ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке в соответствии с 35 U.S.C. §119 испрашивается приоритет по временной заявке США №62/623276, поданной 29 января 2018 года, описание которой включено в настоящее описание в качестве ссылки.
ПРАВИТЕЛЬСТВЕННЫЕ ПРАВА
[0002] Настоящее изобретение создано при поддержке правительства по гранту № HL088093, присужденного Национальным институтом здравоохранения США. Правительство обладает некоторыми правами на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ
[003] Настоящее изобретение относится к способам и лечению TLR2-опосредованных заболеваний и нарушений, включающим введение антитела, фрагмента антитела или полипептида, связывающего и ингибирующего биологическую активность окисленных фосфолипидов.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ В КАЧЕСТВЕ ССЫЛКИ
[0004] Вместе с заявкой подан список последовательностей, названный "Sequence-Listing_ST25.txt", созданный 29 января 2019 года и имеющий размер 34203 байт и машиночитаемый формат для IBM-PC-совместимой операционной системы MS-Windows. Таким образом, список последовательностей включен в настоящее описание в качестве ссылки в полном объеме для всех целей.
УРОВЕНЬ ТЕХНИКИ
[0005] Фосфолипиды, содержащие полиненасыщенные жирные кислоты, очень склонны к модификации под действием активных форм кислорода. Такие фосфолипиды подвержены перекисному окислению с образованием окисленных фосфолипидов (OxPL), индуцирующих цитотоксичность и апоптоз и играющих значительную роль в воспалении. Показано, что OxPL играют роль в транскрипции интерлейкинов, переключении фенотипа гладкомышечных клеток и апоптотических механизмах модифицированных фосфолипидов. Таким образом, перекисное окисление существенно изменяет физико-химические свойства липидных бислоев мембраны и, таким образом, индуцирует передачу сигнала в зависимости от образования или распознавания мембранных доменов или связывания молекул. Различные типы OxPL могут взаимодействовать со специфическими участками связывания и рецепторами, что приводит к активации отдельных путей передачи сигнала.
[0006] Атеросклероз коронарных сосудов человека является хроническим воспалительным заболеванием, возникающим по причине липидных аномалий. Предполагают, что провоспалительный окисленный липопротеин низкой плотности (OxLDL) является связующим звеном между накоплением липидов и воспалением стенок сосудов. Более того, повышенные уровни продуктов окисления фосфолипидов определяют в различных органах и при различных патологических состояниях, включая сосуды при атеросклерозе, воспаленные легкие, неалкогольный стеатогепатоз, плазму пациентов с ишемической болезнью сердца, а также в апоптотических клетках, инфицированных вирусом клетках и клетках, стимулированных воспалительными агонистами. Проведены исследования двух HDL-ассоциированных ферментов: сывороточной параоксоназы (PON1) и PAF-ацетилгидролазы (PAF-AH); обе из которых отвечают за гидролиз окисленных фосфолипидов плазмы, что, таким образом, является доказательством их роли в атеросклерозе. Другим важным маркером оксидативного стресса является связывание OxPL с частицей аполипопротеина B-100 (OxPL/apoB) LDL. Повышенные уровни OxPL/apoB ассоциированы с ишемической болезнью сердца, прогрессированием атеросклероза сонных и бедренных артерий и прогнозированием сердечно-сосудистых явлений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Настоящее изобретение относится к способам и лечению TLR2-опосредованных заболеваний и нарушений, включающим введение антитела, фрагмента антитела или полипептида, связывающего и ингибирующего биологическую активность окисленного фосфолипида. Как показано в исследованиях, представленных в настоящем описании, нейтрализация OxPL с помощью эндогенной экспрессии in vivo антитела EO6 (при использовании EO6-scFv-трансгенной мыши) в значительной степени ингибирует развитие атеросклероза, вызванное агонизмом TLR2. Инъекции агониста TLR2 PAM3CSK4 мышам Ldlr-/- с питанием с высоким содержанием холестерина приводят к значительному усилению атеросклероза. Схожий набор инъекций EO6-scFv-трансгенным мышам (с фоном Ldlr-/-) приводил к значительному ингибированию образования очагов повреждения. В других исследованиях, представленных в настоящем описании, показано, что нейтрализация OxPL может защищать от прогрессирования заболевания в TLR2-регулируемой модели болезни Кавасаки на мышах. Показано, что введение патогена Lactobaccilus casei вызывает подобное болезни Кавасаки заболевание у мышей, приводящее к усиленному атеросклерозу, артерииту коронарных артерий и аневризмам брюшной аорты. Оно зависит от TLR2, т.к. введение L. Casei мышам с дефицитом TLR-2 не приводит к возникновению заболевания. Экспрессия ИЛ-1 также сильно ассоциирована с болезнью Кавасаки. OxPL также являются мощными индукторами высвобождения ИЛ-1 и воспаления. Инъекция L. casei EO6-трансгенным мышам (с фоном Ldlr-/-) по идентичному протоколу приводит к значительному снижению не только атеросклероза, но и, что имеет важное значение, артериита коронарных артерий по сравнению с инъекциями мышам Ldlr-/-. Антитело EO6 не связывается с L. casei напрямую и, таким образом, вероятно, нейтрализует воспалительные эффекты OxPL, вызванные воспалительными эффектами, ассоциированными с TLR2-опосредованным агонизмом.
[0008] Развитие артериита коронарных артерий, а затем и аневризм коронарных артерий является фатальным осложнением у детей, страдающих болезнью Кавасаки, по разным оценкам возникающим у 25% детей, несмотря на существующую терапию, в основном, представляющую собой лечение внутривенным иммуноглобулином (IVIg), полученным из смешанной и очищенной плазмы человека, и аспирином, представляющим собой общее, неспецифическое противовоспалительное терапевтическое средство. Благодаря способности любых антител против OxPL уменьшать воспалительные процессы, включая снижение продукции ИЛ-1B, а также их способности обеспечивать защиту в TLR2-опосредованной модели синдрома Кавасаки на мышах, инъекции гуманизированного или человеческого эквивалента антитела против OxPL, модифицированного для повышения его биологической эффективности, могут обеспечивать защиту против TLR2-ассоциированных заболеваний, включая болезнь Кавасаки. Т.к. такие антитела против OxPL присутствуют в репертуаре B-клеток человека, направленное рекомбинантное терапевтическое средство может обеспечивать клиническую пользу при таком заболевании без побочных эффектов иммуносупрессии или терапевтических средств, полученных из плазмы.
[0009] Таким образом, экспериментальные данные свидетельствуют о том, что атеросклероз и воспалительный артериит, вызванные TLR2-опосредованным агонизмом in vivo у мышей, можно предотвращать посредством нейтрализации OxPL. Разумеется, агонизм TLR2 связан с многочисленными бактериальными заболевания, но при этом и с различными так называемыми аутоиммунно-опосредованными заболеваниями, такими как системная красная волчанка, ревматоидный артрит и др. В целом, данные свидетельствуют о том, что посредством нейтрализации OxPL с использованием антител, фрагментов антител или других связывающих доменов против PC OxPL можено улучшать или предотвращать развитие многих заболеваний, усиливаемых или ухудшающихся в результате активации TLR2-опосредованных путей передачи сигнала.
В некоторых вариантах осуществления настоящее изобретение относится к способу лечения индивидуума с опосредованным Толл-подобным рецептором 2 (TLR2) заболеванием или нарушением, включающему введение терапевтически эффективного количества антитела, фрагмента антитела или полипептида, специфически связывающегося с окисленным фосфолипидом (OxPL), где антитело, фрагмент антитела или полипептид ингибирует биологическую активность OxPL. В дополнительном варианте осуществления способ дополнительно включает введение индивидууму дополнительного терапевтического средства, которое можно использовать для лечения TLR2-опосредованного заболевания или нарушения. Неограничивающие примеры TLR2-опосредованных заболеваний или нарушений включают болезнь Кавасаки, диабет 2 типа, ревматоидный артрит, дерматологические заболевания, рассеянный склероз, системную красную волчанку, язвенный колит, болезнь Грейвса, синдром Шегрена, аутоиммунные заболевания щитовидной железы или васкулит.В конкретном варианте осуществления TLR2-опосредованное заболевание или нарушение является болезнью Кавасаки. В дополнительном варианте осуществления способ дополнительно включает введение индивидууму внутривенного иммуноглобулина (IVIG) и/или салицилатов. В дополнительном варианте осуществления изобретения индивидуум является человеком, возраст которого составляет менее пяти лет.В другом варианте осуществления биологическая активность OxPL включает активацию пути апоптоза CD36-TLR2. В еще одном варианте осуществления антитело, фрагмент антитела или полипептид является одноцепочечным вариабельным фрагментом (ScFv). В некоторых вариантах осуществления антитело или фрагмент антитела распознает и связывает фосфохолиновую концевую группу окисленного фосфолипида, где антитело или фрагмент антитела содержит вариабельный домен тяжелой цепи (VH) и/или вариабельный домен легкой цепи (VL), и где (a) домен VH содержит аминокислотную последовательность, включающую одну, две или три определяющие комплементарность области (CDR), выбранные из группы, состоящей из: SEQ ID NO: 6 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 6; SEQ ID NO: 7 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 7; и SEQ ID NO: 8 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 8; и (b) домен VL содержит аминокислотную последовательность, включающую одну, две или три определяющие комплементарность области (CDR), выбранные из группы, состоящей из: SEQ ID NO: 9 или 12 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 9 или 12; SEQ ID NO: 10 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 10; и SEQ ID NO: 11 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 11. В дополнительном варианте осуществления антитело, фрагмент антитела или полипептид вводят внутрисосудисто. В дополнительном варианте осуществления изобретения домен VH содержит аминокислотную последовательность, включающую CDR, содержащие SEQ ID NO: 6, 7 и 8, и/или домен VL содержит аминокислотную последовательность, включающую CDR, содержащие SEQ ID NO: 9, 10 и 11 или SEQ ID NO: 10, 11 и 12. В другом варианте осуществления антитело или фрагмент антитела выбраны из группы, состоящей из: (a) антитела или scFv с доменами тяжелой и легкой цепи, содержащими определяющие комплементарность области SEQ ID NO: 6, 7, 8, 9, 10 и 11; и (b) антитела или scFv с доменами тяжелой и легкой цепи, содержащими определяющие комплементарность области SEQ ID NO: 6, 7, 8, 10, 11 и 12. В другом варианте осуществления домены тяжелой и легкой цепи соединены с Fc или областью FC2. В еще одном варианте осуществления фрагмент антитела содержит одноцепочечный вариабельный фрагмент ("scFv"), распознающий фосфохолиновую концевую группу окисленного фосфолипида. В конкретном варианте осуществления scFv является растворимым в физиологических условиях. Можно использовать другие OxPL-связывающие средства, ингибирующие биологическую активность OxPL (см., например, международную публикацию №WO 2013/020995, включенную в настоящее описание в качестве ссылки для всех целей).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0010] На фигуре 1A-B показана (A) диаграмма способа, который можно использовать для получения одноцепочечного вариабельного фрагмента ("scFv"). Как указано, можно использовать сайт-специфический мутагенез для мутагенеза вариабельного домена тяжелой цепи ("VH") двухцепочечного иммуноглобулинового антитела для повышения растворимости scFv (слева). Также указаны линкер, лидерная и эффекторная области scFv (справа). (B) Показана общая схема, на которой представлен план генетических компонентов, кодирующих фрагмент антитела scFv EO6 (вверху); и общая схема вектора, где показана кодирующая последовательность для фрагмента антитела EO6-scFv относительно других элементов вектора, используемых для получения трансгенных мышей (внизу).
[0011] На фигуре 2 показана нуклеотидная (SEQ ID NO: 1) и аминокислотная (SEQ ID NO: 2) последовательности и аннотация scFv.
[0012] На фигуре 3 показаны диаграммы модели in vivo на мышах для исследования функций scFv по изобретению у мышей с провоспалительными условиями и питанием с высоким содержанием холестерина, когда мышей подвергали воздействию экзогенного агониста Толл-подобного рецептора (TLR2) Pam3CSK4 (PAM3).
[0013] На фигуре 4A-C показаны эффекты в отношении потребления и массы тела мышей при использовании модели in vivo, показанной на фиг.3. (A) Не наблюдали значимых различий в потреблении пищи у мышей Ldlr-/- по сравнению с мышами Ldlr-/- EO6scFv+/+при введении носителя. (B) Когда мышам вводили Pam3, мыши Ldlr-/- потребляли меньше пищи по сравнению с мышами Ldlr-/- EO6scFv+/+. (C) Через 12 недель масса тела мышей Ldlr-/- EO6scFv+/+была значимо выше, чем у мышей Ldlr-/-. На неделе 12 не наблюдали значимых различий массы тела между мышами Ldlr-/- EO6scFv+/+и мышами Ldlr-/- при введении носителя.
[0014] На фигуре 5A-B показано, что не наблюдали значимых различий (A) холестерина или (B) триглицеридов в плазме крови мышей EO6scTg Ldlr-/-(LDLrKO (нокаут)) по сравнению с мышами Ldlr-/-.
[0015] На фигуре 6A-B показано, что не наблюдали значимых различий (A) профиля холестерина липопротеинов или (B) профиля триглицеридов липопротеинов в плазме крови мышей Ldlr-/-EO6scFv+/+по сравнению с мышами Ldlr-/-, например, уровни липопротеинов были схожи у обеих линий мышей.
[0016] На фигуре 7A-E показано, что наблюдали меньший измеримый атеросклероз у мышей Ldlr-/-EO6scFv+/+по сравнению с мышами Ldlr-/-. (A) Степень общего атеросклероза аорты была выше у мышей Ldlr-/- по сравнению с мышами Ldlr-/-EO6scFv+/+. (B) В частности, более высокую степень атеросклероза определяли в брюшной аорте (ниже диафрагмы) у мышей Ldlr-/- по сравнению с мышами Ldlr-/-EO6scFv+/+. (C) На фигурах (A) и (B) выше показан двухмерный анализ степени атеросклероза посредством планиметрии. Точная масса иссеченных и очищенных аорт является лучшим показателем общего атеросклероза, т.к. она включает измерение толщины. Масса аорт мышей Ldlr-/- была значимо выше, чем у мышей Ldlr-/-EO6scFv+/+. (D) При контроле массы тела соотношение массы аорты и тела мышей Ldlr-/- было значимо выше, чем соотношении массы аорты и тела мышей Ldlr-/-EO6scFv+/+. (E) Степень атеросклероза в корне аорты не отличалась между двумя группами.
[0017] На фигуре 8A-H показаны результаты количественной ПЦР (qPCR) для анализа экспрессии генов системы воспаления в жировой ткани мышей Ldlr-/-EO6scFv+/+по сравнению с мышами Ldlr-/-. В частности, экспрессия (A) ИЛ-1b, (B) ИЛ-6, (C) ФНОα, (E) MCP1, (F) MIP1α, (G) MIP1β и (H) ИЛ-10, как правило, была более низкой у мышей Ldlr-/-EO6scFv+/+по сравнению с мышами Ldlr-/-, в то время как экспрессия ИЛ-12 была немного более высокой (D).
[0018] На фигуре 9A-G показаны результаты твердофазного иммуноферментного анализа (ELISA) для измерения уровней цитокинов в экстрактах жировой ткани мышей Ldlr-/-EO6scFv+/+по сравнению с мышами Ldlr-/-. В частности, измеренные уровни цитокинов (A) ИЛ-1b, (B) ИЛ-6, (C) ФНОα, (D) MCP1, (E) MIP1α, (F) MIP1β и (G) ИЛ-10 отражали результаты анализа экспрессии генов, показанные на фиг.8.
[0019] На фигуре 10A-H показано, что полученные из костного мозга клетки Ldlr-/-EO6scFv+/+при дифференцировке в макрофагальные клетки M1 или M2 и стимуляции PAM3 демонстрировали более низкую экспрессию (A) ИЛ-1β, (B) ИЛ-6, (C) ИЛ-12, (D) ФНОα, (E) MCP1, (F) MIP1α, (G) MIP1β и (H) RANTES по сравнению с дифференцированными макрофагами мышей Ldlr-/-. Приведенные данные представляют собой сравнение клеток M1, полученных из мышей Ldlr-/-EO6scFv+/+или Ldlr-/-. Схожие данные получали в случае клеток M2, например, определяли более низкую экспрессию в клетках M2 Ldlr-/-EO6scFv+/+по сравнению с Ldlr-/-, за исключением того, что абсолютные уровни экспрессии цитокинов были более низкими.
[0020] На фигуре 11 показана устойчивая экспрессия мРНК EO6-scFv в разной жировой ткани, полученной из мышей Ldlr-/-EO6scFv+/+. Это является результатом инфильтрации макрофагами. Макрофаги экспрессируют промотор apoE и, таким образом, экспрессируют трансген EO6-scFv.
[0021] На фигуре 12 показаны изображения флуоресцентной микроскопии, где показано, что обработка макрофагов (RAW264.7) PAM3 индуцировала продукцию OxPL. Т.к. макрофаги мышей Ldlr-/-EO6scFv+/+меньше отвечали на стимуляцию TLR2, и т.к. макрофаги экспрессируют и секретируют EO6-scFv в культуру, предполагают, что стимулированные TLR2 макрофаги образовывали OxPL, и что это, в свою очередь, активируют экспрессию генов системы воспаления аутокринным образом. Для проверки этой гипотезы макрофаги стимулировали PAM3: левая панель, обработка носителем; правая панель, обработка агонистом TLR2 PAM3. Культуры RAW264.7 инкубировали с Pam3 (1 мкг/мл) или контрольным носителем в течение 18 ч. и поверхностно окрашивали на OxPL с помощью антитела EO6 IgM и конъюгата антитела козы против msIgM с FITC. Это свидетельствует о том, что стимулированные TLR2 макрофаги образовывали OxPL.
[0022] На фигуре 13A-F показано, что наблюдали меньший измеримый атеросклероз у мышей Ldlr-/-EO6scFv+/+RAG1-/- по сравнению с мышами Ldlr-/-RAG1-/-. Т.к. мыши RAG1 KO не имеют B- или T-клеток, все атеросклеротические явления, связанные с иммунными клетками, напрямую приписывают действию макрофагов. Примечательно, что абсолютный уровень атеросклероза у мышей Ldlr-/- RAG1-/- был снижен на половину по сравнению с мышами Ldlr-/-. (A) Брюшные аорты, выделенные из мышей Ldlr-/-RAG1-/- (слева) и мышей Ldlr-/-EO6scFv+/+RAG1-/- (справа). Брюшные аорты мышей Ldlr-/-EO6scFv+/+RAG1-/-, очевидно, имели меньше атеросклеротических повреждений, чем у мышей Ldlr-/-RAG1-/-. (B) Размер повреждений в синусе аорты был значимо меньше у мышей Ldlr-/-EO6scFv+/+RAG1-/- по сравнению с мышами Ldlr-/-RAG1-/-. (C) Мыши Ldlr-/- RAG1-/- имели более высокий процент общих повреждений всего организма по сравнению с мышами Ldlr-/-EO6scFv+/+RAG1-/-. (D) Мыши Ldlr-/-RAG1-/- имели более высокий процент повреждений брюшной аорты по сравнению с мышами Ldlr-/-EO6scFv+/+RAG1-/-. (E) Масса аорт мышей Ldlr-/-RAG1-/- была значимо выше, чем у мышей Ldlr-/-EO6scFv+/+RAG1-/-. (F) При контроле массы тела соотношение массы аорты и тела мышей Ldlr-/-RAG1-/- было значимо выше, чем соотношение массы аорты и тела мышей Ldlr-/-EO6scFv+/+RAG1-/-. Т.к. у мышей RAG1 KO отсутствуют B- и T-клетки, основным типом иммунных клеток, способствующих развитию атеросклероза у этих мышей, является макрофаг.Таким образом, эти данные свидетельствуют о том, что основным механизмом, посредством которого макрофаги вносят вклад в развитие атеросклероза в этой TLR2-индуцированной модели, является ответ на OxPL.
[0023] На фигуре 14 показаны изображения детей с болезнью Кавасаки и, кроме того, аневризм коронарных и брюшных артерий, ассоциированных с болезнью Кавасаки.
[0024] На фигуре 15 показана модель на мышах для исследования влияния LCWE и питания HFC на атеросклероз у мышей Ldlr-/-, мышей Ldlr-/-EO6Tg, мышей Ldlr-/-IK17Tg, мышей TLR2-/-Ldlr-/-.
[0025] На фигуре 16 показан анализ анфасных изображений повреждений аорты у мышей с питанием HFC в течение 12 недель и TLR2, активированным посредством LCWE. EO6scFv-Tg значительно снижало анфасные повреждения аорты.
[0026] На фигуре 17 показан анализ площади повреждений брюшной аорты у различных мышей Ldlr-/-, которым инъецировали LCWE и которых кормили HFC в течение 12 недель. Данные выражены как проценты атеросклеротического повреждения, измеренного в брюшной аорте при окрашивании суданом IV. Обнаружено, что EO6Tg и IK17Tg вызывали статистически значимый протективный эффект.Также обнаружено, что TLR2 статистически значимо снижался по сравнению с мышами Ldlr-/-.
[0027] На фигуре 18 показаны поперечные срезы корней аорты мышей Ldlr-/- и мышей EO6-Tg Ldlr-/-, которым вводили LCWE. EO6scFv-Tg уменьшало повреждения корня аорты, размер некротического ядра и, что наиболее важно в отношении манифестации болезни Кавасаки, артериита коронарных артерий. Стрелками указаны коронарные артерии в поперечном сечении. Обширный артериит (крупная клеточная масса) присутствовал у мышей Ldlr-/-, но отсутствовал у мышей EO6-Tg Ldlr-/-.
[0028] На фигуре 19 показаны дополнительные поперечные срезы корней аорты мышей Ldlr-/- и мышей EO6-Tg Ldlr-/-, которым вводили LCWE. Стрелками указаны коронарные артерии в поперечном сечении. Обширный артериит (крупная клеточная масса) присутствовал у мышей Ldlr-/-, но отсутствовал у мышей EO6-Tg Ldlr-/-.
[0029] На фигуре 20 показаны поперечные срезы корней аорты мышей Ldlr-/- TLR2-/- и мышей IK17-Tg+/+Ldlr-/-, которым вводили LCWE. Стрелками указаны коронарные артерии в поперечном сечении. И EO6scFv, и IK17scFv уменьшали артериит коронарных артерий. EO6 (антитело против OxPL), но не IK17 (антитело против MDA), уменьшало повреждения корня аорты.
[0030] На фигуре 21 показаны дополнительные поперечные срезы корней аорты мышей Ldlr-/-TLR2-/- и мышей IK17-Tg+/+Ldlr-/-, которым вводили LCWE. Стрелками указаны коронарные артерии в поперечном сечении. Отсутствие артериита в коронарных артериях у мышей Ldlr-/-TLR2-/- свидетельствует о важности активации TLR2 в модели болезнь Кавасаки на мышах.
[0031] На фигуре 22 показаны уровни воспалительных цитокинов в плазме у контрольных мышей LDLr-/- и мышей EO6-Tg LDLr-/-, которым вводили LCWE и которых кормили HFC в течение 12 недель. Цитокины в плазме измеряли одновременно посредством мультиплексных анализов цитокинов с использованием набора для анализа цитокинов мыши Bio-Plex Pro (Bio-Rad Laboratories, USA). Наблюдали значительное снижение (p<0,04) уровней белков ФНОα, RANTES, MCP-1, CXCL1, ИЛ-6 и ИЛ-12 в сыворотке у мышей EO6scFv-Tg LDLr-/- по сравнению с контрольными мышами.
ПОДРОБНОЕ ОПИСАНИЕ
[0032] В рамках изобретения и формулы изобретения термины в единственном числе включают ссылку на множественное число, если контекст четко не указывает на иное. Таким образом, например, ссылка на "одноцепочечный вариабельный фрагмент" или "scFv" включает множество одноцепочечных вариабельных фрагментов, и ссылка на "окисленный фосфолипид" включает ссылку на один или более окисленных фосфолипидов и их эквивалентов, известных специалистам в этой области, и т.д.
[0033] Если не указано иначе, все технические и научные термины, используемые в настоящем описании, обладают значением, общепринято понятным специалисту в области, к которой принадлежит изобретение. Хотя в практическом осуществлении описываемых способов и композиций можно использовать любые способы и реагенты, схожие или эквивалентные представленным в настоящем описании, примеры способов и материалов описаны далее.
[0034] Все публикации, упомянутые в настоящем описании, включены в него в качестве ссылки в полном объеме в целях описания методологии, представленной в публикациях, которую можно использовать вместе с настоящим описанием. Кроме того, что касается любого термина, представленного в одной или более публикациях, схожего или идентичного термину, конкретно определенного в настоящем описании, определение термина, конкретно представленного в настоящем описании, будет обладать приоритетом во всех смыслах.
[0035] Кроме того, использование союза "и" означает "и/или", если не указано иначе. Аналогично, термины "содержат", "содержит", "содержащий", "включают", "включает" и "включающий" используют взаимозаменяемо, и не являются ограничивающими.
[0036] Если в описании различных вариантов осуществления используют термин "содержащий", специалистам в этой области также следует понимать, что в некоторых конкретных случаях вариант осуществления можно описывать альтернативно с использованием выражений "состоящий, по существу, из" или "состоящий из".
[0037] Термины "антитело" и "иммуноглобулин" используют взаимозаменяемо в самом широком смысле, и они включают моноклональные антитела (например, полноразмерные или интактные моноклональные антитела), поликлональные антитела, мультивалентные антитела, мультиспецифические антитела (например, биспецифические антитела, при условии, что они проявляют желательную биологическую активность), а также могут включать некоторые фрагменты антител. Антитело может являться человеческим, гуманизированным и/или аффинно зрелым.
[0038] В зависимости от аминокислотной последовательности константного домена их тяжелых цепей, иммуноглобулины можно приписывать к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно дополнительно разделять на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, соответствующие разным классам иммуноглобулинов, названы α, δ, ε, γ и μ, соответственно. Структуры и трехмерные конфигурации субъединиц разных классов иммуноглобулинов хорошо известны.
[0039] "Фрагменты антител" содержат только часть интактного антитела, где часть, как правило, сохраняет по меньшей мере одну, но, в основном, большинство или все из функций, обычно ассоциированных с этой частью при ее наличии в интактном антителе. Примеры фрагментов антител включают Fab-, Fab'-, F(ab')2- и Fv-фрагменты; диатела; линейные антитела; молекулы одноцепочечных антител и мультиспецифические антитела, образованные из фрагментов антител. В одном из вариантов осуществления фрагмент антитела содержит антигенсвязывающий участок интактного антитела и, таким образом, сохраняет способность связываться с антигеном. В другом варианте осуществления фрагмент антитела, например, фрагмент, содержащий Fc-область, сохраняет по меньшей мере одну из биологических функций, обычно ассоциированных с Fc-областью при ее наличии в интактном антителе, таких как связывание FcRn, модуляция времени полужизни антитела, функция ADCC и связывание антитела. В одном из вариантов осуществления фрагмент антитела является моновалентным антителом, имеющим время полужизни in vivo, по существу, схожее с таковым у интактного антитела. Например, такой фрагмент антитела может содержать антигенсвязывающее плечо, связанное с последовательностью Fc, способной придавать фрагменту стабильность in vivo. Однако следует понимать, что большое время полужизни антитела не является обязательным в некоторых случаях (например, при лечении острого повреждения при ишемии/реперфузии).
[0040] "Антиген" является заранее определенным антигеном, с которым антитело может селективно связываться. Целевой антиген может являться полипептидом, углеводом, нуклеиновой кислотой, липидом, гаптеном или другим природным или синтетическим соединением. В одном из вариантов осуществления настоящего изобретения антиген является OxPL.
[0041] Термин "антитело против OxPL" или "антитело, связывающееся с OxPL" относится к антителу, которое может связываться с OxPL с достаточной аффинностью таким образом, что антитело можно использовать в качестве диагностического и/или терапевтического средства для направленного воздействия на OxPL. В некоторых вариантах осуществления настоящего изобретения антитело против OxPL имеет такую же или схожую специфичность связывания и Kd, что и антитело EO6 или антитело QX5. В еще одном варианте осуществления антитело против OxPL связывается с концевой группой PC OxPL.
[0042] "Блокирующее" антитело или "антагонистическое" антитело является антителом, ингибирующим или снижающим биологическую активность антигена, с которым оно связывается. Некоторые блокирующие антитела или антагонистические антитела существенно или полностью ингибируют биологическую активность антигена.
[0043] Термин "аффинность связывания", как правило, относится к силе общей суммы нековалентных взаимодействий между отдельным участком связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иначе, в рамках изобретения термин "аффинность связывания" относится к характерной аффинности связывания, отражающей взаимодействие 1:1 между членами связывающейся пары (например, антитела и антигена). Аффинность молекулы X к ее партнеру Y, как правило, может быть представлена константой диссоциации (Kd). Аффинность можно измерять известными в этой области способами, включая способы, представленные в настоящем описании. Низкоаффинные антитела, как правило, медленно связываются с антигеном и имеют склонность к быстрой диссоциации, в то время как высокоаффинные антитела, как правило, быстрее связывают антиген и имеют склонность дольше оставаться связанными. В этой области известно множество способов измерения аффинности связывания, любые из которых можно использовать в целях по настоящему изобретению.
[0044] Термин "биологический образец" включает различные типы образцов, полученные из индивидуума, и их можно использовать в диагностическом анализе или мониторинге. Определение включает кровь и другие жидкие образцы биологического происхождения, образцы солидных тканей, таких как биоптаты, или культуры тканей, или клетки, полученные из них, и их потомство. Определение также включает образцы, обработанные каким-либо образом после их получения, например, посредством обработки реагентами, солюбилизации или обогащения некоторыми компонентами, такими как белки или полинуклеотиды, или погружения в полутвердую или твердую матрицу для получения срезов. Термин "биологический образец" включает клинический образец, а также клетки в культуре, супернатанты клеток, лизаты клеток, сыворотку, плазму, биологическую жидкость и образцы ткани. Источник биологического образца может являться солидной тканью в виде свежего, замороженного и/или консервированного образца органа или ткани, или биоптата, или аспирата; кровью или любым компонентом крови; физиологическими жидкостями, такими как спинномозговая жидкость, амниотическая жидкость, перитонеальная жидкость или интерстициальная жидкость; клетками с любой стадии развития эмбриона или индивидуума. В некоторых вариантах осуществления биологический образец получают из первичной или метастазирующей опухоли. Биологический образец может содержать соединения, в природе не смешанные с тканью, такие как консерванты, антикоагулянты, буферы, фиксативы, питательные вещества, антибиотики или т.п.
[0045] Термин "диатела" относится к небольшим фрагментам антител с двумя антигенсвязывающими участками, где фрагменты содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) в одной полипептидной цепи (VH-VL). Используя линкер, слишком короткий для спаривания двух доменов на одной цепи, домены заставляют спариваться с комплементарными доменами другой цепи и получают два антигенсвязывающих участка. Диатела более подробно описаны, например, в EP 404,097; WO 93/11161; и Hollinger et al., Proc. Natl. Acad. Sci. USA, 90:6444-6448 (1993). Триатела и тетратела также описаны в Hudson et al., Nat. Med. 9:129-134 (2003).
[0046] "Нарушение" или "заболевание" является любым состоянием, при котором будут получать пользу от лечения с помощью вещества/молекулы или способа по настоящему изобретению. Этот термин включает TLR2-опосредованное заболевание или нарушение, такое как болезнь Кавасаки.
[0047] Термин "эффективное количество" относится к количеству, в необходимых дозах и в течение необходимых периодов времени эффективному для достижения желаемого терапевтического или профилактического результата.
[0048] В рамках изобретения термин "Fc-область" относится к C-концевой области тяжелой цепи иммуноглобулина, включая нативную последовательность Fc-области и вариант Fc-области.
[0049] "Функциональная Fc-область" обладает эффекторной функцией нативной последовательности Fc-области. Для таких эффекторных функций, как правило, необходимо комбинирование Fc-области и связывания с доменом (например, вариабельным доменом антитела), и их можно оценивать с использованием различных анализов, как описано, например, в определениях в настоящем описании.
[0050] "Нативная последовательность Fc-области" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. Нативные последовательности Fc-области человека включают нативную последовательность Fc-области IgG1 человека (аллотипы не-A и A), нативную последовательность Fc-области IgG2 человека, нативную последовательность Fc-области IgG3 человека и нативную последовательность Fc-области IgG4 человека, а также их природные варианты.
[0051] Термин "Fc-рецептор" или "FcR" означает рецептор, связывающийся с Fc-областью антитела. В некоторых вариантах осуществления FcR является нативным FcR человека. В других вариантах осуществления FcR является рецептором, связывающимся с антителом IgG (гамма-рецептор), и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы FcγRII включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), имеющие схожие аминокислотные последовательности, отличающиеся, главным образом, своими цитоплазматическими доменами. Активирующий рецептор FcγRIIA содержит иммунорецепторный тирозиновый активирующий мотив (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный тирозиновый ингибирующий мотив (ITIM) в своем цитоплазматическом домене (см., например, Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). Обзор FcR представлен, например, в Ravetch and Kinet, Annu. Rev. Immunol9:457-92 (1991); Capel et al., Immunomethods 4:25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, включая FcR, которые будут идентифицированы в будущем, включены в термин "FcR" в настоящем описании.
[0052] Fc-рецептор также включает неонатальный рецептор FcRn, отвечающий за транспорт материнских IgG к плоду (Guyeret al., J. Immunol. 117:587 (1976) и Kim et al., J. Immunol. 24:249 (1994)) и регуляцию гомеостаза иммуноглобулинами. Известны способы измерения связывания с FcRn (см., например, Ghetie and Ward.,Immunol. Today 18(12):592-598 (1997); Ghetieet al., Nature Biotechnology, 15(7):637-640 (1997); Hinton et al., J. Biol. Chem. 279(8):6213-6216 (2004); WO 2004/92219 (Hinton et al.).
[0053] Связывание FcRn человека in vivo и время полужизни в сыворотке высокоаффинных FcRn-связывающих полипептидов человека можно анализировать, например, в трансгенных мышах или трансфицированных линиях клеток человека, экспрессирующих FcRn человека, или в приматах, которым вводят полипептиды с вариантом Fc-области. В WO 2000/42072 (Presta) описывают варианты антител с улучшенным или сниженным связыванием с FcR. Также см., например, Shields et al., J. Biol. Chem. 9(2):6591-6604 (2001).
[0054] "Fv" является минимальным фрагментом антитела, содержащим полный антигенраспознающий и антигенсвязывающий участок. В двухцепочечных видах Fv эта область состоит из димера сильно, нековалентно связанных одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи. В одноцепочечных видах Fv один вариабельный домен тяжелой цепи и один вариабельный домен легкой цепи можно ковалентно связывать с помощью гибкого пептидного линкера таким образом, что легкие и тяжелые цепи могут связываться в "димерной" структуре, аналогичной структуре двухцепочечных видов Fv. Они находятся в такой конфигурации, что три HVR каждого вариабельного домена взаимодействуют с образованием антигенсвязывающего участка на поверхности димера VH-VL. В совокупности, шесть HVR придают антителу специфичность связывания антигена. Однако, даже один вариабельный домен (или половина Fv, содержащая только три HVR, специфические для антигена) обладает способностью распознавать и связывать антиген, хотя и с более низкой аффинностью, чем целый участок связывания.
[0055] Fab-фрагмент также содержит константный домен легкой цепи и первый константный домен (CH1) тяжелой цепи. Фрагменты Fab' отличаются от Fab-фрагментов добавлением нескольких остатков на карбокси-конец домена CH1 тяжелой цепи, включая один или более цистеинов из шарнирной области антитела. В настоящем описании "Fab'-SH" является обозначением Fab', в котором остатки цистеина константных доменов содержат тиоловую группу. F(ab')2-фрагменты антител исходно получали как пары Fab'-фрагментов, содержащих цистеины шарнирной области между собой. Также известны другие химические соединения фрагментов антител.
[0056] Расщепление антител папаином приводит к получению двух идентичных антигенсвязывающих фрагментов, названных "Fab"-фрагментами, каждый из которых содержит один антигенсвязывающий участок, и остаточного "Fc"-фрагмента, название которого отражает его способность легко кристаллизоваться. Обработка пепсином приводит к получению F(ab')2-фрагмента, содержащего два антигенсвязывающих участка и все равно способного перекрестно связывать антиген.
[0057] "Каркасные" остатки или "FR" являются остатками вариабельного домена, иными, чем остатки гипервариабельной области, определенные в настоящем описании.
[0058] "Гуманизированные" формы не принадлежащих человеку (например, мышиных) антител являются химерными антителами, содержащими минимальную последовательность, полученную из не принадлежащего человеку иммуноглобулина. В одном из вариантов осуществления гуманизированное антитело является иммуноглобулином человека (реципиентное антитело), в котором остатки HVR реципиента заменяют остатками из HVR не являющихся человеком видов (донорное антитело), таких как мышь, крыса, кролик или не являющийся человеком примат, имеющих желаемую специфичность, аффинность и/или емкость. В некоторых случаях, каркасные остатки иммуноглобулина человека заменяют соответствующими не принадлежащими человеку остатками. Кроме того, гуманизированные антитела могут содержать остатки, не обнаруживаемые в реципиентном антителе или донорном антителе. Эти модификации можно осуществлять для дальнейшего улучшения свойств антитела. В основном, гуманизированное антитело будет содержать, по существу, все из по меньшей мере одного и, как правило, двух вариабельных доменов, в которых все или, по существу, все из гипервариабельных петель соответствуют петлям не принадлежащего человеку иммуноглобулина, и все или, по существу, все из FR являются FR последовательности иммуноглобулина человека. Гуманизированное антитело, необязательно, также будет содержать по меньшей мере часть константной области иммуноглобулина (Fc), как правило, из иммуноглобулина человека. Более подробно, см., например, Jones et al., Nature 321:522-525 (1986); Riechmannet al., Nature 332:323-329 (1988); и Presta, Curr. Op.Struct. Biol. 2:593-596 (1992). Также см., например, Vaswani and Hamilton, Ann. Allergy, Asthma & Immunol. 1:105-115 (1998); Harris, Biochem. Soc. Transactions 23:1035-1038 (1995); Hurle and Gross, Curr. Op.Biotech, 5:428-433 (1994); и патенты США №№6982321 и 7087409.
[0059] "Антитело человека" является антителом, имеющим аминокислотную последовательность, соответствующую последовательности антитела, продуцируемого человеком и/или полученного любым из способов получения антител человека, представленных в настоящем описании. Это определение антитела человека конкретно исключает гуманизированное антитело, содержащее не принадлежащие человеку антигенсвязывающие остатки. Антитела человека можно получать различными способами, известными в этой области, включая библиотеки фагового дисплея. Hoogenboom and Winter, J. Mol. Biol., 227:381 (1991); Marks et al., J. Mol. Biol., 222:581 (1991). Также для получения моноклональных антител человека доступны способы, описанные в Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985); Boerner et al., J. Immunol., 147(1):86-95 (1991). Также см. van Dijk and van de Winkel, Curr. Opin. Pharmacol., 5: 368-74 (2001). Антитела человека можно получать посредством введения антигена трансгенному животному, модифицированному для продукции таких антител в ответ на стимуляцию антигеном, но эндогенные локусы которых инактивированы, например, иммунизированным ксеномышам (см., например, патенты США №№6075181 и 6150584). Также см., например, Li et al., Proc. Natl. Acad. Sci. USA, 103:3557-3562 (2006), касающуюся антител человека, полученных с помощью технологии B-клеточной гибридомы человека. Необходимо отметить, что термин "антитело человека" не включает природные антитела, продуцируемые человеком, скорее он относится к антителам, несодержащим какой-либо эпитоп или антигенный фрагмент, который человек не будет распознавать как "чужеродный".
[0060] "Эффекторные клетки человека" являются лейкоцитами, экспрессирующими один или более FcR и осуществляющими эффекторные функции. В некоторых вариантах осуществления клетки экспрессируют по меньшей мере FcγRIII и осуществляют эффекторные функции ADCC. Примеры лейкоцитов человека, опосредующих ADCC, включают мононуклеарные клетки периферической крови (PBMC), естественные киллеры (NK), моноциты, цитотоксические T-клетки и нейтрофилы. Эффекторные клетки можно выделять из нативного источника, например, крови.
[0061] Термин "гипервариабельная область", "HVR" или "HV" при использовании в настоящем описании относится к областям вариабельного домена антитела, являющимся гипервариабельными по своей последовательности и/или образующими структурно определенные петли. Как правило, антитела содержат шесть HVR: три в домене VH (H1, H2, H3) и три в домене VL (L1, L2, L3). В нативных антителах, H3 и L3 демонстрируют самое высокое разнообразие среди шести HVR, и, в частности, считают, что H3 играет уникальную роль в придании антителам точной специфичности. Смотри, например, Xu et al., Immunity 13:37-45 (2000); Johnson and Wu, в Methods in Molecular biology 248:1-25 (Lo, ed., Human Press, Totowa, N.J., 2003). Фактически, природные антитела Верблюжьих, состоящие только из тяжелой цепи, являются функциональными и стабильными в отсутствие легкой цепи. См., например, Hamers-Castermanet al., Nature 363:446-448 (1993); Sheriff et al., Nature Struct. Biol. 3:733-736 (1996).
[0062] "Индивидуум" или "пациент" является позвоночным. В некоторых вариантах осуществления позвоночное представляет собой млекопитающего. Млекопитающие включают, в качестве неограничивающих примеров, сельскохозяйственных животных (таких как коровы), спортивных животных, домашних питомцев (таких как кошки, собаки и лошади), приматов, мышей и крыс. В некоторых вариантах осуществления млекопитающее является человеком.
[0063] "Выделенное" антитело или фрагмент антитела является антителом или фрагментом антитела, идентифицированным и отделенным от компонента его природного окружения. Контаминирующие компоненты природного окружения являются материалами, мешающими диагностическому или терапевтическому использованию антитела, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах осуществления антитело будут очищать (1) до более чем 95% по массе антитела, что определяют способом Лоури, и, как правило, более чем 99% по массе, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности с использованием секвенатор с вращающимся стаканом, или (3) до гомогенность посредством электрофореза в ПААГ в присутствии SDS в восстановительных или невосстановительных условиях с использованием Кумасси синего или окрашивания серебром. Выделенное антитело включает антитело in situ в рекомбинантных клетках, т.к. по меньшей мере один компонент природного окружения антитела будет отсутствовать. Однако, как правило, выделенное антитело будут получать с помощью по меньшей мере одной стадии очистки.
[0064] "Выделенная" молекула нуклеиновой кислоты является молекулой нуклеиновой кислоты, идентифицированной и отделенной от по меньшей мере одной контаминирующей молекулы нуклеиновой кислоты, с которой она, как правило, ассоциирована в природном источнике нуклеиновой кислоты антитела. Выделенная молекула нуклеиновой кислоты находится в иной форме или условиях, чем обнаруживаемые в природе. Таким образом, выделенные молекулы нуклеиновой кислоты отличаются от молекулы нуклеиновой кислоты, существующей в природных клетках. Однако, выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, как правило, экспрессирующих антитело, где, например, молекула нуклеиновой кислоты находится в иной хромосомной локализации, чем в природных клетках.
[0065] В рамках изобретения термин "метка" относится к соединению или композиции, конъюгированным или слитым прямо или косвенно с реагентом, таким как зонд нуклеиновой кислоты или антитело, и облегчающим детекцию реагента, с которым они конъюгированы или слиты. Метка сама может являться детектируемой (например, радиоактивные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать детектируемое химическое изменение субстратного соединения или композиции.
[0066] "Легкие цепи" антител (иммуноглобулинов) любого вида позвоночных можно приписывать к одному из двух четко различаемых типов, названных каппа (κ) и лямбда (λ), на основе аминокислотных последовательностей их константных доменов.
[0067] В рамках изобретения термин "моноклональное антитело" относится к антителу, полученному из популяции, по существу, гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных мутаций, например, природных мутаций, которые могут присутствовать в незначительных количествах. Таким образом, определение "моноклональное" означает природу антитела, как неявляющегося смесью отличающихся антител. В некоторых вариантах осуществления такое моноклональное антитело, как правило, включает антитело, содержащее полипептидную последовательность, связывающуюся с мишенью, где полипептидную последовательность, связывающую мишень, получали способом, включающим селекцию одной связывающей мишень полипептидной последовательности из множества полипептидных последовательностей. Например, способ селекции может представлять собой селекцию уникального клона из множества клонов, таких как совокупность клонов гибридом, фаговых клонов или клонов рекомбинантной ДНК. Следует понимать, что выбранную связывающую мишень последовательность можно дополнительно изменять, например, для улучшения аффинности к мишени, для гуманизации связывающей мишень последовательности, для улучшения ее продукции в культуре клеток, для снижения ее иммуногенности in vivo, для получения мультиспецифического антитела и т.д., и что антитело, содержащее измененную связывающую мишень последовательность, также является моноклональным антителом для целей по настоящему изобретению. В отличие от препаратов поликлональных антител, которые, как правило, включают разные антитела против разных детерминант (эпитопов), каждое моноклональное антитело из препарата моноклональных антител направлено против одной и той же детерминанты на антигене. В дополнение к своей специфичности, препараты моноклональных антител являются предпочтительными, т.к. они, как правило, не загрязнены другими иммуноглобулинами.
[0068] Определение "моноклональное" свидетельствует о природе антитела как получаемого из, по существу, гомогенной популяции антител, и не следует истолковывать его как требующего получения антитела каким-либо конкретным способом. Например, моноклональные антитела для использования в настоящем изобретении можно получать различными способами, включая, например, гибридомный способ (например, Kohler and Milstein, Nature, 256:495-97 (1975); Hongo et al., Hybridoma, 14 (3): 253-260 (1995), Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., в: Monoclonal Antibodies and T-Cell Hybridomas 563-681 (Elsevier, N.Y., 1981)), способы рекомбинантной ДНК (см., например, патент США №4816567), технологии фагового дисплея (см., например, Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol. 222: 581-597 (1992); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Natl. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al., J. Immunol. Methods 284(1-2): 119-132 (2004) и технологии получения антител человека или подобных им антител в животных, содержащих части или все из иммуноглобулиновых локусов человека или генов, кодирующих последовательности иммуноглобулинов человека (см., например, WO 1998/24893; WO 1996/34096; WO 1996/33735; WO 1991/10741; Jakobovits et al., Proc. Natl. Acad. Sci. USA 90: 2551 (1993); Jakobovits et al., Nature 362: 255-258 (1993); Bruggemann et al., Year in Immunol. 7:33 (1993); патенты США №№5545807; 5545806; 5569825; 5625126; 5633425 и 5661016; Marks et al., Bio/Technology 10: 779-783 (1992); Lonberg et al., Nature 368: 856-859 (1994); Morrison, Nature 368: 812-813 (1994); Fishwild et al., Nature Biotechnol. 14: 845-851 (1996); Neuberger, Nature Biotechnol. 14: 826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol. 13: 65-93 (1995).
[0069] Моноклональные антитела в настоящем описании конкретно включают "химерные" антитела, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретных биологических видов или принадлежащих к конкретному классу или подклассу антител, в то время как остальная часть цепей идентична или гомологична соответствующим последовательностям в антителах, полученных из других биологических видов или принадлежащих к другому классу или подклассу антител, а также фрагментам таких антител, при условии, что они проявляют желательную биологическую активность (см., например, патент США №4816567; и Morrison et al., Proc. Natl. Acad. Sci. USA 81:6851-6855 (1984)). Химерные антитела включают антитела, в которых антигенсвязывающую область антитела получают из антитела, полученного, например, посредством иммунизации макак интересующим антигеном.
[0070] В рамках изобретения термины "полинуклеотид" или "нуклеиновая кислота" относятся к полимерам нуклеотидов любой длины, и включают ДНК и РНК. Нуклеотиды могут являться дезоксирибонуклеотидами, рибонуклеотидами, модифицированными нуклеотидами или основаниями и/или их аналогами, которые могут включаться в полимер под действием ДНК- или РНК-полимеразы или посредством синтетической реакции. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если они есть, модификацию в структуру нуклеотида можно вносить до или после сборки полимера. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Полинуклеотид можно дополнительно модифицировать после синтеза, например, посредством конъюгации с меткой. Другие типы модификаций включают, например, "кэпирование", замену одного или более природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, модификации с незаряженными связями (например, метилфосфонаты, фосфотриэфиры, фосфоамидаты, карбаматы и т.д.) и с заряженными связями (например, фосфотиоаты, фосфодитиоаты и т.д.), модификации, включающие "подвешенные" молекулы, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), модификации с интеркаляторами (например, акридином, псораленом и т.д.), модификации, включающие хелаторы (например, металлы, радиоактивные металлы, бор, металлы-окислители и т.д.), модификации, включающие алкиляторы, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т.д.), а также немодифицированные формы полинуклеотидов. Кроме того, любые из гидроксильных групп, как правило, присутствующих в сахарах, можно заменять, например, фосфонатными группами, фосфатными группами, защищенными стандартными защитными группами или активированными для получения дополнительных связей с дополнительными нуклеотидами, или их можно конъюгировать с твердыми или полутвердыми подложками. 5'- и 3'-концевой OH можно фосфорилировать или заменять аминами или органическими кэпирующими группами из 1-20 атомов углерода. Другие гидроксилы также можно дериватизировать до стандартных защитных групп.Полинуклеотиды также могут содержать аналогичные формы рибозных или дезоксирибозных сахаров, как правило, известных в этой области, включая, например, 2'-O-метил-, 2'-O-аллил-, 2'-фтор- или 2'-азидо-рибозу, карбоциклические аналоги сахаров, альфа-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и основные аналоги нуклеозидов, такие как метилрибозид. Одну или более фосфодиэфирных связей можно заменять альтернативными связывающими группами. Эти альтернативные связывающие группы включают, в качестве неограничивающих примеров, варианты осуществления, где фосфат заменяют P(O)S ("тиоатом"), P(S)S ("дитиоатом"), "(O)NR2 ("амидатом"), P(O)R, P(O)OR', CO или CH2 ("формацеталем"), в которых каждый R или R' независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно, содержащий эфирную связь (--O--), арил, алкенил, циклоалкил, циклоалкенил или аралдил. Не все связи в полинуклеотиде должны быть идентичными. Предшествующее описание относится ко всем полинуклеотидам, приведенным в настоящем описании, включая РНК и ДНК.
[0071] "Одноцепочечный Fv" или "scFv"-фрагменты антител содержат домены VH и VL антитела, где эти домены находятся в одной полипептидной цепи. Как правило, полипептид scFv дополнительно содержит полипептидный линкер между доменами VH и VL, позволяющий scFv образовывать желаемую структуру для связывания антигена. На фиг.1 показана структура антитела и scFv. Обзор scFv см. в Pluckthun, The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994).
[0072] В рамках изобретения термин "по существу, схожий" или "по существу, тот же" означает достаточно высокую степень сходства между двумя числовыми значениями (например, одно из которых ассоциировано с антителом по настоящему изобретению, а другое ассоциировано с референсным/компараторным антителом), таким образом, что специалисту в этой области будет понятно, что различия между двумя значениями являются небольшими или не имеют биологической и/или статистической значимости в контексте биологической характеристики, измеряемой с помощью значений (например, значений Kd). Различие между указанными двумя значениями составляет, например, менее приблизительно 50%, менее приблизительно 40%, менее приблизительно 30%, менее приблизительно 20% и/или менее приблизительно 10% как функция референсного/компараторного значения.
[0073] В рамках изобретения фраза "значимо сниженный" или "значимо отличающийся" означает значимо высокую степень различия между двумя числовыми значениями (как правило, одно из которых ассоциировано с молекулой, и а другое ассоциировано с референсной/компараторной молекулой) таким образом, что специалисту в этой области будет понятно, что различие между двумя значениями имеет статистическую значимость в контексте биологической характеристики, измеряемой с помощью указанных значений (например, значений Kd). Различие между указанными двумя значениями составляет, например, более приблизительно 10%, более приблизительно 20%, более приблизительно 30%, более приблизительно 40% и/или более приблизительно 50% как функция значения для референсной/компараторной молекулы.
[0074] "TLR2-связанное заболевание и нарушения" включает, в качестве неограничивающих примеров, аутоиммунные заболевания, включая ревматоидный артрит, системную красную волчанку, системный склероз, синдром Шегрена, псориаз, рассеянный склероз и аутоиммунный диабет.TLR-связанные состояния (например, напрямую и/или косвенно ассоциированные с TLR, такие как TLR2, и т.д.) могут включать любой один или более из: диабета, ожирения, сепсиса, воспалительного заболевания (например, болезни Крона), иммунных нарушений, метаболического заболевания (например, состояний, ассоциированных с метаболическим синдромом), эндокринного заболевания, атеросклероза, астмы, сердечно-сосудистого заболевания, иммунологических состояний и/или любых других подходящих состояний. Например, TLR2-опосредованное заболевание или нарушение может быть выбрано из группы, состоящей из болезни Кавасаки, диабета 2 типа, ревматоидного артрита, дерматологического заболевания, рассеянного склероза, системной красной волчанки, язвенного колита, болезни Грейвса, синдрома Шегрена, аутоиммунных заболеваний щитовидной железы, васкулита и любой их комбинации.
[0075] В рамках изобретения термин "лечение" или "обработка" относится к клиническому вмешательству в попытке изменить природное состояние индивидуума, подвергаемого лечению, или клетки, подвергаемой обработке, и это можно осуществлять для профилактики или в ходе развития клинической патологии. Желаемые эффекты лечения включают профилактику возникновения или рецидивирования заболевания, облегчение симптомов, уменьшение любых прямых или косвенных патологических последствий заболевания, снижение скорости прогрессирования заболевания, улучшения или временного облегчения состояния заболевания и ремиссию или улучшение прогноза. В некоторых вариантах осуществления антитело (гуманизированное или негуманизированное), фрагмент антитела или полипептид по настоящему изобретению или гуманизированное антитело по настоящему изобретению используют для задержки развития заболевания или нарушения.
[0076] Термин "вариабельный" относится к тому факту, что части вариабельных доменов значительно отличаются по своей последовательности среди антитела и вносят вклад в связывание и специфичность каждого конкретного антитела к конкретному антигену. Однако вариабельность распределяется по вариабельным доменам антител неравномерно. Она сконцентрирована в трех сегментах, названных определяющими комплементарность областями или гипервариабельными областями (CDR или HVR, как взаимозаменяемо используют в настоящем описании) в вариабельных доменах легкой и тяжелой цепи. Более консервативные части вариабельных доменов называют каркасными (FR). Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит четыре области FR, в значительной степени имеющие конфигурацию β-листа, соединенные тремя HVR, образующими петли, соединяющие, и в некоторых случаях образующие часть, структуры β-листа. HVR в каждой цепи удерживаются в непосредственной близости областями FR и, вместе с HVR другой цепи, участвуют в образовании антигенсвязывающего участка антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, Md. (1991)). Константные домены напрямую не участвуют в связывании антитела с антигеном, но демонстрируют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности.
[0077] В рамках изобретения термин "вектор" предназначен для обозначения молекулы нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой он связан. Одним из типов вектора является "плазмида", являющаяся замкнутой кольцевой двухцепочечной ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другим типом вектора является фаговый вектор. Другим типом вектора является вирусный вектор, где дополнительные сегменты ДНК можно лигировать в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую их встраивают (например, бактериальные векторы, содержащие бактериальный участок начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут интегрироваться в геном клетки-хозяина мосле введения в клетку-хозяина и реплицироваться вместе с геномом хозяина. Кроме того, некоторые векторы могут направлять экспрессию генов, с которыми они функционально связаны. Такие векторы обозначают в настоящем описании как "экспрессирующие векторы". В основном, экспрессирующие векторы, подходящие для способов рекомбинантной ДНК, находятся в форме плазмид.
[0078] "Вариант Fc-области" содержит аминокислотную последовательность, отличающуюся от нативной последовательности Fc-области в силу по меньшей мере одной модификации аминокислоты, как правило, одной или более замен аминокислот.Как правило, вариант Fc-области содержит по меньшей мере одну замену аминокислоты по сравнению с нативной последовательностью Fc-области или Fc-областью родительского полипептида, например, от приблизительно одной до приблизительно десяти замен аминокислот и, как правило, от приблизительно одной до приблизительно пяти замен аминокислот в нативной последовательности Fc-области или в Fc-области родительского полипептида. Вариант Fc-области по настоящему изобретению имеет по меньшей мере приблизительно 80% гомологии в отношении нативной последовательности Fc-области и/или Fc-области родительского полипептида, по меньшей мере приблизительно 90% гомологии и, как правило, по меньшей мере приблизительно 95% гомологии.
[0079] Термин "окисленные фосфолипиды" (OxPL) относится к фосфолипидам с фосфохолиновой (PC) концевой группой. OxPL являются высоко провоспалительными и проатерогенными. Фосфорилхолин, полярная концевая группа на некоторых фосфолипидах, в значительной степени связана с развитием сердечно-сосудистых заболеваний. Активные формы кислорода, образующиеся во время коронарного воспаления, вызывают окисление липопротеинов низкой плотности (LDL) с образованием окисленных LDL (oxLDL). Фактически, показано, что сердечно-сосудистые заболевания (CVD), такие как атеросклероз, нестабильная стенокардия или острый коронарный синдром, ассоциированы с повышенными уровнями oxLDL в плазме. LDL представляют собой циркулирующую липопротеиновую частицу, содержащую липиды с полярной концевой группой PC и белки, в частности, белок apoB100.
[0080] Во время окисления LDL образуется PC, содержащий неоэпитопы, отсутствующие в немодифицированных LDL. Образовавшийся экспонируемый PC на oxLDL распознается скавенджер-рецепторами на макрофагах, таких как CD36, и образующиеся поглощенные макрофагами oxLDL способствуют образованию провоспалительных пенистых клеток в стенке сосуда. Окисленные LDL также распознаются рецепторами на поверхности эндотелиальной клетки и, как показано, стимулируют ряд ответов, включая дисфункцию эндотелия, апоптоз и ответ на несвернутые белки. Неоэпитопы PC также экспонируются на LDL после модификации под действием фосфолипазы A2 или аминореактивных метаболитов, связанных с заболеванием, таких как альдегиды, образующиеся при окислении гликированных белков. Эти альтернативно модифицированные частицы LDL также являются провоспалительными факторами при CVD. Показано, что антитела против фосфорилхолина (PC) связываются с окисленными или иначе модифицированными LDL и блокируют провоспалительную активность oxLDL в моделях in vivo или исследованиях in vitro.
[0081] Глицерофосфолипиды представляют собой распространенный класс липидов, важных для целостности клеточных мембран. Окисление этерифицированных ненасыщенных жирных кислот значительно изменяет биологическую активность фосфолипидов. Помимо нарушения структурной функции, окисление делает окисленные фосфолипиды (OxPL) маркерами "самомодифицированного" типа, распознаваемыми растворимыми и клеточно-ассоциированными рецепторами врожденного иммунитета, включая скавенджер-рецепторы, природные (кодируемые зародышевой линией) антитела и C-реактивный белок, таким образом, направляя удаление стареющих и апоптотических клеток или окисленных липопротеинов. Кроме того, OxPL приобретают новые виды биологической активности, нехарактерные для их неокисленных предшественников, включая способность регулировать врожденный и адаптивный иммунные ответы. Эффекты OxPL, описанные in vitro и in vivo, позволяют предполагать их потенциальную важность при разных патологиях, включая атеросклероз, острое воспаление, повреждение легких и многие другие состояния.
[0082] Глицерофосфолипиды содержат обширный класс липидов, состоящих из остова глицерина, фосфат-содержащей полярной концевой группы и двух остатков жирной кислоты. PL-связанные полиненасыщенные жирные кислоты (PUFA) представляют собой основную мишень неферментативного или ферментативного окисления, не связанную с образованием метаболической энергии. Окислительная фрагментация молекулы PL приводит к образованию нескольких биологически активных продуктов, включая небольшие реакционноспособными фрагментами PUFA, такими как неэтерифицированные окисленные жирные кислоты (например, гидропероксиды и изопростаны) и лизофосфолипиды. Эти продукты демонстрируют множество видов биологической активности. Доступные доказательства свидетельствуют о том, что неферментативное окисление PL-PUFA происходит в соответствии с теми же основными механизмами окисления свободных (неэтерифицированных) PUFA. Это предположение подтверждено определением схожих классов молекул, образующихся при окислении свободных и PL-связанных PUFA, представленных в настоящем описании. В отличие от неферментативного окисления, окисление PL-PUFA под действием ферментов значительно отличается от окисления неэтерифицированных PUFA. В то время как свободные PUFA могут окисляться множеством ферментов, принадлежащих к разным семействам белков и встраивающих различные окисленные группы, только одна группа липоксигеназ (12/15 липокисгеназы) использует PL-PUFA в качестве субстратов, продуцируя PL-гидропероксиды. Дальнейшее окисление и перестройка продолжаются без участия ферментов, и, таким образом, окисление, инициируемое ферментативными и неферментативными механизмами, приводит к образованию множества схожих продвинутых продуктов окисления PL.
[0083] Толл-подобный рецептор 2, также известный как TLR2, является белком, который у людей кодируется геном TLR2. TLR2 также обозначают как CD282 (кластер дифференцировки 282). TLR2 играет роль в иммунной системе. TLR2 является мембранным рецептором, экспрессирующимся на поверхности некоторых клеток, распознающим чужеродные вещества и передающим соответствующие сигналы в клетки иммунной системы. TLR2 играет фундаментальную роль в распознавании патогена и активации врожденного иммунитета. Толл-подобные рецепторы (TLR) являются высококонсервативными от дрозофилы до людей и обладают структурным и функциональным сходством. Они распознают патоген-ассоциированные молекулярные паттерны (PAMP), экспрессирующиеся на возбудителе инфекции, и опосредуют продукцию цитокинов, необходимых для развития эффективного иммунитета. Различные TLR имеют разные профили экспрессии. Этот ген экспрессируется больше всего в лейкоцитах периферической крови и опосредует ответ организма-хозяина на грамположительные бактерии и дрожжи посредством стимуляции NF-κB. TLR2 определяет широкий спектр микробных компонентов, таких как липотейхоевая кислота грамположительных бактерий, бактериальные липопротеины и зимозан. Из 11 охарактеризованных TLR, TLR2 является уникальным в силу своей способности гетеродимеризоваться с TLR1 или TLR6, что приводит к относительно широкой специфичности к лигандам.
[0084] CD36, являясь корецептором TLR2, позволяет предполагать, что существует провоспалительный путь между эндогенно образующимися липидами и активацией врожденного иммунитета. Исследования также показали, что повышенная эндотелиальная экспрессия и активация TLR2, возникающая в областях нарушения кровотока, таких как области предрасположенности к повреждению в стволе аорты и сердце. Таким образом, экспрессия TLR2 может способствовать атеросклерозу в клетках, не происходящих из BM, таких как эндотелиальные клетки, и, таким образом, может вносить вклад в провоспалительный фенотип активированных эндотелиальных клеток.
[0085] У предрасположенных к атеросклерозу мышей с дефицитом рецепторов липопротеинов низкой плотности (Ldlr-/-) полный дефицит TLR2 приводит к уменьшению атеросклероза. Однако утрата экспрессии TLR2 в клетках, происходящих из BM, не имеет эффекта в отношении прогрессирования заболевания. Данные позволяют предполагать, что неизвестный эндогенный агонист TLR2 влиял на прогрессирование повреждений посредством активации TLR2 в клетках, не происходящих из BM. Как показано в настоящем описании, при интраперитонеальном введении синтетического агониста TLR2/TLR1 Pam3CSK4 бремя заболевания значительно повышалось у мышей Ldlr-/-. Полный дефицит TLR2 у мышей Ldlr-/-, а также дефицит TLR2 только в полученных из BM клетках у мышей Ldlr-/- уменьшали Pam3CSK4-опосредованный атеросклероз, что позволяет предполагать роль экспрессии TLR2 клетками, полученными из BM, в трансдукции эффектов экзогенного агониста TLR2.
[0086] OxPL могут активировать передачу сигнала TLR2-опосредованными путями, что приводит к провоспалительной передаче сигнала в клетке. Кроме того, OxPL-опосредованная активация TLR2 может приводить к апоптозу и гибели клеток, если действует совместно с путями передачи сигнала, способствующему ER стрессу. OxPL индуцирует передачу сигнала ИЛ-8 из эндотелиальных клеток и индуцирует передачу сигнала ИЛ-1β и ФНОα в макрофагах через TLR2-зависимый путь передачи сигнала. Как показано в настоящем описании, активация макрофагов посредством синтетического агониста TLR2 PAM3CSK4 напрямую стимулирует макрофаги с образованием OxPL. Также показано, что активация TLR4 с помощью агонистов, таких как LPS, также будет приводить к образованию макрофагами OxPL (Popat et al., JCI, 2017). В целом, данные, представленные в настоящем описании, свидетельствуют о том, что OxPL может напрямую активировать макрофаги через TLR2 (или TLR4) для индуцирования провоспалительного сигнала и/или апоптоза, и напротив, активация макрофагов через передачу сигнала TLR2 или TLR4, в свою очередь, будет заставлять макрофаги образовывать OxPL. В случае последнего, если макрофаги стимулируют агонистами TLR2/4, локально образованные OxPL потенциально могут амплифицировать и усиливать воспалительный путь с помощью аутокринных эффектов. Таким образом, описанные исследования позволяют предполагать, что OxPL могут напрямую стимулировать пути TLR2, а также действовать паракринным образом для амплификации передачи сигнала провоспалительного агониста TLR2/4. Эти идеи позволяют объяснить, почему нейтрализация OxPL антителом против OxPL in vivo в различных воспалительных условиях обеспечивает такие выраженные противовоспалительные эффекты, которые проявляются в снижении развития заболевания.
[0087] Данные позволяют предполагать, что антитела или их фрагменты, связывающиеся с OxPL, включая EO6, или другие антитела, сконструированные для связывания с фосфохолиновыми (PC) концевыми группами PC-содержащих окисленных фосфолипидов (OxPL), можно использовать в улучшении неблагоприятных эффектов агонизма TLR2, происходящего при различных заболеваниях. Они включают атеросклероз, аутоиммунные нарушения и, в частности, болезнь Кавасаки, заболевание детей неизвестной этиологии, при котором TLR2-опосредованный агонизм, как считают, способствует артерииту коронарных сосудов, приводящему к аневризмам коронарных артерий, тяжелой кальцификации коронарных артерий, нарушению коронарного кровотока, острому тромбозу и значительной заболеваемости и смерти. Заболевание также может поражать молодежь, когда бессимптомные аневризмы коронарных артерий преобразуются в острый тромбоз, вызывая острый инфаркт миокарда. Болезнь Кавасаки также может быть ассоциирована с миокардитом, сердечной недостаточностью и необходимостью трансплантации сердца.
[0088] Врожденные природные антитела (NAb) представляют собой первую линию защиты организма-хозяина против распространенных окислительно-специфических эпитопов (OSE) на эндогенных неоэпитопах (OxLDL и апоптотических клетках) и экзогенных эпитопах патогенов и поддерживают гомеостаз организма-хозяина. OSE являются убиквитарными, образуются во многих воспалительных тканях, включая атеросклеротические повреждения, и являются основной мишенью IgM NAb. Типичное IgM NAb EO6 связывается с фосфохолиновой (PC) концевой группой в окисленных фосфолипидах (OxPL) и блокирует захват OxLDL макрофагами. Клонировано и охарактеризовано природное антитело IgM мыши против OxPL, связывающееся с фосфорилхолиновой ("PC") концевой группой OxPL, но не с нативными неокисленными фосфолипидами ("PL"). Однако, антитела, подобные IgM Nab EO6, имеют ограниченную растворимость, и их нелегко синтезировать.
[0089] Родительское антитело EO6 является антителом IgM мыши, клонированным, охарактеризованным и являющимся объектом патента США №6225070, включенного в настоящее описание в качестве ссылки. В патентной публикации США №20150376268 A1 описывают полностью функциональное одноцепочечное антитело и гуманизированные антитела, связывающиеся с OxPL. В ней описывают многочисленные уникальные молекулярные изменения в последовательности ДНК каркасных областей родительского антитела, тяжелых и легких цепей и линкерных последовательностей, которые определяли с помощью повторных раундов экспериментов, что привело к разработке полностью функционального EO6-scFv. Когда эту последовательность встраивают в соответствующий вектор, полученный scFc экспрессируется в растворенной форме и обладает всеми иммунологическими связывающими свойствами родительского антитела в отношении его идентифицированных антигенов-мишеней, включая способность связываться с уникальным антиидиотипическим антителом AB1-2, эпитопы которого состоят из тяжелых и легких цепей родительского антитела. Описание той заявки также относится к одноцепочечным вариабельным фрагментам антител ("scFv") VH, VL и определяющим комплементарность областям, селективно связывающимся с окисленными фосфолипидами. ScFv по изобретению являются растворимыми, и их легко синтезировать. Описание патентной публикации США №20150376268 A1 включено в настоящее описание в качестве ссылки для всех целей.
[0090] В исследованиях, представленных в настоящем описании, нейтрализация OxPL посредством эндогенной экспрессии in vivo антитела EO6 (с использованием EO6-scFv-трансгенной мыши) значительно ингибировала образование атеросклероза, вызванное агонизмом TLR2. В частности, инъекции агониста TLR2 PAM3CSK4 мышам Ldlr-/-, которых держали на питании с высоким содержанием холестерина, приводит к значительному повышению атеросклероза. Схожий набор инъекций EO6-scFv-трансгенным мышам (на фоне Ldlr-/-) приводил к значительному ингибированию образования повреждений.
[0091] В других исследованиях, представленных в настоящем описании, нейтрализация OxPL может защищать в модели болезни Кавасаки на мышах. Показано, что введение патогена Lactobaccilus casei выбывает заболевание, подобное болезни Кавасаки, у мышей с образующимся повышением атеросклероза, артериита коронарных артерий и аневризм брюшной аорты. Это зависит от TLR2, т.к. введение L. casei мышам с дефицитом TLR2 не вызывает заболевания. Важно, что, показано, что в этом участвует ИЛ-1, и как отмечают, OxPL также являются мощным индуктором высвобождения ИЛ-1. Инъекция L. casei EO6-трансгенным мышам (с фоном Ldlr-/-) по идентичному протоколу приводила к значительному снижению не только атеросклероза, но, что еще важнее, артериита коронарных артерий по сравнению с инъекциями мышей Ldlr-/-. Антитело EO6 не связывается с L. casei напрямую и, таким образом, нейтрализует OxPL, образование которых вызвано воспалительными эффектами, ассоциированными с TLR2-опосредованным агонизмом. Развитие артериита коронарных артерий, а затем аневризм коронарных артерий является основным и очень опасным осложнением у детей, у которых развивается болезнь Кавасаки, и, по разным оценкам, оно происходит у до 25% детей, несмотря на существующую терапию. Как правило, болезнь Кавасаки лечат с использованием внутривенного иммуноглобулина (IVIG), полученного из смешанной и очищенной плазмы человека, и аспирина (общего, но неспецифического противовоспалительного терапевтического средства). Инъекции высоких титров гуманизированных или эквивалентных человеческим антител против OxPL, модифицированных для повышения биологической эффективности, затем могут обеспечивать защиту без каких-либо ожидаемых побочных эффектов, т.к. такие антитела против OxPL присутствует в репертуаре B-клеток человека.
[0092] Таким образом, экспериментальные данные свидетельствуют о том, что атеросклероз и воспалительный артериит, вызванные TLR2 опосредованным агонизмом in vivo у мышей, можно предотвращать посредством нейтрализации OxPL. Разумеется, агонизм TLR2 вносит вклад во множество бактериальных заболеваний, но также и в различные так называемые аутоиммунно-опосредованные заболевания, такие как системная красная волчанка, ревматоидный артрит и др. Данные свидетельствуют о том, что нейтрализация OxPL с помощью антител против PC OxPL может улучшать или предотвращать многие заболевания, усиливаемые или подвергающиеся влиянию активации TLR-опосредованных путей передачи сигнала.
[0093] Настоящее изобретение относится к применению антител, фрагментов антител и гуманизированных антител, связывающихся с OxPL и, в некоторых случаях, имеющих ту же или схожую специфичность связывания с антителом EO6. Фрагменты антител можно получать общепринятыми способами, такими как ферментативное расщепление, или рекомбинантными способами. В некоторых случаях существуют преимущества использования фрагментов антител, а не целых антител. Меньший размер фрагментов делает возможным быстрый клиренс и может приводить к лучшему доступу в ткань. В острых условиях время полужизни фрагментов антител не является критическим. Обзор некоторых фрагментов антител, см. Hudson et al. (2003) Nat. Med. 9:129-134.
[0094] Хотя в настоящем описании представлены конкретные последовательности антител и последовательности фрагментов антител, имеющих биологическую активность, в нем дополнительно описано, что эти последовательности можно использовать для получения улучшенных вариантов. Таким образом, в некоторых случаях антитело или фрагмент антитела может иметь процент идентичности в отношении последовательности по настоящему изобретению.
[0095] В некоторых вариантах осуществления предусмотрены модификации аминокислотной последовательности антител, представленных в настоящем описании. Например, это может быть желательным для улучшения аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела можно получать посредством внесения соответствующих изменений в нуклеотидную последовательность, кодирующую антитело, или посредством пептидного синтеза. Такие модификации включают, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. Для получения конечного продукта можно осуществлять любую комбинацию делеции, инсерции и замены при условии, что конечная конструкция обладает желаемыми характеристиками. Изменения аминокислот можно вносить в аминокислотную последовательность антитела во время получения последовательности.
[0096] Настоящее изобретение относится к антителу или фрагменту антитела, способному связываться с OxPL, или фосфорилхолином, и/или конъюгатом фосфорилхолина, где антитело или фрагмент антитела содержит вариабельный домен тяжелой цепи (VH) и/или вариабельный домен легкой цепи (VL), и где
(a) домен VH содержит аминокислотную последовательность, включающую одну, две или три определяющие комплементарность области (CDR), выбранные из группы, состоящей из:
SEQ ID NO: 6 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 6;
SEQ ID NO: 7 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 7; и
SEQ ID NO: 8 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 8;
(b) домен VL содержит аминокислотную последовательность, включающую одну, две или три определяющие комплементарность области (CDR), выбранные из группы, состоящей из:
SEQ ID NO: 9 или 12 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 9 или 12;
SEQ ID NO: 10 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 10; и
SEQ ID NO: 11 и последовательности, являющейся по меньшей мере на 95%, 96%, 97%, 98%, 99% или 99,9% идентичной SEQ ID NO: 11.
[0097] В одном из вариантов осуществления антитело или фрагмент антитела содержит домен VH, содержащий аминокислотную последовательность, включающую CDR, содержащие SEQ ID NO: 6, 7 и 8, и/или домен VL, содержащий аминокислотную последовательность, включающую CDR, содержащие SEQ ID NO: 9, 10 и 11 или SEQ ID NO: 10, 11 и 12.
[0098] В одном из вариантов осуществления настоящее изобретение относится к антителу или scFv, выбранному из группы, состоящей из: (a) антитела или scFv с доменами тяжелой и легкой цепи, содержащими определяющие комплементарность области SEQ ID NO: 6, 7, 8, 9, 10 и 11; и (b) антитело или scFv с доменами тяжелой и легкой цепи, содержащими определяющие комплементарность области SEQ ID NO: 6, 7, 8, 10, 11 и 12. В одном из вариантов осуществления любые из (a) или (b) соединены с Fc-областью.
[0099] В одном из вариантов осуществления настоящее изобретение относится к антителу, содержащему вариабельную область легкой цепи, приведенную в SEQ ID NO: 2 от аминокислоты 1 до приблизительно аминокислоты 146. В другом варианте осуществления настоящее изобретение относится к антителу с гуманизированной вариабельной областью легкой цепи, содержащей последовательность SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 135. В другом варианте осуществления настоящее изобретение относится к антителу, содержащему вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO: 2 от приблизительно аминокислоты 162 до приблизительно аминокислоты 269. В другом варианте осуществления настоящее изобретение относится к антителу, содержащему гуманизированную вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO: 4 от приблизительно аминокислоты 152 до приблизительно аминокислоты 258.
[00100] В другом варианте осуществления настоящее изобретение относится к химерному антителу, содержащему, например, VH и/или VL мыши и Fc-область человека. Например, SEQ ID NO: 14 включает последовательность химерного антитела по настоящему изобретению. В SEQ ID NO: 14 аминокислоты 1-33 содержат лидерную последовательность каппа-цепи Ig для секреции антитела; аминокислоты 34-146 содержат вариабельную область легкой цепи EO6; аминокислоты 147-161 включают гибкую линкерную последовательность; аминокислоты 162-284 содержат вариабельную область тяжелой цепи EO6 с тройной мутацией P201A, S224A и A225D относительно антитела EO6 мыши дикого типа; аминокислоты 285-517 содержат Fc-область, в SEQ ID NO: 14 Fc-область является IgG1-Fc человека с модификацией C290S и H294Y для повышения активности ADCC. SEQ ID NO: 14 также включает дополнительный линкер и последовательность His-метки, которые, как очевидно специалисту в этой области, необязательны (например, SEQ ID NO: 14 от аминокислоты 518 до аминокислоты 528). Настоящее изобретение также включает кодирующую последовательность SEQ ID NO: 14, содержащую SEQ ID NO: 13. Специалист в этой области легко может определять последовательность нуклеиновой кислоты, соответствующую различным доменам, определенным выше. Настоящее изобретение также включает последовательность химерного антитела, т.е. по меньшей мере на 90%, 95%, 96%, 97%, 98%, 99% или 99,9% идентичного SEQ ID NO: 14 от аминокислоты 1 до аминокислоты 284, связанным с Fc-областью другого иммуноглобулина (например, IgA, IgD, IgE, IgG и IgM или любого из подклассов (изотипов), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2).
[00101] В одном из вариантов осуществления настоящее изобретение относится к scFv, содержащему линкер между вариабельной областью легкой цепи и вариабельной областью тяжелой цепи. Линкер может представлять собой любое количество общеупотребительных пептидных линкеров. В одном из вариантов осуществления линкер содержит повторяющуюся единицу GGGS (SEQ ID NO: 5). GGGS (SEQ ID NO: 5) может повторяться 2, 3, 4 или более раз.
[00102] В другом варианте осуществления настоящее изобретение включает scFv, содержащий вариабельную область легкой цепи SEQ ID NO: 2 от аминокислоты 1 до аминокислоты 146, связанные пептидным линкером с вариабельной областью тяжелой цепи SEQ ID NO: 2 от аминокислоты 162 до приблизительно аминокислоты 269. В конкретном варианте осуществления scFv содержит последовательность SEQ ID NO: 2 от аминокислоты 1 до аминокислоты 269. В другом варианте осуществления настоящее изобретение относится к scFv, имеющему полипептидную последовательность SEQ ID NO: 2 от аминокислоты 1 до приблизительно аминокислоты 269 или от аминокислоты 1 до приблизительно аминокислоты 303. В дополнительном варианте осуществления настоящее изобретение относится к scFv, имеющему полипептидную последовательность, имеющую по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности в отношении SEQ ID NO: 2 от аминокислоты 1 до приблизительно аминокислоты 303, и селективно связывающуюся с окисленным фосфолипидом.
[00103] В дополнительных вариантах осуществления слитые конструкции содержат первый домен, содержащий SEQ ID NO: 2 от аминокислоты 1 до приблизительно аминокислоты 269 или от аминокислоты 1 до приблизительно аминокислоты 303 или ее вариант, функционально связанныый со вторым доменом, содержащим (i) детектируемую метку или (ii) интересующий полипептид. Специалисту в этой области будет понятно, что такие слитые конструкции можно получать химическими способами или способами молекулярной биологии, с помощью которых соединяют кодирующую последовательность, содержащую последовательность SEQ ID NO: 1 или ее вариант, с кодирующей последовательностью, например, интересующего полипептида. Кодирующие последовательности и домены можно разделять линкером или соединять напрямую.
[00104] В еще одном варианте осуществления настоящее изобретение включает scFv, содержащий вариабельную область легкой цепи SEQ ID NO: 4 от аминокислоты 1 до аминокислоты 135, связанные посредством пептидного линкера с вариабельной областью тяжелой цепи SEQ ID NO: 4 от аминокислоты 152 до приблизительно аминокислоты 258. В конкретном варианте осуществления scFv содержит последовательность SEQ ID NO: 4 от аминокислоты 1 до аминокислоты 258. В другом варианте осуществления настоящее изобретение относится к scFv, имеющему полипептидную последовательность SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 258 или от аминокислоты 1 до приблизительно аминокислоты 263. В дополнительном варианте осуществления настоящее изобретение относится к scFv, имеющему полипептидную последовательность, имеющую по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 258, и селективно связывающемуся с окисленным фосфолипидом.
[00105] В дополнительных вариантах осуществления слитые конструкции содержат первый домен, содержащий SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 258 или от аминокислоты 1 до приблизительно аминокислоты 264 или ее вариант, функционально связанный со вторым доменом, содержащим (i) детектируемую метку или (ii) интересующий полипептид. Специалисту в этой области будет понятно, что такие слитые конструкции можно получать химическими способами или способами молекулярной биологии, с помощью которых соединяют кодирующую последовательность, содержащую последовательность SEQ ID NO: 3 или ее вариант, с кодирующей последовательность, например, интересующего полипептида. Кодирующие последовательности и домены можно разделять линкером или соединять напрямую.
[00106] Молекулы нуклеиновой кислоты, кодирующие аминокислотные последовательности антител, фрагментов антител и вариантов антител, получают различными, известными в этой области способами. В случае получения вариантов такие способы включают, в качестве неограничивающих примеров, выделение из природного источника (в случае вариантов природной аминокислотной последовательности) или получение посредством олигонуклеотид-опосредованного (или сайт-специфического) мутагенеза, ПЦР-мутагенеза и кассетного мутагенеза ранее полученного варианта или невариантной версии антитела.
[00107] В конкретном варианте осуществления настоящее изобретение относится к scFv мыши, кодируемому полинуклеотидной последовательностью SEQ ID NO: 1. В дополнительном варианте осуществления настоящее изобретение относится к scFv мыши, кодируемому полинуклеотидной последовательностью, имеющей по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к SEQ ID NO: 1 и приводящей к образованию полипептида, селективно связывающегося с окисленными фосфолипидами.
[00108] В другом варианте осуществления настоящее изобретение относится к scFv, содержащему линкер между вариабельной областью легкой цепи и вариабельной областью тяжелой цепи. Линкер может представлять собой любое количество общеупотребительных пептидных линкеров. В одном из вариантов осуществления линкер содержит повторяющуюся единицу GGGS (SEQ ID NO: 5). GGGS (SEQ ID NO: 5) может повторяться 2, 3, 4 или более раз.
[00109] Настоящее изобретение также включает гуманизированные антитела. В этой области известны различные способы гуманизации не принадлежащих человеку антител. Например, гуманизированное антитело может содержать один или более аминокислотных остатков, встроенных в него из источника, не принадлежащего человеку. Эти непринадлежащие человеку аминокислотные остатки зачастую обозначают как "импортные" остатки, как правило, взятые из "импортного" вариабельного домена. Гуманизацию, по существу, можно осуществлять способом Winter и соавт.(Jones et al. (1986) Nature 321:522-525; Riechmann et al. (1988) Nature 332:323-327; Verhoeyen et al. (1988) Science 239:1534-1536) посредством замены последовательностей гипервариабельной области соответствующими последовательностями антитела человека. Таким образом, такие "гуманизированные" антитела являются химерными антителами (патент США №4816567), где, по существу, меньший, чем интактный вариабельный домен человека, заменяют соответствующей последовательностью не являющихся человеком видов. На практике, гуманизированные антитела, как правило, являются антителами человека, в которых некоторые остатки гипервариабельной области и, вероятно, некоторые остатки FR заменяют остатками из аналогичных участков в антителах грызуна.
[00110] При выборе вариабельных доменов человека легких и тяжелых цепей для использования в получении гуманизированных антител может быть важным снижение антигенности. В соответствии с так называемым способом "лучшего соответствия", последовательность вариабельного домена антитела грызуна подвергают скринингу по всей библиотеке известных последовательностей вариабельных доменов человека. Затем последовательность человека, наиболее близкую к последовательности грызуна, принимают в качестве каркаса человека для гуманизированного антитела. См., например, Sims et al. (1993) J. Immunol. 151:2296; Chothia et al. (1987) J. Mol. Biol. 196:901. В другом способе используют конкретный каркас, полученный из консенсусной последовательности всех антител человека из конкретной подгруппы легких или тяжелых цепей. Тот же каркас можно использовать для нескольких других гуманизированных антител. См., например, Carter et al. (1992) Proc. Natl. Acad. Sci. USA, 89:4285; Presta et al. (1993) J. Immunol., 151:2623.
[00111] Как правило, также желательно гуманизировать антитела с сохранением высокой аффинности к антигену и других благоприятных биологических свойств. Для достижения этой цели одним способом гуманизированные антитела получают посредством анализа родительских последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и хорошо известны специалистам в этой области. Доступны компьютерные программы, с помощью которых иллюстрируют и отображают возможные трехмерные конформационные структуры выбранных иммуноглобулиновых последовательностей-кандидатов. Изучение этих отображений позволяет анализировать возможную роль остатков в функционировании иммуноглобулиновой последовательности-кандидата, т.е. анализ остатков, влияющих на способность иммуноглобулина-кандидата связываться с его антигеном. Таким образом, остатки FR можно выбирать и комбинировать из реципиента и импортированных последовательностей так, чтобы достигать желаемой характеристики антитела, такой как повышенная аффинность для антигенов-мишеней. В основном, остатки гипервариабельной области напрямую и наиболее существенно влияют на связывание антигена.
[00112] В конкретном варианте осуществления настоящее изобретение относится к гуманизированному scFv, кодируемому полинуклеотидной последовательностью SEQ ID NO: 3. В дополнительном варианте осуществления настоящее изобретение относится к гуманизированному scFv, кодируемому полинуклеотидной последовательностью, имеющей по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к SEQ ID NO: 3 и приводящей к получению полипептида, селективно связывающегося с окисленными фосфолипидами.
[00113] Настоящее изобретение также относится к scFv, представленному в настоящем описании, дополнительно содержащему кристаллизуемый фрагмент ("Fc") антитела. В конкретном варианте осуществления Fc-область получают из антитела человека или гуманизированного антитела. Fc-область представляет собой хвостовую область антитела, взаимодействующего с рецепторами поверхности клетки, названными Fc-рецепторами, и некоторыми белками системы комплемента. Это свойство позволяет антителам активировать иммунную систему. В изотипах антител IgG, IgA и IgD Fc-область состоит из двух идентичных белковых фрагментов, полученных из второго и третьего константных доменов двух тяжелых цепей антитела; Fc-области IgM и IgE содержат три константных домена тяжелой цепи (домены CH 2-4) в каждой полипептидной цепи. Fc-области IgG несут высококонсервативные участки N-гликозилирования. Гликозилирование Fc-фрагмента необходимо для опосредованной Fc-рецептором активности. N-гликаны, присоединенные к этому участку, преимущественно являются кор-фукозилированными биантненнарными структурами комплексного типа. Кроме того, небольшие количества этих N-гликанов также несут бисектный GlcNAc и α-2,6-связанные остатки сиаловой кислоты. Другая часть антитела, названная Fab-областью, содержит вариабельные секции, определяющие специфическую мишень, которую может связывать антитело. scFv по настоящему изобретению состоят из элементов Fab-области. В отличие от этого, Fc-области всех антител в классе являются одинаковыми для каждого биологического вида; они являются константными, а не вариабельными. Таким образом, Fc-область иногда обозначают как "фрагмент-константная область". Таким образом, полинуклеотидные и полипептидные последовательности, кодирующие Fc-области для бесконечного количества биологических видов, уже определены и известны специалисту в этой области. В частности, в варианте осуществления настоящее изобретение относится к полинуклеотидной последовательности scFv, представленной в настоящем описании, дополнительно содержащей полинуклеотидную последовательность, кодирующую Fc-область из антитела IgG (например, антитела IgG человека). В дополнительном варианте осуществления настоящее изобретение относится к полипептидной последовательности scFv, представленной в настоящем описании, дополнительно содержащей полипептидную последовательность Fc-области антитела IgG.
[00114] В частности, в варианте осуществления настоящее изобретение относится к полинуклеотидной последовательности scFv, представленной в настоящем описании, дополнительно содержащей полинуклеотидную последовательность, кодирующую Fc-область антитела IgG (например, антитела IgG человека). В дополнительном варианте осуществления настоящее изобретение относится к полипептидной последовательности scFv, представленной в настоящем описании, дополнительно содержащей полипептидную последовательность Fc-области из антитела IgG. В одном из вариантов осуществления кодирующая последовательность Fc-области содержит последовательность, приведенную в SEQ ID NO: 3 от приблизительно нуклеотида 790 до приблизительно нуклеотида 1518.
[001115] В дополнительном варианте осуществления настоящее изобретение относится к вектору, содержащему полинуклеотидную последовательность, кодирующую scFv, приведенный выше со ссылкой на SEQ ID NO: 1 и 3, или последовательности, имеющие идентичность последовательности по меньшей мере 99%, по меньшей мере 95%, по меньшей мере 90%, по меньшей мере 85%, по меньшей мере 80%, по меньшей мере 75% или по меньшей мере 70% идентичности по отношению к SEQ ID NO: 1 или SEQ ID NO: 3.
[00116] Настоящее изобретение также относится к гуманизированному антителу, имеющему специфичность связывания антитела EO6. Гуманизированное антитело содержит (i) последовательность, приведенную в SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 506, или (ii) последовательность, являющуюся по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,8% идентичной по отношению к SEQ ID NO: 4 от аминокислоты 1 до приблизительно аминокислоты 506.
[00117] Настоящее изобретение также относится к полинуклеотиду, кодирующему гуманизированное антитело по настоящему изобретению. Полинуклеотид содержит последовательность, выбранную из группы, состоящей из (i) полинуклеотида, кодирующего SEQ ID NO: 4, (ii) полинуклеотида, гибридизующегося в строгих условиях с полинуклеотидом, состоящим из SEQ ID NO: 3 и кодирующим гуманизированное антитело, связывающееся с OxPL со специфичностью, достаточно схожей с антителом EO6, (iii) полинуклеотида, являющегося по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,5% или 99,8% идентичным SEQ ID NO: 3 и кодирующим антитело, связывающееся OxPL со специфичностью, достаточно схожей с антителом EO6; (iv) полинуклеотида, приведенного в SEQ ID NO: 3; (v) полинуклеотида по любому из пп. (i)-(iv), где полинуклеотид содержит РНК.
[00118] Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела или фрагментов антител по настоящему изобретению, можно получать стандартными рекомбинантными способами. Желаемые полинуклеотидные последовательности можно выделять и секвенировать из продуцирующих антитела клеток, таких как гибридомные клетки. Альтернативно, полинуклеотиды можно синтезировать с использованием нуклеотидного синтезатора или способов ПЦР. После получения, последовательности, кодирующие полипептиды, встраивают в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерологичные полинуклеотиды в прокариотических клетках-хозяевах. Многие векторы, доступные и известные в этой области, можно использовать для целей по настоящему изобретению. Выбор подходящего вектора будет зависеть, в основном, от размера нуклеиновых кислот, подлежащих встраиванию в вектор, и конкретной клетки-хозяина, подлежащей трансформации с использованием вектора. Каждый вектор содержит различные компоненты в зависимости от своей функции (амплификации, или экспрессии гетерологичного полинуклеотида, или и того, и другого) и совместимости с конкретной клеткой-хозяином, в которой он находится. Компоненты вектора, как правило, включают, в качестве неограничивающих примеров: участок начала репликации, маркерный ген для селекции, промотор, участок связывания рибосомы (RBS), сигнальную последовательность, вставку гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции.
[00119] В основном, плазмидные векторы, содержащие репликон и контрольные последовательности, полученные из видов, совместимых с клеткой-хозяином, используют вместе с этими хозяевами. Вектор, как правило, несет участок начала репликации, а также маркерные последовательности, с помощью которых можно осуществлять фенотипическую селекции трансформированных клеток. Например, E.coli, как правило, трансформируют с использованием pBR322, плазмиды, полученной из вида E. Coli. pBR322 содержит гены, кодирующие резистентность к ампициллину (Amp) и тетрациклину (Tet), и, таким образом, обеспечивает простоту идентификации трансформированных клеток. pBR322, ее производные или другие микробные плазмиды или бактериофаги также могут содержать, или их можно модифицировать так, чтобы они содержали, промоторы, которые микроорганизм может использовать для экспрессии эндогенных белков. Примеры производных pBR322, используемые для экспрессии конкретных антител, подробно описаны в Carter et al., патенте США №5648237.
[00120] Кроме того, фаговые векторы, содержащие репликон и контрольные последовательности, совместимые с микроорганизмом-хозяином, можно использовать в качестве трансформирующих векторов вместе с этими хозяевами. Например, бактериофаговые векторы можно использовать в получении рекомбинантного вектора, который можно использовать для трансформации восприимчивых клеток-хозяев, таких как E.coli LE392.
[00121] Экспрессирующий вектор по настоящему изобретению может содержать две или более пары промотор-цистрон, кодирующие каждый из полипептидных компонентов. Промотор является нетранслируемой регуляторной последовательностью, находящейся выше (5') цистрона и модулирующей его экспрессию. Прокариотические промоторы, как правило, относятся к двум классам, индуцибельные и конститутивные. Индуцибельный промотор является промотором, инициирующим повышенные уровни транскрипции цистрона под своим контролем в ответ на изменения условий культивирования, например, наличие или отсутствие питательного вещества или изменение температуры.
[00122] Хорошо известно большое количество промоторов, распознаваемых различными потенциальными клетками-хозяевами. Выбранный промотор может быть функционально связан с цистроном ДНК, кодирующим легкую или тяжелую цепь посредством удаления промотора из источника ДНК посредством расщепления ферментами рестрикции и встраивания промоторной последовательности в вектор по изобретению. Для регуляции амплификации и/или экспрессии генов-мишеней можно использовать нативную промоторную последовательность и многие гетерологичные промоторы. В некоторых вариантах осуществления используют гетерологичные промоторы, т.к. они, как правило, делают возможной более высокую транскрипцию и более высокие выходы экспрессируемого гена-мишени по сравнению с нативным промотором полипептида-мишени.
[00123] Промоторы, подходящие для использования с прокариотическими хозяевами, включают промотор PhoA, системы промотора β-галактамазы и лактозы, систему триптофанового (trp) промотора и гибридные промоторы, такие как промотор tac или trc. Однако также подходят другие промоторы, функционирующие в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, что позволяет специалисту в этой области функционально лигировать их с цистронами, кодирующими целевые легкие и тяжелые цепи (Siebenlist et al., (1980) Cell 20: 269) с использованием линкеров или адапторов для обеспечения любых необходимых участков рестрикции.
[00124] В другом варианте осуществления продукция иммуноглобулинов по настоящему изобретению может происходить в цитоплазме клетки-хозяина и, таким образом, не требует наличия секреторных сигнальных последовательностей в каждом цистроне. В связи с этим, легкие и тяжелые цепи иммуноглобулинов экспрессируются, сворачиваются и собираются с образованием функциональных иммуноглобулинов в цитоплазме. Некоторые штаммы клеток-хозяев (например, штаммы trxB E.coli) обеспечивают цитоплазматические условия, благоприятные для образования дисульфидной связи, таким образом, делая возможным правильный фолдинг и сборку экспрессируемых белковых субъединиц. Proba and Pluckthun Gene, 159:203 (1995).
[00125] Прокариотические клетки-хозяева, подходящие для экспрессии антител по изобретению, включают Archaebacteria и Eubacteria, такие как грамотрицательные или грамположительные организмы. Примеры бактерий, которые можно использовать, включают Escherichia (например, E.coli), Bacilli (например, B. subtilis), Enterobacteria, виды Pseudomonas (например, P. aeruginosa), Salmonella typhimurium, Serratia marcescans, Klebsiella, Proteus, Shigella, Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов осуществления используют грамотрицательные клетки. В одном из вариантов осуществления в качестве клеток-хозяев по настоящему изобретению используют клетки E.coli. Примеры штаммов E.coli включают штамм W3110 (Bachmann, Cellular and Molecular biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp.1190-1219; ATCC, депозит №27,325) и их производные, включая штамм 33D3 (патент США №5639635). Также подходят другие штаммы и их производные, такие как E.coli 294 (ATCC 31,446), E.coli B, E.coli X 1776 (ATCC 31,537) и E.coli RV308. Эти примеры являются иллюстративными, а не ограничивающими. Способы конструирования производных любых из указанных выше бактерий, имеющих определенные генотипы, известны в этой области и описаны, например, в Bass et al., Proteins, 8:309-314 (1990). Как правило, необходимо выбирать подходящие бактерии с учетом реплицируемости репликона в клетках бактерии. Например, E.coli, виды Serratia или Salmonella можно соответствующим образом использовать в качестве организма-хозяина, когда для доставки репликона используют хорошо известные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Как правило, клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и желательным может являться включение дополнительных ингибиторов протеаз в культуру клеток.
[00126] Клетки-хозяева трансформируют с использованием описанных выше экспрессирующих векторов и культивируют в общепринятых питательных средах, модифицированных, при необходимости, для индуцирования промоторов, селекции трансформантов или амплификации генов, кодирующих желаемые последовательности.
[00127] Термин "трансформация" означает встраивание ДНК в прокариотического хозяина таким образом, что ДНК реплицируется как экстрахромосомный элемент или посредством встраивания в хромосому. В зависимости от используемой клетки-хозяина трансформацию осуществляют стандартными способами, подходящими для таких клеток. Обработку кальцием с помощью хлорида кальция, как правило, используют для бактериальных клеток, содержащих значительные клеточные стенки. В другом способе для трансформации используют полиэтиленгликоль/DMSO. Другим используемым способом является электропорация.
[00128] Прокариотические клетки, используемые для получения полипептидов по настоящему изобретению, выращивают в средах, известных в этой области и подходящих для культивирования выбранных клеток-хозяев. Примеры подходящих сред включают бульон Луриа (LB) с необходимыми добавками питательных веществ. В некоторых вариантах осуществления среды также содержат средство для селекции, выбранное с учетом конструкции экспрессирующего вектора, для селективного роста прокариотических клеток, содержащих экспрессирующий вектор. Например, ампициллин добавляют в среду для роста клеток, экспрессирующих ген резистентности к ампициллину.
[00129] Любые необходимые дополнительные вещества, помимо источников углерода, азота и неорганического фосфата, также можно включать в подходящих концентрациях в отдельности или смеси с другим дополнительным веществом или средой, таким как комплексный источник азота. Необязательно, среда для культивирования может содержать один или более восстановителей, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритрита и дитиотреитола. Прокариотические клетки-хозяева культивируют при подходящих температурах.
[00130] В одном из вариантов осуществления экспрессируемые полипептиды секретируются в периплазму клеток-хозяев, и их выделяют из нее. Выделение белков, как правило, включает разрушение микроорганизма, как правило, посредством осмотического шока, обработки ультразвуком или лизиса. После разрушения клеток, клеточный детрит или цельные клетки можно удалять посредством центрифугирования или фильтрации. Белки можно дополнительно очищать, например, с помощью смолы для аффинной хроматографии. Альтернативно, белки могут транспортироваться в среды для культивирования, и их можно выделять из нее. Можно удалять клетки из культуры и супернатант культуры можно фильтровать и концентрировать для дальнейшей очистки продуцируемого белка. Экспрессируемые полипептиды можно дополнительно выделять и идентифицировать с использованием общеизвестных способов, таких как электрофорез в полиакриламидном геле (ПААГ) и вестерн-блоттинг. Специалист в этой области может использовать и оптимизировать крупномасштабную или маломасштабную ферментацию.
[00131] Можно использовать стандартные способы очистки белков, известные в этой области. Следующие способы являются примерами подходящих способов очистки: фракционирование на иммуноаффинных или ионообменных колонках, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на диоксиде кремния или катионообменной смоле, такой как DEAE, хроматофокусирование, электрофорез в ПААГ в присутствии SDS, осаждение сульфатом аммония и гель-фильтрация.
[00132] Настоящее изобретение также относится к экспрессирующему вектору, кодирующему антитело, фрагмент антитела или полипептид, представленные в настоящем описании, переносимому в подходящий организм-хозяина. Подходящий организм-хозяин является микроорганизмом, дрожжами или системой клеток млекопитающего. Как правило, система клеток млекопитающего представляет собой клетки моноцитарного происхождения (например, макрофаги, моноциты и нейтрофилы), клетки лимфоцитарного происхождения (например, миелома, гибридома и нормальная иммортализованная B-клетка), паренхимные (например, гепатоциты) и непаренхимные клетки (например, звездчатые клетки).
[00133] Настоящее изобретение также относится к фармацевтическим композициям или составам, содержащим терапевтически эффективное количество антитела, фрагмента антитела или полипептида по настоящему изобретению. Фармацевтические композиции или составы могут дополнительно содержать носители, эксципиенты, дилюенты, солюбилизаторы, стабилизаторы, буферы, модификаторы тоничности, объемообразующие средства, средства, повышающие/снижающие вязкость, поверхностно-активные вещества, хелатирующие средства и адъюванты.
[00134] "Терапевтически эффективное количество" вещества/молекулы по настоящему изобретению, агониста или антагониста могут варьироваться в зависимости от таких факторов, как состояние заболевания, возраст, пол и масса индивидуума, и способности вещества/молекулы, агониста или антагониста вызывать желаемый ответ у индивидуума. Терапевтически эффективное количество также является количеством, при котором любые токсические или неблагоприятные эффекты вещества/молекулы, агониста или антагониста перевешиваются терапевтически благоприятными эффектами.
[00135] Термин "профилактически эффективное количество" относится к количеству, эффективному в необходимых дозах и в течение необходимых периодов времени в достижении желаемого профилактического результата. Как правило, но не обязательно, т.к. профилактическую дозу используют для индивидуумов до развития заболевания или на его ранних стадиях, профилактически эффективное количество будет меньше терапевтически эффективного количества.
[00136] Терапевтические составы, содержащие антитело или его фрагмент по настоящему изобретению, подготавливают для хранения посредством смешивания антитела или фрагмента, имеющего желаемую степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington: The Science and Practice of Pharmacy 20th edition (2000)) в форме водных растворов, лиофилизированных или других высушенных составов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают буферы, такие как фосфат, цитрат, гистидин и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и m-крезол); низкомолекулярные (менее приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие средства, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексные соединения с металлами (например, Zn-белковые комплексы); и/или неионные поверхностно-активные средства или полиэтиленгликоль (PEG).
[00137] Фармацевтические композиции, представленные в настоящем описании, можно вводить парентерально посредством инъекции, инфузии или имплантации, для местного или системного введения. В рамках изобретения парентеральное введение включает внутривенное, внутриартериальное, интраперитонеальное, интратекальное, внутрижелудочковое, внутриуретральное, интрастернальное, внутричерепное, внутримышечное, интрасиновиальное и подкожное введение.
[00138] Фармацевтические композиции, представленные в настоящем описании, можно составлять в любых лекарственных формах, подходящих для парентерального введения, включая растворы, суспензии, эмульсии и твердые формы, подходящие для получения растворов или суспензий в жидкости перед инъекцией. Такие лекарственные формы можно получать общепринятыми способами, известными специалистам в области фармацевтики (см. Remington: The Science and Practice of Pharmacy, выше).
[00139] Фармацевтические композиции, предназначенные для парентерального введения, могут включать один или более фармацевтически приемлемых носителей и эксципиентов, включая, в качестве неограничивающих примеров, водные носители, смешиваемые с водой носители, неводные носители, противомикробные средства или консерванты против роста микроорганизмов, стабилизаторы, усилители растворимости, изотонические средства, буферные средства, антиоксиданты, местные анестетики, суспендирующие и диспергирующие средства, увлажнители или эмульгаторы, комплексообразователи, секвестранты или хелатирующие средства, криопротекторы, лиопротекторы, загустители, pH-корректирующие средства и инертные газы.
[00140] Подходящие водные носители включают, в качестве неограничивающих примеров, воду, солевой раствор, физиологический раствор или фосфатно-солевой буфер (PBS), хлорид натрия для инъекций, раствор Рингера для инъекций, изотонический раствор декстрозы для инъекций, стерильную воду для инъекций, раствор декстрозы и лактат Рингера для инъекций. Неводные носители включают, в качестве неограничивающих примеров, жирные масла растительного происхождения, касторовое масло, кукурузное масло, хлопковое масло, оливковое масло, арахисовое масло, масло перечной мяты, сафлоровое масло, сезамовое масло, соевое масло, гидрогенизированные растительные масла, гидрогенизированное соевое масло, среднецепочечные триглицериды кокосового масла и пальмовое масло. Смешиваемые с водой носители включают, в качестве неограничивающих примеров, этанол, 1,3-бутандиол, жидкий полиэтиленгликоль (например, полиэтиленгликоль 300 и полиэтиленгликоль 400), пропиленгликоль, глицерин, N-метил-2-пирролидон, диметилацетамид и диметилсульфоксид.
[00141] В одном из вариантов осуществления фармацевтические композиции, представленные в настоящем описании, составляют в виде стерильных растворов, готовых к использованию. В другом варианте осуществления фармацевтические композиции, представленные в настоящем описании, составляют в виде стерильных сухих растворимых продуктов, включая порошки и гиподермальные таблетки, которые, при необходимости, можно восстанавливать с помощью носителя перед использованием. В еще одном варианте осуществления фармацевтические композиции описывают как стерильные суспензии, готовые к использованию. В еще одном варианте осуществления фармацевтические композиции описывают как стерильные сухие нерастворимые продукты для восстановления с помощью носителя перед использованием. В другом варианте осуществления фармацевтические композиции описывают как стерильные эмульсии, готовые к использованию.
[00142] Фармацевтические композиции можно составлять в виде суспензии, твердого, полутвердого вещества или тиксотропной жидкости для введения в виде имплантируемого депо-препарата. В одном из вариантов осуществления фармацевтические композиции, представленные в настоящем описании, диспергируют в твердой внутренней матрице, окруженной внешней полимерной мембраной, нерастворимой в физиологических жидкостях, но позволяющей активному ингредиенту в фармацевтических композициях диффундировать через нее.
[00143] Подходящие внутренние матрицы включают полиметилметакрилат, полибутилметакрилат, пластифицированный или непластифицированный поливинилхлорид, пластифицированный нейлон, пластифицированный полиэтилентерефталат, натуральный каучук, полиизопрен, полиизобутилен, полибутадиен, полиэтилен, сополимеры этиленвинилацетата, кремнийорганический каучук, полидиметилсилоксан, сополимеры карбоната кремния, гидрофильные полимеры, такие как гидрогели сложных эфиров акриловой и метакриловой кислоты, коллаген, перекрестно сшитый поливиниловый спирт и перекрестно сшитый, частично гидролизованный поливинилацетат.
[00144] Подходящие внешние полимерные мембраны включают полиэтилен, полипропилен, сополимеры этилена/пропилена, сополимеры этилена/этилакрилата, сополимеры этилена/винилацетата, кремнийорганический каучук, полидиметилсилоксаны, неопреновый каучук, хлорированный полиэтилен, поливинилхлорид, сополимеры винилхлорида с винилацетатом, винилиденхлорид, этилен и пропилен, иономер полиэтилентерефталата, бутилкаучук, эпихлоргидриновый каучук, сополимер этилена/винилового спирта, терполимер этилена/винилацетата/винилового спирта и сополимер этилена/винилоксиэтанола.
[00145] Состав по настоящему изобретению также может содержать более одного активного соединения, при необходимости, для конкретного показания, подвергаемого лечению, предпочтительно, с взаимодополняющими активностями, не влияющими друг на друга неблагоприятно. Такие молекулы присутствуют в комбинации в количествах, эффективных для предполагаемой цели.
[00146] Активные ингредиенты также можно заключать в микрокапсулу, полученную, например, способами коацервации или интерфазной полимеризации, например, микрокапсулу из гидроксиметилцеллюлозы или желатина и микрокапсулу полиметилметакрилата, соответственно, в коллоидных системах для доставки лекарственных средств (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах) или макроэмульсиях. Такие способы описывают в Remington: The Science and Practice of Pharmacy 20th edition (2000).
[00147] Составы для использования для введения in vivo должны быть стерильными. Этого легко достигают посредством фильтрации через мембраны для стерильной фильтрации.
[00148] Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих иммуноглобулин по изобретению, где матрицы находятся в форме профилированных изделий, например, пленок или микрокапсулы. Примеры матриц с замедленном высвобождением включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, недеградируемый этилен-винилацетат, деградируемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOT (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и лейпролида ацетата) и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этилен-винилацетат и молочная кислота-гликолевая кислота, делают возможным высвобождение молекул в течение более 100 дней, некоторые гидрогели высвобождают белки в течение меньшего периода времени. Когда инкапсулированные иммуноглобулины остаются в организме в течение длительного периода времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к утрате биологической активности и возможным изменениям иммуногенности. Можно разрабатывать рациональные стратегии для стабилизации в зависимости от затрагиваемого механизма. Например, если обнаруживают, что механизмом агрегации является образование межмолекулярной S--S связи посредством тиодисульфидного обмена, стабилизации можно достигать посредством модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля содержания влаги, использования подходящих добавок и разработки специальных композиций полимерных матриц.
[00149] Антитела, фрагменты антител и полипептиды, представленные в настоящем описании, связываются с OxPL и блокируют их провоспалительные эффекты. Ожидают, что использование in vivo антитела, фрагмента антитела или полипептида по настоящему изобретению будет приводить к блокированию биологических эффектов OxPL во множестве разных ситуаций. Например, показано, что OxPL образуются макрофагами и другими клетками с помощью TLR2-опосредованного механизма. Эти высвободившиеся OxPL способствуют нежелательным вазоактивным эффектам по всему организму пациента. Таким образом, инъекция/инфузия антитела, фрагмента антитела или полипептида по настоящему изобретению в случае неотложного и/или хронического использования может блокировать эти неблагоприятные эффекты и/или, альтернативно, блокировать или уменьшать схожие воспалительные явления, являющиеся результатом активации TLR2. Аналогично, антитело, фрагмент антитела или полипептид по настоящему изобретению также можно инфузировать индивидууму для блокирования провоспалительных эффектов, опосредованных OxPL, образующихся при различных патологических состояниях, таких как вирусные или бактериальные инфекции или аутоиммунные нарушения. Таким образом, антитело, фрагмент антитела или полипептид по настоящему изобретению будут эффективными в качестве противовоспалительных средств при других системных воспалительных условиях, опосредованных активацией TLR2, таких как ревматоидный артрит. Таким образом, антитело, фрагмент антитела или полипептид по настоящему изобретению можно использовать во многих клинических обстоятельствах или условиях, когда противовоспалительные и/или противоатеросклеротические средства необходимо вводить временно и/или хронически.
[00150] Настоящее изобретение относится к способам лечения с использованием антитела, фрагмента антитела и полипептида по настоящему изобретению для лечения индивидуума с TLR2-опосредованным заболеванием или нарушением. В конкретном варианте осуществления настоящее изобретение относится к лечению TLR2-опосредованного заболевания или нарушения с помощью терапевтически эффективного количества антитела, фрагмента антитела или полипептида по настоящему изобретению. Неограничивающие примеры TLR2-опосредованных заболеваний или нарушений включают болезнь Кавасаки (Kang et al., Korean J Pediatr 60(7):208-215 (2017)), диабет 2 типа (Sepehriet al., Cell MolBiol Lett 21:2 (2016)), ревматоидный артрит (McGarryet al., Arthritis Res Ther 17:153 (2015)), дерматологические заболевания (Kang et al., Journal of American Academy of Dermatology, 54(6):951-983 (2006)), рассеянный склероз (Hossain et al., Oncotarget 6(34):35131-35132 (2015)), системную красную волчанку (Liu et al., European Journal of Immunology, 45(9):2683-2693 (2015)), язвенный колит (Folova et al., Journal of Histochemistry & Cytochemistry 56(3):267-274 (2008), болезнь Грейвса (Peng et al., Front Immunol, 7:578 (2016)), синдром Шегрена (Sisto et al., ClinExp Med 17(3):341-350 (2017), аутоиммунные заболевания щитовидной железы (Peng et al., Front Immunol 7:547 (2016) и васкулит (Summers et al., Arthritis Rheum 63(4):1124-35 (2011)). В конкретном варианте осуществления настоящее изобретение относится к лечению индивидуума с болезнью Кавасаки с помощью терапевтически эффективного количества антитела, фрагмента антитела или полипептида по настоящему изобретению.
[00151] Для терапевтического применения, представленного в настоящем описании, в настоящем описании также представлены наборы и промышленные изделия. Такие наборы могут содержать носитель, упаковку или контейнер, компартментализированный для вмещения одного или более контейнеров, таких как флаконы, пробирки и т.п., где каждый из контейнеров содержит один из отдельных элементов для использования в способе, представленном в настоящем описании. Подходящие контейнеры включают, например, бутыли, флаконы, шприцы и пробирки. Контейнеры можно получать из различных материалов, таких как стекло или пластик.
[00152] Например, контейнеры могут содержать одно или более антител, фрагментов антител или полипептидов, представленных в настоящем описании, необязательно, в композиции или в комбинации с другим средством, как представлено в настоящем описании. Контейнеры, необязательно, имеют стерильное входное отверстие (например, контейнер может являться мешком для внутривенного раствора или флаконом с пробкой, прокалываемой иглой для подкожных инъекций). Такие наборы, необязательно, содержат описание, или ярлык, или инструкции, относящиеся к использованию в способах, представленных в настоящем описании.
[00153] Набор, как правило, будет содержать один или более дополнительных контейнеров, каждый из которых содержит один или более из различных материалов (таких как реагенты, необязательно, в концентрированной форме и/или устройства), желательных с коммерческой точки зрения и точки зрения пользователя для использования соединения, представленного в настоящем описании. Неограничивающие примеры таких материалов включают буферы, дилюенты, фильтры, иглы, шприцы; носитель, упаковку, контейнер, ярлык на флакон и/или пробирку, где указано содержимое и/или инструкции по использованию, и вкладыши в упаковку с инструкциями по использованию. Как правило, также будут включать набор инструкций.
[00154] Ярлык может находиться на контейнере или быть связанным с контейнером. Ярлык может находиться на контейнере, когда буквы, числа или другие знаки, образующие ярлык, прикрепляют, прессуют или гравируют на самом контейнере, ярлык можно связывать с контейнером, когда он находится во вместилище или носителе, также удерживающем контейнер, например, в виде вкладыша в упаковку. Ярлык можно использовать для указания того, что содержимое будут использовать для конкретного терапевтического применения. На ярлыке также можно указывать инструкции по использованию содержимого, например, в способах, представленных в настоящем описании. Эти другие терапевтические средства можно использовать, например, в количестве, указанном в Настольном справочнике врача (PDR) или иначе определенном специалистом в этой области.
[00155] Следующие примеры представлены в иллюстративных целях, а не для ограничения изобретения. Хотя можно использовать типичные способы, альтернативно можно использовать другие способы, известные специалистам в этой области.
ПРИМЕРЫ
[00156] Материалы. Синтетические стандарты 1,2-динаноил-sn-глицеро-3-фосфохолин (DNPC), 1-пальмитоил-2-(5-окосвалероил)-sn-глицеро-3-фосфохолин (POVPC), 1-пальмитоил-2-глутароил-sn-глицеро-3-фосфохолин (PGPC), 1-пальмитоил-2-азелаоил-sn-глицеро-3-фосфохолин (PAzPC), 1-пальмитоил-2-(9-оксо)наноил-sn-глицеро-3-фосфохолин (PONPC) и природное антитело мыши IgM EO6, несодержащее LPS, получали в Avanti Polar Lipids (Alabaster, AL). 1-(пальмитоил)-2-(5-кето-6-октен-диоил)-3-фосфохолин (KOdiAPC) и 1-пальмитоил-2-(4-кето-додец-3-ен-диоил)-sn-глицеро-3-фосфохолин (KDdiAPC) приобретали в Cayman Chemicals (Ann Arbor, MI). Все растворители имели категорию ВЭЖХ.
[00157] Получение и характеризация EO6-scFv-трансгенных мышей. Получение трансгенных мышей C57BL/6, экспрессирующих идиотип T15/EO6 в виде одноцепочечного вариабельного фрагмента антитела, обозначенного как EO6-scFv-Tg. В кратком изложении, кДНК, кодирующую вариабельные области EO6 тяжелых и легких цепей, соединяли с 15-аминокислотным пептидом посредством ПЦР с удлинением перекрывающихся фрагментов и клонировали в экспрессирующий вектор pSecTag2A (Invitrogen), содержащий лидерную последовательность каппа-цепи Ig мыши для секреции и c-myc и поли-His в качестве эпитопных меток. Трансфицировали клетки HEK293 и обнаруживали, что связывающие свойства EO6-scFv, секретируемого в супернатант культуры, напоминали свойства интактного EO6. Затем ту же конструкцию клонировали в печень-специфический экспрессирующий вектор pLiv7 и использовали для получения трансгенных (Tg) мышей с фоном C57BL/6, экспрессирующих трансген EO6-scFv, регулируемый промотором apoE. Потомство подвергали скринингу на титр EO6-scFv в плазме и встраивание трансгена посредством ПЦР-амплификации ДНК-хвоста. Исходные EO6-scFv-трансгенные линии скрещивали друг с другом для получения "гомозиготных" трансгенных мышей и, в свою очередь, их скрещивали для получения мышей Ldl-/- с фоном C57BL/6. Всех животных генотипировали по EO6-scFv и Ldlr-/-, соответственно, и анализировали плазму для подтверждения экспрессии EO6-scFv посредством иммунологического анализа. мРНК EO6-scFv выраженно экспрессировалась в печени, перитонеальных макрофагах и селезенке и, в меньшей степени, в сердце. В этом исследовании уровни EO6-scFv в плазме в среднем составляли 20-30 мкг/мл.
[00158] Масс-спектрометрия OxPL. PC-содержащие фосфолипиды выделяли из NNCM. Удаляли клеточные среды и промывали клетки PBS. Содержимое каждой лунки соскребали в 1 мл раствора метанола/уксусной кислоты (3% об./об.), содержащего 0,01% BHT, переносили в стеклянную коническую пробирку 10 мл и закрывали ее крышкой в атмосфере N2 (газ.). В каждый образец с целью количественного анализа добавляли десять нанограммов DNPC в качестве внутреннего стандарта. В пробирку добавляли два миллилитра гексана, содержащего BHT, закрывали ее крышкой в атмосфере N2 (газ.), перемешивали на центрифуге типа вортекс в течение пяти секунд, а затем центрифугировали в течение 5 мин при 3500 об./мин. и 4°C. Затем верхний гексановый слой откачивали с использованием стеклянной пастеровской пипетки и выбрасывали. Промывку гексаном/BHT повторяли трижды, закрывая крышкой в атмосфере N2 (газ.), перемешивали на центрифуге типа вортекс в течение пяти секунд и центрифугировали после каждой промывки. После конечной промывки гексаном/BHT в пробирку добавляли 2 мл хлороформа, содержащего BHT, и 750 мкл PBS, а затем перемешивали на центрифуге типа вортекс и центрифугировали, как описано выше. Нижний органический слой удаляли с использованием стеклянной пастеровской пипетки и переносили в чистую стеклянную коническую пробирку 15 мл, где раствор аспирировали с использованием испарителя жидкого азота, а затем переразводили в 300 мкл хлороформа/метанола (2:1 об./об.) для хранения при -80°C.
[00159] Отделение OxPL осуществляли с использованием обращенно-фазовой (RP) хроматографии. Экстракты сердец переразводили в элюенте RP, состоящем из ацетонитрила:воды 60:40, 10 мМ формиата аммония и 0,1% муравьиной кислоты, непосредственно перед инъекцией. Тридцать микролитров образца впрыскивали на колонку Ascentis Express C18 ВЭЖХ (15 см × 2,1 мм, 2,7 мкм; Supelco Analytical, Bellefonte, Pennsylvania, USA) с разделением с помощью системы Prominence UFLC от Shimadzu Corporation (Canby, Oregon, USA). Элюцию осуществляли с использованием линейного градиента растворителя A (ацетонитрил/вода, 60:40 об./об.) и растворителя B (изопропанол/ацетонитрил, 90:10, об./об.), при этом оба растворителя содержат 10 мМ формиата аммония и 0,1% муравьиной кислоты. Композиция используемой подвижной фазы являлась следующей: начальный растворитель B при 32% до 4,00 мин; переход на 45% B; 5,00 мин 52% B; 8,00 мин 58% B; 11,00 мин 66% B; 14,00 мин 70% B; 18,00 мин 75% B; 21,00 мин 97% B; 25,00 мин 97% B; 25,10 мин 32% B. Для анализа использовали скорость потока 260 мкл/мин, и лоток для проб и колоночный термостат держали при 4 и 45°C, соответственно.
[00160] Детекцию OxPL осуществляли посредством масс-спектрометрии в режиме положительной полярности. Сканирование MRM осуществляли с 6 переходами с использованием ионного продукта 184,3 m/z, соответствующего расщепленному фосфохолиновому остатку. Шесть коммерчески доступных стандартов PONPC, POVPC, PGPC, PAzPC, KOdiAPC и KDdiAPC впрыскивали и точное приписывание пиков зависело от времени удержания и массовых переходов. Условия масс-спектрометрии являлись следующими: газовая завеса, 26 фунт/дюйм2; газ для соударений, средний; напряжение ионораспыления, 5500 В; температура, 500,0°C; газ источника ионизации 1, 40,0 фунт/дюйм2; газ источника ионизации 2, 30,0 фунт/дюйм2; потенциал декластеризации, 125 В, потенциал поступления, 10 В; энергия соударения, 53 В; потенциал на выходе ячейки соударений, 9 В; и время выдержки, 50 мсек. Внешнюю калибровку массы осуществляли с регулярными интервалами. Для количественного анализа получали калибровочные кривые мониторинга множественных реакций (MRM) для каждого из 6 коммерчески доступных стандартов OxPL и пики нормализовали по их относительным ответам. Во время экстракции во все образцы добавляли десять нанограммов внутреннего стандарта. Систему тройного квадрупольного масс-спектрометра 4000 QTRAP® с источником электрораспыления ионов Turbo V от AB Sciex (Framingham, Massachusetts, USA) соединяли с системой жидкостной хроматографии.
[00161] Развитие аневризм брюшной аорты и дилатации в модели LCWE-индуцированного васкулита KD на мышах. Болезнь Кавасаки вызывает персистирующий артериит коронарных артерий (CA) у детей младшего возраста, и в настоящее время его считают основной причиной приобретенного заболевания сердца у детей в экономически развитых странах. В модели на животных CA, индуцируемая экстрактом клеточной стенки Lactobacillus casei (LCWE), у мышей точно имитирует патогенез KD у людей. L. casei группы B (ATCC 11578) выращивали в бульоне MRS Lactobacillus, собирали посредством центрифугирования во время экспоненциальной фазы роста и промывали PBS при pH 7,40. После сбора клетки обрабатывали 4% SDS в течение ночи, а затем последовательно инкубировали с 250 мкг/мл РНКазы, ДНКазы I и трипсина. Затем конечный осадок обрабатывали ультразвуком (5 г уплотненного сырого веса в 15 мл PBS) в течение 2 ч. в импульсных условиях 9 сек импульс/5 сек пауза с частотой 20 кГц (Vibra Cell, Sonics & Materials Inc., Newtown, CT). После центрифугирования в течение 1 ч. при 20000×g определяли концентрацию супернатанта с учетом содержания рамнозы по результатам колориметрического анализа с использованием фенол-серной кислоты (Dubois et al. 1956). Концентрация эндотоксина в этом препарате составляла <1,5 пг/мкг, что определяли посредством реакции с лизатом амебоцитов мечехвоста (Associates of Cape Cod Inc., East Falmouth, MA).
[00162] Мышам C57/BL6 возрастом четыре недели инъецировали 250 мкг LCWE в PBS или только физиологический раствор. Мышей умерщвляли в 4 момента времени - 7, 14, 21 и 28 дней. Абдоминальные и коронарные артерии идентифицировали на серийных срезах (7 мкм), фиксировали формалином и окрашивали гематоксилином и эозином. Для иммуногистохимического анализа срезы предварительно обрабатывали 0,3% пероксидом водорода в PBS в течение 30 мин. Антитела против маркеров воспаления или изотипические контрольные антитела наносили в 0,5% бычьем сывороточном альбумине в PBS при 1:100 на 1 ч. Затем стекла промывали и наносили биотинилированное, конъюгированное с пероксидазой хрена вторичное антитело (Vector Lab, Burlingame, CA) при 1:500 в течение 30 мин, промывали и окрашивали с помощью конъюгированной с стрептавидином пероксидазой хрена при 1:1000 в течение 30 мин. Иммуногистохимическое окрашивание определяли с использованием набора SK-4100 DAB по инструкциям производителя (Vector Lab). Данные свидетельствуют о том, что образование LCWE-индуцированных AAA и гистологическая картина интенсивного воспаления у мышей EO6-Tg значительно снижались по сравнению с контрольными мышами (фиг.17-21).
[00163] Анализ воспалительных цитокинов в сыворотке. Для одновременной детекции различных цитокинов в плазме мышей Ldlr-/- или Ldlr-/-/EO6scFv-Tg, которым вводили LCWE и держали на питании HFC в течение 12 недель, использовали 23-плексный иммунологический анализ цитокинов мыши Bio-Plex Pro (Bio-Rad Laboratories, Inc.). Измерения и анализ данных всех анализов осуществляли на основе протокола системы Bio-Plex в комбинации с программным обеспечением Bio-Plex Manager. Результаты показаны на фиг.10. По сравнению с мышами Ldlr-/- (n=7-10) у мышей Ldlr-/-EO6-scFv-Tg (n=7-10) наблюдали, что уровни некоторых провоспалительных цитокинов/хемокинов в плазме (ФНОα, CCL2, CCL5, CXCL1, ИЛ-6 и ИЛ-12) значимо снижались по результатам мультиплексных анализов Bio-Plex (Bio-Rad), что свидетельствует о генерализованном снижении системного воспаления у мышей EO6scFv-TG Ldlr-/-.
[00164] В настоящем описании представлен ряд вариантов осуществления изобретения. Несмотря на это, следует понимать, что можно осуществлять различные модификации без отклонения от объема и сущности настоящего изобретения. Таким образом, другие варианты осуществления входят в объем следующей формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>The Regents of the University of California
<120>ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ
ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ
<130>00015-348WO1
<140>Еще не приписано
<141>2019-01-29
<150>US 62/623,276
<151>2018-01-29
<160>14
<170>PatentIn version 3.5
<210>1
<211>930
<212>ДНК
<213>Искусственная псоедовательность
<220>
<223>Сконструированный домен антитела scFV E06
<220>
<221>CDS
<222>(1)..(930)
<400>1
atg gag aca gac aca ctc ctg cta tgg gta ctg ctg ctc tgg gtt cca 48
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
ggt tcc act ggt gac gcg gcc cag ccg gcc agg cgc gcc gta cga agc 96
Gly Ser Thr Gly Asp Ala Ala Gln Pro Ala Arg Arg Ala Val Arg Ser
20 25 30
tta gac att gtg atg act cag tct cca tct tcc ctt tct gtg tca gca 144
Leu Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala
35 40 45
ggt aag aag gtc acc att agt tgc acg gcc agt gag agc ctt tat tca 192
Gly Lys Lys Val Thr Ile Ser Cys Thr Ala Ser Glu Ser Leu Tyr Ser
50 55 60
agc aaa cac aag gtg cac tac ttg gct tgg tac cag aag aaa cca gag 240
Ser Lys His Lys Val His Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Glu
65 70 75 80
caa tct cct aaa ctg ctg ata tac ggg gca tcc aac cga tac att ggg 288
Gln Ser Pro Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Ile Gly
85 90 95
gtc cct gat cgc ttc aca ggc agt gga tct ggg aca gat ttc act ctg 336
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
100 105 110
acc atc agc agt gta cag gtt gaa gac ctc aca cat tat tac tgt gca 384
Thr Ile Ser Ser Val Gln Val Glu Asp Leu Thr His Tyr Tyr Cys Ala
115 120 125
cag ttt tac agc tat ccg ctc acg ttc ggt gct ggg acc aag ctg gaa 432
Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
130 135 140
atc aaa ggt ggt gga gga tca ggt gga ggt ggt tca gga ggt ggc gga 480
Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
tcc gag gtg aag ctg gtg gag tct gga gga ggc ttg gta cag cct ggg 528
Ser Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
165 170 175
ggt tct ctg aga ctc tcc tgt gca act tct ggg ttc acc ttc agt gat 576
Gly Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp
180 185 190
ttc tac atg gag tgg gtc cgc cag gct cca ggg aag aga ctg gag tgg 624
Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp
195 200 205
att gct gca agt aga aac aaa gct aat gat tat aca aca gag tac gct 672
Ile Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr Thr Thr Glu Tyr Ala
210 215 220
gac tct gtg aag ggt cgg ttc atc gtc tcc aga gac act tcc caa agc 720
Asp Ser Val Lys Gly Arg Phe Ile Val Ser Arg Asp Thr Ser Gln Ser
225 230 235 240
atc ctc tac ctt cag atg aat gcc ctg aga gcc gag gac act gcc att 768
Ile Leu Tyr Leu Gln Met Asn Ala Leu Arg Ala Glu Asp Thr Ala Ile
245 250 255
tat tac tgt gca aga gat tac tac ggt agt agc tac tgg tac ttc gat 816
Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp
260 265 270
gtc tgg ggc gca ggg acc acg gtc acc gtc tcc tct cga gga ggg ccc 864
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Arg Gly Gly Pro
275 280 285
gaa caa aaa ctc atc tca gaa gag gat ctg aat agc gcc gtc gac cat 912
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Ser Ala Val Asp His
290 295 300
cat cat cat cat cat tga 930
His His His His His
305
<210>2
<211>309
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>Синтетическая конструкция
<400>2
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ala Ala Gln Pro Ala Arg Arg Ala Val Arg Ser
20 25 30
Leu Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala
35 40 45
Gly Lys Lys Val Thr Ile Ser Cys Thr Ala Ser Glu Ser Leu Tyr Ser
50 55 60
Ser Lys His Lys Val His Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Glu
65 70 75 80
Gln Ser Pro Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Ile Gly
85 90 95
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
100 105 110
Thr Ile Ser Ser Val Gln Val Glu Asp Leu Thr His Tyr Tyr Cys Ala
115 120 125
Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
130 135 140
Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
Ser Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
165 170 175
Gly Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp
180 185 190
Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp
195 200 205
Ile Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr Thr Thr Glu Tyr Ala
210 215 220
Asp Ser Val Lys Gly Arg Phe Ile Val Ser Arg Asp Thr Ser Gln Ser
225 230 235 240
Ile Leu Tyr Leu Gln Met Asn Ala Leu Arg Ala Glu Asp Thr Ala Ile
245 250 255
Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp
260 265 270
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Arg Gly Gly Pro
275 280 285
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu Asn Ser Ala Val Asp His
290 295 300
His His His His His
305
<210>3
<211>1554
<212>ДНК
<213>Искусственная псоедовательность
<220>
<223>Гуманизированное одноцепочечное антитело E06 с IgG1-Fc (E06scFv-Fc)
<220>
<221>CDS
<222>(1)..(1554)
<400>3
atg gag acc gac aca ctg ttg ttg tgg gtg ttg ctg ctc tgg gtg cca 48
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
gga agc aca ggt gac gct gct gac atc gtc atg acc cag agc ccc gac 96
Gly Ser Thr Gly Asp Ala Ala Asp Ile Val Met Thr Gln Ser Pro Asp
20 25 30
tct ctc gcg gtt tct ctg gga gag cgg gca aca atc aac tgc aca gca 144
Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Thr Ala
35 40 45
agc gaa tcc ctg tac tca tcc aag cac gtg cat tac ctc gct tgg tac 192
Ser Glu Ser Leu Tyr Ser Ser Lys His Val His Tyr Leu Ala Trp Tyr
50 55 60
cag cag aaa cca ggg caa cca cca aag ctc ctc att tat ggg gcc agc 240
Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Gly Ala Ser
65 70 75 80
aac aga tat att gga gtc cca gat cga ttc agc ggt tcc ggc tcc gga 288
Asn Arg Tyr Ile Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly
85 90 95
aca gac ttt acc ctc acg ata agc agc ctg cag gcg gaa gat gtg gcc 336
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala
100 105 110
gtg tat tac tgc gca caa ttc tac agc tat cct ctg acc ttc gga gga 384
Val Tyr Tyr Cys Ala Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Gly
115 120 125
gga aca aaa gtg gag atc aaa ggc gga ggt gga tcc gga ggg ggt gga 432
Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
tct gga ggt ggc ggt agt gaa gtg cag ctg gtg gaa agt gga ggc ggc 480
Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly
145 150 155 160
ctg gtg caa cca ggt ggc tct ctg agg ctg tca tgc gct gcc tct gga 528
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
165 170 175
ttt acc ttc tca gat ttc tac atg gaa tgg gtc aga caa gcc cct gga 576
Phe Thr Phe Ser Asp Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly
180 185 190
aag ggg ctc gag tgg gtg gcc gct tcc agg aac aag gct aat gac tac 624
Lys Gly Leu Glu Trp Val Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr
195 200 205
acc aca gag tac gcc gca agt gtt aaa ggc cgc ttt ata atc tcc cgc 672
Thr Thr Glu Tyr Ala Ala Ser Val Lys Gly Arg Phe Ile Ile Ser Arg
210 215 220
gat gac tct aag aac tcc ttg tac ctt caa atg aat agt ctc aag aca 720
Asp Asp Ser Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr
225 230 235 240
gaa gat aca gcg gta tac tac tgc gcc cgc gac tac tac gga tca agt 768
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser
245 250 255
tat tgg tac ttc gat gtt tgg aga gct ggc aca ctt gtg act gtc agc 816
Tyr Trp Tyr Phe Asp Val Trp Arg Ala Gly Thr Leu Val Thr Val Ser
260 265 270
agt ctt gat cct aaa tcc tct gac aag acc tat acc tgc cca cct tgt 864
Ser Leu Asp Pro Lys Ser Ser Asp Lys Thr Tyr Thr Cys Pro Pro Cys
275 280 285
ccc gcc cca gaa ctt ctg ggt ggc cca tcc gtg ttt ctg ttc cca cca 912
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
290 295 300
aag cca aag gat aca ctc atg atc tct cgc act ccg gaa gtc acg tgc 960
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
305 310 315 320
gtc gtg gtt gat gtg tca cac gag gac ccg gag gtc aaa ttc aat tgg 1008
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
325 330 335
tac gtg gac gga gtc gag gtg cac aac gcc aag aca aag cca cgc gaa 1056
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
340 345 350
gag cag tac aac agc acg tat aga gta gtg agc gtg ctg aca gtg ctc 1104
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
355 360 365
cac cag gat tgg ctt aac ggt aag gaa tac aag tgt aag gtc tcc aac 1152
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
370 375 380
aaa gct ctt cct gct cca ata gaa aag acc att tca aag gcc aag ggg 1200
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
385 390 395 400
caa cct cga gaa ccc cag gtg tac acg ctg cct ccc agc cga gag gag 1248
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
405 410 415
atg acc aag aac caa gta agt ctg aca tgc ctt gtc aaa ggg ttc tac 1296
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
420 425 430
ccc tca gac atc gcc gtg gaa tgg gaa agc aac ggt caa ccc gaa aac 1344
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
435 440 445
aat tac aag aca acg cca ccg gta ctc gat tcc gat ggt tcc ttt ttt 1392
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
450 455 460
ctg tac tcc aaa ctc acg gtg gac aag agt cga tgg cag cag gga aac 1440
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
465 470 475 480
gtt ttc tcc tgt tcc gtg atg cac gaa gca ctg cac aat cac tat acc 1488
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
485 490 495
cag aag tca ctg agt ttg agc cct ggc aaa gga ggg ggc gga tca cat 1536
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser His
500 505 510
cat cac cat cac cat taa 1554
His His His His His
515
<210>4
<211>517
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>Синтетическая конструкция
<400>4
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ala Ala Asp Ile Val Met Thr Gln Ser Pro Asp
20 25 30
Ser Leu Ala Val Ser Leu Gly Glu Arg Ala Thr Ile Asn Cys Thr Ala
35 40 45
Ser Glu Ser Leu Tyr Ser Ser Lys His Val His Tyr Leu Ala Trp Tyr
50 55 60
Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Gly Ala Ser
65 70 75 80
Asn Arg Tyr Ile Gly Val Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly
85 90 95
Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala Glu Asp Val Ala
100 105 110
Val Tyr Tyr Cys Ala Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Gly
115 120 125
Gly Thr Lys Val Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly
145 150 155 160
Leu Val Gln Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly
165 170 175
Phe Thr Phe Ser Asp Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly
180 185 190
Lys Gly Leu Glu Trp Val Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr
195 200 205
Thr Thr Glu Tyr Ala Ala Ser Val Lys Gly Arg Phe Ile Ile Ser Arg
210 215 220
Asp Asp Ser Lys Asn Ser Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr
225 230 235 240
Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser
245 250 255
Tyr Trp Tyr Phe Asp Val Trp Arg Ala Gly Thr Leu Val Thr Val Ser
260 265 270
Ser Leu Asp Pro Lys Ser Ser Asp Lys Thr Tyr Thr Cys Pro Pro Cys
275 280 285
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
290 295 300
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
305 310 315 320
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
325 330 335
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
340 345 350
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
355 360 365
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
370 375 380
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
385 390 395 400
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
405 410 415
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
420 425 430
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
435 440 445
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
450 455 460
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
465 470 475 480
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
485 490 495
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser His
500 505 510
His His His His His
515
<210>5
<211>5
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>Пептидный линкер
<400>5
Gly Gly Gly Gly Ser
1 5
<210>6
<211>7
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR1 VH
<400>6
Gly Phe Thr Phe Ser Asp Phe
1 5
<210>7
<211>8
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR2 VH
<400>7
Arg Asn Lys Ala Asn Asp Tyr Thr
1 5
<210>8
<211>16
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR3 VH
<400>8
Cys Ala Arg Asp Tyr Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp Val Trp
1 5 10 15
<210>9
<211>17
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR1 VL
<400>9
Thr Ala Ser Glu Ser Leu Tyr Ser Ser Lys His Lys Val His Tyr Leu
1 5 10 15
Ala
<210>10
<211>7
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR2 VL
<400>10
Gly Ala Ser Asn Arg Tyr Ile
1 5
<210>11
<211>10
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR3 VL
<400>11
Cys Ala Gln Phe Tyr Ser Tyr Pro Leu Thr
1 5 10
<210>12
<211>16
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>CDR1 VL-гуманизированная
<400>12
Thr Ala Ser Glu Ser Leu Tyr Ser Ser Lys His Val His Tyr Leu Ala
1 5 10 15
<210>13
<211>1587
<212>ДНК
<213>Искусственная псоедовательность
<220>
<223>Последовательность химерного антитела E06scFv с IgG1-Fc человека
<220>
<221>CDS
<222>(1)..(1587)
<400>13
atg gag aca gac aca ctc ctg cta tgg gta ctg ctg ctc tgg gtt cca 48
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
ggt tcc act ggt gac gcg gcc cag ccg gcc agg cgc gcc gta cga agc 96
Gly Ser Thr Gly Asp Ala Ala Gln Pro Ala Arg Arg Ala Val Arg Ser
20 25 30
tta gac att gtg atg act cag tct cca tct tcc ctt tct gtg tca gca 144
Leu Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala
35 40 45
ggt aag aag gtc acc att agt tgc acg gcc agt gag agc ctt tat tca 192
Gly Lys Lys Val Thr Ile Ser Cys Thr Ala Ser Glu Ser Leu Tyr Ser
50 55 60
agc aaa cac aag gtg cac tac ttg gct tgg tac cag aag aaa cca gag 240
Ser Lys His Lys Val His Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Glu
65 70 75 80
caa tct cct aaa ctg ctg ata tac ggg gca tcc aac cga tac att ggg 288
Gln Ser Pro Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Ile Gly
85 90 95
gtc cct gat cgc ttc aca ggc agt gga tct ggg aca gat ttc act ctg 336
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
100 105 110
acc atc agc agt gta cag gtt gaa gac ctc aca cat tat tac tgt gca 384
Thr Ile Ser Ser Val Gln Val Glu Asp Leu Thr His Tyr Tyr Cys Ala
115 120 125
cag ttt tac agc tat ccg ctc acg ttc ggt gct ggg acc aag ctg gaa 432
Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
130 135 140
atc aaa ggt ggt gga gga tca ggt gga ggt ggt tca gga ggt ggc gga 480
Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
tcc gag gtg aag ctg gtg gag tct gga gga ggc ttg gta cag cct ggg 528
Ser Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
165 170 175
ggt tct ctg aga ctc tcc tgt gca act tct ggg ttc acc ttc agt gat 576
Gly Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp
180 185 190
ttc tac atg gag tgg gtc cgc cag gct cca ggg aag aga ctg gag tgg 624
Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp
195 200 205
att gct gca agt aga aac aaa gct aat gat tat aca aca gag tac gct 672
Ile Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr Thr Thr Glu Tyr Ala
210 215 220
gac tct gtg aag ggt cgg ttc atc gtc tcc aga gac act tcc caa agc 720
Asp Ser Val Lys Gly Arg Phe Ile Val Ser Arg Asp Thr Ser Gln Ser
225 230 235 240
atc ctc tac ctt cag atg aat gcc ctg aga gcc gag gac act gcc att 768
Ile Leu Tyr Leu Gln Met Asn Ala Leu Arg Ala Glu Asp Thr Ala Ile
245 250 255
tat tac tgt gca aga gat tac tac ggt agt agc tac tgg tac ttc gat 816
Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp
260 265 270
gtc tgg ggc gca ggg acc acg gtc acc gtc tcc tct ctg gac ccg aag 864
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Leu Asp Pro Lys
275 280 285
tct tct gac aaa act tac aca tgc cca ccg tgc cca gca cct gaa ctc 912
Ser Ser Asp Lys Thr Tyr Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
290 295 300
ctg ggg gga ccg tca gtc ttc ctc ttc ccc cca aaa ccc aag gac acc 960
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
305 310 315 320
ctc atg atc tcc cgg acc cct gag gtc aca tgc gtg gtg gtg gac gtg 1008
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
325 330 335
agc cac gaa gac cct gag gtc aag ttc aac tgg tac gtg gac ggc gtg 1056
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
340 345 350
gag gtg cat aat gcc aag aca aag ccg cgg gag gag cag tac aac agc 1104
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
355 360 365
acg tac cgt gtg gtc agc gtc ctc acc gtc ctg cac cag gac tgg ctg 1152
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
370 375 380
aat ggc aag gag tac aag tgc aag gtc tcc aac aaa gcc ctc cca gcc 1200
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
385 390 395 400
ccc atc gag aaa acc atc tcc aaa gcc aaa ggg cag ccc cga gaa cca 1248
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
405 410 415
cag gtg tac acc ctg ccc cca tcc cgg gag gag atg acc aag aac cag 1296
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
420 425 430
gtc agc ctg acc tgc ctg gtc aaa ggc ttc tat ccc agc gac atc gcc 1344
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
435 440 445
gtg gag tgg gag agc aat ggg cag ccg gag aac aac tac aag acc acg 1392
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
450 455 460
cct ccc gtg ctg gac tcc gac ggc tcc ttc ttc ctc tac agc aag ctc 1440
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
465 470 475 480
acc gtg gac aag agc agg tgg cag cag ggg aac gtc ttc tca tgc tcc 1488
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
485 490 495
gtg atg cac gag gct ctg cac aac cac tac acg cag aag agc ctc tcc 1536
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
500 505 510
ctg tct ccg ggt aaa ggt gga ggt gga tca cat cat cat cat cat cat 1584
Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser His His His His His His
515 520 525
taa 1587
<210>14
<211>528
<212>БЕЛОК
<213>Искусственная псоедовательность
<220>
<223>Синтетическая конструкция
<400>14
Met Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro
1 5 10 15
Gly Ser Thr Gly Asp Ala Ala Gln Pro Ala Arg Arg Ala Val Arg Ser
20 25 30
Leu Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Val Ser Ala
35 40 45
Gly Lys Lys Val Thr Ile Ser Cys Thr Ala Ser Glu Ser Leu Tyr Ser
50 55 60
Ser Lys His Lys Val His Tyr Leu Ala Trp Tyr Gln Lys Lys Pro Glu
65 70 75 80
Gln Ser Pro Lys Leu Leu Ile Tyr Gly Ala Ser Asn Arg Tyr Ile Gly
85 90 95
Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
100 105 110
Thr Ile Ser Ser Val Gln Val Glu Asp Leu Thr His Tyr Tyr Cys Ala
115 120 125
Gln Phe Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
130 135 140
Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
Ser Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
165 170 175
Gly Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Asp
180 185 190
Phe Tyr Met Glu Trp Val Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp
195 200 205
Ile Ala Ala Ser Arg Asn Lys Ala Asn Asp Tyr Thr Thr Glu Tyr Ala
210 215 220
Asp Ser Val Lys Gly Arg Phe Ile Val Ser Arg Asp Thr Ser Gln Ser
225 230 235 240
Ile Leu Tyr Leu Gln Met Asn Ala Leu Arg Ala Glu Asp Thr Ala Ile
245 250 255
Tyr Tyr Cys Ala Arg Asp Tyr Tyr Gly Ser Ser Tyr Trp Tyr Phe Asp
260 265 270
Val Trp Gly Ala Gly Thr Thr Val Thr Val Ser Ser Leu Asp Pro Lys
275 280 285
Ser Ser Asp Lys Thr Tyr Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
290 295 300
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
305 310 315 320
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
325 330 335
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
340 345 350
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
355 360 365
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
370 375 380
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
385 390 395 400
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
405 410 415
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
420 425 430
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
435 440 445
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
450 455 460
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
465 470 475 480
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
485 490 495
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
500 505 510
Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser His His His His His His
515 520 525
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ И СПОСОБЫ | 2016 |
|
RU2770001C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ИЗБИРАТЕЛЬНОЙ ЭКСПРЕССИИ БЕЛКА | 2017 |
|
RU2795467C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) ПРОТИВ CD123 ДЛЯ ИСПОЛЬЗОВАНИЯ В ЛЕЧЕНИИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2015 |
|
RU2724999C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ B-КЛЕТОК | 2013 |
|
RU2766608C2 |
| ЛЕЧЕНИЕ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ С ИСПОЛЬЗОВАНИЕМ ГУМАНИЗИРОВАННОГО ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА ПРОТИВ ВСМА | 2015 |
|
RU2751660C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
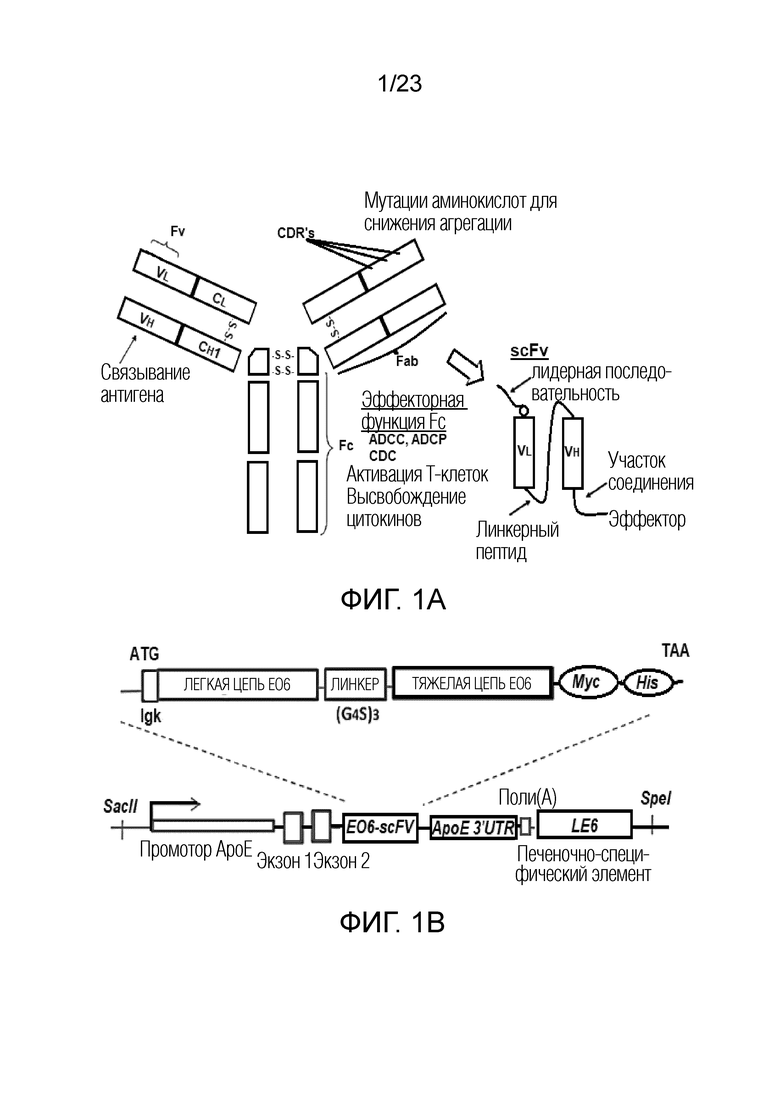


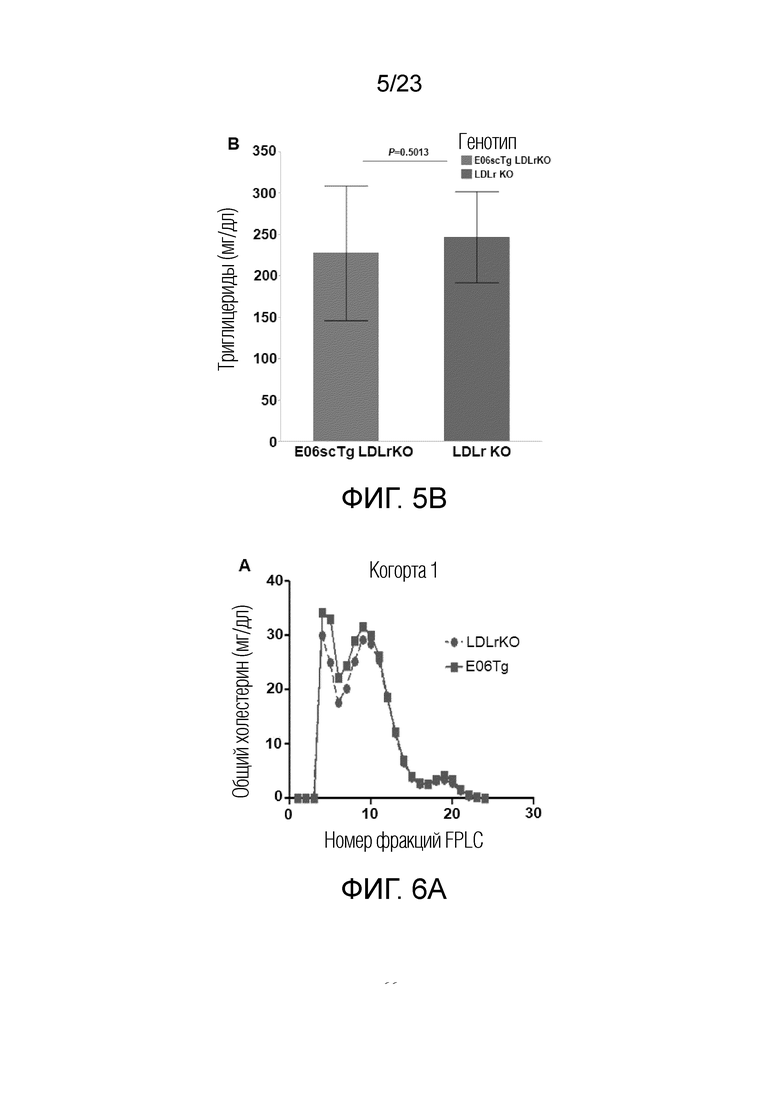
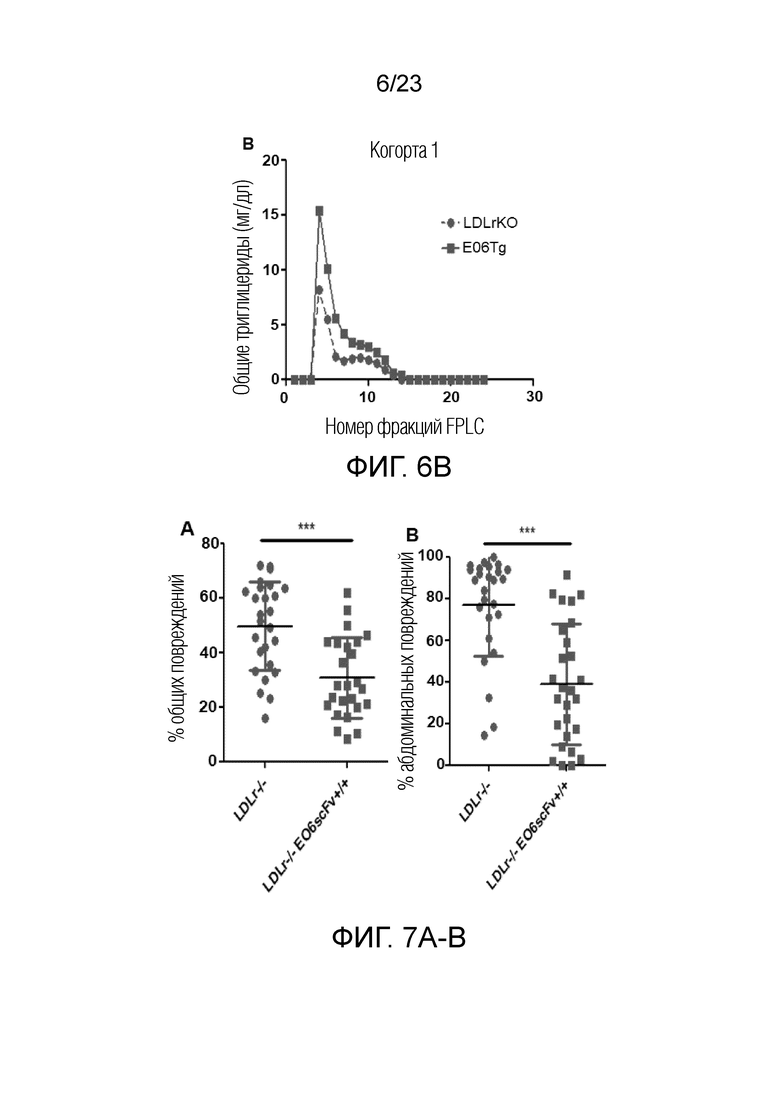
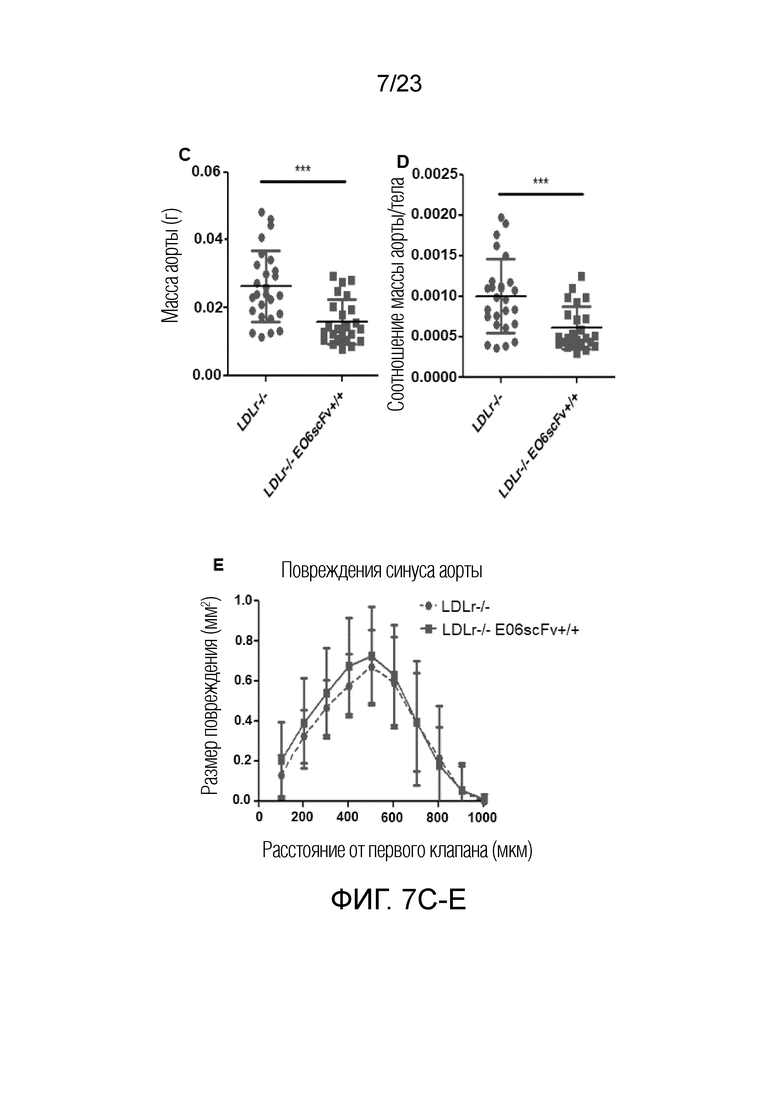
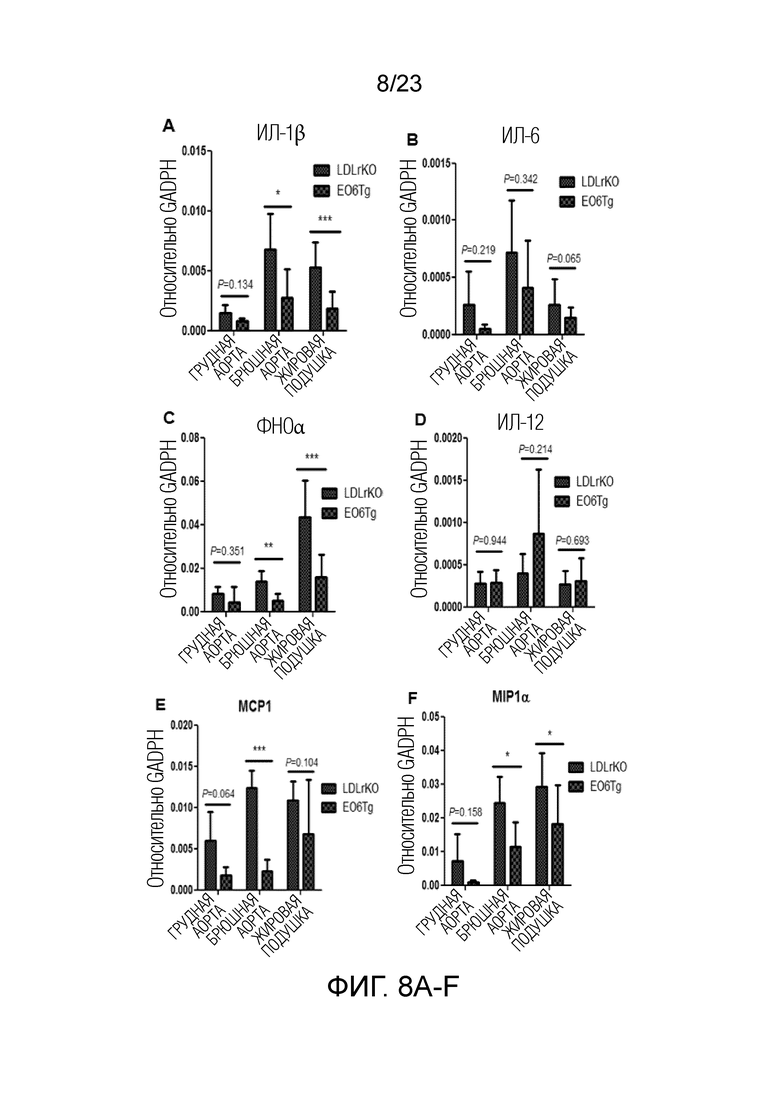
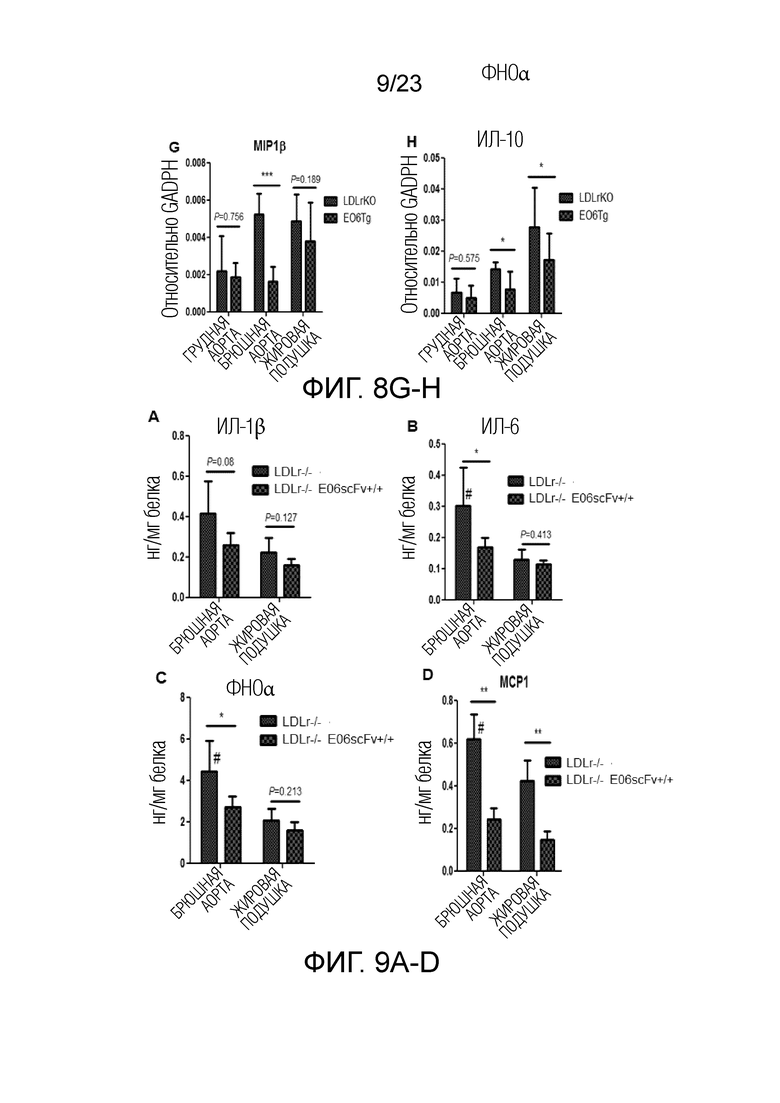
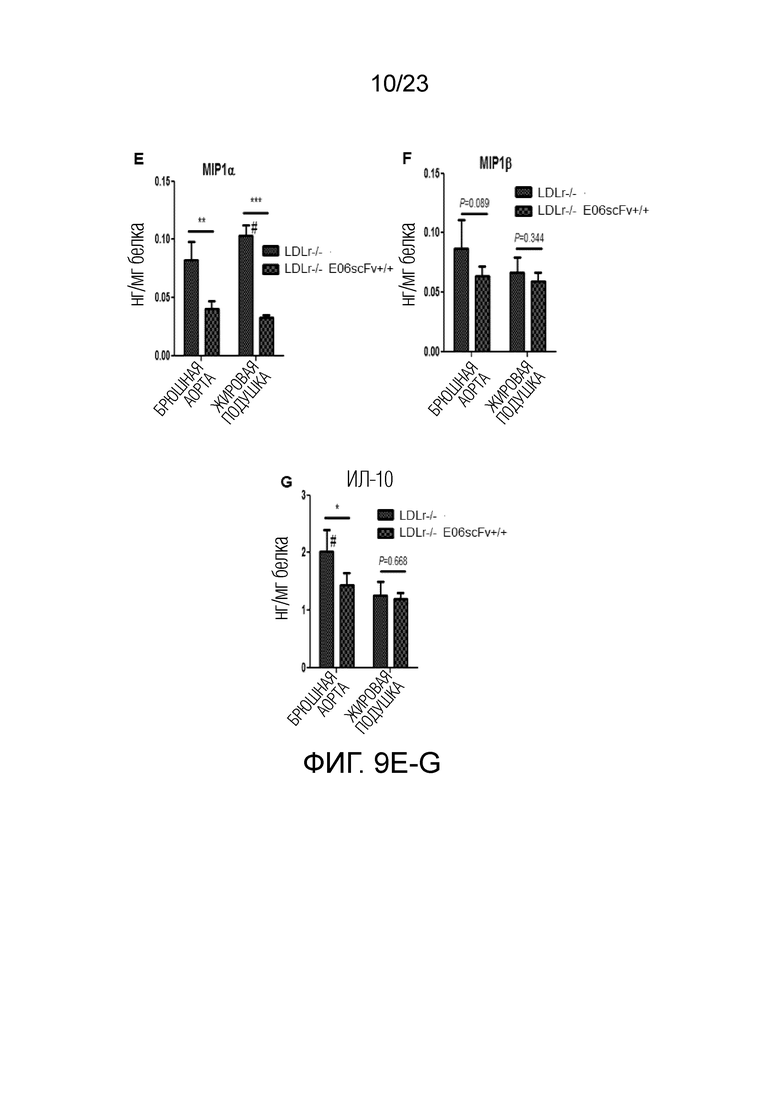
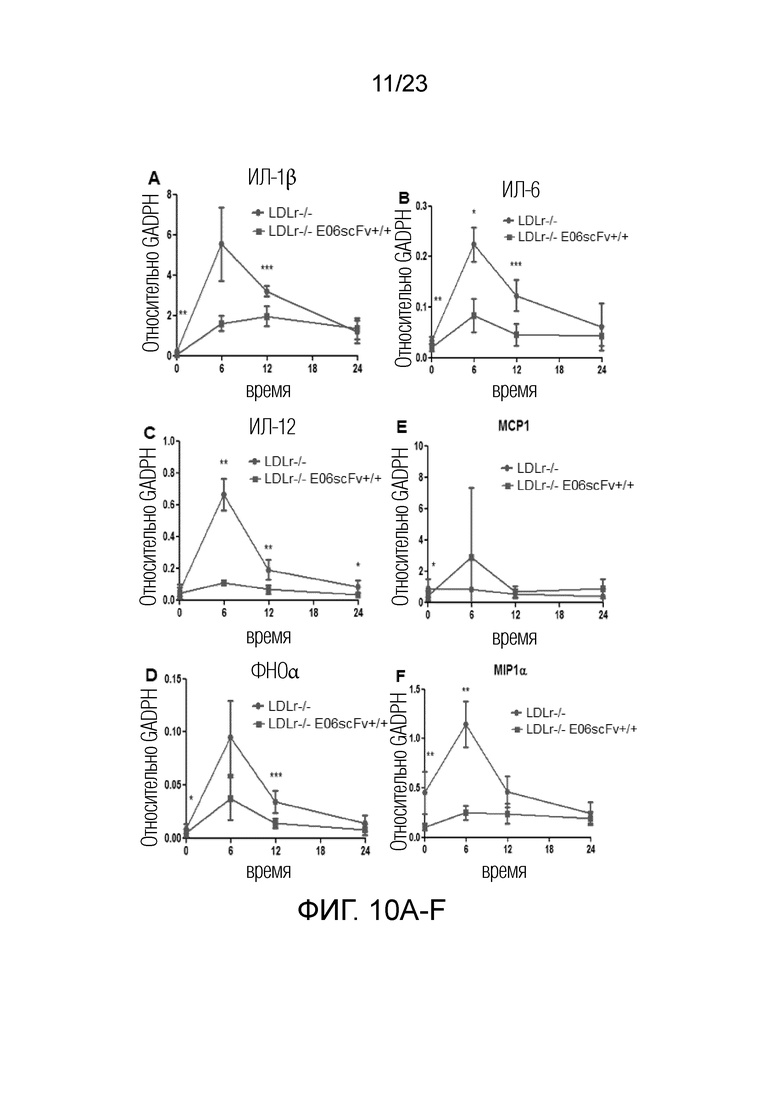
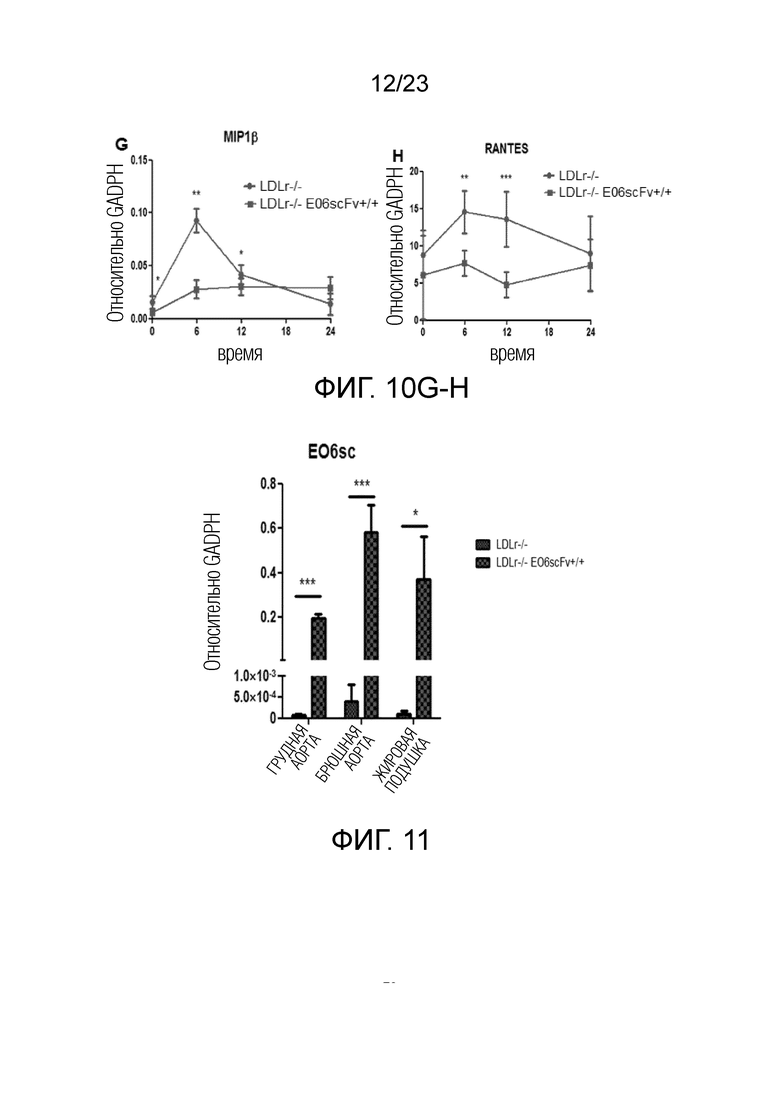
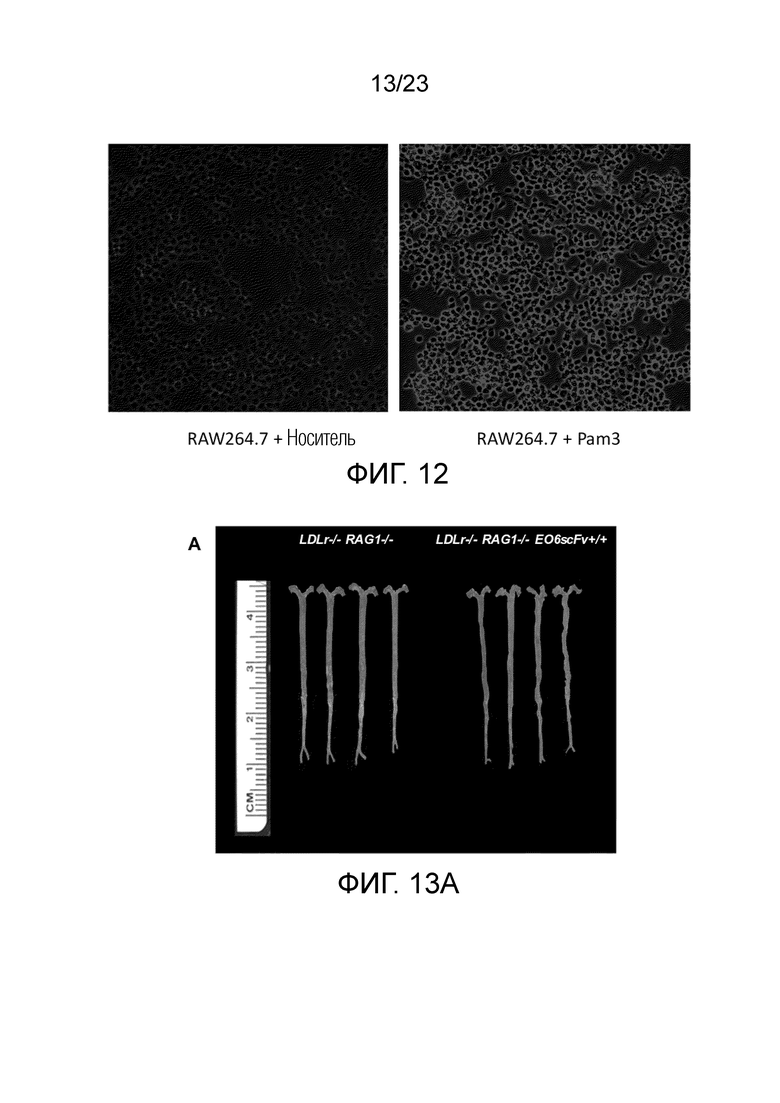
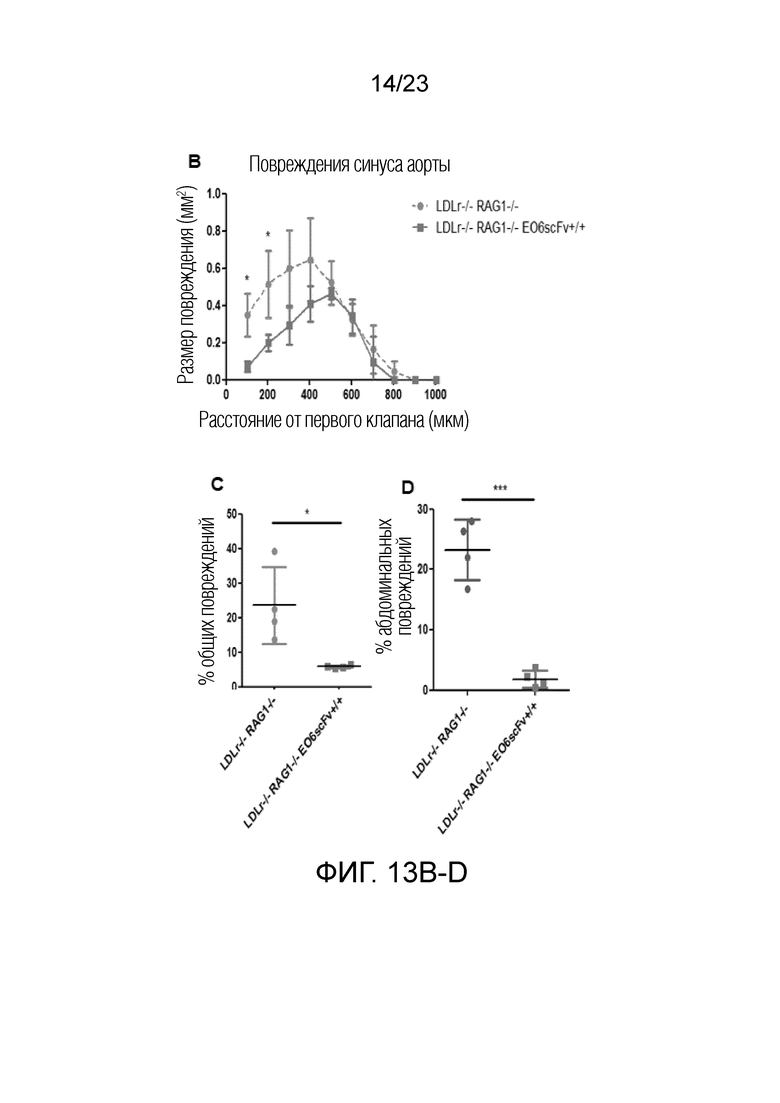
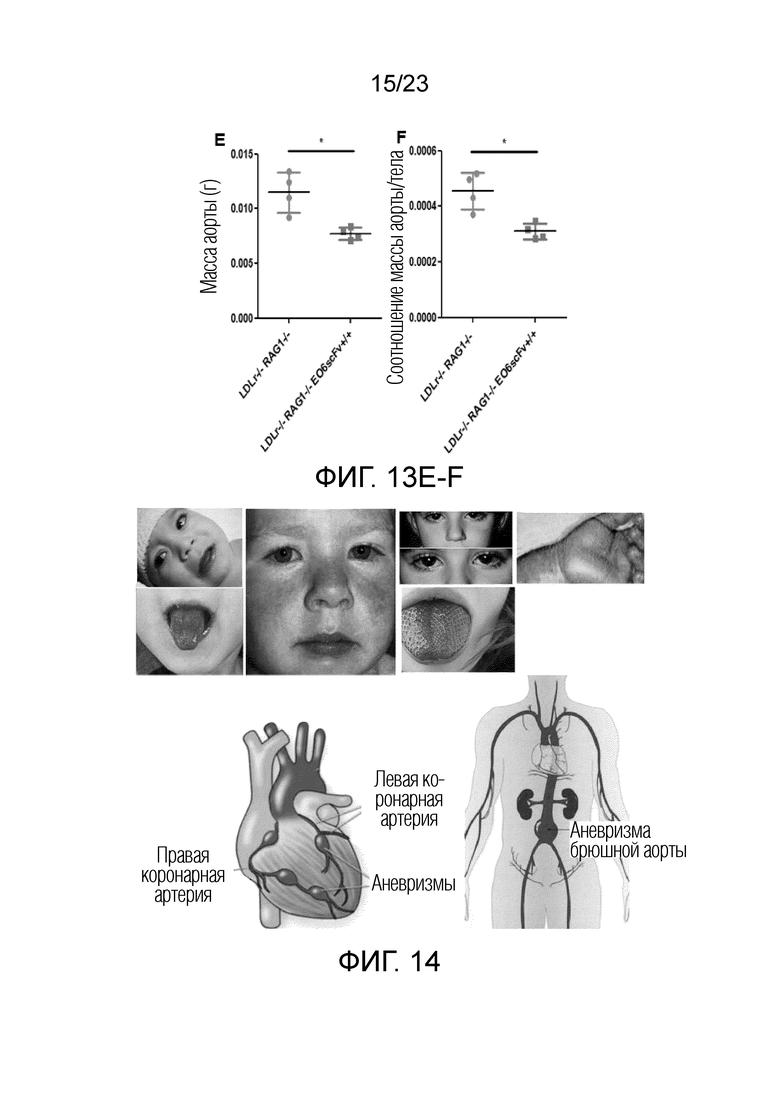
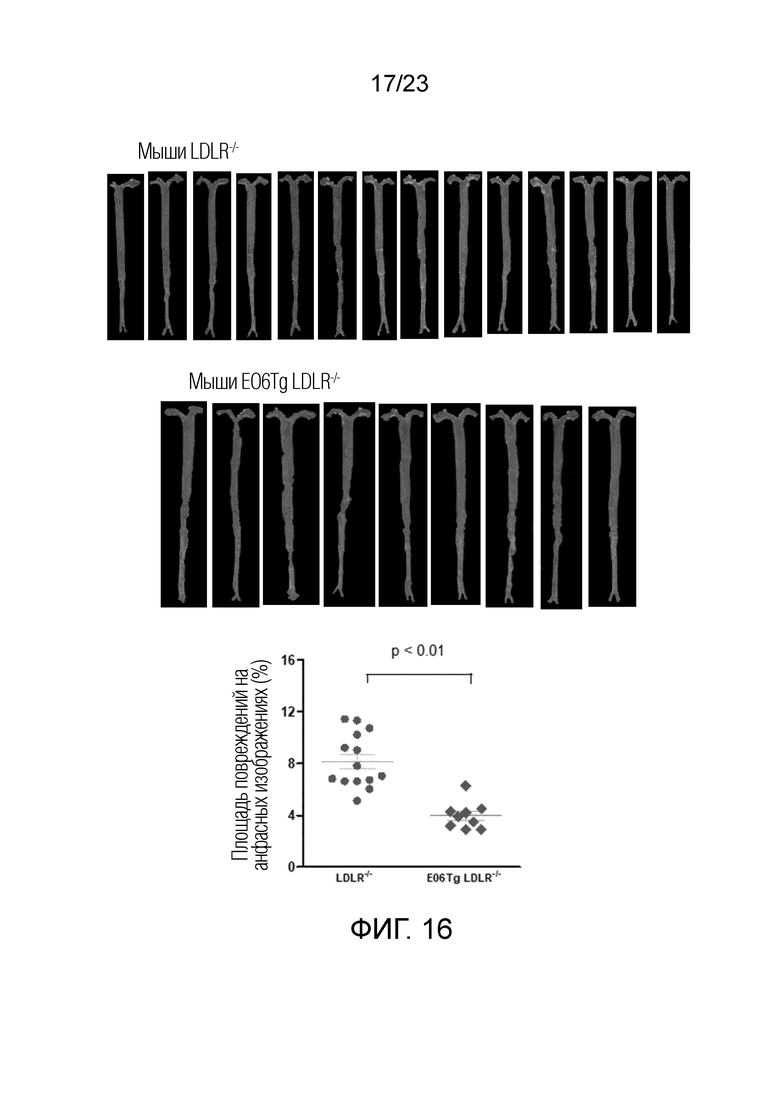
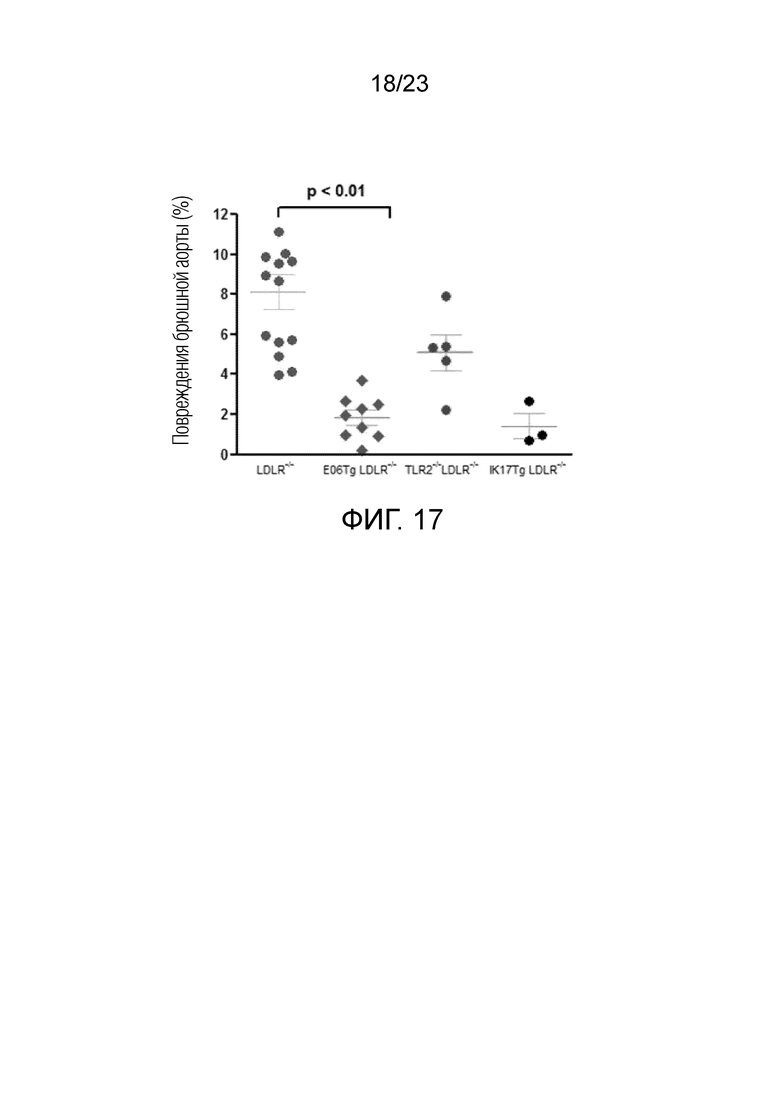
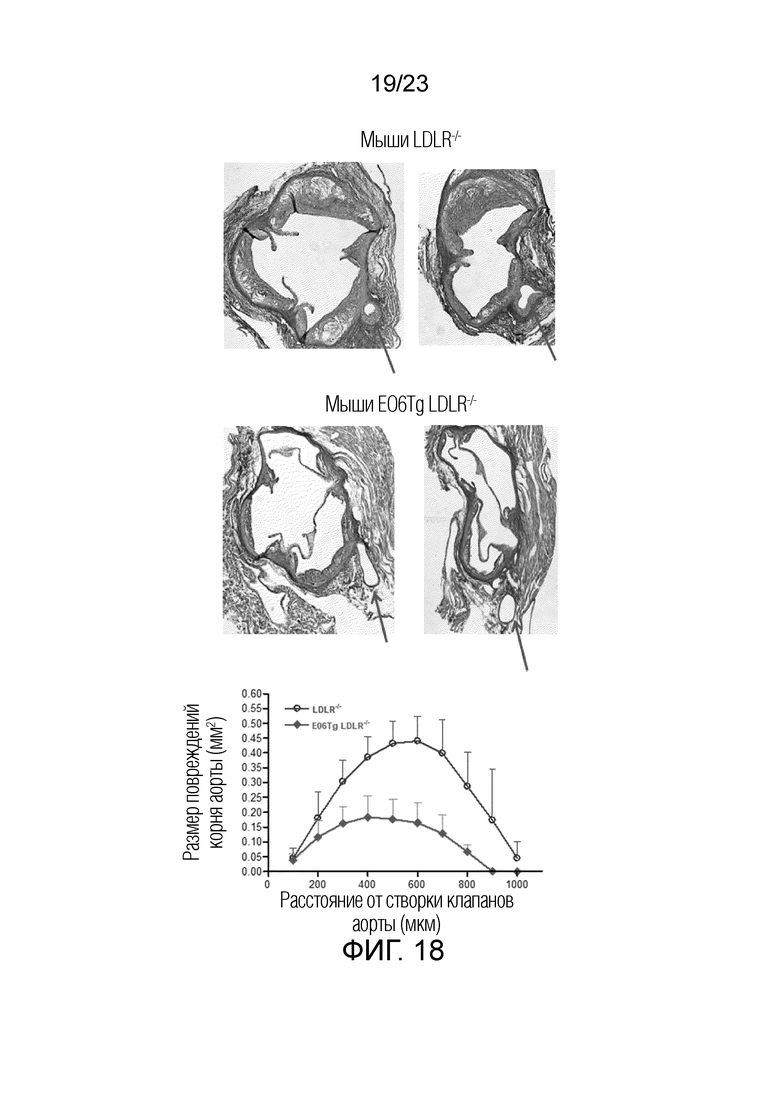
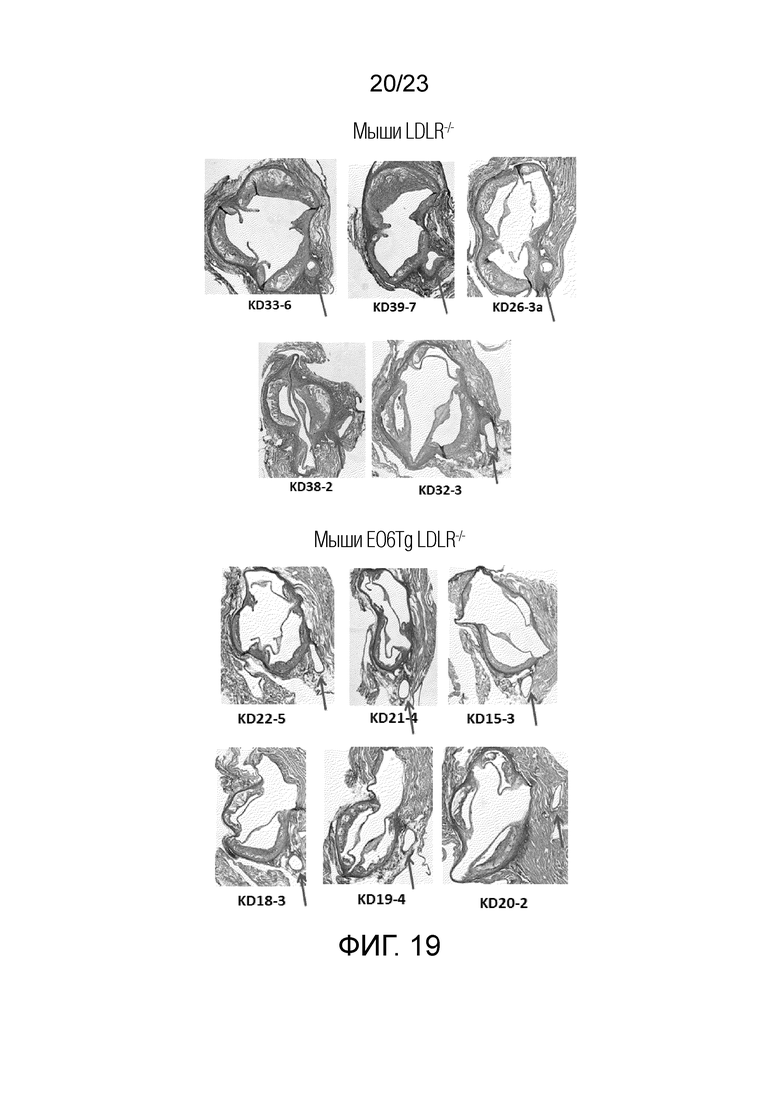
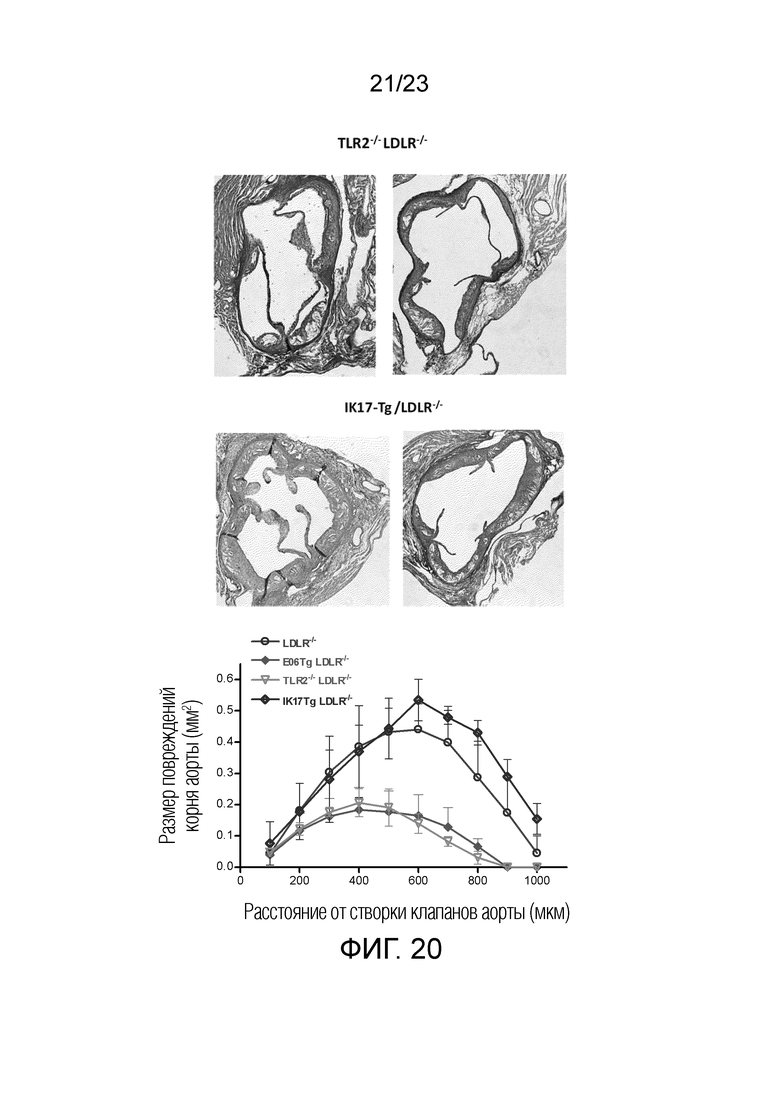
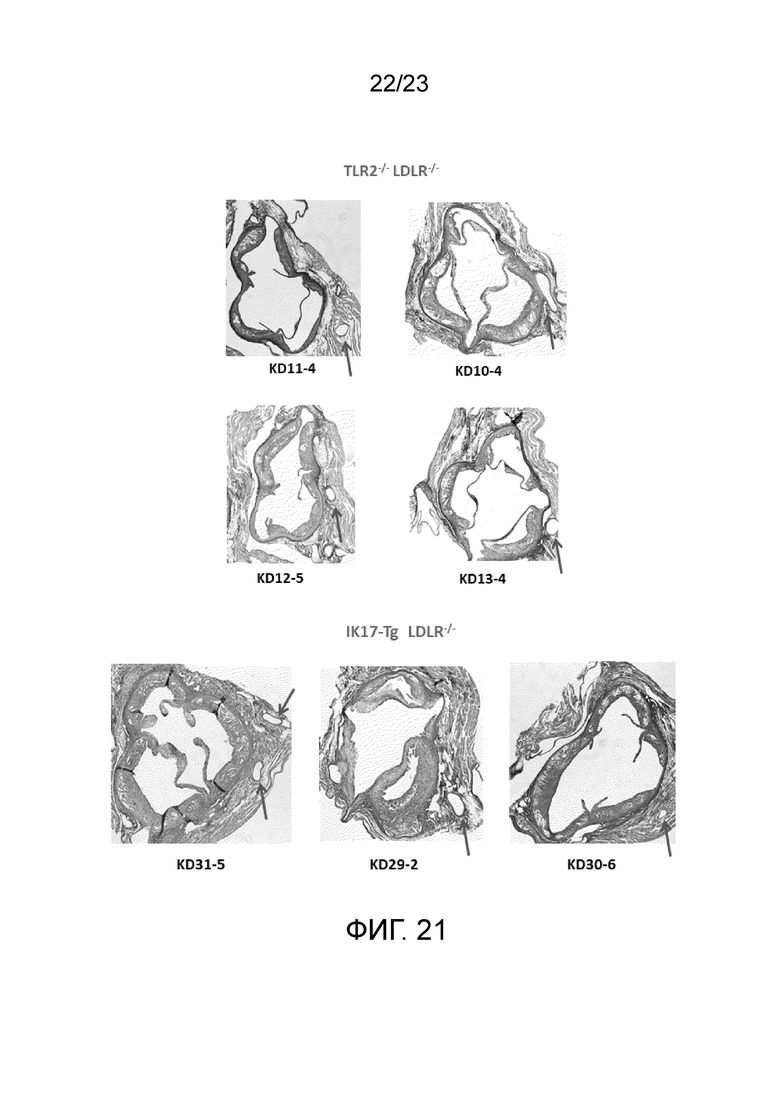
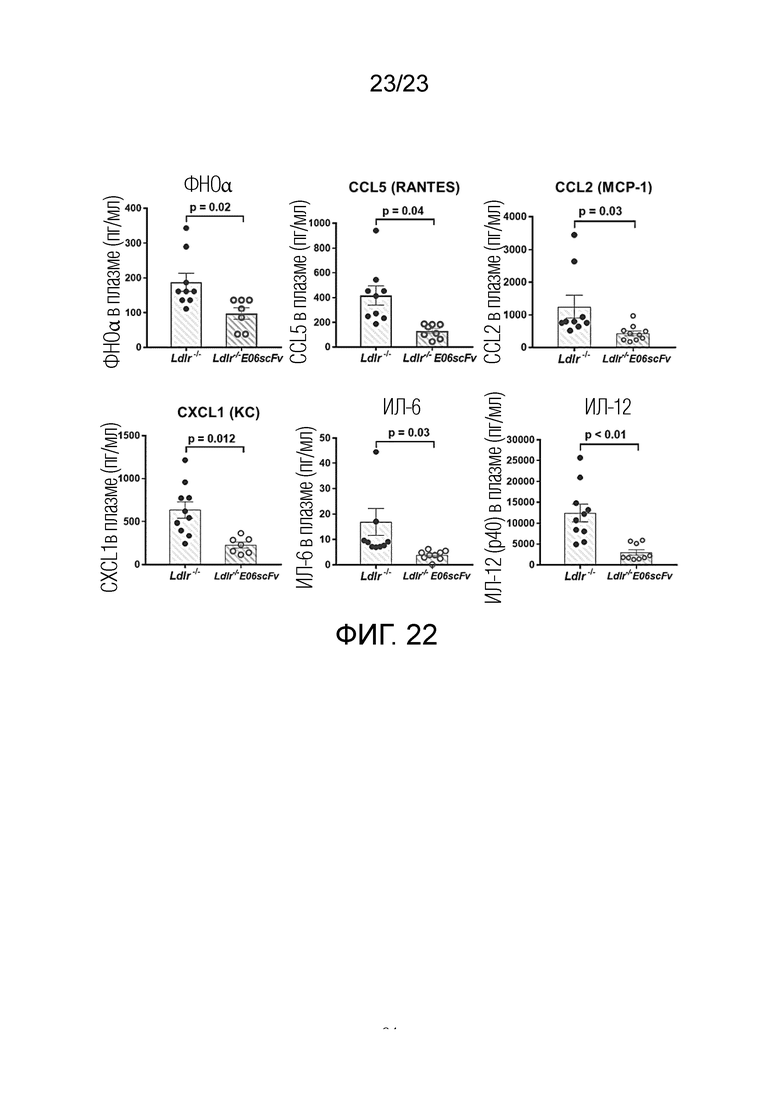
Изобретение относится к биотехнологии. Раскрыто применение полипептида, специфически связывающегося с окисленным фосфолипидом (OxPL), в лечении болезни Кавасаки, где полипептид ингибирует биологическую активность OxPL. При этом полипептид содержит: вариабельный домен тяжелой цепи (VH), содержащий определяющие комплементарность области (CDR) аминокислотной последовательности VH, представленной в SEQ ID NO: 2, и вариабельный домен легкой цепи (VL), содержащий CDR аминокислотной последовательности VL, представленной в SEQ ID NO: 2 или 4. Изобретение позволяет применять его для эффективного лечения болезни Кавасаки у пациента. 14 з.п. ф-лы, 22 ил., 1 пр.
1. Применение полипептида, специфически связывающегося с окисленным фосфолипидом (OxPL), в лечении болезни Кавасаки,
где полипептид ингибирует биологическую активность OxPL и
где полипептид содержит: вариабельный домен тяжелой цепи (VH), содержащий определяющие комплементарность области (CDR) аминокислотной последовательности VH, представленной в SEQ ID NO: 2, и вариабельный домен легкой цепи (VL), содержащий CDR аминокислотной последовательности VL, представленной в SEQ ID NO: 2 или 4.
2. Применение по п.1, дополнительно включающее введение индивидууму дополнительного терапевтического средства, пригодного для лечения болезни Кавасаки.
3. Применение по п.1, дополнительно включающее введение индивидууму внутривенного иммуноглобулина и/или салицилата.
4. Применение по любому из пп.1-3, где индивидуум является человеком возрастом менее пяти лет.
5. Применение по любому из пп.1-4, где биологическая активность OxPL включает активацию пути апоптоза CD36-TLR2.
6. Применение по любому из пп.1-5, где полипептид содержит одноцепочечный вариабельный фрагмент (ScFv).
7. Применение по любому из пп.1-6, где полипептид вводят внутрисосудисто.
8. Применение по любому из пп.1-7, где VH и VL соединены с Fc или областью FC2.
9. Применение по п.6, где scFv связывается с фосфохолиновой концевой группой окисленного фосфолипида и/или где scFv является растворимым в физиологических условиях.
10. Применение по п.1, где VH содержит аминокислотные последовательности CDR, представленные в SEQ ID NO: 6, 7 и 8, и VL содержит аминокислотные последовательности CDR, представленные в SEQ ID NO: 9, 10 и 11.
11. Применение по п.10, где VH содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности VH, представленной в SEQ ID NO: 2, и VL содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности VL, представленной в SEQ ID NO: 2.
12. Применение по п.10, где VH содержит аминокислотную последовательность VH, представленную в SEQ ID NO: 2, и VL содержит аминокислотную последовательность VL, представленную в SEQ ID NO: 2.
13. Применение по п.1, где VH содержит аминокислотные последовательности CDR, представленные в SEQ ID NO: 6, 7 и 8, и VL содержит аминокислотные последовательности CDR, представленные в SEQ ID NO: 12, 10 и 11.
14. Применение по п.13, где VH содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности VH, представленной в SEQ ID NO: 4, и VL содержит аминокислотную последовательность, которая по меньшей мере на 95% идентична аминокислотной последовательности VL, представленной в SEQ ID NO: 4.
15. Применение по п.13, где VH содержит аминокислотную последовательность VH, указанную в SEQ ID NO: 4, и VL содержит аминокислотную последовательность VL, представленную в SEQ ID NO: 4.
| KADL et al., "Oxidized phospholipid-induced inflammation is mediated by Toll-like receptor 2" Free Radlc Biol Med, 15 November 2011, Vol | |||
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Приспособление для присоединения стойки полевого колеса плуга к главной оси передка | 1924 |
|
SU1903A1 |
| WO 201431034 A2, 28.08.2014 | |||
| MORTAZAVI et al., "Down-regulation of TLR2, 3, 9 and Signaling Mediators, MyD88 and TRIF, Gene Transcript Levels in Patients with Kawasaki Disease Treated | |||

