Область техники
Настоящее изобретение относится к области биотехнологии, в частности к аминоацил-тРНК синтетазе, которая эффективно вводит в белок производное лизина.
Уровень техники
Интегральный белок неприродной аминокислоты (цельный белок), полученный путем замены аминокислотного остатка в требуемом месте белка аминокислотами, отличающимися от тех 20 типов, которые участвуют в синтезе обычного белка (неприродные аминокислоты), можно использовать в качестве эффективного средства для анализа структуры и функции белка. Путем применения пар аминоацил-тРНК синтетаза (aaRS)/тРНК, происходящих от различных биологических видов, было синтезировано более 30 типов цельных белков. Системой с наиболее длительной историей применения для введения многих полезных неприродных аминокислот является пара мутантной тирозил-тРНК синтетазы (TyrRS) и ген амбер-супрессора тРНКTyr. Что касается данного способа, ключевым является его ортогональное соотношение, то есть aaRS в двух группах эубактерий, архей и эукариот аминоацилирует тРНК в своих соответствующих группах, но при этом не могут быть аминоацилированы тРНК других групп.
С другой стороны, пирролизил-тРНК синтетаза (PylRS) и ген амбер-супрессора tRNAPyl, полученный из Methanosarcina mazei, в клетках Е. coli функционируют как ортогональная пара aaRS/тРНК. Пирролизин представляет собой производное лизина с объемным метилпирролиновым фрагментом в боковой цепи. PylRS дикого типа в Е. coli может связывать Nε-Boc-L-лизин с tRNAPyl. Однако, поскольку LysRS имеет точное распознавание лизина, было затруднительно сайт-специфичным образом вводить в белки производные лизина, имеющие функциональные группы различного размера и формы.
Следовательно, специалистам в данной области необходимо модифицировать лизил-тРНК-синтетазу дикого типа и разработать аминоацил-тРНК-синтетазу, которая эффективно вводит производные лизина в белок.
Сущность изобретения
Целью настоящего изобретения является предложение аминоацил-тРНК синтетазы и способа эффективного введения производных лизина в белки.
В первом объекте настоящего изобретения предложена мутантная лизил-тРНК-синтетаза, причем такая мутантная лизил-тРНК-синтетаза имеет мутацию в позиции 19 аргинина (R) и/или позиции 29 гистидина (Н) в аминокислотной последовательности, соответствующей лизил-тРНК синтетазе дикого типа.
В другом предпочтительном варианте осуществления лизил-тРНК-синтетаза дикого типа получена из Methanosarcina mazei, Methanosarcina barken или Methanosarcina acetivorans, относящихся к метаногенным археям.
В другом предпочтительном варианте осуществления аминокислотная последовательность лизил-тРНК синтетазы дикого типа показана в SEQ ID NO: 1.
В другом предпочтительном варианте осуществления аминокислотная последовательность лизил-тРНК синтетазы дикого типа показана в SEQ ID NO: 2.
В другом предпочтительном варианте осуществления аргинин (R) в позиции 19 мутирован на гистидин (Н) или лизин (K); и/или
гистидин (Н) в позиции 29 мутирован на аргинин (R) или лизин (K).
В другом предпочтительном варианте осуществления мутация мутантной лизил-тРНК синтетазы выбрана из группы, состоящей из R19H, R19K, H29R, Н29K и их комбинации.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза дополнительно включает в себя мутацию на сайте, выбранном из группы, состоящей из изолейцина (I) в позиции 26, треонина (Т) в позиции 122, лейцина (L) в позиции 309, цистеина (С) в позиции 348, тирозина (Y) в позиции 384 и их комбинации.
В другом предпочтительном варианте осуществления сайт мутации мутантной лизил-тРНК синтетазы дополнительно включает в себя изолейцин (I) в позиции 26; предпочтительно, изолейцин (I) в позиции 26 мутирован на валин (V).
В другом предпочтительном варианте осуществления сайт мутации мутантной лизил-тРНК синтетазы дополнительно включает в себя треонин (Т) в позиции 122; предпочтительно, треонин (Т) в позиции 122 мутирован на триптофан (S).
В другом предпочтительном варианте осуществления сайт мутации мутантной лизил-тРНК синтетазы дополнительно включает в себя лейцин (I) в позиции 309; предпочтительно, лейцин (L) в позиции 309 мутирован на аланин (А).
В другом предпочтительном варианте осуществления сайт мутации мутантной лизил-тРНК синтетазы дополнительно включает в себя цистеин (С) в позиции 348; предпочтительно, цистеин (С) в позиции 348 мутирован на триптофан (S).
В другом предпочтительном варианте осуществления сайт мутации мутантной лизил-тРНК синтетазы дополнительно включает в себя тирозин (Y) в позиции 384; предпочтительно, тирозин (Y) в позиции 384 мутирован на фенилаланин (F).
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза дополнительно включает в себя мутацию, выбранную из группы, состоящей из: I26V, T122S, L309A, C348S, Y384F и их комбинаций.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19H и H29R.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19K и H29R.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19H и Н29К.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19K и Н29К.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19H, I26V и H29R.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19H, H29R, T122S и Y384F.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза включает в себя мутацию, выбранную из группы, состоящей из R19H, H29R, L309A и C348S.
В другом предпочтительном варианте осуществления, за исключением мутации (такой как в позиции 19 и/или 29 и, необязательно, позиции 26, 122, 309, 348 и/или 384), другие аминокислоты мутантной лизил-тРНК синтетазы являются такими же или практически такими же, как последовательность, показанная в SEQ ID NO: 1 или SEQ ID NO: 2.
В другом предпочтительном варианте осуществления «практически такие же» означает, что отличаются самое большее 50 (предпочтительно 1-20, более предпочтительно 1-10) аминокислот, причем такое отличие включает в себя замещение, делецию или добавление аминокислоты, а мутантный белок все еще имеет активность лизил-тРНК синтетазы.
В другом предпочтительном варианте осуществления по сравнению с SEQ ID NO. 1 или SEQ ID NO: 2, последовательность аминокислоты мутантной лизил-тРНК синтетазы имеет, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, 80%), 85%о, 90% и, более предпочтительно, по меньшей мере, 95%о, 96%о, 97%о, 98%, 99% идентичность последовательности.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза образована путем мутирования лизил-тРНК-синтетазы дикого типа, как показано в SEQ ID NO.: 1 или SEQ ID NO: 2.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза выбрана из группы, состоящей из:
(1) полипептида, имеющего аминокислотную последовательность, показанную в любой из SEQ ID NO.: 3-8; или
(2) полипептида, образованного замещением, делецией или добавлением к аминокислотной последовательности одного или нескольких (предпочтительно 1-20, более предпочтительно 1-15, более предпочтительно 1-10, более предпочтительно 1-8, более предпочтительно 1-3, наиболее предпочтительно 1) аминокислотных остатков, как показано в любой из SEQ ID NO.: 3-8, и полученного из полипептида, имеющего аминокислотную последовательность, показанную в любой из SEQ ID NO.: 3-8, имеющей функцию полипептида, как описано в (1).
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантной лизил-тРНК синтетазы показана в любой из SEQ ID NO.: 3-8.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза представляет собой неприродный белок.
В другом предпочтительном варианте осуществления мутантную лизил-тРНК синтетазу используют для введения в целевой белок производных лизина.
В другом предпочтительном варианте осуществления мутантная лизил-тРНК синтетаза имеет следующие характеристики:
по сравнению с лизил-тРНК синтетазой дикого типа, в белки могут быть введены производные лизина с большими функциональными группами.
Во втором объекте настоящего изобретения предложен изолированный полинуклеотид, кодирующий мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения.
В другом предпочтительном варианте осуществления такой полинуклеотид кодирует полипептид, как показано в любой из SEQ ID NO.: 3-8.
В другом предпочтительном варианте осуществления такой полинуклеотид включает в себя последовательность ДНК, последовательность РНК и их комбинации.
В другом предпочтительном варианте осуществления такой полинуклеотид дополнительно содержит вспомогательный элемент, выбранный из группы, состоящей из сигнального пептида, секреторного пептида, последовательности метки (такой как 6His) и их комбинации на фланкировании открытой рамки считывания (ОРС) мутантной лизил-тРНК синтетазы.
В третьем объекте настоящего изобретения предложен вектор, содержащий полинуклеотид согласно второму объекту настоящего изобретения.
В другом предпочтительном варианте осуществления такой вектор включает в себя вектор экспрессии, челночный вектор и интегративный вектор.
В другом предпочтительном варианте осуществления такой вектор выбран из группы, состоящей из рЕТ, pCW, pUC, pPIC9k, рМА5 и их комбинаций.
В другом предпочтительном варианте осуществления такой вектор представляет собой вектор pEvol и/или вектор pBAD.
В другом предпочтительном варианте осуществления такой вектор используется для экспрессирования мутантной лизил-тРНК синтетазы.
В третьем объекте настоящего изобретения предложена клетка-хозяин, которая содержит вектор согласно третьему объекту настоящего изобретения, или ее геном, который интегрирует полинуклеотид согласно второму объекту настоящего изобретения.
В другом предпочтительном варианте осуществления, клетка-хозяин выбрана из группы, состоящей из прокариотической клетки, эукариотической клетки и их комбинации.
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой эукариотическую клетку, такую как дрожжевая клетка, растительная клетка или клетка животного происхождения (такая как клетка млекопитающего).
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой прокариотическую клетку, такую как Escherichia coli.
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой Тор10 или BL21 Е. coli.
В другом предпочтительном варианте осуществления клетка-хозяин содержит:
(a) мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения; а также
(b) ортогональную тРНК, способную связываться с производным лизина в присутствии мутантной лизил-тРНК синтетазы; и, не обязательно
(c) последовательность целевой нуклеиновой кислоты, кодирующая целевой белок, которая включает в себя кодон, распознаваемый ортогональной тРНК в позиции для введения производного лизина.
В другом предпочтительном варианте осуществления клетка-хозяин дополнительно содержит:
(d) производное лизина.
В другом предпочтительном варианте осуществления производное лизина представляет собой субстрат мутантной лизил-тРНК синтетазы.
В другом предпочтительном варианте осуществления производное лизина выбрано из группы, состоящей из производного алкинилоксикарбониллизина, производного ВОС-лизина (трет-бутилоксикарборил-L-лизин), лизина, ацилированного с помощью жирных кислот, и их комбинации.
В другом предпочтительном варианте осуществления структура алкинилоксикарбониллизина показана в Формуле I:
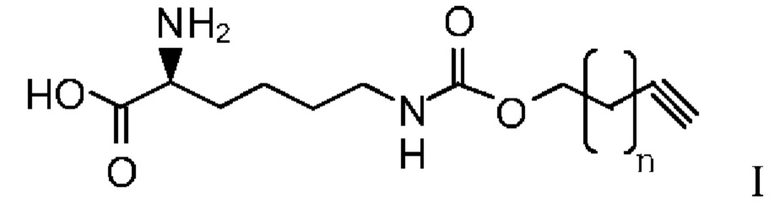
где n представляет собой 0-8.
В другом предпочтительном варианте осуществления ортогональная тРНК является супрессором тРНК, предпочтительно амбер-супрессором тРНК.
В другом предпочтительном варианте осуществления кодирующая аминокислотная последовательность ортогональной тРНК показана в SEQ ID NO.: 9.
GGAAACCTGATCATGTAGATCGAATGGACTCTAAATCCGTTCAGCCGGGTTAGATTCCCGGGGTTTCCGCCA (SEQ ID NO.: 9)
В другом предпочтительном варианте осуществления кодон, распознаваемый ортогональной тРНК, представляет собой амбер-кодон.
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой прокариотическую клетку или эукариотическую клетку, предпочтительно Escherichia coli.
Пятый объект настоящего изобретения предлагает систему экспрессии, при этом такая система экспрессии содержит:
(a) мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения; а также
(b) ортогональную тРНК, способную связываться с производным лизина в присутствии мутантной лизил-тРНК синтетазы; и, не обязательно
(c) последовательность целевой нуклеиновой кислоты, кодирующая целевой белок, которая включает в себя кодон, распознаваемый ортогональной тРНК в позиции для введения производного лизина.
В другом предпочтительном варианте осуществления система экспрессии дополнительно содержит:
(d) производное лизина.
В другом предпочтительном варианте осуществления производное лизина выбрано из группы, состоящей из производного алкинилоксикарбониллизина, производного ВОС-лизина (трет-бутилоксикарборил-L-лизин), лизина, ацилированного с помощью жирных кислот, и их комбинации.
В другом предпочтительном варианте осуществления структура алкинилоксикарбониллизина показана в Формуле I:
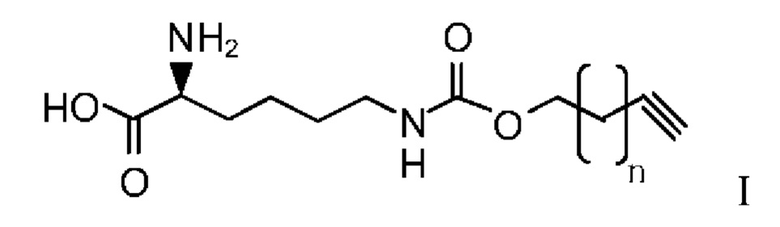
где n представляет собой 0-8.
В другом предпочтительном варианте осуществления ортогональная тРНК является супрессором тРНК, предпочтительно амбер-супрессором тРНК.
В другом предпочтительном варианте осуществления аминокислотная последовательность ортогональной тРНК показана в SEQ ID NO.: 9.
В другом предпочтительном варианте осуществления кодон, распознаваемый ортогональной тРНК, представляет собой амбер-кодон.
В другом предпочтительном варианте осуществления система экспрессии представляет собой клетку или клеточный экстракт.
В другом предпочтительном варианте осуществления система экспрессии применяется для введения алкинилоксикарбониллизина в целевой белок или для получения целевого белка, содержащего неприродную аминокислоту.
В шестом объекте настоящего изобретения предложен способ введения неприродной аминокислоты в целевой белок, включающий в себя следующие этапы:
(1) обеспечение клетки-хозяина согласно четвертому объекту настоящего изобретения или системы экспрессии согласно пятому объекту настоящего изобретения; и
(2) в присутствии производного лизина целевой белок экспрессируется в клетке-хозяине или системе экспрессии.
В другом предпочтительном варианте осуществления целевой белок выбран из группы, состоящей из: белка-предшественника человеческого инсулина, белка-предшественника лиспро-инсулина, белка-предшественника инсулина гларгина, паратироидного гормона, кортикорелина, кальцитонина, бивалирудина, глюкагоноподобного пептида и его производных, экзенатида и лираглутида, зиконотида, серморелина, соматорелина, секретина, тедуглутида, гирудина, гормона роста, фактор роста, фактора высвобождения гормона роста, адренокортикотропного гормона, фактора высвобождения, дезлорелина, десмопрессина, элкатонина, глюкагона, лейпрорелина, лютеинизирующего гормона-гормона высвобождения, соматостатина, тиротропин-
высвобождающего гормона, трипторелина, вазоактивного кишечного пептида, интерферона, паратироидного гормон, пептида ВН3, амилоидного пептида или фрагмента указанного выше пептида, и их комбинации.
В другом предпочтительном варианте осуществления неприродная аминокислота представляет собой производное лизина.
В другом предпочтительном варианте осуществления способ включает в себя этапы:
(1) обеспечение клетки или клеточного экстракта, экспрессирующего мутантную лизил-тРНК синтетазу, ортогональную тРНК и последовательность целевой нуклеиновой кислоты согласно первому объекту настоящего изобретения; и
(2) культивирование клетки или клеточного экстракта в присутствии производного лизина, и, как следствие, введение производного лизина в целевой белок посредством пары ортогональной тРНК-синтетазы и ортогональной тРНК.
В седьмом объекте настоящего изобретения предложен набор, содержащий (а) контейнер и (б) мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения или полинуклеотид согласно второму объекту настоящего изобретения, содержащиеся в контейнере.
В другом предпочтительном варианте осуществления такой набор включает в себя:
(i) первую последовательность нуклеиновой кислоты, содержащую первую кассету экспрессии для экспрессирования мутантной лизил-тРНК синтетазы; и/или
(ii) вторую последовательность нуклеиновой кислоты, содержащую вторую кассету экспрессии для экспрессирования ортогональной тРНК, причем ортогональная тРНК может быть объединена с производным лизина в присутствии мутантной лизил-тРНК синтетазы; и/или
(iii) третью последовательность нуклеиновой кислоты, содержащую третью кассету экспрессии для экспрессирования целевого белка, и кодирующую последовательность целевого белка, которая включает в себя кодон, распознаваемый ортогональной тРНК в позиции для введения производного лизина.
В другом предпочтительном варианте осуществления такой набор дополнительно включает в себя клеточный экстракт.
В другом предпочтительном варианте осуществления соответствующие последовательности нуклеиновой кислоты находятся в одном или разных контейнерах.
В другом предпочтительном варианте осуществления каждая последовательность нуклеиновой кислоты расположена на другом векторе.
В восьмом объекте настоящего изобретения предложена система трансляции, при этом такая система трансляции содержит:
(a) мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения;
(b) ортогональную тРНК, способную связываться с производным лизина в присутствии мутантной лизил-тРНК синтетазы; и
(c) производное лизина.
В девятом объекте настоящего изобретения предлагается применение мутантной лизил-тРНК синтетазы в соответствии с первым объектом настоящего изобретения, или клетки-хозяина в соответствии с четвертым объектом настоящего изобретения, или системы экспрессии в соответствии с пятый объектом настоящего изобретения или набора в соответствии с седьмым объектом настоящего изобретения, для внедрения неприродной аминокислоты в целевой белок или для получения целевого белка, содержащего неприродную аминокислоту.
В десятом объекте настоящего изобретения предлагается способ получения целевого белка, содержащего неприродную аминокислоту, включающий в себя этапы:
(1) обеспечение клетки-хозяина согласно четвертому объекту настоящего изобретения или системы экспрессии согласно пятому объекту настоящего изобретения; и
(2) в присутствии производного лизина целевой белок экспрессируется в клетке-хозяине или системе экспрессии.
В одиннадцатом объекте настоящего изобретения предложен способ получения мутантной лизил-тРНК синтетазы согласно первому объекту настоящего изобретения, содержащий этапы: (i) культивирование клетки-хозяина согласно четвертому объекту настоящего изобретения, и, посредством этого, экспрессирование мутантной лизил-тРНК синтетазы.
В двенадцатом объекте настоящего изобретения предложен ферментный препарат, содержащий мутантную лизил-тРНК синтетазу согласно первому объекту настоящего изобретения.
В другом предпочтительном варианте осуществления лекарственная форма фармацевтического препарата включает в себя: лиофилизированный препарат, жидкий препарат и их комбинацию.
Следует понимать, что в пределах объема настоящего изобретения вышеупомянутые технические признаки настоящего изобретения и технические признаки, конкретно описанные ниже (такие как варианты осуществления), могут быть объединены друг с другом для образования нового или предпочтительного технического решения. Из-за ограничения по объему я не буду повторять их в данном документе один за другим.
Описание чертежей
На Фиг. 1 показана карта плазмиды pEvol-pylRs(R19H, H29R)-pylT.
Подробное описание
После обширных и углубленных исследований и обширного скрининга изобретатели неожиданно получили мутантную лизил-тРНК синтетазу. По сравнению с лизил-тРНК-синтетазой дикого типа мутантная лизил-тРНК-синтетаза по настоящему изобретению может значительно увеличить количество инсерций неприродных аминокислот и количество целевого белка, содержащего неприродные аминокислоты. Кроме того, мутантная лизил-тРНК синтетаза по настоящему изобретению также может улучшать стабильность целевого белка так, что его не удается легко разрушить. На этом основании изобретатели совершили настоящее изобретение.
Термины
Чтобы облегчить понимание настоящего описания, сначала определим некоторые термины. При использовании в данной заявке, если явно не указано иное, каждый из следующих терминов имеет значение, указанное ниже. Другие определения приводятся по мере применения.
Термин «около» может относиться к значению или составу в пределах допустимого диапазона погрешности конкретного значения или состава, определенного специалистом в данной области техники, которые частично будут зависеть от того, каким образом такое значение или состав измеряют или определяют. Например, используемое здесь выражение «около 100» включает в себя все значения между 99 и 101 (такие как 99,1; 99,2; 99,3; 99,4 и т.д.).
Используемый в данном документе термин «содержащий» или «включающий в себя» могут подразумевать открытое, полуоткрытое и закрытое множество. Другими словами, такой термин также включает в себя «по существу, состоящий из» или «состоящий из».
Идентичность последовательности (или ее гомологичность) определяют путем сравнения двух выровненных последовательностей в пределах заданного окна сравнения (это может быть 50%, 60%, 70%, 80%, 90%, 95% или 100% длины эталонной нуклеотидной последовательности или белка) и определения количества позиций, в которых появляется один и тот же остаток. Как правило, этот показатель выражают в процентах. Измерение идентичности нуклеотидных последовательностей является способом, который хорошо известен специалистам в данной области.
В данном документе термины «аминоацил-тРНК-синтетаза» и «лизил-тРНК-синтетаза» используются как взаимозаменяемые.
Лизил-тРНК-синтетаза дикого типа
Используемый в данном документе термин «лизил-тРНК-синтетаза дикого типа» относится к встречающейся в природе аминоацил-тРНК-синтетазе, которая не была модифицирована искусственным путем, а ее нуклеотиды могут быть получены с помощью технологий генной инженерии, таких как секвенирование генома, полимеразная цепная реакция (ПЦР) и т.д., при этом из нуклеотидной последовательности может быть выведена аминокислотная последовательность. Источник лизил-тРНК синтетазы дикого типа не имеет конкретного ограничения. Предпочтительным источником являются организмы Methanosarcina mazei, Methanosarcina barkeri и Methanosarcina acetivorans из метаногенных архей, но не ограничен только ими.
В предпочтительном варианте осуществления настоящего изобретения аминокислотная последовательность лизил-тРНК синтетазы дикого типа показана в SEQ ID NO: 1.
MDKKPLNTLISATGLWMSRTGTIHKIKHHEVSRSKIYIEMACGDHLVVNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSATRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDESLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 1)
В предпочтительном варианте осуществления настоящего изобретения аминокислотная последовательность лизил-тРНК синтетазы дикого типа показана в SEQ ID NO: 2.
MDKKPLNTLISATGLWMSRTGTIHKIKHHEVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSGSYYNGISTNL (SEQ ID NO.: 2)
Мутантная лизил-тРНК синтетаза
Используемые в данном документе термины «мутантный белок», «мутантный белок по настоящему изобретению», «мутантная аминоацил-тРНК-синтетаза по настоящему изобретению», «мутантная лизил-тРНК-синтетаза», «мутантный фермент», «мутанты аминоил-тРНК синтаза» могут использоваться взаимозаменяемо, и все относятся к неприродной мутантной аминоацил-тРНК-синтетазе, при этом мутантная аминоацил-тРНК-синтетаза представляет собой белок, искусственно модифицированный полипептидом, как показано в SEQ ID NO.: 1 или SEQ ID NO: 2. В частности, мутантная аминоацил-тРНК синтетаза представляет собой ту, которая описана в первом объекте настоящего изобретения.
Следует понимать, что нумерация аминокислот в мутантной лизил-тРНК-синтетазе по настоящему изобретению основана на лизил-тРНК-синтетазе дикого типа (предпочтительно, SEQ ID NO.: 1 или SEQ ID NO: 2). Когда конкретный мутантный белок имеет 80% или более гомологии с последовательностью, показанной в SEQ ID NO.: 1 или SEQ ID NO: 2, номер аминокислоты мутантного белка может быть смещен относительно нумерации аминокислоты с SEQ ID NO.: 1 или SEQ ID NO: 2, как, например, несовпадение позиций 1-5 с N-концом или С-концом аминокислоты с использованием традиционных в данной области методик выравнивания последовательностей, при этом специалисты в данной области техники в целом должны понимать, что такое несовпадение находится в разумных пределах и мутеины с 80%) гомологией и аналогичной или подобной активностью гликозилтрансферазы не следует исключать из охвата мутантного белка по настоящему изобретению из-за неправильной нумерации аминокислот.
Мутантный белок по настоящему изобретению представляет собой синтетический белок или рекомбинантный белок, то есть он может быть химически синтезированным продуктом или может быть получен от прокариотического или эукариотического хозяина (например, бактерий, дрожжей и растений) с использованием рекомбинантной технологии. В зависимости от хозяина, используемого в протоколе рекомбинантного получения, мутеин по настоящему изобретению может быть гликозилированным или негликозилированным. Мутеин по настоящему изобретению может также включать в себя исходный остаток метионина или не включать его.
Настоящее изобретение также включает в себя фрагменты, производные и аналоги мутеина. Используемые в данном документе термины «фрагмент», «производное» и «аналог» относятся к белку, который по существу сохраняет ту же биологическую функцию или активность, что и мутеин.
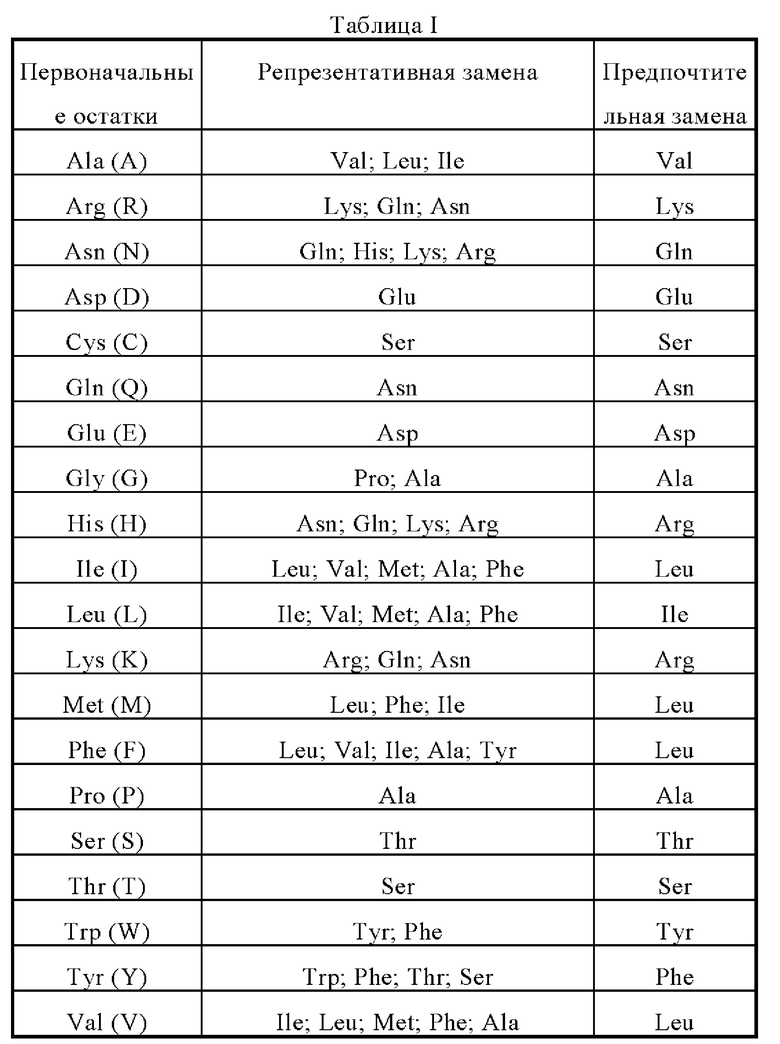
Фрагмент, производное или аналог мутеина по настоящему изобретению может быть (i) мутеином, в котором заменены один или несколько консервативных или неконсервативных аминокислотных остатков (предпочтительно консервативных аминокислотных остатков), причем такие замещенные аминокислотные остатки могут быть кодированы или не кодированы генетическим кодоном, или (ii) мутеином с замещающей группой в одном или нескольких аминокислотных остатках, или (iii) мутеином, образованным путем слияния зрелого мутеина с другим соединением (таким как соединение, которое продлевает длительность периода полураспада мутеина, таким как полиэтиленгликоль), или (iv) мутеином, образованным путем слияния дополнительной аминокислотной последовательности с данной последовательностью мутеина (такой как лидерная последовательность или секреторная последовательность, или последовательность, используемая для очистки данного мутантного белка или последовательности пробелка, или белком слияния, образованным с фрагментом антигена IgG). Согласно изложенному в данном документе принципу, эти фрагменты, производные и аналоги находятся в пределах компетенции специалистов в данной области. В настоящем изобретении консервативно замещенные аминокислоты предпочтительно получают путем аминокислотных замещений согласно таблице I.
Распознание аминокислотного субстрата PylRS связано со стереохимической структурой каталитически активного функционального домена. Величина производных лизина, которые могут быть активированы PylRS дикого типа, ограничена, и производные лизина с большими функциональными группами не могут быть введены в белки. Следовательно, улучшить эффект можно путем мутации сайта PylRS, избегая стерических несоответствий связывающего субстрата или взаимодействия мутантной аминокислоты с аминокислотой субстрата или частью основной цепи.
Предпочтительно, мутеин представляет собой такой, как показан в любой
из SEQ ID NO.: 3-8.
MDKKPLNTLISATGLWMSHTGTIHKVKHREVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 3)
MDKKPLNTLISATGLWMSHTGTIHKIKHREVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENSEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVFGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 4)
MDKKPLNTLISATGLWMSHTGTIHKIKHREVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYARKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFSQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 5)
MDKKPLNTLISATGLWMSHTGTIHKIKHREVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 6)
MDKKPLNTLISATGLWMSHTGTIHKIKHHEVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 7)
MDKKPLNTLISATGLWMSRTGTIHKIKHREVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID NO.: 8)
Следует понимать, что по сравнению с последовательностью, показанной в SEQ ID NO.: 1 или SEQ ID NO: 2, мутеин по настоящему изобретению обычно имеет более высокую гомологию (идентичность). Предпочтительно гомология между мутантным белком и последовательностью, показанной в SEQ ID NO.: 1 или SEQ ID NO: 2, составляет, по меньшей мере, 80%, предпочтительно, по меньшей мере, 85-90%), более предпочтительно, по меньшей мере, 95%, более предпочтительно, по меньшей мере, 98%) и, наиболее предпочтительно, по меньшей мере, 99%.
Кроме того, мутантный белок по настоящему изобретению также может быть модифицирован. Модифицированные (обычно не изменяющие первичную структуру) формы включают в себя: формы мутеина in vivo или in vitro, полученные химическим способом, таким как ацетилирование или карбоксилирование. Модификации также включают в себя гликозилирование, такое как то, с помощью которого производят модификации гликозилирования во время синтеза и процессинга мутантного белка или во время следующих этапов процессинга. Эта модификация может быть осуществлена путем воздействия на мутеин фермента, который выполняет гликозилирование (например, гликозилазы или дегликозилазы млекопитающих).
Модифицированные формы также включают в себя последовательности с фосфорилированными аминокислотными остатками (такими как фосфотирозин, фосфосерин, фосфотреонин). Они также включают в себя мутантные белки, которые были модифицированы для повышения их устойчивости к протеолизу или оптимизации их растворимости.
Термин «полинуклеотид, кодирующий мутантную лизил-тРНК-синтетазу» может включать в себя полинуклеотид, кодирующий мутантную лизил-тРНК-синтетазу по настоящему изобретению, или также может быть полинуклеотидом, который включает в себя дополнительные кодирующие и/или некодирующие последовательности.
Настоящее изобретение также относится к вариантам вышеуказанных полинуклеотидов, которые кодируют фрагменты, аналоги и производные полипептидов или мутеинов, имеющих такую же аминокислотную последовательность, что и в настоящем изобретении. Эти варианты нуклеотидов включают в себя варианты замещения, варианты делеции и варианты инсерции. Как известно в данной области, аллельный вариант представляет собой альтернативную форму полинуклеотида. Это может быть замещение, делеция или инсерция одного или нескольких нуклеотидов, но не приводящие к существенному изменению функции кодируемого мутантного белка.
Настоящее изобретение также относится к полинуклеотидам, которые гибридизуются с вышеупомянутыми последовательностями и имеют, по меньшей мере, 50%, предпочтительно, по меньшей мере, 70% и, более предпочтительно, по меньшей мере, 80%) идентичность между двумя последовательностями. Настоящее изобретение, в частности, относится к полинуклеотидам, которые могут гибридизоваться с полинуклеотидом по настоящему изобретению в строгих условиях (или обязательных условиях). В настоящем изобретении «строгие условия» относятся к: (1) гибридизации и элюции при пониженной ионной силе и более высокой температуре, такой как 0,2 × SSC, 0,1% SDS, 60°С; или (2) добавлению денатурирующего агента во время гибридизации, такого как 50%) (об./ об.) формамида, 0,1% телячьей сыворотки/0,1% фиколла, 42°С и т.д.; или (3) гибридизации, происходящей только тогда, когда идентичность между двумя последовательностями составляет, по меньшей мере, 90% или более, а более предпочтительно 95% или более.
Мутеины и полинуклеотиды по настоящему изобретению предпочтительно предоставляют в изолированной форме и, более предпочтительно, очищенными до гомогенности.
Полноразмерную последовательность полинуклеотида по настоящему изобретению обычно можно получить способом ПЦР-амплификации, способом рекомбинации или способом искусственного синтеза. Для способа ПЦР-амплификации праймеры могут быть сконструированы в соответствии с соответствующей нуклеотидной последовательностью, описанной в настоящем изобретении, особенно с последовательностью открытой рамки считывания, и с использованием в качестве шаблона для амплификации соответствующей последовательности библиотеки кДНК, имеющейся в коммерческом доступе, или библиотеки кДНК, подготовленной обычным способом, который известен специалистам, в данной области техники. Если последовательность является длинной, часто необходимо выполнить две или более ПЦР-амплификаций, а затем все амплифицированные фрагменты сплайсируют вместе в правильном порядке.
Как только соответствующая последовательность получена, для получения соответствующей последовательности в больших количествах можно использовать способ рекомбинации. Обычно это делается путем клонирования в вектор с последующим переносом в клетку и последующим выделением соответствующей последовательности из пролиферированной клетки-хозяина обычными способами.
Кроме того, способы искусственного синтеза также можно использовать для синтеза родственных последовательностей, особенно при незначительной длине фрагмента. Обычно сначала синтезируют несколько небольших фрагментов, а затем лигируют их для получения фрагментов с очень длинными последовательностями.
В настоящее время последовательность ДНК, кодирующая белок (или его фрагмент или производное) по настоящему изобретению, может быть полностью получена с помощью химического синтеза. Затем последовательность ДНК может быть введена в разнообразные существующие молекулы ДНК (или такие, как векторы) и клетки, известные в данной области. Кроме того, мутации также могут быть введены в последовательность белка по настоящему изобретению путем химического синтеза.
Способ использования технологии ПЦР для амплификации ДНК/РНК предпочтительно используют для получения полинуклеотида по настоящему изобретению. В особенности в тех случаях, когда затруднительно получить из библиотеки полноразмерную кДНК, предпочтительно может быть использован способ RACE (способ быстрой амплификации конца кДНК), а праймеры, используемые для ПЦР, могут быть выбраны надлежащим образом в соответствии с информацией о последовательности по настоящему изобретению, содержащейся в данном описании, и могут быть синтезированы обычными способами. Амплифицированные фрагменты ДНК/РНК могут быть разделены и очищены обычными способами, такими как гель-электрофорез.
Вектор экспрессии
Настоящее изобретение также относится к вектору, содержащему полинуклеотид по настоящему изобретению, к клетке-хозяину, полученной с помощью генной инженерии с использованием вектора по настоящему изобретению или кодирующей последовательности мутантного белка по настоящему изобретению, и к способу получения полипептида по настоящему изобретению с помощью рекомбинантной технологии.
Посредством общепринятой технологии рекомбинантной ДНК полинуклеотидная последовательность по настоящему изобретению может быть использована для экспрессии или получения рекомбинантного мутеина. В общем случае, предусмотрены следующие этапы:
(1) применение полинуклеотида (или варианта) по настоящему изобретению, кодирующего мутантный белок по настоящему изобретению, или применение рекомбинантного вектора экспрессии, содержащего полинуклеотид, для трансформации или трансдукции подходящей клетки-хозяина;
(2) клетка-хозяин, культивируемая в подходящей среде;
(3) отделение и очистка белка от среды культивирования или клеток.
В настоящем изобретении полинуклеотидная последовательность, кодирующая мутантный белок, может быть вставлена в рекомбинантный вектор экспрессии. Термин «рекомбинантный вектор экспрессии» относится к бактериальным плазмидам, бактериофагам, дрожжевым плазмидам, вирусам растительных клеток, вирусам клеток млекопитающих, таким как аденовирус, ретровирус или другим векторам, хорошо известным в данной области. Могут быть использованы любая плазмида или вектор, если они могут быть реплицированы и стабилизированы в хозяине. Важной особенностью вектора экспрессии является то, что он обычно содержит точку начала репликации, промотор, маркерный ген и элементы управления трансляцией.
Для конструирования вектора экспрессии, содержащего последовательность ДНК, кодирующую мутеин по настоящему изобретению, и соответствующие сигналы управления транскрипцией/трансляцией, могут быть использованы способы, хорошо известные специалистам в данной области. Эти способы включают в себя технологию рекомбинантной ДНК in vitro, технологию синтеза ДНК и технологию рекомбинации in vivo. Для направления синтеза мРНК последовательность ДНК может быть эффективно связана в векторе экспрессии с подходящим промотором. Типичными примерами таких промоторов являются: Промотор Escherichia coli lac или trp; промотор фага лямбда PL; эукариотические промоторы, включая промотор немедленного ответа CMV, промотор тимидинкиназы HSV, ранний и поздний промоторы SV40, ретровирусные LTR и некоторые другие известные промоторы, которые могут управлять экспрессией генов в прокариотических или эукариотических клетках или вирусах. Вектор экспрессии также включает в себя сайт связывания рибосомы для инициации трансляции и терминатор транскрипции.
Кроме того, вектор экспрессии предпочтительно содержит один или несколько селективных маркерных генов для обеспечения фенотипических признаков отбора трансформированных клеток-хозяев, таких как дигидрофолатредуктаза для культуры эукариотических клеток, устойчивость к неомицину и зеленый флуоресцентный белок (GFP), или устойчивость к тетрациклину или ампициллину для Е. coli.
Вектор, содержащий вышеупомянутую подходящую последовательность ДНК и соответствующий промотор или управляющую последовательность, можно использовать для трансформации подходящей клетки-хозяина так, чтобы она могла экспрессировать упомянутый белок.
Клетка-хозяин может представлять собой прокариотическую клетку, такую как бактериальная клетка; или низшую эукариотическую клетку, такую как дрожжевая клетка; или высшую эукариотическую клетку, такую как клетка млекопитающего. Типичные примеры включают в себя: Escherichia coli, Streptomyces; бактериальные клетки Salmonella typhimurium; грибковые клетки, такие как дрожжевые и растительные клетки (например, клетки женьшеня).
Когда полинуклеотид по настоящему изобретению экспрессируют в клетках высших эукариот, вставка в вектор энхансерной последовательности обеспечит усиление транскрипции. Энхансеры - это цис-действующие факторы ДНК, обычно имеющие от 10 до 300 пар оснований, воздействующие на промоторы для усиления транскрипции генов. Примеры включают от 100 до 270 пар оснований энхансера SV40 на поздней стороне точки инициации репликации, энхансера полиомы на поздней стороне точки инициации репликации, энхансеры аденовируса и т.п.
Специалистам в данной области известно, как выбрать подходящие векторы, промоторы, энхансеры и клетки-хозяева.
Трансформация клеток-хозяев рекомбинантной ДНК может осуществляться обычными способами, хорошо известными специалистам в данной области. Когда хозяином является прокариот, такой как Escherichia coli, компетентные клетки, которые способны абсорбировать ДНК, могут быть собраны после фазы экспоненциального роста и обработаны способом СаСl2. Применяемые этапы хорошо известны в данной области техники. Другим способом является использование MgCl2. При необходимости трансформация также может быть выполнена путем электропорации. Если хозяином является эукариот, могут быть выбраны следующие способы трансфекции ДНК: способ совместного осаждения фосфатом кальция, обычные механические способы, такие как микроинъекция, электропорация, липосомная упаковка и т.д.
Для экспрессии полипептида, кодируемого геном по настоящему изобретению, полученные трансформанты можно культивировать обычными способами. В зависимости от используемой клетки-хозяина среда, используемая для культивирования среда может быть выбрана из разнообразных обычных сред. Культивирование проводят в условиях, подходящих для роста клетки-хозяина. После того, как клетки-хозяева вырастут до приемлемой плотности клеток, выбранный промотор индуцируют подходящим способом (таким как температурное преобразование или химическая индукция), а клетки культивируют в течение определенного периода времени.
Рекомбинантный полипептид в указанном выше способе может быть экспрессирован в клетке или на клеточной мембране или секретирован из клетки. При необходимости, используя физические, химические и другие характеристики, можно отделить и очистить рекомбинантный белок с помощью различных способов разделения. Эти способы хорошо известны специалистам в этой области техники. Примеры этих способов включают в себя, но не ограничиваются перечисленным: обычная обработка ренатурацией, обработка агентом осаждения белка (способ высаливания), центрифугирование, разрушение бактерий посредством осмоса, ультраобработка, ультрацентрифугирование, хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, высокоэффективная жидкостная хроматография (ВЭЖХ) и различные другие технологии жидкостной хроматографии и их комбинации.
По сравнению с предшествующим уровнем техники настоящее изобретение имеет следующие преимущества:
После большого количества скринингов и модификаций в настоящем изобретении впервые было обнаружено, что мутация аргинина (R) в позиции 19 и/или гистидина (Н) в позиции 29, соответствующая лизил-тРНК синтетазе дикого типа (такой как SEQ ID NO.: 1 или 2), полученная мутантная лизил-тРНК синтетаза способна вводить в белки производные лизина с большими функциональными группами.
По сравнению с лизил-тРНК-синтетазой дикого типа мутантная лизил-тРНК-синтетаза по настоящему изобретению может значительно увеличить количество инсерций неприродных аминокислот и количество целевого белка, содержащего неприродные аминокислоты. Кроме того, мутантная лизил-тРНК синтетаза по настоящему изобретению также может улучшать стабильность целевого белка так, что его не удается легко разрушить.
В настоящем изобретении также оказалось, что мутация одной или нескольких аминокислот в других позициях лизил-тРНК-синтетазы, таких как позиции 26, 122, 309, 348 и/или 384 аллелей, может дополнительно повысить величину инсерций неприродных аминокислот, количество целевого белка, содержащего неприродные аминокислоты, и/или стабильность целевого белка.
Настоящее изобретение будет дополнительно разъяснено ниже на конкретных вариантах осуществления. Следует понимать, что эти варианты осуществления предназначены только для иллюстрирования данного изобретения, а не для ограничения его объема. В следующих примерах не указаны подробно условия экспериментальных способов, обычно соответствующих общепринятым условиям, такими как условия, описанные в «Руководстве по лабораторному молекулярному клонированию» (перевод Хуанг Пейтанг и др., Пекин: Science Press, 2002) издательства Sambrook. J et al., или в соответствии с условиями, рекомендованными производителем (например, в технических описаниях продуктов). Если не указано иное, процентные доли и части рассчитывают по массе.
Материалы и реагенты для экспериментов, используемые в последующих примерах, могут быть получены по коммерческим каналам, если не указано иное.
Пример 1 Конструирование мутантов
В соответствии с аминокислотной последовательностью 1 (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, она была клонирована в сайт Spel-Sall ниже промотора araBAD плазмиды вектора экспрессии pEvol-pBpF (приобретена у компании NTCC, устойчива к хлорамфениколу), причем сайт рестрикции Spel был увеличен с помощью ПЦР, а сайт SalI принадлежал самому вектору. Сохранение исходного промотора глутамина glnS плазмиды вектора экспрессии pEvol-pBpF. Ниже промотора рroK плазмиды вектора экспрессии pEvol-pBpF последовательность ДНК (SEQ ID NO.: 9) тРНК (pylTcua) лизил-тРНК синтетазы вставляли с помощью ПЦР. Эта плазмида была названа pEvol-pylRs-pylT.
В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутацию R19H для получения мутантной лизил-тРНК синтетазы pylRs (R19H) с аминокислотной последовательностью, как показано в SEQ ID NO.: 7. В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутацию H29R для получения мутантной лизил-тРНК синтетазы pylRs (H29R) с аминокислотной последовательностью, как показано в SEQ ID NO.: 8. В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутации R19H, H29R и получали мутантную лизил-тРНК синтетазу pylRs (R19H, H29R), как показано в SEQ ID NO.: 6. В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутации R19H, I26V, H29R для получения мутантной лизил-тРНК синтетазы pylRs (R19H, 126V, H29R), как показано в SEQ ID NO.: 3. В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутации R19H, H29R, T122S, Y384F и получали аминокислотную последовательность мутантной лизил-тРНК синтетазы pylRs (R19H, H29R, T122S, Y384F), как показано в SEQ ID NO.: 4. В соответствии с аминокислотной последовательностью (SEQ ID NO.: 1) лизил-тРНК синтетазы дикого типа pylRs, вводили мутации R19H, H29R, L309A, C348S и получали аминокислотную последовательность мутантной лизил-тРНК синтетазы pylRs (R19H, H29R, L309A, C348S), как показано в SEQ ID NO.: 5.
Используя плазмиду pEvol-pylRs-pylT в качестве шаблона, конструировали праймеры, см. SEQ ID NO.: 10 и SEQ ID NO.: 11, и вводили сайт-направленную мутацию R19H. Продукты ПЦР обрабатывали Dpnl, разделяли с помощью электрофореза нуклеиновых кислот, экстрагировали с помощью набора для восстановления ДНК из агарозного геля, лигировали ДНК-лигазой Т4 и трансформировали в большие компетентные клетки E.coli Тор10 химическим способом (способ СаСl2). Трансформированные клетки культивировали на агаровой среде LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl, 1,5% агар), содержащей хлорамфеникол, при температуре 37°С в течение ночи. Отбирали единичную живую колонию и культивировали в жидкой среде LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl), содержащей хлорамфеникол, при 37°С и при 220 об/мин в течение ночи. Полученную плазмиду экстрагировали с помощью набора Plasmid Mini Extraction Kit и назвали pEvol-pylRs(R19H)-pylT;
Используя плазмиду pEvol-pylRs-pylT в качестве шаблона, конструировали праймеры, см. SEQ ID NO.: 12 и SEQ ID NO.: 13, вводили сайт-направленную мутацию H29R, а результирующая плазмида была названа pEvol-pylRs(H29R)-pylT;
Используя плазмиду pEvol-pylRs(R19H)-pylT в качестве шаблона, конструировали праймеры, см. SEQ ID NO.: 12 и SEQ ID NO.: 13, вводили сайт-направленную мутацию H29R, а результирующая плазмида была названа pEvol-pylRs(R19H, H29R)-pylT. Карта плазмиды показана на Фиг. 1.
5'-ATACCGGCACCATCCACAAG-3' (SEQ ID NO.: 10)
5'-GGCTCATCCACAGACCAGTT-3' (SEQ ID NO.: 11)
5'-GCGAGGTTTCCCGTAGCAAA-3' (SEQ ID NO.: 12)
5'-GGTGTTTGATCTTGTGGATGGT-3' (SEQ ID NO.: 13)
Использовали описанные выше способы для раздельного конструирования pEvol-pylRs (R19H, I26V, H29R)-pylT, плазмиды pEvol-pylRs (R19H, H29R, T122S, Y384F)-pylT и плазмиды pEvol-pylRs (R19H, H29R, L309A, C348S)-pylT.
Пример 2 Конструирование штамма и экспрессия высокой плотности белка слияния, модифицированного трет-бутоксикарбонилом (Вое)
Плазмида pEvol-pylRs-pylT, плазмида pEvol-pylRs(R19H)-pylT, плазмида pEvol-pylRs (H29R)-pylT, плазмида pEvol-pylRs(R19H, H29R)-pylT, плазмида pEvol-pylRs(R19H, I26V, H29R)-pylT и плазмида pEvol-pylRs(R19H, H29R, T122S, Y384F)-pylT были соответственно совместно трансформированы вектором экспрессии белка слияния инсулина pBAD-INS (устойчив к канамицину) в компетентные клетки E.coli Тор10 (компетентные клетки приобретены у компании Thermo) химическим способом (способ СаСl2). Трансформированные клетки в течение ночи культивировали на агаровой среде LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl, 1,5% агар), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при температуре 37°С. Отбирали единичную живую колонию и в течение ночи культивировали в жидкой среде LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С и при 220 об/мин. К окончательному концентрату добавляли 20% глицерол для сохранения штамма.
Каждый штамм инокулировали в жидкую среду LB при 37°С и 220 об/мин в течение ночи, инокуляцию осуществляли в емкости с 1% (об./об.) ферментационной средой (12 г/л дрожжевого пептона, 24 г/л порошка дрожжевого экстракта, 4 мл/л глицерина, 12,8 г/л двузамещенного фосфорнокислого натрия, 3 г/л однозамещенного фосфорнокислого калия и 0,3%о пеногасителя), культивировали при 35 (±3)°С, при 200-1000 об/мин и расходе воздуха 2-6 л/мин. После культивирования в течение 3-10 часов до конца ферментации со ступенчатым расходом подавали питательную среду, содержащую глицерин и дрожжевой пептон. После культивирования, пока показатель OD600 не достигал 25-80, для индукции добавляли конечную концентрацию 0,25% L-ara и конечную концентрацию 5 ммоль/л Boc-Lys. Продолжали культивировать, пока показатель OD600 не достигал 180-220, после чего емкость извлекали. Затем штамм собирали центрифугированием (5000 об/мин, 30 мин, 25°С). Для выявления экспрессии Вос-модифицированных лизинсодержащих белков слияния в целых клетках различных штаммов использовали электрофорез в SDS-полиакриламиде.
Белок слияния экспрессирован в форме нерастворимого «внутриклеточного тельца». Чтобы высвободить внутриклеточное тельце, клетки Е. coli разрушали с помощью гомогенизатора высокого давления. Нуклеиновые кислоты, остатки клеток и растворимые белки удаляли центрифугированием при 10000 g. Внутриклеточные тельца, содержащие белок слияния, промывали чистой водой, и полученный осадок внутриклеточного тельца использовали в качестве исходного материала для укладки.
Уровни экспрессии белков слияния различных мутантных ферментов являются следующими: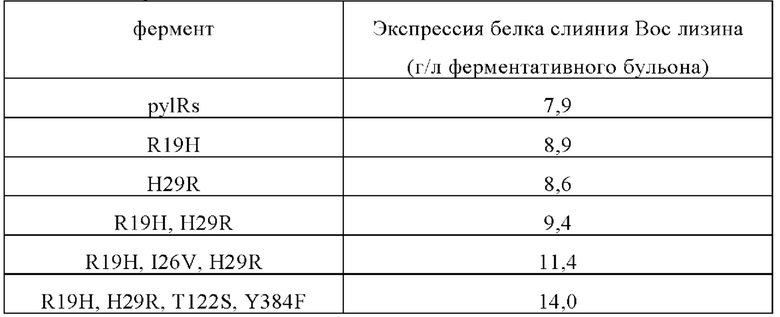
Для повторной укладки белка слияния внутриклеточные тельца растворяли в 7,5 моль/л растворе мочевины с рН 10,5, содержащем 2-10 ммоль/л меркаптоэтанола так, чтобы общая концентрация белка после растворения составляла 10-25 мг/мл. Образец разбавляли в 5-10 раз, и обычную укладку выполняли в течение 16-30 часов при температуре от 4 до 8°С и рН от 10,5 до 11,7. При температуре 18-25°С значение рН поддерживали на уровне 8,0-9,5 и использовали трипсин и карбоксипептидазу В для гидролиза гибридной протеазы в течение 10-20 часов, а затем для прекращения реакции ферментативного гидролиза добавляли 0,45 моль/л сульфата аммония. Результаты анализа обращенно-фазовой ВЭЖХ показывают, что выход на этой стадии ферментативного гидролиза превышает 90%. Аналог инсулина, полученный после разложения с помощью трипсина и карбоксипептидазы В, называется ВОС-лизиновый инсулин. Вос-лизиновый инсулин не может быть гидролизирован с помощью ферментов в указанных выше условиях. Образец осветляли мембранной фильтрацией, используя в качестве буфера раствор сульфата аммония 0,45 ммоль/л, и первоначально очищали гидрофобной хроматографией. Чистота после электрофореза в полиакриламидных гелях с использованием додецилсульфата натрия (SDS) достигает 90%, а полученный Вос-инсулин человека проанализировали с помощью времяпролетной масс-спектрометрии с матрично-активированной лазерной десорбцией/ионизацией (MALDI-TOF), и было обнаружено, что его молекулярная масса соответствует теоретической молекулярной массе в 5907,7 Да. Образцы собирали путем элюирования гидрофобной хроматографией и добавляли соляную кислоту для проведения реакции снятия защитных групп с Вос-инсулина человека. Для регулирования рН до уровня 2,8-3,2 и для прекращения реакции добавляли раствор гидроксида натрия. После двух этапов обращенно-фазовой хроматографии под высоким давлением выход рекомбинантного инсулина человека превышает 85%.
Уровни экспрессии рекомбинантного инсулина человека для различных мутантных ферментов являются следующими:
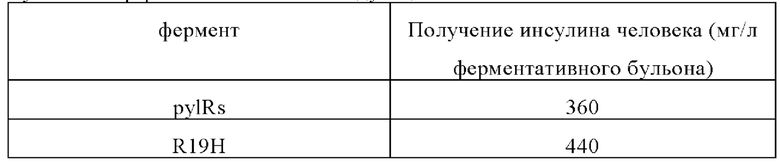

Результаты показывают, что использование мутантного фермента по настоящему изобретению для получения целевого белка, содержащего Вос-лизин, может значительно увеличить объем инсерций неприродных аминокислот и количество целевого белка, содержащего неприродные аминокислоты.
Пример 3 Конструирование штамма и экспрессия высокой плотности белка слияния, модифицированного бутиноксикарбонилом
Плазмида pEvol-pylRs-pylT, плазмида pEvol-pylRs(R19H)-pylT, плазмида pEvol-pylRs(H29R)-pylT, плазмида pEvol-pylRs(R19H, H29R)-pylT, плазмида pEvol-pylRs(R19H29, I26V, H29R))-pylT и плазмида pEvol-pylRs(R19H, H29R, L309A, C348S)-pylT, были соответственно совместно трансформированы вектором экспрессии белка слияния инсулина pBAD-INS (плазмида, сконструированная нашей компанией, устойчивая к канамицину) химическим способом (способ СаСl2) в большие компетентные клетки E.coli Тор10 (компетентные клетки приобретены у компании Thermo). Трансформированные клетки в течение ночи культивировали на агаровой среде LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl, 1,5% агар), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при температуре 37°С. Отбирали единичную живую колонию и помещали на ночь в жидкую среду LB (10 г/л дрожжевого пептона, 5 г/л порошка дрожжевого экстракта, 10 г/л NaCl), содержащую 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С и при 220 об/мин. К окончательному концентрату добавляли 20% глицерол для сохранения штамма.
Каждый штамм инокулировали в жидкую среду LB при 37°С и 220 об/мин в течение ночи, инокуляцию осуществляли в емкости с 1% (об./об.) ферментационной средой (12 г/л дрожжевого пептона, 24 г/л порошка дрожжевого экстракта, 4 мл/л глицерина, 12,8 г/л двузамещенного фосфорнокислого натрия, 3 г/л однозамещенного фосфорнокислого калия и 0,3% пеногасителя), культивировали при 35 (±3)°С, при 200-1000 об/мин и расходе воздуха 2-6 л/мин. После культивирования в течение 3-10 часов до конца ферментации со ступенчатым расходом подавали питательную среду, содержащую глицерин и дрожжевой пептон. После культивирования, пока показатель OD600 не достигал 25-80, для индукции добавляли конечную концентрацию 0,25% L-ara и конечную концентрацию 5 ммоль/л бутинилоксикарбонила-Lys. Продолжали культивировать, пока показатель OD600 не достигал 180-220, после чего емкость извлекали. Затем штамм собирали центрифугированием (5000 об/мин, 30 мин, 25°С). Для выявления экспрессии модифицированного бутинилоксикарбонилом белка слияния лизина в целых клетках различных штаммов использовали электрофорез в SDS-полиакриламиде
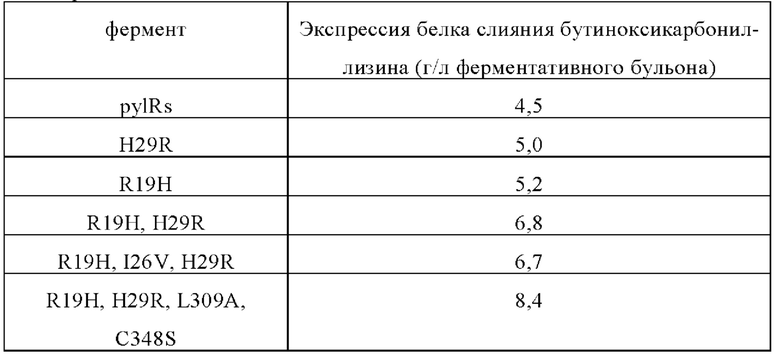
Белок слияния экспрессирован в форме нерастворимого «внутриклеточного тельца». Чтобы высвободить внутриклеточное тельце, клетки Е. coli разрушали с помощью гомогенизатора высокого давления. Нуклеиновые кислоты, остатки клеток и растворимые белки удаляли центрифугированием при 10000 g. Внутриклеточные тельца, содержащие белок слияния, промывали чистой водой, и полученный осадок внутриклеточного тельца использовали в качестве исходного материала для укладки. Для повторной укладки белка слияния внутриклеточные тельца растворяли в 7,5 моль/л растворе мочевины с рН 10,5, содержащем 2-10 ммоль/л меркаптоэтанола так, чтобы общая концентрация белка после растворения составляла 10-25 мг/мл. Образец разбавляли в 5-10 раз, и обычную укладку выполняли в течение 16-30 часов при температуре от 4 до 8°С и рН от 10,5 до 11,7. При температуре 18-25°С значение рН поддерживали на уровне 8,0-9,5 и использовали трипсин и карбоксипептидазу В для гидролиза гибридной протеазы в течение 10-20 часов, а затем для прекращения реакции ферментативного гидролиза добавляли 0,45 моль/л сульфата аммония. Результаты анализа обращенно-фазовой ВЭЖХ показывают, что выход на этой стадии ферментативного гидролиза превышает 90%. Аналог инсулина, полученный после разложения с помощью трипсина и карбоксипептидазы В, называется бутинилоксикарбонил-лизиновый инсулин. Бутинилоксикарбонил-лизиновый инсулин не может быть гидролизирован с помощью ферментов в указанных выше условиях. Образец осветляли мембранной фильтрацией, используя в качестве буфера раствор сульфата аммония 0,45 ммоль/л, и первоначально очищали гидрофобной хроматографией. Чистота после электрофореза в полиакриламидных гелях с использованием додецилсульфата натрия (SDS) достигает 90%, а полученный бутиноксикарбонил-инсулин человека проанализировали с помощью времяпролетной масс-спектрометрии с матрично-активированной лазерной десорбцией/ионизацией (MALDI-TOF), и было обнаружено, что его молекулярная масса соответствует теоретической молекулярной массе в 5907,7 Да.
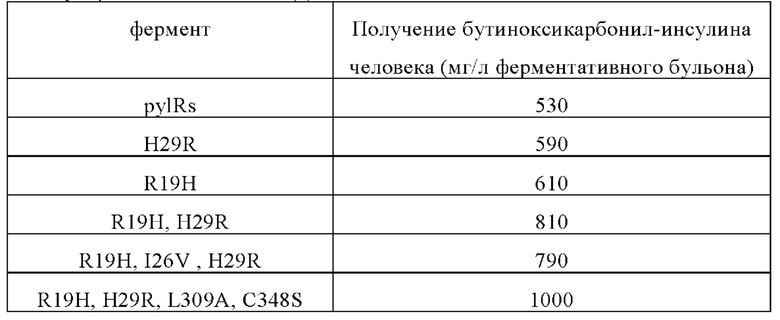
Результаты показывают, что использование мутантного фермента по настоящему изобретению для получения модифицированного бутиноксикарбонилом целевого белка может значительно увеличить объем инсерций неприродной аминокислоты и количество целевого белка, содержащего неприродную аминокислоту.
Все документы, упомянутые в настоящем изобретении, включены в данный документ посредством ссылки, как если бы каждый документ был включен с помощью отдельной ссылки. Кроме того, следует понимать, что после изучения вышеизложенного описания изобретения специалистами в данной области в изобретение могут быть внесены различные изменения или модификации, и что эти аналоги в равной степени попадают в пределы объема формулы изобретения, приложенной к этой заявке.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> НИНГБО КУНПЕНГ БИОТЕКХ КО, ЛТД.
<120> АМИНОАЦИЛ-тРНК СИНТЕТАЗА ДЛЯ ЭФФЕКТИВНОГО ВВЕДЕНИЯ ПРОИЗВОДНОГО
ЛИЗИНА В
БЕЛОК
<130> P2020-0085
<150> CN201910209626.6
<151> 2019-03-19
<160> 13
<170> PatentIn version 3.5
<210> 1
<211> 454
<212> ПРТ
<213> Methanosarcina mazei
<400> 1
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 2
<211> 454
<212> ПРТ
<213> Methanosarcina mazei
<400> 2
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Gly Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 3
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК-синтетаза
<400> 3
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser His Thr Gly Thr Ile His Lys Val Lys His Arg Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 4
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК-синтетаза
<400> 4
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser His Thr Gly Thr Ile His Lys Ile Lys His Arg Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Ser Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Phe
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 5
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК-синтетаза
<400> 5
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser His Thr Gly Thr Ile His Lys Ile Lys His Arg Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Ala Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Ser Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 6
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК-синтетаза
<400> 6
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser His Thr Gly Thr Ile His Lys Ile Lys His Arg Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 7
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК-синтетаза
<400> 7
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser His Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 8
<211> 454
<212> ПРТ
<213> искусственная последовательность
<220>
<223> мутантная лизил-тРНК синтетаза
<400> 8
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His Arg Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 9
<211> 72
<212> ДНК
<213> искусственная последовательность
<220>
<223> кодирующая аминокислотная последовательность ортогональной тРНК
<400> 9
ggaaacctga tcatgtagat cgaatggact ctaaatccgt tcagccgggt tagattcccg
60
gggtttccgc ca
72
<210> 10
<211> 20
<212> ДНК
<213> искусственная последовательность
<220>
<223> праймер
<400> 10
ataccggcac catccacaag
20
<210> 11
<211> 20
<212> ДНК
<213> искусственная последовательность
<220>
<223> праймер
<400> 11
ggctcatcca cagaccagtt
20
<210> 12
<211> 20
<212> ДНК
<213> искусственная последовательность
<220>
<223> праймер
<400> 12
gcgaggtttc ccgtagcaaa
20
<210> 13
<211> 22
<212> ДНК
<213> искусственная последовательность
<220>
<223> праймер
<400> 13
ggtgtttgat cttgtggatg gt
22
<---
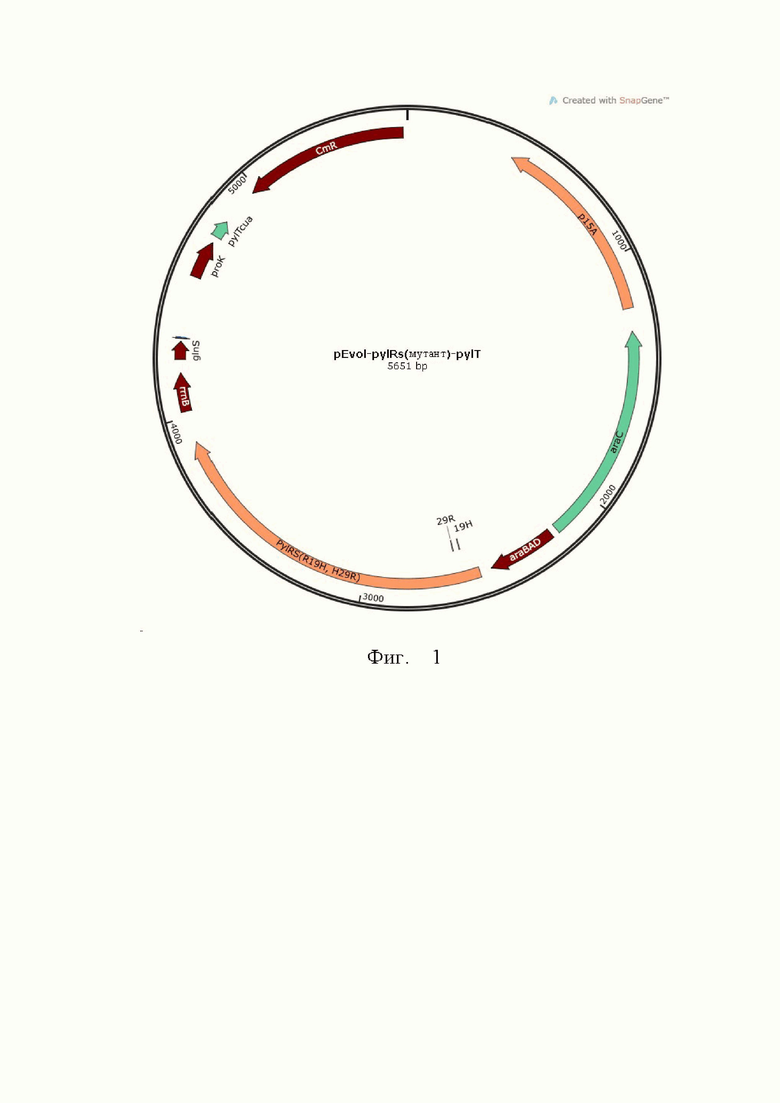
Группа изобретений относится к биотехнологии. Предложена мутантная лизил-тРНК-синтетаза, имеющая мутацию в позиции 19 аргинина (R) и/или позиции 29 гистидина (H) в аминокислотной последовательности, соответствующей лизил-тРНК-синтетазе дикого типа, причем такая мутантная лизил-тРНК-синтетаза необязательно включает в себя мутацию на сайте, выбранном из группы, состоящей из изолейцина (I) в позиции 26, треонина (T) в позиции 122, лейцина (L) в позиции 309, цистеина (C) в позиции 348, тирозина (Y) в позиции 384 и их комбинации. Кроме того, предложены: изолированный полинуклеотид, кодирующий мутантную лизил-тРНК-синтетазу; вектор для экспрессии мутантной лизил-тРНК-синтетазы; клетка-хозяин для экспрессии мутантной лизил-тРНК-синтетазы; система экспрессии; набор для получения целевого белка, содержащего неприродную аминокислоту, а также их применения для получения целевого белка, содержащего неприродную аминокислоту. Изобретения позволяют значительно увеличить количество инсерций производных лизина и количество целевого белка, содержащего производные лизина. 7 н. и 2 з.п. ф-лы, 1 ил., 1 табл., 3 пр.
1. Мутантная лизил-тРНК-синтетаза, где мутантная лизил-тРНК-синтетаза имеет мутацию в позиции 19 аргинина (R) и/или позиции 29 гистидина (H) в аминокислотной последовательности, соответствующей лизил-тРНК-синтетазе дикого типа, причем такая мутантная лизил-тРНК-синтетаза необязательно включает в себя мутацию на сайте, выбранном из группы, состоящей из изолейцина (I) в позиции 26, треонина (T) в позиции 122, лейцина (L) в позиции 309, цистеина (C) в позиции 348, тирозина (Y) в позиции 384 и их комбинации.
2. Мутантная лизил-тРНК-синтетаза по п. 1, причем такая мутантная лизил-тРНК-синтетаза представляет собой полипептид, имеющий аминокислотную последовательность, показанную в любой из SEQ ID NO: 3-8.
3. Изолированный полинуклеотид, кодирующий мутантную лизил-тРНК-синтетазу по п. 1.
4. Вектор для экспрессии мутантной лизил-тРНК-синтетазы, содержащий полинуклеотид по п. 3.
5. Клетка-хозяин для экспрессии мутантной лизил-тРНК-синтетазы, содержащая вектор по п. 4.
6. Клетка-хозяин по п. 5, причем такая клетка-хозяин содержит:
(а) мутантную лизил-тРНК-синтетазу по п. 1; и
(b) ортогональную тРНК, способную связываться с производным лизина в присутствии мутантной лизил-тРНК-синтетазы; и, необязательно,
(c) последовательность целевой нуклеиновой кислоты, кодирующую целевой белок, которая включает в себя кодон, распознаваемый ортогональной тРНК в позиции для введения производного лизина.
7. Система экспрессии, содержащая:
(а) мутантную лизил-тРНК-синтетазу по п. 1; и
(b) ортогональную тРНК, способную связываться с производным лизина в присутствии мутантной лизил-тРНК-синтетазы; и, необязательно,
(c) последовательность целевой нуклеиновой кислоты, кодирующую целевой белок, которая включает в себя кодон, распознаваемый ортогональной тРНК в позиции для введения производного лизина.
8. Набор для получения целевого белка, содержащего неприродную аминокислоту, содержащий (a) контейнер и (b) мутантную лизил-тРНК-синтетазу по п. 1, или полинуклеотид по п. 3, или вектор по п. 4, находящиеся в контейнере.
9. Применение мутантной лизил-тРНК-синтетазы по п. 1, или клетки-хозяина по п. 5, или системы экспрессии по п. 7, или набора по п. 8 для получения целевого белка, содержащего неприродную аминокислоту.
| WO 2019010164 A1, 10.01.2019 | |||
| WO 9739016 A1, 23.10.1997 | |||
| YANAGISAWA T | |||
| ET AL | |||
| Multistep Engineering of Pyrrolysyl-tRNA Synthetase to Genetically Encode Nɛ-(o-Azidobenzyloxycarbonyl) lysine for Site-Specific Protein Modification | |||
| CHEM BIOL | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| NEUMANN H | |||
| ET AL | |||
| Genetically encoding Nε-acetyllysine in recombinant | |||

