Область техники, к которой относится изобретение
Данное изобретение относится к фармацевтическим композициям, содержащим фермент, зависимый от пиридоксальфосфата, например метиониназу, и при необходимости его кофактор (пиридоксальфосфат) и/или фосфатный или нефосфатный предшественник пиридоксальфосфата; к их применение в качестве лекарственного средства; к способу их получения и к соответствующему терапевтическому способу лечения.
Уровень техники
Пиридоксальфосфат (PLP) - одна из форм витамина В6; он служит кофактором для многих ферментов. Эта разнородная группа ферментов, которых насчитывается около 145, в настоящем документе называется ферментами, зависимыми от пиридоксальфосфата, или PLP-ферментами; они в большинстве своем участвуют в метаболических путях превращений аминокислот. Реакции, катализируемые этими ферментами, включают декарбоксилирование, переаминирование или иные реакции с отщеплением функциональных групп (Percudani and Perrachi, EMBO reports Vol. 4 no. 9, 2003).
Поскольку зависимых от пиридоксальфосфата ферментов и катализируемых ими реакций много, исследовались потенциальные возможности их терапевтического использования применительно к человеку. Среди различных вариантов терапевтического применения, связанного с ферментами, зависимыми от пиридоксальфосфата, множество работ были посвящены использованию таких ферментов для лечения рака и сердечнососудистых расстройств (El-Sayed and Shindia “Targets in gene therapy” Prof. Yongping You ed., 2011). В частности, вызывает интерес метиониназа для снижения уровня метионина в плазме крови и индукции апоптоза опухолевых клеток, не способных к синтезу этой аминокислоты. Было показано, что у человека многие опухолевые клетки не могут пролиферировать, когда метионин заменен его предшественником гомоцистеином, тогда как нормальные клетки вполне могут размножаться в такой среде. Эта зависимость от метионина выражена у клеточных линий, происходящих из молочной железы, легких, толстой кишки, почек, мочевого пузыря, а также у меланомных и глиобластомных клеток (Durando et al., Bull Cancer 2008; 95 (1): 69-76).
Хотя зависимые от пиридоксальфосфата ферменты представляют терапевтический интерес, разработка методов лечения на основе системного введения таких ферментов сталкивается со значительными ограничениями:
- зависимые от пиридоксальфосфата ферменты получают в основном из прокариотических организмов, а такие ферментные препараты весьма иммуногенны при введении человеку;
- время полужизни указанных ферментов в плазме крови очень невелико, так что для того, чтобы получить достаточную ферментативную активность, требуются частые введения или большие дозы
- из-за низкой биологической доступности пиридоксальфосфата как кофактора в плазме крови фактическая ферментативная активность быстро падает после введения.
В случае метиониназы эти ограничения подробно описаны. В работе Sun et al. получали рекомбинантную метиониназу в клетках бактерий Escherichia coli, используя кодирующий этот фермент ген, выделенный из других бактерий - Pseudomonas putida. Полученный в результате фермент (rMETase) вводили внутривенно мышам с иммунодефицитом. Через 24 часа после введения метиониназная активность в плазме крови, определенная in vitro без добавления пиридоксальфосфата не выявлялась, что свидетельствует о краткости периода действия (Sun et al. Cancer Research 63, 8377-8383, 2003).
Год спустя те же исследователи опубликовали результаты введения рекомбинантной метиониназы макакам (Yang et al. Clinical Cancer Research Vol. 10, 2131-2138, 2004). В этой работе шести обезьянам вводили внутривенно указанный фермент в дозах 1000, 2000 и 4000 единиц на килограмм массы тела. Через 28 суток после первого введения делали вторую инъекцию, что вызвало у двух особей анафилактический шок, от которого одна из этих двух обезьян умерла. Вследствие иммуногенности рекомбинантной метиониназы в организме вырабатываются антитела против этого белка; у большинства подопытных животных (у четырех из шести особей) эти антитела являлись в основном иммуноглобулинами типа G (IgG), а также типа М (IgM). Была продемонстрирована (in vitro) нейтрализующая способность этих антител.
Для того, чтобы преодолеть ограничения из-за короткого времени полужизни и иммуногенности полученных ферментных препаратов, указанные авторы предложили прибегнуть к «пэгилированию» фермента, то есть присоединить к нему молекулы полиэтиленгликоля (PEG). «Пэгилирование» - известный в данной области техники метод увеличения времени полужизни и снижения иммуногенности белков, используемых в терапевтических целях. Для получения «пэгилированной» рекомбинантной метиониназы использовали активированные производные полиэтиленгликоля. Такая модификация фермента привела к увеличению его времени полужизни у мышей с двух часов (фермент без PEG) до 38 часов («пэгилированная» рекомбинантная метиониназа). И это значительное продление полужизни сопровождалось снижением иммуногенности белка-фермента (Sun et al., Cancer Research 63, 8377-8383, 2003).
Хотя «пэгилирование» отчасти решает проблему времени полужизни и иммуногенности, но основная трудность для применения ферментов, зависимых от пиридоксальфосфата, а именно низкая биологическая доступность кофактора в плазме крови, остается. Ферменты, зависимые от пиридоксальфосфата, проявляют свою каталитическую активность только в форме холофермента, то есть в комплексе со своим кофактором. Будучи введен в организм, холофермент быстро превращается в апофермент - неактивную форму, не содержащую пиридоксальфосфата.
Экзогенный пиридоксальфосфат быстро становится недоступным для ферментов, так как время полужизни свободного пиридоксальфосфата в плазме крови составляет всего лишь около 15 минут. Это было продемонстрировано в случае тирозин-фенол-лиазы, принадлежащей к группе ферментов, зависимых от пиридоксальфосафта. Elmer et al. (Cancer Research 38; 3663-3667, 1978) получили очищенную тирозин-фенол-лиазу, ввели ее здоровым мышам, через 5 часов после введения брали у подопытных животных кровь и определяли в ней активность этого фермента в двух вариантах условий: в одних образцах крови активность тирозин-фенол-лиазы определяли без добавления пиридоксальфосфата, а в других - пиридоксальфосфат добавляли в реакционную смесь в оптимальном для данного метода определения количестве (эти два варианта условий отражают реально определяемую активность данного фермента в плазме крови и его потенциальную активность при наличии доступного кофактора - пиридоксальфосфата). Сравнение полученных результатов показало, что реально в плазме крови определяется только 7% потенциальной тирозин-фенол-лиазной активности. Такой же анализ проводили с группой мышей, которым наряду с тирозин-фенол-лиазой вводили большое количество пиридоксальфосфата, причем введение кофактора повторяли каждый час. В таких условиях сравнение результатов определения ферментативной активности показало, что в плазме крови достигается 37% потенциальной тирозин-фенол-лиазной активности. Таким образом, одновременное введение пиридоксальфосфата позволяло в некоторой степени увеличить тирозин-фенол-лиазную активность в плазме крови. Однако экзогенный пиридоксальфосфат быстро становится недоступным для тирозин-фенол-лиазы, поскольку время полужизни свободного пиридоксадьфосфата в плазме крови составляет около 15 минут. Следовательно, повышение уровня пиридоксальфосфата в плазме крови путем повторных его введений в растворенном виде нецелесообразно. Авторы описанной выше работы предлагают обеспечить нужный уровень пиридоксальфосфата путем длительного его высвобождения из имплантата, состоящего из спермацета и арахисового масла и вводимого путем внутримышечной инъекции в бедро. Однако такое решение проблемы представляется неубедительным: оно не позволяет достигать реальной тирозин-фенол-лиазной активности в плазме крови более 25% от потенциальной и не усиливает (статистически значимым образом) противоопухолевый эффект пиридоксальфосфата у мышей с имплантированной меланомой В-16. Сходные наблюдения имеются в отношении метиониназы (Sun et al., Cancer Research 63, 8377-8383, 2003). По данным этих исследователей, холофермент PLP-rMETase относительно стабилен in vitro, но in vivo комплекс фермента с кофактором быстро диссоциирует, что ведет к потере активности рекомбинантной метиониназы. Авторы указанной работы показали, что продолжительность снижения уровня метионина в случае как самой рекомбинантной метиониназы, так и ее комплекса с полиэтиленгликолем (PEG-rMETase) может быть увеличена с помощью имплантата, непрерывно выделяющего пиридоксальфосфат. Однако эффективности такого имплантируемого устройства с непрерывным высвобождением активного агента неизбежно противостоит низкая биологическая доступность пиридоксальфосфата в плазме крови.
Итак, несмотря на то, что терапевтическим возможностям использования ферментов, зависимых от пиридоксальфосфата, посвящено множество исследований, в том числе есть работы, в которых применение метиониназы доведено до пробных клинических испытаний, клиническая эффективность таких ферментов до сих пор не продемонстрирована.
Таким образом, с целью использовать терапевтические возможности ферментов, зависимых от пиридоксальфосфата, предпочтительно было бы найти средства/способ обеспечения этих ферментов доступным пиридоксальфосфатом в оптимальной для них концентрации.
Описаны различные способы включения активных ингредиентов в эритроциты. В их числе наиболее широко распространен метод лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны. Этот метод предполагает применение одного из трех вариантов лизиса: гипотонический диализ, предварительное набухание клеток в гипотонической среде и разбавление гипотоническим раствором; все эти способы основаны на разнице осмотического давления между внутренним содержимом эритроцитов и окружающей клетки средой. Во всех трех вариантах осуществляются следующие пять этапов: эритроцитарную массу промывают физиологическим буферным раствором и центрифугируют один или несколько раз; обеспечивают контактирование эритроцитов с гипотонической жидкой средой, что приводит к открыванию пор в клеточной мембране; активный ингредиент проникает внутрь эритроцитов; с помощью гипертонического буферного раствора восстанавливается закрытое состояние пор (поры «запечатываются»), так что активный ингредиент оказывается заключенным внутри эритроцитов, которые затем суспендируют в консервирующем растворе. Наибольший интерес представляет метод с гипотоническим диализом, который стал объектом промышленных разработок. В настоящее время чаще всего прибегают к способу, описанному в ЕР 1 773 452: он обладает преимуществами хорошей воспроизводимости и повышенной степени включения активного ингредиента в клетки.
В опубликованных исследовательских работах уже предлагалось заключение ферментов в эритроциты с точки зрения ограничения риска, связанного с иммуногенностью вводимых ферментов, и продления времени их полужизни в организме. В ЕР 1 773 452 описано включение в эритроциты L-аспарагиназы, а в ЕР 1 874 341 - аргининдезаминазы.
Но проведенные до сегодняшнего дня исследования не касались ферментов, которым требуется кофактор, и не рассматривали сложность кинетики ферментов, зависимых от пиридоксальфосфата, и их кофактора - пиридоксальфосфата.
Раскрытие изобретения
Цель данного изобретения - предложить фармацевтическую композицию, содержащую фермент, зависимый от пиридоксальфосфата, которая позволяет ограничить риск, связанный с иммуногенностью фермента, продлить время его полужизни в организме, при этом обеспечивая фермент доступным кофактором (пиридоксальфосфатом) в оптимальной для данного фермента концентрации.
Таким образом в задачи данного изобретения входит получение суспензии эритроцитов в фармацевтически приемлемом носителе или фармацевтической композиции, содержащей эритроциты и фармацевтически приемлемый носитель, причем в эритроцитах заключен фермент, зависимый от пиридоксальфосфата. Далее в настоящем документе о них говорится как о композиции, равно как о суспензии и фармацевтической композиции. Термин «заключение внутрь/инкапсулирование» означает, что активный ингредиент (фермент и при необходимости кофактор и/или другие вещества) полностью или в основном находится внутри клеток. Выражение «в основном» означает, что небольшая доля активного ингредиента может, тем не менее, быть обнаружена в клеточной мембране.
Конкретно говоря, указанная композиция может содержать от 0,01 мг дo 30 мг, предпочтительно от 0,05 мг дo 10 мг фермента, зависимого от пиридоксальфосфата, в 1 мл препарата эритроцитов.
В первом воплощении данного изобретения фермент, зависимый от пиридоксальфосфата, является метиониназой, называемой далее также L-метиониназой или метионин-гамма-лиазой (MGL; номер по Классификации и номенклатуре ферментов 4.4.1.11, регистрационный номер CAS 42616-25-1). Что касается источников метиониназы, которые можно использовать по данному изобретению, то упомянем публикацию El Sayed A, Applied Microbiol. Biotechnol. (2010) 86: 445-467.
Во втором воплощении данного изобретения фермент, зависимый от пиридоксальфосфата, является тирозин-фенол-лиазой (TPL; номер по Классификации и номенклатуре ферментов 4.1.99.2, регистрационный номер CAS 9059-31-8). Можно сослаться на работу H. Kumagai et al., J. Biol. Chem. 245, 7: 1767-72 и 245, 7: 1773-7.
В третьем воплощении данного изобретения фермент, зависимый от пиридоксальфосфата, является тирозинаминотрансферазой (hTATase; номер по Классификации и номенклатуре ферментов 2.6.1.5, регистрационный номер CAS 9014-55-5). Сошлемся на работу R. Rettenmeier et al., Nucleic Acids Res. 1990, 18, 13: 3583-61.
В четвертом воплощении данного изобретения фермент, зависимый от пиридоксальфосфата, является цистатионин-бета-синтетазой (номер по Классификации и номенклатуре ферментов 4.2.1.22, регистрационный номер CAS 9023-99-8). Можно сослаться на работу J. Kraus et al., J. Biol. Chem. 1978, 253, 18: 6523-8.
Композиция по данному изобретению может содержать также кофактор фермента, то есть пиридоксальфосфат, и/или его предшественник, которым может быть нефосфатный предшественник (например, нефосфатная форма витамина В6) и/или фосфатный предшественник, например пиридоксинфосфат (PNP).
Витамин В6 существует в различных формах - фосфатных либо нефосфатных. К фосфатным формам принадлежат пиридоксинфосфат (PNP), пиридоксальфосфат (PLP) и пиридоксаминфосфат (PMP). Соответствующие нефосфатные формы - это пиридоксин (PN), пиридоксаль (PL), и пиридоксамин (PM). Нефосфатные формы витамина В6 могут проникать через мембрану эритроцитов, а фосфатные формы пересекают ее лишь с трудом. Преобладает метаболический путь, описанный в работе Anderson et al., J. Clin. Invest. 1971, Vol. 50, 1901-1909), в котором пиридоксин (PN) внутри эритроцитов превращается в пиридоксинфосфат (PNP) при участии фермента пиридоксин-киназы; пиридокиснфосфат затем преобразуется в пиридоксальфосфат под действием фермента пиридоксинфосфат-оксидазы. Пиридоксальфосфат может потом быть превращен в пиридоксаль (PL) под действием фермента пиридоксальфосфат-фосфатазы, а пиридоксаль способен выйти из эритроцитов. Понятно, что в процессе получения или во время хранения композиции предлагаемый предшественник в эритроцитах может претерпевать превращения.
В настоящем документе под нефосфатной формой витамина В6 понимается один из трех его «витамеров»: пиридоксаля, пиридоксина и пиридоксамина - либо смесь двух или трех этих витамеров. Предпочтительной формой является пиридоксин. Он может быть также в виде соли.
Композиция по данному изобретению содержит пиридоксальфосфат, заключенный внутри эритроцитов. Он может попасть туда в результате процедуры включения (инкапсулирования) или же быть полностью или частично образован в эритроцитах из своего предшественника. Пиридоксальфосфат - изначально присутствующий или образованный - может быть ассоциирован с ферментом. Композиция по данному изобретению может, таким образом, содержать соответствующий холофермент, например комплекс метиониназа-пиридоксальфосфат. В ассоциации со своим кофактором активный фермент обладает значительно более продолжительным временем полужизни, как можно наблюдать, например, по времени исчезновения его субстрата из плазмы крови. Композиция по данному изобретению дает возможность, в частности, сохранения ферментативной активности долее 24 часов после введения, в частности в течение 1, 5, 10 или 15 суток. Под ферментативной активностью имеется в виду снижение концентрации субстрата в плазме крови более чем на 20%, 30%, 40% или 50%.
В одном из воплощений данного изобретения предлагаемая композиция содержит пиридоксальфосфат (PLP) и/или нефосфатную форму витамина B6 и/или фосфатный предшественник, пиридоксинфосафт (PNP) и/или пиридоксаминфосфат (PMP).
По одному из вариантов данного изобретения пиридоксинфосфат и/или пиридоксаминфосфат заключены внутри эритроцитов в составе композиции по данному изобретению. Этот предшественник может быть заключен в клетки вместе с ферментом либо он может полностью или частично образовываться в эритроцитах из своего предшественника.
Композиция по данному изобретению, в частности, содержит от около 0,05 мкмоль до около 600 мкмоль, в том числе от около 0,5 мкмоль до около 100 мкмоль, предпочтительно от около 5 мкмоль до около 50 мкмоль пиридоксальфосфата и/или пиридоксинфосфата и/или пиридоксаминфосфата в 1 л.
По одному из вариантов данного изобретения предлагаемая композиция содержит эритроциты, внутри которых заключен фермент, зависимый от пиридоксальфосфата, и пиридоксальфосфат, а также нефосфатный предшественник пиридоксальфосфата, заключенный в эритроцитах, находящийся внутри эритроцитов или находящийся внутри и снаружи эритроцитов. Этим нефосфатным предшественником может быть пиридоксин. пиридоксаль или пиридоксамин (предпочтительно пиридоксин), или смесь двух или трех из этих соединений. Нефосфатный предшественник может находиться внутри и/или снаружи эритроцитов. Присутствие нефосфатного предшественника позволяет достичь значительно более высокого уровня пиридоксальфосфата внутри эритроцитов, чем это возможно в отсутствие нефосфатного предшественника.
В одном из воплощений данного изобретения предлагаемая композиция содержит эритроциты, внутри которых заключен фермент, зависимый от пиридоксальфосфата, и в дополнение к нему пиридоксальфосфат и один из его фосфатных предшественников - пиридоксинфосфат, пиридоксальфосфат и/или пиридоксаминфосфат. Эта же композиция может также содержать предпочтительно нефосфатный предшественник, в частности пиридоксин, что было описано выше.
Композиция по данному изобретению предпочтительно характеризуется гематокритом, составляющим 35%, 40% или 45% либо превышающим эти значения.
В одном из воплощений данного изобретения предлагаемая композиция содержит эритроциты и фармацевтически приемлемый носитель, причем в эритроцитах заключен, с одной стороны, фермент, зависимый от пиридоксальфосфата, например метиониназа, и с другой стороны, витамин В6 в нефосфатной форме, предпочтительно в виде пиридоксина, для одновременного, раздельного или последовательного введения. Композиция по данному изобретению может, в частности, быть представлена набором, содержащим отдельно эритроциты (суспензию) и витамин В6 в нефосфатной форме, предпочтительно в виде пиридоксина (раствор). В одном из воплощений данного изобретения фармацевтически приемлемый носитель является консервирующим раствором для эритроцитов, то есть раствором, в котором эритроциты, заключающие в себе активный ингредиент, суспендированы в форме, пригодной для хранения вплоть до введения пациенту. Консервирующий раствор предпочтительно содержит по меньшей мере один агент, способствующий сохранению эритроцитов, а именно выбираемый из глюкозы, декстрозы, аденина и маннита. Предпочтительно консервирующий раствор содержит неорганический фосфат, что позволяет подавить активность содержащейся внутри эритроцитов пиридоксальфосфат-фосфатазы.
Консервирующий раствор по данному изобретению может быть водным раствором, содержащим хлорид натрия, аденин и по меньшей мере одно соединение из следующих: глюкоза, декстроза и маннит. По одному из признаков данного изобретения этот раствор также содержит неорганический фосфат.
Консервирующий раствор по данному изобретению может содержать хлорид натрия, аденин и декстрозу, предпочтительно это среда AS3. По одному из признаков данного изобретения этот раствор также содержит неорганический фосфат.
Консервирующий раствор по данному изобретению может содержать хлорид натрия, аденин, глюкозу и маннит, предпочтительно SAG-маннит или среду ADsol.
По одному из вариантов данного изобретения этот раствор также содержит неорганический фосфат.
В частности, композиция или суспензия (в консервирующем растворе) по данному изобретению отличается содержанием внеклеточного гемоглобина на уровне, равном или меньшим 0,5 г/дл, в частности 0,3 г/дл, в том числе 0,2 г/дл, предпочтительно 0,15 г/дл, более предпочтительно 0,1 г/дл через 72 часа хранения при температуре от 2 до 8°C.
В частности, композиция или суспензия (в консервирующем растворе) по данному изобретению отличается содержанием внеклеточного гемоглобина на уровне, равном или меньшим 0,5 г/дл, в частности 0,3 г/дл, в том числе 0,2 г/дл, предпочтительно 0,15 г/дл, более предпочтительно 0,1 г/дл в течение периода времени, составляющего от 24 часов до 20 суток, в частности от 24 часов до 72 часов, причем хранение осуществляется при температуре от 2 до 8°C.
Определение уровня внеклеточного гемоглобина проводится предпочтительно вручную стандартным методом, описанным в работе G. B. Blakney and A. J. Dinwoodie, Clin. Biochem. 8, 96-102, 1975. Существуют также автоматические устройства, позволяющие осуществлять это определение с чувствительностью, свойственной данному устройству.
В частности, композиция или суспензия (в консервирующем растворе) по данному изобретению отличается степенью гемолиза, держащейся на уровне, равном или меньшим 2%, в том числе 1,5%, предпочтительно 1% через 72 часа хранения при температуре от 2 до 8°C.
В частности, композиция или суспензия (в консервирующем растворе) по данному изобретению отличается степенью гемолиза, держащейся на уровне, равном или меньшим 2%, в том числе 1,5%, предпочтительно 1% в течение периода времени, составляющего от 24 часов до 20 суток, в частности от 24 часов до 72 часов, причем хранение осуществляется при температуре от 2 до 8°C.
В частности гематокрит суспензии по данному изобретению составляет или превышает 35%, 40%, 45%.
Определенным способом метаболизм витамина В6 в эритроцитах модифицируется таким образом, чтобы повысить внутриклеточную концентрацию пиридоксальфосфата путем увеличения внутриклеточных уровней пиридоксин-киназы и пиридоксинфосфат- оксидазы и/или уменьшения внутриклеточного уровня пиридоксальфосфат-фосфатазы.
Согласно отличительному признаку данного изобретения предлагаемая композиция помимо фермента, зависимого от пиридоксальфосфата, например метиониназы, и пиридоксальфосфата или его предшественника, содержит пиридоксин-киназу и/или пиридоксинфосфат-оксидазу и/или агент, ингибирующий пиридоксальфосфат-фосфатазу. Эти ферменты или агенты могут быть заключены внутри эритроцитов или находиться вне и внутри эритроцитов.
Указанные ферменты или агенты можно вводить в организм по отдельности, в частности путем смешивания с препаратом нефосфатной формы витамина В6, отдельным от суспензии эритроцитов.
Целью данного изобретения является такие композиции для применения в качестве лекарственного средства.
Целью данного изобретения является, в частности, лекарственное средство, позволяющее обеспечит нуждающегося в том индивида ферментом, зависимым от пиридоксальфосфата, и его кофактором в условиях хорошей биологической доступности, что подразумевает доступность указанных фермента и кофактора друг для друга и наличие их в эффективном количестве, так что фермент активен и действует нужным образом при терапевтическом применении. Такое лекарственное средство предназначено для снижения или ликвидации концентрации субстрата этого фермента в плазме крови, в циркулирующей крови и/или в рассматриваемом органе.
В рамках первой подзадачи по данному изобретению предлагаемое лекарственное средство содержит метиониназу и позволяет снизить или ликвидировать концентрацию метионина в плазме крови или в кровотоке у нуждающегося в том индивида. Это лекарственное средство является противораковым, позволяющим лечить рак, в частности раковые заболевания, при которых опухолевые клетки ауксотрофны по метионину, а именно рак молочной железы, легких, толстого кишечника, почек, мочевого пузыря, а также меланому и глиобластому.
В рамках второй подзадачи по данному изобретению предлагаемое лекарственное средство содержит метиониназу и позволяет снизить или ликвидировать концентрацию гомоцистеина в плазме крови, кровотоке или в печени у нуждающегося в том индивида. Это лекарственное средство позволяет лечить гомоцистеинурию и/или гипергомоцистеинемию и/или связанные с ними патологические состояния, например сердечнососудистые заболевания, расстройства центральной нервной системы, органов зрения и/или скелета (El-Sayed and Shindia Targets in gene therapy Prof. Yongping You ed., 2011).
В рамках третьей подзадачи по данному изобретению предлагаемое лекарственное средство содержит тирозин-фенол-лиазу и позволяет снизить или ликвидировать концентрацию тирозина в плазме крови или в кровотоке у нуждающегося в том индивида. Это лекарственное средство является противораковым, позволяющим лечить рак, в частности раковые заболевания, при которых опухолевые клетки ауксотрофны по тирозину, в частности меланому.
В рамках четвертой подзадачи по данному изобретению предлагаемое лекарственное средство содержит тирозинаминотрансферазу и позволяет снизить или ликвидировать концентрацию тирозина в плазме крови, кровотоке или в печени у нуждающегося в том индивида. Это лекарственное средство позволяет лечить редкое заболевание, связанное с дефицитом указанного фермента, зависимого от пиридоксальфосфата, а именно синдром Рихнера-Ханхарта (тирозинемию типа II).
В рамках пятой подзадачи по данному изобретению предлагаемое лекарственное средство содержит цистатионин-бета-синтазу и позволяет снизить или ликвидировать концентрацию гомоцистеина в плазме крови, кровотоке или в печени у нуждающегося в том индивида. Это лекарственное средство позволяет лечить гомоцистеинурию и/или гипергомоцистеинемию и/или связанные с ними патологические состояния, например сердечнососудистые заболевания, расстройства центральной нервной системы, органов зрения и/или скелета.
Данное изобретение относится также к способу получения фармацевтических композиций, содержащих эритроциты, внутри которых заключен фермент, зависимый от пиридоксальфосфата (например, метиониназа), фармацевтически приемлемый носитель и пиридоксальфосфат (PLP) и при необходимости фосфатный или нефосфатный предшественник пиридоксальфосфата, причем этот способ включает следующие этапы: при необходимости и предпочтительно осадок эритроцитов промывают и центрифугируют один или несколько раз с физиологическим буферным раствором; суспензия эритроцитов контактирует с гипотонической жидкой средой, что приводит к открыванию пор в клеточной мембране; суспензия эритроцитов контактирует с ферментом, зависимым от пиридоксальфосфата, например метиониназой, до и после открывания пор; фермент, зависимый от пиридоксальфосфата, например метиониназа, проникает внутрь эритроцитов; поры в клеточной мембране закрываются в результате воздействия изотонического или гипертонического, предпочтительно гипертонического, буферного раствора, и собирают суспензию эритроцитов с восстановленным закрытым состоянием пор клеточной мембраны, заключающих в себе фермент, зависимый от пиридоксальфосфата, например метиониназу; при необходимости суспензию эритроцитов инкубируют для удаления наиболее хрупких клеток; суспензию эритроцитов промывают и клетки помещают в консервирующий раствор; в этом способе
- пиридоксальфосфат и/или фосфатный предшественник пиридоксальфосфата (если он используется) заключаются в эритроциты вместе с ферментом, зависимым от пиридоксальфосфата, например, метиониназой,
- нефосфатный предшественник пиридоксальфосфата (если он используется) добавляется в суспензию эритроцитов до и/или после открывания пор, и/или
- нефосфатный предшественник пиридоксальфосфата (если он используется) добавляется в ходе инкубации или в консервирующий раствор.
Предпочтительно некоторое количество пиридоксальфосфата заключается внутрь эритроцитов вместе с ферментом, зависимым от пиридоксальфосфата, и по меньшей мере один нефосфатный предшественник, например пиридоксин, пиридоксаль и/или пиридоксамин добавляют в суспензию эритроцитов до и/или после открывания пор клеточной мембраны, и/или в ходе инкубации и/или в консервирующий раствор. Предпочтительно нефосфатным предшественником является пиридоксин.
Обеспечивается контакт суспензии эритроцитов с гипотонической жидкой средой, приводящий к открыванию пор в клеточной мембране. Существуют три варианта применения метода лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны: гипотонический диализ, предварительное набухание клеток в гипотонической среде и разбавление гипотоническим раствором; все эти способы основаны на разнице осмотического давления между внутренним содержимом эритроцитов и окружающей клетки средой. Предпочтителен гипотонический диализ.
Суспензию эритроцитов, внутри которых заключен фермент, зависимый от пиридоксальфосфата, например метиониназа, и при необходимости пиридоксальфосфат и/или предшественник пиридоксальфосфата, можно получить, в частности, следующим способом:
1 - суспендирование осадка эритроцитов в изотоническом растворе при гематокрите, равном или выше 65%; охлаждение при температуре от +1°C до +8°C,
2 - процедура лизиса при температуре от +1°C до +8°C, включающая перенос суспензии эритроцитов с гематокритом, равным или превышающим 65%, и охлажденного гипотонического раствора для лизиса с температурой от +1°C до +8°C, в устройство для диализа, например в змеевик или картридж (предпочтителен картридж);
3 - процедура заключения агентов внутрь эритроцитов (инкапсулирование), состоящая в добавлении, предпочтительно постепенном, предназначенного для включения внутрь клеток активного ингредиента, в частности в растворе, приготовленном заранее, в суспензию эритроцитов до или во время лизиса при температуре от +1°C до +8°C; и
4 - процедура восстановления закрытого состояния пор клеточной мембраны, осуществляемая в присутствии изотонического или гипертонического, предпочтительно гипертонического раствора, при более высокой температуре, а именно при температуре от +30°C до +42°C.
Предпочтительную альтернативу представляет способ, описанный в публикации WO-A-2006/016247 (EР 1 773 452):
1 - суспендирование осадка эритроцитов в изотоническом растворе при гематокрите, равном или выше 65%; охлаждение при температуре от +1°C до +8°C,
2 - определение осмотической хрупкости клеток в образце эритроцитов в образце эритроцитов из того же осадка,
3 - процедура лизиса при температуре от +1°C до +8°C, включающая перенос суспензии эритроцитов с гематокритом, равным или превышающим 65%, и охлажденного гипотонического раствора для лизиса с температурой от +1°C до +8°C, в устройство для диализа, например в змеевик или картридж (картридж предпочтителен); параметры лизиса эритроцитов берут соответственно их осмотической хрупкости, определенной ранее, а именно в зависимости от измеренной осмотической хрупкости устанавливается поток суспензии эритроцитов через устройство для диализа или осмолярность раствора для лизиса; и
4 - процедура заключения агентов внутрь эритроцитов (инкапсулирование), состоящая в добавлении, предпочтительно постепенном, предназначенного для включения внутрь клеток активного ингредиента, в частности в растворе, приготовленном заранее, в суспензию эритроцитов до или во время лизиса при температуре от +1°C до +8°C; и
5 - процедура восстановления закрытого состояния пор клеточной мембраны, осуществляемая в присутствии изотонического или гипертонического, предпочтительно гипертонического раствора, при более высокой температуре, а именно при температуре от +30°C до +42°C.
Говоря конкретно, для диализа осадок эритроцитов суспендируют в изотоническом растворе с высоким гематокритом - равным или превышающим 65%, предпочтительно равным или превышающим 70% - и полученную суспензию охлаждают до температуры от +1°C до +8°C, предпочтительно от +2°C до +6°C, обычно при температуре около +4°C. По одной из конкретных методик гематокрит составляет от 65% до 80%, предпочтительно от 70% до 80%.
Если определяют осмотическую хрупкость эритроцитов, то предпочтительно это делается непосредственно перед этапом лизиса в присутствии или в отсутствие, предпочтительно в присутствии, одного или более активных ингредиентов, подлежащих заключению внутрь эритроцитов. Эритроциты или содержащая их суспензия предпочтительно имеют температуру, близкую или равную той, при которой предполагается осуществлять лизис. Согласно другому предпочтительному признаку данного изобретения проведенные измерения осмотической хрупкости быстро используются, то есть процедуру лизиса осуществляют через короткое время после взятия образца. Предпочтительно этот промежуток времени между забором образца и началом лизиса не превышает 30 минут, желательно не превышает 25 минут или даже 20 минут.
Относительно подробностей проведения процедуры лизиса эритроцитов с последующим восстановлением закрытого состояния пор клеточной мембраны, определения и учета осмотической хрупкости клеток, специалист в данной области техники может обратиться к публикации WO-A-2006/016247, которая включается в настоящий документ путем отсылки.
Усовершенствование методик включения нужных агентов в эритроциты (инкапсулирования) описано в патенте FR 1 354 204, поданном 07.05.2013, к которому можно адресовать специалистов в данной области техники и который включен в настоящий документ путем отсылки. Таким образом, в одном из воплощений данного изобретения эритроциты, несущие внутри себя активные ингредиенты, то есть фермент, зависимый от пиридоксальфосфата, например метиониназу, и при необходимости один или несколько других активных ингредиентов, например пиридоксальфосфат и/или его предшественник, получают способом, включающим инкапсулирование активного ингредиента в эритроциты в результате лизиса с последующим восстановление закрытого состояния пор клеточной мембраны; получение суспензии эритроцитов или осадка эритроцитов, содержащих эритроциты с заключенным в них активным ингредиентом и раствор с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг; инкубацию осадка или суспензии эритроцитов как есть или после добавления раствора для инкубации при осмоляльности, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг. Инкубацию проводят в частности в течение периода времени, составляющем или превышающем 30 минут, в частности, составляющем или превышающем 1 час. После этого удаляют жидкую среду раствора для инкубации и полученные эритроциты суспендируют в растворе, пригодном для введения путем инъекции пациентам, предпочтительно в консервирующем растворе, пригодном для введения путем инъекции этой суспензии пациентам. Указанная осмоляльность - это осмоляльность раствора, в котором суспендированы эритроциты, или в осадке в соответствующий момент времени.
Согласно одной из конкретных методик, нефосфатный предшественник пиридоксальфосфата - нефосфатная форма витамина В6 - включается в препарат в процессе получения или хранения или же на конечной стадии. Это соединение можно, например, включать в состав раствора для инкубации или в консервирующий раствор, или также в препарат перед его введением пациенту путем инъекции в том случае, когда перед введением препарат разводят.
По одному из вариантов данного изобретения пиридоксин и/или пиридоксаль и/или пиридоксамин включается в препарат в процессе получения или хранения или же на конечной стадии в концентрации от 0,1 мМ до 250 мМ, предпочтительно от 1 мМ до 50 мМ. Как описано выше, в эритроцитах эти нефосфатные производные витамина В6 частично превращаются в пиридоксальфосфат.
Термин «стабилизированная суспензия эритроцитов» в настоящем документе означает суспензию, в которой внеклеточное содержание гемоглобина остается меньше или равно 0,2 г/дл вплоть до ее использования применительно к человеку, осуществляемого через 1-72 часа после получения партии эритроцитов, в которых заключен активный ингредиент.
Термин «готовая для использования стабилизированная суспензия эритроцитов» в настоящем документе означает стабилизированную суспензию клеток в растворе, пригодном для введения путем инъекции пациенту, в частности в консервирующем растворе. Гематокрит этого препарата составляет или превышает 35%, 40% или 45%.
Термин «осадок эритроцитов» в нестоящем документе означает концентрат эритроцитов, полученный после отделения этих клеток от жидкой среды, в которой они были суспендированы до того. Отделение клеток от среды может достигаться путем фильтрации или центрифугирования. Обычно для указанного отделения применяется центрифугирование. Осадок эритроцитов содержит некоторое количество жидкой среды. Обычно гематокрит осадка эритроцитов составляет от 70% до 85%.
Термин «раствор для инкубации» в настоящем документе означает раствор, в котором находятся эритроциты с заключенным внутри них активным ингредиентом в ходе этапа инкубации. Инкубацию можно проводить в широком диапазоне значений гематокрита, конкретно - от 10% до 85%.
Термин «хрупкие эритроциты» в настоящем документе означает эритроциты, полученные в результате процедуры включения в них активного ингредиента, которые. будучи суспендированы в консервирующем растворе, могут претерпевать лизис спустя 1-72 часа после хранения эритроцитной суспензии при температуре от 2°C до 8°C.
Термин «исходный гематокрит» в настоящем документе означает значение гематокрита до потери клеток из-за лизиса хрупких эритроцитов в процессе инкубации.
Способ по изобретению включает, в частности, следующие стадии:
(a) инкапсулирование - заключение в эритроциты одного или более нужных ингредиентов (фермента, зависимого от пиридоксальфосфата, например метиониназы, и при необходимости пиридкосальфосфата и/или его предшественника), состоящее в том, что эритроциты контактируют с гипотонической средой (что дает возможность открыться порам в клеточной мембране) и затем с активным ингредиентом (что дает возможность активному ингредиенту проникнуть внутрь эритроцитов), после чего восстанавливается закрытое состояние пор клеточной мембраны, конкретно говоря - с помощью изотонической или гипертонической среды, предпочтительно - гипертонической;
(b) получение или приготовление суспензии или осадка, содержащих эритроциты, внутри которых заключен активный ингредиент, и раствор с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг;
(c) инкубация осадка или суспензии эритроцитов, полученных на этапе (b), как есть или после добавления раствора для инкубации, при осмоляльности, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг, в течение промежутка времени, составляющего или превышающего 30 минут, в частности, составляющего или превышающего 1 час;
(d) удаление жидкой среды инкубированной суспензии, полученной на этапе (c);
(e) суспендирование эритроцитов, полученных на этапе (d), в растворе, пригодном для введения путем инъекции указанной суспензии пациенту, предпочтительно в консервирующем растворе, пригодном для введения путем инъекции указанной суспензии пациенту.
Витамин B6 в нефосфатной форме можно добавлять в ходе этапа инкапсулирования на этапе (а) или в процессе инкубации на этапе (с), или также в консервирующем растворе.
По первому способу этап, осуществляемый после включения активного ингредиента в эритроциты путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны, а именно этап (b), включает по меньшей мере один цикл промывания, предпочтительно два или три цикла промывания, которое осуществляется путем разбавления суспензии или осадка, полученных на этапе лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны или на этапе (а) раствором с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг, и затем получение осадка или суспензии эритроцитов. Этот осадок или эта суспензия содержит эритроциты, внутри которых заключен активный ингредиент, и раствор с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг. Затем осуществляются следующие этапы, например, (c), (d) и (e).
По второму способу на этапе лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны, или этапе (a) в результате восстановления закрытого состояния пор клеточной мембраны с помощью изотонической или гипертонической среды получается суспензия эритроцитов, которые затем могут быть инкубированы, например суспензия этапа (b), в растворе с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг. Другими словами, этап лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны, или этап (a) включает этап для восстановления закрытого состояния пор клеточной мембраны эритроцитов, когда суспендированные эритроциты, в которых заключен активный ингредиент, смешивают с изотоническим или гипертоническим, предпочтительно гипертоническим, «восстанавливающим» раствором, в результате чего получают суспензию эритроцитов с осмоляльностью, составляющей или превышающей 280 мосмоль/кг, в частности от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг. В этом способе этап инкубации, или этап (с) включает инкубацию суспензии эритроцитов, полученной в результате восстановления закрытого состояния пор клеточной мембраны. Инкубация осуществляется в течение промежутка времени, составляющего или превышающего 30 минут, в частности составляющего или превышающего 1 час. Затем осуществляются следующие этапы, например (d) и (e).
Этапы, осуществляемые после лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны, например этапы с (b) пo (e), проводятся в условиях, вызывающих лизис хрупких эритроцитов или большинства из них, в частности больше 50%, 60%, 70%, 80% или 90% или более. Для этого можно варьировать продолжительность периода инкубации, температуру инкубации и осмоляльность раствора, в котором суспендированы эритроциты. Чем выше осмоляльность, тем продолжительнее будет инкубация. Чем меньше осмоляльность, тем короче инкубация с тем же результатом. Также чем выше температура, тем меньше длится инкубация, и наоборот. После этого делают один или несколько циклов промывания, что позволяет удалить обломки клеток и внеклеточный гемоглобин, а также внеклеточный активный ингредиент.
По данному изобретению цикл промывания включает разбавление суспензии или осадка эритроцитов и затем отделение эритроцитов от промывающего раствора. Предпочтительно этап промывания включает два или три цикла разбавления-отделения. Отделение клеток от раствора достигается любым подходящим способом, например фильтрацией и центрифугированием. Предпочтительно центрифугирование.
Инкубацию не ограничивает гематокрит суспензии эритроцитов. Таким образом, можно инкубировать клеточную суспензию с исходным гематокритом, составляющим от10% до 85%, в частности от 40% до 80%. При гематокрите выше 70% следует говорить скорее об осадке эритроцитов, а суспензия характеризуется значениями ниже указанного.
Целью этапа удаления, или этапа (d) является удаление жидкой части суспензии или инкубированного осадка для того, чтобы, в частности, удалить клеточные обломки и внеклеточный гемоглобин, а также, соответственно, внеклеточный активный ингредиент.
По первому способу на этапе удаления, или этапе (d) осуществляют отделение, а именно центрифугирование, что применимо, в частности к суспензии. После такого отделения могут следовать один или несколько, например два или три, циклов промывания путем разбавления изотоническим раствором с последующим отделением в результате центрифугирования.
По второму способу на этапе удаления, или этапе (d), перед разделением клеток и жидкости путем центрифугирования осуществляют разбавление, что применимо как к суспензии, так и к осадку эритроцитов. Для разбавления берут изотонический промывающий раствор или консервирующий раствор.
Конечный этап, или этап (е) состоит в получении окончательной суспензии, которую можно вводить пациентам без дальнейшей обработки.
По первому способу на этом этапе осадок эритроцитов, полученный на этапе удаления, или этапе (d), разбавляют раствором, пригодным для инъекций, а именно консервирующим раствором.
По второму способу на этом этапе для одного или нескольких циклов промывания осадка эритроцитов, полученного в результате этапа удаления, или этапа (d) используется раствор, пригодный для инъекций, а именно консервирующий раствор; промывание состоит в разбавлении им с последующим отделением клеток от жидкости. После промывания эритроциты ресуспендируют в растворе, пригодном для инъекций, а именно в консервирующем растворе.
Способ по изобретению может также включать один, несколько или все из следующих признаков:
- этап инкубации, или этап (c) осуществляется при температуре от около 2°C до около 39°C, в течение промежутка времени, достаточного для того, чтобы произошел лизис хрупких эритроцитов;
- этап инкубации, или этап (c) осуществляется при низкой температуре, а именно составляющей от около 2°C до около 10°C, в частности, от около 2°C до около 8°C, и продолжается в течение от около 1 часа до около 72 часов, в частности от около 6 часов до около 48 часов, предпочтительно от около 19 часов до около 30 часов;
- этап инкубации, или этап (c) осуществляется при более высокой температуре, составляющей от около 20°C до около 39°C, в частности при комнатной температуре (25°C ± 5°C) и продолжается в течение от около 30 минут до около 10 часов, в частности от около 1 часа до около 6 часов, предпочтительно от около 2 часов до около 4 часов; возможно действовать и при температуре даже выше комнатной, но это может отрицательно сказываться на выходе клеток, P50 и/или содержании 2,3-дифосфоглицерата (2,3-DPG);
- на этапе инкубации, или этапе (c) используется суспензия эритроцитов, характеризующаяся исходным гематокритом от 10% до 85%, в частности от 40% до 80%; осадок, полученный в результате отделения эритроцитов от жидкости, с гематокритом, например от 70% до около 85%, или разведенный клеточный осадок с гематокритом от около 40% до 70%;
- этап инкубации включает перемешивание суспензии эритроцитов;
- этап инкубации не включает никакого перемешивания;
- для достижения желаемой осмоляльности в качестве раствора для промывания и\или инкубации используется водный раствор хлорида натрия известной концентрации; например, этот раствор может содержать 0,9% NaCl; помимо хлорида натрия указанный раствор может содержать глюкозу, а именно моногидрат глюкозы, однозамещенного фосфата натрия дигидрат, двузамещенного фосфата натрия додекагидрат; например, может использоваться такой состав: 0,9% NaCl; 0,2% глюкозы моногидрат; 0,034% однозамещенного фосфата натрия дигидарат; 0,2% двузамещенного фосфата натрия додекагидрат;
- для промывания на конечном этапе, или этапе (е) используется консервирующий раствор;
- осмоляльность раствора (жидкой части) суспензии эритроцитов, которая готова к использованию или может быть введена путем инъекции пациенту, составляет от около 280 мосмоль/кг до около 380 мосмоль/кг, предпочтительно от около 290 мосмоль/кг до около 330 мосмоль/кг;
- гематокрит суспензии эритроцитов, которая готова к использованию или может быть введена путем инъекции пациенту, составляет или превышает 35%, 40% или 45%;
- для всех этапов промывания, инкубации используется консервирующий раствор;
- на этапе (b) и/или на этапе (e) раствор для промывания и консервирующий раствор имеют одинаковый состав и содержат одно или более веществ, способствующих сохранению эритроцитов;
- консервирующий раствор (и растворы для промывания или для инкубации, если нужно) является водным раствором, содержащим хлорид натрия, аденин и по меньшей мере одно из следующих веществ: глюкоза, декстроза и маннит;
- консервирующий раствор (и растворы для промывания или для инкубации, если нужно) содержит хлорид натрия, аденин и декстрозу, предпочтительно в виде среды AS3;
- консервирующий раствор (и растворы для промывания или для инкубации, если нужно) содержит хлорид натрия, аденин, глюкозу и маннит, предпочтительно в виде среды SAG-маннит или ADsol.
Способы по изобретению содержат, в частности, следующий этап:
(a) заключение активного ингредиента внутрь эритроцитов (инкапсулирование), предполагающее контактирование эритроцитов с гипотонической средой, что позволяет открыться порам в их клеточной мембране; контактирование с активным ингредиентом, чтобы он проник внутрь эритроцитов; восстановление закрытого состояния пор в клеточной мембране эритроцитов с помощью изотонической или гипертонической среды. Следует отметить, что активный ингредиент может присутствовать в суспензии эритроцитов до их лизиса или его можно также добавлять в процессе лизиса или после лизиса, но непременно до восстановления закрытого состояния пор клеточной мембраны. В одном из воплощений этого этапа (а) способ по данному изобретению включает следующие подпункты этап (а):
(a1) получение суспензии эритроцитов с гематокритом, составляющим или превышающим 60% или 65%;
(a2) определение осмотической хрупкости эритроцитов в этой суспензии;
(a3) процедура для лизиса эритроцитов и включения в них одного или нескольких активных ингредиентов, предполагающая перенос суспензии эритроцитов в устройство для диализа, а именно в диализный картридж против раствора для лизиса; установление скорости потока эритроцитарной суспензии или скорости потока раствора для лизиса, или осмолярности раствора для лизиса в зависимости от осмотической хрупкости. определенной в подпункте (а2);
(a4) процедура для восстановления закрытого состояния пор клеточной мембраны эритроцитов.
Другой целью изобретения является способ терапевтического лечения, предназначенный для обеспечения нуждающегося в том индивида ферментом, зависимым от пиридоксальфосфата, и его кофактором в условиях хорошей биологической доступности, что предполагает возможность взаимодействия фермента со своим кофактором и наличие эффективных количеств того и другого, так чтобы фермент обладал нужной активностью и эффективностью при терапевтическом применении. Этот способ, в частности. предназначен для ликвидации или уменьшения концентрации субстрата указанного фермента в плазме крови или в кровотоке и/или в органе-мишени. Этот способ включает введение эффективного количества композиции по данному изобретению или использование набора по данному изобретению.
По первому подпункту этой цели изобретения ею является терапевтический способ лечения, позволяющий ликвидировать или снизить уровень метионина в плазме крови или в кровотоке у нуждающегося в том индивида. Этот способ включает введение указанному индивиду эффективного количества композиции по данному изобретению или использование набора по данному изобретению, содержащего метиониназу и ее кофактор. Этот способ является способом лечения рака, в частности раковых заболеваний, при которых опухолевые клетки ауксотрофны по метионину, а именно рака молочной железы, легких, толстой кишки, почки, мочевого пузыря, а также меланомы и глиобластомы.
По второму подпункту этой цели изобретения ею является терапевтический способ лечения, позволяющий ликвидировать или снизить уровень гомоцистеина в плазме крови или в кровотоке или в печени у нуждающегося в том индивида. Этот способ включает введение указанному индивиду эффективного количества композиции по данному изобретению или использование набора по данному изобретению, содержащего метиониназу и ее кофактор. Этот способ является способом лечения гомоцистеинурии и/или гипергомоцистеинемии и/или патологических состояний, связанных с гипергомоцистеинемией, например сердечнососудистых заболевании, заболеваний центральной нервной системы, органов зрения и/или скелета.
По третьему подпункту этой цели изобретения ею является терапевтический способ лечения, позволяющий ликвидировать или снизить уровень тирозина в плазме крови или в кровотоке у нуждающегося в том индивида. Этот способ включает введение указанному индивиду эффективного количества композиции по данному изобретению или использование набора по данному изобретению, содержащего тирозин-фенол-лиазу и ее кофактор. Этот способ является способом лечения рака, в частности раковых заболеваний, при которых опухолевые клетки ауксотрофны по тирозину, а именно меланомы.
По четвертому подпункту этой цели изобретения ею является терапевтический способ лечения, позволяющий ликвидировать или снизить уровень тирозина в плазме крови или в кровотоке и/или в печени у нуждающегося в том индивида. Этот способ включает введение указанному индивиду эффективного количества композиции по данному изобретению или использование набора по данному изобретению, содержащего тирозинаминотрансферазу и ее кофактор. Этот способ является способом лечения редкого заболевания, связанного с этим ферментом, зависимым от пиридоксальфосфата, а именно синдрома Рихнера-Ханхарта (тирозинемии типа II).
По пятому подпункту этой цели изобретения ею является терапевтический способ лечения, позволяющий ликвидировать или снизить уровень гомоцистеина в плазме крови или в кровотоке и/или в печени у нуждающегося в том индивида. Этот способ включает введение указанному индивиду эффективного количества композиции по данному изобретению или использование набора по данному изобретению, содержащего цистатионин-бета-синтазу и ее кофактор. Этот способ является способом лечения гомоцистеинурии и/или гипергомоцистеинемии и/или патологических состояний, связанных с гипергомоцистеинемией, например сердечно-сосудистых заболеваний, заболеваний центральной нервной системы, органов зрения и/или скелета.
Композиция по изобретению, используемая в указанных выше терапевтических применениях, может также содержать кофактор соответствующего фермента, зависимого от пиридоксальфосфата, то есть собственно пиридоксальфосфат и/или его предшественник, который может быть нефосфатным, например нефосфатной формой витамина В6, и/или фосфатным, например пиридоксинфосфатом (PNP). Эта композиция может также содержать пиридоксинкиназу, пиридоксинфосфат-оксидазу и агент, ингибирующий пиридоксальфосфат-фосфатазу. В более широком смысле способ лечения, о котором идет речь, может включать введение композиции или набора по данному изобретению, как описано выше.
В ходе лечения по изобретению пациенту вводят одну или несколько доз в месяц, а именно одну или две, представляющих от 50 мл до 300 мл суспензии эритроцитов или композиции по данному изобретению с гематокритом, составляющим или превышающим 35%, 40% или 45%; введение осуществляется в один или несколько приемов путем внутривенной или интраартериальной инъекции, в частности путем перфузии.
Или же пациенту вводят раздельно эффективное количество композиции, содержащей эритроциты с заключенным внутри них ферментом, зависимым от пиридоксальфосфата (например, метиониназой), и эффективное количество раствора, содержащего нефосфатную форму витамина В6, предпочтительно пиридоксин. Нефосфатная форма витамина В6 может быть ведена путем инъекции одновременно с суспензией эритроцитов или отдельно от нее или любым другим путем, в частности пероральным.
В первом воплощении изобретения пациенту вводят путем инъекции суспензию эритроцитов с заключенными в них одним или более активными ингредиентами, полученную за 1-72 часа, в частности за 10-72 часа до введения. Эта суспензия характеризуется гематокритом, составляющим или превышающим 35%, 40% или 45%. Эта суспензия находится в консервирующем растворе. Уровень внеклеточного гемоглобина в ней составляет 0,5 г/дл или менее, в частности 0,3 г/дл или менее, в том числе 0,2 г/дл или менее, предпочтительно 0,15 г/дл или менее, еще предпочтительнее 0,1 г/дл или менее и/или степень гемолиза составляет 2% или менее, в частности 1,5% или менее, предпочтительно 1% или менее. Эту суспензию перед введением пациенту не промывают и не подвергают никаким сходным манипуляциям.
Другой целью изобретения является способ получения метиониназы в очищенном виде с высоким выходом, включающий следующие этапы:
(a) культивирование бактерий, трансформированных таким образом, чтобы они продуцировали метиониназу; центрифугирование этой культуры и отделение осадка клеток;
(b) суспендирование полученного клеточного осадка в буферном растворе для лизиса; лизис бактериальных клеток, центрифугирование и отделение супернатанта;
(c) обработка супернатанта осаждающим агентом, осаждение и отделение осадка;
(d) с полученным осадком проводят два цикла кристаллизации или осаждения с использованием полиэтиленгликоля (PEG) при температуре от около 25°C до около 40°C и отделяют полученный осадок.
(e) суспендируют полученный осадок в солюбилизирующем буферном растворе (например, [25 мM Tris; 0,5 мM пиридоксаль-5-фосфат (P5P); 0,5 мг/мл бета-меркаптоэтанол; pH 7.5]) и проводят два цикла анионобменной хроматографии, выделяя раствор метиониназы
(f) полученный раствор метиониназы подвергают еще одному этапу очистки путем хроматографии; в результате получают раствор очищенной метиониназы.
В одном из предпочтительных воплощений изобретения используется нуклеотидная последовательность, кодирующая метиониназу, из бактерий Pseudomas putida. Эту последовательность можно оптимизировать, чтобы она подходила к последовательностям продуцирующего штамма бактерий, которым служит предпочтительно штамм HMS174 бактерий E. coli. Для трансформирования штамма, предназначенного для продуцирования нужного фермента, используется экспрессионный вектор, содержащий нуклеотидную последовательность (предпочтительно оптимизированную), кодирующую метиониназу; отбирают продуцирующий клон. Продуцирование метиониназы отобранным клоном осуществляется в ферментере при обычных для подобных задач условиях.
Предпочтительно клеточный осадок, полученный на этапе (a) ресуспендируют в буферном растворе для лизиса (например [100 мM фосфат натрия; 4,4 мM этилендиаминтетраацетат (EDTA); 3,3 мM пиридоксаль-5-фосфат (P5P); 1 мM дитиотреитол (DTT); pH 7,6]) (7 мл на 1 г сырого веса). Предпочтительно лизис достигается путем гомогенизации высокого давления, предпочтительно в несколько приемов, предпочтительно в три приема. Как правило, перед каждой гомогенизацией температура поддерживается около 10°C (от 9°C до 12°C). Предпочтительно после лизиса и перед центрифугированием лизат клеток осветляют, используя катионный коагулянт, предпочтительно полиэтиленимин (PEI). Как правило. концентрация полиэтиленимина берется от около 0,05% до около 0,5% (объем/объем), в частности от около 0,1% до около 0,3%; предпочтительно концентрация полиэтиленимина составляет около 0,2%.
Осаждение на этапе (с) можно осуществлять с помощью сульфата аммония, как правило при 60%-ном насыщении. Предпочтительно перед осаждением супернатант фильтруют, используя мембрану с порами диаметром 0,2 мкм.
На этапе (d) полиэтиленгликоль берут предпочтительно с молекулярной массой 6000 (PEG-6000). Его конечная концентрация составляет от около 5% до около 25% (масса/объем), в частности от около 5% до около 15%. Первый цикл осуществляют предпочтительно в присутствии сульфата аммония. Как правило, сульфат аммония берут в насыщении около 10% (от 9% до 11%). Как правило, конечная концентрация полиэтиленгликоля составляет около 10%. Второй цикл осуществляют предпочтительно в присутствии неорганической соли; как правило, берут соль щелочного металла, например хлорид натрия или хлорид калия, предпочтительно хлорид натрия. Конечная концентрация указанной соли составляет около 0,20 М (от около 0,19 М до около 0,21 М). Как правило, конечная концентрация полиэтиленгликоля составляет около 12%. Температура при этом составляет от около 25°C до около 35 °C, в частности от около 28°C до около 32°C, как правило около 30°C.
На этапе (e) проводят хроматографию с использованием диэтиламиноэтил-сефарозы (DEAE-сефарозы). Предпочтительно перед проведением хроматографии ресуспендированный осадок пропускают через фильтр с порами диаметром 0,45 мкм.
На этапе (f), удаляют оставшиеся примеси, например эндотоксины, белки клетки-хозяина (HCP) и ДНК. Для этого проводят мембранную хроматографию (типа Q).
Очищенную метиониназу концентрируют и проводит диафильтрацию. Для консервации препарата применяют лиофилизацию и хранят полученный продукт при температуре около -80°C.
Далее изобретение описывается подробнее в его воплощениях, взятых в качестве не ограничивающих примеров и сопровождающихся иллюстрациями, в числе которых
Фигура 1 представляет схемы способов очистки метионин-гамма-лиазы (MGL): слева - описанный в Европейском патенте № 0 978 560, справа - усовершенствованный, описанный в настоящей заявке. Изменения, внесенные в способ, описанный в Европейском патенте № 0 978 560 B1, касаются этапов, осуществляемых после этапа осаждения сульфатом аммония.
Фигуры 2 и 3 представляет сравнение внутриклеточных (фиг. 2) и внеклеточных (фиг. 3) концентраций пиридоксальфосфата после инкубации суспензии эритроцитов с заключенными в них метионин-гамма-лиазой и пиридоксальфосфатом (RC-MGL-PLP) с пиридоксином (PN) в различных концентрациях. В суспензии RC-MGL-PLP, инкубированной в течение 3 и 24 часов при комнатной температуре в отсутствие пиридоксина (0 мМ) исходный уровень пиридоксальфосфата составлял 3,9 мкM. В результате инкубации этой суспензии с пиридоксином в концентрациях 2 мМ и 4 мМ внутриклеточная концентрация пиридоксальфосфата возрастала до 8 мкМ через 3 часа инкубации (светло-серые столбики) и до еще более высокого уровня (11 мкМ и 14 мкМ соответственно) через 24 часа инкубации (темно-серые столбики).
Фигура 4. Фармакокинетика эритроцитов (RC), нагруженных метионин-гамма-лиазой с пиридоксальфосфатом (MGL-PLP). Продукт RC-MGL-PLP2 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~30 мкM PLP. Продукт RC-MGL-PLP3 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~125 мкM PLP. Продукт RC-MGL-PLP4 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 5 мг/мл MGL и 33 мкM PLP. Продукт RC-MGL-PLP5 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 6 мг/мл MGL и 100 мкM PLP. Флуоресцентное мечение (CFSE) указанных продуктов позволяло проследить эритроциты in vivo. Указанные продукты, введенные путем внутривенной инъекции мышам линии CD1 (8 мл/кг массы тела в случае продуктов RC-MGL-PLP2, RC-MGL-PLP3 и RC-MGL-PLP5; 10 мл/кг массы тела в случае продукта RC-MGL-PLP4), обладали превосходной стабильностью: выживаемость введенных эритроцитов через 120 часов (то есть 5 суток) после введения превышала 75%. В случае продукта RC-MGL-PLP4 выживаемость эритроцитов становилась менее 75% спустя ~10 суток.
Фигура 5. Фармакодинамика свободной метионин-гамма-лиазы (MGL). Препарат фермента разводили буферным раствором фосфата калия, содержащим 10 мкМ пиридоксаль-5-фосфата (Р5Р), чтобы получить два продукта для введения путем инъекции (MGL-L1 и MGL-L2). Эти продукты были такими, что обеспечивалась 1) такая же концентрация MGL, как в случае продукта RC-MGL-PLP2, то есть 0,45 мг/мл, и 2) концентрация MGL, вдвое большая, чем в случае продукта RC-MGL-PLP2, то есть 0,90 мг/мл. Эти два продукта вводили путем внутривенной инъекции (IV) мышам линии CD1 в дозе 8 мл/кг массы тела с добавлением (также путем внутривенной инъекции) пиридоксина через 6 часов после введения. Определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS). Уровень L-метионина в плазме крови у контрольных (не получавших указанных введений) мышей линии CD1 составлял 68 мкM. Введение как продукта MGL-L1, так и продукта MGL-L2 приводило к быстрому (через 15 минут или менее после введения) снижению содержания L-метионина в плазме крови, но пониженный уровень держался недолго.
Фигура 6. Фармакодинамика эритроцитов с заключенными в них метионин-гамма-лиазой и пиридоксальфосфатом (RC-MGL-PLP) за короткий период времени. Продукт RC-MGL-PLP2 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~30 мкM PLP. Продукт RC-MGL-PLP3 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~125 мкM PLP. Эти продукты вводили путем внутривенной инъекции (IV) мышам линии CD1 в дозе 8 мл/кг массы тела; в случае животных, получивших RC-MGL-PLP2, - с добавлением (также путем внутривенной инъекции) пиридоксина через 6 часов после введения. Определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS). Уровень L-метионина в плазме крови у контрольных (не получавших указанных введений) мышей линии CD1 составлял 82 мкM. Введение как продукта RC-MGL-PLP2, так и продукта RC-MGL-PLP3 приводило к быстрому (через 15 минут или менее после введения) снижению содержания L-метионина в плазме крови до уровней 15,0 ± 3,6 мкM и 22,7 ± 1,5 мкМ соответственно, после чего в период от 48 часов до 120 часов после введения продукта сохранялся более умеренно пониженный уровень (35 мкМ)
Фигура 7. Фармакодинамика эритроцитов с заключенными в них метионин-гамма-лиазой и пиридоксальфосфатом (RC-MGL-PLP) за продолжительный период времени. Продукт RC-MGL-PLP4 (0,5 мг/мл) получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 5 мг/мл MGL и 33 мкM PLP. Этот продукт вводили путем внутривенной инъекции (IV) мышам линии CD1 в дозе 10 мл/кг массы тела с добавлением (также путем внутривенной инъекции) пиридоксина через 6 часов после введения. Определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS). Уровень L-метионина в плазме крови у контрольных (не получавших указанных введений) мышей линии CD1 составлял 68 мкM. Введение продукта RC-MGL-PLP4 приводило к быстрому (через 15 минут или менее после введения) снижению содержания L-метионина в плазме крови до уровня ~10 мкМ, после чего в период от 24 часов до 48 часов после введения продукта сохранялся более умеренно пониженный уровень (~25 мкМ); затем уровень L-метионина постепенно возвращался к наблюдавшемуся у контрольных животных (через 12 суток после введения).
Фигура 8. Остаточная активность ферментов, зависимых от пиридоксальфосфата, в циркулирующей крови. Препарат метионин-гамма-лиазы (MGL) разводили буферным раствором фосфата калия, содержащим 10 мкМ пиридоксаль-5-фосфата (Р5Р), чтобы получить продукт для введения путем инъекции MGL-L2. Продукт RC-MGL-PLP2 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~30 мкM PLP. Продукт RC-MGL-PLP3 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 3 мг/мл MGL и ~125 мкM PLP. Продукт RC-MGL-PLP4 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 5 мг/мл MGL и 33 мкM PLP. Продукт RC-MGL-PLP5 получали путем лизиса с последующим восстановлением закрытого состояния пор клеточной мембраны в суспензии эритроцитов, содержащей 6 мг/мл MGL и 100 мкM PLP. Указанные продукты вводили путем внутривенной инъекции мышам линии CD1 (8 мл/кг массы тела в случае продуктов MGL-L2, RC-MGL-PLP2, RC-MGL-PLP3 и RC-MGL-PLP5; 10 мл/кг массы тела в случае продукта RC-MGL-PLP4) Определяли остаточную активность введенной метионин-гамма-лиазы (эритроциты суммарно) по образованию аммиака в катализируемой этим ферментом реакции способом, описанным в примере 4.
Пример 1. Способ получения и свойства метионин-гамма-лиазы (MGL)
Получение штамма и выделение клона-суперпродуцента.
Природную нуклеотидную последовательность, кодирующую метионин-гамма-лиазу Pseudomonas putida (GenBank: D88554.1), оптимизировали путем модификации редких кодонов (чтобы приспособить указанную последовательность из P. putida к штамму-продуценту Escherichia coli). Были также сделаны другие изменения, чтобы улучшить условия инициации трансляции. Наконец, создали «молчащие» мутации, чтобы удалить три элемента, являющиеся частью предполагаемого бактериального промотора в кодирующей нуклеотидной последовательности (бокс-35, бокс-10 и сайт связывания фактора транскрипции в положении 56). Штамм-продуцент E. coli HMS174 (DE3) трансформировали экспрессионным вектором pGTPc502_MGL (промотор T7), содержащим оптимизированную последовательность; в итоге был отобран клон для продуцирования нужного белка. Клон-продуцент предварительно культивировали в течение 6-8 часов (предварительное культивирование 1) и 16 часов (предварительное культивирование 2) в среде GY с добавлением 0,5% глюкозы и канамицина при температуре 37°C.
Ферментация
Продуцирование осуществлялось после предварительного культивирования 2 при оптической плотности 0,02 в ферментере со средой GY при перемешивании с контролем давления и рН. Фаза роста (при температуре 37°C) длилась до достижения оптической плотности 10; экспрессию индуцировали при температуре 28°C путем добавления в культуральную среду изопропилтиогалактозида (IPTG; 1 мМ). Через 20 часов после индукции экспрессии собирали клеточный осадок в два приема: среду с клетками пропускали через полые волокна с порогом отсечения по молекулярной массе 500 кДа и затем концентрировали в 5-10 раз; получали клеточный осадок путем центрифугирования при 15 900 g, после чего хранили при температуре -20°C.
Очистка
Клеточный осадок оттаивали и суспендировали в буферном растворе для лизиса (7 объем/масса). Лизис проводили при температуре 10°C в три приема путем гомогенизации высокого давления (один раз при давлении 1000 бар, затем два раза при давлении 600 бар). Лизат клеток осветляли при температуре 10°C путем добавления 0,2%-ного полиэтиленимина (PEI) и центрифугирования при 15 900 g. Растворимую фракцию стерилизовали (0,2 мкм), затем осаждали сульфатом аммония (60%-ное насыщение) при температуре 6°Cв течение 20 часов. Проводили два раза кристаллизацию (осадок растворяли в солюбилизирующем буферном растворе): для этого первый раз добавляли политэтиленгликоль с молекулярной массой 6000 (PEG-6000) в конечной концентрации 10% и сульфат аммония (10%-ное насыщение), второй раз добавляли PEG-6000 в конечной концентрации 12% и хлорид натрия в конечной концентрации 0,2 M при температуре 30°C. Каждый раз после центрифугирования при 15 900 g собирали осадок, содержащий желаемый белок - метионин-гамма-лиазу (MGL). Осадок, содержащий белок MGL, ресуспендировали в солюбилизирующем буферном растворе и пропускали через фильтр с порами диаметром 0,45 мкм, после чего проводили два раза анионобменную хроматографию (DEAE-сефароза FF). Очищенный белок подвергали дополнительной тонкой очистке путем мембранной хроматографии (капсулы типа Q) для удаления различных примесей (эндотоксинов, белков клетки-хозяина, остаточной ДНК). Наконец, очищенный белок MGL концентрировали до 40 мг/мл и проводили диафильтрацию в буферном растворе для белкового препарата, используя кассету для тангенциальной поточной фильтрации с порогом отсечения по молекулярной массе 10 кДа. Полученный продукт разливали на аликвоты по ~ 50 мг белка на один флакон, лиофилизировали при контролируемых давлении и температуре и хранили при температуре -80°C.
Свойства
Удельную активность фермента определяли, измеряя количество образующегося NH3, как описано в Примере 4. Чистоту полученного препарата белка определяли путем электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE). Уровень пиридоксальфосфата определяли после разведения водой способом, описанным в Примере 5. Осмолярность определяли при помощи осмометра (Micro-Osmometer Loser Type 15) .
В приведенной ниже таблице представлены основные свойства одной из полученных партий метионин-гамма-лиазы.
Свойства после разведения водой (625 мкл):
78,7 мг/мл; ~622 мкМ PLP; 50 мM фосфата натрия; pH 7,2; Осмолярность 300 мосмоль/кг
Обсуждение способа получения
Способ очистки метионин-гамма-лиазы, описанный в примере 1, базируется на способе, подробно описанном в Европейском патенте № 0 978 560 B1 и связанной с ним публикации (Takakura et al., Appl Microbiol Biotechnol 2006). Такой выбор объясняется простотой и надежностью этапа кристаллизации, который в описании авторов предстает как особенно практичный и легко приспосабливаемый к получению белка в больших масштабах. Этот способ основывается на использовании полиэтиленгликоля с молекулярной массой 6000 (PEG6000) и сульфата аммония после нагревания раствора фермента (метионин-гамма-лиазы), полученного в результате лизиса, осветления и удалении примесей на добавочных этапах с использованием PEG6000/сульфата аммония. Важным моментом этого этапа также является возможность быстро достигнуть высокого уровня чистоты в ходе этапа удаления примесей путем центрифугирования после обработки раствора метионин-гамма-лиазы полиэтиленгликолем. В результате центрифугирования примеси оказываются в осадке, а большая часть метионин-гамма-лиазы - в растворенном виде в супернатанте. Благодаря достигаемой таким образом степени чистоты, для получения очищенного искомого белка достаточно одного этапа хроматографии, в котором метионин-гамма-лиазу пропускают через анионообменную колонку (DEAE), и очистки путем гель-фильтрации на колонке с сефакрилом S200 HR.
Про опробовании этого патентованного способа в небольшом масштабе обнаружилось, что получаемые результаты не воспроизводятся. Согласно Европейскому патенту № 0 978 560 B1, в конце этапа удаления примесей (обработка PEG6000/сульфатом аммония и центрифугирование) большая часть метионин-гамма-лиазы содержится в растворимой фракции, причем примеси в результате центрифугирования удаляются с осадком. Однако в ходе получения метионин-гамма-лиазы способом, описанным в Европейском патенте № 0 978 560 B1, искомый белок - метионин-гамма-лиаза после центрифугирования по большей части (~80%) оказывался в осадке. В приведенной ниже таблице приведены значения доли метионин-гамма-лиазы (в процентах) в растворимой фракции, определенные денситометрически по гелям SDS-PAGE.
Этот неожиданный результат побудил усовершенствовать указанный патентованный способ следующим образом: 1) обрабатывать получаемый в результате центрифугирования осадок, содержащий метионин-гамма-лиазу; 2) проводить два последовательных этапа кристаллизации для лучшего удаления примесей после нанесения на колонку с DEAE; 3) оптимизировать хроматографическую очистку на колонке с DEAE.
Что касается последнего пункта, то было обнаружено, что смола DEAE-сефароза FF недостаточно сильный ионообменный материал в использовавшихся условиях (состав и рН буферного раствора). После ряда различных пробных опытов было в конце концов решено 1) заменить фосфатный буферный раствор, используемый в исходном способе, на буферный раствор Tris с pH 7,6 для повышения надежности способа и 2) пропускать белковый препарат через DEAE-колонку второй раз, чтобы существенно снизить уровень эндотоксинов и повысит чистоту получаемого белкового продукта без потери метионин-гамма-лиазы (0,8 эндотоксиновых единиц/мг по работе Takakura et al., по сравнению с 0,57 ЭЕ/мг по модифицированному способу).
Наконец, чтобы добиться соответствия способа получения белка требованиям, связанным с его применением в больших масштабах согласно стандартам надлежащей производственной практики (GMP), был добавлен этап дополнительной тонкой очистки путем мембранной хроматографии (типа Q) с целью уменьшить уровни остаточных эндотоксинов и белков клетки-хозяина (HCP). Для этого этапа окончательной тонкой очистки не требуется гель-фильтрации c использованием S200, которую трудно осуществлять при получении продукта в промышленных масштабах (этот вид хроматографии предполагает существенные затраты времени и денег).
Различные этапы очистки способа по Европейскому патенту № 0 978 560 B1 и способа, предлагаемого в настоящей заявке, представлены на фиг. 1.
В приведенной ниже таблице представлены данные, позволяющие убедиться, что предлагаемые изменения способа получения метионин-гамма-лиазы обеспечивают очистку фермента с выходом по меньшей мере таким же, какой, по описанию, достигается исходным способом.
$ после очистки с сефакрилом S-200 HR (способ по Европейскому патенту № 978 560) или после мембранной хроматографии типа Q (способ по данному изобретению).
Пример 2. Заключение метионин-гамма-лиазы в мышиные эритроциты вместе с пиридоксальфосфатом
Цельную кровь мышей линии CD1 (от Charles River Laboratories, Inc.) центрифугировали при 1000 g в течение 10 минут при температуре 4°C, чтобы удалить плазму и светлый слой лейкоцитов. Эритроциты промывали три раза 0,9%-ным раствором хлорида натрия (объем/объем). Лиофилизованную метионин-гамма-лиазу ресуспендировали в воде в концентрации 78,7 мг/мл и добавляли этот раствор в суспензию эритроцитов с таким расчетом, чтобы гематокрит окончательной суспензии составлял 70% при различных концентрациях метионин-гамма-лиазы и пиридоксальфосфата. Эту суспензию загружали в устройство для гемодиализа при скорости потолка 120 мл/ч и проводили диализ против гипотонического раствора при скорости потока 15 мл/мин (с противотоком). Затем в суспензии восстанавливали закрытое состояние пор в клеточной мембране эритроцитов с помощью гипертонического раствора, после чего инкубировали в течение 30 минут при температуре 37°C. Суспензию промывали три раза 0,9%-ным раствором хлорида натрия, содержащим 0,2% глюкозы, после чего клетки помещали в консервирующий раствор SAG-маннит, содержащий 6% бычьего сывороточного альбумина (BSA). Определяли свойства полученных продуктов в день 0 (D0) не позже чем через 2 часа после получения и в день 1 (D1), то есть через ~18-24 часа хранения при температуре 2-8°C). Гематологические показатели определяли с помощью автоматического гематологического анализатора для ветеринарии Sysmex (PocH-100iV).
Результаты
В описываемых далее различных экспериментах активность метионин-гамма-лиазы (MGL) в конечных продуктах определяли способом, описанным в Примере 4, используя для сравнения независимую калибровочную кривую активности MGL в водном растворе. Полученные результаты в сочетании с поясняющими экспериментами, показывают, что активность MGL в конечных продуктах тем больше, чем больше количество фермента, с которым манипулируют способом по данному изобретению, и что нетрудно получить конечный продукт, содержащий внутри эритроцитов MGL в количестве до 32 МЕ/мл и при этом вполне стабильный.
В другом эксперименте были получены три мышиных конечных продукта (RC-MGL-PLP1, RC-MGL-PLP2 и RC-MGL-PLP3) следующими способами:
- RC-MGL-PLP1. Достигалось включение в эритроциты MGL вместе с PLP в суспензии, содержащей 3 мг/мл MGL и ~30 мкМ PLP. Конечный продукт помещали в раствор SAG-маннит с 6% BSA с добавлением PLP в конечной концентрации 10 мкМ.
- RC-MGL-PLP2. Достигалось включение в эритроциты MGL вместе с PLP в суспензии, содержащей 3 мг/мл MGL и ~30 мкМ PLP. Конечный продукт помещали в раствор SAG-маннит с 6% BSA
- RC-MGL-PLP3. Достигалось включение в эритроциты MGL вместе с PLP в суспензии, содержащей 3 мг/мл MGL и ~124 мкМ PLP. Конечный продукт помещали в раствор SAG-маннит с 6% BSA.
В третьем эксперименте получали мышиный конечный продукт RC-MGL-PLP4 из новой партии метионин-гамма-лиазы следующим способом:
- RC-MGL-PLP4. Достигалось включение в эритроциты MGL вместе с PLP в суспензии, содержащей 5 мг/мл MGL и ~35 мкМ PLP. Конечный продукт помещали в раствор SAG-маннит с 6% BSA.
Наконец, в четвертом эксперименте получали мышиный конечный продукт RC-MGL-PLP5 из третьей партии метионин-гамма-лиазы следующим способом:
- RC-MGL-PLP5. Достигалось включение в эритроциты MGL вместе с PLP в суспензии, содержащей 6 мг/мл MGL и ~100 мкМ PLP. Конечный продукт помещали в раствор SAG-маннит с 6% BSA.
В приведенной ниже таблице подробно представлены гематологические и биохимические свойства трех конечных продуктов в день 0 (D0), то есть после их получения. Выход (включение в эритроциты, выраженное в процентах) в этих трех случаях был вполне удовлетворительным - от 18,6% дo 30,5%.
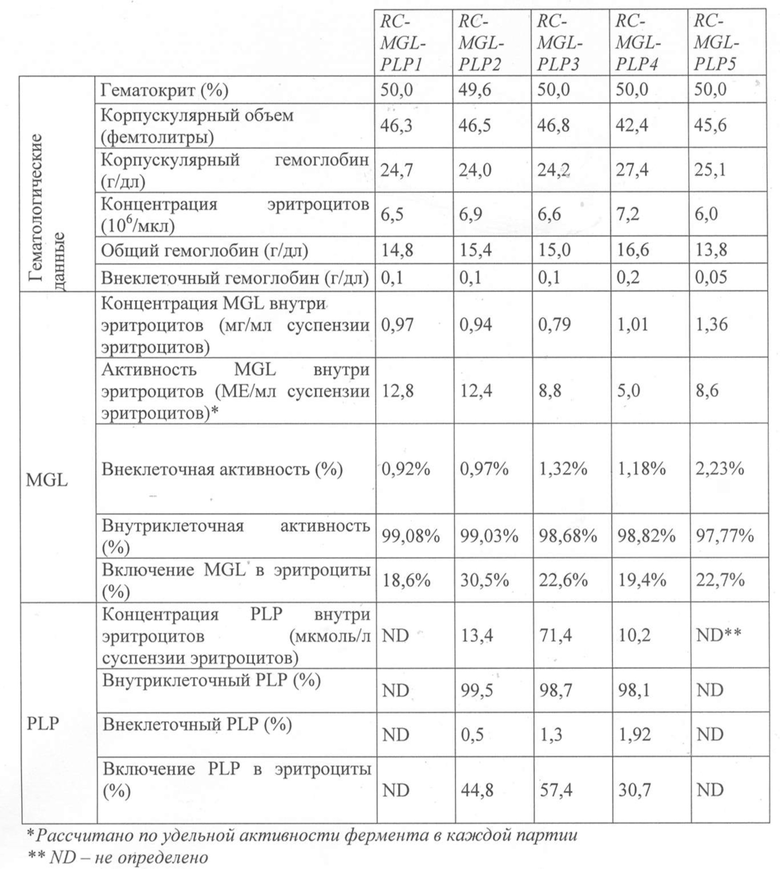
Пример 3. Получение человеческих эритроцитов с заключенными внутри них метионин-гамма-лиазой и пиридоксальфосфатом промышленным способом
Содержимое мешка с человеческой эритроцитарной массой, обедненной лейкоцитами (предоставлено “Etablissement Français du Sang”), промывали (один цикл трехкратного промывания в клеточном процессоре Cobe 2991) 0,9%-ным раствором хлорида натрия. Лиофилизованный препарат метионин-гамма-лиазы (MGL) ресуспендировали в 0,7%-ном растворе хлорида натрия и добавляли полученный раствор к суспензии эритроцитов так, чтобы получить конечную суспензию с гематокритом 70%, содержащую 3 мг/мл MGL и около 30 мкМ пиридоксальфосфата (из препарата MGL). Эту суспензию гомогенизировали и проводили инкапсулирование способом, описанным в Европейском патенте № 1 773 452. После восстановления закрытого состояния пор в клеточной мембране эритроцитов суспензию инкубировали в течение 3 часов при комнатной температуре, чтобы избавиться от наиболее хрупких эритроцитов. Затем суспензию промывали три раза 0,9%-ным раствором хлорида натрия, содержащим 0,2% глюкозы (с помощью клеточного процессора Cobe 2991) и ресуспендировали в 80 мл консервирующего раствора (AS-3). Уровень заключенной внутри эритроцитов метионин-гамма-лиазы определяли, как описано в примере 4.
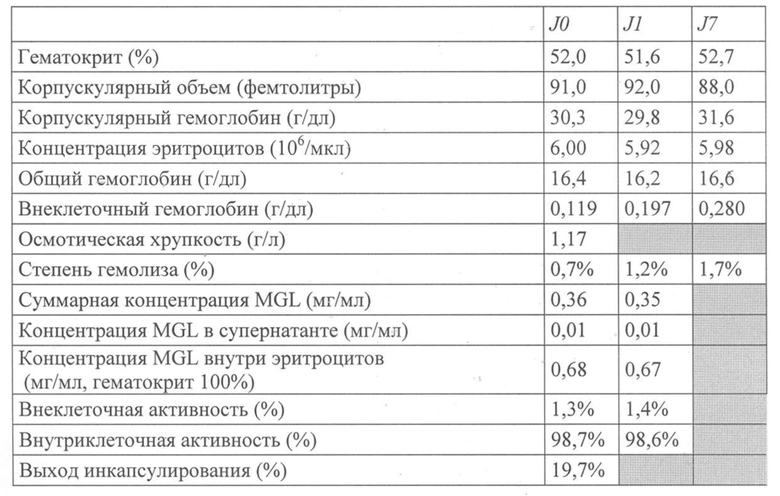
Пример 4. Определение метионин-гамма-лиазы, заключенной внутри эритроцитов
Определение метионин-гамма-лиазной активности в клеточной суспензии (суммарные эритроциты) и в супернатанте основывается на измерении количества NH3, образующегося в результате действия этого фермента. Аммиак определяют непосредственно путем ферментативной реакции (с помощью глутаматдегидрогеназы), используя набор производства Roche Diagnostics (11877984).
Получение стандартных кривых
Определяли активность метионин-гамма-лиазы при различных ее концентрациях в составе эритроцитов (суммарную в клеточной суспензии и в супернатанте) либо в водном растворе.
- Для получения стандартной кривой ферментативной активности в водном растворе готовили растворы метионин-гамма-лиазы концентрациями от 0 до 12 мкг/мл в фосфатном буферном растворе (100 мМ; рН 7,2) в присутствии 20 мкМ пиридоксальфосфата;
- Для получения стандартной кривой ферментативной активности метионин-гамма-лиазы в суммарной суспензии эритроцитов 10 мкл эритроцитарной массы, обедненной лейкоцитами, подвергали лизису в 90 мкл раствора, содержащего 260 мкМ пиридоксальфосфата и метионин-гамма-лиазу в концентрации от 0 to 100 мкг/мл. Затем полученную «стандартную» суммарную суспензию разводили в 20 раз фосфатным буферным раствором (100 мМ; рН 7,2).
- Для получения стандартной кривой ферментативной активности метионин-гамма-лиазы в супернатанте суспензии эритроцитов 10 мкл супернатанта эритроцитарной массы, обедненной лейкоцитами, подвергали лизису в 50 мкл раствора, содержащего 6,4 мкМ пиридоксальфосфата и метионин-гамма-лиазу в концентрации от 0 до 20 мкг/мл.
Предварительная обработка образцов
Предназначенные к определению ферментативной активности образцы (10 мкл) предварительно обрабатывали так же, как стандартные (добавляли пиридоксальфосфат и разводили так же, но не добавляли метионин-гамма-лиазу).
Определение метионин-гамма-лиазы
Стандартные (STD) либо анализируемые образцы (7,5 мкл) вносили в лунки планшета, прозрачного для ультрафиолетового излучения. Для того, чтобы удалить эндогенный аммиак в образцах в лунки добавляли 94 мкл реагента R1 и 56 мкл реагента R2 из указанного выше набора производства Roche Diagnostics, содержащие α-кетоглутарат в буферном растворе, восстановленный никотинамидадениндинуклеотидфосфат (NADPH) и глутаматдегидрогеназу (GLDH). Инкубировали 10 минут, после чего прибавляли 75 мкл L-метионина в концентрации 78,3 мM и реакционную смесь инкубировали в течение 30 минут. Превращение NADPH в NADP+ непрерывно отслеживали, измеряя оптическую плотность на длине волны 340 нм. Для стандартных и анализируемых образцов рассчитывали величину ΔOD/мин по линейному участку кривых оптической плотности при 340 нм. Строили калибровочную кривую ΔOD/мин = f (концентрация или активность MGL в стандартных образцах). Используя параметры регрессии, определяли концентрацию метионин-гамма-лиазы в анализируемых образцах. Результат выражали в мг/мл или в МЕ/мл (удельную активность MGL определяли для каждой партии). Уровень MGL внутри эритроцитов рассчитывали по формуле
[MGL]внутри эритроцитов =
= ([MGL]суммарная - ([MGL]в супернатанте x (1- гематокрит/100)) / (гематокрит/100).
Пример 5. Определение пиридоксальфосфата в образцах крови методом высокоэффективной жидкостной хроматографии
Определение пиридоксальфосфата в суммарной клеточной суспензии и в ее супернатанте проводили способом, представляющим собой модификацию метода, описанного в работе Van de Kamp et al., Nutrition Research 15, 415-422, 1995. Определение осуществляли методом обращено-фазовой высокоэффективной жидкостной хроматографии (RP-HPLC), используя хроматографическую систему высокого давления Shimadzu UFLC) с флуориметрической детекцией (флуориметр RF-10AX; длина волны возбуждения 300 нм, длина волны испускания 400 нм). Содержащийся в образцах пиридоксальфосфат экстрагировали трихлоруксусной кислотой (TCA) в конечной концентрации 6%. Центрифугировали при 15 000 g в течение 10 минут, собирали супернатант и разводили в подвижной фазе А. Наносили 50 мкл образца на колонку 5 мкм C-18 Gemini 250x4,6 мм (Phenomenex). Подвижная фаза А состояла из : 33 мМ однозамещенного фосфата калия, 8 мМ 1-октансульфоната натрия с добавлением бисульфита натрия (0,5 г/л) для усиления сигнала от пиридоксальфосфата; подвижная фаза В состояла из: 33 мМ однозамещенного фосфата калия, 17% (объем/объем) 2-пропанола. Использовали градиент от 100% подвижной фазы А с постепенным увеличением доли подвижной фазы В (доля В увеличивалась от 0% до 8% за промежуток времени 8 минут). Скорость потока через колонку поддерживали на уровне 1 мл/мин. Концентрацию пиридоксальфосфата в анализируемых образцах определяли с внешним стандартом (пиридоксальфосфат в различных концентрациях подвергали такой же обработке трихлоруксусной кислотой, что и анализируемые образцы). Время удерживания для пиридоксальфосфата около 3,4 минут. Уровень пиридоксальфосфата внутри эритроцитов рассчитывали по формуле
[PLP]внутри эритроцитов =
= ([PLP]суммарная - ([PLP]в супернатанте x (1- гематокрит/100)) / (гематокрит/100).
Пример 6. Возрастание уровня пиридоксальфосфата внутри эритроцитов в результате включения в них вместе с метионин-гамма-лиазой
С суспензией мышиных эритроцитов проводили включение в них метионин-гамма-лиазы и пиридоксальфосфата, как описано в Примере 2. Определение внутриклеточного пиридоксальфосфата осуществляли способом, описанным в Примере 5.
С суспензией человеческих эритроцитов проводили включение в них метионин-гамма-лиазы и пиридоксальфосфата, как описано в Примере 3. Перед этапом инкубации при комнатной температуре отбирали в качестве образца часть человеческих эритроцитов, содержащих метионин-гамма-лиазу и пиридоксальфосфат (RC-MGL-PLP), чтобы определить уровень внутриклеточного пиридоксальфосфата способом, описанным в Примере 5. В приведенной ниже таблице сравниваются физиологические уровни пиридоксальфосфата в человеческих или мышиных эритроцитах с уровнями, достигнутыми в результате заключение пиридоксальфосфата внутрь эритроцитов вместе с метионин-гамма-лиазой.
эритроциты
эритроциты
(Natta &
Reynolds)
(Fonda)
RC-MGL-PLP1 или
RC-MGL-PLP2
• 3 мг/мл MGL
~• 30 µM PLP
в RC-MGL-PLP3
• 3 мг/мл MGL
~• 125 мкM PLP
* Рассчитывали следующим образом: 7,5 нмоль/г гемоглобина (Hb) ~2,4 мкM (при средней корпускулярной концентрации гемоглобина 32 г/дл).
Пример 7. Увеличение концентрации пиридоксальфосфата в эритроцитах с заключенной в них метионин-гамма-лиазой при инкубации с пиридоксином in vitro
Суспензию человеческих эритроцитов подвергали процедуре заключения в них метионин-гамма-лиазы способом, описанным в примере 3. Перед этапом 3-часовой инкубации отбирали часть эритроцитов, разделяли это количество на три порции и инкубировали с пиридоксином (объем/объем) в разных концентрациях (0 мM, 2 мM и 4 мM). После гомогенизации эти суспензии инкубировали при комнатной температуре. Через 3 часа и через 24 часа инкубации образцы клеточных суспензий и супернатантов (полученных путем центрифугирования суспензий при 1000 g при температуре 4°C в течение 10 минут) замораживали для определения концентрации пиридоксальфосфата путем высокоэффективной жидкостной хроматографии, как описано в Примере 5.
Полученные результаты представлены на фиг. 2 и 3.
В отсутствие пиридоксина внутриклеточный уровень пиридоксальфосфата составляет 3,9 мкM (этот пиридоксальфосфат оказывается в клетках в результате включения в них метионин-гамма-лиазы вместе с пиридоксальфосфатом). Эта концентрация пиридоксальфосфата не изменяется спустя 3 часа и 24 часа инкубации. Некоторое снижение концентрации пиридоксальфосфата отмечается через 24 часа и сопровождается наличием внеклеточного пиридоксальфосфата, что можно объяснить гемолизом в конце инкубации.
В присутствии пиридоксина (при его концентрации 2 мМ или 4 мМ) эритроциты, содержащие метионин-гамма-лиазу, обогащаются пиридоксальфосфатом: через 3 часа инкубации его внутриклеточная концентрация возрастает в 2 раза, достигая около 8 мкМ, а через 24 часа инкубации - почти в 3 раза, причем зависимым от концентрации пиридоксина образом (11 мкМ пиридоксальфосфата при концентрации пиридоксина 2 мМ и 14 мкМ - при 4 мМ). Эти результаты показывают, что инкубация с пиридоксином суспензии эритроцитов, содержащих инкапсулированный фермент, зависимый от пиридоксальфосфата, приводит к увеличению внутриклеточного уровня пиридоксальфосфата и этот увеличенный уровень сохраняется продолжительное время.
Пример 8. Фармакокинетические показатели для эритроцитов с заключенными в них метионин-гамма-лиазой и пиридоксальфосфатом у мышей
В мышиные продукты RC-MGL-PLP2, RC-MGL-PLP3, RC-MGL-PLP4 и RC-MGL-PLP5 включали интернализируемую флуоресцентную метку CFSE и вводили их внутривенно мышам линии CD1. Спустя различные промежутки времени (D0 + 15 минут, D1, D2, D5 для трех из указанных продуктов, для продукта RC-MGL-PLP4 еще дополнительно D14 и D28, для продукта RC-MGL-PLP5 дополнительно D14), животных умерщвляли и собирали кровь в пробирки с гепаринатом лития, которые держали в темноте при температуре +4°C для определения фармакокинетических показателей. Долю эритроцитов, помеченных CFSE, в цельной крови определяли методом проточной цитометрии. Цельную кровь в количестве 5 мкл разводили в 1 мл PBS, содержащего 0,5% BSA, и каждый образец анализировали в трех параллельных пробах (10 000 клеток, канал FL-1; цитометр FC500, Beckman Coulter). Определяли выживаемость эритроцитов с заключенной в них метионин-гамма-лиазой, складывая долю эритроцитов, меченных CFSE в различные моменты времени, с долей эритроцитов, меченных CFSE в момент T0 + 15 минут (100% контроль). По полученным значениям (в процентах) строили график (см. фиг. 4), который иллюстрирует изменение доли эритроцитов с заключенной в них метионин-гамма-лиазой в кровотоке с течением времени.
Результаты определения доли эритроцитов, меченных CFSE, в кровотоке в различные моменты времени показывают, что четыре испытывавшихся продукта обладают превосходной стабильностью in vivo (у мышей): до 120 часов после введения в организм (выживаемость составила 83,5 ± 0,6 %; 94,7 ± 0,6 %; 87,3 ± 5,6 % и 76,8 ± 1,3 % соответственно). Для продукта RC-MGL-PLP4 фармакокинетическое исследование, проведенное через 29 суток, показало, что время полужизни эритроцитов с заключенной в них метионин-гамма-лиазой составляет около 12,6 суток.
Пример 9. Снижение уровня L-метионина через 24 часа
Мышиные продукты RC-MGL-PLP1, RC-MGL-PLP2 и RC-MGL-PLP3, полученные и охарактеризованные, как описано в примере 2, вводили внутривенно мышам линии CD1 в дозе 8 мл/кг массы тела. Через 6 часов после инъекции животным, получившим RC-MGL-PLP2, вводили около 0,09 мг пиридоксина (150 мкл раствора пиридоксина гидрохлорида концентрацией 2,9 мМ). Через 24 часа определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS) (Piraud M. et al., Rapid Commun. Mass Spectrum. 19, 3287-97, 2005). В приведенной ниже таблице представлены данные по снижению уровня L-метионина у различных групп мышей.
L-метионина в плазме крови (мкM)
• PLP в конечном продукте
(~5 мкмоль/л эритроцитов)
• PLP в консервирующем растворе конечного продукта (10 мкM)
пиридоксин
• PLP, заключенный в конечном продукте
(~13,4 мкмоль/л эритроцитов)
• Внутривенная инъекция ~0,09 мг пиридоксина
• PLP, заключенный в конечном продукте (~71,4 мкмоль/л эритроцитов)
У контрольных особей уровень L-метионина в плазме крови составил 82,7 ± 22,5 мкМ. Продукт RC-MGL-PLP1, содержащий метионин-гамма-лиазу с низкой концентрацией пиридоксальфосфата, вызывал снижение уровня L-метионина в плазме крови на 44% (через 24 часа после введения). Как мы полагаем, пиридоксальфосфат, добавленный в консервирующий раствор продукта, не доступен для метионин-гамма-лиазы, поскольку 1) он по большей части связан с бычьим сывороточным альбумином, содержащимся в консервирующем растворе, и 2) он не может пройти через клеточную мембрану эритроцитов.
Полученные результаты показывают, что более целенаправленное обеспечение эритроцитов пиридоксальфосфатом - путем внутривенного введения пиридоксина (RC-MGL-PLP2) или путем включения пиридоксальфосфата в них в большей концентрации дает возможность достигнуть в ~1,5 раза большего снижения уровня L-метионина (на 73% и на 64% соответственно).
Пример 10. Фармакодинамические показатели для эритроцитов с заключенной в них метионин-гамма-лиазой
Мышам линии CD1 вводили внутривенно метионин-гамма-лиазу в свободном виде в дозе 8 мл/кг массы тела. Было сделано две серии инъекций: в первой серии концентрация фермента была 0,45 мг/мл (продукт MGL-L1), во второй - вдвое выше (0,90 мг/мл; продукт MGL-L2). Через 6 часов после введения фермента животным, получившим MGL-L1 и MGL-L2, вводили (также путем инъекции) около 0,09 мг пиридоксина (150 мкл раствора пиридоксина гидрохлорида концентрацией 2,9 мМ). Определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS) через 15 минут, 24 часа, 48 часов и 120 часов после введения MGL-L1 и через 15 минут, 24 часа, 48 часов, 120 часов и 144 часа после введения MGL-L2. На фиг. 5 представлены результаты этого определения для разных групп подопытных мышей.
Полученные результаты показывают, что в обеих группах животных имело место значительное (≈4 мкM) и быстрое (через 15 минут после введения) снижение уровня L-метионина. Однако это снижение преходящее и не сохраняется во времени: уровни L-метионина возвращались к контрольным значениям через 24 часа после введения продукта несмотря на изначальное присутствие пиридоксаль-5-фосфата (присутствовавшего в разбавляющем буферном растворе, а также в препарате фермента, разведенном водой) и добавку витамина В6, как в случае продукта RC-MGL-PLP2. Таким образом, активность свободной метионин-гамма-лиазы утрачивается в период начиная через 15 минут и кончая 24 часами после введения - вероятно, из-за быстрого исчезновения фермента из кровотока.
На втором этапе эксперимента мышам линии CD1 вводили внутривенно мышиные продукты RC-MGL-PLP1, RC-MGL-PLP2 и RC-MGL-PLP3, полученные и охарактеризованные, как описано в Примере 2, в дозе 8 мл/кг массы тела. Через 6 часов после инъекции животным, получившим RC-MGL-PLP2, вводили около 0,09 мг пиридоксина (150 мкл раствора пиридоксина гидрохлорида концентрацией 2,9 мМ). Через 15 минут, 24, 48 и 120 часов определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS). Результаты этого определения у разных групп подопытных мышей представлены на фиг. 6 и в приведенной ниже таблице.
Полученные результаты показывают, что в обеих группах мышей снижение уровня L-метионина в плазме крови стабилизировалось на уровне ~35 мкM и не изменялось со временем (в период от 48 дo 120 часов после введения продукта). Это свидетельствует, что у мышей добавка пиридоксальфосфата или его предшественника (витамина В6) позволяет поддерживать активность метионин-гамма-лиазы, заключенной в эритроцитах, в течение по меньшей мере 120 часов после введения. В приведенной ниже таблице представлены значения концентрации L-метионина в плазме крови через 24 и 120 часов после введения различных продуктов (свободной метионин-гамма-лиазы либо заключенной внутри эритроцитов с пиридоксальфосфатом).
Чтобы получить картину фармакодинамики за промежуток времени более 120 часов, мышам линии CD1 вводили внутривенно мышиный продукт RC-MGL-PLP4 с концентрацией метионин-гамма-лиазы, заключенной в эритроцитах, 0,5 мг/мл конечного продукта в дозе 10 мл/кг массы тела. Животным, получившим RC-MGL-PLP4, через 6 часов после этой инъекции вводили около 0,09 мг пиридоксина (150 мкл раствора пиридоксина гидрохлорида концентрацией 2,9 мМ). Определяли уровень L-метионина в плазме крови путем высокоэффективной жидкостной хроматографии-тандемной масс-спектрометрии (HPLC-MS-MS). На фиг. 7 представлены данные, полученные через 15 минут и в моменты D1, D2, D5, D14 и D28 после инъекции. Полученные результаты демонстрируют значительное (≈10 мкM по сравнению с 68 мкМ в контроле) и быстрое (через 15 минут после введения) снижение уровня L-метионина. Однако это снижение в период от 24 до 48 часов после инъекции несколько стабилизировалось на величине около 25 мкМ и возрастало до около 40 мкМ через 5 суток, достигая в конце концов контрольных значения через около 12 суток после введения RC-MGL-PLP4.
Наконец, определяли остаточную активность введенной метионин-гамма-лиазы способом, описанным в примере 4, в присутствии пиридоксальфосфата. На фиг. 8 представлены данные об остаточной активности в зависимости от времени для свободного фермента MGL-L2, различных продуктов RC-MGL-PLP и для тирозин-фенол-лиазы (по литературным источникам). Эти результаты показывают, что путем включения метионин-гамма-лиазы в эритроциты возможно сохранить высокую ферментативную активность в течение 24 часов (остаточная активность составляет от около 60% до 100%). Остаточная активность фермента несколько снижается через 48 часов (составляет от около 35% до 100%) и на протяжении 120 часов, то есть пяти суток после введения фермента, держится на уровне от около 20% до около 65%. Остаточная активность свободной метионин-гамма-лиазы резко падает в первые минуты после введения, так что через 24 часа она практически нулевая (остаточная активность <10%). Для сравнения заметим, что остаточная активность тирозин-фенол-лиазы, введенной в свободном виде, отмечается на графике и через 5 часов после введения, составляя самое большее 37% (Elmer et al., 1978). Определение остаточной ферментативной активности с очевидностью демонстрирует преимущество заключения ферментов, зависимых от пиридоксальфосфата, внутрь эритроцитов для сохранения их активности.
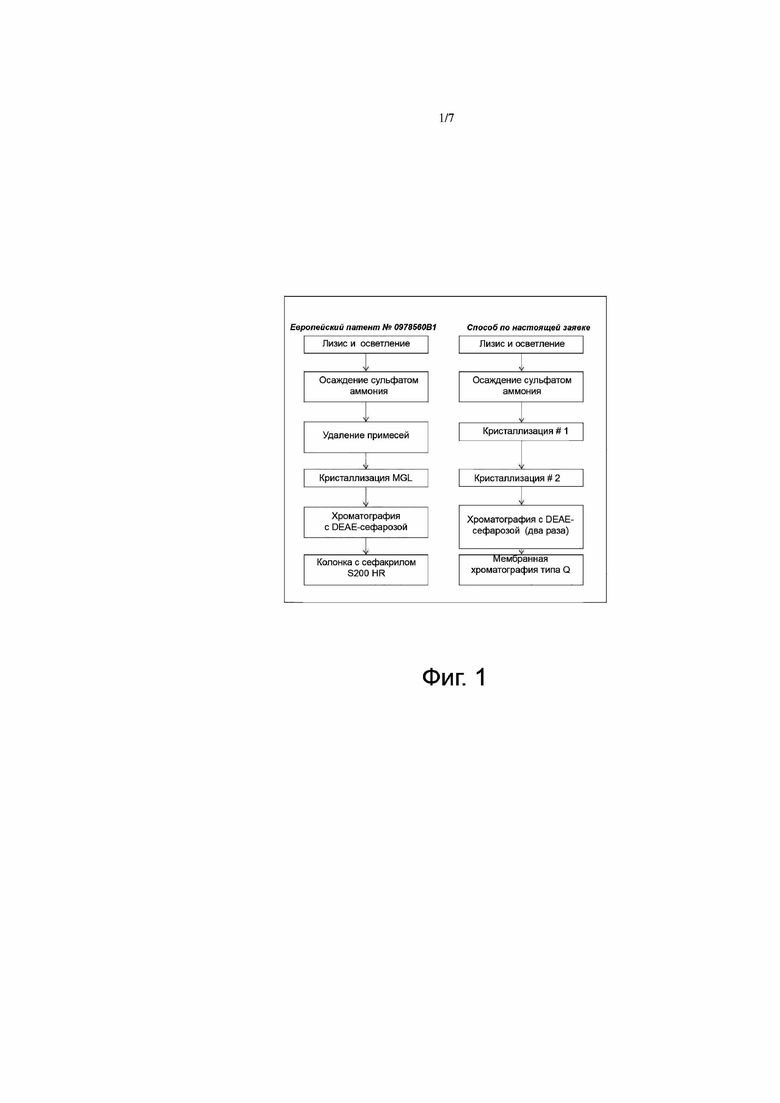

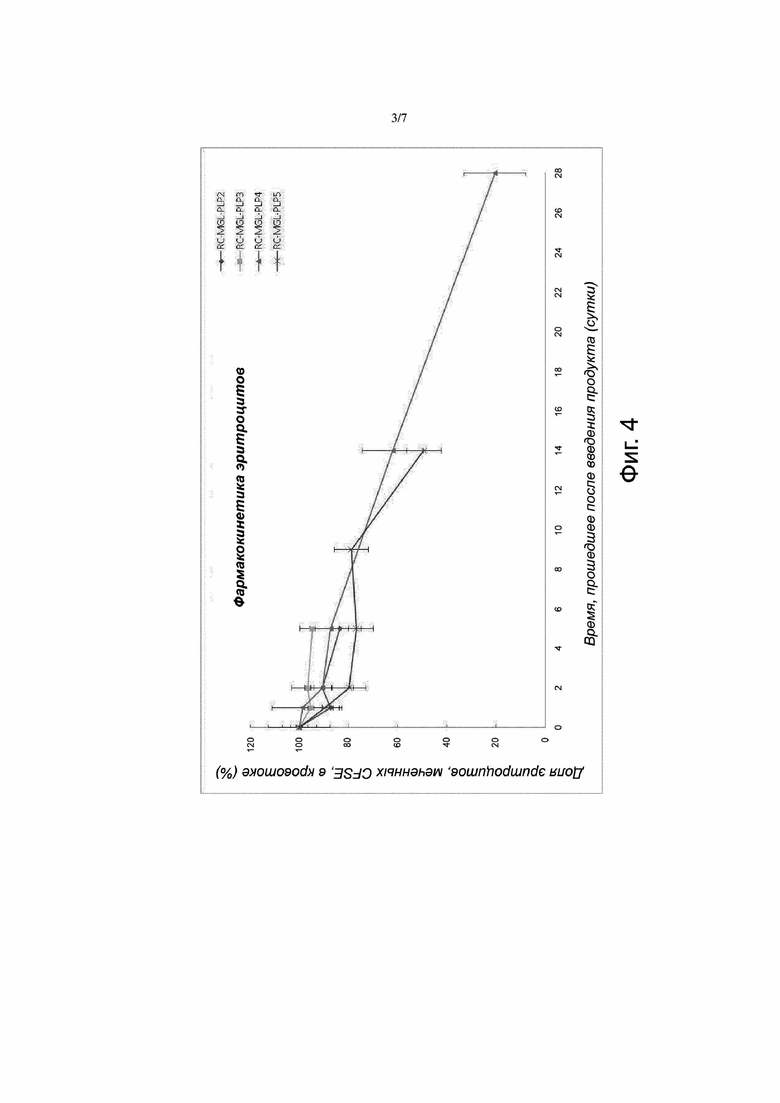
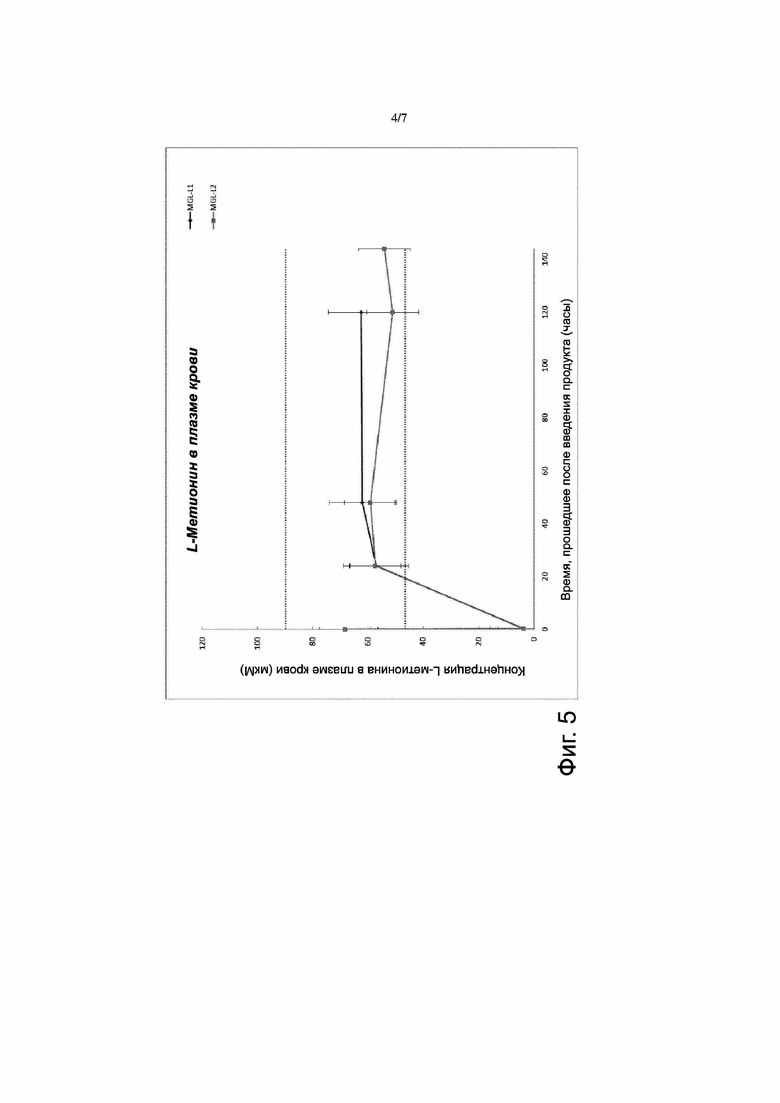
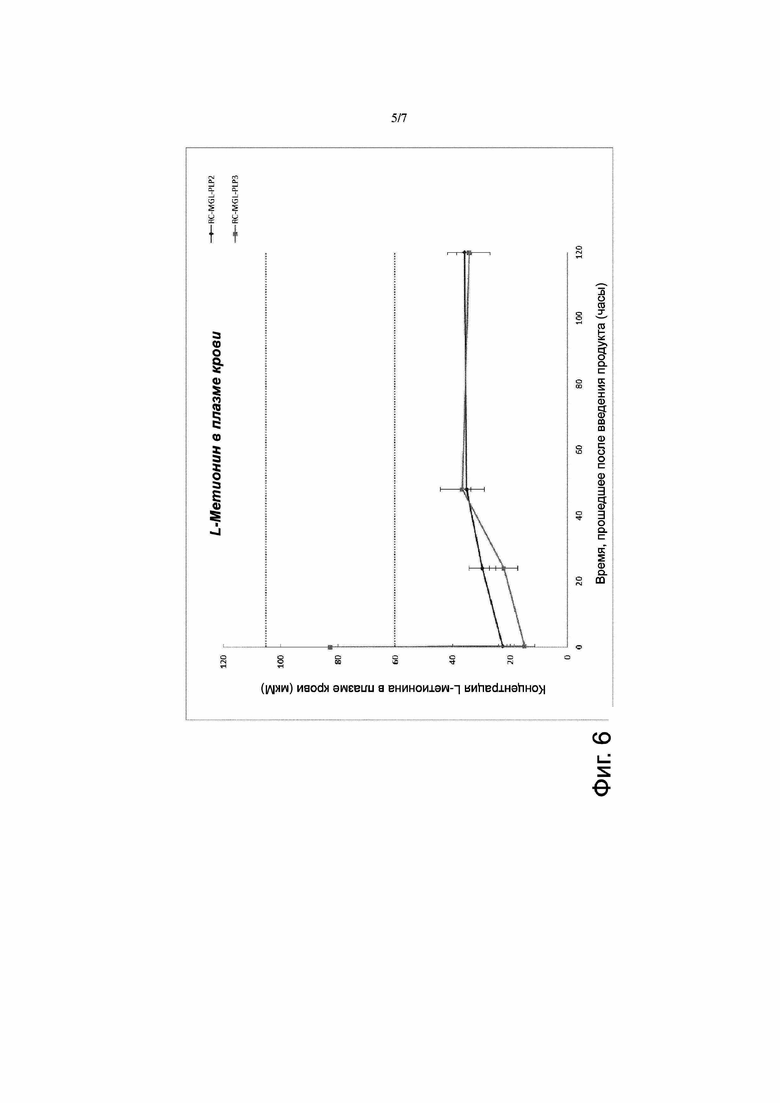
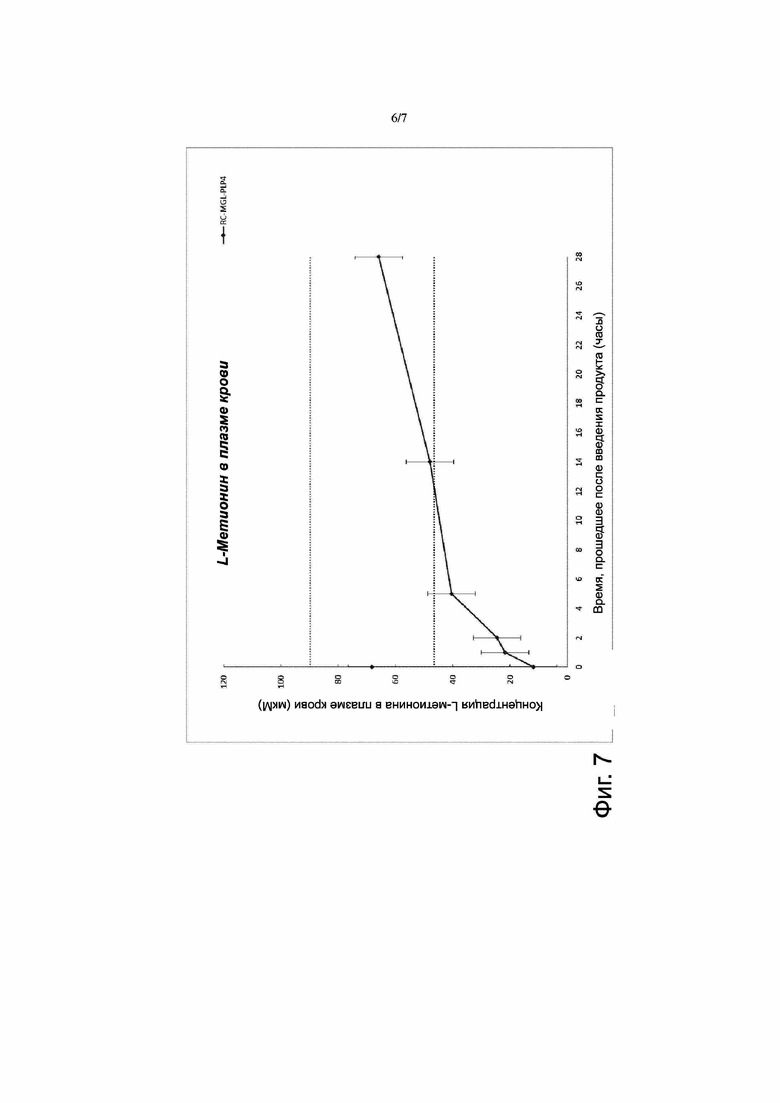
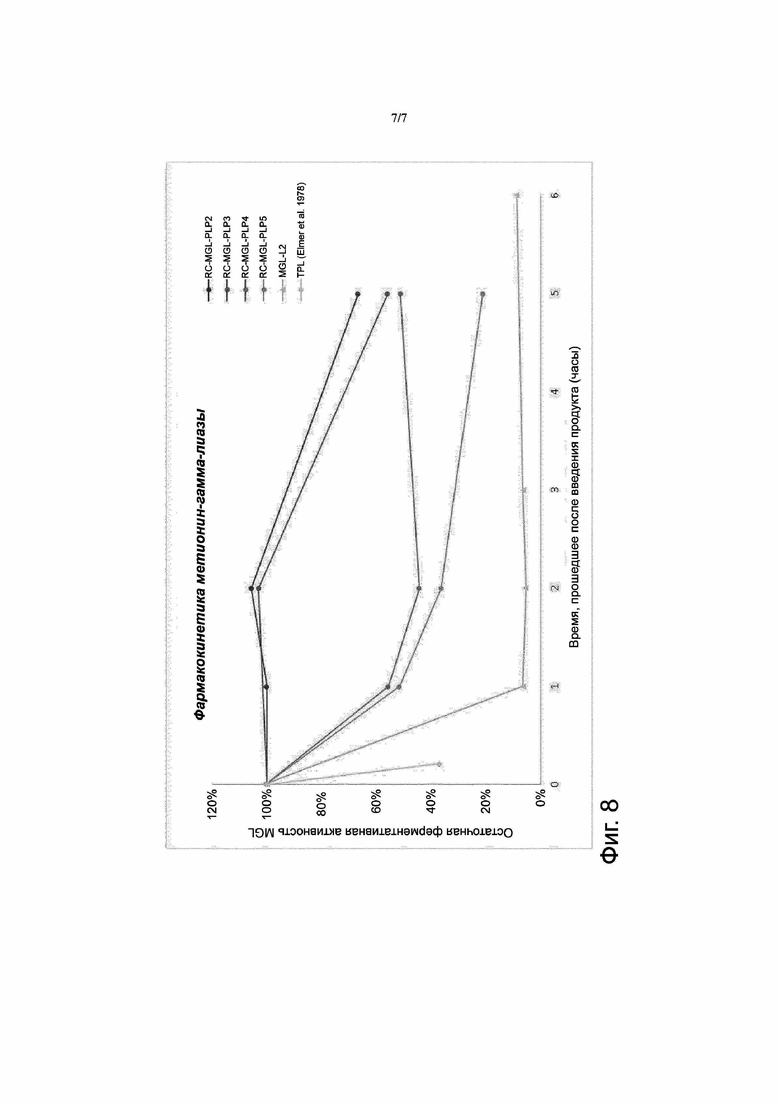
Группа изобретений относится к фармацевтике. Комбинация для уменьшения в плазме или кровотоке метионина, гомоцистеина или тирозина у пациента содержит инъецируемую фармацевтическую суспензию, обеспечивающую длительную ферментативную активность, зависимую от пиридоксальфосфата (PLP), содержащую эритроциты и фармацевтически приемлемый носитель; где в эритроцитах инкапсулировано множество молекул PLP-зависимого фермента и содержится PN-киназа и PNP-оксидаза в количестве, достаточном для производства пиридоксальфосфата (PLP) из предшественника PLP, присутствующего в кровотоке, в количестве, достаточном для поддержания достаточной части молекул PLP-зависимого фермента в его голоферментных формах для сохранения ферментативной активности через 24 ч после введения; композицию, содержащую достаточное количество фармацевтически приемлемого витамина В6, предшественника PLP, подходящего для введения пациенту, где комбинация представлена в форме набора или композиции. Также раскрыт терапевтический способ лечения для уменьшения в кровотоке метионина, гомоцистеина или тирозина у пациента. Группа изобретений обеспечивает ферменты доступным пиридоксальфосфатом в оптимальной для них концентрации. 2 н. и 19 з.п. ф-лы, 8 табл., 10 пр., 8 ил.
1. Комбинация для истощения или уменьшения в плазме или кровотоке субстрата, выбранного из метионина, гомоцистеина и тирозина у пациента или субъекта, нуждающегося в этом, содержащая:
(а) инъецируемую фармацевтическую суспензию, которая обеспечивает длительную ферментативную активность, зависимую от пиридоксальфосфата (PLP), пациенту или субъекту, нуждающемуся в этом, содержащую эритроциты и фармацевтически приемлемый носитель, причем в эритроцитах инкапсулировано множество молекул PLP-зависимого фермента и содержится PN-киназа и PNP-оксидаза в количестве, достаточном для производства пиридоксальфосфата (PLP) из предшественника PLP, присутствующего в кровотоке пациента или субъекта, в количестве, достаточном для поддержания достаточной части молекул PLP-зависимого фермента в его голоферментных формах для сохранения ферментативной активности через 24 часа после введения; где суспензия необязательно содержит консервирующий раствор, содержащий неорганический фосфат, для ингибирования ферментативной активности PLP-фосфатазы внутри эритроцитов;
(б) композицию, содержащую достаточное количество фармацевтически приемлемого витамина В6, предшественника PLP, подходящего для введения пациенту или субъекту,
где комбинация представлена в форме набора или композиции.
2. Комбинация по п.1, которая необязательно содержит PLP, инкапсулированный в эритроцитах вместе с молекулами PLP-зависимого фермента.
3. Комбинация по любому из пп.1 и 2, где суспензия дополнительно содержит PLP или предшественник PLP, инкапсулированный в эритроцитах и/или присутствующий вне указанных эритроцитов, и где, когда он присутствует, количество инкапсулированного предшественника PLP является эффективным для поддержания инкапсулированного PLP-зависимого фермента в его голоферментной форме дольше относительно того же PLP-зависимого фермента, не инкапсулированного в эритроцитах.
4. Комбинация по п.3, где предшественник PLP представляет собой нефосфатный предшественник, выбранный из пиридоксаля, пиридоксина, пиридоксамина и их комбинаций.
5. Комбинация по п.3, в которой предшественник PLP, присутствующий в суспензии, является предшественником фосфата.
6. Комбинация по п.5, в которой предшественник фосфата, присутствующий в суспензии, содержит пиридоксинфосфат (PNP) и/или пиридоксаминфосфат (PMP).
7. Комбинация по любому из пп.1 и 2, включающая от 0,01 до 30 мг PLP-зависимого фермента на мл эритроцитов.
8. Комбинация по п.7, включающая от 0,05 до 10 мг PLP-зависимого фермента на мл эритроцитов.
9. Комбинация по п.3, включающая от 0,05 до 600 мкмоль PLP и/или PNP и/или PMP на литр эритроцитов.
10. Комбинация по любому из пп.1 и 2, где суспензия дополнительно содержит пиридоксинкиназу (PN-киназу), пиридоксинфосфатоксидазу (PNP оксидазу) и/или агент, ингибирующий PLP-фосфатазу.
11. Комбинация по любому из пп.1 и 2, где PLP-зависимый фермент представляет собой метиониназу (METase), тирозин-феноллиазу (TPL), тирозин-аминотрансферазу (TATase) или цистатионин-бета-синтазу (CBS).
12. Комбинация по любому из пп.1 и 2, в которой суспензия содержит консервирующий раствор, содержащий неорганический фосфат для ингибирования ферментативной активности PLP-фосфатазы внутри эритроцитов.
13. Комбинация по любому из пп.1 и 2, в которой суспензию получают путем открытия пор в мембранах эритроцитов, контактирования эритроцитов с PLP-зависимым ферментом до, во время и/или после открытия пор, обеспечения проникновения фермента в эритроциты, закрытия пор эритроцитов, содержащих фермент, введения энзимсодержащих эритроцитов в буфер для образования первой суспензии, необязательно инкубации первой суспензии для обеспечения лизиса наиболее хрупких эритроцитов, удаления лизированных эритроцитов и суспендирования эритроцитов в консервирующем растворе, получая таким образом суспензию.
14. Комбинация по п.13, где предшественником PLP является пиридоксин.
15. Терапевтический способ лечения для истощения или уменьшения в плазме крови или в кровотоке субстрата, выбранного из метионина, гомоцистеина и тирозина, у пациента или субъекта, нуждающегося в этом, включающий введение указанному пациенту или субъекту по меньшей мере одной дозы эффективного количества комбинации по любому из пп.1-14.
16. Способ по п.15, включающий введение эффективного количества нефосфатной формы витамина В6, где суспензию и нефосфатную форму витамина В6 вводят отдельно или последовательно; и где введение 1) эффективного количества витамина B6 и эффективного количества инкапсулированного PLP-зависимого фермента обеспечивает значительно большую остаточную активность PLP-зависимого фермента, чем 2) введение того же количества витамина B6 и того же количества неинкапсулированной свободной формы PLP-зависимого фермента.
17. Способ по п.15, в котором PLP-зависимый фермент представляет собой метиониназу (METазу), тирозинфеноллиазу (TPL), тирозин аминотрансферазу (TATазу) или цистатионин бета-синтазу (CBS); и
при этом, когда фермент представляет собой METазу, способ истощает или снижает гомоцистеин в плазме крови, или в кровотоке, или в печени у пациента или субъекта, нуждающегося в этом;
при этом, когда фермент представляет собой TPL или TATазу, способ с помощью TPL истощает или снижает тирозин в плазме крови или в кровотоке у пациента или субъекта, нуждающегося в этом; а также
при этом, когда фермент представляет собой CBS, способ истощает или снижает гомоцистеин в плазме крови, или в кровотоке, и/или в печени у пациента, страдающего от гомоцистинурии и/или гипергомоцистинурии и/или связанных патологий.
18. Способ по п.17, в котором пациент или субъект, нуждающийся в этом, страдает от рака.
19. Способ по п.18, где указанный рак включает опухолевые клетки, ауксотрофные по метионину или тирозину, и PLP-зависимый фермент представляет собой METазу, TPL, TATазу или комбинации TPL и TATазы.
20. Способ по п.15, в котором пациент или субъект страдает синдромом Ричнера-Ханхарта, а PLP-зависимый фермент представляет собой ТАТазу.
21. Способ по п.15, где пациент или субъект страдает от гомоцистинурии и/или гипергомоцистинурии, и PLP-зависимый фермент представляет собой CBS.
| Agrawal V | |||
| et al | |||
| Targeting methionine auxotrophy in cancer: discovery & exploration // Expert Opin Biol Ther | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| СПОСОБ СНИЖЕНИЯ КОНЦЕНТРАЦИИ АММИАКА В КРОВИ С ПОМОЩЬЮ АММОЦИТОВ И ИНКАПСУЛИРОВАННОЙ ГЛУТАМИНСИНТЕТАЗЫ | 2007 |
|
RU2362572C2 |
| WO 2013139906 A1, 26.09.2013 | |||
| Рукавный фильтр с импульсной регенерацией | 1990 |
|
SU1773452A1 |

