или 22 нуклеотида. Таким образом, для супрессии та-миРНК обеспечивается рекомбинантная молекула, конструкция или вектор ДНК, включающие транскрибируемую последовательность ДНК или элемент супрессии, кодирующий молекулу та-миРНК для целевой супрессии гена(ов) оксидазы GA. Такая транскрибируемая последовательность ДНК и элемент супрессии могут иметь длину по меньшей мере 20 нуклеотидов и иметь последовательность, соответствующую одному или более гену(ам) оксидазы GA, и/или последовательность, комплементарную одному или более гену(ам) оксидазы GA. Относительно способов конструирования подходящих каркасов та-миРНК, см., например, патент США № 9,309,512, который включен в данное описание посредством ссылки во всей своей полноте.
[94] В соответствии с вариантами реализации данного изобретения предлагается рекомбинантная молекула, вектор или конструкция ДНК, включающие транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая связывается или гибридизуется с целевой мРНК в клетке растения, при этом молекула целевой мРНК кодирует ген оксидазы GA20 или GA3, и при этом транскрибируемая последовательность ДНК функционально связана с конститутивным или тканеспецифичным, или тканепредпочтительным промотором. В дополнение к нацеливанию на зрелую последовательность мРНК, некодирующая молекула РНК может вместо этого нацеливаться на интронную последовательность гена оксидазы GA или транскрипта мРНК, или на последовательность мРНК оксидазы GA, перекрывающую кодирующие и некодирующие последовательности. В соответствии с другими вариантами реализации изобретения предлагается рекомбинантная молекула, вектор или конструкция ДНК, включающие транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК (предшественник), которая расщепляется или процессируется в зрелую некодирующую молекулу РНК, которая связывается или гибридизуется с целевой мРНК в клетке растения, при этом молекула целевой мРНК кодирует белок оксидазы GA20 или GA3, и при этом транскрибируемая последовательность ДНК функционально связана с конститутивным или тканеспецифичным, или тканепредпочтительным промотором. Для целей данного изобретения "некодирующая молекула РНК" представляет собой молекулу РНК, которая не кодирует белок. Неограничивающие примеры некодирующей молекулы РНК включают микро РНК (микроРНК), предшественника микроРНК, малую интерферирующую РНК (миРНК), предшественника миРНК, малую РНК (длиной 18-26 нуклеотидов) и предшественника, кодирующих то же самое, гетерохроматическую миРНК (гх-миРНК), взаимодействующую с Piwi РНК (piРНК), двухцепочечную РНК, образующую шпильки (дцРНК, образующая шпильки), транс-активную миРНК (та-миРНК), встречающуюся в природе антисмысловую миРНК (nat-миРНК - naturally occurring antisense siRNA) CRISPR РНК (crРНК), tracer (транс-активирующую CRISPR) РНК (tracrРНК), направляющую РНК (нРНК) и одиночную направляющую РНК (онРНК - single-guide RNA).
[95] В соответствии с вариантами реализации данного изобретения подходящие тканеспецифичные или тканепредпочтительные промоторы для экспрессии элемента супрессии оксидазы GA20 или GA3 могут включать в себя те промоторы, которые управляют или вызывают экспрессию связанного с ним элемента или последовательности супрессии, по меньшей мере, в сосудистой и/или листовой ткани кукурузы или злаковых растений, или, возможно, в другой ткани в случае оксидазы GA3. Экспрессия элемента или конструкции супрессии оксидазы GA, с помощью тканеспецифического или тканепредпочтительного промотора также может происходить в других тканях злаковых или кукурузных растений за пределами сосудистых и листовых тканей, но уровни активной GA в развивающихся репродуктивных тканях растения (особенно в женском репродуктивном органе или початке) предпочтительно не подвергаются значительному уменьшению или воздействию (по сравнению с диким типом или контрольными растениями), так что развитие женского органа или початка может происходить нормально в трансгенном растении без каких-либо отклонений в початке и потери потенциальной урожайности.
[96] Любые сосудистые промоторы, известные в данной области техники, могут потенциально применяться в качестве тканеспецифического или тканепредпочтительного промотора. Примеры сосудистых промоторов включают в себя промотор RTBV (см., например, SEQ ID NO: 65), известный промотор гена сахарозосинтазы, такой как промотор кукурузной сахарозосинтазы-1 (Sus1 или Sh1) (см., например, SEQ ID NO: 67), промотор паралога гена кукурузы Sh1, промотор сахарозосинтазы (Ss1) ячменя, промотор рисовой сахарозосинтазы-1 (RSs1) (см., например, SEQ ID NO: 68) или промотор рисовой сахарозосинтазы-2 (RSs2) (см., например, SEQ ID NO: 69), известный промотор гена транспортера сахарозы, такой как промотор транспортера рисовой сахарозы (SUT1) (см., например, SEQ ID NO: 70), или различные известные вирусные промоторы, такие как промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) гемивируса полосы кукурузы (MSV - maize streak geminivirus) или рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или OsYSL2 (SEQ ID NO: 71), и любой участок функциональной последовательности или усечение любого из вышеуказанных промоторов со сходным профилем экспрессии, такой как усеченный промотор RTBV (см., например, SEQ ID NO: 66).
[97] Любые листовые промоторы, известные в данной области техники, могут потенциально применяться в качестве тканеспецифического или тканепредпочтительного промотора. Примеры листовых промоторов включают в себя промотор пираватфосфатдикиназы кукурузы или промотор PPDK (pyruvate phosphate dikinase) (см., например, SEQ ID NO: 72), промотор фруктозо-1,6-бисфосфат альдолазы кукурузы или FDA (см., например, SEQ ID NO: 73) и рисовый промотор Nadh-Gogat (см., например, SEQ ID NO: 74) и любую часть функциональной последовательности или усечение любого из вышеуказанных промоторов со сходным профилем экспрессии. Другие примеры листовых промоторов генов однодольных растений включают в себя промотор рибулозофосфаткарбоксилазы (Рубиско) или малой субъединицы Рубиско (RBCS - RuBisCO small subunit), промотор гена, связывающего хлорофилл а/b белка, промотор фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы) и промотор гена Myb, и любую часть функциональной последовательности или усечение любого из этих промоторов со сходным профилем экспрессии.
[98] Любые другие сосудистые и/или листовые промоторы, известные в данной области техники, также могут быть применены, включая промоторные последовательности из родственных генов (например, последовательности промоторов сахарозосинтазы, транспортера сахарозы и вирусных генов) одного и того же или разных видов растений или вирусов, которые имеют сходный профиль экспрессии. Кроме того, представлены промоторные последовательности с высокой степенью гомологии к любому из вышеперечисленного. Например, сосудистый промотор может содержать последовательность ДНК, которая является, по меньшей мере, на 70%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 65, 66, 67, 68, 69, 70 и 71, любой части функциональной последовательности или ее усечению, и/или любую последовательность, комплементарную любой из вышеуказанных последовательностей; листовой промотор может содержать, например, последовательность ДНК, которая является, по меньшей мере, на 70%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 72, 73 и 74, любой части функциональной последовательности или ее усечению, и/или любой последовательности, комплементарной любой из вышеуказанных последовательностей; и конститутивный промотор может содержать последовательность ДНК, которая является, по меньшей мере, на 70%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 75, 76, 77, 78, 79, 80, 81, 82 и 83, любая часть функциональной последовательности или усечение, и/или любой последовательности, комплементарной любой из вышеуказанных последовательностей. Примеры сосудистых и/или листовых промоторов могут дополнительно включать другие известные, сконструированные и/или идентифицированные впоследствии промоторные последовательности, которые, как показано, имеют профиль экспрессии в сосудистой и/или листовой ткани(нях) злакового или кукурузного растения. Кроме того, любой известный или идентифицированный впоследствии конститутивный промотор также может быть применен для экспрессии элемента супрессии оксидазы GA20 или GA3. Общие примеры конститутивных промоторов приведены ниже.
[99] Как понятно в данной области техники, термин "промотор" может, как правило, относиться к последовательности ДНК, которая содержит сайт связывания РНК-полимеразы, сайт инициации транскрипции и/или ТАТА-бокс и способствует или стимулирует транскрипцию и экспрессию ассоциированной транскрибируемой полинуклеотидной последовательности и/или гена (или трансгена). Промотор может быть синтетическим или искусственным и/или сконструированным, измененным или полученным из известной или встречающейся в природе последовательности промотора. Промотор может представлять собой химерный промотор, содержащий комбинацию двух или более гетерологичных последовательностей. Таким образом, промотор по данному изобретению может включать варианты последовательностей промотора, которые сходны по составу, но не идентичны другим промоторным последовательностям, известным или представленным в данном документе. Промотор может быть классифицирован в соответствии со множеством критериев, касающихся характера профиля экспрессии, ассоциированной кодирующей или транскрибируемой последовательности, или гена (включая трансген), функционально связанных с промотором, таким как конститутивный, специфический к стадии развития, тканеспецифический, индуцибельный и тому подобное. Промоторы, которые управляют экспрессией во всех или почти во всех тканях растения, называют "конститутивными" промоторами. Однако уровень экспрессии с "конститутивным промотором" не обязательно является одинаковым для разных типов тканей и клеток. Промоторы, которые управляют экспрессией в определенные периоды или стадии развития, называются "промоторами, специфическими к стадии развития". Промоторы, которые управляют усиленной экспрессией в определенных тканях растения по сравнению с другими растительными тканями, называют промоторами, "усиленными в определенной ткани" или "тканепредпочтительными промоторами". Таким образом, "тканепредпочтительный" промотор вызывает относительно более высокую или преимущественную, или преобладающую экспрессию в конкретной ткани(нях) растения, но с более низкими уровнями экспрессии в другой ткани(нях) растения. Промоторы, которые экспрессируют в определенной ткани(нях) растения, практически не экспрессируясь в других тканях растения, называют "тканеспецифическими" промоторами. Тканеспецифический или тканепредпочтительный промотор также может быть определен относительно специфической или предпочтительной ткани(ей), в которой он управляет экспрессией ассоциированной транскрибируемой последовательности ДНК или элемента супрессии. Например, промотор, который вызывает специфическую экспрессию в сосудистых тканях, может упоминаться как "сосудисто-специфический промотор", тогда как промотор, который вызывает преимущественную или преобладающую экспрессию в сосудистых тканях, может упоминаться как "сосудисто-предпочтительный промотор". Аналогично, промотор, который вызывает специфическую экспрессию в тканях листьев, может упоминаться как "промотор, специфический к листовой ткани", тогда как промотор, который вызывает преимущественную или преобладающую экспрессию в тканях листьев, может упоминаться как "промотор, предпочтительный к листовой ткани". "Индуцибельный" промотор представляет собой промотор, который инициирует транскрипцию в ответ на стимул окружающей среды, такой как холод, засуха или свет, или другие стимулы, такие как ранение или химическое воздействие. Промотор также может быть классифицирован по своему происхождению, например, гетерологичный, гомологичный, химерный, синтетический и тому подобное. "Гетерологичный" промотор представляет собой промоторную последовательность, имеющую иное происхождение по сравнению с ассоциированной с ним транскрибируемой последовательностью, кодирующей последовательность или ген (или трансген), и/или не встречающуюся в природе у видов растений, подлежащих трансформации, как определено выше.
[100] Некоторые из оксидаз GA в злаковых растениях состоят из семейства родственных генов оксидазы GA. Например, кукуруза имеет семейство из по меньшей мере девяти генов оксидазы GA20, которые включают оксидазу_1 GA20, оксидазу_2 GA20, оксидазу_3 GA20, оксидазу_4 GA20, оксидазу_5 GA20, оксидазу_6 GA20, оксидазу_7 GA20, оксидазу_8 GA20 и оксидазу_9 GA20. Однако в кукурузе есть только две оксидазы GA3: оксидаза_1 GA3 и оксидаза_2 GA3. Последовательности SEQ ID NO ДНК и белка для каждого из этих генов оксидазы GA20 представлены в Таблице 1, а последовательности SEQ ID NO ДНК и белка для каждого из этих генов оксидазы GA3 представлены в Таблице 2.
Таблица 1. Последовательности ДНК и белка по идентификатору последовательности для генов оксидазы GA20 в кукурузе.
Таблица 2. Последовательности ДНК и белка по идентификатору последовательности для генов оксидазы GA3 в кукурузе.
[101] Последовательность геномной ДНК оксидазы_3 GA20 представлена в SEQ ID NO: 34, а последовательность геномной ДНК оксидазы_5 GA20 представлена в SEQ ID NO: 35. Для гена оксидазы_3 GA20, SEQ ID NO: 34 предоставляет 3000 нуклеотидов в 3'-5' направлении от 5'-НТО оксидазы_3 GA20; нуклеотиды 3001-3096 соответствуют 5'-НТО; нуклеотиды 3097-3665 соответствуют первому экзону; нуклеотиды 3666-3775 соответствуют первому интрону; нуклеотиды 3776-4097 соответствуют второму экзону; нуклеотиды 4098-5314 соответствуют второму интрону; нуклеотиды 5315-5584 соответствуют третьему экзону; и нуклеотиды 5585-5800 соответствуют 3'-НТО. SEQ ID NO: 34 также обеспечивает 3000 нуклеотидов в 5'-3' направлении от конца 3'-НТО (нуклеотиды 5801-8800). Для гена оксидазы_5 GA20, SEQ ID NO: 35 обеспечивает 3000 нуклеотидов в 3'-5' направлении от стартового кодона оксидазы_5 GA20 (нуклеотиды 1-3000); нуклеотиды 3001-3791 соответствуют первому экзону; нуклеотиды 3792-3906 соответствуют первому интрону; нуклеотиды 3907-4475 соответствуют второму экзону; нуклеотиды 4476-5197 соответствуют второму интрону; нуклеотиды 5198-5473 соответствуют третьему экзону; и нуклеотиды 5474-5859 соответствуют 3'-НТО. SEQ ID NO: 35 также обеспечивает 3000 нуклеотидов в 5'-3' направлении от конца 3'-НТО (нуклеотиды 5860-8859).
[102] Последовательность геномной ДНК оксидазы_1 GA3 представлена в SEQ ID NO: 36, а последовательность геномной ДНК оксидазы_2 GA3 представлена в SEQ ID NO: 37. Для гена оксидазы_1 GA3, нуклеотиды 1-29 из SEQ ID NO: 36 соответствуют 5'-НТО; нуклеотиды 30-514 из SEQ ID NO: 36 соответствуют первому экзону; нуклеотиды 515-879 из SEQ ID NO: 36 соответствуют первому интрону; нуклеотиды 880-1038 из SEQ ID NO: 36 соответствуют второму экзону; нуклеотиды 1039-1158 из SEQ ID NO: 36 соответствуют второму интрону; нуклеотиды 1159-1663 из SEQ ID NO: 36 соответствуют третьему экзону; и нуклеотиды 1664-1788 из SEQ ID NO: 36 соответствуют 3'-НТО. Для гена оксидазы_2 GA3, нуклеотиды 1-38 из SEQ ID NO: 37 соответствуют 5-НТО; нуклеотиды 39-532 из SEQ ID NO: 37 соответствуют первому экзону; нуклеотиды 533-692 из SEQ ID NO: 37 соответствуют первому интрону; нуклеотиды 693-851 из SEQ ID NO: 37 соответствуют второму экзону; нуклеотиды 852-982 из SEQ ID NO: 37 соответствуют второму интрону; нуклеотиды 983-1445 из SEQ ID NO: 37 соответствуют третьему экзону; и нуклеотиды 1446-1698 из SEQ ID NO: 37 соответствуют 3'-НТО.
[103] В дополнение к фенотипическим наблюдениям с нацеливанием на ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20, или ген(ы) оксидазы_1 GA3 и/или оксидазы_2 GA3, с целью супрессии, также наблюдается полукарликовый фенотип с супрессией гена оксидазы_4 GA20. Последовательность геномной ДНК оксидазы_4 GA20 представлена в SEQ ID NO: 38. Для гена оксидазы_4 GA, SEQ ID NO: 38 предоставляет нуклеотиды 1-1416 в 3'-5' направлении от 5'-НТО; нуклеотиды 1417-1543 из SEQ ID NO: 38 соответствуют 5'-НТО; нуклеотиды 1544-1995 из SEQ ID NO: 38 соответствуют первому экзону; нуклеотиды 1996-2083 из SEQ ID NO: 38 соответствуют первому интрону; нуклеотиды 2084-2411 из SEQ ID NO: 38 соответствуют второму экзону; нуклеотиды 2412-2516 из SEQ ID NO: 38 соответствуют второму интрону; нуклеотиды 2517-2852 из SEQ ID NO: 38 соответствуют третьему экзону; нуклеотиды 2853-3066 из SEQ ID NO: 38 соответствуют 3'-НТО; и нуклеотиды 3067-4465 из SEQ ID NO: 38 соответствуют геномной последовательности в 5'-3' направлении от 3'-НТО.
[104] В соответствии с вариантами реализации данного изобретения предложена молекула, вектор или конструкция рекомбинантной ДНК, содержащие транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, сегменту или части молекулы мРНК (i) экспрессирующиеся из эндогенного гена оксидазы GA и/или (ii) кодирующие эндогенный белок оксидазы GA в растении, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, и при этом растение представляет собой злаковое или кукурузное растение.
[105] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацелена на ген(ы) оксидазы GA20, такой как ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20, с целью супрессии, и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из одной или более из SEQ ID NO: 7, 8, 13 и 14. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным одной или более из SEQ ID NO: 9 и 15. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным одной или более из SEQ ID NO: 9 и 15. В дополнение к нацеливанию на зрелую последовательность мРНК (включая одну или обе из нетранслируемых или экзонных последовательностей), некодирующая молекула РНК может дополнительно нацеливаться на интронные последовательности гена или транскрипта оксидазы GA20.
[106] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацелена, с целью супрессии, на ген(ы) оксидазы GA3, и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из одной или более из SEQ ID NO: 28, 29, 31 и 32. В соответствии с другими вариантами реализации изобретения, некодирующая молекула РНК является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным одному или обоим из SEQ ID NO: 30 и 33. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным одному или обоим из SEQ ID NO: 30 и 33. В дополнение к нацеливанию на зрелую последовательность мРНК (включая одну или обе из нетранслируемых или экзонных последовательностей), некодирующая молекула РНК может дополнительно нацеливаться на интронные последовательности гена или транскрипта оксидазы GA3.
[107] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацелена, с целью супрессии, на ген оксидазы_4 GA20, и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из одной или обоих из SEQ ID NO: 10 и 11. В соответствии с другими вариантами реализации изобретения, некодирующая молекула РНК является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 12. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 12. В дополнение к нацеливанию на зрелую последовательность мРНК (включая одну или обе из нетранслируемых или экзонных последовательностей), некодирующая молекула РНК может дополнительно нацеливаться на интронные последовательности гена или транскрипта оксидазы GA20.
[108] В соответствии со многими вариантами реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК молекулы, вектора или конструкции рекомбинантной ДНК, может представлять собой предшественника микроРНК или миРНК, который процессируется или расщепляется в растительной клетке с образованием зрелой микроРНК или миРНК, которая нацеливается на ген оксидазы GA20 или оксидазы GA3.
[109] В соответствии с вариантами реализации данного изобретения уровни GA могут быть снижены в стебле злакового или кукурузного растения путем нацеливания только на ограниченную подгруппу генов в семействе оксидазы GA с целью супрессии. Не ограничиваясь какой-либо теорией, предлагается, что нацеливание на ограниченное число генов в семействе оксидаз GA с целью супрессии может привести к низкорослому фенотипу и устойчивости к полеганию в трансгенных растениях, но без отклонений в репродуктивных или початковых тканях растения из-за дифференциальной экспрессии среди генов оксидаз GA, достаточной компенсации для супрессии гена(ов) оксидазы GA другим геном(ами) оксидазы GA в этих репродуктивных тканях и/или неполной супрессии целевого гена(ов) оксидазы GA. Таким образом, путем ограничения экспрессии или супрессии гена(ов) оксидазы GA с помощью тканеспецифического или тканепредпочтительного промотора можно избежать не только отклонений, предлагается, что ограниченное подмножество генов оксидазы GA (например, ограниченное количество генов оксидазы GA20) может быть направлено на супрессию, так что другие гены оксидаз GA в пределах того же семейства генов (например, другие гены оксидазы GA20) могут компенсировать потерю экспрессии гена(ов) супрессированной оксидазы GA в этих тканях. Неполная супрессия целевого гена(ов) оксидазы GA также может обеспечить достаточный уровень экспрессии целевого гена(ов) оксидазы GA в одной или более тканях, чтобы избежать отклонений или нежелательных признаков в растении, которые могут негативно повлиять на урожайность, таких как репродуктивные отклонения или чрезмерное сокращение высоты растений. В отличие от мутаций полной потери функции в гене, супрессия может сохранить частичную активность целевого гена. Поскольку разные гены оксидазы GA20 имеют разные профили экспрессии в растениях, нацеливание на ограниченное подмножество генов оксидазы GA20 с целью супрессии может позволить модифицировать определенные признаки, избегая при этом отклонений, ранее связанных с мутантами GA в злаковых растениях. Другими словами, признаки роста, развития и репродуктивные признаки или отклонения, ранее связанные с мутантами GA в кукурузе и других злаковых культурах, могут быть разделены путем нацеливания только на ограниченное количество или подмножество (то есть один или более, но не всех) генов оксидазы GA20 или GA3 и/или неполной супрессии целевого гена оксидазы GA. Посредством трансгенного нацеливания на подмножество одного или более эндогенных генов оксидазы GA3 или GA20 с целью супрессии в растении, можно применять более первазивный профиль экспрессии (например, с конститутивным промотором) для получения полукарликовых растений без значительных репродуктивных отклонений и/или других нежелательных признаков в растении, даже с экспрессией трансгенной конструкции в репродуктивной ткани(нях). В самом деле, в данном документе представлены элементы и конструкции супрессии, которые избирательно нацелены на гены оксидазы_3 GA20 и/или оксидазы_5 GA20 (идентифицированные в приведенной выше Таблице 1) с целью супрессии, которые могут быть функционально связаны с сосудистым, листовым и/или конститутивным промотором.
[110] С конструкцией супрессии, которая нацелена только на ограниченное подмножество генов оксидазы GA20, таких как ген(ы) оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20, или которая нацелена на ген(ы) оксидазы оксидазы_1 GA3 и/или оксидазы_2 GA3, ограничение профиля экспрессии элемента супрессии может быть менее важным для достижения нормального репродуктивного развития злаковых или кукурузных растений и предотвращения отклонений в женском органе или початке из-за компенсации и тому подобного, от других генов оксидазы GA20 и/или GA3. Таким образом, экспрессия конструкции и элемента супрессии, селективно или предпочтительно нацеленных, например, на ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20, ген оксидазы_4 GA20 и/или ген оксидазы_1 GA3, и/или оксидазы_2 GA3 в кукурузе, или на аналогичные гены и гомологи в других растениях злаков, может быть вызван различными типами промоторов, экспрессирующимся в растениях, включая конститутивные и тканеспецифичные или тканепредпочтительные промоторы, такие как сосудистый или листовой промотор, которые могут включать в себя, например, промотор RTBV, приведенный выше (например, промотор, содержащий последовательность RTBV (SEQ ID NO: 65) или усеченную последовательность RTBV (SEQ ID NO: 66)), и любые другие промоторы, которые управляют экспрессией в тканях, охватывающих большую часть или всю сосудистую и/или листовую ткань(ни) растения. Любой известный или впоследствии идентифицированный конститутивный промотор с достаточно высоким уровнем экспрессии также может быть применен для экспрессии супрессирующей конструкции, нацеленной на подмножество генов оксидазы GA20 и/или GA3 в кукурузе, в частности на ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20, ген оксидазы_4 GA20 и/или ген(ы) оксидазы_1 GA3 и/или оксидазы_2 GA3, или подобные гены и гомологи в других злаковых растениях.
[111] Примеры конститутивных промоторов, которые могут быть применены в однодольных растениях, таких как злаковые или кукурузные растения, включают в себя, например, различные промоторы гена актина, такие как промотор Актина 1 риса (см., например, Патент США № 5,641,876; см., также SEQ ID NO: 75 или SEQ ID NO: 76) и промотор Актина 2 риса (см., например, Патент США № 6,429,357; см., также, например, SEQ ID NO: 77 или SEQ ID NO: 78), промотор CaMV 35S или 19S (см., например, Патент США № 5,352,605; см., также, например, SEQ ID NO: 79 для CaMV 35S), промотор юбиквитина кукурузы (см., например, Патент США № 5,510,474), полиюбиквитиновый промотор Coix lacryma-jobi (см., например, SEQ ID NO: 80), промотор риса или кукурузы Gos2 (см., например, Pater и соавт., The Plant Journal, 2(6): 837-44 1992 год; см., также, например, SEQ ID NO: 81 для рисового промотора Gos2), промотор FMV 35S (см., например, Патент США № 6,372,211), двойной улучшенный промотор CMV (см., например, Патент США № 5,322,938), промотор MMV (см., например, Патент США № 6,420,547; см., также, например, SEQ ID NO: 82), промотор PCLSV (см., например, Патент США № 5,850,019; см., также, например, SEQ ID NO: 83), промотор Emu (см., например, Last и соавт., Theor. Appl. Genet. 81:581 (1991 год); и Mcelroy и соавт., Mol. Gen. Genet. 231:150 (1991 год)), тубулиновый промотор кукурузы, риса или других видов, промотор нопалинсинтазы (nos), промотор октопинсинтазы (ocs), промотор маннопинсинтазы (mas) или промотор алкогольдегидрогеназы растения (например, Adh1 маиса), любые другие промоторы, включая вирусные промоторы, известные или идентифицированные в данной области техники для обеспечения конститутивной экспрессии в злаковом или кукурузном растении, любые другие конститутивные промоторы, известные в данной области техники, которые можно применять в однодольных или злаковых растениях, и любую часть функциональной последовательности или усечение любого из вышеуказанных промоторов.
[112] Достаточный уровень экспрессии транскрибируемой последовательности ДНК, кодирующей некодирующую молекулу РНК, нацеленную на ген оксидазы GA с целью супрессии, может быть необходим для получения низкорослого, полукарликового фенотипа, который является устойчивым к полеганию, поскольку более низкие уровни экспрессии могут быть недостаточными для снижения уровней активной GA в растении в достаточной степени, чтобы повлечь за собой существенный фенотип. Таким образом, могут быть предпочтительными тканеспецифичные и тканепредпочтительные промоторы, которые управляют и тому подобное, умеренным или сильным уровнем экспрессии их ассоциированной транскрибируемой последовательности ДНК в ткани(нях), продуцирующей активную GA растения. Кроме того, такие тканеспецифичные и тканепредпочтительные (промоторы) должны стимулировать и тому подобное, экспрессию ассоциированной с ними транскрибируемой последовательности ДНК в течении одной или более вегетативной стадии(стадий) развития растений, когда растение растет и/или удлиняется, включая одну или более из следующей вегетативной стадии(стадий): VE, V1, V2, V3, V4, V5, V6, V7, V8, V9, V10, V11, V12, V13, V14, Vn, VT, например, экспрессия по меньшей мере в течении V3-V12, V4-V12, V5-V12, V6-V12, V7-V12, V8-V12, V3-V14, V5-V14, V6-V14, V7-V14, V8-V14, V9-V14, V10-V14, и тому подобное, или в течении любого другого диапазона вегетативных стадий, когда происходит рост и/или удлинение растения.
[113] В соответствии со многими вариантами реализации изобретения, экспрессируемый растением промотор может предпочтительно стимулировать экспрессию конститутивно или, по меньшей мере, в части сосудистой и/или листовой ткани растения. Различные промоторы, управляющие экспрессией элемента супрессии, нацеленного на эндогенный ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20, ген оксидазы_4 GA20, ген(ы) оксидазы_1 GA3 и/или оксидазы_2 GA3 в кукурузе, или подобные гены и гомологи в других злаковых растениях, могут быть эффективными для уменьшения высоты растения и повышения устойчивости к полеганию в различной степени в зависимости от их конкретного профиля и силы экспрессии в растении. Однако некоторые тканеспецифичные и тканепредпочтительные промоторы, управляющие экспрессией элемента супрессии оксидазы GA20 или GA3 в растении, могут не приводить к фенотипу значительно низкого роста или устойчивости к полеганию из-за пространственно-временного профиля экспрессии промотора во время развития растения, и/или количество или сила экспрессии промотора являются слишком низкими или слабыми. Дополнительно, некоторые конструкции супрессии могут только уменьшать и не устранять экспрессию целевого гена(ов) оксидазы GA20 или GA3 при экспрессии в растении и, таким образом, в зависимости от профиля и силы экспрессии данного промотора, профиль и уровень экспрессии конструкции супрессии оксидазы GA20 или GA3 с таким промотором могут быть недостаточными для получения наблюдаемого фенотипа высоты растений и устойчивости к полеганию у растений.
[114] В соответствии с данными вариантами реализации изобретения предлагается рекомбинантная молекула, вектор или конструкция ДНК, для супрессии одного или более эндогенного гена(ов) оксидазы GA20 или GA3 в растении, содержащие транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, сегменту или части молекулы мРНК, экспрессирующиеся из эндогенного гена оксидазы GA и кодирующие эндогенный белок оксидазы GA в растении, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, и при этом растение представляет собой злаковое или кукурузное растение. Как сказано выше, помимо нацеливания на зрелую последовательность мРНК, некодирующая молекула РНК может дополнительно нацеливаться на интронную последовательность(и) гена или транскрипта оксидазы GA. В соответствии со многими вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться, для супрессии, на ген оксидазы_3 GA20, и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 7 или SEQ ID NO: 8. Согласно некоторым вариантам реализации изобретения некодирующая молекула РНК, нацеленная на ген оксидазы_3 GA20 для супрессии, может быть комплементарной к, по меньшей мере, 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из SEQ ID NO: 7 или SEQ ID NO: 8. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться на ген оксидазы GA20 для супрессии и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 9. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 9.
[115] Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA вместо или в дополнение к экзонному, 5'-НТО или 3'-НТО гена оксидазы GA. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_3 GA20, может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 34 и/или нуклеотидам 3666-3775 или 4098-5314 из SEQ ID NO: 34. Важно отметить, что представленные в данном документе последовательности для гена оксидазы_3 GA20 могут варьироваться в зависимости от разнообразия растений кукурузы, линий и зародышевых плазм из-за полиморфизмов и/или наличия различных аллелей гена. Дополнительно, ген оксидазы_3 GA20 может быть экспрессирован в виде альтернативно сплайсированных изоформ, которые могут приводить к различным мРНК, кДНК и кодирующим последовательностям, которые могут влиять на конструкцию супрессирующей конструкции и некодирующей молекулы РНК. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_3 GA20, может быть более широко определена как содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 34.
[116] В соответствии с вариантами реализации данного изобретения предложена молекула, вектор или конструкция рекомбинантной ДНК для супрессии эндогенного гена оксидазы_5 GA20 в растении, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК, нацеленная на ген оксидазы_5 GA20 для супрессии, содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 13 или SEQ ID NO: 14. Согласно некоторым вариантам реализации изобретения некодирующая молекула РНК, нацеленная на ген оксидазы_5 GA20 для супрессии, может быть комплементарной к, по меньшей мере, 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из SEQ ID NO: 13 или SEQ ID NO: 14. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться на ген оксидазы GA20 для супрессии и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 15. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 15.
[117] Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA вместо или в дополнение к экзонной или нетранслируемой области зрелой мРНК гена оксидазы GA. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_5 GA20, может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 35 и/или нуклеотидам 3792-3906 или 4476-5197 из SEQ ID NO: 35. Представленные в данном документе последовательности для оксидазы_5 GA20 могут варьироваться в зависимости от разнообразия растений кукурузы, линий и зародышевых плазм из-за полиморфизмов и/или наличия различных аллелей гена. Дополнительно, ген оксидазы_5 GA20 может быть экспрессирован в виде альтернативно сплайсированных изоформ, которые могут приводить к различным мРНК, кДНК и кодирующим последовательностям, которые могут влиять на конструкцию супрессирующей конструкции и некодирующей молекулы РНК. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_3 GA20, может быть определена более широко как содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 35.
[118] В соответствии с дополнительными вариантами реализации изобретения, предлагается рекомбинантная молекула, вектор или конструкция рекомбинантной ДНК для совместной супрессии эндогенных генов оксидазы_3 GA20 и оксидазы_5 GA20 в растении, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК нацелена на гены оксидазы_3 GA20 и оксидазы_5 GA20, для супрессии, содержит последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 7 и/или SEQ ID NO: 8, и (ii) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 13 и/или SEQ ID NO: 14. Согласно некоторым из этих вариантов реализации изобретения, некодирующая молекула РНК, совместно нацеленная на гены оксидазы_3 GA20 и оксидазы_5 GA20 для супрессии, может быть комплементарной по меньшей мере 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из (i) SEQ ID NO: 7 (и/или SEQ ID NO: 8) и (ii) SEQ ID NO: 13 (и/или SEQ ID NO: 14). Согласно многим вариантам реализации изобретения, некодирующая молекула РНК, совместно нацеленная на гены оксидазы_3 GA20 и оксидазы_5 GA20 для супрессии, содержит последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 9, и (ii) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 15. Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA. Таким образом, некодирующая молекула РНК может нацеливаться на интронную последовательность(ти) одного или обоих гена(ов) оксидазы_3 GA20 и/или оксидазы_5 GA20, как указано выше.
[119] Согласно конкретным вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, содержит (i) последовательность, которая является, по меньшей мере, на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной SEQ ID NO: 39, 41, 43 или 45, и/или (ii) последовательность или элемент супрессии, кодирующие некодирующую молекулу РНК, содержащую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичной SEQ ID NO: 40, 42, 44 или 46. Согласно некоторым вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность с одним или более ошибочными спариваниями, например, 1, 2, 3, 4, 5 или более комплементарных ошибочных спариваний, относительно последовательности целевого сайта или сайта узнавания целевой мРНК гена оксидазы GA20, такой как последовательность, которая почти комплементарна SEQ ID NO: 40, но с одним или более комплементарными ошибочными спариваниями по отношению к SEQ ID NO: 40. Согласно конкретному варианту реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, содержит последовательность, которая на 100% идентична SEQ ID NO: 40, которая на 100% комплементарна целевой последовательности в кДНК и кодирующих последовательностях оксидазы_3 GA20 (то есть, SEQ ID NO: 7 и 8, соответственно) и/или соответствующей последовательности мРНК, кодируемой эндогенным геном оксидазы_3 GA20. Однако последовательность некодирующей молекулы РНК, кодируемой транскрибируемой последовательностью ДНК, которая на 100% идентична SEQ ID NO: 40, 42, 44 или 46, может быть не полностью комплементарной целевой последовательности в кДНК и кодирующих последовательностях ген оксидазы_5 GA20 (то есть SEQ ID NO: 13 и 14, соответственно) и/или соответствующей последовательности мРНК, кодируемой эндогенным геном оксидазы_5 GA20. Например, наибольшее комплементарное совпадение между некодирующей молекулой РНК или последовательностью микроРНК в SEQ ID NO: 40 и кДНК, и кодирующими последовательностями гена оксидазы_5 GA20 может включать в себя одно ошибочное спаривание в первом положении SEQ ID NO: 39 (то есть, "C" в первом положении SEQ ID NO: 39 заменяется на "G"; то есть GTCCATCATGCGGTGCAACTA). Однако некодирующая молекула РНК или последовательность микроРНК в SEQ ID NO: 40 могут по-прежнему связываться и гибридизоваться с мРНК, кодируемой эндогенным геном оксидазы_5 GA20, несмотря на это небольшое несовпадение.
[120] В соответствии с вариантами реализации данного изобретения, предлагается рекомбинантная молекула, вектор или конструкция ДНК, для супрессии одного или более эндогенного гена(ов) оксидазы GA3 в растении, содержащие транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, сегменту или части молекулы мРНК, экспрессирующиеся из эндогенного гена оксидазы GA3 и кодирующие эндогенный белок оксидазы GA3 в растении, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, и при этом растение представляет собой злаковое или кукурузное растение. Помимо нацеливания на зрелую последовательность мРНК, некодирующая молекула РНК может дополнительно нацеливаться на интронные последовательности гена или транскрипта оксидазы GA3.
[121] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться, для супрессии, на ген оксидазы_1 GA3, и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 28 или SEQ ID NO: 29. Согласно некоторым вариантам реализации изобретения некодирующая молекула РНК, нацеленная на ген оксидазы GA3 для супрессии, может быть комплементарной к, по меньшей мере, 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из SEQ ID NO: 28 или SEQ ID NO: 29. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацеливается на ген оксидазы GA3 для супрессии и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 30. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 30.
[122] Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA3 вместо или в дополнение к экзонному, 5'-НТО или 3'-НТО гена оксидазы GA. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_1 GA3, может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 36 и/или нуклеотидам 515-879 или 1039-1158 из SEQ ID NO: 36. Представленные в данном документе последовательности для оксидазы_1 GA3 могут варьироваться в зависимости от разнообразия растений кукурузы, линий и зародышевых плазм из-за полиморфизмов и/или наличия различных аллелей гена. Дополнительно, ген оксидазы_1 GA3 может быть экспрессирован в виде альтернативно сплайсированных изоформ, которые могут приводить к различным мРНК, кДНК и кодирующим последовательностям, которые могут влиять на конструкцию супрессирующей конструкции и некодирующей молекулы РНК. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_1 GA3, может быть определена более широко как содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 36.
[123] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться, для супрессии, на ген оксидазы_2 GA3, и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 31 или SEQ ID NO: 32. Согласно некоторым вариантам реализации изобретения некодирующая молекула РНК, нацеленная на ген оксидазы GA3 для супрессии, может быть комплементарной к, по меньшей мере, 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из SEQ ID NO: 31 или SEQ ID NO: 32. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацеливается на ген оксидазы GA3 для супрессии и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 33. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 33.
[124] Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA3 вместо или в дополнение к экзонному, 5'-НТО или 3'-НТО гена оксидазы GA3. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_2 GA3, может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 37 и/или нуклеотидам 533-692 или 852-982 из SEQ ID NO: 37. Представленные в данном документе последовательности для оксидазы_2 GA3 могут варьироваться в зависимости от разнообразия растений кукурузы, линий и зародышевых плазм из-за полиморфизмов и/или наличия различных аллелей гена. Дополнительно, ген оксидазы_2 GA3 может быть экспрессирован в виде альтернативно сплайсированных изоформ, которые могут приводить к различным мРНК, кДНК и кодирующим последовательностям, которые могут влиять на конструкцию супрессирующей конструкции и некодирующей молекулы РНК. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_2 GA3, может быть определена более широко как содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 37.
[125] Согласно конкретным вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК для нацеливания на ген оксидазы GA3, содержит (i) последовательность, которая является, по меньшей мере, на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной SEQ ID NO: 57 или 59, и/или (ii) последовательность или элемент супрессии, кодирующие некодирующую молекулу РНК, содержащую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичной SEQ ID NO: 58 или 60. Согласно некоторым вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность с одним или более ошибочными спариваниями, например, 1, 2, 3, 4, 5 или более комплементарных ошибочных спариваний, относительно последовательности целевого сайта или сайта узнавания целевой мРНК гена оксидазы GA3, такой как последовательность, которая почти комплементарна SEQ ID NO: 57 или 59, но с одним или более комплементарными ошибочными спариваниями по отношению к SEQ ID NO: 57 или 59. Согласно конкретному варианту реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, содержит последовательность, которая на 100% идентична SEQ ID NO: 58 или 60, которая на 100% комплементарна целевой последовательности в кДНК и кодирующих последовательностях гена оксидазы_1 GA3 или оксидазы_2 GA3 в кукурузе (то есть, SEQ ID NO: 28, 29, 31 и/или 32) и/или соответствующей последовательности мРНК, кодируемой эндогенным геном оксидазы_1 GA3 или оксидазы_2 GA3.
[126] В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК может нацеливаться, для супрессии, на ген оксидазы_4 GA20, и содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 10 или SEQ ID NO: 11. Согласно некоторым вариантам реализации изобретения некодирующая молекула РНК, нацеленная на ген оксидазы_4 GA20 для супрессии, может быть комплементарной к, по меньшей мере, 19 последовательным нуклеотидам, но не более чем 27 последовательным нуклеотидам, например, комплементарной 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из SEQ ID NO: 10 или SEQ ID NO: 11. В соответствии с некоторыми вариантами реализации изобретения, некодирующая молекула РНК нацеливается на ген оксидазы GA20 для супрессии и содержит последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 12. В соответствии с дополнительными вариантами реализации изобретения, некодирующая молекула РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным SEQ ID NO: 12.
[127] Как упомянуто выше, некодирующая молекула РНК может нацеливаться на интронную последовательность гена оксидазы GA20 вместо или в дополнение к экзонному, 5'-НТО или 3'-НТО гена оксидазы GA20. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_4 GA20, может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 38 и/или нуклеотидам 1996-2083 или 2412-2516 из SEQ ID NO: 38. Представленные в данном документе последовательности для оксидазы_4 GA20 могут варьироваться в зависимости от разнообразия растений кукурузы, линий и зародышевых плазм из-за полиморфизмов и/или наличия различных аллелей гена. Дополнительно, ген оксидазы_4 GA20 может быть экспрессирован в виде альтернативно сплайсированных изоформ, которые могут приводить к различным мРНК, кДНК и кодирующим последовательностям, которые могут влиять на конструкцию супрессирующей конструкции и некодирующей молекулы РНК. Таким образом, некодирующая молекула РНК нацеленная, для супрессии, на ген оксидазы_4 GA20, может быть определена более широко как содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 38.
[128] Согласно конкретным вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК для нацеливания на ген оксидазы_4 GA20, содержит (i) последовательность, которая является, по меньшей мере, на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной SEQ ID NO: 61, и/или (ii) последовательность или элемент супрессии, кодирующие некодирующую молекулу РНК, содержащую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичной SEQ ID NO: 62. Согласно некоторым вариантам реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность с одним или более ошибочными спариваниями, например, 1, 2, 3, 4, 5 или более комплементарных ошибочных спариваний, относительно последовательности целевого сайта или сайта узнавания целевой мРНК гена оксидазы GA20, такой как последовательность, которая почти комплементарна SEQ ID NO: 61, но с одним или более комплементарными ошибочными спариваниями по отношению к SEQ ID NO: 61. Согласно конкретному варианту реализации изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, содержит последовательность, которая на 100% идентична SEQ ID NO: 62, которая на 100% комплементарна целевой последовательности в кДНК и кодирующих последовательностях оксидазы_4 GA20 в кукурузе (то есть SEQ ID NO: 10 или 11), и/или соответствующей последовательности мРНК, кодируемой эндогенным геном оксидазы_4 GA20.
[129] В соответствии с вариантами реализации данного изобретения предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_3 GA20 и/или ген(ы) оксидазы_5 GA20 для супрессии, при этом последовательность транскрибируемой ДНК является функционально связанной с конститутивным, тканеспецифичным или тканепредпочтительным промотором, и при этом транскрибируемая последовательность ДНК вызывает снижение или уменьшение уровня экспрессии эндогенного гена(ов) оксидазы_3 GA20 и/или оксидазы_5 GA20 в одной или более ткани(нях) растения, трансформированного транскрибируемой последовательностью ДНК. Такая некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 9, и/или (ii) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 15.
[130] В соответствии с вариантами реализации данного изобретения предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_1 GA3 и/или ген(ы) оксидазы_2 GA3 для супрессии, при этом последовательность транскрибируемой ДНК является функционально связанной с конститутивным, тканеспецифичным или тканепредпочтительным промотором, и при этом транскрибируемая последовательность ДНК вызывает снижение или уменьшение уровня экспрессии эндогенного гена(ов) оксидазы_1 GA3 и/или оксидазы_2 GA3 в одной или более ткани(нях) растения, трансформированного транскрибируемой последовательностью ДНК. Такая некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 30, и/или (ii) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 33.
[131] В соответствии с вариантами реализации данного изобретения предложена конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы_4 GA20 для супрессии, при этом последовательность транскрибируемой ДНК является функционально связанной с конститутивным, тканеспецифичным или тканепредпочтительным промотором, и при этом транскрибируемая последовательность ДНК вызывает снижение или уменьшение уровня экспрессии эндогенного гена оксидазы_4 GA20 в одной или более ткани(нях) растения, трансформированного транскрибируемой последовательностью ДНК. Такая некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, может содержать последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 12.
[132] В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20 для супрессии, и/или имеет эндогенный ген оксидазы_3 GA20 и/или оксидазы_5 GA20, отредактированный с помощью целевых методов редактирования генома, как предложено в данном документе, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень экспрессии эндогенного гена(ов) оксидазы_3 GA20 и/или оксидазы_5 GA20 устраняется, уменьшается или понижается в одной или более растительной ткани(нях), такой как одна или более сосудистая и/или листовая ткань(и) модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением. В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20 для супрессии, и/или имеет эндогенный ген оксидазы_3 GA20 и/или оксидазы_5 GA20, отредактированный с помощью целевых методов редактирования генома для снижения или устранения уровня его экспрессии и/или активности, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень одной или более активных GA, таких как GA1, GA3, GA4 и/или GA7, уменьшается или понижается в одной или более растительной ткани(нях), таких как одна или более стеблевая, междоузловая, сосудистая и/или листовая ткань(и) или одна или более стеблевая и/или междоузловая ткань(и), модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением.
[133] В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_1 GA3 и/или оксидазы_2 GA3 для супрессии, и/или имеет эндогенный ген оксидазы_1 GA3 или оксидазы_2 GA3, отредактированный с помощью целевых методов редактирования генома, как предложено в данном документе, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень экспрессии эндогенного гена(ов) оксидазы_1 GA3 и/или оксидазы_2 GA3 устраняется, уменьшается или понижается в одной или более растительной ткани(нях), такой как одна или более сосудистая и/или листовая ткань(и) модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением. В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_1 GA3 и/или оксидазы_2 GA3 для супрессии, и/или имеет эндогенный ген оксидазы_1 GA3 и/или оксидазы_2 GA3, отредактированный с помощью целевых методов редактирования генома для снижения или устранения уровня его экспрессии и/или активности, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень одной или более активных GA, таких как GA1, GA3, GA4 и/или GA7, уменьшается или понижается в одной или более растительной ткани(нях), таких как одна или более стеблевая, междоузловая, сосудистая и/или листовая ткань(и) или одна или более стеблевая и/или междоузловая ткань(и), модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением.
[134] В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы_4 GA20 для супрессии, и/или имеет эндогенный ген оксидазы_4 GA20, отредактированный с помощью целевых методов редактирования генома, как предложено в данном документе, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень экспрессии эндогенного гена(ов) оксидазы_4 GA20 устраняется, уменьшается или понижается в одной или более растительной ткани(нях), такой как одна или более сосудистая и/или листовая ткань(и) модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением. В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформируется рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_4 GA20 для супрессии, и/или имеет эндогенный ген оксидазы_4 GA20, отредактированный с помощью целевых методов редактирования генома для снижения или устранения уровня его экспрессии и/или активности, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, таким как сосудистый промотор или листовой промотор, и при этом уровень одной или более активных GA, таких как GA1, GA3, GA4 и/или GA7, уменьшается или понижается в одной или более растительной ткани(нях), таких как одна или более стеблевая, междоузловая, сосудистая и/или листовая ткань(и) или одна или более стеблевая и/или междоузловая ткань(и), модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением.
[135] В соответствии со многими вариантами реализации изобретения предлагается модифицированное или трансгенное растение, которое трансформировано рекомбинантной конструкцией ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 для супрессии, трансформировано конструкцией рекомбинантной ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, нацеленную на эндогенный ген(ы) оксидазы_1 GA3, оксидазы_2 GA3 для супрессии, и/или имеет эндогенный ген оксидазы_3 GA20, оксидазы_4 GA20 или оксидазы_5 GA20, отредактированный с помощью целевых методов редактирования генома, для снижения или устранения уровня его экспрессии и/или активности, как представлено в данном документе документе, при этом транскрибируемая последовательность ДНК функционально связана с конститутивным промотором или тканеспецифичным или тканепредпочтительным промотором, такой как сосудистый промотор или листовой промотор, и при этом модифицированное или трансгенное растение имеет один или более следующих признаков: полукарликовость или уменьшенную высоту, или рост растения, уменьшенную длину междоузлия стебля, повышенную устойчивость к полеганию и/или увеличенный диаметр стебля. Такое модифицированное или трансгенное растение может не иметь каких-либо значительных репродуктивных отклонений. Модифицированное или трансгенное растение может иметь один или более из следующих дополнительных признаков: уменьшенный излом стебля, более глубокие корни, увеличенную площадь листьев, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, повышение эффективности использования воды, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение количества зерна, увеличение массы зерна, увеличение урожайности и/или увеличенный уборочный индекс. Согласно многим из этих вариантов реализации изобретения уровень экспрессии и/или активности эндогенного гена(ов) оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 или эндогенного гена(ов) оксидазы_1 GA3 и/или оксидазы_2 GA3, может быть устранен, уменьшен или понижен в одной или более растительных ткани(нях), такой как одна или более сосудистая и/или листовая ткань(и), модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением, и/или уровень одной или более активных GA, таких как GA1, GA3, GA4 и/или GA7, уменьшается или понижается в одной или более растительной ткани(нях), таких как одна или более стеблевая, междоузловая, сосудистая и/или листовая ткань(и), или одна или более стеблевая и/или междоузловая ткань(и), модифицированного или трансгенного растения, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 40%, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 90% или 100% по сравнению с диким типом или контрольным растением.
[136] В соответствии со многими вариантами реализации изобретения, описанными в приведенных выше абзацах, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК молекулы, вектора или конструкции рекомбинантной ДНК, может представлять собой предшественника микроРНК или миРНК, который впоследствии может быть процессирован или расщеплен в растительной клетке с образованием зрелой микроРНК или миРНК.
[137] Молекула, конструкция или вектор рекомбинантной ДНК по данному изобретению может содержать транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, таким как конститутивный или сосудистый и/или листовой промотор. Для целей данного изобретения, некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, может включать зрелую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, и/или молекулу-предшественник РНК, которая может процессироваться в растительной клетке в зрелую некодирующую молекулу РНК, такую как микроРНК или миРНК, которая нацелена на эндогенный ген оксидазы GA для супрессии. В дополнение к связанному с ней промотору последовательность транскрибируемой ДНК, кодирующая некодирующую молекулу РНК для супрессии эндогенного гена оксидазы GA, также может быть функционально связана с одним или более дополнительными регуляторными элементами, такими как энхансер(ы), лидер, сайт инициации транскрипции (TSS - transcription start site), линкер, 5' и 3' нетранслируемая область(и) (НТО), интрон(ы), сигнал полиаденилирования, область терминации или стоп-кодон и тому подобное, которые являются подходящими, необходимыми или предпочтительными для усиления, регуляции или обеспечения возможности экспрессии транскрибируемой последовательности ДНК в клетке растения. Такой дополнительный регуляторный элемент(ы) может быть необязательным и/или применяться для усиления или оптимизации экспрессии последовательности трансгена или транскрибируемой ДНК. Как указано в данном документе, "энхансер" отличается от "промотора" тем, что энхансер обычно не имеет сайта инициации транскрипции, TATA-бокса или эквивалентной последовательности и, следовательно, его недостаточно для запуска транскрипции. В данном документе термин "лидер" может быть определен, как правило, как последовательность ДНК 5'-НТО гена (или трансгена) между сайтом инициации транскрипции (TSS) и 5'-концом транскрибируемой последовательности ДНК, или сайтом инициации кодирующей белок последовательности трансгена.
[138] В соответствии с дополнительными вариантами реализации изобретения предложены способы трансформации растительной клетки, ткани или эксплантата молекулой, или конструкцией рекомбинантной ДНК, содержащей транскрибируемую последовательность ДНК или трансген, функционально связанный с экспрессируемым в растении промотором, для получения трансгенного растения. Последовательность транскрибируемой ДНК может кодировать некодирующую молекулу РНК, которая нацелена на ген(ы) оксидазы GA для супрессии, или предшественника РНК, который процессируется в зрелую молекулу РНК, такую как микроРНК или миРНК, которая нацелена на один или более ген(ов) оксидазы GA для супрессии. В данной области техники известны многочисленные способы трансформации хромосом или пластид в растительной клетке с помощью молекулы или конструкции рекомбинантной ДНК, которые можно применять в соответствии с вариантами реализации способа по данному изобретению для получения трансгенной растительной клетки и растения. Любой подходящий способ или метод трансформации растительной клетки, известный в данной области техники, может быть применен в соответствии с данными способами. Эффективные способы трансформации растений включают бактериально-опосредованную трансформацию, такую как трансформация, опосредованная Agrobacterium или Rhizobium, и трансформация, опосредованная бомбардировкой частицами или бомбардировкой микрочастицами. В данной области техники известно множество способов трансформации эксплантов с помощью вектора для трансформации посредством бактериально-опосредованной трансформации или бомбардировки микрочастицами, или частицами и последующего культивирования, и тому подобного, данных эксплантов для регенерации или развития трансгенных растений. В данной области техники также известны другие способы трансформации растений, такие как микроинъекция, электропорация, вакуумная инфильтрация, давление, соникация, перемешивание карбиднокремниевого волокна, ПЭГ-опосредованная трансформация и тому подобное.
[139] Способы трансформации растительных клеток и эксплантов хорошо известны специалистам в данной области техники. Способы трансформации растительных клеток путем бомбардировки микрочастицами с частицами, покрытыми рекомбинантной ДНК, представлены, например, в патентах США № 5,550,318; 5,538,880 6,160,208; 6,399,861; и 6,153,812, а опосредованная Agrobacterium трансформация описана, например, в патентах США № 5,159,135; 5,824,877; 5,591,616; 6,384,301; 5,750,871; 5,463,174; и 5,188,958, все из которых включены в данный документ посредством ссылки. Дополнительные способы трансформации растений можно найти, например, в Compendium of Transgenic Crop Plants (2009 год) Blackwell Publishing. Любой подходящий способ трансформации растения, известный или разработанный в данной области техники, может быть применен для трансформации растительной клетки или эксплантата с помощью любой из молекул нуклеиновой кислоты, конструкций или векторов, представленных в данном документе.
[140] Трансгенные растения, полученные способами трансформации, могут быть химерными или нехимерными для события трансформации в зависимости от применяемых способов и эксплантов. Дополнительно предложены способы экспрессии некодирующей молекулы РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии в одной или более растительных клетках или тканях под контролем экспрессируемого в растении промотора, такого как конститутивный, тканеспецифический, тканепредпочтительный, сосудистый и/или листовой промотор, как представлено в данном документе. Такие способы могут быть применены для создания трансгенных растений злаков или кукурузы, имеющих более низкий рост, полукарликовость, уменьшенную длину междоузлия, увеличенный диаметр стебля и/или улучшенную устойчивость к полеганию. Такие трансгенные злаковые или кукурузные растения могут также иметь другие признаки, которые могут быть полезными для урожайности, например, уменьшенный излом стебля, более глубокие корни, увеличенную площадь листьев, более раннюю сомкнутость полога, повышение устойчивости к засухе, повышение эффективности использования азота, повышение эффективности использования воды, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, снижение содержания антоцианов и/или площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение количества зерна, увеличение массы зерна, увеличение урожайности и/или увеличенный уборочный индекс по сравнению с диким типом или контрольным растением. В данном документе термин "уборочный индекс" относится к массе собранного зерна, деленной на общую массу надземной биомассы растения на уборочной площади.
[141] Трансгенные растения, экспрессирующие трансгенную или некодирующую молекулу РНК оксидазы GA, которая нацелена на эндогенный ген оксидазы GA для супрессии, могут иметь более раннюю сомкнутость полога (например, приблизительно на один день более раннюю или на 12-48 часов, на 12-36 часов, на 18-36 часов или на около 24 часа более раннюю сомкнутость полога), чем растение дикого типа или контрольное растение. Хотя трансгенные растения, экспрессирующие трансгенную или некодирующую молекулу РНК оксидазы GA, которая нацелена на эндогенный ген оксидазы GA для супрессии, могут иметь более низкую высоту початка, чем растение дикого типа или контрольное растение, высота початка обычно может быть не менее 18 сантиметров над землей. Трансгенные растения, экспрессирующие некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, могут иметь большую биомассу и/или площадь листьев на одной или более поздних вегетативных стадиях (например, V8-V12), чем растение дикого типа или контрольное растение. Трансгенные растения, экспрессирующие трансгенную или некодирующую молекулу РНК оксидазы GA, которая нацелена на эндогенный ген оксидазы GA для супрессии, могут иметь более глубокие корни на более поздних вегетативных стадиях при выращивании в поле, чем растение дикого типа или контрольное растение, что может быть связано с повышенной скоростью роста корня. Эти трансгенные растения могут достигать глубины 90 см под землей раньше (например, на 10-25 дней раньше, на 15-25 дней раньше или на около 20 дней раньше), чем растение дикого типа или контрольное растение, что может происходить в результате вегетативного и репродуктивного перехода растения (например, на V16/R1 через около 50 дней после посадки, в отличие от через около 70 дней после посадки для контрольных растений).
[142] Клетка(и) реципиента или цели эксплантата или клеточные цели для трансформации включают, но без ограничений, клетку семени, клетку плода, клетку листа, семядольную клетку, клетку гипокотиля, клетку меристемы, клетку эмбриона, клетку эндосперма, корневую клетку, клетку проростка, стволовую клетку, клетку стручка, клетку цветка, клетку соцветия, клетку стебля, клетку цветоножки, клетку пестика, клетку стигмы, клетку рецептакула, клетку лепестка, клетку чашелистника, пыльцевую клетку, клетку пыльника, клетку тычиночной нити, клетку завязи, клетку семезачатка, клетку перикарпа, клетку флоэмы, клетку почки, клетку каллуса, хлоропласт, устьичную клетку, клетку трихомы, клетку корневого волоска, клетку корнеплода или клетку сосудистой ткани, семя, эмбрион, меристему, семядоль, гипокотиль, эндосперм, корень, побег, стебель, узел, каллус, клеточную суспензию, протопласт, цветок, лист, пыльцу, пыльник, завязь, семезачаток, перикарп, почку и/или сосудистую ткань или любую трансформируемую часть любого из вышеперечисленного. Для трансформации растений любые целевые клетки, ткань(и), эксплантат(ы) и тому подобное, которые могут быть применены для получения вектора с целью трансформации рекомбинантной ДНК или молекулы по данному изобретению, могут в совокупности называться "эксплантат" для трансформации. Предпочтительно трансформируемая или трансформированная клетка, или ткань эксплантата может быть дополнительно развита или регенерирована в растение. Любая клетка или эксплантат, из которых можно выращивать или регенерировать плодородное растение, рассматривается в качестве полезной клетки-реципиента или эксплантата для практического применения данного описания (то есть, в качестве целевого эксплантата для трансформации). Каллус может быть инициирован или создан из различных тканевых источников, включая, но без ограничения, эмбрионы или части эмбрионов, неэмбриональные ткани семян, апикальные меристемы проростков, микроспоры и тому подобное. Любые клетки, которые способны пролиферировать как каллус, могут служить клетками-реципиентами для трансформации. Способы трансформации и материалы для получения трансгенных растений (например, различные среды и реципиентные целевые клетки или эксплантаты и способы трансформации и последующая регенерация в трансгенные растения) известны в данной области техники.
[143] Трансформация целевого растительного материала или эксплантата может быть осуществлена в культуре ткани на питательной среде, например, смеси питательных веществ, которые позволяют клеткам расти in vitro или клеточной культуре. Как известно в данной области техники, трансформированные эксплантаты, клетки или ткани могут быть подвергнуты дополнительным этапам культивирования, таким как индукция каллуса, отбор, регенерация и тому подобное. Трансформация также может осуществляться без создания или применения ткани каллуса. Трансформированные клетки, ткани или эксплантаты, содержащие вставку или событие рекомбинантной последовательности ДНК, можно выращивать, развивать или регенерировать в трансгенные растения в культуре, пробках или почве в соответствии со способами, известными в данной области техники. Трансгенные растения могут быть затем скрещены с собой или другими растениями для получения трансгенных семян и потомства. Трансгенное растение также может быть получено путем скрещивания первого растения, содержащего последовательность рекомбинантной ДНК или событие трансформации, со вторым растением, в котором отсутствует вставка. Например, конструкция или последовательность рекомбинантной ДНК может быть введена в первую линию растений, которая поддается трансформации, которая затем может быть скрещена со второй линией растений с целью интрогрессии конструкции или последовательности рекомбинантной ДНК во вторую линию растений. Потомство этих скрещиваний может быть затем обратно скрещено в более желательную линию множество раз, например, через 6-8 поколений или обратных скрещиваний, чтобы получить потомства растения с по существу тем же генотипом, что и у исходной родительской линии, но для введения конструкции или последовательности рекомбинантной ДНК.
[144] Представленное в данном документе трансгенное или отредактированное растение, часть растения, клетка или эксплантат могут относиться к элитному сорту или элитной линии. Элитный сорт или элитная линия относится к сорту, полученному в результате селекции и отбора для обеспечения превосходных агрономических характеристик. Представленное в данном документе трансгенное или отредактированное растение, клетка или эксплантат может представлять собой гибридное растение, клетку или эксплантат. Как подразумевается в данном документе "гибрид" создается путем скрещивания двух растений из разных сортов, линий, инбредов или видов, так что потомство содержит генетический материал от каждого родителя. Специалисты в данной области признают, что могут быть получены гибриды более высокого порядка. Например, первый гибрид может быть получен путем скрещивания сорта A с сортом B, чтобы создать гибрид A x B, и второй гибрид может быть получен путем скрещивания сорта C с сортом D, чтобы создать гибрид C x D. Первый и второй гибриды могут быть далее скрещены для создания гибрида высшего порядка (A x B) x (C x D), содержащего генетическую информацию от всех четырех родительских сортов.
[145] В соответствии с вариантами реализации данного изобретения предлагается модифицированное растение, содержащее элемент супрессии оксидазы GA, который нацелен на два или более генов оксидазы GA для супрессии, или комбинацию двух или более элемента(ов) супрессии оксидазы GA и/или редактирования гена(ов). Конструкция или вектор рекомбинантной ДНК может содержать одну кассету или элемент супрессии, содержащий транскрибируемую последовательность ДНК, сконструированную или выбранную для кодирования некодирующей молекулы РНК, которая является комплементарной распознаванию мРНК или целевой последовательности двух или более генов оксидазы GA, включая по меньшей мере первый ген оксидазы GA и второй ген оксидазы GA, то есть мРНК целевых генов оксидазы GA имеют идентичную или почти идентичную (или подобную) последовательность, так что один элемент супрессии и кодированная некодирующая молекула РНК могут нацеливаться на каждый из целевых генов оксидазы GA для супрессии. Например, экспрессионная кассета и конструкция супрессии представлены в данном документе, включают транскрибируемую последовательность ДНК, которая кодирует одну некодирующую молекулу РНК, которая нацелена на гены оксидазы_3 GA20 и оксидазы_5 GA20 для супрессии.
[146] Согласно другим вариантам реализации изобретения конструкция рекомбинантной ДНК или вектора могут содержать два или более элемента супрессии или последовательности, которые могут быть объединены вместе в конструкции или векторе либо в тандеме в одной экспрессионной кассете, либо по отдельности в двух или более экспрессионных кассетах. Конструкция или вектор рекомбинантной ДНК может содержать одну экспрессионную кассету или элемент супрессии, содержащие последовательность транскрибируемой ДНК, которая кодирует некодирующую молекулу РНК, содержащую две или более целевые последовательности, расположенные в тандеме, включая, по меньшей мере, первую целевую последовательность и вторую целевую последовательность, при этом первая целевая последовательность является комплементарной распознаванию мРНК или целевому сайт первого гена оксидазы GA, а вторая целевая последовательность комплементарна распознаванию мРНК или целевого сайту второго гена оксидазы GA, и при этом транскрибируемая последовательность ДНК является функционально связанной с экспрессируемым в растении промотором. Промотор, экспрессируемый в растении может быть конститутивным промотором или тканеспецифичным или тканепредпочтительным промотором, как представлено в данном документе. Некодирующая молекула РНК может быть экспрессирована в виде пре-микроРНК, которая процессируется в две или более зрелые микроРНК, включая, по меньшей мере, первую зрелую микроРНК и вторую микроРНК, при этом первая микроРНК содержит целевую последовательность, которая комплементарна распознаванию мРНК или целевому сайту первого гена оксидазы GA, а вторая микроРНК содержит целевую последовательность, которая комплементарна распознаванию мРНК или целевому сайту второго гена оксидазы GA.
[147] В соответствии с другими вариантами реализации изобретения конструкция рекомбинантной ДНК или вектор могут содержать две или более экспрессионные кассеты, включая первую экспрессионную кассету и вторую экспрессионную кассету, при этом первая экспрессионная кассета содержит первую транскрибируемую последовательность ДНК, функционально связанную с первым промотором, экспрессируемым в растении, и вторая экспрессионная кассета содержит вторую транскрибируемую последовательность ДНК, функционально связанную со вторым промотором, экспрессируемым в растении, при этом первая транскрибируемая последовательность ДНК кодирует первую некодирующую молекулу РНК, содержащую целевую последовательность, которая комплементарна распознаванию мРНК или целевому сайту первого гена оксидазы GA, и вторая транскрибируемая последовательность ДНК кодирует вторую некодирующую молекулу РНК, содержащую целевую последовательность, которая комплементарна распознаванию мРНК или целевому сайту второго гена оксидазы GA. Каждый из первого и второго промоторов, экспрессируемых в растении, может быть конститутивным промотором или тканеспецифичным, или тканепредпочтительным промотором, как предусмотрено в данном документе, и первый и второй промоторы, экспрессируемые в растении могут быть одинаковыми или разными промоторами.
[148] В соответствии с другими вариантами реализации изобретения два или более элемента супрессии или конструкции, нацеленную на ген(ы) оксидазы GA и/или редактирование(я) гена оксидазы GA, могут быть объединены в модифицированном растении путем скрещивания двух или более растений вместе в одном или более поколениях для получения модифицированного растения, имеющего желаемую комбинацию элемента(ов) супрессии и/или редактирование(я) генов. В соответствии с этими вариантами реализации изобретения первое модифицированное растение, содержащее элемент или конструкцию супрессии, нацеленную на ген(ы) оксидазы GA (или редактирование гена оксидазы GA), может быть скрещено со вторым модифицированным растением, содержащим элемент или конструкцию супрессии, нацеленную на ген(ы) оксидазы GA (или редактирование гена оксидазы GA), так что может быть получено модифицированное растение потомства, содержащее первый элемент или конструкцию супрессии и второй элемент или конструкцию супрессии, элемент или конструкцию супрессии и редактирование гена оксидазы GA, или редактирование первого гена оксидазы GA и редактирование второго гена оксидазы GA. В альтернативном варианте, модифицированное растение, содержащее два или более элементов или конструкций супрессии, нацеленных на ген(ы) оксидазы GA и/или редактирование гена(ов) оксидазы GA, может быть осуществлено посредством (i) котрансформации первого элемента или конструкции супрессии и второго элемента или конструкции супрессии (каждый из которых нацелен на ген оксидазы GA для супрессии), (ii) трансформации модифицированного растения вторым элементом или конструкцией супрессии, при этом модифицированное растение уже содержит первый элемент или конструкцию супрессии, (iii) трансформации модифицированного растения с помощью элемента или конструкции супрессии, при этом модифицированное растение уже содержит отредактированный ген оксидазы GA, (iv) трансформации модифицированного растения конструкцией(ями) для внесения одного или более изменений в ген(ы) оксидазы GA, при этом модифицированное растение уже содержит элемент или конструкцию супрессии или (v) трансформации с помощью конструкции(ий) для внесения двух или более изменений в ген(ы) оксидазы GA.
[149] В соответствии с вариантами реализации данного изобретения предложены модифицированные растения, содержащие две или более конструкций, нацеленных на ген(ы) оксидазы GA для супрессии, включая первую конструкцию рекомбинантной ДНК и вторую конструкцию рекомбинантной ДНК, при этом первая конструкция рекомбинантной ДНК содержит первую транскрибируемую последовательность ДНК, кодирующую первую некодирующую молекулу РНК, которая является комплементарной распознаванию мРНК или целевой последовательности первого гена оксидазы GA, и вторая конструкция рекомбинантной ДНК содержит вторую транскрибируемую последовательность ДНК, кодирующую вторую молекулу некодирующей РНК, которая является комплементарной распознаванию мРНК или целевой последовательность второго гена оксидазы GA. Первая и вторая конструкции рекомбинантной ДНК могут быть помещены в один вектор и трансформированы в растение как одно событие трансформации или представлены в отдельных векторах или конструкциях, которые могут быть трансформированы в виде отдельных событий трансформации. В соответствии с этими вариантами реализации изобретения первый ген оксидазы GA может представлять собой ген оксидазы_3 GA20, оксидазы_5 GA20, оксидазы_4 GA20, оксидазы_1 GA3 или оксидазы_2 GA3, первая некодирующая молекула РНК является комплементарной последовательности распознавания или целевой последовательности мРНК, экспрессируемой из такого гена оксидазы GA, а второй ген оксидазы GA может представлять собой ген оксидазы_3 GA20, оксидазы_5 GA20, оксидазы_4 GA20, оксидазы_1 GA3 или оксидазы_2 GA3. В соответствии с некоторыми вариантами реализации изобретения, первый и второй гены оксидазы GA могут быть одинаковыми или разными генами оксидазы GA. В альтернативном варианте, второй ген оксидазы GA может представлять собой другой ген оксидазы GA, такой как ген оксидазы_1 GA20, оксидазы_2 GA20, оксидазы_6 GA20, оксидазы_7 GA20, оксидазы_8 GA20 или оксидазы_9 GA20, а вторая некодирующая молекула РНК является комплементарной к последовательности распознавания или целевой последовательности мРНК, экспрессируемой из такого гена оксидазы GA.
[150] В соответствии с вариантами реализации данного изобретения предложены модифицированные растения, содержащие конструкцию рекомбинантной ДНК, нацеленную на гены оксидазы GA для супрессии, включающую транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая содержит две или более нацеливающих последовательностей, расположенных в тандеме, включая, по меньшей мере, первую нацеливающую последовательность, которая комплементарна последовательности распознавания мРНК или целевой последовательности первого гена оксидазы GA, и вторую нацеливающую последовательность, которая комплементарна последовательности распознавания мРНК или целевой последовательности второго гена оксидазы GA. Некодирующая молекула РНК может быть экспрессирована в виде пре-микроРНК, которая процессирует в две или более зрелые микроРНК, включая, по меньшей мере, первую зрелую микроРНК и вторую микроРНК, при этом первая микроРНК содержит первую нацеливающую последовательность, которая комплементарна распознаванию мРНК или целевому сайту первого гена оксидазы GA, а вторая микроРНК содержит вторую нацеливающую последовательность, которая комплементарна распознаванию мРНК или целевому сайту второго гена оксидазы GA. В соответствии с этими вариантами реализации изобретения первый ген оксидазы GA может представлять собой ген оксидазы_3 GA20, оксидазы_5 GA20, оксидазы_4 GA20, оксидазы_1 GA3 или оксидазы_2 GA3, первая некодирующая молекула РНК является комплементарной последовательности распознавания или целевой последовательности мРНК, экспрессируемой из такого гена оксидазы GA, а второй ген оксидазы GA может представлять собой ген оксидазы_3 GA20, оксидазы_5 GA20, оксидазы_4 GA20, оксидазы_1 GA3 или оксидазы_2 GA3. В соответствии с некоторыми вариантами реализации изобретения, первый и второй гены оксидазы GA могут быть одинаковыми или разными генами оксидазы GA. В альтернативном варианте, второй ген оксидазы GA может представлять собой другой ген оксидазы GA, такой как ген оксидазы_1 GA20, оксидазы_2 GA20, оксидазы_6 GA20, оксидазы_7 GA20, оксидазы_8 GA20 или оксидазы_9 GA20, а вторая некодирующая молекула РНК является комплементарной к последовательности распознавания или целевой последовательности мРНК, экспрессируемой из такого гена оксидазы GA.
[151] В описанных выше сценариях расположения и независимо от того, объединены ли целевые последовательности в тандем в одной транскрибируемой последовательности ДНК (или в экспрессионной кассете) или в отдельных транскрибируемых последовательностях ДНК (или экспрессионных кассетах), второй ген оксидазы GA может быть геном оксидазы GA, отличным от гена оксидазы_3 GA20, оксидазы_5 GA20, оксидазы_4 GA20, оксидазы_1 GA3 или гена оксидазы_2 GA3, такого как ген оксидазы_1 GA20, оксидазы_2 GA20, оксидазы_6 GA20, оксидазы_7 GA20, оксидазы_8 GA20 или оксидазы_9 GA20. В соответствии с этими вариантами реализации изобретения вторая нацеливающая последовательность некодирующей молекулы РНК может быть, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из любого одного или более из SEQ ID NO: 1, 2, 4, 5, 16, 17, 19, 20, 22, 23, 25, и/или 26. Согласно некоторым вариантам реализации изобретения вторая нацеливающая последовательность некодирующей молекулы РНК может быть комплементарной, по меньшей мере 19 последовательным нуклеотидам, но не более 27 последовательным нуклеотидам, например, комплементарна 19, 20, 21, 22, 23, 24, 25, 26 или 27 последовательным нуклеотидам из любого одного или более из SEQ ID NO: 1, 2, 4, 5, 16, 17, 19, 20, 22, 23, 25 и/или 26. В соответствии с некоторыми вариантами реализации изобретения, вторая нацеливающая последовательность некодирующей молекулы РНК является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA в растении, который является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным любому одной или более из SEQ ID NO: 3, 6, 18, 21, 24 и/или 27. В соответствии с дополнительными вариантами реализации изобретения, вторая нацеливающая последовательность некодирующей молекулы РНК может содержать последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% подобным одной или более из SEQ ID NO: 3, 6, 18, 21, 24 и/или 27.
[152] Молекула или конструкция рекомбинантной ДНК по данному изобретению может содержать или быть включена в вектор для трансформации ДНК для применения в трансформации целевой клетки растения, ткани или эксплантата. Такой вектор для трансформации может обычно содержать последовательности или элементы, необходимые или полезные для эффективной трансформации, в дополнение к по меньшей мере одной последовательности трансгена, экспрессионной кассеты и/или транскрибируемой ДНК, кодирующей ген оксидазы GA или некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы GA для супрессии. Для Agrobacterium-опосредованной, Rhizobia-опосредованной или другой бактериально-опосредованной трансформации вектор для трансформации может содержать участок или область сконструированной транспортной ДНК (или Т-ДНК), имеющие две граничные последовательности, левую границу (LB- left border) и правую границу (RB - right border), фланкирующих по меньшей мере транскрибируемую последовательность ДНК или трансген, так что вставка Т-ДНК в геном растения создает событие трансформации для транскрибируемой последовательности ДНК, трансгена или экспрессионной кассеты. Таким образом, транскрибируемая последовательность ДНК, трансген или экспрессионная кассета, кодирующая некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы GA для супрессии, может быть расположена между левой и правой границами Т-ДНК, возможно, вместе с дополнительным трансгеном(ами) или экспрессионной кассетой(ами), такими как трансген селектируемого растением маркера и/или другой ген(ы) агрономического интереса, который может придавать растению признак или фенотип агрономического интереса. Согласно альтернативным вариантам реализации изобретения транскрибируемая последовательность ДНК, трансген или экспрессионная кассета, кодирующая некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы GA для супрессии, и трансген селектируемого растением маркера (или другой ген агрономического интереса) могут присутствовать в отдельных сегментах Т-ДНК на одной и той же или другой молекуле(ах) рекомбинантной ДНК, например, для котрансформации. Вектор или конструкция для трансформации может дополнительно содержать прокариотические поддерживающие элементы, которые могут быть расположены в векторе вне области(ей) Т-ДНК.
[153] Трансген селектируемого растением маркера в векторе или конструкции для трансформации по данному изобретению может быть применен для помощи в селекции трансформированных клеток или ткани вследствие присутствия селекционного агента, такого как антибиотик или гербицид, при этом трансген селектируемого растением маркера обеспечивает толерантность или устойчивость к селективному агенту. Таким образом, селектируемый агент может оказывать влияние или способствовать выживанию, развитию, росту, пролиферации и тому подобного, трансформированных клеток, экспрессирующих трансген селектируемого растением маркера, например, для увеличения доли трансформированных клеток или тканей в растении R0. Обычно применяемые гены селектируемого растением маркера включают в себя, например, гены, обеспечивающие толерантность или устойчивость к антибиотикам, таким как канамицин и паромомицин (nptII), гигромицин B (aph IV), стрептомицин или спектиномицин (aadA) и гентамицин (aac3 и aacC4) или те, которые обеспечивают толерантность или устойчивость к гербицидам, таким как глюфосинат (bar или pat), дикамба (DMO) и глифосат (aroA или EPSPS). Можно также применять гены скрининга растений, которые обеспечивают возможность визуального скрининга трансформантов, такие как люцифераза или зеленый флуоресцентный белок (ЗФБ), или ген, экспрессирующий бета-глюкуронидазу или ген (GUS) uidA, для которого известны различные хромогенные субстраты. В некоторых вариантах реализации изобретения вектор или полинуклеотид, представленный в данном документе, содержит по меньшей мере один селектируемый маркерный ген, выбранный из группы, состоящей из nptII, aph IV, aadA, aac3, aacC4, bar, pat, DMO, EPSPS, aroA, ЗФБ и GUS. Трансформация растений может также проводиться без селекции во время одного или более этапов, или стадий культивирования, развития или регенерации трансформированных эксплантатов, тканей, растений и/или частей растения.
[154] Согласно данным вариантам реализации изобретения способы трансформации растительной клетки, ткани или эксплантата с помощью молекулы или конструкции рекомбинантной ДНК могут дополнительно включать сайт-направленную или целевую интеграцию. В соответствии с этими способами часть молекулы донорной матрицы рекомбинантной ДНК (то есть, последовательности вставки) может быть вставлена или интегрирована в желаемом сайте или локусе в геноме растения. Последовательность вставки донорной матрицы может содержать трансген или конструкцию, такую как последовательность трансгена или транскрибируемой ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии. Донорская матрица также может иметь один или два плеча гомологии, фланкирующих последовательность вставки, чтобы способствовать целевому событию вставки посредством гомологичной рекомбинации и/или репарации, направляемой гомологией. Каждое гомологичное плечо может являться, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК в геноме однодольного или злакового растения. Таким образом, молекула рекомбинантной ДНК по данному изобретению может содержать донорную матрицу для сайт-направленной или нацеленной интеграции трансгена или конструкции, такой как последовательность трансгена или транскрибируемой ДНК, кодирующая некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии в геноме растения.
[155] Любой сайт или локус в геноме растения могут потенциально быть выбраны для сайт-направленной интеграции последовательности трансгена, конструкции или транскрибируемой ДНК, представленной в данном документе. Для сайт-направленной интеграции сначала можно сделать двухцепочечный разрыв (ДЦР) или одноцепочечный разрыв в выбранном геномном локусе с помощью сайт-специфической нуклеазы, такой как, например, нуклеаза с цинковыми пальцами, сконструированная или нативная мегануклеаза, TALE-эндонуклеаза или РНК-направленная эндонуклеаза (например, Cas9 или Cpf1). Для сайт-направленной интеграции может быть применен любой способ, известный в данной области техники. В присутствии молекулы донорной матрицы с последовательностью вставки, ДЦР или одноцепочечный разрыв может затем быть восстановлен с помощью гомологичной рекомбинации между гомологичным плечом(ами) донорной матрицы и геномом растения или с помощью негомологичного соединения концов (НГСК), что приводит к сайт-направленной интеграции последовательности вставки в геном растения для создания события целевой вставки в сайт ДЦР или одноцепочечного разрыва. Таким образом, может быть достигнута сайт-специфическая вставка или интеграция трансгена, конструкции или последовательности.
[156] Введение ДЦР или одноцепочечного разрыва также может быть применено для введения целевых мутаций в геном растения. В соответствии с этим подходом мутации, такие как делеции, вставки, инверсии и/или замены, могут быть введены в целевой сайт посредством несовершенного восстановления ДЦР или одноцепочечного разрыва с целью вызвать нокаут или нокдаун гена оксидазы GA. Такие мутации могут быть получены путем несовершенного восстановления целевого локуса даже без применения молекулы донорной матрицы. "Нокаут" гена оксидазы GA может быть достигнут путем индукции ДЦР или одноцепочечного разрыва в эндогенном локус гена оксидазы GA или около него, что приводит к отсутствию экспрессии белка оксидазы GA или экспрессии нефункционального белка, тогда как "нокдаун" гена оксидазы GA может быть достигнут аналогичным образом путем индукции ДЦР или одноцепочечного разрыва в эндогенном локусе гена оксидазы GA или около него, который несовершенно восстанавливается в сайте, который не влияет на кодирующую последовательность гена оксидазы GA таким образом, чтобы исключить функцию кодируемого белка оксидазы GA. Например, сайт ДЦР или одноцепочечного разрыва внутри эндогенного локуса может находиться в 3'-5' направлении или 5'-области гена оксидазы GA (например, последовательности промотора и/или энхансера), с целью влияния или уменьшения его уровеня экспрессии. Подобным образом, такие целевые нокаут или нокдаун мутации гена оксидазы GA могут создаваться с помощью молекулы донорной матрицы для направления конкретной или желаемой мутации в целевой сайт или около него, посредством репарации ДЦР или одноцепочечного разрыва. Молекула донорной матрицы может содержать гомологичную последовательность с последовательностью вставки или без нее и содержать одну или более мутаций, таких как одна или более делеций, вставок, инверсий и/или замен, относительно целевой геномной последовательности в сайте ДЦР или одноцепочечного разрыва или около него. Например, целевые нокаут мутации гена оксидазы GA могут быть достигнуты путем удаления или инвертирования по меньшей мере части гена или путем введения сдвига рамки считывания или преждевременного стоп-кода в кодирующую последовательность гена. Удаление части гена оксидазы GA также может быть осуществлено путем создания ДЦР или одноцепочечного разрыва в двух целевых сайтах, что приводит к делеции промежуточной целевой области, фланкированной целевыми сайтами.
[157] Сайт-специфическая нуклеаза, представленная в данном документе, может быть выбрана из группы, состоящей из нуклеазы c цинковыми пальцами (ZFN), мегануклеазы, РНК-направленной эндонуклеазы, TALE-эндонуклеазы (TALEN), рекомбиназы, транспозазы или любой их комбинации. См., например, Khandagale K., и соавт., "Genome editing for targeted improvement in plants,", Plant Biotechnol Rep 10: 327-343 (2016 год); и Gaj T., и соавт., "ZFN, TALEN and CRISPR/Cas-based methods for genome engineering", Trends Biotechnol. 31(7): 397-405(2013), содержание и описание которых включены в данный документ посредством ссылки. Рекомбиназа может представлять собой сериновую рекомбиназу, присоединенную к мотиву распознавания ДНК, тирозиновую рекомбиназу, присоединенную к мотиву распознавания ДНК или другой фермент рекомбиназы, известный в данной области техники. Рекомбиназа или транспозаза может представлять собой ДНК-транспозазу или рекомбиназу, присоединенную к ДНК-связывающему домену. Тирозиновая рекомбиназа, присоединенная к мотиву распознавания ДНК, может быть выбрана из группы, состоящей из рекомбиназы Cre, рекомбиназы Flp и рекомбиназы Tnp1. Согласно некоторым вариантам реализации изобретения рекомбиназа Cre или рекомбиназа Gin, представленные в данном документе, связаны с ДНК-связывающим доменом цинкового пальца. В другом варианте реализации изобретения сериновая рекомбиназа, присоединенная к мотиву распознавания ДНК, представленному в данном документе, выбрана из группы, состоящей из интегразы PhiC31, интегразы R4 и интегразы TP-901. В другом варианте реализации изобретения транспозаза ДНК, присоединенная к ДНК-связывающему домену, предоставленному в данном документе, выбрана из группы, состоящей из TALE-piggyBac и TALE-Mutator.
[158] В соответствии с вариантами реализации данного описания РНК-направленная эндонуклеаза может быть выбрана из группы, состоящей из Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9 (также известных как Csn1 и Csx12), Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, Cpf1, CasX, CasY, их гомологов или модифицированных версий, Argonaute (неограничивающие примеры белков Argonaute, включают Thermus thermophilus Argonaute (TtAgo), Pyrococcus furiosus Argonaute (PfAgo), Natronobacterium gregoryi Argonaute (NgAgo) и их гомологи или модифицированные версии. Согласно некоторым вариантам реализации изобретения РНК-направленная эндонуклеаза может представлять собой фермент Cas9 или Cpf1.
[159] В одном аспекте сайт-специфическая нуклеаза, предоставленная в данном документе, выбрана из группы, состоящей из нуклеазы с цинковыми пальцами, мегануклеазы, РНК-направленной нуклеазы, нуклеазы TALE, рекомбиназы, транспозазы или любой их комбинации. В другом аспекте сайт-специфическая нуклеаза, представленная в данном документе, выбрана из группы, состоящей из Cas9 или Cpf1. В другом аспекте сайт-специфическая нуклеаза, предоставленная в данном документе, выбрана из группы, состоящей из Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, Cpf1, CasX, CasY их гомолога или их модифицированной версии. В другом аспекте предоставленная в данном документе РНК-направленная нуклеаза выбрана из группы, состоящей из Cas9 или Cpf1. В другом аспекте предоставленная в данном документе РНК-направленная нуклеаза выбрана из группы, состоящей из Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, Cpf1, CasX, CasY их гомолога или их модифицированной версии. В другом аспекте способ и/или композиция, представленная в данном документе, содержит, по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять сайт-специфических нуклеаз. В еще одном аспекте способ и/или композиция, представленная в данном документе, содержит, по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять полинуклеотидов, кодирующих по меньшей мере одну, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять или по меньшей мере десять сайт-специфических нуклеазы.
[160] Для РНК-направленных эндонуклеаз дополнительно предоставлена молекула направляющей РНК (нРНК), которая направляет эндонуклеазу в целевой сайт в геноме растения посредством спаривания оснований или гибридизации, чтобы вызвать ДЦР или одноцепочечный разрыв в целевом сайте или около него. нРНК может трансформироваться или вводиться в растительную клетку или ткань (возможно, вместе с нуклеазой или кодирующей нуклеазу молекулой, конструкцией или вектором ДНК) в виде молекулы нРНК или в виде молекулы, конструкции или вектора рекомбинантной ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую направляющую РНК функционально связанную с экспрессируемым в растении промотором. Как понятно в данной области техники, "направляющая РНК" может включать в себя, например, CRISPR РНК (crРНК), одноцепочечную направляющую РНК (sgРНК) или любую другую молекулу РНК, которая может направлять или устремлять эндонуклеазу к конкретному целевому сайту в геноме. "Одноцепочечная направляющая РНК" (или "sgРНК") представляет собой молекулу РНК, содержащую crРНК, ковалентно связанную с tracrРНК линкерной последовательностью, которая может быть экспрессирована в виде одного транскрипта или молекулы РНК. Направляющая РНК содержит направляющую или нацеливающую последовательность, которая идентична или комплементарна целевому сайту в геноме растения, например, в гене оксидазы GA или около него. Мотив, прилегающий к протоспейсеру (PAM - protospacer-adjacent motif) может присутствовать в геноме, непосредственно примыкающим и расположенным выше 5'-конца последовательности геномного целевого сайта, комплементарно нацеливающей последовательности, направляющей РНК, то есть, непосредственно в 5'-3' направлении (3') к смысловой (+) цепи геномного целевого сайта (относительно целевой последовательности направляющей РНК), как известно в данной области техники. См., например, Wu, X. и соавт., ʺTarget specificity of the CRISPR-Cas9 system, ʺQuant Biol. 2(2): 59-70 (2014 год), содержание и описание которых включено в данный документ посредством ссылки. Геномная последовательность PAM на смысловой (+) цепи, прилегающей к целевому сайту (относительно нацеливающей последовательности направляющей РНК), может содержать 5'-NGG-3'. Тем не менее, соответствующая последовательность направляющей РНК (то есть, непосредственно в 5'-3' направлении (3') к нацеливающей последовательности направляющей РНК), как правило, может быть не комплементарной геномной последовательности PAM. Направляющая РНК обычно может представлять собой некодирующую молекулу РНК, которая не кодирует белок. Направляющая последовательность направляющей РНК может иметь длину по меньшей мере 10 нуклеотидов, например, 12-40 нуклеотидов, 12-30 нуклеотидов, 12-20 нуклеотидов, 12-35 нуклеотидов, 12-30 нуклеотидов, 15-30 нуклеотидов, 17- 30 нуклеотидов или 17-25 нуклеотидов, или около 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более нуклеотидов в длину. Направляющая последовательность может быть, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14 по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам последовательность ДНК в геномном целевом сайте.
[161] Для редактирования генома в гене оксидазы_3 GA20 или около него с помощью РНК-направленной эндонуклеазы можно применять направляющую РНК, содержащую направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам из SEQ ID NO: 34 или последовательности, комплементарной ей (например, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам из SEQ ID NO: 34 или последовательности, комплементарной ей). Для редактирования генома в гене оксидазы_5 GA20 или около него с помощью РНК-направленной эндонуклеазы можно применять направляющую РНК, содержащую направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам из SEQ ID NO: 35 или последовательности, комплементарной ей (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам из SEQ ID NO: 35 или последовательности, комплементарной ей). В данном документе термин "последовательный" в отношении полинуклеотидной или белковой последовательности означает отсутствие делеций или пробелов в последовательности.
[162] Для мутаций нокдауна (и, возможно, нокаута) посредством редактирования генома, РНК-направленная эндонуклеаза может быть нацелена на последовательность, расположенную в 3'-5' направлении или в 5'-3' направлении, такую как последовательность промотора и/или энхансера, или интрон, последовательность 5'НТО и/или 3'НТО гена оксидазы_3 GA20 или оксидазы_5 GA20, для мутации одной или более промоторных и/или регуляторных последовательностей гена и влияет на уровень его экспрессии или снижает его. Для нокдауна (и, возможно, нокаута) гена оксидазы_3 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-3096 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 3666-3775 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 4098-5314 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 5585-5800 из SEQ ID NO: 34 или диапазоне нуклеотидных последовательностей 5801-8800 из SEQ ID NO: 34, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-3096, 3666-3775, 4098-5314, 5585-5800, 5801-8800, или 5585-8800 из SEQ ID NO: 34, или последовательности, комплементарной им).
[163] Для нокдауна (и, возможно, нокаута) гена оксидазы_5 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-3000 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 1-3000 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 3792-3906 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 4476-5197 из SEQ ID NO: 35 или диапазоне нуклеотидных последовательностей 5860-8859 из SEQ ID NO: 35, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-3000, 3792-3906, 4476-5197 или 5860-8859 из SEQ ID NO: 35, или последовательности, комплементарной им).
[164] Для мутаций нокдауна (и, возможно, нокаута) посредством редактирования генома, РНК-направленная эндонуклеаза может быть нацелена на кодирующую и/или интронную последовательность гена оксидазы_3 GA20 или оксидазы_5 GA20, чтобы потенциально устранить экспрессию и/или активность функционального белка оксидазы GA из гена. Однако в некоторых случаях может быть достигнут нокаут экспрессии гена оксидазы GA путем нацеливания на последовательность(и) гена в 3'-5' направлении и/или 5'НТО или на другие последовательности в геномном локусе гена или около него. Таким образом, нокаут экспрессии гена оксидазы GA может быть достигнут путем нацеливания на геномную последовательность в сайте или локусе, или около него, целевого гена оксидазы_3 GA20 или оксидазы_5 GA20, в 3'-5' направлении или в 5'-3' направлении последовательности, такой как последовательность промотора и/или энхансера или последовательность интрона, 5'НТО и/или 3'НТО гена оксидазы_3 GA20 или оксидазы_5 GA20, как описано выше для нокдауна гена оксидазы_3 GA20 или оксидазы_5 GA20.
[165] Для нокаута (и, возможно, нокдауна) гена оксидазы_3 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 3097-5584 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 3097-3665 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 3776-4097 из SEQ ID NO: 34, диапазоне нуклеотидных последовательностей 5315-5584 из SEQ ID NO: 34, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 3097-5584, 3097-3665, 3097-3775, 3665-4097, 3776-4097, 3776-5314, 4098-5584 или 5315-5584 из SEQ ID NO: 34, или последовательности, комплементарной им).
[166] Для нокаута (и, возможно, нокдауна) гена оксидазы_5 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 3001-5473 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 3001-3791 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 3907-4475 из SEQ ID NO: 35, диапазоне нуклеотидных последовательностей 5198-5473 из SEQ ID NO: 35, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 3001-5473, 3001-3791, 3001-3906, 3792-4475, 3907-4475, 3907-5197, 4476-5473 или 5198-5473 из SEQ ID NO: 35, или последовательности, комплементарной им).
[167] В соответствии с некоторыми вариантами реализации изобретения предложена направляющая РНК для нацеливания на эндогенный ген оксидазы_3 GA20 и/или оксидазы_5 GA20, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21 или более последовательным нуклеотидам из любого одного или более из SEQ ID NO: 138-167.
[168] Для редактирования генома в гене оксидазы_4 GA20 или около него с помощью РНК-направленной эндонуклеазы можно применять направляющую РНК, содержащую направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам из SEQ ID NO: 38 или последовательности, комплементарной ей (например, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам из SEQ ID NO: 38 или последовательности, комплементарной ей).
[169] Для мутаций нокдауна (и, возможно, нокаута) посредством редактирования генома, РНК-направленная эндонуклеаза может быть нацелена на кодирующую и/или интронную последовательность гена оксидазы_4 GA20, чтобы потенциально устранить экспрессию и/или активность функционального белка оксидазы_4 GA20 из гена. Для гена оксидазы_4 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1544-2852 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 1544-1995 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 2084-2411 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 2517-2852 из SEQ ID NO: 38, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1544-2852, 1544-1995, 1544-2083, 1996-2411, 2084-2411, 2084-2516, 2412-2852 или 2517-2852 из SEQ ID NO: 38, или последовательности, комплементарной им).
[170] Для мутаций нокдауна (и, возможно, нокаута) посредством редактирования генома, РНК-направленная эндонуклеаза может быть нацелена на последовательность, расположенную в 3'-5' направлении или в 5'-3' направлении, такую как последовательность промотора и/или энхансера, или интрон, последовательность 5'НТО и/или 3'НТО гена оксидазы_4 GA20, для мутации одной или более промоторных и/или регуляторных последовательностей гена и влияет на уровень его экспрессии или снижает его. Для нокдауна гена оксидазы_3 GA20 в кукурузе может быть применена направляющая РНК, содержащая направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-1416 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 1417-1543 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 1996-2083 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 2412-2516 из SEQ ID NO: 38, диапазоне нуклеотидных последовательностей 2853-3066 из SEQ ID NO: 38 или диапазоне нуклеотидных последовательностей 3067-4465 из SEQ ID NO: 38, или последовательности, комплементарной им (например, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более последовательным нуклеотидам в диапазоне нуклеотидных последовательностей 1-1416, 1417-1543, 1-1543, 1996-2083, 2412-2516, 2853-3066, 3067-4465 или 2853-4465 из SEQ ID NO: 38, или последовательности, комплементарной им).
[171] В дополнение к направляющей последовательности направляющая РНК может дополнительно содержать одну или более другую структурную или каркасную последовательность(и), которые могут связываться или взаимодействовать с РНК-направленной эндонуклеазой. Такие каркасные или структурные последовательности могут дополнительно взаимодействовать с другими молекулами РНК (например, tracrРНК). В данной области техники известны способы и методы конструирования нацеливающих конструкций и направляющих РНК для редактирования генома и сайт-направленной интеграции в целевом сайте в геноме растения с применением РНК-направленной эндонуклеазы.
[172] В соответствии с некоторыми вариантами реализации изобретения предложены конструкции и векторы рекомбинантной ДНК, содержащие полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу, такую как нуклеаза с цинковыми пальцами (ZFN), мегануклеаза, РНК-направленная эндонуклеаза, TALE-эндонуклеаза (TALEN) рекомбиназа или транспозаза, при этом кодирующая последовательность функционально связана с промотором, экспрессируемым в растении. Для РНК-направленных эндонуклеаз, дополнительно предложены конструкции и векторы рекомбинантной ДНК, содержащие полинуклеотидную последовательность, кодирующую направляющую РНК, при этом направляющая РНК содержит направляющую последовательность достаточной длины, имеющую процентную идентичность или комплементарность к целевому сайту в геноме растения, например, в целевом гене оксидазы GA или около него. В соответствии с некоторыми вариантами реализации изобретения полинуклеотидная последовательность конструкции и вектора рекомбинантной ДНК, которая кодирует сайт-специфическую нуклеазу или направляющую РНК, может быть функционально связана с экспрессируемым в растении промотором, таким как индуцибельный промотор, конститутивный промотор, тканеспецифический промотор и тому подобное.
[173] Согласно некоторым вариантам реализации изобретения, конструкция или вектор рекомбинантной ДНК могут содержать первую полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу, и вторую полинуклеотидную последовательность, кодирующую направляющую РНК, которые могут быть введены в клетку растения вместе с помощью методов трансформации растения. В альтернативном варианте могут быть предоставлены две конструкции или векторы рекомбинантной ДНК, включая первую конструкцию или вектор рекомбинантной ДНК и вторую конструкцию или вектор рекомбинантной ДНК, которые могут быть введены в клетку растения вместе или последовательно с помощью методов трансформации растений, при этом первая конструкция или вектор рекомбинантной ДНК содержит полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу, а вторая конструкция или вектор рекомбинантной ДНК содержит полинуклеотидную последовательность, кодирующую направляющую РНК. Согласно некоторым вариантам реализации изобретения конструкция или вектор рекомбинантной ДНК, содержащая полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу, может быть введена с помощью методов трансформации растений в клетку растения, которая уже содержит (или трансформирована ею) конструкцию или вектор рекомбинантной ДНК, содержащую полинуклеотидную последовательность, кодирующую направляющую РНК. В альтернативном варианте, конструкция или вектор рекомбинантной ДНК, содержащая полинуклеотидную последовательность, кодирующую направляющую РНК, может быть введена с помощью методов трансформации растения в клетку растения, которая уже содержит (или трансформирована ею) конструкцию или вектор рекомбинантной ДНК, содержащую полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу. В соответствии с еще одним дополнительным вариантом реализации изобретения первое растение, содержащее (или трансформированное) конструкцию или вектор рекомбинантной ДНК, содержащую полинуклеотидную последовательность, кодирующую сайт-специфическую нуклеазу, может быть скрещено со вторым растением, содержащим (или трансформированным) конструкцию или вектор рекомбинантной ДНК, содержащую полинуклеотидную последовательность, кодирующую направляющую РНК. Такие конструкции или векторы рекомбинантной ДНК могут быть временно трансформированы в клетку растения или стабильно трансформированы, или интегрированы в геном клетки растения.
[174] В одном аспекте векторы, содержащие полинуклеотиды, кодирующие сайт-специфическую нуклеазу, и, необязательно, одну или более, две или более, три или более, или четыре или более нРНК предоставляются растительной клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией). В одном аспекте векторы, содержащие полинуклеотиды, кодирующие нуклеазу Cas9, и, необязательно, одну или более, две или более, три или более, или четыре или более нРНК предоставляются растительной клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией). В другом аспекте векторы, содержащие полинуклеотиды, кодирующие Cpf1, и, необязательно, одну или более, две или более, три или более, или четыре или более crРНК предоставляются клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, вирусной трасфекцией, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией).
[175] Некоторые сайт-специфические нуклеазы, такие как рекомбиназы, нуклеазы с цинковыми пальцами (ZFN), мегануклеазы и TALEN, не являются РНК-направленными и вместо этого полагаются на свою белковую структуру для определения своего целевого сайта для того, чтобы вызвать ДЦР или одноцепочечный разрыв, или они слиты, связаны или присоединены к ДНК-связывающему белковому домену или мотиву. Структура белка сайт-специфической нуклеазы (или слитого/присоединенного/связанного ДНК-связывающего домена) может нацеливать сайт-специфическую нуклеазу на целевой сайт. Согласно многим из этих вариантов реализации изобретения, не-РНК-направленные сайт-специфические нуклеазы, такие как рекомбиназы, нуклеазы с цинковыми пальцами (ZFN), мегануклеазы и TALEN, могут быть спроектированы, сконструированы и изготовлены в соответствии с известными способами для нацеливания и связывания с целевым сайтом в геномном локусе эндогенного гена оксидазы GA кукурузы или злакового растения или около него, такого как ген оксидазы_3 GA20 или ген оксидазы_5 GA20 в кукурузе, с целью создания ДЦР или одноцепочечного разрыва в таком геномном локусе для нокаута или нокдауна экспрессии гена оксидазы GA путем восстановления ДЦР или одноцепочечного разрыва. Например, сконструированная сайт-специфическая нуклеаза, такая как рекомбиназа, нуклеаза с цинковыми пальцами (ZFN), мегануклеаза или TALEN, может быть сконструирована для нацеливания и связывания с (i) целевым сайтом в геноме растения, соответствующим последовательности из SEQ ID NO: 34 или его комплементарной последовательности, с целью создания ДЦР или одноцепочечного разрыва в геномном локусе гена оксидазы_3 GA20, (ii) целевым сайтом в геноме растения, соответствующим последовательности из SEQ ID NO: 35 или его комплементарной последовательности, с целью создания ДЦР или одноцепочечного разрыва в геномном локусе гена оксидазы_5 GA20, или (iii) целевым сайтом в геноме растения, соответствующим последовательности из SEQ ID NO: 38 или его комплементарной последовательности, с целью создания ДЦР или одноцепочечного разрыва в геномном локусе гена оксидазы_4 GA20, что может затем привести к созданию мутации или вставке последовательности в сайт ДЦР или одноцепочечного разрыва через механизмы клеточной репарации, которые могут направляться донорной молекулой или матрицей.
[176] В одном аспекте описанная в данном документе методика целевого редактирования генома может включать применение рекомбиназы. В некоторых вариантах реализации изобретения тирозиновая рекомбиназа, присоединенная, и тому подобное, к мотиву или домену распознавания ДНК, может быть выбрана из группы, состоящей из рекомбиназы Cre, рекомбиназы Flp и рекомбиназы Tnp1. В одном аспекте Cre-рекомбиназа или Gin-рекомбиназа, представленные в данном документе, могут быть связаны с ДНК-связывающим доменом с цинковыми пальцами. Система сайт-направленной рекомбинации Flp-FRT может происходить из 2-микронной плазмиды пекарских дрожжей Saccharomyces cerevisiae. В этой системе рекомбиназа Flp (флиппаза) может рекомбинировать последовательности между целевыми сайтами распознавания флиппазой (FRT- flippase recognition target). FRT-сайты содержат 34 нуклеотида. Flp может связываться с "плечами" сайтов FRT (одно плечо находится в обратной ориентации) и расщепляет сайт FRT с любого конца промежуточной последовательности нуклеиновой кислоты. После расщепления Flp может рекомбинировать последовательности нуклеиновых кислот между двумя сайтами FRT. Cre-lox представляет собой систему сайт-направленной рекомбинации, полученную из бактериофага P1, которая аналогична системе рекомбинации Flp-FRT. Cre-lox можно применять для инвертирования последовательности нуклеиновой кислоты, удаления последовательности нуклеиновой кислоты или транслокации последовательности нуклеиновой кислоты. В этой системе рекомбиназа Cre может рекомбинировать пару последовательностей нуклеиновой кислоты lox. Lox-сайты содержат 34 нуклеотида, причем первый и последний 13 нуклеотиды (плечи) являются палиндромными. Во время рекомбинации белок Cre рекомбиназы связывается с двумя сайтами lox на разных нуклеиновых кислотах и расщепляется в сайтах lox. Расщепленные нуклеиновые кислоты сплайсируются вместе (реципрокно транслоцируются) и рекомбинация завершается. В другом аспекте предоставленный в данном документе сайт lox представляет собой сайт loxP, lox 2272, loxN, lox 511, lox 5171, lox71, lox66, M2, M3, M7 или M11.
[177] ZFN представляют собой синтетические белки, состоящие из сконструированного ДНК-связывающего домена цинкового пальца, слитого с доменом расщепления (или полудоменом расщепления), который может быть получен из эндонуклеазы рестрикции (например, FokI). ДНК-связывающий домен может быть каноническим (C2H2) или неканоническим (например, C3H или C4). ДНК-связывающий домен может содержать один или более цинковых пальцев (например, 2, 3, 4, 5, 6, 7, 8, 9 или более цинковых пальцев) в зависимости от целевого сайта. Несколько цинковых пальцев в ДНК-связывающем домене могут быть разделены линкерной последовательностью(ями). ZFN могут быть сконструированы для расщепления практически любого участка двухцепочечной ДНК путем модификации ДНК-связывающего домена цинкового пальца. ZFN образуют димеры из мономеров, состоящих из неспецифического домена расщепления ДНК (например, полученного из нуклеазы FokI), слитого с ДНК-связывающим доменом, содержащим набор цинковых пальцев, сконструированных для связывания последовательности ДНК целевого сайта. ДНК-связывающий домен ZFN обычно может состоять из 3-4 (или более) цинковых пальцев. Аминокислоты в положениях -1, +2, +3 и +6 относительно начала α-спирали цинкового пальца, которые способствуют сайт-специфическому связыванию с целевым сайтом, могут быть изменены и настроены для соответствия конкретной целевой последовательности. Другие аминокислоты могут образовывать основный консенсусный каркас для генерации ZFN с различной специфичностью последовательности. Способы и правила конструирования ZFN для нацеливания и связывания с конкретными целевыми последовательностями известны в данной области техники. См., например, заявки на патент США № 2005/0064474, 2009/0117617 и 2012/0142062, содержание и описание которых включены в данный документ посредством ссылки. Для нуклеазного домена FokI может потребоваться димеризация для расщепления ДНК, и поэтому необходимы два ZFN с их С-концевыми областями для связывания противоположных цепей ДНК сайта расщепления (разделенных 5-7 п.о.). Мономер ZFN может разрезать целевой сайт, если два ZF-связывающих сайта являются палиндромными. Применяемый в данном документе ZFN является широким и включает мономерный ZFN, который может расщеплять двухцепочечную ДНК без помощи другого ZFN. Термин ZFN может также применяться для обозначения одного или обоих членов пары ZFN, которые сконструированы для совместной работы по расщеплению ДНК в одном и том же сайте.
[178] Не ограничиваясь какой-либо научной теорией, поскольку специфичность ДНК-связывающих доменов цинкового пальца может быть изменена с применением одного из различных способов, теоретически могут быть созданы индивидуальные ZFN для нацеливания практически на любую целевую последовательность (например, на ген оксидазы GA или около него в геноме растения). Общедоступные способы конструирования доменов с цинковыми пальцами включают в себя Context-dependent Assembly (CoDA), Oligomerized Pool Engineering (OPEN) или Modular Assembly. В одном аспекте способ и/или композиция, предоставленные в данном документе, включают одну или более, две или более, три или более, четыре или более, или пять или более ZFN. В другом аспекте ZFN, предоставленный в данном документе, способен генерировать целевой ДЦР или одноцепочечный разрыв. В одном аспекте векторы, содержащие полинуклеотиды, кодирующие один или более, два или более, три или более, или четыре или более, или пять или более ZFN предоставляются клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, вирусной трасфекцией, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией). ZFN могут быть введены в виде белков ZFN, в виде полинуклеотидов, кодирующих белки ZFN, и/или в виде комбинаций белков и полинуклеотидов, кодирующих белок.
[179] Мегануклеазы, которые обычно идентифицируются у микробов, такие как семейство хоуминг-эндонуклеаз LAGLIDADG, являются уникальными ферментами с высокой активностью и длинными последовательностями распознавания (>14 п.о.), что приводит к сайт-специфическому расщеплению целевой ДНК. Сконструированные версии встречающихся в природе мегануклеаз обычно имеют удлиненные последовательности распознавания ДНК (например, от 14 до 40 п.о.). Согласно некоторым вариантам реализации изобретения, мегануклеаза может содержать каркасный или основной фермент, выбранный из группы, состоящей из I-CreI, I-CeuI, I-MsoI, I-SceI, I-AniI и I-DmoI. Конструирование мегануклеаз может быть более сложным, чем ZFN и TALEN, потому что функции распознавания и расщепления ДНК мегануклеазами тесно связаны в одном домене. Специализированные способы мутагенеза и скрининга высокой производительности были применены для создания новых вариантов мегануклеазы, которые распознают уникальные последовательности и обладают улучшенной нуклеазной активностью. Таким образом, мегануклеаза может быть выбрана или сконструирована для связывания с геномной целевой последовательностью в растении, например, в геномном локусе гена оксидазы GA или около него. В одном аспекте способ и/или композиция, предоставленные в данном документе, включают одну или более, две или более, три или более, четыре или более, или пять или более мегануклеаз. В другом аспекте мегануклеаза, предоставленная в данном документе, способна генерировать целевой ДЦР. В одном аспекте векторы, содержащие полинуклеотиды, кодирующие одну или более, две или более, три или более, или четыре или более, или пять или более мегануклеаз предоставляются клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, вирусной трасфекцией, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией).
[180] TALEN представляют собой искусственные ферменты рестрикции, полученные путем слияния ДНК-связывающего домена эффектора, подобного активаторам транскрипции (TALE) с нуклеазным доменом (например, FokI). Когда каждый член пары TALEN связывается с сайтами ДНК, фланкирующими целевой сайт, мономеры FokI димеризуются и вызывают разрыв двухцепочечной ДНК в целевом сайте. Помимо домена расщепления FokI дикого типа, варианты домена расщепления FokI с мутациями были разработаны для улучшения специфичности расщепления и активности расщепления. Домен FokI функционирует как димер, требуя двух конструкций с уникальными ДНК-связывающими доменами для сайтов в целевом геноме с правильной ориентацией и расстоянием. Как количество аминокислотных остатков между ДНК-связывающим доменом TALEN и доменом расщепления FokI, так и количество оснований между двумя отдельными сайтами связывания TALEN являются параметрами для достижения высоких уровней активности.
[181] TALEN представляют собой искусственные ферменты рестрикции, полученные путем слияния ДНК-связывающего домена эффектора, подобного активаторам транскрипции (TALE), с нуклеазным доменом. В некоторых аспектах нуклеаза выбрана из группы, состоящей из PvuII, MutH, TevI, FokI, AlwI, MlyI, SbfI, SdaI, StsI, CleDORF, Clo051 и Pept071. Когда каждый член пары TALEN связывается с сайтами ДНК, фланкирующими целевой сайт, мономеры FokI димеризуются и вызывают разрыв двухцепочечной ДНК в целевом сайте. Термин TALEN, применяемый в данном документе, является широким по значению и включает мономерный TALEN, который может расщеплять двухцепочечную ДНК без помощи другого TALEN. Термин TALEN также относится к одному или обоим членам пары TALEN, которые работают вместе для расщепления ДНК в одном и том же сайте.
[182] Эффекторы, подобные активаторам транскрипции (TALE - transcription activator-like effector), могут быть сконструированы так, чтобы связывать практически любую последовательность ДНК, например, в геномном локусе гена оксидазы GA в растении или около него. TALE имеет центральный ДНК-связывающий домен, состоящий из 13-28 повторных мономеров 33-34 аминокислот. Аминокислоты каждого мономера высококонсервативны, за исключением гипервариабельных аминокислотных остатков в положениях 12 и 13. Две вариабельные аминокислоты называются повторяющимися вариабельными диостатками (RVD - repeat-variable diresidue). Аминокислотные пары NI, NG, HD и NN RVD предпочтительно распознают аденин, тимин, цитозин и гуанин/аденин соответственно, а модуляция RVD может распознавать последовательные основания ДНК. Эта простая связь между аминокислотной последовательностью и распознаванием ДНК позволила создать специфические ДНК-связывающие домены путем выбора комбинации повторяющихся сегментов, содержащих соответствующие RVD.
[183] Помимо домена расщепления FokI дикого типа, варианты домена расщепления FokI с мутациями были разработаны для улучшения специфичности расщепления и активности расщепления. Домен FokI функционирует как димер, требуя двух конструкций с уникальными ДНК-связывающими доменами для сайтов в целевом геноме с правильной ориентацией и расстоянием. Как количество аминокислотных остатков между ДНК-связывающим доменом TALEN и доменом расщепления FokI так и количество оснований между двумя отдельными сайтами связывания TALEN являются параметрами для достижения высоких уровней активности. Домены расщепления PvuII, MutH и TevI cleavage являются полезными альтернативами вариантам FokI и FokI для применения с TALE. PvuII функционирует как высокоспецифичный домен расщепления, когда соединен с TALE (см., Yank и соавт., 2013 год. PLoS One. 8: e82539). MutH способен вводить специфические для цепи одноцепочечные разрывы в ДНК (см., Gabsalilow и соавт., 2013 год. Nucleic Acids Research. 41: e83). TevI вводит двухцепочечные разрывы в ДНК в целевых сайтах (см., Beurdeley и соавт., 2013 год. Nature Communications. 4: 1762).
[184] Взаимосвязь между аминокислотной последовательностью и распознаванием ДНК связывающего домена TALE позволяет конструировать белки. Программные обеспечения, такие как DNA Works, могут применяться для разработки конструкций TALE. Другие способы конструирования конструкций TALE известны специалистам в данной области техники. См., Doyle и соавт., Nucleic Acids Research (2012 год) 40: W117-122.; Cermak и соавт., Nucleic Acids Research (2011 год). 39:e82; и tale-nt.cac.cornell.edu/about. В одном аспекте способ и/или композиция, предоставленные в данном документе, включают один или более, два или более, три или более, четыре или более, или пять или более TALEN. В другом аспекте TALEN, предоставленный в данном документе, способен генерировать целевой ДЦР. В одном аспекте векторы, содержащие полинуклеотиды, кодирующие один или более, два или более, три или более, или четыре или более, или пять или более TALEN предоставляются клетке с помощью способов трансформации, известных в данной области техники (например, без ограничений, вирусной трасфекцией, бомбардировкой частицами, ПЭГ-опосредованной трансфекцией протопласта или Agrobacterium-опосредованной трансформацией). См., например, заявки на патент США № 2011/0145940, 2011/0301073 и 2013/0117869, содержание и описание которых включены в данный документ посредством ссылки.
[185] В данном документе термин "целевой метод редактирования генома" относится к любому способу, протоколу или методу, которые позволяют осуществлять точное и/или целевое редактирование определенного местоположения в геноме растения (то есть, редактирование в значительной степени или полностью не случайным образом) с применением сайт-специфической нуклеазы, такой как мегануклеаза, нуклеаза с цинковыми пальцами (ZFN), РНК-направленная эндонуклеаза (например, система CRISPR/Cas9), эндонуклеаза TALE (TALEN), рекомбиназа или транспозаза. В данном документе термин "редактирование" или "редактирование генома" относится к созданию целевой мутации, делеции, инверсии или замены по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 75, по меньшей мере 100, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500, по меньшей мере 5000, по меньшей мере 10000 или по меньшей мере 25000 нуклеотидов последовательности нуклеиновой кислоты эндогенного генома растения. В данном документе термин "редактирование" или "редактирование генома" также охватывает целевую вставку или сайт-направленную интеграцию по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 75, по меньшей мере 100, по меньшей мере 250, по меньшей мере 500, по меньшей мере 750, по меньшей мере 1000, по меньшей мере 1500, по меньшей мере 2000, по меньшей мере 2500, по меньшей мере 3000, по меньшей мере 4000, по меньшей мере 5000, по меньшей мере 10000 или по меньшей мере 25 000 нуклеотидов в эндогенном геноме растения. "Редактирование" или "геномное редактирование" в единственном числе относится к одной такой целевой мутации, делеции, инверсии, замене или вставке, тогда как "редактирования" или "геномные редактирования" относится к двум или более целевой мутации(ям), делеции(ям), инверсии(ям), замене(ам) и/или вставке(ам), причем каждое "редактирование" вводится с помощью целевого метода редактирования генома.
[186] Учитывая, что супрессия генов оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 в кукурузе приводит к образованию растений с меньшей высотой и длиной междоузлия в дополнение к другим полезным признакам, предполагается, что экспрессия одного или обоих этих генов может быть уменьшена или устранены путем геномного редактирования одного или более этого гена(ов) с целью обеспечения сходных полезных признаков для растений кукурузы. Дополнительно, учитывая, что конститутивная экспрессия конструкций супрессии, нацеленных на эти гены оксидазы GA20, продуцирует растения кукурузы, обладающие полезными признаками низкого роста без отклонений в початке, и что экспрессия непосредственно в репродуктивных тканях початка также не приводит к появлению репродуктивных отклонений, предполагается, что один или оба из этих генных локусов могут быть отредактированы с целью нокдауна или нокаута их экспрессии для получения сходных эффектов у растений кукурузы. Подходы редактирования целевых генов могут быть применены для модификации последовательности промотора и/или регуляторной области(ей) одного или более генов оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 с целью нокдауна или нокаута экспрессии этого гена(ов), например, посредством целевых делеций, вставок, мутаций или других изменений последовательности. Действительно, промотор и/или регуляторная область(и) или последовательность(и), или 5'-НТО, 3'-НТО, и/или интронная последовательность(и) одного или более генов оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 могут быть в значительной степени удалены или мутированы. В альтернативном варианте, все или часть кодирующей (экзон), 5'-НТО, 3'-НТО и/или интронной последовательности(ей) одного или более генов оксидазы_3 GA20, оксидазы_4 GA20 и/или оксидазы_5 GA20 могут быть редактированы, удалены, мутированы или иным образом модифицированы для нокдауна или нокаута экспрессии, или активности этого гена(ов). Такие целевые модификации в локусах генов оксидазы_3 GA20 оксидазы_4 GA20 и/или оксидазы_5 GA20 могут быть достигнуты с применением любой подходящей технологии редактирования генома, известной в данной области техники, например, путем восстановления двухцепочечного разрыва (ДЦР) или одноцепочечного разрыва, введенного сайт-специфической нуклеазой, такой как, например, нуклеаза с цинковыми пальцами, сконструированная или нативная мегануклеаза, TALE-эндонуклеаза или РНК-направленная эндонуклеаза (например, Cas9 или Cpf1). Такое восстановление ДЦР или одноцепочечного разрыва может привести к спонтанным или стохастическим делециям, добавлениям, мутациям и тому подобному, в целевом сайте, где был введен ДЦР или одноцепочечный разрыва, или репарация сайта может включать в себя применение молекулы донорной матрицы с целью направить или вызвать предпочтительную или специфическую делецию, добавление, мутацию и тому подобное, в целевом сайте.
[187] Как указано в данном документе, растение, трансформированное молекулой рекомбинантной ДНК или вектором для трансформации, содержащими трансген, кодирующий транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, может включать в себя различные однодольные растения или злаковые растения, такие как кукуруза и другие однодольные или злаковые растения, которые имеют отдельные мужские и женские цветки (аналогично кукурузе) и, таким образом, могут быть восприимчивы к отклонениям в женских репродуктивных органах, структурах или тканях с мутациями в пути GA.
[188] Данные композиции и способы могут быть дополнительно применимы к другим злаковым растениям, которые выиграли бы от уменьшенной высоты растения и/или повышенной устойчивости к полеганию. Такие растения могут быть трансформированы молекулами или конструкциями рекомбинантной ДНК для супрессии одного или более эндогенных генов оксидазы GA20 и/или GA3 в растении в соответствии со способами и подходами, представленными в данном документе, с целью получения злакового растения, которое может быть короче и/или устойчивым к полеганию. Действительно, злаковое растение, эктопически экспрессирующее транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA для супрессии, может иметь множество полезных признаков, таких как более низкий рост или высоту растения, более короткую длину междоузлия, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, в дополнение к другим улучшенным признакам, связанным с урожайностью и/или устойчивостью к засухе, как предусмотрено в данном документе, относительно растения дикого типа или контрольного растения, не имеющего последовательности трансгена или транскрибируемой ДНК. Как описано дополнительно ниже, растения злаковых культур, которые уже были модифицированы с целью увеличения урожайности и устойчивости к полеганию посредством мутаций в пути GA, такие как пшеница, рис, просо, ячмень и сорго, могут вместо этого трансформироваться молекулой или конструкцией рекомбинантной ДНК как предусмотрено в данном документе. В отличие от многих мутаций пути GA в этих культурах, которые могут быть рецессивными, трансгенные конструкции, экспрессирующие элемент супрессии, нацеленный на эндогенный биосинтетический ген оксидазы GA в этих культурах, могут быть доминантными, даже если они гемизиготны или присутствуют в растении в виде единственной копии. Таким образом, растения, которые могут быть трансформированы молекулой рекомбинантной ДНК или конструкцией, экспрессирующей конструкцию супрессии, могут потенциально включать различные однодольные или злаковые культуры. Наличие доминантного трансгенного локуса, вызывающего полукарликовый, устойчивый к полеганию фенотип может быть преимущественным и предпочтительным по сравнению с рецессивным мутантным аллелем для того же фенотипа из-за преимуществ в селекции и интеграции признаков.
[189] В соответствии с вариантами реализации данного изобретения дополнительно предлагается, чтобы гены оксидазы GA в других растениях злаковых имеющие наибольшую идентичность/сходство последовательностей с генами оксидазы_3 GA20, оксидазы_4 GA20, оксидазы_5 GA20, оксидазы_1 GA3 и/или оксидазы_2 GA3 в кукурузе, которые как показано данном документе, вызывают фенотип короткого роста, полукарликовость и другие полезные признаки при супрессированы с помощью конструкции рекомбинантной ДНК для супрессии, также могут быть целями для супрессии с целью получения трансгенных растений злаков, имеющих сходные фенотипы полукарликовости и/или устойчивости к полеганию. В Таблице 3 представлен список генов оксидазы GA из других злаковых растений (сорго - Sorghum bicolor; рис - Oryza sativa; просо итальянское - Setaria italica; пшеница - Triticum aestivum; и ячмень - Hordeum vulgare), имеющих высокую степень идентичности последовательности с одним из генов оксидазы GA в кукурузе, который при супрессии приводит к низкорослому, полукарликовому фенотипу.
Таблица 3. Гомологи генов оксидазы GA кукурузы из других растений злаковых культур.
(SEQ ID NO)
(SEQ ID NO)
(SEQ ID NO)
(SEQ ID NO)
оксидаза 2
оксидаза 2-подобная
italica
оксидаза 2
sativa
оксидаза-D2
italica
sativa
диоксигеназа 2-2
диоксигеназа 2-2-подобная
italica
диоксигеназа 2-3
sativa
гидроксилаза
D2 белок
[190] В соответствии с другим аспектом данного изобретения, молекула, вектор или конструкция рекомбинантной ДНК предоставлена для супрессии эндогенного гена оксидазы GA (или оксидаза-подобного GA) в злаковом растении, причем молекула, вектор или конструкция рекомбинантной ДНК, содержат транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является (i) по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам любого одного или более из SEQ ID NO: 84, 85, 87, 88, 89, 91, 92, 93, 95, 96, 98, 99, 100, 102, 103, 105, 106, 107, 109, 110, 111, 113, 114, 115, 119, 120, 122, 123, 124, 126, 127, 128, 130, 131, 132, 134, 135 и/или 137, и/или (ii) по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей белок в злаковом растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным любому одной или более из SEQ ID NO: 86, 90, 94, 97, 101, 104, 108, 112, 116, 118, 121, 125, 129, 133 и/или 136. Аналогично, некодирующая молекула РНК может нацеливаться на эндогенный ген оксидазы GA (или оксидаза-подобного GA) в злаковом растении, имеющий процентную идентичность с геном(ами) оксидазы GA, который, как показано, влияет на высоту растения кукурузы. Таким образом, дополнительно предоставляется некодирующая молекула РНК, содержащая последовательность, которая является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок в злаковом растении, который является, по меньшей мере, на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным любому одной или более из SEQ ID NO: 9, 12, 15, 30 и/или 33. Как упомянуто выше, некодирующая молекула РНК может нацеливаться на последовательность экзона, интрона и/или НТО гена оксидазы GA (или оксидаза-подобного GA).
[191] Кроме того, предоставлены способы введения или трансформации в растение зерна, часть растения или клетку растения любой из вышеупомянутых конструкций, векторов или конструкций в соответствии с любым из способов, описанных в данном документе, которые могут быть сконструированы любым подходящим способом, описанным в данном документе, включая различные укладки или совместные нацеливающие расположения, а также модифицированные злаковые растения, части растений, ткани растений и клетки растений, полученные таким образом и/или содержащие любую такую молекулу, вектор или конструкцию рекомбинантной ДНК. Поскольку некодирующая молекула РНК, экспрессируемая из вышеуказанных конструкций, будет сконструирована для нацеливания на эндогенный ген оксидазы GA, растение зерна, трансформированное такими молекулами, векторами или конструкциями рекомбинантной ДНК, должно предпочтительно соответствовать виду происхождения для целевой последовательности или близкородственным видам, штаммам, зародышевым плазмам, линиям и тому подобному. Например, конструкция супрессии, комплементарная SEQ ID NO: 84, должна применяться для трансформации растения сорго, такого как растение Sorghum bicolor или, возможно, родственных видов сорго, штаммов и тому подобного, которые, как ожидается, будут иметь близкородственную или сходную последовательность гена оксидазы GA (или оксидаза-подобного GA).
[192] Геномные последовательности для каждого из указанных выше генов из растений злаков дополнительно представлены в Таблице 3, и могут быть применены для нацеливания этих генов для редактирования генома в соответствии с любым известным методом. Любая сайт-специфическая нуклеаза и способ могут быть применены, как описано в данном документе, для создания ДЦР или одноцепочечного разрыва для гена в геномном локусе или около него, который может быть репарирован неидеально или посредством опосредованной матрицей рекомбинации с целью создания мутаций и тому подобного, в гене или около него. Подходящие нуклеазы могут быть выбраны из группы, состоящей из нуклеазы c цинковыми пальцами (ZFN), мегануклеазы, РНК-направленной эндонуклеазы, TALE-эндонуклеазы (TALEN), рекомбиназы, транспозазы или любой их комбинации. Для РНК-направленной эндонуклеазы может быть применена конструкция или вектор рекомбинантной ДНК, содержащие направляющую РНК, для направления нуклеазы в целевой сайт. Соответственно, направляющая РНК для редактирования гена оксидазы GA (или оксидаза-подобного GA) в злаковой культуре может содержать направляющую последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25 или более последовательным нуклеотидам любого одного или более из SEQ ID NO: 84, 85, 87, 88, 89, 91, 92, 93, 95, 96, 98, 99, 100, 102, 103, 105, 106, 107, 109, 110, 111, 113, 114, 115, 119, 120, 122, 123, 124, 126, 127, 128, 130, 131, 132, 134, 135 и/или 137. Для сайт-специфических нуклеаз, которые не являются РНК-направленными, таких как нуклеаза с цинковыми пальцами (ZFN), мегануклеаза, TALE-эндонуклеаза (TALEN), рекомбиназа и/или транспозаза, специфичность геномной цели для редактирования определяется структурой белка, в частности, его ДНК-связывающим доменом. Такие сайт-специфические нуклеазы могут быть выбраны, разработаны или сконструированы так, чтобы связывать и разрезать желаемый целевой сайт в любом из вышеуказанных генов оксидазы (или оксидаза-подобного GA) в геноме злакового растения. Подобно трансформации с помощью конструкции супрессии, злаковое растение, трансформированное определенной направляющей РНК или молекулой, вектором или конструкцией рекомбинантной ДНК, кодирующими направляющую РНК, предпочтительно должно быть видом, в котором существует целевая геномная последовательность или близкородственным видом, штаммом, зародышевой плазмой, линией и тому подобным, так что направляющая РНК способна распознавать и связываться с нужным целевым сайтом разрезания.
[193] Дополнительно предоставлены способы введения или трансформации в злаковом растении, части растения или клетке растения любой направляющей РНК, описанной выше, или любой конструкции, вектора или конструкции, кодирующей такую направляющую РНК, возможно, в дополнение к РНК-направленной нуклеазе, согласно любому из описанных в данном документе способов, а также модифицированным растениям злаков, частям растений, тканям растений и клеткам растений, полученным таким образом и/или содержащим любую такую молекулу, вектор или конструкцию рекомбинантной ДНК и/или отредактированный ген оксидазы GA (или оксидаза-подобного GA). Модифицированные растения злаков, имеющие отредактированный ген оксидазы GA (или оксидаза-подобного GA) и/или элемент супрессии, нацеленный на ген оксидазы GA (или оксидаза-подобного GA), могут иметь один или более полезных признаков, представленных в данном документе, таких как более низкая высота растения, более короткая длина междоузлия, увеличенный диаметр стебля, улучшенная устойчивость к полеганию и/или устойчивость к засухе по сравнению с растением дикого типа или контрольным растением, не имеющим какого-либо такого элемента редактирования или супрессии. В дополнение к редактированию генома мутации в гене оксидазы GA (или оксидаза-подобного GA) могут быть введены с помощью других методов мутагенеза, как описано в данном документе.
[194] В соответствии с другим аспектом данного изобретения предоставляется трансгенное растение(я), растительная клетка(и), семя(ена) и часть(и) растения, содержащее событие трансформации или вставку в геном, по меньшей мере, одной его растительной клетки, при этом событие трансформации или вставка содержит последовательность, конструкцию или экспрессионную кассету рекомбинантной ДНК, содержащую транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA с целью супрессии, при этом последовательность транскрибируемой ДНК функционально связана с промотором, экспрессируемым в растении, таким как конститутивный, сосудистый и/или листовой промотор. Такое трансгенное растение может быть получено любым подходящим способом трансформации, как указано выше, для получения трансгенного растения R0, которое затем может быть самоопылено или скрещено с другими растениями, чтобы производить семена R1 и последующие поколения потомства и семян посредством дополнительных скрещиваний и тому подобного. Варианты реализации данного описания дополнительно включают в себя растительную клетку, ткань, эксплантат, часть растения и тому подобное, содержащую одну или более трансгенных клеток, имеющих событие трансформации или геномную вставку рекомбинантной ДНК или полинуклеотидной последовательности, содержащей транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, которая нацелена на эндогенный ген оксидазы GA с целью супрессии.
[195] Трансгенные растения, клетки растений, семена и части растений по данному изобретению могут быть гомозиготными или гемизиготными для трансгенного события или вставки транскрибируемой последовательности ДНК с целью супрессии гена оксидазы GA в геноме, по меньшей мере, одной его растительной клетки, или событие целевого редактирования генома, таким образом растения, клетки растения, семена и части растения в соответствии с данными вариантами реализации изобретения могут содержать любое количество копий такого трансгенного события(ий), вставки(ок) и/или редактирования(ий). Дозу или количество экспрессии трансгенной или транскрибируемой последовательности ДНК можно изменять по ее зиготности и/или количеству копий, что может влиять на уровень или степень фенотипических изменений в трансгенном растении и тому подобное. Как указано выше, трансгенные растения, представленные в данном документе, могут включать в себя различные однодольные или злаковые растения и даже сельскохозяйственные растения, такие как пшеница, рис и сорго, уже имеющие повышенный урожай и/или устойчивость к полеганию вследствие предшествующих селекционных усилий и мутаций пути GA в этих растениях. Преимущества применения трансгенной или транскрибируемой последовательности ДНК для экспрессии элемента супрессии, нацеленного на ген биосинтетической оксидазы GA, включают не только способность ограничивать экспрессию тканеспецифическим или тканепредпочтительным образом, но также и потенциальное доминирование (например, доминантно-негативные эффекты) одиночной или гемизиготной копии транскрибируемой последовательности ДНК, что приводит к полезным признакам или фенотипам низкорослости, полукарликовости у сельскохозяйственных растений. Таким образом, молекулы или конструкции рекомбинантной ДНК по данному изобретению могут быть применены для создания полезных признаков у множества однодольных или злаковых растений без отклонений с применением только одной копии трансгенного события, вставки или конструкции. В отличие от ранее описанных мутаций или аллелей в пути GA, которые являются рецессивными и требуют, чтобы растения были гомозиготными по мутантному аллелю, растения, трансформированные GA-модифицирующими трансгенами и супрессирующими конструкциями по данному изобретению, могут улучшать признаки, урожайность и усилия по селекции, облегчая производство гибридных злаковых растений, поскольку им требуется только одиночная или гемизиготная копия трансгенной или супрессирующей конструкции.
[196] В соответствии с некоторыми вариантами реализации изобретения трансгенное или модифицированное злаковое или кукурузное растение, содержащее трансген гена оксидазы GA или последовательность транскрибируемой ДНК для супрессии эндогенного гена оксидазы GA или геномно редактированного гена оксидазы GA, может быть дополнительно охарактеризовано как имеющее один или более полезных признаков, таких как более низкий рост или высота полукарликового растения, уменьшенная длина междоузлия, увеличенный диаметр стебля, улучшенная устойчивость к полеганию, уменьшенный излом стебля, более глубокие корни, увеличенная площадь листьев, более ранняя сомкнутость полога, повышенное содержание воды в листьях и/или более высокая устьичная проводимость в условиях ограничения воды, снижение содержания и/или площади антоцианов в листьях в нормальных или стрессовых условиях, ограничивающих азот или воду, улучшенные признаки, связанные с урожайностью, включая более крупный женский репродуктивный орган или початок, увеличение массы початка, индекса сбора урожая, урожайности, увеличение количества зерна и/или увеличение массы зерна, по отношению к растению дикого типа или контрольному растению. Такое трансгенное злаковое или кукурузное растение может дополнительно иметь повышенную устойчивость к стрессу, такую как повышенная устойчивость к засухе, повышенное использование азота и/или устойчивость к посадке с высокой плотностью.
[197] Для целей данного описания термин "растение" включает в себя эксплантат, часть растения, рассаду, проросток или целое растение на любой стадии регенерации или развития. В данном документе термин "трансгенное растение" относится к растению, геном которого был изменен в результате интеграции или вставки молекулы, конструкции или последовательности рекомбинантной ДНК. Трансгенное растение включает в себя растение R0 развитое или регенерированное из первоначально трансформированной растительной клетки(ок), а также потомство трансгенных растений в более поздних поколениях или скрещивание с трансгенным растением R0. В данном документе термин "часть растения" может относиться к любому органу или неизмененной ткани растения, таким как меристема, орган/структура побега (например, лист, стебель или узел), корень, цветок или цветковый орган/структура (например, верховой лист, чашелистник, лепесток, тычинка, плодолистик, пыльник и семезачаток), семена (например, эмбрион, эндосперм и оболочка семени), плод (например, зрелая завязь), росток или другие ткани растения (например, сосудистая ткань, эпидермис, основная паренхима и тому подобное) или любой их части. Части растений по данному изобретению могут быть жизнеспособными, нежизнеспособными, регенерируемыми и/или нерегенерируемыми. "Ростком" может быть любая часть растения, которая может прорасти в целое растение.
[198] Согласно данным вариантам реализации изобретения растительная клетка, трансформированная конструкцией или молекулой, содержащей транскрибируемую последовательность ДНК с целью супрессии эндогенного гена оксидазы GA, или конструкцией, применяемой для редактирования генома, может включать в себя любую растительную клетку, которая является компетентной для трансформации, как понятно в данной области техники, основанной на способе трансформации, такой как меристемная клетка, эмбриональная клетка, каллусная клетка и тому подобное. В данном документе "трансгенная растительная клетка" просто относится к любой растительной клетке, которая трансформирована стабильно интегрированной молекулой, конструкцией или последовательностью рекомбинантной ДНК. Трансгенная растительная клетка может включать в себя первоначально трансформированную растительную клетку, трансгенную растительную клетку регенерированного или развитого растения R0, трансгенную растительную клетку, культивируемую из другой трансгенной растительной клетки, или трансгенную растительную клетку из любого потомственного растения или потомка трансформированного растения R0, включая клетку(и) семени растения или эмбриона, или культивируемую растительную клетку, клетку каллуса и тому подобное.
[199] Варианты реализации данного описания дополнительно включают в себя способы создания или получения трансгенных или модифицированных растений, такие как трансформация, редактирование генома, скрещивание и тому подобное, при этом способ включает в себя введение молекулы, конструкции или последовательности рекомбинантной ДНК, содержащей трансген оксидазы GA или транскрибируемую последовательность ДНК для супрессии эндогенного гена оксидазы GA в клетке растения или редактирования геномного локуса эндогенного гена оксидазы GA, а затем регенерацию или развитие трансгенного или модифицированного растения из трансформированной или редактированной растительной клетки, что может быть выполнено под давлением отбора в пользу трансгенного события. Такие способы могут включать в себя трансформацию клетки растения молекулой, конструкцией или последовательностью рекомбинантной ДНК, содержащей транскрибируемую последовательность ДНК для супрессии эндогенного гена оксидазы GA и отбор растения, имеющего один или более измененных фенотипов или признаков, таких как один или более из следующих признаков на одной или более стадиях развития: более низкий рост или высота полукарликового растения, уменьшенная длина междоузлия в одном или более междоузлии(ях), увеличенный диаметр стебля, улучшенная устойчивость к полеганию, уменьшенный излом стебля, более глубокие корни, увеличенная площадь листьев, более ранняя сомкнутость полога, повышенное содержание воды в листьях и/или более высокая устьичная проводимость в условиях ограничения воды, снижение содержания и/или площади антоцианов в листьях в нормальных или стрессовых условиях, ограничивающих азот или воду, улучшенные признаки, связанные с урожайностью, включая более крупный женский репродуктивный орган или початок, увеличение массы початка, индекса сбора урожая, урожайности, увеличение количества зерна и/или увеличение массы зерна, повышенная устойчивость к стрессу, такая как повышенная устойчивость к засухе, повышенное использование азота и/или повышенная устойчивость к посадке с высокой плотностью по сравнению с растением дикого типа или контрольным растением.
[200] В соответствии с другим аспектом данного изобретения предложены способы для посадки модифицированного или трансгенного растения(ий), представленных в данном документе, с нормальной/стандартной или высокой плотностью посадки в поле. В соответствии с некоторыми вариантами реализации изобретения урожай культурного растения на гектар (или на площадь земли) может быть увеличен путем посадки модифицированного или трансгенного растения(ий) по данному изобретению с более высокой плотностью посадки в поле. Как описано в данном документе, модифицированные или трансгенные растения, экспрессирующие транскрибируемую последовательность ДНК, которая кодирует некодирующую молекулу РНК, нацеленную на эндогенный ген оксидазы GA с целью супрессии, или имеющие редактированный ген оксидазы GA, могут иметь уменьшенную высоту растения, более короткое междоузлие(лия), увеличенный диаметр стебля и/или повышенную устойчивость к полеганию. Предполагается, что модифицированные или трансгенные растения могут переносить условия посадки с высокой плотностью, поскольку увеличение диаметра стебля может противостоять полеганию, а более короткая высота растения может обеспечивать повышенную проницаемость света для нижних листьев в условиях посадки с высокой плотностью. Таким образом, модифицированные или трансгенные растения, представленные в данном документе, могут быть посажены с более высокой плотностью с целью увеличения урожайности с гектара (или площади земли) в поле. Для пропашных культур более высокая плотность посадки может быть достигнута путем высадки большего количества семян/растений на длину ряда и/или путем уменьшения расстояния между рядами.
[201] Согласно некоторым вариантам реализации изобретения модифицированное или трансгенное растение может быть посажено с плотностью в поле (количество растений на площадь земли/поля), которая является, по меньшей мере на 5%, 10%, 15%, 20%, 25%, 50%, 75%, 100%, 125%, 150%, 175%, 200%, 225% или 250% выше, чем нормальная плотность посадки для этого сельскохозяйственного растения в соответствии со стандартной агрономической практикой. Модифицированное или трансгенное растение может быть посажено с плотностью посадки в поле по меньшей мере 93 480 растений на гектар, по меньшей мере 98 400 растений на гектар, по меньшей мере 103 320 растений на гектар, по меньшей мере 108 240 растений на гектар, по меньшей мере 110 700 растений на гектар, по меньшей мере 113 160 растений на гектар, по меньшей мере 118 080 растений на гектар, по меньшей мере 123 000 растений на гектар, по меньшей мере 127 920 растений на гектар, по меньшей мере 132 840 растений на гектар или по меньшей мере 137 760 растений на гектар. Например, растения кукурузы могут быть посажены с более высокой плотностью посадки, например, в диапазоне от около 93 480 растений на гектар до около 147 600 растений на гектар или от около 98 400 растений на гектар до около 142 680 растений на гектар, или от около 103 320 растений на гектар до около 142 680 растений на гектар, или от около 98 400 растений на гектар до около 110 700 растений на гектар, или от около 110 700 растений на гектар до около 123 000 растений на гектар, или от около 123 000 растений на гектар до около 142 680 растений на гектар, или от около 127 920 растений на гектар до около 137 760 растений на гектар, или около 93 480 растений на гектар, около 103 320 растений на гектар, около 113 160 растений на гектар или около 118 080 растений на гектар, около 123 000 растений на гектар или около 127 920 растений на гектар, или около 132 840 растений на гектар, в отличие от стандартного диапазона плотности посадки, например, от около 44 280 растений на гектар до около 93 480 растений на гектар.
[202] В соответствии с вариантами реализации данного описания предлагается модифицированное растение(я) кукурузы, которые включают в себя (i) высоту растения менее чем 2000 мм, менее чем 1950 мм, менее чем 1900 мм, менее чем 1850 мм, менее чем 1800 мм, менее чем 1750 мм, менее чем 1700 мм, менее чем 1650 мм, менее чем 1600 мм, менее чем 1550 мм, менее чем 1500 мм, менее чем 1450 мм, менее чем 1400 мм, менее чем 1350 мм, менее чем 1300 мм, менее чем 1250 мм, менее чем 1200 мм, менее чем 1150 мм, менее чем 1100 мм, менее чем 1050 мм или менее чем 1000 мм и/или (ii) средний диаметр стебля по меньшей мере 18 мм, по меньшей мере 18,5 мм, по меньшей мере 19 мм, по меньшей мере 19,5 мм, по меньшей мере 20 мм, по меньшей мере 20,5 мм, по меньшей мере 21 мм, по меньшей мере 21,5 мм и по меньшей мере 22 мм. Другими словами, предлагаются модифицированное растение(я) кукурузы, которые имеют высоту растений менее чем 2000 мм, менее чем 1950 мм, менее чем 1900 мм, менее чем 1850 мм, менее чем 1800 мм, менее чем 1750 мм, менее чем 1700 мм, менее чем 1650 мм, менее чем 1600 мм, менее чем 1550 мм, менее чем 1500 мм, менее чем 1450 мм, менее чем 1400 мм, менее чем 1350 мм, менее чем 1300 мм, менее чем 1250 мм, менее чем 1200 мм, менее чем 1150 мм, менее чем 1100 мм, менее чем 1050 мм или менее чем 1000 мм и/или средний диаметр стебля более 18 мм, более 18,5 мм, более 19 мм, более 19,5 мм, более 20 мм, более 20,5 мм, более 21 мм, более 21,5 мм или более 22 мм. Любой такой признак или диапазон высоты растения, выражается в миллиметрах (мм), может быть преобразован в другую единицу измерения на основе известных преобразований (например, один дюйм равен 2,54 см или 25,4 миллиметра, а миллиметры (мм), сантиметры (см) и метры (м) отличаются только на одну или более степеней от десяти). Таким образом, любое измерение, представленное в данном документе, дополнительно описано в терминах любых других сопоставимых единиц измерения в соответствии с известными и установленными преобразованиями. Однако точная высота растения и/или диаметр стебля модифицированного растения кукурузы могут зависеть от окружающей среды и генетического фона. Таким образом, изменение высоты растения и/или диаметра стебля модифицированного растения кукурузы вместо этого может быть описано в терминах минимальной разницы или процентного изменения относительно контрольного растения. Модифицированное растение кукурузы может дополнительно содержать, по меньшей мере, один початок, который по существу не содержит мужских репродуктивных тканей или структур, или других отклонений.
[203] В соответствии с вариантами реализации данного изобретения предложены модифицированные растения кукурузы, которые имеют высоту растения на поздних вегетативных и и/или репродуктивных стадиях развития (например, на стадии R3) от 1000 мм до 1800 мм, от 1000 мм до 1700 мм, от 1050 до 1700 мм, от 1100 до 1700 мм, от 1150 до 1700 мм, от 1200 до 1700 мм, от 1250 до 1700 мм, от 1300 до 1700 мм, от 1350 до 1700 мм, от 1400 мм и 1700 мм, от 1450 мм до 1700 мм, от 1000 мм до 1500 мм, от 1050 мм до 1500 мм, от 1100 мм до 1500 мм, от 1150 мм до 1500 мм, от 1200 мм до 1500 мм, от 1250 мм до 1500 мм, от 1300 до 1500 мм, от 1350 до 1500 мм, от 1400 до 1500 мм, от 1450 до 1500 мм, от 1000 до 1600 мм, от 1100 до 1600 мм, от 1200 мм до 1600 мм, от 1300 до 1600 мм, от 1350 до 1600 мм, от 1400 до 1600 мм, от 1450 мм до 1600 мм, от 1000 мм до 2000 мм, от 1200 мм до 2000 мм, от 1200 мм до 1800 мм, от 1300 мм до 1700 мм, от 1400 мм до 1700 мм, от 1400 мм до 1600 мм, от 1400 мм до 1700 мм, от 1400 до 1800 мм, от 1400 до 1900 мм, от 1400 до 2000 мм или от 1200 до 2500 мм, и/или средний диаметр стебля от 17,5 до 22 мм, от 18 мм до 22 мм, от 18,5 до 22 мм, от 19 до 22 мм, от 19,5 до 22 мм, от 20 до 22 мм, от 20,5 до 22 мм, от 21 до 22 мм, от 21,5 до 22 мм, от 17,5 до 21 мм, от 17,5 до 20 мм, от 17,5 до 19 мм, от 17,5 до 18 мм, от 18 до 21 мм, от 18 до 20 мм или от 18 мм до 19 мм. Модифицированное растение кукурузы может быть по существу не содержащим отклонений, таких как мужские репродуктивные ткани или структуры в одном или более початках модифицированного растения кукурузы.
[204] Согласно вариантам реализации данного изобретения представлены модифицированные растения кукурузы, которые имеют (i) высоту растения, которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70% или по меньшей мере на 75% меньше, чем высота растения дикого типа или контрольного растения и/или (ii) диаметр стебля, который является, по меньшей мере на 5%, меньшей мере на 10%, меньшей мере на 15%, меньшей мере на 20%, меньшей мере на 25%, меньшей мере на 30%, меньшей мере на 35%, меньшей мере на 40%, меньшей мере на 45%, меньшей мере на 50%, меньшей мере на 55%, меньшей мере на 60%, меньшей мере на 65%, меньшей мере на 70%, меньшей мере на 75%, меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или, по меньшей мере, на 100% больше диаметра стебля растения дикого типа или контрольного растения. В соответствии с вариантами реализации данного изобретения модифицированное растение кукурузы может иметь уменьшенную высоту растения, которая является не более чем на 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55% или 60% короче, чем высота растения дикого типа или контрольного растения, и/или диаметр стебля, который является менее чем (или не более чем) на 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100% больше, чем диаметр стебля растения дикого типа или контрольного растения. Например, модифицированное растение может иметь (i) высоту растения, которая является по меньшей мере на 10%, по меньшей мере на 15% или по меньшей мере на 20% меньше или короче (то есть короче более чем или ровно на 10%, 15% или 20%), но не более чем на 50% короче, чем у растения дикого типа или контрольного растения, и/или (ii) диаметр стебля, который является, по меньшей мере на 5%, по меньшей мере на 10% или по меньшей мере на 15% больше, но не более чем на 30%, 35% или 40% больше, чем у растения дикого типа или контрольного растения. По определению, фразы "по меньшей мере на 20% короче" и "короче более чем или ровно на 20%" исключают, например, на 10% короче. Аналогично, по определению, фраза "не более чем на 50% короче" исключает на 60% короче; фраза "по меньшей мере на 5% больше" исключает на 2% больше; и фраза "не более чем на 30% больше" исключает на 40% больше.
[205] В соответствии с вариантами реализации данного изобретения представлены модифицированные растения кукурузы, которые имеют высоту от 5% до 75%, от 5% до 50%, от 10% до 70%, от 10% до 65%, от 10% до 60%, от 10% до 55%, от 10% до 50%, от 10% до 45%, от 10% до 40%, от 10% до 35%, от 10% до 30%, от 10% до 25%, от 10% до 20%, от 10% до 15%, от 10% до 10%, от 10% до 75%, от 25% до 75%, от 10% до 50%, от 20% до 50%, от 25% до 50%, от 30% до 75%, от 30% до 50%, от 25% до 50%, от 15% до 50%, от 20% до 50%, от 25% до 45% или от 30% до 45% меньше чем высота растения дикого типа или контрольного растения и/или диаметр стебля от 5% до 100%, от 5% до 95%, от 5% до 90%, от 5% до 85%, от 5% до 80%, от 5% до 75%, от 5% до 70%, от 5% до 65%, от 5% до 60%, от 5% до 55%, от 5% до 50%, от 5% до 45%, от 5% до 40%, от 5% до 35%, от 5% до 30%, от 5% до 25%, от 5% до 20%, от 5% до 15%, от 5% до 10%, от 10% до 100%, от 10% до 75%, от 10% до 50%, от 10% до 40%, от 10% до 30%, от 10% до 20%, от 25% до 75%, от 25% до 50%, от 50% до 75%, от 8% до 20% или от 8% до 15% больше чем диаметр стебля растения дикого типа или контрольного растения.
[206] В соответствии с вариантами реализации данного изобретения представлены модифицированные растения кукурузы, которые содержат среднюю длину междоузлия (или длину междоузлия минус-2 и/или длину междоузлия минус-4 относительно положения початка), которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70% или по меньшей мере на 75% меньше той же или средней длины междоузлия растения дикого типа или контрольного растения. "Минус-2 междоузлия" растения кукурузы относится ко второму междоузлию под початком растения, а "минус-4 междоузлия" растения кукурузы относится к четвертому междоузлию ниже початка растения. В соответствии со многими вариантами реализации изобретения предлагаются модифицированные растения кукурузы, которые имеют среднюю длину междоузлия (или длину минус-2 междоузлия и/или минус-4 длину междоузлия относительно положения початка), которая на от 5% до 75%, от 5% до 50%, от 10% до 70%, от 10% до 65%, от 10% до 60%, от 10% до 55%, от 10% до 50%, от 10% до 45%, от 10% до 40%, от 10% до 35%, от 10% до 30%, от 10% до 25%, от 10% до 20%, от 10% до 15%, от 10% до 10%, от 10% до 75%, от 25% до 75%, от 10% до 50%, от 20% до 50%, от 25% до 50%, от 30% до 75%, от 30% до 50%, от 25% до 50%, от 15% до 50%, от 20% до 50%, от 25% до 45% или от 30% до 45% меньше, чем одинаковая или средняя длина междоузлия растения дикого типа или контрольного растения.
[207] В соответствии с вариантами реализации данного изобретения представлены модифицированные растения кукурузы, которые содержат массу початка (индивидуально или в среднем), которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% большей, чем масса початка растения дикого типа или контрольного растения. Модифицированное растение кукурузы, представленное в данном документе, может содержать массу початка, которая на от 5% до 100%, от 5% до 95%, от 5% до 90%, от 5% до 85%, от 5% до 80%, от 5% до 75%, от 5% до 70%, от 5% до 65%, от 5% до 60%, от 5% до 55%, от 5% до 50%, от 5% до 45%, от 5% до 40%, от 5% до 35%, от 5% до 30%, от 5% до 25%, от 5% до 20%, от 5% до 15%, от 5% до 10%, от 10% до 100%, от 10% до 75%, от 10% до 50%, от 25% до 75%, от 25% до 50% или от 50% до 75% больше, чем масса початка растения дикого типа или контрольного растения.
[208] В соответствии с вариантами реализации данного изобретения предлагаются модифицированные растения кукурузы или злаковые растения, которые имеют уборочный индекс, по меньшей мере 0,57, по меньшей мере 0,58, по меньшей мере 0,59, по меньшей мере 0,60, по меньшей мере 0,61, по меньшей мере 0,62, по меньшей мере 0,63 по меньшей мере 0,64 или по меньшей мере 0,65 (или выше). Модифицированное растение кукурузы может иметь уборочный индекс от 0,57 до 0,65, от 0,57 до 0,64, от 0,57 до 0,63, от 0,57 до 0,62, от 0,57 до 0,61, от 0,57 до 0,60, от 0,57 до 0,59, от 0,57 до 0,58, от 0,58 до 0,65, от 0,59 до 0,65 или от 0,60 до 0,65. Модифицированное растение кукурузы может иметь уборочный индекс, который является, по меньшей мере на 1%, по меньшей мере на 2%, по меньшей мере на 3%, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8% по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45% или по меньшей мере на 50% больше, чем уборочный индекс растения дикого типа или контрольного растения. Модифицированные растения кукурузы могут иметь уборочный индекс, который от 1% до 45%, от 1% до 40%, от 1% до 35%, от 1% до 30%, от 1% до 25%, от 1% до 20%, от 1% до 15%, от 1% до 14%, от 1% до 13%, от 1% до 12%, от 1% до 11%, от 1% до 10%, от 1% до 9%, от 1% до 8%, от 1% до 7%, от 1% до 6%, от 1% до 5%, от 1% до 4%, от 1% до 3%, от 1% до 2%, от 5% до 15%, от 5% до 20%, от 5% до 30% или от 5% до 40% больше, чем уборочный индекс растения дикого типа или контрольного растения.
[209] В соответствии с вариантами реализации данного изобретения предложены модифицированные растения кукурузы или злаковые растения, которые имеют урожайность, повышенную по меньшей мере на 0,0627 тонн на гектар, по меньшей мере 0,1254 тонн на гектар, по меньшей мере 0,1881 тонн на гектар, по меньшей мере 0,2508 тонн на гектар не менее 0,3135 тонн на гектар, по меньшей мере 0,3762 тонн на гектар, по меньшей мере 0,4389 тонн на гектар, по меньшей мере 0,5016 тонн на гектар, по меньшей мере 0,5643 тонн на гектар или по меньшей мере 0,627 тонн на гектар относительно растения дикого типа или контрольного растения. Модифицированные растения кукурузы могут иметь увеличение урожайности от 0,0627 до 0,627, от 0,0627 до 0,5016, от 0,1254 до 0,5016, от 0,1254 до 0,3762, от 0,1254 до 0,3135, от 0,15675 до 0,28215 или от 0,1881 до 0,2508 тонн на гектар. Модифицированное растение кукурузы может иметь урожайность, повышенную по меньшей мере на 1%, по меньшей мере на 2%, по меньшей мере на 3%, по меньшей мере на 4%, по меньшей мере на 5%, по меньшей мере на 6%, по меньшей мере на 7%, по меньшей мере на 8%, по меньшей мере на 9%, по меньшей мере на 10%, по меньшей мере на 11%, по меньшей мере на 12%, по меньшей мере на 13%, по меньшей мере на 14%, по меньшей мере на 15%, по меньшей мере на 20% или по меньшей мере на 25%, чем урожайность растения дикого типа или контрольного растения. Модифицированная кукуруза может иметь урожайность от 1% до 25%, от 1% до 20%, от 1% до 15%, от 1% до 14%, от 1% до 13%, от 1% до 12%, от 1% до 11%, от 1% до 10%, от 1% до 9%, от 1% до 8%, от 1% до 7%, от 1% до 6%, от 1% до 5%, от 1% до 4%, от 1% до 3%, от 1% до 2%, от 5% до 15%, от 5% до 20%, от 5% до 25%, от 2% до 10%, от 2% до 9%, от 2% до 8%, от 2% до 7%, от 2% до 6%, от 2% до 5% или от 2% до 4% больше, чем урожайность растения дикого типа или контрольного растения.
[210] В соответствии с вариантами реализации данного изобретения предложено модифицированное злаковое или кукурузное растение, которое имеет частоту полегания, которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или на 100% меньше или ниже, чем у растения дикого типа или контрольного растения. Модифицированные злаковые или кукурузные растения могут иметь частоту полегания от 5% до 100%, от 5% до 95%, от 5% до 90%, от 5% до 85%, от 5% до 80%, от 5% до 75%, от 5% до 70%, от 5% до 65%, от 5% до 60%, от 5% до 55%, от 5% до 50%, от 5% до 45%, от 5% до 40%, от 5% до 35%, от 5% до 30%, от 5% до 25%, от 5% до 20%, от 5% до 15%, от 5% до 10%, от 10% до 100%, от 10% до 75%, от 10% до 50%, от 10% до 40%, от 10% до 30%, от 10% до 20%, от 25% до 75%, от 25% до 50% или от 50% до 75% меньше или ниже, чем у растения дикого типа или контрольного растения. Кроме того, представлены популяции злаковых или кукурузных растений, обладающие повышенной устойчивостью к полеганию и пониженной частотой полегания. Предложены популяции модифицированных злаковых или кукурузных растений, имеющие частоту полегания, которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или на 100% меньше или ниже, чем у популяции растений дикого типа или контрольных растений. Популяция модифицированных растений кукурузы может иметь частоту полегания от 5% до 100%, от 5% до 95%, от 5% до 90%, от 5% до 85%, от 5% до 80%, от 5% до 75%, от 5% до 70%, от 5% до 65%, от 5% до 60%, от 5% до 55%, от 5% до 50%, от 5% до 45%, от 5% до 40%, от 5% до 35%, от 5% до 30%, от 5% до 25%, от 5% до 20%, от 5% до 15%, от 5% до 10%, от 10% до 100%, от 10% до 75%, от 10% до 50%, от 10% до 40%, от 10% до 30%, от 10% до 20%, от 25% до 75%, от 25% до 50% или от 50% до 75% меньше или ниже, чем у популяции растений дикого типа или контрольных растений, что может быть выражено как среднее значение для определенного числа растений или посевной площади равной плотности.
[211] Согласно вариантам реализации данного изобретения предлагаются модифицированные растения кукурузы, имеющие значительно уменьшенную или пониженную высоту растения (например, 2000 мм или менее) и значительно увеличенный диаметр стебля (например, 18 мм или более) по сравнению с растением дикого типа или контрольным растением. В соответствии с этими вариантами реализации изобретения уменьшение или понижение высоты растения и увеличение диаметра стебля могут находиться в пределах любого из диапазонов высоты, диаметра или процента, указанных в данном документе. Такие модифицированные растения кукурузы, имеющие уменьшенную высоту растения и увеличенный диаметр стебля относительно растения дикого типа или контрольного растения, могут быть трансформированы транскрибируемой последовательностью ДНК, кодирующей некодирующую молекулу РНК, которая нацелена по меньшей мере на один ген оксидазы GA20 и/или по меньшей мере один ген оксидазы GA3 с целью супрессии. Модифицированные растения кукурузы, имеющие значительно уменьшенную высоту растения и/или значительно увеличенный диаметр стебля по сравнению с растением дикого типа или контрольным растением, могут дополнительно иметь по меньшей мере один початок, который по существу не содержит мужских репродуктивных тканей или структур и/или других отклонений. Модифицированные растения кукурузы, имеющие значительно уменьшенную высоту растения и/или увеличенный диаметр стебля относительно растения дикого типа или контрольного растения, могут иметь пониженную активность одного или более гена(ов) оксидазы GA20 и/или оксидазы GA3 в одной или более ткани(нях) растения, такой как одна или более сосудистая и/или листовая ткань(ни) растения, относительно одной и той же ткани(ней) растения дикого типа или контрольного растения. Согласно многим вариантам реализации изобретения модифицированные растения кукурузы могут содержать, по меньшей мере, одну полинуклеотидную или транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, функционально связанную с промотором, который может быть конститутивным, тканеспецифичным или тканепредпочтительным промотором, при этом некодирующая молекула РНК направлена, по меньшей мере, на один ген(ы) оксидазы GA20 и/или оксидазы GA3 с целью супрессии, как предусмотрено в данном документе. Некодирующая молекула РНК может представлять собой микроРНК или миРНК или молекулу-предшественника микроРНК или миРНК. В соответствии с некоторыми вариантами реализации изобретения модифицированные растения кукурузы, имеющие значительно уменьшенную высоту растения и/или увеличенный диаметр стебля относительно растения дикого типа или контрольного растения, могут дополнительно иметь повышенный уборочный индекс и/или повышенную устойчивость к полеганию по сравнению с растением дикого типа или контрольным растением.
[212] Модифицированные растения кукурузы или злаковые растения, имеющие значительно уменьшенную высоту растения и/или значительно увеличенный диаметр стебля по сравнению с растением дикого типа или контрольным растением, могут содержать мутацию (например, вставку, делецию, замену и тому подобное) в гене оксидазы GA, введенную с помощью технологии редактирования генов или другой техники мутагенеза, при этом экспрессия гена оксидазы GA снижена или устранена в одной или более тканях модифицированного растения. Такие модифицированные растения кукурузы, имеющие уменьшенную высоту растения и/или увеличенный диаметр стебля относительно растения дикого типа или контрольного растения, могут дополнительно иметь повышенный уборочный индекс и/или повышенную устойчивость к полеганию относительно растения дикого типа или контрольного растения. Такие модифицированные растения кукурузы могут быть практически свободны от отклонений, таких как мужские репродуктивные ткани или структуры и/или другие отклонения в, по меньшей мере, одном початке модифицированных растений. Методы мутагенеза растения (исключая редактирование генома) могут включать в себя химический мутагенез (то есть, обработку химическим мутагеном, таким как азид, гидроксиламин, азотистая кислота, акридин, аналог нуклеотидного основания или алкилирующий агент - например, EMS (этилметансульфонат), MNU (N-метил-N-нитрозомочевина) и тому подобное), физический мутагенез (например, гамма-лучи, рентгеновское излучение, ультрафиолетовое излучение, ионный пучок, другие формы излучения и тому подобное) и инсерционный мутагенез (например, транспозон или вставка Т-ДНК). Растения или различные части растения, ткани растения или клетки растения могут подвергаться мутагенезу. Обработанные растения могут быть воспроизведены для сбора семян или получения потомства растения, а обработанные части растения, ткани растения или клетки растения могут развиваться или регенерироваться в растения или другие ткани растения. Мутации, полученные с помощью методов химического или физического мутагенеза, могут включать в себя мутации сдвига рамки считывания, миссенс- или нонсенс-мутации, приводящие к потере функции или экспрессии целевого гена, такого как ген оксидазы GA3 или GA20.
[213] Один способ мутагенеза гена называется "TILLING" (для нацеливания на индуцированные локальные поражения в геномах), при котором мутации создаются в растительной клетке или ткани, предпочтительно в семени, репродуктивной ткани или зародышевой линии растения, например, применяя мутаген, такой как обработка EMS. Полученные растения выращивают и самоопыляют, а потомство применяют для приготовления образцов ДНК. ПЦР-амплификация и сиквенирование последовательности нуклеиновой кислоты гена оксидазы GA могут быть применены для определения того, имеет ли мутированное растение мутацию в гене оксидазы GA. Растения, имеющие мутации в гене оксидазы GA, могут затем быть проверены на измененный признак, такой как уменьшенная высота растения. В альтернативном варианте, подвергнутые мутагенезу растения могут быть проверены на измененный признак, такой как уменьшенная высота растения, и затем ПЦР-амплификация и сиквенирование последовательности нуклеиновой кислоты гена оксидазы GA могут быть применены для определения того, имеет ли растение с измененным признаком также мутацию в гене оксидазы GA. См., например, Colbert и соавт., 2001 год, Plant Physiol 126:480-484; и McCallum и соавт., 2000 год, Nature Biotechnology 18:455-457. TILLING может применяться для идентификации мутаций, которые изменяют экспрессию гена или активность белков, кодируемых геном, которые могут применяться для введения и отбора целевой мутации в гене оксидазы GA растения кукурузы или злакового растения.
[214] Кукурузные или злаковые растения, которые были подвергнуты мутагенезу или обработке с целью редактирования генома, могут быть подвергнуты скринингу и отобраны на основе наблюдаемого фенотипа (например, любого фенотипа, описанного в данном документе, такого как более короткая высота растения, увеличенный диаметр стебля и тому подобное), или применения селекционного агента с селектируемым маркером (например, гербицидом и тому подобным), скринируемым маркером или молекулярным методом (например, более низкие уровни GA, более низкие уровни транскрипта или оксидазы GA, наличие трансгена или транскрибируемой последовательности и тому подобное). Такие методы скрининга и/или отбора могут быть применены для идентификации и отбора растений, имеющих мутацию в гене оксидазы GA, которая приводит к желаемому фенотипу растений.
[215] В соответствии с вариантами реализации данного описания предлагается популяция модифицированных растений кукурузы или злаковых растений, при этом популяция модифицированных растений кукурузы или злаковых растений имеет среднюю высоту растения, которая является значительно меньшей, и/или средний диаметр стебля, который является значительно большим, чем популяция растений дикого типа или контрольных растений. Популяция модифицированных растений кукурузы или злаковых растений может иметь общую родословную с одним модифицированным растением кукурузы или злаковых растений и/или иметь одну совместную вставку, событие или редактирование конструкции для супрессии трансгенной оксидазы GA. Модифицированные растения кукурузы в популяции модифицированных растений кукурузы, как правило, могут содержать, по меньшей мере, один початок, который по существу не содержит мужских репродуктивных тканей или структур и/или других отклонений. Популяция модифицированных растений кукурузы или злаковых растений может иметь повышенную устойчивость к полеганию в среднем или на число растений или полевых площадей, чем популяция растений дикого типа или контрольных растений. Популяция модифицированных растений кукурузы или злаковых растений может иметь частоту полегания, которая является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 30%, по меньшей мере на 40%,по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 90% или на 100% меньше (или ниже), чем у популяции контрольных растений кукурузы или злаковых. Популяция модифицированных растений кукурузы может иметь уборочный индекс по меньшей мере 0,57 или более.
[216] В соответствии с вариантами реализации данного изобретения предлагаются модифицированные растения кукурузы или злаковых, имеющие пониженное содержание гиббереллина (в активной форме), по меньшей мере, в ткани(ях) стебля и междоузлия, таких как ткань(и) стебля, междоузлия, листа и/или сосудистая ткань(и), по сравнению с той же тканью(ями) растений дикого типа или контрольных растений. В соответствии со многими вариантами реализации изобретения предлагаются модифицированные растения кукурузы или злаковые растения, имеющие значительно уменьшенную высоту растения и/или значительно увеличенный диаметр стебля по сравнению с растениями дикого типа или контрольными растениями, при этом модифицированные кукурузные или злаковые растения дополнительно имеют значительно сниженный или пониженный уровень(и) активных гиббереллинов или активных GA (например, одного или более из GA1, GA3, GA4 и/или GA7) в одной или более ткани(ей) стебля, междоузлия, листа и/или сосудистой ткани(ях) относительно одной и той же ткани(ей) растений дикого типа или контрольных растений. Например, уровень одной или более активных GA в ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях) модифицированного кукурузного или злакового растения может являться, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% меньше или ниже, чем в той же ткани(нях) растения дикого типа или контрольного растения.
[217] Согласно некоторым вариантам реализации изобретения модифицированное кукурузное или злаковое растение может содержать уровень(и) активного гиббереллина (GA) (например, одна или более из GA1, GA3, GA4 и/или GA7) в одной или более ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях), который на от 5% до 50%, от 10% до 100%, от 20% до 100%, от 30% до 100%, от 40% до 100%, от 50% до 100%, от 60% до 100%, от 70% до 100%, от 80% до 100%, от 80% до 90%, от 10% до 90%, от 10% до 80%, от 10% до 70%, от 10% до 60%, от 10% до 50%, от 10% до 40%, от 10% до 30%, от 10% до 20%, от 50% до 100%, от 20% до 90%, от 20% до 80%, от 20% до 70%, от 20% до 60%, от 20% до 50%, от 20% до 40%, от 20% до 40%, от 20% до 30%, от 30% до 90%, от 30% до 80%, от 30% до 70%, от 30% до 60%, от 30% до 50%, от 30% до 40%, от 40% до 90% от 40% до 80%, от 40% до 70%, от 40% до 60%, от 40% до 50%, от 50% до 90%, от 50% до 80%, от 50% до 70%, от 50% до 60%, от 60% до 90%, от 60% до 80%, от 60% до 70%, от 70% до 90% или от 70% до 80% меньше или (или ниже), чем в той же ткани(нях) растения кукурузы дикого типа или контрольного растения. Модифицированное кукурузное или злаковое растение, имеющее пониженный уровень(и) активного гиббереллина (GA) в одной или более ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях), может также по существу не содержать отклонений, таких как мужские репродуктивные ткани или структуры и/или другие отклонения по меньшей мере в одном початке модифицированного растения кукурузы.
[218] В соответствии с вариантами реализации данного изобретения предлагаются модифицированные кукурузные или злаковые растения, имеющие значительно пониженный или устраненный уровень экспрессии одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA3 и/или GA20 в одной или более ткани(нях), такой как одна или более ткань(и) стебля, междоузлия, листа и/или сосудистая ткань(и) модифицированных растений, по сравнению с той же тканью(ми) растений дикого типа или контрольных растений. Согласно многим вариантам реализации изобретения предлагается модифицированное кукурузное или злаковое растение, содержащее значительно уменьшенную высоту растения и/или значительно увеличенный диаметр стебля по сравнению с растениями дикого типа или контрольными растениями, при этом модифицированное кукурузное или злаковое растение имеет значительно сниженный или устраненный уровень экспрессии одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA20 и/или оксидазы GA3 в одной или более ткани(нях), такой как одна или более ткань(и) стебля, междоузлия, листа и/или сосудистая ткань(и) модифицированного растения, по сравнению с той же тканью(ми) растений дикого типа или контрольных растений. Например, модифицированное кукурузное или злаковое растение имеет значительно сниженный или устраненный уровень экспрессии транскрипта(ов) и/или белка(ов) гена оксидазы_3 GA20 и/или оксидазы_5 GA20, и/или значительно сниженный или устраненный уровень экспрессии транскрипта(ов) и/или белка(ов) гена оксидазы_1 GA3 и/или оксидазы_2 GA3 в одной или более ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях) модифицированного растения, по сравнению с той же тканью(ми) растений дикого типа или контрольных растений. Например, уровень одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA3 и/или оксидазы GA20 или одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA (или оксидаза-подобного GA) в одной или более ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях) модифицированного растения кукурузы может являться, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35%, по меньшей мере на 40%, по меньшей мере на 45%, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 100% меньше или ниже, чем в той же ткани(нях) растения дикого типа или контрольного кукурузного или злакового растения.
[219] Согласно некоторым вариантам реализации изобретения модифицированное кукурузное или злаковое растение может содержать уровень(и) одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA3 и/или GA20, одного или более транскрипта(ов) и/или белка(ов) гена оксидазы GA (или оксидаза-подобного GA) в одной или более ткани(нях) стебля, междоузлия, листа и/или сосудистой ткани(нях), который на от 5% до 50%, от 10% до 100%, от 20% до 100%, от 30% до 100%, от 40% до 100%, от 50% до 100%, от 60% до 100%, от 70% до 100%, от 80% до 100%, от 80% до 90%, от 10% до 90%, от 10% до 80%, от 10% до 70%, от 10% до 60%, от 10% до 50%, от 10% до 40%, от 10% до 30%, от 10% до 20%, от 50% до 100%, от 20% до 90%, от 20% до 80%, от 20% до 70%, от 20% до 60%, от 20% до 50%, от 20% до 40%, от 20% до 40%, от 20% до 30%, от 30% до 90%, от 30% до 80%, от 30% до 70%, от 30% до 60%, от 30% до 50%, от 30% до 40%, от 40% до 90% от 40% до 80%, от 40% до 70%, от 40% до 60%, от 40% до 50%, от 50% до 90%, от 50% до 80%, от 50% до 70%, от 50% до 60%, от 60% до 90%, от 60% до 80%, от 60% до 70%, от 70% до 90% или от 70% до 80% меньше или ниже, чем в той же ткани(нях) растения дикого типа или контрольного кукурузного или злакового растения. Модифицированное кукурузное или злаковое растение, имеющее пониженный или устраненный уровень экспрессии по меньшей мере одного гена(ов) оксидазы GA20 и/или оксидазы GA3 в одной или более ткани(нях), может также по существу не содержать отклонений, таких как мужские репродуктивные ткани или структуры и/или другие отклонения по меньшей мере в одном початке модифицированного растения кукурузы.
[220] Согласно некоторым вариантам реализации изобретения предлагаются способы, включающие в себя снижение или устранение экспрессии по меньшей мере одного гена оксидазы GA20 и/или по меньшей мере одного гена оксидазы GA3 в злаковом растении, например в одной или более ткани стебля, междоузлия, сосудистой и/или листовой ткани злакового растения, при этом экспрессия по меньшей мере одного гена оксидазы GA20 и/или по меньшей мере одного гена(ов) оксидазы GA3 существенно не изменяется или изменяется по меньшей мере в одной репродуктивной ткани растения, и/или при этом уровень(и) одной или более активных GA существенно не изменен или изменен по меньшей мере в одной репродуктивной ткани растения по сравнению с растением дикого типа или контрольным растением. Согласно многим вариантам реализации изобретения уровень(и) экспрессии по меньшей мере одного гена оксидазы GA20 или оксидазы GA3 снижается или устраняется по меньшей мере в одной ткани модифицированного растения с помощью конструкции рекомбинантной ДНК, содержащей транскрибируемую последовательность ДНК, кодирующую элемент супрессии гена оксидазы GA20 или оксидазы GA3, такой как по меньшей мере одна зрелая микроРНК или предшественник микроРНК, который процессируется в зрелую микроРНК, при этом микроРНК способна снижать или супрессировать уровень экспрессии по меньшей мере одного гена оксидазы GA20 или оксидазы GA3, и при этом транскрибируемая последовательность ДНК функционально связана с конститутивным, тканеспецифичным или тканепредпочтительным промотором.
[221] Предлагаются способы и методы для скрининга и/или идентификации клеток или растений и тому подобного, на предмет наличия целевых изменений или трансгенов и отбора клеток или растений, содержащих целевые изменения или трансгены, которые могут основываться на одном или более фенотипах, или признаках, или на наличие или отсутствие молекулярного маркера или полинуклеотидной или белковой последовательности в клетках или растениях. Нуклеиновые кислоты могут быть выделены и обнаружены с применением методов, известных в данной области техники. Например, нуклеиновые кислоты могут быть выделены и обнаружены с применением, без ограничения, технологии рекомбинантных нуклеиновых кислот и/или полимеразной цепной реакции (ПЦР). Общие методы ПЦР описаны, например, в PCR Primer: A Laboratory Manual, Dieffenbach & Dveksler, Ред., Cold Spring Harbor Laboratory Press, 1995 год. Методы рекомбинантных нуклеиновых кислот включают в себя, например, расщепление эндонуклеазой рестрикции и лигирование, которые могут быть применены для выделения нуклеиновой кислоты. Выделенные нуклеиновые кислоты также могут быть химически синтезированы либо в виде одной молекулы нуклеиновой кислоты, либо в виде ряда олигонуклеотидов. Полипептиды могут быть очищены из природных источников (например, биологического образца) известными способами, такими как ионообменная ДЭАЭ (диэтиламиноэтил), гельфильтрационная хроматография и хроматография на гидроксиапатите. Полипептид также может быть очищен, например, путем экспрессии нуклеиновой кислоты в экспрессионном векторе. Кроме того, очищенный полипептид может быть получен химическим синтезом. Степень чистоты полипептида может быть измерена с применением любого подходящего способа, например, колоночной хроматографии, электрофореза в полиакриламидном геле или анализа с помощью ВЭЖХ. Любой способ, известный в данной области техники, может применяться для скрининга и/или идентификации клеток, растений и тому подобного, имеющих трансген или редактирование генома в своем геноме, способ может быть основан на любой подходящей форме визуального наблюдения, отбора, молекулярного метода и тому подобного.
[222] В некоторых вариантах реализации изобретения предлагаются способы обнаружения рекомбинантных нуклеиновых кислот и/или полипептидов в клетках растений. Например, нуклеиновые кислоты могут быть обнаружены с применением гибридизационных зондов или путем производства ампликонов с применением ПЦР с праймерами, как известно в данной области техники. Гибридизация между нуклеиновыми кислотами обсуждается в Sambrook и соавт. (1989 год, Molecular Cloning: A Laboratory Manual, 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, штат Нью-Йорк). Полипептиды могут быть обнаружены с применением антител. Методы обнаружения полипептидов с применением антител включают твердофазные иммуноферментные анализы (ELISA), вестерн-блоты, иммунопреципитацию, иммунофлуоресценцию и тому подобное. Антитело, представленное в данном документе, может представлять собой поликлональное антитело или моноклональное антитело. Антитело, обладающее специфической аффинностью связывания с полипептидом, представленным в данном документе, может быть получено с применением способов, известных в данной области техники. Антитело или гибридизационный зонд могут быть прикреплены к твердой подложке, такой как пробирка, планшет или лунка, с применением способов, известных в данной области техники.
[223] Обнаружение (например, продукта амплификации, комплекса гибридизации, полипептида) может быть осуществлено с применением детектируемых меток, которые могут быть присоединены или связаны с гибридизационным зондом или антителом. Термин "метка" предназначен для применения как прямых меток, так и непрямых меток. Детектируемые метки включают ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы и радиоактивные материалы.
[224] Скрининг и отбор модифицированных, редактированных или трансгенных растений или клеток растений могут проводиться с помощью любых методологий, известных специалистам в области молекулярной биологии. Примеры методик скрининга и отбора включают, но без ограничений, Саузерн-анализ, ПЦР-амплификацию для выявления полинуклеотида, нозерн-блоттинг, защиту от РНКазы, достройку праймера, ОТ-ПЦР-амплификацию для обнаружения РНК-транскриптов, сиквенирование по Сенгеру, технологии сиквенирования следующего поколения (например, Illumina®, PacBio®, Ion Torrent™ и тому подобные), ферментные анализы для определения ферментативной или рибозимной активности полипептидов и полинуклеотидов, а также гель-электрофорез белков, вестерн-блоттинг, иммунопреципитацию и иммуноферментный анализ для выявления полипептидов. Другие методы, такие как гибридизация in situ, окрашивание ферментов и иммуноокрашивание, также могут быть применены для обнаружения присутствия или экспрессии полипептидов и/или полинуклеотидов. Способы выполнения всех упомянутых методов известны в данной области техники.
ВАРИАНТЫ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
[225] В следующих абзацах перечислено подмножество иллюстративных вариантов реализации изобретения.
[226] Вариант реализации изобретения 1. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в злаковом растении или клетке растения, эндогенный белок оксидазы GA20 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 9, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[227] Вариант реализации изобретения 2. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 7 или SEQ ID NO: 8.
[228] Вариант реализации изобретения 3. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или растительной клетке, причем эндогенный белок оксидазы GA20 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 15.
[229] Вариант реализации изобретения 4. Конструкция рекомбинантной ДНК по варианту реализации изобретения 3, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 13 или SEQ ID NO: 14.
[230] Вариант реализации изобретения 5. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[231] Вариант реализации изобретения 6. Конструкция рекомбинантной ДНК по варианту реализации изобретения 5, отличающаяся тем, что сосудистый промотор содержит один из следующих: промотор сахарозосинтазы, промотор транспортера сахарозы, промотор Sh1, промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) гемивируса полосы кукурузы (MSV - maize streak geminivirus), рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или рисовый промотор желтой полосы 2 (OsYSL2).
[232] Вариант реализации изобретения 7. Конструкция рекомбинантной ДНК по варианту реализации изобретения 5, отличающаяся тем, что сосудистый промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичной одной или более из SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70 или SEQ ID NO: 71, или ее функциональной части.
[233] Вариант реализации изобретения 8. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[234] Вариант реализации изобретения 9. Конструкция рекомбинантной ДНК по варианту реализации изобретения 8, отличающаяся тем, что промотор, экспрессируемый в растении содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 65 или SEQ ID NO: 66, или ее функциональной части.
[235] Вариант реализации изобретения 10. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[236] Вариант реализации изобретения 11. Конструкция рекомбинантной ДНК по варианту реализации изобретения 10, отличающаяся тем, что листовой промотор включает один из следующего: промотор Рубиско, промотор PPDK, промотор FDA, промотор Nadh-Gogat, промотор гена белка, связывающего хлорофилл а/b, промотор фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы) или промотор гена Myb.
[237] Вариант реализации изобретения 12. Конструкция рекомбинантной ДНК по варианту реализации изобретения 10, отличающаяся тем, что листовой промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 72, SEQ ID NO: 73 или SEQ ID NO: 74, или ее функциональной части.
[238] Вариант реализации изобретения 13. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[239] Вариант реализации изобретения 14. Конструкция рекомбинантной ДНК по варианту реализации изобретения 13, отличающаяся тем, что конститутивный промотор выбран из группы, состоящей из: промотора актина, промотора 35S или 19S CaMV, растительного промотора убиквитина, растительного промотора Gos2, промотора FMV, промотора CMV, промотора MMV, промотора PCLSV, промотора Emu, промотора тубулина, промотора нопалинсинтазы, промотора октопинсинтазы, промотора маннопинсинтазы или алкогольдегидрогеназы маиса, или их функциональной части.
[240] Вариант реализации изобретения 15. Конструкция рекомбинантной ДНК по варианту реализации изобретения 13, отличающаяся тем, что конститутивный промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83, или ее функциональной части.
[241] Вариант реализации изобретения 16. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, отличающаяся тем, что некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК представляет собой предшественника микроРНК или миРНК, который процессирован или расщеплен в растительной клетке с образованием зрелой микроРНК или миРНК.
[242] Вариант реализации изобретения 17. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 1.
[243] Вариант реализации изобретения 18. Трансгенное злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 1.
[244] Вариант реализации изобретения 19. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что трансгенное растение имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[245] Вариант реализации изобретения 20. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[246] Вариант реализации изобретения 21. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения дикого типа.
[247] Вариант реализации изобретения 22. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что диаметр стебля трансгенного растения на одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем диаметр стебля на одном или более междоузлиях контрольного растения дикого типа.
[248] Вариант реализации изобретения 23. Трансгенное злаковое растение по любому из вариантов реализации изобретения 18, отличающееся тем, что трансгенное злаковое растение представляет собой растение кукурузы, и при этом диаметр стебля трансгенного растения кукурузы на одном или более из первого, второго, третьего и/или четвертого междоузлия, расположенного ниже початка, является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у того же междоузлия контрольного растения дикого типа.
[249] Вариант реализации изобретения 24. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[250] Вариант реализации изобретения 25. Трансгенное злаковое растение по варианту реализации изобретения 18, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения дикого типа.
[251] Вариант реализации изобретения 26. Трансгенное кукурузное растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 1.
[252] Вариант реализации изобретения 27. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по варианту реализации изобретения 1 и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
[253] Вариант реализации изобретения 28. Способ по варианту реализации изобретения 25, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[254] Вариант реализации изобретения 29. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA20 является, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98%, по меньшей мере, на 99%, по меньшей мере, на 99,5% или на 100% идентичным SEQ ID NO: 15, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[255] Вариант реализации изобретения 30. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 13 или SEQ ID NO: 14.
[256] Вариант реализации изобретения 31. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[257] Вариант реализации изобретения 32. Конструкция рекомбинантной ДНК по варианту реализации изобретения 31, отличающаяся тем, что сосудистый промотор содержит один из следующих: промотор сахарозосинтазы, промотор транспортера сахарозы, промотор Sh1, промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) гемивируса полосы кукурузы (MSV - maize streak geminivirus), рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или рисовый промотор желтой полосы 2 (OsYSL2).
[258] Вариант реализации изобретения 33. Конструкция рекомбинантной ДНК по варианту реализации изобретения 31, отличающаяся тем, что сосудистый промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70 или SEQ ID NO: 71, или ее функциональной части.
[259] Вариант реализации изобретения 34. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[260] Вариант реализации изобретения 35. Конструкция рекомбинантной ДНК по варианту реализации изобретения 34, отличающаяся тем, что промотор, экспрессируемый в растении содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 65 или SEQ ID NO: 66, или ее функциональной части.
[261] Вариант реализации изобретения 36. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[262] Вариант реализации изобретения 37. Конструкция рекомбинантной ДНК по варианту реализации изобретения 36, отличающаяся тем, что листовой промотор включает в себя один из следующего: промотор Рубиско, промотор PPDK, промотор FDA, промотор Nadh-Gogat, промотор гена белка, связывающего хлорофилл а/b, промотор фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы) или промотор гена Myb.
[263] Вариант реализации изобретения 38. Конструкция рекомбинантной ДНК по варианту реализации изобретения 36, отличающаяся тем, что листовой промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 72, SEQ ID NO: 73 или SEQ ID NO: 74, или ее функциональной части.
[264] Вариант реализации изобретения 39. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[265] Вариант реализации изобретения 40. Конструкция рекомбинантной ДНК по варианту реализации изобретения 39, отличающаяся тем, что конститутивный промотор выбран из группы, состоящей из: промотора актина, промотора 35S или 19S CaMV, растительного промотора юбиквитина, растительного промотора Gos2, промотора FMV, промотора CMV, промотора MMV, промотора PCLSV, промотора Emu, промотора тубулина, промотора нопалинсинтазы, промотора октопинсинтазы, промотора маннопинсинтазы или алкогольдегидрогеназы маиса, или их функциональной части.
[266] Вариант реализации изобретения 41. Конструкция рекомбинантной ДНК по варианту реализации изобретения 39, отличающаяся тем, что конститутивный промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83, или ее функциональной части.
[267] Вариант реализации изобретения 42. Конструкция рекомбинантной ДНК по варианту реализации изобретения 29, отличающаяся тем, что некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК представляет собой предшественника микроРНК или миРНК, который процессирован или расщеплен в растительной клетке с образованием зрелой микроРНК или миРНК.
[268] Вариант реализации изобретения 43. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 29.
[269] Вариант реализации изобретения 44. Трансгенное злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 29.
[270] Вариант реализации изобретения 45. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что трансгенное растение имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[271] Вариант реализации изобретения 46. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[272] Вариант реализации изобретения 47. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения дикого типа.
[273] Вариант реализации изобретения 48. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что диаметр стебля трансгенного растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем диаметр стебля в одном или более междоузлиях контрольного растения дикого типа.
[274] Вариант реализации изобретения 49. Трансгенное злаковое растение по любому из вариантов реализации изобретения 44, отличающееся тем, что трансгенное злаковое растение представляет собой растение кукурузы, и при этом диаметр стебля трансгенного растения кукурузы в одном или более из первого, второго, третьего и/или четвертого междоузлия, расположенного ниже початка, является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у того же междоузлия контрольного растения дикого типа.
[275] Вариант реализации изобретения 50. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[276] Вариант реализации изобретения 51. Трансгенное злаковое растение по варианту реализации изобретения 44, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения дикого типа.
[277] Вариант реализации изобретения 52. Трансгенное кукурузное растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 29.
[278] Вариант реализации изобретения 53. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по варианту реализации изобретения 29, и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
[279] Вариант реализации изобретения 54. Способ по варианту реализации изобретения 29, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[280] Вариант реализации изобретения 55. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA3 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA3 является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 30 или 33, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[281] Вариант реализации изобретения 56. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 28, 29, 31 или 32.
[282] Вариант реализации изобретения 57. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[283] Вариант реализации изобретения 58. Конструкция рекомбинантной ДНК по варианту реализации изобретения 57, отличающаяся тем, что сосудистый промотор содержит один из следующих: промотор сахарозосинтазы, промотор транспортера сахарозы, промотор Sh1, промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) геминивируса полосы маиса (MSV - maize streak geminivirus), рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или рисовый промотор желтой полосы 2 (OsYSL2).
[284] Вариант реализации изобретения 59. Конструкция рекомбинантной ДНК по варианту реализации изобретения 57, отличающаяся тем, что сосудистый промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70 или SEQ ID NO: 71, или ее функциональной части.
[285] Вариант реализации изобретения 60. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[286] Вариант реализации изобретения 61. Конструкция рекомбинантной ДНК по варианту реализации изобретения 60, отличающаяся тем, что промотор, экспрессируемый в растении содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 65 или SEQ ID NO: 66, или ее функциональной части.
[287] Вариант реализации изобретения 62. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[288] Вариант реализации изобретения 63. Конструкция рекомбинантной ДНК по варианту реализации изобретения 62, отличающаяся тем, что листовой промотор включает в себя один из следующего: промотор Рубиско, промотор PPDK, промотор FDA, промотор Nadh-Gogat, промотор гена белка, связывающего хлорофилл а/b, промотор фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы) или промотор гена Myb.
[289] Вариант реализации изобретения 64. Конструкция рекомбинантной ДНК по варианту реализации изобретения 62, отличающаяся тем, что листовой промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 72, SEQ ID NO: 73 или SEQ ID NO: 74, или ее функциональной части.
[290] Вариант реализации изобретения 65. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[291] Вариант реализации изобретения 66. Конструкция рекомбинантной ДНК по варианту реализации изобретения 65, отличающаяся тем, что конститутивный промотор выбран из группы, состоящей из: промотора актина, промотора 35S или 19S CaMV, растительного промотора юбиквитина, растительного промотора Gos2, промотора FMV, промотора CMV, промотора MMV, промотора PCLSV, промотора Emu, промотора тубулина, промотора нопалинсинтазы, промотора октопинсинтазы, промотора маннопинсинтазы или алкогольдегидрогеназы маиса, или их функциональной части.
[292] Вариант реализации изобретения 67. Конструкция рекомбинантной ДНК по варианту реализации изобретения 65, отличающаяся тем, что конститутивный промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83, или ее функциональной части.
[293] Вариант реализации изобретения 68. Конструкция рекомбинантной ДНК по варианту реализации изобретения 55, отличающаяся тем, что некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК представляет собой предшественника микроРНК или миРНК, который процессирован или расщеплен в растительной клетке с образованием зрелой микроРНК или миРНК.
[294] Вариант реализации изобретения 69. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 55.
[295] Вариант реализации изобретения 70. Трансгенное злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 55.
[296] Вариант реализации изобретения 71. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что трансгенное растение имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[297] Вариант реализации изобретения 72. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[298] Вариант реализации изобретения 73. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения дикого типа.
[299] Вариант реализации изобретения 74. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что диаметр стебля трансгенного растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем диаметр стебля в одном или более междоузлиях контрольного растения дикого типа.
[300] Вариант реализации изобретения 75. Трансгенное злаковое растение по любому из вариантов реализации изобретения 70, отличающееся тем, что трансгенное злаковое растение представляет собой растение кукурузы, и при этом диаметр стебля трансгенного растения кукурузы в одном или более из первого, второго, третьего и/или четвертого междоузлия, расположенного ниже початка, является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у того же междоузлия контрольного растения дикого типа.
[301] Вариант реализации изобретения 76. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[302] Вариант реализации изобретения 77. Трансгенное злаковое растение по варианту реализации изобретения 70, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения дикого типа.
[303] Вариант реализации изобретения 78. Трансгенное кукурузное растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 55.
[304] Вариант реализации изобретения 79. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по варианту реализации изобретения 55, и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
[305] Вариант реализации изобретения 80. Способ по варианту реализации изобретения 79, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[306] Вариант реализации изобретения 81. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA20 в однодольном или злаковом растении или клетке растения, эндогенный белок оксидазы GA20 является, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90%, по меньшей мере, на 95%, по меньшей мере, на 96%, по меньшей мере, на 97%, по меньшей мере, на 98%, по меньшей мере, на 99%, по меньшей мере, на 99,5% или на 100% идентичным SEQ ID NO: 12, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[307] Вариант реализации изобретения 82. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 10 или 11.
[308] Вариант реализации изобретения 83. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[309] Вариант реализации изобретения 84. Конструкция рекомбинантной ДНК по варианту реализации изобретения 83, отличающаяся тем, что сосудистый промотор содержит один из следующих: промотор сахарозосинтазы, промотор транспортера сахарозы, промотор Sh1, промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) геминивируса полосы маиса (MSV - maize streak geminivirus), рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или рисовый промотор желтой полосы 2 (OsYSL2).
[310] Вариант реализации изобретения 85. Конструкция рекомбинантной ДНК по варианту реализации изобретения 83, отличающаяся тем, что сосудистый промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70 или SEQ ID NO: 71, или ее функциональной части.
[311] Вариант реализации изобретения 86. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[312] Вариант реализации изобретения 87. Конструкция рекомбинантной ДНК по варианту реализации изобретения 86, отличающаяся тем, что промотор, экспрессируемый в растении содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 65 или SEQ ID NO: 66, или ее функциональной части.
[313] Вариант реализации изобретения 88. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[314] Вариант реализации изобретения 89. Конструкция рекомбинантной ДНК по варианту реализации изобретения 88, отличающаяся тем, что листовой промотор включает в себя один из следующего: промотор Рубиско, промотор PPDK, промотор FDA, промотор Nadh-Gogat, промотор гена белка, связывающего хлорофилл а/b, промотор фосфоенолпируваткарбоксилазы (ФЕП-карбоксилазы) или промотор гена Myb.
[315] Вариант реализации изобретения 90. Конструкция рекомбинантной ДНК по варианту реализации изобретения 88, отличающаяся тем, что листовой промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 72, SEQ ID NO: 73 или SEQ ID NO: 74, или ее функциональной части.
[316] Вариант реализации изобретения 91. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[317] Вариант реализации изобретения 92. Конструкция рекомбинантной ДНК по варианту реализации изобретения 91, отличающаяся тем, что конститутивный промотор выбран из группы, состоящей из: промотора актина, промотора 35S или 19S CaMV, растительного промотора юбиквитина, растительного промотора Gos2, промотора FMV, промотора CMV, промотора MMV, промотора PCLSV, промотора Emu, промотора тубулина, промотора нопалинсинтазы, промотора октопинсинтазы, промотора маннопинсинтазы или алкогольдегидрогеназы маиса, или их функциональной части.
[318] Вариант реализации изобретения 93. Конструкция рекомбинантной ДНК по варианту реализации изобретения 91, отличающаяся тем, что конститутивный промотор содержит последовательность ДНК, которая является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% одной или более из SEQ идентичной одной или более из SEQ ID NO: 75, SEQ ID NO: 76, SEQ ID NO: 77, SEQ ID NO: 78, SEQ ID NO: 79, SEQ ID NO: 80, SEQ ID NO: 81, SEQ ID NO: 82 или SEQ ID NO: 83, или ее функциональной части.
[319] Вариант реализации изобретения 94. Конструкция рекомбинантной ДНК по варианту реализации изобретения 81, отличающаяся тем, что некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК представляет собой предшественника микроРНК или миРНК, который процессирован или расщеплен в растительной клетке с образованием зрелой микроРНК или миРНК.
[320] Вариант реализации изобретения 95. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 81.
[321] Вариант реализации изобретения 96. Трансгенное злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 81.
[322] Вариант реализации изобретения 97. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что трансгенное растение имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[323] Вариант реализации изобретения 98. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[324] Вариант реализации изобретения 99. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения дикого типа.
[325] Вариант реализации изобретения 100. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что диаметр стебля трансгенного растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем диаметр стебля в одном или более междоузлиях контрольного растения дикого типа.
[326] Вариант реализации изобретения 101. Трансгенное злаковое растение по любому из вариантов реализации изобретения 96, отличающееся тем, что трансгенное злаковое растение представляет собой растение кукурузы, и при этом диаметр стебля трансгенного растения кукурузы в одном или более из первого, второго, третьего и/или четвертого междоузлия, расположенного ниже початка, является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у того же междоузлия контрольного растения дикого типа.
[327] Вариант реализации изобретения 102. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[328] Вариант реализации изобретения 103. Трансгенное злаковое растение по варианту реализации изобретения 96, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения дикого типа.
[329] Вариант реализации изобретения 104. Трансгенное кукурузное растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 81.
[330] Вариант реализации изобретения 105. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по варианту реализации изобретения 81, и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
[331] Вариант реализации изобретения 106. Способ по варианту реализации изобретения 105, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[332] Вариант реализации изобретения 107. Конструкция рекомбинантной ДНК по варианту реализации изобретения 1, 29, 55 или 81, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок оксидазы GA в однодольном или злаковом растении или растительной клетке, причем эндогенный белок оксидазы GA является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным одной или более из SEQ ID NO: 3, 6, 9, 12, 15, 18, 21, 24, 27, 30 и 33.
[333] Вариант реализации изобретения 108. Конструкция рекомбинантной ДНК по варианту реализации изобретения 107, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из одного или более SEQ ID NO: 1, 2, 4, 5, 7, 8, 10, 11, 13, 14, 16, 17, 19, 20, 22, 23, 25, 26, 28, 29, 31, и 32.
[334] Вариант реализации изобретения 109. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% комплементарной по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей эндогенный белок в однодольном или злаковом растении или клетке растения, при этом эндогенный белок является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентичным SEQ ID NO: 86, 90, 94, 97, 101, 104, 108, 112, 116, 118, 121, 125, 129, 133 или 136, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[335] Вариант реализации изобретения 110. Конструкция рекомбинантной ДНК по варианту реализации изобретения 109, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам из SEQ ID NO: 84, 85, 87, 88, 89, 91, 92, 93, 95, 96, 98, 99, 100, 102, 103, 105, 106, 107, 109, 110, 111, 113, 114, 115, 119, 120, 122, 123, 124, 126, 127, 128, 130, 131, 132, 134, 135 или 137.
[336] Вариант реализации изобретения 111. Конструкция рекомбинантной ДНК по варианту реализации изобретения 109, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[337] Вариант реализации изобретения 112. Конструкция рекомбинантной ДНК по варианту реализации изобретения 109, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[338] Вариант реализации изобретения 113. Конструкция рекомбинантной ДНК по варианту реализации изобретения 109, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[339] Вариант реализации изобретения 114. Конструкция рекомбинантной ДНК по варианту реализации изобретения 109, отличающаяся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[340] Вариант реализации изобретения 115. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 81.
[341] Вариант реализации изобретения 116. Трансгенное злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 109.
[342] Вариант реализации изобретения 117. Трансгенное злаковое растение по варианту реализации изобретения 116, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[343] Вариант реализации изобретения 118. Трансгенное злаковое растение по варианту реализации изобретения 116, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[344] Вариант реализации изобретения 119. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по варианту реализации изобретения 116, и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
[345] Вариант реализации изобретения 120. Способ по варианту реализации изобретения 119, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[346] Вариант реализации изобретения 121. Способ снижения уровня по меньшей мере одной активной молекулы GA в стебле кукурузы или злакового растения, включающий в себя: супрессию одного или более генов оксидазы GA3 или оксидазы GA20 рекомбинантной конструкцией ДНК в одной или более тканях трансгенного злакового или кукурузного растения.
[347] Вариант реализации изобретения 122. Способ по варианту реализации изобретения 121, отличающийся тем, что конструкция рекомбинантной ДНК кодирует некодирующую молекулу РНК, которая нацелена на один или более генов оксидазы GA3 или GA20 с целью супрессии, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[348] Вариант реализации изобретения 123. Способ по варианту реализации изобретения 122, отличающийся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[349] Вариант реализации изобретения 124. Способ по варианту реализации изобретения 122, отличающийся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[350] Вариант реализации изобретения 125. Способ по варианту реализации изобретения 122, отличающийся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[351] Вариант реализации изобретения 126. Способ по варианту реализации изобретения 122, отличающийся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[352] Вариант реализации изобретения 127. Способ по варианту реализации изобретения 121, отличающийся тем, что трансгенное растение кукурузы или зерна представляет собой растение кукурузы.
[353] Вариант реализации изобретения 128. Трансгенное растение кукурузы или зерна, содержащее конструкцию рекомбинантной ДНК, при этом конструкция рекомбинантной ДНК содержит последовательность транскрибируемой ДНК, кодирующую некодирующую молекулу РНК, которая нацелена по меньшей мере на один эндогенный ген оксидазы GA20 или GA3 с целью супрессии, при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении, и при этом трансгенное однодольное растение или злаковое растение имеет меньшую высоту растения по сравнению с контрольным растением дикого типа.
[354] Вариант реализации изобретения 129. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что трансгенное растение имеет один или более из следующих дополнительных признаков относительно контрольного растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и увеличение плодовитости.
[355] Вариант реализации изобретения 130. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения.
[356] Вариант реализации изобретения 131. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что диаметр стебля трансгенного растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у контрольного растения.
[357] Вариант реализации изобретения 132. Трансгенное кукурузное или злаковое растение по любому из вариантов реализации изобретения 128, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения.
[358] Вариант реализации изобретения 133. Трансгенное кукурузное или злаковое растение по любому из вариантов реализации изобретения 128, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения.
[359] Вариант реализации изобретения 134. Трансгенное кукурузное или злаковое растение по любому из вариантов реализации изобретения 128, отличающееся тем, что трансгенное растение не имеет каких-либо значительных отклонений в по меньшей мере одном женском органе или початке.
[360] Вариант реализации изобретения 135. Трансгенное кукурузное или злаковое растение по любому из вариантов реализации изобретения 128, отличающееся тем, что трансгенное злаковое растение представляет собой растение кукурузы, и при этом некодирующая молекула РНК нацелена на эндогенный ген(ы) оксидазы_3 GA20 и/или оксидазы_5 GA20 с целью супрессии.
[361] Вариант реализации изобретения 136. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что промотор, экспрессируемый в растении представляет собой сосудистый промотор.
[362] Вариант реализации изобретения 137. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что промотор, экспрессируемый в растении представляет собой промотор RTBV.
[363] Вариант реализации изобретения 138. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что промотор, экспрессируемый в растении представляет собой конститутивный промотор.
[364] Вариант реализации изобретения 139. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что промотор, экспрессируемый в растении представляет собой листовой промотор.
[365] Вариант реализации изобретения 140. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 128, отличающееся тем, что трансгенное растение имеет один или более из следующих дополнительных признаков относительно контрольного растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и увеличение плодовитости.
[366] Вариант реализации изобретения 141. Злаковое растение, содержащее мутацию в эндогенном гене оксидазы GA или около него, введенную методом мутагенеза, при этом уровень экспрессии эндогенного гена оксидазы GA снижается или отсутствует в злаковом растении, и при этом злаковое растение имеет меньшую высоту по сравнению с контрольным растением дикого типа.
[367] Вариант реализации изобретения 142. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что злаковое растение, содержащее мутацию, имеет один или более из следующих дополнительных признаков относительно контрольного растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и увеличение плодовитости.
[368] Вариант реализации изобретения 143. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что высота злакового растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения.
[369] Вариант реализации изобретения 144. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что диаметр стебля злакового растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у контрольного растения.
[370] Вариант реализации изобретения 145. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля злакового растения ниже, чем в той же ткани междоузлия контрольного растения.
[371] Вариант реализации изобретения 146. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля злакового растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения.
[372] Вариант реализации изобретения 147. Злаковое растение по варианту реализации изобретения 141, отличающееся тем, что злаковое растение не имеет каких-либо существенных отклонений в по меньшей мере одном женском органе или початке.
[373] Вариант реализации изобретения 148. злаковое растение по варианту реализации изобретения 141, отличающееся тем, что злаковое растение представляет собой кукурузное растение.
[374] Вариант реализации изобретения 149. Кукурузное или злаковое растение, содержащие геномное редактирование, введенное с помощью целевого метода редактирования генома в локус эндогенного гена оксидазы GA или около него, при этом уровень экспрессии эндогенного гена оксидазы GA снижен или отсутствует по сравнению с контрольным растением, и при этом редактированное злаковое растение имеет меньшую высоту по сравнению с контрольным растением.
[375] Вариант реализации изобретения 150. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что редактированное растение имеет один или более из следующих дополнительных признаков относительно контрольного растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и увеличение плодовитости.
[376] Вариант реализации изобретения 151. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что высота редактированного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения.
[377] Вариант реализации изобретения 152. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что диаметр стебля редактированного растения в одном или более междоузлиях стебля является, по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% больше, чем у контрольного растения.
[378] Вариант реализации изобретения 153. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля редактированного растения ниже, чем в той же ткани междоузлия контрольного растения.
[379] Вариант реализации изобретения 154. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля редактированного растения является по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже, чем у той же ткани междоузлия контрольного растения.
[380] Вариант реализации изобретения 155. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что редактированное растение не имеет каких-либо существенных отклонений в по меньшей мере одном женском органе или початке.
[381] Вариант реализации изобретения 156. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что геномное редактирование вводят с применением мегануклеазы, нуклеазы c цинковыми пальцами (ZFN), РНК-направленной эндонуклеазы, TALE-эндонуклеазы (TALEN), рекомбиназы или транспозазы.
[382] Вариант реализации изобретения 157. Редактированное кукурузное или злаковое растение по варианту реализации изобретения 149, отличающееся тем, что геномное редактирование включает замену, делецию, вставку или инверсию одного или более нуклеотидов относительно последовательности эндогенного гена оксидазы GA в контрольном растении.
[383] Вариант реализации изобретения 158. Композиция, содержащая направляющую РНК, при этом направляющая РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичной или комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере, 25 последовательным нуклеотидам целевой последовательности ДНК в геномном локусе эндогенного гена оксидазы GA растения зерна или около него.
[384] Вариант реализации изобретения 159. Композиция по варианту реализации изобретения 158, отличающаяся тем, что молекула направляющей РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере 25 последовательным нуклеотидам из SEQ ID NO: 34, 35 или 38 или последовательности, комплементарной им.
[385] Вариант реализации изобретения 160. Композиция по варианту реализации изобретения 158, отличающаяся тем, что молекула направляющей РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%,по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере 25 последовательным нуклеотидам из SEQ ID NO: 87, 91, 95, 98, 105, 109, 113, 117, 122, 126, 130 или 137 или последовательности, комплементарной им.
[386] Вариант реализации изобретения 161. Композиция по варианту реализации изобретения 158, дополнительно включающая в себя эндонуклеазу, направляемую РНК.
[387] Вариант реализации изобретения 162. Композиция по варианту реализации изобретения 161, отличающаяся тем, что РНК-направленная эндонуклеаза в присутствии молекулы направляющей РНК вызывает двухцепочечный разрыв или одноцепочечный разрыв в целевой последовательности ДНК или около него в геноме злакового растения.
[388] Вариант реализации изобретения 163. Композиция по варианту реализации изобретения 161, отличающаяся тем, что РНК-направленная эндонуклеаза выбрана из группы, состоящей из Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn1, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx12, Csx15, Csf1, Csf2, Csf3, Csf4, Cpf1, CasX, CasY, Argonaute и любых их гомологов или модифицированных версий, имеющих активность РНК-направленной эндонуклеазы.
[389] Вариант реализации изобретения 164. Композиция по варианту реализации изобретения 158, дополнительно включающая в себя донорную матрицу рекомбинантной ДНК содержащую, по меньшей мере, одну гомологичную последовательность или гомологичное плечо, при этом, по меньшей мере, одна гомологичная последовательность или гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК, при этом последовательность целевой ДНК представляет собой геномную последовательность в геномном локусе эндогенного гена оксидазы GA кукурузы или злакового растения или около него.
[390] Вариант реализации изобретения 165. Конструкция рекомбинантной ДНК, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу направляющей РНК, при этом молекула направляющей РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или по меньшей мере 25 последовательным нуклеотидам целевой последовательности ДНК в геномном локусе эндогенного гена оксидазы GA кукурузы или злакового растения или около него.
[391] Вариант реализации изобретения 166. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что направляющая РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере 25 последовательным нуклеотидам из SEQ ID NO: 34, 35 или 38 или последовательности, комплементарной им.
[392] Вариант реализации изобретения 167. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что молекула направляющей РНК содержит направляющую последовательность, которая является, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% комплементарной по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или, по меньшей мере 25 последовательным нуклеотидам из SEQ ID NO: 87, 91, 95, 98, 105, 109, 113, 117, 122, 126, 130 или 137 или последовательности, комплементарной им.
[393] Вариант реализации изобретения 168. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что последовательность транскрибируемой ДНК функционально связана с промотором, экспрессируемым в растении.
[394] Вариант реализации изобретения 169. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что молекула направляющей РНК представляет собой CRISPR РНК (crРНК) или одноцепочечную направляющую РНК (онРНК).
[395] Вариант реализации изобретения 170. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что направляющая РНК содержит последовательность, комплементарную последовательности мотива, прилегающего к протоспейсеру (PAM), присутствующую в геноме злакового растения, непосредственно прилегающую к последовательности целевой ДНК в геномном локусе эндогенного гена оксидазы GA или около него.
[396] Вариант реализации изобретения 171. Конструкция рекомбинантной ДНК по любому варианту реализации изобретения 165, отличающаяся тем, что последовательность PAM содержит каноническую последовательность 5'-NGG-3'.
[397] Вариант реализации изобретения 172. Конструкция рекомбинантной ДНК по варианту реализации изобретения 165, отличающаяся тем, что ген эндогенной оксидазы GA кодирует белок, который является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 9, 12 или 15.
[398] Вариант реализации изобретения 173. Молекула ДНК, содержащая конструкцию рекомбинантной ДНК по варианту реализации изобретения 165.
[399] Вариант реализации изобретения 174. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по варианту реализации изобретения 165.
[400] Вариант реализации изобретения 175. Бактериальная клетка, содержащая конструкцию рекомбинантной ДНК по варианту реализации изобретения 165.
[401] Вариант реализации изобретения 176. Кукурузное или злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 165.
[402] Вариант реализации изобретения 177. Композиция, содержащая конструкцию рекомбинантной ДНК по варианту реализации изобретения 165.
[403] Вариант реализации изобретения 178. Композиция по варианту реализации изобретения 177, дополнительно включающая в себя эндонуклеазу, направляемую РНК.
[404] Вариант реализации изобретения 179. Композиция по варианту реализации изобретения 177, отличающаяся тем, что РНК-направленная эндонуклеаза выбрана из группы, состоящей из Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Csm3, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, Cpf1, Argonaute и их гомологов или модифицированных версий, имеющих активность РНК-направленной эндонуклеазы.
[405] Вариант реализации изобретения 180. Композиция по варианту реализации изобретения 177, дополнительно включающая в себя вторую конструкцию рекомбинантной ДНК, содержащую вторую транскрибируемую последовательность ДНК, кодирующую эндонуклеазу, направляемую РНК.
[406] Вариант реализации изобретения 181. Композиция по варианту реализации изобретения 177, включающая в себя молекулу ДНК или вектор, содержащие конструкцию рекомбинантной ДНК и вторую конструкцию рекомбинантной ДНК.
[407] Вариант реализации изобретения 182. Композиция по варианту реализации изобретения 177, включающая в себя первую молекулу ДНК или вектор и вторую молекулу ДНК или вектор, при этом первая молекула или вектор ДНК содержит конструкцию рекомбинантной ДНК, кодирующую молекулу направляющей РНК, а вторая молекула или вектор ДНК содержит вторую конструкцию рекомбинантной ДНК, кодирующую эндонуклеазу, направляемую РНК.
[408] Вариант реализации изобретения 183. Композиция по варианту реализации изобретения 177, дополнительно включающая в себя донорную матрицу рекомбинантной ДНК содержащую, по меньшей мере, одну гомологичную последовательность или гомологичное плечо, при этом, по меньшей мере, одна гомологичная последовательность или гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК, при этом последовательность целевой ДНК представляет собой геномную последовательность в геномном локусе эндогенного гена оксидазы GA кукурузы или злакового растения или около него.
[409] Вариант реализации изобретения 184. Донорная матрица рекомбинантной ДНК содержащая, по меньшей мере, одну гомологичную последовательность, при этом по меньшей мере одна гомологичная последовательность является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК, при этом последовательность целевой ДНК представляет собой геномную последовательность в геномном локусе эндогенного гена оксидазы GA кукурузы или злакового растения или около него.
[410] Вариант реализации изобретения 185. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 184, отличающаяся тем, что, по меньшей мере, одна гомологичная последовательность содержит, по меньшей мере, одну мутацию относительно комплементарной цепи последовательности целевой ДНК в геномном локусе эндогенного гена оксидазы GA или около него.
[411] Вариант реализации изобретения 186. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 185, отличающаяся тем, что, по меньшей мере, одна мутация включает замену, делецию, вставку или инверсию одного, или более нуклеотидов относительно комплементарной цепи последовательности целевой ДНК.
[412] Вариант реализации изобретения 187. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 184, отличающаяся тем, что по меньшей мере одна гомологичная последовательность является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичной или комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам SEQ ID NO: 34, 35 или 38, или последовательности, комплементарной им.
[413] Вариант реализации изобретения 188. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 184, отличающаяся тем, что по меньшей мере одна гомологичная последовательность является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичной или комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам SEQ ID NO: 87, 91, 95, 98, 105, 109, 113, 117, 122, 126, 130 или 137, или последовательности, комплементарной им.
[414] Вариант реализации изобретения 189. Донорная матрица рекомбинантной ДНК содержащая два гомологичных плеча, включая первое гомологичное плечо и второе гомологичное плечо, при этом первое гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам первой фланкирующей последовательности ДНК, при этом второе гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам второй фланкирующей последовательности ДНК, и при этом первая фланкирующая последовательность ДНК и вторая фланкирующая последовательность ДНК представляют собой геномные последовательности в геномном локусе эндогенного гена оксидазы GA кукурузного или злакового растения или около него.
[415] Вариант реализации изобретения 190. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 189, дополнительно содержащая последовательность вставки, расположенную между первым гомологичным плечом и вторым гомологичным плечом.
[416] Вариант реализации изобретения 191. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 189, отличающаяся тем, что последовательность вставки содержит, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 400, по меньшей мере 500, по меньшей мере 750, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 нуклеотидов.
[417] Вариант реализации изобретения 192. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 189, отличающаяся тем, что каждое гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичным или комплементарным, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам из SEQ ID NO: 34, 35 или 38, или последовательности, комплементарной им.
[418] Вариант реализации изобретения 193. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 189, отличающаяся тем, что каждое гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичным или комплементарным, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам из SEQ ID NO: 87, 91, 95, 98, 105, 109, 113, 117, 122, 126, 130 или 137, или последовательности, комплементарной им.
[419] Вариант реализации изобретения 194. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 189, отличающаяся тем, что один или более нуклеотидов, присутствующих в геноме однодольного или злакового растения между первой фланкирующей последовательностью ДНК и второй фланкирующей последовательностью ДНК, отсутствуют в молекуле донорной матрицы рекомбинантной ДНК между первым гомологичным плечом и вторым гомологичным плечом.
[420] Вариант реализации изобретения 195. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 194, отличающаяся тем, что, по меньшей мере 1, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 200, по меньшей мере 300, по меньшей мере 400, по меньшей мере 500, по меньшей мере 750, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 нуклеотидов, присутствующих в геноме однодольного или злакового растения между первой и второй фланкирующими последовательностями ДНК, отсутствуют в молекуле донорной матрицы рекомбинантной ДНК между первым и вторым гомологичными плечами.
[421] Вариант реализации изобретения 196. Молекула или вектор ДНК, содержащие донорную матрицу рекомбинантной ДНК по варианту реализации изобретения 189.
[422] Вариант реализации изобретения 197. Бактериальная клетка или клетка-хозяин, содержащая донорную матрицу рекомбинантной ДНК по варианту реализации изобретения 189.
[423] Вариант реализации изобретения 198. Кукурузное или злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 189.
[424] Вариант реализации изобретения 199. Сконструированная сайт-специфическая нуклеаза, которая связывается с целевым сайтом в геномном локусе эндогенного гена оксидазы GA кукурузного или злакового растения, или рядом с ним и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте.
[425] Вариант реализации изобретения 200. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 199, отличающаяся тем, что сайт-специфическая нуклеаза представляет собой мегануклеазу или хоуминг-эндонуклеазу.
[426] Вариант реализации изобретения 201. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 200, отличающаяся тем, что сконструированная мегануклеаза или хоуминг-эндонуклеаза включает фермент остова или основной фермент, выбранный из группы, состоящей из I-CreI, I-CeuI, I-MsoI, I-SceI, I-AniI и I-DmoI.
[427] Вариант реализации изобретения 202. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 199, отличающаяся тем, что сайт-специфическая нуклеаза представляет собой нуклеазу с цинковыми пальцами (ZFN), содержащую ДНК-связывающий домен и домен расщепления.
[428] Вариант реализации изобретения 203. Сконструированная нуклеаза с цинковыми пальцами по варианту реализации изобретения 202, отличающаяся тем, что домен расщепления представляет собой домен нуклеазы FokI.
[429] Вариант реализации изобретения 204. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 199, отличающаяся тем, что сайт-специфическая нуклеаза представляет собой эффекторную нуклеазу, подобную активаторам транскрипции (TALEN), содержащую ДНК-связывающий домен и домен расщепления.
[430] Вариант реализации изобретения 205. Сконструированный TALEN по варианту реализации изобретения 204, отличающийся тем, что домен расщепления выбран из группы, состоящей из нуклеазного домена PvuII, нуклеазного домена MutH, нуклеазного домена TevI, нуклеазного домена FokI, нуклеазного домена AlwI, нуклеазного домена MlyI, нуклеазного домена SbfI, нуклеазного домена SdaI, нуклеазного домена StsI, нуклеазного домена CleDORF, нуклеазного домена Clo051 и нуклеазного домена Pept071.
[431] Вариант реализации изобретения 206. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 199, отличающаяся тем, что целевой сайт, связанный сайт-специфической нуклеазой, является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичным или комплементарным, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам из SEQ ID NO: 34, 35 или 38, или последовательности, комплементарной им.
[432] Вариант реализации изобретения 207. Сконструированная сайт-специфическая нуклеаза по варианту реализации изобретения 199, отличающаяся тем, что целевой сайт, связанный сайт-специфической нуклеазой, является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% идентичным или комплементарным, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам из SEQ ID NO: 87, 91, 95, 98, 105, 109, 113, 117, 122, 126, 130 или 137, или последовательности, комплементарной им.
[433] Вариант реализации изобретения 208. Конструкция рекомбинантной ДНК, содержащая трансген, кодирующий сайт-специфическую нуклеазу, при этом сайт-специфическая нуклеаза связывается с целевым сайтом в геномном локусе эндогенного гена оксидазы GA однодольного или злакового растения, или около него и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте.
[434] Вариант реализации изобретения 209. Конструкция рекомбинантной ДНК по варианту реализации изобретения 208, отличающаяся тем, что трансген функционально связан с промотором, экспрессируемым в растении.
[435] Вариант реализации изобретения 210. Конструкция рекомбинантной ДНК по варианту реализации изобретения 208, отличающаяся тем, что сайт-специфическая нуклеаза представляет собой мегануклеазу или хоуминг-эндонуклеазу, нуклеазу с цинковыми пальцами или эффекторную нуклеазу, подобную активаторам транскрипции (TALEN).
[436] Вариант реализации изобретения 211. Молекула или вектор ДНК, содержащая конструкцию рекомбинантной ДНК по варианту реализации изобретения 208.
[437] Вариант реализации изобретения 212. Бактериальная клетка или клетка-хозяин, содержащая конструкцию рекомбинантной ДНК по варианту реализации изобретения 208.
[438] Вариант реализации изобретения 213. Кукурузное или злаковое растение, часть растения или клетка растения, содержащее конструкцию рекомбинантной ДНК по варианту реализации изобретения 208.
[439] Вариант реализации изобретения 214. Донорная матрица рекомбинантной ДНК, содержащая по меньшей мере одно гомологичное плечо и последовательность вставки, при этом по меньшей мере одно гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарным, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности геномной ДНК кукурузного или злакового растения, и при этом последовательность вставки содержит конструкцию рекомбинантной ДНК, содержащую транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК нацелена, с целью супрессии, на один или более эндогенных генов оксидазы GA20 или GA3 в однодольном или злаковом растении или в растительной клетке, и при этом транскрибируемая последовательность ДНК функционально связана с промотором, экспрессируемым в растении.
[440] Вариант реализации изобретения 215. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 214, отличающаяся тем, что по меньшей мере одно гомологичное плечо содержит два гомологичных плеча, включая в себя первое гомологичное плечо и второе гомологичное плечо, при этом первое гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам первой фланкирующей последовательности ДНК, а второе гомологичное плечо содержит последовательность, которая является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам второй фланкирующей последовательности ДНК, при этом первая фланкирующая последовательность ДНК и вторая фланкирующая последовательность ДНК представляют собой геномные последовательности в одном и том же геномном локусе однодольного или злакового растения или около него, и при этом последовательность вставки расположена между первым гомологичным плечом и вторым гомологичным плечом и содержит конструкцию рекомбинантной ДНК, содержащую транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК.
[441] Вариант реализации изобретения 216. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 215, отличающаяся тем, что последовательность транскрибируемой ДНК функционально связана с промотором, экспрессируемым в растении.
[442] Вариант реализации изобретения 217. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 215, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей белок оксидазы GA, который является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 9, 12, 15, 30 или 33.
[443] Вариант реализации изобретения 218. Донорная матрица рекомбинантной ДНК по варианту реализации изобретения 215, отличающаяся тем, что некодирующая молекула РНК содержит последовательность, которая является, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% комплементарной, по меньшей мере, 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26 или по меньшей мере 27 последовательным нуклеотидам молекулы мРНК, кодирующей белок оксидазы GA, который является, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или 100% идентичным SEQ ID NO: 86, 90, 94, 97, 101, 104, 108, 112, 116, 118, 121, 125, 129, 133 или 136.
[444] Вариант реализации изобретения 219. Композиция, содержащая донорную матрицу рекомбинантной ДНК по варианту реализации изобретения 214.
[445] Вариант реализации изобретения 220. Бактериальная клетка или клетка-хозяин, содержащая донорную матрицу рекомбинантной ДНК по варианту реализации изобретения 214.
[446] Вариант реализации изобретения 221. Трансгенное кукурузное или злаковое растение, часть растения или клетка растения, содержащие последовательность вставки донорной матрицы рекомбинантной ДНК по варианту реализации изобретения 214.
[447] Вариант реализации изобретения 222. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 214, отличающееся тем, что трансгенное растение имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[448] Вариант реализации изобретения 223. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 222, отличающееся тем, что трансгенное растение имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
[449] Вариант реализации изобретения 224. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 222, отличающееся тем, что высота трансгенного растения является, по меньшей мере на 10%, по меньшей мере на 20%, по меньшей мере на 25%, по меньшей мере на 30%, по меньшей мере на 35% или по меньшей мере на 40% ниже чем у контрольного растения.
[450] Вариант реализации изобретения 225. Трансгенное кукурузное или злаковое растение по варианту реализации изобретения 222, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения ниже, чем в той же ткани междоузлия контрольного растения.
[451] Вариант реализации изобретения 226. Способ получения трансгенной кукурузы или злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата с помощью указанной донорной матрицы рекомбинантной ДНК по варианту реализации изобретения 215, и (b) регенерацию или развитие трансгенной кукурузы или злакового растения из трансформированного эксплантата, при этом трансгенное кукурузное или злаковое растение содержит последовательность вставки донорной матрицы рекомбинантной ДНК.
[452] Вариант реализации изобретения 227. Способ по варианту реализации изобретения 226, отличающийся тем, что однодольное или злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
[453] Вариант реализации изобретения 228. Способ получения кукурузного или злакового растения, имеющего геномное редактирование в эндогенном гене оксидазы GA или около него, включающий в себя: (а) введение по меньшей мере в одну клетку эксплантата кукурузного или злакового растения сайт-специфической нуклеазы или молекулы рекомбинантной ДНК, содержащей трансген, кодирующий сайт-специфическую нуклеазу, при этом сайт-специфическая нуклеаза связывается с целевым сайтом в геномном локусе эндогенного гена оксидазы GA или около него и вызывает двухцепочечный или одноцепочечный разрыв в целевом сайте и (b) регенерацию или развитие редактированной кукурузного или злакового растения по меньшей мере из одной клетки эксплантата, содержащей геномное редактирование в эндогенном гене оксидазы GA или около него отредактированного однодольного или злакового растения.
[454] Вариант реализации изобретения 229. Способ по варианту реализации изобретения 228, отличающийся тем, что этап введения (a) дополнительно содержит введение донорной матрицы ДНК, содержащей, по меньшей мере, одну гомологичную последовательность или гомологичное плечо, при этом, по меньшей мере, одна гомологичная последовательность или гомологичное плечо является, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 99% или на 100% комплементарной, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90, по меньшей мере 100, по меньшей мере 150, по меньшей мере 200, по меньшей мере 250, по меньшей мере 500, по меньшей мере 1000, по меньшей мере 2500 или по меньшей мере 5000 последовательным нуклеотидам последовательности целевой ДНК, при этом последовательность целевой ДНК представляет собой геномную последовательность в геномном локусе эндогенного гена оксидазы GA однодольного или злакового растения или около него.
[455] Вариант реализации изобретения 230. Способ по варианту реализации изобретения 228, дополнительно включающий в себя: (c) отбор редактированного кукурузного или злакового растения.
[456] Вариант реализации изобретения 231. Способ по варианту реализации изобретения 230, отличающийся тем, что этап отбора (c) включает в себя определение, был ли редактирован локус эндогенного гена оксидазы GA с применением молекулярного анализа.
[457] Вариант реализации изобретения 232. Способ по варианту реализации изобретения 230, отличающийся тем, что этап отбора (c) включает в себя определение, был ли редактирован локус эндогенного гена оксидазы GA путем наблюдения фенотипа растения.
[458] Вариант реализации изобретения 233. Способ по варианту реализации изобретения 231, отличающийся тем, что фенотип растения представляет собой уменьшение высоты растения относительно контрольного растения.
[459] Вариант реализации изобретения 234. Способ по варианту реализации изобретения 228, отличающийся тем, что этап введения (а) создает по меньшей мере одну мутацию в геномном локусе эндогенного гена оксидазы GA или около него, и при этом мутация включает замену, делецию, вставку или инверсию одного, или более нуклеотидов относительно последовательности геномной ДНК контрольного растения.
[460] Вариант реализации изобретения 235. Модифицированное растение кукурузы, имеющее высоту растения менее чем 2000 мм, менее чем 1950 мм, менее чем 1900 мм, менее чем 1850 мм, менее чем 1800 мм, менее чем 1750 мм, менее чем 1700 мм, менее чем 1650 мм, менее чем 1600 мм, менее чем 1550 мм, менее чем 1500 мм, менее чем 1450 мм, менее чем 1400 мм, менее чем 1350 мм, менее чем 1300 мм, менее чем 1250 мм, менее чем 1200 мм, менее чем 1150 мм, менее чем 1100 мм, менее чем 1050 мм или менее чем 1000 мм и либо (i) средний диаметр стебля более 18 мм, более 18,5 мм, более 19 мм, более 19,5 мм, более 20 мм, более 20,5 мм, более 21 мм, более 21,5 мм или более 22 мм, (ii) улучшение устойчивости к полеганию по сравнению с контрольным растением дикого типа, либо (iii) повышение устойчивости к засухе по сравнению с контрольным растением дикого типа.
[461] Вариант реализации изобретения 236. Модифицированное кукурузное растение по варианту реализации изобретения 235, отличающееся тем, что кукурузное растение имеет один или более из следующих признаков относительно контрольного растения дикого типа: увеличенный диаметр стебля, улучшенную устойчивость к полеганию, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
[462] Вариант реализации изобретения 237. Модифицированное кукурузное растение по варианту реализации изобретения 235, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля кукурузного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
[463] Вариант реализации изобретения 238. Модифицированное злаковое растение, имеющее уменьшенную высоту растения по сравнению с контрольным растением дикого типа, и (i) увеличенный диаметр стебля по сравнению с контрольным растением дикого типа, (ii) улучшенную устойчивость к полеганию по сравнению с контрольным растением дикого типа, или (iii) улучшенную устойчивость к засухе по сравнению с контрольным растением дикого типа.
[464] Вариант реализации изобретения 239. Модифицированное злаковое растение по варианту реализации изобретения 238, отличающееся тем, что уровень одной или более активных GA в стебле злакового растения ниже, чем в контрольном растении дикого типа.
ПРИМЕРЫ
Пример 1. Уменьшение высоты растений у инбредных линий кукурузы через события трансформации по элементу супрессии оксидазы GA20.
[465] Линия инбредных растений кукурузы была трансформирована с помощью опосредованной Agrobacterium трансформации вектором трансформации, имеющим экспрессионную конструкцию, включающую транскрибируемую последовательность ДНК с последовательностью (SEQ ID NO: 39), кодирующей нацеливающую последовательность (SEQ ID NO: 40) микроРНК под контролем промотора палочковидного вируса риса тунгро (RTBV) (SEQ ID NO: 65), который, как известно, вызывает экспрессию в сосудистых тканях растений. МикроРНК, кодируемая данной конструкцией, содержит последовательность РНК, нацеленную на гены оксидазы_3 GA20 и оксидазы_5 GA20 в растениях кукурузы с целью супрессии. С помощью данной конструкции было проведено несколько событий трансформации, и эти трансформанты были протестированы в теплице, чтобы определить, не уменьшилась ли у них высота растений по сравнению с нетрансгенными контрольными растениями дикого типа. Как проиллюстрировано на Фиг. 1, значительное уменьшение высоты растений регулярно наблюдалось у трансгенных растений, экспрессирующих конструкцию супрессии, через несколько событий трансформации (см. 1-8 события) по сравнению с контрольными растениями дикого типа (ДТ). Высота растений для каждого из событий трансформации была рассчитана как среднее значение для приблизительно 10 растений для каждого события и сравнена со средней высотой контрольных растений. Среднеквадратические ошибки, представленные в виде планок погрешностей на Фиг. 1, были рассчитаны для каждого события и контрольных растений. Более того, развитие початка у каждого из этих трансформантов было нормальным.
[466] Как видно из результатов данного эксперимента, средняя высота растений у растений, экспрессирующих микроРНК, нацеленную на супрессию генов оксидазы_3 GA20 и оксидазы_5 GA20, последовательно снижалась в течение нескольких событий до 35% по сравнению с контрольными растениями. Эти данные подтверждают вывод о том, что эффекты, наблюдаемые с данной конструкцией супрессии, не обусловлены вставкой конструкции в какой-либо один локус в геноме растения.
[467] Эти данные дополнительно указывают на то, что экспрессия данной конструкции для супрессии оксидазы GA20 при использовании сосудистого промотора RTBV эффективна для создания данных фенотипов высоты растений. Кроме того, ранние данные на растениях кукурузы R0, конститутивно экспрессирующих ту же конструкцию супрессии оксидазы GA20 под контролем различных конститутивных промоторов, также дают низкорослые растения (см. Пример 15 ниже). Таким образом, экспрессия конструкции супрессии, нацеленной на оксидазу GA20, может быть эффективной для уменьшения высоты растения и обеспечения других полезных признаков, связанных с урожайностью и препятствующих полеганию, описанных в данном документе, учитывая, что различные профили экспрессии, включая сосудистую и конститутивную экспрессию, обеспечивают сходные фенотипы высоты растений без видимых отклонений початков.
Пример 2. Уменьшенная высота растений у гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20.
[468] Гибридные растения кукурузы, имеющие конструкцию супрессии оксидазы GA20, описанную в Примере 1, также показали уменьшенную высоту растений по сравнению с контрольными растениями дикого типа при выращивании в полевых условиях. Вычисляли среднюю высоту растения трансгенных гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20, на 10 микроучастках и сравнивали со средней высотой растения контрольных гибридных растений кукурузы дикого типа (нетрансгенных) на 32 микроучастках. Каждая микроучасток для трансгенного и нетрансгенного контроля включала приблизительно 6 растений, хотя фактическое количество растений на участок могло варьироваться в зависимости от количества растений, которые прорастают и развиваются в растения, имеющие початки. Как проиллюстрировано Фиг. 2А, значительное уменьшение средней высоты растений наблюдалось у трансгенных гибридных растений, экспрессирующих конструкцию супрессии (гибрид SUP-GA20ox), по сравнению с гибридными растениями кукурузы дикого типа (контроль). Среднеквадратические ошибки, представленные в виде планок погрешностей на Фиг. 2А, были рассчитаны для трансгенных гибридов и контрольных растений. Изображение гибридного контрольного растения (слева) рядом с трансгенным гибридным растением, экспрессирующим элемент супрессии оксидазы GA20 (справа), дополнительно приведено на Фиг. 2B.
[469] В данном эксперименте средняя высота выращенных в поле гибридных растений кукурузы, экспрессирующих микроРНК, нацеленных на гены оксидазы_3 GA20 и оксидазы_5 GA20, уменьшилась примерно на 40% по сравнению с контрольными гибридными растениями дикого типа. Эти данные свидетельствуют, что фенотип высоты растений присутствует у гибридных растений кукурузы в дополнении к инбредным линиям. Тем не менее, общая биомасса в данном эксперименте оказалась нейтральной у полукарликовых растений кукурузы по сравнению с контролями.
Пример 3. Увеличенный диаметр стебля у гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20.
[470] Гибридные растения кукурузы, имеющие конструкцию супрессии оксидазы GA20, описанную в Примере 1, также продемонстрировали увеличенный диаметр стебля по сравнению с контрольными растениями дикого типа при выращивании в полевых условиях. Диаметр стебля измеряли на втором междоузлие ниже первичного початка. Рассчитывали средний диаметр стебля трансгенных гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20, на 8 микроучастках и сравнивали со средним диаметром стебля контрольных гибридных растений кукурузы дикого типа (нетрансгенных) на 8 микроучастках. Каждая микроучасток включала около 6 растений. Как проиллюстрировано на Фиг. 3А, значительное увеличение среднего диаметра стебля наблюдалось у трансгенных гибридных растений, экспрессирующих конструкцию супрессии (гибрид SUP-GA20ox), по сравнению с гибридными растениями кукурузы дикого типа (контроль). Среднеквадратические ошибки, представленные в виде планок погрешностей на Фиг. 3А, были рассчитаны для трансгенных гибридов и контрольных растений. Изображение поперечного среза стебля гибридного контрольного растения (контроль; слева) приведено рядом с поперечным срезом стебля трансгенного гибридного растения, экспрессирующего элемент супрессии оксидазы GA20 (SUP_GA20ox; справа), далее изображено на Фиг. 3B.
[471] В данном эксперименте средний диаметр стебля выращенных в поле гибридных растений кукурузы, экспрессирующих микроРНК, нацеленные на гены оксидазы_3 GA20 и оксидазы_5 GA20, увеличился примерно на 13% по сравнению с контрольными гибридными растениями дикого типа. Эти данные демонстрируют, что гибридные растения кукурузы, экспрессирующие микроРНК оксидазы GA20, могут иметь более толстые стебли в дополнение к фенотипу с уменьшенной высотой растений.
Пример 4. Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, имели увеличенный сырой вес початка.
[472] Гибридные растения кукурузы, содержащие конструкцию супрессии оксидазы GA20, описанную в Примере 1, также продемонстрировали увеличенный сырой вес початка по сравнению с контрольными растениями дикого типа при выращивании в полевых условиях. Рассчитывали средний вес сырого початка на участок трансгенных гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20, на 24 микроучастках и сравнивали со средним весом сырого початка гибридных контрольных растений кукурузы (нетрансгенных) на 8 микроучастках. Опять же, каждая микроучасток включала около 6 растений. Как изображено на Фиг. 4, увеличение среднего веса сырого початка на участок наблюдалось у трансгенных гибридных растений, экспрессирующих конструкцию супрессии (гибрид SUP-GA20ox), по сравнению с гибридными растениями кукурузы дикого типа (контроль), и развитие початка и зерна было нормальным. Стандартные отклонения для данного эксперимента, представленные в виде планок погрешностей на Фиг. 4, были рассчитаны для трансгенных гибридов и контрольных растений. Как проиллюстрировано на Фиг. 5, аналогичные результаты были получены на другом сайте для полевых испытаний, который также подвергся повреждению от ветра.
[473] В данном эксперименте средний вес сырого початка выращенных в поле гибридных растений кукурузы, экспрессирующих микроРНК, нацеленные на гены оксидазы_3 GA20 и оксидазы_5 GA20, был увеличен по сравнению с контрольными гибридными растениями дикого типа, что указывает на то, что эти трансгенные растения могут дополнительно иметь улучшенные признаки, связанные с урожайностью. Тем не менее, эти результаты основаны на данных наблюдений без крупномасштабного статистического сравнения с контролями, и показатели урожайности следует испытывать в условиях выращивания на обширных полях.
Пример 5. Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, проявляли повышенную устойчивость к полеганию.
[474] В месте проведения полевых испытаний повреждение ветром гибридных растений кукурузы перед цветением показало повышенную устойчивость к полеганию растений, экспрессирующих конструкцию супрессии оксидазы GA20, описанную в Примере 1, по сравнению с контрольными гибридными растениями дикого типа. В то время как гибридные контрольные растения дикого типа (нетрансгенные) были визуально положенными в ответ на данное сильное ветровое воздействие, трансгенные гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, на соседнем поле, показали устойчивость к полеганию. Чтобы оценить эффекты устойчивости к полеганию гибридными растениями кукурузы, экспрессирующими конструкцию супрессии оксидазы GA20, средний вес сырых початков на участок у трансгенных гибридных растений кукурузы, супрессирующих оксидазу GA20, в двух местах полевых испытаний, подверженных полеганию, сравнивали со средним весом сырых початков гибридных контрольных растений дикого типа. Данные, собранные в этих двух исследованиях, показали, что гибридные контрольные растения имели сниженный средний вес сырого початка примерно на 57% и 81%, соответственно для двух исследований, по сравнению с гибридными растениями, экспрессирующими конструкцию супрессии оксидазы GA20.
[475] Визуальное наблюдение того, что гибридные трансгенные растения кукурузы, супрессирующие оксидазу GA20, имели повышенную устойчивость к полеганию в сравнении с нетрансгенными контрольными растениями, наряду с увеличением среднего веса сырого початка в данных испытаниях с трансгенными растениями, супрессирующими оксидазу GA20, показывают, что повышенная устойчивость к полеганию может объяснять увеличение среднего веса сырого початка. Таким образом, повышенная устойчивость к полеганию у растений, супрессирующих оксидазу GA20, может дополнительно увеличить потенциал урожайности/стабильность этих трансгенных растений кукурузы благодаря противостоянию эффектам полегания во время суровых погодных явлений.
Пример 6. Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, показали увеличение индекса урожайности.
[476] Гибридные растения кукурузы, содержащие конструкцию супрессии оксидазы GA20, описанную в Примере 1, дополнительно продемонстрировали увеличение индекса урожайности по сравнению с контрольными растениями дикого типа при выращивании в полевых условиях. Индекс урожайности трансгенных гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20, определяли для растений, выращенных на 8 микроучастках, и сравнивали с нетрансгенными гибридными контрольными растениями кукурузы. Каждая микроучасток включала около 6 растений. Как проиллюстрировано на Фиг. 6, значительное увеличение индекса урожайности наблюдалось у трансгенных гибридных растений, экспрессирующих конструкцию супрессии (гибрид SUP-GA20ox), по сравнению с гибридными растениями кукурузы дикого типа (контроль). Среднеквадратические ошибки, представленные в виде планок погрешностей на Фиг. 6, были рассчитаны для трансгенных гибридов и контрольных растений.
[477] В данном эксперименте индекс урожайности выращенных в поле гибридных растений кукурузы, экспрессирующих микроРНК, нацеленные на гены оксидазы_3 GA20 и оксидазы_5 GA20, увеличился примерно на 11% по сравнению с контрольными гибридными растениями дикого типа. Данное увеличение индекса урожайности было дополнительно связано с уменьшением веса соломы по сравнению с контрольными растениями дикого типа, однако у трансгенных растений не наблюдалось различий в общем сухом весе биомассы.
Пример 7. Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, показали увеличение средней оценки урожайности зерна.
[478] Средняя оценка урожайности зерна для гибридных растений кукурузы, экспрессирующих элемент супрессии оксидазы GA20 (идентифицированный в Примере 1), была рассчитана для 16 микроделянок на поле (с приблизительно 6 растениями на участок). Рассчитанная средняя оценка урожайности зерна для этих трансгенных гибридных растений кукурузы, супрессирующих оксидазу GA20, увеличилась примерно на 15% по сравнению с контрольными гибридными растениями кукурузы (Фиг. 7). Оценка урожайности зерна являет собой показатель, который представляет общую оценку ожидаемой урожайности, основанную на показателях признаков початка. Оценка урожайности зерна получена из початков, собранных вручную на небольших участках, и единицами измерения являются кг/га (вместо буш/акр). Оценка урожайности зерна (кг/га) рассчитывается по формуле (количество зерна на единицу площади (зерен/м2) × вес одного зерна (мг) × 15,5%/100).
Пример 8. Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20, имели увеличение среднего показателя плодовитости.
[479] Гибридные растения кукурузы, экспрессирующие элемент супрессии оксидазы GA20 (идентифицированный в Примере 1), также показали в эксперименте на микроучастке, что они обладают повышенной плодовитостью и вторичными початками по сравнению с нетрансгенными гибридными контрольными растениями. Оценка плодовитости была определена по 10 микроучастках трансгенных гибридных растений кукурузы в поле (приблизительно 6 растений на участок). Как проиллюстрировано на Фиг. 8, средний показатель плодовитости трансгенных гибридных растений кукурузы, супрессирующих оксидазу GA20, составлял 3, тогда как средний показатель плодовитости контрольных растений составлял 1. Для определения оценки плодовитости растения анализировали на развитие вторичных початков на стадии развития R1. Растения оценивали по следующей шкале: 1=образование небольшого вторичного початка или его отсутствие; 2=выметывания пестичных столбиков видно на вторичном початке; 3=развитый вторичный початок возникал из листового влагалища початка; и 4=хорошее развитие вторичного початка, подобного первичному початку. Сбор урожая в конце сезона также показал, что по крайней мере некоторые вторичные початки были продуктивными с нормально развитыми зернами.
Пример 9. Обширные исследования урожайности и признаков на поле с гибридными растениями кукурузы, трансформированными конструкцией супрессии оксидазы GA20.
[480] Конструкция супрессии оксидазы GA20, описанная в Примере 1, была трансформирована в коммерческую женскую инбредную линию кукурузы, и был создан ряд событий трансформации. Трансформированные растения выращивали и самоопыляли для накопления достаточного количества семян, а затем скрещивали с различными коммерческими мужскими инбредными линиями кукурузы для получения гибридных растений кукурузы. Каждая отдельная мужская инбредная линия, применяемая для получения мужского-женского гибрида, называется тестером. Затем гибридные растения кукурузы с различными тестерами выращивали на обширных полях в полевых условиях в соответствии со стандартной агрономической практикой (SAP - standard agronomic practice). Плотность посадки для SAP составляла 83 640 растений на гектар с интервалом 76 сантиметров.
[481] Для полевых испытаний, четыре различных события трансформации, экспрессирующих конструкцию супрессии оксидазы GA20, были скрещены с двумя различными линиями коммерческого тестера. Затем гибридные растения кукурузы были протестированы в 16 географических точках в 6 штатах США на Среднем Западе. Урожай трансгенных гибридных растений кукурузы в этих местах рассчитывали и сравнивали с урожаем нетрансгенных гибридных контрольных растений кукурузы. В Таблице 4 представлена разница урожайности в тоннах/гектар между трансгенными гибридными растениями кукурузы для каждого события по сравнению с нетрансгенным контролем. Отрицательное число указывает на снижение урожая. Различия по урожайности со статистическим значением p менее 0,2 указаны в Таблице 4 жирным шрифтом и курсивом. Эта запись также применяется для указания статистической значимости для оставшихся таблиц в этих примерах, если не указано иное. Как показано в Таблице 4 под заголовком SAP, значительное увеличение урожайности наблюдалось у трансгенных гибридных растений кукурузы, экспрессирующих супрессирующую конструкцию (трансгенные растения) в условиях SAP, относительно гибридных растений кукурузы дикого типа (Контроль). Значительное увеличение урожайности наблюдалось по 4 трансгенным событиям и 2 контрольным линиям.
[482] Сравнимое обширное исследование урожайности было проведено в условиях высокой плотности (HD), с 103 320 растений на гектар и расстоянием между рядами 76 сантиметров, и в сравнении со стандартной агрономической практикой (SAP). Различия в урожайности в условиях HD представлены в Таблице 4 под заголовком HD. Смешанные результаты были получены в этих условиях высокой плотности с урожаем, варьирующимся в зависимости от событий и тестеров. Тем не менее увеличение урожая наблюдалось для двух событий с одним из двух тестеров, и остается возможность для более высокого урожая через большее количество зародышевой плазмы в различных условиях высокой плотности.
[483] Испытания признаков также проводились в полях с целью измерения ряда признаков развития и репродуктивных признаков. Эти испытания проводились с нормальной плотностью (SAP), как описано выше, и в условиях посадки сверхвысокой плотности (UHD) 132 840 растений на гектар с расстоянием между рядами 50 сантиметров. Испытания проводились на гибридных растениях кукурузы с 7 событиями трансформации и 3 тестерами, и данные для каждого тестера были объединены по 7 событиям.
[484] Таблица 5 суммирует результаты испытаний признаков на гибридных растениях кукурузы. Измерением является либо разница в процентах, либо разница в днях или количестве листьев между трансгенными растениями и контролем. Где это уместно, стадия развития, такая как R3 и тому подобное, на которой было выполнено измерение, указана в скобках под столбцом "Название Признака". Сбрасывание пыльцы измеряется с точки зрения количества дней от прорастания до 50% растений, сбрасывающих пыльцу. Появление выметывания пестичных столбиков измеряют по количеству дней от прорастания до 50% растений, выметывающих пестичные столбики. Интервал между сбрасыванием пыльцы и выметыванием пестичных столбиков представляет собой измерение количества дней от 50% растений, сбрасывающих пыльцу до выметывания пестичных столбиков. Сила стебля представляет собой меру силы, при которой сегмент стебля ломается в поперечном направлении с применением специального инструмента для ломания стебля. Индекс площади листьев (LAI - leaf area index) представляет собой безразмерную величину, характеризующую протяженность полога растения, определяемую как площадь одностороннего зеленого листа на единицу площади поверхности земли в пределах пространства широколиственного полога.
Таблица 5. Различия в признаках между трансгенными и контрольными растениями в SAP и UHD.
[485] Как показано в Таблице 5, у трансгенных растений по сравнению с контролем наблюдалось значительное уменьшение высоты растения, высоты початка и длины междоузлия. Трансгенные растения постоянно демонстрируют высоту растений ниже 1,82 метра и высоту початков выше 45 сантиметров, что позволяет собирать урожай без внесения изменений в оборудование. В этом эксперименте наблюдалось увеличение диаметра стебля, особенно в условиях с более высокой плотностью посадки.
[486] Таблица 6 суммирует результаты испытания признаков початка для гибридной кукурузы. Испытания проводились на гибридных растениях кукурузы с 7 событиями трансформации и 3 тестерами, и данные для каждого тестера были объединены по 7 событиям. Измерения представляют собой разницу в процентах дельты между трансгенными растениями и контролем. Где это уместно, стадия развития, такая как R3 и тому подобное, на которой было выполнено измерение, указана в скобках под столбцом "Название Признака". Площадь початка представляет собой средний размер початка на графике с точки зрения площади с двухмерного изображения, полученного при визуализации початка, включая зерна и свободное пространство. Диаметр початка представляет собой средний размер диаметра початка, измеренный как максимальная "широкая" ось початка по его самому широкому сечению. Длина початка представляет собой средний размер длины початка, измеренный от кончика початка по прямой линии до основания узла початка. Кончик початка void_pct представляет собой средний размер процента площади свободного пространства в верхней 30% области початка, с двухмерного изображения, полученного при визуализации початка, включая зерна и свободное пространство. Свободное пространство початка представляет собой средний размер процента площади свободного пространства на початке, с двухмерного изображения, измеренного при визуализации початка, включая зерна и свободное пространство. Оценка урожайности зерна определена в Примере 7. Зерна на единицу площади измеряются как среднее значение числа зерен на единицу площади поля. Початки собирали с заданной длины ряда, обычно одного метра, и обмолачивали и объединяли для подсчета зерен, а затем количество пересчитывали в общее количество зерен на единицу площади поля. Вес одного зерна измеряли как среднее значение веса на зерно. Это рассчитывалось как соотношение (масса образца зерна отрегулированная до 15,5% влажности)/(количество образца зерна). Зерна на початок представляют собой среднее значение количества зерен на початок. Это рассчитывалось как (общий вес зерна/(единичный вес зерна* общее количество початков), где общий вес зерна и общее количество початков измеряются по образцам початка на площади от 0,19 до 10 квадратных метров.
Признака
[487] Как показано в Таблице 6, наблюдалось значительное увеличение площади початка и длины початка в этих экспериментах для трансгенных растений по сравнению с контролем. Также было заметно уменьшение диаметра початка. В этом эксперименте оценка урожайности зерна была в основном нейтральной между трансгенными растениями и контролем.
[488] Дополнительные данные были собраны в полевых условиях со стандартной плотностью по 8 событиям, скрещенным с одним тестером, что свидетельствует об уменьшении высоты растения, высоты початка и длины междоузлия, а также увеличении диаметра стебля и уборочного индекса по сравнению с контролем (данные не показаны). Высоту растений измеряли от земли до самого верхнего листа с язычком на стадии R3. Высота початка измерялась от земли до узла початка на стадии R3. Диаметр стебля измеряли в середине 2-го узла стебля ниже початка, если не указано иное. Эти данные продемонстрировали высокую пенетрантность признаков высоты растений и стебля по событиям, хотя увеличение плодовитости (или количества вторичных початков) не было значительным или выраженным в этих исследованиях.
[489] В отдельном эксперименте рост высоты растения измеряли от стадии V11 до стадии R1 и далее. На Фиг. 9 проиллюстрированы различия в высоте растений между трансгенными растениями и контролем за этот период времени. На фигуре нарисованы пунктирные линии для высоты 5 футов и 6 футов для справки.
Пример 10. Трансгенные растения проявляли улучшенные признаки в стрессовых условиях, вызванных недостатком азота и воды в условиях контролируемой среды.
[490] Этот пример иллюстрирует усиленную реакцию на стресс, вызванный недостатком воды и азота у трансгенных растений кукурузы, имеющих конструкцию супрессии оксидазы GA20, описанную в Примере 1, по сравнению с контролем в автоматизированной теплице (АТ) или в поле, как указано. Устройство и способы автоматического фенотипического анализа растений в АТ описаны, например, в публикации патента США № 2011/0135161, которая включена в данный документ посредством ссылки во всей своей полноте.
[491] В условиях АТ растения кукурузы тестировали в пяти различных условиях, включая отсутствие стресса, стрессовые условия слабого и умеренного дефицита азота и умеренного дефицита воды. Растения кукурузы выращивали в особых стрессовых условиях, приведенных в Таблице 7.
[492] Дефицит воды определяются как удельное объемное содержание воды (ОСВ), которое ниже, чем ОСВ растения, не подверженного стрессу. Например, растение, не подверженное стрессу можно поддерживать при 50% ОСВ, а ОСВ для анализа дефицита воды можно определить между 35% и 40% ОСВ. Данные были собраны с применением визуализации в видимом свете и гиперспектральной визуализации, а также непосредственного измерения массы горшка и количества воды и питательных веществ, ежедневно вносимых к отдельным растениям. Дефицит азота определяется (частично) как удельная концентрация азота в мМ, которая ниже концентрации азота в растении, не подверженном стрессу. Например, растение, не подверженное стрессу может поддерживаться при 8 мМ азота, тогда как концентрация азота, применяемая в анализе дефицита азота, может поддерживаться в концентрации от 4 до 6 мМ.
[493] Для проверки было измерено до десяти параметров. Измерения на основе визуализации в видимом свете: высота растения, биомасса и площадь полога. Высота растения (PlntH) относится к расстоянию от вершины горшка до самой высокой точки растения, полученному из бокового изображения (мм). Биомасса (Bmass) определяется как расчетный сырой вес побегов (г) растения, полученная из изображений, полученных с разных углов. Площадь полога (Cnop) определяется как площадь листа, если смотреть сверху вниз (мм2). Оценка и площадь антоциана, оценка и концентрация хлорофилла и оценка содержания воды были измерены с помощью гиперспектральной визуализации. Оценка антоцианов (AntS) представляет собой оценку антоцианов в листовом пологе, полученную из гиперспектральной визуализации сверху вниз. Область антоцианов (AntA) представляет собой оценку антоцианов в стебле, полученную из гиперспектральной визуализации при виде сбоку. Оценка хлорофилла (ClrpS) и концентрация хлорофилла (ClrpC) являются измерениями хлорофилла в листовом пологе, полученными из гиперспектральной визуализации сверху вниз, где оценка хлорофилла измеряется в относительных единицах, а концентрация хлорофилла измеряется в единицах частей на миллион (чнм). Содержание воды в листве (FlrWtrCt) представляет собой измерение воды в листовом пологе, полученное из гиперспектральной визуализации сверху вниз. Коэффициент полезного использования воды (WUE - Water Use Efficiency) определяется из граммов растительной биомассы на литр добавленной воды. Внесенная вода (WtrAp) представляет собой прямое измерение воды, добавленной в горшок (горшок без отверстия) в ходе эксперимента. Эти физиологические эксперименты были организованы таким образом, чтобы тестируемые трансгенные растения сравнивались с контролем. Трансгенные растения с двумя событиями трансформации измеряли по сравнению с контролем. Все данные представлены в процентном различии дельты трансгенного растения по отношению к контролю. Точка измерения со статистическим p-значением <0,1 показана жирным курсивом. Другие точки измерений имеют p-значение >0,1.
[494] В Таблице 8 приведены результаты испытания признаков в АТ, измеренных через 21 день после посадки на вегетативной стадии, тогда как в Таблице 9 приведены результаты испытаний признаков в АТ, измеренных через 55 дней после посадки на репродуктивной стадии, у трансгенных растений, имеющих одно из двух событий конструкции супрессии оксидазы GA20, описанной в Примере 1, относительно контрольных растений.
Таблица 8. Трансгенные растения против контрольных растений в теплице в нормальных и стрессовых условиях, через 21 день после посадки.
[495] Как показано в Таблице 8, по сравнению с контролем трансгенные растения проявляли некоторые улучшенные признаки, связанные со стрессоустойчивостью, и сохраняли другие положительные признаки в условиях стресса. Высота растений значительно уменьшилась во всех обработках и не была затронута стрессовым состоянием. Площадь биомассы и полога была нейтральной в условиях отсутствия стресса, но увеличилась в более тяжелых стрессовых условиях. Содержание влаги в листве значительно увеличилось в условиях отсутствия стресса и стрессовых условиях, что указывает на то, что трансгенные растения удерживают больше воды в тканях листьев. Площадь антоцианов значительно уменьшилась в условиях отсутствия стресса и стрессовых условиях, что указывает на отсутствие дефицита азота у трансгенных растений.
[496] Как показано в Таблице 9, по сравнению с контролем, у трансгенных растений наблюдалось значительное уменьшение в признаках внесенной воды, WUE, биомассы и массы стебля, что указывает на то, что у трансгенных растений была улучшена эффективность использования воды, причем растениям с более низкой биомассой требуется меньше воды. Уборочный индекс значительно возрос в условиях отсутствия стресса и стрессовых условиях.
Пример 11. Трансгенные растения проявляли повышенную устойчивость к засухе, устьичную проводимость и скорость роста корня на репродуктивных стадиях как при стандартной, так и при высокой плотности посадки в поле.
[497] Прямые наблюдения были сделаны по уменьшению скручивания листьев в трансгенных растениях кукурузы, имеющих конструкцию супрессии оксидазы GA20 из Примера 1, в условиях засухи в полевых условиях по сравнению с контрольными растениями. Скручивание листьев кукурузы происходит, когда потенциал воды в листьях падает ниже порога примерно -1,1 МПа. Устьичная проводимость также уменьшается при водном стрессе. Стабильные соотношения изотопов кислорода (δ18O) применялись в качестве показателя устьичной проводимости, которые обратно пропорциональны устьичной проводимости. Значительное снижение δ18O, и таким образом, значительное увеличение устьичной проводимости трансгенных растений по сравнению с контролем наблюдалось в образцах листьев початка, собранных на стадии R5 (см. Фиг. 10). Данные были взяты из трансгенных растений за два события трансформации и усреднены по 10 тестерам с 2 повторениями на тестер. Увеличение δ18O в листьях контрольных растений указывает на то, что устьичная проводимость была ниже для контроля. В сочетании с уменьшением скручивания листьев, наблюдаемым в полевых условиях, значительное увеличение устьичной проводимости в листьях трансгенных растений в результате испытаний на урожайность в 15 из 16 полевых местоположений указывает на улучшение состояния влаги в листьях во время позднего вегетационного роста трансгенных растений.
[498] Эффективное поглощение воды корнями является важным фактором роста растений. Чтобы измерить прогресс в развитии глубины укоренения, емкостные датчики влажности почвы Sentek® SOLO были установлены на стадии V4 в ряду между растениями в одном полевом местоположении. Влажность почвы ежечасно измеряли емкостными датчиками на глубинах 10, 20, 30, 50, 70, 90, 120 и 150 см от уровня земли. Глубина фронта укоренения определялась наличием суточных структур при истощении влаги в почве, зафиксированных датчиками. Корневая активность уже присутствовала на глубине 10, 20 и 30 см во время установки на стадии V4. Мы обнаружили первое появление истощения влаги в почве на глубинах 50, 70 и 90 см. Почва на глубине 120 и 150 см была насыщена в течение всего периода вегетации. Хотя рост корней, возможно, достиг этих глубин, мы не смогли обнаружить активность корней на этих глубинах для этого эксперимента из-за невозможности обнаружить истощение влаги в почве в насыщенной зоне. На Фиг. 11 проиллюстрировано время (дни после посадки) для фронтального корня растения, необходимое для достижения различных глубин на оси Y. Линии с кружками предназначены для растений с шагом междурядий 76 сантиметров и густотой посадки 83 640 растений на гектар, а линии с квадратами - для растений с шагом междурядий 50 сантиметров и густотой посадки 135 300 растений на гектар. Стадии роста показаны на оси X.
[499] Как проиллюстрировано на Фиг. 11, рост корня в этом эксперименте был сходным между трансгенными и контрольными растениями вплоть до V12, причем корни достигали глубины 50 и 70 см через около 30 и 36 дней после посадки, соответственно. Однако корни трансгенных растений достигли глубины 90 см в R1 или раньше (то есть, через около 50 дней после посадки), или на около 20 дней раньше, чем корни контрольных растений. Трансгенные растения проявляли повышенную скорость фронта укоренения после стадии V11/V12, что может привести к усилению предотвращения засухи в критический период развития растения во время цветения. Это увеличение скорости фронта укоренения может позволить трансгенным растениям использовать более глубокие запасы почвенной воды в течение критического периода на стадии R1, что позволяет избежать, уменьшить или минимизировать влияние засухи на цветение и опыление. Улучшенное опыление в условиях засухи может улучшить набор зерен и потенциал урожайности.
[500] Чтобы дополнить приведенный выше полевой эксперимент с датчиками влажности, скорость корневого фронта для трансгенных растений кукурузы, имеющих конструкцию супрессии оксидазы GA20 из Примера 1 (n=10), была измерена в эксперименте с корневым ящиком и сравнена с контрольными растениями дикого типа (n=9). Корневые ящики из плексигласа (1,5 метра в высоту и пятнадцать на двадцать сантиметров в поперечном сечении; толщина стенок 1,25 сантиметра) были заполнены смесью # 10 полевая почва/вермикулит/перлит (соотношение 1:1:1) и применялись для визуализации корней каждого растения. Максимальная глубина укоренения в каждом ящике измерялась через равные промежутки времени после посадки (примерно каждые два дня). В этом эксперименте средняя глубина фронтового корня трансгенных растений была последовательно больше или глубже, чем у контрольных растений ДТ, начиная с около 21-го дня после посадки (то есть примерно на стадии V4) и продолжая, по крайней мере, до 34 дня после посадки, когда измерения были остановлены (данные не показаны). Это наблюдение в корневых ящиках с контролируемой средой согласуется с увеличенной глубиной корней, наблюдаемой с помощью датчиков влажности в полевых условиях, и показывает, что более глубокие корни могут возникать на более ранних стадиях развития, хотя различия в глубине корней не были обнаружены в полевом эксперименте до окончания стадии V11/V12.
[501] Хотя признаки корня, измеренные в экспериментах с контролируемой средой, описанных в Примере 14 ниже, как правило, не показывают существенной разницы в глубине корня (или показывают только минимальную разницу), эксперимент с вермикулитом в Примере 14 проводился на стадии V3 до того, как различие в глубине корня наблюдалось в эксперименте с корневым ящиком в этом Примере 11 (то есть, начиная со стадии V4), и хотя эксперимент с аэропонным аппаратом в Примере 14 проводился на стадии V5, аэропоническая система не имеет какого-либо взаимодействия растения с почвой (в отличие от эксперимента с вермикулитом), которое может повлиять на нормальный (или более естественный) рост и развитие корней.
Пример 12. Трансгенные растения имеют более высокую устьичную проводимость в нормальных условиях и в условиях засухи и поддерживают более высокую способность фотосинтеза в условиях стресса от засухи.
[502] В теплице также измеряли уровни устьичной проводимости и фотосинтеза в листьях при нормальных условиях и условиях засухи. Для этого эксперимента трансгенные растения с конструкцией супрессии оксидазы GA20 из Примера 1 и контрольные растения дикого типа подвергали хорошему поливу (1500 мл воды в день) или воздействию ограниченного количества воды/хронической засухи (1000 мл воды в день). Двадцать (20) повторений контрольных растений дикого типа и десять (10) повторений на событие (всего два события) для конструкции супрессии оксидазы GA20 подвергали хорошему поливу, и сто сорок (140) повторений контрольных растений дикого типа и семьдесят (70) повторений на событие (всего два события) для конструкции супрессии оксидазы GA20 подвергали воздействию ограниченного количества воды/хронической засухи. Пограничные растения соответствующей высоты (гибриды для растений ДТ и инбреды для трансгенных растений) были размещены по периметру экспериментальных растений в теплице для нормализации эффектов затенения. Суточные измерения устьичной проводимости и фотосинтеза проводились утром и днем с помощью устройства LI-COR® на стадии V12 в соответствии с инструкциями производителя. Как проиллюстрировано на Фиг. 12A, было установлено, что устьичная проводимость была постоянно выше для трансгенных растений как в условиях хорошего полива, так и в условиях засухи в оба момента времени дня. Также наблюдалось, что у трансгенных растений было меньше скручивание листьев в условиях засухи. Как дополнительно проиллюстрировано на Фиг. 12B, более высокая скорость фотосинтеза также наблюдалась в ответ на условия засухи, которая существенно не реагировала на увеличение солнечного света во второй половине дня, в отличие от контрольных растений, которые показали снижение скорости фотосинтеза днем, особенно в условиях засухи.
[503] Эти результаты (в сочетании с отдельными полевыми наблюдениями выше) демонстрируют, что трансгенные растения с конструкцией супрессии оксидазы GA20 не только имели более высокий газообмен и фотосинтез в листе, но поддерживали более высокий газообмен и фотосинтез в листе в ответ на условия ограничения воды/хронической засухи. Дополнительно было отмечено, что у трансгенных растений температура листьев была ниже, чем у контрольных растений (данные не показаны). Таким образом, предсказано, что трансгенные растения, экспрессирующие конструкцию супрессии оксидазы GA20, могут обладать большей устойчивостью к засухе и способностью поддерживать фотосинтез в условиях ограничения воды по сравнению с контролями. Не ограничиваясь теорией, дополнительно предлагается, что более глубокие корни, наблюдаемые для трансгенных растений с конструкцией супрессии оксидазы GA20 (особенно на поздних вегетативных и ранних репродуктивных стадиях), могут способствовать устойчивости к засухе этих трансгенных растений.
Пример 13. Трансгенные растения проявляли репродуктивные признаки, сравнимые с контрольными в тепличных условиях.
[504] Трансгенные растения кукурузы, имеющие конструкцию супрессии оксидазы GA20, описанную в Примере 1, а также контрольные растения выращивали в горшках в теплице до репродуктивной стадии R1, а репродуктивные признаки измеряли на стадиях V8 и R1. Данные были взяты для трансгенных растений двух событий трансформации (Таблица 10). Данные представлены либо с точки зрения разницы в количестве дней, либо в виде процентной разницы для трансгенных растений по сравнению с контролем дикого типа, причем существенные изменения выделены жирным шрифтом. Названия признаков определены в Примерах 9 и 10 выше. Конкретные наблюдения за признаками и классами признаков цветения, незрелого початка, зрелого початка и султана суммированы в таблице. В целом, репродуктивное развитие у трансгенных растений было почти эквивалентно контрольным растениям с небольшими или незначительными изменениями.
1
Вдоль
Пример 14. Корневые признаки трансгенных и контрольных растений в тепличных условиях.
[505] Трансгенные растения, имеющие конструкцию супрессии оксидазы GA20, описанную в Примере 1, и контрольные растения выращивали в теплице на среде с вермикулитом до стадии V3 или в аэропонном аппарате до стадии V5. Растения были извлечены, а корни промыты для непосредственного или оптического измерения корневых признаков. Трансгенные растения с 4 событиями трансформации были протестированы в сравнении с контролем. Результаты измерений суммированы в Таблицах 11 и 12 для растений, выращенных на вермикулитовой среде, или в аэропонном аппарате для роста, соответственно. Количество точек ветвления корня измеряет количество концевых точек ветвления корня растения посредством визуализации корня растения. Изображение корневой системы было представлено в виде наброска для измерения длины корня. Было получено до 40 изображений под разными углами вокруг вертикальной оси корня, и измерения были усреднены по изображениям. Общая длина корня измеряет совокупную длину корней растения, как если бы все корни были выстроены в ряд, путем визуализации корневой системы растения. Изображение корневой системы было представлено в виде наброска для измерения длины корня. Было получено до 40 изображений под разными углами вокруг вертикальной оси корня, и измерения были усреднены по изображениям. Данные в Таблицах 11 и 12 представлены в процентном различии дельты трансгенных растений по сравнению с контролем со значительными изменениями, представленными жирным шрифтом.
[506] Как показано в Таблицах 11 и 12, у трансгенных растений наблюдалось значительное уменьшение высоты растений на стадиях V3 и V5, но в этих экспериментах между трансгенными и контрольными растениями наблюдались только незначительные изменения в общей структуре корней.
Пример 15. Фенотипические наблюдения трансгенных растений с альтернативными промоторами.
[507] В Примерах 1-14 трансгенные растения содержали элемент супрессии оксидазы GA20, функционально связанный с промотором RTBV. Растения кукурузы также трансформировали тем же элементом супрессии, функционально связанным с различными другими промоторами, чтобы проверить, как различные профили экспрессии элемента супрессии оксидазы GA20 могут влиять на высоту растения и другие фенотипы.
[508] Трансгенные растения (растения R0), регенерированные из эксплантов, трансформированных конструкциями, функционально связанными с различными промоторами, наблюдали на стадии роста R5 в теплице, а початки наблюдали после того, как их снимали для просушки. Различные протестированные промоторы указаны в Таблице 13. Наблюдения проводились для растений с множественными событиями трансформации для каждой конструкции, содержащей разные промоторы по сравнению с контрольными растениями той же линии скрещивания без конструкции супрессии оксидазы GA20. Результаты этих наблюдений суммированы в Таблице 13 по событиям трансформации для каждой конструкции.
Таблица 13. Обобщенные результаты наблюдений R0 трансгенных растений с конструкцией супрессии микроРНК для оксидазы GA20 под контролем различных промоторов.
растений R0
без отклонений
без отклонений
без отклонений
без отклонений
без отклонений
без отклонений
сосудисто усиленный
без отклонений
без отклонений
без отклонений
без отклонений
[509] Как показано в Таблице 13, по сравнению с контролями, трансгенные растения R0 с конструкцией супрессии оксидазы GA20 не демонстрировали каких-либо существенных отклонений при наблюдении за всеми протестированными промоторами. Даже экспрессия непосредственно в репродуктивных тканях початка не вызывала заметных отклонений. Высота растений была явно уменьшена не только для конструкции промотора RTBV (в предыдущих Примерах), но также и у трансгенных растений, имеющих одинаковую конструкцию супрессии оксидазы GA20, функционально связанную с различными конститутивными промоторами, листовыми промоторами на разных уровнях экспрессии, некоторыми сосудистыми промоторами и корневым промотором с высоким уровнем экспрессии. Был протестирован сконструированный промотор с конститутивной экспрессией (С1), связанный с конструкцией супрессии оксидазы GA20, и также было обнаружено, что он вызывает фенотип короткого роста. Аналогично, было обнаружено, что, по меньшей мере, один сконструированный промотор с сосудистой экспрессией (V1), связанный с конструкцией для супрессии оксидазы GA20, вызывает фенотип короткого роста в дополнение к сосудистому промотору Cyp2 риса, хотя растения с двумя другими сконструированными сосудистыми промоторами (V2, V3) и тремя сконструированными промоторами стебля (S1, S2, S3) не имели уменьшенной высоты растения. Однако изменение последовательности терминатора транскрипции для конструкции супрессии оксидазы GA20 под контролем промотора RTBV не изменило фенотип короткого роста (не показано в Таблице 11). Применяемый в данном документе термин "средне-короткий" относится к умеренному уменьшению высоты растения (относительно уменьшения высоты растения, наблюдаемого с промотором RTBV), а наблюдение "некоторых коротких" означает, что были некоторые различия в степени уменьшения высоты растения.
[510] Эти результаты показывают, что экспрессия элемента супрессии оксидазы GA20 конститутивными промоторами неизменно приводила к фенотипу короткого роста, хотя с этими конститутивными промоторами наблюдалась некоторая изменчивость в фенотипах высоты растений. Аналогичным образом, комбинация промотора RTBV с энхансерным элементом для преобразования профиля экспрессии из сосудистого в конститутивный по-прежнему вызывает фенотип короткого роста (что указывает на достаточность промотора RTBV). Несколько сосудистых промоторов, включая промотор RTBV, продуцировали фенотип с коротким ростом, но пара других сконструированных сосудистых промоторов не продуцировала фенотип с коротким ростом, что может быть связано с более низким уровнем экспрессии с этими промоторами. Ни один из промоторов стебля не продуцировал фенотип короткого роста, что указывало на то, что экспрессии конструкции супрессии GA20 в стебле было недостаточно для получения этого фенотипа. Удивительно, но экспрессия конструкции супрессии GA20 в листе постоянно приводила к фенотипам короткого роста с различными уровнями экспрессии, хотя результаты были в определенной степени вариабельными. Эти данные указывают на то, что производство активных GA в ткани листьев способствует росту растений и, в конечном итоге, высоте растений, даже если такой вертикальный рост происходит в стебле растения. Экспрессия конструкции супрессии оксидазы GA20 с различными корневыми промоторами, как правило, не давала фенотипа короткого роста, хотя один корневой промотор действительно продуцировал умеренный фенотип, что может быть связано с дополнительной экспрессией в надземных растительных тканях.
[511] Затем растения R0 самоопылялись и полученные семена выращивали в рассаднике для получения гомозиготного инбредного потомства растений (растения R1). Наблюдения за потомками трансгенных растений R1 с некоторыми промоторными конструкциями (по меньшей мере, 4 события трансформации на конструкцию) проводили на стадии развития R1 по сравнению с контрольными растениями той же линии скрещивания без конструкции супрессии оксидазы GA20. Как и растения R0, было обнаружено, что растения потомков R1, экспрессирующие конструкцию супрессии оксидазы GA20 с каждым из промоторов RTBV, CAMV e35S и полиюбиквитиновым промотором Coix lacryma-jobi имеют низкорослый, полукарликовый фенотип, без каких-либо существенных отклонений.
Пример 16. Фенотипические наблюдения трансгенных растений кукурузы с конструкциями, нацеленными на различные гены оксидазы GA.
[512] Приведенные выше Примеры демонстрируют, что микроРНК-экспрессирующая конструкция, нацеленная на гены оксидазы_3 GA20 и оксидазы_5 GA20 для супрессии, и функционально связанная с экспрессируемым в растении сосудистым, конститутивным и/или листовым промотором, может быть применена для создания короткорослого, полукарликового кукурузного растения. Чтобы проверить, как нацеливание на различные гены оксидазы GA20 или GA3, или разные части генов оксидазы_3 GA20 и/или оксидазы_5 GA20 для супрессии, может повлиять на высоту растения, было создано несколько конструкций и трансформировано в растения кукурузы. Конструкции также были созданы с той же нацеливающей последовательностью, что и в приведенных выше Примерах, но с другой последовательностью остова микроРНК (две из микроРНК кукурузы, одна из микроРНК сои и одна из микроРНК хлопка - конструкция в приведенных выше Примерах применяла рисовую последовательность остова микроРНК). В Таблице 14 приведен список этих дополнительных конструкций супрессии, а также наблюдения за трансгенными растениями R0 в теплице, содержащими эти конструкции (по сравнению с контрольными растениями дикого типа). Конструкции, нацеленные на (i) оксидазу_1 GA20/оксидазу_2 GA20, (ii) оксидазу_3 GA20/оксидазу_9 GA20, (iii) оксидазу_7 GA20/оксидазу_8 GA20 и (iv) оксидазу_3 GA20/оксидазу_5 GA20 (с разными остовами микроРНК), каждая из которых кодируется микроРНК с одной последовательность нацеливания, комплементарной обоим целевым генам, в то время как наборы (i) отдельных последовательностей, нацеленных на оксидазу_3 GA20 и оксидазу_5 GA20, (ii) отдельных последовательностей, нацеленных на оксидазу_4 GA20 и оксидазу_6 GA20, и (iii) отдельных последовательностей, нацеленных на оксидазу_4 и оксидазу_7/8 GA20, каждая экспрессировалась в виде одной пре-микроРНК с двумя последовательностями нацеливания, расположенными в тандеме, которые расщепляются и разделяются на две зрелые микроРНК. В Таблице 14 представлена последовательность, нацеленная на микроРНК, и последовательность кДНК, комплементарная последовательности, нацеленной на микроРНК. Для конструкции оксидаза_1 GA20/оксидазы_2 GA20 звездочка (*) указывает на то, что длина выравнивания между последовательностью нацеливания микроРНК и целью мРНК или сайтом узнавания была короче (17 против 20 нуклеотидов) для оксидазы_1 GA20, чем для оксидазы_2 GA20. Аналогично для конструкции оксидазы_3 GA20/оксидазы_9 GA20 звездочка (*) указывает на то, что длина выравнивания между последовательностью нацеливания микроРНК и целью мРНК или сайтом узнавания была короче (17 против 20 нуклеотидов) для оксидазы_9 GA20, чем для оксидазы_3 GA20. Для каждой из конструкций, перечисленных в Таблице 14, значительных отклонений не наблюдалось, за исключением наблюдений, представленных в таблице.
Таблица 14. Обобщенные результаты наблюдений R0 трансгенных растений кукурузы с конструкциями супрессии микроРНК, нацеленными на различные гены оксидазы GA.
(Конструкция/Промотор)
Целевая область
Оксидаза_2 GA20
(промотор RTBV)
2: экзон
Оксидаза_9 GA20
(промотор RTBV)
9: экзон*
Оксидаза_8 GA20
(промотор RTBV)
(Отдельный; промотор RTBV
и 35S)
(~ 6 сантиметров против ДТ)
(Отдельный; промотор RTBV
и 35S)
Оксидаза_5 GA20
(Отдельный; тандемный набор)
Оксидаза_5 GA20
(Разные остова микроРНК)
(промотор RTBV)
(промотор RTBV)
(наблюдалось только 3 события)
(промотор CAMV e35S )
(промотор RTBV)
(более темные зеленые листья)
(промотор CAMV e35S )
Оксидаза_6 GA20
(Отдельный; тандемный набор)
умеренно короче (~ 20%)
Оксидаза_7/8 GA20
(Отдельный; тандемный набор)
умеренно короче (~ 20%)
[513] Наблюдения, обобщенные в Таблице 14, демонстрируют, что нацеливание на некоторые другие гены оксидазы GA20 не приводило к низкорослому, полукарликовому фенотипу. Ни одна из конструкций, нацеленных на (i) родственные гены оксидазы_1 GA20 и оксидазы_2 GA20, (ii) родственные гены оксидазы_3 GA20 и оксидазы_9 GA20, (iii) родственные гены оксидазы_7 GA20 и оксидазы_8 GA20 или (iv) только ген оксидазы_9 GA20 не приводила к наблюдаемому низкорослому, полукарликовому фенотипу у растений R0. Напротив, те конструкции, которые кодируют одну микроРНК, совместно нацеленную на гены оксидазы_3 GA20 и оксидазы_5 GA20 в трансгенных растениях R0 и R1, действительно давали низкорослый, полукарликовый фенотип, даже если применялась другая последовательность терминации транскрипции или разные остова микроРНК (всего из 5 протестированных последовательностей остова микроРНК). Кроме того, нацеливание на различные последовательности генов оксидазы_3 GA20 и оксидазы_5 GA20 все еще приводит к получению полукарликовых растений. Интересно, что конструкции супрессии, которые были сконструированы для нацеливания на каждый из генов оксидазы_3 GA20 и оксидазы_5 GA20 по отдельности, не продуцировали низкорослый, полукарликовый фенотип, в отличие от конструкций, совместно нацеленных на гены оксидазы_3 GA20 и оксидазы_5 GA20, хотя конструкция, отдельно нацеленная на ген оксидазы_3 GA20, действительно вызывала небольшое уменьшение высоты растения. Тем не менее, трансгенные растения, имеющие тандемный векторный набор конструкций супрессии, отдельно нацеленных на гены оксидазы_3 GA20 и оксидазы_5 GA20, действительно продуцировали низкорослый, полукарликовый фенотип, подобно конструкциям, кодирующим одну микроРНК, совместно нацеленную на гены оксидазы_3 GA20 и оксидазы_5 GA20. Эти данные демонстрируют, что низкорослый, полукарликовый фенотип наблюдается с конструкциями, нацеленными как на гены оксидазы_3 GA20, так и на гены оксидазы_5 GA20, но полный полукарликовый фенотип не наблюдается при нацеливании на гены оксидазы_3 GA20 и оксидазы_5 GA20 по отдельности для супрессии (только небольшое уменьшение высоты при нацеливании на оксидазу_3 GA20 и отсутствие фенотипа высоты растения при нацеливании на оксидазу_5 GA20). Кроме того, как описано, фенотип высоты растения не наблюдался при нацеливании на ген(ы) оксидазы_1 GA20, оксидазы_2 GA20, оксидазы_6 GA20, оксидазы_7 GA20, оксидазы_8 GA20 и/или оксидазы_9 GA20.
[514] Помимо генов оксидазы_3 GA20 и оксидазы_5 GA20, наблюдалось умеренное уменьшение высоты растений в трансгенных растениях R0 с конструкцией супрессии, включающей две целевые последовательности в тандеме, комплементарном совместной цели: (i) гены оксидазы_4 GA20 и оксидазы_6 GA20 или (ii) гены оксидазы_4 GA20, оксидазы_7 GA20 и оксидазы_8 GA20 - одна из двух целевых последовательностей направлена как на гены оксидазы_7 GA20, так и на гены оксидазы_8 GA20. Учитывая, что отдельная конструкция, которая нацелена на гены оксидазы_7 GA20 и оксидазы_8 GA20, не продуцировала фенотип высоты растения, а конструкция супрессии, нацеленная на гены оксидазы_4 GA20 и ген оксидазы_6 GA20, продуцировала фенотип высоты растения, который был сходен с конструкцией супрессии нацеленной на гены оксидазы_4 GA20, оксидазы_7 GA20 и оксидазы_8 GA20, предполагается, что нацеливание на ген оксидазы_4 GA20 в значительной степени (если не полностью) отвечает за фенотип высоты растений, наблюдаемый у этих трансгенных растений. Кроме того, трансгенные растения кукурузы с конструкциями, нацеленными на гены оксидазы_1 GA3 или оксидазы_2 GA3, также продемонстрировали снижение высоты растения, хотя в этом фенотипе была некоторая изменчивость в зависимости от конститутивного промотора. Таким образом, в дополнение к генам оксидазы_3 GA20 и оксидазы_5 GA20, гены оксидазы_4 GA20, оксидазы_1 GA3 и оксидазы_2 GA3 также могут быть целью для супрессии с целью получения низкорослых, полукарликовых растений.
Пример 17. Фенотипические наблюдения растений кукурузы, имеющих редактированный ген оксидазы_3 GA20 или оксидазы_5 GA20.
[515] В дополнение к вышеупомянутым конструкциям супрессии в эндогенных генах оксидазы_3 GA20 и оксидазы_5 GA20 в растениях кукурузы были созданы несколько мутаций путем редактирования генома для проверки фенотипического эффекта нокаута каждого из этих генов. Для каждого из генов оксидазы_3 GA20 и оксидазы_5 GA20 была создана серия из десяти одноцепочечных направляющих РНК (sgРНК), кодирующих нацеливающие конструкции, которые нацелены на разные позиции вдоль геномной последовательности для каждого гена. Была создана дополнительная серия из десяти sgРНК, каждая из которых нацелена на оба гена оксидазы_3 GA20 и оксидазы_5 GA20 в одинаковых или разных положениях вдоль геномной последовательности для каждого гена. Целевые изменения в геноме были сделаны путем доставки sgРНК вместе с экспрессией белка Cas9 эксплантам кукурузы, чтобы вызвать появление ДЦР или одноцепочечный разрыв в геномного целевом сайте для нRNA или около него, который затем может быть неправильно репарирован с целью введения мутации в целевом сайте или около него. Присутствие мутации было впоследствии подтверждено с помощью анализа ПДРФ (полиморфизма длин рестрикционных фрагментов) и/или сиквенированием растений. В приведенной ниже Таблице 15 представлен список протестированных конструкций направляющих РНК (нРНК), которые могут быть применены для редактирования генома одного или обоих гена(ов) оксидазы_3 GA20 и оксидазы_5 GA20 с помощью РНК-направленной эндонуклеазы. Эти конструкции направляющих РНК, как правило, предназначены для нацеливания на кодирующие последовательности генов оксидазы_3 GA20 и оксидазы_5 GA20, но некоторые из комбинированных целевых конструкций могут вместо этого нацеливаться на последовательность НТО одного из двух генов. Эти нРНК могут быть применены с подходящей эндонуклеазой для получения двухцепочечного разрыва (ДЦР) или одноцепочечного разрыва в геноме в геномном целевом сайте, или около него, соответствующей нРНК, который может быть несовершенно репарирован, с целью создания мутации (например, вставки, делеции, замены и тому подобного). Трансгенные растения, которые были гомозиготными по редактированному гену оксидазы_3 GA20 или гомозиготными по редактированному гену оксидазы_5 GA20, были получены из нескольких конструкций (см., жирный текст). События также генерировались из конструкций, предназначенных для редактирования обоих генов. Для конструкций, совместно нацеленных на гены оксидазы_3 GA20 и оксидазы_5 GA20, координаты кодирующей последовательности (CDS) приведены в отношении одного из двух генов, указанных в скобках. Таблица 15 далее показывает, какие конструкции произвели события редактирования генов, были ли эти события гомозиготными или гетерозиготными в растениях R0, а числа±в скобках указывают вероятное изменение последовательности с мутацией (например, +1 означает вставку 1 нуклеотида и тому подобное, и более крупные или более сложные инделы помечены как "дел." или "вставка"). Для комбинированного нацеливания на оксидазу_3 GA20 и оксидазу_5 GA20 идентичность мутированного гена также указана в скобках. В соответствии с результатами для конструкций супрессии, трансгенные растения, гомозиготные по редактированному гену оксидазы_3 GA20 или оксидазы_5 GA20, не имели низкорослого, полукарликового фенотипа, а имели нормальную высоту растения относительно контрольных растений (см., конструкции оксидазы_3-D GA20 и оксидазы_3-G GA20, а также конструкции оксидазы_5-B GA20 и оксидазы_5-G GA20 в Таблице 15), что указывает на то, что нокаут только одного из этих генов недостаточен для получения полукарликового фенотипа.
Таблица 15. Направляющие РНК (нРНК) нацеленные на гены оксидазы_3 GA20 и оксидазы_5 GA20 с целью редактирования.
(SEQ ID NO)
2. гетерозиготный (-1)
3. биаллельный (-2, +1)
2. гетерозиготный (-1)
3. биаллельный (+1, -1)
2. гетерозиготный (+1)
3. гетерозиготный (+1, -7)
4. гетерозиготный (-3, -1)
2. гомозиготный (-1)
(Ox_3 GA20)
(Ox_3 GA20)
(Ox_5 GA20)
(Ox_3 GA20)
(Ox_5 GA20)
(Ox_3 GA20)
гетеро (Ox5, +1, дел.)
2. гетеро (Ox3, +1, дел.)
гетеро (Ox5, +1, вставка)
(Ox_3 GA20)
(Ox_5 GA20)
(Ox_5 GA20)
(Ox_5 GA20)
Пример 18. Конструкция супрессии, нацеленная на гены оксидазы_3 GA20 и оксидазы_5 GA20, снижает уровни транскриптов оксидазы GA20 и уровни активной GA в растении.
[516] Чтобы определить, как уровни транскрипта оксидазы GA20 были затронуты в трансгенных растениях с помощью конструкции супрессии, нацеленной на гены оксидазы_3 GA20 и оксидазы_5 GA20, целые ткани из различных частей трансгенных растений, выращенных в теплице, были взяты на разных вегетативных стадиях (V3, V8 и V14), а уровни транскриптов мРНК для каждого из генов оксидазы GA20 анализировали с применением анализа TaqMan®. Для этих экспериментов суммарную РНК экстрагировали с применением набора для экстракции РНК от Zymo Research™ и обрабатывали ДНКазой Turbo™ для уменьшения загрязнения геномной ДНК. Затем РНК подвергали обратной транскрипции с целью получения двухцепочечной кДНК. Количественную ПЦР с обратной транскрипцией проводили с применением геноспецифических праймеров и меченных FAM зондов TaqMan® в Bio-Rad® CFX96 Real Time System. Метрики контроля качества рассчитывались с применением тканеспецифических стандартов для определения эффективности кПЦР и суммарной РНК, которая не подвергалась обратной транскрипции для учета остаточного загрязнения геномной ДНК. Разница между пороговыми значениями цикла для генов, представляющих интерес против нормализующих генов определяла относительное количество каждого транскрипта гена в каждой ткани. Эта относительная величина была рассчитана с применением одного (18S) или среднего геометрического двух (18S и ELF1A) нормализующих генов.
[517] В этом эксперименте уровень транскрипта оксидазы_3 GA20 был снижен в большинстве вегетативных тканей на этих стадиях, включая ткань листьев и стеблей в V3, ткань междоузлия в V8 и ткань листьев и междоузлия в V14, хотя уровень транскрипта оксидазы_3 GA20 в корне V3 и листе V8 оказался неизменным (данные не показаны). Кроме того, уровень транскрипта оксидазы_5 GA20 для этого эксперимента в целом оставался неизменным в тестируемых вегетативных тканях (данные не показаны), хотя уровень экспрессии транскрипта оксидазы_5 GA20 был относительно низким в этих тканях. Ни оксидаза_3 GA20, ни оксидаза_5 GA20 не были значительно снижены в образцах тканей корня трансгенных растений. Уровень транскриптов каждого из других генов оксидазы GA20 (то есть подтипы 1, 2, 4 и 6-9), как правило, не изменялся или увеличивался в некоторых тканях трансгенных растений.
[518] Аналогичный эксперимент был проведен с репродуктивными тканями трансгенных растений, экспрессирующих ту же самую конструкцию супрессии. Цельные ткани из различных частей трансгенных растений, выращенных в теплице, отбирали на разных репродуктивных стадиях (R1 и R3), а уровни транскриптов мРНК для каждого из генов оксидазы GA20 анализировали с применением анализа TaqMan®. В этом эксперименте уровни транскриптов оксидазы_3 GA20 и оксидазы_5 GA20 были в основном неизменными в R1 листьев, початков, султанов и междоузлий и в R3 листьев и междоузлий по сравнению с контролями (данные не представлены). Результаты для других генов оксидазы GA20 были в основном смешанными или нейтральными (данные не показаны).
[519] Эти данные показывают, что уровень транскриптов оксидазы_3 GA20 в трансгенных растениях кукурузы во время вегетативных стадий, как правило, снижены в этом эксперименте, но, по-видимому, практически не изменялся по сравнению с контрольными тканями растений на более поздних репродуктивных стадиях. Хотя явного снижения уровня транскриптов гена оксидазы_5 GA20 в этих трансгенных растительных тканях обычно не наблюдалось, уровень экспрессии этого гена был относительно низким. Таким образом, изменения в уровне его экспрессии, возможно, было трудно обнаружить с помощью этого способа. Кроме того, конструкция супрессии, по-видимому, специфична для целевых генов оксидазы GA20, поскольку в этом эксперименте не наблюдалось последовательного снижения уровня экспрессии ни для одного из других генов оксидазы GA20.
[520] В отдельном эксперименте уровни экспрессии оксидазы GA20 были определены в тканях стебля трансгенных растений, экспрессирующих конструкцию супрессии из предыдущих Примеров (нацеленную на гены оксидазы_3 GA20 и оксидазы_5 GA20 для супрессии под контролем промотора RTBV), по сравнению с контролем дикого типа. Образцы тканей отбирали из стеблей V3-V6, и части этих стеблей дополнительно рассекали для отделения сосудистых и несосудистых тканей с целью определения уровней дифференциальной экспрессии среди этих тканей. Уровни экспрессии транскрипта определяли с применением метода РНК-сиквенирования (RNA-Seq) для количественного сравнения между тканями трансгенного растения и растения дикого типа. Данные, представленные на Фиг. 13А получены из трансгенных растений, имеющих одно из двух событий, и контрольных растений дикого типа, имеющих одну из двух зародышевых плазм, причем каждый столбец на Фиг. 13А, представляет одно из двух трансгенных событий или гермоплазм, соответственно. Для этих экспериментов отдельные сосудистые пучки отделяли от остальной ткани стебля образцов и подвергали отдельному анализу. Как проиллюстрировано на Фиг. 13А, микроРНК, экспрессируемая конструкцией супрессии, была обнаружена в общей стеблевой ткани растения ("общая", то есть, без разделения сосудистой и несосудистой ткани), а также в отделенной сосудистой ("Сосуд") и несосудистой ("Несосуд") тканях из общего образца стебля. Однако уровень экспрессии микроРНК был значительно выше в сосудистой, чем в несосудистой ткани, что указывает на сосудистый профиль экспрессии промотора RTBV.
[521] Общие образцы стебля и отделенные сосудистые и несосудистые образцы также анализировали в аналогичном эксперименте RNA-Seq для измерения и сравнения уровней транскриптов гена оксидазы_3 GA20 и оксидазы_5 GA20 в трансгенных растениях по сравнению с контрольными растениями дикого типа (наряду с другими генами оксидазы GA20), хотя для каждого типа ткани показан только один образец дикого типа. Для этих экспериментов ткань стебля из контрольных или трансгенных растений (два события) разрезали на отдельные сосудистые пучки и несосудистые ткани, как описано выше. Общая sРНК и мРНК были сиквенированы для каждого образца, и данные были проанализированы и сравнены с применением анализа главных компонентов.
[522] Как проиллюстрировано на Фиг. 13B, уровни транскрипта гена оксидазы_3 GA20 были значительно снижены в общей стволовой ткани (Общая) и в отдельных сосудистых тканях стебля (Сосуд) трансгенных растений (ТР) по сравнению с контролями дикого типа (ДТ), но оставались неизменными в отделенной несосудистой (Несосуд) ткани. Однако уровни транскриптов гена оксидазы_5 GA20 были значительно снижены в общей ткани стебля (Общая), но относительно не изменились в отделенных сосудистых (Сосуд) и несосудистых (Несосуд) тканях трансгенных растений, хотя наблюдалась тенденция к снижению для транскрипта оксидазы_5 GA20 в образцах сосудистой (Сосуд) ткани из трансгенных растений. Уровень экспрессии гена оксидазы_5 GA20 был низким, особенно в несосудистых тканях. Все другие гены оксидазы GA20 не показали значительного снижения уровней транскриптов в анализируемых тканях трансгенных растений, хотя пара генов оксидазы GA20 показала небольшую тенденцию к повышению уровня их экспрессии. Эти данные дополнительно демонстрируют, что уровни экспрессии генов оксидазы_3 GA20 и оксидазы_5 GA20 снижаются в разной степени в одной или более тканях трансгенных растений, имеющих конструкцию супрессии относительно контролей. Действительно, более высокая экспрессия микроРНК и более сильная супрессия эндогенного гена оксидазы_3 GA20 в сосудистых тканях согласуется с сосудистым профилем экспрессии промотора RTBV и, возможно, более высокие уровни экспрессии гена оксидазы_3 GA20 в сосудистых против несосудистых тканей растений дикого типа. Аналогичный профиль наблюдается также для гена оксидазы_5 GA20, хотя и не настолько выраженный, как у оксидазы_3 GA20 между сосудистыми и несосудистыми тканями.
[523] Низкорослый, полукарликовый феномен, наблюдаемый с супрессией оксидазы GA20 в трансгенных растениях, вероятно, опосредован снижением уровня активных GA, присутствующих в тканях стебля или междоузлия и/или в тканях растений, которые продуцируют активные GA. Чтобы определить уровни активных GA (особенно G1, G3 и G4) относительно других неактивных форм гормона, уровни GA измеряли в различных образцах ткани, взятых у контрольных растений трансгенного и дикого типа на разных стадиях развития. Для этих экспериментов свежезамороженные образцы для каждой ткани измельчали и распределяли в 96-луночные стеклянные пробирки вместе с внутренними стандартами. Образцы экстрагировали с применением растворителя метанол:вода:уксусная кислота (80:19:1 об./об.) два раза в течение 4 часов при 4°С. Растворитель выпаривали из экстракта почти до сухости, применяя многоканальный SPE с азотом. Образцы дополнительно очищали с применением картриджа SPE. После очистки образцы обрабатывали с применением стандартного способа ЖХ-МС/МС с масс-спектр измерительными приборами Shimadzu® Nexera® UPLC и SCIEX® triple quad 5500. Хроматографы были проанализированы и определены количественно с применением внутренних стандартов.
[524] Два набора экспериментов были выполнены с образцами, взятыми из различных тканей вегетативной стадии растения. Как показано в Таблице 16 для одного эксперимента в теплице, сниженные уровни активных GA (GA1, GA3 и GA4) наблюдались в различных тканях трансгенных растений на разных вегетативных стадиях. Данные в Таблице 16 представлены в виде числа трансгенных растений, имеющих значительное изменение количества каждого гормона GA для данной ткани ("U"=вверх или увеличен; "D"=вниз или уменьшен; "N"=нейтральный или без изменений и "Т"=общее количество растений). GA, которые демонстрировали, по крайней мере, частичное уменьшение в образцах ткани, выделены жирным шрифтом. Активная GA1 была снижен в тканях листьев и междоузлий на стадии V8 и в ткани междоузлий на стадии V14, а активная GA4 была уменьшена в стебле V3 и междоузлиях V8 и V14. Однако активная GA3 не была заметно уменьшена в этом эксперименте. Другие неактивные формы GA были изменены в различных тканях трансгенных растений, как показано в Таблице 16. В целом, GA, которые находятся в 5'-3' направлении генов оксидазы GA20 в пути гиббереллиновой кислоты (например, GA9, GA20 и GA34), имеют тенденцию к снижению, тогда как GA, которые находятся в 3'-5' направлении генов оксидазы GA20, имеют тенденцию к повышению (например, GA12 и GA53), что может быть связано с более низкой активностью генов оксидазы GA, вызывающих накопление предшественников GA в 3'-5' направлении. Эти данные согласуются с супрессией активности оксидазы GA20 в этих тканях и более низким уровнем активных гормонов GA в стебле и листьях трансгенных растений.
[525] В отдельном эксперименте аналогичные измерения гормонов GA были взяты из различных тканей растений во время вегетативных стадий развития. Как показано в Таблице 17 для эксперимента с применением тканей, взятых у растений в теплице и поле, пониженные уровни одной или более активных GA (GA1, GA3 и GA4) наблюдались в листьях и междоузлиях трансгенных растений на стадиях V3 и V8. Образцы листьев на стадии V8 для этого эксперимента были взяты у растений в поле, в отличие от других образцов, взятых у растений в теплице. Данные в Таблице 17 отображаются аналогично тому, как описано в Таблице 16. Другие неактивные формы GA были изменены в различных тканях трансгенных растений, как показано в Таблице 17. Подобно наблюдениям, изложенным выше, GA, которые находятся в 5'-3' направлении генов оксидазы GA20 в пути гиббереллиновой кислоты (например, GA9, GA20 и GA34), имеют тенденцию к снижению, тогда как GA, которые находятся в 3'-5' направлении генов оксидазы GA20, имеют тенденцию к повышению (например, GA12 и GA53). Эти данные снова согласуются с супрессией активности оксидазы GA20 в этих тканях и более низким уровнем активных гормонов GA в стебле и листьях трансгенных растений.
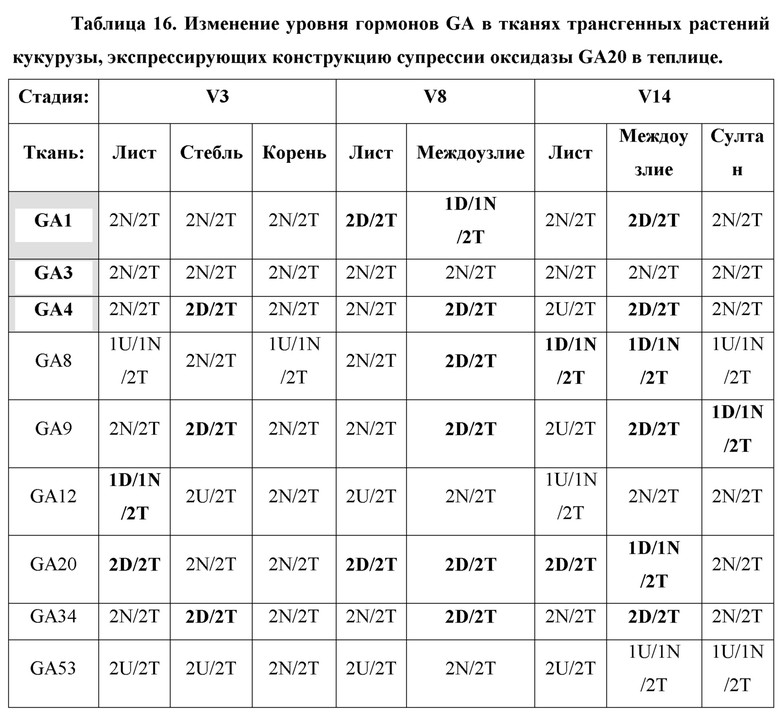
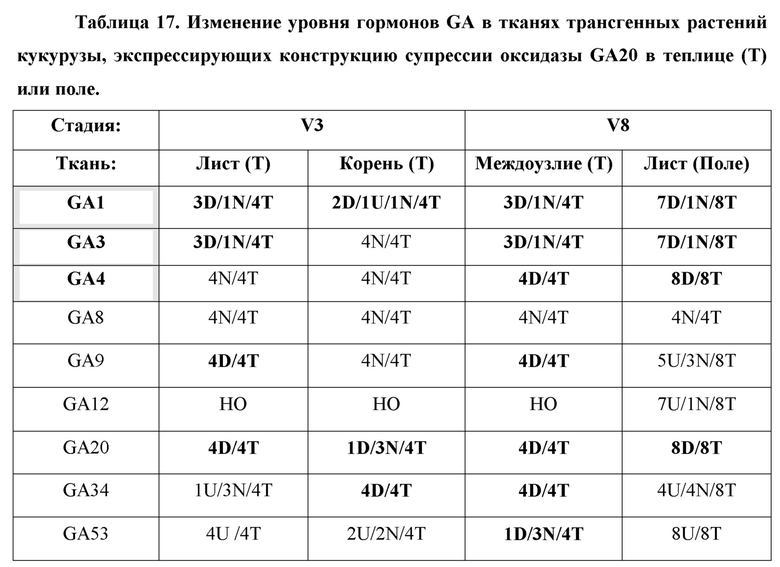
[526] Супрессия генов оксидазы оксидазы_3 GA20 и оксидазы_5 GA20 в трансгенных растениях кукурузы снижает уровни целевых транскриптов оксидазы GA в различных тканях, включая стебель, междоузлие сосудистые ткани и листья, и супрессия этих генов оксидазы GA20 также дополнительно связана со снижением уровней активных GA в тканях трансгенного растения, включая стебель и междоузлия, которые являются местом воздействия на рост растений на вегетативных стадиях и, в конечном счете, на высоту растений на более поздних вегетативных и репродуктивных стадиях. Аналогично наблюдениям, что уровни транскрипта оксидазы GA20 в тканях репродуктивной стадии в основном не изменяются или смешиваются, уровни гормонов GA, включая активные GA, также в основном не изменяются или смешиваются в тканях репродуктивной стадии (данные не показаны).
[527] После подробного изложения данного описания будет очевидно, что возможны модификации, варианты и эквивалентные варианты реализации изобретения без отклонения от сущности и объема данного описания, как описано в данном документе и в прилагаемой формуле изобретения. Кроме того, следует понимать, что все примеры в данном описании представлены в качестве неограничивающих примеров.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Monsanto Technology LLC
Allen, Edwards M.
Boddu, Jayanand
Dietrich, Charles R.
Goldshmidt, Alexander
Howell, Miya
Kosola, Kevin R.
Neelam, Anil
Slewinski, Thomas L.
Venkatesh, Tyamagondlu V.
Wang, Huai
<120> СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УВЕЛИЧЕНИЯ УРОЖАЙНОСТИ НИЗКОРОСЛЫХ
РАСТЕНИЙ ПУТЕМ МАНИПУЛЯЦИИ МЕТАБОЛИЗМА ГИББЕРЕЛЛИНА
<130> P34494WO00
<150> США 62/376,298
<151> 2016-08-17
<150> США 62/442,377
<151> 2017-01-04
<150> США 62/502,313
<151> 2017-05-05
<160> 167
<170> PatentIn версия 3,5
<210> 1
<211> 1741
<212> ДНК
<213> Zea mays
<400> 1
gacggtagtt ttcatctaaa gtttattctt cgtcacatgg gatggccgtt tgcttgtttg 60
ttgcttccgg gaggcggtgg tgaattgaag cagatcgaca agcatggctg cccactggtc 120
tcgatcgatc ggcctgccat gccatgccat gccactagag tccgtcctga ctggccgccc 180
gttcccccgt ataaaaaggc aggcaggcag gcagagcggg gacgagcaag caagcagttg 240
cagttgcagc ggcctcctcc tctgcttcct ccctcctcct cctcaccatg gtgctggctg 300
cgcacgatcc ccctcccctt gtgttcgacg ctgcccgcct gagcggcctc tccgacatcc 360
cgcagcagtt catctggccg gcggacgaga gccccacccc ggactccgcc gaggagctgg 420
ccgtgccgct catcgacctc tccggggacg ccgccgaggt ggtccggcag gtccggcgcg 480
cctgcgacct gcacggcttc ttccaggtgg tggggcacgg catcgacgcg gcgctgacgg 540
cggaggccca ccgctgcatg gacgccttct tcacgctgcc gctcccggac aagcagcgcg 600
cgcagcgccg ccagggggac agctgcggct acgccagcag cttcacgggc cggttcgcgt 660
ccaagctgcc ctggaaggag acgctgtcgt tccgctacac cgacgacgac gacggcgaca 720
agtccaagga cgtcgtggcg tcctacttcg tggacaagct gggcgagggg taccggcacc 780
acggggaggt gtacgggcgc tactgctctg agatgagccg tctgtcgctg gagctcatgg 840
aggtgctagg cgagagcctg ggcgtgggcc ggcgccactt ccggcgcttc ttccagggga 900
acgactccat catgcgcctc aactactacc cgccgtgcca gcggccctac gacacgctgg 960
gcacggggcc gcattgcgac cccacgtcgc tcaccatcct gcaccaggac gacgtgggcg 1020
gactccaggt gttcgacgcc gccacgctcg cgtggcgctc catcaggccc cgcccgggcg 1080
ccttcgtcgt caacatcggc gacaccttca tggcgctctc caacgggcgc tacaggagct 1140
gcctccaccg cgccgtcgtc aacagccggg tggcacgccg ctcgctcgcc ttcttcctgt 1200
gcccggagat ggacaaggtg gtcaggccgc ccaaggagct ggtggacgac gccaacccga 1260
gggcgtaccc ggacttcacg tggaggacgc tgctggactt caccatgagg cactacaggt 1320
cggacatgag gacgctcgag gccttctcca actggctcag caccagtagc aatggcggac 1380
agcacctgct ggagaagaag taggcatgct atttgggtat ggaagatggt ggatgtaagc 1440
aaacaaagcc aaattaagca gagtaggtta attaaggttg gctgatgatc catttaggga 1500
aggagctgat ctccctgact ccctcctcca attttctcaa ccaaatttat atagtataat 1560
aataataata aaatagcaag taatagttgt atcgtattat tattaattaa tttattagct 1620
ggtaggcaag tagtattaaa taccatttgt agtacgatgg gcgtatttct attttggcgt 1680
tttgctctgt gttttttgac gtttcctttg gatttggggg gacctcagat cagctcggcc 1740
t 1741
<210> 2
<211> 1116
<212> ДНК
<213> Zea mays
<400> 2
atggtgctgg ctgcgcacga tccccctccc cttgtgttcg acgctgcccg cctgagcggc 60
ctctccgaca tcccgcagca gttcatctgg ccggcggacg agagccccac cccggactcc 120
gccgaggagc tggccgtgcc gctcatcgac ctctccgggg acgccgccga ggtggtccgg 180
caggtccggc gcgcctgcga cctgcacggc ttcttccagg tggtggggca cggcatcgac 240
gcggcgctga cggcggaggc ccaccgctgc atggacgcct tcttcacgct gccgctcccg 300
gacaagcagc gcgcgcagcg ccgccagggg gacagctgcg gctacgccag cagcttcacg 360
ggccggttcg cgtccaagct gccctggaag gagacgctgt cgttccgcta caccgacgac 420
gacgacggcg acaagtccaa ggacgtcgtg gcgtcctact tcgtggacaa gctgggcgag 480
gggtaccggc accacgggga ggtgtacggg cgctactgct ctgagatgag ccgtctgtcg 540
ctggagctca tggaggtgct aggcgagagc ctgggcgtgg gccggcgcca cttccggcgc 600
ttcttccagg ggaacgactc catcatgcgc ctcaactact acccgccgtg ccagcggccc 660
tacgacacgc tgggcacggg gccgcattgc gaccccacgt cgctcaccat cctgcaccag 720
gacgacgtgg gcggactcca ggtgttcgac gccgccacgc tcgcgtggcg ctccatcagg 780
ccccgcccgg gcgccttcgt cgtcaacatc ggcgacacct tcatggcgct ctccaacggg 840
cgctacagga gctgcctcca ccgcgccgtc gtcaacagcc gggtggcacg ccgctcgctc 900
gccttcttcc tgtgcccgga gatggacaag gtggtcaggc cgcccaagga gctggtggac 960
gacgccaacc cgagggcgta cccggacttc acgtggagga cgctgctgga cttcaccatg 1020
aggcactaca ggtcggacat gaggacgctc gaggccttct ccaactggct cagcaccagt 1080
agcaatggcg gacagcacct gctggagaag aagtag 1116
<210> 3
<211> 371
<212> ПРТ
<213> Zea mays
<400> 3
Met Val Leu Ala Ala His Asp Pro Pro Pro Leu Val Phe Asp Ala Ala
1. 5 10 15
Arg Leu Ser Gly Leu Ser Asp Ile Pro Gln Gln Phe Ile Trp Pro Ala
20 25 30
Asp Glu Ser Pro Thr Pro Asp Ser Ala Glu Glu Leu Ala Val Pro Leu
35 40 45
Ile Asp Leu Ser Gly Asp Ala Ala Glu Val Val Arg Gln Val Arg Arg
50 55 60
Ala Cys Asp Leu His Gly Phe Phe Gln Val Val Gly His Gly Ile Asp
65 70 75 80
Ala Ala Leu Thr Ala Glu Ala His Arg Cys Met Asp Ala Phe Phe Thr
85 90 95
Leu Pro Leu Pro Asp Lys Gln Arg Ala Gln Arg Arg Gln Gly Asp Ser
100 105 110
Cys Gly Tyr Ala Ser Ser Phe Thr Gly Arg Phe Ala Ser Lys Leu Pro
115 120 125
Trp Lys Glu Thr Leu Ser Phe Arg Tyr Thr Asp Asp Asp Asp Gly Asp
130 135 140
Lys Ser Lys Asp Val Val Ala Ser Tyr Phe Val Asp Lys Leu Gly Glu
145 150 155 160
Gly Tyr Arg His His Gly Glu Val Tyr Gly Arg Tyr Cys Ser Glu Met
165 170 175
Ser Arg Leu Ser Leu Glu Leu Met Glu Val Leu Gly Glu Ser Leu Gly
180 185 190
Val Gly Arg Arg His Phe Arg Arg Phe Phe Gln Gly Asn Asp Ser Ile
195 200 205
Met Arg Leu Asn Tyr Tyr Pro Pro Cys Gln Arg Pro Tyr Asp Thr Leu
210 215 220
Gly Thr Gly Pro His Cys Asp Pro Thr Ser Leu Thr Ile Leu His Gln
225 230 235 240
Asp Asp Val Gly Gly Leu Gln Val Phe Asp Ala Ala Thr Leu Ala Trp
245 250 255
Arg Ser Ile Arg Pro Arg Pro Gly Ala Phe Val Val Asn Ile Gly Asp
260 265 270
Thr Phe Met Ala Leu Ser Asn Gly Arg Tyr Arg Ser Cys Leu His Arg
275 280 285
Ala Val Val Asn Ser Arg Val Ala Arg Arg Ser Leu Ala Phe Phe Leu
290 295 300
Cys Pro Glu Met Asp Lys Val Val Arg Pro Pro Lys Glu Leu Val Asp
305 310 315 320
Asp Ala Asn Pro Arg Ala Tyr Pro Asp Phe Thr Trp Arg Thr Leu Leu
325 330 335
Asp Phe Thr Met Arg His Tyr Arg Ser Asp Met Arg Thr Leu Glu Ala
340 345 350
Phe Ser Asn Trp Leu Ser Thr Ser Ser Asn Gly Gly Gln His Leu Leu
355 360 365
Glu Lys Lys
370
<210> 4
<211> 1517
<212> ДНК
<213> Zea mays
<400> 4
caggaataaa ataagcctcc gcccggcttc gttgcatcca cgcacgcagc aagcgatcgg 60
atttcgccag catggcggcg gcggccgtgg tgttcgacgc cgaggcgctg agccgggagg 120
agcacatccc ggcgcagttc gtgtggccca ccgaggagcg ggcgccggcg ggcggcgtgg 180
aggaggtcgc catccccgtg gtcgacctcg gcgagttcct ccgccgcggg gtgctcccgc 240
gcggcgtggc ggaggcgtgc gagcgccacg gcgtcttcca ggtggtgaac cacggcgtgg 300
gcgccgcgct gctcgccgag gcctaccgct gttgcgacgc cttttacgcg ctcccgctcg 360
cggacaagca gcgcgcgcag cgccggcacg gggagaacca cggctacgcc agcagcttca 420
cgggccgctt ccactgctgc ctgccgtgga aggagacgct gtccttcaac tgccccgccg 480
gtgccgggac tgcgcgcgcc gtcgtcggct acttcgtcga cgtcctcggc gaggactacc 540
gccacatggg ggaggtgtac caggagtact gcgacgcgat gacgcgtctg gcgctggacg 600
tgacggaggt gctggcggca gcgctggggc tggaccgcgg cgcactgcgc ggcttcttcg 660
agggcggcga ctccgtcatg cggctgaacc actacccggc gtgccggcag ccgcacctga 720
cgctggggac gggcccgcac cgggacccga cgtcgctgac gctgctgcac caggacgacg 780
tgggcgggct gcaggtgcgc gccggcggcg ggccgtggcg cgcggtgcgg ccccgcgcgg 840
acgcgttcgt ggtcaacatt ggcgacacct tcgccgcgct caccgacggg cgtcacacca 900
gctgcctgca ccgcgccgtg gtgaccggcg gcggctcccg ccggtcgctc gccttcttcc 960
tcaacccgcc gctggaccgc gtcgtccgcc cgccgggcgc gctcctccag gagaacaagc 1020
aggcgggccg cccgcgcgcg ttcccggact tcacgtggcg cgagttcctc gagttcacgc 1080
agaagcacta ccggtcggac gcgggcacca tggacgcctt cgtgtcgtgg atcgcgggag 1140
gccgccgcca ccatggcgga caggaggagg gcaactgaga tcgatgcatc tctagctgta 1200
ggcagcagcg cagcagctac caagaataat ggccggcgac ggagatgcag ctacgacgca 1260
caaataaatt gagtgtttgt ggtacaataa ggacgaggac gatcaatggc gacctgtaac 1320
cggtgcagtt ttagttaatc tttcatggcg atatggcatt aaccaatcgt tggtgtaaaa 1380
tgcgtgcatg ctttgcatgc caatgttggc catgtgatgg cacagcgtga gtgtagctca 1440
cccaccgtga caacgtgcta atttcgtgtg gtcctagata ccaaggtcgt ctaatgaact 1500
tgatggattg atgattt 1517
<210> 5
<211> 1107
<212> ДНК
<213> Zea mays
<400> 5
atggcggcgg cggccgtggt gttcgacgcc gaggcgctga gccgggagga gcacatcccg 60
gcgcagttcg tgtggcccac cgaggagcgg gcgccggcgg gcggcgtgga ggaggtcgcc 120
atccccgtgg tcgacctcgg cgagttcctc cgccgcgggg tgctcccgcg cggcgtggcg 180
gaggcgtgcg agcgccacgg cgtcttccag gtggtgaacc acggcgtggg cgccgcgctg 240
ctcgccgagg cctaccgctg ttgcgacgcc ttttacgcgc tcccgctcgc ggacaagcag 300
cgcgcgcagc gccggcacgg ggagaaccac ggctacgcca gcagcttcac gggccgcttc 360
cactgctgcc tgccgtggaa ggagacgctg tccttcaact gccccgccgg tgccgggact 420
gcgcgcgccg tcgtcggcta cttcgtcgac gtcctcggcg aggactaccg ccacatgggg 480
gaggtgtacc aggagtactg cgacgcgatg acgcgtctgg cgctggacgt gacggaggtg 540
ctggcggcag cgctggggct ggaccgcggc gcactgcgcg gcttcttcga gggcggcgac 600
tccgtcatgc ggctgaacca ctacccggcg tgccggcagc cgcacctgac gctggggacg 660
ggcccgcacc gggacccgac gtcgctgacg ctgctgcacc aggacgacgt gggcgggctg 720
caggtgcgcg ccggcggcgg gccgtggcgc gcggtgcggc cccgcgcgga cgcgttcgtg 780
gtcaacattg gcgacacctt cgccgcgctc accgacgggc gtcacaccag ctgcctgcac 840
cgcgccgtgg tgaccggcgg cggctcccgc cggtcgctcg ccttcttcct caacccgccg 900
ctggaccgcg tcgtccgccc gccgggcgcg ctcctccagg agaacaagca ggcgggccgc 960
ccgcgcgcgt tcccggactt cacgtggcgc gagttcctcg agttcacgca gaagcactac 1020
cggtcggacg cgggcaccat ggacgccttc gtgtcgtgga tcgcgggagg ccgccgccac 1080
catggcggac aggaggaggg caactga 1107
<210> 6
<211> 368
<212> ПРТ
<213> Zea mays
<400> 6
Met Ala Ala Ala Ala Val Val Phe Asp Ala Glu Ala Leu Ser Arg Glu
1. 5 10 15
Glu His Ile Pro Ala Gln Phe Val Trp Pro Thr Glu Glu Arg Ala Pro
20 25 30
Ala Gly Gly Val Glu Glu Val Ala Ile Pro Val Val Asp Leu Gly Glu
35 40 45
Phe Leu Arg Arg Gly Val Leu Pro Arg Gly Val Ala Glu Ala Cys Glu
50 55 60
Arg His Gly Val Phe Gln Val Val Asn His Gly Val Gly Ala Ala Leu
65 70 75 80
Leu Ala Glu Ala Tyr Arg Cys Cys Asp Ala Phe Tyr Ala Leu Pro Leu
85 90 95
Ala Asp Lys Gln Arg Ala Gln Arg Arg His Gly Glu Asn His Gly Tyr
100 105 110
Ala Ser Ser Phe Thr Gly Arg Phe His Cys Cys Leu Pro Trp Lys Glu
115 120 125
Thr Leu Ser Phe Asn Cys Pro Ala Gly Ala Gly Thr Ala Arg Ala Val
130 135 140
Val Gly Tyr Phe Val Asp Val Leu Gly Glu Asp Tyr Arg His Met Gly
145 150 155 160
Glu Val Tyr Gln Glu Tyr Cys Asp Ala Met Thr Arg Leu Ala Leu Asp
165 170 175
Val Thr Glu Val Leu Ala Ala Ala Leu Gly Leu Asp Arg Gly Ala Leu
180 185 190
Arg Gly Phe Phe Glu Gly Gly Asp Ser Val Met Arg Leu Asn His Tyr
195 200 205
Pro Ala Cys Arg Gln Pro His Leu Thr Leu Gly Thr Gly Pro His Arg
210 215 220
Asp Pro Thr Ser Leu Thr Leu Leu His Gln Asp Asp Val Gly Gly Leu
225 230 235 240
Gln Val Arg Ala Gly Gly Gly Pro Trp Arg Ala Val Arg Pro Arg Ala
245 250 255
Asp Ala Phe Val Val Asn Ile Gly Asp Thr Phe Ala Ala Leu Thr Asp
260 265 270
Gly Arg His Thr Ser Cys Leu His Arg Ala Val Val Thr Gly Gly Gly
275 280 285
Ser Arg Arg Ser Leu Ala Phe Phe Leu Asn Pro Pro Leu Asp Arg Val
290 295 300
Val Arg Pro Pro Gly Ala Leu Leu Gln Glu Asn Lys Gln Ala Gly Arg
305 310 315 320
Pro Arg Ala Phe Pro Asp Phe Thr Trp Arg Glu Phe Leu Glu Phe Thr
325 330 335
Gln Lys His Tyr Arg Ser Asp Ala Gly Thr Met Asp Ala Phe Val Ser
340 345 350
Trp Ile Ala Gly Gly Arg Arg His His Gly Gly Gln Glu Glu Gly Asn
355 360 365
<210> 7
<211> 1522
<212> ДНК
<213> Zea mays
<400> 7
gcacactcgc agctcgcaca tctcatggtg tcctaagaac ggcaagagcc agctctgcct 60
agcagcagcg cacagccaca tccatggacg ccagcccgac cccaccgctc cccctccgcg 120
ccccaactcc cagcattgac ctccccgctg gcaaggacag ggccgacgcg gcggctaaca 180
aggccgcggc tgtgttcgac ctgcgccggg agcccaagat cccggagcca ttcctgtggc 240
cgcacgaaga ggcgcggccg acctcggccg cggagctgga ggtgccggtg gtggacgtgg 300
gcgtgctgcg caatggcgac ggcgcggggc tccgccgcgc cgcggcgcaa gtggcggcgg 360
cgtgcgcgac gcacgggttc ttccaggtgt gcgggcacgg cgtggacgcg gcgctggggc 420
gcgccgcgct ggacggcgcc agcgacttct tccggctgcc gctggctgag aagcagcggg 480
cccggcgcgt ccccggcacc gtgtccgggt acacgagcgc gcacgccgac cggttcgcgt 540
ccaagctccc ctggaaggag accctgtcct tcggcttcca cgacggcgcc gcggcgcccg 600
tcgtcgtgga ctacttcacc ggcaccctcg gccaagattt cgagccagtg gggcgggtgt 660
accagaggta ctgcgaggag atgaaggagc tgtcgctgac gatcatggag ctgctggagc 720
tgagcctggg cgtggagcgc ggctactacc gggagttctt cgaggacagc cgctccatca 780
tgcggtgcaa ctactacccg ccgtgcccgg tgccggagcg cacgctgggc acgggcccgc 840
actgcgaccc cacggcgctg accatcctcc tgcaggacga cgtcggcggg ctggaggtcc 900
tggtggacgg cgagtggcgc cccgtccggc ccgtcccagg cgccatggtc atcaacatcg 960
gcgacacctt catggcgctg tccaacgggc ggtacaagag ctgcctgcac cgcgcggtgg 1020
tgaaccggcg gcaggagcgg caatcgctgg ccttcttcct gtgcccgcgc gaggaccggg 1080
tggtgcgccc gccggccagc gccgcgccgc ggcagtaccc ggacttcacc tgggccgacc 1140
tcatgcgctt cacgcagcgc cactaccgcg ccgacacccg cacgctggac gccttcaccc 1200
gctggctctc ccacggcccg gcggcggcgg ctccctgcac ctaacgagcc ggccgtctct 1260
ttcgccgggg cccgcgcggg gttcgcccac gtggtgatca ggtggcagac atgtggccca 1320
cgggccccgc gccgccttcc ccatttttgg acgaccctac tgctactact actagtgtac 1380
atatgcaaaa aaatacatat atatataggt actttctcta atatttttat atataagcaa 1440
ggcggcctgg tgttcttttc tttgttttgt cgacaactgt ttgatcccat cctatggacg 1500
atggatagtt caatgtttgt ac 1522
<210> 8
<211> 1161
<212> ДНК
<213> Zea mays
<400> 8
atggacgcca gcccgacccc accgctcccc ctccgcgccc caactcccag cattgacctc 60
cccgctggca aggacagggc cgacgcggcg gctaacaagg ccgcggctgt gttcgacctg 120
cgccgggagc ccaagatccc ggagccattc ctgtggccgc acgaagaggc gcggccgacc 180
tcggccgcgg agctggaggt gccggtggtg gacgtgggcg tgctgcgcaa tggcgacggc 240
gcggggctcc gccgcgccgc ggcgcaagtg gcggcggcgt gcgcgacgca cgggttcttc 300
caggtgtgcg ggcacggcgt ggacgcggcg ctggggcgcg ccgcgctgga cggcgccagc 360
gacttcttcc ggctgccgct ggctgagaag cagcgggccc ggcgcgtccc cggcaccgtg 420
tccgggtaca cgagcgcgca cgccgaccgg ttcgcgtcca agctcccctg gaaggagacc 480
ctgtccttcg gcttccacga cggcgccgcg gcgcccgtcg tcgtggacta cttcaccggc 540
accctcggcc aagatttcga gccagtgggg cgggtgtacc agaggtactg cgaggagatg 600
aaggagctgt cgctgacgat catggagctg ctggagctga gcctgggcgt ggagcgcggc 660
tactaccggg agttcttcga ggacagccgc tccatcatgc ggtgcaacta ctacccgccg 720
tgcccggtgc cggagcgcac gctgggcacg ggcccgcact gcgaccccac ggcgctgacc 780
atcctcctgc aggacgacgt cggcgggctg gaggtcctgg tggacggcga gtggcgcccc 840
gtccggcccg tcccaggcgc catggtcatc aacatcggcg acaccttcat ggcgctgtcc 900
aacgggcggt acaagagctg cctgcaccgc gcggtggtga accggcggca ggagcggcaa 960
tcgctggcct tcttcctgtg cccgcgcgag gaccgggtgg tgcgcccgcc ggccagcgcc 1020
gcgccgcggc agtacccgga cttcacctgg gccgacctca tgcgcttcac gcagcgccac 1080
taccgcgccg acacccgcac gctggacgcc ttcacccgct ggctctccca cggcccggcg 1140
gcggcggctc cctgcaccta a 1161
<210> 9
<211> 386
<212> ПРТ
<213> Zea mays
<400> 9
Met Asp Ala Ser Pro Thr Pro Pro Leu Pro Leu Arg Ala Pro Thr Pro
1. 5 10 15
Ser Ile Asp Leu Pro Ala Gly Lys Asp Arg Ala Asp Ala Ala Ala Asn
20 25 30
Lys Ala Ala Ala Val Phe Asp Leu Arg Arg Glu Pro Lys Ile Pro Glu
35 40 45
Pro Phe Leu Trp Pro His Glu Glu Ala Arg Pro Thr Ser Ala Ala Glu
50 55 60
Leu Glu Val Pro Val Val Asp Val Gly Val Leu Arg Asn Gly Asp Gly
65 70 75 80
Ala Gly Leu Arg Arg Ala Ala Ala Gln Val Ala Ala Ala Cys Ala Thr
85 90 95
His Gly Phe Phe Gln Val Cys Gly His Gly Val Asp Ala Ala Leu Gly
100 105 110
Arg Ala Ala Leu Asp Gly Ala Ser Asp Phe Phe Arg Leu Pro Leu Ala
115 120 125
Glu Lys Gln Arg Ala Arg Arg Val Pro Gly Thr Val Ser Gly Tyr Thr
130 135 140
Ser Ala His Ala Asp Arg Phe Ala Ser Lys Leu Pro Trp Lys Glu Thr
145 150 155 160
Leu Ser Phe Gly Phe His Asp Gly Ala Ala Ala Pro Val Val Val Asp
165 170 175
Tyr Phe Thr Gly Thr Leu Gly Gln Asp Phe Glu Pro Val Gly Arg Val
180 185 190
Tyr Gln Arg Tyr Cys Glu Glu Met Lys Glu Leu Ser Leu Thr Ile Met
195 200 205
Glu Leu Leu Glu Leu Ser Leu Gly Val Glu Arg Gly Tyr Tyr Arg Glu
210 215 220
Phe Phe Glu Asp Ser Arg Ser Ile Met Arg Cys Asn Tyr Tyr Pro Pro
225 230 235 240
Cys Pro Val Pro Glu Arg Thr Leu Gly Thr Gly Pro His Cys Asp Pro
245 250 255
Thr Ala Leu Thr Ile Leu Leu Gln Asp Asp Val Gly Gly Leu Glu Val
260 265 270
Leu Val Asp Gly Glu Trp Arg Pro Val Arg Pro Val Pro Gly Ala Met
275 280 285
Val Ile Asn Ile Gly Asp Thr Phe Met Ala Leu Ser Asn Gly Arg Tyr
290 295 300
Lys Ser Cys Leu His Arg Ala Val Val Asn Arg Arg Gln Glu Arg Gln
305 310 315 320
Ser Leu Ala Phe Phe Leu Cys Pro Arg Glu Asp Arg Val Val Arg Pro
325 330 335
Pro Ala Ser Ala Ala Pro Arg Gln Tyr Pro Asp Phe Thr Trp Ala Asp
340 345 350
Leu Met Arg Phe Thr Gln Arg His Tyr Arg Ala Asp Thr Arg Thr Leu
355 360 365
Asp Ala Phe Thr Arg Trp Leu Ser His Gly Pro Ala Ala Ala Ala Pro
370 375 380
Cys Thr
385
<210> 10
<211> 1457
<212> ДНК
<213> Zea mays
<400> 10
taatcacctc atcacaggtc cccccagcct cactctcgcg ccggctcaag gtacattgcg 60
tgtcctagcc aagacacgca gctcatctca gcctcacacg cacagcaaga gcgaggcgtg 120
attcgccatg ggcggcctca ctatggacca ggccttcgtg caggcccccg agcaccgccc 180
caagcccatc gtcaccgagg ccaccggcat ccctctcatc gacctctcgc ctctggccgc 240
cagcggcggc gccgtggacg cgctggccgc cgaggtgggc gcggcgagcc gggactgggg 300
cttcttcgtg gtcgtgggcc acggcgtgcc cgcagagacc gtggcgcgcg cgacggaggc 360
gcagcgagcg ttcttcgcgc tgccggcaga gcggaaggcc gccgtgcgga ggaacgaggc 420
ggagccgctc gggtactacg agtcggagca caccaagaac gtgagggact ggaaggaggt 480
gtacgacctc gtgccgcgcg agccgccgcc gccggcagcc gtggccgacg gcgagcttgt 540
gttcgataac aagtggcccc aggatctacc gggcttcaga gaggcgctgg aggagtacgc 600
gaaagcgatg gaagagctgg cgttcaagct gctggagctg atcgcccgga gcctgaagct 660
gaggcccgac cggctgcacg gcttcttcaa ggaccagacg accttcatcc ggctgaacca 720
ctaccctcct tgcccgagcc ccgacctggc cctcggcgtg gggcggcaca aggacgccgg 780
cgccctgacc atcctgtacc aggacgacgt cggggggctc gacgtccggc ggcgctccga 840
cggcgagtgg gtccgcgtca ggcccgtgcc cgactcgttc atcatcaacg tcggcgacct 900
catccaggta cgagagcgcg gagcaccggg tgtcggtgaa ctcggcgagg gagaggttct 960
ccatgcccta cttcttcaac ccggcgacct acaccatggt ggagccggtg gaggagctgg 1020
tgagcaagga cgatccgccc aggtacgacg cctacaactg gggcgacttc ttcagcacca 1080
ggaagaacag caacttcaag aagctcaacg tggagaacat tcagatcgcg catttcaaga 1140
agagcctcgt cctcgcctaa ctactgctac tgctaggatc catgccattg ccatgtcgtc 1200
ttcagattca gagcacgcca tgtcgtcgct agcttcgtgg tagaacaaat aatgatgtgc 1260
gtgctgtgtg taagcatgga tatggatgtg aatatgtaat atgatgagca ctcctacttt 1320
ggtatgtttg ggaataacag acttgtgttg gtctggttca ttatttgtaa gaaaatcaaa 1380
aagagttagt agggcaggag gctaaccaca gtcatgctgc accacatccc tggtggaaag 1440
ctggccgggt tacgcta 1457
<210> 11
<211> 1116
<212> ДНК
<213> Zea mays
<400> 11
atgggcggcc tcactatgga ccaggccttc gtgcaggccc ccgagcaccg ccccaagccc 60
atcgtcaccg aggccaccgg catccctctc atcgacctct cgcctctggc cgccagcggc 120
ggcgccgtgg acgcgctggc cgccgaggtg ggcgcggcga gccgggactg gggcttcttc 180
gtggtcgtgg gccacggcgt gcccgcagag accgtggcgc gcgcgacgga ggcgcagcga 240
gcgttcttcg cgctgccggc agagcggaag gccgccgtgc ggaggaacga ggcggagccg 300
ctcgggtact acgagtcgga gcacaccaag aacgtgaggg actggaagga ggtgtacgac 360
ctcgtgccgc gcgagccgcc gccgccggca gccgtggccg acggcgagct tgtgttcgat 420
aacaagtggc cccaggatct accgggcttc agagaggcgc tggaggagta cgcgaaagcg 480
atggaagagc tggcgttcaa gctgctggag ctgatcgccc ggagcctgaa gctgaggccc 540
gaccggctgc acggcttctt caaggaccag acgaccttca tccggctgaa ccactaccct 600
ccttgcccga gccccgacct ggccctcggc gtggggcggc acaaggacgc cggcgccctg 660
accatcctgt accaggacga cgtcgggggg ctcgacgtcc ggcggcgctc cgacggcgag 720
tgggtccgcg tcaggcccgt gcccgactcg ttcatcatca acgtcggcga cctcatccag 780
gtacgagagc gcggagcacc gggtgtcggt gaactcggcg agggagaggt tctccatgcc 840
ctacttcttc aacccggcga cctacaccat ggtggagccg gtggaggagc tggtgagcaa 900
ggacgatccg cccaggtacg acgcctacaa ctggggcgac ttcttcagca ccaggaagaa 960
cagcaacttc aagaagctca acgtggagaa cattcagatc gcgcatttca agaagagcct 1020
cgtcctcgcc taactactgc tactgctagg atccatgcca ttgccatgtc gtcttcagat 1080
tcagagcacg ccatgtcgtc gctagcttcg tggtag 1116
<210> 12
<211> 371
<212> ПРТ
<213> Zea mays
<400> 12
Met Gly Gly Leu Thr Met Asp Gln Ala Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Pro Lys Pro Ile Val Thr Glu Ala Thr Gly Ile Pro Leu Ile Asp
20 25 30
Leu Ser Pro Leu Ala Ala Ser Gly Gly Ala Val Asp Ala Leu Ala Ala
35 40 45
Glu Val Gly Ala Ala Ser Arg Asp Trp Gly Phe Phe Val Val Val Gly
50 55 60
His Gly Val Pro Ala Glu Thr Val Ala Arg Ala Thr Glu Ala Gln Arg
65 70 75 80
Ala Phe Phe Ala Leu Pro Ala Glu Arg Lys Ala Ala Val Arg Arg Asn
85 90 95
Glu Ala Glu Pro Leu Gly Tyr Tyr Glu Ser Glu His Thr Lys Asn Val
100 105 110
Arg Asp Trp Lys Glu Val Tyr Asp Leu Val Pro Arg Glu Pro Pro Pro
115 120 125
Pro Ala Ala Val Ala Asp Gly Glu Leu Val Phe Asp Asn Lys Trp Pro
130 135 140
Gln Asp Leu Pro Gly Phe Arg Glu Ala Leu Glu Glu Tyr Ala Lys Ala
145 150 155 160
Met Glu Glu Leu Ala Phe Lys Leu Leu Glu Leu Ile Ala Arg Ser Leu
165 170 175
Lys Leu Arg Pro Asp Arg Leu His Gly Phe Phe Lys Asp Gln Thr Thr
180 185 190
Phe Ile Arg Leu Asn His Tyr Pro Pro Cys Pro Ser Pro Asp Leu Ala
195 200 205
Leu Gly Val Gly Arg His Lys Asp Ala Gly Ala Leu Thr Ile Leu Tyr
210 215 220
Gln Asp Asp Val Gly Gly Leu Asp Val Arg Arg Arg Ser Asp Gly Glu
225 230 235 240
Trp Val Arg Val Arg Pro Val Pro Asp Ser Phe Ile Ile Asn Val Gly
245 250 255
Asp Leu Ile Gln Val Arg Glu Arg Gly Ala Pro Gly Val Gly Glu Leu
260 265 270
Gly Glu Gly Glu Val Leu His Ala Leu Leu Leu Gln Pro Gly Asp Leu
275 280 285
His His Gly Gly Ala Gly Gly Gly Ala Gly Glu Gln Gly Arg Ser Ala
290 295 300
Gln Val Arg Arg Leu Gln Leu Gly Arg Leu Leu Gln His Gln Glu Glu
305 310 315 320
Gln Gln Leu Gln Glu Ala Gln Arg Gly Glu His Ser Asp Arg Ala Phe
325 330 335
Gln Glu Glu Pro Arg Pro Arg Leu Thr Thr Ala Thr Ala Arg Ile His
340 345 350
Ala Ile Ala Met Ser Ser Ser Asp Ser Glu His Ala Met Ser Ser Leu
355 360 365
Ala Ser Trp
370
<210> 13
<211> 1733
<212> ДНК
<213> Zea mays
<400> 13
atgaggccgc gcctccctcc aaatgttccc tccctgcctt cgtctttgtc gttgctcgca 60
aactccctgt cctcccctgt tacaaatacc cccacccgcc cggacagctt ccctgcatac 120
ttgcagctcg cacatctcat ggtgtcgcag gaacgacaag agccagctgt gcctagcagc 180
agcagcagca gcgccaagcg cgcagccacg tccatggacg ccagcccggc cccgccgctc 240
ctcctccgcg cccccactcc cagccccagc attgacctcc ccgctggcaa ggacaaggcc 300
gacgcggcgg ccagcaaggc cggcgcggcc gtgttcgacc tgcgccggga gcccaagatc 360
cccgcgccat tcctgtggcc gcaggaagag gcgcggccgt cctcggccgc ggagctggag 420
gtgccgatgg tggacgtggg cgtgctgcgc aatggcgacc gcgcggggct gcggcgcgcc 480
gcggcgcagg tggccgcggc gtgcgcgacg cacgggttct tccaggtgtg cgggcacggc 540
gtggacgcgg cgctggggcg cgccgcgctg gacggcgcca gcgacttctt ccggctgccg 600
ctcgccgaga agcagcgcgc ccggcgcgtc cccggcaccg tgtccgggta cacgagcgcg 660
cacgccgacc ggttcgcggc caagctcccc tggaaggaga ccctgtcgtt cggctaccac 720
gacggcgccg cgtcgcctgt cgtcgtggac tacttcgtcg gcaccctcgg ccaggatttc 780
gagccaatgg ggtgggtgta ccagaggtac tgcgaggaga tgaaggagct gtcgctgacg 840
atcatggagc tgctggagct gagcctgggc gtggagctgc gcggctacta ccgggagttc 900
ttcgaggaca gccggtccat catgcggtgc aactactacc cgccgtgccc ggagccggag 960
cgcacgctgg gcacgggccc gcactgcgac cccacggcgc tcaccatcct cctgcaggac 1020
gacgtgggcg ggctggaggt gctggtggac ggtgagtggc gccccgtccg gcccgtcccg 1080
ggcgccatgg tcatcaacat cggcgacacc ttcatggcgc tgtcgaacgg gaggtacaag 1140
agctgcctgc accgcgcggt ggtgaaccag cggcgggcgc ggcggtcgct ggccttcttc 1200
ctgtgcccgc gcgaggaccg ggtggtgcgc ccgccggcca gtgctgcgcc gcggcgctac 1260
ccggacttca cctgggccga cctcatgcgc ttcacgcagc gccactaccg cgccgacacc 1320
cgcacgctgg acgccttcac ccgctggctc tcccacggcc cggcccaggc ggcggcgcct 1380
ccctgcacct agcgagccgg gccaaggccg tctctttcgc cccacgtgcg cgcccagctg 1440
ggcaggtggc cagacacgcg gcccgcgggc cccgcgccgc cttgccattt tttgacgctg 1500
gccctactgc tgtgctacta gtgtacatat gcaagagtac atatatatat atatatatac 1560
gtattttcta tatattatat ataaaagcaa ggcggcccgg tgcccttctc ttgttttgtc 1620
cacaactgtt tgatcccatt attctatgga ccatggatac ttcaatgttt gtactaagac 1680
cgtgaacgtg ggattctttt ccttcctctg tgttttttct gagaaaaatt aaa 1733
<210> 14
<211> 1392
<212> ДНК
<213> Zea mays
<400> 14
atgaggccgc gcctccctcc aaatgttccc tccctgcctt cgtctttgtc gttgctcgca 60
aactccctgt cctcccctgt tacaaatacc cccacccgcc cggacagctt ccctgcatac 120
ttgcagctcg cacatctcat ggtgtcgcag gaacgacaag agccagctgt gcctagcagc 180
agcagcagca gcgccaagcg cgcagccacg tccatggacg ccagcccggc cccgccgctc 240
ctcctccgcg cccccactcc cagccccagc attgacctcc ccgctggcaa ggacaaggcc 300
gacgcggcgg ccagcaaggc cggcgcggcc gtgttcgacc tgcgccggga gcccaagatc 360
cccgcgccat tcctgtggcc gcaggaagag gcgcggccgt cctcggccgc ggagctggag 420
gtgccgatgg tggacgtggg cgtgctgcgc aatggcgacc gcgcggggct gcggcgcgcc 480
gcggcgcagg tggccgcggc gtgcgcgacg cacgggttct tccaggtgtg cgggcacggc 540
gtggacgcgg cgctggggcg cgccgcgctg gacggcgcca gcgacttctt ccggctgccg 600
ctcgccgaga agcagcgcgc ccggcgcgtc cccggcaccg tgtccgggta cacgagcgcg 660
cacgccgacc ggttcgcggc caagctcccc tggaaggaga ccctgtcgtt cggctaccac 720
gacggcgccg cgtcgcctgt cgtcgtggac tacttcgtcg gcaccctcgg ccaggatttc 780
gagccaatgg ggtgggtgta ccagaggtac tgcgaggaga tgaaggagct gtcgctgacg 840
atcatggagc tgctggagct gagcctgggc gtggagctgc gcggctacta ccgggagttc 900
ttcgaggaca gccggtccat catgcggtgc aactactacc cgccgtgccc ggagccggag 960
cgcacgctgg gcacgggccc gcactgcgac cccacggcgc tcaccatcct cctgcaggac 1020
gacgtgggcg ggctggaggt gctggtggac ggtgagtggc gccccgtccg gcccgtcccg 1080
ggcgccatgg tcatcaacat cggcgacacc ttcatggcgc tgtcgaacgg gaggtacaag 1140
agctgcctgc accgcgcggt ggtgaaccag cggcgggcgc ggcggtcgct ggccttcttc 1200
ctgtgcccgc gcgaggaccg ggtggtgcgc ccgccggcca gtgctgcgcc gcggcgctac 1260
ccggacttca cctgggccga cctcatgcgc ttcacgcagc gccactaccg cgccgacacc 1320
cgcacgctgg acgccttcac ccgctggctc tcccacggcc cggcccaggc ggcggcgcct 1380
ccctgcacct ag 1392
<210> 15
<211> 463
<212> ПРТ
<213> Zea mays
<400> 15
Met Arg Pro Arg Leu Pro Pro Asn Val Pro Ser Leu Pro Ser Ser Leu
1. 5 10 15
Ser Leu Leu Ala Asn Ser Leu Ser Ser Pro Val Thr Asn Thr Pro Thr
20 25 30
Arg Pro Asp Ser Phe Pro Ala Tyr Leu Gln Leu Ala His Leu Met Val
35 40 45
Ser Gln Glu Arg Gln Glu Pro Ala Val Pro Ser Ser Ser Ser Ser Ser
50 55 60
Ala Lys Arg Ala Ala Thr Ser Met Asp Ala Ser Pro Ala Pro Pro Leu
65 70 75 80
Leu Leu Arg Ala Pro Thr Pro Ser Pro Ser Ile Asp Leu Pro Ala Gly
85 90 95
Lys Asp Lys Ala Asp Ala Ala Ala Ser Lys Ala Gly Ala Ala Val Phe
100 105 110
Asp Leu Arg Arg Glu Pro Lys Ile Pro Ala Pro Phe Leu Trp Pro Gln
115 120 125
Glu Glu Ala Arg Pro Ser Ser Ala Ala Glu Leu Glu Val Pro Met Val
130 135 140
Asp Val Gly Val Leu Arg Asn Gly Asp Arg Ala Gly Leu Arg Arg Ala
145 150 155 160
Ala Ala Gln Val Ala Ala Ala Cys Ala Thr His Gly Phe Phe Gln Val
165 170 175
Cys Gly His Gly Val Asp Ala Ala Leu Gly Arg Ala Ala Leu Asp Gly
180 185 190
Ala Ser Asp Phe Phe Arg Leu Pro Leu Ala Glu Lys Gln Arg Ala Arg
195 200 205
Arg Val Pro Gly Thr Val Ser Gly Tyr Thr Ser Ala His Ala Asp Arg
210 215 220
Phe Ala Ala Lys Leu Pro Trp Lys Glu Thr Leu Ser Phe Gly Tyr His
225 230 235 240
Asp Gly Ala Ala Ser Pro Val Val Val Asp Tyr Phe Val Gly Thr Leu
245 250 255
Gly Gln Asp Phe Glu Pro Met Gly Trp Val Tyr Gln Arg Tyr Cys Glu
260 265 270
Glu Met Lys Glu Leu Ser Leu Thr Ile Met Glu Leu Leu Glu Leu Ser
275 280 285
Leu Gly Val Glu Leu Arg Gly Tyr Tyr Arg Glu Phe Phe Glu Asp Ser
290 295 300
Arg Ser Ile Met Arg Cys Asn Tyr Tyr Pro Pro Cys Pro Glu Pro Glu
305 310 315 320
Arg Thr Leu Gly Thr Gly Pro His Cys Asp Pro Thr Ala Leu Thr Ile
325 330 335
Leu Leu Gln Asp Asp Val Gly Gly Leu Glu Val Leu Val Asp Gly Glu
340 345 350
Trp Arg Pro Val Arg Pro Val Pro Gly Ala Met Val Ile Asn Ile Gly
355 360 365
Asp Thr Phe Met Ala Leu Ser Asn Gly Arg Tyr Lys Ser Cys Leu His
370 375 380
Arg Ala Val Val Asn Gln Arg Arg Ala Arg Arg Ser Leu Ala Phe Phe
385 390 395 400
Leu Cys Pro Arg Glu Asp Arg Val Val Arg Pro Pro Ala Ser Ala Ala
405 410 415
Pro Arg Arg Tyr Pro Asp Phe Thr Trp Ala Asp Leu Met Arg Phe Thr
420 425 430
Gln Arg His Tyr Arg Ala Asp Thr Arg Thr Leu Asp Ala Phe Thr Arg
435 440 445
Trp Leu Ser His Gly Pro Ala Gln Ala Ala Ala Pro Pro Cys Thr
450 455 460
<210> 16
<211> 1510
<212> ДНК
<213> Zea mays
<400> 16
aaagagcgcg cgacggcggc ccctgggaga gccatgcgag actggaggcg gaaccgcgca 60
cgacaccaag ctgccgcgcc ggactgctgc acgcaagcgc agcgcaggac cgaccgacct 120
ccgtaggcac gcacggcgcc ggcggcatgg cggagcacct cctgtcgacg gccgtgcacg 180
acacgctgcc ggggagctac gtgcggccgg agccggagcg cccgcgcctc gcggaggtcg 240
tgaccggcgc gcgcatcccc gtcgtggacc tgggcagccc cgaccgcggc gcggtcgtgg 300
ccgccgtcgg cgacgcctgc cgctcgcacg gcttcttcca ggtcgtcaac cacgggatac 360
acgccgccct ggtcgcggcg gtgatggccg cggggcgcgg cttcttccgg ctgccccccg 420
aggagaaggc caagctctac tccgacgacc ccgccaggaa gatccggctg tccaccagct 480
tcaacgtgcg caaggagacg gtgcacaact ggcgcgacta cctccgcctg cactgccatc 540
ccctcgacga gttcctgccc gattggccgt ccaacccgcc cgatttcaag gagaccatgg 600
gcacctactg caaggaggtc cgggagctcg ggttcaggct gtacgccgcg atctcggaga 660
gcctgggcct agaggcgagc tacatgaagg aagcgctggg ggagcaggag cagcacatgg 720
cggtcaactt ctacccgccg tgcccggagc cggagctcac ctacggcctc ccggcgcaca 780
ccgaccccaa cgcgctcacc atcctgctca tggacccgga cgtcgccggc ctgcaggtgc 840
tccacgccgg ccagtgggtc gccgtcaacc cgcagcccgg cgcgctcatc atcaacatcg 900
gcgaccagct gcaggcgctg agcaacgggc agtaccggag cgtgtggcac cgcgcggtgg 960
tgaactcgga ccgggagcgc atgtccgtgg cgtcgttcct gtgcccgtgc aaccacgtcg 1020
tgctcggccc cgcgcggaag ctcgtcaccg aggacacccc ggccgtgtac aggaactaca 1080
cgtacgacaa gtactacgcc aagttctgga gcaggaacct ggaccaggag cactgcctcg 1140
agctcttcag aacctagcga atcggatacg gatggatgga tacattacat acgcgccctc 1200
tgtttttctc catgacgtta gaagaacacg ttctgcaatg tttgtccatt caaggtggta 1260
tcaatcaagg ctgtggtcgt tgcaattctt ccgctccata tacatgatta aatgctttga 1320
aagaaaaaga aaaaaaagaa acacaagtat tatggcacta ctagtgtttt taggaacaag 1380
gaaagagggg ttgcccctgc tggctatata tattaaatat aaataaaggt aaggctgtag 1440
acattggtga ataagagaaa gtatttgagt ttctctattg tcactccaga acagactcct 1500
ttgcctcgat 1510
<210> 17
<211> 1011
<212> ДНК
<213> Zea mays
<400> 17
atggcggagc acctcctgtc gacggccgtg cacgacacgc tgccggggag ctacgtgcgg 60
ccggagccgg agcgcccgcg cctcgcggag gtcgtgaccg gcgcgcgcat ccccgtcgtg 120
gacctgggca gccccgaccg cggcgcggtc gtggccgccg tcggcgacgc ctgccgctcg 180
cacggcttct tccaggtcgt caaccacggg atacacgccg ccctggtcgc ggcggtgatg 240
gccgcggggc gcggcttctt ccggctgccc cccgaggaga aggccaagct ctactccgac 300
gaccccgcca ggaagatccg gctgtccacc agcttcaacg tgcgcaagga gacggtgcac 360
aactggcgcg actacctccg cctgcactgc catcccctcg acgagttcct gcccgattgg 420
ccgtccaacc cgcccgattt caaggagacc atgggcacct actgcaagga ggtccgggag 480
ctcgggttca ggctgtacgc cgcgatctcg gagagcctgg gcctagaggc gagctacatg 540
aaggaagcgc tgggggagca ggagcagcac atggcggtca acttctaccc gccgtgcccg 600
gagccggagc tcacctacgg cctcccggcg cacaccgacc ccaacgcgct caccatcctg 660
ctcatggacc cggacgtcgc cggcctgcag gtgctccacg ccggccagtg ggtcgccgtc 720
aacccgcagc ccggcgcgct catcatcaac atcggcgacc agctgcaggc gctgagcaac 780
gggcagtacc ggagcgtgtg gcaccgcgcg gtggtgaact cggaccggga gcgcatgtcc 840
gtggcgtcgt tcctgtgccc gtgcaaccac gtcgtgctcg gccccgcgcg gaagctcgtc 900
accgaggaca ccccggccgt gtacaggaac tacacgtacg acaagtacta cgccaagttc 960
tggagcagga acctggacca ggagcactgc ctcgagctct tcagaaccta g 1011
<210> 18
<211> 336
<212> ПРТ
<213> Zea mays
<400> 18
Met Ala Glu His Leu Leu Ser Thr Ala Val His Asp Thr Leu Pro Gly
1. 5 10 15
Ser Tyr Val Arg Pro Glu Pro Glu Arg Pro Arg Leu Ala Glu Val Val
20 25 30
Thr Gly Ala Arg Ile Pro Val Val Asp Leu Gly Ser Pro Asp Arg Gly
35 40 45
Ala Val Val Ala Ala Val Gly Asp Ala Cys Arg Ser His Gly Phe Phe
50 55 60
Gln Val Val Asn His Gly Ile His Ala Ala Leu Val Ala Ala Val Met
65 70 75 80
Ala Ala Gly Arg Gly Phe Phe Arg Leu Pro Pro Glu Glu Lys Ala Lys
85 90 95
Leu Tyr Ser Asp Asp Pro Ala Arg Lys Ile Arg Leu Ser Thr Ser Phe
100 105 110
Asn Val Arg Lys Glu Thr Val His Asn Trp Arg Asp Tyr Leu Arg Leu
115 120 125
His Cys His Pro Leu Asp Glu Phe Leu Pro Asp Trp Pro Ser Asn Pro
130 135 140
Pro Asp Phe Lys Glu Thr Met Gly Thr Tyr Cys Lys Glu Val Arg Glu
145 150 155 160
Leu Gly Phe Arg Leu Tyr Ala Ala Ile Ser Glu Ser Leu Gly Leu Glu
165 170 175
Ala Ser Tyr Met Lys Glu Ala Leu Gly Glu Gln Glu Gln His Met Ala
180 185 190
Val Asn Phe Tyr Pro Pro Cys Pro Glu Pro Glu Leu Thr Tyr Gly Leu
195 200 205
Pro Ala His Thr Asp Pro Asn Ala Leu Thr Ile Leu Leu Met Asp Pro
210 215 220
Asp Val Ala Gly Leu Gln Val Leu His Ala Gly Gln Trp Val Ala Val
225 230 235 240
Asn Pro Gln Pro Gly Ala Leu Ile Ile Asn Ile Gly Asp Gln Leu Gln
245 250 255
Ala Leu Ser Asn Gly Gln Tyr Arg Ser Val Trp His Arg Ala Val Val
260 265 270
Asn Ser Asp Arg Glu Arg Met Ser Val Ala Ser Phe Leu Cys Pro Cys
275 280 285
Asn His Val Val Leu Gly Pro Ala Arg Lys Leu Val Thr Glu Asp Thr
290 295 300
Pro Ala Val Tyr Arg Asn Tyr Thr Tyr Asp Lys Tyr Tyr Ala Lys Phe
305 310 315 320
Trp Ser Arg Asn Leu Asp Gln Glu His Cys Leu Glu Leu Phe Arg Thr
325 330 335
<210> 19
<211> 1387
<212> ДНК
<213> Zea mays
<400> 19
gttttctttt tgaacgtaac tgacagaagc tatctgccta gctacggcgt gtcggttgct 60
tgtctcacca aagcagcgac atggaagcct gacagctcgt cgcgtcgcgc catttccacc 120
caacaaagcg gcggcgccag cacgcactgc ttctgcttgt gcgtgctcct ccgttccggg 180
cacgcctcta aagtctatac agcctcgaat ccatcccggc cgccgctcct gggggatact 240
acagcgagcc gaagcgggga tggcggagat ccctgtgatc gacctgcgcg tcgccggctc 300
ggcggccgag gagtccgcgc ggctgcgggc cgcgtgcgag cgcctgggct gcttccgggt 360
gaccggccac ggcgtgccct cggtgctcct ggcagagatg aaggccgccg tgcgcgcgct 420
cttcgacctc cccgacgacg ccaagcgccg caacgccgac gtcatcaccg gcagcggcta 480
cgtcgccccc agcccgacca acccgctcta cgaggccttc gggctcctcg acgccgccgt 540
gcccaccgac gtcgacgcct tttgcgcgct cctcgacgcg ccgcccaaca tcagggagac 600
cgtcaaggcc tacgcggaga agatgcacga tgtgatcgtt ggcgtcgccc gcgagctggc 660
gtctagcctg gggctagtcg aggagcactc gttccaggac tggccgtgcc agttccgcat 720
caacaggtac aactacacgc gggagacggt gggctcctcc ggcgtgcaga cccacacgga 780
ctcgggcttc ctcaccgtgc tccatgagga cgagtgtgtc ggcggcctcg aggtcctgga 840
cccgggcacc ggcgagttcg tgcccgtgga ccccgtcgcg ggctcctttc tcgtaaacat 900
cggcgacgtc ggcacggcgt ggagcaacgg gaggctgcac aacgtgaagc accgggtgcg 960
gtgcgtcgca cccgtgccgc gcatctccat cgccatgttc ctgctcgcac ccaaggacga 1020
cagcgtgagc gcaccggcgg cgttcgtgga cgcggaccac ccgcgcaggt acaaggtgtt 1080
caactacaac gactatcgga ggctcagact gtccaccggc gagcacgcag gcgaggcgct 1140
cgcacggatg gcggcgtgac gtggctggag ctgcaaattg gattggaagc cgagacaagc 1200
cgttagttat ttaccatgcc cgtgcgttca ccgcacacaa tcatattcaa aagccataaa 1260
ataaaaaata attttaatat cagtcaacat atggtttaaa tatcatatgg agtacaatat 1320
tccgaatttt tttttgtaat ttagtctgtc ttttgaaaaa aatgcacatc tagacctccg 1380
gatgact 1387
<210> 20
<211> 900
<212> ДНК
<213> Zea mays
<400> 20
atggcggaga tccctgtgat cgacctgcgc gtcgccggct cggcggccga ggagtccgcg 60
cggctgcggg ccgcgtgcga gcgcctgggc tgcttccggg tgaccggcca cggcgtgccc 120
tcggtgctcc tggcagagat gaaggccgcc gtgcgcgcgc tcttcgacct ccccgacgac 180
gccaagcgcc gcaacgccga cgtcatcacc ggcagcggct acgtcgcccc cagcccgacc 240
aacccgctct acgaggcctt cgggctcctc gacgccgccg tgcccaccga cgtcgacgcc 300
ttttgcgcgc tcctcgacgc gccgcccaac atcagggaga ccgtcaaggc ctacgcggag 360
aagatgcacg atgtgatcgt tggcgtcgcc cgcgagctgg cgtctagcct ggggctagtc 420
gaggagcact cgttccagga ctggccgtgc cagttccgca tcaacaggta caactacacg 480
cgggagacgg tgggctcctc cggcgtgcag acccacacgg actcgggctt cctcaccgtg 540
ctccatgagg acgagtgtgt cggcggcctc gaggtcctgg acccgggcac cggcgagttc 600
gtgcccgtgg accccgtcgc gggctccttt ctcgtaaaca tcggcgacgt cggcacggcg 660
tggagcaacg ggaggctgca caacgtgaag caccgggtgc ggtgcgtcgc acccgtgccg 720
cgcatctcca tcgccatgtt cctgctcgca cccaaggacg acagcgtgag cgcaccggcg 780
gcgttcgtgg acgcggacca cccgcgcagg tacaaggtgt tcaactacaa cgactatcgg 840
aggctcagac tgtccaccgg cgagcacgca ggcgaggcgc tcgcacggat ggcggcgtga 900
<210> 21
<211> 299
<212> ПРТ
<213> Zea mays
<400> 21
Met Ala Glu Ile Pro Val Ile Asp Leu Arg Val Ala Gly Ser Ala Ala
1. 5 10 15
Glu Glu Ser Ala Arg Leu Arg Ala Ala Cys Glu Arg Leu Gly Cys Phe
20 25 30
Arg Val Thr Gly His Gly Val Pro Ser Val Leu Leu Ala Glu Met Lys
35 40 45
Ala Ala Val Arg Ala Leu Phe Asp Leu Pro Asp Asp Ala Lys Arg Arg
50 55 60
Asn Ala Asp Val Ile Thr Gly Ser Gly Tyr Val Ala Pro Ser Pro Thr
65 70 75 80
Asn Pro Leu Tyr Glu Ala Phe Gly Leu Leu Asp Ala Ala Val Pro Thr
85 90 95
Asp Val Asp Ala Phe Cys Ala Leu Leu Asp Ala Pro Pro Asn Ile Arg
100 105 110
Glu Thr Val Lys Ala Tyr Ala Glu Lys Met His Asp Val Ile Val Gly
115 120 125
Val Ala Arg Glu Leu Ala Ser Ser Leu Gly Leu Val Glu Glu His Ser
130 135 140
Phe Gln Asp Trp Pro Cys Gln Phe Arg Ile Asn Arg Tyr Asn Tyr Thr
145 150 155 160
Arg Glu Thr Val Gly Ser Ser Gly Val Gln Thr His Thr Asp Ser Gly
165 170 175
Phe Leu Thr Val Leu His Glu Asp Glu Cys Val Gly Gly Leu Glu Val
180 185 190
Leu Asp Pro Gly Thr Gly Glu Phe Val Pro Val Asp Pro Val Ala Gly
195 200 205
Ser Phe Leu Val Asn Ile Gly Asp Val Gly Thr Ala Trp Ser Asn Gly
210 215 220
Arg Leu His Asn Val Lys His Arg Val Arg Cys Val Ala Pro Val Pro
225 230 235 240
Arg Ile Ser Ile Ala Met Phe Leu Leu Ala Pro Lys Asp Asp Ser Val
245 250 255
Ser Ala Pro Ala Ala Phe Val Asp Ala Asp His Pro Arg Arg Tyr Lys
260 265 270
Val Phe Asn Tyr Asn Asp Tyr Arg Arg Leu Arg Leu Ser Thr Gly Glu
275 280 285
His Ala Gly Glu Ala Leu Ala Arg Met Ala Ala
290 295
<210> 22
<211> 1496
<212> ДНК
<213> Zea mays
<400> 22
gtcggtctct tgtctcacca aaccggcgac atggtacatg gaggccagcc cgtcgcttgg 60
cgccacaagt ctcggtgccg tccgtccgac aagcggcgcc agcgcacgct ggctgctcgt 120
gcacgcctct aaatacggcc ccggacccgc caccaagcga aggccaatcc cgtccgccgc 180
cccccaccaa ccacgaacca cgcaagcgaa cccggccggc gcggggcagc ggcgatggcg 240
gagatcccgg tgatcgacct gcgcctcgcc ggctcgtcgc ccgacgagtc ggcgcggctg 300
cgcgacgcgt gcgagcgcct gggctgcttt cgggtgaccg gccacggcgc gcccgcgggg 360
ctcctggccg acatgaaggc cgccgtgcgc gcgctcttcg acctccccga cgacgccaag 420
cgccgcaacg ccgacgtcat ccccggcagc ggctacgtcg cgccctgccc cgccaacccg 480
ctctacgagg ccttcgggct cctcgacgcc gccgcgcccg ccgacgtcga cgccttctgc 540
gcgcgcctcg acgcgccgcc caaagtcagg gagaccgtca agacctacgc ggagaagatg 600
cacgacgtga tcgtcggcgt cgccggcgag ctggccacca gcctggggct gggcctggag 660
gagcactcgt tccaggactg gccgtgccag ttccgcatca acaggtacaa ctacacgcag 720
gagacggtgg gctcctccgg cgtgcagacc cacacggact cgggcttcct caccgtgctc 780
caggaggacg agtgcgtcgg cggcctcgag gtgctggacc ccgccgccgg tgagttcgtg 840
cccgtggacc ccgtcgccgg ctccttcctc gtcaacatcg gcgacgtcgg cacggcgtgg 900
agcaacggga ggctccacaa cgtgaagcac cgggtgcggt gcgtcgcgcc cgtgccgcgc 960
atctccatcg ccatgttcct gctggcgccc aaggacgacc gcgtgagcgc cccggaggcg 1020
ttggtcgacg cgggccaccc gcgtcggtac aagccgttca actacgacga ctaccggagg 1080
ctccggctgt ccaccggcga gcgcgcaggc gaggcgctcg cgcggatggc ggcgtgatgt 1140
cgtcacgcac gtgcaagccg ttaattatag gctcgcgcat gcatacgcct acacgagagg 1200
ttgtctcgtt aagccgttct attaaaatgt gtgggggaga aagatgacta ccgtggtgcc 1260
atgtggattg ctatcgggtc tgatcaataa aatcttgcaa cacttgcacg tgcgattcca 1320
tatcctagca cgggtgggcg ccacgctagt aggtagagac cggagcggcc aaaaaatggc 1380
tacagcacca gtaggtgaac tctcaagcaa cactggctat cccacttctg acgttgtctc 1440
tctcatcact atgtatgacc agcgaatgaa gtgtttaaaa atctgacgcc gtgaaa 1496
<210> 23
<211> 903
<212> ДНК
<213> Zea mays
<400> 23
atggcggaga tcccggtgat cgacctgcgc ctcgccggct cgtcgcccga cgagtcggcg 60
cggctgcgcg acgcgtgcga gcgcctgggc tgctttcggg tgaccggcca cggcgcgccc 120
gcggggctcc tggccgacat gaaggccgcc gtgcgcgcgc tcttcgacct ccccgacgac 180
gccaagcgcc gcaacgccga cgtcatcccc ggcagcggct acgtcgcgcc ctgccccgcc 240
aacccgctct acgaggcctt cgggctcctc gacgccgccg cgcccgccga cgtcgacgcc 300
ttctgcgcgc gcctcgacgc gccgcccaaa gtcagggaga ccgtcaagac ctacgcggag 360
aagatgcacg acgtgatcgt cggcgtcgcc ggcgagctgg ccaccagcct ggggctgggc 420
ctggaggagc actcgttcca ggactggccg tgccagttcc gcatcaacag gtacaactac 480
acgcaggaga cggtgggctc ctccggcgtg cagacccaca cggactcggg cttcctcacc 540
gtgctccagg aggacgagtg cgtcggcggc ctcgaggtgc tggaccccgc cgccggtgag 600
ttcgtgcccg tggaccccgt cgccggctcc ttcctcgtca acatcggcga cgtcggcacg 660
gcgtggagca acgggaggct ccacaacgtg aagcaccggg tgcggtgcgt cgcgcccgtg 720
ccgcgcatct ccatcgccat gttcctgctg gcgcccaagg acgaccgcgt gagcgccccg 780
gaggcgttgg tcgacgcggg ccacccgcgt cggtacaagc cgttcaacta cgacgactac 840
cggaggctcc ggctgtccac cggcgagcgc gcaggcgagg cgctcgcgcg gatggcggcg 900
tga 903
<210> 24
<211> 300
<212> ПРТ
<213> Zea mays
<400> 24
Met Ala Glu Ile Pro Val Ile Asp Leu Arg Leu Ala Gly Ser Ser Pro
1. 5 10 15
Asp Glu Ser Ala Arg Leu Arg Asp Ala Cys Glu Arg Leu Gly Cys Phe
20 25 30
Arg Val Thr Gly His Gly Ala Pro Ala Gly Leu Leu Ala Asp Met Lys
35 40 45
Ala Ala Val Arg Ala Leu Phe Asp Leu Pro Asp Asp Ala Lys Arg Arg
50 55 60
Asn Ala Asp Val Ile Pro Gly Ser Gly Tyr Val Ala Pro Cys Pro Ala
65 70 75 80
Asn Pro Leu Tyr Glu Ala Phe Gly Leu Leu Asp Ala Ala Ala Pro Ala
85 90 95
Asp Val Asp Ala Phe Cys Ala Arg Leu Asp Ala Pro Pro Lys Val Arg
100 105 110
Glu Thr Val Lys Thr Tyr Ala Glu Lys Met His Asp Val Ile Val Gly
115 120 125
Val Ala Gly Glu Leu Ala Thr Ser Leu Gly Leu Gly Leu Glu Glu His
130 135 140
Ser Phe Gln Asp Trp Pro Cys Gln Phe Arg Ile Asn Arg Tyr Asn Tyr
145 150 155 160
Thr Gln Glu Thr Val Gly Ser Ser Gly Val Gln Thr His Thr Asp Ser
165 170 175
Gly Phe Leu Thr Val Leu Gln Glu Asp Glu Cys Val Gly Gly Leu Glu
180 185 190
Val Leu Asp Pro Ala Ala Gly Glu Phe Val Pro Val Asp Pro Val Ala
195 200 205
Gly Ser Phe Leu Val Asn Ile Gly Asp Val Gly Thr Ala Trp Ser Asn
210 215 220
Gly Arg Leu His Asn Val Lys His Arg Val Arg Cys Val Ala Pro Val
225 230 235 240
Pro Arg Ile Ser Ile Ala Met Phe Leu Leu Ala Pro Lys Asp Asp Arg
245 250 255
Val Ser Ala Pro Glu Ala Leu Val Asp Ala Gly His Pro Arg Arg Tyr
260 265 270
Lys Pro Phe Asn Tyr Asp Asp Tyr Arg Arg Leu Arg Leu Ser Thr Gly
275 280 285
Glu Arg Ala Gly Glu Ala Leu Ala Arg Met Ala Ala
290 295 300
<210> 25
<211> 1614
<212> ДНК
<213> Zea mays
<400> 25
accacacgaa ttgcacatct ccacagctca cgattccaac actagctaca tatatatgta 60
gctttctagg ctactatata cactcaccac caagtgtgaa gtgtgtatat atagtgacag 120
ctactgcaat atatacatac gcgtcaccta tatattagcc aagctagcta tatgagcttg 180
gttgcggcgc caatggcgat cgtcgacgtg gccaacgccc agctgcagca agcagcagca 240
gcagctgcca agaaagacga ggacggccat gagcagcagg agtcgtccta cgactacggc 300
gcgctgatga aaggcgtgag gcacctgtcg gacagcggca ttaccaggct gcccgacagg 360
tacgtcctgc ccgcgtccga ccgccccggc gtccttgccg tctcgtcgtc cgtggcgggc 420
agcggcaggg tcaagctccc tgtcgtcaac ctcgccggcc tccgcgaccc ctgccagcgc 480
gccgccgtgc tggccacgct cgacgccgcg tgccgggagt acggcttctt tcaggtggta 540
aaccacgggt tcgggagcga cgtgagcggc gggatgctgg acgtggcgca gcgcttcttc 600
gagctgccgc tggccgagcg agcgcggcac atgtcggcgg acgtgcgggc gccggtgcgc 660
tacggcacca gcttcaacca ggccaaggac gacgtgctct gctggcgcga cttcctcaag 720
ctcgtctgcc agccgctgca ggcggtgctc ccgtactggc cccagcagcc ggcggacctc 780
agggacgtgg ccaccaggta cgccacggcg agccaccggc tgttcatgga ggtcatggag 840
gcggcgctgg aggccctggg catccccacg gccggcggcg tgctcgggga gctggcagcg 900
tcgtcgtcgc acatgatgac ggtgaactgc tacccggcgt gcccgcagcc tgagctcacg 960
ctggggatgc cctcgcactc ggactacggc ctcttcacgt tcgtcctgca ggaccacgtc 1020
gagggcctcc aggtcatgca cgacggccgc tggctcacca tcgaccccat cccgggatcg 1080
ttcgtcgtca acgtcggcga ccacctagag atctacagca acgggcggta caagagcgcg 1140
ctgcaccggg tgcacgtgaa ctccacgcgg ccgcgcatct cggtggcgtc gttccacagc 1200
ctgccggcgg agcgagtgat cgggccggcg ccggagctgg tggacgacga ggccggcaac 1260
ccgcggcggt acatggacac cgacttcgct accttcctcg cctacctcgc atccgcggac 1320
ggcaagaaca agaccttcct ccagtcaagg aagctgcctg ctgctgctcc tccatgcctc 1380
tagctaacta gatagctgct tattaatctg acagaataaa attaatcagt tcagcgcaca 1440
attccacaag cgaaaacaaa cctggatttg ttttaattag ctctgccctt cattattaca 1500
ttcaagctag ctcttggtca acgcatgcac acaagcttga gcattgactg gtcccttttc 1560
aatcggttgc attgtactcc ctccgtacca aaattggttg tcgctatagt attt 1614
<210> 26
<211> 1212
<212> ДНК
<213> Zea mays
<400> 26
atgagcttgg ttgcggcgcc aatggcgatc gtcgacgtgg ccaacgccca gctgcagcaa 60
gcagcagcag cagctgccaa gaaagacgag gacggccatg agcagcagga gtcgtcctac 120
gactacggcg cgctgatgaa aggcgtgagg cacctgtcgg acagcggcat taccaggctg 180
cccgacaggt acgtcctgcc cgcgtccgac cgccccggcg tccttgccgt ctcgtcgtcc 240
gtggcgggca gcggcagggt caagctccct gtcgtcaacc tcgccggcct ccgcgacccc 300
tgccagcgcg ccgccgtgct ggccacgctc gacgccgcgt gccgggagta cggcttcttt 360
caggtggtaa accacgggtt cgggagcgac gtgagcggcg ggatgctgga cgtggcgcag 420
cgcttcttcg agctgccgct ggccgagcga gcgcggcaca tgtcggcgga cgtgcgggcg 480
ccggtgcgct acggcaccag cttcaaccag gccaaggacg acgtgctctg ctggcgcgac 540
ttcctcaagc tcgtctgcca gccgctgcag gcggtgctcc cgtactggcc ccagcagccg 600
gcggacctca gggacgtggc caccaggtac gccacggcga gccaccggct gttcatggag 660
gtcatggagg cggcgctgga ggccctgggc atccccacgg ccggcggcgt gctcggggag 720
ctggcagcgt cgtcgtcgca catgatgacg gtgaactgct acccggcgtg cccgcagcct 780
gagctcacgc tggggatgcc ctcgcactcg gactacggcc tcttcacgtt cgtcctgcag 840
gaccacgtcg agggcctcca ggtcatgcac gacggccgct ggctcaccat cgaccccatc 900
ccgggatcgt tcgtcgtcaa cgtcggcgac cacctagaga tctacagcaa cgggcggtac 960
aagagcgcgc tgcaccgggt gcacgtgaac tccacgcggc cgcgcatctc ggtggcgtcg 1020
ttccacagcc tgccggcgga gcgagtgatc gggccggcgc cggagctggt ggacgacgag 1080
gccggcaacc cgcggcggta catggacacc gacttcgcta ccttcctcgc ctacctcgca 1140
tccgcggacg gcaagaacaa gaccttcctc cagtcaagga agctgcctgc tgctgctcct 1200
ccatgcctct ag 1212
<210> 27
<211> 403
<212> ПРТ
<213> Zea mays
<400> 27
Met Ser Leu Val Ala Ala Pro Met Ala Ile Val Asp Val Ala Asn Ala
1. 5 10 15
Gln Leu Gln Gln Ala Ala Ala Ala Ala Ala Lys Lys Asp Glu Asp Gly
20 25 30
His Glu Gln Gln Glu Ser Ser Tyr Asp Tyr Gly Ala Leu Met Lys Gly
35 40 45
Val Arg His Leu Ser Asp Ser Gly Ile Thr Arg Leu Pro Asp Arg Tyr
50 55 60
Val Leu Pro Ala Ser Asp Arg Pro Gly Val Leu Ala Val Ser Ser Ser
65 70 75 80
Val Ala Gly Ser Gly Arg Val Lys Leu Pro Val Val Asn Leu Ala Gly
85 90 95
Leu Arg Asp Pro Cys Gln Arg Ala Ala Val Leu Ala Thr Leu Asp Ala
100 105 110
Ala Cys Arg Glu Tyr Gly Phe Phe Gln Val Val Asn His Gly Phe Gly
115 120 125
Ser Asp Val Ser Gly Gly Met Leu Asp Val Ala Gln Arg Phe Phe Glu
130 135 140
Leu Pro Leu Ala Glu Arg Ala Arg His Met Ser Ala Asp Val Arg Ala
145 150 155 160
Pro Val Arg Tyr Gly Thr Ser Phe Asn Gln Ala Lys Asp Asp Val Leu
165 170 175
Cys Trp Arg Asp Phe Leu Lys Leu Val Cys Gln Pro Leu Gln Ala Val
180 185 190
Leu Pro Tyr Trp Pro Gln Gln Pro Ala Asp Leu Arg Asp Val Ala Thr
195 200 205
Arg Tyr Ala Thr Ala Ser His Arg Leu Phe Met Glu Val Met Glu Ala
210 215 220
Ala Leu Glu Ala Leu Gly Ile Pro Thr Ala Gly Gly Val Leu Gly Glu
225 230 235 240
Leu Ala Ala Ser Ser Ser His Met Met Thr Val Asn Cys Tyr Pro Ala
245 250 255
Cys Pro Gln Pro Glu Leu Thr Leu Gly Met Pro Ser His Ser Asp Tyr
260 265 270
Gly Leu Phe Thr Phe Val Leu Gln Asp His Val Glu Gly Leu Gln Val
275 280 285
Met His Asp Gly Arg Trp Leu Thr Ile Asp Pro Ile Pro Gly Ser Phe
290 295 300
Val Val Asn Val Gly Asp His Leu Glu Ile Tyr Ser Asn Gly Arg Tyr
305 310 315 320
Lys Ser Ala Leu His Arg Val His Val Asn Ser Thr Arg Pro Arg Ile
325 330 335
Ser Val Ala Ser Phe His Ser Leu Pro Ala Glu Arg Val Ile Gly Pro
340 345 350
Ala Pro Glu Leu Val Asp Asp Glu Ala Gly Asn Pro Arg Arg Tyr Met
355 360 365
Asp Thr Asp Phe Ala Thr Phe Leu Ala Tyr Leu Ala Ser Ala Asp Gly
370 375 380
Lys Asn Lys Thr Phe Leu Gln Ser Arg Lys Leu Pro Ala Ala Ala Pro
385 390 395 400
Pro Cys Leu
<210> 28
<211> 1863
<212> ДНК
<213> Zea mays
<400> 28
tgccaccata ccactagtgc aaggtcctag atttacactt ggtgctacac cttgcttcgc 60
ccccttcctt ccttccttcc ttccttccct ccttccttgg tctctaggca gctagcagtg 120
tggtgctgct gccggccgcc tattggccgc ctgggactgg gatccattaa ttactgcgcg 180
cgcgcggcta accaaccaat cccagcgtgc gtaatctatt gcccacatgc cgacgccgtc 240
gcacctcaac aagaacccgc gctacctgga cttccgggcg gcgcggcggg tgccggagtc 300
gcacgcctgg ccgggcctgc acgaccaccc cgtcgtggac ggcggcgcgc cgggccccga 360
cgccgtgccg gtggtggacc tgggcgccgc ggacccggcg ccggcgccgg cggcggcggt 420
ggcccgcgcc gccgagcaat ggggcgcgtt cctgctcacg ggccacggcg tccccgcgga 480
cctgctggcg cgcgtggagg accggatcgc caccatgttc gcgctgccgg ccgacgacaa 540
gatgcgcgcc gtgcgcgggc ccggcgacgc ctgcggctac ggctccccgc ccatctcctc 600
cttcttctcc aagtgcatgt ggtccgaggg ctacaccttc tcgccggcct ccctccgcgc 660
cgacctccgc aagctctggc ccaaggccgg cgacgactac accagcttct gtgatgtgat 720
ggaggagttc cacaagcaca tgcgcgccct cgcggacaag ctgctggagc tgttcctcat 780
ggcgctgggg ctcaccgacg agcaggccag cgccgtcgag gccgagcgga ggatcgccga 840
gacgatgacc gccaccatgc atctcaactg gtacccgagg tgcccggacc cgcggcgcgc 900
gctggggctg atcgcgcaca ccgactcggg cttcttcacc ttcgtgatgc agagcctcgt 960
gcccgggctg cagctcttcc gccacgcccc ggaccggtgg gtggcggtgc cggccgtgcc 1020
gggcgccttc gtcgtcaacg tgggcgacct cttccacatc ctcaccaacg gccggttcca 1080
cagcgtgtac caccgcgccg tcgtgaaccg ggacctcgac aggatctcgc tcggctactt 1140
cctcggcccg ccgccgcacg ccaaggtggc gccgctgcgc gaggccgtgc cgcccggccg 1200
ggcccccgcg taccgcgccg tcacgtggcc cgagtacatg ggcgtccgca agaaggcctt 1260
caccaccggc gcctccgcgc tcaagatggt cgccctcgcc gccgccgccg acctcgacga 1320
cgacggcgac gccgccgtcg tccatcagca gcagcagcta gtcgtctcgt cgtagccgag 1380
accgatcgcc ggagactgat gctgatgatg atgcatatat acatgagaga aatcgtcgag 1440
tagactagcc gattgcaaaa gcaaccccag ctgccgaaac ctggcatatc gatcccattc 1500
tctgctgcgc acatgtatgc atgcatgcgc ttcgtccgtt cgactcgtgt gtgcttgctt 1560
gcttgcgcgt gcagcagaac taattccgtt ccgcagctag ctgctctgct ctgctctgct 1620
ggaatgtaat taagtagtag tatatggtag tagagaaaag attagctagg cgatcgatat 1680
agatgacggg ccggggaaga agacgaatta attaagatcg atcgacgacg acgagctgtg 1740
cgtggctggc tgtgttcttc tctagcctag ttacagaggc cggctgctgc tgcttccaat 1800
cgggctgctt gtcgctactg acgatcgtta gtggatccat taactaatct ggaattctgg 1860
att 1863
<210> 29
<211> 1149
<212> ДНК
<213> Zea mays
<400> 29
atgccgacgc cgtcgcacct caacaagaac ccgcgctacc tggacttccg ggcggcgcgg 60
cgggtgccgg agtcgcacgc ctggccgggc ctgcacgacc accccgtcgt ggacggcggc 120
gcgccgggcc ccgacgccgt gccggtggtg gacctgggcg ccgcggaccc ggcgccggcg 180
ccggcggcgg cggtggcccg cgccgccgag caatggggcg cgttcctgct cacgggccac 240
ggcgtccccg cggacctgct ggcgcgcgtg gaggaccgga tcgccaccat gttcgcgctg 300
ccggccgacg acaagatgcg cgccgtgcgc gggcccggcg acgcctgcgg ctacggctcc 360
ccgcccatct cctccttctt ctccaagtgc atgtggtccg agggctacac cttctcgccg 420
gcctccctcc gcgccgacct ccgcaagctc tggcccaagg ccggcgacga ctacaccagc 480
ttctgtgatg tgatggagga gttccacaag cacatgcgcg ccctcgcgga caagctgctg 540
gagctgttcc tcatggcgct ggggctcacc gacgagcagg ccagcgccgt cgaggccgag 600
cggaggatcg ccgagacgat gaccgccacc atgcatctca actggtaccc gaggtgcccg 660
gacccgcggc gcgcgctggg gctgatcgcg cacaccgact cgggcttctt caccttcgtg 720
atgcagagcc tcgtgcccgg gctgcagctc ttccgccacg ccccggaccg gtgggtggcg 780
gtgccggccg tgccgggcgc cttcgtcgtc aacgtgggcg acctcttcca catcctcacc 840
aacggccggt tccacagcgt gtaccaccgc gccgtcgtga accgggacct cgacaggatc 900
tcgctcggct acttcctcgg cccgccgccg cacgccaagg tggcgccgct gcgcgaggcc 960
gtgccgcccg gccgggcccc cgcgtaccgc gccgtcacgt ggcccgagta catgggcgtc 1020
cgcaagaagg ccttcaccac cggcgcctcc gcgctcaaga tggtcgccct cgccgccgcc 1080
gccgacctcg acgacgacgg cgacgccgcc gtcgtccatc agcagcagca gctagtcgtc 1140
tcgtcgtag 1149
<210> 30
<211> 382
<212> ПРТ
<213> Zea mays
<400> 30
Met Pro Thr Pro Ser His Leu Asn Lys Asn Pro Arg Tyr Leu Asp Phe
1. 5 10 15
Arg Ala Ala Arg Arg Val Pro Glu Ser His Ala Trp Pro Gly Leu His
20 25 30
Asp His Pro Val Val Asp Gly Gly Ala Pro Gly Pro Asp Ala Val Pro
35 40 45
Val Val Asp Leu Gly Ala Ala Asp Pro Ala Pro Ala Pro Ala Ala Ala
50 55 60
Val Ala Arg Ala Ala Glu Gln Trp Gly Ala Phe Leu Leu Thr Gly His
65 70 75 80
Gly Val Pro Ala Asp Leu Leu Ala Arg Val Glu Asp Arg Ile Ala Thr
85 90 95
Met Phe Ala Leu Pro Ala Asp Asp Lys Met Arg Ala Val Arg Gly Pro
100 105 110
Gly Asp Ala Cys Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe Ser
115 120 125
Lys Cys Met Trp Ser Glu Gly Tyr Thr Phe Ser Pro Ala Ser Leu Arg
130 135 140
Ala Asp Leu Arg Lys Leu Trp Pro Lys Ala Gly Asp Asp Tyr Thr Ser
145 150 155 160
Phe Cys Asp Val Met Glu Glu Phe His Lys His Met Arg Ala Leu Ala
165 170 175
Asp Lys Leu Leu Glu Leu Phe Leu Met Ala Leu Gly Leu Thr Asp Glu
180 185 190
Gln Ala Ser Ala Val Glu Ala Glu Arg Arg Ile Ala Glu Thr Met Thr
195 200 205
Ala Thr Met His Leu Asn Trp Tyr Pro Arg Cys Pro Asp Pro Arg Arg
210 215 220
Ala Leu Gly Leu Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe Val
225 230 235 240
Met Gln Ser Leu Val Pro Gly Leu Gln Leu Phe Arg His Ala Pro Asp
245 250 255
Arg Trp Val Ala Val Pro Ala Val Pro Gly Ala Phe Val Val Asn Val
260 265 270
Gly Asp Leu Phe His Ile Leu Thr Asn Gly Arg Phe His Ser Val Tyr
275 280 285
His Arg Ala Val Val Asn Arg Asp Leu Asp Arg Ile Ser Leu Gly Tyr
290 295 300
Phe Leu Gly Pro Pro Pro His Ala Lys Val Ala Pro Leu Arg Glu Ala
305 310 315 320
Val Pro Pro Gly Arg Ala Pro Ala Tyr Arg Ala Val Thr Trp Pro Glu
325 330 335
Tyr Met Gly Val Arg Lys Lys Ala Phe Thr Thr Gly Ala Ser Ala Leu
340 345 350
Lys Met Val Ala Leu Ala Ala Ala Ala Asp Leu Asp Asp Asp Gly Asp
355 360 365
Ala Ala Val Val His Gln Gln Gln Gln Leu Val Val Ser Ser
370 375 380
<210> 31
<211> 1439
<212> ДНК
<213> Zea mays
<400> 31
gacctccatt ttgattatct ctatcctgta cgtgccgaga gtccttcaaa gccgacgacg 60
agacgacgat gcagtcgtcg tcgtcatcag cctcgacgcc ggctgccgct tccggcctcg 120
tcttcgatct cgggtctgcg gcgggcgtgc cggagacaca cgcgtggccg ggggtgaacg 180
agtacccgtc ggtggagtcc gctggccgcg acgtggtccc ggtggtggac atgggggtgg 240
cctgcccgga cgcgacgcgg gcgttggcgc gcgccgcaga cgagtggggc gtgtttctgc 300
tcgtcggcca cggcgtgccc cgggaagtgg cggcgcgtgc cgaggagcag gtcgcgcgcc 360
tgttcgtgct cccggctcct gacaaggccc gcgcggggcg ccgccccggg gagcccacgg 420
ccaccggcta cggcaggccg cccctggcac tccgcttctc caagctcatg tggtccgagg 480
ggtacacgtt ccgcgccgcc accgtccgcg aagagttccg ccgcgtctgg cccgacggcg 540
gcgacgacta cctccgcttc tgcgacgtga tggaggagta cgacagagag atgagggctc 600
tcggtggcag gctgctcgac ctcttcttca tggcgctcgg cctcaccgac gtccagttcg 660
ccaccggcga gacggagcgg aggatccgcg agacctggac ggcgacgatg cacccaatcc 720
tgtgtccgga accggagcgc gccatcgggc tgacggcgca cacggactcg ggcttcatca 780
cgctcatcat gcagagcccc gtgcccgggc tgcagctgct ccgccgcggg ccggaccggt 840
gggtgacggt gccggcgccg ccgggcgcgc tcatcgtcat gctcggcgac ctgttccagg 900
tgctcacgaa cggccgcttc cggagcccta tccaccgcgc cgtcgtaagc cgagagcgcg 960
agcggatctc cgtgccctac ttcctctgcc cgccggagga catgacggtg gcgccgctcg 1020
cgtccgctct gctgccgggg aggaaggccg tgttccgggc cgtgacgtgg ccagagtaca 1080
tggaggtcaa gcacaaggtg ttcggcacgg atgcgccggc cctggagatg ctgcagctgc 1140
aggtggatga ggaagaacaa ggtgaaaggg ccgccaccac ctaagcccta aggaactact 1200
agctgaatcc ataaactaat aaagaattcg tgaataaggg cgttggaaga ctggacacaa 1260
cacaagagag ttgctatata tcgtatttct gaaatttaag gcaaatatct tagttaaaaa 1320
actggtatat ttaaatagac aatatatatc taaaataaag atagttcacc atttttacgg 1380
tcgaacaatg ataaagttat atattgtctg aatagtaaca aattaaagat ttccaggag 1439
<210> 32
<211> 1116
<212> ДНК
<213> Zea mays
<400> 32
atgcagtcgt cgtcgtcatc agcctcgacg ccggctgccg cttccggcct cgtcttcgat 60
ctcgggtctg cggcgggcgt gccggagaca cacgcgtggc cgggggtgaa cgagtacccg 120
tcggtggagt ccgctggccg cgacgtggtc ccggtggtgg acatgggggt ggcctgcccg 180
gacgcgacgc gggcgttggc gcgcgccgca gacgagtggg gcgtgtttct gctcgtcggc 240
cacggcgtgc cccgggaagt ggcggcgcgt gccgaggagc aggtcgcgcg cctgttcgtg 300
ctcccggctc ctgacaaggc ccgcgcgggg cgccgccccg gggagcccac ggccaccggc 360
tacggcaggc cgcccctggc actccgcttc tccaagctca tgtggtccga ggggtacacg 420
ttccgcgccg ccaccgtccg cgaagagttc cgccgcgtct ggcccgacgg cggcgacgac 480
tacctccgct tctgcgacgt gatggaggag tacgacagag agatgagggc tctcggtggc 540
aggctgctcg acctcttctt catggcgctc ggcctcaccg acgtccagtt cgccaccggc 600
gagacggagc ggaggatccg cgagacctgg acggcgacga tgcacccaat cctgtgtccg 660
gaaccggagc gcgccatcgg gctgacggcg cacacggact cgggcttcat cacgctcatc 720
atgcagagcc ccgtgcccgg gctgcagctg ctccgccgcg ggccggaccg gtgggtgacg 780
gtgccggcgc cgccgggcgc gctcatcgtc atgctcggcg acctgttcca ggtgctcacg 840
aacggccgct tccggagccc tatccaccgc gccgtcgtaa gccgagagcg cgagcggatc 900
tccgtgccct acttcctctg cccgccggag gacatgacgg tggcgccgct cgcgtccgct 960
ctgctgccgg ggaggaaggc cgtgttccgg gccgtgacgt ggccagagta catggaggtc 1020
aagcacaagg tgttcggcac ggatgcgccg gccctggaga tgctgcagct gcaggtggat 1080
gaggaagaac aaggtgaaag ggccgccacc acctaa 1116
<210> 33
<211> 371
<212> ПРТ
<213> Zea mays
<400> 33
Met Gln Ser Ser Ser Ser Ser Ala Ser Thr Pro Ala Ala Ala Ser Gly
1. 5 10 15
Leu Val Phe Asp Leu Gly Ser Ala Ala Gly Val Pro Glu Thr His Ala
20 25 30
Trp Pro Gly Val Asn Glu Tyr Pro Ser Val Glu Ser Ala Gly Arg Asp
35 40 45
Val Val Pro Val Val Asp Met Gly Val Ala Cys Pro Asp Ala Thr Arg
50 55 60
Ala Leu Ala Arg Ala Ala Asp Glu Trp Gly Val Phe Leu Leu Val Gly
65 70 75 80
His Gly Val Pro Arg Glu Val Ala Ala Arg Ala Glu Glu Gln Val Ala
85 90 95
Arg Leu Phe Val Leu Pro Ala Pro Asp Lys Ala Arg Ala Gly Arg Arg
100 105 110
Pro Gly Glu Pro Thr Ala Thr Gly Tyr Gly Arg Pro Pro Leu Ala Leu
115 120 125
Arg Phe Ser Lys Leu Met Trp Ser Glu Gly Tyr Thr Phe Arg Ala Ala
130 135 140
Thr Val Arg Glu Glu Phe Arg Arg Val Trp Pro Asp Gly Gly Asp Asp
145 150 155 160
Tyr Leu Arg Phe Cys Asp Val Met Glu Glu Tyr Asp Arg Glu Met Arg
165 170 175
Ala Leu Gly Gly Arg Leu Leu Asp Leu Phe Phe Met Ala Leu Gly Leu
180 185 190
Thr Asp Val Gln Phe Ala Thr Gly Glu Thr Glu Arg Arg Ile Arg Glu
195 200 205
Thr Trp Thr Ala Thr Met His Pro Ile Leu Cys Pro Glu Pro Glu Arg
210 215 220
Ala Ile Gly Leu Thr Ala His Thr Asp Ser Gly Phe Ile Thr Leu Ile
225 230 235 240
Met Gln Ser Pro Val Pro Gly Leu Gln Leu Leu Arg Arg Gly Pro Asp
245 250 255
Arg Trp Val Thr Val Pro Ala Pro Pro Gly Ala Leu Ile Val Met Leu
260 265 270
Gly Asp Leu Phe Gln Val Leu Thr Asn Gly Arg Phe Arg Ser Pro Ile
275 280 285
His Arg Ala Val Val Ser Arg Glu Arg Glu Arg Ile Ser Val Pro Tyr
290 295 300
Phe Leu Cys Pro Pro Glu Asp Met Thr Val Ala Pro Leu Ala Ser Ala
305 310 315 320
Leu Leu Pro Gly Arg Lys Ala Val Phe Arg Ala Val Thr Trp Pro Glu
325 330 335
Tyr Met Glu Val Lys His Lys Val Phe Gly Thr Asp Ala Pro Ala Leu
340 345 350
Glu Met Leu Gln Leu Gln Val Asp Glu Glu Glu Gln Gly Glu Arg Ala
355 360 365
Ala Thr Thr
370
<210> 34
<211> 4095
<212> ДНК
<213> Zea mays
<400> 34
taaatttgtg atccttgtga agttgttata tcatgaattg tgaacttgtt gcatttgtga 60
tcttttgtca actttgttgt attgtgaagt ttgatatgtt taccgatcgt attttagatt 120
tcgatcgtta ccggtgtatt ttccgcacca aacttttgtt tccgatgttt tcgaaatacc 180
gatatcgttt ccgtttctat agttaccctt ttcaatttta tttccgatta aaaatatgaa 240
aacggtaatg gttttagtgt ttatcgaccg ttttcatctc taatcatccc tgccggtgaa 300
gtttaatttt tcccttggct aaagagatgc aagctgctgt aaaatacgtt aaaacaggca 360
aggcagcccc agcagccagc atcgcgtgcc cgtctatgta catcagtgga tacgtagcat 420
ctctagtgag taatataacg attgcatttg gctggaggac gtatgttata taagtatgtc 480
atttaccagt tgcattagta tcttccctaa ctcctataat aactctcttc gtggaatgga 540
cgtagacgta tgctatataa gtattaaaaa atagtttttt aagctggtgt cctcaatttt 600
gctattgttc tcgtttttat ctttagttgt gtcacaaatt taatccgtac aacaaatcaa 660
aaataccata cccttcttat attaattttc taacataaca tttgtttaga tattttcagt 720
cgtgaaaata caattctaat tctaacgtcg tagtatcaaa tcaaaccatc cagaatttga 780
ccaagcttaa ttataaaaaa tataaaattt atgatactga atagatagca ttagatttgt 840
tatataatat atttttataa aataccattt ttatggtata aatattggta ctcctttact 900
ttaaactata gatagttttg actaaggatg caactagaat tgcatcctct tttcactgca 960
ccttcattag ttttaatatt tatttagatg ggcccttgca aactgtagat atcatctctt 1020
gcaacattct ttctatagca ccacgaaaat gtattgcggc tttgaaatta taattgaatt 1080
agttgtatca tttctttcac cgatgcgtta aattcaaaat taagtgttat atttcttcat 1140
aatttgttaa atatatagac cctataatcc accattattt actataatag catacattaa 1200
cattggtttt agcctacact acgacactcg aggcattgaa ttttcctcta tcaaagaatt 1260
atatgtgtag tagtattgtt cttgacaaaa agggggatta aaattaaact accaatattg 1320
atacttatct tatcacatcc atgaatacaa tcaacactct tacaaaagat aagatacaag 1380
attaaaaagt accatgataa tacattaaga ttattagcaa tgcattaaat taaataaatg 1440
tgcaagtgaa tcatgatttt agttttatct attttacttt taaaatatga tattctctga 1500
ctacttctaa gcataaatgt gattctaagt catgaccgat cgtgcttatt cagaaaaatg 1560
aaggagacac agatttctat aaaaaaaggt tgtcatggga ctattgggtc aaccatctta 1620
ttcatttggg aaaataagtt tagaacacat caacccattt tagatgttga gtttggccct 1680
aatggtccat tgaccttact tttgtgggtt gacatagacc atctatccca agttattgtt 1740
gtgtcacatt ccctgatatc atgaatctat attttagctt tccgttttca tatttttagt 1800
cgttacatat tttttatccg cgtactagat taaaactcta gttgttgcaa tacattttgt 1860
tcattttttt ctatttcttc tttactaaca acatattcta gttcctagct acattcttaa 1920
gtaccatagt gctataaaca ttttttatcc tacattattc cacttaagaa attgaatttt 1980
ctgcataaaa aaattatatg tccagtagtg ttgtcttata aaagcataaa gtgattaaaa 2040
ttaaaaccat tattgatatc ttatttttca aaaaaaaata taagcttata gaaagtgaat 2100
taatttcatg gtaaattaat atagtttaaa ttgaattatt agtgttatta ctatgtttat 2160
tatcaatgaa acatttttca tggttgatat aacttagtgt tacttatttt agtatttttt 2220
atataattct agttaacttt tagtttttga tttaaaaaaa cgagaattgt gtccttttgt 2280
ggagtgagta taaagaaagt aatatctgtt catcataatt tggtttttta aggtacgtga 2340
aacttgcttt atatttggac tcaagctatg tctaaataca tagtaaaaaa gcaatatttc 2400
tagaaaagac aaaacatctt ataatttaga atcaaggaaa tatatagatt ttatgtgcag 2460
tgagaagcca tttacaatgg aacgttcaac gttgggccaa tagatatttt gcgatatgat 2520
gatgggcata tttttgcatg gttgtccctc cactagctat agtttgatga tacgatacgc 2580
tgcacacacc attgggttgt accatgttag tgtagcaaca gtagaaaccc aattgtggcc 2640
gtgaaccatg ataatactag gtagagtgct agctagaggt ttcaggctat tgatgcgtga 2700
attaaacttt ctgttgtgtt gcgaggaaac gagtattgtg aaatatttga aacggttttt 2760
tttgtgaaag atttgaaacg gtatttttgt tgtgaaataa agatcaaggc taaataaatt 2820
caaactaata aaacatatta attgacggcc tgaagccccc gcccccatgg ccccatgcca 2880
tagcatcagg tcccacatga catgaggccg cgcctccctc tatgttggct ccctgccttc 2940
gccgttgtcg tcgctcccga actccctctc ctcccctgtt acaaataccc ccacccgccc 3000
ggacagcttc cctgcacact cgcagctcgc acatctcatg gtgtcctaag aacggcaaga 3060
gccagctctg cctagcagca gcgcacagcc acatccatgg acgccagccc gaccccaccg 3120
ctccccctcc gcgccccaac tcccagcatt gacctccccg ctggcaagga cagggccgac 3180
gcggcggcta acaaggccgc ggctgtgttc gacctgcgcc gggagcccaa gatcccggag 3240
ccattcctgt ggccgcacga agaggcgcgg ccgacctcgg ccgcggagct ggaggtgccg 3300
gtggtggacg tgggcgtgct gcgcaatggc gacggcgcgg ggctccgccg cgccgcggcg 3360
caagtggcgg cggcgtgcgc gacgcacggg ttcttccagg tgtgcgggca cggcgtggac 3420
gcggcgctgg ggcgcgccgc gctggacggc gccagcgact tcttccggct gccgctggct 3480
gagaagcagc gggcccggcg cgtccccggc accgtgtccg ggtacacgag cgcgcacgcc 3540
gaccggttcg cgtccaagct cccctggaag gagaccctgt ccttcggctt ccacgacggc 3600
gccgcggcgc ccgtcgtcgt ggactacttc accggcaccc tcggccaaga tttcgagcca 3660
gtggggtgag taaagaagaa gatggcgccg aatttacatt tataagtagg accagcagaa 3720
gcccctgccc ctgggggcct tagcattgca ttcgactgat gaatacgcat ggcaggcggg 3780
tgtaccagag gtactgcgag gagatgaagg agctgtcgct gacgatcatg gagctgctgg 3840
agctgagcct gggcgtggag cgcggctact accgggagtt cttcgaggac agccgctcca 3900
tcatgcggtg caactactac ccgccgtgcc cggtgccgga gcgcacgctg ggcacgggcc 3960
cgcactgcga ccccacggcg ctgaccatcc tcctgcagga cgacgtcggc gggctggagg 4020
tcctggtgga cggcgagtgg cgccccgtcc ggcccgtccc aggcgccatg gtcatcaaca 4080
tcggcgacac cttca 4095
<210> 35
<211> 7404
<212> ДНК
<213> Zea mays
<400> 35
cctattttgt gtctaatact cttcttatat taattgtttg gtcaaacttt agataaattt 60
gactaatgat gcaattaaaa ctgcatcacc tttactaagg tactgcttta tatgtttcga 120
caaaattttc aattattctc tatgtgtttt aatctttgcg ctacacctcc attgatttaa 180
atactcattt attttaaacc ataacttaaa ttatatcgga tctttgcatc ctttctatgg 240
caccatacat gaatcgatat tttggctgca aatttttaat catgttagtt ttagcatttt 300
ttcatatcca tgtgttaagt ttgaatcatg tgttgttttt atataattta ttgaaaatat 360
agatcctaaa cttcactaat acttacaaca atagcatcat catgtgtttt aatccacgcc 420
acaacactca aggcattgaa ttttcttcta ccaaagagtt gtatgtgtgt attgttcttt 480
aaaaaataga gtgattataa ttaaactacc agtattcata tgtaaaatgt atagacatct 540
aaaataaaat ttgcaaaaaa cattgttgca gactttcaat ataattaaga atgggtttta 600
gggtcatgat atatggtttg ttaaagaaac ttgttttttt ttgcaattga taaactataa 660
aatacatttt cactattgtg tgcatatgta cttggtatac atagtggcat atatcatttt 720
tgtttacttt gaggtttgaa ttatctatgt taaaattgga taacatagat acattggtgt 780
gcgtcctttg gcccatttac ttgactgagg agcaatacta taaagtaaaa catatttgga 840
tattttatct taaactccta gcataatatt gatttaatta tgaacaaata tatgtttagg 900
tgatagtttc atgggtggta aactatataa gaaggcttac catgatcttt gcaaactcta 960
ggctatgaaa gagttccatg atttgtctta gaagcataga caaaacagtg ataatgatct 1020
aaatcacact tatggcactg atgaccatat atgcaaagct aaatgcatgt taagttgtat 1080
tatatcatat gtttacaatg actatcgcat ataacgagga atacattgtc tatatagata 1140
gctattactg tagtagtgcc aaatgttgga caacatgaat cataatcttc aaacctagag 1200
aaattgtagt cagtcgtaca catatcgtct agtaagttgt ctatactttt tatttattgt 1260
atcaaatttt attgttatct tgcttgcttg tttgtttgta ccatagacac aatatggtca 1320
aaaagtggtc aatcgattcg aagaagattg caattgacga gtgctaacag ttgatccttt 1380
tgttgtgcac gctagcggag tagcatgaaa agagtaaaat atgaaattag cgttctaaac 1440
tgtttgtgct ataggtactt cgtatttaat ggagtgacta actataggaa ggtgagagct 1500
cagaagtcag caccctcaca cagagttcta gagttagtgg tcatcgaacc acgacaaact 1560
acatgatgag cagaagaggc aacatcaaga ctatgatcaa tagtttcggg tcaatgaatg 1620
acatcgtgat gagtatttat ctaactatat agaacaacaa cacatgatgt tttaagtaag 1680
ttcaactgat cttctattgc tatctttaag tatttaacgt agcgaataat gttttatcta 1740
tttcattcat aaataatgtt gtgacaaaag gggataacca tcacttttac catgttctag 1800
ataccacaac catctccacc atcataatgg gttcttcatt ggtgcttgga cctcaaataa 1860
tcatatctat agccaactta gctcaattct aataaaatta ggcaacttgg cttcattgta 1920
gcaaaaatag ccaacttagc tcaattttat ctaaacttag ctaatctagc acaacttaga 1980
tcaatattag gaaaaactaa tcaatctaat ctagctcaac tatagcgaaa gatagatatt 2040
gtagcataac ttagtagatc tatctcaaat tttagcaaaa actaatcaat ttagataaac 2100
tctataaaat tttaatcatt atgacttatt tccaactaat tgtaacttgc atgattttta 2160
tgttccttct ttataattag caacacctaa agacacgaat gatgaggggt ctaacgcatt 2220
cattaaccag ttgttaaata atactctagg tagatgataa gaactctaat tattctatga 2280
atctaagcta aaagatgttt aatatttaag tattggtgtt tattatgtta tttagaacga 2340
ttcatgttac ttaaagattt gttatgattt ttaaatatga ttatgataat ttatgtggtg 2400
tggattaact tgtgaacata tgtgatgtag atgaatatgt atgttgtgga tggaaccata 2460
tgaatatata tacacactca tatactattc gttggtgtag gtaaagcttc atccatcggt 2520
aattactaaa tggtcttcag tcattaccac taggtgaagc ttcacacgac cgataattat 2580
tgaagaacgc tcattaattt ccggtaatgg cttattggcc ttcactagtc ggtgaaaatt 2640
agctattttt ataccaataa aaattagcta atatatgtaa accaggtcta atttttatgg 2700
gcctcttacc gaccaaaatt gattagatta ttgttacaat agttttagtc aaaagctagc 2760
tatgctataa aaattttgaa ttaaagtgag tttcgtaata aaaattgcat acttttaaaa 2820
taaaataatt aaaaaacagt ttttagaaat acaatcaaac accttatgct ataaaaaaat 2880
tgtaatgtac ctacaaatat ataatacttt actttaaaat aggcctgtgc cttctcggct 2940
ctatatgggc tgcctccaac gaagcgccat ggccatgggc tccactgtgt cgggtcccac 3000
atgaggccgc gcctccctcc aaatgttccc tccctgcctt cgtctttgtc gttgctcgca 3060
aactccctgt cctcccctgt tacaaatacc cccacccgcc cggacagctt ccctgcatac 3120
ttgcagctcg cacatctcat ggtgtcgcag gaacgacaag agccagctgt gcctagcagc 3180
agcagcagca gcgccaagcg cgcagccacg tccatggacg ccagcccggc cccgccgctc 3240
ctcctccgcg cccccactcc cagccccagc attgacctcc ccgctggcaa ggacaaggcc 3300
gacgcggcgg ccagcaaggc cggcgcggcc gtgttcgacc tgcgccggga gcccaagatc 3360
cccgcgccat tcctgtggcc gcaggaagag gcgcggccgt cctcggccgc ggagctggag 3420
gtgccgatgg tggacgtggg cgtgctgcgc aatggcgacc gcgcggggct gcggcgcgcc 3480
gcggcgcagg tggccgcggc gtgcgcgacg cacgggttct tccaggtgtg cgggcacggc 3540
gtggacgcgg cgctggggcg cgccgcgctg gacggcgcca gcgacttctt ccggctgccg 3600
ctcgccgaga agcagcgcgc ccggcgcgtc cccggcaccg tgtccgggta cacgagcgcg 3660
cacgccgacc ggttcgcggc caagctcccc tggaaggaga ccctgtcgtt cggctaccac 3720
gacggcgccg cgtcgcctgt cgtcgtggac tacttcgtcg gcaccctcgg ccaggatttc 3780
gagccaatgg ggtaagtaag gtagtaagaa ggagcgccgg tttacattta ccgcacgtcg 3840
gcgtgcggtc gagtcgggac tcgggagacg tatgaacccc cgtcccgtcc catgcatgtg 3900
tggcaggtgg gtgtaccaga ggtactgcga ggagatgaag gagctgtcgc tgacgatcat 3960
ggagctgctg gagctgagcc tgggcgtgga gctgcgcggc tactaccggg agttcttcga 4020
ggacagccgg tccatcatgc ggtgcaacta ctacccgccg tgcccggagc cggagcgcac 4080
gctgggcacg ggcccgcact gcgaccccac ggcgctcacc atcctcctgc aggacgacgt 4140
gggcgggctg gaggtgctgg tggacggtga gtggcgcccc gtccggcccg tcccgggcgc 4200
catggtcatc aacatcggcg acaccttcat ggtaacgaaa cgaaagcgct cgctcctctg 4260
ttttccttgg ccgctcttgt cctgtgtgta tattcagttg agctctctct gtgctgttat 4320
ttcccgaatc ctagtggacc taaacgggca ggttattaca gcacgcacac gtaggcatgt 4380
catgtagcta gtacatacat agcgatgccg atgcaaatgc aatagagaca tgcgttcgag 4440
ttggttccta tctcggcggg ctacggcagg tacacgcggc cgcggcgcgc tctctctagt 4500
ctatccgcgg ccgcgcccag gccgatcgag gcttccgggg gagagttgcg acaagagaac 4560
ggaccgaggg ggtcggctag cggtagcaag ttccctgttg gtttgtggcg ttggagcgtt 4620
gcggagaggc ttgcgcggcg gcggggacgt cgacggggac gtggcgggga gacgatacga 4680
tgggtgccgg gcaggtttcc gaattccaaa cgtttttgtg gcgtgcgtcc atggggcgcc 4740
cccaaacttc ggacgtttcc ggcgctccaa caaatcttct cgcttcacac gtcaccgtcg 4800
tcccggattc atttgcctcg tcgctccacc attcgctgct ctcctctcca cgtactctta 4860
ccctgacctt tgggaaagaa ctgaacattc gagatgcaca acagttcaaa tataacatat 4920
gcagcacaag atcgttcgac tgctatccga caagccaaca acgtgcccag tagaactgaa 4980
tgtacctgtg atttccagca ctaacttaca gcaacgttgt gaaaaaacaa aaacgaaaac 5040
aaacggcaga aaaaacagat gtattgttct acagttacac caaatatttt ctggtccttt 5100
cagcaccaac aagagccata cgcatatcta gaagacaaaa ttcctctaat ttcaccccta 5160
cgtggtagca gttcctcctc aacacagttc acgtgctagc gtcgagttct ttgggccgcc 5220
acatcgactt ctcgacgcag agcaggccct cgctgccctt ggtgtaggtc atccgcacct 5280
cccactgcac ggacttggcc atgctctcca gctcatttat cgtgtccgcg gtgtccctca 5340
cgatcagctt gccctgtggc ctcagtacac ggtcgacctc ggcgaaaact gcagccagtt 5400
tgcatctgta aacaggcaac acagattttt agtatctaaa acactgcagg caaacgccac 5460
aggttttagt cgcaagaagc aataaaagca tgcaaacaat gctacgtgta cgtatcaaag 5520
gaacatgtca aaactcgttg catgaacgat cattgatgtt tccttgctga actagtcaca 5580
tcagtctgct tcaacttctg ggtttcacta gtagatatac cagaagggta gaataatgtg 5640
aagagcaaga aatacagacc tctttctgag ctttgagaac agatggtccg cgtgcagaag 5700
gtcatacgtt cttgggtaag tgctgaaaga ctcgcaccag tcatggtaca tgccaaacaa 5760
accgcgctcg tagatgatgg gcagcgtgtc tggtgaatcg atcggcacga tattcatgac 5820
ccagaccttt tggtccctca gagctgcagc aaaactgcca tgcaacaatg taaagcatta 5880
gtcaagaaga aggtgtacag tgcatttctc cttgtcaaca gtcttcagta acaaaaaaaa 5940
agtgttatgc ttgactgaat ctttcaaaga aatatgcttg atgacttatg gtggacaagt 6000
tgcctgttat agtgttatgt tttaattaac tatgtgccag cttgggtaac tagtagttat 6060
gtagtgtgat ctgaattacc aaaatataaa taaataaata aacatgccca agaaactacg 6120
aaaaccattt acttaccctc catagacagc tctcatgtcc atgacatttc tcactttgga 6180
ccagtcaatt cccatgccat tcacatacga tttacttaca acccgtttcc agtgggcatt 6240
atctgcctca aaatcttcat ttgcaggctt tccatagaca ccaaccttgg aaccatcaat 6300
ccagaaaggg gtcttctcaa gcctttgcgg ccataactct ggccattttg atcctcggac 6360
ttttgagcca ccaggcagtt tgtgcatgca tgcttccaac ggtacattcc tgcaaatcaa 6420
aaggctgtgt aagcaaagca gagaagcact tttctccatt gaaaatatac tcttctcaaa 6480
gaaccgaaac cataccaagc agcatctgca tcatcagatt ccttgcacaa tggcgggctg 6540
ttttcagatc ttttctcata gcaaatattg tccattggtt tctgatatat gaccatacca 6600
acttggttta acttatcctt agtcttgttg accatcttcc agcacatgga ctttgtcaaa 6660
gtagacatgg ctgaaaaggg tatgtggcca catgttatgt tagaaataaa attcaatttt 6720
gaacagttgg tccatagcat gtattttgaa caaatgcaat ccttctccat ccatgaaaga 6780
agttgaccct tcatacttag gattattcag tactttcact catgtctgct gaatttgttc 6840
tcttggtagt tgctatacaa gaaaggggga agtacagagt agctaaactt atacaagcta 6900
tagtctgata tttgtatgaa acataaattt tggtatggat gtcttattaa aatgggaggt 6960
tgtataatat ttttctagcc tacctcaact tgcttgagac taaaaggctt tgttgttgtt 7020
gttgaggctg tatggtgctt tgactttaca aatcaagtta tcagctaccc tacttatgga 7080
tatacacctc tcataaaatg atggtaagaa gtttcgatat gtcacattaa cataagaact 7140
tcattcagtt agggtacaac gaagttaagt agttacggaa ataccattcc aaatctcaac 7200
atcctctggg agcttttggt aaacaggagt ggcagaccag acaaagtaac caccagggcg 7260
taacaagcgg ttcaattcca gcaaaagcat gccacctaaa agtagcgagc cagcaataag 7320
attcagttct atagcaaatc aataaatgaa aggaggacat gtcaatatgt aaccagcagg 7380
acaaaccttc gatgtgccaa ggga 7404
<210> 36
<211> 1788
<212> ДНК
<213> Zea mays
<400> 36
aatcccagcg tgcgtaatct attgcccaca tgccgacgcc gtcgcacctc aacaagaacc 60
cgcgctacct ggacttccgg gcggcgcggc gggtgccgga gtcgcacgcc tggccgggcc 120
tgcacgacca ccccgtcgtg gacggcggcg cgccgggccc cgacgccgtg ccggtggtgg 180
acctgggcgc cgcggacccg gcgccggcgc cggcggcggc ggtggcccgc gccgccgagc 240
aatggggcgc gttcctgctc acgggccacg gcgtccccgc ggacctgctg gcgcgcgtgg 300
aggaccggat cgccaccatg ttcgcgctgc cggccgacga caagatgcgc gccgtgcgcg 360
ggcccggcga cgcctgcggc tacggctccc cgcccatctc ctccttcttc tccaagtgca 420
tgtggtccga gggctacacc ttctcgccgg cctccctccg cgccgacctc cgcaagctct 480
ggcccaaggc cggcgacgac tacaccagct tctggtacgt tgcgttgcgt gcttgtgtgc 540
gcgcacacct gccgaccgcg gccacaccgt acgcaaccca cgcgtacgta cgtgcgctag 600
ctacctgctt cgctcgcttc gctcctctcg cctcgccatg catatgcacg tacggccgta 660
caggtacagc agcaggtcac acgcacgaac gcacgcacgc accagcaccg atatgataca 720
tcatcgacgt gtcgtccccc cgtctaaggc catgcatgca tgcaagcacg cctagctagc 780
ccttttggct tgctagctga cgaggggagc taggacgagc atacttactg tgcgcgtcat 840
gctcaattgc tcacactata ctactacttg ttactacagt gatgtgatgg aggagttcca 900
caagcacatg cgcgccctcg cggacaagct gctggagctg ttcctcatgg cgctggggct 960
caccgacgag caggccagcg ccgtcgaggc cgagcggagg atcgccgaga cgatgaccgc 1020
caccatgcat ctcaactggt gggtatatat tattgtctgt catgttgtcg tcgtcgtacg 1080
cgttgcggtt gggtgtacat gtatataaca caaacaacaa aaaactaacg ccgtgccgac 1140
gacgacgacg atcatcaggt acccgaggtg cccggacccg cggcgcgcgc tggggctgat 1200
cgcgcacacc gactcgggct tcttcacctt cgtgatgcag agcctcgtgc ccgggctgca 1260
gctcttccgc cacgccccgg accggtgggt ggcggtgccg gccgtgccgg gcgccttcgt 1320
cgtcaacgtg ggcgacctct tccacatcct caccaacggc cggttccaca gcgtgtacca 1380
ccgcgccgtc gtgaaccggg acctcgacag gatctcgctc ggctacttcc tcggcccgcc 1440
gccgcacgcc aaggtggcgc cgctgcgcga ggccgtgccg cccggccggg cccccgcgta 1500
ccgcgccgtc acgtggcccg agtacatggg cgtccgcaag aaggccttca ccaccggcgc 1560
ctccgcgctc aagatggtcg ccctcgccgc cgccgccgac ctcgacgacg acggcgacgc 1620
cgccgtcgtc catcagcagc agcagctagt cgtctcgtcg tagccgagac cgatcgccgg 1680
agactgatgc tgatgatgat gcatatatac atgagagaaa tcgtcgagta gactagccga 1740
ttgcaaaagc aaccccagct gccgaaacct ggcatatcga tcccattc 1788
<210> 37
<211> 1698
<212> ДНК
<213> Zea mays
<400> 37
cgtgccgaga gtccttcaaa gccgacgacg agacgacgat gcagtcgtcg tcgtcatcag 60
cctcgacgcc ggctgccgct tccggcctcg tcttcgatct cgggtctgcg gcgggcgtgc 120
cggagacaca cgcgtggccg ggggtgaacg agtacccgtc ggtggagtcc gctggccgcg 180
acgtggtccc ggtggtggac atgggggtgg cctgcccgga cgcgacgcgg gcgttggcgc 240
gcgccgcaga cgagtggggc gtgtttctgc tcgtcggcca cggcgtgccc cgggaagtgg 300
cggcgcgtgc cgaggagcag gtcgcgcgcc tgttcgtgct cccggctcct gacaaggccc 360
gcgcggggcg ccgccccggg gagcccacgg ccaccggcta cggcaggccg cccctggcac 420
tccgcttctc caagctcatg tggtccgagg ggtacacgtt ccgcgccgcc accgtccgcg 480
aagagttccg ccgcgtctgg cccgacggcg gcgacgacta cctccgcttc tggtacgtac 540
gagcgccatg tcacgtgctt gtgctttcat gcctcgtacc gtcgtcgtgc tgtacgtgtt 600
atgtttatcg gccggtacgt cacgcgtgct acactggtta acgacgtgag cgtgcccacg 660
ttgactgcat gcatgtgcat gcgcgcgccc agcgacgtga tggaggagta cgacagagag 720
atgagggctc tcggtggcag gctgctcgac ctcttcttca tggcgctcgg cctcaccgac 780
gtccagttcg ccaccggcga gacggagcgg aggatccgcg agacctggac ggcgacgatg 840
cacccaatcc tgtacgtacg tcaaaaacga atatctgacc aatgcaaacg tttttctgca 900
atgccagtca tccactcatc ctgtacgtac ctctggactc tgcttgtcca tctactgatg 960
acacgtatgg taggtacccc aggtgtccgg aaccggagcg cgccatcggg ctgacggcgc 1020
acacggactc gggcttcatc acgctcatca tgcagagccc cgtgcccggg ctgcagctgc 1080
tccgccgcgg gccggaccgg tgggtgacgg tgccggcgcc gccgggcgcg ctcatcgtca 1140
tgctcggcga cctgttccag gtgctcacga acggccgctt ccggagccct atccaccgcg 1200
ccgtcgtaag ccgagagcgc gagcggatct ccgtgcccta cttcctctgc ccgccggagg 1260
acatgacggt ggcgccgctc gcgtccgctc tgctgccggg gaggaaggcc gtgttccggg 1320
ccgtgacgtg gccagagtac atggaggtca agcacaaggt gttcggcacg gatgcgccgg 1380
ccctggagat gctgcagctg caggtggatg aggaagaaca aggtgaaagg gccgccacca 1440
cctaagccct aaggaactac tagctgaatc cataaactaa taaagaattc gtgaataagg 1500
gcgttggaag actggacaca acacaagaga gttgctatat atcgtatttc tgaaatttaa 1560
ggcaaatatc ttagttaaaa aactggtata tttaaataga caatatatat ctaaaataaa 1620
gatagttcac catttttacg gtcgaacaat gataaagtta tatattgtct gaatagtaac 1680
aaattaaaga tttccagg 1698
<210> 38
<211> 4095
<212> ДНК
<213> Zea mays
<400> 38
cggtctaagt gaccgtttga gagaggaaaa gggttgaaag agacccggtc tttgtgacca 60
cctcaacggg gagtaggttt ataagaaccg aacctcggta aaacgaatca ccgtgtcatc 120
cgccttattt gcttgtgatt tgttttcgcc ctctctttcg gactcgttta tatttctaac 180
gctaaccccg acttgtagtt gtgcttaaag tttgtaaatt tcagattcgc cctattcacc 240
ccctctaggc gactttcata taaatattgg gagaaatatg aaaaacaaat gaaggtcgaa 300
cgagtcagag acaccataaa aaagaggtcg tcttaactag ggtgctaaac ctcaacattg 360
tagtagatct tagtactgag tttgacatct ttgacaccaa caagatggtg atacgttact 420
ttctacgtta acttgggtag gtatatcgac tatagtggcc tataacacta ggctatgtaa 480
tatgatattg tgttgagtct ttataaacat gatttttttt aaaaaaaaga gctaaaataa 540
aaaatagaaa tcgacggtac gatgcaagtt cttctcaaga caaccaaacg cacccttgcc 600
cctttattga aattgaagta tgtgctttat caaatgttta aatactaatt ataagtatta 660
aatataattt aattataata ctaattatat agataaagac taaataacaa gacaaattta 720
ttaaatataa ttaattcatt attaacaaat acttaatgta gcacgatcga atcatggact 780
aattagtctt gatagactcg tcttaccatt taatcataat tagttttgta tactgtttat 840
aatatttcta actagctagt attaaacttt tgatgtaacc taactaaagt ttagtcacgc 900
caatacataa ggactcggat cgttcgatca cccatgacat cacgtatact aagagcatct 960
ccaaaagctc tccagaagtc tcccctaaat ctattttttt gggaaaaaca caaaaacatg 1020
tctccaacag ttcccttaaa gcgcccccaa ctttttcata gcccttaaaa ctccctcatt 1080
tgtagctaca aatgaggggt tttttgggct ccccagaaac aaactgttga tttaagggat 1140
ctgttggaga aaggattaaa atttaccctc acttattatt tagatgtccc ttaaaactga 1200
ttttgaggag tcgttttatg tagagctctt ggagatgctc taacacaccg agcacaaccg 1260
catcatcaat caaaacaacc caaagtttgt tcggtacaag tcatcagcct gtgtacacac 1320
atcagcctcg gccccgggag aagcgctagc aaacaaggtt cacctaaaaa tccatccaga 1380
ttcattgaat ccaaccagca caaacgtccc atttattaat cacctcatca caggtccccc 1440
cagcctcact ctcgcgccgg ctcaaggtac attgcgtgtc ctagccaaga cacgcagctc 1500
atctcagcct cacacgcaca gcaagagcga ggcgtgattc gccatgggcg gcctcactat 1560
ggaccaggcc ttcgtgcagg cccccgagca ccgccccaag cccatcgtca ccgaggccac 1620
cggcatccct ctcatcgacc tctcgcctct ggccgccagc ggcggcgccg tggacgcgct 1680
ggccgccgag gtgggcgcgg cgagccggga ctggggcttc ttcgtggtcg tgggccacgg 1740
cgtgcccgca gagaccgtgg cgcgcgcgac ggaggcgcag cgagcgttct tcgcgctgcc 1800
ggcagagcgg aaggccgccg tgcggaggaa cgaggcggag ccgctcgggt actacgagtc 1860
ggagcacacc aagaacgtga gggactggaa ggaggtgtac gacctcgtgc cgcgcgagcc 1920
gccgccgccg gcagccgtgg ccgacggcga gcttgtgttc gataacaagt ggccccagga 1980
tctaccgggc ttcaggtgac gaaattaact atatatccct ttcgatcata gttgcgttaa 2040
taaattaagg gaatcgtgag cgtacgtacg taagtttccg cagagaggcg ctggaggagt 2100
acgcgaaagc gatggaagag ctggcgttca agctgctgga gctgatcgcc cggagcctga 2160
agctgaggcc cgaccggctg cacggcttct tcaaggacca gacgaccttc atccggctga 2220
accactaccc tccttgcccg agccccgacc tggccctcgg cgtggggcgg cacaaggacg 2280
ccggcgccct gaccatcctg taccaggacg acgtcggggg gctcgacgtc cggcggcgct 2340
ccgacggcga gtgggtccgc gtcaggcccg tgcccgactc gttcatcatc aacgtcggcg 2400
acctcatcca ggtacgtgcc cacctgatga actgagctga acgtaggttg catgcactgc 2460
atgtgtatag gcttctcaga tcgcttcgtg tggcgtaagg tgtggagcaa cgacaggtac 2520
gagagcgcgg agcaccgggt gtcggtgaac tcggcgaggg agaggttctc catgccctac 2580
ttcttcaacc cggcgaccta caccatggtg gagccggtgg aggagctggt gagcaaggac 2640
gatccgccca ggtacgacgc ctacaactgg ggcgacttct tcagcaccag gaagaacagc 2700
aacttcaaga agctcaacgt ggagaacatt cagatcgcgc atttcaagaa gagcctcgtc 2760
ctcgcctaac tactgctact gctaggatcc atgccattgc catgtcgtct tcagattcag 2820
agcacgccat gtcgtcgcta gcttcgtggt agaacaaata atgatgtgcg tgctgtgtgt 2880
aagcatggat atggatgtga atatgtaata tgatgagcac tcctactttg gtatgtttgg 2940
gaataacaga cttgtgttgg tctggttcat tatttgtaag aaaatcaaaa agagttagta 3000
gggcaggagg ctaaccacag tcatgctgca ccacatccct ggtggaaagc tggccgggtt 3060
acgctacgct cgtgcagcca gattactgca gggccgggat atgcttccgg tggaaggaag 3120
gggacggtgg ctgaggacca tggggctgga gcctgggaga gaggtcgagc tagaagaaag 3180
ggggagagag aagacgcaca acgaagatgg gtcagccagg gatttcgacc caagggggag 3240
ctagtggatt ttgggagaaa acagaaaaga gaaaagagaa aagaagaaaa atttgttggt 3300
gtgaacacaa ggttgatttg tcttttctta tttggattga tgatgagtcg tggactaacc 3360
gacccgtgag ctattgtgtc gtataatcat gtctctcggt ttctggtgtg caggtttgaa 3420
gcacagagac ggtggtcgac gcaaaggtga acgtcatgca ggttcgtgcc gatggaccgg 3480
gagcagtgaa agacgagcgt tgggacttga acaagggacc agagtcgccg gatgactagc 3540
cgcagtggct gacgcctgga acacgcatag acgtgaggac gtggtagagc aggtgaaaat 3600
cgcctagagg gggggggggt gaatagacaa aacctaaaaa ttataaactt tgaacacaaa 3660
ctttacctga ggttaccgtt agaacgagta ttaatgaaat cggagtgcgg aaggcaagtt 3720
cttcttgcta cgagttgctt aatcaatatt gataactttg ggagtcaact caaaatgatc 3780
acaagcaaaa gaactagaga gagaggagag gaagaatcaa ctcgcaaagt aatgatcaac 3840
acaaatgaac acaatgattt atttctcgag gtttggttcc gaagaaccta ctccccgttc 3900
aggagtccac ataggacatg tctctttcaa ccctttctct ctctcaaatg gtcacataga 3960
ctggttcagt tgagagcacc tagagggggg tgaataggtg atcttgtaaa atcaaacact 4020
aatagccaca aaacttagtt taaagtgtta gtacggctaa gtagctttga agcgagttat 4080
tgtgaacaca acaat 4095
<210> 39
<211> 21
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 39
ctccatcatg cggtgcaact a 21
<210> 40
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 40
uaguugcacc gcaugaugga g 21
<210> 41
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 41
ggtactgcga ggagatgaa 19
<210> 42
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 42
uucaucuccu cgcaguaccu a 21
<210> 43
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 43
caggcgccat ggtcatcaa 19
<210> 44
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 44
uugaugacca uggcgccugg a 21
<210> 45
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 45
tcatgcggtg caactacta 19
<210> 46
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 46
uaguaguugc accgcaugau a 21
<210> 47
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 47
tcgctcgcct tcttcctca 19
<210> 48
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 48
ugaggaagaa ggcgagcgac a 21
<210> 49
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 49
tccaacgggc ggtacaaga 19
<210> 50
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 50
ucuuguaccg cccguuggac c 21
<210> 51
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 51
gcatcaacag gtacaacta 19
<210> 52
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 52
uaguuguacc uguugaugcg a 21
<210> 53
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 53
tggacgatgg atagttcaa 19
<210> 54
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 54
uugaacuauc caucguccau c 21
<210> 55
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 55
tggaccatgg atacttcaa 19
<210> 56
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 56
uugaaguauc caugguccau c 21
<210> 57
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 57
gcaaggtcct agatttaca 19
<210> 58
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 58
uguaaaucua ggaccuugca a 21
<210> 59
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 59
cagagtacat ggaggtcaa 19
<210> 60
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 60
uugaccucca uguacucugg a 21
<210> 61
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 61
ccatgcccta cttcttcaa 19
<210> 62
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 62
uugaagaagu agggcaugga a 21
<210> 63
<211> 19
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 63
acatggcggt caacttcta 19
<210> 64
<211> 21
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 64
uagaaguuga ccgccaugug a 21
<210> 65
<211> 726
<212> ДНК
<213> Палочковидный вирус риса тунгро
<400> 65
tcctacaaaa gggagtagta atatttaatg agcttgaagg aggatatcaa ctctctccaa 60
ggtttattgg agacctttat gctcatggtt ttattaaaca aataaacttc acaaccaagg 120
ttcctgaagg gctaccgcca atcatagcgg aaaaacttca agactataag ttccctggat 180
caaataccgt cttaatagaa cgagagattc ctcgctggaa cttcaatgaa atgaaaagag 240
aaacacagat gaggaccaac ttatatatct tcaagaatta tcgctgtttc tatggctatt 300
caccattaag gccatacgaa cctataactc ctgaagaatt tgggtttgat tactacagtt 360
gggaaaatat ggttgatgaa gacgaaggag aagttgtata catctccaag tatactaaga 420
ttatcaaagt cactaaagag catgcatggg cttggccaga acatgatgga gacacaatgt 480
cctgcaccac atcaatagaa gatgaatgga tccatcgtat ggacaatgct taaagaagct 540
ttatcaaaag caactttaag tacgaatcaa taaagaagga ccagaagata taaagcggga 600
acatcttcac atgctaccac atggctagca tctttacttt agcatctcta ttattgtaag 660
agtgtataat gaccagtgtg cccctggact ccagtatata aggagcacca gagtagtgta 720
atagat 726
<210> 66
<211> 165
<212> ДНК
<213> Палочковидный вирус риса тунгро
<400> 66
acgaatcaat aaagaaggac cagaagatat aaagctggaa catcttcaca tgctaccaca 60
tggctagcat ctttacttta gcatctctat tattgtaaga gtgtataatg accagtgtgc 120
ccctggactc cagtatataa ggagcaccag agtagtgtaa tagat 165
<210> 67
<211> 2000
<212> ДНК
<213> Zea mays
<400> 67
ctgcaatata tacaccaaaa gtattataaa ctgtcatata tatgaccaaa acctttttat 60
tttagaaaag tatattaatc atggtatatt aatcaaagtt gttgttgggg ctgcaaaaat 120
catacccttc ttccacaagc tgttccttga actgcaggta ctcaggaact ctcagctcct 180
caacagcgag ctcactgacg ttgaccctca catactccca gacaccaggc ctagggcgga 240
tggcaagtgc aacccatggg gggatgacaa tcgcctcctg taagataata gagctagaat 300
gattaaagaa ggtgcacact acaaaaggaa cagtgctgtc cagcgagatc tgaatctgat 360
gcaaacctga gctgccctca ggacatcctc aaaagcacca tccttgagct tctcgcgctc 420
agcctcaggg atcgcattgt tgtactcggc aatgatctgg tggggctgca gcataccctt 480
tccaaggttt ttcagcctgc gcaaaacgat gtgccaaata acatcagact atgccagatc 540
tataaactca tcaaacatat acaatttcaa gaaatagttt agacgtatga tcagcagtca 600
gtagcgtggg aacatatgca acatagcgaa gaggcacaac agcaaattca ttcgaaaaaa 660
tgaaaacaaa gattcctctc ttttaactga acttctcgaa acccctttca tgcctacaca 720
tccgatctag tcagatgcct atgcgttcat gctgaacaga acgtgtcaga actaagcata 780
aactggttag caagcattat cgtattcgat agacccttta gtaacaagct atacattggg 840
taagttcaga ctccaatcat tctgttcaga aacatcgtat tgaatataaa actaaagaac 900
acacatgcag gtgcagccag atctaacagc agtttacagt cggtactaaa aaaagcatgg 960
tgtatgtatg tatcatcagt atccagtact aggtttcgac aaaatcctgg atgctaatta 1020
aatactcatc ttattaggga acacaggaac attatgtcta cagcattgaa tgatggccac 1080
atcatgctag atctaacaat acataatatg atggaactgg tcttaaaaag tcgcattcgc 1140
tcaaataata cccgtagcaa aataaatgta aacttgcaga cgaagcgggg gaaatgaggg 1200
cagacctggt gaagacggcg acaagctcat tggggtgggc agagagtgag tcgccaatgc 1260
gctccctgac gctgtggagg cggctcagga cacggtcacc tgcaccttcc cccattgctg 1320
tcctcttcct ggatcctcag gcctgcacag cgaaaccgaa acggaagcgg aagcttcagt 1380
cagcagagaa aactgaaacc gaaaaacggt tcagatccgt tgacataaaa gctgcgatga 1440
catcctaaaa ctaaaacccc tccagcaaga cataaaccca actgccaaca accagtcttt 1500
taagtctcga cacacccttg acgctgcgcc acgaaactat attgcaggca agaaaccaac 1560
agaacctaac tctggaaggg gggaaagaaa cggcagacag gagcaagacc caaaaaaaaa 1620
cgactcagat cctggtacta tagtcctagt acctagacca gaaagaagaa acaaccaata 1680
caacaagagg catacaagaa ctgaatcgat gaactgaaac gcttcagagg accgaggaat 1740
ggcggagaag ggaggcgcct atttatacag atctgacgag agaaccgaac aaaaacacat 1800
cgatgggaac catggagaag aaaagggctg gccgcatggc accaatggcc tcggcctcca 1860
aaaagccgtt gaatccaaag caggcgagga cgaagcgtga cgcggcaggg tacttctcta 1920
gaaaagcacg gcatcagcaa ggtggggggg ctggggttcc ttattgcagg caatcacgag 1980
gtgattagca caaacggaag 2000
<210> 68
<211> 2000
<212> ДНК
<213> Oryza sativa
<400> 68
ctgaaatata catcagagat attacaatga catatatatc gataaaagaa aaataataaa 60
attaagtttt aaattttaag aatatatgtt tttagtatcc caattatgca gatttcatac 120
ccttcttcca caagctgttc cttgaactgc aagtactcgg ggactgtcag caactcaaca 180
gcgagctcgc tcacattgac cctcacatac tcccagacac cgggcctcgg gcggatggca 240
agggcaaccc atggggagat aacaatcccc tcctgcatga taaaaaacaa ttacaagtta 300
agttagagca agcggtagag taaagatgga tctctgtgat gcaatgaaat ctgaatctga 360
ttcaaacctg tgcactcctc aggacatcct caaaagcacc gtccttcagc ttctcacgat 420
cagcctcaga gattgcgttg ttgtactcag caatgatctg gtgggcctga agcattccct 480
ttccgaggtt aaccagcctg cgcaaataac agtgtcaaca aaaatatcag gccagatcta 540
tcaactcagc ctataaatat ctcaataaga taattttagc acttgagcat ttgcgcataa 600
taagaaaatt tgctattagc cacttaaaaa gaccatatat gatctgtttg cattgagatg 660
aattaaaaat ttcattgtag atatgaaatg attagttttg accatttaat tggacttaat 720
gaaatatgcg cgataatcag atctacgcgc tcgcgccaat agatctagta agatgtaggt 780
tttttatttt ttttgtgaaa ctttgctacc acaacaagca tctgtaccag tgcagaattc 840
attacttgta ttcagtttgt aaaccgtata tataatataa ataacatgca catgcagtca 900
gatctagcac taccagtcca cagtaatcca aaactacatt tgtatatttc atcattattc 960
agtagtacta ggtttgtaca aaatcttggc tgcagaaggc cgcacttaaa tattcattct 1020
aatcagaaac ttaaaaaaaa agtgactaca aaatgattgc atccaattca gtaaatatga 1080
gccattcctg gccagatcta acaatctcaa caacaaagat cctatatgaa catctccttc 1140
taaaagaaaa tacagtaaca tctgaaggca gtagactaga aaccaacaaa atctaatgct 1200
gggaaatcac taaatcagca cgaacctggt gaagacggcg acgagctcat tggggtgggc 1260
ggagagggag tcgccgatgc gctccctgac gctgtggagg cggctcagga cgcggtcgcc 1320
ggcagcttcc cccattgctc tcctcttcct cttggctcct caagcctgcg tgcacaacca 1380
accaccatca tcagatacat ccagacccag tcaacacaat cactccagga aaaaaaaaag 1440
tcaagccata aaccccaacc aaaaaccacg cctttgacaa acactggaag aaaaagaaaa 1500
tcgcagcttt ttcacaagca atctagaaga aaagaaaaag aaaagactac atagcagcta 1560
taattgactg agaagcatac aggaatcaaa caatggagaa ggggagggag gaagaacaat 1620
gatgctccag gctgaggacc gaggaactgg gtgaagcggg gtaggcgcgt atttatgcag 1680
atctgaggag agaaaccacc aaaacaatcc gatggtttca acgaaaaaga tcgtcgcttc 1740
ttgctgcacc agctcaccca tagccgttga gatcgaagct aagctagcag cagcaaagct 1800
ggaacgaaga gtgacgcatt caagctcctc tcctctcctc tcctctcctc cggagcacga 1860
ggccagcatg ggatggattg gggtttcttg ttggccatgg caaaggagga ggtcattaac 1920
gttgacacgg cgtaatttaa ttaaatctta tcttaaaata tgatttaagt ggtagtaaca 1980
aggaagatta atactatgaa 2000
<210> 69
<211> 2000
<212> ДНК
<213> Oryza sativa
<400> 69
gctgtttaag aaaaacagaa gtaaaattca gtcactgtta ttttgcttca gttataatct 60
gcaaatcgtc gttctggtac ttactgtcca tcaacaagct gttccttgaa tgccaagtac 120
tcagaaacac tcagctcttc cactgccaac tcacttacat tcacccgaat gtagtcccag 180
acaccaggcc ttggcctgat ggccagtgca acccagggcg gcagcacaat ggcttcctac 240
atacagtcaa ggaagtaagt tagaaagact ggtatttgac tttgagttga ctatcataac 300
catcctggct cattgccaaa tttacctgag cagcccggag aatgtcttca aagggagcat 360
atttctcttt gtcagcttcg atcaaggcat cgaactccgc aagcagctgg tgacgctgga 420
gcattccctt tccctggtta acatacctgc atagagtgat atttaagaaa tagaaccaat 480
gcttagatct cacatccttt ctgcggctga actatgttaa tggcactacc acataaacct 540
gatttttact tcttattttt aagaccacat gatctgtact taatctagct atgaacaaac 600
aatatttcaa catcatctaa gattcatgac tcaagacaaa aatgttagag ctcatcacag 660
attattatag ataccatcat taaaactaaa gagatgcata accttgtcag ctaagaattt 720
gtaacatact aacatgttat cgtttcacat ctgggttgac taagaactaa ccaactgtat 780
ggataaaatc attgaaaact caaaacaatt agtagcaggt tccaagaaga cacaagatat 840
tatattgaga tcttcaccta gagaagagtg caatcaactc attgggatga gacgagaagg 900
tggcaccgag gcgttcgcgg agactgtgga ggcgagctag cttggcagcc atgactcaat 960
ttcaggaact gcaaagaaag gttacactta gcaacacgta ccaaaaccac tcacttgcac 1020
aagaataatt agtcaacagc catcactaag cattgcaaga ctatctctga acaggaaagc 1080
catgctaaat caacactaat aacatcacac aaaagcattg gaagatcaaa acataactaa 1140
aaacagctgt ttcatctaca caactgaaag catctatggt ttacgaagca gagtgcgagt 1200
actgattcaa aataaatcaa cctgaaccaa tatactctga caatgttttc aaagggataa 1260
aagaaccagc tttatcaaat ggatttgttg ggttttagta agtatcattg agataccgat 1320
ggcatatctc aaactttgca aaattataat ggcatggttc caaattaccc tttagtatta 1380
gcaccagtta gatcctaatt cctaaatccg ataggacaga gcgaaagatc cctggagata 1440
tgaagatttg gctacagatt aagcagagcc aacatgaagt tccgaatatt atgaatccgc 1500
aagcggggag atcaaagaga agaatacgga aggtcgcgac tccatgaaag aatccaacca 1560
aaaacccaaa gatttttctc agttcaaaaa aaaaaaaccc ttcatttttg gttcgccatc 1620
caccgacagg caccaagaca ttcctcagga agcaaaaaag attaagcaga acaagtgata 1680
agcaagacac agtatcaacg gactacgagt cgagaaaatc actgaggcgc gattcttact 1740
gcaccaagta aaaaaaaatt tgggggcaaa aagaactctg caatggggcg gagcaacgtg 1800
gcagcaaaac taaaggtcga ggatttgagg ttttttgccg gttttcctcg aaaccccgaa 1860
tccgctcata gtaaacccac taaactgcag cagaaacccc cctcttggtt cagatttacc 1920
gaaagcagta aacccaagaa catgtcagca aaaactcctg caagattcag ctgacgaccc 1980
accaaagaat cgcaagaaat 2000
<210> 70
<211> 2000
<212> ДНК
<213> Oryza sativa
<400> 70
tggcggacgc gccacgcaca aacacaaacc tgcacacccc tgtgtcagag gaggagaggc 60
caagaaagga aatcgagtgg aggaagtgag gagcggcgga gacgtaggag gaggaggggg 120
agatggaaat ggaaagccgc gcgagagagg aggcgcgtgc tggatgggag gaggaggagg 180
aggtggtggg tttgtgtttg gagagacgag cgagagaggc gaagcattta aagggaggaa 240
gagggggaga gagagagaga gagagagaga gagagagaga gagagagaaa ggaggaatat 300
aataaagggt ggtgcacctg ccaactgcta tgctcaccaa cactttgtac acacccagtt 360
acacccccct gcctttatta tttccagtgc agtaataact tcaacaatta ttgaaatgaa 420
aatggaatta atggagttag tatcggatta gcgacacgct tgccgagctt ctagacggtg 480
cgattatttc agcgggaacg actttctgta ggtgaattta atagaggagt gttttaaatc 540
cactcgacgt tgtaatagct ggtttaattc gtttgtactg tcgagtagtt atccaaaatc 600
aattttggat atttaaaaga aaaaaaaaca gatccgaagt attggaccta ctggcaaata 660
ggaattttgc tatatatagg tgtgcgttca tttataatgg agtagcatgg agttttatta 720
atccagtaaa tgttttcatt gatttaatta atataacgaa tttcgcttga ggccatattt 780
gttaaacgct tttatctcta tcatcattca tcctaccagt aaagagcacc ggagatcgca 840
cttcatttaa atatatgtcc atgttggata aaccatagtt tattatagtg ttcttttata 900
tgttttgtgg ggaatttaga ttgtttaata tggcatacat atccatccat cattattata 960
ttctaacaca actggataag tgttctaaac tattgtagaa taactttgta gtatgatcga 1020
tcttgtggaa taaaaaaagt ctgacaataa cctttcataa aggaatatga atacccgtaa 1080
tcaacgcatc aaatcattca cggtgtacgc ctagcgaatt cgttggcgag tgctcgtgcg 1140
gccgtgggct cgctgtgatg catgcatggc tctctggcta cgtcgagata gcgattagta 1200
gcaaaattaa gcaagccact tattaattaa tctttggaga tatcatatga ttaaggcatt 1260
aattcgtacg tactcgtcgt cagcgttttc tgcaaagtcc actacagttt tttctttctt 1320
tgctgaaaat gctgatgtgt tggagatgga gtgacgtgca caacctgccg ccacgtggat 1380
ggttgctgga gcctacgtgt catcttaatt tgaacaaaaa aaaaagagga ataatacatc 1440
aatacatttt cgaatttcag ttctgccatt gaccagtaat acacatgtcg gcctcacatt 1500
ttaccctgat cttagtaacg ggtggtcgcc tggtcggtca ctgaaaaaag ttcaggaaat 1560
tatagtcaaa ctgaaacgaa catattcact ccttaaaaaa actaaatctt tttatatatt 1620
tgtgatattg taaaatagct acgggataat gatatagata tatatagtga taagggatag 1680
atggatcgag atatggagtt gtgctttctt taatttccac tacttgggct accatattat 1740
ggtagttggt atgaaaagat acacagcagt atagtgatgt gatcaatgac atgtatatct 1800
cacatgctcc catgttggag tcaaattttg ctagactaaa atccaattcc aagcagtccc 1860
tagccaagaa caaacaaaat tcagtgaggt cactgctgca ccaaggactg catgcatgca 1920
ggagaagggc attttctctt ttttcttttg gagactcgat tcaattcggt cggtcggtcg 1980
caatggtcag cttaattaaa 2000
<210> 71
<211> 2000
<212> ДНК
<213> Oryza sativa
<400> 71
tgtgaaaggt ggcggcacca gcttagccgc agcttctctc gtcgtctccc tgaaacgaga 60
gggaggaagt tggtagcgtg atatatttag gcatgtcatc tcttgtataa gaagtcttat 120
ctgtgctaat tcacacggtt ctctaatctc tctccattct gtttttgtaa attggttcag 180
tagatagcgt agggttatgc ttatatatac tccgtgaagt atatatttaa aaattagtca 240
cacgtaaagt actatacatg ttttatcgtc taataacaat aaaaacacta atcataaaat 300
ttttttaaat aatacgaatg gttaaacgtt gaatatgaac cgtgcaaaac tatatttatt 360
ttgtaacaga gaaaatattt cacattaatt agattgttgt tttatggaag gttggagagc 420
tgcgccgccg ttgcgcagac ctaggaggct gcttataagt tataatcaat caattcacgg 480
atgccggctg ggacgcggcc catcgtccgg gaagacgaca actcaacgca aaaagccgat 540
atgcctccaa attgccattg ccacctctac ggctgtttat actgctccaa atcaaaagcg 600
tccatggaag aatctagtat ttcccgcaaa gacgatgatg atatgcagga ttggatatat 660
agggggttgt tgcatgattg ctagaactcc cgtttccgaa gttgttcgtc catttttaaa 720
gctgccaaat aggaatttat tttgttttca agtgtaatag agttctgtcc agatgagtga 780
attataattt ggttcacatt ttatttgcta agtttcagtt tgaacattct caaataactt 840
ttttcttcac tttttaaccg agtaacttag ttattttttc cgtttggacc acccaacaat 900
ttgttgctaa gtgcatctca cccgtcaaat aattcctttg aatccaaatt caattatatc 960
ccaaaaataa aaaacttctg aattccacat caattcaaac cccaaccatt ttaatttctc 1020
tccatatttt ccatttctct atttttacct ttctcttttt tccatctatt tatttttttc 1080
cttttctatt tctttctttc tccttccttt ctctgtttcc ttcttcttct cctcggctag 1140
gcccgagcca gcccgtgccg cctcgcgcca accctgtgcc gccttacgcc gcgcttgcgt 1200
gcgctcgcgc ccacctcgtg cccaacccgc gcacgccaca cgcacacacg aggacgatcg 1260
acggacgaat gcaatcatat ccccttcctt actcagctag aaggctcaag aaccgcaact 1320
ttgatctctt ccaccctctc aaatccgccc caacccctgc tgactcaatc gccattaccg 1380
gaggaaaaat ccccgaaacc ctattaccgg cgccactaac agagctccaa aattcgtcgc 1440
ataattcgaa aatattctga aattgaaggt aaaaatggaa tctacatgcg aagtactccc 1500
tttcccctcc aatccgtcac tggaacgccg ccggcgccgc ctcccgctgc cactgccctg 1560
tttggccgcc gacagccgca cggcgcgccg ctgctccagg ccgccctagc ttcaaccacc 1620
gccacctttg gctccgcctc cctcctctta tgctcaccaa gcccgcctcc ctcgccggag 1680
atcgccggaa ccaccgccgc catggccgcc accgcctcct gcttctggcc gccgccgcca 1740
gcctcgccac cggcgcctat gccaccgccg accacggaaa cggagtccct acaccttggg 1800
gaccacaaaa ccggcggcat ccctcccaaa accggcctcc tccaccgccg gcgttcgtgg 1860
gattccggcc agttctgtgc agagcgagag aagaagagga aaaatagatt ttcctattga 1920
aagataaatc agaaaattcc tttttctttt cctatcaagt tgaccatccg tttgacctca 1980
aaatcaaaat ctgagaccta 2000
<210> 72
<211> 786
<212> ДНК
<213> Zea mays
<400> 72
gacatggagg tggaaggcct gacgtagata gagaagatgc tcttagcttt cattgtcttt 60
cttttgtagt catctgattt acctctctcg tttatacaac tggtttttta aacactcctt 120
aacttttcaa attgtctctt tctttaccct agactagata attttaatgg tgattttgct 180
aatgtggcgc catgttagat agaggtaaaa tgaactagtt aaaagctcag agtgataaat 240
caggctctca aaaattcata aactgttttt taaatatcca aatattttta catggaaaat 300
aataaaattt agtttagtat taaaaaattc agttgaatat agttttgtct tcaaaaatta 360
tgaaactgat cttaattatt tttccttaaa accgtgctct atctttgatg tctagtttga 420
gacgattata taattttttt tgtgcttaac tacgacgagc tgaagtacgt agaaatacta 480
gtggagtcgt gccgcgtgtg cctgtagcca ctcgtacgct acagcccaag cgctagagcc 540
caagaggccg gaggtggaag gcgtcgcggc actatagcca ctcgccgcaa gagcccaaga 600
gaccggagct ggaaggatga gggtctgggt gttcacgaat tgcctggagg caggaggctc 660
gtcgtccgga gccacaggcg tggagacgtc cgggataagg tgagcagccg ctgcgatagg 720
ggcgcgtgtg aaccccgtcg cgccccacgg atggtataag aataaaggca ttccgcgtgc 780
aggatt 786
<210> 73
<211> 1160
<212> ДНК
<213> Zea mays
<400> 73
atgtgctggt gccccataag gtaggcacct aggtctgtgt ttgaagcatc gacagatttg 60
taaacatgtt cctatgaacc tatttctgat tgataatttg tcaaaactca tcatttgtct 120
tcatccttgc ctgcttgcgt tcacgtgaca aagtacgtgt atgtcttcgg cctttgctgt 180
gtatgtttcg cattgcttag atgtggtgaa agaacatcag aagatgcatt gatggcgtgc 240
ttaaaccagt gatgtgctcc aggtgttcct gcagtctgca gagatattta ctcttgtagt 300
cttgttgaca gcacagttgt atgtgatttc ttggatgtaa tgtaaaccaa atgaaagata 360
ggaacagttc gtcctcttcc gtatacgaag gtcactgtat catttgtcgt ggcacaagat 420
gatctgcagg caggactgca acatggtttc ttggactgtc ctgaatgccc gttcttgttc 480
tttagttgag ccagagcagc agcctggtgt cggtgcctga gacctgacga agcacacggc 540
aaacaaacaa gtcgcagcag ctagcagggg cgttgccatc gccacaagcc cccaagagac 600
ccgccgagga aaagaaaaaa aaactacggc cgccgttgcc aagccgagcg tgcgaaccga 660
tccacggatg ggagatcaga gatcacccac cgcaggcggg cggcagtggc tggcgaggtg 720
cgtccacaga acctgctgca ggtccctgtc cgtcccggcg accccttttc taggcgagca 780
actccccatg gcagagctgc acgcagcagg gcccgtcgtt ggttgcagct ttaacccttt 840
ttgttttaac catacaatgc agagtcgcag aggtgaaaca ggacggaaat tacagaaaag 900
atggtggtgt gccagcagcc ccagcatgaa gaagatcagg acaaaagaaa agcttgtgat 960
tggtgacagc aacaggattg gattggagcc aagctaggca gtgagaggca ggcagcaaga 1020
cgcgtcagcc actgaaatcc agagggcaac ctcggcctca caactcatat ccccttgtgc 1080
tgttgcgcgc cgtggttagc caggtgtgct gcaggcctcc tccttgttta tatatgggag 1140
atgctctcac cctctaaggt 1160
<210> 74
<211> 1532
<212> ДНК
<213> Oryza sativa
<400> 74
tagtcctcta atatatgaaa ttttgatata ggtaaagaag ggtattgcaa ggataagaat 60
gtaaaaagaa ataagagtaa tccttaccga taatagtatt ccttctctac cgttaaaagt 120
taaacctgtg cgtgtagcat tttaatccag gatctatcga atccgtccct cgttggcgtg 180
ggcgacgaac acgtgcagaa gaagctttcc ccagaaagca cctcaccgcc tcgccgtctg 240
gcagactggc acgcggggcc ctaccctcgc tgcgcctggg cccgtccgcc ttctgcacac 300
tgtcacgccc ccacccgctc gccgcctcgc gcctctctct ccgcctccgc cgcggccgcc 360
cgacgtgata gcgacacgta ggactcgcca aacacaaaaa atccatcgcg atttttggaa 420
ttttgttaca aaccaaatcc cgcattagag atttaatttg atttaattta attacgtagg 480
agtaccagat aaggagatcg agttaaaaaa gctaacggcg cggcgtggtt atctccgaat 540
cggctgtggc tccccgcgtc ggcgtcggcg cggcggcggc gcgccggccg aaccctggcc 600
gtcggatcgg gcgtcgtcct gggccccacg cgccacgggc ggctgtcgtt tgctcctcgg 660
agcggggtgg gcccaccatg gccaccacca caggtcgcgg tcgcggctga cctggcggtg 720
gtcccgtgct cgcggtgttt tttttttttc actctctttc tctcggtgga cagtagcggg 780
ggccgcggcc cgcgggggca gagattgcaa aaacagcgga aacggaagat tgcaaaattg 840
caactgcttt cctgttttta attcgggatc aaaaagattc tttcgtcggg gtccccgtgc 900
cattgttgta ttgcgcgtag gtccttgctt gtaaaagata atctccttaa ttttttcttt 960
gtactactag tgtatatgca gtaagaatat accatgagta aaatgaacca caaaactaat 1020
tacgatatac cattctcatg tagacgttct cttttctttt gctagtcata cgtgcatata 1080
taaccaaaca aaaaaatgtt tgaagtactc ctatccaatt tattactcca gtagacaaca 1140
aaagaaaatg tttgaagtaa taactgatcc atggtacagt agggttgtcg tcaatcttgt 1200
gtttctttca ttccattgta cttacaatcg tactccagct agcacagcac aatgggctta 1260
agctttggac cccaaattct gatcttgtcg gggacccgta cgaaaatact cccgtagaga 1320
tgcagatacc gtcacaacct acaaccaacg aatgttaaga aaacaaaggg aaaaaaaaag 1380
aggcgaattc ggaggagaaa aaacggtggc taaaatatag tgcgggtgtg gggacgcgac 1440
gcgagcgacg aaagaggaga gaggatgggt tggcctgccc ccccctcccc tgtctataaa 1500
tgcagaggcg ccgagtgccc tagtcgccgc tc 1532
<210> 75
<211> 841
<212> ДНК
<213> Oryza sativa
<400> 75
tcgaggtcat tcatatgctt gagaagagag tcgggatagt ccaaaataaa acaaaggtaa 60
gattacctgg tcaaaagtga aaacatcagt taaaaggtgg tataaagtaa aatatcggta 120
ataaaaggtg gcccaaagtg aaatttactc ttttctacta ttataaaaat tgaggatgtt 180
tttgtcggta ctttgatacg tcatttttgt atgaattggt ttttaagttt attcgctttt 240
ggaaatgcat atctgtattt gagtcgggtt ttaagttcgt ttgcttttgt aaatacagag 300
ggatttgtat aagaaatatc tttagaaaaa cccatatgct aatttgacat aatttttgag 360
aaaaatatat attcaggcga attctcacaa tgaacaataa taagattaaa atagctttcc 420
cccgttgcag cgcatgggta ttttttctag taaaaataaa agataaactt agactcaaaa 480
catttacaaa aacaacccct aaagttccta aagcccaaag tgctatccac gatccatagc 540
aagcccagcc caacccaacc caacccaacc caccccagtc cagccaactg gacaatagtc 600
tccacacccc cccactatca ccgtgagttg tccgcacgca ccgcacgtct cgcagccaaa 660
aaaaaaaaga aagaaaaaaa agaaaaagaa aaaacagcag gtgggtccgg gtcgtggggg 720
ccggaaacgc gaggaggatc gcgagccagc gacgaggccg gccctccctc cgcttccaaa 780
gaaacgcccc ccatcgccac tatatacata cccccccctc tcctcccatc cccccaaccc 840
t 841
<210> 76
<211> 1392
<212> ДНК
<213> Oryza sativa
<400> 76
ctcgaggtca ttcatatgct tgagaagaga gtcgggatag tccaaaataa aacaaaggta 60
agattacctg gtcaaaagtg aaaacatcag ttaaaaggtg gtataagtaa aatatcggta 120
ataaaaggtg gcccaaagtg aaatttactc ttttctacta ttataaaaat tgaggatgtt 180
ttgtcggtac tttgatacgt catttttgta tgaattggtt tttaagttta ttcgcgattt 240
tggaaatgca tatctgtatt tgagtcgggt tttaagttcg tttgcttttg taaatacaga 300
gggatttgta taagaaatat ctttaaaaaa acccatatgc taatttgaca taatttttga 360
gaaaaatata tattcaggcg aattctcaca atgaacaata ataagattaa aatagcttgc 420
ccccgttgca gcgatgggta ttttttctag taaaataaaa gataaactta gactcaaaac 480
atttacaaaa acaaccccta aagtcctaaa gcccaaagtg ctatgcacga tccatagcaa 540
gcccagccca acccaaccca acccaaccca ccccagtgca gccaactggc aaatagtctc 600
cacaccccgg cactatcacc gtgagttgtc cgcaccaccg cacgtctcgc agccaaaaaa 660
aaaaaaagaa agaaaaaaaa gaaaaagaaa aaacagcagg tgggtccggg tcgtgggggc 720
cggaaaagcg aggaggatcg cgagcagcga cgaggccggc cctccctccg cttccaaaga 780
aacgcccccc atcgccacta tatacatacc cccccctctc ctcccatccc cccaacccta 840
ccaccaccac caccaccacc tcctcccccc tcgctgccgg acgacgagct cctcccccct 900
ccccctccgc cgccgccggt aaccaccccg cgtccctctc ctctttcttt ctccgttttt 960
tttttccgtc tcgtctcgat ctttggcctt ggtagtttgg gggcgagagg cggcttcgtc 1020
gcccagatcg gtgcgcggga ggggcgggat ctcgcggctg ggtctcggcg tgcggccgga 1080
tcctcgcggg gaatggggct ctcggatgta gatctgatcc gccgttgttg ggggagatga 1140
tggggcgttt aaaatttcgc catgctaaac aagatcagga agaggggaaa agggcactat 1200
ggtttatatt tttatatatt tctgctgctg ctcgtcaggc ttagatgtgc tagatctttc 1260
tttcttcttt ttgtgggtag aatttgaatc cctcagcatt gttcatcggt agtttttctt 1320
ttcatgattt gtgacaaatg cagcctcgtg cggagctttt ttgtaggtag aagatggctg 1380
acgccgagga ta 1392
<210> 77
<211> 743
<212> ДНК
<213> Oryza sativa
<400> 77
gaattcccgg acctccatgc ctacatcaac taatttgatt ccttgagttt acgtttagtg 60
atatgtctat ttttagagct tgttggggct tcggcctcag ctctagccag ccaaacatgt 120
tctaccaagt accctatgtt ggcatgatat agtgatgcat tataacaata aatgagcgag 180
ggattgctgg ctgaaaaagc tatactagct gcatttggtt atagttaacc gaactattaa 240
ttgcgtgtac aacaaaataa aaaaaatgca tgttgcacat tctttcatta acattatgtt 300
ttggtagtgt gaattagaaa tttgattgac agtagatcga caaacatagt ttcaatatgc 360
ttaagttagt tatgacttta acatatcagt ctccttgata ttttcgtttt agattcgtct 420
ctctactagt gtgtatgtcc accttccata gcagtgaagg gttccattcc atccctggta 480
aaaaaaaatc aaccactact atttatttcc taaaaagcaa aatgataaaa tatcattttt 540
ttaataaaaa taaaaaaatt ttggggtaca taattgatgt tgccccttgg gattaacctt 600
aaaaaagggc gaattttcta gggtttggcc aagttttgca atgcaccaaa ttattcccct 660
tgggccggcc gccaccccaa aaaaaacccc aacccccaac tttccattga aggccgggcc 720
cccttaaatc ctcatccccc caa 743
<210> 78
<211> 144
<212> ДНК
<213> Oryza sativa
<400> 78
taaaaaaggg cgaattttct agggtttggc caagttttgc aatgcaccaa attattcccc 60
ttgggccggc cgccacccca aaaaaaaccc caacccccaa ctttccattg aaggccgggc 120
ccccttaaat cctcatcccc ccaa 144
<210> 79
<211> 612
<212> ДНК
<213> Вирус мозаики цветной капусты
<400> 79
ggtccgattg agacttttca acaaagggta atatccggaa acctcctcgg attccattgc 60
ccagctatct gtcactttat tgtgaagata gtggaaaagg aaggtggctc ctacaaatgc 120
catcattgcg ataaaggaaa ggccatcgtt gaagatgcct ctgccgacag tggtcccaaa 180
gatggacccc cacccacgag gagcatcgtg gaaaaagaag acgttccaac cacgtcttca 240
aagcaagtgg attgatgtga tggtccgatt gagacttttc aacaaagggt aatatccgga 300
aacctcctcg gattccattg cccagctatc tgtcacttta ttgtgaagat agtggaaaag 360
gaaggtggct cctacaaatg ccatcattgc gataaaggaa aggccatcgt tgaagatgcc 420
tctgccgaca gtggtcccaa agatggaccc ccacccacga ggagcatcgt ggaaaaagaa 480
gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg atatctccac tgacgtaagg 540
gatgacgcac aatcccacta tccttcgcaa gacccttcct ctatataagg aagttcattt 600
catttggaga gg 612
<210> 80
<211> 837
<212> ДНК
<213> Coix lacryma-jobi
<400> 80
agcagactcg cattatcgat ggagctctac caaactggcc ctaggcatta acctaccatg 60
gatcacatcg taaaaaaaaa accctaccat ggatcctatc tgttttcttt ttgccctgaa 120
agagtgaagt catcatcata tttaccatgg cgcgcgtagg agcgcttcgt cgaagaccca 180
taggggggcg gtactcgcac cgtggttgtt tcctgttatg taatatcgga tgggggagca 240
gtcggctagg ttggtcccat cggtactggt cgtcccctag tgcgctagat gcgcgatgtt 300
tgtcctcaaa aactcttttc ttcttaataa caatcatacg caaatttttt gcgtattcga 360
gaaaaaaaga agattctatc tgtttttttt ttgaaatggc tccaatttat aggaggagcc 420
cgtttaacgg cgtcgacaaa tctaacggac accaaccagc gaatgagcga acccaccagc 480
gccaagctag ccaagcgaag cagacggccg agacgctgac acccttgcct tggcgcggca 540
tctccgtcgc tggctcgctg gctctggccc cttcgcgaga gttccggtcc acctccacct 600
gtgtcggttt ccaactccgt tccgccttcg cgtgggactt gttccgttca tccgttggcg 660
gcatccggaa attgcgtggc gtagagcacg gggccctcct ctcacacggc acggaaccgt 720
cacgagctca cggcaccggc agcacggcgg ggattccttc cccaccaccg ctccttccct 780
ttcccttcct cgcccgccat cataaatagc cacccctccc agcttccttc gccacat 837
<210> 81
<211> 947
<212> ДНК
<213> Oryza sativa
<400> 81
aatccgaaaa gtttctgcac cgttttcacg tcctaactaa caatataggg aacgtgtgct 60
aaatataaaa tgagacctta tatatgtagc gctgataact agaactatgt aagaaaaact 120
catccaccta ctttagtggc aatcgggcta aataaaaaag agtcgctaca ctagtttcgt 180
tttccttagt aattaagtgg gaaaatgaaa tcattattgc ttagaatata cgttcacatc 240
tctgtcatga agttaaatta ttcgaggtag ccataattgt catcaaactc ttcttgaata 300
aaaaaatctt tctagctgaa ctcaatgggt aaagagagat attttttttt aaaaaaaaat 360
agaatgaaga tattctgaac gtatcggcaa agatttaaac atataattat ataattttat 420
agtttgtgca ttcgttatat cgcacgtcat taaggacatg tcttactcca tctcaatttt 480
tatttagtaa ttaaagacaa ttgacttatt tttattattt atcttttttc gattagatgc 540
aaggtactta cgcacacact ttgtgctcat gtgcatgtgt gagtgcacct cctcaataca 600
cgttcaacta gcgacacatc tccaatatca ctcgcctatt taatacattt aggtagcaat 660
atctgaattc aagcactcca ccatcaccag accactttta ataatatcta aaatacaaaa 720
aataatttta cagaatagca tgaaaagtat gaaacgaact atttaggttt ttcacataca 780
aaaaaaaaaa gaattttgct cgtgcgcgag cgccaatctc ccatattggg cacacaggca 840
acaacagagt ggctgcccac agaacaaccc acaaaaaacg atgatctaac ggaggacagc 900
aagtccgcaa caacctttta acagcaggct ttgcggccag gagagag 947
<210> 82
<211> 721
<212> ДНК
<213> Каулимовирус мозаики Mirabilis
<400> 82
tggagattca gaaaaatctc catcaacaaa taatccaagt aaggattaat ggattgatca 60
acatccttac cgctatgggt aagattgatg aaaagtcaaa aacaaaaatc aattatgcac 120
accagcatgt gttgatcacc agctattgtg ggacaccaat ttcgtccaca gacatcaaca 180
tcttatcgtc ctttgaagat aagataataa tgttgaagat aagagtggga gccaccacta 240
aaacattgct ttgtcaaaag ctaaaaaaga tgatgcccga cagccacttg tgtgaagcat 300
gtgaagccgg tccctccact aagaaaatta gtgaagcatc ttccagtggt ccctccactc 360
acagctcaat cagtgagcaa caggacgaag gaaatgacgt aagccatgac gtctaatccc 420
acaagaattt ccttatataa ggaacacaaa tcagaaggaa gagatcaatc gaaatcaaaa 480
tcggaatcga aatcaaaatc ggaatcgaaa tctctcatct ctctctacct tctctctaaa 540
aaacacttag atgtgtgagt aatcacccac ttggggttgt aatatgtagt agtaaataag 600
ggaaccttag ggtataccat tgttgtaata ttattttcag tatcaataaa ataatctttc 660
agtttatctt atattcattt gtgtgacacc gtattcccat aaaaccgatc ctaatctctc 720
c 721
<210> 83
<211> 352
<212> ДНК
<213> Peanut chlorotic streak caulimovirus
<400> 83
acagagggat ttctctgaag atcatgtttg ccagctatgc gaacaatcat cgggagatct 60
tgagccaatc aaagaggagt gatgtagacc taaagcaata atggagccat gacgtaaggg 120
cttacgccat tacgaaataa ttaaaggctg atgtgacctg tcggtctctc agaaccttta 180
ctttttatat ttggcgtgta tttttaaatt tccacggcaa tgacgatgtg acctgtgcat 240
ccgctttgcc tataaataag ttttagtttg tattgatcga cacgatcgag aagacacggc 300
catttggacg atcatttgag agtctaaaag aacgagtctt gtaatatgtt tt 352
<210> 84
<211> 1648
<212> ДНК
<213> Sorghum bicolor
<400> 84
cactcgcaca tctcatggtg tcccaagaac ggcaagagcc agcactgcct ctgcctagca 60
acagcagcag cgccaagcga gcagccgcgt ccatggacgc cagcagcccg gccccgccgc 120
tcctcctccg cgcccccact cccagtccca gcattgacct ccccgctgcc gctggcaagg 180
ccgcggccgt gttcgacctg cggcgggagc ccaagatccc ggcgccattc ctgtggccgc 240
acgaggaggc gcgcccgacc tcggccgcgg agctggaggt tccggtggtg gacgtgggcg 300
tgctgcgcaa tggcgaccgc gcggggctgc ggcgcgccgc ggcgcaggtg gcctcggcgt 360
gcgcgacgca cgggttcttc caggtgtgcg ggcacggcgt ggacgcggcc ctggggcgcg 420
ccgcgctgga cggcgccagc gacttcttcc ggctgccgct ggccgacaag cagcgcgccc 480
ggcgcgtccc cggcaccgtg tccgggtaca cgagcgcgca cgccgaccgg ttcgcgtcca 540
agctcccctg gaaggagacc ctgtccttcg gcttccacga cggcgccgcg tcgcccgtcg 600
tcgtggacta cttcaccggc accctcggcc aagatttcga gccaatgggg cgggtgtacc 660
agaggtactg cgagaagatg aaggagctgt cgctgacgat catggagctg ctggagctga 720
gcctgggcgt ggagcgcggc tactaccggg agttcttcga ggacagccgc tccatcatgc 780
ggtgcaacta ctacccgccg tgcccggagc cggagcgcac gctgggcacg ggcccgcact 840
gcgaccctac ggcgctgacc atcctcctgc aggacgacgt cggcgggctg gaggtgctgg 900
tggacggcga gtggcgcccc gtccggcccg tcccaggcgc catggtcatc aacatcggcg 960
acaccttcat ggcgctgtcg aacgggcggt acaagagctg cctgcaccgc gcggtggtga 1020
accagcggca ggagcggcgg tcgctggcct tcttcctgtg cccgcgcgag gaccgggtgg 1080
tgcggccgcc ggccagcagc gccacgccgc ggcagtaccc ggacttcacc tgggccgacc 1140
tcatgcgctt cacgcagcgc cactaccgcg ccgacacccg cacgctggac gccttcaccc 1200
gctggctctc ccacggccca gtcccagccc aggaggcggc ggctccctgc acctagcgag 1260
cgagcgagcc gggccaaaca aacaaggggc aaaggccatc tctttcgccg gggcccgcgc 1320
gcggggttcg cccacgtgcg cgcccaggtg ggcgctggcc gcgggcaggt ggcggacatg 1380
tggcctgcgg gccccgcgcc gccttcccat ttttggacgc tgccgcgcat gccgcatgcg 1440
tgcgtcgacg gccctactac ttctactact gctactgcga ctactagtgt acatacgcaa 1500
aaatacatat atacgtattt tctatatata tatatataag caaggcggcc ccccggtgac 1560
cttttctttg tttttgtcga caactgtgtt ttgatcccat tctagctgtt ctatggacca 1620
tggatggttc gttcaatgtt tgtacgta 1648
<210> 85
<211> 1242
<212> ДНК
<213> Sorghum bicolor
<400> 85
atggtgtccc aagaacggca agagccagca ctgcctctgc ctagcaacag cagcagcgcc 60
aagcgagcag ccgcgtccat ggacgccagc agcccggccc cgccgctcct cctccgcgcc 120
cccactccca gtcccagcat tgacctcccc gctgccgctg gcaaggccgc ggccgtgttc 180
gacctgcggc gggagcccaa gatcccggcg ccattcctgt ggccgcacga ggaggcgcgc 240
ccgacctcgg ccgcggagct ggaggttccg gtggtggacg tgggcgtgct gcgcaatggc 300
gaccgcgcgg ggctgcggcg cgccgcggcg caggtggcct cggcgtgcgc gacgcacggg 360
ttcttccagg tgtgcgggca cggcgtggac gcggccctgg ggcgcgccgc gctggacggc 420
gccagcgact tcttccggct gccgctggcc gacaagcagc gcgcccggcg cgtccccggc 480
accgtgtccg ggtacacgag cgcgcacgcc gaccggttcg cgtccaagct cccctggaag 540
gagaccctgt ccttcggctt ccacgacggc gccgcgtcgc ccgtcgtcgt ggactacttc 600
accggcaccc tcggccaaga tttcgagcca atggggcggg tgtaccagag gtactgcgag 660
aagatgaagg agctgtcgct gacgatcatg gagctgctgg agctgagcct gggcgtggag 720
cgcggctact accgggagtt cttcgaggac agccgctcca tcatgcggtg caactactac 780
ccgccgtgcc cggagccgga gcgcacgctg ggcacgggcc cgcactgcga ccctacggcg 840
ctgaccatcc tcctgcagga cgacgtcggc gggctggagg tgctggtgga cggcgagtgg 900
cgccccgtcc ggcccgtccc aggcgccatg gtcatcaaca tcggcgacac cttcatggcg 960
ctgtcgaacg ggcggtacaa gagctgcctg caccgcgcgg tggtgaacca gcggcaggag 1020
cggcggtcgc tggccttctt cctgtgcccg cgcgaggacc gggtggtgcg gccgccggcc 1080
agcagcgcca cgccgcggca gtacccggac ttcacctggg ccgacctcat gcgcttcacg 1140
cagcgccact accgcgccga cacccgcacg ctggacgcct tcacccgctg gctctcccac 1200
ggcccagtcc cagcccagga ggcggcggct ccctgcacct ag 1242
<210> 86
<211> 413
<212> ПРТ
<213> Sorghum bicolor
<400> 86
Met Val Ser Gln Glu Arg Gln Glu Pro Ala Leu Pro Leu Pro Ser Asn
1. 5 10 15
Ser Ser Ser Ala Lys Arg Ala Ala Ala Ser Met Asp Ala Ser Ser Pro
20 25 30
Ala Pro Pro Leu Leu Leu Arg Ala Pro Thr Pro Ser Pro Ser Ile Asp
35 40 45
Leu Pro Ala Ala Ala Gly Lys Ala Ala Ala Val Phe Asp Leu Arg Arg
50 55 60
Glu Pro Lys Ile Pro Ala Pro Phe Leu Trp Pro His Glu Glu Ala Arg
65 70 75 80
Pro Thr Ser Ala Ala Glu Leu Glu Val Pro Val Val Asp Val Gly Val
85 90 95
Leu Arg Asn Gly Asp Arg Ala Gly Leu Arg Arg Ala Ala Ala Gln Val
100 105 110
Ala Ser Ala Cys Ala Thr His Gly Phe Phe Gln Val Cys Gly His Gly
115 120 125
Val Asp Ala Ala Leu Gly Arg Ala Ala Leu Asp Gly Ala Ser Asp Phe
130 135 140
Phe Arg Leu Pro Leu Ala Asp Lys Gln Arg Ala Arg Arg Val Pro Gly
145 150 155 160
Thr Val Ser Gly Tyr Thr Ser Ala His Ala Asp Arg Phe Ala Ser Lys
165 170 175
Leu Pro Trp Lys Glu Thr Leu Ser Phe Gly Phe His Asp Gly Ala Ala
180 185 190
Ser Pro Val Val Val Asp Tyr Phe Thr Gly Thr Leu Gly Gln Asp Phe
195 200 205
Glu Pro Met Gly Arg Val Tyr Gln Arg Tyr Cys Glu Lys Met Lys Glu
210 215 220
Leu Ser Leu Thr Ile Met Glu Leu Leu Glu Leu Ser Leu Gly Val Glu
225 230 235 240
Arg Gly Tyr Tyr Arg Glu Phe Phe Glu Asp Ser Arg Ser Ile Met Arg
245 250 255
Cys Asn Tyr Tyr Pro Pro Cys Pro Glu Pro Glu Arg Thr Leu Gly Thr
260 265 270
Gly Pro His Cys Asp Pro Thr Ala Leu Thr Ile Leu Leu Gln Asp Asp
275 280 285
Val Gly Gly Leu Glu Val Leu Val Asp Gly Glu Trp Arg Pro Val Arg
290 295 300
Pro Val Pro Gly Ala Met Val Ile Asn Ile Gly Asp Thr Phe Met Ala
305 310 315 320
Leu Ser Asn Gly Arg Tyr Lys Ser Cys Leu His Arg Ala Val Val Asn
325 330 335
Gln Arg Gln Glu Arg Arg Ser Leu Ala Phe Phe Leu Cys Pro Arg Glu
340 345 350
Asp Arg Val Val Arg Pro Pro Ala Ser Ser Ala Thr Pro Arg Gln Tyr
355 360 365
Pro Asp Phe Thr Trp Ala Asp Leu Met Arg Phe Thr Gln Arg His Tyr
370 375 380
Arg Ala Asp Thr Arg Thr Leu Asp Ala Phe Thr Arg Trp Leu Ser His
385 390 395 400
Gly Pro Val Pro Ala Gln Glu Ala Ala Ala Pro Cys Thr
405 410
<210> 87
<211> 12906
<212> ДНК
<213> Sorghum bicolor
<400> 87
cactcgcaca tctcatggtg tcccaagaac ggcaagagcc agcactgcct ctgcctagca 60
acagcagcag cgccaagcga gcagccgcgt ccatggacgc cagcagcccg gccccgccgc 120
tcctcctccg cgcccccact cccagtccca gcattgacct ccccgctgcc gctggcaagg 180
ccgcggccgt gttcgacctg cggcgggagc ccaagatccc ggcgccattc ctgtggccgc 240
acgaggaggc gcgcccgacc tcggccgcgg agctggaggt tccggtggtg gacgtgggcg 300
tgctgcgcaa tggcgaccgc gcggggctgc ggcgcgccgc ggcgcaggtg gcctcggcgt 360
gcgcgacgca cgggttcttc caggtgtgcg ggcacggcgt ggacgcggcc ctggggcgcg 420
ccgcgctgga cggcgccagc gacttcttcc ggctgccgct ggccgacaag cagcgcgccc 480
ggcgcgtccc cggcaccgtg tccgggtaca cgagcgcgca cgccgaccgg ttcgcgtcca 540
agctcccctg gaaggagacc ctgtccttcg gcttccacga cggcgccgcg tcgcccgtcg 600
tcgtggacta cttcaccggc accctcggcc aagatttcga gccaatgggg taagcgaagc 660
accgatttac atttaccgcg cgtcggcccc tgaggcctgg gtcttagtct tagcactgca 720
tatacggtcg gtagctctgg atatgatacg tatatatgaa accccgttcc aatcccatgc 780
acggtgtaca caggcgggtg taccagaggt actgcgagaa gatgaaggag ctgtcgctga 840
cgatcatgga gctgctggag ctgagcctgg gcgtggagcg cggctactac cgggagttct 900
tcgaggacag ccgctccatc atgcggtgca actactaccc gccgtgcccg gagccggagc 960
gcacgctggg cacgggcccg cactgcgacc ctacggcgct gaccatcctc ctgcaggacg 1020
acgtcggcgg gctggaggtg ctggtggacg gcgagtggcg ccccgtccgg cccgtcccag 1080
gcgccatggt catcaacatc ggcgacacct tcatggtaac ccctgctctg ttttttcttg 1140
tcctcctctt gtcctgtgtg tgtgtatatt cacttctctc tgtttttttg ccccgaatcc 1200
tagtggacct aactggacgg attacagcac gcacacgtag gcatgtcatg tagcagcagt 1260
ctgcagcact gtagtactta gcgatgcaat agagacatgc gttccagtcg gttccatctc 1320
ggtgggctac agctacagtc ctacacggac gcggctcgta gtcgtaggga cgggcgcgtt 1380
ctctgtatcc acacacggct gcgcccaggc cgaggcttcc gccgcgggaa agttgcgaca 1440
acagaacggg gtttgtgccg ttggagcgtt gcggagaggc agaggcttgg ggggacgggg 1500
gcgcgatacg ctgcgatggg tgggtgaccg aggcgacgct ttcggcgggg gcccgggcct 1560
gcccaggtgc gcgcggcctc gtcgccttcc cctgtttttt tgatgccgcc gctcggtcct 1620
cggtgttctg gctccgcccg cccgctcgct gggtgcccat cccatctgat ccgatccgct 1680
ccgctccgcg gtggcggtcc tatgcgatgc cgccgcacga gcgcgggggg ccgcccgtgg 1740
aggagtagaa agtggtacaa ggttggttgg aacttggaat tgtggggggt tactgctgct 1800
ggtggctgct gctttgcaac ttgccaggct gctgcctgtt gccccccgcg ttttctagcc 1860
gtttccgctc gcgatccggc acgcggcgcc cacaccgggg ctccagctcg gccccttggc 1920
cgtgtaggta gcaggcactt gcatctgtcc gttcgacacg atgattcttg tgcactgtgt 1980
acgtatgtac taaccctttc tggtatgatg tacgcatggc atgcaggcgc tgtcgaacgg 2040
gcggtacaag agctgcctgc accgcgcggt ggtgaaccag cggcaggagc ggcggtcgct 2100
ggccttcttc ctgtgcccgc gcgaggaccg ggtggtgcgg ccgccggcca gcagcgccac 2160
gccgcggcag tacccggact tcacctgggc cgacctcatg cgcttcacgc agcgccacta 2220
ccgcgccgac acccgcacgc tggacgcctt cacccgctgg ctctcccacg gcccagtccc 2280
agcccaggag gcggcggctc cctgcaccta gcgagcgagc gagccgggcc aaacaaacaa 2340
ggggcaaagg ccatctcttt cgccggggcc cgcgcgcggg gttcgcccac gtgcgcgccc 2400
aggtgggcgc tggccgcggg caggtggcgg acatgtggcc tgcgggcccc gcgccgcctt 2460
cccatttttg gacgctgccg cgcatgccgc atgcgtgcgt cgacggccct actacttcta 2520
ctactgctac tgcgactact agtgtacata cgcaaaaata catatatacg tattttctat 2580
atatatatat ataagcaagg cggccccccg gtgacctttt ctttgttttt gtcgacaact 2640
gtgttttgat cccattctag ctgttctatg gaccatggat ggttcgttca atgtttgtac 2700
gtactccacg taaccaaact actctagtgg actagtagat cgggctcatg tgatgaaact 2760
ggaccgacgc ggacgtcacg tgcgtcaccc gcgtctggta gcggtagcgc acgagcgccg 2820
aatgtttcct gggcccgcaa gagaatcgct tctcatctcc tctcaccatg aatggggaaa 2880
aatgctgcgt cgaaagttcc agacgtttcc aaattccaaa cggttttgtg gcgtccgatc 2940
catggggcgc cccaaacttc caagacgttt tcaggttcca aatcttcgtg ctccacatca 3000
ccttcttccc agattcattt gcctcgtcgc ttgctctcct gtgttattca cgggtcccac 3060
tgttgccccg tctgcgagaa agaaatttat tagagttgaa gcattcgaca tttcgactga 3120
ctgattgtta gtatcactaa attttgtgca catgtttctt tggtcattca tctctggata 3180
ttttttttag ataatggata taaatatcgg gcctctacat ctgaggaagt acacagccaa 3240
ttattttcat ctctggacat gggacgatgg aagaggcaga tagatttagg agacccttca 3300
attcagaatt tcaggtgcac aaggcctgcc tggcttgccc ggattcttgt ttcggacatg 3360
accaactagg ccgcactact tgcactgata gctggagaaa aaacaaaact ttgcaaacag 3420
caggattatc tacaagggaa actccatcca cgtgaaccag catttcaggg agagatgcga 3480
caaaaaaaaa gaggcggcaa caaaaaaatc ttactgcaat tttatctctg cattgaacct 3540
cttccaacca tgccgcatcc tgtactgttt tgtatctttc ccggtggtcc gttgcgttct 3600
cacgcagttg ataacatgca gtcacgcacc accgaatcca gtgtactagg ggtagtgact 3660
tgtcacgcgg aacaacaggt cggtagcacc aagcaagtcg ctgtagactt gggcgtttaa 3720
caacgacttg cacaacagtt caaatatagc atatgcaatt atgcacaaga ttgttcgact 3780
gctatccgac aaactgaaga agctgcccaa ttgaacagaa tgtaccagtg atttccagca 3840
cactatctta cagcagcgtt gagaatgaaa caacaaatgg gggaaaacag atgtgtatta 3900
ttctacagtt acaccaaaga gtttgtcctt tcagcatcaa caagaatcat atgcatatct 3960
agtgacaaaa attcctctaa ttttacccta cttggtaaca gttctcttca acacatatat 4020
ttcacgtgct tgcatcgagt tccttgggcc gccacatcga cttctcgacg caaagcaagc 4080
cctcgttgcc cttggtgtag gtcattcgca cctcccactg cagggacttg gccatgcttt 4140
ccagttcgtt tattgtgtcc gcagtgtccc tcacaatcag tttgccttgg ggcctcagta 4200
cacgatcaac ctcggcaaaa actgccatca atttgcatct gtaaacaagc aacacagatt 4260
tagcatctgt aaacaccaca ggtttcattg caagaagcat aaagcatgca aacatgctac 4320
ttgtacatgt caaagaaaca tgtcaaactc aaacacatga aaatcattat tattgttttc 4380
ttgctgaact gatcacatta gttggtttca atttctgagt tccactagta atctatacca 4440
gaaggataga ataatgtcaa gaacaagaga tacaaacctc tttgtgagct ttgagaatag 4500
atggtccgcg tgcagaaggt cataagttct tgggtaagtg ctcaaagact cgcaccagtc 4560
atggtacata ccaaacaaac cacgctcgta aatgatgggc agtgtgtctg gtgaatcaat 4620
cggcacaata ttcatgaccc agaccttttg gtccctcaga gctgcagcaa aactgccatg 4680
caacgatgta aagcattagt aaaaatattg ggttttttaa accaaaacca agaaagataa 4740
ttcctccagc ttaactgaaa gaaagaaaga aaaaaactgc ttaatgactt atggtggaca 4800
agttgcctgt tatgttttat gatagctatg tgccagcttg gctaactggt agttatgtag 4860
tgtgatctga attaccaaaa aagagaagaa aaaaaaatca tgcccaagaa actgagaaag 4920
acacccattt acttaccctc catacacagc tctcatgtcc attacatttc tcactttgga 4980
ccagtcaatt cccatgccat tcacatacga tttacttaca acccttttcc agtgagcatt 5040
atctgcctcg aaatcttcat ttgcaggctt tccatagacc ccaaccttgg aaccatcaat 5100
ccagaaagga gtcttctcaa gcctttgtgg ccaaaactct ggccattttg accctcgaac 5160
tttcgagcca acaggcagtt tgtgcatgca tgcttccaaa ggtacattcc tgcaaatcaa 5220
aagattgtgt aagcaaagca gaggaagcac ttcgccgcat tgaaaatacg ttcttctcaa 5280
agaaacaaaa ccataccaag ctgcatctgc atcatcagat tccttgcaca atggtgggtt 5340
gttttcggat cttttctcat agcaaatgtt gtccattggt ttctgaaata tgaccatacc 5400
aacttgattt aacttatcct tagtcttgtt gaccatcttc cagcacatgg actttgtcaa 5460
agtggacatc gctgaaaaga ttaaggggtc atatgttatg atagaaataa aattcaattt 5520
tgcactgttg gtacatagca tctgttttga acaaatgcaa tccttcctta tccatgaaag 5580
aagttaaccc ctgatactta ggattattca gtactttcac tcatgaactg ctgaatttgt 5640
tctgccagta gttgctatac tagaaatgtt cagtgtacca aacataaatt tggtacgggt 5700
tccttattaa agatgggagg ctgtatggta tttcgacgta acaaatcaag ttagcagcta 5760
ccctacttat ggatatacac ttctcaaaat gaatatacat agttttgata ggtgacatta 5820
attaatataa gaacttcatg cagttagggt gaaactaaac taagcagtta cggaaatacc 5880
attccaaatc tcaacatcct ctgggagctt ttggtaaaca ggagtggcag accagacaaa 5940
gtaaccacca gggcgtaaca agcggttcaa ttccagcaaa agcatgccac ctaaaaggag 6000
tcagtaataa gattcagttc tatagcaaat caataaatga aaggaagaca tgtcaccaac 6060
aagacaaacc ttcaatgtgc caagggaccc tgcagcgagc gcaatgaatg acatcaaaga 6120
ctctgctggg gtatggaagt ctcttggtgc ccatcacagc tgatattgct ggaattcccc 6180
tttctaatgc aaattgtact tgagcttcat gctcatcttt cggagcaaaa gacatggtaa 6240
gcacatctct atcaaacatg tagcctccaa agctggcaac tccacaaccg acatctagaa 6300
tgacgcggct tcgtttgccc catgcaatat caggcagtgc ctgtgaatga cagtttaatc 6360
agcatatgat gaaagcaagt gtgataatat caagttcaaa gatgcaacat gaaactttca 6420
taatcatgga cagtactaag cttgcttgat agattaatgt atggatgaga ctaaaaaaaa 6480
ggaaagttgt atccatcaga acgagaggct gaaaacacat ggctggctgt gaaagcctga 6540
tgtcgtttag tctagcataa acaaactgtc ctcagcatgt agatttccat agggtggcat 6600
ttgacaaatt atgattgtgg actagcgaat caatcactga ttctcaaaag tgtgagacag 6660
atgagttcaa gtctaagggg tgactaatat gggatgctgg gatgatgatg atgatatata 6720
cctgctgaat agtatcaata tagtggaggg caccattctt gaactgagtc ccacccccag 6780
ggaacaggag gtagtcacct gatactttaa cccaattttg atgtcccttg tactctgcga 6840
gcctagtgtg aggaacattg ctgtaccata cctgcaaaaa gcagcacaag atggtaataa 6900
gtaaacagag atcttggtca gctaaagatg attcagtgtc gtacaattta gaatagacag 6960
aatcaccttg tccctgctcc ttggccactc aattgggcgt ttatatcctt ctgggagtgg 7020
aacaaggcag gtaggaggct cctcagggca atgcctctca cgatgttcat aatgtttagt 7080
agttcgaagc ttcttgatag ccttctcgtt gtcaaggcaa ggtatgtaat ctgtcgaggc 7140
actgctatta catagtttcc aggaatagct agtcgcatca cctgaagact ttgatgacgc 7200
ttggacttcc ttttcattct tggactctgc agcctgtgtg gggaatgaac cattctgggt 7260
atttgactcc ttcagaagct ctgattgggc cccatcagga aatacctcgt tggagtttga 7320
gctctgatcc ttctctccat tttcttccac cttctcttct atctgaggtt gctcctcctg 7380
agtggcatca ccttcaggct tctcttcttg atcatccttg ctctctccat caggtttttc 7440
atcaccactc tcatttgtga tttcatcatc tttcttctcc ccactcttct ccccgtcacc 7500
atcgttcttc atgtcatctg accgcccttc tgattttcca tttgcatcat caaacatatc 7560
cttggtctca gctttctctg tcggcacttc cggctccttc tcttcaggct tctcctctgg 7620
cttctcagtg aacttctctt ccatcgtggc atccttgtta ttcggctcct ccggcatcgt 7680
ggcatcattg ttgtcggtgt cctcaaattt ctcagaacct tcaccagcat tgtcctgtga 7740
ggccccaaaa ttgacaggcg caggctgctg cttcaccacc ggcttcttat tcgaggagat 7800
ctccagcggg aagacagtgg acgaggtcat catccacgcg ccgaccaggc agagcgccac 7860
aaagacgacg accgtggtgg tcgtgcagaa cgacgacgac gtcgaggacg gccggcggcc 7920
gtccatcttc ccacctcggc caaatgccat tagtgcctgg cgaacatgta ccagagcacc 7980
gaccttcacg cgatttatct ccaccaacta ctgctggacc aagaaccccc aaaaaaatcg 8040
cacctttgtc tgctttgtgc tgctacagcc gcgcggcacc tgaagcaaac cacaaaaaaa 8100
acttaaatcg ccgcggacat aaatcaaggt gctggatcta aagaacaaac gctggatcta 8160
ctcaagcaac aacggaagga agatccgcta ttggtgctag tattagcttc ttgtttccta 8220
gtactacagc ggctctttcc cagtataaga acacgggaaa acgcggagaa atcccccttc 8280
gtggccaaac atggaaagaa aattagtaaa gcgtgtgctt taaaaccccc tcgttctgtt 8340
ccttccgcgg agagctaccg catcttccaa ttgagctggt tctcagctgg gcgcaaaacg 8400
cgcactaatc aatgtccgat tccatccaca aagaaaaaaa agacgggaac agctaatcca 8460
gcagctcgct cgctagctag ctagctcatc ggcggaagga cggaaccagc tttgctggat 8520
ccaggacagc aagagtgtgc aaggagaaag aacggagcag caatgcggat tgcggaggcg 8580
gtggattggt acctcgccgg aaccgaccgg agtggtcgcg gtggccctcc gcgcggatct 8640
cgaagaggag cgaggaaggg gaaggcggat gcgcgtcctt gggttctctg ccaccgcact 8700
gggcctcgcc gcgttataaa ggcgggcggg cgggcgggca gcgcagtgtg agtggagtgc 8760
aatctgttgt gtagtgtgtg aagaggcgga agcggaagcg gaggagatgg gttcgcatta 8820
gacgaccgta cgtaattata cgctatacta gtacttgggt tagattactc gggagatctt 8880
ggccaaaatg tccggtctga gtgtttggta gttttatgga tttgcccttt taagatgttg 8940
gtatttctcc gggagcttag aaagaagaaa tggcgatgct ttaggccttg tttagatgcg 9000
aaaaaaattt ggatttcgct actgtggcat ttttatttgt ttgtagcaaa tattgtccaa 9060
acacggacta actaagattc atctcgcgat ttacagttaa actgtacaat tagtttttat 9120
tttcatttat atttaatgtt tcatggatgt gtcgaaagat acgatatgat agaaaatttt 9180
gaaaactttt tagttattga ggttaactaa acaatgcctt aattgagaat ttactcgagc 9240
aaaaagagtt aggtcagtct cagtggagag tttcatggtg ttgtttccaa gactgccata 9300
tcatgtgaaa tgaaatgaaa cttggttgaa acactcactc tcaatggaga gtttcatttt 9360
atagtttcat gggcatttaa tttcaatact catagagagt tgatatcgtg ccaactcatt 9420
tcttctctct cttcttaaat acacagtcat atcatcaaaa aaaatcctat gtagcaacat 9480
atttaatgca aataaaactc atatggtgga ctgtaggagt agcattaggc caagggcaca 9540
cacacggtca cggtgtgagt gcgacggtgc gagtgggccc gcggcggtag taagtgcgtg 9600
cgcgcccggc gcccccctcc gcggcgacga cgcagcggca gcgcgtcgtc cagtgcaccg 9660
tctgctgttc ggcgctgcgg gtcctccgcg ccacggcgca gtgaaccggg cgcgtgcatc 9720
ccgggagcgg cggcttggca ctcccctgct tgtcggtggc ggccgtcggc atcgctcggc 9780
cccggagcgt cacgaggctg ctgattggga gcgagagcga gtagtggggc tggttgggga 9840
caatcccatt cccacccggc ccaccaggct gggactggcc cactagtcac tagtgggtgg 9900
ctcatgggtg tgggtgggct ggctaatgcc gcctgcccaa caaccaaccc aacccctgtg 9960
gacgctggta ccggtagttg ccgcgccatg gtggactgct gccgcctgat gcctttgcct 10020
gccacgctcc acgagttgag gcgcaccaaa ctgtgctgtg ctcctgattt gtgctaatcg 10080
gccgacgcgt accattcttt ctttctttcg tctacgcgca gagaggccgg ttgactgttt 10140
cttcgttgga gggccatgtt gactcgtact aataataaaa ataataatac taggttgact 10200
ttttcaattc caacgcagca gtgcaaagct gcccacctat gagcacaggt ccttttttaa 10260
ctccgttttt gtacgtacac acgtactgtc cagcctgtgt ctaataatct taccaaaaac 10320
ctgtcatctc actatcaacc aatcaggctc tccgcctgtt cgtcgaggaa cagcagttgt 10380
tttccctact ccaacataga gtacactatg gacgcacatt accatgccag cttgagctta 10440
gcattgccca ccgttggata actgccatgc cattctcagg ccctgtttag ttcccatcta 10500
aaaatttttc atccattcca tcgaatcttt ggacacatgc atggaacatt aaatgtagat 10560
aaaaaaataa actaattaca cagtttagtt gagaatcgcg agacgaatct tttaagtcta 10620
gttactccat aattagcctt aagtgctaca gtaatccaca tatactaatg acagattaat 10680
tatgcttaat aaatttgtct tacagtttcc tgacgagcta tgtaatttgt ttttttatta 10740
gtttctaaaa acccctcccg acatccttcc gacatatccg atgtgacaac caaaaaattt 10800
tcatcttcaa tctaaacagg ccctcactct catcatctca tgccggggca gcaggtccgt 10860
cgtcaggtct gtcgtcccgt cccgtgccgt ctgaagcaac aggcgagaga aacgccgttc 10920
catcggtttg ccgagcgtgc agaggataga gctatactcg atccggagag gattgtgaaa 10980
cgaagcacgg ttaagcagtg ccgcgcacgt gctgctctgc tcctggatcc gatccagatc 11040
gactcggggc gtctcggcct cagcggcgat ggcaatcatc gcgcgcgctg ctggagctgg 11100
acgttttcgt cttgcattgc aggaggcgga acagaacgga gaaagccacg gcgcgctttg 11160
ccgacgccac gcgctgacac gagggacccg ttcagcggcc agcacgcagc ctaatcatgc 11220
ctgtcggggg gagctcatcc gttcctgaat ttgggtcatg ctccagtatc aggtattcag 11280
gtactagtac tcctgagcca tgtgctgcga caaaaaagcg aggctcctgt agtagagcct 11340
tgtttactta caaaattttt tacattctca gttatattaa atcttgtgac acatgcataa 11400
agcattaaat atacataaaa gaaataatta tttacacagt tacttataat ttgcgaaacg 11460
aatcttttaa gactggttag tttatgatta gataatattt attaaataca aatgaaagta 11520
atattattta tattttgcaa aaagtaaata agacctaggt agctaggcca acgtgagcat 11580
gtcggacccg gaccggttcg ttctacggcg cgtcccgcaa acctgcagcc aggtagtagt 11640
agtacaccgt gcacgggaga ggtgcgccat gcatgctcgg gcaaaagatc atagagaaag 11700
gtgcagcgtt tcagttgcac acctgaccga gtgacgcctc gccttgtttg gctttgttcc 11760
caaaattttt taaaattcct catcacatta aatctttgaa cgaatatatg gagcattaaa 11820
tataaataaa agaaataatt aatcatacaa tttgtctgta atttgcgaga cgaatctttt 11880
gagcctagtt agtttataat taaataatat ttgttaaata caaacgaaaa tgctacgtta 11940
gccaaaacta aaatttttct ccaaacgtga cccagcacct tccgatcaat catcactcag 12000
cgggtcacgt cagaagatca gatggacctt gccgtccggg cctgtctctc ggcctcctcc 12060
ccatctggaa cgaacagagg tccagtcctg tttcgagtcg agctgagtcg atcagatggg 12120
cctaaatagg ccgaagacgt aggcaaaggg cccgctgatt tatctgattc ttctaggacc 12180
gtgcatgcgc ggatgggcct aggtggaaac ccaacagatg tgaggcttca aagaggaaga 12240
agtccgttac acatggagag ttagtctata atgggataat atttaccaca aacaaataaa 12300
aatactacag tagcgaaatc caaaattttt cacatctaaa caaggcccta gatgttttgt 12360
cagtgccaga ccagagaaaa tctcgtcttc tgctgtcaat agctttgatg attcctggcg 12420
gcagaggtaa agcttgcctg ggccttgttt agttccgaaa agtgaaaagt tttcggtact 12480
gtagcacttt tgtttgttcg tgacaaatat catccaatta tggactaact agaattaaaa 12540
gattcgtctc gtgatctaca gctaaactgt gtaattagtt tttgttttcg tctatattta 12600
atgtttcatg catgtgccac aagattcgat gtgacggaga attttgaaaa ttttttggtt 12660
ttcagagtga actaaacaag gcccagatgt aattgaccat gccatcgagc gcgagttgac 12720
tagagtgagt cggccctgat ggttaagtag tgcagactgc caagtggaca accgtctatc 12780
aactttgcag agtggggcga atgcactgag gatgttggag aggggcaagc caaggtaaac 12840
ttgaggaaag atgcttgttg acactgtagt atgtgaacaa tcctgtttaa ttttgtgtcc 12900
tcgacg 12906
<210> 88
<211> 1790
<212> ДНК
<213> Setaria italica
<400> 88
tctcatggtg tcccaagcac agcaagagcc agctctgcct cacagcagca gcaccgccaa 60
gcgcgcagcc gcgtcactca tggacgcccg cccggcccag cctctcctcc tccgcgcccc 120
gactcccagc attgacctcc ccgcgtccaa gccggacagg gccgccgcgg cggccggcaa 180
ggccgccgcc gcctccgtgt tcgacctgcg gcgggagccc aagatcccgg cgccattcgt 240
gtggccgcac gacgacgcgc ggccggcgtc ggcggcggag ctggacgtgc cgttggtgga 300
cgtgggcgtg ctgcgcaatg gcgaccgcgc ggggctgcgg cgcgctgcgg cgcaggtggc 360
cgcggcgtgc gcgacgcacg ggttcttcca ggtgtgcggg cacggcgtgg gcgcggacct 420
ggcgcgcgcg gcgctggacg gcgccagtga cttcttccgg ctgccgctgg cggagaagca 480
gcgcgcccgg cgcgtcccgg ggaccgtgtc cgggtacacg agcgcgcacg ccgaccggtt 540
cgcgtccaag ctcccctgga aggagaccct ctccttcggg ttccacgacg gcgccgcgtc 600
gcccgtcgtc gtcgactact tcgccggcac cctcgggcag gacttcgagg cagtggggcg 660
ggtgtaccag aggtactgcg aggagatgaa ggctctgtcg ctgacgatca tggagctcct 720
ggagctgagc ctgggcgtgg agcgcggcta ctaccgcgac ttcttcgagg acagccgctc 780
catcatgcgg tgcaactact acccgccgtg cccggagccg gagcgcacgc tgggcacggg 840
cccgcactgc gaccccaccg cgctgaccat cctcctccag gacgacgtcg gcgggctcga 900
ggtcctcgtc gacggcgact ggcgccccgt ccgccccgtc cccggcgcca tggtcatcaa 960
catcggcgac accttcatgg ctctgtccaa cgggcggtac aagagctgcc tgcaccgggc 1020
ggtggtgaac cagcggcagg agcggcggtc gctggccttc ttcctgtgcc cgcgcgagga 1080
ccgggtggtg cgcccgccgg ccagcggcgc cgtcggcgag gcgccccgcc gctacccgga 1140
cttcacctgg gccgacctca tgcgcttcac gcagcgccac taccgcgccg acacccgcac 1200
gctggacgcc ttcacacgct ggctctccca cggcccggcc caggacgcgc cagtggcggc 1260
ggcggcttcc acctagctag cggcgcggat ccgaccgagc ccattgacga cgccgtccct 1320
ttccgccgcc gccggggccc gcgcgggggt tcaccccacg tgcgcgccca ggtgggcgag 1380
gtggcggcct cgtggcccgc gggccccgcg ccgccttccc atttttgggc gctgccgccc 1440
cgcgcgcatg ccggatgcgt gcgtccacgg cctactgctg ctactagtgt acatatacaa 1500
acatacatat atacgtagta taaatatata agcaagcggc ccggtgcccc ttttcgtttt 1560
cttgttttgt cgatcacaat ctctggattc gatggatgga taaatgtttg tacgcatgca 1620
tgtagatggg ctcatgaaat ttcagaatct ggaacggacg aggagctcac gtgcctcttc 1680
cgtgtctggt agcggtagct gcgtgccaaa tgtctggtgg gcccaaagaa attctagtgc 1740
cacccgtccg gatccggcat ccgaaagttc ccgacggttc gacacccgaa 1790
<210> 89
<211> 1272
<212> ДНК
<213> Setaria italica
<400> 89
atggtgtccc aagcacagca agagccagct ctgcctcaca gcagcagcac cgccaagcgc 60
gcagccgcgt cactcatgga cgcccgcccg gcccagcctc tcctcctccg cgccccgact 120
cccagcattg acctccccgc gtccaagccg gacagggccg ccgcggcggc cggcaaggcc 180
gccgccgcct ccgtgttcga cctgcggcgg gagcccaaga tcccggcgcc attcgtgtgg 240
ccgcacgacg acgcgcggcc ggcgtcggcg gcggagctgg acgtgccgtt ggtggacgtg 300
ggcgtgctgc gcaatggcga ccgcgcgggg ctgcggcgcg ctgcggcgca ggtggccgcg 360
gcgtgcgcga cgcacgggtt cttccaggtg tgcgggcacg gcgtgggcgc ggacctggcg 420
cgcgcggcgc tggacggcgc cagtgacttc ttccggctgc cgctggcgga gaagcagcgc 480
gcccggcgcg tcccggggac cgtgtccggg tacacgagcg cgcacgccga ccggttcgcg 540
tccaagctcc cctggaagga gaccctctcc ttcgggttcc acgacggcgc cgcgtcgccc 600
gtcgtcgtcg actacttcgc cggcaccctc gggcaggact tcgaggcagt ggggcgggtg 660
taccagaggt actgcgagga gatgaaggct ctgtcgctga cgatcatgga gctcctggag 720
ctgagcctgg gcgtggagcg cggctactac cgcgacttct tcgaggacag ccgctccatc 780
atgcggtgca actactaccc gccgtgcccg gagccggagc gcacgctggg cacgggcccg 840
cactgcgacc ccaccgcgct gaccatcctc ctccaggacg acgtcggcgg gctcgaggtc 900
ctcgtcgacg gcgactggcg ccccgtccgc cccgtccccg gcgccatggt catcaacatc 960
ggcgacacct tcatggctct gtccaacggg cggtacaaga gctgcctgca ccgggcggtg 1020
gtgaaccagc ggcaggagcg gcggtcgctg gccttcttcc tgtgcccgcg cgaggaccgg 1080
gtggtgcgcc cgccggccag cggcgccgtc ggcgaggcgc cccgccgcta cccggacttc 1140
acctgggccg acctcatgcg cttcacgcag cgccactacc gcgccgacac ccgcacgctg 1200
gacgccttca cacgctggct ctcccacggc ccggcccagg acgcgccagt ggcggcggcg 1260
gcttccacct ag 1272
<210> 90
<211> 423
<212> ПРТ
<213> Setaria italica
<400> 90
Met Val Ser Gln Ala Gln Gln Glu Pro Ala Leu Pro His Ser Ser Ser
1. 5 10 15
Thr Ala Lys Arg Ala Ala Ala Ser Leu Met Asp Ala Arg Pro Ala Gln
20 25 30
Pro Leu Leu Leu Arg Ala Pro Thr Pro Ser Ile Asp Leu Pro Ala Ser
35 40 45
Lys Pro Asp Arg Ala Ala Ala Ala Ala Gly Lys Ala Ala Ala Ala Ser
50 55 60
Val Phe Asp Leu Arg Arg Glu Pro Lys Ile Pro Ala Pro Phe Val Trp
65 70 75 80
Pro His Asp Asp Ala Arg Pro Ala Ser Ala Ala Glu Leu Asp Val Pro
85 90 95
Leu Val Asp Val Gly Val Leu Arg Asn Gly Asp Arg Ala Gly Leu Arg
100 105 110
Arg Ala Ala Ala Gln Val Ala Ala Ala Cys Ala Thr His Gly Phe Phe
115 120 125
Gln Val Cys Gly His Gly Val Gly Ala Asp Leu Ala Arg Ala Ala Leu
130 135 140
Asp Gly Ala Ser Asp Phe Phe Arg Leu Pro Leu Ala Glu Lys Gln Arg
145 150 155 160
Ala Arg Arg Val Pro Gly Thr Val Ser Gly Tyr Thr Ser Ala His Ala
165 170 175
Asp Arg Phe Ala Ser Lys Leu Pro Trp Lys Glu Thr Leu Ser Phe Gly
180 185 190
Phe His Asp Gly Ala Ala Ser Pro Val Val Val Asp Tyr Phe Ala Gly
195 200 205
Thr Leu Gly Gln Asp Phe Glu Ala Val Gly Arg Val Tyr Gln Arg Tyr
210 215 220
Cys Glu Glu Met Lys Ala Leu Ser Leu Thr Ile Met Glu Leu Leu Glu
225 230 235 240
Leu Ser Leu Gly Val Glu Arg Gly Tyr Tyr Arg Asp Phe Phe Glu Asp
245 250 255
Ser Arg Ser Ile Met Arg Cys Asn Tyr Tyr Pro Pro Cys Pro Glu Pro
260 265 270
Glu Arg Thr Leu Gly Thr Gly Pro His Cys Asp Pro Thr Ala Leu Thr
275 280 285
Ile Leu Leu Gln Asp Asp Val Gly Gly Leu Glu Val Leu Val Asp Gly
290 295 300
Asp Trp Arg Pro Val Arg Pro Val Pro Gly Ala Met Val Ile Asn Ile
305 310 315 320
Gly Asp Thr Phe Met Ala Leu Ser Asn Gly Arg Tyr Lys Ser Cys Leu
325 330 335
His Arg Ala Val Val Asn Gln Arg Gln Glu Arg Arg Ser Leu Ala Phe
340 345 350
Phe Leu Cys Pro Arg Glu Asp Arg Val Val Arg Pro Pro Ala Ser Gly
355 360 365
Ala Val Gly Glu Ala Pro Arg Arg Tyr Pro Asp Phe Thr Trp Ala Asp
370 375 380
Leu Met Arg Phe Thr Gln Arg His Tyr Arg Ala Asp Thr Arg Thr Leu
385 390 395 400
Asp Ala Phe Thr Arg Trp Leu Ser His Gly Pro Ala Gln Asp Ala Pro
405 410 415
Val Ala Ala Ala Ala Ser Thr
420
<210> 91
<211> 2888
<212> ДНК
<213> Setaria italica
<400> 91
tctcatggtg tcccaagcac agcaagagcc agctctgcct cacagcagca gcaccgccaa 60
gcgcgcagcc gcgtcactca tggacgcccg cccggcccag cctctcctcc tccgcgcccc 120
gactcccagc attgacctcc ccgcgtccaa gccggacagg gccgccgcgg cggccggcaa 180
ggccgccgcc gcctccgtgt tcgacctgcg gcgggagccc aagatcccgg cgccattcgt 240
gtggccgcac gacgacgcgc ggccggcgtc ggcggcggag ctggacgtgc cgttggtgga 300
cgtgggcgtg ctgcgcaatg gcgaccgcgc ggggctgcgg cgcgctgcgg cgcaggtggc 360
cgcggcgtgc gcgacgcacg ggttcttcca ggtgtgcggg cacggcgtgg gcgcggacct 420
ggcgcgcgcg gcgctggacg gcgccagtga cttcttccgg ctgccgctgg cggagaagca 480
gcgcgcccgg cgcgtcccgg ggaccgtgtc cgggtacacg agcgcgcacg ccgaccggtt 540
cgcgtccaag ctcccctgga aggagaccct ctccttcggg ttccacgacg gcgccgcgtc 600
gcccgtcgtc gtcgactact tcgccggcac cctcgggcag gacttcgagg cagtggggta 660
agtatgtagg aatgaacttg gcacgcattg catccacatg gcgtgctgat cgaacgagct 720
gagccaaccg gcatgcacac atggcgtggc aggcgggtgt accagaggta ctgcgaggag 780
atgaaggctc tgtcgctgac gatcatggag ctcctggagc tgagcctggg cgtggagcgc 840
ggctactacc gcgacttctt cgaggacagc cgctccatca tgcggtgcaa ctactacccg 900
ccgtgcccgg agccggagcg cacgctgggc acgggcccgc actgcgaccc caccgcgctg 960
accatcctcc tccaggacga cgtcggcggg ctcgaggtcc tcgtcgacgg cgactggcgc 1020
cccgtccgcc ccgtccccgg cgccatggtc atcaacatcg gcgacacctt catggtacgg 1080
ccgccgctaa tccatccttt tgttgctctt atctcctctg gcgagtgcga gtaacgaaag 1140
cgctagctcc cctgctcctt gtcctgctct gtttcccaag tcctaatgga gctaaccggg 1200
cagactgcaa cacgcacgcg taggcatgtc acgtagccac cacttgcact gtgctgcgca 1260
gcgacgacgc aacgcggacg tgcgttcgag tcggttccat ctcggcgccg ctacacgcgg 1320
ccgcggctcc tagcctccta gggctccctg atccctatcc ccgagccctt ccgcgggaaa 1380
agttcgttgg cgacggcaga ggagagccga cgggtccgtg ccgttggagc gtggcggcag 1440
gagaggccgg gagggtgttt tgttgcgttg cgcggcggcg cggaggatgc gatggcgcgg 1500
gcgggcggcg ctttcggcgg tggcccccgc gacccacgtg cgcgcgcggt ctcgtcgcct 1560
tccctgtttt ggtgccacct ctctgtgtcc gggaatgggt tggcttagcg gcgaccgaga 1620
ccgggcggtg gtctggcctg ctcccggcgc ccatcccgcc tggtctctca tcctgctcct 1680
cctatgcgcg agggggcctg tagcggctgg agtacaagca gattggttgg gttgggttgc 1740
tgctgcttgg ctgttgcccg cccgctttct agccgtttcc gctcgccatc cggcacgcgg 1800
cgcccacgcc ggggctccag ctcggcccct ttggccgtgt gggtggcagg cacccctgca 1860
tcgtctcgtg cgtccggttt ccgcgcctgg ccccccgcct tgaggtttcc ctgtgctttt 1920
gacaagactt tcgtagatat atgtgtgtgt atgtgtgtgt gtgcgtgcgc gcgtgtgtgt 1980
atatatatat ataaataaat aacatctgtg aatgatggat tacacgtgta gctgaccggc 2040
tgattgtgtt cgcgtgtgtg tcttcgatgc attgcaggct ctgtccaacg ggcggtacaa 2100
gagctgcctg caccgggcgg tggtgaacca gcggcaggag cggcggtcgc tggccttctt 2160
cctgtgcccg cgcgaggacc gggtggtgcg cccgccggcc agcggcgccg tcggcgaggc 2220
gccccgccgc tacccggact tcacctgggc cgacctcatg cgcttcacgc agcgccacta 2280
ccgcgccgac acccgcacgc tggacgcctt cacacgctgg ctctcccacg gcccggccca 2340
ggacgcgcca gtggcggcgg cggcttccac ctagctagcg gcgcggatcc gaccgagccc 2400
attgacgacg ccgtcccttt ccgccgccgc cggggcccgc gcgggggttc accccacgtg 2460
cgcgcccagg tgggcgaggt ggcggcctcg tggcccgcgg gccccgcgcc gccttcccat 2520
ttttgggcgc tgccgccccg cgcgcatgcc ggatgcgtgc gtccacggcc tactgctgct 2580
actagtgtac atatacaaac atacatatat acgtagtata aatatataag caagcggccc 2640
ggtgcccctt ttcgttttct tgttttgtcg atcacaatct ctggattcga tggatggata 2700
aatgtttgta cgcatgcatg tagatgggct catgaaattt cagaatctgg aacggacgag 2760
gagctcacgt gcctcttccg tgtctggtag cggtagctgc gtgccaaatg tctggtgggc 2820
ccaaagaaat tctagtgcca cccgtccgga tccggcatcc gaaagttccc gacggttcga 2880
cacccgaa 2888
<210> 92
<211> 1567
<212> ДНК
<213> Oryza sativa
<400> 92
tgcccagaca gctcgccctg cacacacaca cacactcaca ctcacacacg ctctcaactc 60
actcccgctc aacacagcgc tcacttctca tctccaatct catggtggcc gagcacccca 120
cgccaccaca gccgcaccaa ccaccgccca tggactccac cgccggctct ggcattgccg 180
ccccggcggc ggcggcggtg tgcgacctga ggatggagcc caagatcccg gagccattcg 240
tgtggccgaa cggcgacgcg aggccggcgt cggcggcgga gctggacatg cccgtggtcg 300
acgtgggcgt gctccgcgac ggcgacgccg aggggctgcg ccgcgccgcg gcgcaggtgg 360
ccgccgcgtg cgccacgcac gggttcttcc aggtgtccga gcacggcgtc gacgccgctc 420
tggcgcgcgc cgcgctcgac ggcgccagcg acttcttccg cctcccgctc gccgagaagc 480
gccgcgcgcg ccgcgtcccg ggcaccgtgt ccggctacac cagcgcccac gccgaccgct 540
tcgcctccaa gctcccatgg aaggagaccc tctccttcgg cttccacgac cgcgccgccg 600
cccccgtcgt cgccgactac ttctccagca ccctcggccc cgacttcgcg ccaatgggga 660
gggtgtacca gaagtactgc gaggagatga aggagctgtc gctgacgatc atggaactcc 720
tggagctgag cctgggcgtg gagcgaggct actacaggga gttcttcgcg gacagcagct 780
caatcatgcg gtgcaactac tacccgccat gcccggagcc ggagcggacg ctcggcacgg 840
gcccgcactg cgaccccacc gccctcacca tcctcctcca ggacgacgtc ggcggcctcg 900
aggtcctcgt cgacggcgaa tggcgccccg tcagccccgt ccccggcgcc atggtcatca 960
acatcggcga caccttcatg gcgctgtcga acgggaggta taagagctgc ctgcacaggg 1020
cggtggtgaa ccagcggcgg gagcggcggt cgctggcgtt cttcctgtgc ccgcgggagg 1080
acagggtggt gcggccgccg ccgagcgccg ccacgccgca gcactacccg gacttcacct 1140
gggccgacct catgcgcttc acgcagcgcc actaccgcgc cgacacccgc acgctcgacg 1200
ccttcacgcg ctggctcgcg ccgccggccg ccgacgccgc cgcgacggcg caggtcgagg 1260
cggccagctg atcgccgaac ggaacgaaac ggaacgaaca gaagccgatt tttggcgggg 1320
cccacgccca cgtgaggccc cacgtggaca gtgggcccgg gcggaggtgg cacccacgtg 1380
gaccgcgggc cccgcgccgc cttccaattt tggaccctac cgctgtacat attcatatat 1440
tgcaagaaga agcaaaacgt acgtgtgggt tgggttgggc ttctctctat tactaaaaaa 1500
aatataatgg aacgacggat gaatggatgc ttatttattt atctaaattg aattcgaatt 1560
cggctca 1567
<210> 93
<211> 1170
<212> ДНК
<213> Oryza sativa
<400> 93
atggtggccg agcaccccac gccaccacag ccgcaccaac caccgcccat ggactccacc 60
gccggctctg gcattgccgc cccggcggcg gcggcggtgt gcgacctgag gatggagccc 120
aagatcccgg agccattcgt gtggccgaac ggcgacgcga ggccggcgtc ggcggcggag 180
ctggacatgc ccgtggtcga cgtgggcgtg ctccgcgacg gcgacgccga ggggctgcgc 240
cgcgccgcgg cgcaggtggc cgccgcgtgc gccacgcacg ggttcttcca ggtgtccgag 300
cacggcgtcg acgccgctct ggcgcgcgcc gcgctcgacg gcgccagcga cttcttccgc 360
ctcccgctcg ccgagaagcg ccgcgcgcgc cgcgtcccgg gcaccgtgtc cggctacacc 420
agcgcccacg ccgaccgctt cgcctccaag ctcccatgga aggagaccct ctccttcggc 480
ttccacgacc gcgccgccgc ccccgtcgtc gccgactact tctccagcac cctcggcccc 540
gacttcgcgc caatggggag ggtgtaccag aagtactgcg aggagatgaa ggagctgtcg 600
ctgacgatca tggaactcct ggagctgagc ctgggcgtgg agcgaggcta ctacagggag 660
ttcttcgcgg acagcagctc aatcatgcgg tgcaactact acccgccatg cccggagccg 720
gagcggacgc tcggcacggg cccgcactgc gaccccaccg ccctcaccat cctcctccag 780
gacgacgtcg gcggcctcga ggtcctcgtc gacggcgaat ggcgccccgt cagccccgtc 840
cccggcgcca tggtcatcaa catcggcgac accttcatgg cgctgtcgaa cgggaggtat 900
aagagctgcc tgcacagggc ggtggtgaac cagcggcggg agcggcggtc gctggcgttc 960
ttcctgtgcc cgcgggagga cagggtggtg cggccgccgc cgagcgccgc cacgccgcag 1020
cactacccgg acttcacctg ggccgacctc atgcgcttca cgcagcgcca ctaccgcgcc 1080
gacacccgca cgctcgacgc cttcacgcgc tggctcgcgc cgccggccgc cgacgccgcc 1140
gcgacggcgc aggtcgaggc ggccagctga 1170
<210> 94
<211> 389
<212> ПРТ
<213> Oryza sativa
<400> 94
Met Val Ala Glu His Pro Thr Pro Pro Gln Pro His Gln Pro Pro Pro
1. 5 10 15
Met Asp Ser Thr Ala Gly Ser Gly Ile Ala Ala Pro Ala Ala Ala Ala
20 25 30
Val Cys Asp Leu Arg Met Glu Pro Lys Ile Pro Glu Pro Phe Val Trp
35 40 45
Pro Asn Gly Asp Ala Arg Pro Ala Ser Ala Ala Glu Leu Asp Met Pro
50 55 60
Val Val Asp Val Gly Val Leu Arg Asp Gly Asp Ala Glu Gly Leu Arg
65 70 75 80
Arg Ala Ala Ala Gln Val Ala Ala Ala Cys Ala Thr His Gly Phe Phe
85 90 95
Gln Val Ser Glu His Gly Val Asp Ala Ala Leu Ala Arg Ala Ala Leu
100 105 110
Asp Gly Ala Ser Asp Phe Phe Arg Leu Pro Leu Ala Glu Lys Arg Arg
115 120 125
Ala Arg Arg Val Pro Gly Thr Val Ser Gly Tyr Thr Ser Ala His Ala
130 135 140
Asp Arg Phe Ala Ser Lys Leu Pro Trp Lys Glu Thr Leu Ser Phe Gly
145 150 155 160
Phe His Asp Arg Ala Ala Ala Pro Val Val Ala Asp Tyr Phe Ser Ser
165 170 175
Thr Leu Gly Pro Asp Phe Ala Pro Met Gly Arg Val Tyr Gln Lys Tyr
180 185 190
Cys Glu Glu Met Lys Glu Leu Ser Leu Thr Ile Met Glu Leu Leu Glu
195 200 205
Leu Ser Leu Gly Val Glu Arg Gly Tyr Tyr Arg Glu Phe Phe Ala Asp
210 215 220
Ser Ser Ser Ile Met Arg Cys Asn Tyr Tyr Pro Pro Cys Pro Glu Pro
225 230 235 240
Glu Arg Thr Leu Gly Thr Gly Pro His Cys Asp Pro Thr Ala Leu Thr
245 250 255
Ile Leu Leu Gln Asp Asp Val Gly Gly Leu Glu Val Leu Val Asp Gly
260 265 270
Glu Trp Arg Pro Val Ser Pro Val Pro Gly Ala Met Val Ile Asn Ile
275 280 285
Gly Asp Thr Phe Met Ala Leu Ser Asn Gly Arg Tyr Lys Ser Cys Leu
290 295 300
His Arg Ala Val Val Asn Gln Arg Arg Glu Arg Arg Ser Leu Ala Phe
305 310 315 320
Phe Leu Cys Pro Arg Glu Asp Arg Val Val Arg Pro Pro Pro Ser Ala
325 330 335
Ala Thr Pro Gln His Tyr Pro Asp Phe Thr Trp Ala Asp Leu Met Arg
340 345 350
Phe Thr Gln Arg His Tyr Arg Ala Asp Thr Arg Thr Leu Asp Ala Phe
355 360 365
Thr Arg Trp Leu Ala Pro Pro Ala Ala Asp Ala Ala Ala Thr Ala Gln
370 375 380
Val Glu Ala Ala Ser
385
<210> 95
<211> 3140
<212> ДНК
<213> Oryza sativa
<400> 95
tgcccagaca gctcgccctg cacacacaca cacactcaca ctcacacacg ctctcaactc 60
actcccgctc aacacagcgc tcacttctca tctccaatct catggtggcc gagcacccca 120
cgccaccaca gccgcaccaa ccaccgccca tggactccac cgccggctct ggcattgccg 180
ccccggcggc ggcggcggtg tgcgacctga ggatggagcc caagatcccg gagccattcg 240
tgtggccgaa cggcgacgcg aggccggcgt cggcggcgga gctggacatg cccgtggtcg 300
acgtgggcgt gctccgcgac ggcgacgccg aggggctgcg ccgcgccgcg gcgcaggtgg 360
ccgccgcgtg cgccacgcac gggttcttcc aggtgtccga gcacggcgtc gacgccgctc 420
tggcgcgcgc cgcgctcgac ggcgccagcg acttcttccg cctcccgctc gccgagaagc 480
gccgcgcgcg ccgcgtcccg ggcaccgtgt ccggctacac cagcgcccac gccgaccgct 540
tcgcctccaa gctcccatgg aaggagaccc tctccttcgg cttccacgac cgcgccgccg 600
cccccgtcgt cgccgactac ttctccagca ccctcggccc cgacttcgcg ccaatggggt 660
aattaaaacg atggtggacg acattgcatt tcaaattcaa aacaaattca aaacacaccg 720
accgagatta tgctgaattc aaacgcgttt gtgcgcgcag gagggtgtac cagaagtact 780
gcgaggagat gaaggagctg tcgctgacga tcatggaact cctggagctg agcctgggcg 840
tggagcgagg ctactacagg gagttcttcg cggacagcag ctcaatcatg cggtgcaact 900
actacccgcc atgcccggag ccggagcgga cgctcggcac gggcccgcac tgcgacccca 960
ccgccctcac catcctcctc caggacgacg tcggcggcct cgaggtcctc gtcgacggcg 1020
aatggcgccc cgtcagcccc gtccccggcg ccatggtcat caacatcggc gacaccttca 1080
tggtaaacca tctcctattc tcctctcctc tgttctcctc tgcttcgaag caacagaaca 1140
agtaattcaa gctttttttt ctctctcgcg cgaaattgac gagaaaaata agatcgtggt 1200
aggggcgggg ctttcagctg aaagcgggaa gaaaccgacc tgacgtgatt tctctgttcc 1260
aatcacaaac aatggaatgc cccactcctc catgtgttat gatttatctc acatcttata 1320
gttaatagga gtaagtaaca agctactttt ttcatattat agttcgtttg attttttttt 1380
tttaaagttt ttttagtttt atccaaattt attgaaaaac ttagcaacgt ttataatacc 1440
aaattagtct catttagttt aatattgtat atattttgat aatatattta tgttatatta 1500
aaaatattac tatatttttc tataaacatt attaaaagcc atttataata taaaatggaa 1560
ggagtaatta atatggatct cccccgacat gagaatattt tccgatggtg tgacgacgcc 1620
atgtaagctt cggtgggcct ggacggccag aggtgccaac agccacgtcc aacaacccct 1680
gggtcccccc ctaacactcc aaacagtagt gagtagtgtc tcgtcgcgtt ttagtatttg 1740
atgacaaaca aagtgtgagt tgagttagcc accaccaact tgcacacgag cacatacatt 1800
tgtgtccatt ctcgccagtc acttccatct ctagtcctaa ctcctatcta gcgatgtaag 1860
cggataattt catcatccgt atataaacct gtttgttata gttaatttcc tatataatac 1920
tataacagta tacattttaa aagaaaacaa aattaggata aacaggccct gctcctatcc 1980
atccatggca cttggaagga ccagactcgg tcatgccatg ccaagccaag atatgggtta 2040
tggaagagta gagaagagga gagatgagag ataagcatgc gttctcctcc tcgttggatg 2100
tgtattttgg agggatttgt gtagtagtag cagcggcgcc gcggggacgg atgcggatgg 2160
tggcgctttc ggtggcgttt tcccgggggg gttttggttt ggcgcttggg ggggatggca 2220
tggcgcggcg tgcggctgca cgccacacac acgcgcgcgc acgcacgtac gtcgtcgtcg 2280
ccgcgggcgg acggtagctt agggtggtgt gttccgcgcg cgggcgcgga ttgttccatg 2340
ccgatcgatt tggcgccacc ctcgccgcgg ctcttgtcgc gtcgtgcgcc tctctcgcgc 2400
ggtttgtcct tgtcgcgttg ctcagccggc gacgggggca cggacattgg cgatgtagcc 2460
ctgcacgtgt cggcctctcc gttgatgaat gatgatgtat gtatgtattt ttttttgtct 2520
gaaggaattt gtggggaatt gttgtgtgtg caggcgctgt cgaacgggag gtataagagc 2580
tgcctgcaca gggcggtggt gaaccagcgg cgggagcggc ggtcgctggc gttcttcctg 2640
tgcccgcggg aggacagggt ggtgcggccg ccgccgagcg ccgccacgcc gcagcactac 2700
ccggacttca cctgggccga cctcatgcgc ttcacgcagc gccactaccg cgccgacacc 2760
cgcacgctcg acgccttcac gcgctggctc gcgccgccgg ccgccgacgc cgccgcgacg 2820
gcgcaggtcg aggcggccag ctgatcgccg aacggaacga aacggaacga acagaagccg 2880
atttttggcg gggcccacgc ccacgtgagg ccccacgtgg acagtgggcc cgggcggagg 2940
tggcacccac gtggaccgcg ggccccgcgc cgccttccaa ttttggaccc taccgctgta 3000
catattcata tattgcaaga agaagcaaaa cgtacgtgtg ggttgggttg ggcttctctc 3060
tattactaaa aaaaatataa tggaacgacg gatgaatgga tgcttattta tttatctaaa 3120
ttgaattcga attcggctca 3140
<210> 96
<211> 1170
<212> ДНК
<213> Triticum aestivum
<400> 96
atggacacca gccctgcaac tcccctgctc ctccagcctc ctgctcccag cattgacccg 60
ttcgccgcca aggcggccgt caacaagggc ggcggcgcgg caaccgcggt gtacgacctc 120
cggagggagc cgaagatccc cgccccgttc gtgtggccgc acgccgaggt gcgccccacc 180
acggcccagg agctggccgt gccggtggtg gacgtgggcg tgctgcgcaa tggcgacgcc 240
gcggggctcc gccgcgccgt ggcgcaggtg gccgcggcgt gcgccacgca cgggttcttc 300
caggtgtccg ggcacggcgt ggacgaggcc ctggcgcgcg cggcgctgga cggcgcgagc 360
ggcttcttcc ggctgccgct ggccgagaag cagcgcgcgc ggcgcgtccc ggggaccgtg 420
tccgggtaca cgagcgcgca cgccgaccgg ttcgcctcca agctcccctg gaaggagacc 480
ctctccttcg gcttccacga ccgcgccggc gccgcgcccg tcgtggtgga ctacttcacc 540
agcaccctcg ggccggacta cgagccaatg gggagggtgt accaggagta ctgcgggaag 600
atgaaggagc tgtcgctgag gatcatggag ctgctggagc tgagccaggg cgtggagaag 660
cgcgggtact accgggagtt cttcgcggac agcagctcca tcatgcggtg caactactac 720
ccgccgtgcc cggagccgga gcgcacgctg ggcacgggcc cgcactgcga ccccacggcg 780
ctcaccatcc tactgcagga cgacgtgggc gggctggagg tcctcgtcga cggcgactgg 840
cgccccgtcc gccccgtccc cggcgccatg gtcatcaaca tcggcgacac cttcatggcg 900
ctgtcgaacg ggcggtacaa gagctgcctg caccgcgcgg tggtgaaccg gcggcaggag 960
cggcggtcgc tggccttctt cctgtgcccg cgcgaggacc gcgtggtgcg gccgccgccg 1020
ggcctgagga gcccgcggcg gtacccggac ttcacctggg ctgacctcat gcgcttcacg 1080
cagcgccact accgcgccga cacgcgcacc ctcgacgcct tcacccagtg gttctcctcc 1140
tcctcctcct cggcccagga ggcggcctga 1170
<210> 97
<211> 389
<212> ПРТ
<213> Triticum aestivum
<400> 97
Met Asp Thr Ser Pro Ala Thr Pro Leu Leu Leu Gln Pro Pro Ala Pro
1. 5 10 15
Ser Ile Asp Pro Phe Ala Ala Lys Ala Ala Val Asn Lys Gly Gly Gly
20 25 30
Ala Ala Thr Ala Val Tyr Asp Leu Arg Arg Glu Pro Lys Ile Pro Ala
35 40 45
Pro Phe Val Trp Pro His Ala Glu Val Arg Pro Thr Thr Ala Gln Glu
50 55 60
Leu Ala Val Pro Val Val Asp Val Gly Val Leu Arg Asn Gly Asp Ala
65 70 75 80
Ala Gly Leu Arg Arg Ala Val Ala Gln Val Ala Ala Ala Cys Ala Thr
85 90 95
His Gly Phe Phe Gln Val Ser Gly His Gly Val Asp Glu Ala Leu Ala
100 105 110
Arg Ala Ala Leu Asp Gly Ala Ser Gly Phe Phe Arg Leu Pro Leu Ala
115 120 125
Glu Lys Gln Arg Ala Arg Arg Val Pro Gly Thr Val Ser Gly Tyr Thr
130 135 140
Ser Ala His Ala Asp Arg Phe Ala Ser Lys Leu Pro Trp Lys Glu Thr
145 150 155 160
Leu Ser Phe Gly Phe His Asp Arg Ala Gly Ala Ala Pro Val Val Val
165 170 175
Asp Tyr Phe Thr Ser Thr Leu Gly Pro Asp Tyr Glu Pro Met Gly Arg
180 185 190
Val Tyr Gln Glu Tyr Cys Gly Lys Met Lys Glu Leu Ser Leu Arg Ile
195 200 205
Met Glu Leu Leu Glu Leu Ser Gln Gly Val Glu Lys Arg Gly Tyr Tyr
210 215 220
Arg Glu Phe Phe Ala Asp Ser Ser Ser Ile Met Arg Cys Asn Tyr Tyr
225 230 235 240
Pro Pro Cys Pro Glu Pro Glu Arg Thr Leu Gly Thr Gly Pro His Cys
245 250 255
Asp Pro Thr Ala Leu Thr Ile Leu Leu Gln Asp Asp Val Gly Gly Leu
260 265 270
Glu Val Leu Val Asp Gly Asp Trp Arg Pro Val Arg Pro Val Pro Gly
275 280 285
Ala Met Val Ile Asn Ile Gly Asp Thr Phe Met Ala Leu Ser Asn Gly
290 295 300
Arg Tyr Lys Ser Cys Leu His Arg Ala Val Val Asn Arg Arg Gln Glu
305 310 315 320
Arg Arg Ser Leu Ala Phe Phe Leu Cys Pro Arg Glu Asp Arg Val Val
325 330 335
Arg Pro Pro Pro Gly Leu Arg Ser Pro Arg Arg Tyr Pro Asp Phe Thr
340 345 350
Trp Ala Asp Leu Met Arg Phe Thr Gln Arg His Tyr Arg Ala Asp Thr
355 360 365
Arg Thr Leu Asp Ala Phe Thr Gln Trp Phe Ser Ser Ser Ser Ser Ser
370 375 380
Ala Gln Glu Ala Ala
385
<210> 98
<211> 3050
<212> ДНК
<213> Triticum aestivum
<400> 98
ctcatggtgc tccagaccgc tcagcaagaa ccatccctga cgcgtccgcc tcactgcagc 60
gtcgccagcg cgcgctcgcc ggcggccatg gacaccagcc ctgcaactcc cctgctcctc 120
cagcctcctg ctcccagcat tgacccgttc gccgccaagg cggccgtcaa caagggcggc 180
ggcgcggcaa ccgcggtgta cgacctccgg agggagccga agatccccgc cccgttcgtg 240
tggccgcacg ccgaggtgcg ccccaccacg gcccaggagc tggccgtgcc ggtggtggac 300
gtgggcgtgc tgcgcaatgg cgacgccgcg gggctccgcc gcgccgtggc gcaggtggcc 360
gcggcgtgcg ccacgcacgg gttcttccag gtgtccgggc acggcgtgga cgaggccctg 420
gcgcgcgcgg cgctggacgg cgcgagcggc ttcttccggc tgccgctggc cgagaagcag 480
cgcgcgcggc gcgtcccggg gaccgtgtcc gggtacacga gcgcgcacgc cgaccggttc 540
gcctccaagc tcccctggaa ggagaccctc tccttcggct tccacgaccg cgccggcgcc 600
gcgcccgtcg tggtggacta cttcaccagc accctcgggc cggactacga gccaatgggg 660
taatatatcc acccgcccac acccctatcc ggccagcacg aatccatccc cgccactgca 720
tttttttcct tttgtttccg cgcgaccgta cgttcgatcg gcgcccacgt acgtacgtgc 780
gtacgcagta gcagtacttg aagccgccgt actacgtgct gagtagtgac aactgaacac 840
gtgcaggagg gtgtaccagg agtactgcgg gaagatgaag gagctgtcgc tgaggatcat 900
ggagctgctg gagctgagcc agggcgtgga gaagcgcggg tactaccggg agttcttcgc 960
ggacagcagc tccatcatgc ggtgcaacta ctacccgccg tgcccggagc cggagcgcac 1020
gctgggcacg ggcccgcact gcgaccccac ggcgctcacc atcctactgc aggacgacgt 1080
gggcgggctg gaggtcctcg tcgacggcga ctggcgcccc gtccgccccg tccccggcgc 1140
catggtcatc aacatcggcg acaccttcat ggtaattact cctctctcag cgttgctttc 1200
gctgattaat tgcagaaaca gtagtcaact acccatgctc tgttccgctg tgctctgctt 1260
cccaacgagc gaaccggccc ataaaaactg ccttgctgtc ttggaaccaa gaggaaaggg 1320
accgtgggag cctaccgaca cgacgtgatt gcactctgct tcctaacaag cgagccgccg 1380
gtagggctat caccgtaagg gctcctttga ttcaaaggaa tttcttagga tttctgaagg 1440
attgaaatcc ttaggatttt ttcctatgtt ggtacttcga ttcataggat tgaatcccat 1500
aggatttttt tcctatgaaa tcttctgtac tacatttcat aggaaatcta acatccactc 1560
caaccttttt ttatatttcc tttgtttttc atgtgccatc aaacactcct tgttaatcct 1620
ataggattca agtgggcatg ccactccaat cctatacttt tcccattcct acgttttcaa 1680
aatcctacga atcaaagagg ccctaaagct gctgacatga cgtgattttt tttttctttt 1740
ctttctttct ttctcagctc caatcaacgc tggttattag atcattagag tggacaggtt 1800
gaattaacat gcagtagtta gtagttagca gccacaaacg ggtcccgttc tctgaagtct 1860
gaactgacat aagtcctgat catcgaccat tctttgcttc ctaggacgat gcctgttgga 1920
acttgcgtcc aatgcccgtt agggagtggt aattgtcatc acttttagac tcgtcgattc 1980
cactgatgaa gacgtagcac atggatgagc caacgtatcc gtttctagtg gtctcgaaaa 2040
gtagggtttc attcattcta tctatctatc cgtccgtcca aaagggctgc gatgcgagca 2100
cttgagtcgg agccaatcag agcgcgagaa aagatagggg gggtagcaag ccatgtcgga 2160
ggggcgtttg cttccggcag gtttggattc ttgtggtagg cgggcggctc tgtacagtag 2220
cggcggtgac ggtgaggtgg cggcgctttc ggtggcgggc caacccaggt gcatgcacgc 2280
gcgctcgtcg ttttcccgcc tgaatctgcc gctgcgccca tggcaagggg gtgggtgctg 2340
ccgccgggcg atggagtaga tcacggtcgc cgtcgggctc ggccagttga tcacggttcg 2400
ttcgtgcggt actaggttcc cccacggcac tgtgactgca tcgttccggc cctcgccatt 2460
ggcgatcggg caatctcctg ttcatccgtc gctgttgatt cctcggccac gatagaccat 2520
gcgcgtgccg gtcgtcgccc cgtcgcgctc gcttcacgtg ctcgtcgcgt ggctcccgtc 2580
ccacacgagg ccgccgcttt ctgacccagt ggagcgcgtg atttacagtt tatatatgtc 2640
gctgcatttt tctttttgtg tgctgctcat tttgcttgga cggagaccgg gaacgattag 2700
ccacggatct aacgcgttgt tgcttgtttt caatgcatgc atgcaggcgc tgtcgaacgg 2760
gcggtacaag agctgcctgc accgcgcggt ggtgaaccgg cggcaggagc ggcggtcgct 2820
ggccttcttc ctgtgcccgc gcgaggaccg cgtggtgcgg ccgccgccgg gcctgaggag 2880
cccgcggcgg tacccggact tcacctgggc tgacctcatg cgcttcacgc agcgccacta 2940
ccgcgccgac acgcgcaccc tcgacgcctt cacccagtgg ttctcctcct cctcctcctc 3000
ggcccaggag gcggcctgat tctgctctgc cacgaaacga tcggtccaca 3050
<210> 99
<211> 1486
<212> ДНК
<213> Hordeum vulgare
<400> 99
gaccagtagc atatagtttt tcttgtgttt gccatggtgg acgtgtcgaa ctttgtagaa 60
gccaatggca atgcagcagt atcgattcct gccatggaag ttgctgggag tcctcacgtc 120
ccgttcgttc ctcgggacgc gaacgcgaca gacagcaaga atgccaagga cgtcctcgac 180
ctctggcggc agcagaaaca aatcccggct cccttcatct ggccccacgc cgacgcgcgg 240
ccgtcgtcga tcttggagct ggacgtgccc gtggtcgaca tcggcgcggc cctgcacagc 300
gccgccggga tggcccgcgc cgcggcgcag gtggccgagg catgcgcgag ccacggcttc 360
ttccaggtga ccgggcacgg cgtcgacccc gcgctggccc aagcagcgct cgacggcgca 420
gcggacttct tccgcctgcc gctcgccacc aagcagcgcg cccgccgatc cccggggacc 480
gtcaaagggt acgcctccgc ccacgccgac cgcttcgccg ccaagcttcc ctggaaggag 540
actctctcct tcatccacaa ccacgtccac gaggacgtcg gcgcccgcgc aagcagtcac 600
gtcgtcgact acttcacctc cgcccttggc gacgacttca tgcacctagg ggaggtgtac 660
caggagtact gtgaggcgat ggaggacgcg tcgctggcga taatggaggt gctgggggtg 720
agcctggggc tggggagagg gtactacagg gacttcttcg ccgacggcag ctccatcatg 780
aggtgcaact actacccgcg gtgcccggag ccggaccgga cgctggggac ggggccgcac 840
tgcgacccgt cggcgctgac catcctgctg caggacggcg aggtggacgg gctccaggtg 900
ctcgtcgacg gcgcatggcg ctccgtgcgg cccaagcccg gcgagctcgt cgtaaacatc 960
ggcgacacct tcatggcgct gtcgaacggc cggtacaaga gctgcctcca ccgcgcggtg 1020
gtgcaccggg agaaggagcg ccggtcgctg gcctacttcc tcgccccgcg ggaggaccgg 1080
gtggttcgcc cgccgccttc gccggcgccg gcgccgcggc tctacccgga cttcacctgg 1140
gcggagctca tgcgattcac gcagcgccac taccgcgccg acgcccgcac gctcgacgcc 1200
ttcgcgtgct ggctcgacct gcccagctgc cccaccacgc cccaggccca agggactgtc 1260
tagtgtctgt gatgtatcat ctgtctcagc tgttgtatac gaccacttgt gtctgctagc 1320
tctgcgcttg tgtttcttat gtgagctaac taactaaata gtgtgtatat ttcttgccgc 1380
gccttatgca agccctagtc tagaacatgt aataattaac ttaagcatat acgttgatct 1440
ttggtgtatt tttcatattt ccttcataat gaataatcta ttatgc 1486
<210> 100
<211> 1230
<212> ДНК
<213> Hordeum vulgare
<400> 100
atggtggacg tgtcgaactt tgtagaagcc aatggcaatg cagcagtatc gattcctgcc 60
atggaagttg ctgggagtcc tcacgtcccg ttcgttcctc gggacgcgaa cgcgacagac 120
agcaagaatg ccaaggacgt cctcgacctc tggcggcagc agaaacaaat cccggctccc 180
ttcatctggc cccacgccga cgcgcggccg tcgtcgatct tggagctgga cgtgcccgtg 240
gtcgacatcg gcgcggccct gcacagcgcc gccgggatgg cccgcgccgc ggcgcaggtg 300
gccgaggcat gcgcgagcca cggcttcttc caggtgaccg ggcacggcgt cgaccccgcg 360
ctggcccaag cagcgctcga cggcgcagcg gacttcttcc gcctgccgct cgccaccaag 420
cagcgcgccc gccgatcccc ggggaccgtc aaagggtacg cctccgccca cgccgaccgc 480
ttcgccgcca agcttccctg gaaggagact ctctccttca tccacaacca cgtccacgag 540
gacgtcggcg cccgcgcaag cagtcacgtc gtcgactact tcacctccgc ccttggcgac 600
gacttcatgc acctagggga ggtgtaccag gagtactgtg aggcgatgga ggacgcgtcg 660
ctggcgataa tggaggtgct gggggtgagc ctggggctgg ggagagggta ctacagggac 720
ttcttcgccg acggcagctc catcatgagg tgcaactact acccgcggtg cccggagccg 780
gaccggacgc tggggacggg gccgcactgc gacccgtcgg cgctgaccat cctgctgcag 840
gacggcgagg tggacgggct ccaggtgctc gtcgacggcg catggcgctc cgtgcggccc 900
aagcccggcg agctcgtcgt aaacatcggc gacaccttca tggcgctgtc gaacggccgg 960
tacaagagct gcctccaccg cgcggtggtg caccgggaga aggagcgccg gtcgctggcc 1020
tacttcctcg ccccgcggga ggaccgggtg gttcgcccgc cgccttcgcc ggcgccggcg 1080
ccgcggctct acccggactt cacctgggcg gagctcatgc gattcacgca gcgccactac 1140
cgcgccgacg cccgcacgct cgacgccttc gcgtgctggc tcgacctgcc cagctgcccc 1200
accacgcccc aggcccaagg gactgtctag 1230
<210> 101
<211> 409
<212> ПРТ
<213> Hordeum vulgare
<400> 101
Met Val Asp Val Ser Asn Phe Val Glu Ala Asn Gly Asn Ala Ala Val
1. 5 10 15
Ser Ile Pro Ala Met Glu Val Ala Gly Ser Pro His Val Pro Phe Val
20 25 30
Pro Arg Asp Ala Asn Ala Thr Asp Ser Lys Asn Ala Lys Asp Val Leu
35 40 45
Asp Leu Trp Arg Gln Gln Lys Gln Ile Pro Ala Pro Phe Ile Trp Pro
50 55 60
His Ala Asp Ala Arg Pro Ser Ser Ile Leu Glu Leu Asp Val Pro Val
65 70 75 80
Val Asp Ile Gly Ala Ala Leu His Ser Ala Ala Gly Met Ala Arg Ala
85 90 95
Ala Ala Gln Val Ala Glu Ala Cys Ala Ser His Gly Phe Phe Gln Val
100 105 110
Thr Gly His Gly Val Asp Pro Ala Leu Ala Gln Ala Ala Leu Asp Gly
115 120 125
Ala Ala Asp Phe Phe Arg Leu Pro Leu Ala Thr Lys Gln Arg Ala Arg
130 135 140
Arg Ser Pro Gly Thr Val Lys Gly Tyr Ala Ser Ala His Ala Asp Arg
145 150 155 160
Phe Ala Ala Lys Leu Pro Trp Lys Glu Thr Leu Ser Phe Ile His Asn
165 170 175
His Val His Glu Asp Val Gly Ala Arg Ala Ser Ser His Val Val Asp
180 185 190
Tyr Phe Thr Ser Ala Leu Gly Asp Asp Phe Met His Leu Gly Glu Val
195 200 205
Tyr Gln Glu Tyr Cys Glu Ala Met Glu Asp Ala Ser Leu Ala Ile Met
210 215 220
Glu Val Leu Gly Val Ser Leu Gly Leu Gly Arg Gly Tyr Tyr Arg Asp
225 230 235 240
Phe Phe Ala Asp Gly Ser Ser Ile Met Arg Cys Asn Tyr Tyr Pro Arg
245 250 255
Cys Pro Glu Pro Asp Arg Thr Leu Gly Thr Gly Pro His Cys Asp Pro
260 265 270
Ser Ala Leu Thr Ile Leu Leu Gln Asp Gly Glu Val Asp Gly Leu Gln
275 280 285
Val Leu Val Asp Gly Ala Trp Arg Ser Val Arg Pro Lys Pro Gly Glu
290 295 300
Leu Val Val Asn Ile Gly Asp Thr Phe Met Ala Leu Ser Asn Gly Arg
305 310 315 320
Tyr Lys Ser Cys Leu His Arg Ala Val Val His Arg Glu Lys Glu Arg
325 330 335
Arg Ser Leu Ala Tyr Phe Leu Ala Pro Arg Glu Asp Arg Val Val Arg
340 345 350
Pro Pro Pro Ser Pro Ala Pro Ala Pro Arg Leu Tyr Pro Asp Phe Thr
355 360 365
Trp Ala Glu Leu Met Arg Phe Thr Gln Arg His Tyr Arg Ala Asp Ala
370 375 380
Arg Thr Leu Asp Ala Phe Ala Cys Trp Leu Asp Leu Pro Ser Cys Pro
385 390 395 400
Thr Thr Pro Gln Ala Gln Gly Thr Val
405
<210> 102
<211> 1423
<212> ДНК
<213> Sorghum bicolor
<400> 102
cctctcatca caggccccag cctcactctt ctcacagcaa gacatcgcag cctcacaacc 60
acacagcaac gtgatcgcca tgggcgggct caccatggag caggccttcg tgcaggcccc 120
cgagcaccgc cccaagccca ccgtcaccga ggccaccggc atcctggtca tcgacctctc 180
gcctctcacc gccagcgaca ccgacgcggc cgcggtggac gcgctggccg ccgaggtggg 240
cgcggcgagc cgggactggg gcttcttcgt ggtggttggc cacggcgtgc ccgcggagac 300
cgtggcgcgc gcgacggcgg cgcagcgcgc gttcttcgcg ctgccggcgg agcggaaggc 360
cgccgtgcgg aggagcgagg cggagccgct cgggtactac gagtcggagc acaccaagaa 420
cgtcagggac tggaaggagg tgttcgacct cgtcccgcgc gatccgccgc cgccagcagc 480
cgtggccgac ggcgagctcg tcttcaagaa caagtggccc caggatctgc cgggcttcag 540
agaggcgctg gaggagtacg cggcagcgat ggaggagctg tcgttcaagc tgctggagct 600
gatcgcccgg agcttgaagc tgaggcccga ccggctgcac ggcttcttca aggaccagac 660
gacgttcatc cggctgaacc actaccctcc atgcccgagc ccggacctgg cgctgggagt 720
ggggcggcac aaggacgcgg gggcgctgac catcctgtac caggacgaag tgggcgggct 780
ggacgtccgg cggcgctcct ccgacggcgg cggcggcgag tgggtgcggg tgaggcccgt 840
gccggagtcg ttcgtcatca acgtcggcga cctcgtccag gtgtggagca acgacaggta 900
cgagagcgcg gagcaccggg tgtcggtgaa ctcggcgagg gagaggttct ccatgcccta 960
cttcttcaac ccggcgagct acaccatggt ggagccggtg gaggagctgg tgagcgacga 1020
cgacccgccc aggtacgacg cctacagctg gggcgagttc ttcagcacca ggaagaacag 1080
caacttcaag aagctcagcg tggagaacat tcagatcgcg catttcaaga agaccctcgt 1140
cctcgcctag ataagcagca ggatactaca ggtctacagg actaggacaa gccgatcgag 1200
gtgaccggcc gtcgtcttca gattcagtat atgcgtgtcg ccgttcgtgt tagaacaaat 1260
taataatgtg cgcgctgtgt gctgtgtgtg tggagtaaaa aaaaactaaa catggatgtg 1320
catgttcaaa aaaaaaaaca tggatgcgag tatgtttggg aataataaca ggcttgtgac 1380
ggtctggttt atttgcaaat tcaaaccgaa ttggttgatc ttc 1423
<210> 103
<211> 1071
<212> ДНК
<213> Sorghum bicolor
<400> 103
atgggcgggc tcaccatgga gcaggccttc gtgcaggccc ccgagcaccg ccccaagccc 60
accgtcaccg aggccaccgg catcctggtc atcgacctct cgcctctcac cgccagcgac 120
accgacgcgg ccgcggtgga cgcgctggcc gccgaggtgg gcgcggcgag ccgggactgg 180
ggcttcttcg tggtggttgg ccacggcgtg cccgcggaga ccgtggcgcg cgcgacggcg 240
gcgcagcgcg cgttcttcgc gctgccggcg gagcggaagg ccgccgtgcg gaggagcgag 300
gcggagccgc tcgggtacta cgagtcggag cacaccaaga acgtcaggga ctggaaggag 360
gtgttcgacc tcgtcccgcg cgatccgccg ccgccagcag ccgtggccga cggcgagctc 420
gtcttcaaga acaagtggcc ccaggatctg ccgggcttca gagaggcgct ggaggagtac 480
gcggcagcga tggaggagct gtcgttcaag ctgctggagc tgatcgcccg gagcttgaag 540
ctgaggcccg accggctgca cggcttcttc aaggaccaga cgacgttcat ccggctgaac 600
cactaccctc catgcccgag cccggacctg gcgctgggag tggggcggca caaggacgcg 660
ggggcgctga ccatcctgta ccaggacgaa gtgggcgggc tggacgtccg gcggcgctcc 720
tccgacggcg gcggcggcga gtgggtgcgg gtgaggcccg tgccggagtc gttcgtcatc 780
aacgtcggcg acctcgtcca ggtgtggagc aacgacaggt acgagagcgc ggagcaccgg 840
gtgtcggtga actcggcgag ggagaggttc tccatgccct acttcttcaa cccggcgagc 900
tacaccatgg tggagccggt ggaggagctg gtgagcgacg acgacccgcc caggtacgac 960
gcctacagct ggggcgagtt cttcagcacc aggaagaaca gcaacttcaa gaagctcagc 1020
gtggagaaca ttcagatcgc gcatttcaag aagaccctcg tcctcgccta g 1071
<210> 104
<211> 356
<212> ПРТ
<213> Sorghum bicolor
<400> 104
Met Gly Gly Leu Thr Met Glu Gln Ala Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Pro Lys Pro Thr Val Thr Glu Ala Thr Gly Ile Leu Val Ile Asp
20 25 30
Leu Ser Pro Leu Thr Ala Ser Asp Thr Asp Ala Ala Ala Val Asp Ala
35 40 45
Leu Ala Ala Glu Val Gly Ala Ala Ser Arg Asp Trp Gly Phe Phe Val
50 55 60
Val Val Gly His Gly Val Pro Ala Glu Thr Val Ala Arg Ala Thr Ala
65 70 75 80
Ala Gln Arg Ala Phe Phe Ala Leu Pro Ala Glu Arg Lys Ala Ala Val
85 90 95
Arg Arg Ser Glu Ala Glu Pro Leu Gly Tyr Tyr Glu Ser Glu His Thr
100 105 110
Lys Asn Val Arg Asp Trp Lys Glu Val Phe Asp Leu Val Pro Arg Asp
115 120 125
Pro Pro Pro Pro Ala Ala Val Ala Asp Gly Glu Leu Val Phe Lys Asn
130 135 140
Lys Trp Pro Gln Asp Leu Pro Gly Phe Arg Glu Ala Leu Glu Glu Tyr
145 150 155 160
Ala Ala Ala Met Glu Glu Leu Ser Phe Lys Leu Leu Glu Leu Ile Ala
165 170 175
Arg Ser Leu Lys Leu Arg Pro Asp Arg Leu His Gly Phe Phe Lys Asp
180 185 190
Gln Thr Thr Phe Ile Arg Leu Asn His Tyr Pro Pro Cys Pro Ser Pro
195 200 205
Asp Leu Ala Leu Gly Val Gly Arg His Lys Asp Ala Gly Ala Leu Thr
210 215 220
Ile Leu Tyr Gln Asp Glu Val Gly Gly Leu Asp Val Arg Arg Arg Ser
225 230 235 240
Ser Asp Gly Gly Gly Gly Glu Trp Val Arg Val Arg Pro Val Pro Glu
245 250 255
Ser Phe Val Ile Asn Val Gly Asp Leu Val Gln Val Trp Ser Asn Asp
260 265 270
Arg Tyr Glu Ser Ala Glu His Arg Val Ser Val Asn Ser Ala Arg Glu
275 280 285
Arg Phe Ser Met Pro Tyr Phe Phe Asn Pro Ala Ser Tyr Thr Met Val
290 295 300
Glu Pro Val Glu Glu Leu Val Ser Asp Asp Asp Pro Pro Arg Tyr Asp
305 310 315 320
Ala Tyr Ser Trp Gly Glu Phe Phe Ser Thr Arg Lys Asn Ser Asn Phe
325 330 335
Lys Lys Leu Ser Val Glu Asn Ile Gln Ile Ala His Phe Lys Lys Thr
340 345 350
Leu Val Leu Ala
355
<210> 105
<211> 1499
<212> ДНК
<213> Sorghum bicolor
<400> 105
cctctcatca caggccccag cctcactctt ctcacagcaa gacatcgcag cctcacaacc 60
acacagcaac gtgatcgcca tgggcgggct caccatggag caggccttcg tgcaggcccc 120
cgagcaccgc cccaagccca ccgtcaccga ggccaccggc atcctggtca tcgacctctc 180
gcctctcacc gccagcgaca ccgacgcggc cgcggtggac gcgctggccg ccgaggtggg 240
cgcggcgagc cgggactggg gcttcttcgt ggtggttggc cacggcgtgc ccgcggagac 300
cgtggcgcgc gcgacggcgg cgcagcgcgc gttcttcgcg ctgccggcgg agcggaaggc 360
cgccgtgcgg aggagcgagg cggagccgct cgggtactac gagtcggagc acaccaagaa 420
cgtcagggac tggaaggagg tgttcgacct cgtcccgcgc gatccgccgc cgccagcagc 480
cgtggccgac ggcgagctcg tcttcaagaa caagtggccc caggatctgc cgggcttcag 540
gtgacgaaat caacttatct tttcgatcat attttaccat ttaatagttt aacaataatt 600
gaactttttt ttgcagagag gcgctggagg agtacgcggc agcgatggag gagctgtcgt 660
tcaagctgct ggagctgatc gcccggagct tgaagctgag gcccgaccgg ctgcacggct 720
tcttcaagga ccagacgacg ttcatccggc tgaaccacta ccctccatgc ccgagcccgg 780
acctggcgct gggagtgggg cggcacaagg acgcgggggc gctgaccatc ctgtaccagg 840
acgaagtggg cgggctggac gtccggcggc gctcctccga cggcggcggc ggcgagtggg 900
tgcgggtgag gcccgtgccg gagtcgttcg tcatcaacgt cggcgacctc gtccaggtgt 960
ggagcaacga caggtacgag agcgcggagc accgggtgtc ggtgaactcg gcgagggaga 1020
ggttctccat gccctacttc ttcaacccgg cgagctacac catggtggag ccggtggagg 1080
agctggtgag cgacgacgac ccgcccaggt acgacgccta cagctggggc gagttcttca 1140
gcaccaggaa gaacagcaac ttcaagaagc tcagcgtgga gaacattcag atcgcgcatt 1200
tcaagaagac cctcgtcctc gcctagataa gcagcaggat actacaggtc tacaggacta 1260
ggacaagccg atcgaggtga ccggccgtcg tcttcagatt cagtatatgc gtgtcgccgt 1320
tcgtgttaga acaaattaat aatgtgcgcg ctgtgtgctg tgtgtgtgga gtaaaaaaaa 1380
actaaacatg gatgtgcatg ttcaaaaaaa aaaacatgga tgcgagtatg tttgggaata 1440
ataacaggct tgtgacggtc tggtttattt gcaaattcaa accgaattgg ttgatcttc 1499
<210> 106
<211> 1490
<212> ДНК
<213> Setaria italica
<400> 106
accccacaca cacacccgca ctgcatgcgg cgtcctagct aatcagtcgc tgctggcagc 60
ctcacaagtc acacaactcc gacgcaggaa agctcgatcc atcgccatgg gcggcttctc 120
catggatcag tccttcgtgc aggcccccga gcaccgcccc aagcccaccg tcaccgaggc 180
cacgggcatc ccgctcatcg acctctcgcc actcaccggc ggtggcggcg gcgacgcggc 240
cgccgtggac gcgctggccg ccgaggtggg cgcggcgagc cgggactggg gcttcttcgt 300
ggtggtgggg cacggtgtgc cggcggagac cgtggcgcgc gccacggagg cgcagcgcgc 360
gttcttcgcc ctgccggcgg agcggaaagc cgccgtgcgg aggagcgagg cggagccgct 420
cgggtactac gagtcggagc acaccaagaa cgtcagggac tggaaggagg tgtacgacct 480
cgtcccgggc gggcttcagc cgccgatagc cgtggccgac ggcgaggtcg tgttcgaaaa 540
caagtggccc gaagacctgc cgggattcag agaggcgttg gaggagtaca tgcaagcgat 600
ggaagagctg gcattcaaga tactggagct gatcgcccgg agcctgaacc tgaggcctga 660
cagactgcac ggcttcttca aggaccagac caccttcatc cggctcaacc actaccctcc 720
ctgcccgagc cccgacctcg ccctcggcgt cggccggcac aaggacgccg gagcactgac 780
catcctctac caggacgacg tcggcgggct cgacgtccgg cgccgttccg acggcgattg 840
ggtccgcgtc aagcctgtcc ccgactcctt catcatcaac gtcggcgacc tcatccaggt 900
ttggagcaac gacaggtacg agagcgcgga gcaccgggtt acggtgaact cggccaagga 960
gaggttctcc aggccctact tcttcaaccc ggcgggctac accatggtgg agccggtgga 1020
ggagctggtg agcgaggagg acccgccccg gtacgacgcc tacaactggg gcaacttctt 1080
cagcaccagg aagaacagca acttcaagaa gctgagcgtg gagaacatcc agatcgcgca 1140
tttcaagagg agcgtcgccg cctaggatac gcacagaaag atcccatatg ctgacttgct 1200
gatgaggcga caggcggccg tgtcgtcttc agattcagag actgggagta aacatttgtg 1260
cggtgttctg taatcgtgat gtgacgagaa ctttagatat atgtttggaa ataacagcct 1320
tgtgttggtc tggcttatcc gcaaagtcaa gattttcttc tacattttgg gattattgtt 1380
ggtaagcatt aagcaacgtc cagttcttac ttcttagctc gatcagtgga cgtaggaccg 1440
gcctctgatg acaagggtga tttatgagaa atgtcatgta tatatgttcc 1490
<210> 107
<211> 1059
<212> ДНК
<213> Setaria italica
<400> 107
atgggcggct tctccatgga tcagtccttc gtgcaggccc ccgagcaccg ccccaagccc 60
accgtcaccg aggccacggg catcccgctc atcgacctct cgccactcac cggcggtggc 120
ggcggcgacg cggccgccgt ggacgcgctg gccgccgagg tgggcgcggc gagccgggac 180
tggggcttct tcgtggtggt ggggcacggt gtgccggcgg agaccgtggc gcgcgccacg 240
gaggcgcagc gcgcgttctt cgccctgccg gcggagcgga aagccgccgt gcggaggagc 300
gaggcggagc cgctcgggta ctacgagtcg gagcacacca agaacgtcag ggactggaag 360
gaggtgtacg acctcgtccc gggcgggctt cagccgccga tagccgtggc cgacggcgag 420
gtcgtgttcg aaaacaagtg gcccgaagac ctgccgggat tcagagaggc gttggaggag 480
tacatgcaag cgatggaaga gctggcattc aagatactgg agctgatcgc ccggagcctg 540
aacctgaggc ctgacagact gcacggcttc ttcaaggacc agaccacctt catccggctc 600
aaccactacc ctccctgccc gagccccgac ctcgccctcg gcgtcggccg gcacaaggac 660
gccggagcac tgaccatcct ctaccaggac gacgtcggcg ggctcgacgt ccggcgccgt 720
tccgacggcg attgggtccg cgtcaagcct gtccccgact ccttcatcat caacgtcggc 780
gacctcatcc aggtttggag caacgacagg tacgagagcg cggagcaccg ggttacggtg 840
aactcggcca aggagaggtt ctccaggccc tacttcttca acccggcggg ctacaccatg 900
gtggagccgg tggaggagct ggtgagcgag gaggacccgc cccggtacga cgcctacaac 960
tggggcaact tcttcagcac caggaagaac agcaacttca agaagctgag cgtggagaac 1020
atccagatcg cgcatttcaa gaggagcgtc gccgcctag 1059
<210> 108
<211> 352
<212> ПРТ
<213> Setaria italica
<400> 108
Met Gly Gly Phe Ser Met Asp Gln Ser Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Pro Lys Pro Thr Val Thr Glu Ala Thr Gly Ile Pro Leu Ile Asp
20 25 30
Leu Ser Pro Leu Thr Gly Gly Gly Gly Gly Asp Ala Ala Ala Val Asp
35 40 45
Ala Leu Ala Ala Glu Val Gly Ala Ala Ser Arg Asp Trp Gly Phe Phe
50 55 60
Val Val Val Gly His Gly Val Pro Ala Glu Thr Val Ala Arg Ala Thr
65 70 75 80
Glu Ala Gln Arg Ala Phe Phe Ala Leu Pro Ala Glu Arg Lys Ala Ala
85 90 95
Val Arg Arg Ser Glu Ala Glu Pro Leu Gly Tyr Tyr Glu Ser Glu His
100 105 110
Thr Lys Asn Val Arg Asp Trp Lys Glu Val Tyr Asp Leu Val Pro Gly
115 120 125
Gly Leu Gln Pro Pro Ile Ala Val Ala Asp Gly Glu Val Val Phe Glu
130 135 140
Asn Lys Trp Pro Glu Asp Leu Pro Gly Phe Arg Glu Ala Leu Glu Glu
145 150 155 160
Tyr Met Gln Ala Met Glu Glu Leu Ala Phe Lys Ile Leu Glu Leu Ile
165 170 175
Ala Arg Ser Leu Asn Leu Arg Pro Asp Arg Leu His Gly Phe Phe Lys
180 185 190
Asp Gln Thr Thr Phe Ile Arg Leu Asn His Tyr Pro Pro Cys Pro Ser
195 200 205
Pro Asp Leu Ala Leu Gly Val Gly Arg His Lys Asp Ala Gly Ala Leu
210 215 220
Thr Ile Leu Tyr Gln Asp Asp Val Gly Gly Leu Asp Val Arg Arg Arg
225 230 235 240
Ser Asp Gly Asp Trp Val Arg Val Lys Pro Val Pro Asp Ser Phe Ile
245 250 255
Ile Asn Val Gly Asp Leu Ile Gln Val Trp Ser Asn Asp Arg Tyr Glu
260 265 270
Ser Ala Glu His Arg Val Thr Val Asn Ser Ala Lys Glu Arg Phe Ser
275 280 285
Arg Pro Tyr Phe Phe Asn Pro Ala Gly Tyr Thr Met Val Glu Pro Val
290 295 300
Glu Glu Leu Val Ser Glu Glu Asp Pro Pro Arg Tyr Asp Ala Tyr Asn
305 310 315 320
Trp Gly Asn Phe Phe Ser Thr Arg Lys Asn Ser Asn Phe Lys Lys Leu
325 330 335
Ser Val Glu Asn Ile Gln Ile Ala His Phe Lys Arg Ser Val Ala Ala
340 345 350
<210> 109
<211> 1886
<212> ДНК
<213> Setaria italica
<400> 109
accccacaca cacacccgca ctgcatgcgg cgtcctagct aatcagtcgc tgctggcagc 60
ctcacaagtc acacaactcc gacgcaggaa agctcgatcc atcgccatgg gcggcttctc 120
catggatcag tccttcgtgc aggcccccga gcaccgcccc aagcccaccg tcaccgaggc 180
cacgggcatc ccgctcatcg acctctcgcc actcaccggc ggtggcggcg gcgacgcggc 240
cgccgtggac gcgctggccg ccgaggtggg cgcggcgagc cgggactggg gcttcttcgt 300
ggtggtgggg cacggtgtgc cggcggagac cgtggcgcgc gccacggagg cgcagcgcgc 360
gttcttcgcc ctgccggcgg agcggaaagc cgccgtgcgg aggagcgagg cggagccgct 420
cgggtactac gagtcggagc acaccaagaa cgtcagggac tggaaggagg tgtacgacct 480
cgtcccgggc gggcttcagc cgccgatagc cgtggccgac ggcgaggtcg tgttcgaaaa 540
caagtggccc gaagacctgc cgggattcag gtgaatcaac ttgcgcatat tgttgtttct 600
ggcattgcat atgatcgtcg tgccagtatg ttttgacaat atttttgttt tcatattttt 660
ggtgaagatg ggaaaatctt tgttgaaata atcagggaat tttcacatct ttttttaatc 720
aaagatagaa taggttcggt tactgaattt tgatgatgga cagaaaaagc tgtgttttca 780
ctttccatct cagcgatgtt tttttgtgga tgaattctcc taaatttttg tcttttcatg 840
ttaaaacttg aacgggaatt ctcgcagaga ggcgttggag gagtacatgc aagcgatgga 900
agagctggca ttcaagatac tggagctgat cgcccggagc ctgaacctga ggcctgacag 960
actgcacggc ttcttcaagg accagaccac cttcatccgg ctcaaccact accctccctg 1020
cccgagcccc gacctcgccc tcggcgtcgg ccggcacaag gacgccggag cactgaccat 1080
cctctaccag gacgacgtcg gcgggctcga cgtccggcgc cgttccgacg gcgattgggt 1140
ccgcgtcaag cctgtccccg actccttcat catcaacgtc ggcgacctca tccaggtaca 1200
acaaacaaaa acacacgtca ttctcaaatc ttttcgtgct gttaatgctc attcacgaat 1260
tgatatctta catgaacgac tgagactttt tcaggtttgg agcaacgaca ggtacgagag 1320
cgcggagcac cgggttacgg tgaactcggc caaggagagg ttctccaggc cctacttctt 1380
caacccggcg ggctacacca tggtggagcc ggtggaggag ctggtgagcg aggaggaccc 1440
gccccggtac gacgcctaca actggggcaa cttcttcagc accaggaaga acagcaactt 1500
caagaagctg agcgtggaga acatccagat cgcgcatttc aagaggagcg tcgccgccta 1560
ggatacgcac agaaagatcc catatgctga cttgctgatg aggcgacagg cggccgtgtc 1620
gtcttcagat tcagagactg ggagtaaaca tttgtgcggt gttctgtaat cgtgatgtga 1680
cgagaacttt agatatatgt ttggaaataa cagccttgtg ttggtctggc ttatccgcaa 1740
agtcaagatt ttcttctaca ttttgggatt attgttggta agcattaagc aacgtccagt 1800
tcttacttct tagctcgatc agtggacgta ggaccggcct ctgatgacaa gggtgattta 1860
tgagaaatgt catgtatata tgttcc 1886
<210> 110
<211> 1379
<212> ДНК
<213> Oryza sativa
<400> 110
aagccacacg cacacacaca cacacgctga cacacgagac gaacacttgt gctacagctt 60
ctcgccacca gctactgatc gaccatgggc ggcctctcca tggaccaggc gttcgtgcag 120
gcccccgagc accgccccaa ggcgtccgtc gccgaggccg acggcatccc ggtcatcgac 180
ctctcccctc tcctcgccgc cggcgatggc gacgccgacg gggtggacgc gctcgcggcg 240
gaggtcggga gggcgagccg ggactggggc ttcttcgtgg tggtgcgcca cggtgtgccc 300
gcggaggcgg tggcgcgcgc ggcggaggcg cagaggacgt tcttcgcgct gccgccggag 360
cggagggcgg ccgtggcgcg gagcgaggcg gcgccgatgg ggtactacgc gtccgagcac 420
accaagaacg tcagggactg gaaggaggtg ttcgacctcg tcccgcgcca gacgccgccg 480
ccgccgacga ccgccgtggc cgacggcgac ctggtgttcg acaacaagtg gcccgacgac 540
ctgccgggat tcagggaggc aatggaggag tacggcgaag cggtggagga gctggcgttc 600
aagctgctgg agctgatcgc caggagcctc ggcctgagac ccgaccgcct ccatggcttc 660
ttcaaggacg accagaccac cttcatccgg ctcaaccact accctccctg cccgagcccc 720
gacctcgccc tcggcgtcgg ccgccacaag gacgccggcg cgctcaccgt gctctaccag 780
gacgatgtcg gcggcctcga cgtccgccgc cgatccgacg gcgagtgggt gcgcgtcagg 840
cccgtccctc actccttcat catcaacgtc ggcgacatca tccaggtgtg gagcaatgac 900
aggtacgaga gcgcggagca ccgggtggcg gtgaacgtgg agaaggagag gttctccatc 960
cctttcttct tcaacccggc gggccacacc atggtggagc cactggagga ggtcgtgagc 1020
gacgagagcc cggccaggta caacccctac aactggggcg aattcttcag caccaggaag 1080
aacagcaact tcaagaagct ggacgtggag aacgtccaga tcacgcattt caggaagaat 1140
taacgcgccg gctagatcat gttcagtaaa ttttcagatg atgatgcgtg gacaaccata 1200
tagcctttgc gtcataagtt aataatgtct gtgacagtat atcatgtaaa caatcgtatg 1260
atgtggcttc tctatctgcc ggtgatggta atgtgacatt gtagaagagg gtttgtgaga 1320
tacttccttc acttaacttt tacgaatgaa tatagacaac cacaacatcc ttgtcgtga 1379
<210> 111
<211> 1059
<212> ДНК
<213> Oryza sativa
<400> 111
atgggcggcc tctccatgga ccaggcgttc gtgcaggccc ccgagcaccg ccccaaggcg 60
tccgtcgccg aggccgacgg catcccggtc atcgacctct cccctctcct cgccgccggc 120
gatggcgacg ccgacggggt ggacgcgctc gcggcggagg tcgggagggc gagccgggac 180
tggggcttct tcgtggtggt gcgccacggt gtgcccgcgg aggcggtggc gcgcgcggcg 240
gaggcgcaga ggacgttctt cgcgctgccg ccggagcgga gggcggccgt ggcgcggagc 300
gaggcggcgc cgatggggta ctacgcgtcc gagcacacca agaacgtcag ggactggaag 360
gaggtgttcg acctcgtccc gcgccagacg ccgccgccgc cgacgaccgc cgtggccgac 420
ggcgacctgg tgttcgacaa caagtggccc gacgacctgc cgggattcag ggaggcaatg 480
gaggagtacg gcgaagcggt ggaggagctg gcgttcaagc tgctggagct gatcgccagg 540
agcctcggcc tgagacccga ccgcctccat ggcttcttca aggacgacca gaccaccttc 600
atccggctca accactaccc tccctgcccg agccccgacc tcgccctcgg cgtcggccgc 660
cacaaggacg ccggcgcgct caccgtgctc taccaggacg atgtcggcgg cctcgacgtc 720
cgccgccgat ccgacggcga gtgggtgcgc gtcaggcccg tccctcactc cttcatcatc 780
aacgtcggcg acatcatcca ggtgtggagc aatgacaggt acgagagcgc ggagcaccgg 840
gtggcggtga acgtggagaa ggagaggttc tccatccctt tcttcttcaa cccggcgggc 900
cacaccatgg tggagccact ggaggaggtc gtgagcgacg agagcccggc caggtacaac 960
ccctacaact ggggcgaatt cttcagcacc aggaagaaca gcaacttcaa gaagctggac 1020
gtggagaacg tccagatcac gcatttcagg aagaattaa 1059
<210> 112
<211> 352
<212> ПРТ
<213> Oryza sativa
<400> 112
Met Gly Gly Leu Ser Met Asp Gln Ala Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Pro Lys Ala Ser Val Ala Glu Ala Asp Gly Ile Pro Val Ile Asp
20 25 30
Leu Ser Pro Leu Leu Ala Ala Gly Asp Gly Asp Ala Asp Gly Val Asp
35 40 45
Ala Leu Ala Ala Glu Val Gly Arg Ala Ser Arg Asp Trp Gly Phe Phe
50 55 60
Val Val Val Arg His Gly Val Pro Ala Glu Ala Val Ala Arg Ala Ala
65 70 75 80
Glu Ala Gln Arg Thr Phe Phe Ala Leu Pro Pro Glu Arg Arg Ala Ala
85 90 95
Val Ala Arg Ser Glu Ala Ala Pro Met Gly Tyr Tyr Ala Ser Glu His
100 105 110
Thr Lys Asn Val Arg Asp Trp Lys Glu Val Phe Asp Leu Val Pro Arg
115 120 125
Gln Thr Pro Pro Pro Pro Thr Thr Ala Val Ala Asp Gly Asp Leu Val
130 135 140
Phe Asp Asn Lys Trp Pro Asp Asp Leu Pro Gly Phe Arg Glu Ala Met
145 150 155 160
Glu Glu Tyr Gly Glu Ala Val Glu Glu Leu Ala Phe Lys Leu Leu Glu
165 170 175
Leu Ile Ala Arg Ser Leu Gly Leu Arg Pro Asp Arg Leu His Gly Phe
180 185 190
Phe Lys Asp Asp Gln Thr Thr Phe Ile Arg Leu Asn His Tyr Pro Pro
195 200 205
Cys Pro Ser Pro Asp Leu Ala Leu Gly Val Gly Arg His Lys Asp Ala
210 215 220
Gly Ala Leu Thr Val Leu Tyr Gln Asp Asp Val Gly Gly Leu Asp Val
225 230 235 240
Arg Arg Arg Ser Asp Gly Glu Trp Val Arg Val Arg Pro Val Pro His
245 250 255
Ser Phe Ile Ile Asn Val Gly Asp Ile Ile Gln Val Trp Ser Asn Asp
260 265 270
Arg Tyr Glu Ser Ala Glu His Arg Val Ala Val Asn Val Glu Lys Glu
275 280 285
Arg Phe Ser Ile Pro Phe Phe Phe Asn Pro Ala Gly His Thr Met Val
290 295 300
Glu Pro Leu Glu Glu Val Val Ser Asp Glu Ser Pro Ala Arg Tyr Asn
305 310 315 320
Pro Tyr Asn Trp Gly Glu Phe Phe Ser Thr Arg Lys Asn Ser Asn Phe
325 330 335
Lys Lys Leu Asp Val Glu Asn Val Gln Ile Thr His Phe Arg Lys Asn
340 345 350
<210> 113
<211> 2027
<212> ДНК
<213> Oryza sativa
<400> 113
aagccacacg cacacacaca cacacgctga cacacgagac gaacacttgt gctacagctt 60
ctcgccacca gctactgatc gaccatgggc ggcctctcca tggaccaggc gttcgtgcag 120
gcccccgagc accgccccaa ggcgtccgtc gccgaggccg acggcatccc ggtcatcgac 180
ctctcccctc tcctcgccgc cggcgatggc gacgccgacg gggtggacgc gctcgcggcg 240
gaggtcggga gggcgagccg ggactggggc ttcttcgtgg tggtgcgcca cggtgtgccc 300
gcggaggcgg tggcgcgcgc ggcggaggcg cagaggacgt tcttcgcgct gccgccggag 360
cggagggcgg ccgtggcgcg gagcgaggcg gcgccgatgg ggtactacgc gtccgagcac 420
accaagaacg tcagggactg gaaggaggtg ttcgacctcg tcccgcgcca gacgccgccg 480
ccgccgacga ccgccgtggc cgacggcgac ctggtgttcg acaacaagtg gcccgacgac 540
ctgccgggat tcaggtcagg tcaccacatc gatcgatcgt cttcttcatc ctcgcatcaa 600
ttcagttcaa cctcatcgaa ttcttgagca gggaggcaat ggaggagtac ggcgaagcgg 660
tggaggagct ggcgttcaag ctgctggagc tgatcgccag gagcctcggc ctgagacccg 720
accgcctcca tggcttcttc aaggacgacc agaccacctt catccggctc aaccactacc 780
ctccctgccc gagccccgac ctcgccctcg gcgtcggccg ccacaaggac gccggcgcgc 840
tcaccgtgct ctaccaggac gatgtcggcg gcctcgacgt ccgccgccga tccgacggcg 900
agtgggtgcg cgtcaggccc gtccctcact ccttcatcat caacgtcggc gacatcatcc 960
aggtactttt ttttttgagc agctacatat ttatcaacaa attttcttct aacaatttat 1020
cggacataaa tatattacaa tgaaagaata attgtatcat aacttgtgtg tccttatatg 1080
taagttttag aaatcctata gtaacatggt attttcgcga aagcggagat tgtgagaccg 1140
tatcttttca cccatgcgcg tcatatgatt tttttttctt gccaacttaa ataaatttca 1200
aagtaaatct aatagattaa aattatgtga aacttacata taagttttct acggtaacac 1260
gctattttca cgaaacggag gtcgttccaa gttgaatgaa tcttgaagta aatctaacga 1320
tttaaaatta tgtgcataca cgttatatta cagttatata caagttataa tataattaca 1380
ctacaattat aacggtattc atagttgaca aacttttaaa agagaattag ttaataaata 1440
tataacaaca ttgtagttta attgttacta tttgacatca tttttatttg cattttgaat 1500
ttgactgaaa aaattgagag tgcgcttgtc caggtgtgga gcaatgacag gtacgagagc 1560
gcggagcacc gggtggcggt gaacgtggag aaggagaggt tctccatccc tttcttcttc 1620
aacccggcgg gccacaccat ggtggagcca ctggaggagg tcgtgagcga cgagagcccg 1680
gccaggtaca acccctacaa ctggggcgaa ttcttcagca ccaggaagaa cagcaacttc 1740
aagaagctgg acgtggagaa cgtccagatc acgcatttca ggaagaatta acgcgccggc 1800
tagatcatgt tcagtaaatt ttcagatgat gatgcgtgga caaccatata gcctttgcgt 1860
cataagttaa taatgtctgt gacagtatat catgtaaaca atcgtatgat gtggcttctc 1920
tatctgccgg tgatggtaat gtgacattgt agaagagggt ttgtgagata cttccttcac 1980
ttaactttta cgaatgaata tagacaacca caacatcctt gtcgtga 2027
<210> 114
<211> 1747
<212> ДНК
<213> Triticum aestivum
<400> 114
tcactcaagg ccacaacaca ctcgccagtc catcgccacc atacgtgaca acttgagtta 60
cttgatctgt tgctcatcga tctcgacatc gccatgggcg gcctctccat ggaccaggcc 120
ttcgtgcagg cccccgagca tcgcaccaag gcgaacctcg ccgacgcggc cggcatcccg 180
gtcatcgacc tctcccctct cgccgccggc gacaaggccg gcctggacgc cctcgcggcc 240
gaggtgggca gggcgagccg tgactggggg ttcttcgtgg tggtgcgcca cggcgtgccg 300
gcggagacgg tggcgcgggc gctggaggcg cagagggcct tcttcgcgct gcccgcggac 360
cggaaggcgg ccgtgcggag ggacgaggcg gcgccgctgg ggtactacga gtcggagcac 420
accaagaacg tcagggactg gaaggaggtg ttcgacctcg tcccccgcga gccgccgccg 480
cctgccgcgg ttgccgacgg cgagctcatg ttcgagaaca agtggcccga ggacctgccg 540
gggttcagag aggctctcga agagtacgag aaagcgatgg aggagctggc gttcaagctg 600
ctggagctga tcgcccggag cctgggactg agaccggacc ggctgcacgg cttcttcaag 660
gaccagacca ccttcatccg gctgaaccac tacccgccct gccccagccc cgacctcgcc 720
ctcggcgtcg gtcgccacaa ggacgccggc gcgctcacca tcctctacca ggacgacgtc 780
ggcgggctcg acgtccggcg ccgctccgac ggcgagtggg tgcgcgtcag gcctgtcccg 840
gactcctacg tcatcaacgt cggcgacatc atccaggtgt ggagcaacga caggtacgag 900
agcgcggagc acagggtgtc ggtgaactcg cacaaggaga ggttctccat gccctacttc 960
ttcgaccccg ggagcgacgc catgatcgag ccgttggagg agatggtgag cgacgaaagg 1020
ccggccaggt acgacgccta caactggggc aacttcttca gcaccaggaa gaacagcaac 1080
ttcaggaagc tcgccgtcga aaacgtccag atcgcacact tcagaaagga ccgaccttaa 1140
atgaaggatc cctcatgaat tcatgatcct tccgctctcc tcagtgatcc tagtgctaca 1200
actacaagca tctccccgtt tgtagtaatc atatataaat aagtattccc tccgtaaact 1260
aatataagag catttaaaac actactctag tgatctaaat gctcttatat tagtttacag 1320
agagagtatt gtgtattaat aatgactttc tctgtttcaa aataagtgat gacgtggttt 1380
tagttcaatt ttttttagag aggaggcatc tgacgggcct taaactgagg accttagagt 1440
acaaacaagg ttcgacgaaa gtaagtttaa gggatacaag gccgtagcca acaaaacgcg 1500
acgcagcgcg caatctaaaa tcagcgtgct gtcaaggtag ctggagacgt ccatgccgtt 1560
aatctctctc aagaagctcg ccgaagctca gtgcaccttg cgtgcactct tgtgaagagc 1620
accttcacgt gtcctttgtc ctgagatttt gtcaacagtt tccatgactg caagaaaaac 1680
actagtttgt ataatagctc agcgggatgt cgaatgaatt gcccctcaat caaagcttta 1740
tttctag 1747
<210> 115
<211> 1047
<212> ДНК
<213> Triticum aestivum
<400> 115
atgggcggcc tctccatgga ccaggccttc gtgcaggccc ccgagcatcg caccaaggcg 60
aacctcgccg acgcggccgg catcccggtc atcgacctct cccctctcgc cgccggcgac 120
aaggccggcc tggacgccct cgcggccgag gtgggcaggg cgagccgtga ctgggggttc 180
ttcgtggtgg tgcgccacgg cgtgccggcg gagacggtgg cgcgggcgct ggaggcgcag 240
agggccttct tcgcgctgcc cgcggaccgg aaggcggccg tgcggaggga cgaggcggcg 300
ccgctggggt actacgagtc ggagcacacc aagaacgtca gggactggaa ggaggtgttc 360
gacctcgtcc cccgcgagcc gccgccgcct gccgcggttg ccgacggcga gctcatgttc 420
gagaacaagt ggcccgagga cctgccgggg ttcagagagg ctctcgaaga gtacgagaaa 480
gcgatggagg agctggcgtt caagctgctg gagctgatcg cccggagcct gggactgaga 540
ccggaccggc tgcacggctt cttcaaggac cagaccacct tcatccggct gaaccactac 600
ccgccctgcc ccagccccga cctcgccctc ggcgtcggtc gccacaagga cgccggcgcg 660
ctcaccatcc tctaccagga cgacgtcggc gggctcgacg tccggcgccg ctccgacggc 720
gagtgggtgc gcgtcaggcc tgtcccggac tcctacgtca tcaacgtcgg cgacatcatc 780
caggtgtgga gcaacgacag gtacgagagc gcggagcaca gggtgtcggt gaactcgcac 840
aaggagaggt tctccatgcc ctacttcttc gaccccggga gcgacgccat gatcgagccg 900
ttggaggaga tggtgagcga cgaaaggccg gccaggtacg acgcctacaa ctggggcaac 960
ttcttcagca ccaggaagaa cagcaacttc aggaagctcg ccgtcgaaaa cgtccagatc 1020
gcacacttca gaaaggaccg accttaa 1047
<210> 116
<211> 348
<212> ПРТ
<213> Triticum aestivum
<400> 116
Met Gly Gly Leu Ser Met Asp Gln Ala Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Thr Lys Ala Asn Leu Ala Asp Ala Ala Gly Ile Pro Val Ile Asp
20 25 30
Leu Ser Pro Leu Ala Ala Gly Asp Lys Ala Gly Leu Asp Ala Leu Ala
35 40 45
Ala Glu Val Gly Arg Ala Ser Arg Asp Trp Gly Phe Phe Val Val Val
50 55 60
Arg His Gly Val Pro Ala Glu Thr Val Ala Arg Ala Leu Glu Ala Gln
65 70 75 80
Arg Ala Phe Phe Ala Leu Pro Ala Asp Arg Lys Ala Ala Val Arg Arg
85 90 95
Asp Glu Ala Ala Pro Leu Gly Tyr Tyr Glu Ser Glu His Thr Lys Asn
100 105 110
Val Arg Asp Trp Lys Glu Val Phe Asp Leu Val Pro Arg Glu Pro Pro
115 120 125
Pro Pro Ala Ala Val Ala Asp Gly Glu Leu Met Phe Glu Asn Lys Trp
130 135 140
Pro Glu Asp Leu Pro Gly Phe Arg Glu Ala Leu Glu Glu Tyr Glu Lys
145 150 155 160
Ala Met Glu Glu Leu Ala Phe Lys Leu Leu Glu Leu Ile Ala Arg Ser
165 170 175
Leu Gly Leu Arg Pro Asp Arg Leu His Gly Phe Phe Lys Asp Gln Thr
180 185 190
Thr Phe Ile Arg Leu Asn His Tyr Pro Pro Cys Pro Ser Pro Asp Leu
195 200 205
Ala Leu Gly Val Gly Arg His Lys Asp Ala Gly Ala Leu Thr Ile Leu
210 215 220
Tyr Gln Asp Asp Val Gly Gly Leu Asp Val Arg Arg Arg Ser Asp Gly
225 230 235 240
Glu Trp Val Arg Val Arg Pro Val Pro Asp Ser Tyr Val Ile Asn Val
245 250 255
Gly Asp Ile Ile Gln Val Trp Ser Asn Asp Arg Tyr Glu Ser Ala Glu
260 265 270
His Arg Val Ser Val Asn Ser His Lys Glu Arg Phe Ser Met Pro Tyr
275 280 285
Phe Phe Asp Pro Gly Ser Asp Ala Met Ile Glu Pro Leu Glu Glu Met
290 295 300
Val Ser Asp Glu Arg Pro Ala Arg Tyr Asp Ala Tyr Asn Trp Gly Asn
305 310 315 320
Phe Phe Ser Thr Arg Lys Asn Ser Asn Phe Arg Lys Leu Ala Val Glu
325 330 335
Asn Val Gln Ile Ala His Phe Arg Lys Asp Arg Pro
340 345
<210> 117
<211> 1863
<212> ДНК
<213> Triticum aestivum
<400> 117
tcactcaagg ccacaacaca ctcgccagtc catcgccacc atacgtgaca acttgagtta 60
cttgatctgt tgctcatcga tctcgacatc gccatgggcg gcctctccat ggaccaggcc 120
ttcgtgcagg cccccgagca tcgcaccaag gcgaacctcg ccgacgcggc cggcatcccg 180
gtcatcgacc tctcccctct cgccgccggc gacaaggccg gcctggacgc cctcgcggcc 240
gaggtgggca gggcgagccg tgactggggg ttcttcgtgg tggtgcgcca cggcgtgccg 300
gcggagacgg tggcgcgggc gctggaggcg cagagggcct tcttcgcgct gcccgcggac 360
cggaaggcgg ccgtgcggag ggacgaggcg gcgccgctgg ggtactacga gtcggagcac 420
accaagaacg tcagggactg gaaggaggtg ttcgacctcg tcccccgcga gccgccgccg 480
cctgccgcgg ttgccgacgg cgagctcatg ttcgagaaca agtggcccga ggacctgccg 540
gggttcaggt acggtcatca actcaatcaa ttctgcgacc ccgagagaaa tggttcacta 600
ttattcgtgg ttcatacgta tgattcagac gttaatctcg atgcaaattg atttgtgcat 660
gcagagaggc tctcgaagag tacgagaaag cgatggagga gctggcgttc aagctgctgg 720
agctgatcgc ccggagcctg ggactgagac cggaccggct gcacggcttc ttcaaggacc 780
agaccacctt catccggctg aaccactacc cgccctgccc cagccccgac ctcgccctcg 840
gcgtcggtcg ccacaaggac gccggcgcgc tcaccatcct ctaccaggac gacgtcggcg 900
ggctcgacgt ccggcgccgc tccgacggcg agtgggtgcg cgtcaggcct gtcccggact 960
cctacgtcat caacgtcggc gacatcatcc aggtgtggag caacgacagg tacgagagcg 1020
cggagcacag ggtgtcggtg aactcgcaca aggagaggtt ctccatgccc tacttcttcg 1080
accccgggag cgacgccatg atcgagccgt tggaggagat ggtgagcgac gaaaggccgg 1140
ccaggtacga cgcctacaac tggggcaact tcttcagcac caggaagaac agcaacttca 1200
ggaagctcgc cgtcgaaaac gtccagatcg cacacttcag aaaggaccga ccttaaatga 1260
aggatccctc atgaattcat gatccttccg ctctcctcag tgatcctagt gctacaacta 1320
caagcatctc cccgtttgta gtaatcatat ataaataagt attccctccg taaactaata 1380
taagagcatt taaaacacta ctctagtgat ctaaatgctc ttatattagt ttacagagag 1440
agtattgtgt attaataatg actttctctg tttcaaaata agtgatgacg tggttttagt 1500
tcaatttttt ttagagagga ggcatctgac gggccttaaa ctgaggacct tagagtacaa 1560
acaaggttcg acgaaagtaa gtttaaggga tacaaggccg tagccaacaa aacgcgacgc 1620
agcgcgcaat ctaaaatcag cgtgctgtca aggtagctgg agacgtccat gccgttaatc 1680
tctctcaaga agctcgccga agctcagtgc accttgcgtg cactcttgtg aagagcacct 1740
tcacgtgtcc tttgtcctga gattttgtca acagtttcca tgactgcaag aaaaacacta 1800
gtttgtataa tagctcagcg ggatgtcgaa tgaattgccc ctcaatcaaa gctttatttc 1860
tag 1863
<210> 118
<211> 349
<212> ПРТ
<213> Hordeum vulgare
<400> 118
Met Gly Gly Leu Ser Met Gly Gln Ala Phe Val Gln Ala Pro Glu His
1. 5 10 15
Arg Thr Lys Pro Thr Leu Ala Asp Ala Asp Gly Ile Pro Val Ile Asp
20 25 30
Leu Ser Pro Leu Ala Ala Gly Asp Glu Ala Gly Val Asp Ala Leu Ala
35 40 45
Ala Glu Val Gly Arg Ala Ser Arg Asp Trp Gly Phe Phe Val Val Val
50 55 60
Arg His Gly Val Pro Ala Glu Thr Val Ala Arg Ala Leu Glu Ala Gln
65 70 75 80
Arg Ala Phe Phe Ala Leu Pro Ala Glu Arg Lys Ala Ala Val Arg Arg
85 90 95
Asp Glu Ala Ala Pro Leu Gly Tyr Tyr Glu Ser Glu His Thr Lys Asn
100 105 110
Val Arg Asp Trp Lys Glu Val Phe Asp Phe Val Pro Arg Glu Pro Pro
115 120 125
Pro Pro Ala Ala Val Ala Asp Gly Glu Leu Val Phe Glu Asn Lys Trp
130 135 140
Pro Glu Asp Leu Pro Gly Phe Arg Val Ala Phe Glu Glu Tyr Ala Lys
145 150 155 160
Ala Met Glu Glu Leu Ala Phe Lys Leu Leu Glu Leu Ile Ala Arg Ser
165 170 175
Leu Gly Leu Thr Pro Asp Arg Leu Asn Gly Phe Phe Lys Asp His Gln
180 185 190
Thr Thr Phe Ile Arg Leu Asn His Tyr Pro Pro Cys Pro Ser Pro Asp
195 200 205
Leu Ala Leu Gly Val Gly Arg His Lys Asp Ala Gly Ala Leu Thr Val
210 215 220
Leu Tyr Gln Asp Asp Val Gly Gly Leu Asp Val Arg His Arg Ser Asp
225 230 235 240
Gly Glu Trp Val Arg Val Arg Pro Val Pro Asp Ser Tyr Val Ile Asn
245 250 255
Val Gly Asp Ile Ile Gln Val Trp Ser Asn Asp Arg Tyr Glu Ser Ala
260 265 270
Glu His Arg Val Ser Val Asn Ser Asp Lys Glu Arg Phe Ser Met Pro
275 280 285
Tyr Phe Phe Asn Pro Gly Ser Asp Ala Met Val Glu Pro Leu Glu Glu
290 295 300
Met Val Ser Asp Glu Arg Pro Ala Arg Tyr Asp Ala Tyr Asn Trp Gly
305 310 315 320
His Phe Phe Ser Thr Arg Lys Asn Ser Asn Phe Lys Lys Leu Asp Val
325 330 335
Glu Asn Val Gln Ile Ala His Phe Arg Lys Leu His Leu
340 345
<210> 119
<211> 1963
<212> ДНК
<213> Sorghum bicolor
<400> 119
tataaatacc acgccatgta cttctctgct tctacacttc tccagcttct ctcatgccat 60
accactagtg caaggtccta gatttacact tggtgctaca gcttcttcct ccctccctcc 120
cctctctagg cagctagcac gcagcgcagc acacgaaaca tctattgacc ggccgcctcc 180
gccggggatc cataattact atactaccaa tcggccagcg tcatgccgac gccgtcgcac 240
ctcgcgaacc cgcgctactt cgacttccgt gcggcgcggc gggtgccgga gacgcacgcc 300
tggccggggc tgcacgacca ccccgtcgtg gacggcggcg cgccggggcc agacgccgtc 360
cccgtggtgg acctcgcggg ggcggcggac gagccgagag ccgcggtggt ggcccaagtg 420
gcgcgcgccg ccgagcaatg gggcgcgttc ctgctcacgg ggcacggcgt ccccgcggag 480
ctgctggcgc gcgtcgagga ccggatcgcc accatgttcg cgctgccagc ggacgacaag 540
atgcgcgccg tgcgcgggcc tggcgacgcc tgcggctacg gctccccgcc catctcctcc 600
ttcttctcca agtgcatgtg gtcggaggga tacaccttct cgccggccaa cctccgcgcc 660
gacctccgca agctctggcc taaggccggc gacgactaca ccagcttctg tgatgtgatg 720
gaggagttcc acaagcacat gcgtgccctc gcggacaagc tgctggagct gttcctcatg 780
gcgctggggc tcaccgacga gcaggtcggc ggcgtggagg cggagcggag gatcgccgag 840
acgatgaccg ccaccatgca cctcaactgg taccctcggt gcccggaccc gcgccgcgcg 900
ctggggctga tcgcgcacac cgactcgggc ttcttcacct tcgtgctgca gagcctcgtc 960
ccggggctgc agctcttccg ccacgccccg gaccggtggg tggcggtgcc ggcggtaccg 1020
ggcgccttcg tcgtcaacgt gggcgacctc ttccacatcc tcaccaacgg ccggttccac 1080
agcgtgtacc accgcgccgt cgtgaaccgg gacctcgaca ggatatctct cggctacttc 1140
ctcggcccgc cgccgcacgc caaggtggcg ccgctaaggg aggccgtgcc gcccggccgc 1200
acccccgcgt accgcgccgt cacgtggccc gagtacatgg gcgtccgcaa gaaggccttc 1260
accaccggcg catccgcgct caagatggtc gccctcgccg ccgccgccgc cgccgccgac 1320
ctcgacgatg acgccggtgc tggcgccgcc gccgaacctg tcgtccatca gcagctactc 1380
gtctcgtcgt agccgatcga tcgccggatc ggtcgagact gatgatgatg atgcatatat 1440
actcgtcgat ggagtagaca gactaatcaa gcaaccctga aactatgaat gcatgcgtgc 1500
gcttcgtgct tgcttgcgca tgcagctagc aggcttcatt ccgttccgca gctgctctgc 1560
tccaacctgc tctgctggat tgatgtatat ggtagaagaa ttaagagatc gatggatgac 1620
ggaggaagaa gaagacgaag acgacgatga ggaaaaggac acgctgtacg tagctggttc 1680
ttctagtcta gtttacagca ggccgggcgg ccggctgctg cttccaatcg agtttgtcgt 1740
tactgacgat tgttagtgga tcgattaact aatctggaat tctggattat taatataatg 1800
catgtggttt ggcatctggc gtaaagcagg taatggtacc tagccagtag ccagtagcca 1860
ggctggtcaa tgataggtct ataccctgat cctgtactgt tgtttctttc ggtctttctg 1920
agagagaaaa aaaacgaata tatggcgtac tcaattcatc aaa 1963
<210> 120
<211> 1170
<212> ДНК
<213> Sorghum bicolor
<400> 120
atgccgacgc cgtcgcacct cgcgaacccg cgctacttcg acttccgtgc ggcgcggcgg 60
gtgccggaga cgcacgcctg gccggggctg cacgaccacc ccgtcgtgga cggcggcgcg 120
ccggggccag acgccgtccc cgtggtggac ctcgcggggg cggcggacga gccgagagcc 180
gcggtggtgg cccaagtggc gcgcgccgcc gagcaatggg gcgcgttcct gctcacgggg 240
cacggcgtcc ccgcggagct gctggcgcgc gtcgaggacc ggatcgccac catgttcgcg 300
ctgccagcgg acgacaagat gcgcgccgtg cgcgggcctg gcgacgcctg cggctacggc 360
tccccgccca tctcctcctt cttctccaag tgcatgtggt cggagggata caccttctcg 420
ccggccaacc tccgcgccga cctccgcaag ctctggccta aggccggcga cgactacacc 480
agcttctgtg atgtgatgga ggagttccac aagcacatgc gtgccctcgc ggacaagctg 540
ctggagctgt tcctcatggc gctggggctc accgacgagc aggtcggcgg cgtggaggcg 600
gagcggagga tcgccgagac gatgaccgcc accatgcacc tcaactggta ccctcggtgc 660
ccggacccgc gccgcgcgct ggggctgatc gcgcacaccg actcgggctt cttcaccttc 720
gtgctgcaga gcctcgtccc ggggctgcag ctcttccgcc acgccccgga ccggtgggtg 780
gcggtgccgg cggtaccggg cgccttcgtc gtcaacgtgg gcgacctctt ccacatcctc 840
accaacggcc ggttccacag cgtgtaccac cgcgccgtcg tgaaccggga cctcgacagg 900
atatctctcg gctacttcct cggcccgccg ccgcacgcca aggtggcgcc gctaagggag 960
gccgtgccgc ccggccgcac ccccgcgtac cgcgccgtca cgtggcccga gtacatgggc 1020
gtccgcaaga aggccttcac caccggcgca tccgcgctca agatggtcgc cctcgccgcc 1080
gccgccgccg ccgccgacct cgacgatgac gccggtgctg gcgccgccgc cgaacctgtc 1140
gtccatcagc agctactcgt ctcgtcgtag 1170
<210> 121
<211> 389
<212> ПРТ
<213> Sorghum bicolor
<400> 121
Met Pro Thr Pro Ser His Leu Ala Asn Pro Arg Tyr Phe Asp Phe Arg
1. 5 10 15
Ala Ala Arg Arg Val Pro Glu Thr His Ala Trp Pro Gly Leu His Asp
20 25 30
His Pro Val Val Asp Gly Gly Ala Pro Gly Pro Asp Ala Val Pro Val
35 40 45
Val Asp Leu Ala Gly Ala Ala Asp Glu Pro Arg Ala Ala Val Val Ala
50 55 60
Gln Val Ala Arg Ala Ala Glu Gln Trp Gly Ala Phe Leu Leu Thr Gly
65 70 75 80
His Gly Val Pro Ala Glu Leu Leu Ala Arg Val Glu Asp Arg Ile Ala
85 90 95
Thr Met Phe Ala Leu Pro Ala Asp Asp Lys Met Arg Ala Val Arg Gly
100 105 110
Pro Gly Asp Ala Cys Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe
115 120 125
Ser Lys Cys Met Trp Ser Glu Gly Tyr Thr Phe Ser Pro Ala Asn Leu
130 135 140
Arg Ala Asp Leu Arg Lys Leu Trp Pro Lys Ala Gly Asp Asp Tyr Thr
145 150 155 160
Ser Phe Cys Asp Val Met Glu Glu Phe His Lys His Met Arg Ala Leu
165 170 175
Ala Asp Lys Leu Leu Glu Leu Phe Leu Met Ala Leu Gly Leu Thr Asp
180 185 190
Glu Gln Val Gly Gly Val Glu Ala Glu Arg Arg Ile Ala Glu Thr Met
195 200 205
Thr Ala Thr Met His Leu Asn Trp Tyr Pro Arg Cys Pro Asp Pro Arg
210 215 220
Arg Ala Leu Gly Leu Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe
225 230 235 240
Val Leu Gln Ser Leu Val Pro Gly Leu Gln Leu Phe Arg His Ala Pro
245 250 255
Asp Arg Trp Val Ala Val Pro Ala Val Pro Gly Ala Phe Val Val Asn
260 265 270
Val Gly Asp Leu Phe His Ile Leu Thr Asn Gly Arg Phe His Ser Val
275 280 285
Tyr His Arg Ala Val Val Asn Arg Asp Leu Asp Arg Ile Ser Leu Gly
290 295 300
Tyr Phe Leu Gly Pro Pro Pro His Ala Lys Val Ala Pro Leu Arg Glu
305 310 315 320
Ala Val Pro Pro Gly Arg Thr Pro Ala Tyr Arg Ala Val Thr Trp Pro
325 330 335
Glu Tyr Met Gly Val Arg Lys Lys Ala Phe Thr Thr Gly Ala Ser Ala
340 345 350
Leu Lys Met Val Ala Leu Ala Ala Ala Ala Ala Ala Ala Asp Leu Asp
355 360 365
Asp Asp Ala Gly Ala Gly Ala Ala Ala Glu Pro Val Val His Gln Gln
370 375 380
Leu Leu Val Ser Ser
385
<210> 122
<211> 2321
<212> ДНК
<213> Sorghum bicolor
<400> 122
tataaatacc acgccatgta cttctctgct tctacacttc tccagcttct ctcatgccat 60
accactagtg caaggtccta gatttacact tggtgctaca gcttcttcct ccctccctcc 120
cctctctagg cagctagcac gcagcgcagc acacgaaaca tctattgacc ggccgcctcc 180
gccggggatc cataattact atactaccaa tcggccagcg tcatgccgac gccgtcgcac 240
ctcgcgaacc cgcgctactt cgacttccgt gcggcgcggc gggtgccgga gacgcacgcc 300
tggccggggc tgcacgacca ccccgtcgtg gacggcggcg cgccggggcc agacgccgtc 360
cccgtggtgg acctcgcggg ggcggcggac gagccgagag ccgcggtggt ggcccaagtg 420
gcgcgcgccg ccgagcaatg gggcgcgttc ctgctcacgg ggcacggcgt ccccgcggag 480
ctgctggcgc gcgtcgagga ccggatcgcc accatgttcg cgctgccagc ggacgacaag 540
atgcgcgccg tgcgcgggcc tggcgacgcc tgcggctacg gctccccgcc catctcctcc 600
ttcttctcca agtgcatgtg gtcggaggga tacaccttct cgccggccaa cctccgcgcc 660
gacctccgca agctctggcc taaggccggc gacgactaca ccagcttctg gtacgtgcac 720
ccgccggccg cgcgccgcca cacaccgtac ccacacacgt gcgcgctcgc gcctagctac 780
tagtagctgc tttgctttgc ttacctttga ttctcgcctt tgccatgcat atgcatgatg 840
cacgtacagg tactgcaggt acaacatgtc acacgcacgc acgcacgcac aacccatagt 900
ccgatacgat acatcatcga tcgacgtgtc gtcaccgtct aaggccatgc atgcatgcaa 960
gcacacgcct agaccttttt agcatgctgg ctgacgagga gtatactagc taataagcta 1020
cttgtcactg cgcgtcttgc ttaattacac tagtgcatat ttctacagtg atgtgatgga 1080
ggagttccac aagcacatgc gtgccctcgc ggacaagctg ctggagctgt tcctcatggc 1140
gctggggctc accgacgagc aggtcggcgg cgtggaggcg gagcggagga tcgccgagac 1200
gatgaccgcc accatgcacc tcaactggta ccctcggtgc ccggacccgc gccgcgcgct 1260
ggggctgatc gcgcacaccg actcgggctt cttcaccttc gtgctgcaga gcctcgtccc 1320
ggggctgcag ctcttccgcc acgccccgga ccggtgggtg gcggtgccgg cggtaccggg 1380
cgccttcgtc gtcaacgtgg gcgacctctt ccacatcctc accaacggcc ggttccacag 1440
cgtgtaccac cgcgccgtcg tgaaccggga cctcgacagg atatctctcg gctacttcct 1500
cggcccgccg ccgcacgcca aggtggcgcc gctaagggag gccgtgccgc ccggccgcac 1560
ccccgcgtac cgcgccgtca cgtggcccga gtacatgggc gtccgcaaga aggccttcac 1620
caccggcgca tccgcgctca agatggtcgc cctcgccgcc gccgccgccg ccgccgacct 1680
cgacgatgac gccggtgctg gcgccgccgc cgaacctgtc gtccatcagc agctactcgt 1740
ctcgtcgtag ccgatcgatc gccggatcgg tcgagactga tgatgatgat gcatatatac 1800
tcgtcgatgg agtagacaga ctaatcaagc aaccctgaaa ctatgaatgc atgcgtgcgc 1860
ttcgtgcttg cttgcgcatg cagctagcag gcttcattcc gttccgcagc tgctctgctc 1920
caacctgctc tgctggattg atgtatatgg tagaagaatt aagagatcga tggatgacgg 1980
aggaagaaga agacgaagac gacgatgagg aaaaggacac gctgtacgta gctggttctt 2040
ctagtctagt ttacagcagg ccgggcggcc ggctgctgct tccaatcgag tttgtcgtta 2100
ctgacgattg ttagtggatc gattaactaa tctggaattc tggattatta atataatgca 2160
tgtggtttgg catctggcgt aaagcaggta atggtaccta gccagtagcc agtagccagg 2220
ctggtcaatg ataggtctat accctgatcc tgtactgttg tttctttcgg tctttctgag 2280
agagaaaaaa aacgaatata tggcgtactc aattcatcaa a 2321
<210> 123
<211> 1796
<212> ДНК
<213> Setaria italica
<400> 123
actagtgcaa ggtcctagat ttacacttgg tgcttgcttg tttcttccta gttgctactg 60
gtagcacgca gtggctggct ggccgtaatc tattgtctgg gctcgatcgg tgattaggaa 120
gtagccaaag caagctaagg ccgccgccgc cgccgccatg ccgacgccgt cgcacctcaa 180
gaacccgctc tacttcgact tccgcgccgc gcggcgggtg ccggagtccc acgcctggcc 240
ggggctcgac gaccaccccg tggtggacgg cggcggcgcg ccggggtccc cggacgccgt 300
gccggtggtg gacctgcgcg agccgggcgc cgcggcggtg gcccgcgtgg cgcgcgccgc 360
cgagcagtgg ggcgcgttcc tgctcaccgg ccacggcgtc cccgcggagc tcctggcgcg 420
cgtcgaggac cgcgtcgcgt gcatgttcgc gctgccggcc gccgacaaga tgcgcgccgt 480
gcgcgggccg ggggacgcct gcggctacgg ctcgccgccc atctcctcct tcttctccaa 540
gtgcatgtgg tccgagggct acaccttctc gccggcctcc ctccgccgcg acctccgcaa 600
gctctggccc aaggccggcg acgactacga cagcttctgt gacgtgatgg aggagttcca 660
caaggagatg cgcgccctcg ccgacaggct cctggagctg ttcctcaggg cgctcgggct 720
caccggcgag caggtcggcg ccgtcgaggc ggagcggagg atcggcgaga cgatgaccgc 780
caccatgcac ctcaactggt atccgaggtg cccggacccg cggcgcgcgc tggggctgat 840
cgcgcacacg gactcgggct tcttcacctt cgtgctgcag agcctcgtgc cggggctgca 900
gctgttccgg cacggcccca accggtgggt ggcggtgccg gccgtgccgg gcgccttcgt 960
cgtcaacgtc ggcgacctct tccacatcct cacgaacggc cgcttccaca gcgtgtacca 1020
ccgcgccgtc gtcaaccggg acctcgaccg gatatcgctc ggctacttcc tcggcccgcc 1080
gccccacgcc aaggtggcgc cgctccggga ggtcgtgccg ccgggccggg cccccgccta 1140
ccgcgccgtc acgtggcccg agtacatggg cgtccgcaag aaggccttca ccaccggcgc 1200
ctccgcgctc aagatggtcg ccgccgccgc cgccgccacc gaatccgacg acaccgacgc 1260
agccgccgcc gccgttcacc agccgccggt cgtcgtctca tcgtagccga tcgatcgccg 1320
gaaacacaga cgatgcatac cgtaccccga gcaatctaat caaaacaagg catccattct 1380
cgcgcgcatg cagcggccag ccgggcttcc gcagctgctc ggcctcctct gctggctgtg 1440
gaaatggaaa attttaatct gagatgaaga cgaagacgaa gacgaaacgg agaggaaaag 1500
gacatgctgt agctgtttct tctagttgcg caggccgctc ccagtcgagt ttgtcgttac 1560
tgacgattat tactctgatg aaaactaatc tgaattaatg catgtagttt ggcaatttgg 1620
tactaaaggt aggcacctag ccaggctggt caatgatagg tctataacct gatcctgttc 1680
tctgttgttt tcctttgtct gagaaaaaat ggaaataatt gatccggccg gacgggtgta 1740
ctgataggtg atgctgaatt gctgatgcaa gaggttgcga gctgcagtga gcagca 1796
<210> 124
<211> 1149
<212> ДНК
<213> Setaria italica
<400> 124
atgccgacgc cgtcgcacct caagaacccg ctctacttcg acttccgcgc cgcgcggcgg 60
gtgccggagt cccacgcctg gccggggctc gacgaccacc ccgtggtgga cggcggcggc 120
gcgccggggt ccccggacgc cgtgccggtg gtggacctgc gcgagccggg cgccgcggcg 180
gtggcccgcg tggcgcgcgc cgccgagcag tggggcgcgt tcctgctcac cggccacggc 240
gtccccgcgg agctcctggc gcgcgtcgag gaccgcgtcg cgtgcatgtt cgcgctgccg 300
gccgccgaca agatgcgcgc cgtgcgcggg ccgggggacg cctgcggcta cggctcgccg 360
cccatctcct ccttcttctc caagtgcatg tggtccgagg gctacacctt ctcgccggcc 420
tccctccgcc gcgacctccg caagctctgg cccaaggccg gcgacgacta cgacagcttc 480
tgtgacgtga tggaggagtt ccacaaggag atgcgcgccc tcgccgacag gctcctggag 540
ctgttcctca gggcgctcgg gctcaccggc gagcaggtcg gcgccgtcga ggcggagcgg 600
aggatcggcg agacgatgac cgccaccatg cacctcaact ggtatccgag gtgcccggac 660
ccgcggcgcg cgctggggct gatcgcgcac acggactcgg gcttcttcac cttcgtgctg 720
cagagcctcg tgccggggct gcagctgttc cggcacggcc ccaaccggtg ggtggcggtg 780
ccggccgtgc cgggcgcctt cgtcgtcaac gtcggcgacc tcttccacat cctcacgaac 840
ggccgcttcc acagcgtgta ccaccgcgcc gtcgtcaacc gggacctcga ccggatatcg 900
ctcggctact tcctcggccc gccgccccac gccaaggtgg cgccgctccg ggaggtcgtg 960
ccgccgggcc gggcccccgc ctaccgcgcc gtcacgtggc ccgagtacat gggcgtccgc 1020
aagaaggcct tcaccaccgg cgcctccgcg ctcaagatgg tcgccgccgc cgccgccgcc 1080
accgaatccg acgacaccga cgcagccgcc gccgccgttc accagccgcc ggtcgtcgtc 1140
tcatcgtag 1149
<210> 125
<211> 382
<212> ПРТ
<213> Setaria italica
<400> 125
Met Pro Thr Pro Ser His Leu Lys Asn Pro Leu Tyr Phe Asp Phe Arg
1. 5 10 15
Ala Ala Arg Arg Val Pro Glu Ser His Ala Trp Pro Gly Leu Asp Asp
20 25 30
His Pro Val Val Asp Gly Gly Gly Ala Pro Gly Ser Pro Asp Ala Val
35 40 45
Pro Val Val Asp Leu Arg Glu Pro Gly Ala Ala Ala Val Ala Arg Val
50 55 60
Ala Arg Ala Ala Glu Gln Trp Gly Ala Phe Leu Leu Thr Gly His Gly
65 70 75 80
Val Pro Ala Glu Leu Leu Ala Arg Val Glu Asp Arg Val Ala Cys Met
85 90 95
Phe Ala Leu Pro Ala Ala Asp Lys Met Arg Ala Val Arg Gly Pro Gly
100 105 110
Asp Ala Cys Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe Ser Lys
115 120 125
Cys Met Trp Ser Glu Gly Tyr Thr Phe Ser Pro Ala Ser Leu Arg Arg
130 135 140
Asp Leu Arg Lys Leu Trp Pro Lys Ala Gly Asp Asp Tyr Asp Ser Phe
145 150 155 160
Cys Asp Val Met Glu Glu Phe His Lys Glu Met Arg Ala Leu Ala Asp
165 170 175
Arg Leu Leu Glu Leu Phe Leu Arg Ala Leu Gly Leu Thr Gly Glu Gln
180 185 190
Val Gly Ala Val Glu Ala Glu Arg Arg Ile Gly Glu Thr Met Thr Ala
195 200 205
Thr Met His Leu Asn Trp Tyr Pro Arg Cys Pro Asp Pro Arg Arg Ala
210 215 220
Leu Gly Leu Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe Val Leu
225 230 235 240
Gln Ser Leu Val Pro Gly Leu Gln Leu Phe Arg His Gly Pro Asn Arg
245 250 255
Trp Val Ala Val Pro Ala Val Pro Gly Ala Phe Val Val Asn Val Gly
260 265 270
Asp Leu Phe His Ile Leu Thr Asn Gly Arg Phe His Ser Val Tyr His
275 280 285
Arg Ala Val Val Asn Arg Asp Leu Asp Arg Ile Ser Leu Gly Tyr Phe
290 295 300
Leu Gly Pro Pro Pro His Ala Lys Val Ala Pro Leu Arg Glu Val Val
305 310 315 320
Pro Pro Gly Arg Ala Pro Ala Tyr Arg Ala Val Thr Trp Pro Glu Tyr
325 330 335
Met Gly Val Arg Lys Lys Ala Phe Thr Thr Gly Ala Ser Ala Leu Lys
340 345 350
Met Val Ala Ala Ala Ala Ala Ala Thr Glu Ser Asp Asp Thr Asp Ala
355 360 365
Ala Ala Ala Ala Val His Gln Pro Pro Val Val Val Ser Ser
370 375 380
<210> 126
<211> 2146
<212> ДНК
<213> Setaria italica
<400> 126
actagtgcaa ggtcctagat ttacacttgg tgcttgcttg tttcttccta gttgctactg 60
gtagcacgca gtggctggct ggccgtaatc tattgtctgg gctcgatcgg tgattaggaa 120
gtagccaaag caagctaagg ccgccgccgc cgccgccatg ccgacgccgt cgcacctcaa 180
gaacccgctc tacttcgact tccgcgccgc gcggcgggtg ccggagtccc acgcctggcc 240
ggggctcgac gaccaccccg tggtggacgg cggcggcgcg ccggggtccc cggacgccgt 300
gccggtggtg gacctgcgcg agccgggcgc cgcggcggtg gcccgcgtgg cgcgcgccgc 360
cgagcagtgg ggcgcgttcc tgctcaccgg ccacggcgtc cccgcggagc tcctggcgcg 420
cgtcgaggac cgcgtcgcgt gcatgttcgc gctgccggcc gccgacaaga tgcgcgccgt 480
gcgcgggccg ggggacgcct gcggctacgg ctcgccgccc atctcctcct tcttctccaa 540
gtgcatgtgg tccgagggct acaccttctc gccggcctcc ctccgccgcg acctccgcaa 600
gctctggccc aaggccggcg acgactacga cagcttctgg tacgtcgtcg tctatagcta 660
gtagctagcc gccggcacac gtgcgcctga cctgctccgc catgcatggt gcacgtatgc 720
agatcgatca cacgcaccga tcgatcgacg tgtcccggtc aaggccatgc atgcatgcaa 780
gcaaccaaca gcacgcctcc tgatactgct tgttgcttac accgttggta tgtgcctgtt 840
gcctacagtg acgtgatgga ggagttccac aaggagatgc gcgccctcgc cgacaggctc 900
ctggagctgt tcctcagggc gctcgggctc accggcgagc aggtcggcgc cgtcgaggcg 960
gagcggagga tcggcgagac gatgaccgcc accatgcacc tcaactggta tgtgccatgc 1020
catgaccacc tgcgtctatg aactaacgga agcttccatc gcgtgtccat gacgatttag 1080
aagctgtagt ccagagcttg agacaaacga aacgaagctt acatggtggc gtgacgtgtc 1140
gcgtgcaggt atccgaggtg cccggacccg cggcgcgcgc tggggctgat cgcgcacacg 1200
gactcgggct tcttcacctt cgtgctgcag agcctcgtgc cggggctgca gctgttccgg 1260
cacggcccca accggtgggt ggcggtgccg gccgtgccgg gcgccttcgt cgtcaacgtc 1320
ggcgacctct tccacatcct cacgaacggc cgcttccaca gcgtgtacca ccgcgccgtc 1380
gtcaaccggg acctcgaccg gatatcgctc ggctacttcc tcggcccgcc gccccacgcc 1440
aaggtggcgc cgctccggga ggtcgtgccg ccgggccggg cccccgccta ccgcgccgtc 1500
acgtggcccg agtacatggg cgtccgcaag aaggccttca ccaccggcgc ctccgcgctc 1560
aagatggtcg ccgccgccgc cgccgccacc gaatccgacg acaccgacgc agccgccgcc 1620
gccgttcacc agccgccggt cgtcgtctca tcgtagccga tcgatcgccg gaaacacaga 1680
cgatgcatac cgtaccccga gcaatctaat caaaacaagg catccattct cgcgcgcatg 1740
cagcggccag ccgggcttcc gcagctgctc ggcctcctct gctggctgtg gaaatggaaa 1800
attttaatct gagatgaaga cgaagacgaa gacgaaacgg agaggaaaag gacatgctgt 1860
agctgtttct tctagttgcg caggccgctc ccagtcgagt ttgtcgttac tgacgattat 1920
tactctgatg aaaactaatc tgaattaatg catgtagttt ggcaatttgg tactaaaggt 1980
aggcacctag ccaggctggt caatgatagg tctataacct gatcctgttc tctgttgttt 2040
tcctttgtct gagaaaaaat ggaaataatt gatccggccg gacgggtgta ctgataggtg 2100
atgctgaatt gctgatgcaa gaggttgcga gctgcagtga gcagca 2146
<210> 127
<211> 1933
<212> ДНК
<213> Oryza sativa
<400> 127
actactcatt ccactattgt aaagtcatag aaaaaattta tatagagaga aaaaattagt 60
gttgttattg ttactggctt tctgccagac gagacgagcg agcgcgcgag tgtgttgctc 120
tctggtcatc gtcgtcgtcg tcgcgatgcc gacgccgtcg cacttgaaga acccgctctg 180
cttcgacttc cgggcggcga ggcgggtgcc ggagacgcac gcgtggccgg ggctggacga 240
ccacccggtg gtggacggcg gcggcggcgg cggcgaggac gcggtgccgg tggtggacgt 300
cggggcgggc gacgcggcgg cgcgggtggc gcgggcggcg gagcagtggg gcgcgttcct 360
tctggtcggg cacggcgtgc cggcggcgct gctgtcgcgc gtcgaggagc gcgtcgcccg 420
cgtgttctcc ctgccggcgt cggagaagat gcgcgccgtc cgcggccccg gcgagccctg 480
cggctacggc tcgccgccca tctcctcctt cttctccaag ctcatgtggt ccgagggcta 540
caccttctcc ccttcctccc tccgctccga gctccgccgc ctctggccca agtccggcga 600
cgactacctc ctcttctgtg acgtgatgga ggagtttcac aaggagatgc ggcggctagc 660
cgacgagttg ctgaggttgt tcttgagggc gctggggctc accggcgagg aggtcgccgg 720
agtcgaggcg gagaggagga tcggcgagag gatgacggcg acggtgcacc tcaactggta 780
cccgaggtgc ccggagccgc ggcgagcgct ggggctcatc gcgcacacgg actcgggctt 840
cttcaccttc gtgctccaga gcctcgtccc ggggctgcag ctgttccgtc gagggcccga 900
ccggtgggtg gcggtgccgg cggtggcggg ggccttcgtc gtcaacgtcg gcgacctctt 960
ccacatcctc accaacggcc gcttccacag cgtctaccac cgcgccgtcg tgaaccgcga 1020
ccgcgaccgg gtctcgctcg gctacttcct cggcccgccg ccggacgccg aggtggcgcc 1080
gctgccggag gccgtgccgg ccggccggag ccccgcctac cgcgctgtca cgtggccgga 1140
gtacatggcc gtccgcaaga aggccttcgc caccggcggc tccgccctca agatggtctc 1200
caccgacgcc gccgccgccg ccgacgaaca cgacgacgtc gccgccgccg ccgacgtcca 1260
cgcataagct atagctacta gctacctcga tctcacgcaa aaaaaaaaag aaacaattaa 1320
tagagcaaaa aaaaaaagaa gagaaaatgg tggtacttgt gtttaaggtt tcctccatgc 1380
aaaatggttt gcatgcatgc atgcaaagct agcatctgca gctgcaagaa ttacaagagc 1440
agagaagcag acagctagat ggagataatt aattaattaa ttaatctaat taagcatgca 1500
ataattaaga ttattattct gatttcagaa ctgaaaaaaa aagtgtggtt aattaattat 1560
tggttaggct taattttatc tagatgtaga aaaagaatca agatcttcaa gcaagagaga 1620
agaggatcga agaagaagga aaagaaaacg aaaaggacat gctgtgttgt ctcttctagt 1680
tgtaccctgg ctgctgatta agtgctttgt tttgttgctg caagcttgtc gttactgatt 1740
attagttagt tatgcatcta attgattaaa ctaatctgtt tggcattttg gctcgagcta 1800
agctatagcc aggctggtca atgataggaa cttgtacaat ttaagcaatt gaacctgatc 1860
ctgtactggc atgtatgtat atatgcaagt gatgagaacc actagctagt atagctagac 1920
atgtatttgt ata 1933
<210> 128
<211> 1122
<212> ДНК
<213> Oryza sativa
<400> 128
atgccgacgc cgtcgcactt gaagaacccg ctctgcttcg acttccgggc ggcgaggcgg 60
gtgccggaga cgcacgcgtg gccggggctg gacgaccacc cggtggtgga cggcggcggc 120
ggcggcggcg aggacgcggt gccggtggtg gacgtcgggg cgggcgacgc ggcggcgcgg 180
gtggcgcggg cggcggagca gtggggcgcg ttccttctgg tcgggcacgg cgtgccggcg 240
gcgctgctgt cgcgcgtcga ggagcgcgtc gcccgcgtgt tctccctgcc ggcgtcggag 300
aagatgcgcg ccgtccgcgg ccccggcgag ccctgcggct acggctcgcc gcccatctcc 360
tccttcttct ccaagctcat gtggtccgag ggctacacct tctccccttc ctccctccgc 420
tccgagctcc gccgcctctg gcccaagtcc ggcgacgact acctcctctt ctgtgacgtg 480
atggaggagt ttcacaagga gatgcggcgg ctagccgacg agttgctgag gttgttcttg 540
agggcgctgg ggctcaccgg cgaggaggtc gccggagtcg aggcggagag gaggatcggc 600
gagaggatga cggcgacggt gcacctcaac tggtacccga ggtgcccgga gccgcggcga 660
gcgctggggc tcatcgcgca cacggactcg ggcttcttca ccttcgtgct ccagagcctc 720
gtcccggggc tgcagctgtt ccgtcgaggg cccgaccggt gggtggcggt gccggcggtg 780
gcgggggcct tcgtcgtcaa cgtcggcgac ctcttccaca tcctcaccaa cggccgcttc 840
cacagcgtct accaccgcgc cgtcgtgaac cgcgaccgcg accgggtctc gctcggctac 900
ttcctcggcc cgccgccgga cgccgaggtg gcgccgctgc cggaggccgt gccggccggc 960
cggagccccg cctaccgcgc tgtcacgtgg ccggagtaca tggccgtccg caagaaggcc 1020
ttcgccaccg gcggctccgc cctcaagatg gtctccaccg acgccgccgc cgccgccgac 1080
gaacacgacg acgtcgccgc cgccgccgac gtccacgcat aa 1122
<210> 129
<211> 373
<212> ПРТ
<213> Oryza sativa
<400> 129
Met Pro Thr Pro Ser His Leu Lys Asn Pro Leu Cys Phe Asp Phe Arg
1. 5 10 15
Ala Ala Arg Arg Val Pro Glu Thr His Ala Trp Pro Gly Leu Asp Asp
20 25 30
His Pro Val Val Asp Gly Gly Gly Gly Gly Gly Glu Asp Ala Val Pro
35 40 45
Val Val Asp Val Gly Ala Gly Asp Ala Ala Ala Arg Val Ala Arg Ala
50 55 60
Ala Glu Gln Trp Gly Ala Phe Leu Leu Val Gly His Gly Val Pro Ala
65 70 75 80
Ala Leu Leu Ser Arg Val Glu Glu Arg Val Ala Arg Val Phe Ser Leu
85 90 95
Pro Ala Ser Glu Lys Met Arg Ala Val Arg Gly Pro Gly Glu Pro Cys
100 105 110
Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe Ser Lys Leu Met Trp
115 120 125
Ser Glu Gly Tyr Thr Phe Ser Pro Ser Ser Leu Arg Ser Glu Leu Arg
130 135 140
Arg Leu Trp Pro Lys Ser Gly Asp Asp Tyr Leu Leu Phe Cys Asp Val
145 150 155 160
Met Glu Glu Phe His Lys Glu Met Arg Arg Leu Ala Asp Glu Leu Leu
165 170 175
Arg Leu Phe Leu Arg Ala Leu Gly Leu Thr Gly Glu Glu Val Ala Gly
180 185 190
Val Glu Ala Glu Arg Arg Ile Gly Glu Arg Met Thr Ala Thr Val His
195 200 205
Leu Asn Trp Tyr Pro Arg Cys Pro Glu Pro Arg Arg Ala Leu Gly Leu
210 215 220
Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe Val Leu Gln Ser Leu
225 230 235 240
Val Pro Gly Leu Gln Leu Phe Arg Arg Gly Pro Asp Arg Trp Val Ala
245 250 255
Val Pro Ala Val Ala Gly Ala Phe Val Val Asn Val Gly Asp Leu Phe
260 265 270
His Ile Leu Thr Asn Gly Arg Phe His Ser Val Tyr His Arg Ala Val
275 280 285
Val Asn Arg Asp Arg Asp Arg Val Ser Leu Gly Tyr Phe Leu Gly Pro
290 295 300
Pro Pro Asp Ala Glu Val Ala Pro Leu Pro Glu Ala Val Pro Ala Gly
305 310 315 320
Arg Ser Pro Ala Tyr Arg Ala Val Thr Trp Pro Glu Tyr Met Ala Val
325 330 335
Arg Lys Lys Ala Phe Ala Thr Gly Gly Ser Ala Leu Lys Met Val Ser
340 345 350
Thr Asp Ala Ala Ala Ala Ala Asp Glu His Asp Asp Val Ala Ala Ala
355 360 365
Ala Asp Val His Ala
370
<210> 130
<211> 2040
<212> ДНК
<213> Oryza sativa
<400> 130
actactcatt ccactattgt aaagtcatag aaaaaattta tatagagaga aaaaattagt 60
gttgttattg ttactggctt tctgccagac gagacgagcg agcgcgcgag tgtgttgctc 120
tctggtcatc gtcgtcgtcg tcgcgatgcc gacgccgtcg cacttgaaga acccgctctg 180
cttcgacttc cgggcggcga ggcgggtgcc ggagacgcac gcgtggccgg ggctggacga 240
ccacccggtg gtggacggcg gcggcggcgg cggcgaggac gcggtgccgg tggtggacgt 300
cggggcgggc gacgcggcgg cgcgggtggc gcgggcggcg gagcagtggg gcgcgttcct 360
tctggtcggg cacggcgtgc cggcggcgct gctgtcgcgc gtcgaggagc gcgtcgcccg 420
cgtgttctcc ctgccggcgt cggagaagat gcgcgccgtc cgcggccccg gcgagccctg 480
cggctacggc tcgccgccca tctcctcctt cttctccaag ctcatgtggt ccgagggcta 540
caccttctcc ccttcctccc tccgctccga gctccgccgc ctctggccca agtccggcga 600
cgactacctc ctcttctggt atatatacat atatactctc ccatgcattc catgcacata 660
cactctacgt atatatctac ctctacgtat atatctacgt attgatctac gtataatata 720
cgcagtgacg tgatggagga gtttcacaag gagatgcggc ggctagccga cgagttgctg 780
aggttgttct tgagggcgct ggggctcacc ggcgaggagg tcgccggagt cgaggcggag 840
aggaggatcg gcgagaggat gacggcgacg gtgcacctca actggtaccc gaggtgcccg 900
gagccgcggc gagcgctggg gctcatcgcg cacacggact cgggcttctt caccttcgtg 960
ctccagagcc tcgtcccggg gctgcagctg ttccgtcgag ggcccgaccg gtgggtggcg 1020
gtgccggcgg tggcgggggc cttcgtcgtc aacgtcggcg acctcttcca catcctcacc 1080
aacggccgct tccacagcgt ctaccaccgc gccgtcgtga accgcgaccg cgaccgggtc 1140
tcgctcggct acttcctcgg cccgccgccg gacgccgagg tggcgccgct gccggaggcc 1200
gtgccggccg gccggagccc cgcctaccgc gctgtcacgt ggccggagta catggccgtc 1260
cgcaagaagg ccttcgccac cggcggctcc gccctcaaga tggtctccac cgacgccgcc 1320
gccgccgccg acgaacacga cgacgtcgcc gccgccgccg acgtccacgc ataagctata 1380
gctactagct acctcgatct cacgcaaaaa aaaaaagaaa caattaatag agcaaaaaaa 1440
aaaagaagag aaaatggtgg tacttgtgtt taaggtttcc tccatgcaaa atggtttgca 1500
tgcatgcatg caaagctagc atctgcagct gcaagaatta caagagcaga gaagcagaca 1560
gctagatgga gataattaat taattaatta atctaattaa gcatgcaata attaagatta 1620
ttattctgat ttcagaactg aaaaaaaaag tgtggttaat taattattgg ttaggcttaa 1680
ttttatctag atgtagaaaa agaatcaaga tcttcaagca agagagaaga ggatcgaaga 1740
agaaggaaaa gaaaacgaaa aggacatgct gtgttgtctc ttctagttgt accctggctg 1800
ctgattaagt gctttgtttt gttgctgcaa gcttgtcgtt actgattatt agttagttat 1860
gcatctaatt gattaaacta atctgtttgg cattttggct cgagctaagc tatagccagg 1920
ctggtcaatg ataggaactt gtacaattta agcaattgaa cctgatcctg tactggcatg 1980
tatgtatata tgcaagtgat gagaaccact agctagtata gctagacatg tatttgtata 2040
<210> 131
<211> 1332
<212> ДНК
<213> Hordeum vulgare
<400> 131
acactcactc ctcaatccat ccgtctccac cattgctcgc tagctcgagc tctactagct 60
agcactgcaa agtcagccgg gccggagttg atttggtcct tgttagcttg accgatcgta 120
tacgtatcgc caggatgccg acgccgtcgc acctgagcaa ggacccgcac tacttcgact 180
tccgggcggc gcggcgggtg ccggagacac acgcgtggcc ggggctgcac gaccacccgg 240
tggtggacgg cggcggcgcg ggcggagggc cggacgcggt gccggtggtg gacatgcgcg 300
acccgtgcgc cgcggaggcg gtggcgctgg ccgcgcagga ctggggcgcc ttcctcttgc 360
agggccacgg cgtcccgttg gagctgctgg cccgcgtgga ggccgcgata gcgggcatgt 420
tcgcgctgcc ggcgtcggag aagatgcgcg ccgtgcggcg gcccggcgac tcgtgcggct 480
acgggtcgcc gcccatctcc tccttcttct ccaagtgcat gtggtccgag ggctacacct 540
tctccccggc caacctccgc tccgacctcc gcaagctctg gcccaaggcc ggccacgact 600
accgccactt ctgtgccgtg atggaggagt tccacaggga gatgcgcgtt ctggccgaca 660
agctgctgga gctgttcctg gtggccctcg ggctcaccgg cgagcaggtc gccgccgtcg 720
agtcggagca caagatcgcc gagaccatga ccgccacaat gcacctcaac tggtacccca 780
agtgcccgga cccgaagcga gcgctgggcc tgatcgcgca cacggactcg ggcttcttca 840
ccttcgtgct ccagagcctg gtgcccgggc tgcagctgtt ccggcacggc cccgaccgtt 900
gggtgacggt gcccgccgtg ccgggcgcca tggtcgtcaa cgtcggcgac ctcttccaca 960
tcctcaccaa tggccgcttc cacagcgtct accaccgcgc cgtcgtcaac cgcgacagcg 1020
accggatatc gctggggtac ttcctcggcc cgcccgccca cgttaaggtg gcgccgctca 1080
gggaggccct cgccggcacg cccgctgcct accgcgccgt cacgtggccc gagtacatgg 1140
gcgtgcgcaa gaaggccttc accaccggcg cctccgcgct caagatggtc gccatctcca 1200
ccgacgacgc cgccgacgtc ctccccgacg tcctctcgtc gtagatcggc gccggccatc 1260
acccggccgg ccaagagacc gatctataca aacaattagt gaacaaaaaa aaaaaaaaaa 1320
aaaaaaaaaa aa 1332
<210> 132
<211> 1110
<212> ДНК
<213> Hordeum vulgare
<400> 132
atgccgacgc cgtcgcacct gagcaaggac ccgcactact tcgacttccg ggcggcgcgg 60
cgggtgccgg agacacacgc gtggccgggg ctgcacgacc acccggtggt ggacggcggc 120
ggcgcgggcg gagggccgga cgcggtgccg gtggtggaca tgcgcgaccc gtgcgccgcg 180
gaggcggtgg cgctggccgc gcaggactgg ggcgccttcc tcttgcaggg ccacggcgtc 240
ccgttggagc tgctggcccg cgtggaggcc gcgatagcgg gcatgttcgc gctgccggcg 300
tcggagaaga tgcgcgccgt gcggcggccc ggcgactcgt gcggctacgg gtcgccgccc 360
atctcctcct tcttctccaa gtgcatgtgg tccgagggct acaccttctc cccggccaac 420
ctccgctccg acctccgcaa gctctggccc aaggccggcc acgactaccg ccacttctgt 480
gccgtgatgg aggagttcca cagggagatg cgcgttctgg ccgacaagct gctggagctg 540
ttcctggtgg ccctcgggct caccggcgag caggtcgccg ccgtcgagtc ggagcacaag 600
atcgccgaga ccatgaccgc cacaatgcac ctcaactggt accccaagtg cccggacccg 660
aagcgagcgc tgggcctgat cgcgcacacg gactcgggct tcttcacctt cgtgctccag 720
agcctggtgc ccgggctgca gctgttccgg cacggccccg accgttgggt gacggtgccc 780
gccgtgccgg gcgccatggt cgtcaacgtc ggcgacctct tccacatcct caccaatggc 840
cgcttccaca gcgtctacca ccgcgccgtc gtcaaccgcg acagcgaccg gatatcgctg 900
gggtacttcc tcggcccgcc cgcccacgtt aaggtggcgc cgctcaggga ggccctcgcc 960
ggcacgcccg ctgcctaccg cgccgtcacg tggcccgagt acatgggcgt gcgcaagaag 1020
gccttcacca ccggcgcctc cgcgctcaag atggtcgcca tctccaccga cgacgccgcc 1080
gacgtcctcc ccgacgtcct ctcgtcgtag 1110
<210> 133
<211> 369
<212> ПРТ
<213> Hordeum vulgare
<400> 133
Met Pro Thr Pro Ser His Leu Ser Lys Asp Pro His Tyr Phe Asp Phe
1. 5 10 15
Arg Ala Ala Arg Arg Val Pro Glu Thr His Ala Trp Pro Gly Leu His
20 25 30
Asp His Pro Val Val Asp Gly Gly Gly Ala Gly Gly Gly Pro Asp Ala
35 40 45
Val Pro Val Val Asp Met Arg Asp Pro Cys Ala Ala Glu Ala Val Ala
50 55 60
Leu Ala Ala Gln Asp Trp Gly Ala Phe Leu Leu Gln Gly His Gly Val
65 70 75 80
Pro Leu Glu Leu Leu Ala Arg Val Glu Ala Ala Ile Ala Gly Met Phe
85 90 95
Ala Leu Pro Ala Ser Glu Lys Met Arg Ala Val Arg Arg Pro Gly Asp
100 105 110
Ser Cys Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe Ser Lys Cys
115 120 125
Met Trp Ser Glu Gly Tyr Thr Phe Ser Pro Ala Asn Leu Arg Ser Asp
130 135 140
Leu Arg Lys Leu Trp Pro Lys Ala Gly His Asp Tyr Arg His Phe Cys
145 150 155 160
Ala Val Met Glu Glu Phe His Arg Glu Met Arg Val Leu Ala Asp Lys
165 170 175
Leu Leu Glu Leu Phe Leu Val Ala Leu Gly Leu Thr Gly Glu Gln Val
180 185 190
Ala Ala Val Glu Ser Glu His Lys Ile Ala Glu Thr Met Thr Ala Thr
195 200 205
Met His Leu Asn Trp Tyr Pro Lys Cys Pro Asp Pro Lys Arg Ala Leu
210 215 220
Gly Leu Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe Val Leu Gln
225 230 235 240
Ser Leu Val Pro Gly Leu Gln Leu Phe Arg His Gly Pro Asp Arg Trp
245 250 255
Val Thr Val Pro Ala Val Pro Gly Ala Met Val Val Asn Val Gly Asp
260 265 270
Leu Phe His Ile Leu Thr Asn Gly Arg Phe His Ser Val Tyr His Arg
275 280 285
Ala Val Val Asn Arg Asp Ser Asp Arg Ile Ser Leu Gly Tyr Phe Leu
290 295 300
Gly Pro Pro Ala His Val Lys Val Ala Pro Leu Arg Glu Ala Leu Ala
305 310 315 320
Gly Thr Pro Ala Ala Tyr Arg Ala Val Thr Trp Pro Glu Tyr Met Gly
325 330 335
Val Arg Lys Lys Ala Phe Thr Thr Gly Ala Ser Ala Leu Lys Met Val
340 345 350
Ala Ile Ser Thr Asp Asp Ala Ala Asp Val Leu Pro Asp Val Leu Ser
355 360 365
Ser
<210> 134
<211> 1653
<212> ДНК
<213> Triticum aestivum
<220>
<221> прочий_признак
<222> (1594)..(1600)
<223> n представляет собой a, c, g или t
<220>
<221> прочий_признак
<222> (1641)..(1641)
<223> n представляет собой a, c, g или t
<400> 134
cacgagatcc atccgtctcc accattgctc gctagctcga gctcctagct agtactgcaa 60
agtcagccgg ggagttgatt tggtccttct tggcttgacc gatcgtacgt gccgccagga 120
tgccgacgcc ggcgcacctg agcaaggacc cgcgctactt cgacttccgg gcggcgcggc 180
gggtgccgga gacgcacgcg tggcccgggc tgcacgacca ccccgtggtg gacggcagcg 240
gcgcgggcgg agggccggac gcggtgccgg tggtggacat gcgcgacccg tgcgcggcgg 300
aggcggtggc gctggcggcg caggactggg gcgccttcct cctggagggc cacggcgtcc 360
cgttggagct gctggcgcgc gtggaggccg cgatcgcggg catgttcgcg ctgccggcgt 420
cggagaagat gcgcgccgtg cggcggcccg gcgactcgtg cggctacggg tcgccgccca 480
tctcctcctt cttctccaag tgcatgtggt ccgagggcta caccttctcc ccggccaacc 540
tccgctccga cctccgcaag ctctggccca aggccggcca cgactaccgc cacttctgcg 600
ccgtgatgga ggagttccac agggagatgc gcgcgctggc cgacaagctg ctggagctgt 660
tcctggtggc cctcgggctc accggcgagc aggtcgccgc cgtcgagtcc gagcagaaga 720
tcgccgagac catgaccgcc acaatgcacc tcaactggta ccccaagtgc ccggacccga 780
agcgggcgct gggcctgatc gcgcacacgg actcgggctt cttcaccttc gtgctgcaga 840
gccttgtgcc cgggctgcag ctgttccggc acggccccga ccggtgggtg acggtgcccg 900
ccgtgccggg ggccatggtc gtcaacgtcg gcgacctctt ccagatcctc accaacggcc 960
gcttccacag cgtctaccac cgcgccgtcg tcaaccgcga cagcgaccgg atatcgctcg 1020
gctacttcct cggcccgccc gcccacgtca aggtggcgcc gctcagggag gccctggccg 1080
gcacgcccgc cgcctaccgc gccgtcacgt ggcccgagta catgggcgtg cgcaagaagg 1140
ccttcaccac cggcgcctcc gcgctcaaga tggtcgccat ctccactgac aacgacgccg 1200
ccaaccacac ggacgacctg atctcgtcgt agatcggcgc cggccatcac cggccggcca 1260
agggatcgat ctacacacac aattagtgaa caaaaaaatg ccagagatgg tgcatggtgg 1320
gctggtagct tagctgaggt agctaggagg aagagcgcgc gtgcggctgt cgttcgtgcg 1380
gctgttcccg caaaaaaaaa aaaggtttcc tccatatatg tctccatgca gaactgcaga 1440
tgctggtggt ggatgcgtcc atgcagcagg gaacgaacta attgtaagaa aatcaagcaa 1500
acttagttct acatctgtaa ttaagtatgc atgccacttg gtttaattca attcaagtgc 1560
agaaaaaatt atgatgggaa aaaaaaagac atgnnnnnnn aaaaaaaaaa aaaaaaaaaa 1620
aaaaaaaaaa aaaaaaaaaa naaaaaaaaa aaa 1653
<210> 135
<211> 1113
<212> ДНК
<213> Triticum aestivum
<400> 135
atgccgacgc cggcgcacct gagcaaggac ccgcgctact tcgacttccg ggcggcgcgg 60
cgggtgccgg agacgcacgc gtggcccggg ctgcacgacc accccgtggt ggacggcagc 120
ggcgcgggcg gagggccgga cgcggtgccg gtggtggaca tgcgcgaccc gtgcgcggcg 180
gaggcggtgg cgctggcggc gcaggactgg ggcgccttcc tcctggaggg ccacggcgtc 240
ccgttggagc tgctggcgcg cgtggaggcc gcgatcgcgg gcatgttcgc gctgccggcg 300
tcggagaaga tgcgcgccgt gcggcggccc ggcgactcgt gcggctacgg gtcgccgccc 360
atctcctcct tcttctccaa gtgcatgtgg tccgagggct acaccttctc cccggccaac 420
ctccgctccg acctccgcaa gctctggccc aaggccggcc acgactaccg ccacttctgc 480
gccgtgatgg aggagttcca cagggagatg cgcgcgctgg ccgacaagct gctggagctg 540
ttcctggtgg ccctcgggct caccggcgag caggtcgccg ccgtcgagtc cgagcagaag 600
atcgccgaga ccatgaccgc cacaatgcac ctcaactggt accccaagtg cccggacccg 660
aagcgggcgc tgggcctgat cgcgcacacg gactcgggct tcttcacctt cgtgctgcag 720
agccttgtgc ccgggctgca gctgttccgg cacggccccg accggtgggt gacggtgccc 780
gccgtgccgg gggccatggt cgtcaacgtc ggcgacctct tccagatcct caccaacggc 840
cgcttccaca gcgtctacca ccgcgccgtc gtcaaccgcg acagcgaccg gatatcgctc 900
ggctacttcc tcggcccgcc cgcccacgtc aaggtggcgc cgctcaggga ggccctggcc 960
ggcacgcccg ccgcctaccg cgccgtcacg tggcccgagt acatgggcgt gcgcaagaag 1020
gccttcacca ccggcgcctc cgcgctcaag atggtcgcca tctccactga caacgacgcc 1080
gccaaccaca cggacgacct gatctcgtcg tag 1113
<210> 136
<211> 370
<212> ПРТ
<213> Triticum aestivum
<400> 136
Met Pro Thr Pro Ala His Leu Ser Lys Asp Pro Arg Tyr Phe Asp Phe
1. 5 10 15
Arg Ala Ala Arg Arg Val Pro Glu Thr His Ala Trp Pro Gly Leu His
20 25 30
Asp His Pro Val Val Asp Gly Ser Gly Ala Gly Gly Gly Pro Asp Ala
35 40 45
Val Pro Val Val Asp Met Arg Asp Pro Cys Ala Ala Glu Ala Val Ala
50 55 60
Leu Ala Ala Gln Asp Trp Gly Ala Phe Leu Leu Glu Gly His Gly Val
65 70 75 80
Pro Leu Glu Leu Leu Ala Arg Val Glu Ala Ala Ile Ala Gly Met Phe
85 90 95
Ala Leu Pro Ala Ser Glu Lys Met Arg Ala Val Arg Arg Pro Gly Asp
100 105 110
Ser Cys Gly Tyr Gly Ser Pro Pro Ile Ser Ser Phe Phe Ser Lys Cys
115 120 125
Met Trp Ser Glu Gly Tyr Thr Phe Ser Pro Ala Asn Leu Arg Ser Asp
130 135 140
Leu Arg Lys Leu Trp Pro Lys Ala Gly His Asp Tyr Arg His Phe Cys
145 150 155 160
Ala Val Met Glu Glu Phe His Arg Glu Met Arg Ala Leu Ala Asp Lys
165 170 175
Leu Leu Glu Leu Phe Leu Val Ala Leu Gly Leu Thr Gly Glu Gln Val
180 185 190
Ala Ala Val Glu Ser Glu Gln Lys Ile Ala Glu Thr Met Thr Ala Thr
195 200 205
Met His Leu Asn Trp Tyr Pro Lys Cys Pro Asp Pro Lys Arg Ala Leu
210 215 220
Gly Leu Ile Ala His Thr Asp Ser Gly Phe Phe Thr Phe Val Leu Gln
225 230 235 240
Ser Leu Val Pro Gly Leu Gln Leu Phe Arg His Gly Pro Asp Arg Trp
245 250 255
Val Thr Val Pro Ala Val Pro Gly Ala Met Val Val Asn Val Gly Asp
260 265 270
Leu Phe Gln Ile Leu Thr Asn Gly Arg Phe His Ser Val Tyr His Arg
275 280 285
Ala Val Val Asn Arg Asp Ser Asp Arg Ile Ser Leu Gly Tyr Phe Leu
290 295 300
Gly Pro Pro Ala His Val Lys Val Ala Pro Leu Arg Glu Ala Leu Ala
305 310 315 320
Gly Thr Pro Ala Ala Tyr Arg Ala Val Thr Trp Pro Glu Tyr Met Gly
325 330 335
Val Arg Lys Lys Ala Phe Thr Thr Gly Ala Ser Ala Leu Lys Met Val
340 345 350
Ala Ile Ser Thr Asp Asn Asp Ala Ala Asn His Thr Asp Asp Leu Ile
355 360 365
Ser Ser
370
<210> 137
<211> 1884
<212> ДНК
<213> Triticum aestivum
<400> 137
tatatataca gctccttgta cttctctcgt tcttacactc actcctcaat ccatccgtct 60
ccaccattgc tcgctagctc gagctcctag ctagtactgc aaagtcagcc ggggagttga 120
tttggtcctt cttggcttga ccgatcgtac gtgccgccag gatgccgacg ccggcgcacc 180
tgagcaagga cccgcgctac ttcgacttcc gggcggcgcg gcgggtgccg gagacgcacg 240
cgtggcccgg gctgcacgac caccccgtgg tggacggcag cggcgcgggc ggagggccgg 300
acgcggtgcc ggtggtggac atgcgcgacc cgtgcgcggc ggaggcggtg gcgctggcgg 360
cgcaggactg gggcgccttc ctcctggagg gccacggcgt cccgttggag ctgctggcgc 420
gcgtggaggc cgcgatcgcg ggcatgttcg cgctgccggc gtcggagaag atgcgcgccg 480
tgcggcggcc cggcgactcg tgcggctacg ggtcgccgcc catctcctcc ttcttctcca 540
agtgcatgtg gtccgagggc tacaccttct ccccggccaa cctccgctcc gacctccgca 600
agctctggcc caaggccggc cacgactacc gccacttctg gtacgtacgc cggccgccga 660
tgcgcatata cacgtcatag tacggcacct acctaactgg ctctggccaa ccgtccgtac 720
acacgtgaag gggcgacgtg tccgactccg accatgcatg catgcacgcg cgcgaaactt 780
gttactcctg ttctgctatg gcagcagcta gccgcgtgtg tccgttcgta ggagtagtta 840
cttacacagt tacacttacg ccgtccgtcg tgttcctcga cgtgcagcgc cgtgatggag 900
gagttccaca gggagatgcg cgcgctggcc gacaagctgc tggagctgtt cctggtggcc 960
ctcgggctca ccggcgagca ggtcgccgcc gtcgagtccg agcagaagat cgccgagacc 1020
atgaccgcca caatgcacct caactggtac gttccactac tactccagta gtacaagtac 1080
aatatataga atacaaatgg cagcagccac gacgacacgt actccaccat gcagcaaagc 1140
atatattgtc ggtgcggcgg ttgacacgga gttgtgtcgt gtcgttgatt cacaggtacc 1200
ccaagtgccc ggacccgaag cgggcgctgg gcctgatcgc gcacacggac tcgggcttct 1260
tcaccttcgt gctgcagagc cttgtgcccg ggctgcagct gttccggcac ggccccgacc 1320
ggtgggtgac ggtgcccgcc gtgccggggg ccatggtcgt caacgtcggc gacctcttcc 1380
agatcctcac caacggccgc ttccacagcg tctaccaccg cgccgtcgtc aaccgcgaca 1440
gcgaccggat atcgctcggc tacttcctcg gcccgcccgc ccacgtcaag gtggcgccgc 1500
tcagggaggc cctggccggc acgcccgccg cctaccgcgc cgtcacgtgg cccgagtaca 1560
tgggcgtgcg caagaaggcc ttcaccaccg gcgcctccgc gctcaagatg gtcgccatct 1620
ccactgacaa cgacgccgcc aaccacacgg acgacctgat ctcgtcgtag atcggcgccg 1680
gccatcaccg gccggccaag ggatcgatct acacacacaa ttagtgaaca aaaaaatgcc 1740
agagatggtg catggtgggc tggtagctta gctgaggtag ctaggaggaa gagcgcgcgt 1800
gcggctgtcg ttcgtgcggc tgttcccgca aaaaaaaaaa ggtttcctcc atatakgtcc 1860
ccakscaaaa tsgmaawgct gggg 1884
<210> 138
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 138
acggguucuu ccaggugugc 20
<210> 139
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 139
cacggguucu uccaggugug 20
<210> 140
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 140
cauugaccuc cccgcuggca 20
<210> 141
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 141
ccagcgggga ggucaaugcu 20
<210> 142
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 142
cccagcauug accuccccgc 20
<210> 143
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 143
cgcgcucgug uacccggaca 20
<210> 144
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 144
cucccggcgc aggucgaaca 20
<210> 145
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 145
guguacccgg acacggugcc 20
<210> 146
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 146
ugcagggaag cuguccgggc 20
<210> 147
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 147
uucuuccagg ugugcgggca 20
<210> 148
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 148
agauccccgc gccauuccug 20
<210> 149
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 149
augcagggaa gcuguccggg 20
<210> 150
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 150
auuccugugg ccgcaggaag 20
<210> 151
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 151
cagcggggag gucaaugcug 20
<210> 152
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 152
caggaauggc gcggggaucu 20
<210> 153
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 153
gacuacuucg ucggcacccu 20
<210> 154
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 154
gccaggauuu cgagccaaug 20
<210> 155
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 155
ggaacauuug gagggaggcg 20
<210> 156
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 156
gggaggucaa ugcuggggcu 20
<210> 157
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 157
uuggcucgaa auccuggccg 20
<210> 158
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 158
acggguucuu ccaggugugc 20
<210> 159
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 159
cacggguucu uccaggugug 20
<210> 160
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 160
cauugaccuc cccgcuggca 20
<210> 161
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 161
ccagcgggga ggucaaugcu 20
<210> 162
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 162
cccagcauug accuccccgc 20
<210> 163
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 163
cgcgcucgug uacccggaca 20
<210> 164
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 164
cucccggcgc aggucgaaca 20
<210> 165
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 165
guguacccgg acacggugcc 20
<210> 166
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 166
ugcagggaag cuguccgggc 20
<210> 167
<211> 20
<212> РНК
<213> Искусственная Последовательность
<220>
<223> олигонуклеотид супрессии
<400> 167
uucuuccagg ugugcgggca 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ УВЕЛИЧЕНИЯ УРОЖАЙНОСТИ НИЗКОРОСЛЫХ РАСТЕНИЙ ПУТЕМ МАНИПУЛЯЦИИ МЕТАБОЛИЗМА ГИББЕРЕЛЛИНА | 2017 |
|
RU2837282C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ РЕДУЦИРУЮЩИХ САХАРОВ В РАСТЕНИИ | 2019 |
|
RU2801948C2 |
| СНИЖЕНИЕ ПРЕВРАЩЕНИЯ НИКОТИНА В НОРНИКОТИН В РАСТЕНИЯХ | 2015 |
|
RU2733837C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2817591C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2816526C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗМЕНЕНИЯ ЦВЕТЕНИЯ И АРХИТЕКТУРЫ РАСТЕНИЙ ДЛЯ УЛУЧШЕНИЯ ПОТЕНЦИАЛЬНОЙ УРОЖАЙНОСТИ | 2017 |
|
RU2775653C2 |
| КОМПОЗИЦИИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2015 |
|
RU2745322C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2740312C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| МОДУЛИРОВАНИЕ СОДЕРЖАНИЯ АМИНОКИСЛОТ В РАСТЕНИИ | 2019 |
|
RU2799785C2 |
Изобретение относится к области биохимии, в частности к конструкции рекомбинантной ДНК. Также раскрыты вектор для трансформации, трансгенное растение кукурузы, содержащие указанную конструкцию рекомбинантной ДНК. Раскрыт способ получения трансгенного злакового растения, с помощью указанной молекулы ДНК. Изобретение позволяет эффективно получать трансгенное растение, имеющее уменьшенную высоту растения по сравнению с контрольным растением дикого типа. 5 н. и 24 з.п. ф-лы, 17 табл., 18 пр.
1. Конструкция рекомбинантной ДНК для супрессии гена оксидазы GA в кукурузе, содержащая транскрибируемую последовательность ДНК, кодирующую некодирующую молекулу РНК, при этом некодирующая молекула РНК содержит нацеливающую последовательность, которая:
(a) по меньшей мере на 90% комплементарна по меньшей мере 21 последовательному нуклеотиду первой молекулы мРНК, кодирующей первый эндогенный белок оксидазы GA в растении кукурузы или клетке растения, где первый эндогенный белок оксидазы GA по меньшей мере на 90% идентичен SEQ ID NO: 9; и
(b) по меньшей мере на 90% комплементарна по меньшей мере 21 последовательному нуклеотиду второй молекулы мРНК, кодирующей второй эндогенный белок оксидазы GA в растении кукурузы или клетке растения, где второй эндогенный белок оксидазы GA по меньшей мере на 90% идентичен SEQ ID NO: 15;
где указанная транскрибируемая последовательность ДНК функционально связана с сосудистым промотором.
2. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что нацеливающая последовательность некодирующей молекулы РНК по меньшей мере на 90% комплементарна по меньшей мере 21 последовательному нуклеотиду последовательности, выбранной из группы, состоящей из SEQ ID NO: 7, 8, 13 и 14.
3. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что нацеливающая последовательность некодирующей молекулы РНК содержит последовательность, которая на 100% комплементарна по меньшей мере 21 последовательному нуклеотиду первой молекулы мРНК, кодирующей первый эндогенный белок оксидазы GA20.
4. Конструкция рекомбинантной ДНК по п. 3, отличающаяся тем, что нацеливающая последовательность некодирующей молекулы РНК содержит последовательность, которая на 100% комплементарна по меньшей мере 21 последовательному нуклеотиду SEQ ID NO: 7 или 8.
5. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что сосудистый промотор содержит один из следующих: промотор сахарозосинтазы, промотор транспортера сахарозы, промотор Sh1, промотор пятнистого вируса желтой коммелины (CoYMV - Commelina yellow mottle virus), промотор большого межгенного региона (LIR - large intergenic region) геминивируса карликовой пшеницы (WDV - wheat dwarf geminivirus), промотор белка оболочки (СР - coat protein) гемивируса полосы кукурузы (MSV - maize streak geminivirus), рисовый промотор, подобный желтой полосе 1 (YS1 - yellow stripe 1) или рисовый промотор желтой полосы 2 (OsYSL2).
6. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что сосудистый промотор содержит последовательность ДНК, которая является по меньшей мере на 90% идентичной одной или более из SEQ ID NO: 67, SEQ ID NO: 68, SEQ ID NO: 69, SEQ ID NO: 70 или SEQ ID NO: 71, или ее функциональной части.
7. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что промотор, экспрессируемый в растении, представляет собой промотор RTBV.
8. Конструкция рекомбинантной ДНК по п. 7, отличающаяся тем, что промотор, экспрессируемый в растении, содержит последовательность ДНК, которая является по меньшей мере на 90% идентичной одной или более из SEQ ID NO: 65 или SEQ ID NO: 66, или ее функциональной части.
9. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что некодирующая молекула РНК, кодируемая транскрибируемой последовательностью ДНК, представляет собой предшественника микроРНК или миРНК, который процессируется или расщепляется в растительной клетке с образованием зрелой микроРНК или миРНК.
10. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что нацеливающая последовательность некодирующей молекулы РНК содержит последовательность, которая на 100% комплементарна по меньшей мере 21 последовательному нуклеотиду второй молекулы мРНК, кодирующей второй эндогенный белок оксидазы GA20.
11. Конструкция рекомбинантной ДНК по п. 10, отличающаяся тем, что нацеливающая последовательность некодирующей молекулы РНК содержит последовательность, которая на 100% комплементарна по меньшей мере 21 последовательному нуклеотиду SEQ ID NO: 13 или SEQ ID NO: 14.
12. Вектор для трансформации, содержащий конструкцию рекомбинантной ДНК по п. 1.
13. Трансгенное растение кукурузы, имеющее уменьшенную высоту растения по сравнению с контрольным растением дикого типа, при этом трансгенное растение кукурузы содержит конструкцию рекомбинантной ДНК по п. 1.
14. Трансгенное растение кукурузы по п. 13, отличающееся тем, что трансгенное растение кукурузы имеет один или более из следующих признаков относительно контрольного растения: более короткую высоту растения, увеличенный диаметр стебля, улучшенную устойчивость к полеганию, уменьшенный излом стебля, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
15. Трансгенное растение кукурузы по п. 13, отличающееся тем, что указанное трансгенное растение кукурузы имеет более короткую высоту растения и/или улучшенную устойчивость к полеганию.
16. Трансгенное растение кукурузы по п. 13, отличающееся тем, что высота трансгенного растения кукурузы по меньшей мере на 10% ниже чем у контрольного растения дикого типа.
17. Трансгенное растение кукурузы по п. 13, отличающееся тем, что диаметр стебля трансгенного растения кукурузы в одном или более междоузлиях стебля по меньшей мере на 5% больше, чем диаметр стебля в одном или более междоузлиях контрольного растения дикого типа.
18. Трансгенное растение кукурузы по п. 13, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения кукурузы ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
19. Трансгенное растение кукурузы по п. 13, отличающееся тем, что уровень одной или более активной GA в по меньшей мере одной ткани междоузлия стебля трансгенного растения кукурузы по меньшей мере на 5% ниже, чем у той же ткани междоузлия контрольного растения дикого типа.
20. Трансгенное растение кукурузы по п. 13, отличающееся тем, что трансгенное растение кукурузы не имеет каких-либо значительных отклонений в по меньшей мере одном женском органе или початке.
21. Способ получения трансгенного злакового растения, включающий в себя: (а) трансформацию по меньшей мере одной клетки эксплантата конструкцией рекомбинантной ДНК по п. 1 и (b) регенерацию или развитие трансгенного злакового растения из трансформированного эксплантата.
22. Способ по п. 21, отличающийся тем, что злаковое растение трансформируется с помощью опосредованной Agrobacterium трансформации или бомбардировки частицами.
23. Модифицированное растение кукурузы, имеющее высоту растения менее чем 2000 мм, которое имеет (i) средний диаметр стебля более 18 мм, (ii) улучшение устойчивости к полеганию по сравнению с контрольным растением дикого типа, либо (iii) повышение устойчивости к засухе по сравнению с контрольным растением дикого типа, где модифицированное растение кукурузы содержит конструкцию рекомбинантной ДНК по п.1.
24. Модифицированное растение кукурузы по п. 23, отличающееся тем, что модифицированное растение кукурузы имеет один или более из следующих признаков относительно контрольного растения дикого типа: увеличенный диаметр стебля, улучшенную устойчивость к полеганию, уменьшенный излом стебля, более глубокие корни, увеличенную площадь листа, более раннюю сомкнутость полога, более высокую устьичную проводимость, более низкую высоту початка, повышенное содержание влаги в листве, повышение устойчивости к засухе, повышение эффективности использования азота, снижение содержания антоцианов и площади антоцианов в листьях в нормальных условиях или стрессовых условиях, вызванных ограниченным количеством азота или воды, увеличение массы початка, увеличение уборочного индекса, увеличение урожая, увеличение количества зерна, увеличение массы зерна и/или увеличение плодовитости.
25. Модифицированное растение кукурузы по п. 23, отличающееся тем, что уровень одной или более активных GA в по меньшей мере одной ткани междоузлия стебля кукурузного растения ниже, чем в той же ткани междоузлия контрольного растения дикого типа.
26. Модифицированное растение кукурузы, имеющее уменьшенную высоту растения по сравнению с контрольным растением дикого типа, которое имеет (i) увеличенный диаметр стебля по сравнению с контрольным растением дикого типа, (ii) улучшенную устойчивость к полеганию по сравнению с контрольным растением дикого типа, или (iii) улучшенную устойчивость к засухе по сравнению с контрольным растением дикого типа, где модифицированное растение кукурузы содержит конструкцию рекомбинантной ДНК по п.1.
27. Модифицированное растение кукурузы по п. 26, отличающееся тем, что уровень одной или более активных GA в стебле злакового растения ниже, чем в контрольном растении дикого типа.
28. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что некодирующая молекула РНК снижает уровни экспрессии первой и второй молекул мРНК по меньшей мере в одной ткани трансгенного растения кукурузы, содержащего конструкцию рекомбинантной ДНК, по сравнению с контрольным растением, когда некодирующая молекула РНК экспрессируется в трансгенном растении кукурузы.
29. Конструкция рекомбинантной ДНК по п. 1, отличающаяся тем, что некодирующая молекула РНК снижает уровни экспрессии первого и второго эндогенных белков оксидазы GA20 по меньшей мере в одной ткани трансгенного растения кукурузы, содержащего конструкцию рекомбинантной ДНК, по сравнению с контрольным растением, когда некодирующая молекула РНК экспрессируется в трансгенном растении кукурузы.
| WO 2011023537 A1, 03.03.2011 | |||
| RU 2013151447 A, 27.05.2015 | |||
| RU 2013135491 A, 10.02.2015. |

