ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к области медицины и биологии, в частности к способам создания лекарственных форм для различных ферментных препаратов, а также других биологически активных компонентов (БАК), на основе эритроцитов, которые могут быть использованы как в лечении ряда заболеваний, так и в целях диагностики.
УРОВЕНЬ ТЕХНИКИ
Ферменты, включенные в эритроциты, могут быть использованы как в фермент-заместительной и противоопухолевой терапии, так и для удаления из кровотока ряда низкомолекулярных токсичных соединений таких как аммиак, цианид, этиловый и метиловый спирт, ацетальдегид и т.д. [1]. В настоящее время фермент L-аспарагиназа (аспарагиназа) является неотъемлемой частью комплексной терапии острого лимфобластного лейкоза (ОЛЛ) у детей и взрослых [2], а также некоторых других видов онкологических заболеваний [3]. Аспарагиназа это фермент, гидролизующий аминокислоту аспарагин до аспарагиновой кислоты и аммиака. Механизм ее противоопухолевого действия связан с уменьшением концентрации аспарагина в плазме, так как эта аминокислота необходима для синтеза белка и ее отсутствие приводит к замедлению деления опухолевых клеток из-за невозможности такого синтеза. Особенно эффективное действие аспарагиназа оказывает на клетки некоторых опухолей, которые, в отличие от нормальных клеток, не могут самостоятельно синтезировать аспарагин (например, при ОЛЛ) [4]. Однако внутривенное введение аспарагиназы ограничено существованием многих побочных эффектов, основными из которых являются возникновение сильных аллергических реакций, а также короткое время жизни аспарагиназы в кровотоке [5,6]. Создание новой лекарственной формы аспарагиназы (аспарагиназы, включенной в эритроциты) улучшает ее фармакологические свойства, т.к. сильно увеличивает время жизни аспарагиназы в кровотоке [7,8]. Это происходит потому, что аспарагиназа проводит свою реакцию, находясь внутри эритроцитов, что защищает ее от находящихся в плазме антител и протеаз, способных ее разрушить. Субстрат, с которым работает аспарагиназа - аспарагин, способен проникать из плазмы внутрь эритроцита через эритроцитарную мембрану [9]. Аспарагиназу для терапевтического применения получают из бактерий (Esherichia coli, либо из фитопатогенной Erwinia chryzanthemi), т.е. она является для организма человека чужеродным белком. Однако если такая аспарагиназа спрятана внутри собственных клеток организма, это предупреждает развитие аллергических реакций на введенный чужеродный белок. Аналогично обстоит дело и с другими ферментными препаратами, которые включены в эритроциты и могут быть использованы для терапии ряда заболеваний.
Включение в эритроциты других БАК, например, низкомолекулярного антрациклинового антибиотика доксорубицина или терпено-индольных алкалоидов винкристина и винбластина, которые также широко используются в терапии ряда онкологических заболеваний [10-12], позволяет создать в эритроцитах депо этих лекарственных препаратов, которые постепенно выходят из эритроцитов-носителей, поддерживая в кровотоке достаточную терапевтическую концентрацию лекарства в течение долгого времени. Это важно, т.к., несмотря на высокую эффективность этих препаратов при прямом внутривенном введении, дозы препаратов, которые могут быть введены в организм, лимитируются их высокой токсичностью, например, их супрессивным действием на костный мозг [12], кардио- и нефротоксичностью [13]. При введении этих препаратов в эритроциты удается избавиться от их высоких пиковых концентраций в плазме в момент введения, которые и определяют, в большой степени, токсичность препаратов. Таким образом, включение ряда лекарственных ферментов и других биологически активных компонентов в эритроциты может обеспечивать улучшение фармакологических свойств данных БАК (их фармакокинетики и фармакодинамики), а также снижение их токсических эффектов.
В настоящее время наиболее мягкими, эффективными и щадящими способами, которые позволяют получить эритроциты-носители лекарственных препаратов со свойствами близкими к свойствам исходных эритроцитов, являются способы, основанные на обратимом гипоосмотическом воздействии на клетки.
Из современного уровня техники известны различные вариации способов включения аспарагиназы и других ферментов, а также низкомолекулярных БАК, в эритроциты, в основе которых лежит обратимое гипоосмотическое воздействие на клетки:
1. Ropars С., Nicolau Y.C., Chassaigne М. Encapsulating biological active substances into erythrocytes, US Patent 4,652,449, March 14, 1987, A61K 35/18 [14].
2. Pages E., Ropars С., Bailleul С. Apparatus for causing medical products to penetrate into red blood cells. US Patent 5,589,389, December 31, 1996, C12M 1/12, C12M 3/06) [15].
3. a) Magnani M., Panzani I., Bigi L., Zanella A. Method of encapsulating biologically active agents within erythrocytes and apparatus thereof (Dideco Sri, Italy). Европейский патент ЕР 0882448, дата публикации 12.01.2005, бюллетень 2005/02, А61К 9/50) [16];
б). Мамбрини Д., Серафини С. Устройство и набор для инкапсуляции в эритроцитах по меньшей мере одного соединения для терапевтического и диагностического применения (EryDel SpA, Italy). Патент России RU 2595844, дата публикации заявки РСТ 03.11.2011, А61К 9/50, А61М 1/36, B01J 4/00) [17];
в). Мамбрини Д., Бенатти Л., Капогросси Д., Мандолини М. Способ получения эритроцитов, нагруженных одним или несколькими веществами, представляющими фармацевтический интерес, и полученные таким образом эритроциты (EryDel SpA, Italy). Патент России RU 2670070, дата публикации заявки РСТ 13.11.2014, А61К9/50) [18].
4. Godfrin Y. Lisis/resealing process and device for incorporating an active ingredient, in particular asparaginase or inositol hexaphosphate, in erythrocytes (EryTech Pharma, France). US Patent 8,617,840, data of patent 31.12.2013, C12Q 1/24, C12N 13/00, C12M 1/33, C12M 3/08 [19].
Все указанные способы используют для включения ферментов и других БАК в эритроциты различные варианты гипоосмотического способа.
Общий принцип данной технологии состоит в том, что эритроциты приводят в контакт с гипоосмотическим раствором (т.е. раствором, имеющим осмоляльность ниже физиологической, составляющей 300 мОсм/кг). В результате этого возникает разница в осмотическом давлении по обе стороны эритроцитарной мембраны. Для уравновешивания осмотического давления внутри и снаружи эритроцита в него начинает поступать вода. Эритроцит постепенно набухает, в результате чего, после достижения максимально возможного объема (при сохранении постоянной площади поверхности эритроцита), в его мембране появляются разрывные дефекты (поры) диаметром от 8 до 50 нм, через которые из эритроцита может выходить гемоглобин и другие соединения, а внутрь эритроцита могут поступать компоненты (в том числе аспарагиназа или другие ферменты и низкомолекулярные соединения, а также нагруженные БАК наночастицы), присутствующие во внешнем растворе. Диаметр пор был измерен экспериментально и составлял от 8 до 50 нм [20-23], при этом было установлено, что для выхода гемоглобина из эритроцита достаточно образования пор диаметром около 10 нм [23,24].
Способы, раскрытые в патенте US 4,652,449 (Ropars С., et al.) [14], позволяют работать с широким диапазоном объемов упакованных эритроцитов (от 200 мл до нескольких мл), которые должны быть сначала выделены из соответствующего объема исходной крови (от 450 мл до 10 мл).
Вещество, которое должно быть включено в эритроциты, вводится с помощью насоса в трубку, через которую суспензия эритроцитов подается в диализный элемент (до прохождения этой суспензии через теплообменное устройство, обеспечивающее нужную для диализа клеток температуру).
Общими чертами всех вариантов проведения процедуры включения биологически активного компонента в эритроциты является использование для лизиса эритроцитов диализного элемента (диализатора), состоящего из двух отсеков (компартментов), разделенных диализной мембраной. Устройство диализных элементов может быть различным. Они могут содержать 2 отсека, разделенных только одной плоской полупроницаемой мембраной, или множество полых волокон из полупроницаемой мембраны, окруженных внешним отсеком. В первом отсеке циркулирует суспензия эритроцитов, а во втором - гипоосмотический раствор для обеспечения лизиса эритроцитов.
Запечатывание полученных эритроцитов-носителей может быть проведено путем запечатывания пор в мембранах этих эритроцитов, при восстановлении нормальной осмотичности в суспензии лизированных в присутствии включаемого препарата эритроцитов, путем их инкубации при температуре от 20°С до 40°С с гиперосмотическим раствором (т.е. раствором с осмоляльностью выше физиологической, составляющей 300 мОсм/кг) как в отдельном резервуаре, так и во втором диализном элементе, в первом отсеке которого циркулирует суспензия лизированных эритроцитов, а во втором гиперосмотический раствор.
В отдельных частных случаях выполнения способа [14] для целей лизиса и запечатывания эритроцитов может быть использован один и тот же диализный элемент, после соответствующей замены буфера во втором отсеке и установления температуры, соответствующей каждой из стадий (примерно от 0°С до 10°С для лизиса и примерно от 20°С до 40°С для запечатывания эритроцитов).
Способ включения биологически активных веществ в эритроциты, описанный в патенте [14], отличается тем, что в первый отсек диализного элемента непрерывно подают суспензию эритроцитов в изотоническом растворе, содержащую вещество (или вещества) с биологической активностью, а во второй отсек диализного элемента - водный солевой раствор, который является гипотоническим по отношению к суспензии эритроцитов и предназначен для лизиса эритроцитов. В этом случае лизат эритроцитов контактирует с одним или более веществом, обладающим биологической активностью. Далее проводят повышение осмотичности лизата эритроцитов, что позволяет повторно запечатать мембрану эритроцитов, в которые оказывается включен один или более биологически активный компонент.
В качестве недостатка данного способа следует отметить, что в качестве исходного компонента может быть использована лишь суспензия отмытых эритроцитов, что в свою очередь приводит к необходимости предварительного отмывания эритроцитов из цельной крови на внешнем устройстве. Следующим недостатком является возможность работы только с большими объемами суспензии отмытых эритроцитов (полученных из объемов крови от 200 до 450 мл), тогда как работа с малыми объемами суспензии эритроцитов (в несколько мл) осложнена, т.к. способ не предусматривает концентрирования клеток после лизиса, а кинетика процесса изменяется при существенном разбавлении исходной суспензии эритроцитов, которое происходит в этом случае. Требуется строгий подбор параметров диализаторов (площади диализной мембраны и пропускной способности диализатора, которая определяется, в том числе, скоростью протекания суспензии эритроцитов и лизирующего раствора через отдельные компартменты диализного элемента), а также строгий выбор отношений площадей мембран диализных элементов для лизиса клеток и для их запечатывания, которые не всегда могут быть обеспечены существующими на сегодняшний день моделями диализных элементов.
В способе, раскрытом в патенте US 5,589,389 (Pages Е., Ropars С. и Bailleul С., 1996), который фактически является вариантом выполнения способа по патенту US 4,652,449, для включения БАК (инозитолгексафосфата, ИГФ) используют большой объем исходной крови (1 трансфузионная единица, т.е. ~450 мл крови) [15]. Способ включает стадию получения суспензии отмытых эритроцитов из цельной крови. Данный способ также основан на обратимом гипоосмотическом лизисе исходных эритроцитов, отмытых при 4°С, их инкубации с включаемым веществом (при 4°С) и последующем запечатывании полученных эритроцитов-носителей путем возвращения эритроцитарной суспензии нормальной осмоляльности при инкубации с гиперосмотическим раствором (при 37°С). Пример патента содержит информацию о включении в эритроциты инозитолгексафосфата, который добавляют в среду уже на второй и третьей стадии отмывания исходных эритроцитов. На стадии лизиса эритроцитов поддерживается температура ниже 10°С (оптимально 4°С), стадия запечатывания полученных эритроцитов-носителей, происходит при поддержании температуры выше 20°С (оптимально 37°С).
Способы, раскрытые в патентах ЕР 0882448 (Dideco Sri, Italy), RU 2595844 и RU 2 670070C2 (EryDel SpA, Italy) позволяют работать с небольшими объемами (обычно 50 мл, реже до 100 мл) цельной аутологичной крови пациента [16,17,18]. Исходную кровь центрифугируют, чтобы отделить эритроциты от плазмы, и отмывают эритроциты изоосмотическим (т.е. имеющим физиологическую осмоляльность равную 300 мОсм/кг) солевым раствором с помощью центрифужного колокола для отмывания эритроцитов. Для достижения открытия пор в эритроцитарной мембране используют способ предварительного постепенного набухания (pre-swelling), когда эритроциты помещают последовательно в два гипоосмотических раствора (причем осмоляльность второго раствора ниже осмоляльности первого из них). В патентах [16,17] осмоляльности гипоосмотических растворов выбраны так, что после контакта с первым раствором эритроциты набухают и сферулируются, а после добавления второго гипоосмотического раствора эритроциты лизируются (или частично лизируются), с образованием в эритроцитарной мембране временных пор. При этом суспензия эритроцитов каждый раз сильно разбавляется гипоосмотическим раствором (до финального гематокрита около 4%-2.5%), поэтому после стадии инкубации с гипоосмотическими растворами суспензию лизированных эритроцитов концентрируют. Для этого используют диализное устройство (например, картридж с полыми волокнами из полупроницаемой мембраны), в котором суспензия лизированных (или частично лизированных) эритроцитов прокачивается по внутреннему контуру диализатора, а из внешнего контура осуществляется частичная откачка лизирующего раствора во внешнее приемное устройство. Затем в сконцентрированную суспензию лизированных клеток добавляют водный раствор включаемого БАК, который начинает поступать внутрь эритроцитов через образовавшиеся в мембране эритроцита поры. После окончания процесса инкубации осмоляльность внешней среды восстанавливают до нормальной величины, добавляя в суспензию клеток гиперосмотический раствор, в результате чего поры в мембране эритроцита закрываются и часть включаемого компонента остается внутри эритроцитов-носителей. Большинство стадий процесса регулируется автоматически с помощью специального электронного блока.
Недостатком способа по изобретению является большая потеря внутриклеточного содержимого эритроцитов, обусловленная сильным разбавлением эритроцитов при инкубации и лизисе в присутствии гипоосмотических растворов, что, в свою очередь, приводит к снижению содержания внутриклеточных веществ в эритроцитах после того, как их мембрана становится частично лизированной. И хотя макромолекулы не могут уйти из гемофильтра (диализатора), в котором затем проводят концентрирование, содержание низкомолекулярных метаболитов гликолиза (с молекулярным весом меньше 10000 дальтон) должно снижаться при откачке водного раствора из внешнего контура диализатора, т.к. эти метаболиты сначала выходят в большой объем гипоосмотических растворов на стадии лизиса, а затем легко проходят через диализную мембрану на стадии концентрирования эритроцитов. Кроме того, стадии набухания и лизиса эритроцитов идут при комнатной температуре без охлаждения растворов и суспензии до примерно 4°С, что вызывает снижение выхода включения препарата, т.к. известно, что время жизни образующихся в мембране пор уменьшается при повышении температуры до комнатной.
В патенте RU 2670070 [18], описан вариант способа включения БАК в эритроциты, при котором потеря внутриклеточного содержимого снижена, а эффективность включения БАК увеличена относительно ранее описанного в патентах [16,17] способа за счет того, что осмотичность второго гипоосмотического раствора выбрана такой, что лизиса набухших эритроцитов при контакте со вторым гипоосмотическим раствором не происходит. Эритроциты, набухшие после двух последовательных ступеней обработки гипоосмотическими растворами, где осмоляльность второго раствора ниже, чем осмоляльность первого гипоосмотического раствора, сначала концентрируются, а потом к ним добавляется водный раствор включаемого вещества, что и приводит к лизису набухших эритроцитов (уже в присутствии БАК). При этом осмотичность суспензии набухших эритроцитов после добавления первого гипоосмотического раствора составляет 250-200 мОсм/кг, осмотичность суспензии набухших эритроцитов после добавления второго гипоосмотического раствора составляет 200-170 мОсм/кг, а осмотичность суспензии эритроцитов после двух ступеней набухания и добавления включаемого биологически активного компонента составляет примерно 150-110 мОсм/кг.
В известном способе, раскрытом в патенте US 8,617,840 (фирмы EryTech Pharma, France) [19], выбранном нами в качестве ближайшего аналога, для снижения осмоляльности среды снаружи эритроцитов используют процесс диализа суспензии эритроцитов против гипоосмотического раствора с помощью диализатора, в котором суспензия эритроцитов протекает по внутреннему контуру, а гипоосмотический раствор протекает противотоком по его внешнему контуру. БАК, который должен быть включен в эритроциты, при этом помещают либо в исходную суспензию отмытых эритроцитов (т.е. во внутренний контур), либо в суспензию уже лизированных эритроцитов после их выхода из диализатора. Затем загруженные БАК эритроциты запечатывают, восстанавливая, как и в других методах, осмоляльность внешней среды.
Особенности данного способа таковы, что в качестве исходного компонента может быть использована только отмытая эритромасса, работа с цельной кровью не предусматривается. При этом объем этой эритромассы может быть от 10 мл до 250 мл упакованных эритроцитов.
Незадолго до стадии лизиса проводят измерение осмотической хрупкости эритроцитов, на основании результатов которого выбирают условия лизиса (осмолярность лизирующего раствора и/или скорость протекания суспензии клеток через диализатор).
Измерение хрупкости клеток проводят на внешнем устройстве на образце, начальная температура которого составляет от +1 до +8°С. Процесс занимает 20-30 минут.
На основании полученных данных рассчитывают скорость потока клеток в диализаторе или осмолярность диализующего раствора с применением следующих формул:
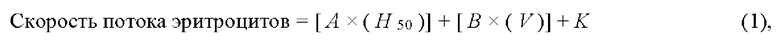
где Н50 - осмолярность, при которой лизировано 50% эритроцитов, V - объем суспензии эритроцитов, А и В - переменные, которые регулируются в зависимости от параметров диализатора и осмолярности лизирующего раствора; K - регулируемая константа.
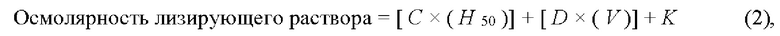
где С и D - переменные, которые регулируются в зависимости от параметров диализатора и скорости потока эритроцитов в диализаторе; K - регулируемая константа.
Рассчитанные параметры процесса лизиса в диализаторе (скорость протекания суспензии эритроцитов по внутреннему контуру диализатора и/или осмолярность лизирующего раствора) устанавливаются оператором вручную, т.к. коэффициенты в уравнениях, связывающих различные параметры проведения лизиса с отдельными показателями осмотической хрупкости исходных эритроцитов должны быть установлены для каждого вида используемых диализаторов отдельно, т.к. зависят от параметров самого диализатора.
Такой способ корректировки и установления параметров процесса лизиса является длительным и трудоемким, и не может быть легко использован в любой лаборатории. Кроме того, его использование повышает вероятность установления некорректных значений ввиду вероятности ошибки оператора.
РАСКРЫТИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Техническая проблема, которую решает настоящее изобретение, заключается в устранении недостатков, присущих известным способам.
Технической задачей, решаемой настоящим изобретением, является создание высокоэффективного и воспроизводимого способа включения в эритроциты биологически активных компонентов методом проточного гипоосмотического диализа.
Техническим результатом настоящего изобретения является обеспечение высокой и воспроизводимой эффективности (степени) включения различных биологически активных компонентов в эритроциты, снижение потери внутриклеточного содержимого. Дополнительным техническим результатом является возможность использовать в качестве исходного материала как цельную кровь, так и предварительно приготовленную эритромассу без потери эффективности включения биологически активных компонентов в эритроциты.
Эффективность включения биологически активных компонентов в эритроциты, согласно настоящему способу, превосходит аналогичную эффективность для известных способов.
Указанный технический результат достигается посредством создания способа включения в эритроциты, по меньшей мере, одного биологически активного компонента методом проточного гипоосмотического диализа, содержащего следующие стадии:
- добавление, по меньшей мере, одного биологически активного компонента к суспензии сконцентрированных отмытых из цельной крови эритроцитов или к предварительно полученной эритромассе;
- лизис эритроцитов в присутствии, по меньшей мере, одного биологически активного компонента с помощью гипоосмотического проточного диализа;
- инкубация суспензии эритроцитов, лизированных в присутствии, по меньшей мере, одного биологически активного компонента, с гиперосмотическим раствором с образованием запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- отмывание запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- концентрирование с помощью диализатора запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- вымывание из диализатора оставшихся там запечатанных эритроцитов-носителей и добавление их к ранее полученным запечатанным эритроцитам-носителям, содержащим, по меньшей мере, один биологически активный компонент;
при этом, стадию лизиса эритроцитов в присутствии, по меньшей мере, одного биологически активного компонента, проводят при поддержании постоянного трансмембранного давления в диализаторе 160-180 мм рт.ст. и давлении на входе во внутренний контур диализатора, не превышающем атмосферное более, чем на 120 мм рт.ст., а концентрирование эритроцитов осуществляют при поддержании постоянного трансмембранного давления в диализаторе 90-120 мм рт.ст. с учетом измеряемых показателей давления во внешнем и внутреннем контурах диализатора.
Все приведенные выше стадии процесса являются необходимыми для достижения заявленного технического результата. Параметры проведения каждой стадии также были выбраны так, чтобы обеспечить достижение этого технического результата.
Концентрирование суспензии эритроцитов позволяет откачать из суспензии лишнюю жидкость, максимально сохранив число целых клеток в образце. Для выбора трансмембранного давления на стадии концентрирования суспензии эритроцитов было исследовано, как повышение этого давления влияет на гемолиз эритроцитов, находящихся в диализаторе. На фиг. 1 представлены результаты экспериментов на эритроцитах из двух различных образцов крови. Как видно на представленной фигуре, при трансмембранном давлении выше 120 мм рт.ст. уже наблюдается увеличение содержания гемоглобина в сливаемой из диализатора жидкости, что соответствует увеличению гемолиза в ходе процесса концентрирования. С другой стороны, трансмембранное давление ниже 90 мм рт.ст. допустимо, но сильно увеличивает длительность процедуры концентрирования. Таким образом, оптимальным для стадии концентрирования суспензии эритроцитов был выбран интервал трансмембранного давления 90-120 ммрт.ст. По той же причине, чтобы не терять эритроциты в результате их гемолиза, давление на входе во внутренний контур диализатора, по которому проходит суспензия эритроцитов, не должно превышать атмосферное более, чем на 120 мм. рт.ст.
Так как лизированные в результате гипоосмотического диализа эритроциты имеют в своей мембране временные поры, они способны выдержать более высокие трансмембранные давления без полного разрушения клеток. Чтобы выбрать оптимальный интервал трансмембранного давления в диализаторе на стадии лизиса, были проведены опыты, в которых был определен процент инкапсуляции аспарагиназы при различном трансмембранном давлении (фиг. 2). На основании проведенных экспериментов было установлено, что поддержание во время диализа трансмембранного давления в диализаторе 160-180 мм рт.ст. обеспечивает стабильно высокий процент включения вещества в эритроциты. Более низкое давление недостаточно для эффективного включения, а слишком высокое давление приводит к уменьшению эффективности включения, по-видимому, в результате более сильного гемолиза клеток. Таким образом, как оптимальный интервал трансмембранного давления в ходе гипоосмотического диализа эритроцитов был выбран диапазон 160-180 мм рт.ст.
Биологически активный компонент (или компоненты) добавляется в исходную эритромассу или, в случае, когда в качестве исходного материала использовали цельную кровь, в отмытые из исходной цельной крови и затем сконцентрированные эритроциты.
Если в качестве исходного материала использована цельная кровь, стадии добавления, по меньшей мере, одного биологически активного компонента к суспензии сконцентрированных отмытых из цельной крови эритроцитов предшествуют стадия отмывания эритроцитов из исходной цельной крови и стадия их последующего концентрирования. В частности, концентрирование суспензии эритроцитов, отмытых из цельной крови, позволяет избежать их сильного разбавления перед добавлением БАК и, таким образом, проводить процесс последующего гипоосмотического лизиса при более высоком гематокрите и концентрации БАК, что дополнительно повышает эффективность включения БАК и снижает потерю внутриклеточного содержимого в ходе диализа.
Таким образом, дополнительным преимуществом заявленного способа является возможность работать не только с отмытой эритроцитарной массой, но и цельной кровью пациента или донора.
Схема последовательных стадий осуществления способа представлена на фиг. 3, где используются следующие обозначения:
ПН1 - перистальтический насос, посредством которого на разных стадиях процесса во внутренний контур диализатора поступает суспензия эритроцитов или физиологический раствор; ПН2 и ПН3 перистальтические насосы, посредством которых физиологический раствор или гипоосмотический раствор из соответствующих мешков поступают во внешний контур диализатора и откачиваются из этого контура, соответственно; ПН1 и ПН2 - шприцевые насосы для подачи включаемого биологически активного компонента (или компонентов) и гиперосмотического раствора, соответственно; ЭН - полученные эритроциты-носители, содержащие БАК.
Каждый этап способа обозначен отдельным индексом. В различных частных случаях выполнения способа в качестве исходного материала используется либо цельная кровь, либо предварительно отмытая эритромасса (указано, как этапы a1 или а2 на фиг. 3, соответственно, которые соответствуют введению в систему исходного материала). Перемещения суспензии эритроцитов по магистралям устройства на каждом этапе осуществления способа обозначены следующим образом: a1 и а2 введение исходного материала (исходной крови или исходных отмытых эритроцитов, соответственно) в систему; b - подача цельной крови из исходного мешка на отмывание; с - сбор отмытых эритроцитов в мешок для суспензии отмытых эритроцитов; d - концентрирование отмытых эритроцитов; е гипоосмотический диализ и сбор лизированных в присутствии БАК эритроцитов в мешок, находящийся в блоке для запечатывания, с последующей инкубацией суспензии лизированных ЭН в этом мешке с гиперосмотическим раствором для из запечатывания; f - подача запечатанных ЭН на отмывание; g - сбор запечатанных эритроцитов-носителей после их отмывания в мешок для суспензии ЭН; h - концентрирование полученных ЭН после их отмывания.
Когда в качестве исходного материала используют цельную кровь, то после этапа a1 последовательно осуществляются этапы b-h; в то время как когда в качестве исходного материала используют предварительно отмытую эритромассу, то после этапа а2 осуществляются этапы e-h.
Для способа по изобретению предпочтительны представленные ниже температурные режимы.
Стадию отмывания эритроцитов осуществляют при комнатной температуре.
Стадию лизиса и концентрирования суспензии эритроцитов осуществляют при поддержании температуры от +2°С до +10°С, предпочтительно, от +4°С до +6°С.
Стадию запечатывания лизированных в присутствии биологически активного компонента или компонентов эритроцитов осуществляют при температуре от +25°С до +40°С.
Такие диапазоны температур стандартны для большинства способов включения БАК в эритроциты, и продиктованы физиологическими особенностями мембраны эритроцита. Известно, что деформируемость эритроцита меняется с температурой [25]. Для создания стабильных пор в мембране на стадии диализа необходимо обеспечить минимальную текучесть мембраны эритроцита, чему способствует низкая температура. Однако, слишком низкая температура может приводить к агрегации эритроцитов [26], поэтому оптимальным считается диапазон, приведенный выше. На стадии запечатывания лизированных эритроцитов необходимо обеспечить оптимальную текучесть мембраны эритроцита для быстрого закрытия пор в мембране. Так как текучесть мембраны эритроцитов повышается с повышением температуры, для восстановления формы эритроцита логично создавать температуру близкую к физиологической. Температура выше 40°С может приводить к денатурации белков в мембране клетки и, как следствие, к гемолизу эритроцитов.
Предпочтительными, но не ограничивающими, являются следующие варианты реализации способа.
В частном варианте реализации способа на стадии лизиса эритроцитов с помощью гипоосмотического диализа поддерживают постоянное трансмембранное давление в диализаторе 170 мм рт.ст., а на стадии концентрирования поддерживают постоянное трансмембранное давление 100 мм рт.ст.
В частном варианте реализации способа стадию концентрирования отмытых эритроцитов проводят до достижения величины гематокрита суспензии эритроцитов 75%-80%.
Способ проводят в стерильных условиях.
В качестве биологически активного компонента (БАК) для включения в эритроциты может быть использован целый ряд веществ, обладающих какой-либо лекарственной или биологической активностью. В эритроциты могут быть включены самые разнообразные соединения, относящиеся к различным классам веществ, и микрочастицы, обладающие биологической активностью и способные быть удержанными внутри эритроцита. Приведенные здесь конкретные примеры молекул не являются ограничивающими способ по изобретению, а лишь демонстрируют возможности конкретных вариантов реализации. Ряд используемых БАК включает (но не ограничивается) следующие вещества и классы соединений: различные белки, в том числе ферменты (например, аспарагиназа, глутаматдегидрогиназа, аланинаминотрансфераза или их комбинации); пептиды, олигопептиды и полипептиды, аминокислоты; нуклеиновые кислоты, нуклеотиды, олигонуклеотиды, фосфаты нуклеотидов, в том числе ди- и трифосфаты; аналоги нуклеозидов, используемые как терапевтические агенты (иммуносупрессанты и ингибиторы роста опухолевых клеток), такие как 6-меркаптопурин, азатиопурин, флударабинфосфат; противовирусные агенты, такие как фосфорилированный азидотимидин, (AZT), дидезоксицитозин (ddC), природные и синтетические иммуномодуляторы (активаторы и супрессоры), например, производные мурамилдипептида (MDP); вещества с антикарциногенной активностью (метатрексат, винкристин, винбластин, доксорубицин и т.п.); гормоны (например, кортикостероиды и глюкокортикостероиды); нестероидные противовоспалительные агенты; аллостерические регуляторы гемоглобина (например, инозитолгексафосфат или пиридоксальфосфат); вещества, защищающие гемоглобин или эритроциты (например, криопротекторы); простагландины; лейкотриены (например, лейкотриен II); цитокины; антитела; α-, β- и γ-интерфероны; глутатион; токсины; и другие вещества, имеющие фармакологическую активность.
Кроме того, в качестве биологически активного компонента в эритроциты могут быть включены различные искусственные наночастицы, с включенными биологически активными препаратами или соединениями. Это позволяет, например, использовать эритроцит-носитель как депо-форму в случае наночастиц, которые из-за состава своей поверхности обладают коротким временем циркуляции в кровотоке. Кроме того, такие наночастицы могут обладать, например, сильной флюоресценцией или магнитными свойствами, которые могут быть использованы для создания биосенсоров, концентрирования лекарственного компонента в нужном органе с использованием магнитов или для усиления контрастности при создании биологических изображений, например, при магнитно-резонансной томографии (МРТ) и т.п.
В качестве раствора для отмывания эритроцитов можно использовать физиологический раствор, раствор Bio-Wash или другие изоосмотические растворы для отмывания эритроцитов. Для лизиса эритроцитов и последующего их запечатывания используют гипоосмотический и гиперосмотический растворы, соответственно.
Для реализации заявленного способа могут использоваться различные варианты установки, содержащие устройство для отмывания эритроцитов и диализный элемент, предназначенный для лизиса эритроцитов. В частности, может быть использован диализный элемент, предназначенный для концентрирования и гипоосмотического лизиса эритроцитов. Установка может включать различные насосы для перекачки растворов и суспензий, а также соответствующие емкости для растворов и суспензий эритроцитов и соединяющие их магистрали, на которых установлены клапаны, управляющие движением жидкостей по магистралям. В качестве устройства для отмывания эритроцитов может быть использовано любое известное устройство для такого отмывания, например, устройство фирмы Haemonetics АСР-215 (Haemonetics SA, Швейцария), Cell Saver® и др., или специально сконструированный встроенный в установку узел для отмывания, содержащий центрифугу, сменную центрифугируемую емкость, необходимые насосы и мешки с исходным и отработанным раствором для отмывания эритроцитов. В качестве диализного элемента могут быть использованы стандартные диализные патроны, например, стандартные педиатрические диализаторы фирмы Fresenius (FX Paed) объемом 18 мл и т.п.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Заявляемый способ может быть проиллюстрирован следующими фигурами:
На фиг. 1 представлена зависимость концентрации гемоглобина в суспензии эритроцитов, прошедших через внутренний контур диализатора в ходе стадии концентрирования суспензии эритроцитов, от уровня трансмембранного давления во внешнем и внутреннем контурах диализатора. Показаны результаты экспериментов на эритроцитах из двух различных образцов крови, в которых концентрирование 10% суспензии эритроцитов проводили в течение 8 мин при различных трансмембранных давлениях в диализаторе.
На фиг. 2 представлена зависимость степени инкапсуляции аспарагиназы от уровня трансмембранного давления в диализаторе на стадии гипоосмотического диализа эритроцитов в присутствии аспарагиназы.
На фиг. 3 представлена схема последовательных стадий осуществления заявляемого способа.
На фиг. 4 представлены электронные микрофотографии исходных эритроцитов, а также полученных из них с помощью предлагаемого способа эритроцитов-носителей, содержащих либо аспарагиназу, либо совместно включенные глутаматдегидрогеназу и аланинаминотрансферазу.
На фиг. 5 представлены гематологические показатели (эритроцитарные индексы) исходных эритроцитов и эритроцитов-носителей, нагруженных различными БАК, полученных с помощью заявляемого способа.
На фиг. 6 представлена осмотическая резистентность исходных эритроцитов и эритроцитов-носителей, нагруженных различными БАК, полученных с помощью заявляемого способа.
На фиг. 7 представлены параметры, характеризующие фильтруемость исходных эритроцитов и эритроцитов-носителей различных БАК, полученных с помощью заявляемого способа.
Термины и определения
БАК биологически активный компонент;
Комнатная температура - температура от 20°С до 25°С;
Рабочие растворы - растворы, используемые на разных стадиях осуществления способа включения биологически активных компонентов в эритроциты, к которым относятся физиологический раствор, раствор Bio-Wash или другие изоосмотические растворы для отмывания эритроцитов, гипоосмотический и гиперосмотический растворы и т.п.;
Исходный материал - цельная кровь пациента или соответствующая по группе цельная кровь донора (исходная кровь), или предварительно отмытые эритроциты (исходные эритроциты);
Эритроциты - красные клетки крови;
Эритроцитарная масса (эритромасса, упакованные эритроциты) - концентрированная суспензия отмытых эритроцитов с высоким гематокритом (60-80%);
Эритроциты-носители (ЭН) - эритроциты, содержащие включенный биологически активный компонент (или компоненты);
Лизированные эритроциты - эритроциты, полученные в результате обратимого гипоосмотического лизиса, например, путем диализа суспензии эритроцитов против гипоосмотического буфера с помощью диализного устройства (диализатора), в мембране которых образовались временные поры;
Лизированные эритроциты-носители - эритроциты, лизированные в присутствии БАК, например, путем диализа против гипоосмотического буфера, содержащего БАК, в мембране которых образовались временные поры и часть БАК поступила внутрь эритроцитов;
Эритроциты-биореакторы - частный случай эритроцитов, нагруженных БАК. Эритроциты, нагруженные ферментным препаратом, способным проводить свою реакцию внутри клеток, в которые он включен;
Запечатанные эритроциты-носители - эритроциты, загруженные БАК, с восстановленной целостностью мембраны клетки, получаемые путем восстановления осмотичности в суспензии лизированных эритроцитов-носителей при добавлении к ней гиперосмотического раствора;
Эффективность (степень или выход) включения БАК (Е) - процент от общего количества введенного в систему БАК, оказавшийся запечатанным внутри эритроцитов-носителей в результате проведения процедуры включения БАК в эритроциты (процент или выход инкапсуляции);
Относительная эффективность включения БАК (R) - процент, который составляет реально полученная в эритроцитах-носителях концентрация (или, в случае включения ферментов, активность) БАК (в расчете на 1 мл эритроцитов-носителей при гематокрите 100%) от максимально возможной в данных условиях, за которую принимают концентрацию (или активность) БАК в суспензии отмытых из цельной крови или исходных предварительно отмытых эритроцитов;
Выход клеток (С) - процент клеток, сохранившихся после проведения процедуры включения БАК в эритроциты, от исходного числа клеток;
Фармакоциты - эритроциты с включенным биологически активным компонентом;
Магистрали - пластиковые или силиконовые гибкие трубки, соединяющие различные узлы устройства;
PBS - физиологический раствор, содержащий 10 мМ фосфатного буфера, имеющий рН 7.4.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Представленные ниже сведения приведены для лучшего понимания принципа заявленного изобретения. Эти сведения не являются ограничивающими, а лишь иллюстрируют общие принципы осуществления способа.
Для получения эритроцитов-носителей биологически активного компонента (компонентов) в качестве исходного материала может быть использована как цельная кровь пациента или донора с аналогичной группой крови, так и предварительно отмытые упакованные эритроциты (эритроцитарная масса, эритромасса).
Ниже приведены примеры включения биологически активных компонентов в эритроциты заявляемым способом гипоосмотического проточного диализа.
Пример 1
Включение в эритроциты L-аспарагиназы
Аспарагиназа (L-аспарагин-амидогидролаза, АСП) - фермент класса гидролаз, катализирующий гидролиз L-аспарагина с образованием L-аспарагиновой кислоты и иона аммония согласно реакции (3):
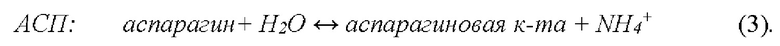
Аспарагиназа является неотъемлемой частью комплексного курса терапии при ряде онкологических заболеваний, например, ОЛЛ у взрослых и детей. Так как прямое внутривенное введение свободного фермента имеет серьезные недостатки (главные из которых, сильные аллергические реакции, короткое время жизни фермента в кровотоке, снижение эффективности лекарства при повторных введениях из-за образования в плазме антител к данному ферменту, влияние высоких пиковых концентраций фермента в момент введения на систему свертывания крови, поджелудочную железу, печень и ряд других систем организма, и т.п.), в качестве лекарственной формы аспарагиназы, имеющей улучшенные фармакологические свойства, используют аспарагиназу, включенную в эритроциты [7].
Для включения аспарагиназы в эритроциты предлагаемым способом в данном примере в качестве исходного материала была использована цельная кровь доноров, которую забирали с помощью пункции локтевой вены в стандартный раствор дигидрата трехосновного цитрата натрия (3.2%, 0.109 М), при соотношении цитрат : кровь составляющем 1:9, а также лиофилизованная L-аспарагиназа из Е. coli Веро-аспарагиназа (ВероФарм ООО, Россия) по 10000 МЕ/флакон, в состав субстанции которой входят дополнительно маннитол 50 мг и глицин 50 мг (на 10000 ME). Для проведения проточного диализа были использованы стандартные педиатрические диализаторы фирмы Fresenius (FX Paed) объемом 18 мл. Раствор аспарагиназы готовили путем разведения лиофилизата ферментного препарата буфером (0.015 М Трис-HCl, 0.015% бычьего сывороточного альбумина (BSA), рН 7.3) до конечной активности 4000 МЕ/мл.
В проведенных экспериментах (n=11) объем исходной крови составлял 100 мл, а объем добавляемого раствора аспарагиназы - 0.4 мл (с активностью 4000 МЕ/мл).
В каждом эксперименте 100 мл исходной цельной крови донора (с гематоКритом от 39.4% до 45.2%) было помещено в находящийся при комнатной температуре мешок, который стерильно соединяли (припаивали) с устройством для отмывания эритроцитов. В качестве устройства для отмывания эритроцитов было использовано устройство АСР-215 (Haemonetics, SA, Швейцария) с объемом центрифужного колокола 275 мл.
Для удаления из системы магистралей воздуха сначала была проведена промывка диализатора. Промывка проводилась в течение 2 мин, путем прокачивания посредством работы насосов через внутренний и внешний контур диализатора со скоростью 100 мл/мин по 200 мл физиологического раствора из мешков для физиологического раствора в мешки для сбора отработанных растворов. Для проведения отмывания эритроцитов объем суспензии эритроцитов в мешке с исходным материалом должен быть равен объему центрифужного колокола устройства, используемого для отмывания эритроцитов (примерно 275 мл). Чтобы увеличить объем материала в мешке с исходной кровью до этого объема, после окончания промывки диализатора, когда клапаны и насосы на внешнем контуре диализатора прекращали работу, открывался клапан, через который физиологический раствор из мешков для физиологического раствора посредством насоса на внутреннем контуре диализатора прокачивался в мешок с исходной кровью со скоростью 100 мл/мин в течение 2 мин.
Далее для осуществления стадии отмывания эритроцитов из исходной цельной крови, кровь из мешка с исходным материалом посредством работы внутреннего насоса устройства для отмывания эритроцитов поступала в центрифужный колокол, куда посредством работы другого внутреннего насоса этого устройства поступал также раствор для отмывания эритроцитов из мешков для растворов, используемых для отмывания эритроцитов. В качестве раствора для отмывания был использован стандартный раствор Bio-Wash, который представляет собой физиологический раствор с добавкой глюкозы (0.2%). Отработанный раствор для отмывания поступал в мешок для отходов, а отмытые из крови эритроциты поступали в мешок для суспензии отмытых эритроцитов, находящийся при температуре +4°С. Стадия отмывания эритроцитов продолжалась до тех пор, пока датчик наличия жидкости в магистрали не зафиксировал окончание перемещения суспензии из отмывающего устройства в мешок для отмытых эритроцитов.
Так как объем полученной суспензии отмытых эритроцитов равен объему центрифужного колокола устройства для отмывания эритроцитов, т.е. полученная суспензия отмытых эритроцитов была достаточно сильно разбавлена, перед началом стадии диализа проводилась стадия концентрирования отмытых эритроцитов. Для этого суспензия эритроцитов из мешка для отмытых эритроцитов, находящегося на шейкере и постоянно перемешиваемая, посредством работы насоса, через открытый клапан перемещалась по внутреннему контуру диализатора со скоростью 20 мл/мин и собиралась вновь в том же мешке. Одновременно, посредством работы двух насосов, по внешнему контуру диализатора, прокачивали физиологический раствор из мешка для физиологического раствора в мешок для сбора отработанных растворов. Насос, подающий физиологический раствор во внешний контур диализатора, работал с постоянной скоростью 20 мл/мин, а скорость второго насоса, выкачивающего раствор из внешнего контура диализатора, была исходно выше, но менялась таким образом, чтобы в диализаторе поддерживалось постоянное трансмембранное давление 100 мм рт.ст., которое определялось по разности давлений, зафиксированных датчиками давления на внутреннем и внешнем контуре диализатора. При приближении величины гематокрита, фиксируемой датчиком гематокрита, к величине порядка 70% 75%, снижение скорости насоса на внешнем контуре диализатора уже не могло поддерживать постоянное трансмембранное давление в диализаторе на уровне 100 мм рт.ст. При этом давление на входе во внутренний контур диализатора начинало расти. Когда разность давлений на входе во внутренний контур диализатора и выходе из него (где давление равно атмосферному) становилась выше 120 мм рт.ст., насос на внутреннем контуре диализатора начинал снижать скорость, чтобы сохранить уровень трансмембранного давления 100 мм рт.ст. Суспензию отмытых эритроцитов прокачивали через диализатор до тех пор, пока скорости работы насосов на внешнем контуре диализатора не становились равными. При этом датчик измерения гематокрита фиксировал на выходе из внутреннего контура диализатора величину гематокрита суспензии эритроцитов равную 75%-80%. После выравнивания скоростей работы насосов на внешнем контуре диализатора стадия концентрирования завершалась, насосы на внешнем контуре диализатора прекращали работу, а через внутренний контур диализатора посредством работы насоса на внутреннем контуре диализатора, в мешок с суспензией эритроцитов из мешков для физиологического раствора поступало еще 35 мл физиологического раствора, чтобы собрать в нем оставшиеся в диализаторе эритроциты. Скорость работы насоса на внутреннем контуре диализатора при этом автоматически поддерживалась максимально возможной, при которой давление на входе во внутренний контур диализатора не превышало атмосферное более, чем на 120 мм рт.ст.
Одновременно посредством шприцевого насоса в магистраль, ведущую в мешок с суспензией отмытых эритроцитов, поступал раствор аспарагиназы (0.4 мл, 4000 МЕ/мл). Затем клапаны перекрывались.
Далее осуществлялась стадия лизиса, при этом сначала через внешний контур диализатора посредством работы насосов на внешнем контуре диализатора в течение 1 мин прогоняли гипоосмотический раствор (5 мМ KH2PO4/K2HPO4, 2 мМ MgCl2, 5 мМ глюкозы, 37 мМ NaCl, 60 мОсм/кг, рН 7.4) со скоростью 30 мл/мин из мешка для гипоосмотического раствора в мешок для сбора отработанных растворов. Затем из мешка с отмытыми эритроцитами (постоянно мягко перемешиваемого шейкером) суспензия эритроцитов с аспарагиназой посредством работы насоса прокачивалась через внутренний контур диализатора и собиралась в мешке для сбора и последующего запечатывания лизированных эритроцитов-носителей, содержащих БАК (примерно за 10-20 мин). Одновременно, во внешнем контуре диализатора в противоток суспензии эритроцитов с аспарагиназой, поступающей из мешка с эритроцитами, посредством работы насосов на внешнем контуре диализатора прокачивали гипоосмотический раствор из мешка для гипоосмотического раствора в мешок для сбора отработанных растворов. Величина скорости работы насосов на внешнем контуре диализатора была одинаковой и составляла 30 мл/мин. Во время диализа поддерживалось постоянное трансмембранное давление 170 мм рт.ст. Это достигалось в автоматическом режиме регулировкой скорости насоса на внутреннем контуре диализатора, которая сначала была установлена как 6.4 мл/мин, но потом насос на внутреннем контуре диализатора изменял скорость, с учетом показаний датчиков давления, чтобы поддерживать трансмембранное давление в диализаторе на выбранном уровне 170 мм рт.ст. при максимальной возможной скорости работы насоса на внутреннем контуре диализатора, при которой давление на входе во внутренний контур диализатора не превышало атмосферное более, чем на 120 мм рт.ст.
Предпочтительно чтобы скорость подачи гипоосмотического раствора во внешний контур диализатора была близкой или равной скорости откачки этого раствора из данного контура, что не позволяет избыточной жидкости входить во внутренний контур диализатора и разбавлять находящуюся там суспензию эритроцитов. Как следствие достигается дополнительное увеличение эффективности включения БАК и снижение потери внутриклеточного содержимого за счет снижения разбавления суспензии эритроцитов в ходе диализа. Близость скоростей подразумевает расхождение в пределах 1 отн. %, предпочтительно в пределах 0.5 отн. %.
После полного прохождения суспензии эритроцитов через диализатор, остатки эритроцитов из диализатора были собраны в мешок для сбора и последующего запечатывания лизированных эритроцитов-носителей путем прокачивания через внутренний контур диализатора с помощью насоса еще 30 мл физиологического раствора. Насос на внутреннем контуре диализатора при этом работал со скоростью 30 мл/мин примерно 1 мин. Посредством шприцевого насоса в магистраль, ведущую в мешок для сбора и последующего запечатывания лизированных эритроцитов-носителей, было введено 9 мл гиперосмотического раствора (1 М NaCl, 50 мМ KH2PO4/K2HPO4, 5 мМ АТФ, 50 мМ глюкозы, 50 мМ натриевой соли пирувата, рН 7.4, 2240 мОсм/кг). Расчет необходимого объема гиперосмотического раствора был сделан заранее в соответствии с формулой (4):
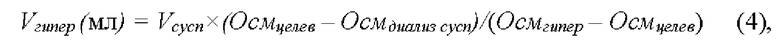
где Vгипер - объем гиперосмотического раствора, который необходимо добавить, Vcycn - объем диализированной суспензии эритроцитов, Осмдиализ cycn - исходная осмоляльность диализированной суспензии, Осмцелев - конечная осмоляльность суспензии, которой требуется достичь (равная примерно 300 мОсм/кг) и Осмгипер. - осмоляльность гиперосмотического раствора (2240 мОсм/кг).
Чтобы рассчитать объем необходимой добавки гиперосмотического раствора, в проведенных экспериментах, где объем исходной цельной крови составлял 100 мл, предварительно были измерены получаемые в аналогичных условиях объемы суспензии эритроцитов после диализа (Vcycn) и осмоляльность этой суспензии (Осмдиализ cycn) при осмоляльности гипоосмотического раствора 60 мОсм/кг, которые составляли в среднем 92.8±14.5 мл, и 112.11±5.64 мОсм/кг, соответственно (n=9). На основании этих измерений было рассчитано, что для восстановления нормальной осмоляльности в суспензии лизированных в присутствии аспарагиназы клеток необходимо использовать добавку гиперосмотического раствора (с осмоляльностью 2240 мОсм/кг) равную 9 мл на каждые 100 мл объема исходной крови.
Только после того, как вся суспензия лизированных эритроцитов-носителей была собрана в мешке для сбора и последующего запечатывания лизированных эритроцитов-носителей, был включен термостат, который обеспечил температуру равную 37°С, и мешок с суспензией лизированных в присутствии аспарагиназы эритроцитов был проинкубирован при 37°С и постоянном перемешивании с помощью шейкера в течение 30 мин, чтобы обеспечить запечатывание полученных эритроцитов-носителей.
Далее полученные запечатанные эритроциты-носители, содержащие аспарагиназу, были отмыты с помощью устройства для отмывания эритроцитов раствором Bio-Wash и собраны в чистый мешок для полученных запечатанных и отмытых эритроцитов-носителей. Для этого сначала они посредством работы внутреннего насоса АСР-215 поступали в центрифужный колокол устройства для отмывания эритроцитов (АСР-215). Одновременно посредством работы насоса на внутреннем контуре диализатора из мешков для физиологического раствора через внутренний контур диализатора в мешок для сбора и последующего запечатывания лизированных эритроцитов-носителей поступили дополнительные 200 мл физиологического раствора (скорость работы насоса на внутреннем контуре диализатора составляла 100 мл/мин). Запечатанные эритроциты-носители были отмыты, и собраны в чистый мешок для полученных запечатанных и отмытых эритроцитов-носителей. Полученная сильно разбавленная суспензия отмытых запечатанных эритроцитов-носителей была затем сконцентрирована с помощью повторения стадии, описанной выше для концентрирования эритроцитов, отмытых из исходной крови, за исключением того, что сконцентрированные эритроциты-носители были собраны в мешок для полученных запечатанных и отмытых эритроцитов-носителей. Насос прокачивал суспензию через внутренний контур диализатора со скоростью 20 мл/мин, а насосы на внешнем контуре диализатора прокачивали физиологический раствор из мешка для физиологического раствора в мешок для сбора отработанных растворов (скорость работы одного насоса на внешнем контуре диализатора была постоянна и составляла 20 мл/мин, а скорость другого насоса на внешнем контуре диализатора была переменная и устанавливалась на уровне, который обеспечивал постоянное трансмембранное давление 100 мм рт.ст.).
Во всех проведенных экспериментах были рассчитаны выходы инкапсуляции фермента (Е) и выход клеток после проведения процедуры включения ферментов (С), согласно формулам (5) и (6), соответственно:
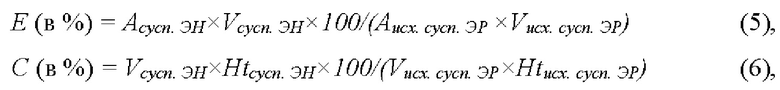
где Аисх.cycn.ЭР и Acycn.ЭН - активности фермента в исходной суспензии отмытых эритроцитов и в конечной суспензии эритроцитов-носителей с включенным ферментом, соответственно, Vисх.cycn.ЭР и Vcycn.ЭН - объемы, a Htисх.cycn.ЭР и Htcycn.ЭН - гематокриты этих суспензий соответственно.
Кроме того, еще один показатель эффективности включения - относительная эффективность включения (R), был оценен как процент, который полученная в эритроцитах удельная активность фермента составляет от максимально возможной удельной активности фермента, которая может быть получена в данных условиях (формула (7)). Под удельной активностью понимается величина активности фермента в расчете на мл клеток-носителей (при гематокрите 100%). При этом за максимально возможную включенную удельную активность принимали активность включаемого вещества в суспензии исходных эритроцитов сразу после внесения аспарагиназы в суспензию отмытых из исходной крови эритроцитов (до начала процедуры включения), т.к. именно такая активность должна установиться в клетках и окружающей среде, если вещество полностью уравновешивается между этими фазами.
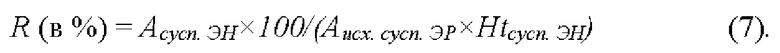
Активность аспарагиназы измеряли индооксиновым методом, используя в качестве субстрата аспарагиназы аспартат-β-гидроксамат (AHA) и определяя образовавшийся при этом гидроксиламин спектрофотометрически на длине волны 705 нм после его превращения в индооксин в присутствии 8-гидроксихинолина [27]. Для измерения активности аспарагиназы, включенной в эритроциты, клетки сначала лизировали, разбавляя водой в 200 раз, а затем определяли активность фермента в лизатах (для удобного измерения, активность аспарагиназы в растворе должна составлять примерно 100 МЕ/л). Полученные результаты представлены в Таблице 1.
Пример 2
Совместное включение в эритроциты глутаматдегидрогеназы и аланинаминотрансферазы
Состояние гипераммониемии (повышение концентрации аммония в крови) является опасным осложнением многих патологических состояний, связанных с дефицитами ферментов цикла мочевины, хроническими и острыми патологиями печени (рак, цирроз и т.д.), а также некоторыми инфекциями желудочно-кишечного тракта.
Избыточный аммоний обладает сильным нейротоксическим действием, что может вызывать ряд когнитивных расстройств и даже летальный исход [28]. Поскольку избыточный аммоний очень токсичен, его необходимо быстро убирать из кровотока, однако существующие медикаментозные средства недостаточно эффективны. Чтобы вернуть концентрацию аммония к нормальному уровню (≤60 μM), требуется от 2 до 7 дней. Ранее было установлено [29], что совместное включение в эритроциты двух ферментов - глутаматдегидрогеназы (ГДГ) и аланинаминотрансферазы (ААТ), позволяет создать эритроциты-биореакторы, достаточно эффективно убирающие из кровотока избыточный аммоний, согласно реакциям (8) и (9):
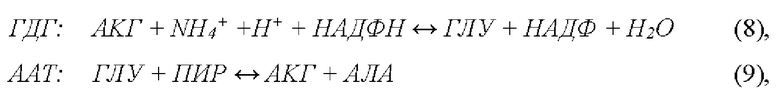
где АКГ - α-кетоглутарат, ГЛУ - глутамат, ПИР - пируват, АЛА - аланин, 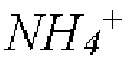 - ион аммония, а НАДФ и НАДФН - окисленная и восстановленная формы никотинамидадениндинуклеотидфосфата, соответственно.
- ион аммония, а НАДФ и НАДФН - окисленная и восстановленная формы никотинамидадениндинуклеотидфосфата, соответственно.
Для получения эритроцитов-носителей, нагруженных совместно ГДГ и ААТ, в качестве исходного материала были использованы предварительно отмытые упакованные эритроциты, полученные стандартным образом на станции переливания крови, а также растворы с различными активностями бактериальной ГДГ из Proteus, sp. (Toyobo Co. Ltd., Osaka, Japan) и ААТ из сердца свиньи (Sigma-Aldrich, St. Louis, MO, USA). Исходный водный раствор ГДГ имел активность 1945 МЕ/мл (при рН 7.4). Объем его добавки составлял от 0.065 до 0.784 мл. Исходная ААТ была лиофильно высушена и имела активность примерно 75 МЕ/мг белка. Для добавления в систему этот фермент был предварительно разбавлен буфером (0.1 М Трис-HCl, рН 7.4) для получения раствора с активностью 250 МЕ/мл. Объем добавки этого фермента в суспензию эритроцитов в разных экспериментах составлял от 0.319 до 14.84 мл. Таким образом, суммарный объем добавки смеси ферментов варьировал в разных экспериментах, в результате чего варьировал и суммарный объем суспензии эритроцитов с добавками ферментов.
Во всех экспериментах (n=17) для включения ферментов в эритроциты использовали способ проточного гипоосмотического диализа, описанный в Примере 1 для аспарагиназы, за исключением первых стадий способа, т.к. в качестве исходного материала в данном примере использовали предварительно отмытую эритроцитарную массу, описанную выше.
В мешок помещали от 52.5 до 60 мл суспензии эритроцитов в физиологическом растворе с гематокритом 80%, предварительно отмытых путем стандартного центрифугирования. Мешок затем был стерильно присоединен (припаян) к магистралям, ведущим к трем каналам одноразовой системы магистралей устройства в блоке запечатывания, ведущим к датчику для определения гематокрита, датчику наличия жидкости в магистрали и клапану, открывающему доступ в мешок из внутреннего контура диализатора.
В течение 2 мин была проведена промывка диализатора, для этого посредством работы насосов через внутренний и внешний контуры диализатора пропустили по 200 мл физиологического раствора со скоростью 100 мл/мин. В конце промывки посредством шприцевого насоса в магистраль, ведущую в мешок с эритромассой была введена смесь растворов включаемых ферментов (в разных экспериментах от 0.384 мл до 15.624 мл), а после этого, посредством работы насоса, в течение 15 сек со скоростью 20 мл/мин через внутренний контур диализатора в мешок пропустили еще 5.0 мл физиологического раствора, чтобы полностью смыть в мешок введенную смесь ферментов. После этого клапаны были перекрыты. Расчетный гематокрит полученной после этого в мешке суспензии эритроцитов с добавленными ферментами колебался от 59.0 до 70% (в среднем 66.6±4.6%, n=17). Активности ферментов, добавленных в исходную суспензию эритроцитов, тоже были различны и составляли от 2.11 до 18.74 МЕ/мл суспензии для ГДГ и от 1.33 до 45.6 МЕ/мл суспензии для ААТ.
Все последующие стадии способа (гипоосмотический диализ эритроцитов в присутствии добавленных ферментов, запечатывание полученной суспензии лизированных в присутствии ферментов эритроцитов, отмывание запечатанных эритроцитов-носителей и их последующее концентрирование) были проведены строго по протоколу, описанному в Примере 1 и при тех же условиях.
Как и в Примере 1, были определены эффективности включения Е и R (раздельно для каждого фермента) по формулам (5) и (7), а также выход клеток в результате процедуры включения ферментов (по формуле (6)). Полученные результаты также представлены в Таблице 1.
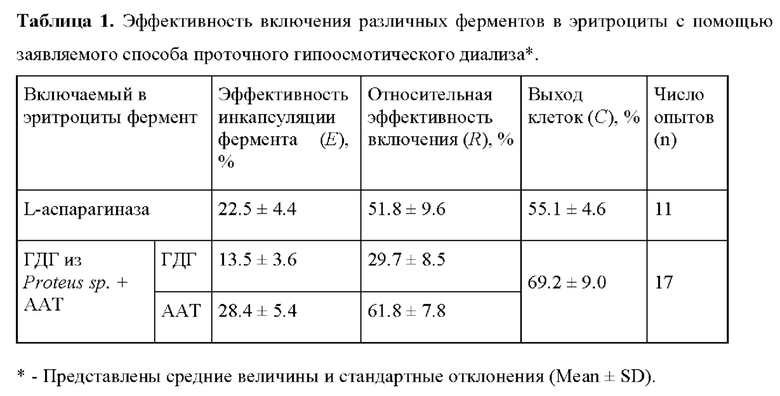
Сравнение результатов, полученных в Примерах 1 и 2, показало, что выход клеток при использовании в качестве исходного материала отмытой эритромассы оказался выше. Это можно объяснить тем, что все старые и хрупкие клетки, присутствующие в исходной крови, в этом случае были отмыты еще на стадии приготовления эритромассы. Второй факт, который можно наблюдать в Примере 2, состоит в том, что процент включения двух использованных ферментов различен. ГДГ из Proteus sp. похожа на хорошо изученную ГДГ из печени быка, про которую известно, что ее эллипсоидальная по форме молекула имеет молекулярный вес 340 кДа, длину 13.6 нм и диаметр 4.3 нм [30,31]. Молекула ГДГ из Proteus sp. также имеет вытянутую форму, но немного меньший молекулярный вес (300 кДа) [32]. В тоже время глобулярная молекула ААТ имеет молекулярный вес только 150 кДа и диаметр около 4 нм [33]. Таким образом размер молекулы ГДГ гораздо больше, чем у ААТ, что снижает скорость проникновения этого фермента внутрь клетки через поры эритроцитарной мембраны, имеющие диаметр около 8-10 нм.
Эффективность включения БАК в эритроциты способом проточного гипоосмотического диализа по настоящему изобретению была также сравнена с эффективностью включения БАК в эритроциты с помощью обратимого гипоосмотического воздействия в соответствие с другими существующими способами. В Таблице 2 представлены такие показатели, приведенные в опубликованных работах или рассчитанные нами самостоятельно из опубликованных данных по включению БАК, с помощью каждого из известных способов.
В качестве показателя включения в разных работах авторы рассчитывали либо относительную эффективность включения (К) (т.е. какой процент включенная в клетки удельная активность составляет от максимально возможной в данных условиях), либо суммарный процент инкапсуляции вещества (Е) (т.е. суммарный процент вещества, включенный в эритроциты с помощью использованного способа), рассчитанные по формулам (7) и (5).
Представленные в Таблице 2 данные демонстрируют, что эффективность включения БАК с помощью заявляемого способа превосходит аналогичные эффективности, получаемые в других способах.
Чтобы оценить качество эритроцитов-носителей, полученных заявляемым способом, были исследованы их свойства, которые были сравнены с аналогичными свойствами исходных эритроцитов.
Свойства эритроцитов-носителей, полученных заявленным способом
Кроме эффективности включения БАК в эритроциты, для проверки качества эритроцитов-носителей БАК по сравнению со свойствами исходных эритроцитов, были исследованы такие свойства, как: форма клеток после включения БАК, эритроцитарные индексы клеток, их осмотическая резистентность и способность деформироваться.
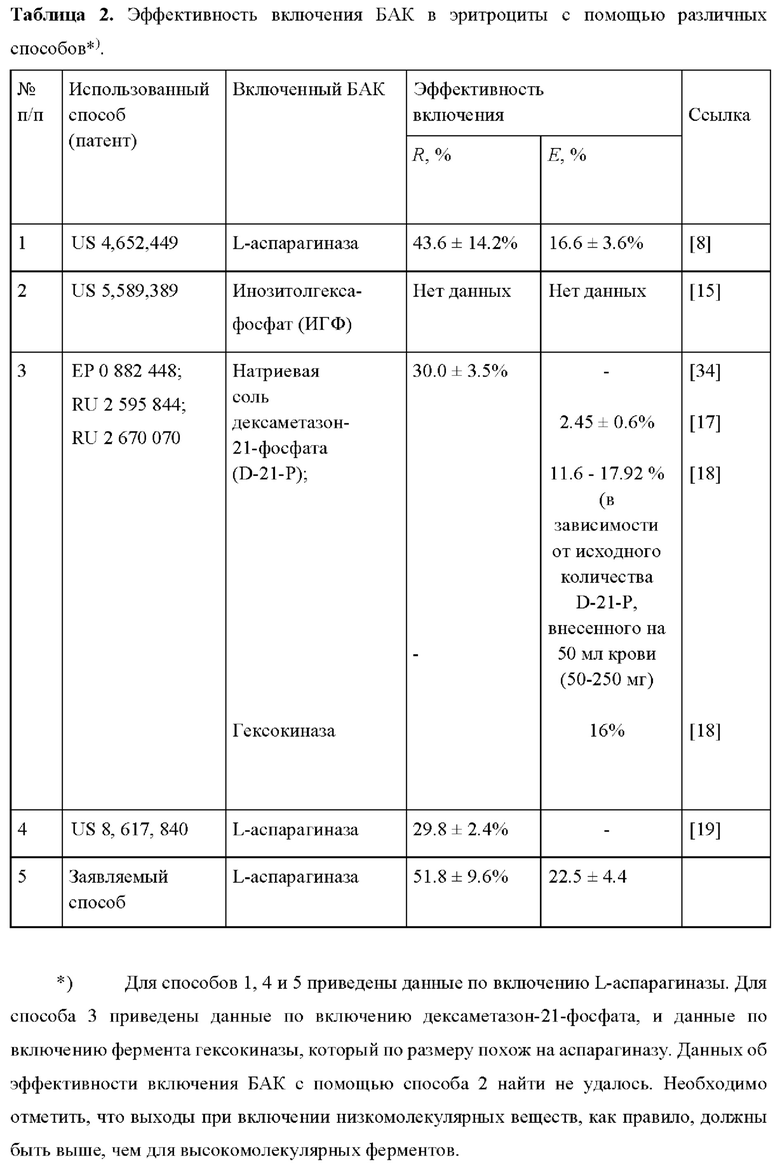
А. Форма клеток после проведения процедуры включения фермента в эритроциты
О физическом состоянии клеток после включения фермента может говорить форма полученных эритроцитов-носителей. В настоящей работе эта форма была исследована методом дифференциальной интерференционно-контрастной (конфокальной) микроскопии. Для получения микрофотографий эритроциты сначала фиксировали. Для этого их инкубировали в PBS с альбумином (10 мг/мл) и глутаровым альдегидом (2.5%) 1 час при комнатной температуре. Гематокрит суспензии клеток при фиксации составлял 25%. Затем данную суспензию разводили до гематокрита около 0.1%. Фотографии были получены с помощью микроскопа Zeiss Axio Observer Z.1 (Carl Zeiss, Йена, Германия), иммерсионный объектив 100х, 1,3 NA, камера QuantEm 512sc.
На фиг. 4 представлены микрофотографии исходных эритроцитов (А), а также эритроцитов-носителей с включенной аспарагиназой (Б) или совместно включенными ГДГ из Proteus sp. и ААТ из сердца свиньи (В).
Из анализа фотографий можно сделать вывод, что для обоих видов эритроцитов-носителей, содержащих различные БАК, большинство полученных клеток возвращает после восстановления осмоляльности среды свою двояковогнутую форму, которая благоприятствует способности эритроцитов деформироваться и проходить сквозь узкие капиллярные сосуды в организме. Размер большей части эритроцитов с включенными ГДГ и ААТ практически не отличался от размера исходных эритроцитов, тогда как для эритроцитов, нагруженных аспарагиназой, он очень незначительно уменьшился.
Б. Гематологические показатели эритроцитов-носителей, полученных заявленным способом
Основные стандартные гематологические показатели клеток (эритроцитарные индексы) были исследованы для исходных эритроцитов (n=9), а также эритроцитов-носителей, нагруженных глутаматдегидрогеназой из Proteus sp. совместно с аланинаминотрансферазой из сердца свиньи (ГДГ+ААТ) (n=6) или аспарагиназой (n=9). В качестве основных показателей были исследованы: средний объем эритроцита (MCV, в фл), среднее содержание гемоглобина в эритроците (МСН, в пг) и средняя концентрация гемоглобина в эритроците (МСНС, в г/дл). Измерение гематологических показателей проводили на гематологическом анализаторе Micros ОТ (ABX-France, Франция).
Полученные результаты представлены на фиг. 5. После включения различных ферментов в эритроциты наблюдалось умеренное изменение эритроцитарных индексов. Средний объем клеток, среднее содержание и средняя концентрация в них гемоглобина всегда достоверно снижались (ANOVA, р<0.05). Это полностью согласуется с данными, полученными ранее в других работах [35]. Несмотря на то, что концентрация и содержание гемоглобина в эритроцитах-носителях снижены по сравнению с исходными эритроцитами, а, следовательно, снижена и их способность переносить кислород, это не влияет критически на выполнение функции газообмена эритроцитами в целом, т.к. вводимый объем эритроцитов-носителей не будет превышать 10% от общего объема эритроцитов в крови человека. Поэтому, даже если концентрация гемоглобина в них будет снижена на 50%, суммарное содержание гемоглобина в организме снизится несущественно (примерно на 5%).
Поскольку размер молекул гликолитических ферментов близок к размеру молекул гемоглобина, можно предполагать, что концентрация этих ферментов в ходе гипоосмотического диализа и открытия пор в клеточной мембране снизится также, как и для Hb не более чем на 30%. Такое снижение не может повлиять на скорость гликолитических реакций в эритроците, т.к. все необходимые ферменты присутствуют в нем в существенном избытке. В результате, можно полагать, что уровень энергообеспечения клетки существенно не изменится, что позволит эритроцитам-носителям выживать в кровотоке достаточно долго [36].
В. Осмотическая резистентность эритроцитов-носителей
Для измерения осмотической резистентности различных эритроцитов, суспензии исходных эритроцитов или полученных эритроцитов-носителей разводили в PBS до гематокрита 5%. Измерения проводили в растворах с разной осмоляльностью по методике, описанной в [37]. Кривую осмотической резистентности строили как зависимость процента нелизированных эритроцитов от осмоляльности использованных растворов. Кривые осмотической резистентности исходных эритроцитов (n=13) и эритроцитов-носителей, полученных методом проточного гипоосмотического диализа, с включенными совместно ГДГ из Proteus sp. и ААТ из сердца свиньи (n=9), а также с включенной аспарагиназой (n=13) представлены на фиг. 6 (для каждой точки представлены средние величины ± средняя ошибка среднего, Mean ± SEM).
Вид кривой осмотической резистентности для эритроцитов-носителей обоих видов достаточно сильно отличался от вида кривой для исходных эритроцитов, что согласуется с результатами, полученными в других работах по измерению осмотической резистентности эритроцитов-носителей [35,38]. Кривые осмотической резистентности для эритроцитов-носителей стали более пологими и сместились в сторону низких осмоляльностей. Таким образом, эритроциты-носители более устойчивы чем исходные эритроциты к изменениям осмоляльности среды в области осмоляльностей ниже 150 мОсм/кг. Лизис 50% исходных (нативных) клеток происходит уже при осмоляльности 135 мОсм/кг, тогда как у эритроцитов-носителей при этой осмоляльности наблюдается только 25 30% лизированных клеток. Это можно объяснить тем, что во время процедуры включения БАК на стадии отмывания клеток были удалены неустойчивые к осмотическому стрессу эритроциты, а также тем, что эритроциты-носители имеют сниженное содержание гемоглобина, что и повлияло на общую кривую осмотической резистентности. С другой стороны, в области осмоляльностей близких к физиологическим, осмотическая резистентность эритроцитов-носителей почти не отличается от осмотической резистентности исходных эритроцитов.
Г. Деформируемость полученных эритроцитов-носителей БАК
Жизнеспособность эритроцитов-носителей БАК (фармакоцитов) в кровотоке во многом определяется их способностью деформироваться и легко проходить через узкие капиллярные сосуды. Деформируемость эритроцитов можно определить с помощью фильтрационных методов, т.к. скорость фильтрации суспензии эритроцитов или эритроцитов-носителей через искусственный мембранный фильтр с порами диаметром 3-5 мкм, близкими к диаметру эритроцитов (~3-5 мкм), и длиной 10 мкм, прямо зависит от этой способности клеток. Фильтруемость клеток была оценена по следующим параметрам: процентное содержание не фильтрующихся эритроцитов в суспензии (Z) и индекс фильтруемости (F), которые определяли методом последовательного пропускания через искусственный полиэтилентерфталатный фильтр (Объединенный Институт ядерных исследований, Дубна, РФ) с порами диаметром 3.5 мкм фиксированного объема (250 мкл) сначала буфера PBS, а затем суспензии клеток с гематокритом 0.1%. Затем, отмыв фильтр после пропускания суспензии, через него повторно пропускали тот же объем PBS. Расчет проводили по формулам (10,11) в соответствии с работой [39]:
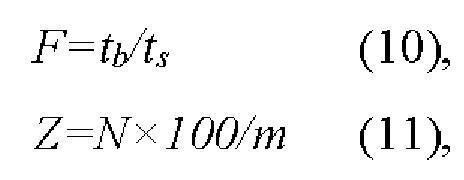
где tb - время протекания через фильтр 250 мкл буфера (PBS), ts - время протекания через тот же фильтр 250 мкл 0.1% суспензии клеток (исходных эритроцитов или эритроцитов-носителей, содержащих БАК), т - общее число клеток, помещенных на фильтр, а N - число пор, которые были блокированы нефильтрующимися клетками при пропускании суспензии через фильтр.
Величина N была определена согласно формуле (12):
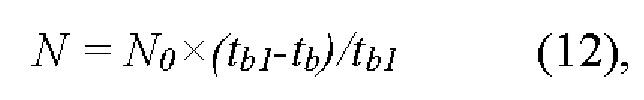
где N0 - известное общее число пор на фильтре, a tb1 - время повторного протекания 250 мкл буфера через фильтр, который был отмыт после пропускания через него суспензии эритроцитов.
Параметры фильтруемости исходных эритроцитов и эритроцитов-носителей представлены на фиг. 7, где представлено процентное содержание нефильтрующихся эритроцитов (Z) и индекс фильтруемости (F) для исходных эритроцитов (1) (n=8), свежеполученных эритроцитов-носителей, нагруженных совместно ГДГ из Proteus sp. и ААТ (2) (n=5), свежеполученных эритроцитов-носителей, нагруженных L-аспарагиназой (3) (n=3), и эритроцитов-носителей, нагруженных L-аспарагиназой, после их инкубации в буфере для хранения в течение 2 часов при комнатной температуре (4) (n=3). Во всех случаях приведены средние величины ± SD.
Процентное содержание нефильтрующихся клеток в суспензии свежеполученных эритроцитов-носителей обоих типов было достоверно выше по сравнению с данным параметром нативных (исходных) эритроцитов (ANOVA, p<0.05). Чтобы понять, насколько необратимы эти изменения, для нагруженных аспарагиназой эритроцитов была измерена фильтруемость не только свежеприготовленных эритроцитов-носителей, но и их фильтруемость после 2 часовой инкубации при комнатной температуре в буфере для хранения клеток (137 мМ NaCl, 2.7 мМ KCl, 10 мМ Na2HPO4, 2 мМ KH2PO4, 1.3 мМ CaCl2, 5 мМ MgCl2, 10 мМ глюкозы, 5% (по массе) BSA, 30 мМ HEPES, 0.28 мМ аденина и 0.02 мг/мл ампициллина (рН 7.4)). Было показано, что после инкубации процентное содержание нефильтрующихся клеток в суспензии эритроцитов-носителей понижается (относительно суспензии свежеполученных эритроцитов-носителей) и не отличается достоверно от значений данного параметра для исходных эритроцитов (ANOVA, p<0.05).
Литература
1. Koleva L.D., Bovt E.A., Ataullakhanov F.I., Sinauridze E.I. Erythrocytes as carriers: from drug delivery to biosensors. Pharmaceutics 2020, 12(3), No 276 (44 pp.). DOI: 10.3 3 90/pharmaceutics 12030276.
2. Halfon-Domenech C, Thomas X., Chabaud S., Baruchel A., Gueyffier F., Mazingue F., Auvrignon A., Corm S., Dombret H., Chevallier P., et al. L-asparaginase loaded red blood cells in refractory or relapsing acute lymphoblastic leukaemia in children and adults: results of the GRASPALL 2005-01 randomized trial. Br. J. Haematol. 2011, 153(1), 58-65. DOI: 10.1111/j.365-2141.2011.08588.x.
3. Hammel P., Berardi R., Van Cutsem E., Feliu J., Greil R., Wasan H.S., Metges J.-P., Nygren P., Osterlund P.J., Parner V., et al. Trybeca-1: a randomized, phase 3 study of eryaspase in combination with chemotherapy versus chemotherapy alone as second-line treatment in patients with pancreatic adenocarcinoma (NCT03665441). J. Clin. Oncol. 2019, 37(4), suppl. TPS471. DOI: 10.1200/JCO.2019.37.4_suppl.TPS471.
4. Batool Т., Makky E. A., Jalal M., Yusoff M.M. A Comprehensive review on L-asparaginase and its applications. Appl. Biochem. Biotechnol. 2016, 178(5), 900 923. DOI: 10.1007/s 12010-015-1917-3.
5. Van Den Berg H. Asparaginase revisited. Leuk. Lymphoma 2011, 52(2), 168-178. DOI: 10.3109/10428194.2010.537796.
6. Mtiller H., Boos J. Use of L-asparaginase in childhood ALL. Crit. Rev. Oncol.Hematol. 1998; 28(2), 97-113. DOI: 10.1016/sl040-8428(98)00015-8.
7. Корсакова Д.В., Синауридзе Е.И. L-аспарагиназа: новые подходы к улучшению фармакологических свойств. Вопросы гематол., онкол. и иммунопатол. в педиатрии. 2018, 17(4), 80-97. DOI: 10.24287/1726-1708-2018-17-4-80-97.
8. Kravtzoff R., Desbois I., Lamagnere J.P., Muh J.P., Valat C, Chassaigne M., Colombat P., Ropars, C. Improved pharmacodynamics of L-asparaginase loaded in human red blood cells. Eur. J. Clin. Pharmacol. 1996, 49(6), 465 470. DOI: 10.1007/BF00195932.
9. Атауллаханов Ф.И., Витвицкий B.M., Жаботинский A.M., Пичугин A.B. Проницаемость эритроцитов человека для аспарагина. Биохимия 1985, 50(10), 1733-1737.
10. Booser, D.J.; Hortobagyi, G.N. Anthracycline antibiotics in cancer therapy. Focus on drug resistance. Drugs 1994, 47(2), 223-258. DOI: 10.2165/00003495-199447020-00002.
11. Moudi M., Go R., Yong C, Nazre M. Vinca alkaloids. Int. J. Prev. Med. 2013, 4(11), 1231-1235. DOI: 10.2165/00128415-200711380-00080.
12. Arora R., Malhotra P., Mathur A., Mathur A., Govil СМ., Ahuja P.S. Anticancer alkaloids of Catharanthus roseus: transition from traditional to modern medicine. In: Herbal Medicine: A Cancer Chemopreventive and Therapeutic Perspective, 1st ed., Arora, R.Ed., Jaypee Brothers Medical Publishers: New Delhi, India, 2010, Chapter: 21, pp.292 309. ISBN 9788184488418.
13. Sawyer D.B., Peng X., Chen В., Pentassuglia L., Lim C.C. Mechanisms of anthracycline cardiac injury: can we identify strategies for cardioprotection? Prog. Cardiovasc. Dis. 2010, 53(2), 105 113. DOI: 10.1016/j.pcad.2010.06.007.
14. Ropars C, Nicolau Y.C., Chassaigne M. Encapsulating biological active substances into erythrocytes, US Patent 4,652,449, March 14, 1987, A61K 35/18
15. Pages E., Ropars C, Bailleul C. Apparatus for causing medical products to penetrate into red blood cells. US Patent 5,589,389, December 31, 1996, C12M 1/12, C12M 3/06.
16. Magnani M., Panzani I., Bigi L., Zanella A. Method of encapsulating biologically active agents within erythrocytes and apparatus thereof (Dideco Sri, Italy). EP 0 882 448 B1, data of publication 12.01.2005, bulletin 2005/02, A61K 9/50.
17. Мамбрини Д., Серафини С.Устройство и набор для инкапсуляции в эритроцитах по меньшей мере одного соединения для терапевтического и диагностического применения (EryDel SpA, Italy). Патент России RU 2595844, дата публикации заявки РСТ 03.11.2011, А61К 9/50, А61М 1/36, B01J 4/00.
18. Мамбрини Д., Бенатти Л., Капогросси Д., Мандолини М. Способ получения эритроцитов, нагруженных одним или несколькими веществами, представляющими фармацевтический интерес, и полученные таким образом эритроциты (EryDel SpA, Italy). Патент России RU 2670070, дата публикации заявки РСТ 13.11.2014, А61К9/50.
19. Godfrin Y. Lisis/resealing process and device for incorporating an active ingredient, in particular asparaginase or inositol hexaphosphate, in erythrocytes (EryTech Pharma, France). US Patent 8,617,840, data of patent 31.12.2013, C12Q 1/24, C12N 13/00, C12M 1/33, C12M 3/08
20. Linderkamp O., Meiselman H.J. Geometric, osmotic, and membrane mechanical properties of density-separated human red cells. Blood 1982, 59(6), 1121-1127.
21. Ponder E. Diffractometric measurements of the tonicity volume relations of human red cells in hypotonic systems. J. Gen. Physiol. 1951, 34 (5), 567-571. DOI: 10.1085/jgp.34.5.567.
22. Evans J., Gratzer W., Mohandas N., Parker K., Sleep J. Fluctuations of the red blood cell membrane: Relation to mechanical properties and lack of ATP dependence. Biophys. J. 2008, 94(10), 4134-4144. DOI: 10.1529/biophysj. 107.117952.
23. Seeman P. Transient holes in the erythrocyte membrane during hypotonic hemolysis and stable holes in the membrane after lysis by saponin and lysolecithin. J. Cell Biol. 1967, 32(1), 55-70. DOI: 10.1083/jcb.32.1.55.
24. Borsakova D.V., Protasov E.S., Nazarenko S.V., Alexandrovich Y.G., Butylin A.A., Ataullakhanov F.I., Sinauridze E.I. Ways to increase the activity of glutamate dehydrogenase in erythrocyte-bioreactors for the ammonium removal. Biochemistry (Moscow), Supplement Series A: Membrane and Cell Biology, 2019, 13(3), 212-224. DOI: 10.1134/S0233475519030046.
25. Williamson J.R., Shanahan M.O., Hochmuth R.M. The influence of temperature on red cell deformability. Blood 1975; 46(4), 611-624. Доступно на: https://doi.Org/10.l182/blood.V46.4.611.611.
26. Wolkers W.F., Crowe L.M., Isvetkova N.M., Tablin F., Crowe J.H. In situ assessment of erythrocyte membrane properties during cold storage. Molec. Membrane Biol., 2002, 19(1), 59-65. DOI: 10.1080/09687680110103613.
27. Lanvers C, Pinheiro J., Hempel G. et al. Analytical validation of a microplate reader-based method for the therapeutic drug monitoring of L-asparaginase in human serum. Anal. Biochem. 2002; 309(1): 117-126. DOI: 10.1016/S0003-2697(02)00232-4.
28. Auron A., Brophy P.D. 2012. Hyperammonemia in review: Pathophysiology, diagnosis, and treatment. Pediatr. Nephrol. 2012; 27(2), 207-222. DOI: 10.1007/s00467-011-1838-5.
29. Protasov, E.S.; Borsakova, D. V.; Alexandravich, Y.G.; Korotkov, A.V.; Kosenko, E.A.; Butylin, A. A.; Ataullakhanov, F.I.; Sinauridze, E.I. Erythrocytes as bioreactors to decrease excess ammonium concentration in blood. Sci. Rep.2019; 9, 1455. DOI: 10.1038/s41598-018-37828-5.
30. Santera, E. Glutamate Dehydrogenases: Enzymology, Physiological Role and Biotechnological Relevance. In: Dehydrogenases (A. B. Hervas, Ed.) (Ch. 12). Rijeka: IntechOpen. 2012, pp.289-318. DOI: https://doi.org/10.5772/47767" 10.5772/47767.
31. Colman R.F. Dehydrogenases-structure and catalytic mechanisms: Glutamate dehydrogenase (bovine liver). In: A study of enzymes. Ed. S.A. Kuby S.A. Ed. Vol.II, Mechanism of enzyme action. Ch. 7. Boston, Roca Raton, Ann Arbor: CRC Press, 1990, pp.173-192.
32. L-Glutamic Dehydrogenase (NADP) from Proteus sp.Доступно на сайте: https://www.sigmaaldrich.com/catalog/product/sigma/g4387.
33. Saier M.H., Jenkins W.T. Alanine aminotransferase. I. Purification and properties. J. Biol. Chem. 1967; 242(1), 91-100.
34. Mangani M., Rossi L., D'ascenzo M., Panzani I., Bigi L., Zanella A. Erythrocyte engineering for drug delivery and targeting. Biotechnol. Appl. Biochem. 1998; 28(1): 1-6. DOI: 10.1111/j. 1470-8744.1998.tb00505.x.
35. Sanz S., Pinilla M., Garin M., Tipton K.F., Luque J. The influence of enzyme concentration on the encapsulation of glutamate dehydrogenase and alcohol dehydrogenase in red blood cells. Biotechnol. Appl. Biochem. 1995; 22(2):223-231. DOI: 10.1111/j.l470-8744.1995.tb00348.x.
36. Martinov M.V., Plotnikov A.G., Vitvitsky V.M., Ataullakhanov F.I. Deficiencies of glycolytic enzymes as a possible cause of hemolytic anemia. Biochim. Biophys. Acta - Gen. Subject 2000; 1474(l):75-87. DOI: 10.1016/S0304-4165(99)00218-4.
37. Shcherbachenko I., Lisovskaya I., Tikhonov V. Oxidation-induced calcium-dependent dehydration of normal human red blood cells. Free Radic. Res. 2007; 41(5):536-545. DOI: 10.1080/10715760601161452.
38. Kosenko E.A., Venediktova N.I., Kudryavtsev A.A., Ataullakhanov F.I., Kaminsky Y.G., Felipo V., Montoliu C. Encapsulation of glutamine synthetase in mouse erythrocytes: a new procedure for ammonia detoxification. Biochem. Cell Biol. 2008; 86(6):469-476. DOI: 10.1139/008-134.
39. Lisovskaya I.L., Shurkhina E.S., Nesterenko V.M., Rozenberg Y.M, Ataullakhanov F.I. Determination of the content of nonfilterable cells in erythrocyte suspensions as a function of the medium osmolality, Biorheology 1998; 35(2): 141-153. DOI: 10.1016/S0006-355X(99)80004-3.
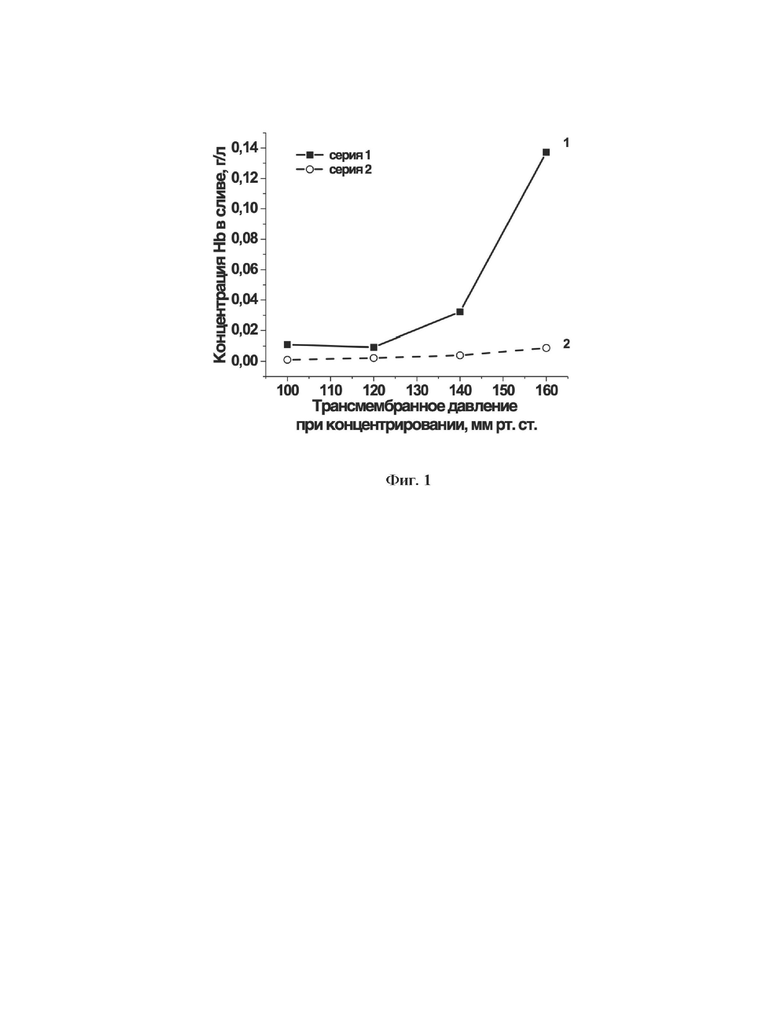
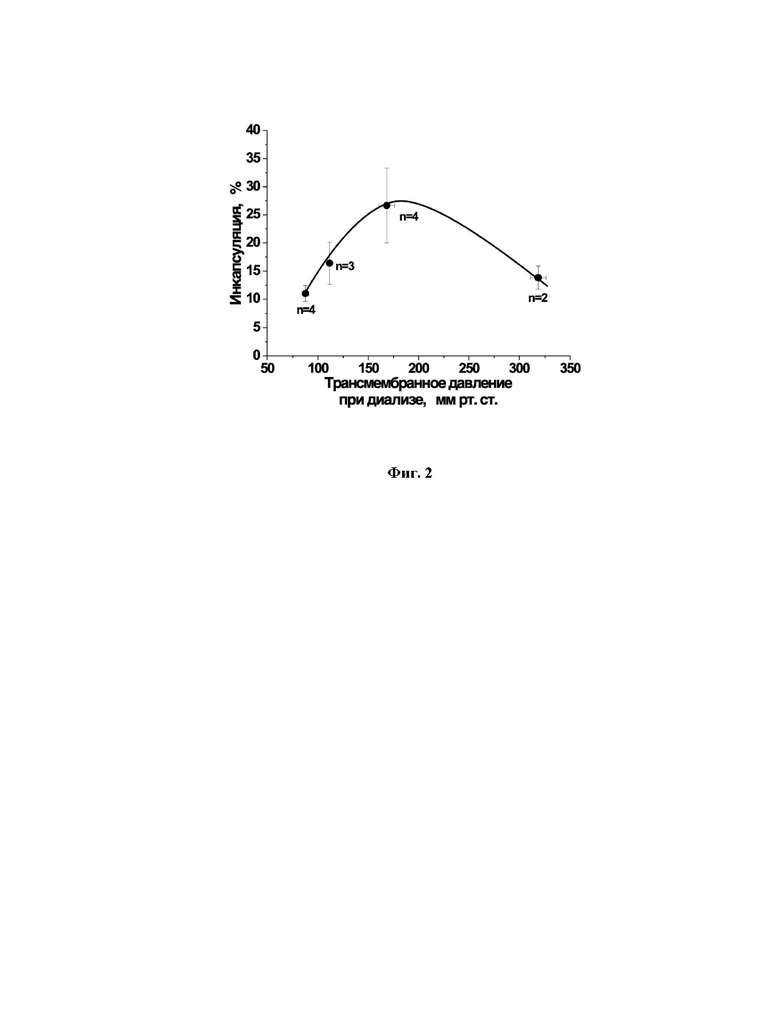
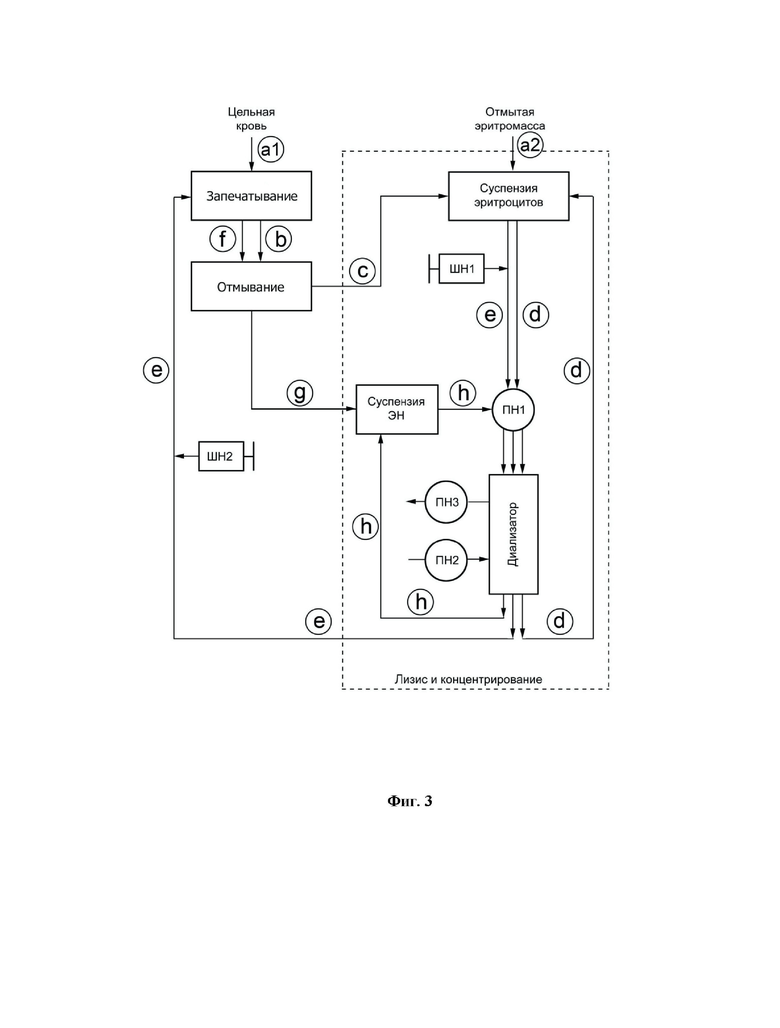

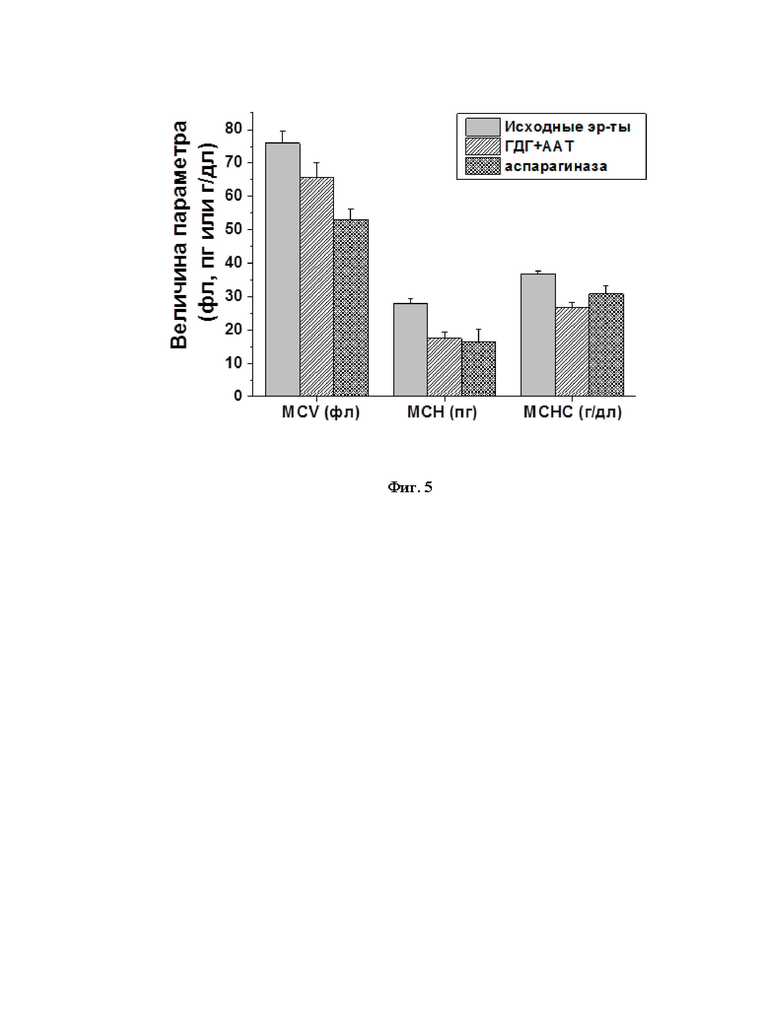
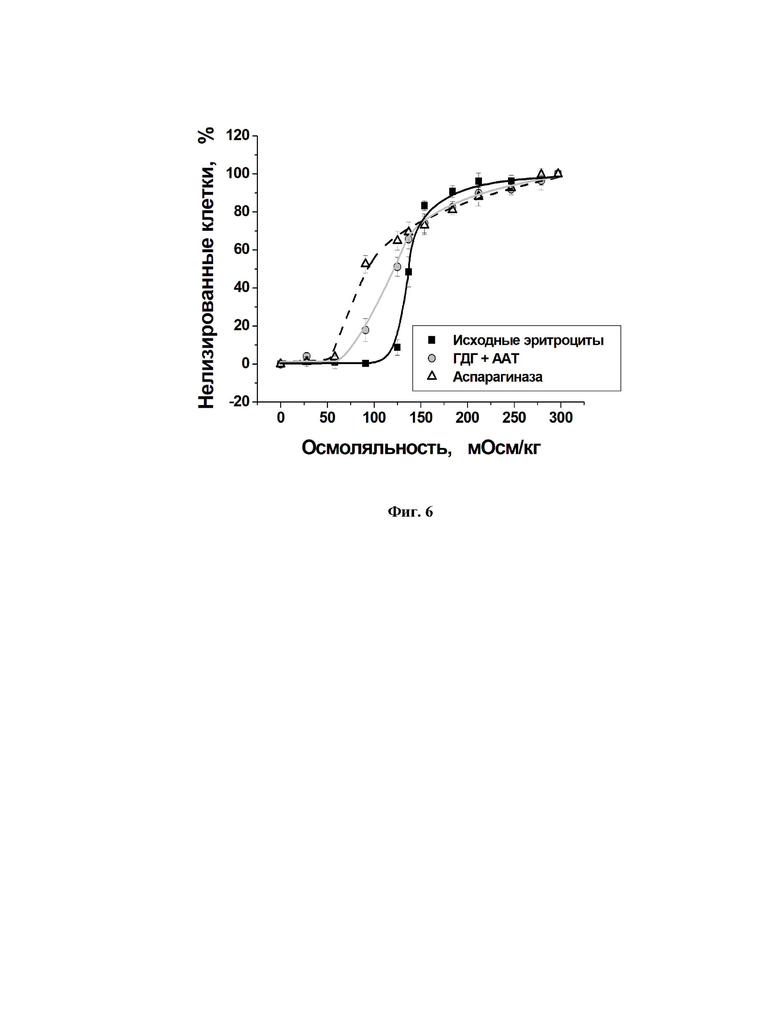
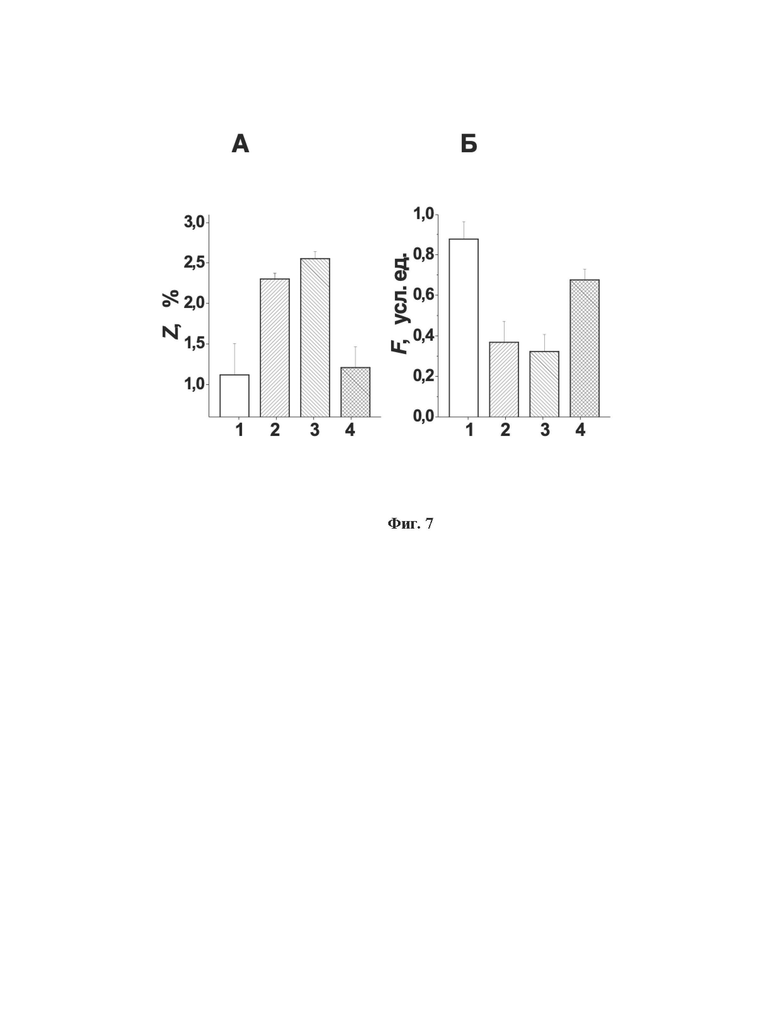
Изобретение относится к фармацевтической промышленности, а именно к способу включения в эритроциты биологически активного компонента. Способ включения в эритроциты биологически активного компонента методом проточного гипоосмотического диализа содержит следующие стадии: добавление, биологически активного компонента к суспензии сконцентрированных отмытых из цельной крови эритроцитов или к предварительно полученной эритромассе; лизис эритроцитов в присутствии, по меньшей мере, одного биологически активного компонента с помощью гипоосмотического проточного диализа; инкубацию суспензии эритроцитов, лизированных в присутствии биологически активного компонента, с гиперосмотическим раствором с образованием запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент; отмывание запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент; концентрирование с помощью диализатора запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент; вымывание из диализатора оставшихся там запечатанных эритроцитов-носителей и добавление их к ранее полученным запечатанным эритроцитам-носителям, содержащим, по меньшей мере, один биологически активный компонент; при этом стадию лизиса эритроцитов в присутствии биологически активного компонента проводят при поддержании постоянного трансмембранного давления в диализаторе 160-180 мм рт. ст. и давлении на входе во внутренний контур диализатора, не превышающем атмосферное более чем на 120 мм рт. ст., и поддержании температуры от +2°С до +10°С; отмывание эритроцитов осуществляют при комнатной температуре; концентрирование эритроцитов осуществляют при поддержании постоянного трансмембранного давления в диализаторе 90-120 мм рт. ст. с учетом измеряемых показателей давления во внешнем и внутреннем контурах диализатора и поддержании температуры от +2°С до +10°С; стадию запечатывания лизированных в присутствии биологически активного компонента или компонентов эритроцитов осуществляют при температуре от +25°С до +40°С; причем стадию концентрирования отмытых из цельной крови эритроцитов и стадию концентрирования запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент, проводят до достижения величины гематокрита суспензии эритроцитов 75–80%. Вышеописанный способ позволяет обеспечить высокую эффективность и воспроизводимую степень включения различных биологически активных компонентов в эритроциты, снижение потери внутриклеточного содержимого. 15 з.п. ф-лы, 7 ил., 2 табл., 2 пр.
1. Способ включения в эритроциты, по меньшей мере, одного биологически активного компонента методом проточного гипоосмотического диализа, содержащий следующие стадии:
- добавление, по меньшей мере, одного биологически активного компонента к суспензии сконцентрированных отмытых из цельной крови эритроцитов или к предварительно полученной эритромассе;
- лизис эритроцитов в присутствии, по меньшей мере, одного биологически активного компонента с помощью гипоосмотического проточного диализа;
- инкубация суспензии эритроцитов, лизированных в присутствии, по меньшей мере, одного биологически активного компонента, с гиперосмотическим раствором с образованием запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- отмывание запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- концентрирование с помощью диализатора запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент;
- вымывание из диализатора оставшихся там запечатанных эритроцитов-носителей и добавление их к ранее полученным запечатанным эритроцитам-носителям, содержащим, по меньшей мере, один биологически активный компонент;
при этом, стадию лизиса эритроцитов в присутствии, по меньшей мере, одного биологически активного компонента, проводят при поддержании постоянного трансмембранного давления в диализаторе 160-180 мм рт. ст. и давлении на входе во внутренний контур диализатора, не превышающем атмосферное более чем на 120 мм рт. ст., и поддержании температуры от +2°С до +10°С; отмывание эритроцитов осуществляют при комнатной температуре; концентрирование эритроцитов осуществляют при поддержании постоянного трансмембранного давления в диализаторе 90-120 мм рт. ст. с учетом измеряемых показателей давления во внешнем и внутреннем контурах диализатора и поддержании температуры от +2°С до +10°С; стадию запечатывания лизированных в присутствии биологически активного компонента или компонентов эритроцитов осуществляют при температуре от +25°С до +40°С; причем стадию концентрирования отмытых из цельной крови эритроцитов и стадию концентрирования запечатанных эритроцитов-носителей, содержащих, по меньшей мере, один биологически активный компонент, проводят до достижения величины гематокрита суспензии эритроцитов 75–80%.
2. Способ по п.1, дополнительно включающий стадии отмывания эритроцитов из цельной крови и их последующего концентрирования, предшествующие стадии добавления, по меньшей мере, одного биологически активного компонента к суспензии сконцентрированных отмытых из цельной крови эритроцитов.
3. Способ по п.1 или 2, в котором на стадии лизиса эритроцитов путем гипоосмотического диализа через внешний контур диализатора пропускают гипоосмотический раствор.
4. Способ по п.1 или 2, в котором на стадии лизиса эритроцитов путем гипоосмотического диализа через внутренний контур диализатора пропускают суспензию эритроцитов с, по меньшей мере, одним биологически активным компонентом.
5. Способ по п.1 или 2, в котором на стадии лизиса эритроцитов поддерживают постоянное трансмембранное давление в диализаторе 170 мм рт. ст.
6. Способ по п.1 или 2, в котором на стадии концентрирования суспензии эритроцитов поддерживают постоянное трансмембранное давление в диализаторе 100 мм рт. ст.
7. Способ по п.1 или 2, в котором, по меньшей мере, один биологически активный компонент выбран из группы:
- белков, в том числе ферментов, пептидов, олигопептидов, полипептидов, аминокислот; или
- нуклеиновых кислот, нуклеотидов, олигонуклеотидов, фосфатов нуклеотидов, аналогов нуклеозидов; или
- противовирусных агентов, природных или синтетических иммуномодуляторов, веществ с антикарциногенной активностью; гормонов; нестероидных противовоспалительных агентов; аллостерических регуляторов гемоглобина, веществ, защищающих гемоглобин или эритроциты, простагландинов, лейкотриенов; цитокинов; антител, α-, β- или γ-интерферонов, глутатиона, токсинов; или
- наночастиц, содержащих компоненты, имеющие фармакологическую или диагностическую значимость.
8. Способ по п. 7, в котором включаемый фермент представляет собой аспарагиназу, или пары совместно включенных ферментов глутаматдегидрогеназы и аланинаминотрансферазы, или алкогольдегидрогеназы и ацетальдегиддегидрогеназы.
9. Способ по п. 7, в котором аналог нуклеозида представляет собой 6-меркаптопурин, азатиопурин или флударабинфосфат.
10. Способ по п. 7, в котором противовирусный агент представляет собой фосфорилированный азидотимидин или дидезоксицитозин.
11. Способ по п. 7, в котором природный или синтетический иммуномодулятор представляет собой производное мурамилдипептида.
12. Способ по п. 7, в котором вещество с антикарциногенной активностью представляет собой метатрексат, винкристин, винбластин или антрациклиновый антибиотик.
13. Способ по п. 7, в котором антрациклиновый антибиотик представляет собой доксорубицин, даунорубомицин или митоксантрон.
14. Способ по п. 7, в котором гормон представляет собой кортикостероид или глюкокортикостероид.
15. Способ по п. 7, в котором аллостерический регулятор гемоглобина представляет собой инозитолгексафосфат или пиридоксальфосфат.
16. Способ по п. 7, в котором вещество, защищающее гемоглобин или эритроциты, представляет собой криопротектор.
| БОРСАКОВА Д.В | |||
| и др | |||
| Способы повышения активности глутаматдегидрогеназы в эритроцитах-биореакторах для удаления аммония // Биологические мембраны | |||
| Т | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| С | |||
| Вагонный распределитель для воздушных тормозов | 1921 |
|
SU192A1 |
| Mangani M., Rossi L., D’ascenzo M., Panzani I., Bigi L., Zanella A | |||
| Erythrocyte engineering for drug delivery and targeting | |||
| Biotechnol | |||
| Appl | |||
| Biochem | |||

