Изобретение относится к области биотехнологии, в частности к генетической инженерии высших растений, и касается способа получения низконикотиновых нетрансгенных мутантных растений табака путем нокаута генов семейства путресцин-N-метилтрансферазы (РМТ).
Табак является перспективной культурой в качестве системы синтеза рекомбинантных белков для фармацевтики, промышленности и ветеринарии, а никотин является загрязняющим веществом для таких систем. Снижение содержания никотина в растениях табака является актуальной задачей, поскольку в настоящее время нарастает тренд на отказ от никотина в курительном табаке, что порождает спрос на низконикотиновые сорта.
Известен способ получения растений табака со сниженным содержанием никотина путем сайленсинга генов семейства путресцин-N-метилтрансферазы [1]. Недостатком способа является повышение содержания анатабина в листьях модифицированных растений.
Наиболее близким к заявленному способу - прототипом, является способ получения растений табака со сниженным содержанием никотина путем РНК-интерференции [2]. В качестве мишени для подавления РНК был выбран фрагмент длиной 540 п.н. Фрагменты ДНК лигировали в вектор pHANNIBAL в прямой и обратной ориентации под контролем промотора 35S вируса мозаики цветной капусты (CaMV) и терминатора октопинсинтазы (OCS). Экспрессионная кассета переносилась в вектор pCAMBIA1301, которая затем переносилась в штамм Agrobacterium tumefaciens ЕНА105, который затем использовали для трансформации листовых эксплантов табака.
К недостаткам РНК-интерференции методом агробактериальной трансформации относится получение регенерантов с очень широким спектром содержания никотина и нестабильностью супрессии.
Задачей настоящего изобретения является получение генетически-стабильных линий растений табака с нокаутом генов семейства путресцин-N-метилтрансферазы путем использования технологии геномного редактирования.
Технический результат: повышение эффективности способа получения табака со сниженным содержанием никотина за счет нокаута генов семейства РМТ, обеспечивающего получение стабильных линий.
Поставленная задача достигается предлагаемым способом, заключающимся в следующем.
Осуществляют подбор (дизайн) специфической направляющей РНК (нРНК), имеющей нуклеотидную последовательность SEQ ID NO 3.
Фрагмент ДНК, кодирующий нРНК, синтезируют с помощью гибридизации пары специфических F (прямой) и R (обратный) олигонуклеотидов: PMT_RGEN_F, имеющего последовательность SEQ ID NO 1 и PMT_RGEN_R, имеющего последовательность SEQ ID NO 2.
В табл. 1 (см. в графической части) представлена структура используемых олигонуклеотидов и направляющей РНК. В перечне последовательностей представлены нуклеотидные последовательности используемых олигонуклеотидов и направляющей РНК.
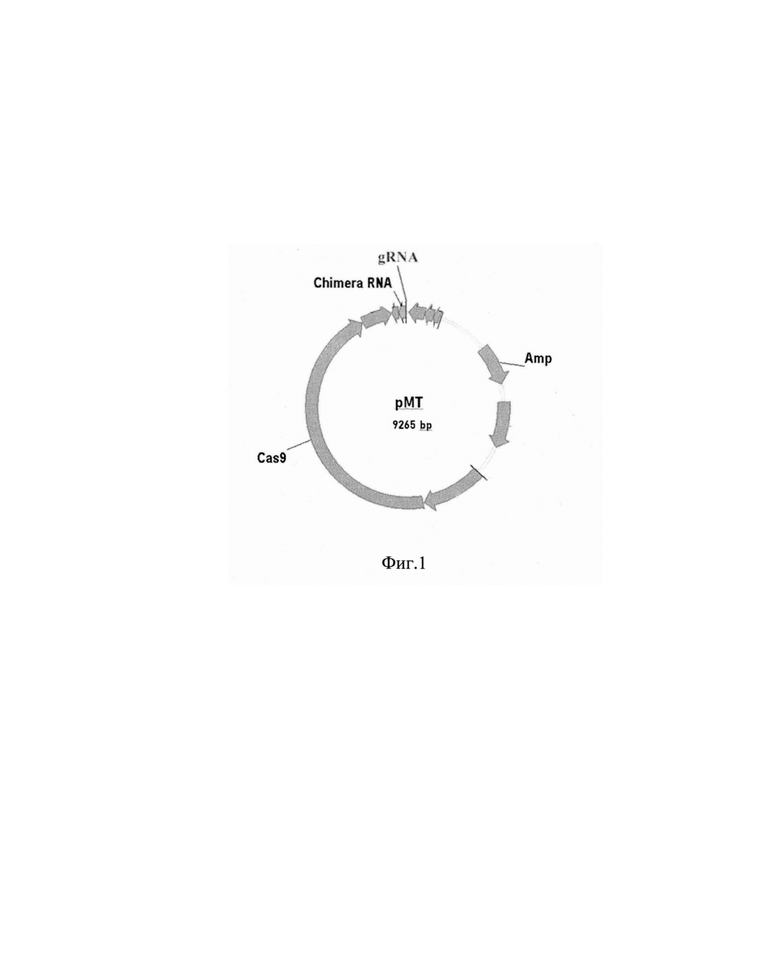
Далее конструируют рекомбинантную плазмидную ДНК (рМТ) путем клонирования фрагментов ДНК, кодирующих специфичную направляющую РНК (нРНК) в вектор, содержащий систему для редактирования генома, pSI57 [3]. В результате получают рекомбинантную плазмидную ДНК рМТ (размером 9265 п.н.), обеспечивающую мутагенез.
Плазмида состоит из следующих элементов (фиг. 1):
- фрагмента ДНК векторной плазмиды (pSI57) размером 9244 п.н.;
- фрагмента ДНК, содержащего последовательность направляющей РНК размером 21 п.н. (нРНК).
Созданную генетическую конструкцию используют для мутагенеза генов семейства путресцин-N-метилтрансферазы. Для этого смешивают полученную плазмиду рМТ с плазмидой pBi 121, несущей ген устойчивости к канамицину и ген-репортер β-глюкуронидазы (GUS) Е. coli. Далее иммобилизуют смесь плазмид на золотых частицах и проводят трансформацию листовых эксплантов табака (Nicotiana tabacum) путем бомбардировки золотыми частицами при помощи генной пушки. Получают растения-регенеранты путем селекции на средах с добавлением канамицина. Отбор трансформантов осуществляют при помощи метода гистохимического окрашивания с реактивом X-Gluc, а отбор мутантных растений-регенерантов осуществляют при помощи секвенирования по Сэнгеру [4].
Определяющими отличительными признаками заявляемого способа, по сравнению с прототипом, являются:
- вместо интерферирующей РНК для подавления функции генов путресцин-N-метилтрансферазы используют мутагенез методом геномного редактирования, что позволяет получить генетически-стабильные низконикотиновые растения табака.
- трансформацию растений табака осуществляют при помощи генной пушки (вместо агробактериальной трансформации), что позволяет использовать сразу несколько плазмид (рМТ для внесения направленных мутаций и pBi121 для осуществления селективного отбора растений-регенерантов);
- отбор растений-регенерантов осуществляют при помощи метода гистохимического окрашивания с реактивом X-Gluc, что позволяет быстро обнаружить функциональную встройку трансгена.
- конструкции для селективного отбора и конструкции для геномного редактирования собирают в разных плазмидах, что позволяет существенно снизить вероятность получения генетических химер, и развития нежелательных побочных эффектов за счет того, что при использовании данного метода, конструкции для геномного редактирования экспрессируются транзиентно.
Предлагаемый способ позволяет в поколениях от самоопыления получить нетрансгенные стабильные мутантные линии табака со стабильно сниженным содержанием никотина.
Изобретение поясняется следующими примерами конкретного выполнения способа.
Пример 1. Конструирование генетической конструкции рМТ.
Клонирование фрагментов ДНК, кодирующих сайт-специфические последовательности нРНК, в составе вектора pSI57 осуществляли в один этап посредством лигирования.
В конструкцию собирали два фрагмента: фрагмент ДНК, кодирующий специфичную нРНК, и фрагмент плазмиды pSI57, содержащий ген нуклеазы Cas9, ген каркаса нРНК и ген устойчивости к ампициллину. Полученную плазмиду обозначали как рМТ.
Для синтеза фрагментов ДНК, кодирующих специфичную нРНК, производили реакцию гибридизации пары F и R олигонуклеотидов, структура которых представлена в таблице 1.
Программа: Денатурация (95°С, 5 мин), отжиг (95-85°С - 2°С/сек, 85-25°С - 0,1°С/сек). Параллельно с гибридизацией олигонуклеотидов обрабатывали плазмиду pSI57 рестриктазой BstV2I (2 ч при температуре 55°С). Очищали набором для очистки ДНК от реакционной смеси (БиоСилика).
Следующим этапом проводили реакцию лигирования. Состав реакционной смеси: 50 нг порезанной и очищенной плазмиды рМТ, 10 пмоль гибридизованных олигонуклеотидов (см. Таблицу 1), 1 мкл 10х лигазного буфера (СибЭнзим), 0,5 мкл Т4 лигазы, вода (H2O) до V=10 мкл. Инкубировали смесь 3 ч при температуре 16°С.
Полученной лигазной смесью трансформировали хемокомпетентные клетки Е. coli DH10B стандартным способом [5].
Трансформированные бактерии выращивали в чашках Петри на агаризованной среде LB с добавлением антибиотика ампициллина (200 мкг/мл) в течение 12 ч при температуре 37°С. Далее полученные колонии переносили в жидкую среду LB (с ампициллином 200 мкг/мл) и выращивали ночную культуру в термостатируемом шейкере при температуре 37°С.
После выделения плазмиды рМТ из ночной культуры клеток проводили их скрининг на наличие встройки методом рестрикционного анализа. Для рестрикционного анализа проводили гидролиз исходного вектора и всех выделенных плазмид по сайтам рестрикции BstV2I и Sfr274I. После рестрикционного анализа проверяли наличие встройки секвенированием по Сэнгеру.
Пример 2. Получение нетрансгенных мутантных растений табака со сниженным содержанием никотина.
Полученную по примеру 1 конструкцию рМТ, несущую фрагмент, кодирующий направляющую РНК, которая нацеливает нуклеазу Cas9 на гены семейства путресцин-N-метилтрансферазы, использовали для трансформации растений табака (Nicotiana tabacum L. сорта Petit Havana линии SRI) с помощью генной пушки. Для биобаллистики брали крупные, здоровые листья растений табака из культуры in vitro без признаков некроза. Готовили смесь из двух генетических векторов, содержащую плазмиду рМТ и вектор pBi121, несущий ген устойчивости к канамицину и ген-репортер β-глюкуронидазы (GUS) Е. coli. Полученную смесь наносили на золотые частицы и доставляли в клетки эпидермиса нижней стороны листьев растений табака при помощи генной пушки (Bio-Rad PDS-1000 Не) при следующих условиях: размер золотых частиц 1 мкм, разрывной диск 1100 пси, расстояние 6 см, вакуум при выстреле 27 мм рт.ст.
Трансформированные экспланты инкубировали на каллусообразующей питательной среде (MS), содержащей 1 мг/л 6-бензиламинопурина (БАП) (Sigma-Aldrich) и 0,1 мг/л α-нафтилуксусной кислоты (НУК) (Sigma-Aldrich) с добавлением селектирующего агента канамицина (100 мг/л) до каллусообразования. Полученные каллусы перемещали на среду MS с добавлением 0,1 мг/л БАП без канамицина для органогенеза и регенерации, затем получали первичные растения-регенеранты. Полученные растения были пересажены на среду MS (Murashige and Skoog) без гормонов и проанализированы на наличие мутаций в целевых локусах генов семейства путресцин-N-метилтрансферазы. Трансгенность полученных каллусов и регенерантов проверяли методом гистохимического окрашивания с реактивом X-Gluc для выявления активности гена β-глюкуронидазы. Ткани каллусов и регенерантов помещали в раствор, содержащий 950 мкл фосфатно-солевого буфера (PBS) и 50 мкл X-Gluc и осуществляли инфильтрацию путем вакуумирования. Далее инкубировали образцы (ткани каллусов) в течение 24 ч при температуре 37°С. После инкубирования образцы промывали 70% этанолом 3-кратно и проводили визуальную оценку окрашивания тканей каллусов и регенерантов.
Было получено два органогенных каллуса, один из которых, обозначенный РМТ(1), дал начало популяции клонов-регенерантов, поддерживаемых in vitro. Данная линия была проанализирована методом ПЦР-секвенирования по Сэнгеру на предмет мутаций в генах РМТ, и показано присутствие мутаций, приводящих к нокауту генов РМТ в полученной линии. Клоны-регенеранты выращивали в гидропонном комплексе до формирования соцветий, созревания и достижения максимального размера. Полученные из семян mut PMT поколения Т1 выращивали in vitro и отбирали нетрансгенные растения. Нетрансгенные мутантные растения поколения Т1 были проанализированы на содержание никотина в листовой ткани методом высокоэффективной жидкостной хроматографии (ВЭЖХ).
Для измерения использовали растения, выращенные в условиях гидропонного комплекса, одного возраста при достижении максимального размера, с каждого растения отбирали созревшие листья на трех уровнях: первый уровень - 2-й лист снизу; второй уровень - средний лист на стебле и третий уровень - 2-й лист вниз от соцветия). Собранные листья растений табака мелко нарезали, удаляли жилки, помещали во флаконы и замораживали в жидком азоте. Высушивание листьев табака проводили в лиофильной сушилке ИНЕЙ-4, производительность 40 г воды/час, вакуум 7 Па, температура - 30°С. Время высушивания составляло от 6 до 16 часов. Лиофильно высушенное сырье измельчали, навеску (0.2÷0.5 г) заливали 0.1% HCl в 40% водном метаноле и экстрагировали на качалке с перемешиванием в течение 24 ч при температуре 25°С. Осадок отделяли центрифугированием, экстракт анализировали с использованием системы ВЭЖХ Agilent Infinity 1260. Результаты измерений приведены в таблице 2 (см. в графической части).
В качестве контрольных растений использовалась исходная линия табака Nicotiana tabacum L. сорта Petit Havana линии SRI.
Из представленных в таблице 2 данных видно, что полученные нетрансгенные мутантные растения табака имеют пониженный в 2 раза уровень содержания никотина, по сравнению с контрольными растениями.
Таким образом, предлагаемый способ позволяет получать стабильные линии нетрансгенных мутантных растений табака со сниженным содержанием никотина, что упрощает использование табака в качестве модельного растения в биоинженерии и в производстве рекомбинантных белков, где никотин и подобные ему алкалоиды нежелательны.
Источники информации
1. Y. Chintapakorn and J.D. Hamil. (2003) Antisense-mediated down-regulation of putrescine N-methyltransferase activity in transgenic Nicotiana tabacum L. can lead to elevated levels of anatabine at the expense of nicotine. Plant Molecular Biology 53: 87-105.
2. Wang, P., Liang, Z., Zeng, J., Li, W., Sun, X., Miao, Z., and Tang, K. (2008) Generation of tobacco lines with widely different reduction in nicotine levels via RNA silencing approaches. J. Biosci., 33, 177-184.
3. Костина H.E., и др. Создание линии табака со сниженными антифидантными свойствами по отношению к колорадскому жуку. Биотехнология и селекция растений. 2020;3(1):24-30).
4. Sanger F., S. Nicklen, and A.R. Coulson. 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 12:5463-5467.
5. Мазин A.B. Методы молекулярной генетики и генной инженерии: научное издание / А.В. Мазин, К.Д. Кузнеделов, А.С. Краев: АН СССР. Сиб. отд-е. Ин-т цитологии и генетики. - Новосибирск: Наука. Сиб. отд-е, 1990. - 247 с.
Перечень последовательностей
attgacctcaaagagcatgacatct
aaacagatgtcatgctctttgaggt
acctcaaagagcatgacatct
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения мутантных растений табака со сниженным содержанием никотина | 2021 |
|
RU2803332C2 |
| РАСТИТЕЛЬНЫЕ КЛЕТКИ И ОРГАНИЗМЫ РАСТЕНИЙ С МОДИФИЦИРОВАННЫМ КЛЕТОЧНЫМ РОСТОМ, РАЗВИТИЕМ И ДИФФЕРЕНЦИРОВКОЙ | 2004 |
|
RU2349642C2 |
| МОЛЕКУЛА РНК-ПРОВОДНИКА ДЛЯ ГЕНОМНОГО РЕДАКТИРОВАНИЯ ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ОДНОДОЛЬНЫХ ЗЕРНОВЫХ С ПРИМЕНЕНИЕМ СИСТЕМЫ CRISPR/CAS9 | 2020 |
|
RU2762831C1 |
| ЭКСПРЕССИЯ ПРОМОТОРА СИНТАЗЫ АЦЕТООКСИКИСЛОТ В ИНТРОДУЦИРОВАННЫХ ГЕНАХ РАСТЕНИЙ | 1995 |
|
RU2197527C2 |
| Молекула РНК-проводника sgRNA для внесения мутаций в консервативный участок промоторной области гена PPD-D1 мягкой пшеницы с применением системы редактирования генома CRISPR/Cas9 | 2024 |
|
RU2822358C1 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С НУКЛЕОТИДНОЙ ДЕЛЕЦИЕЙ В ПРОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772577C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
| ТРЕОНИНСИНТАЗА ИЗ NICOTIANA TABACUM И ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2689719C2 |
| СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ ПШЕНИЦЫ С БИАЛЛЕЛЬНЫМИ МУТАЦИЯМИ В ПРОТОМОТОРНОЙ ОБЛАСТИ ГЕНА VRN-A1 ПРИ ПОМОЩИ ТЕХНОЛОГИИ РЕДАКТИРОВАНИЯ ГЕНОМА РАСТЕНИЙ CRISPR/CAS9 | 2020 |
|
RU2772575C2 |
| МОДУЛИРОВАНИЕ БЕТА-ДАМАСЦЕНОНА У РАСТЕНИЙ | 2012 |
|
RU2681497C2 |

Изобретение относится к области биотехнологии, а именно к генетической инженерии высших растений, в частности к способу получения нетрансгенных мутантных растений табака со сниженным содержанием никотина путем нокаута генов семейства путресцин-N-метилтрансферазы (РМТ). Данный способ позволяет в поколениях от самоопыления получать нетрансгенные стабильные мутантные линии табака со сниженным содержанием никотина. 2 з.п. ф-лы, 1 ил., 2 табл., 2 пр.
1. Способ получения нетрансгенных мутантных растений табака со сниженным содержанием никотина путем мутагенеза генов семейства путресцин-N-метилтрансферазы, включающий дизайн специфической РНК, клонирование фрагмента ДНК, кодирующего специфическую РНК, в вектор, последующую трансформацию растений табака полученной плазмидой, регенерацию их на селективной среде и отбор трансформантов, отличающийся тем, что проводят прямой мутагенез генов семейства путресцин-N-метилтрансферазы методом геномного редактирования посредством нуклеазы Cas9, при этом трансформацию листовых эксплантов табака осуществляют путем бомбардировки золотыми частицами при помощи генной пушки, для чего готовят смесь из двух генетических векторов, содержащую плазмиду рМТ и вектор pBi121, несущий ген устойчивости к канамицину и ген-репортер β-глюкуронидазы Е. coli, полученную смесь наносят на золотые частицы и доставляют в клетки эпидермиса нижней стороны листьев растений табака при помощи генной пушки, а отбор трансформантов выполняют при помощи гистохимического окрашивания с реактивом X-Gluc, при этом плазмиду рМТ конструируют с использованием специфической направляющей РНК (нРНК), имеющей нуклеотидную последовательность SEQ ID NO 3, а фрагмент ДНК, кодирующий нРНК, синтезируют с помощью гибридизации пары специфических олигонуклеотидов, имеющих последовательности SEQ ID NO 1 и SEQ ID NO 2.
2. Способ по п. 1, отличающийся тем, что бомбардировку проводят золотыми частицами размером 1 мкм, разрывной диск 1100 пси, расстояние 6 см, вакуум при выстреле 27 мм рт.ст.
3. Способ по п. 1, отличающийся тем, что отбор мутантных растений-регенерантов осуществляют при помощи секвенирования по Сэнгеру.
| WANG P | |||
| et al., Generation of tobacco lines with widely different reduction in nicotine levels via RNA silencing approaches | |||
| J Biosci | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Vol | |||
| Способ сопряжения брусьев в срубах | 1921 |
|
SU33A1 |
| No | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Pp | |||
| Кулисный парораспределительный механизм | 1920 |
|
SU177A1 |
| STEPPUHN A | |||
| et al., Nicotine's defensive function in nature | |||
| PLoS Biol | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Vol | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| No | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Art | |||
| Искусственный двухслойный мельничный жернов | 1921 |
|
SU217A1 |
| КОСТИНА Н.Е | |||
| и др., Создание линии табака со сниженными антифидантными | |||

