Ссылка на родственные заявки
[0001] Согласно настоящей заявке испрашивается преимущество в соответствии с заявкой на выдачу патента США №62/592905, поданной 30 ноября 2017 г., и заявкой на выдачу патента США №62/661373, поданной 23 апреля 2018 г., каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей.
Ссылка на перечень последовательностей, представленный в виде текстового файла посредством файловой системы EFS
[0002] Перечень последовательностей, представленный в файле 523380SEQLIST.txt, характеризуется размером, составляющим 154 КБ, был создан 30 ноября 2018 г. и включен в настоящий документ посредством ссылки.
Предшествующий уровень техники настоящего изобретения
[0003] Тропомиозин-рецепторная киназа В (TRKB) является многообещающей мишенью для нейропротекции при нейродегенеративных заболеваниях, таких как глаукома. TRKB является одним из наиболее широко распространенных нейротрофических рецепторов (NTR) в головном мозге, который в высокой степени представлен в неокортексе, гиппокампе, стриатуме и стволе мозга. Связывание нейротрофического фактора головного мозга (BDNF) с рецептором TRKB запускает его димеризацию посредством конформационных изменений и аутофосфорилирования остатков тирозина во внутриклеточном домене, что приводит к активации сигнальных путей с участием митоген-активируемой протеинкиназы (MAPK), фосфатидилинозитол-3-киназы (PI3K) и фосфолипазы С-γ (PLC-γ).
[0004] TRKB важна для выживаемости, дифференцировки и функционирования нейронов, и агонисты TRKB могут характеризоваться терапевтическим потенциалом при многочисленных неврологических, психических и метаболических нарушениях. Однако по-прежнему существует потребность в подходящих животных, отличных от человека, обеспечивающих истинную человеческую мишень или близкую к истинной человеческой мишени для реагентов, нацеленных на TRKB человека, что позволяет проводить испытание эффективности и способа действия таких агентов на живых животных, а также фармакокинетические и фармакодинамические исследования.
Краткое раскрытие настоящего изобретения
[0005] Предусмотрены не являющиеся человеком животные, содержащие гуманизированный локус TRKB, а также способы применения таких не являющихся человеком животных. Кроме того, предусмотрены геномы или клетки не являющегося человеком животного, содержащие гуманизированный локус TRKB.
[0006] Согласно одному аспекту предусмотрены геномы не являющегося человеком животного, клетки не являющегося человеком животного или не являющиеся человеком животные, содержащие гуманизированный локус TRKB. Такие геномы не являющегося человеком животного, клетки не являющегося человеком животного или не являющиеся человеком животные могут содержать генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, причем белок TRKB содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и весь или часть внеклеточного домена кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
[0007] Согласно одному аспекту предусмотрены не являющиеся человеком животные, содержащие гуманизированный локус TrkB. Такие не являющиеся человеком животные могут содержать генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, причем белок TRKB содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и весь или часть внеклеточного домена кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
[0008] Согласно другому аспекту предусмотрены клетки не являющегося человеком животного, содержащие в своем геноме генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, причем белок TRKB содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и весь или часть внеклеточного домена кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
[0009] Согласно другому аспекту предусмотрены геномы не являющегося человеком животного, содержащие генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, причем белок TRKB содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и весь или часть внеклеточного домена кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
[0010] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB содержит внеклеточный домен TRKB человека. Необязательно внеклеточный домен содержит последовательность, представленную в SEQ ID NO: 60. Необязательно весь внеклеточный домен кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека, необязательно при этом кодирующая последовательность для внеклеточного домена содержит последовательность, представленную в SEQ ID NO: 72.
[0011] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB содержит эндогенный сигнальный пептид. Необязательно сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 51 или 55. Необязательно весь сигнальный пептид кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 63 или 67.
[0012] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB содержит эндогенный трансмембранный домен TRKB. Необязательно трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 53 или 57. Необязательно весь трансмембранный домен кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ Ш NO: 65 или 69.
[0013] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB содержит эндогенный цитоплазматический домен TRKB. Необязательно цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 54 или 58. Необязательно весь цитоплазматический домен кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 66 или 70.
[0014] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB содержит эндогенный сигнальный пептид TRKB, эндогенный трансмембранный домен TRKB и эндогенный цитоплазматический домен TRKB. Необязательно сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 51, трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 53, и цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 54. Необязательно сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 55, трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 57, и цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 58. Необязательно весь сигнальный пептид, весь транс мембранный домен и весь цитоплазматический домен кодируются эндогенной последовательностью TrkB. Необязательно кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 63, кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ ID NO: 65, и кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 66. Необязательно кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 67, кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ ID NO: 69, и кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 70.
[0015] В некоторых таких геномах не являющегося человеком животного, клетках не являющегося человеком животного или не являющихся человеком животных белок TRKB представляет собой химерный белок TRKB не являющегося человеком животного/человека. Необязательно внеклеточный домен представляет собой внеклеточный домен TRKB человека, трансмембранный домен представляет собой трансмембранный домен эндогенного белка TRKB, и цитоплазматический домен представляет собой цитоплазматический домен эндогенного белка TRKB. Необязательно белок TRKB содержит последовательность, представленную в SEQ ID NO: 4 или 5. Необязательно кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 12 или 13.
[0016] Некоторые такие геномы не являющегося человеком животного, клетки не являющегося человеком животного или не являющиеся человеком животные являются гетерозиготными по генетически модифицированному эндогенному локусу TrkB. Некоторые такие геномы не являющегося человеком животного, клетки не являющегося человеком животного или не являющиеся человеком животные являются гомозиготными по генетически модифицированному эндогенному локусу TrkB.
[0017] Некоторые такие не являющиеся человеком животные представляют собой млекопитающих. Необязательно не являющееся человеком животное представляет собой грызуна. Необязательно грызун представляет собой крысу или мышь.
[0018] Некоторые такие не являющиеся человеком животные представляют собой крыс. Необязательно белок TRKB содержит последовательность, представленную в SEQ ID NO: 5. Необязательно кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 13.
[0019] Некоторые такие не являющиеся человеком животные представляют собой мышей. Необязательно белок TRKB содержит последовательность, представленную в SEQ ID NO: 4. Необязательно кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 12.
[0020] Согласно другому аспекту предусмотрены способы оценки активности нацеленного на TRKB человека реагента in vivo с использованием представленных выше не являющихся человеком животных. Некоторые такие способы предусматривают следующее: (а) введение нацеленного на TRKB человека реагента не являющемуся человеком животному; и (b) оценка активности нацеленного на TRKB человека реагента у не являющегося человеком животного.
[0021] Согласно некоторым таким способам оцениваемая активность представляет собой нейропротекторную активность.
[0022] Согласно некоторым таким способам стадия (а) предусматривает инъекцию нацеленного на TRKB человека реагента не являющемуся человеком животному.
[0023] Согласно некоторым таким способам стадия (b) предусматривает оценку изменений одного или нескольких или всех из следующего: масса тела, состав тканей организма, метаболизм и локомоция, по сравнению с контрольным не являющимся человеком животным. Необязательно оценка изменений в составе тканей организма предусматривает оценку массы нежировых тканей и/или массы жировой ткани по сравнению с контрольным не являющимся человеком животным. Необязательно оценка изменений в метаболизме предусматривает оценку изменений в потреблении пищи и/или потреблении воды.
[0024] Согласно некоторым таким способам стадия (b) предусматривает оценку фосфорилирования TRKB и/или активации путей MAPK/ERK и PI3K/Akt по сравнению с контрольным не являющимся человеком животным.
[0025] Согласно некоторым таким способам стадия (b) предусматривает оценку нейропротекторной активности. Согласно некоторым таким способам стадия (b) предусматривает оценку нейропротекторной активности, и не являющееся человеком животное представляет собой крысу. Согласно некоторым таким способам стадия (b) предусматривает оценку жизнеспособности ганглиозных клеток сетчатки. Необязательно оценка жизнеспособности ганглиозных клеток сетчатки предусматривает оценку плотности ганглиозных клеток сетчатки. Необязательно плотность ганглиозных клеток сетчатки измеряют в иссеченных сетчатках, окрашенных в отношении ганглиозных клеток сетчатки. Необязательно жизнеспособность ганглиозных клеток сетчатки оценивают в модели полного пересечения зрительного нерва после повреждения зрительного нерва. Необязательно жизнеспособность ганглиозных клеток сетчатки оценивают в модели раздавливания зрительного нерва.
[0026] Согласно некоторым таким способам нацеленный на TRKB человека реагент представляет собой антигенсвязывающий белок. Необязательно антигенсвязывающий белок представляет собой антитело - агонист TRKB человека. Согласно некоторым таким способам нацеленный на TRKB человека реагент представляет собой малую молекулу. Необязательно малая молекула представляет собой агонист TRKB человека.
[0027] Согласно другому аспекту предусмотрены нацеливающие векторы для создания генетически модифицированного эндогенного локуса TrkB, кодирующего белок TRKB, причем белок TRKB содержит цитоплазматический домен, трансмембранный домен и внеклеточный домен, и весь или часть внеклеточного домена кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека, и причем нацеливающий вектор содержит нуклеиновую кислоту - вставку, содержащую ортологичную последовательность TRKB человека, фланкированную 5' гомологичным плечом, нацеленным на 5' целевую последовательность на эндогенном локусе TrkB, и 3' гомологичным плечом, нацеленным на 3' целевую последовательность на эндогенном локусе TrkB.
[0028] Согласно другому аспекту предусмотрены способы получения любого из не являющихся человеком животных, описанных выше. Некоторые такие способы могут предусматривать следующее: (а) введение в плюрипотентную клетку не являющегося человеком животного, которая не является эмбрионом на стадии одной клетки, следующего:
(i) экзогенная матрица репарации, содержащая нуклеиновую кислоту - вставку, фланкированную 5' гомологичным плечом, которое гибидизуется с 5' целевой последовательностью на эндогенном локусе TrkB, и 3' гомологичным плечом, которое гибидизуется с 3' целевой последовательностью на эндогенном локусе TrkB, причем нуклеиновая кислота - вставка содержит ортологичную последовательность TRKB человека; и
(ii) нуклеазный агент, нацеленный на целевую последовательность в пределах эндогенного локуса TrkB, причем геном модифицируют для включения в него генетически модифицированного эндогенного локуса TrkB; (b) введение модифицированной плюрипотентной клетки не являющегося человеком животного в эмбрион-хозяин; и (с) имплантация эмбриона-хозяина в организм суррогатной матери для получения генетически модифицированного не являющегося человеком животного поколения F0, содержащего генетически модифицированный эндогенный локус TrkB. Необязательно плюрипотентная клетка представляет собой эмбриональную стволовую (ES) клетку. Необязательно нуклеазный агент представляет собой белок Cas9 и гидовую РНК, которая нацелена на целевую последовательность для гидовой РНК в пределах эндогенного локуса TrkB. Необязательно стадия (а) дополнительно предусматривает введение в плюрипотентную клетку не являющегося человеком животного второй гидовой РНК, которая нацелена на вторую целевую последовательность для гидовой РНК в пределах эндогенного локуса TrkB. Необязательно экзогенная матрица репарации представляет собой большой нацеливающий вектор, длина которого составляет по меньшей мере 10 т.п.н., или при этом экзогенная матрица репарации представляет собой большой нацеливающий вектор, в котором общая сумма 5' гомологичного плеча и 3' гомологичного плеча составляет по меньшей мере 10 т.п.н. в длину.
[0029] Некоторые такие способы предусматривают следующее: (а) введение в эмбрион на стадии одной клетки не являющегося человеком животного следующего: (i) экзогенная матрица репарации, содержащая нуклеиновую кислоту - вставку, фланкированную 5' гомологичным плечом, которое гибидизуется с 5' целевой последовательностью на эндогенном локусе TrkB, и 3' гомологичным плечом, которое гибидизуется с 3' целевой последовательностью на эндогенном локусе TrkB, причем нуклеиновая кислота - вставка содержит ортологичную последовательность TRKB человека; и (ii) нуклеазный агент, нацеленный на целевую последовательность в пределах эндогенного локуса TrkB, причем геном модифицируют для включения в него генетически модифицированного эндогенного локуса TrkB; и (b) имплантация модифицированного эмбриона на стадии одной клетки не являющегося человеком животного в организм суррогатной матери для получения генетически модифицированного не являющегося человеком животного поколения F0, содержащего генетически модифицированный эндогенный локус TrkB. Необязательно нуклеазный агент представляет собой белок Cas9 и гидовую РНК, которая нацелена на целевую последовательность для гидовой РНК в пределах эндогенного локуса TrkB. Необязательно стадия (а) дополнительно предусматривает введение в не относящийся к человеку эмбрион на стадии одной клетки второй гидовой РНК, которая нацелена на вторую целевую последовательность для гидовой РНК в пределах эндогенного локуса TrkB.
Краткое описание фигур
[0030] На фигуре 1 (не в масштабе) показано схематическое изображение схемы нацеливания для гуманизации области локуса TrkB мыши (Ntrk2 мыши), кодирующей внеклеточный домен TRKB. В верхней части фигуры показан эндогенный локус TrkB мыши (Ntrk2 мыши), а в нижней части фигуры показан большой нацеливающий вектор.
[0031] На фигуре 2 (не в масштабе) показано схематическое изображение анализов TAQMAN® для скрининга гуманизации локуса TrkB мыши (Ntrk2 мыши). Анализы на приобретение аллеля (GOA) включают в себя 7138hU и 7138hD. Анализы на потерю аллеля (LOA) включают в себя 7138U и 7138D.
[0032] На фигуре 3 показаны вестерн-блоты, оценивающие уровни общего TRKB и уровни фосфо-TRKB у гомозиготных гуманизированных в отношении TRKB мышей через 1 час, 4 часа и 18 часов после прямой гиппокампальной инъекции антитела - агониста TRKB Н4Н9816Р2 или антитела изотипического контроля.
[0033] На фигуре 4 (не в масштабе) показано схематическое изображение схемы нацеливания для гуманизации области локуса TrkB крысы (Ntrk2 крысы), кодирующей внеклеточный домен TRKB. В верхней части фигуры показан эндогенный локус TrkB крысы (Ntrk2 крысы), а в нижней части фигуры показан большой нацеливающий вектор.
[0034] На фигуре 5 (не в масштабе) показано схематическое изображение анализов TAQMAN® для скрининга гуманизации локуса TrkB крысы (Ntrk2 крысы) и положения гидовой РНК (целевые последовательности для гидовой РНК, как представлено в SEQ ID NO: 41-44) для нацеливания на локус TrkB крысы (Ntrk2 крысы). Анализы на приобретение аллеля (GOA) включают в себя 7138hU и 7138hD. Анализы на потерю аллеля (LOA) включают в себя rnoTU, rnoTM и rnoTD. Анализы CRISPR, предназначенные для охвата области, которая нарушается при нацеливании CRISPR/Cas9, включают в себя rnoTGU и rnoTGD. Анализы удерживания включают в себя rnoTAU2 и rnoTAD2.
[0035] На фигуре 6 показано выравнивание белков TRKB (NTRK2) мыши, крысы и человека.
[0036] На фигуре 7 показаны вестерн-блоты фосфо-TrkB, общего TrkB, фосфо-Akt, общего АКТ, фосфо-ERK и общего ERK через 15 минут и 2 часа после обработки первичных кортикальных нейронов, выделенных из детенышей первого дня после рождения гомозиготных гуманизированных в отношении TRKB мышей различными антителами -агонистами TrkB или BDNF.
[0037] На фигуре 8 показаны фармакокинетические профили антитела к TRKB Н4Н9816Р2 у гомозиготных TrkBhu/hu мышей и мышей дикого типа.
[0038] На фигуре 9 показана клеточная выживаемость в дифференцированных клетках SH-SY5Y нейробластомы человека, обработанных различными дозами антител - агонистов TrkB или BDNF. mAb1 к TrkB представляет собой Н4Н9816Р2; mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB крысы и TrkB мыши. Антитело изотипического контроля человека использовали в качестве отрицательного контроля. Данные нормировали к бессывороточной среде без антител.
[0039] На фигуре 10 показана клеточная выживаемость в первичных ганглиозных клетках сетчатки мыши, обработанных различными дозами антитела - агониста TrkB или BDNF. mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB крысы и TrkB мыши. Данные нормировали к бессывороточной среде без антител.
[0040] На фигурах 11А и 11В показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у крыс и мышей дикого типа, соответственно, после пересечения зрительного нерва и лечения с помощью BDNF, антитела - агониста TrkB, антитела изотипического контроля или контроля -носителя. Крысам вводили BDNF (5 мкг), антитело - агонист TrkB (18 мкг), антитело изотипического контроля (18 мкг) или контроль - носитель интравитреально через 3 дня и 10 дней после пересечения зрительного нерва. Мышам вводили BDNF (2,5 мкг), антитело -агонист TrkB (10 мкг), антитело изотипического контроля (10 мкг) или контроль - носитель интравитреально через 3 дня и 10 дней после пересечения зрительного нерва. mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB крысы и TrkB мыши.
[0041] На фигурах 12А и 12В показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у мышей и крыс дикого типа, соответственно, после пересечения зрительного нерва или раздавливания зрительного нерва и лечения различными дозами BDNF. На фигуре 12А показана зависимость ответа от дозы BDNF в модели раздавливания зрительного нерва (ONC) у мышей WT. На фигуре 12 В показана зависимость ответа от дозы BDNF в модели пересечения зрительного нерва у крыс WT от 0,13 мкг до 30 мкг.
[0042] На фигуре 13А показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у гомозиготных, гетерозиготных или крыс дикого типа в отношении TrkB, которым вводили либо антитело -агонист TrkB, либо антитело изотипического контроля интравитреально через 3 и 10 дней после пересечения зрительного нерва (**** = р <0,0001; *** р <0,001; двухфакторный дисперсионный анализ). Сетчатки иссекали через 14 дней после пересечения. mAb1 к TrkB представляет собой Н4Н9816Р2.
[0043] На фигуре 13 В показана плотность ганглиозных клеток сетчатки в неповрежденных глазах, иссеченных из гомозиготных, гетерозиготных или крыс дикого типа в отношении TrkB.
[0044] На фигуре 13С показана масса тела гомозиготных мышей в отношении TRKB человека, которым вводили антитело - агонист TrkB (Н4Н9816Р2; TrkB) или антитело изотипического контроля (REGN1945; контроль).
[0045] На фигуре 14 показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у гомозиготных в отношении TRKB человека крыс, которым вводили либо антитело - агонист TrkB (hTrkB; Н4Н9816Р2), либо антитело изотипического контроля (REGN1945) интравитреально через 3 и 10 дней после пересечения зрительного нерва. Сетчатки иссекали через 14 дней после пересечения.
[0046] На фигурах 15А и 15В показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у гомозиготных в отношении TRKB человека крыс, которым вводили различные антитела -агонисты TrkB (H4H9816P2-L9, H4H9814P-L9, H4H9780P-L5 или комбинацию всех трех) или антитело изотипического контроля (REGN1945) интравитреально через 3 и 10 дней после пересечения зрительного нерва (** р <0,01; критерий Краскела-Уоллиса по сравнению с антителом изотипического контроля). Сетчатки иссекали через 14 дней после пересечения. Фигура 15А включает в себя не получивший лечение контроль (неповрежденный контра латеральный глаз), а фигура 15В - нет.
[0047] На фигуре 16 показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у крыс дикого типа, которым вводили различные антитела - агонисты TrkB (Н4Н9780Р и Н4Н9814Р) или антитело изотипического контроля (REGN1945) интравитреально через 3 и 10 дней после пересечения зрительного нерва. Сетчатки иссекали через 14 дней после пересечения.
[0048] На фигурах 17А и 17В показана плотность ганглиозных клеток сетчатки в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки у гомозиготных в отношении TRKB человека мышей, которым вводили антитело - агонист TrkB (Н4Н9780Р) или антитело изотипического контроля (REGN1945) интравитреально через 3 и 10 дней после пересечения зрительного нерва. Сетчатки иссекали через 14 дней после пересечения. Фигура 17А включает в себя нормальный контроль (неповрежденный контра латеральный глаз), а фигура 17В - нет.
[0049] На фигуре 17С показана масса тела гомозиготных в отношении TRKB человека мышей, которым вводили антитело - агонист TrkB (Н4Н9780Р) или антитело изотипического контроля (REGN1945).
Определения
[0050] Используемые взаимозаменяемо в настоящем документе термины «белок», «полипептид» и «пептид» включают в себя полимерные формы аминокислот любой длины, включая в себя кодированные и некодированные аминокислоты и химически или биохимически модифицированные или дериватизированные аминокислоты. Термины также включают в себя полимеры, которые были модифицированы, такие как полипептиды, характеризующиеся модифицированными пептидными каркасами. Термин «домен» относится к любой части белка или полипептида, характеризующейся конкретной функцией или структурой.
[0051] Считается, что белки имеют «N-конец» и «С-конец». Термин «N-конец» относится к началу белка или полипептида, оканчивающемуся аминокислотой со свободной аминогруппой (-NH2). Термин «С-конец» относится к концу аминокислотной цепи (белка или полипептида), оканчивающейся свободной карбоксильной группой (-СООН).
[0052] Используемые взаимозаменяемо в настоящем документе термины «нуклеиновая кислота» и «полинуклеотид» включают в себя полимерные формы нуклеотидов любой длины, включая в себя рибонуклеотиды, дезоксирибонуклеотиды или их аналоги или модифицированные версии. Они включают в себя одно-, двух- и многоцепочечные ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК и полимеры, содержащие пуриновые основания, пиримидиновые основания или другие природные, химически модифицированные, биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
[0053] Считается, что нуклеиновые кислоты характеризуются наличием «5' концов» и «3' концов», потому что мононуклеотиды реагируют с образованием олигонуклеотидов таким образом, что 5'-фосфат одного мононуклеотидпентозного кольца прикреплен к 3'-кислороду его соседа в одном направлении через фосфодиэфирную связь. Конец олигонуклеотида называют «5'-концом», если его 5'-фосфат не связан с 3'-кислородом мононуклеотидного пентозного кольца. Конец олигонуклеотида называют «3 '-концом», если его 3 '-кислород не связан с 5'-фосфатом другого мононуклеотидного пентозного кольца. Можно сказать, что последовательность нуклеиновой кислоты, даже если она находится внутри более крупного олигонуклеотида, характеризуются наличием 5' и 3' концов. В линейной или кольцевой молекуле ДНК дискретные элементы называются «расположенными выше по отношению к ходу транскрипции» или 5' относительно «расположенных ниже» или 3' элементов.
[0054] Термин «геномно интегрированный» относится к нуклеиновой кислоте, которая была введена в клетку так, что нуклеотидная последовательность интегрируется в геном клетки и способна наследоваться ее потомством. Любой протокол можно использовать для стабильного включения нуклеиновой кислоты в геном клетки.
[0055] Термин «нацеливающий вектор» относится к рекомбинантной нуклеиновой кислоте, которую можно ввести путем гомологичной рекомбинации, лигирования, опосредованного негомологичным соединением концов, или любым другим способом рекомбинации в отношении целевого положения в геноме клетки.
[0056] Термин «вирусный вектор» относится к рекомбинантной нуклеиновой кислоте, которая включает в себя по меньшей мере один элемент вирусного происхождения и включает в себя элементы, достаточные для или разрешающие упаковку в частицу вирусного вектора. Вектор и/или частицу можно использовать с целью переноса ДНК, РНК или других нуклеиновых кислот в клетки либо ex vivo, либо in vivo. Известны многочисленные формы вирусных векторов.
[0057] Термин «дикий тип» включает в себя объекты, характеризующиеся структурой и/или активностью, обнаруживаемой в нормальном (в отличие от мутантного, болезненного, измененного и т.д.) состоянии или контексте. Гены и полипептиды дикого типа часто существуют в нескольких различных формах (например, аллели).
[0058] Термин «эндогенная» относится к последовательности нуклеиновой кислоты, которая встречается в природе внутри клетки или животного, не являющегося человеком. Например, эндогенная последовательность TrkB не являющегося человеком животного относится к нативной последовательности TrkB, которая естественным образом встречается в локусе TrkB у не являющегося человеком животного.
[0059] «Экзогенные» молекулы или последовательности включают в себя молекулы или последовательности, которые в норме не присутствуют в клетке в этой форме. Нормальное присутствие включает в себя присутствие на конкретной стадии развития и при конкретных условиях окружающей среды клетки. Например, экзогенная молекула или последовательность может включать в себя мутантную версию соответствующей эндогенной последовательности в клетке, такую как гуманизированная версия эндогенной последовательности, или может включать в себя последовательность, соответствующую эндогенной последовательности в клетке, но в другой форма (т.е. не в пределах хромосомы). Напротив, эндогенные молекулы или последовательности включают в себя молекулы или последовательности, которые в норме присутствуют в такой форме в конкретной клетке на определенной стадии развития в определенных условиях окружающей среды.
[0060] Термин «гетерологичный» при использовании в контексте нуклеиновой кислоты или белка указывает на то, что нуклеиновая кислота или белок содержит по меньшей мере две части, которые в природе не встречаются вместе в одной и той же молекуле. Например, термин «гетерологичный» при использовании в отношении частей нуклеиновой кислоты или частей белка указывает, что нуклеиновая кислота или белок содержит две или более подпоследовательности, которые не находятся в таком же отношении друг к другу (например, объединены) в природе. В качестве одного примера, «гетерологичная» область нуклеиновокислотного вектора представляет собой сегмент нуклеиновой кислоты в пределах или прикрепленный к другой молекуле нуклеиновой кислоты, которая не обнаружена в ассоциации с другой молекулой в природе. Например, гетерологичная область нуклеиновокислотного вектора может включать в себя кодирующую последовательность, фланкированную последовательностями, которые не обнаружены в природе в ассоциации с кодирующей последовательностью. Аналогично, «гетерологичная» область белка представляет собой сегмент аминокислот в пределах или прикрепленный к другой пептидной молекуле, который не обнаружен в природе в ассоциации с другой пептидной молекулой (например, слитый белок или белок с меткой). Аналогично, нуклеиновая кислота или белок могут содержать гетерологичную метку или гетерологичную последовательность секреции или гетерологичную последовательность локализации.
[0061] «Оптимизация ко донов» использует преимущества вырожденности ко донов, что проявляется в множественности комбинаций ко донов с тремя парами оснований, которые определяют аминокислоту, и, как правило, включает в себя процесс модификации последовательности нуклеиновой кислоты для усиленной экспрессии в конкретных клетках-хозяевах путем замены по меньшей мере одного кодона нативной последовательности на ко дон, который чаще или наиболее часто используется в генах клетки-хозяина при сохранении нативной аминокислотной последовательности. Например, нуклеиновую кислоту, кодирующую белок Cas9, можно модифицировать для замены кодонов, характеризующихся более высокой частотой использования в данной прокариотической или эукариотической клетке, включая в себя бактериальную клетку, дрожжевую клетку, клетку человека, клетку, не относящуюся к человеку, клетку млекопитающего, клетку грызуна, клетку мыши, клетку крысы, клетку хомяка или любую другую клетку-хозяина по сравнению с встречающейся в природе последовательностью нуклеиновой кислоты. Таблицы использования кодонов легко доступны, например, в «Базе данных использования кодонов» («Codon Usage Database»). Эти таблицы можно адаптировать несколькими способами. См. Nakamura et al. (2000) Nucleic Acids Research 28:292, полностью включенный в настоящий документ посредством ссылки для всех целей. Также доступны компьютерные алгоритмы для оптимизации кодонов конкретной последовательности для экспрессии в конкретном хозяине (см., например, Gene Forge).
[0062] Термин «локус» относится к конкретному местоположению гена (или значимой последовательности), последовательности ДНК, кодирующей полипептид последовательности или положению в хромосоме генома организма. Например, «локус TrkB» может относиться к конкретному местоположению гена TrkB, последовательности ДНК TrkB, последовательности, кодирующей TrkB, или положению TrkB в хромосоме генома организма, который был идентифицирован относительно того, где такая последовательность расположена. «Локус TrkB» может содержать регуляторный элемент гена TrkB, включая в себя, например, энхансер, промотор, 5'- и/или З'-нетранслируемую область (UTR) или их комбинацию.
[0063] Термин «ген» относится к последовательности ДНК в хромосоме, которая кодирует продукт (например, продукт РНК и/или полипептидный продукт) и включает в себя кодирующую область, прерываемую некодирующими нитронами, и последовательность, расположенную смежно с кодирующей областью на 5' и 3' концах так, что ген соответствует полноразмерной мРНК (включая в себя 5' и 3' нетранслируемые последовательности). Термин «ген» также включает в себя другие некодирующие последовательности, включая в себя регуляторные последовательности (например, промоторы, энхансеры и сайты связывания транскрипционных факторов), сигналы полиаденилирования, участки внутренней посадки рибосомы, сайленсеры, изолирующую последовательность и области прикрепления матрицы. Эти последовательности могут быть близки к кодирующей области гена (например, в пределах 10 т.п.н.) или могут быть расположены на удаленных сайтах, и они влияют на уровень или скорость транскрипции и трансляции гена.
[0064] Термин «аллель» относится к вариантной форме гена. Некоторые гены имеют множество различных форм, которые расположены в одном и том же положении или генетическом локусе в хромосоме. Диплоидный организм содержит два аллеля в каждом генетическом локусе. Каждая пара аллелей представляет генотип определенного генетического локуса. Генотипы описаны как гомозиготные, если в конкретном локусе есть два идентичных аллеля, и как гетерозиготные, если два аллеля различаются.
[0065] «Промотор» представляет собой регуляторную область ДНК, как правило, содержащую ТАТА-бокс, способный направлять РНК-полимеразу II для инициации синтеза РНК в соответствующем сайте инициации транскрипции для конкретной полинуклеотидной последовательности. Промотор может дополнительно содержать другие области, которые влияют на скорость инициации транскрипции. Раскрытые в настоящем документе промоторные последовательности модулируют транскрипцию функционально связанного полинуклеотида. Промотор может являться активным в одном или нескольких типах клеток, раскрытых в настоящем документе (например, эукариотическая клетка, клетка млекопитающего, не являющегося человеком, клетка человека, клетка грызуна, плюрипотентная клетка, эмбрион на стадии одной клетки, дифференцированная клетка или их комбинация). Промотор может являться, например, конститутивно активным промотором, условным промотором, индуцируемым промотором, ограниченным по времени промотором (например, промотором, регулируемым стадией развития) или пространственно ограниченным промотором (например, клеточноспецифическим или тканеспецифическим промотором). Примеры промоторов можно найти, например, в международной патентной публикации WO 2013/176772, полностью включенной в настоящий документ посредством ссылки для всех целей.
[0066] Термин «функциональная связь» или «функционально связанный» включает в себя смежное положение двух или более компонентов (например, промотора и другого элемента последовательности) так, что оба компонента функционируют нормально и обеспечивают возможность того, что по меньшей мере один из компонентов может опосредовать функцию, на которую действует по крайней мере один из других компонентов. Например, промотор может быть функционально связан с кодирующей последовательностью, если промотор контролирует уровень транскрипции кодирующей последовательности в ответ на присутствие или отсутствие одного или нескольких транскрипционных регуляторных факторов. Функциональная связь может включать в себя такие последовательности, которые являются смежными друг с другом или действуют in trans (например, регуляторная последовательность может действовать на расстоянии для управления транскрипцией кодирующей последовательности).
[0067] Термин «вариант» относится к нуклеотидной последовательности, отличающейся от последовательности, наиболее распространенной в популяции (например, одним нуклеотидом), или последовательности белка, отличающейся от последовательности, наиболее распространенной в популяции (например, одной аминокислотой).
[0068] Термин «фрагмент» применительно к белку означает белок, который является более коротким или содержит меньше аминокислот, чем полноразмерный белок. Термин «фрагмент» в отношении нуклеиновой кислоты означает нуклеиновую кислоту, которая является более короткой или содержит меньше нуклеотидов, чем полноразмерная нуклеиновая кислота. Фрагмент может являться, например, N-концевым фрагментом (т.е. удаление части С-концевого конца белка), С-концевым фрагментом (т.е. удаление части N-концевого конца белка) или внутренним фрагментом.
[0069] Термин «идентичность последовательности» или «идентичность» в контексте двух полинуклеотидных или полипептидных последовательностей относится к остаткам в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия в указанном окне сравнения. Когда процент идентичности последовательности используют в отношении белков, положения остатков, которые не являются идентичными, часто отличаются консервативными аминокислотными заменами, где аминокислотные остатки заменены другими аминокислотными остатками со сходными химическими свойствами (например, заряд или гидрофобность) и, следовательно, не меняют функциональные свойства молекулы. Когда последовательности отличаются по консервативным заменам, процентную идентичность последовательности можно повысить для корректировки консервативного характера замены. Говорят, что последовательности, которые отличаются такими консервативными заменами, характеризуются «сходством последовательностей» или «сходством». Средства для такой корректировки хорошо известны. Как правило, это включает в себя оценку консервативной замены как частичного, а не полного несовпадения, что увеличивает процентную идентичность последовательности. Таким образом, например, когда идентичная аминокислота получает балл 1, а неконсервативная замена получает оценку 0, консервативная замена получает балл от 0 до 1. Рассчитывают балл консервативных замен, например, как это реализовано в программе PC/GENE (Intelligenetics, Маунтин-Вью, Калифорния США).
[0070] «Процентное отношение идентичности последовательности» включает в себя значение, определенное путем сравнения двух оптимально выровненных последовательностей (наибольшее количество идеально совпадающих остатков) в окне сравнения, причем часть полинуклеотидной последовательности в окне сравнения может содержать добавления или удаления (т.е. пропуски) по сравнению с эталонной последовательностью (которая не содержит добавления или удаления) для оптимального выравнивания двух последовательностей. Процентное отношение рассчитывают путем определения количества положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях, для получения числа совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения и умножение результата на 100 для получения процентного отношения идентичности последовательности. Если не указано иное (например, более короткая последовательность включает в себя связанную гетерологичную последовательность), окно сравнения представляет собой полную длину более короткой из двух сравниваемых последовательностей.
[0071] Если не указано иное, значения идентичности/сходства последовательности включают в себя значение, полученное с использованием GAP версии 10 с использованием следующих параметров: % идентичности и % сходства для нуклеотидной последовательности с использованием штрафа за внесение пропуска согласно GAP, составляющего 50, и штрафа за удлинение пропуска, составляющего 3, и матрицы замен nwsgapdna.cmp; % идентичности и % сходства для аминокислотной последовательности с использованием штрафа за внесение пропуска согласно GAP, составляющего 8, и штрафа за удлинение пропуска, составляющего 2, и матрицы замен BLOSUM62; или любой эквивалентной им программы. «Эквивалентная программа» включает в себя любую программу сравнения последовательностей, которая для любых двух рассматриваемых последовательностей генерирует выравнивание, характеризующееся идентичными совпадениями нуклеотидных или аминокислотных остатков и идентичным процентным отношением идентичности последовательности по сравнению с соответствующим выравниванием, сгенерированным с помощью GAP версии 10.
[0072] Термин «консервативная аминокислотная замена» относится к замене аминокислоты, которая обычно присутствует в последовательности, на другую аминокислоту схожего размера, заряда или полярности. Примеры консервативных замен включают в себя замену неполярного (гидрофобного) остатка, такого как изолейцин, валин или лейцин, на другой неполярный остаток. Аналогичным образом, примеры консервативных замен включают в себя замену одного полярного (гидрофильного) остатка на другой, например, замена между аргинином и лизином, между глутамином и аспарагином или между глицином и серином. Кроме того, замена основного остатка, такого как лизин, аргинин или гистидин, на другой, или замена одного кислотного остатка, такого как аспарагиновая кислота или глутаминовая кислота, на другой кислотный остаток являются дополнительными примерами консервативных замен. Примеры неконсервативных замен включают в себя замену неполярного (гидрофобного) аминокислотного остатка, такого как изолейцин, валин, лейцин, аланин или метионин, на полярный (гидрофильный) остаток, такой как цистеин, глутамин, глутаминовая кислота или лизин, и/или полярного остатка на неполярный остаток. Типичные классификации аминокислот приведены в таблице ниже.

[0073] «Гомологичная» последовательность (например, последовательность нуклеиновой кислоты) включает в себя последовательность, которая либо является идентичной, либо по существу сходной известной эталонной последовательности, так что она является, например, по меньшей мере на 50%, по меньшей мере на 55%, по меньшей мере на 60%, по меньшей мере на 65%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичной известной эталонной последовательности. Гомологичные последовательности могут включать в себя, например, ортологичную последовательность и паралогичные последовательности. Гомологичные гены, например, как правило, происходят от общей предковой последовательности ДНК, либо посредством события видообразования (ортологичные гены), либо события генетической дупликации (паралогичные гены). «Ортологичные» гены включают в себя гены у различных видов, которые произошли от общего предкового гена путем видообразования. Ортологи, как правило, сохраняют ту же функцию в ходе эволюции. «Паралогичные» гены включают в себя гены, связанные с дупликацией в геноме. Паралоги могут развивать новые функции в ходе эволюции.
[0074] Термин «in vitro» включаете себя искусственные среды и относится к процессам или реакциям, которые происходят в искусственной среде (например, в пробирке). Термин «in vivo» включает в себя природные среды (например, клетку, организм или тело) и относится к процессам или реакциям, которые происходят в естественной среде. Термин «ех vivo» включает в себя клетки, которые были удалены из организма человека, а также относится к процессам или реакциям, которые происходят в таких клетках.
[0075] Термин «репортерный ген» относится к нуклеиновой кислоте с последовательностью, кодирующей продукт гена (как правило, фермент), который легко и количественно анализируется, когда конструкт, содержащий последовательность репортерного гена, функционально связан с эндогенным или гетерологичным промоторным и/или энхансерным элементом, введен в клетки, содержащие (или можно сделать так, чтобы они содержали) факторы, необходимые для активации промоторных и/или энхансерных элементов. Примеры репортерных генов включают в себя без ограничения гены, кодирующие бета-галактозидазу (lacZ), гены бактериальной хлорамфениколацетилтрансферазы (кошка), гены люциферазы светлячка, гены, кодирующие бета-глюкуронидазу (GUS), и гены, кодирующие флуоресцентные белки. «Репортерный белок» относится к белку, кодируемому репортерным геном.
[0076] Используемый в настоящем документе термин «флуоресцентный репортерный белок» означает репортерный белок, который можно обнаружить на основе флуоресценции, где флуоресценция может быть либо непосредственно от репортерного белка, активности репортерного белка на флуорогенном субстрате, либо белка с аффинностью в отношении связывания с флуоресцентным меченым соединением. Примеры флуоресцентных белков включают в себя зеленые флуоресцентные белки (например, GFP, GFP-2, tagGFP, turboGFP, eGFP, изумрудно-зеленый, Azami Green, мономерный Azami Green, CopGFP, AceGFP и ZsGreenl), желтые флуоресцентные белки (например, YFP, eYFP, цитрин, Venus, YPet, PhiYFP и ZsYellowl), синие флуоресцентные белки (например, BFP, eBFP, eBFP2, азурит, mKalamal, GFPuv, сапфир и Т-сапфир), голубые флуоресцентные белки (например, CFP, eCFP, лазурь, CyPet, AmCyanl и Midoriishi-Cyan), красные флуоресцентные белки (например, RFP, mKate, mKate2, mPlum, DsRed-мономер, mCherry, mRFPl, DsRed-Express, DsRed2, DsRed-мономер, HcRed-Tandem, HcRedl, AsRed2, eqFP611, mRaspberry, mStrawberry и Jred), оранжевые флуоресцентные белки (например, mOrange, mKO, Kusabira-Orange, мономерный Kusabira-Orange, mTangerine и tdTomato) и любой другой подходящий флуоресцентный белок, присутствие которого в клетках можно обнаружить с помощью способов проточной цитометрии.
[0077] Термин «рекомбинация» включает в себя любой процесс обмена генетической информацией между двумя полинуклеотидами и может происходить по любому механизму. Рекомбинация в ответ на двухцепочечные разрывы (DSB) происходит главным образом посредством двух консервативных путей репарации ДНК: негомологичного соединения концов (NHEJ) и гомологичной рекомбинации (HR). См. Kasparek & Humphrey (2011) Seminars in Cell & Dev. Biol. 22: 886-897, полностью включенный в настоящий документ посредством ссылки для всех целей. Аналогичным образом, репарация целевой нуклеиновой кислоты, опосредованное экзогенной донорной нуклеиновой кислотой, может включать в себя любой процесс обмена генетической информацией между двумя полинуклеотидами.
[0078] NHEJ включает в себя репарацию двухцепочечных разрывов в нуклеиновой кислоте путем прямого лигирования концов разрывов друг с другом или с экзогенной последовательностью без необходимости гомологичной матрицы. Лигирование несмежных последовательностей с помощью NHEJ часто может приводить к делециям, вставкам или транслокациям вблизи сайта двухцепочечного разрыва. Например, NHEJ также может приводить к целенаправленной интеграции экзогенной донорной нуклеиновой кислоты посредством прямого лигирования концов разрыва с концами экзогенной донорной нуклеиновой кислоты (т.е. захват на основе NHEJ). Такая NHEJ-опосредованная целевая интеграция может быть предпочтительной для вставки экзогенной донорной нуклеиновой кислоты, когда пути гомологичной репарации (HDR) нелегко использовать (например, в неделящихся клетках, первичных клетках и клетках, которые плохо выполняют репарацию ДНК на основе гомологии). Кроме того, в отличие от гомологичной репарации, знание относительно больших областей идентичности последовательностей, фланкирующих сайт расщепления, не требуется, что может быть благоприятным при попытке целенаправленной вставки в организмы, имеющие геномы, для которых знания о геномной последовательности ограничены. Интеграция может происходить посредством лигирования тупых концов между экзогенной донорной нуклеиновой кислотой и расщепленной геномной последовательностью или посредством лигирования липких концов (т.е. характеризующихся 5' или 3' свисающими концами) с использованием экзогенной донорной нуклеиновой кислоты, которая фланкирована свисающими концами, которые совместимы с таковыми, созданными нуклеазным агентом в расщепленной геномной последовательности. См., например, документы US 2011/020722, WO 2014/033644, WO 2014/089290 и Maresca et al. (2013) Genome Res. 23(3):539-546, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Если тупые концы лигированы, может потребоваться вырезание мишени и/или донора для создания областей микрогомологии, необходимых для соединения фрагментов, что может привести к нежелательным изменениям в целевой последовательности.
[0079] Рекомбинация может происходить посредством гомологичной репарации (HDR) или гомологичной рекомбинации (HR). HDR или HR включают в себя форму репарации нуклеиновой кислоты, которая может требовать гомологии нуклеотидной последовательности, использует «донорную» молекулу в качестве матрицы для репарации «целевой» молекулы (т.е. той, которая испытала двухцепочечный разрыв) и приводит к переносу генетической информации от донора к мишени. Не желая быть связанными какой-либо конкретной теорией, такой перенос может включать в себя коррекцию несовпадения гетеродуплексной ДНК, которая образуется между поврежденной мишенью и донором, и/или отжиг в зависимости от синтеза нитей, при котором донора используют для повторного синтеза генетической информации, которая станет частью мишени, и/или связанные процессы. В некоторых случаях донорный полинуклеотид, часть донорного полинуклеотида, копия донорного полинуклеотида или часть копии донорного полинуклеотида интегрируется в целевую ДНК. См. Wang et al. (2013) Cell 153:910-918; Mandalos et al. (2012) PLOS ONE7:e45768:l-9; и Wang et al. (2013) Nat Biotechnol. 31:530-532, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[0080] Термин «антигенсвязывающий белок» включает в себя любой белок, который связывается с антигеном. Примеры антигенсвязывающих белков включают в себя антитело, антигенсвязывающий фрагмент антитела, мультиспецифическое антитело (например, биспецифическое антитело), scFV, би-scFV, диатела, триатела, тетратела, V-NAR, VHH, VL, F(ab), F(ab)2, DVD (антигенсвязывающий белок с двойным вариабельным доменом), SVD (антигенсвязывающий белок с одинарным вариабельным доменом), привлекающий Т-клетки биспецифический активатор (BiTE) или Davisbody (патент США №8586713, полностью включенный в настоящий документ посредством ссылки для всех целей).
[0081] Используемое в настоящем документе выражение «антитело к TRKB» включает в себя как моновалентные антитела с одинарной специфичностью, а также биспецифические антитела, содержащие первое плечо, которое связывается с TRKB, и второе плечо, которое связывается со вторым (целевым) антигеном, причем анти-TRKB плечо содержит, например, любую из последовательностей HCVR/LCVR или CDR, как изложено в таблице 22 в настоящем документе. Выражение «антитело к TRKB» также включает в себя конъюгаты антитела с лекарственным средством (ADC), содержащие антитело к TRKB или его антигенсвязывающую часть, конъюгированные с лекарственным средством или токсином (т.е. цитотоксическим агентом). Выражение «антитело к TRKB» также включает в себя конъюгаты антитела с радионуклидом (ARC), содержащие антитело к TRKB или его антигенсвязывающую часть, конъюгированные с радионуклидом.
[0082] Используемый в настоящем документе термин «антитело к TRKB» означает любую антигенсвязывающую молекулу или молекулярный комплекс, содержащие по меньшей мере одну определяющую комплементарность область (CDR), которая специфически связывается или взаимодействует с TRKB или частью TRKB. Термин «антитело» включает в себя молекулы иммуноглобулина, содержащие четыре полипептидные цепи, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями, а также их мультимеры (например, IgM). Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (сокращенно в настоящем документе HCVR или VH) и константную область тяжелой цепи. Константная область тяжелой цепи содержит три домена, CH1, СН2 и СН3. Каждая легкая цепь содержит вариабельную область легкой цепи (сокращенно в настоящем документе «LCVR или «VL») и константную область легкой цепи (CL). Области VH и VL можно дополнительно подразделить на области гипервариабельности, называемые определяющими комплементарность областями (CDR), чередующиеся с областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбоксиконца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Согласно различным вариантам осуществления FR антитела к TRKB (или его антигенсвязывающей части) могут являться идентичными последовательностям зародышевой линии человека или могут быть естественным образом или искусственно модифицированы. Аминокислотную консенсусную последовательность можно определить на основании параллельного сравнительного анализа двух или более CDR.
[0083] Используемый в настоящем документе термин «антитело» также включает в себя антигенсвязывающие фрагменты молекул полноразмерных антител. Используемые в настоящем документе термины «антигенсвязывающая часть» антитела, «антигенсвязывающий фрагмент» антитела и тому подобное включают в себя любой полученный ферментативно, синтетический или генетически сконструированный полипептид или гликопротеин, который специфически связывается с антигеном с образованием комплекса. Антигенсвязывающие фрагменты антитела можно получить, например, из полных молекул антитела с использованием любых подходящих стандартных техник, таких как протеолитическое расщепление или рекомбинантные техники генной инженерии, предусматривающие манипуляции с ДНК и экспрессию ДНК, кодирующую вариабельные и необязательно константные домены. Такая ДНК известна и/или легко доступна, например, из коммерческих источников, библиотек ДНК (включая в себя, например, фаговые библиотеки антител), или же ее можно синтезировать. ДНК можно секвенировать и воздействовать на нее химически или с использованием техник молекулярной биологии, например, для расположения одного или нескольких вариабельных и/или константных доменов в подходящую конфигурацию или для введения кодонов, создания цистеиновых остатков, модификации, добавления или удаления аминокислот и т.д.
[0084] Неограничивающие примеры антигенсвязывающих фрагментов включают в себя следующее: (i) фрагменты Fab; (ii) фрагменты F(ab')2; (iii) фрагменты Fd; (iv) фрагменты Fv; (v) одноцепочечные молекулы Fv (scFv); (vi) фрагменты dAb; и (vii) минимальные распознающие звенья, состоящие из аминокислотных остатков, которые имитируют гипервариабельную область антитела (например, выделенная определяющая комплементарность область (CDR), такая как пептид CDR3) или пептид с ограниченной конформационной свободой FR3-CDR3-FR4. Другие сконструированные молекулы, такие как домен-специфические антитела, однодоменные антитела, антитела с удаленными доменами, химерные антитела, CDR-привитые антитела, диатела, триатела, тетратела, миниантитела, наноантитела (например моновалентные наноантитела, бивалентные наноантитела и т.д.), иммунофармацевтические средства на основе модульного белка малого размера (SMIP) и вариабельные домены IgNAR акулы, также предусмотрены используемым в настоящем документе выражением «антигенсвязывающий фрагмент».
[0085] Антигенсвязывающий фрагмент антитела, как правило, будет содержать по меньшей мере один вариабельный домен. Вариабельный домен может характеризоваться любым размером или аминокислотным составом и, как правило, будет содержать по меньшей мере одну CDR, которая является смежной или расположена в одной рамке считывания с одной или несколькими каркасными последовательностями. В антигенсвязывающих фрагментах, содержащих домен VH, ассоциированный с доменом VL, домены VH и VL могут быть расположены относительно друг друга в любом подходящем расположении. Например, вариабельная область может являться димерной и может содержать димеры VH-VH, VH-VL или VL-VL. Альтернативно, антигенсвязывающий фрагмент антитела может содержать мономерный домен VH или VL.
[0086] Согласно определенным вариантам осуществления антигенсвязывающий фрагмент антитела может содержать по меньшей мере один вариабельный домен, ковалентно связанный по меньшей мере с одним константным доменом. Неограничивающие иллюстративные конфигурации вариабельных и константных доменов, которые можно обнаружить в пределах антигенсвязывающего фрагмента антитела согласно настоящему изобретению, включают в себя следующее: (i) VH-CH1; (ii) VH-CH2; (iii) VH-CH3; (iv) VH-CH1-CH2; (v) VH-CH1-CH2-CH3; (vi) VH-CH2-CH3; (vii) VH-CL; (viii) VL-CH1; (ix) VL-CH2; (x) VL-CH3; (xi) VL-CH1-CH2; (xii) VL-CH1-CH2-CH3; (xiii) VL-CH2-CH3; и (xiv) VL-CL. В любой конфигурации вариабельных и константных доменов, включая в себя любую из иллюстративных конфигураций, перечисленных выше, вариабельные и константные домены могут быть либо напрямую связаны друг с другом, либо могут быть связаны с помощью полной или частичной шарнирной или линкерной области. Шарнирная область может состоять по меньшей мере из 2 (например, 5, 10, 15, 20, 40, 60 или больше) аминокислот, что дает в результате гибкое или полугибкое соединение между смежными вариабельными и/или константными доменами в одной полипептидной молекуле. Более того, антигенсвязывающий фрагмент антитела согласно настоящему изобретению может содержать гомодимер или гетеродимер (или другой мультимер) любой из конфигураций вариабельных и константных доменов, перечисленных выше, в нековалентной ассоциации друг с другом и/или с одним или несколькими мономерными доменами VH или VL (например, с помощью дисульфидной(ых) связи(ей)).
[0087] Как и в случае с полными молекулами антитела, антигенсвязывающие фрагменты могут являться моноспецифическими или мультиспецифическими (например, биспецифическими). Мультиспецифический антигенсвязывающий фрагмент антитела, как правило, будет содержать по меньшей мере два различных вариабельных домена, причем каждый вариабельный домен способен специфически связываться с отдельным антигеном или с другим эпитопом на одном и том же антигене. Любой формат мультиспецифических антител, включая в себя иллюстративные форматы биспецифических антител, раскрытые в настоящем документе, можно адаптировать для применения в контексте антигенсвязывающего фрагмента антитела согласно настоящему изобретению с использованием стандартных техник.
[0088] Термин «эпитоп» относится к антигенной детерминанте, которая взаимодействует со специфическим сайтом связывания с антигеном в вариабельной области молекулы антитела, известной как паратоп. Отдельный антиген может содержать больше одного эпитопа. Таким образом, различные антитела могут связываться с различными областями на антигене и могут характеризоваться различными биологическими эффектами. Эпитопы могут являться либо конформационными, либо линейными. Конформационный эпитоп образован пространственно смежными аминокислотами из различных сегментов неразветвленной полипептидной цепи. Линейный эпитоп образован смежными аминокислотными остатками в полипептидной цепи. В определенных обстоятельствах эпитоп может включать в себя фрагменты сахаридов, фосфорильных групп или сульфонильных групп на антигене.
[0089] Термин «специфически связывает» или «специфически связывается с» или тому подобное означает, что антитело или его антигенсвязывающий фрагмент образует комплекс с антигеном, который является относительно стабильным в физиологических условиях. Специфическое связывание может характеризоваться равновесной константой диссоциации, равной по меньшей мере приблизительно 1×10-6 М или меньше (например, меньшее значение KD обозначает более плотное связывание). Способы определения того, являются ли две молекулы специфически связывающимися, хорошо известны в настоящей области техники и включают в себя, например, равновесный диализ, поверхностный плазмонный резонанс и тому подобное. Как описано в настоящем документе, с помощью поверхностного плазмонного резонанса, например, BIACORE™, были идентифицированы антитела, которые связываются специфически с TRKB. Более того, мультиспецифические антитела, которые связываются с белком TRKB и одним или несколькими дополнительными антигенами, или биспецифические, которые связываются с двумя различными областями TRKB, тем не менее считаются антителами, которые «связываются специфически», как используется в настоящем документе.
[0090] Раскрытые в настоящем документе антитела к TRKB могут содержать одну или несколько аминокислотных замен, вставок и/или делеции в каркасных областях и/или областях CDR вариабельных доменов тяжелой и легкой цепей. Такие мутации можно легко установить путем сравнения аминокислотных последовательностей, раскрытых в настоящем документе, с последовательностями, доступными, например, из общедоступных баз данных последовательностей антител. После получения антитела и антигенсвязывающие фрагменты, которые содержат одну или несколько мутаций, можно легко испытать в отношении одного или нескольких требуемых свойств, таких как улучшенная специфичность связывания, увеличенная аффинность связывания, улучшенные или усиленные антагонистические или агонистические биологические свойства (в зависимости от конкретной ситуации), сниженная иммуногенность и т.д. Антитела и антигенсвязывающие фрагменты, полученные этим общим способом, предусмотрены настоящим изобретением.
[0091] Кроме того, предусмотрены антитела к TRKB, содержащие варианты любой из аминокислотных последовательностей HCVR, LCVR и/или CDR, раскрытых в настоящем документе, с одной или несколькими консервативными заменами. Например, согласно настоящему изобретению предусмотрены антитела к TRKB с аминокислотными последовательностями HCVR, LCVR и/или CDR, например, с 10 или меньше, 8 или меньше, 6 или меньше, 4 или меньше и т.д. консервативными аминокислотными заменами по отношению к любой из аминокислотных последовательностей HCVR, LCVR и/или CDR, раскрытых в таблице 22.
[0092] Термин «существенная идентичность» или «по существу идентичный», когда он относится к нуклеиновой кислоте или ее фрагменту в контексте антител к TRKB, указывает на то, что при оптимальном выравнивании с соответствующими нуклеотидными вставками или делециями с другой нуклеиновой кислотой (или ее комплементарной цепью) существует идентичность нуклеотидной последовательности по меньшей мере приблизительно в 90% и более предпочтительно по меньшей мере приблизительно в 95%, 96%, 97%, 98% или 99% нуклеотидных оснований, как измерено с помощью любого хорошо известного алгоритма измерения идентичности последовательностей, такого как FASTA, BLAST или GAP, как обсуждается ниже. Молекула нуклеиновой кислоты с существенной идентичностью по отношению к эталонной молекуле нуклеиновой кислоты в некоторых случаях может кодировать полипептид, характеризующийся такой же или по существу сходной аминокислотной последовательностью, что и полипептид, кодируемый эталонной молекулой нуклеиновой кислоты.
[0093] Применительно к полипептидам в контексте антител к TRKB термин «существенное сходство» или «по существу сходный» означает, что две пептидные последовательности, при оптимальном выравнивании, например, с помощью программ GAP или BESTFIT с использованием штрафов за внесение пропусков по умолчанию, характеризуются по меньшей мере 95% идентичностью последовательностей, еще более предпочтительно по меньшей мере 98% или 99% идентичностью последовательностей. Предпочтительно положения остатков, которые не являются идентичными, отличаются консервативными аминокислотными заменами. «Консервативная аминокислотная замена» применительно к полипептидам в контексте антител к TRKB представляет собой замену, при которой аминокислотный остаток заменен другим аминокислотным остатком, содержащим боковую цепь (группу R) со сходными химическими свойствами (например, заряд или гидрофобность). В целом, консервативная аминокислотная замена существенно не изменит функциональные свойства белка. В случаях, когда две или более аминокислотных последовательностей отличаются друг от друга консервативными заменами, процентное отношение или степень сходства можно скорректировать в сторону увеличения, чтобы отрегулировать консервативный характер замены. Средства для осуществления этой регулировки хорошо известны специалистам в настоящей области техники. См., например, Pearson (1994) Methods Mol. Biol. 24: 307-331, которая включена в настоящий документ посредством ссылки. Примеры групп аминокислот, которые содержат боковые цепи со сходными химическими свойствами, включают в себя следующее: 1) алифатические боковые цепи: глицин, аланин, валин, лейцин и изолейцин; 2) алифатически-гидроксильные боковые цепи: серии и треонин; 3) амидсодержащие боковые цепи: аспарагин и глутамин; 4) ароматические боковые цепи: фенилаланин, тирозин и триптофан; 5) основные боковые цепи: лизин, аргинин и гистидин; 6) кислотные боковые цепи: аспартат и глутамат и 7) серосодержащие боковые цепи: цистеин и метионин. Предпочтительные группы консервативных аминокислотных замен представляют собой: валин-лейцин-изолейцин, фенилаланин-тирозин, лизин-аргинин, аланин-валин, глутамат-аспартат и аспарагин-глутамин. Альтернативно, консервативной заменой является любое изменение, характеризующееся положительным значением в матрице логарифмического правдоподобия РАМ250, раскрытой в Gonnet et al. (1992) Science 256: 1443 45, включенной в настоящий документ посредством ссылки. «Умеренно консервативная» замена представляет собой любое изменение, характеризующееся неотрицательным значением в матрице логарифмического правдоподобия РАМ250.
[0094] Композиции или способы, «предусматривающие» или «включающие в себя» один или несколько перечисленных элементов, могут включать в себя другие элементы, которые не указаны конкретно. Например, композиция, которая «содержит» или «включает в себя» белок, может содержать белок отдельно или в комбинации с другими ингредиентами. Переходная фраза «состоящий по существу из» означает, что объем формулы изобретения должен интерпретироваться как охватывающий указанные элементы, перечисленные в формуле изобретения, и элементы, которые не оказывают существенного влияния на основную(ые) и новую(ые) характеристику(и) заявленного изобретения. Таким образом, не предусмотрено, что термин «состоящий по существу из» при использовании в формуле изобретения следует интерпретировать как эквивалент «содержащий».
[0095] «Необязательно» или «необязательно» означает, что впоследствии описанное событие или обстоятельство может произойти или может не произойти, и что описание включает в себя случаи, в которых происходит событие или обстоятельство, и случаи, в которых оно не происходит.
[0096] Обозначение диапазона значений включает в себя все целые числа в пределах диапазона или определяющие диапазон и все поддиапазоны, определенные целыми числами в пределах диапазона.
[0097] Если иное не очевидно из контекста, термин «приблизительно» охватывает значения в пределах стандартного предела погрешности измерения (например, SEM) установленного значения.
[0098] Термин «и/или» относится и охватывает любые возможные комбинации одного или нескольких связанных перечисленных пунктов, а также отсутствие комбинаций при интерпретации в альтернативе («или»).
[0099] Термин «или» относится к любому одному представителю конкретного перечня, а также включает в себя любую комбинацию представителей этого перечня.
[00100] Формы единственного числа включают в себя формы множественного числа, если контекст явно не предписывает иное. Например, термин «белок» или «по меньшей мере один белок» может включать в себя множество белков, включая в себя их смеси.
[00101] Статистически значимый означает р ≤0,05.
Подробное раскрытие настоящего изобретения
I. Обзор
[00102] В настоящем документе раскрыты геномы не являющегося человеком
животного, клетки не являющегося человеком животного и не являющиеся человеком животные, содержащие гуманизированный локус TRKB, и способы применения таких клеток не являющегося человеком животного и не являющихся человеком животных. Клетки не являющегося человеком животного или не являющиеся человеком животные, содержащие гуманизированный локус TRKB, экспрессируют белок TRKB человека или химерный белок TRKB, содержащий один или несколько фрагментов белка TRKB человека (например, весь или часть внеклеточного домена TRKB человека).
[00103] Гуманизированный аллель TRKB (например, полученный в результате замены всей или части геномной ДНК не являющегося человеком животного «один к одному» ортологичной геномной ДНК человека) обеспечит истинную человеческую мишень или близкое приближение к истинной человеческой мишени нацеленных на TRKB человека реагентов (например, антитела - агонисты или малые молекулы-агонисты, предназначенные для нацеливания на TRKB человека), что позволяет испытывать эффективность и способ действия таких агентов у живых животных, а также проводить исследования фармакокинетики и фармакодинамики. Например, как показано в рабочих примерах, раскрытых в настоящем документе, интравитреальное введение агонистов - антител к TRKB обладает значительным нейропротекторным эффектом после повреждения зрительного нерва у гуманизированных в отношении TrkB крыс.
II. Не являющиеся человеком животные, содержащие гуманизированный локус TRKB
[00104] Геномы не являющегося человеком животного, клетки не являющегося человеком животного и не являющиеся человеком животные, раскрытые в настоящем документе, содержат гуманизированный локус TRKB. Клетки или не являющиеся человеком животные, содержащие гуманизированный локус TRKB, экспрессируют белок TRKB человека или частично гуманизированный, химерный белок TRKB, в котором один или несколько фрагментов нативного белка TRKB были заменены соответствующими фрагментами из TRKB человека (например, весь или часть внеклеточного домена).
A. TRKB
[00105] Клетки и не являющиеся человеком животные, описанные в настоящем
документе, содержат гуманизированный локус TRKB. TRKB (также известный как рецептор факторов роста BDNF-NT-3, GP145-TrkB, Trk-B, TrkB, рецептор нейротрофической тирозинкиназы 2 типа, тирозинкиназа TrkB, связанная с тропомиозином киназа В, тропомиозин-рецепторная киназа В, нейротрофическая рецепторная тирозинкиназа 2 и NTRK2) кодируется геном TRKB (также известным как NTRK2, OBHD, TRK-B и GP145-TRKB). TRKB является рецепторной тирозинкиназой, участвующей в развитии и созревании центральной и периферической нервной системы посредством регуляции выживаемости, пролиферации, миграции, дифференцировки нейронов, а также образования и пластичности синапсов. TRKB является рецептором для BDNF/нейротрофического фактора головного мозга и NTF4/нейротрофина-4. Альтернативно, TRKB также может связывать NTF/нейротрофин-3, который является менее эффективным в активации рецептора, но регулирует выживаемость нейронов посредством TRKB. После связывания лиганда TRKB подвергается гомодимеризации, аутофосфорилированию и активации. Каноническая изоформа TRKB экспрессируется в центральной и периферической нервной системе. В центральной нервной системе (ЦНС) экспрессия наблюдается в коре головного мозга, гиппокампе, таламусе, сосудистом сплетении, зернистом слое мозжечка, стволе головного мозга и спинном мозге. В периферической нервной системе он экспрессируется во многих черепных ганглиях, зрительном нерве, вестибулярной системе, множественных структурах лица, подчелюстных железах и дорсальных корешковых ганглиях.
[00106] TRKB человека соответствует 9q21.33 человека на хромосоме 9 (регистрационный номер гена согласно RefSeq NCBI 4915; сборка GRCh38.p7; положение NC 000009.12 (84668368..85027070)). Сообщалось, что ген характеризуется 23 экзонами. Белку TRKB дикого типа человека присвоен регистрационный номер UniProt Q16620. Известны по меньшей мере семь изоформ (Q16620-1 - Q16620-7). Последовательность для одной изоформы, Q16620-4 (идентичной регистрационному номеру NCBI NP 006171.2), представлена в SEQ ID NO: 3. мРНК (кДНК), кодирующей каноническую изоформу, присвоен регистрационный номер NCBI AF410899.1 и она представлена в SEQ ID NO: 8. Другому примеру мРНК (кДНК), кодирующей изоформу человека TRKB, присвоен регистрационный номер RefSeq мРНК NM 006180.4. Иллюстративная кодирующая последовательность (CDS) представлена в SEQ ID NO: 11. Полноразмерный белок TRKB человека, представленный в SEQ TD NO: 3, содержит 838 аминокислоты, включая в себя сигнальный пептид (аминокислоты 1-31), внеклеточный домен (аминокислоты 32-430), трансмембранный домен (аминокислоты 431-454) и цитоплазматический домен (аминокислоты 455-838). Разграничения между этими доменами являются такими, как обозначено в UniProt. Ссылка на TRKB человека включает в себя канонические формы (дикого типа), а также все аллельные формы и изоформы. Любые другие формы TRKB человека содержат аминокислоты, пронумерованные для максимального выравнивания с формой дикого типа, при этом выровненные аминокислоты обозначены тем же номером. Пример другой изоформы TRKB человека представляет собой Q16620-1 (идентичный регистрационному номеру NCBI NP 001018074.1), представленный в SEQ ID NO: 75. мРНК (кДНК), кодирующей эту изоформу, присвоен регистрационный номер NCBI NM 001018064.2 и она представлена в SEQ ID NO: 76. Иллюстративная кодирующая последовательность (CDS) для этой изоформы (CCDS ГО CCDS35050.1) представлена в SEQ ID NO: 77.
[00107] TrkB крысы соответствует 17р14 крысы на хромосоме 17 (регистрационный номер гена согласно RefSeq NCBI 25054; сборка Rnor 6.0; положение NC 005116.4 (5934651..6245778, комплементарная цепь)). Сообщалось, что ген характеризуется 23 экзонами. Белку TRKB крысы дикого типа присвоен регистрационный номер UniProt Q63604. Известны по меньшей мере три изоформы (Q63604-1 - Q63604-3). Последовательность для канонической изоформы, Q63604-1 (идентичной регистрационному номеру NCBI NP 036863.1), представлена в SEQ ID NO: 2. мРНК (кДНК), кодирующей каноническую изоформу присвоен регистрационный номер NCBI NM 012731.2 и она представлена в SEQ ID NO: 7. Другому примеру мРНК (кДНК), кодирующей изоформу TRKB крысы, присвоен RefSeq мРНК М55291. Иллюстративная кодирующая последовательность (CDS) представлена в SEQ ID NO: 10. Канонический полноразмерный белок TRKB крысы, представленный в SEQ ID NO: 2, содержит 821 аминокислоту, включая в себя сигнальный пептид (аминокислоты 1-31), внеклеточный домен (аминокислоты 32-429), трансмембранный домен (аминокислоты 430-453) и цитоплазматический домен (аминокислоты 454-821). Разграничения между этими доменами являются такими, как обозначено в UniProt. Ссылка на TRKB крысы включает в себя канонические формы (дикого типа), а также все аллельные формы и изоформы. Любые другие формы TRKB крысы содержат аминокислоты, пронумерованные для максимального выравнивания с формой дикого типа, при этом выровненные аминокислоты обозначены тем же номером.
[00108] TrkB мыши соответствует 13 В1; 13 31.2 сМ мыши на хромосоме 12 (регистрационный номер гена согласно RefSeq NCBI 18212; сборка GRCm38.p4 (GCF 000001635.24); положение NC 000079.6 (58806569..59133970)). Сообщалось, что ген характеризуется 23 экзонами. Белку TRKB дикого типа мыши присвоен регистрационный номер UniProt PI 5209. Известны по меньшей мере четыре изоформы (Р15209-1 - Р15209-4). Последовательность для канонической изоформы, Р15209-1 (идентичной регистрационным номерам NCBI NP 001020245.1 и NP 001269890.1), представлена в SEQ ID NO: 1. Иллюстративной изоформе мРНК (кДНК), кодирующей каноническую изоформу, присвоен регистрационный номер NCBI NM 001025074.2 и она представлена в SEQ ID NO: 6. Иллюстративная кодирующая последовательность (CDS) (CCDS ID CCDS26 573.1) представлена в SEQ ID NO: 9. Канонический полноразмерный белок TRKB мыши, представленный в SEQ ID NO: 1, содержит 821 аминокислоту, включая в себя сигнальный пептид (аминокислоты 1-31), внеклеточный домен (аминокислоты 32-429), трансмембранный домен (аминокислоты 430-453) и цитоплазматический домен (аминокислоты 454-821). Разграничения между этими доменами являются такими, как обозначено UniProt. Ссылка на TRKB мыши включает в себя канонические формы (дикого типа), а также все аллельные формы и изоформы. Любые другие формы TRKB мыши содержат аминокислоты, пронумерованные для максимального выравнивания с формой дикого типа, при этом выровненные аминокислоты обозначены тем же номером.
В. Гуманизированные локусы TRKB
[00109] Гуманизированный локус TRKB может представлять собой локус TrkB, в котором весь ген TrkB заменен соответствующей ортологичной последовательностью TRKB человека, или представлять собой локус TrkB, в котором только часть гена TrkB заменена соответствующей ортологичной последовательностью TRKB человека (т.е. гуманизированная). Необязательно соответствующая ортологичная последовательность TRKB человека модифицируют для оптимизации кодонов на основании частоты использования кодона у не являющегося человеком животного. Замененные (т.е. гуманизированные) области могут включать в себя кодирующие области, такие как экзон, некодирующие области, такие как интрон, нетранслируемую область или регуляторную область (например, промотор, энхансер или элемент, связывающий транскрипционный репрессор) или любую их комбинацию. В качестве одного примера экзоны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22 или всем 23 экзонам гена TRKB человека, могут являться гуманизированными. Например, экзоны, соответствующие экзонам 3-10 гена TRKB человека, могут являться гуманизированными, включая в себя сегмент экзона 2 (кодирующий экзон 1) от кодона, кодирующего аминокислоту 33, начиная сразу после сигнального пептида. Альтернативно, область TrkB, кодирующая эпитоп, распознаваемый антигенсвязывающим белком к TRKB человека, или область, на которую нацелен нацеленный на TRKB человека реагент (например, малая молекула), может являться гуманизированной. Аналогично, нитроны, соответствующие 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или всем 22 интронам гена TRKB человека, могут являться гуманизированными или могут оставаться эндогенными. Например, интроны, соответствующие интронам между экзонами 2 и 10 (т.е. интроны 2-9, между кодирующим экзоном 1 и экзоном 10) гена TRKB человека, могут являться гуманизированными, необязательно включая в себя часть нитрона после экзона 10 (т.е. интрон 10). Фланкирующие нетранслируемые области, включая в себя регуляторные последовательности, также могут являться гуманизированными или оставаться эндогенными. Например, 5' нетранслируемая область (UTR), 3'UTR или как 5' UTR, так и 3' UTR могут являться гуманизированными, или 5' UTR, 3'UTR или как 5' UTR, так и 3' UTR могут оставаться эндогенными. Согласно конкретному примеру как 5' UTR, так и 3' UTR остаются эндогенными. В зависимости от степени замены на ортологичные последовательности, регуляторные последовательности, такие как промотор, могут являться эндогенными или снабжаться заменяющей ортологичной последовательностью человека. Например, гуманизированный локус TRKB может включать в себя эндогенный промотор TrkB не являющегося человеком животного.
[00110] Одна или несколько или все из областей, кодирующих сигнальный пептид, цитоплазматический домен, трансмембранный домен или внеклеточный, могут являться гуманизированными, или одна или несколько таких областей могут оставаться эндогенными. Иллюстративные кодирующие последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB мыши представлены в SEQ ID NO: 63-66 соответственно. Иллюстративные кодирующие последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB крысы представлены в SEQ ID NO: 67-70 соответственно. Иллюстративные кодирующие последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB человека представлены в SEQ ID NO: 71-74 соответственно.
[00111] Например, вся или часть области локуса TrkB, кодирующей сигнальный пептид, может являться гуманизированной, и/или вся или часть области локуса TrkB, кодирующей внеклеточный домен, может являться гуманизированной, и/или вся или часть области локуса TrkB, кодирующей трансмембранный домен, может являться гуманизированной, и/или вся или часть области локуса TrkB, кодирующей цитоплазматический домен, может являться гуманизированной. Согласно одному примеру вся или часть области локуса TrkB, кодирующей внеклеточный домен, является гуманизированной. Необязательно CDS внеклеточного домена TRKB человека содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной по отношению к SEQ ID NO: 72 (или ее вырожденным последовательностям). Белок TRKB может сохранять активность нативного TRKB (например, сохраняет способность фосфорилироваться, сохраняет способность активировать нижележащие сигнальные пути, такие как пути PI3K/AKT и MAPK/ERK, или сохраняет способность регулировать выживаемость, пролиферацию, миграцию, дифференцировку нейронов или образование синапсов и их пластичность или производить любой из фенотипов, раскрытых в другом месте в настоящем документе). Например, область локуса TrkB, кодирующая внеклеточный домен, может являться гуманизированной так, что химерный белок TRKB производиться с эндогенным сигнальным пептидом, эндогенным цитоплазматический доменом, эндогенным трансмембранным доменом и гуманизированным внеклеточным доменом.
[00112] Одна или несколько из областей, кодирующих сигнальный пептид, цитоплазматический домен, трансмембранный домен или внеклеточный, могут оставаться эндогенными. Например, область, кодирующая сигнальный пептид и/или цитоплазматический домен и/или трансмембранный домен, может оставаться эндогенной. Необязательно CDS эндогенного сигнального пептида TRKB содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 63 или 67 (или их вырожденным последовательностям). Необязательно CDS эндогенного трансмембранного домена TRKB содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 65 или 69 (или их вырожденным последовательностям). Необязательно CDS эндогенного цитоплазматического домена TRKB содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 66 или 70 (или их вырожденным последовательностям). В каждом случае белок TRKB может сохранять активность нативного TRKB.
[00113] Белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать один или несколько доменов, которые происходят из белка TRKB человека, и/или один или несколько доменов, которые происходят эндогенного (т.е. нативного) белка TRKB. Иллюстративные аминокислотные последовательности сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB мыши представлены в SEQ ID NO: 51-54 соответственно. Иллюстративные аминокислотные последовательности сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB крысы представлены в SEQ ID NO: 55-58 соответственно. Иллюстративные аминокислотные последовательности сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB человека представлены в SEQ ID NO: 59-62 соответственно.
[00114] Белок TRKB может содержать один или несколько или все из следующего: сигнальный пептид TRKB человека, внеклеточный домен TRKB человека, трансмембранный домен TRKB человека и цитоплазматический домен TRKB человека. В качестве одного примера белок TRKB может содержать внеклеточный домен TRKB человека.
[00115] Белок TRKB, кодируемый гуманизированным локусом TRKB, также может содержать один или несколько доменов, которые происходят из эндогенного (т.е. нативного) белка TRKB не являющегося человеком животного. В качестве одного примера белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать сигнальный пептид из эндогенного (т.е. нативного) белка TRKB не являющегося человеком животного и/или цитоплазматический домен из эндогенного (т.е. нативного) белка TRKB не являющегося человеком животного и/или трансмембранный домен из эндогенного (т.е. нативного) белка TRKB не являющегося человеком животного.
[00116] Домены в химерном белке TRKB, которые происходят из белка TRKB человека, могут кодироваться полностью гуманизированной последовательностью (т.е. вся последовательность, кодирующая этот домен, заменена ортологичной последовательностью TRKB человека) или могут кодироваться частично гуманизированной последовательностью (т.е. некоторая часть последовательности, кодирующей этот домен, заменена ортологичной последовательностью TRKB человека, а остальная эндогенная (т.е. нативная) последовательность, кодирующая этот домен, кодирует те же аминокислоты, что и ортологичная последовательность TRKB человека, так что кодируемый домен является идентичным таковому домену в белке TRKB человека). Аналогично, домены в химерном белке, которые происходят из эндогенного белка TRKB, могут кодироваться полностью эндогенной последовательностью (т.е. вся последовательность, кодирующая этот домен, представляет собой эндогенную последовательность TrkB) или могут кодироваться частично гуманизированной последовательностью (т.е. некоторая часть последовательности, кодирующей этот домен, заменена ортологичной последовательностью TRKB человека, но ортологичная последовательность TRKB человека кодирует те же аминокислоты, что и замененная эндогенная последовательность TrkB, так что кодируемый домен является идентичным таковому домену в эндогенном белке TRKB). Например, часть области локуса TrkB, кодирующей трансмембранный домен (например, кодирующей N-концевую область трансмембранного домена), можно заменить ортологичной последовательностью TRKB человека, причем аминокислотная последовательность области трансмембранного домена, кодируемая ортологичной последовательностью TRKB человека, является идентичной соответствующей эндогенной аминокислотной последовательности.
[00117] В качестве одного примера белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать внеклеточный домен TRKB человека. Необязательно внеклеточный домен TRKB человека содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 60. Белок TRKB сохранять активность нативного TRKB (например, сохраняет способность фосфорилироваться, сохраняет способность активировать нижележащие сигнальные пути, такие как пути PI3K/AKT и MAPK/ERK, или сохраняет способность регулировать выживаемость, пролиферацию, миграцию, дифференцировку нейронов или образование синапсов и их пластичность или производить любой из фенотипов, раскрытых в другом месте в настоящем документе). В качестве другого примера белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать эндогенный цитоплазматический домен TRKB не являющегося человеком животного (например, цитоплазматический домен TRKB мыши или цитоплазматический домен TRKB крысы). Необязательно цитоплазматический домен TRKB не являющегося человеком животного содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 54 или 58. В качестве другого примера белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать эндогенный трансмембранный домен TRKB не являющегося человеком животного (например, трансмембранный домен TRKB мыши или трансмембранный домен TRKB крысы). Необязательно трансмембранный домен TRKB не являющегося человеком животного содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 53 или 57. В качестве другого примера белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать эндогенный сигнальный пептид TRKB не являющегося человеком животного (например, сигнальный пептид TRKB мыши или TRKB сигнальный пептид TRKB крысы). Необязательно сигнальный пептид TRKB не являющегося человеком животного содержит, состоит по существу из или состоит из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 51 или 55. В каждом случае белок TRKB может сохранять активность нативного TRKB. Например, белок TRKB, кодируемый гуманизированным локусом TRKB, может содержать, состоять по существу из или состоять из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 4 или 5. Необязательно CDS TRKB, кодируемая гуманизированным локусом TRKB, может содержать, состоять по существу из или состоять из последовательности, которая является по меньшей мере на 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной SEQ ID NO: 12 или 13 (или их вырожденным последовательностям). В каждом случае белок TRKB может сохранять активность нативного TRKB.
[00118] Необязательно гуманизированный локус TRKB может содержать другие элементы. Примеры таких элементов могут включать в себя селекционные кассеты, репортерные гены, сайты распознавания рекомбиназы или другие элементы. Альтернативно, в гуманизированном локусе TRKB могут отсутствовать другие элементы (например, могут отсутствовать маркер селекции или кассета селекции). Примеры подходящих репортерных генов и репортерных белков раскрыты в другом месте в настоящем документе. Примеры подходящих маркеров селекции включают в себя неомицинфосфотрансферазу (пеог), гигромицин В-фосфотрансферазу (hygr), пуромицин-N-ацетилтрансферазу (puror), бластицидин S-дезаминазу (bsrr), ксантин/гуанинфосфорибозилтрансферазу (gpt) и тимидинкиназу вируса простого герпеса (HSV-k). Примеры рекомбиназ включают в себя рекомбиназы Cre, Flp и Dre. Одним примером гена Cre-рекомбиназы является Crei, в котором два экзона, кодирующие Cre-рекомбиназу, разделены интроном, чтобы предотвратить его экспрессию в прокариотической клетке. Такие рекомбиназы могут дополнительно содержать сигнал ядерной локализации для облегчения локализации в ядре (например, NLS-Crei). Сайты распознавания рекомбиназы включают в себя нуклеотидные последовательности, которые распознаются сайт-специфической рекомбиназой и могут служить в качестве субстрата для события рекомбинации. Примеры сайтов распознавания рекомбиназы включают в себя FRT, FRT11, FRT71, attp, att, rox и сайты lox, такие как loxP, lox511, lox2272, lox66, lox71, loxM2 и lox5171.
[00119] Другие элементы, такие как репортерные гены или кассеты селекции, могут представлять собой самоудаляющиеся кассеты, фланкированные сайтами распознавания рекомбиназы. См., например, US 8697851 и US 2013/0312129, причем каждый из документов включен полностью включен в настоящий документ посредством ссылки для всех целей. В качестве примера, самоудаляющаяся кассета может содержать ген Crei (содержит два экзона, кодирующих рекомбиназу Cre, которые разделены интроном), функционально связанный с промотором Prml мыши, и ген устойчивости к неомицину, функционально связанный с промотором убиквитина человека. При использовании промотора Prml, самоудаляющаяся кассета может быть специфически удалена в мужских половых клетках животных F0. Полинуклеотид, кодирующий маркер селекции, может быть функционально связан с промотором, активным в целевой клетке. Примеры промоторов представлены в другом месте в настоящем документе. В качестве другого конкретного примера самоудаляющаяся кассета селекции может содержать кодирующую последовательность гена устойчивости к гигромицину, функционально связанную с одним или несколькими промоторами (например, как убиквитин человека и промоторы ЕМ7), за которыми следует сигнал полиаденилирования, за которым следует кодирующая последовательность Crei, функционально связанная с одним или несколькими промоторами (например, промотором mPrml), за которым следует другой сигнал полиаденилирования, причем вся кассета фланкирована сайтами loxP.
[00120] Гуманизированный локус TRKB также может представлять собой условный аллель. Например, условный аллель может представлять собой многофункциональный аллель, как описано в US 2011/0104799, полностью включенной в настоящий документ посредством ссылки для всех целей. Например, условный аллель может содержать следующее: (а) пусковая последовательность в смысловой ориентации относительно транскрипции целевого гена; (b) кассета для отбора по чувствительности к лекарственному средству (DSC) в смысловой или антисмысловой ориентации; (с) представляющая интерес нуклеотидная последовательность (NSI) в антисмысловой ориентации; и (d) модуль условной инверсии (COIN, который использует экзон-расщепляющий интрон и обратимый модуль, подобный ловушке гена) в обратной ориентации. См., например, US 2011/0104799. Условный аллель может дополнительно содержать рекомбинируемые звенья, которые рекомбинируют при воздействии первой рекомбиназы с образованием условного аллеля, который (i) не содержит пусковую последовательность и DSC; и (ii) содержит NSI в смысловой ориентации и COIN в антисмысловой ориентации. См., например, US 2011/0104799.
[00121] Один иллюстративный гуманизированный локус TRKB (например, гуманизированный локус TrkB мыши или гуманизированный локус TrkB крысы) представляет собой локус, в котором область в экзоне 2/кодирующем экзоне 1 от кодона, кодирующего аминокислоту 33, начиная сразу после сигнального пептида (или кодона, соответствующего кодону, кодирующему аминокислоту 33 в TrkB мыши, TrkB крысы или TRKB человека при оптимальном выравнивании с CDS TrkB мыши, TrkB крысы или TRKB человека соответственно) до экзона 10 (или экзона, соответствующего экзону 10 TrkB мыши, TrkB крысы или TRKB человека TRKB при оптимальном выравнивании с CDS TrkB мыши, TrkB крысы или TRKB человека соответственно), необязательно включая в себя часть нитрона 10, заменена соответствующей последовательностью человека. Замененная область кодирует внеклеточный домен TRKB. См. фигуры 1 и 4 и SEQ ID NO: 4 и 5.
С. Геномы не являющегося человеком животного, клетки не являющегося человеком животного и не являющиеся человеком животные, содержащие гуманизированный локус TRKB
[00122] Предусмотрены геномы не являющегося человеком животного, клетки не являющегося человеком животного и не являющиеся человеком животные, содержащие гуманизированный локус TRKB, как описано в другом месте в настоящем документе. Геномы, клетки или не являющиеся человеком животные могут являться мужскими ил женскими. Геномы, клетки или не являющиеся человеком животные могут являться гетерозиготными или гомозиготными в отношении гуманизированного локуса TRKB. Диплоидный организм имеет два аллеля на каждом генетическом локусе. Каждая пара аллелей представляет генотип конкретного генетического локуса. Генотипы описывают как гомозиготные, если присутствуют два идентичных аллеля на конкретном локусе, и как гетерозиготные, если два аллеля отличаются.
[00123] Предусмотренные в настоящем документе геномы или клетки не
являющегося человеком животного могут представлять собой, например, любой геном или клетку не являющегося человеком животного, содержащие локус TrkB или геномный локус, гомологичный или ортологичный локусу TRKB человека. Геномы могут происходить из следующего или клетки могут представлять собой, например, следующее: эукариотические клетки, которые включают в себя, например, клетки грибов (например, дрожжи), клетки растений, клетки животных, клетки млекопитающих, клетки млекопитающих, не являющихся человеком, и клетки человека. Термин «животное» включает в себя любого представителя царства животных, включая в себя, например, млекопитающих, рыб, рептилий, амфибий, птиц и червей. Клеткой млекопитающего может являться, например, клетка млекопитающего, не являющегося человеком, клетка грызуна, клетка крысы, клетка мыши или клетка хомяка. Другие млекопитающие, не являющиеся человеком, включают в себя, например, не являющихся человеком приматов, низших обезьян, человекообразных обезьян, орангутангов, кошек, собак, кроликов, лошадей, быков, оленей, бизонов, домашний скот (например, виды крупного рогатого скота, такие как коровы, быки и т.д.; виды овечьих, такие как овцы, козы и т.д.; и виды свинообразных, такие как свиньи и кабаны). Птицы включают в себя, например, кур, индеек, страусов, гусей, уток и так далее. Домашние животные и сельскохозяйственные животные также предусмотрены. Термин «не являющийся человеком» исключает людей.
[00124] Клетки также могут находиться в недифференцированном или дифференцированном состоянии любого типа. Например, клетка может представлять собой тотипотентную клетку, плюрипотентную клетку (например, плюрипотентную клетку человека или плюрипотентную клетку, не относящуюся к человеку, такую как эмбриональная стволовая клетка мыши (ES) или ES клетка крысы), или не плюрипотентную клетку. Тотипотентные клетки включают в себя недифференцированные клетки, которые могут давать клетки любого типа, а плюрипотентные клетки включают в себя недифференцированные клетки, которые обладают способностью развиваться в более чем один тип дифференцированных клеток. Такими плюрипотентными и/или тотипотентными клетками могут являться, например, ES клетки или ES-подобные клетки, такие как индуцированные плюрипотентные стволовые (iPS) клетки. ES клетки включают в себя происходящие из эмбриона тотипотентные или плюрипотентные клетки, которые способны вносить вклад в любую ткань развивающегося эмбриона при введении в эмбрион. ES клетки могут быть получены из внутренней клеточной массы бластоцисты и способны дифференцироваться в клетки любого из трех зародышевых слоев позвоночных (энтодерма, эктодерма и мезодерма).
[00125] Клетки, предусмотренные в настоящем документе, также могут представлять собой половые клетки (например, сперматозоиды или ооциты). Клетки могут быть митотически компетентными клетками или митотически неактивными клетками, мейотически компетентными клетками или мейотически неактивными клетками. Аналогично, клетки также могут являться первичными соматическими клетками или клетками, которые не являются первичными соматическими клетками. Соматические клетки включают в себя любую клетку, которая не является гаметой, зародышевой клеткой, гаметоцитом или недифференцированной стволовой клеткой. Например, клетки могут представлять собой нейроны, такие как гиппокампальные нейроны или кортикальные нейроны.
[00126] Подходящие клетки, предусмотренные в настоящем документе, также включают в себя первичные клетки. Первичные клетки включают в себя клетки или культуры клеток, которые были выделены непосредственно из организма, органа или ткани. Первичные клетки включают в себя клетки, которые не являются ни трансформированными, ни бессмертными. Они включают в себя любую клетку, полученную из организма, органа или ткани, которая ранее не была перенесена в культуру ткани или ранее была перенесена в культуру ткани, но не способна бесконечно переноситься в культуру ткани. Такие клетки могут быть выделены с помощью общепринятых техник и включают в себя, например, гиппокампальные нейроны или кортикальные нейроны.
[00127] Другие подходящие клетки, предусмотренные в настоящем документе, включают в себя иммортализованные клетки. Иммортализованные клетки включают в себя клетки многоклеточного организма, которые в норме не размножаются бесконечно долго, но из-за мутации или изменения избежали нормального клеточного старения и вместо этого могут продолжать подвергаться делению. Такие мутации или изменения могут происходить естественным путем или быть преднамеренно вызванными. Конкретный пример линии иммортализованных клеток представляет собой клеточную линию нейробластомы, такую как N18TG2 или Т48, или клеточную линию, такую как клеточная линия NIH-3T3. Многочисленные типы иммортализованных клеток хорошо известны. Иммортализованные или первичные клетки включают в себя клетки, которые, как правило, используют для культивирования или для экспрессии рекомбинантных генов или белков.
[00128] Клетки, предусмотренные в настоящем документе, также включают в себя эмбрионы на стадии одной клетки (т.е. оплодотворенные ооциты или зиготы). Такие одноклеточные эмбрионы могут происходить из любого генетического фона (например, BALB/c, C57BL/6, 129 или их комбинации для мышей), могут являться свежими или замороженными и могут быть получены в результате естественного разведения или оплодотворения in vitro.
[00129] Клетки, предусмотренные в настоящем документе, могут являться нормальными, здоровыми клетками или могут являться патологическими или содержащими мутации клетками.
[00130] Не являющиеся человеком животные, содержащие гуманизированный локус TRKB, как описано в настоящем документе, могут быть получены с помощью способов, описанных в другом месте в настоящем документе. Термин «животное» включает в себя любого представителя царства животных, включая в себя, например, млекопитающих, рыб, рептилий, амфибий, птиц и червей. Согласно конкретному примеру не являющееся человеком животное представляет собой не являющееся человеком млекопитающее. Не являющиеся человеком млекопитающие включают в себя, например, не являющихся человеком приматов, низших обезьян, человекообразных обезьян, орангутангов, кошек, собак, лошадей, быков, оленей, бизонов, овец, кроликов, грызунов (например, мышей, крыс, хомяков и морских свинок) и домашний скот (например, виды крупного рогатого скота, такие как коровы и быки; виды овечьих, такие как овцы и козы; и виды свинообразных, такие как свиньи и кабаны). Птицы включают в себя, например, кур, индеек, страусов, гусей и уток. Домашние животные и сельскохозяйственные животные также предусмотрены. Термин «не являющееся человеком животное» исключает людей. Предпочтительные не являющиеся человеком животные включают в себя, например, грызунов, таких как мыши и крысы.
[00131] Не являющиеся человеком животные могут быть любого генетического происхождения. Например, подходящими мышами могут являться линия 129, линия C57BL/6, смесь 129 и C57BL/6, линия BALB/c или линия Swiss Webster. Примеры линий 129 включают в себя 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129S1/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1 и 129Т2. См., например, Festing et al. (1999)Mammalian Genome 10:836, полностью включенный в настоящий документ посредством ссылки для всех целей. Примеры линий C57BL включают в себя C57BL/A, C57BL/An, C57BL/GrFa, C57BL/Kal wN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/Ola. Подходящие мыши также могут быть из смеси вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6 (например, 50% 129 и 50% C57BL/6). Аналогично, подходящие мыши могут быть из смеси вышеупомянутых линий 129 или смеси вышеупомянутых линий BL/6 (например, линия 129S6 (129/SvEvTac)).
[00132] Аналогичным образом, крысы могут быть из любой линии крыс, включая в себя, например, линию крыс ACI, линию крыс Dark Agouti (DA), линию крыс Wistar, линию крыс LEA, линию крыс Sprague Dawley (SD) или линию крыс Fischer, такой как Fisher F344 или Fisher F6. Крысы также могут быть получены из линии, полученного из смеси двух или более линий, указанных выше. Например, подходящая крыса может быть из линии DA или линии ACI. Линия крыс ACI характеризуется тем, что имеет окрас шерсти «черный агути», белый живот и лапы и гаплотип RT1av1. Такие линии доступны из различных источников, включая в себя Harlan Laboratories. Линия крыс Dark Agouti (DA) характеризуется тем, что имеет окрас шерсти «агути» и гаплотип RT1av1. Такие крысы доступны из различных источников, включая в себя Charles River и Harlan Laboratories. В некоторых случаях подходящими крысами могут являться инбредные крысы. См., например, US 2014/0235933, которая полностью включена в настоящий документ посредством ссылки для всех целей.
III. Способы применения не являющихся человеком животных, содержащих гуманизированный локус TRKB, для оценки эффективности нацеленных на TRKB человека реагентов in vivo или ex vivo
[00133] Предусмотрены различные способы применения не являющихся человеком животных, содержащих гуманизированный локус TRKB, как описано в другом месте в настоящем документе, для оценки или оптимизации доставки или эффективности нацеленных на TRKB человека реагентов (например, терапевтических молекул - агонистов) in vivo или ex vivo. Поскольку не являющиеся человеком животные содержат гуманизированный локус TRKB, не являющиеся человеком животные будут более точно отражать эффективность нацеленного на TRKB человека реагента.
А. Способы испытания эффективности нацеленных на TRKB человека реагентов in vivo или ex vivo
[00134] Предусмотрены различные способы оценки доставки или эффективности нацеленных на TRKB человека реагентов in vivo с использованием не являющихся человеком животных, содержащих гуманизированный локус TRKB, как описано в другом месте в настоящем документе. Такие способы могут предусматривать следующее: (а) введение не являющемуся человеком животному нацеленного на TRKB человека реагента; и (b) оценка активности нацеленного на TRKB человека реагента.
[00135] Нацеленный на TRKB человека реагент может представлять собой нацеленное на TRKB человека антитело или антигенсвязывающий белок или любую другую большую молекулу или малую молекулу, которая нацелена на человека TRKB. Альтернативно, нацеленный на TRKB человека реагент может представлять собой любой биологический или химический агент, который нацелен на локус TRKB человека (ген TRKB человека), мРНК TRKB человека или белок TRKB человека. Примеры нацеленных на TRKB человека реагентов раскрыты в другом месте в настоящем документе.
[00136] Такие нацеленные на TRKB человека реагенты можно вводить в клетку или не являющемуся человеком животному посредством любого способа доставки (например, AAV, LNP, или HDD), как раскрыто в другом месте в настоящем документе, и любого пути введения. Средства доставки терапевтических молекул и пути введения раскрыты более подробно в другом месте в настоящем документе. Согласно конкретным способам реагенты доставляют посредством инъекции (например, прямая гиппокампальная инъекция, подкожная инъекция или интравитреальная инъекция).
[00137] Способы оценки активности нацеленного на TRKB человека реагента являются хорошо известными и предусмотрены в другом месте в настоящем документе. Согласно некоторым способам оценка активности нацеленного на TRKB человека реагента (например, агонистической активности или ингибирующей активности) предусматривает оценку активности TRKB (например, фосфорилирование TRKB, TRKB-опосредованная активация нижележащих сигнальных путей или TRKB-индуцированные фенотипы), как раскрыто в другом месте в настоящем документе. Оценку активности можно проводить в любом типе клеток, любом типе тканей или любом типе органов, как раскрыто в другом месте в настоящем документе. Согласно некоторым способам оценку активности проводят в ткани головного мозга (например, гиппокампе или стриатуме) или нейронах (например, ганглиозных клетках сетчатки, гиппокампальных нейронах или кортикальных нейронах).
[00138] Если нацеленный на TRKB реагент представляет собой реагент для редактирования генома (например, нуклеазный агент), такие способы могут предусматривать оценку модификации гуманизированного локуса TRKB. Например, оценка может предусматривать секвенирование гуманизированного локуса TRKB в одной или нескольких клетках, выделенных из организма не являющегося человеком животного (например, секвенирование следующего поколения). Оценка могут включать в себя выделение целевого органа (например, головного мозга) или ткани из организма не являющегося человеком животного и оценку модификации гуманизированного локуса TRKB в целевом органе или ткани. Оценка также может включать в себя оценку модификации гуманизированного локуса TRKB в двух или более различных типах клеток в пределах целевого органа или ткани. Аналогично, оценка может включать в себя выделение нецелевого органа или ткани (например, двух или более нецелевых органов или тканей) из организма не являющегося человеком животного и оценку модификации гуманизированного локуса TRKB в нецелевом органе или ткани.
[00139] Такие способы также могут содержать измерение уровней экспрессии мРНК, произведенной гуманизированным локусом TRKB, или измерение уровней экспрессии белка, кодируемого гуманизированным локусом TRKB. Например, содержания белка можно измерить в конкретном типе клеток, тканей или органов (например, головной мозг) или секретированные уровни можно измерить в сыворотке. Способы оценки экспрессии мРНК TRKB или белка, экспрессированного из гуманизированного локуса TRKB, предусмотрены в другом месте в настоящем документе и являются хорошо известными.
[00140] Различные способы, предусмотренные выше для оценки активности in vivo, также можно использовать для оценки активности нацеленных на TRKB человека реагентов ex vivo, как описано в другом месте в настоящем документе.
В. Способы оптимизации доставки или эффективности нацеленного на TRKB человека реагента in vivo или ex vivo
[00141] Предусмотрены различные способы оптимизации доставки нацеленных на TRKB человека реагентов в клетку или не являющемуся человеком животному или оптимизации активности или эффективности доставки нацеленных на TRKB человека реагентов in vivo. Такие способы могут предусматривать, например, следующее: (а) проведение способа испытания эффективности нацеленного на TRKB человека реагента, как описано выше, в первый раз у первого не являющегося человеком животного или в первой клетке; (b) изменение переменной и проведение способа во второй раз у второго не являющегося человеком животного (т.е. того же вида) или во второй клетке с измененной переменной; и (с) сравнение активности нацеленного на TRKB человека реагента на стадии (а) с активностью нацеленного на TRKB человека реагента на стадии (b), и выбор способа, приводящего к большей эффективности или активности.
[00142] Способы измерения доставки, эффективности или активности нацеленных на TRKB человека реагентов раскрыты в другом месте в настоящем документе. Более высокая эффективность может означать разные вещи в зависимости от требуемого эффекта в организме не являющегося человеком животного или клетке. Например, более высокая эффективность может означать более высокую активность и/или более высокую специфичность. Более высокой активностью может являться, например, активность в активации TRKB или активность в ингибировании TRKB. Это может относиться к более высокому проценту клеток, подлежащих нацеливанию, в конкретном целевом типе клеток (например, нейронах, таких как ганглиозные клетки сетчатки) или в конкретной целевой ткани или органе (например, головном мозге). Более высокая специфичность может относиться к более высокой специфичности в отношении TRKB по сравнению с нецелевыми эффектами, более высокой специфичности в отношении целевого типа клеток или более высокой специфичности в отношении целевого типа ткани или органа.
[00143] Переменная, которая изменяется, может представлять собой любой параметр. В качестве одного примера измененная переменная может представлять собой способ упаковки или доставки, с помощью которого нацеленный на TRKB человека реагент или реагенты вводят в клетку или не являющемуся человеком животному. Примеры способов доставки раскрыты в настоящем документе в другом месте. В качестве другого примера измененная переменная может представлять собой путь введения для введения нацеленного на TRKB человека реагента или реагентов в клетку или не являющемуся человеком животному. Примеры способов введения раскрыты в другом месте в настоящем документе.
[00144] В качестве другого примера измененной переменной может являться концентрация или количество введенного нацеленного на TRKB человека реагента или реагентов. В качестве другого примера, измененная переменная может представлять собой время введения нацеленного на TRKB человека реагента или реагентов относительно времени проведения оценки активности или эффективности реагентов. В качестве другого примера, измененной переменной может являться число раз или частота, с которой вводят нацеленный на TRKB человека реагент или реагенты. В качестве другого примера, измененной переменной может являться нацеленный на TRKB человека реагент или реагенты, которые вводят (например, сравнение одного реагента с другим реагентом).
С. Нацеленные на TRKB человека реагенты
[00145] Нацеленный на TRKB человека реагент может представлять собой любой реагент, который нацелен на белок TRKB человека, ген TRKB человека или мРНК TRKB человека. Нацеленный на TRKB человека реагент может представлять собой, например, агонист (т.е. молекула, которая опосредованно или прямо активирует TRKB человека) или может представлять собой антагонист (т.е. ингибитор или ингибирующий реагент, который блокирует активность TRKB человека). Согласно конкретному примеру нацеленный на TRKB человека реагент представляет собой агонист TRKB. Нацеленные на TRKB человека реагенты в способах, раскрытых в настоящем документе, могут представлять собой известные нацеленные на TRKB человека реагенты, могут представлять собой предполагаемые нацеленные на TRKB человека реагенты (например, кандидатные реагенты, предназначенные для нацеливания на TRKB человека) или могут представлять собой реагенты, подвергаемые скринингу в отношении нацеленной на TRKB человека активности.
[00146] Например, нацеленный на TRKB человека реагент может представлять собой антигенсвязывающий белок (например, антитело - агонист), нацеленный на эпитоп белка TRKB человека. Пример такого реагента представляет собой антитело - агонист TRKB, Н4Н9816Р2. Другие антитела к TRKB раскрыты в другом месте в настоящем документе. В некоторых случаях антитела к TRKB связываются с TRKB человека со значением Kd, составляющим меньше чем приблизительно 200 нМ, как измерено с помощью поверхностного плазмонного резонанса при 25°С или при 37°С. В других случаях антитела к TRKB связываются с TRKB человека со значением Kd, составляющим меньше чем приблизительно 600 пМ, меньше чем приблизительно 300 пМ, меньше чем приблизительно 200 пМ, меньше чем приблизительно 150 пМ, меньше чем приблизительно 100 пМ, меньше чем приблизительно 80 пМ, меньше чем приблизительно 50 пМ, меньше чем приблизительно 40 пМ, меньше чем приблизительно 30 пМ, меньше чем приблизительно 20 пМ, меньше чем приблизительно 10 пМ, меньше чем приблизительно 5 пМ, меньше чем приблизительно 3 пМ или меньше чем приблизительно 1 пМ. В некоторых случаях антитела к TRKB связываются с TRKB человека со значением диссоциативного периода полужизни (t½) составляющим больше чем приблизительно 10 минут, как измерено с помощью поверхностного плазмонного резонанса при 25°С или 37°С. В других случаях антитела к TRKB связываются с TRKB человека со значением t½, составляющим больше чем приблизительно 20 минут, больше чем приблизительно 50 минут, больше чем приблизительно 100 минут, больше чем приблизительно 120 минут, больше чем приблизительно 150 минут, больше чем приблизительно 300 минут, больше чем приблизительно 350 минут, больше чем приблизительно 400 минут, больше чем приблизительно 450 минут, больше чем приблизительно 500 минут, больше чем приблизительно 550 минут, больше чем приблизительно 600 минут, больше чем приблизительно 700 минут, больше чем приблизительно 800 минут, больше чем приблизительно 900 минут, больше чем приблизительно 1000 минут, больше чем приблизительно 1100 минут или больше чем приблизительно 1200 минут. В качестве конкретного примера антитело к TRKB может содержать набор из шести CDR (HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3), выбранных из группы, представленной в таблице 22, или по существу сходных последовательностей, характеризующихся по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, или по меньшей мере 99% идентичности последовательности по отношению к ним.
[00147] Другие нацеленные на TRKB человека реагенты включают в себя малые молекулы (например, агонисты), нацеленные на белок TRKB человека. Примеры малых молекул - агонистов TRKB включают в себя 7,8-дигидроксифлавон (7,8-DHF), дезоксигедунин, LM22A-4 (N,N',N'',-трис(2-гидроксиэтил)-1,3,5-бензолтрикарбоксамид) и LM22B-10 (2-[[4-[[4-[бис-(2-гидроксиэтил)амино]фенил]-(4-хлорфенил)метил]фенил]-(2-гидроксиэтил)амино]этанол). См., например, Liu et al. (2015) Translational Neurodegeneration 5:2; Massa et al. (2010) j. Clin. Invest. 120(5):1774-1785; и Yang et al. (2016) Neuropharmacology 110:343-361, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Примером нацеленного на TRKB реагента, который представляет собой ингибитор, является K252a. См., например, Yang et al. (2016) Neuropharmacology 110:343-361, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00148] Другие нацеленные на TRKB человека реагенты включают в себя пептиды или пептидные миметики (например, агонисты), нацеленные на белок TRKB человека. Примеры пептидных миметиков, которые служат в качестве агонистов TRKB человека, раскрыты, например, в O'Leary et al. (2003) J. Biol. Chem. 278(28):25738-25744, полностью включенном в настоящий документ посредством ссылки для всех целей.
[00149] Другие нацеленные на TRKB человека реагенты могут включать в себя реагенты для редактирования генома, такие как нуклеазный агент (например, нуклеаза системы сгруппированных регулярно перемежающихся коротких палиндромных повторов (CRISPR)/CRISPR-ассоциированных белков, нуклеаза домена «цинковые пальцы» (ZFN) и эффекторная нуклеаза, подобная активатору транскрипции (TALEN)), которая расщепляет сайт распознавания в пределах гена TRKB человека. Аналогично, нацеленный на TRKB человека реагент может представлять собой экзогенную донорную нуклеиновую кислоту (например, нацеливающий вектор или одноцепочечный олигодезоксинуклеотид (ssODN)), предназначенную для рекомбинации с геном TRKB человека).
[00150] Другие нацеленные на TRKB человека реагенты могут включать в себя антисмысловые олигонуклеотиды (например, миРНК (малые интерферирующие РНК) или миРНК (малые шпилечные РНК)), нацеленные на мРНК TRKB человека. Антисмысловые олигонуклеотиды (ASO) или антисмысловые РНК представляют собой короткие синтетические последовательности нуклеотидов, предназначенные для предотвращения экспрессии целевого белка путем селективного связывания с РНК, которая кодирует целевой белок и тем самым предотвращая трансляцию. Эти соединения связываются с РНК с высокой аффинностью и селективностью посредством хорошо охарактеризованного спаривания Уотсона-Крика (гибридизации). РНК-интерференция (РНКи) представляет собой эндогенный клеточный механизм для контроля экспрессии генов, при котором малые интерферирующие РНК (миРНК), которые связаны с индуцированным РНК комплексом сайленсинга (RISC), опосредуют расщепление целевой матричной РНК (мРНК).
[00151] Активность любого другого известного или предполагаемого нацеленного на TRKB человека реагента также можно оценить с использованием не являющихся человеком животных, раскрытых в настоящем документе. Аналогично, любую другую молекулу можно подвергнуть скринингу в отношении нацеленной на TRKB человека активности с использованием не являющихся человеком животных, раскрытых в настоящем документе.
D. Введение нацеленных на TRKB человека реагентов не являющимся человеком животным или клеткам
[00152] Раскрытые в настоящем документе способы могут предусматривать введение в организм не являющегося человеком животного или клетку различных молекул (например, нацеленных на TRKB человека реагентов, таких как антитела или малые молекулы), включая в себя нуклеиновые кислоты, белки, комплексы нуклеиновой кислоты и белка, пептидные миметики, антигенсвязывающие белка или малые молекулы. «Введение» включает в себя представление клетке или не являющемуся человеком животному молекулы (например, нуклеиновой кислоты или белка или малой молекулы) таким образом, чтобы она получила доступ к внутренней части клетки или внутренней части клеток в организме не являющегося человеком животного. Введение можно осуществлять любыми средствами. Если вводят несколько компонентов, их можно вводить одновременно или последовательно в любой комбинации. Кроме того, два или больше компонентов можно вводить в клетку или организм не являющегося человеком животного с помощью одного и того же способа доставки или с помощью различных способов доставки. Аналогично, два или больше компонентов можно вводить не являющемуся человеком животному одним и тем же путем введения или различными путями введения.
[00153] Молекулы, введенные в организм или клетку не являющегося человеком животного, могут быть предусмотрены в композициях, содержащих носитель, увеличивающий стабильность введенных молекул (например, продлевая период при данных условиях хранения (например, -20°С, 4°С или температура окружающей среды), для которых продукты разложения остаются ниже порогового значения, такого как ниже 0,5% по массе исходной нуклеиновой кислоты или белка, или увеличение стабильности in vivo). Неограничивающие примеры таких носителей включают в себя микросферы поли(молочной кислоты) (PLA), микросферы, микросферы сополимера D,L-молочной и гликолевой кислот (PLGA), липосомы, мицеллы, обратные мицеллы, липидные кохлеаты и липидные микротрубочки.
[00154] В настоящем документе предусмотрены различные способы и композиции, позволяющие вводить нацеленный на TRKB человека реагент в клетку или животному, не являющемуся человеком. Способы введения нуклеиновых кислот в клетки различных типов известны в настоящей области техники и включают в себя, например, способы стабильной трансфекции, способы транзиентной трансфекции и способы, опосредованные вирусом.
[00155] Протоколы трансфекции, а также протоколы для введения последовательностей нуклеиновых кислот в клетки могут различаться. Неограничивающие способы трансфекции включают в себя химические способы трансфекции; способы с использованием липосом; наночастиц; фосфата кальция (Graham et al. (1973) Virology 52 (2): 456-67, Bacchetti et al. (1977) Proc. Natl. Acad. Sci. USA 74 (4): 1590-4, и Kriegler, M (1991). Transfer and Expression: A Laboratory Manual. New York: W. H. Freeman and Company, pp.96-97); дендримеров или катионных полимеров, таких как DEAE-декстран или полиэтиленимин. Нехимические способы включают в себя электропорацию, сонопорацию и оптическую трансфекцию. Трансфекция на основе частиц включает в себя использование генной пушки или магнитной трансфекции (Bertram (2006) Current Pharmaceutical Biotechnology 7, 277-28). Вирусные техники также можно использовать для трансфекции.
[00156] Введение нацеленных на TRKB человека реагентов в клетку также может быть опосредовано электропорацией, внутрицитоплазматической инъекцией, вирусной инфекцией, аденовирусом, аденоассоциированным вирусом, лентивирусом, ретровирусом, трансфекцией, липид-опосредованной трансфекцией или нуклеофекцией. Нуклеофекция представляет собой усовершенствованную технологию электропорации, которая позволяет доставлять нуклеиновокислотные субстраты не только в цитоплазму, но также через ядерную мембрану и в ядро. Кроме того, использование нуклеофекции в раскрытых в настоящем документе способах, как правило, требует гораздо меньше клеток, чем обычная электропорация (например, только приблизительно 2 миллионов по сравнению с 7 миллионами при обычной электропорации). В одном примере нуклеофекцию выполняют с использованием системы LONZA® NUCLEOFECTOR™.
[00157] Введение нацеленных на TRKB человека реагентов в клетку (например, зиготу) также можно осуществить путем микроинъекции. В зиготах (т.е. эмбрионах на стадии одной клетки) микроинъекцию можно осуществлять в пронуклеус матери и/или отца или в цитоплазму. Если микроинъекция происходит только в одном пронуклеусе, отцовский пронуклеус предпочтительнее из-за его большего размера. Микроинъекцию мРНК предпочтительно осуществляют в цитоплазму (например, для доставки мРНК напрямую в аппарату трансляции), при этом микроинъекцию белка или полинуклеотида, кодирующего белок или кодирующего РНК, предпочтительно осуществляют в ядро/пронуклеус. Альтернативно, микроинъекцию можно осуществлять путем инъекции как в ядро/пронуклеус, так и в цитоплазму: сначала можно ввести иглу в ядро/пронуклеус и можно ввести первое количество, при удалении иглы из эмбриона на стадии одной клетки второе количество может быть введено инъекцией в цитоплазму. Способы проведения микроинъекции хорошо известны. См., например, Nagy et al. (Nagy A, Gertsenstein M, Vintersten K, Behringer R., 2003, Manipulating the Mouse Embryo. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); см. также Meyer et al. (2010) Proc. Natl. Acad. Sci. USA 107:15022-15026 и Meyer et al. (2012)Proc. Natl. Acad. Sci. USA 109:9354-9359.
[00158] Другие способы введения нацеленных на TRKB человека реагентов в клетку или животному, не являющемуся человеком, могут включать в себя, например, векторную доставку, опосредованную частицами доставку, опосредованную экзосомами доставку, опосредованную липидными наночастицами доставку, опосредованную проникающим в клетку пептидом доставку или опосредованную имплантируемыми устройствами доставку. В качестве конкретных примеров, нуклеиновую кислоту или белок можно вводить в клетку или животному, не являющемуся человеком, в носителе, таком как микросфера поли(молочной кислоты) (PLA), микросфера сополимера D,L-молочной и гликолевой кислот) (PLGA), липосома, мицелла, обратная мицелла, липидный кохлеат или липидная микротрубочка. Некоторые конкретные примеры доставки животному, не являющемуся человеком, включают в себя гидродинамическую доставку, опосредованную вирусом доставку (например, опосредованную аденоассоциированным вирусом (AAV) доставку) и опосредованную липидными наночастицами доставку.
[00159] Введение нацеленных на TRKB человека реагентов в клетки или организм животных, не являющихся человеком, можно осуществить с помощью гидродинамической доставки (HDD). Гидродинамическая доставка появилась как способ внутриклеточной доставки ДНК in vivo. Для доставки генов в паренхиматозные клетки через выбранный кровеносный сосуд необходимо вводить только необходимые последовательности ДНК, что устраняет проблемы безопасности, связанные с современными вирусными и синтетическими векторами. При введении в кровоток ДНК способна достигать клеток в различных тканях, доступных для крови. Гидродинамическая доставка использует силу, создаваемую быстрой инъекцией большого объема раствора в несжимаемую кровь в кровотоке, чтобы преодолеть физические барьеры эндотелия и клеточных мембран, которые препятствуют проникновению крупных и непроницаемых для мембран соединений в паренхиматозные клетки. В дополнение к доставке ДНК этот способ применим для эффективной внутриклеточной доставки РНК, белков и других небольших соединений in vivo. См., например, Bonamassa et al. (2011) Pharm. Res. 28(4):694-701, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00160] Введение нацеленных на TRKB человека реагентов также можно осуществить путем опосредованной вирусом доставки, такой как AAV-опосредованная доставка или опосредованная лентивирусом доставка. Другие иллюстративные вирусы/вирусные векторы включают в себя ретровирусы, аденовирусы, вирусы коровьей оспы, поксвирусы и вирусы простого герпеса. Вирусы могут инфицировать делящиеся клетки, неделящиеся клетки или как делящиеся, так и неделящиеся клетки. Вирусы могут интегрироваться в геном хозяина или, альтернативно, не интегрироваться в геном хозяина. Такие вирусы также могут быть разработаны так, чтобы получить сниженный иммунитет. Вирусы могут являться способными к репликации или могут являться дефектными по репликации (например, дефектными в одном или нескольких генах, необходимых для дополнительных циклов репликации и/или упаковки вириона). Вирусы могут вызывать транзиентную экспрессию, длительную экспрессию (например, в течение по меньшей мере 1 недели, 2 недель, 1 месяца, 2 месяцев или 3 месяцев) или постоянную экспрессию. Иллюстративные титры вирусов (например, титры AAV) включают в себя 1012, 1013, 1014, 1015 и 1016 векторных геномов/мл.
[00161] Геном оцДНК AAV состоит из двух открытых рамок считывания, Rep и Сар, фланкированных двумя инвертированными концевыми повторами, которые позволяют синтезировать цепь комплементарной ДНК. При конструировании переносящей плазмиды AAV трансген помещают между двумя ITR, и Rep и Сар могут доставляться in trans. В дополнение к Rep и Cap, AAV может потребовать наличие плазмиды - помощника, содержащей гены аденовируса. Эти гены (Е4, Е2а и VA) опосредуют репликацию AAV. Например, переносящая плазмида, Rep/Cap, и плазмида - помощник могут быть трансфицированы в клетки HEK293, содержащие ген аденовируса Е1+, для получения инфекционных частиц AAV. Альтернативно, Rep, Сар и гены - помощники аденовируса могут быть объединены в одну плазмиду. Подобные упаковывающие клетки и способы можно использовать для других вирусов, таких как ретровирусы.
[00162] Идентифицированы многочисленные серотипы AAV. Эти серотипы различаются по типам клеток, которые они инфицируют (т.е. их тропизм), что позволяет осуществлять преимущественную трансдукцию определенных типов клеток. Серотипы ткани ЦНС включают в себя AAV1, AAV2, AAV4, AAV5, AAV8 и AAV9. Серотипы для ткани сердца включают в себя AAV1, AAV8 и AAV9. Серотипы почечной ткани включают в себя AAV2. Серотипы легочной ткани включают в себя AAV4, AAV5, AAV6 и AAV9. Серотипы ткани поджелудочной железы включают в себя AAV8. Серотипы для фоторецепторных клеток включают AAV2, AAV5 и AAV8. Серотипы ткани пигментного эпителия сетчатки включают в себя AAV1, AAV2, AAV4, AAV5 и AAV8. Серотипы скелетной мышечной ткани включают в себя AAV1, AAV6, AAV7, AAV8 и AAV9. Серотипы ткани печени включают в себя AAV7, AAV8 и AAV9 и, в частности, AAV8.
[00163] Тропизм можно дополнительно уточнить с помощью псевдотипирования, которое представляет собой смешивание капсида и генома из различных вирусных серотипов. Например, AAV2/5 указывает на вирус, содержащий геном серотипа 2, упакованный в капсид из серотипа 5. Использование псевдотипированных вирусов может повысить эффективность трансдукции, а также изменить тропизм. Гибридные капсиды, полученные из разных серотипов, также можно использовать для изменения вирусного тропизма. Например, AAV-DJ содержит гибридный капсид из восьми серотипов и демонстрирует высокую инфекционность в широком диапазоне типов клеток in vivo. AAV-DJ8 является еще одним примером, который проявляет свойства AAV-DJ, но с улучшенным поглощением тканями головного мозга. Серотипы AAV также можно модифицировать с помощью мутаций. Примеры мутационных модификаций AAV2 включают в себя Y444F, Y500F, Y730F и S662V. Примеры мутационных модификаций AAV3 включают в себя Y705F, Y731F и T492V. Примеры мутационных модификаций AAV6 включают в себя S663V и T492V. Другие псевдотипированные/модифицированные варианты AAV включают в себя AAV2/1, AAV2/6, AAV2/7, AAV2/8, AAV2/9, AAV2.5, AAV8.2 и AAV/SASTG.
[00164] Для ускорения экспрессии трансгена можно использовать самокомплементарные варианты AAV (scAAV). Поскольку AAV зависит от механизма репликации ДНК клетки, чтобы синтезировать комплементарную цепь генома одноцепочечной ДНК AAV, экспрессия трансгена может быть отсрочена. Чтобы устранить эту задержку, можно использовать scAAV, содержащие комплементарные последовательности, которые способны к самопроизвольному отжигу при заражении, что устраняет необходимость синтеза ДНК клетки - хозяина.
[00165] Чтобы увеличить емкость упаковки, более длинные трансгены можно разделить между двумя переносящими плазмидами AAV, первая с 3'-донорным сайтом сплайсинга, а вторая с 5'-акцепторным сайтом сплайсинга. При коинфекции клетки эти вирусы образуют конкатемеры, сплайсируются вместе, и можно экспрессировать полноразмерный трансген. Хотя это позволяет увеличить экспрессию трансгена, экспрессия является менее эффективной. В подобных способах увеличения емкости используют гомологичную рекомбинацию. Например, трансген может быть разделен между двумя переносящими плазмидами, но с существенным перекрытием последовательности, так что коэкспрессия индуцирует гомологичную рекомбинацию и экспрессию полноразмерного трансгена.
[00166] Введение нацеленных на TRKB человека реагентов также можно осуществить путем доставки, опосредованной липидными наночастицами (LNP). Липидные составы могут защищать биологические молекулы от разложения при одновременном улучшении их клеточного поглощения. Липидные наночастицы представляют собой частицы, содержащие множество молекул липидов, физически связанных друг с другом межмолекулярными силами. Они включают в себя микросферы (включая в себя однослойные и многослойные везикулы, например, липосомы), дисперсную фазу в эмульсии, мицеллы или внутреннюю фазу в суспензии. Такие липидные наночастицы можно использовать для инкапсуляции одной или нескольких нуклеиновых кислот или белков для доставки. Составы, которые содержат катионные липиды, являются применимыми для доставки полианионов, таких как нуклеиновые кислоты. Другими липидами, которые могут быть включены, являются нейтральные липиды (т.е. незаряженные или цвиттерионные липиды), анионные липиды, вспомогательные липиды, которые усиливают трансфекцию, и липиды-невидимки, которые увеличивают продолжительность времени, в течение которого наночастицы могут существовать in vivo. Примеры подходящих катионных липидов, нейтральных липидов, анионных липидов, вспомогательных липидов и липидов-невидимок можно найти в международной патентной публикации WO 2016/010840 А1, полностью включенной в настоящий документ посредством ссылки для всех целей. Иллюстративная липидная наночастица может содержать катионный липид и один или несколько других компонентов. В одном примере другой компонент может содержать вспомогательный липид, такой как холестерин. В другом примере другие компоненты могут содержать вспомогательный липид, такой как холестерин, и нейтральный липид, такой как DSPC. В другом примере другие компоненты могут содержать вспомогательный липид, такой как холестерин, необязательный нейтральный липид, такой как DSPC, и липид-невидимку, такой как S010, S024, S027, S031 или S033.
[00167] Способ доставки можно выбрать для снижения иммуногенности. Например, если несколько компонентов, их можно доставлять разными способами (например, бимодальной доставкой). Эти различные способы могут придавать различные фармакодинамические или фармакокинетические свойства изучаемой доставляемой молекуле. Например, разные способы могут привести к разному распределению в ткани, разному периоду полужизни или разному распределению по времени. Некоторые способы доставки (например, доставка нуклеиновокислотного вектора, который сохраняется в клетке посредством автономной репликации или геномной интеграции) приводят к более постоянной экспрессии и присутствию молекулы, тогда как другие способы доставки являются временными и менее постоянными (например, доставка РНК или белка).
[00168] Введение in vivo можно осуществлять любым подходящим путем, включая в себя, например, парентеральный, внутривенный, пероральный, подкожный, внутриартериальный, внутричерепной, интратекальный, интраперитонеальный, местный, интраназальный или внутримышечный. Способы системного введения включают в себя, например, пероральный и парентеральный пути. Примеры парентеральных путей включают в себя внутривенный, внутриартериальный, внутрикостный, внутримышечный, внутрикожный, подкожный, интраназальный и интраперитонеальный пути. Конкретным примером является внутривенная инфузия. Закапывание в нос и интравитреальная инъекция являются другими конкретными примерами. Способы местного введения включают в себя, например, интратекальный, интрацеребровентрикулярный, интрапаренхиматозный (например, локализованная интрапаренхиматозная доставка в следующее: стриатум (например, в хвостатое ядро или в путамен), кора головного мозга, прецентральная извилина, гиппокамп (например, зубчатая извилина или область СА3), височная кора, миндалина, лобная кора, таламус, мозжечок, продолговатый мозг, гипоталамус, крыша среднего мозга, покрышка среднего мозга или черная субстанция), интраокулярный, интраорбитальный, субконъюктивальный, интравитреальный, субретинальный и транссклеральный пути. Значительно меньшие количества компонентов (по сравнению с системными подходами) могут оказывать эффект при местном введении (например, интрапаренхиматозном или интравитреальном) по сравнению с системным введением (например, введением внутривенно). Местные способы введения также могут уменьшать или устранять возникновение потенциально токсических побочных эффектов, которые могут возникнуть, когда терапевтически эффективные количества компонента вводятся системно. Согласно конкретному примеру нацеленные на TRKB человека реагенты вводят посредством прямой гиппокампальной инъекции, подкожной инъекции или интравитреальной инъекции.
[00169] Композиции, содержащие нацеленные на TRKB человека реагенты можно составить с использованием одного или нескольких физиологически и фармацевтически приемлемых носителей, разбавителей, вспомогательных веществ или добавок. Состав может зависеть от выбранного пути введения. Термин «фармацевтически приемлемый» означает, что носитель, разбавитель, вспомогательное вещество или добавка совместимы с другими ингредиентами состава и по существу не вредны для их реципиента.
[00170] Частота введения и количество дозировок могут зависеть от периода полужизни нацеленных на TRKB человека реагентов и пути введения среди других факторов. Введение нацеленных на TRKB человека реагентов в клетку или животному, не являющемуся человеком, можно выполнить один раз или несколько раз в течение периода времени. Например, введение можно выполнить по меньшей мере два раза в течение периода времени, по меньшей мере три раза в течение периода времени, по меньшей мере четыре раза в течение периода времени, по меньшей мере пять раз в течение периода времени, по меньшей мере шесть раз в течение периода времени, по меньшей мере семь раз в течение периода времени, по меньшей мере восемь раз в течение периода времени, по меньшей мере девять раз в течение периода времени, по меньшей мере в десять раз в течение периода времени, по меньшей мере одиннадцать раз, по меньшей мере двенадцать раз в течение периода времени, по меньшей мере тринадцать раз в течение периода времени, по меньшей мере четырнадцать раз в течение периода времени, по меньшей мере пятнадцать раз в течение периода времени, по меньшей мере шестнадцать раз в течение периода времени, по меньшей мере семнадцать раз в течение периода времени, по меньшей мере восемнадцать раз в течение периода времени, по меньшей мере девятнадцать раз в течение периода времени или по меньшей мере двадцать раз в течение периода времени.
E. Измерение доставки, активности или эффективности нацеленных на TRKB человека реагентов in vivo или ex vivo
[00171] Раскрытые в настоящем документе способы могут дополнительно предусматривать обнаружение или измерение активности нацеленных на TRKB человека реагентов. Измерение активности таких реагентов (например, активность агониста или активность ингибитора) может включать в себя измерение активности TRKB. Активность TRKB можно измерить с помощью любых известных средств. Например, можно оценить фосфорилирование TRKB (например, в головном мозге или нейронах), можно оценить активацию нижележащих путей, таких как PI3K/AKT и MAPK/ERK за счет TRKB (например, в головном мозге или нейронах, таких как первичные кортикальные нейроны) или можно оценить клеточную выживаемость (например, клеточная выживаемость нейронов, такая как выживаемость ганглиозных клеток сетчатки). Например, фосфорилирование или активацию нижележащих сигнальных путей можно оценить через 15 минут, 30 минут, 1 час, 2 часа, 4 часа или 18 часов после введения дозы. Увеличение фосфорилирования TRKB, активации нижележащих сигнальных путей или клеточной выживаемости может являться признаком активации TRKB, тогда как их уменьшение может указывать на ингибирование TRKB.
[00172] У не являющихся человеком животных оценка может предусматривать оценку одного или нескольких или всех из следующего: масса тела, состав тканей организма, метаболизм и локомоция, по сравнению с контрольным не являющимся человеком животным (например, через 12 часов, 24 часа, 48 часов, 72 часа, 96 часов или 120 часов после введения дозы). См., например, Lin et al. (2008) PLoS ONE 3(4):e1900; Rios et al. (2013) Trends in Neurosciences 36(2):83-90; и Zomer et al. (2003) Biol. Psychiatry 54:972-982, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Оценка изменений состава тканей организма может предусматривать, например, оценку массы нежировых тканей и/или массы жировой ткани. Оценка изменений метаболизма может предусматривать, например, оценку изменений в потреблении пищи и/или потреблении воды. Уменьшение массы тела, массы жировой ткани, массы нежировых тканей, потребления пищи и потребления воды может являться признаком активации TRKB, тогда как уменьшение этих показателей может являться признаком ингибирования TRKB. Увеличение локомоции может являться признаком активации TRKB, тогда как уменьшение этого показателя может являться признаком ингибирования TRKB.
[00173] Оценка может предусматривать оценку нейропротекторной активности. В качестве одного примера клеточную выживаемость можно оценить у не являющихся человеком животных. Например, ганглиозных клеток сетчатки грызуна (RGC) часто используют для исследования нейродегенеративных процессов, ассоциированных с поражением аксона, а также для оценки нейропротекторной терапии. См., например,  (2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. Выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить (например, в модели полного пересечения зрительного нерва после повреждения зрительного нерва) после лечения с помощью нацеленного на TRKB человека реагента по сравнению с контрольным не являющимся человеком животным. Например, выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить в модели полного пересечения зрительного нерва после повреждения зрительного нерва. См., например, (2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. В качестве другого примера выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить в модели раздавливания зрительного нерва. В этой модели повреждение зрительного нерва раздавливанием приводит к постепенному апоптозу ганглиозных клеток сетчатки. См., например, Tang et al. (2011) J. Vis. Exp. 50:2685, полностью включенный в настоящий документ посредством ссылки для всех целей. Выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить, например, путем измерения плотности ганглиозных клеток сетчатки (например, в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки). Увеличенная выживаемость/жизнеспособность может являться показателем активации TRKB, тогда как уменьшенная выживаемость/жизнеспособность может являться показателем ингибирования TRKB.
(2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. Выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить (например, в модели полного пересечения зрительного нерва после повреждения зрительного нерва) после лечения с помощью нацеленного на TRKB человека реагента по сравнению с контрольным не являющимся человеком животным. Например, выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить в модели полного пересечения зрительного нерва после повреждения зрительного нерва. См., например, (2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. В качестве другого примера выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить в модели раздавливания зрительного нерва. В этой модели повреждение зрительного нерва раздавливанием приводит к постепенному апоптозу ганглиозных клеток сетчатки. См., например, Tang et al. (2011) J. Vis. Exp. 50:2685, полностью включенный в настоящий документ посредством ссылки для всех целей. Выживаемость/жизнеспособность ганглиозных клеток сетчатки можно оценить, например, путем измерения плотности ганглиозных клеток сетчатки (например, в сетчатках, иссеченных и окрашенных в отношении ганглиозных клеток сетчатки). Увеличенная выживаемость/жизнеспособность может являться показателем активации TRKB, тогда как уменьшенная выживаемость/жизнеспособность может являться показателем ингибирования TRKB.
[00174] Если нацеленный на TRKB человека реагент представляет собой реагент для редактирования генома, измерение может содержать оценку гуманизированного локуса TRKB в отношении модификаций. Различные способы можно использовать для идентификации клеток с целевой генетической модификацией. Скрининг может включать в себя количественный анализ для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ можно выполнить с помощью количественной ПЦР, такой как ПНР в режиме реального времени (кПЦР). ПНР в режиме реального времени может использовать первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может содержать флуоресцентный зонд, который распознает амплифицированную последовательность. Другие примеры подходящих количественных анализов включают в себя флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды TAQMAN® Molecular Beacon или технологию зондов ECLIPSE™ (см., например, US 2005/0144655, полностью включенную в настоящий документ посредством ссылки для всех целей). Секвенирование следующего поколения (NGS) также можно использовать для скрининга. Секвенирование следующего поколения может также носить название «NGS» или «массовое параллельное секвенирование» или «высокопроизводительное секвенирование». NGS можно использовать в качестве инструмента скрининга в дополнение к анализам МОА для определения точного характера целевой генетической модификации и того, является ли это согласованным по типам клеток или типов тканей или типов органов.
[00175] Оценку у животного, не являющегося человеком, можно проводить в любом типе клеток из любой ткани или органа. Например, оценку можно осуществить в нескольких типах клеток из одной и той же ткани или органа (например, головной мозг) или в клетках из нескольких мест в ткани или органе (например, гиппокамп и стриатум). Это может предоставить информацию о том, какие типы клеток в целевой ткани или органе модифицируются или какие участки ткани или органа достигаются нацеленным на TRKB человека реагентом. В качестве другого примера оценку можно осуществить в нескольких типах тканей или в нескольких органах. В способах, в которых нацеленное воздействие производят на конкретную ткань или орган, это может предоставить информацию о том, насколько эффективно происходит нацеленное воздействие на эту ткань или орган и есть ли нецелевые эффекты в других тканях или органах.
[00176] Если реагент предназначен для инактивации гуманизированного локуса TRKB, воздействия на экспрессию гуманизированного локуса TRKB или предотвращения трансляции мРНК гуманизированного TRKB, измерение может включать в себя оценку экспрессии мРНК гуманизированного TRKB или белка. Это измерение можно проводить в головном мозге или конкретных типах клеток (например, нейронах, таких как ганглиозные клетки сетчатки).
IV. Способы получения не являющихся человеком животных, содержащих гуманизированный локус TRKB
[00177] Предусмотрены различные способы получения генома не являющегося человеком животного, клетки не являющегося человеком животного или не являющегося человеком животного, содержащих гуманизированный локус TRKB, как раскрыто в другом месте в настоящем документе. Любой общепринятый способ или протокол получения генетически модифицированного организма является подходящим для получения такого генетически модифицированного не являющегося человеком животного. См., например, Cho et al. (2009) Current Protocols in Cell Biology 42:19.11:19.11.1-19.11.22 и Gama Sosa et al. (2010) Brain Struct. Fund. 214(2-3):91-109, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Такие генетически модифицированный не являющиеся человеком животные могут быть получены, например, посредством нокина гена в целевом локусе TrkB.
[00178] Например, способ получения не являющегося человеком животного, содержащего гуманизированный локус TRKB, может предусматривать следующее: (1) модификация генома плюрипотентной клетки для включения гуманизированного локуса TRKB; (2) идентификация или отбор генетически модифицированной плюрипотентной клетки, содержащей гуманизированный локус TRKB; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион-хозяин не являющегося человеком животного; и (4) имплантация и вынашивание эмбриона-хозяина в организме суррогатной матери. Например, способ получения не являющегося человеком животного, содержащего гуманизированный локус TRKB, может предусматривать следующее: (1) модификация генома плюрипотентной клетки для включения гуманизированного локуса TRKB; (2) идентификация или отбор генетически модифицированной плюрипотентной клетки, содержащей гуманизированный локус TRKB; (3) введение генетически модифицированной плюрипотентной клетки в эмбрион-хозяин не являющегося человеком животного; и (4) вынашивание эмбриона-хозяина в организме суррогатной матери. Необязательно эмбрион-хозяин, содержащий модифицированную плюрипотентную клетку (например, ES клетку, не относящуюся к человеку), можно инкубировать до стадии бластоцисты перед имплантацией и гестацией в организме суррогатной матери для получения не являющегося человеком животного F0. Затем суррогатная мать может произвести не являющееся человеком животное поколения F0, содержащее гуманизированный локус TRKB.
[00179] Способы могут дополнительно предусматривать идентификацию клетки или животного с модифицированным целевым геномным локусом. Различные способы можно использовать для идентификации клеток и животных с целевой генетической модификацией.
[00180] На стадии модификации генома можно использовать, например, экзогенные матрицы репарации (например, нацеливающие векторы) для модификации локуса TrkB так, чтобы он содержал гуманизированный локус TRKB, раскрытый в настоящем документе. В качестве одного примера нацеливающий вектор может быть предусмотрен для получения гуманизированного гена TRKB на эндогенном локусе TrkB (например, эндогенный локус TrkB не являющегося человеком животного), причем нацеливающий вектор содержит 5' гомологичное плечо, нацеленное на 5' целевую последовательность на эндогенном локусе TrkB, и 3' гомологичное плечо, нацеленное на 3' целевую последовательность на эндогенном локусе TrkB. Экзогенные матрицы репарации также могут содержать нуклеиновые кислоты - вставки, включая в себя сегменты ДНК, подлежащие интеграции в локус TrkB. Интеграция нуклеиновой кислоты - вставки в локус TrkB может приводить к добавлению представляющей интерес последовательность нуклеиновой кислоты в локус TrkB, делеции представляющей интерес последовательности нуклеиновой кислоты в локус TrkB или замене представляющей интерес последовательности нуклеиновой кислоты в локус TrkB (т.е. делеция и вставка). Гомологичные плечи могут фланкировать нуклеиновую кислоту - вставку, содержащую последовательность TRKB человека, для создания гуманизированного локуса TRKB (например, для делеции сегмента эндогенного локуса TrkB и замены ортологичной последовательностью TRKB человека).
[00181] Экзогенные матрицы репарации могут быть предусмотрены для опосредованной негомологичным соединением концов (NHEJ) вставки или гомологичной репарации. Экзогенные матрицы репарации могут содержать дезоксирибонуклеиновую кислоту (ДНК) или рибонуклеиновую кислоту (РНК), они могут являться одноцепочечными или двухцепочечными, и они могут быть в линейной или кольцевой форме. Например, матрица репарации может представлять собой одноцепочечный олигодезоксинуклеотид (ssODN).
[00182] Экзогенные матрицы репарации также могут содержать гетерологичную последовательность, которая не присутствует на ненацеленном эндогенном локусе TrkB. Например, экзогенная матрица репарации может содержать кассету селекции, такую как кассета селекции, фланкированная сайтами распознавания рекомбиназы.
[00183] Некоторые экзогенные матрицы репарации содержат гомологичные плечи. Если экзогенная матрица репарации также содержит нуклеиновую кислоту - вставку, гомологичные плечи могут фланкировать нуклеиновую кислоту - вставку. Для удобства пользования гомологичные плечи в настоящем документе носят название 5' и 3' (т.е. расположенные выше и ниже относительно хода транскрипции) гомологичные плечи. Эта терминология относится к относительному положению гомологичных плеч по отношению к нуклеиновой кислоте - вставке в пределах экзогенной матрицы репарации. 5' и 3' гомологичные плечи соответствуют областям в пределах локуса TrkB, которые в настоящем документе носят название «5' целевая последовательность» и «3' целевая последовательность» соответственно.
[00184] Гомологичное плечо и целевая последовательность «соответствуют» или являются «соответствующими» друг другу, когда две области обладают достаточным уровнем идентичности последовательности относительно друг друга, чтобы действовать в качестве субстратов для реакции гомологичной рекомбинации. Термин «гомология» включает в себя последовательности ДНК, которые либо являются идентичными, либо обладают идентичностью последовательности относительно соответствующей последовательности. Идентичность последовательности между данной целевой последовательностью и соответствующим гомологичным плечом, встречающимся в экзогенной матрице репарации, может быть любой степени идентичности последовательности, которая позволяет происходить гомологичной рекомбинации. Например, количество идентичности последовательности, которой обладают гомологичное плечо экзогенной матрицы репарации (или его фрагмент) и целевая последовательность (или ее фрагмент), может составлять по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности, так что последовательности подвергаются гомологичной рекомбинации. Более того, соответствующая область гомологии между гомологичным плечом и соответствующей целевой последовательностью может быть любой длины, которая является достаточной для стимулирования гомологичной рекомбинации. В некоторых нацеливающих векторах предусмотренная мутация в эндогенном локусе TrkB включена в нуклеиновую кислоту - вставку, фланкированную гомологичными плечами.
[00185] В клетках, отличных от эмбрионов на стадии одной клетки, экзогенная матрица репарации может представлять собой «большой нацеливающий вектор» или «LTVEC», который включает в себя нацеливающие векторы, которые содержат гомологичные плечи, которые соответствуют и происходят из последовательностей нуклеиновой кислоты, больших чем те, которые, как правило, используют в других подходах, предусмотренных для проведения гомологичной рекомбинации в клетках. LTVEC также включают в себя нацеливающие векторы, содержащие нуклеиновые кислоты - вставки, характеризующиеся последовательностями нуклеиновой кислоты, большими чем те, которые, как правило, используют в других подходах, предусмотренных для проведения гомологичной рекомбинации в клетках. Например, LTVEC делают возможным модификацию больших локусов, которые не могут поместиться в общепринятые нацеливающие векторы на основе плазмиды из-за их ограничений по размеру. Например, целевой локус может представлять собой (т.е. 5' и 3' гомологичные плечи могут соответствовать локусу) локус клетки, который не может быть нацелен с использованием общепринятого способа или который может быть нацелен только неправильно или только со значительно низкой эффективностью при отсутствии одно- или двухцепочечного разрыва, вызванного нуклеазным агентом (например, белком Cas). LTVEC могут быть любой длины и, как правило, их длина составляет по меньшей мере 10 т.п.н. Общая сумма 5' гомологичного плеча и 3' гомологичного плеча в LTVEC, как правило, составляет по меньшей мере 10 т.п.н.
[00186] Стадия скрининга может предусматривать, например, количественный анализ для оценки модификации аллеля (МОА) родительской хромосомы. Например, количественный анализ можно выполнить с помощью количественной ПНР, такой как ПНР в режиме реального времени (кПЦР). В ПНР в режиме реального времени можно использовать первый набор праймеров, который распознает целевой локус, и второй набор праймеров, который распознает нецелевой эталонный локус. Набор праймеров может содержать флуоресцентный зонд, который распознает амплифицированную последовательность
[00187] Другие примеры подходящих количественных анализов включают в себя флуоресцентно-опосредованную гибридизацию in situ (FISH), сравнительную геномную гибридизацию, изотермическую амплификацию ДНК, количественную гибридизацию с иммобилизованным(и) зондом(ами), зонды INVADER®, зонды TAQMAN® Molecular Beacon или технологию зондов ECLIPSE™ (см., например, US 2005/0144655, полностью включенную в настоящий документ посредством ссылки для всех целей).
[00188] Примером подходящей плюрипотентной клетки является эмбриональная стволовая (ES) клетка (например, ES клетка мыши или ES клетка крысы). Модифицированную плюрипотентную клетку можно создать, например, путем следующего: (а) введение в клетку одной или нескольких экзогенных донорных нуклеиновых кислот (например, нацеливающих векторов), содержащих нуклеиновую кислоту - вставку, фланкированную, например, 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, причем нуклеиновая кислота - вставка содержит последовательность TRKB человека для создания гуманизированного локуса TRKB; и (b) идентификация по меньшей мере одной клетки, содержащей в своем геноме нуклеиновую кислоту - вставку, интегрированную в эндогенный локусе TrkB (т.е. идентификация по меньшей мере одной клетки, содержащей гуманизированный локус TRKB). Модифицированную плюрипотентную клетку можно создать, например, посредством рекомбинации путем следующего: (а) введение в клетку одного или нескольких нацеливающих векторов, содержащих нуклеиновую кислоту - вставку, фланкированную 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, причем нуклеиновая кислота - вставка содержит гуманизированный локус TRKB; и (b) идентификация по меньшей мере одной клетки, содержащей в своем геноме нуклеиновую кислоту - вставку, интегрированную в целевом геномном локусе.
[00189] Альтернативно, модифицированную плюрипотентную клетку можно получить следующим образом: (а) введение в клетку следующего: (i) нуклеазный агент, причем нуклеазный агент индуцирует одноцепочечный разрыв или двух цепочечный разрыв на целевом сайте в пределах эндогенного локуса TrkB; и (ii) одна или несколько экзогенных донорных нуклеиновых кислот (например, нацеливающих векторов), содержащих нуклеиновую кислоту - вставку, фланкированную, например, 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, расположенным в достаточной близости от целевого сайта нуклеазы, причем нуклеиновая кислота - вставка содержит последовательность TRKB человека для создания гуманизированного локуса TRKB; и (с) идентификация по меньшей мере одной клетки, содержащей в своем геноме нуклеиновую кислоту - вставку, интегрированную в эндогенном локусе TrkB (т.е. идентификация по меньшей мере одной клетки, содержащей гуманизированный локус TRKB). Альтернативно, модифицированную плюрипотентную клетку можно получить следующим образом: (а) введение в клетку следующего: (i) нуклеазный агент, причем нуклеазный агент индуцирует одноцепочечный разрыв или двух цепочечный разрыв на целевом сайте в пределах целевого геномного локуса; и (ii) один или несколько нацеливающих векторов, содержащих нуклеиновую кислоту - вставку, фланкированную 5' и 3' гомологичными плечами, соответствующими 5' и 3' целевым сайтам, расположенным в достаточной близости от сайт распознавания, причем нуклеиновая кислота - вставка содержит гуманизированный локус TRKB; и (с) идентификация по меньшей мере одной клетки, содержащей модификацию (например, интеграцию нуклеиновой кислоты - вставки) в целевом геномном локусе. Можно использовать любой нуклеазный агент, который вызывает одноцепочечный разрыв или разрыв двухцепочечной цепи в требуемый сайт распознавания. Примеры подходящих нуклеаз включают в себя эффекторную нуклеазу, подобную активатору транскрипции (TALEN), нуклеазу домена «цинковые пальцы» (ZFN), мегануклеазу и системы сгруппированных регулярно перемежающихся коротких палиндромных повторов (CRISPRyCRISPR-ассоциированных белков (Cas) или компоненты таких системы (например, CRISPR/Cas9). См., например, US 2013/0309670 и US 2015/0159175, каждая из которых полностью включена в настоящий документ посредством ссылки для всех целей.
[00190] Донорную клетку можно вводить в эмбрион-хозяин на любой стадии, такой как стадия бластоцисты или стадия предморула (т.е. стадия 4 клеток или стадия 8 клеток). Создается потомство, способное передавать генетическую модификацию через зародышевую линию. См., например, патент США №7294754, полностью включенный в настоящий документ посредством ссылки для всех целей.
[00191] Альтернативно, способ получения не являющихся человеком животных, описанных в другом месте в настоящем документе, может предусматривать следующее: (1) модификация генома эмбриона на стадии одной клетки для включения в него гуманизированного локуса TRKB с использованием способов, описанных выше, для модификации плюрипотентных клеток; (2) отбор генетически модифицированных эмбрионов; и (3) имплантация и вынашивание генетически модифицированного эмбриона в организме суррогатной матери. Альтернативно, способ получения не являющихся человеком животных, описанных в другом месте в настоящем документе, может предусматривать следующее: (1) модификация генома эмбриона на стадии одной клетки для включения в него гуманизированного локуса TRKB с использованием способов, описанных выше, для модификации плюрипотентных клеток; (2) отбор генетически модифицированных эмбрионов; и (3) вынашивание генетически модифицированного эмбриона в организме суррогатной матери. Создается потомство, способное передавать генетическую модификацию через зародышевую линию.
[00192] Техники ядерного переноса также можно использовать для получения не являющихся человеком млекопитающих. Вкратце, способы переноса ядра могут предусматривать следующие стадии: (1) энуклеация ооцита или получение энуклеированного ооцита; (2) выделение или получение донорной клетки или ядра для объединения с энуклеированным ооцитом; (3) вставка клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки; (4) имплантация реконструированной клетки в матку животного с образованием эмбриона; и (5) обеспечение развития эмбриона. В таких способах ооциты, как правило, извлекают у умерших животных, хотя они могут быть выделены также из яйцеводов и/или яичников живых животных. Ооциты могут созревать в различных хорошо известных средах до энуклеации. Энуклеацию яйцеклетки можно выполнить с помощью ряда хорошо известных способов. Вставку донорной клетки или ядра в энуклеированный ооцит с образованием реконструированной клетки можно осуществить путем микроинъекции донорной клетки под вителлиновый слой перед слиянием. Слияние может быть индуцировано приложением электрического импульса постоянного тока через плоскость контакта/слияния (электрослияние), воздействием на клетки химических веществ, способствующих слиянию, таких как полиэтиленгликоль, или посредством инактивированного вируса, такого как вирус Сендай. Реконструированная клетка может быть активирована электрическими и/или неэлектрическими способами до, во время и/или после слияния донора - ядра и реципиента - ооцита. Способы активации включают в себя электрические импульсы, химически индуцированный шок, проникновение с помощью сперматозоидов, повышение содержания двухвалентных катионов в ооците и снижение фосфорилирования клеточных белков (как с помощью ингибиторов киназы) в ооците. Активированные реконструированные клетки или эмбрионы можно культивировать в хорошо известных средах и затем переносить в матку животного. См., например, документы US 2008/0092249, WO 1999/005266, US 2004/0177390, WO 2008/017234 и патент США №7612250, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[00193] Различные способы, предусмотренные в настоящем документе, позволяют создавать генетически модифицированное не являющееся человеком животное F0, причем клетки генетически модифицированного животного F0 содержат гуманизированный локус TRKB. Признано, что в зависимости от способа, используемого для получения животного F0, число клеток в животном F0, которые содержат гуманизированный локус TRKB, будет варьироваться. Введение донорных ES клеток в эмбрион на стадии предморула из соответствующего организма (например, эмбриона мыши на стадии 8 клеток), например, с помощью способа VELOCIMOUSE® позволяет получать больший процент клеточной популяции животного F0, которое содержит клетки, содержащие представляющую интерес нуклеотидную последовательность, содержащую целевую генетическую модификацию. Например, по меньшей мере 50%, 60%, 65%, 70%, 75%, 85%, 86%, 87%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% клеточного вклада животного F0, не являющегося человеком, может содержать клеточную популяцию, содержащую целевую модификацию.
[00194] Клетки генетически модифицированного животного F0 могут являться гетерозиготными по гуманизированному локусу TRKB или могут являться гомозиготными по гуманизированному локусу TRKB.
[00195] Все патентные заявки, веб-сайты, другие публикации, регистрационные номера и тому подобное, указанные выше или ниже, полностью включены посредством ссылки для всех целей в той же степени, как если бы было специально и индивидуально указано, что каждый отдельный документ включен в него посредством ссылки. Если разные версии последовательности связаны с регистрационным номером в разное время, подразумевается версия, связанная с регистрационным номером на дату подачи настоящей заявки. Действительная дата подачи означает более раннюю из действительной даты подачи или даты подачи приоритетной заявки со ссылкой на регистрационный номер, в соответствующих случаях. Аналогичным образом, если разные версии публикации, веб-сайта или тому подобного публикуются в разное время, подразумевается последняя версия, опубликованная на действительную дату подачи, если не указано иное. Любой признак, стадию, элемент, вариант осуществления или аспект настоящего изобретения можно использовать в сочетании с любым другим, если специально не указано иное. Хотя настоящее изобретение было описано более подробно с помощью иллюстрации и примера для ясности и понимания, будет очевидно, что определенные изменения и модификации можно осуществить в рамках объема прилагаемой формулы изобретения.
Краткое описание последовательностей
[00196] Нуклеотидные и аминокислотные последовательности, перечисленные в прилагаемом перечне последовательностей, показаны с использованием стандартных буквенных сокращений для нуклеотидных оснований и трехбуквенного кода для аминокислот. Нуклеотидные последовательности показаны в соответствии со стандартным соглашением, начиная с 5' конца последовательности и продвигаясь вперед (т.е. слева направо в каждой строке) до 3' конца. Показана только одна цепь каждой нуклеотидной последовательности, но считается, что комплементарная цепь включена в любую ссылку на отображаемую цепь. Когда представлена нуклеотидная последовательность, кодирующая аминокислотную последовательность, подразумевается, что также представлены ее кодон-вырожденные варианты, которые кодируют ту же аминокислотную последовательность. Аминокислотные последовательности показаны в соответствии со стандартным соглашением, начиная с амино конца последовательности и продвигаясь вперед (т.е. слева направо в каждой строке) к карбокси-концу.
[00197]



ПРИМЕРЫ
Пример 1. Получение мышей, содержащих гуманизированный локус TRKB
[00198] Большой нацеливающий вектор (LTVEC), содержащий 5' гомологичное плечо, содержащее 41,6 т.п.н., локус TrkB мыши и 3' гомологичное плечо, содержащее 62,4 т.п.н. локуса TrkB мыши, создавали для замены области, составляющей 65,7 т.п.н., из гена TrkB мыши, кодирующего внеклеточный домен TRKB мыши, на 74,4 т.п.н. соответствующей последовательности TRKB человека. Информация о TRKB мыши и человека представлена в таблице 2. Описание создания большого нацеливающего вектора представлено в таблице 3. Создание и применение больших нацеливающих векторов (LTVEC), полученных из ДНК бактериальной искусственной хромосомы (ВАС) посредством реакций бактериальной гомологичной рекомбинации (BHR) с использованием технологии генной инженерии VELOCIGENE®, описаны, например, в патенте США №6586251 и Valenzuela et al. (2003) Nat. Biotechnol. 21(6):652-659, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Создание LTVEC посредством способов сборки in vitro описано, например, в документах US 2015/0376628 и WO 2015/200334, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей.
[00199]

[00200]

[00201] В частности, область, начиная с экзона 2 (кодирующий экзон 1; от аминокислоты 32, сохраняя сигнальный пептид) до экзона 10, включая в себя первые 137 пар оснований интрона 10 и все интроны между экзонами 2 и 10 (т.е. между кодирующим экзоном 1 и экзоном 10) подвергали делеции из локуса TrkB мыши (сохраняя трансмембранный домен мыши, кодируемый экзонами 10 и 11). Область, включая в себя экзон 2/кодирующий экзон 1 (от аминокислоты 32, начиная после сигнального пептида) до экзона 10, включая в себя первые 177 пар оснований интрона 10 и все интроны между экзонами 2 и 10 (т.е. между кодирующим экзоном 1 и экзоном 10) вставляли вместо удаленной области крысы (сохраняя трансмембранный домен крысы, кодируемый экзонами 10 и 11).
[00202] Последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB мыши представлены в SEQ ID NO: 51-54, соответственно, с соответствующей кодирующей последовательностью, представленной в SEQ ID NO: 63-66 соответственно. Последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB человека представлены в SEQ ID NO: 59-62 соответственно, с соответствующей кодирующей последовательностью, представленной в SEQ ID NO: 71-74 соответственно. Предполагаемый кодируемый химерный белок TRKB содержит трансмембранный и внутриклеточный домены TRKB мыши, сигнальный пептид TRKB мыши и внеклеточный домен TRKB человека. См. фиг. 1. Выравнивание белков TRKB мыши и человека в фиг. 6. Кодирующие последовательности TrkB/TRKB мыши и человека представлены в SEQ ID NO: 9 и 11 соответственно. Последовательности белка TRKB мыши и человека представлены в SEQ ID NO: 1 и 3 соответственно. Последовательности для предполагаемой химерной кодирующей последовательности TRKB мыши/человека и предполагаемого химерного белка TRKB мыши/человека представлены в SEQ ID NO: 12 и 4 соответственно.
[00203] Для создания мутантного аллеля большой нацеливающий вектор вводили в эмбриональные стволовые клетки F1H4 мыши. После отбора с использованием антибиотика колонии отбирали, размножали и подвергали скринингу с помощью TAQMAN®. См. фиг. 2. Анализы на потерю аллеля проводили для обнаружения потери эндогенного аллеля крысы, и анализы на приобретение аллеля проводили для обнаружения приобретения гуманизированного аллеля с использованием праймеров и зондов, представленных в таблице 4.
[00204]

[00205] Анализы на модификацию аллеля (МОА), включая в себя анализы на потерю аллеля (LOA) и приобретение аллеля (GOA) описаны, например, в US 2014/0178879; US 2016/0145646; WO 2016/081923; и Frendewey et al. (2010) Methods Enzymol. 476:295-307, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Анализ на потерю аллеля (LOA) инвертирует традиционную логику скрининга и количественно определяет количество копий в образце геномной ДНК нативного локуса, на который была направлена мутация. В правильно нацеленном клоне гетерозиготных клеток анализ LOA обнаруживает один из двух нативных аллелей (для генов, не находящихся в хромосоме X или Y), причем другой аллель разрушается в результате целевой модификации. Тот же принцип может быть применен в обратном порядке, как и анализ усиления аллеля (GOA), для количественного определения количества копий вставленного нацеливающего вектора в образец геномной ДНК.
[00206] Мышей F0 создавали с использованием способа VELOCFMOUSE®. См., например, патенты США №№7576259; 7659442; 7294754; документы US 2008/0078000; и Poueymirou et al. (2007) Nat. Biotechnol. 25(1):91-99, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. В способе VELOCIMOUSE® целевые мышиные эмбриональные стволовые (ES) клетки вводят посредством лазерной инъекции в эмбрионы на стадии до образования морулы, например, в эмбрионы на стадии восьми клеток, которые эффективно дают мышей поколения F0, которые полностью происходят из ES-клеток. Все эксперименты, выполненные на гуманизированных в отношении TRKB мышах, как описано ниже, проводили на мышах, у которых самоудаляющаяся кассета селекции удалялась самостоятельно.
Пример 2. In vivo сравнение эффекта Н4Н9816 и REGN1945 антител изотипического контроля на массу тела и метаболизм у TrkBhu/hu мышей (MAID7139)
Экспериментальная процедура
[00207] Для определения эффекта антитела - агониста TRKB, Н4Н9816Р2, на массу тела и состав тканей тела метаболическое исследование мышей, гомозиготных в отношении экспрессии рецептора TRKB человека вместо рецептора TRKB мыши (TrkBhu/hu мыши) проводили после однократной подкожной инъекции антитела. Эти исследования предприняли частично на основе предыдущих исследований агонистов TrkB и мышей с «выключенным» TrkB. См., например, Lin et al. (2008) PLoS ONE 3(4):e1900; Rios et al. (2013) Trends in Neurosciences 36(2):83-90; и Zorner et al. (2003) Biol. Psychiatry 54:972-982, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. TrkBhu/hu мышей (самцы, возраст - 20 недель) вначале переносили из групповой клетки в индивидуальную клетку, в которой из размещали на две недели для акклиматизации. После окончания этого периода мышей переносили в метаболический клетки (CLAMS, Columbus Instruments) для оценки изменений в потреблении пищи и воды, локомоции, расходе энергии и дыхании после введения антител. Обычный порошкообразный корм хранили в напольной камере на весах с пружинным нажимом (Mettler Toledo, PL602E) для измерения потребления пищи посредством изменения общей массы корма. Вода была доступна через поилку, расположенную вверху клетки, и потребление измеряли путем отслеживания изменений в объеме насосной линии (Oxymax®/CLAMS Liquid Unit). Метаболические клетки CLAMS измеряли каждый из этих параметров в непрерывных 16-18-минутных интервалах на протяжении всего исследования. Метаболические данные анализировали в единичных измерениях и суммировали в 24-часовых интервалах, содержащих один полный цикл темноты и света, с использованием программного обеспечения OXYMAX®/CLAMS (Columbus instruments, v5.35). После акклиматизации к клеткам в течение двух недель мыши TrkBhu/hu получали однократную подкожную дозу 50 мг/кг либо антитела - агониста TRKB, Н4Н9816Р2, либо антитела изотипического контроля IgG4 в PBS при рН 7,2. Группа не получивших лечение контрольных мышей TrkBhu/hu не получала инъекцию. Мышей взвешивали непосредственно перед введением дозы и через 24, 48, 72, 96 и 120 часов после введения дозы. Чтобы измерить состав тканей организма каждой мыши, с помощью анализатора EchoMRI™-500 (EchoMRI LLC) проводили ядерно-магнитную резонансную релаксометрию, также называемую количественным магнитным резонансом. Перед введением дозы мышей помещали в прозрачный пластиковый держатель и вставляли в устройство ЯМР-МРТ для измерения массы нежировых тканей, массы жировой ткани и состояния гидратации каждого субъекта. Измерения выполняли в течение 0,5-3,2 минуты на мышь и снова проводили приблизительно через 120 часов после введения дозы.
Результаты и выводы
[00208] Ежедневный мониторинг массы тела проводили для определения того, вызывает ли единичная подкожная инъекция Н4Н9816Р2 потерю массы тела у мышей TrkBhu/hu. До введения дозы не было значительных различий в средней массе тела в трех группах лечения, так как каждая из них характеризовалась средней массой тела до введения дозы, составляющей 28,39-29,85 г (таблица 5). Однако через 48 часов после введения дозы мыши TrkBhu/hu, получившие лечение с помощью Н4Н9816Р2, потеряли в среднем 1,70 г, или 5,96% от массы тела до введения дозы. В то же время не получившие лечение и мыши TrkBhu/hu, получившие лечение с помощью антител изотипического контроля, набрали от 1,79 до 2,37% их массы тела до введения дозы. Мыши TrkBhu/hu, получившие лечение с помощью Н4Н9816Р2, продолжали терять массу тела на протяжении всего курса исследования, и через 72 и 96 часов после введения дозы эти мыши теряли в среднем 8,42% и 11,80% от массы тела до введения дозы соответственно. Через 120 часов после введения дозы мыши TrkBhu/hu, получившие лечение с помощью Н4Н9816Р2, потеряли в среднем 12,67% от массы тела до введения дозы. Наоборот, у не получивших лечение и мышей TrkBhu/hu, получивших лечение с помощью антител изотипического контроля, не наблюдалось потери массы тела до введения дозы в течение всего исследования. Поскольку масса тела у мышей TrkBhu/hu, получивших лечение с помощью Н4Н9816Р2, была значительно снижена по сравнению как с не получившими лечение, так и с изотипическими контролями через 48, 72, 96 и 120 часов после введения дозы, было установлено, что антитело - агонист TRKB Н4Н9816Р2 вызывает значительную потерю массы тела у TrkBhu/hu мышей.
[00209]

[00210] Эффект инъекции антитело - агонист TRKB Н4Н9816Р2 на состав тканей организма также измеряли путем проведения ЯМР-МРТ на каждом субъекте до и после введения дозы. Перед введением дозы три группы лечения TrkBhu/hu мышей не проявляли каких-либо значимых различий в массе жировой ткани или массе нежировых тканей, при этом каждая группа характеризовалась в среднем 4,19-4,75 г массы жировой ткани и 21,32-21,70 г массы нежировых тканей (таблица 6). После введения антител, тем не менее, TrkBhu/hu мыши, которым вводили дозу Н4Н9816Р2, теряли в среднем 48,90% своей общей массы жировой ткани тела в течение всего исследования (таблица 6). Не получившие лечение и получившие лечение с помощью антитела изотипического контроля TrkBhu/hu мыши теряли в среднем 8,49% и 9,48% своей массы жировой ткани перед введением дозы, соответственно, что было значимо меньшей, чем у получивших лечение с помощью Н4Н9816Р2 субъектов (таблица 6). Более того, получившие лечение с помощью Н4Н9816Р2 TrkBhu/hu мыши теряли в среднем 7,84% своей массы нежировых тканей в течение всего исследования, что было значимо больше, чем 2,41% и 1,75% средней потери массы нежировых тканей перед введением дозы у групп, не получивших лечение и получивших лечение с помощью антитела изотипического контроля, соответственно (таблица 6). Таким образом, описанная потеря массы тела можно объяснить значительной потерей массы жировой ткани и умеренной потерей массы нежировых тканей после инъекции антитела - агониста TRKB Н4Н9816Р2 у TrkBhu/hu мышей.
[00211]


[00212] В дополнение к оценке эффектов инъекции антитела - агониста TRKB Н4Н9816Р2 на массу тела и состав тканей тела у мышей TrkBhu/hu, непрерывно измеряли потребление пищи, питьевую активность и двигательную активности с помощью метаболической клетки. Перед введением дозы TrkBhu/hu мыши потребляли в среднем от 3,49 до 3,73 г корма в день. Однако через 24 часа после введения дозы получившие лечение с помощью Н4Н9816Р2 TrkBhu/hu мыши значительно снижали потребление пищи до 2,20 г корма в день. Средний уровень потребления пищи у получивших лечение с помощью Н4Н9816Р2 TrkBhu/hu мышей не превышал 2,49 г корма в день в течение оставшейся части исследования, в то время как не получившие лечение и получившие лечение с помощью изотипического антитела TrkBhu/hu мыши постоянно потребляли в среднем 3,62-4,07 г корма в день (таблица 7).
[00213] Аналогично, не было значительных различий в ежедневном потреблении воды между группами лечения до введения дозы. TrkBhu/hu мыши потребляли в среднем 4,67-5,55 мл воды в день в каждой группе лечения (таблица 8). После введения дозы получившие лечение с помощью Н4Н9816Р2 TrkBhu/hu мыши снижали потребление воды до 2,05-3,24 мл воды в день. Это было значительно ниже, чем у не получивших лечение и получивших лечение с помощью антитела изотипического контроля TrkBhu/hu мышей, которые постоянно потребляли 4,50-5,77 мл воды в день в течение всего исследования (таблица 8). Таким образом, инъекция антитела - агониста TRKB, Н4Н9816Р2, по-видимому, приводила к значительному снижению потребления пищи и воды у TrkBhu/hu мышей по сравнению как с не получившими лечение мышами, так и с изотипическими контролями.
[00214]

[00215]


[00216] Для определения эффектов лечения с помощью антитела на активность анализировали локомоцию с помощью программного обеспечения OXYMAX®/CLAMS (Columbus instruments, v5.35), которое непрерывно измеряло общее количество передвижений в х-плоскости каждой мыши. Одна мышь демонстрировала гиперактивность до введения дозы и была исключена из статистического анализа после введения дозы. В то время как у не получивших лечение и получивших лечение с помощью изотипического антитела субъектов постоянно регистрировали в среднем 11000-15000 передвижений в день в течение всего исследования, у получивших лечение с помощью Н4Н9816Р2 TrkBhu/hu мышей регистрировали 28260 передвижений в течение 24-48 часов после введения дозы и регистрировали 21193 и 27028 передвижений с 48-72 до 72-96 часов после введения дозы, соответственно (таблица 9). У получивших лечение с помощью Н4Н9816Р2 TrkBhu/hu мышей регистрировали большее общее количество передвижений в каждый момент времени после введения антитела, что указывает на гиперактивность как дополнительную активность при инъекции Н4Н9816Р2. В сочетании эти эффекты позволяют предположить, что однократная подкожная инъекция антитела - агониста TRKB, Н4Н9816Р2, вызывала значительные изменения в массе тела, составе тканей организма, метаболизме и локомоции у TrkBhu/hu мышей.
[00219]

Пример 3. In vivo сравнение эффекта антитела - агониста TRKB Н4Н9816Р2 и изотипического контроля IgG4 REGN1945 на фосфорилирование TRKB в головном мозге после стереотаксической инъекции у TrkBhu/hu мышей (MAID7139)
Экспериментальная процедура
[00218] Тирозин-рецепторная киназа В (TRKB) активируется посредством связывания со своим лигандом нейротрофический фактор головного мозга (BDNF) на внеклеточном домене рецептора, которое индуцирует димеризацию и аутофосфорилирование тирозиновых остатков в внутриклеточном домене рецептора и последующую активацию цитоплазматических сигнальных путей. См., например, Haniu et al. (1997) J. Biol. Chem. 272(40):25296-25303 и Rogalski et al. (2000) J. Biol. Chem. 275(33):25082-25088, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Чтобы определить влияние антитела - агониста TRKB, Н4Н9816Р2, на кинетику активации TRKB, проводили исследование временной динамики фосфорилирования TRKB после прямой гиппокампальной инъекции у мышей, гомозиготных по химерному рецептору TRKB мыши/человека, у которых внеклеточный домен был гуманизирован (MAID7139) (которые называются TrkBhu/hu мыши). Мыши TrkBhu/hu (N=48) получали двусторонние стереотаксические инъекции либо с 2 мкл любой носителя (PBS), либо REGN1945, обозначенного как антитело изотипического контроля IgG4 (конечная концентрация 27,5 мг/мл), или антитела - агониста TRKB Н4Н9816Р2 (конечная концентрация 27,5 мг/мл) в гиппокамп, -2 мм сзади и +1,5 мм латеральнее брегмы. Чтобы свести к минимуму повреждение ткани, инъекцию и удаление иглы выполняли постепенно в течение 5-минутных интервалов. Затем мышей TrkBhu/hu умерщвляли путем эвтаназии с помощью СО2 приблизительно через 30 минут, 1 час, 4 часа или 18 часов после инъекции. Терминальное кровотечение выполняли с помощью пункции сердца для сбора крови, и мышей затем транскардиально перфузировали холодным гепаринизированным физиологическим раствором. Головной мозг осторожно удаляли из черепа, и срез ткани размером 2 мм3, окружающий место инъекции, иссекали, собирали в пробирку Эппендорфа и хранили на льду. Затем срез головного мозга лизировали в 300 мкл буфера для лизиса RIPA (ThermoFisher Scientific, № по кат. 89901), содержащего 2× ингибиторы протеазы и фосфатазы (ThermoFisher Scientific, № по кат. 78444), и хранили на льду. Затем лизированную ткань гомогенизировали для дальнейшей обработки, аликвотировали и хранили при -80°С.
[00219] Для оценки фосфорилирования TRKB в ткани головного мозга проводили иммуноосаждение и вестерн-блоттинг. Антитело к TRKB человека H4H10108N, которое не конкурирует за связывание с Н4Н9816Р2, связывали с гранулами сефарозы, активированными NHS (приготовленными с использованием протокола производителя; GE Healthcare, № по кат. 17-0906), и трижды промывали DPBS для удаления остатков консервирующего раствора. Гомогенизированные лизаты головного мозга оттаивали на льду и разбавляли до концентрации 1 мг/мл (от массы мозга к объему буфера) в буфере, состоящем из 1% NP-40, 0,1% Tween-20, ингибиторов протеазы и фосфатазы в TBST. Концентрацию белка в гомогенизированном лизате головного мозга определяли количественно, выполняя стандартный анализ ВСА в соответствии с инструкциями производителя (Thermo Scientific Pierce, № по кат. 23225). На каждые 100 мкг белка в раствор лизата головного мозга добавляли 15 мкл активированных NHS шариков сефарозы с антителом к TRKB человека (H4H10108N) и смесь инкубировали в течение ночи при 4°С с легким встряхиванием 20 об/мин (шейкер Thermo). На следующий день образцы центрифугировали при 1000 × g в течение одной минуты, а затем супернатант осторожно удаляли. Затем шарики дважды промывали 400 мкл трис-буферного солевого раствора (Bio-Rad, № по кат. 1706435) с 1% Tween-20 (Sigma Aldrich, № по кат. Р9416) (TBST). После тщательной аспирации промывочного буфера к каждому образцу добавляли 60 мкл 0,1% трифторуксусной кислоты (TFA; Sigma-Aldrich, Т62200) в воде при рН 3,0. Раствор перемешивали и оставляли на две минуты, затем собирали и переносили в отдельную пробирку. Этот процесс повторяли с другими 60 мкл 0,1% TFA при рН 3,0. Два 0,1% раствора TFA для каждого образца затем объединяли и добавляли 2 мкл 1М Трис-HCl (ThermoFisher Scientific, № по кат. 15567-027) при рН 8,5.
[00220] Раствор сушили с использованием высокоскоростного вакуума, а затем повторно суспендировали и восстанавливали смесью из 20 мкл 1-кратного буфера Лэммли (Bio-Rad, № по кат. 1610737) вместе с 355 нМ 2-меркаптоэтанола (ВМЕ; Gibco, № по кат. 21985-023). Образцы кипятили при 95°С в течение 10 минут и загружали на 10-луночный 4-15% трис-глициновый гель Mini-Protean (Bio-Rad, № по кат. 4561086). После электрофореза образцы белка переносили из трис-глицинового геля на мембрану PVDF (Bio-Rad, № по кат. 170-4156) через систему Trans-Blot Turbo Transfer System (Bio-Rad, № по кат. 1704156) в течение 30 минут с постоянной скоростью 1,3А и 25В. После переноса мембрану блокировали 2,5% молоком (Bio-Rad, № по кат. 170-6406) в TBST в течение одного часа при комнатной температуре, а затем исследовали в течение ночи любым из следующего: антитело к фосфо-TRKB (Novus, № по кат. NB100-92656), разведенное 1:1000 в растворе 2,5% BSA или первичное антитело к TRKB (Cell Signaling, № по кат. 4603), разведенное до 1:1000 в 2,5% молоке TBST при 4°С на шейкере при 30 об/мин. На следующий день блоты промывали TBST и инкубировали с антителом к IgG кролика, конъюгированным с пероксидазой хрена (Jackson, № по кат. 111-035-144) при разведении 1:1000 в 1% молоке в TBST в течение 1 часа при комнатной температуре. Затем блоты снова промывали, проявляли в растворе ECL (PerkinElmer, Inc. № по кат. RPN2106), и последующие изображения делали каждые 30 секунд.
Результаты и выводы
[00221] Иммунопреципитация и последующий вестерн-блоттинг белка, полученного из лизатов головного мозга мыши TrkBhu/hu, продемонстрировали, что фосфорилирование TRKB гиппокампа было обнаруживаемым у мышей, которым вводили инъекцию антитела - агониста TRKB, Н4Н9816Р2, но не у мышей, получивших носитель или антитело изотипического контроля, как показано на фигуре 3. Среди оцененных моментов времени фосфорилирование TRKB достигло пика через 4 часа после стереотаксической инъекции мышам, которым вводили дозу Н4Н9816Р2. Фосфорилирование TRKB было также обнаружено вестерн-блоттингом через 18 часов после введения дозы у некоторых, но не у всех мышей. И наоборот, инъекция носителя и антитела изотипического контроля IgG4, не вызывала фосфорилирования TRKB ни в какой момент времени. Вестерн-блоттинг также показал, что общие уровни рецептора TRKB были снижены у некоторых, но не у всех мышей TrkBhu/hu, которым вводили дозу Н4Н9816Р2, относительно мышей, которым вводили дозу носителя и изотипического контроля. Общие уровни TRKB, по-видимому, слегка снижены у субъектов, получивших лечение с помощью Н4Н9816Р2, через 18 часов после введения дозы. Таким образом, эти результаты показывают, что прямая инъекция антитела - агониста TRKB, Н4Н9816Р2, индуцирует фосфорилирование рецепторов TRKB гиппокампа у TrkBhu/hu мышей.
Пример 4. Активация нижележащих сигнальных путей с помощью антител -агонистов TrkB в первичных кортикальных нейронах от TrkBhu/hu мышей первого дня после рождения
Экспериментальная процедура
[00222] Все процедуры проводили в соответствии с Предписанием ARVO об использовании животных в исследованиях офтальмологии и зрения и IACUC (Институциональный комитет по содержанию и использованию животных) Regeneron Pharmaceutical Inc. Первичные кортикальные нейроны мыши выделяли и культивировали у гуманизированных в отношении TrkB мышей (МАШ 7139). См., например, Beaudoin et al. (2012) Nat. Protoc. 7(9):1741-1754, полностью включенный в настоящий документ посредством ссылки для всех целей. Вестерн-блоттинг проводили для определения эффектов антител - агонистов TrkB на нижележащие пути Akt и Erk (p-Akt, p-Erk1/2). Первичные кортикальные нейроны от гуманизированных в отношении TrkB детенышей мышей первого дня после рождения (Р1) культивировали в течение 4 дней (DIV-4) в базальной среде NeuralQ (Global Stem, № по кат. GSM-9420) с добавлением добавки для нейронов GS21 (Global Stem, № по кат. GSM-3100), Glutamax (Invitrogen, № по кат. 35050-061) и пенициллина/стрептомицина. Клетки обрабатывали антителом - агонистом TrkB Н4Н9816Р-L1 (10 мкг/мл), антителом - агонистом TrkB H4H9780P-L1 (10 мкг/мл), антителом - агонистом TrkB H4H9814P-L1 (10 мкг/мл), изотипическим контролем IgG4 REGN1945 (10 мкг/мл), контрольным антителом H1M8037C-L1 (10 мкг/мл) или BDNF (1 мкг/мл) в течение 15 минут или 2 часов. Вестерн-блоттинг проводили для определения того, имеют ли агонисты разницу в поддержании и активности нижележащих сигналов. Обработанные клетки промывали и очищали в холодном PBS, содержащем 1% ингибиторов протеазы и фосфатазы (Sigma). Концентрацию белка определяли с помощью анализа белка методом Бредфорда (Pierce). Образцы (50 мкг) разделяли с помощью SDS-PAGE в 3-8% восстановленных трис-ацетатом гелях (Novex) и переносили на нитроцеллюлозную мембрану (Bio-Rad).
[00223] Мембрану инкубировали в течение 1 часа в блокирующем растворе, содержащем 5% молока и 0,1% Tween-20, рН 7,6. За этим следовала инкубация в течение ночи при 4°С в блокирующем буфере, содержащем 5% BSA, 0,1% Tween-20 и кроличье антитело к фосфо-Trk (Cell Signaling, кат. №9141, 1:500), кроличье антитело к фосфо-Akt (Cell Signaling, кат. №9271, 1:1000) или кроличье антитело к фосфо-ERK1/2 (Sigma, кат. № Е7028, 1:5000). Затем меченые белки визуализировали путем инкубации с конъюгированным с пероксидазой хрена (HRP) антителом к IgG козы, мыши или кролика с последующим проявлением с помощью хемилюминесцентного субстрата для HRP (Pierce). Для определения количества общего TrkB, МАРК или Akt, присутствующих в каждой полосе, нитроцеллюлозные мембраны очищали путем стрипирования от антител в буфере для стрипирования (Pierce) в течение 20 минут и инкубировали с кроличьим антителом к TrkB (Cell Signaling, кат. №4603, 1:1000), кроличьим антитело к Erk1/2 (Cell Signaling, кат. №06-182, 1:1000) или кроличьим антителом к Akt (Cell Signaling, кат. №9272, 1:1000) и затем визуализировали, как описано выше. Бета-актин (Sigma, кат. № А5316, 1:20000 и GAPDH (Sigma, кат. № G9295) исследовали в качестве контроля загрузки образца.
Материалы
[00224]

[00225]

[00226]

[00227]

[00228]

Результаты и выводы
[00229] Как показано на фигуре 7, хотя все антитела - агонисты TrkB демонстрировали активацию путей MAPK/ERK и PI3K/Akt через 15 минут после инкубации, только BDNF и Н4Н9814Р демонстрировали фосфорилирование TrkB. Через 2 часа после инкубации все антитела - агонисты TrkB показали активацию TrkB.
Пример 5. Фармакокинетическая оценка антитела к TRKB у гуманизированных в отношении TrkB мышей и мышей дикого типа
Экспериментальная процедура
[00230] Оценку фармакокинетики антитела к TrkB, Н4Н9816Р2 (серия H4H9816P2-L7), проводили у гуманизированных в отношении TrkB (мыши, гомозиготные по экспрессии химерного TrkB мыши/человека, TrkBhu/hu) (MAID7139) мышей и мышей дикого типа (WT). Когорты содержали 5 мышей на линию мыши. Все мыши получали однократную подкожную (SC) дозу, составляющую 10 мг/кг. Образцы крови собирали через 6 часов и 1, 2, 3, 6, 9, 16, 21 и 30 дней после введения дозы. Кровь обрабатывали до сыворотки и замораживали при -80°С до анализа.
[00231] Концентрации антител в кровотоке определяли путем анализа общего антитела IgG4/hIgG1 человека с использованием GyroLab xPlore™ (Gyros, Упсала, Швеция). Вкратце, биотинилированное мышиное моноклональное антитело, специфическое к IgG4/IgG1 человека (REGN2567; серия RSCH15088)), разведенное до 100 мкг/мл в буфере для разведения антител (0,05% Tween-20 + PBS), захватывали на CD Gyrolab Bioaffy 200, который содержали аффинные колонки, предварительно загруженные гранулами, покрытыми стрептавидином (Dynospheres™). Стандарт, используемый для калибровки в этом анализе, представлял собой Н4Н9816Р в концентрациях от 0,488 до 2000 нг/мл в буфере для разведения (0,5% BSA+PBS), содержащем 0,1% нормальной мышиной сыворотки (NMS). Образцы сыворотки разводили 1:100 в буфере для разведения антител. IgG человека, захваченный на аффинных колонках, покрытых антителом к REGN2567, на CD, исследовали при комнатной температуре, обнаруживали путем добавления 0,5 мкг/мл конъюгированного с Alexa-647 мышиного моноклонального антитела к каппа-цепи человека (REGN654; серия RSCH13067), разведенного в буфер для обнаружения (буфер Rexxip F); и полученный флуоресцентный сигнал регистрировали в единицах ответа (RU) с помощью прибора GyroLab xPlore. Концентрации образцов определяли путем интерполяции по стандартной кривой, которую подбирали с использованием подбора логистической кривой с 5 параметрами с использованием программного обеспечения Gyrolab Evaluator. Средние концентрации из двух повторных экспериментов использовали для последующего анализа PK.
[00232] Параметры PK определяли некомпартментным анализом (NCA) с использованием программного обеспечения Phoenix®WinNonlin® версии 6.3 (Certara, L.P., Принстон, Нью-Джерси) и модели внесосудистого введения дозы. Используя соответствующие средние значения концентрации для каждого антитела, все параметры РК, включая в себя наблюдаемую максимальную концентрацию в сыворотке (Стих), предполагаемый наблюдаемый период полужизни (t1/2) и площадь под кривой зависимости концентрации от времени вплоть до последней измеряемой концентрации (AUClast), определяли с использованием линейного правила трапеций с линейной интерполяцией и равномерным взвешиванием.
Результаты и выводы
[00233] После 10 мг/кг подкожного введения антитела к TrkB, Н4Н9816Р2, аналогичные максимальные концентрации (Cmax) антитела наблюдали к 1 или 2 дню как у мышей TrkBhu/hu, так и у мышей WT (135 и 131 мкг/мл соответственно). К 9 дню Н4Н9816Р2 проявляло более выраженное удаление лекарственного средства у мышей TrkBhu/hu, чем у мышей WT, что указывает на опосредованный мишенью эффект. На 30-й день концентрации антител были приблизительно в 35 раз меньше у мышей TrkBhu/hu. Воздействие антител (AUClast) для Н4Н9816Р2 у мышей WT было в ~ 1,7 раза выше, чем у мышей TrkBhu/hu (1730 и 1020 д*мкг/мл соответственно). У мышей WT также наблюдали увеличение времени полужизни приблизительно в 3 раза (Т1/2) по сравнению с мышами TrkBhu/hu (8,4 и 2,9 дня соответственно).
[00234] Обобщение данных для общих концентраций антител к TrkB представлено в таблице 15, средние параметры PK описаны в таблице 16, а средние общие концентрации антител в зависимости от времени показаны на фигуре 8. На фигуре 8 мышам вводили разовую подкожную дозу, составляющую 10 мг/кг в день 0. Концентрации общего Н4Н9816Р2 в сыворотке крови измеряли с помощью иммуноанализа Gyros. Точки данных после введения дозы через 6 часов, 1, 2, 3, 6, 9, 16, 21 и 30 дней указывают на среднюю концентрацию антитела. Общие концентрации антител Н4Н9816Р2 представлены сплошными кружками для мышей TrkBhu/hu и сплошными квадратами для мышей дикого типа. Данные представлены как среднее значение ±SD.
[00235]

[00236]

Пример 6. Получение крыс, содержащих гуманизированный локус TRKB
[00237] Большой нацеливающий вектор, содержащий 5' гомологичное плечо, содержащее 7 т.п.н. локуса TrkB крысы, и 3' гомологичное плечо, содержащее 47 т.п.н. локуса TrkB крысы, создавали для замены области 68,5 т.п.н. из гена TrkB крысы, кодирующего внеклеточный домен TRKB крысы, на 74,4 т.п.н. соответствующей последовательности TRKB человека. Создание и применение больших нацеливающих векторов (LTVEC), полученных из ДНК бактериальной искусственной хромосомы (ВАС) посредством реакций бактериальной гомологичной рекомбинации (BHR) с использованием технологии генной инженерии VELOCIGENE®, описаны, например, в патенте США №6586251 и Valenzuela et al. (2003) Nat. Biotechnol. 21(6):652-659, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Создание LTVEC посредством способов сборки in vitro описано, например, в документах US 2015/0376628 и WO 2015/200334, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Информация о TRKB крысы и человека представлена в таблице 17. Описание создания большого нацеливающего вектора представлена в таблице 18.
[00238]

[00239]

[00240] В частности, область, начиная с экзона 2 (кодирующий экзон 1; от аминокислоты 32, сохраняя сигнальный пептид) до экзона 10, включая в себя первые 50 пар оснований интрона 10 и все интроны между экзонами 2 и 10 (т.е. между кодирующим экзоном 1 и экзоном 10) подвергали делеции из локуса TrkB крысы (сохраняя трансмембранный домен крысы, кодируемый экзонами 10 и 11). Область, включая в себя экзон 2/кодирующий экзон 1 (от аминокислоты 32, начиная после сигнального пептида) до экзона 10, включая в себя первые 66 пар оснований интрона 10 и все интроны между экзонами 2 и 10 (т.е. между кодирующим экзоном 1 и экзоном 10) вставляли вместо удаленной области крысы (сохраняя трансмембранный домен крысы, кодируемый экзонами 10 и 11).
[00241] Последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB крысы представлены в SEQ ID NO: 55-58 соответственно, с соответствующей кодирующей последовательностью, представленной в SEQ ID NO: 67-70 соответственно. Последовательности для сигнального пептида, внеклеточного домена, трансмембранного домена и цитоплазматического домена TRKB человека представлены в SEQ ID NO: 59-62 соответственно, с соответствующей кодирующей последовательностью, представленной в SEQ ID NO: 71-74 соответственно. Предполагаемый кодируемый химерный белок TRKB содержит трансмембранный и внутриклеточный домены TRKB крысы, сигнальный пептид TRKB крысы и внеклеточный домен TRKB человека. См. фиг. 4. Выравнивание белков TRKB крысы и человека на фигуре 6. Кодирующие последовательности TrkB/TRKB крысы и человека представлены в SEQ ID NO: 10 и 11 соответственно. Последовательности белка TRKB крысы и человека представлены в SEQ ID NO: 2 и 3 соответственно. Последовательности для предполагаемой химерной кодирующей последовательности TRKB крысы/человека и предполагаемого химерного белка TRKB крысы/человека представлены в SEQ ID NO: 13 и 5 соответственно.
[00242] Для создания мутантного аллеля компоненты CRISPR/Cas9, включая в себя четыре гидовые РНК (целевые последовательности для гидовых РНК, приведенных в SEQ ID NO: 41-44), вводили в эмбриональные стволовые клетки крысы вместе с большим нацеливающим вектором. Конкретно, 4×106 крысиных ES-клеток (линия Dark Agouti DA2B) подвергали электропорации со следующим: 2 мг TrkB LTVEC; 5 мг экспрессирующей Cas9 плазмиды; и 5 мг каждая из гРНК: gU, gU2, gD и gD2. Условия электропорации: напряжение 400 В; емкость 100 мФ; и сопротивление 0 Вт.Отбор с помощью антибиотиков проводили с использованием G418 в концентрации 75 мг/мл. Колонии отбирали, размножали и подвергали скринингу с помощью TAQMAN®. См. фиг. 5. Анализы на потерю аллеля проводили для обнаружения потери эндогенного аллеля крысы, и анализы на приобретение аллеля проводили для обнаружения приобретения гуманизированного аллеля и анализы CRISPR и анализы удерживания проводили с использованием праймеров и зондов, представленных в таблице 19.
[00243]



[00244] Анализы на модификацию аллеля (МОА), включая в себя анализы на потерю аллеля (LOA) и приобретение аллеля (GOA) описаны, например, в US 2014/0178879; US 2016/0145646; WO 2016/081923; и Frendewey et al. (2010) Methods Enzymol. 476:295-307, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. Анализ на потерю аллеля (LOA) инвертирует традиционную логику скрининга и количественно определяет количество копий в образце геномной ДНК нативного локуса, на который была направлена мутация. В правильно нацеленном клоне гетерозиготных клеток анализ LOA обнаруживает один из двух нативных аллелей (для генов, не находящихся в хромосоме X или Y), причем другой аллель разрушается в результате целевой модификации. Тот же принцип может быть применен в обратном порядке, как и анализ усиления аллеля (GOA), для количественного определения количества копий вставленного нацеливающего вектора в образец геномной ДНК.
[00245] Анализы удерживания описаны в US 2016/0145646 и WO 2016/081923, каждый из которых полностью включен в настоящий документ посредством ссылки для всех целей. В анализах удерживания проводят различие между правильными целевыми вставками нуклеиновой кислоты - вставки в целевой геномный локус и случайными трансгенными вставками нуклеиновой кислоты - вставки в места генома за пределами целевого геномного локуса путем оценки количества копий матриц ДНК из 5' и 3' целевых последовательностей, соответствующих 5' и 3' гомологическим плечам нацеливающего вектора, соответственно. В частности, в анализах удерживания определяют количество копий в образце геномной ДНК 5' матрицы ДНК целевой последовательности, предназначенной для сохранения в модифицированном целевом геномном локусе, и/или 3' матрицы ДНК целевой последовательности, предназначенной для сохранения в модифицированном целевом геномном локусе. В диплоидных клетках правильно нацеленные клоны сохранят число копий, равное двум. Число копий больше двух, как правило, указывает на трансгенную интеграцию нацеливающего вектора случайным образом вне целевого геномного локуса, а не в целевом геномном локусе. Число копий меньше, чем обычно, указывает на большие делеции, выходящие за пределы области, нацеленной для делеции.
[00246] Анализы CRISPR представляют собой анализы TAQMAN®, предназначенные для охвата области, которая нарушается из-за гРНК CRISPR. Когда гРНК CRISPR разрезает и создает индел (вставку или делецию), анализ TAQMAN® не сможет амплифицироваться и, таким образом, сообщает о расщеплении CRISPR.
[00247] Положительный клон СВ1 оттаивали, размножали и повторно подтверждали с помощью TAQMAN®. СВ1 также подтверждали с помощь успешной ПЦР от 5'-конца последовательности замены человека до фланкирующей геномной последовательности крысы за пределами конца 5' гомологичного плеча. Ампликон ПЦР подтверждали как правильный путем секвенирования концов.
[00248] Крыс F0 и F1 получали с использованием способов, описанных в документах US 2014/0235933, US 2014/0310828, WO 2014/130706 и WO 2014/172489, каждый из которых полностью включен в настоящий документ п посредством ссылки для всех целей. В таких способах подтвержденные целевые клоны ES-клеток крысы (например, клоны ES-клеток Dark Agouti) подвергают микроинъекции в бластоцисты (например, бластоцисты Sprague Dawley (SD)), которые затем переносят псевдобеременным самкам-реципиентам (например, самкам-реципиентам SD) для вынашивания с использованием стандартных техник. Химеры идентифицируют (например, по цвету шерсти), и химеры самцов F0 скрещивают с самками крыс дикого типа той же линии (например, самки SD). Затем детенышей зародышевой линии (например, Agouti) F1 генотипируют на наличие целевого аллеля. Все эксперименты, выполненные на гуманизированных в отношении TRKB крысах, как описано ниже, проводили на крысах, у которых самоудаляющаяся кассета селекции удалялась самостоятельно.
Пример 7. In vivo сравнение эффекта антитела - агониста TRKB Н4Н9816Р2 и изотипического контроля IgG4 REGN1945 на выживаемость ганглиозных клеток сетчатки (RGC) крыс TrkBhu/hu
Экспериментальная процедура
[00249] Все процедуры проводили в соответствии с Предписанием ARVO об использовании животных в исследованиях офтальмологии и зрения и IACUC (Институциональный комитет по содержанию и использованию животных) Regeneron Pharmaceutical Inc. Использовали взрослых гуманизированных в отношении TrkB крыс (MAID100010) в возрасте 8-10 недель, каждая массой 200-250 г. Все хирургические вмешательства на крысах проводили под общим наркозом с использованием внутрибрюшинной инъекции кетамина (63 мг/кг) и ксилазина (6,0 мг/кг). Для защиты роговицы применяли глазную мазь, содержащую эритромицин (0,5%, Bausch & Lomb).
[00250] Интраорбитальная аксотомия зрительного нерва и интравитреальная инъекция. На левый зрительный нерв (ON) воздействовали интраорбитально, его твердую оболочку вскрывали. ON пересекали приблизительно на 1,5 мм позади глазного яблока. Были приняты меры, чтобы не повредить кровоснабжение сетчатки. Интравитреальные инъекции выполняли непосредственно позади плоской части ресничного тела с помощью вытянутой стеклянной пипетки, соединенной с шприцем Гамильтона объемом 50 мкл. Были приняты меры, чтобы не повредить хрусталик. Крысы с любыми значительными послеоперационными осложнениями (например, ишемия сетчатки, катаракта) были исключены из дальнейшего анализа. Животных распределяли по разным экспериментальным группам. Одна контрольная группа получала интравитреальные инъекции 3 мкл изотипического контроля REGN1945 (46,6 мкг/мкл); другая группа получала инъекцию 3 мкл антитела к TRKB человека Н4Н9816Р2 (45,7 мкг/мкл) через 3 и 10 дней после ON-аксотомии.
[00251] Иммуногистохимическое окрашивание и подсчет жизнеспособных ганглиозных клеток сетчатки (RGC). BRN3A (специфический для головного мозга гомеобокс/белок домена РОА ЗА) использовали в качестве маркера для выживших ганглиозных клеток сетчатки (RGC), поскольку было показано, что он является эффективным и надежным способом селективного мечения жизнеспособных RGC в тотальных препаратах сетчатки после повреждения зрительного нерва. См., например,  (2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. Для иммуноокрашивания BRN3A сетчатки блокировали в 10% нормальной сыворотке осла и 0,5% Triton Х-100 в течение 1 часа, затем инкубировали в той же среде с антителом к BRN3A (1:400; № по кат.: sc-31984, Santa Cruz) 2 ч при комнатной температуре. После дальнейшей промывки сетчатки инкубировали с конъюгированным с Alexa594 ослиным вторичным антителом к антителу козла (1:400; № по кат.: А-11058, mvitrogen) в течение ночи при 4°С.
(2009) Invest. Ophthalmol. Vis. Sic. 50(8):3860-3868, полностью включенный в настоящий документ посредством ссылки для всех целей. Для иммуноокрашивания BRN3A сетчатки блокировали в 10% нормальной сыворотке осла и 0,5% Triton Х-100 в течение 1 часа, затем инкубировали в той же среде с антителом к BRN3A (1:400; № по кат.: sc-31984, Santa Cruz) 2 ч при комнатной температуре. После дальнейшей промывки сетчатки инкубировали с конъюгированным с Alexa594 ослиным вторичным антителом к антителу козла (1:400; № по кат.: А-11058, mvitrogen) в течение ночи при 4°С.
Результаты и выводы
[00252] Для оценки эффект антитела - агониста TRKB на выживаемость RGC in vivo авторы настоящего изобретения использовали модель полного пересечения зрительного нерва. Антитело - агонист TRKB (Н4Н9816Р2) или антитело изотипического контроля применяли через 3 и 10 дней после операции. Животных умерщвляли через 14 дней после аксотомии. Плотность RGC в неповрежденном контралатеральном глазе схожа для трех генотипов TRKB (гомозиготный гуманизированный, гетерозиготный гуманизированный и дикий тип), в среднем составляя около 1600 на мм2, как показано в таблице 20. Плотность выживших RGC оценивали в тотальных препаратах сетчатки с использованием окрашивания BRN3A. Авторы настоящего изобретения обнаружили, что у гомозиготных TrkBhu/hu гуманизированных крыс антитело - агонист TRKB (Н4Н9816Р2) значительно (р<0,01, критерий Манна-Уитни) увеличивало выживаемость RGC по сравнению с контролями (685±106 по сравнению с 255±66 RGC на мм2). У гетерозиготных TrkBhu/+ гуманизированных крыс также наблюдали значительный (р<0,05, критерий Манна-Уитни) эффект на выживаемость со стороны антитела - агониста TrkB (444±90 по сравнению с 208±50 RGC на мм2). У крыс дикого типа наблюдали небольшое, но незначительное увеличение количества RGC у крыс, получивших антитело - агонист TRKB, по сравнению с изотипическим контролем (таблица 21). В заключение, антитело - агонист TRKB (Н4Н9816Р2) значительно увеличивало выживаемость RGC у гуманизированных крыс TrkBhu/hu.
[00253]

[00254]
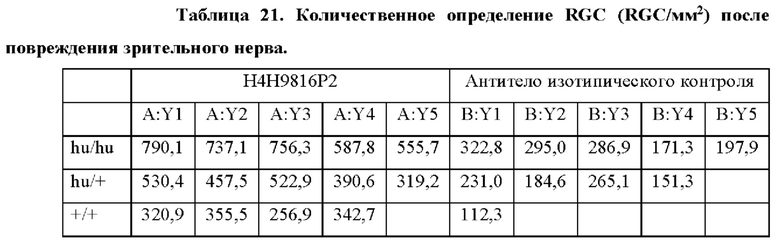
Пример 8. Аминокислотные последовательности вариабельной области тяжелой и легкой цепей антител к TRKB, используемых в примерах.
[00255] Несколько полностью человеческих антител к TRKB (т.е. антител, обладающих вариабельными доменами человека и константными доменами человека) испытывали в примерах, включая в себя те, которые обозначены как Н4Н9780Р, Н4Н9814Р и Н4Н9816Р2. В таблице 22 представлены идентификаторы аминокислотных последовательностей вариабельных областей тяжелой и легкой цепей и CDR выбранных антител к TRKB, используемых в примерах. В таблице 23 представлены идентификаторы последовательностей нуклеиновых кислот вариабельных областей тяжелой и легкой цепей и CDR выбранных антител к TRKB, используемых в примерах. Указанные антитела описаны более подробно в заявке на выдачу патента США №16/202881, поданной 28 ноября 2018 г., которая полностью включена в настоящий документ посредством ссылки для всех целей.
[00256]

[00257]
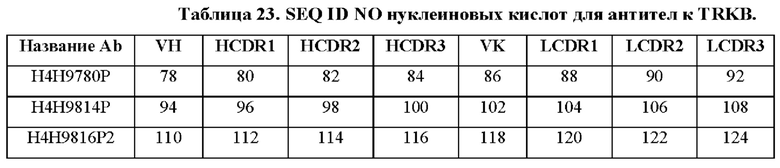
[00258] Как правило, антитела в настоящем документе называют согласно следующей номенклатуре: приставка Fc (например, «Н4Н») с последующим числовым идентификатором (например, «9780», «9816» и т.д., как показано в таблице 22), за которым следует суффикс «Р» или «Р2». Приставки Н4Н в обозначениях антител, используемых в настоящем документе, указывают на конкретный изотип Fc-области антитела. Таким образом, в соответствии с этой номенклатурой антитело в настоящем документе может называться, например, «Н4Н9780Р» и т.д., что указывает на Fc IgG4 человека. Вариабельные области являются полностью человеческими, что обозначено первой «Н» в обозначении антитела. Среднему специалисту в настоящей области техники понятно, что антитело с конкретным изотипом Fc можно превратить в антитело с другим изотипом Fc (например, антитело с Fc IgG1 мыши можно превратить в антитело с IgG4 человека и т.д.), но в любом случае вариабельные домены (включая в себя CDR), которые указаны с помощью числовых идентификаторов, показанных в таблице 22, будут оставаться прежними, и, как предполагают, свойства связывания с антигеном являются идентичными или по существу сходными независимо от природы Fc-домена.
Пример 9. Нейропротекторный эффект агонистов - антитела к TrkB человека у гуманизированных в отношении TrkB крыс
[00259] Приведенные ниже эксперименты предприняли для оценки нейропротекторного действия эндогенного агониста TRKB, нейротрофического фактора головного мозга (BDNF) и моноклонального антитела - агониста TRKB (mAb) у мышей и крыс дикого типа (WT) и у гуманизированных в отношении TrkB мышей и крыс.
[00260] Эффекты in vitro BDNF и антитела (Ab) к TRKB количественно определяли с помощью анализов клеточной выживаемости с использованием дифференцированной клеточной линии нейробластомы человека SH-SY5Y. In vitro BDNF или Ab к TRKB значительно увеличивали клеточную выживаемость в дифференцированных по ретиноевой кислоте клетках SH-SY5Y. Эффекты показали колоколообразные кривые зависимости ответов от дозы с оптимальной дозой, составляющей 1 мкг/мл для BDNF или 10 мкг/мл для Ab к TRKB. Клеточную линию нейробластомы SH-SY5Y культивировали в среде для дифференцировки, содержащей 10 мкМ полностью транс-ретиноевой кислоты, в течение 4 дней. Культуру заменяли на бессывороточную среду для дифференцировки, содержащую разные дозы антител (0,01-100 мкг/мл). Через два дня добавляли реагент ССК8, планшеты инкубировали в течение 3-4 часов и измеряли OD450 для определения процента выживших клеток. Данные нормализовали по отношению к бессывороточной среде без антител. Как показано на фигуре 9, mAb к TRKB (mAb1 к TrkB представляет собой Н4Н9816Р2; mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB) в зависимости от дозы увеличивало выживаемость клеток SH-SY5Y. Изотипический контроль человека не влиял на выживаемость клеток SH-SY5Y. Бессывороточная среда без антител приводила к 100% выживаемости.
[00261] Затем сетчатки от мышей Р2 C57BL/6J иссекали и диссоциировали. Ганглиозные клетки сетчатки очищали с помощью иммуно-пэннинга и культивировали в 96-луночном планшете с обработкой или без обработки. Через 24 часа в каждую лунку добавляли МТТ (3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолийбромид) для расчета выживаемости клеток для каждой группы. Как показано на фигуре 10, BDNF имел колоколообразную кривую ответа и оптимальную дозу при 1 мкг/мл. mAb2 к TrkB (контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB мыши и TrkB крысы) также может характеризоваться колоколообразной кривой при более высоких дозах, но проявляет нейропротекторный эффект.
[00262] Для испытания нейропротекторного эффекта in vivo использовали WT и гуманизированных в отношении TrkB мышей и крыс. Животные получали интравитреальные (IVT) инъекции BDNF или mAb к TRKB на 3 и 10 день после пересечения зрительного нерва (ONT). Количество ганглиозных клеток сетчатки (RGC) определяли количественно с использованием программного обеспечения HALO (Indica Labs) через 1 неделю для мыши или 2 недели для крысы после пересечения зрительного нерва с помощью ИГХ (IHC) Brn3a на плоских держателях сетчатки.
[00263] Гибель RGC у мышей TrkBhu/hu являлась сходной с гибелью у мышей WT через 1 или 2 недели после пересечения зрительного нерва. BDNF или Ab к TRKB обладали небольшим или отсутствием значимого нейропротекторного эффекта у мышей WT или TrkBhu/hu. В отличие от этого, у крыс TrkBhu/hu наблюдали значительную нейро протекция RGC с помощью IVT Ab к TRKB. Уменьшение массы тела наблюдали у мышей TrkBhu/hu, но не у крыс после лечения с помощью IVT Ab к TRKB. BDNF не влиял на массу тела ни у мышей, ни у крыс.
[00264] На фигурах 11А и 11В показаны результаты эксперимента по оценке нейропротекции на модели пересечения зрительного нерва у мышей и крыс WT. На фигуре 11А крысам линии Dark Agouti в возрасте 8-9 недель вводили BDNF (5 мкг), mAb2 к TrkB (18 мкг), антитело изотипического контроля (18 мкг) или контроль - носитель интравитреально через 3 дня и 10 дней после пересечения. mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB мыши и TrkB крысы. Сетчатки иссекали и окрашивали на ганглиозные клетки сетчатки через 14 дней после пересечения. BDNF и mAb к TRKB показали значительную нейропротекцию, измеренную по плотности ганглиозных клеток сетчатки (RGC). На фигуре 11В 8-недельным мышам C57BL/6J WT вводили BDNF (2,5 мкг), mAb2 к TrkB (10 мкг), антитело изотипического контроля (10 мкг) или контроль - носитель интравитреально через 3 дня и 10 дней после пересечения. mAb2 к TrkB представляет собой контрольное антитело - агонист TrkB с аффинностью к TrkB человека, TrkB мыши и TrkB крысы. Не наблюдали какой-либо значительной нейропротекции. Таким образом, лечение с помощью BDNF и mAb к TRKB приводило к значительному увеличению плотности RGC в иссеченных сетчатках у крыс дикого типа после пересечения зрительного нерва, тогда как в той же модели у мышей дикого типа не наблюдалось значительного влияния на плотность RGC.
[00265] На фигурах 12А и 12В показана зависимость ответа от дозы BDNF у мышей и крыс WT. На фигуре 12А зависимость ответа от дозы BDNF в модели раздавливания зрительного нерва (ONC) у мышей WT показывает небольшое окно нейропротекции. На фигуре 12 В показана зависимость ответа от дозы BDNF в модели пересечения зрительного нерва у крыс WT от 0,13 мкг до 30 мкг. Существует колоколообразный ответ, аналогичный данным in vitro, с оптимальной дозой, составляющей 0,8 мкг. Сетчатки иссекали и окрашивали в отношении ганглиозных клеток сетчатки через 14 дней после пересечения. Таким образом, лечение BDNF привело к гораздо более выраженной кривой зависимости ответа от дозы, измеренной по плотности RGC в иссеченных сетчатках у крыс дикого типа после пересечения зрительного нерва, по сравнению с гораздо менее выраженной кривой зависимости ответа от дозы BDNF при измерении по плотности RGC в иссеченных сетчатках мышей дикого типа после раздавливания зрительного нерва.
[00266] Нейропротекторный эффект Ab к TRKB затем испытывали на гуманизированных в отношении TrkB крысах. Результаты на фигурах 13А и 13В показывают, что интравитреальная инъекция mAb к TRKB у гуманизированных в отношении TrkB крыс с пересеченным зрительным нервом, демонстрирует значительную нейропротекцию ганглиозных клеток сетчатки. Гомозиготным по TRKB человека, гетерозиготным по TRKB человека или крысам с TrkB дикого типа вводили либо mAb1 к TrkB, либо антитело изотипического контроля интравитреально (3 мкл) через 3 и 10 дней после пересечения зрительного нерва. Через четырнадцать дней после пересечения сетчатки иссекали и окрашивали в отношении RGC. Крысы являлись самками в возрасте 17-19 недель. Как показано на фигуре 13А, крысы, получившие лечение с помощью mAb1 к TrkB (Н4Н9816Р2), показали нейропротекцию во всех трех генотипах по сравнению с соответствующими крысами, получившими антитело изотипического контроля. Гомозиготные и гетерозиготные по TRKB человека крысы, получившие изотипический контроль, характеризовались более высокой плотностью RGC, чем крысы дикого типа, получившие изотипический контроль. На фигуре 13 В показано отсутствие различий количества RGC в необработанных глазах между генотипами. На фигуре 13С показана масса тела гомозиготных по TRKB человека мышей, которым вводили либо антитело - агонист TrkB (Н4Н9816Р2), либо антитело изотипического контроля (REGN1945) через 14 дней после пересечения.
[00267] Затем создавали карты изоплотности RGC тотального препарата для сетчатки крысы, показывающие меченные Brn3a клетки неповрежденных и обработанных поврежденных глаз в трех генотипах (данные не показаны). Реконструкцию тотального препарата получали с помощью столика с электроприводом на флуоресцентном микроскопе (Nikon Eclipse Ti). RGC подсчитывали с использованием программного обеспечения для анализа изображений (HALO®; mdica Labs, Корралес, Нью-Мексико, США). Карты изоплотности получали с помощью Matlab. Более высокую плотность RGC наблюдали у гуманизированных в отношении TrkB крыс, получивших лечение с помощью mAb1 к TrkB (Н4Н9816Р2), по сравнению с крысами, получившими лечение с помощью изотипического контроля (данные не показаны).
[00268] Взятые вместе, данные, показанные на фигурах 11А, 11В, 12А, 12В и 13А-13С, демонстрируют, что интравитреальное введение mAb - агониста TRKB обладает значительным нейропротекторным эффектом после повреждения зрительного нерва у гуманизированных в отношении TrkB крыс, в отличие от небольшого или отсутствия значимого нейропротекторного эффекта, наблюдаемого после повреждения зрительного нерва у гуманизированных в отношении TrkB мышей.
[00269] Чтобы дополнительно оценить влияние антител - агонистов TRKB на выживаемость RGC у крыс в модели пересечения зрительного нерва (ONT), предприняли исследование зависимости ответа от дозы. Использовали гомозиготных по TRKB человека крыс (МАГО100010; 75% SD, 25% DA) в возрасте 1-9 месяцев. Шесть крыс использовали в каждой группе. Гомозиготным по TRKB человека крысам вводили различные дозы либо mAb1 к TrkB, либо антитела изотипического контроля (REGN1945) интравитреально (3 мкл) через 3 и 10 дней после пересечения зрительного нерва. Через четырнадцать дней после пересечения сетчатки иссекали и окрашивали в отношении RGC. Как показано на фигуре 14, mAb1 к TrkB в зависимости от дозы увеличивало выживаемость RGC у гуманизированных в отношении TrkB крыс.
[00270] Затем нейропротекторный эффект различных антител - агонистов TrkB сравнивали у гомозиготных по TRKB человека крыс на модели пересечения зрительного нерва (ONT). Использовали гуманизированных в отношении TrkB крыс (MAID100010; 75% SD, 25% DA) в возрасте 8-10 недель. В каждой группе использовали от пяти до шести крыс. Гомозиготным по TRKB человека крысам вводили либо H4H9816P2-L9 (10 мкг), Н4Н9814Р-L9 (10 мкг), H4H9780P-L5 (10 мкг), комбинацию всех трех (3,3 мкг каждая), либо антитело изотипического контроля (REGN1945; 10) мкг) интравитреально (3 мкл) через 3 и 10 дней после пересечения зрительного нерва. Через четырнадцать дней после пересечения сетчатки иссекали и окрашивали в отношении RGC. Результаты показаны на фигурах 15А и 15В. Каждое антитело - агонист TrkB обладает нейропротекторным эффектом по сравнению с антителом изотипического контроля. Масса тела в каждой группе была одинаковой (данные не показаны).
[00271] Напротив, антитела - агонисты TrkB, Н4Н9780Р и Н4Н9814Р, не оказывали нейропротекторного эффекта у крыс дикого типа. Нейропротекторный эффект оценивали у крыс дикого типа с использованием модели пересечения зрительного нерва (ONT). Использовали самок крыс дикого типа в возрасте 8-10 недель. В каждой группе использовали от пяти до шести крыс. Крысам дикого типа вводили интравитреально (3 мкл) Н4Н9780Р (120 мкг), Н4Н9814Р (120 мкг) или антитело изотипического контроля (REGN1945; 120 мкг) через 3 и 10 дней после пересечения зрительного нерва. Через четырнадцать дней после пересечения сетчатки иссекали и окрашивали в отношении RGC. Как показано на фигуре 16, ни одно из антител - агонистов TrkB не оказывало значительного нейропротекторного эффекта у крыс дикого типа.
[00272] Кроме того, антитело - агонист TrkB (Н4Н9780Р) не оказывало нейропротекторного эффекта у гомозиготных по TRKB человека мышей. Использовали гомозиготных по TRKB мышей-самцов (MAID7139; 75% C57BL/6, 25% 129), которым было 5 месяцев. В каждой группе использовали от пяти до шести мышей. Гомозиготным по TRKB человека мышам вводили либо Н4Н9780Р (40 мкг на глаз), либо антитело изотипического контроля (REGN1945; 40 мкг на глаз) интравитреально (1 мкл) через 3 и 10 дней после пересечения зрительного нерва. Через четырнадцать дней после пересечения сетчатки иссекали и в отношении RGC. Как показано на фигурах 17А и 17В, антитело - агонист TrkB не оказывало нейропротективного эффекта у гомозиготных по TRKB человека мышей в отличие от нейропротективного эффекта, наблюдаемого у гомозиготных по TRKB человека крыс. На фигуре 17С показана масса тела гомозиготных по TRKB человека мышей, которым вводили либо Н4Н9780Р, либо антитело изотипического контроля через 14 дней после пересечения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Регенерон Фармасьютикалз, Инк.
<120> НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННЫЙ
ЛОКУС TrkB
<130> 57766-523380
<150> US 62/592905
<151> 2017-11-30
<150> US 62/661373
<151> 2018-04-23
<160> 125
<170> PatentIn версии 3.5
<210> 1
<211> 821
<212> PRT
<213> Mus musculus
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (32)..(429)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (430)..(453)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (454)..(821)
<223> Цитоплазматический домен
<400> 1
Met Ser Pro Trp Leu Lys Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ser Ala Arg Ile Trp Cys Thr Glu Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Leu Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala Tyr Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Arg His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Arg His Phe Arg His Leu Asp Leu Ser Asp Leu Ile
130 135 140
Leu Thr Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Leu Lys
145 150 155 160
Thr Leu Gln Glu Thr Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Met Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Arg Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Val Thr Leu Ser Cys Ser Val Gly Gly Asp Pro
210 215 220
Leu Pro Thr Leu Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Arg Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Met Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Arg Gln Ile Ser Ala His Phe Met Gly Arg Pro Gly Val
370 375 380
Asp Tyr Glu Thr Asn Pro Asn Tyr Pro Glu Val Leu Tyr Glu Asp Trp
385 390 395 400
Thr Thr Pro Thr Asp Ile Gly Asp Thr Thr Asn Lys Ser Asn Glu Ile
405 410 415
Pro Ser Thr Asp Val Ala Asp Gln Ser Asn Arg Glu His Leu Ser Val
420 425 430
Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu Val
435 440 445
Met Leu Leu Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met Lys
450 455 460
Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro Leu
465 470 475 480
His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly Gly
485 490 495
Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu Asn
500 505 510
Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr Phe
515 520 525
Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu Gly
530 535 540
Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu Cys
545 550 555 560
Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp Ala
565 570 575
Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu Thr
580 585 590
Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val Glu
595 600 605
Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp Leu
610 615 620
Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala Glu
625 630 635 640
Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile Ala
645 650 655
Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe Val
660 665 670
His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu Leu
675 680 685
Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr Asp
690 695 700
Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met Pro
705 710 715 720
Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val Trp
725 730 735
Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln Pro
740 745 750
Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln Gly
755 760 765
Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu Leu
770 775 780
Met Leu Gly Cys Trp Gln Arg Glu Pro His Thr Arg Lys Asn Ile Lys
785 790 795 800
Ser Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val Tyr
805 810 815
Leu Asp Ile Leu Gly
820
<210> 2
<211> 821
<212> PRT
<213> Rattus norvegicus
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (32)..(429)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (430)..(453)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (454)..(821)
<223> Цитоплазматический домен
<400> 2
Met Ser Pro Trp Pro Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala Cys
20 25 30
Pro Met Ser Cys Lys Cys Ser Thr Thr Arg Ile Trp Cys Thr Glu Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Ile Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Leu Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Lys Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala Tyr Lys Ala Phe Leu
100 105 110
Lys Asn Gly Asn Leu Arg His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Arg His Phe Arg His Leu Asp Leu Ser Asp Leu Ile
130 135 140
Leu Thr Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Leu Lys
145 150 155 160
Thr Leu Gln Glu Thr Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Thr Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Arg Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Val Thr Ile Ser Cys Ser Val Gly Gly Asp Pro
210 215 220
Leu Pro Thr Leu Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Arg Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Met Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Arg Gln Ile Ser Ala His Phe Met Gly Arg Pro Gly Val
370 375 380
Asp Tyr Glu Thr Asn Pro Asn Tyr Pro Glu Val Leu Tyr Glu Asp Trp
385 390 395 400
Thr Thr Pro Thr Asp Ile Gly Asp Thr Thr Asn Lys Ser Asn Glu Ile
405 410 415
Pro Ser Thr Asp Val Ala Asp Gln Thr Asn Arg Glu His Leu Ser Val
420 425 430
Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu Val
435 440 445
Met Leu Leu Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met Lys
450 455 460
Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro Leu
465 470 475 480
His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly Gly
485 490 495
Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu Asn
500 505 510
Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr Phe
515 520 525
Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu Gly
530 535 540
Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu Cys
545 550 555 560
Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp Ala
565 570 575
Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu Thr
580 585 590
Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val Glu
595 600 605
Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp Leu
610 615 620
Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala Glu
625 630 635 640
Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile Ala
645 650 655
Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe Val
660 665 670
His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu Leu
675 680 685
Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr Asp
690 695 700
Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met Pro
705 710 715 720
Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val Trp
725 730 735
Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln Pro
740 745 750
Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln Gly
755 760 765
Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu Leu
770 775 780
Met Leu Gly Cys Trp Gln Arg Glu Pro His Thr Arg Lys Asn Ile Lys
785 790 795 800
Asn Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val Tyr
805 810 815
Leu Asp Ile Leu Gly
820
<210> 3
<211> 838
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (32)..(430)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (431)..(454)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (455)..(838)
<223> Цитоплазматический домен
<400> 3
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Phe Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Asp Phe Ser Trp Phe Gly Phe Gly Lys Val Lys Ser Arg Gln Gly
465 470 475 480
Val Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
485 490 495
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
500 505 510
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
515 520 525
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
530 535 540
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
545 550 555 560
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
565 570 575
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
580 585 590
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
595 600 605
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
610 615 620
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
625 630 635 640
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
645 650 655
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
660 665 670
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
675 680 685
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
690 695 700
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
705 710 715 720
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
725 730 735
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
740 745 750
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
755 760 765
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
770 775 780
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
785 790 795 800
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Met Arg Lys Asn Ile
805 810 815
Lys Gly Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
820 825 830
Tyr Leu Asp Ile Leu Gly
835
<210> 4
<211> 822
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Последовательность мыши
<220>
<221> MISC_FEATURE
<222> (32)..(430)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (32)..(432)
<223> Последовательность человека
<220>
<221> MISC_FEATURE
<222> (431)..(454)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (433)..(822)
<223> Последовательность мыши
<220>
<221> MISC_FEATURE
<222> (455)..(822)
<223> Цитоплазматический домен
<400> 4
Met Ser Pro Trp Leu Lys Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Leu Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
465 470 475 480
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
485 490 495
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
500 505 510
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
515 520 525
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
530 535 540
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
545 550 555 560
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
565 570 575
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
580 585 590
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
595 600 605
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
610 615 620
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
625 630 635 640
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
645 650 655
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
660 665 670
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
675 680 685
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
690 695 700
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
705 710 715 720
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
725 730 735
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
740 745 750
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
755 760 765
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
770 775 780
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Thr Arg Lys Asn Ile
785 790 795 800
Lys Ser Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
805 810 815
Tyr Leu Asp Ile Leu Gly
820
<210> 5
<211> 822
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Последовательность крысы
<220>
<221> MISC_FEATURE
<222> (32)..(430)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (32)..(432)
<223> Последовательность человека
<220>
<221> MISC_FEATURE
<222> (431)..(454)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (433)..(822)
<223> Последовательность крысы
<220>
<221> MISC_FEATURE
<222> (455)..(822)
<223> Цитоплазматический домен
<400> 5
Met Ser Pro Trp Pro Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Leu Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
465 470 475 480
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
485 490 495
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
500 505 510
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
515 520 525
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
530 535 540
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
545 550 555 560
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
565 570 575
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
580 585 590
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
595 600 605
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
610 615 620
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
625 630 635 640
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
645 650 655
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
660 665 670
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
675 680 685
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
690 695 700
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
705 710 715 720
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
725 730 735
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
740 745 750
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
755 760 765
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
770 775 780
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Thr Arg Lys Asn Ile
785 790 795 800
Lys Asn Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
805 810 815
Tyr Leu Asp Ile Leu Gly
820
<210> 6
<211> 8744
<212> DNA
<213> Mus musculus
<400> 6
gtctggaggg tgctatgcta tgcgtgtgtg cgtgtgtgtg cgcgcgcgcg tgtgtgagcg 60
tgtgtgtttt tggatttcat actaattttc tggagtttct gcccctgctc tgcgtcagcc 120
ctcacgtcac ttcgccagca gtagcagagg cggcggcggc ggcggccgcc ggttagagcc 180
cagtcgctgc ttcagctgct gttgctgctt ctgcagcgct ctgctccctg cgcttgctac 240
gggaggccgg ggaagccgcg cggacagtcc tcggtggcct gggccggcac tgtcctgcta 300
ccgcagttgc tccccagccc tgaggtgcgc accgatatcg atattcgtgc cggtttagcg 360
gttctgcgac ccaaagagtc cagggagatc caccgagtgg tgcctggcgt ataggactat 420
gcagccgcct tgtggctcgg agcagcggcc cgcgatgtcc cagccactgt gaaccatttg 480
gtcagcgcca acctgctcag ccccagcacc gacaggctca gcctctggta cgctccactc 540
cgcgggaggc caccagcacc aagcagcaag agggcgcagg gaaggcctcc cccctccggc 600
gggggacgcc tggctcagcg tagggacacg cactccgact gactggcact ggcagctcgg 660
gatgtcgccc tggctgaagt ggcatggacc cgccatggcg cggctctggg gcttatgcct 720
gctggtcttg ggcttctgga gggcctctct cgcctgcccg acgtcctgca aatgcagttc 780
cgctaggatt tggtgtactg agccttctcc aggcatcgtg gcattcccga ggttggaacc 840
taacagcgtt gacccggaga acatcacgga aattctcatt gcaaaccaga aaaggctaga 900
aatcatcaat gaagatgacg ttgaagctta cgtggggctg agaaacctta caattgtgga 960
ttccggctta aagtttgtgg cttacaaagc gtttctgaaa aacagcaacc tgcggcacat 1020
aaatttcaca cgaaacaagc tgacgagttt gtccaggaga catttccgcc accttgactt 1080
gtctgacctg atcctgacgg gtaatccgtt cacgtgctcc tgcgacatca tgtggctcaa 1140
gactctccag gagactaaat ccagccccga cactcaggat ttgtactgcc tcaatgagag 1200
cagcaagaac atgcccctgg cgaacctgca gatacccaat tgtggtctgc catctgcacg 1260
tctggctgct cctaacctca ccgtggagga aggaaagtct gtgacccttt cctgcagtgt 1320
ggggggtgac ccactcccca ccttgtactg ggacgttggg aatttggttt ccaagcacat 1380
gaatgaaaca agccacacac agggctcctt aaggataacg aacatttcat ctgatgacag 1440
tggaaagcaa atctcttgtg tggcagaaaa ccttgtagga gaagatcaag attctgtgaa 1500
cctcactgtg cattttgcgc caactatcac gtttctcgag tctccaacct cagatcacca 1560
ctggtgcatt ccattcactg tgagaggcaa ccccaagcct gcgcttcagt ggttctacaa 1620
tggggccata ctgaatgagt ccaagtacat ctgtactaag atccacgtca ccaatcacac 1680
ggagtaccat ggctgcctcc agctggataa ccccactcat atgaataacg gagactacac 1740
cctgatggcc aagaacgagt atgggaagga tgagagacag atctccgctc acttcatggg 1800
ccggcctgga gtcgactacg agacaaaccc aaattaccct gaagtcctct atgaagactg 1860
gaccacgcca actgacattg gggatactac gaacaaaagt aatgaaatcc cctccacgga 1920
tgttgctgac caaagcaatc gggagcatct ctcggtctat gccgtggtgg tgattgcatc 1980
tgtggtggga ttctgcctgc tggtgatgtt gctcctgctc aagttggcga gacattccaa 2040
gtttggcatg aaaggcccag cttcggtcat cagcaacgac gatgactctg ccagccccct 2100
ccaccacatc tccaatggga gtaacactcc atcttcttcg gagggcggtc ccgacgctgt 2160
cattattgga atgaccaaga ttcctgttat tgaaaacccc cagtactttg gcatcaccaa 2220
cagtcagctc aagccagaca catttgttca gcatatcaag agacacaaca tcgttctgaa 2280
gagggaactt ggggaaggag ccttcgggaa agttttcctt gccgagtgct acaacctctg 2340
cccagagcag gataagatcc tggtggctgt gaagacgctg aaggacgcca gcgacaatgc 2400
acgcaaggac tttcatcggg aagctgagct gctgaccaac ctccagcacg agcacattgt 2460
caagttctac ggtgtctgtg tggagggcga cccactcatc atggtctttg agtacatgaa 2520
gcacggggac ctcaacaagt tccttagggc acacgggccc gacgcagtgc tgatggcaga 2580
gggtaacccg cccacagagc tgacgcagtc gcagatgctg cacatcgctc agcaaatcgc 2640
agcaggtatg gtctacctgg cgtcccaaca ctttgtgcac cgtgacctgg ccacccggaa 2700
ctgcctggtg ggagagaacc tgctggtgaa aattggggac tttgggatgt cccgagatgt 2760
gtacagcacc gactactatc gggtcggtgg ccacacaatg ttgcccatcc gatggatgcc 2820
tccagagagc atcatgtaca ggaaattcac caccgagagc gacgtctgga gcctgggcgt 2880
tgtgttgtgg gagatcttca cctacggcaa gcagccctgg tatcagctat cgaacaatga 2940
ggtgatagag tgcatcaccc agggaagagt ccttcagcgg cctcgaacgt gtccccagga 3000
ggtgtatgag ctcatgcttg gatgctggca gcgggaacca cacacccgga agaacatcaa 3060
gagcatccac accctccttc agaacttggc caaggcatct cccgtctacc tggatatcct 3120
aggctagggt cctccttctg cccagaccgt ccttcccaag gccctcctca gactggccta 3180
cacgacgaac ctcttgactg ccgctgacgt catgaccttg ctgtccttcg ctctgacagt 3240
gttgacagga ccaggagcgg ctctttgggg gaggcagtgt gtgcttctcc atccacagac 3300
agtattaact cgcttctggc attgtctctt tctctccctt gggtttgttt ctttcttttg 3360
ccccttcccc ttttatcatt atttattcat ttatttattt tctggtcttc accgcttcac 3420
ggccctcagt ctctccttga ccaatctggc ttctgcattc ctattaactg tacatagaca 3480
aaggccttaa caaacctaat ttgttatatc agcagacact ccagtttgcc caccacaact 3540
aacaatgcct tgttgtattc ctgcctttga cgtggatgaa aaaaagggaa aaaaaaatca 3600
aacatctgac ttaagctgtc acttccgatg tacagacgtg gggcgtttct atggattcac 3660
ttctatttat tatttattaa tttatttatt tatcactctt cttattgttt tctggtggtt 3720
ttaacctatg tgtgagaagg aaaagttgtg tacaatctgg gaaaacttta tcagtgggaa 3780
atgaaaacga gagcgagcaa gcgagcaaga gagggagaga gagagagaag cgttaccata 3840
aaccacggca tgagcgagac agagacaagc catgggatca gtcgggagtc cgttgtgctt 3900
aggaaaaccc agcagccatt agctggggga gcatgttcgg ctctgtcccc caagcacctt 3960
tctgaggagg acacaggatg ttgaactctg cttcacgggc agagcttcta atgacagata 4020
ctggcttgca ctggaaagac agttcccacg ggacctggac ggacaacaca tcctacattc 4080
agacattgtg gtcgggcacg gtgacagagt tgatccgttt ctcaagtgtt atctaccaag 4140
cttttgtgaa gttccatcga aggaggtaga ttcttgctca gatataattt caggaaaacc 4200
cgagtccttg acaaagacag gagacgctct caatttggag gcaagtttct cttaccttga 4260
actttttcag acagcaactc cgcccagccc ccatcttcca ctctcacctg tcttgtaact 4320
gtgcaaacaa aagtgtgcat ggtctttgtc aattgatacc tatgtgcacg tgtgcagaaa 4380
ctgttgttcc agctggggtg tctgattagg agggcagatc cataaaaggt ctaacctagg 4440
caacttcggg aaaggagacc agatcagtag ctggaggcac tctccagtag gcggtgaggg 4500
gtttactgag taggcatgct gaagcccgga tattcaccca tctcaaaccc cccgggctgc 4560
aggacaggca caggccatcc ctgaggagaa ggggagccct tttgggatac cacctgaggt 4620
tatgttcagt gtgctctggt caagtccctt gctcggggct ctgtttgggg agagtggttt 4680
cattccaagg tactcattat tagtatgctg ttttgttaac tatactccat taaaaagtta 4740
aaaaaaaaaa gaattaagcc ttgacactgt atggctgaca ggaggctgtg cccagactga 4800
gcctggagat ttgcgcccgc acatggtcat tggttttccg aaaagagagg gtaaatttat 4860
atagaaattt acaggtattt gggtagtcat ttagaccgag ggagaccagt gtttccattt 4920
ctctgcgccc cctctgtgag gggaagttcg atacacttga cacctttata aacggagcca 4980
gataggaagg gagtgactta attcacctta gaacatttca tttggtgttt atttctgaag 5040
gtgcaagagc tctgtgtagg tttcatttgt gcccgagcat ttctggagca ctgtgttttc 5100
tagcagaaac tccgagagcc agttctcaca atgaaacttt aaaacctgtg taaattgaca 5160
gagagcagaa ggcgatccaa gaggcccact caagtgagtg gtggcacgag gcacatgcgg 5220
tggccctttc tgttgtgctg gcccatgaga gatgggagct attttgtcct cttcgtccat 5280
taaaacaaac ccctccagaa tacctatagt aatataatga aagccatatc tctgtgatct 5340
ataccgtcag gtatagatca ttaaaggacc catggtactg tcaggcactg tggaaccgtg 5400
aggcagcgga aaggcaaggg cacatttgta catgttcctc tagcttccgc cagccgtgac 5460
ctatgaactc acataggccg gttgctccta gtctgacggg ctgccctggg aggaaaagct 5520
gcaagatgct cagcagagag caaaggagag gatagtcttg gaaaaggcca aggttgggga 5580
tgccatacag gattacacca aagggctgta ctgagaatgt gggagcattc cattccaggt 5640
ttgtattttc ctcagcaaga aaaaagagca gatagaaggg cagaggagga tagagaggaa 5700
gagaaaggag agggagaaga gagcagagaa gagagagaga gagagagaga gagagaatct 5760
ttatactttt tggcaagtcc tcgaaggtct caaaatgaaa gtgtctatgc aagtgcaaat 5820
ttttacagtt atttatacta attattatta ttgttattaa ttattattat tactagtctg 5880
ttgtctcaag aatatgtgca gatttcagag cattaaagag tatttggttt cttttaaagt 5940
agttgggtga gccaaggaca ccttaaaata attgtcactg ttcatctgct atcgcctttc 6000
tcatggtatt ttaagtttta gaaaagaaac attcttgttc taaaacatat ccctctccat 6060
agatgacaaa aaaaaatggt gttagcaaac cgaaacctcc cccgattcct gtctgacatc 6120
cgatccattt ctcaactgct tgatgatttc cgagcgcttt ttgcatatgt tgtcaaaata 6180
ttcatattat tttggtgagg gaatggagac tcagaaactc agaggacaaa cataagcttc 6240
accgaagtct gtctgccaga tacaaactaa catacaaaca agacagccgg gaaggaaatg 6300
tgctggtagc gtgtctgtgg agcagggtgg gggttagaac agaaggagac ctgggcagtg 6360
cacgcagaag agatgttaca tcatgtgctt ctgctcctgg cttctgctga ctcccaactt 6420
tgcgctcagc ctcatttaga aaacgcttca aacactgctc ttcagggttc tgtcagaact 6480
cacacacacg tgctcacacc agccccgctt gtgacgtttt tatgtacttc tatataaata 6540
tatacataat atatgttata aacacttgat ttatacatat acaaatgcac acatgtagtg 6600
tgtttgtgtg tttatgtata aacatatagg ctcatggtaa tagaataatt ataagtaggg 6660
tgcaatatga ataatttgct taatatttgc taagtaacca aaactttcca acgtcatgtg 6720
gcagttcata tttccactct cagtgtgggt aaggatgggc tacaccacac tttcagctct 6780
gtgcagccct tggatggaag atgtgttgaa aaaaaaatca catcttccct gaaacaactg 6840
actgaaagcc atcctctaca ttgaatctgt tctctggctc tttgcaaagg agaaatgcag 6900
acaaatggtc tccttgtgtc ccagtcctgg gaaggagaag tgggttttct cttcttgcac 6960
atttgggccg tgctcacctt tcctggctct atgcccttta cactgggctc agagatgaag 7020
cagtctagac ccaagctgca gggctgtagg gaggcaaatg tcttcgaggc tgatagcaaa 7080
gcatccaaaa atctatagca agatgcgcag cttgaagtca ctcctcatcc acacggcaag 7140
caagagccag ggcatgcagt gctgtccatc tctaggactg ggcagtgggg tccccggtca 7200
agggcagtct ttccctaagc ctctcctttt gcaaatggca ctgccagcag tacctggcat 7260
taagtcacga aaaatttcta aaatggcagc acagacccgt gtgccctcat tgctctgccc 7320
ccacatgcct catcattaga gggctccagt tttcaaactt gactttccat ttggccctgt 7380
atcgtggtta aaggaaatct caacagaggg atggccaatt gacctaaatc cccagatgtc 7440
caagtttgca gtgaaagcca gttcttccca aatgtataac tggaatgtag ggactcaggt 7500
tgtgatactt catacaggaa tctcagatgt tattgtaaag aactgggttg ggagggattc 7560
atcactaagc caacaataat gcaggagggc gtggctgatc gttaagccaa tagtagtgca 7620
ggagggcgtg gctgctgtga gggctggaat ccatctctgc tgctaaagtt cagggttctc 7680
aactcttttc agctgttgca gcccttcact tctttccctt cggcgttatt tatttgtgta 7740
tttatttatt tatttattta tttatctatc tatctatcaa cttttggtcc attcatcacc 7800
aacataaagc aaaaacaatt ttttacatat atgtaaatgt gttgcaagtg aaacactgtg 7860
aatctcacaa ccacctccta gcaactatgc tgccatcttg acgtgcgtct caggagtcga 7920
gacgggaaaa gacgaggacg tcattttgca gcctacgcat tctgggccag gtccgtgttc 7980
gttttatttc tttagtctgt gagttaagaa aacaatccga gtggagggtg acggatgttc 8040
ggcaagttcg gtgaagcagc acgagctctt tgcagctata ggttcgagca agaagaaaaa 8100
ccatgggaga tggagaaggg aaatttgatg gaggggtggg gtggggtggg ggctctgtct 8160
tacagctcgt gtagcttact gttgcttaaa aatgtataca acagctggaa atgttttcaa 8220
cacaaggtat ttgaaataaa tgtgaatctt aaatatgtac tcccttaagg aatgaacata 8280
ttatggtgaa atattgctcc cccgcgtccc ctacaaatct gcctagagat gtggatacct 8340
acatttgctt aagcatcaga ccagtgtttg tatatacccg ggccatacac actcatctcg 8400
acgtctctct gcggatgtat ttccccttca ctggtgacct ggtatttaga actccgtctt 8460
ttcagttggt ttatttccct ttaatgtgat gtctctgtgc cgattattac cggttcttac 8520
ttgtttttgc aatccgtttt gaggtccagt gttttactga gactcattgc atcttggctg 8580
atttcaaagt gacacccgaa tacagtgttt aaaaaaaaag tttgtttgta aatcatgtga 8640
ccagcttctc tcaccctgac atggaatgtc tcttgtacta cagtgtattt aataataaaa 8700
gaaaaaaatg atgtcttaca ataaataaca tcttccacaa gagg 8744
<210> 7
<211> 4750
<212> DNA
<213> Rattus norvegicus
<400> 7
tgtgcgtgcg tgcgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg tgtgtgtgtg 60
tgagcgtgtg tgtttttgga tttcatacta attttctgga gtttctgccc ctgctctgcg 120
tcagccctca cgtcacttcg ccagcagtag cagaggcggc ggcggccgcc ggttagagcc 180
cagtcgctgc ttcagctgct gttgctgctt ctgcggcgct ctgctccctg cgctggctac 240
gggaggccgg gggagccgcg ccgacagtcc tctgtggcca gggccggcac tgtcctgcta 300
ccgcagttgc tccccagccc tgaggtgcgc accgatatcg atatccgtgc cggtttagcg 360
gttctgcgac ccaaagagtc cagggagagc caccgagtgg cgcctggcgt ataggaccat 420
gcagccgcct tgtggcttgg agcagcggcc cgtgatgttc cagccactgt gaaccatttg 480
gtcagcgcca acctgctcag ccccagcacc gacaggctca gcctctggta cgctcctctc 540
ggcgggaggc catcagcacc aagcagcaag agggctcagg gaaggcctcc cccctccggc 600
gggggacgcc tggctcagcg tagggacacg cactctgact gactggcact ggcagctcgg 660
gatgtcgccc tggccgaggt ggcatggacc cgccatggcg cggctctggg gcttatgctt 720
gctggtcttg ggcttctgga gggcttctct tgcctgcccc atgtcctgca aatgcagcac 780
cactaggatt tggtgtaccg agccttctcc tggcatcgtg gcatttccga ggttggaacc 840
taacagcatt gacccagaga acatcaccga aattctcatt gcaaaccaga aaaggttaga 900
aatcatcaat gaagatgatg tcgaagctta cgtggggctg aaaaacctta caattgtgga 960
ttccggctta aagtttgtgg cttacaaggc gtttctgaag aacggcaacc tgcggcacat 1020
caatttcact cgaaacaagc tgacgagttt gtccaggaga catttccgcc accttgactt 1080
gtctgacctg atcctgacgg gtaatccgtt cacgtgttcc tgtgacatca tgtggctcaa 1140
gactctccag gagacgaaat ccagccccga cactcaggat ttgtattgcc tcaatgagag 1200
cagcaagaat acccctctgg cgaacctgca gattcccaat tgtggtctgc cgtctgcacg 1260
tctggccgct cctaacctca cggtggagga agggaagtct gtgaccattt cctgcagcgt 1320
cgggggtgac ccgctcccca ccttgtactg ggacgttggg aatttggttt ccaaacacat 1380
gaatgaaaca agccacacac agggctcctt aaggataaca aacatttcat cggatgacag 1440
tgggaaacaa atctcttgtg tggcagaaaa cctcgttgga gaagatcaag actctgtgaa 1500
cctcactgtg cattttgcac caaccatcac atttctcgaa tctccaacct cagaccacca 1560
ctggtgcatc ccattcactg tgagaggcaa ccccaagcca gcacttcagt ggttctacaa 1620
cggagccata ctgaatgaat ccaagtacat ctgtaccaaa atacacgtca ccaatcacac 1680
ggagtaccac ggctgcctcc agctggataa ccccactcat atgaataatg gagactacac 1740
cctaatggcc aagaatgaat atgggaagga cgagagacag atttctgctc acttcatggg 1800
ccggcctgga gttgactatg agacaaaccc aaattaccct gaagtcctct atgaagactg 1860
gaccacgcca actgacatcg gggatactac aaacaaaagt aatgagatcc cctccacgga 1920
tgttgctgac caaaccaatc gggagcatct ctcggtctat gccgtggtgg tgattgcctc 1980
tgtggtagga ttctgcctgc tggtgatgct gcttctgctc aagttggcga gacattccaa 2040
gtttggcatg aaaggcccag cttccgtcat cagcaacgac gatgactctg ccagccctct 2100
ccaccacatc tccaacggga gcaacactcc gtcttcttcg gagggcgggc ccgatgctgt 2160
catcattggg atgaccaaga tccctgtcat tgaaaacccc cagtacttcg gtatcaccaa 2220
cagccagctc aagccggaca catttgttca gcacatcaag agacacaaca tcgttctgaa 2280
gagggagctt ggagaaggag cctttgggaa agttttccta gcggagtgct ataacctctg 2340
ccccgagcag gataagatcc tggtggccgt gaagacgctg aaggacgcca gcgacaatgc 2400
tcgcaaggac tttcatcgcg aagccgagct gctgaccaac ctccagcacg agcacattgt 2460
caagttctac ggtgtctgtg tggagggcga cccactcatc atggtctttg agtacatgaa 2520
gcacggggac ctcaacaagt tccttagggc acacgggcca gatgcagtgc tgatggcaga 2580
gggtaacccg cccaccgagc tgacgcagtc gcagatgctg cacatcgctc agcaaatcgc 2640
agcaggcatg gtctacctgg catcccaaca cttcgtgcac cgagacctgg ccacccggaa 2700
ctgcttggta ggagagaacc tgctggtgaa aattggggac ttcgggatgt cccgggatgt 2760
atacagcacc gactactacc gggttggtgg ccacacaatg ttgcccatcc gatggatgcc 2820
tccagagagc atcatgtaca ggaaattcac caccgagagt gacgtctgga gcctgggagt 2880
tgtgttgtgg gagatcttca cctacggcaa gcagccctgg tatcagctat caaacaacga 2940
ggtgatagaa tgcatcaccc agggcagagt ccttcagcgg cctcgcacgt gtccccagga 3000
ggtgtacgag ctgatgctgg gatgctggca gcgggaacca cacacaagga agaacatcaa 3060
gaacatccac acactccttc agaacttggc gaaggcgtcg cccgtctacc tggacatcct 3120
aggctagact ccctcttctc ccagacggcc cttcccaagg cacccctcag acctcttaac 3180
tgccgctgat gtcaccacct tgctgtcctt cgctctgaca gtgttaacaa gacaaggagc 3240
ggctctccgg ggtgaggcag tgcgcacttc cccatccaca gacagtatcg actcgcttct 3300
ggctttgtcg ctttctctcc ctttggtttg tttctttctt ttgcccattc tccatttatt 3360
tatttattta tttatttatt tatttattta tttatctatc tatctatcta tctatctatc 3420
tatttattta tttattggtc ttcactgctt catggtcctc ggcctctctc cttgaccgat 3480
ctggcttctg tactcctatt cactgtacat agacaaaggc cttaacaaac ctgatttgtt 3540
atatcagcag acactccagt ttgcccacca caactaacaa tgccttgttg tattcctgcc 3600
tttgatgtgg atgaaaaaaa gggaaaaaaa aataatcaaa catctgactt aaaccgtcac 3660
ttccgatgta cagacacggg gcgtttctat ggattcactt ctatctatct atttatttat 3720
ttatctattt atttatttct cttctttgtt gttttccggt ggttttagcc tgtgtatgag 3780
aagggaaagt catgtacagt ctgggaaaac tttatctgtg ggaaatggaa accagaaagg 3840
gaaagaagct ttaccataaa gcacagcagg agtgagacac agaaaagcca ttggatcagc 3900
cagagtccgt cctgcatagg aaaacccagc agccatcagg ctggaggatc atgttcggca 3960
ctgacccccg aggacctttc tgaggaggac acagaatgtt aaactctgca tcatggacac 4020
agtttccgat cacagatact ggccttcaat ggaaaaaaaa aaaaacccag atagttcttg 4080
tgagacctgg acagcacgtc caacatccag acattgtggt cgggcacagt gacagagttg 4140
atgcatttct cacgggttat tctacagagc ttttgtcaag tccaatggaa ggaggtagat 4200
tcttgttcag atatgatttc gggaaaaacc gagtccttga caaagacagg agacaccctc 4260
agttgggagg caagtttctc ttaccttgga ctttctcaca cagcaattct cacccccacc 4320
ccctccactc tcacctgtct tgtaactgtg caaacaaaag tgtgcatggt ctttgtcagt 4380
tgataccttt gtgcacctct gtgcagaaac tgctgtctgt cccggctgtg gtacccgatc 4440
agtggggtag atccacgaaa ggtctcattt taggccgctt tgggaaggta accagatcgg 4500
tagctggaag cactctccag taggtggcga agggtgagtg ggtctgctga agcctgcata 4560
tcttcaccca cctcaaaccc accgggctgc acaggggaca ggcacaggcc acccctgagg 4620
gacagggaag ctctcttggg ataccacctg agtttacatt cagtgtgctc aggtcaagtc 4680
tctcgctcgg ggctctgttt cggggagaat ggtttcattc caacgcactc attatcagga 4740
ttctgttttc 4750
<210> 8
<211> 4057
<212> DNA
<213> Homo sapiens
<400> 8
gggagcagga gcctcgctgg ctgcttcgct cgcgctctac gcgctcagtc cccggcggta 60
gcaggagcct ggacccaggc gccggcggcg ggcgtgaggc gccggagccc ggcctcgagg 120
tgcataccgg acccccattc gcatctaaca aggaatctgc gccccagaga gtcccggacg 180
ccgccggtcg gtgcccggcg cgccgggcca tgcagcgacg gccgccgcgg agctccgagc 240
agcggtagcg cccccctgta aagcggttcg ctatgccggg accactgtga accctgccgc 300
ctgccggaac actcttcgct ccggaccagc tcagcctctg ataagctgga ctcggcacgc 360
ccgcaacaag caccgaggag ttaagagagc cgcaagcgca gggaaggcct ccccgcacgg 420
gtgggggaaa gcggccggtg cagcgcgggg acaggcactc gggctggcac tggctgctag 480
ggatgtcgtc ctggataagg tggcatggac ccgccatggc gcggctctgg ggcttctgct 540
ggctggttgt gggcttctgg agggccgctt tcgcctgtcc cacgtcctgc aaatgcagtg 600
cctctcggat ctggtgcagc gacccttctc ctggcatcgt ggcatttccg agattggagc 660
ctaacagtgt agatcctgag aacatcaccg aaattttcat cgcaaaccag aaaaggttag 720
aaatcatcaa cgaagatgat gttgaagctt atgtgggact gagaaatctg acaattgtgg 780
attctggatt aaaatttgtg gctcataaag catttctgaa aaacagcaac ctgcagcaca 840
tcaattttac ccgaaacaaa ctgacgagtt tgtctaggaa acatttccgt caccttgact 900
tgtctgaact gatcctggtg ggcaatccat ttacatgctc ctgtgacatt atgtggatca 960
agactctcca agaggctaaa tccagtccag acactcagga tttgtactgc ctgaatgaaa 1020
gcagcaagaa tattcccctg gcaaacctgc agatacccaa ttgtggtttg ccatctgcaa 1080
atctggccgc acctaacctc actgtggagg aaggaaagtc tatcacatta tcctgtagtg 1140
tggcaggtga tccggttcct aatatgtatt gggatgttgg taacctggtt tccaaacata 1200
tgaatgaaac aagccacaca cagggctcct taaggataac taacatttca tccgatgaca 1260
gtgggaagca gatctcttgt gtggcggaaa atcttgtagg agaagatcaa gattctgtca 1320
acctcactgt gcattttgca ccaactatca catttctcga atctccaacc tcagaccacc 1380
actggtgcat tccattcact gtgaaaggca accccaaacc agcgcttcag tggttctata 1440
acggggcaat attgaatgag tccaaataca tctgtactaa aatacatgtt accaatcaca 1500
cggagtacca cggctgcctc cagctggata atcccactca catgaacaat ggggactaca 1560
ctctaatagc caagaatgag tatgggaagg atgagaaaca gatttctgct cacttcatgg 1620
gctggcctgg aattgacgat ggtgcaaacc caaattatcc tgatgtaatt tatgaagatt 1680
atggaactgc agcgaatgac atcggggaca ccacgaacag aagtaatgaa atcccttcca 1740
cagacgtcac tgataaaacc ggtcgggaac atctctcggt ctatgctgtg gtggtgattg 1800
cgtctgtggt gggattttgc cttttggtaa tgctgtttct gcttaagttg gcaagacact 1860
ccaagtttgg catgaaagat ttctcatggt ttggatttgg gaaagtaaaa tcaagacaag 1920
gtgttggccc agcctccgtt atcagcaatg atgatgactc tgccagccca ctccatcaca 1980
tctccaatgg gagtaacact ccatcttctt cggaaggtgg cccagatgct gtcattattg 2040
gaatgaccaa gatccctgtc attgaaaatc cccagtactt tggcatcacc aacagtcagc 2100
tcaagccaga cacatttgtt cagcacatca agcgacataa cattgttctg aaaagggagc 2160
taggcgaagg agcctttgga aaagtgttcc tagctgaatg ctataacctc tgtcctgagc 2220
aggacaagat cttggtggca gtgaagaccc tgaaggatgc cagtgacaat gcacgcaagg 2280
acttccaccg tgaggccgag ctcctgacca acctccagca tgagcacatc gtcaagttct 2340
atggcgtctg cgtggagggc gaccccctca tcatggtctt tgagtacatg aagcatgggg 2400
acctcaacaa gttcctcagg gcacacggcc ctgatgccgt gctgatggct gagggcaacc 2460
cgcccacgga actgacgcag tcgcagatgc tgcatatagc ccagcagatc gccgcgggca 2520
tggtctacct ggcgtcccag cacttcgtgc accgcgattt ggccaccagg aactgcctgg 2580
tcggggagaa cttgctggtg aaaatcgggg actttgggat gtcccgggac gtgtacagca 2640
ctgactacta cagggtcggt ggccacacaa tgctgcccat tcgctggatg cctccagaga 2700
gcatcatgta caggaaattc acgacggaaa gcgacgtctg gagcctgggg gtcgtgttgt 2760
gggagatttt cacctatggc aaacagccct ggtaccagct gtcaaacaat gaggtgatag 2820
agtgtatcac tcagggccga gtcctgcagc gaccccgcac gtgcccccag gaggtgtatg 2880
agctgatgct ggggtgctgg cagcgagagc cccacatgag gaagaacatc aagggcatcc 2940
ataccctcct tcagaacttg gccaaggcat ctccggtcta cctggacatt ctaggctagg 3000
gcccttttcc ccagaccgat ccttcccaac gtactcctca gacgggctga gaggatgaac 3060
atcttttaac tgccgctgga ggccaccaag ctgctctcct tcactctgac agtattaaca 3120
tcaaagactc cgagaagctc tcgagggaag cagtgtgtac ttcttcatcc atagacacag 3180
tattgacttc tttttggcat tatctctttc tctctttcca tctcccttgg ttgttccttt 3240
ttcttttttt aaattttctt tttcttcttt tttttcgtct tccctgcttc acgattctta 3300
ccctttcttt tgaatcaatc tggcttctgc attactatta actctgcata gacaaaggcc 3360
ttaacaaacg taatttgtta tatcagcaga cactccagtt tgcccaccac aactaacaat 3420
gccttgttgt attcctgcct ttgatgtgga tgaaaaaaag ggaaaacaaa tatttcactt 3480
aaactttgtc acttctgctg tacagatatc gagagtttct atggattcac ttctatttat 3540
ttattattat tactgttctt attgtttttg gatggcttaa gcctgtgtat aaaaaagaaa 3600
acttgtgttc aatctgtgaa gcctttatct atgggagatt aaaaccagag agaaagaaga 3660
tttattatga accgcaatat gggaggaaca aagacaacca ctgggatcag ctggtgtcag 3720
tccctactta ggaaatactc agcaactgtt agctgggaag aatgtattcg gcaccttccc 3780
ctgaggacct ttctgaggag taaaaagact actggcctct gtgccatgga tgattctttt 3840
cccatcacca gaaatgatag cgtgcagtag agagcaaaga tggcttccgt gagacacaag 3900
atggcgcata gtgtgctcgg acacagtttt gtcttcgtag gttgtgatga tagcactggt 3960
ttgtttctca agcgctatcc acagaacctt tgtcaacttc agttgaaaag aggtggattc 4020
atgtccagag ctcatttcgg ggtcaggtgg gaaagcc 4057
<210> 9
<211> 2466
<212> DNA
<213> Mus musculus
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (94)..(1287)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (1288)..(1359)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1360)..(2463)
<223> Цитоплазматический домен
<400> 9
atgtcgccct ggctgaagtg gcatggaccc gccatggcgc ggctctgggg cttatgcctg 60
ctggtcttgg gcttctggag ggcctctctc gcctgcccga cgtcctgcaa atgcagttcc 120
gctaggattt ggtgtactga gccttctcca ggcatcgtgg cattcccgag gttggaacct 180
aacagcgttg acccggagaa catcacggaa attctcattg caaaccagaa aaggctagaa 240
atcatcaatg aagatgacgt tgaagcttac gtggggctga gaaaccttac aattgtggat 300
tccggcttaa agtttgtggc ttacaaagcg tttctgaaaa acagcaacct gcggcacata 360
aatttcacac gaaacaagct gacgagtttg tccaggagac atttccgcca ccttgacttg 420
tctgacctga tcctgacggg taatccgttc acgtgctcct gcgacatcat gtggctcaag 480
actctccagg agactaaatc cagccccgac actcaggatt tgtactgcct caatgagagc 540
agcaagaaca tgcccctggc gaacctgcag atacccaatt gtggtctgcc atctgcacgt 600
ctggctgctc ctaacctcac cgtggaggaa ggaaagtctg tgaccctttc ctgcagtgtg 660
gggggtgacc cactccccac cttgtactgg gacgttggga atttggtttc caagcacatg 720
aatgaaacaa gccacacaca gggctcctta aggataacga acatttcatc tgatgacagt 780
ggaaagcaaa tctcttgtgt ggcagaaaac cttgtaggag aagatcaaga ttctgtgaac 840
ctcactgtgc attttgcgcc aactatcacg tttctcgagt ctccaacctc agatcaccac 900
tggtgcattc cattcactgt gagaggcaac cccaagcctg cgcttcagtg gttctacaat 960
ggggccatac tgaatgagtc caagtacatc tgtactaaga tccacgtcac caatcacacg 1020
gagtaccatg gctgcctcca gctggataac cccactcata tgaataacgg agactacacc 1080
ctgatggcca agaacgagta tgggaaggat gagagacaga tctccgctca cttcatgggc 1140
cggcctggag tcgactacga gacaaaccca aattaccctg aagtcctcta tgaagactgg 1200
accacgccaa ctgacattgg ggatactacg aacaaaagta atgaaatccc ctccacggat 1260
gttgctgacc aaagcaatcg ggagcatctc tcggtctatg ccgtggtggt gattgcatct 1320
gtggtgggat tctgcctgct ggtgatgttg ctcctgctca agttggcgag acattccaag 1380
tttggcatga aaggcccagc ttcggtcatc agcaacgacg atgactctgc cagccccctc 1440
caccacatct ccaatgggag taacactcca tcttcttcgg agggcggtcc cgacgctgtc 1500
attattggaa tgaccaagat tcctgttatt gaaaaccccc agtactttgg catcaccaac 1560
agtcagctca agccagacac atttgttcag catatcaaga gacacaacat cgttctgaag 1620
agggaacttg gggaaggagc cttcgggaaa gttttccttg ccgagtgcta caacctctgc 1680
ccagagcagg ataagatcct ggtggctgtg aagacgctga aggacgccag cgacaatgca 1740
cgcaaggact ttcatcggga agctgagctg ctgaccaacc tccagcacga gcacattgtc 1800
aagttctacg gtgtctgtgt ggagggcgac ccactcatca tggtctttga gtacatgaag 1860
cacggggacc tcaacaagtt ccttagggca cacgggcccg acgcagtgct gatggcagag 1920
ggtaacccgc ccacagagct gacgcagtcg cagatgctgc acatcgctca gcaaatcgca 1980
gcaggtatgg tctacctggc gtcccaacac tttgtgcacc gtgacctggc cacccggaac 2040
tgcctggtgg gagagaacct gctggtgaaa attggggact ttgggatgtc ccgagatgtg 2100
tacagcaccg actactatcg ggtcggtggc cacacaatgt tgcccatccg atggatgcct 2160
ccagagagca tcatgtacag gaaattcacc accgagagcg acgtctggag cctgggcgtt 2220
gtgttgtggg agatcttcac ctacggcaag cagccctggt atcagctatc gaacaatgag 2280
gtgatagagt gcatcaccca gggaagagtc cttcagcggc ctcgaacgtg tccccaggag 2340
gtgtatgagc tcatgcttgg atgctggcag cgggaaccac acacccggaa gaacatcaag 2400
agcatccaca ccctccttca gaacttggcc aaggcatctc ccgtctacct ggatatccta 2460
ggctag 2466
<210> 10
<211> 2466
<212> DNA
<213> Rattus norvegicus
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (94)..(1287)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (1288)..(1359)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1360)..(2463)
<223> Цитоплазматический домен
<400> 10
atgtcgccct ggccgaggtg gcatggaccc gccatggcgc ggctctgggg cttatgcttg 60
ctggtcttgg gcttctggag ggcttctctt gcctgcccca tgtcctgcaa atgcagcacc 120
actaggattt ggtgtaccga gccttctcct ggcatcgtgg catttccgag gttggaacct 180
aacagcattg acccagagaa catcaccgaa attctcattg caaaccagaa aaggttagaa 240
atcatcaatg aagatgatgt cgaagcttac gtggggctga aaaaccttac aattgtggat 300
tccggcttaa agtttgtggc ttacaaggcg tttctgaaga acggcaacct gcggcacatc 360
aatttcactc gaaacaagct gacgagtttg tccaggagac atttccgcca ccttgacttg 420
tctgacctga tcctgacggg taatccgttc acgtgttcct gtgacatcat gtggctcaag 480
actctccagg agacgaaatc cagccccgac actcaggatt tgtattgcct caatgagagc 540
agcaagaata cccctctggc gaacctgcag attcccaatt gtggtctgcc gtctgcacgt 600
ctggccgctc ctaacctcac ggtggaggaa gggaagtctg tgaccatttc ctgcagcgtc 660
gggggtgacc cgctccccac cttgtactgg gacgttggga atttggtttc caaacacatg 720
aatgaaacaa gccacacaca gggctcctta aggataacaa acatttcatc ggatgacagt 780
gggaaacaaa tctcttgtgt ggcagaaaac ctcgttggag aagatcaaga ctctgtgaac 840
ctcactgtgc attttgcacc aaccatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcatcc cattcactgt gagaggcaac cccaagccag cacttcagtg gttctacaac 960
ggagccatac tgaatgaatc caagtacatc tgtaccaaaa tacacgtcac caatcacacg 1020
gagtaccacg gctgcctcca gctggataac cccactcata tgaataatgg agactacacc 1080
ctaatggcca agaatgaata tgggaaggac gagagacaga tttctgctca cttcatgggc 1140
cggcctggag ttgactatga gacaaaccca aattaccctg aagtcctcta tgaagactgg 1200
accacgccaa ctgacatcgg ggatactaca aacaaaagta atgagatccc ctccacggat 1260
gttgctgacc aaaccaatcg ggagcatctc tcggtctatg ccgtggtggt gattgcctct 1320
gtggtaggat tctgcctgct ggtgatgctg cttctgctca agttggcgag acattccaag 1380
tttggcatga aaggcccagc ttccgtcatc agcaacgacg atgactctgc cagccctctc 1440
caccacatct ccaacgggag caacactccg tcttcttcgg agggcgggcc cgatgctgtc 1500
atcattggga tgaccaagat ccctgtcatt gaaaaccccc agtacttcgg tatcaccaac 1560
agccagctca agccggacac atttgttcag cacatcaaga gacacaacat cgttctgaag 1620
agggagcttg gagaaggagc ctttgggaaa gttttcctag cggagtgcta taacctctgc 1680
cccgagcagg ataagatcct ggtggccgtg aagacgctga aggacgccag cgacaatgct 1740
cgcaaggact ttcatcgcga agccgagctg ctgaccaacc tccagcacga gcacattgtc 1800
aagttctacg gtgtctgtgt ggagggcgac ccactcatca tggtctttga gtacatgaag 1860
cacggggacc tcaacaagtt ccttagggca cacgggccag atgcagtgct gatggcagag 1920
ggtaacccgc ccaccgagct gacgcagtcg cagatgctgc acatcgctca gcaaatcgca 1980
gcaggcatgg tctacctggc atcccaacac ttcgtgcacc gagacctggc cacccggaac 2040
tgcttggtag gagagaacct gctggtgaaa attggggact tcgggatgtc ccgggatgta 2100
tacagcaccg actactaccg ggttggtggc cacacaatgt tgcccatccg atggatgcct 2160
ccagagagca tcatgtacag gaaattcacc accgagagtg acgtctggag cctgggagtt 2220
gtgttgtggg agatcttcac ctacggcaag cagccctggt atcagctatc aaacaacgag 2280
gtgatagaat gcatcaccca gggcagagtc cttcagcggc ctcgcacgtg tccccaggag 2340
gtgtacgagc tgatgctggg atgctggcag cgggaaccac acacaaggaa gaacatcaag 2400
aacatccaca cactccttca gaacttggcg aaggcgtcgc ccgtctacct ggacatccta 2460
ggctag 2466
<210> 11
<211> 2517
<212> DNA
<213> Homo sapiens
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (94)..(1290)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (1291)..(1362)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1363)..(2514)
<223> Цитоплазматический домен
<400> 11
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgctgtggt ggtgattgcg 1320
tctgtggtgg gattttgcct tttggtaatg ctgtttctgc ttaagttggc aagacactcc 1380
aagtttggca tgaaagattt ctcatggttt ggatttggga aagtaaaatc aagacaaggt 1440
gttggcccag cctccgttat cagcaatgat gatgactctg ccagcccact ccatcacatc 1500
tccaatggga gtaacactcc atcttcttcg gaaggtggcc cagatgctgt cattattgga 1560
atgaccaaga tccctgtcat tgaaaatccc cagtactttg gcatcaccaa cagtcagctc 1620
aagccagaca catttgttca gcacatcaag cgacataaca ttgttctgaa aagggagcta 1680
ggcgaaggag cctttggaaa agtgttccta gctgaatgct ataacctctg tcctgagcag 1740
gacaagatct tggtggcagt gaagaccctg aaggatgcca gtgacaatgc acgcaaggac 1800
ttccaccgtg aggccgagct cctgaccaac ctccagcatg agcacatcgt caagttctat 1860
ggcgtctgcg tggagggcga ccccctcatc atggtctttg agtacatgaa gcatggggac 1920
ctcaacaagt tcctcagggc acacggccct gatgccgtgc tgatggctga gggcaacccg 1980
cccacggaac tgacgcagtc gcagatgctg catatagccc agcagatcgc cgcgggcatg 2040
gtctacctgg cgtcccagca cttcgtgcac cgcgatttgg ccaccaggaa ctgcctggtc 2100
ggggagaact tgctggtgaa aatcggggac tttgggatgt cccgggacgt gtacagcact 2160
gactactaca gggtcggtgg ccacacaatg ctgcccattc gctggatgcc tccagagagc 2220
atcatgtaca ggaaattcac gacggaaagc gacgtctgga gcctgggggt cgtgttgtgg 2280
gagattttca cctatggcaa acagccctgg taccagctgt caaacaatga ggtgatagag 2340
tgtatcactc agggccgagt cctgcagcga ccccgcacgt gcccccagga ggtgtatgag 2400
ctgatgctgg ggtgctggca gcgagagccc cacatgagga agaacatcaa gggcatccat 2460
accctccttc agaacttggc caaggcatct ccggtctacc tggacattct aggctag 2517
<210> 12
<211> 2469
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (1)..(93)
<223> Последовательность мыши
<220>
<221> misc_feature
<222> (94)..(1290)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (94)..(1296)
<223> Последовательность человека
<220>
<221> misc_feature
<222> (1291)..(1362)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1297)..(2466)
<223> Последовательность мыши
<220>
<221> misc_feature
<222> (1363)..(2466)
<223> Цитоплазматический домен
<400> 12
atgtcgccct ggctgaagtg gcatggaccc gccatggcgc ggctctgggg cttatgcctg 60
ctggtcttgg gcttctggag ggcctctctc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgccgtggt ggtgattgca 1320
tctgtggtgg gattctgcct gctggtgatg ttgctcctgc tcaagttggc gagacattcc 1380
aagtttggca tgaaaggccc agcttcggtc atcagcaacg acgatgactc tgccagcccc 1440
ctccaccaca tctccaatgg gagtaacact ccatcttctt cggagggcgg tcccgacgct 1500
gtcattattg gaatgaccaa gattcctgtt attgaaaacc cccagtactt tggcatcacc 1560
aacagtcagc tcaagccaga cacatttgtt cagcatatca agagacacaa catcgttctg 1620
aagagggaac ttggggaagg agccttcggg aaagttttcc ttgccgagtg ctacaacctc 1680
tgcccagagc aggataagat cctggtggct gtgaagacgc tgaaggacgc cagcgacaat 1740
gcacgcaagg actttcatcg ggaagctgag ctgctgacca acctccagca cgagcacatt 1800
gtcaagttct acggtgtctg tgtggagggc gacccactca tcatggtctt tgagtacatg 1860
aagcacgggg acctcaacaa gttccttagg gcacacgggc ccgacgcagt gctgatggca 1920
gagggtaacc cgcccacaga gctgacgcag tcgcagatgc tgcacatcgc tcagcaaatc 1980
gcagcaggta tggtctacct ggcgtcccaa cactttgtgc accgtgacct ggccacccgg 2040
aactgcctgg tgggagagaa cctgctggtg aaaattgggg actttgggat gtcccgagat 2100
gtgtacagca ccgactacta tcgggtcggt ggccacacaa tgttgcccat ccgatggatg 2160
cctccagaga gcatcatgta caggaaattc accaccgaga gcgacgtctg gagcctgggc 2220
gttgtgttgt gggagatctt cacctacggc aagcagccct ggtatcagct atcgaacaat 2280
gaggtgatag agtgcatcac ccagggaaga gtccttcagc ggcctcgaac gtgtccccag 2340
gaggtgtatg agctcatgct tggatgctgg cagcgggaac cacacacccg gaagaacatc 2400
aagagcatcc acaccctcct tcagaacttg gccaaggcat ctcccgtcta cctggatatc 2460
ctaggctag 2469
<210> 13
<211> 2469
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (1)..(93)
<223> Последовательность крысы
<220>
<221> misc_feature
<222> (94)..(1290)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (94)..(1296)
<223> Последовательность человека
<220>
<221> misc_feature
<222> (1291)..(1362)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1297)..(2466)
<223> Последовательность крысы
<220>
<221> misc_feature
<222> (1363)..(2466)
<223> Цитоплазматический домен
<400> 13
atgtcgccct ggccgaggtg gcatggaccc gccatggcgc ggctctgggg cttatgcttg 60
ctggtcttgg gcttctggag ggcttctctt gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgccgtggt ggtgattgcc 1320
tctgtggtag gattctgcct gctggtgatg ctgcttctgc tcaagttggc gagacattcc 1380
aagtttggca tgaaaggccc agcttccgtc atcagcaacg acgatgactc tgccagccct 1440
ctccaccaca tctccaacgg gagcaacact ccgtcttctt cggagggcgg gcccgatgct 1500
gtcatcattg ggatgaccaa gatccctgtc attgaaaacc cccagtactt cggtatcacc 1560
aacagccagc tcaagccgga cacatttgtt cagcacatca agagacacaa catcgttctg 1620
aagagggagc ttggagaagg agcctttggg aaagttttcc tagcggagtg ctataacctc 1680
tgccccgagc aggataagat cctggtggcc gtgaagacgc tgaaggacgc cagcgacaat 1740
gctcgcaagg actttcatcg cgaagccgag ctgctgacca acctccagca cgagcacatt 1800
gtcaagttct acggtgtctg tgtggagggc gacccactca tcatggtctt tgagtacatg 1860
aagcacgggg acctcaacaa gttccttagg gcacacgggc cagatgcagt gctgatggca 1920
gagggtaacc cgcccaccga gctgacgcag tcgcagatgc tgcacatcgc tcagcaaatc 1980
gcagcaggca tggtctacct ggcatcccaa cacttcgtgc accgagacct ggccacccgg 2040
aactgcttgg taggagagaa cctgctggtg aaaattgggg acttcgggat gtcccgggat 2100
gtatacagca ccgactacta ccgggttggt ggccacacaa tgttgcccat ccgatggatg 2160
cctccagaga gcatcatgta caggaaattc accaccgaga gtgacgtctg gagcctggga 2220
gttgtgttgt gggagatctt cacctacggc aagcagccct ggtatcagct atcaaacaac 2280
gaggtgatag aatgcatcac ccagggcaga gtccttcagc ggcctcgcac gtgtccccag 2340
gaggtgtacg agctgatgct gggatgctgg cagcgggaac cacacacaag gaagaacatc 2400
aagaacatcc acacactcct tcagaacttg gcgaaggcgt cgcccgtcta cctggacatc 2460
ctaggctag 2469
<210> 14
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 14
aggtgggtag gtcctggaag tg 22
<210> 15
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 15
aatgctgtcc caagagtggg 20
<210> 16
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 16
gtcctgcatc ccttgtcttt g 21
<210> 17
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 17
atgtgggcgt tgtgcagtct c 21
<210> 18
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 18
cgctgcagtg cattgaactc agca 24
<210> 19
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 19
ctgtggaggg acgtgaccag 20
<210> 20
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 20
tccgctagga tttggtgtac tg 22
<210> 21
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 21
agccttctcc aggcatcgtg gcat 24
<210> 22
<211> 19
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 22
tccgggtcaa cgctgttag 19
<210> 23
<211> 18
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 23
tcctgcgagg gttctgac 18
<210> 24
<211> 29
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 24
tgggtgctca tatgccagag aaattgtca 29
<210> 25
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 25
cgatctgtga tggcctgctt ac 22
<210> 26
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 26
gggctcaggc aggtatatgt tg 22
<210> 27
<211> 27
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 27
acagatgctg tcccaaacat agcaaga 27
<210> 28
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 28
ccaaccctaa gccagtgaaa cag 23
<210> 29
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 29
gtgctggaga ccaggagact 20
<210> 30
<211> 28
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 30
tgccatactc agtttatacg gtgctgac 28
<210> 31
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 31
gcctggtggc tcagtcaatg 20
<210> 32
<211> 19
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 32
gcagacactg gatgggtca 19
<210> 33
<211> 26
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 33
ccattcgcga gttatgagaa gctgca 26
<210> 34
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 34
acagggttag ctggtgaatg ga 22
<210> 35
<211> 19
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 35
tcggagcaca ggactacag 19
<210> 36
<211> 25
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 36
caagaggaac tgtgtccagg aaagc 25
<210> 37
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 37
agcgtgcctc acctaacctc ta 22
<210> 38
<211> 19
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 38
gcacagcact gtaaaggca 19
<210> 39
<211> 29
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 39
acggaactcg aaggaattgg tattgttgt 29
<210> 40
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 40
acacagctat gggagaaaga ctg 23
<210> 41
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 41
ccggttggcg agtgcgcatg cac 23
<210> 42
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 42
ccttccggtt ggcgagtgcg cat 23
<210> 43
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 43
ccaagggtgc gttgatggat cta 23
<210> 44
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 44
gaatgccaag ggtgcgttga tgg 23
<210> 45
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 45
ctgggtgatt gggactgaga aag 23
<210> 46
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 46
cagccttgaa agtatggctt gggc 24
<210> 47
<211> 18
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 47
gcactcgcca accggaag 18
<210> 48
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 48
gaccagctca cccttactta tgg 23
<210> 49
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 49
actgaatgcc aagggtgcgt tga 23
<210> 50
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 50
tcttggaaat ccgctgaaga gtt 23
<210> 51
<211> 31
<212> PRT
<213> Mus musculus
<400> 51
Met Ser Pro Trp Leu Lys Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala
20 25 30
<210> 52
<211> 398
<212> PRT
<213> Mus musculus
<400> 52
Cys Pro Thr Ser Cys Lys Cys Ser Ser Ala Arg Ile Trp Cys Thr Glu
1 5 10 15
Pro Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val
20 25 30
Asp Pro Glu Asn Ile Thr Glu Ile Leu Ile Ala Asn Gln Lys Arg Leu
35 40 45
Glu Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn
50 55 60
Leu Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala Tyr Lys Ala Phe
65 70 75 80
Leu Lys Asn Ser Asn Leu Arg His Ile Asn Phe Thr Arg Asn Lys Leu
85 90 95
Thr Ser Leu Ser Arg Arg His Phe Arg His Leu Asp Leu Ser Asp Leu
100 105 110
Ile Leu Thr Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Leu
115 120 125
Lys Thr Leu Gln Glu Thr Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr
130 135 140
Cys Leu Asn Glu Ser Ser Lys Asn Met Pro Leu Ala Asn Leu Gln Ile
145 150 155 160
Pro Asn Cys Gly Leu Pro Ser Ala Arg Leu Ala Ala Pro Asn Leu Thr
165 170 175
Val Glu Glu Gly Lys Ser Val Thr Leu Ser Cys Ser Val Gly Gly Asp
180 185 190
Pro Leu Pro Thr Leu Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His
195 200 205
Met Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile
210 215 220
Ser Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu
225 230 235 240
Val Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro
245 250 255
Thr Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile
260 265 270
Pro Phe Thr Val Arg Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr
275 280 285
Asn Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His
290 295 300
Val Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro
305 310 315 320
Thr His Met Asn Asn Gly Asp Tyr Thr Leu Met Ala Lys Asn Glu Tyr
325 330 335
Gly Lys Asp Glu Arg Gln Ile Ser Ala His Phe Met Gly Arg Pro Gly
340 345 350
Val Asp Tyr Glu Thr Asn Pro Asn Tyr Pro Glu Val Leu Tyr Glu Asp
355 360 365
Trp Thr Thr Pro Thr Asp Ile Gly Asp Thr Thr Asn Lys Ser Asn Glu
370 375 380
Ile Pro Ser Thr Asp Val Ala Asp Gln Ser Asn Arg Glu His
385 390 395
<210> 53
<211> 24
<212> PRT
<213> Mus musculus
<400> 53
Leu Ser Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys
1 5 10 15
Leu Leu Val Met Leu Leu Leu Leu
20
<210> 54
<211> 368
<212> PRT
<213> Mus musculus
<400> 54
Lys Leu Ala Arg His Ser Lys Phe Gly Met Lys Gly Pro Ala Ser Val
1 5 10 15
Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro Leu His His Ile Ser Asn
20 25 30
Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly Gly Pro Asp Ala Val Ile
35 40 45
Ile Gly Met Thr Lys Ile Pro Val Ile Glu Asn Pro Gln Tyr Phe Gly
50 55 60
Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr Phe Val Gln His Ile Lys
65 70 75 80
Arg His Asn Ile Val Leu Lys Arg Glu Leu Gly Glu Gly Ala Phe Gly
85 90 95
Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu Cys Pro Glu Gln Asp Lys
100 105 110
Ile Leu Val Ala Val Lys Thr Leu Lys Asp Ala Ser Asp Asn Ala Arg
115 120 125
Lys Asp Phe His Arg Glu Ala Glu Leu Leu Thr Asn Leu Gln His Glu
130 135 140
His Ile Val Lys Phe Tyr Gly Val Cys Val Glu Gly Asp Pro Leu Ile
145 150 155 160
Met Val Phe Glu Tyr Met Lys His Gly Asp Leu Asn Lys Phe Leu Arg
165 170 175
Ala His Gly Pro Asp Ala Val Leu Met Ala Glu Gly Asn Pro Pro Thr
180 185 190
Glu Leu Thr Gln Ser Gln Met Leu His Ile Ala Gln Gln Ile Ala Ala
195 200 205
Gly Met Val Tyr Leu Ala Ser Gln His Phe Val His Arg Asp Leu Ala
210 215 220
Thr Arg Asn Cys Leu Val Gly Glu Asn Leu Leu Val Lys Ile Gly Asp
225 230 235 240
Phe Gly Met Ser Arg Asp Val Tyr Ser Thr Asp Tyr Tyr Arg Val Gly
245 250 255
Gly His Thr Met Leu Pro Ile Arg Trp Met Pro Pro Glu Ser Ile Met
260 265 270
Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val Trp Ser Leu Gly Val Val
275 280 285
Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln Pro Trp Tyr Gln Leu Ser
290 295 300
Asn Asn Glu Val Ile Glu Cys Ile Thr Gln Gly Arg Val Leu Gln Arg
305 310 315 320
Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu Leu Met Leu Gly Cys Trp
325 330 335
Gln Arg Glu Pro His Thr Arg Lys Asn Ile Lys Ser Ile His Thr Leu
340 345 350
Leu Gln Asn Leu Ala Lys Ala Ser Pro Val Tyr Leu Asp Ile Leu Gly
355 360 365
<210> 55
<211> 31
<212> PRT
<213> Rattus norvegicus
<400> 55
Met Ser Pro Trp Pro Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Leu Cys Leu Leu Val Leu Gly Phe Trp Arg Ala Ser Leu Ala
20 25 30
<210> 56
<211> 398
<212> PRT
<213> Rattus norvegicus
<400> 56
Cys Pro Met Ser Cys Lys Cys Ser Thr Thr Arg Ile Trp Cys Thr Glu
1 5 10 15
Pro Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Ile
20 25 30
Asp Pro Glu Asn Ile Thr Glu Ile Leu Ile Ala Asn Gln Lys Arg Leu
35 40 45
Glu Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Lys Asn
50 55 60
Leu Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala Tyr Lys Ala Phe
65 70 75 80
Leu Lys Asn Gly Asn Leu Arg His Ile Asn Phe Thr Arg Asn Lys Leu
85 90 95
Thr Ser Leu Ser Arg Arg His Phe Arg His Leu Asp Leu Ser Asp Leu
100 105 110
Ile Leu Thr Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Leu
115 120 125
Lys Thr Leu Gln Glu Thr Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr
130 135 140
Cys Leu Asn Glu Ser Ser Lys Asn Thr Pro Leu Ala Asn Leu Gln Ile
145 150 155 160
Pro Asn Cys Gly Leu Pro Ser Ala Arg Leu Ala Ala Pro Asn Leu Thr
165 170 175
Val Glu Glu Gly Lys Ser Val Thr Ile Ser Cys Ser Val Gly Gly Asp
180 185 190
Pro Leu Pro Thr Leu Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His
195 200 205
Met Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile
210 215 220
Ser Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu
225 230 235 240
Val Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro
245 250 255
Thr Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile
260 265 270
Pro Phe Thr Val Arg Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr
275 280 285
Asn Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His
290 295 300
Val Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro
305 310 315 320
Thr His Met Asn Asn Gly Asp Tyr Thr Leu Met Ala Lys Asn Glu Tyr
325 330 335
Gly Lys Asp Glu Arg Gln Ile Ser Ala His Phe Met Gly Arg Pro Gly
340 345 350
Val Asp Tyr Glu Thr Asn Pro Asn Tyr Pro Glu Val Leu Tyr Glu Asp
355 360 365
Trp Thr Thr Pro Thr Asp Ile Gly Asp Thr Thr Asn Lys Ser Asn Glu
370 375 380
Ile Pro Ser Thr Asp Val Ala Asp Gln Thr Asn Arg Glu His
385 390 395
<210> 57
<211> 24
<212> PRT
<213> Rattus norvegicus
<400> 57
Leu Ser Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys
1 5 10 15
Leu Leu Val Met Leu Leu Leu Leu
20
<210> 58
<211> 368
<212> PRT
<213> Rattus norvegicus
<400> 58
Lys Leu Ala Arg His Ser Lys Phe Gly Met Lys Gly Pro Ala Ser Val
1 5 10 15
Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro Leu His His Ile Ser Asn
20 25 30
Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly Gly Pro Asp Ala Val Ile
35 40 45
Ile Gly Met Thr Lys Ile Pro Val Ile Glu Asn Pro Gln Tyr Phe Gly
50 55 60
Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr Phe Val Gln His Ile Lys
65 70 75 80
Arg His Asn Ile Val Leu Lys Arg Glu Leu Gly Glu Gly Ala Phe Gly
85 90 95
Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu Cys Pro Glu Gln Asp Lys
100 105 110
Ile Leu Val Ala Val Lys Thr Leu Lys Asp Ala Ser Asp Asn Ala Arg
115 120 125
Lys Asp Phe His Arg Glu Ala Glu Leu Leu Thr Asn Leu Gln His Glu
130 135 140
His Ile Val Lys Phe Tyr Gly Val Cys Val Glu Gly Asp Pro Leu Ile
145 150 155 160
Met Val Phe Glu Tyr Met Lys His Gly Asp Leu Asn Lys Phe Leu Arg
165 170 175
Ala His Gly Pro Asp Ala Val Leu Met Ala Glu Gly Asn Pro Pro Thr
180 185 190
Glu Leu Thr Gln Ser Gln Met Leu His Ile Ala Gln Gln Ile Ala Ala
195 200 205
Gly Met Val Tyr Leu Ala Ser Gln His Phe Val His Arg Asp Leu Ala
210 215 220
Thr Arg Asn Cys Leu Val Gly Glu Asn Leu Leu Val Lys Ile Gly Asp
225 230 235 240
Phe Gly Met Ser Arg Asp Val Tyr Ser Thr Asp Tyr Tyr Arg Val Gly
245 250 255
Gly His Thr Met Leu Pro Ile Arg Trp Met Pro Pro Glu Ser Ile Met
260 265 270
Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val Trp Ser Leu Gly Val Val
275 280 285
Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln Pro Trp Tyr Gln Leu Ser
290 295 300
Asn Asn Glu Val Ile Glu Cys Ile Thr Gln Gly Arg Val Leu Gln Arg
305 310 315 320
Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu Leu Met Leu Gly Cys Trp
325 330 335
Gln Arg Glu Pro His Thr Arg Lys Asn Ile Lys Asn Ile His Thr Leu
340 345 350
Leu Gln Asn Leu Ala Lys Ala Ser Pro Val Tyr Leu Asp Ile Leu Gly
355 360 365
<210> 59
<211> 31
<212> PRT
<213> Homo sapiens
<400> 59
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala
20 25 30
<210> 60
<211> 399
<212> PRT
<213> Homo sapiens
<400> 60
Cys Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp
1 5 10 15
Pro Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val
20 25 30
Asp Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu
35 40 45
Glu Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn
50 55 60
Leu Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe
65 70 75 80
Leu Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu
85 90 95
Thr Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu
100 105 110
Ile Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile
115 120 125
Lys Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr
130 135 140
Cys Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile
145 150 155 160
Pro Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr
165 170 175
Val Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp
180 185 190
Pro Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His
195 200 205
Met Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile
210 215 220
Ser Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu
225 230 235 240
Val Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro
245 250 255
Thr Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile
260 265 270
Pro Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr
275 280 285
Asn Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His
290 295 300
Val Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro
305 310 315 320
Thr His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr
325 330 335
Gly Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly
340 345 350
Ile Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp
355 360 365
Tyr Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn
370 375 380
Glu Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His
385 390 395
<210> 61
<211> 24
<212> PRT
<213> Homo sapiens
<400> 61
Leu Ser Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys
1 5 10 15
Leu Leu Val Met Leu Phe Leu Leu
20
<210> 62
<211> 384
<212> PRT
<213> Homo sapiens
<400> 62
Lys Leu Ala Arg His Ser Lys Phe Gly Met Lys Asp Phe Ser Trp Phe
1 5 10 15
Gly Phe Gly Lys Val Lys Ser Arg Gln Gly Val Gly Pro Ala Ser Val
20 25 30
Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro Leu His His Ile Ser Asn
35 40 45
Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly Gly Pro Asp Ala Val Ile
50 55 60
Ile Gly Met Thr Lys Ile Pro Val Ile Glu Asn Pro Gln Tyr Phe Gly
65 70 75 80
Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr Phe Val Gln His Ile Lys
85 90 95
Arg His Asn Ile Val Leu Lys Arg Glu Leu Gly Glu Gly Ala Phe Gly
100 105 110
Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu Cys Pro Glu Gln Asp Lys
115 120 125
Ile Leu Val Ala Val Lys Thr Leu Lys Asp Ala Ser Asp Asn Ala Arg
130 135 140
Lys Asp Phe His Arg Glu Ala Glu Leu Leu Thr Asn Leu Gln His Glu
145 150 155 160
His Ile Val Lys Phe Tyr Gly Val Cys Val Glu Gly Asp Pro Leu Ile
165 170 175
Met Val Phe Glu Tyr Met Lys His Gly Asp Leu Asn Lys Phe Leu Arg
180 185 190
Ala His Gly Pro Asp Ala Val Leu Met Ala Glu Gly Asn Pro Pro Thr
195 200 205
Glu Leu Thr Gln Ser Gln Met Leu His Ile Ala Gln Gln Ile Ala Ala
210 215 220
Gly Met Val Tyr Leu Ala Ser Gln His Phe Val His Arg Asp Leu Ala
225 230 235 240
Thr Arg Asn Cys Leu Val Gly Glu Asn Leu Leu Val Lys Ile Gly Asp
245 250 255
Phe Gly Met Ser Arg Asp Val Tyr Ser Thr Asp Tyr Tyr Arg Val Gly
260 265 270
Gly His Thr Met Leu Pro Ile Arg Trp Met Pro Pro Glu Ser Ile Met
275 280 285
Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val Trp Ser Leu Gly Val Val
290 295 300
Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln Pro Trp Tyr Gln Leu Ser
305 310 315 320
Asn Asn Glu Val Ile Glu Cys Ile Thr Gln Gly Arg Val Leu Gln Arg
325 330 335
Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu Leu Met Leu Gly Cys Trp
340 345 350
Gln Arg Glu Pro His Met Arg Lys Asn Ile Lys Gly Ile His Thr Leu
355 360 365
Leu Gln Asn Leu Ala Lys Ala Ser Pro Val Tyr Leu Asp Ile Leu Gly
370 375 380
<210> 63
<211> 93
<212> DNA
<213> Mus musculus
<400> 63
atgtcgccct ggctgaagtg gcatggaccc gccatggcgc ggctctgggg cttatgcctg 60
ctggtcttgg gcttctggag ggcctctctc gcc 93
<210> 64
<211> 1194
<212> DNA
<213> Mus musculus
<400> 64
tgcccgacgt cctgcaaatg cagttccgct aggatttggt gtactgagcc ttctccaggc 60
atcgtggcat tcccgaggtt ggaacctaac agcgttgacc cggagaacat cacggaaatt 120
ctcattgcaa accagaaaag gctagaaatc atcaatgaag atgacgttga agcttacgtg 180
gggctgagaa accttacaat tgtggattcc ggcttaaagt ttgtggctta caaagcgttt 240
ctgaaaaaca gcaacctgcg gcacataaat ttcacacgaa acaagctgac gagtttgtcc 300
aggagacatt tccgccacct tgacttgtct gacctgatcc tgacgggtaa tccgttcacg 360
tgctcctgcg acatcatgtg gctcaagact ctccaggaga ctaaatccag ccccgacact 420
caggatttgt actgcctcaa tgagagcagc aagaacatgc ccctggcgaa cctgcagata 480
cccaattgtg gtctgccatc tgcacgtctg gctgctccta acctcaccgt ggaggaagga 540
aagtctgtga ccctttcctg cagtgtgggg ggtgacccac tccccacctt gtactgggac 600
gttgggaatt tggtttccaa gcacatgaat gaaacaagcc acacacaggg ctccttaagg 660
ataacgaaca tttcatctga tgacagtgga aagcaaatct cttgtgtggc agaaaacctt 720
gtaggagaag atcaagattc tgtgaacctc actgtgcatt ttgcgccaac tatcacgttt 780
ctcgagtctc caacctcaga tcaccactgg tgcattccat tcactgtgag aggcaacccc 840
aagcctgcgc ttcagtggtt ctacaatggg gccatactga atgagtccaa gtacatctgt 900
actaagatcc acgtcaccaa tcacacggag taccatggct gcctccagct ggataacccc 960
actcatatga ataacggaga ctacaccctg atggccaaga acgagtatgg gaaggatgag 1020
agacagatct ccgctcactt catgggccgg cctggagtcg actacgagac aaacccaaat 1080
taccctgaag tcctctatga agactggacc acgccaactg acattgggga tactacgaac 1140
aaaagtaatg aaatcccctc cacggatgtt gctgaccaaa gcaatcggga gcat 1194
<210> 65
<211> 72
<212> DNA
<213> Mus musculus
<400> 65
ctctcggtct atgccgtggt ggtgattgca tctgtggtgg gattctgcct gctggtgatg 60
ttgctcctgc tc 72
<210> 66
<211> 1104
<212> DNA
<213> Mus musculus
<400> 66
aagttggcga gacattccaa gtttggcatg aaaggcccag cttcggtcat cagcaacgac 60
gatgactctg ccagccccct ccaccacatc tccaatggga gtaacactcc atcttcttcg 120
gagggcggtc ccgacgctgt cattattgga atgaccaaga ttcctgttat tgaaaacccc 180
cagtactttg gcatcaccaa cagtcagctc aagccagaca catttgttca gcatatcaag 240
agacacaaca tcgttctgaa gagggaactt ggggaaggag ccttcgggaa agttttcctt 300
gccgagtgct acaacctctg cccagagcag gataagatcc tggtggctgt gaagacgctg 360
aaggacgcca gcgacaatgc acgcaaggac tttcatcggg aagctgagct gctgaccaac 420
ctccagcacg agcacattgt caagttctac ggtgtctgtg tggagggcga cccactcatc 480
atggtctttg agtacatgaa gcacggggac ctcaacaagt tccttagggc acacgggccc 540
gacgcagtgc tgatggcaga gggtaacccg cccacagagc tgacgcagtc gcagatgctg 600
cacatcgctc agcaaatcgc agcaggtatg gtctacctgg cgtcccaaca ctttgtgcac 660
cgtgacctgg ccacccggaa ctgcctggtg ggagagaacc tgctggtgaa aattggggac 720
tttgggatgt cccgagatgt gtacagcacc gactactatc gggtcggtgg ccacacaatg 780
ttgcccatcc gatggatgcc tccagagagc atcatgtaca ggaaattcac caccgagagc 840
gacgtctgga gcctgggcgt tgtgttgtgg gagatcttca cctacggcaa gcagccctgg 900
tatcagctat cgaacaatga ggtgatagag tgcatcaccc agggaagagt ccttcagcgg 960
cctcgaacgt gtccccagga ggtgtatgag ctcatgcttg gatgctggca gcgggaacca 1020
cacacccgga agaacatcaa gagcatccac accctccttc agaacttggc caaggcatct 1080
cccgtctacc tggatatcct aggc 1104
<210> 67
<211> 93
<212> DNA
<213> Rattus norvegicus
<400> 67
atgtcgccct ggccgaggtg gcatggaccc gccatggcgc ggctctgggg cttatgcttg 60
ctggtcttgg gcttctggag ggcttctctt gcc 93
<210> 68
<211> 1194
<212> DNA
<213> Rattus norvegicus
<400> 68
tgccccatgt cctgcaaatg cagcaccact aggatttggt gtaccgagcc ttctcctggc 60
atcgtggcat ttccgaggtt ggaacctaac agcattgacc cagagaacat caccgaaatt 120
ctcattgcaa accagaaaag gttagaaatc atcaatgaag atgatgtcga agcttacgtg 180
gggctgaaaa accttacaat tgtggattcc ggcttaaagt ttgtggctta caaggcgttt 240
ctgaagaacg gcaacctgcg gcacatcaat ttcactcgaa acaagctgac gagtttgtcc 300
aggagacatt tccgccacct tgacttgtct gacctgatcc tgacgggtaa tccgttcacg 360
tgttcctgtg acatcatgtg gctcaagact ctccaggaga cgaaatccag ccccgacact 420
caggatttgt attgcctcaa tgagagcagc aagaataccc ctctggcgaa cctgcagatt 480
cccaattgtg gtctgccgtc tgcacgtctg gccgctccta acctcacggt ggaggaaggg 540
aagtctgtga ccatttcctg cagcgtcggg ggtgacccgc tccccacctt gtactgggac 600
gttgggaatt tggtttccaa acacatgaat gaaacaagcc acacacaggg ctccttaagg 660
ataacaaaca tttcatcgga tgacagtggg aaacaaatct cttgtgtggc agaaaacctc 720
gttggagaag atcaagactc tgtgaacctc actgtgcatt ttgcaccaac catcacattt 780
ctcgaatctc caacctcaga ccaccactgg tgcatcccat tcactgtgag aggcaacccc 840
aagccagcac ttcagtggtt ctacaacgga gccatactga atgaatccaa gtacatctgt 900
accaaaatac acgtcaccaa tcacacggag taccacggct gcctccagct ggataacccc 960
actcatatga ataatggaga ctacacccta atggccaaga atgaatatgg gaaggacgag 1020
agacagattt ctgctcactt catgggccgg cctggagttg actatgagac aaacccaaat 1080
taccctgaag tcctctatga agactggacc acgccaactg acatcgggga tactacaaac 1140
aaaagtaatg agatcccctc cacggatgtt gctgaccaaa ccaatcggga gcat 1194
<210> 69
<211> 72
<212> DNA
<213> Rattus norvegicus
<400> 69
ctctcggtct atgccgtggt ggtgattgcc tctgtggtag gattctgcct gctggtgatg 60
ctgcttctgc tc 72
<210> 70
<211> 1104
<212> DNA
<213> Rattus norvegicus
<400> 70
aagttggcga gacattccaa gtttggcatg aaaggcccag cttccgtcat cagcaacgac 60
gatgactctg ccagccctct ccaccacatc tccaacggga gcaacactcc gtcttcttcg 120
gagggcgggc ccgatgctgt catcattggg atgaccaaga tccctgtcat tgaaaacccc 180
cagtacttcg gtatcaccaa cagccagctc aagccggaca catttgttca gcacatcaag 240
agacacaaca tcgttctgaa gagggagctt ggagaaggag cctttgggaa agttttccta 300
gcggagtgct ataacctctg ccccgagcag gataagatcc tggtggccgt gaagacgctg 360
aaggacgcca gcgacaatgc tcgcaaggac tttcatcgcg aagccgagct gctgaccaac 420
ctccagcacg agcacattgt caagttctac ggtgtctgtg tggagggcga cccactcatc 480
atggtctttg agtacatgaa gcacggggac ctcaacaagt tccttagggc acacgggcca 540
gatgcagtgc tgatggcaga gggtaacccg cccaccgagc tgacgcagtc gcagatgctg 600
cacatcgctc agcaaatcgc agcaggcatg gtctacctgg catcccaaca cttcgtgcac 660
cgagacctgg ccacccggaa ctgcttggta ggagagaacc tgctggtgaa aattggggac 720
ttcgggatgt cccgggatgt atacagcacc gactactacc gggttggtgg ccacacaatg 780
ttgcccatcc gatggatgcc tccagagagc atcatgtaca ggaaattcac caccgagagt 840
gacgtctgga gcctgggagt tgtgttgtgg gagatcttca cctacggcaa gcagccctgg 900
tatcagctat caaacaacga ggtgatagaa tgcatcaccc agggcagagt ccttcagcgg 960
cctcgcacgt gtccccagga ggtgtacgag ctgatgctgg gatgctggca gcgggaacca 1020
cacacaagga agaacatcaa gaacatccac acactccttc agaacttggc gaaggcgtcg 1080
cccgtctacc tggacatcct aggc 1104
<210> 71
<211> 93
<212> DNA
<213> Homo sapiens
<400> 71
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcc 93
<210> 72
<211> 1197
<212> DNA
<213> Homo sapiens
<400> 72
tgtcccacgt cctgcaaatg cagtgcctct cggatctggt gcagcgaccc ttctcctggc 60
atcgtggcat ttccgagatt ggagcctaac agtgtagatc ctgagaacat caccgaaatt 120
ttcatcgcaa accagaaaag gttagaaatc atcaacgaag atgatgttga agcttatgtg 180
ggactgagaa atctgacaat tgtggattct ggattaaaat ttgtggctca taaagcattt 240
ctgaaaaaca gcaacctgca gcacatcaat tttacccgaa acaaactgac gagtttgtct 300
aggaaacatt tccgtcacct tgacttgtct gaactgatcc tggtgggcaa tccatttaca 360
tgctcctgtg acattatgtg gatcaagact ctccaagagg ctaaatccag tccagacact 420
caggatttgt actgcctgaa tgaaagcagc aagaatattc ccctggcaaa cctgcagata 480
cccaattgtg gtttgccatc tgcaaatctg gccgcaccta acctcactgt ggaggaagga 540
aagtctatca cattatcctg tagtgtggca ggtgatccgg ttcctaatat gtattgggat 600
gttggtaacc tggtttccaa acatatgaat gaaacaagcc acacacaggg ctccttaagg 660
ataactaaca tttcatccga tgacagtggg aagcagatct cttgtgtggc ggaaaatctt 720
gtaggagaag atcaagattc tgtcaacctc actgtgcatt ttgcaccaac tatcacattt 780
ctcgaatctc caacctcaga ccaccactgg tgcattccat tcactgtgaa aggcaacccc 840
aaaccagcgc ttcagtggtt ctataacggg gcaatattga atgagtccaa atacatctgt 900
actaaaatac atgttaccaa tcacacggag taccacggct gcctccagct ggataatccc 960
actcacatga acaatgggga ctacactcta atagccaaga atgagtatgg gaaggatgag 1020
aaacagattt ctgctcactt catgggctgg cctggaattg acgatggtgc aaacccaaat 1080
tatcctgatg taatttatga agattatgga actgcagcga atgacatcgg ggacaccacg 1140
aacagaagta atgaaatccc ttccacagac gtcactgata aaaccggtcg ggaacat 1197
<210> 73
<211> 72
<212> DNA
<213> Homo sapiens
<400> 73
ctctcggtct atgctgtggt ggtgattgcg tctgtggtgg gattttgcct tttggtaatg 60
ctgtttctgc tt 72
<210> 74
<211> 1152
<212> DNA
<213> Homo sapiens
<400> 74
aagttggcaa gacactccaa gtttggcatg aaagatttct catggtttgg atttgggaaa 60
gtaaaatcaa gacaaggtgt tggcccagcc tccgttatca gcaatgatga tgactctgcc 120
agcccactcc atcacatctc caatgggagt aacactccat cttcttcgga aggtggccca 180
gatgctgtca ttattggaat gaccaagatc cctgtcattg aaaatcccca gtactttggc 240
atcaccaaca gtcagctcaa gccagacaca tttgttcagc acatcaagcg acataacatt 300
gttctgaaaa gggagctagg cgaaggagcc tttggaaaag tgttcctagc tgaatgctat 360
aacctctgtc ctgagcagga caagatcttg gtggcagtga agaccctgaa ggatgccagt 420
gacaatgcac gcaaggactt ccaccgtgag gccgagctcc tgaccaacct ccagcatgag 480
cacatcgtca agttctatgg cgtctgcgtg gagggcgacc ccctcatcat ggtctttgag 540
tacatgaagc atggggacct caacaagttc ctcagggcac acggccctga tgccgtgctg 600
atggctgagg gcaacccgcc cacggaactg acgcagtcgc agatgctgca tatagcccag 660
cagatcgccg cgggcatggt ctacctggcg tcccagcact tcgtgcaccg cgatttggcc 720
accaggaact gcctggtcgg ggagaacttg ctggtgaaaa tcggggactt tgggatgtcc 780
cgggacgtgt acagcactga ctactacagg gtcggtggcc acacaatgct gcccattcgc 840
tggatgcctc cagagagcat catgtacagg aaattcacga cggaaagcga cgtctggagc 900
ctgggggtcg tgttgtggga gattttcacc tatggcaaac agccctggta ccagctgtca 960
aacaatgagg tgatagagtg tatcactcag ggccgagtcc tgcagcgacc ccgcacgtgc 1020
ccccaggagg tgtatgagct gatgctgggg tgctggcagc gagagcccca catgaggaag 1080
aacatcaagg gcatccatac cctccttcag aacttggcca aggcatctcc ggtctacctg 1140
gacattctag gc 1152
<210> 75
<211> 822
<212> PRT
<213> Homo sapiens
<220>
<221> MISC_FEATURE
<222> (1)..(31)
<223> Сигнальный пептид
<220>
<221> MISC_FEATURE
<222> (32)..(430)
<223> Внеклеточный домен
<220>
<221> MISC_FEATURE
<222> (431)..(454)
<223> Трансмембранный домен
<220>
<221> MISC_FEATURE
<222> (455)..(822)
<223> Цитоплазматический домен
<400> 75
Met Ser Ser Trp Ile Arg Trp His Gly Pro Ala Met Ala Arg Leu Trp
1 5 10 15
Gly Phe Cys Trp Leu Val Val Gly Phe Trp Arg Ala Ala Phe Ala Cys
20 25 30
Pro Thr Ser Cys Lys Cys Ser Ala Ser Arg Ile Trp Cys Ser Asp Pro
35 40 45
Ser Pro Gly Ile Val Ala Phe Pro Arg Leu Glu Pro Asn Ser Val Asp
50 55 60
Pro Glu Asn Ile Thr Glu Ile Phe Ile Ala Asn Gln Lys Arg Leu Glu
65 70 75 80
Ile Ile Asn Glu Asp Asp Val Glu Ala Tyr Val Gly Leu Arg Asn Leu
85 90 95
Thr Ile Val Asp Ser Gly Leu Lys Phe Val Ala His Lys Ala Phe Leu
100 105 110
Lys Asn Ser Asn Leu Gln His Ile Asn Phe Thr Arg Asn Lys Leu Thr
115 120 125
Ser Leu Ser Arg Lys His Phe Arg His Leu Asp Leu Ser Glu Leu Ile
130 135 140
Leu Val Gly Asn Pro Phe Thr Cys Ser Cys Asp Ile Met Trp Ile Lys
145 150 155 160
Thr Leu Gln Glu Ala Lys Ser Ser Pro Asp Thr Gln Asp Leu Tyr Cys
165 170 175
Leu Asn Glu Ser Ser Lys Asn Ile Pro Leu Ala Asn Leu Gln Ile Pro
180 185 190
Asn Cys Gly Leu Pro Ser Ala Asn Leu Ala Ala Pro Asn Leu Thr Val
195 200 205
Glu Glu Gly Lys Ser Ile Thr Leu Ser Cys Ser Val Ala Gly Asp Pro
210 215 220
Val Pro Asn Met Tyr Trp Asp Val Gly Asn Leu Val Ser Lys His Met
225 230 235 240
Asn Glu Thr Ser His Thr Gln Gly Ser Leu Arg Ile Thr Asn Ile Ser
245 250 255
Ser Asp Asp Ser Gly Lys Gln Ile Ser Cys Val Ala Glu Asn Leu Val
260 265 270
Gly Glu Asp Gln Asp Ser Val Asn Leu Thr Val His Phe Ala Pro Thr
275 280 285
Ile Thr Phe Leu Glu Ser Pro Thr Ser Asp His His Trp Cys Ile Pro
290 295 300
Phe Thr Val Lys Gly Asn Pro Lys Pro Ala Leu Gln Trp Phe Tyr Asn
305 310 315 320
Gly Ala Ile Leu Asn Glu Ser Lys Tyr Ile Cys Thr Lys Ile His Val
325 330 335
Thr Asn His Thr Glu Tyr His Gly Cys Leu Gln Leu Asp Asn Pro Thr
340 345 350
His Met Asn Asn Gly Asp Tyr Thr Leu Ile Ala Lys Asn Glu Tyr Gly
355 360 365
Lys Asp Glu Lys Gln Ile Ser Ala His Phe Met Gly Trp Pro Gly Ile
370 375 380
Asp Asp Gly Ala Asn Pro Asn Tyr Pro Asp Val Ile Tyr Glu Asp Tyr
385 390 395 400
Gly Thr Ala Ala Asn Asp Ile Gly Asp Thr Thr Asn Arg Ser Asn Glu
405 410 415
Ile Pro Ser Thr Asp Val Thr Asp Lys Thr Gly Arg Glu His Leu Ser
420 425 430
Val Tyr Ala Val Val Val Ile Ala Ser Val Val Gly Phe Cys Leu Leu
435 440 445
Val Met Leu Phe Leu Leu Lys Leu Ala Arg His Ser Lys Phe Gly Met
450 455 460
Lys Gly Pro Ala Ser Val Ile Ser Asn Asp Asp Asp Ser Ala Ser Pro
465 470 475 480
Leu His His Ile Ser Asn Gly Ser Asn Thr Pro Ser Ser Ser Glu Gly
485 490 495
Gly Pro Asp Ala Val Ile Ile Gly Met Thr Lys Ile Pro Val Ile Glu
500 505 510
Asn Pro Gln Tyr Phe Gly Ile Thr Asn Ser Gln Leu Lys Pro Asp Thr
515 520 525
Phe Val Gln His Ile Lys Arg His Asn Ile Val Leu Lys Arg Glu Leu
530 535 540
Gly Glu Gly Ala Phe Gly Lys Val Phe Leu Ala Glu Cys Tyr Asn Leu
545 550 555 560
Cys Pro Glu Gln Asp Lys Ile Leu Val Ala Val Lys Thr Leu Lys Asp
565 570 575
Ala Ser Asp Asn Ala Arg Lys Asp Phe His Arg Glu Ala Glu Leu Leu
580 585 590
Thr Asn Leu Gln His Glu His Ile Val Lys Phe Tyr Gly Val Cys Val
595 600 605
Glu Gly Asp Pro Leu Ile Met Val Phe Glu Tyr Met Lys His Gly Asp
610 615 620
Leu Asn Lys Phe Leu Arg Ala His Gly Pro Asp Ala Val Leu Met Ala
625 630 635 640
Glu Gly Asn Pro Pro Thr Glu Leu Thr Gln Ser Gln Met Leu His Ile
645 650 655
Ala Gln Gln Ile Ala Ala Gly Met Val Tyr Leu Ala Ser Gln His Phe
660 665 670
Val His Arg Asp Leu Ala Thr Arg Asn Cys Leu Val Gly Glu Asn Leu
675 680 685
Leu Val Lys Ile Gly Asp Phe Gly Met Ser Arg Asp Val Tyr Ser Thr
690 695 700
Asp Tyr Tyr Arg Val Gly Gly His Thr Met Leu Pro Ile Arg Trp Met
705 710 715 720
Pro Pro Glu Ser Ile Met Tyr Arg Lys Phe Thr Thr Glu Ser Asp Val
725 730 735
Trp Ser Leu Gly Val Val Leu Trp Glu Ile Phe Thr Tyr Gly Lys Gln
740 745 750
Pro Trp Tyr Gln Leu Ser Asn Asn Glu Val Ile Glu Cys Ile Thr Gln
755 760 765
Gly Arg Val Leu Gln Arg Pro Arg Thr Cys Pro Gln Glu Val Tyr Glu
770 775 780
Leu Met Leu Gly Cys Trp Gln Arg Glu Pro His Met Arg Lys Asn Ile
785 790 795 800
Lys Gly Ile His Thr Leu Leu Gln Asn Leu Ala Lys Ala Ser Pro Val
805 810 815
Tyr Leu Asp Ile Leu Gly
820
<210> 76
<211> 8498
<212> DNA
<213> Homo sapiens
<400> 76
cctcgaggtg cataccggac ccccattcgc atctaacaag gaatctgcgc cccagagagt 60
cccgggagcg ccgccggtcg gtgcccggcg cgccgggcca tgcagcgacg gccgccgcgg 120
agctccgagc agcggtagcg cccccctgta aagcggttcg ctatgccggg gccactgtga 180
accctgccgc ctgccggaac actcttcgct ccggaccagc tcagcctctg ataagctgga 240
ctcggcacgc ccgcaacaag caccgaggag ttaagagagc cgcaagcgca gggaaggcct 300
ccccgcacgg gtgggggaaa gcggccggtg cagcgcgggg acaggcactc gggctggcac 360
tggctgctag ggatgtcgtc ctggataagg tggcatggac ccgccatggc gcggctctgg 420
ggcttctgct ggctggttgt gggcttctgg agggccgctt tcgcctgtcc cacgtcctgc 480
aaatgcagtg cctctcggat ctggtgcagc gacccttctc ctggcatcgt ggcatttccg 540
agattggagc ctaacagtgt agatcctgag aacatcaccg aaattttcat cgcaaaccag 600
aaaaggttag aaatcatcaa cgaagatgat gttgaagctt atgtgggact gagaaatctg 660
acaattgtgg attctggatt aaaatttgtg gctcataaag catttctgaa aaacagcaac 720
ctgcagcaca tcaattttac ccgaaacaaa ctgacgagtt tgtctaggaa acatttccgt 780
caccttgact tgtctgaact gatcctggtg ggcaatccat ttacatgctc ctgtgacatt 840
atgtggatca agactctcca agaggctaaa tccagtccag acactcagga tttgtactgc 900
ctgaatgaaa gcagcaagaa tattcccctg gcaaacctgc agatacccaa ttgtggtttg 960
ccatctgcaa atctggccgc acctaacctc actgtggagg aaggaaagtc tatcacatta 1020
tcctgtagtg tggcaggtga tccggttcct aatatgtatt gggatgttgg taacctggtt 1080
tccaaacata tgaatgaaac aagccacaca cagggctcct taaggataac taacatttca 1140
tccgatgaca gtgggaagca gatctcttgt gtggcggaaa atcttgtagg agaagatcaa 1200
gattctgtca acctcactgt gcattttgca ccaactatca catttctcga atctccaacc 1260
tcagaccacc actggtgcat tccattcact gtgaaaggca accccaaacc agcgcttcag 1320
tggttctata acggggcaat attgaatgag tccaaataca tctgtactaa aatacatgtt 1380
accaatcaca cggagtacca cggctgcctc cagctggata atcccactca catgaacaat 1440
ggggactaca ctctaatagc caagaatgag tatgggaagg atgagaaaca gatttctgct 1500
cacttcatgg gctggcctgg aattgacgat ggtgcaaacc caaattatcc tgatgtaatt 1560
tatgaagatt atggaactgc agcgaatgac atcggggaca ccacgaacag aagtaatgaa 1620
atcccttcca cagacgtcac tgataaaacc ggtcgggaac atctctcggt ctatgctgtg 1680
gtggtgattg cgtctgtggt gggattttgc cttttggtaa tgctgtttct gcttaagttg 1740
gcaagacact ccaagtttgg catgaaaggc ccagcctccg ttatcagcaa tgatgatgac 1800
tctgccagcc cactccatca catctccaat gggagtaaca ctccatcttc ttcggaaggt 1860
ggcccagatg ctgtcattat tggaatgacc aagatccctg tcattgaaaa tccccagtac 1920
tttggcatca ccaacagtca gctcaagcca gacacatttg ttcagcacat caagcgacat 1980
aacattgttc tgaaaaggga gctaggcgaa ggagcctttg gaaaagtgtt cctagctgaa 2040
tgctataacc tctgtcctga gcaggacaag atcttggtgg cagtgaagac cctgaaggat 2100
gccagtgaca atgcacgcaa ggacttccac cgtgaggccg agctcctgac caacctccag 2160
catgagcaca tcgtcaagtt ctatggcgtc tgcgtggagg gcgaccccct catcatggtc 2220
tttgagtaca tgaagcatgg ggacctcaac aagttcctca gggcacacgg ccctgatgcc 2280
gtgctgatgg ctgagggcaa cccgcccacg gaactgacgc agtcgcagat gctgcatata 2340
gcccagcaga tcgccgcggg catggtctac ctggcgtccc agcacttcgt gcaccgcgat 2400
ttggccacca ggaactgcct ggtcggggag aacttgctgg tgaaaatcgg ggactttggg 2460
atgtcccggg acgtgtacag cactgactac tacagggtcg gtggccacac aatgctgccc 2520
attcgctgga tgcctccaga gagcatcatg tacaggaaat tcacgacgga aagcgacgtc 2580
tggagcctgg gggtcgtgtt gtgggagatt ttcacctatg gcaaacagcc ctggtaccag 2640
ctgtcaaaca atgaggtgat agagtgtatc actcagggcc gagtcctgca gcgaccccgc 2700
acgtgccccc aggaggtgta tgagctgatg ctggggtgct ggcagcgaga gccccacatg 2760
aggaagaaca tcaagggcat ccataccctc cttcagaact tggccaaggc atctccggtc 2820
tacctggaca ttctaggcta gggccctttt ccccagaccg atccttccca acgtactcct 2880
cagacgggct gagaggatga acatctttta actgccgctg gaggccacca agctgctctc 2940
cttcactctg acagtattaa catcaaagac tccgagaagc tctcgaggga agcagtgtgt 3000
acttcttcat ccatagacac agtattgact tctttttggc attatctctt tctctctttc 3060
catctccctt ggttgttcct ttttcttttt ttaaattttc tttttctttt ttttttcgtc 3120
ttccctgctt cacgattctt accctttctt ttgaatcaat ctggcttctg cattactatt 3180
aactctgcat agacaaaggc cttaacaaac gtaatttgtt atatcagcag acactccagt 3240
ttgcccacca caactaacaa tgccttgttg tattcctgcc tttgatgtgg atgaaaaaaa 3300
gggaaaacaa atatttcact taaactttgt cacttctgct gtacagatat cgagagtttc 3360
tatggattca cttctattta tttattatta ttactgttct tattgttttt ggatggctta 3420
agcctgtgta taaaaaagaa aacttgtgtt caatctgtga agcctttatc tatgggagat 3480
taaaaccaga gagaaagaag atttattatg aaccgcaata tgggaggaac aaagacaacc 3540
actgggatca gctggtgtca gtccctactt aggaaatact cagcaactgt tagctgggaa 3600
gaatgtattc ggcaccttcc cctgaggacc tttctgagga gtaaaaagac tactggcctc 3660
tgtgccatgg atgattcttt tcccatcacc agaaatgata gcgtgcagta gagagcaaag 3720
atggcttccg tgagacacaa gatggcgcat agtgtgctcg gacacagttt tgtcttcgta 3780
ggttgtgatg atagcactgg tttgtttctc aagcgctatc cacagaacct ttgtcaactt 3840
cagttgaaaa gaggtggatt catgtccaga gctcatttcg gggtcaggtg ggaaagccaa 3900
gaacttggaa aagataagac aagctataaa ttcggaggca agtttctttt acaatgaact 3960
tttcagatct cacttccctc cgacccctaa cttccatgcc cacccgtcct tttaactgtg 4020
caagcaaaat tgtgcatggt cttcgtcgat taataccttg tgtgcagaca ctactgctcc 4080
agacgtcgtt tccctgatag gtagagcaga tccataaaaa ggtatgactt atacaattag 4140
gggaagctaa tggagtttat tagctgagta tcaatgtctc tgcgttgtac ggtggtgatg 4200
ggttttaatg aatatggacc ctgaagcctg gaaatcctca tccacgtcga acccacagga 4260
ctgtgggaag ggcagaatca atccctaagg gaaaggaaac ctcaccctga gggcatcaca 4320
tgcactcatg ttcagtgtac acaggtcaag tcccttgctc tgggctctag ttgggagagt 4380
ggtttcattc caagtgtact ccattgtcag tatgctgttt ttgtttcctt cactccattc 4440
aaaaagtcaa aatacaaaat ttggcacagc atgccaacgg gaggctgtgc ccagaccaag 4500
cactggaagt gtgcttctag gcatagtcat tggttttgca aaaagagggc tcaaatttaa 4560
atagaaattt acagctattt gaatggtcag atataccaag aaagaaaaat atttctgttc 4620
ctcaagaaaa cttgctaccc tctgtgaggg gaattttgct aaacttgaca tctttataac 4680
atgagccaga ttgaaaggga gtgattttca ttcatcttag gtcatgttat ttcatatttg 4740
tttctgaagg tgcgatagct ctgttttagg ttttgcttgc gcctgttaat tactggaaca 4800
ccttattttt cattaaaggc tttgaaagcc aattctcaaa aattcaaaag tgcaaattaa 4860
cagaacaaaa ggaaatccag tagcaactgc agtcaagcga gggagttgac aagataaacc 4920
ttacgtccat tcaagttata tgctggccta tgagagatga gagttgggtc gtttgttctc 4980
tttgttgatg attttaaaaa aaccctctag aatacacata ataacataat gaaagccata 5040
tctccatgat atatatgtgc acatatatat acatacatgt gcatgtatgt atcatattaa 5100
ggacccatgg tactcttaaa acactgtaga actctgtgac gcagtaagga aggggcagat 5160
ttgtacaaaa acttttctag attccatcag caaaaaccaa cacaggtttg tcacgctgca 5220
tgtctggcca gctaatctcg ggggaaaagc tacaagttat ttattttatt ttaagagaat 5280
aaagtaggta ataatttaag ggatcaaatt caaggaggaa tgtgcaattt tagagcaaag 5340
atttgtttaa ggcaaatgag actttgggag catcccattc cagttttgtc ttttttttct 5400
ctgaaagaaa aaagcaaaaa ataaaataaa attccactta taccttctga caagtcccta 5460
aaggtcttga aataaaaggt tctatgcaag tgcaaagttt tatagttatt tttattgctg 5520
attattacta ttactatctc tgttgtctta agagtatgtg ctgatttcag agacatctca 5580
aattgaaaga atatcagatt gcttttaaag tagctgaacg agccacagaa tatctgaaat 5640
tattcattgt tgttcctcca ccaccccctt tctcatggtc tgatttttag aagagtggca 5700
tcctcgttct aaaatgtaat gatcaccaaa tacggccttc catcaaattt gtgaaaacta 5760
caacagtata acagtgacaa acctaattct ctagcccaaa cctggtctga caatcatttc 5820
catttagaag tcattgaata gttttccaaa cactttccat gtgtgttagc aaattattcc 5880
tattttgtgt agatgaggac gttgagactc agagacattc agaggcacgc tagaggtctc 5940
cagcctagct tccagcacca ttgggactga atccaagtac tctcactctg aacttcgtgg 6000
ttctgtccac tagagactct aatatgcaaa caagcagttc aggaaagaaa gcatgctaac 6060
acattcatga agcagtatat gaagttagaa gaacaaaagg aaatacagga gatgacaagc 6120
aactgagata ttgtgatata taatcatgct cttagcttca gctaaattca gctaaattct 6180
tgtacactga accaatgtca taatcaggct tatttagaaa acactttgaa ctatgctata 6240
aaagattata tcagaattca tattatacat gtgttcacat cagcgctacc tgtgatgttt 6300
tcatgtattt atgtatgtgt tataaatact tgatttatac atatacaaat gcacatacgt 6360
agtgtgtttg tgtgtttatg tataaattta taggcacaca ataatagagg taattataag 6420
taggatgcgg tatgaataat ttgcttaaaa tatgctaaat aaccaaaact gtttaacgtc 6480
atgttgctgt tagtgcttcc atactccacg tgggtaggac tacatcacac ttttcaactc 6540
tgtgcagtac tgcatgggtg gaagacatat ttaagataat gtgcttccca aaacaactga 6600
ataaaagcca tcccactaca ttgagtgctt tctctggctc cttgcaaaga aagatacttt 6660
ttgtaatggt ccaggaaagg aacattgctt tctttttgtc tttcagcaca tttgtattat 6720
gctcaccttg tctctgtctc actgtgaccc ctttacactt gagttcagag ttcaagcatt 6780
ccaaatataa attggaatgt tggcagccca gtggcttgaa ggccaatgat gagcagtcca 6840
agaccccaca gcgagatgag caactcttag gaattcccac atcctagagt gaatgcacca 6900
actaacagta tagaatgctg tccttttcaa agcgtcctaa cagcaggatt acctggtcaa 6960
gtatggactt tctttgaatc tttcttttca caaattggac tgcctgtaat accaataaca 7020
ttgttgtatc taactaaata aatgactgca tatacacaca taccctcaat tctcttgctt 7080
ccccattttc tttttcatcc cctgtctcag gacttttatt ttcaatgttg acctttggtt 7140
tggccatata tcactgttat aggaaatctc atgagaggaa tggctagtga cccaactctc 7200
caaatgtcta agttagtagt tacagctgat tttttatgat gcataattgg aatgtggagc 7260
ctctgaggtt gtgatagctt gtacatgaat ttcaaatgtc attctaaaga atgaggggtg 7320
ggagggattt atagttagaa acgacagtgc aggaaggggt attttcttgt tgtcagggct 7380
ggaatgaatc actgctgctc aagtcaaagg ttcttgaata tccttagttt ttgcatttcc 7440
cctccttttc ctttgacctt tatttattta attatgtatt tatttattta tttatatact 7500
tttgctccat tcagcacaaa cacaaagcaa agcaaaaaaa aaaatatata tatatatatc 7560
tgtatatgtg ttgtaggcaa aacactgtga atttcacaac aaccaccacc aagcaactat 7620
tttgccatct taacatacat ctcaggagac gaaatgagaa aagatgggga tgtcattttt 7680
tagtctatgc gtttgaggcc aggtccatgt ttatttattt ctttagtcta tgcattaatg 7740
aaaatgatcc tgagtggagg ttagctgaac gttcaatgta ctggagcaag catcataaaa 7800
gctgctagta gccatgtgtt tgaacaggaa aaatattaca gaaaatgaaa tgtaaaggcc 7860
tatatcttgc agcttgtata tcttactatt gcttaaaaaa tgtataaagc agctggaaat 7920
gttttaaata caaggtcttt gaattaaatg tggattttaa atatgtaatc ccttgacaaa 7980
tgaccaaatt atggtgaact attgctccct gcgttctttg atcattacct atgacttaca 8040
aatctgcctg gagatgtgga cattctgcat ttgcttctgt atctggagag atgtttgtat 8100
atatccaggc cgtatacaca cacatttcca tatctctcta cagatatatt tccccttcaa 8160
tcgtgacctg gtatttggaa ctctcctttt catttggctt atcttccttt taatgtgatg 8220
tctctgtgct aatacttacc agttcttgtt ttgcaatctg ttttgaggtc cattgcttta 8280
ctaagaccca ctgcatcttg gctgatttca aagtgacacc tgaatacagt gtttaaaaaa 8340
aaaaaagttt tgtttgtaaa tcatgtgacc agcttctctc aacctgacat ggaaagtctc 8400
ttgtactaca gtgtatttaa taaaaatgat gtcttacaat aaataacata ctccaaaaga 8460
gagactaaaa atgaaaaaaa aaaaaaaaaa aaaaaaaa 8498
<210> 77
<211> 2469
<212> DNA
<213> Homo sapiens
<220>
<221> misc_feature
<222> (1)..(93)
<223> Сигнальный пептид
<220>
<221> misc_feature
<222> (94)..(1290)
<223> Внеклеточный домен
<220>
<221> misc_feature
<222> (1291)..(1362)
<223> Трансмембранный домен
<220>
<221> misc_feature
<222> (1363)..(2466)
<223> Цитоплазматический домен
<400> 77
atgtcgtcct ggataaggtg gcatggaccc gccatggcgc ggctctgggg cttctgctgg 60
ctggttgtgg gcttctggag ggccgctttc gcctgtccca cgtcctgcaa atgcagtgcc 120
tctcggatct ggtgcagcga cccttctcct ggcatcgtgg catttccgag attggagcct 180
aacagtgtag atcctgagaa catcaccgaa attttcatcg caaaccagaa aaggttagaa 240
atcatcaacg aagatgatgt tgaagcttat gtgggactga gaaatctgac aattgtggat 300
tctggattaa aatttgtggc tcataaagca tttctgaaaa acagcaacct gcagcacatc 360
aattttaccc gaaacaaact gacgagtttg tctaggaaac atttccgtca ccttgacttg 420
tctgaactga tcctggtggg caatccattt acatgctcct gtgacattat gtggatcaag 480
actctccaag aggctaaatc cagtccagac actcaggatt tgtactgcct gaatgaaagc 540
agcaagaata ttcccctggc aaacctgcag atacccaatt gtggtttgcc atctgcaaat 600
ctggccgcac ctaacctcac tgtggaggaa ggaaagtcta tcacattatc ctgtagtgtg 660
gcaggtgatc cggttcctaa tatgtattgg gatgttggta acctggtttc caaacatatg 720
aatgaaacaa gccacacaca gggctcctta aggataacta acatttcatc cgatgacagt 780
gggaagcaga tctcttgtgt ggcggaaaat cttgtaggag aagatcaaga ttctgtcaac 840
ctcactgtgc attttgcacc aactatcaca tttctcgaat ctccaacctc agaccaccac 900
tggtgcattc cattcactgt gaaaggcaac cccaaaccag cgcttcagtg gttctataac 960
ggggcaatat tgaatgagtc caaatacatc tgtactaaaa tacatgttac caatcacacg 1020
gagtaccacg gctgcctcca gctggataat cccactcaca tgaacaatgg ggactacact 1080
ctaatagcca agaatgagta tgggaaggat gagaaacaga tttctgctca cttcatgggc 1140
tggcctggaa ttgacgatgg tgcaaaccca aattatcctg atgtaattta tgaagattat 1200
ggaactgcag cgaatgacat cggggacacc acgaacagaa gtaatgaaat cccttccaca 1260
gacgtcactg ataaaaccgg tcgggaacat ctctcggtct atgctgtggt ggtgattgcg 1320
tctgtggtgg gattttgcct tttggtaatg ctgtttctgc ttaagttggc aagacactcc 1380
aagtttggca tgaaaggccc agcctccgtt atcagcaatg atgatgactc tgccagccca 1440
ctccatcaca tctccaatgg gagtaacact ccatcttctt cggaaggtgg cccagatgct 1500
gtcattattg gaatgaccaa gatccctgtc attgaaaatc cccagtactt tggcatcacc 1560
aacagtcagc tcaagccaga cacatttgtt cagcacatca agcgacataa cattgttctg 1620
aaaagggagc taggcgaagg agcctttgga aaagtgttcc tagctgaatg ctataacctc 1680
tgtcctgagc aggacaagat cttggtggca gtgaagaccc tgaaggatgc cagtgacaat 1740
gcacgcaagg acttccaccg tgaggccgag ctcctgacca acctccagca tgagcacatc 1800
gtcaagttct atggcgtctg cgtggagggc gaccccctca tcatggtctt tgagtacatg 1860
aagcatgggg acctcaacaa gttcctcagg gcacacggcc ctgatgccgt gctgatggct 1920
gagggcaacc cgcccacgga actgacgcag tcgcagatgc tgcatatagc ccagcagatc 1980
gccgcgggca tggtctacct ggcgtcccag cacttcgtgc accgcgattt ggccaccagg 2040
aactgcctgg tcggggagaa cttgctggtg aaaatcgggg actttgggat gtcccgggac 2100
gtgtacagca ctgactacta cagggtcggt ggccacacaa tgctgcccat tcgctggatg 2160
cctccagaga gcatcatgta caggaaattc acgacggaaa gcgacgtctg gagcctgggg 2220
gtcgtgttgt gggagatttt cacctatggc aaacagccct ggtaccagct gtcaaacaat 2280
gaggtgatag agtgtatcac tcagggccga gtcctgcagc gaccccgcac gtgcccccag 2340
gaggtgtatg agctgatgct ggggtgctgg cagcgagagc cccacatgag gaagaacatc 2400
aagggcatcc ataccctcct tcagaacttg gccaaggcat ctccggtcta cctggacatt 2460
ctaggctag 2469
<210> 78
<211> 354
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 78
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt cagcttcagt agctttggca tgcactgggt ccgccaggct 120
ccaggcaagg gactggagtg ggtgtcagtt atatcatatg atggaattaa tacatactat 180
acagactccg tgaagggccg attcaccatc tccagagaca attccaagaa cacgctgtat 240
ctgcaaatga acggcctgag agctgaggac acggctcttt attactgtgt gcaagggtca 300
attggaaccg tttttgaata ctggggccag ggaaccctgg tcaccgtctc ctca 354
<210> 79
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 79
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Ser Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Ser Tyr Asp Gly Ile Asn Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Gly Leu Arg Ala Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Val Gln Gly Ser Ile Gly Thr Val Phe Glu Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 80
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 80
ggattcagct tcagtagctt tggc 24
<210> 81
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 81
Gly Phe Ser Phe Ser Ser Phe Gly
1 5
<210> 82
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 82
atatcatatg atggaattaa taca 24
<210> 83
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 83
Ile Ser Tyr Asp Gly Ile Asn Thr
1 5
<210> 84
<211> 33
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 84
gtgcaagggt caattggaac cgtttttgaa tac 33
<210> 85
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 85
Val Gln Gly Ser Ile Gly Thr Val Phe Glu Tyr
1 5 10
<210> 86
<211> 321
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 86
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcgagtca gggcattagc aattatttag cctggtatca gcagaaacca 120
gggaaagttc ctaaactcct gatctatgct gcatccactt tacaatcagg ggtcccatct 180
cggttcggtg gcagtggatc tgggacagat ttcactctca ccatcagcag cctgcagcct 240
gaagatgttg caacttatta ctgtcaaaag tataccagtg ccccattcac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 87
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Gly Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Tyr Thr Ser Ala Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 88
<211> 18
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 88
cagggcatta gcaattat 18
<210> 89
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 89
Gln Gly Ile Ser Asn Tyr
1 5
<210> 90
<211> 9
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 90
gctgcatcc 9
<210> 91
<211> 3
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 91
Ala Ala Ser
1
<210> 92
<211> 27
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 92
caaaagtata ccagtgcccc attcact 27
<210> 93
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 93
Gln Lys Tyr Thr Ser Ala Pro Phe Thr
1 5
<210> 94
<211> 366
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 94
gaggtgcagc tggtggagtc tgggggaggc ttggtccagc ctggggggtc cctaaaactc 60
tcctgtacag cctctgggtt caccttcagt ggctctgtta ttcactgggt ccgccaggct 120
tccgggaaag ggctggagtg gattggccgt attagaaaca aggctaacag ttacgcgaca 180
gcatatggtg cgtcggtgac aggcaggttc accatctcca gagatgattc aaagaacacg 240
gcgtatctgc aaatgaacag cctgaaaacc gaggacacgg ccgtttatta ctgtactttc 300
ccgggtgtag tgggacgagg aggttttgac tactggggcc agggcaccct ggtcaccgtc 360
tcctca 366
<210> 95
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 95
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Thr Ala Ser Gly Phe Thr Phe Ser Gly Ser
20 25 30
Val Ile His Trp Val Arg Gln Ala Ser Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Arg Asn Lys Ala Asn Ser Tyr Ala Thr Ala Tyr Gly Ala
50 55 60
Ser Val Thr Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Ala Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Thr Phe Pro Gly Val Val Gly Arg Gly Gly Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 96
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 96
gggttcacct tcagtggctc tgtt 24
<210> 97
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 97
Gly Phe Thr Phe Ser Gly Ser Val
1 5
<210> 98
<211> 30
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 98
attagaaaca aggctaacag ttacgcgaca 30
<210> 99
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 99
Ile Arg Asn Lys Ala Asn Ser Tyr Ala Thr
1 5 10
<210> 100
<211> 39
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 100
actttcccgg gtgtagtggg acgaggaggt tttgactac 39
<210> 101
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 101
Thr Phe Pro Gly Val Val Gly Arg Gly Gly Phe Asp Tyr
1 5 10
<210> 102
<211> 339
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 102
gacatcgtga tgacccagtc tccagactcc ctggctgtgt ctctgggcga gggggccacc 60
atcaactgca tgtccagcca gagtgtttta ttcagctcca acaataagaa ctacttagct 120
tggtaccaac agaaaccagg acagcctcct aagttgctct tttactgggc atctacccgg 180
gaatccgggg tccctgaccg attcggtggc agcgggtctg ggacagattt ctctctcacc 240
atcaacagcc tgcagactga agatgtggca gtttattact gtctccaata ttatagtatt 300
ccgtggacgt tcggccaagg gaccaaggtg gaaatcaaa 339
<210> 103
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 103
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Gly Ala Thr Ile Asn Cys Met Ser Ser Gln Ser Val Leu Phe Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Phe Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Gly Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Thr
65 70 75 80
Ile Asn Ser Leu Gln Thr Glu Asp Val Ala Val Tyr Tyr Cys Leu Gln
85 90 95
Tyr Tyr Ser Ile Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys
<210> 104
<211> 36
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 104
cagagtgttt tattcagctc caacaataag aactac 36
<210> 105
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 105
Gln Ser Val Leu Phe Ser Ser Asn Asn Lys Asn Tyr
1 5 10
<210> 106
<211> 9
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 106
tgggcatct 9
<210> 107
<211> 3
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 107
Trp Ala Ser
1
<210> 108
<211> 27
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 108
ctccaatatt atagtattcc gtggacg 27
<210> 109
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 109
Leu Gln Tyr Tyr Ser Ile Pro Trp Thr
1 5
<210> 110
<211> 381
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 110
gaggtgcagc tggtggagtc tgggggaggc ttggtacagc ctggagggtc cctgacactc 60
tcctgtgcag cctctggatt caatttccgt gattatgaaa tgatctgggt ccgccagact 120
ccagggaagg ggctggagtg gatttcatac attagtaata gtggttatac catatactac 180
gcagactctg tgaagggccg attcaccatc tccagagaca acgccaggaa ctcaatatat 240
ctgcaagtga acagcctgag agccgaggac acggctgttt attactgttc gagacgtact 300
actatgattc ggggcattag ggcgtactac tattacggtc tggacgtctg gggccaaggg 360
accacggtca ccgtctcctc a 381
<210> 111
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 111
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Thr Leu Ser Cys Ala Ala Ser Gly Phe Asn Phe Arg Asp Tyr
20 25 30
Glu Met Ile Trp Val Arg Gln Thr Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Ser Tyr Ile Ser Asn Ser Gly Tyr Thr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Arg Asn Ser Ile Tyr
65 70 75 80
Leu Gln Val Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Arg Thr Thr Met Ile Arg Gly Ile Arg Ala Tyr Tyr Tyr Tyr
100 105 110
Gly Leu Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 112
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 112
ggattcaatt tccgtgatta tgaa 24
<210> 113
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 113
Gly Phe Asn Phe Arg Asp Tyr Glu
1 5
<210> 114
<211> 24
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 114
attagtaata gtggttatac cata 24
<210> 115
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 115
Ile Ser Asn Ser Gly Tyr Thr Ile
1 5
<210> 116
<211> 60
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 116
tcgagacgta ctactatgat tcggggcatt agggcgtact actattacgg tctggacgtc 60
<210> 117
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 117
Ser Arg Arg Thr Thr Met Ile Arg Gly Ile Arg Ala Tyr Tyr Tyr Tyr
1 5 10 15
Gly Leu Asp Val
20
<210> 118
<211> 324
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 118
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagagtcacc 60
atcacttgcc gggcaagtca gagcattagc agctatttaa attggtatca gcagaaacca 120
gggaaagccc ctaagctcct gatctatgct gcatccagtt tgcaaagtgg ggtcccgtca 180
aggttcagtg gcagtggatc tgggacagat ttcactctca ccatcagcag tctgcaacct 240
gaagattttg caacttacta ctgtcaacag agttacagta cccctccgat caccttcggc 300
caagggacac gactggagat taaa 324
<210> 119
<211> 108
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 119
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Pro
85 90 95
Ile Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys
100 105
<210> 120
<211> 18
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 120
cagagcatta gcagctat 18
<210> 121
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 121
Gln Ser Ile Ser Ser Tyr
1 5
<210> 122
<211> 9
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 122
gctgcatcc 9
<210> 123
<211> 3
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 123
Ala Ala Ser
1
<210> 124
<211> 30
<212> DNA
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 124
caacagagtt acagtacccc tccgatcacc 30
<210> 125
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Синтетическая
<400> 125
Gln Gln Ser Tyr Ser Thr Pro Pro Ile Thr
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ БЕТА-АМИЛОИДУ/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2730682C1 |
| КРИОКОНСЕРВИРОВАННЫЕ КЛЕТКИ-ЕСТЕСТВЕННЫЕ КИЛЛЕРЫ, ПРЕДВАРИТЕЛЬНО НАГРУЖЕННЫЕ КОНСТРУКЦИЕЙ АНТИТЕЛА | 2019 |
|
RU2819927C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ PSMA | 2019 |
|
RU2794450C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ CD20/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2753390C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ PD-L1 и LAG-3 | 2017 |
|
RU2784388C2 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| Терапевтические антитела к CD47 | 2016 |
|
RU2748401C2 |
| АНТИТЕЛО К ЦИТОМЕГАЛОВИРУСУ ЧЕЛОВЕКА И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2817217C1 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С ROR2 | 2018 |
|
RU2784586C2 |







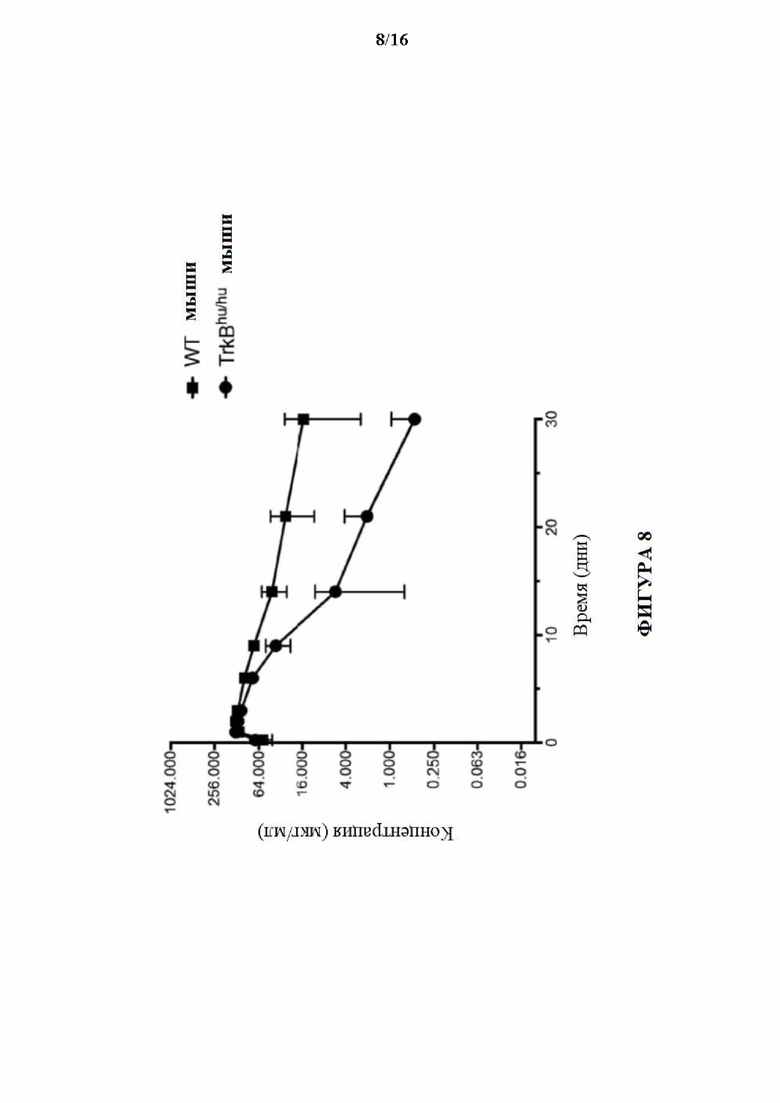








Изобретение относится к области биотехнологии, в частности к не являющемуся человеком животному для экспрессии химерного белка тропомиозин-рецепторной киназы В (TRKB), содержащему генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, его клетке, а также к способу получения вышеуказанного животного. Изобретение эффективно для оценки активности нацеленного на TRKB человека реагента in vivo. 4 н. и 18 з.п. ф-лы, 17 ил., 23 табл., 9 пр.
1. Не являющееся человеком животное для экспрессии химерного белка тропомиозин-рецепторной киназы В (TRKB), содержащего один или несколько фрагментов белка TRKB человека, причем не являющееся человеком животное содержит генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB, где не являющееся человеком животное представляет собой мышь или крысу, причем белок TRKB содержит эндогенный сигнальный пептид TRKB, эндогенный цитоплазматический домен TRKB, эндогенный трансмембранный домен TRKB и внеклеточный домен TRKB человека, и весь внеклеточный домен кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
2. Не являющееся человеком животное по п. 1, причем внеклеточный домен содержит последовательность, представленную в SEQ ID NO: 60, необязательно при этом кодирующая последовательность для внеклеточного домена содержит последовательность, представленную в SEQ ID NO: 72.
3. Не являющееся человеком животное по п. 1 или 2, причем:
(I) сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 51 или 55; или
(II) весь сигнальный пептид кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 63 или 67.
4. Не являющееся человеком животное по любому из предыдущих пунктов, причем:
(I) трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 53 или 57; или
(II) весь трансмембранный домен кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ ID NO: 65 или 69.
5. Не являющееся человеком животное по любому из предыдущих пунктов, причем:
(I) причем цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 54 или 58; или
(II) весь цитоплазматический домен кодируется эндогенной последовательностью TrkB, необязательно при этом кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 66 или 70.
6. Не являющееся человеком животное по любому из предыдущих пунктов, причем сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 51, трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 53, и цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 54, или
причем сигнальный пептид содержит последовательность, представленную в SEQ ID NO: 55, трансмембранный домен содержит последовательность, представленную в SEQ ID NO: 57, и цитоплазматический домен содержит последовательность, представленную в SEQ ID NO: 58.
7. Не являющееся человеком животное по любому из предыдущих пунктов, причем весь сигнальный пептид, весь трансмембранный домен и весь цитоплазматический домен кодируются эндогенной последовательностью TrkB,
необязательно при этом кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 63, кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ ID NO: 65, и кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 66, или
необязательно при этом кодирующая последовательность для сигнального пептида содержит последовательность, представленную в SEQ ID NO: 67, кодирующая последовательность для трансмембранного домена содержит последовательность, представленную в SEQ ID NO: 69, и кодирующая последовательность для цитоплазматического домена содержит последовательность, представленную в SEQ ID NO: 70.
8. Не являющееся человеком животное по любому из предыдущих пунктов, причем белок TRKB содержит последовательность, представленную в SEQ ID NO: 4 или 5.
9. Не являющееся человеком животное по п. 8, причем кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 12 или 13.
10. Не являющееся человеком животное по любому из предыдущих пунктов, причем крыса или мышь является гомозиготной по генетически модифицированному эндогенному локусу TrkB.
11. Не являющееся человеком животное по любому из предыдущих пунктов, причем не являющееся человеком животное представляет собой крысу, причем необязательно белок TRKB содержит последовательность, представленную в SEQ ID NO: 5, и необязательно кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 13.
12. Не являющееся человеком животное по любому из пп. 1-10, причем не являющееся человеком животное представляет собой мышь, причем необязательно белок TRKB содержит последовательность, представленную в SEQ ID NO: 4, и необязательно кодирующая последовательность генетически модифицированного локуса TrkB, кодирующего белок TRKB, содержит последовательность, представленную в SEQ ID NO: 12.
13. Способ оценки активности нацеленного на TRKB человека реагента in vivo, предусматривающий следующее:
(a) введение нацеленного на TRKB человека реагента не являющемуся человеком животному по любому из пп. 1-12; и
(b) оценка активности нацеленного на TRKB человека реагента у не являющегося человеком животного.
14. Способ по п. 13, при котором стадия (а) предусматривает инъекцию нацеленного на TRKB человека реагента не являющемуся человеком животному.
15. Способ по п. 13 или 14, при котором стадия (b) предусматривает оценку изменений одного или нескольких или всего из следующего: масса тела, состав тканей организма, метаболизм и локомоция, по сравнению с контрольным не являющимся человеком животным, причем необязательно оценка изменений в составе тканей организма предусматривает оценку массы нежировых тканей и/или массы жировой ткани по сравнению с контрольным не являющимся человеком животным, и необязательно оценка изменений в метаболизме предусматривает оценку изменений в потреблении пищи и/или потреблении воды.
16. Способ по любому из пп. 13-15, при котором стадия (b) предусматривает оценку фосфорилирования TRKB и/или активации путей MAPK/ERK и PI3K/Akt.
17. Способ по любому из пп. 13-16, при котором стадия (b) предусматривает оценку нейропротекторной активности.
18. Способ по любому из пп. 13-17, при котором стадия (b) предусматривает оценку жизнеспособности ганглиозных клеток сетчатки, причем необязательно жизнеспособность ганглиозных клеток сетчатки оценивают в модели полного пересечения зрительного нерва после повреждения зрительного нерва или оценивают в модели раздавливания зрительного нерва.
19. Способ по любому из пп. 13-18, при котором нацеленный на TRKB человека реагент представляет собой антигенсвязывающий белок, причем необязательно антигенсвязывающий белок представляет собой антитело - агонист TRKB человека.
20. Способ по любому из пп. 13-18, при котором нацеленный на TRKB человека реагент представляет собой малую молекулу, причем необязательно малая молекула представляет собой агонист TRKB человека.
21. Способ получения не являющегося человеком животного по любому из пп. 1-12, предусматривающий следующее:
(I) (а) модификацию генома плюрипотентной клетки мыши или крысы для включения генетически модифицированного эндогенного локуса TrkB, кодирующего белок TRKB, причем белок TRKB содержит эндогенный сигнальный пептид TRKB, эндогенный цитоплазматический домен TRKB, эндогенный трансмембранный домен TRKB и внеклеточный домен TRKB человека, и весь внеклеточный домен кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека;
(b) идентификацию или отбор генетически модифицированных плюрипотентных клеток мыши или крысы, содержащих генетически модифицированный эндогенный локус TrkB;
(c) введение генетически модифицированной плюрипотентной клетки мыши или крысы в эмбрион мыши или крысы-хозяина; и
(d) имплантацию и вынашивание эмбриона-хозяина мыши или крысы суррогатной матерью мышью или крысой; или
(II) (а) модификацию генома мышиного или крысиного эмбриона на стадии одной клетки для включения генетически модифицированного эндогенного локуса TrkB, кодирующего белок TRKB, причем белок TRKB содержит эндогенный сигнальный пептид TRKB, эндогенный цитоплазматический домен TRKB, эндогенный трансмембранный домен TRKB и внеклеточный домен TRKB человека, и весь внеклеточный домен кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека;
(b) отбор генетически модифицированного эмбриона мыши или крысы на стадии одной клетки, содержащего генетически модифицированный эндогенный локус TrkB; и
(c) имплантацию и вынашивание генетически модифицированного эмбриона мыши или крысы на стадии одной клетки суррогатной матерью мышью или крысой.
22. Клетка не являющегося человеком животного для экспрессии химерного белка тропомиозин-рецепторной киназы В (TRKB), содержащего один или несколько фрагментов белка TRKB человека, причем не являющееся человеком животное содержит генетически модифицированный эндогенный локус TrkB, кодирующий белок TRKB,
причем не являющееся человеком животное представляет собой мышь или крысу, причем белок TRKB содержит эндогенный сигнальный пептид TRKB, эндогенный цитоплазматический домен TRKB, эндогенный трансмембранный домен TRKB и внеклеточный домен TRKB человека, и весь внеклеточный домен кодируется сегментом эндогенного локуса TrkB, который был подвергнут делеции и заменен ортологичной последовательностью TRKB человека.
| MYLENE PERREAULT et al | |||
| Деревянное стыковое устройство | 1920 |
|
SU163A1 |
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| EIJA KOPONEN et al | |||
| Transgenic mice overexpressing the full-length neurotrophin receptor trkB exhibit increased activation of the trkB-PLC; pathway, reducedanxiety, and facilitated | |||

