Изобретение относится к медицине, а именно к иммунологии и онкологии, клеточным биотехнологиям, способам усиления противоопухолевого Т-клеточного иммунитета при раке легкого.
Одна из важнейших функций иммунной системы заключается в том числе, в уничтожении опухолевых клеток. Несмотря на то, что каждый тип опухолей имеет существенные особенности развития, в патогенезе многих, особенно злокачественных видов опухолей, отмечен такой общий механизм развития и прогрессии как подавление противоопухолевой активности иммунной системы. В частности, под влиянием опухоли и опухолевого микроокружения снижается функциональная активность и количество Т-лимфоцитов. Так, у больных раком легкого количество и пролиферативная активность CD8+ Т-лимфоцитов обычно снижены по сравнению со здоровыми людьми [Sathaliyawala Т., et al., 2013]. В этой связи, повышение уровня CD8+ Т-клеток после лечения рассматривается как потенциальный предиктор лучшей выживаемости у пациентов с раком легкого [Liu Q. et al., 2021]. Кроме того, у CD8 Т-лимфоцитов при взаимодействии с опухолью снижается активность метаболических путей, и они приобретают гипореактивные фенотипы, которые не могут быть восстановлены простой стимуляцией [Speiser D.E. et al., 2016].
Итак, истощение эффекторных Т-лимфоцитов с одновременным снижением их цитостатической активности лежит в основе снижения противоопухолевого иммунного ответа при раке легкого. Для повышения его эффективности необходима разработка подходов, которые могут преодолеть ограничения, возникающие у Т-лимфоцитов.
Адекватного прототипа в проанализируемой нами литературе не обнаружено.
Новый технический результат - повышение противоопухолевого иммунитета и подавление роста опухоли легкого за счет увеличения специфичной цитотоксичности и устойчивости к действию опухолевых клеток CD8+ Т-лимфоцитов, индуцированных метаболическим репрограммированием.
Для достижения нового технического результата в способе усиления противоопухолевого Т-клеточного иммунитета при раке легкого, реципиенту внутривенно вводят репрограммированные CD8 позитивные Т-лимфоциты, которые готовят in vitro следующим образом: к наивным CD8 позитивным Т-лимфоцитам в концентрации 1×108/мл добавляют ингибитор протеинкиназ MEK дозе 1 μM совместно с 50 мкл антигенпрезентирующей смеси, полученной на основе лизата опухолевых клеток, культивируют в течение 48 часов, далее добавляют ингибитор контрольных точек PD1 в концентрации 10 мкг/мл и культивируют в течение 2 часов.
Новым в предлагаемом изобретении является то, что предварительно до внутривенного введения проводят in vitro репрограммирование популяции наивных CD8 позитивных Т-лимфоцитов, для этого к культуре наивных CD8 позитивных Т-лимфоцитов с концентрацией 1×108 /мл добавляют смесь ингибитора протеинкиназ MEK в дозе 1 μM и 50 мкл антигенпрезентирующей смеси, полученной на основе лизата опухолевых клеток, культивируют в течение 48 часов, далее CD8 позитивные Т-лимфоциты культивируют с ингибитором контрольных точек PD1 в концентрации 10 мкг/мл в течение 2 часов.
Отличительные признаки проявили в заявляемой совокупности новые свойства явным образом не вытекающие из уровня техники в данной области и неочевидные для специалиста.
Идентичной совокупности признаков не обнаружено в проанализированной патентной и научно-медицинской литературе. Способ апробировании в экпериментальной практике.
Использование предлагаемого способа позволило обеспечить более высокую устойчивость формирующегося фенотипа CD8+ Т-лимфоцитов к цитотоксическому действию самой опухоли и целевую элиминацию опухолевых клеток.
Исходя из вышеизложенного, заявляемое изобретение соответствует критериям патентоспособности изобретения «Новизна» и «Изобретательский уровень» и «Промышленная применимость».
Рак легкого является вторым наиболее распространенным видом рака и основной причиной смертности от рака во всем мире: в 2020 году было диагностировано 2,2 миллиона новых случаев и 1,8 миллиона случаев смерти [Петрова Г.В. и др., 2019]. Высокая смертность онкологических больных обусловлена невосприимчивостью метастатических опухолевых клеток, в том числе популяции стволовых опухолевых клеток, к современным методам лечения. Поэтому необходимы новые методы лечения больных раком легкого, чтобы снизить смертность и улучшить качество их жизни.
Известно, что опухоли содержат небольшую субпопуляцию стволовых опухолевых клеток, которые поддерживают первичные опухоли и метастазы [Hochmair M., et al., 2020]. Обычная химиотерапия уничтожает большую часть опухолевых клеток, но стволовые опухолевые клетки выживают и восстанавливают опухоль, а также они ответственны за метастазы [Hochmair, M, et al., 2020]. Таким образом, возможно терапевтическое влияние на стволовые опухолевые клетки в сочетании со стандартной химиотерапией может улучшить исходы у онкологических больных и обеспечить долгосрочный ответ. Стволовые опухолевые клетки были идентифицированы в большинстве солидных опухолей. Для большинства солидных опухолей в качестве селективных маркеров была предложена экспрессия CD44, CD117, EGR, Ax1, Sox2 [Hochmair M., et al., 2020].
Популяция CD8 Т-лимфоцитов является неоднородной и состоит из популяций клеток, различающихся по продолжительности жизни, функциям и анатомическому расположению. Популяции CD8 Т-лимфоцитов различают по экспрессии поверхностных маркеров, в частности, CCR7 [Sallusto F. et al., 2004]. CD8 Т-лимфоциты, относящиеся к клеткам «центральной» Т-памяти, характеризуются повышенной экспрессией CCR7 и продукцией большого количества IL-2, устойчивым пролиферативным ответом на вторичную инфекцию, большей способностью к самообновлению и гомеостатической пролиферации (базальная пролиферация) по сравнению с клетками «эффекторной» Т-памяти [Jung Y.W. et al., 2016].
Ингибирование MEK (mitogen-activated protein kinase/митоген-активируемая протеинкиназа) представляет собой фармакологически осуществимый метод, который можно использовать для улучшения различных иммунных терапевтических подходов, включая комбинацию с праймирующими агентами, иммуномодуляторами или повышение эффективности клеточной терапии. Ингибирование пути MEKi (mitogen-activated protein kinase inhibition/ингибирование митоген-активируемой протеинкиназы) одобрено Федеральным управлением по лекарственным средствам США и используются для лечения рака (NCT02407405) [Verma, V., et al. 2021]. Признается, однако, недостаточная специфичность и частые побочные эффекты, наблюдаемые после системного введения ингибитора MEK [Herrick W.G., et al., 2021].
Одним из представителей препаратов ингибиторов контрольных точек PD1 является ниволумаб, который представляет собой иммуноглобулин G4 (IgG4) с молекулярной массой 146 кДа, он блокирует взаимодействие между рецептором программированной смерти (PD-1) и его лигандами (PD-L1 и PD-L2). PD-1 рецептор является негативным регулятором активности Т-лимфоцитов. Связывание PD-1 с лигандами PD-L1 и PD-L2, которые экспрессируют клетки опухоли или клетки микроокружения опухоли, приводит к ингибированию пролиферации Т-лимфоцитов и секреции цитокинов. Для повышения противоопухолевой активности Т-клеток нами предложено использовать моноклональные антитела к PD1 рецептору.
Способ может быть использован для создания популяции репрограммированных CD8 позитивных Т-лимфоцитов, обладающих противоопухолевой и антиметастатической активностью. Исходя из вышеизложенного, заявляемое изобретение соответствует критериям патентоспособности изобретения «Новизна» и «Изобретательский уровень» и «Промышленная применимость».
Предлагаемый способ был изучен в экспериментах на 8-10 недельных мышах линии C57BL/6 в количестве 100 штук. Животные первой категории, инбредные мыши, получены из питомника отдела экспериментального биомедицинского моделирования НИИФиРМ им. Е.Д. Гольдберга Томского НИМЦ РАН. Содержание животных и дизайн экспериментов были одобрены биоэтическим комитетом НИИФиРМ им. Е.Д. Гольдберга Томского НИМЦ.
Изобретение будет понятно из следующего описания и приложенных к нему фигур 1, 2, 3 Приложения.
На фигуре 1 изображены Т-лимфоциты, полученные с использованием клеточного мультиридера Cytation 5 и окрашенных: Hoechst (синий) для идентификации ядер клеток; CD8 FITC (зеленый) для идентификации CD8+ лимфоцитов; CCR7 AF555 (красный); (Hoechst+CD8+CCR7) составное изображение с использованием всех трех цветов. Определение процента клеток CD8+CCR7+ производят по отношению клеток, подсчитанных по зеленому и красному каналам, к общему количеству клеток, подсчитанных по синему (DAPI) каналу. Все масштабные линейки имеют размер 1000 мкм. Увеличение ×4.
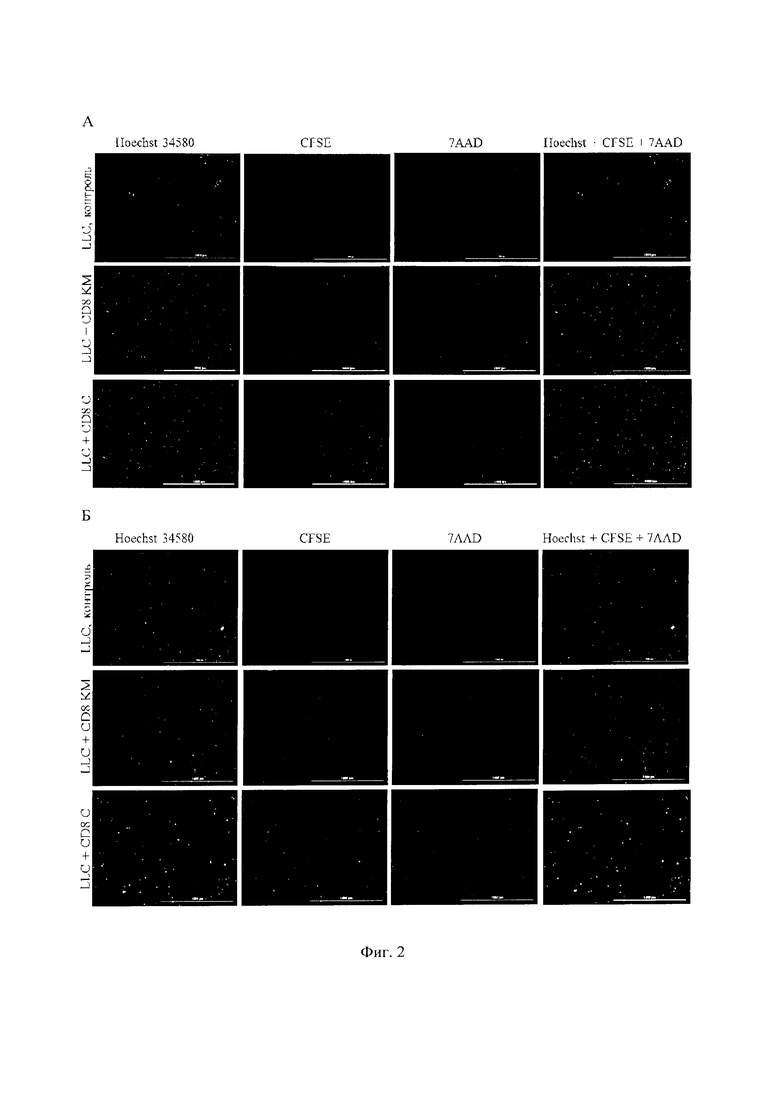
На фигуре 2 изображены наивные (А) и репрограммированные (Б) CD8+ Т-лимфоциты костного мозга (CD8 KM) и селезенки (CD8 С) в культуре клеток карциномы легкого Льюис (LLC) в соотношении 2,5:1, окрашенных: Hoechst (синий) для идентификации ядер клеток; CD8+ CFSE (зеленый) для идентификации CD8+ Т-лимфоцитов; 7ААД (красный); (Hoechst+CFSE+7AAD) составное изображение с использованием всех трех цветов. Определение процента погибших Т-клеток Hoechst+CFSE+производят по отношению клеток, подсчитанных в синем и зеленом каналах, к общему количеству клеток. Все масштабные линейки имеют размер 1000 мкм. Увеличение ×4.
На фигуре 3 изображена гистологическая картина легкого мыши интактного контроля (А, Д), мыши с карциномой легкого Льюис (Б, Е), мыши с карциномой легкого Льюис, леченные репрограммированными CD8+ Т-лимфоцитами, изолированными из костного мозга мышей-доноров (В, Ж) и мыши с карциномой легкого Льюис, леченые CD8 репрограммированными Т-лимфоцитами, изолированными из селезенки мышей-доноров (Г, З). Окраска гематоксилином и эозином, ув. ×100 (А, Б, В, Г) и ×400 (Д, Е, Ж, 3). Окраска гематоксилином и эозином, ув. ×100. Препараты изготовлены на 7-е сутки эксперимента.
Способ осуществляют следующим образом:
Проводят репрограммирование CD8+ Т-лимфоцитов, выделенных из костного мозга. Для подтверждения полученного результата проводят анализ стабильности экспрессии маркера CCR7 на CD8+ Т-лимфоцитах в культуре Т-лимфоцитов in vitro. Исследуют выживаемость и цитотоксическую активность репрограммированных CD8+ Т-лимфоцитов в культуре клеток карциномы легкого Льюис in vitro. Наконец, in vivo оценивают противоопухолевую и антиметастастическую активности репрограммированных CD8+ Т-лимфоцитов.
Для достижения наиболее оптимального результата более выраженной цитотоксичности и повышения выживаемости репрограммированных Т-лимфоцитов используют предварительную подготовку лимфоцитов для репрограммирования.
Для получения мононуклеаров костный мозг выделяют из бедренной кости мыши 0,5 мл препаровочной среды, следующего состава: 95% RPMI-1640, 5% эмбриональной телячьей сыворотки, ресуспендируют. Полученную суспензию наслаивают на градиент плотности HISTOPAQUE-1077 («Sigma», США) в соотношении суспензия-градиент 2:1 и центрифугируют в течение 25 минут при 1500 об/мин. Клетки интерфазного кольца собирают и разбавляют 5-кратным объемом фосфатного буфера Хэнкса с 2% инактивированной эмбриональной телячьей сывороткой и дважды отмывают центрифугированием при 1500 об/мин в течение 7-10 мин.
Для выделения мононуклеаров из селезенки, предварительно селезенку выделяют из брюшной полости мыши, помещают в чашку Петри диаметром 90 мм с фосфатным буфером Хэнкса («Sigma», США) и измельчают, используя одноразовые скальпели, до получения однородной клеточной взвеси. Полученную клеточную взвесь собирают в центрифужные пробирки и фильтруют через нейлоновый фильтр диаметром 70 мкм для удаления стромы и крупных агрегатов. После центрифугирования взвеси клеток в течение 10 мин при 1500 об/мин надосадочную жидкость сливают. Осадок клеток осторожно ресуспендируют, используя 2 мл холодной среды следующего состава: 90% DMEM-LG («Sigma», США), 10% инактивированной эмбриональной телячьей сывороткой («Sigma», США), 2 мМ L-глютамина («Sigma», США), раствора антибиотиков (пенициллин/стрептомицин 100 Ед/мл и 100 мкг/мл («Sigma», США). Полученный аспират переносят в пробирку с равным объемом фосфатного буфера Хэнкса с 2%, инактивированной эмбриональной телячьей сывороткой и далее наслаивают на градиент плотности Histopaque-1077 («Sigma», США) в соотношении суспензия-градиент 2:1, после чего центрифугируют в течение 25 минут при 1500 об/мин. Клетки интерфазного кольца собирают и разбавляют 5-кратным объемом фосфатного буфера Хэнкса с 2% инактивированной эмбриональной телячьей сывороткой и дважды отмывают центрифугированием при 1500 об/мин в течение 7-10 мин. Количество мононуклеаров подсчитывают в камере Горяева
Проводят обогащение клеточной суспензии, полученной из мононуклеаров костного мозга или селезенки, Т-лимфоцитами заданного иммунофенотипа, используя магнитную сепарацию. Обогащение проводят по стандартному протоколу с использованием набора для мышей (EasySep™ Mouse Naïve CD8+ T Cell Isolation Kit) в соответствии с инструкцией производителя (Stemcell Technologies, Ванкувер, Канада). В результате популяция мононуклеаров костного мозга или селезенки обогащается наивными CD8 позитивными Т-лимфоцитами (CD3+CD8+CD44-CD62L+). Далее клеточную суспензию, обогащенную наивными CD8 позитивными Т-лимфоцитами, инкубируют в течение 2-3 ч в рекомендуемой для Т-лимфоцитов среде, содержащей: раствор фосфатно-солевого буфера, 2% инактивированной эмбриональной телячьей сыворотки и 1 мМ EDTA без Са2+, Mg2+ и биотина, при температуре 37°С и 5% СО2. По окончании культивирования проводится оценка количества CD8 позитивных Т-лимфоцитов и оценка чистоты полученной субпопуляции на проточном цитофлюориметре FACS Canto II (BD Biosciences, США).
Для активации Т-лимфоцитов готовят антигенпрезентирующую смесь, представляющую собой лизат опухолевых клеток (смесь опухолевых антигенов). Лизат опухолевых клеток карциномы легкого Льюис получают с использованием цикла замораживания-оттаивания в 0,85% растворе NaCl. Цикл повторяют пять раз в быстрой последовательности, от -70°С до 37°С, а затем повторно замораживают и хранят при -70°С до использования [Dong В. et al., 2014]. Для усиления иммунного ответа в лизат опухолевых клеток добавляют адъювант. Адъювант для антигенпрезентирующей смеси готовят по стандартному протоколу производителя, в нашем случае это полный адъювант Фрейнда («Sigma», США). Раствор адъюванта Фрейнда смешивают с лизатом опухолевых клеток (3×104/мл) в соотношении 1:1 с образованием густой эмульсии.
Для репрограммирования CD8 позитивных Т-лимфоцитов во флакон с Т-лимфоцитами в концентрации 1×108/мл и объемом среды во флаконе не менее 5 мл вносят 50 мкл антигенпрезентирующей смеси с 1 μM ингибитора MEK. MEK представляет собой MAPK/ERK киназу, фермент сигнального пути ERK (ERK - extracellular signal-regulated kinase). Полученную клеточную взвесь инкубируют 48 ч при температуре 37°С и 5% CO2. В целях защиты от гуморального действия опухолевых клеток (в нашем случае клеток карциномы легкого Льюис) репрограммированные Т-лимфоциты, инкубируют в течение 2 ч в присутствии человеческого моноклонального антитела (МАТ) ниволумаб в концентрации 10 мкг/мл. По окончании цикла инкубации проводят 2-х кратную отмывку рекомендуемой для Т-лимфоцитов средой. Репрограммированные Т-лимфоциты готовы к использованию.
Для подтверждения устойчивости и эффективности полученной популяции репрограммированных CD8 позитивных Т-лимфоцитов проводят исследования in vitro и in vivo.
Для оценки устойчивости полученной популяции проводится процедура истощения репрограммированных Т-лимфоцитов in vitro [Dunsford L.S., et al., 2020]. Репрограммированные Т-лимфоциты в количестве 1-3×106/мл в рекомендованной для Т-лимфоцитов культуральной среде с добавлением IL-2 (30 МЕ/мл) («Stemcell Technologies», Канада) стимулируют T-Activator CD3/CD28 Dynabeads® («Thermo Fisher Scientific Baltics», Литва) в соотношении частиц и клеток 1:1. Далее каждые 48 ч проводят ре-стимуляцию, суммарно 3-4 раза. После чего Dynabeads удаляют. Определяют иммунофенотип, проводят анализ стабильности и цитотоксичности репрограммированных Т-лимфоцитов in vitro.
Анализ стабильности экспрессии CCR7 на CD8+ Т-лимфоцитах проводят в культуре Т-лимфоцитов in vitro. Культивирование наивных, репрограммированных и репрограммированных истощенных Т-лимфоцитов проводят в среде RPMI 1640 («Sigma», США) с добавлением 10% эмбриональной телячьей сыворотки (ЭТС, «Sigma», США), 2 мМ L-глутамина («Sigma», США), 10 мМ HEPES («Sigma», США) и 55 мкМ β-меркаптоэтанола («Thermo Scientific», США) при температуре 37°С и 5% СО2 в течение 1 ч. Концентрация Т-лимфоцитов - 1×108/ мл. Объем среды во флаконе - не менее 5 мл. Далее проводят окрашивание Т-лимфоцитов: добавляют первичные антитела CCR7 в концентрации 100 мкл на лунку, окрашивают 60 мин при 37°С, убирают надосадок, промывают фосфатным буфером 3 раза, далее инкубируют со вторичным антителом AF555 в течение 60 мин при 37°С, убирают надосадок, промывают фосфатным буфером 3 раза, добавляют красители Hoechst 34580 для идентификации ядер и CD8 FITC для идентификации CD8 позитивных Т-лимфоцитов по 1 мкл на 1 мл среды каждого, инкубируют 30 мин при 37°С, убирают надосадок, промывают фосфатным буфером 3 раза. После завершения окрашивания планшеты помещают в мультимодальный ридер клеточного имиджинга Cytation 5. С помощью Cytation 5, оснащенного кубами: DAPI (синий), GFP (зеленый), RFP (желтый), оценивают количество CD8+CCR7+ Т-лимфоцитов. Подсчитывают количество CD8+CCR7+ Т-лимфоцитов в культуре наивных, репрограммированных, истощенных репрограммированных CD8+ Т-лимфоцитов, выделенных из костного мозга или селезенки. Изображения, полученные на мультиридере Cytation 5 (увеличение 4× или 20×), анализируют с использованием программного обеспечения для анализа данных Gen5™ (Biotek, Bad Friedrichshall, Германия). Все изображения предварительно обрабатывают для выравнивания фона перед выполнением анализа. Определение процента клеток CD8+CCR7+ производят по отношению клеток, подсчитанных по зеленому и красному каналам, к общему количеству клеток, подсчитанных по синему (DAPI) каналу.
Анализ цитотоксичности и выживаемости репрограммированных и наивных Т-лимфоцитов in vitro проводят на культуре клеток карциномы легкого Льюис (LLC). На первом этапе готовят культуру клеток карциномы легкого Льюис. Для этого 2×103 клеток карциномы легкого Льюис вносят в 0,2 мл культуральной среды, состоящей из RPMI 1640 с добавлением 2 мМ L-глутамина и 10% эмбриональной телячьей сыворотки, и высевают в 96-луночные планшеты с оптическим дном и в течение 24 ч дают возможность клеткам прикрепиться к пластику.
На втором этапе готовят Т-лимфоциты. Т-лимфоциты переносят в коническую пробирку на 15 мл и добавляют витальный краситель сукцинимидиловый эфир карбоксифлуоресцеина (CFSE) в концентрации 1 мкМ. Взвесь Т-лимфоцитов с CFSE инкубируют при 37°С, 5% СО2 в течение 45 минут. Затем пробирки центрифугируют в течение 15 минут при 2000 об/мин. Среду, содержащую избыток красителя, удаляют и заменяют свежей культуральной средой (RPMI 1640 с добавлением 2 мМ L-глутамина и 10% ЭТС). Окрашенные CFSE Т-лимфоциты разводят в среде RPMI, содержащей 10 мкл/мл красителя 7AAD.
На третьем этапе окрашенные CFSE Т-лимфоциты вносят в 96-луночные планшеты к клеткам-мишеням (клетки карциномы легкого Льюис). Соотношение Т-лимфоцитов к клеткам-мишеням и компоновка клеток в культуральном планшете указано в таблице 1.

Группа отрицательного контроля (ряд G, лунки с культуральной средой); группа положительного контроля (ряд А, лунки с клетками карциномы легкого Льюис); группа с Т-лимфоцитарным контролем (ряд Н), с 1 по 6 столбец к клеткам-мишеням добавляются репрограммированные CD8+ Т-лимфоциты, а с 7 по 12 столбец - наивные CD8+ Т-лимфоциты соответственно.
После завершения цикла культивирования планшеты переносят в Cytation 5 для светлопольного и флуоресцентного изображения тестовых лунок через 24 часа. Для культур клеток карциномы легкого Льюис в лунках было сделано одно изображение с 4-х кратным увеличением для каждого канала, чтобы захватить репрезентативную популяцию клеток на лунку. Изображения CD8+ Т-лимфоцитов и клеток карциномы легкого Льюис получают с помощью системы визуализации клеток Cytation 5 (4-кратное или 20-кратное увеличение). Анализ клеток проводят с использованием программного обеспечения для анализа данных Gen5™. (Biotek, Bad Friedrichshall, Германия). Перед анализом изображения предварительно обрабатывают для выравнивания фона.
Выживаемость наивных и репрограммированных CD8+ Т-лимфоцитов костного мозга или селезенки в культуре клеток карциномы легкого Льюис оценивают, анализируя составное изображение с использованием всех трех цветов. Определение процента погибших Т-лимфоцитов Hoechst+CFSE+производят по отношению клеток, подсчитанных в синем и зеленом каналах, к общему количеству клеток.
Цитотоксичность наивных и репрограммированных CD8+ Т-лимфоцитов костного мозга или селезенки в культуре клеток карциномы легкого Льюис оценивают, анализируя по отношению клеток, подсчитанных в синем и красном каналах, к общему количеству клеток карциномы легкого Льюис (процент погибших клеток карциномы легкого Льюис Hoechst+7AAD+). Определяют количество апоптотических опухолевых клеток карциномы легкого Льюис после совместного культивирования с CD8+ Т-лимфоцитами костного мозга или селезенки (% от добавленных клеток).
В экспериментах in vivo и in vitro используется линия клеток карциномы легкого Льюис, штамм C57BL (CLS Cell Lines. Service, Германия).
Для оценки противоопухолевой и антиметастатической активности мышам-реципиентам с карциномой легкого Льюис репрограммированные CD8+ Т-лимфоциты, выделенные из костного мозга или селезенки, вводят внутривенно в дозе 1×106 клеток/мышь в 0,1 мл стерильного фосфатного буфера на 4-й и 6-й дни эксперимента.
Противоопухолевую активность репрограммированных CD8 Т-лимфоцитов исследуют в ортотопической модели карциномы легкого Льюис [Justilien V., Fields А.Р., 2013]. Перед введением опухолевых клеток карциномы легкого Льюис мышь наркотизируют, используя для анестезии изофлуран, вводимый ингаляционно с помощью аппарата для ингаляционного наркоза UGO BASIELE модель 21050 (Италия). Далее животным под наркозом в стерильных условиях удаляют волосяной покров на левой половине грудной клетки выше нижней линии ребер и чуть ниже нижней границы лопатки. 50 мкл суспензии опухолевых клеток карциномы легкого Льюис штамма C57BL (1,5×106 клеток) вводят инсулиновым шприцом, снабженным иглой для подкожных инъекций 30G, на глубину 5 мм в левое легкое между 6-м и 7-м ребром.
Рост опухоли контролируют после эвтаназии животных на 7-е сутки после введения клеток карциномы легкого Льюис. Проводят исследование размера опухоли, морфологическое и гистологическое исследование легких.
Для оценки роста опухоли у животных исследуют линейные размеры опухолевых узлов в ортогональных плоскостях и рассчитывают их объем в эллиптическом приближении [Tomayko М.М., Reynolds С.Р., 1989]. Опухоль измеряют с помощью штангенциркуля и объем опухолей рассчитывают по формуле: V = π/6 × длина × ширина × высота.
Специфическое влияние клеточной терапии на опухолевый процесс изучают по росту опухоли. Влияние клеточной терапии на рост карциномы легкого Льюис оценивают путем статистического сравнения объема опухолевых узлов в контрольных и подопытных группах при разных сроках наблюдения, по длительности задержки роста опухоли и индексу торможения роста опухоли (ТРО):
TPO=(VK-VO)/VK×100%;
где VK и VO - средние объемы узлов в контрольной и опытной группах.
Для гистологического исследования легкие фиксируют в 10% растворе нейтрального формалина, проводят через спирты восходящих концентраций до ксилола и заливают в парафин по стандартной методике [Cardiff R.D. et al., 2014]. Депарафинизированные срезы толщиной 5 мкм окрашивают гематоксилином и эозином.
Для оценки экспрессии маркеров стволовых опухолевых клеток выделяют мононуклеары из легких. Для этого легкие промываются стерильным фосфатно-солевым буфером (PBS, «Sigma», США) через правый желудочек сердца и после удаления трахеи тщательно измельчаются стерильными ножницами до получения однородной клеточной взвеси. Клеточная взвесь инкубируется в растворе 0,2% коллагеназы 2 типа («Sigma», США), разведенной стерильным фосфатно-солевым буфером при 37°С в течение 30 мин с периодическим перемешиванием. Для удаления стромы и крупных агрегатов клеточную взвесь фильтруют через нейлоновый фильтр диаметром 70 мкм («BD Falcon», США). После центрифугирования в течение 10 мин при 1500 об/мин надосадочную жидкость сливают. Осадок клеток осторожно ресуспендируют, используя 2 мл холодной среды следующего состава: 90% DMEM-LG («Sigma», США), 10% инактивированной ЭТС («Sigma», США), 2 мМ L-глютамина («Sigma», США), раствора антибиотиков (пенициллин/стрептомицин 100 Ед/мл и 100 мкг /мл («Sigma», США)). Полученный аспират переносят в пробирку с равным объемом PBS с 2% инактивированной ЭТС и далее наслаивают на градиент плотности Histopaque-1077 в соотношении суспензия-градиент 2:1 и центрифугируют в течение 25 минут при 1500 об/мин. Клетки интерфазного кольца собирают и разбавляют 5-кратным объемом фосфатно-солевого буфера с 2% инактивированной ЭТС и дважды отмывают центрифугированием при 1500 об/мин в течение 7-10 мин. Количество мононуклеаров подсчитывают в камере Горяева.
Влияние репрограммированных CD8 Т-лимфоцитов на содержание стволовых опухолевых клеток исследуют, оценивая экспрессию мембранных и внутриклеточных рецепторов стволовых и прогениторных клеток с использованием мышиных моноклональных антител по стандартному протоколу для проточной цитометрии. Суспензию клеток преинкубируют с CD16/CD32 мыши (BD FcBlock™) в течение 5 минут. Затем полученные суспензии клеток суспендируют в концентрации 1×106/100 мкл в фосфатно-солевом буфере, содержащем 1% ЭТС, и инкубируют в течение 30 минут при 4°С с соответствующими антителами: CD8 FITC, CD44 АРС-Су™7, CD117 FITC, EGR Alexa Fluor® 647, Ax1 BV421. После инкубации образцы дважды отмывают при 300 g в течение 5 мин. После окрашивания поверхностными маркерами для окрашивания внутриклеточным антителом Sox2 образцы инкубируют со свежеприготовленным рабочим раствором фиксирующего пермеабилизирующего буфера (Fix/Perm Buffer, «BD Biosciences)), США) при 4°С в течение 40 минут в защищенном от света месте. Далее образцы дважды отмывают специализированным промывающим буфером (Perm/Wash Buffer, «BD Biosciences)), США) при 350 g при 4°С в течение 6 минут. Внутриклеточное окрашивание Sox2 проводят при инкубации образцов в течение 40 минут в защищенном от света месте при 4°С. Затем образцы дважды отмывают при 350 g в течение 6 минут. Добавляют 500 мкл фосфатного буфера и анализируют при помощи прибора FACS Canto II с программным обеспечением FACS Diva («BD Biosciences)), США).
Математическую обработку результатов производят с применением стандартных методов вариационной статистики. Достоверность различий оценивают с использованием параметрического t критерия Стьюдента или непараметрического U критерия Манна-Уитни.
Пример 1.
Для оценки эффективности проведенного репрограммирования in vitro изучают экспрессию хемокинового рецептора CCR7 CD8+ Т-лимфоцитами костного мозга или селезенки (Фигура 1 Приложения). Репрограммированные CD8+ Т-лимфоциты, выделенные из костного мозга или селезенки, показывают большую экспрессию хемокинового рецептора CCR7, чем наивные CD8+ Т-лимфоциты, независимо от места их выделения, в культуре CD8+ Т-лимфоцитов (Таблица 2). В целях проверки стойкости изменений, вызванных ингибитором MEK и ниволумабом, проведена процедура истощения репрограммированных CD8+ Т-лимфоцитов. Истощение не вызывает изменений в экспрессии CCR7 репрограммированными CD8+ Т-лимфоцитами, что говорит о стабильности индуцированных MEKi и ниволумабом изменений (Таблица 2). Сравнение экспрессии CCR7 клетками костного мозга и селезенки не выявляет тканевых различий параметра.

Таким образом, использование ингибиторов протеинкиназ MEK, ингибиторов контрольных точек PD1 и предварительное обучение антигенпрезентирующей смесью вызывают повышение экспрессии хемокинового рецептора CCR7 CD8+ Т-лимфоцитами.
Исследование апоптоза репрограммированных и наивных CD8+ Т-лимфоцитов, выделенных из костного мозга или селезенки, в культуре клеток карциномы легкого Льюис показывает, что в сравнении с наивными лимфоцитами наиболее стойкой к цитотоксическому действию опухолевых клеток является популяция репрограммированных лимфоцитов, независимо от места выделения (Фигура 2, Таблица 3). По окончании цикла культивирования наивных CD8+ Т-лимфоцитов в стандартной среде процент мертвых клеток от общего числа клеток составляет 1,92% у Т-лимфоцитов, выделенных из костного мозга, и 2,48% у Т-лимфоцитов, выделенных из селезенки. Репрограммированные CD8+ Т-лимфоциты более стабильны при культивировании: в состоянии апоптоза находится 0,69% Т-лимфоцитов, выделенных из костного мозга, и 0,79% Т-лимфоцитов, выделенных из селезенки (Таблица 3).

При разведении Т-лимфоциты и опухолевые клетки 0,25:1 мы наблюдаем увеличение апоптоза наивных CD8+ Т-лимфоцитов в культуре клеток карциномы легкого Льюис. Уровень апоптоза выше в группе наивных CD8+ Т-лимфоцитов более чем в 2 раза. Повышение концентрации наивных CD8+ Т-лимфоцитов в культуре клеток карциномы легкого Льюис вызывает снижение уровня апоптоза до уровня группы «контроль» лимфоцитов. Репрограммированные CD8+ лимфоциты костного мозга или селезенки более устойчивы к цитотоксическому действию опухолевых клеток по сравнению с наивными CD8+ Т-лимфоцитами. На это указывает снижение уровня апоптоза репрограммированных CD8+ Т-лимфоцитов, выделенных из костного мозга, по сравнению с наивными CD8+ Т-лимфоцитами в разведении 0,25:1; 1:1 и 2,5:1 (Таблица 3). При сравнении уровня апоптоза репрограммированных CD8+ Т-лимфоцитов, выделенных из селезенки, в культуре клеток карциномы легкого Льюис выявляется большая гибель клеток лимфоцитами в разведении 1:1 и 2,5:1 (Таблица 3).
Таким образом, in vitro репрограммированные CD8+ Т-лимфоциты демонстрируют большую устойчивость к цитотоксическому действию клеток карциномы легкого Льюис, чем наивные CD8+ Т-лимфоциты.
Исследование цитотоксичности репрограммированных CD8+ Т-лимфоцитов проводят при совместном культивировании с клетками карциномы легкого Льюис. В культуре клеток карциномы легкого Льюис соотношение между CD8+ Т-лимфоцитами и опухолевыми клетками составляет 0:1, 0,25:1, 1:1, 2,5:1, 5:1 и 10:1. Наивные и репрограммированные CD8+ Т-лимфоциты, независимо от места выделения, вызывают апоптоз опухолевых клеток уже в соотношении 0,25:1. Максимальный уровень апоптоза опухолевых клеток наблюдается в соотношении клеток 10:1 (CD8+ Т-лимфоциты : опухолевые клетки). Цитотоксичность репрограммированных CD8+ Т-лимфоцитов превосходит таковую у наивных CD8+ Т-лимфоцитов (Таблица 4). Достоверных различий в цитотоксичности между репрограммированными клетками костного мозга и селезенки не выявляется.

Таким образом, в культуре карциномы легкого Льюис репрограммированные CD8+ Т-лимфоциты, выделенные из костного мозга или селезенки, демонстрируют большую цитотоксическую активность, чем наивные CD8+ Т-лимфоциты.
Пример 2.
Морфологическое и гистологическое исследование легких мышей-реципиентов линии C57BL/6 с карциномой легкого Льюис после клеточной терапии репрограммированными CD8+ Т-лимфоцитами проводят на 7-е сутки после моделирования ортотопической модели. В легких мышей с карциномой легкого Льюис без лечения выявляются хорошо васкуляризированные крупные опухолевые узлы, состоящие из атипичных клеток. Для этой группы клеток был характерен клеточный и ядерный полиморфизм. В общей популяции атипичных клеток выявляются гигантские многоядерные клетки, множество клеток в состоянии митоза. В опухолевой ткани встречаются многочисленные мелкие очаги некроза (Фигура 3 Б, Е Приложения). Клеточная терапия репрограммированными CD8+ Т-лимфоцитами, выделенными из костного мозга или селезенки, не изменяет общую гистологическую картину легких мышей с карциномой легкого Льюис (Фигура 3 В, Г, Ж, 3 Приложения). На фоне терапии отмечается уменьшение количества опухолевых эмболов в сосудах, периваскулярных и перибронхиальных метастазов.
Индекс торможения роста опухоли (ТРО) на модели карциномы легкого Льюис после клеточной терапии репрограммированными CD8+ Т-лимфоцитами костного мозга или селезенки составляет 57% и 54,7% соответственно (таблица 5). Индекс ингибирования процесса метастазирования составляет после клеточной терапии репрограммированными CD8+ Т-лимфоцитами костного мозга 91,665% и селезенки 98%. На фоне введения репрограммированных CD8+ Т-лимфоцитов выявляются уменьшение объема опухоли, частоты метастазирования, степени поражения легких метастазами и среднего числа метастазов.

Таким образом, репрограммированные CD8+ Т-лимфоциты, выделенные из костного мозга или селезенки оказывают, противоопухолевое и антиметастическое действие на модели карциномы легкого Льюис.
На фоне развития карциномы легкого Льюис в легких мышей на 7-е сутки эксперимента наблюдается увеличение количества опухолевых и стволовых опухолевых клеток с различными фенотипами: Ах1+, Ax1+CD117+, EGF+CD44+Sox2+, EGF+Sox2+, CD44+Sox2+, CD117+Sox2+, CD117+EGF+CD44+Sox2+(Таблица 6). Введение репрограммированных CD8+ Т-лимфоцитов, как из костного мозга, так и из селезенки, уменьшает количество всех исследованных популяций опухолевых и стволовых опухолевых клеток в легких мышей-реципиентов.
Таким образом, метаболическое репрограммирование CD8+ Т-лимфоцитов, основанное на совместном применении ингибиторов протеинкиназ MEK и антигенпрезентирующей смеси, ингибитора контрольных точек PD1, приводит к формированию популяции репрограммированных CD8+ Т-лимфоцитов, устойчивых к действию опухолевых клеток, обладающих цитотоксической активностью in vitro, противоопухолевым и антиметастическим действием. Следует отметить, активность репрограммированных CD8+ Т-лимфоцитов не зависит от места их выделения, что в перспективе предполагает возможность использования для выделения Т-лимфоцитов периферическую кровь пациентов.

Источники информации, принятые во внимание при составлении описания:
1. Liu, Q., Zhao, С., Jiang, P., Liu, D. Circulating tumor cells counts are associated with CD8+ T cell levels in programmed death-ligand 1-negative non-small cell lung cancer patients after radiotherapy: A retrospective study. // Medicine. - 2021. - 100(29), e26674.
2. Sathaliyawala, Т., Kubota, M., Yudanin, N., Turner, D., Camp, P., Thome, J. J., Bickham, K. L., Lerner, H., Goldstein, M., Sykes, M., Kato, Т., Farber, D. L. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. // Immunity. - 2013. - 38(1), 187-197.
3. Speiser DE, Ho P-C, Verdeil G. Regulatory circuits of T cell function in cancer. // Nat. Rev. Immunol. - 2016. - 16, 599. doi: 10.1038/nri.2016.80
4. Петрова Г.В., Грецова О.П., Старинский В.В. Сравнение данных государственной онкологической статистики и ракового регистра России // Сибирский онкологический журнал. - 2019. - T. 18, №5. - С. 5-17.
5. Hochmair, M., Rath, В., Klameth, L., Ulsperger, E., Weinlinger, С., Fazekas, A., Plangger, A., Zeillinger, R., Hamilton, G. Effects of salinomycin and niclosamide on small cell lung cancer and small cell lung cancer circulating tumor cell lines. // Investigational new drugs. - 2020. - 38(4), 946-955.
6. Sallusto F., Geginat J., Lanzavecchia A. Central memory and effector memory T cell subsets: function, generation, and maintenance // Annu. Rev. Immunol. - 2004. - Vol. 22. P. 745-763. doi: 10.1146/annurev.immunol.22.012703.104702Sallusto F. et al., 2004
7. Jung Y.W., Kim H.G., Perry C.J., Kaech S.M. CCR7 expression alters memory CD8 T-cell homeostasis by regulating occupancy in IL-7- and IL-15-dependent niches // Proc. Natl. Acad. Sci. USA. - 2016. - Vol. 113, No. 29. P. 8278-8283. doi: 10.1073/pnas.l602899113
8. Verma, V., Jafarzadeh, N., Boi, S. et al. MEK inhibition reprograms CD8+ T lymphocytes into memory stem cells with potent antitumor effects. // Nat Immunol. - 2021. - 22, 53-66. doi:. 10.1038/s41590-020-00818-9
9. Herrick, W.G., Kilpatrick, C.L., Hollingshead, M.G., Esposito, D., O'Sullivan Coyne, G., et al. Isoform- and Phosphorylation-specific Multiplexed Quantitative Pharmacodynamics of Drugs Targeting PI3K and MAPK Signaling in Xenograft Models and Clinical Biopsies. // Molecular cancer therapeutics. - 2021. - 20(4), 749-760. doi: 10.1158/1535-7163.MCT-20-0566
10. Dong В., Minze L.J., Xue W., Chen W. Molecular insights into the development of T cell-based immunotherapy for prostate cancer // Expert Rev. Clin. Immunol. - 2014. - Vol. 10, No. 11. P. 1547-1557. doi: 10.1586/1744666X.2014.962515
11. Dunsford L.S., Thoirs R.H., Rathbone E., Patakas A. A Human In Vitro T Cell Exhaustion Model for Assessing Immuno-Oncology Therapies // Immuno-Oncology. Methods in Pharmacology and Toxicology. - 2020. - P. 89-101. doi: 10.1007/978-1-0716-0171-6_6
12. Justilien V., Fields A.P. Utility and applications of orthotopic models of human non-small cell lung cancer (NSCLC) for the evaluation of novel and emerging cancer therapeutics // Curr. Protoc. Pharmacol. - 2013. - Vol. 62. P. 14.27.1-14.27.17. doi: 10.1002/0471141755.ph1427s6
13. Tomayko M.M., Reynolds CP. Determination of subcutaneous tumor size in athymic (nude) mice // Cancer Chemother. Pharmacol. - 1989. - Vol. 24, No. 3. P. 148-154. doi: 10.1007/BF00300234
14. Cardiff R.D., Miller H., Munn R.J. Manual hematoxylin and eosin staining of mouse tissue sections // Cold Spring Harb. Protoc. - 2014. - Vol. 6. P. 655-658. doi: 10.1101/pdb.prot073411
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2012 |
|
RU2688185C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2540490C2 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2013 |
|
RU2700765C2 |
| ПЕПТИД CDCA1 И ВКЛЮЧАЮЩЕЕ ЕГО ФАРМАЦЕВТИЧЕСКОЕ СРЕДСТВО | 2008 |
|
RU2486195C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2478400C2 |
| СПОСОБ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2010 |
|
RU2489169C2 |
| Способ усиления роста меланомы В16/F10 по сравнению с ростом меланомы В16/F10 при самостоятельной перевивке и замедления роста LLC (карциномы Льюиса) по сравнению с ростом LLC при самостоятельной перевивке при первично-множественных злокачественных опухолях на фоне первичного иммунодефицита | 2021 |
|
RU2759487C1 |
| ЧАСТИЦЫ PM21 ДЛЯ УЛУЧШЕНИЯ ХОМИНГА NK-КЛЕТОК В КОСТНОМ МОЗГЕ | 2018 |
|
RU2777993C2 |
| УСТРОЙСТВА И СПОСОБЫ ДЛЯ ОТБОРА КЛЕТОК, УСТОЙЧИВЫХ К АПОПТОТИЧЕСКИМ СИГНАЛАМ, И ОБЛАСТИ ИХ ПРИМЕНЕНИЯ | 2013 |
|
RU2630301C2 |
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |


Изобретение относится к медицине, а именно к иммунологии и клеточной биотехнологии, в частности к способу усиления противоопухолевого Т-клеточного иммунитета при раке легкого. Изобретение позволяет подавить рост опухоли легкого за счет повышения специфичной цитотоксичности и устойчивости к действию опухолевых клеток CD8+ Т-лимфоцитов, индуцированных метаболическим репрограммированием. 3 ил., 6 табл., 2 пр.
Способ усиления противоопухолевого Т-клеточного иммунитета при раке легкого, характеризующийся тем, что реципиенту внутривенно вводят репрограммированные CD8 позитивные Т-лимфоциты, которые готовят in vitro следующим образом: к наивным CD8 позитивным Т-лимфоцитам в концентрации 1×108/мл добавляют ингибитор протеинкиназ MEK в дозе 1 μΜ совместно с 50 мкл антигенпрезентирующей смеси, полученной на основе лизата опухолевых клеток, культивируют в течение 48 часов, далее добавляют ингибитор контрольных точек PD1 в концентрации 10 мкг/мл и культивируют в течение 2 часов.
| WO2014195852 A1, 11.12.2014 | |||
| СПОСОБ ГЕНЕРАЦИИ АНТИГЕН-СПЕЦИФИЧЕСКИХ ЦИТОТОКСИЧЕСКИХ КЛЕТОК С ПРОТИВООПУХОЛЕВОЙ АКТИВНОСТЬЮ ПРИ РАКЕ МОЛОЧНОЙ ЖЕЛЕЗЫ | 2013 |
|
RU2521506C1 |
| VERMA V | |||
| et al | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Nat Immunol | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Vol | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| No | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Pp | |||
| Веникодробильный станок | 1921 |
|
SU53A1 |
| WO2021025177 A1, 11.02.2021. | |||

